Способ получения клетки, способной продуцировать гетеропротеины с высоким выходом - RU2486245C2
Код документа: RU2486245C2
Чертежи













Описание
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к способу получения клетки, способной продуцировать гетеропротеины с высоким выходом.
УРОВЕНЬ ТЕХНИКИ
Если белки, пригодные в качестве фармацевтических средств, получают с помощью технологии рекомбинантных ДНК, то применение клеток животных позволяет осуществить их сложную посттрансляционную модификацию и сворачивание, к чему прокариотические клетки не способны. Поэтому в качестве клеток-хозяев для продуцирования рекомбинантных белков часто используют клетки животных.
Недавно было разработано множество биопрепаратов, таких как антитела и физиологически активные белки. Методики, которые позволяют осуществить эффективную продукцию рекомбинантных белков клетками животных, способствуют снижению стоимости биопрепаратов и сулят их стабильную поставку пациентам.
В этих обстоятельствах желателен способ продуцирования белков, обладающий повышенной эффективностью продуцирования.
Было известно, что под воздействием метотрексата (MTX) в клетках амплифицируется (амплификация гена) некоторое число копий гена дигидрофолатредуктазы (DHFR), в результате чего клетки становятся устойчивыми к MTX. В промышленном производстве широко применяется способ количественного повышения продуцирования полезного белка, при котором плазмиды с геном белка, расположенным непосредственно ниже гена DHFR, вводят в клетки животных, а затем эти клетки культивируют в питательной среде с добавкой MTX для того, чтобы индуцировать амплификацию гена (патентный документ 1).
[Патентный документ 1]
Японская патентная публикация №6-30588
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
ПРОБЛЕМА, РЕШАЕМАЯ ИЗОБРЕТЕНИЕМ
Целью настоящего изобретения является создание клетки, способной продуцировать белки с высоким выходом, и обеспечение способа получения такой клетки.
СРЕДСТВА ДЛЯ РЕШЕНИЯ ПРОБЛЕМЫ
Авторы настоящего изобретения направили все свои усилия на решение вышеуказанной проблемы. В результате они обнаружили, что клетки-хозяева приобретали устойчивость к MTX при сильной экспрессии в этих клетках переносчика таурина (TauT), а также что клетки, которые стали продуцировать антитело после введения в них гена антитела, приобретали устойчивость к MTX при сильной экспрессии в этих клетках TauT и, кроме того, что способность к продукции антитела клетками, продуцирующими антитело, в которых одновременно экспрессировались TauT и декарбоксилаза цистеинсульфиновой кислоты (CSAD), улучшалась при обработке клеток высокой концентрацией MTX. Эти открытия привели к завершению настоящего изобретения.
Настоящее изобретение в целом можно описать следующим образом.
(1) Способ получения клетки, способной продуцировать желаемый полипептид с высоким выходом, в котором клетку с сильной экспрессией переносчика таурина, куда была введена ДНК, кодирующая желаемый полипептид, культивируют в присутствии высокой концентрации метотрексата, а из числа выживших клеток отбирают клетку, способную продуцировать желаемый полипептид с высоким выходом.
(2) Способ согласно (1) выше, в котором в клетку с сильной экспрессией переносчика таурина, куда была введена ДНК, кодирующая желаемый полипептид, также вводят ДНК, кодирующую дигидрофолатредуктазу (DHFR).
(3) Способ согласно (2) выше, в котором клетка с сильной экспрессией переносчика таурина, куда была введена ДНК, кодирующая желаемый полипептид, представляет собой клетку, котрансформированную одной молекулой, содержащей ДНК, кодирующую желаемый полипептид, и ДНК, кодирующую дигидрофолатредуктазу (DHFR).
(4) Способ согласно (3) выше, в котором молекула, содержащая ДНК, кодирующую желаемый полипептид, и ДНК, кодирующую дигидрофолатредуктазу (DHFR), представляет собой вектор.
(5) Способ согласно любому (1)-(4) выше, в котором клетка с сильной экспрессией переносчика таурина также сильно экспрессирует декарбоксилазу цистеинсульфиновой кислоты.
(6) Клетка, полученная способом согласно любому из (1)-(5) выше.
(7) Способ получения желаемого полипептида, в котором культивируют клетку согласно (6) выше.
(8) Способ согласно (7) выше, в котором желаемый полипептид представляет собой антитело.
(9) Способ получения фармацевтического продукта, содержащего полипептид, полученный способом согласно (7) или (8).
(10) Способ количественного увеличения продукции полипептида клеткой с сильной экспрессией переносчика таурина, в которую была введена ДНК, кодирующая желаемый полипептид, причем указанный способ включает обработку клетки высокой концентрацией метотрексата.
(11) Способ согласно (10) выше, в котором в клетку с сильной экспрессией переносчика таурина, куда была введена ДНК, кодирующая желаемый полипептид, также вводят ДНК, кодирующую дигидрофолатредуктазу.
(12) Способ согласно (11) выше, в котором клетка с сильной экспрессией переносчика таурина, куда была введена ДНК, кодирующая желаемый полипептид, представляет собой клетку, котрансформированную одной молекулой, содержащей ДНК, кодирующую желаемый полипептид, и ДНК, кодирующую дигидрофолатредуктазу.
(13) Способ согласно любому из (10)-(12) выше, в котором клетка с сильной экспрессией переносчика таурина, также сильно экспрессирует декарбоксилазу цистеинсульфиновой кислоты.
(14) Способ получения клетки, способной продуцировать желаемый полипептид с высоким выходом, в котором клетку с сильной экспрессией переносчика таурина, куда была введена ДНК, кодирующая желаемый полипептид, культивируют в присутствии высокой концентрации метотрексата, а из числа выживших клеток отбирают клетку, способную продуцировать желаемый полипептид с высоким выходом.
(15) Способ количественного увеличения продукции полипептида клеткой с сильной экспрессией переносчика таурина, в которую была введена ДНК, кодирующая желаемый полипептид, причем указанный способ включает обработку клетки метотрексатом.
ЭФФЕКТ ИЗОБРЕТЕНИЯ
Настоящее изобретение сделало возможной продукцию желаемого полипептида, включая антитело, с высоким выходом.
Настоящее описание изобретения охватывает содержание, раскрытое в описании изобретения и/или чертежах японской патентной заявки №2007-267384, приоритет которой испрашивается по настоящей патентной заявке.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
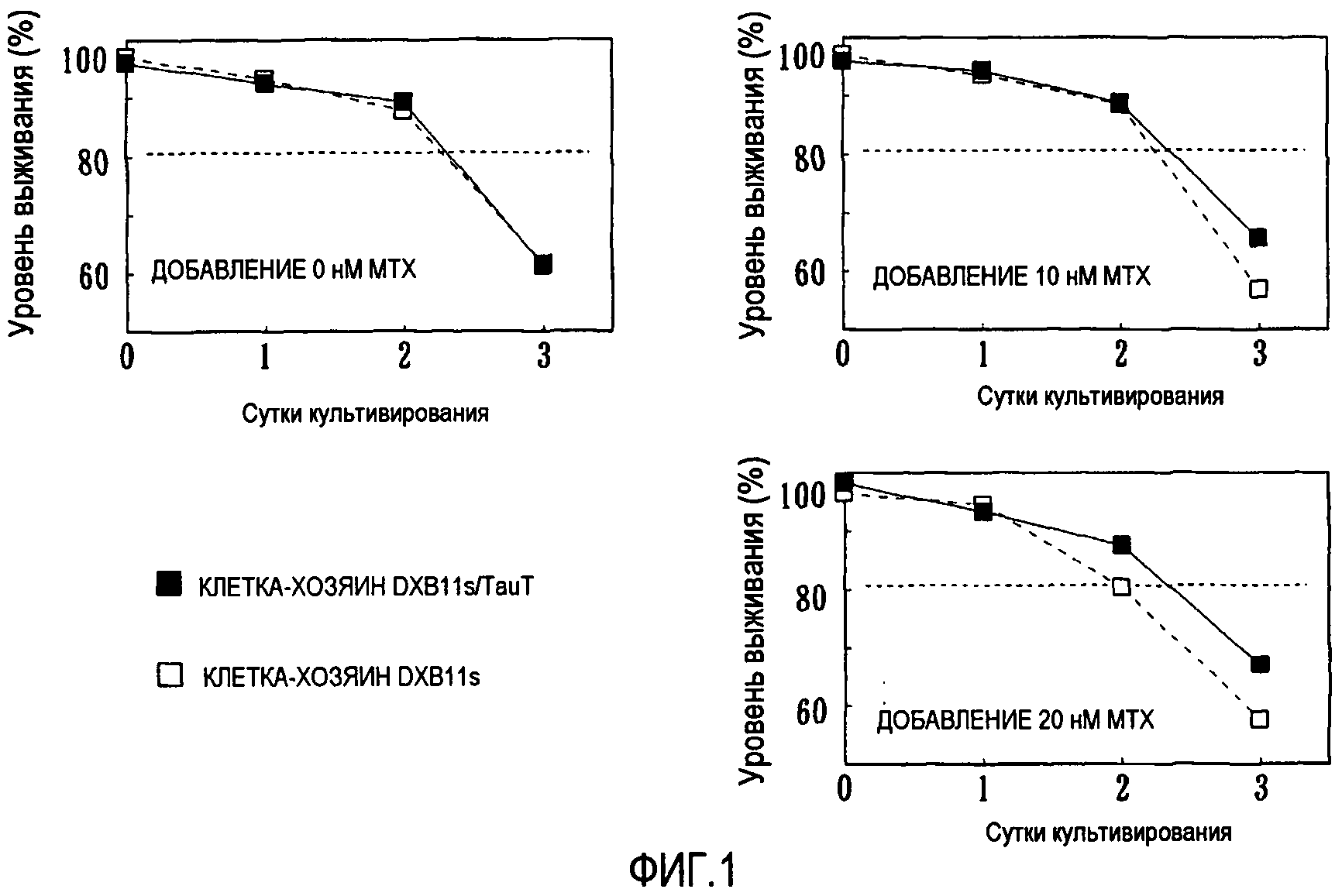
На фиг.1 представлены клетки-хозяева, в которых сильно экспрессируется TauT, что характеризует устойчивость к MTX.
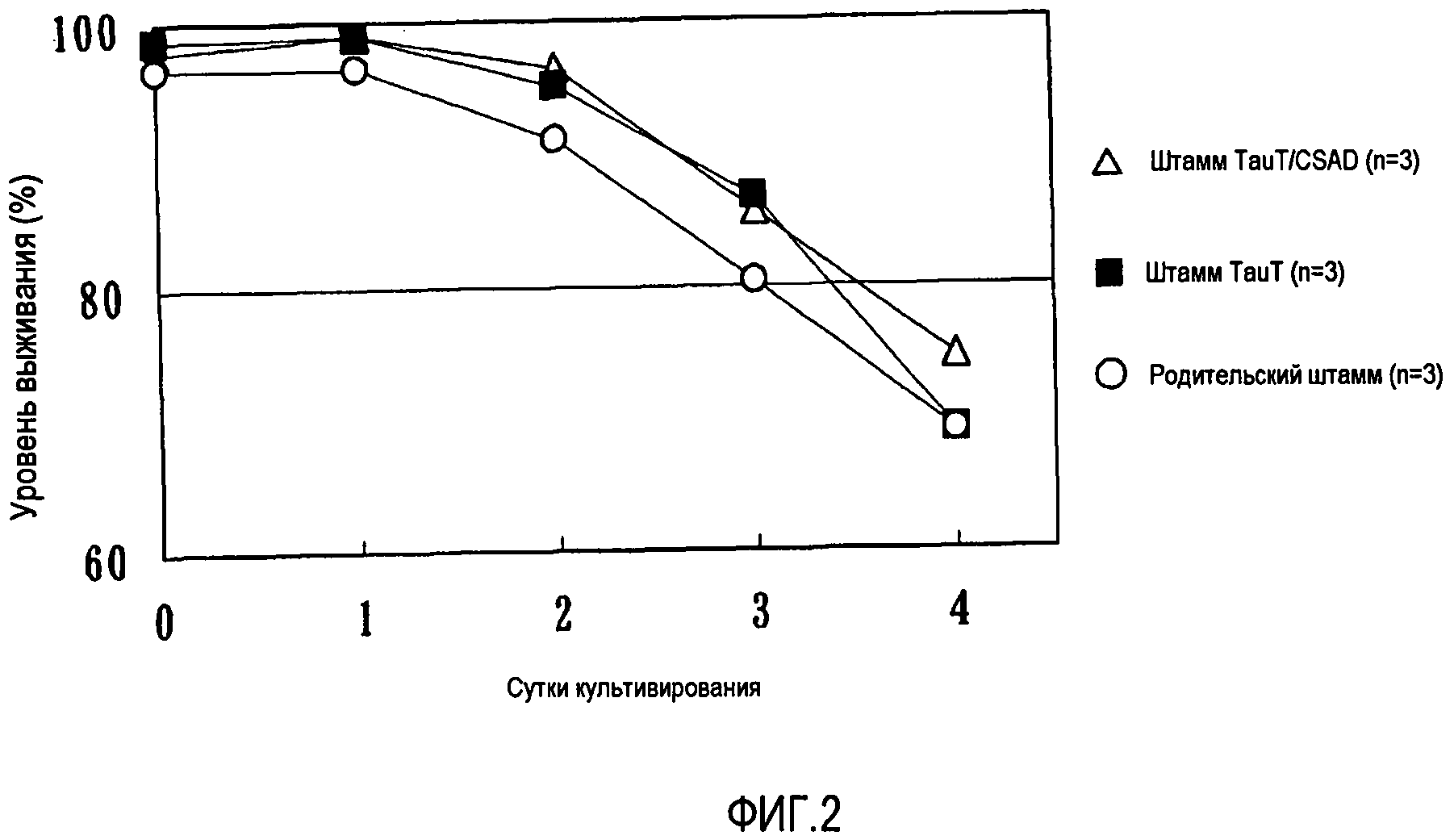
На фиг.2 показано, что штамм TauT, относящийся к клеткам, продуцирующим антитело к глипикану-3, в которых сильно экспрессируется TauT, и штамм TauT/CSAD, относящийся к вышеуказанным клеткам, продуцирующим антитело, в которых также сильно экспрессируется CSAD, обладают большей устойчивостью к MTX по сравнению с родительским штаммом.
На фиг.3 представлено изменение количества клеток по времени при непрерывном субкультивировании клеток в присутствии высокой концентрации MTX. На 28-ые сутки после инициации культуры условия были восстановлены до начального состояния 20 нМ MTX с целью повторного начала пролиферации.
На фиг.4 представлена усиленная способность клеток к продукции антител после их обработки высокой концентрацией MTX. Наибольшую величину продукции в расчете на клетку демонстрировали клетки, обработанные 200 нМ MTX.
На фиг.5 представлена плазмида, в которой был экспрессирован TauT хомяка (622 аминокислоты), для селекции с гигромицином.
На фиг.6 представлена плазмида, в которой был экспрессирован CSAD хомяка (493 аминокислоты), для селекции с пуромицином.
На фиг.7 представлена нуклеотидная последовательность вновь клонированного происходящего из клеток CHO гена переносчика таурина хомяка, и установленная на ее основе аминокислотная последовательность.
На фиг.8 представлена топология на мембране переносчика таурина, которая была воссоздана на основе трансмембранных участков и ориентаций, предсказанных программой TMpred по аминокислотной последовательности вновь клонированного происходящего из клеток CHO TauT хомяка, со ссылкой на ФИГ. 5 из публикации Shinichi Uchida et al., Proc. Natl. Acad. Sci. USA Vol.89, pp.8230-8234, September 1992. Знак

На фиг.9 представлена нуклеотидная последовательность вновь клонированного происходящего из клеток CHO гена CSAD хомяка и установленная на ее основе аминокислотная последовательность.
На фиг.10 представлена структура phGC33CAG1, которая представляет собой экспрессирующую плазмиду для гуманизированного антитела к глипикану-3 человека.
НАИЛУЧШИЙ СПОСОБ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Ниже в настоящем документе более подробно описаны различные варианты осуществления настоящего изобретения.
Настоящее изобретение относится к способу получения клетки, способной продуцировать желаемый полипептид с высоким выходом, в котором клетку с сильной экспрессией переносчика таурина, куда была введена ДНК, кодирующая желаемый полипептид, культивируют в присутствии высокой концентрации MTX, а из числа выживших клеток отбирают клетку, способную продуцировать желаемый полипептид с высоким выходом.
В соответствии со способом по настоящему изобретению клетку с сильной экспрессией TauT, в которую была введена ДНК, кодирующая желаемый полипептид, культивируют в присутствии высокой концентрации MTX.
ДНК, кодирующую желаемый полипептид, вводят в клетку, сильно экспрессирующую TauT.
В способе по настоящему изобретению желаемый полипептид конкретно не ограничен. Полипептид может представлять собой любой полипептид, такой как антитело (например, антитело к рецептору IL-6, антитело к IL-6, антитело к глипикану-3, антитело к CD3, антитело к CD20, антитело к GPIIb/IIIa, антитело к TNF, антитело к CD25, антитело к EGFR, антитело к Her2/neu, антитело к RSV, антитело к CD33, антитело к CD52, антитело к IgE, антитело к CD11a, антитело к VEGF, антитело к VLA4 и т.д.) или физиологически активный белок (например, гранулоцитарный колониестимулирующий фактор (G-CSF), гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF), эритропоэтин, интерферон, интерлейкины, такие как IL-1 или IL-6, t-PA, урокиназу, сывороточный альбумин, фактор свертывающей системы крови, парагормон [PTH] и т.д.). Особенно предпочтительным является антитело, и этим антителом может быть любое антитело, например природное антитело, антитело с низкой молекулярной массой (например, Fab, scFv, sc(Fv)2), химерное антитело, гуманизированное антитело и т.д.
Известно, что переносчик таурина представляет собой мембранный белок, обладающий функцией захвата аминокислот (таких как таурин и β-аланин) в клетки.
Клетка, которая сильно экспрессирует переносчик таурина, конкретно не ограничена, при условии, что эта клетка обладает повышенным уровнем экспрессии переносчика таурина по сравнению с соответствующей природной клеткой. Природная клетка также конкретно не ограничена. Можно использовать клетку, которую используют в качестве хозяина при продукции рекомбинантного белка (например, клетки CHO).
Наряду с тем, что клетка, сильно экспрессирующая TauT, может представлять собой любую клетку, которая может являться эукариотической клеткой, включая клетки животных, клетки растений и клетки дрожжей, или прокариотической клеткой, включая Escherichia coli и Bacillus subtilis, пригодной является культивируемая клетка, используемая в качестве хозяина при продукции рекомбинантного белка. Предпочтительными являются клетки животных, такие как клетка CHO и клетка COS, и среди них особенно предпочтительной является клетка CHO. Для продукции желаемого полипептида также особенно предпочтительной является клетка CHO с дефицитом dhfr (например, клеточная линия DXB11 из клеток CHO или клеточная линия DG44 из клеток CHO). Поскольку клетка CHO с дефицитом dhfr является ауксотрофной по гипоксантину и тимидину, такая клетка не может расти в среде, лишенной гипоксантина и тимидина (далее называемой в этом документе как "среда без HT"); однако клетка приобретает способность к росту в среде без HT после трансформации ее рекомбинантным вектором, содержащим ген DHFR. В соответствии с этим в качестве хозяина удобно использовать клетку CHO с дефицитом dhfr, поскольку селекцию трансформированной клетки можно проводить, основываясь на ауксотрофности клетки по гипоксантину и тимидину.
В качестве клетки, сильно экспрессирующей переносчик таурина, может быть приведена клетка, в которую искусственно перенесен ген переносчика таурина. Клетка, в которую был искусственно перенесен ген переносчика таурина, может быть получена способами, известными специалисту в данной области. Например, такую клетку можно получить встраиванием гена переносчика таурина в вектор и трансформацией этого вектора в клетку. Кроме того, понятие "клетки, в которые был искусственно перенесен ген TauT", охватывает в этом документе клетки, в которых был активирован эндогенный ген TauT с помощью технологии активации генов (см., например, международную публикацию WO 94/12650), так чтобы осуществлялась сильная экспрессия TauT.
В качестве TauT, подлежащего сильной экспрессии в клетке, можно использовать TauT, происходящий из любого организма. Более конкретно, можно использовать TauT, выделенный из организма человека или грызуна (такого как мышь, крыса или хомяк). Предпочтительно использовать TauT, выделенный из организма человека, грызуна или того же биологического вида, к которому принадлежит клетка-хозяин. Например, если клетка, которой позволяют сильно экспрессировать TauT, представляет собой клетку яичника китайского хомяка (клетку CHO), то предпочтительно использовать TauT, происходящий из человека или хомяка.
Кроме того, в качестве гена TauT, подлежащего сильной экспрессии в клетке, можно использовать любую из перечисленных ниже ДНК, от (a) до (e), кодирующих TauT.
(a) ДНК, кодирующая полипептид с аминокислотной последовательностью, которая представлена в SEQ ID NO: 2, или с аминокислотной последовательностью SC6A6_RAT (P31643), SC6A6_MOUSE (O35316), SC6A6_HUMAN (P31641), SC6A6_BOVIN (Q9MZ34) или SC6A6_CANFA (Q00589) в UniProt Knowledgebase;
(b) ДНК, кодирующая полипептид с аминокислотной последовательностью, происходящей из аминокислотной последовательности, представленной в SEQ ID NO: 2, или с аминокислотной последовательностью SC6A6_RAT (P31643), SC6A6_MOUSE (O35316), SC6A6_HUMAN (P31641), SC6A6_BOVIN (Q9MZ34) или SC6A6_CANFA (Q00589) в UniProt Knowledgebase, полученная путем замены, делеции, вставки и/или инсерции одного или более аминокислотных остатков, и, тем не менее, обладающая активностью TauT;
(с) ДНК, кодирующая полипептид, обладающий 70% или более гомологией аминокислотной последовательности с аминокислотной последовательностью, представленной в SEQ ID NO: 2, или с аминокислотной последовательностью SC6A6_RAT (P31643), SC6A6_MOUSE (O35316), SC6A6_HUMAN (P31641), SC6A6_BOVIN (Q9MZ34) или SC6A6_CANFA (Q00589) в UniProt Knowledgebase при сохранении активности TauT;
(d) ДНК, имеющая нуклеотидную последовательность, представленную в SEQ ID NO: 1 или нуклеотидную последовательность M96601, L03292, Z18956, AF260239 или M95495 в банке данных GenBank;
(e) ДНК, которая гибридизируется с ДНК, комплементарной ДНК, имеющей нуклеотидную последовательность, представленную в SEQ ID NO: 1, или нуклеотидную последовательность M96601, L03292, Z18956, AF260239 или M95495 в банке данных GenBank при строгих условиях, и которая, тем не менее, кодирует полипептид, обладающий активностью TauT.
ДНК согласно (a) представляет собой ДНК, кодирующую TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки, иллюстрируемую ДНК согласно (d).
ДНК согласно (b) представляет собой ДНК, кодирующую полипептид, функционально эквивалентный TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки, иллюстрируемый полипептидом, имеющим аминокислотную последовательность, происходящую из аминокислотной последовательности TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки в результате делеции одной или нескольких аминокислот, предпочтительно 1-30 аминокислот, более предпочтительно 1-10 аминокислот; полипептидом, имеющим аминокислотную последовательность, происходящую из аминокислотной последовательности TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки в результате вставки одной или нескольких аминокислот, предпочтительно 1-30 аминокислот, более предпочтительно 1-10 аминокислот; и полипептидом, имеющим аминокислотную последовательность, происходящую из аминокислотной последовательности TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки в результате замены одной или нескольких аминокислот, предпочтительно 1-30 аминокислот, более предпочтительно 1-10 аминокислот, другими аминокислотами.
Выражение "функционально эквивалентный TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки" означает наличие видов активности, сходных с видами активности TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки, например активности связывания таурина, активности транспортировки таурина в клетки и т.д. К таким полипептидам относятся, например, мутанты TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки. Этот термин охватывает TauT, происходящий из организмов других биологических видов.
Аминокислотные остатки, подлежащие мутации, конкретно не ограничены. Предпочтительно аминокислотные остатки подвергаются мутации в другие аминокислотные остатки, в которых сохраняются свойства исходной боковой цепи аминокислот. Конкретные примеры свойств боковой цепи аминокислот включают гидрофобные аминокислоты (A, I, L, M, F, P, W, Y и V), гидрофильные аминокислоты (R, D, N, C, E, Q, G, H, K, S и T), аминокислоты с алифатической боковой цепью (G, A, V, L, I и P), аминокислоты с боковой цепью, содержащей гидрофильную группу (S, T и Y), аминокислоты с боковой цепью, содержащей атом серы (C и M), аминокислоты с боковой цепью, содержащей карбоновую кислоту и амид (D, N, E и Q), аминокислоты с боковой цепью, содержащей основание (R, K и H) и аминокислоты с боковой цепью, содержащей ароматическую группу (H, F, Y и W) (В скобках приведены однобуквенные коды аминокислот).
Сообщалось о том, что полипептид, имеющий аминокислотную последовательность, происходящую из первоначальной аминокислотной последовательности в результате модификации (такой как делеция, вставка и/или замещение одной или нескольких аминокислот), сохраняет биологическую активность исходного полипептида (Mark, D. F. et al., Proc. Natl. Acad. Sci. USA (1984) 81, 5662-5666; Zoller, M. J. & Smith, M. Nucleic Acids Research (1982) 10, 6487-6500; Wang, A. et al., Science 224, 1431-1433; Dalbadie-McFarland, G. et al., Proc. Natl. Acad. Sci. USA (1982) 79, 6409-6413).
ДНК согласно (c) кодирует полипептид, функционально эквивалентный TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки, который представляет собой полипептид, обладающий по меньшей мере 70%, предпочтительно 97% или более, более предпочтительно 98% или более, более предпочтительно 99% или более гомологией с аминокислотной последовательностью TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки. Для определения гомологии полипептидов можно следовать алгоритму, описанному в Wilbur, W. J. and Lipman, D. J., Proc. Natl. Acad. Sci. USA (1983) 80, 726-730.
ДНК можно получить способами, известными специалисту в данной области. Например, ДНК можно получать получением библиотеки кДНК из клетки, экспрессирующей TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки, и проведением гибридизации с использованием части последовательности ДНК TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки (например, SEQ ID NO: 1) в качестве зонда. Библиотеку кДНК можно получать, например, способом, описанным в Sambrook, J. et al., Molecular Cloning, Cold Spring Harbor Laboratory Press (1989). Альтернативно можно использовать коммерческую библиотеку кДНК. Также ДНК TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки можно получать выделением РНК из клетки, экспрессирующей TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки, синтезом олигонуклеотидных молекул ДНК на основе последовательности ДНК, кодирующей TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки (например, SEQ ID NO: 1), и проведением ПЦР с использованием олигонуклеотидных молекул ДНК в качестве праймеров, тем самым амплифицируя кДНК, кодирующую TauT.
Кроме того, путем определения нуклеотидной последовательности полученной кДНК, можно определить участок трансляции, кодирующий полипептид, и получить аминокислотную последовательность TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки или полипептида, функционально эквивалентного ему. Более того, путем скрининга геномной библиотеки с применением полученной кДНК в качестве зонда можно выделить геномную ДНК.
Конкретно, можно использовать следующие способы. Во-первых, из клеток, тканей или сходных с ними, экспрессирующих TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки, выделяют мРНК. Для выделения мРНК получают тотальную РНК известными способами, например способом ультрацентрифугирования с гуанидином (Chirgwin, J. M. et al., Biochemistry (1979) 18, 5294-5299), способом AGPC (Chomczynski, P. and Sacchi, N., Anal. Biochem. (1987) 162, 156-159) или сходными с ними, а затем мРНК очищают от тотальной РНК, применяя набор для очистки мРНК (Pharmacia) и т.д. Альтернативно, мРНК можно получить прямо, используя набор QuickPrep для очистки мРНК (Pharmacia).
Из полученной мРНК с использованием обратной транскриптазы синтезируют кДНК. Альтернативно кДНК можно синтезировать с использованием коммерческого набора, такого как AMV Reverse Transcriptase First-Strand cDNA Synthesis (SEIKAGAKU CORPORATION). Также можно синтезировать и амплифицировать кДНК способом 5'-RACE (Frohman, M. A. et al., Proc. Natl. Acad. Sci. USA (1988) 85, 8998-9002; Belyavsky, A. et al., Nucleic Acids Res. (1989) 17, 2919-2932) с использованием набора 5'-Ampli FINDER RACE (Clontech) и полимеразной цепной реакции (ПЦР) с праймерами. Далее можно использовать точечный мутагенез для внесения мутаций в полученную таким образом ДНК. Затем мутированную таким способом ДНК можно амплифицировать с помощью ПЦР.
Из полученного в результате ПЦР продукта извлекают представляющий интерес фрагмент ДНК и лигируют его в векторную ДНК с получением тем самым рекомбинантного вектора. Вектор вводят в хозяина (например, E.coli), а затем проводят селекцию полученных колоний, с получением тем самым требуемого рекомбинантного вектора. Нуклеотидную последовательность представляющей интерес ДНК можно подтвердить известным способом, таким как дидезоксинуклеотидный способ обрыва цепи.
Кроме того, для ДНК можно сконструировать нуклеотидную последовательность с повышенной эффективностью экспрессии, принимая во внимание частоту применения кодонов в хозяине, подлежащем применению для экспрессии (Grantham, R. et al., Nucleic Acids Research (1981) 9, p.43-74). Кроме того, ДНК можно модифицировать с использованием коммерчески доступных наборов или известных способов. Примеры таких модификаций включают, но не ограничиваются ими, расщепление ферментами рестрикции, вставку синтетических олигонуклеотидов или соответствующих фрагментов ДНК, добавление линкеров и вставку кодона инициации (ATG) и/или стоп-кодона (TAA, TGA или TAG).
ДНК согласно (e) представляет собой ДНК, которая гибридизируется с ДНК, комплементарной ДНК, имеющей нуклеотидную последовательность, представленную в SEQ ID NO: 1, или нуклеотидную последовательность M96601, L03292, Z18956, AF260239 или M95495 в GenBank, в строгих условиях и кодирует полипептид, функционально эквивалентный TauT хомяка, крысы, мыши, человека, крупного рогатого скота или собаки. Строгие условия, включая, например, условия низкой строгости, может правильно выбрать специалист в данной области. К условиям низкой строгости относятся, например, 42°C, 2×SSC и 0,1% SDS, предпочтительно 50°C, 2×SSC и 0,1% SDS. Более предпочтительно выбирать условия высокой строгости. К условиям высокой строгости относятся, например, 65°C, 2×SSC и 0,1% SDS. В этих условиях по мере повышения температуры гибридизации можно получить ДНК с повышенной гомологией. Описанная выше ДНК, которая гибридизуется, предпочтительно представляет собой ДНК, полученную из природных источников, например кДНК или хромосомную ДНК. Эти ДНК, выделенные способами гибридизации, обычно обладают высокой идентичностью нуклеотидной последовательности с нуклеотидной последовательностью, представленной в SEQ ID NO: 1, или нуклеотидной последовательностью M96601, L03292, Z18956, AF260239 или M95495 в GenBank. Термин "высокая идентичность" обычно относится к гомологии 96% или более, предпочтительно к гомологии 98% или более, более предпочтительно к гомологии 99% или более. Идентичность нуклеотидных последовательностей можно определить при помощи алгоритма BLAST (Karlin and Altschul, Proc. Natl. Acad. Sci. USA 90:5873-5877, 1993). На основе этого алгоритма были разработаны такие компьютерные программы как BLASTN и BLASTX (Altschul et al. J. Mol. Biol. 215:403-410, 1990). Если нуклеотидные последовательности анализируют в программе BLASTN, основанной на алгоритме BLAST, можно установить, например, такие параметры как score = 100 и wordlength = 12. Известны конкретные процедуры для этих способов анализа (.).
Когда используется клетка, в которую искусственно перенесен ген TauT, порядок переноса гена TauT и переноса ДНК, кодирующей желаемый полипептид, конкретно не ограничен. ДНК, кодирующую желаемый полипептид, можно переносить после переноса гена TauT. Альтернативно ген TauT можно переносить после переноса ДНК, кодирующей желаемый полипептид. Также можно переносить ген TauT и ДНК, кодирующую желаемый полипептид, одновременно.
Клетка, сильно экспрессирующая TauT, может также сильно экспрессировать CSAD.
В качестве клетки, сильно экспрессирующей CSAD, может быть взята клетка, в которую был искусственно перенесен ген CSAD. Клетка, в которую был искусственно перенесен ген CSAD, может быть получена способами, известными специалисту в данной области. Например, такую клетку можно получить, встраивая ген CSAD в вектор и трансформируя этим вектором клетки. Кроме того, понятие "клетки, в которые был искусственно перенесен ген CSAD", охватывает в этом документе клетки, в которых был активирован эндогенный ген CSAD посредством технологии активации генов (см., например, международную публикацию WO 94/12650), так чтобы осуществлялась сильная экспрессия CSAD.
В качестве CSAD, подлежащего сильной экспрессии в клетке, можно использовать CSAD, происходящий из любого организма. Конкретно, можно использовать CSAD, выделенный из организма человека, грызуна (такого как мышь, крыса или хомяк), иглобрюха (такого как тигровая рыба фугу) или асцидии (такой как Ciona intestnalis). Предпочтительно можно использовать CSAD, выделенный из организма человека, грызуна или того же биологического вида, к которому принадлежит клетка-хозяин. Например, если клетка, которой дают возможность сильно экспрессировать CSAD, представляет собой клетку яичника китайского хомяка (клетку CHO), то предпочтительно использовать CSAD, происходящий из организма человека или хомяка.
Кроме того, в качестве гена CSAD, подлежащего сильной экспрессии в клетке, можно использовать любую из перечисленных ниже ДНК от (a1) до (e1).
(a1) ДНК, кодирующая полипептид с аминокислотной последовательностью, которая представлена в SEQ ID NO: 4, или с аминокислотной последовательностью CSAD_RAT (Q64611), CSAD_MOUSE (Q9DBE0) или CSAD_HUMAN (Q9Y600) в UniProt Knowledgebase;
(b1) ДНК, кодирующая полипептид с аминокислотной последовательностью, происходящей из аминокислотной последовательности, представленной в SEQ ID NO: 4, или с аминокислотной последовательностью CSAD_RAT (Q64611), CSAD_MOUSE (Q9DBE0) или CSAD_HUMAN (Q9Y600) в UniProt Knowledgebase путем замены, делеции, вставки и/или инсерции одного или более аминокислотных остатков, но с сохранением активности CSAD;
(c1) ДНК, кодирующая полипептид, обладающий гомологией аминокислотной последовательности 70% или более с аминокислотной последовательностью, представленной в SEQ ID NO: 4, или с аминокислотной последовательностью CSAD_RAT (Q64611), CSAD_MOUSE (Q9DBE0) или CSAD_HUMAN (Q9Y600) в UniProt Knowledgebase, и тем не менее сохраняющий активность CSAD.
(d1) ДНК, имеющая нуклеотидную последовательность, представленную в SEQ ID NO: 3, или нуклеотидную последовательность M64755, AK005015 или AF116546 (GenBank);
(e1) ДНК, которая гибридизируется с ДНК, комплементарной ДНК, имеющей нуклеотидную последовательность, представленную в SEQ ID NO: 3, или нуклеотидную последовательность M64755, AK005015 или AF116546 (GenBank) в строгих условиях, и тем не менее кодирует полипептид, обладающий активностью CSAD.
ДНК согласно (a1) представляет собой ДНК, кодирующую CSAD хомяка, крысы, мыши или человека. Например, она может представлять собой ДНК согласно (d1). Ферментативная активность CSAD крупного рогатого скота идентифицирована, однако информация о его последовательности недоступна.
ДНК согласно (b1) представляет собой ДНК, кодирующую полипептид, функционально эквивалентный CSAD хомяка, крысы, мыши или человека, иллюстрируемый полипептидом, имеющим аминокислотную последовательность, происходящую из аминокислотной последовательности CSAD хомяка, крысы, мыши или человека, путем делеции одной или более аминокислот, предпочтительно 1-30 аминокислот, более предпочтительно 1-10 аминокислот; полипептидом, имеющим аминокислотную последовательность, происходящую из аминокислотной последовательности CSAD хомяка, крысы, мыши или человека, путем вставки одной или более аминокислот, предпочтительно 1-30 аминокислот, более предпочтительно 1-10 аминокислот; и полипептидом, имеющим аминокислотную последовательность, происходящую из аминокислотной последовательности CSAD хомяка, крысы, мыши или человека, путем замены одной или нескольких аминокислот, предпочтительно 1-30 аминокислот, более предпочтительно 1-10 аминокислот, другими аминокислотами.
Выражение "полипептид, который функционально эквивалентен CSAD хомяка, крысы, мыши или человека" используется для обозначения того, что полипептид обладает декарбоксилирующей активностью, которая эквивалентна активности, присущей CSAD хомяка, крысы, мыши или человека, такой как ферментативная активность в отношении синтеза гипотаурина из 3-сульфиновокислого аланина, ферментативная активность в отношении синтеза таурина из цистеиновой кислоты, и ферментативная активность по типу глутаматдекарбоксилазы в отношении синтеза β-аланина из аспарагиновой кислоты. Такой полипептид охватывает, например, мутанты CSAD хомяка, крысы, мыши или человека. Он также охватывает CSAD, выделенный из других биологических видов (например, CSAD крупного рогатого скота и т.п.).
Аминокислотные остатки, подлежащие мутации, конкретно не ограничены. Предпочтительно аминокислотные остатки подвергаются мутации в другие аминокислотные остатки, в которых сохраняются свойства исходной боковой цепи аминокислот. Конкретные примеры свойств боковой цепи аминокислот включают гидрофобные аминокислоты (A, I, L, M, F, P, W, Y и V), гидрофильные аминокислоты (R, D, N, C, E, Q, G, H, K, S и T), аминокислоты с алифатической боковой цепью (G, A, V, L, I и P), аминокислоты с боковой цепью, содержащей гидрофильную группу (S, T и Y), аминокислоты с боковой цепью, содержащей атом серы (C и M), аминокислоты с боковой цепью, содержащей карбоновую кислоту и амид (D, N, E и Q), аминокислоты с боковой цепью, содержащей основание (R, K и H) и аминокислоты с боковой цепью, содержащей ароматическую группу (H, F, Y и W) (В скобках приведены однобуквенные коды аминокислот).
ДНК согласно (c1) кодирует полипептид, функционально эквивалентный CSAD хомяка, крысы, мыши или человека, который представляет собой полипептид, обладающий, по меньшей мере, 70%, предпочтительно 97% или более, более предпочтительно 98% или более, еще предпочтительнее 99% или более гомологией с аминокислотной последовательностью CSAD хомяка, крысы, мыши или человека. Для определения гомологии полипептидов можно следовать алгоритму, описанному в Wilbur, W. J. and Lipman, D. J., Proc. Natl. Acad. Sci. USA (1983) 80, 726-730.
ДНК можно получить способами, известными специалисту в данной области. Например, ДНК можно получать получением библиотеки кДНК из клетки, экспрессирующей CSAD хомяка, крысы, мыши или человека, и проведением гибридизации с использованием части последовательности ДНК CSAD хомяка, крысы, мыши или человека (например, SEQ ID NO: 3) в качестве зонда. Библиотеку кДНК можно получать, например, способом, описанным в Sambrook, J. et al., Molecular Cloning, Cold Spring Harbor Laboratory Press (1989). Альтернативно можно использовать коммерческую библиотеку кДНК. Также ДНК CSAD хомяка, крысы, мыши или человека можно получать выделением РНК из клетки, экспрессирующей CSAD хомяка, крысы, мыши или человека, синтезом олигонуклеотидных молекул ДНК на основе последовательности ДНК, кодирующей CSAD хомяка, крысы, мыши или человека (например, SEQ ID NO: 3), и проведением ПЦР с использованием олигонуклеотидных молекул ДНК в качестве праймеров, тем самым амплифицируя кДНК, кодирующую CSAD.
Кроме того, путем определения нуклеотидной последовательности полученной кДНК, можно определить участок трансляции, кодирующий полипептид, и получить аминокислотную последовательность CSAD хомяка, крысы, мыши или человека или полипептида, функционально эквивалентного ему. Более того, путем скрининга геномной библиотеки с применением полученной кДНК в качестве зонда можно выделить геномную ДНК.
Конкретно, можно использовать следующие способы. Во-первых, из клеток, тканей или сходных с ними экспрессирующих CSAD хомяка, крысы, мыши или человека выделяют мРНК. Для выделения мРНК получают тотальную РНК известными способами, например способом ультрацентрифугирования с гуанидином (Chirgwin, J. M. et al., Biochemistry (1979) 18, 5294-5299), способом AGPC (Chomczynski, P. and Sacchi, N., Anal. Biochem. (1987) 162, 156-159) или сходными с ними, а затем мРНК очищают от тотальной РНК, применяя набор для очистки мРНК (Pharmacia) и т.д. Альтернативно, мРНК можно получить прямо, используя набор QuickPrep для очистки мРНК (Pharmacia).
Из полученной мРНК с использованием обратной транскриптазы синтезируют кДНК. Альтернативно кДНК можно синтезировать с использованием коммерческого набора, такого как AMV Reverse Transcriptase First-Strand cDNA Synthesis (SEIKAGAKU CORPORATION). Также можно синтезировать и амплифицировать кДНК способом 5'-RACE (Frohman, M. A. et al., Proc. Natl. Acad. Sci. USA (1988) 85, 8998-9002; Belyavsky, A. et al., Nucleic Acids Res. (1989) 17, 2919-2932) с использованием набора 5'-Ampli FINDER RACE (Clontech) и полимеразной цепной реакции (ПЦР) с праймерами. Далее можно использовать точечный мутагенез для внесения мутаций в полученную таким образом ДНК. Затем мутированную таким способом ДНК можно амплифицировать с помощью ПЦР.
Из полученного в результате ПЦР продукта извлекают представляющий интерес фрагмент ДНК и лигируют его в векторную ДНК с получением тем самым рекомбинантного вектора. Вектор вводят в хозяина (например, E.coli), а затем проводят селекцию полученных колоний, с получением тем самым требуемого рекомбинантного вектора. Нуклеотидную последовательность представляющей интерес ДНК можно подтвердить известным способом, таким как дидезоксинуклеотидный способ обрыва цепи.
Кроме того, для ДНК можно сконструировать нуклеотидную последовательность с повышенной эффективностью экспрессии, принимая во внимание частоту применения кодонов в хозяине, подлежащем применению для экспрессии (Grantham, R. et al., Nucleic Acids Research (1981) 9, p.43-74). Кроме того, ДНК можно модифицировать с использованием коммерчески доступных наборов или известных способов. Примеры таких модификаций включают, но не ограничиваются ими, расщепление ферментами рестрикции, вставку синтетических олигонуклеотидов или соответствующих фрагментов ДНК, добавление линкеров и вставку кодона инициации (ATG) и/или стоп-кодона (TAA, TGA или TAG).
ДНК согласно (e1) представляет собой ДНК, которая гибридизируется с ДНК, комплементарной ДНК, имеющей нуклеотидную последовательность, представленную в SEQ ID NO: 3, или нуклеотидную последовательность M64755, AK005015 или AF116546 в GenBank, в строгих условиях и кодирует полипептид, функционально эквивалентный CSAD хомяка, крысы, мыши или человека. Строгие условия, включая, например, условия низкой строгости, может правильно выбрать специалист в данной области. К условиям низкой строгости относятся, например, 42°C, 2×SSC и 0,1% SDS, предпочтительно 50°C, 2×SSC и 0,1% SDS. Более предпочтительно выбирать условия высокой строгости. К условиям высокой строгости относятся, например, 65°C, 2×SSC и 0,1% SDS. В этих условиях по мере повышения температуры гибридизации можно получить ДНК с повышенной гомологией. Описанная выше ДНК, которая гибридизуется, предпочтительно представляет собой ДНК, полученную из природных источников, например кДНК, или хромосомную ДНК. Эти ДНК, выделенные способами гибридизации, обычно обладают высокой идентичностью нуклеотидной последовательности с нуклеотидной последовательностью, представленной в SEQ ID NO: 3, или с нуклеотидной последовательностью M64755, AK005015 или AF116546 в GenBank. Термин "высокая идентичность" обычно относится к гомологии 96% или более, предпочтительно к гомологии 98% или более, более предпочтительно к гомологии 99% или более. Идентичность нуклеотидных последовательностей можно определить при помощи алгоритма BLAST (Karlin and Altschul, Proc. Natl. Acad. Sci. USA 90:5873-5877, 1993). На основе этого алгоритма были разработаны такие компьютерные программы как BLASTN и BLASTX (Altschul et al. J. Mol. Biol. 215:403-410, 1990). Если нуклеотидные последовательности анализируют в программе BLASTN, основанной на алгоритме BLAST, можно установить, например, такие параметры как score = 100 и wordlength = 12. Известны конкретные процедуры для этих способов анализа (.).
Ген DHFR можно ввести в клетку, сильно экспрессирующую TauT, вместе с ДНК, кодирующей желаемый полипептид. Под воздействием MTX в клетках амплифицируется некоторое число копий гена DHFR (амплификация гена), вследствие чего клетки становятся устойчивыми к MTX. Клетку, сильно экспрессирующую TauT, можно получить котрансформацией одной молекулы (например, вектором), содержащей ДНК, кодирующую желаемый полипептид, и ДНК, кодирующую DHFR. Когда в клетки вводят рекомбинантный вектор, в котором ДНК, кодирующая желаемый полипептид, связана с геном DHFR, и полученные таким образом клетки культивируют в среде в присутствии MTX, то можно получить клетки, способные продуцировать желаемый полипептид с высоким выходом за счет амплификации гена. Источником гена DHFR могут быть самые разные организмы, и их последовательности ДНК общеизвестны (мышь: GenBank V00734, крыса: GenBank AF318150, человек: GenBank J00140). Следовательно, можно получить ген DHFR в соответствии с этой доступной информацией и встроить его в вектор. Кроме того, также можно использовать коммерчески доступный экспрессирующий вектор, в который встроен ген DHFR (pOptiVEC™-TOPO (зарегистрированная торговая марка), продукт Invitrogen Corporation).
Когда в качестве хозяина используется клетка, в которую искусственно введен ген TauT, ДНК, кодирующую желаемый полипептид, и ДНК, кодирующую DHFR, можно вводить в клетку после введения в нее гена TauT или, наоборот, ген TauT можно вводить в клетку после введения в нее ДНК, кодирующей желаемый полипептид, и ДНК, кодирующей DHFR. Альтернативно ген TauT, ДНК, кодирующую желаемый полипептид, и ДНК, кодирующую DHFR, можно вводить в клетку одновременно.
Ген TauT (а также в некоторых случаях ген CSAD) и ДНК, кодирующую желаемый полипептид, можно вводить одновременно в одном векторе или независимо друг от друга, используя несколько векторов.
Кроме того, ДНК, кодирующую желаемый полипептид, и ген DHFR можно встраивать в один вектор или в разные векторы. Для того чтобы эффективно создать штамм, способный к продукции с высоким выходом за счет амплификации гена, предпочтительно вводить ДНК, кодирующую желаемый полипептид, и ген DHFR в одном векторе. Для того чтобы повысить эффективность амплификации гена, предпочтительно ген DHFR встраивать ниже промотора, обладающего низкой эффективностью транскрипции (например, промотора SV40), а ДНК, кодирующую желаемый полипептид, встраивать ниже промотора, обладающего высокой эффективностью транскрипции (например, промотора CMV, промотора SRα или промотора EF-1α). Когда ДНК, кодирующую желаемый полипептид, и ген DHFR встраивают в разные векторы, векторы можно вводить в клетку хозяина посредством котрансфекции. В этом случае вектор, в который встроена ДНК, кодирующая желаемый полипептид, предпочтительно вводить в клетку-хозяина в избыточном количестве (обычно, в избытке, составляющем приблизительно от двух до 40 раз) относительно вектора, в который встроен ген DHFR.
Для конструирования вектора, экспрессирующего полипептид в клетках CHO, может быть предусмотрено использование предраннего участка энхансера/промотора CMV, типичной последовательности Козака (-6 GCCR (R=A/G) CCAUGG+4) (SEQ ID NO: 5) и маркера устойчивости к лекарственному средству (например, к неомицину, гигромицину или пуромицину), а также добавление на N-концевой стороне сигнального пептида для секреции (такого как MGWSCIILFLVATATGVHS (SEQ ID NO: 6)).
Перед введением в клетки-хозяева экспрессирующий вектор экспрессии можно расщеплять соответствующим ферментом рестрикции, переводя его в линейную форму. Экспрессирующий элемент представляющего интерес гена можно легко встроить в хромосому клетки-хозяина, получив вектор в линейной форме. Способ переноса гена конкретно не ограничен, и он может представлять собой любой способ, например способ с фосфатом кальция, способ с DEAE декстраном, способ липофекции или способ электропорации. Множество копий можно вводить проведением генетического переноса с использованием системы NUCLEOFECTOR (продукт Amaxa Biosystems).
После осуществления переноса гена в клетки-хозяева такие клетки культивируют на селективной среде, в результате чего можно отобрать клетки, в которые был введен представляющий интерес ген. Например, если ДНК, кодирующую желаемый полипептид, встраивают в вектор, имеющий ген устойчивости к лекарственному средству, и клетки-хозяева трансформируют полученным таким образом вектором, а затем культивируют в среде, содержащей лекарственное средство, то выживающие клетки можно отбирать в качестве трансформированных клеток. Кроме того, если ДНК, кодирующую желаемый полипептид, встраивают в вектор, имеющий ген DHFR, и dhfr-дефицитные клетки CHO, используемые в качестве клеток-хозяев, трансформируют полученным таким образом вектором, а затем культивируют в среде без HT, то выживающие клетки можно отбирать в качестве трансформированных клеток. В качестве среды можно использовать смешанную среду CHO-S-SFMII/CD-CHO (продукт Invitrogen Corporation) и сходные с ней. Трансформированные клетки непрерывно культивируют, определяя количество продуцированного представляющего интерес полипептида, через определенное время (обычно приблизительно в интервале с 14 по 21 сутки). Затем отбирают клетки, способные к продукции с высоким выходом, для обработки MTX. Предпочтительно, чтобы трансформированные клетки, подлежащие обработке MTX, были способны не только к продукции полипептида с высоким выходом, но и к быстрой пролиферации. Скорости пролиферации можно сравнивать, измеряя количество выживших клеток, которые подвергали субкультивированию.
Обработка MTX относится, например, к культивированию клеток (предварительное субкультивирование) в среде, в которую добавлена высокая концентрация MTX. К высокой концентрации относится такая концентрация, которая по меньшей мере в два раза превышает нормальную концентрацию MTX, при которой проводят селекцию клеток (что составляет приблизительно 20 нМ для клеток CHO); например, она представляет собой концентрацию, при которой 90% или более штамма, в который не был введен TauT, погибает через три недели после субкультивирования. Хотя концентрация варьирует в зависимости от клеток, в случае, когда для продуцирования рекомбинантного белка используются клетки CHO, такие как CHO DXB11s, обычно подходящей считается концентрация 50 нМ или более; предпочтительно она составляет 80 нМ или более и более предпочтительно 100 нМ или более. Подходящий период культивирования составляет от семи до 35 суток, предпочтительно от 14 до 28 суток и более предпочтительно от 21 до 28 суток.
Когда трансформированные клетки культивируют в среде, в которую добавляют MTX в высокой концентрации, то концентрацию MTX можно увеличивать постепенно. Например, клетки от 14 до 21 суток культивируют в среде, содержащей MTX в концентрации 10 нМ, а затем от 14 до 28 суток культивируют в среде, содержащей MTX в концентрации 100 нМ.
Штамм, способный к продукции с высоким выходом, можно отобрать на любой стадии культивирования с изменением концентрации MTX. Также, если клеточная пролиферация более не наблюдается вследствие культивирования в среде с добавлением высокой концентрации MTX, то клеточную пролиферацию можно восстановить, вернув клетки в среду с добавлением низкой концентрации MTX и продолжая культивирование в этой среде.
В качестве питательной среды с добавлением высокой концентрации MTX можно использовать смешанную среду CHO-S-SFMII/CD-CHO (продукт корпорации Invitrogen) и сходные с ней.
Даже если трансформированный штамм перед обработкой MTX почти однороден, после обработки MTX он может утратить эту однородность, поскольку в ходе обработки он может приобрести разнообразие. Общее количество полипептида, продуцируемого группой однородных клеток, увеличивается по сравнению с количеством, продуцируемым почти однородным штаммом перед обработкой MTX. Из группы клеток, приобретших разнообразие, можно клонировать только тот штамм, который способен к продукции с высоким выходом. Например, эффективными являются лимитирующее разведение с использованием 96-луночного планшета и клонирование единичных клеток, сопровождаемое клеточной сортировкой. Для любого из этих способов можно использовать общедоступный способ.
Клетка по настоящему изобретению, сильно экспрессирующая TauT, обладает превосходной устойчивостью к MTX, что показано в описанных ниже примерах, следовательно, селекцию таких клеток можно проводить при более высокой концентрации MTX, чем нормальная концентрация, при которой проводят селекцию.
Из группы клеток по настоящему изобретению, полученных в результате обработки высокой концентрацией MTX, как описано выше, можно эффективно и в большом количестве выделить штамм, способный продуцировать желаемый полипептид с высоким выходом, который невозможно или очень трудно получить при нормальной обработке MTX. Таким образом, клетка по настоящему изобретению, сильно экспрессирующая TauT, крайне полезна в качестве трансформированной клетки для селекции с помощью MTX.
Таким образом, настоящее изобретение также относится к клетке, способной продуцировать желаемый полипептид с высоким выходом, который продуцируется способами, описанными выше. Клетка может относиться к группе неоднородных клеток или к клонированному однородному штамму.
Настоящее изобретение относится к способу продукции полипептида, в котором культивируют клетку, полученную описанными выше способами. Кроме того, желаемый полипептид можно получить с использованием клетки, в которой был активирован эндогенный ген с помощью технологии активации генов (см., например, международную публикацию WO 94/12650), чтобы продуцировался желаемый полипептид.
Для культивирования клетки можно использовать среды, используемые в общепринятых клеточных культурах (предпочтительно культурах клеток животных). Эти среды обычно содержат аминокислоты, витамины, липидные факторы, источники энергии, регуляторы осмотического давления, источники железа и регуляторы pH. Содержание этих компонентов обычно является следующим: аминокислоты 0,05-1500 мг/л, витамины 0,001-10 мг/л, липидные факторы 0-200 мг/л, источники энергии 1-20 г/л, регуляторы осмотического давления 0,1-10000 мг/л, источники железа 0,1-500 мг/л, регуляторы pH 1-10000 мг/л, микроэлементы металлов 0,00001-200 мг/л, поверхностно-активные вещества 0-5000 мг/л, кофакторы роста 0,05-10000 мкг/л и нуклеозиды 0,001-50 мг/л. Однако содержание не ограничивается этими диапазонами и его можно соответствующим образом выбирать, в зависимости от типа клетки, подлежащей культивированию, типа требуемого полипептида и т.д.
В дополнение к этим компонентам можно добавлять микроэлементы металлов, поверхностно-активные вещества, кофакторы роста, нуклеозиды и т.п.
Конкретные примеры таких компонентов включают аминокислоты, такие как L-аланин, L-аргинин, L-аспарагин, L-аспарагиновая кислота, L-цистеин, L-цистин, L-глутамин, L-глутаминовая кислота, глицин, L-гистидин, L-изолейцин, L-лейцин, L-лизин, L-метионин, L-орнитин, L-фенилаланин, L-пролин, L-серин, L-треонин, L-триптофан, L-тирозин и L-валин, предпочтительно L-аланин, L-аргинин, L-аспарагин, L-аспарагиновая кислота, L-цистин, L-глутамин, L-глутаминовая кислота, глицин, L-гистидин, L-изолейцин, L-лейцин, L-лизин, L-метионин, L-фенилаланин, L-пролин, L-серин, L-треонин, L-триптофан, L-тирозин и L-валин; витамины, такие как i-инозитол, биотин, фолиевая кислота, липоевая кислота, никотинамид, никотиновая кислота, п-аминобензойная кислота, пантотенат кальция, пиридоксаля гидрохлорид, гидрохлорид пиридоксина, рибофлавин, гидрохлорид тиамина, витамин B12 и аскорбиновая кислота, предпочтительно биотин, фолиевая кислота, липоевая кислота, никотинамид, пантотенат кальция, пиридоксаля гидрохлорид, рибофлавин, гидрохлорид тиамина, витамин B12 и аскорбиновая кислота; липидные факторы, такие как хлорид холина, тартрат холина, линолеиновая кислота, олеиновая кислота и холестерин, предпочтительно хлорид холина; источники энергии, такие как глюкоза, галактоза, манноза и фруктоза, предпочтительно глюкоза; регуляторы осмотического давления, такие как хлорид натрия, хлорид калия и нитрат калия, предпочтительно хлорид натрия; источники железа, такие как железо-ЭДТА, цитрат трехвалентного железа, хлорид двухвалентного железа, хлорид трехвалентного железа, сульфат двухвалентного железа, сульфат трехвалентного железа и нитрат трехвалентного железа, предпочтительно хлорид трехвалентного железа, железо-ЭДТА и цитрат трехвалентного железа; и регуляторы pH, такие как гидрокарбонат натрия, хлорид кальция, дигидрофосфат натрия, HEPES и MOPS, предпочтительно гидрокарбонат натрия. В качестве примеров могут быть приведены культуральные среды, содержащие любые из этих компонентов.
Помимо указанных выше компонентов можно добавлять микроэлементы металлов, такие как сульфат меди, сульфат марганца, сульфат цинка, сульфат магния, хлорид никеля, хлорид олова, хлорид магния и субсиликат натрия, предпочтительно сульфат меди, сульфат цинка и сульфат магния; поверхностно-активные вещества, такие как Tween 80 и Pluronic F68; кофакторы роста, такие как рекомбинантный инсулин, рекомбинантный IGF-1, рекомбинантный EGF, рекомбинантный FGF, рекомбинантный PDGF, рекомбинантный TGF-α, гидрохлорид этаноламина, селенит натрия, ретиноевая кислота и дигидрохлорид путресцина, предпочтительно селенит натрия, гидрохлорид этаноламина, рекомбинантный IGF-1 и дигидрохлорид путресцина; и нуклеозиды, такие как дезоксиаденозин, дезоксицитидин, дезоксигуанозин, аденозин, цитидин, гуанозин и уридин. В предпочтительных примерах указанные выше среды могут содержать антибиотики, такие как стрептомицин, пенициллин-G калий и гентамицин, и индикаторы pH, такие как феноловый красный.
pH среды варьирует, в зависимости от клетки, подлежащей культивированию. Как правило, подходящим является pH 6,8-7,6. Во многих случаях подходящим является pH 7,0-7,4.
Также можно использовать коммерческую среду для культивирования клеток животных, например D-MEM (модифицированная способом Игла среда Дульбекко), смесь 1:1 D-MEM/F-12 (модифицированная способом Игла среда Дульбекко: смесь питательных веществ F-12), RPMI1640, CHO-S-SFMII (Invitrogen), CHO-SF (Sigma-Aldrich), EX-CELL 301 (JRH Biosciences), CD-CHO (Invitrogen), IS CHO-V (Irvine Scientific), PF-ACF-CHO (Sigma-Aldrich) или сходные с ними.
Альтернативно, среда может представлять собой бессывороточную среду.
Когда клетка представляет собой клетку CHO, клетку CHO можно культивировать способами, известными специалистам в данной области. Например, клетки CHO обычно можно культивировать в атмосфере с концентрацией CO2 в газообразной фазе от 0 до 40%, предпочтительно от 2 до 10%, при 30-39°C, предпочтительно при приблизительно 37°C.
Как будет ясно из приведенных ниже примеров, продукция побочных продуктов (таких как лактат), которые становятся веществами, ингибирующими рост клетки, может подавляться в клетке, сильно экспрессирующей TauT. В результате этого клетка демонстрирует эффект поддержания высокого коэффициента выживания. Клетка способна к культивированию в течение трех месяцев или более длительного периода.
Более того, в случае, когда требуемый полипептид, такой как антитело, продуцируется клеточной культурой, на поздней стадии культивирования расположение клеток становится высоко плотным (приблизительно 1×107 клеток/мл), и эффект продуктов их жизнедеятельности, таких как лактат, становится крайне высоким. Если требуемый полипептид продуцируется клетками, на высоком уровне экспрессирующими TauT, поддерживается высокий уровень выживаемости даже на поздней стадии культивирования, и также можно ожидать повышения выхода требуемого полипептида.
Надлежащий период культивирования для продукции желаемого полипептида с использованием такой клетки, как правило, составляет от 1 суток до 3 месяцев, предпочтительно от 1 суток до 2 месяцев, более предпочтительно от 1 суток до 1 месяца.
Что касается различных устройств для культивирования клеток животных, то можно использовать резервуарное устройство типа ферментера (биореактора), культуральное устройство с перемешиванием струей воздуха, культуральное устройство типа колбы с культурой, культуральное устройство типа вращающейся колбы, культуральное устройство типа микроносителя, культуральное устройство типа реактора кипящего слоя, культуральное устройство типа системы полых волокон, культуральное устройство типа вращающегося флакона, культуральное устройство типа биореактора с фильтрующим наполнителем и т.п.
Культивирование можно проводить любым способом культивирования, таким как периодическая культура, культура с подпиткой или непрерывная культура. Предпочтительно используют культуру с подпиткой или непрерывную культуру. Более предпочтительной является культура с подпиткой.
Когда культивируют клетку, в среду можно добавлять таурин для обеспечения захвата таурина в клетки. Концентрация таурина, подлежащая добавлению в среду, конкретно не ограничена. Обычно концентрация составляет 0-100 г/л, предпочтительно 0-20 г/л и более предпочтительно 0-10 г/л.
Когда полипептид, продуцируемый способом по настоящему изобретению, обладает биологической активностью, полезной в качестве фармацевтического средства, можно продуцировать фармацевтическое средство смешиванием этого полипептида с фармацевтически приемлемым носителями или добавками и изготовлением из них препарата.
Конкретные примеры фармацевтически приемлемых носителей и добавок включают воду, органические растворители, которые являются фармацевтически приемлемыми, коллаген, поливиниловый спирт, поливинилпирролидон, карбоксивиниловый полимер, карбоксиметилцеллюлозу натрия, полиакрилат натрия, альгинат натрия, растворимый в воде декстран, карбоксиметилкрахмал натрия, пектин, метилцеллюлозу, этилцеллюлозу, ксантановую смолу, гуммиарабик, казеин, агар-агар, полиэтиленгликоль, диглицерин, глицерин, пропиленгликоль, вазелин, парафин, стеариловый спирт, стеариновую кислоту, сывороточный альбумин человека (HSA), маннит, сорбит, лактозу и поверхностно-активные вещества, которые являются приемлемыми в качестве фармацевтических добавок.
Фактические добавки можно выбирать из указанных выше добавок по отдельности или в сочетании в соответствии с дозированной формой лекарственного средства по настоящему изобретению, но они не ограничиваются добавками, приведенными выше. Например, когда полипептид используют в качестве инъецируемого состава, очищенный полипептид можно растворять в растворителе, таком как физиологический раствор, буфер или раствор глюкозы, а затем в раствор можно добавлять ингибитор адсорбции, такой как Tween 80, Tween 20, желатин или сывороточный альбумин человека. Альтернативно, можно использовать лиофилизированное средство для получения дозированной формы, которую растворяют и разбавляют перед применением. Примеры эксципиента, пригодного для лиофилизации, включают спирты сахаров и сахариды, такие как маннит и глюкоза.
Эффективные дозы полипептида можно надлежащим образом выбирать, в зависимости от типа полипептида, тапа заболевания, подлежащего лечению или профилактике, возраста пациента, тяжести заболевания, и т.д. Например, когда полипептид представляет собой антитело против глипикана, эффективную дозу антитела против глипикана выбирают из диапазона от 0,001 мг до 1000 мг на кг массы тела на введение. Альтернативно можно выбирать дозу 0,01-100000 мг/на организм пациента. Однако эффективная доза не ограничивается этими диапазонами.
Полипептид можно вводить либо перорально, либо парентерально, однако предпочтительным является парентеральное введение. Конкретно, могут быть приведены инъекция (например, системное или местное введение путем внутривенной инъекции, внутримышечная инъекция, внутрибрюшинная инъекция, подкожная инъекция и т.д.), введение через нос, введение через легкие, чрескожное введение и сходные с ними.
В другом варианте осуществления настоящего изобретения предусмотрен способ увеличения величины продукции полипептида клеткой, сильно экспрессирующей TauT, в которую была введена ДНК, кодирующая желаемый полипептид, причем указанный способ включает обработку клетки высокой концентрацией MTX.
В настоящем изобретении понятие "клетки, в которые была перенесена ДНК", охватывает не только клетки, в которые была встроена экзогенная ДНК с помощью технологии генетической рекомбинации, но и клетки, в которых была активирована эндогенная ДНК при помощи технологии активации генов (см., например, международную публикацию WO94/12650), так чтобы инициировалась или повышалась экспрессия белка, соответствующего эндогенной ДНК, или транскрипция этой ДНК.
ПРИМЕРЫ
Далее в этом документе настоящее изобретение более подробно описано с помощью следующих примеров. Следует отметить, что эти примеры приведены только в целях иллюстрирования настоящего изобретения, но не для ограничения объема настоящего изобретения.
[Пример приготовления 1] Получение экспрессирующей плазмиды для гуманизированного антитела к глипикану-3 человека
Ген H-цепи гуманизированного антитела к глипикану-3 человека получили следующим образом. Мышь (MRL/1pr, продукт Charles River Laboratories Japan, Inc.) иммунизировали фрагментами глипикана-3 (которые были получены в результате экспрессии гена слитого с GST белка, полученного с помощью ПЦР). Затем получали гибридому с использованием клеток селезенки мыши. Гибридому подвергали скринингу способом ELISA с использованием глипикана-3 в качестве антитела, посредством чего был отобран клон, способный продуцировать антитело, связывающее глипикан-3. Затем из гибридомы экстрагировали мРНК и при помощи реакции обратной транскрипции с использованием обратной транскриптазы получили кДНК. С помощью ПЦР с использованием праймера (CAGGGGCCAGTGGATAGACCGATG) (SEQ ID NO: 7) амплифицировали ген вариабельной области H-цепи антитела мыши к глипикану-3, который имел кДНК и последовательность оснований, комплементарную гену вариабельной области H-цепи мыши, и ген получали с помощью связывания с pGEM-T Easy (продукт Promega Corporation). Ген вариабельной области H-цепи антитела человека, который обладал гомологией с каркасной областью гена вариабельной области H-цепи антитела мыши к глипикану-3, идентифицировали с помощью поиска по базе данных Kabat. Конструировали, а затем синтезировали с помощью ПЦР последовательность оснований гена вариабельной области H-цепи гуманизированного антитела к глипикану-3, в котором каждая каркасная часть гена вариабельной области H-цепи антитела человека, идентифицированная таким способом, была соединена с каждой CDR-частью гена вариабельной области H-цепи антитела мыши к глипикану-3. Затем ген вариабельной области H-цепи гуманизированного антитела к глипикану-3 связывали с геном константной области IgG1 человека и получали ген H-цепи гуманизированного антитела к глипикану-3 посредством оптимизации путем замены аминокислот (см. WO 06/06693). Затем ген H-цепи гуманизированного антитела к глипикану-3 присоединяли ниже промотора CAG, а ниже него присоединяли сигнальную последовательность поли(A) β-глобина мыши, с получением в результате единицы экспрессии H-цепи. Единицу экспрессии H-цепи можно вырезать рестриктазами BamHI и HindIII в участке, расположенном выше нее, и рестриктазой XhoI в участке, расположенном ниже нее.
Затем получали ген L-цепи гуманизированного антитела к глипикану-3 человека следующим образом. Мышь иммунизировали фрагментами глипикана-3. Далее получали гибридому с использованием клеток селезенки мыши. Гибридому подвергали скринингу способом ELISA с применением глипикана-3 в качестве антитела, посредством чего был отобран клон, способный к продукции антитела, связывающего глипикан-3. Затем из гибридомы экстрагировали мРНК и при помощи реакции обратной транскрипции с использованием обратной транскриптазы получали кДНК. Ген вариабельной области H-цепи антитела мыши к глипикану-3 амплифицировали с помощью ПЦР с использованием праймера (GCTCACTGGATGGTGGGAAGATG) (SEQ ID NO: 8), который имел кДНК и последовательность оснований, комплементарную гену вариабельной области L-цепи мыши, и ген получали с помощью связывания с pGEM-T Easy (продукт Promega Corporation). Ген вариабельной области L-цепи антитела человека, который обладал гомологией с каркасной областью гена вариабельной области L-цепи антитела мыши к глипикану-3, идентифицировали с помощью поиска по базе данных Kabat. Конструировали, а затем синтезировали с помощью ПЦР, последовательность оснований гена вариабельной области L-цепи гуманизированного антитела к глипикану-3, в котором каждая каркасная часть гена вариабельной области L-цепи антитела человека, идентифицированная таким способом, была связана с каждой CDR-частью гена вариабельной области H-цепи антитела мыши к глипикану-3. Затем ген вариабельной области L-цепи гуманизированного антитела к глипикану-3 связывали с геном константной области κ IgG человека, и получали ген L-цепи гуманизированного антитела к глипикану-3 с помощью оптимизации путем замены аминокислот (см. WO 06/06693). Затем ген L-цепи гуманизированного антитела к глипикану-3 присоединяли ниже промотора CAG и ниже его присоединяли сигнальную последовательность поли(A) β-глобина мыши с получением в результате единицы экспрессии L-цепи. Единицу экспрессии L-цепи можно вырезать рестриктазой HindIII.
I69NPEP4, плазмиду, созданную IDEC Corporation, разрезали рестриктазами BamHI и XhoI и присоединяли к единице экспрессии H-цепи для продукции pINP-CG33-H1. Затем соединяли pINP-CG33-H1, расщепленную рестриктазой HindIII, и единицу экспрессии L-цепи, вырезанную рестриктазой HindIII. Посредством действия, описанного выше, получили phGC33CAG1, 2-копийную экспрессирующую плазмиду для L-цепи, содержащую две копии экспрессирующей единицы L-цепи и одну копию экспрессирующей единицы H-цепи на плазмиду (фиг.10).
[Пример 1] Приобретение устойчивости к MTX путем введения переносчика таурина хомяка (TauT)
В клетки CHO DXB11s, служившие клетками-хозяевами, вводили экспрессирующие плазмиды pHyg-TauT (см. пример получения 2, приведенный ниже, фиг.5) способом электропорации, с получением клеток-хозяев DXB11s/TauT, которые сильно экспрессировали TauT. После этого проводили сравнение чувствительности к MTX между клетками-хозяевами DXB11s/TauT и клетками-хозяевами DXB11s, которые использовали в качестве родительского штамма. Поскольку как клетки-хозяева DXB11s/TauT, так и клетки-хозяева DXB11s, были дефицитными по гену DHFR (ауксотрофными по HT), их уровни выживания снижались одинаково при культивировании в смешанной среде CHO-S-SFM II/CD-CHO без HT при отсутствии MTX (фиг.1, "добавление 0 нМ MTX"). При добавлении 10 нМ или 20 нМ MTX наблюдалась тенденция к ускоренному снижению уровня выживания клеток-хозяев DXB11s вследствие цитотоксичности MTX. С другой стороны, у клеток-хозяев DXB11s/TauT снижение уровня выживания подавлялось до такого уровня, который соответствовал уровню выживания при отсутствии MTX (фиг.1, "добавление 10 нМ, 20 нМ MTX"). Этот результат свидетельствует о том, что клетки-хозяева DXB11s/TauT обладали устойчивостью к MTX по сравнению с клетками-хозяевами DXB11s.
Затем в клетки CHO, способные продуцировать антитела к глипикану-3, которые служили родительским штаммом (dhfr-дефицитные клетки CHO, содержащие phGC33CAG1, экспрессирующий вектор, полученный по примеру получения 1, который содержал встроенные в него промотор CAG, ген антитела к глипикану-3, промотор β-глобина мыши и ген DHFR), способом электропорации вводили экспрессирующие плазмиды pHyg-TauT с получением штамма GC33/DXB11s/TauT, в котором сильно экспрессировался TauT. Затем в указанный выше штамм GC33/DXB11s/TauT были дополнительно введены экспрессирующие плазмиды pPur-CSAD (см. ссылочный пример 4, приведенный ниже, фиг.6) с получением штамма GC33/DXB11s/TauT/CSAD, в котором сильно экспрессировался CSAD. Позднее проводили сравнение штамма GC33/DXB11s/TauT и штамма GC33/DXB11s/TauT/CSAD с родительским штаммом GC33/DXB11s по чувствительности к MTX. Поскольку родительский штамм представлял собой штамм, способный продуцировать антитела, в котором ген антитела амплифицировался до 20 копий или более (относительно величины перед обработкой MTX) посредством обработки 20 нМ MTX, указанные выше штаммы могли стабильно пролиферировать в смешанной среде CHO-S-SFM II/CD-CHO, в которую был добавлен 20 нМ MTX. Однако вследствие цитотоксичности MTX уровни выживания всех указанных выше штаммов снижались в смешанной среде CHO-S-SFM II/CD-CHO при повышении в ней концентрации MTX до 200 нМ, что в 10 раз превышало начальную концентрацию. Эта экспериментальная система также показала, что два штамма, в которых сильно экспрессировался TauT, были в большей степени устойчивыми к MTX, чем родительский штамм (фиг.2).
[Пример 2] Количественное увеличение продукции антитела при использовании устойчивого к MTX штамма, сильно экспрессирующего TauT
Штамм GC33/DXB11s/TauT/CSAD, который был наиболее устойчив к MTX согласно примеру 1, субкультивировали в ротационной культуре в течение 28 дней с применением смешанной среды CHO-S-SFM II/CD-CHO, в которую была добавлена высокая концентрация (100 нМ или 200 нМ) в условиях 2×105 клеток/мл на начальной стадии с заменой среды через каждые три-пять суток. Как представлено на фиг.3, поскольку клеточная пролиферация не наблюдалась даже на 28-ые сутки после инициации культуры, концентрацию MTX возвращали к начальному уровню 20 нМ, после чего продолжали субкультивирование по типу ротационной культуры. Через 14 суток после этого, т.е. на 42-й сутки, клетки начали пролиферировать. Указанные выше два штамма, уровни выживания которых восстанавливались при субкультивировании в разбавленной среде (клетки, обработанные 100 нМ или 200 нМ MTX), культивировали в 15-мл пробирках с использованием смешанной среды CHO-S-SFM II/CD-CHO в присутствии 20 нМ MTX в условиях 1×105 клеток/мл на начальной стадии. В результате уровни клеточной пролиферации штамма GC33/DXB11s/TauT/CSAD до и после обработки высокой концентрацией MTX различались, и штамм, обработанный 200 нМ MTX, обладал наибольшей продуктивностью (количество жизнеспособных клеток: 18,0×105 клеток/мл, количественная продукция антитела: 144 мг/л), в то время как штамм, обработанный 100 нМ MTX (количество жизнеспособных клеток: 22,3×105 клеток/мл, велиина продукции антитела: 162 мг/л), также демонстрировал потенциал к продукции большего количества антитела, чем штамм TauT/CSAD перед обработкой высокой концентрацией MTX (количество жизнеспособных клеток: 13,5×105 клеток/мл, величина продукции антитела: 66 мг/л) (фиг.4). Кроме того, количество антитела, продуцированного штаммом TauT/CSAD перед обработкой высокой концентрацией MTX, составляло 86 мг/л, даже если количество жизнеспособных клеток возрастало до 19,6×105 клеток/мл. Таким образом, было показано, что штамм TauT/CSAD перед обработкой высокой концентрацией MTX имел наименьшую продуктивность.
Полученные результаты, как описано выше, позволяют предположить, что клетки приобретают устойчивость к MTX при сильной экспрессии переносчика таурина (TauT) искусственным образом, а клетки, способные к продукции большего количества антитела, можно получить при обработке высокой концентрацией MTX, если в качестве клеток-хозяев используются клетки, сильно экспрессирующие TauT.
Настоящее изобретение применимо к клеткам, способным продуцировать все типы желаемых полипептидов (предпочтительно антител).
[Справочный пример 1] Клонирование гена переносчика таурина хомяка, происходящего из клетки CHO
Тотальную РНК извлекали из клеток, продуцирующих антитело к рецептору IL-6 (линия клеток CHO DXB11, в которую был перенесен ген антитела против рецептора IL-6) (нерассмотренная японская патентная публикация № Hei 8-99902), а затем из нее синтезировали кДНК поли(A)-зависимым способом. Ген переносчика таурина хомяка (TauT) получали в ПЦР с применением матрицы кДНК, фрагментированной тремя рестрикционными ферментами, SalI, XhoI и EcoRI. В качестве праймеров для ПЦР были сконструированы фрагменты ДНК, содержащие 5'-концевую и 3'-концевую последовательность, консервативную для TauT крысы и мыши. Определяли нуклеотидную последовательность клонированного гена. По ее гомологии с другими генами TauT известных биологических видов было подтверждено, что клонированный ген кодирует TauT хомяка (фиг.7). Аминокислотная последовательность TauT хомяка имеет высокую гомологию с TauT мыши (идентичность 96%), TauT крысы (идентичность 96%) и TauT человека (идентичность 93%). Было предсказано, что TauT хомяка является переносчиком с 12 трансмембранными участками (фиг.8). Нуклеотидная последовательность TauT хомяка представлена в SEQ ID NO: 1. Аминокислотная последовательность TauT хомяка представлена в SEQ ID NO: 2.
[Справочный пример 2] Увеличение плотности жизнеспособных клеток, ингибирование продукции лактата и увеличение выхода антител, вызванное переносом переносчика таурина хомяка
Экспрессирующую плазмиду промотора CMV pHyg/TauT (фиг.5) конструировали, добавляя последовательность Козака к гену TauT хомяка (далее TauT), полученному клонированием согласно справочному примеру 1. Контрольную плазмиду pHyg без гена pHyg/TauT или TauT вводили способом элетропорации в родительский штамм клеток CHO, продуцирующих антитело к глипикану-3 (см. WO 2006/006693). После селекции клеток со встроенной экспрессирующей плазмидой в присутствии гигромицина (400 мкг/мл) все стабильно растущие клеточные штаммы размножали (pHyg/TauT: 8 штаммов, pHyg: 7 штаммов). Получали мРНК TauT. Позднее способом TaqMan было подтверждено, что 7 штаммов экспрессируют TauT более сильно, чем родительский штамм, и они были отобраны в качестве клеток с перенесенным pHyg/TauT. Средний уровень экспрессии мРНК в этих трансформированных клетках (7 штаммов) был приблизительно в 40 раз выше по сравнению с контролем (7 штаммов). Клетки всех 14 штаммов выращивали в периодической культуре и подпитываемой культуре в 50-мл встряхиваемых колбах при начальной плотности клеток 2×105 клеток/мл. На 7-ые сутки культивирования (поздняя стадия) проводили сравнение этих штаммов по плотности жизнеспособных клеток, выходу лактата и выходу антитела к глипикану-3. По мере роста клеток в периодической культуре в культуральном бульоне накапливаются ингибиторные вещества, такие как лактат, и рост клеток подавляется. Однако плотность жизнеспособных клеток (9,28±3,27×105 клеток/мл) и выход лактата (1,54±0,20 г/л) в клетках, трансформированных pHyg/TauT, превосходили соответствующие показатели в клетках, трансформированных pHyg (плотность жизнеспособных клеток: 5,69±2,09×105 клеток/мл, выход лактата: 1,75±0,15 г/л) (t-критерий; p<0,05). Что касается выхода антитела к глипикану-3, то 4 из 7 клеточных штаммов, трансформированных pHyg/TauT (средний выход антитела: 440,6 мг/л), демонстрировали выход антитела, превышающий самый большой выход в клетках, трансформированных pHyg (389,6 мг/л). Далее, поскольку превосходство клеток, трансформированных pHyg/TauT, с точки зрения выхода антитела к глипикану-3 становилось более очевидным в подпитываемой культуре (t-критерий; P<0,01, фиг.7), штамм T10, трансформированный pHyg/TauT (который демонстрировал наибольшую способность к росту среди указанных выше 4 штаммов), и родительский штамм выращивали в подпитываемой культуре в сосуде емкостью 1 л. В результате коэффициент жизнеспособности T10 поддерживался на уровне 80% или более даже на 32 сутки культивирования при подавлении продукции лактата. Вследствие этого выход антитела к глипикану-3 в указанном штамме достигал 2,9 г/л на 35-ые сутки культивирования. Проточным цитометрическим анализом было подтверждено, что клетки T10, трансформированные TauT, экспрессировали молекулы TauT на клеточной мембране. Эти результаты позволяют предположить, что при искусственной экспрессии Taut хомяка можно повысить потенциал клеток, продуцирующих антитело, и создать штаммы, способные к усиленной продукции антитела.
[Справочный пример 3] Клонирование гена декарбоксилазы цистеинсульфиновой кислоты (CSAD) хомяка, полученного из клетки CHO
Из клеток, продуцирующих антитело к рецептору IL-6 (линия клеток CHO DXB11, в которую был перенесен ген антитела против рецептора IL-6) (японская нерассмотренная патентная публикация № Hei 8-99902), экстрагировали тотальную РНК, а затем из нее синтезировали кДНК поли(A)-зависимым способом. Ген CSAD хомяка получали с помощью ПЦР с использованием матрицы кДНК, фрагментированной тремя ферментами рестрикции, SalI, XhoI и EcoRI. В качестве праймеров для ПЦР были сконструированы фрагменты ДНК, содержащие 5'-концевую и 3'-концевую последовательность, консервативную для CSAD крысы и мыши. Определяли нуклеотидную последовательность клонированного гена. По ее гомологии с другими генами CSAD известных биологических видов было подтверждено, что клонированный ген кодирует CSAD хомяка (фиг.9). Аминокислотная последовательность CSAD хомяка имела высокую гомологию с аминокислотными последовательностями CSAD мыши (идентичность 96%), CSAD крысы (идентичность 96%) и CSAD человека (идентичность 91%). Было предсказано, что CSAD представляет собой фермент, обладающий такой же активностью. Нуклеотидная последовательность CSAD хомяка представлена в SEQ ID NO: 3. Аминокислотная последовательность CSAD хомяка представлена в SEQ ID NO: 4.
[Справочный пример 4] Конструирование экспрессирующей плазмиды CSAD хомяка для селекции с помощью пуромицина
Экспрессирующую плазмиду с промотором CMV pPur/CSAD (фиг.6) конструировали, добавляя последовательность Козака к гену CSAD хомяка (далее CSAD), полученному клонированием согласно справочному примеру 3.
Все публикации, патенты и патентные заявки, процитированные в этом документе, включены в настоящий документ в качестве ссылок в полном объеме.
ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ
Настоящее изобретение применимо для производства белков.
СВОБОДНЫЙ ТЕКСТ СПИСКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ
В SEQ ID NO: 1 представлена нуклеотидная последовательность гена, кодирующего TauT хомяка.
В SEQ ID NO: 2 представлена аминокислотная последовательность TauT хомяка.
В SEQ ID NO: 3 представлена нуклеотидная последовательность гена, кодирующего CSAD хомяка.
В SEQ ID NO: 4 представлена аминокислотная последовательность CSAD хомяка.
В SEQ ID NO: 5 представлена последовательность Козака.
В SEQ ID NO: 6 представлена последовательность секреторного сигнального пептида.
В SEQ ID NO: 7 представлена последовательность праймера, имеющего последовательность оснований, комплементарную гену вариабельной области H-цепи мыши.
В SEQ ID NO: 8 представлена последовательность праймера, имеющего последовательность оснований, комплементарную гену вариабельной области L-цепи мыши.
Реферат
Изобретение относится к области биотехнологии, а именно к способу получения клетки СНО, способной продуцировать желаемый полипептид с высоким выходом, клетке, полученной данным способом, способу получения желаемого полипептида, способу количественного увеличения продукции полипептида клеткой СНО с сильной экспрессией переносчика таурина. Способ включает культивирование клетки СНО, в которую был искусственно перенесен ген переносчика таурина (TauT), была введена ДНК, кодирующая желаемый полипептид, и ДНК, кодирующая дигидрофолатредуктазу (DHFR), в присутствии концентрации метотрексата, при которой 90% или более клеток, в которые не был введен TauT, погибают в течение 3 недель после субкультивирования. Отбирают из числа выживших клеток СНО, клетку СНО, способную продуцировать желаемый полипептид с высоким выходом. Клетка, полученная данным способом, продуцирует желаемый полипептид с более высоким выходом, чем клетка СНО, трансфицированная ДНК, кодирующей желаемый полипептид, и ДНК, кодирующей DHFR, но не геном переносчика таурина. Способ получения желаемого полипептида включает культивирование вышеуказанной клетки и выделение желаемого пептида. Предложенное изобретение позволяет продуцировать желаемый полипептид с высоким выходом. 4 н. и 5 з.п. ф-лы, 10 ил., 6 пр.
Комментарии