Клетка, продуцирующая с высокой эффективностью активный белок арилсульфатазу в, и способ получения этой клетки - RU2763990C2
Код документа: RU2763990C2
Чертежи







Описание
Основой изобретения явилось открытие того факта, что адаптированная к суспензионному культивированию производная клеточной линии СНО-K1, экспрессирующая арилсульфатазу В человека, значительно увеличивает свою продуктивность в случае введения в нее конструкций, экспрессирующих рекомбинантный модифицирующий сульфатазу фактор 1. Результатом таких модификаций являются клетки, продуцирующие с высоким выходом активные рекомбинантные лизосомальные ферменты сульфатазы (в 10 раз выше, чем аналогичные клетки без SUMF1). Изобретение относится также к применению любой производной клеточной линии СНО-K1, ко-экспрессирующей рекомбинантный SUMF1 человека и рекомбинантный лизосомальный фермент сульфатазу, и может быть пригодным для изготовления активных лизосомальных сульфатаз для терапии болезней лизосомального накопления путем фермент-заместительной терапии.
Краткое описание чертежей.
Фиг. 1. Продуктивность клеток во время скрининга 96-ти луночных планшетов каждой группы. ARSB+SUMF1 - клетки, трансфицированные двумя плазмидами, несущими гены ARSB и SUMF1; ARSB - клетки, трансфицированные одной плазмидой, несущей ген ARSB. Для статистического обсчета данных использовали Unpaired t test (программа GraphPad) Pvalue<0,0001.
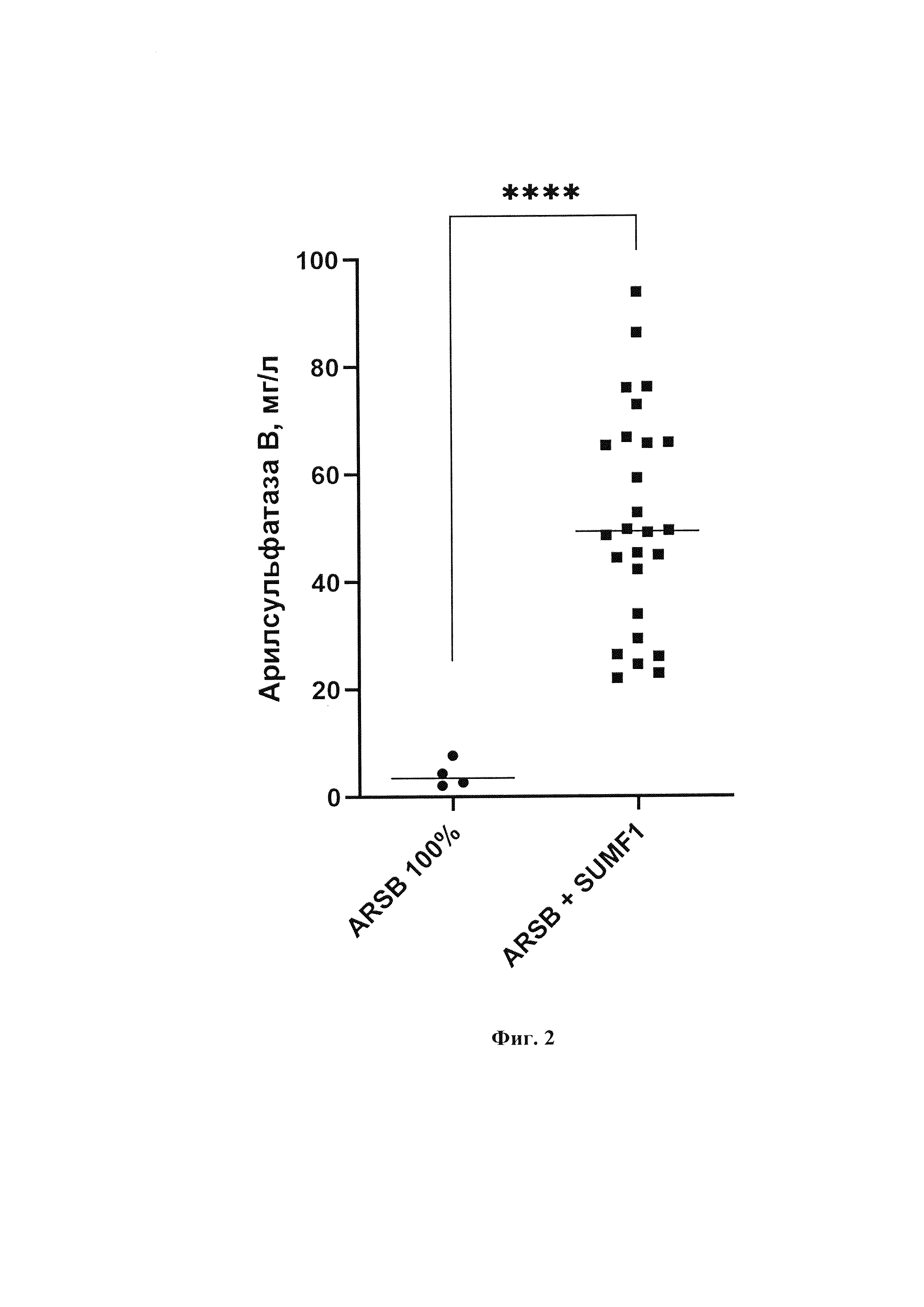
Фиг. 2. Продуктивность клеток на 6-е сутки процесса batch-культивирования. ARSB+SUMF1 - клетки, трансфицированные двумя плазмидами, несущими гены ARSB и SUMF1; ARSB - клетки, трансфицированные одной плазмидой, несущей ген ARSB. Для статистического обсчета данных использовали Unpaired t test (программа GraphPad)
Pvalue<0,0001.
Фиг. 3. Специфическая продуктивность клеток. ARSB+SUMF1 - клетки, трансфицированные двумя плазмидами, несущими гены ARSB и SUMF1; ARSB - клетки, трансфицированные одной плазмидой, несущей ген ARSB. Для статистического обсчета данных использовали Unpaired t test (программа GraphPad) Pvalue=0,0001.
Фиг. 4. Электрофореграмма (слева) и вестерн-блот (справа) КЖ клеток. Пробы КЖ клеток нанесены по 20 мкл на трек. Ladder - маркер Protein Ladder (Thermo Fisher Scientific, США). Стандарт - рекомбинантный белок галсульфаза, препарат «Наглазим» (BIOMARIN, Англия). ARSB+SUMF1 - клетки, трансфицированные двумя плазмидами, несущими гены ARSB и SUMF1; ARSB - клетки, трансфицированные одной плазмидой, несущей ген ARSB.
Фиг. 5. Электрофореграмма (слева) и вестерн-блот (справа) клеточных лизатов. Пробы нанесены по 8×104 клеток на трек. Ladder - маркер Protein Ladder, 10 мкл. ASB - рекомбинантный белок галсульфаза, препарат «Наглазим» (BIOMARIN, Англия). ARSB+SUMF1 - клетки, трансфицированные двумя плазмидами, несущими гены ARSB и SUMF1; ARSB - клетки, трансфицированные одной плазмидой, несущей ген ARSB.
Фиг. 6. Продуктивность клеток CHO-K1-V и CHO-K1-GEN после супер-трансфекции плазмидой, несущей ген SUMF1 на 6 день batch-культивирования. ARSB+SUMF1 - клетки, трансфицированные двумя плазмидами, несущими гены ARSB и SUMF1; ARSB - клетки, трансфицированные одной плазмидой, несущей ген ARSB.
Для статистического обсчета данных использовали Unpaired t test (программа GraphPad). Unpaired t test for CHO-K1-V: Pvalue<0,0001, Unpaired t test for CHO-K1-GEN: Pvalue<0,0001.
Фиг. 7. Электрофореграмма (слева) и вестерн-блот (справа) клеточных лизатов после супер-трансфекции геном SUMF1. Пробы нанесены по 8×104клеток на трек. Ladder - маркер Protein Ladder, 10 мкл на трек. ARSB+SUMF1 - клетки, трансфицированные двумя плазмидами, несущими гены ARSB и SUMF1; ARSB - клетки, трансфицированные одной плазмидой, несущей ген ARSB.
Фиг. 8. Электрофореграмма (слева) и вестерн-блот (справа) проб КЖ клеток после супер-трансфекции геном SUMF1 на 6 день batch-культивирования. Пробы нанесены по 25 мкл на трек. Ladder - маркер Protein Ladder, 10 мкл. Стандарт - «Наглазим». ARSB+SUMF1 - клетки, трансфицированные двумя плазмидами, несущими гены ARSB и SUMF1; ARSB - клетки, трансфицированные одной плазмидой, несущей ген ARSB.
Список используемых сокращений


Настоящее изобретение относится к новому высокоэффективному модифицированному штамму-продуценту клеток СНО-K1 или ее производных для получения активных рекомбинантных лизосомальных ферментов сульфатаз человека или их биологически активных фрагментов, мутантов, вариантов в количествах, которые обеспечивают их терапевтическое применение. В общем случае, предложенная клетка-продуцент содержит (а) первый экспрессирующий вектор для рекомбинантного модифицирующего сульфатазу фактора 1 человека, содержащий последовательности CMVe/EF1a pro, гена SUMF1, BGH рА, и регуляторного элемента, повышающие экспрессию гена, и (b) второй экспрессирующий вектор для рекомбинантного лизосомального фермента арилсульфатазы В, содержащий последовательности CMVe/EF1a pro, гена ARSB, BGH рА, и регуляторного элемента, повышающие экспрессию гена. Получение штамма-продуцента включает стадии: (а) культивирования СНО-K1 клетки или ее производного; (б) получения первого экспрессирующего вектора млекопитающих, способного экспрессировать активный рекомбинантный лизосомальный фермент сульфатазу человека или его биологически активный фрагмент, мутант, вариант или производное в происходящей из СНО-K1 клетке или ее производном; (в) получения второго экспрессирующего вектора млекопитающих, способного экспрессировать рекомбинантный модифицирующий сульфатазу фактор 1 (SUMF1) человека или его биологически активный фрагмент, мутант, вариант или производное в СНО-K1 клетке или ее производной линии, недефектной по эндосомальному закислению; (г) трансфекции СНО-K1 клетки или ее производного первым и вторым экспрессирующими векторами; (д) селекции и клонирования трансфектанта СНО-K1 клетки или ее производного, которая экспрессирует активный рекомбинантный лизосомальный фермент сульфатазу человека или его биологически активный фрагмент, мутант, вариант или производное; (е) оптимизации способа культивирования клеток для изготовления рекомбинантного лизосомального фермента сульфатазы человека или его биологически активного фрагмента, мутанта, варианта или производного.
Предпочтительно, рекомбинантная лизосомальная сульфатаза человека представляет собой арилсульфатазу В (ген ARSB). В других вариантах осуществления с помощью клетки по изобретению могут быть получены также арилсульфатаза А (ген ARSA), идуронат-2-сульфатаза (ген IDS), сульфамидаза/гепарин-N-сульфатаза (ген SGSH), N-ацетилглюкозамин-сульфатаза (ген GNS) и N-ацетилгалактозамин-6-сульфатаза (ген GALNS). При получении клетки используют стадии трансфекции кДНК, кодирующей весь лизосомальный фермент сульфатазу или его часть, и кДНК, кодирующей весь SUMF1 человека или его часть, в СНО-K1 клетку или ее производное, не дефектную по эндосомальному закислению. В некоторых вариантах осуществления, первый и второй экспрессирующие векторы, которые способны экспрессировать кДНК, кодирующую активный рекомбинантный лизосомальный фермент сульфатазу человека и SUMF1 человека, соответственно, трансфицируют в клетку одновременно. В некоторых вариантах осуществления, первый и второй экспрессирующие векторы трансфицируют в клетку последовательно. В некоторых вариантах осуществления, используют кДНК, кодирующую полноразмерный лизосомальный фермент сульфатазу человека, в то время как в других вариантах осуществления используют кДНК, кодирующую его биологически активный фрагмент, мутант, вариант или производное. В некоторых вариантах осуществления, используют кДНК, кодирующую полноразмерный SUMF1 человека, в то время как в других вариантах осуществления используют кДНК, кодирующую его биологически активный фрагмент, мутант, вариант или производное. В некоторых вариантах осуществления, для одновременного переноса кДНК лизосомального фермента сульфатазы человека и SUMF1 или последовательно в СНО-K1 клетку или ее производное используют несколько экспрессирующих векторов. В некоторых вариантах осуществления, для одновременного переноса кДНК лизосомального фермента сульфатазы и SUMF1 человека в СНО-K1 клетку или ее производное используют один экспрессирующий вектор. В предпочтительном варианте осуществления, СНО-K1 клетка или ее производное представляет собой клеточную линию СНО-K1. В предпочтительном варианте осуществления, способ включает продукцию активного рекомбинантного лизосомального фермента арилсульфатазы В (ASB) клеточной линией СНО-K1 или ее производным. Изобретение относится также к модифицированной клеточной линии СНО-K1, характеризующейся ее способностью продуцировать активные рекомбинантные лизосомальные ферменты сульфатазы человека в количествах, которые дают возможность терапевтического применения лизосомального фермента сульфатазы. В различных вариантах осуществления, изобретение относится также к происходящим из СНО-K1 клеточным линиям, или к их производным, которые способны продуцировать с высоким выходом активные рекомбинантные лизосомальные ферменты сульфатазы человека, тем самым позволяя крупномасштабную продукцию таких терапевтических лизосомальных ферментов сульфатаз. В одном варианте осуществления, клеточная линия СНО-K1 содержит (а) экспрессирующий вектор для рекомбинантного модифицирующего сульфатазу фактора 1 (SUMF1) человека, нуклеотидная последовательность которого включает в себя полностью последовательность SEQ ID#1 и (b) экспрессирующий вектор для рекомбинантного лизосомального фермента арилсульфатазы В (ARSB), нуклеотидная последовательность которого включает в себя полностью последовательность SEQ ID#2.
Известен способ получения различных сульфатаз, в т.ч. арилсульфатазы В, при их ко-эксперессии с геном SUMF1 (патенты RU 2510820 и RU 2607376). Изобретения относятся к клеточной линии G71, адаптированной для роста в бессывороточной суспензионной культуре, обозначаемой G71S. Клеточная линия G71S, дефицитная по эндосомальному закислению, экспрессирующая модифицирующий сульфатазу фактор 1 (SUMF1) человека, продуцирует лизосомальные ферменты сульфатазы с более высокими уровнями активности (т.е. превращения остатка цистеина в активном центре арилсульфатазы В в Сα-формилглицин (FGly)). Известный способ включает стадии культивирования клеток линии G71S группы комплементации END3 или ее производного; получения вектора, содержащего рекомбинантный лизосомальный фермент сульфатазу человека, получения второго экспрессирующего вектора, содержащего рекомбинантный модифицирующий сульфатазу фактор 1 (SUMF1); затем трансфекции указанной клеточной линии G71S первым и вторым экспрессирующими векторами. Селективный маркер представлял собой ген фосфотрансферазы неомицина, который несет точечную мутацию, снижающую эффективность фермента. Данный маркер экспрессировали под слабым промотором HSV-tk. Продуктивность составляла 0,6-1,2 пкг/клетка/день, при активности фермента 1000-1400 нг/мл. Для данной линии клеток показано положительное влияние SUMF1 как на продуктивность, так и на активность полученного фермента.
Известен также способ получения арилсульфатазы В в недефектной по системе комплементации END3 клеточной линии СНО, при этом показано влияние ко-экспрессии SUMF1 на активность сульфатаз, но не на продуктивность клетки (WO 2004072275, данный способ является наиболее близким к заявленному изобретению).
Таким образом, из уровня техники в настоящее время неизвестен способ, позволяющий получать арилсульфатазу В с высокой продуктивностью и активностью в какой-либо иной клеточной линии, помимо дефектной линии G71S.
В заявленном изобретении, как и в прототипе WO 2004072275, для продукции сульфатазы была использована недефектная по эндосомному закислению линия СНО-K1, при этом, за счет введения в клетку собственных экспрессирующих конструкций, имеющих последовательности SEQ ID#1 и SEQ ID#2, была достигнута более высокая продуктивность фермента по сравнению с протототипом и другими известными способами, составляющая 1-3,5 пг/клетку/день (фиг. 3). Арилсульфатаза В содержит остаток цистеина в каталитическом центре; этот остаток цистеина посттрансляционно модифицируется в Cα-формилглицин (FGly) для активации фермента. Посттрансляционная активация фермента, которая катализируется вспомогательным белком FGE (ген SUMF1), происходит сразу после трансляции на несвернутых сульфатазах в эндоплазматической сети, после чего молекула арилсульфатаза В выводится с помощью сигнального пептида из клетки. Поскольку клетка экспрессирует избыточное количество арилсульфатазы В, то достаточного эндогенного количества FGE нет в эндоплазматической сети, чтобы конвертировать каждую молекулу арилсульфатазы В. При ко-экспрессии генами ARSB (целевой фермент, арилсульфатаза В) и SUMF1 (вспомогательный белок, FGly-generating enzyme, FGE), который способствует правильному фолдингу арилсульфатазы В, клеточному аппарату легче проводить биосинтез избыточной арилсульфатазы В, в результате увеличивается выход белка. Использование при трансфекции клеток линии СНО-K1 комбинаций ДНК, содержащих кодирующие последовательности белков в предложенном способе получения сульфатаз привела к неожиданному значительному усилению уровня экспрессии целевой сульфатазы, не наблюдаемому ранее в уровне техники. Хотя различные варианты получения сульфатаз в различных клеточных линях известны, ранее не было показано, что введение в клетку экспрессирующей конструкции SUMF1 на порядок повышает экспрессию целевой сульфатазы. Данный технический результат является новым и неочевидным из уровня техники. Ключевым фактором, по нашему мнению, являлось использование линии клеток СНО-K1, недефектной по эндосомальному закислению, в сочетании с экспрессией белков FGE и ASB в составе оригинальных экспрессирующих конструкций, обеспечивающих оптимальный уровень продукции обоих белков, что всегда является нетривиальной задачей при совместной экспрессии двух и более белков (см. пример 2, фиг. 2, 6). Плазмида для экспрессии по существу содержит следующие элементы, по порядку: ген, обеспечивающий устойчивость бактерии к ампициллину (селективный маркер, SV40 pro - промотор вируса sv40, puro - ген, обеспечивающий устойчивость бактерии к пуромицину или неомицину (селективный маркер), CMVe/EF1a pro - энхансер вируса CMV и промотор транскрипции гена фактора элонгации альфа, ген экспрессируемого белка - ARSB или SUMF1, BGH рА - сигнал полиаденилирования бычьего гормона роста, дополнительно может быть введен UCOE - универсальный элемент разворачивания хроматина. При этом для обеспечения оптимального уровня продукции обоих белков в клетке необходимо присутствие двух кодирующих последовательностей, одна из которых содержит CMVe/EF1a pro, ген, кодирующий ARSB, BGH рА, и другая, которая содержит CMVe/EF1a pro, ген, кодирующий FGE, BGH рА.
Дополнительный вклад к продуктивности может быть получен введением в состав плазмиды нуклеотидной последовательности, которая специфически связывается с изолированным ядерным матриксом в условиях in vitro. Применение подобных последовательностей для усиления экспрессии трансгена известно с 1990-х годов. BODE et al. BIOLOGICAL SIGNIFICANCE OF UNWINDING CAPABILITY OF NUCLEAR MATRIX-ASSOCIATING DNAs SCIENCE. - 1992, p. 195-197. J Biol Chem. 1995 Oct 13; 270(41):24010-8. Mutually exclusive interaction of a novel matrix attachment region binding protein and the NF-muNR enhancer repressor. Implications for regulation of immunoglobulin heavy chain expression. Zong R et al. J Biol Chem. 1995 Oct 13; 270(41):24010-8. J Immunol. 2002 Sep 1; 169(5):2477-87. High frequency of matrix attachment regions and cut-like protein x/CCAAT-displacement protein and В cell regulator of IgH transcription binding sites flanking Ig V region genes. Goebel Pat al., J Biol. Chem. 1997 Jul 18; 272(29):18440-52. Interaction of the nuclear matrix-associated region (MAR)-binding proteins, SATB1 and CDP/Cux., with a MAR element (L2a) in an upstream regulatory region of the mouse CD8a gene. Banan et al., Crit Rev Eukaryot Gene Expr. 1999; 9(3-4):311-8. Origin and roles of nuclear matrix proteins. Specific functions of the MAR-binding protein MeCP2/ARBP. Strätling WH, Yu F. Construction of a chromosome specific library of human MARs and mapping of matrix attachment regions on human chromosome 19 JOURNAL Nucleic Acids Res. 24 (7), 1330-1336 (1996) В патенте RU 2235786 (C2), опубл. 22.01.1999, также раскрыто применение так называемой "области присоединения матрикса", являющейся регуляторным элементом высшего порядка структурной организации хроматина, с помощью которых хроматин разделяется на топологически независимые "петли", что позволяет осуществлять независимую пространственную и временную регуляцию экспрессии гена и репликации ДНК. Помимо S/MAR и MAR, другими известными регуляторными элементами, повышающими экспрессию гена, являются, например, элементы UCOE, STAR (Регуляторные элементы в векторах для эффективного получения клеточных линий-продуцентов целевых белков. Acta Naturae (русскоязычная версия), 2015 Максименко О.Г. Гасанов Н.Б. Георгиев П.Г.).
Осуществление изобретения
1. Получение экспрессирующих векторов
кДНК полноразмерного модифицирующего сульфатазу фактора 1 (SUMF1) человека, кодирующая полипептид из 374 аминокислот, клонировали в экспрессирующий вектор pGNR2, который содержит энхансер вируса CMV и промотор транскрипции гена фактора элонгации альфа, сигнал полиаденилирования бычьего гормона роста и универсальный элемент открытия хроматина - UCOE. Селективный маркер представлял собой ген устойчивости к неомицину под контролем промотора sv40. Полученная плазмида была обозначена как pGNR-071-007.
кДНК полноразмерной N-acetylgalactosamine-4-sulfatase (Arylsulfatase В) человека, кодирующая полипептид из 515 аминокислот, включающий сигнальный пептид из 20 аминокислот, клонировали в экспрессирующий вектор pGNR1, который содержит энхансер вируса CMV и промотор транскрипции гена фактора элонгации альфа, сигнал полиаденилирования бычьего гормона роста и универсальный элемент открытия хроматина - UCOE. Селективный маркер представлял собой ген устойчивости к пуромицину под контролем промотора sv40. Полученная плазмида была обозначена как pGNR-071-006.
2. Получение клеточных линий, ко-экспрессирующих ASB и FGE
Клетки линии СНО-K1, были адаптированы к суспензионному культивированию в среде, не содержащей сыворотку или другие компоненты животного происхождения. Клетки были адаптированы к бессывороточной среде путем постепенного уменьшения объема содержащей сыворотку среды (100%, 50%, 20% и 0%) и увеличения объема бессывороточной среды (0%, 50% 80% и 100%) BalanCD (IrvineScientific, США). Клетки культивировали при 37°С, 5% СО2 и стандартных условиях увлажнения.
Трансфекцию клеток-хозяев проводили на системе трансфекций Neon Transfection System (Thermo Fisher Scientific, США) по стандартному протоколу для CHO клеток. Для этого клетки СНО смешивали с парными комбинациями плазмид, кодирующих гены SUMF1 и ARSB. После трансфекции пулы-трансфектанты помещали в среду BalanCD. Контрольную электропорацию проводили без добавления плазмиды. Жизнеспособность и плотность жизнеспособных клеток подсчитывались через 24 и 48 часов после трансфекции. Затем пулы-трансфектанты культивировали в среде BalanCD с селективными антибиотиками: пуромицин 5 мкг/мл (Sigma, Япония) и/или неомицин 400 мкг/мл (Gibco, США). Наличие целевого фермента экспрессируемого в КЖ измеряли с помощью ИФА.
Максимальная продуктивность клеток во время проведения скрининга 96-ти луночных планшетов со вспомогательным геном SUMF1 составляла 3,5 мг/л, максимальная продуктивность клеток без вспомогательного белка - 1,5 мг/л (фиг. 1)
3. Проведение иммуноферментного анализа
Для сорбции на твердой фазе разводили очищенные поликлональные антитела anti-galsulfase (МБЦ «Генериум») в карбонатно-бикарбонатном буфере рН 9,6 до концентрации 5 мкг/мл и вносили по 50 мкл на лунку 96-луночного сорбционного планшета (Nunc MaxiSorp, ThermoFisher). Разведенные образцы КЖ в буфере PBS-Ta вносили в лунки по 50 мкл. Стандарт (галсульфаза, препарат «Наглазим» 1,0 мг/мл) разводили в буфере PBS-Ta до концентрации 40, 20, 10, 5, 2,5 1,25, 0,625 нг/мл и вносили по 50 мкл на лунку. Разведенные образцы КЖ и стандарта инкубировали 1 час при +37°С, 300 об/мин. Промывку проводили буфером PBS-T в 3-х повторах по 200 мкл на лунку. Затем добавляли конъюгированные антитела (anti-galsulfaze HRP) разведенные в PBS-Ta. Промывку проводили буфером PBS-T в 3-х повторах по 200 мкл на лунку. Затем в лунки добавляли раствор ТМБ по 100 мкл. Для остановки реакции добавляли СТОП-раствор (0,5М H2SO4) по 50 мкл на лунку. Измерения проводили на спектрофотометре Benchmark Plus (Bio-Rad, США) при OD 450 нм.
Оценка специфической продуктивности клеточных линий экспрессирующих арилсульфатазу В с FGE и без вспомогательного белка показала, что максимальная продуктивность клеток с SUMF1 в процессе batch-культивирования составила около 90 мг/л. Продуктивность арилсульфатазы В у клеток с дополнительным геном SUMF1 в среднем выше в 10 раз (фиг. 2). Клеточные линии без вспомогательного белка FGE имеют самую низкую продуктивность около 5 мг/л (фиг. 2). Специфическая продуктивность клеточных линий, экспрессирующих только ASB и ABS+FGE также представлена на фиг. 3.
4. Оценка активности целевого белка
Целевой белок может быть выделен и очищен из КЖ продуцентов любым стандартным методом. Удельная активность полученной по изобретению арилсульфатазы В имеет сравнимое значение с препаратом-стандартом «Наглазим».

5. Супер-трансфекция геном SUMF1 низкопродуктивных клеточных линий арилсульфатазы В
Для супер-трансфекции использовали клеточные линии CHO-K1-GEN и CHO-K1-V, которые экспрессировали арилсульфатазу В с очень низким выходом (около 1 мг/л). Клетки линий CHO-K1-GEN и CHO-K1-V смешивали с плазмидами, несущее гены SUMF1. Трансфекцию проводили на Neon Transfection System (Thermo Fisher Scientific, США) по стандартному протоколу для CHO. Далее клетки помещали в среду с селективным антибиотиком, и проводили стабилизацию пула-трансфектанта.
Примеры
Пример 1. Конструирование экспрессирующей плазмиды, несущей ген ARSB.
Для создания целевых конструкций были использованы готовые минипрепы плазмид с целевыми генами pGNR-071-002 (ген ARSB), pGNR-071-001 (ген SUMF1) и вектора pGNR2 и pGNRl соответственно, с рестрикцией по сайтам HindIII+XbaI. Нужные фрагменты бэкбонов и вставок были очищены из агарозного геля на микроколонках по стандартному протоколу. Лигазные смеси трансформировали в компетентные клетки Е. coli XL10 Gold. После ночной инкубации на чашках Петри, скололи по 2 колонии для каждой конструкции и перенесли их в индивидуальные пробирки с 3 mL среды 2-YT+Amp.После ночной инкубации в шейкере по 2 мл каждой культуры использовали для очистки ДНК согласно стандартной процедуре на силикатных колонках. Полученные препараты валидировали аналитической рестрикцией. Во всех клонах размеры полученных фрагментов соответствовали теоретическим. Плазмиды наработали из 100 мл ночной культуры, перепроверили аналитической рестрикцией по сайтам PstI и линеаризовали по сайтам PvuI.
Пример 2. Оценка продуктивности клеточных линий в условиях batch-культивирования.
Для оценки влияния вспомогательного белка на продуктивность по арилсульфатазе В было получено 26 клеточных линий, ко-экспрессирующих целевой белок (ASB) и вспомогательный (FGE) и 4-е линии клеток, экспрессирующие только ASB. Анализ клеточных линий проводили в режиме batch: посевная плотность клеток 0,2×106 клеток/мл; температурный режим +37°С; скорость перемешивания 200 об/мин; содержание CO2 5%. Анализ полученных результатов во время batch культивирования показал, что ко-трансфекция двух генов положительно влияет на выход целевого белка ASB (фиг. 2).
Пример 3. Электрофорез в восстанавливающих условиях и вестерн-блот КЖ клеточных линий, экспрессирующих арилсульфатазу В с FGE и без вспомогательного белка.
Анализ клеточных линий, ко-экспрессирующих целевой белок (ASB) и вспомогательный (FGE) также проводили с помощью электрофореза в восстанавливающих условиях и вестерн-блота.
Для получения проб культуральной жидкости клеток, КЖ смешивали 1X денатурирующим буфером, содержащим 2-меркаптоэтанолом, после чего образцы прогревали 10 мин при +95°С. Для визуализации белков в SDS-PAGE добавлен трихлорэтанол.
Для получения проб лизатов клеток, суспензию клеток (1×106 клеток) осаждали центрифугированием при 1000 об/мин 5 минут, надосадок декантировали, осадок промывали в PBS в объеме равном удаленному надосадку. После промывки клетки осаждали центрифугированием при 1000 об/мин 5 минут, надосадок удаляли и клетки ресуспендировали в 100 мкл 1X денатурирующего буфера с 2-меркаптоэтанолом, после чего пробы прогревали 5 минут при +95°С.
Для вестерн-блот анализа белки из геля переносили на нитроцеллюлозную мембрану (Thermo Scientific, США). Затем переносили мембрану в блокировочный раствор (5% сухое обезжиренное молоко для микробиологии (Merck, Германия) в буфере PBS-Ta). Мембрану инкубировали 1 час; мембрану переносили в раствор первичных антител в течение 1 часа; отмывали мембрану в PBS-T 3 раза по 5 минут; мембрану переносили в раствор вторичных антител в течение 1 час; отмывали мембрану в PBS-T 4 раза по 5 минут; наносили проявочный раствор Western blotting detection Kit (Advanstal, США) на мембрану; использовали систему детекции ChemDoc (Bio-Rad, США) для визуализации мест специфического связывания белков. Для детекции арилсульфатазы В использовали в качестве первичных антител крысиные поликлональные антитела против галсульфазы (МБЦ «Генериум»), в качестве окрашивающих антител использовали кроличьи антитела против иммуноглобулинов крысы (p-RAQ-Iss, ИМТЕК, Россия). Для детекции продукта гена SUMF1 использовали кроличьи поликлональные антитела Anti-SUMFl (ab91479, Abeam, Англия), в качестве окрашивающих антител использовали кроличьи антитела к иммуноглобулинам козы (P-RAG Iss, ИМТЕК, Россия).
Электрофоретическая подвижность целевого белка ASB схожа с используемым стандартом - препаратом «Наглазим», однако, можно наблюдать более интенсивный сигнал в КЖ клеток, которые были ко-трансфицированы генами ARSB+SUMF1 (фиг. 4, треки 1-2), что подтверждает данные ИФА batch-культивирования (фиг. 2). Клетки, которые были трансфицированы одной плазмидой, содержащие ген ARSB, имеют очень слабый специфический сигнал (фиг. 4, треки 4-7).
Пример 4. Получение продукта гена SUMF1 - вспомогательного белка FGE. Экспрессия FGE в клетках с высокой продуктивностью арилсульфатазы В.
Белок FGE находится в эндоплазматическом ретикулуме. Молекулярный вес по аминокислотной последовательности FGE составляет 37-42 kD. Для проверки экспрессии вспомогательного белка FGE был проведен электрофорез в денатурирующих восстанавливающих условиях и вестерн-блот клеточных лизатов. На фиг. 5 справа можно наблюдать сигнал, полученный благодаря взаимодействию специфических антител к FGE. Все клеточные линии, которые имеют высокую продуктивность арилсульфатазы В (в среднем 50 мг/л) были ко-трансфицированы генами SUMF1 и ARSB и имеют вспомогательный белок FGE (фиг. 5, треки 1-3). Для сравнения были показаны клеточные линии без вспомогательного белка (фиг. 5, трек 5-8), для них специфического сигнала FGE не наблюдали. Полученные данные подтверждают вклад FGE в увеличение выхода арилсульфатазы В во время биосинтеза.
Пример 5. Супер-трансфекция геном SUMF1 низко продуктивных клеточных линий, экспрессирующих арилсульфатазы В.
После стабилизации пулов-трансфектантов в присутствии двух селективных антибиотиков клеточные линии были посеяны на batch культивирование. Продуктивность клеточных линий была измерена с помощью ИФА. В результате супер-трансфекции низко продуктивных клеточных линий CHO-K1-V и CHO-K1-GEN плазмидой, несущей ген SUMF1, удалось увеличить выход целевого продукта в 10 раз (фиг. 6).
Увеличение выхода целевого фермента низко продуктивной клеточной линии CHO-K1-GEN - 2G2 с помощью супер-трансфекции вспомогательным геном SUMF1 также подтверждается результатами (фиг. 8). На треках 1-2 были нанесены пробы КЖ клеточной линии 2G2+SUMF1, специфический сигнал виден отчетливо и сравним со стандартом в 5-м треке. Специфический сигнал к галсульфазе в КЖ клеточной линии 2G2 (4-й трек, фиг. 8), без вспомогательного гена SUMF1, не наблюдали, что связано с очень низкой продуктивностью клеточной линии без SUMF1. Для подтверждения наличия в клетках FGE с дополнительной супер-трансфекцией SUMF1, у клеток с большей продуктивностью, был проведен электрофорез и вестерн-блот лизатов клеточной линии 2G2 (фиг. 7). На мембране отчетливо можно наблюдать специфический сигнал к SUMF1, в клеточном лизате, у продуцентов с дополнительной трансфекцией геном SUMF1 (фиг. 7, треки 2-3). Это коррелирует с результатами электрофореза и вестерн-блота КЖ клеток (фиг. 7, треки 1-2). Таким образом, продуктивность арилсульфатазы В увеличилась после супер-трансфекции SUMF1.
--->
Список нуклеотидных последовательностей
SEQ ID#1 CMVe/EF1a pro – энхансер вируса CMV и промотор
транскрипции гена фактора элонгации альфа
CTAGTTATTAATAGTAATCAATTACGGGGTCATTAGTTCATAGCCCATATATGGAG
TTCCGCGTTACATAACTTACGGTAAATGGCCCGCCTGGCTGACCGCCCAACGACCCCCGC
CCATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAATAGGGACTTTCCATTGA
CGTCAATGGGTGGAGTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCAT
ATGCCAAGTACGCCCCCTATTGACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCC
CAGTACATGACCTTATGGGACTTTCCTACTTGGCAGTACATCTACGTATTAGTCATCGCT
ATTACCATGGGTGAGGCTCCGGTGCCCGTCAGTGGGCAGAGCGCACATCGCCCACAGTCC
CCGAGAAGTTGGGGGGAGGGGTCGGCAATTGAACCGGTGCCTAGAGAAGGTGGCGCGGGG
TAAACTGGGAAAGTGATGTCGTGTACTGGCTCCGCCTTTTTCCCGAGGGTGGGGGAGAAC
CGTATATAAGTGCAGTAGTCGCCGTGAACGTTCTTTTTCGCAACGGGTTTGCCGCCAGAA
CACAGGTAAGTGCCGTGTGTGGTTCCCGCGGGCCTGGCCTCTTTACGGGTTATGGCCCTT
GCGTGCCTTGAATTACTTCCACCTGGCTGCAGTACGTGATTCTTGATCCCGAGCTTCGGG
TTGGAAGTGGGTGGGAGAGTTCGAGGCCTTGCGCTTAAGGAGCCCCTTCGCCTCGTGCTT
GAGTTGAGGCCTGGCCTGGGCGCTGGGGCCGCCGCGTGCGAATCTGGTGGCACCTTCGCG
CCTGTCTCGCTGCTTTCGATAAGTCTCTAGCCATTTAAAATTTTTGATGACCTGCTGCGA
CGCTTTTTTTCTGGCAAGATAGTCTTGTAAATGCGGGCCAAGATCTGCACACTGGTATTT
CGGTTTTTGGGGCCGCGGGCGGCGACGGGGCCCGTGCGTCCCAGCGCACATGTTCGGCGA
GGCGGGGCCTGCGAGCGCGGCCACCGAGAATCGGACGGGGGTAGTCTCAAGCTGGCCGGC
CTGCTCTGGTGCCTGGCCTCGCGCCGCCGTGTATCGCCCCGCCCTGGGCGGCAAGGCTGG
CCCGGTCGGCACCAGTTGCGTGAGCGGAAAGATGGCCGCTTCCCGGCCCTGCTGCAGGGA
GCTCAAAATGGAGGACGCGGCGCTCGGGAGAGCGGGCGGGTGAGTCACCCACACAAAGGA
AAAGGGCCTTTCCGTCCTCAGCCGTCGCTTCATGTGACTCCACGGAGTACCGGGCGCCGT
CCAGGCACCTCGATTAGTTCTCGAGCTTTTGGAGTACGTCGTCTTTAGGTTGGGGGGAGG
GGTTTTATGCGATGGAGTTTCCCCACACTGAGTGGGTGGAGACTGAAGTTAGGCCAGCTT
GGCACTTGATGTAATTCTCCTTGGAATTTGCCCTTTTTGAGTTTGGATCTTGGTTCATTC
TCAAGCCTCAGACAGTGGTTCAAAGTTTTTTTCTTCCATTTCAGGTGTCGTGAGGAATTA
GCTTGGTACTAATACGACTC
SEQ ID#2 Galsulfase – ген арилсульфатазы B со стоп-кодоном
ATGGTGAGCCAGGCCCTGAGACT
GCTGTGCCTGCTGCTGGGCCTGCAGGGCTGCCTGGCCGCTGGCGCCAGCAGACCTCCCCA
CCTGGTGTTCCTGCTGGCCGACGACCTGGGCTGGAACGACGTGGGCTTCCACGGCAGCAG
AATCAGAACCCCCCACCTGGACGCCCTGGCTGCTGGAGGCGTGCTGCTGGACAACTACTA
CACCCAGCCCCTGTGCACCCCCAGCAGAAGCCAGCTCCTGACAGGAAGATACCAGATCAG
AACCGGCCTGCAGCACCAGATCATCTGGCCCTGCCAGCCCAGCTGCGTGCCTCTGGACGA
GAAGCTGCTGCCCCAGCTGCTGAAGGAGGCCGGCTACACCACCCACATGGTGGGCAAGTG
GCACCTGGGCATGTACAGAAAGGAGTGCCTGCCCACCAGAAGAGGCTTCGACACCTACTT
CGGCTACCTGCTGGGCAGCGAGGACTACTACAGCCACGAGAGATGCACCCTGATCGACGC
CCTGAACGTGACCAGATGTGCCCTGGACTTCAGAGATGGCGAGGAGGTGGCCACCGGCTA
CAAGAACATGTACAGCACCAACATCTTCACCAAGAGAGCCATCGCCCTGATCACCAACCA
CCCCCCCGAGAAGCCCCTGTTCCTGTACCTGGCCCTGCAGAGCGTGCACGAGCCCCTGCA
GGTGCCCGAGGAGTACCTGAAGCCCTACGACTTCATCCAGGACAAGAACAGACACCACTA
CGCCGGCATGGTGAGCCTGATGGACGAGGCCGTGGGCAACGTGACCGCCGCCCTGAAGTC
CAGCGGCCTGTGGAACAACACCGTGTTCATCTTCAGCACCGACAATGGCGGCCAGACCCT
GGCTGGAGGCAACAATTGGCCCCTGAGAGGCAGAAAGTGGAGCCTGTGGGAGGGCGGCGT
GAGAGGCGTGGGCTTCGTGGCCAGCCCCCTGCTGAAGCAGAAGGGCGTGAAGAACAGAGA
GCTGATCCACATCAGCGACTGGCTGCCCACCCTGGTGAAGCTGGCCAGAGGCCACACCAA
TGGCACCAAGCCCCTGGACGGCTTCGACGTGTGGAAGACCATCAGCGAGGGCAGCCCCAG
CCCCAGAATCGAGCTGCTGCACAACATCGACCCCAACTTCGTGGACAGCAGCCCCTGCCC
CAGAAACAGCATGGCCCCCGCCAAGGACGACAGCAGCCTGCCCGAGTACAGCGCCTTCAA
CACCAGCGTGCACGCCGCCATCAGACACGGCAACTGGAAGCTGCTGACCGGCTACCCCGG
CTGCGGCTACTGGTTCCCTCCTCCCAGCCAGTACAACGTGAGCGAGATCCCCAGCAGCGA
TCCCCCTACCAAGACCCTGTGGCTGTTCGACATCGACAGAGACCCCGAGGAGAGACACGA
CCTGAGCAGAGAGTACCCCCACATCGTGACCAAGCTGCTGAGCAGACTGCAGTTCTACCA
CAAGCACAGCGTGCCCGTGTACTTCCCCGCCCAGGACCCCAGATGCGACCCCAAGGCCAC
CGGCGTGTGGGGCCCTTGGATGTGA
SEQ ID#3 BGH pA - сигнал полиаденилирования бычьего
гормона роста
GCTCGCTGAT
CAGCCTCGACTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTT
CCTTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCAT
CGCATTGTCTGAGTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGG
GGGAGGATTGGGAAGACAATAGCAGGCATGCTGGG
SEQ ID#4 Amp – ген β-лактамазы
TTACCAATGCTTAATCAGTGAGGCACCTATCTCA
GCGATCTGTCTATTTCGTTCATCCATAGTTGCCTGACTCCCCGTCGTGTAGATAACTACG
ATACGGGAGGGCTTACCATCTGGCCCCAGTGCTGCAATGATACCGCGAGACCCACGCTCA
CCGGCTCCAGATTTATCAGCAATAAACCAGCCAGCCGGAAGGGCCGAGCGCAGAAGTGGT
CCTGCAACTTTATCCGCCTCCATCCAGTCTATTAATTGTTGCCGGGAAGCTAGAGTAAGT
AGTTCGCCAGTTAATAGTTTGCGCAACGTTGTTGCCATTGCTACAGGCATCGTGGTGTCA
CGCTCGTCGTTTGGTATGGCTTCATTCAGCTCCGGTTCCCAACGATCAAGGCGAGTTACA
TGATCCCCCATGTTGTGCAAAAAAGCGGTTAGCTCCTTCGGTCCTCCGATCGTTGTCAGA
AGTAAGTTGGCCGCAGTGTTATCACTCATGGTTATGGCAGCACTGCATAATTCTCTTACT
GTCATGCCATCCGTAAGATGCTTTTCTGTGACTGGTGAGTACTCAACCAAGTCATTCTGA
GAATAGTGTATGCGGCGACCGAGTTGCTCTTGCCCGGCGTCAATACGGGATAATACCGCG
CCACATAGCAGAACTTTAAAAGTGCTCATCATTGGAAAACGTTCTTCGGGGCGAAAACTC
TCAAGGATCTTACCGCTGTTGAGATCCAGTTCGATGTAACCCACTCGTGCACCCAACTGA
TCTTCAGCATCTTTTACTTTCACCAGCGTTTCTGGGTGAGCAAAAACAGGAAGGCAAAAT
GCCGCAAAAAAGGGAATAAGGGCGACACGGAAATGTTGAATACTCAT
SEQ ID#5 sv40 pro – промотор вируса sv40
TGCATCTCAATTAGTCA
GCAACCATAGTCCCGCCCCTAACTCCGCCCATCCCGCCCCTAACTCCGCCCAGTTCCGCC
CATTCTCCGCCCCATCGCTGACTAATTTTTTTTATTTATGCAGAGGCCGAGGCCGCCTCG
GCCTCTGAGCTATTCCAGAAGTAGTGAGGAGGCTTTTTTGGAGGCCTAGGCTTTTGCAAA
AAGCT
SEQ ID#6 puro - ген устойчивости к пуромицину
ATGACCGAGTACAAGCCCACGGTGCGCCTCGCCACCCGCGACGACGTC
CCCAGGGCCGTACGCACCCTCGCCGCCGCGTTCGCCGACTACCCCGCCACGCGCCACACC
GTCGATCCGGACCGCCACATCGAGCGGGTCACCGAGCTGCAAGAACTCTTCCTCACGCGC
GTCGGGCTCGACATCGGCAAGGTGTGGGTCGCGGACGACGGCGCCGCGGTGGCGGTCTGG
ACCACGCCGGAGAGCGTCGAAGCGGGGGCGGTGTTCGCCGAGATCGGCCCGCGCATGGCC
GAGTTGAGCGGTTCCCGGCTGGCCGCGCAGCAACAGATGGAAGGCCTCCTGGCGCCGCAC
SEQ ID#7 UCOE – универсальный элемент открытия хроматина
GGGCGGCCGCACGCGTGGCCCTCCGCGCCTACAGCTCAAGCCACATCCGAA
GGGGGAGGGAGCCGGGAGCTGCGCGCGGGGCCGCCGGGGGGAGGGGTGGCACCGCCCACG
CCGGGCGGCCACGAAGGGCGGGGCAGCGGGCGCGCGCGCGGCGGGGGGAGGGGCCGGCGC
CGCGCCCGCTGGGAATTGGGGCCCTAGGGGGAGGGCGGAGGCGCCGACGACCGCGGCACT
TACCGTTCGCGGCGTGGCGCCCGGTGGTCCCCAAGGGGAGGGAAGGGGGAGGCGGGGCGA
GGACAGTGACCGGAGTCTCCTCAGCGGTGGCTTTTCTGCTTGGCAGCCTCAGCGGCTGGC
GCCAAAACCGGACTCCGCCCACTTCCTCGCCCGCCGGTGCGAGGGTGTGGAATCCTCCAG
ACGCTGGGGGAGGGGGAGTTGGGAGCTTAAAAACTAGTACCCCTTTGGGACCACTTTCAG
CAGCGAACTCTCCTGTACACCAGGGGTCAGTTCCACAGACGCGGGCCAGGGGTGGGTCAT
TGCGGCGTGAACAATAATTTGACTAGAAGTTGATTCGGGTGTTTCCGGAAGGGGCCGAGT
CAATCCGCCGAGTTGGGGCACGGAAAACAAAAAGGGAAGGCTACTAAGATTTTTCTGGCG
GGGGTTATCATTGGCGTAACTGCAGGGACCACCTCCCGGGTTGAGGGGGCTGGATCTCCA
GGCTGCGGATTAAGCCCCTCCCGTCGGCGTTAATTTCAAACTGCGCGACGTTTCTCACCT
GCCTTCGCCAAGGCAGGGGCCGGGACCCTATTCCAAGAGGTAGTAACTAGCAGGACTCTA
GCCTTCCGCAATTCATTGAGCGCATTTACGGAAGTAACGTCGGGTACTGTCTCTGGCCGC
AAGGGTGGGAGGAGTACGCATTTGGCGTAAGGTGGGGCGTAGAGCCTTCCCGCCATTGGC
GGCGGATAGGGCGTTTACGCGACGGCCTGACGTAGCGGAAGACGCCTTAGTGGGGGGGAA
GGTTCTAGAAAAGCGGCGGCAGCGGCTCTAGCGGCAGTAGCAGCAGCGCCGGGTCCCGTG
CGGAGGTGCTCCTCGCAGAGTTGTTTCTCCAGCAGCGGCAGTTCTCACTACAGCGCCAGG
ACGAGTCCGGTTCGTGTTCGTCCGCGGAGATCTCTCTCATCTCGCTCGGCTGCGGGAAAT
CGGGCTGAAGCGACTGAGTCCGCGATGGAGGTAACGGGTTTGAAATCAATGAGTTATTGA
AAAGGGCATGGCGAGGCCGTTGGCGCCTCAGTGGAAGTCGGCCAGCCGCCTCCGTGGGAG
AGAGGCAGGAAATCGGACCAATTCAGTAGCAGTGGGGCTTAAGGTTTATGAACGGGGTCT
TGAGCGGAGGCCTGAGCGTACAAACAGCTTCCCCACCCTCAGCCTCCCGGCGCCATTTCC
CTTCACTGGGGGTGGGGGATGGGGAGCTTTCACATGGCGGACGCTGCCCCGCTGGGGTGA
AAGTGGGGCGCGGAGGCGGGACTTCTTATTCCCTTTCTAAAGCACGCTGCTTCGGGGGCC
ACGGCGTCTCCTCGGACGGCCGGGCGCGCCG
SEQ ID#8 SUMF – ген SUMF со стоп-кодоном
ATGGCCGCCCCTGCCCTGGGCCT
GGTGTGTGGCAGATGCCCTGAGCTGGGACTGGTGCTGCTCCTGCTGTTGCTCAGCCTGCT
GTGTGGCGCCGCCGGCAGCCAGGAGGCCGGCACCGGCGCTGGAGCCGGCAGCCTGGCCGG
CAGCTGCGGCTGTGGCACCCCCCAGAGACCTGGCGCCCACGGCAGCTCTGCCGCTGCCCA
CAGATACAGCAGAGAGGCCAATGCCCCTGGCCCTGTGCCTGGCGAGAGACAGCTGGCCCA
CAGCAAGATGGTGCCCATTCCTGCCGGCGTGTTCACCATGGGCACCGACGATCCTCAGAT
CAAGCAGGATGGCGAAGCCCCTGCCAGAAGAGTGACCATCGACGCCTTCTACATGGACGC
CTATGAGGTGAGCAACACCGAGTTCGAGAAGTTCGTGAATAGCACCGGCTACCTGACCGA
AGCCGAGAAGTTTGGCGACAGCTTCGTGTTCGAGGGCATGCTGAGCGAGCAGGTGAAGAC
CAACATCCAGCAGGCCGTGGCCGCTGCCCCTTGGTGGCTGCCCGTGAAAGGCGCCAATTG
GAGACACCCCGAGGGCCCCGACAGCACCATCCTGCACAGACCCGACCACCCTGTGCTGCA
CGTGAGCTGGAACGACGCCGTGGCCTACTGCACCTGGGCCGGCAAGAGACTGCCCACCGA
GGCCGAGTGGGAGTACAGCTGCAGAGGCGGCCTGCACAATAGACTGTTTCCCTGGGGCAA
CAAGCTGCAGCCCAAAGGCCAGCACTATGCCAATATCTGGCAGGGCGAGTTTCCCGTGAC
CAATACCGGCGAGGACGGCTTCCAGGGCACCGCCCCTGTGGACGCCTTCCCTCCCAATGG
CTATGGCCTGTACAACATTGTGGGCAATGCCTGGGAGTGGACCAGCGACTGGTGGACCGT
GCACCACAGCGTGGAGGAGACCCTGAATCCCAAAGGCCCTCCCAGCGGCAAGGACAGAGT
GAAGAAAGGCGGCAGCTACATGTGCCACAGAAGCTACTGCTACAGATACAGATGTGCCGC
CAGAAGCCAGAACACCCCCGACAGCAGCGCCAGCAATCTGGGCTTTAGATGCGCTGCCGA
CAGACTGCCCACCATGGACTGA
SEQ ID#9 Neo - ген устойчивости к неомицину
ATGATTGAACAAGATGGATTGCACGCAGGTTCTCCGGCCGCTTGGGTGGAG
AGGCTATTCGGCTATGACTGGGCACAACAGACAATCGGCTGCTCTGATGCCGCCGTGTTC
CGGCTGTCAGCGCAGGGGCGCCCGGTTCTTTTTGTCAAGACCGACCTGTCCGGTGCCCTG
AATGAACTGCAGGACGAGGCAGCGCGGCTATCGTGGCTGGCCACGACGGGCGTTCCTTGC
GCAGCTGTGCTCGACGTTGTCACTGAAGCGGGAAGGGACTGGCTGCTATTGGGCGAAGTG
CCGGGGCAGGATCTCCTGTCATCTCACCTTGCTCCTGCCGAGAAAGTATCCATCATGGCT
GATGCAATGCGGCGGCTGCATACGCTTGATCCGGCTACCTGCCCATTCGACCACCAAGCG
AAACATCGCATCGAGCGAGCACGTACTCGGATGGAAGCCGGTCTTGTCGATCAGGATGAT
CTGGACGAAGAGCATCAGGGGCTCGCGCCAGCCGAACTGTTCGCCAGGCTCAAGGCGCGC
ATGCCCGACGGCGAGGATCTCGTCGTGACACATGGCGATGCCTGCTTGCCGAATATCATG
GTGGAAAATGGCCGCTTTTCTGGATTCATCGACTGTGGCCGGCTGGGTGTGGCGGACCGC
TATCAGGACATAGCGTTGGCTACCCGTGATATTGCTGAAGAGCTTGGCGGCGAATGGGCT
GACCGCTTCCTCGTGCTTTACGGTATCGCCGCTCCCGATTCGCAGCGCATCGCCTTCTAT
CGCCTTCTTGACGAGTTCTTCTGA
<---
Реферат
Группа изобретений относится к биотехнологии. Предложена трансформированная клетка СНО-K1, не являющаяся дефектной по эндосомальному закислению, продуцирующая арилсульфатазу В с высоким выходом, достигаемым за счет ее коэкспрессии с рекомбинантным модифицирующим сульфатазу фактором 1 в составе экспрессирующего вектора под EF1a промотором. Раскрыта также векторная экспрессирующая конструкция для получения трансформированной клетки, которая по существу содержит последовательности: CMVe/EF1a pro, BGH рА, регуляторный элемент, повышающий экспрессию гена, и ген, кодирующий экспрессируемый белок, выбранный из ARSB и SUMF1. Изобретение может быть использовано для изготовления активных лизосомальных арилсульфатаз для терапии болезней лизосомального накопления путем фермент-заместительной терапии. 2 н. и 5 з.п. ф-лы, 8 ил., 1 табл., 5 пр.
Формула
Документы, цитированные в отчёте о поиске
Производство активной высокофосфорилированной n-ацетилгалактозамин-6-сульфатазы человека и ее применение
Комментарии