Рекомбинантные hvt-векторы, экспрессирующие множественные антигены патогенов птиц, и содержащие их вакцины - RU2752836C2
Код документа: RU2752836C2
Чертежи






























Описание
Перекрестная ссылка на связанные заявки
Эта заявка испрашивает приоритет по предварительной заявке США 62/433842, поданной 14 декабря 2016 года.
Область техники, к которой относится изобретение
Изобретение относится к рекомбинантным вирусным векторам для вставки и экспрессии чужеродных генов для применения в качестве безопасных средств иммунизации для защиты от различных патогенов. Оно также относится к поливалентной композиции или вакцине, содержащей один или несколько рекомбинантных вирусных векторов, для защиты от различных патогенов. Настоящее изобретение относится к способам получения и применения рекомбинантных вирусных векторов.
Предшествующий уровень техники
Вакцинация домашней птицы широко используется для защиты стай домашней птицы от разрушительных болезней, включая ньюкаслскую болезнь (ND), инфекционный бурсит птиц (IBD), болезнь Марека (MD), инфекционный бронхит (IB), инфекционный ларинготрахеит (ILT) и птичий грипп (AI). ND вызывается птичьим парамиксовирусом 1 (APMV-1), также обозначаемым вирусом ND (NDV), принадлежащим к семейству Paramyxoviridae. MD вызывается герпесивирусом кур 2 (семейство Herpesviridae), также обозначаемым как вирус MD серотипа 1 (MDV1). IB вызывается вирусом IB (IBV), принадлежащим к семейству Coronaviridae, ILT вызывается вирусом герпеса 1 (семейство Herpesviridae), также называемым вирусом ILT (ILTV), и AI вызывается вирусом AI (AIV), принадлежащим к семейству Orthomyxoviridae.
С целью вакцинации птиц от этих птичьих патогенов был предложен ряд рекомбинантных птичьих вирусных векторов. Используемые вирусные векторы включают авипоксвирусы, особенно оспы кур (ЕР-А-0,517,292), вирус Марека, такой как серотипы 1, 2 и 3 (HVT) (WO87/04463; WO2013/082317) или, в ином случае, ITLV, NDV и птичий аденовирус. Когда некоторые из этих рекомбинантных птичьих вирусных векторов использовались для вакцинации, они демонстрировали различные уровни защиты.
Было разработано несколько рекомбинантных вирусов герпеса индеек (HVT, также обозначаемых векторами Meleagrid herpesvirus 1 или MDV серотипа 3), экспрессирующих антигены из различных патогенов (патенты США №№ 5980906, 5853733, 6183753, 5187087), включая IBDV, NDV, ILTV и лицензированные AIV. Особый интерес представляет защитный ген IBDV VP2, экспрессируемый HVT-вектором, который продемонстрировал явные преимущества по сравнению с классическими вакцинами против IBD (Bublot et al. J.Comp. Path.2007, Vol.137, S81-S84; US 5980906). Другие представляющие интерес HVT-векторы представляют собой те, которые экспрессируют защитный(ые) ген(ы) либо NDV (Morgan et al 1992, Avian dis. 36, 858-70; US 6866852; US 5650153), ILTV (Johnson et al., 2010 Avian Dis 54, 1251-1259; US 6299882; US5,853,733, EP 1801204) либо NDV и IBDV (US9,114,108; WO2016102647, WO2013/057235, WO2015032910, WO20131445. В US2016/0158347 сообщалось об использовании олигодезоксинуклеотидного агониста TLR21 для усиления иммунного ответа против антигена, который экспрессируется вектором HVT.
Одной из практических проблем совместного использования нескольких рекомбинантных вакцин на основе HVT является их интерференция. Снижение защиты, по меньшей мере, от одного заболевания происходит, когда смешиваются два рекомбинанта HVT, экспрессирующих разные антигены (Rudolf Heine 2011; Issues of the Poultry Recombinant Viral Vector Vaccines which May Cause an Effect on the Economic Benefits of those Vaccines; работа представлена на XVII конгрессе Всемирной ассоциации специалистов по ветеринарии домашней птицы (WVPA) в Канкуне, Мексика, 14-18 августа 2011 года; Slacum G, Hein R. and Lynch P., 2009, The compatibility of HVT recombinants with other Marek’s disease vaccines, 58th Western Poultry Disease Conference, Sacramento, CA, USA, March 23rd-25th, p 84).
Учитывая потенциальное влияние патогенов животных, таких как NDV и IBDV, на ветеринарное здравоохранение и экономику, необходимы эффективные способы профилактики инфекции и защиты животных. Существует потребность в комбинированных эффективных векторных вакцинах и подходящем способе создания вакцины, которая могла бы облегчить проблему интерференции, наблюдаемой между двумя векторными вакцинами на основе HVT.
Сущность изобретения
Настоящее изобретение показало неожиданный результат, когда поливалентные композиции или вакцины, содержащие рекомбинантный HVT-вектор, были эффективными при защите животных от различных птичьих патогенов без интерференции. Неожиданные результаты также наблюдались, когда различные комбинации промоторов/линкеров, кодон-оптимизированного гена, polyA-хвостов и сайтов вставки придают разные уровни эффективности и стабильности экспрессии одного или нескольких гетерологичных генов in vivo и in vitro. Настоящее изобретение обеспечивает стабильные HVT-векторы, которые способны эффективно экспрессировать несколько генов, и преодолевает известную проблему, заключающуюся в том, что HVT-векторы с несколькими вставками являются менее стабильными.
Настоящее изобретение относится к рекомбинантному HVT-вектору, содержащему один, два или более гетерологичных полинуклеотидов, кодирующих и экспрессирующих, по меньшей мере, один антиген птичьего патогена.
Настоящее изобретение относится к композиции или вакцине, содержащей один или несколько рекомбинантных HVT-векторов, содержащих один или несколько гетерологичных полинуклеотидов, кодирующих и экспрессирующих, по меньшей мере, один антиген птичьего патогена.
Настоящее изобретение относится к способу вакцинации животного или индукции иммуногенного или защитного ответа у животного, включающему, по меньшей мере, одно введение композиции или вектора по настоящему изобретению.
Краткое описание чертежей
Следующее подробное описание, данное в качестве примера и не предназначенное для ограничения изобретения конкретными описанными воплощениями, может быть понято в сочетании с прилагаемыми фигурами, включенными в настоящий документ ссылкой, в которых:
фиг. 1 представляет собой таблицу, показывающую SEQ ID NO, присвоенные каждой последовательности ДНК и белка;
фиг. 2 отображает структуру генома HVT и сайтов вставки в нем;
фиг. 3 отображает плазмидную карту pFSV40VP2;
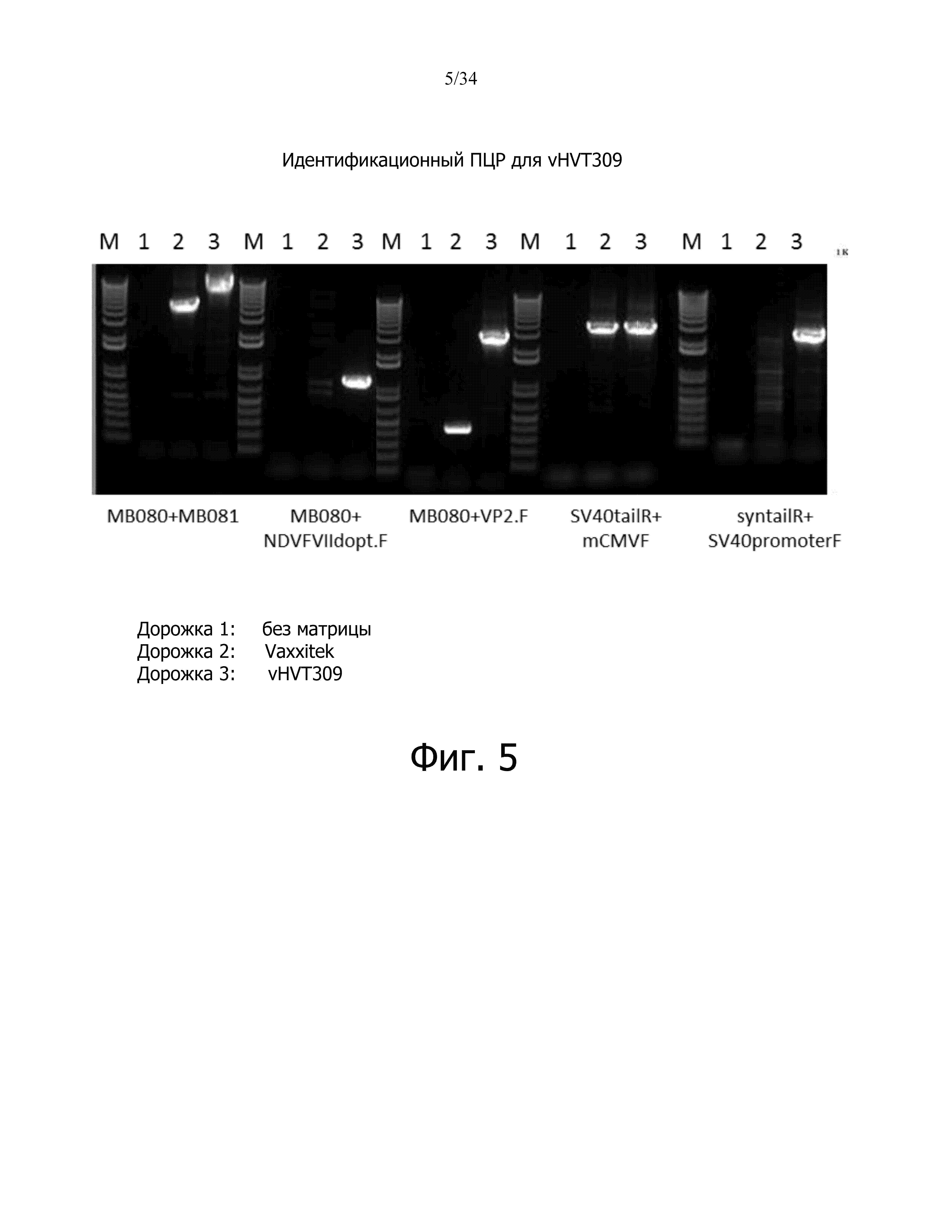
фиг. 4 отображает схематическое представление сайтов связывания праймеров для vHVT309;
фиг. 5 отображает результат идентификации ПЦР для vHVT309;
фиг. 6 отображает плазмидную карту pFIRESVP2;
фиг. 7 отображает схематическое представление сайтов связывания праймеров для vHVT310;
фиг. 8 отображает результат идентификации ПЦР для vHVT310;
фиг. 9 отображает плазмидную карту pFP2AVP2;
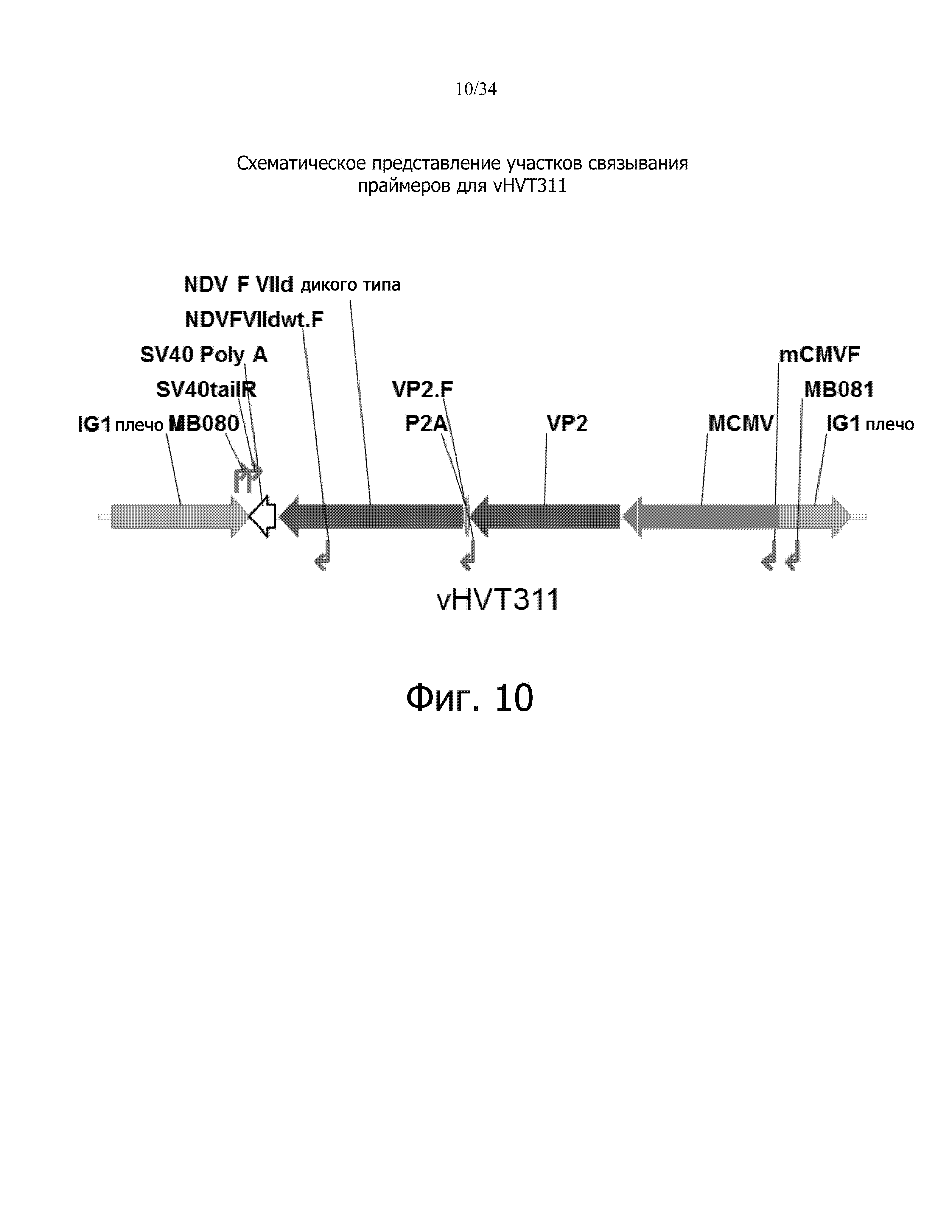
фиг. 10 отображает схематическое представление сайтов связывания праймеров для vHVT311;
фиг. 11 отображает результат идентификации ПЦР для vHVT311;
фиг. 12 отображает плазмидную карту pVP2IRESgD;
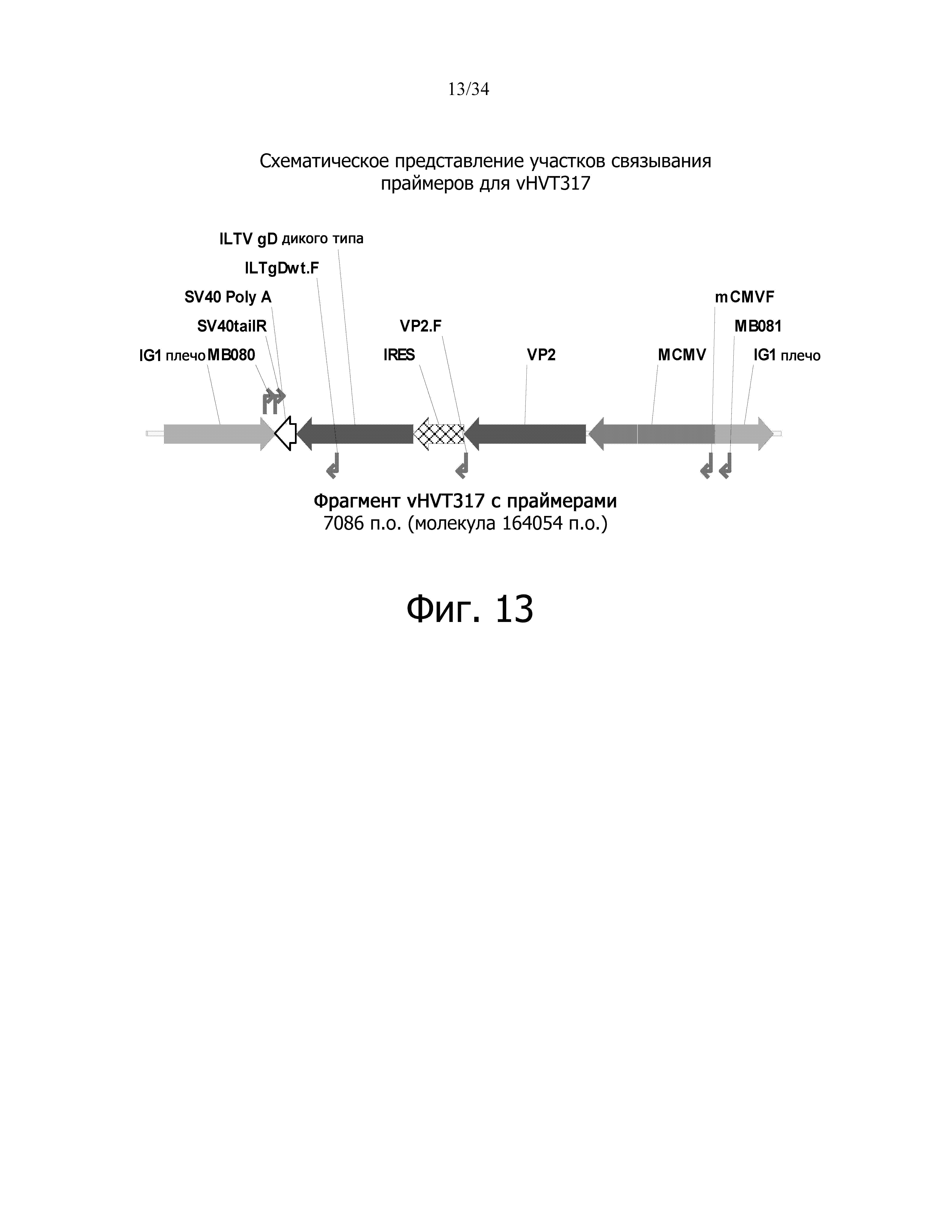
фиг. 13 отображает схематическое представление сайтов связывания праймеров для vHVT317;
фиг. 14 отображает результат идентификации ПЦР для vHVT317;
фиг. 15 отображает плазмидную карту pFwtSV40VP2;
фиг. 16 отображает схематическое представление сайтов связывания праймеров для vHVT313;
фиг. 17 отображает результат идентификации ПЦР для vHVT313;
фиг. 18 отображает плазмидную карту pVP2IRESFwt;
фиг. 19 отображает схематическое представление сайтов связывания праймеров для vHVT316;
фиг. 20 отображает результат идентификации ПЦР для vHVT316;
фиг. 21А-21D отображает выравнивания последовательностей ДНК и белка;
фиг. 22 отображает плазмидную карту HVT US2SVgDwtsyn;
фиг. 23 отображает плазмидную карту pHVTIG1gDCaFopt;
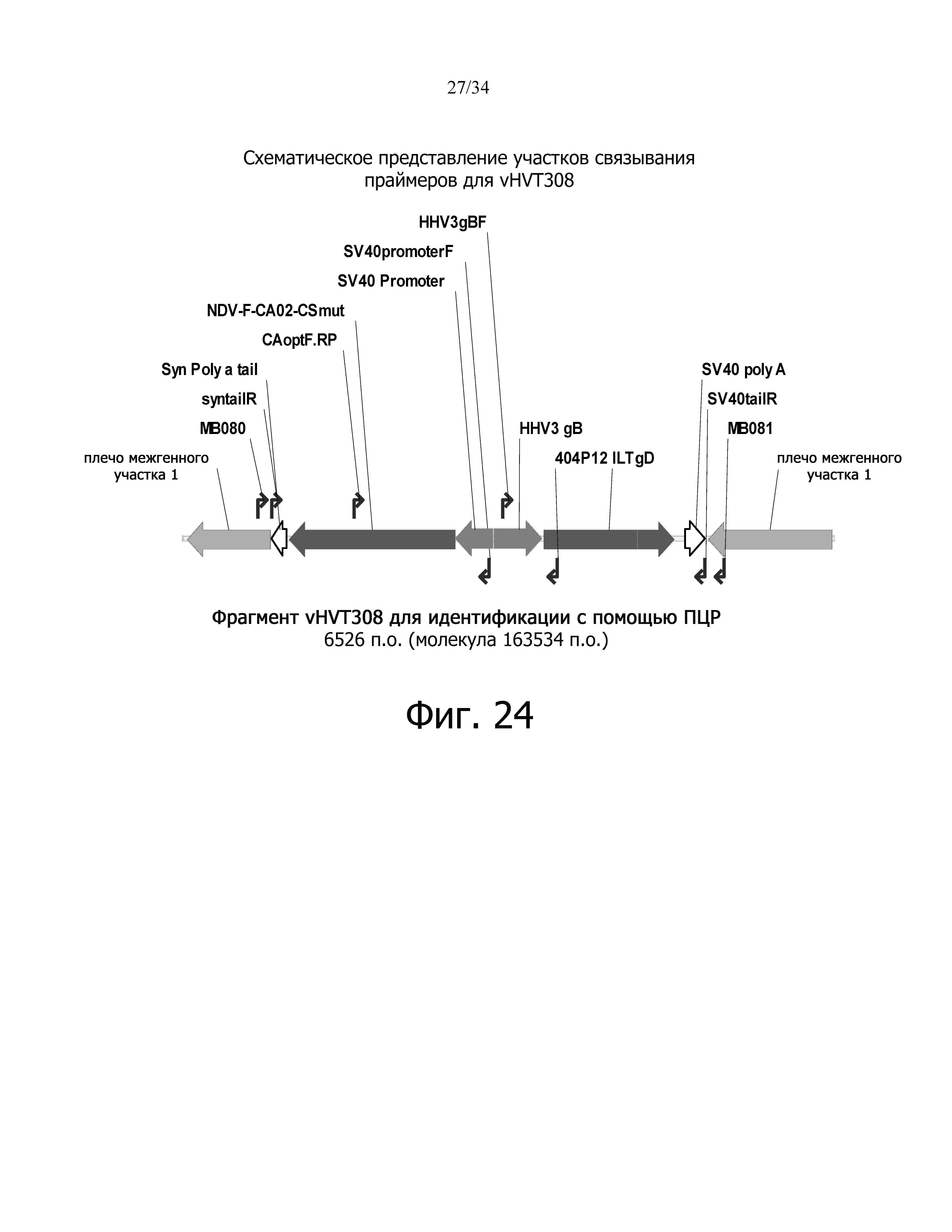
фиг. 24 отображает схематическое представление сайтов связывания праймеров для vHVT308;
фиг. 25 отображает результат идентификации ПЦР для vHVT308;
фиг. 26 отображает плазмидную карту pFwtIRESgD;
фиг. 27 отображает схематическое представление сайтов связывания праймеров для vHVT322;
фиг. 28 отображает результат идентификации ПЦР для vHVT322;
фиг. 29 отображает плазмидную карту pHVTUS2SVgDwtsyn;
фиг. 30 отображает схематическое представление сайтов связывания праймеров для vHVT406;
фиг. 31 отображает результат идентификации ПЦР для vHVT406.
Подробное описание изобретения
Хотя изобретение было описано в связи с его конкретными воплощениями, следует понимать, что оно способно к дальнейшим модификациям, и эта заявка предназначена для охвата любых вариантов, применений или адаптаций изобретения в соответствии, в общем, с принципами изобретения и включая такие отступления от настоящего раскрытия, которые соответствуют известной или обычной практике в области техники, к которой относится изобретение, и которые могут быть применены к существенным признакам, изложенным выше, и как указано далее в объеме прилагаемой формулы изобретения. Данное изобретение включает все модификации и эквиваленты объекта изобретения, изложенного в аспектах или формуле изобретения, представленных в данном документе, в максимальной степени, допускаемой применимым законодательством.
Следует отметить, что в этом раскрытии и, в частности, в формуле изобретения такие термины, как «включает», «включенный», «включающий» и тому подобное, могут иметь значение, приписанное ему в патентном законодательстве США; например, они могут означать «содержит», «содержащийся», «содержащий» и тому подобное; и что такие термины, как «состоящий в основном из» и «состоит в основном из», имеют значение, приписываемое им в Патентном законе США, например, они допускают элементы, которые не указаны явно, но исключают элементы, которые встречаются в предшествующем уровне техники или которые влияют на основную или новую характеристику изобретения.
Термины, указанные в единственном числе, включают множественное число, если контекст явно не указывает иное. Точно так же, слово «или» предназначено для включения «и», если контекст явно не указывает иное. Слово «или» означает любой элемент определенного списка, а также включает любую комбинацию элементов этого списка.
Также следует понимать, что хотя термины «первый», «второй» и т.д. могут использоваться в данном документе для описания различных элементов, эти элементы не должны ограничиваться этими терминами. Эти термины используются только для различения одного элемента от другого. Например, первый жест может быть назван вторым жестом, и, аналогично, второй жест может быть назван первым жестом, не выходя за пределы объема настоящего изобретения. Все описанные в данном документе способы могут быть выполнены в любом подходящем порядке, если в данном документе не указано иное или иное явно противоречит контексту.
Термин «животное» используется в данном документе для включения всех млекопитающих, птиц и рыб. Используемое в данном документе животное может быть выбрано из группы, состоящей из лошадиных (например, лошади), собачьих (например, собаки, волки, лисы, койоты, шакалы), кошачьих (например, львы, тигры, домашние кошки, дикие кошки, другие крупные кошки и другие кошачьи, включая гепардов и рысь), крупного рогатого скота (например, коровы), свиней (например, домашние свиньи), овец (например, овцы, козы, ламы, бизоны), птиц (например, курица, утка, гусь, индейка, перепел, фазан, попугай, зяблик, ястреб, ворона, страус, эму и казуар), приматов (например, полуобезьяна, долгопят, мартышка, гиббон, человекообразная обезьяна), людей и рыб. Термин «животное» также включает отдельное животное на всех стадиях развития, включая эмбриональную и фетальную стадии.
Используемый в данном документе термин «около» означает приблизительно, в области, ориентировочно или приближенно. Когда термин «около» используется в сочетании с численным интервалом, то он модифицирует этот интервал путем расширения границ выше и ниже указанных числовых значений. В общем, термин «около» используется в данном документе для изменения числового значения выше и ниже указанного значения посредством варьирования на 10%. В одном аспекте термин «около» означает плюс или минус 20% от значения числа, с которым он используется. Следовательно, около 50% означает в диапазоне от 45 до 55%. Численные диапазоны, указанные в данном документе конечными точками, включают все числа и дроби, включенные в этот диапазон (например, от 1 до 5 включает 1, 1,5, 2, 2,75, 3, 3,90, 4 и 5). Также следует понимать, что все числа и их дроби, как предполагается, модифицируются термином «около».
Термины «полипептид» и «белок» используются в данном документе взаимозаменяемо для обозначения полимера из последовательных аминокислотных остатков.
Термины «нуклеиновая кислота», «нуклеотид» и «полинуклеотид» используются взаимозаменяемо и относятся к РНК, ДНК, кДНК или кРНК и их производным, таким как те, которые содержат модифицированные каркасы. Следует понимать, что изобретение обеспечивает полинуклеотиды, содержащие последовательности, комплементарные описанным в данном документе. «Полинуклеотид», рассматриваемый в настоящем изобретении, включает как прямую цепь (от 5' к 3'), так и обратную комплементарную цепь (от 3' к 5'). Полинуклеотиды согласно изобретению могут быть получены различными способами (например, химическим синтезом, клонированием генов и т.д.) и могут принимать различные формы (например, линейные или разветвленные, одноцепочечные или двухцепочечные или их гибриды, праймеры, зонды и т.д.).
Термин «геномная ДНК» или «геном» используется взаимозаменяемо и относится к наследственной генетической информации организма-хозяина. Геномная ДНК включает ДНК ядра (также известная как хромосомная ДНК), но и ДНК пластид (например, хлоропластов) и других клеточных органелл (например, митохондрий). Геномная ДНК или геном, рассматриваемый в настоящем изобретении, также относится к РНК вируса. РНК может быть РНК с положительной полярностью или отрицательной полярностью. Термин «геномная ДНК», рассматриваемый в настоящем изобретении, включает геномную ДНК, содержащую последовательности, комплементарные описанным в данном документе. Термин «геномная ДНК» также относится к матричной РНК (мРНК), комплементарной ДНК (кДНК) и комплементарной РНК (кРНК).
Термин «ген» используется в широком смысле для обозначения любого сегмента полинуклеотида, связанного с биологической функцией. Таким образом, гены или полинуклеотиды включают интроны и экзоны, как в геномной последовательности, или только кодирующие последовательности, как в кДНК, такие как открытая рамка считывания (ORF), начиная от стартового кодона (метионинового кодона) и заканчивая сигналом терминации (стоп кодон). Гены и полинуклеотиды могут также включать области, которые регулируют их экспрессию, такие как инициация транскрипции, трансляция и терминация транскрипции. Таким образом, также включены промоторы и районы связывания рибосом (в общем, эти регуляторные элементы лежат приблизительно между 60 и 250 нуклеотидами выше от стартового кодона кодирующей последовательности или гена; Doree SM et al.; Pandher K et al.; Chung JY et al.), терминаторы транскрипции (обычно терминатор расположен в пределах приблизительно 50 нуклеотидов ниже стоп-кодона кодирующей последовательности или гена; Ward CK et al.). Ген или полинуклеотид также относится к фрагменту нуклеиновой кислоты, который экспрессирует мРНК или функциональную РНК или кодирует конкретный белок и который включает регуляторные последовательности.
Используемый в данном документе термин «гетерологичная ДНК» относится к ДНК, полученной из другого организма, такого как другой тип клеток или отличный от реципиента вид. Термин также относится к ДНК или ее фрагменту в том же геноме ДНК-хозяина, где гетерологичная ДНК встроена в область генома, которая отличается от ее первоначального расположения.
Используемый в данном документе термин «антиген» или «иммуноген» означает вещество, которое индуцирует специфический иммунный ответ у животного-хозяина. Антиген может включать целый организм, убитый, ослабленный или живой; субъединицу или часть организма; рекомбинантный вектор, содержащий вставку с иммуногенными свойствами; часть или фрагмент ДНК, способных индуцировать иммунный ответ при представлении животному-хозяину; полипептид, эпитоп, гаптен, или любые их комбинации. С другой стороны, иммуноген или антиген может включать токсин или антитоксин.
Термин «иммуногенный белок или пептид», используемый в данном документе, включает полипептиды, которые иммунологически активны в том смысле, что, как только их вводят хозяину, они способны вызывать иммунный ответ гуморального и/или клеточного типа, направленный против белка. Предпочтительно, если белковый фрагмент является таким, что имеет по существу такую же иммунологическую активность, что и общий белок. Таким образом, фрагмент белка согласно изобретению содержит или состоит по существу или состоит, по меньшей мере, из одного эпитопа или антигенной детерминанты. «Иммуногенный» белок или полипептид, при использовании в данном документе, включает последовательность полной длины белка, их аналоги, или их иммуногенные фрагменты. Под термином «иммуногенный фрагмент» понимается фрагмент белка, который включает один или несколько эпитопов, и, таким образом, вызывает иммунологический ответ, как описано выше. Такие фрагменты могут быть идентифицированы с использованием любого числа методик картирования эпитопов, хорошо известных в данной области. Например, линейные эпитопы могут быть определены, например, параллельным синтезом большого числа пептидов на твердых подложках, которые соответствуют частям белковой молекулы, и взаимодействием пептидов с антителами, когда пептиды все еще прикреплены к подложкам. Точно так же, конформационные эпитопы легко идентифицируются путем определения пространственной конформации аминокислот, например, путем рентгеновской кристаллографии и 2-мерного ядерного магнитного резонанса.
Термин «иммуногенный белок или пептид» дополнительно рассматривает делеции, добавления и замены последовательности, при условии, что полипептид функционирует для получения иммунологического ответа, как определено в данном документе. Термин «консервативная вариация» означает замену аминокислотного остатка другим биологически аналогичным остатком, или замены нуклеотидов в последовательности нуклеиновой кислоты, таким образом, что закодированный остаток аминокислоты не изменяется или является другим биологически сходным остатком. В связи с этим, в частности, предпочтительные замены, как правило, консервативны по своей природе, т.е. являются заменами внутри семейства аминокислот. Так, например, аминокислоты, как правило, делятся на четыре семейства: (1) кислые - аспартат и глутамат; (2) основные - лизин, аргинин, гистидин; (3) неполярные - аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан; и (4) незаряженные полярные - глицин, аспарагин, глутамин, цистин, серин, треонин, тирозин. Фенилаланин, триптофан и тирозин иногда классифицируют как ароматические аминокислоты. Примеры консервативных вариаций включают замену одного гидрофобного остатка, такого как изолейцин, валин, лейцин или метионин на другой гидрофобный остаток, или замены одного полярного остатка на другой полярный остаток, например замены лизина на аргинин, глутаминовой кислоты на аспарагиновую кислоту, или глутамина на аспарагин, и тому подобное; или подобная консервативная замена аминокислоты на структурно родственную аминокислоту, которая не будет иметь большое влияние на биологическую активность. Белки, имеющие по существу одну и ту же аминокислотную последовательность, в качестве исходной молекулы, но имеющие незначительные аминокислотные замены, которые не оказывают существенного влияния на иммуногенность белка, таким образом, находятся в пределах определения эталонного полипептида. Все полипептиды, полученные этими изменениями, включены в данный документ. Термин «консервативная вариация» также включает применение замещенной аминокислоты вместо незамещенной родительской аминокислоты при условии, что антитела, индуцированные полипептидом с заменой также иммунореагируют с незамещенным полипептидом.
Термин «эпитоп» относится к участку на антигене или к гаптену, на который отвечают специфические В-клетки и/или Т-клетки. Термин также используется взаимозаменяемо с термином «антигенная детерминанта» или «участок антигенной детерминанты». Антитела, которые распознают тот же эпитоп, могут быть идентифицированы простым иммуноанализом, показывающим способность одного антитела блокировать связывание другого антитела с антигеном-мишенью.
«Иммунологический ответ» к композиции или вакцине представляет собой развитие в хозяине клеточного и/или антитело-опосредованного иммунного ответа на представляющую интерес композицию или вакцину. Обычно, «иммунологический ответ» включает, без ограничения перечисленным, одним или несколькими из следующих эффектов: продуцирование антител, В-клеток, Т-хелперов, и/или цитотоксических Т-клеток, специфически направленных на антиген или антигены, включенные в представляющую интерес композицию или вакцину. Предпочтительно, если хозяин будет демонстрировать либо терапевтическую, либо защитную иммунологическую реакцию таким образом, что устойчивость к новой инфекции будет усилена и/или клиническая тяжесть заболевания снижена. Такая защита будет продемонстрирована либо снижением, либо отсутствием симптомов, обычно демонстрируемых зараженным хозяином, более быстрым временем восстановления и/или пониженным титром вируса в зараженном хозяине.
Термины «рекомбинантный» и «генетически модифицированный» используются взаимозаменяемо и относятся к любой модификации, изменению или конструированию полинуклеотида или белка в его нативной форме или структуре или любой модификации, изменению или конструированию полинуклеотида или белка в его естественной среде или окружении. Модификация, изменение или конструирование полинуклеотида или белка может включать, без ограничения указанным, делецию одного или нескольких нуклеотидов или аминокислот, делецию целого гена, кодоновую оптимизацию гена, консервативную аминокислотную замену, вставку одного или нескольких гетерологичных полинуклеотидов.
Термины «поливалентная вакцина или композиция», «комбинированная или комбо вакцина или композиция» и «поливалентная вакцина или композиция» используются взаимозаменяемо для обозначения композиции или вакцины, содержащей более одной композиции или вакцины. Поливалентная вакцина или композиция может содержать две, три, четыре или более композиций или вакцин. Поливалентная вакцина или композиция может содержать рекомбинантные вирусные векторы, активные или аттенуированные или убитые вирусы дикого типа или смесь рекомбинантных вирусных векторов и вирусов дикого типа в активной или аттенуированной или убитой формах.
В другом воплощении изобретения предлагается вирусный вектор рекомбинантного поксвируса, содержащий один или несколько гетерологичных полинуклеотидов, кодирующих и экспрессирующих, по меньшей мере, один антиген или полипептид птичьего патогена. Штаммы HVT, используемые для рекомбинантного вирусного вектора, может представлять собой любые штаммы HVT, в том числе, без ограничения указанным, штамм HVT FC126 (Igarashi T. et al., J. Gen. Virol. 70, 1789-1804, 1989).
Гены, кодирующие антиген или полипептид, могут представлять собой гены, кодирующие слитый белок вируса ньюкаслской болезни (NDV-F), гемагглютинин-нейраминидазу вируса ньюкаслской болезни (NDV-HN), гликопротеин C вируса болезни Марека (gC), гликопротеин B вируса болезни Марека (gB), гликопротеин E вируса болезни Марека (gE), гликопротеин I вируса болезни Марека (gI), гликопротеин H вируса болезни Марека H (gH) или гликопротеин L вируса болезни Марека (gL), VP2 вируса инфекционного бурсита (IBDV), IBDV VP3, IBDV VP4, гликопротеин B ILTV, гликопротеин I ILTV, гликопротеин D ILTV, гликопротеин E ILTV, гликопротеин ILTV, гликопротеин C ILTV, гемагглютинин гриппа (HA), нейраминидазу гриппа (NA), защитные гены, полученные из Mycoplasma gallisepticum или Mycoplasma synoviae (MS) или их комбинации. Антиген или полипептид может быть любым антигеном из патогена птицы, выбранного из группы, состоящей из вируса птичьего энцефаломиелита, птичьего реовируса, птичьего парамиксовируса, птичьего метапневмовируса, вируса птичьего гриппа, птичьего аденовируса, вируса оспы кур, птичьего коронавируса, птичьего ротавируса, вируса анемии цыплят, птичьего астровируса, птичьего ретровируса, птичьего парвовируса, птичьего ретровируса, птичьего пикорнавируса, кокцидиоза (Eimeria sp.), Campylobacter sp., Salmonella sp., Pasteurella sp., Avibacterium sp., Mycoplasma gallisepticum, Mycoplasma synoviae, Clostridium sp., и Escherichia coli.
Кроме того, предполагается, что гомологи вышеупомянутых антигенов или полинуклеотидов входят в объем настоящего изобретения. Используемый в данном документе термин «гомологи» включает ортологи, аналоги и паралоги. Термин «аналоги» относится к двум полинуклеотидам или полипептидам, которые имеют одинаковую или сходную функцию, но которые развиваются отдельно у неродственных организмов. Термин «ортологи» относится к двум полинуклеотидам или полипептидам от разных видов, эволюционировавших из общего предкового гена при видообразовании. Обычно ортологи кодируют полипептиды, имеющие одинаковые или сходные функции. Термин «паралоги» относится к двум полинуклеотидам или полипептидам, которые родственны из-за дублирования в геноме. Паралоги обычно имеют разные функции, но эти функции могут быть родственными. Аналоги, ортологи и паралоги полипептида дикого типа могут отличаться от полипептида дикого типа посттрансляционными модификациями, различиями аминокислотных последовательностей или и тем, и другим. В частности, гомологи по изобретению, как правило, будут демонстрировать, по меньшей мере, 80-85%, 85-90%, 90-95% или 95%, 96%, 97%, 98%, 99% идентичности последовательности, с частью или полными полинуклеотидными или полипептидными последовательностями антигенов, описанных выше, и будут проявлять аналогичную функцию.
В одном воплощении настоящее изобретение относится к рекомбинантному вирусному HVT-вектору, содержащему один, два или более гетерологичных полинуклеотидов, кодирующих и экспрессирующих антиген или полипептид NDV-F, антиген или полипептид IBDV VP2, антиген или полипептид ILD gD или их комбинацию. В одном аспекте воплощения антиген или полипептид NDV-F имеет, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 5 или 22, или консервативным вариантом, аллельным вариантом, гомологом или иммуногенным фрагментом, включающим, по меньшей мере, восемь или, по меньшей мере, десять последовательных аминокислот одного из этих полипептидов, или комбинацию этих полипептидов. В другом аспекте воплощения гетерологичный полинуклеотид кодирует антиген или полипептид NDV-F, имеющий, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 5. В еще одном аспекте воплощения гетерологичный полинуклеотид имеет, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полинуклеотидом, имеющим последовательность, представленную в SEQ ID NO: 3, 4 или 21.
В другом аспекте воплощения антиген или полипептид IBDV VP2 имеет, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 2, или консервативным вариантом, аллельным вариантом, гомологом или иммуногенным фрагментом, включающим, по меньшей мере, восемь или, по меньшей мере, десять последовательных аминокислот одного из этих полипептидов или комбинацию этих полипептидов. В другом аспекте воплощения гетерологичный полинуклеотид кодирует антиген или полипептид IBDV VP2, имеющий, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 2. В еще одном аспекте воплощения гетерологичный полинуклеотид имеет, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полинуклеотидом, имеющим последовательность, представленную в SEQ ID NO: 1.
В другом аспекте воплощения антиген или полипептид ILD gD имеет, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом с последовательностью, представленной в SEQ ID NO: 17, или с консервативным вариантом, аллельным вариантом, гомологом или иммуногенным фрагментом, включающим, по меньшей мере, восемь или, по меньшей мере, десять последовательных аминокислот одного из этих полипептидов или комбинацию этих полипептидов. В другом аспекте воплощения гетерологичный полинуклеотид кодирует антиген или полипептид ILTV gD, имеющий, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, имеющем последовательность, представленную в SEQ ID NO: 17. В еще одном аспекте воплощения гетерологичный полинуклеотид имеет, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полинуклеотидом, имеющим последовательность, представленную в SEQ ID NO: 16.
Варианты включают аллельные варианты. Термин «аллельный вариант» относится к полинуклеотиду или полипептиду, содержащему полиморфизм, которые приводят к изменениям в аминокислотных последовательностях белка и которые существуют в пределах естественной популяции (например, видов или разновидностей вируса). Такие естественные аллельные вариации обычно могут приводить к дисперсии 1-5% в полинуклеотиде или полипептиде. Аллельные варианты могут быть идентифицированы путем секвенирования последовательности нуклеиновой кислоты, представляющей интерес, для ряда различных видов, которые могут быть легко осуществлены с применением гибридизационных зондов для идентификации того же генетического локуса генов у этих видов. Любые и все такие вариации нуклеиновой кислоты и полученные аминокислотные полиморфизмы или вариации, которые являются результатом естественных аллельных вариаций и которые не изменяют функциональную активность гена, представляющего интерес, подразумеваются охваченными изобретением.
Термин «идентичность» в отношении последовательностей может относиться, например, к количеству положений с идентичными нуклеотидами или аминокислотами, деленному на число нуклеотидов или аминокислот в более короткой из двух последовательностей, где выравнивание двух последовательностей может быть определено в соответствии с алгоритмом Wilbur и Lipman (Wilbur and Lipman). Идентичность последовательности или сходство последовательностей двух аминокислотных последовательностей или идентичность последовательности между двумя нуклеотидными последовательностями можно определить с помощью программного пакета Vector NTI (Invitrogen, 1600 Faraday Ave., Карлсбад, Калифорния). Когда говорят, что последовательности РНК являются сходными или имеют степень идентичности последовательности или гомологию с последовательностями ДНК, тимидин (Т) в последовательности ДНК считается равным урацилу (U) в последовательности РНК. Таким образом, последовательности РНК входят в объем изобретения и могут быть получены из последовательностей ДНК, причем тимидин (Т) в последовательности ДНК считается равным урацилу (U) в последовательностях РНК.
Полинуклеотиды раскрытия включают последовательности, которые являются вырожденными из-за вырожденности генетического кода, например, в случае оптимизированного использования кодонов для конкретного хозяина. Используемый в данном документе термин «оптимизированный» относится к полинуклеотиду, который генетически модифицирован так, чтобы увеличить его экспрессию у данного вида. Чтобы обеспечить оптимизированные полинуклеотиды, кодирующие полипептиды gD NDV-F, IBDV VP2 или ILTV, последовательность ДНК этих генов может быть модифицирована так, чтобы: 1) включать кодоны, предпочтительные для высокоэкспрессируемых генов в конкретном виде; 2) включать содержание A + T или G + C в составе нуклеотидных оснований сопоставимое с содержанием, по существу обнаруженным у указанных видов; 3) сформировать последовательность инициации указанных видов; или 4) устранить последовательности, которые вызывают дестабилизацию, несоответствующее полиаденилирование, деградацию и терминацию РНК или которые образуют шпильки вторичной структуры или сайты сплайсинга РНК. Повышенная экспрессия белка NDV F, IBDV VP2 или ILTV gD у указанных видов может быть достигнута путем использования частоты распределения использования кодонов у эукариот и прокариот или у конкретного вида. Термин «частота предпочтительного использования кодонов» относится к предпочтению, проявляемому конкретной клеткой-хозяином в использовании нуклеотидных кодонов, для указания данной аминокислоты. Есть 20 природных аминокислот, большинство из которых указаны более чем одним кодоном. Следовательно, все вырожденные нуклеотидные последовательности включены в раскрытие, пока аминокислотная последовательность полипептида NDV-F, IBDV VP2 или ILTV gD, кодируемого нуклеотидной последовательностью, функционально не изменяется.
Успешная экспрессия гетерологичных полинуклеотидов рекомбинантным/модифицированным инфекционным вирусом требует двух условий. Во-первых, гетерологичные полинуклеотиды должны быть вставлены или введены в область генома вируса, чтобы модифицированный вирус оставался жизнеспособным. Вторым условием для экспрессии встроенных гетерологичных полинуклеотидов является наличие регуляторных последовательностей, позволяющих экспрессировать ген в вирусном фоне (например, промотор, энхансер, донорный и акцепторный сайты сплайсинга и интрон, консенсусная последовательность инициации трансляции Kozak, сигналы полиаденилирования, нетранслируемые элементы последовательности).
Сайт вставки может быть любой несущественной областью генома HVT, включая, без ограничения указанным, область между STOP-кодоном ORF UL55 и местом соединения UL с соседней областью повторов (межгенная область 1, локус IG1, US 5980906), локус IG2 (межгенная область 2), локус IG3 (межгенная область 3), локус UL43, локус US10, локус US2, локус SORF3/US2 (см. фиг. 2).
Как правило, выгодно использовать сильный промоторный функциональный в эукариотических клетках. Промоторы включают, без ограничения указанным, немедленно ранний промотор цитомегаловируса (CMV), промотор CMV IE мыши (mCMV), промотор CMV морской свинки, промотор SV40, промоторы вируса псевдобешенства, такие как промотор гликопротеина X, промотор вируса простого герпеса-1, такого как промотор альфа-4, промотор вируса болезни Марека (включая MDV-1, MDV-2 и HVT), такие как те, которые управляют экспрессией гликопротеинов gC, gB, g или gI, промотор HHV3gB (промотор гликопротеина B герпесвируса человека типа III), промоторы вируса инфекционного ларинготрахеита, такие как гены гликопротеинов gB, gE, gI, gD или другие промоторы герпесвируса.
В одном воплощении изобретения предложен рекомбинантный HVT-вектор, включающий первый гетерологичный полинуклеотид, кодирующий и экспрессирующий антиген или полипептид IBDV VP2, и второй полинуклеотид, кодирующий и экспрессирующий антиген или полипептид NDV-F. В одном аспекте воплощения антиген или полипептид NDV-F имеет, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 5. В другом аспекте воплощения антиген или полипептид IBDV VP2 имеет, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 2. В другом аспекте полинуклеотид, кодирующий полипептид NDV-F, функционально связан с промотором SV40, имеющим последовательность, представленную в SEQ ID NO: 7, и экспрессия антигена или полипептида NDV-F регулируется промотором SV40. В еще одном аспекте экспрессия антигена или полипептида NDV-F регулируется polyA-сигналом SV40, имеющим последовательность, представленную в SEQ ID NO: 8, или сигналом синтетического polyA, имеющим последовательность, представленную в SEQ ID NO: 9. В другом аспекте экспрессия антигена или полипептида IBDV VP2 регулируется промотором mCMV-IE, имеющим последовательность, представленную в SEQ ID NO: 6, и polyA-сигналом SV40, имеющим последовательность, представленную в SEQ ID NO: 8, или синтетическим polyA-сигналом, имеющим последовательность, представленную в SEQ ID NO: 9.
В другом воплощении изобретения предложен рекомбинантный HVT-вектор, включающий первый гетерологичный полинуклеотид, кодирующий и экспрессирующий антиген или полипептид IBDV VP2, и второй полинуклеотид, кодирующий и экспрессирующий антиген или полипептид NDV-F, и дополнительно включающий последовательность, которая регулирует экспрессию второго полинуклеотида. Регуляторные последовательности или линкеры могут представлять собой участок внутренней посадки рибосомы (IRES), последовательность РНК, полученную из вируса энцефаломиокардита (EMCV), или последовательность, кодирующую саморасщепляющийся пептид 2A свиного тешовируса-1 или пептид вируса ящура (FMDV) (P2A).
В одном аспекте воплощения рекомбинантный HVT-вектор содержит первый полинуклеотид, кодирующий антиген IBDV VP2, и второй полинуклеотид, кодирующий антиген NDV-F, и дополнительно содержит IRES, имеющий последовательность, представленную в SEQ ID NO: 10. В другом аспекте воплощения рекомбинантный HVT содержит первый полинуклеотид, кодирующий антиген IBDV VP2, и второй полинуклеотид, кодирующий антиген NDV-F, и дополнительно содержит полинуклеотид, кодирующий P2A, имеющий последовательность, представленную в SEQ ID NO: 11.
Одно воплощение изобретения обеспечивает рекомбинантный HVT-вектор, включающий первый гетерологичный полинуклеотид, кодирующий и экспрессирующий антиген или полипептид NDV F, и второй полинуклеотид, кодирующий и экспрессирующий антиген или полипептид ILTV gD, и дополнительно включающий последовательность, которая регулирует экспрессию второго полинуклеотида. Регуляторные последовательности или линкеры могут представлять собой участок внутренней посадки рибосомы (IRES), последовательность РНК, полученную из вируса энцефаломиокардита (EMCV), или последовательность, кодирующую саморасщепляющийся пептид 2A свиного тешовируса-1 или вируса ящура (FMDV) (P2A). В одном аспекте воплощения антиген или полипептид ILD gD имеет, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 17. В другом аспекте воплощения антиген или полипептид NDV F имеет, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 5 или 22. В еще одном аспекте воплощения рекомбинантный HVT-вектор содержит первый полинуклеотид, кодирующий антиген NDV F, и второй полинуклеотид, кодирующий антиген ILTV gD, и дополнительно содержит IRES, имеющий последовательность, представленную в SEQ ID NO: 10.
В другом воплощении изобретения предложен рекомбинантный HVT-вектор, включающий первый гетерологичный полинуклеотид, кодирующий и экспрессирующий антиген или полипептид NDV F, и второй полинуклеотид, кодирующий и экспрессирующий антиген или полипептид ILTV gD. В одном аспекте воплощения антиген или полипептид ILD gD имеет, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 17. В другом аспекте воплощения антиген или полипептид NDV F имеет, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 5 или 22. В одном аспекте полинуклеотид, кодирующий полипептид NDV F, функционально связан с промотором SV40, и экспрессия антигена или полипептида NDV F регулируется промотором SV40. В другом аспекте полинуклеотид, кодирующий полипептид ILTV gD, функционально связан с промотором HHV3gB, и экспрессия антигена или полипептида ILTV gD регулируется промотором HHV3gB. В еще одном аспекте промотор HHV3gB находится в обратном направлении. В еще одном аспекте экспрессии антигена NDV F и антигена ILTV gD регулируются промотором SV40 и обратным промотором HHV3gB и направлены в разные стороны.
В другом воплощении изобретения предложен рекомбинантный HVT-вектор, включающий первый гетерологичный полинуклеотид, кодирующий и экспрессирующий антиген или полипептид IBDV VP2, и второй полинуклеотид, кодирующий и экспрессирующий антиген или полипептид ILTV gD. В одном аспекте воплощения антиген или полипептид ILD gD имеет, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 17. В другом аспекте воплощения антиген или полипептид IBDV VP2 имеет, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 2. В еще одном аспекте воплощения рекомбинантный HVT-вектор содержит первый полинуклеотид, кодирующий антиген IBDV VP2, и второй полинуклеотид, кодирующий антиген ILTV gD, и дополнительно содержит IRES, имеющий последовательность, представленную в SEQ ID NO: 10.
В другом воплощении изобретения предложен рекомбинантный HVT-вектор, включающий гетерологичный полинуклеотид, кодирующий и экспрессирующий антиген или полипептид ILTV gD. В одном аспекте воплощения антиген или полипептид ILD gD имеет, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 17. В другом аспекте воплощения полинуклеотид, кодирующий полипептид ILTV gD, функционально связан с промотором SV40, и экспрессия антигена или полипептида ILTV gD регулируется промотором SV40.
В одном воплощении полинуклеотиды, кодирующие антиген IBDV VP2, и/или антиген NDV-F, и/или антиген ILD gTV, могут быть вставлены в одну или несколько областей локуса, выбранных из группы, состоящей из IG1, IG2, US10, US2, SORF3 -US2 и gD генома HVT. В другом воплощении полинуклеотиды, кодирующие антиген IBDV VP2, и/или антиген NDV-F, и/или антиген ILTV gD, встроены в один и тот же локус, такой как IG1 генома HVT.
В одном воплощении настоящее изобретение относится к фармацевтической композиции или вакцине, содержащей один или несколько рекомбинантных HVT-векторов по настоящему изобретению и фармацевтически или ветеринарно приемлемый носитель, наполнитель, носитель или адъювант. HVT-вектор может содержать два гетерологичных полинуклеотида, где первый полинуклеотид содержит полинуклеотид, кодирующий полипептид, выбранный из группы, состоящей из антигена VP2 вируса инфекционного бурсита (IBDV), антигена гликопротеина D (gD) вируса инфекционного ларинготрахеита (ILTV) и антигена F вируса ньюкаслской болезни (NDV-F), и где второй полинуклеотид содержит полинуклеотид, кодирующий полипептид, выбранный из группы, состоящей из антигена VP2 вируса инфекционного бурсита (IBDV), антигена гликопротеина D (gD) вируса инфекционного ларинготрахеита (ILTV) и антигена F вируса ньюкаслской болезни (NDV-F).
В другом воплощении настоящее изобретение относится к композиции или вакцине, содержащей вирусный HVT-вектор, включающий: i) первый гетерологичный полинуклеотид, кодирующий и экспрессирующий антиген IBDV VP2 или антиген NDV-F; ii) второй полинуклеотид, кодирующий и экспрессирующий антиген NDV-F или антиген IBDV VP2; и iii) возможно, фармацевтически или ветеринарно приемлемый носитель, наполнитель, носитель или адъювант. В еще одном воплощении настоящее изобретение относится к композиции или вакцине, содержащей вирусный HVT-вектор, включающий: i) первый гетерологичный полинуклеотид, кодирующий и экспрессирующий антиген IB2 VP2 или антиген ILTV gD; ii) второй полинуклеотид, кодирующий и экспрессирующий антиген ILTV gD или IBDV VP2; и iii) возможно, фармацевтически или ветеринарно приемлемый носитель, наполнитель, носитель или адъювант. В еще одном воплощении настоящее изобретение относится к композиции или вакцине, содержащей вирусный HVT-вектор, включающий: i) первый гетерологичный полинуклеотид, кодирующий и экспрессирующий антиген NDV-F или антиген ILTV gD; ii) второй полинуклеотид, кодирующий и экспрессирующий антиген ILTV gD или антиген NDV-F; и iii) возможно, фармацевтически или ветеринарно приемлемый носитель, наполнитель, носитель или адъювант. В еще одном воплощении настоящее изобретение относится к композиции или вакцине, содержащей вирусный HVT-вектор, включающий гетерологичный полинуклеотид, кодирующий и экспрессирующий антиген ILTV gD, и, необязательно, фармацевтически или ветеринарно приемлемый носитель, наполнитель, носитель или адъювант. В еще одном воплощении настоящее изобретение относится к композиции или вакцине, содержащей HVT, включающую полинуклеотид, имеющий, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99 % идентичности последовательности с полинуклеотидом, имеющим последовательность, представленную в SEQ ID NO: 1, 3, 4, 12, 13, 14, 15, 16, 18, 19, 20, 21, 25, 26 или 27. В одном воплощении показано, что вставка двух или более гетерологичных полинуклеотидов в один локус обеспечивает лучшую защиту и эффективность, чем вставка в несколько локусов. В другом воплощении показано, что экспрессия более одного гетерологичного полинуклеотида с одной мРНК через IRES или P2A обеспечивает лучшую защиту и эффективность против болезней птиц. В еще одном воплощении экспериментальные данные, представленные в настоящем изобретении, раскрывают, что конструкции, содержащие элементы IRES, обеспечивали лучшую защиту, чем конструкции, содержащие элементы P2A.
Фармацевтически или ветеринарно приемлемые носители или адъюванты или несущие среды или эксципиенты хорошо известны специалистам в данной области. Например, фармацевтически или ветеринарно приемлемый носитель или адъювант или несущая среда или эксципиент могут представлять собой разбавитель вакцины болезни Марека, используемый для вакцин против MD. Другие фармацевтически или ветеринарно приемлемые носитель или адъювант или несущая среда или эксципиент, которые можно использовать для способов по данному изобретению, включают, без ограничения указанным, 0,9% раствор NaCl (например, физиологический раствор) или фосфатный буфер, поли- (L-глутамат), разбавитель раствора Рингера с лактатом для инъекций (хлорид натрия, лактат натрия, хлорид калия и хлорид кальция) или поливинилпирролидон. Фармацевтически или ветеринарно приемлемый носитель или адъювант или несущая среда или эксципиент могут представлять собой любое соединение или комбинацию соединений, облегчающих введение вектора (или белка, экспрессированного из вектора изобретения) in vitro, или облегчающих трансфекцию или инфекцию и/или улучшающих сохранение вектора (или белка). Дозы и дозовые объемы в данном документе обсуждаются в общем описании, и могут также быть определены специалистами в данной области из этого описания в сочетании с информацией в данной области, без проведения излишних экспериментов.
Необязательно другие соединения могут быть добавлены в качестве фармацевтически или ветеринарно приемлемых носителей или адъювантов или носителей или наполнителей, включая, без ограничения указанным, квасцы; CpG-олигонуклеотиды (ODN), в частности ODN 2006, 2007, 2059 или 2135 (Pontarollo RA et al., Vet. Immunol. Immunopath, 2002, 84: 43-59; Wernette C.M. et al., Vet. Immunol. Immunopath, 2002, 84: 223-236; Mutwiri G. et al., Vet. Immunol. Immunopath, 2003, 91: 89-103); polyA-polyU, диметилдиоктадециламмонийбромид (DDA) (“Vaccine Design The Subunit and Adjuvant Approach», edited by Michael F. Powell and Mark J. Newman, Pharmaceutical Biotechnology, 6: p.03, p.157); N, N-диоктадецил-N',N'-бис (2-гидроксиэтил) пропандиамин (такой как AVRIDINE®) (там же, с. 148); карбомер, хитозан (см. патент США № 5980912).
Фармацевтические композиции и вакцины согласно изобретению могут содержать или состоять по существу из одного или нескольких адъювантов. Подходящими адъювантами для практического применения настоящего изобретения являются (1) полимеры акриловой или метакриловой кислоты, малеинового ангидрида и алкенильных производных полимеров, (2) иммуностимулирующие последовательности (ISS), такие как олигодезоксирибонуклеотидные последовательности, имеющие один или несколько неметилированных CpG (Klinman et al., 1996; WO 98/16247), (3) эмульсия масло-в-воде, такая как эмульсия SPT, описанная на стр. 147 «Vaccine Design, The Subunit and Adjuvant Approach», опубликованная M. Powell, M. Newman, Plenum Press 1995, и эмульсия MF59, описанная на стр. 183 той же работы, (4) катионные липиды, содержащие четвертичную аммониевую соль, например DDA (5) цитокины, (6) гидроксид алюминия или фосфат алюминия, (7) сапонин или (8) другие адъюванты, обсуждаемые в любом документе, приведенном и включенном ссылкой в настоящую заявку, или (9) любые комбинации или их смеси.
В одном воплощении адъювант может включать TS6, TS7, TS8 и TS9 (US 7371395), LR2, LR3 и LR4 (US 7691368), TSAP (US 20110129494), TRIGEN™ (Newport Labs), синтетические дцРНК (например, поли-IC), поли-ICLC [HILTONOL®]), а также вспомогательные вещества MONTANIDE™ (W/O W/O/W, O/W, IMS и гель, все производства SEPPIC).
В другом воплощении изобретение предусматривает введение терапевтически эффективного количества вакцины или композиции для доставки рекомбинантных HVT-векторов в целевую клетку. Определение терапевтически эффективного количества представляет собой обычные экспериментальные работы для специалиста в данной области.
Другой аспект изобретения относится к способу индукции иммунологического ответа у животного против одного или нескольких антигенов или защитного ответа у животного против одного или нескольких птичьих патогенов, причем этот способ включает инокуляцию животного, по меньшей мере, один раз вакциной или фармацевтической композицией по настоящему изобретению. Еще один аспект изобретения относится к способу индукции иммунологического ответа у животного на один или несколько антигенов или защитного ответа у животного против одного или нескольких птичьих патогенов в режиме введения «прайм-буст», которое состоит, по меньшей мере, из одного первичного введения и, по меньшей мере, одно бустерного введение с использованием, по меньшей мере, одного общего полипептида, антигена, эпитопа или иммуногена. Иммунологическая композиция или вакцина, используемые при первичном введении, могут быть одинаковыми, могут отличаться по природе от тех, которые используются в качестве бустера.
Птичьими патогенами могут быть вирус ньюкаслской болезни (NDV), вирус инфекционного бурсита (т.е. вирус IBDV или болезнь Гумборо), вирус болезни Марека (MDV), вирус инфекционного ларинготрахеита (ILTV), вирус птичьего энцефаломиелита, вирус птичьего энцефаломиелита, птичий реовирус, птичий парамиксовирус, птичий метапневмовирус, вирус птичьего гриппа, птичий аденовирус, вирус оспы кур, птичий коронавирус, птичий ротавирус, птичий парвовирус, птичий астровирус и вирус анемии цыплят, кокцидиоз (Eimeria sp.), Campylobacter sp., Salmonella sp., Mycoplasma gallisepticum, Mycoplasma synoviae, Pasteurella sp., Avibacterium sp., E. coli или Clostridium sp.
Обычно одно введение вакцины птице проводится в возрасте одного дня подкожным или внутримышечным путем или in ovo у 17-19-дневного эмбриона. Повторное введение может быть сделано в течение 5-30 дней после первого введения.
Могут быть использованы различные способы введения суточным цыплятам, например, подкожно или внутримышечно, внутрикожно, трансдермально. In ovo вакцинации может быть осуществлены в амниотической мешок и/или эмбрион. Для вакцинации можно использовать имеющиеся в продаже устройства для введения in ovo и SC.
Композиция или вакцина может содержать дозу от около 102 до около 1020, от около 103 до около 1018, от около 104 до около 1016, от около 105 до около 1012 VLP (вирусоподобных частиц), продуцируемых in vitro или in vivo из вирусного вектора, плазмиды или бакуловируса. Вирусный вектор может быть титрован на основании любых способов титрования вируса, включая, без ограничения указанным, FFA (анализ образования фокусов) или FFU (фокусобразующая единица), TCID50 (50%-ная инфекционная доза для тканевой культуры), PFU (бляшкообразующая единица), и FAID50 (50% -ная инфекционная доза флуоресцентного антитела), и VLP, полученные in vitro, можно титровать с помощью анализа гемагглютинации, ELISA и электронной микроскопии. Другие способы также могут быть применимы в зависимости от типа VLP.
Композиция или вакцина может содержать от около 102,0 до около 107,0 TCID50 или PFU/дозу, от около 102,0 до около 107,0 TCID50 или PFU/дозу и от около 102,0 до около 106,5 TCID50 или PFU/дозу.
Объем доз может составлять от около 0,1 до около 10 мл, от около 0,2 до около 5 мл.
Далее раскрытие будет описано посредством следующих неограничивающих примеров.
Примеры
Конструирование ДНК-вставок, плазмид и рекомбинантных вирусных векторов проводили с применением стандартных методов молекулярной биологии, описанных J. Sambrook et al. (Molecular Cloning: A Laboratory Manual, 4th edition, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 2014).
Пример 1. Конструирование рекомбинантных HVT-векторов, экспрессирующих два гена.
Пример 1.1 Конструирование рекомбинантного vHVT309, экспрессирующего IBDV-VP2 и NDV-F
Цель исследования заключалась в создании рекомбинантного HVT, в котором экспрессирующая кассета, содержащая промотор цитомегаловируса мыши (mCMV), ген, кодирующий вирусный белок 2 вируса инфекционного бурсита (VP2), polyA-хвост вируса обезьян 40 (SV40 polyA), промотор вируса обезьян 40 (промотор SV40), ген, кодирующий слитый белок вируса ньюкаслской болезни (NDV-F) и синтетический polyA-хвост (syn polyA-хвост), интегрирована в межгенный сайт 1 (IG1).
Родительским вирусом, используемым в конструкции, является vHVT13 (HVT-вектор, экспрессирующий ген IBDV VP2, активный ингредиент вакцины Merial VAXXITEK® (HVT + IBD), также известный как vHVT17 в US 5980906). Вектор vHVT13 содержит экспрессирующую кассету, состоящую из промотора mCMV IE (SEQ ID NO: 6), гена IBDV VP2 (SEQ ID NO: 1, кодирующая SEQ ID NO: 2), и polyA-хвоста SV40 (SEQ ID NO: 8), вставленного в сайт вставки IG1. Слитый белок вируса ньюкаслской болезни (NDV-F), соответствующий последовательности генотипа VIId, был синтезирован химически и оптимизирован по кодонам (GenScript). Сайт расщепления F-белка этого синтетического гена был изменен так, чтобы соответствовать лентогенной последовательности сайта расщепления F, и результирующая последовательность гена NDV-F имеет 99% идентичность аминокислотной последовательности с последовательностью NDV-F, депонированной в GenBank (AY337464). Мышиный промотор CMV IE использовали для IBD-VP2, а промотор SV40 использовали для NDV-F. Локус вставки является межгенным сайтом 1 (IG1) в HVT (фиг. 2). Донорную плазмиду pFSV40VP2 (вставочную плазмиду, содержащую polyA VP2/SV40 и фланкирующее плечо промотора IG1 + SV40 + синтетический polyA NDV-F) конструировали, как описано ниже. Клетки фибробластов куриных эмбрионов (CEF) использовали для рекомбинации in vitro.
Конструирование донорной плазмиды
Синтетическая ДНК в pUC57, содержащая ген IBDV VP2 (SEQ ID NO: 1, кодирующая SEQ ID NO: 2), polyA-хвост SV40 (SEQ ID NO: 8), промотор SV40 (SEQ ID NO: 7), ген NDV-F (SEQ ID NO: 3, кодирующий SEQ ID NO: 5), и синтетический polyA-хвост (SEQ ID NO: 9) была синтезирована с помощью GeneScript (фиг. 3). Плазмиду pFSV40VP2 трансформировали с использованием набора Top10 Oneshot (кат. № C404002, Invitrogen), выращивали крупномасштабную культуру, и осуществляли экстракцию плазмиды с использованием набора Qiagens Maxi Prep. Транзиторная экспрессия maxi prep была подтверждена с использованием реагента для трансфекции Fugene в клетках фибробластов куриных эмбрионов (CEF) и поликлональной куриной сыворотки против NDV.
Получение рекомбинантов
За стандартной процедурой гомологичной рекомбинации следовала совместная электропорация вторичных клеток CEF с использованием плазмиды pFSV40VP2 и вирусной ДНК, выделенной из вакцины vHVT13. Совместную электропорацию проводили с использованием 1×107 2 CEF в 300 мкл Opti-MEM и шокового воздействия при 150 В с ёмкостным сопротивлением 950 в 2 мм кювете для электропорации. Трансфицированные клетки высевали в 96-луночный планшет и инкубировали в течение 4 дней. Клетки, выращенные в 96-луночном планшете, затем дублировали в два 96-луночных планшета и инкубировали еще 3 дня. Один набор 96-луночных планшетов использовали для IFA с использованием куриных поликлональных сывороток против NDV-F для идентификации положительных лунок, содержащих рекомбинанты, а другой набор 96-луночных планшетов использовали для извлечения инфицированных клеток из положительных лунок.
Способы очистки рекомбинантных вирусов выполняли сначала путем дупликации 96-луночного планшета и отбора IFA для лунок, содержащих большинство IFA-положительных бляшек с наименьшим количеством IFA-негативных бляшек. Затем лунки, соответствующие этим критериям, собирали и доводили до 1 мл в DMEM + 2% FBS. Из 1 мл исходного материала 5-20 мкл удаляли и смешивали с 1×107 CEF в 10 мл DMEM + 2% FBS и разделяли на аликвоты на новый 96-луночный планшет, чтобы иметь по одной вирусной бляшке на лунку. 96-луночные планшеты были продублированы после 5 дней инкубации, и лунки, которые содержали бляшки, были проверены на присутствие двойного рекомбинанта и отсутствие родительского вируса vHVT13 с помощью IFA и ПЦР. Опять же лунки, которые, как оказалось, содержат больше рекомбинантного вируса, путем сравнения результатов ПЦР-полос, собирали и доводили до 1 мл и аликвотили в новые 96-луночные планшеты. После двух циклов очистки инфицированных вирусом клеток выделяли рекомбинантный вирус, экспрессирующий белок NDV-F, и чистоту рекомбинантного вируса проверяли с помощью IFA и ПЦР для подтверждения отсутствия родительского вируса.
Анализ рекомбинантов методом ПЦР
ДНК выделяли из исходного вируса экстракцией фенолом/хлороформом, осаждением этанолом, и ресуспендировали в 20 мМ HEPES. Праймеры для ПЦР (таблица 1) были сконструированы для специфической идентификации гена IBDV-VP2 и NDV-F VIId, промоторов, polyA, а также чистоты рекомбинантного вируса из родительского вируса Vaxxitek. Расположение сайтов связывания праймеров показано на фиг. 4. ПЦР проводили с использованием 200 мкг ДНК-матрицы вместе с указанными парами праймеров, указанными в таблице 1. Условия циклов ПЦР следующие: 94°C - 2 мин; 30 циклов: 94°C - 30 с, 60°C - 45 с, 68°C - 3 мин (5 мин для набора праймеров MB080 + MB081); 68°C - 5 минут (7 минут для набора праймеров MB080 + MB081).
Таблица 1. Ожидаемые полосы ПЦР с использованием определенных наборов праймеров

Анализ экспрессии
Для иммунофлуоресцентного тестирования рекомбинантный материал разводили в среде 1: 100. Приблизительно 50 мкл разведенного вируса добавляли к 20 мл DMEM + 2% FBS с 2 × 107 CEF и затем разделяли на аликвоты в двух 96-луночных планшетах (100 мкл/лунку). Планшеты инкубировали в течение 4 дней при 37°C + 5% CO2, пока вирусные бляшки не становились видны. Планшеты фиксировали 95% ледяным ацетоном в течение трех минут, давали высохнуть на воздухе в течение десяти минут и трижды промывали водой. Двойное иммунофлуоресцентное окрашивание проводили на планшете #1, используя куриную антисыворотку против вируса ньюкаслской болезни (NDV Pab) (лот # C0117A, Charles Rivers Laboratories) при 1: 500 и моноклонального антитела HVT L78 (HVT Mab) (Lee et al. 1983, J. Immunol. 130 (2) 1003-6; Мериал) (1: 3000) и инкубировали планшет при 37°C в течение 1 часа. Двойной иммунофлуоресцентный анализ проводили на планшете #2 с использованием куриной антисыворотки против вируса инфекционного бурсита (IBDV Pab) в разведении 1: 500 (лот # G0117, Charles Rivers Laboratories) и моноклонального антитела HVT L78 (HVT Mab) (Merial) в разведении 1: 3000 и инкубировали планшет при 37°C в течение 1 часа. После одного часа инкубации планшеты трижды промывали PBS. На оба планшета # 1 и # 2 добавляли меченые FITC антитела против куриных IgG (кат. № F8888, Sigma) в разведении 1: 500 и меченые TRITC ослиные антитела против мышиных антител Alex Fluor (кат. № A10037, Invitrogen) в разведении 1: 300. Планшеты опять инкубировали при 37°C в течение 1 часа. После одного часа инкубации клетки трижды промывали PBS и визуализировали с помощью флуоресцентного микроскопа, используя фильтр изотиоцианата флуоресцеина (FITC) и фильтр изотиоцианата тетраметил родамина (TRITC).
Результаты
Нуклеотидной и аминокислотной последовательностям донорной плазмиды pFSV40VP2 присвоены SEQ ID NO, как показано на фиг. 1.
Получение рекомбинантов и анализ экспрессии
Геномную ДНК вируса vHVT13 совместно электропорировали с донорной плазмидой pFSV40VP2 для получения рекомбинанта с использованием метода гомологичной рекомбинации. Рекомбинантный вирус отделяли от родительского вируса Vaxxitek с помощью отбора иммунофлуоресцентно положительных лунок и ПЦР-скрининга в нескольких раундах очистки бляшек. Очищенный из бляшек рекомбинантный вирус, экспрессирующий белок NDV-F, обозначенный vHVT309, масштабировали из колб для тканевых культур в 5 × 850 см2 роллерные флаконы. Через около 72 часа после инфекции инфицированные CEF собирали. Аликвоты замораживали в жидком азоте, каждая аликвота содержала 10% FBS и 10% ДМСО. Титрование проводили в трех повторах на CEF, и для vHVT309 был получен титр 1,5 × 105 PFU/мл.
Двойное иммунофлуоресцентное окрашивание проводили с использованием куриной антисыворотки (Pab) в разведении 1: 500 и моноклонального антитела HVT L78 (Mab) в разведении 1: 3000 с последующим FITC-меченными антителами против куриных IgG в разведении 1: 500 и TRITC-мечеными ослиными антителами против мышиных антител Alex Fluor в разведении 1: 300. На планшете #1 сравнивали экспрессию вируса ньюкаслской болезни с HVT, а на планшете #2 сравнивали экспрессию вируса инфекционного бурсита с HVT. Было обнаружено, что все исследованные HVT-TRITC-положительные бляшки vHVT309 экспрессируют белки NDV-F и IBDV-VP2.
ПЦР-анализ vHVT309
Чистота рекомбинантного вируса была подтверждена с помощью ПЦР с использованием пар праймеров, которые специфичны для фланкирующих плеч HVT, промоторов, генов NDV-F и IBDV-VP2 и polyA-хвостов. Результаты ПЦР показывают, что рекомбинантный вирус vHVT309 несет предполагаемую экспрессирующую кассету, а сток вируса не содержит определяемых количеств родительского вируса Vaxxitek (таблица 1 и фиг. 5).
Заключение
На основании анализа ПЦР и иммунофлуоресцентного анализа, vHVT309 представляет собой рекомбинантный вирус, содержащий ген IBDV-VP2 под контролем промотора mCMV и ген NDV-F под контролем промотора SV40. Вновь полученный vHVT309 не содержит обнаруживаемого родительского вируса vHVT13.
Пример 1.2. Конструирование рекомбинантного vHVT310, экспрессирующего IBDV-VP2 и NDV-F
Цель исследования заключалась в конструировании рекомбинантного HVT, в котором экспрессирующая кассета, содержащая промотор цитомегаловируса мыши (mCMV), ген, кодирующий вирусный белок 2 вируса инфекционного бурсита (VP2), участок внутренней посадки рибосомы (IRES), ген кодирующий слитый белок вируса ньюкаслской болезни (NDV-F) и polyA-хвост вируса обезьян 40 (SV40-polyA), интегрирована в межгенный сайт 1 (IG1) (фиг. 2).
В конструкции используется родительский вирус vHVT13. Слитый белок вируса ньюкаслской болезни (NDV-F), соответствующий последовательности генотипа VIId, был синтезирован химически и оптимизирован по кодонам (GenScript).Сайт расщепления F-белка этого синтетического гена был изменен так, чтобы соответствовать лентогенной последовательности сайта расщепления F, и результирующая последовательность гена NDV-F имеет 99% идентичность аминокислотной последовательности с последовательностью NDV-F, депонированной в GenBank (AY337464). Мышиный промотор CMV IE использовали для IBD-VP2 (в родительском вирусе Vaxxitek). IRES, последовательность РНК, происходящая из вируса энцефаломиокардита (EMCV), которая позволяет инициировать трансляцию в мРНК непосредственно ниже от того места, где находится IRES, была вставлена в конец гена VP2 для инициации трансляции нижележащего гена NDV-F. Это был первый раз, когда IRES использовался в векторе HVT.
Локус вставки является межгенным сайтом 1 (IG1) в HVT (фиг. 2). Донорная плазмида pFIRESVP2 (вставочная плазмида, содержащая ген VP2 + IRES + NDV-F и SV40 polyA/фланкирующее плечо IG1) была сконструирована, как описано ниже. Клетки фибробластов куриных эмбрионов (CEF) использовали для рекомбинации in vitro.
Конструирование донорной плазмиды
Синтетическая ДНК в pUC57, содержащая ген IBDV VP2 (SEQ ID NO: 1, кодирующий SEQ ID NO: 2), IRES (SEQ ID NO: 10), ген NDV-F (SEQ ID NO: 3, кодирующий SEQ ID NO: 5), и SV40 polyA-хвост (SEQ ID NO: 8), была синтезирована с помощью GeneScript (фиг. 6). Плазмиду pFIRESVP2 трансформировали с использованием набора Top10 Oneshot (кат. № C404002, Invitrogen) и выращивали крупномасштабную культуру, и осуществляли экстракцию плазмиды с использованием набора Qiagens Maxi Prep.
Получение рекомбинантов
Процедуру гомологичной рекомбинации, описанную в Примере 1.1, использовали для получения рекомбинантного vHVT310.
Анализ рекомбинантов методом ПЦР
Процедура ПЦР-анализа, описанную в Примере 1.1, была выполнена для проверки vHVT310.
Анализ экспрессии
Анализ экспрессии, описанный в Примере 1.1, был выполнен для анализа экспрессии vHVT310.
Результаты
Нуклеотидной и аминокислотной последовательностям донорной плазмиды pFIRESVP2 присвоены SEQ ID NO, как показано на фиг. 1.
Получение рекомбинантов и анализ экспрессии
Геномную ДНК вируса Vaxxitek совместно электропорировали с донорной плазмидой pFIRESVP2 для получения рекомбинантного вируса с использованием метода гомологичной рекомбинации. Рекомбинантный вирус отделяли от родительского вируса HVT с помощью отбора иммунофлуоресцентно положительных лунок и ПЦР-скрининга в нескольких раундах очистки бляшек. Очищенный из бляшек рекомбинантный вирус, экспрессирующий белок NDV-F, обозначенный как vHVT310, масштабировали из колб для тканевых культур во флаконы 5 × 850 см2. Через около 72 часа после инфекции инфицированные CEF собирали. Аликвоты замораживали в жидком азоте, каждая аликвота содержала 10% FBS и 10% ДМСО. Титрование проводили в трех повторах на CEF, и для vHVT310 был получен титр 2,0 × 106 PFU/мл.
Двойное иммунофлуоресцентное окрашивание осуществляли с использованием куриной антисыворотки (Pab) в разведении 1: 500 и моноклонального антитела HVT L78 (Mab) в разведении 1: 3000 с последующим FITC-меченными антителами против куриных IgG в разведении 1: 500 и TRITC-мечеными ослиными антитителами против мышиных антител Alex Fluor в разведении 1: 300. На планшете #1 сравнивали экспрессию вируса ньюкаслской болезни с помощью HVT, а на планшете #2 сравнивали экспрессию вируса инфекционного бурсита с помощью HVT. Было обнаружено, что все исследованные HVT-TRITC-положительные бляшки vHVT310 экспрессируют белки NDV-F и IBDV-VP2.
ПЦР-анализ vHVT310
Чистота рекомбинантного вируса была подтверждена с помощью ПЦР с использованием пар праймеров, которые специфичны для фланкирующих плеч HVT, промотора, генов NDV-F и IBDV-VP2 и polyA-хвоста. Результаты ПЦР показывают, что рекомбинантный вирус vHVT310 несет предполагаемую экспрессирующую кассету, а сток вируса не содержит определяемых количеств родительского вируса Vaxxitek (таблица 2 и фиг. 7-8).
Таблица 2. Ожидаемые полосы ПЦР с использованием определенных наборов праймеров

Заключение
На основании ПЦР-тестирования и иммунофлуоресцентного анализа vHVT310 представляет собой рекомбинантный вирус, содержащий ген IBDV-VP2 и NDV-F под контролем промотора mCMV, где трансляция гена NDV-F инициируется IRES из EMCV. Вновь созданный рекомбинантный vHVT310 не содержит какого-либо обнаруживаемого родительского вируса vHVT13.
Пример 1.3. Конструирование рекомбинантного vHVT311, экспрессирующего IBDV-VP2 и NDV-F
Цель исследования заключалась в конструировании рекомбинантного HVT, в котором экспрессирующая кассета, содержащая промотор цитомегаловируса мыши (mCMV), ген, кодирующий вирусный белок 2 вируса инфекционного бурсита (VP2), саморасщепляющийся пептид 2A свиного тешовируса-1 (P2A), ген, кодирующий слитый белок вируса ньюкаслской болезни (NDV-F) и polyA-хвост вируса обезьян 40 (SV40-polyA), интегрирована в межгенный сайт 1 (IG1) (фиг. 2).
Родительский вирус, используемый в конструкции, представляет собой vHVT13 (HVT-вектор, экспрессирующий ген IBDV VP2, вакцину Merial VAXXITEK® (HVT + IBD)). Полинуклеотид, соответствующий последовательности слитого белка вируса ньюкаслской болезни генотипа VIId дикого типа (NDV-F), был синтезирован химически (GenScript). Сайт расщепления F-белка этого синтетического гена был изменен так, чтобы соответствовать лентогенной последовательности сайта расщепления F, и результирующая последовательность гена NDV-F имеет 99% идентичность аминокислотной последовательности с последовательностью NDV-F, депонированной в GenBank (AY337464). Мышиный промотор CMV IE использовали для IBD-VP2 (в родительском вирусе Vaxxitek). В конец гена VP2 был встроен саморасщепляющийся пептид 2A свиного тешовируса-1 (P2A), который делает возможным ко-трансляционное «расщепление» вышележащих и нижележащих генов, VP2 и F, соответственно, с одного промотора mCMV, соответственно. Это первый раз, когда P2A был использован в векторах HVT.
Локус вставки является межгенным сайтом 1 (IG1) в HVT (фиг. 2). Донорную плазмиду pFP2AVP2 (вставочную плазмиду, содержащую VP2 + P2A + NDV-F и SV40 polyA/фланкирующее плечо IG1) конструировали, как описано ниже. Клетки фибробластов куриных эмбрионов (CEF) использовали для рекомбинации in vitro.
Конструирование донорной плазмиды
Синтетические. ДНК в pUC57, содержащая ген IBDV VP2 (SEQ ID NO: 1, кодирующий SEQ ID NO: 2), P2A-кодирующую ДНК (SEQ ID NO: 11), ген NDV-F (SEQ ID NO: 4, кодирующий SEQ ID NO: 5) и polyA-хвост SV40 (SEQ ID NO: 8), синтезировали с помощью GeneScript (фиг. 9). Плазмиду pFP2AVP2 трансформировали с использованием набора Top10 Oneshot (кат. № C404002, Invitrogen), выращивали крупномасштабную культуру, и осуществляли экстракцию плазмиды с использованием набора Qiagens Maxi Prep.
Получение рекомбинантов
Процедуру гомологичной рекомбинации, описанную в Примере 1.1, использовали для получения рекомбинантного vHVT311.
Анализ рекомбинантов методом ПЦР
Процедура ПЦР-анализа, описанную в Примере 1.1, была выполнена для проверки vHVT311.
Анализ экспрессии
Анализ экспрессии, описанный в Примере 1.1, был выполнен для анализа экспрессии vHVT311.
Результаты
Нуклеотидной и аминокислотной последовательностям донорной плазмиды pFP2AVP2 присвоены SEQ ID NO, как показано на фиг. 1.
Получение рекомбинантов и анализ экспрессии
Геномную ДНК вируса Vaxxitek совместно электропорировали с донорной плазмидой pFP2AVP2 для получения рекомбинантного вируса с использованием метода гомологичной рекомбинации. Рекомбинантный вирус отделяли от родительского вируса Vaxxitek с помощью отбора иммунофлуоресцентно положительных лунок и ПЦР-скрининга в нескольких раундах очистки бляшек. Очищенный из бляшек рекомбинантный вирус, экспрессирующий белок NDV-F, обозначенный как vHVT311, масштабировали из колб для тканевых культур во флаконы 5 × 850 см2. Через около 72 часа после инфекции инфицированные CEF собирали. Аликвоты замораживали в жидком азоте, каждая аликвота содержала 10% FBS и 10% ДМСО. Титрование проводили в трех повторностях на CEF, и для vHVT311 был получен титр 2,5 × 106 PFU/мл.
Двойной иммунофлуоресцентный анализ проводили с использованием куриной антисыворотки (Pab) в разведении 1: 500 и моноклонального антитела (Mab) в разведении 1: 3000 с последующим введением FITC-меченых антител против куриных IgG в разведении 1: 500 и TRITC-меченых ослиных антител против мышиных антител Alex Fluor, в разведении 1: 300. На планшете #1 сравнивали экспрессию вируса ньюкаслской болезни с помощью HVT, а на планшете #2 сравнивали экспрессию вируса инфекционного бурсита с вирусом ньюкаслской болезни. Было обнаружено, что все исследованные HVT-TRITC-положительные бляшки vHVT311 экспрессируют NDV-F, а все NDV-TRITC-положительные бляшки экспрессируют белки IBDV-VP2.
ПЦР-анализ vHVT311
Чистота рекомбинантного вируса была подтверждена с помощью ПЦР с использованием пар праймеров, которые специфичны для фланкирующих плеч HVT, промотора, генов NDV-F и IBDV-VP2 и polyA-хвоста. Результаты ПЦР показывают, что рекомбинантный вирус vHVT311 несет предполагаемую экспрессирующую кассету, а сток вируса не содержит определяемых количеств родительского вируса Vaxxitek (таблица 3 и фиг. 10-11).
Таблица 3. Ожидаемые полосы ПЦР с использованием определенных наборов праймеров

Заключение
На основании ПЦР-тестирования и иммунофлуоресцентного анализа, vHVT311 представляет собой рекомбинантный вирус, содержащий ген IBDV-VP2 и NDV-F под контролем промотора mCMV, в котором опосредованное пептидом 2A расщепление приводит к коэкспрессии белков VP2 и F. Вновь созданный рекомбинантный vHVT311 не содержит какого-либо обнаруживаемого родительского вируса vHVT13.
Пример 1.4. Конструирование рекомбинантного vHVT317, экспрессирующего IBDV-VP2 и ILTV-gD
Цель исследования заключалась в конструировании рекомбинантного HVT, в котором экспрессирующая кассета, содержащая промотор цитомегаловируса мыши (mCMV), ген, кодирующий вирусный белок 2 вируса инфекционного бурсита (VP2), участок внутренней посадки рибосомы (IRES), ген кодирующий белок гликопротеина D инфекционного ларинготрахеита (ILTV-gD) и polyA-хвост вируса обезьян 40 (SV40-polyA) интегрирована в межгенный сайт 1 (IG1) (фиг. 2).
В конструкции используется родительский вирус vHVT13. В конструкции используется последовательность гликопротеина D вируса инфекционного ларинготрахеита (ILTV gD), которая была синтезирована химически (GenScript). Мышиный промотор CMV IE использовали для IBD-VP2 (в родительском вирусе vHVT13). Последовательность РНК (IRES), полученная из вируса энцефаломиокардита (EMCV), которая позволяет инициировать трансляцию в мРНК непосредственно ниже от того места, где находится IRES, была вставлена в конец гена VP2 для инициации трансляции нижестоящего гена ILTV-gD.
Локус вставки является межгенным сайтом 1 (IG1) в HVT (фиг. 2). Донорную плазмиду pVP2IRESgD (вставочную плазмиду, содержащую ген VP2 + IRES + ILTV-gD и polyA SV40/фланкирующее плечо IG1) конструировали, как описано ниже. Клетки фибробластов куриных эмбрионов (CEF) использовали для рекомбинации in vitro.
Конструирование донорной плазмиды
Синтетическую ДНК в pUC57, содержащую ген IBDV VP2 (SEQ ID NO: 1, кодирующий SEQ ID NO: 2), IRES (SEQ ID NO: 10), ген ILTV-gD (SEQ ID NO: 16, кодирующий SEQ ID NO: 17), и polyA-хвост SV40 (SEQ ID NO: 8) синтезировали с помощью GenScript. Плазмиду pFIRESVP2 трансформировали в dcm-/dam-компетентные клетки (New England Biolabs, cat # C2925I), затем расщепляли Hind III/Sal I. Фрагмент 5kb экстрагировали из геля. Синтетическая ДНК в pUC57, содержащая частичный IRES, ILTV-gD дикого типа и polyA-хвост SV40, синтезировали с помощью GenScript. Плазмиду Sal-Fse gD-IRES расщепляли Hind III/Sal I. Фрагмент 1,9 т.п.н. экстрагировали из геля. Два фрагмента лигировали и трансформировали с использованием набора Top10 Oneshot (кат. № C404002, Invitrogen). Колонии были проверены Hind III/Sbf I на корректный паттерн. Конечная донорная плазмида была секвенирована и обозначена как pVP2IRESgD (см. фиг. 12).
Получение рекомбинантов
Процедуру гомологичной рекомбинации, описанную в Примере 1.1, использовали для получения рекомбинантного vHVT317.
Анализ рекомбинантов методом ПЦР
Процедура ПЦР-анализа, описанную в Примере 1.1, была выполнена для проверки vHVT317.
Анализ экспрессии
Анализ экспрессии, описанный в Примере 1.1, был выполнен для анализа экспрессии vHVT317.
Результаты
Нуклеотидной и аминокислотной последовательностям донорной плазмиды pVP2IRESgD присвоены SEQ ID NO, как показано на фиг. 1.
Получение рекомбинантов и анализ экспрессии
Двойной иммунофлуоресцентный анализ проводили с использованием куриной антисыворотки (поликлональное антитело) в разведении 1: 500 и моноклонального антитела (Mab) в разведении 1: 3000 с последующим введением FITC-меченых антител против куриных IgG в разведении 1: 500 и TRITC-меченых ослиных антител против мышиных антител Alex Fluor в разведении 1: 300. Было обнаружено, что все исследованные бляшки vHVT317 экспрессируют белки IBDV-VP2 по сравнению с HVT-положительными бляшками, и все бляшки экспрессируют белки ILTV-gD при сравнении с IBDV-положительными бляшками.
ПЦР-анализ vHVT317
Чистота рекомбинантного вируса была подтверждена с помощью ПЦР с использованием пар праймеров, которые специфичны для фланкирующих плеч HVT, промотора, генов ILTV-gD и IBDV-VP2 и polyA-хвоста. Результаты ПЦР показывают, что рекомбинантный вирус vHVT317 несет предполагаемую экспрессирующую кассету, а сток вируса не содержит определяемых количеств родительского вируса Vaxxitek (таблица 4 и фиг. 13-14).
Таблица 4. Ожидаемые полосы ПЦР с использованием определенных наборов праймеров

Заключение
Основываясь на ПЦР-тестировании и иммунофлуоресцентном анализе, vHVT317 представляет собой рекомбинантный вирус, содержащий ген IBDV-VP2 и ILTV-gD под контролем промотора mCMV, где трансляция гена ILTV-gD инициируется IRES из EMCV. Вновь созданный рекомбинантный vHVT317 не содержит какого-либо обнаруживаемого родительского вируса vHVT13.
Пример 1.5. Конструирование рекомбинантного vHVT313, экспрессирующего IBDV-VP2 и NDV-F
Цель исследования заключается в создании рекомбинантного HVT, в котором экспрессирующая кассета, содержащая промотор цитомегаловируса мыши (mCMV), ген, кодирующий вирусный белок 2 вируса инфекционного бурсита (VP2), polyA-хвост вируса полиомиелита вируса обезьян 40 (SV40 polyA), промотор вируса обезьян 40 (промотор SV40), ген, кодирующий слитый белок вируса ньюкаслской болезни дикого типа (NDV-F) и синтетический polyA-хвост (syn polyA-хвост), интегрирована в межгенный сайт 1 (IG1) (фиг. 2).
В конструкции используется родительский вирус vHVT13. Белок вируса ньюкаслской болезни дикого типа (NDV-F), соответствующий последовательности дикого типа генотипа VIId, был синтезирован химически (GenScript). Сайт расщепления F-белка этого синтетического гена был изменен так, чтобы соответствовать лентогенной последовательности сайта расщепления F, и результирующая последовательность гена NDV-F имеет 99% идентичность аминокислотной последовательности с последовательностью NDV-F, депонированной в GenBank (AY337464). Использовали мышиный промотор CMV IE для IBD-VP2 (в родительском вирусе Vaxxitek) и промотор SV40 для NDV-F.
Локус вставки является межгенным сайтом 1 (IG1) (фиг. 2). Донорную плазмиду pFwtSV40VP2 (вставочную плазмиду, содержащую VP2/SV40 polyA и фланкирующее плечо IG1 + SV40 промотор + NDV-F + синтетический polyA) конструировали, как описано ниже. Клетки фибробластов куриных эмбрионов (CEF) использовали для рекомбинации in vitro.
Конструирование донорной плазмиды
Синтетическую ДНК в pUC57, содержащую ген IBDV VP2 (SEQ ID NO: 1, кодирующая SEQ ID NO: 2), polyA-хвост SV40 (SEQ ID NO: 8), промотор SV40 (SEQ ID NO: 7), ген NDV-F SEQ ID NO: 4, кодирующая SEQ ID NO: 5), и синтетический polyA-хвост (SEQ ID NO: 9), синтезировали с помощью GeneScript.
Плазмиду, pFSV40VP2 затем гидролизовали с помощью Sbf I/AvrII и фрагмент 5.6 тыс. н. экстрагировали из геля. Плазмиду, pHM103NDVFwtsyn также расщепляли SbfI/Avr II и фрагмент 1,9 тыс. н. экстрагировали из геля. Затем фрагменты лигировали вместе и трансформировали, используя набор Top10 Oneshot (кат. № C404002, Invitrogen). Колонии подвергали скринингу с помощью PstI на предмет корректного паттерна. Транзиторная экспрессия maxi prep была подтверждена с использованием реагента для трансфекции Fugene в клетках фибробластов куриных эмбрионов (CEF) и поликлональной куриной сыворотки против NDV. Конечная донорная плазмида была секвенирована и обозначена как pFwtSV40VP2 (см. фиг. 15).
Получение рекомбинантов
Процедуру гомологичной рекомбинации, описанную в Примере 1.1, использовали для получения рекомбинантного vHVT313.
Анализ рекомбинантов методом ПЦР
Процедура ПЦР-анализа, описанную в Примере 1.1, была выполнена для проверки vHVT313.
Анализ экспрессии
Анализ экспрессии, описанный в Примере 1.1, был выполнен для анализа экспрессии vHVT313.
Результаты
Нуклеотидной и аминокислотной последовательностям донорной плазмиды pFwtSV40VP2 присвоены SEQ ID NO, как показано на фиг. 1.
Получение рекомбинантов и анализ экспрессии
Двойной иммунофлуоресцентный анализ проводили с использованием куриной антисыворотки (Pab) и моноклонального антитела против HVT (Mab), за которым следовали FITC-меченные антитела против куриных IgG, и TRITC-меченные ослиные антитела против мышиных антител Alex Fluor. Было обнаружено, что все исследованные TRITC-положительные бляшки vHVT313 экспрессируют белки NDV-F и IBDV-VP2.
ПЦР-анализ vHVT313
Чистота рекомбинантного вируса была подтверждена с помощью ПЦР с использованием пар праймеров, которые специфичны для фланкирующих плеч HVT, промоторов, генов NDV-F и IBDV-VP2 и polyA-хвостов. Результаты ПЦР показывают, что рекомбинантный вирус vHVT313 несет предполагаемую экспрессирующую кассету, а сток вируса не содержит определяемых количеств родительского вируса Vaxxitek (таблица 5 и фиг. 16-17).
Таблица 5. Ожидаемые полосы ПЦР с использованием определенных наборов праймеров

Заключение
На основании ПЦР-тестирования и иммунофлуоресцентного анализа, vHVT313 представляет собой рекомбинантный вирус, содержащий ген IBDV-VP2 под контролем промотора mCMV и ген дикого типа NDV-F под контролем промотора SV40. Вновь созданный vHVT313 не содержит обнаруживаемых родительских вирусов Vaxxitek.
Пример 1.6. Конструирование рекомбинантного vHVT316, экспрессирующего IBDV-VP2 и NDV-F
Цель исследования заключалась в конструировании рекомбинантного HVT, в котором экспрессирующая кассета, содержащая промотор цитомегаловируса мыши (mCMV), ген, кодирующий вирусный белок 2 вируса инфекционного бурсита (VP2), участок внутренней посадки рибосомы (IRES), ген кодирующий слитый белок вируса ньюкаслской болезни дикого типа (NDV-F) и polyA-хвост вируса обезьяны 40 (SV40-polyA), интегрированный в локус IG1 (фиг. 2).
В конструкции используется родительский вирус vHVT13. Белок вируса ньюкаслской болезни дикого типа (NDV-F), соответствующий последовательности дикого типа генотипа VIId, был синтезирован химически (GenScript). Сайт расщепления F-белка этого синтетического гена был изменен так, чтобы соответствовать лентогенной последовательности сайта расщепления F, и результирующая последовательность гена NDV-F имеет 99% идентичность аминокислотной последовательности с последовательностью NDV-F, депонированной в GenBank (AY337464). Мышиный промотор CMV IE использовали для IBD-VP2 (в родительском вирусе Vaxxitek). IRES был вставлен в конец гена VP2 для инициации трансляции нижележащего гена NDV-F.
Локус вставки - IG1 (фиг. 2). Донорная плазмида pVP2IRESFwt (вставочная плазмида, содержащая ген VP2 + IRES + NDV-F и SV40 polyA/фланкирующее плечо IG1) была сконструирована, как описано ниже. Клетки фибробластов куриных эмбрионов (CEF) использовали для рекомбинации in vitro.
Конструирование донорной плазмиды
Синтетическая ДНК в pUC57, содержащая ген IBDV VP2 (SEQ ID NO: 1, кодирующий SEQ ID NO: 2), IRES (SEQ ID NO: 10), ген NDV-F (SEQ ID NO: 4, кодирующий SEQ ID NO: 5), и polyA-хвост SV40 (SEQ ID NO: 8), была синтезирована с помощью GenScript. Плазмиду pFIRESVP2 трансформировали в dcm-/dam-компетентные клетки (New England Biolabs, cat # C2925I), затем расщепляли Hind III/Sal I. Фрагмент 5 тыс. н. экстрагировали из геля. Синтетическую ДНК в pUC57, содержащую частичный IRES, NDV-F дикого типа и SV40 polyA-хвост, синтезировали с помощью GenScript. Плазмиду Sal-Hind-Fwt + расщепляли с помощью Hind III/Sal I. Фрагмент в 2,2 тыс. н. экстрагировали из геля. Два фрагмента лигировали и трансформировали с использованием набора Top10 Oneshot (кат. № C404002, Invitrogen). Конечная донорная плазмида была секвенирована и обозначена как pVP2IRESFwt (см. фиг. 18).
Получение рекомбинантов
Процедуру гомологичной рекомбинации, описанную в Примере 1.1, использовали для получения рекомбинантного vHVT316.
Анализ рекомбинантов методом ПЦР
Процедура ПЦР-анализа, описанную в Примере 1.1, была выполнена для проверки vHVT316.
Анализ экспрессии
Анализ экспрессии, описанный в Примере 1.1, был выполнен для анализа экспрессии vHVT316.
Результаты
Нуклеотидной и аминокислотной последовательностям донорной плазмиды pVP2IRESFwt присвоены SEQ ID NO, как показано на фиг. 1.
Получение рекомбинантов и анализ экспрессии
Двойное иммунофлуоресцентное окрашивание проводили с использованием куриной антисыворотки (Pab) и моноклонального антитела (Mab) с последующими FITC-меченными антителами против куриных IgG и TRITC-мечеными ослиными антителами против мышиных антител Alex Fluor. Было обнаружено, что все исследованные бляшки vHVT316 экспрессируют белки IBDV-VP2 по сравнению с HVT-положительными бляшками, и все бляшки экспрессируют белки IBDV-VP2 по сравнению с NDV-положительными бляшками.
ПЦР-анализ vHVT316
Чистота рекомбинантного вируса была подтверждена с помощью ПЦР с использованием пар праймеров, которые специфичны для фланкирующих плеч HVT, промотора, генов NDV-F и IBDV-VP2 и polyA-хвоста. Результаты ПЦР показывают, что рекомбинантный вирус vHVT316 несет предполагаемую экспрессирующую кассету, а сток вируса не содержит определяемых количеств родительского вируса Vaxxitek (таблица 6 и фиг. 19-20).
Таблица 6. Ожидаемые полосы ПЦР с использованием специфических наборов праймеров

Заключение
Основываясь на ПЦР-тестировании и иммунофлуоресцентном анализе, vHVT316 представляет собой рекомбинантный вирус, содержащий ген IBDV-VP2 и NDV-F под контролем промотора mCMV, где трансляция гена NDV-F инициируется IRES из EMCV. Вновь созданный рекомбинантный vHVT316 не содержит какого-либо обнаруживаемого родительского вируса Vaxxitek.
Пример 1.7. Конструирование рекомбинантного vHVT407, экспрессирующего IBDV-VP2 и ILTV-gD
Цель исследования заключалась в конструировании рекомбинантного HVT, в котором экспрессирующая кассета, содержащая промотор SV40, гликопротеин D ILTV и синтетический поли A в сайте SORF3-US2 из vHVT13.
В конструкции использовали родительский вирус vHVT13. В конструкции использовали последовательность гликопротеина D вируса инфекционного ларинготрахеита (ILTV gD), которая была химически синтезирована (GenScript). Промотор SV40 использовали для ILTV gD. Локус вставки представляет собой SORF3-US2 для ILTV gD и IG1 для IBDV VP2 из vVHT13 (фиг. 2). Была сконструирована донорная плазмида HVT US2SVgDwtsyn, содержащая плечи SORF3-US2, промотор SV40 (SEQ ID NO: 7), ген, кодирующий ILTV gD дикого типа (SEQ ID NO: 16, кодирующий SEQ ID NO: 17), и синтетический polyA (SEQ ID NO: 9) (см. фиг. 22). Клетки фибробластов куриных эмбрионов (CEF) использовали для рекомбинации in vitro.
Получение рекомбинантов
Процедуру гомологичной рекомбинации, описанную в Примере 1.1, использовали для получения рекомбинантного vHVT407.
Анализ рекомбинантов методом ПЦР
Процедура ПЦР-анализа, описанную в Примере 1.1, была выполнена для проверки vHVT407.
Анализ экспрессии
Анализ экспрессии, описанный в Примере 1.1, был выполнен для анализа экспрессии vHVT407.
Результаты
Нуклеотидной и аминокислотной последовательностям донорной плазмиды HVT US2SVgDwtsyn присвоены SEQ ID NO, как показано на фиг. 1.
Получение рекомбинантов и анализ экспрессии
Двойное иммунофлуоресцентное окрашивание проводили с использованием куриной антисыворотки (Pab) и моноклонального антитела (Mab) с последующими FITC-меченными антителами против куриных IgG и TRITC-мечеными ослиными антителами против мышиных антител Alex Fluor. Было обнаружено, что все исследованные бляшки vHVT407 экспрессируют белки IBDV-VP2 и ILTV gD.
ПЦР-анализ vHVT407
Чистота рекомбинантного вируса была подтверждена с помощью ПЦР с использованием пар праймеров, специфичных для фланкирующих плеч HVT, промотора, генов ILTV gD и IBDV-VP2 и polyA-хвоста. Результаты ПЦР демонстрируют, что рекомбинантный вирус vHVT407 несет предполагаемую экспрессирующую кассету, и сток вируса не содержит определяемых количеств родительского вируса vHVT13.
Заключение
Основываясь на ПЦР-тестировании и иммунофлуоресцентном анализе, vHVT407 является рекомбинантным вирусом, содержащим гены IBDV-VP2 и ILTV gD. Вновь созданный рекомбинантный vHVT407 не содержит какого-либо обнаруживаемого родительского вируса vHVT13.
Пример 1.8. Конструирование рекомбинантного vHVT308, экспрессирующего NDV-F и ILTV-gD в противоположных направлениях
Цель исследования заключалась в создании вставочной плазмиды для сайта межгенной области I, которая будет содержать синтетический polyA-хвост, NDV F, промотор SV40, промотор HHV3gB, ILTV gD и polyA-хвост SV40 для гомологичной рекомбинации в HVT FC126.
Родительским вирусом, использованным в конструкции, является HVT FC126. Синтетический слитый белок вируса ньюкаслской болезни (NDV-F) (SEQ ID NO: 21, кодирующий SEQ ID NO: 22), соответствующий последовательности генотипа V, был синтезирован химически и оптимизирован по кодонам (GenScript). Сайт расщепления F-белка этого синтетического гена был изменен так, чтобы соответствовать лентогенной последовательности сайта расщепления F. Синтетический гликопротеин D ILTV дикого типа (SEQ ID NO: 16, кодирующий SEQ ID NO: 17) был синтезирован химически. Была сконструирована донорная плазмида pHVTIG1gDCaFopt, содержащая промотор HHV3gB (промотор гликопротеина B герпесвируса человека 3 типа) в обратной ориентации, управляющий экспрессией ILTV-gD + полиА-хвост SV40, и промотор SV40, управляющий экспрессией слитого белка вируса ньюкаслской болезни + синтетический polyA-хвост (см. фиг. 23). Клетки фибробластов куриных эмбрионов (CEF) использовали для рекомбинации in vitro.
Получение рекомбинантов
Для получения рекомбинантного vHVT308 использовали процедуру гомологичной рекомбинации, описанную в Примере 1.1. Последовательные пассажи были выполнены до пре-MSV + 13.
Анализ рекомбинантов методом ПЦР
Для проверки рекомбинантного vHVT308 проводили процедуру ПЦР-анализа, описанную в Примере 1.1.
Анализ экспрессии
Для анализа экспрессии рекомбинантного vHVT308 проводили анализ экспрессии, описанный в Примере 1.1.
Результаты
Нуклеотидной и аминокислотной последовательностям донорной плазмиды pHVTIG1gDCaFopt присвоены SEQ ID NO, как показано на фиг. 1.
Получение рекомбинантов и анализ экспрессии
Двойное иммунофлуоресцентное окрашивание проводили с использованием куриной антисыворотки (Pab) и моноклонального антитела (Mab) с последующими FITC-меченными антителами против куриных IgG и TRITC-мечеными ослиными антителами против мышиных антител Alex Fluor. Было обнаружено, что все исследованные бляшки vHVT308 экспрессируют белки NDV-F и ILTV-gD.
ПЦР-анализ vHVT308
Чистоту рекомбинантного вирусу подтверждали с помощью ПЦР с использованием пар праймеров, которые специфичны для фланкирующих плеч HVT, промоторов, генов NDV-F и ILTV-gD и polyA-хвостов. Результаты ПЦР показывают, что рекомбинантный вирус vHVT308 несет предполагаемую экспрессирующую кассету, а сток вируса не содержит определяемых количеств родительского вируса HVT (таблица 6.1 и фиг. 24 и 25).
Таблица 6.1. Ожидаемые полосы ПЦР с использованием определенных наборов праймеров

Реакции ПЦР со всеми парами праймеров привели к ожидаемым продуктам ПЦР и паттерну полос. Как показано выше, нет никаких свидетельств родительского вируса HVT в vHVT308, и vHVT308 стабилен при пассажах до пре-MSV + 13.
Заключение
На основании ПЦР-тестирования и иммунофлуоресцентного анализа, vHVT308 представляет собой рекомбинантный вирус HVT, содержащий ген NDV-F под контролем промотора SV40 и ген ILTV-gD под контролем промотора HHV3gB. vHVT308 не содержит обнаруживаемого родительского вируса HVT.
Пример 1.9. Конструирование рекомбинантного vHVT322, экспрессирующего NDV-F и ILTV-gD
Цель исследования заключается в создании рекомбинантного HVT, в котором экспрессирующая кассета, содержащая промотор mCMV, слитый белок вируса ньюкаслской болезни (NDV-F), участок внутренней посадки рибосомы (IRES), гликопротеин D инфекционного ларинготрахеита (ILTV-gD), и poly-A-хвост вируса обезьян 40 (поли А SV40) будет гомологично рекомбинирована с фланкирующими плечами в межгенной области 1 (IG1) vHVT13 (HVT + IBD).
В конструкции использовался родительский вирус vHVT13. Был химически синтезирован (GenScript) слитый белок вируса ньюкаслской болезни (NDV-F), соответствующий последовательности VIId генотипа дикого типа (SEQ ID NO: 4, кодирующая SEQ ID NO: 5). Сайт расщепления F-белка этого синтетического гена был изменен для соответствия лентогенной последовательности сайта расщепления F, и результирующая последовательность гена NDV-F имеет 99% идентичность нуклеотидной, а также аминокислотной последовательности NDV-F, депонированной в GenBank (AY337464). Синтетический гликопротеин D ILTV дикого типа (SEQ ID NO: 16, кодирующий SEQ ID NO: 17) был синтезирован химически. Донорная плазмида pFwtIRESgD содержала левое фланкирующее плечо IG1, mCMV (мышиный CMV IE), NDV-F, IRES, ILTV-gD, SV40 polyA и правое фланкирующее плечо IG1 (см. фиг. 26). Клетки фибробластов куриных эмбрионов (CEF) использовали для рекомбинации in vitro.
Получение рекомбинантов
Процедуру гомологичной рекомбинации, описанную в Примере 1.1, использовали для получения рекомбинантного vHVT322. Последовательный пассаж был выполнен до пре-MSV + 13.
Анализ рекомбинантов методом ПЦР
Процедуру ПЦР-анализа, описанную в Примере 1.1, проводили для проверки рекомбинантного vHVT322.
Анализ экспрессии
Анализ экспрессии, описанный в Примере 1.1, проводили для анализа экспрессии рекомбинантного vHVT322.
Результаты
Нуклеотидной и аминокислотной последовательностям донорной плазмиды pFwtIRESgD присвоены SEQ ID NO, как показано на фиг. 1.
Получение рекомбинантов и анализ экспрессии
Двойное иммунофлуоресцентное окрашивание проводили с использованием куриной антисыворотки (Pab) и моноклонального антитела (Mab) с последующими FITC-меченными антителами против куриных IgG и TRITC-мечеными ослиными антителами против мышиных антител Alex Fluor. Было обнаружено, что все исследованные TRITC-положительные бляшки vHVT322 экспрессируют белки NDV-F и ILTV-gD.
ПЦР-анализ vHVT322
Чистота рекомбинантного вируса была подтверждена с помощью ПЦР с использованием пар праймеров, которые специфичны для фланкирующих плеч HVT, промотора, генов NDV-F и ILTV-gD и polyA-хвоста. Результаты ПЦР показывают, что рекомбинантный вирус vHVT322 несет предполагаемую экспрессирующую кассету, и сток вируса не содержит определяемых количеств родительского vHVT13 (таблица 6.2 и фиг. 27 и 28).
Таблица 6.2. Ожидаемые полосы ПЦР с использованием наборов специфических праймеров

Реакции ПЦР со всеми парами праймеров привели к ожидаемым продуктам ПЦР и паттерну полос. Как показано выше, нет никаких признаков родительского вируса vHVT13 в vHVT322.
Заключение
Основываясь на ПЦР-тестировании и иммунофлуоресцентном анализе, vHVT322 представляет собой рекомбинантный вирус HVT, содержащий ген NDV-F и ILTV-gD под контролем промотора mCMV. vHVT322 не содержит обнаруживаемого родительского вируса vHVT13.
Пример 1.10. Конструирование рекомбинантного vHVT406, экспрессирующего ILT-gDwt
Цель исследования заключается в создании рекомбинантного HVT, сайт которого SORF3-US2 содержит промотор SV40, gD инфекционного лариготрахеита и синтетический polyA-хвост для гомологичной рекомбинации в HVT FC126.
Родительским вирусом, использованным в конструкте, является HVT FC126. Синтезированный гликопротеин D дикого типа (gDwt) синтетического вируса инфекционного ларинготрахеита (ILTV) был синтезирован химически. Донорная плазмида pHVTUS2SVgDwtsyn содержала плечи SORF3 и US2 HVT FC126, промотор SV40, gDwt ILTV (SEQ ID NO: 16, кодирующий SEQ ID NO: 17) и синтетический пpolyA (см. фиг. 29). Клетки фибробластов куриных эмбрионов (CEF) использовали для рекомбинации in vitro.
Получение рекомбинантов
Процедуру гомологичной рекомбинации, описанную в Примере 1.1, использовали для получения рекомбинантного vHVT406. Последовательный пассаж проводили до пре-MSV + 13 (x + 12).
Анализ рекомбинантов методом ПЦР
Процедуру ПЦР-анализа, описанную в Примере 1.1, проводили для проверки рекомбинантного vHVT406.
Анализ экспрессии
Анализ экспрессии, описанный в Примере 1.1, проводили для анализа экспрессии рекомбинантного vHVT406.
Результаты
Нуклеотидной и аминокислотной последовательностям донорной плазмиды pHVTUS2SVgDwtsyn присвоены SEQ ID NO, как показано на фиг. 1.
Получение рекомбинантов и анализ экспрессии
Геномную ДНК вируса HVT совместно электропорировали с донорной плазмидой pHVTUS2SVgDwtsyn для получения рекомбинантного HVT с использованием метода гомологичной рекомбинации. Рекомбинантный вирус отделяли от родительского вируса HVT с помощью отбора иммунофлуоресцентно положительных лунок и ПЦР-скрининга в нескольких раундах очистки бляшек. Очищенный из бляшек рекомбинантный вирус HVT, экспрессирующий белок ILTV-gD, был обозначен vHVT406.
Рекомбинантные вирусные бляшки vHVT406 визуализировали с использованием фильтров TRITC и FITC для двойного окрашивания. FITC показал экспрессию ILTV-gDwt, а TRITC показал экспрессию HVT. Из-за небольших лунок 96-луночных планшетов каждую лунку регистрировали по бляшкам, которые сначала подсчитывали с помощью фильтра TRITC, а затем пересчитывали с помощью фильтра FITC. Объединенные 600+ бляшек подсчитывали между пассажами от пре-MSV и до пре-MSV + 13. Все бляшки были положительными как по FITC, так и по TRITC для обоих пассажей.
ПЦР-анализ vHVT406
ПЦР-анализ vHVT406 проводили с использованием праймеров для ПЦР, перечисленных в таблице 6.3 (см. фиг. 30). Как показано на фиг. 31, размеры продуктов ПЦР после гель-электрофореза хорошо соответствуют ожидаемым размерам и паттернам полос. Нет никаких свидетельств родительского вируса HVT FC126 в vHVT406.
Таблица 6.3. Ожидаемые полосы ПЦР с использованием специальных наборов праймеров

Заключение
Основываясь на ПЦР-тестировании и иммунофлуоресцентном анализе, vHVT406 представляет собой рекомбинантный вирус HVT, содержащий промотор SV40, ген ILTV-gDwt и синтетический polyA-хвост в сайте SOrf3-US2. vHVT406 не содержит обнаруживаемого родительского вируса HVT.
Пример 1.11. Исследование стабильности HVT-векторов in vitro
HVT-векторы, сконструированные выше, тестировали на стабильность генома/экспрессии после множественных пассажей in vitro в клетках фибробластов куриного эмбриона (CEF). HVT-векторы, экспрессирующие два гена, были стабильными после многократных пассажей. Вопреки общему мнению, что HVT с несколькими вставками менее стабильны, результаты неожиданно продемонстрировали, что HVT-векторы по настоящему изобретению стабильны и эффективно экспрессируют два гена.
Пример 2. Эффективность от ньюкаслской болезни (ND), индуцированная в D28 с помощью vHVT306, vHVT309, vHVT310 и vHVT311 у цыплят SPF
Цель данного исследования заключалась в оценке эффективности четырех рекомбинантных конструкций HVT (vHVT306, vHVT309, vHVT310 и vHVT311), экспрессирующих ген IBDV VP2 и ген NDV F, вводимых односуточным цыплятам SPF против заражения ньюкаслской болезнью (штамм Texas GB) в D28.
Характеристики этих вакцин-кандидатов описаны в таблице 7 ниже.
Таблица 7. Характеристики векторов, использованных в контрольном исследовании

vHVT306*: вектор vHVT, экспрессирующий IBDV VP2 и NDV F (см. US 9114108), используемый в качестве контроля.
vHVT13** является активным ингредиентом лицензированной вакцины VAXXITEK HVT-IBD на основе вектора HVT, экспрессирующего ген IBDV VP2 (описан как vHVT17 в US 5980906 и EP 0 719 864).
Девяносто пять безпатогенных цыплят (SPF) в возрасте одного дня были распределены на 5 групп, как показано в таблице 8. Всех птиц от 1 до 4 групп (20 птиц/группу) вакцинировали подкожным (SC) путем с использованием 0,2 мл различных конструкций HVT-IBD + ND в указанной дозе (см. таблицу 8). 15 птиц из 5 группы остались невакцинированными. Через двадцать восемь (D28) дней после вакцинации птиц в каждой группе заражали штаммом NDV Texas GB внутримышечным (IM) путем (104,0 50% инфекционной дозы в яйце (EID50) по 0,1 мл/особь). Птиц наблюдали на предмет клинических признаков в течение 14 дней после заражения. Птицы, у которых в течение 14 дней после заражения не было выявлено клинических признаков ND (включая признаки центральной нервной системы или респираторных заболеваний и/или смерть), считались защищенными.
Результаты защиты приведены в таблице 8. Все контрольные птицы группы 5 погибли после заражения. Защита в вакцинированных группах достигла не менее 90%.
Таблица 8. Эффективность от ND, индуцированная различными двойными конструкциями HVT-IBD + ND у цыплят SPF

vHVT306*: используется в качестве контроля
Пример 3 Эффективность от IBD, индуцированная vHVT309, vHVT310, vHVT311 и vHVT407 против стандартного заражения IBDV в D35
Цель данного исследования заключалась в оценке эффективности трех рекомбинантных конструкций HVT (vHVT309, vHVT310 и vHVT311), экспрессирующих ген IBDV VP2 и ген NDV F, и одной конструкции (vHVT407), экспрессирующей ген IBDV VP2 и ген ILTV gD, вводимых односуточным цыплятам SPF против стандартного заражения IBDV в D35.
Цыплят односуточного возраста, не имеющих специфических патогенов (SPF), распределяли по 4 группам, как показано в таблице 9. Всех птиц от 1 до 4 групп (20 птиц/группу) вакцинировали подкожным (SC) путем с использованием 0,2 мл различных конструкций HVT-IBD + ND или HVT-IBD + ILT в указанной дозе. 20 птиц из группы 5 остались невакцинированными. Через тридцать пять дней после вакцинации (в D35) всех птиц заражали классическим штаммом STC вируса инфекционного бурсита (IBDV) внутриглазным (IO) путем (102,0 EID50 по 0,03 мл/птицу). Через четыре дня после заражения (в день 39) все птицы были умерщвлены и вскрыты для исследования грубых поражений фабрициевой сумки.
Результаты защиты приведены в таблице 9. Все вакцинированные птицы (кроме двух птиц, вакцинированных vHVT311) были защищены от IBD, тогда как ни одна из контрольных птиц не была защищена.
Таблица 9. Эффективность от IBD, индуцированная различными двойными конструкциями HVT-IBD + ND или HVT-IBD + ILT у цыплят SPF после заражения в D35 штаммом STC IBDV

Пример 4. Эффективность от IBD, индуцированная vHVT309, vHVT310, vHVT311 и vHVT407 против заражения вариантом IBDV в D35
Цель данного исследования заключалась в оценке эффективности трех рекомбинантных конструкций HVT (vHVT309, vHVT310 и vHVT311), экспрессирующих ген IBDV VP2 и ген NDV F, и одной конструкции (vHVT407), экспрессирующей ген IBDV VP2 и ген ILTV gD, вводимых односуточным цыплятам SPF против заражения вариантом (Delaware E) IBDV, проведенного в D35.
Цыплят односуточного возраста, не имеющих специфических патогенов (SPF), распределяли по 6 группам, как показано в таблице 10. Всех птиц с 1 по 4 группу (19-20 птиц/группу) вакцинировали подкожным (SC) введением 0,2 мл различных конструкций HVT-IBD + ND или HVT-IBD + ILT в указанной дозе. Птицы из группы 5 (19 птиц) и группы 6 (18 птиц) были оставлены невакцинированными. В D35, всех птиц из групп с 1 по 5 заражали штаммом варианта Delaware E вируса инфекционного бурсита (IBDV) внутриглазным (МО) путем (103,0 EID50 в 0,03 мл/особь). Птицы из группы 6 остались незараженными. В D46 измеряли массу тела и массу фабрициевой сумки всех птиц. Соотношения масс Ф/Т (отношение массы фабрициевой сумки к массе тела х100) рассчитывали для всех групп.
Результаты защиты приведены в таблице 10. Вакцинированные птицы из групп 1 и 2 имели среднее значение соотношение масс Ф/Т, такое же, как и у невакцинированных не подвергнутых заражению контролей (группа 6), и больше, чем у невакцинированных зараженных контролей (группа 5). Птицы группы 3 не были защищены, а птицы группы 4 были частично защищены. Удивительно, но vHVT310, который содержит IRES, обеспечил лучшую защиту, чем vHVT311, который содержит P2A.
Таблица 10. Эффективность от IBD, индуцированная различными двойными конструкциями HVT-IBD + ND или HVT-IBD + ILT у цыплят SPF после заражения в D35 штаммом варианта E IBDV

Пример 5. Эффективность от IBD, индуцированная vHVT306, vHVT309 и vHVT310 против заражения vvIBDV в D28 у бройлеров.
Цель исследования заключалась в оценке эффективности трех рекомбинантных конструкций HVT (vHVT306, vHVT309 и vHVT310), экспрессирующих ген IBDV VP2 и ген NDV F, введенных односуточным цыплятам-бройлерам, против заражения vvIBDV, проведенного в D28.
Семьдесят односуточных цыплят-бройлеров (линия Hubbard JA957) были распределены на 5 групп, как показано в таблице 11. Всех птиц из 2-5 групп (около 15 птиц/группу) вакцинировали подкожным (SC) путем с использованием 0,2 мл различных конструкций HVT-IBD + ND в указанной дозе. Десять птиц из группы 1 остались невакцинированными. Через двадцать восемь дней после вакцинации (в день 28) всех птиц заражали сильновирулентным штаммом IBDV (vvIBDV) 91-168 внутриглазным (IO) путем (104,3 EID50 в 0,05 мл на птицу). Через десять дней после заражения (в D38) все птицы были умерщвлены и вскрыты для исследования грубых поражений фабрициевой сумки. Фабрициеву сумку и тело взвешивали, и на фабрициевой сумке проводили гистопатологию. Гистологические поражения фабрициевой сумки оценивались от 0 до 5 по следующей шкале: 0 - поражения нет, нормальная фабрициева сумка; От 1 до 1% от 25% фолликулов показывают лимфоидное истощение (т.е. менее 50% истощения в 1 пораженном фолликуле), приток гетерофилов в очагах поражения; 2 - от 26% до 50% фолликулов показывают почти полное истощение лимфоидных клеток (то есть с более чем 75% истощения в 1 пораженном фолликуле), пораженные фолликулы демонстрируют некротические повреждения и может быть обнаружен сильный приток гетерофилов; 3 - от 51% до 75% фолликулов показывают лимфоидное истощение; пораженные фолликулы имеют очаги некроза и обнаруживается сильный приток гетерофилов; 4 - от 76% до 100% фолликулов показывают почти полное лимфоидное истощение; обнаружены гиперплазия и кистозные структуры; пораженные фолликулы имеют очаги некроза и обнаруживается сильный приток гетерофилов; и 5 - 100% фолликулов показывают почти полное лимфоидное истощение; полная потеря фолликулярной структуры; утолщенный и свернутый эпителий; фиброз ткани фабрициевой сумки. Птицы считались защищенными, если они не проявляли клинических признаков после заражения и если их гистологический показатель составлял ≤2.
В течение первой недели в этой партии бройлеров было несколько ранних случаев смерти, вероятно, из-за колибактериоза. Доза тестируемых вакцин была ниже ожидаемой (2000 PFU). Результаты защиты приведены в таблице 11. Частичная защита была вызвана вакцинацией, которая показывает, что у vHVT310 она была выше, чем vHVT306 и vHVT309.
Таблица 11. Эффективность от IBD, индуцированная различными двойными конструкциями HVT-IBD + ND у цыплят-бройлеров после заражения в D28 штаммом vvIBDV
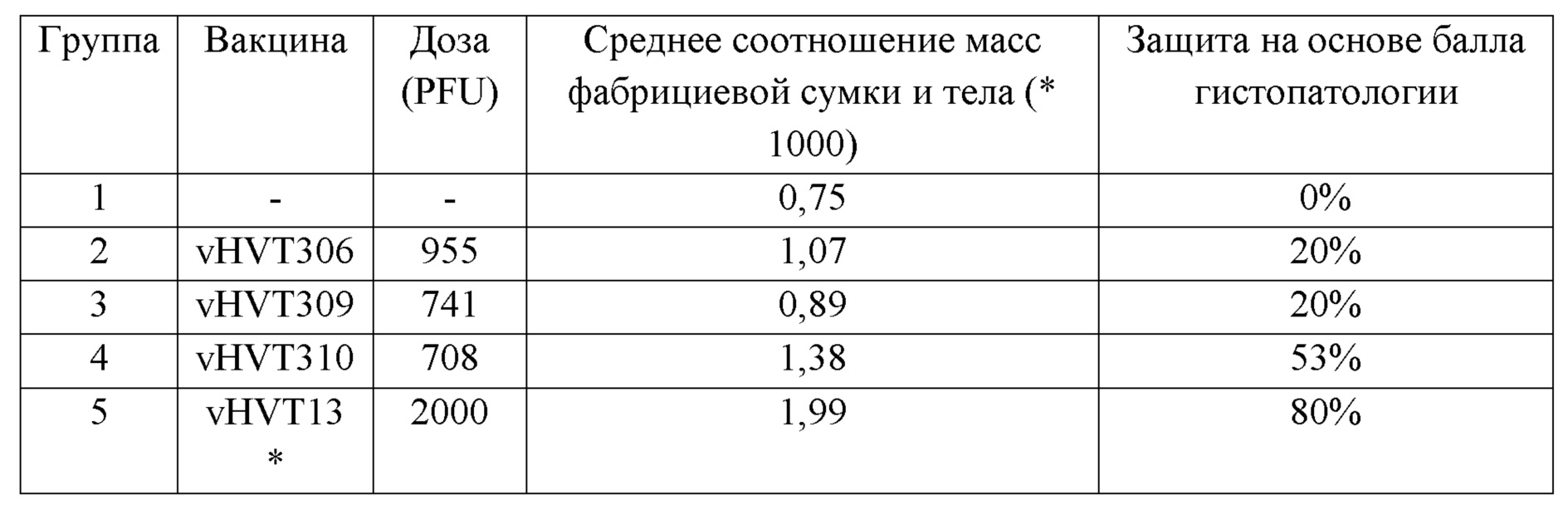
vHVT13*: используется в качестве контроля.
Пример 6. Эффективность от ND, индуцированная vHVT306, vHVT309 и vHVT310, против велогенного заражения NDV в D42 у бройлеров.
Цель исследования заключалась в оценке эффективности трех рекомбинантных конструкций HVT (vHVT306, vHVT309 и vHVT310), экспрессирующих ген IBDV VP2 и ген NDV F, введенных односуточным цыплятам-бройлерам, против велогенного заражения NDV, проведенного в D42.
Однодневные цыплята-бройлеры (линия Hubbard JA957) были распределены на 4 группы, как показано в таблице 12. Всех птиц со 2 по 4 группу (16-20 птиц/группу) вакцинировали подкожным (SC) путем с использованием 0,2 мл различных конструкций HVT-IBD + ND в указанной дозе. Двенадцать птиц из группы 1 остались невакцинированными. Сорок два дня после вакцинации (в D42), всех птиц заражали велогенным штаммом NDV Herts 33 внутримышечным (IM) путем (105,0 EID50 в 0,2 мл/особь). Всех птиц наблюдали на предмет клинических признаков в течение 14 дней после заражения. Птицы считались защищенными, если они не умирали и не демонстрировали клинических признаков ND.
В течение первой недели в этой партии бройлеров было несколько ранних случаев смерти, вероятно, из-за колибактериоза. Доза тестируемых вакцин была ниже ожидаемой (2000 PFU). Результаты защиты приведены в таблице 12. Лучшие защиты были индуцированы вакцинацией vHVT309 и vHVT310, а затем vHVT306.
Таблица 12. Эффективность от ND, индуцированная различными двойными конструкциями HVT-IBD + ND у цыплят-бройлеров после заражения в D42 велогенным штаммом NDV

Пример 7. Эффективность от ND, индуцированная vHVT306, vHVT309 и vHVT310, против велогенного заражения NDV в D42 у бройлеров.
Цель исследования заключалась в оценке эффективности трех рекомбинантных конструкций HVT (vHVT306, vHVT309 и vHVT310), экспрессирующих ген IBDV VP2 и ген NDV F, введенных односуточным цыплятам-бройлерам, против велогенного заражения NDV, выполненного в D42.
Однодневные цыплята-бройлеры (линия Hubbard JA957) были распределены на 4 группы, как показано в таблице 13. Всех птиц из групп со 2 по 4 (16-20 птиц/группу) вакцинировали подкожным (SC) путем с использованием 0,2 мл различных конструкций HVT-IBD + ND при 2000 PFU. Девятнадцать птиц из группы 1 остались невакцинированными. Через сорок два дня после вакцинации (в D42), всех птиц заражали велогенным штаммом NDV Herts 33 внутримышечным (IM) путем (105,0 EID50 в 0,2 мл/особь). Всех птиц наблюдали на предмет клинических признаков в течение 14 дней после заражения. Птицы считались защищенными, если они не умирали и не демонстрировали клинических признаков ND.
Результаты защиты приведены в таблице 13. В целом, уровни защиты были выше, чем в предыдущем исследовании (см. Пример 6), но они следуют той же тенденции: лучшие защиты были вызваны вакцинацией vHVT309 и vHVT310, а затем vHVT306.
Результаты показали, что vHVT309 более эффективен, чем vHVT306, против заражения ND у SPF, а также у бройлеров (таблицы 12 и 13), что позволяет предположить, что вставка гетерологичных полинуклеотидов в один локус оказывает меньшее отрицательное влияние на общую приспособленность вируса, чем вставка в несколько локусов.
Таблица 13. Эффективность от ND, индуцированная различными двойными конструкциями HVT-IBD + ND, у цыплят-бройлеров после заражения в D42 велогенным штаммом NDV

Пример 8. Эффективность от IBD, индуцированная vHVT306, vHVT309 и vHVT310 против стандартного заражения IBDV в D14 у цыплят SPF.
Цель исследования заключалась в оценке эффективности трех рекомбинантных конструкций HVT (vHVT306, vHVT309 и vHVT310), экспрессирующих ген IBDV VP2 и ген NDV F, введенных односуточным цыплятам SPF, против стандартного заражения IBDV, выполненного в D14.
Цыплят односуточного возраста, не имеющих специфических патогенов (SPF), распределяли по 4 группам, как показано в таблице 14. Всех птиц из групп 1-3 (21-22 птицы/группу) вакцинировали подкожным (SC) путем с использованием 0,2 мл различных конструкций HVT-IBD + ND в указанной дозе. 22 птицы из группы 4 остались невакцинированными. Через четырнадцать дней после вакцинации (в D14) всех птиц заражали классическим штаммом STC вируса инфекционного бурсита (IBDV) внутриглазным (IO) путем (101,4 EID50 в 0,03 мл/птицу). Через четыре дня после заражения (в день D18) всех птиц умерщвляли и вскрывали для исследования грубых поражений фабрициевой сумки.
Результаты защиты приведены в таблице 14. 3 экспериментальные вакцины индуцировали схожие уровни защиты от IBD, в то время как все контрольные птицы, кроме одной, оказались инфицированными.
Таблица 14. Эффективность от IBD, индуцированная различными двойными конструкциями HVT-IBD + ND у цыплят SPF после заражения в D14 штаммом STC IBDV

Пример 9. Эффективность от IBD, индуцированная vHVT306, vHVT309 и vHVT310 у цыплят SPF после заражения вариантом IBD в D14.
Цель данного исследования заключалась в оценке эффективности трех рекомбинантных конструкций HVT (vHVT306, vHVT309 и vHVT310), экспрессирующих ген IBDV VP2 и ген NDV F, введенных односуточным цыплятам SPF, против вариантного заражения IBDV, проведенного в D14.
Цыплят односуточного возраста, не имеющих специфических патогенов (SPF), распределяли по 5 группам, как показано в таблице 15. Всех птиц из групп 1-3 (20 птиц/группу) вакцинировали подкожным (SC) путем с использованием 0,2 мл различных конструкций HVT-IBD + ND в указанной дозе. Птицы из группы 4 и группы 5 (19-20 птиц/группу) были оставлены невакцинированными. В D14 все птицы из групп 1-4 подвергались заражению штаммом варианта Delaware E вируса инфекционного бурсита (IBDV) внутриглазным (IO) путем (102,2 EID50 по 0,03 мл/птицу). Птиц из группы 5 не заражали. На D25 измеряли массу тела и вес фабрициевой сумки всех птиц. Соотношения масс Ф/Т (соотношение массы фабрициевой сумки к массе тела х100) рассчитывали для всех групп.
Результаты защиты приведены в таблице 15. Частичная защита была индуцирована в D14 3 вакцинами, причем защита была выше для vHVT309 и vHVT310.
Рекомбинантные vHVT306 и vHVT309 имеют две независимые экспрессирующие кассеты (две мРНК). Конструкции, экспрессирующие два гена через IRES или P2A (например, vHVT310, vHVT317, vHVT311, vHVT316, vHVT322), находятся не только в одном сайте вставки, но также гены экспрессируются из одной мРНК. Сравнение всех данных, представленных в таблицах 11-19, показывает, что рекомбинанты с одним сайтом вставки vHVT309 и vHVT310 более эффективны, чем рекомбинант с двумя сайтами вставки vHVT306, что указывает на то, что рекомбинанты HVT, несущие более одного гетерологичного полинуклеотида в одном локусе вставки, биологически более пригодны, чем рекомбинанты HVT, несущие гетерологичные полинуклеотиды в нескольких локусах вставки. Кроме того, неожиданно оказалось, что экспрессия более чем одного гетерологичного полинуклеотида из одной мРНК, экспрессируемой через IRES, оказывает меньшее отрицательное влияние на эффективность от IBD, особенно у бройлеров (см. результаты в таблице 11).
Таблица 15. Эффективность от IBD, индуцированная различными двойными конструкциями HVT-IBD + ND у цыплят SPF после заражения в D14 штаммом варианта E IBDV

Пример 10. Эффективность от IBD, индуцированная vHVT306, vHVT309 и vHVT310 против заражения стандартным IBDV в D28 у цыплят SPF.
Цель исследования заключалась в оценке эффективности трех рекомбинантных конструкций HVT (vHVT306, vHVT309 и vHVT310), экспрессирующих ген IBDV VP2 и ген NDV F, введенных односуточным цыплятам SPF, против заражения стандартным IBDV, выполненного в D28.
Цыплят односуточного возраста, не имеющих специфических патогенов (SPF), распределяли на 4 группы, как показано в таблице 16. Всех птиц от 1 до 3 групп (20-22 птицы/группу) вакцинировали подкожным (SC) введением 0,2 мл различных конструкций HVT-IBD + ND в указанной дозе. 22 птицы из группы 4 остались невакцинированными. Через двадцать восемь дней после вакцинации (в D28), всех птиц заражали классическим штаммом STC вируса инфекционного бурсита (IBDV) внутриглазным (IO) путем (102.0 EID50 в 0,03 мл/птиц). Через четыре дня после заражения (в день D32) всех птиц умерщвляли и вскрывали для исследования грубых поражений фабрициевой сумки.
Результаты защиты приведены в таблице 16. Полная защита была вызвана vHVT310, при этом только несколько птиц не были защищены другими вакцинами-кандидатами.
Таблица 16. Эффективность от IBD, индуцированная различными двойными конструкциями HVT-IBD + ND у цыплят SPF после заражения в D28 штаммом STC IBDV

Пример 11. Эффективность от IBD, индуцированная vHVT306, vHVT309 и vHVT310 у цыплят SPF после заражения вариантом IBD в D28.
Цель исследования заключалась в оценке эффективности трех рекомбинантных конструкций HVT (vHVT306, vHVT309 и vHVT310), экспрессирующих ген IBDV VP2 и ген NDV F, введенных односуточным цыплятам SPF, против заражения вариантом IBDV, проведенного в D28.
Односуточные цыплята, не имеющие специфических патогенов (SPF), были распределены на 5 групп, как показано в таблице 17. Всех птиц из групп 1-3 (20 птиц/группу) вакцинировали подкожным (SC) путем с использованием 0,2 мл различных конструкций HVT-IBD + ND в указанной дозе. Птицы из группы 4 и группы 5 (18-19 птиц/группу) были оставлены невакцинированными. В D28 все птицы из групп 1-4 подвергались заражению штаммом варианта Delaware E вируса инфекционного бурсита (IBDV) внутриглазным (IO) путем (102,2 EID50 в 0,03 мл/птицу). Птицы из группы 5 оставались незараженными. В D39 измеряли массу тела и массу фабрициевой сумки всех птиц. Соотношения масс Ф/Т (соотношение массы фабрициевой сумки к массе тела х100) рассчитывали для всех групп.
Результаты защиты приведены в таблице 17. Соотношение масс Ф/Т для группы 5 (незараженная группа) не могло быть получено, поскольку эта группа неожиданно была инфицирована штаммом STC IBDV. Защита, индуцированная vHVT310, была выше, чем защита, индуцированная vHVT306 и vHVT309.
Таблица 17. Эффективность от IBD, индуцированная различными двойными конструкциями HVT-IBD + ND у цыплят SPF после заражения в D28 штаммом варианта E IBDV

*Не сделано из-за воздействия на эту группу стандартного IBDV
Пример 12. Эффективность от ньюкаслской болезни (ND), индуцированная в D21 и D28, с помощью vHVT306, vHVT309 и vHVT310 у цыплят SPF.
Цель исследования заключалась в оценке эффективности трех рекомбинантных конструкций HVT (vHVT309, vHVT310 и vHVT311), экспрессирующих ген IBDV VP2 и ген NDV F, введенных односуточным цыплятам SPF, против заражения ньюкаслской болезнью (штамм Texas GB) в D21 и D28.
Цыплят односуточного возраста, не имеющих специфических патогенов (SPF), распределяли по 4 группам, как показано в таблице 18. Всех птиц из групп 1-3 (50 птиц/группу) вакцинировали подкожным (SC) путем с использованием 0,2 мл различных конструкций HVT-IBD + ND в указанной дозе. 30 птиц из группы 4 остались невакцинированными. Через двадцать один (D21) день после вакцинации 20 птиц из групп 1-3 и 15 птиц из группы 4 заражали штаммом NDV Texas GB внутримышечным (IM) путем (104,2 50% инфекционная доза в яйце (EID50) по 0,1 мл/птицу). Через двадцать восемь (D28) дней после вакцинации 30 птиц из групп 1-3 и 15 птиц из группы 4 заражали штаммом NDV Texas GB внутримышечным (IM) путем (104,3 50% инфекционная доза в яйце (EID50) по 0,1 мл/птицу). Птиц наблюдали на предмет клинических признаков в течение 14 дней после заражения. Птицы, у которых в течение 14 дней после заражения не было выявлено клинических признаков ND (включая признаки центральной нервной системы или респираторных заболеваний и/или смерть), считались защищенными.
Результаты защиты приведены в таблице 18. Все контрольные птицы группы 4 погибли после заражения. Защита, вызванная vHVT310 была наилучшей, за ней следовали vHVT306 и vHVT309.
Таблица 18. Эффективность от ND в D21 и D28, индуцированная различными двойными конструкциями HVT-IBD + ND у цыплят SPF

vHVT306*: использовался в качестве контроля
Пример 13. Эффективность от болезни Марека (MD), индуцированная vHVT306, vHVT309 и vHVT310 у цыплят SPF
Цель исследования заключалась в оценке эффективности трех рекомбинантных конструкций HVT (vHVT309, vHVT310 и vHVT311), экспрессирующих ген IBDV VP2 и ген NDV F, введенных односуточным цыплятам SPF, против болезней Марека (штамм GA, 2 партии и 2 разведения).
Односуточные цыплята, не имеющие специфических патогенов (SPF), были распределены на 4 группы, как показано в таблице 19. Все птицы из групп с 1 по 3 (20 птиц/группу) были вакцинированы подкожным (SC) путем 0,2 мл различных конструкций HVT-IBD + ND в указанной дозе. 20 птиц из группы 4 остались невакцинированными. Через четыре дня после вакцинации (D4) 18-20 птиц из групп 1-4 заражали SC-путем двумя разведениями (1: 5 и 1: 640) двух разных партий (# 1 и # 2) штамма vMDV GA22. Птиц наблюдали на предмет клинических признаков болезни Марека в течение 46-50 дней после вылупления. В D46-D50 все оставшиеся птицы были вскрыты и проверены на наличие поражений при болезни Марека. Птицы, у которых не было клинических признаков или повреждений MD, считались защищенными.
Результаты защиты приведены в таблице 19. Инфекция у контрольных птиц группы 4 варьировала в пределах 75-90%. В целом, защита, индуцированная vHVT310, была наилучшей, за ней следовали vHVT306, а затем vHVT309.
Таблица 19. Эффективность MD, индуцированная различными двойными конструкциями HVT-IBD + ND у цыплят SPF против заражения 2 разными партиями GA22, разведенного 1: 5 или 1: 640
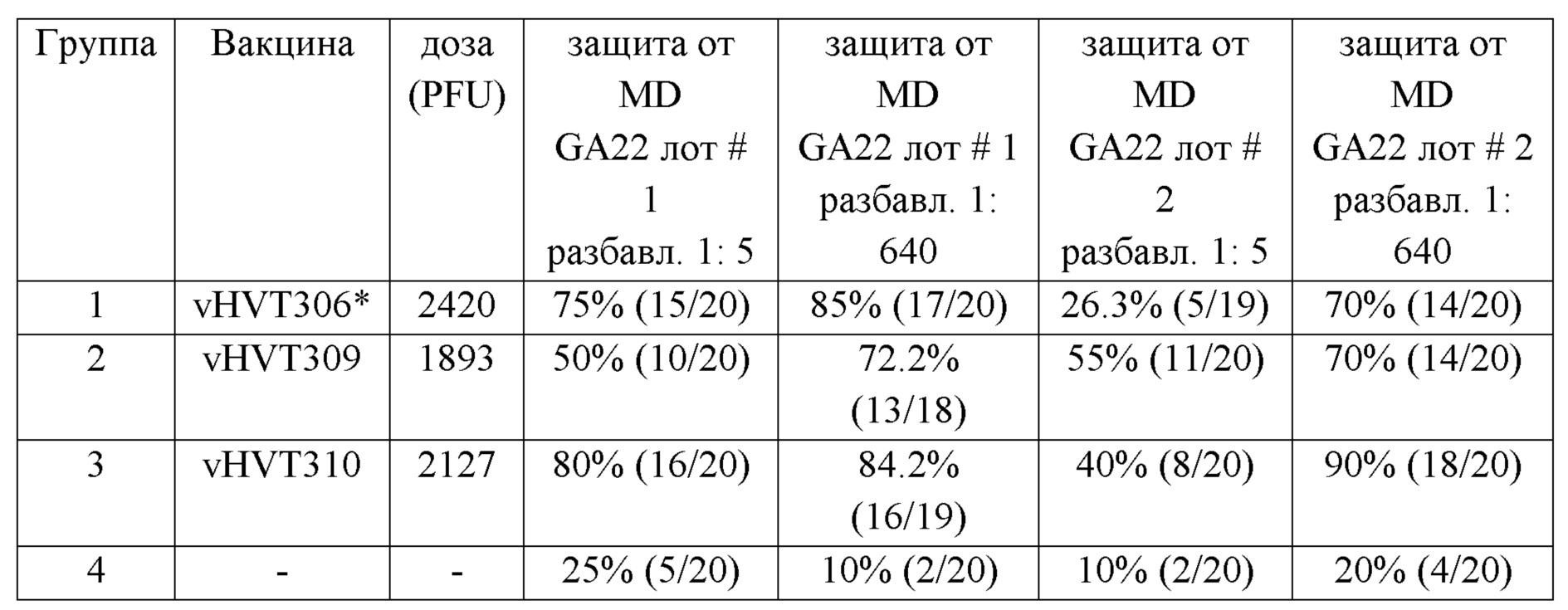
vHVT306*: использовался в качестве контроля
Пример 14. Эффективность от IBD, индуцированная vHVT306 и vHVT407 против заражения классическим IBDV в D21 у цыплят SPF.
Цель данного исследования заключалась в оценке эффективности двух рекомбинантных конструкций HVT, одна из которых, (vHVT306), экспрессирует ген IBDV VP2 и ген NDV F, а другая (vHVT407), экспрессирует ген IBDV VP2 и ген ILD gD, вводимых односуточным цыплятам SPF против заражения классическим IBDV, проведенным в D21.
Сорок односуточных цыплят SPF (белый леггорн) распределяли на 3 группы, как показано в таблице 20. Всех птиц из групп 2 и 3 (около 15 птиц/группу) вакцинировали подкожным (SC) путем 0,2 мл конструкции vHVT306 или vHVT407 в указанной дозе. Десять птиц из группы 1 остались невакцинированными. Через двадцать один день после вакцинации (в D21), всех птиц заражали классическим штаммом 52/70 Faragher IBDV внутриглазным (IO) путем (102,0 EID50 в 0,05 мл/птицу). Через одиннадцать дней после заражения (в день D32) всех птиц умерщвляли и вскрывали для выявления грубых поражений фабрициевой сумки. Фабрициева сумка и тело были взвешены для расчета соотношения массы фабрициевой сумки к массе тела. Птицы считались защищенными, если у них не было клинических признаков или поражения фабрициевой сумки после заражения.
Результаты защиты приведены в таблице 20. Вакцинация vHVT306 или vHVT407 вызывала полную защиту от IBD.
Таблица 20. Эффективность от IBD, индуцированная двумя конструкциями HVT, экспрессирующими два гена, у цыплят SPF после заражения в D21 штаммом Faragher IBDV

vHVT306*: использовался в качестве контроля. **среднее ± стандартное отклонение
Пример 15. Эффективность от ILT, индуцированная vHVT407 против заражения ILTV в D21 у цыплят SPF
Цель данного исследования заключалась в оценке эффективности двух рекомбинантных конструкций vHVT407, экспрессирующих ген IBDV VP2 и ген ILTV gD, введенных односуточным цыплятам SPF против заражения ILTV, проведенного в D21.
Двадцать четыре односуточных цыпленка SPF (белый леггорн) были распределены на 2 группы, как показано в таблице 21. Всех птиц (около 12 птиц на группу) вакцинировали подкожным (SC) путем с использованием 0,2 мл vHVT13 (используемого в качестве отрицательного контроля) или конструкции vHVT407 в указанной дозе. Через двадцать один день после вакцинации (в день D21) всех птиц заражали штаммом ILTV ILT-96-3 интратрахеальным (IT) путем (103,6 EID50 в 0,5 мл/птицу). Птиц наблюдали на предмет клинических признаков в течение 11 дней после заражения. В дни исследования 25-29 и 32 у всех птиц наблюдали на предмет клинических признаков, включая характер дыхания, конъюнктивит, депрессию и смертность. В день изучения 32 все оставшиеся птицы были умерщвлены. Птицы считались защищенными, если у них не было клинических признаков ILT, таких как респираторный дистресс, связанный с кашлем, чиханием, хрипами, депрессией, удушьем и/или кровянистыми слизистыми эксудатами, включая смертность.
Результаты защиты приведены в таблице 21. Значительная защита ILT была вызвана вакцинацией vHVT407 в этих условиях заражения.
Таблица 21. Эффективность от ILT, индуцированная конструкцией vHVT407 у цыплят SPF после заражения в D21 штаммом ILTV ILT-96-3

vHVT13*: использовали в качестве отрицательного контроля.
Пример 16. Эффективность от ILT, индуцированная vHVT407, коммерческой вакциной HVT-ILT и коммерческой вакциной из эмбрионов кур (CEO) против заражения ILTV в D21 у цыплят-бройлеров.
Цель исследования заключалась в оценке эффективности рекомбинантной конструкции vHVT407, экспрессирующей ген IBDV VP2 и ген ILTV gD, вводимой односуточным цыплятам-бройлерам, по сравнению с коммерческой вакциной HVT-ILT (INNOVAX® ILT) против заражения ILTV, проведенного в D21.
Сорок восемь односуточных коммерческих цыплят-бройлеров распределяли на 3 группы, как показано в таблице 22. Всех птиц (около 12 птиц/группу) 1-3 групп вакцинировали подкожным (SC) путем по 0,2 мл vHVT13 (используемого в качестве отрицательного контроля), vHVT407 или INNOVAX® ILT (используемого в качестве положительного контроля) в указанной дозе. Через двадцать один день после вакцинации (в день D21) всех птиц заражали штаммом ILTV ILT-96-3 интратрахеальным (IT) путем (104,2 EID50 в 0,5 мл/птицу). Птиц наблюдали на предмет клинических признаков в течение 1 2 дней после заражения. В дни исследования 25-29 и 32-33 все птицы наблюдались и оценивались по клиническим признакам, включая характер дыхания, конъюнктивит, депрессию и смертность. В день исследования 34 все оставшиеся птицы были умерщвлены. Птицы считались защищенными, если у них не было клинических признаков ILT, таких как респираторный дистресс, связанный с кашлем, чиханием, хрипами, депрессией, удушьем и/или кровянистыми слизистыми эксудатами, включая смертность.
Результаты защиты приведены в таблице 22. Защита от ILT была вызвана вакцинацией vHVT407, которая была выше, чем индуцированная INNOVAX ILT.
Таблица 22. Эффективность от ILT, индуцированная конструкциями vHVT407 и INNOVAX ILT у цыплят-бройлеров после заражения в D21 штаммом ILTV ILT-96-3

vHVT13*: использовали в качестве отрицательного контроля. **INNOVAX® ILT использовали в качестве положительного контроля
Пример 17. Эффективность от ньюкаслской болезни (ND), индуцированная в D14, D21 и D32 с помощью vHVT310 и vHVT316 у цыплят SPF
Цель исследования заключалась в сравнении возникновения иммунитета к ND для двух рекомбинантных конструкций HVT (vHVT310 и vHVT316), экспрессирующих ген IBDV VP2 и ген NDV F, вводимых односуточным цыплятам SPF против заражения ньюкаслской болезнью (штамм Texas GB), проведенного в D14, D21 и D32.
Односуточных цыплят, не имеющих специфических патогенов (SPF), распределяли по 3 группам, как показано в таблице 23. Всех птиц с 1 по 2 группу (59-70 птиц/группу; см. таблицу) вакцинировали подкожным (SC) путем с использованием 0,2 мл различных конструкций HVT-IBD + ND в указанной дозе. 45 птиц из группы 3 были оставлены невакцинированными. В D14, D21 и D32, 15-30 птиц (см. таблицу) из групп 1-3 заражали штаммом NDV Texas GB внутримышечным (IM) путем (104,0 EID50/особь) по 0,1 мл/особь). Птиц наблюдали на предмет клинических признаков в течение 14 дней после заражения. Птицы, у которых в течение 14 дней после заражения не было выявлено клинических признаков ND (включая признаки центральной нервной системы или респираторных заболеваний и/или смерти), считались защищенными.
Результаты защиты приведены в таблице 23. Все контрольные птицы группы 3 погибли после заражения. Уровни защиты, индуцированные как vHVT310, так и vHVT316, были сходными, с возможным более ранним началом иммунитета, индуцированного vHVT316.
Таблица 23. Эффективность от ND при D14, D21 и D32, индуцированных различными двойными конструкциями HVT-IBD + ND у цыплят SPF

Пример 18. Эффективность от ILTV, индуцированная векторами HVT, экспрессирующими ILTV gD и IBDV VP2 или экспрессирующими ILTV gD и NDV F
Цель данного исследования заключалась в оценке эффективности рекомбинантных конструкций HVT, экспрессирующих гены ILTV gD и IBDV VP2 (таких как vHVT317 и vHVT407) или экспрессирующих гены ILTV gD и NDV F (таких как vHVT308 и vHVT322), вводимых цыплятам против заражения ILTV.
Цыплят распределяли по разным группам. Птиц вакцинировали подкожным (SC) путем по 0,2 мл различных конструкций HVT. Птицы из одной группы оставались невакцинированными. Птиц заражали ILTV внутритрахеальным (IT) путем или через инфраорбитальный синус. Птиц наблюдали по клиническим признакам в течение 11-14 дней после заражения. Птицы, у которых отсутствовали какие-либо клинические признаки ILTV (включая респираторный дистресс, связанный с кашлем, чихание, хрипы, депрессия, удушье и/или кровянистые слизистые эксудаты и/или смерть) в течение 14 дней после заражения, считались защищенными.
Результаты показывают, что HVT-векторы обеспечивают защиту от инфекции ILTV.
Пример 19. Эффективность от IBD, индуцированная векторами HVT, экспрессирующими ILTV gD и IBDV VP2
Цель данного исследования заключалась в оценке эффективности рекомбинантных конструкций HVT, экспрессирующих ILTV gD и IBDV VP2 (таких как vHVT317 и vHVT407), вводимых цыплятам против заражения IBD.
Цыплята распределяли по разным группам. Птиц вакцинировали подкожным (SC) путем с использованием 0,2 мл различных конструкций HVT. Птицы из одной группы оставляли невакцинированными. Птиц заражали IBD внутриглазным (IO) путем. Птиц наблюдали на предмет клинических признаков в течение 4-10 дней после заражения. Птицы, у которых отсутствовали клинические признаки IBD (включая депрессию и/или смерть) и которые не демонстрировали поражений фабрициевой сумки и/или атрофии в течение 10 дней после заражения, считались защищенными.
Результаты показывают, что HVT-векторы обеспечивают защиту от инфекции IBD.
Пример 20. Эффективность от NDV, индуцируемая векторами HVT, экспрессирующими ILTV gD и NDV F
Цель исследования заключалась в оценке эффективности рекомбинантных конструкций HVT, экспрессирующих гены ILD gD и NDV F (таких как vHVT308 и vHVT322), вводимых цыплятам против заражения NDV.
Цыплята распределяли по разным группам. Птиц вакцинировали подкожным (SC) путем по 0,2 мл различных конструкций HVT. Птицы из одной группы остались невакцинированными. Птиц заражали NDV внутримышечно (IM). Птицы наблюдали на предмет клинических признаков в течение 14 дней после заражения. Птицы, которые не демонстрировали клинических признаков ND (включая признаки центральной нервной системы или дыхания и/или смерть) в течение 14 дней после заражения, считались защищенными.
Результаты показывают, что HVT-векторы обеспечивают защиту от инфекции NDV.
Пример 21. Эффективность от IBD, индуцированная vHVT316 и vHVT317 против заражения стандартным IBDV в D28 у цыплят SPF.
Цель исследования заключалась в оценке эффективности двух рекомбинантных конструкций HVT (vHVT316 и vHVT317), экспрессирующих либо ген VPD IBDV и ген NDV F (vHVT316), либо ген IBDV VP2 и ген ILTV gD (vHVT317), введенных односуточным цыплятам SPF, против заражения стандартным BDV, проведенного в D28.
Односуточные цыплята, не имеющие специфических патогенов (SPF), были распределены на 3 группы, как показано в Таблице 24. Всех птиц из групп 1 и 2 (15 птиц/группу) вакцинировали подкожным (SC) путем введения 0,2 мл vHVT316 и vHVT317 в указанной дозе. 15 птиц из группы 3 оставили невакцинированными. Через двадцать восемь дней после вакцинации (в D28), всех птиц заражали классическим штаммом STC вируса инфекционного бурсита (IBDV) внутриглазным (IO) путем (102,0 EID50 в 0,03 мл/птиц). Через четыре дня после заражения (в день D32) все птицы были убиты и вскрыты для исследования грубых поражений фабрициевой сумки.
Результаты защиты показаны в таблице 24. vHVT316 и vHVT317 вызвали 100% и 80% защиту, соответственно; однако доза, вводимая vHVT317, была почти в 3 раза ниже, чем доза vHVT316.
Таблица 24. Эффективность от IBD, индуцированная двойными конструкциями HVT-IBD + ND (vHVT316) и HVT-IBD + ILT (vHVT317) у цыплят SPF после заражения в D28 штаммом STC IBDV

Пример 22. Эффективность от IBD, индуцированная vHVT310, vHVT316 и vHVT317 у цыплят SPF после заражения вариантом IBD в D28.
Цель исследования заключалась в оценке эффективности трех рекомбинантных конструкций HVT (vHVT310, vHVT316 и vHVT317), экспрессирующих либо ген VPD IBDV и ген NDV F (ген vHVT310 & vHVT316), либо ген IBDV VP2 и ген ILTV gD (vHVT317), вводимых односуточным цыплятам SPF против заражения вариантом IBDV, выполненного в D28.
Односуточные цыплята, не имеющие специфических патогенов (SPF), были распределены на 5 групп, как показано в таблице 25. Всех птиц из групп 1-3 (15 птиц/группу) вакцинировали введением подкожным (SC) путем 0,2 мл vHVT310, vHVT316 и vHVT317 в указанной дозе. Птицы из группы 4 и группы 5 (15 птиц/группу) были оставлены невакцинированными. В D28 всех птиц из групп 1-4 заражали штаммом варианта Delaware E вируса инфекционного бурсита IBDV, внутриглазным (IO) путем (103,0 EID50 по 0,03 мл/особь). Птицы из группы 5 оставались незараженными. В D39 измеряли массу тела и массу фабрициевой сумки всех птиц. Соотношения масс Ф/Т (отношение массы фабрициевой сумки к массе тела х100) рассчитывали для всех групп.
Результаты защиты приведены в таблице 25. Защита наблюдалась во всех вакцинированных группах. Защита с помощью vHVT317 была несколько выше, чем защита, вызванная vHVT310 и vHVT316, несмотря на его более низкую дозу.
Таблица 25. Эффективность от IBD, индуцированная различными конструкциями HVT с двойными вставками у цыплят SPF после заражения в D28 штаммом варианта E IBDV

Пример 23. Эффективность от ILT, индуцированная vHVT317 у цыплят SPF против заражения ILTV в D28
Цель данного исследования заключалась в оценке эффективности рекомбинантной конструкции vHVT317, экспрессирующей ген IBDV VP2 и ген ILTV gD, введенных односуточным цыплятам SPF против заражения ILTV, проведенного в D28.
Тридцать шесть односуточных цыплят SPF (белый леггорн) были распределены на 2 группы, как показано в таблице 26. Всех птиц (около 18 птиц/группу) либо вакцинировали подкожным (SC) путем введения 0,2 мл vHVT317, либо оставляли невакцинированным. Двадцать восемь дней после вакцинации (в D28), всех птиц заражали штаммом ILT-96-3 ILTV интратрахеальным (IT) путем (103,0 EID50 по 0,2 мл/особь). Птиц наблюдали на предмет клинических признаков и смертности в D32, D36 и D39. Клинические признаки включали дыхание, конъюнктивит, депрессию и смертность. В день изучения 32 все оставшиеся птицы были умерщвлены. Оценку защиты осуществляли по 3 различным критериям: (1) любая птица, проявляющая какие-либо клинические признаки в течение трех последовательных дней или погибшая после заражения, считается ilt-положительной; (2) любая птица, имеющая какие-либо умеренные или тяжелые клинические признаки в любой категории за любой день или умершая после заражения, считается ilt-положительной; и (3) любая птица, проявляющая какие-либо умеренные или тяжелые клинические признаки в любой категории в течение двух дней подряд или погибшая после заражения, считается ILT-положительной.
Результаты защиты, основанные на 3 различных критериях, показаны в Таблице 26. Заражение ILT было тяжелым, так как оно убило (или птицы были подвергнуты эвтаназии по этическим причинам, когда они демонстрировали очень тяжелые клинические признаки) 86,7% невакцинированных птиц. В этих условиях заражения высокие уровни защиты от ILT были вызваны вакцинацией vHVT317.
Таблица 26. Эффективность от ILT, индуцированная конструкцией vHVT317 у цыплят SPF после заражения в D28 штаммом ILTV ILT-96-3

Пример 24. Эффективность от ILT, индуцированная vHVT317, коммерческой HVT-ILT и коммерческой вакциной из куриных эмбрионов (CEO) против заражения ILTV в D21 у цыплят-бройлеров.
Цель исследования заключалась в оценке эффективности рекомбинантной конструкции vHVT317, экспрессирующей ген IBDV, VP2 и ген ILTV gD, вводимой односуточным цыплятам - бройлерам по сравнению с коммерческой вакциной HVT-ILT (INNOVAX® ILT, Merck Animal Health) против заражения ILTV, проведенного в D28.
Пятьдесят один коммерческий цыпленок-бройлер односуточного возраста были распределены на 3 группы, как показано в таблице 27. Всех птиц (17 птиц/группу) вакцинировали либо подкожным (SC) путем по 0,2 мл vHVT317, либо INNOVAX® ILT (использовали в качестве положительного контроля) в указанной дозе, либо оставляли невакцинированными. В день D26 количество птиц в группе было уменьшено до 15, и каждая птица была взвешена. Через двадцать восемь дней после вакцинации (в день 28) всех птиц заражали штаммом 63140 ILTV инфраорбитальным путем (104,3 EID50 в 0,2 мл на птицу). В дни исследования 31-35 и день исследования 38 всех птиц индивидуально осматривали на предмет наличия клинических признаков. В день исследования 38 исследования все оставшиеся птицы были взвешены по отдельности и убиты. Оценку защиты осуществляли про 3 различным критериям: (1) любая птица, проявляющая какие-либо клинические признаки в течение трех последовательных дней или погибшая после заражения, считается ilt-положительной; (2) любая птица, имеющая какие-либо умеренные или тяжелые клинические признаки в любой категории за любой день или умершая после заражения, считается ilt-положительной; и (3) любая птица, проявляющая какие-либо умеренные или тяжелые клинические признаки в любой категории в течение двух дней подряд или погибшая после заражения, считается ILT-положительной. Массу тела также сравнивали в D26 и D38.
Результаты защиты с использованием 3 критериев приведены в таблице 27. Все контроли были сочтены незащищенными по 3 критериям. Обе испытанные вакцины вызывали высокую и сходную защиту от ILT. Не было значительной разницы между массой тела в D26 (до заражения); однако после заражения масса тела вакцинированных птиц была значительно (р <0,0001) выше, чем у невакцинированных птиц, что указывает на защиту от потери массы.
Таблица 27. Эффективность от ILT, индуцированная конструкциями vHVT317 и INNOVAX ILT (положительный контроль) у цыплят-бройлеров после заражения в D28 штаммом 63140 ILTV

*среднее ± стандартное отклонение в г
Пример 25. Эффективность от IBD, индуцированной введением in ovo vHVT317, после заражения вариантом IBD цыплят SPF в возрасте 28 дней.
Цель исследования заключалась в оценке эффективности vHVT317, экспрессирующего ген IBDV VP2 и ген ILD gTV, вводимых in ovo 18-19 дневным эмбрионам кур SPF против заражения вариантом IBDV, проведенного в возрасте 28 дней (31 день после вакцинации).
18-19 дневные эмбрионы кур, не имеющих патогенов (SPF), были распределены на 3 группы, как показано в Таблице 28. Все птицы из групп 1 и 2 (около 30 яиц на группу) были вакцинированы in ovo (SC) путем по 0,05 мл vHVT317 в указанной дозе. Яйца с эмбрионами из группы 3 иммунизировали 0,05 мл разбавителя вакцины Марека. При вылуплении оставляли 22 птенца на группу, и до заражения все 3 группы были сокращены до 20 птиц. Через тридцать один день после вакцинации (в возрасте 28 дней) птиц из всех трех групп заражали штаммом варианта Delaware E вируса инфекционного бурсита (IBDV) интраокулярным (IO) путем (целевая доза 103,0 EID50 в 0,03 мл/птицу). Птицы из группы 5 подвергались фиктивному заражению TPB (триптозофосфатный бульон, 0,03 мл/птицу). Через одиннадцать дней после заражения измеряли массу тела и массу фабрициевой сумки всех птиц. Соотношения масс Ф/Т (соотношение массы фабрициевой сумки к массе тела х100) рассчитывали для всех групп.
Результаты защиты приведены в таблице 28. Четкая атрофия фабрициевой сумки наблюдалась у всех невакцинированных зараженных птиц. Защита наблюдалась в группах, вакцинированных vHVT317 2 тестируемыми дозами.
Таблица 28. Эффективность от IBD, индуцированная vHVT317, вводимой in ovo после заражения цыплят SPF в возрасте 28 дней с использованием штамма варианта Е IBDV

Пример 26. Эффективность от ILT, индуцированная vHVT317, введенной in ovo цыплятам SPF, против заражения ILTV в D28.
Цель исследования заключалась в оценке эффективности рекомбинантной конструкции vHVT317, экспрессирующей ген IBDV VP2 и ген ILTV gD, вводимой in ovo 18-19 дневным эмбрионам кур SPF, против заражения ILTV, проведенного в D28 (в 25 дневном возрасте).
18-19 дневные эмбрионы кур, не имеющих патогенов (SPF), были распределены на 2 группы, как показано в таблице 29. Всех птиц из группы 1 (около 30 яиц на группу) вакцинировали in ovo (SC) 0,05 мл vHVT317 в указанной дозе. Яйца с эмбрионами из группы 2 иммунизировали 0,05 мл разбавителя вакцины Марека. При вылупливании оставляли 22 птенца на группу, и за день до заражения обе группы сокращали до 20 птиц. Через двадцать пять дней после вакцинации (в день 28) птиц обеих групп заражали штаммом 63140 ILTV, введенным в инфраорбитальный синус (104,7 EID50 в 0,2 мл на птицу). Птиц наблюдали на предмет клинических признаков и смертности с D28 по D38. В день исследования 38 все оставшиеся птицы были умерщвлены. Эффективность в отношении заражения ILT определяли по отсутствию типичных клинических признаков ILT, таких как: депрессия, респираторный дистресс, связанный с кашлем, чиханием, хрипами, затрудненное дыхание с расширенной шеей, с кровянистыми и/или слизистыми выделениями или без них; одышка и дыхание через рот; отек подглазничных пазух, с дренажем или без него; и/или опухшая конъюнктива с частичным или полным закрытием глаз. Любая смертность после заражения, за исключением травмы или какого-либо явного состояния, исключающего птицу из исследования, считалась ILT-положительной.
Результаты защиты от ILT приведены в таблице 29. Результаты показали, что большинство вакцинированных птиц vHVT317 были защищены.
Таблица 29. Эффективность от ILT, индуцированная vHVT317, вводимой in ovo цыплятам SPF после инфраорбитального заражения в D28 штаммом 63140 ILTV

Пример 27. Эффективность от ND, индуцированная vHVT309 и vHVT310 у цыплят SPF против велогенного заражения NDV в D21
Цель исследования заключалась в оценке эффективности двух рекомбинантных конструкций HVT (vHVT309 и vHVT310), экспрессирующих ген IBDV VP2 и ген NDV F, введенных односуточным цыплятам SPF, против велогенного заражения NDV, проведенного в D21.
Односуточных цыплят SPF (белый леггорн) распределяли по 3 группам птиц, как показано в таблице 30. Всех птиц из групп 2 и 3 вакцинировали подкожным (SC) путем по 0,2 мл vHVT309 (14 птиц; группа 2) или vHVT310 (15 птиц; группа 3) в целевой дозе 2000 PFU. Птицы из группы 1 (5 птиц) были оставлены невакцинированными. Две птицы группы 2 погибли в D5 по неизвестной причине. Через двадцать один день после вакцинации (в день 21) кровь 10 птиц из группы 3 собирали для серологии; затем всех птиц из всех 3 групп заражали велогенным штаммом NDV Herts 33 внутримышечным (IM) путем (105,0 EID50 по 0,2 мл на птицу). Всех птиц наблюдали на предмет клинических признаков в течение 14 дней после заражения. Птицы считались защищенными, если они не умирали и не демонстрировали клинических признаков ND.
Результаты защиты приведены в таблице 30. Все невакцинированные птицы группы 1 умерли после заражения; все вакцинированные птицы были защищены. Конструкция vHVT310 индуцировала значимое анти-NDV (среднее значение 3,7 ± 0,3 (стандартное отклонение) log10 по ELISA (набор для непрямого скрининга «ID Screen Newcastle Disease Indirect» от ID-VET) и среднее значение 3,9 ± 0,7 log2 с помощью теста HI) и анти-IBDV (среднее значение 3,7 log10 ± 0,2 log10 по ELISA (набор ProFLOK IBD Plus ELISA от Zoetis) во всех 10 сыворотках G3-птиц, отобранных в D21.
Таблица 30. Эффективность от ND, индуцированная различными двойными конструкциями HVT-IBD + ND, у цыплят-бройлеров после заражения в D21 велогенным штаммом NDV

Пример 28. Эффективность от IBD, индуцированная vHVT309 и vHVT310 у цыплят SPF против заражения классическим IBDV в D21
Цель исследования заключалась в оценке эффективности двух рекомбинантных конструкций HVT (vHVT309 и vHVT310), экспрессирующих ген IBDV VP2 и ген NDV F, введенных односуточным цыплятам SPF, против заражения классическим IBDV, проведенного в D21.
Односуточные цыплята SPF (белый леггорн) были распределены на 3 группы, как показано в таблице 31. Все птицы из групп 2 и 3 (около 15 птиц/группу) были вакцинированы подкожным (SC) введением по 0,2 мл vHVT30 9 (группа 2) или vHVT310 (группа 3) в целевой дозе 2000 PFU. Десять птиц из группы 1 остались невакцинированными. В группе 2 были зарегистрированы две неспецифические ранние смерти. Через двадцать один день после вакцинации (в D21), всех птиц заражали классическим штаммом 52/70 Faragher IBDV внутриглазным (IO) путем (102,0 EID50 в 0,05 мл/птицу). Через одиннадцать дней после заражения (в день D32) всех птиц умерщвляли и вскрывали для выявления грубых поражений фабрициевой сумки. Фабрициеву сумку и тело взвешивали для расчета соотношения фабрициевой сумки к массе тела. Фабрициеву сумку затем хранили в формальдегиде для гистологии. Гистологические поражения фабрициевой сумки оценивали по шкале, представленной в таблице 32. Тяжесть заражения была подтверждена, если (1), по меньшей мере, 50% контролей заражения умерли или имели характерные признаки заболевания, особенно апатию/взъерошенные перья в течение более 2 дней или прострацию, и (2) 100% выживших контролей заражения показали гистологические баллы фабрициевой сумки ≥3. Эффективность вакцин-кандидатов была продемонстрирована, если, по меньшей мере, 90% цыплят были защищены. Цыплят считали защищенными, если (1) они выжили и не продемонстрировали заметных клинических признаков заболевания, особенно отсутствие апатии/взъерошенных перьев в течение более 2 дней или отсутствие прострации, и (2) они показали гистологический балл фабрициевой сумки <3.
Результаты защиты приведены в таблице 31. Все контроли были положительными на инфекцию IBD. Полная защита IBD была вызвана вакцинацией vHVT309 или vHVT310.
Таблица 31. Эффективность от IBD, индуцированная двумя конструкциями HVT, экспрессирующими два гена, у цыплят SPF после заражения в D21 штаммом Faragher IBDV

** среднее ± стандартное отклонение
Таблица 32. Балльная шкала гистологических поражений фабрициевой сумки*

Пример 29. Влияние введения vHVT317 in ovo на выводимость цыплят SPF.
Цель исследования заключалась в оценке безопасности vHVT317 в отношении выводимости при введении in ovo.
Результаты представляют собой обобщение данных нескольких исследований, в том числе описанных в Примерах 23, 24, 25 и 26. Яйца с эмбрионом на 18-19 день инкубации инокулировали либо vHVT317 в целевой дозе 2000 или 3000 PFU, либо разбавителем вакцины от болезни Марека. Процент выводимости оценивали для каждой группы. Результаты суммированы в таблице 33 и показали отличные уровни выводимости в вакцинированных яйцах.
Таблица 33. Выводимость после введения in ovo vHVT317

Пример 30. Эффективность от ILT, индуцированная vHVT406, против заражения ILTV.
Пример 30.1. Эффективность от ILT, индуцированная vHVT406, против заражения ILTV в D28
Цель исследования заключалась в оценке и сравнении эффективности рекомбинантной конструкции vHVT406, экспрессирующей ген ILTV gD, и коммерческой векторной вакцины HVT-ILT против заражения ILT.
Двенадцать (12) односуточных цыплят SPF распределяли в каждую группу. Птиц в группах 1-2 вакцинировали SQ по 0,2 мл на птицу. После вакцинации все птицы были помещены в их соответствующие секции. На 28-й день всех птиц заражали внутритрахеальным (IT) путем с помощью вируса инфекционного ларинготрахеита (ILT), ILT-93-3 EP2. Всех птиц наблюдали в течение 11 дней после заражения на предмет клинических признаков заражения. На 32 день у всех оставшихся птиц были взяты мазки трахеи и конъюнктивы. Мазки были обработаны для анализа q-PCR. На 39 день все оставшиеся птицы были умерщвлены.
Результаты показаны в таблице 34 ниже. Результаты показали, что все вакцинированные птицы vHVT406 были защищены. Удивительно, но результаты также показали, что хорошая защита (100% защита) была достигнута в группе vHVT406, когда использовалась более низкая доза (6 960 PFU/0,2 мл) по сравнению с более высокой дозой (10 340 PFU/0,2 мл), используемой для коммерческого продукта Innovax HVT-ILT.
Таблица 34. Количество птиц, положительных по ILT, и процент положительных по группам1

1 Птицы считались положительными, если они демонстрировали клинические признаки в течение трех последовательных дней, включая смертность или смертность после мазка.
2 Бляшкообразующие единицы (pfu) - подкожное введение (SQ); 0,20 мл на дозу.
3 Одна птица в группе vHVT406 была исключена из исследования из-за паралича.
4 Коммерческий продукт MSD Animal Health
Пример 30.2. Эффективность от ILT, индуцированная vHVT406 против заражения ILTV в D21
Цель исследования заключалась в оценке и сравнении эффективности vHVT406 и двух коммерческих вакцин против HVT-ILT против заражения ILT.
Двенадцать (12) односуточных птиц SPF были отнесены к каждой группе. В рандомизации также были определены изолирующие секции, в которые были помещены птицы (12 птиц на секцию, одна секция на группу). Птиц в группах 1-3 вакцинировали SQ по 0,2 мл на птицу. На 21-й день всех птиц в группах 1-2 заражали интратрахеальным (IT) вирусом инфекционного ларинготрахеита (ILT), ILT-96-3 EP2. Птиц наблюдали в течение 11 дней после заражения на предмет клинических признаков заражения. На 25 день у всех оставшихся птиц были собраны мазки из трахеи и конъюнктивы. Образцы мазка были обработаны для q-PCR. На 32 день все оставшиеся птицы были умерщвлены.
Результаты показаны в таблице 35 ниже. Результаты показали, что все, кроме одной вакцинированной птицы vHVT406, были защищены. Удивительно, но результаты также показали, что хорошая защита (91,7% защита) была достигнута в группе vHVT406, когда использовалась более низкая доза (810 PFU/0,2 мл) по сравнению с более высокой дозой (1590 PFU/0,2 мл), используемой для коммерческого продукта Innovax HVT- ILT для достижения того же уровня защиты (91,7%). Кроме того, vHVT406 обеспечивает лучшую защиту (91,7%) при использовании в более низкой дозе, чем коммерческий продукт Vectormune HVT-ILT, который обеспечивает защиту только на 75%.
Таблица 35. Количество птиц, положительных по ILT, и процент положительных по группе1
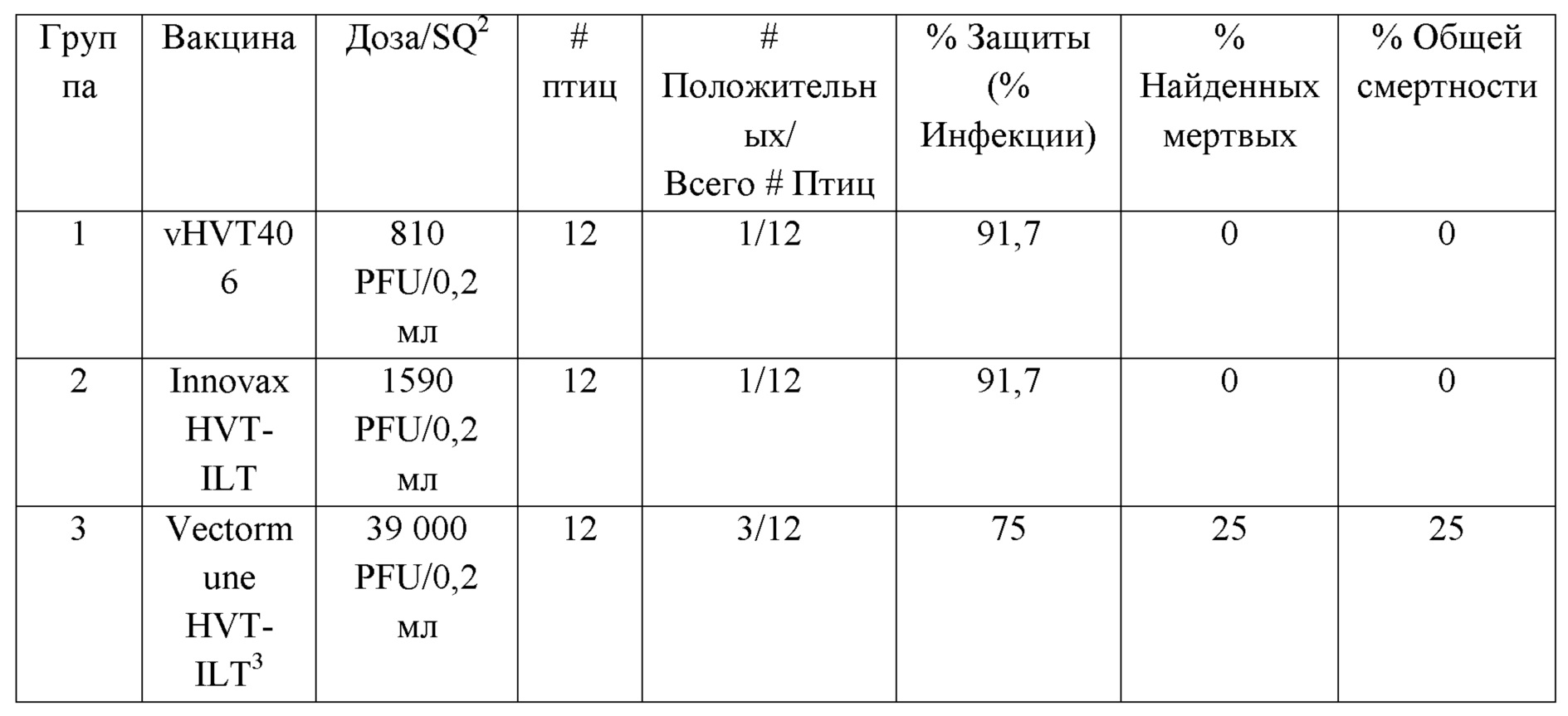
1 Птицы считались положительными, если они демонстрировали клинические признаки в течение трех последовательных дней, включая смертность или смертность после мазка.
2 бляшкообразующие единицы (pfu) - подкожное введение (SQ); 0,20 мл на дозу.
3 Коммерческий продукт Ceva
***
Опираясь, таким образом, на подробные воплощения настоящего раскрытия, следует понимать, что раскрытие, определенное вышеприведенными примерами, не должно ограничиваться конкретными деталями, изложенными в вышеприведенном описании, поскольку многие его возможные варианты возможны без отступления от духа или объема настоящего раскрытия.
Все процитированные или упоминаемые в данном документе документы («документы, процитированные в данном документе») и все документы, цитируемые или упоминаемые в процитированных документах, вместе с инструкциями изготовителя, описаниями, спецификациями продукта и технологическими картами для любых продуктов, упомянутых в данном документе или в любом документе, включенном ссылкой в данный документ, включены в данный документ ссылкой и могут быть использованы при практическом применении данного изобретения.
Реферат
Настоящее изобретение относится к биотехнологии, а именно к вакцине для иммунизации в целях защиты от птичьих патогенов. Вакцина содержит один или несколько рекомбинантных векторов вируса герпеса индеек (HVT). Вектор содержит по меньшей мере два гетерологичных полинуклеотида. Каждый из полинуклеотидов кодирует и экспрессирует антиген птичьего патогенна. Гетерологичные полинуклеотиды связаны IRES или P2A. Рекомбинантный HVT-вектор для экспрессии антигенов патогенов птиц содержит по меньшей мере два гетерологичных полинуклеотида, каждый из которых кодирует и экспрессирует антиген птичьего патогена. Два гетерологичных полинуклеотида кодируют два антигена птичьего патогена. Первый гетерологичный полинуклеотид содержит полинуклеотид, кодирующий полипептид, выбранный из группы, состоящей из антигена VP2 вируса инфекционного бурсита (IBDV), антигена гликопротеина D вируса инфекционного ларинготрахеита (ILTV) и антигена F вируса ньюкаслской болезни (NDV-F). Второй гетерологичный полинуклеотид содержит полинуклеотид, кодирующий полипептид, выбранный из группы, состоящей из антигена VP2 вируса инфекционного бурсита (IBDV), антигена гликопротеина D вируса инфекционного ларинготрахеита (ILTV) и антигена F вируса ньюкаслской болезни (NDV-F). Или вектор содержит гетерологичные полинуклеотиды, кодирующие полипептиды, выбранные из группы, состоящей из антигена VP2 вируса инфекционного бурсита (IBDV), антигена гликопротеина D (gD) вируса инфекционного ларинготрахеита (ILTV) и антигена F вируса ньюкаслской болезни (NDV-F). Способ вакцинации или индукции иммунного или защитного ответа у животного против одного или нескольких птичьих патогенов включает по меньшей мере одно введение вакцины или вектора. Неожиданно вакцины, содержащие рекомбинантный HVT-вектор, были эффективными при защите животных от различных птичьих патогенов без интерференции. 3 н. и 22 з.п. ф-лы, 38 табл., 34 ил., 30 пр.
Формула
Документы, цитированные в отчёте о поиске
Рекомбинантный непатогенный mdv-вектор, обеспечивающий полиспецифический иммунитет
Комментарии