Регуляторные элементы растений и их применение - RU2675524C2
Код документа: RU2675524C2
Чертежи























































Описание
ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка заявляет приоритет по предварительной заявке США 61/785245, поданной 14 марта 2013 года, которая включена в данное описание посредством ссылки в полном объеме.
ВКЛЮЧЕНИЕ ПЕРЕЧНЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Перечень последовательностей, которые содержатся в файле с именем "MONS331WO.txt", размером 54,4 Кб (измерено в Microsoft Windows®), который был создан 12 марта 2014 года, подается при этом в электронной форме и включен в данный документ посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
[0003] Настоящее изобретение относится к области молекулярной биологии растений, генной инженерии растений и молекул ДНК, пригодных для модуляции экспрессии генов в растениях.
УРОВЕНЬ ТЕХНИКИ
[0004] Регуляторные элементы представляют собой генетические элементы, которые регулируют активность генов путем модуляции транскрипции функционально связанной транскрибируемой молекулы ДНК. Такие элементы включают промоторы, лидеры, интроны и 3'-нетранслируемые области и могут быть применены в области молекулярной биологии растений и генетической инженерии растений.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0005] В данном изобретении предложены новые регуляторные элементы для использования в растениях и генетические конструкции, содержащие регуляторные элементы. В данном изобретении также предложены трансгенные растения, клетки растений, части растений и семена, содержащие регуляторные элементы. В одном варианте реализации изобретение предлагает регуляторные элементы, описанные в данном документе, функционально связанные с транскрибируемой молекулой ДНК. В некоторых вариантах реализации изобретения транскрибируемая молекула ДНК является гетерологической по отношению к последовательности регуляторного элемента, предложенного в данном документе. В данном документе также предложены способы получения и применения регуляторных элементов, описанных в данном документе, включая генетические конструкции, содержащие регуляторные элементы, и трансгенные растения, клетки растений, части растений и семена, содержащие регуляторные элементы, функционально связанные с транскрибируемой молекулой ДНК, гетерологической по отношению к регуляторному элементу.
[0006] Таким образом, в одном аспекте в изобретении предложена рекомбинантная молекула ДНК, содержащая последовательность ДНК, выбранную из группы, состоящей из: а) последовательности ДНК с по меньшей мере около 85 процентами идентичности последовательности с любой из SEQ ID NOs: 1-20; б) последовательности ДНК, содержащей любую из SEQ ID NOs: 1-20; и в) фрагмента любой из SEQ ID NOs: 1-20, причем данный фрагмент обладает ген-регуляторной активностью; в котором последовательность ДНК функционально связана с гетерологической транскрибируемой молекулой ДНК. Под «гетерологической транскрибируемой молекулой ДНК» подразумевается, что транскрибируемая молекула ДНК является гетерологической по отношению к последовательности ДНК. В конкретных вариантах реализации изобретения рекомбинантная молекула ДНК содержит последовательность ДНК, имеющую по меньшей мере около 85 процентов, по меньшей мере около 86 процентов, по меньшей мере около 87 процентов, по меньшей мере около 88 процентов, по меньшей мере около 89 процентов, по меньшей мере около 90 процентов, по меньшей мере около 91 процента, по меньшей мере около 92 процентов, по меньшей мере около 93 процентов, по меньшей мере около 94 процентов, по меньшей мере около 95 процентов, по меньшей мере около 96 процентов, по меньшей мере около 97 процентов, по меньшей мере около 98 процентов или по меньшей мере 99 процентов идентичности последовательности с последовательностью ДНК любой из SEQ ID NOs: 1-20. В конкретных вариантах реализации изобретения гетерологическая транскрибируемая молекула ДНК содержит ген, который представляет агрономический интерес, такой как ген способный придавать растениям устойчивость к гербицидам или вредителям. В еще других вариантах реализации изобретения в изобретении предложена генетическая конструкция, содержащая рекомбинантную молекулу ДНК, как предложено в данном документе.
[0007] В другом аспекте в данном документе предлагаются трансгенные клетки растений, содержащие рекомбинантную молекулу ДНК, которая включает последовательность ДНК, выбранную из группы, состоящей из: а) последовательности ДНК с по меньшей мере около 85 процентами идентичности последовательности с любой из SEQ ID NOs: 1-20; б) последовательности ДНК, которая содержит любую из SEQ ID NOs: 1-20; и в) фрагмента любой из SEQ ID NOs: 1-20, причем данный фрагмент обладает ген-регуляторной активностью; при этом последовательность ДНК функционально связана с гетерологической транскрибируемой молекулой ДНК. В некоторых вариантах реализации изобретения трансгенная клетка растения представляет собой клетку однодольного растения. В других вариантах реализации изобретения трансгенная клетка растения представляет собой клетку двудольного растения.
[0008] В еще другом аспекте в данном документе ниже приведено трансгенное растение, или его часть, содержащее рекомбинантную молекулу ДНК, которая содержит последовательность ДНК, выбранную из группы, состоящей из: а) последовательности ДНК с по меньшей мере около 85 процентами идентичности последовательности с любой из SEQ ID NOs 1-20; б) последовательности ДНК, которая содержит любую из SEQ ID NOs: 1-20; и в) фрагмента любой из SEQ ID NOs: 1-20, причем данный фрагмент обладает ген-регуляторной активностью; при этом последовательность ДНК функционально связана с гетерологической транскрибируемой молекулой ДНК. В конкретных вариантах реализации изобретения трансгенное растение представляет собой дочернее растение любого поколения по отношению к исходному трансгенному растению и содержит молекулу рекомбинантной ДНК. Трансгенное семя, содержащее рекомбинантную молекулу ДНК, которое порождает такое трансгенное растение при выращивании, также предлагается в изобретении.
[0009] В другом аспекте в изобретении предложен способ получения товарного продукта из трансгенного растения, которое содержит рекомбинантную молекулу ДНК согласно настоящему изобретению. Товарная продукция согласно данному изобретению содержит определяемое количество SEQ ID NOs: 1-20. Используемый в данном документе термин "товарный продукт" относится к любой композиции или продукту, который состоит из материала, полученного из трансгенного растения, части растения, растительной клетки, или семени, содержащих рекомбинантную молекулу ДНК по изобретению. Товарная продукция, включает, но не ограничивается этим, обработанные семена, зерна, части растений и пищу. Трансгенные растения, которые содержат рекомбинантную молекулу ДНК согласно данному изобретению, могут быть использованы для изготовления любого товарного продукта, как правило, полученного из растения. Товарный продукт согласно данному изобретению будет содержать определяемое количество ДНК, которое соответствует рекомбинантной молекуле ДНК согласно данному изобретению. Обнаружение в образце этой одной или более рекомбинантных молекул ДНК может быть использовано для определения содержимого или источника товарного продукта. Любой стандартный способ обнаружения молекул ДНК может быть использован, включая способы детекции, описанные в данном документе.
[00010] Еще в одном аспекте в данном изобретении предложен способ экспрессии транскрибируемой молекулы ДНК, например, гена, представляющего агрономический интерес, в трансгенном растении путем получения трансгенного растения, которое содержит рекомбинантную молекулу ДНК согласно данному изобретению, и культивирования растения.
[00011] В данном документе также предложен способ получения трансгенного растения путем трансформации растительной клетки рекомбинантной молекулой ДНК согласно данному изобретению, чтобы получить трансформированную растительную клетку, и регенерирования трансформированной растительной клетки, чтобы получить трансгенное растение.
[00012] Также настоящим изобретением предложен модифицированный ко дон кишечной палочки (E.coli), кодирующей последовательности β-глюкуронидазы (GUS); причем модифицированный кодон GUS-кодирующей последовательности демонстрирует более высокую экспрессию в трансгенном растении, чем нативная GUS-кодирующая последовательность Е. coll. В одном варианте реализации изобретения модифицированный кодон GUS кодирующей последовательности может быть выбран из группы, состоящей из SEQ ID NOs: 29 и 30. Трансгенное растение может быть однодольным растением. В одном варианте реализации изобретения однодольное растение может быть выбрано из группы, которая включает кукурузу (Zea mays), рис (Oryza sativa), пшеницу (Triticum), ячмень (Hordeum vulgare), сорго (Sorghum spp.), просо, африканское просо (Pennisetum glaucum), даггус (Eleusine coracana), просо обыкновенное (Panicum miliaceum), щетинник итальянский (Setaria italica), овес (Avena sativa), тритикале, рожь (Secale cereale), росичку (Digitaria), лук (Allium spp.), ананас (Ananas spp.), газонную траву, сахарный тростник (Saccharum spp.), пальмы (Arecaceae), бамбук (Bambuseae), бананы (Musaceae), имбирные (Zingiberaceae), лилии (Lilium), нарциссы (Narcissus), ирисы (Iris), амариллисы, орхидеи (Orchidaceae), канны, колокольчики (Hyacinthoides) и тюльпаны (Tulipa). Трансгенные растения также могут быть двудольными растениями. В одном варианте реализации изобретения двудольные растения выбраны из группы, которая включает сою (Glycine max), дикую сою (Glycine soja), хлопок (Gossypium), томаты (Solanum Lycopersicum), перец (Piper), тыкву (Cucurbita), горох (Pisum sativum), люцерну (Medicago sativa), Medicago truncatula, фасоль (Phaseolus), нут (Cicer arietinum), подсолнечник (Helianthus annuus), картофель (Solanum tuberosum), арахис (Arachis hypogaea), кинву, гречиху посевную (Fagopyrum esculentum), рожковое дерево (Onia siliqua), свеклу (Beta vulgaris), шпинат (Spinacia olerácea) и огурец (Cucumis sativus).
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[00013] Фигуры 1а-1в иллюстрируют выравнивание между кодирующей последовательностью (CR-Ec.uidA-1:1:4, SEQ ID NO: 31) β-глюкуронидазы (GUS) нативной Ε. coli, и последовательностью модифицированных кодонов GUS кодирующей последовательности Е. coli (CR-Ec.uidA_nno-1:1:1, SEQ ID NO: 30). Одинаковые нуклеотиды при выравнивании указаны звездочкой.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[00014] SEQ ID NOs: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21 и 23 представляют собой промоторные последовательности.
[00015] SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22 и 24 представляют собой лидерные последовательности.
[00016] SEQ ID NO: 25-28 представляют собой последовательности праймеров амплификации.
[00017] SEQ ID NO: 29 и 30 представляют собой модифицированные кодоны GUS кодирующей последовательности. SEQ ID NO: 29 содержит вырезаемый интрон, а последовательность SEQ ID NO: 30 представляет собой непрерывную кодирующую последовательность.
[00018] SEQ ID NO: 31 представляет собой кодирующую последовательность β-глюкуронидазы нативной Escherichia coli.
[00019] SEQ ID NO: 32 представляет собой GUS кодирующую последовательность с вырезаемым интроном на основании SEQ ID NO: 31 β-глюкуронидазы нативной Ε. coli.
[00020] SEQ ID NO: 33, 39 и 40 представляет собой последовательность 3'-нетранслируемой области (3'-НТО).
[00021] SEQ ID NO: 34-37, 41 и 44 представляют собой последовательности групп транскрипционных регуляторных элементов (ЕХР), содержащие либо промоторную последовательность, функционально связанную 5'-лидерной последовательностью, которая функционально связана 5'-последовательностью интрона, либо в случае SEQ ID 44, промоторную последовательность, функционально связанную 5'-лидерной последовательностью.
[00022] SEQ ID NO: 38 представляет собой последовательность интрона.
[00023] SEQ ID NOs: 42 и 44 представляют собой последовательности, кодирующие белки люциферазы, полученные из Photinus pyralis и Renilla reniformis, соответственно.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[00024] Данное изобретение относится к молекулам ДНК, имеющим ген-регуляторную активность в растениях. Нуклеотидные последовательности этих молекул ДНК приводятся в SEQ ID NOs: 1-20. Эти молекулы ДНК, например, способны влиять на экспрессию функционально связанной транскрибируемой молекулы ДНК в тканях растений и, следовательно, регулировать экспрессию функционально связанного трансгена в трансгенных растениях. Изобретение также относится к способам их модификации, создания и применения. Данное изобретение также относится к композициям, которые содержат трансгенные клетки растений, растения, части растений и семена, содержащие рекомбинантные молекулы ДНК согласно данного изобретения, и к способам их получения и применения.
[00025] Следующие определения и способы предлагаются для того, чтобы лучше обозначить данное изобретение и направлять к специалистам в данной области техники при применении изобретения. Если не указано иное, термины следует понимать в соответствии с общепринятым использованием специалистами в данной области техники.
Молекулы ДНК
[00026] Используемый в данном документе термин «ДНК» или «молекула ДНК» относится к двухцепочечной молекуле ДНК клеточного или синтетического происхождения, т.е. к полимеру дезоксирибонуклеотидных оснований. Используемый в данном документе термин «последовательность ДНК» относится к нуклеотидной последовательности молекулы ДНК. Номенклатура, использованная в данном документе, соответствует таковой по разделу 37 Свода федеральных нормативных актов Соединенных Штатов § 1.822 и изложена в таблицах WIPO Standard ST.25 (1998), приложение 2, таблицы 1 и 3.
[00027] Как используется в данном документе «рекомбинантная молекула ДНК» представляет собой молекулу ДНК, содержащую комбинацию молекул ДНК, которые бы не объединились естественным путем без вмешательства человека. Например, рекомбинантная молекула ДНК может быть молекулой ДНК, которая состоит по меньшей мере из двух гетерологических по отношению друг к другу молекул ДНК; молекулой ДНК, содержащей последовательность ДНК, которая отклоняется от существующих в природе последовательностей ДНК; или молекулой ДНК, которая была встроена в ДНК клетки-хозяина путем генетической трансформации.
[00028] Как используется в данном документе, термин «идентичность последовательностей» относится к степени, в которой две оптимально выровненные последовательности ДНК являются идентичными. Оптимальное выравнивание последовательностей создается путем выравнивания двух последовательностей ДНК вручную, например, референсной последовательности и другой последовательности ДНК, для создания максимального количества соответствий между нуклеотидами при выравнивании последовательностей с соответствующими внутренними нуклеотидными инсерциями, делециями или разрывами. Используемый в данном документе термин «референсная последовательность» относится к последовательности ДНК, приводимой в SEQ ID NOs: 1-20.
[00029] Используемый в данном документе термин «процент идентичности последовательностей», или «процент идентичности», или «% идентичности» представляет собой долю идентичности, умноженную на 100. «Доля идентичности» для последовательности ДНК, оптимально выровненной с начальной последовательностью, представляет собой число соответствий между нуклеотидами при оптимальном выравнивании, деленное на общее число нуклеотидов в референсной последовательности, например, на общее число нуклеотидов во всей полноразмерной референсной последовательности. Таким образом, в одном варианте реализации данное изобретение относится к молекуле ДНК, содержащей последовательность ДНК, которая при оптимальном выравнивании с референсной последовательностью, представленной в данном документе как SEQ ID NOs: 1-20 имеет по меньшей мере около 85 процентов идентичности, по меньшей мере около 86 процентов идентичности, по меньшей мере около 87 процентов идентичности, по меньшей мере около 88 процентов идентичности, по меньшей мере около 89 процентов идентичности, по меньшей мере около 90 процентов идентичности, по меньшей мере около 91 процента идентичности, по меньшей мере около 92 процентов идентичности, по меньшей мере около 93 процентов идентичности, по меньшей мере около 94 процентов идентичности, по меньшей мере около 95 процентов идентичности, по меньшей мере около 96 процентов идентичности, по меньшей мере около 97 процентов идентичности, по меньшей мере около 98 процентов идентичности, по меньшей мере около 99 процентов идентичности или по меньшей мере около 100 процентов идентичности с референсной последовательностью.
Регуляторные элементы
[00030] Регуляторные элементы, такие как промоторы, лидеры, энхансеры, интроны и области терминации транскрипции (или 3' НТО), играют существенную роль во всеобщей экспрессии генов в живых клетках. Термин «регуляторный элемент», используемый в данном документе, относится к молекулам ДНК, имеющим ген-регуляторную активность. Термин «ген-регуляторная активность», используемый в данном документе, относится к способности влияния на экспрессию функционально связанной транскрибируемой молекулы ДНК, например, путем воздействия на транскрипцию и/или трансляцию функционально связанной транскрибируемой молекулы ДНК. Регуляторные элементы, такие как промоторы, лидеры, энхансеры и интроны, 3'-НТО, функционирующие в растениях, являются, таким образом, полезными для модификации фенотипов растений с помощью генетической инженерии.
[00031] Используемая в данном документе «группа регуляторных элементов экспрессии» или «ЕХР» последовательность может относиться к группе функционально связанных регуляторных элементов, таких как энхансеры, промоторы, лидеры и интроны. Таким образом, группа регуляторных элементов экспрессии может состоять, например, из промотора, функционально связанного с 5'-лидерной последовательностью, которая, в свою очередь, функционально связана с 5'-последовательностью интрона.
[00032] Регуляторные элементы могут быть охарактеризованы по своему характеру экспрессии генов, например, по положительным и/или отрицательным воздействиям, таким как конститутивная экспрессия или временная, пространственная, зависящая от стадии развития, тканевая, зависящая от окружения, физиологическая, патологическая, клеточного цикла и/или химически чувствительная экспрессия, и любая из их комбинаций, а также по количественным или качественным показателям. Используемый в данном документе термин «характер экспрессии гена» представляет собой любую особенность транскрипции функционально связанной молекулы ДНК в транскрибируемую молекулу РНК. Транскрибируемая молекула РНК может быть транслирована для получения молекулы белка или может образовать антисмысловую или другую регуляторную молекулу РНК, такую как двухцепочечная РНК (дцРНК), транспортная РНК (тРНК), рибосомальная РНК (рРНК), микроРНК (микроРНК) и тому подобные.
[00033] Используемый в данном документе термин «экспрессия белка» относится к любому паттерну трансляции транскрибируемой молекулы РНК в молекулу белка. Экспрессия белка может характеризоваться своими временными, пространственными, морфологическими свойствами или свойствами развития, а также количественными или качественными показателями.
[00034] Промотор является полезным в качестве регуляторного элемента для модуляции экспрессии функционально связанной транскрибируемой молекулы ДНК. Как используется в данном документе, термин «промотор» относится в общем смысле к молекуле ДНК, которая участвует в распознавании и связывании РНК-полимеразы II и других белков, таких как транс-действующие факторы транскрипции, для инициации транскрипции. Промотор может происходить из 5'-нетранслируемой области (5'-НТО) гена. В альтернативном варианте промоторы могут быть получены синтетическим путем или в результате манипулирования молекулой ДНК. Промоторы также могут быть химерными. Химерные промоторы получают путем слияния двух или более гетерологических молекул ДНК. Промоторы, полезные в практической реализации данного изобретения, включают SEQ ID NOs: 1, 3, 5, 7, 9, 11, 13, 15, 17 и 19, включая их фрагменты или варианты. В конкретных вариантах реализации данного изобретения такие молекулы ДНК и любые их варианты или производные, описанные в данном документе, далее определяются как обладающие промоторной активностью, т.е. способны действовать в качестве промотора в клетке хозяина, такого как трансгенное растение. Еще в дополнительных конкретных вариантах реализации изобретения фрагмент может быть определен как проявляющий промоторную активность, которой обладает исходная молекула промотора, из которого он был получен, или фрагмент может содержать «минимальный промотор», обеспечивающий основной уровень транскрипции и включающий TATA бокс или эквивалентную последовательность ДНК для распознавания и связывания комплекса РНК-полимеразы II с целью инициации транскрипции.
[00035] В одном варианте реализации данного изобретения фрагменты снабжены промоторной последовательностью, описанной в данном документе. Фрагменты промоторов могут включать промоторную активность, как описано выше, и могут быть полезными по отдельности или в комбинации с другими промоторами и фрагментами промоторов, например, при создании химерных промоторов. В конкретных вариантах реализации данного изобретения предлагаются фрагменты промотора, содержащие по меньшей мере около 50, по меньшей мере около 75, по меньшей мере около 95, по меньшей мере около 100, по меньшей мере около 125, по меньшей мере около 150, по меньшей мере около 175, по меньшей мере около 200, по меньшей мере около 225, по меньшей мере около 250, по меньшей мере около 275, по меньшей мере около 300, по меньшей мере около 500, по меньшей мере около 600, по меньшей мере около 700, по меньшей мере около 750, по меньшей мере около 800, по меньшей мере около 900 или по меньшей мере около 1000 непрерывных нуклеотидов, или больше, молекулы ДНК, имеющей промоторную активность, описанную в данном документе. Способы получения таких фрагментов из исходной молекулы промотора хорошо известны в данной области техники.
[00036] Композиции, полученные из любых промоторов, представленных в SEQ ID NOs: 1, 3, 5, 7, 9, 11, 13, 15, 17 и 19, такие как внутренние или 5'-делеции, например, могут быть получены с использованием хорошо известных в данной области техники способов для улучшения или изменения экспрессии, включая удаление элементов, имеющих положительные или отрицательные воздействия на экспрессию; удвоение элементов, имеющих положительные или отрицательные воздействия на экспрессию; и/или удвоение или удаление элементов, имеющих ткане- или клеточно-специфические воздействия на экспрессию. Композиции, полученные из любых промоторов, представленных в SEQ ID NOs: 1, 3, 5, 7, 9, 11, 13, 15, 17 и 19, содержащие 3'-делеции, в которых элемент ТАТА-бокса или его эквивалентная последовательность и нижележащая последовательность удалены, могут быть использованы, например, для создания энхансерных элементов. Дополнительные делеции могут быть внесены для удаления любых элементов, имеющих положительные или отрицательные, тканево-специфичные, клеточно-специфичные или время-специфичные (такие как, но не ограничиваясь этим, суточные ритмы) воздействия на экспрессию. Любой из промоторов, представленный в SEQ ID NOs: 1, 3, 5, 7, 9, 11, 13, 15, 17 и 19, и фрагменты или энхансеры, полученные из них, могут быть использованы для получения композиций химерных регуляторных элементов, содержащих любой из промоторов, представленных в SEQ ID NOs: 1, 3, 5, 7, 9, 11, 13, 15, 17 и 19, и фрагменты или энхансеры, полученные из них, функционально связанные с другими энхансерами и промоторами.
[00037] В соответствии с настоящим изобретением, промотор или фрагмент промотора могут быть проанализированы на наличие известных промоторных элементов, т.е. характеристик последовательности ДНК, таких как ТАТА-бокс и других известных мотивов сайтов связывания транскрипционных факторов. Идентификация таких известных промоторных элементов может быть использована специалистом в данной области техники для разработки вариантов промотора, имеющих подобный характер экспрессии, как и исходный промотор.
[00038] Используемый в данном документе термин «лидер» относится к молекуле ДНК из 5'-нетранслируемой области (5'-НТО) гена и определяется в общем смысле как сегмент ДНК между сайтом инициации транскрипции (СИТ) и начальным сайтом последовательности, кодирующей белок. В альтернативном варианте, лидеры могут быть получены синтетическим путем или в результате манипулирования элементами ДНК. Лидер может быть использован в качестве 5'-регуляторного элемента для модуляции экспрессии функционально связанной транскрибируемой молекулы ДНК. Молекулы лидеров могут быть использованы с гетерологическим промотором или с их нативным промотором. Молекулы промоторов по изобретению, таким образом, могут быть функционально связанными с их нативным лидером или могут быть функционально связанными с гетерологическим лидером. Лидеры, полезные в практической реализации изобретения, включают SEQ ID NOs: 2, 4, 6, 8, 10, 12, 14, 16, 18 и 20 или их фрагменты или варианты. В конкретных вариантах реализации данного изобретения такие последовательности ДНК могут быть определены как способные действовать в качестве лидера в клетке-хозяине, включая, например, трансгенную клетку растения. В одном варианте реализации настоящего изобретения такие последовательности ДНК могут быть расшифрованы как включающие лидерную активность.
[00039] Лидерные последовательности (5'-НТО), представленные в SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18 и 20, могут состоять из регуляторных элементов или могут принимать вторичные структуры, оказывающие влияние на транскрипцию или трансляцию функционально связанной молекулы ДНК. Лидерные последовательности, представленные в SEQ ID NOs: 2, 4, 6, 8, 10, 12, 14, 16, 18 и 20, могут быть использованы, в соответствии с данным изобретением, для создания химерных регуляторных элементов, которые влияют на транскрипцию или трансляцию функционально связанной молекулы ДНК. В дополнение, лидерные последовательности представлены в SEQ ID NOs: 2, 4, 6, 8, 10, 12, 14, 16, 18 и 20 могут быть использованы для создания химерных лидерных последовательностей, которые влияют на транскрипцию или трансляцию функционально связанной молекулы ДНК.
[00040] Используемый в данном документе термин «интрон» относится к молекуле ДНК, которая может быть выделена или идентифицирована из геномной копии гена и может быть определена в общем смысле как область, которая вырезается во время процессинга информационной РНК (мРНК) до трансляции. В альтернативном варианте интрон может быть получен синтетическим путем или в результате манипулирования элементом ДНК. Интрон может содержать энхансерные элементы, влияющие на транскрипцию функционально связанных генов. Интрон может быть использован в качестве регуляторного элемента для модуляции экспрессии функционально связанной транскрибируемой молекулы ДНК. Генетическая конструкция может содержать интрон, и интрон может быть или не быть гетерологическим по отношению к транскрибируемой молекуле ДНК. Примеры интронов в данной области техники включают интрон актина риса и интрон HSP70 кукурузы.
[00041] По сравнению с генетическими конструкциями без интрона, включение некоторых интронов в генетические конструкции приводит к увеличению накопления мРНК и белка в растениях. Этот эффект был назван как «интрон опосредованное усиление» (IME) экспрессии генов (Mascarenhas et al, Plant Mol. Biol. 15:913-920, 1990). Интроны, стимулирующие экспрессию в растениях, были идентифицированы в генах кукурузы (например, tubA1, Adh1, Sh1 и Ubi1), в генах риса (например, tpi) и в генах двудольных растений, таких как петуния (например, rbcS), картофель (например, st-ls1) и Arabidopsis thaliana (например, ubq3 и pat1). Было показано, что делеции или мутации в пределах сайтов сплайсинга интрона снижают экспрессию генов, указывая, что сплайсинг может быть необходим для IME. Однако сплайсинг сам по себе не требуется, так как IME было показано для двудольных растений, с помощью точечных мутаций в пределах сайтов сплайсинга pat1 гена из A. thaliana. Было показано, что многократное использование одного и того же интрона в одном растении проявляет недостатки. В таких случаях необходимо иметь коллекцию основных элементов контроля для создания соответствующих рекомбинантных элементов ДНК.
[00042] Используемый в данном документе термин «3'-молекула терминации транскрипции», «3'-нетранслируемая область» или «3'-НТО» относится к молекуле ДНК, которая используется во время транскрипции с нетранслируемой области 3'- части молекулы мРНК. 3'-нетранслируемая область молекулы мРНК может быть получена путем специфического расщепления и 3'-полиаденилированием, также известным как поли(А)-хвост. 3'-НТО может быть функционально связанной с транскрибируемой молекулой ДНК и располагаться ниже от нее, и может включать сигнал полиаденилирования и другие регуляторные сигналы, способные воздействовать на транскрипцию, процессинг мРНК или экспрессию генов. Полагают, что поли(А)-хвосты выполняют функции стабилизации мРНК и инициации трансляции. Примеры 3'- молекул терминации транскрипции в данной области техники представляют собой 3'- область нопалинсинтазы, 3'-область hspl7 пшеницы, 3' - область малой субъединицы РуБисКО гороха, 3'-область Е6 хлопка и 3'-НТО бусенника.
[00043] 3'-НТО, как правило, находят полезное применение для рекомбинантной экспрессии специфических молекул ДНК. Слабая 3'-НТО может генерировать сквозное пропитывание, что может повлиять на экспрессию молекулы ДНК, расположенной в соседних экспрессионных кассетах. Подходящий контроль для терминации транскрипции может предотвратить сквозное прочитывание в последовательностях ДНК (например, других экспрессионных кассет), локализованных ниже, и может дополнительно обеспечить эффективное повторное использование РНК-полимеразы, для улучшения экспрессии генов. Эффективная терминация транскрипции (освобождение РНК-полимеразы II от ДНК) является предпосылкой для следующей инициации транскрипции и, таким образом, непосредственно влияет на общий уровень транскрипта. После терминации транскрипции зрелая мРНК освобождается от места синтеза, и матрица транспортируется в цитоплазму. Эукариотические мРНК накапливаются в формах поли(А) in vivo, что затрудняет обнаружение сайтов терминации транскрипции общепринятыми методами. Однако предсказание функциональных и эффективных 3'-НТО с помощью биоинформатических методов затруднено, так как отсутствуют консервативные последовательности ДНК, позволяющие легко предсказывать эффективную 3'-НТО.
[00044] С практической точки зрения, как правило, полезно, чтобы 3'-НТО, использованная в экспрессионной кассете, обладала следующими характеристиками. 3'-НТО должна быть способной рационально и эффективно завершать транскрипцию ДНК и предотвращать сквозное прочитывание транскрипта в любой из соседних последовательностей ДНК, которая может содержаться в другой экспрессионной кассете, как и в случае нескольких экспрессионных кассет, принадлежащих одной трансферной ДНК (Т-ДНК), или соседней хромосомной ДНК, в которую Т-ДНК была встроена. 3'-НТО не должна вести к снижению транскрипционной активности, наделяемой промотором, лидером, энхансерами и нитронами, которые используются для контролирования экспрессии молекулы ДНК. В биотехнологии растений 3'-НТО часто используется для затравки реакций амплификации обратно транскрибируемой РНК, выделенной из трансформированных растений, и используется для: (1) оценки транскрипционной активности или экспрессии экспрессионной кассеты, интегрированной в хромосому растения; (2) оценки количества копий вставок в ДНК растения; и (3) оценки зиготности полученного семени после размножения. 3'-НТО также используется в реакциях амплификации ДНК, экстрагированной из трансформированных растений, для определения неиспользованной встроенной кассеты.
[00045] Используемый в данном документе термин "энхансер" или "энхансерный элемент" относится к цис-действующему регуляторному элементу, называемому цис-элемент, который придает аспект общего характера экспрессии, но, как правило, недостаточный сам по себе для контролирования транскрипции, функционально связанной последовательности ДНК. В отличие от промоторов, энхансерные элементы обычно не включают сайт инициации транскрипции (СИТ) или ТАТА-бокс, или эквивалентную последовательность ДНК. Промотор или фрагмент промотора может по природе содержать один или более энхансерных элементов, влияющих на транскрипцию функционально связанной последовательности ДНК. Энхансерный элемент также может быть объединен с промотором для получения химерного промоторного цис-элемента, придающего аспект общей модуляции экспрессии генов.
[00046] Полагают, что многие энхансерные элементы промотора связывают ДНК-связывающие белки и/или влияют на топологию ДНК, производя локальные конформации, которые избирательно позволяют или ограничивают доступ РНК-полимеразы к ДНК-матрице или которые облегчают избирательное открытие двойной спирали на участке инициации транскрипции. Энхансерный элемент также может выполнять функцию связывания транскрипционных факторов, регулирующих транскрипцию. Некоторые энхансерные элементы связывают более чем один фактор транскрипции, и транскрипционные факторы могут взаимодействовать с различной аффинностью с более чем одним доменом энхансера. Энхансерные элементы могут быть идентифицированы с помощью ряда методов, включая анализ делеций, т.е. удаления одного или более нуклеотидов с 5'-конца или внутри промотора; анализ ДНК-связывающего белка, используя футпринтинг ДНКазы I, интерференцию метилирования, анализы сдвига электрофоретической мобильности, in vivo геномный футпринтинг путем опосредованной лигированием полимеразной цепной реакции (ПЦР) и других общепринятых анализов; или путем анализа подобия последовательностей ДНК, используя известные мотивы цис-элементов или энхансерных элементов, как целевую последовательность или целевой мотив, с общепринятыми способами сравнения последовательностей ДНК, таких как BLAST. Тонкая структура домена энхансера может быть дополнительно исследована путем мутагенеза (или замещения) одного или более нуклеотидов или другими общепринятыми способами, известными в данной области техники. Энхансерные элементы могут быть получены путем химического синтеза или путем выделения из регуляторных элементов, которые включают такие элементы, и они могут быть синтезированы с дополнительными фланкирующими нуклеотидами, включающими применимые сайты рестрикции для облегчения последующих манипуляций. Таким образом, разработка, создание и использование энхансерных элементов, в соответствии со способами, описанными в данном документе, для модуляции экспрессии функционально связанных транскрибируемых молекул ДНК, охватываемых согласно данному изобретению.
[00047] Используемый в данном документе термин «химерный» относится к отдельной молекуле ДНК, полученной путем слияния первой молекулы ДНК со второй молекулой ДНК, причем ни первая, ни вторая молекула ДНК не будут по обыкновению содержаться в такой конфигурации, т.е. не будут объединены друг с другом. Химерная молекула ДНК представляет, таким образом, новую молекулу ДНК, иначе говоря, не содержащуюся обыкновенно в природе. Используемый в данном документе термин «химерный промотор» относится к промотору, полученному посредством такого манипулирования молекулами ДНК. Химерный промотор может объединять два или более фрагмента ДНК, например, слияние промотора с энхансерным элементом. Таким образом, разработка, создание и использование химерных промоторов, в соответствии со способами, описанными в данном документе, для модуляции экспрессии функционально связанных транскрибируемых полинуклеотидных молекул, охватываются согласно изобретению.
[00048] Используемый в данном документе термин «вариант» относится ко второй молекуле ДНК, такой как регуляторный элемент, которая аналогична по композиции, но не идентична первой молекуле ДНК, и причем вторая молекула ДНК все еще сохраняет общую функциональность, т.е. такой же или аналогичный характер экспрессии, например, через большую или меньшую, или эквивалентную транскрипционную или трансляционную активность по сравнению с первой молекулой ДНК. Вариант может быть укороченной или усеченной версией первой молекулы ДНК и/или измененной версией последовательности ДНК первой молекулы ДНК, такой как вариант с различными сайтами рестрикции и/или внутренними делециями, заменами и/или вставками. «Варианты» регуляторного элемента также охватывают варианты, возникающие в результате мутаций, которые происходят во время или в результате трансформации клеток бактерий и растений. В данном изобретении последовательность ДНК, приводимая в SEQ ID NOs: 1-20, может быть использована для создания вариантов, которые аналогичны по композиции, но не идентичны последовательности ДНК исходного регуляторного элемента, хотя все еще сохраняют общую функциональность, т.е. такой же или аналогичный характер экспрессии, как и исходный регуляторный элемент. Получение таких вариантов по изобретению находится в пределах обычной квалификации в данной области техники в свете описания и осуществлено в пределах объема данного изобретения.
[00049] Химерные регуляторные элементы могут быть разработаны для включения различных составных элементов, которые могут быть функционально связаны различными способами, известными в данной области техники, такими как расщепление ферментами рестрикции и лигирования, клонирование независимое от лигирования, модульная сборка продуктов ПЦР в процессе амплификации или прямой химический синтез регуляторного элемента, а также другими способами, известными в данной области техники. Полученные различные химерные регуляторные элементы могут состоять из тех же или вариантов тех же составных элементов, но отличатся по последовательности ДНК, или последовательностей ДНК, включающих связывающую последовательность ДНК, или последовательностей, которые позволяют составным частям быть функционально связанными. В данном изобретении последовательность ДНК, приводимая в SEQ ID NOs: 1-20, может обеспечить референсную последовательность регуляторного элемента, в которой составные элементы, включающие референсную последовательность, могут быть соединены способами, известными в данной области техники, и могут включать замены, делеции и/или вставки одного или более нуклеотидов или мутации, которые по обыкновению встречаются при трансформации клеток бактерий и растений.
[00050] Действенность модификаций, публикаций или делеций, описанных в данном документе, на желаемые аспекты экспрессии конкретного трансгена может быть проверена эмпирически в стабильных и транзиентных тестах растений, таких как те, которые описаны в рабочих примерах в данном документе, для того чтобы подтвердить результаты, которые могут варьировать в зависимости от произведенных изменений и цели изменения исходной молекулы ДНК.
Генетические конструкции
[00051] Используемый в данном документе термин «генетическая конструкция» означает любую рекомбинантную молекулу ДНК, такую как плазмида, космида, вирус, фаг или линейную, или кольцевую молекулу ДНК, или РНК, полученную из любого источника, способную к геномной интеграции или автономной репликации, включающую молекулу ДНК, причем по меньшей мере одна молекула ДНК связана с другой молекулой ДНК функционально действующим способом, т.е. они функционально связаны между собой. Используемый в данном документе термин «вектор» означает любую генетическую конструкцию, которая может быть использована для целей трансформации, т.е. введения гетерологической ДНК или РНК в клетку хозяина. Генетическая конструкция, как правило, включает одну или более экспрессионных кассет. Как используется в данном документе «экспрессионная кассета» относится к молекуле ДНК, содержащей по меньшей мере транскрибируемую молекулу ДНК, функционально связанную с одним или более регуляторных элементов, как правило, по меньшей мере промотором и 3'-НТО.
[00052] Используемый в данном документе термин «функционально связанный» относится к первой молекуле ДНК, соединенной со второй молекулой ДНК, причем первая и вторая молекулы ДНК расположены так, что первая молекула ДНК влияет на функцию второй молекулы ДНК. Две молекулы ДНК могут быть или не быть частью единой непрерывной молекулы ДНК и могут быть или не быть смежными. Например, промотор является функционально связанным с транскрибируемой молекулой ДНК, если промотор модулирует транскрипцию целевой транскрибируемой молекулы ДНК в клетке. Лидер, например, является функционально связанным с последовательностью ДНК, когда он способен воздействовать на транскрипцию или трансляцию последовательности ДНК.
[00053] Генетические конструкции по данному изобретению могут быть предложены в одном варианте реализации изобретения как генетические конструкции, состоящие из двух граничных областей опухолеиндуцирующей плазмиды, (Ti), имеющие области правого пограничного района (RB или AGR.tu.RB) и левого пограничного района (LB или AGRtu.LB) Ti-плазмиды, изолированной из Agrobacterium tumefaciens, содержащей Т-ДНК, которая, наряду с переносом молекул, который обеспечивается клетками A. tumefaciens, позволяет интеграцию Т-ДНК в геном клетки растения (см., например, патент США 6603061). Генетические конструкции также могут содержать сегменты плазмидного каркаса ДНК, обеспечивающие функцию репликации и селекцию на антибиотике в клетках бактерий, например, начало репликации Escherichia coli, такой как ori322, начало репликации для широкого спектра хозяев, такой как oriV или oriRi, и кодирующую область селективного маркера, такого как Spec/Strp, кодирующего Тп7 аминогликозид аденилтрансферазу (aadA), придающую устойчивость спектиномицину или стрептомицину, или селективный маркерный ген гентамицина (Gm, Gent). Для трансформации растений часто используют бактериальный штамм хозяина ABI, С58 или LBA4404 A. tumefaciens; однако другие штаммы, известные специалистам в области трансформации растений, могут функционировать в данном изобретении.
[00054] В данной области техники известны способы сборки и введения генетических конструкций в клетку таким образом, что транскрибируемая молекула ДНК транскрибируется в функциональную молекулу мРНК, которая транслируется и экспрессируется в виде белка. Для осуществления на практике настоящего изобретения, общепринятые композиции и способы получения и использования генетических конструкций и клеток хозяина хорошо известны специалисту в данной области техники. Типичные векторы, используемые для экспрессии нуклеиновых кислот в высших растениях хорошо известны в данной области техники и включают векторы, полученные из Ti-плазмиды Agrobacterium tumefaciens и pCaMVCN контрольного вектора для перенесения.
[00055] Различные регуляторные элементы могут быть включены в генетическую конструкцию, включая любой из тех, что предложен в данном документе. Любые такие регуляторные элементы могут быть предложены в сочетании с другими регуляторными элементами. Такие комбинации могут быть разработаны или модифицированы для получения желаемых регуляторных функций. В одном варианте реализации настоящего изобретения генетические конструкции по изобретению содержат по меньшей мере один регуляторный элемент, функционально связанный с транскрибируемой молекулой ДНК, функционально связанной с 3'-НТО.
[00056] Генетические конструкции по изобретению могут включать любой промотор или лидер, предложенные в данном документе или известные в данной области техники. Например, промотор по изобретению может быть функционально связан с гетерологическим нетранслируемым 5'-лидером, таким как лидер, происходящий от гена белка теплового шока. В альтернативном варианте лидер по изобретению может быть функционально связан с гетерологическим промотором, таким как транскрипционный промотор Cauliflower Mosaic Virus 35S.
[00057] Экспрессионные кассеты также могут включать кодирующую последовательность транзитного пептида, которая кодирует пептид, полезный для внутриклеточного направления функционально связанного белка, в частности, к хлоропласту, лейкопласту или другой пластидной органелле, митохондрии, пероксисоме, вакуоле или внеклеточному пространству. Многие белки, локализованные в хлоропластах, экспрессируются генами ядра как предшественники и направляются в хлоропласт с помощью транзитного пептида хлоропласта (ТПХ). Примеры таких белков, изолированных из хлоропластов, включают, но не ограничиваются этим, белки, ассоциированные с малой субъединицей (МСЕ) рибулозо-1,5,-бифосфат карбоксилазы, ферредоксин, ферредоксин оксидоредуктазу, белок I и белок II фотосенсибилизирующего комплекса, тиоредоксин F и энолпирувил шикимат фосфатсинтазу (ЭПШФС). Транзитные пептиды хлоропластов описаны, например, в патенте США 7193133. Было показано, что нехлоропластные белки могут быть направлены в хлоропласт при экспрессии гетерологического ТПХ, функционально связанного с трансгеном, кодирующим нехлоропластные белки.
Транскрибируемая молекула ДНК
[00058] Используемый в данном документе термин «транскрибируемая молекула ДНК» относится к любой молекуле ДНК, способной транскрибироваться в молекулу РНК, включая, но не ограничиваясь этим, те, которые имеют последовательности, кодирующие белок, и те, которые производят молекулы РНК, имеющие последовательности, полезные для супрессии гена. Тип молекулы ДНК может включать, но не ограничивается этими, молекулу ДНК из того же растения, молекулу ДНК из другого растения, молекулу ДНК из другого организма или синтетическую молекулу ДНК, такую как молекула ДНК, содержащая антисмысловую последовательность гена, или молекула ДНК, кодирующая искусственную, синтетическую или иным образом модифицированную версию трансгена. Типовая транскрибируемая молекула ДНК для включения в генетические конструкции по изобретению включает, например, молекулы ДНК или гены из другого вида, чем тот, в который молекула ДНК встроена, или гены, происходящие из или присутствующие в том же виде, но которые включены в клетки-реципиенты способами генной инженерии, а не классическими методами размножения.
[00059] «Трансген» относится к транскрибируемой молекуле ДНК, гетерологической для клетки-хозяина по меньшей мере в отношении ее расположения в геноме клетки-хозяина и/или транскрибируемой молекуле ДНК, искусственно включенной в геном клетки-хозяина в текущем или любом предыдущем поколении клеток.
[00060] Регуляторный элемент, такой как промотор согласно данному изобретению, может быть функционально связанным с транскрибируемой молекулой ДНК, которая гетерологична по отношению к регуляторному элементу. Используемый в данном документе термин «гетерологический» относится к комбинации из двух или более молекул ДНК, когда такая комбинация по обыкновению не встречается в природе. Например, две молекулы ДНК могут быть получены из разных видов и/или две молекулы ДНК могут быть получены из разных генов, например, разных генов одного и того же вида или же одних и тех же генов из разных видов. Регуляторный элемент является, таким образом, гетерологическим по отношению к функционально связанной транскрибируемой молекуле ДНК, если такая комбинация по обыкновению не встречается в природе, т.е. транскрибируемая молекула ДНК по обыкновению не встречается функционально связанной с регуляторным элементом.
[00061] Транскрибируемая молекула ДНК в общем смысле может быть любой молекулой ДНК, для которой желательна экспрессия транскрипта. Такая экспрессия транскрипта может приводить к трансляции полученной молекулы мРНК и, соответственно, к экспрессии белка. В альтернативном варианте, например, транскрибируемая молекула ДНК может быть разработана, чтобы в итоге может вызвать уменьшение экспрессии конкретного гена или белка. В одном варианте реализации настоящего изобретения это может быть достигнуто с помощью транскрибируемой молекулы ДНК, ориентированной в антисмысловом направлении. Специалист в данной области техники хорошо знаком с использованием такой антисмысловой технологии. Любой ген может негативно регулироваться таким способом, и в одном варианте реализации настоящего изобретения транскрибируемая молекула ДНК может быть разработана для супрессии конкретного гена через экспрессию молекулы дцРНК, миРНК или микроРНК.
[00062] Таким образом, в одном варианте реализации настоящего изобретения представляется рекомбинантная молекула ДНК, содержащая регуляторный элемент по изобретению, такой как те, что предложены в SEQ ID NOs: 1-20, функционально связанный с гетерологической транскрибируемой молекулой ДНК для того, чтобы модулировать транскрипцию транскрибируемой молекулы ДНК на желаемом уровне или по желаемому характеру, когда генетическая конструкция интегрирована в геном клетки трансгенного растения. В одном варианте реализаций настоящего изобретения транскрибируемая молекула ДНК содержит область гена, кодирующую белок, а в другом варианте реализации данного изобретения транскрибируемая молекула ДНК содержит антисмысловую область гена.
Гены, которые представляют агрономический интерес
[00063] Транскрибируемая молекула ДНК может быть геном, представляющим агрономический интерес. Используемый в данном документе термин «ген, представляющий агрономический интерес» относится к транскрибируемой молекуле ДНК, которая при экспрессии в конкретной ткани растений, клетке или типе клетки придает желаемую характеристику. Продукт гена, представляющего агрономический интерес, может действовать внутри растения для того, чтобы влиять на морфологию растения, физиологию, рост, развитие, урожайность, композицию зерна, питательный профиль, устойчивость к болезням или вредителям и/или толерантность к факторам окружающей среды или химическим факторам, или может действовать в качестве пестицидного агента в рационе вредителя, который питается растением. В одном варианте реализации данного изобретения регуляторный элемент по изобретению включен в генетическую конструкцию таким образом, что регуляторный элемент функционально связан с транскрибируемой молекулой ДНК, которая является геном, представляющим агрономический интерес. В трансгенном растении, содержащем такую генетическую конструкцию, экспрессия гена, представляющего агрономический интерес, может придать полезную агрономическую черту. Полезная агрономическая черта может включать, но не ограничиваться этими, например, толерантность к гербицидам, борьбу с насекомыми, модифицирование урожайности, устойчивость к болезням, устойчивость к патогену, модифицирование роста и развития растения, модифицированное содержание крахмала, модифицированное содержание масла, модифицированное содержание жирных кислот, модифицированное содержание белка, модифицированное созревание плодов, улучшенное питание животных и человека, производства биополимеров, устойчивость к стрессовым факторам окружающей среды, фармацевтические пептиды, улучшенные свойства для переработки, улучшение вкуса, полезное производство гибридных семян, улучшенное производство волокна и желательное производство биотоплива.
[00064] Примеры генов, представляющих агрономический интерес, известных в данной области техники, включают гены для устойчивости к гербицидам (патенты США №№6803501, 6448476; 6248876; 6225114; 6107549; 5866775; 5804425; 5633435 и 5463175), повышения урожайности (патенты США №№USRE38446, 6716474; 6663906; 6476295; 6441277; 6423828; 6399330; 6372211; 6235971; 6222098 и 5716837), борьбы с насекомыми (патенты США №№6809078; 6713063; 6686452; 6657046; 6645497; 6642030; 6639054; 6620988; 6593293; 6555655; 6538109; 6537756; 6521442; 6501009; 6468523; 6326351; 6313378; 6284949; 6281016; 6248,536; 6242241; 6221649; 6177615; 6156573; 6153814; 6110464; 6093695; 6063756; 6063597; 6023013; 5959091; 5942664; 5942658; 5880275; 5763245 и 5763241), устойчивости к грибковым заболеваниям (патенты США №№6653280; 6573361; 6506962; 6316407; 6215048; 5516671; 5773696; 6121436; 6316407 и 6506962), устойчивости к вирусам (патенты США №№6617496; 6608241; 6015940; 6013864; 5850023 и 5304730), устойчивости к нематоде (патент США №6228992), устойчивости к бактериальным заболеваниям (патент США №5516671), роста и развития растений (патенты США №№6723897 и 6518488), накопления крахмала (патенты США №№6538181; 6538179; 6538178; 5750876; 6476295), производства модифицированных масел (патенты №№США 6444876; 6426447 и 6380462), повышенной продукции масел (патенты США №№6495739; 5608149; 6483008 и 6476295), модифицированного содержания жирных кислот (патенты США №№6828475; 6822141; 6770465; 6706950; 6660849; 6596538; 6589767; 6537750; 6489461 и 6459018), повышенной продукции белков (патент США №6380466), созревания плодов (патент США №5512466), улучшенного питания животных и человека (патенты США №№6723837; 6653530; 6541259; 5985605 и 6171640), биополимеров (патенты США №№USRE37543 6228623 и 5958745 и 6946588), устойчивости к стрессовым факторам окружающей среды (патент США №6072103), фармацевтических пептидов и секретируемых пептидов (патенты США №№6812379; 6774283; 6140075 и 6080560), улучшенных свойств для переработки (патент США №6476295), улучшенной усвояемости (патент США №6531648), низкого содержания рафинозы (патент США №6166292), промышленного производства фермента (патент США №5543576), улучшенного вкуса (патент США №6011199), фиксации азота (патент США №5229114), производства гибридных семян (патент США №5689041), производства волокна (патенты США №№6576818; 6271443, 5981834 и 5869720) и производства биотоплива (патент США №5998700).
[00065] В альтернативном варианте ген, представляющий агрономический интерес, может повлиять на вышеуказанные характеристики растения или фенотипы, посредством кодирования молекулы РНК, которая причиняет направленную модуляцию экспрессии эндогенного гена, например, через антисмысловые (см., например, патент США 5107065), ингибиторные РНК («РНК-интерференции», включающие модуляцию экспрессии генов с помощью микроРНК-, миРНК-, транс-действующих миРНК- и фазовых малых РНК-опосредованных механизмов, например, как описано в опубликованных заявках США 2006/0200878 и 2008/0066206 и в заявке на патент США 11/974469); или косупрессорно-опосредованные механизмы. РНК также может быть каталитической молекулой РНК (например, рибозимом или рибосвитчем, см., например, опубликованную заявку США 2006/0200878), разработанной для отщепления желаемого эндогенного продукта мРНК. В данной области техники известны способы для разработки и введения генетических конструкций в клетку, таким образом, что транскрибируемая молекула ДНК транскрибируется в молекулу, способную вызвать супрессию гена.
[00066] Экспрессия транскрибируемой молекулы ДНК в клетке растения также может быть использована для супрессии вредителей растений, питающихся клетками растения, например, композиции, выделенные из жесткокрылых насекомых - вредителей, и композиций, выделенных из нематодных вредителей. Вредители растений включают, но не ограничиваются этими, членистоногих вредителей, нематодных вредителей и грибковых или микробных вредителей.
Селективные маркеры
[00067] Селективные маркерные трансгены также могут быть использованы с регуляторными элементами по изобретению. Используемый в данном документе термин «селективный маркерный трансген» относится к любой транскрибируемой молекуле ДНК, чья экспрессия в трансгенном растении, ткани или клетке, или отсутствие таковых, может быть подвергнута скринингу или подсчитана каким-нибудь способом. Селективные маркерные гены и, связанные с ними, методы отбора и скрининга, предназначены для использования на практике изобретения, известны в данной области техники и включают, но не ограничиваются этим, транскрибируемые молекулы ДНК, кодирующие бета-глюкуронидазу (GUS), зеленый флуоресцентный белок (GFP), белки, придающие устойчивость к антибиотикам, и белки, придающие устойчивость к гербицидам.
β-глюкуронидаза
[00068] Ген β-глюкуронидазы (GUS), который выделен из Escherichia coli К-12, представляет собой один из наиболее широко используемых генов для отчетов по биотехнологии растений. Ген GUS Е. coli, uidA, представляет собой часть оперона GUS на бактериальной хромосоме. Его индуцируют широким спектром β-D-глюкуронидов. Фермент GUS представляет собой экзогидролазу, которая катализирует гидролиз β-D-глюкуронидов на D-глюкуроновую кислоту и агликон. Е. coli живет в пищеварительном тракте позвоночных, включая человека. Позвоночные используют путь глюкуронизации для детоксикации ксенобиотиков и эндогенных соединений, таких как отходы, стероиды, алифатические спирты, фенолы, карбоновые кислоты, сахара и другие различные метаболиты. Глюкуронидация вовлекает в конъюгирование с D-глюкуроновой кислотой. Это происходит главным образом в печени, но также происходит в других тканях и органах, таких как почки, надпочечники, и желудочно-кишечный тракт.Глюкуроновая кислота, может быть использована E.coli, в качестве основного источника углерода и энергии. Следовательно, белок GUS E.coli предоставляет средство, с помощью которого бактерии могут разрушать продукты путем глюкуронизации в пищеварительном тракте позвоночных с получением глюкуроновой кислоты в качестве источника углерода и энергии. Агликоны, которые также высвобождаются GUS ферментом, как правило, не разлагаются бактериями, но используется в качестве трансфера для D-глюкуроновой кислоты (Gilissen et ai, Transgenic Research, 7: 157-163, 1998).
[00069] Использование гена β-глюкуронидазы Ε. coli в качестве репортера было впервые описано Jefferson et al. (Proc. Natl. Acad. Sci., 83: 8447-8451, 1986) и был использован во многом таким же образом, как впервые описано с момента его введения. Ген GUS используется для мониторинга экспрессии генов растений и часто используется для описания промоторов или других экспрессионных элементов. Тем не менее, некоторые растительные промоторы экспрессируют на очень низком уровне, и могут быть недетектируемыми с использованием анализа на основе GUS. Эти более слабоэкспрессирующие промоторы могут быть полезны для развития трансгенньгх культур с желаемыми признаками, такими как повышение урожайности.
[00070] В самом начале развития трансгенных растений сельскохозяйственных культур самыми желаемыми были промоторы, которые обеспечивали высокую конститутивную экспрессию. Такие конститутивные промоторы, которые были получены из геномов вирусов растений, таких как вирус мозаики цветной капусты и вирус мозаики норичника, были использованы для контроля трансгенов, на которые возложена толерантность к гербицидам или устойчивость к насекомым. В области биотехнологии растений по своей сложности увеличивается разработка новых трансгенных черт, которые требуют более специфических паттернов экспрессии или более низких уровней экспрессии. Избыточная экспрессия в неподходящих растительных тканях может привести к нежелательным эффектам в трансформированном растении. Например, смещенная экспрессия (экспрессия гена в аномальном месте в организме) генов ферментов в растениях может привести к уменьшению желаемого конечного продукта из-за нехватки предшественника в точке разветвления метаболического пути (Iwase et al., Plant Biotech. 26: 29-38, 2009).
[00071] Так как транскрипционные факторы (TFs), в норме, выступают в качестве основных регуляторов клеточных процессов, они, как ожидается, будут отлично подходить для модификации сложных признаков культурных растений, и TF-технология, вероятно, будут заметной частью следующего поколения эффективных биотехнологических культур. TF-технологии часто требуют оптимизации, как снижения нежелательных побочных эффектов, таких как задержка роста, так и повышение желаемых признаков до уровня, при котором они имеют коммерческую ценность. Оптимизация часто достигается изменением экспрессии трансгена TF. Тканеспецифичные, этапо-специфичные, или индуцируемые промоторы, а не обычные конститутивных промоторы, могут быть использованы для ограничения экспрессии трансгена в соответствующие ткани или при условиях окружающей среды (Century et al, Plant Physiology, 147: 20-29, 2008).
[00072] Отчасти из-за этих изменений, существует необходимость в более чувствительном анализе характера экспрессии элементов для идентификации элементов экспрессии, которые обеспечивают требуемый уровень и характер экспрессии. Настоящее изобретение предлагает улучшение модифицированного кодона GUS-кодирующей последовательности, функционально связанной с промотором, которые экспрессирует лучше, чем GUS-кодирующая последовательность нативной Е. coli, используемой обычно в данной области техники. Улучшенный модифицированный кодон GUS-кодирующей последовательности может быть использован для обеспечения большей чувствительности качественного и количественного анализа и позволяет характеризовать промоторы и другие элементы экспрессии, которые невозможны с GUS-кодирующей последовательностью нативной Е. coli. Улучшенная модифицированная кодон GUS кодирующая последовательность может быть использована для характеристики элементов экспрессии в однодольных и двудольных растениях. Однодольные растения, используемые в практической реализации данного изобретения, включают, но не ограничиваются ими, кукурузу (Zea mays), рис (Oryza sativa), пшеницу (Triticum), ячмень (Hordeum vulgaré), сорго (Sorghum spp.), просо, африканское просо (Pennisetum glaucum), даггус (Eleusine coracana), просо обыкновенное (Panicum miliaceum), щетинник итальянский (Setaria italica), овес (Avena sativa), тритикале, рожь (Secale cereale), росичку (Digitaria), лук (Allium spp.), ананас (Ananas spp.), газонную траву, сахарный тростник (Saccharum spp.), пальмы (Arecaceae), бамбук (Bambuseae), бананы (Musaceae), имбирные (Zingiberaceae), лилии (Lilium), нарциссы (Narcissus), ирисы (Iris), амариллисы, орхидеи (Orchidaceae), канны, колокольчики (Hyacinthoides) и тюльпаны (Tulipa). Двудольные растения, используемые в практической реализации изобретения, включают, но не ограничиваются ими, сою (Glycine max), дикую сою (Glycine soja), хлопок (Gossypium), томаты (Solanum Lycopersicum), перец (Piper), тыкву (Cucurbita), горох (Pisum sativum), люцерну (Medicago sativa), Medicago truncatula, фасоль (Phaseolus), нут (Cicer arietinum), подсолнечник (Helianthus annuus), картофель (Solanum tuberosum), арахис (Arachis hypogaea), кинву, гречиху посевную (Fagopyrum esculentum), рожковое дерево (Onia siliqua), свеклу (Beta vulgaris), шпинат (Spinacia olerácea) и огурец (Cucumis sativus).
Трансформация клеток
[00073] Изобретение также направляется к способу получения трансформированных клеток и растений, содержащих один или более регуляторных элементов, функционально связанных с транскрибируемой молекулой ДНК.
[00074] Термин «трансформация» относится к введению молекулы ДНК в реципиентного хозяина. Используемый в данном документе термин «хозяин» относится к бактериям, грибам или растениям, включая любые клетки, ткани, органы, или к потомству бактерий, грибов или растений. Ткани и клетки растений, представляющие собой особый интерес, включают протопласты, каллус, корни, клубни, семена, стебли, листья, проростки, завязи и пыльцу.
[00075] Используемый в данном документе термин «трансформированный» относится к клетке, ткани, органу или организму, в который была введена молекула чужеродной ДНК, такая как генетическая конструкция. Введенная молекула ДНК может быть интегрирована в геномную ДНК клетки, ткани, органа или организма реципиента так, что введенная молекула ДНК наследуется в дальнейшем потомстве. «Трансгенная» или «трансформированная» клетка или организм также может включать потомство клетки или организма и потомство, полученное в результате программы размножения с использованием такого трансгенного организма в качестве родителя при скрещивании и проявляющее измененный фенотип как результат присутствия чужеродных молекул ДНК. Введенная молекула ДНК также может быть временно введенной в клетку-реципиент так, что введенная молекула ДНК не наследуется в дальнейшем потомстве. Термин «трансгенный» относится к бактерии, грибу или растению, содержащим одну или более гетерологических молекул ДНК.
[00076] Существует много способов, хорошо известных специалистам в данной области техники, для введения молекулы ДНК в клетки растений. Процесс, как правило, включает этапы выбора подходящей клетки-хозяина, трансформации клетки-хозяина вектором и получения трансформированной клетки-хозяина. Способы и материалы для трансформации клеток растений, посредством введения растительной генетической конструкции в геном растений, в реализации на практике изобретения могут включать любой из известных и показанных способов. Подходящие способы включают, но не ограничиваются этими, бактериальную инфекцию (например, Agrobacterium), бинарные БИХ векторы, прямой перенос ДНК (например, ПЭГ-опосредованную трансформацию, поглощение ДНК, опосредованное высушиванием/ингибированием, электропорацию, перемешивание с волокнами карбида кремния и ускорение частиц, покрытых ДНК) из числа других.
[00077] Клетки-хозяева могут представлять собой любую клетку или организм, например, клетку растения, клетку водоросли, водоросли, клетку гриба, грибы, бактериальную клетку или клетку насекомых. В конкретных вариантах реализации изобретения клетки-хозяева и трансформированные клетки могут включать клетки культурных растений.
[00078] Трансгенное растение впоследствии может быть регенерировано из клетки трансгенного растения по изобретению. С использованием общепринятых методов размножения или самоопыления можно получить семена из этого трансгенного растения. Такое семя и полученное в результате растение-потомок, выращенное из такого семени, будут содержать рекомбинантную молекулу ДНК согласно изобретению, и, следовательно, будут трансгенными.
[00079] Трансгенные растения по изобретению могут быть самоопылены, для предоставления семян гомозиготных трансгенных растений по изобретению (гомозиготных по рекомбинантной молекуле ДНК), или скрещены с нетрансгенными растениями или другими трансгенными растениями, для предоставления семян гомозиготных трансгенных растений по изобретению (гомозиготных по рекомбинантной молекуле ДНК). Как гомозиготные, так и гетерозиготные трансгенные растения упоминаются в данном документе как «растения-потомки». Растения потомки представляют собой трансгенные растения, произошедшие от исходного трансгенного растения и содержащие рекомбинантную молекулу ДНК по изобретению. Семена, полученные с использованием трансгенного растения по изобретению, могут быть собраны и использованы для выращивания поколений трансгенных растений, т.е. растений-потомков по изобретению, включающих генетическую конструкцию настоящего изобретения и экспрессию гена, представляющего агрономический интерес. Описания способов размножения, которые обычно используются для различных культур, могут быть найдены в одной из нескольких книжных ссылок, см., например, Allard, Principles of Plant Breeding, John Wiley & Sons, NY, U. of CA, Davis, CA, 50-98 (1960); Simmonds, Principles of Crop Improvement, Longman, Inc., NY, 369-399 (1979); Sneep and Hendriksen, Plant breeding Perspectives, Wageningen (ed), Center for Agricultural Publishing and Documentation (1979); Fehr, Soybeans: Improvement, Production and Uses, 2nd Edition, Monograph, 16:249 (1987); Fehr, Principles of Variety Development, Theory and Technique, (Vol.1) и Crop Species Soybean (Vol.2), Iowa State Univ., Macmillan Pub. Co., NY, 360-376 (1987).
[00080] Трансформированные растения могут быть проанализированы на наличие целевого гена или генов и по уровню экспрессии и/или профилю, предоставленному регуляторными элементами по изобретению. Специалистам в данной области техники известны многочисленные способы, доступные для анализа трансформированных растений. Например, способы анализа растений включают, но не ограничиваются ими, саузерн-блоттинги или нозерн-блоттинги, подходы на основе ПЦР, биохимические анализы, способы фенотипического скрининга, полевые апробации и иммунодиагностические анализы. Экспрессия транскрибируемой молекулы ДНК может быть измерена с использованием реагентов TaqMan® (Applied Biosystems, Foster City, CA) и по способам, описанным производителем, и по количеству циклов ПЦР, определенному с использованием TaqMan® Testing Matrix. В альтернативном варианте реагенты Invader® (Third Wave Technologies, Madison, WI) и способы, описанные производителем, могут быть использованы для оценки экспрессии трансгена.
[00081] Изобретение также относится к частям растения по изобретению. Части растений включают листья, стебли, корни, клубни, семена, эндосперм, семязачаток и пыльцу, но не ограничиваются этим. Части растений по изобретению могут быть жизнеспособными, нежизнеспособными, способными и/или не способными к регенерации. В изобретении также включены и предложены трансформированные клетки растения, содержащие молекулу ДНК по изобретению. Трансформированные клетки или клетки трансгенного растения по изобретению включают клетки способные и/или не способные к регенерации.
[00082] Изобретение может быть более легко понятным посредством ссылки на следующие примеры, которые предложены в качестве иллюстрации, и не подразумевают ограничения изобретения, если это не установлено. Специалистам в данной области техники должно быть понятно, что способы, описанные в следующих примерах, представляют собой способы, обнаруженные авторами изобретения, для нормального функционирования при применении изобретения. Однако специалистам в данной области техники должно быть понятно, в свете настоящего описания, что многие изменения могут быть совершены в конкретных описанных вариантах реализации изобретения, и по-прежнему получится похожий или аналогичный результат без отступления от сущности и объема изобретения, поэтому все имеющее значение размещается далее или показывается в прилагаемых графических материалах и должно быть интерпретировано в иллюстративном, а не в ограничивающем смысле.
ПРИМЕРЫ
Пример 1
Идентификация и клонирование регуляторных элементов
[00083] Новые RCc3 промоторы и лидеры были идентифицированы и клонированы из геномной ДНК однодольных видов: бусенника обыкновенного (Coix lacryma-jobi), росички кроваво-красной (Digitaria sanguinalis (L.) Scop.), мискантуса китайского (Miscanthus sinensis f. gracillimus), трипсакума двудомного (Tripsacum dactyloides) и сахарного тросника (Saccharum offwinarum). Белок RCc3 принадлежит надсемейству проламина, которое получило свое название от спирторастворимого пролина и богатого на глютамин белка зерновых. Надсемейство проламина (также называемое семейством ингибитор протеазы/липидо-транспортирующего белка/семян 2S альбумина; Pfam ID: PF00234) представляет собой одно из самых распространенных над семейств белков в геноме растения. Членами надсемейства проламина изобилуют фрукты, орехи, семена и овощи у разнообразных растений. Они, как известно, проявляют разнообразные функции: хранение и защита семян, связывание или передача липидов, и ингибирование ферментов. Липидо-транспортирующие белки (LTPs) принадлежат к надсемейству проламинов и экспрессируются в различных растительных тканях. Белок риса RCc3 является LTP, что экспрессируется в корнях риса, хотя и не все белки LTP корень-специфичны.
[00084] Праймеры амплификации ДНК (представленные как SEQ ID NO: 25-28) были разработаны с использованием кодирующих последовательностей двадцати четырех (24) LTP белков из Zea mays, Oryza sativa, Sorghum bicolor и Brachypoium distachyon. Праймеры амплификации были использованы с GenomeWalker™ (Clontech Laboratories, Inc, Mountain View, CA) библиотеками, построенными для клонирования 5' области соответствующей последовательности геномной ДНК.
[00085] Биоинформационный анализ был проведен, чтобы определить регуляторные элементы внутри амплифицированной ДНК. Используя результаты этого анализа, были определены регуляторные элементы в последовательности ДНК и праймеры для амплификации регуляторных элементов. Соответствующую молекулу ДНК по каждому регуляторному элементу амплифицировали с использованием стандартных условий полимеразной цепной реакции (ПЦР) с применением праймеров, содержащих уникальные сайты рестрикции фермента и геномной ДНК, выделенной из С. lacryma-jobi, D. sanguinalis (L.) Scop., M. sinensis f. gracillimus, T. dactyloides и S. officinarum. Полученные фрагменты ДНК были лигированы в векторы и секвенированы.
[00086] Последовательности ДНК идентифицированных промоторов и лидеров RCc3 перечислены в таблице 1. Промоторные последовательности приведены в данном документе в качестве SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17 и 19. Лидерные последовательности представлены в данном документе как SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18 и 20.


Пример 2
Анализ регуляторных элементов, контролирующих GUS в трансгенной кукурузе
[00087] Растения кукурузы были преобразованы с использованием векторов, специальных бинарных плазмидных конструкций, содержащих промотор RCc3, функционально связанный с его нативной лидерной последовательностью RCc3, управляющей экспрессией трансгенной (GUS) β-глюкуронидазы. Полученные трансформированные растения анализировали на GUS-экспрессию белка.
[00088] Векторы, используемые в данных экспериментах, были сконструированы с использованием способов клонирования, известных в данной области техники. Полученные векторы, содержащие правый пограничный участок от A. tumefaciens; первую трансгенную экспрессионную кассету для анализа промоторной/лидерной последовательности RCc3, функционально связанную с ко доном переработанной кодирующей последовательностью GUS, обладающий смысловым интроном GOI-Ec.uidA+St.LSl.nno:3 (SEQ ID NO: 29), функционально связанным 5'-концом с 3'-НТО из гена S-аденозилметионин синтетазы 1 проса итальянского (T-SETit.Amsl-1: 1:1, SEQ ID NO: 159); вторую трансгенную экспрессионную кассету, используемую для отбора трансформированных растительных клеток, что проявляют устойчивость к глифосату (управляемую промотором Actin 1 риса); и левый пограничный участок от A. tumefaciens. Полученные плазмиды были использованы для трансформации растений кукурузы с использованием методов, известных в данной области техники. Экспрессия GUS, обусловленная новыми промоторами и лидерами RCc3, была сравнена с экспрессией, управляемой гомологами промоторов и лидеров RCc3 проса итальянского и риса. В таблице 2 приведены плазмидные конструкции, промоторные и лидерные последовательности RCc3, и SEQ ID NOs.

[00089] В некоторых случаях растения были преобразованы с помощью агробактериальных способов трансформации, известных в данной области техники и описанных в заявке на патент США 2009/0138985.
[00090] Гистохимический анализ GUS был использован для качественного анализа экспрессии генов в трансформированных растениях. Образцы тканей инкубировали с GUS-окрашивающим раствором X-Gluc (5-бромо-4-хлоро-3-индолил-L-глюкуронид) (1 мг/мл) в течение соответствующего периода времени, промывали и визуально исследовали на наличие синего окрашивания. GUS активность качественно определяется путем прямого визуального осмотра или осмотра под микроскопом, используя выбранные органы и ткани растений. Ro растения были проверены на экспрессию в корнях и листьях, а также пыльниках, пестиках, и развивающихся семенах и эмбрионах через 21 день после опыления (21 ДПО).
[00091] Для количественного анализа общий белок экстрагировали из выбранных тканей трансформированных растений кукурузы. Один микрограмм общего белка был использован с флюорогенным субстратом 4-митилумбрелиферил-β-D-глюкуронид (МУГ) в общем реакционном объеме 50 мкл. Продукт реакции, 4-митилумбрелиферил (4-МУ), максимально флуоресцирует при высоком рН, когда гидроксильная группа ионизирована. Добавление щелочного раствора карбоната натрия одновременно останавливает анализ и регулирует рН для подсчета флуоресцентного продукта. Флуоресценцию измеряли с возбуждением при 365 нм, эмиссией при 445 нм, используя Fluoromax-3 (Horiba; Киото, Япония) с Micromax Reader, с шириной щели, установленной на возбуждение 2 нм, 3 нм на излучение. Средние значения экспрессии были предоставлены в пмоль/мкг 4МУ белка/час.
[00092] Среднее значение экспрессии R0 GUS для каждого преобразования было записано, и средний уровень экспрессии и стандартную ошибку определяли на основании измерений образцов, полученных из нескольких событий трансформации.
Пример 3
Энхансеры, полученные из регулярных элементов
[00093] Энхансеры могут быть получены из промоторных элементов, представленных в данном изобретении, таких как SEQ ID NOs: 1, 3, 5, 7, 9, 11, 13, 15, 17 и 19. Эти энхансерные элементы могут состоять из одного или нескольких цис-регуляторных элементов, которые, когда функционально связанны с 5' или с 3' промотором элемента или функционально связанны с 5' или с 3' дополнительных элементов энхансера, которые функционально связаны с промотором, может повысить или модулировать экспрессию транскрибируемой молекулы ДНК, или предоставить экспрессию транскрибируемой молекулы ДНК в определенном типе клеток или органе растения, или в конкретной временной точке развития или циркадного ритма. Энхансеры изготовлены путем удаления ТАТА-бокса или функционально подобных элементов и любой последующей последовательности ДНК из промоторов, которые позволяют транскрибировать промоторы, представленные в данном документе, как описано выше, в том числе их фрагменты, в которых ТАТА-бокс или функционально аналогичные элементы и последовательности ДНК, расположенные после ТАТА-бокса, удаляются.
[00094] Энхансерные элементы могут быть получены из промоторных элементов, представленных в данном изобретении, и клонированы с использованием способов, известных в данной области техники, будут функционально связаны 5'-концом или 3'-концом с промоторным элементом или функционально связаны 5'-концом или 3'-концом с дополнительными энхансерными элементами,, которые функционально связаны с промотором. Энхансерные элементы могут быть клонированы, чтобы быть функционально связанными 5'-концом или 3'-концом с промоторным элементом, полученным от организма того же или другого рода; или функционально связанным 5'-концом или 3'-концом с дополнительными энхансерными элементами, полученными от организма того же или другого рода; или функционально связанными с промотором, полученным от организма того же или другого рода, полученного с использованием химерного регуляторного элемента. Вектор экспрессии GUS может быть сконструирован с использованием способов, известных в данной области техники, подобных конструкциям, описанным в предыдущих примерах, в которых в результате векторы экспрессии растений содержат правый пограничный участок от A. tumefaciens; первую трансгенную экспрессионную кассету для регуляторного или химерного регуляторного элемента, состоящую из регуляторного или химерного регуляторного элемента, функционально связанного с интроном, полученным из белка теплового шока HSP70 Ζ mays (I-Zm.DnaK-1:1:1, SEQ ID NO: 38) или любой из интронов, представленных в данном документе, или любой другой интрон, функционально связанный с последовательностью, кодирующей GUS, который либо обладает процессируемым интроном (GUS-2, SEQ ID NO: 32), либо не обладает интроном (CR-EC.uidA-1: 1: 4 (GUS.nat), SEQ ID NO: 31), функционально связанным с нопалинсинтазным 3'-НТО из A. tumefaciens (T-AGRtu.nos-1: 1:13, SEQ ID NO: 39), либо обладает 3'-НТО из гена белка-переносчика липидов риса (Т-Os.LTP-1: 1: 1, SEQ ID NO: 40); вторую трансгенную экспрессионную кассету, используемую для отбора трансформированных растительных клеток, что проявляют устойчивость к глифосату (управляемую промотором Actin 1 риса) или же, в альтернативном варианте, к антибиотику канамицину (управляемую промотором Actin 1 риса); и левый пограничный участок от A. tumefaciens. Полученные плазмиды могут быть использованы для трансформации растений кукурузы или растений других родов путем описанных выше способов или других способов, известных в данной области. В качестве альтернативы, протопласты, полученные из кукурузы или растений других родов, могут быть трансформированы с использованием способов, известных в данной области техники, для проведения временных тестов.
[00095] Экспрессия GUS управляемая регуляторными элементами, содержащими один или более энхансер может быть оценена в стабильных или временных тестах растений, чтобы определить влияние энхансерного элемента на экспрессию трансгена. Модификации одного или более трансгенных элементов или дупликация одного или более энхансерных элементов может выполняться на основании эмпирических экспериментов и в результате конечной регуляции экспрессии генов, наблюдаемой с использованием композиции каждого регулирующего элемента. Изменение относительных положений одного или нескольких энхансеров в результате регулирующего или химерного регуляторного элемента может влиять на активность транскрипции или специфичность регулирующего или химерного регуляторного элемента и определяется эмпирически, с целью определения наилучших энхансеров для требуемого профиля экспрессии трансгена для кукурузы или растений другого рода.
Пример 4
Увеличение аналитической чувствительности при использовании кодон-модифицированной β-глюкоронидазы (GUS)
[00096] Промоторы растений часто экспрессируют на уровнях, которые ниже нормального порога обнаружения многих количественных анализов, но их характеристики экспрессии могут быть очень ценными для экспрессии определенных трансгенов. Ранее в биотехнологии растений промоторы, которые давали высокую конститутивную экспрессию являлись желательными и были использованы для регуляции транскрипции молекул ДНК, которые производятся конкретной признак, требующий высокой конститутивной экспрессии, такой как устойчивость к гербицидам или устойчивость к насекомым. Эти высококонститутивные промоторы часто создаются из геномов вирусов растений, а не геномов растений, например, промоутеры 35S, полученные из вируса мозаики цветной капусты и вирус мозаики норичника. Примечательно, что в некоторых случаях, высокая конститутивная экспрессия генов определенных транскрибируемы молекул ДНК может привести к негативным последствиям, таким как сайленсинг трансгенов off-phenotypes или yield drag. Например, высокий уровень экспрессии гена GUS в трансгенных растениях сахарного тростник,а в которых используются два различных промотора убиквитина, полученные из сахарного тростника, а также из кукурузы, приводит к посттранскрипционному сайленсингу гена GUS (Wei et al., J. Plant Physiol. 160: 1241-1251, 2003).
[00097] Кроме того, в последнее время необходимы промоторы, которые демонстрируют конкретные паттерны экспрессии или повышают экспрессию в конкретных тканях растения. Например, эктопическая экспрессия генов ферментов в растениях может привести к снижению желаемого конечного продукта из-за нехватки предшественника в точке разветвления метаболического пути (Iwase et al., Plant Biotech. 26:29-38, 2009). В этих случаях желательно использовать промотор, который экспрессирует функционально связанную к транскрибируемую молекулу ДНК в правильных типах тканей или клеток, или в определенном окне развития. Промоторы, полученные из геномов растений, часто демонстрируют экспрессию в конкретных клетках, тканях или на необходимом этапе. Из-за более низких уровней экспрессии этих растительных промоторов, анализы экспрессия часто требуют использования энхансеров, для повышения уровня экспрессии, чтобы обеспечить обнаружение в количественном анализе. Тем не менее, использование таких энхансеров часто изменяется общий характер экспрессии промотора растений.
[00098] Повышение экспрессии гена-репортера, используемого в анализе, устраняет необходимость в усилении промотора растительного происхождения и, таким образом, обеспечивает более точную оценку характера экспрессии, предоставляемой промотором. Этот пример демонстрирует использование модифицированного кодона GUS кодирующей последовательности, чтобы улучшить количественную чувствительность анализа при исследовании нескольких различных ЕХР, состоящих из промоторной последовательности, функционально связанной 5'-концом с лидерной последовательностью, функционально связанной 5'-концом с последовательностью интрона.
[00099] Растения кукурузы были трансформированы с использованием растительных векторов, содержащих ЕХР последовательности, управляющие экспрессией как родного трансгена β-глюкоронидазы (GUS) Escherichia coli, так и кодон-модифицированного трансгена β-глюкуронидазы (GUS.nno), и полученные растения были проанализированы на экспрессию белка GUS. Кодирующие последовательности ЕХР и GUS клонировали в бинарной плазмиды конструкций с использованием способов, известных в данной области техники.
[000100] Полученные экспрессионные растительные конструкции содержат правый пограничный участок от A. tumefaciens; первую трансгенную для анализа чувствительности двух последовательностей, кодирующих GUS, состоящих из ЕХР, функционально связанной с любой нативной последовательностью Е. Coli, кодирующей GUS (CPv-Ec.uidA-1:1:4 (GUS.nat), SEQ ID NO: 31) или кодон-модифицированной последовательностью, кодирующей GUS (CR-Ec.uidA_nno-1:1:1 (GUS.nno), SEQ ID NO: 30), функционально связанную 5'-концом с 3'-НТО гена белка-переносчика липидов риса (T-Os.LTP-1:1:1, SEQ ID NO: 40); вторую трансгенную экспрессионную кассету, используемую для отбора трансформированных растительных клеток, что проявляют устойчивость к глифосату (управляемую промотором Actin 1 риса); и левый пограничный участок от A. tumefaciens. Фиг. 1А-1С иллюстрируют выравнивание между нативной последовательностью, кодирующей GUS (CR-Ec.uidA-1:1:4), и кодон-модифицированной последовательностью, кодирующей GUS (CR-Ec.uidA_nno-1:1:1). Идентичные нуклеотиды в выравнивании отмечены звездочкой. Кодон-модифицированная последовательность, кодирующая GUS, на 77,9% идентична нативной последовательности, кодирующей GUS, и была разработана для лучшей экспрессии в растении.
[000101] Три (3) различных класса ЕХР были использованы, при этом каждый проявлял специфический паттерн экспрессии. EXP EXP-SETit.Cab3+Zm.DnaK: 1:1 (SEQ ID NO: 34) и EXP-SETit.Cab3+Zm.DnaK: 1:2 (SEQ ID NO: 35) проявляли профиль экспрессии в листьях кукурузы и по существу идентичны, за исключением вставки в пять нуклеотидов 5'-CCGGA-3' в позиции 1408-1412 EXP-SETit.Cab3+Zm.DnaK: 1:2. Последовательность EXP EXP-CaMV.35S-enh+Os.Rcc3+Zm.DnaK:1:5 (SEQ ID NO: 36) обеспечивает повышенный профиль экспрессии в корнях кукурузы. Последовательность EXP EXP-Zm.UbqM1:1:2 (SEQ ID NO: 37) обеспечивает высококонститутивный профиль экспрессии у кукурузы. Полученные плазмиды были использованы для трансформации растений кукурузы с использованием способов, известных в данной области техники. В таблице 3 приведены обозначения плазмидных конструктов и соответствующие последовательности ЕХР и GUS.

[000102] В некоторых случаях растения были трансформированы с помощью агробактериалъных способов трансформации, известных в данной области техники и описанных в заявке на патент США 2009/0138985 публикации.
[000103] Гистохимический анализ GUS был использован для качественного анализа экспрессии генов в трансформированных растениях. Образцы тканей инкубировали с GUS-окрашивающим раствором X-Gluc (5-бром-4-хлор-3-индолил-b-глюкуронид) (1 мг/мл) в течение соответствующего периода времени, промывали и визуально исследовали на наличие синего окрашивания. GUS активность качественно определяется путем прямого визуального осмотра или осмотра под микроскопом, используя выбранные органы и ткани растений. R0 растения были проверены на экспрессию в корнях и листьях, а также пыльниках, пестиках, и развивающихся семенах и эмбрионах через 21 день после опыления (21 ДПО).
[000104] Для количественного анализа общий белок экстрагировали из выбранных тканей трансформированных растений кукурузы. Один микрограмм общего белка был использован с флюорогенным субстратом 4-метилумбеллиферил-β-D-глюкуронид (МУГ) в общем реакционном объеме 50 мкл. Продукт реакции, 4-метилумбеллиферил (4-МУ), максимально флуоресцирует при высоком рН, когда гидроксильная группа ионизирована. Добавление щелочного раствора карбоната натрия одновременно останавливает анализ и регулирует рН для подсчета флуоресцентного продукта. Флуоресценцию измеряли с возбуждением при 365 нм, эмиссией при 445 нм, используя Fluoromax-3 (Horiba; Киото, Япония) с Micromax Reader, с шириной щели, установленной на возбуждение 2 нм, 3 нм на излучение.
[000105] Среднее значение экспрессии GUS для поколения трансформантов R0 представлено в таблицах 4, 5 и 6.

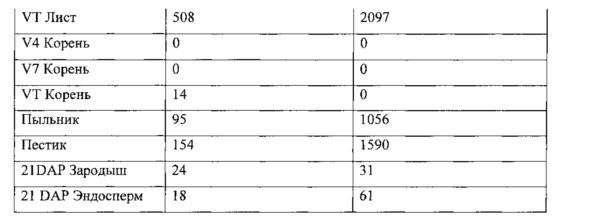
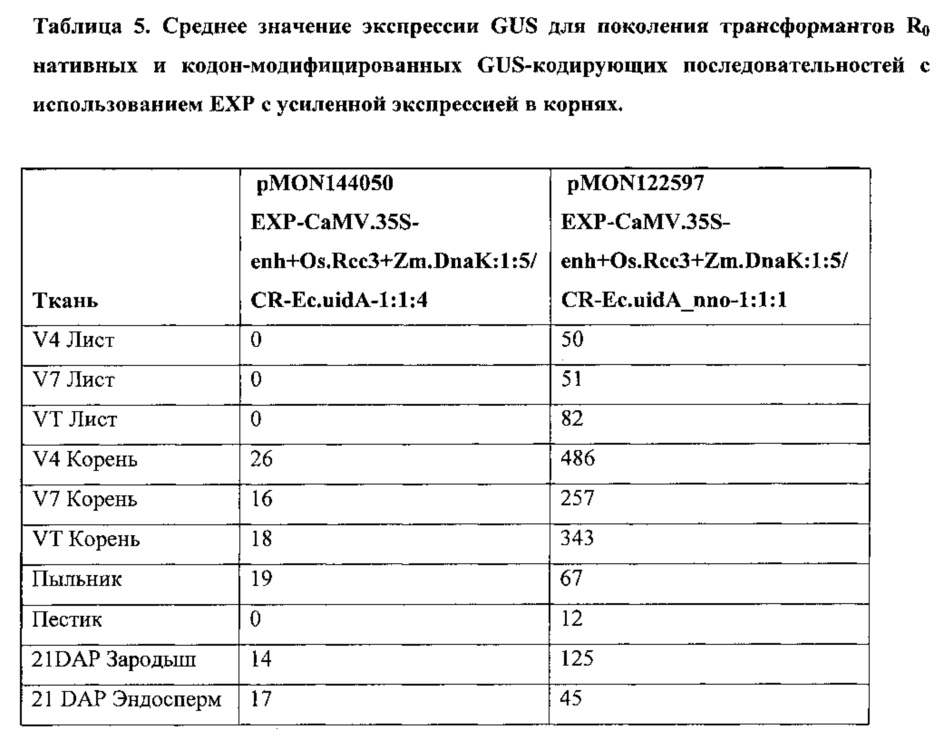


[000106] Как видно из таблиц 4, 5 и 6, существует возможность повысить чувствительность количественного анализа с использованием кодон-модифицированной GUS-кодирующей последовательности по сравнению с нативной кодирующей GUS последовательностью. Некоторые изменения между популяциями GUS.rmo и GUS.nat следует ожидать, поскольку экспрессия может зависеть от вставки участков Т-ДНК; однако общая тенденция чувствительности показывает гораздо больше чувствительности, при использовании GUS.rmo. GUS, управляемый EXP-SETit.Cab3 + Zm.DnaK: 1:1 (SEQ ID NO: 34) и EXP-SETit.Cab3 + Zm.DnaK: 1:2 (SEQ ID NO: 35), продемонстрировали в 2,26-8,1 раза большую чувствительность с применением GUS.nno, чем в случае с GUS.nat. Аналогичным образом, обеспечивается усиленная экспрессия в корне EXP-CaMV.35S-Enh + Os.Rcc3 + Zm.DnaK: 1:5 (SEQ ID NO: 36) в 16,06-19,06 раза с помощью GUS.nno, чем GUS.nat, благодаря чему кодон-модифицированная GUS-кодирующая последовательность идеально подходит для скрининга корневых промоторов, особенно тех, которые экспрессируют на низком уровне, и могут демонстрировать уровень GUS на уровне или ниже фоновых уровней при использовании нативной GUS последовательности, кодирующей 8.1 кратно большую чувствительность с помощью GUS.nno. Высококонститутивный профиль экспрессии, обеспечиваемый EXP-Zm.UbqMl: 1:2 (SEQ ID NO: 37), был продемонстрирован в 1,42-4,09 раза более высокий уровень с использованием GUS.nno, по сравнению с GUS.nat.
[000107] Качественная GUS-реакция была более чувствительна и лучше наблюдалась при использовании кодон-модифицированной GUS-кодирующей последовательности. Как правило, качественные реакции окрашивания менее чувствительны, чем количественные анализы. Использование кодон-модифицированной GUS-кодирующей последовательности обеспечивает лучшие и более однородные микроскопические осмотры окрашенных тканей. Например, в ткани корней, где GUS управлялся ЕХР-CaMV.35S-Enh + Os.Rcc3 + Zm.DnaK:1:5 (SEQ ID NO: 36), гистохимическое окрашивание тканей, трансформированных кодон-модифицированной GUS-кодирующей последовательностью, был более выраженным и видимым во всех образцах V7 корня: коре, эпидермисе, эндодерме, корневых волосках и кончиках боковых корней. В противоположность этому, GUS окрашивание качественно не наблюдалось в соответствующих V7 тканях корня, когда нативную последовательность, кодирующую GUS, управляли при помощи EXP-CaMV.35S-Enh + Os.Rcc3 + Zm.DnaK:1:5. Улучшенная кодон-модифицированная GUS-кодирующая последовательность, (CR-Ec.uidA_nno-1:1:1, SEQ ID NO: 30), обеспечивает большую чувствительность анализа и является особенно ценной при измерении экспрессия промоторов, которые экспрессируют на низком уровне.
Пример 5
Анализ регуляторных элементов, контролирующих GUS в протопластах в корнях и листьях кукурузы
[000108] Протопласты листьев и корней кукурузы были трансформированы с помощью векторов, содержащих RCc3 промотор, функционально связанный с нативным лидером RCc3, контролирующим экспрессию трансгена β-глюкуронидазы (GUS), и в результате трансформированные протопласты были проанализировали на экспрессию белка GUS. RCc3 промоторные и лидерные последовательности были клонированы в бинарные плазмидные конструкции с использованием способов, известных в данной области техники и описанных ранее в Примере 2.
[000109] Также были построены две плазмиды для использования при ко-трансформации и нормировании данных, с использованием способов, известных в данной области техники. Каждая плазмида содержала специфическую последовательность, кодирующую люциферазу, под контролем конститутивной последовательности ЕХР. Растительный вектор pMON19437 содержит экспрессионную кассету с конститутивным промотором, функционально связанным с 5'-интроном, (EXP-CaMV.35S-enh+Zm.DnaK:1:1, SEQ ID NO: 41), функционально связанным с 5'- кодирующей последовательностью люциферазы светляка (Photinus pyralis) (LUCIFERASE:1:3, SEQ ID NO: 42), функционально связанной с 5'-3'-НТО из гена нопалинсинтазы Agrobacterium tumefaciens (T-AGRtu.nos-1:1:13, SEQ ID NO: 39). Растительный вектор pMON63934 содержит экспрессионную кассету с конститутивной последовательностью ЕХР (ЕХР-CaMV.35S-enh-Lhcb1, SEQ ID NO: 44), функционально связанной с 5'-кодирующей последовательностью люциферазы морской губки (Renilla reniformis) (CR-Ren.hRenilla Lucife-0:0:1, SEQ ID NO: 43), функционально связанной с 5'-3'-HTO из гена нопалинсинтазы Agrobacterium tumefaciens (T-AGRtu.nos-1:1:13, SEQ ID NO: 39).
[000110] Протопласты листьев и корней кукурузы были трансформированы с использованием способа трансформации на основе ПЭГ, который хорошо известен в данной области техники. Клетки протопластов трансформировали плазмидными ДНК pMON19437 и pMON63934, и плазмидами, представленными в таблице 7. После трансформации трансформированные протопласты инкубировали в течение ночи в полной темноте. В дальнейшем измерения как GUS, так и люциферазы проводили путем помещения аликвот лизированных препаратов клеток, трансформированных как описано ранее, в два различных планшета с мелкими лунками. Один планшет использовали для измерения GUS, а второй планшет использовали для выполнения двойного анализа люциферазы с использованием системы двойного анализа репортера люциферазы (Promega Corp., Madison, WI; см., например, Promega Notes Magazine, No: 57, 1996, p.02).
[000111] Четыре трансформации были выполнены для каждого ЕХР или промотора + лидера + последовательности интрона. Были определены средние значения экспрессии для каждой последовательности ЕХР или промотора + лидера + последовательности интрона из нескольких образцов каждого эксперимента по трансформации. Измерения образцов были проведены с использованием четырех повторностей для каждой трансформации генетической конструкции (плазмиды) с последовательностью ЕХР или промотором + лидером + последовательностью интрона. Фоновая экспрессия GUS была определена с использованием отрицательного контроля плазмидной конструкции, в которой отсутствовал трансген GUS. Средние уровни экспрессии GUS и люциферазы приведены в таблицах 7 (лист) и 8 (корень). В этих таблицах значения люциферазы светлячков (например, экспрессия pMON19437) приведены в столбце "FLUC", а значения люциферазы синих анютиных глазок (например, экспрессия pMON63934) приведены в столбце "RLUC." Также в таблицах 7 и 8 представлены средние показатели GUS/FLUC и GUS/RLUC, которые представляют собой меру относительной разницы показателей силы экспрессии в анализе протопластов.



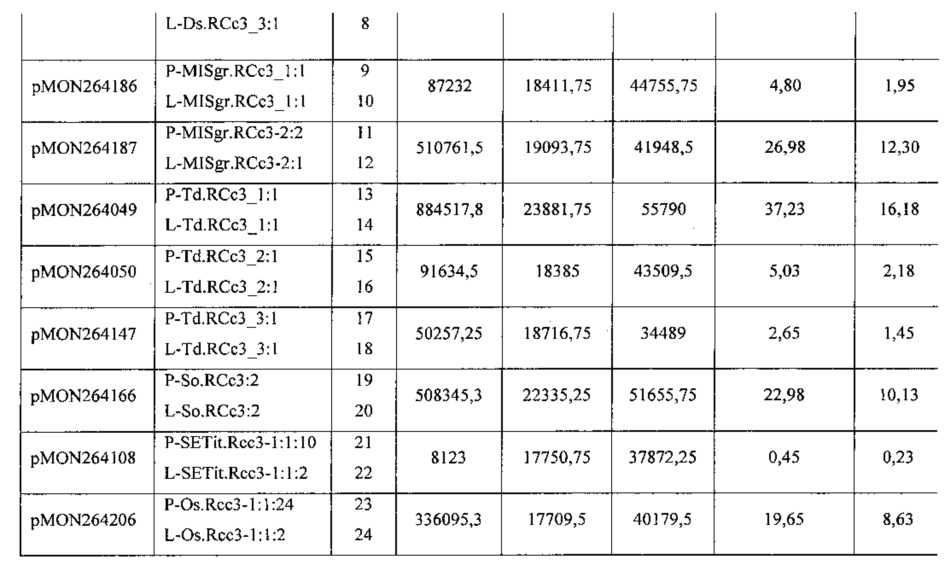
[000112] Как показано в таблице 7, все гомологи RCc3 промотора продемонстрировали способность контролировать экспрессию трансгена в протопластах листьев кукурузы. Некоторые из гомологов RCc3 промотора контролируют экспрессию лучше, чем другие в этом тесте, на основании показателей GUS/FLUC и GUS/RLUC. В дальнейшем, как показано в таблице 8 выше, все гомологи RCc3 промотора продемонстрировали способность контролировать экспрессию трансгена в протопластах корня кукурузы в разной степени.
Пример 6
Анализ регуляторных элементов, контролирующих GUS в трансгенной кукурузе
[000113] Растения кукурузы были трансформированы векторами, содержащими RCc3 промотор, функционально связанный с его нативным RCc3 лидером, контролирующим экспрессию трансгена β-глюкуронидазы (GUS). Полученные трансформированные растения были проанализированы экспрессией GUS белка.
[000114] RCc3 промоторные и лидерные последовательности были клонированы в бинарные плазмидные конструкции с использованием способов, известных в данной области техники и таких, как описано ранее в Примере 2. Полученные бинарные плазмидные конструкции: pMON264146, pMON264148, pMON264088, pMON264107, pMON264186, pMON264187, pMON264049, pMON264050, pMON264147 и pMON264166. Растения кукурузы были стабильно трансформированы pMON264108 и pMON264206. Качественный и количественный анализ экспрессии GUS проводили, как описано в Примере 2. Растения анализировали на V4, V7 и VT стадии развития. На R1 и R3 показан отбор проб. Таблица 9 показывает средние количественные показатели экспрессии GUS для стабильно трансформированных растений кукурузы.
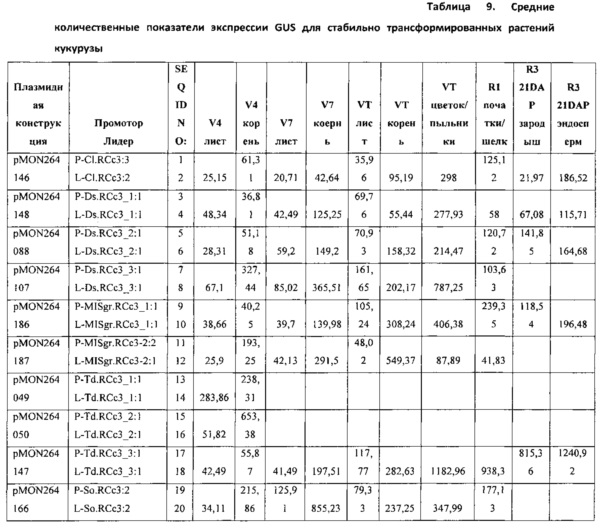
[000115] Как показано в таблице 9, все гомологи RCc3 промотора контролировали экспрессию трансгена GUS в стабильно трансформированных растениях кукурузы. Кроме того, каждый промотор имел уникальный характер экспрессии. Например, экспрессия VT цветок/пыльник отличалась среди гомологов RCc3 промотора. Экспрессия, контролируемая P-Td.RCc3_3:1 (SEQ ID NO: 17), была наиболее высокой для всех наблюдаемых промоторов, тогда как экспрессия, контролируемая P-MISgr.RCc3-2: 2 (SEQ ID NO: 11) была самой низкой. В отношении экспрессии R1 початки/шелк, P-Td.RCc3_3:1 (SEQ ID NO: 17) показала самый высокий уровень экспрессии в этих тканях и Р-MISgr.RCc3-2:2 (SEQ ID NO: 11) показала самый низкий уровень экспрессии. Экспрессия, контролируемая P-Td.RCc3 3:1 (SEQ ID NO: 17) увеличена в тканях, которые развиваются позже. Уровень экспрессии увеличивался в корне от стадии V4 к стадии VT и даже превысил VT цветок/пыльник, R1 початки/шелк и R3 21ДПО эмбриона и эндосперма. Экспрессия, контролируемая P-Td.RCc3_3:1, была самой высокой среди гомологов RCc3 промотора в VT цветы/пыльники, R1 початки/шелк и R3 21ДПО эмбриона и эндосперма.
[000116] В отношении экспрессии в листе и корне, некоторые из гомологов RCc3 промотора продемонстрировали более высокую экспрессию в корне по отношению к экспрессии в листе. Таблица 10 показывает соотношение экспрессии в корне к экспрессии в листе для всех проанализированных промоторов RCc3.


[000117] Как показано в Таблице 10, каждый гомолог RCc3 промотора продемонстрировал различные коэффициенты экспрессии корня к листу и различный ее характер от V4 стадии к стадии VT. Например, P-C1.RCc3:3 (SEQ ID NO: 1) поддерживает аналогичное соотношение экспрессии от V4 стадии до стадии VT с небольшим снижением, которое происходит на стадии V7. Как показано в таблице 9, экспрессия в корне слегка снизилась от V4 до V7, а затем увеличилась на этапе VT. Промотор Р-Ds.RCc3_3:1 (SEQ ID NO: 7) продемонстрировал изменение соотношения экспрессии от стадии V4 до стадии VT с более высокой экспрессией в корне по отношению к листу в стадиях V4 и V7, а затем сдвиг, который приблизил экспрессию в листе по отношению к корню на стадии VT (1,25). Как показано в таблице 9, этот промотор демонстрирует увеличение экспрессии в листе от стадии V4 до VT, тогда как экспрессия в корне снижается от стадии V7 до VT. Промотор P-So.RCc3:2 (SEQ ID NO: 19) поддерживает соотношение экспрессии в корне к экспрессии в листе на уровне 6,33 на стадии V4 и на уровне 6,79 на стадии V7, но затем снижает до значения 2,99 на стадии VT. Вместе с тем, экспрессия с этим промотором увеличилась в 3,69 и 3,96 раза в листе и корне, соответственно, от стадии V4 до V7, а затем снизилась до 2,33 и 1,10 по отношению к V4 на стадии VT.
[000118] Примечательно, что не все промоторы имели более высокий коэффициент отношения корня к листу. Например, промоторы P-Ds.RCc3_1:1 (SEQ ID NO: 3) и P-Td.RCc3_1:1 (SEQ ID NO: 13) имели соотношение корня к листу ниже одного на стадии V4. Вместе с тем, экспрессия, контролируемая P-Td.RCc3_1:1, была 6,6 раза больше, чем P-Ds.RCc3_1:1 в корне на стадии V4. Наибольшее соотношение корня и листа на этапе V4 было достигнуто с помощью P-Td.RCc3_2:1 (SEQ ID NO: 15). Соотношение экспрессии корня и листа с промотором P-Ds.RCc3_1:1 увеличилось с V4 (0,76) до V7 (2.95), а затем вернулось к соотношению, аналогичному на V4 (0,79).
[000119] Промотор P-MISgr.RCc3-2:2 (SEQ ID NO: 11) продемонстрировал увеличение экспрессии как в листе, так и в корне со стадии V4 по стадию VT. Этот промотор имел соотношение корня к листу выше 6,9 на протяжении всех трех стадий, но соотношение изменилось с 7,46 на стадии V4 до 6,92 на стадии V7, а затем увеличилось до 11,44 на стадии VT. Экспрессия, контролируемая P-MISgr.RCc3-2:2, увеличилась в листе и корне от стадии V4 до стадии VT.
[000120] Каждый из гомологов RCc3 промотора продемонстрировал паттерны экспрессии в стабильно трансформированной кукурузе, которые не могут быть предсказаны в силу того, что получены из гомологичных генов, особенно при использовании для трансформации гетерологичных видов, таких как кукуруза. Большинство промоторов продемонстрировали более высокую экспрессию в корне по отношению к листу в некоторых точках или на стадиях V4, V7 или VT, или на всех этапах анализа. Следует отметить, что величина экспрессии широко отличалась между промоторами. Уникальные свойства экспрессии каждого из гомологов RCc3 промотора делают некоторые из них более подходящим для определенных типов экспрессии транскрибируемой молекулы ДНК. Например, экспрессия транскрибируемой молекулы ДНК, которая может иметь решающее значение для усвоения питательных веществ в почве и которая лучше всего экспрессируется в более поздней стадии развития, когда растение собирается начать размножение и производство семян, может извлечь наибольшую выгоду с таким промотором, как P-MISgr.RCc3-2:2 (SEQ ID NO: 11), который увеличивает экспрессию в корне на стадии VT.

[000121] Из иллюстраций и описания принципов изобретения специалистам в данной области техники будет очевидно, что форма и детали данного изобретения могут быть модифицированы без отступления от этих принципов. Мы заявляем все модификации, находящиеся в пределах сущности и объема формулы изобретения. Все публикации и опубликованные патентные документы, приведенные в данном документе, включены посредством ссылки в той же степени, как если бы каждая отдельная публикация или заявка на патент конкретно и индивидуально указывались как включенные посредством ссылки.
Реферат
Изобретение относится к области биохимии, в частности к рекомбинантной молекуле ДНК для экспрессии гетерологической транскрибируемой полинуклеотидной молекулы, а также к клетке трансгенного растения, части трансгенного растения, семени трансгенного растения и трансгенному растению, ее содержащим. Также раскрыт способ получения товарного продукта с использованием вышеуказанного растения или его части. Изобретение также относится к способу получения трансгенного растения с помощью вышеуказанной молекулы ДНК. Изобретение позволяет эффективно осуществлять экспрессию гетерологической транскрибируемой полинуклеотидной молекулы в растении. 7 н. и 7 з.п. ф-лы, 1 ил., 10 табл., 6 пр.
Формула
Документы, цитированные в отчёте о поиске
Экспрессия промотора синтазы ацетооксикислот в интродуцированных генах растений
Комментарии