Конъюгаты квантовой точки с пептидом-носителем, пригодные для визуализации и доставки молекул в растения - RU2609637C2
Код документа: RU2609637C2
Чертежи







Описание
Заявление о праве на приоритет
Настоящая заявка притязает на преимущество приоритета предварительной заявки на патент № 61/466804, поданной 23 марта 2011 г.
Область техники, к которой относится изобретение
Настоящее изобретение относится к способам введения представляющей интерес молекулы в растительную клетку, имеющую клеточную оболочку, при помощи QD-пептидного конъюгата, содержащего квантовую точку (QD) и один или несколько проникающих в клетку пептидов (СРР).
Уровень техники
Наночастицы обладают уникальными свойствами, используемыми для доставки ДНК в клетки. Наночастицы металлов, такие как наночастицы золота (Au), используются для доставки ДНК благодаря низкой цитотоксичности и легкому взаимодействию с разными биологически значимыми лигандами. Помимо наночастиц металлов, в качестве носителей для доставки молекул в клетки используются наночастицы полупроводников (например, квантовые точки) (”QD”) размером 3-5 нм. ДНК и белки могут быть связаны с лигандом, присоединенным к поверхности QD (см., например, публикацию F. Patolsky et al., J. Am. Chem. Soc. 125, 13918 (2003)).
Наночастицы используются для доставки плазмидной ДНК в разные животные клетки. Установлено, что при инкубации наночастиц, покрытых ДНК, с клетками, не имеющими клеточной оболочки, указанные клетки поглощают такие наночастицы и начинают экспрессировать любые гены, кодированные ДНК. Однако доставка генов в растения затруднена из-за наличия у растительных клеток оболочек, что требует применения инвазивных средств доставки для генетической трансформации растений. Поэтому для опосредованной наночастицами доставки молекулы в клетки, имеющие клеточную оболочку, до введения частиц в протопласты растения удаляют клеточную оболочку (см. публикацию F. Torney et al., Nature Nanotechnol. 2 (2007)). Клеточная оболочка в растительных клетках является барьером, препятствующим доставке экзогенных молекул. Для доставки генов и мелких молекул в растительные клетки, имеющие оболочку, существуют многочисленные инвазивные методы, такие как генное ружье (биолистика), микроинъекция, электропорация и агробактерии, но доставка белков может быть осуществлена только с помощью микроинъекции. Доставка мелких молекул и белков при наличии оболочки у растительной клетки до сих пор не была исследована, при этом было бы желательно разработать методы, позволяющие манипулировать интактными растительными клетками, тканями или органами in vitro и in vivo.
Проникающие в клетки пептиды (СРР) являются новым и быстро растущим классом коротких пептидов, которые, как известно, играют важную роль в транслокации целого ряда несущих комплексов, включающих белки и ДНК, через биологические мембраны в линиях клеток млекопитающих и человека.

Хотя СРР облегчают доставку молекул в клетки млекопитающих, использование СРР для трансфекции растительных клеток ограничено рядом факторов. Главным препятствием для применения данной технологии при работе с растениями является то, что в отличие от животных клеток растительные клетки имеют двойной барьер (клеточную оболочку и плазматическую мембрану), препятствующий интернализации СРР и переносимых ими молекул. Поэтому СРР должны преодолеть два указанных барьера для эффективной транслокации. СРР используются в растительных клетках, но обычно благодаря применению агентов и методов, обеспечивающих проникновение СРР в растительные клетки. Домен, способствующий трансдукции белка ТАТ из ВИЧ-1, (PTD) является одним из наиболее хорошо исследованных перемещающихся пептидов. В недавно опубликованных отчетах была показана возможность использования ТАТ-PTD и его олигомеров для доставки в клетки млекопитающих плазмидной ДНК путем образования комплекса с отрицательно заряженной ДНК.

Другие пептиды, которые, как известно, обладают свойствами транслокации, включают pVEC, транспортан, пенетратин, пептиды рер-1 и их фрагменты.
Нанесение пептидов на QD является одним методом, широко применяемым при создании наночастиц для разных биотехнологических процессов. Например, присоединение проникающих в клетку пептидов, таких как полиаргинин и ТАТ-выделенные пептиды, к поверхности QD позволяет переносить QD в животные клетки. СРР широко используются в качестве носителей для доставки в клетки молекул при выполнении фундаментальных и прикладных биомедицинских исследований. С их помощью теперь можно вводить в клетки млекопитающих вещества, не способные проникать через мембрану, такие как нуклеиновые кислоты пепсина (PNA), белки, олигонуклеотиды или наночастицы. Биологи, работающие с растениями, считают весьма привлекательным использование СРР для доставки и временной экспрессии биомолекул в клетках. Таким образом, по-прежнему существует потребность в методе устойчивого введения генов и других представляющих интерес молекул в растения с помощью наночастиц.
Сущность изобретения
Нижеследующие варианты осуществления изобретения описаны наряду с системами, средствами и способами, которые поясняют и иллюстрируют изобретение, но не ограничивают его объем.
Один вариант осуществления изобретения относится к способу введения представляющей интерес молекулы в растительную клетку, имеющую клеточную оболочку, для достижения устойчивой трансформации растения и семян. Указанный способ включает получение растительной клетки, имеющей клеточную оболочку, осуществление взаимодействия квантовой точки (QD) с одним или несколькими проникающими в клетку пептидами (СРР) с образованием QD-пептидного конъюгата и присоединение одной или нескольких представляющих интерес молекул к одному или нескольким СРР с образованием активированного QD-пептидного конъюгата. Клетку и активированный QD-пептидный конъюгат вводят в соприкосновение друг с другом в условиях, обеспечивающих поглощение указанного конъюгата клеткой, имеющей клеточную оболочку.
Другой вариант осуществления изобретения относится к способу проведения устойчивой экспрессии гена. Указанный способ включает получение растительной клетки, имеющей клеточную оболочку, осуществление взаимодействия квантовой точки (QD) с одним или несколькими проникающими в клетку пептидами (СРР) с образованием QD-пептидного конъюгата и присоединение одного или нескольких генов к одному или нескольким СРР с образованием активированного QD-пептидного конъюгата. Растительную клетку, имеющую клеточную оболочку, и активированный QD-пептидный конъюгат вводят в соприкосновение друг с другом в условиях, обеспечивающих поглощение QD-пептидного конъюгата и одного ли нескольких генов растительной клеткой, имеющей клеточную оболочку. Указанный ген экспрессируется в потомстве растения, содержащего данную растительную клетку.
Другой вариант осуществления изобретения относится к способу переноса молекулярного вещества в растительную клетку. Указанный способ включает осуществление взаимодействия квантовой точки (QD) с одним или несколькими проникающими в клетку пептидами (СРР) с образованием QD-пептидного конъюгата и осуществление взаимодействия QD-пептидного конъюгата с плазмидной ДНК с образованием структуры активированного QD-пептидного конъюгата. Структуру активированного QD-пептидного конъюгата вводят в соприкосновение с интактной растительной клеткой, имеющей клеточную оболочку, в условиях, обеспечивающих поглощение растительной клеткой одного или нескольких СРР и одного или нескольких генов из плазмидной ДНК.
Другой вариант осуществления изобретения относится к способу скрининга и идентификации трансформации растения. Указанный способ включает получение растительной клетки, имеющей клеточную оболочку, осуществление взаимодействия квантовой точки (QD) с одним или несколькими проникающими в клетку пептидами (СРР) с образованием QD-пептидного конъюгата и присоединение одной или нескольких представляющих интерес молекул к одному или нескольким СРР с образованием активированного QD-пептидного конъюгата. Клетку, имеющую клеточную оболочку, и активированный QD-пептидный конъюгат вводят в соприкосновение друг с другом в условиях, обеспечивающих поглощение QD-пептидного конъюгата и представляющей интерес молекулы растительной клеткой, имеющей клеточную оболочку. Растительную клетку, имеющую клеточную оболочку, затем визуализируют.
Помимо вышеуказанных объектов и вариантов осуществления изобретения, из нижеследующего описания изобретения станут очевидными другие объекты и варианты осуществления изобретения.
Краткое описание чертежей
На фиг. 1 показан вариант конъюгата квантовой точки/пептида.
На фиг. 2 показана плазмидная карта pDAB3831.
Варианты осуществления изобретения
В нижеследующем описании изобретения и таблицах использован ряд терминов. Для достижения ясного и правильного понимания описания изобретения и формулы изобретения далее приведены следующие определения терминов.
Обратное скрещивание. Обратное скрещивание означает процесс, в соответствии с которым селекционер повторно скрещивает гибридное потомство с одним из родителей, например, гибрид F1 первого поколения скрещивают с одним из родительских генотипов гибрида F1.
Зародыш. Зародыш является маленьким растением, находящимся в созревшем зерне.
Устойчивость к гербициду. Устойчивость к гербициду означает способность растения выживать (то есть растение не может быть уничтожено) при внесении данной дозы гербицида. В некоторых случаях толерантные растения могут временно пожелтеть или каким-либо другим образом отреагировать на повреждение гербицидом (например, избыточным кущением и/или замедлением роста), но затем восстанавливаются.
Стабилизированный. Стабилизированный означает характеристики растения, которые воспроизводимо передаются одним поколением следующему поколению инбредных растений того же сорта.
Поглощение. Поглощение означает транслокацию частицы, такой как квантовая точка, несущие пептиды, проникающие в клетку пептиды и хоминг-пептиды, через клеточную оболочку или клеточную мембрану, при этом транслокация происходит не только под воздействием стимула, сообщаемого частице самой клеткой, поглощающей данную частицу. Неограничивающие примеры устройств или методов, обеспечивающих транслокацию частицы через клеточную оболочку или клеточную мембрану под воздействием стимула, сообщаемого частице, включают биолистику, генное ружье, микроинъекцию и/или прокалывание.
В некоторых вариантах осуществления изобретения в одном или нескольких пептидах может быть создано несколько сайтов присоединения или заполнения “добавляемой” или “гостевой” молекулой. Указанное свойство может быть использовано, например, для специфического направленного воздействия и изменения сайтов молекул в клетках в таких областях, как биомиметика, направленная доставка, с целью создания негенетически модифицированных организмов и временной трансформации древесных или растительных культур для сообщения им определенных признаков и устойчивости к болезням. Варианты осуществления изобретения также относятся к созданию приемлемых биосенсоров. Кроме того, в способах по настоящему изобретению могут быть использованы искусственные хромосомы (ACES) в качестве альтернативы современным эукариотическим векторам для точного воздействия и гомологичной рекомбинации.
Конкретные варианты осуществления изобретения относятся к использованию многофункциональных флуоресцирующих наночастиц, пригодных для доставки отрицательно заряженных молекул, таких как, например, ДНК/РНК. Несущие и проникающие в клетку пептиды (СРР)/хоминг-пептиды (НР) (совместно определяемые в настоящем описании изобретения как ”СРР”), такие как R9, TAT, MPG и γ-Zein, были присоединены к поверхности люминесцентных квантовых точек (QD). QD-пептидные конъюгаты были использованы для эффективной доставки ДНК в растение Arabidopsis. QD-пептидные биоконъюгаты не оказывали токсического воздействия на рост осей цветков Arabidopsis и образование семян. Было идентифицировано несколько устойчивых трансформантов Т1 и был произведен анализ рассады. Установлено, что доставка ДНК в растения произведена с помощью носителя и достигнута устойчивая трансформация. Могут быть созданы интеллектуальные системы для доставки биомолекул и направленного воздействия на клетки и компартменты клеток. В конкретных вариантах осуществления изобретения такие автофлуоресцентные QD могут быть использованы для визуализации признаков в растениях.
Определенные варианты осуществления изобретения относятся к способу введения представляющей интерес молекулы в растительную клетку, имеющую клеточную оболочку, для достижения устойчивой трансформации растения и семян. Указанный способ включает получение растительной клетки, имеющей клеточную оболочку, осуществление взаимодействия квантовой точки (QD) с одним или несколькими проникающими в клетку пептидами (СРР) с образованием QD-пептидного конъюгата и присоединение одной или нескольких представляющих интерес молекул к одному или нескольким СРР с образованием активированного QD-пептидного конъюгата. Клетку и активированный QD-пептидный конъюгат вводят в соприкосновение друг с другом в условиях, обеспечивающих поглощение указанного конъюгата клеткой, имеющей клеточную оболочку.
В некоторых вариантах осуществления изобретения несколько пептидов были ковалентно связаны с наночастицами QD и доставлены в растительные клетки. Наночастицы с проникающими в клетку пептидами R9, γ-Zein и MPG были успешно доставлены в растения для достижения устойчивой трансформации растений. В других вариантах осуществления изобретения была использована меченая биомолекула для отслеживания введенных молекул в цитоплазму. Было достигнуто эффективное поглощение указанных QD-пептид-ДНК конъюгатов, при этом комплекс ДНК с QD-пептидным конъюгатом был устойчивым, о чем свидетельствует устойчивая трансформация, передаваемая семенами, и получение устойчивой рассады Т1.
Другие объекты изобретения относятся к использованию QD-пептидных конъюгатов для многофункциональных вариантов интеллектуальной доставки биофункциональных биомолекул (например, для доставки ДНК/РНК и ферментов), визуализации и разных биотехнологических диагностических функций. Современная методика может способствовать развитию химии поверхностных взаимодействий и инкапсуляции, что позволит облегчить синтез целого ряда молекул, обладающих разными функциональными свойствами. Основные свойства указанных веществ с точки зрения их возможного использования для доставки биомолекул и генов определяются большой плотностью концевых групп в таких системах. Данное явление улучшает поверхностные характеристики молекул, позволяет создать много сайтов присоединения (например, для конъюгации сигнальных или направленно воздействующих частей) и определяет молекулярный объем, который имеет важное значение для включения других молекул в указанный комплекс. Конъюгированные несущие пептиды одновременно обеспечивают доставку QD и присоединенных молекул. Кроме того, в результате образования комплекса переносимой молекулы непосредственно с несущим пептидом можно устранить трэйдоф в целом ряде присоединенных видов на поверхности QD. Так как отрицательно заряженные олигонуклеотиды не могут проникать через барьеры, создаваемые клеточной оболочкой и клеточной мембраной и сами клеточные мембраны, настоящее изобретение наряду с прочим обеспечивает создание эффективных систем доставки для встраивания ДНК, регуляции генов и корреляции.
В соответствии с вариантами осуществления настоящего изобретения растительная клетка, имеющая клеточную оболочку, может быть любой растительной клеткой, имеющей интактную и целую клеточную оболочку. Примеры клеток, имеющих клеточную оболочку, включают, не ограничиваясь ими, водоросли, табак, морковь, кукурузу, канолу, рапс, хлопчатник, пальму, арахис, сою, сахарный тростник, Oryza sp., Arabidopsis sp. и Ricinus sp., предпочтительно табак, морковь, кукурузу, хлопчатник, канолу, сою и сахарный тростник, более предпочтительно табак и морковь. Варианты осуществления настоящего изобретения относятся к клеткам, имеющим клеточную оболочку, из любой ткани или органа, которые включают, не ограничиваясь ими, зародыши, меристематические клетки, каллюс, пыльцу, листья, пыльники, корни, корневые отростки, цветки, семена, бобы, стебли и культуру ткани.
В вариантах осуществления настоящего изобретения представляющая интерес молекула может быть любой молекулой, доставляемой в растительную клетку по настоящему изобретению. Представляющие интерес молекулы или компоненты таких молекул могут включать, не ограничиваясь ими, нуклеиновые кислоты, ДНК, РНК, иРНК, гены, плазмиды, космиды, YAC, BAC, искусственные хромосомы растений, минихромосомы растений, генетически созданные ДНК локусов признаков, полипептиды, ферменты, гормоны, гликопептиды, сахара, жиры, сигнальные пептиды, антибиотики, витамины, мессенджеры, вторичные мессенджеры, аминокислоты, сАМР, лекарственные средства, гербициды, фунгициды, антибиотики и/или их комбинации.
Варианты осуществления настоящего изобретения относятся к способам профилактики или лечения заболевания. Неограничивающие примеры вариантов осуществления изобретения включают доставку фунгицидов, антибиотиков и/или других лекарственных средств в клетки, нуждающиеся в таких средствах, способами по настоящему изобретению.
В соответствии с объектами настоящего изобретения QD-пептидный конъюгат может быть поглощен разными частями клетки. Примеры частей клетки, способных поглощать QD-пептидный конъюгат, включают, не ограничиваясь ими, цитозоль, ядро, тонопласты, пластиды, этиопласты, хромопласты, лейкопласты, элайопласты, протеинопласты, амилопласты, хлоропласты и полость двойной мембраны. В других вариантах осуществления изобретения QD-пептидный конъюгат может быть поглощен клеткой, имеющей клеточную оболочку, по симпластическому или апопластическому пути.
Дополнительные варианты осуществления изобретения относятся к генетически модифицированным растительным клеткам и способам создания таких клеток, в соответствии с которыми в растительные клетки вводят одну или несколько нуклеиновых кислот способами по настоящему изобретению. В одном варианте осуществления изобретения в растительную клетку, имеющую клеточную оболочку, с помощью QD-пептидного комплекса по настоящему изобретению вводят плазмиду, содержащую представляющий интерес ген и селектируемый маркер. В других вариантах осуществления изобретения могут быть выбраны устойчивые трансформанты, в которые устойчиво встроен представляющий интерес ген и/или селектируемый маркер. В альтернативных вариантах осуществления изобретения растительная клетка, включающая представляющий интерес ген, может быть размножена с образованием других клеток, включающих представляющую интерес молекулу. В других вариантах осуществления изобретения растительные клетки, содержащие представляющую интерес молекулу, могут быть регенерируемыми клетками, используемыми для регенерации всего растения, включающего представляющую интерес молекулу.
Другим объектом настоящего изобретения являются способы создания регенерируемых растительных клеток, включающих представляющую интерес молекулу, которые предназначены для использования в культуре ткани. Культура ткани предпочтительно способна регенерировать растения, относящиеся к такому же генотипу, что и регенерируемые клетки. Регенерируемые клетки в таких культурах ткани могут представлять собой зародыши, протопласты, меристематические клетки, каллюс, пыльцу, листья, пыльники, корни, корневые отростки, цветки, семена, бобы или стебли. Кроме того, вариант осуществления изобретения относится к растениям, регенерированным из культур ткани по настоящему изобретению.
Альтернативно, настоящее изобретение относится к способу сообщения желаемого признака растительной клетке, имеющей клеточную оболочку, который включает введение QD-пептидного конъюгата и представляющей интерес молекулы, сообщающей желаемый признак растительной клетке, в соприкосновение с указанной клеткой в условиях, обеспечивающих проникновение QD-пептидного конъюгата через клеточную оболочку. Примеры желаемых признаков включают, не ограничиваясь ими, признаки, выбираемые из мужской стерильности, устойчивости к гербициду, устойчивости к насекомым, устойчивости к бактериальному заболеванию, грибковому заболеванию и/или вирусному заболеванию.
Другими объектами настоящего изобретения являются способы создания устойчивых линий растений, включающих желаемый признак или представляющую интерес молекулу, в соответствии с которыми растительной клетке сначала сообщают желаемый признак или вводят представляющую интерес молекулу, обеспечивая проникновение QD-пептидного конъюгата через клеточную оболочку растения. Методы создания устойчивых линий растений хорошо известны специалисту с данной области и могут представлять собой методы, которые включают, не ограничиваясь ими, самоопыление, обратное скрещивание, создание гибридов, скрещивание с популяциями и тому подобные. В объем настоящего изобретения входят все растения и растительные клетки, включающие желаемый признак или представляющую интерес молекулу, которые были введены в растительную клетку (или ее предшественники) путем поглощения QD-пептидного конъюгата через клеточную оболочку. Растительные клетки, включающие желаемый признак или представляющую интерес молекулу, которые были введены в растение или клетку (или ее предшественники) путем поглощения QD-пептидного конъюгата через клеточную оболочку, могут быть использованы для скрещивания с другими растительными клетками с целью получения гибридных клеток первого поколения (F1), семян и/или растений, обладающих лучшими характеристиками.
В вариантах осуществления изобретения, в которых представляющая интерес молекула содержит один или несколько генов, указанные гены могут быть доминантными или рецессивными аллелями. В качестве примера можно отметить, что указанные гены могут сообщать растению такие признаки, как устойчивость к гербициду, устойчивость к насекомым, устойчивость к бактериям, устойчивость к грибам, устойчивость к вирусному заболеванию, мужская фертильность, мужская стерильность, более высокая питательная ценность и промышленная применимость.
Благодаря созданию методов молекулярной биологии, которые позволили выделять и исследовать гены, кодирующие определенные белковые продукты или РНК (например, иРНК), ученые, работающие в области биологии растений, стали проявлять повышенный интерес к созданию генома клеток, содержащего и экспрессирующего чужеродные гены, дополнительные или модифицированные варианты нативных или эндогенных генов (возможно, стимулируемых разными промоторами), с целью соответствующего изменения признаков клетки. Такие чужеродные дополнительные и/или модифицированные гены определяются в настоящем описании изобретения как “трансгены”. На протяжении последних пятнадцати-двадцати лет было разработано несколько методов создания трансгенных клеток, и конкретные варианты осуществления настоящего изобретения относятся к трансформированным вариантам клеток и способам создания таких клеток путем введения трансгена в клетку, имеющую клеточную оболочку, в результате поглощения QD-пептидного конъюгата через клеточную оболочку. В вариантах осуществления настоящего изобретения трансген может находиться в экспрессирующем векторе.
Трансформация клетки может включать создание экспрессирующего вектора, функционирующего в конкретной клетке. Такой вектор может содержать ДНК, включающую ген, контролируемый или функционально связанный с регуляторным элементом (таким как, например, промотор). Экспрессирующий вектор может содержать одну или нескольких таких функционально связанных комбинаций гена/регуляторного элемента. Вектор может быть плазмидой и может быть использован отдельно или в комбинации с другими плазмидами с образованием клеток, трансформированных методами, обеспечивающими введение трансгена в генетический материал растительной клетки, имеющей клеточную оболочку.
Использование QD-пептидных конъюгатов в соответствии со способами по настоящему изобретению позволило получить устойчиво трансформированные растения и продемонстрировало наличие экспрессии устойчиво трансформированного гена, относящегося к фенотипу, сообщающего трансгенному растению Т1 высокую устойчивость к гербициду. Было показано, что данное растение является плодородным, так как позволило получить семена Т2.
Экспрессирующие векторы, применяемые для поглощения молекул с помощью QD-пептидного конъюгата: гены-маркеры
Экспрессирующие векторы могут включать по меньшей мере один генетический маркер, функционально связанный с регуляторным элементом (таким как, например, промотор), который позволяет выявить трансформированные клетки, содержащие маркер, путем отбора по отрицательному признаку (то есть ингибирование роста клеток, не содержащих селектируемый ген-маркер) или путем отбора по положительному признаку (то есть скрининг продукта, кодированного генетическим маркером). В области трансформации клеток хорошо известны многие селектируемые гены-маркеры, которые включают, например, гены, кодирующие ферменты, метаболически детоксифицирующие селективный химический агент, который может быть антибиотиком или гербицидом, или гены, кодирующие измененную мишень, которая может быть невосприимчива к ингибитору. В данной области также известны несколько методов отбора по положительному признаку.
Одним широко используемым селектируемым геном-маркером, пригодным для трансформации растений, является ген неомицин-фосфотрансферазы II (nptII), контролируемый регуляторными сигналами растения, который сообщает устойчивость к канамицину. См., например, публикацию Fraley et al., Proc. Natl. Acad. Sci. U.S.A. 80:4803 (1983). Другим широко используемым селектируемым геном-маркером является ген гигромицин-фосфотрансферазы, сообщающий устойчивость к антибиотику гигромицину. См., например, публикацию Vanden Elzen et al., Plant Mol. Biol. 5:299 (1985).
Дополнительные селектируемые гены-маркеры бактериального происхождения, сообщающие устойчивость к антибиотикам, включают гентамицин-ацетилтрансферазу, стрептомицин-фосфотрансферазу, аминогликозид-3'-аденилтрансферазу и детерминанту устойчивости к блеомицину. См. публикации Hayford et al., Plant Physiol. 86:1216 (1988); Jones et al., Mol. Gen. Genet. 210:86 (1987); Svab et al., Plant Mol. Biol. 14:197 (1990); Hille et al., Plant Mol. Biol. 7:171 (1986). Другие селектируемые гены-маркеры сообщают устойчивость к гербицидам, таким как глифозат, глюфозинат или бромоксинил. См. публикации Comai et al., Nature 317:741-744 (1985); Gordon-Kamm et al., Plant Cell 2:603-618 (1990); и Stalker et al., Science 242:419-423 (1988).
Другие селектируемые гены-маркеры, пригодные для трансформации растений, имеют происхождение, отличное от бактериального. Указанные гены включают, например, мышиную дигидрофолат-редуктазу, растительную 5-енолпирувилшикимат-3-фосфат-синтазу и растительную ацетолактат-синтазу. См. публикации Eichholtz et al., Somatic Cell Mol. Genet. 13:67 (1987); Shah et al., Science 233:478 (1986); Charest et al., Plant Cell Rep. 8:643 (1990).
Другой класс генов-маркеров, пригодных для трансформации растений, требует скрининга предположительно трансформированных растительных клеток, а не прямого генетического отбора трансформированных клеток, обладающих устойчивостью к токсическому веществу, такому как антибиотик. Указанные гены особенно пригодны для количественного определения или визуализации пространственного паттерна экспрессии гена в определенных тканях и часто определяются как гены-репортеры, так как указанные гены могут быть гибридизированы с геном или регуляторной последовательностью гена для исследования экспрессии гена. Гены, обычно используемые для скрининга трансформированных клеток, включают β-глюкуронидазу (GUS), β-галактозидазу, люциферазу и хлорамфеникол-ацетилтрансферазу. См. публикации R.A. Jefferson, Plant Mol. Biol. Rep. 5:387 (1987); Teeri et al., EMBO J. 8:343 (1989); Koncz et al., Proc. Natl. Acad. Sci. U.S.A. 84:131 (1987), DeBlock et al., EMBO J. 3:1681 (1984).
Недавно была разработаны методы визуализации активности GUS in vivo, не требующие разрушения растительной ткани. См. публикации Molecular Probes, 2908, Imagene Green™, p. 1-4 (1993) и Naleway et al., J. Cell Biol. 115:151a (1991). Однако указанные методы визуализации активности GUS in vivo оказались непригодными для выявления трансформированных клеток из-за низкой чувствительности, высокой фоновой флуоресценции и ограничений, связанных с использованием генов люфицеразы в качестве селектируемых маркеров.
Недавно в качестве маркеров экспрессии генов в прокариотических и эукариотических клетках были использованы гены, кодирующие флуоресцирующие белки (например, GFP, EGFP, EBFP, ECFP и YFP). См. публикацию Chalfie et al., Science 263:802 (1994). В качестве селектируемых маркеров могут быть использованы флуоресцирующие белки и мутации флуоресцирующих белков.
Экспрессирующие векторы, применяемые для поглощения молекул с помощью QD-пептидных конъюгатов: промоторы
Гены, введенные в экспрессирующие векторы, должны стимулироваться нуклеотидной последовательностью, включающей регуляторный элемент, например, промотор. В настоящее время в области трансформации клеток хорошо известны несколько типов промоторов наряду с другими регуляторными элементами, которые могут быть использованы отдельно или в комбинации с промоторами.
В использованном здесь значении термин “промотор” означает область ДНК, расположенную вверху от начала транскрипции, которая участвует в узнавании и связывании РНК-полимеразы и других белков для инициации транскрипции. “Растительный промотор” может быть промотором, способным инициировать транскрипцию в растительных клетках. В качестве примеров создаваемых в настоящее время промоторов можно привести промоторы, которые предпочтительно инициируют транскрипцию в определенных тканях, таких как листья, корни, семена, волокна, сосуды ксилемы, трахеиды или склеренхима. Такие промоторы определяются как “тканепредпочтительные”. Промоторы, которые инициируют транскрипцию только в определенных тканях, определяются как “тканеспецифичные”. Промотор, специфичный к “типу клетки”, стимулирует главным образом экспрессию в определенных типах клеток в одном или нескольких органах, например, в сосудистых клетках корней или листьев. “Индуцибельный” промотор может быть промотором, активность которого регулируется окружающими условиями. Примеры окружающих условий, которые могут влиять на транскрипцию, вызываемую индуцибельными промоторами, включают анаэробные условия или наличие света. Тканеспецифичные, тканепредпочтительные, специфичные к типу клетки и индуцибельные промоторы составляют класс “неконститутивных” промоторов. “Конститутивный” промотор является промотором, который может быть активным в большинстве окружающих условий.
А. Индуцибельные промоторы
Индуцибельный промотор может быть функционально связан с геном для экспрессии в клетке. Индуцибельный промотор может быть необязательно функционально связан с нуклеотидной последовательностью, кодирующей сигнальную последовательность, которая может быть функционально связана с геном для экспрессии в клетке. При наличии индуцибельного промотора скорость транскрипции повышается под воздействием индуцирующего агента.
В настоящем изобретении может быть использован любой индуцибельный промотор. См. публикацию Ward et al., Plant Mol. Biol. 22:361-366 (1993). Типичные индуцибельные промоторы включают, не ограничиваясь ими, промоторы из системы ACEI, реагирующие на медь (Mett et al., PNAS 90:4567-4571 (1993)); ген In2 кукурузы, который реагирует на защитные средства от бензолсульфонамидного гербицида (Hershey et al., Mol. Gen. Genetics 227:229-237 (1991); and Gatz et al., Mol. Gen. Genetics 243:32-38 (1994)); и Tet-репрессор из Tn10 (Gatz et al., Mol. Gen. Genetics 227:229-237 (1991)). Особенно пригодным индуцибельным промотором может быть промотор, реагирующий на индуцирующий агент, на который растения обычно не реагируют. Типичным индуцибельным промотором является индуцибельный промотор гена стероидного гормона, транскрипционная активность которого может быть вызвана глюкокортикостероидным гормоном. Schena et al., Proc. Natl. Acad. Sci. U.S.A. 88-0421 (1991).
В. Конститутивные промоторы
Конститутивный промотор может быть функционально связан с геном для экспрессии в клетке, или конститутивный промотор может быть функционально связан с нуклеотидной последовательностью, кодирующей сигнальную последовательность, которая может быть функционально связана с геном для экспрессии в клетке.
В настоящем изобретении могут быть использованы разные конститутивные промоторы. Типичные конститутивные промоторы включают, не ограничиваясь ими, промоторы из растительных вирусов, такие как промотор 35S из CaMV (Odell et al., Nature 313:810-812 (1985)); промоторы из генов актина риса (McElroy et al., Plant Cell 2:163-171 (1990)); убиквитин (Chrictensen et al., Plant Mol. Biol. 12:619-632 (1989) and Christensen et al., Plant Mol. Biol. 18:675-689 (1992)); pEMU (Last et al., Theor. Appl. Genet. 81:581-588 (1991)); MAS (Velten et al., EMBO J. 3:2723-2730 (1984)) и гистон Н3 кукурузы (Lepetit et al., Mol. Gen. Genetics 231:276-285 (1992) и Atanassova et al., Plant Journal 2(3):291-300 (1992)). Особенно пригодным конститутивным промотором является промотор ALS, 5'-концевой фрагмент Xba1/NcoI структурного гена ALS3 Brassica napus (или нуклеотидная последовательность, подобная указанному фрагменту Xba1/NcoI). См. заявку на патент РСТ WO 96/30530.
С. Тканеспецифичные или тканепредпочтительные промоторы
Тканеспецифичный промотор может быть функционально связан с геном для экспрессии в клетке. Тканеспецифичный промотор может быть необязательно функционально связан с нуклеотидной последовательностью, кодирующей сигнальную последовательность, которая может быть функционально связана с геном для экспрессии в клетке. Растения, трансформированные представляющим интерес геном, функционально связанным с тканеспецифичным промотором, могут продуцировать белковый продукт трансгена исключительно или предпочтительно в определенной ткани.
В настоящем изобретении может быть использован любой тканеспецифичный или тканепредпочтительный промотор. Типичные тканеспецифичные или тканепредпочтительные промоторы включают, не ограничиваясь ими, промотор, предпочтительно инициирующий транскрипцию в корнях, такой как ген фазеолина (Murai et al., Science 23:476-482 (1983) и Sengupta-Gopalan et al., Proc. Natl. Acad. Sci. U.S.A. 82:3320-3324 (1985); промотор, специфичный к листьям и индуцируемый светом, такой как промотор из cab или rubisco (Simpson et al., EMBO J. 4(11):2723-2729 (1985) и Timko et al., Nature 318:579-582 (1985)); промотор, специфичный к пыльнику, такой как промотор из LAT52 (Twell et al., Mol. Gen. Genetics 217:240-245 (1989)); промотор, специфичный к пыльце, такой как промотор из Zm13 (Guerrero et al., Mol. Gen. Genetics 244:161-168 (1993)), или промотор, предпочитающий микроспоры, такой как промотор из apg (Twell et al., Sex. Plant Reprod. 6: 217-2245 (1993)).
Белок, продуцированный трансгенами, может быть перенесен в субклеточный компартмент, такой как хлоропласт, вакуоль, пероксисома, глиоксисома, клеточная оболочка или митохондрион, для секреции в апопласт, путем функционального связывания нуклеотидной последовательности, кодирующей сигнальную последовательность, с 5'- и/или 3'-концевой областью гена, кодирующего представляющий интерес белок. Направленно воздействующие последовательности, расположенные у 5'- и/или 3'-конца структурного гена, могут определять во время синтеза и процессинга белка окончательную локализацию кодированного белка в компартменте. Альтернативно, такие белки, определяющие субклеточный компартмент, могут быть непосредственно связаны с QD-пептидным конъюгатом для направления указанного конъюгата, содержащего представляющую интерес молекулу, в требуемый субклеточный компартмент.
Сигнальная последовательность направляет полипептид во внутриклеточную органеллу, в субклеточный компартмент или для секреции в апопласт. В данной области известны многие сигнальные последовательности. См., например, публикации

Гены чужеродных белков и агрономические гены
Трансгенные растения по настоящему изобретению способны продуцировать чужеродный белок в коммерческих количествах. Таким образом, методы отбора и размножения трансформированных растений, хорошо известные в данной области, позволяют получить множество трансгенных растений, которые могут быть собраны обычным способом и использованы для экстракции чужеродного белка из представляющей интерес ткани или общей биомассы. Белок может быть экстрагирован из растительной биомассы известными методами, рассмотренными, например, в публикации Heney and Orr, Anal. Biochem. 114:92-6 (1981).
В соответствии с объектами настоящего изобретения трансгенное растение, предназначенное для коммерческого продуцирования чужеродного белка, может быть клеткой или растением. В соответствии с другими объектами изобретения представляющей интерес биомассой могут быть семена. Для относительно небольшого количества трансгенных растений, характеризующихся более высокими уровнями экспрессии, может быть создана генетическая карта в результате выполнения стандартных анализов методами RFLP, ПЦР и SSR, которые позволяют идентифицировать примерную локализацию в хромосоме встроенной молекулы ДНК. Для ознакомления с типичными методами, применяемыми для указанной цели, см. публикацию Glick and Thompson, Methods in Plant Molecular Biology and Biotechnology CRC Press, Boca Raton 269:284 (1993). Содержащаяся на карте информация о локализации в хромосоме может быть использована для соответствующей защиты трансгенного растения. При несанкционированном размножении и скрещивании с другой зародышевой плазмой карту области встраивания можно сравнить с аналогичными картами неустановленных растений для определения общего происхождения с эталонным растением. Стандартными методами, применяемыми для сравнения карт, являются гибридизация, RFLP, ПЦР, SSR и секвенирование.
Аналогичным образом в трансформированных клетках или их потомстве могут быть экспрессированы агрономические гены. В частности, при помощи способов по настоящему изобретению могут быть генетически созданы растения, экспрессирующие разные фенотипы, интересные в агрономическом отношении. Типичные гены, которые могут быть использованы в данной связи, включают, не ограничиваясь ими, гены, рассмотренные ниже в настоящем описании изобретения.
1. Гены, сообщающие устойчивость к вредителям или болезням
А) Гены устойчивости к болезням растений. Защитные функции растения часто активируются в результате специфического взаимодействия между продуктом гена устойчивости к заболеванию (R) в растении и продуктом соответствующего авирулентного (Avr) гена в патогенном организме. Растение может быть трансформировано клонированными генами устойчивости для создания растений, устойчивых к определенным штаммам патогенных организмов. См., например, публикации Jones et al., Science 266:789 (1994) (клонирование гена Cf-9 томатов, сообщающего устойчивость к Cladosporum fulvum); Martin et al., Science 262:1432 (1993) (ген Pro томатов, сообщающий устойчивость к Pseudomonas syringae, кодирует протеинкиназу); Mindrinos et al., Cell 78:1089 (1994) (ген RSP2 Arabidopsis, сообщающий устойчивость к Pseudomonas syringae).
В) Ген, сообщающий устойчивость к вредителю, такому как соевая нематода. См., например, заявку на патент РСТ WO 96/30517; заявку на патент PCT WO 93/19181.
С) Белок Bacillus thuringiensis, его производное или синтетический полипептид, смоделированный с указанного белка. См., например, публикацию Geiser et al., Gene 48:109 (1986), в которой описано клонирование и нуклеотидная последовательность гена δ-эндотоксина Bt. Кроме того, молекулы ДНК, кодирующие гены δ-эндотоксина, могут быть приобретены в Американской коллекции типовых культур, Manassas, Va., например, под номерами доступа АТСС 40098, 67136, 31995 и 31998.
D) Лектин. См., например, публикацию Van Damme et al., Plant Molec. Biol. 24:25 (1994), в которой описаны нуклеотидные последовательности нескольких генов лектина, связывающихся с маннозой Clivia miniata.
Е) Белок, связывающийся с витаминами, такой как авидин. См. заявку на патент РСТ US93/06487. В указанной заявке на патент рассмотрено применение авидина и гомологов авидина в качестве ларвицидов против насекомых-вредителей.
F) Ингибитор фермента, например, ингибитор протеазы или протеиназы либо ингибитор амилазы. См., например, публикации Abe et al., J. Biol. Chem. 262:16793 (1987) (нуклеотидная последовательность ингибитора цистеин-протеиназы риса), Huub et al., Plant Molec. Biol. 21:985 (1993) (нуклеотидная последовательность кДНК, кодирующая ингибитор протеиназы табака I), Sumitani et al., Biosci. Biotech. Biochem. 57:1243 (1993) (нуклеотидная последовательность ингибитора альфа-амилазы Strepromyces nitrosporeus) и патент США № 5494813 (Hepher and Atkinson, выдан 27 февраля 1996 г.).
G) Специфичный к насекомым гормон или феромон, такой как экдистероидный или ювенильный гормон, его вариант, миметик на его основе, его антагонист или агонист. См., например, публикацию Hammock et al., Nature 344:458 (1990), в которой описана экспрессия в бакуловирусе клонированного ювенильного гормона эстеразы, инактиватора ювенильного гормона.
Н) Специфичный к насекомым пептид или нейропептид, который, будучи экспрессированным, нарушает физиологию зараженного насекомого. См., например, публикации Regan, J. Biol. Chem. 269:9 (1994) (в результате экспрессии образуется ДНК, кодирующая рецептор диуретического гормона насекомых) и Pratt et al., Biochem. Biophys. Res. Comm. 163:1243 (1989) (аллостатин может быть идентифицирован в Diploptera puntata). См. также патент США № 5266317, выданный Tomalski et al., в котором описаны гены, кодирующие специфичные к насекомым паралитические нейротоксины.
I) Специфичный к насекомым яд, продуцируемый в природе змеей, осой или любым другим организмом. См., например, публикацию Pang et al., Gene 116:165 (1992), в которой описана гетерологичная экспрессия в растениях гена, кодирующего токсичный для насекомых пептид скорпиона.
J) Фермент, отвечающий за сверхнакопление монотерпена, сесквитерпена, стероида, гидроксамовой кислоты, фенилпропаноидное производное или другая небелковая молекула, обладающая инсектицидной активностью.
K) Фермент, участвующий в модификации, включая посттрансляционную модификацию биологически активной молекулы, например, природный или синтетический гликолитический фермент, протеолитический фермент, липолитический фермент, нуклеаза, циклаза, трансаминаза, эстераза, гидролаза, фосфатаза, киназа, форсфорилаза, полимераза, эластаза, хитиназа и глюканаза. См. заявку на патент РСТ WO 93/02197 на имя Scott et al., в которой описана нуклеотидная последовательность гена каллазы. Молекулы ДНК, содержащие последовательности, кодирующие хитиназу, могут быть получены, например, в АТСС под номерами доступа 39637 и 67152. См. также публикацию Kramer et al., Insect Biochem. Molec. Biol. 23:691 (1993), в которой рассмотрена нуклеотидная последовательность кДНК, кодирующая хитиназу табачного бражника, и публикацию Kawalleck et al., Plant Molec. Biol. 21:673 (1993), в которой представлена нуклеотидная последовательность гена полиубиквитина ubi4-2 огородной петрушки.
L) Молекула, стимулирующая сигнальную трансдукцию. См., например, публикацию Botella et al., Plant Molec. Biol. 24:757 (1994), в которой описаны нуклеотидные последовательности клонов кДНК кальмодулина золотистой фасоли, и публикацию Griess et al., Plant Physiol. 104:1467 (1994), в которой представлена нуклеотидная последовательность клона кДНК кальмодулина кукурузы.
М) Гидрофобный пептид. См. заявку на патент РСТ WO 95/16776 (в которой описаны производные пептидов тахиплезина, ингибирующие грибные патогены растений) и заявку на патент РСТ WO 95/18855 (в которой описаны синтетические антимикробные пептиды, сообщающие устойчивость к болезням).
N) Мембранная пермеаза, образователь или блокатор каналов. См., например, публикацию Jaynes et al., Plant Sci. 89:43 (1993), в которой описана гетерологичная экспрессия аналога литического пептида β-цекропина, сообщающего трансгенным растениям табака устойчивость к Pseudomonas solanacearum.
О) Проникающий в вирусы белок или выделенный из него комплексный токсин. Аккумуляция белков вирусной оболочки в трансформированных растительных клетках сообщает устойчивость к вирусной инфекции и/или заболеванию, вызываемому вирусом, из которого может быть выделен ген белка оболочки, а также к родственным вирусам. См. публикацию Beachy et al., Ann. Rev. Phytopathol. 28:451 (1990). Белок оболочки вируса сообщает трансформированным растениям устойчивость к вирусу мозаики люцерны, вирусу мозаики огурцов, вирусу полосатости табака, вирусу Х картофеля, вирусу Y картофеля, вирусу гравировки табака, вирусу дробления табака и вирусу мозаики табака. Там же.
Р) Антитело, специфичное к насекомым, или выделенный из него иммунотоксин. Таким образом, антитело, направленно воздействующее на важную функцию обмена веществ в кишечнике насекомого, инактивирует пораженный фермент, убивая насекомое. Сравни публикацию Taylor et al., Abstract #497, Seventh Int'l Symposium on Molecular Plant-Microbe Interactions (Edinburgh, Scotland) (1994) (инактивация ферментов в трансгенном табаке при помощи одноцепочечных фрагментов антитела).
Q) Антитело, специфичное к вирусу. См., например, публикацию Tavladoraki et al., Nature 366:469 (1993), в которой показано, что трансгенные растения, экспрессирующие гены рекомбинантного антитела, защищены от воздействия вируса.
R) Блокирующий развитие белок, продуцируемый в природных условиях патогенным организмом или паразитом. Например, грибные эндо-α-1,4-D-полигалактуроназы облегчают колонизацию растения грибами и высвобождение питательных веществ растения путем солюбилизации гомо-α-1,4-D-галактуроназы в клеточной оболочке растения. См. публикацию Lamb et al., Bio/Technology 10:1436 (1992). Клонирование и исследование гена, кодирующего белок, ингибирующий эндополигалактуроназу бобов, описано в публикации Toubart et al., Plant J. 2:367 (1992).
S) Блокирующий развитие белок, продуцируемый в природных условиях растением. Например, в публикации Logemann et al., Bio/Technology 10:305 (1992) показано, что трансгенные растения, экспрессирующие ген, инактивирующий рибосому ячменя, обладают повышенной устойчивостью к грибковому заболеванию.
2. Гены, сообщающие устойчивость к гербициду
А) Гербицид, ингибирующий точку роста или меристему, такой как имидазолинон, сульфонамид или сульфонилмочевина. Типичные гены, относящиеся к данной категории и кодирующие мутантные ферменты ALS и AHAS, описаны соответственно в публикациях Lee et al., EMBO J. 7:1241 (1988) и Miki et al., Theor. Appl. Genet. 80:449 (1990).
В) Глифозат (устойчивость, сообщаемая, например, мутантными генами 5-енолпирувилшикимат-3-фосфат-синтазы (EPSP) (в результате введения рекомбинантных нуклеиновых кислот и/или выполнения разных форм мутагенеза in vivo нативных генов EPSP), генами aroA и генами глифозат-ацетилтрансферазы (GAT)), другие фосфоносоединения, такие как глюфозинат (гены фосфинотрицин-ацетилтрансферазы (РАТ) из вида Strepromyces, включая Streptomyces hygroscopicus и Strepromyces viridichromogenes), пиридинокси- или феноксипропионовые кислоты и циклогексоны (гены, кодирующие ингибитор АССазы). См., например, патент США № 4940835, выданный Shah et al., и патент США № 6248876, выданный Barry et al., в которых описаны нуклеотидные последовательности форм EPSP, способные сообщать растению устойчивость к глифозату. Молекула ДНК, кодирующая мутантный ген aroA, может быть приобретена в АТСС под номером доступа 39256, и нуклеотидная последовательность мутантного гена описана в патенте США № 4769061, выданном Comai. В заявке на европейский патент № 0333033 на имя Kumada et al. и в патенте США № 4975374, выданном Goodman et al., описаны нуклеотидные последовательности генов глутаминсинтетазы, сообщающие устойчивость к гербицидам, таким как L-фосфинотрицин. Нуклеотидная последовательность гена РАТ представлена в заявке на европейский патент № 0242246 на имя Leemans et al., и в публикации DeGreef et al., Bio/Technology 7:61 (1989) описано создание трансгенных растений, экспрессирующих химерные гены, кодирующие активность РАТ. Типичные гены, сообщающие устойчивость к феноксипропионовым кислотам и циклогексонам, таким как сетоксидим и галоксифоп, включают гены Acc1-S1, Acc1-S2 и Acc1-S3, описанные в публикации Marshall et al., Theor. Appl. Genet. 83:435 (1992). Гены GAT, способные сообщать устойчивость к глифозату, описаны в заявке на патент WO 2005012515 на имя Castle et al. Гены, сообщающие устойчивость к гербицидам на основе 2,4-D-феноксипропионовой кислоты и пиридилоксиауксина, описаны в заявке на патент WO 2005107437 компании Dow AgroSciences LLC.
С) Гербицид, ингибирующий фотосинтез, такой как триазин (гены psbA и gs+) или бензонитрил (ген нитрилазы). В публикации Przibila et al., Plant Cell 3:169 (1991) описана трансформация Chlamydomonas плазмидами, кодирующими мутантные гены psbA. Нуклеотидные последовательности для генов нитрилазы представлены в патенте США № 4810648, выданном Stalker, и молекулы ДНК, содержащие указанные гены, могут быть приобретены в АТСС под номерами доступа 53435, 67441 и 67442. Клонирование и экспрессия ДНК, кодирующей глутатион-S-трансферазу, описаны в публикации Hayes et al., Biochem. J. 285:173 (1992).
3. Гены, сообщающие лучший признак или способствующие улучшению признака
а) Изменение обмена жирных кислот, например, путем трансформации растения антисмысловым геном стеарил-АСР-дезатуразы, с целью увеличения содержания стеариновой кислоты в растении. См. публикацию Knultzon et al., Proc. Natl. Acad. Sci. U.S.A. 89:2624 (1992).
В) Уменьшение содержания фитата. 1) Введение гена, кодирующего фитазу, усиливает разрушение фитата, в результате чего образуется больше свободного фосфата в трансформированном растении. См., например, публикацию Van Hartingsveldt et al., Gene 127:87 (1993), в которой представлена нуклеотидная последовательность гена фитазы Aspergillus niger. 2) Введение гена, уменьшающего содержание фитата. В случае кукурузы содержание фитата может быть уменьшено путем клонирования и последующего повторного введения ДНК, ассоциированной с единичным аллелем, отвечающим за мутанты кукурузы, характеризующиеся низкими уровнями фитиновой кислоты. См. публикацию Raboy et al., Maydica 35:383 (1990).
С) Изменение состава углеводов, достигаемое, например, путем трансформации растений геном, кодирующим фермент, изменяющий паттерн разветвления цепи крахмала. См. публикации Shiroza et al., J. Bacteol. 170:810 (1988) (нуклеотидная последовательность гена фруктозилтрансферазы мутантов Streprococcus), Steinmetz et al., Mol. Gen. Genet. 20:220 (1985) (нуклеотидная последовательность гена левансахаразы Bacillus subtilis), Pen et al., Bio/Technology 10:292 (1992) (создание трансгенных растений, экспрессирующих 4-амилазу Bacillus lichenifonn), Elliot et al., Plant Molec. Biol. 21:515 (1993) (нуклеотидные последовательности генов инвертазы томатов), Sogaard et al., J. Biol. Vhem. 268:22480 (1993) (сайтнаправленный мутагенез гена α-амилазы ячменя) и Fisher et al., Plant Physiol. 102:1045 (1993) (фермент II разветвления цепи крахмала эндоспермы кукурузы).
Примеры
Настоящее изобретение далее описано с помощью следующих примеров, которые являются иллюстративными и не ограничивают объем изобретения.
Пример 1
Синтез пептидов
В таблице 1 приведены последовательности следующих проникающих в клетку пептидов (СРР): R9 (Futaki et al., 2001, Suzuki et al., 2002), MPG (Morris, 1997 and Morris, 1999) и γ-ZEIN (Kogan et al., 2001 and 2002). Указанные пептиды были синтезированы в компании American Peptide Company (Sunnyvale, CA) в виде С-концевых амидов. Целостность образцов была исследована при помощи масс-спектрофотометра методами, известными в данной области.
Получение конъюгатов квантовой точки-пептида
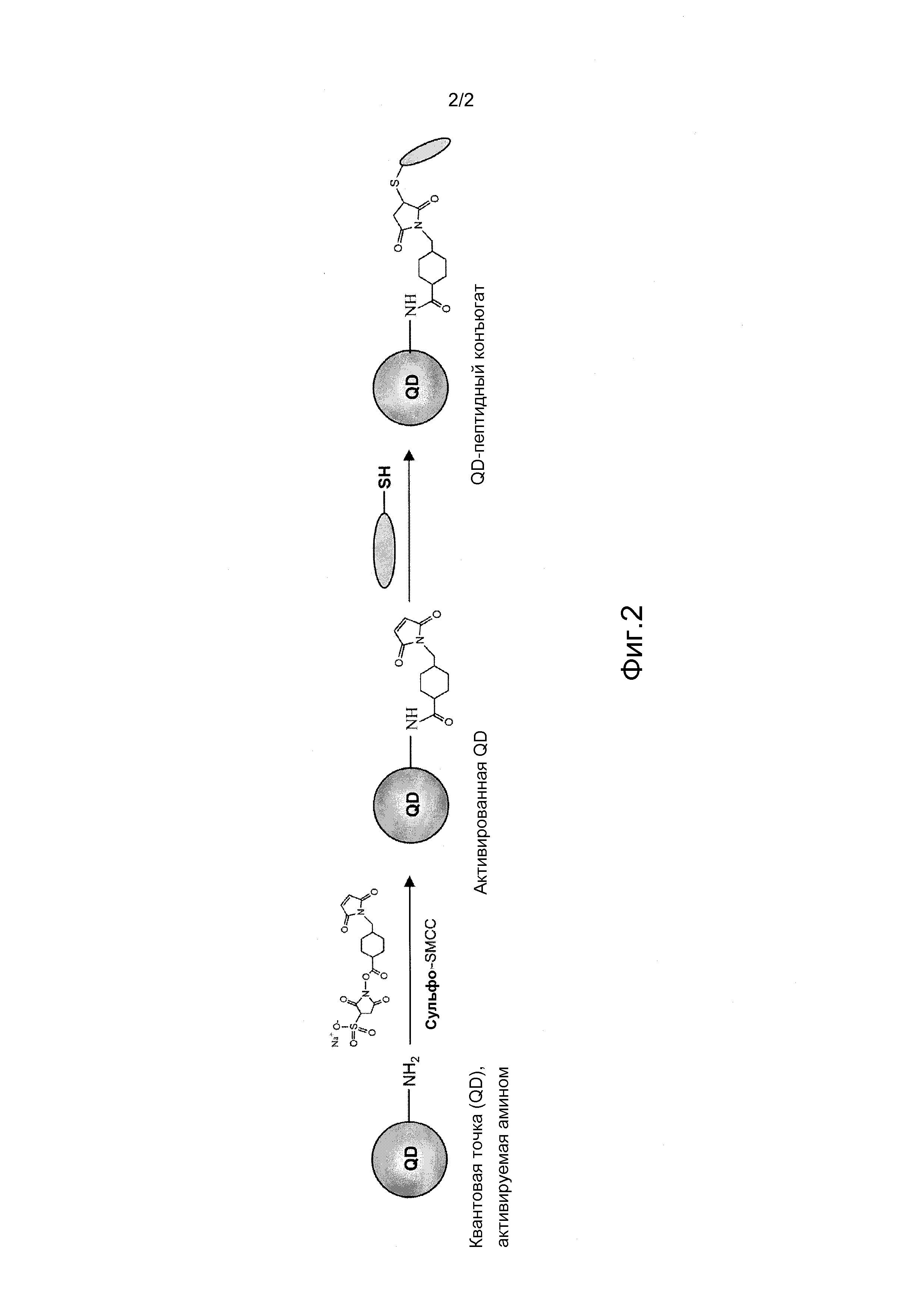
Были получены конъюгаты квантовой точки (QD) и СРР.
Квантовые точки с функциональным амином, приобретенные в компании EvidentTech (Troy, NY), активировали, добавляя примерно 1 мг сульфо-SMCC к 200 мкл квантовых точек с амином (QD514; n=2,3 нмоль) в 200 мкл 50 мМ фосфата натрия, рН 7,4). СРР независимо смешивали с квантовой точкой, активированной имидом малеиновой кислоты, в буфере для конъюгации (1 мМ EDTA, 0,1 М фосфата, 0,15 М NaCl, рН 7,2) и инкубировали при 4°С в течение ночи. Полученные QD-CPP конъюгаты центрифугировали со скоростью 90000 оборотов/мин в течение 3 часов, осадок растворяли в растворе PBS (физиологический раствор с фосфатным буфером). Были получены QD-CPP конъюгаты с молярным соотношением QD и СРР в интервале от 1:100 до 1:300.
Доставка ДНК на QD, опосредованная проникающим в клетку пептидом
Был создан комплекс плазмидной ДНК, pDAB3831 (фигура 1) с QD-CPP конъюгатом. До образования комплекса ДНК с QD-CPP конъюгатом плазмидную ДНК денатурировали и оставляли для повторного отжига. Затем ДНК разводили до конечного объема, равного 10 мкл, в воде, не содержащей ДНКазы. Раствор денатурировали, нагревая до 70°С в течение 5 минут, после чего раствор оставляли медленно охлаждаться до комнатной температуры. Комплекс ДНК с QD-CPP конъюгатом был получен при конечном соотношении CPP-ДНК и QD, равном 1:100. Образование комплекса QD-CPP-ДНК происходило в течение 1 часа при 37°С. Затем добавляли 3% стерильную сахарозу до достижения конечного объема, равного 10 мл. Раствор комплекса QD-CPP-ДНК использовали для трансформации цветочных почек Arabidopsis thaliana.
Трансформация Arabidopsis thaliana комплексом QD-CPP-ДНК
Растительный материал для трансформации in planta
Синхронизированное прорастание семян имеет важное значение для однородного развития цветков у растений Т0. Семена Arabidopsis thaliana cv. Columbia суспендировали в 0,1% растворе агара и инкубировали при 4°С в течение 48 часов до полной стратификации. 60 мг семян взвешивали и переносили в 15 мл пробирку. Добавляли 13 мл 0,1% раствора агара и интенсивно перемешивали до равномерного диспергирования семян. Концентрация семян была равна 4,6 мг/1 мл раствора (или примерно 230 семян/мл). Было получено 6 пробирок (72 мл раствора) для засева 4 плоских поддонов, содержащих 18 (3½ дюйма; 8,89 см) вегетационных сосудов на каждом лотке. Семена инкубировали при 4°С в течение 48 часов до полной стратификации. Каждый вегетационный сосуд засевали отдельно 1,0 мл стратифицированных семян. После засева всех сосудов лотки закрывали куполообразной крышкой для сохранения влаги в почве. Куполообразные крышки удаляли через 15 дней после посева. После прорастания семян растения выращивали в устройстве Conviron (модели СМР4030 и СМР3244, Controlled Environments Limited, Winnipeg, Manitoba, Canada) в условиях длинного светового дня (16-часовой светлый период/8-часовой темный период) при интенсивности света 120-150 мкмоль/м2сек. при постоянной температуре (22°С) и влажности (40-50%). Растения поливали через 10-14 дней после посева раствором Хоагланда и затем деионизированной водой так, чтобы почва была влажной, но не сырой. Через 4 недели после посева цветы срезали для более равномерного роста последующих цветов. На 5-ю неделю после посева были получены растения для трансформации.
Трансформация in planta и скрининг устойчивых растений Т1
Arabidopsis thaliana сорта Columbia трансформировали в соответствии с измененной методикой, разработанной Clough и Bent (S.J. Clough and A.F. Bent, 1998, Plant J. 16:735-43). Из раствора QD-CPP-ДНК было получено 10 мл суспензии, которую использовали для обработки растений Arabidopsis (в основном несозревшие рацемозные соцветия с некоторыми оплодотворенными стручками). До погружения растений в раствор QD-CPP-ДНК добавляли Silwet L-77 до концентрации 0,05% (250 мкл/500 мл) - 0,005% и тщательно смешивали. Надземные части растения погружали в раствор QD-CPP-ДНК на 2-30 секунд при осторожном перемешивании. Обработанные растения выдерживали под пластиковой куполообразной крышкой в течение 16-24 часа при 22-24°С. Растения переносили в устройство Conviron и продолжали выращивать до зрелого состояния и сбора семян. Селекционные лотки (10,5”×21”×1”) использовали для скрининга семян растений Т0 при наличии примерно 10000 семян на каждом лотке. Были использованы два контрольных образца для гарантии правильного выполнения опрыскивания, отрицательный контрольный образец трансформации Co1-0 и гомозиготные семена Columbia для селектируемого маркера РАТ (фосфинотрицин-ацетилтранрсфераза) в качестве положительного контрольного образца трансформации. Для синхронизации роста семена стратифицировали в 0,1% растворе агара в течение 48 часов до посева. Для получения 10000 семян на селекционном лотке 200 мг семян вводили в 0,1% раствор агара и интенсивно перемешивали до равномерного распределения семян. Стратифицированные семена высевали на селекционные лотки, заполненные смесью Sunshine LP5, и слегка поливали раствором Хоагланда. Чтобы селекционное опрыскивание было эффективным, необходимо равномерно посеять 40 мл суспендированных семян на селекционный лоток. После посева все селекционные лотки покрывали куполообразными крышками и выращивали растения для селекции. Куполообразные крышки удаляли примерно через 5 дней после посева.
Кроме того, был выполнен контрольный эксперимент. При выполнении данного эксперимента для трансформации Arabidopsis thaliana использовали раствор, содержащий только ДНК, не образующую комплекс с QD-CPP конъюгатом. Трансформацию отдельно использованной ДНК в качестве контрольного образца выполняли в соответствии с вышеуказанной методикой.
Отбор трансформированных растений
Только что собранные семена Т1 сушили в течение 7 дней при комнатной температуре. Семена Т1 высевали в лотки для прорастания размером 26,5×51 см, в каждый из которых высевали 200 мг аликвоты стратифицированных семян Т1 (~10000 семян), предварительно суспендированных в 40 мл 0,1% раствора агарозы, и хранили при 4°С в течение 2 дней для соответствия требованиям покоя растений и гарантии синхронного прорастания семян.
Смесь Sunshine LP5 покрывали тонкоизмельченным вермикулитом, слегка поливали раствором Хоагланда до влажного состояния и оставляли раствор стекать под действием силы тяжести. 40-мл аликвоту стратифицированных семян равномерно высевали на вермикулит с помощью пипетки и покрывали сохраняющими влажность куполообразными крышками на 4-5 дней. Куполообразные крышки удаляли за 1 день до первоначального отбора трансформантов, который производили, выполняя послевсходовое опрыскивание глюфозинатом.
Через семь дней после посева (DAP) растения Т1 (стадия образования семядоли и 2-4 листьев) в течение пяти дней пять раз опрыскивали 0,2% раствором гербицида Liberty (200 г аэ/л глюфозината, Bayer Crop Sciences, Kansas City, MO) в распыляемом объеме, равном 10 мл/лоток (703 л/га), используя пневматический распылитель DeVilbiss для внесения эффективной нормы глюфозината, равной 280 г аэ/га за одно применение. Выжившие растения (активно растущие растения) выявляли через 4-7 дней после последнего опрыскивания и пересаживали по одному в 3-дюймовые (7,62 см) горшки, содержащие горшечную среду (Metro Mix 360). Пересаженные растения покрывали сохраняющими влажность куполообразными крышками на 3-4 дня и помещали в камеру для выращивания при 22°С или переносили в теплицу. Затем куполообразные крышки удаляли и растения возвращали в теплицу (22±5°С, 50±30% относительная влажность, 14 часов света: 10 часов темноты, минимум 500 мкЕ/м2сек-1 естественного+дополнительного света).
Молекулярные анализы
Геномную ДНК трансгенных растений Arabidopsis экстрагировали из всех листьев 6-недельных растений, используя набор для растений DNAZOL (Invitrogen, Carlsbad, CA) в соответствии с инструкциями производителя. Затравки для ПЦР были предназначены для обнаружения трансгенов yfp и pat. Затравки для yfp представлены в виде SEQ ID NO:4 и SEQ ID NO:5. Затравки для pat представлены в виде SEQ ID NO:6 и SEQ ID NO:7.
SEQ ID NO:4: 5'-TGTTCCACGGCAAGATCCCCTACG-3'
SEQ ID NO:5: 5'-TATTCATCTGGGTGTGATCGGCCA-3'
SEQ ID NO:6: 5'-GGAGAGGAGACCAGTTGAGATTAG-3'
SEQ ID NO:7: 5'-AGATCTGGGTAACTGGCCTAACTG-3'
Реакции амплификации методом ПЦР для pat и yfp выполняли, используя набор TaKaRa Ex Taq (Takara, Otsu, Shiga, Japan). Генные продукты амплифицировали в общем объеме реакционной смеси, равном 50 мкл. Реакционная смесь для ПЦР содержала 100 нг матрицы геномной ДНК, однократный реакционный буфер ExTaq, 0,2 мМ dNTP, 10 пмоль каждой затравки и 0,25 единиц/мкл ExTaq. ПЦР выполняли в следующих условиях: 1 цикл при 96°С в течение 5 минут и 31 цикл в следующих условиях: 94°С в течение 15 секунд, 65°С в течение 30 секунд, 72°С в течение 1 минуты и конечное удлинение при 72°С в течение 7 минут. Продукт амплификации ПЦР анализировали при помощи электрофореза в 0,8% агарозном геле ТАЕ и визуализировали путем окрашивания бромидом этидия. В таблице 2 приведены результаты амплификации продуктов, полученных в результате выполнения указанных реакций.
Пример 2
Витальная визуализация in planta при помощи конъюгатов квантовой точки с проникающим в клетку пептидом
Витальная визуализация при помощи DHLA-кэппированной квантовой точки (QD) и QD-CPP конъюгата
Гибридные белки, состоящие из проникающих в клетку пептидов (СРР) и желтого флуоресцирующего белка (YFP), были получены и выделены в соответствии с описанием, приведенным в предварительной заявке на патент США № 61/319764 и публикации Chen et al., 2007. Разные проникающие в клетку пептиды субклонировали вверху от кодирующей последовательности YFP на однозначных сайтах рестрикции NcoI-SpeI в бактериальном экспрессирующем векторе рЕТ280. Индуцировали экспрессию белков, которые выделяли и очищали в соответствии с описанием, приведенным в предварительной заявке на патент США № 61/319764 и публикации Chen et al., 2007. Последовательности гибридов СРР-YFP приведены в таблице 3.





DHLA-кэппированные квантовые точки (QD) с максимальным излучением при 620 нм были синтезированы в результате выполнения поэтапных реакций металлоорганических предшественников в горячих смесях координирующих растворителей ранее описанными методами. См. публикации Aron et al., 2006; Lu et al., 2007; Doyon et al., 2006; Collins et al., 2003; и Lanio et al., 2000. Нанокристаллы делали гидрофильными путем замены нативной кэппирующей оболочки, состоящей главным образом из триоктилфосфина (ТОР) и оксида триоктилфосфина (ТОРО), бифункциональными лигандами, как было описано ранее. См. публикации Lie et al., 2002; Mani et al., 2006; Desjarlais and Berg, 1993. Были использованы два набора гидрофильных квантовых точек: (1) нанокристаллы, кэппированные дигидролипоевой кислотой, и (2) нанокристаллы, кэппированные смесью дигидролипоевой кислоты с присоединенным полиэтиленгликолем (молекулярная масса ~600) (DHLA-PEG) и DHLA-PEG с концевым биотином (молекулярная масса ~400) (DHLA-PEG-биотин) при молярном соотношении лигандов, равном 9:1. Полученные квантовые точки были соответственно определены как DHLA-QD и DHLA-PEG-биотин-QD.
Помимо вышеописанных молекул СРР (γ-ZEIN, MPG и R9), с DHLA-кэппированными квантовыми точками были связаны две дополнительные молекулы, РЕР1 и ТАТ (таблица 4), ранее описанным методом (Aron et al., 2006). QD-CPP конъюгаты с вышеуказанными молярными соотношениями добавляли к 0,3 мкМ DHLA-кэппированных квантовых точек с излучением 510-620 нм в 10 мМ трис-буфера с хлором, рН 8,0, и инкубировали при комнатной температуре в течение 30 минут. Конъюгаты исследовали при помощи гель-электрофореза, при этом было обнаружено изменение электрофоретической подвижности QD, связанных с СРР. Образцы разводили в однократном буфере ТВЕ (0,09 М трис-буфера, 0,002 М Na2-EDTA, 0,09 М борной кислоты, рН 8,3) и исследовали в 1% или 2% агарозных гелях. Эффект изменения числа молекул СРР в квантовой точке исследовали, наблюдая за флуоресценцией комплекса. Изображения в геле были получены в результате возбуждения QD и/или белка и обнаружения изображений для отдельных флуоресцентных полос в гелях. Кроме того, образование конъюгата было подтверждено в результате исследования изменений в переносе энергии между квантовыми точками (QD) и проникающими в клетку пептидами (СРР) в процессе самосборки.
Поглощение и субклеточная локализация конъюгата квантовой точки с проникающим в клетку пептидом в растительных клетках
Биоконъюгаты квантовой точки разводили полной культуральной средой и добавляли к культурам кластерообразующих клеток Arabidopsis, культурам единичных клеток табака JTNT1 и моркови с интактными оболочками (предварительная заявка на патент США № 61/319764). Раствор инкубировали в количестве 40-150 мкг/мл при 37°С в течение 1-4 часов. Смешанные конъюгаты квантовой точки, состоящие из QD/CPP с соотношением 1:5 или 1:10 при наличии 50 молекул СРР на одну QD, инкубировали с культурами клеток. Избыток несвязанных конъюгатов QD удаляли, промывая культуру по меньшей мере три раза однократным физиологическим раствором с фосфатным буфером (PBS). Затем клетки инкубировали в течение 30 минут при комнатной температуре и дважды промывали PBS.
Эпифлуоресцентные изображения были получены с помощью конфокального микроскопа Leica. Разделенные последовательные флуоресцентные изображения подвергали количественному определению с помощью сдвоенной системы просмотра, оснащенной 565 нм дихроичным фильтром. Для квантовых точек с длиной волны 620 нм был визуализирован комплекс QD-CPP. Для визуализации клеток образцы возбуждали при 488 нм и излучения регистрировали/отделяли с помощью 565 нм дихроичного фильтра и подвергали обратному преобразованию свертки. Флуоресценцию квантовой точки регистрировали при λ<620 нм, и хвост флуоресценции YFP регистрировали при λ>537 нм, если метка гибрида СРР была использована отдельно без квантовых точек. Утечку YFP в окно QD вычитали в виде части деконволюции. Используемые отдельно квантовые точки с излучением 620 нм возбуждали при 488 нм, и их излучения отделяли с помощью 565 нм дихроичного фильтра и подвергали обратному преобразованию свертки. Флуоресценцию красителей DAPI и Calcuofluor возбуждали при помощи ксеноновой (Хе) лампы, и излучение регистрировали, используя кубическое значение DAPI (D350/50X для возбуждения, 400DCLP для дихроичного фильтра, De460/50m для обнаружения). AF647-TF возбуждали при помощи ксеноновой лампы и флуоресценцию обнаруживали, используя кубическое значение Су5 (HQ620/60X для возбуждения, Q660LP для дихроичного фильтра, HQ700/75m для излучения). Кубические значения возбуждения/обнаружения были предоставлены компанией Chroma Technology (Bellows Falls, VT). Дифференциальные интерферирующие контрастные (DIC) изображения были получены при использовании источника яркого света.
Таким образом, функциональные квантовые точки, содержащие разные проникающие в клетку пептиды, предназначенные для отслеживания локализации в единичных клетках с оболочкой Arabidopsis, моркови и табака JTNT1, были обнаружены в результате визуализации с помощью конфокального микроскопа LSM710 Zeiss. Квантовые точки обладают более высокой устойчивостью к разрушению в процессе обмена веществ и более высокой устойчивостью к фотообесцвечиванию. Амины QD, образующие комплекс с СРР, такими как R9, MPG, γ-ZEIN, РЕР1 и ТАТ, инкубировали с единичными клетками Arabidopsis, моркови и табака JTNT1 в течение 30 минут, после чего клетки промывали средой и визуализировали при помощи конфокального микроскопа LSM710 Zeiss. Конфокальный сканер Zeiss LSM710, оснащенный инвертированным микроскопом Axio Observer Z1, был использован с длиной волны возбуждения 514 нм в 3-часовом эксперименте по поглощению и с длиной волны возбуждения 561 нм в 5-часовом эксперименте по поглощению.
Полученные результаты показывают, что живые взвешенные клетки Arabidopsis, табака JTNT1 и моркови с интактными оболочками не флуоресцируют при λ=620 нм. Однако при введении в клетки QD620 была обнаружена интернализация QD в растительные клетки. Изображения каналов, полученные в синем и красном излучении красителя Calcofluor при окрашивании клеточной оболочки и в диапазоне флуоресценции QD620, показали наличие синей флуоресценции, указывающей на присутствие клеточной оболочки, или красной флуоресценции, указывающей на присутствие QD620 в ядре, где была локализована QD620 в результате направленного проникновения в ядро. Наложение всех изображений показало, что интернализированные квантовые точки были сконцентрированы в цитоплазме и ядре.
Было продемонстрировано направленное проникновение СРР-конъюгированной QD620 в ядро взвешенных клеток Arabidopsis, табака JTNT1 и моркови. Направленное проникновение в ядро было подтверждено контрастным окрашиванием ядра ядерным красителем DAPI. DAPI является витальным ядерным красителем, широко используемым в живых растительных клетках. Указанный краситель является флуоресцентным красителем, возбуждаемым ультрафиолетовым светом, для которого характерна синяя флуоресценция при связывании с ДНК в ядре. Хотя диапазон излучения красителя Calcofluor находится в синей части спектра, такое излучение характерно только для оболочки. Наложение изображений QD620-СРР конъюгатов и изображений с окрашиванием DAPI позволило выявить совместную локализацию QD-MPG конъюгатов в ядре.
Интернализированные QD620, конъюгированные с проникающими в клетки пептидами MPG, R9, γ-ZEIN, РЕР1 и ТАТ, характеризовались поверхностными изменениями, и значения их зета-потенциала были измерены в диапазоне 9,5466-10,1586 мВ. Гидродинамические значения конъюгатов находились в диапазоне 122-342 нм. Конъюгаты, инкубированные с взвешенными клетками Arabidopsis, табака JTNT1 и моркови, были интернализированы в интактные клетки и локализованы в ядре, что свидетельствует о направленном проникновении конъюгатов QD в ядро живых клеток в отличие от обработки конъюгатом амина QD620, когда частицы были обнаружены главным образом в цитоплазме и изредка в ядре. Транслокация в ядро клеток Arabidopsis, табака JTNT1 и моркови была обнаружена при использовании комплекса квантовых точек с проникающими в клетку пептидами (MPG, TAT, PEP1, R9 и γ-zein).
Данный пример иллюстрирует возможность применения квантовых точек, меченных проникающими в клетку пептидами в качестве флуоресцирующих носителей, в исследованиях по отслеживанию веществ в живых клетках. Квантовые точки действуют в качестве устойчивых маркеров.
Пример 3
Витальная визуализация in planta при помощи конъюгатов полистирола с проникающими в клетку пептидами
Была произведена доставка и локализация в клетке наночастиц полистирола, гибридизированных с СРР. Наночастицы комплекса полистирола/СРР были введены в живые растительные клетки и направлены в определенные компартменты клетки.
Наночастицы полистирола, меченные проникающим в клетку пептидом, ТАТ, были интернализированы в единичные клетки табака JTNT1.
Конъюгация пептида ТАТ с карбоксилированными частицами FluoSphere
Поглощение флуоресцирующих наночастиц полистирола FluoSphere (диаметр 20 нм) единичными клетками табака JTNT1, имеющими оболочку, анализировали при наличии и отсутствии проникающего в клетку пептида ТАТ.
Наночастицы FluoSphere (Invitrogen, Carlsbad, CA) были получены после хранения при температуре 4°С и приготовлены путем добавления гидрохлорида 1-этил-3-[3-диметиламинопропил]карбодиимида (EDCL) и N-гидроксисукцинимида (NHS). Полученную смесь инкубировали в течение 1 часа для достижения взаимодействия EDCL с наночастицами FluoSphere. Проникающий в клетку пептид ТАТ (карбоксильный конец которого защищен трет-бутанолом) был использован для образования комплекса с наночастицами FluoSphere. Комплекс ТАТ-FluoSphere инкубировали в течение ночи при комнатной температуре. Полученный комплекс очищали при помощи фильтрующей центрифуги Ultra-4 Amicon с пределом отсечения молекулярной массы, равной 50000. Полученный комплекс ТАТ-FluoSphere переносили в чистую пробирку. Комплекс ТАТ-FluoSphere хранили при 4°С до использования в экспериментах.
Исследования поглощения комплекса ТАТ-FluoSphere и неконъюгированных наночастиц Fluosphere взвешенными клетками табака JTNT1
В взвеси единичных клеток JTNT1, полученные в среде NT1B, вводили 3% глицерина. 20 мкл обработанного ультразвуком и интенсивно перемешанного комплекса ТАТ-FluoSphere и не образующих комплекс наночастиц FluoSphere добавляли к клеткам JTNT1 и инкубировали в течение 30, 60 или 120 минут. Единичные клетки JTNT1 выделяли путем центрифугирования пробирок со скоростью 700 оборотов/минуту в течение 5 минут. Супернатант удаляли и клетки дважды промывали 2 мл порциями NT1B. Промытые клетки JTNT1 повторно ресуспендировали в 1 мл среды NT1B, содержащей глицерин, и несколько клеток пипетировали на отдельные предметные стекла, визуализировали при помощи вертикального флуоресцентного микроскопа в режиме яркого поля, и регистрировали флуоресцентное излучение при 580 нм и кубическом значении фильтра. Микроскоп был использован для визуализации клеток JTNT1, обработанных комплексом ТАТ-FluoSphere. Программное обеспечение ImageJ было использовано для отображения и наложения (стековая обработка) изображений в ярком поле и флуоресцентных изображений для более легкого определения локализации наночастиц FluoSphere в клетках табака JTNT1.
В результате обработки клеток табака JTNT1 комплексом ТАТ-FluoSphere в течение 120 минут произошла интернализация комплекса ТАТ-FluoSphere и направленная доставка в ядра клеток. Обработка в течение 30 минут не вызвала значительного поглощения клеткой комплекса ТАТ-FluoSphere. Было отмечено, что некоторое количество комплекса ТАТ-FluoSphere было ассоциировано с оболочками единичных клеток JTNT1. При обработке комплексом ТАТ-FluoSphere в течение 60 минут произошла интернализация некоторого количества комплекса в единичных клетках JTNT1. Большое количество комплекса ТАТ-FluoSphere было обнаружено на периферии клеток, однако, лишь небольшое количество комплекса находилось в непосредственной близости от ядра клеток табака JTNT1. После обработки в течение 120 минут значительное количество комплекса ТАТ-FluoSphere было поглощено клетками табака JTNT1 и направленно доставлено в ядра указанных клеток.
Интернализация наночастиц полистирола была отмечена проникающими в клетку пептидами (СРР) ТАТ, MPG и γ-ZEIN во взвешенных клетках Arabidopsis.
Конъюгация проникающих в клетку пептидов с карбоксилированными наночастицами FluoSphere и поглощение указанных частиц взвешенными клетками Arabidopsis
СРР разных типов метили для оценки проникновения наночастиц через клеточные оболочки и клеточные мембраны в цитоплазму и ядра живых взвешенных клеток Arabidopsis при помощи конфокальной микроскопии. Комплекс наночастиц FluoSphere с проникающими в клетку пептидами (ТАТ, MPG и γ-zein) был получен в соответствии с приведенным выше описанием. Полученный комплекс СРР-FluoSphere смешивали с 0,1 мл взвеси агрегированных клеток Arabidopsis и инкубировали при комнатной температуре в темноте в течение 3 часов (первый эксперимент) или 5 часов (второй эксперимент). Взвеси клеток центрифугировали и удаляли супернатант. Клетки ресуспендировали в свежей культуральной среде. Каплю ресуспендированной взвеси клеток пипетировали на покровное стекло, используя вакуумную смазку для очерчивания периметра вокруг капли взвеси клеток до наложения второго предметного стекла поверх взвеси клеток с образованием сэндвича клеток между двух предметных стекол. Для отображения агрегированных клеток Arabidopsis через 3 часа и 5 часов после воздействия комплекса СРР-FluoSphere использовали конфокальный микроскоп (был использован конфокальный сканер Zeiss LSM710, оснащенный инвертированным микроскопом Axio Observer Z1 с длиной волны возбуждения 514 нм в трехчасовом эксперименте по поглощению и с длиной волны возбуждения 561 нм в пятичасовом эксперименте по поглощению).
В агрегированных клетках Arabidopsis не было обнаружено видимой автофлуоресценции при возбуждении лазером с длиной волны 561 нм и 514 нм в конфокальном микроскопе. Однако в протопластах Arabidopsis была обнаружена некоторая автофлуоресценция при указанной длине волны. Фоновая автофлуоресценция клеток не препятствовала визуализации в экспериментах по поглощению комплекса СРР-FluoSphere.
Протопласты Arabidopsis визуализировали через 5 часов после воздействия конъюгатов СРР-FluoSphere и затем подвергали контрастному окрашиванию красителем Calcofluor. Конъюгаты СРР-FluoSphere были интернализированы протопластами несмотря на то, что протопласты регенерировали материал клеточной оболочки. Жизнеспособность клеток была определена благодаря наличию активных тяжей протопластов, наблюдаемых при помощи конфокальной микроскопии. Кроме того, поглощенный клеткой комплекс проникающего в клетку пептида ТАТ с наночастицами FluoSphere был обнаружен в ядре клетки. Было установлено, что агрегированные клетки Arabidopsis через 5 часов после воздействия конъюгатов проникающего в клетку пептида γ-zein с наночастицами FluoSphere содержат наночастицы FluoSphere в ядре клетки. На полученных изображениях видно, что проникающий в клетку пептид γ-zein опосредует перенос наночастиц FluoSphere в ядра клеток.
Через 6 часов после воздействия немодифицированными наночастицами FluoSphere и контрастного окрашивания красителем Calcofluor указанные наночастицы не были интернализированы в протопласты Arabidopsis и не были перенесены в ядра клеток. Были обнаружены разрушенные и мертвые протопласты, ассоциированные с немодифицированными наночастицами FluoSphere. Таким образом, было продемонстрировано, что немодифицированные наночастицы FluoSphere не были доставлены в клетку живых протопластов Arabidopsis. Однако разрушенные или мертвые клетки интернализировали немодифицированные наночастицы FluoSphere. Полученные данные показывают, что при нарушении целостности мембраны клеток происходит интернализация немодифицированных наночастиц FluoSphere, при этом немодифицированные наночастицы FluoSphere не проникают в живые клетки, имеющие интактную клеточную оболочку и клеточную мембрану.
Приведенные примеры показывают, что гибридизация с СРР обеспечивает эффективную доставку и направленное воздействие на клетку. Кроме того, в зависимости от типа использованного СРР может быть произведено направленное воздействие на ядра. Использование конъюгатов СРР-FluoSphere облегчает поглощение макромолекул, не способных проникать в клетку. Несмотря на то, что данный метод позволяет метить клетки меньшим количеством QD или наночастиц полистирола, по-прежнему существует зависимость от способности клеток к включению чужеродных веществ, так как проникновение в клетки отсутствовало при температуре инкубации 4°С или инкубации с ингибитором, таким как вортманнин.
Конъюгаты TRITC-меченного дендримера РАМАМ, использованные в качестве новых зондов для визуализации in vivo субклеточных компартментов в растительных клетках Arabidopsis
Маркер на основе наночастиц, используемый для количественного определения эндоцитоза в интактных живых клетках, имеющих оболочку, позволяет визуализировать органеллы, опосредующие эндоцитоз растения in vivo. Кроме того, такая визуализация производится без выцветания или обесцвечивания красителя. И наконец, с помощью такого маркера на основе наночастиц можно отслеживать доставку частиц или вещества в живые растительные клетки.
В данном примере рассмотрена визуализация эндоцитозного компартмента в живой растительной клетке с помощью TRITC-меченных частиц дендримера РАМАМ. Указанные частицы используются для витальной визуализации вместо стирильных красителей, таких как FM4-64, которые, как известно, применяются для отслеживания растительных вакуолей и в исследованиях эндоцитоза. Известно, что указанные красители перемещаются из одного компартмента в другой с течением времени, позволяя эффективно визуализировать эндоцитозные компартменты в живых растительных клетках. Однако для отслеживания эндоцитоза можно использовать флуоресцирующие частицы дендримера РАМАМ, при этом биомолекулы могут быть связаны с дендримером или иметь метки для отслеживания их перемещения в живых растительных клетках (в отличие от стирильных красителей, которые окрашивают только пузырьки). Введение вортманнина ингибировало эндоцитоз в данном исследовании, из чего следует, что частицы дендримера РАМАМ обволакивают пузырьки эндосомы и могут быть использованы в качестве маркера в исследованиях по отслеживанию пузырьков. Была продемонстрирована возможность использования TRITC-меченных частиц дендримера РАМАМ для отслеживания эндоцитоза, при этом эндоцитоз был ингибирован при использовании вортманнина. Таким образом, в данном примере было продемонстрировано новое применение указанных частиц в качестве маркера при визуализации живых клеток в исследованиях по отслеживанию пузырьков в растительных клетках.
Мечение TRITC и дендример РАМАМ
TRITC-меченный дендример РАМАМ был получен в соответствии с описанием, приведенным в публикации Pasupathy ert al., 2008. TRITC-меченный дендример РАМАМ добавляли к 0,5-мл аликвоте взвешенных клеток Arabidopsis thaliana, полученных из 7-дневных культур. Кроме того, за 5 минут до визуализации к смеси добавляли краситель Calcofluor. Культуры растений инкубировали в течение 30 минут с комплексами TRITC-дендримера, после чего культуры растений сразу же исследовали при помощи конфокального микроскопа. Для нескольких контрольных образцов к культуре добавляли 25 мкл 10 мкМ вортманнина (MP Biomedicals, Solon, OH) за 30 минут до добавления комплексов TRITC-дендримера. Затем образцы снова инкубировали в течение 30 минут. Живые клетки и кластеры визуализировали при помощи конфокального микроскопа LSM710 в соответствии с вышеописанной методикой. В контрольных клетках, которые не инкубировали с дендримерами, отсутствовала автофлуоресценция при рассмотрении под конфокальным микроскопом.
Конфокальная лазерная сканирующая микроскопия (CLSM)
Для визуализации ассоциации комплексов TRITC-РАМАМ в обработанных клетках и исследования необработанных контрольных клеток с учетом соответствующего времени инкубации клетки дважды промывали культуральной средой или буфером PBS (рН 7,4), добавляли краситель Calcofluor для окрашивания клеточной оболочки в течение 5 минут и сразу же исследовали под конфокальным микроскопом. Клетки исследовали при помощи CLSM (Carl Zeiss LSM-710, Germany), используя аргоновый лазер с длиной волны 600-620 нм для визуализации живых клеток. Внутриклеточное распределение TRITC-РАМАМ исследовали в одной плоскости, а также в виде z-срезов.
Результаты, полученные для клеток, не обработанных вортманнином, показали, что мембраны клеток и эндосомы были окрашены TRITC-меченным дендримером РАМАМ. TRITC-меченные дендримеры РАМАМ проникали в эндосомы. С помощью конфокального лазерного сканирующего микроскопа можно было отслеживать проникновение дендримера РАМАМ в эндосомы клеток через интактную клеточную оболочку. Дендример РАМАМ проникал в клетку в соответствии с процессом эндоцитоза. В присутствии вортманнина TRITC-меченный дендример РАМАМ был локализован в мембране клетки. Так как процесс эндоцитоза был ингибирован, комплекс дендример РАМАМ-СРР не проник в цитозоль клеток Arabidopsis.
Несмотря на то что настоящее изобретение было описано на примере определенных вариантов осуществления изобретения, в данное изобретение могут быть внесены изменения, не выходящие за пределы существа и объема изобретения. Поэтому данная заявка на патент охватывает любые варианты, применения или адаптации изобретения, соответствующие общим принципам. Кроме того, настоящая заявка на патент охватывает такие отступления от настоящего описания изобретения, которые являются известной или общепринятой практикой в той области, к которой относится настоящее изобретение, и которые входят в объем прилагаемой формулы изобретения и ее эквивалентов.
Реферат
Изобретение относится к области биохимии, в частности к способу введения представляющей интерес нуклеиновой кислоты в растительную клетку, имеющую клеточную стенку, который включает взаимодействие растительной клетки, имеющей клеточную стенку, с активированным QD-пептидным конъюгатом. Изобретение также относится к способу проведения устойчивой экспрессии гена, способу переноса молекулярного вещества в растительную клетку, а также к способу идентификации трансформанта растения. Изобретение позволяет эффективно осуществлять введения представляющей интерес нуклеиновой кислоты в растительную клетку, имеющую клеточную стенку. 4 н. и 15 з.п. ф-лы, 2 ил., 4 табл., 3 пр.
Формула
Документы, цитированные в отчёте о поиске
Трансформация хлоропластов ряски
Комментарии