Простатоассоциированные антигены и иммунотерапевтические схемы на основе вакцин - RU2609651C2
Код документа: RU2609651C2
Чертежи



















































































































































































































































Описание
ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Эта заявка притязает на приоритет предварительной заявки США №61/642844, поданной 4 мая 2012, которая во всей полноте включена в данный документ путем ссылки.
ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Это заявка подается вместе с перечнем последовательностей в электронной форме. Перечень последовательностей предоставлен в виде файла в текстовом формате под названием «РС71854А SEQ LISTING_ST25.TXT», созданного 4 апреля 2013 года и имеющего размер 257Кб. Перечень последовательностей, содержащихся в текстовом формате, является частью заявки и включен в данное описание посредством ссылки во всей полноте.
ОБЛАСТЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Настоящее изобретение в целом относится к иммунотерапии и особенно к вакцинам и способам лечения и предотвращения неопластических нарушений.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Рак является ведущей причиной смертности во всем мире. Традиционные схемы лечения рака добились успеха в лечении селективной группы циркулирующих и солидных раков. Тем не менее, многие опухоли устойчивы к традиционным подходам. В последние годы была исследована иммунотерапия для лечения рака, которая включает генерацию активного системного опухолеспецифического иммунного ответа хозяина, возникающего вследствие введения вакцинной композиции в участок, удаленный от опухоли. Были предложены различные типы вакцин, в том числе те, которые содержат выделенные опухолевые антигены.
Рак предстательной железы является вторым, наиболее часто диагностируемым раком и четвертой по значимости причиной смерти вследствие рака у мужчин в развитых странах по всему миру. Различные ассоциированные с простатой антигены (РАА), такие как простатоспецифический антиген (PSA), простатоспецифический мембранный антиген (PSMA) и антиген стволовой клетки простаты (PSCA), как было показано, избыточно экспрессируются клетками рака простаты по сравнению с нормальными аналогами. Поэтому, эти антигены представляют собой возможные цели для индукции специфических иммунных ответов против раков, экспрессирующих антигены, посредством использования иммунотерапии, основанной на вакцинах, (смотри, например, Marrari, Α., M. lero, et al. (2007). "Vaccination therapy in prostate cancer." Cancer Immunol Immunother 56(4):429-45.)
PSCA является 123-аминокислотным мембранным белком. Аминокислотная последовательность непроцессированного человеческого PSCA состоит из аминокислот 4-123 SEQ ID №21. PSCA имеет высокую тканевую специфичность и экспрессируется более чем 85% образцами рака простаты при уровнях экспрессии, повышающихся с более высокими индексами Глиссона и андрогенной независимостью. Он экспрессируется в 80-100% костных метастазов у больных раком предстательной железы.
PSA является калликреиноподобной сериновой протеазой, которая продуцируется исключительно столбчатыми эпителиальными клетками, выстилающими ацинус и протоки предстательной железы. Матричная РНК PSA транслируется в виде 261-аминокислотного препроPSA предшественника. ПрепроРБА имеет 24 дополнительных остатков, которые составляют предварительную область (сигнальный полипептид) и прополипептид. Высвобождение прополипептида приводит к 237-аминокислоте, зрелой внеклеточной форме, которая является ферментативно активной. Аминокислотная последовательность человеческого непроцессированного PSA представлена в SEQ ID №15. PSA является органоспецифическим и, как результат, его продуцируют эпителиальные клетки доброкачественной гиперпластической (ВРН) ткани, ткани первичного рака простаты или ткани метастатического рака простаты.
PSMA, также известный как фолатгидролаза 1 (FOLH1), состоит из 750 аминокислот. Аминокислотная последовательность непроцессированного PSMA представлена в SEQ ID NO: 1. PSMA включает цитоплазматический домен (аминокислоты 1-19), трансмембранный домен (аминокислоты 20-43) и внеклеточный домен (аминокислоты 44-750). PSMA представляет собой тип II димерного трансмембранного белка, экспрессируемого на поверхности клеток рака простаты и на неоваскулатуре. Он также экспрессируется на нормальных клетках простаты, головного мозга, слюнной железы и панкреатическое дерево. Однако, было обнаружено, что в клетках рака простаты он экспрессируется при в 1000 раз высших уровнях, чем в нормальных тканях. Он с избытком экспрессируется на неоваскулатуре других разных солидных опухолей, таких как опухоли толстой кишки, молочной железы, печени, мочевого пузыря, поджелудочной железы, легких, почек, а также меланомы и саркомы. Таким образом, PSMA считается мишенью не только специфической для клеток рака простаты, но также панкарциномной мишенью для других видов рака. Экспрессирование PSMA является универсальной характеристикой карциномы простаты и его повышенная экспрессия коррелирует с агрессивностью опухоли. Экспрессия PSMA является повышенной при опухолях высокой степени злокачественности, метастатических поражениях и андрогенонезависимом заболевании.
Несмотря на то, что было выявлено большое количество опухолевых антигенов и многие из этих антигенов были изучены в качестве вакцин на основе белков или на основе ДНК для лечения или профилактики рака, большинство клинических испытаний до сих пор не привели к терапевтическому продукту. Одна из проблем при разработке вакцин рака состоит в том, что раковые антигены, как правило, получаются сами по себе и, следовательно, являются слабо иммуногенными, потому что иммунная система является саморегулируемой и не распознает собственные белки. Соответственно, существует потребность в способе повышения иммуногенности или терапевтического эффекта противораковых вакцин.
Для повышения иммуногенности или повышения противоопухолевой эффективности вакцин рака были исследованы многочисленные подходы. Один из таких подходов включает использование различных иммунных модуляторов, таких как агонисты TLR, агонисты TNFR, ингибиторы CTLA-4 и ингибиторы протеинкиназы.
Toll-подобные рецепторы (TLR) относятся к типу 1 мембранных рецепторов, которые экспрессируются на кроветворных и некроветворных клетках. По меньшей мере, 11 представителей были определены в семействе TLR. Эти рецепторы характеризуются по их способности распознавать патоген-ассоциированные молекулярные паттерны (РАМР), экспрессируемые патогенными организмами. Было обнаружено, что инициирование TLR вызывает серьезные воспалительные реакции путем повышения продукции цитокинов, экспрессии рецепторов хемокинов (CCR2, CCR5 и CCR7) и костимулирующей молекулярной экспрессии. Как таковые, эти рецепторы во врожденных иммунных системах осуществляют контроль над полярностью последующего приобретенного иммунного ответа. Среди TLR TLR9 широко исследуется по поводу его функций в иммунных реакциях. Стимуляция рецепторов TLR9 направляет антиген-представляющие клетки (АРС) в направлении примирующих высокоактивных ТН1-доминирующих Т-клеточных реакций, за счет увеличения продуцирования провоспалительных цитокинов и представления костимулирующих молекул к Т-клеткам. Было обнаружено, что олигонуклеотиды CpG, лиганды для TLR9, представляют класс сильнодействующих иммуностимулирующих факторов. CpG-терапия была испытана против широкого спектра моделей опухолей у мышей, и, как было показано, сообразно способствует ингибированию или регрессии опухоли.
Цитотоксический Т-лимфоцитарный антиген 4 (CTLA-4) является членом суперсемейства иммуноглобулинов и экспрессируется на поверхности Т-хелперов. CTLA-4 является негативным регулятором активации CD28, зависящим от активации Т-клеток, и действует в качестве контрольной точки для ингибирующего адаптивного иммунного ответа. Как и у костимулирующего белка CD28 Т-клеток, CTLA-4 связывается с CD80 и CD86 на антиген-представляющих клетках. CTLA-4 передает ингибирующий сигнал Т-клеток, в то время как CD28 передает стимулирующий сигнал. Человеческие антитела против человеческого CTLA-4 были описаны как иммуностимулирующие модуляторы в ряде болезненных состояний, таких как лечение или профилактика вирусной и бактериальной инфекции и для лечения рака (WO 01/14424 и WO 00/37504). Различные доклинические исследования показали, что блокада CTLA-4 моноклональными антителами усиливает иммунный ответ против иммуногенных опухолей и может даже отвергнуть раковые опухоли. Два полностью человеческие моноклональные антитела к человеческому CTLA-4 (mAb), ипилимумаб (MDX-010) и тремелимумаб (также известный как СР-675206), были исследованы в клинических испытаниях при лечении различных видов солидных опухолей.
Суперсемейство фактора некроза опухоли (TNF) представляет собой группу цитокинов, которые привлекают специфические родственные рецепторы клеточной поверхности, суперсемейство рецепторов TNF (TNFR) надсемейство. Члены суперсемейства факторов некроза опухоли действуют через лиганд-опосредованную тримеризацию, вызывая рекрутинг нескольких внутриклеточных адаптеров для активации нескольких путей сигнальной трансдукции, таких как апоптоз, путь NF-KB, путь JNK, а также иммунных и воспалительных реакций. Примеры суперсемейства TNF включают лиганды CD40, лиганды ОХ40, лиганды 4-1 ВВ, лиганды CD27, лиганд CD30 лиганд (CD153), лиганды TNF-альфа, TNF-бета, RANK, LT-альфа, LT-бета, GITR и LIGHT. Суперсемейство TNFR включает, например, CD40, ОХ40, 4-1 ВВ, CD70 (лиганд CD27), CD30, TNFR2, RANK, LT-бета R, HVEM, GITR, TROY и RELT. CD40 обнаруживается на поверхности В-лимфоцитов, дендритных клеток, фолликулярных дендритных клеток, гемопоэтических клеток-предшественников, эпителиальных клеток и карцином. CD40 связывается с лигандом (CD40-L), который представляет собой гликопротеин и экспрессируется на активированных Т-клетках, в основном CD4+, но и на некоторых CD8+, а также базофилах/тучных клетках. Из-за роли CD40 во врожденном и адаптивном иммунных ответах, агонисты CD40, включая различные агонистические антитела CD40, такие как полностью человеческое моноклональное антитело СР870893 к CD40, были изучены для использования в качестве адъювантов вакцин, а также в терапии.
Протеинкиназы представляют собой семейство ферментов, которые катализируют фосфорилирование специфических остатков в белках. Протеинкиназы являются ключевыми элементами в сигнальных путях, ответственных за трансдукцию внеклеточных сигналов, включая действие цитокинов на их рецепторы, к ядрам, вызывая различные биологические события. Многие роли протеинкиназ в нормальной физиологии клеток включают контроль клеточного цикла и рост клеток, дифференцировку, апоптоз клеток, подвижность и митогенез. Киназы, такие как C-Src, С-Аbl, митоген-активируемая протеинкиназа (MAP), фосфотидилинозитол-3-киназа (PI3K) АКТ и рецептор эпидермального фактора роста (EGF), как правило, активируются в раковых клетках, и, как известно, способствуют к туморогенезу. Логически, в настоящее время разрабатывается ряд ингибиторов киназ для противораковой терапии, в частности ингибиторы тирозинкиназы (TKI): циклин-зависимые ингибиторы киназы, ингибиторы киназы Аврора, ингибиторы контрольных точек клеточного цикла, рецептора эпидермального фактора роста (ингибиторы EGFR), FMS-подобные ингибиторы тирозинкиназы, рецептора тромбоцитарного фактор роста (PDGFR), ингибиторы киназы домена вставки, ингибиторы, ориентированные на путь PI3K/Akt/mTOR, ингибиторы, ориентированные на путь Ras-Raf-MEK-ERK (ERK), киназные ингибиторы рецептора фактора роста эндотелия сосудов (VEGFR), c-kit ингибиторы и киназные ингибиторы серина/треонина. Ряд ингибиторов киназы был исследован в клинических испытаниях по поводу использования в противораковых препаратах, которые включают, например, МК0457, VX-680, ZD6474, MLN8054, AZD2171, СНС-032, РТК787 / ZK222584, сорафениб (BAY43-9006), SU5416, SU6668 AMG706, зактима (ZD6474), МР-412, дазатиниб, КЭП-701, (лестауртиниб), XL647, XL999, тикерб (лапатиниб), MLN518 (ранее известный как СТ53518), РКС412, ST1571, AMN107, АЕЕ 788, OSI-930, OSI-817, сунитиниба малат (сутент; SU1 1248), ваталаниб (PTK787/ZK 222584), SNS-032, SNS-314 и акситиниб (AG-013736). Гефитиниб и эрлотиниб представляют собой перорально доступный EGFR-TKI.
Иммуномодуляторы, которые были изучены, как правило, вводят системно пациентам, например, путем перорального введения, внутривенных инъекций или вливаний, или внутримышечной инъекции. Одним из основных факторов, которые ограничивают эффективное применение некоторых иммунных модуляторов, является токсичность, вызванная высоким системным воздействием вводимых агентов. Например, относительно СО40-агонистов, сообщалось, что 0,3 мг/кг является максимально переносимой дозой для примера антитела к CD40 и что более высокие дозы могут вызывать побочные эффекты, включая тромбоэмболии, головные боли 3 степени, высвобождение цитокинов, вызывающее токсичные эффекты, такие как озноб и тому подобное, и переходящую токсичность печени. (Vanderheide et al., J Clin. Oncol. 25(7):876-8833 (March 2007)). В клиническом испытании для исследования комбинации внутривенного тремелимумаба (антитело к CTLA-4) с пероральным сунитинибом у пациентов с метастатической почечно-клеточной карциномой наблюдалось быстрое начало почечной недостаточности и, как следствие, не рекомендовалось дальнейшее исследование тремелимумаба в дозах более высоких, чем 6 мг/кг плюс сунитиниб в дозе 37,5 мг в день. Смотрите: Brian I. Rini et al.: Phase 1 Dose-Escalation Trial of Tremelimumab Plus Sunitinib in Patients With Metastatic Renal Cell Carcinoma. Cancer 117(4):758-767 (2011). Таким образом, существует потребность в иммунотерапевтических схемах на основе вакцин, где иммуномодуляторы вводят в эффективных дозах, которые не вызывают серьезных побочных эффектов, таких как токсичность печени или почечную недостаточность.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В некоторых аспектах настоящее изобретение предусматривает выделенные иммуногенные полипептиды PSMA и иммуногенные полипептиды PSA, которые являются полезными, например, для индукции иммунного ответа in vivo (например, у животных, включая человека) или in vitro, генерирования антител или для использования в качестве компонента в вакцинах для лечения рака, включая рак простаты. В одном аспекте, настоящее изобретение предусматривает выделенные иммуногенные полипептиды PSMA, которые имеют, по меньшей мере, 90% идентичность аминокислотам 15-11, 12, 13, 14, 15, 16, 17, 18, или 19 из консервативных Т-клеточных эпитопов человеческого PSMA в соответствующих положениях.
В других аспектах настоящее изобретение предусматривает молекулы нуклеиновых кислот, которые кодируют иммуногенные полипептды РАА. В некоторых вариантах осуществления настоящее изобретение предусматривает выделенные молекулы нуклеиновых кислот или их вырожденные варианты, которые содержат нуклеотидную последовательность, кодирующую иммуногенный полипептид PSMA или функциональный вариант указанного полипептида, предусмотренный данным описанием
В некоторых других аспектах настоящее изобретение предусматривает мультиантигенные конструкты нуклеиновых кислот, где каждый кодирует два или более иммуногенных полипептида РАА.
Описание также предусматривает векторы, содержащие одну или несколько молекул нуклеиновых кислот согласно изобретению. Векторы могут быть использованы для клонирования или экспрессии иммуногенных полипептидов РАА, кодируемых молекулами нуклеиновых кислот, или для доставки молекул нуклеиновых кислот в композиции, такой как вакцина, к клетке-хозяину или животному-хозяину, такому как человек.
В некоторых дополнительных аспектах настоящее изобретение предусматривает композиции, содержащие один или более иммуногенных полипептидов РАА, выделенные молекулы нуклеиновых кислот, кодирующие иммуногенные полипептиды РАА, или векторы, или плазмиды, содержащие молекулы нуклеиновых кислот, кодирующие иммуногенные полипептиды РАА. В некоторых вариантах осуществления композиция является иммуногенной композицией, полезной для индукции иммунного ответа против РАА в организме млекопитающего, такого как мышь, собака, обезьяна или человека. В некоторых вариантах осуществления композиция является вакцинной композицией, полезной для иммунизации млекопитающего, такого как человек, для ингибирования аномальной клеточной пролиферации, для обеспечения защиты против развития рака (используется в качестве профилактики) или для лечения нарушений (используется в качестве терапии), связанных с повышенной экспрессией РАА, таких как рак, особенно рак простаты.
В еще других аспектах настоящее изобретение предусматривает способы использования иммуногенных полипептидов РАА, выделенных молекул нуклеиновых кислот и композиций, содержащих иммуногенный полипептид РАА или выделенные молекулы нуклеиновых кислот, описанные в данном документе выше. В некоторых вариантах осуществления настоящее изобретение предусматривает способ индукции иммунного ответа против РАА в организме млекопитающего, в частности человека, включающий введение указанному млекопитающему эффективного количества полипептида, предусмотренного изобретением, который является иммуногенным по отношению к целевому РАА, эффективного количества выделенной молекулы нуклеиновой кислоты, кодирующей такой иммуногенный полипептид, или композиции, содержащей такой иммуногенный полипептид РАА. Вакцины полипептида или нуклеиновой кислоты могут быть использованы вместе с одним или несколькими адъювантами.
В еще других аспектах настоящее изобретение предусматривает схемы иммунотерапии на основе вакцины (или "VBIR"), которые включают совместное введение вакцины, доставляющей разные опухолеассоциированные антигены (ТАА) для индуцирования ТАА-специфических иммунных ответов для лечения разнообразных раков, в комбинации с, по меньшей мере, одним иммуноугнетающим клеточным ингибитором и, по меньшей мере, одним иммуноэффекторным клеточным энхансером. В частности, в некоторых аспектах изобретение предусматривает способ усиления иммуногенности или терапевтического эффекта вакцины для лечения опухолевого заболевания у млекопитающего, включающий введение млекопитающему, получающему вакцину, эффективного количества, по меньшей мере, одного иммунноугнетающего клеточного ингибитора и, по меньшей мере, одного иммунноэффекторного клеточного энхансера. В дополнительном аспекте изобретение предусматривает способ лечения опухолевого заболевания у млекопитающего, включающий введение млекопитающему вакцины, по меньшей мере, одного иммуноугнетающего клеточного ингибитора и, по меньшей мере, одного иммунноэффекторного клеточного энхансера.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
ФИГ. 1. Схематическая иллюстрация вектора PJV7563.
ФИГ. 2. Аминокислотное выравнивание пяти вирусных кассет 2А. Пропущенные глицин-пролиновые связи помечены звездочками.
ФИГ. 3. Последовательность предпочтительного EMCV IRES. Сайт инициации трансляции обозначен звездочкой. Минимальный элемент IRES исключает подчеркнутые первые 5 кодонов белка EMCV L.
ФИГ. 4. Точечные участки, показывающие экспрессию человеческого модифицированного антигена PSMA (аминокислоты 15-750) и непроцессированного человеческого PSCA на поверхности клеток НЕК293, трансфицированных двойными
ФИГ. 5. Изображение вестерн-блоттинга, показывающее экспрессию человеческого модифицированного антигена PSMA (аминокислоты 15-750) и непроцессированного человеческого PSCA в клетках НЕК293, трансфицированных двойными антигенными вакцинными конструктами, как измерено с помощью вестерн-блоттинга со специфическими моноклональными антителами к PSMA и PSCA.
ФИГ. 6. Изображение вестерн-блоттинга, показывающее экспрессию человеческого цитостатического антигена PSA (аминокислоты 25-261) в клетках НЕК293, трансфицированных двойными антигенными вакцинными конструктами, как измерено с помощью вестерн-блоттинга со специфическим моноклональным антителом к PSA. Дорожка 5300 демонстрирует относительно слабую полосу в приблизительно 2 кДа больше, чем PSA, в соответствии с С-концевым слиянием белка 2А.
ФИГУРЫ 7А, 7В. Точечная диаграмма, показывающая экспрессию человеческого модифицированного антигена PSMA (аминокислоты 15-750) и непроцессированного человеческого PSCA на поверхности клеток НЕК293, трансфицированных триплетноантигенным вакцинным конструктом, измеренную с помощью проточной цитометрии. (ФИГ. 7А. Одноантигенные контроли и однопромоторные трехантигенные конструкты. ФИГ. 7В. Двухпромоторные трехантигенные конструкты).
ФИГУРЫ 8А, 8В. Изображения вестерн-блоттинга, показывающие экспрессию человеческого PSA в клетках НЕК293, трансфицированных триплетноантигенными вакцинными конструктами, как измерено с помощью вестерн-блоттинга со специфическими моноклональными антителами PSA. Полосы на дорожках 5259 и 456 выходят за пределы дорожки 5297. Хотя это и не видно в сканированном геле, дорожки 456, 457 и 458 и выставили полосу в приблизительно 2 кДа больше, чем PSA, в соответствии с С-концевым слиянием пептида 2А. (ФИГ. 8А. Однопромоторные триплетноантигенные конструкты. ФИГ. 8В. Двухпромоторные трехантигенные конструкты)
ФИГУРЫ 9A-9D. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность трехантигенных вакцин с помощью анализа IFN-γ ELISPOT.
ФИГУРЫ 10A-10D. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность трехантигенных вакцин с помощью анализа IFN-γ ELISPOT assay.
ФИГ. 11. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность трехантигенных вакцин с помощью титров антител к PSMA.
ФИГ. 12. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность трехантигенных вакцин с помощью титров антител к PSCA.
ФИГ. 13. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность трехантигенных вакцин с помощью поверхностноклеточного связывания антител к PSMA.
ФИГ. 14. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность трехантигенных вакцин с помощью поверхностноклеточного связывания антител к PSCA.
ФИГУРЫ 15А-15С. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью анализа IFN-γ ELISPOT.
ФИГУРЫ 16А-16С. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью анализа IFN-γ ELISPOT.
ФИГ. 17. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью титров антител к PSMA.
ФИГ. 18. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью титров антител к PSCA.
ФИГ. 19. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью поверхностноклеточного связывания антител к PSMA.
ФИГ. 20. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью поверхностноклеточного связывания антител к PSCA.
ФИГУРЫ 21А-21D. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью анализа IFN-γ ELISPOT.
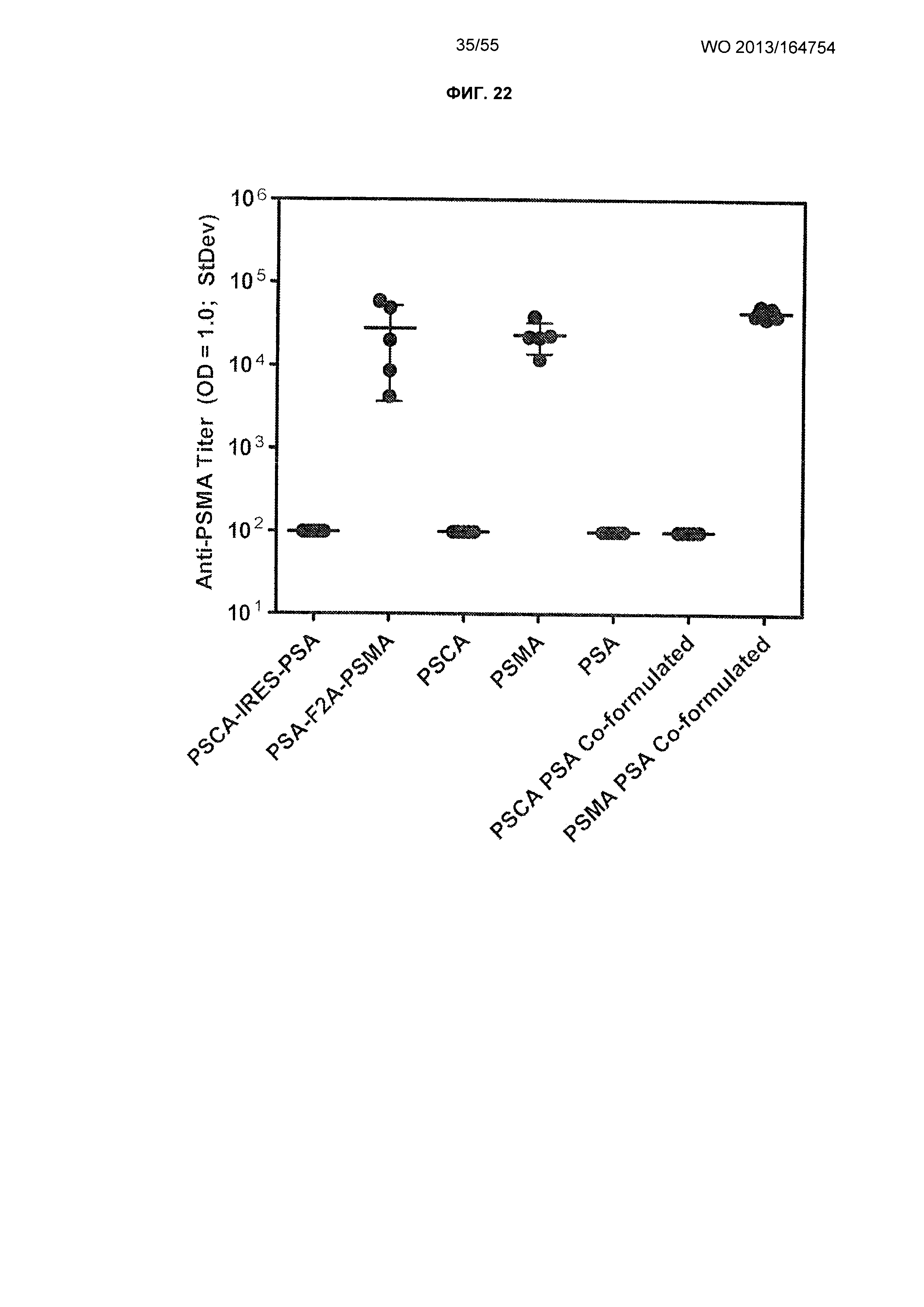
ФИГ. 22. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью титров антител к PSMA.
ФИГ. 23. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью титров антител к PSCA.
ФИГ. 24. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью поверхностноклеточного связывания антител к PSMA.
ФИГ. 25. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью поверхностноклеточного связывания антител к PSCA.
ФИГ. 26. Графики, изображающие результаты репрезентативного исследования, которое оценивает Т-клеточный иммунный ответ, вызванный человеческим модифицированным антигеном PSMA (аа 15-750), по сравнению с непроцессированным PS MA (аа 1-750) у мышей C57BL/6.
ФИГУРЫ 27А, 27В. Графики, изображающие результаты репрезентативного исследования, которое оценивает Т-клеточный иммунный ответ, вызванный человеческим модифицированным антигеном PSMA (аа 15-750) по сравнению с непроцессированным антигеном PSMA (аа 1-750) у трансгенных мышей Пастера (HLA-А2/DR1) с помощью анализа IFN-Y ELISPOT.
ФИГ. 28. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность вакцин человеческого модифицированного и непроцессированного PSMA с помощью титров антител к PSMA.
ФИГ. 29. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуногенность вакцин человеческого модифицированного и непроцессированного PSMA с помощью поверхностноклеточного связывания антител к PSMA.
ФИГ. 30. Графики, изображающие результаты репрезентативного исследования, которое оценивает уровни моноклональных антител к CTLA-4 в крови, измеренные с помощью конкурентного ELISA у индийских макак-резус, инъецированных антителом к CTLA-4 (СР-675, 206) в дозе 10 мг/кг.
ФИГУРЫ 31А и 31В. Графики, изображающие результаты репрезентативного исследования, которое оценивает иммуномодулирующую активность моноклонального антитела к мышиному CTLA-4 (клон 9Н10) по количеству вакцины, вызывающей иммунные реакции, с помощью анализа окрашивания внутриклеточных цитокинов.
ФИГ. 32. Графики, изображающие результаты репрезентативного исследования, которое оценивает и сравнивает скорость роста подкожной опухоли при лечении малатом сунитиниба (Sutent) в качестве монотерапии или в комбинации с контролем (контроль) или вакциной против рака (rHER2).
ФИГУРЫ 33А-33D. Графики, показывающие индивидуальные темпы роста опухоли у мышей из репрезентативного исследования, оценивающего и сравнивающего противоопухолевую эффективность сунитиниба малата (Sutent) в дозе 20 мг/кг с контролем (контроль) или противораковой вакциной (rHER2).
ФИГ. 34. График, показывающий кривые выживаемости Каплана-Мейера групп мышей из исследования, описанного на Фигуре 33, которое оценивает противоопухолевую эффективность сунитиниба малата (Sutent) с контрольной (контроль) или противоопухолевой вакциной (rHER2), которых лечили малатом сунитиниба (Sutent) с контрольной (контроль) или противоопухолевой вакциной (рак).
ФИГУРЫ 35А, 35В. Графики, показывающие изменения в миелоидных супрессорных клетках (Gr1+CD11b+) и Treg-содержащих CD25+CD4+ клеток в периферической крови мышей, лечившихся как на Фигуре 33.
ФИГУРЫ 36А-36С. Графики, показывающие изменения общего количества миелоидных супрессорных клеток (Gr1+CD11b+), клеток Treg (CD4+CD25+Foxp3+) и PD-1+CD8 Т-клеток, выделенных из опухолей мышей.
ФИГ. 37. График, показывающий кривые выживаемости Каплана-Мейера групп мышей из исследования, которое оценивает противоопухолевую эффективность сунитиниба малата (Sutent), моноклонально антитела к мышиному CTLA-4 (клон 9D9), в комбинации с контрольной (контроль) или противораковой вакциной (вакцина) у мышей BALB/neuT, несущих подкожные опухоли TUBO.
ФИГ. 38. График, показывающий кинетику уровней сунитиниба в крови мышей BALB/neuT с подкожными опухолями TUBO.
ФИГ. 39. График, показывающий кривые выживаемости Каплана-Мейера групп мышей из исследования, которое оценивает противоопухолевую эффективность у мышей BALB/neuT с подкожными опухолями TUBO, лечившихся малатом сунитиниба (Sutent) и контрольной или противораковой вакциной.
ФИГ. 40. График, показывающий результаты репрезентативного исследования IFNy ELISPO, которое оценивает амплитуду антиген-специфических Т-клеточных реакций, вызванных вакциной rHER2 в группах мышей при введении с иммунными модуляторами.
ФИГУРЫ 41А, 41В. Графики, показывающие результаты репрезентативного исследования, которое оценивает иммуномодуляторную активность CpG7909 (PF-03512676) по качеству индуцированного вакциной иммунного ответа с помощью анализа внутриклеточного окрашивания от цитокинов.
ФИГУРЫ 42А, 42В. Графики, показывающие результаты репрезентативного исследования, которое оценивает иммуномодуляторную активность агонистического моноклонального антитела к мышиному CD40 по качеству индуцированного вакциной иммунного ответа с помощью анализа внутриклеточного окрашивания от цитокинов.
ФИГ. 43. График, показывающий кривые выживаемости Каплана-Мейера групп мышей из исследования, которое оценивает противоопухолевую эффективность низкой дозы малата сунитиниба (Sutent) и контрольной или противораковой вакцины у мышей BALB/neuT со спонтанными опухолями молочной железы.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
А. ОПРЕДЕЛЕНИЯ
Термин "адъювант" относится к веществу, которое способно повысить, ускорить или продлить иммунный ответ при введении с вакцинным иммуногеном.
Термин "агонист" относится к веществу, которое способствует (индуцирует, вызывает, усиливает или повышает) активность другой молекулы или рецептора. Термин агонист охватывает вещества, которые связывают рецептор (например, антитело, гомолог природного лиганда из других видов), и вещества, которые способствуют рецепторной функции без вещества, которые способствуют функции рецептора без привязки к ней (например, путем активации связанного белка).
Термин "антагонист" или "ингибитор" относится к веществу, которое частично или полностью блокирует, ингибирует или нейтрализует биологическую активность другой молекулы или рецептора.
Термин "совместное введение" относится к введению двух или более средств одному и тому же субъекту в течение периода лечения. Два или более средств могут находиться в одной композиции и, следовательно, вводиться одновременно. Альтернативно, два или более средств могут находиться в разных физических композициях и вводиться раздельно, последовательно или одновременно субъекту. Термин "вводить одновременно" или "одновременное введение" означает, что введение первого средства и второго средства перекрываются по времени друг с другом, в то время как термин "вводить последовательно" или "последовательное введение" означает, что введение первого средства и второго средства не перекрываются по времени друг с другом.
Термин "консервативный Т-клеточный эпитоп" относится к одной из следующих аминокислотных последовательностей человеческого белка PSMA, в порядке, предусмотренном в SEQ ID NO. 1:
аминокислоты 168-176 (GMPEGDLVY),
аминокислоты 347-356 (HSTNGVTRIY),
аминокислоты 557-566 (ETYELVEKFY),
аминокислоты 207-215 (KVFRGNKVK),
аминокислоты 431-440 (STEWAEENSR),
аминокислоты 4-12 (LLHETDSAV),
аминокислоты 27-35 (VLAGGFFLL),
аминокислоты 168-177 (GMPEGDLVYV),
аминокислоты 441-450 (LLQERGVAYI),
аминокислоты 469-477 (LMYSLVHNL),
аминокислоты 711-719 (ALFDIESKV),
аминокислоты 663-671 (MNDQVMFL),
аминокислоты 178-186 (NYARTEDFF),
аминокислоты, 227-235 (LYSDPADYF),
аминокислоты 624-632 (TYSVSFDSL),
аминокислоты 334-348 (TGNFSTQKVKMHIHS),
аминокислоты 459-473 (NYTLRVDCTPLMYSL),
аминокислоты 687-701 (YRHVIYAPSSHNKYA), и
аминокислоты 730-744 (RQIYVAAFTVQAAAE).
Термин "цитозольный" означает, что после нуклеотидной последовательности, кодирующей конкретный полипептид, экспрессирующийся в клетке-хозяине, экспрессированный полипептид удерживается внутри клетки-хозяина.
Термин "вырожденный вариант" относится к последовательностям ДНК, которые имеют замещения оснований, но кодируют тот же самый полипептид.
Термин "эффективное количество" относится к количеству, введенному млекопитающему, которое является достаточным для того, чтобы вызвать требуемый эффект у млекопитающего.
Термин "фрагмент" данного полипептида относится к полипептиду, который короче, чем данный полипептид, и разделяет 100% идентичность с последовательностью данного полипептида.
Термин "идентичный" или процент "идентичности" в контексте двух или более последовательностей нуклеиновых кислот или полипептидов, относится к двум или более последовательностями или подпоследовательностям, которые одинаковы или имеют определенный процент аминокислотных остатков или нуклеотидов, которые одинаковы при сравнении и выровнены для максимального соответствия.
Термин "энхансер иммуноэффекторных клеток" или "IEC энхансер" относится к веществу, способному повышать или усиливать количество, качество или функцию одного или нескольких типов иммунных эффекторных клеток млекопитающего. Примеры иммунных эффекторных клеток включают цитолитические CD8 Т-клетки, CD40 Т-лимфоциты, NK-клетки и В-клетки.
Термин "иммунный модулятор" относится к веществу, способному изменять (например, ингибировать, снижать, повышать, усиливать или стимулировать) работу любого компонента врожденной, гуморальной или клеточный иммунной системы млекопитающих. Таким образом, термин "иммуномодулятор" охватывает "энхансер иммуноэффекторных клеток", как определено в данном документе, и "ингибитор иммуносупрессорных клеток", как определено в данном документе, а также вещество, которое влияет на другие компоненты иммунной системы млекопитающего.
Термин "иммунный ответ" относится к любому обнаруживаемому ответу на конкретное вещество (например, антиген или иммуноген) с помощью иммунной системы хозяина - позвоночного животного, включая врожденные иммунные реакции (например, активацию сигнального каскада Toll-рецептора), клеточноопосредованные иммунные реакции (например, реакции, опосредованные Т-клетками, такими как антигенспецифические Т-клетки, и неспецифическими клетками иммунной системы) и гуморальные иммунные реакции (например, реакции, опосредованные В-клетками, такие как продуцирование и секреция антител в плазме, лимфе и/или тканевых жидкостях), но не ограничиваются ими. Примеры иммунных реакций включают изменение (например, повышение) активации Toll-подобного рецептора, экспрессии или секреции лимфокина (например, цитокина (например, Th1, Th2 или Th17 типов цитокинов) или хемокина), активации макрофагов, активации дендритных клеток, активации Т-клеток (например, CD4+ или CD8+ Т-клеток), активации NK-клеток, активации В-клеток (например, продуцирование или секрецию антител), связывания иммуногена (например, антиген (например, иммуногенного полипептида)) с молекулой МНС, индуцирование реакции цитотоксического Т-лимфоцита ("CTL"), индуцирование реакции В-клеток (например, продуцирование антител), и, распространение (например, рост популяции клеток) клеток иммунной системы (например, Т-клеток или В-клеток), и повышенного процессинга и представления антиген представляющими клетками. Термин "иммунная реакция" также охватывает любой обнаруживаемый ответ на конкретное вещество (такое как антиген или иммуноген) одного или более компонентов иммунной системы позвоночного животного in vitro.
Термин "иммуногенность" относится к способности вещества порождать, вызывать, стимулировать или индуцировать иммунный ответ, либо улучшать, повышать, увеличивать или продлевать уже существующий иммунный ответ на конкретный антиген, самого по себе или когда связано с носителем, в присутствии или в отсутствие адъюванта.
Термин "иммуногенный полипептид PSA" относится к полипептиду, который является иммуногенным против человеческого белка PSA, или против клеток, экспрессирующих человеческий белок PSA.
Термин "иммуногенный полипептид PSCA" относится к полипептиду, который является иммуногенным против человеческого белка PSCA, или против клеток, экспрессирующих человеческий белок PSCA.
Термин "иммуногенный полипептид PSMA" относится к полипептиду, который является иммуногенным против человеческого белка PSMA, или против клеток, экспрессирующих человеческий белок PSMA. Термин "иммуногенный полипептид РАА" относится к "иммуногенному полипептиду PSA", "иммуногенному полипептиду PSCA" или "иммуногенному полипептиду PSMA", как определено в данном документе выше.
Термин "молекула нуклеиновой кислоты иммуногенного PSA" относится к молекуле нуклеиновой кислоты, которая кодирует иммуногенный полипептид PSA, как определено в данном документе.
Термин "молекула нуклеиновой кислоты иммуногенного PSCA" относится к молекуле нуклеиновой кислоты, которая кодирует "иммуногенный полипептид PSCA", как определено в данном документе.
Термин "молекула нуклеиновой кислоты иммуногенного PSMA" относится к молекуле нуклеиновой кислоты, которая кодирует "иммуногенный полипептид PSMA", как определено в данном документе.
Термин "молекула нуклеиновой кислоты иммуногенного РАА" относится к молекуле нуклеиновой кислоты, которая кодирует "иммуногенный полипептид PSA", "иммуногенный полипептид PSCA" или "иммуногенный полипептид PSMA", как определено в данном документе выше.
Термин "ингибитор иммуносупрессивных клеток" или "ингибитор ISC" относится к веществу, способному снижать или подавлять количество или функцию иммуносупрессивных клеток млекопитающего. Примеры иммуносупрессивных клеток включают регуляторные Т-лимфоциты ("Т reg"), миелоидные супрессорные клетки и опухоль-ассоциированные макрофаги.
Термин "внутрикожное введение" или "вводят внутрикожно" в контексте введения вещества, например в качестве терапевтического агента или иммуномодулятора, млекопитающему, включая человека, относится к доставке вещества в дермальный слой кожи млекопитающего. Кожа млекопитающих состоит из трех слоев - эпидермиса, дермы и подкожного слоя. Эпидермис относительно тонкий, жесткий внешний слой кожи. Большинство клеток в эпидермисе являются кератиноцитами. Дерма, следующий слой кожи, является толстым слоем волокнистой и эластичной ткани (в основном из коллагена, эластина и фибриллина), который придает коже ее гибкость и прочность. Дерма содержит нервные окончания, потовые железы и масло (сальных) желез, волосяные фолликулы и кровеносные сосуды. Дерма изменяется по толщине в зависимости от места кожи. В организме человека она составляет приблизительно 0,3 мм на веке и приблизительно 3,0 мм на спине. Подкожный слой состоит из жира и соединительной ткани, где находятся более крупные кровеносные сосуды и нервы. Толщина этого слоя изменяется по всему телу и от человека к человеку. Термин "внутрикожное введение" относится к доставке вещества к внутренней стороне дермального слоя. В отличие от этого, "подкожное введение" относится к введению вещества в подкожный слой, а "местное введение" относится к введению вещества на поверхность кожи. Термин "местное введение" или "введенный местно" охватывает "местное введение", "внутрикожное введение" и "подкожное введение", как определено в данном документе выше, этот термин охватывает также "внутриопухолевое введение", которое относится к введению вещества во внутрь опухоли. Местное введение предназначено для обеспечения высоких локальных концентраций вокруг места введения в течение периода времени, пока не будет достигнуто системное биораспределение введенного вещества, в то время как "системное введение" предназначено для введения вещества для абсорбирования в кровь и достижения быстрого системного воздействия путем распространения через кровеносную систему к органам или тканям по всему телу.
Термин "млекопитающее" относится к любым видам животных класса млекопитающих. Примеры млекопитающих включают: людей; нечеловекообразных приматов, таких как обезьяны; лабораторных животных, таких как крысы, мыши, морские свинки; домашних животных, таких как кошки, собаки, кролики, крупный рогатый скот, овцы, козы, лошади и свиньи; и в неволе диких животных, таких как львы, тигры, слоны и подобные.
Термин "мембранно-связанный" означает, что после нуклеотидной последовательности, кодирующей конкретный полипептид, экспрессирующийся в клетке-хозяине, экспрессированный полипептид связывается с, прикрепляется к или иным образом соединяется с мембраной клетки.
Термин "опухолевое заболевание" относится к состоянию, при котором клетки размножаются с аномально высокой и неконтролируемой скоростью, в размере, превышающем и несогласованным с теми, что в окружающих нормальных тканях. Это обычно приводит к солидному поражению или комку, известному как "опухоль". Этот термин охватывает доброкачественные и злокачественные опухолевые расстройства. Термин "злокачественное опухолевое заболевание", которое используется как синоним термина "рак" в данном описании, относится к опухолевым заболеваниям, характеризующимся способностью опухолевых клеток распространяться на другие участки тела (известный как "метастаз"). Термин "доброкачественное опухолевое заболевание" относится к опухолевым заболеванием, при которых опухолевые клетки не обладают способностью метастазировать.
Термин "функционально связанный" относится к смежному положению, при котором компоненты, также описанные, находятся во взаимосвязи, позволяющей им функционировать предназначенным им способом. Контрольная последовательность, "функционально связанная" с трансгеном, лигирована таким образом, что экспрессия трансгена достигается в условиях, совместимых с контрольными последовательностями.
Термин "ортологические" относится к генам различных видов, которые похожи друг на друга и возникли от общего предка. Термин "фармацевтически приемлемый наполнитель" относится к веществу в иммуногенной композиции или вакцине, кроме активных ингредиентов (например, антиген, антигенкодирующая нуклеиновая кислота, иммуномодулятор или адъювант), который совместим с активными ингредиентами и не вызывает значительного неблагоприятного воздействия на субъектов, которым его вводят.
Термины "пептид", "полипептид" и "белок" используются в данном документе взаимозаменяемо и относятся к полимерной форме аминокислот любой длины, которая может включать кодированные и некодированные аминокислоты, химически или биохимически модифицированные или дериватизированные аминокислоты и полипептиды, имеющие модифицированные полипептидные скелеты.
Термин "предотвращение" или "предотвратить" относится к (а) удерживанию расстройства от появления или (б) более позднее начало заболевания или появления симптомов расстройства.
Термин "простатоассоциированный антиген" (или РАА) относится к ТАА (как определено в данном документе), который специфически экспрессируется на опухолевых клетках предстательной железы или экспрессируется с более высокой частотой или плотностью опухолевыми клетками, чем неопухолевыми клетками того же типа ткани. Примеры РАА включают PSA, PSCA и PSMA.
Термин "секретируется" в контексте полипептида означает, что после нуклеотидной последовательности, кодирующей полипептид, экспрессирующийся в клетке-хозяине, экспрессированный полипептид секретируется вне клетки-хозяина.
Термин "субоптимальная доза" при использовании для описания количества иммуномодулятора, такого как ингибитор протеинкиназы, относится к дозе иммунного модулятора, которая ниже минимального количества, необходимого для получения желаемого терапевтического эффекта для заболевания, подлежащего лечению, когда иммуномодулятор сам по себе вводят пациенту.
Термин "лечение" или "лечить" относится к упразднению расстройства, снижению тяжести расстройства или снижению тяжести или частоте встречаемости симптомом расстройства.
Термин "опухолеассоциированный антиген" или "ТАА" относится к антигену, который специфически экспрессируется в опухолевых клетках или экспрессируется с более высокой частотой или плотностью опухолевыми клетками, чем неопухолевыми клетками одного и того же типа ткани. Опухолевые антигены могут быть антигенами, обычно неэкспрессируемыми хозяином; они могут быть мутированными, усеченными, неправильно упакованными или другими неприятными проявлениями молекул, обычно экспрессируемых хозяином; они могут быть идентичными обычно экспрессируемым молекулам, но экспрессированными при аномально высоких уровнях; или они могут быть экспрессированными в контексте или среде, которые являются ненормальными. Опухолеассоциированные антигены могут быть, например, белками или фрагментами белков, сложными углеводами, ганглиозидами, гаптенами, нуклеиновыми кислотами или любой комбинацией этих или других биологических молекул. Термин "вакцина" относится к иммуногенной композиции для введения млекопитающему для индукции иммунного ответа против конкретного антигена.
Термин "вариант" данного полипептида относится к полипептиду, который разделяет менее 100%, но более 80% идентичность с аминокислотной последовательностью данного полипептида и демонстрирует, по меньшей мере, некоторую иммуногенную активность этого данного полипептида.
Термин "вектор" относится к молекуле нуклеиновой кислоты, способной транспортировать или передавать чужеродную молекулу нуклеиновой кислоты. Термин охватывает и векторы экспрессии, и векторы транскрипции. Термин "вектор экспрессии" относится к вектору, способному экспрессировать вставку в клетку-мишень, и обычно содержит регуляторные последовательности, такие как энхансерные, промоторные и терминирующие последовательности, которые управляют экспрессией вставки. Термин "вектор транскрипции" относится к вектору, способному к транскрипции, но не трансляции. Транскрипционные векторы используются для усиления их вставки. Молекулу чужеродной нуклеиновой кислоты называют "вставкой" или "трансгеном". Вектор обычно состоит из вставки и более длинной последовательности, которая служит в качестве основы вектора. Исходя из структуры или происхождения векторов, основные типы векторов включают плазмидные векторы, космидные векторы, фаговые векторы, такие как фаг лямбда, вирусные векторы, такие как аденовирусные (Ad) векторы, а также искусственные хромосомы.
В. ИММУНОГЕН ЯВЛЯЕТСЯ ПРОСТАТОАССОЦИИРОВАННЫМ АНТИГЕНОМ (РАА)
ПОЛИПЕПТИДЫ
В некоторых аспектах настоящее изобретение предусматривает выделенные иммуногенные полипептиды PSA и полипептиды PSMA, полезные, например, для вызова иммунной реакции in vivo (например, у животного, включая людей) или in vitro, активируя эффекторные Т-клетки или продуцирование антител, специфических к PSA и PSMA, соответственно, или для использования в качестве компонента в вакцинах для лечения рака, особенно рака простаты. Эти полипептиды могут быть получены с помощью методов, известных из уровня техники в свете настоящего описания. Способность полипептидов вызвать иммунную реакцию может быть измерена в in vitro анализах или в in vivo анализах. In vitro анализы определения способности полипептида или ДНК конструкта вызывать иммунную реакцию известны в данной области техники. Одним из примеров таких in vitro анализов является измерение способности полипептида или нуклеиновой кислоты, экспрессирующей полипептид, стимулировать реакцию Т-клеток, как описано в патенте США №7387882, описание которого включено в эту заявку. Метод количественного определения включает этапы: (1) контактирования антигенпрезентирующих клеток в культуре с антигеном таким образом, что антиген может быть принятым и обработанным с помощью антиген-представляющих клеток, продуцирующих один или более обработанных антигенов; (2) контактирование антиген-представляющих клеток с Т-клетками в условиях, достаточных для реакции Т-клеток в ответ на один или более из обработанных антигенов; (3) определения того, реагируют ли Т-клетки на один или более из обработанных антигенов. Использованные Т-клетки могут быть CD8+ Т-клетками или CD4+ Т-клетками. Т-клеточную реакцию можно определить путем измерения высвобождения одного из нескольких цитокинов, таких как интерферон-гамма и интерлейкин-2, лизиса антигенпрезентирующих клеток (опухолевые клетки), и продуцирования антител В-клетками.
В-1. Иммуногенный полипептид PSMA
В одном аспекте, настоящее изобретение предусматривает выделенный иммуногенный полипептид PSMA, который имеет, по меньшей мере, 90% идентичность с аминокислотами 15-750 человеческого PSMA из SEQ ID NO: 1 и содержит аминокислоты, по меньшей мере, 10, 11, 12, 13, 14, 15, 16, 17, 18 или 19, консервативных Т-клеточных эпитопов человеческого PSMA в соответствующих положениях.
В некоторых вариантах осуществления иммуногенный полипептид PSMA содержит аминокислоты, по меньшей мере, 15, 16, 17, 18 или 19, консервативных Т-клеточных эпитопов человеческого PSMA.
В некоторых вариантах осуществления настоящее изобретение предусматривает иммуногенный полипептид PSMA, состоящий из аминокислотной последовательности SEQ ID NO: 9, или иммуногенный полипептид PSMA, имеющий 93%-99%, 94%-98% или 94%-97% идентичность с аминокислотной последовательностью SEQ ID NO: 9.
Примеры некоторых отдельных иммуногенных полипептидов PSMA включают:
1) полипептид, состоящий из аминокислот 15-750 из SEQ ID NO: 1;
2) полипептид, содержащий аминокислоты 4-739 из SEQ ID NO: 3;
3) полипептид, содержащий аминокислоты 4-739 из SEQ ID NO: 5;
4) полипептид, содержащий аминокислоты 4-739 из SEQ ID NO: 7;
2) полипептид, содержащий аминокислотную последовательность SEQ ID NO: 3;
3) полипептид, содержащий аминокислотную последовательность SEQ ID NO: 5; и
4) полипептид, содержащий аминокислотную последовательность SEQ ID NO: 7.
В других вариантах осуществления настоящее изобретение предусматривает иммуногенный полипептид PSMA, выбранный из группы, состоящей из:
1) полипептида, состоящего из аминокислотной последовательности SEQ ID NO: 11
2) полипептида, состоящего из аминокислотной последовательности SEQ ID NO: 13;
3) полипептида, содержащего аминокислотную последовательность SEQ ID NO: 13.
В некоторых других вариантах осуществления настоящее изобретение предусматривает выделенные иммуногенные полипептиды PSMA, которые являются вариантами любого из следующих полипептидов:
2) полипептида, содержащего аминокислоты 4-739 из SEQ ID NO: 3;
3) полипептида, содержащего аминокислоты 4-739 из SEQ ID NO: 5; и
4) полипептида, содержащего аминокислоты 4-739 из SEQ ID NO: 7,
причем аминокислотная последовательность варианта имеет 93%-99% идентичность с последовательностью SEQ ID NO: 1 и разделяет, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95% или, по меньшей мере, 99% идентичность с аминокислотной последовательностью SEQ ID NO: 3, 5 или 7.
Варианты полипептида данного РАА могут быть получены путем удаления, вставки или замещения одной или более аминокислот в родительском иммуногенном полипептиде РАА. Примером получения таких вариантов является консервативное замещение отдельных аминокислот полипептидов, то есть, путем замены одной аминокислоты другой, имеющей аналогичные свойства.
Иммуногенный полипептид PSMA согласно изобретению может быть получен путем сохранения некоторых или всех консервативных эпитопов Т-клеток человеческого PSMA в SEQ ID NO: 1, не смотря на замещения определенных аминокислот в остальных областях человеческого PSMA аминокислотами, обнаруженными в одном или нескольких ортологах человеческого PSMA, в соответствующих положениях. Последовательности различных ортологов PSMA которые могут быть использованы для получения иммуногенных полипептидов PSMA, доступны из базы данных GeneBank. Эти ортологи вместе с их идентификационными номерами NCBI приведены в Таблице 18. Замещения аминокислот человеческого PSMA аминокислотами из одного или более ортологов могут быть консервативными замещениями или неконсервативными замещениями, или и тем и другим, и могут выбираться, исходя из ряда факторов, известных в данной области, включая дивергенцию, необходимую для достижения МНС-связывания, присутствие ортологичных аминокислот на участке замещения, воздействие поверхности и поддержание 3-D структуры белка для оптимальной обработки и представления.
B-2. Иммуногенные полипептиды PSA
В другом аспекте, настоящее изобретение предусматривает выделенные иммуногенные полипептиды PSA. В одном варианте осуществления выделенный иммуногенный полипептид PSA является полипептидом, состоящим из аминокислотной последовательности SEQ ID NO: 15 или аминокислот 4-263 из SEQ ID NIO: 15, или их вариантом. В другом варианте осуществления выделенный иммуногенный полипептид PSA является полипептидом, состоящим из аминокислотной последовательности SEQ ID NO: 17 или аминокислот 4-240 из SEQ ID NO: 17, или их вариантом. В дополнительном варианте осуществления выделенный иммуногенный полипептид PSA является полипептидом, состоящим из аминокислотной последовательности SEQ ID NO: 19 или аминокислот 4-281 из SEQ ID NO: 19, или их вариантом.
С.МОЛЕКУЛЫ НУКЛЕИНОВЫХ КИСЛОТ, КОДИРУЮЩИЕ ИММУНОГЕННЫЙ РАА
ПОЛИПЕПТИДЫ
В некоторых аспектах настоящее изобретение предусматривает молекулы нуклеиновых кислот, которые кодируют иммуногенные полипептиды РАА. Молекулы нуклеиновых кислот могут быть дезоксинуклеотидами (ДНК) или рибонуклеотидами (РНК). Таким образом, молекула нуклеиновой кислоты может содержать нуклеотидную последовательность, раскрытую в данном документе, где тимидин (Т) может быть также урацилом (U), что отражает различия между химическими структурами ДНК и РНК. Молекулы нуклеиновой кислоты могут быть модифицированными формами, одно- или двухцепочечными формами или линейными или кольцевыми формами. Молекулы нуклеиновой кислоты могут быть получены с использованием способов, известных в данной области свете настоящего описания.
С-1. Молекулы нуклеиновых кислот, кодирующие иммуногенные полипептиды PSMA
В одном аспекте настоящее изобретение предусматривает молекулы выделенных нуклеиновых кислот или их вырожденные варианты, которые содержат нуклеотидную последовательность, кодирующую иммуногенный полипептид PSMA, включая иммуногенные полипептиды PSMA, предусмотренные настоящим описанием, или их функциональные фрагменты.
В некоторых вариантах осуществления нуклеотидная последовательность кодирует мембраносвязанный иммуногенный полипептид PSMA. В некоторых отдельных вариантах осуществления молекула выделенной нуклеиновой кислоты содержит нуклеотидную последовательность или ее вырожденный вариант, выбранные из группы, состоящей из:
1) нуклеотидной последовательности, кодирующей аминокислотную последовательность SEQ ID NO: 9;
2) нуклеотидной последовательности, кодирующей аминокислоты 4-739 из SEQ ID NO: 3;
3) нуклеотидной последовательности, кодирующей аминокислоты 4-739 из SEQ ID NO: 5; и
4) нуклеотидной последовательности, кодирующей аминокислоты 4-739 из SEQ ID NO: 7.
В некоторых других отдельных вариантах осуществления нуклеотидная последовательность кодирует вариант иммуногенного полипептида PSMA из SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7 или SEQ ID NO: 9, причем вариант имеет аминокислотную последовательность, которая имеет (а) от 93% до 99% идентичность с аминокислотной последовательностью SEQ ID NO: 1 и (b) по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95% или, по меньшей мере, 99% идентичность с аминокислотной последовательностью SEQ ID NO: 3, 5 или 7.
В еще некоторых других отдельных вариантах осуществления молекула выделенной нуклеиновой кислоты содержит нуклеотидную последовательность или ее вырожденный вариант, выбранные из группы, состоящей из:
1) нуклеотидной последовательности, содержащей нуклеотиды 10-2217 из SEQ ID NO: 4;
2) нуклеотидной последовательности, содержащей нуклеотиды 10-2217 из SEQ ID NO: 6;
3) нуклеотидной последовательности, содержащей нуклеотиды 10-2217 из SEQ ID NO: 8; и
4) нуклеотидной последовательности, содержащей нуклеотиды 10-2217 из SEQ ID NO: 10.
C-2. Молекулы нуклеиновых кислот, кодирующие иммуногенные полипептиды PSA
В другом аспекте настоящее изобретение предусматривает молекулы выделенных нуклеиновых кислот или их вырожденные варианты, которые кодируют иммуногенный полипептид PSA, включая иммуногенные полипептиды PSA, предусмотренные настоящим описанием.
В некоторых вариантах осуществления молекула выделенной нуклеиновой кислоты содержит или состоит из нуклеотидной последовательности, кодирующей цитозольный иммуногенный полипептид PSA. В одном варианте осуществления нуклеотидная последовательность кодирует цитозольный иммуногенный полипептид PSA, состоящий из следующих аминокислотных остатков 4-240 из SEQ ID NO: 17. В другом варианте осуществления нуклеотидная последовательность кодирует цитозольный иммуногенный полипептид PSA, содержащий аминокислотную последовательность SEQ ID NO: 17. В еще другом варианте осуществления нуклеотидная последовательность кодирует цитозольный иммуногенный полипептид PSA, состоящий из аминокислотной последовательности SEQ ID NO: 17. В другом варианте осуществления нуклеотидная последовательность кодирует функциональный вариант любого из указанных цитозольных иммуногенных полипептидов PSA, предусмотренных в данном документе выше.
В некоторых других вариантах осуществления молекула выделенной нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую полипептид мембраносвязанного иммуногенного PSA. В одном варианте осуществления нуклеотидная последовательность кодирует полипептид мембраносвязанного иммуногенного PSA, содержащий следующие аминокислотные остатки 4-281 из SEQ ID NO: 19. В другом варианте осуществления нуклеотидная последовательность кодирует полипептид мембраносвязанного иммуногенного PSA, содержащий аминокислотную последовательность SEQ ID NO: 19. В еще другом варианте осуществления нуклеотидная последовательность кодирует полипептид мембраносвязанного иммуногенного PSA, состоящий из аминокислотной последовательности SEQ ID NO: 19. В еще других вариантах осуществления нуклеотидная последовательность кодирует функциональный вариант любого из указанных полипептидов мембраносвязанного иммуногенного PSA, предусмотренных в данном документе выше.
Примеры молекул отдельных нуклеиновых кислот, предусмотренных настоящим описанием, включают:
1) молекулу нуклеиновой кислоты, состоящую из нуклеотидной последовательности SEQ ID NO: 18;
2) молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность SEQ ID NO: 18;
3) молекулу нуклеиновой кислоты, состоящую из нуклеотидной последовательности SEQ ID NO: 20;
4) молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность SEQ ID NO: 20; и
5) вырожденный вариант любой из молекул нуклеиновых кислот 1) - 4) выше.
С-3. Молекулы нуклеиновых кислот, кодирующие два или более иммуногенных полипептидов РАА
В другом аспекте настоящее изобретение предусматривает молекулу нуклеиновой кислоты, которая кодирует более чем один иммуногенный полипептид РАА, например, по меньшей мере, два, по меньшей мере, три или, по меньшей мере, четыре полипептида иммуногенных РАА. Такие молекулы нуклеиновых кислот также называют "мультиантигенными конструктами", "мультиантигенной вакциной", "мультиантигенной плазмидой" и т.п. в настоящем описании. Таким образом, в некоторых аспектах одна молекула нуклеиновой кислоты несет две кодирующие нуклеотидные последовательности, причем каждая из кодирующих последовательностей экспрессирует отдельный иммуногенный полипептид РАА. Такую молекулу нуклеиновой кислоты также называют "биантигенным конструктом", "биантигенной вакциной" или "биантигенной плазмидой" и т.п. в этом описании. В некоторых других аспектах одна молекула нуклеиновой кислоты несет три кодирующие нуклеотидные последовательности, причем каждая из кодирующих последовательностей экспрессирует отдельный иммуногенный полипептид РАА. Такую молекулу нуклеиновой кислоты также называют "триантигенным конструктом", "триантигенной вакциной" или "триантигенной плазмидой" в этом описании. Отдельные полипептиды РАА, кодируемые мультиантигенным конструктом, могут быть иммуногенными против одного и того же антигена, такого как PSMA, PSA или PSCA. Отдельные полипептиды РАА, кодируемые мультиантигенным конструктом, могут быть иммуногенными против разных антигенов, например, одного полипептида РАА, являющегося полипептидом PSMA, и другого одного полипептида PSA. В частности, один мультиантигенный конструкт может кодировать два или более иммуногенных полипептидов РАА в любой одной из следующих комбинаций:
1) по меньшей мере, один иммуногенный полипептид PSMA и, по меньшей мере, один иммуногенный полипептид PSA;
2) по меньшей мере, один иммуногенный полипептид PSMA и, по меньшей мере, один полипептид PSCA;
3) по меньшей мере, один иммуногенный полипептид PSA и, по меньшей мере, один полипептид PSCA; и
4) по меньшей мере, один иммуногенный полипептид PSMA, по меньшей мере, один иммуногенный полипептид PSA, и, по меньшей мере, один полипептид PSCA.
Полипептиды иммуногенных PSMA, кодируемые мультиантигенным конструктом, могут быть цитозольными, секретируемыми или мембраносвязанными, но преимущественно мембраносвязанными. Подобным образом, иммуногенный полипептид PSA, кодируемого мультиантигенным конструктом, могут быть цитозольными, секретируемыми или мембраносвязанными, но преимущественно цитозольным. В некоторых вариантах осуществления настоящее изобретение предусматривает мультиантигенный конструкт, который кодирует, по меньшей мере, один мембраносвязанный иммуногенный полипептид PSMA и, по меньшей мере, один мембраносвязанный полипептид PSA.
В некоторых других вариантах осуществления настоящее изобретение предусматривает мультиантигенный конструкт, который кодирует, по меньшей мере, один мембраносвязанный иммуногенный полипептид PSMA, по меньшей мере, один цитозольный полипептид PSA и, по меньшей мере, один полипептид PSCA, причем, по меньшей мере, один цитозольный полипептид PSA содержит аминокислоты 4-240 из SEQ ID NO: 17 и где, по меньшей мере, один иммуногенный полипептид PSMA выбран из группы, состоящей из:
1) полипептида, содержащего аминокислоты 15-750 из SEQ ID NO: 1;
2) полипептида, содержащего аминокислотную последовательность SEQ ID NO: 3;
3) полипептида, содержащего аминокислотную последовательность SEQ ID NO: 5;
4) полипептида, содержащего аминокислотную последовательность SEQ ID NO: 7;
5) полипептида, содержащего аминокислоты 4-739 из SEQ ID NO: 9;
6) полипептида, содержащего аминокислоты 4-739 из SEQ ID NO: 3;
7) полипептида, содержащего аминокислоты 4-739 из SEQ ID NO: 5;
8) полипептида, содержащего аминокислоты 4-739 из SEQ ID NO: 7; и
9) полипептида, содержащего аминокислотную последовательность SEQ ID NO: 9.
В некоторых отдельных вариантах осуществления настоящее изобретение предусматривает мультиантигенный конструкт, содержащий, по меньшей мере, одну нуклеотидную последовательность, кодирующую иммуногенный полипептид PSMA, по меньшей мере, одну нуклеотидную последовательность, кодирующую иммуногенный полипептид PSA, и, по меньшей мере, одну нуклеотидную последовательность, кодирующую полипептид человеческого PSCA, причем нуклеотидная последовательность кодирует иммуногенный полипептид PSA из SEQ ID NO: 17 аминокислоты 4-240 из SEQ ID NO. 17, и где нуклеотидную последовательность, кодирующую иммуногенный полипептид PSMA выбирают из группы, состоящей из:
1) нуклеотидной последовательности SEQ ID NO: 2;
2) нуклеотидной последовательности SEQ ID NO: 4;
3) нуклеотидной последовательности SEQ ID NO: 6;
4) нуклеотидной последовательности SEQ ID NO: 8;
5) нуклеотидной последовательности SEQ ID NO: 10;
6) нуклеотидной последовательности, содержащей нуклеотиды 10-2217 из SEQ ID NO: 4;
7) нуклеотидной последовательности, содержащей нуклеотиды 10-2217 из SEQ ID NO: 6;
8) нуклеотидной последовательности, содержащей нуклеотиды 10-2217 из SEQ ID NO: 8; и
9) нуклеотидной последовательности, содержащей нуклеотиды 10-2217 из SEQ ID NO: 10.
Примеры специфических мультиантигенных конструктов, предусмотренных настоящим описанием, включают молекулы нуклеиновых кислот, которые содержат нуклеотидную последовательность, представленную в SEQ ID NO: 23-36.
Мультиантигенные конструкты, предусмотренные настоящим описанием, могут быть получены с использованием различных технологий, известных в уровне техники и в свете описания. Например, мультиантигенный конструкт можно получить путем внедрения множества независимых промоторов в одну плазмиду (Huang, Υ., Ζ. Chen, et al. (2008). "Design, construction, and characterization of a dual-promoter multigenic DNA vaccine directed against an HIV-1 subtype С/В' recombinant." J Acquir Immune Defic Syndr 47(4):403-411; Xu, Κ., Ζ.Y. Ling, et al. (2011). "Broad humoral and cellular immunity elicited by a bivalent DNA vaccine encoding HA and NP genes from an H5N1 virus." Viral Immunol 24(1):45-56). Плазмида может быть сконструирована для того, чтобы нести несколько кассет экспрессии, каждая из которых состоит из а) эукариотического промотора для инициирования РНК-полимеразной зависимой транскрипции, с усиливающим элементом или без него, б) гена, кодирующего антиген-мишень, и в) терминирующей транскрипцию последовательности. При доставке плазмиды в ядра трансфицированных клеток транскрипция будет инициироваться от каждого промотора, приводя к продуцированию отдельных мРНК, кодирующих один из антигенов-мишеней. Матричные РНК будут независимо транслироваться, тем самым продуцируя желаемые антигены.
Мультиантигенные конструкты, предусмотренные настоящим описанием, могут также быть получены с использованием одного вектора посредством применения вирусных 2А-подобных полипептидов (Szymczak, A. L. and D.A. Vignali (2005). "Development of 2А peptide-based strategies in the design of multicistronic vectors." Expert Opin Biol Ther 5(5):627-638; de Felipe, P., G. A. Luke, et al. (2006). "E unum pluribus: multiple proteins from a self-processing polyprotein." Trends Biotechnol 24(2):68-75; Luke, G.Α., P. de Felipe, et al. (2008). "Occurrence, function and evolutionary origins of '2A-like' sequences in virus genomes." J Gen Virol 89(Pt 4): 1036-1042; Ibrahimi, Α., G. Vande Velde, et al. (2009). "Highly efficient multicistronic lentiviral vectors with peptide 2A sequences." Hum Gene Ther 20(8):845-860; Kim, J.H., S.R. Lee, et al. (2011). "High cleavage efficiency of a 2A peptide derived from porcine teschovirus-1 in human cell lines, zebrafish and mice." PLoS One 6(4):e18556). Эти полипептиды, также названные кассетами расщепления или CHYSEL (действующие в цис-положении гидролазные элементы), составляют приблизительно 20 аминокислот в длину с высококонсервативным карбоксиконцевым мотивом D-V/I-EXNPGP (Фигура 2). Кассеты редки в природе, наиболее часто встречаются у вирусов, таких как вирус ящура (FMDV), вирус А лошадиного ринита (ERAV), вирус энцефаломиокардита (EMCV), Porcine teschovirus (PTV) и вирус Thosea asigna (TAV) (Luke, G.Α., P. de Felipe, et al. (2008). "Occurrence, function and evolutionary origins of '2A-like' sequences in virus genomes." J Gen Virol 89(Pt 4):1036-1042). С мультиантигенной стратегией экспрессии на основе 2А гены, кодирующие несколько целевых антигенов, могут быть связаны друг с другом в одной открытой рамке считывания, разделенные 2А-кассетами. Вся открытая рамка считывания может быть клонирована в вектор с одним промотором и терминатором. При доставке конструктов в клетку-хозяин мРНК, кодирующая несколько антигенов, будет транскрибироваться и транслироваться в виде одного полипротеина. В течение трансляции кассет 2А рибосомы пропускают связь между С-концевым глицином и пролином. Рибосомный пропуск действует подобно котрансляционному автокаталитическому "расщеплению", которое высвобождает вышележащие от нижележащих белков. Внедрение кассеты 2А между двумя белковыми антигенами приводит к добавлению ~20 аминокислот на С-концах вышележащего полипептида и одну аминокислоту (пролин) к N-концу нижележащего белка. В адаптации этой методологии, протеазные сайты расщепления могут быть внедрены на N-конце кассеты 2А таким образом, что протеазы будут повсеместно расщеплять кассету от вышележащего белка (Fang, J., S. Yi, et al. (2007). "An antibody delivery system for regulated expression of therapeutic levels of monoclonal antibodies in vivo." Mol Ther 15(6):1153-1159).
Другая стратегия построения мультиантигенных конструктов, предусмотренных настоящим изобретением, включает использование участка внутренней посадки рибосомы или 1RES. Участки внутренней посадки рибосомы являются РНК-элементами (Фигура 3), обнаруженными в 5' нетранслируемых участках молекул некоторых РНК (Bonnal, S., С. Boutonnet, et al (2003). "IRESdb: the IntePHKI Ribosome Entry Site database." Nucleic Acids Res 31(1):427-428). Они привлекают эукариотические рибосомы к РНК для облегчения прямой трансляции открытых рамок считывания. В отличие от обычной клеточной 7-метилгуанозиновой кэп-зависимой трансляции, IRES-опосредованная трансляция может инициироваться у AUG-кодонов далеко в молекуле РНК. Высокоэффективный процесс может быть использован для применения в мультицистронных векторах экспрессии (Bochkov, Y.A. and А.С. Palmenberg (2006). "Translational efficiency of EMCV IRES in bicistronic vectors is dependent upon IRES sequence and gene location." Biotechniques 41(3):283-284, 286, 288). Как правило, два трансгена вставляются в вектор между промотором и терминатором транскрипции, как две отдельные открытые рамки считывания, разделенные IRES. При доставке конструктов в клетку-хозяин будет транскрибироваться один длинный транскрипт, кодирующий оба трансгена. Первая ORF будет транслироваться традиционным кэп-зависимым образом, терминируя у стоп-кодона выше от 1RES.Вторая ORF будет транслироваться кэп-независимым способом с помощью 1RES. Таким образом, два независимых белка могут быть получены из одной мРНК, транскрибированной из вектора с одной экспрессионной кассетой.
Хотя мультиантигенная стратегия экспрессии описана в данном документе в контексте ДНК-вакцинного конструкта, принципы применимы так же в контексте вирусных векторных генетических вакцин.
D. ВЕКТОРЫ, СОДЕРЖАЩИЕ МОЛЕКУЛУ НУКЛЕИНОВОЙ КИСЛОТЫ, КОДИРУЮЩУЮ ИММУНОГЕННЫЙ ПОЛИПЕПТИД РАА
Другой аспект изобретения касается векторов, содержащих одну или более молекул нуклеиновых кислот согласно изобретению. Векторы полезны для клонирования или экспрессирования иммуногенных полипептидов РАА, кодируемых молекулами нуклеиновых кислот, или для доставки молекулы нуклеиновой кислоты в составе композиции, такой как вакцина, в клетку-хозяин или животное-хозяин, такое как человек. Может быть получено широкое разнообразие векторов, содержащих или экспрессирующих молекулу нуклеиновой кислоты согласно изобретению, таких как плазмидные векторы, космидные векторы, фаговые векторы и вирусные векторы.
В некоторых вариантах осуществления описание предусматривает плазмидный вектор, содержащий молекулу нуклеиновой кислоты согласно изобретению. Репрезентативные примеры подходящих плазмидных векторов включают pBR325, pUC18, pSKF, pET23D и pGB-2. Другие репрезентативные примеры плазмидных векторов, а также способы конструирования таких векторов, описаны в патентах США №5580859, 5589466, 5688688, 5814482 и 5580859.
В других вариантах осуществления настоящее изобретение предусматривает векторы, которые сконструированы из вирусов, таких как ретровирусы, альфавирусы, аденовирусы. Репрезентативные примеры ретровирусных векторов описаны более детально в ЕР 0415731; публикациях РСТ WO 90/07936; WO 91/0285, WO 93/11230; WO 93/10218, WO 9403622; WO 93/25698; WO 93/25234; и патентах США №5219740, 5716613, 5851529, 5591624, 5716826, 5716832 и 5817491. Репрезентативные примеры векторов, которые могут быть получены из альфавирусов, описаны в патентах США №5091309 и 5217879, 5843723 и 5789245. В некоторых отдельных вариантах осуществления настоящее изобретение предусматривает аденовирусные векторы, полученные из аденовирусов нечеловекообразных приматов, таких как аденовирусы обезьян. Примеры таких аденовирусных векторов, а также их получение, раскрыты в публикациях заявок РСТ WO 2005/071093 и WO 2010/086189, и включают нерепликационные векторы, такие как ChAd3, ChAd4, ChAd5, ChAd7, ChAd8, ChAd9, ChAd10, ChAd11, ChAd16, ChAd17, ChAd19, ChAd20, ChAd22, ChAd24, ChAd26, ChAd30, ChAd31, ChAd37, ChAd38, ChAd44, ChAd63, ChAd68, ChAd82, ChAd55, ChAd73, ChAd83, ChAd146, ChAd147, Pan Ad1, Pan Ad2 и Pan Ad3, и репликационно компетентные векторы, такие как векторы Ad4 и Ad7. Предпочтительно, чтобы при построении аденовирусных векторов из аденовирусов обезьян один или более из ранних генов из геномной области вируса, выбранного из Е1А, В, Е1, Е2А, Е2В, Е3 и Е4, были либо удалены, либо оказались нефункциональными из-за делеции или мутации. В отдельном варианте осуществления вектор сконструирован из ChAd3 или ChAd68. Подходящие векторы также могут быть получены из других вирусов, таких как: вирусы оспы, таких как поксвирус канареек или вирус коровьей оспы (Fisher-Hoch et al., PNAS 86:317-321, 1989; Flexner et al., Ann. N.Y. Acad. Sci. 569:86-103, 1989; Flexner et al., Vaccine 8:17-21, 1990; патенты США №4603112, 4769330 и 5017487; WO 89/01973); аденоассоциированные векторы (смотри, например, патент США №5872005); SV40 (Mulligan et al., Nature 277:108-114, 1979); герпеса (Kit, Adv. Exp.Med. Biol. 215:219-236, 1989; патент США №5288641); и лентивируса, такого как HIV (Poznansky, J. Virol. 65:532-536, 1991).
Способы конструирования векторов хорошо известны в данной области техники. Векторы экспрессии обычно включают один или несколько элементов управления, которые функционально связаны с последовательностью нуклеиновой кислоты, подлежащей экспрессированию. Термин "элементы контроля" относится коллективно к промоторным областям, сигналам полиаденилирования, терминирующим транскрипцию последовательностям, входным регуляторным доменам, точкам начала репликации, участкам внутренней посадки рибосомы ("1RES"), энхансерам и т.п., которые в совокупности обеспечивают репликацию, транскрипцию и трансляцию кодирующей последовательности в клетку-реципиент. Не все из этих элементов управления должны всегда присутствовать, поскольку выбранная кодирующая последовательность способна к репликации, транскрипции и трансляции в соответствующую клетку-хозяин. Элементы управления выбираются, исходя из ряда факторов, известных специалистам в этой области техники, таких как конкретные клетки-хозяева и источник или структуры других векторных компонентов. Для усиления экспрессии иммуногенного полипептида РАА последовательность Козака может быть предоставлена выше последовательности, кодирующей иммуногенный полипептид РАА. Для позвоночных, известная последовательность Козака представляет собой (GCC) NCCATGG, в котором N является А или G и GCC является менее консервативной. Примеры последовательностей Козака, которые могут быть использованы, включают ACCAUGG и ACCATGG.
Е. КОМПОЗИЦИИ, СОДЕРЖАЩИЕ ИМУНОГЕННЫЙ РАА ПОЛИПЕПТИД (ПОЛИПЕПТИДНЫЕ КОМПОЗИЦИИ)
В другом аспекте, настоящее изобретение предусматривает композиции, содержащие один или более полипептидов выделенных иммуногенных РАА, предусмотренных настоящим описанием ("полипептидная композиция"). В некоторых вариантах осуществления полипептидная композиция является иммуногенной композицией, полезной для вызова иммунной реакции против белка РАА у млекопитающего, такого как мышь, собака, нечеловекообразный примат или человек. В некоторых других вариантах осуществления полипептидная композиция представляет собой вакцинную композицию, полезную для иммунизации млекопитающего, такого как человек, для ингибирования анормальной клеточной пролиферации, для обеспечения защиты против развития рака (используется в качестве профилактики) или для лечения нарушений (используется в качестве терапии), ассоциированных с надмерной экспрессией РАА, таких как раковые заболевания, особенно рак простаты.
Полипептидная композиция, предусмотренная настоящим описанием, может содержать один тип иммуногенного полипептида РАА, такой как иммуногенный полипептид PSMA, иммуногенный полипептид PSA или иммуногенный полипептид PSCA. Композиция также может содержать комбинацию двух или более разных типов иммуногенных полипептидов РАА. Например, полипептидная композиция может содержать иммуногенные полипептиды РАА в любой из следующих комбинаций:
1) иммуногенный полипептид PSMA и иммуногенный полипептид PSA;
2) иммуногенный полипептид PSMA и полипептид PSCA; или
3) иммуногенный полипептид PSMA, иммуногенный полипептид PSA и полипептид PSCA.
Иммуногенная композиция или вакцинная композиция, предусмотренные настоящим описанием, могут дополнительно содержать фармацевтически приемлемый эксципиент. Фармацевтически приемлемые эксципиенты для иммуногенных или вакцинных композиций известны из уровня техники. Примеры подходящих эксципиентов включают биосовместимые масла, такие как рапсовое масло, подсолнечное масло, арахисовое масло, хлопковое масло, масло жожоба, сквалан, сквален, физиологический солевой раствор, консерванты и осмотические агенты регулирования давления, газы-носители, рН-регулирующие агенты, органические растворители, гидрофобные агенты, ингибиторы ферментов, поглощающие воду полимеры, поверхностно-активные вещества, промоторы абсорбции, модификаторы рН и антиокислительные агенты.
Иммуногенный полипептид РАА в композиции, особенно иммуногенной композиции или вакцинной композиция, может быть связан с, конъюгирован с или иным образом включен в качестве носителя для введения реципиенту. Термин "носитель" относится к веществу или структуре, к которой иммуногенный полипептид может быть присоединен или иным образом связан, для доставки иммуногенного полипептида к рецепиенту (например, пациенту). Сам носитель может быть иммуногенным. Примеры носителей включают иммуногенные полипептиды, иммунные островки CpG, гемоцианин лимфы улитки (KLH), столбнячный анатоксин (ТТ), субъединицу В холерного токсина (СТВ), бактерии или бактериальные "тени", липосомы, хитосомы, виросомы, микросферы, дендритные клетки или им подобные. Одна или более молекул иммуногенного полипептида РАА может быть соединенной с одной молекулой носителя. Способы присоединения иммуногенного полипептида к носителю известны из уровня техники. Вакцинная композиция или иммуногенная композиция, предусмотренные настоящим описанием, могут быть использованы в сочетании с одним или более иммунных модуляторов или адъювантов. Иммунные модуляторы или адъюванты могут быть оформлены отдельно от вакцинной композиции или они могут быть частью той же вакцинной композиции. Таким образом, в одном варианте осуществления, вакцинная композиция дополнительно содержит один или более иммунных модуляторов или адъювантов. Примеры иммунных модуляторов или адъювантов предусмотрены в данном документе ниже.
Полипептидные композиции, в том числе и иммуногенные вакцинные композиции, могут быть получены в любой подходящей лекарственной форме, такой как жидкие формы (например, растворы, суспензии или эмульсии) и твердые формы (например, капсулы, таблетки или порошки), а также с помощью способов, известных специалистам в данной области техники.
F. КОМПОЗИЦИИ, СОДЕРЖАЩИЕ НУКЛЕИНОВУЮ КИСЛОТУ ИММУНОГЕННОГО РАА МОЛЕКУЛА (КОМПОЗИЦИИ НУКЛЕИНОВЫХ КИСЛОТ)
Настоящее описание также предусматривает композицию, содержащую молекулу выделенной нуклеиновой кислоты или вектор, предусмотренные настоящим описанием, (в данном документе "композиция нуклеиновой кислоты"). Композиции нуклеиновых кислот полезны для вызова иммунной реакции против белка РАА in vitro или in vivo у млекопитающего, включая человека.
В некоторых отдельных вариантах осуществления, композиция нуклеиновой кислоты представляет собой ДНК-вакцинную композицию для введения людям для ингибирования анормальной клеточной пролиферации, обеспечения защиты против развития рака (используется в качестве профилактики) или для лечения рака (используется в качестве терапии), ассоциированного с надмерной экспрессией РАА, или для вызова иммунной реакции на конкретный человеческий РАА, такой как PSMA, PSA и PSCA. Молекула нуклеиновой кислоты в композиции может быть "оголенной" молекулой нуклеиновой кислоты, т.е. просто в виде выделенной ДНК, свободной от элементов, которые способствуют трансфекции или экспрессии. Альтернативно, молекула нуклеиновой кислоты в композиции может быть включена в вектор.
Композиция нуклеиновой кислоты, предусмотренная настоящим описанием, может содержать отдельные выделенные молекулы нуклеиновых кислот, каждая из которых кодирует только один тип иммуногенного полипептида РАА, такой как иммуногенный полипептид PSMA, иммуногенный полипептид PSA или иммуногенный полипептид PSCA.
Композиция нуклеиновой кислоты может содержать мультиантигенный конструкт, предусмотренный настоящим описанием, который кодирует два или более типов иммуногенных полипептидов РАА. Мультиантигенный конструкт может кодировать два или более иммуногенных полипептидов РАА в любой из следующих комбинаций:
1) иммуногенный полипептид PSMA и иммуногенный полипептид PSA;
2) иммуногенный полипептид PSMA и иммуногенный полипептид PSCA;
3) иммуногенный полипептид PSA и иммуногенный полипептид PSCA; и
4) иммуногенный полипептид PSMA, иммуногенный полипептид PSA, и иммуногенный полипептид PSCA.
Композиции нуклеиновых кислот, включая ДНК-вакцинные композиции, могут дополнительно содержать фармацевтически приемлемый эксципиент. Примеры подходящих фармацевтически приемлемых эксципиентов для композиций нуклеиновых кислот, в том числе ДНК-вакцинных композиций, хорошо известны специалистам в данной области и включают сахара, и т.д. Такие эксципиенты могут быть водными или неводными растворами, суспензиями и эмульсиями. Примеры неводных эксципиентов включают пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и инъецируемые органические сложные эфиры, такие как этилолеат. Примеры водного эксципиента включают воду, спиртовые / водные растворы, эмульсии или суспензии, в том числе солевые и буферные среды. Подходящие эксципиенты также включают агенты, которые помогают клеточному поглощению полинуклеотидной молекулы. Примерами таких агентов являются (I) химические вещества, которые изменяют клеточную проницаемость, такие как бупивакаин, (II) липосомы или вирусные частицы для инкапсуляции полинуклеотида, или (III) катионные липиды или микрочастицы кремнезема, золота или вольфрама, которые присоединяются к полинуклеотидам. Анионные и нейтральные липосомы хорошо известны в данной области техники (смотрите, например, Liposomes: A Practical Approach, RPC New Ed, IRL press (1990), для подробного описания способов получения липосом) и полезны для доставки большого ассортимента продуктов, включая полинуклеотиды. Катионные липиды также известны в данной области и обычно используются для доставки генов. Такие липиды включают Лиофектин™, также известный как DOTMA (N-[1-(2,3-диолеилокси)пропилы, Ν,Ν,Ν-триметиламмония хлорид), DOTAP (1,2-бис(олеилокси)-3(триметиламмонио)пропан), DDAB (диметилдиоктадецилбромид), DOGS (диоктадесиламидологлицил спермин) и производные холестерина, такие как DCChol (3-6eTa-(N-(N',N'-диметиламинометан)карбамоил)холестерин). Описание этих катионных липидов могут быть найдены в ЕР 187702, WO 90/11092, патенте США №5283185, WO 91/15501, WO 95/26356 и патенте США №5527928. Особенно полезным катионным липидным препаратом, который может быть использован с нуклеиновой вакциной, предусмотренной описанием, является Vaxfectin, который представляет собой смесь катионного липида (GAP-DMORIE) и нейтрального фосфолипида (DPyPE), которые при комбинировании с водным носителем самоорганизовываются с образованием липосом.
Катионные липиды для доставки генов предпочтительно используют в сочетании с липидом, таким как DOPE (диолеилфосфатидилэтаноламин), как описано в WO 90/11092 в качестве примера. Кроме того, ДНК-вакцина также может быть приготовлена с неионным блоксополимером, таким как CRL1005.
G. ИСПОЛЬЗОВАНИЕ ИММУНОГЕННЫХ ПОЛИПЕПТИДОВ РАА, МОЛЕКУЛ НУКЛЕИНОВЫХ КИСЛОТ И КОМПОЗИЦИЙ
В других аспектах настоящее изобретение предусматривает способы использования иммуногенных полипептидов РАА, молекул выделенных нуклеиновых кислот и композиций, содержащих иммуногенный полипептид РАА или молекулу выделенной нуклеиновой кислоты, описанные в данном документе выше.
В одном аспекте настоящее изобретение предусматривает способ вызова иммунной реакции против РАА у млекопитающего, особенно человека, включающий введение млекопитающему эффективного количества (1) иммуногенного полипептида РАА, предусмотренного описанием, который является иммуногенным против целевого РАА, (2) молекулы выделенной нуклеиновой кислоты, кодирующей такой иммуногенный полипептид РАА, (3) композиции, содержащей такой иммуногенный полипептид РАА, или (4) композиции, содержащей молекулу выделенной нуклеиновой кислоты, кодирующей такой иммуногенный полипептид РАА.
В некоторых вариантах осуществления описание изобретения предусматривает способ вызова иммунной реакции против PSMA у человека, включающий введение человеку эффективного количества композиции иммуногенного PSMA, предусмотренной настоящим описанием, где композицию иммуногенного PSMA выбирают из: (1) иммуногенного полипептида PSMA, (2) молекулы выделенной нуклеиновой кислоты, кодирующей иммуногенный полипептид PSMA, (3) композиции, содержащей иммуногенный полипептид PSMA, или (4) композиции, содержащей молекулу выделенной нуклеиновой кислоты, кодирующей иммуногенный полипептид PSMA.
В некоторых других вариантах осуществления описание изобретения предусматривает способ вызова иммунной реакции против PSA у человека, включающий введение человеку эффективного количества композиции иммуногенного PSA, предусмотренной настоящим описанием, где композицию иммуногенного PSA выбирают из: (1) иммуногенного полипептида PSA, (2) молекулы выделенной нуклеиновой кислоты, кодирующей иммуногенный полипептид PSA, (3) композиции, содержащей иммуногенный полипептид PSA, или (4) композиции, содержащей молекулу выделенной нуклеиновой кислоты, кодирующей иммуногенный полипептид PSA.
В другом аспекте настоящее изобретение предусматривает способ ингибирования анормальной клеточной пролиферации у человека, причем анормальная клеточная пролиферация ассоциирована с надмерной экспрессией РАА. Способ включает введение человеку эффективного количества композиции иммуногенного РАА, предусмотренной настоящим описанием, которая иммуногенна против надмерно экспрессированного РАА. Композиция иммуногенного РАА может быть (1) иммуногенным полипептидом РАА, (2) молекулой выделенной нуклеиновой кислоты, кодирующей один или более иммуногенных полипептидов РАА, (3) композицией, содержащей иммуногенный полипептид РАА, или (4) композицией, содержащей молекулу выделенной нуклеиновой кислоты кодирующей один или более иммуногенных полипептидов РАА. В некоторых вариантах осуществления, способ предусмотрен для ингибирования анормальной клеточной пролиферации в простате у человека. В конкретном варианте осуществления настоящее описание изобретения предусматривает способ ингибирования анормальной клеточной пролиферации в простате, надмерно экспрессирующей PSMA, включающий введение человеку эффективного количества (1) иммуногенного полипептида PSMA, (2) молекулы выделенной нуклеиновой кислоты, кодирующей один или более иммуногенных полипептидов PSMA, (3) композиции, содержащей иммуногенный полипептид PSMA, или (4) композиции, содержащей молекулу выделенной нуклеиновой кислоты, кодирующей один или более иммуногенных полипептидов PSMA.
В другом аспекте настоящее изобретение предусматривает способ лечения рака у человека, где рак ассоциирован с надмерной экспрессией РАА. Способ включает введение человеку эффективного количества композиции иммуногенного РАА, способного вызывать иммунный ответ против надмерно экспрессированного РАА. Композиция иммуногенного РАА может быть (1) иммуногенным полипептидом РАА, (2) молекулой выделенной нуклеиновой кислоты, кодирующей один или более иммуногенных полипептидов РАА, (3) композицией, содержащей иммуногенный полипептид РАА, или (4) композицией, содержащей молекулу выделенной нуклеиновой кислоты, кодирующей один или более иммуногенных полипептидов РАА. Примеры раковых заболеваний, которые можно лечить с помощью способа, включают рак молочной железы, рак желудка, рак яичников, рак легких, рак мочевого пузыря, колоректальный рак, рак почки, рак поджелудочной железы и рак простаты.
В некоторых вариантах осуществления описание изобретения предусматривает способ лечения рака простаты у человека, включающий введение человеку эффективного количества композиции нуклеиновой кислоты, предусмотренной в данном документе выше. Нуклеиновые кислоты в композиции могут кодировать только один конкретный Τ иммуногенный полипептид РАА, такой как иммуногенный полипептид PSMA, иммуногенный полипептид PSA или иммуногенный полипептид PSCA. Нуклеиновые кислоты в композиции могут также кодировать два или более разных иммуногенных полипептидов РАА, таких как: (1) иммуногенный полипептид PSMA и иммуногенный полипептид PSA; (2) иммуногенный полипептид PSMA и иммуногенный полипептид PSCA; (3) иммуногенный полипептид PSA ииммуногенный полипептид PSCA; (4) иммуногенный полипептид PSMA, иммуногенный полипептид PSA и иммуногенный полипептид PSCA. Каждая отдельная молекула нуклеиновой кислоты в композиции может кодировать только один конкретный иммуногенный полипептид РАА, такой как полипептид PSMA, полипептид PSA или полипептид PSCA. Альтернативно, отдельная молекула нуклеиновой кислоты в композиции может быть мультиантигенными конструктами, кодирующими два разных типа иммуногенны полипептидов РАА, таких как: (1) иммуногенный полипептид PSMA и иммуногенный полипептид PSA; (2) иммуногенный полипептид PSMA и иммуногенный полипептид PSCA; (3) иммуногенный полипептид PSCA и иммуногенный полипептид PSA; или (4) иммуногенный полипептид PSMA, иммуногенный полипептид PSA и иммуногенный полипептид PSCA. В некоторых отдельных вариантах осуществления композиция нуклеиновой кислоты содержит мультиантигенный конструкт t, который кодирует, по меньшей мере, (4) иммуногенный полипептид PSMA, иммуногенный полипептид PSA и иммуногенный полипептид PSCA. Иммуногенный полипептид PSCA, содержащийся в вакцинных композициях или экспрессирующийся нуклеиновой кислотой в вакцинных композициях для лечения рака простаты у человека, предпочтительно является человеческим непроцессированным белком PSCA.
Полипептидные композиции и композиции нуклеиновых кислот можно вводить в организм животного, включая человека, с помощью ряда способов, известных в данной области техники. Примеры подходящих способов включают: (1) внутримышечное, внутрикожное, внутриэпидермальное, внутривенное, внутриартериальное, подкожное, внутрибрюшинное введение, или (2) пероральное введение, и (3) местное применение (например, глазное, интраназальное и наружное применение). Один конкретный метод внутрикожного или внутриэпидермального введения вакцинной композиции нуклеиновой кислоты, который может быть применен для доставки с помощью генной пушки, использует устройство для доставки вакцины Particle Mediated Epidermal Delivery (PMED™), продаваемое PowderMed. PMED представляет собой безыгольный метод введения вакцин животным или людям. Система PMED включает преципитацию ДНК на микроскопические частицы золота, которые затем нагнетают с помощью гелиевого газа в эпидермис. Покрытые ДНК частицы золота доставляются к АРС и кератиноцитам эпидермиса и, оказавшись внутри ядер этих клеток, ДНК элюируется из золота и становится транскрипционно активной, продуцируя закодированный белок. Этот белок затем представляется с помощью АРС в лимфоциты для индуцирования Т-клеточных иммунных реакций. Другим конкретным методом внутримышечного введения вакцины нуклеиновой кислоты, предусмотренной настоящим описанием, является электропорация. Электропорация использует контролируемые электрические импульсы для создания временных пор в клеточной мембране для облегчения клеточного поглощения вакцины нуклеиновой кислоты, введенной в мышцу. Когда CpG используется в комбинации с вакциной нуклеиновой кислоты, то предпочтительно, чтобы CpG и вакцина нуклеиновой кислоты были одной композицией, а препарат вводили внутримышечно путем электропорации. Эффективное количество иммуногенного полипептида РАА или нуклеиновой кислоты, кодирующей иммуногенный полипептид РАА, в композиции для введения в данном способе, предусмотренном настоящим описанием, может быть легко определено специалистом в данной области и будет зависеть от ряда факторов. В способе лечения рака, такого как рак простаты, факторы, которые могут быть рассмотрены при определении эффективного количества иммуногенного полипептида РАА или нуклеиновой кислоты включают: (1) субъект, подлежащий лечению, включая иммунный статус и состояние здоровья субъекта, (2) степень тяжести и стадии рака, подлежащие лечению, (3) конкретные иммуногенные полипептиды РАА, использованные или экспрессированные, (4) желательную степень защиты или лечения (5) способ введения и график, и (6) другие используемые терапевтические агенты (такие как адъюванты или иммуномодуляторы). В случае вакцинных композиций нуклеиновых кислот, включая мультиантигенные вакцинные композиции, способ разработки и доставки являются одними из ключевых факторов для определения дозы нуклеиновой кислоты, необходимой для вызова эффективного иммунного ответа. Например, эффективные количества нуклеиновой кислоты может находиться в диапазоне от 2 мкг/дозу - 10 мг/дозу, когда вакцинная композиция нуклеиновой кислоты сформулирована в виде водного раствора и вводиться путем подкожной инъекций или пневматической инъекции, тогда как только 16 нг/дозу - 16 мкг/дозу могут быть необходимыми, когда нуклеиновую кислоту получают в виде покрытых золотых шариков и доставляют с использованием технологии генной пушки. Диапазон доз для вакцины нуклеиновой кислоты, вводимой путем электропорации, как правило, находится в диапазоне от 0,5-10 мг/дозу. В случае, когда вакцину нуклеиновой кислоты вводят вместе с CpG путем электропорации в совместной композиции, доза вакцины нуклеиновой кислоты может находиться в пределах от 0,5-5 мг/дозу, а доза CpG обычно находится в диапазон 0,05 мг - 5 мг/дозу, например, 0,05, 0,2, 0,6, 0,2 или 1 мг/дозу на человека.
Вакцинная композиция нуклеиновой кислоты или полипептида согласно настоящему изобретению может быть использована в стратегии "прайм-буст" для вызова надежной и длительной иммунной реакции. Протоколы вакцинации с примированием и реиммунизацией, базирующиеся на повторных инъекциях одного и того же иммуногенного конструкта, хорошо известны. Обычно, первая доза может не дать защитный иммунитет, но только "зарядить" ("прайм") иммунную систему. Защитный иммунный ответ развивается после второй или третьей дозы ("бусты"). Бусты осуществляют в соответствии с обычными методами, и могут быть дополнительно оптимизированы эмпирически по графику введения, путям введения, выбору адъювантов, дозы и потенциальной последовательности при введении с другой вакциной. В одном варианте осуществления, вакцины нуклеиновых кислот или полипептидов согласно настоящему изобретению используются в обычной стратегии "прайм-буст", при которой одну и ту же вакцину вводят животному в виде нескольких доз. В другом варианте осуществления, вакцинные композиции нуклеиновых кислот или полипептидов используются в гетерологичной вакцинации "прайм-буст", при которой различные типы вакцин, содержащие те же антигены, вводят через заданные интервалы времени. Например, конструкт нуклеиновой кислоты может быть введен в форме плазмиды в начальной дозе ("прайм") и в составе вектора в последующих дозах ("буст"), или наоборот.
Для лечения рака предстательной железы, вакцины полипептидов или нуклеиновых кислот согласно настоящему изобретению могут быть использованы вместе с вакцинами против рака простаты, исходя из других антигенов, таких как антигены на основе кислой фосфатазы предстательной железы и андрогенных рецепторов.
Полипептид или нуклеиновая кислота, Вакцинная композиция полипептида или нуклеиновой кислоты согласно настоящему изобретению может быть использована вместе с одним или более вспомогательными веществами. Примеры подходящих адъювантов включают: (1) эмульсионные композиции масло-в-воде (с или без других специфических иммуностимулирующих агентов, таких как мурамиловые полипептиды или компоненты бактериальной клеточной стенки), такие как, например (a) MF59™ (публикация РСТ №WO 90/14837; Chapter 10 in Vaccine design: the subunit and adjuvant approach, eds. Powell & Newman, Plenum Press 1995), содержащая 5% сквалена, 0,5% твина 80 (полиоксиэтиленсорбитана моноолеат) и 0,5% спана 85 (сорбитана триолеат) приготовленная в виде субмикронных частиц с использованием микрофлюидизатора, (б) SAF, содержащий 10% сквалена, 0,4% твина 80, 5% плюроник-блокполимер L121 и Thr-MDP, микрофлюидизаторованный в субмикронную эмульсию или путем встряхивания сформированный в эмульсию с большим размером частиц и (с) адъювантная система RIBI™ (RAS) (Ribi Immunochem, Hamilton, МТ), содержащая 2% сквалена, 0,2% твина 80 иодин или более компонентов бактериальной стенки, таких как монофосфорил-липид А (MPL), димиколат трегалозы (TDM) и скелет клеточной стенки (CWS); (2) сапониновые адъюванты, такие как QS21, STIMULON™ (Cambridge Bioscience, Worcester, MA), Abisco® (Isconova, Sweden) или Iscomatrix® (Commonwealth Serum Laboratories, Australia); (3) полный адъювант Фрейнда (CFA) и неполный адъювант Фрейнда (IFA); (4) цитокины, такие как интерлейкины (например IL-1, IL-2, IL-4, IL-5, IL-6, IL-7, IL-12 (публикация РСТ №. WO 99/44636), и т.п.), интерфероны (например гамма-интерферон), макрофагальный колониестимулирующий фактор (M-CSF), фактор некроза опухоли (TNF), и т.п.; (5) монофосфорил-липид A (MPL) или 3-О-деацитилированный MPL (3dMPL), необязательно, практически в отсутствие квасцов при использовании пневмококковых сахаридов [например GB-2220221, ЕР-А-0689454, WO 00/56358); (6) комбинации 3dMPL с, например, QS21 и/или эмульсий масло-в-воде (например ЕР-А-0835318, ЕР-А-0735898, ЕР-А-0761231); (7) олигонуклеотиды, содержащие мотивы CpG, т.е. содержащие, по меньшей мере, один динуклеотид CG, когда цитозин неметилирован (например, Krieg, Vaccine (2000) 19:618-622; Krieg, Curr Opin Mol Ther (2001) 3:15-24; WO 98/40100, WO 98/55495, WO 98/37919 и WO 98/52581); (8) полиоксиэтиленовый этер или полиоксиэтиленовый эстер (например WO 99/52549); (9) поверхностно-активный эстер полиоксиэтиленсорбитана в комбинации с октоксинолом (например, WO 01/21207) или поверхностно-активный олиоксиэтиленалкиловый этер или эстер в комбинации с, по меньшей мере, одним дополнительным неионым поверхностно-активным веществом, таким как октоксинол (например, WO 01/21152); (10) сапонин и иммуностимулирующий олигонуклеотид (например олигонуклеотид CpG) (например, WO 00/62800); (11) металлическая соль, включая соли алюминия (такие как квасцы, фосфат алюминия, гидроксид алюминия); (12) сапонин и эмульсия масло-в-воде (например WO 99/11241); (13) сапонин (например QS21)+3dMPL+IM2 (необязательно + стерол) (например WO 98/57659); (14) другие вещества, которые действуют как иммуностимулирующие агенты для повышения эффективности композиции, такие как мурамиловые полипептиды, включая N-ацетилмурамил-L-треонил-D-изоглутамин (thr-MDP), N-ацетил-25 ацетилнормурамил-L-аланил-D-изоглутамин (nor-MDP), N-ацетилмурамил-L-аланил-D-изоглутаминил-L-аланин-2-(1'-2'-дипальмитоил-sn-глицеро-3-гидроксифосфорилокси)-этиламин МТР-РЕ), (15) лиганды для Toll-подобных рецепторов (TLR), природных или синтезированных (например, Kanzler et al., Nature Med. 13:1552-1559 (2007)), включая лиганды TLR3, такие как полил:С и подобные соединения, такие как гилтонол и амплиген.
Вакцинные композиции полипептидов и нуклеиновых кислот согласно настоящему изобретению могут быть использованы вместе с одним или более иммуномодуляторов. Примеры подходящих иммуномодуляторов включают белковые ингибиторы тирозинкиназы (например, афатиниб, акситиниб, цедираниб, эрлотинибом, гефитинибом, грандинин, лапатиниб, лестауртиниб, нератиниб, пазопаниб, хузартиниб, регорафениб, семаксаниб, сорафениб, сунитиниб, тивозаниб, тоцераниб, босутиниб и вандетаниб), агонисты CD40 (например, агонист антитела CD40), агонисты ОХ40 (например, агонист антитела ОХ40), ингибиторы CTLA-4 и (например, антитело к CTLA-4 ипилимумаб и тремелимумаб), агонисты TLR, агонисты 4-1 ВВ, антагонисты Tim-1, антагонисты LAGE-3 и антагонисты PD-L1 и PD-1.
H. ИММУНОТЕРАПЕВТИЧЕСКИЕ СХЕМЫ, ОСНОВАННЫЕ НА ВАКЦИНАХ (VBIR)
В дополнительном аспекте, настоящее изобретение предусматривает способ повышения иммуногенности или терапевтического эффекта вакцины для лечения опухолевого заболевания у млекопитающего, особенно человека. Способ включает введение млекопитающему, принимающему вакцину для лечения опухолевого заболевания, (1) эффективного количества, по меньшей мере, одного иммуносупрессивного клеточного ингибитора (ингибитор ISC) и (2) эффективного количества, по меньшей мере, одного иммуноэффекторного клеточного энхансера (энхансер IEC). Способ может быть использован в комбинации с вакциной в любой форме или составе, например, субъединичной вакциной, белковой вакциной, пептидной вакциной или вакцинами на основе нуклеиновых кислот, таких как ДНК-вакцина, РНК-вакцина, плазмидная вакцина или векторная вакцина. Кроме того, способ не ограничивается каким-либо конкретным типом вакцин или каким-либо конкретным типом рака. Предпочтительно, способ может быть использован в комбинации с любой вакциной, предназначенной для лечения опухолевого заболевания, включая доброкачественные, предзлокачественные и злокачественные неопластические нарушения. Например, способ может быть использован в комбинации с вакциной, которая предназначена для лечения любого из следующих неопластических нарушений: карциномы, включая карциному мочевого пузыря (включая прогрессирующий и метастатический рак мочевого пузыря), молочной железы, толстой кишки (включая рак ободочной и прямой кишки), почек, печени, легких (включая мелкоклеточный и немелкоклеточный рак легкого и легочную аденокарциному), яичников, простаты, семенников, мочеполового тракта, лимфатической системы, прямой кишки, гортани, поджелудочной железы (включая экзокринную панкреатическую карциному), пищевода, желудка, желчного пузыря, шейки матки, щитовидной железы и кожи (включая плоскоклеточный рак); гематобластозы лимфоидной линии, включая лейкоз, острый лимфобластный лейкоз, острый лимфобластный лейкоз, В-клеточную лимфому, Т-клеточную лимфому, лимфому Ходжкина, неходжкинскую лимфому, волосатоклеточную лимфому, лимфому гистиоцитарную и лимфому Беркитта; гемопоэтические опухоли миелоидной линии, включая острые и хронические миелогенные лейкозы, миелодиспластический синдром, миелоидный лейкоз и промиелоцитарную лейкемию; опухоли центральной и периферической нервной системы, включая астроцитомы, глиомы, нейробластомы и шванномы; опухоли мезенхимального происхождения, включая фибросаркому, рабдомиосаркому и остеосаркому; другие опухоли, включая меланому, пигментную ксеродерму, роговую кератому, семиному, фолликулярный рак щитовидной железы и тератокарциному; меланому, неоперабельной стадии III или IV злокачественную меланому неоперабельной стадии III или IV, плоскоклеточный рак, мелкоклеточный рак легких, немелкоклеточный рак легкого, глиомы, рак желудочно-кишечного тракта, рак почки, рак яичника, рак печени, рак прямой кишки, рак эндометрия, рак почки, рак предстательной железы, рак щитовидной железы, нейробластомы, рак поджелудочной железы, глиобластомы, рак шейки матки, рак желудка, рак мочевого пузыря, гепатомы, рак молочной железы, рак толстой кишки и рака головы и шеи, рак желудка, опухоль половых клеток, рак кости, опухоли костей, злокачественная фиброзная гистиоцитома кости взрослых; злокачественная фиброзная гистиоцитома кости детей, саркому, детскую саркому, синоназальных природных киллеров, новообразования, новообразования клеток плазмы; миелодиспластические синдромы; нейробластому; опухоль зародышевых тестикулярных клеток, внутриглазную меланому, миелодиспластические синдромы; миелодиспластические/миелопролиферативные заболевания, синовиальные саркомы, хронический миелоидный лейкоз, острый лимфобластный лейкоз, положительный острый лимфобластный лейкоз филадельфийской хромосомы (Ph+ALL), множественную миелому, острый миелобластный лейкоз, хронический лимфолейкоз и мастоцитоз. В некоторых вариантах осуществления, настоящее изобретение обеспечивает способ повышения иммуногенности или терапевтического эффекта вакцины для лечения рака предстательной железы у человека. Вводимая вакцина способна вызывать иммунный ответ против любого человеческого РАА, такого как PSMA, PSA или PSCA. В некоторых отдельных вариантах осуществления, вводимая вакцина содержит молекулу нуклеиновой кислоты, кодирующую антиген, способный вызывать иммуногенность против человеческого РАА, такого как PSMA, PSA или PSCA. Примеры конкретных молекул нуклеиновых кислот, которые могут содержаться в вакцине, включают следующее, предусмотренные настоящим описанием:
1) молекулу нуклеиновой кислоты, кодирующую иммуногенный полипептид PSMA, иммуногенный полипептид PSA или иммуногенный полипептид PSCA;
2) молекулу нуклеиновой кислоты, кодирующую два иммуногенных полипептида РАА, предусмотренные настоящим описанием, такие как а) иммуногенный полипептид PSMA и иммуногенный полипептид PSA; b) иммуногенный полипептид PSMA и иммуногенный полипептид PSCA; или с) иммуногенный полипептид PSA и иммуногенный полипептид PSCA; и
3) молекулу нуклеиновой кислоты, кодирующую три иммуногенных полипептида РАА, которые представляют собой иммуногенный полипептид PSMA, иммуногенный полипептид PSA и иммуногенный полипептид PSCA.
В другом дополнительном аспекте, настоящее изобретение предусматривает способ лечения опухолевых заболеваний у млекопитающего, особенно человека. Способ включает введение млекопитающему (1) эффективного количества вакцины, способной вызывать иммунный ответ против ТАА, ассоциированного с опухолевым заболеванием, (2) эффективного количества, по меньшей мере, одного иммуносупрессивного клеточного ингибитора (ингибитор ISC) и (3) эффективного количества, по меньшей мере, одного иммуноэффекторного энхансера (энхансер IEC). Любая вакцина, которая способна вызывать иммунный ответ против конкретного ТАА, может быть использована в способе. Многие ТАА известны из уровня техники. Кроме того, для простатоассоциированных антигенов ниже приведены примеры ТАА, которые известны из уровня техники: CEA, MUC-1, Ер-САМ, 5Т4, ХГЧ-В, К-RAS и TERT для колоректального рака; CEA, MUC-1, р53, мезотелин, сьюрвивин и NY-ESO-1 для рака яичников; Muc-1, 5Т4, WT-1, TERT, CEA, EGF-R и MAGE-А3 для немелкоклеточного рака легкого; 5Т4 для почечно-клеточной карциномы; и Muc-1, мезотелин, K-Ras, Annexin А2, TERT и CEA для рака поджелудочной железы. Новые ТАА продолжают идентифицироваться. Вакцина, которая способна вызывать иммунный ответ против любого из известных или новых ТАА, может быть использована в способе. Кроме того, вводимая вакцина может находиться в любой форме или составе, например, субъединичной вакцины, пептидных вакцин или вакцин на основе нуклеиновых кислот, таких как ДНК-вакцина, РНК-вакцина, плазмидных вакцин или векторных вакцин.
В некоторых вариантах осуществления настоящее изобретение предусматривает способ лечения рака простаты у человека, способ, включающий введение человеку вакцины, способной вызывать иммунный ответ против любого человеческого РАА, такого как PSMA, PSA или PSCA. В некоторых отдельных вариантах осуществления, вводимая вакцина содержит молекулу нуклеиновой кислоты, кодирующую антиген, способный вызывать иммунный ответ против человеческого РАА, такого как PSMA, PSA или PSCA. Примеры конкретных молекул нуклеиновых кислот, которые могут содержаться в вакцине, включают следующее, предусмотренные настоящим описанием:
1) молекулу нуклеиновой кислоты, кодирующую иммуногенный полипептид PSMA, иммуногенный полипептид PSA или иммуногенный полипептид PSCA;
2) молекулу нуклеиновой кислоты, кодирующую два иммуногенных полипептида РАА, предусмотренные настоящим описанием, такие как а) иммуногенный полипептид PSMA и иммуногенный полипептид PSA; b) иммуногенный полипептид PSMA и иммуногенный полипептид PSCA; или с) иммуногенный полипептид PSA и иммуногенный полипептид PSCA; и
3) молекулу нуклеиновой кислоты, кодирующую три иммуногенных полипептида РАА, которые представляют собой иммуногенный полипептид PSMA, иммуногенный полипептид PSA и иммуногенный полипептид PSMA.
Способ лечения опухолевого заболевания у млекопитающего и способ повышения иммуногенности или терапевтического эффекта вакцины для лечения опухолевого заболевания у млекопитающего, описанного в данном документе выше, альтернативно называют "иммунотерапевтическими схемами на основе вакцин" (или "VBIR").
В иммунотерапевтических схемах на основе вакцин энхансеры IEC и ингибиторы ISC могут вводиться с помощью любого пригодного метода и пути, включая (1) системное введение, такое как внутривенное, внутримышечное или пероральное введение, и (2) местное введение, такое как внутрикожное и подкожное введение. Где это уместно или подходит, местное введение, как правило, является более предпочтительным, чем системное введение. Местное введение любого энхансера IEC и ингибитора ISC может осуществляться в любом месте тела млекопитающего, которое подходит для местного введения лекарственных средств; тем не менее, более предпочтительно, чтобы эти иммуномодуляторы вводились местно в непосредственной близости от вакцинодренирующего лимфатического узла. Два или более конкретных энхансера IEC из одного класса энхансеров IEC (например, два CTLA-агониста) можно вводить в сочетании с ингибитором ISC. Кроме того, два или более конкретных энхансера IEC из двух или более различных классов энхансеров IEC (например, один антагонист CTLA-4 и один агонист TLR) можно вводить вместе. Аналогичным образом, два или более конкретных ингибитора ISC из одного класса ингибиторов ISC (например, два или более ингибиторов протеинкиназы) могут быть введены в комбинации с энхансером IEC. Кроме того, два или более конкретных ингибиторов ISC из двух или более различных классов ингибиторов ISC (например, один ингибитор протеинкиназы и один ингибитор СОХ-2) могут быть введены вместе.
В иммунотерапевтических схемах на основе вакцин вакцину можно вводить одновременно или последовательно с любым или всеми используемыми иммуномодуляторами (например, ингибиторами ISC и энхансерами IEC). Точно так же, когда используют два или более иммуномодуляторов, то они могут быть введены одновременно или последовательно относительно друг друга. В некоторых вариантах осуществления, вакцину вводят одновременно (например, в смеси) относительно одного иммуномодулятора, но последовательно относительно одного или более дополнительных иммунных модуляторов. Совместное введение вакцины и иммуномодуляторов в иммунотерапевтической схеме, основанной на вакцине, может включать случаи, в которых вакцина и, по меньшей мере, один иммуномодулятор вводят таким образом, что каждый присутствует в месте введения, таком как вакцинодренирующий лимфатический узел, в одно и то же время, несмотря на то, что антиген и иммуномодуляторы не вводятся одновременно. Совместное введение вакцины и иммуномодуляторов также может включать случаи, при которых вакцина или иммуномодулятор удаляются из места введения, но, по меньшей мере, один клеточный эффект удаленной вакцины или иммуномодулятора сохраняется в месте введения, таком как вакцинодренирующий лимфатический узел, по меньшей мере, до тех пор, пока один или более дополнительных иммуномодуляторов не будут введены в это место. В случаях, когда вакцину нуклеиновой кислоты вводят в комбинации с вакциной CpG, то вакцина и CpG могут содержаться в одной композиции и вводиться вместе любым подходящим способом. В некоторых вариантах осуществления, вакцину нуклеиновой кислоты и CpG в совместном препарате (смесь) вводят внутримышечно в сочетании с электропорацией.
Любые ингибиторы ISC могут быть использованы в иммунотерапевтических схемах на основе вакцин. Примеры классов ингибиторов SIC включают ингибиторы протеинкиназы, ингибиторы циклооксигеназы-2 (СОХ-2), ингибиторы фосфодиэстеразы типа 5 (PDE5) и ДНК-сшивающие агенты. Примеры ингибиторов СОХ-2 включают целекоксиб и рофекоксиб. Примерами ингибиторов PDE5 являются аванафил, лоденафил, мироденафил, силденафил, тадалафил, варденафил, уденафил и запринаст. Примером ДНК-сшивающих агентов является циклофосфамид. Примеры специфических ингибиторов протеинкиназы подробно описаны ниже.
Термин "ингибитор протеинкиназы" относится к любому веществу, которое действует как селективный или неселективный ингибитор протеинкиназы. Термин "протеинкиназы" относится к ферментам, которые катализируют перенос концевого фосфата аденозинтрифосфата к тирозиновым, сериновым или треониновым остаткам в белковых веществах. Протеинкиназы включают рецепторные тирозинкиназы и нерецепторные тирозинкиназы. Примеры рецепторных тирозинкиназ включают EGFR (например, EGFR/HER1/ErbB1, HER2/Neu/ErbB2, HER3/ЕrbВ3, HER4/ErbB4), INSR (инсулиновый рецептор), IGF-IR, IGF-II1 R, IRR (рецептор, связанный с инсулиновым рецептором), PDGFR (например, PDGFRA, PDGFRB), c-KIT/SCFR, VEGFR-1/FLT-1, VEGFR-2/FLK-1/KDR, VEGFR-3/FLT-4, FLT-3/FLK-2, CSF-1 R, FGFR 1-4, ССК4, TRK А-С, MET, RON, ЕРНА 1-8, ЕРНВ 1-6, AXL, MER, TYR03, TIE, ТЕК, RYK, DDR 1-2, RET, с-ROS, LTK (лейкоцитная тирозинкиназа), ALK (киназа анапластической лимфомы), ROR 1-2, MUSK, AATYK 1-3 и RTK 106. Примеры нерецепторных тирозинкиназ включают BCR-ABL, Src, Frk, Btk, Csk, Abl, Zap70, Fes/Fps, Fak, Jak, Ack и LIMK. В иммунотерапевтических схемах на основе вакцин, предусмотренных настоящим описанием, ингибиторы тирозинкиназы вводят млекопитающему в субоптимальной дозе. Термин "субоптимальная доза" относится к количеству дозы, которая ниже минимальной эффективной дозы, когда ингибитор тирозинкиназы вводят в виде монотерапии (т.е., где ингибитор протеинкиназы вводят в сам по себе без других терапевтических агентов) для заданного опухолевого заболевания.
Примеры специфических ингибиторов протеинкиназ, пригодных для использования в иммунотерапевтических схемах на основе вакцин, включают лапатиниб, Azd 2171, ЕТ180СН 3, индирубин-3'-оксим, NSC-154020, PD 169316, кверцетин, росковитин, трицирибин, ZD 1839, 5-лодотуберцидин, адафостин, алоизин, алстерпаулон, аминогенистеин, API-2, апигенин, арктигенин, арри-334543, акситиниб (AG-013736), AY-22989, AZD 2171, бисиндолилмалеимид IX, CCI-779, хелеритрин, DMPQ, DRB, едельфосин, ENMD-981693, аналог эрбстатина, ерлотиниб, фасудил, гефитиниб (ZD1839), Н-7, Н-8, Н-89, НА-100, НА-1004, НА-1077, НА-1100, гидроксифасудил, кенпаулон, KN-62, KY12420, ЛЧМ-А13, лютеолин, LY294002, LY-294002, малотоксин, ML-9, MLN608, NSC-226080, NSC-231634, NSC-664704, NSC-680410, NU6102, оломоуцин, оксиндол I, PD 153035, PD 98059, флоридзин, пицеатанол, пикроподофилин, PKI, РР1, РР2, РТК787/ZK222584, РТК787/ZK-222584, пурваланол А, рапамун, рапамыцин, Ro 31-8220, ротлерин, SB202190, SB203580, сиролимус, SL327, SP600125, стауроспорин, STI-571, SU1498, SU4312, SU5416, SU5416 (семаксантиб), SU6656, SU6668, ингибитор Syk-киназы, ТВВ, TCN, тирфостин AG 1024, тирфостин AG 490, Τ тирфостин AG 825, тирфостин AG 957, U0126, W-7, вортманин, Y-27632, зактима (ZD6474), ΖΜ 252868. гефитиниб (Iressa.RTM.), сунитиниба малат (Sutent.RTM.; SU11248), эрлотиниб (Tarceva.RTM.; OSI-1774), лапатиниб (GW572016; GW2016), канертиниб (Cl 1033), семаксиниб (SU5416), ваталаниб (РТК787 / ZK222584), сорафениб (BAY 43-9006), иматиниб (Gleevec.RTM.; STI571), дасатиниб (BMS-354825), лефлуномид (SU101), вандетаниб (Zactima. RTM.; ZD6474) и нилотиниб. Дополнительные ингибиторы протеинкиназы, пригодные для использования в настоящем изобретении, описаны в, например, патенты США №5618829, 5639757, 5728868, 5804396, 6100254, 6127374, 6245759, 6306874, 6313138, 6316444, 6329380, 6344459, 6420382, 6479512, 6498165, 6544988, 6562818, 6586423, 6586424, 6740665, 6794393, 6875767, 6927293 и 6958340.
В некоторых вариантах осуществления ингибитор протеинкиназы может представлять собой мультикиназный ингибитор, который является ингибитором, который действует на более чем одну конкретную киназы. Примеры мультикиназных ингибиторов включают иматиниб, сорафениб, лапатиниб, BIRB-796 и AZD-1152, AMG706, зактима (ZD6474), МР-412 сорафениб (BAY 43-9006), дазатиниб, СЕР-701 (лестауриниб), XL647, XL999, тукерб (лапатиниб), MLN518 (ранее известный как СТ53518), РКС412, ST1571, АЕЕ 788, OSI-930, OSI-817, сунитиниба малат (сутент), акситиниб (AG-013736), эрлотиниб, гефитиниб, акситиниб, босутиниб, темсиролисмус и нилотиниб (AMN107). В некоторых отдельных вариантах осуществления ингибитор тирозинкиназы представляет собой сунитиниб, сорафениб или фармацевтически приемлемые соли или производные (такие как малат или тозилат) сунитиниба или сорафениба.
Сунитиниба малат, который продается компанией Pfizer Inc. под торговым названием Сутент, описан химически как бутандиовая кислота, гидрокси-, (2S)-, соединение с N-[2-(диэтиламино)этил]-5-[(2)-(5-фтор-1,2-дигидро-2-оксо-3Н-индол-3-илидин)метил]-2,4-диметил-1Н-пиррол-3-карбоксамидом (1:1). Соединение, его синтез и конкретные полиморфные описаны в патенте США №6573293, патентных публикациях США №2003-0229229, 2003-0069298 и 2005-0059824, и у J.M. Manley, M.J. Kalman, В.G. Conway, С.С. Ball, J.L Havens and R. Vaidyanathan, "Early Amidation Approach to 3-[(4-amido)pyrrol-2-yl]-2-indolinones," J. Org. Chew. 68, 6447-6450 (2003). Композиции сунитиниба и его L-малатной соли описаны в публикации РСТ № WO 2004/024127. Сунитиниба малат был одобрен в США для лечения желудочно-кишечной стромальной опухоли, распространенной почечно-клеточной карциномы и прогрессирующих, высокодифференцированных нейроэндокринных опухолей поджелудочной железы у больных с неоперабельным местно-распространенным или метастатическим заболеванием. Рекомендуемая доза сунитиниба малата для желудочно-кишечной стромальной опухоли (GIST) и распространенного почечно-клеточного рака (RCC) для человека составляет 50 мг для приема внутрь один раз в день, по схеме 4 недели лечения, а затем 2 недели отдыха (схема 4/2). Рекомендуемая доза сунитиниба малата для нейроэндокринных опухолей поджелудочной железы (pNET) составляет 37,5 мг, принимаемые перорально один раз в день.
В иммунотерапевтических схемах на основе вакцин, сунитиниба малат можно вводить перорально в виде разовой дозы или нескольких доз. Как правило, сунитиниба малеат доставляется в течение двух, трех, четырех или более последовательных еженедельных доз, за которыми следует период "отдыха", составляющий приблизительно 1 или 2 недели, или более, в течение которых сунитиниба малат не доставляется. В одном варианте осуществления дозы доставляются в течение 4 недель с 2 неделями отдыха. В другом варианте осуществления сунитиниба малат доставляется в течение двух недель с 1 неделей отдыха. Кроме того, он также может быть доставлен без периода "отдыха" в течение всего периода лечения. Эффективное количество сунитиниба малеата, вводимого перорально человеку в иммунотерапевтических схемах на основе вакцин, составляет, как правило, ниже 40 мг на человека на дозу. Например, его можно вводить перорально в количестве 37,5, 31,25, 25, 18,75, 12,5, 6,25 мг на человека в день. В некоторых вариантах осуществления сунитиниба малеат вводят перорально в диапазоне 1-25 мг на человека на дозу. В некоторых других вариантах осуществления сунитиниба малеат вводят перорально в диапазоне от 6,25, 12,5 или 18,75 мг на человека на дозу. Другие лекарственные схемы и вариации предвидятся заранее и будут определяться под руководством врача.
Сорафениба тозилат, продаваемый под торговым названием Nexavar, также является мультикиназным ингибитором. Его химическое название 4-(4-{3-[4-хлор-3-(трифторметил)фенил]уреидо}фенокси)-N-метилпиридин-2-карбоксамид. Он одобрен в США для лечения первичного рака почек (распространенной почечно-клеточной карциномы) и распространенного первичного рака печени (гепатоцеллюлярной карциномы). Рекомендуемая суточная доза составляет 400 мг, принимаемые перорально дважды в день. В иммунотерапевтических схемах на основе вакцин, предусмотренных настоящим описанием, эффективное количество вводимого перорально сорафениба тозилата составляет ниже 400 мг на человека в день. В некоторых вариантах осуществления эффективное количество вводимого перорально сорафениба тозилата находится в диапазоне 10-300 мг на человека в день. В некоторых других вариантах осуществления эффективное количество вводимого перорально сорафениба тозилата находится между 10-200 мг на человека в день и составляет 10, 20, 60, 80, 100, 120, 140, 160, 180 или 200 мг на человека в день.
Акситиниб, продаваемый под торговым названием INLYTA, является селективным ингибитором VEGF-рецепторов 1, 2 и 3. Его химическое название (N-метил-2-[3-((Е)-2-пиридин-2-ил-винил)-1-индазол-6-илсульфанил]бензамид. Он одобрен для лечения распространенной почечно-клеточной карциномы после безуспешности одной предварительной системной терапии. Начальная доза составляет 5 мг перорально дважды в день. Дозу можно регулировать, исходя из индивидуальной безопасности и переносимости. В иммунотерапевтических схемах на основе вакцин, предусмотренных настоящим описанием, эффективное количество вводимого перорально акситиниба, как правило, составляет ниже 5 мг дважды в день. В некоторых других вариантах осуществления эффективное количество вводимого перорально акситиниба находится между 1-5 мг дважды в день. В некоторых других вариантах осуществления, эффективное количество вводимого перорально акситиниба находится между 1, 2, 3, 4 или 5 мг дважды в день.
В иммунотерапевтических схемах на основе вакцин может быть использованы любые энхансеры IEC. Они могут быть небольшими молекулами и большими молекулами (например, белками, полипептидами, ДНК, РНК и антителами). Примеры энхансеров IEC, которые могут быть использованы, включают агонисты TNFR, антагонисты CTLA-4, агонисты TLR, антагонисты белка 1 (PD-1) запрограммированной гибели клеток (например, BMS-936558), антитело СТ-011 к PD-1) и антагонисты лиганда 1 белка 1 запрограммированной гибели клеток (PD-L1) (например, BMS-936559), антагонисты гена 3 активации лимфоцитов (LAG3) и антагонисты Т-клеточной молекулы-3, содержащей иммуноглобулин и муциновый домен (TIM- 3). Примеры конкретных агонистов TNFR, антагонистов CTLA-4, агонистов TLR представлены в деталях ниже.
Агонисты TNFR.
Примеры агонистов TNFR включают агонисты ОХ40, 4-1 ВВ (такой как BMS-663513), GITR (такой как TRX518) и CD40. Примеры специфических агонистов CD40 подробно описаны в данном документе ниже.
Агонисты CD40 представляют собой вещества, которые связываются с рецептором CD40 на клетке и повышают одну или более связанных с CD40 или CD40L активностей. Таким образом, "агонист" CD40 охватывает "лиганды" CD40.
Примеры агонистов CD40 включают агонистические антитела CD40, фрагменты агонистических антител CD40, лиганды CD40 (CD40L) и фрагменты и производные CD40L, такие как содержащие олигомерные (например, бивалентные, тримерные CD40L), слитые белки, и их варианты.
Лиганды CD40 для использования в настоящем изобретении включают любой пептид, полипептид или белок, или нуклеиновую кислоту, кодирующую пептид, полипептид или белок, которые могут связываться с и активировать один или более рецепторов CD40 на клетке. Пригодные лиганды CD40 описаны, например, в патенте США №6482411, 6410711; патенте США №6391637; и патенте США №5981724, все эти патенты и заявки и последовательности CD40L, описанные в них, включены во всей полноте в данный документ путем ссылки. Хотя человеческие лиганды CD40 являются предпочтительными для использования в терапии человека, в настоящем изобретении могут быть использованы лиганды CD40 от любых видов. Для использования в других видах животных, например, в ветеринарных вариантах, вид лиганда CD40, соответствующий животному, которое лечат, будет предпочтительным. В некоторых вариантах осуществления, лиганд CD40 представляет собой пептид или белковый олигомер gp39, включая природным путем образованные пептид, полипептид или белковый олигомер gp39, а также пептиды, полипептиды или белки др39 (и кодирующие нуклеиновые кислоты), которые содержат олигомеризационную последовательность. В то время как олигомеры, такие как димеры, тримеры и тетрамеры, являются предпочтительными в некоторых аспектах изобретения, в других аспектах изобретения для использования рассматриваются крупные олигомерные структуры, при условии, что олигомерные структуры сохраняют способность связываться и активировать один или более рецепторов CD40.
В других вариантах осуществления агонист CD40 представляет собой антитело к CD40 или его антигенсвязывающий фрагмент. Антитело может быть человеческим, гуманизированным или частично человеческим химерным антителом к CD4. Примеры специфических моноклональных антител к CD40 включают моноклональное антитело G28-5, гпАЬ89, ЕА-5 или S2C6 и СР870893. В конкретном варианте осуществления, агонистичным антителом к CD40 является СР870893 или десетузумаб (SGN-40).
СР-870893 является полностью человеческим агонистичным моноклональным антителом к CD40 (mAb), которое было исследовано клинически в качестве противоопухолевой терапии. Структура и получение СР870893 описана в WO 2003/041070 (где антитело идентифицируется по внутреннему идентификатору "21.4.1"). Аминокислотные последовательности тяжелой цепи и легкой цепи СР-870893 приведены в SEQ ID NO: 40 и SEQ ID NO: 41, соответственно. В ходе клинических испытаний СР870893 вводили путем внутривенной инфузии, как правило, в дозах в диапазонах 0,05-0,25 мг/кг на инфузию. В фазе I клинических исследований максимально переносимая доза (MTD) СР-870893 была определена как 0,2 мг/кг, а ограничивающая дозу токсичность включала CRS 3-ей степени и крапивницу 3-й степени. [Jens Ruter et al.: Immune modulation with weekly dosing of an agonist CD40 antibody in a phase I study of patients with advanced solid tumors. Cancer Biology & Therapy 10:10, 983-993; November 15, 2010]. В иммунотерапевтических схемах на основе вакцин, предусмотренных настоящим описанием, СР-870893 можно вводить внутрикожно, подкожно или местно. Предпочтительно, чтобы его вводили внутрикожно. Эффективное количество для введения СР870893 в схеме, как правило, составляет ниже 0,2 мг/кг, как правило, находится в диапазоне 0,01 мг - 0,15 мг/кг или 0,05-0,1 мг/кг.
Дацетузумаб (также известный как SGN-40 или huS2C6; CAS №88-486-59-9) является другим агонистичным антителом к CD40, которое исследовалось в клинических испытаниях для лимфом, диффузных крупных В-клеточных лимфом и множественной миеломы. В клинических испытаниях, дацетузумаб вводили внутривенно в недельных дозах от 2 мг/кг до 16 мг/кг. В иммунотерапевтических схемах на основе вакцин, предусмотренных настоящим описанием, дацетузумаб можно вводить внутрикожно, подкожно или местно. Предпочтительно, чтобы его вводили внутрикожно. Эффективное количество дацетузумаюа, вводимого в иммунотерапевтических схемах на основе вакцин, как правило, составляет ниже 16 мг/кг, как правило, находится в диапазоне 0,2-14 мг/кг, или 0,5-8 мг/кг, или 1-5 мг/кг.
Ингибиторы CTLA-4
Пригодные антагонистические агенты против CTLA-4 для использования в иммунотерапевтических схемах на основе вакцин, предусмотренных описанием изобретения, включают без ограничения антитела к CTLA-4 (такие как человеческие антитела к CTLA-4, мышиные антитела к CTLA-4, антитела млекопитающих к CTLA-4, гуманизированные антитела к CTLA-4, моноклональные антитела к CTLA-4, поликлональные антитела к CTLA-4, химерные антитела к CTLA-4, доменные антитела к CTLA-4), фрагменты антител к CTLA-4 (такие как (одноцепочечные фрагменты антител к CTLA-4, фрагменты тяжелой цепи антител к CTLA-4 и фрагменты легкой цепи антител к CTLA-4), и ингибиторы CTLA-4, которые выступают в роли агонистов ко-стимулирующего пути. В некоторых вариантах осуществления, ингибитором CTLA-4 является ипилимумаб или тремелимумаб.
Ипилимумаб (также известный как МЕХ-010 или MDX-101), продаваемый как YERVOY, является человеческим антителом к человеческому CTLA-4. Ипилимумаб известен под регистрационным номером CAS 477202-00-9 и описан как антитело 10DI в патентной публикации РСТ № WO 01/14424, включенной в данное описание посредством ссылки во всей своей полноте и для всех целей. Примеры фармацевтической композиции, содержащей ипилимумаб, приведены в патентной публикации РСТ № WO 2007/67959. Ипилимумаб утверждается в США для лечения неоперабельной или метастатической меланомы. Рекомендуемая доза ипилимумаба в качестве монотерапии составляет 3 мг/кг при внутривенном введении каждые 3 недели в течение в общей сложности 4 доз. В способах, предусмотренных настоящим изобретением, ипилимумаб вводят местно, в частности внутрикожно или подкожно. Эффективное количество ипилимумаба, вводимого внутрикожно или подкожно, как правило, находится в диапазоне 5-200 мг/дозу на человека. В некоторых вариантах осуществления, эффективное количество ипилимумаба находится в диапазоне 10-150 мг/дозу на человека на дозу. В некоторых отдельных вариантах осуществления, эффективное количество ипилимумаба составляет приблизительно 10, 25, 50, 75, 100, 125, 150, 175 или 200 мг/дозу на человека.
Тремелимумаб (также известный как СР-675206) является полностью человеческим моноклональным антителом IgG2 и имеет номер CAS 745013-59-6. Тремелимумаб описан как антитело 11.2.1 в патенте США №6682736, включенном в настоящее описание посредством ссылки во всей своей полноте и для всех целей. Аминокислотные последовательности тяжелой цепи и легкой цепи тремилимаба изложены в SEQ IND NO: 42 и 43, соответственно. Тремелимумаб был исследован в клинических испытаниях для лечения различных опухолей, включая меланомы и рак молочной железы; в которых тремелимумаб вводили внутривенно либо в виде одной дозы или нескольких доз каждые 4 или 12 недель при диапазоне доз от 0,01 до 15 мг/кг. В схемах, предусмотренных в соответствии с настоящим изобретением, тремелимумаб вводят местно, в частности внутрикожно или подкожно. Эффективное количество тремелимумаба, вводимого внутрикожно или подкожно, как правило, находится в диапазоне 5-200 мг/дозу на человека. В некоторых вариантах осуществления, эффективное количество тремелимумаба находится в диапазоне 10-150 мг/дозу на человека на дозу. В некоторых отдельных вариантах осуществления, эффективное количество тремелимумаба составляет приблизительно 10, 25, 50, 75, 100, 125, 150, 175 или 200 мг/дозу на человека.
Агонисты Toll-подобного рецептора (TLR).
Термин "агонист toll-подобного рецептора" или "агонист TLR" относится к соединению, которое действует как агонист toll-подобного рецептора (TLR). Он включает агонисты TLR1, TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9, TLR10 и TLR11 или их комбинацию. Если не указано иное, то соединение-агонист TLR может включать соединение в любой фармацевтически приемлемой форме, включая любой изомер (например, диастереомер или энантиомер), соль, сольват, полиморф и подобное. В частности, если соединение является оптически активным, то ссылка на соединение может включать каждый из энантиомеров соединения, а также рацемические смеси энантиомеров. Кроме того, соединение может быть идентифицировано в качестве агониста одного или нескольких конкретных TLR (например, агонист TLR7, агонист TLR8 или агонист TLR7/8).
Агонизм TLR для конкретного соединения можно оценить любым подходящим способом, известным в данной области техники. Независимо от конкретного используемого анализа, соединение может быть идентифицировано в качестве агониста конкретного TLR, если выполнение анализа с соединением приводит, по меньшей мере, к пороговому увеличению некоторой биологической активности, опосредованной конкретным TLR. С другой стороны, соединение может быть идентифицировано как не действующее в качестве агониста указанного TLR, если при использовании для выполнения анализа, предназначенного для обнаружения биологической активности, опосредованной указанным TLR, соединение не вызывает пороговое увеличение биологической активности. Если не указано иное, то увеличение биологической активности относится к увеличению той же биологической активности, которая наблюдалось в соответствующем контроле. Анализ может проводиться с соответствующим контролем или без него. С опытом, любой специалист в данной области может развить достаточное знание с конкретным анализом (например, диапазон значений, наблюдаемых в соответствующем контроле при определенных условиях анализа), что выполнение контроля не всегда может быть необходимым для определения агонизма TLR соединения в конкретном анализе. Некоторые агонисты TLR, используемые в способе согласно настоящему изобретению, представляют собой небольшие органические молекулы, в отличие от крупных биологических молекул, таких как белки, пептиды, и тому подобное. Примеры небольших молекул антагонистов TLR включают молекулы, раскрытые, например, в патентах США №4689338; 4929624, 4988815; 5037986; 5175296; 5238944; 5266575; 5268376; 5346905; 5352784; 5367076; 5389640; 5395937; 5446153; 5482936; 5693811; 5741908; 5756747; 5939090; 6039969; 6083505; 6110929; 6194425; 6245776; 6331539; 6376669; 6451810; 6525064; 6545016; 6545017; 6558951 и 6573273. Примеры конкретных низкомолекулярных агонистов TLR, применимые в способах, предусмотренных в соответствии с настоящим изобретением, включают 4-аминоальфа, альфа, 2-триметил-1Н-имидазо[4,5-с]хинолин-1-этанол, N-(2-{2-[4-амино-2-(2-метоксиэтил)-1Н-имидазо[4,5-с]хинолин-1-ил]этокси}этил)-N-метилморфолин-4-карбоксамид, 1-(2-амино-2-метилпропил)-2-(этоксиметил)-1Н-имидазо[4,5-с]хинолин-4-амин, N-[4-(4-амино-2-этил-1Н-имидазо[4,5-с]хинолин-1-ил)бутил]метансульфонамид, N-[4-(4-амино-2-пропил-1Н-имидазо[4,5-с]хинолин-1-ил)бутил]метансульфонамид и имиквимод. Некоторые агонисты TLR, особенно полезные в методах или схеме, предусмотренных настоящим описанием, обсуждаются в обзорной статье: Folkert Steinhagen, et al.: TLR-based immune adjuvants. Vaccine 29 (2011):3341-3355.
В некоторых вариантах осуществления агонисты TLR представляют собой агонисты TLR9, в частности олигонуклеотиды CpG (или CpG.ODN). Олигонуклеотид CpG представляет собой короткую молекулу нуклеиновой кислоты, содержащую цитозин, за которым следует гуанин, связанные фосфатной связью, в которой пиримидиновое кольцо цитозина неметилировано. Мотив CpG представляет собой множество оснований, которые включают неметилированный центральный CpG, окруженный, по меньшей мере, одним основанием, фланкирующим (на 3' и 5' стороне) центральный CpG. Олигонуклеотиды CpG включают как D так и К олигонуклеотиды. Весь CpG-олигонуклеотид может быть неметилированным или может быть частично неметилированным. Примеры CpG-олигонуклеотидов, применимые в способах, предусмотренных настоящим описанием, включают олигонуклеотиды, раскрытые в патентах США №6194388, 6207646, 6214806, 628371, 6239116 и 6339068.
Олигонуклеотиды CpG могут включать различные химические модификации и замещения по сравнению с природной РНК и ДНК, включая фосфодиэфирный межнуклеозидный мост, единицу бета-D-рибозы (дезоксибозу) и/или природное нуклеозидное основание (аденин, гуанин, цитозин, тимин, урацил). Примеры химических модификаций известны специалистам в данной области и описаны, например у Uhlmann Ε. et al. (1990), Chem. Rev. 90:543; "Protocols for Oligonucleotides and Analogs", Synthesis and Properties and Synthesis and Analytical Techniques, S. Agrawal, Ed., Humana Press, Totowa, USA 1993; Crooke, ST. et al. (1996) Annu. Rev. Pharmacol. Toxicol. 36:107-129; и Hunziker J. et al., (1995), Mod. Synth. Methods 7:331-417. В частности, олигонуклеотид CpG может содержать модифицированный цитозин. Модифицированный цитозин является природным или не встречающиеся в природе пиримидиновым основным аналогом цитозина, который может заменить это основание без ущерба иммуностимулирующей активности олигонуклеотида. Модифицированные цитозины включают 5-замещенные цитозины (например 5-метилцитозин, 5-фторцитозин, 5-хлорцитозин, 5-бромцитозин, 5-йодцитозин, 5-гидроксицитозин, 5-гидроксиметилцитозин, 5-дифторметилцитозин и незамещенный или замещенный 5-алкинилцитозин), 6-замещенные цитозины, N4-замещенные цитозины (например N4-этилцитлозин), 5-азацитозин, 2-меркаптоцитозин, изоцитозин, псевдоизоцитозин, аналоги цитозина с конденсированными кольцевыми системами (например Ν,Ν'-пропиленцитозин или феноксазин) и уроцил и его производные (например 5-фторурацил, 5-бромурацил, 5-бромовинилурацил, 4-тиоурацил, 5-гидроксиурацил, 5-пропинилурацил), но не ограничиваются ими. Некоторые предпочтительные цитозины включают 5-метилцитозин, 5-фторцитозин, 5-гидроксицитозин, 5-гидроксиметилцитозин и N4-этилцитозин.
Олигонуклеотид CpG может также содержать модифицированный гуанин. Модифицированный гуанин является природным или не встречающимся в природе пуриновым основным аналогом гуанина, который может заменить это основание без ущерба иммуностимулирующей активности олигонуклеотида. Модифицированные гуанины включают 7-деазагуанин, 7-деаза-7-замещенный гуанин, гипоксантин, N2-замещенные гуанины (например, N2-метилгуанин), 5-амино-3-метил-3Н,6Н-тиазоло[4,5-d]пиримидин-2,7-дион, 2,6-диаминопурин, 2-аминопурин, пурин, индол, аденин, замещенные аденины (например, Ν6-метиладенин, 8-оксоаденин), 8-замещенный гуанин (например, 8-гидроксигуанин и 8-бромгуанин) и 6-тиогуанин, но не ограничиваются ими. В некоторых вариантах осуществления описания изобретения гуаниновое основание замещено универсальным основанием (например, 4-метилиндолом, 5-нитроиндолом и К-основанием), ароматической кольцевой системой (например, бензимидазолом или дихлорбензимидазолом, амидом 1-метил-1Н-[1,2,4]триазол-3-карбоновой кислоты) или атомом водорода.
В некоторых аспектах олигонуклеотиды CpG включают модифицированные скелеты. Было показано, что модификация скелета нуклеиновой кислоты обеспечивает повышенную активность нуклеиновых кислот при введении in vivo. Вторичные структуры, такие как "петля-на-стебле", могут стабилизировать нуклеиновые кислоты от деградации. Альтернативно, стабилизация нуклеиновой кислоты может быть осуществлена с помощью фосфатной модификации скелета. Предпочтительная стабилизированная нуклеиновая кислота имеет, по меньшей мере, частично фосфоротиоатно модифицированный скелет. Фосфоротиоаты могут быть синтезированы с использованием автоматизированных методик, использующих химию фосфороамидатов или Н-фосфонатов. Арил- и алкилфосфонаты могут быть получены, например, как описано в патенте США №4469863; а алкилфосфотриэфиры (в которых заряженный фрагмент кислорода алкилируется, как описано в патенте США №5023243 и европейском патенте №092574) могут быть получены с использованием коммерчески доступных реагентов с помощью автоматизированного твердофазного синтеза. Способы получения других модификаций и замещений скелета ДНК уже описаны (Uhlmann, Ε. and Peyman, Α. (1990) Chem. Rev. 90:544; Goodchild, J. (1990) Bioconjugate Chem. 1:165). 2'-O-метилнуклеиновые кислоты с мотивами CpG также вызывают иммунную активацию, так же как этоксимодифицированные нуклеиновые кислоты CpG. На самом деле не были обнаружены модификации скелета, которые полностью уничтожают эффект CpG, хотя он значительно снижается вследствие замены С на 5-метил С.Конструкции, имеющие фосфоротиоатные связи, обеспечивают максимальную активность и защиту нуклеиновой кислоты от деградации внутриклеточными экзо- и эндонуклеазами. Другие модифицированные олигонуклеотиды включают фосфодиэфирные модифицированные олигонуклеотиды, комбинации фосфодиэфира и фосфоротиоатных олигонуклеотидов, метилфосфоната, метилфосфоротиоата, фосфородитиоата, п-этокси и их комбинации. Каждая из этих комбинаций и их конкретные воздействия на клетки иммунной системы обсуждаются более подробно со ссылкой на нуклеиновые кислоты CpG в патентных публикациях РСТ WO 96/02555 и WO 98/18810 и в патентах США №6194388 и 6239116.
Олигонуклеотиды CpG могут иметь один или два доступных 5' конца. Можно создать модифицированные олигонуклеотиды, имеющие два таких 5' конца, например, путем присоединения двух олигонуклеотидов через 3'-3' связь для создания олигонуклеотида, имеющего один или два доступных 5' конца. Связь 3'-3' может быть фосфодиэфирным, фосфоротиоатным или любым другим модифицированным межнуклеозидным мостом. Методы создания таких связей известны из уровня техники. Например, такие связи описаны у Seliger, H. et al., Oligonucleotide analogs with terminal 3'-3'- and 5'-5'-internucleotidic linkages as antisense inhibitors of viral gene expression, Nucleosides and Nucleotides (1991), 10(1-3), 469-77 and Jiang, et al., Pseudo-cyclic oligonucleotides: in vitro and in vivo properties, Bioorganic and Medicinal Chemistry (1999), 7(12), 2727-2735.
Кроме того, 3'-3'-связанные олигонуклеотиды, где связь между 3'-концевыми нуклеозидами не является фосфодиэфирным, фосфоротиоатным или другим модифицированным мостом, могут быть получены с использованием дополнительного спейсера, такого как три- или тетраэтиленгликоль фосфатного фрагмента (Durand, M. et al., Triple-helix formation by an oligonucleotide containing one (dA)12 and two (dT)12 sequences bridged by two hexaethylene glycol chains, Biochemistry (1992), 31(38), 9197-204, патенты США №5658738 и 5668265). Альтернативно, ненуклеотидный линкер может быть получен из этандиола, пропандиола или из лишенной азотистого основания единицы (d-спейсера) (Fontanel, Marie Laurence et al., Nucleic Acids Research (1994), 22(11), 2022-7) с использованием стандартной фосфорамидитной химии. Ненуклеотидные линкеры могут быть включены один раз или несколько раз, либо в сочетании друг с другом, допуская любое желаемое расстояние между 3'-концами двух олигонуклеотидов, которые будут связаны между собой.
Фосфодиэфирный межнуклеозидный мост расположен на 3' и/или 5'-конце нуклеозида может быть заменен на модифицированный межнуклеозидный мост, где модифицированный межнуклеозидный мост, например, выбран из фосфортиоатного, фосфородитиоатного, NR1R2-фосфорамидатного, боранофосфатного, а-гидроксибензилфосфонатного моста, фосфат-(С-С21)-O-алкилового эфира, фосфат-[(С6-С21)-арил- (С-С21)-O-алкил]сложного эфира, (С1-С6)-алкилфосфонатного и/или (С6-С12)арилфосфонатного моста, (C7-C12) -а-гидроксиметиларильного (например, раскрытых в публикации РСТ № WO 95/01363), где (С6-С12)арил, (С6-С20)арил и (С6-С1)арил, необязательно, замещены галогеном, алкилом, алкокси, нитро, циано и где R1 и R2 представляют собой независимо друг от друга водород, (С С18)-алкил, (С6-С20)-арил, (С6-С4)-арил, (С С8)-алкил, предпочтительно водород, (С-С8)-алкил, предпочтительно (С-С4)-алкил и/или метоксиэтил, или R1 и R2 образуют вместе с атомом азота, несущим их, 5-6-членное гетероциклическое кольцо, которое может дополнительно содержать дополнительный гетероатом, выбранный из группы О, S и N.
Замена фосфодиэфирного моста, расположенного на 3' и/или 5' конце нуклеозида, дефосфомостом (дефосфомосты описаны, например, у Uhlmann Ε. and Peyman A. in "Methods in Molecular Biology", Vol. 20, "Protocols for Oligonucleotides and Analogs", S. Agrawal, Ed., Humana Press, Totowa 1993, Chapter 16, pp. 355 и далее), где дефосфомост, например, выбирают из дефосфомостов формацеталя, 3'-тиоформацеталя, метилгидроксиламина, оксима, метилендиметилгидразо, диметиленсульфона и/или силильных групп.
Олигонуклеотиды CpG для использования в способах или схеме, предусмотренных описанием изобретения, могут необязательно иметь химерные скелеты. Химерный скелет является скелетом, который содержит больше одного типа связи. В одном варианте осуществления, химерный скелет может быть представлен формулой: 5'1 N1 ZN2Y2 3'. Y1 и Y2 представляют собой молекулы нуклеиновой кислоты, имеющие от 1 до 10 нуклеотидов. Y1 и Y2 включают, по меньшей мере, одну модифицированную межнуклеотидную связь. Поскольку, по меньшей мере, два нуклеотида химерных олигонуклеотидов включают скелетные модификации, то эти нуклеиновые кислоты являются примером одного типа "стабилизированных иммуностимулирующих нуклеиновых кислот".
Что касается химерных олигонуклеотидов, Y1 и Y2 рассматриваются независимо друг от друга. Это означает, что каждый из Y1 и Y2 могут иметь или не иметь разные последовательности и отличающиеся друг от друга скелетные связи в одной и той же молекуле. В некоторых вариантах осуществления, Y1 и/или Y2 имеют от 3 до 8 нуклеотидов. N1 и N2 являются молекулами нуклеиновых кислот, содержащими от 0 до 5 нуклеотидов, тогда как N1 ZN2 имеет, по меньшей мере, 6 нуклеотидов, в общей сложности. Нуклеотиды N1 ZN2 имеют фосфодиэфирный скелет, и не включают нуклеиновые кислоты, имеющие модифицированный скелет. Ζ является иммуностимулирующим мотивом нуклеиновой кислоты, предпочтительно выбранным из приведенных в настоящем документе.
Центральные нуклеотиды (N1 ZN2) формулы Y1 N1 ZN2Y2 имеют фосфодиэфирные межнуклеотидные связи и Y1 и Y2 имеют, по меньшей мере, один, но могут иметь более чем один или, возможно, даже все измененные межнуклеотидные связи. В предпочтительных вариантах Y1 и/или Y2 имеют, по меньшей мере, два или от двух до пяти модифицированных межнуклеотидных связей или Y1 имеет пять модифицированных межнуклеотидных связей и Y2 имеет две модифицированные межнуклеотидные связи. Модифицированная межнуклеотидная связь, в некоторых вариантах осуществления, является фосфоротиоатной модифицированной связью, фосфородитиоатной связью или р-этокси модифицированной связью.
Примеры конкретных олигонуклеотидов CpG, полезных в способах, предусмотренных настоящим описанием, включают:
5' TCGTCGTTTTGTCGTTTTGTCGTT3' (CpG 7909);
5' TCGTCGTTTTTCGGTGCTTTT3' (CpG 24555); и
5' TCGTCGTTTTTCGGTCGTTTT3' (CpG 10103).
CpG7909, синтетический одноцепочечный 24 мер, широко исследован для лечения рака как в виде монотерапии, так и в сочетании с химиотерапевтическими агентами, а также в качестве адъюванта для вакцины против рака и инфекционных заболеваний. Сообщалось, что однократная внутривенная доза CpG 7909 хорошо переносится без каких-либо клинических эффектов и высокой токсичности вплоть до 1,05 мг/кг, в то время как однократная доза подкожного CpG 7909 была максимально переносимой дозой (MTD), составляющей 0,45 мг/кг с ограничивающей дозу токсичностью миалгии и конституционных эффектов. [Смотрите Zent, Clive S, et al: Phase I clinical trial of CpG oligonucleotide 7909 (PF-03512676) in patients with previously treated chronic lymphocytic leukemia. Leukemia and Lymphoma, 53(2):211-217(7) (2012)]. В схемах, предусмотренных настоящим описанием, CpG7909 можно вводить путем инъекции в мышцу или любыми другими подходящими способами. Предпочтительно, чтобы его вводили местно в непосредственной близости от вакцинного дренирующего лимфатического узла, в частности, путем внутрикожного или подкожного введения. Для использования с вакциной на основе нуклеиновой кислоты, такой как ДНК-вакцина, предпочтительно формировать CpG совместно с вакциной в одной композиции и вводить внутримышечно в сочетании с электропорацией. Эффективное количество CpG7909 для внутримышечного, внутрикожного или подкожного введения, как правило, находится в диапазоне 10 мкг/дозу - 10 мг/дозу. В некоторых вариантах осуществления эффективное количество CpG7909 находится в диапазоне 0,05 мг - 14 мг/дозу. В некоторых отдельных вариантах осуществления эффективное количество CpG7909 составляет приблизительно 0,05; 0,1; 0,2; 0,3; 0,4; 0,5; 1 мг/дозу. Другие олигонуклеотиды CpG, включая CpG 24555 и CpG 10103, могут быть введены аналогичным образом и при аналогичных уровнях доз.
В некоторых отдельных вариантах осуществления настоящее изобретение предусматривает способ повышения иммуногенности или терапевтического эффекта вакцины для лечения опухолевого заболевания у человека, включающий введение человеку (1) эффективного количества, по меньшей мере, одного ингибитора ISC и (2) эффективного количества, по меньшей мере, одного энхансера IEC, причем, по меньшей мере, один ингибитор ISC является ингибитором протеинкиназы, выбранным из сорафениба тозилата, сунитиниба малата, акситиниба, эрлотиниба, гефитиниба, акситиниба, босутиниба, темсиролимуса или нилотиниба, и где, по меньшей мере, один энхансер IEC выбирают из ингибитора CTLA-4, агониста TLR или агониста CD40. В некоторых предпочтительных вариантах осуществления схема включает введение человеку (1) эффективного количества, по меньшей мере, одного ингибитора ISC и (2) эффективного количества, по меньшей мере, одного энхансера IEC, причем, по меньшей мере, один ингибитор ISC является ингибитором протеинкиназы, выбранным из акситиниба, сорафениба тозилата или сунитиниба малата, и где, по меньшей мере, один энхансер IEC выбирают из ингибитора CTLA-4, выбранного из ипилимумаба или тремелимумаба. В некоторых дополнительных предпочтительных вариантах осуществления схема включает введение человеку (1) эффективного количества, по меньшей мере, одного ингибитора ISC и (2) эффективного количества, по меньшей мере, двух энхансеров IEC, причем, по меньшей мере один ингибитор ISC является ингибитором протеинкиназы, выбранным из сунитиниба или акситиниба, и где, по меньшей мере, два энхансера IEC представляют собой тремелимумаб и агонист TLR, выбранный из CpG7909, CpG2455 или CpG10103.
В некоторых других вариантах осуществления настоящее изобретение предусматривает способ лечения рака простаты у человека, включающий введение человеку (1) эффективного количества вакцины, способной вызывать иммунную реакцию против человеческого РАА, (2) эффективного количества, по меньшей мере, одного ингибитора ISC, и (3) эффективного количества, по меньшей мере, одного энхансера IEC, причем, по меньшей мере, один ингибитор ISC является ингибитором протеинкиназы, выбранным из сорафениба тозилата, сунитиниба малата, акситиниба, эрлотиниба, гефитиниба, акситиниба, босутиниба, темсиролимуса или нилотиниба, и где, по меньшей мере, один энхансер IEC выбирают из ингибитора CTLA-4, агониста TLR или агониста CD40. В некоторых предпочтительных вариантах осуществления способ включает введение человеку (1) эффективного количества вакцины, способной вызывать иммунную реакцию против человеческого РАА, (2) эффективного количества, по меньшей мере, одного ингибитора ISC, и (3) эффективного количества, по меньшей мере, одного энхансера IEC, причем, по меньшей мере, один ингибитор ISC является ингибитором протеинкиназы, выбранным из сорафениба тозилата, сунитиниба малата или акситиниба, и где, по меньшей мере, один энхансер IEC является ингибитором CTLA-4, выбранным из ипилимумаба или тремелимумаба
В некоторых дополнительных специфических вариантах осуществления способ включает введение человеку (1) эффективного количества, по меньшей мере, одного ингибитора ISC и (2) эффективного количества, по меньшей мере, двух энхансеров IEC, причем, по меньшей мере, один ингибитор ISC является ингибитором протеинкиназы, выбранным из сунитиниба или акситиниба, и где, по меньшей мере, два энхансера IEC представляют собой тремелимумаб и агонист TLR, выбранный из CpG7909, CpG2455 или CpG10103.
Дополнительные терапевтические агенты.
Иммунотерапевтическая схема, основанная на вакцинах, предусмотренная настоящим описанием, может дополнительно содержать дополнительный терапевтический агент. Может быть использовано широкое разнообразие онкологических терапевтических агентов, включая химиотерапевтические агенты и гормональные терапевтические агенты. Специалисту в данной области техники будет понятно наличие и развитие других методов лечения рака, которые могут быть использованы в VBIR, предусмотренной настоящим описанием, и лечение не будет ограничено теми формами терапии, которые изложены в данном документе.
Термин "химиотерапевтическое средство" относится к химическим или биологическим веществам, которые могут вызвать гибель раковых клеток, или препятствовать росту, делению, восстановлению и/или функции раковых клеток. Примеры химиотерапевтических агентов включают те, которые раскрыты в WO 2006/088639, WO 2006/129163 и US 20060153808, описания которых включены в данный документ в качестве ссылки. Примеры конкретных химиотерапевтических агентов включают: (1) алкилирующие агенты, такие как хлорамбуцил (LEUKERAN), циклофосфамид (CYTOXAN), ифосфамид (IFEX), мехлорэтамингидрохлорид (MUSTARGEN), тиотепа (THIOPLEX), стрептозотоцин (ZANOSAR), кармустин (BICNU, GLIADEL WAFER), ломустин (CEENU) и дакарбазин (DTIC-DOME); (2) алкалоиды или алкалоиды барвинка, в том числе цитотоксические антибиотики, такие как доксорубицин (адриамицин), эпирубицин (ELLENCE, PHARMORUBICIN), даунорубицин (Cerubidine, DAUNOXOME), неморубицин, идарубицин IDAMYCIN PFS, ZAVEDOS), митоксантрон (DHAD, новантрон), дактиномицин (актиномицин D, COSMEGEN), пликамицин (MITHRACIN), митомицин (MUTAMYCIN) и блеомицин (BLENOXANE), винорелбина тартрата (NAVELBINE), винбластин (VELBAN), винкристин (ONCOVIN) и виндезин (ELDISINE); (3) антиметаболиты, такие как капецитабин (XELODA), цитарабином (CYTOSAR-U), флударабин (FLUDARA), гемцитабин (GEMZAR), гидроксимочевина (HYDRA), метотрексат (FOLEX, MEXATE, TREXALL), неларабин (ARRANON), триметрексат (NEUTREXIN) и пеметрексед (ALIMTA); (4) пиримидиновые антагонисты, такие как 5-фторурацил (5-FU); капецитабин (XELODA), ралтитрексед (TOMUDEX), тегафурурацил (UFTORAL) и гемцитабин (GEMZAR); (5) таксаны, такие как доцетаксел (TAXOTERE), паклитаксел (TAXOL); (6) препараты платины, такие как цисплатин (PLATINOL) и карбоплатин (PARAPLATIN) и оксалиплатин (ELOXATIN); (7) ингибиторы топоизомеразы, такие как иринотекан (CAMPTOSAR), топотекан (HYCAMTIN), этопозид (ETOPOPHOS, VEPESSID, TOPOSAR) и тенипозид (VUMON); (8) эпиподофиллотоксины (производные подофиллотоксина), такие как этопозид (ETOPOPHOS, VEPESSID, TOPOSAR); (9) производные фолиевой кислоты, такие как лейковорин (WELLCOVORIN); (10) нитрозомочевины, такие как кармустин (BiCNU), ломустин (CeeNU); (11) ингибиторы рецептора тирозинкиназы, включая рецептор эпидермального фактора роста (EGFR), фактора роста эндотелия сосудов (VEGF), рецептор инсулина, рецептор инсулиноподобного фактора роста (IGFR), рецептор фактора роста гепатоцитов (HGFR) и рецептор фактора роста тромбоцитов (PDGFR), такие как гефитиниб (IRESSA), эрлотиниб (TARCEVA), бортезомибом (VELCADE), мезилат иматиниба (GLEEVEC), генефитиниб, лапатиниб, сорафениб, талидомид, сунитиниб (SUTENT), акситиниб, ритуксимаб (RITUXAN, MABTHERA), трастузумаб (HERCEPTIN), цетуксимаб (ERBITUX), бевацизумаб (AVASTIN) и ранибизумаб (LUCENTIS), lym-1 (ONCOLYM), антитела к рецептору инсулиноподобного фактора роста-1 (IGF-1R), которые описаны в WO 2002/053596); (12) ингибиторы ангиогенеза, такие как бевацизумаб (AVASTIN), сурамина (GERMANIN), ангиостатин, SU5416, талидомид и ингибиторы матриксной металлопротеиназы (например, батимастат и маримастат), и те, которые раскрыты в WO 2002/055106; и (13) ингибиторы протеасом, такие как бортезомиб (VELCADE).
Термин "гормональный терапевтический агент" относится к химическому или биологическому веществу, которое ингибирует или исключает продуцирование гормона или ингибирует или нейтрализует действие гормона на рост и/или выживаемость клеток рака. Примеры таких агентов, пригодных для VBIR, включают те, которые раскрыты в US 20070117809. Примеры конкретных гормональных терапевтических агентов включают тамоксифен (NOLVADEX), торемифен (Fareston), фулвестрант (FASLODEX), анастрозол (ARIMIDEX), эксеместан (AROMASIN), летрозол (FEMARA), мегестролацетат (MEGACE), госерелин (ZOLADEX), леупролид (LUPRON), абиратерон и MDV3100.
VBIR, предусмотренная этим описанием изобретения, также может быть использована в комбинации с другими видами терапии, включая (1) хирургические методы, которые удаляют все или часть органов или желез, которые участвуют в продуцировании гормона, таких как яичники, тестикулы, надпочечники и гипофиз, и (2) облучение, при котором органы или железы пациента подвергают облучению в количестве, достаточном, чтобы ингибировать или исключить продуцирование целевого гормона.
I. ПРИМЕРЫ
Для иллюстрации некоторых вариантов осуществления изобретения предусмотрены следующие примеры. Они никоим образом не должны толковаться как ограничивающие объем изобретения. Исходя из изложенного выше описания и этих примеров, специалист в данной области техники сможет определить существенные характеристики изобретения, и, не отступая от его сущности и объема, сможет осуществить разные изменения и модификации изобретения, чтобы приспособить его для разных использований и условий.
ПРИМЕР 1. АНТИГЕНЫ В ЦИТОЗОЛЬНОМ, СЕКРЕТИРУЕМОМ И МЕМБРАНОСВЯЗАННОМ ФОРМАТАХ, ПОЛУЧЕННЫЕ ИЗ БЕЛКА ЧЕЛОВЕЧЕСКОГО PSMA
Пример 1 иллюстрирует конструкцию трех иммуногенных полипептидов PSMA, относящихся к "цитозольный антиген человеческого PSMA", "секретируемый антиген человеческого PSMA" и "мембраносвязанный антиген человеческого PSMA", соответственно, и биологические свойства этих полипептидов.
1А. Конструкция полипептидов иммуногенного PSMA
ДНК-конструкты, кодирующие полипептиды иммуногенного PSMA в цитозольных, секретируемых и модифицированных форматах, были созданы, исходя из белковой последовательности природного человеческого PSMA и исследованы по поводу их способности вызывать противоопухолевые эффекторные иммунные реакции. Структура и получение каждого из антигенных форматов человеческого PSMA предусмотрены, как изложено ниже.
1А1. Цитозольный антиген человеческого PSMA. Полипептид иммуногенного PSMA в цитозольной форме был задуман для удержания иммуногенного полипептида внутри клетки, в которой он экспрессируется. Цитоплазматический домен (аминокислоты 1-19) и трансмембранный домен (аминокислоты 20-43) человеческого PSMA были удалены, что привело к цитозольному полипептиду PSMA, который состоит из аминокислот 44-750 (внеклеточный домен или ECD) человеческого PSMA из SEQ ID ΝΟ: 1. К N-концу полипептида для повышения экспрессии может быть добавлена оптимальная последовательность Козака "MAS".
1А2. Секретируемый антиген человеческого PSMA. Полипептид иммуногенного PSMA в секретируемой форме был задуман для секретирования полипептида наружу клетки, в которой он экспрессируется. Секретируемый полипептид создан с аминокислотами 44-750 (ECD) человеческого PSMA из SEQ ID NO: 1 и секреторного элемента каппа lg, который имеет аминокислотную последовательность ETDTLLLWVLLLWVPGSTGD и два аминокислотных линкера (АА) на N-конце, чтобы максимизировать секрецию антигена PSMA как только он экспрессируется.
1А3. Мембраносвязанный антиген человеческого PSMA. Иммуногенный мембраносвязанный полипептид PSMA был задуман, чтобы стабилизировать полипептид на клеточной поверхности. Первые 14 аминокислот белка человеческого PSMA были удалены, а полученный иммуногенный полипептид состоит из аминокислот 15 - 750 белка человеческого PSMA из SEQ ID N0:1. Иммуногенный полипептид, который состоит из аминокислот 15 - 750 белка природного человеческого PSMA из SES ID N0:1 и разделяет 100% идентичность последовательности с белком природного человеческого PSMA, в настоящем описании также называют антигеном "модифицированного человеческого PSMA", "модифицированного hPSMA" или "hPSMAmod".
IB. Получение ДНК-плазмид для экспрессирования антигенов PSMA ДНК-конструкты, кодирующие цитозольные PSMA-антигены, секретируемые PSMA-
антигены и модифицированные PSMA-антигены, клонировали индивидуально в вектор PJV7563, который пригоден для in vivo исследования на животных (Фигура 1). Обе цепи ДНК в векторах PJV7563 сиквенировали для поддержания задуманной работоспособности.
Крупномасштабное получение ДНК-плазмид (Qiagen/CsCl) продуцировалось из последовательности подтвержденного клона. Качество плазмидной ДНК было подтверждено высоким отношением 260/280, высоким отношением в высшей степени скрученной в спираль ДНК и ДНК с одноцепочечным разрывом, низкими уровнями эндотоксинов (<10 Ед./мгДНК) и негативного биогруза.
IC. Экспрессия конструктов PSMA в млекопитающих клетках
Экспрессию цитозольных, секретируемых и модифицированных антигенов PSMA определяли с помощью FACS. Млекопитающие клетки 293 трансфицировали векторами PJV7563 PMED, кодирующими разные полипептиды иммуногенного PSMA. Через три дня клетки 293 окрашивали с мышиным антителом к PSMA, после с флюоресцентным конъюгированным (FITC) крысиным антимышиным вторичным антителом. Данные, представленные ниже как средняя интенсивность флюоресценции (MFI) через негативные контроли, подтверждают, что модифицированный антиген человеческого PSMA экспрессируется на клеточной поверхности.

1D. Композиции плазмид PSMA на частицах золота (для ND10X15)
Технология кожной доставки с помощью частиц (Particle Mediated Epidermal Delivery technology (PMED)) представляет собой безыгольный способ введения вакцин животным или людям. Система PMED включает преципитацию ДНК на микроскопические частицы золота, которые затем Система PMED включает преципитацию ДНК на микроскопические частицы золота, которые затем нагнетают с помощью гелиевого газа в эпидермис.ND10, одно применяемое устройство, использует находящийся под давлением гелий из внутреннего цилиндра для доставки частиц золота, и Х15, дублирующее доставляющее устройство, использует внешний гелиевый резервуар, который соединен с Х15 через шланг высокого давления для доставки частиц золота. Оба этих устройства были использованы в исследованиях доставки ДНК-плазмид PSMA. Частицы золота обычно составляли 1-3 мкм в диаметре и частицы были сформированы таким образом, чтобы содержать 2 мкг плазмид PSMA на 1 мг частиц золота. (Sharpe, M. et al.: P. Protection of mice from H5N1 influenza challenge by prophylactic DNA vaccination using particle mediated epidermal delivery. Vaccine, 2007, 25(34): 6392-98: Roberts LK, et al.: Clinical safety and efficacy of a powdered Hepatitis В nucleic acid vaccine delivered to the epidermis by a commercial prototype device. Vaccine, 2005; 23(40):4867-78).
IE. Трансгенные мыши, использованные для исследований in vivo
Использовали две трансгенные мышиные модели для человеческих HLA для оценки презентации разных антигенов PSMA разными HLA, а трансгенную мышиную модель для человеческого PSMA использовали для оценки нарушения иммунной толерантности к человеческому PSMA. В первой трансгенной мышиной модели HLA использовали мышей HLA А2/DR1 (из института Пастера, Париж, Франция; названные также "мыши Пастера"). Мыши Пастера «накаутированы» по мышиному β-2-микроглобулину и не экспрессируют молекулы функционального Н-2b; поэтому эта модель, как предполагают, репрезентирует презентацию антигена в человеческом HLA А2 и контексте DR1 (Pajot, Α., M.-L. Michel, N. Faxilleau, V. Pancre, С. Auriault, D.M. Ojcius, F.A. Lemonnier, and Y.-C. Lone. A mouse model of human adaptive immune functions: HLA-A2.1/HLA-DR1-transgenic H-2 class I-/class ll-knockout mice. Eur. J. Immunol. 2004, 34:3060-69.). Во второй трансгенной мышиной модели HLA используют мышей, которые «накаутированы» человеческим HLA А24, который ковалентно связан с человеческим β-2-микроглобулином у локуса H2bk. Эти мыши лишены мышиного β-2-микроглобулина и не экспрессируют молекулы функционального Н-2b. Эта модель позволяет оценивать антигенную презентацию в контексте человеческих HLA А24.
IF. Иммуногенность белков человеческого PSMA в цитозольном, секретируемом и модифицированном форматах
Схема исследования. Восьми-десятинедельных мышей иммунизировали, используя метод PMED, разными ДНК-конструктами PSMA в режиме прайм/буст/буст/, с интервалом в две недели между каждой вакцинацией. Альтернативно, мышей примировали аденовирусным вектором, кодирующим антиген PSMA при концентрации 1×109 вирусных частиц в 50 мкл (PBS) путем внутримышечной инъекции. Аденовирусный вектор (вектор pShuttle-CMV от Stratagene) модифицировали, чтобы он содержал сайты рестрикции Nhel и Bglll в пределах множественного сайта клонирования. ДНК, клонирующую модифицированный человеческий PSMA, затем выделяли, обрабатывая Nhel и Bglll, лигировали в этот вектор и закрепляли последовательность. Вектор pShuttle модифицированного человеческого PSMA затем рекомбинировали с вектором pAdEasy-1, а вирус размножали согласно системе AdEasy (Stratagene). Через двадцать дней их бустировали PMED, как описано выше. В каждой использованной схеме антиген-специфическую Т-клеточную реакцию измеряли через 7 дней после последней иммунизации в анализе ELISPOT с интерфероном-гамма (IFN-y). Анализ ELISPOT подобен твердофазному иммуноферментному сэндвич-анализу (ELISA). Кратко, иммобилизованное антитело, специфическое к IFN-y (BD Bioscience, # 51-2525kc) наносили на поливинилиденфторидную (ПВДФ) мембрану в микропланшете в течение ночи при 4°C. Планшет блокировали сывороткой/белком, чтобы предотвратить неспецифическое связывание с антителом. После блокирования эффекторные клетки (например, спленоциты, выделенные из мышей, иммунизированных PSMA) и мишени (например, пептиды PSMA из пептидной библиотеки, активированные клетки-мишени с пептидами или опухолевые клетки, экспрессирующие соответствующие антигены) или митоген (который будет стимулировать спленоциты неспецифически продуцировать IFN-у) добавляли в лунки и инкубировали в течение ночи при 37°C в 5% CO2 инкубаторе. Цитокины, секретируемый эффекторными клетками, захватываются антителом покрытия на поверхности PVDF-мембраны. После удаления клеток и питательные среды, 100 мкл биотинилированного поликлонального антитела к мышиному IFN-y (0,5 мг/мл BD Bioscience, # 51-1818kz) добавляли в каждую из лунок для обнаружения. Пятна визуализировали добавлением стрептавидина/пероксидазы хрена (HRP, BD Bioscience, # 557630) и преципитата субстрата, 3-амино-9-этилкарбазола (АЕС), получая пятна красного цвета. Каждое пятно представляет собой один цитокин, продуцирующий Т-клетку. В общем, в исследованиях, раскрытых в данном документе, анализ ELISpot проводили следующим образом: 5×105 спленоцитов от мышей, иммунизированных PSMA, культивировали (1)в присутствии конкретных пептидов PSMA, полученных из библиотеки пептидов PSMA (см. Таблицу 16), созданной из 15-аминокислотных пептидов, перекрывающихся 11 аминокислотами, (2) с известными HLA А2.1-рестриктированными специфическими пептидами PSMA, или (3) с опухолевыми клетками. Для измерения распознавания эндогенной презентации антигена, спленоциты культивировали с клетками рака простаты человека HLA А2 (т.е. LNCaP, доступные от АТСС), которые, естественно, экспрессируют PSMA, или культивировали с опухолевыми клетками HLA А2, трансдуцированными с аденовирусом, кодирующим и, таким образом, экспрессирующим модифицированный антиген человеческого PSMA. Кроме того, к анализу ELISPOT добавили белок ECD человеческого PSMA для измерения определенных CD4 IFN-y-продуцирующих клеток. Для контроля в случае необходимости, HLA А2-рестриктированный HER-2 специфический пептид р168-175 или опухолевые клетки, не экспрессирующие PSMA, или иррелевантный белок, такой как BSA, были использованы в качестве отрицательного контроля в анализе ELISPOT IFN-y. Данные результатов приводятся в нормализованной форме для числа образующих пятно клеток (SFC), которые секретируют IFN-y в 1×106 спленоцитов. По меньшей мере, было проведено по три исследования для каждого тестируемого антигенного пептида PSMA.
Результаты. Данные анализа ELISPOT с спленоцитами мышей Пастера, культивированными с пептидами, полученными из библиотеки пептидов PSMA, представлены в Таблице 1. Положительный ответ определяли как имеющий SFC>100. Как показано в Таблице 1, иммуногенные полипептиды PMSA, созданные со всеми тремя антигенными форматами, цитозольными, секретируемыми и модифицированными антигенами человеческого PSMA, описанные в Примере 1 выше, способны индуцировать Т-клеточный ответ. Модифицированный антигенный формат человеческого PSMA измененный формат антиген человеческого индуцировал лучшую широту и амплитуду ответных реакций Т-клеток.
Таблица 1. Т-клеточный ответ, вызванный цитозольными, секретируемыми и модифицированными антигенами человеческого PSMA, у мышей Пастера


Данные анализа ELISpot на Т-клеточные реакции, индуцированные различными вакцинными форматами PSMA у мышей Пастера (которые распознали HLA А2.1-рестриктированные пептиды PSMA активированных клеток-мишеней, а также PSMA + опухолевые клетки HLA А2.1 LNCaP) представлены в Таблице 2. Клетки РС3, являющиеся клеточной линией рака простаты человека, которая не экспрессируют PSMA, были использованы в данном исследовании в качестве негативного контроля. Положительный ответ определяется как имеющий SFC>50. Как показано в Таблице 2, различные тестируемые конструкты PSMA способны индуцировать Т-клетки, которые распознают известный HLA А2, рестриктированный эпитопами PSMA, а также белок PSMA и клетки LNCaP рака простаты человека. Тем не менее, модифицированный конструкт PSMA индуцирует наилучший по широте и амплитуде ответ Т-клеток.
Таблица 2. Т-клеточные реакции, вызванные цитозольными, секретируемыми и модифицированными антигенами человеческого PSMA, у мышей Пастера, которые распознали HLA А2.1-рестриктированные пептиды PSMA активированных клеток, а также PSMA + опухолевые клетки HLA А2.1 LNCaP.

1G. Humoral immune response measured in Pasteur mice or nonhuman primates
1G1. Сэндвич-анализ ELISA. Стандартный сэндвич-анализ ELISA осуществляли с помощью автоматизированной системы Biotek. Планшеты покрывали 25 мкл природного белка PSMA с концентрацией 1,0 мкг/мл в PBS в течение ночи, планшеты промывали и блокировали 35 мкл/лунку 5% FBS 1 × PBS-T 0,05% и инкубировали в течение 1 часа при комнатной температуре на шейкере при 600 оборотов в минуту. Блокирующую среду декантировали, серийно разводили вакцинированную мышиную сыворотку с половинными разведениями в 5% FBS 1 × PBS-T 0,05%, начиная с 1:100 или 1:500, а образцы по 25 мкл разбавленной сыворотки добавляли в каждую лунку 96-луночных планшет и инкубировали в течение 1 часа при комнатной температуре на шейкере при 600 оборотов в минуту. Планшеты промывали 3 раза 75 мкл/лунку 1 × PBS-T 0,05% с использованием Biotek ELx405 и по 25 мкл/лунку разведенного 1:30000 вторичного антитела к мышиному IgG HRP (Abeam Cat # ab20043) (разведенного в 1 × PBS-T 0,05%) добавляли в каждую лунку 96-луночных планшет и инкубировали в течение 1 часа при комнатной температуре на шейкере при 600 оборотов в минуту. Планшеты промывали 5 раз 75 мкл/лунку 1 × PBS-T 0,05% с использованием Biotek Elx405. Субстрат ТМВ разводили в соотношении 1:10 и по 25 мкл добавляли в каждую лунку и инкубировали при комнатной температуре в течение 30 минут. Реакцию останавливали добавлением 12,5 мкл/лунку 1М H2SO4. Планшеты считывали с помощью Spectramax Plus при длине волны 450 нм. Данные считывали как титры и они могут быть представлены как первый положительный (среднее и оба значения выше 5% FBS PBS+3 временное стандартное отклонение) и/или в качестве расчетных титров при OD 0,5 или 1,0. Сыворотку от иррелевантных вакцинированных мышей использовали в качестве негативного контроля.

Результаты. Данные, представленные в Таблице 3, показывает, что цитозольный антиген человеческого PSMA не вызывает каких-либо реакций против PSMA, в то время как измененный антиген человеческого PSMA последовательно индуцировал хорошие ответные реакции антител против PSMA у всех мышей.
Данные, представленные в Таблице 3, показывают, что антитела, индуцированные антигенам человеческого PSMA, отреагировали на множественные пептидные эпитопы в библиотеке PSMA. Сыворотку от каждой мыши в каждой группе собирали в равных количествах и разводили 1:500 в анализе ELISA. Группу мышей негативного контроля, вакцинированных противодифтерийным (CRM) токсоидом, исследовали параллельно. Каждую лунку в 96-луночной планшете ELISA покрывали 0,03 мг одним 15аа пептидом, полученным из библиотеки пептидов PSMA. значение OD выше 0,10 считается позитивным.
1G2. Анализ клеточного связывания FACS. Различные линии раковых клеток предстательной железы были использованы для данного анализа. LNCaP (АТСС) использовали в качестве клеток рака простаты человека, экспрессирующих PSMA, а РС3 (АТСС) использовали в качестве негативных клеток рака простаты человека, которые не экспрессируют PSMA. В некоторых анализах клеточную линию TRAMP-C2, созданную для стабильной экспрессии природного непроцессированного человеческого PSMA, и клеточную линию родительской TRAMP-C2, которая не экспрессирует PSMA (негативный контроль), использовали для анализа связывания клеток. Анализ на связывание клеток проводили следующим образом: клетки LNCaP и РСЗ (или TRAMP-C2PSMA и TRAMP С2) высевали в отдельные лунки в количестве 2×105 клеток/лунку (50 мкл) в 96-луночный планшет. Сыворотку от мышей, вакцинированных PSMA, как описано в 1F, разводили 1:50 с буфером FACS (PBS, рН 7,4, 1% FBS, 25 мМ HEPES и 1 мМ ЭДТА). Пятьдесят микролитров разбавленного J591-A антитела (мышиного антитела против человеческого PSMA, клон J591-A из АТСС) добавляли к разбавленной тест-сыворотке или буферу FACS (неокрашенные образцы) для достижения соответствующего количества клеток на лунку в окрашивающей планшете. Все смешивали с помощью пипетки, а затем выдерживали на льду в течение 20 мин. Клетки дважды промывали буфером FACS; каждую промывку центрифугировали при 1200 оборотах в минуту при 5°C в течение 5 минут. Добавляли пятьдесят микролитров вторичного окрашивающего раствора, содержащего разведение 1:200 РЕ-меченного козьего антитела к lg мыши (Sigma, cat Р9670-5) и 0,25 мкл водного красителя "живой / мертвый" (Invitrogen, Cat # L34957) в каждую содержащую клетки лунку и выдерживали на льду в течение 20 мин. Клетки промывали дважды, как описано выше. Промытые осадки клеток ресуспендировали в 125 мкл FACS-буфера, а затем в каждую лунку добавляли по 75 мкл 4% раствора параформальдегида, чтобы зафиксировать клетки. Образцы хранили на льду в защищенном от света месте в течение, по меньшей мере, 15 мин. Образцы прогоняли на FACS Canto II. Было зарегистрировано десять тысяч событий живых клеток для каждого образца. Контрольными образцами для каждого типа клеток были 1) неокрашенные клетки, 2) клетки только с вторичными антителами, 3) клетки с J591 плюс вторичное антитело, и 4) клетки с интактной сывороткой плюс вторичное антитело. Данные сообщались как средняя интенсивность флуоресценции (MFI) относительно негативных контролей.
Результаты анализа клеточного связывания FACS. Таблица 4 показывает, что антитела, индуцированные как секретируемыми, так и модифицированными антигенами человеческого PSMA, способны связываться с позитивными клетками рака простаты (LNCaP), а не с негативными клетками рака простаты (РС3). Модифицированный антиген PSMA последовательно индуцировал хорошие ответные реакции антител к PSMA у всех мышей.


1G3. Анализ антителозависимой клеточноопосредованной цитотоксичности (ADCC)
Схема исследования. Индийских резус макак иммунизировали нуклеиновой кислотой, кодирующей модифицированный антиген человеческого PSMA, доставляемый аденовирусом (1011 V.P. внутримышечно), с последующими 2 PMED иммунизациями (8 запусков/иммунизация, по 4 запуска на правую и левую стороны нижней части живота) с 8 и 6 недельными интервалами соответственно. Животное также получало внутрикожные инъекции 3 мг CpG (PF-03512676) в непосредственной близости от каждой пахового дренирующего лимфатического узла во время второй иммунизации PMED. Антителозависимую клеточноопосредованную цитотоксичность определяли из плазмы, собранной из крови перед любой иммунизацией (преиммунная плазма) и через 8 дней после последней иммунизации PMED (иммунная плазма).
Анализ антителозависимой клеточноопосредованной цитотоксичности. Антителозависимую клеточноопосредованную цитотоксичность определяли с использованием стандартного теста с радиоактивным хромом 51. В качестве клеток-мишеней использовали клеточные линии рака простаты человека LNCaP и РС3. В качестве эффекторных клеток использовали клетки человеческой РВМС. Эффекторы к клеткам-мишеням были установлены на уровне 30:1. Кратко, для одной реакции мечения в 200 мкл инкубировали 1,5×106 клеток-мишеней с 200 пКи 51Сr (37°C, 5% CO2 в течение 1 часа). Клетки промывали три раза, концентрацию клеток доводили до 2×105 клеток/мл. Контрольные моноклональные антитела (mAb) или исследуемую плазму (1:50) готовили при концентрации 2Х и по 175 мкл из каждого разбавления mAb/плазма (в зависимости от размера анализа) добавляли к 175 мкл клеток-мишеней. Смесь инкубировали в течение 30 минут при 4°C в пробирке Эппендорфа. Клетки промывали один раз до свободных несвязанных антител. В это время, по 100 мкл свежевыделенных эффекторных клеток добавляли в каждую лунку 96-луночного планшета вместе с 100 мкл моноклональных антител или исследуемой плазмы, связывающих клетки-мишени, и инкубировали при 37°C и 5% С02 в течение 4 ч. Образцы испытывали в двух повторностях. По 100 мкл 2N HCI добавляли в целевые лунки для максимального высвобождения и по 100 мкл среды добавляли в целевые лунки для спонтанного высвобождения. Специфический лизис рассчитывали следующим образом: процент высвобождения = (ER-SR)/(MR-SR)×100, где ER (эффекторы + высвобождение клеток-мишеней) - экспериментальное высвобождение, SR (клетки-мишени, инкубированные только со средой) - спонтанное высвобождение и MP (клетки-мишени, инкубированные только с 2N HCl) - максимальное высвобождение. Процент специфического лизиса вычисляли путем вычитания высвобождения иррелевантной мишени (РС3) от высвобождения антиген-специфической мишени (LNCaP).

Результаты. Данные анализа антителозависимой цитотоксичности представлены в Таблице 6. LNCaP, линия PSMA + раковых клеток простаты человека, покрытых иммунной плазмой, полученной от иммунизированного hPSMA животного, лизировали под действием эффекторных клеток, тогда как РС3, линия PSMA - раковых клеток простаты человека, покрытых той же иммунной сывороткой, не лизируются эффекторными клетками. Аналогичным образом, LNCaP, покрытые преиммунной плазмой, не лизируются эффекторными клетками. Герцептин, моноклональное антитело против HER-2, использовали в качестве позитивного контроля, поскольку клетки LNCaP, как известно, экспрессируют HER-2 (Li, Cozzi et al. 2004). Ритуксан, моноклональное антитело против В-клеточного антигена (CD20), использовали в качестве негативного контрольного антитела, так LNCaP клетки не экспрессируют CD20. Оба моноклональных антитела, как сообщается, имеют ADCC-активность (Dall'Ozzo, Tartas et al. 2004; Collins, O'Donovan et al. 2011).
ПРИМЕР 2. СОЗДАНИЕ ПЕРЕТАСОВАННЫХ АНТИГЕНОВ PSMA
Этот пример иллюстрирует конструирование и некоторые биологические свойства различных иммуногенных полипептидов PSMA, которые являются вариантами модифицированного антигена человеческого PSMA (SEQ ID NO: 9), как описано в Примере 1.
2А. Дизайн перетасованных антигенов PSMA
Были разработаны различные иммуногенные полипептиды PSMA, которые являются вариантами модифицированного антигена человеческого PSMA (SEQ ID NO: 9), как описано в Примере 1. Эти варианты были созданы путем введения мутаций, выбранных из ортологов человеческого PSMA, в последовательность модифицированного антигена человеческого PSMA. Эти варианты в описании называют, взаимозаменяемо, как "перетасованные антигены PSMA" или "перетасованные модифицированные антигены PSMA". Принцип и методика, использованные при создании этих вариантов, представлены ниже.
Для отбора точечных мутаций перетасованного варианта был написан вычислительный алгоритм. Во-первых, множественное выравнивание последовательностей PSMA и 12 ортологов (Приложение 2а) было собрано с использованием NCBI PSI-BLAST. Выход из PSI-BLAST включал склонности для каждого остатка в каждом положении PSMA среди ортологов. Perl-скрипт затем использовал эти склонности, чтобы выбрать точечные мутации, как указано ниже:
1) Среди всех положений выбирают наиболее часто наблюдаемый остаток, что не соответствует идентичности природного человеческого PSMA.
2) Убедитесь, что это положение мутации не совпадает с идентифицированным Классом I или II эпитопов человеческого PSMA для того, чтобы точечная мутация не находилась в пределах консервативного Т-клеточного эпитопа, как определено в данном документе выше (Таблица 19).
3) Рассчитайте сходство мутации с человеческим остатком через матрицу BLOSUM62, чтобы убедиться, что оценка сходства BLOSUM62 для замещения остатков находится в диапазоне 0-1 (включительно).
Эта итерационная процедура продолжается пока не будет достигнут определенный процент идентичности последовательности (ниже 100) по отношению к человеческому PSMA.
Чтобы служить в качестве ввода в этом алгоритме были собраны ортологи PSMA для построения положение специфической матрицы с использованием PSI-BLAST от NCBI. Кроме того, выявленные эпитопные участки PSMA были перечислены в файле, который также был предусмотрен для алгоритма перетасовки. Неперетасовочные участки также были расширены до цитозольных и трансмембранных областей белка, чтобы избежать проблем мембраносвязывающей функциональности. Ортологичные белковые последовательности PSMA, матрица BLOSUM62 и программа PSI-BLAST были загружены с сайта NCBI.
Затем сценарий перетасовки был запущен с помощью этих входных данных и дал вариант человеческого PSMA с 94% идентичностью последовательности с оригинальным человеческим PSMA. Кроме того, были введены три мутации для улучшения HLA-A2-связывания, исходя из их производительности в алгоритме Epitoptimizer (Houghton, Engelhorn et al. 2007). Эти мутации представляют собой M664L (эпитоп: 663-671), I676V (эпитоп: 668-676) и N76L (эпитоп: 75-83). Полученный антиген называют "перетасованный модифицированный антиген PSMA 1", "перетасованный модифицированный PSMA 1" или "перетасованный антиген PSMA 1".
Результаты, основанные на эпитопах с консенсусным рангом <1%, и IC50, с помощью нейронной сети (единственный лучший метод), <500, показали, что предсказанные эпитопы из HLA А2.1, HLA A3, HLA А11, HLA А24 и HLA В7 были высоко консервативными в этом перетасованном антигене. Два дополнительных варианта модифицированного антигена человеческого PSMA, описанные в Примере 1, были разработаны с более высокими идентичностями последовательностей и более рестрективным баллом запирания BLOSUM, составляющим 1, для удаления всех неконсервативных замен. Эти два варианта также упоминаются как "перетасованный модифицированный антиген PSMA 2" и "перетасованный модифицированный антиген PSMA 3", соответственно. Процент идентичности перетасованных модифицированных антигенов PSMA 1-3 по отношению к модифицированному конструкту человеческого PSMA (например, аминокислоты 15-750 человеческого PSMA) составляет приблизительно 93,6%, 94,9% и 96,4%, соответственно.
Перетасованный модифицированный антиген PSMA 1 имеет аминокислотную последовательность SEQ ID NO: 3 и имеет следующие мутации относительно модифицированного антигена человеческого PSMA: N47S, T53S, K55Q, M58V, L65M, N76L, S98A, Q99E, К122Е, N132D, V154I, 1157V, F161Y, D191Ε, M192L, V201L, V225I, I258V, G282E, I283L, R320K, L362I, S380A, E408K, L417I, H475Y, K482Q, M509V, S513N, E542K, M583L, N589D, R598Q, S613N, 1614L, S615A, Q620E, M622L, S647N, E648Q, S656N, I659L, V660L, L661V, M664L, I676V
Перетасованный модифицированный антиген PSMA 2 имеет аминокислотную последовательность SEQ ID NO: 5 и имеет следующие мутации относительно модифицированного антигена человеческого PSMA: мутации:
N47S, K55Q, M58V, Q91Е, S98A, А111S, К122Е, N132D, V154I, 1157V, F161Y, V201L, V225I, I258V, S312A, R320K, K324Q, R363K, S380A, Е408К, H475Y, K482Q, Y494F, E495D, К499Е, M509L, N540D, Е542К, N544S, M583I, 1591V, R598Q, R605K, S613N, S647N, E648Q, S656N, V660L
Перетасованный модифицированный антиген PSMA 3 имеет аминокислотную последовательность SEQ ID NO: 7 и имеет следующие мутации относительно модифицированного антигена человеческого PSMA: мутации:
Т339А, V342L, M344L, T349N, N350T, Е351К, S401Т, Е408К, M470L, Y471Н, H475Y, F506L, M509L, А531S, N540D, Е542К, N544S, G548S, V555I, E563V, V603A, R605K, K606N, Y607H, D609E, K610N, 1611L
2B. Иммунные реакции, измеренные после вакцинации у мышей Пастера
Схема исследования. Восьми-десятинедельных мышей Пастера иммунизировали, используя метод PMED, разными плазмидными ДНК, экспрессирующими перетасованные модифицированные антигены PSMA, в схеме прайм/буст/буст с интервалом в две недели между каждой вакцинацией, как описано в Примере 1F. Антиген-специфические Т и В-клеточные реакции измеряли через 7 дней после последней иммунизации в Антиген-специфические Т и В-клеточные реакции измеряли через 7 дней после последней иммунизации в анализе ELISPOT с гамма-интерфероном (IFN-y) ELISPOT и сэндвич-ELISA соответственно.


Результаты. Данные ELISpot, представленные в Таблице 7, показывают, что, в целом, перетасованные модифицированные антигены PSMA способны индуцировать Т-клеточный ответ по широте и амплитуде очень похожий на ответ модифицированного антигена человеческого PSMA. Показатель SFC>100 считается позитивным. Символ "*" представляет слишком много, чтобы точно подсчитать.

Как показано в Таблице 8, все перетасованные модифицированные антигены PSMA способны индуцировать Т-клетки, которые распознают известные HLA А2-рестриктированные эпитопы PSMA, а также человеческие опухолевые клетки HLA А2, трансдуцированные аденовирусом, несущим трансген PSMA для экспрессирования PSMA. Опухолевые клетки, которые не экспрессируют PSMA, служили в качестве негативного контроля и не распознавались. Показатель SFC>50 считается позитивным.


Данные ELISpot, представленные в Таблице 9, были получены со спленоцитами, которые были истощены из CD8; поэтому они представляют Т-клеточные реакции на специфичные к CD4 Т-клетки. Эти данные показывают, что ответ CD4, вызванный перетасованным модифицированным антигеном PSMA 2 очень похож на ответ, вызванный модифицированным антигеном человеческого PSMA. Показатель SFC>50 считается позитивным.

Данные в Таблице 10 демонстрируют, что все перетасованные модифицированные антигены PSMA способны вызывать реакции антител к человеческому PSMA. Перетасованный модифицированный антиген PSMA 2 и модифицированный антиген человеческого PSMA индуцировали последовательные реакции антител у всех мышей.


Данные ELISpot, представленные в Таблице 11, демонстрируют, что в целом Т-клеточная реакция, индуцированная перетасованным модифицированным антигеном PSMA у мышей HLA А24 по широте и амплитуде очень похожа на ответ модифицированного антигена человеческого PSMA. Показатель SFC>100 считается позитивным.
2С. Подавление иммунной толерантности к человеческому PSMA перетасованными модифицированными антигенами PSMA
Схема исследования. Трансгенная мышиная модель человеческого PSMA t использует мышей, которые были созданы с использованием минимального крысиного пробазинового промотора, запускающего экспрессию PSMA, особенно в предстательной железе (Zhang, Thomas et al. 2000) Endocrinology 141(12):4698-4710. Эти мыши были созданы на основе C57BL/6. Данные RT-PCR иммунного гистохимического окрашивания Данные подтвердили экспрессию PSMA в вентральных и дорсолатеральных корнях железы у этих PSMA-трансгенных мышей. Эндогенная экспрессия человеческого белка PSMA у этих мышей, как ожидалось, генерирует иммунную толерантность.
Результаты. Как показано в Таблице 12, только 20% из трансгенных мышей PSMA смогли поднять Т-клеточный ответ на человеческий PSMA с использованием модифицированного антигена человеческого PSMA. Тем не менее, 67% из трансгенных мышей PSMA были способны поднять PSMA-специфический Т-клеточный ответ, используя перетасованный модифицированный антиген PSMA 2. Данные показывают, что включение "не своих" аминокислотных последовательностей в перетасованный модифицированный антиген PSMA 2 улучшило подавление толерантности к "своему" антигену человеческого PSMA. Показатель SFC>50 считается позитивным.

ПРИМЕР 3. ДИЗАЙН РАЗЛИЧНЫХ ИММУНОГЕННЫХ ПОЛИПЕПТИДОВ PSA
Пример 3 иллюстрирует конструкцию и некоторые биологические свойства иммуногенных полипептидов PSA в цитозольной, секретируемой и мембраносвязанной формах.
3А. Создание различных антигенных форм PSA
Подобно тому, как описано в Примере 1 для трех различных полипептидных форм иммуногенного PSMA (например, цитозольной, мембраносвязанный и секретируемой формы), иммуногенные полипептиды PSA в трех формах были также разработаны на основе последовательности человеческого PSA. Иммуногенный полипептид PSA в цитозольной форме, который состоит из аминокислот 25-261 природного человеческого PSA, строится путем удаления секреторного сигнала и продомена (аминокислоты 1-24). Аминокислотная последовательность этого цитозольного полипептида иммуногенного PSA представлена в SEQ ID NO: 17. Секретируемая форма полипептида PSA представляет собой природный непроцессированный человеческий PSA (аминокислоты 1-261). Полипептид иммуногенного PSA в мембраносвязанной форме строится путем связывания цитозольной формы полипептида иммуногенного PSA (аминокислоты 25-261 природного человеческого PSA) с трансмембранным доменом человеческого PSMA (аминокислоты 15-54 человеческого PSMA).
3В. Иммунные реакции у мышей Пастера и мышей HLA А24
Схема исследования. Восьми-десятинедельных мышей Пастера HLA А2 или мышей HLA А24 иммунизировали ДНК, экспрессирующей разные антигены PSA, используя PMED, предусмотренный в Примере ЗА, по схеме прайм/буст/буст с двухнедельными интервалами между каждой вакцинацией, как описано в Примере 1. Антиген-специфические Т и В-клеточные реакции измеряли через 7 дней после последней иммунизации в анализе ELISPOT с интерфероном-гамма и в сэндвич-ELISA.

Результаты. Таблица 13 показывает данные ELISpot, полученные от спленоцитов, выделенных из мышей Пастера HLA А2 или мышей HLA А24, культивированных с пептидами, полученными из библиотеки пептидов PSA. Т-клеточные реакции могут быть обнаружены у обоих HLA А2 и HLA-A24 мышей. Показатель SFC>100 считается позитивным.

Данные ELISpot, показанные в Таблице 14, указывают, что полипептиды иммуногенного PSA в цитозольной и мембраносвязанной форме способны индуцировать Т-клетки, которые распознают человеческие опухолевые клетки, трансдуцированные с аденовирусом для экспрессирования цитозольного антигена PSA (SKmel5-Ad-PSA), но не клетки, трансдуцированные с аденовирусом для экспрессирования eGFP (SKmel5-Ad-eGFP). Эти два антигена также вызывают реакцию на белок PSA. Секретируемый антиген PSA не способен индуцировать Т-клетки к SKmel5-Ad-PSA или белку PSA. Показатель SFC>50 считается позитивным.

Данные в Таблице 15 показывают, что полипептиды иммуногенного PSA в секретируемой и мембраносвязанной форме способны индуцировать реакцию антитела к PSA.







ПРИМЕР 4. СОЗДАНИЕ МУЛЬТИАНТИГЕННЫХ ВАКЦИННЫХ КОНСТРУКТОВ
В этом Примере описано несколько стратегий экспрессирования множественных антигенов из однокомпонентного ДНК-вакцинного конструкта. Эти мультиантигенные ДНК-вакцинные конструкты разделяют тот же основной плазмидный скелет, что и pPJV7563. Хотя стратегии мультиантигенной экспрессии описаны в данном документе в контексте ДНК-вакцин, их принципы могут применяться подобным образом в контексте вирусных векторных генетических вакцин (таких как аденовирусные векторы). Если специально не указано иное, то гены, включенные в состав мультиантигенных конструктов, кодируют модифицированный антиген человеческого PSMA (указанный как PSMA), непроцессированный человеческий PSCA (указанный как PSCA) и цитозольный антиген человеческого PSA (указанный как PSA), как описано в примерах данного документа выше.
ПРИМЕР 4А. ДВУХАНТИГЕННЫЕ КОНСТРУКТЫ
4А1. СОЗДАНИЕ ДВУХАНТИГЕННЫХ КОНСТРУКТОВ С ИСПОЛЬЗОВАНИЕМ НЕСКОЛЬКИХ ПРОМОТОРОВ
Основная стратегия. Одна из стратегий для создания поливалентных нуклеинокислотных вакцинных конструктов включает несколько независимых промоторов в одну плазмиду (Huang, Υ., Ζ. Chen, et al. (2008). "Design, construction, and characterization of a dual-promoter multigenic DNA vaccine directed against an HIV-1 subtype С / В'recombinant." J Acquir Immune Defic Syndr 47(4):403-411; Xu, Κ., Ζ. Y. Ling, et al. (2011). "Broad humoral and cellular immunity elicited by a bivalent DNA vaccine encoding HA and NP genes from an H5N1 virus." Viral Immunol 24(1):45-56). Плазмида может быть сконструирована, чтобы нести несколько кассет экспрессии, каждая из которых состоит из а) эукариотического промотора для инициирования РНК полимеразой зависимой транскрипции при наличии или без энхансерного элемента, б) гена, кодирующего антиген-мишень, и в) последовательности терминации транскрипции. При доставке плазмиды в трансфицированные клеточные ядра, транскрипция будет инициироваться от каждого промотора, приводя к продуцированию отдельных мРНК, каждая из которых кодирует один из антигенов-мишеней. Матричная РНК будет транслироваться независимо, тем самым продуцируя желательные антигены.
Плазмида 460 (двойной промотор PSMA / PSCA). Плазмиду 460 сконструировали с использованием методики сайт-направленного мутагенеза, ПЦР и вставки рестрикционных фрагментов. Во-первых, сайт рестрикции Κρη I вводили выше промотора CMV в плазмиду 5259 с помощью сайт-направленного мутагенеза с праймерами MD5 и MD6 в соответствии с протоколом производителя (Quickchange kit, Agilent Technologies, Santa Clara, CA). Во-вторых, кассету экспрессии, состоящую из минимального промотора CMV, человеческого PSMA и терминатора транскрипции кроличьего глобулина В, амплифицировали с помощью ПЦР из плазмиды 5166 с использованием праймеров, которые несут сайты рестрикции Kpn I (MD7 и MD8). Ампликон ПЦР расщепляли Κpn I и вставляли в только что введенный сайт Κpn I плазмиды 5259, обработанной телячьей кишечной щелочной фосфатазой.
4А2. СОЗДАНИЕ ДВУХАНТИГЕННЫХ КОНСТРУКТОВ С ИСПОЛЬЗОВАНИЕМ пептидов 2А
Основная стратегия. Мультибелковые антигены также можно экспрессировать из одного вектора, используя вирусные 2А-подобные пептиды (Szymczak, A.L. and D.А. Vignali (2005). "Development of 2А peptide-based strategies in the design of multicistronic vectors." Expert Opin Biol Ther 5(5):627-638; de Felipe, P., G.A. Luke, et al. (2006). "Ε unum pluribus: multiple proteins from a self-processing polyprotein." Trends Biotechnol 24(2):68-75; Luke, G.Α., P. de Felipe, et al. (2008). Occurrence, function and evolutionary origins of '2A-like' sequences in virus genomes." J Gen Virol 89(Pt 4):1036-1042; Ibrahimi, Α., G. Vande Velde, et al. (2009). "Highly efficient multicistronic lentiviral vectors with peptide 2A sequences." Hum Gene Ther 20(8):845-860; Kim, J.H., S.R. Lee, et al. (2011). "High cleavage efficiency of a 2A peptide derived from porcine teschovirus-1 in human cell lines, zebrafish and mice." PLoS One 6(4):e18556). Эти пептиды, называемые также кассетами расщепления или CHYSEL (цис-действующие гидролазные элементы), примерно 20 аминокислот в длину с высоко консервативным карбоксиконцевым мотивом D-V/I-EXNPGP (Фигура 2). Кассеты редки в природе, наиболее часто встречается у вирусов, таких как вирус ящура (FMDV), вирус ринита лошадей (ERAV), вирус энцефаломиокардита (EMCV), свинной teschovirus (PTV) и вирус Thosea asigna (TAV) (Luke, G.Α., P. de Felipe, et al. (2008). "Occurrence, function and evolutionary origins of '2A-like' sequences in virus genomes." J Gen Virol 89(Pt 4):1036-1042). С мультиантигенной стратегией экспрессии на основе 2А, гены, кодирующие несколько целевых антигенов, можно связывать друг с другом в одной открытой рамке считывания, разделенной кассетами 2А. Вся открытая рамка считывания может быть клонирована в вектор с одним промотором и терминатором. При доставке генетической вакцины в клетку, мРНК, кодирующая несколько антигенов, будет транскрибироваться и транслироваться как единый полипротеин. Во время трансляции кассет 2А рибосомы пропускают связь между С-концевым глицином и пролином. Рибосомный пропуск действует подобно котрансляционному автокаталитическому "расщеплению", которое высвобождает вышележащие от нижележащих белков. Включение кассеты 2А между двумя белковыми антигенами приводит к добавлению аминокислот -20 на С-конце вышележащего полипептида и одной аминокислоты (пролина) к N-концу нижележащему белку. При адаптации этой методологии, протеазные сайты расщепления могут быть включены в N-конец кассеты 2А таким образом, что повсеместно протеазы будут расщеплять кассету от вышележащего белка (Fang, J., S. Yi, et al. (2007). "An antibody delivery system for regulated expression of therapeutic levels of monoclonal antibodies in vivo." Mol Ther 15(6):1153-1159).
Плазмида 451 (PSMA-T2A-PSCA). Плазмиду 451 сконструировали с использованием методики ПЦР с перекрывающимися праймерами и обмена рестрикционными фрагментами. Во-первых, ген, кодирующий аминокислоты 15-750 человеческого PSMA, амплифицировали с помощью ПЦР с использованием плазмиды 5166 в качестве матрицы с праймерами 119 и 117. Ген, кодирующий непроцессированный человеческий PSCA, амплифицировали с помощью ПЦР с использованием плазмиды 5259 в качестве матрицы с праймерами 118 и 120. ПЦР привело к добавлению перекрытия последовательностей TAV 2А (t2A) на 3' конце PSMA и 5' конце PSCA. Ампликоны были смешаны вместе и амплифицированы с помощью ПЦР с использованием праймеров 119 и 120. Ампликон PSMA-T2A-PSCA обрабатывали Nhe I и Bgl II, и вставляли в подобным образом расщепленную плазмиду 5166. Глицин-сериновый линкер был включен между PSMA и кассетой Т2А для способствования высокой эффективности расщепления.
Плазмида 454 (PSCA-F2A-PSMA). Плазмиду 454 создали, используя методики ПЦР и обмена рестрикционными фрагментами. Во-первых, ген, кодирующий непроцессированный человеческий PSCA, амплифицировали с помощью ПЦР, используя плазмиду 5259 в качестве матрицы с праймерами 42 и 132. Ампликон обрабатывали BamH I и вставляли в подобным образом расщепленную, CIP-обработанную плазмиду 5300. Глицин-сериновый линкер был включен между PSCA и кассетой FMDV 2А (F2A) для способствования высокой эффективности расщепления.
Плазмида 5300 (PSA-F2A-PSMA). Плазмиду 5300 сконструировали с использованием методики ПЦР с перекрывающимися праймерами и обмена рестрикционными фрагментами. Во-первых, ген, кодирующий аминокислоты 25-261 PSA амплифицировали с помощью ПЦР из плазмиды 5297 с праймерами MD1 и MD2. Ген, кодирующий аминокислоты 15-750 человеческого PSMA амплифицировали с помощью ПЦР из плазмиды 5166 с праймерами MD3 и MD4. ПЦР привело к добавлению перекрытия последовательностей F2A на 3' конце PSA и 5' конце PSMA. Ампликоны смешивали вместе и удлиняли с помощью ПЦР. Ампликон PSA-F2A-PSMA обрабатывали Nhe I и Bgl II и вставляли в подобным образом расщепленную плазмиду pPJV7563.
4А3. Двухантигенные конструкты, использующие внутренние сайты связывания рибосомы
Основная стратегия: Третья стратегия экспрессии нескольких белковых антигенов из одной плазмиды или вектора включает использование внутреннего сайта связывания рибосомы или 1RES. Внутренние сайты связывания рибосом являются РНК-элементами (Фигура 3), обнаруженными в 5' нетранслируемых участках некоторых молекул РНК (Bonnal, S., С. Boutonnet, et al. (2003). "IRESdb: the Internal Ribosome Entry Site database." Nucleic Acids Res 31(1):427-428). Они привлекают эукариотические рибосомы к РНК для облегчения трансляции нижележащих открытых рамок считывания. В отличие от обычной клеточной 7-метилгуанозиновой кэп-зависимой трансляции, IRES-опосредованная трансляция может инициировать в кодонах AUG далеко в пределах молекулы РНК. Может быть использован высокоэффективный процесс для применения в мультицитронных векторах экспрессии (Bochkov, Y.A. and А.С. Palmenberg (2006). "Translational efficiency of EMCV 1RES in bicistronic vectors is dependent upon IRES sequence and gene location." Biotechniques 41(3):283-284, 286, 288). Как правило, в вектор вставляют два трансгена между промотором и терминатором транскрипции в виде двух отдельных открытых рамок считывания, разделенных IRES. При доставке генетической вакцины в клетку, будет транскрибироваться один длинный транскрипт, кодирующий оба трансгена. Первая ORF будет транслироваться традиционным кэп-зависимым образом, терминируя на вышележащем стоп-кодоне IRES. Вторая ORF будет транслироваться кэп-независимым способом с помощью IRES. Таким образом, два независимых белка могут быть получены из одной мРНК транскрипции из вектора с одной экспрессионной кассетой.
Плазмида 449 (PSMA-mlRES-PSCA). Плазмиду 449 сконструировали с использованием методики ПЦР с перекрывающимися праймерами и обмена рестрикционными фрагментами. Во-первых, ген, кодирующий непроцессированный человеческий PSCA амплифицировали с помощью ПЦР из плазмиды 5259 с праймерами 124 и 123. Минимальный EMCV 1RES амплифицировали с помощью PCR из pShuttle-IRES с праймерами 101 и 125. Перекрывающиеся ампликоны смешивали вместе и амплифицировали с помощью ПЦР с праймерами 101 и 123. Ампликон IRES-PSCA обрабатывали Bgl II и BamH I и вставляли в Bgl II-расщепленную, CIP-обработанную плазмиду 5166. Для того, чтобы зафиксировать спонтанную мутацию в пределах IRES, IRES, содержащий AVR II к последовательности Κpn I, был заменен эквивалентным фрагментом из pShuttle-IRES.
Плазмида 603 (PSCA-pIRES-PSMA). Плазмиду 603 сконструировали, используя методы ПЦР и непрерывного клонирования. Ген, кодирующий непроцессированный человеческий PSCA, присоединенный на своем 3'-конце к предпочтительному EMCV 1RES, амплифицировали из плазмиды 455 с помощью ПЦР с праймерами SD546 и SD547. Ген, кодирующий аинокислоты 15-750 человеческого PSMA амплифицировали с помощью ПЦР из плазмиды 5166, используя праймеры SD548 и SD550. Два перекрывающихся апликона ПЦР вставили в Nhe I и Bgl II-расщепленную pPJV7563 с помощью непрерывного клонирования, согласно инструкциям производителя (Invitrogen, Carlsbad, СА).
Плазмида 455 (PSCA-mlRES-PSA). Плазмиду 455 сконструировали с использованием методики ПЦР с перекрывающимися праймерами и обмена рестрикционными фрагментами. Во-первых, ген, кодирующий аинокислоты 25-261 человеческого PSA амплифицировали с помощью ПЦР из плазмиды 5297 с праймерами 115 и 114. Минимальный EMCV 1RES амплифицировали с помощью ПЦР из pShuttle-IRES с праймерами 101 и 116. Перекрывающиеся ампликоны смешивали вместе и амплифицировали с помощью ПЦР с праймерами 101 и 114. IRES-PSA обрабатывали Bgl II и BamH I и вставляли в Bgl II-расщепленную, CIP-обработанную плазмиду 5259. Для того, чтобы зафиксировать спонтанную мутацию в пределах этого клона, последовательность от Bgl II до BstE II заменили эквивалентным фрагментом из свежей реакции ПЦР с перекрывающимися праймерами.
ПРИМЕР 4В. ТРЕХАНТИГЕННЫЕ ДНК-КОНСТРУКТЫ
Основная стратегия. Способности антител двухантигенных векторов экспрессии направлять экспрессию PSMA, PSCA, и/или PSA были охарактеризованы в трансфицированных клетках НЕК293 (Фигуры 4-6). Двухантигенные кассеты экспрессии, включая PSA-F2A-PSMA, PSMA-mlRES-PSCA, PSMA-T2A-PSCA, PSA-T2A-PSCA, PSCA-F2A-PSMA, PSCA-pIRES-PSMA и PSMA-m IRES-PSA, были выбраны для включения в разных комбинациях в трехантигенные векторы экспрессии. Во всех случаях векторы базировались на скелете родительской плазмиды pPJV7563. Четыре вектора (плазмиды 456, 457, 458 и 459) использовали один полный CMV-промотор с терминатором транскрипции кроличьего В-глобулина для управления экспрессией всех трех антигенов. Два других вектора (плазмиды 846 и 850) включены в стратегию двойных промоторов в комбинации с 1RES или 2А для управления экспрессией трех антигенов. Векторы с несколькими кассетами 2А были разработаны, чтобы нести разные кассеты для сведения к минимуму вероятности рекомбинации между первой и второй кассетами во время амплификации плазма id/вектор. Экспрессия антигена была продемонстрирована с помощью проточной цитометрии (Фигура 7) и вестерн-блоттинга (Фигура 8).
Плазмида 456 (PSA-F2A-PSMA-mlRES-PSCA). Плазмиду 456 сконструировали с помощью обмена рестрикционными фрагментами. Плазмиду 5300 обработали Nhe I и Нра I и вставку ~1,8 т.п.н. лигировали в подобным образом расщепленную плазмиду 449.
Плазмида 457 (PSA-F2A-PSMA-T2A-PSCA). Плазмиду 457 сконструировали с помощью обмена рестрикционными фрагментами. Плазмиду 5300 обработали Nhe I и Нра I и вставку ~1,8 т.п.н. лигировали в подобным образом расщепленную плазмиду 451.
Плазмида 458 (PSA-T2A-PSCA-F2A-PSMA). Плазмиду 458 сконструировали, используя методику ПЦР и обмена рестрикционными фрагментами. Ген, кодирующий аминокислоты 25 - 261 человеческого PSA амплифицировали с помощью ПЦР из плазмиды 5297 с праймерами 119 и 139, приводя к добавлению последовательности Т2А и сайта рестрикции Nhe I к 3'-концу. Ампликон обрабатывали Nhe I и вставляли в подобным образом расщепленную плазмиду 454.
Плазмида 459 (PSCA-F2A-PSMA-mlRES-PSA). Плазмиду 459 сконструировали с помощью обмена рестрикционными фрагментами. Плазмиду 454 обработали Nhe I и Bgl II, a PSCA-F2A-PSMA, содержащую вставку, лигировали в подобным образом расщепленную плазмиду 455.
Плазмида 846 (CBA-PSA, CMV-PSCA-pIRES-PSMA). Плазмиду 846 сконструировали, используя методы ПЦР и непрерывного клонирования. Во-первых, экспрессионную кассету синтезировали так, что она состоит из 1) промотора и 5'-нетранслируемой области из гена куриного бета-актина (СВА) 2) гибридного интрона куриного бета-актина / кроличьего бета-глобина, 3) гена, кодирующего аминокислоты 25 -26 человеческого PSA, и 4) терминатора бычьего гормона роста. Эту PSA-экспрессирующую кассету амплифицировали с помощью ПЦР из плазмиды 796 с праймерами 3SallCBA и 5SallBGH. Ампликон клонировали в сайт Sail плазмиды 603, используя GeneArt Seamless Cloning and Assembly Kit (Invitrogen, Carlsbad, CA). При доставке этой плазмиды в клетку, экспрессия PSA будет зависеть от промотора СВА, тогда как экспрессия PSCA и PSMA будет зависеть от CMV-промотора.
Плазмида 850 (CBA-PSA, CMV-PSCA-F2A-PSMA). Плазмиду 850 сконструировали, используя методы ПЦР и непрерывного клонирования. Во-первых, СВА-промоторно управляемую PSA-экспрессирующую кассету амплифицировали с помощью ПЦР из плазмиды 796 с праймерами 3SallCBA и 5SallBGH. Ампликон клонировали в сайт Sail плазмиды 454, используя GeneArt Seamless Cloning and Assembly Kit (Invitrogen, Carlsbad, CA). При доставке этой плазмиды в клетку экспрессия PSA будет управляться СВА-промотором, тогда как экспрессия PSCA и PSMA будет управляться CMV-промотором.



ПРИМЕР 4С. ТРЕХАНТИГЕННЫЕ АДЕНОВИРУСНЫЕ КОНСТРУКТЫ
Основная стратегия. Как и ДНК плазмиды, вирусные вакцинные векторы могут быть разработаны для доставки нескольких антигенов рака простаты. Три мультиантигенные стратегии экспрессии, описанные выше для ДНК-вакцин - двойные промоты, пептиды 2А и внутренние сайты связывания рибосомы - были включены в разных комбинациях для создания трехантигенных аденовирусных векторов. Кратко, мультиантигенные кассеты экспрессии клонировали в плазмиду pShuttle-CMV, модифицированную для того, чтобы нести две копии тетрациклиновой операторной последовательности (ТеtO2). Рекомбинантные векторы серотипа 5 аденовируса создали, используя векторную систему AdEasy согласно инструкциям производителя (Agilent Technologies, Inc., Santa Clara, CA). Вирусы амплифицировали в клетки НЕК293 и очищали с помощью двойного связывания с хлоридом цезия согласно стандартным протоколам. Перед исследованием in vivo вирусные штаммы тщательно охарактеризованы по поводу концентрации вирусных частиц, титра инфекционности, стерильности, эндотоксина, геномной и трансгенной целостности, трансгенной идентичности и экспрессии.
Аденовирус-733 (PSA-F2A-PSMA-T2A-PSCA). Ad-733 является вирусным эквивалентом плазмиды 457. Экспрессией управляет единичный CMV-промотор с тетрациклиновым оператором для подавления экспрессии трансгена во время крупномасштабного продуцирования в линиях НЕК293, экспрессирующих Tet-penpeccop. Стратегии мультиантигенной экспрессии включают две разные последовательности 2А.
Аденовирус-734 (PSA-T2A-PSCA-F2A-PSMA). Ad-734 является вирусным эквивалентом плазмиды 458. Экспрессией управляет единичный CMV-промотор с тетрациклиновым оператором для подавления экспрессии трансгена во время крупномасштабного продуцирования в линиях НЕК293, экспрессирующих Tet-penpeccop. Стратегии мультиантигенной экспрессии включают две разные последовательности 2А.
Аденовирус-735 (PSCA-F2A-PSMA-mlRES-PSA). Ad-735 является вирусным эквивалентом плазмиды 459. Экспрессией управляет единичный CMV-промотор с тетрациклиновым оператором для подавления экспрессии трансгена во время крупномасштабного продуцирования в линиях НЕК293, экспрессирующих Tet-репрессор. Стратегии мультиантигенной экспрессии включают последовательность 2А и IRES.
Аденовирус-796 (CBA-PSA, CMV-PSCA-plRES-PSMA). Ad-796 является вирусным эквивалентом плазмиды 846. Экспрессией PSA управляет промотор куриного бета-актина, тогда как экспрессией PSCA и PSMA управляет промотор CMV-TetO2. Стратегии мультиантигенной экспрессии включают два промотора и 1RES.
Аденовирус-809 (CBA-PSA, CMV-PSCA-F2A-PSMA). Ad-809 является вирусным эквивалентом плазмиды 850. Экспрессией PSA управляет промотор куриного бета-актина, тогда как экспрессией PSCA и PSMA управляет промотор CMV-TetO2. Стратегии мультиантигенной экспрессии включают два промотора и последовательность 2А.
ПРИМЕР 5. ИММУНОГЕННОСТЬ ТРЕХАНТИГЕННЫХ ДНК-ВАКЦИН
Пример 5 иллюстрирует способность трехантигенных нуклеинокислотных вакцинных продуктов, экспрессирующих PSMA, PSCA и PSA, вызывать антиген-специфические Τ и В-клеточные ответные реакции на все три закодированные антигены простаты.
Исследование клеточной иммунной реакции. Иммуногенность трехантигенных конструктов, содержащих PSMA, PSCA и PSA, как описано в Примере 5, изучали у мышей C57BL/6, согласно процедуре, описанной ниже. Самки мышей C57BL/6 примировали в день 0 и бустировали в дни 14, 28 и 49 ДНК-вакцинными конструктами, кодирующими антигены человеческих PSMA, PSCA и PSA путем PMED-введения. В целом оценили четыре разные стратегии трехантигенной вакцинации, которые включали три ДНК-вакцины, соэкспрессирующие белки-мишени, и один совместный подход. Для соэкспрессии использовали единичные ДНК-плазмиды, кодирующие все три антигена простаты, соединенные пептидами 2А или внутренний сайты связывания рибосомы (IRES): PSA-F2A-PSMA-T2A-PSCA (плазмида ID#457), PSA-T2A-PSCA- F2A-PSMA (плазмида ID#458) и PSCA-F2A-PSMA-IRES-PSA (плазмида ID#459). Для формирования совместного подхода три разные ДНК-плазмиды, каждая из которых индивидуально кодирует PSMA, PSCA или PSA, совместно сформировали на одну золотую частицу для PMED-доставки. За исключением совместного формирования, элементы ДНК, которые контролируют соэкспрессию (2а и IRES), отличаются по длине, эффективности экспрессии трансгена и наличию чужеродного генетического материала, прикрепленного к трансгенам-мишеням. В качестве контроля, мышей C57BL/6 вакцинировали ДНК, экспрессирующей один антигена простаты, или PSMA, или PSCA, или PSA. Для соэкспрессии триплетных или одноантигенных ДНК-вакцин дозу в 2 мкг ДНК-вакцинной плазмиды давали на PMED-введение, в то время как по 1 мкг каждой из совместно сформированных трехантигенных ДНК-вакцин (всего 3 мкг) вводили на PMED-введение. Клеточную иммунную реакцию на три- и одноантигенные вакцины измеряли путем изъятия селезенок от каждого животного в день 56 через семь дней после последней PMED-вакцинации. Спленоциты выделили и подвергли анализу IFN-y ELISPOT для измерения PSMA, PSCA и PSA-специфических Т-клеточных реакций. Кратко, 2×105 спленоцитов от отдельных животных помещали в каждую лунку с 5×104 на лунку клеток TRAMP-C2 (трансгенная аденокарцинома мышиной простаты), стабильно экспрессирующих один антиген простаты человека или PSMA, PSCA и PSA вместе, или с индивидуальным пептидом или пулом человеческих PSMA, PSCA и PSA-специфических пептидов в дозе 10 мкг/мл (смотри Таблицу 22 для пептидов и композиции пептидного пула), или только среду в качестве контроля. Каждое условие осуществляли в трех повторностях. Планшеты инкубировали в течение 20 ч при 37°C и 5% СO2, промывали и после инкубации разрабатывали в соответствии с инструкциями изготовителя. Количество IFN-пятнообразующих клеток (SFC) подсчитали с помощью ридера Technology Ltd. (CTL). Результаты представлены на Фигурах 9 и 10, которые показывают среднее количество PSMA, PSCA или PSA-специфических SFC+/ - стандартное отклонение пяти мышей в каждой группе, нормированное к 1×106 спленоцитов.


Изучение реакции антител. Реакции антител на три- и одноантигенные вакцины измеряли путем забора сыворотки от каждого животного в день 56, на седьмой день после последней PMED-вакцинации. Сыворотку подвергали иммуноферментному твердофазному анализу (ELISA) для определения титров антител к PSMA и PSCA. Кратко, ELISA-планшеты покрывали 1 мкг/мл человеческого PSMA или PSCA и инкубировали в течение ночи при 4°C. Планшеты затем блокировали и инкубировали при комнатной температуре в течение 1 ч с 1% бычьего сывороточного альбумина (BSA). Каждый образец сыворотки разводили серийно в двух экземплярах, начиная с 1:100 разбавления и инкубировали в течение 1 ч при комнатной температуре. После промывания козье антимышиное поликлональное IgG-антитело, конъюгированное с пероксидазой хрена (HRP), инкубировали при комнатной температуре в течение 1 ч. После промывания ТМВ пероксидазный EIA-субстрат инкубировали при комнатной температуре в течение 30 мин. Колориметрическую реакцию останавливали добавлением 1N серной кислоты, а затем считывали поглощение при 450 нм. Кривые титрования строили для каждого образца сыворотки (разведения образцов по сравнению с оптической плотностью). Титр сыворотки (в дальнейшем преобразованный во взаимодействующий титр) затем принимали в качестве наиболее разведенного образца сыворотки, тестированного со значением оптической плотности (OD) выше нижнего предела обнаружения (LLOD; фон плюс 3 стандартных отклонения), или разведение сыворотки, рассчитанное по достижению значения OD 1,0. Результаты представлены на Фигурах 11 и 12, которые показывают средние титры +/- стандартное отклонение пяти мышей в каждой группе.
Сыворотку также подвергали сортировке клеток с активированной флуоресценцией (FACS) для измерения связывания антитела с человеческим PSMA или PSCA, экспрессированных на клеточной поверхности пригодной клеточной линии, тем самым определяя будут ли антитела, вызванные мультиантигенными вакцинами, способны распознавать природные конформации PSMA и PSCA, соответственно. Клетки LNCaP (аденокарциномы простаты человека) использовали для измерения связывания антител с природным PSMA. Клетки РС3 (рака простаты человека) служили контролем в анализе FACS, так как эти клетки не экспрессируют человеческий PSMA. Клетки MIA-РаСа-2 (карциномы поджелудочной железы человека), трансдуцированые аденовирусом, экспрессирующим человеческий PSCA (Ad-PSCA), использовали для измерения связывания антител с природным PSCA. Нетрансдуцированные клетки MIA-PaCa-2 служили контролем. Вкратце, для измерения связывания антитела к PSMA по 2×105 клеток LNCaP или РС3 инкубировали с 1:100 разведением мышиной сыворотки или 15 мкг/мл контрольного мышиного моноклонально антитела (mAb) к человеческому PSMA (клон J591-A) в течение 20 мин при 4°C. Для измерения связывания антитела к PSCA, по 2×105 Ad-PSCA трансдуцированных и нетрансдуцированных MIA-PaCa-2 клеток инкубировали с 1:30 разведением сыворотки мыши или 4 мкг/мл контрольного мышиного mAb к человеческому PSCA (клон 7F5) в течение 20 мин при 4°C. Затем клетки промывали и инкубировали со вторичным фикоэритрин (РЕ)-конъюгированным козьим антимышиным lgG-антителом с красителем "живой / мертвый" еще в течение 20 мин при 4°C. После инкубации клетки промывали и ресуспендировали в 1,5% параформальдегида, было обнаружено 10000 живых клеток и на FACS Canto II. Результаты представлены на Фигурах 13 и 14, которые показывают среднее изменение при средней интенсивности флуоресценции (MFI) в сыворотке мыши по сравнению с вторичным антимышиным антителом самим по себе +/- стандартное отклонение пяти мышей в каждой группе. Титры антител не измеряли, поскольку PSA экспрессировался в виде цитоплазматического белка мультиантигенными вакцинами, изученными в этом исследовании.
Результаты:
На Фигурах 9А-9D представлены результаты репрезентативного исследования, которое оценивает клеточные иммунные реакции трехантигенных вакцин с помощью анализа IFN-γ ELISPO. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14, 28 и 49. В день 56, семь дней после последней PMED-вакцинации, распознание эндогенных антигенов простаты оценивали, изучая Т-клеточные реакции на (A) TRAMP C2-PSMA, (В) TRAMP C2-PSCA, (С) TRAMP C2-PSA, и (D) TRAMP C2-PSMA-PSA-PSCA клетки с помощью анализа IFN-γ ELISPOT. Клетки TRAMP С2 служили фоновым контролем для анализа. Для IFN-γ Т-клеточной реакции на эндогенный PSMA, значительная реакция на TRAMP C2-PSMA наблюдалась после одной PSMA PMED-вакцинации, которая согласовывалась с реакциями, наблюдаемыми в других исследованиях. Подобная PSMA-специфическая IFN-γ Т-клеточная реакция на TRAMP C2-PSMA была обнаружена после трехантигенных вакцинаций. В отличие от этого, полная абляция реакции наблюдалась после сформированной PSMA, PSCA и PSA вакцинации (* указывает p<0,05 в двухфакторном дисперсионном анализе ANOVA). Для IFN-γ Т-клеточной реакции на эндогенный PSCA, не наблюдалось существенной разницы в реакции на клетки TRAMP C2-PSCA при сравнении единичной PSCA-вакциной с четырьмя разными трехантигенными вакцинами. Для IFN-γ Т-клеточной реакции на эндогенный PSA, было обнаружено значительное снижение амплитуды реакции на TRAMP C2-PSA при сравнении с иммуногенностью единичной PSA-вакцины к PSCA-F2A-PSMA-IRES-PSA (*** указывает p<0,001 в двухфакторном дисперсионном анализе ANOVA) или соформированной вакцины (* указывает p<0,05 в двухфакторном дисперсионном анализе ANOVA). При исследовании реакции на TRAMP C2-PSMA-PSCA-PSA, наиболее высокая амплитуда IFN-γ Т-клеточной реакции наблюдалась после вакцинации PSA-F2A-PSMA-T2A-PSCA. Взятые вместе, эти данные демонстрируют распознание эндогенных PSMA, PSCA и PSA и генерирование антиген-специфических IFN-γ Т-клеточных реакций на все три антигена простаты с использованием стратегии соэкспрессионной ДНК-вакцинации, особенно вакцинным конструктом PSA-F2A-PSMA-T2A-PSCA. Кроме того, стратегия соформированной ДНК-вакцинации привела к потере антиген-специфических IFN-γ Т-клеточных реакций на PSMA и PSA.
На Фигурах 10A-10D представлены результаты репрезентативного исследования, которое оценивает иммуногенность трехантигенных вакцин с помощью анализа IFN-γ ELISPOT. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14, 28 и 49. В день 56, Т-клеточные реакции на (А) пептиды PSMA, (В) пулы пептидов PSMA, (С) пептиды PSCA и (D) пептиды PSA (смотри Таблицу 22) оценивали с помощью анализа IFN-γ ELISPOT. Среда сама по себе служила фоновым контролем для анализа. Для IFN-γ Т-клеточных реакций на индивидуальные пептиды и пулы пептидов PSMA при сравнении с единичной PSMA-вакциной PSMA более высокая амплитуда реакции наблюдалась после введения трехантигенной вакцины PSA-F2A-PSMA-T2A-PSCA. Подобным образом, более высокая амплитуда IFN-γ Т-клеточных реакций на PSCA и PSA-специфические пептиды после введения вакцины PSA-F2A-PSMA-T2A-PSC. Совместно сформированная вакцина PSMA, PSCA и PSA не вызвала или вызвала низкие Т-клеточные реакции на PSMA-специфические пептиды и низкую амплитуду реакций на PSCA и PSA-специфические пептиды. Эти данные также демонстрируют генерирование Т-клеточных реакций на PSMA, PSCA и PSA, соэкспрессированных из одного вакцинного конструкта. Наблюдались последовательные и сильные IFN-γ Т-клеточные реакции на все три антигена простаты после вакцинации PSA-F2A-PSMA-T2A-PSCA, и значительное снижение амплитуды IFN-γ Т-клеточных реакций на антигены простаты после вакцинации PSA-T2A-PSCA-F2A-PSMA, PSMA-F2A-PSMA-I RES-PSA и вакцинации совместно сформированными PSMA, PSCA и PSA.
На Фигуре 11 представлены результаты репрезентативного исследования, которое оценивает иммуногенность трехантигенных вакцин с помощью титров антител к PSMA. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14, 28 и 49. В день 56, титры антител к PSMA в сыворотке оценивали с помощью ELISA. Всех животных вакцинировали PSMA, генерируя значительные титры антител к PSMA. Между титрами не было существенных отличий, хотя вакцинация PSA-F2A-PSMA-T2A-PSCA привела к более низким титрам по сравнению с другими группами, вакцинированными PSMA. Эти данные демонстрируют генерирование антител, специфических к PSMA, после трехантигенной вакцинации с использованием стратегий соэкспрессионной вакцины и совместно сформированной вакцины.
На Фигуре 12 представлены результаты репрезентативного исследования, которое оценивает иммуногенность трехантигенных вакцин с помощью титров антител к PSCA. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14, 28 и 49. В день 56, титры антител к РСМА в сыворотке оценивали с помощью ELISA. Титры антител обнаружили у мышей, вакцинированных только PSCA и совместно сформированными PSMA, PSCA и PSA. Эти результаты указывают на то, что совместно сформированные PSMA, PSCA и PSA вызывают обнаруживаемый титр антитела к PSCA по сравнению со стратегией соэкспрессионной ДНК-вакцинации.
На Фигуре 13 представлены результаты репрезентативного исследования, которое оценивает иммуногенность трехантигенных вакцин с помощью поверхностноклеточного связывания антител к PSMA. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14, 28 и 49. В день 56, распознание клеточноповерхностного природного PSMA оценивали путем связывания антител в сыворотке с клетками LNCaP и РС3. Клетки РС3 служили фоновым контролем для анализа. Вакцинация PSA-F2A-PSMA-T2A-PSCA дала антитела к PSMA с значительно более низкой связывающей способностью с клетками LNCaP по сравнению с мышами, вакцинированными PSA-T2A-PSCA-F2A-PSMA или только PSMA (* указывает значение p<0,05 в однофакторном дисперсионном анализе ANOVA). Все остальные PSMA-вакцинированные группы не показали существенных отличий в связывании антител к PSMA. Кратность изменения в отношении вторичного антитела для J591-A mAb составила 45,3 (данные не показаны). В целом, эти данные демонстрируют генерирование PSMA-специфических антител, которые распознают природный PSMA после трехантигенной вакцинации с использованием стратегий соэкспрессионной и совместно сформированной ДНК-вакцинации.
На Фигуре 14 представлены результаты репрезентативного исследования, которое оценивает иммуногенность трехантигенных вакцин с помощью поверхностноклеточного связывания антител к PSCA. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14, 28 и 49. В день 56, распознание поверхностноклеточного природного PSCA оценивали путем связывания антител в сыворотке с клетками MIA-РаСа-2, трандуцированными Ad-PSCA и нетрансдуцированными. Нетрандуцированные парентеральные клетки служили фоновым контролем для анализа. За исключением одинарной PSMA и одинарной PSA цитовакцин, все вакцинные схемы с PSCA привели к значительному связыванию антител к PSCA с клетками MIA-PaCa-2, трандуцированными Ad-PSCA, по сравнению с парентеральными клетками. Не было существенных отличий в связывание антител к PSCA с клетками MIA-PaCa-2, трандуцированными Ad-PSCA, среди PSCA-вакцинированных групп (однофакторный дисперсионный анализ ANOVA, значение p>0,05). Кратность изменения в отношении вторичного антитела для 7F5 mAb составила 18,7 (данные не показаны). В целом, эти данные демонстрируют генерирование PSCA-специфических антител, которые распознают природный PSCA после трехантигенной вакцинации с использованием стратегий соэкспрессионной и совместно сформированной ДНК-вакцинации.
ПРИМЕР 6. ИММУНОГЕННОСТЬ ДВУХАНТИГЕННЫХ ВАКЦИН
Следующие примеры предусмотрены для иллюстрации способности двухантигенных вакцин экспрессировать два антигена простаты для вызова антиген-специфических Τ и В-клеточных реакций на два закодированных антигена простаты.
6А. Иммуногенность двухантигенных вакцин, содержащих PSMA и PSCA, у мышей C57BU6:
Процедура исследования.
Исследование клеточных иммунных реакций. Самок мышей C57BL/6 примировали в день 0 и бустировали в дни 14, 28, 42 и 70 ДНК, экспрессирующей человеческий PSMA и PSCA, с помощью эпидермальной инъекции PMED. В целом, оценили пять разных стратегий двухантигенных ДНК-вакцинаций, которые включали четыре ДНК-вакцины, которые соэкспрессировали антигены, и один подход совместного формирования. Для соэкспрессии единичные ДНК-вакцинные плазмиды, кодирующие два антигена простаты - PSMA и PSCA, соединили с помощью двойного промотора, вводили пептиды 2А или 1RES. Они включали двойной промотор PSMA-PSCA (плазмида Ю# 460), PSMA-T2A-PSCA (плазмида ID#451), PSCA-F2A-PSMA (плазмида Ю#454) и PSCA-IRES-PS MA (плазмида ID#603). Для совместного формирования две разные ДНК-плазмиды, каждая из которых индивидуально кодирует PSMA и PSCA, сформировали на одной частице золота для PMED-доставки. За исключением совместного формирования, ДНК-элементы, которые контролируют совместную экспрессию (двойной промотор, 2А и IRES), отличались по длине, эффективности экспрессии трансгена и присутствию чужеродного генетического материала, присоединенного к трансгенам-мишеням. В качестве контроля мышей C57BL/6 вакцинировали ДНК, экспрессирующей один антиген простаты - PSMA или PSCA. Для соэкспрессирующих двойных или единичных ДНК-вакцин давали суммарную дозу, составляющую 2 мкг ДНК-вакцины на PMED-введение, тогда как по 2 мкг каждой ДНК-вакцинной плазмиды (в сумме 4 мкг ДНК на введение) давали для совместной композиции. Клеточные иммунные реакции двойных и единичных антигенных вакцин измеряли путем изъятия селезенок у каждого животного в день 77, семь дней после последней PMED-вакцинации. Выделяли спленоциты и подвергали анализу IFN-γ ELISPOT для измерения PSMA и PSCA-специфических Т-клеточных реакций. Кратко, 2×105 спленоцитов от отдельных животных засевали в лунки с 2×104 на лунку клетками TRAMP-C2, экспрессирующими один эндогенный антиген простаты человека или PSMA, PSCA и PSA вместе, или с индивидуальными пептидами или пулом пептидов, специфических к человеческим PSMA и PSCA, в концентрации 10 мкг/мл (смотри Таблицу 22 для пептидов и композиции пептидного пула), или только среду в качестве контроля. Каждое условие осуществляли в трех повторностях. Планшеты инкубировали в течение 20 ч при 37°C и 5% CO2, промывали и после инкубации разрабатывали в соответствии с инструкциями изготовителя. Количество of IFN-γ SFC подсчитали с помощью ридера CTL. Результаты представлены на Фигурах 15 и 16, которые показывают среднее количество PSMA или PSCA-специфических SFCs +/- стандартное отклонение пяти мышей в каждой группе, нормированное к 1×106 спленоцитов.
Изучение реакции антител. Реакции антител на двух- и одноантигенные вакцины измеряли путем забора сыворотки от каждого животного в день 77, на седьмой день после последней PMED-вакцинации. Сыворотку подвергали иммуноферментному твердофазному анализу (ELISA) для определения титров антител к PSMA и PSCA, как описано в Примере 5. Результаты представлены на Фигурах 17 и 18, которые показывают средние титры +/- стандартное отклонение пяти мышей в каждой группе.
Сыворотку также подвергали сортировке клеток с активированной флуоресценцией (FACS) для измерения связывания антитела с человеческим PSMA или PSCA, экспрессированных на клеточной поверхности пригодной клеточной линии, тем самым определяя будут ли антитела, вызванные мультиантигенными вакцинами, способны распознавать природные конформации PSMA и PSCA, соответственно. Связывание антител с поверхностноклеточным природным PSA не измеряли, поскольку PSA экспрессируется в виде цитоплазматичного белка мультиантигенными вакцинами, изученными в этом исследовании. Анализ FACS проводили согласно процедуре, описанной в Примере 5. Результаты, представленные на Фигурах 19 и 20, показывают среднее изменение при средней интенсивности флуоресценции (MFI) в сыворотке мыши по сравнению с вторичным антимышиным антителом самим по себе +/- стандартное отклонение пяти мышей в каждой группе. Титры антител и связывание с поверхностноклеточным природным PSA не измеряли, поскольку PSA экспрессировался в виде цитоплазматического белка мультиантигенными вакцинами, изученными в этом исследовании.
Результаты.
На Фигурах 15А-165С представлены результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин в анализе IFN-γ ELISPOT. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14, 28, 42 и 70. В день 77, распознавание эндогенных PSMA и PSCA оценивали, изучая Т-клеточные реакции на клетки (A) TRAMP C2-PSMA, (В) TRAMP C2-PSCA и (С) TRAMP С2-PSMA-PSA-PSCA в анализе IFN-γ ELISPOT. Клетки TRAMP С2 служили фоновым контролем для анализа. Для IFN-γ Т-клеточных реакций на эндогенный PSMA, амплитуда реакции TRAMP C2-PSMA значительно снижалась после вакцинации с двойным промотором PSMA-PSCA, PS MA-T2A-PSCA, PSCA-IRES-PSMA и совместно сформированными PSMA и PSCA по сравнению с вакцинацией только PSMA (** и*** указывают значение ps<0,01 и <0,001, соответственно, в двухфакторном дисперсионном анализе ANOVA). Однако, вакцинный конструкт PSCA-F2A-PSMA вызвал подобную амплитуду IFN-γ Т-клеточной реакции на клетки TRAMP C2-PSMA в качестве единичной PSMA-вакцины. Для IFN-γ Т-клеточных реакций на эндогенный PSCA, наблюдалось значительное повышение ответных реакций после вакцинации несколькими двухантигенными вакцинами, включающими двойной промотор PSMA-PSCA, PSCA-F2A-PSMA, PSCA-IRES-PSMA и совместно сформированные PSMA и PSCA, по сравнению с PSCA-вакциной (*,** и*** указывают значение ps <0,05, 0,01 и 0,001, соответственно, в двухфакторном дисперсионном анализе ANOVA). Вакцинный конструкт PSCA-T2A-PSMA вызывает подобную амплитуду IFN-γ Т-клеточной реакции на клетки TRAMP C2-PSCA в качестве единичной PSCA-вакцины. При сравнении IFN-γ Т-клеточных реакций на TRAMP C2-PSMA-PSA-PSCA, не наблюдалось существенных отличий между группами, вакцинированными разными двухантигенными вакцинами. Взятые вместе, эти данные демонстрируют генерирование PSMA и PSCA-специфических Т-клеточных ответов после двухантигенной вакцинации с использованием стратегий соэкспрессионной и совместно сформированной ДНК-вакцинации.
На Фигурах 16А-16С представлены результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин в анализе IFN-γ ELISPOT. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14, 28, 42 и 70. В день 77, Т-клеточные реакции на (А) пептиды PSMA, (В) пулы пептидов PSMA и (С) пептиды PSCA (смотри Таблицу 22) оценивали с помощью анализа IFN-γ ELISPOT. Среда сама по себе служили фоновым контролем для анализа. Для IFN-γ Т-клеточных реакций на индивидуальные пептиды и пулы пептидов PSMA наблюдалась более высокая амплитуда реакций по сравнению с единичной PSMA-вакциной после двухантигенных вакцинаций PSMA-T2A-PSCA и PSCA-F2A-PSMA. Значительное снижение IFN-γ Т-клеточной реакции на индивидуальные пептиды PSMA наблюдалось после вакцинации двойным промотором PSMA-PSCA, PSCA-IRES-PSMA и совместно сформированными PSMA и PSCA. IFN-y Т-клеточная реакция на PSCA-специфический пептид была похожей среди групп, вакцинированных разными двухантигенными вакцинами. Эти данные также демонстрируют генерирование Т-клеточных реакций на PSMA и PSCA, соэкспрессированные на одном ДНК-вакцинном конструкте, или доставляемые в виде совместно сформированной композиции.
На Фигуре 17 представлены результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью титров антител к PSMA. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14, 28, 42 и 70. В день 77, титры антител к PSMA в сыворотке оценивали с помощью ELISA. Все животные, вакцинированные PSMA, генерировали значительные титры антител к PSMA. Мыши, вакцинированные двойным вакцинным конструктом - PSCA-F2A-PSMA и единичной PSMA-вакциной, генерировали значительно более высокие титры антител по сравнению со всеми другими группами мышей, вакцинированных PSMA (однофакторный дисперсионный анализ ANOVA, значение p<0,05). Вакцинация двойным промотором PSMA-PSCA и совместно сформированными PSMA и PSCA привела к более высоким титрам антител по сравнению с мышами, которые получили вакцину PSMA-T2A-PSCA. Взятые вместе, эти данные демонстрируют генерирование PSMA-специфических антител после двухантигенной ДНК-вакцинации с PSMA и PSCA с использованием стратегий соэкспрессионной и совместно сформированной вакцинации.
На Фигуре 18 представлены результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью титров антител к PSCA. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14, 28, 42 и 70. В день 77, титры антител к PSCA в сыворотке оценивали с помощью. Мыши, вакцинированные совместно сформированными PSMA и PSCA и единичной PSCA-вакциной, генерировали значительно более высокие титры антител по сравнению со всеми другими группами мышей, вакцинированных PSCA (однофакторный дисперсионный анализ ANOVA). Вакцинация двойным промотором PSMA-PSCA привела к более высоким титрам антител по сравнению с вакцинацией PSMA-T2A-PSCA, PSCA-F2A-PSMA и PSCA-IRES-PSMA. Эти результаты указывают на то, что соэкспрессированные и совместно сформированные PSMA и PSCA вызывают антитела к PSCA.
На Фигуре 19 представлены результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью поверхностноклеточного связывания антител к PSMA. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14, 28, 42 и 70. В день 77, распознание поверхностноклеточного природного PSMA оценивали путем связывания антител в сыворотке с клетками LNCaP и РС3. Клетки РС3 служили фоновым контролем для анализа. За исключением единичной PSCA-вакцины, все вакцинные схемы с PSMA привели к значительному связыванию антител к PSMA с клетками LNCaP по сравнению с контрольными клетками РС3. Не было существенных отличий в связывании антител к PSMA с клетками LNCaP между PSMA-вакцинированными группами (однофакторный дисперсионный анализ ANOVA, значение p>0,05). Кратность изменения в отношении вторичного антитела для J591-A mAb составила 45,3 (данные не показаны). Эти данные демонстрируют генерирование PSMA-специфических антител, которые распознают природный PSMA, после двухантигенной ДНК-вакцинации с использованием стратегий соэкспрессионной и совместно сформированной вакцинации.
На Фигуре 20 представлены результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью поверхностноклеточного связывания антител к PSCA. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14, 28, 42 и 70. В день 77, распознание поверхностноклеточного природного PSCA оценивали путем связывания антител в сыворотке с клетками MIA-PaCa-2, трансдуцированными Ad-PSCA и нетрансдуцированными. Нетрансдуцированные парентеральные клетки служили фоновым контролем для анализа. За исключением единичной PSMA-вакцины, все вакцинные схемы с PSCA привели к значительному связыванию антител к PSCA с клетками MIA-PaCa-2, трансдуцированными Ad-PSCA, по сравнению с контрольными клетками. Не было существенных отличий в связывании антител к PSCA с клетками MIA-РаСа-2, трансдуцированными Ad-PSCA, между PSCA-вакцинированными группами (однофакторный дисперсионный анализ ANOVA, значение p>0,05). Кратность изменения в отношении вторичного антитела для 7F5 mAb составила 18,7 (данные не показаны). В целом, эти данные демонстрируют генерирование PSCA-специфических антител, которые распознают природный PSCA, после двухантигенной ДНК-вакцинации с использованием стратегий соэкспрессионной и совместно сформированной вакцинации.
6 В. Иммуногенность двухантигенных вакцин, содержащих PSMA и PSA или PSCA и PSA, у мышей C57BU6
Процедура исследования.
Исследование клеточных иммунных реакций. Самок мышей C57BL/6 примировали в день 0 и бустировали в дни 14 и 28 ДНК, экспрессирующей человеческий PSMA, PSCA и PSA, с помощью эпидермальной инъекции PMED. В целом, оценили четыре разных стратегий двухантигенных ДНК-вакцинаций, которые включали две соэкспрессионных подхода и две стратегии совместного формирования. Для соэкспрессии вводили единичные ДНК-вакцинные плазмиды, кодирующие два антигена простаты - PSMA и PSA, соединенные пептидом 2А (плазмида ID# 5300), или PSCA и PSA, соединенные IRES (плазмида ID#455). Для совместного формирования плазмиды плазмиды, индивидуально кодирующие PSMA, PSCA или PSA, совместно формировали на одной частице золота для PMED-доставки. В частности, они включали совместно сформированные PSMA и PSA и совместно сформированные PSCA и PSA. В качестве контролей, мышей C57BL/6 вакцинировали ДНК, экспрессирующей единичный антиген простаты - PSMA, PSCA или PSA. Для соэкспрессии двух- или одноантигенных вакцин на PMED-введение давали дозу в 2 мкг ДНК, тогда как по 2 мкг каждой ДНК-вакцинной плазмиды (в сумме 4 мкг ДНК на введение) давали для совместного формулирования. Клеточные иммунные реакции двух- и одноантигенных вакцин измеряли путем извлечения селезенки от каждого животного в день 35. Выделяли спленоциты и подвергали анализу IFN-γ ELISPOT для измерения PSMA, PSCA и PSA-специфических Т-клеточных реакций. Кратко, по 2×105 спленоцитов от индивидуального животного помещали в лунку с 2×104 клеток TRAMP-C2, экспрессирующих единственный эндогенный антиген простаты человека или PSMA, PSCA и PSA вместе, или с индивидуальным пептидом или пулами пептидов, специфических к человеческим PSMA, PSCA и PSA, в концентрации 10 мкг/мл (смотри Таблицу 22 для пептидов и пептидных пуловых композиций), или только средой в качестве контроля. Каждое событие осуществляли в трех повторностях. Планшеты инкубировали в течение 20 часов при 37°C и 5% CO2, промывали и разрабатывали после инкубации в соответствии с указаниями изготовителя. Количество IFN-γ SFC подсчитывали с помощью ридера CTL. Результаты представлены на Фигурах 21 и 22, которые показывают среднее количество PSMA, PSCA и PSA-специфических SFC +/- стандартное отклонение пяти мышей в каждой группе, нормированное к 1×106 спленоцитов.
Исследование реакции антител. Самок мышей C57BL/6 примировали в день 0 и бустировали в дни 14, 28 и 49 ДНК, экспрессирующим PSMA, PSCA и PSA, путем PMED. Реакции антител на двух- и одноантигенные вакцины измеряли путем сбора сыворотки от каждого животного в день 56, семь дней после последней PMED-вакцинации. Титры антител к PSMA и PSCA в сыворотке определяли, используя анализ ELISA, как описано в Примере 5. Результаты представлены на Фигурах 23 и 24, которые показывают средние титры +/- стандартное отклонение пяти мышей в каждой группе.
Сыворотку также подвергали анализу FACS для измерения связывания антитела с человеческим PSMA или PSCA, экспрессированным на клеточной поверхности соответствующих клеточных линий, тем самым определения, являются ли антитела, полученные с помощью мультиантигенных вакцин, способными распознавать конформации природных PSMA и PSCA, соответственно. Связывание антител с поверхностноклеточным природным PSA не измеряли, поскольку PSA экспрессировались в виде цитоплазматического белка мультиантигенными вакцинами, изученными в данном исследовании. Анализ FACS проводили в соответствии с процедурой, описанной в Примере 5. Результаты представлены на Фигурах 25 и 26, которые показывают среднюю кратность изменения в MFI в мышиной сыворотке по сравнению с вторичным антимышиным антителом самим по себе +/- стандартное отклонение пяти мышей в каждой группе. Титры антител не измеряли, поскольку PSA экспрессировался в виде цитоплазматического белка мультиантигенными вакцинами, изученными в этом исследовании.
Результаты.
На Фигуре 21 представлены результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин в анализе IFN-γ ELISPOT. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14 и 28. В день 35, распознавание эндогенных PSMA, PSCA и PSA оценивали, изучая Т-клеточные реакции на клетки (A) TRAMP C2-PSMA, (В) TRAMP C2-PSCA, (С) TRAMP C2-PSA и (D) TRAMP C2-PSMA-PSA-PSCA в анализе IFN-γ ELISPOT. Клетки TRAMP С2 служили фоновым контролем для анализа. Для IFN-γ Т-клеточных реакций на эндогенно экспрессированный PSMA на клетках не наблюдалось значительных отличий между реакциями на TRAMP C2-PSMA после вакцинации двойными антигенами, содержащими PSMA (PSA-F2A-PSMA и совместно сформированные PSMA и PSA), и PSMA самим по себе. Также, для IFN-γ Т-клеточных реакций на эндогенный PSCA, не наблюдалось значительных отличий в амплитуде реакции на TRAMP C2-PSCA между вакцинами двойных PSCA-IRES-PSA и совместно сформированных PSCA и PSA по сравнению с единичной PSCA-вакциной. Для IFN-γ Т-клеточных реакций на эндогенный PSA, было обнаружено существенное повышение амплитуды реакции на TRAMP C2-PSA при сравнении иммуногенности единичной PSA-вакцины к PSA-F2A-PSMA (*** указывает p<0,001 в двухфакторном дисперсионном анализе ANOVA) и совместно сформированных PSMA и PSA (** указывает p<0,01 в двухфакторном дисперсионном анализе ANOVA). Не наблюдалось отличий в амплитуде реакции на TRAMP C2-PSA при сравнении животных, которые получали двойную вакцину PSCA и PSA и единичную вакцину PSA. При рассмотрении IFN-γ Т-клеточного ответа на TRAMP C2-PSMA-PSA-PSCA не было существенных отличий в реакции между группами, вакцинированными разными двухантигенными вакцинами. Взятые вместе, эти данные демонстрируют генерирование PSMA и PSA-специфических Т-клеточных реакций, а также PSCA и PSA-специфических Т-клеточных реакций после двухантигенной ДНК-вакцинации с использованием стратегий совместной экспрессии и совместного формирования вакцин.
На Фигуре 22 представлены результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью титров антител к PSMA. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14, 28 и 49. В день 56, титры антител к PSMA в сыворотке оценили с помощью ELISA. Все животные, вакцинированные PSMA, генерировали значительные титры антител к PSMA. Не наблюдалось существенных отличий в титрах антител между мышами, вакцинированными PSA-F2A-PSMA, совместно сформированными PSMA и PSA и PSMA самим по себе (однофакторный дисперсионный анализ ANOVA, значение p>0,05). Взятые вместе, эти данные демонстрируют генерирование PSMA-специфических антител после двухантигенной ДНК-вакцинации с PSMA и PSA с использованием стратегий совместной экспрессии и совместного формирования вакцин.
На Фигуре 23 представлены результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью титров антител к PSCA. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14, 28 и 49. В день 56, титры антител к PSCA в сыворотке оценили с помощью ELISA. Все животные, вакцинированные PSCA, генерировали значительные титры антител к PSCA. Не наблюдалось существенных отличий в титрах антител между мышами, вакцинированными PSCA-IRES-PSA, совместно сформированными PSCA и PSA и PSCA самим по себе (однофакторный дисперсионный анализ ANOVA, значение p>0,05), хотя титры антител, генерированные после вакцинации PSCA-IRES-PSA, изменялись меньше, чем в других группах, вакцинированных PSCA. Эти результаты указывают на то, что соэкспрессия PSCA и PSA или совместно сформированные PSCA и PSA вызывали антитела к PSCA
На Фигуре 24 представлены результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью поверхностноклеточного связывания антител к PSMA. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14, 28 и 49. В день 56, распознание поверхностноклеточного природного PSMA оценивали путем связывания антител в сыворотке с клетками LNCaP и РСЗ. Клетки РСЗ служили фоновым контролем для анализа. Не было существенных отличий в связывании антител к PSMA с клетками LNCaP между PSMA-вакцинированными группами (однофакторный дисперсионный анализ ANOVA, p-значение >0,05). Кратность изменения в отношении вторичного антитела для J591-A mAb составила 45,3 (данные не показаны). В целом, эти данные демонстрируют генерирование PSMA-специфических антител, которые распознают природный PSMA после двухантигенной вакцинации с использованием стратегий совместной экспрессии и совместного формирования вакцин.
На Фигуре 25 представлены результаты репрезентативного исследования, которое оценивает иммуногенность двухантигенных вакцин с помощью поверхностноклеточного связывания антител к PSCA. Кратко, по 5 мышей в группе примировали в день 0 и бустировали с PMED в дни 14, 28 и 49. В день 56, распознание поверхностноклеточного природного PSCA оценивали путем связывания антител в сыворотке с клетками MIA-РаСа-2, трансдуцированными Ad-PSCA и нетрансдуцированными. Нетрансдуцированные парентеральные клетки служили фоновым контролем для анализа. Все группы мышей, вакцинированные PSCA, демонстрировали очень низкое связывание антител к PSCA клетками MIA-PaCa-2, трансдуцированными Ad-PSCA. Вакцинация PSCA-IRES-PSA привела к значительному снижению связывания с клетками MIA-PaCa-2, трансдуцированными Ad-PSCA, по сравнению с мышами, вакцинированными только PSCA (* указывает p<0,05 в однофакторном дисперсионном анализе ANOVA). Взятые вместе, эти данные демонстрируют, что соэкспрессия PSCA и PSA и совместно сформированные PSCA и PSA приводят к очень низкому распознаванию природного PSCA антителом, специфическим к PSCA.
ПРИМЕР 7. ИММУНОГЕННОСТЬ МОДИФИЦИРОВАННЫХ АНТИГЕНОВ ЧЕЛОВЕЧЕСКОГО PSMA
Схема исследования. Иммунные реакции, индуцированные ДНК-вакцинацией с использованием конструкта, кодирующего полипептид иммуногенного PSMA ("модифицированный антиген человеческого PSMA" или "модифицированный hPSMA"), содержащий 15-750 аминокислоты(ах) белка природного человеческого PSMA из SEQ ID NO: 1, сравнивали с реакциями, индуцированными белком природного человеческого непроцессированного PSMA (непроцессированный hPSMA). Группы самок мышей C57BL/6 или самок трансгенных мышей Пастера (HLA-A2 / DR1) примировали в день 0 и бустировали в дни 14, 28 путем PMED-введения 2 мкг ДНК-вакцины, кодирующей белок непроцессированного hPSMA или модифицированного hPSMA. Мышей обескравливали и умерщвляли в день 35 (через 7 дней после третьей вакцинации) и определяли Т-клеточные иммунные реакции на белок непроцессированного hPSMA в спленоцитах в анализе IFN-γ ELISPOT. Для мышей C57BL/6, одноклеточные суспензии по 2×105 спленоцитов от отдельных животных помещали в лунки с 10 мкг очищенного белка hPSMA, 2×104 клетками TRAMP-C2 самими по себе или клетками TRAMP-C2, экспрессирующими hPSMA или слитый белок PSMA-PSA-PSCA. Для трансгенных мышей Пастера (HLA-A2/DR1), одноклеточные суспензии по 2×105 спленоцитов от отдельных животных помещали в лунки с 2×104 клетками К562, экспрессирующими человеческие HLA-A2, которые были активированы известными НLА-А2-рестриктированными CD8+ Т-клеточными эпитопами, полученными из белковой последовательности человеческого PSMA (Таблица 23). Реакции у мышей Пастера также обнаруживали, используя 10 мкг/мкл очищенного белка PSMA protein или 2×104 клеток SK-Mel5, которые были трансдуцированы аденовирусными векторами, экспрессирующими контрольный белок (Ad-eGFP) или белок непроцессированного человеческого PSMA (Ad-hPSMA). Каждое событие осуществляли в трех повторностях. Планшеты инкубировали в течение 20 ч при 37°C и 5% СO2, промывали и разрабатывали после инкубации в соответствии с инструкцией производителя. Количество IFN-γ SFC подсчитывали с помощью ридера CTL. Результаты представлены на Фигурах 19 и 20, которые показывают среднее количество PSMA-специфических SFC/млн спленоцитов +/- стандартное отклонение в каждой группе.
Анализ ELISA. Реакции антител, вызванные вакцинами модифицированного и непроцессированного PSMA, измеряли в сыворотке от каждого животного, собранной в день 35. Сыворотку подвергли ELISA для определения титров антител к PSMA, как описано в Примере 5. Результаты представлены на Фигуре 26, которые показывают средние титры +/- стандартное отклонение количества мышей в группе.
Анализ FACS. Сыворотку также подвергли анализу FACS для измерения связывания антитела с человеческими PSMA, экспрессированными на клеточной поверхности приемлемых клеточных линий, тем самым определяя, способны ли антитела, генерируемые вакцинами модифицированного и непроцессированного PSMA, распознавать конформацию природного PSMA. Анализ FACS проводили согласно процедуре, описанной в Примере 5. Результаты представлены на Фигуре 29, которые показывают среднюю кратность изменения в MFI в мышиной сыворотке по сравнению с вторичным антимышиным антителом самим по себе +/- стандартное отклонение количества мышей в группе.

Результаты. На Фигуре 25 представлены результаты репрезентативного исследования для оценки Т-клеточной иммунной реакции, вызванной модифицированным человеческим PSMA (ак 15-750) по сравнению с непроцессированным человеческим PSMA (ак 1-750) у мышей C57BL/6, определенные с помощью анализа IFN-γ ELISPOT. По пять (5) мышей в группе примировали в день 0 и бустировали PMED ДНК-вакцинами, экспрессирующими белки модифицированного hPSMA или непроцессированного hPSMA в дни 14 и 28. В день 35, реакцию, вызванную непроцессированным белком hPSMA, сравнивали, определяя Т-клеточные реакции на TRAMP C2-PSMA или белок ECD очищенного человеческого PSMA ECD (называют белком очищенного hPSMA на Фиг. 26) с помощью анализа IFN-γ ELISPOT. Клетки TRAMP С2 служили фоновым контролем для анализа. Амплитуда IFN-Y Т-клеточных реакций, вызванных TRAMP C2-PSMA или белком очищенного hPSMA, не отличалась существенно (двухфакторный дисперсионный анализ ANOVA, значение p>0,05) среди групп. Эти результаты указывают на то, что ДНК-вакцины, экспрессирующие белки модифицированного hPSMA и непроцессированного hPSMA, вызывают эквивалентные Т-клеточные реакции у мышей C57BL/6.
На Фигурах 27А и 27В представлены результаты репрезентативного исследования для оценки Т-клеточной иммунной реакции антигена модифицированного человеческого PSMA (ак 15-750) по сравнению с антигеном непроцессированного человеческого PSMA (ак 1-750) у трансгенных мышей Пастера (HLA-A2/DR1) с помощью анализа IFN-γ ELISPOT. Десятерых (10) мышей в группе примировали в день 0 и бустировали PMED ДНК-вакцинами, кодирующими белок модифицированного hPSMA или непроцессированного hPSMA в дни 14 и 28. В день 35, Т-клеточную реакцию, вызванную на белок непроцессированного hPSMA, определяли с помощью анализа IFN-γ ELISPOT, используя (A) PSMA, полученный из HLA-А2-рестриктированных пептидов, репрезентирующих известные CD8+ эпитопы, и (В) клетки SK-Mel5, трансдуцированные Ad-hPSMA или непроцессированным белком очищенного hPSMA. Белок hHER2 106 и SK-Mel5 Ad-eGFP служили негативными контролями в анализах. Вакцина модифицированного hPSMA вызвала более высокую амплитуда IFN-γ Т-клеточных реакций на НLА-А2-рестриктированные CD8+ Т-клеточные эпитопы, хотя отличия между группами не были существенными (двухфакторный дисперсионный анализ ANOVA, значение p>0,05). Подобным образом, вакцина модифицированного hPSMA вызвала более высокую амплитуду иммунной реакции на клетки SK-Mel5, трансдуцированные Ad-hPSMA, и значительно (двухфакторный дисперсионный анализ ANOVA, значение P>0,05) более высокую частоту IFN-γ SFC к белку очищенного hPSMA. Эти результаты демонстрируют, что ДНК-вакцина, экспрессирующая белок модифицированного hPSMA, является более сильной относительно индукции Т-клеточной реакции на белок hPSMA, чем на белок непроцессированного hPSMA у трансгенных мышей Пастера (HLA-A2/DR1).
На Фигуре 28 представлены результаты репрезентативного исследования, которое оценивает иммуногенность вакцин человеческого модифицированного и непроцессированного PSMA с помощью титров антител к PSMA. Кратко, мышей примировали в день 0 и бустировали с PMED в дни 14 и 28. Девять мышей Пастера вакцинировали модифицированным PSMA, 10 мышей Пастера вакцинировали непроцессированным PSMA и по 5 мышей C57BL / 6 в каждой группе вакцинировали или модифицированным PSMA, или непроцессированным PSMA. В день 35, титры антител к PSMA в сыворотке оценивали с помощью ELISA. Как и ожидалось, мыши C57BL/6 генерировали значительно более высокие титры антител к PSMA по сравнению с мышами Пастера (однофакторный дисперсионный анализ ANOVA). При сравнение титров антител между теми же штаммами мышей не было обнаружено существенных отличий в титрах антител у мышей, вакцинированных модифицированным или непроцесированным PSMA (однофакторный дисперсионный анализ ANOVA, значение p>0,05). В целом, эти результаты демонстрируют, что вакцинация непроцессированной версией человеческого PSMA генерирует эквивалентный титр антител к PSMA по сравнению с вакцинной человеческого модифицированного PSM.
На Фигуре 29 представлены результаты репрезентативного исследования, которое оценивает иммуногенность вакцин человеческого модифицированного и непроцессированного PSMA с помощью поверхностноклеточного связывания антител к PSMA. Кратко, по 5 мышей C57BL/6 в группе примировали в день 0 и бустировали с PMED в дни 14 и 28. В день 35, распознание поверхностноклеточного природного PSMA оценивали путем связывания антител в сыворотке с клетками LNCaP и РС3. Клетки РС3 служили фоновым контролем для анализа. Не было существенных отличий в связывании антител к PSMA с клетками LNCaP между мышами, вакцинированными модифицированным или непроцессированным PSMA (однофакторный дисперсионный анализ ANOVA, значение p>0,05). Кратность изменения в отношении вторичного антитела для J591-A mAb составила 14,3 (данные не показаны). В целом, эти данные демонстрируют, что возможна генерация PSMA-специфических антител, которые распознают природный PSMA после вакцинации модифицированным или непроцессированным PSMA.
ПРИМЕР 8. ДЕЙСТВИЕ АНТИТЕЛА К CTLA-4 НА ИНДУЦИРОВАННУЮ ВАКЦИНОЙ ИММУННУЮ РЕАКЦИЮ
Действие местного введения моноклонального антитела к CTLA-4 (СР-675, 206) на иммунные реакции, вызванные молекулой нуклеиновой кислоты человеческого PSMA, предусмотренной изобретением, было изучено в исследовании на обезьянах, в котором иммунную реакцию оценивают путем измерения PSMA-специфических Т-клеточных реакций, используя анализ IFN-y ELISPOT.
Лечение животных и взятие проб. Три группы самок индийского макака резус, от пяти до шести (#1 до 5 или 6) на каждую тестируемую группу, иммунизировали гуклеиновой кислотой (SEQ ID NO: 10), которая кодирует модифицированный антиген человеческого PSMA (SEQ ID NO: 9), доставляемый аденовирусом (1011 V.P., инъецированных внутримышечно) после 2 ДНК-иммунизаций (8 нажатий/иммунизацию, по 4 нажатия на каждую правую и левую стороны нижней части живота) с помощью PMED с 6 и 9-недельными интервалами, соответственно. Животные в группах 2 и 3 дополнительно получали билатеральные интрадермальные инъекции по 3 мг CpG (PF-03512676) после PMED-иммунизации в непосредственной близости от каждого пахового дренирующего лимфатического узла. Группа 2 также получала внутривенные инъекции моноклонального антитела к CTLA-4 (СР-675, 206) в дозе 10 мг/кг, а группа 3 получала интрадермальные инъекции моноклонального антитела к CTLA-4 (СР-675, 206) в дозе 5 мг/кг в непосредственной близости от каждого левого и правого вакцинодренирующего лимфатического узла во время второй PMED-иммунизации.
Образцы периферической крови брали от каждого животного через шесть дней после последней PMED-иммунизации. Из образцов выделили мононуклеарные клетки периферической крови (РВМС) и подвергли анализу IFN-y ELISPOT для измерения PSMA-специфических Т-клеточных реакций. Кратко, по 4×105 РВМС от каждого животного помещали в лунки планшет для IFN-y ELISPOT с пулами PSMA-специфических пептидов, по 2 мкг/мл каждого, белком ECD hPSMA в дозе 10 мкг/мл, белком ECD резусного PSMA в дозе 10 мкг/мл или неспецифическими контрольными белками (пептидный пул человеческого HER2) по 2 мкг/мл каждого. Композиции каждого пула PSMA-специфических белков представлены в Таблице 24А. Планшеты инкубировали в течение 16 часов при 37°C и 5% CO2 промывали и разрабатывали после инкубации в соответствии с инструкцией производителя. Количество IFN-y - пятнообразующих клеток (SFC) подсчитывали с помощью ридера CTL. Каждое условие осуществляли в двух повторностях. Результаты представлены в Таблице 24В, которые показывают среднее количество PSMA-специфических SFC из трех повторностей за вычетом среднего количества SFC от неспецифических контрольных пептидов, нормализированное до 106 РВМС. Значек ^ указывает, что количество не является точным, потому что количество пятен было слишком большим для подсчета.
Процедура анализа IFN-y ELISPOT. Иммобилизированное антитело, специфическое к IFN-y (BD Bioscience, #51-2525kc) наносили на поливинилиденфторидную мембрану (PVDF) в микропланшете в течение ночи при 4°C. Планшет блокировали сывороткой/белком, чтобы предотвратить неспецифическое связывание с антителом. После блокирования эффекторные клетки (такие как спленоциты, выделенные от иммунизированных мышей, или РВМС, выделенные от макак резус) и мишени (такие как пептиды PSMA из пептидной библиотеки, клетки-мишени, активированные антиген-специфическими пептидами, или опухолевые клетки, экспрессирующие релевантные антигены) добавляли в лунки и инкубировали в течение ночи при 37°C в 5% CO2 инкубаторе. Цитокины, секретируемые эффекторными клетками, захватывались антителом покрытия на поверхности PVDF-мембраны. После удаления клеток и культуральной среды, по 100 мкл биотинилированного поликлонального антитела к человеческому IFNy добавляли в каждую лунку для определения. Пятна визуализировали добавлением стрептавидина-пероксидазы хрена и преципитата субстрата, 3-амино-9-этилкарбазола (АЕС), для получения красного цвета пятна, как в протоколе производителя (Mabtech). Каждое пятно представляет собой одну Т-клетку, продуцирующую цитокины.
Результаты. Таблица 24В показывает результаты репрезентативного анализа IFN-y ELISPOT, который оценивает и сравнивает Т-клеточные реакции, вызванные вакциной без моноклонального антитела (группа 1) или с моноклональным антителом к CTLA-4 (СР-675, 206), введенной системно путем внутривенных инъекций (группа 2) или местно внутрикожными инъекциями в непосредственной близости от вакцинодренирующего лимфатического узла (группа 3). Как показано в Таблице 1В, вакцина PSMA индуцировала измеримые IFN-y Т-клеточные реакции на несколько PSMA-специфические пептиды и белки в отсутствие CpG (PF-03512676) и моноклонального антитела к CTLA-4 (СР-675, 206). Реакции слегка повышались при добавлении CpG (PF-03512676) и системной доставке антитела к CTLA-4 (СР-675, 206; группа 2). В то же время, наблюдалась более мощная и значительно усиленная реакция на несколько пептидов PSMA и белок PSMA, когда моноклональное антитело к CTLA-4 (СР-675, 206) доставлялось местно внутрикожными инъекциями в непосредственной близости от вакцинодренирующего лимфатического узла (группа 3).



ПРИМЕР 9. СИСТЕМНОЕ ВОЗДЕЙСТВИЕ АНТИТЕЛА CTLA-4 ПОСЛЕ ВВЕДЕНИЯ У ОБЕЗЬЯН
Уровни в крови тремелимумаба - антитела к CTLA-4 (СР675206) были исследованы у индийских макак резус после введения антитела внутрикожной или внутривенной инъекциями.
Лечение животных и взятие проб. Трех животных на группу лечения инъецировали антителом к CTLA-4 - тремелимумабом в дозе 10 мг/кг одной внутривенной инъекцией в подкожную вену или несколькими внутрикожными билатеральными инъекциями по 0,2 мл в верхнюю часть бедра в непосредственно близости к паховым дренирующим лимфатическим узлам. Образцы крови брали через 0, 1, 2, 4, 8, 12, 24 и 48 часов после инъекции и помещали в вакуумные пробирки (вакутайнер-пробирки) на 2,0 мл, содержащие литий-гепарин в качестве антикоагулянта. Плазму собирали из супернатанта в вакутайнер-пробирках после центрифугирования при 1500 оборотах в минуту при 4°C в течение 10 минут. Уровни тремелимумаба в плазме измеряли с помощью количественного анализа ELISA согласно процедуре, описанной ниже.
Процедура количественного анализа тремелимумаба с помощью ELISA. Аналитические 384-луночные планшеты для высокого связывания (VWR-Greiner Bio-One Cat# 82051-264) покрывали 25 мкл/лунку CD-152 (CTLA-4; Ancell Immunology Research Products Cat# 501-020) в 1 г/мл 100 мМ карбонатно-бикарбонатного покрывающего буфера и инкубировали в течение ночи при 4°C. Планшеты промывали шесть раз 1 × PBS-Tween (0,01 М PBS рН 7,4/0,05% Tween 20) и блокировали, используя 40 мкл/лунку 5% FBS/1 × PBS-Tween, и инкубировали, встряхивая при 600 оборотах в минуту при комнатной температуре в течение 1 часа. Стандарты получали путем следующих разведений тремелимумаба: 200, 67, 22, 7,4, 2,5, 0,82, 0,27, 0,09 и 0,03 нг/мл. Образцы разводили до 1:100, 1:1000 и 1:10,000. Разбавитель состоял из 1% «необученной» сыворотки яванского макака и 5% FBS в 1 × PBS-Tween (0,01 М PBS рН 7,4/0,05% Tween 20). По 25 мкл/лунку стандарта, образца и контрольного разбавителя переносили в двух повторностях в планшет и инкубировали, встряхивая при 600 оборотах в минуту при комнатной температуре в течение 1 часа. После шестикратного промывания 1 × PBS-Tween, добавляли по 25 мкл/лунку вторичного антитела (козье антитело к человеческому IgG HRP, Southern Biotech Cat# 9042-05) при разведении 1:5,000 с 1 × PBS-Tween, а затем инкубировали, встряхивая при 600 оборотах в минуту при комнатной температуре в течение 1 часа. После шестикратного промывания с 1 × PBS-Tween, добавляли по 25 мкл/лунку ТМВ пероксидазного EIA-субстрата (раствор А+В) (Bio-Rad Cat# 172-1067), планшеты инкубировали при комнатной температуре в течение 4 минут. Колориметрическую реакцию останавливали добавлением 12,5 мкл/лунку 1 N серной кислоты, а поглощение затем считывали при 450 нм. Количество тремелимумаба в каждом образце определяли, используя стандартную кривую с концентрациями от 0,27 до 67 нг/мл, используемыми в качестве количественного диапазона.
Результаты. Уровни антител к CTLA-4 в плазме из репрезентативного исследования представлены на Фигуре 30. Как показано, внутрикожные инъекции антитела к CTLA-4 тремелимумаба отображают более медленную кинетику высвобождения антитела в крови и низкий профиль системного воздействия (AUCo-24=4,9×106 нг ч/мл) чем внутривенные инъекции (AUCo-24=7,2×106 нг ч/мл).
ПРИМЕР 10. ВОЗДЕЙСТВИЕ АНТИТЕЛА CTLA-4 НА ИММУННЫЕ РЕАКЦИИ, ВЫЗВАННЫЕ ВАКЦИНАМИ У МЫШЕЙ
Процедура исследования. Самок мышей BALB/с, по 6 на группу, примировали и бустировали ДНК, экспрессирующей rHer, с помощью PMED, разделенного четырехнедельным интервалом. По 150 мкг моноклонального антитела, специфического к мышиному CTLA-4 (клон 9Н10, Bioxcell or #ВЕ0131), или изотипных контрольных моноклональных антител (Bioxcell # ВЕ0091) вводили в дни приведения в действие PMED, и по 100 мкг в дни после PMED путем местных внутрикожных или системных внутрибрюшинных инъекций, как указано в рецептах. Полифункциональные (мультицитокинпозитивные) Т-клеточные реакции измеряли из спленоцитов, изъятых от отдельных мышей на 7 после финальных PMED-иммунизаций, с помощью анализа ICS. После 5-ти часовой стимуляции вакциноспецифическим эпитопным пептидом (rHer-2 специфический антиген-специфический CD8 (рбб), эпитоп CD4 (р169) или иррелевантный пептид HBV (коровый антиген р87)) в дозе 10 мкг/мл, спленоциты первыми окрашивались по CD4, CD3 и CD8, за которыми последовала пермеабилизация и окрашивание экспрессии IFNA, TNFa и IL-2, которую анализировали с помощью проточной цитометрии. Общее количество антиген-специфических одно-, двух- или трехцитокино положительных Т-клеток в селезенке каждого животного вычисляли путем вычитания реакций на иррелевантный пептид HBV от вакциноспецифических реакций и нормировали на общее количество спленоцитов, выделенных из селезенки.
Результаты. На Фигурах 31А и 31В представлены результаты репрезентативного исследования, которое оценивает иммуномодуляторную активность моноклонального антитела к CTLA-4 (клон 9Н10) от качества вакциноиндуцированных иммунных ответов с помощью анализа внутриклеточного цитокинового окрашивания. Через семь дней после последнего PMED наблюдалось значительное повышение антиген-специфических одно- и двухцитокиноположительных реакций CD8 Т-клеток при местной внутрикожной доставке антитела к CTLA-4 и двух- и трехцитокиноположительных реакций CD8 Т-клеток при системной доставке. Кроме того, наблюдалось значительное повышение антиген-специфических одноцитокиноположительных CD4 Т-клеток при внутрикожной доставке и двухцитокиноположительных клеток при системной доставке антитела к CTLA-4 (указывает p<0,05 по Т-критерию Стьюдента).
ПРИМЕР 11. СИНЕРГИТИЧЕСКИЙ ЭФФЕКТ СУНИТИНИБА В КОМБИНАЦИИ С ПРОТИВОРАКОВОЙ ВАКЦИНОЙ
Процедура исследования. Противоопухолевую активность сунитиниба в комбинации с антираковой ДНК-вакциной исследовали у самок трансгенных мышей BALB/neuT.
Самкам трансгенных гетерозиготных мышей BALB/neuT, которые экспрессируют крысиный опухолеасоциированный антиген, подкожно имплантировали 106 клеток TUBO, экспрессирующих rHer-2, которые происходят от спонтанных млекопитающих опухолей мышей BALB/neu. Через 7 дней после имплантации опухолевых клеток, мышам один раз в день перорально давали носитель или малат сунитиниба в дозах, указанных в рецепте. Через три дня после начала терапии малатом сунитиниба мышей иммунизировали в режиме, содержащем (а) контрольную вакцину, которая экспрессирует антиген, который не экспрессируется в опухоли или мышью, или (b) ДНК раковый вакцинный конструкт, который экспрессирует крысиный антиген Her-2 из SEQ ID NO: 54 (rHER2), который экспрессируется в опухоли или мышью. Скорость роста опухоли анализировали путем измерения длинного (а) и короткого диметра (b) подкожных опухолей TUBO дважды в неделю и рассчитывали объем как «а×b2x 0,5 мм3». Среднее значение и стандартную ошибку среднего объема опухолей от 10 мышей из каждой группы наносили на график в зависимости от дней после имплантации опухоли.
Результаты. На Фигуре 32 показаны результаты репрезентативного исследования, которое оценивает и сравнивает скорость роста подкожной опухоли с момента лечения малатом сунитиниба в качестве монотерапии или в комбинации с противораковой ДНК-вакциной. Не смотря на то, что опухоли от мышей, которые получали противораковую ДНК-вакцину (rHER2: внутримышечная инъекция 109 V.P. аденовируса, экспрессирующего rHer-2, после двух активаций ДНК, экспрессирующей rHer-2, дважды в неделю с помощью PMED), продолжали быстро расти, опухоли от мышей, которые получали малат сунетиниба в дозах 20 мг/к или 40 мг/к с контрольными вакцинами (контроль: внутримышечная инъекция 109 V.P. аденовируса, экспрессирующего eGFP, после двух активаций ДНК, экспрессирующей коровый антиген HBV, дважды в неделю с помощью PMED) значительно снизили скорость роста опухоли с дозой 20 мг/кг, показывающей субоптимальную эффективность, по сравнению с дозой 40 мг/кг. В то же время, когда противоопухолевую вакцину вводили совместно с субоптимальной дозой малата сунитиниба, составляющей 20 мг/кг, опухоли росли с более медленной скоростью, чем у мышей, лечившихся той же дозой малата сунитиниба, введенного совместно с контрольной вакциной, и подобно тем мышам, которые лечились малатом сунитиниба в более высоких дозах. Противоопухолевая вакцина обеспечивает дополнительный терапевтический эффект у мышей, получивших субоптимальные дозы малата сунитиниба.
На Фигурах 33А-33D показаны скорости роста отдельных опухолей у мышей из репрезентативного исследования, которое оценивает и сравнивает противоопухолевую эффективность малата сунитиниба в дозе 20 мг/кг с контролем (контроль) или в противораковой ДНК-вакциной (rHER2). Кратко, через 7 дней после имплантации опухолевых клеток, десяти мышам в группе лечения перорально вводили носитель или 20 мг/кг малата сунитиниба (Сутент) в течение 34 дней. Через три дня после начала дозирования Сутента осуществляли серии иммунизаций, примированных аденовирусом согласно PMED, которые продолжались после прекращения терапии Сутентом. В частности мышей иммунизировали контрольной вакциной, состоящей из внутримышечной инъекции 109 V.P. аденовируса, экспрессирующего eGFP, далее с последующими двумя активациями дважды в неделю, двумя активациями каждые 9 дней и четырьмя активациями еженедельно ДНК, экспрессирующей коровые и поверхностные антигены HBV, путем PMED, или противораковой вакциной, состоящей из аденовируса, экспрессирующего rHer-2, взамен ДНК. Опухоли животных, которые получали носитель с контрольной вакциной, становились измеряемыми приблизительно на 7 день и продолжали расти, достигнув 2000 мм3 через 50 дней после имплантирования опухоли. Рост опухоли у мышей, которые получали Сутент с контрольной вакциной, был значительно ослабленным, пока терапия Сутентом не прекратилась. Опухоли демонстрировали быстрый рост непосредственно после прекращения лечения сутентом, большинство достигло 2000 мм3 через 50 дней после имплантации опухоли. Рост опухоли у мышей, которые получали только протираковые вакцинации, был умеренно ниже, чем у мышей, которые не получали противоопухолевой вакцины или Сутента. Комбинация противоопухолевой вакцины и Сутента не только угнетала рост опухоли во время терапии сутентом (Фигура 33), но также значительно ухудшала прогрессирование опухоли у 60% животных после прекращения лечения Сутентом.
На Фигуре 34 показана Кривая выживаемости Каплана-Мейера для групп мышей из исследования, описанного на Фигуре 2В, которое оценивает противоопухолевую эффективность Сутента с контролем (контроль) или противораковой вакциной (rHER2). Мышей умерщвляли после того, как объем опухоли достигал 2000 мм3, согласно руководству IAUCUC. Только мыши, лечившиеся Сутентом (в дозе 20 мг/кг) и противораковой вакциной, демонстрировали существенное продление выживаемости по сравнению с ившами, лечившимися носителем или контрольной вакциной, противоопухолевой вакциной без Сутента или Сутентом без противоопухолевой вакцины (*P<0,01 согласно логарифмическому ранговому критерию). На Фигурах 35А-35D показан процент миелоидных супрессорных клеток (Gr1+CD11b+) и Treg, содержащих клетки CD25+CD4+ в периферической крови мышей, лечившихся в ФИГ. 2В. Кратко, РВМС окрашивали и анализировали с помощью проточной цитометрии по поводу экспрессии Gr1, CD11b, CD3, CD4 и CD25 из подчелюсного кровопускания у пяти мышей в каждой группе в день d27 (20 дней после начала лечения Сутентом или носителем). Показана средняя и стандартная ошибка среднего в каждой группе лечения. Статистически значимое снижение процента % миелоидных супрессорных клеток наблюдалось у мышей, которые лечились Сутентом с контролем или с противоопухолевой вакциной, по сравнению с мышами, которые не получали Сутент или противоопухолевую вакцину (носитель + контроль). В то же время, существенно белее низкое количество миелоидных супрессорных клеток наблюдалось у мышей, лечившихся комбинацией Сутента с противоопухолевой вакциной (Сутент + rHER2), по сравнению с мышами, которые лечились противоопухолевой вакциной без Сутента (носитель + rHER2) или Сутентом без противоопухолевой вакцины (Сутент + контроль). Статистически значимое снижение Treg, содержащих CD25+CD4+ Т-клетки в популяции CD4, наблюдалось для Сутента с противораковой вакциной. Эти мыши имели значительно более низкий процент Treg, содержащих CD25+CD4+ Τ-клетки в популяции CD4, у мышей, которые лечились противоопухолевой вакциной без Сутента или Сутентом без противоопухолевой вакцины, в их крови. Значек * указывает Ρ<0,05 согласно параметрическому t-критерию Стьюдента.
На Фигурах 36А-36С показано суммарное количество миелоидных супрессорных клеток (Gr1+CD11b+), Treg (CD4+CD25+Foxp3+) и PD-1+CD8 Т-клетки, выделенные из опухолей мышей. Кратко, мыши получали одноразовую суточную пероральную дозу носителя или сутента в количестве 20 мг/кг через три дня после имплантации клеток TUBO в течение 28 дней. Тех же мышей иммунизировали контрольной вакциной, состоящей из внутримышечной инъекции 109 V.P. аденовируса, экспрессирующего eGFP, далее с последующими двумя активациями дважды в неделю ДНК, экспрессирующей коровый антиген HBV, доставляемой с помощью PMED, или протираковой вакциной, состоящей из аденовируса, экспрессирующего rHer-2, и ДНК. При внутрикожной инъекции 50 мкг CpG (PF-03512676) давали путем PMED-введений в непосредственной близости к правостороннему паховому дренирующему лимфатическому узлу. Через семь дней после второго PMED и CpG-введения у шести мышей из группы лечения забирали отдельные опухоли. Одноклеточную суспензию, полученную из выделенных подкожных опухолей, окрашивали антителами, специфическими к Gr1, Cd11b, CD3, CD8, CD25, FoxP3 и PD-1 и анализировали с помощью проточной цитометрии. Среднее значение и стандартную ошибку среднего суммарного количества специфических клеток, как указано на Фигурах, на мкг опухоли из каждой группы лечения наносили на график (значок ^ указывает Ρ<0,05 согласно параметрическому t-критерию Стьюдента). Несмотря на то, что не наблюдалось существенных отличий в частоте обнаружения иммуносупрессивных Treg или MDSC в опухолях, когда мыши получали противораковую вакцину (А и В), по сравнению с мышами, которые получали контрольную вакцину (А и В), значительное снижение наблюдалось, когда мышей лечили Сутентом (А и В), по сравнению с мышами, которые получали только противораковую вакцину. Снижение PD-1+CD8 Т-клеток также наблюдалось у мышей, которые лечились Сутентом (С), по сравнению с мышами, которые получали только противораковую вакцину (С). Взятые вместе, эти данные демонстрируют, что агенты, которые снижают Treg, MDSC или CD8+PD-1+ Т-клетки, в комбинации с вакциной будут полезными для снижения опухолевой нагрузки у животных, несущих опухоли.
ПРИМЕР 12. ПРОТИВОРАКОВАЯ ЭФФЕКТИВНОСТЬ ВАКЦИНЫ В КОМБИНАЦИИ С СУНИТИНИБОМ И АНТИТЕЛОМ К CTLA-4
Противоопухолевую эффективность противораковой вакцины в комбинации с моноклональным антителом к CTLA-4 (клон 9D9) исследовали у мышей BALB/neuT, несущих подкожные опухоли TUBO.
Процедура исследования. Кратко, десяти мышам в каждой группе ежедневно перорально вводили носитель или малат сунитиниба в дозе 20 мг/кг, начиная с 10-го дня после имплантации опухоли, и до дня 64. Вакцинацию ДНК-конструктами, которые не кодируют антиген (контрольная вакцина) или кодируют крысиный антиген Her-2 из SEQ ID NO: 54 (противораковая вакцина), в виде аденовирусных векторов в день 13, затем следовали две еженедельные иммунизации, две иммунизации раз в две недели и семь еженедельных иммунизации соответствующими антигенами (антигенами HBV или rHer-2) с помощью ДНК. Группам мышей (закрашенный кружок и открытый треугольник), которые лечились моноклональным антителом к мышиному CTLA-4, внутрибрюшинно вводили 250 мкг антитела в день 20, 27, 41, 55, 62, 69, 76, 83, 90 и 97 сразу после инъекций PMED.
Результаты. На Фигуре 37 показана кривая выживаемости Каплана-Мейера для групп мышей из репрезентативного исследования, оценивающего противоопухолевую эффективность сунитиниба и моноклонального антитела к мышиному CTLA-4 (клон 9D9) в комбинации с противораковой вакциной. Увеличение продолжительности жизни наблюдалось у мышей, лечившихся Сутентом с контрольной вакциной (не закрашенный кружок), моноклональным антителом к мышиному CTLA-4 (не закрашенный треугольник) или противораковой вакциной (закрашенный треугольник). Дальнейшее увеличение выживаемости наблюдалось у мышей, лечившихся Сутентом и противораковой вакциной в комбинации с моноклональным антителом к мышиному CTLA-4 (закрашенный кружок). Значения Ρ рассчитывали с помощью логарифмического рангового критерия.
ПРИМЕР 13. СИСТЕМНОЕ ВОЗДЕЙСТВИЕ СУНИТИНИБА И ПРОТИВОРАКОВАЯ ЭФФЕКТИВНОСТЬ ПРОТИВОРАКОВОЙ ВАКЦИНЫ В КОМБИНАЦИИ С НИЗКОЙ ДОЗОЙ СУНИТИНИБА
Исследование системного воздействия Сунитиниба.
Кинетику сунитиниба в крови исследовали у мышей BALB/neuT с подкожными опухолями TUBO. Кратко, 20 мышам в каждой группе лечения давали Сутент перорально в дозе 20 мг кг один раз в день (SID) или 10 мг/кг два раза в день (BID) с 6-ти часовыми интервалами через 6 дней после имплантации опухоли. Подчелюстную кровь собирали от 2-3 мышей в пробирки с гепарин-литием в разные моменты времени после введения Сутента, дозированного как указано (0, 2, 4, 6, 8, 10, 12 и 24 часа). Надосадочную плазму удаляли из пробирок после центрифугирования при ×1000 оборотах в течение 15 мин., уровни сунитиниба в образцах плазмы измеряли с помощью LC/MS/MS. Среднее значение и стандартная ошибка среднего значения каждой группы в каждый момент времени наносились на график.
Результаты представлены на Фигуре 38. Среднее значение и стандартная ошибка среднего значения каждой группы в каждый момент времени наносились на график. Пунктирная горизонтальная линия обозначает минимальный уровень сунитиниба в крови, 50 нг/мл, который необходим для эффективного ингибирования роста опухоли при монотерапии (Mendel, D., et al.: "In vivo antitumor activity of SU11248, a novel tyrosine kinase inhibitor targeting vascular endothelial growth factor and platelet-derived growth factor receptors: determination of a pharmacokinetic/pharmacodynamic relationship". Clinical Cancer Research, 203, 9:327-337). Как показано, уровни сунитиниба в крови у мышей, которые получали 20 мг/кг SID или 10 мг/кг BID, только поддерживают целевую эффективную дозу 50 нг/мл, которая временно эффективно ингибирует рост опухоли в течение 24 ч. Уровни в крови в группе, получавшей 20 мг/кг SID, достигли максимума выше 50 нг/мл через 2 часа, упали до 50 нг/мл через 6 часов, а кровь очистилась через 12 ч после дозирования Сутента, его уровни в группе, получавшей 10 мг/кг BID, достигли максимума выше 50 нг/мл через 2 часа, но быстро упали ниже 50 нг/мл через 4 часа и достигли своего пика снова через 2 часа после второй дозы. Уровни быстро упали до 50 нг/мл через 4 часа и кровь очистилась через 18 часов после второй дозы. Несмотря на режим дозирования два раза в сутки, животные, получавшие 10 мг/кг, продолжали демонстрировать более низкую продолжительность воздействия на целевую концентрацию, чем животные, получавшие 20 мг/кг в режиме дозирования раз в сутки.
Исследование противоопухолевой эффективности.
Противоопухолевую эффективность длительного применения низкой дозы сунитиниба в комбинации с противораковой вакциной исследовали у мышей BALB/neuT с подкожными опухолями TUBO. Кратко, мыши получали малат сунитиниба (Сутент) в течение 31 дня в дозе 20 мг/кг SID или в течение 104 дней в дозе 10 мг/кг BID и получали контрольную вакцину или противораковую вакцину. Контрольную вакцину, которая не доставляет антиген, и противораковую вакцину, которая доставляет крысиный антиген Her-2 (rHer-2) из SEQ ID NO: 54, вводили с помощью аденовируса в день 9, затем следуют пять введений ДНК один раз в две недели, доставляющие с помощью PMED антигены HBV или rHer-2, соответственно.
Результаты представлены на Фигуре 39. Помимо того, что противораковая вакцина улучшила выживаемость мышей, получавших Сутент в дозе 20 мг/кг, даже имелось значительное улучшение выживаемости мышей, получавших Сутент в дозе 10 мг/кг (*P=0,05, рассчитанное с помощью логарифмического рангового критерия).
ПРИМЕР 14. ВОЗДЕЙСТВИЕ АГОНИСТА CPG ИЛИ CD40 НА ИММУННЫЕ РЕАКЦИИ, ВЫЗВАННЫЕ ПРОТИВОРАКОВОЙ ВАКЦИНОЙ
Исследование иммуногенности у мышей BALB/с
Воздействие местного введения иммунных модуляторов на амплитуду и качество антиген-специфических иммунных реакций, вызванных раком, исследовали у мышей BALB/с, у которых иммунную реакцию оценивали, измеряя гНЕР2-специфические Т-клеточные реакции, используя анализ IFN-y ELISPOT или анализ окрашивания внутриклеточных цитокинов. Кратко, от 4 до 6 самок мышей BALB/с в группе, как указано, иммунизировали экспрессионным ДНК-плазмидным конструктом, кодирующим антигенную последовательность rHER2 (SEQ ID NO: 54), с помощью системы доставки PMED. Иммунные модуляторы, CpG7909 (PF-03512676) и CD40 моноклональное агонистичное антитело к CD40, вводили местно внутрикожными инъекциями в непосредственной близости к вакцинодренирующему лимфатическому узлу сразу после PMED-активаций. Антиген-специфические Т-клеточные реакции измеряли с помощью IFN-y ELISPOT или анализа окрашивания внутриклеточных цитокинов в соответствии с процедурой, описанной ниже.
Анализ окрашивания внутриклеточных цитокинов (ICS)
rHer-2-специфические полифункциональные (мультицитокинопозитивные) Т-клеточные иммунные ответы измеряли у спленоцитов или РВМС, выделенных от отдельных животных, с помощью анализа ICS. Обычно 106 спленоцитов инкубировали с брефелдином А в концентрации 1 мкг/мл и пептидным стимулятором (rHer-2-специфический CD8 рбб, rHer-2-специфический CD4 р169 или иррелевантный HBV р87) в концентрации 10 нг/мл в течение 5 часов при 37°C в 5% CO2 инкубаторе. После стимуляции спленоциты промывали и блокировали Fey-блоком (антимышиный CD16/CD32) в течение 10 мин. при 4°C с последующим 20-минутным окрашиванием водным красителем «живой/мертвый», антимышиным CD3ePE-Cy7, антимышиным CD8a тихоокеанским голубым и антимышиным CD45R/В220 РеrСР-Су5.5. Клетки промывали, фиксировали в 4% параформальдегиде в течение ночи при 4°C, пермеабилизировали с BD фикс./перм. Раствором в течение 30 мин. при комнатной температуре и инкубировали с антимышиным IFN-y APC, антимышиным TNFa Alexa488 и антимышиным IL-2 РЕ в течение 30 минут при комнатной температуре. Клетки промывали и 20000 CD4 или CD8 Τ клеток отобрали для анализа с помощью проточной цитометрии. Общее количество антиген-специфических одно-, двух- или трехцитокиноположительных Т-клеток в селезенке каждого животного подсчитывали путем вычитания rHer-2-специфических реакций на иррелевантный пептид HBV от вакциноспецифических реакций и нормализовали к общему количеству спленоцитов, выделенных из селезенки.
Результаты анализа IFN-y ELISPOT
На Фигуре 40 показаны результаты IFN-y ELISPOT групп мышей из репрезентативного исследования, оценивающего амплитуду антиген-специфических Т-клеточных реакций, вызванных rHER2-вакциной, вводимой с иммунными модулятороми, как показано. Кратко, каждую мышь в группе лечения (n=4) иммунизировали ДНК-плазмидными экспрессионными конструктами, кодирующими антигенные последовательности rHER2 (SEQ ID NO: 54), с помощью PMED, после чего сразу следует введение 100 мкг контрольного крысиного IgG моноклонального антитела (Bioxcell # ВЕ0089: контрольное mAb) или 50 мкг CpG7909 или 100 мкг моноклонального антитела к CD40 (Bioxcell #ВЕ0016-2: a-CD40 mAb), как показано. Антиген-специфические иммунные реакции измеряли с помощью анализа IFN-y ELISPOT у 505 спленоцитов, смешанных с контрольным или rHer-2-специфическим рбб пептидами в концентрации 10 мкг 7 через 7 дней после активизации PMED. Общее количество клеток, секретирующих IFN-y, из спленоцитов, содержащих 105 CD8 Т-клеток, были подсчитано по результатам ELISPOT от отдельных животных, а % от CD8 Т-клеток в спленоцитах и среднее значение и стандартная средняя погрешность каждой группы нанесены на график. Как показано, CpG7909 и моноклональное антитело к CD40 значительно повышали амплитуду антиген-специфических иммунных реакций, вызванных rHer-2 DNA, по сравнению с мышами, которые получали контрольные антитела.
Результаты анализа окрашивания внутриклеточных цитокинов (ICS). На Фигурах 41А и 41В представлены результаты репрезентативного исследования, которое оценивает иммуномодуляторную активность CpG 7909 по качеству вакциноиндуцированных иммунных реакции с помощью анализа окрашивания внутриклеточных цитокинов. Кратко, каждое животное иммунизировали дважды ДНК-плазмидными экспрессионными конструктами, кодирующими антигенную последовательность rHER2 (SEQ ID NO: 54), доставляемыми PMED с 4-недельным интервалом. Мышам в каждой группе (n=5) вводили внутрикожные инъекции PBS (группа PMED) или 50 мкг CpG 7909 (группа PMED+CpG) в непосредственной близости к правому вакцинодренирующему лимфатическому узлу сразу после двух ДНК-иммунизаций с помощью PMED. Через семь дней после последней иммунизации с помощью PMED, анализ ICS осуществляли с спленоцитами, выделенными от каждой мыши, для обнаружения антиген-специфических полифункциональных CD8 или CD4 Т-клеток, которые секретируют IFN-y, TNFa и/или IL-2. Значительное повышение rHer-2-специфических мультицитокиноположительных CD8 и CD4 Т-клеточных реакций обнаружили у мышей, лечившихся местной доставкой CpG 7909 по сравнению с PBS. Повышение в одноцитокиноположительной CD8 популяции наблюдалось у животных, которые получали местное введение CpG7909, по сравнению с PBS (указывает Ρ<0,05 согласно параметрическому t-критерию Стьюдента).
На Фигурах 42А и 42В представлены результаты репрезентативного исследования, которое оценивает иммуномодуляторную активность агонистического моноклонального антитела к CD40 по качеству вакциноиндуцированных иммунных реакции с помощью анализа окрашивания внутриклеточных цитокинов. Кратко, каждое животное иммунизировали дважды ДНК-плазмидными экспрессионными конструктами, кодирующими антигенные последовательности rHER2 (SEQ ID NO: 54) доставляемыми PMED с 4-недельным интервалом. Мышам в каждой группе (n=6) вводили внутрикожные инъекции в дозе 100 мкг контрольного изотипа IgG (PMED с IgG) или моноклонально антитела к CD40 (PMED с aCD40) в непосредственной близости к правому вакцинодренирующему лимфатическому узлу через день после первой иммунизации, введенной с помощью PMED. Через семь дней после последней иммунизации PMED анализ ICS осуществляли с спленоцитами, выделенными от каждой мыши, для обнаружения rHer-2-специфических полифункциональных CD8 или CD4 Т-клеток, которые секретируют IFN-y, TNFa и/или IL-2. Значительное повышение rHer-2-специфический трицитокинопозитивных CD8 и CD4 Т-клеточных реакций обнаружили у мышей, лечившихся местной доставкой моноклонально антитела к CD40 по сравнению с контрольным изотипом IgG. Имелись также значительные повышения rHer-2-специфических одно- и двухцитокинопозитивных CD4 Т-клеток в ответ на моноклональное антитело к CD40, введенное местно (указывает Ρ<0,05, согласно параметрическому t-критерию Стьюдента).
ПРИМЕР 15. ПРОТИВОРАКОВАЯ ЭФФЕКТИВНОСТЬ ПРОТИВОРАКОВОЙ ВАКЦИНЫ В КОМБИНАЦИИ С НИЗКОЙ ДОЗОЙ СУНИТИНИБА
Противоопухолевую эффективность противораковой вакцины в комбинации с низкой дозой сунитиниба исследовали у мышей BALB/neuT с спонтанными опухолями молочной железы.
Лечение животных. Кратко, 13-14-недельным самкам мышей перорально давали малат сунитиниба (Сутент) а дозе 5 мкг/кг в течение 112 дней дважды в сутки. Контрольную вакцину, которая не доставляет антиген, и противораковую вакцину, которая доставляет крысиный антиген Неr-2 из SEQ ID NO: 54 (rНеr-2), вводили с помощью аденовирусных инъекций в день 3 в качестве прайма с последующими 7 введениями два раз в неделю ДНК, доставляющей антигены HBV (контрольная вакцина) или rHer-2 (противораковая вакцина), соответственно, с помощью PMED. Конечную точку выживаемости определяли, когда все десять молочных желез становились опухолепозитивными или когда объем опухолей достигал 2000 мм3. Результаты представлены на Фигуре 43.
Результаты. По сравнению с ранее опубликованным фармакокинетическим профилем Сутента (Mendel, D., Laird, D., et al.: "In vivo antitumor activity of SU1 1248, a novel tyrosine kinase inhibitor targeting vascular endothelial growth factor and platelet-derived growth factor receptors: determination of a pharmacokinetic/pharmacodynamic relationship". Clinical Cancer Research, 203, 9:327-337) и предыдущими данными (Фигура 38), СМах Сутента у мышей, получавших дважды в сутки по 5 мг/кг, как ожидалось, значительно ниже минимальных уровней в крови, необходимых для достижения эффективного противоопухолевого действия у мышей и человека. Данные показывают быстрое и временное улучшение выживаемости мышей, лечившихся монотерапией низкой дозой Сутента, тогда как при введении с противораковой вакциной наблюдалось более стойкое и значительное улучшение выживания (Р<0,0001, согласно логарифмическому ранговому критерию).
ПРИМЕР 16. ПОВЫШЕНИЕ ВЫЗВАННЫХ ВАКЦИНОЙ ИММУННЫХ РЕАКЦИЙ ПРИ МЕСТНОМ ВВЕДЕНИИ CPG
Повышение иммунных реакций, вызванных нуклеиновой кислотой человеческого PSMA, предусмотренной изобретением, при местном введении CpG (PF-03512676) изучали в исследовании с обезьянами, у которых иммунную реакцию оценивали, измеряя PSMA-специфические Т-клеточные реакции, используя анализ IFN-y ELISPOT.
Лечение животных и сбор образцов. Шесть групп китайских макак крабоедов, по шесть (# 1 до 6) в каждой тестируемой группе, иммунизировали плазмидной ДНК, кодирующей модифицированный антиген человеческого PSMA (аминокислоты 15-750 из SEQ ID NO: 1), доставляемой электропорацией. Кратко, все животные получили билатеральные внутримышечные инъекции по 5 сг плазмидной ДНК с последующей электропорацией (DNA ЕР) в день 0. Сразу после электропорации группа 2 получила билатеральные внутримышечные инъекции по 2 мг CpG, смешанного с 1 мг квасцами в непосредственной близости к месту введения ДНК. Группа 3 и 4 получила билатеральные внутримышечные инъекции по 2 мг CpG, доставленного без квасцов, в непосредственной близости к месту введения ДНК в день 0 или день 3, соответственно. Группа 5 получила билатеральные внутрикожные инъекции по 2 мг CpG, доставленные в непосредственной близости к вакцинодренирующим лимфатическим узлам в день 3. Группа 6 получила билатеральные инъекции по 200 мкг CpG, смешанного с раствором ДНК, которые совместно ввели электропорацией в мышцу в день 0.
Процедура анализа IFN-y ELISPOT. Образцы периферической крови брали у каждого животного через пятнадцать дней после иммунизации ДНК. Мононуклеарные клетки периферической крови (РВМС) выделяли из образцов крови и подвергли анализу IFN-y ELISPOT для измерения PSMA-специфических Т-клеточных реакций. Кратко, по 405 РВМС от каждого животного помещали в лунки с пулом PSMA-специфических пептидов или неспецифических контрольных пептидов (пептидный пул человеческого HER2), каждый в дозе 2 мкг/мл, в планшеты для анализа IFN-y ELISPOT. Композиция каждого PSMA-специфического пептидного пула предоставлена в Таблице 1А. Планшеты инкубировали в течение 16 часов при 37°C и 5% CO2, промывали и разрабатывали после инкубации в соответствии с инструкцией производителя. Количество IFN-y - пятнообразующих клеток (SFC) подсчитывали с помощью ридера CTL. Каждое условие осуществляли в двух повторностях. Результат репрезентативного эксперимента представлен в Таблице 1В. Зарегистрированную PSMA-специфическую реакцию рассчитывали путем вычитания среднего количества SFC с неспецифическими контрольными пептидами (пептидный пул человеческого HER2) от среднего количества SFC пулов пептидов PSMA и нормировали к SFC, наблюдаемым с 106 РВМС. Значок А указывает, что количество не является точным, потому что количество пятен было слишком большим для подсчета. НО означает «не определяли».
Результаты. Таблица 28 показывает результат репрезентативного анализа IFN-y ELISPOT, который оценивает и сравнивает IFN-y Т-клеточные реакции, вызванные вакциной без CpG (группа 1) и с CpG (PF-03512676), введенного местно внутримышечными (группы 2, 3, 4 и 5) и внутрикожными инъекциями (группа 6). Результаты Таблицы 1В представлены на Фигуре 1. Как показано в Таблице 1В и на ФИГ. 1, PSMA-специфические IFN-y Т-клеточные реакции были обнаружены на несколько PSMA-специфических пептидных пулов в отсутствие CpG (PF-03512676) у всех шести животных (группа 1). Суммарный измеренный ответ на пептиды PSMA был незначительно выше у нескольких животных, которые дополнительно получали CpG (PF-03512676) внутримышечными (группа 4: 3/6) или внутрикожными (группа 5: 2/6) инъекциями, через 3 дня после электропорации ДНК. Однако, когда CpG доставляли сразу после электропорации в тот же день (группы 2 и 3), было несколько животных, которые не испытывали высоких ответов (группа 2: 4/6 и группа 3: 3/6), будь то смесь с квасцами или нет. В то же время, более высокие реакции были обнаружены у 4/6 животных, когда десятикратно более низкую дозу CpG вводили совместной электропорацией с раствором ДНК в мышцу (группа 6) со статистически более высоким ответом (Р<0,05) на пептидные пулы Н1 и R1 по сравнению с животными, которые не получали CpG (группа 1). Данные показывают, что низкая доза CpG может эффективно повысить IFN-Y Т-клеточные реакции, вызванные ДНК-вакциной, когда вводится путем совместной электропорации в мышцы.


ПРИМЕР 17. ПОВЫШЕНИЕ ВЫЗВАННЫХ ВАКЦИНОЙ ИММУННЫХ РЕАКЦИЙ ПРИ МЕСТНОМ ВВЕДЕНИИ АНТИТЕЛА К CTLA-4
Действие низкой дозы подкожного введения моноклонального антитела к CTLA-4 (СР-675, 206) на иммунные реакции, вызванные нуклеиновой кислотой PSMA макака резус, изучали в исследовании с обезьянами, у которых иммунную реакцию оценивали, измеряя PSMA-специфические Т-клеточные реакции, используя анализ IFN-y ELISPOT. Нуклеиновая кислота PSMA макака резус, использованная в исследовании, имеет последовательность, указанную в SEQ ID NO: 56, и кодирует полипептид иммуногенного PSMA из SEQ ID NO: 55.
Лечение животных и сбор образцов. Пять групп самок индийского макак резус, по семь (# 1 до 7) в каждой тестируемой группе, иммунизировали аденовирусом, кодирующим модифицированный полипептид PSMA макака резус, доставляемым билатеральными внутримышечными (2×510 V.P.), сразу после аденовирусных инъекций группа 1 получала носитель, а группы 2-4 получали подкожные билатеральные инъекции антитела к CTLA-4 (СР-675, 206) в дозах 2×25 мг, 2×16,7 мг и 2×8,4 мг, соответственно, в непосредственной близости от вакцинодренирующего лимфатического узла.
Через девять дней после иммунизации мононуклеарные клетки периферической крови (РВМС) брали у каждого животного и подвергли анализу IFN-y ELISPOT для измерения PSMA-специфических Т-клеточные реакции. Кратко, 405 РВМС от каждого животного помещали в лунки с пулом PSMA-специфических пептидов (Р1, Р2, Р3 или R1+R2, определенных в Таблице 24А) или неспецифическими контрольными пептидами (пептидный пул человеческого HER2), каждый по 2 мкг/мл, в планшеты для IFN-y ELISPOT. Планшеты инкубировали в течение 16 часов при 37°C и 5% CO2, промывали и разрабатывали после инкубации в соответствии с инструкцией производителя. Количество IFN-y - пятнообразующих клеток (SFC) подсчитывали с помощью ридера CTL. Каждое условие осуществляли в двух повторностях. Среднее двух повторностей на фоне кучных SFC PSMA-специфических пептидных пулов нормализовали к ответу 106 РВМС. Индивидуальный и суммарные ответы на пептидные пулы от каждого отдельного животного представлены в Таблице 29.
Процедура анализа IFN-y ELISPOT. Иммобилизированные антитела, специфические к IFN-y (BD Bioscience, #51-2525kc), наносили на поливинилиденфторидную (PVDF) мембрану в микропланшете в течение ночи при 4°С.Планшет блокировали сывороткой / белком, чтобы предотвратить неспецифическое связывание с антителом. После блокирования эффекторные клетки (такие как спленоциты, выделенные от иммунизированных мышей, или РВМС, выделенные от макак резус) и мишени (такие как пептиды PSMA из пептидной библиотеки, клетки-мишени, активированные антиген-специфическими пептидами, или опухолевые клетки, экспрессирующие релевантные антигены) добавляли в лунки, инкубировали в течение ночи при 37°C в 5% CO2 инкубаторе. Цитокины, секретируемый эффекторными клетками, захватывались антителом покрытия на поверхности PVDF-мембраны. После удаления клеток и культуральной среды, по 100 мкл биотинилированного поликлонального антитела к человеческому IFNy добавили в каждую лунку для определения. Пятна визуализировали добавлением стрептавидин-пероксидазы хрена и преципитата субстрата, 3-амино-9-этилкарбазола (АЕС), для получения красного цвета пятна, как в протоколе производителя (Mabtech). Каждое пятно представляет собой одну Т-клетку, продуцирующую цитокины.
Результаты. Таблица 29. показывает результаты репрезентативного анализа IFN-у ELISPOT, который сравнивает Т-клеточные реакции, вызванные вакциной без моноклонального антитела (группа 1) или с моноклональным антителом к CTLA-4 (СР-675, 206) (группы 2-4), введенной местно подкожными инъекциями в непосредственной близости от вакцинодренирующего лимфатического узла. Вакцина генерировала иммунную реакцию (группа 1), который значительно повышался местным введением антитела к CTLA-4 (СР-675, 206) в дозе 50 мг (группа 2, Ρ=0,001 по Т-критерию Стьюдента с использованием заниженных значений). Реакция также значительно повышалась благодаря малым дозам антитела к CTLA-4, составляющим 33,4 мг (группа 3: Ρ=0,004 по Т-критерию Стьюдента с использованием заниженных значений) и 16,7 мг (группа 4: Ρ=0,05 по Т-критерию Стьюдента), соответственно. Данные свидетельствуют о том, что низкие дозы антитела к CTLA-4, доставляемые в виде подкожных инъекций, могут значительно повысить вызванные вакциной иммунные реакции.

ПРИМЕР 18. ИММУНОМОДУЛИРОВАНИЕ МИЕЛОИДНЫХ СУПРЕССОРНЫХ КЛЕТОК НИЗКИМИ ДОЗАМИ СУНИТИНИБА
Следующий пример предусмотрен для иллюстрации иммуномодулирующего действия низкой дозы сунитиниба на миелоидные супрессорные клетки (MDSC) in vivo в неопухолевой мышиной модели.
Процедура исследования. Чтобы получить обогащенные MDSC спленоциты, клетки TUBO (1×106) имплантировали в бока 5 мышам 5 BALB/neuT и оставили на, приблизительно, 20-30 дней, пока объем опухоли не достиг 1000-1500 мм3. Мышей затем умерщвляли, удаляли селезенки и выделяли спленоциты, обогащенные MDSC. Спленоциты метили в течение 10 минут 5 мкМ CFSE, промывали с PBS и подсчитывали. Меченые клетки впоследствии ресуспендировали по 2×107 спленоцитов/мл в растворе PBS и через капельницу инъекцией в хвостовую вену адаптивно перенесли интактным реципиентным мышам BALB/с. За три дня до адаптивного переноса мышам-реципиентам стали дважды в сутки вводить носитель или малат сунитиниба (Сутент) в дозе 5 мг/кг, 10 мг/кг и 20 мг/кг. После адоптивного переноса, мыши-реципиенты продолжали получать бисуточную дозу носителя или сунитиниба еще в течение двух дней, после чего мышей умерщвляли, селезенки удаляли, спленоциты выделяли и обрабатывали для фенотипического анализа.
Спленоциты подсчитывали и ресуспендировали по 2×106 клеток/мл в окрашивающем буфере FACS (PBS, 0,2% (вес/объем) бычьего сывороточного альбумина и 0,02% (вес/объем) азида натрия). Для окрашивания спленоцитов проточной цитометрией, 2,2×106 клеток сначала инкубировали с антителами к CD16/CD32 в течение 10 минут при 4°C, чтобы блокировать рецепторы Fe и свести к минимуму неспецифическое связывание. Спленоциты затем окрашивали в течение 20 минут при 4°C соответствующими флуорофорными конъюгированными антителами (BioLegend) к мышиным маркерам клеточных поверхностей. Для Т-клеток (анти-CD3 (Pacific Blue), клон 17А2) и для MDSC (анти-GR-1 (APC), клон RB6-8C5 и анти-CD11b (РеrСр Су5.5), клон М1/70) также был включен краситель «живой/мертвый». После инкубации антитела, окрашенные спленоциты промывали 2 мл в буфере FACS, осаждали центрифугированием и ресуспендировали в 0,2 мл буфера FACS перед сбором данных на проточном цитрометре BD CANTO II. Для контроля влияния сунитиниба или носителя на выживание адаптивно перенесенных MDSC, мы рассчитали процент CFSE+, CD3-, GR1+, CD11b+ в живых синглетных вратах. Затем мы определили количество адаптивно перенесенных MDSC на селезенку путем расчета, какое фактическое количество клеток в процентах репрезентировало общее количество спленоцитов. Данные анализировали с помощью программного обеспечения «FloJo and Graph pad».
Результаты. Данные, представленные в Таблице 31, репрезентируют среднее количество адаптивно перенесенных клеток CSFE+, CD3-, GR1+, CD11b+, регенерированных из селезенки (п=7/группу), через 2 дня после адаптивного переноса из мышей, дважды в сутки получавших носитель или по 5 мг/кг, 10 мг/кг и 20 мг/кг сунитиниба. Данные показывают, что Сунитиниб, дозированный два раза в сутки in vivo обладает иммуномодулирующим действием на MDSC, даже когда вводится в дозе всего лишь 5 мг/кг, приводя к статистически значимому снижению количества, регенерированного по сравнению с контрольной группой, получавшей носитель.

ПРИМЕР 19. ИММУНОГЕННОСТЬ ТРЕХАНТИ ГЕННЫХ АДЕНОВИРУСНОГО КОНСТРУКТА И ДНК-КОНСТРУКТА
Следующий пример предусмотрен для иллюстрации способности тирантигенных вакцинных конструктов (в виде аденовирусного вектора или ДНК-плазмиды), экспрессирующих три антигена PSMA, PSCA и PSA, предусмотренных изобретением, вызывать специфические Т-клеточные реакции на все три закодированных антигена у нечеловекообразных приматов.
Процедура исследования in vivo. Т-клеточную иммуногенность пяти аденовирусных векторов, каждый из которых экспрессирует три антигена (PSMA, PSCA и PSA; Ad-733, Ad-734, Ad-735, Ad-796 и Ad-809), предусмотренные изобретением, сравнивали со смесью трех аденовирусных векторов, каждый из которых экспрессирует только один антиген (PSMA, PSA или PSCA), через 9 дней после примирования. Реакцию на единичный аденовирус, экспрессирующий один антиген (группы 1-3), оценивали, чтобы продемонстрировать специфичность. Кратко, индийских макак резус (n=6 для групп 1 и 3, n=7 для группы 2 и n=8 для групп 4-9) внутримышечно инъецировали в сумме 1011 V.P. с последующими внутрикожными инъекциями антитела к CTLA-4 в дозе 10 мг/кг в тот же день. Через девять дней после инъекций, мононуклеарные клетки периферической крови (PBMCs) выделяли от каждого животного и подвергали анализу IFN-y ELISPOT для измерения PSMA, PSA и PSCA специфических Т-клеточных реакций. Через тринадцать недель после инъекций аденовируса и антитела к CTLA-4, когда Т-клеточные реакции снижаются, обезьяны получали буст-вакцинации ДНК (Группа 1: PSMA, плазмида 5166; Группа 2: PSA, плазмида 5297; Группа 3: PSCA, плазмида 5259; Группа 4: смесь PSMA, PSA и PSCA, плазмиды 5166, 5259 и 5297; Группа 4: плазмида 457; Группа 6: плазмида 458; Группа 7: плазмида 459; Группа 8: плазмида 796 и Группа 9: плазмида 809), доставляемые электропорацией. В общих словах, каждое животное получало по 5 мг плазмидной ДНК, предусмотренной изобретением, которая доставляет ту же кассету экспрессии, которая кодируется аденовирусом, использованным для примирования. Через девять после буст-вакцинации мононуклеарные клетки периферической крови (PBMCs) выделяли от каждого животного и подвергали анализу IFN-y ELISPOT.
Анализ IFN-y ELISPOT. Кратко, по 405 РВМС от каждого животного помещали в лунки с пулами PSMA-специфических пептидов Р1, Р2, Р3 или Н1 и Н2 (Таблица 24А), PSA-специфическим пулом 1 или 2 (Таблица 25), PSCA-специфическим пулом (Таблица 26) или неспецифическими контрольными пептидами (пептидный пул человеческого HER2), каждый по 2 мкг/мл, в планшеты для IFN-y ELISPOT. Планшеты инкубировали в течение 16 часов при 37°C и 5% CO2, промывали и разрабатывали после инкубации в соответствии с инструкцией производителя. Количество IFN-y - пятнообразующих клеток (SFC) подсчитывали с помощью ридера CTL. Каждое условие осуществляли в двух повторностях. Среднее двух повторностей из фона, урегулированного SFC из антиген-специфических пептидных пулов, нормировали к ответу в 106 РВМС. Антиген-специфические реакции в таблицах представляют сумму реакций на соответствующие антиген-специфические пептиды или пептидные пулы.
Результаты: Таблица 27 репрезентирует исследование IFN-y ELISPOT, которое оценивает Т-клеточную иммуногенность пяти разных аденовирусов, каждый из которых экспрессирует все три антигена, по сравнению со смесью трех аденовирусов, каждый из которых экспрессирует один антиген у индийских макак резус. У большинства животных, которые получали только инъекции Ad-PSMA (группа 1), были индуцированы специфические реакции на PSMA, но не на PSA или PSCA (Т-критерий Стьюдента, Ρ<0,03). Одно животное (# 4), которое предпочтительно индуцировало реакции на PSCA, удалили из статистического анализа). У животных, которые получали только инъекции Ad-PSA (группа 2), были индуцированы специфические реакции на PSA, но не PSMA или PSCA (Т-критерий Стьюдента, Ρ<0,02). У животных, которые получали только инъекции Ad-PSCA (группа 3), были индуцированы специфические реакции на PSCA, но не на PSMA или PSA (Т-критерий Стьюдента, Ρ<0,03). Все пять триантигеноэкспрессирующих аденовирусных вектора (группы 5-9) индуцировали IFN-y Т-клеточные реакции на все три антигена, амплитуда которых изменялась животным. Амплитуда реакций на PSCA, индуцированных триантигеноэкспрессирующими аденовирусными векторами, была подобной смеси отдельных векторов (группа 4). Однако, амплитуда реакций на PSMA, индуцированных Ad-809 (группа 9), и реакций на PSA, индуцированных Ad-796 (группа 8), значительно превосходила смесь (Т-критерий Стьюдента, Р=0,04 и Р=0,02), соответственно. Эти результаты указывают на то, что вакцинация аденовирусом, экспрессирующим три антигена одновременно, может вызывать Т-клеточные иммунные реакции, которые эквивалентны или превосходят реакции, которые возникают в ответ на вакцинацию смесью отдельных аденовирусов, у нечеловекообразных приматов.
Таблица 28 показывает результаты IFN-y ELISPOT, репрезентирующие исследование, которое оценивает иммуногенность пяти разных трехантигенных экспрессионных кассет, предусмотренных изобретением, доставляемых аденовирусным примированием в комбинации с антителом к CTLA-4 с последующей буст-электропорацией соответствующей плазмидной ДНК. Иммунные реакции сравнивали со смесью трех конструктов, экспрессирующих по одному антигену, доставляемых подобно аденовирусному примированию с антителом к CTLA-4 и буст-электропорационными инъекциям ДНК.
Все животные, которые получали только Ad-PSMA с антителом к CTLA-4 с последующими иммунизациями PSMA-плазмидой (группа 1), вызывали специфические реакции на PSMA, но не на PSA или PSCA. Подобным образом, все животные, которые получали только Ad-PSA с антителом к CTLA-4 с последующими иммунизациями PSA-плазмидой (группа 2), вызывали специфические реакции на PSA, но не на PSMA или PSCA и, наконец, все животные, которые получали только с антителом к CTLA-4 с последующими иммунизациями PSCA-плазмидой (группа 3), вызывали специфические реакции на PSCA, но не на PSMA или PSA (Т-критерий Стьюдента, Ρ<0,01).
Все животные, которых иммунизировали трехантигенными экспрессирующими векторами (группы 5-9) или смесью (группа 4), индуцировали IFN-y Т-клеточные реакции на все три антигена. Частота обнаружения IFN-y Т-клеток, специфических к PSCA или PSA, была похожей во всех трех группах (группы 4-9), соответственно. Однако, группы 7 и 9, которые получали трехантигенные экспрессирующие векторы, продуцировали значительно более высокую частоту реакций на PSMA, чем смесь трех одноантигенных экспрессирующих конструктов (группа 4). Эти результаты указывают на то, что аденовирусная вакцина и ДНК-вакцина, экспрессирующие три антигена одновременно в одной кассете, могут вызывать IFN-y Т-клеточные реакции, которые эквивалентны или превосходят реакции, которые возникают в ответ на смесь аденовирусов и ДНК, экспрессирующих единичные антигены, у нечеловекообразных приматов.









ПРИМЕР 20. СНИЖЕНИЕ ФОСФОРИЛИРОВАНИЯ STAT3 СУНИТИНИБОМ
Следующий пример предусмотрен для иллюстрирования способности сунитиниба непосредственно ингибировать фосфорилирование STAT3 (сигнальный трансдуктор активатора транскрипции 3), ключевого медиатора иммунной супрессии в селезенке.
Процедура исследования. Сильное влияние Сутента на статус фосфорилирования STAT3 в селезенке исследовали в мышиной модели подкожной опухоли для оценки прямого иммуномодулирующего действия соединения. Кратко, 10-12-недельным самкам мышей BALB/neuT имплантировали 106 клеток TUBO подкожно в правый бок. Через сорок один день после имплантации Сутент вводили через желудочный зонд в дозе 20 мг/кг. Через 0, 1, 3, 7 и 24 после введения Сутента, по три животных на временную точку умерщвляли согласно руководству IAUCUC и селезенки сразу же замораживали в жидком азоте, чтобы сохранить статус фосфорилирования. Селезенки самок мышей BALB/c замораживали для использования в качестве здоровых мышиных контролей.
Процедура анализа STAT3. Мгновенно замороженные селезенки гомогенизировали в количестве 100 мг ткани на 500 мл лизисного буфера (70 мМ NaCl, 50 мМ В-глицеролфосфата, 10 мМ HEPES, 1% Triton Х-100, 100 мМ Na3V04, 100 мМ PMSF, 1 мг/мл леупептина), используя политронный гомогенизатор тканей. Полученные гидролезаты центрифугировали в количестве 10000 г в течение 15 минут. Супернатант выделяли, а концентрации белка определяли, используя набор для анализа белка ВСА (Pierce, Rockford, Illinois). По сорок мкг белка добавляли в каждую лунку общего STAT3 (eBioscience, cat no. 85-86101-11) или фосфор-STAT3 (eBioscience, по кат.85-86102-11) из набора ELISA. Относительные уровни белков сравнивали со стандартами, предусмотренными в наборе, и с стандартами, приобретенными самостоятельно (Signaling Technologies, cat no. 9333-S).
Результаты. Таблица 29 показывает результаты репрезентативного анализа STAT, который оценивает действие Сутента на статус фосфорилирования STAT3 в селезенке. Оба экстракта селезенки от здоровых мышей или мышей, несущих опухоль, демонстрировали аналогичные уровни белка STAT3 в анализе ELISA (Всего STAT3). Однако, по сравнению со здоровыми BALB/c, экстракты от мышей, несущих опухоль, имели значительно более высокие уровни фосфорилированного STAT3 (Т-критерий Стьюдента, p<0,001). Уровни фосфорилирования резко сократилось до уровней здоровых животных только через 1 час после обработки Сутентом и поддерживались на более низких уровнях, чем у необработанных мышей до 7 часов. Через 24 часа уровни фосфорилирования STAT3 полностью восстановились до уровней перед лечением Сутентом. Кинетика фосфорилирования отражает уровень циркулирующего Сутента в крови. Быстрая реакция фосфорилирования STAT3 в селезенке, отражающая фармакокинетический профиль Сутента, предполагает прямую иммуномодулирующую функцию Сутента у животных, несущих опухоль.


Групповой перечень последовательностей представлен после формулы изобретения.
Реферат
Изобретение относится к области биотехнологии, конкретно к получению мультиантигенных ДНК-конструктов, векторов экспрессии, содержащих указанные конструкты, и фармацевтических композиций, что может быть использовано в медицине. Получают мультиантигенные ДНК-конструкты для экспрессии иммуногенных полипептидов простатоспецифических антигенов РSА, PSCA, PSMA, которые используют в составе композиции для ингибирования аномальной клеточной пролиферации или индуцирования иммунного ответа против простатоассоциированного антигена у млекопитающих. Изобретение позволяет с высокой эффективностью лечить рак предстательной железы у человека. 7 н. и 17 з.п. ф-лы, 43 ил., 33 табл., 20 пр.
Формула

Документы, цитированные в отчёте о поиске
Выделенное антитело против специфического мембранного антигена простаты (psma) и способ ингибирования роста клеток, экспрессирующих psma
Комментарии