Полинуклеотид, кодирующий гомолог ацил-соа-синтетазы, и его применение - RU2528248C2
Код документа: RU2528248C2
Чертежи












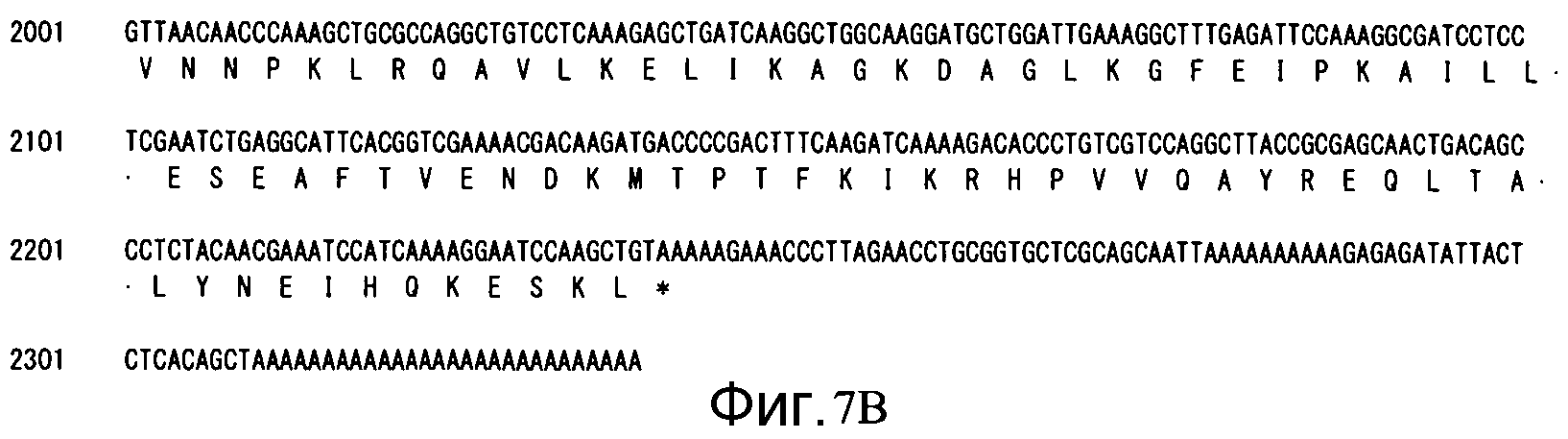



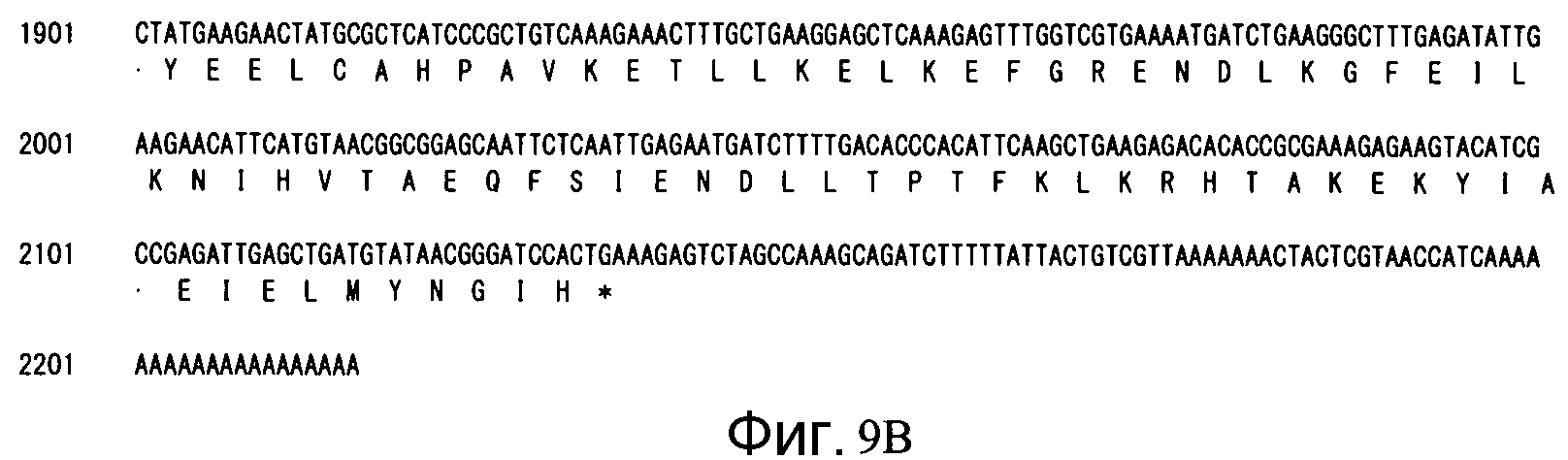



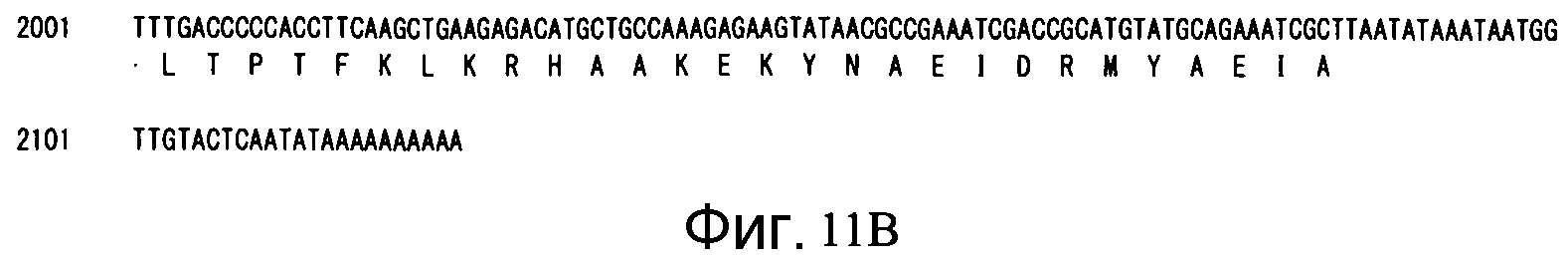



















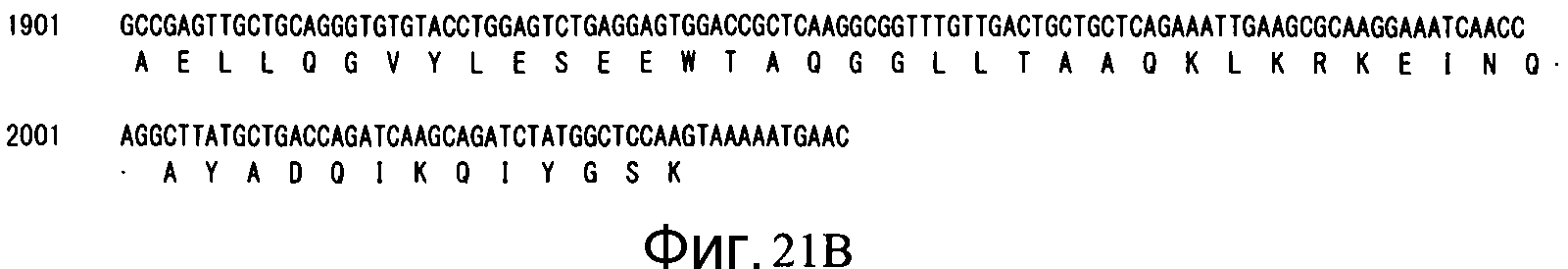











Описание
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Данное изобретение относится к полинуклеотиду, кодирующему гомолог ацил-СоА-синтетазы, и его применению.
УРОВЕНЬ ТЕХНИКИ
Жирные кислоты, содержащие две или более ненасыщенные связи, вместе называют полиненасыщенными жирными кислотами (PUFA), и известно, что они в частности включают в себя арахидоновую кислоту (ARA), дигомо-γ-линоленовую кислоту (DGLA), эйкозапентаеновую кислоту (EPA), докозагексаеновую кислоту (DHA) и т.д. Некоторые из этих полиненасыщенных жирных кислот не могут быть синтезированы в организме животного. Таким образом, необходимо компенсировать эти полиненасыщенные жирные кислоты в виде незаменимых аминокислот из пищи.
Полиненасыщенные жирные кислоты широко распространены; например, арахидоновая кислота может быть выделена из липидов, экстрагированных из надпочечников и печени животных. Однако полиненасыщенные жирные кислоты, содержащиеся в органах животных, составляют лишь малое количество и не могут быть получены в достаточном количестве для больших подач при простом экстрагировании или выделении из органов животных. По этой причине были разработаны микробиологические способы для получения полиненасыщенных жирных кислот культивированием различных микроорганизмов. Кроме того, известно, что микроорганизмы рода Mortierella продуцируют липиды, содержащие полиненасыщенные жирные кислоты, такие как арахидоновая кислота и т.п.
Были предприняты другие попытки получения полиненасыщенных жирных кислот в растениях. Полиненасыщенные жирные кислоты входят в состав запасных липидов, таких как триацилглицериды, и известно, что они накапливаются в мицелии микроорганизмов или семенах растений.
Ацил-СоА-синтетаза (ACS) является ферментом, катализирующим тиоэстерификацию жирных кислот, и кофермент А (CoA) катализирует следующую реакцию.
Жирная кислота+CoASH+АТФ→Ацил-CoA+АМФ+PPi
Ацил-СоА, продуцируемый ACS, участвует в различных явлениях живой природы, включающих в себя биосинтез и ремоделирование липидов, продуцирование энергии β-окислением, ацилирование белков, регуляцию экспрессии жирными кислотами и т.д. Кроме того, сообщалось, что ACS ассоциирована с внеклеточным поглощением жирных кислот, внутриклеточным транспортом жирных кислот и т.д. (Непатентные Документы 1 и 2). В связи с этим считается, что он регулирует активность ACS при продуцировании полиненасыщенных жирных кислот или т.п. утилизацией микроорганизмами или растениями.
В дрожжах Saccharomyces cerevisiae, используемых в качестве эукариота-модели, известны шесть (6) генов ацил-CoA-синтетазы (ScFAA1, ScFAA2, ScFAA3, ScFAA4, ScFAT1 и ScFAT2) (Непатентный Документ 1). Белки, кодируемые этими генами, являются разными по субстратной специфичности, таймингу экспрессии, внутриклеточной локализации и функции.
Патентный Документ 1 описывает девять (9) генов ацил-CoA-синтетазы (ScACS), полученных из Schizochytrium sp. Патентный Документ 1 описывает также увеличенное продуцирование DPA (n-6) (докозапентановой кислоты (n-6)) или DHA при коэкспрессии гена, кодирующего систему PUFA-синтаз Schizochytrium sp., с ScACS, в сравнении со случаем, когда не участвует коэкспрессия с ScACS.
Кроме того, сообщалось также о генах ацил-CoA-синтетаз, полученных из животных и растений (Непатентный Документ 2 и Патентный документ 2).
[Патентный Документ 1] Japanese Unexamined Patent Application Publication (Translation of PCT Application) No. 2009-529890
[Патентный Документ 2] PCT International Publication Pamphlet WO 0209295
[Непатентный Документ 1] B. B. A. 1771, 286-298, 2007
[Непатентный Документ 2] Exp. Biol. Med., 233 (5), 507-521, 2008
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
При описанных выше обстоятельствах было желательным выделение нового гена, который увеличивает количество жирных кислот, продуцируемых в клетке-хозяине, или изменяет состав продуцируемых жирных кислот, при экспрессии этого гена в этой клетке-хозяине.
В результате обширных исследований авторам данного изобретения удалось клонировать ген, кодирующий гомолог ACS липидпродуцирующего гриба Mortierella alpina (далее "M. alpina") (MaACS), и осуществить данное изобретение. То есть, данное изобретение обеспечивает следующие полинуклеотиды, белки, экспрессирующие векторы, трансформанты и способ получения липидов или липидных композиций и пищевых продуктов, и т.д. с использованием этих трансформантов, а также пищевые продукты, получаемые при помощи этого способа, и т.д.
Таким образом, данное изобретение характеризуется следующим образом.
[1] Полинуклеотид по любому из пунктов, выбранных из группы, состоящей из (a)-(e), ниже:
(a) полинуклеотид, содержащий любую нуклеотидную последовательность, выбранную из группы, состоящей из нуклеотидных последовательностей, представленных в SEQ ID NO: 1, 6, 11, 16, 21, 26, 31, 36, 41, 46, 51 и 56;
(b) полинуклеотид, кодирующий белок, состоящий из любой аминокислотной последовательности, выбранной из группы, состоящей из аминокислотных последовательностей, представленных в SEQ ID NO: 2, 7, 12, 17, 22, 27, 32, 37, 42, 47, 52 и 57;
(c) полинуклеотид, кодирующий белок, состоящий из аминокислотной последовательности, где 1-100 аминокислот делетированы, заменены, инсертированы и/или добавлены в любую аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей, представленных в SEQ ID NO: 2, 7, 12, 17, 22, 27, 32, 37, 42, 47, 52 и 57, и имеющий ацил-CoA-синтетазную активность или активность увеличения количества или изменения состава жирных кислот, продуцируемых в клетке-хозяине при экспрессии в этой клетке-хозяине;
(d) полинуклеотид, кодирующий белок, имеющий аминокислотную последовательность, имеющую по меньшей мере 60% идентичность любой аминокислотной последовательности, выбранной из группы, состоящей из аминокислотных последовательностей, представленных в SEQ ID NO: 2, 7, 12, 17, 22, 27, 32, 37, 42, 47, 52 и 57, и имеющий ацил-CoA-синтетазную активность или активность увеличения количества или изменения состава жирных кислот, продуцируемых в клетке-хозяине при экспрессии в этой клетке-хозяине; и
(e) полинуклеотид, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной любой нуклеотидной последовательности, выбранной из группы, состоящей из нуклеотидных последовательностей, представленных в SEQ ID NO: 1, 6, 11, 16, 21, 26, 31, 36, 41, 46, 51 и 56, при строгих условиях, и который кодирует белок, имеющий ацил-CoA-синтетазную активность или активность увеличения количества или изменения состава жирных кислот, продуцируемых в клетке-хозяине при экспрессии в этой клетке-хозяине.
[2] Полинуклеотид по п. 1, который представляет собой полинуклеотид, определенный либо в (f), либо в (g) ниже:
(f) полинуклеотид, кодирующий белок, состоящий из аминокислотной последовательности, где 1-10 аминокислот делетированы, заменены, инсертированы и/или добавлены в любую из аминокислотных последовательностей, выбранных из группы, состоящей из аминокислотных последовательностей, представленных в SEQ ID NO: 2, 7, 12, 17, 22, 27, 32, 37, 42, 47, 52 и 57, и имеющий ацил-CoA-синтетазную активность или активность увеличения количества или изменения состава жирных кислот, продуцируемых в клетке-хозяине при экспрессии в этой клетке-хозяине; и
(g) полинуклеотид, кодирующий белок, имеющий аминокислотную последовательность, имеющую по меньшей мере 90% идентичность любой аминокислотной последовательности, выбранной из группы, состоящей из аминокислотных последовательностей, представленных в SEQ ID NO: 2, 7, 12, 17, 22, 27, 32, 37, 42, 47, 52 и 57, и ацил-CoA-синтетазную активность или активность увеличения количества или изменения состава жирных кислот, продуцируемых в клетке-хозяине при экспрессии в этой клетке-хозяине.
[3] Полинуклеотид по [1] выше, содержащий любую нуклеотидную последовательность, выбранную из группы, состоящей из нуклеотидных последовательностей, представленных в SEQ ID NO: 1, 6, 11, 16, 21, 26, 31, 36, 41, 46, 51 и 56.
[4] Полинуклеотид по [1] выше, кодирующий белок, состоящий из любой аминокислотной последовательности, выбранной из группы, состоящей из аминокислотных последовательностей, представленных в SEQ ID NO: 2, 7, 12, 17, 22, 27, 32, 37, 42, 47, 52 и 57.
[5] Полинуклеотид по любому из [1]-[4] выше, который представляет собой ДНК.
[6] Белок, кодируемый полинуклеотидом по любому из [1]-[5] выше.
[7] Вектор, содержащий полинуклеотид по любому из [1]-[5] выше.
[8] Трансформант, не являющийся человеком, в который введен полинуклеотид по любому из [1]-[5] выше, или вектор по [7] выше.
[9] Способ получения композиции липидов или жирных кислот, который предусматривает сбор композиции липидов или жирных кислот из культуры трансформанта по [8] выше.
[10] Способ по [9] выше, где этим липидом является триацилглицерин.
[11] Способ по [9] выше, где этой жирной кислотой является полиненасыщенная жирная кислота, имеющая по меньшей мере 18 атомов углерода.
[12] Пищевой продукт, фармацевтический препарат, косметический препарат или мыло, содержащие композицию липидов или жирных кислот, полученную способом получения по [9] выше.
Полинуклеотид по данному изобретению может быть использован для трансформации подходящей клетки-хозяина. Полученный таким образом трансформант может быть использован для получения композиций жирных кислот, пищевых продуктов, косметических препаратов, фармацевтических препаратов, мыла и т.д.
Более конкретно, трансформант по этому изобретению обеспечивает чрезвычайно высокую эффективность продуцирования липидов и жирных кислот. Таким образом, данное изобретение может быть эффективно использовано для приготовления фармацевтических препаратов или диетических пищевых продуктов, которые требуют большого количества липидов или жирных кислот.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Фиг. 1 показывает соответствие между кДНК-последовательностью и предположительной аминокислотной последовательностью MaACS-1.
Фиг. 2A показывает сопоставление между геномной последовательностью и последовательностью CDS MaACS-1.
Фиг. 2В является продолжением Фиг. 2А.
Фиг. 3A показывает соответствие между кДНК-последовательностью и предположительной аминокислотной последовательностью MaACS-2.
Фиг. 3В является продолжением Фиг. 3А.
Фиг. 4А показывает сопоставление между геномной последовательностью и последовательностью CDS MaACS-2.
Фиг. 4В является продолжением Фиг. 4А.
Фиг. 4С является продолжением Фиг. 4В.
Фиг. 5 показывает соответствие между кДНК-последовательностью и предположительной аминокислотной последовательностью MaACS-3.
Фиг. 6А показывает сопоставление между геномной последовательностью и последовательностью CDS MaACS-3.
Фиг. 6B является продолжением Фиг. 6A.
Фиг. 7A показывает соответствие между кДНК-последовательностью и предположительной аминокислотной последовательностью MaACS-4.
Фиг. 7B является продолжением Фиг. 7A.
Фиг. 8A показывает сопоставление между геномной последовательностью и последовательностью CDS MaACS-4.
Фиг. 8B является продолжением Фиг. 8A.
Фиг. 8C является продолжением Фиг. 8B.
Фиг. 9А показывает соответствие между кДНК-последовательностью и предположительной аминокислотной последовательность MaACS-5.
Фиг. 9B является продолжением Фиг. 9A.
Фиг. 10A показывает сопоставление между геномной последовательностью и последовательностью CDS MaACS-5.
Фиг. 10B является продолжением Фиг. 10A.
Фиг. 11A показывает соответствие между кДНК-последовательностью и предположительной аминокислотной последовательностью MaACS-6.
Фиг. 11B является продолжением Фиг. 11A.
Фиг. 12A показывает сопоставление между геномной последовательностью и последовательностью CDS MaACS-6.
Фиг. 12B является продолжением Фиг. 12A.
Фиг. 13 показывает соответствие между кДНК-последовательностью и предположительной аминокислотной последовательностью MaACS-7.
Фиг. 14A показывает сопоставление между геномной последовательностью и последовательностью CDS MaACS-7.
Фиг. 14B является продолжением Фиг. 14A.
Фиг. 15A показывает соответствие между кДНК-последовательностью и предположительной аминокислотной последовательностью MaACS-8.
Фиг. 15B является продолжением Фиг 15A.
Фиг. 16A показывает сопоставление между геномной последовательностью и последовательностью CDS MaACS-8.
Фиг. 16B является продолжением Фиг. 16A.
Фиг. 16C является продолжением Фиг. 16B.
Фиг. 17 показывает соответствие между кДНК-последовательностью и предположительной аминокислотной последовательностью MaACS-9.
Фиг. 18A показывает сопоставление между геномной последовательностью и последовательностью CDS MaACS-9.
Фиг. 18B является продолжением Фиг. 18A.
Фиг. 19A показывает соответствие между кДНК-последовательностью и предположительной аминокислотной последовательностью MaACS-10.
Фиг. 19B является продолжением Фиг. 19A.
Фиг. 20A показывает сопоставление между геномной последовательностью и последовательностью CDS MaACS-10.
Фиг. 20B является продолжением Фиг. 20A.
Фиг. 20C является продолжением Фиг. 20B.
Фиг. 21A показывает соответствие между кДНК-последовательностью и предположительной аминокислотной последовательностью MaACS-11.
Фиг. 21B является продолжением Фиг. 21A.
Фиг. 22A показывает сопоставление между геномной последовательностью и последовательностью CDS MaACS-11.
Фиг. 22B является продолжением Фиг. 22A.
Фиг. 23A показывает соответствие между кДНК-последовательностью и предположительной аминокислотной последовательностью MaACS-12.
Фиг. 23B является продолжением Фиг. 23A.
Фиг. 24A показывает сопоставление между геномной последовательностью и последовательностью CDS MaACS-12.
Фиг. 24B является продолжением Фиг. 24A.
Фиг. 25A показывает сопоставление между MaACS, имеющими относительно высокую гомологию аминокислотной последовательности с полученным из S. cerevisiae FAA-белком (FAA: активация жирных кислот), и этим FAA-белком. Подчеркнутые одной линией и подчеркнутые двойной линией последовательности обозначают мотив АТФ-АМФ и мотив FACS/VLACS-FATP, соответственно.
Фиг. 25В является продолжением Фиг. 25A.
Фиг. 25C является продолжением Фиг. 25B.
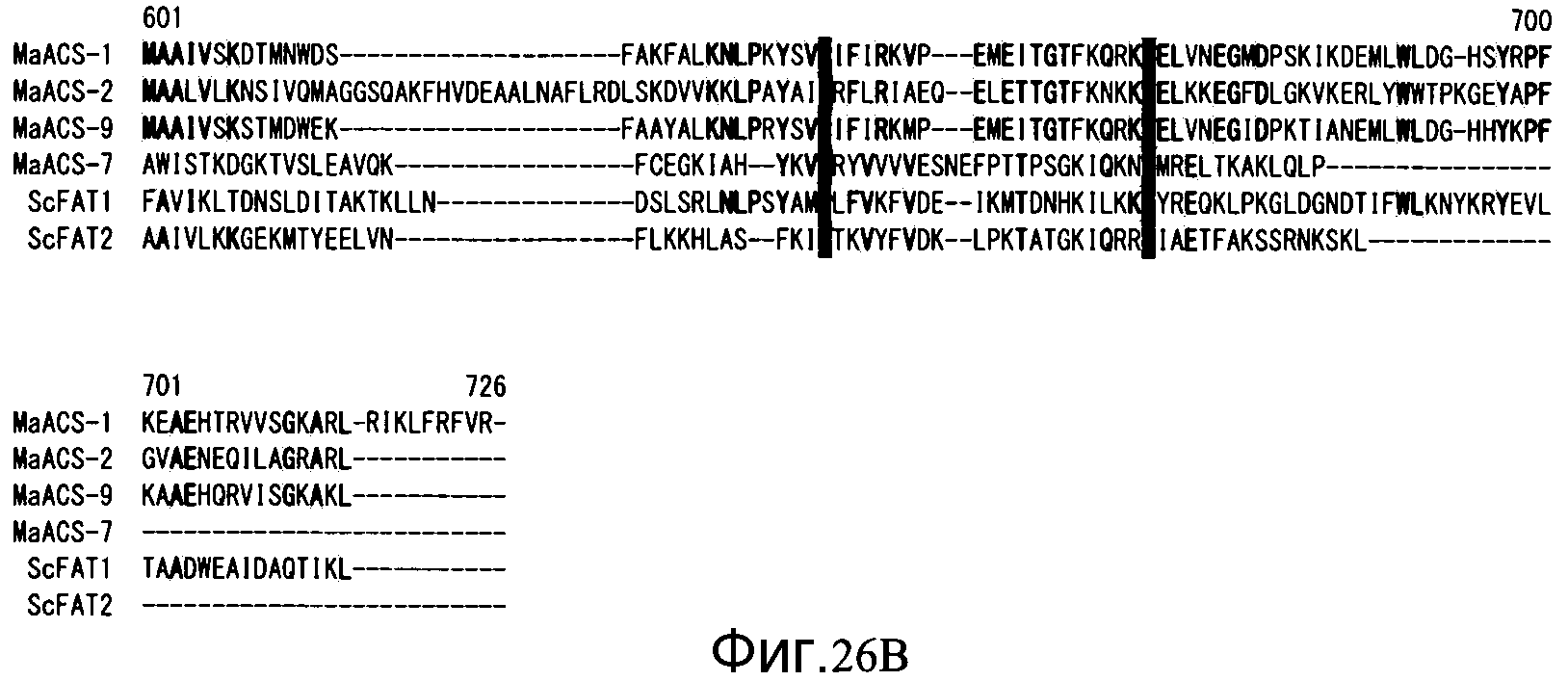
Фиг. 26A показывает сопоставление между MaACS, имеющими относительно высокую гомологию последовательности с полученным из S. cerevisiae FAT-белком (FAT: трансфераза жирных кислот), и этим FAT-белком. Подчеркнутые одной линией и подчеркнутые двойной линией последовательности обозначают мотив АТФ-АМФ и мотив FACS/VLACS-FATP, соответственно.
Фиг. 26B является продолжением Фиг. 26A.
Фиг. 27 показывает изменения на протяжении времени в продуцировании липидов (Фиг. 27A) и продуцировании арахидоновой кислоты (Фиг. 27B) в расчете на мицелий в сверхэкспрессирующем MaACS-10 M. alpina.
Фиг. 28 показывает изменения на протяжении времени в продуцировании липидов (Фиг. 28A) и продуцировании арахидоновой кислоты (Фиг. 28B) в расчете на мицелий в сверхэкспрессирующем MaACS-11 M. alpina.
НАИЛУЧШИЙ СПОСОБ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Данное изобретение подробно описывается далее. Предполагается, что описанные ниже варианты осуществления предназначены только для описания изобретения, но не для ограничения изобретения только следующими вариантами осуществления. Данное изобретение может осуществляться различными путями без отклонения от сущности этого изобретения.
Все публикации, опубликованные заявки на патент, патенты и другие патентные документы, цитируемые в этой заявке, включены здесь посредством ссылки в их полном объеме. Таким образом, эта заявка включает в себя ссылку на содержания описания и фигур в Японской заявке на патент (№ 2010-19967), поданной 1 февраля 2010 года, приоритет которой был заявлен.
Как будет описано позднее подробно в ПРИМЕРАХ, приведенных ниже, авторам данного изобретения впервые удалось клонирование полноразмерной кДНК генов гомолога ACS продуцирующего липиды гриба M. alpina (MaACS-1~12). Авторы данного изобретения идентифицировали также нуклеотидные последовательности геномных ДНК MaACS-1~12 из M. alpina и их предположительные аминокислотные последовательности. ORF-последовательности, предположительные аминокислотные последовательности, CDS-последовательности, кДНК-последовательности и геномные последовательности MaACS-1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 и 12, представлены как последовательности SEQ ID NO: 1, 6, 11, 16, 21, 26, 31, 36, 41, 46, 51 и 56 (далее эти последовательности вместе называются "ORF-последовательностями MaACS-1~12"), SEQ ID NO: 2, 7, 12, 17, 22, 27, 32, 37, 42, 47, 52 и 57 (далее эти последовательности вместе называются "аминокислотными последовательностями MaACS-1~12"), SEQ ID NO: 3, 8, 13, 18, 23, 28, 33, 38, 43, 48, 53 и 58 (далее эти последовательности вместе называются "CDS-последовательностями MaACS-1~12"), SEQ ID NO: 4, 9, 14, 19, 24, 29, 34, 39, 44, 49, 54 и 59 (далее эти последовательности называются "кДНК-последовательностями MaACS-1~12") и SEQ ID NO: 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55 и 60 (далее эти последовательности вместе называются "геномными последовательностиями MaACS-1~12"), соответственно. Эти полинуклеотиды и белки могут быть получены способами, описанными в ПРИМЕРАХ ниже, известными способами генной инженерии, известными способами синтеза и т.д.
1. Полинуклеотид по данному изобретению
Сначала данное изобретение обеспечивает полинуклеотид, описанный в любом полинуклеотиде, выбранном из группы, состоящей из (a)-(g), ниже:
(a) полинуклеотид, содержащий нуклеотидную последовательность, выбранную из группы, состоящей из ORF-последовательностей MaACS-1~12;
(b) полинуклеотид, содержащий любую нуклеотидную последовательность, выбранную из группы, состоящей из кДНК-последовательностей MaACS-1~12;
(c) полинуклеотид, кодирующий белок, состоящий из любой аминокислотной последовательности, выбранной из группы, состоящей из аминокислотных последовательностей MaACS-1~12;
(d) полинуклеотид, кодирующий белок, состоящий из аминокислотной последовательности, где 1-100 аминокислот делетированы, заменены, инсертированы и/или добавлены в любую аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей MaACS-1~12, и имеющий ацил-CoA-синтетазную активность или активность увеличения количества и/или изменения состава жирных кислот, продуцируемых в клетке-хозяине при экспрессии в этой клетке-хозяине;
(e) полинуклеотид, кодирующий белок, имеющий аминокислотную последовательность, имеющую по меньшей мере 60% идентичность любой аминокислотной последовательности, выбранной из группы, состоящей из аминокислотных последовательностей MaACS-1~12, и имеющий ацил-CoA-синтетазную активность или активность увеличения количества и/или изменения состава жирных кислот, продуцируемых в клетке-хозяине при экспрессии в этой клетке-хозяине; и
(f) полинуклеотид, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной любой нуклеотидной последовательности, выбранной из группы, состоящей из ORF-последовательностей MaACS-1~12, при строгих условиях, и который кодирует белок, имеющий ацил-CoA-синтетазную активность или активность увеличения количества и/или изменения состава жирных кислот, продуцируемых в клетке-хозяине при экспрессии в этой клетке-хозяине; и
(g) полинуклеотид, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной любой нуклеотидной последовательности, выбранной из группы, состоящей из кДНК-последовательностей MaACS-1~12, при строгих условиях, и который кодирует белок, имеющий ацил-CoA-синтетазную активность или активность увеличения количества и/или изменения состава жирных кислот, продуцируемых в клетке-хозяине при экспрессии в этой клетке-хозяине.
В данном контексте термин “полинуклеотид" обозначает ДНК или РНК.
В данном контексте термин "полинуклеотид, который гибридизуется при строгих условиях", относится к полинуклеотиду, полученному при помощи способа гибридизации колоний, способа гибридизации бляшек, способа гибридизации по Саузерну или т.п., с использованием в качестве зонда, например полинуклеотида, состоящего из нуклеотидной последовательности, комплементарной любой нуклеотидной последовательности, выбранной из группы, состоящей из ORF-последовательностей MaACS-1~12, или любой нуклеотидной последовательности, выбранной из группы, состоящей из кДНК-последовательностей MaACS-1~12, или целого или части полинуклеотида, состоящего из нуклеотидной последовательности, выбранной из группы, состоящей из аминокислотных последовательностей MaACS-1~12. Для способов гибридизации используются способы, описанные, например, в "Sambrook & Russell, Molecular Cloning: A Laboratory Manual Vol. 3, Cold Spring Harbor, Laboratory Press 2001", "Ausubel, Current Protocols in Molecular Biology, John Wiley & Sons 1987-1997", и т.д.
В данном контексте термин "условия строгости" может обозначать любое из условий низкой строгости, условий средней строгости и условий высокой строгости. Термин “условия низкой строгости” обозначает, например, 5x SSC, 5x раствор Денхардта, 0,5% SDS, 50% формамид при 32°C. Термин "условия средней строгости" обозначает, например, 5x SSC, 5x раствор Денхардта, 0,5% SDS, 50% формамид при 42°C или 5x SSC, 1% SDS, 50 мМ Трис-HCl (pH 7,5), 50% формамид при 42°C. Термин "условия высокой строгости" обозначает, например, 5x SSC, 5x раствор Денхардта, 0,5% SDS, 50% формамид при 50°C или 0,2×SSC, 0,1% SDS при 65°C. При этих условиях ожидается, что ДНК с более высокой идентичностью получают эффективно при более высоких температурах, хотя в строгости гибридизации участвуют многие факторы, включающие в себя температуру, концентрацию зонда, длину зонда, ионную силу, время, концентрацию соли и другие, и специалист с квалификацией в этой области может удобным образом выбирать эти факторы для достижения подобной строгости.
При использовании коммерчески доступных наборов для гибридизации, может быть использована, например, система прямого мечения и детектирования Alkphos (GE Healthcare). В этом случае, согласно прилагаемому протоколу, после культивирования с меченым зондом в течение ночи, мембрану промывают первичным промывочным буфером, содержащим 0,1% (масс./об.) SDS (ДСН), при 55°C для детектирования гибридизованной ДНК. Альтернативно, в получении зонда на основе нуклеотидной последовательности, комплементарной любой нуклеотидной последовательности, выбранной из группы, состоящей из ORF-последовательностей MaACS-1~12, или любой нуклеотидной последовательности, выбранной из группы, состоящей из кДНК-последовательностей MaACS-1~12, или на основе всей или части нуклеотидной последовательности, кодирующей любую аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей MaACS-1~12, гибридизация может быть детектирована с набором детектирования нуклеиновых кислот DIG (Roche Diagnostics) при мечении этого зонда дигоксигенином (DIG) с использованием коммерчески доступного реагента (например, смеси для ПЦР-мечения (Roche Diagnostics) и т.д.).
Кроме полинуклеотидов, описанных выше, другие полинуклеотиды, которые могут быть гибридизованы, включают в себя ДНК, имеющую 50% или более высокую, 51% или более высокую, 52% или более высокую, 53% или более высокую, 54% или более высокую, 55% или более высокую, 56% или более высокую, 57% или более высокую, 58% или более высокую, 59% или более высокую, 60% или более высокую, 61% или более высокую, 62% или более высокую, 63% или более высокую, 64% или более высокую, 65% или более высокую, 66% или более высокую, 67% или более высокую, 68% или более высокую, 69% или более высокую, 70% или более высокую, 71% или более высокую, 72% или более высокую, 73% или более высокую, 74% или более высокую, 75% или более высокую, 76% или более высокую, 77% или более высокую, 78% или более высокую, 79% или более высокую, 80% или более высокую, 81% или более высокую, 82% или более высокую, 83% или более высокую, 84% или более высокую, 85% или более высокую, 86% или более высокую, 87% или более высокую, 88% или более высокую, 89% или более высокую, 90% или более высокую, 91% или более высокую, 92% или более высокую, 93% или более высокую, 94% или более высокую, 95% или более высокую, 96% или более высокую, 97% или более высокую, 98% или более высокую, 99% или более высокую, 99,1% или более высокую, 99,2% или более высокую, 99,3% или более высокую, 99,4% или более высокую, 99,5% или более высокую, 99,6% или более высокую, 99,7% или более высокую, 99,8% или более высокую или 99,9% или более высокую идентичность с ДНК для любой нуклеотидной последовательности, выбранной из группы, состоящей из ORF-последовательностей MaACS-1~12, или для любой нуклеотидной последовательности, выбранной из группы, состоящей из кДНК-последовательностей MaACS-1~12, или с ДНК, кодирующей любую аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей MaACS-1~12, как рассчитано программой поиска гомологии, такой как FASTA, BLAST и т.д., с использованием параметров по умолчанию.
Идентичность между аминокислотными последовательностями или нуклеотидными последовательностями может быть определена с использованием FASTA (Science 227 (4693): 1435-1441, (1985)), алгоритма BLAST (Basic Local Alignment Search Tool) согласно Karlin and Altschul (Proc. Natl. Acad. Sci. USA, 87: 2264-2268, 1990; Proc. Natl. Acad. Sci. USA, 90: 5873, 1993). Были разработаны программы, названные blastn, blastx, blastp, tblastn и tblastx, на основе алгоритма BLAST (Altschul S. F. et al., J. Mol. Biol. 215: 403, 1990). При секвенировании нуклеотидной последовательности с использованием blastn, параметрами являются, например, оценка (счет)=100 и длина слова=12. При секвенировании нуклеотидной последовательности с использованием blastp, параметрами являются, например, оценка (счет)=50 и длина слова=3. При использовании программ BLAST и Gapped BLAST, для каждой из этих программ используются параметры по умолчанию.
Полинуклеотиды по данному изобретению, описанные выше, могут быть получены известными способами генетической инженерии или известными способами синтеза.
2. Белок по данному изобретению
Данное изобретение обеспечивает белки, показанные ниже.
(i) Белок, кодируемый полинуклеотидом любого из (a)-(g) выше.
(ii) Белок, содержащий любую аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей MaACS-1~12.
(iii) Белок, состоящий из аминокислотной последовательности, в которой одна или несколько аминокислот делетированы, заменены, инсертированы и/или добавлены в любую аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей MaACS-1~12, и имеющий ацил-CoA-синтетазную активность или активность увеличения количества и/или изменения состава жирных кислот, продуцируемых в клетке-хозяине при экспрессии в этой клетке-хозяине.
(iv) Белок, имеющий аминокислотную последовательность, имеющую по меньшей мере 90% идентичность любой аминокислотной последовательности, выбранной из группы, состоящей из аминокислотных последовательностей MaACS-1~12, и имеющий ацил-CoA-синтетазную активность или активность увеличения количества и/или изменения состава жирных кислот, продуцируемых в клетке-хозяине при экспрессии в этой клетке-хозяине.
Белки, описанные в (iii) или (iv) выше, являются обычно природно-встречающимися мутантами белка, состоящего из любой аминокислотной последовательности, выбранной из группы, состоящей из аминокислотных последовательностей MaACS-1~12, и включают в себя белки, которые могут быть искусственно получены с использованием сайт-направленного мутагенеза, описанного, например, в "Sambrook & Russell, Molecular Cloning: A Laboratory Manual Vol. 3, Cold Spring Harbor Laboratory Press 2001", "Ausubel, Current Protocols in Molecular Biology, John Wiley & Sons 1987-1997", "Nuc. Acids. Res., 10, 6487 (1982)", "Proc. Natl. Acad. Sci. USA, 79, 6409 (1982)", "Gene, 34, 315 (1985)", "Nuc. Acids. Res., 13, 4431 (1985)", "Proc. Natl. Acad. Sci. USA, 82, 488 (1985)", и т.д.
В данном контексте "белок, состоящий из аминокислотной последовательности, в которой одна или несколько аминокислот делетированы, заменены, инсертированы и/или добавлены в любой аминокислотной последовательности, выбранной из группы, состоящей из аминокислотных последовательностей MaACS-1~12, и имеющий ацил-CoA-синтетазную активность или активность увеличения количества и/или изменения состава жирных кислот, продуцируемых в клетке-хозяине при экспрессии в этой клетке-хозяине" включает в себя белки, состоящие из аминокислотной последовательности, в которой, например, 1-100, 1-90, 1-80, 1-70, 1-60, 1-50, 1-40, 1-39, 1-38, 1-37, 1-36, 1-35, 1-34, 1-33, 1-32, 1-31, 1-30, 1-29, 1-28, 1-27, 1-26, 1-25, 1-24, 1-23, 1-22, 1-21, 1-20, 1-19, 1-18, 1-17, 1-16, 1-15, 1-14, 1-13, 1-12, 1-11, 1-10, 1-9 (1-несколько), 1-8, 1-7, 1-6, 1-5, 1-4, 1-3, 1-2 или одна аминокислота является/являются делетированными, замененными, инсертированными и/или добавленными в любую аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей MaACS-1~12, и имеющий ацил-CoA-синтетазную активность или активность увеличения количества и/или изменения состава жирных кислот, продуцируемых в клетке-хозяине при экспрессии в этой клетке-хозяине. В общем, количество делеций, замен, инсерций и/или добавлений является предпочтительно меньшим.
Такие белки включают в себя белок, имеющий идентичность приблизительно 60% или более высокую, 61% или более высокую, 62% или более высокую, 63% или более высокую, 64% или более высокую, 65% или более высокую, 66% или более высокую, 67% или более высокую, 68% или более высокую, 69% или более высокую, 70% или более высокую, 71% или более высокую, 72% или более высокую, 73% или более высокую, 74% или более высокую, 75% или более высокую, 76% или более высокую, 77% или более высокую, 78% или более высокую, 79% или более высокую, 80% или более высокую, 81% или более высокую, 82% или более высокую, 83% или более высокую, 84% или более высокую, 85% или более высокую, 86% или более высокую, 87% или более высокую, 88% или более высокую, 89% или более высокую, 90% или более высокую, 91% или более высокую, 92% или более высокую, 93% или более высокую, 94% или более высокую, 95% или более высокую, 96% или более высокую, 97% или более высокую, 98% или более высокую, 99% или более высокую, 99,1% или более высокую, 99,2% или более высокую, 99,3% или более высокую, 99,4% или более высокую, 99,5% или более высокую, 99,6% или более высокую, 99,7% или более высокую, 99,8% или более высокую или 99,9% или более высокую относительно любой аминокислотной последовательности, выбранной из группы, состоящей из аминокислотных последовательностей MaACS-1~12, и имеющий диацилглицерол-ацилтрансферазную активность. Когда описанный выше процент идентичности является более высоким, этот белок является обычно предпочтительным.
Предполагается, что термин делеция, замена, инсерция и/или добавление одного или нескольких аминокислотных остатков в аминокислотной последовательности белка по данному изобретению означает, что один или несколько аминокислотных остатков делетированы, заменены, инсертированы и/или добавлены в произвольных и одном или нескольких положениях в одной и той же последовательности. Одновременно могут встречаться два или более типов делеций, замен, инсерций и добавлений.
Примеры аминокислотных остатков, которые являются взаимозаменяемыми, приведены ниже. Аминокислотные остатки в одной и той же группе являются взаимозаменяемыми. Группа А: лейцин, изолейцин, норлейцин, валин, норвалин, аланин, 2-аминобутановая кислота, метионин, о-метилсерин, трет-бутилглицин, трет-бутилаланин и циклогексилаланин; Группа B: аспарагиновая кислота, глутаминовая кислота, изоаспарагиновая кислота, изоглутаминовая кислота, 2-аминоадипиновая кислота и 2-аминосубериновая кислота; Группа C: аспарагин и глутамин; Группа D: лизин, аргинин, орнитин, 2,4-диаминобутановая кислота и 2,3-диаминопропионовая кислота; Группа E: пролин, 3-гидроксипролин и 4-гидроксипролин; Группа F: серин, треонин и гомосерин; и Группа G: фенилаланин и тирозин.
Белок по данному изобретению может быть также получен способами химического синтеза, такими как Fmoc-способ (фторенилметилоксикарбонильный способ), tBoc-способ (трет-бутилоксикарбонильный способ) и т.д. Кроме того, для химического синтеза могут быть также использованы пептидные синтезаторы, доступные из Advanced Automation Peptide Protein Technologies, Perkin Elmer, Protein Technologies, PerSeptive, Applied Biosystems, SHIMADZU Corp., etc.
Белок, кодируемый полинуклеотидом по этому изобретению, и белок по изобретению являются, оба, белками-гомологами ACS, и считается, что они имеют ацил-CoA-синтетазную активность, так как мотив АТР-АМР (АТФ-АТФ) и мотив FACS/VLACS-FATP, которые являются важными для ацил-CoA-синтетазной активности, являются сохраненными. Предполагается, что в данном контексте ATP, AMP, FACS, VLACS и FATP обозначают аденозинтрифосфат, аденозинмонофосфат, (ацил жирных кислот)-CoA-синтетазу, ацил-CoA синтетазу с очень длинной цепью и белок транспорта жирных кислот, соответственно. Конкретные аминокислотные последовательности мотива ATP-AMP и мотива FACS/VLACS-FATP, содержащихся в белке по данному изобретению, показаны на фиг. 25 и 26 в подчеркнутой одной линией и подчеркнутых двойной линией последовательностях, соответственно. Что касается репрезентативных аминокислотных последовательностей мотива ATP-AMP и мотива FACS/VLACS-FATP, может быть сделана ссылка на базы данных, включающие в себя pfam (http://pfam.sanger.ac.uk/), и т.д.
В данном контексте термин "ацил-CoA-синтетазная активность (ACS-активность)" предназначен для обозначения активности, стимулирующей реакцию образования ацил-СоА посредством образования тиоэфирной связи между жирной кислотой и коферментом А (уравнение химической реакции ниже).
Жирная кислота+Кофермент A→Ацил-CoA+H2O
Ацил-CoA-синтетазная активность может быть количественно подтверждена, например, культивированием в течение определенного периода времени клеток-хозяев, в которые введен полипептид по данному изобретению, получением лизата клеток-хозяев, смешиванием этого клеточного лизата с меченой жирной кислотой (например, полиненасыщенной жирной кислотой, меченной радиоактивным изотопом и т.д.) и кофермента А, реакцией их в течение некоторого периода времени, затем экстракцией свободных жирных кислот н-гептаном и определением количества (ацил жирной кислоты)-СоА, который образуется во время вышеописанной реакции и остается в водной фракции, с использованием сцинтилляционного счетчика. В отношении деталей этого способа для подтверждения ацил-CoA-синтетазной активности, может быть сделана ссылка на Black P. N., et al. (J. B. C., 272 (8), 4896-4903, 1997). Альтернативно, активность ацил-СоА-синтетазы может быть также определена способом, описанным в разделе "Оценивание активности ACS" ПРИМЕРА 2, который не включает в себя использование радиоактивной метки.
Термин "активность увеличения количества жирных кислот, продуцируемых в клетке-хозяине при экспрессии в этой клетке-хозяине" предназначен для обозначения активности, которая, при введении (трансформации) полинуклеотида по данному изобретению или полинуклеотида, кодирующего белок по данному изобретению, в клетку-хозяина и экспрессии в этой клетке-хозяине, увеличивает общую продукцию жирных кислот в сравнении со ссылочной клеткой (контролем), полученной из того же самого штамма, что и клетка-хозяин, в которую не вводят вышеописанный полинуклеотид.
Термин "активность изменения состава жирных кислот, продуцируемых в клетке-хозяине при экспрессии в этой клетке-хозяине" предназначен для обозначения активности, которая, при введении полинуклеотида по данному изобретению или полинуклеотида, кодирующего белок по данному изобретению, вводят (трансформируют) в клетку-хозяина и экспрессируют в этой клетке-хозяине, изменяет количество или соотношение различных продуцируемых жирных кислот в сравнении со ссылочной клеткой (контролем), полученной из того же самого штамма, что и клетка-хозяин, в которую не вводят вышеописанный полинуклеотид.
В данном контексте термин "жирная кислота" предназначен для обозначения алифатической монокарбоновой кислоты (карбоновой кислоты, имеющей один карбоксильный остаток и атомы углерода, соединенные друг с другом в цепи), представленной общей формулой RCOOH (где R обозначает алкил). Эта жирная кислота включает в себя насыщенную жирную кислоту, не имеющую двойной связи, и ненасыщенную жирную кислоту, содержащую двойную связь (двойные связи) в этой углеводородной цепи. Эта жирная кислота является предпочтительно ненасыщенной жирной кислотой и, более предпочтительно, полиненасыщенной жирной кислотой, содержащей множество двойных связей в этой углеводородной цепи. Эта полиненасыщенная жирная кислота включает в себя предпочтительно ненасыщенную жирную кислоту, имеющую атомы углерода в количестве 18 или более, например, ненасыщенную жирную кислоту, имеющую 18 или 20 атомов углерода, и примеры включают в себя, но не ограничиваются ими, олеиновую кислоту, линолевую кислоту, линоленовую кислоту (γ-линоленовую кислоту, дигомо-γ-линоленовую кислоту и т.д.), арахидоновую кислоту и т.п. Этими полиненасыщенными жирными кислотами являются особенно предпочтительно линолевая кислота, γ-линоленовая кислота, дигомо-γ-линоленовая кислота и арахидоновая кислота, более предпочтительно, линолевая кислота, дигомо-γ-линоленовая кислота и арахидоновая кислота и, наиболее предпочтительно, линолевая кислота, дигомо-γ-линоленовая кислота и арахидоновая кислота.
В данном изобретении "клетка-хозяин" не ограничивается особо, пока эта клетка способна экспрессировать полинуклеотид по изобретению при введении этого полинуклеотида. Эти клетки включают в себя клетки, полученные из млекопитающих (за исключением человека), насекомых, растений, грибов, бактерий и т.д., предпочтительно клетки из растений и грибов и наиболее предпочтительно липидпродуцирующих грибов или дрожжей.
Липидпродуцирующими грибами, которые могут быть использованы, являются липидпродуцирующие грибы, описанные, например, в MYCOTAXON, Vol. XLIV, No. 2, pp. 257-265 (1992). Конкретные примеры включают в себя, но не ограничиваются ими, микроорганизмы, принадлежащие к роду Mortierella, в том числе подроду Mortierella, например, Mortierella elongata IFO8570, Mortierella exigua IFO8571, Mortierella hygrophila IFO5941, Mortierella alpina IFO8568, ATCC16266, ATCC32221, ATCC42430, CBS 219.35, CBS224.37, CBS250.53, CBS343.66, CBS527.72, CBS528.72, CBS529.72, CBS608.70 и CBS754.68 и т.д., или микроорганизмы, принадлежащие к подроду Micromucor, например, Mortierella isabellina CBS194.28, IFO6336, IFO7824, IFO7873, IFO7874, IFO8286, IFO8308 и IFO7884, Mortierella nana IFO8190, Mortierella ramanniana IFO5426, IFO8186, CBS112.08, CBS212.72, IFO7825, IFO8184, IFO8185 и IFO8287, Mortierella vinacea CBS236.82 и т.д. Среди других, предпочтительной является Mortierella alpina.
Конкретные примеры дрожжей включают в себя род Saccharomyces, род Candida, род Zygosaccharomyces, род Pichia и род Hansenula и предпочтительно Saccharomyces cerevisiae в роде Saccharomyces. В диких штаммах дрожжей, таких как Saccharomyces cerevisiae и т.д., насыщенные жирные кислоты или моновалентные жирные кислоты, имеющие в основном 18 или менее атомов углерода, могут быть синтезированы в этих клетках, но полиненасыщенные жирные кислоты не могут в них синтезироваться. По этой причине, при использовании дрожжей, таких как Saccharomyces cerevisiae, и т.д. в качестве клетки-хозяина предпочтительным является придание способности синтезировать полиненасыщенные жирные кислоты клеткам дрожжей генетической инженерией и т.д. Способность синтезировать полиненасыщенные жирные кислоты может быть придана введением гена, кодирующего белок, полученного из организма, который уже имеет способность синтезировать полиненасыщенные жирные кислоты и участвует в синтезе жирных кислот.
"Организм, который уже имеет способность синтезировать полиненасыщенные жирные кислоты" включает в себя, например, липидпродуцирующие грибы. Конкретные примеры этих липидпродуцирующих грибов являются те же самыми, что и описанные здесь выше.
Примеры гена, кодирующего белок, полученный из организма, который уже имеет способность синтезировать полиненасыщенные жирные кислоты, и "гена, кодирующего белок, который участвует в синтезе жирных кислот" включают в себя, но не ограничиваются ими, ген Δ12-десатуразы жирных кислот, ген Δ6-десатуразы жирных кислот, ген GLELO-элонгазы жирных кислот и ген Δ5-десатуразы жирных кислот, и т.д. Нуклеотидные последовательности гена Δ12-десатуразы жирных кислот, гена Δ6-десатуразы жирных кислот, гена GLELO-элонгазы жирных кислот и гена Δ5-десатуразы жирных кислот являются доступными посредством наличия доступа к базам данных, в том числе GenBank, и т.д. Например, в GenBank для доступа к соответствующим последовательностям вводятся номера доступа Accession No. AB020033, No. AB020032, No. AB193123 и No. AB188307.
Гены для белков, связанных с синтезом жирных кислот, описанных выше, инсертируют в подходящие векторы (например, pESC (Stratagene), pYES (Invitrogen) и т.д.), которые затем вводят в дрожжи способом электропорации, способом сферопластов (Proc. Natl. Acad. Sci. USA, 75 p1929 (1978)), литийацетатным способом (J. Bacteriology, 153, p163 (1983)) и способами, описанными в Proc. Natl. Acad. Sci. USA, 75 p1929 (1978), Methods in Yeast Genetics, 2000 Edition: A Cold Spring Harbor Laboratory Course Manual, и т.д.
Жирные кислоты могут быть экстрагированы из клеток-хозяев, трансформированных полинуклеотидом по данному изобретению или полинуклеотидом, кодирующим белок по данному изобретению, следующим образом. Клетку-хозяина культивируют и затем обрабатывают общепринятым образом, например, центрифугированием, фильтрованием и т.д., для получения культивируемых клеток. Эти клетки тщательно промывают водой и предпочтительно высушивают. Высушивание может выполняться лиофилизацией, высушиванием на воздухе и т.д. В зависимости от необходимости, эти высушенные клетки разрывают с использованием Dynomil или обработки ультразвуком и затем экстрагируют органическим растворителем, предпочтительно в токе азота. Примеры органического растворителя включают в себя эфир, гексан, метанол, этанол, хлороформ, дихлорметан, петролейный эфир и т.д. Альтернативно, хорошие результаты могут быть получены чередующейся экстракцией метанолом и петролейным эфиром или экстракцией системой однофазного растворителя хлороформ-метанол-вода. Удаление органического растворителя из этого экстракта дистилляцией при пониженном давлении может давать содержащие жирную кислоту липиды. Эти экстрагированные жирные кислоты могут быть превращены в метиловые эфиры способом, использующим хлористоводородную кислоту-метанол и т.д.
Количество или соотношение различных жирных кислот может быть определено анализом жирных кислот, экстрагированных, как описано выше, с использованием различных хроматографических способов. Примеры хроматографических способов включают в себя, но не ограничиваются ими, жидкостную хроматографию высокого разрешения и газовую хроматографию и особенно предпочтительно газовую хроматографию.
3. Вектор по данному изобретению и трансформанты с введенным вектором
В другом варианте осуществления данное изобретение дополнительно обеспечивает экспрессирующий вектор, содержащий полинуклеотид по изобретению.
Вектор по изобретению обычно конструируют для содержания экспрессионной кассеты, содержащей:
(i) промотор, который может транскрибироваться в клетке-хозяине;
(ii) любой из полинуклеотидов, определенных в (a)-(g) выше, который связан с этим промотором; и
(iii) экспрессионную кассету, содержащую в качестве компонента сигнал, который функционирует в этой клетке-хозяине в отношении терминации транскрипции и полиаденилирования РНК-молекулы.
Вектор, сконструированный таким образом, вводят в клетку-хозяина. Примеры клеток-хозяев, которые могут быть подходящим образом использованы в данном изобретении, являются теми же, что и описанные выше.
В этих клетках-хозяевах, трансформированных вектором по данному изобретению, активность ACS является более увеличенной, жирные кислоты являются более продуцируемыми, или количество или соотношение различных жирных кислот, содержащихся в этих клетках, являются измененными в сравнении с клетками-хозяевами, которые не трансформированы вектором по данному изобретению.
Примеры векторов, доступных для введения в липидпродуцирующие грибы, включают в себя, но не ограничиваются им, pDura5 (Appl. Microbiol. Biotechnol., 65, 419-425, (2004)).
Любой вектор является доступным в качестве вектора, используемого для введения в дрожжи, и не ограничивается особо, пока он является вектором, способным экспрессировать этот инсерт в клетках дрожжей. Этот вектор включает в себя, например, pYE22m (Biosci. Biotech. Biochem., 59, 1221-1228, 1995).
Могут быть использованы промоторы/терминаторы для регуляции экспрессии генов в клетках-хозяевах в произвольной комбинации, пока они функционируют в этих клетках-хозяевах. Например, могут быть использованы промотор гена гистона Н4.1, промотор глицеральдегид-3-фосфатдегидрогеназы и т.д.
В качестве селектируемых маркеров, используемых для трансформации, могут быть использованы ауксотрофные маркеры (ura5, niaD), устойчивый к гигромицину ген, устойчивый к зеоцину ген, устойчивый к генецитину ген (G418r), устойчивый к меди ген (CUP1) (Marin et al., Proc. Natl. Acad. Sci. USA, 81, 337 1984), устойчивый к церуленину ген (fas2m, PDR4) (Junji Inokoshi, et al., Biochemistry, 64, 660, 1992; и Hussain et al., Gene, 101: 149, 1991, соответственно) и т.п.
Для трансформации клеток-хозяев могут быть использованы обычно известные способы. В липидпродуцирующих грибах трансформация может выполняться, например, способом электропорации (Mackenzie, D. A. et al., Appl. Environ. Microbiol., 66, 4655-4661, 2000) и способом доставки частиц (способом, описанным в JPA 2005-287403 "Method of Breeding Lipid-Producing Fungus"). С другой стороны, для трансформации дрожжей могут быть использованы способ электропорации, сферопластный способ (Proc. Natl. Acad. Sci. USA, 75 p1929 (1978)) и литийацетатный способ (J. Bacteriology, 153 p163 (1983)), а также способы, описанные в Proc. Natl. Acad. Sci. USA, 75 p1929 (1978), Methods in yeast genetics, 2000 Edition: A Cold Spring Harbor Laboratory Course Manual, etc). Однако способ трансформации не ограничивается описанными выше способами.
В отношении общих способов клонирования может быть сделана ссылка на "Sambrook & Russell, Molecular Cloning: A Laboratory Manual Vol. 3, Cold Spring Harbor Laboratory Press 2001", "Methods in Yeast Genetics, A laboratory manual (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY)" и т.д.
4. Способ получения композиции липидов или жирных кислот по изобретению
В другом варианте осуществления данное изобретение дополнительно обеспечивает способ получения композиции липидов или жирных кислот, который предусматривает использование вышеописанного трансформанта.
В данном контексте термин "липид" предназначен для обозначения простого липида, включающего в себя соединение (например, глицерид), которое состоит из жирной кислоты и спирта, соединенных через сложноэфирную связь, или его аналога (например, сложного эфира холестерина) и т.д.; сложного липида, в котором фосфорная кислота, аминокислота (аминокислоты), сахарид (сахариды) или т.п. связаны с частью простого липида; или производного липида, которое является гидролизатом этого липида и является нерастворимым в воде.
В данном контексте термин "масло и жир" предназначен для обозначения сложного эфира глицерина и жирной кислоты (глицерида).
Термин "жирная кислота" является тем же, что и определенный выше.
Способ экстрагирования композиции липидов или жирных кислот по данному изобретению является таким же, что и способ для экстрагирования жирных кислот, описанный выше.
Жирные кислоты могут быть выделены из вышеописанных содержащих жирные кислоты липидов в состоянии смешанных жирных кислот или смешанных эфиров жирных кислот концентрированием и разделением общепринятым образом (например, добавлением мочевины, разделением при охлаждении, колоночной хроматографией и т.д.).
Липиды, полученные способом по данному изобретению, включают в себя предпочтительно ненасыщенные жирные кислоты и, более предпочтительно, полиненасыщенные жирные кислоты. Предпочтительными примерами полиненасыщенных жирных кислот являются ненасыщенные жирные кислоты, имеющие 18 или более атомов углерода, например, полиненасыщенные жирные кислоты, имеющие 18-20 атомов углерода, и включают в себя, но не ограничиваются ими, олеиновую кислоту, линолевую кислоту, линоленовую кислоту (γ-линоленовую кислоту и дигомо-γ-линоленовую кислоту и т.д.), арахидоновую кислоту и т.д. Особенно предпочтительными полиненасыщенными кислотами являются линолевая кислота, γ-линолевая кислота, дигомо-γ-линолевая кислота и арахидоновая кислота, более предпочтительно линолевая кислота, дигомо-γ-линолевая кислота и арахидоновая кислота, и наиболее предпочтительно дигомо-γ-линоленовая кислота и арахидоновая кислота.
Липиды, полученные способом по данному изобретению, и состав (композиция) жирных кислот, содержащихся в этих липидах, могут быть подтверждены способом экстракции липидов или способом разделения жирных кислот, описанных выше, или их комбинацией.
Композиция липидов или жирных кислот, полученная способом получения по данному изобретению, может быть обеспечена для применения в получении, например, пищевых продуктов, фармацевтических препаратов, промышленных материалов (сырьевых материалов для косметических изделий, мыла и т.д.), которые содержат масла и жиры, общепринятым образом.
Еще в одном варианте осуществления данное изобретение обеспечивает способ приготовления пищевых продуктов, косметических изделий, мыла и т.д. с применением трансформанта по данному изобретению. Этот способ предусматривает стадию образования липидов или жирных кислот с использованием трансформанта по данному изобретению.
Пищевые продукты, косметические изделия, фармацевтические препараты, мыло и т.д., содержащие полученные липиды или жирные кислоты, получают общепринятым образом. Таким образом, эти пищевые продукты, косметические изделия, фармацевтические препараты, мыло и т.д., получаемые способом по данному изобретению, содержат липиды или жирные кислоты, полученные с использованием трансформанта по данному изобретению. Это изобретение дополнительно обеспечивает пищевые продукты, косметические изделия, фармацевтические препараты, мыло и т.д., полученные таким способом.
Форма косметического изделия (композиции) или фармацевтического препарата (композиции) по этому изобретению не ограничивается особо и может быть любой формой, включающей в себя состояние раствора, пасты, геля, твердого вещества или порошка. Эта косметическая композиция или фармацевтическая композиция по данному изобретению может также использоваться в качестве косметических изделий или агентов для локального нанесения на кожу, включающих в себя масло, лосьон, крем, эмульсию, гель, шампунь, ополаскиватель для волос, кондиционер для волос, эмаль, крем-основу, губную помаду, пудру для лица, маску для лица, мазь, отдушку, порошок, одеколон, зубную пасту, мыло, аэрозоль, очищающую пену для лица и т.д., агент для ухода за кожей, предупреждающий старение кожи, противовоспалительный агент для кожи, агент для косметического ухода при принятии ванн, содержащий лекарственное средство тоник, эссенцию для поддержания естественной красоты кожи, защищающий от солнца агент или защитный или улучшающий агент для страданий, вызываемых повреждениями кожи, растрескавшейся кожей или имеющей порезы кожей и т.д.
Косметическая композиция по данному изобретению может быть дополнительно приготовлена подходящим образом с другими маслами и жирами и/или красителями, отдушками, консервантами, поверхностно-активными веществами, пигментами, антиоксидантами и т.д., если необходимо. Соотношение этих материалов в композиции может быть удобным образом определено квалифицированными в данной области специалистами, в зависимости от цели (например, масла и жиры могут содержаться в этой композиции в количестве 1-99,99 масс.%, предпочтительно 5-99,99 масс.% и более предпочтительно 10-99,9 масс.%). Если необходимо, фармацевтическая композиция по данному изобретению может также содержать другие фармацевтически активные компоненты (например, противовоспалительные компоненты) или вспомогательные компоненты (например, смягчители или компоненты-носители). Примеры этих других компонентов, обычно используемых в косметическом или кожном препарате для наружного применения, включают в себя агент против угрей, агент для предотвращения перхоти или зуда, антиперспирант и дезодорирующий агент, агент для обработки ожогов, агент против клещей и вшей, агент для смягчения кератина, агент для ксеродермии, антивирусный агент, усиливающий чрескожную абсорбцию агент и т.п.
Пищевой продукт по данному изобретению включает в себя диетическую добавку, диетические пищевые продукты, функциональный пищевой продукт, пищевой продукт для маленьких детей, продукт детского питания, модифицированное молоко для младенцев, модифицированное молоко для недоношенных младенцев, пищевой продукт для людей пожилого и старого возраста и т.д. В данном контексте термины пища или пищевой продукт предназначены для обозначения твердой, жидкой и текучей пищи, а также их смеси и вместе обозначают годный в пищу материал.
Термин диетическая добавка относится к пищевым продуктам, обогащенным специфическими питательными ингредиентами. Термин диетические пищевые продукты относится к пищевым продуктам, которые являются полезными или важными для здоровья и включают в себя диетические добавки, природные пищевые продукты, диетические продукты и т.д. Термин функциональный пищевой продукт относится к пищевому продукту для восполнения питательных ингредиентов, которые помогают регулировать функции организма, и является синонимом пищевого продукта для указанного, полезного для здоровья применения. Термин пищевой продукт для маленьких детей относится к пищевому продукту, предназначенному для детей до приблизительно 6 лет. Термин пищевой продукт для людей пожилого и старого возраста относится к пищевому продукту, обработанному для облегчения переваривания и абсорбции в сравнении с необработанными пищевыми продуктами. Термин модифицированное молоко для младенцев относится к модифицированному молоку, предназначенному для детей до приблизительно одного года. Термин модифицированное молоко для недоношенных младенцев относится к модифицированному молоку, предназначенному для недоношенных младенцев до приблизительно 6 месяцев после рождения.
Форма этих пищевых продуктов включает в себя природные пищевые продукты (обработанные жирами и маслами), такие как мясо, рыба и орехи; пищевые продукты, дополненные жирами и маслами во время варки, например, китайские пищевые продукты, китайские макаронные изделия, супы и т.д.; пищевые продукты, приготовленные с использованием жиров и масел в качестве теплоносителя, например, рыба и овощи, запеченные вместе, или зажаренные во фритюре рыба и овощи, зажаренные во фритюре пищевые продукты, жареный соевый творог, китайский жареный рис, пончики, японские жареные булочки или каринто; пищевые продукты на основе жира и масла или подвергнутые переработке пищевые продукты, дополненные жирами и маслами во время переработки, например, маслом, маргарином, майонезом, заправкой, шоколадом, быстро разваривающиеся макаронные изделия, карамель, бисквиты, печенье, кексы, мороженое; и пищевые продукты, опрысканные или покрытые жирами и маслами после окончательной (финишной) обработки, например, рисовые крекеры, сухие бисквиты, хлеб с добавлением пасты из гледичии (sweet bean paste bread) и т.д. Однако пищевой продукт не ограничивается продуктами, содержащими жиры и масла, и другие примеры включают в себя сельскохозяйственные пищевые продукты, такие как мучные кондитерские изделия, макаронные продукты, вареный рис, сладости (например, конфеты, жевательные резинки, гумми, сладкие таблетки, яЯпонские сладости), соевый творог или тофу и их подвергнутые технологической обработке продукты; ферментированные пищевые продукты, такие как японское рисовое вино или сакэ (японская рисовая водка), содержащий лекарство напиток, сладкий херес или сладкая водка, уксус, соевый соус и ферментированная соевая паста из вареных соевых бобов и риса или мисо и т.д.; пищевые продукты, получаемые от скота, такие как йогурт, ветчина, бекон, колбаса и т.д.; морепродукты, такие как измельченный и пропаренный рыбный брикет (рыбная паста) или камабоко, жареный во фритюре рыбный брикет или агетен и слоистый рыбный брикет или ханпен, и т.д.; а также фруктовые напитки, безалкогольные напитки, спортивные напитки, алкогольные напитки, чай и т.д.
Пищевой продукт по данному изобретению может также быть в форме фармацевтических препаратов, таких как капсулы и т.д., или в форме подвергнутого обработке пищевого продукта, такого как природные жидкие пищевые рационы (диеты), диеты с определенной смесью и элементные смеси (элементные диеты), приготовленные с маслом и жиром по данному изобретению вместе с белками, сахарами, микроэлементами, витаминами, эмульгаторами, химическими ароматизаторами и т.д., полезные для здоровья напитки, энтеральные нутриенты и т.п.
Как описано выше, жирные кислоты могут эффективно продуцироваться экспрессией гена гомолога ACS по данному изобретению в клетках-хозяевах.
Кроме того, этот уровень экспрессии может быть использован в качестве индикатора для исследования условий для культивирования, контроля культивирования и т.д. для эффективного продуцирования жирных кислот.
ПРИМЕРЫ
Далее данное изобретение будет описано более подробно со ссылкой на примеры, но должно быть понятно, что это изобретение не предназначено для ограничения объема изобретения этими примерами.
[ПРИМЕР 1]
Анализ генома M. alpina
Штамм M. alpina 1S-4 высевали на 100 мл среды GY2:1 (2% глюкоза и 1% дрожжевой экстракт, pH 6,0) с последующим культивированием со встряхиванием при 28°C в течение 2 дней. Мицелиальные клетки собирали фильтрованием и геномную ДНК получали с использованием DNeasy (QIAGEN). Нуклеотидную последовательность этой геномной ДНК, описанную выше, определяли с использованием Roche 454 GS FLX Standard. В этом случае секвенирование нуклеотидов библиотеки фрагментов выполняли в двух экспериментах и секвенирование нуклеотидов конъюгационно спаренной библиотеки выполняли в трех экспериментах. Полученные нуклеотидные последовательности собирали с получением 300 суперконтигов.
Синтез кДНК и конструирование кДНК-библиотеки
Штамм M. alpina 1S-4 высевали на 100 мл среды (1,8% глюкоза, 1% дрожжевой экстракт pH 6,0) и предварительно культивировали в течение 3 дней при 28°C. Культуральный сосуд на 10 л (Able Co., Tokyo) загружали 5 л среды (1,8% глюкоза, 1% порошок сои, 0,1% оливковое масло, 0,01% Адеканол, 0,3% KH2PO4, 0,1% Na2SO4, 0,05% CaCl2.2H2O и 0,05% MgCl2.6H2O, pH 6,0), и все количество предварительно культивируемого продукта высевали на нее, с последующим аэробным спин-культивированием (культивированием с постоянным перемешиванием) при условиях 300 об/мин, 1 vvm и 26°C в течение 8 дней. В дни 1, 2 и 3 культивирования добавляли глюкозу в количестве, соответствующем 2%, 2% и 1,5%, соответственно. Мицелиальные клетки собирали на каждой стадии в дни 1, 2, 3, 6 и 8 культивирования для получения общей РНК количественным способом с гуанидингидрохлоридом/CsCl. С использованием набора для очистки мРНК Oligotex-dT30
Поиск на гомолог ACS
С использованием в качестве запроса аминокислотных последовательностей ScFAA1 (YOR317W), ScFAA2 (YER015W), ScFAA3 (YIL009W), ScFAA4 (YMR246W), ScFAT1 (YBR041W) и ScFAT2 (YBR222C), которые являются ACS из дрожжей, выполняли поиск tblastn против геномной нуклеотидной последовательности штамма M. alpina 1S-4. В результате были обнаружены совпадения в двенадцати (12) последовательностях. Таким образом, было обнаружено совпадение на суперконтигах, содержащих последовательность, представленную в SEQ ID NO: 5, SEQ ID NO: 10, SEQ ID NO: 15, SEQ ID NO: 20, SEQ ID NO: 25, SEQ ID NO: 30, SEQ ID NO: 35, SEQ ID NO: 40, SEQ ID NO: 45, SEQ ID NO: 50, SEQ ID NO: 55 или SEQ ID NO: 60. Гены, несущие SEQ ID NO: 5, SEQ ID NO: 10, SEQ ID NO: 15, SEQ ID NO: 20, SEQ ID NO: 25, SEQ ID NO: 30, SEQ ID NO: 35, SEQ ID NO: 40, SEQ ID NO: 45, SEQ ID NO: 50, SEQ ID NO: 55 и SEQ ID NO: 60, были обозначены как MaACS-1, MaACS-2, MaACS-3, MaACS-4, MaACS-5, MaACS-6, MaACS-7, MaACS-8, MaACS-9, MaACS-10, MaACS-11 и MaACS-12.
Клонирование гомолога ACS
Для клонирования кДНК, соответствующих генам MaACS-1~12, выполняли скрининг кДНК-библиотеки, описанной выше. Мечение зонда выполняли при помощи ПЦР с использованием ExTaq (Takara Bio Inc.). То есть, готовили дигоксигенин-(DIG)-меченные амплифицированные ДНК-зонды с использованием смеси для мечения ПЦР (Roche Diagnostics) вместо смеси dNTP, присоединенной к ExTaq.
Условия для гибридизации устанавливали следующим образом.
Буфер: 5x SSC, 1% SDS, 50 мМ Трис-HCl (pH 7,5), 50% формальдегид;
Температура: 42°C (в течение ночи);
Условия промывки: 0,2x SSC, в 0,1% растворе SDS (65°C) в течение 20 минут×3.
Детектирование выполняли с использованием набора для детектирования DIG нуклеиновой кислоты (Roche Diagnositcs). Фаговые клоны получали скринингом, и плазмиды вырезали из фаговых клонов вырезанием in vivo с получением соответствующих плазмидных ДНК.
Праймеры для получения зондов, используемых для скрининга соответствующих генов, количество нуклеотидов в CDS соответствующих генов, количество аминокислот в аминокислотных последовательностях, расшифрованных из этих нуклеотидных последовательностей CDS, и количество экзонов и интронов посредством сравнения последовательностей геномной ДНК с последовательностями CDS приведены ниже.
(1) MaACS-1
Праймер ACS-1-1F: 5'-GTCGGCTCCAAGCTTGCAATCC-3' (SEQ ID NO: 61)
Праймер ACS-1-2R: 5'-GGACAGCTCCAGCACTGTGGTAAAG-3' (SEQ ID NO: 62)
кДНК (SEQ ID NO: 4)
CDS (SEQ ID NO: 3): 1857 п.н.
ORF (SEQ ID NO: 1): 1854 п.н.
Аминокислотная последовательность (SEQ ID NO: 2): 618 аминокислот (см. Фиг. 1)
Количество экзонов: 5, количество интронов: 4 (см. Фиг. 2)
(2) MaACS-2
Праймер ACS-2-1F: 5'-GACCACGGGATTCCCCAAGGCTGC-3' (SEQ ID NO: 63)
Праймер ACS-2-2R: 5'-CTTGGTCGCGCTTGTTCCTGGCCAC-3' (SEQ ID NO: 64)
кДНК (SEQ ID NO: 9)
CDS (SEQ ID NO: 8): 1929 п.н.
ORF (SEQ ID NO: 6): 1926 п.н.
Аминокислотная последовательность (SEQ ID NO: 7): 642 аминокислоты (см. Фиг. 3)
Количество экзонов: 8, количество интронов: 7 (см. Фиг. 4)
(3) MaACS-3
Праймер ACS-3-1F: 5'-TACAGCTTTGTTGCTGTCCCCATC-3' (SEQ ID NO: 65)
Праймер ACS-3-2R: 5'-GATGATGGGTGTGCTTGCAAAGATC-3' (SEQ ID NO: 66)
кДНК (SEQ ID NO: 14)
CDS (SEQ ID NO: 13): 1653 п.н.
ORF (SEQ ID NO: 11): 1650 п.н.
Аминокислотная последовательность (SEQ ID NO: 12): 550 аминокислот (см. Фиг. 5)
Количество экзонов: 9, количество интронов: 8 (см. Фиг. 6)
(4) MaACS-4
Праймер ACS-4-1F: 5'-AACCCAAAGCTGCGCCAGGCTGTCC-3' (SEQ ID NO: 67)
Праймер ACS-4-2R: 5'-TTACAGCTTGGATTCCTTTTGATGG-3' (SEQ ID NO: 68)
кДНК (SEQ ID NO: 19)
CDS (SEQ ID NO: 18): 2067 п.н.
ORF (SEQ ID NO: 16): 2064 п.н.
Аминокислотная последовательность (SEQ ID NO: 17): 688 аминокислот (см. Фиг. 7)
Количество экзонов: 7, количество интронов: 6 (см. Фиг. 8)
(5) MaACS-5
Праймер ACS-5-1F: 5'-GTCGTGCCCGATGCGGAGACGC-3' (SEQ ID NO: 69)
Праймер ACS-5-2R: 5'-TCAGTGGATCCCGTTATACATCAG-3' (SEQ ID NO: 70)
кДНК (SEQ ID NO: 24)
CDS (SEQ ID NO: 23): 1980 п.н.
ORF (SEQ ID NO: 21): 1977 п.н.
Аминокислотная последовательность (SEQ ID NO: 22): 659 аминокислот (см. Фиг. 9)
Количество экзонов: 6, количество интронов: 5 (см. Фиг. 10)
(6) MaACS-6
Праймер ACS-6-1F: 5'-GCGTCCCCCTCTATGATACATTG-3' (SEQ ID NO: 71)
Праймер ACS-6-2R: 5'-GTGGGATGCAGGACGGCAACATCG-3' (SEQ ID NO: 72)
кДНК (SEQ ID NO: 29)
CDS (SEQ ID NO: 28): 1980 п.н.
ORF (SEQ ID NO: 26): 1977 п.н.
Аминокислотная последовательность (SEQ ID NO: 27): 659 аминокислот (см. Фиг. 11)
Количество интронов: по меньшей мере 5 (см. Фиг. 12)
(7) MaACS-7
Праймер ACS-7-1F: 5'-GGATGCCGAACAACAGCGCGTGG-3' (SEQ ID NO: 73)
Праймер ACS-7-2R: 5'-GCACCCTCCTCAGAAACAGCCCTC-3' (SEQ ID NO: 74)
кДНК (SEQ ID NO: 34)
CDS (SEQ ID NO: 33): 1827 п.н.
ORF (SEQ ID NO: 31): 1824 п.н.
Аминокислотная последовательность (SEQ ID NO: 32): 608 аминокислот (см. Фиг. 13)
Количество экзонов: 5, количество интронов: 4 (см. Фиг. 14)
(8) MaACS-8
Праймер ACS-8-1F: 5'-CAGTCGAGTACATTGTCAACCACG-3 (SEQ ID NO: 75)
Primer ACS-8-2R: 5'-GCGGTTCAAGAGGCGAGGCACAGC-3' (SEQ ID NO: 76)
кДНК (SEQ ID NO: 39)
CDS (SEQ ID NO: 38): 2079 п.н.
ORF (SEQ ID NO: 36): 2076 п.н.
Аминокислотная последовательность (SEQ ID NO: 37): 692 аминокислоты (см. Фиг. 15)
Количество экзонов: 8, количество интронов: 7 (см. Фиг. 16)
(9) MaACS-9
Праймер ACS-9-1F: 5'-GTTCATCTTCTGCTGGCTGGGTCTC-3' (SEQ ID NO: 77)
Праймер ACS-9-2R: 5'-GTTGCGTTGTTCACGCGGCAATCC-3' (SEQ ID NO: 78)
кДНК (SEQ ID NO: 44)
CDS (SEQ ID NO: 43): 1851 п.н.
ORF (SEQ ID NO: 41): 1848 п.н.
Аминокислотная последовательность (SEQ ID NO: 42): 616 аминокислот (см. Фиг. 17)
Количество экзонов: 5: количество интронов: 4 (см. Фиг. 18)
(10) MaACS-10
Праймер ACS-10-1F: 5'-ATGGAAACCTTGGTTAACGGAAAG-3' (SEQ ID NO: 79)
Праймер ACS-10-2R: 5'-TCAGCAAAGATGGCCTTGGGCTGG-3' (SEQ ID NO: 80)
кДНК (SEQ ID NO: 49)
CDS (SEQ ID NO: 48): 2076 п.н.
ORF (SEQ ID NO: 46): 2073 п.н.
Аминокислотная последовательность (SEQ ID NO: 47): 691 аминокислота (см. Фиг. 19)
Количество экзонов: 8, количество интронов: 7 (см. Фиг. 20)
(11) MaACS-11
Праймер ACS-11-1F: 5'-GTCAAGGGCGAGACTCGCATCC-3' (SEQ ID NO: 81)
Праймер ACS-11-2R: 5'-CGGTGACGATGGTCATGGACTGC-3' (SEQ ID NO: 82)
кДНК (SEQ ID NO: 54)
CDS (SEQ ID NO: 53): 2043 п.н.
ORF (SEQ ID NO: 51): 2040 п.н.
Аминокислотная последовательность (SEQ ID NO: 52): 680 аминокислот (см. Фиг. 21)
Количество экзонов: 3, количество интронов: 2 (см. Фиг. 22)
(12) MaACS-12
Праймер ACS-12-1F: 5'-GCGAGACCCGCATCCGCCGCTCC-3' (SEQ ID NO: 83)
Праймер ACS-12-2R: 5'-GACCGTCCTCGCCCAGGGTGTCG-3' (SEQ ID NO: 84)
кДНК (SEQ ID NO: 59)
CDS (SEQ ID NO: 58): 2043 п.н.
ORF (SEQ ID NO: 56): 2040 п.н.
Аминокислотная последовательность (SEQ ID NO: 57): 680 аминокислот (см. Фиг. 23)
Количество экзонов: 3, количество интронов: 2 (см. Фиг. 24)
Анализ секвенирования
Идентичность между нуклеотидными последовательностями 12 гомологов ACS из M. alpina показана в таблице 1 и идентичность между аминокислотными последовательностями показана в таблице 2. MaACS-11 и MaACS-12 показали высокую идентичность 80,2% в нуклеотидной последовательности и 84,3% в аминокислотной последовательности.


С использованием в качестве последовательностей-запросов предположительных аминокислотных последовательностей для CDS-последовательностей MaACS-1~12, выполняли поиск BLASTp против аминокислотных последовательностей, зарегистрированных в GenBank. Белки, имеющие аминокислотную последовательность, которая совпадает с предположительными аминокислотными последовательностями MaACS-1~12 с наибольшей оценкой и идентичностью между этими белками и предположительными аминокислотными последовательностями MaACS-1~12, показаны в таблице 3. Идентичность предположительных аминокислотных последовательностей MaACS-1~12 с аминокислотными последовательностями полученных из S. cerevisiae ацил-CoA-синтетаз также показаны в таблице 4.


Фиг. 25 показывает сопоставление между MaACS из MaACS-1~12, которые имеют относительно высокую гомологию аминокислотной последовательности относительно полученных из S. cerevisiae белков FAA и этих белков FAA. Фиг. 26 показывает сопоставление гомологов ACS, имеющих относительно высокую гомологию аминокислотной последовательности относительно полученных из S. cerevisiae белков FAT. Районы мотива ATP-AMP и мотива FACS/VLACS-FATP, которые являются важными мотивами для активности ACS, являются высоко консервативными в обеих группах, показанных на фигурах 25 и 26.
Конструирование экспрессирующего вектора
Векторы для экспрессии MaACS-1, MaACS-10, MaACS-11, MaACS-6, MaACS-8 и MaACS-9, соответственно, в дрожжах конструировали следующим образом с использованием экспрессирующего вектора pYE22m (Biosci. Biotech. Biochem., 59, 1221-1228, 1995).
Плазмиду, содержащую SEQ ID NO: 29, которая была получена скринингом MaACS-6, расщепляли ферментами рестрикции (рестриктазами) BamHI и XhoI. Полученный ДНК-фрагмент приблизительно 2,1 т.п.н. лигировали с ДНК-фрагментом, полученным расщеплением вектора pYE22m рестриктазами BamHI и SalI с использованием Ligation High (TOYOBO) с получением плазмиды pYE-ACS-6.
С использованием плазмиды, содержащей кДНК MaACS-8 в качестве матрицы, выполняли ПЦР с представленными ниже праймерами с использованием ExTaq (Takara Bio Inc.). Амплифицированный таким образом ДНК-фрагмент клонировали при помощи набора для клонирования TOPO-TA (Invitrogen).
Праймер EcoRI-ACS-8-F: 5'-GGATCCATGCCTTCCTTCAAAAAGTACAACC-3' (SEQ ID NO: 85)
Праймер SmaI-ACS-8-R: 5'-CCCGGGCAAAGAGTTTTCTATCTACAGCTT-3' (SEQ ID NO: 86)
Нуклеотидную последовательность этого инсерта подтверждали, и плазмиду, содержащую правильную нуклеотидную последовательность, расщепляли рестриктазами EcoRI и SmaI. С использованием Ligation High (TOYOBO), полученный ДНК-фрагмент приблизительно 2,1 т.п.н. лигировали с ДНК-фрагментом, полученным расщеплением вектора pYE22m рестриктазами EcoRII и SmaI с получением плазмиды pYE-ACS-8.
С использованием плазмиды, содержащей кДНК MaACS-9 в качестве матрицы, выполняли ПЦР с представленными ниже праймерами с использованием ExTaq (Takara Bio Inc.). Амплифицированный таким образом ДНК-фрагмент клонировали при помощи набора для клонирования TOPO-TA (Invitrogen).
Праймер EcoRI-ACS-9-F: 5'-GAATTCATGGTTGCTCTCCCACTCG-3' (SEQ ID NO: 87)
Праймер BamHI-ACS-9-R: 5'-GGATCCCTACTATAGCTTGGCCTTGCC-3' (SEQ ID NO: 88)
Нуклеотидную последовательность этого инсерта подтверждали, и плазмиду, содержащую правильную нуклеотидную последовательность, расщепляли рестриктазами EcoRI и BamHI. С использованием Ligation High (TOYOBO), этот полученный ДНК-фрагмент приблизительно 2,0 т.п.н. лигировали с ДНК-фрагментом, полученным расщеплением вектора pYE22m рестриктазами EcoRII и BamHI с получением плазмиды pYE-ACS-9
С использованием плазмиды, содержащей кДНК MaACS-1 в качестве матрицы, выполняли ПЦР с представленными ниже праймерами с использованием ExTaq (Takara Bio Inc.). Амплифицированный таким образом ДНК-фрагмент клонировали при помощи набора для клонирования TOPO-TA (Invitrogen).
Праймер EcoRI-ACS-1-F: 5'-GGATCCATGTATGTCGGCTCCAAGCTTGC-3' (SEQ ID NO: 89)
Праймер SalI-ACS-1-R: 5'-GTCGACTCAAAGCCTGGCTTTGCCGCTGACG-3' (SEQ ID NO: 90)
Нуклеотидную последовательность этого инсерта подтверждали, и плазмиду, содержащую правильную нуклеотидную последовательность, расщепляли рестриктазами EcoRI и SalI. С использованием Ligation High (TOYOBO), этот полученный ДНК-фрагмент приблизительно 1,9 т.п.н. лигировали с ДНК-фрагментом, полученным расщеплением вектора pYE22m рестриктазами EcoRI и SalI с получением плазмиды pYE-ACS-1.
С использованием плазмиды, содержащей кДНК MaACS-10 в качестве матрицы, выполняли ПЦР с представленными ниже праймерами с использованием ExTaq (Takara Bio Inc.). Амплифицированный таким образом ДНК-фрагмент клонировали при помощи набора для клонирования TOPO-TA (Invitrogen).
Праймер ACS-10-1F: 5'-GGATCCATGGAAACCTTGGTTAACGGAAAG-3' (SEQ ID NO: 91)
Праймер KpnI-ACS-10-R: 5'-GGTACCTAGAACTTCTTCCACATCTCCTC-3' (SEQ ID NO: 92)
Нуклеотидную последовательность этого инсерта подтверждали, и плазмиду, содержащую правильную нуклеотидную последовательность, расщепляли рестриктазами EcoRI и KpnI. С использованием Ligation High (TOYOBO), этот полученный ДНК-фрагмент приблизительно 2,1 т.п.н. лигировали с ДНК-фрагментом, полученным расщеплением вектора pYE22m рестриктазами EcoRI и KpnI. Плазмиду pYE-ACS-10 получали скринингом на ориентацию, которую промотор GAPDH вектора pYE22m помещал на его 5'-конце CDS MaACS-10.
С использованием плазмиды, содержащей кДНК MaACS-11 в качестве матрицы, выполняли ПЦР с представленными ниже праймерами с использованием ExTaq (Takara Bio Inc.). Амплифицированный таким образом ДНК-фрагмент клонировали при помощи набора для клонирования TOPO-TA (Invitrogen).
Праймер SacI-ACS-11-F: 5'-GAGCTCATGCCAAAGTGCTTTACCGTCAACG-3' (SEQ ID NO: 93)
Праймер BamHI-ACS-11-R: 5'-GGATCCTTACTTGGAGCCATAGATCTGCTTG-3' (SEQ ID NO: 94)
Нуклеотидную последовательность этого инсерта подтверждали, и плазмиду, содержащую правильную нуклеотидную последовательность, расщепляли рестриктазами SacI и BamHI. С использованием Ligation High (TOYOBO), полученный ДНК-фрагмент приблизительно 2,0 т.п.н. лигировали с ДНК-фрагментом, полученным расщеплением вектора pYE22m рестриктазами SacI и BamHI с получением плазмиды pYE-ACS-11.
Экспрессия в дрожжах
Получение трансформантов
Дрожжи штамма S. cerevisiae EH13-15 (trp1, MATα) (Appl. Microbiol. Biotechnol., 30, 515-520, 1989) трансформировали плазмидами pYE22m, pYE-MaACS-6, pYE-MaACS-8 и pYE-MaACS-9, соответственно, литийацетатным способом. Трансформанты подвергали скринингу на способность расти на агаровой среде SC-Trp (2% агар) (на литр, 6,7 г азотистого основания дрожжей без аминокислот (DIFCO), 20 г глюкозы, 1,3 г порошка аминокислот (смеси 1,25 г сульфата аденина, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глутаминовой кислоты, 0,6 г гистидина, 1,8 г лейцина, 0,9 г лизина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина, 6 г треонина и 0,6 г урацила)).
Культивирование дрожжей
Каждый один из трансформантов, полученных с использованием соответствующих плазмид, обеспечивали для следующего эксперимента культивирования.
Одну платиновую петлю этих дрожжей высевали на 10 мл SC-Trp и культивировали при встряхивании для предварительного инкубирования при 30°C в течение одного дня. После добавления 1 мл предварительно инкубированного материала к среде SC-Trp, выполняли основное культивирование культивированием со встряхиванием при 30°C в течение одного дня.
Анализ жирных кислот в мицелии
Культуральный бульон дрожжей центрифугировали для извлечения мицелиальных клеток. После промывки 10 мл стерильной воды, эти мицелиальные клетки опять центрифугировали, извлекали и лиофилизировали. Жирные кислоты в этих мицелиальных клетках превращали в метиловые эфиры способом с использованием смеси хлористоводородная кислота-этанол с последующей экстракцией гексаном. После удаления гексана дистилляцией жирные кислоты анализировали газовой хроматографией.
Продукция жирных кислот в расчете на количество среды показано в таблице 5. В штаммах, трансформированных ppYE-MaACS-6, pYE-MaACS-8 или pYE-MaACS-9, продукция в расчете на количество среды была увеличенной в сравнении с контролем, который трансформировали плазмидой pYE22m.

Экспрессия в продуцирующих арахидоновую кислоту дрожжах
(1) Выведение продуцирующих арахидоновую кислоту штаммов
Для выведения продуцирующего арахидоновую кислоту штамма дрожжей (S. cerevisiae), конструировали следующие плазмиды.
Сначала, с использованием кДНК, полученной из штамма 1S-4 M. alpina в качестве матрицы, выполняли ПЦР с ExTaq с использованием пары праймеров Δ12-f и Δ12-r, Δ6-f и Δ6-r, GLELO-f и GLELO-r или Δ5-f и Δ5-r для амплификации гена Δ12 десатуразы жирных кислот (GenBank Accession No. AB020033) (далее здесь "ген Δ12"), гена Δ6 десатуразы жирных кислот (GenBank Accession No. AB020032) (далее здесь "ген Δ6"), гена GLELO элонгазы жирных кислот (GenBank Accession No. AB193123) (далее здесь ген "GLELO") и гена Δ5 десатуразы жирных кислот (GenBank Accession No. AB188307) (далее здесь "ген Δ5") в штамме M. alpina 1S-4.
Δ12-f: 5'-TCTAGAATGGCACCTCCCAACACTATTG-3' (SEQ ID NO: 95)
Δ12-r: 5'-AAGCTTTTACTTCTTGAAAAAGACCACGTC-3' (SEQ ID NO: 96)
Δ6-f: 5'-TCTAGAATGGCTGCTGCTCCCAGTGTGAG-3' (SEQ ID NO: 97)
Δ6-r: 5'-AAGCTTTTACTGTGCCTTGCCCATCTTGG-3' (SEQ ID NO: 98)
GLELO-f: 5'-TCTAGAATGGAGTCGATTGCGCAATTCC-3' (SEQ ID NO: 99)
GLELO-r: 5'-GAGCTCTTACTGCAACTTCCTTGCCTTCTC-3' (SEQ ID NO: 100)
Δ5-f: 5'-TCTAGAATGGGTGCGGACACAGGAAAAACC-3' (SEQ ID NO: 101)
Δ5-r: 5'-AAGCTTTTACTCTTCCTTGGGACGAAGACC-3' (SEQ ID NO: 102)
Эти гены клонировали с набором TOPO-TA-Cloning Kit. Эти клоны подтверждали их нуклеотидными последовательностями. Клоны, содержащие нуклеотидные последовательности гена Δ12, гена Δ6, гена GLELO и гена Δ5, были обозначены как плазмиды pCR-MAΔ12DS (содержащая нуклеотидную последовательность гена Δ12), pCR-MAΔ6DS (содержащая нуклеотидную последовательность гена Δ6), pCR-MAGLELO (содержащая нуклеотидную последовательность гена GLELO) и pCR-MAΔ5DS (содержащая нуклеотидную последовательность гена Δ5), соответственно.
С другой стороны, плазмиду pURA34 (JPA 2001-120276) расщепляли рестриктазой HindIII. Полученный ДНК-фрагмент приблизительно 1,2 т.п.н. инсертировали в сайт HindIII этого вектора, который получали расщеплением вектора pUC18 (Takara Bio Inc.) рестриктазами EcoRI и SphI, затем затуплением концов и самолигированием указанного вектора. Клон, в котором сайт EcoRI этого вектора был локализован на его 5'-конце URA3, был назван pUC-URA3. Кроме того, ДНК-фрагмент приблизительно 2,2 т.п.н., который получали расщеплением YEp13 рестриктазами SalI и XhoI, инсертировали в сайт SalI вектора pUC18. Клон, в котором сайт EcoRI вектора был локализован на его 5'-конце LEU2, был назван pUC-LEU2.
Затем эту плазмиду pCR-MAΔ12DS расщепляли рестриктазой HindIII, с последующим затуплением концов и дополнительным расщеплением рестриктазой XbaI. Полученный ДНК-фрагмент приблизительно 1,2 т.п.н. лигировали с ДНК-фрагментрм приблизительно 6,6 т.п.н., который был получен расщеплением вектора pESC-URA (STRATAGENE) рестриктазой SacI, затуплением концов и дополнительным расщеплением рестриктазой SpeI. Таким образом, получали плазмиду pESC-U-Δ12. Плазмиду pCR-MAΔ6DS расщепляли рестриктазой XbaI, с последующим затуплением концов и дополнительным расщеплением рестриктазой HindIII. Полученный ДНК-фрагмент приблизительно 1,6 т.п.н. лигировали с ДНК-фрагментом приблизительно 8 т.п.н., который получали расщеплением плазмиды pESC-U-Δ12 растриктазой SalI, затуплением концов и дополнительным расщеплением рестриктазой HindIII, с получением посредством этого плазмиды pESC-U-Δ12:Δ6. Эту плазмиду частично расщепляли рестриктазой PvuII. Полученный фрагмент приблизительно 4,2 т.п.н. инсертировали в сайт SmaI pUC-URA3 с получением плазмиды pUC-URA-Δ12:Δ6.
Плазмиду pCR-MAGLELO также расщепляли рестриктазами XbaI и SacI. Полученный ДНК-фрагмент приблизительно 0,95 т.п.н. лигировали с ДНК-фрагментом приблизительно 7,7 т.п.н., который получали расщеплением вектора pESC-LEU (STRATAGENE) рестриктазами XbaI и SacI. Таким образом, получали плазмиду pESC-L-GLELO. Плазмиду pCR-MAΔ5DS расщепляли рестриктазой XbaI, с последующим затуплением концов и дополнительно расщеплением рестриктазой HindIII. Полученный ДНК-фрагмент приблизительно 1,3 т.п.н. лигировали с ДНК-фрагментрм приблизительно 8,7 т.п.н., который получали расщеплением плазмиды pESC-L-GLELO растиктазой ApaI, затуплением концов и дополнительно расщеплением рестриктазой HindIII, с получением посредством этого плазмиды pESC-L-GLELO:Δ5. Эту плазмиду расщепляли рестриктазой PvuII, и полученный фрагмент приблизительно 3,2 т.п.н. инсертировали в сайт SmaI pUC-LEU2 с получением плазмиды pUC-LEU-GLELO:Δ5. Штамм Saccharomyces cerevisiae YPH499 (STRATAGENE) котрансформировали плазмидой pUC-URA-Δ12:Δ6 и плазмидой pUC-LEU-GLELO:Δ5. Эти трансформанты подвергали скринингу на способность расти на SC-Leu,Ura-агаровой среде. Среди этих трансформантов, один случайный штамм был назван штаммом ARA3-1. При культивированием этого штамма в дополненной галактозой среде, этот штамм стал способным экспрессировать от промотора GAL1/10 ген Δ12-десатуразы жирных кислот, ген Δ6-десатуразы, ген GLELO и ген Δ5-десатуразы жирных кислот.
(2) Трансформация в продуцирующие арахидоновую кислоту дрожжи и анализ
Штамм ARA3-1 трансформировали плазмидами pYE22m, pYE-ACS-1, pYE-ACS-10 и pYE-ACS-11, соответственно. Трансформанты подвергали скринингу на способность расти на SC-Trp,Leu,Ura-агаровой среде (2% агар) (на литр, 6,7 г азотистого основания дрожжей без аминокислот (DIFCO), 20 г глюкозы, 1,3 г порошка аминокислот (смеси 1,25 г сульфата аденина, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глутаминовой кислоты, 0,6 г гистидина, 0,9 г тирозина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина и 6 г треонина)). Произвольные четыре штамма из соответствующих трансформированных плазмидой штаммов использовали для последующего культивирования.
Эти штаммы культивировали при 30°C в течение одного дня в 10 мл SC-Trp,Leu,Ura-жидкой среды, описанной выше. Один миллилитр этой культуры высевали на 10 мл SG-Trp,Leu,Ura-жидкой среды (на литр, 6,7 г азотистого основания дрожжей без аминокислот (DIFCO), 20 г галактозы, 1,3 г порошка аминокислот (смеси 1,25 г сульфата аденина, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глутаминовой кислоты, 0,6 г гистидина, 0,9 г лизина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина и 6 г треонина)) и затем культивировали при 15°C в течение 6 дней. Мицелиальные клетки собирали, промывали водой и затем лиофилизировали. После превращения жирных кислот в высушенных мицелиальных клетках в метиловые эфиры способом с использованием хлористоводородной кислоты-метанола, выполняли анализ жирных кислот газовой хроматографией. Отношение каждой PUFA к общим жирным кислотам в контрольном штамме, трансформированном плазмидой pYE22m, и в штаммах, трансформированных каждым гомологом ACS из Mortierella, показано в таблице 6.

Как показано в таблице 6, это соотношение жирных кислот могло бы быть модифицировано экспрессией гомолога ACS из Mortierella. В частности, в экспрессионном штамме MaACS-11, соотношения арахидоновой кислоты, линолевой кислоты и γ-линоленовой кислоты увеличивались приблизительно в 1,8 раза, приблизительно в 1,5 раз и приблизительно в 2,4 раза, соответственно, в сравнении с контрольным штаммом. В экспрессионном штамме MaACS-1 количество арахидоновой кислоты увеличивалось приблизительно в 1,5 раза в сравнении с контрольным штаммом. Дополнительно в экспрессионном штамме MaACS-10 соотношения линолевой кислоты и γ-линоленовой кислоты увеличивались приблизительно в 2 раза и приблизительно в 4 раза, соответственно, в сравнении с контрольным штаммом.
[ПРИМЕР 2]
Конструирование экспрессирующего вектора
Экспрессирующий вектор для дрожжей
Вектор pYE-ACS-12 для экспрессии MaACS-12 в дрожжах конструировали следующим образом. С использованием плазмиды, содержащей кДНК MaACS-12 в качестве матрицы, выполняли ПЦР со следующими праймерами с использованием KOD-Plus- (TOYOBO).
Праймер Eco-ACS-G-F: 5'-GAATTCATGACAAAGTGCCTCACCGTCG-3' (SEQ ID NO: 103)
Праймер Sma-ACS-G-R: 5'-CCCGGGACTTAGGCCGTTCCATAAAGCTG-3' (SEQ ID NO: 104)
Амплифицированный ДНК-фрагмент клонировали с использованием набора Zero Blunt TOPO PCR Cloning Kit (Invitrogen). Нуклеотидную последовательность этого инсерта подтверждали, и плазмиду, содержащую правильную нуклеотидную последовательность, расщепляли рестриктазами EcoRI и SmaI. С использованием Ligation High (TOYOBO), полученный фрагмент ДНК приблизительно 2 т.п.н. лигировали с ДНК-фрагментом, полученным расщеплением вектора pYE22m рестриктазой BamHI и затем затуплением концов с использованием Blunting Kit (TAKARA Bio), и затем расщеплением EcoRI, с получением плазмиды pYE-ACS-12.
Экспрессирующий вектор для M. alpina
Вектор для экспрессии MaACS-10 и MaACS-11 в M. alpina конструировали следующим образом.
Сначала pUC18 расщепляли рестриктазами EcoRI и HindIII, и адаптор, полученный отжигом олиго ДНК MCS-для-pUC18-F2 с MCS-для-pUC18-R2, инсертировали в нее для конструирования pUC18-RF2.
MCS-для-pUC18-F2:
5'-AATTCATAAGAATGCGGCCGCTAAACTATTCTAGACTAGGTCGACGGCGCGCCA-3' (SEQ ID NO: 105)
MCS-для-pUC18-R2:
5'-AGCTTGGCGCGCCGTCGACCTAGTCTAGAATAGTTTAGCGGCCGCATTCTTATG-3' (SEQ ID NO: 106)
С использованием геномной ДНК M. alpina в качестве матрицы, выполняли ПЦР с праймерами Not1-GAPDHt-F и EcoR1-Asc1-GAPDHt-R с использованием KOD-Plus- (Toyobo). Амплифицированный ДНК-фрагмент приблизительно 0,5 т.п.н. клонировали с использованием набора Zero Blunt TOPO PCR Cloning Kit (Invitrogen). После подтверждения нуклеотидной последовательности этого инсерта, этот ДНК-фрагмент приблизительно 0,9 т.п.н., полученный расщеплением рестриктазами NotI и EcoRI, инсертировали в сайт NotI и EcoRI плазмиды pUC18-RF2 для конструирования плазмиды pDG-1.
Not1-GAPDHt-F: 5'-AGCGGCCGCATAGGGGAGATCGAACC-3' (SEQ ID NO: 107)
EcoR1-Asc1-GAPDHt-R: 5'-AGAATTCGGCGCGCCATGCACGGGTCCTTCTCA-3' (SEQ ID NO: 108)
С использованием генома M. alpina в качестве матрицы, выполняли ПЦР с праймерами URA5g-F1 и URA5g-R1 с использованием KOD-Plus- (Toyobo). Амплифицированный ДНК-фрагмент клонировали с использованием набора Zero Blunt TOPO PCR Cloning Kit (Invitrogen). После подтверждения нуклеотидной последовательности этого инсерта, ДНК-фрагмент приблизительно 2 т.п.н., полученный расщеплением SalI, инсертировали в сайт SalI плазмиды pDG-1. Эта плазмида, которая инсертирована на 5'-конце геном URA5, была ориентирована в направлении EcoRI-стороны этого вектора, и была названа плазмидой pDuraG.
URA5g-F1: 5'-GTCGACCATGACAAGTTTGC-3' (SEQ ID NO: 109)
URA5g-R1: 5'-GTCGACTGGAAGACGAGCACG-3' (SEQ ID NO: 110)
Затем выполняли ПЦР с KOD-Plus- (TOYOBO) с использованием генома M. alpina в качестве матрицы и праймеров hisHp+URA5-F и hisHp+MGt-F. С использованием набора In-Fusion (зарегистрированное товарное название) Advantage PCR Cloning Kit (TAKARA Bio), этот амплифицированный ДНК-фрагмент приблизительно 1,0 т.п.н. лигировали с ДНК-фрагментом приблизительно 5,3 т.п.н., амплифицированным при помощи ПЦР с KOD-Plus- (TOYOBO) с использованием pDuraG в качестве матрицы и праймеров pDuraSC-GAPt-F и URA5gDNA-F, с получением плазмиды pDUra-RhG.
hisHp+URA5-F:
5'-GGCAAACTTGTCATGAAGCGAAAGAGAGATTATGAAAACAAGC-3' (SEQ ID NO: 111)
hisHp+MGt-F:
5'-CACTCCCTTTTCTTAATTGTTGAGAGAGTGTTGGGTGAGAGT-3' (SEQ ID NO: 112)
pDuraSC-GAPt-F:
5'-TAAGAAAAGGGAGTGAATCGCATAGGG-3' (SEQ ID NO: 113)
URA5gDNA-F: 5'-CATGACAAGTTTGCCAAGATGCG-3' (SEQ ID NO: 114)
С использованием плазмиды pDUra-RhG в качестве матрицы, этот ДНК-фрагмент приблизительно 6,3 т.п.н. амплифицировали при помощи ПЦР KOD-Plus- (TOYOBO) с использованием праймеров pDuraSC-GAPt-F и pDurahG-hisp-R.
pDurahG-hisp-R: 5'-ATTGTTGAGAGAGTGTTGGGTGAGAGTG-3' (SEQ ID NO: 115)
С использованием плазмиды, содержащей кДНК MaACS-10, этот ДНК-фрагмент приблизительно 2,1 т.п.н. амплифицировали при помощи ПЦР с KOD-Plus- (TOYOBO), с использованием представленных ниже праймеров.
Праймер ACS-10+hisp-F:
5'-CACTCTCTCAACAATATGGAAACCTTGGTTAACGGAAAGT-3' (SEQ ID NO: 116)
Праймер ACS-10+MGt-R:
5'-CACTCCCTTTTCTTACTAGAACTTCTTCCACATCTCCTCAATATC-3' (SEQ ID NO: 117)
Полученный ДНК-фрагмент лигировали с ДНК-фрагментом 6,3 т.п.н., описанным выше, с использованием In-Fusion (зарегистрированное товарное название) Advantage PCR Cloning Kit (TAKARA BIO) с получением плазмиды pDUraRhG-ACS-10.
С использованием плазмиды, содержащей кДНК MaACS-11 в качестве матрицы, ДНК-фрагмент 2,1 т.п.н. амплифицировали при помощи ПЦР с KOD-Plus- (TOYOBO) с использованием представленных ниже праймеров.
Праймер ACS-11+MGt-R:
5'-CACTCCCTTTTCTTATTACTTGGAGCCATAGATCTGCTTGA-3' (SEQ ID NO: 118)
Праймер ACS-11+hisp-F:
5'-CACTCTCTCAACAATATGCCAAAGTGCTTTACCGTCAAC-3' (SEQ ID NO: 119)
Полученный ДНК-фрагмент лигировали с ДНК-фрагментом 6,3 т.п.н., описанным выше, с использованием In-Fusion (зарегистрированное товарное название) Advantage PCR Cloning Kit (TAKARA BIO) с получением плазмиды pDUraRhG-ACS-11.
Оценивание активности ACS
Дрожжи EH13-15 трансформировали плазмидами pYE22m, pYE-ACS-5, pYE-ACS-8, pYE-ACS-10, pYE-ACS-11 и pYE-ACS-12, соответственно, и два случайных полученных трансформанта культивировали следующим образом. Одну платиновую петлю этих мицелиальных клеток высевали на 10 мл SC-Trp и культивировали при встряхивании для предварительного инкубирования при 30°C в течение одного дня. После добавления 1% предварительно инкубированного материала к 100 мл среды SC-Trp, выполняли основное культивирование культивированием со встряхиванием при 28°C в течение одного дня.
Раствор необработанного фермента готовили следующим образом. Мицелиальные клетки собирали центрифугированием, промывали водой и временно хранили при -80°C. Мицелиальные клетки суспендировали в 5 мл Буфера В (50 мМ раствор натрийсульфатного буфера (рН 6,0), 10% глицерина и 0,5 мМ PMSF). Затем эти мицелиальные клетки разрывали с использованием Френч-пресса (16 кПа, 3 раза). Центрифугирование проводили при 1500×g при 4°C в течение 10 минут и центрифугировали. Полученный супернатант использовали в качестве раствора необработанного фермента.
Активность ACS определяли следующими процедурами на основе описания ссылочной литературы (J.B.C., 272 (8), 1896-4903, 1997). Реакционный раствор содержал 200 мМ Трис-HCl (pH 7,5), 2,5 мМ АТФ, 8 мМ MgCl2, 2 мМ ЭДТА, 20 мМ NaF, 0,1% Тритон Х-100, 50 мкг/мл жирных кислот, 50 мкм CoA и 100 мкл необработанного раствора фермента (подходящим образом разведенного в Буфере B), и его готовили в целом в 500 мкл. Эту реакцию проводили при 28°C в течение 30 минут. После завершения этой реакции, добавляли 2,5 мл стоп-раствора (изопропанол:н-гептан:1 M серная кислота (40:20:1)) и смесь тщательно перемешивали. Кроме того, к ней добавляли 2 мл н-гептана. После их тщательного перемешивании, эту смесь центрифугировали для извлечения верхнего слоя. К более низкому слою добавляли еще 2 мл н-гептана и обрабатывали таким же образом для извлечения верхнего слоя. Эти слои верхнего слоя объединяли и упаривали досуха с использованием центрифужного концентратора. Затем к полученному материалу добавляли 50 мкл 0,2 мг/мл трикозановой кислоты (23:0) в качестве внутреннего стандарта. Эти жирные кислоты превращали в метиловые эфиры способом с использованием смеси хлористоводородная кислота-метанол с последующим анализом жирных кислот с использованием газовой хроматографии. Количество жирных кислот, которые изменялись в ацил-CoA и, следовательно, распределялись в нижний слой описанными выше процедурами, рассчитывали из количества детектируемых жирных кислот. Эти результаты показаны в таблице ниже. Активность ACS выражали в виде количества жирных кислот, распределенных в нижний слой описанными выше процедурами, в расчете на массу белка в необработанном растворе фермента. Контролем является штамм, трансформированный pYE22m, а другие штаммы являются трансформантами, в которые были введены экспрессирующие векторы соответствующих генов.
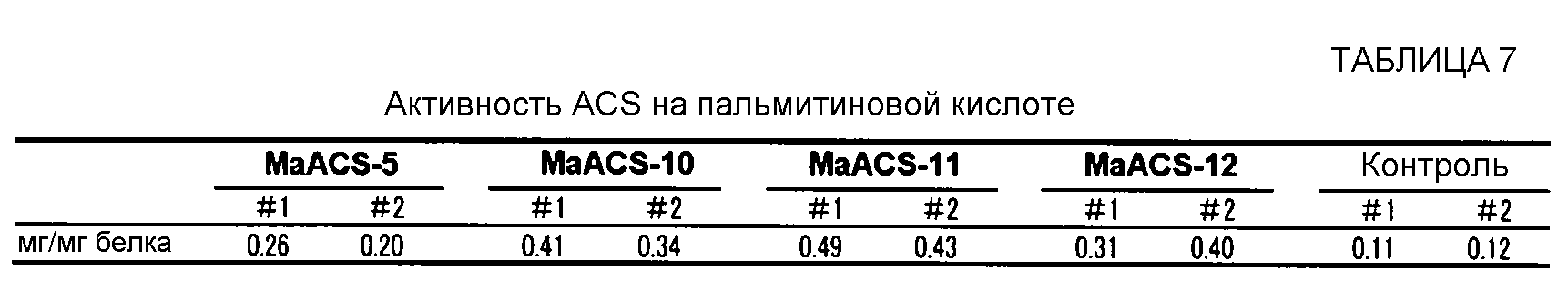
При применении в качестве субстрата пальмитиновой кислоты, MaACS-5, MaACS-10, MaACS-11 и MaACS-12 обнаруживали активность ACS, приблизительно в 2-4 раза более высокую, чем контроль.

При применении олеиновой кислоты в качестве субстрата, MaACS-10, MaACS-11 и MaACS-12 обнаруживали активность ACS, в два раза более высокую, чем контроль.
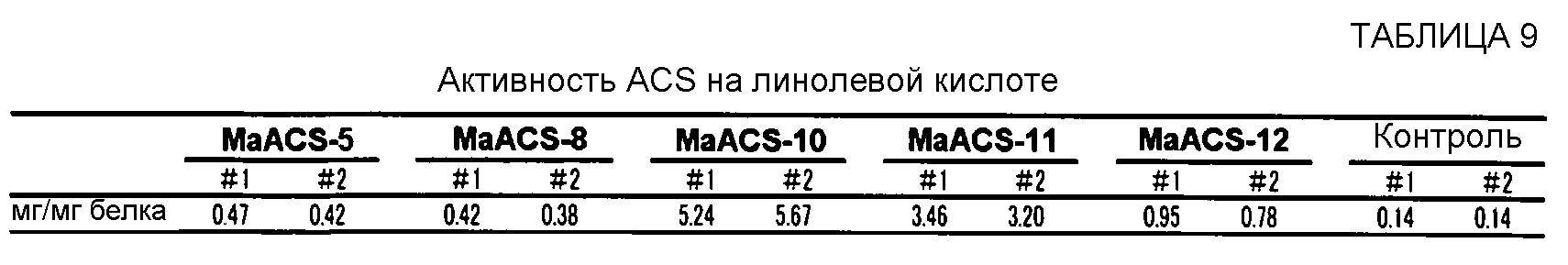
При применении линолевой кислоты в качестве субстрата, MaACS-5, MaACS-8 и MaACS-12 обнаруживали активность ACS, в несколько раз (приблизительно в 3, 3 раза и в 6 раз, соответственно) более высокую, чем контроль, в то время как MaACS-10 и MaACS-11 обнаруживали активность ACS, в несколько десятков раз (приблизительно в 40 раз и в 20 раз, соответственно) более высокую, чем контроль.

При применении γ-линолевой кислоты в качестве субстрата, MaACS-5, MaACS-8, MaACS-10, MaACS-11 и MaACS-12 обнаруживали активность ACS, приблизительно в 2 раза-10 раз более высокую, чем контроль.

При применении дигомо-γ-линолевой кислоты в качестве субстрата, все из MaACS-10, MaACS-11 и MaACS-12 обнаруживали активность ACS, в несколько десятков раз (приблизительно в 60 раз, 40 раз и 30 раз, соответственно) более высокую, чем контроль.

При применении арахидоновой кислоты в качестве субстрата, MaACS-10, MaACS-11 и MaACS-12 обнаруживали активность ACS, в несколько десятков раз (приблизительно в 90 раз, 30 раз и 10 раз, соответственно) более высокую, чем контроль.
Как описано выше, MaACS-10, MaACS-11 и MaACS-12, в частности, обнаруживали более высокую активность на полиненасыщенных жирных кислотах из 20 атомов углерода, таких как дигомо-γ-линолевая или арахидоновая кислота
Активность поглощения арахидоновой кислоты ACS-экспрессируемыми дрожжами
Дрожжи EH13-15 трансформировали плазмидами pYE22m, pYE-ACS-10, pYE-ACS-11 и pYE-ACS-12, соответственно, и полученные случайным образом два трансформанта культивировали следующим образом. Одну платиновую петлю этих клеток высевали на 10 мл SC-Trp и культивировали при встряхивании для предварительного инкубирования при 30°C в течение одного дня. После добавления 100 мкл предварительно инкубированного материала к 10 мл среды SC-Trp, дополненной 50 мкг/мл арахидоновой кислоты, выполняли основное культивирование культивированием со встряхиванием при 25°C в течение одного дня. Эти мицелиальные клетки собирали, лиофилизировали и подвергали анализу жирных кислот. Определяли отношение арахидоновой кислоты, поглощенной в эти мицелиальные клетки, к добавленной арахидоновой кислоте. Эти результаты показаны в таблице 14. Контролем является штамм, трансформированный pYE22m, и другие штаммы являются трансформантами, в которые были индуцированы экспрессирующие векторы соответствующих генов.
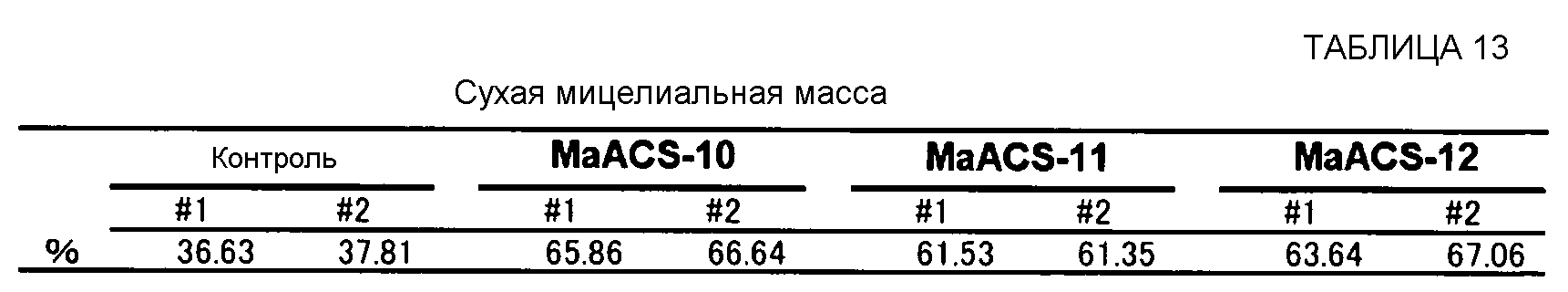
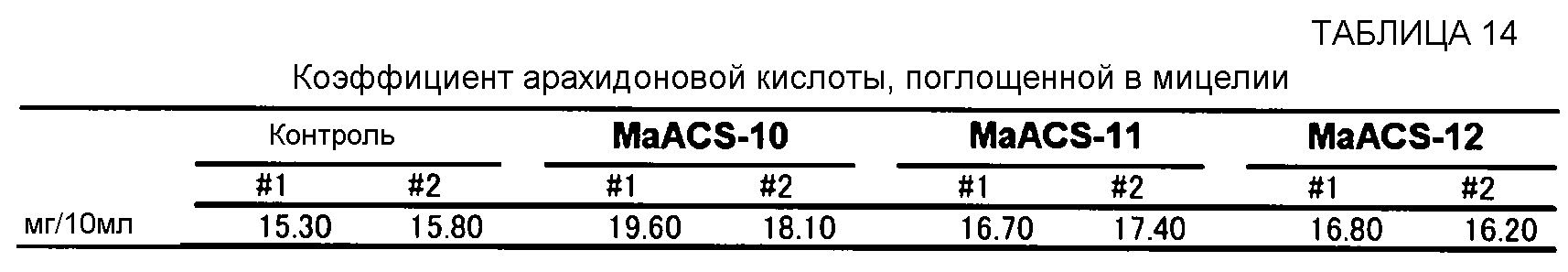
Получение трансформантов M. alpina
С использованием в качестве хозяина урацил-ауксотрофного штамма Δura-3, полученного из штамма M. alpina 1S-4, как описано в PCT International Publication Pamphlet WO 2005/019437 "Method of Breeding Lipid-Producing Fungus", трансформацию выполняли способом доставки частиц с использованием плазмид pDUraRhG-ACS-10 и pDUraRhG-ACS-11, соответственно. Для скрининга этих трансформантов использовали агаровую среду SC (0,5% азотистое основания без аминокислот и сульфата аммония (Difco), 0,17% сульфат аммония, 2% глюкоза, 0,002% аденин, 0,003% тирозин, 0,0001% метионин, 0,0002% аргинин, 0,0002% гистидин, 0,0004% лизин, 0,0004% триптофан, 0,0005% треонин, 0,0006% изолейцин, 0,0006% лейцин, 0,0006% фенилаланин и 2% агар).
Оценивание трансформантов M. alpina
Полученные трансформанты высевали на 4 мл среды GY и культивировали их со встряхиванием при 28°C в течение 2 дней. Мицелиальные клетки собирали фильтрованием, и РНК экстрагировали с набором RNeasy Plant (QIAGEN). Для синтеза кДНК использовали SuperScript First Strand System для ОТ-ПЦР. Для подтверждения экспрессии соответствующих генов из введенных конструктов, выполняли ОТ-ПЦР со следующими парами праймеров.
ACS10-RT1: 5'-GTCCCGAATGGTTCCT-3' (SEQ ID NO: 120)
ACS10-RT2: 5'-AGCGGTTTTCTACTTGC-3' (SEQ ID NO: 121)
ACS11-RT1: 5'-AACTACAACCGCGTCG-3' (SEQ ID NO: 122)
ACS11-RT2: 5'-CGGCATAAACGCAGAT-3' (SEQ ID NO: 123)
В трансформантах, которые подтверждали сверхэкспрессию, один трансформант в каждом случае высевали на 10 мл среды GY (2% глюкоза и 1% дрожжевой экстракт) и культивировали со встряхиванием при 28ºC при 300 об/мин в течение 3 дней. Весь объем этой культуры переносили на 500 мл среды GY (в колбу на 2 л Sakaguchi) и культивировали со встряхиванием при 28ºC и 120 об/мин. При трех, семи, десяти и двенадцати днях после этого дня 5 мл каждой и 10 мл каждой брали и фильтровали. После сушки мицелиальных клеток при 120°C, жирные кислоты превращали в метиловые эфиры способом с использованием смеси хлористоводородная кислота-метанол и анализировали газовой хроматографией. Продуцирование жирных кислот и количество продуцируемой арахидоновой кислоты, в расчете на высушенные мицелиальные клетки, подвергали мониторингу на протяжении времени. Штамм Δura-3 трансформанта-хозяина использовали в качестве контроля. Эти результаты показаны на Фиг. 27 (MaACS-10) и Фиг. 28 (MaACS-11).
Как показано на фигурах 27 и 28, при сверхэкспрессии MaACS-10 и MaACS-11 в M. alpina, как количество жирных кислот, так и количество арахидоновой кислоты в расчете на мицелий увеличивались в сравнении с контролем.
ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ
Полинуклеотид по данному изобретению экспрессируют в подходящей клетке-хозяине для эффективного продуцирования жирных кислот, в частности полиненасыщенных жирных кислот. Эти жирные кислоты, продуцируемые в клетках-хозяевах по данному изобретению, могут быть использованы для получения композиций жирных кислот, пищевых продуктов, косметических средств, фармацевтических средств, мыла и т.д.
Реферат
Изобретение относится к биотехнологии, в частности к белку, имеющему ацил-CoA-синтетазную активность, полученному из микроорганизма рода Mortierella и полинуклеотиду, кодирующему этот белок. Изобретение также относится к экспрессирующему вектору и трансформанту, не являющемся человеком, содержащим заявленные полинуклеотиды, а также к способу получения композиции липидов или жирных кислот с использованием этого трансформанта. Изобретение позволяет за счет использования нового гена увеличивать количество липидов или жирных кислот, а также изменять состав продуцируемых жирных кислот. 5 н. и 5 з.п. ф-лы, 28 ил., 14 табл., 2 пр.
Формула
(a) полинуклеотид, состоящий из любой нуклеотидной последовательности, выбранной из группы, состоящей из нуклеотидных последовательностей, представленных в SEQ ID NO: 46, 51, 56, 36, 21, 1, 6, 11, 16, 26, 31 и 41.
(b) полинуклеотид, кодирующий белок, состоящий из любой аминокислотной последовательности, выбранной из группы, состоящей из аминокислотных последовательностей, представленных в SEQ ID NO: 47, 52, 57, 37, 22, 2, 7, 12, 17, 27, 32 и 42;
(c) полинуклеотид, кодирующий белок, состоящий из аминокислотной последовательности, где 1-10 аминокислот делетированы, заменены, инсертированы и/или добавлены в любую аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей, представленных в SEQ ID NO: 47, 52, 57, 37, 22, 2, 7, 12, 17, 27, 32 и 42, и имеющий ацил-CoA-синтетазную активность;
(d) полинуклеотид, кодирующий белок, имеющий аминокислотную последовательность, имеющую по меньшей мере 90% идентичность любой аминокислотной последовательности, выбранной из группы, состоящей из аминокислотных последовательностей, представленных в SEQ ID NO: 47, 52, 57, 37, 22, 2, 7, 12, 17, 27, 32 и 42, и имеющий ацил-CoA-синтетазную активность; и
(e) полинуклеотид, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной любой нуклеотидной последовательности, выбранной из группы, состоящей из нуклеотидных последовательностей, представленных в SEQ ID NO: 46, 51, 56, 36, 21, 1, 6, 11, 16, 26, 31 и 41, при строгих условиях, и который кодирует белок, имеющий ацил-CoA-синтетазную активность.
Документы, цитированные в отчёте о поиске
Ферменты синтетазы
Комментарии