Антитела, специфичные к cd22 человека, и их применения в терапии и диагностике - RU2342401C2
Код документа: RU2342401C2
Чертежи



















Описание
Данное изобретение относится к молекуле антитела, обладающей специфичностью по отношению к антигенным детерминантам антигена B-лимфоцитов CD22. Данное изобретение также относится к терапевтическим применениям молекулы антитела и способам получения молекулы антитела.
В природной молекуле антитела имеются две тяжелых цепи и две легкие цепи. Каждая тяжелая цепь и каждая легкая цепь имеет на своем N-конце вариабельный домен. Каждый вариабельный домен состоит из четырех каркасных областей (FR), чередующихся с тремя областями, определяющими комплементарность (CDR). Остатки вариабельных доменов традиционно нумеруют согласно системе, разработанной Kabat et al. Данная система указана в Kabat et al, 1987, in Sequences of Proteins of Immunological Interest, US Department of Health and Human Services, NIH, USA (в дальнейшем «Kabat et al. (выше)»). Указанную систему нумерации используют в данном описании, за исключением случаев, оговоренных особо.
Обозначение остатков по Кабату не всегда непосредственно соответствует линейной нумерации аминокислотных остатков. Реальная линейная аминокислотная последовательность может содержать меньше или содержать дополнительные аминокислоты по сравнению со строгой нумерацией Кабата, что соответствует укорочению или инсерции структурного компонента либо в каркасе, либо в CDR основной структуры вариабельного домена. Правильную нумерацию остатков по Кабату можно определить для данного антитела выравниванием остатков гомологии в последовательности антитела со «стандартной» пронумерованной по Кабату последовательностью.
CDR вариабельного домена тяжелой цепи расположены в остатках 31-35 (CDR-H1), остатках 50-65 (CDR-H2) и остатках 95-102 (CDR-H3) согласно нумерации Кабата.
CDR вариабельного домена легкой цепи расположены в остатках 24-34 (CDR-L1), остатках 50-56 (CDR-L2) и остатках 89-97 (CDR-L3) согласно нумерации Кабата.
Конструирование CDR-привитых антител описано в Европейской заявке на выдачу патента EP-A-0239400, в которой заявлен способ, посредством которого CDR мышиного моноклонального антитела прививают в каркасные области вариабельных доменов иммуноглобулина человека сайт-направленным мутагенезом, используя длинные олигонуклеотиды. CDR определяют антигенсвязывающую специфичность антител и представляют собой относительно короткие пептидные последовательности, которые удерживаются каркасными областями вариабельных доменов.
Самую раннюю работу по гуманизации моноклональных антител посредством CDR-прививки осуществляли на моноклональных антителах, узнающих синтетические антигены, такие как NP. Однако описаны примеры, в которых CDC-прививкой гуманизировали мышиное моноклональное антитело, узнающее лизоцим, и крысиное моноклональное антитело, узнающее антиген на T-клетках человека, Verhoeyen et al. (Science, 239, 1534-1536, 1988) и Riechmann et al. (Nature, 332, 323-324, 1988) соответственно.
Riechmann et al. обнаружили, что отдельно перенос CDR (которые определены Kabat (Kabat et al. (выше) и Wu et al., J. Exp. Med., 132, 211-250, 1970)) был недостаточным для обеспечения удовлетворительной антигенсвязывающей активности в CDR-привитом продукте. Обнаружили, что ряд каркасных остатков должны быть изменены так, чтобы они соответствовали остаткам каркасной области донора. Предложенные критерии для выбора каркасных остатков, которые необходимо изменить, описаны в международной заявке на выдачу патента № WO 90/07861.
Опубликован ряд обзоров, в которых обсуждаются CDR-привитые антитела, включая Vaughan et al. (Nature Biotechnology, 16, 535-539, 1998).
Злокачественные лимфомы представляют собой разнообразную группу неоплазм. Большинство случаев встречается у пожилых людей. Неходжкинская лимфома (NHL) является заболеванием, которое в настоящее время поражает от 200000 до 250000 пациентов в США. Это второе наиболее быстро распространяющееся злокачественное опухолевое заболевание в США, распространяющееся со скоростью примерно 55000 новых случаев в год. Частота встречаемости возрастает со скоростью, которая выше, чем можно рассчитать просто из-за увеличения возраста популяции и воздействия известными факторами риска.
Классификация лимфом является сложной, и она была разработана в последние десятилетия. В 1994 г. введена исправленная евро-американская классификация лимфом (REAL). В данной классификации лимфомы B-клеточной (наиболее часто идентифицируемые), T-клеточной и неклассифицируемой природы систематизированы в условные подтипы. В каждодневной практике распределение NHL на категории низкой, промежуточной и высокой степени злокачественности на основании их общего гистологического проявления широко отражает их клиническое поведение.
NHL преимущественно поражает лимфатические узлы, но у отдельных пациентов в опухоль могут быть вовлечены другие анатомические места, такие как печень, селезенка, костный мозг, легкое, кишечник и кожа. Заболевание обычно существует в виде безболезненного увеличения лимфатических узлов. Экстронодальная лимфома наиболее часто поражает кишечник, хотя документально показана первичная лимфома фактически каждого органа. Системные симптомы включают лихорадку, потение, слабость и потерю веса.
До настоящего времени основным определяющим фактором в терапии NHL была система стадирования по Ann Arbor, полностью основанная на анатомической степени заболевания. Данная информация может быть детализирована включением дополнительных прогностических показателей, включая возраст, уровни лактатдегидрогеназы в сыворотке и статус проявления. Но и при этом знание системы стадирования Ann Arbor вместе с гистологическим и иммунологическим подтипом опухоли все еще является основным определяющим фактором лечения.
NHL низкой степени злокачественности имеет вялое течение со средней выживаемостью пациентов от 8 до 10 лет. Имеющееся в настоящее время лечение мало влияет на выживаемость, хотя облучение локального заболевания и химиотерапия в случае системных симптомов улучшает качество жизни пациентов. Комбинированная химиотерапия может быть предназначена для рецидивирующего заболевания. Заболевание промежуточной и особенно заболевание высокой степени злокачественности является чрезвычайно агрессивным и имеет тенденцию к диссеминации. Заболевание такой степени требует неотложного лечения. Лучевая терапия может являться полезным компонентом лечения пациентов с очень массивным заболеванием. Использовано множество различных схем химиотерапии, и долговременная безрецидивная выживаемость может быть получена у более чем половины пациентов. Терапия высокими дозами с поддержкой стволовыми клетками сначала вводилась для пациентов с рецидивирующим или рефрактерным заболеванием, но в настоящее время все больше и больше находит место в терапии первой линии для пациентов с заболеванием высокого риска. В последние годы тенденция ко все более агрессивному терапевтическому подходу должна быть уравновешена в отношении в целом более пожилого возраста и связанной с этим физической слабостью многих пациентов с NHL и необходимостью согласования токсичности лечения с индивидуальным прогнозом каждого случая заболевания пациента.
Необходимы улучшенные схемы лечения, которые являются более эффективными и лучше переносимыми. Недавно введенные агенты включают новые цитотоксические лекарственные средства, постепенно включаемые в комбинации, и введение основанного на антителах лечения.
Неходжкинская лимфома охватывает ряд B-клеточных лимфом. Следовательно, антигены B-клеток представляют собой подходящие мишени для терапии антителами.
CD22 является мембранным гликопротеидом с М.м. 135 кД, относящимся к семейству белков, связывающих сиаловую кислоту, называемых сиалоадгезинами. Он выявляется в цитоплазме на ранней стадии развития B-клеток, появляется на поверхности клеток одновременно с IgD и обнаруживается на большинстве зрелых B-клеток. Экспрессия увеличивается после активации B-клеток. CD22 утрачивается при конечной дифференцировке, и обычно сообщается о том, что он отсутствует в плазматических клетках. Таким образом, указанный интернализующийся антиген присутствует на поверхности пре-B-клеток и зрелых B-клеток, но не стволовых клеток или плазматических клеток.
У человека существуют две изоформы CD22. Преобладающая форма (CD22β) содержит 7 подобных иммуноглобулину (Ig-подобных) доменов во внеклеточной области. У варианта CD22α отсутствует Ig-подобный домен 4 и он может иметь укороченный цитоплазматический домен. Показано, что антитела, которые блокируют адгезию CD22 с моноцитами, нейтрофилами, лимфоцитами и эритроцитами, связываются в первом или втором Ig-подобном домене.
Цитоплазматический домен CD22 фосфорилируется по тирозину при лигировании рецептора B-клеточного антигена и связывается с Lyk, Syk и фосфатидилинозитол-3-киназой. Функция CD22 заключается в понижающем модулировании пороговой величины активации B-клеток. Он также опосредует адгезию клеток через взаимодействие с клетками, несущими соответствующие сиалогликоконъюгаты.
CD22 экспрессируется в случае большинства B-клеточных лейкозов и лимфом, включая NHL, острый лимфобластный лейкоз (B-ALL), хронический лимфоцитарный лейкоз (B-CLL) и особенно острый нелимфоцитарный лейкоз (ANLL).
Моноклональные антитела против CD22 описаны на предшествующем уровне техники. В WO 98/41641 описаны рекомбинантные анти-CD22-антитела с остатками цистеина в VH44 и VL100. В WO 96/04925 описаны области VH и VL анти-CD22-антитела LL2. В патенте США No. 5686072 описаны комбинации анти-CD22 и анти-CD19 иммунотоксинов. В WO 98/42378 описано применение «голых» анти-CD22-антител для лечения B-клеточных злокачественных заболеваний.
Ряд основанных на антителах терапевтических средств либо недавно лицензированы, например ритуксан (немеченая химерная γ1 (+mγ1V-область) человека, специфичная для CD20), либо находятся в стадии клинических испытаний для данного заболевания. Они основаны либо на опосредованной комплементом или ADCC-опосредованной гибели B-клеток, либо на применении радионуклидов, таких как131I или90Y, которые создают связанные с приготовлением и применением проблемы для врачей и пациентов. Существует необходимость в молекуле антитела для лечения NHL, которую можно многократно использовать и можно легко и эффективно получать. Существует также необходимость в молекуле антитела, которая обладает высокой аффинностью в отношении CD22 и низкой иммуногенностью для человека.
Сущность изобретения
В первом аспекте данное изобретение относится к молекуле антитела, обладающей специфичностью по отношению к CD22 человека, содержащей тяжелую цепь, в которой вариабельный домен содержит CDR (которая определена Kabat et al., (выше)), имеющую последовательность, приведенную в виде H1 на фиг.1 (SEQ ID NO: 1) для CDR-H1, в виде H2 на фиг.1 (SEQ ID NO: 2) или H2, из которой был удален потенциальный сайт гликозилирования, или H2, в которой остаток лизина в положении 60 (согласно системе нумерации Kabat) был заменен альтернативной аминокислотой, или H2, в которой и сайт гликозилирования и реакционно-способный лизин в положении 60 были удалены, для CDR-H2 или в виде H3 на фиг.1 (SEQ ID NO: 3) для CDR-H3.
Молекула антитела согласно первому аспекту данного изобретения содержит, по меньшей мере, одну CDR, выбранную из H1, H2 и H3 (SEQ ID NO: 1, SEQ ID NO: 2 и SEQ ID NO:3), в случае вариабельного домена тяжелой цепи. Предпочтительно молекула антитела содержит, по меньшей мере, две или более, предпочтительно все три CDR в вариабельном домене тяжелой цепи.
Во втором аспекте данного изобретения предлагается молекула антитела, обладающая специфичностью по отношению к CD22 человека, содержащая легкую цепь, в которой вариабельный домен содержит CDR (которая определена Kabat et al., (выше)), имеющую последовательность, приведенную в виде L1 на фиг.1 (SEQ ID NO: 4) для CDR-L1, L2 на фиг.1 (SEQ ID NO: 5) для CDR-L2 или L3 на фиг.1 (SEQ ID NO: 6) для CDR-L3.
Молекула антитела согласно второму аспекту данного изобретения содержит, по меньшей мере, одну CDR, выбранную из L1, L2 и L3 (SEQ ID NO: 4; SEQ ID NO: 5 и SEQ ID NO: 6) для вариабельного домена легкой цепи. Предпочтительно молекула антитела содержит, по меньшей мере, две или более, предпочтительно все три CDR в вариабельном домене легкой цепи.
Молекулы антител согласно первому и второму аспектам данного изобретения предпочтительно имеют комплементарную легкую цепь или комплементарную тяжелую цепь соответственно.
Предпочтительно молекула антитела согласно первому или второму аспектам данного изобретения содержит тяжелую цепь, в которой вариабельный домен содержит CDR (которая определена Kabat et al., (выше)), имеющую последовательность, приведенную в виде H1 на фиг.1 (SEQ ID NO: 1) для CDR-H1, в виде H2 на фиг.1 (SEQ ID NO: 2) или H2, из которой был удален потенциальный сайт гликозилирования, или H2, в которой остаток лизина в положении 60 (согласно системе нумерации Kabat) был заменен альтернативной аминокислотой, или H2, в которой и сайт гликозилирования и реакционно-способный лизин в положении 60 были удалены, для CDR-H2 или в виде H3 на фиг.1 (SEQ ID NO: 3) для CDR-H3, и легкую цепь, в которой вариабельный домен содержит CDR (которая определена Kabat et al., (выше)), имеющую последовательность, приведенную в виде L1 на фиг.1 (SEQ ID NO: 4) для CDR-L1, в виде L2 на фиг.1 (SEQ ID NO: 5) для CDR-L2 или в виде L3 на фиг.1 (SEQ ID NO: 6) для CDR-L3.
Вышеуказанные CDR, приведенные в SEQ ID NO: 1-6 и на фиг.1, получены из мышиного моноклонального антитела 5/44.
Полные последовательности вариабельных доменов мышиного антитела 5/44 показаны на фиг.2 (легкая цепь) (SEQ ID NO: 7) и фиг.3 (тяжелая цепь) (SEQ ID NO: 8). Данное мышиное антитело также называют далее «донорное антитело» или «мышиное моноклональное антитело».
Первый альтернативно предпочтительный вариант согласно первому или второму аспектам данного изобретения представляет собой мышиное моноклональное антитело 5/44, имеющее последовательности вариабельных доменов легкой и тяжелой цепей, показанные на фиг.2 (SEQ ID NO: 7) и фиг.3 (SEQ ID NO: 8) соответственно. Константная область легкой цепи 5/44 является каппа типа, а константная область тяжелой цепи является областью IgG1.
Во втором альтернативно предпочтительном варианте антитело согласно любому первому или второму аспектам данного изобретения является химерной молекулой антитела мышь/человек, называемой в данном описании химерной молекулой антитела 5/44. Химерная молекула антитела содержит вариабельные домены мышиного моноклонального антитела 5/44 (SEQ ID NO: 7 и 8) и константные домены человека. Предпочтительно химерная молекула антитела 5/44 содержит C-домен каппа человека (Hieter et al., Cell, 22, 197-207, 1980; номер доступа в Genebank J00241) в легкой цепи и домены гамма 4 человека (Flanagan et al., Nature, 300, 709-713, 1982) в тяжелой цепи, необязательно с заменой остатка серина в положении 241 остатком пролина.
Предпочтительно антитело согласно данному изобретению содержит тяжелую цепь, в которой вариабельный домен содержит в качестве CDR-H2 (которая определена Kabat et al., (выше)) H2', в которой была удалена последовательность потенциального сайта гликозилирования и которая неожиданно увеличивала аффинность химерного антитела 5/44 в отношении антигена CD22, и который предпочтительно имеет в качестве CDR-H2 последовательность, приведенную в виде H2' (SEQ ID NO: 13).
Альтернативно или дополнительно антитело согласно данному изобретению может содержать тяжелую цепь, в которой вариабельный домен содержит в качестве CDR-H2 (которая определена Kabat et al., (выше)) H2'', в которой остаток лизина в положении 60, который расположен в открытом положении в CDR-H2 и который, как полагают, имеет потенциальную возможность взаимодействовать с конъюгирующими агентами, приводя к снижению антигенсвязывающей аффинности, заменен на альтернативную аминокислоту, чтобы в результате получить консервативную замену. Предпочтительно CDR-H2 имеет последовательность, приведенную в виде H2'' (SEQ ID NO: 15).
Альтернативно или дополнительно антитело согласно данному изобретению может содержать тяжелую цепь, в которой вариабельный домен содержит в качестве CDR-H2 (которая определена Kabat et al., (выше)) H2''', в которой и последовательность потенциального сайта гликозилирования и остаток лизина в положении 60 заменены альтернативными аминокислотами. Предпочтительно CDR-H2 имеет последовательность, приведенную в виде H2''' (SEQ ID NO: 16).
В третьем альтернативно предпочтительном варианте антитело согласно любому первому или второму аспектам данного изобретения является CDR-привитой молекулой антитела. Термин «CDR-привитая молекула антитела» в используемом в данном описании смысле относится к молекуле антитела, в которой тяжелая и/или легкая цепь содержит одну или несколько CDR (включая при желании модифицированную CDR) из донорного антитела (например, мышиного моноклонального антитела), привитых в каркас вариабельной области тяжелой и/или легкой цепи акцепторного антитела (например, антитела человека).
Предпочтительно такое CDR-привитое антитело имеет вариабельный домен, содержащий акцепторные каркасные области человека, а также одну или несколько вышеуказанных донорных CDR.
В том случае, когда прививают CDR, можно использовать любую подходящую каркасную последовательность вариабельной области акцептора, относящуюся к классу/типу донорного антитела, из которого получены CDR, включая каркасные области мыши, примата и человека. Примерами каркасных областей человека, которые можно использовать в данном изобретении, являются KOL, NEWM, REI, EU, TUR, TEI, LAY и POM (Kabat et al. (выше)). Например, KOL и NEWM можно использовать для тяжелой цепи, REI можно использовать для легкой цепи и EU, LAY и POM можно использовать как для тяжелой цепи, так и для легкой цепи. Альтернативно можно использовать последовательности зародышевой линии человека. Предпочтительной каркасной областью в случае легкой цепи является последовательность подгруппы зародышевой линии человека (DPK9+JK1), показанная на фиг.5 (SEQ ID NO: 17). Предпочтительной каркасной областью в случае тяжелой цепи является последовательность подгруппы (DP7+JH4) человека, показанная на фиг.6 (SEQ ID NO: 21).
В CDR-привитом антителе согласно данному изобретению предпочтительно использовать в качестве акцепторного антитела антитело, имеющее цепи, которые гомологичны цепям донорного антитела. Тяжелая и легкая цепи акцептора не обязательно должны быть получены из того же самого антитела и при желании могут содержать составные цепи, имеющие каркасные области, полученные из разных цепей.
Также в CDR-привитом антителе согласно данному изобретению каркасные области не должны иметь точно такую же последовательность, как последовательности акцепторного антитела. Например, необычные остатки могут быть заменены на более часто встречающиеся остатки для данного класса или типа акцепторной цепи. Альтернативно выбранные остатки в каркасных областях акцептора могут быть заменены так, чтобы они соответствовали остатку, обнаруженному в таком же положении в донорном антителе, или остатку, который представляет собой консервативную замену остатка, обнаруженного в таком же положении в донорном антителе. Такие замены должны осуществляться по минимуму, необходимому для восстановления аффинности донорного антитела. Протокол выбора остатков в каркасных областях акцептора, которые необходимо изменить, указан в WO 91/09967.
Предпочтительно, если в молекуле CDR-привитого антитела согласно данному изобретению акцепторная легкая цепь имеет последовательность подгруппы DPK9+JK1 человека (показанную на фиг.5) (SEQ ID NO:17 (DPK9) плюс SEQ ID NO:18 (JK1)), то акцепторные каркасные области легкой цепи содержат донорные остатки в положениях 2, 4, 37, 38, 45 и 60 и могут дополнительно содержать донорный остаток в положении 3 (согласно Kabat et al. (выше)).
Предпочтительно, если в молекуле CDR-привитого антитела согласно данному изобретению акцепторная тяжелая цепь имеет последовательность DP7+JH4 человека (показанную на фиг.3) (SEQ ID NO:21), то акцепторные каркасные области тяжелой цепи содержат кроме одной или нескольких донорных CDR донорные остатки в положениях 1, 28, 48, 71 и 93 и могут дополнительно содержать донорные остатки в положениях 67 и 69 (согласно Kabat et al. (выше)).
Донорные остатки представляют собой остатки из донорного антитела, т.е. антитела, из которого исходно получали CDR.
Предпочтительно антитело согласно данному изобретению содержит тяжелую цепь, в которой вариабельный домен содержит в качестве CDR-H2 (которая определена Kabat et al., (выше)) Н2′, в которой последовательность потенциального сайта гликозилирования была удалена, чтобы увеличить аффинность химерного антитела 5/4 4 в отношении антигена CD22, и который предпочтительно имеет в качестве CDR-H2 последовательность, приведенную в виде Н2′ (SEQ ID NO:13).
Альтернативно или дополнительно антитело согласно данному изобретению может содержать тяжелую цепь, в которой вариабельный домен содержит в качестве CDR-H2 (которая определена Kabat et al., (выше)) Н2′′, в которой остаток лизина в положении 60, который расположен в открытом положении в CDR-H2 и который, как полагают, имеет потенциальную возможность взаимодействовать с конъюгирующими агентами, приводя к снижению антигенсвязывающей аффинности, заменяют на альтернативную аминокислоту. Предпочтительно CDR-H2 имеет последовательность, приведенную в виде Н2′′ (SEQ ID NO:15).
Альтернативно или дополнительно антитело согласно данному изобретению может содержать тяжелую цепь, в которой вариабельный домен содержит в качестве CDR-H2 (которая определена Kabat et al., (выше)) H2''', в которой и последовательность потенциального сайта гликозилирования и остаток лизина в положении 60 заменены альтернативными аминокислотами. Предпочтительно CDR-H2 имеет последовательность, приведенную в виде H2''' (SEQ ID NO: 16).
Молекула антитела согласно данному изобретению может содержать полную молекулу антитела, имеющую полноразмерные тяжелые и легкие цепи; его фрагмент, такой как фрагмент Fab, модифицированный Fab, Fab', F(ab')2 или Fv; мономер или димер легкой цепи или тяжелой цепи; одноцепочечное антитело, например одноцепочечный Fv, в котором вариабельные домены тяжелой и легкой цепи связаны пептидным линкером. Подобным образом вариабельные области тяжелой и легкой цепи можно комбинировать с другими доменами антител в зависимости от ситуации.
Молекула антитела согласно данному изобретению может иметь связанную с ней эффекторную или репортерную молекулу. Например, молекула антитела может иметь макроцикл для хелатирования атома тяжелого металла или токсин, такой как рицин, связанный с ней структурой ковалентного мостика. Альтернативно можно использовать способы технологии рекомбинантных ДНК, чтобы получить молекулу антитела, в которой фрагмент Fc (домены CH2, CH3 и шарнирный домен), домены CH2 и CH3 или домен CH3 полной молекулы иммуноглобулина был заменен (заменены) или связан (связаны) пептидной связью с функциональным неиммуноглобулиновым белком, таким как молекула фермента или токсина.
Молекула антитела согласно данному изобретению предпочтительно имеет аффинность связывания, составляющую по меньшей мере 0,85×10-10 М, более предпочтительно по меньшей мере 0,75×10-10 М и наиболее предпочтительно по меньшей мере 0,5×10-10 М.
Предпочтительно молекула антитела согласно данному изобретению содержит вариабельный домен легкой цепи 5/44-gL1 (SEQ ID NO:19) и вариабельный домен тяжелой цепи 5/44-gH7 (SEQ ID NO: 27). Последовательности вариабельных доменов указанных легкой и тяжелой цепей показаны на фиг.5 и 6 соответственно.
Данное изобретение также относится к вариантам молекулы антитела согласно данному изобретению, которые имеют повышенную аффинность по отношению к CD22. Такие варианты можно получить, используя ряд протоколов созревания аффинности, включая мутирование CDR (Yang et al., J. Mol. Biol., 254, 392-403, 1995), перестановку цепей (Marks et al., Bio/Technology, 10, 779-783, 1992), применение мутантных штаммов Е.coli (Low et al., J. Mol. Biol., 250, 359-368, 1996), перестановку ДНК (Patten et al., Curr. Opin. Biotechnol., 8, 724-733, 1997), фаговый дисплей (Thompson et al., J. Mol. Biol., 256, 77-88, 1996) и ПЦР на материале половых клеток (Crameri et al., Nature, 391, 288-291, 1998). Vaughan et al. (выше) обсуждает указанные способы созревания аффинности.
Данное изобретение также относится к последовательности ДНК, кодирующей тяжелую и/или легкую цепь(цепи) молекулы антитела согласно данному изобретению.
Предпочтительно последовательность ДНК кодирует тяжелую или легкую цепь молекулы антитела согласно данному изобретению.
Последовательность ДНК согласно данному изобретению может содержать синтетическую ДНК, полученную, например, химической обработкой, кДНК, геномную ДНК или любую их комбинацию.
Данное изобретение также относится к клонирующему или экспрессирующему вектору, содержащему одну или несколько последовательностей ДНК согласно данному изобретению. Предпочтительно клонирующий или экспрессирующий вектор содержит две последовательности ДНК, кодирующие легкую цепь и тяжелую цепь молекулы антитела согласно данному изобретению соответственно.
Общие способы, посредством которых можно сконструировать векторы, способы трансфекции и способы культивирования хорошо известны специалистам в данной области. В связи с этим приведены ссылка на «Current Protocols in Molecular Biology», 1999, F. M. Ausubel (ed), Wiley Interscience, New York и руководство Maniatis, выпущенное Cold Spring Harbor Publishing.
Последовательности ДНК, которые кодируют молекулу антитела согласно данному изобретению, можно получить способами, хорошо известными специалистам в данной области. Например, последовательности ДНК, кодирующие часть или полностью тяжелую и легкую цепи антитела, могут быть синтезированы в соответствии с требованиями на основе определенных последовательностей ДНК или на основе соответствующих аминокислотных последовательностей.
ДНК, кодирующая акцепторные каркасные последовательности, широко доступна специалистам в данной области и может быть легко синтезирована на основе их известных аминокислотных последовательностей.
Можно использовать стандартные способы молекулярной биологии, чтобы получить последовательности ДНК, кодирующие молекулу антитела согласно данному изобретению. Требуемые последовательности ДНК можно синтезировать полностью или частично, используя методику синтеза олигонуклеотидов. В зависимости от обстоятельств можно использовать способ сайт-направленного мутагенеза и полимеразной цепной реакции (ПЦР).
Любую подходящую систему клетка-хозяин/вектор можно использовать для экспрессии последовательностей ДНК, кодирующих молекулу антитела согласно данному изобретению. Можно использовать бактериальные системы, например E. coli, и другие системы микроорганизмов, в частности для экспрессии фрагментов антител, таких как Fab- и F(ab')2-фрагменты, и особенно фрагменты Fv и одноцепочечные фрагменты антител, например одноцепочечные Fv. Эукариотические системы экспрессии в клетках-хозяевах, например системы млекопитающих, можно использовать для получения более крупных молекул антител, включая полные молекулы антител. Подходящие клетки-хозяева млекопитающих включают CHO, клетки миеломы и гибридомы.
Данное изобретение также относится к способу получения молекулы антитела согласно данному изобретению, включающему культивирование клетки-хозяина, содержащей вектор согласно данному изобретению, в условиях, подходящих для того, чтобы привести к экспрессии белка с ДНК, кодирующей молекулу антитела согласно данному изобретению, и выделение молекулы антитела.
Молекула антитела может содержать только полипептид тяжелой или легкой цепи, и в этом случае для трансфекции клеток-хозяев необходимо использовать последовательность, кодирующую только полипептид тяжелой цепи или легкой цепи. Для получения продуктов, содержащих обе цепи, тяжелую и легкую, линию клеток можно трансфицировать двумя векторами, первым вектором, кодирующим полипептид легкой цепи, и вторым вектором, кодирующим полипептид тяжелой цепи. Альтернативно можно использовать один вектор, при этом вектор содержит последовательности, кодирующие полипептиды легкой цепи и тяжелой цепи.
Данное изобретение также относится к терапевтической или диагностической композиции, содержащей молекулу антитела согласно данному изобретению в комбинации с фармацевтически приемлемым наполнителем, разбавителем или носителем.
Данное изобретение также относится к способу получения терапевтической или диагностической композиции, включающему смешивание молекулы антитела согласно данному изобретению вместе с фармацевтически приемлемым наполнителем, разбавителем или носителем.
Молекула антитела может быть единственным активным ингредиентом в терапевтической или диагностической композиции или может находиться в сопровождении других активных ингредиентов, включая ингредиенты, являющиеся другими антителами, например антителами против T-клеток, антителами против IFNγ или анти-ЛПС-антителами, или ингредиентов, не являющихся антителами, таких как ксантины.
Фармацевтические композиции предпочтительно содержат терапевтически эффективное количество антитела согласно изобретению. Термин «терапевтически эффективное количество» в используемом в данном описании смысле относится к количеству терапевтического средства, необходимого для лечения, ослабления или профилактики заболевания или состояния, являющегося мишенью, или для проявления регистрируемого терапевтического или профилактического эффекта. Для любого антитела терапевтически эффективную дозу сначала можно определить либо в анализах на культуре клеток, либо в моделях на животных, обычно грызунах, кроликах, собаках, свиньях или приматах. Модель на животных также можно использовать для определения соответствующего диапазона концентраций и пути введения. Затем такую информацию можно использовать для определения применимых доз и путей введения для человека.
Точное эффективное количество для человека будет зависеть от тяжести состояния болезни, общего состояния здоровья субъекта, возраста, веса и пола субъекта, питания, времени и частоты введения, комбинации(ций) лекарственных средств, реакционной чувствительности и толерантности/чувствительности к терапии. Данное количество может быть определено обычным экспериментированием, и оно зависит от решения врача. В общем эффективная доза будет составлять от 0,01 мг/кг до 50 мг/кг, предпочтительно от 0,1 мг/кг до 20 мг/кг, более предпочтительно примерно 15 мг/кг.
Композиции можно вводить пациенту отдельно или можно вводить в комбинации с другими агентами, лекарственными средствами или гормонами.
Доза, в которой вводят молекулу антитела согласно данному изобретению, зависит от природы состояния, которое необходимо лечить, степени злокачественности лимфомы или лейкоза и от того, используется ли молекула антитела профилактически или для лечения существующего состояния.
Частота дозы будет зависеть от времени полужизни молекулы антитела и продолжительности его действия. Если молекула антитела имеет короткое время полужизни (например, от 2 до 10 часов), то может быть необходимо давать одну или несколько доз в сутки. Альтернативно, если молекула антитела имеет длительное время полужизни (например, от 2 до 15 дней) может быть необходимо вводить дозу только один раз в сутки, один раз в неделю или даже один раз каждый 1 или 2 месяца.
Фармацевтическая композиция также может содержать фармацевтически приемлемый носитель для введения антитела. Носитель не должен сам по себе индуцировать выработку антител, опасных для индивидуума, получающего композицию, и должен быть нетоксичным. Подходящими носителями могут быть крупные, медленно метаболизируемые макромолекулы, такие как белки, полипептиды, липосомы, полисахариды, полимолочные кислоты, полигликолевые кислоты, полимерные аминокислоты, сополимеры аминокислот и неактивные вирусные частицы.
Могут быть использованы фармацевтически приемлемые соли, например соли неорганических кислот, такие как гидрохлориды, гидробромиды, фосфаты и сульфаты, или соли органических кислот, такие как ацетаты, пропионаты, малонаты и бензоаты.
Фармацевтически приемлемые носители в фармацевтических композициях могут дополнительно содержать жидкости, такие как вода, физиологический раствор, глицерин и этанол. Дополнительно в таких композициях могут присутствовать вспомогательные вещества, такие как увлажняющие или эмульгирующие агенты или вещества для забуферивания pH. Такие носители делают возможным готовить фармацевтические композиции в виде таблеток, пилюль, драже, капсул, жидкостей, гелей, сиропов, взвесей и суспензий для приема пациентом внутрь.
Предпочтительные формы для введения включают формы, подходящие для парентерального введения, например путем инъекции или инфузии, например инъекции болюсов или непрерывной инфузии. В том случае, когда продукт предназначен для инъекции или инфузии, он может принимать форму суспензии, раствора или эмульсии в масляном или водном наполнителе, и он может содержать агенты для приготовления композиции, такие как суспендирующие агенты, консерванты, стабилизаторы и/или диспергирующие агенты. Альтернативно молекула антитела может быть в сухой форме для перерастворения перед применением с использованием соответствующей стерильной жидкости.
После приготовления композиции согласно настоящему изобретению можно вводить непосредственно субъекту. Субъектом, подвергаемым лечению, может быть животное. Однако предпочтительно композиции адаптированы для введения человеку.
Фармацевтические композиции согласно данному изобретению можно вводить любым количеством способов, включая, но не ограничиваясь указанным, пероральный, внутривенный, внутримышечный, внутриартериальный, интрамедуллярный, интратекальный, интравентрикулярный, трансдермальный, чрескожный (например, см. WO 98/20734), подкожный, внутрибрюшинный, интраназальный, энтеральный, местный, подъязычный, внутривагинальный или ректальный пути. Также можно использовать гипоспреи, чтобы ввести фармацевтические композиции согласно настоящему изобретению. Обычно терапевтические композиции могут быть приготовлены в виде инъецируемых препаратов, либо в виде жидких растворов, либо суспензий. Также могут быть приготовлены твердые формы, подходящие для растворения или суспендирования в жидких наполнителях перед инъекцией.
Прямая доставка композиций, как правило, будет осуществляться путем инъекции подкожно, внутрибрюшинно, внутривенно или внутримышечно, или доставляться в интерстициальное пространство ткани. Композиции также можно вводить в место повреждения. Дозированное лечение может представлять собой схему на основе однократной дозы или схему на основе многократных доз.
Будет понятно, что активным ингредиентом в композиции будет молекула антитела. Как таковая она будет чувствительна к деградации в желудочно-кишечном тракте. Таким образом, если композицию необходимо вводить путем с использованием желудочно-кишечного тракта, композиция должна будет содержать агенты, которые защищают антитело от деградации, но которые высвобождают антитело после того, как произойдет всасывание из желудочно-кишечного тракта.
Всестороннее обсуждение фармацевтически приемлемых носителей имеется в Remington's Pharmaceutical Sciences (Mack Publishing Company, N.J. 1991).
Также предусматривается, что антитело согласно данному изобретению будет вводиться с использованием генной терапии. Чтобы этого достичь, последовательности ДНК, кодирующие тяжелую и легкую цепи молекулы антитела, под контролем соответствующих компонентов ДНК вводят пациенту с тем, чтобы цепи антитела экспрессировались с последовательностей ДНК и проходили сборку in situ.
Данное изобретение также относится к молекуле антитела согласно данному изобретению для применения при лечении заболевания, опосредованного клетками, экспрессирующими CD22.
Данное изобретение, кроме того, относится к применению молекулы антитела согласно данному изобретению для производства лекарственного средства для лечения заболевания, опосредованного клетками, экспрессирующими CD22.
Молекулу антитела согласно данному изобретению можно использовать при любой терапии, когда требуется снизить уровень клеток, экспрессирующих CD22, которые присутствуют в организме человека или животного. Указанные CD22-экспрессирующие клетки могут циркулировать в организме или присутствовать на нежелательно высоком уровне локализованными в конкретном месте в организме. Например, повышенные уровни клеток, экспрессирующих CD22, будут присутствовать при B-клеточных лимфомах и лейкозах. Молекулу антитела согласно данному изобретению можно использовать для лечения заболеваний, опосредованных клетками, экспрессирующими CD22.
Молекулу антитела согласно данному изобретению предпочтительно используют для лечения злокачественных лимфом и лейкозов, наиболее предпочтительно NHL.
Данное изобретение также относится к способу лечения человека или животного, страдающего от заболевания или для которого существует риск возникновения заболевания, опосредованного клетками, экспрессирующими CD22, при этом способ включает введение субъекту эффективного количества молекулы антитела согласно данному изобретению.
Молекулу антитела согласно данному изобретению также можно использовать для диагностики, например при диагностике in vivo, и визуализации патологических состояний, в которые вовлечены клетки, которые экспрессируют CD22.
Далее данное изобретение описано только посредством иллюстрации в следующих примерах, в которых даны ссылки на сопровождающие их чертежи.
На фиг.1 показана аминокислотная последовательность CDR мышиного моноклонального антитела 5/44 (SEQ ID NO: 1-6).
На фиг.2 показана полная последовательность вариабельного домена легкой цепи мышиного моноклонального антитела 5/44.
На фиг.3 показана полная последовательность вариабельного домена тяжелой цепи мышиного моноклонального антитела 5/44.
На фиг.4 показана стратегия удаления сайта гликозилирования и реакционно-способного лизина в CDR-H2.
На фиг.5 показана конструкция прививки для последовательности легкой цепи 5/44.
На фиг.6 показана конструкция прививки для последовательности тяжелой цепи 5/44.
На фиг.7 показаны векторы pMRR14 и pMRR10.1.
На фиг.8 показаны результаты анализа Biacore химерных мутантов 5/44.
На фиг.9 показаны олигонуклеотиды для сборки генов gH1 и gL1 5/44.
На фиг.10 показаны промежуточные векторы pCR2.1(544gH1) и pCR2.1(544gL1).
На фиг.11 показаны олигонуклеотидные кассеты, используемые для получения следующих прививок.
На фиг.12 показан анализ конкуренции между флуоресцентно меченым мышиным антителом 5/44 и привитыми вариантами; и
На фиг.13 показана полная последовательность ДНК и белка привитых тяжелой и легкой цепей.
Подробное описание изобретения
Пример 1: Создание отобранных для испытаний вариантов антител
Панель антител против CD22 отбирали из гибридом, используя следующие критерии для отбора: связывание с клетками Дауди, интернализация в клетки Дауди, связывание с периферическими мононуклеарными клетками крови (PBMC), интернализация в PBMC, аффинность (более чем 10-9 М), γ1 мыши и скорость продуцирования. В качестве предпочтительного антитела выбрано антитело 5/44.
Пример 2: Клонирование гена и экспрессия химерной молекулы антитела 5/44
Получение клеток гибридомы 5/44 и получение из них РНК
Гибридому 5/44 создавали, используя обычную методику гибридом после иммунизации мышей белком CD22 человека. РНК из клеток гибридомы 5/44 получали, используя набор RNEasy (Qiagen, Crawley, UK; No. в каталоге 74106). Полученную РНК обратно транскрибировали в кДНК, как описано ниже.
Распределение CD22 на опухолях NHL
Предприняли иммуногистохимическое исследование, чтобы оценить частоту встречаемости и распределение окрашивания с использованием моноклональных антител 5/44 против CD22. В исследование включали контрольные антитела против CD20 и CD79a, чтобы подтвердить B-клеточные области опухолей.
Всего исследовали 50 опухолей и классифицировали их следующим образом, используя систему рабочей формулировки и систему классификации REAL:
- 7 B-лимфобластный лейкоз/лимфома (высокая/I)
- 4 B-CLL/мелкоклеточная лимфоцитарная лимфома (низкая/A)
- 3 лимфоплазмацитарная лимфома/иммуноцитома (низкая/A)
- 1 лимфома мантийных клеток (промежуточная/F)
- 14 лимфома фолликулярного центра (низкая - промежуточная/D)
- 13 диффузная лимфома из крупных клеток (промежуточная - высокая/G,H)
- 6 неклассифицируемые (K)
- 2 T-клеточные лимфомы
40 B-клеточных лимфом были позитивными в отношении антигена CD22 при использовании антитела 5/44 в концентрации 0,1 мкг/мл, а другие 6 становились позитивными, когда концентрацию увеличивали до 0,5 мкг/мл. В случае остальных 2 B-клеточных опухолей, которые были негативными при 0,1 мкг/мл, оставалось недостаточно ткани, чтобы тестировать более высокую концентрацию. Однако параллельное тестирование с другим анти-CD22-антителом Celltech 6/13, которое давало более сильное окрашивание, чем 5/44, дало во всех 48 B-клеточных лимфомах окрашивание, позитивное по отношению к CD22.
Таким образом, можно сделать вывод, что антиген CD22 широко экспрессируется на B-клеточных лимфомах и таким образом представляет собой подходящую мишень для иммунотерапии при NHL.
ПЦР-клонирование VH и VL 5/44
Последовательности кДНК, кодирующие вариабельные домены тяжелой и легкой цепей 5/44, синтезировали, используя обратную транскриптазу, чтобы получить однонитевые кДНК-копии с мРНК, присутствующей в общей массе РНК. Затем их использовали в качестве матрицы для амплификации мышиных последовательностей V-областей, используя специфичные олигонуклеотидные праймеры в полимеразной цепной реакции (ПЦР).
a) Синтез кДНК
кДНК синтезировали в объеме реакции 20 мкл, содержащем следующие реагенты: 50 мМ трис-HCl pH 8,3, 75 мМ KCl, 10 мМ дитиотреитол, 3 мМ MgCl2, 0,5 мМ каждого дезоксирибонуклеозидтрифосфата, 20 единиц RNAsin, 75 нг случайного гексануклеотидного праймера, 2 мкг РНК 5/44 и 200 единиц обратной транскриптазы вируса лейкоза Молони мышей. После инкубации при 42°C в течение 60 минут реакцию останавливали нагреванием при 95°C в течение 5 минут.
b) ПЦР
Аликвоты кДНК подвергали ПЦР, используя комбинации праймеров, специфичных для тяжелой и легкой цепей. В качестве прямых праймеров использовали пулы вырожденных праймеров, сконструированных для отжига с консервативными последовательностями сигнального пептида. Все указанные последовательности содержат по порядку сайт рестрикции (VL SfuI; VH HindIII), начиная с 7 нуклеотидов от их 5'-концов, последовательность GCCGCCACC (SEQ ID NO: 50), чтобы обеспечить оптимальную трансляцию получаемых в результате мРНК, кодон инициации и 20-30 нуклеотидов на основании последовательностей лидерного пептида известных мышиных антител (Kabat et al., Sequences of proteins of immunological interest, 5th Edition, 1991, U.S. Department of Health and Human Services, Public Health Service, National Institutes of Health).
3'-праймеры конструировали так, чтобы они перекрывали соединение каркас 4-J-C антитела и содержали сайт рестрикции для фермента BsiWI, чтобы облегчить клонирование ПЦР-фрагмента VL. 3'-праймеры тяжелой цепи представляют собой смесь, сконструированную так, чтобы они перекрывали соединение J-C антитела. 3'-праймер включает сайт рестрикции ApaI, чтобы облегчить клонирование. 3'-область праймеров содержит смешанную последовательность, основанную на последовательностях, обнаруженных в известных мышиных антителах (Kabat et al., 1991, выше).
Комбинации описанных выше праймеров делают возможным клонирование ПЦР-продуктов для VH и VL непосредственно в подходящем экспрессирующем векторе (см. ниже), чтобы получить химерные (мышь-человек) тяжелые и легкие цепи, и экспрессию данных генов в клетках млекопитающих, чтобы получить химерные антитела требуемого изотипа.
Инкубации (100 мкл) для ПЦР проводили следующим образом. Каждая реакционная смесь содержала 10 мМ трис-HCl pH 8,3, 1,5 мМ MgCl2, 50 мМ KCl, 0,01% мас./об. желатина, 0,25 мМ каждого дезоксирибонуклеозидтрифосфата, 10 пмоль смеси 5'-праймеров, 10 пмоль 3'-праймера, 1 мкл кДНК и 1 единицу полимеразы Taq. Реакционные смеси инкубировали при 95°C в течение 5 минут и затем подвергали циклу 94°C в течение 1 минуты, 55°C в течение 1 минуты и 72°C в течение 1 минуты. После 30 циклов аликвоты каждой реакционной смеси анализировали электрофорезом в агарозном геле.
Для V-области тяжелой цепи амплифицированный продукт ДНК получали только когда пул праймеров, отжигающийся в начале каркаса I, заменял пул праймеров сигнального пептида. Фрагменты клонировали в векторах для секвенирования ДНК. Определяли последовательность ДНК, чтобы получить выведенную аминокислотную последовательность. Данную выведенную последовательность подтверждали, обращаясь к N-концевой последовательности белка, определенной экспериментально. На фиг. 2 и 3 показана последовательность ДНК/белка зрелых V-областей легкой и тяжелой цепи мышиного моноклонального антитела 5/44 соответственно.
c) Молекулярное клонирование ПЦР-фрагментов
Затем последовательности V-области мышей клонировали в экспрессирующих векторах pMRR10.1 и pMRR14 (фиг.7). Указанные векторы представляют собой векторы для экспрессии легкой и тяжелой цепи соответственно, содержащие ДНК, кодирующую константные области легкой цепи каппа человека и тяжелой цепи гамма-4 человека. Область VL субклонировали в экспрессирующий вектор посредством рестрикционного расщепления и лигирования из вектора для секвенирования, используя сайты рестрикции SfuI и BsiWI, создавая плазмиду pMRR10(544cL). ДНК тяжелой цепи амплифицировали в ПЦР, используя 5'-праймер, чтобы ввести сигнальный пептид, так как его не получали в методике клонирования - использовали лидер тяжелой цепи мышиного антитела из другой собственной гибридомы (названной 162). 5'-праймер имел следующую последовательность:

Обратный праймер был идентичен праймеру, используемому в исходном клонировании гена VH. Полученный в результате продукт ПЦР расщепляли ферментами HindIII и ApaI, субклонировали и подтверждали его последовательность ДНК, создавая плазмиду pMRR14(544cH). Временная котрансфекция обоих экспрессирующих векторов в клетки CHO давала химерное антитело c5/44. Котрансфекцию проводили, используя реагент липофектамин согласно протоколам производителя (InVitrogen:Life Technology, Groningen, The Netherlands. No. в каталоге 11668-027).
Удаление сайта гликозилирования и реакционно-способного лизина
Потенциальная N-связанная последовательность сайта гликозилирования обнаружена в CDR-H2, имеющая аминокислотную последовательность N-Y-T (фиг.3). SDS-ПААГ, Вестерн-блоттинг и окрашивание на карбогидраты гелей 5/44 и его фрагментов (включая Fab) показали, что данный сайт был действительно гликозилирован (не показано). Кроме того, обнаружен остаток лизина в открытом положении в CDR-H2, который имел потенциальную возможность уменьшать аффинность связывания антитела, обеспечивая дополнительный сайт для конъюгивания с агентом, с которым антитело может быть конъюгировано.
Использовали методику ПЦР, чтобы ввести аминокислотные замены в последовательность CDR-H2, пытаясь удалить сайт гликозилирования и/или реакционно-способный лизин, как показано на фиг.4. Прямые праймеры, кодирующие мутации N55Q, T57A или T57V, использовали для удаления сайта гликозилирования (фиг.4) и создавали четвертый прямой праймер, содержащий замену K60R, чтобы удалить реакционно-способный остаток лизина (фиг.4). Обратный праймер каркаса 4 использовали в каждой из указанных ПЦР-амплификаций. Продукты ПЦР расщепляли ферментами XbaI и ApaI и встраивали в pMRR14(544cH) (также расщепленный XbaI и ApaI), чтобы создать экспрессирующие плазмиды, кодирующие указанные мутанты. Мутации N55Q, T57A и T57V удаляли сайт гликозилирования за счет изменения аминокислотной последовательности так, что она отличалась от консенсуса N-X-T/S, в то время как мутация K60R заменяла потенциально реакционно-способный лизин сходным образом положительно заряженным остатком аргинина. Полученные в результате варианты плазмид cH котрансфицировали плазмидой cL, чтобы создать экспрессируемые варианты химерных антител.
Оценка активностей химерных генов
Активности химерных генов оценивали после временной трансфекции клеток CHO.
c) Определение констант аффинности анализом BiaCore
Аффинности химерного 5/44 или его вариантов, в которых были удалены их сайт гликозилирования или их реакционно-способный лизин, исследовали, используя методику BIA для связывания с конструкциями CD22-mFc. Результаты показаны на фиг.8. Все измерения связывания осуществляли на приборе BIAcoreTM 2000 (Pharmacia Biosensor AB, Uppsala, Sweden). Анализ осуществляли на основе захвата CD22mFc посредством иммобилизованного анти-Fc мыши. Антитело было в растворимой фазе. Образцы, стандарт и контроли (50 мкл) инъецировали на иммобилизованное анти-Fc мыши, затем антитело в растворенной фазе. После каждого цикла поверхность регенировали 50 мкл 40 мМ HCl со скоростью 30 мкл/мин. Кинетический анализ осуществляли, используя компьютерную программу BIAevaluation 3.1 (Pharmacia).
Удаление сайта гликозилирования в конструкции T57A приводило к несколько более быстрой скорости образования комплекса и немного более медленной скорости распада комплекса по сравнению с химерным 5/44, давая примерно 5-кратное увеличение аффинности. Мутация N55Q не оказывала влияния на аффинность. Данный результат был неожиданным, так как он свидетельствует о том, что удаление карбогидрата само по себе не оказывает влияния на связывание (как в случае с заменой N55Q). Повышенную аффинность наблюдали только в случае замены T57A. Одним из возможных объяснений является то, что независимо от присутствия карбогидрата треонин в положении 57 оказывает негативное влияние на связывание, которое исключается при превращении треонина в аланин. Гипотеза о том, что важное значение имеет малый размер аланина и что негативное действие треонина связано с его размером, подтверждается результатом, полученным с использованием мутации T57V: что замена валином в положении 57 не приносит пользы (результаты не показаны).
Удаление лизина в результате мутации K60R имело нейтральное действие на аффинность, т.е. введение аргинина удаляет потенциально реакционно-способный сайт, не подвергая риску аффинность.
Таким образом обе мутации для удаления сайта гликозилирования и для удаления реакционно-способного лизина включали в конструкцию при гуманизации.
Пример 2: CDR-прививка 5/44
Молекулярное клонирование генов вариабельных областей тяжелой и легкой цепей антитела 5/44 и их применение для получения химерных (мышь/человек) антител 5/44 описано выше. Нуклеотидная и аминокислотная последовательности VL- и VH-доменов 5/44 мыши показаны на фиг. 2 и 3 (SEQ ID NO: 7 и 8) соответственно. В данном примере описана CDR-прививка антитела 5/44 в каркасные области человека, чтобы снизить возможную иммуногенность у человека согласно способу Adair et al., (WO 91/09967).
CDR-прививка легкой цепи 5/44
Выравнивание последовательности белка с консенсусными последовательностями из V-области легкой цепи каппа подгруппы I человека показало 64% идентичность последовательностей. Поэтому выбранные для конструирования CDR-привитой легкой цепи акцепторные каркасные области соответствовали каркасным областям последовательности O12,DPK9 зародышевой линии подгруппы I VK человека. Акцепторную последовательность каркаса 4 получали из последовательности JK1 зародышевой линии J-области человека.
Сравнение аминокислотных последовательностей каркасных областей 5/44 мыши и акцепторной последовательности приведено на фиг.5 и оно показывает, что существует 27 различий между донорными и акцепторными цепями. В каждом положении делали анализ потенциальной возможности мышиного остатка вносить вклад в связывание антигена, либо прямо, либо опосредованно вследствие влияния на укладку или на область контакта VH/VL. Если считали, что мышиный остаток имеет важное значение и в достаточной мере отличается от человеческого остатка по размеру, полярности или заряду, то такой мышиный остаток сохраняли. На основании данного анализа конструировали две версии CDR-привитой легкой цепи, имеющие последовательности, приведенные в SEQ ID NO: 19 и SEQ ID NO: 20 (фиг.5).
CDR-прививка тяжелой цепи 5/44
CDR-прививку тяжелой цепи 5/44 осуществляли, используя такую же методику, которая описана для легкой цепи. Обнаружено, что V-домен тяжелой цепи 5/44 гомологичен тяжелым цепям человека, относящимся к подгруппе I (70% идентичность последовательностей), и поэтому последовательность каркаса VH1-3,DP7 зародышевой линии подгруппы I человека использовали в качестве акцепторного каркаса. Акцепторные последовательности каркаса 4 получали из последовательности JH4 зародышевой линии J-области человека.
Сравнение тяжелой цепи 5/44 с каркасными областями показано на фиг.6, где можно видеть, что тяжелая цепь 5/44 отличается от акцепторной последовательности по 22 положениям. Анализ вклада, который любое из указанных положений может вносить в связывание антигена, привел к 5 вариантам конструируемых CDR-привитых тяжелых цепей, имеющих последовательности, приведенные в SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25, SEQ ID NO: 26 и SEQ ID NO: 27 (фиг.6).
Конструирование генов для привитых последовательностей
Проектировали гены для кодирования привитых последовательностей gH1 и gL1, и проектировали и конструировали серию перекрывающихся олигонуклеотидов (фиг.9). Методику ПЦР-сборки использовали для конструирования генов CDR-привитых V-областей. Готовили реакционные смеси объемом 100 мкл, содержащие 10 мМ трис-HCl pH 8,3, 1,5 мМ MgCl2, 50 мМ KCl, 0,001% желатина, 0,25 мМ каждого дезоксирибонуклеозидтрифосфата, 1 пмоль каждого из «внутренних» праймеров (T1, T2, T3, B1, B2, B3), 10 пмоль каждого из «внешних» праймеров (F1, R1) и 1 единицу полимеразы Taq (AmpliTaq, Applied BioSystems, No. в каталоге N808-0171). Параметры цикла ПЦР представляли собой 94°C в течение 1 минуты, 55°C в течение 1 минуты и 72°С в течение 1 минуты, 30 циклов. Затем продукты реакции разгоняли в 1,5% агарозном геле, вырезали и извлекали, используя центрифужные колонки QIAGEN (набор для экстракции из геля QIAquick, No. в каталоге 28706). ДНК элюировали в объеме 30 мкл. Затем аликвоты (1 мкл) ДНК gH1 и gL1 клонировали в клонирующем векторе InVitrogen TOPO TA pCR2.1 TOPO (No. в каталоге K4500-01) согласно инструкциям производителя. Данный неэкспрессирующий вектор служил в качестве промежуточного продукта клонирования, чтобы облегчить секвенирование большого числа клонов. Секвенирование ДНК с использованием специфичных для вектора праймеров применяли для идентификации правильных клонов, содержащих gH1 и gL1, создавая плазмиды pCR2.1(544gH1) и pCR2.1(544gL1) (фиг.10).
Способ замены олигонуклеотидных кассет использовали для создания гуманизированных прививок gH4,5,6 и 7 и gL2. На фиг.11 показана конструкция олигонуклеотидных кассет. Чтобы сконструировать каждый вариант, вектор (pCR2.1(544gH1) или pCR2.1(544gL1)) разрезали показанными ферментами рестрикции (XmaI/SacII для тяжелой цепи, XmaI/BstEII для легкой цепи). Крупный векторный фрагмент очищали из агарозного геля и использовали для лигирования с олигонуклеотидной кассетой. Указанные кассеты состоят из 2 комплементарных олигонуклеотидов (показанных на фиг.11), смешиваемых в концентрации 0,5 пмоль/мкл в объеме 200 мкл 12,5 мМ трис-HCl pH 7,5, 2,5 мМ MgCl2, 25 мМ NaCl, 0,25 мМ дитиоэритритол. Отжиг осуществляли нагреванием до 95°C в течение 3 минут на водяной бане (объем 500 мл), затем давая возможность реакционной смеси медленно остыть до комнатной температуры. Затем подвергнутую отжигу олигонуклеотидную кассету разбавляли в десять раз в воде перед лигированием в соответствующим образом разрезанный вектор. Секвенирование ДНК использовали для подтверждения правильной последовательности, создавая плазмиды pCR2.1(5/44-gH4-7) и pCR2.1(5/44-gL2). Затем проверенные привитые последовательности субклонировали в экспрессирующих векторах pMRR14 (тяжелая цепь) и pMR10.1 (легкая цепь).
Активность CDR-привитых последовательностей в связывании CD22
Векторы, кодирующие привитые варианты, котрансфицировали в клетки CHO в различных комбинациях вместе с исходными цепями химерного антитела. Активность связывания сравнивали в конкурентном анализе, осуществляя конкуренцию с исходным антителом 5/44 мыши за связывание с клетками Ramos (полученными из ATCC, линия лимфобластных клеток человека лимфомы Беркитта, экспрессирующих поверхностный CD22). Данный анализ считался наилучшим способом сравнения прививок в отношении их способности связываться с CD22 клеточной поверхности. Результаты показаны на фиг.8. Как можно видеть, существует очень незначительное различие между любыми из прививок, при этом все они проявляли себя более эффективно, чем химерное антитело в конкуренции против мышиного родительского антитела. Введение 3 дополнительных остатков человека на конце CDR-H3 (gH5 и gH7) по-видимому не оказывало влияния на связывание.
Была выбрана комбинация прививок с наименьшим количеством мышиных остатков, gL1gH7. Привитая легкая цепь gL1 имела 6 донорных остатков. Остатки V2, V4, L37 и Q45 являются потенциально важными для упаковки остатками. Остаток H38 находится в области контакта VH/VL. Остаток D60 является поверхностным остатком вблизи CDR-L2 и может непосредственно вносить вклад в связывание антигена. Из этих остатков V2, L37, Q45 и D60 обнаружены в последовательностях зародышевой линии каппа-генов человека из других подгрупп. Привитая тяжелая цепь gH7 имеет 4 донорных каркасных остатка (остаток R28 считается частью CDR-H1 согласно структурному определению, используемому при CDR-прививках (см. Adair et al. (1991 WO91/09967)). Остатки E1 и A71 являются поверхностными остатками вблизи CDR. Остаток I48 является потенциальным пакующим остатком. Остаток T93 присутствует в области контакта VH/VL. Из указанных остатков E1 и A71 обнаружены в других генах зародышевой линии подгруппы I человека. Остаток I48 обнаружен в подгруппе 4 зародышевой линии человека, и T73 обнаружен в подгруппе 3 зародышевой линии человека.
Полная последовательность ДНК и белка как легкой цепи, так и тяжелой цепи, включая примерное положение интронов в генах константных областей, предоставляемых векторами, показаны на фиг.13 и приведены в SEQ ID NO: 29 и SEQ ID NO: 28 соответственно для легкой цепи и SEQ ID NO: 31 и SEQ ID NO: 30 соответственно для тяжелой цепи.
ДНК, кодирующую указанные гены легкой и тяжелой цепи, вырезали из указанных векторов. ДНК тяжелой цепи расщепляли в 5'-сайте HindIII, затем обрабатывали фрагментом Кленова ДНК полимеразы I E. coli, чтобы создать тупой 5'-конец. Расщепление в 3'-сайте EcoRI давало в результате фрагмент тяжелой цепи, который очищали из агарозных гелей. Таким же образом получали фрагмент легкой цепи, затупленный в 5'-сайте SfuI и с 3'-сайтом EcoRI. Оба фрагмента клонировали в основанных на DHFR экспрессирующих векторах и использовали для создания стабильных клеточных линий на клетках CHO.














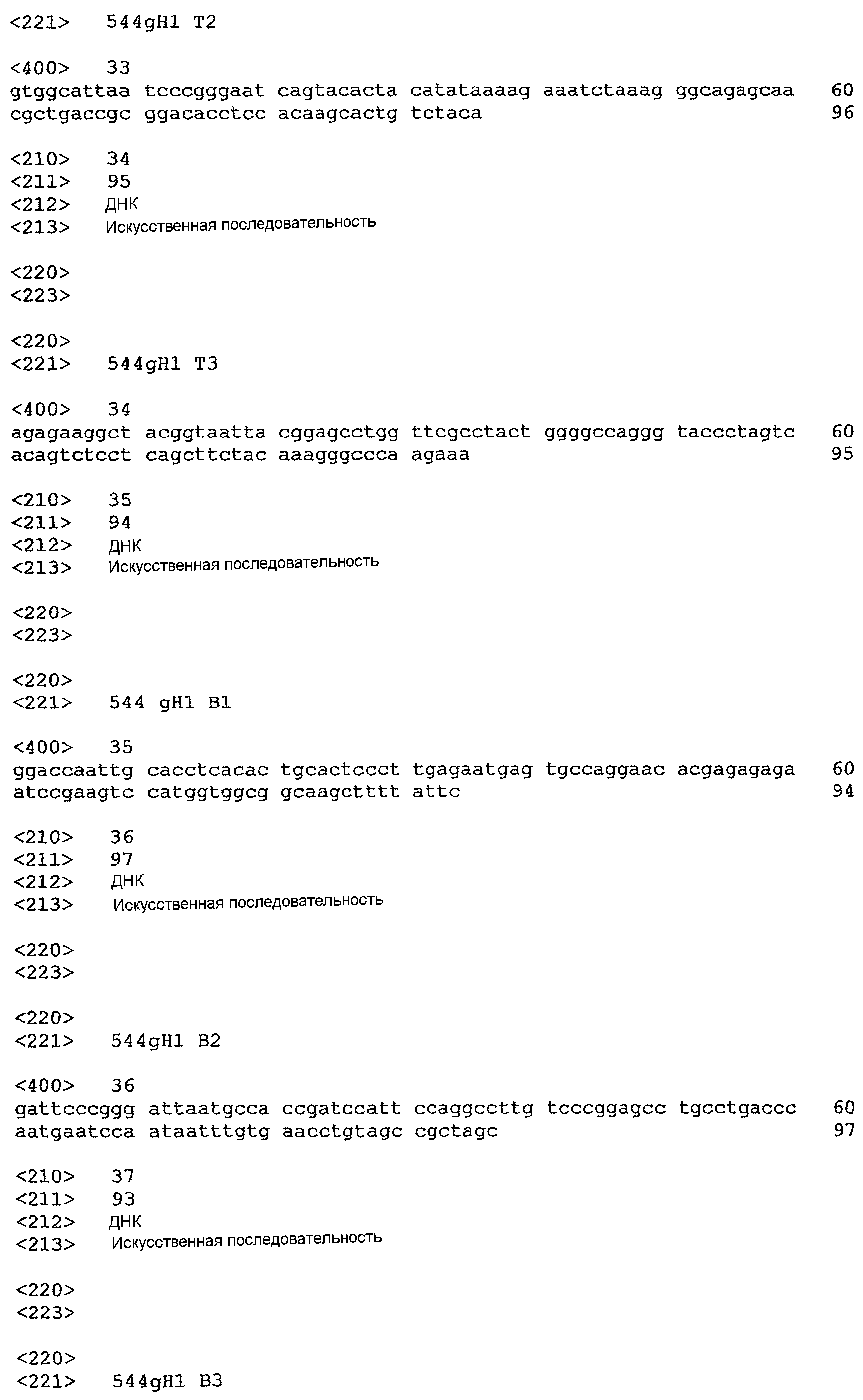




Реферат
Изобретение относится к биотехнологии. Описаны молекулы антител, обладающие специфичностью в отношении CD22 человека, содержащие, по меньшей мере, одну CDR, полученную из мышиного моноклонального антитела. Также раскрыто CDR-привитое антитело, в котором, по меньшей мере, одна из CDR является модифицированной CDR. Описаны последовательности ДНК, кодирующие цепи молекул антител. Предложены векторы, содержащие описанные последовательности ДНК, трансформированные клетки-хозяева, содержащие предложенные векторы. Раскрыто применение молекул антител при лечении заболеваний, опосредованных клетками, экспрессирующими CD22. Изобретение позволяет получать антитело, обладающее высокой аффинностью в отношении CD22 и низкой иммуногенностью для человека. 14 н. и 14 з.п. ф-лы, 13 ил.
Формула
Документы, цитированные в отчёте о поиске
Дизамещенные фураноны, тиазолы и пентеноны в качестве ингибиторов циклооксигеназы, способ их получения, фармацевтическая композиция и способ лечения
Комментарии