Способы генетического контроля поражения растений насекомыми и применяемые для этого композиции - RU2478710C2
Код документа: RU2478710C2
Чертежи




























































































































































































Описание
Приведение в качестве ссылки списка последовательности, представленного на компакт-диске
Список последовательности представлен на одном компакт-диске (Копия 1), вместе с его дубликатом (Копия 2), каждый из которых создан 15 сентября 2006, и каждый из которых содержит один файл размером 669 КБ, имеющий имя "MNDE002.APP.TXT." Материал, содержащийся на компакт-диске, особым образом приведен здесь в качестве ссылки.
Испрашивание приоритета
Эта заявка испрашивает приоритет Предварительной заявки США Сер. № 60/718034, поданной 16 сентября 2005, которая приведена здесь в качестве ссылки в полном объеме.
Уровень техники
1. Область техники, к которой относится изобретение
Настоящее изобретение относится в основном к генетическому контролю поражения паразитами. А именно, настоящее изобретение относится к технологиям рекомбинантной ДНК для посттранскрипционного репрессинга или ингибирования экспрессии заданных кодирующих последовательностей в клетках паразитов для обеспечения защитного эффекта от паразитов.
2. Описание предшествующего уровня техники
Окружающая среда, в которой живут люди, насыщена паразитами. Паразиты, включая насекомых, паукообразных, ракообразных, грибки, бактерии, вирусы, нематоды, плоских червей, круглых червей, острицы, анкилостомы, ленточных червей, трипаносомы, шистосомы, оводов, блох, зудней, клещей и вшей и т.п. распространены в среде, окружающей человека. В попытке контроля над поражениями этими паразитами применяли множество средств. Композиции для контроля над поражениями микроскопическими паразитами, такими как бактерии, грибки и вирусы, предоставляли в форме антибиотических композиций, противовирусных композиций и противогрибковых композиций. Композиции для контроля над поражениями паразитами больших размеров, такими как нематоды, плоские черви, круглые черви, острицы, сердечные черви, солитеры, трипаносомы, шистосомы и т.п. обычно имели форму химических композиций, которые можно наносить на поверхности, на которых присутствуют паразиты, или вводить пораженным животным в форме гранул, порошков, таблеток, паст или капсул и т.п. В области техники существует большая потребность в улучшении этих способов и особенно в способах, которые будут оказывать благоприятное воздействие на окружающую среду по сравнению с предшествующими способами.
Коммерческие зерновые культуры часто являются объектами поражения насекомыми. В последние несколько десятилетий достигнут существенный прогресс в разработке более эффективных способов и композиций для управления поражением растений насекомыми. Химические пестициды были очень эффективны в ликвидации поражений паразитами. Однако, существует ряд недостатков применения химических пестицидных агентов. Химические пестицидные агенты являются неселективными. При применении химических пестицидов, предназначенных для контроля над беспозвоночными паразитами, такими как жесткокрылые насекомые, включая виды кукурузных жуков, которые губительны для различных зерновых культур и других растений, также оказывается воздействие на фауну, не являющуюся мишенью, что часто эффективно стерилизует область на период времени, в течение которого применяются пестицидные агенты. Химические пестицидные агенты персистируют в окружающей среде и в основном медленно подвергаются метаболизму, если это и имеет место. Они аккумулируются в цепи питания и особенно у видов высших хищников. Аккумулирование этих химических пестицидных агентов приводит к развитию устойчивости к агентам, и у видов, находящихся выше на эволюционной лестнице, агенты могут действовать как мутагены и/или канцерогены, вызывая необратимые и губительные генетические модификации. Таким образом, весьма длительно существует потребность в безвредных для окружающей среды способах контроля или искоренения паразитирования насекомых на или в растениях, т.е. в способах, которые являются селективными, инертными к окружающей среде, неустойчивыми, и биоразлагаемыми, и которые хорошо подходят для схем менеджмента устойчивости паразитов.
Композиции, содержащие бактерии Bacillus thuringiensis (Bt), были коммерчески доступны и применялись в качестве экологически безопасных и приемлемых инсектицидов в течение более тридцати лет. Инсектицидный эффект бактерий Bt не сохраняется в окружающей среде, бактерии высокоселективны в отношении пораженных видов-мишеней, проявляют свои эффекты только при приеме внутрь паразитом-мишенью, и, как показано, являются безопасными для растений и других организмов, для которых они не предназначены, включая людей. Трансгенные растения, содержащие один или более генов, кодирующих инсектицидный белок Bt, также доступны в области техники и заметно эффективны в контроле поражения насекомыми паразитами. Реальным результатом применения рекомбинантных растений, экспрессирующих инсектицидные белки Bt, является заметное уменьшение количества химических пестицидных агентов, которые применяются в окружающей среде для контроля поражения паразитами на полях с сельскохозяйственными культурами в областях, в которых применяют подобные трансгенные зерновые культуры. Уменьшение применения химических пестицидных агентов привело к более чистым почвам и более чистой воде, стекающей из почвы в окружающие ручьи, реки, пруды и озера. В дополнение к этим преимуществам в отношении окружающей среды наблюдалось значительное увеличение числа полезных насекомых на полях с сельскохозяйственными культурами, на которых произрастают трансгенные сельскохозяйственные культуры, устойчивые к насекомым, вследствие уменьшения применения химических инсектицидных агентов.
Антисмысловые способы и композиции описаны в области техники и, как полагают, оказывают эффекты посредством синтеза одноцепочечной молекулы РНК, которая, в теории, гибридизируется in vivo с образованием в основном комплементарной смысловой цепочки молекулы РНК. Антисмысловую технологию было трудно применять во многих системах по трем основным причинам. Во-первых, антисмысловая последовательность, экспрессируемая в трансформированной клетке, является неустойчивой. Во-вторых, неустойчивость антисмысловой последовательности, экспрессируемой в трансформированной клетке, одновременно создает затруднение в доставке последовательности хозяину, типу клетки или биологической системе, находящейся на расстоянии от трансгенной клетки. В-третьих, затруднения, связанные с неустойчивостью и доставкой антисенсорной последовательности, создают затруднения в попытке предоставления в пределах рекомбинантной клетки, экспрессирующей антисенсорную последовательность, дозы, которая может эффективно модулировать уровень экспрессии заданной смысловой нуклеотидной последовательности.
Существуют немногочисленные улучшения в технологиях модулирования уровня экспрессии генов в клетках, тканях или организме, и, в частности, не хватает разработанных технологий для задержки, репрессинга или другого ослабления экспрессии специфичных генов с применением технологии рекомбинантной ДНК. Кроме того, как следствие непредсказуемости этих подходов, не доступны никакие коммерчески приемлемые средства для модулирования уровня экспрессии специфичного гена в эукариотическом или прокариотическом организме.
Ранее демонстрировалась двухцепочечная РНК, опосредующая ингибирование специфичных генов у различных паразитов. Подходы к генетическому контролю, опосредованные дцРНК, тестировали на плодовых мушках Drosophila melanogaster (Kennerdell и Carthew, 1998; Kennerdell и Carthew, 2000). Kennerdell и Carthew (1998) описывают способ доставки дцРНК, включающий создание трансгенных насекомых, которые экспрессируют двухцепочечные молекулы РНК, или введение растворов дцРНК в организм насекомого или в защитную оболочку яйца до, или во время эмбрионального развития.
Ранее исследователи доказали, что супрессии генов, опосредованной двухцепочечной РНК, можно достигнуть у нематод либо с помощью кормления, либо с помощью выдерживания нематод в растворах, содержащих двухцепочечные или малые интерферирующие молекулы РНК, и с помощью введения молекул дцРНК. Rajagopal и др. (2002) описали неудачные попытки супрессии эндогенного гена у личинок насекомого паразита Spodoptera litura с помощью кормления или выдерживания новорожденных личинок в растворах, содержащих дцРНК, специфичный для заданного гена, но успешную супрессию после введения в гемолимфу личинкам 5-й возрастной стадии дцРНК, применяя микроаппликатор. Недавно Yadav и др. (2006) сообщили, что произведенная хозяином дцРНК, образующаяся в растениях, может защищать такие растения от инфицирования нематодами. Точно так же в опубликованной заявке на патент США № 2003/0150017 описан предпочтительный локус ингибирования личинок чешуекрылого Helicoverpa armigera с помощью доставки дцРНК в личинки посредством поедания растения, трансформированного для продукции дцРНК. Согласно положениям WO 2005/110068 в пищевом рационе кукурузного жука (CRW) обеспечивается наличие CRW-специфичной дцРНК, направленной на жизненно-важные гены CRW. дцРНК предоставляется в пищевом рационе CRW in vitro и in planta, что в результате приводит к замедлению роста личинок CRW или их гибели после поедания продуктов из пищевого рациона, и этот эффект был продемонстрирован для нескольких различных генов.
Таким образом, существует потребность в выявлении эффективных нуклеотидных последовательностей для применения в улучшенных способах модуляции экспрессии генов с помощью репрессинга, задержки или другого ослабления экспрессии генов у конкретного жесткокрылого паразита с целью контроля поражения паразитом или для привнесения новых фенотипических признаков.
Сущность изобретения
В одном аспекте изобретение предоставляет способ ингибирования экспрессии гена-мишени у жесткокрылого паразита. В некоторых вариантах осуществления способ включает модуляцию или ингибирования экспрессии одного или более генов-мишеней у жесткокрылого паразита, что вызывает прекращение пищевой активности, роста, развития, репродукции и/или инфекционности и в конечном счете приводит к гибели насекомого. Способ включает введение частично или полностью стабилизированной двухцепочечной РНК (дцРНК), включая ее модифицированные формы, такие как последовательности малой интерферирующей РНК (siРНК), в клетки или во внеклеточное пространство, например в среднюю кишку, в организме жесткокрылого паразита, где дцРНК поступает в клетки и ингибирует экспрессию по крайней мере одного или более генов-мишеней, и при этом ингибирование оказывает губительный эффект на жесткокрылого паразита. Способы и сопутствующие композиции могут применяться для ограничения или устранения паразитирования жесткокрылого паразита в организме или на поверхности любого хозяина паразита, симбионта паразита или окружающей среде, в которой присутствует паразит, с помощью обеспечения наличия в диете паразита одной или более композиций, включающих дцРНК молекулы, описанные здесь. Способ будет иметь в частности преимущество в защите растений от поражения насекомыми. В одном варианте осуществления паразит определяется как имеющий pH пищеварительной системы в пределах диапазона от приблизительно 4,5 до приблизительно 9,5, от приблизительно 5 до приблизительно 9, от приблизительно 6 до приблизительно 8, и от приблизительно 7,0.
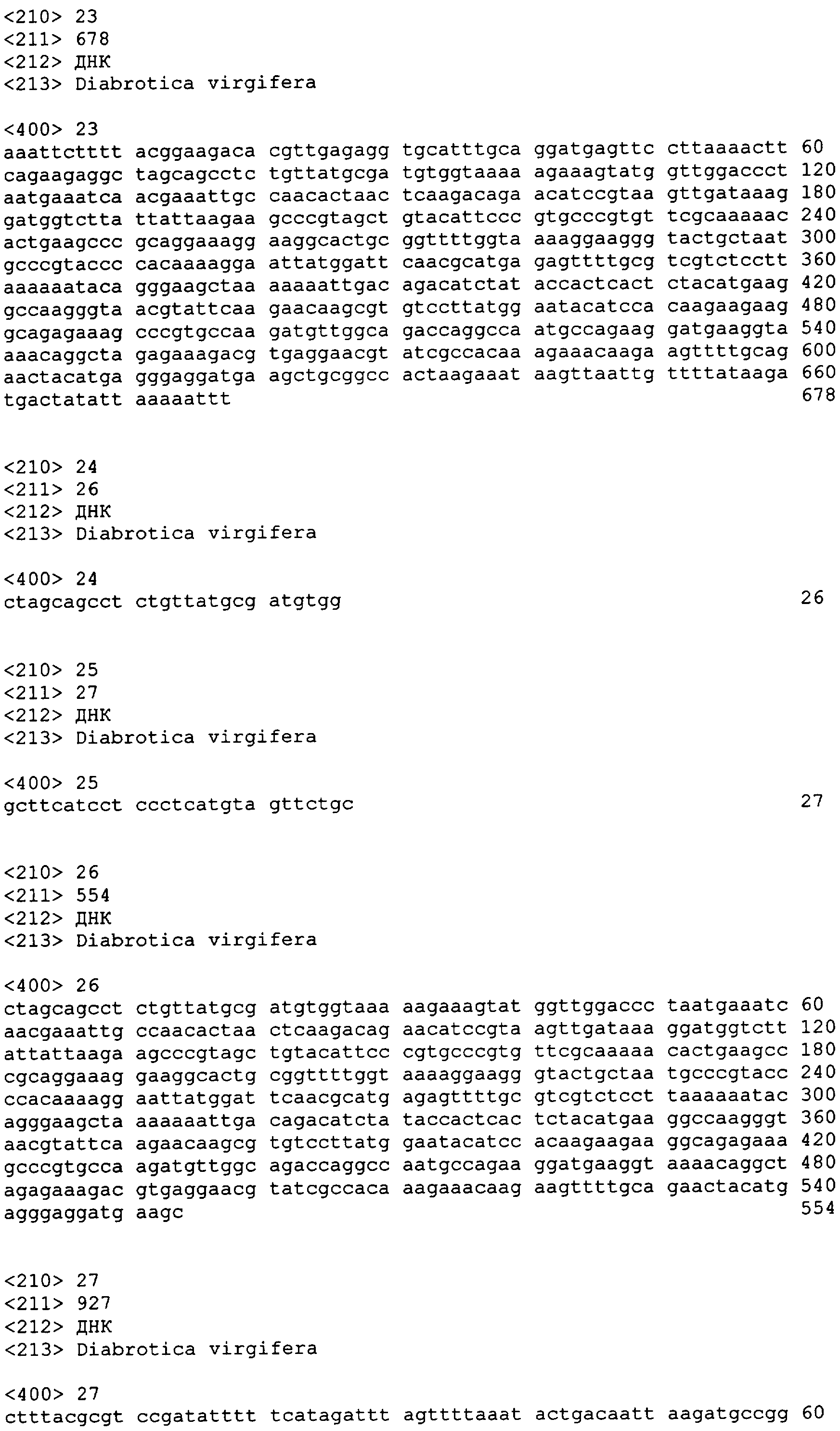
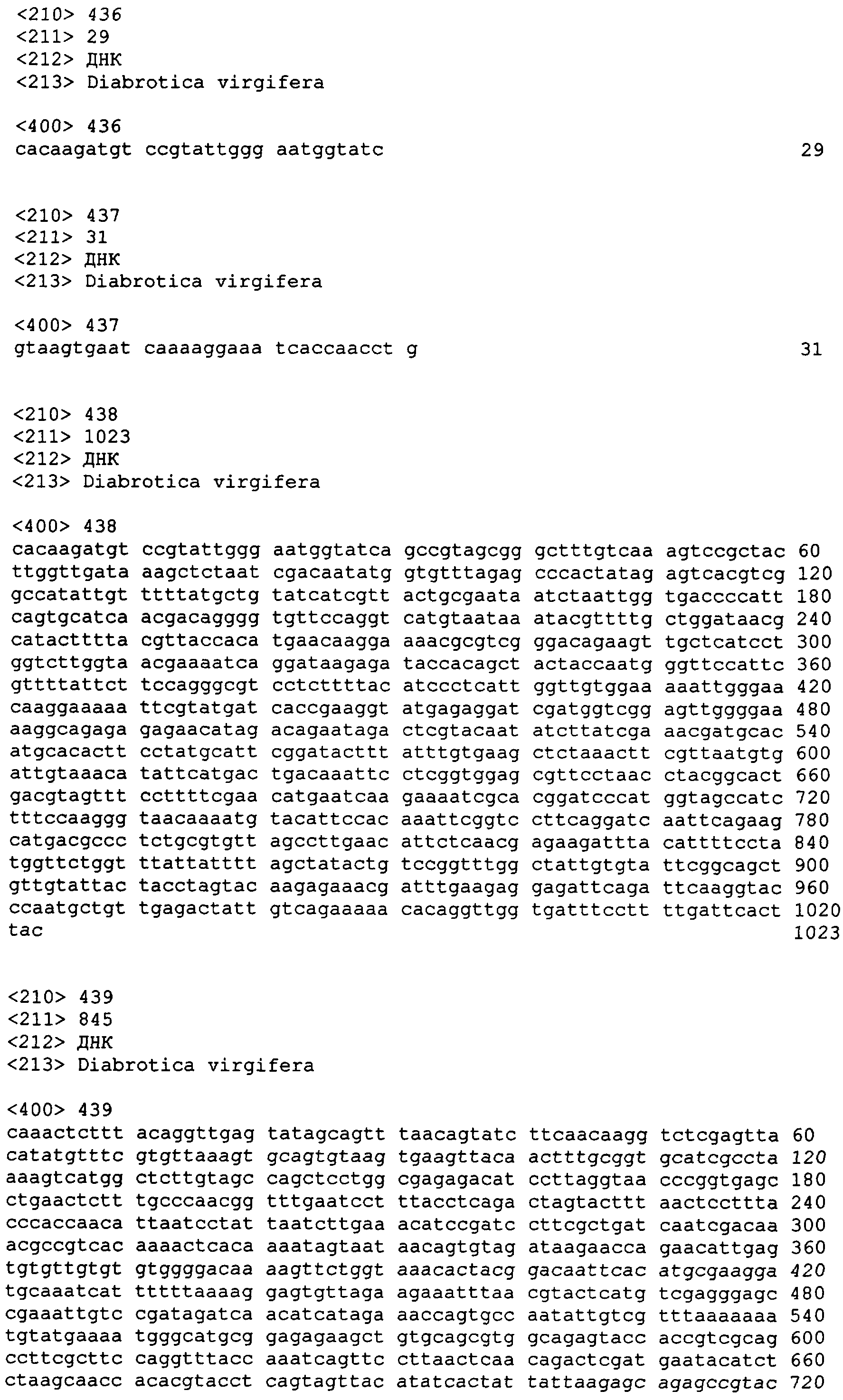
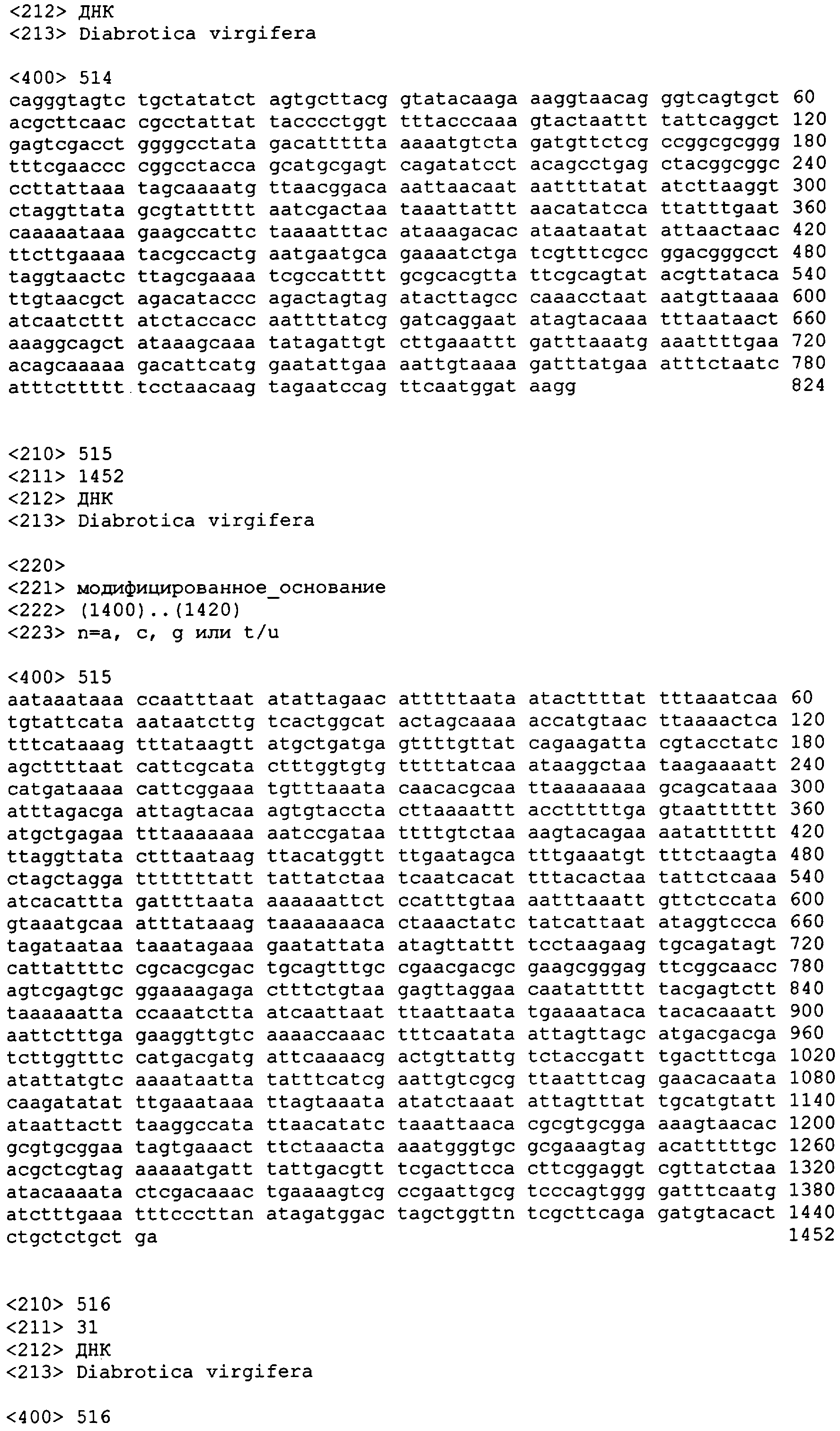
В другом аспекте настоящее изобретение предоставляет примерные композиции нуклеиновой кислоты, которые являются гомологичными по крайней мере части одной или более нативных последовательностей нуклеиновой кислоты у паразита-мишени. В некоторых вариантах осуществления паразит выбран из числа Diabrotica sp., включая западного кукурузного жука (WCR, Diabrotica virgifera или Diabrotica virgifera virgifera), южного кукурузного жука (SCR, Diabrotica undecimpunctata howardi), мексиканского кукурузного жука (MCR, Diabrotica virgifera zeа), бразильского кукурузного жука (BZR, Diabrotica balteata, Diabrotica viridula, Diabrotica speciosa), северного кукурузного жука (NCR, Diabrotica barberi), Diabrotica undecimpunctata; а также картофельного колорадского жука (CPB, Leptinotarsa decemlineata), хрущака каштанового (RFB, Tribolium castaneum) и Мексиканскую зерновку бобовую (Epilachna varivestis). В других вариантах осуществления паразит выбран из числа чешуекрылых насекомых, включая Мотылька кукурузного (ECB, Ostrinia nubilalis], Черную совку (BCW, Agrotis ipsilon), Совку хлопковую (CEW, Helicoverpa zeа), совку травяную (FAW, Spodoptera frugiperda), хлопкового долгоносика (BWV, Anthonomus grandis) тутового шелкопряда (Bombyx mori) и Manduca sexta, и из двукрылых насекомых, включая Drosophila melanogaster, Anopheles gambiae и Aedes aegypti. Специфичные примеры таких нуклеиновых кислот, предоставляемых в соответствии с изобретением, даны в приложенном списке последовательностей с SEQ ID NO:1 по SEQ ID NO:906.
В еще одном аспекте изобретение предоставляет способ супрессии экспрессии гена у жесткокрылого паразита, такого как кукурузный жук или у родственных видов, который включает шаг обеспечения в пищевом рационе паразита в количестве, вызывающем супрессию гена, по крайней мере одной молекулы дцРНК, транскрибированной от нуклеотидной последовательности, как описано здесь, по крайней мере один сегмент которой является комплементарным последовательности мРНК в клетках паразита. Способ может далее включать наблюдение гибели, ингибирования, остановки роста или прекращения пищевой активности паразита. Молекула дцРНК, включая ее модифицированную форму, такую как молекула siРНК, скармливаемая паразиту в соответствии с изобретением, может быть по крайней мере от приблизительно 80, 81, 82, 83, 84, 85, 86, 87, 88 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или приблизительно 100% идентичной молекуле РНК, транскрибированной от нуклеотидной последовательности, выбранной из группы, включающей с SEQ ED NO:1 по SEQ ID NO:906. В отдельных вариантах осуществления нуклеотидная последовательность может быть выбрана из группы, состоящей из SEQ ID NO:697, SEQ ID NO:813-819, SEQ ID NO:841 и SEQ ID NO:874.
Соответственно, в другом аспекте настоящего изобретения предоставляется набор изолированных и очищенных нуклеотидных последовательностей, как указано с SEQ ID NO:1 по SEQ ID NO:906. Настоящее изобретение предоставляет стабилизированную дцРНК молекулу или экспрессию одной или более miРНКs для ингибирования экспрессии гена-мишени у жесткокрылого паразита, экспрессируемого от этих последовательностей и их фрагментов. Стабилизированная молекула дцРНК, включая miРНК или siРНК, может включать по крайней мере две кодирующих последовательности, которые расположены в смысловой и антисмысловой ориентации относительно по крайней мере одного промотора, при этом нуклеотидная последовательность, которая включает смысловую цепь и антисмысловую цепь, связывает или соединяет с помощью спейсерной последовательности по крайней мере от приблизительно пяти до приблизительно одной тысячи нуклеотидов, при этом смысловая цепь и антисмысловая цепь могут быть различной длины, и при этом каждая из двух кодирующих последовательностей имеет по крайней мере 80% идентичность последовательности, по крайней мере 90%, по крайней мере 95%, по крайней мере 98%, или 100% идентичность последовательности любой одной или более нуклеотидной последовательности(ям), указанным с SEQ ID NO:1 по SEQ ID NO:906.
Далее изобретение предоставляет фрагмент или конкатемер последовательности нуклеиновой кислоты, выбранной из группы, включающей от SEQ ID NO:1 до SEQ ID NO:906. В специфических вариантах осуществления нуклеотидная последовательность может включать фрагмент или конкатемер последовательности, выбранной из группы, состоящей из SEQ ID NO:697, SEQ ID NOs:813-819, SEQ ID NO:841, и SEQ ID NO:874.
Фрагмент может быть определен как вызывающий гибель, ингибирование, остановку роста или прекращение пищевой активности паразита при экспрессии в виде дцРНК и предоставлении паразиту. Фрагмент может, например, включать по крайней мере приблизительно 19, 21, 23, 25, 40, 60, 80, 100, 125 или более смежных нуклеотидов из любой одной или более последовательностей с SEQ ID NO:1 по SEQ ID NO:906, или их комплементов. Один полезный сегмент ДНК для применения в настоящем изобретении имеет длину по крайней мере от приблизительно 19 до приблизительно 23, или от приблизительно 23 до приблизительно 100 нуклеотидов, до приблизительно 2000 нуклеотидов или более. Особенно эффективными будут последовательности дцРНК, включающие от приблизительно 23 до приблизительно 300 нуклеотидов, гомологичных заданной последовательности паразита. Изобретение также предоставляет рибонуклеиновую кислоту, экспрессируемую от любой из таких последовательностей, включая дцРНК. Последовательность, выбранная для применения в экспрессии агента супрессии гена, может быть создана из одной последовательности, полученной от одного или более паразитов-мишеней и предназначенной для применения в экспрессии РНК, которая участвует в супрессии одного гена или семейства генов в одном или более паразите-мишени, или эта последовательность ДНК может быть создана в виде химеры из множества последовательностей ДНК.
В еще одном аспекте изобретение предоставляет рекомбинантные конструкты ДНК, включая молекулу нуклеиновой кислоты, кодирующую описанную здесь молекулу дцРНК. дцРНК может быть сформирована с помощью транскрипции одной цепи молекулы дцРНК от нуклеотидной последовательности, которая является по крайней мере от приблизительно 80% до приблизительно 100%, идентичной нуклеотидной последовательности, выбранной из группы, включающей от SEQ ID NO:1 до SEQ ID NO:906. Такие рекомбинантные конструкты ДНК могут быть определены как продуцирующие молекулы дцРНК, способные ингибировать экспрессию эндогенного гена(ов)-мишени в клетке паразита после приема внутрь. Конструкт может включать нуклеотидную последовательность изобретения, операбельно связанную с промоторной последовательностью, которая функционирует в клетке хозяина. Такой промотор может быть тканеспецифичным и может, например, быть специфичным к типу ткани, которая является объектом поражения паразитом. Например, в случае жуков, паразитирующих на корнях растений, может быть желательно применение промотора, обеспечивающего корнеспецифичную экспрессию.
Конструкты нуклеиновой кислоты в соответствии с изобретением могут включать по крайней мере одну нуклеотидную последовательность не природного происхождения, которая может быть транскрибирована в одноцепочечную РНК, способную к образованию молекулы дцРНК in vivo посредством гибридизации. Такие последовательности дцРНК собираются самостоятельно и могут обеспечиваться в пищевом рационе жесткокрылого паразита для достижения желательного ингибирования.
Рекомбинантный конструкт ДНК может включать две различные последовательности не природного происхождения, которые при экспрессии in vivo в виде последовательности дцРНК и нахождении в пищевом рационе жесткокрылого паразита ингибируют экспрессию по крайней мере двух различных генов-мишеней в клетке жесткокрылого паразита. В некоторых вариантах осуществления в клетке или растении, включающем клетку, продуцируются по крайней мере 3, 4, 5, 6, 8 или 10 или более различных дцРНК, которые обладают эффектом ингибирования паразита. дцРНК может экспрессироваться от множества конструктов, включенных в различные объекты трансформации, или может быть включена одиночную молекулу нуклеиновой кислоты. дцРНК могут экспрессироваться с помощью одиночного промотора или множества промоторов. В некоторых вариантах осуществления изобретения продуцируются одиночные дцРНК, которые включают нуклеиновые кислоты, гомологичные множеству локусов у паразита.
В еще одном аспекте изобретение предоставляет рекомбинантное клетки хозяина, имеющие в своем геноме по крайней мере одну рекомбинантную последовательность ДНК, которая транскрибируется для продукции по крайней мере одной молекулы дцРНК, которая после принятия внутрь жесткокрылым паразитом осуществляет ингибирование экспрессии гена-мишени у паразита. Молекула дцРНК может быть закодирована любой из нуклеиновых кислот, описанных здесь и указанных в списке последовательностей. Настоящее изобретение также предоставляет трансформированную клетку растения, имеющую в своем геноме по крайней мере одну описанную здесь рекомбинантную последовательность ДНК. Также предоставляются трансгенные растения, включающие такую трансформированную клетку растения, включая потомство растений любого поколения, семена и продукты растений, каждое из которых включает рекомбинантную ДНК.
Способы и композиции настоящего изобретения могут быть применимы к любому однодольному и двудольному растению, в зависимости от желаемого контроля жесткокрылого паразита. В частности, к растениям относятся, не ограничиваясь ими, люцерна, укроп, яблоко, абрикос, артишок, рукола, спаржа, авокадо, банан, ячмень, бобы, свекла, ежевика, черника, брокколи, брюссельская капуста, капуста кочанная, канола, канталупа, морковь, маниока, цветная капуста, сельдерей, вишня, кинза, цитрусовые, климентины, кофе, кукуруза, хлопок, огурцы, дугласова пихта, баклажан, эндивий, эскариоль, эвкалипт, фенхель, фикус, тыква, виноград, грейпфрут, дыня мускатная белая, хикама, киви, салат-латук, лук-порей, лимон, лайм, сосна ладанная, манго, дыня, грибы, орех, овес, окра, лук, апельсин, декоративные растения, папайя, петрушка, горох, персик, арахис, груша, перец, хурма, сосна, ананас, банан, слива, гранат, тополь, картофель, тыква, айва, сосна лучистая, радиккио, редька, малина, рис, рожь, сорго обыкновенное, южная сосна, соя, шпинат, патиссон, земляника, сахарная свекла, сахарный тростник, подсолнечник, сладкий картофель, амбровое дерево, мандарин, чай, табак, томаты, дерн, виноградная лоза, арбуз, пшеница, ямс и цукини. Таким образом, изобретение также представляет растение, трансформированное с помощью рекомбинантной последовательности ДНК, как указано с SEQ ID NO:1 по SEQ ID NO:906, или ее конкатемера, фрагмента, или комплемента, который транскрибируется для продукции по крайней мере одной молекулы дцРНК, которая после приема внутрь жесткокрылым паразитом приводит к ингибированию экспрессии гена-мишени у паразита. В специфических вариантах осуществления рекомбинантная последовательность ДНК может быть выбрана из группы, состоящей из SEQ ID NO:697, SEQ ID NO:813-819, SEQ ID NO:841, и SEQ ID NO:874, или их фрагмента, комплемента или конкатемера.
Изобретение также предоставляет комбинации способов и композиций для контроля поражения жесткокрылыми паразитами. Тогда как в методах предоставляется способ дцРНК, как описано здесь, для защиты растения от поражения насекомым наряду с одним или более инсектицидными агентами, который обладает свойствами, отличными от свойств, характерных для способов и композиций дцРНК. Например, один или более Bt белков можно включить в пищевой рацион насекомых паразитов в комбинации с одной или более дцРНК, как описано здесь. Композицию, сформулированную для местного нанесения или получаемую с помощью трансгенного подхода, который объединяет способы и композиции дцРНК с Bt, можно применять для обеспечения синергий, которые ранее не были известны в области техники для контроля поражения насекомыми. При этом синергия является сокращением уровня экспрессии, требуемой либо для дцРНК, либо для белка(ов) Bt. При объединении вместе для контроля вредителей может применяться наименьшая эффективная доза каждого агента. Полагают, что инсектицидные белки Bt создают входные поры, через которые молекулы дцРНК способны более эффективно проникать в места, находящиеся на расстоянии от кишечника насекомого-паразита, или более эффективно проникать в клетки, находящиеся вблизи участков повреждения, созданных белками Bt, таким образом, требуется меньшее количество либо Bt, либо дцРНК для достижения желаемого инсектицидного результата или желаемого ингибирования или супрессии заданной биологической функции у паразита-мишени.
Таким образом, настоящее изобретение предоставляет композицию, которая содержит два или более различных пестицидных агента, каждый из которых является токсичным для одного и того же вида паразита или насекомого, по крайней мере один из которых включает описанную здесь дцРНК. В некоторых вариантах осуществления второй агент может быть агентом, выбранным из группы, состоящей из пататина, инсектицидного белка Bacillus thuringiensis, инсектицидного белка Xenorhabdus, инсектицидного белка Photorhabdus, инсектицидного белка Bacillus laterosporous, инсектицидного белка Bacillus sphaericus и лигнина. Инсектицидный белок Bacillus thuringiensis может быть любым из множества инсектицидных белков, включая, но не ограничиваясь ими, Cry1, Cry3, TIC851, CryETTO, Cry22, TIC901, TIC1201, TIC407, TIC417, бинарный инсектицидный белок CryET33 и CryET34, бинарный инсектицидный белок CryETSO и CryET76, бинарный инсектицидный белок TIC100 и TIC101, бинарный инсектицидный белок PS149B1, инсектицидный белок VIP, TIC900 или родственный белок, или комбинации инсектицидных белков ET29 или ET37 с инсектицидными белками TIC810 или TIC812, и инсектицидными химерами любого из вышеуказанных инсектицидных белков.
Рибонуклеиновую кислоту, которую вносят в пищевой рацион, можно вносить в искусственный пищевой субстрат, составленный для обеспечения специфических потребностей в питательных веществах для содержания паразита на таком пищевом субстрате. Пищевой субстрат может быть подкреплен контролирующим паразита количеством РНК, очищенной от системы отдельной экспрессии, для определения количества композиции РНК, контролирующего паразита, или определения степени супрессивной активности после приема внутрь подкрепленного пищевого субстрата паразитом. Пищевой субстрат может также быть рекомбинантной клеткой, преобразованной с помощью последовательности ДНК, созданной для экспрессии агента, РНК, или агента супрессии гена. После приема внутрь паразитом одной или более таких трансформированных клеток наблюдается желаемый фенотипический результат, указывающий на то, что агент функционирует, ингибируя экспрессию заданной нуклеотидной последовательности, которая присутствует в клетках паразита.
Ген, на который направлена супрессия, может кодировать жизненно-необходимый белок, определенная функция которого выбрана из группы, включающей образование мышц, образование ювенильных гормонов, ювенильную гормональную регуляцию, ионную регуляцию и транспорт, синтез и транспорт белка, синтез пищеварительных ферментов, поддержание мембранного потенциала клетки, биосинтез аминокислот, распад аминокислот, образование мужских половых клеток, синтез феромонов, чувствительность к феромонам, образование щупалец, образование крыльев, образование конечностей, развитие и дифференцировку, образование яиц, созревание личинок, образование пищеварительных ферментов, синтез гемолимфы, поддержание гемолимфы, передачу нервных импульсов, деление клетки, энергетический метаболизм, дыхание, неизвестную функцию и апоптоз.
Другой аспект настоящего изобретения также предоставляет способы повышения размера урожая, получаемого от сельскохозяйственной культуры, подвергнутой поражению насекомыми-паразитами, указанный способ включает шаги a) введения полинуклеотида, включающего последовательность, выбранную от SEQ ID NO:1 до SEQ ID NO:906, или ее комплемент, или конкатемер, или фрагмент, в указанную сельскохозяйственную культуру; и b) выращивания сельскохозяйственной культуры для возможности экспрессии указанного полинуклеотида, при этом экспрессия полинуклеотида ингибирует поедание насекомыми-паразитами и потерю урожая вследствие поражения паразитами.
В некоторых вариантах осуществления экспрессия полинуклеотида приводит к образованию молекулы РНК, которая вызывает супрессию по крайней мере первого гена-мишени у насекомого-паразита, который поедал часть указанного сельскохозяйственного растения, при этом ген-мишень выполняет по крайней мере одну существенную функцию, выбранную из группы, включающей пищевую функцию паразита, жизнеспособность паразита, апоптоз клеток паразита, дифференцировку и развитие паразита или любой клетки паразита, половое размножение паразита, образование мышц, возбуждение мышц, сокращение мышц, образование и/или уменьшение содержания ювенильных гормонов, ювенильную гормональную регуляцию, ионную регуляцию и транспорт, поддержание мембранного потенциала клетки, биосинтез аминокислот, распад аминокислот, образование мужских половых клеток, синтез феромонов, чувствительность к феромонам, образование щупалец, образование крыльев, образование конечностей, образование яиц, созревание личинок, образование пищеварительных ферментов, синтез гемолимфы, поддержание гемолимфы, передачу нервных импульсов, переход в личиночную стадию, окукливание, выход из состояния окукливания, деление клеток, энергетический метаболизм, дыхание, синтез и поддержание структуры цитоскелета, метаболизм нуклеотидов, азотистый обмен, потребление воды, задержку воды и сенсорное восприятие.
В других вариантах осуществления насекомое-паразит является кукурузный жуком паразитом, выбранным из группы, включающей Diabrotica undecimpunctata howardi (Южный кукурузный жук (SCR)), Diabrotica virgifera virgifera (Западный кукурузный жук (WCR)), Diabrotica barberi (Северный кукурузный жук (NCR)), Diabrotica virgifera zea (Мексиканский кукурузный жук (MCR)), Diabrotica balteata (Бразильский кукурузный жук (BZR)), Diabrotica viridula (Бразильский кукурузный жук (BZR)) и Diabrotica speciosa (Бразильский кукурузный жук (BZR)).
Также предоставляются способы улучшения устойчивости к засухе сельскохозяйственной культуры, полученной из сельскохозяйственной культуры, подвергнутой поражению насекомым-паразитом, указанный способ включает шаги a) включения полинкулеотидной последовательности, выбранной из SEQ ID NO:1 до SEQ ID NO:906, или их фрагмента, в указанную сельскохозяйственную культуру; и b) культивирования сельскохозяйственной культуры для возможности экспрессии указанного полинуклеотида, при этом экспрессия полинуклеотида ингибирует поедание насекомыми-паразитами и потерю устойчивости к засухе вследствие поражения паразитом.
Еще один аспект изобретения далее предоставляет агрономически и коммерчески важные продукты и/или химические композиции, включая, но не ограничиваясь ими, корм для животных, сырье, продукты и побочные продукты, которые предназначены для применения в качестве пищи для потребления человеком или для применения в композициях и сырьевых продуктах, которые предназначены для потребления человеком, включая, но не ограничиваясь ими, кукурузный порошок, кукурузную муку, кукурузную патоку, кукурузное масло, кукурузный крахмал, попкорн, кукурузный жмых, зерновые продукты и т.п. Такие композиции могут быть определены как содержащие поддающиеся обнаружению количества приведенной здесь нуклеотидной последовательности и, таким образом, также являются диагностическими для любого трансгенного объекта, содержащего такие нуклеотидные последовательности. Эти продукты полезны по крайней мере, потому что их предполагается получать из сельскохозяйственных культур, выращенных с меньшим количеством пестицидов и органофосфатов в результате включения в них нуклеотидов настоящего изобретения, для контроля поражения растений жесткокрылыми паразитами. Такое сырье и сырьевые продукты могут быть получены из семян, полученных от трансгенных растений, при этом трансгенное растение экспрессирует РНК от одного или более смежных нуклеотидов настоящего изобретения или нуклеотидов одного или более жесткокрылых паразитов и их комплементов. Такое сырье и сырьевые продукты могут также быть эффективны в контроле жесткокрылых паразитов такого сырья и сырьевых продуктов, таком как, например, контроль мучного хрущака, благодаря наличию в сырье или сырьевом продукте РНК, подавляющей ген паразита, экспрессируемой из генной последовательности, укзанной в настоящем изобретении.
Также предоставляется способ получения такого сырьевого продукта, включающий получение растения, трансформированного с помощью полинуклеотида, включающего последовательность, выбранную из группы, включающей SEQ ID NO:1 до SEQ ID NO:906, или их конкатемер, или фрагмент, или комплемент, и получения сырьевого продукта из растения или его части. Далее, другим аспектом изобретения является способ изготовления пищевого или кормового продукта, включающий получение растения, трансформированного с помощью полинуклеотида, выбранного из группы, включающей SEQ ID NO:1 до SEQ ID NO:906 или их фрагмент или часть, и изготовление пищевого или кормового продукта из указанного растения или его части.
Изобретение также предоставляет машиночитаемый носитель с записанными на него одной или более нуклеотидными последовательностями, указанной в SEQ ID NO:1 до SEQ ID NO:906, или их комплементами, для применения в множестве компьютерных приложений, включая, но не ограничиваясь, поиск идентичности и подобия ДНК, поиск идентичности и подобия белка, определение характеристик транскрипционного профилирования, проведение сравнения между геномами и исследование искусственной гибридизации.
Краткое описание чертежей
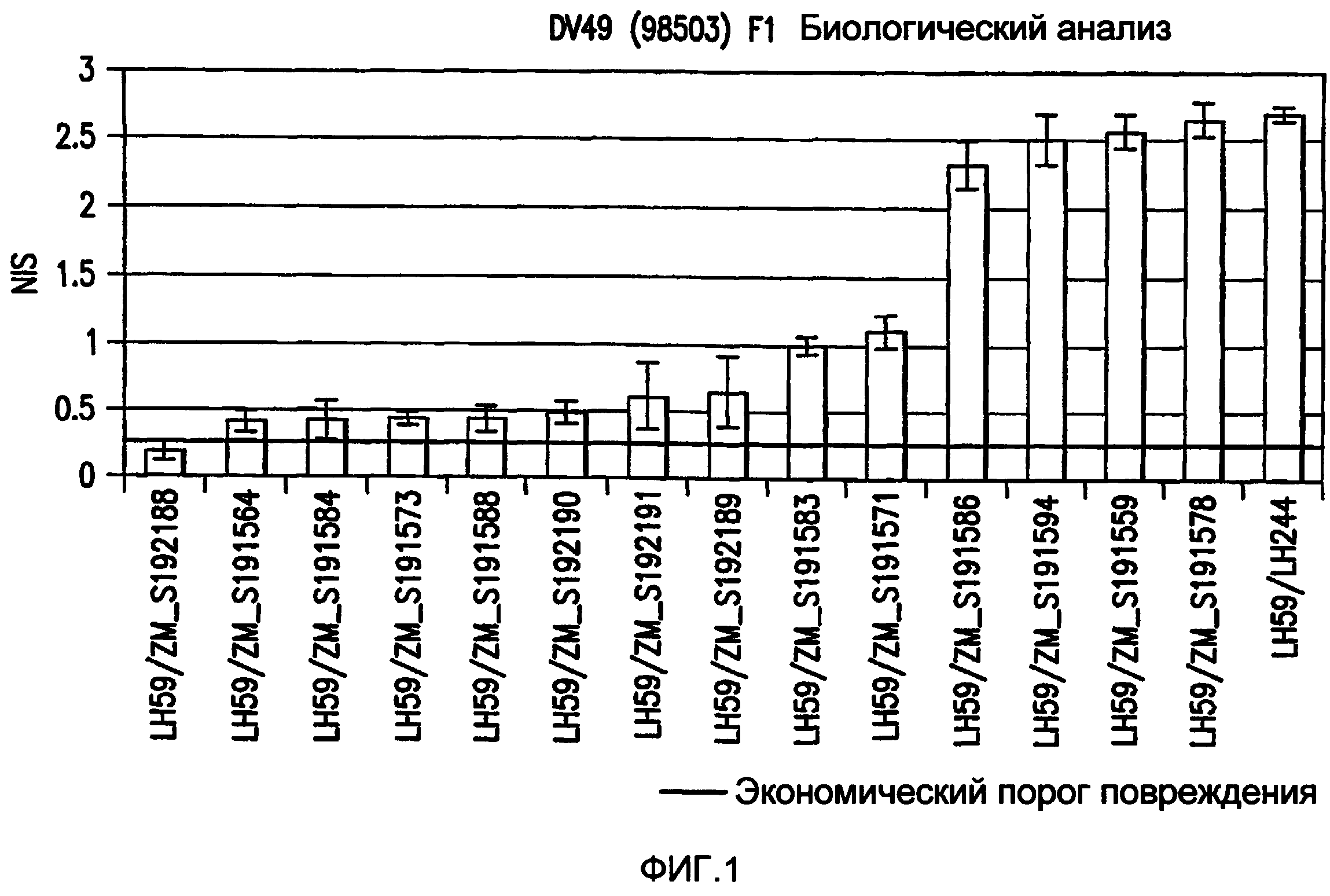
Фиг. 1: Биоанализ растительных объектов кукурузы F1, трансформированных с помощью pMON98503 (SEQ ID NO:820) и обследованных на наличие Западного кукурузного жука (WCR).
Фиг. 2: Биоанализ растительных объектов кукурузы F1, трансформированных с помощью pMON98504, включающих конкатемер C1 (SEQ ID NO:821) и обследованных на наличие Западного кукурузного жука (WCR).
Фиг. 3: Выбор фрагментов Dv49 и Dv248 и схематическая структура конкатемера C38 Dv49-Dv248.
Фиг. 4: дцРНК F1-F13, синтезированные на основе конкатемера C38.
Фиг. 5: Дозозависимая реакция на конкатемер 38 DV49-DV248 (Фрагменты F1-F6).
Фиг. 6: Дозозависмая реакция на конкатемер 38 DV49-DV248 (Фрагменты F7-F10).
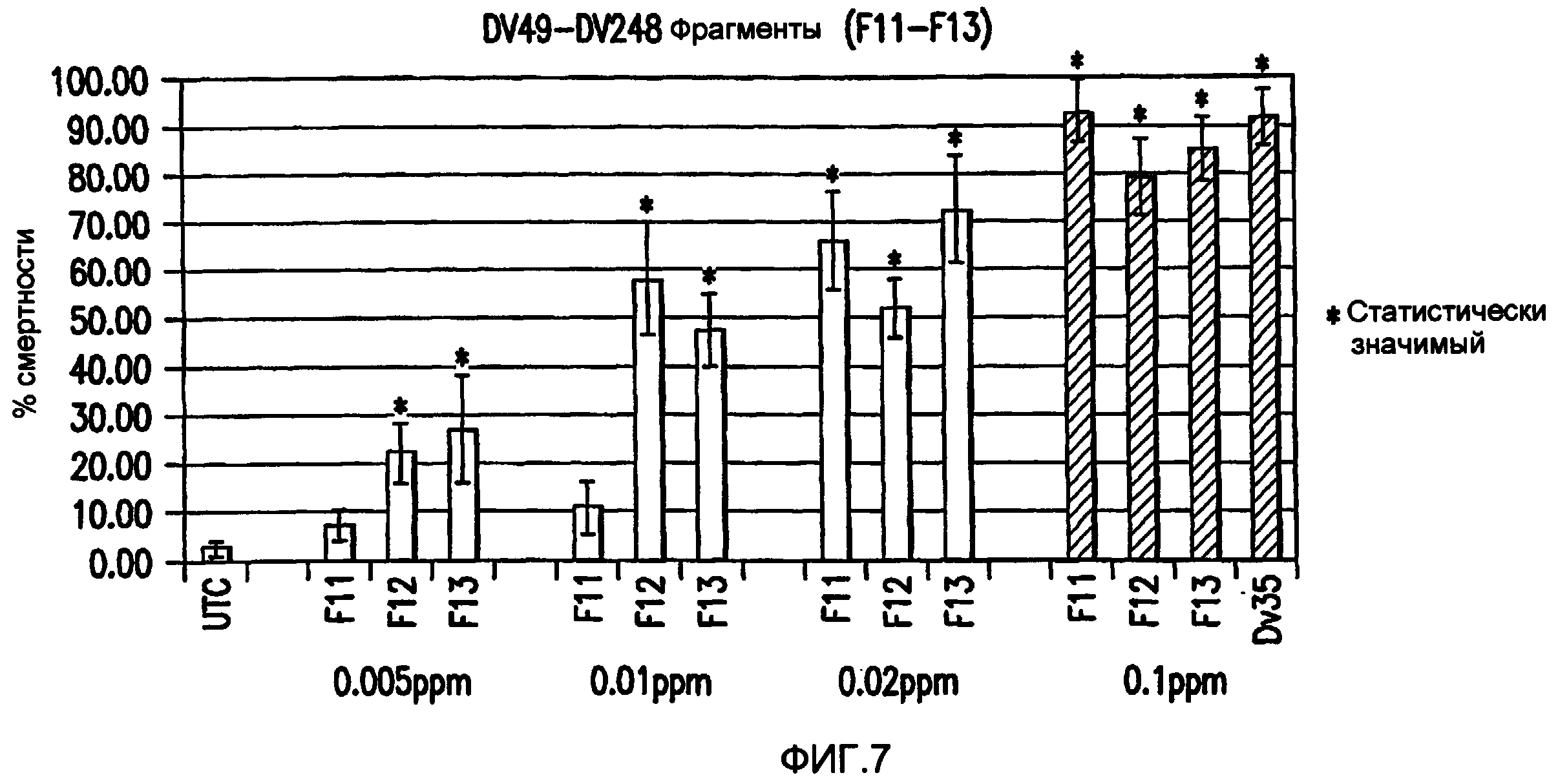
Фиг. 7: Дозозависмая реакция на конкатемер 38 DV49-DV248 (Фрагменты F11-F13).
Подробное описание изобретения
Далее представлено подробное описание изобретения для помощи специалистам в области техники в осуществлении настоящего изобретения. Обычный специалист в области техники может вносить модификации и изменения в описанные здесь варианты осуществления, не отступая от сущности или объема настоящего изобретения.
Настоящее изобретение предоставляет способы и композиции для генетического контроля поражения паразитами. Например, настоящее изобретение предоставляет технологии рекомбинантной ДНК для посттранскрипционного репрессинга или ингибирования экспрессии заданной кодирующей последовательности в клетке паразита для обеспечения защитного эффекта от паразита с помощью скармливания паразиту одной или более молекул двухцепочечных или малых интерферирующих рибонуклеиновых кислот (РНК), считанных со всей или части заданной кодирующей последовательности, таким образом контролируя поражение. Поэтому настоящее изобретение относится к последовательность-специфичному ингибированию экспрессии кодирующих последовательностей с помощью двухспиральной РНК (дцРНК), включая малую интерферирующую РНК (siРНК), для достижения заданных уровней контроля паразита.
Предполагается, что изолированные и в основном очищенные молекулы нуклеиновой кислоты, включая, но не ограничиваясь ими, не встречающиеся в природе нуклеотидные последовательности и рекомбинантные конструкции ДНК, для транскрипции молекул дцРНК настоящего изобретения вызывают супрессию или ингибирование экспрессии эндогенной кодирующей последовательности или заданной кодирующей последовательности у паразита при введении в последнего. Также предоставляются трансгенные растения, которые (a) содержат нуклеотидные последовательности, кодирующие изолированные и в основном очищенные молекулы нуклеиновой кислоты, и не встречающиеся в природе рекомбинантные конструкции ДНК для транскрипции молекул дцРНК для контроля поражения растения паразитами, и (b) проявляют устойчивость и/или повышенную толерантность к поражению насекомыми. Также описаны композиции, содержащие дцРНК нуклеотидные последовательности настоящего изобретения, для применения в виде местного нанесения на растения или на животных, или внесения в среду, окружающую животное, для достижения устранения или уменьшения поражения паразитами.
В данном изобретении изобретатели обнаружили, что вопреки положениям предшествующего уровня техники скармливание композиции, содержащей двухцепочечные молекулы РНК, состоящие из последовательностей, обнаруживаемых в одной или более экспрессируемых нуклеотидных последовательностях вида жесткокрылых, видам, от которых были получены нуклеотидные последовательности, приводит к ингибированию одной или более биологических функций у вида жесткокрылых. В частности, изобретатели обнаружили, что скармливание описанных здесь двухцепочечных молекул РНК видам паразитов сельскохозяйственных культур, таким как кукурузные жуки, приводит к гибели или ингибированию развития и дифференцировки насекомых паразитов, которые поедают эти композиции.
Изобретателями определены нуклеотидные последовательности, описанные здесь, как обеспечивающие эффекты защиты растений от видов жесткокрылых паразитов. Установлены аминокислотные последовательности, закодированные последовательностями кДНК, и произведено сравнение с известными аминокислотными последовательностями. Установлено, что многие последовательности кодируют белки, которые имеют некоторую аннотированную информацию, связанную с ними. Аннотированная информация, которая связана со специфической нуклеотидной последовательностью и закодированной посредством нее последовательностью белка, основана на гомологии или подобии между аминокислотными последовательностями, установленными посредством трансляции описанных здесь кодирующих последовательностей и аминокислотными последовательностями, которые известны в области техники в общедоступных базах данных.
Последовательности кДНК, кодирующие белки или части белков, необходимые для выживания, такие как аминокислотные последовательности, задействованные в различных метаболических или катаболических биохимических путях, делении клетки, репродукции, энергетическом метаболизме, пищеварении, неврологических функциях и т.п. были выбраны для применения в получении двухцепочечных молекул РНК, которые включались в диету жесткокрылых паразитов. Как описано здесь, поедание паразитом-мишенью композиций, содержащих одну или более дцРНК, по крайней мере один сегмент которых соответствует по крайней мере в основном идентичному сегменту РНК, продуцируемой в клетках паразита-мишени, приводит к гибели, остановке роста или другому ингибированию паразита-мишени. Эти результаты указывают, что нуклеотидная последовательность или ДНК или РНК, полученная от жесткокрылого паразита, может быть применена для создания растительных клеток, устойчивых к поражению паразитом. Например, хозяин паразита может быть преобразован таким образом, что будет содержать одну или более нуклеотидных последовательностей, полученных от жесткокрылого паразита. Нуклеотидная последовательность, трансформированная в паразита хозяина или симбионта, может кодировать одну или более РНКs, которые преобразуются в последовательность дцРНК в клетках или биологических жидкостях в транформированном хозяине или симбионте, таким образом, обеспечивая наличие дцРНК в диете паразита, если/когда паразит паразитирует на трансгенном хозяине или симбионте, приводя к супрессии экспрессии одного или более генов в клетках паразита и в конечном счете гибели, остановке роста или другому ингибированию паразита.
Настоящее изобретение относится в основном к генетическому контролю поражения жесткокрылыми паразитами в организме хозяина. Конкретнее, настоящее изобретение включает способы доставки агентов для контроля вредителей жесткокрылому паразиту. Такие агенты для контроля вредителей вызывают, прямо или косвенно, ослабление способности паразита поддерживать свою жизнедеятельность, расти или, иначе, вызывать поражение заданного хозяина или симбионта. Настоящее изобретение предоставляет способы включения стабилизированных молекул дцРНК в пищевой рацион паразита в качестве средства для супрессии генов-мишеней у паразита, таким образом, достигая желаемого контроля над паразитированием паразита внутри, или вокруг хозяина или симбионта, на которого нацелен паразит.
Для осуществления вышеуказанного настоящее изобретение предоставляет способ ингибирования экспрессии гена-мишени у жесткокрылого паразита, включая, например, кукурузных жуков или другие виды жесткокрылых насекомых, что приводит к прекращению пищевой активности, роста, развития, репродукции, инфекционности и, в конечном счете, может привести к гибели паразита. В одном варианте осуществления способ включает включение частично или полностью стабилизированных двухцепочечных нуклеотидных молекул РНК (дцРНК) в пищевую композицию, на которую паразит полагается как на источник пищи, и создание пищевой композиции, пригодной для поедания паразитом. Прием внутрь пищевой композиции, содержащей двухцепочечные или siРНК молекулы, приводит к захвату молекул клетками паразита, приводя к ингибированию экспрессии по крайней мере одного гена-мишени в клетках паразита. Ингибирование гена-мишени оказывает отрицательное воздействие на паразита.
В некоторых вариантах осуществления молекулы дцРНК, предоставляемые изобретением, включают нуклеотидные последовательности, комплементарные последовательности, приведенной в любой с SEQ ID NO:1 по SEQ ID NO:906, ингибирование которых в организме паразита приводит к уменьшению содержания или удалению белка или агента из нуклеотидной последовательности, который необходим для роста и развития или другой биологической функции паразитов. Выбранная нуклеотидная последовательность может иметь от приблизительно 80% по крайней мере до приблизительно 100% идентичность последовательности к одной из нуклеотидных последовательностей, указанных с SEQ ID NO:1 по SEQ ID NO:906, как указано в списке последовательностей, включая их комплементы. Такое ингибирование может быть описано как специфичное в том отношении, что нуклеотидная последовательность от части гена-мишени выбрана из той, с которой считана ингибирующая дцРНК или siРНК. Способ эффективен в ингибировании экспрессии по крайней мере одного гена-мишени и может применяться для ингибирования многих различных типов генов-мишеней у паразита. В конкретных вариантах осуществления нуклеотидная последовательность может быть выбрана из группы, состоящей из SEQ ID NO:697, SEQ ID NО:813-819, SEQ ID NО:841 и SEQ ID NO:874.
Последовательности, идентифицированные как обладающие защитным эффектом от паразита, могут легко экспрессироваться как молекулы дцРНК посредством создания подходящих конструктов для экспрессии. Например, такие последовательности могут экспрессироваться в виде шпильки и стебля и петлевой структуры с помощью использования первого сегмента, соответствующего последовательности, выбранной от SEQ ID NO:1 до SEQ ID NO:906 или их фрагмента, связывания этой последовательности со спейсерной областью второго сегмента, который не является гомологичным или комплементарным первому сегменту, и связывая ее с третьим сегментом, который считывает РНК, при этом, по крайней мере часть третьего сегмента в основном комплементарна первому сегменту. Такая конструкция формирует структуру стебля и петли с помощью гибридизации первого сегмента с третьим сегментом, а структура петли формируется так, что включает второй сегмент (W094/01550, W098/05770, США 2002/0048814A1 и США 2003/0018993A1).
A. Композиции нуклеиновых кислот и конструкты
Изобретение предоставляет рекомбинантные конструкты ДНК для применения в достижении устойчивой трансформации конкретных мишеней паразита-хозяина или симбионта. Трансформированная мишень паразита-хозяина или симбионта может экспрессировать эффективные пестицидные уровни предпочтительных молекул дцРНК или siРНК из рекомбинантных конструктов ДНК и обеспечивать наличие молекул в пищевом рационе паразита. Пары изолированных и очищенных нуклеотидных последовательностей могут быть предоставлены из материалов библиотеки кДНК и/или геномной библиотеки. Пары нуклеотидных последовательностей могут быть получены от любого предпочтительного жесткокрылого паразита для применения в качестве праймеров тепловой амплификации для создания матрицы ДНК для получения молекул дцРНК и siРНК настоящего изобретения.
Применяемый здесь термин "нуклеиновая кислота относится к одно- или двухцепочечному полимеру дезоксирибонуклеотидного или рибонуклеотидного оснований, считываемых от 5' к 3' концу. "Нуклеиновая кислота" может также необязательно включать нуклеотидные основания не природного происхождения или измененные, которые позволяют правильно их считывать с помощью полимеразы и не ослабляют экспрессию полипептида, закодированного этой нуклеиновой кислотой. Термин "нуклеотидная последовательность" или "последовательность нуклеиновой кислоты" относится как к смысловой, так и к антисмысловым цепочкам нуклеиновой кислоты в виде либо отдельных одиночных цепочек, либо двойных спиралей. Термин "рибонуклеиновая кислота" (РНК) включает РНКi (ингибиторная РНК), дцРНК (двухцепочечная РНК), siРНК (малая интерферирующая РНК), мРНК (мессенджерная РНК), miРНК (микро-РНК), tРНК (трансферная РНК, либо нагруженная, либо разгруженная от соответствующей ацилированной аминокислоты), и cРНК (комплементарная РНК), и термин "дезоксирибонуклеиновая кислота" (ДНК) включает кДНК и геномную ДНК и гибриды ДНК-РНК. Словосочетания "сегмент нуклеиновой кислоты", "сегмент нуклеотидной последовательности", или более обобщенно "сегмент" должны пониматься специалистами в области техники как функциональные термины, которые включают как геномные последовательности, так и последовательности рибосомальной РНК, последовательности трансферной РНК, последовательности мессенджерной РНК, оперонные последовательности и меньшие сконструированные нуклеотидные последовательности, которые экспрессируют или могут быть адаптированы для экспрессии белков, полипептидов или пептидов.
Согласно изобретению предоставляются нуклеотидные последовательности, экспрессия которых приводит к последовательности РНК, которая в основном гомологична молекуле РНК направленного гена у насекомого, которая включает последовательность РНК, закодированную нуклеотидной последовательностью в геноме насекомого. Таким образом, после поедания стабилизированной последовательности РНК может быть достигнута понижающая регуляция нуклеотидной последовательности гена-мишени в клетках насекомого, приводящая к вредному воздействию на поддержание жизнедеятельности, жизнеспособность, пролиферацию, репродукцию и способность насекомого к паразитированию.
Применяемый здесь термин "в основном гомологичная" или "значительная гомология", в отношении последовательности нуклеиновой кислоты, включает нуклеотидную последовательность, которая гибридизирует в условиях большой напряженности до кодирующей последовательности, указанной в любой из с SEQ ID NO:1 по SEQ ID NO:906, как указано в списке последовательностей, или ее комплементы. Последовательности, которые гибридизируют в условиях большой напряженности до любой из с SEQ ID NO:1 по SEQ ID NO:906, как указано списке последовательностей, или их комплементы относятся к тем, которые позволяют, чтобы между этими двумя последовательностями происходило антипараллельное выравнивание, и две последовательности способны в условиях большой напряженности формировать водородные связи с основаниями на противоположной цепи для формирования двойной молекулы, которая является достаточно устойчивой в условиях большой напряженности, чтобы поддаваться обнаружению с помощью способов, известных в области техники. В основном гомологичные последовательности имеют предпочтительно приблизительно от 70% до приблизительно 80% идентичность последовательности, или более предпочтительно от приблизительно 80% до приблизительно 85% идентичность последовательности, или наиболее предпочтительно от приблизительно 90% до приблизительно 95% идентичность последовательности, приблизительно до 99% идентичности последовательности сравнительным нуклеотидным последовательностям, указанным в любой из с SEQ ID NO:1 по SEQ ID NO:906, как указано в списке последовательностей, или их комплементам.
Применяемый здесь термин "идентичность последовательности", "подобие последовательности" или "гомология" служит для описания сродства между двумя или более нуклеотидными последовательностями. Процентный показатель "идентичности последовательности" двух последовательностей определяют с помощью сравнения двух оптимально выровненных последовательностей в окне сравнения, при этом часть последовательности в окне сравнения может включать вставки или делеции (то есть, разрывы), в отличие от контрольной последовательности (которая не включает вставки или делеции), для оптимального выравнивания этих двух последовательностей. Процент рассчитывают с помощью определения числа позиций, в которых идентичное основание нуклеиновой кислоты или аминокислотный остаток встречается в обеих последовательностях, для получения числа соответствующих позиций, деления числа соответствующих позиций на общее число позиций в окне сравнения и умножения результата на 100 для получения процентного показателя идентичности последовательности. Последовательность, которая при сравнении с контрольной последовательностью является идентичной ей в каждой позиции, считают идентичной контрольной последовательности и наоборот. Первая нуклеотидная последовательность при рассмотрении в направлении от 5' к 3' называют "комплементом" или комплементарной второй или контрольной нуклеотидной последовательности, рассматриваемой в направлении от 3' к 5', если первая нуклеотидная последовательность полностью комплементарна контрольной или второй последовательности. Применяемый здесь термин молекулы последовательности нуклеиновой кислоты служит для обозначения "полной комплементарности", когда каждый нуклеотид одной из последовательностей, читаемой от 5' к 3', является комплементарным каждому нуклеотиду другой последовательности, читаемой от 3' к 5'. Нуклеотидная последовательность, которая является комплементарной контрольной нуклеотидной последовательности, будет иметь последовательность, идентичную обратной комплементарной последовательности контрольной нуклеотидной последовательности. Эти термины и описания хорошо известны в области техники и легко поняты специалистам в области техники.
Применяемый здесь термин "окно сравнения" относится к концептуальному сегменту по крайней мере из 6 смежных позиций, обычно от приблизительно 50 до приблизительно 100, чаще от приблизительно 100 до приблизительно 150, по которым последовательность сравнивается с контрольной последовательностью из того же числа смежных позиций после оптимального выравнивания этих двух последовательностей. Окно сравнения может включать вставки или делеции (то есть разрывы) в количестве приблизительно 20% или менее по сравнению с контрольной последовательностью (которая не включает вставки или делеции) для оптимального выравнивания этих двух последовательностей. Для подробного обсуждения анализа последовательности специалистам в области техники необходимо обратиться к детализированным способам, применяемым для выравнивания последовательности в Wisconsin Genetics Software Package Release 7.0, Genetics Computer Group, 575 Science Drive Madison, Wis., USA), или обратиться к Ausubel и др. (1998).
Настоящее изобретение предоставляет последовательности ДНК, способные к экспрессии в виде РНК в клетке или микроорганизме, для ингибирования экспрессии гена-мишени в клетке, ткани или органе насекомого. Последовательности включают молекулу ДНК, кодирующую одну или более различных нуклеотидных последовательностей, при этом каждая из различных нуклеотидных последовательностей включает смысловую нуклеотидную последовательность и антисмысловую нуклеотидную последовательность, связанные спейсерной последовательностью, кодирующей молекулу дцРНК настоящего изобретения. Спейсерная последовательность составляет часть смысловой нуклеотидной последовательности или антисмысловой нуклеотидной последовательности и формируется в пределах дцРНК молекулы между смыслововой и антисмысловой последовательностями. Смысловая нуклеотидная последовательность или антисмысловая нуклеотидная последовательность в основном идентичны нуклеотидной последовательности гена-мишени, или его производного, или комплементарной ей последовательности. Молекула дцДНК может быть операбельно помещена под контролем промоторной последовательности, которая функционирует в клетке, ткани или органе хозяина, экспрессирующего дцДНК, для продукции молекул дцРНК. В одном варианте осуществления последовательность ДНК может быть получена из нуклеотидной последовательности, как указано с SEQ ID NO:1 по SEQ ID NO:906 в списке последовательности.
Изобретение также предоставляет последовательность ДНК для экспрессии в клетке растения, которое после экспрессии ДНК до РНК и поедания паразитом-мишенью, вызывает супрессию гена-мишени в клетке, ткани или органе насекомого-паразита. дцРНК включает по крайней мере одну или множество последовательностей структурных генов, в которых каждая из последовательностей структурных генов включает смысловую нуклеотидную последовательность и антисмысловую нуклеотидную последовательность, связанные спейсерной последовательностью, которая формирует петлю в пределах комплементарных и антисмысловых последовательностей. Смысловая нуклеотидная последовательность или антисмысловая нуклеотидная последовательность в основном идентичны нуклеотидной последовательности гена-мишени, его производному или комплементарной ему последовательности. Одну или более последовательностей структурных генов операбельно помещают под контролем одной или более промоторных последовательностей, по крайней мере одна из которых является операбельной, в клетку, ткань или орган прокариотического или эукариотического организма, особенно растения.
Генная последовательность или фрагмент для контроля паразитов согласно изобретению могут быть клонированы между двумя тканеспецифичными промоторами, например двумя корнеспецифичными промоторами, которые являются операбельными в трансгенной растительной клетке, и экспрессированы в ней для продукции мРНК в трансгенной растительной клетке, которые образуют в ней молекулы дцРНК. Молекулы дцРНК, содержащиеся в тканях растения, поглощаются насекомым так, что достигается заданная супрессия экспрессии гена-мишени.
Нуклеотидная последовательность, предоставляемая настоящим изобретением, может включать инвертированный повтор, отделенный "спейсерной последовательностью". Спейсерная последовательность может быть представлена областью, включающей любую последовательность нуклеотидов, которая облегчает формирование вторичной структуры между каждым повтором, где это требуется. В одном варианте осуществления настоящего изобретения спейсерная последовательность является частью смысловой или антисмысловой кодирующей последовательности для мРНК. В альтернативном варианте спейсерная последовательность может включать любую комбинацию нуклеотидов или их гомологов, которые способны ковалентно связываться с молекулой нуклеиновой кислоты. Спейсерная последовательность может включать последовательность нуклеотидов из по крайней мере приблизительно 10-100 нуклеотидов в длину, или в альтернативном варианте из по крайней мере приблизительно 100-200 нуклеотидов в длину, из по крайней мере приблизительно 200-400 нуклеотидов в длину, или из по крайней мере приблизительно 400-500 нуклеотидов в длину.
Молекулы нуклеиновой кислоты, или фрагмент молекул нуклеиновой кислоты, или другие молекулы нуклеиновой кислоты в списке последовательностей способны специфично гибридизироваться с другими молекулами нуклеиновой кислоты при определенных условиях. Считается, что применяемые здесь две молекулы нуклеиновой кислоты способны специфично гибридизироваться друг с другом, если две молекулы способны к образованию антипараллельной, двухцепочечной структуры нуклеиновой кислоты. Считается, что молекула нуклеиновой кислоты является комплементом другой молекулы нуклеиновой кислоты, если они проявляют полную комплементарность. Считается, что две молекулы являются "минимально комплементарными", если они могут гибридизироваться друг с другом с достаточной устойчивостью, что позволяет им оставаться связанными друг с другом по крайней мере при обычных условиях пониженной жесткости. Подобным образом считают, что молекулы являются комплементарными, если они могут гибридизироваться друг с другом с достаточной устойчивостью, что позволяет им оставаться связанными друг с другом при обычных строгих условиях. Обычные строгие условия описаны Sambrook и др. (1989) и Haymes и др. (1985).
Поэтому отступления от полной комплементарности допустимы, пока такие отступления полностью не препятствуют способности молекул формировать двухцепочечную структуру. Таким образом, чтобы выполнять функцию праймера или зонда, молекула нуклеиновой кислоты или фрагмент молекулы нуклеиновой кислоты должен только иметь достаточно комплементарную последовательность, чтобы быть способным формировать устойчивую двухцепочечную структуру в присутствии конкретного растворителя и используемых концентрацииях соли.
Подходящие условия строгости, способствующие гибридизации ДНК, например, 6,0×хлорид натрия/цитрат натрия (SSC) при приблизительно 45°C с последующим промыванием 2,0×SSC при 50°C известны специалистам в области техники или могут быть найдены в Современных Протоколах Молекулярной Биологии (1989). Например, концентрация соли в шаге промывания может быть выбрана от низкой строгости, составляющей приблизительно 2,0×SSC при 50°C до высокой строгости, составляющей приблизительно 0,2×SSC при 50°C. Кроме того, температура в шаге промывания может быть увеличена от условий низкой строгости с комнатной температурой, приблизительно 22°C, до условий высокой строгости с температурой приблизительно 65°C. И температура и соль могут различаться, или либо температура, либо концентрация соли могут поддерживаться на постоянном уровне, тогда как другой показатель будет изменяться. При таких условиях нуклеиновая кислота, применяемая в настоящем изобретении, может специфично гибридизироваться с одной или более молекулами нуклеиновой кислоты от WCR или их комплементами. Предпочтительно, нуклеиновая кислота, применяемая в настоящем изобретении, имеет по крайней мере от приблизительно 80%, или по крайней мере от приблизительно 90%, или по крайней мере от приблизительно 95%, или по крайней мере от приблизительно 98% или даже приблизительно 100% идентичность последовательности с одной или более молекулами нуклеиновой кислоты, указанными с SEQ ID No:1 по SEQ ID NO:906 в списке последовательностей.
Нуклеиновые кислоты настоящего изобретения могут также быть синтезированы, или полностью или частично, особенно когда желательно предоставить предпочтительные для растения последовательности с помощью способов, известных в области техники. Таким образом, все или часть нуклеиновых кислот настоящего изобретения могут быть синтезированы с помощью кодонов, предпочтительных для выбранного хозяина. Предпочтительные для вида кодоны могут быть определены, например, с помощью кодонов, используемых наиболее часто в белках, экспрессируемых у хозяина конкретного вида. Другие модификации нуклеотидных последовательностей могут привести к мутантам, имеющим несколько измененную активность.
Нуклеотидные последовательности дцРНК или siРНК включают двойные цепочки полимеризированного рибонуклеотида и могут включать модификации либо в фосфатно-сахарных боковых цепях, либо в нуклеозидах. Модификации в структуре РНК могут быть рассчитаны для возможности специфического генетического ингибирования. В одном варианте осуществления молекулы дцРНК могут быть изменены в результате ферментативного процесса так, чтобы было возможным создать молекулы siРНК. siРНК может эффективно опосредовать эффект понижающей регуляции для некоторых генов-мишеней у некоторых насекомых. Этот ферментативный процесс может быть осуществлен с помощью фермента РНКаза III или фермента DICER, присутствующих в клетках насекомого, позвоночного животного, грибка или растения в каскаде реакций эукариотической РНКi (Elbashir и др., 2002; Hamilton и Baulcombe, 1999). В этом процессе может также использоваться рекомбинантная DICER или РНКаза III, введенная в клетки насекомого-мишени посредством рекомбинантных технологий ДНК, которые известны специалисту в области техники. И фермент DICER и РНКаза III, образующиеся в природе в насекомом или полученные с помощью рекомбинантных технологий ДНК, расщепляют большие цепи дцРНК на меньшие олигонуклеотиды. Фермент DICER специфично разрезает молекулы дцРНК на части siРНК, каждая из которых имеет длину приблизительно 19-25 нуклеотидов, в то время как фермент РНКаза III обычно расщепляет молекулы дцРНК до siРНК из 12-15 пар оснований. Молекулы siРНК, полученные с помощью любого из ферментов, имеют от 2 до 3 нуклеотидов на 3' "липком", и 5' фосфатном, и 3' гидроксильном концах. Молекулы siРНК, созданные с помощью фермента РНКаза III, сходны с созданными с помощью ферментов DICER в каскаде реакций эукариотической РНКi и, следовательно, затем направляются и распадаются с помощью врожденного клеточного механизма распада РНК после их последовательного раскручивания, разделения на одноцепочечные РНК и гибридизации с последовательностями РНК, транскрибированными геном-мишенью. Этот процесс приводит к эффективному распаду или удалению последовательности РНК, закодированной нуклеотидной последовательностью гена-мишени, у насекомого. В результате подавляется транскрипция направленной нуклеотидной последовательности в организме насекомого. Подробные описания ферментативных процессов могут быть найдены в Hannon (2002).
Нуклеотидная последовательность настоящего изобретения может быть записана на машиночитаемый носитель. Применяемый здесь термин "машиночитаемый носитель" относится к любому материальному носителю экспрессии, который может быть прочитан и доступен непосредственно с помощью компьютера. К таким носителям относятся, но не ограничиваясь ими: магнитные носители данных, такие как гибкие диски, жесткий диск, носитель данных и магнитная лента; оптические носители данных, такие как CD-ROM; электрические носители данных, такие как RAM и ROM; компьютерные файлы в формате оптического распознавания знаков, и гибриды этих категорий, такие как магнитные/оптические носители данных. Специалисту ясно, что любой из известных в настоящее время машиночитаемых носителей может быть применен для создания производства, включающего машиночитаемый носитель с записью на нем нуклеотидной последовательности настоящего изобретения.
Применяемый здесь термин "записанный" относится к процессу сохранения информации на машиночитаемый носитель. Специалист может легко выбрать любой из известных в настоящее время способов записи информации на машиночитаемый носитель для создания носителя, включающего информацию о нуклеотидной последовательности настоящего изобретения. Для создания машиночитаемого носителя с записанной на него нуклеотидной последовательностью настоящего изобретения специалисту доступно множество структур хранения данных. Выбор структуры хранения данных в основном будет основан на средствах, выбранных для доступа к сохраненной информации. Кроме того, для хранения информации о нуклеотидной последовательности настоящего изобретения на машиночитаемом носителе может применяться множество программ процессора и форматов. Информация о последовательности может быть представлена в виде текстового файла, форматированного под коммерчески доступное программное обеспечение, такое как WordPerfect и Microsoft Word, или представлена в виде текстового файла ASCII, сохраненного в приложении для работы с базами данных, таком как DB2, Sybase, Oracle или подобном. Специалист может легко приспособить любой формат структурирования процессорной информации (например, текстовый файл или базу данных) для получения машиночитаемого носителя с записью на нем информации о нуклеотидной последовательности настоящего изобретения.
Программное обеспечение является публично доступным, что позволяет специалисту получать доступ к информации о последовательности, предоставленной на машиночитаемом носителе. Программное обеспечение, которое поддерживает алгоритмы поиска BLAST (Altschul и др., 1990) и BLAZE (Brutlag, и др., 1993) в системе Sybase, может применяться для идентификации открытых рамок считывания (ORFs) в последовательностях, таких как Unigenes и EST's, которые здесь представлены и которые содержат гомологию к ORFs или белкам от других организмов. Такие ORFs являются фрагментами, кодирующими белок, в пределах последовательностей настоящего изобретения и эффективны в продукции коммерчески важных белков, таких как ферменты, применяемые в биосинтезе аминокислот, метаболизме, транскрипции, трансляции, процессинге РНК, распаде нуклеиновых кислот и белка, модификации белка и репликации ДНК, рестрикции, модификации, рекомбинации и восстановлении.
Настоящее изобретение далее предоставляет системы, в частности машинные системы, которые содержат описанную здесь информацию о последовательности. Такие системы разработаны для идентификации коммерчески важных фрагментов молекулы нуклеиновой кислоты настоящего изобретения. Применяемый здесь термин "машинная система" относится к аппаратным средствам, средствам программного обеспечения и средствам хранения данных, применяемым для анализа информации о нуклеотидной последовательности настоящего изобретения. Минимальное аппаратное средство машинных систем настоящего изобретения включает центральный процессор (CPU), средства для ввода данных, средства для вывода данных и средства для хранения данных. Специалист ясно, что любая из доступных в настоящее время машинных систем подходит для применения в настоящем изобретении.
Применяемый здесь термин "структурный мотив-мишень", или "мотив-мишень", относится к любой рационально выбранной последовательности или комбинации последовательностей, в которых последовательности или последовательность(и) выбраны на основе трехмерной конфигурации, которая формируется после укладки мотива-мишени. Существует множество мотивов-мишеней, известных в области техники. К мотивам-мишеням белка относятся, но не ограничиваясь ими, активные сайты ферментов и сигнальные последовательности. К мотивам-мишеням нуклеиновых кислот относятся, но не ограничиваясь ими, промоторные последовательности, цис-элементы, шпилькообразные структуры и элементы индуцируемой экспрессии (последовательности, связывающие белок).
B. Рекомбинантные векторы и трансформация клетки - хозяина
Рекомбинантный вектор ДНК может, например, быть представлен линейной или закрытой кольцевой плазмидой. Векторная система может быть представлена единичным вектором или плазмидой, или двумя или более векторами или плазмидами, которые вместе содержат всю ДНК, которую необходимо ввести в геном бактериального хозяина. Кроме того, бактериальный вектор может быть вектором экспрессии. Молекулы нуклеиновой кислоты, как указано с SEQ ID NО:1 по SEQ ID, NО:906, или их фрагменты и комплементы могут, например, быть соответственно вставлены в вектор под контролем подходящего промотора, который функционирует в одном или более микробном хозяине для запуска экспрессии связанной кодирующей последовательности или другой последовательности ДНК. Для этой цели доступно множество векторов и выбор подходящего вектора будет зависеть главным образом от размера нуклеиновой кислоты, которая будет вставлена в вектор, и конкретной клетки-хозяина, которая будет трансформирована с помощью вектора. Каждый вектор содержит различные компоненты, в зависимости от его функции (амплификация ДНК или экспрессия ДНК), и специфической клетки-хозяина, с которой он совместим. Векторные компоненты для бактериальной трансформации в основном включают, но не ограничиваясь ими, один или более из следующих компонентов: сигнальная последовательность, репликатор, один или более селектируемых маркерных генов и индуцируемый промотор, допускающий экспрессию экзогенной ДНК.
Экспрессирующие и клонирующие векторы в основном содержат ген селекции, также называемый селектируемым маркером. Этот ген кодирует белок, необходимый для выживания или роста трансформированных клеток хозяина, выращенных в селективной культуральной среде. Типичные гены селекции кодируют белки, которые (a) придают устойчивость к антибиотикам или другим токсинам, например ампициллину, неомицину, метотрексату или тетрациклину, (b) восполняют ауксотрофный дефицит или (c) поставляют критические питательные вещества, не доступные в сложных питательных средах, например ген, кодирующий D-аланин рацемазу бацилл. Те клетки, которые успешно трансформированы с гетерологичным белком или его фрагментом, продуцируют белок, придающий устойчивость к лекарственным средствам, что, таким образом, позволяет выживать в режиме селекции.
Вектор экспрессии для продукции мРНК может также содержать индуцируемый промотор, который распознается бактериальным организмом хозяина и операбельно связывается с кодирующей нуклеиновой кислотой, например молекулой нуклеиновой кислоты, кодирующей мРНК или интересующий ее фрагмент у D. v. virgifera. К индуцируемым промоторам, подходящим для применения с бактериальными хозяевами, относятся промотор β-лактамазы, E. coli λ фаг PL и PR промоторы и промотор галактозы E. coli, промотор арабинозы, промотор щелочной фосфатазы, промотор триптофана (концевого), и промотор лактозного оперона и его разновидностей, и гибридные промоторы, такие как tac промотор. Однако, другие известные бактериальные индуцируемые промоторы являются подходящими.
Термин "операбельно связанный", применяемый в ссылке на регуляторную последовательность и структурную нуклеотидную последовательность, означает, что регуляторная последовательность вызывает регулируемую экспрессию связанной структурной нуклеотидной последовательности. Термины "регуляторные последовательности" или "элементы контроля" относятся к нуклеотидным последовательностям, расположенным выше (5' некодирующие последовательности), в пределах или ниже (3' нетранслированные последовательности) структурной нуклеотидной последовательности, и которые влияют на время и уровень или величину транскрипции, процессинг или стабильность РНК, или трансляцию связанной структурной нуклеотидной последовательности. Регулирующие последовательности могут содержать промоторы, лидерные последовательности трансляции, интроны, усилители, структуры стебель-петля, последовательности, связывающие репрессор, и последовательности распознавания полиаденилирования и т.п.
В альтернативном варианте конструкты для экспрессии могут быть интегрированы в бактериальный геном с интегрирующим вектором. Интегрирующие векторы обычно содержат по крайней мере одну последовательность, гомологичную бактериальной хромосоме, которая позволяет вектору интегрироваться. Интеграция является результатом рекомбинаций между гомологичной ДНК в векторе и бактериальной хромосоме. Например, интегрирующие векторы, созданные из ДНК различных штаммов бацилл, интегрируют в хромосому бацилл (Европейский патент 0 127328). Интегрирующие векторы могут также состоять из бактериофага или транспозонных последовательностей. Суицидные векторы также известны в области техники.
Для конструкции подходящих векторов, содержащих один или более вышеупомянутых компонентов, используются стандартные рекомбинантные технологии ДНК. Выделенные плазмиды или фрагменты ДНК расщепляют, настраивают и повторно лигируют в форме, желательной для создания требуемых плазмид. Примеры доступных бактериальных векторов экспрессии включают, но не ограничиваясь ими, многофункциональные векторы клонирования и экспрессии E. coli, такие как Bluescript™ (Stratagene, La Jolla, CA), в которых, например, белок D. v. virgifera или его фрагмент может быть лигирован в вектор в рамке последовательностей для амино-терминали Met и последующих 7 остатков β-галактозидазы так, что получают гибридный белок; pIN векторы (Van Heeke и Schuster, 1989) и т.п.
Дрожжевой рекомбинантный конструкт может обычно содержать один или более следующих компонентов: промоторную последовательность, последовательность для слияния с партнером, лидерную последовательность, последовательность терминации транскрипции, селектируемый маркер. Эти элементы могут быть объединены в полигенный экспрессирующий кластер, который может содержаться в репликоне, таком как внехромосомный элемент (например, плазмиды), способный к устойчивому сохранению в хозяине, таком как дрожжи или бактерии. Репликон может иметь две системы репликации, что, таким образом, позволяет сохраняться ему, например, в дрожжах для экспрессии и в прокариотическом хозяине для клонирования и амплификации. Примеры таких шаттл-векторов бактерий-дрожжей включают YEp24 (Botstein и др., 1979), pCl/1 (Brake и др., 1984) и YRp17 (Stinchcomb и др., 1982). Кроме того, репликон может быть в плазмиде либо с высоким, либо с низким числом копий. Плазмида с высоким числом копий будет в основном иметь число копий в пределах от приблизительно 5 до приблизительно 200, и обычно от приблизительно 10 до приблизительно 150. Хозяин, содержащий плазмиду с высоким числом копий, будет предпочтительно иметь по крайней мере приблизительно 10, и более предпочтительно по крайней мере приблизительно 20 копий.
Эффективная промоторная последовательность дрожжей может быть получена из генов, кодирующих ферменты в метаболическом пути. Примеры таких генов включают алкогольдегидрогеназу (ADH) (EP 0 284044), енолазу, глюкокиназу, глюкозо-6-фосфат-изомеразу, глицеральдегид-3-фосфат-дегидрогеназу (GAP или GAPDH), гексокиназу, фосфофруктокиназу, 3-фосфоглицератмутазу и пируваткиназу (PyK) (EP 0 3215447). Ген PH05 дрожжей, кодирующий кислую фосфатазу, также предоставляет эффективные промоторные последовательности (Myanohara и др., 1983). Кроме того, синтетические промоторы, которые не встречаются в природе, также функционируют как промоторы дрожжей. Примеры таких гибридных промоторов включают регуляторную последовательность ADH, связанную с областью активации транскрипции GAP (Патенты США №№ 4876197 и 4880734). Примеры последовательностей терминатора транскрипции и других распознанных у дрожжей последовательностей терминации, таких как кодирующих гликолитические ферменты, известны специалистам в области техники.
В альтернативном варианте конструкты для экспрессии могут быть интегрированы в геном дрожжей с помощью интегрирующего вектора. Интегрирующие векторы обычно содержат по крайней мере одну последовательность, гомологичную хромосоме дрожжей, которая позволяет вектору интегрироваться, и предпочтительно содержит две гомологичных последовательности, ограничивающие конструкт для экспрессии. Интеграция является результатом рекомбинаций между гомологичной ДНК в векторе и хромосоме дрожжей (Orr-Weaver и др., 1983). Интегрирующий вектор может быть направлен к специфичному локусу в дрожжах с помощью выбора подходящей гомологичной последовательности для включения в вектор (см. Orr-Weaver и др., выше). Один или более конструктов для экспрессии могут интегрировать, возможно изменяя уровни продуцируемого рекомбинантного белка (Rine и др., 1983).
В настоящем изобретении также рассматривается трансформация нуклеотидной последовательности настоящего изобретения в растение для достижения ингибирующих паразита уровней экспрессии одной или более молекул дцРНК. Вектор трансформации может быть легко изготовлен с помощью способов, доступных в области техники. Вектор трансформации включает одну или более нуклеотидных последовательностей, которые способны подвергаться транскрипции до молекулы РНК и которые в основном гомологичны и/или комплементарны одной или более нуклеотидным последовательностям, закодированным геномом насекомого, так что после накопления РНК происходит понижающая регуляция экспрессии по крайней мере одной из соответствующих нуклеотидных последовательностей генома насекомого.
Вектор трансформации может именоваться конструктом дцДНК и может также быть определен как рекомбинантная молекула, агент контроля насекомых, генетическая молекула или химерный генетический конструкт. Химерный генетический конструкт настоящего изобретения может включать, например, нуклеотидные последовательности, кодирующие один или более антисмысловых транскриптов, один или более смысловых транскриптов, один или более каждого из вышеупомянутых, в котором весь или часть соответствующего транскрипта гомологична всей или части молекулы РНК, включающей последовательность РНК, закодированную нуклеотидной последовательностью в геноме насекомого.
В одном варианте осуществления вектор трансформации растения включает выделенную и очищенную молекулу ДНК, включающую промотор, операбельно связанный с одной или более нуклеотидными последовательностями настоящего изобретения. Нуклеотидная последовательность выбрана из группы, включающей от SEQ ID NO:1 до SEQ ID NO:906, как указано в списке последовательностей. Нуклеотидная последовательность содержит сегмент, кодирующий всю или часть РНК, присутствующей в пределах транскрипта РНК паразита-мишени, и может включать инвертированные повторы всей или части РНК паразита-мишени. Молекула ДНК, включающая вектор экспрессии, может также содержать функциональную последовательность интрона, расположенную либо выше кодирующей последовательности, либо даже в пределах кодирующей последовательности, и может также содержать пять первичных (5') нетранслированных лидерных последовательностей (то есть, UTR или 5'-UTR), расположенных между промотором и точкой инициации трансляции.
Вектор трансформации растения может содержать последовательности от более чем одного гена, таким образом, позволяя продуцировать более одной дцРНК для ингибирования экспрессии двух или более генов в клетках паразита-мишени. Специалисту в области техники ясно, что сегменты ДНК, последовательность которых соответствует присутствующей в различных генах, могут быть объединены в единый композитный сегмент ДНК для экспрессии в трансгенном растении. В альтернативном варианте плазмида настоящего изобретения, уже содержащая по крайней мере один сегмент ДНК, может быть изменена с помощью последовательной вставки дополнительных сегментов ДНК между последовательностями усилителя, промотора и терминатора. В агенте контроля насекомых настоящего изобретения, предназначенном для ингибирования многочисленных генов, гены, которые необходимо ингибировать, могут быть получены от тех же видов насекомых, чтобы усилить эффективность агента контроля насекомых. В некоторых вариантах осуществления гены могут быть получены из различных насекомых, чтобы расширить диапазон насекомых, против которых агент является эффективным. Когда предполагается супрессия или комбинация экспрессии и супрессии нескольких генов, полицистронный элемент ДНК может быть изготовлен, как иллюстрировано и раскрыто в Fillatti, Публикация заявки № US 2004-0029283.
Промоторы, которые функционируют в различных видах растений, также известны в области техники. К промоторам, эффективным для экспрессии полипептидов в растениях, относятся те, которые являются индуцируемыми, вирусными, синтетическими или конститутивными, как описано Odell и др. (1985), и/или промоторами, которые подвергаются временной регуляции, пространственной регуляции и пространственно-временной регуляции. Предпочтительные промоторы включают усиленные промоторы CaMVSSS и промотор FMV35S. Для настоящего изобретения, например для оптимального контроля видов, которые питаются корнями, может быть предпочтительным достижение наивысших уровней экспрессии этих генов в корнях растений. Множество промоторов с повышенным содержанием в корнях идентифицировано и известно в области техники (Lu и др., 2000; Патенты США №№ 5837848 и 6489542).
Вектор или конструкт с рекомбинантной ДНК настоящего изобретения будут обычно включать селектируемый маркер, который придает выбираемый фенотип растительным клеткам. Селектируемые маркеры могут также применяться для выбора растений или растительных клеток, которые содержат экзогенные нуклеиновые кислоты, кодирующие полипептиды или белки настоящего изобретения. Маркер может кодировать устойчивость к биоцидам, устойчивость к антибиотикам (например, канамицину, G418 блеомицину, гигромицину и т.д.) или устойчивость к гербицидам (например, глифозат и т.д.). Примеры селектируемых маркеров включают, но не ограничиваясь ими, ген neo, который кодирует устойчивость к канамицину и может быть выбран для применения канамицина, G418 и т.д., ген bar, который кодирует устойчивость к биалафосу; мутантный ген синтазы EPSP, который кодирует устойчивость к глифосфату; ген нитрилазы, который придает устойчивость к бромоксинилу; мутантный ген ацетолактатсинтазы (ALS), который придает устойчивость к имидазолину или сульфонилуреазе; и ген устойчивости к метотрексату DHFR. Примеры таких селектируемых маркеров описаны в патентах США №№ 5550318; 5633435; 5780708 и 6118047.
Рекомбинантный вектор или конструкт настоящего изобретения может также содержать скринируемый маркер. Скринируемые маркеры могут применяться для мониторинга экспрессии. Примеры скринируемых маркеров включают ген β-глюкуронидазы или uidA (GUS), который кодирует фермент, различные хромогенные субстраты которого известны (Jefferson, 1987; Jefferson и др., 1987); ген R-локуса, который кодирует продукт, который регулирует продукцию антоцианиновых пигментов (красный цвет) в тканях растений (Dellaporta и др., 1988); ген β-лактамазы (Sutcliffe и др., 1978), ген, который кодирует фермент, различные хромогенные субстраты которого известны (например, PAD AC, хромогенный цефалоспорин); ген люциферазы (Ow и др., 1986) ген xylE (Zukowsky и др., 1983), который кодирует катехолдиоксигеназу, которая может конвертировать хромогенные катехолы; ген cc-амилазы (Ikatu и др., 1990); ген тирозиназы (Katz и др., 1983), который кодирует фермент, способный окислять тирозин до DOPA и допахинона, который в свою очередь конденсируется до меланина; α-галактозидазу, которая катализирует хромогенный субстрат α-галактозу.
К предпочтительным векторам трансформации растений относятся полученные из Ti плазмиды Agrobacterium tumefaciens (например, Патенты США №№ 4536475, 4693977, 4886937, 5501967 и EP 0122791). Плазмиды Agrobacterium rhizogenes (или "Ri") также эффективны и известны в области техники. К другим предпочтительным векторам трансформации растений относятся те, что раскрыты, например, Herrera-Estrella (1983); Bevan (1983), Klee (1985) и в EP 0 120 516.
В основном предпочтительно включение функциональной рекомбинантной ДНК в неспецифичное местоположение в геном растения. В специальных случаях может быть эффективной вставка рекомбинантного конструкта ДНК с помощью сайт-специфичной интеграции. Существует несколько систем сайт-специфичной рекомбинации, которые известны как функционирующие импланты, и к ним относятся cre-lox, раскрытый в Патенте США 4959317 и FLP-FRT, раскрытый в Патенте США 5527695.
Полагают, что подходящие способы для трансформации клеток хозяина для применения в настоящем изобретении включают фактически любой способ, с помощью которого ДНК может быть введен в клетку, например с помощью непосредственной доставки ДНК, например с помощью PEG-опосредованной трансформации протопластов (Omirulleh и др., 1993), десикационно/ингибиторно-опосредованного захвата ДНК (Potrykus и др., 1985), электропорации (Патент США 5384253), перемешивания с кремний-карбидными волокнами (Kaeppler и др., 1990; Патент США № 5302523; и Патент США № 5464765), Agrobacterium-опосредованной трансформации (Патент США № 5591616 и Патент США № 5563055) и ускорения покрытых ДНК частиц (Патент США № 5550318; Патент США № 5538877; и Патент США № 5538880) и т.д. Применяя технологии, такие как эти, клетки фактически любых видов могут быть устойчиво трансформированы. В случае многоклеточных видов трансгенные клетки могут быть регенерированы в трансгенные организмы.
Способы создания трансгенных растений и экспрессии гетерологичных нуклеиновых кислот в растениях в частности известны и могут применяться с предоставленными здесь нуклеиновыми кислотами для получения трансгенных растений, которые имеют пониженную восприимчивость к поеданию организмом паразита-мишени, таким как кукурузный жук. Векторы для трансформации растения могут быть получены, например, с помощью вставки раскрытых здесь нуклеиновых кислот, продуцирующих дцРНК, в векторы для трансформации растения и введения их в растения. Одна известная векторная система была получена с помощью модификации природной системы переноса генов Agrobacterium tumefaciens. Природная система включает большие Ti(опухоль-индуцирующие)-плазмиды, включающие большой сегмент, известный как T-ДНК, который переносится в трансформированные растения. Другой сегмент Ti-плазмиды, область vir, ответственен за перенос T-ДНК. Область T-ДНК ограничена терминальными повторами. В модифицированных бинарных векторах опухоль-индуцирующие гены удалены, и функции области vir используются для переноса чужеродной ДНК, ограничивающими последовательностями T-ДНК. T-область может также содержать селектируемый маркер для эффективного восстановления трансгенных растений и клеток, и участок многократного клонирования для вставки последовательностей для переноса, таких как кодирующая дцРНК нуклеиновая кислота.
Трансгенное растение, сформированное с помощью способов трансформации Agrobacterium, обычно содержит отдельную простую рекомбинантную последовательность ДНК, вставленную в одну хромосому, и называется трансгенным объектом. Такие трансгенные растения могут являться гетерозиготными для вставленной экзогенной последовательности. Трансгенное растение, гомозиготное относительно трансгена, может быть получено с помощью самоопыления самостоятельного изолированного трансгенного растения для продукции семян F1. Одна четверть полученных семян F1 будет гомозиготными относительно трансгена. После прорастания семян F1 образуются растения, которые могут тестироваться на гетерозиготность или гомозиготность, обычно с помощью анализа SNP или термического анализа амплификации, который учитывает различие между гетерозиготами и гомозиготами (то есть, анализа зиготности).
C. Экспрессия нуклеиновой кислоты и супрессия гена-мишени
Настоящее изобретение предоставляет, в качестве примера, трансформированный организм хозяина или симбионта паразита мишени, трансформированные растительные клетки и трансформированные растения и их потомство. Трансформированные растительные клетки и трансформированные растения могут быть сконструированы для экспрессии одной или более описанных здесь последовательностей дцРНК или siРНК, для обеспечения эффекта защиты от паразита. Эти последовательности могут применяться для генной супрессии в организме паразита, таким образом уменьшая паразитирование паразита на защищенном трансформированном организме хозяина или симбионта. Применяемое здесь выражение "генная супрессия" относится к любому известному способу уменьшения уровней генной транскрипции до мРНК и/или последующей трансляции мРНК.
Генная супрессия также предназначена для обозначения уменьшения экспрессии белка от гена или кодирующей последовательности, включая посттранскрипционную генную супрессию и транскрипционную супрессию. Посттранскрипционная генная супрессия опосредуется гомологией между всей или частью мРНК, транскрибированной от гена или кодирующей последовательности, предназначенной для супрессии, и соответствующей двухцепочечной РНК, применяемой для супрессии, и относится к реальному и измеримому уменьшению количества доступной мРНК, доступной в клетке для связывания с рибосомами. Транскрибированная РНК может иметь смысловую ориентацию для эффекта, называемого ко-супрессией, антисмысловую ориентацию для эффекта, называемого антисмысловой супрессией, или обе ориентации, продуцирующие дцРНК для эффекта, называемого интерференцией РНК (РНКi).
Транскрипционная супрессия опосредуется присутствием в клетке агента супрессии гена дцРНК, имеющего последовательность, в основном идентичную последовательности промотора ДНК или его комплемента, для эффекта, который называется транссупрессией промотора. Генная супрессия может быть эффективной в отношении природного гена растения, связанного с определенным признаком, например, для предоставления растений со сниженными уровнями белка, кодируемого природным геном, или с повышенными или сниженными уровнями метаболита, на который оказывается влияние. Генная супрессия может также быть эффективной в отношении генов-мишеней у паразитов растений, которые могут поедать или контактировать с растительным материалом, содержащим агенты супрессии генов, специфично разработанные для ингибирования или подавления экспрессии одной или более гомологичных или комплементарных последовательностей в клетках паразита. Посттранскрипционная генная супрессия антисмысловой или ориентированной смысловой РНК для регуляции экспрессии генов в растительных клетках раскрыта в Патентах США №№ 5107065, 5759829, 5283184 и 5231020. Применение дцРНК для супрессии генов в растениях раскрыто в WO 99/53050, WO 99/49029, публикации заявки на патент США № 2003/0175965 и 2003/0061626, заявке на патент США № 10/465800 и патентах США №№ 6506559 и 6326193.
В полезном способе посттранскрипционной генной супрессии в растениях используются как ориентированная смысловая, так и ориентированная антисмысловая, транскрибированная РНК, которая является стабилизированной, например, в виде структуры шпильки, стебля и петли. В предпочтительном конструкте ДНК для осуществления посттранскрипционной супрессии генов первый сегмент кодирует РНК, имеющую антисмысловую ориентацию, в основном идентичную сегменту гена, супрессия которого предполагается, который связан со вторым сегментом в смысловой ориентации, кодирующим РНК, в основном комплементарную первому сегменту. Такой конструкт формирует структуру стебель-петля с помощью гибридизации первого сегмента со вторым сегментом и структуру петли из нуклеотидных последовательностей, связывающих два сегмента (см. W094/01550, W098/05770, US 2002/0048814 и US 2003/0018993).
Согласно одному варианту осуществления настоящего изобретения предоставляется нуклеотидная последовательность, in vitro экспрессия которой приводит к транскрипции устойчивой последовательности РНК, которая в основном гомологична молекуле РНК заданного гена у насекомого, которая включает последовательность РНК, закодированную нуклеотидной последовательностью в геноме насекомого. Таким образом, после поедания насекомым устойчивой последовательности РНК, включенной в пищевой рацион или распыленной на поверхности растения, запускается понижающая регуляция нуклеотидной последовательности, соответствующей гену-мишени в клетках насекомого-мишени.
Ингибирование гена-мишени с помощью технологии устойчивой дцРНК настоящего изобретения является последовательность-специфичным, так как нуклеотидные последовательности, соответствующие двухспиральной области РНК, предназначены для генетического ингибирования. РНК, содержащая нуклеотидные последовательности, идентичные части гена-мишени является предпочтительной для ингибирования. Установлено, что последовательности РНК со вставками, делециями и единичными точечными мутациями относительно заданной последовательности также эффективны для ингибирования. При осуществлении настоящего изобретения предпочтительно, чтобы ингибиторная дцРНК и часть гена-мишени имели по крайней мере приблизительно от 80% идентичность последовательности, или приблизительно от 90% идентичность последовательности, или приблизительно от 95% идентичность последовательности, или приблизительно от 99% идентичность последовательности, или даже приблизительно 100% идентичность последовательности. В альтернативном варианте двухспиральная область РНК может быть определена функционально как нуклеотидная последовательность, которая способна гибридизироваться с частью транскрипта гена-мишени. Последовательность с длиной, меньшей чем полная длина, имеющая большую гомологию, компенсирует более длинную, но менее гомологичную последовательность. Длина идентичных нуклеотидных последовательностей может составлять по крайней мере приблизительно 25, 50, 100, 200, 300, 400, 500 или по крайней мере приблизительно 1000 оснований. Обычно, необходимо применять последовательность, состоящую из более 20-100 нуклеотидов, хотя последовательность из более чем приблизительно 200-300 нуклеотидов будет предпочтительнее, а последовательность из более чем приблизительно 500-1000 нуклеотидов будет особенно предпочтительной, в зависимости от размера гена-мишени. Изобретение имеет преимущество в том, что допускает вариации последовательности, которые можно ожидать вследствие генетических мутаций, полиморфизма штамма или эволюционной дивергенции. Включенная молекула нуклеиновой кислоты может не быть абсолютной гомологичной, может не иметь полной длины в сравнении с первичным продуктом транскрипции или полностью обработанной мРНК гена-мишени. Поэтому специалист в области техники должен понимать, что, как раскрыто здесь, 100% идентичность последовательности между РНК и геном-мишенью не является обязательной для осуществления настоящего изобретения.
Ингибирование экспрессии гена-мишени может быть определено количественно с помощью определения либо эндогенной РНК-мишени, либо белка, продуцируемого посредством трансляции РНК-мишени, и последствия ингибирования могут быть подтверждены с помощью изучения внешних свойств клетки или организма. Технологии количественного определения РНК и белков известны специалистам в области техники. Доступны многократные селектируемые маркеры, которые придают устойчивость к ампициллину, блеомицину, хлорамфениколу, гентамицину, гигромицину, канамицину, линкомицину, метотрексату, фосфинотрицину, пуромицину, спектиномицину, рифампицину и тетрациклину и т.п.
В некоторых вариантах осуществления экспрессия гена ингибируется по крайней мере на 10%, предпочтительно по крайней мере на 33%, более предпочтительно по крайней мере на 50%, и еще более предпочтительно по крайней мере на 80%. В особенно предпочтительных вариантах осуществления изобретения экспрессия гена ингибируется по крайней мере на 80%, более предпочтительно по крайней мере на 90%, более предпочтительно по крайней мере на 95%, или по крайней мере на 99% в клетках насекомого, таким образом, имеет место значительное ингибирование. Значительное ингибирование предназначено относится к достаточному ингибированию, которое приводит к поддающемуся обнаружению фенотипическому изменению (например, прекращению роста личинки, параличу или гибели и т.д.) или поддающемуся обнаружению уменьшению РНК и/или белка, соответствующего ингибированному гену-мишени. Несмотря на то, что в некоторых вариантах осуществления изобретения ингибирование происходит в основном во всех клетках насекомого, в других предпочтительных вариантах осуществления ингибирование происходит только в субпопуляции клеток, экспрессирующих ген. Например, если ген, который необходимо ингибировать, играет важную роль в клетках пищеварительного тракта насекомого, ингибирования гена в этих клетках достаточно, чтобы оказать отрицательное воздействие на насекомого.
Молекулы дцРНК могут синтезироваться или in vivo или in vitro. дцРНК может быть сформирован с помощью единичной самокомплементарной цепи РНК или из двух комплементарных цепей РНК. Эндогенная РНК полимераза клетки может опосредовать транскрипцию in vivo, или клонированная РНК полимераза может применяться для транскрипции in vivo или in vitro. Ингибирование может направляться с помощью специфичной транскрипции в органе, ткани или типе клеток; стимуляции внешних условий (например, инфекции, стресса, температуры, химических индукторов); и/или инженерной транскрипции на определенной стадии развития или в определенном возрасте. Цепи РНК могут быть или могут не быть полиаденилированы; цепи РНК могут быть или могут не быть способны к трансляции в полипептид с помощью клеточного аппарата трансляции.
РНК, дцРНК, siРНК или miРНК настоящего изобретения может быть получен химически или ферментативно специалистом в области техники посредством неавтоматических или автоматизированных реакций, или in vivo в другом организме. РНК может также быть получен с помощью частичного или полностью органического синтеза; любой модифицированный рибонуклеотид может быть включен с помощью in vitro ферментного или органического синтеза. РНК может быть синтезирован с помощью клеточной РНК полимеразы или РНК полимеразы бактериофага (например, T3, T7, SP6). Применение и продукция конструкта для экспрессии известны в области техники (см., например, WO 97/32016; Патенты США №№ 5593874, 5698425, 5712135, 5789214 и 5804693). В случае химического синтеза или ферментного синтеза in vitro РНК может быть очищен до введения в клетку. Например, РНК может быть очищен от смеси с помощью экстракции с растворителем или смолой, преципитации, электрофореза, хроматографии или их комбинации. В альтернативном варианте РНК может применяться в отсутствии или с минимальной очисткой, чтобы избежать потерь вследствие процессинга образца. РНК может быть высушена для хранения или растворена в водном растворе. Раствор может содержать буферы или соли для стимуляции ренатурации и/или стабилизации двойных цепей.
Для транскрипции от трансгена in vivo или конструкта для экспрессии может применяться регуляторная область (например, промотор, усилитель, глушитель и область полиаденилирования) для транскрипции цепи РНК (или цепей). Поэтому, в одном варианте осуществления нуклеотидные последовательности, предназначенные для применения в получении молекул РНК, могут быть операбельно связанны с одной или более промоторными последовательностями, функциональными в микроорганизме, грибке или клетке-хозяине растения. В идеале, нуклеотидные последовательности помещают под контролем эндогенного промотора, обычного компонента в геноме хозяина. Нуклеотидная последовательность настоящего изобретения под контролем операбельно связанной промоторной последовательности может далее быть ограничена дополнительными последовательностями, которые преимущественно влияют на транскрипцию и/или стабильность образующегося в результате транскрипта. Такие последовательности в основном располагаются выше операбельно связанного промотора и/или ниже 3' конца конструкта для экспрессии и могут находиться как выше промотора, так и ниже 3' конца конструкта для экспрессии, несмотря на то, что такая последовательность, находящаяся только выше, также предусмотрена.
Применяемый здесь термин "агент контроля насекомых" или "агент супрессии гена" относится к специфической молекуле РНК, включающей первый сегмент РНК и второй сегмент РНК, в которой комплементарность между первым и вторым сегментами РНК приводит к способности двух сегментов гибридизировать in vivo и in vitro с формированием двухцепочечной молекулы. В основном может быть предпочтительным включение третьего сегмента РНК, связывающего и стабилизирующего первую и вторую последовательности так, что все структуры формируют структуры стебля и петли, или еще более сильно гибридизирующие структуры могут формировать узловую структуру стебель-петля. В альтернативном варианте симметричная шпилька может быть сформирована без третьего сегмента, в котором нет никакой сконструированной петли, но по стерическим причинам шпилька будет формировать собственную петлю, если стебель имеет достаточную длину, чтобы стабилизировать. Первый и второй сегменты РНК будут в основном находиться в пределах длины молекулы РНК и будут являться в основном инвертированными повторами друг друга и будут соединены третьим сегментом РНК. Первый и второй сегменты соответствуют неизменно и несравненно смысловой и антисмысловой последовательностям по отношению к РНК-мишени, транскрибируемой от гена-мишени в заданном насекомом паразите, который подвергается супрессии с помощью приема внутрь молекулы дцРНК. Агент контроля насекомых может также быть представлен в основном очищенной (или изолированной) молекулой нуклеиновой кислоты и более конкретно молекулами нуклеиновой кислоты или молекулами фрагментов нуклеиновой кислоты от геномной ДНК (gДНК) или кДНК библиотеки. В альтернативном варианте фрагменты могут включать меньшие олигонуклеотиды, имеющие от приблизительно 15 до приблизительно 250 нуклеотидных остатков, и более предпочтительно, от приблизительно 15 до приблизительно 30 нуклеотидных остатков.
Применяемый здесь термин "геном" в отношении клеток насекомого или хозяина охватывает не только хромосомную ДНК, находящуюся в ядре, но и ДНК органелл, находящихся в пределах субклеточных компонентов клетки. ДНК настоящего изобретения, введенная в клетки растения, поэтому может быть либо интегрирована с хромосомой, либо локализована в органелле. Термин "геном", в отношении бактерий, охватывает и хромосому и плазмиды в пределах бактериальной клетки-хозяина. ДНК настоящего изобретения, введенная в бактериальные клетки-хозяева, поэтому может быть либо интегрирована с хромосомой, либо локализована в плазмиде.
Применяемый здесь термин "паразит" относится к насекомым, паукообразным насекомым, ракообразным, грибам, бактериям, вирусам, нематодам, плоским червям, аскаридам, острицам, анкилостомам, ленточным червям, трипаносомам, шистосомам, оводам, блохам, зудням, клещам, вшам и т.п., которые распространены в среде, окружающей человека, и которые могут поедать или контактировать с одной или более клетками, тканями или жидкостями, продуцируемыми хозяином паразита или симбионтом, трансформированным для экспрессии, или покрытым двухцепочечным агентом супрессии гена, или которые могут поедать растительный материал, содержащий агент супрессии гена. Применяемый здесь признак "устойчивость к паразиту" является характеристикой трансгенного растения, трансгенного животного, трансгенного хозяина или трансгенного симбионта, которая вызывает устойчивость растения, животного, хозяина или симбионта к нападению паразита, который в обычных условиях способен вызывать повреждения или потерю растения, животного, хозяина или симбионта. Такая устойчивость к паразиту может являться результатом спонтанной мутации или более часто инкорпорации рекомбинантной ДНК, которая вызывает устойчивость к паразиту. Для придания устойчивости насекомого к трансгенному растению рекомбинантная ДНК может, например, быть транскрибирована в молекулу РНК, которая формирует молекулу дцРНК в тканях или жидкостях рекомбинантного растения. Молекула дцРНК включает часть сегмента РНК, который является идентичным соответствующему сегменту РНК, закодированному последовательностью ДНК в насекомом паразите, который предпочтительно питается рекомбинантным растением. Экспрессия гена в насекомом паразите-мишени подвергается супрессии с помощью дцРНК, и супрессия экспрессии гена в насекомом паразите мишени приводит к формированию растения, устойчивого к насекомому. Fire и др. (Патент США № 6506599) в общем описал ингибирование поражения паразитами, предоставляя специфические особенности только приблизительно нескольких нуклеотидных последовательностей, которые были эффективны для ингибирования функции генов у нематод вида Caenorhabditis elegans. Подобным образом Plaetinck и др. (US 2003/0061626) описал применение дцРНК для ингибирования функции генов у множества паразитов нематод. Mesa и др. (US 2003/0150017) описал применение последовательности дцДНК для трансформации клеток хозяина для экспрессии соответствующих последовательностей дцРНК, которые в основном идентичны последовательностям-мишеням в специфичных патогенах, и, в частности, описал создание рекомбинантных растений, экспрессирующих такие последовательности дцРНК, для поедания различными вредителями растений, способствуя понижающей регуляции гена в геноме паразита и повышению устойчивости растения к поражению паразитами.
Настоящее изобретение обеспечивает ингибированную экспрессию одного или множества генов-мишеней в паразите-мишени с помощью способов стабилизированной дцРНК. Изобретение особенно эффективно в модуляции экспрессии эукариотических генов, в частности модуляция экспрессии генов, находящихся в паразитах, которые имеют уровень pH в пищеварительной системе от приблизительно 4,5 до приблизительно 9,5, более предпочтительно от приблизительно 5,0 до приблизительно 8,0, и еще более предпочтительно от приблизительно 6,5 до приблизительно 7,5. Для вредителей растений с пищеварительной системой, которая имеет уровень pH вне этих диапазонов, может быть желательным применение способов доставки, которые не требуют приема внутрь молекул дцРНК.
Модулирующий эффект дцРНК применим к множеству генов, экспрессируемых в паразитах, включая, например, эндогенные гены, ответственные за клеточный метаболизм или клеточную трансформацию, включая гены поддержания жилища, факторы транскрипции и другие гены, которые кодируют полипептиды, задействованные в клеточном метаболизме.
Применяемая здесь фраза "ингибирование экспрессии гена" или "ингибированная экспрессия гена-мишени в клетке насекомого" относится к отсутствию (или объективному уменьшению) уровня белка и/или продукта мРНК от гена-мишени. Специфичность относится к способности ингибировать ген-мишень без обнаруживаемых эффектов на другие гены клетки и без любых эффектов на любой ген в клетке, которая продуцирует молекулу дцРНК. Ингибирование экспрессии гена-мишени у насекомого-паразита может привести к новым фенотипическим признакам у насекомого-паразита.
Настоящее изобретение частично предоставляет систему доставки для доставки агентов контроля насекомых к насекомым посредством воздействия на их пищевой рацион, содержащую агенты контроля насекомых настоящего изобретения. В соответствии с одним из вариантов осуществления стабилизированные молекулы дцРНК или siРНК могут быть включены в пищевой рацион насекомого или могут быть нанесены на поверхность поедаемого субстрата для поедания насекомым. Настоящее изобретение также частично предоставляет систему доставки для доставки агентов контроля насекомых к насекомым посредством воздействия на них микроорганизма или хозяина, такого как растение, содержащее агенты контроля насекомых настоящего изобретения, с помощью скармливания клеток микроорганизма или хозяина, или содержимого клеток. В соответствии с другим вариантом осуществления настоящее изобретение включает создание трансгенной растительной клетки или растения, которое содержит рекомбинантный конструкт ДНК, транскрибирующий стабилизированные молекулы дцРНК настоящего изобретения. Применяемая здесь фраза "создание трансгенной растительной клетки или растения" относится к способам использования рекомбинантных технологий ДНК, легко доступных в области техники (например, Sambrook, и др., 1989), для создания вектора трансформации растения, транскрибирующего стабилизированные молекулы дцРНК настоящего изобретения, для трансформации растительной клетки или растения и создания трансгенной растительной клетки или трансгенного растения, которое содержит транскрибированные, стабилизированные молекулы дцРНК.
В еще одном варианте осуществления непатогенные, аттенуированные штаммы микроорганизмов могут применяться в качестве носителя для агентов контроля насекомых и, с этой точки зрения, микроорганизмы, несущие такие агенты, также называются агентами контроля насекомых. Микроорганизмы могут быть сконструированы для экспрессии нуклеотидной последовательности гена-мишени, для продукции молекул РНК, включающих последовательности РНК, гомологичные или комплементарные последовательностям РНК, обычно обнаруживаемым в клетках насекомого. Воздействие микроорганизмов на насекомых приводит к приему внутрь микроорганизмов и понижающей регуляции экспрессии генов-мишеней, опосредованных прямо или косвенно молекулами РНК, или их фрагментами, или производными.
В альтернативном варианте настоящее изобретение предоставляет воздействие на насекомого агентов контроля насекомых настоящего изобретения, включенных в мешалку с распылением и нанесения на поверхность хозяина, такого как растение-хозяин. В примере варианта осуществления поедание насекомым агентов контроля насекомых приводит к доставке агентов контроля насекомых в кишечник насекомого и затем в клетки организма насекомого. В другом варианте осуществления инфицирование насекомого агентами контроля насекомых с помощью других средств, таких как инъекции или другие физические способы, также позволяет доставить агенты контроля насекомых. В еще одном варианте осуществления молекулы непосредственно РНК инкапсулируют в синтетическую матрицу, такую как полимер, и наносят на поверхность хозяина, такого как растение. Поедание насекомым клеток хозяина позволяет доставить агенты контроля насекомых в насекомое и приводит к понижающей регуляции гена-мишени у хозяина.
Предполагается, что композиции настоящего изобретения могут быть включены в семена определенных видов растений либо в виде продукта экспрессии рекомбинантного гена, включенного в геном растительных клеток, либо с помощью включения в покрытие или в средство для обработки семян, которое наносят на семена перед посадкой. Считается, что растительная клетка, содержащая рекомбинантный ген, является здесь трансгенным объектом.
Полагается, что пестицидное средство для обработки семян может предоставить значительные преимущества при объединении с трансгенным объектом, который обеспечивает защиту от поражения жесткокрылыми паразитами, который находится в пределах предпочтительного диапазона эффективности в отношении защиты от паразита-мишени. Кроме того, полагается, что бывают ситуации, известные специалистам в области техники, когда благоприятно иметь такие трансгенные объекты, которые находятся в пределах предпочтительного диапазона эффективности.
Настоящее изобретение частично предоставляет систему доставки для доставки агентов контроля насекомых в насекомых. Стабилизированные молекулы дцРНК или siРНК настоящего изобретения могут быть непосредственно введены в клетки насекомого, или введены во внеклеточное пространство, интерстициальное пространство, лимфатическую систему, пищеварительную систему в кровоток насекомого посредством орального приема внутрь или других способов, которые может использовать специалист в области техники. Способы для орального введения могут включать прямое смешивание РНК с пищей насекомого, а также разработку методов, в которых виды, которые используются в качестве пищи, конструируют для экспрессии дцРНК или siРНК, затем скармливают насекомому для оказания на него воздействия. Например, в одном варианте осуществления молекулы дцРНК или siРНК могут быть включены внутрь или нанесены на поверхность пищевого субстрата насекомого. В другом варианте осуществления РНК может быть распылена на поверхность растения. В еще одном варианте осуществления дцРНК или siРНК могут экспрессироваться микроорганизмами, а микроорганизмы могут наноситься на поверхность растения или вводиться в корень, стебель с помощью физического способа, такого как инъекция. В еще одном варианте осуществления растение может быть генетически сконструировано для экспрессии дцРНК или siРНК в количестве, достаточном, чтобы вызвать гибель насекомых, которые, как известно, инфицируют растение.
В частности, при осуществлении настоящего изобретения в WCR стабилизированная дцРНК или siРНК может быть введена в среднюю кишку насекомого и достигать желательного ингибирования заданных генов. Молекулы дцРНК или siРНК могут быть включены в пищевой рацион или нанесены на пищевой субстрат как обсуждено выше и могут поедаться насекомыми. В любом случае, дцРНК's настоящего изобретения предоставляются в диете паразита-мишени. Паразит-мишень настоящего изобретения будет иметь pH пищеварительного тракта от приблизительно 4,5 до приблизительно 9,5, или от приблизительно 5 до приблизительно 8,5, или от приблизительно 6 до приблизительно 8, или от приблизительно 6,5 до приблизительно 7,7, или приблизительно 7,0. Пищеварительный тракт паразита-мишени определен здесь как орган, локализующийся в организме паразита, в котором пищевой субстрат, который поедается паразитом-мишенью, подвергается воздействию среды, которая является благоприятной для поглощения молекул дцРНК настоящего изобретения, не изменяя pH настолько значительно, что приводило бы к диссоциации водородных связей между двойными цепями дцРНК и формированию одноцепочечных молекул.
Также ожидается, что дцРНК's, продуцируемые с помощью химического или ферментативного синтеза могут быть сформулированы таким образом, что будут соответствовать общепринятой сельскохозяйственной практике и их можно будет применять в виде распыляемых продуктов для контроля поражения насекомыми. Композиции могут включать подходящие связующие вещества и смачиватели, необходимые для эффективного покрытия листвы, а также UV-протектанты, для защиты дцРНК от повреждения под воздействием UV-излучения. Такие добавки обычно применяются в биоинсектицидной промышленности и известны специалистам в области техники. Такое применение можно комбинировать с другими методами распыления инсектицидов, биологически обоснованными или нет для усиления защиты растений от повреждения вследствие поедания насекомыми.
Настоящие изобретатели предполагают, что бактериальные штаммы, продуцирующие инсектицидные белки, можно применять для продукции дцРНК с целью контроля насекомых. Эти штаммы могут иметь улучшенные свойства контроля насекомых. Для создания дцРНК для контроля насекомых можно использовать множество различных бактериальных хозяев. Примеры бактерий могут включать E. coli, B. thuringiensis, Pseudomonas sp., Photorhabdus sp., Xenorhabdus sp., Serratia entomophila и родственные Serratia sp., B. sphaericus, B. cereus, B. laterospams, B. popilliae, Clostridium bifermentans и другие виды Clostridium или другие спорообразующие грамположительные бактерии. В некоторых вариантах осуществления бактерии могут быть сконструированы для контроля паразитов, таких как москиты.
Настоящее изобретение также относится к рекомбинантным конструктам ДНК для экспрессии в микроорганизме. Экзогенные нуклеиновые кислоты, от которых транскрибируется интересующая РНК, могут быть введены в микробную клетку-хозяина, такую как бактериальная клетка или грибковая клетка, применяя способы, известные в области техники.
Нуклеотидные последовательности настоящего изобретения могут быть введены в широкий спектр прокариотических и эукариотических микроорганизмов-хозяев для продукции стабилизированных молекул дцРНК или siРНК. Термин "микроорганизм" включает прокариотические и эукариотические микробные виды, такие как бактерии, грибы и водоросли. Грибы включают дрожжи и нитчатые грибы, и др. К иллюстративным прокариотам, как к грамотрицательным, так и грамположительным, относятся Enterobacteriaceae, такие как Escherichia, Erwinia, Shigella, Salmonella, и Proteus; Bacillaceae; Rhizobiceae, такие как Rhizobium; Spirillaceae, такие как photobacterium, Zymomonas, Serratia, Aeromonas, Vibrio, Desulfovibrio, Spirillum; Lactobacillaceae; Pseudomonadaceae, такие как Pseudomonas и Acetobacter; Azotobacteraceae, Actinomycetales, и Nitrobacteraceae. К эукариотам относятся грибы, такие как Phycomycetes и Ascomycetes, которые включают дрожжи, такие как Saccharomyces и Schizosaccharomyces; и Basidiomycetes, такие как Rhodotorula, Aureobasidium, Sporobolomyces и т.п.
Для защиты растений от насекомых большое количество микроорганизмов, являющихся известными обитателями филлопланы (поверхность листов растения) и/или ризосферы (почва, окружающая корни растения) широкого ряда важных зерновых культур, могут также являться желательными клетками-хозяевами для манипуляции, распространения, хранения, доставки и/или мутагенеза раскрытых рекомбинантных конструктов. К этим микроорганизмам относятся бактерии, водоросли и грибы. Особенно интересны такие микроорганизмы, как бактерии, например, рода Bacillus (включая виды и подвиды B. thuringiensis kurstaki HD-1, B. thuringiensis kurstaki HD-73, B. thuringiensis sotto, B. thuringiensis berliner, B. thuringiensis thuringiensis, B. thuringiensis tolworthi, B. thuringiensis dendrolimus, B. thuringiensis alesti, B. thuringiensis galleriae, B. thuringiensis aizawai, B. thuringiensis subtoxicus, B. thuringiensis entomocidus, B. thuringiensis tenebrionis и B. thuringiensis san diego); Pseudomonas, Erwinia, Serratia, Klebsiella, Zanthomonas, Streptomyces, Rhizobium, Rhodopseudomonas, Methylophilius, Agrobacterium, Acetobacter, Lactobacillus, Arthrobacter, Azotobacter, Leuconostoc, и Alcaligenes; грибы, в частности дрожжи, например, рода Saccharomyces, Cryptococcus, Kluyveromyces, Sporobolomyces, Rhodotorula и Aureobasidium. Особенно интересны такие бактериальные виды фитосферы, как Pseudomonas syringae, Pseudomonas fluorescens, Serratia marcescens, Acetobacter xylinum, Agrobacterium tumefaciens, Rhodobacter sphaeroides, Xanthomonas campestris, Rhizobium melioti, Alcaligenes eutrophus и Azotobacter vinlandii; и виды дрожжей фитосферы, такие как Rhodotorula rubra, R. glutinis, R. marina, R. aurantiaca, Cryptococcus albidus, C. diffluens, C. laurentii, Saccharomyces rosei, S. pretoriensis, S. cerevisiae, Sporobolomyces roseus, S. odorus, Kluyveromyces veronae и Aureobasidium pollulans.
D. Трансгенные растения
Настоящее изобретение предоставляет семена и растения, имеющие один или более трансгенных объектов. Комбинации объектов упоминаются как "многоуровневые" трансгенные объекты. Эти многоуровневые трансгенные объекты могут быть объектами, которые направлены на одного и того же паразита-мишень, или они могут быть направлены на различных паразитов-мишеней. В одном варианте осуществления семя, обладающее способностью экспрессировать предоставляемую здесь нуклеиновую кислоту, также обладает способностью экспрессировать по крайней мере один другой инсектицидный агент, включая, но не ограничиваясь ими, молекулу РНК, последовательность которой получена из последовательности РНК, экспрессируемой в паразите-мишени, и которая образует двухцепочечную структуру РНК после экспрессии в семени или клетках растения, выращенного из семени, при этом прием внутрь одной или более клеток растения паразитом-мишенью приводит к супрессии экспрессии РНК в клетках паразита-мишени.
В некоторых вариантах осуществления семя, обладающее способностью экспрессировать дцРНК, последовательность которой получена от паразита-мишени, также имеет трансгенный объект, который обеспечивает толерантность к гербицидам. Один полезный пример гена толерантности к гербицидам обеспечивает устойчивость к глифосфату, N-(фосфонометил)глицину, включая форму соли изопропиламина такого гербицида.
В настоящем способе комбинация экспрессии инсектицидного количества дцРНК в клетках трансгенного семени или растения, выращенного из семени, с обработкой семени или растения определенными химическими или белковыми пестицидами может применяться для обеспечения неожиданных синергичных преимуществ, включая неожиданно превосходящую эффективность защиты образующегося в результате трансгенного растения от повреждения паразитом-мишенью. В специфических вариантах осуществления обработка трансгенного семени, способного экспрессировать некоторые конструкты, которые формируют молекулы дцРНК, последовательность которых получают из одной или более последовательностей, экспрессируемых в кукурузном жуке, в количестве от приблизительно 100 граммов до приблизительно 400 граммов пестицида на 100 кг семени, обеспечивает неожиданно превосходящую защиту от кукурузного жука. Кроме того, полагают, что такие комбинации также эффективны в защите полупогруженных растений от истребления другими паразитами. Семена настоящего изобретения могут также применяться для уменьшения стоимости использования пестицида, так как для достижения необходимой защиты можно применять меньшее количество пестицида, чем когда такие способы не применяются. Кроме того, вследствие применения меньшего количества пестицида и вследствие его применения до засевания и без нанесения на отдельное поле полагают, что рассматриваемый способ, таким образом, является более безопасным для использующего его человека и для окружающей среды и потенциально менее дорогостоящий, чем обычные способы.
"Синергичный" означает наличие синергичного влияния комбинации на пестицидную активность (или эффективность) комбинации трансгенного объекта и пестицида. Несмотря на то, что не предполагается, что такие синергичные эффекты ограничивают пестицидную активность, они должны также включать такие неожиданные преимущества, как увеличенный диапазон активности, благоприятный профиль активности в части типа и размеров уменьшения повреждения, уменьшенная стоимость пестицида и применения, уменьшенное распределение пестицида в окружающей среде, уменьшенное воздействие пестицида на персонал, который производит, транспортирует и засевает семена кукурузы, и другие преимущества, известные специалистам в области техники.
Пестициды и инсектициды, которые являются эффективными в композициях в комбинации со способами и композициями настоящего изобретения, включая обработку и нанесение покрытия на семена, а также способы применения таких композиций могут быть найдены, например, в Патенте США 6551962, который приведен здесь в качестве ссылки в полном объеме.
Несмотря на то, что полагают, что средства обработки семян могут наноситься на трансгенное семя в любом физиологическом состоянии, может быть предпочтительным, чтобы семя находилось в достаточно устойчивом состоянии, чтобы оно не подвергалось никакому повреждению во время процесса обработки. Как правило, семя является семенем, собранным с нивы; забранным от трансгенного растения; и отделенным от любого другого материала несеменного растения. Семя предпочтительно должно быть биологически устойчивым при условии, что обработка не вызовет никакого биологического повреждения семени. В одном варианте осуществления, например, может быть произведена обработка посевного зерна, которое было собрано, очищено и высушено до влажности менее приблизительно 15% по весу. В альтернативном варианте осуществления семя может быть высушено, а затем залито водой и/или другим материалом и затем повторно высушено до или во время обработки пестицидом. С учетом описанных ограничений полагают, что семя может быть обработано в любое время между сбором семени и засеванием семени. Применяемый здесь термин "незасеянное семя" включает семя в любой период между сбором семени и засеванием семени в почву с целью прорастания и роста растения. Когда сказано, что незасеянное семя "обработано" пестицидом, то это не означает, что такая обработка включает технологию, в которой пестицид наносят на почву, а не на семя. Например, считается, что такая обработка, как нанесение пестицида в виде полос, "T"-полос или бороздок, одновременно с засеванием семени, не включена в настоящее изобретение.
Пестицид или комбинация пестицидов, к которым можно применить понятие "беспримесный", не содержит каких-либо разбавляющих или дополнительных компонентов. Однако, пестицид обычно наносят на семена в форме пестицидной композиции. Эта композиция может содержать один или более других желательных компонентов, включая, но не ограничиваясь ими, жидкие разбавители, связующие вещества, служащие матрицей для пестицида, наполнители для защиты семян в условиях стресса, и пластификаторы для улучшения гибкости, адгезии и/или растекаемости покрытия.
Рассматриваемые пестициды можно наносить на семя в виде компонента оболочки семени. Способы и композиции нанесения оболочки на семя, известные в области техники, являются эффективными, когда они модифицированы с помощью добавления одного из вариантов осуществления комбинации пестицидов настоящего изобретения. Такие способы нанесения оболочки и прибор для их нанесения раскрыты, например, в Патентах США №№ 5918413, 5891246, 5554445, 5389399, 5107787, 5080925, 4759945 и 4465017. Композиции оболочек семян раскрыты, например, в Патентах США №№ 5939356, 5882713, 5876739, 5849320, 5834447, 5791084, 5661103, 5622003, 5580544, 5328942, 5300127, 4735015, 4634587, 4383391, 4372080, 4339456, 4272417 и 4245432 и других.
Пестициды, которые являются эффективными в нанесении оболочки, являются пестицидами, которые описаны здесь. Количество пестицида, которое применяется для обработки семени, варьирует в зависимости от типа семени и типа активных ингредиентов, но обработка будет включать контактирование семян с определенным количеством комбинации пестицидов, которая является пестицидно эффективной. Когда паразитом-мишенью является насекомое, этим количеством будет являться количество инсектицида, которое является инсектицидно эффективным. Применяемый здесь термин "инсектицидно эффективное количество" означает количество инсектицида, которое будет вызывать гибель насекомых-паразитов в состоянии роста личинки или куколки, или будет последовательно уменьшать или замедлять степень повреждения, вызываемого насекомым-паразитом.
В основном, количество пестицида, которое наносится на семена при обработке, составляет от приблизительно 10 г до приблизительно 2000 г активного компонента пестицида на 100 кг веса семян. Предпочтительно, количество пестицида будет в пределах диапазона от приблизительно 50 г до приблизительно 1000 г активного компонента на 100 кг семян, более предпочтительно в пределах диапазона от приблизительно 100 г до приблизительно 600 г активного компонента на 100 кг семян, и еще более предпочтительно в пределах диапазона от приблизительно 200 г до приблизительно 500 г активного компонента на 100 кг веса семян. В альтернативном варианте установлено, что предпочтительно, чтобы количество пестицида составляло от приблизительно 60 г активного компонента пестицида на 100 кг семян и более предпочтительно от приблизительно 80 г на 100 кг семян.
Пестициды, применяемые для обработки, не должны ингибировать прорастание семян и должны быть эффективными в защите семян и/или растения во время жизненного цикла насекомого-мишени, во время которого оно вызывает повреждение семян или растения. В основном, покрытие будет эффективно в течение от приблизительно 0 до 120 дней после засевания. Пестициды рассматриваемого изобретения могут наноситься на семена в форме оболочки.
Выгоды, предоставляемые настоящим изобретением, могут включать, но не ограничиваясь ими: простоту введения дцРНК в клетки насекомого, низкую концентрацию дцРНК, которая может применяться, устойчивость dsRJSFA и эффективность ингибирования. Возможность использования низкой концентрации стабилизированной дцРНК позволяет избежать некоторых недостатков антисмысловой интерференции. Настоящее изобретение не ограничено применением in vitro или композициями специфических последовательностей, конкретным набором генов-мишеней, специфической частью нуклеотидной последовательности гена-мишени, или конкретным трансгеном, или конкретным способом доставки, в противоположность некоторым из доступных технологий, известных в области техники, таким как антисмысловая и ко-супрессия. Кроме того, генетическая манипуляция становится возможной в организмах, которые не являются классическими генетическими моделями.
При осуществлении настоящего изобретения можно осуществить селекцию для гарантии того, что наличие нуклеотидных последовательностей, транскрибированных от рекомбинантного конструкта, не является губительным для клеток, не принадлежащих паразиту. Это может быть достигнуто с помощью направления генов, которые имеют низкую степень идентичности последовательности с соответствующими генами растения или позвоночного животного. Предпочтительная степень идентичности последовательности составляет менее приблизительно 80%. Более предпочтительно степень идентичности последовательности составляет менее приблизительно 70%. Наиболее предпочтительно степень идентичности последовательности составляет менее приблизительно 60%.
В дополнение к прямой трансформации растения с помощью рекомбинантного конструкта ДНК трансгенные растения могут быть получены с помощью скрещивания первого растения, имеющего рекомбинантный конструкт ДНК, со вторым растением, не имеющего конструкта. Например, рекомбинантная ДНК для генной супрессии может быть введена в первую линию растения, которая является доступной для трансформации, для продукции трансгенного растения, которое может быть скрещено со второй линией растения для интрогрессии рекомбинантной ДНК для генной супрессии во вторую линию растения.
При практическом осуществлении настоящее изобретение может быть объединено с другими признаками контроля насекомых в растении для достижения желательных признаков для повышенного контроля поражения насекомыми. Объединение признаков контроля насекомых, которые задействуют различные способы активности, может предоставить защищенные от насекомых трансгенные растения с устойчивостью, превосходящей таковую у растений, содержащих один признак контроля насекомых, вследствие меньшей вероятности развития этой устойчивости на ниве.
Механизм инсектицидной активности кристаллических белков B. thuringiensis был широко изучен в прошлом десятилетии. Показано, что кристаллические белки являются токсичными для личиночной формы насекомого только после приема белка внутрь. В личинках чешуекрылых щелочной pH и протеолитические ферменты в средней кишке насекомого растворяют белки, таким образом, обеспечивая высвобождение компонентов, которые являются токсичными для насекомого. Эти токсичные компоненты разрушают клетки средней кишки, приводят к прекращению пищевой активности насекомого, и, в конечном счете, вызывают гибель насекомого. По этой причине доказано, что токсины B. thuringiensis являются эффективными и экологически безопасными инсектицидами при контакте с различными насекомыми-паразитами. Жесткокрылые и полужесткокрылые насекомые, и, вероятно, двукрылые, lygus и другие колющие и сосущие насекомые имеют слабо кислый pH кишечника, и, таким образом, токсины Bt, эффективные в отношении личинок чешуекрылых, неэффективны против этих паразитов. Также полагают, что слабо кислый pH кишечника этих насекомых является более благоприятным для композиций настоящего изобретения, и без ограничений до конкретной теории, полагают, что щелочной pH кишечника личинок чешуекрылых является значительной причиной, по которой предшествующие попытки демонстрации эффективности дцРНК терпели неудачу (Fire et al., Патент США №6506559; Mesa et al. Публикация патента №US2003/0150017; Rajagopal et al., 2002; Tabara et al, 1998). Поэтому полагают, что раскрытые здесь способы дцРНК необходимо избирательно применять в композициях и на растениях для контроля жесткокрылых, двукрылых, полужесткокрылых, lygus и колющих и сосущих насекомых. Приведенные здесь способы и композиции особенно эффективны для направления генов для супрессии в насекомых, имеющих pH кишки от приблизительно 4,5 до приблизительно 9,5, или от приблизительно 5,0 до приблизительно 9,0, или от приблизительно 5,5 до приблизительно 8,5, или от приблизительно 6,0 до приблизительно 8,0, или от приблизительно 6,5 до приблизительно 7,7, или от приблизительно 6,8 до приблизительно 7,6, или приблизительно 7,0. Однако предполагается, что насекомые и другие виды паразитов, которые имеют pH кишки от приблизительно 7,5 до приблизительно 11,5, или от приблизительно 8,0 до приблизительно 11,0, или от приблизительно 9,0 до приблизительно 10,0, такие как личинки чешуекрылых насекомых, также входят в объем настоящего изобретения. Это особенно верно, когда дцРНК, специфичную для ингибирования гена в личинках чешуекрылых, вносят в диету личинок наряду с одним или более белками Bt, что в сравнении с белком Bt обычно будет токсичным для этих чешуекрылых личинок при применении на пороговом или надпороговом уровнях. Наличие одного или более токсинов Bt, токсичных для одинаковых видов насекомых, будет эффективно снижать pH кишечника, обеспечивая устойчивую окружающую среду для двухцепочечных молекул РНК, для проявления их эффектов подавления гена-мишени у насекомого-паразита.
Ожидается, что комбинация некоторых стабилизированных конструктов дцРНК с одним или более генами белка контроля насекомых приведет к синергиям, которые будут усиливать фенотип контроля насекомых трансгенного растения. Биологические анализы насекомого с применением искусственной диеты или ткани целых растений могут применяться для определения влияния дозы на гибель или ингибирования роста личинок, применяя как белки контроля насекомых, так и дцРНК. Специалист в области техники может тестировать смеси молекул дцРНК и белков контроля насекомых в биологическом анализе для идентификации комбинации активных компонентов, которые являются синергичными и желательными для помещения в защищенные от насекомых растения (Tabashnik, 1992). Сообщалось о синергизме между различными белками контроля насекомых в уничтожении насекомых-паразитов (для обзора см. Schnepf и др., 1998). Ожидается, что синергизм будет наблюдаться между некоторыми дцРНК и между некоторыми дцРНК и некоторыми белками контроля насекомых.
Изобретение также относится к сырьевым продуктам, содержащим одну или более последовательностей настоящего изобретения, и получаемых от рекомбинантного растения или семени, содержащего одну или более нуклеотидных последовательностей настоящего изобретения, которые специфично рассмотрены как варианты осуществления настоящего изобретения. Предполагается, что к сырьевым продуктам, содержащим одну или более последовательностей настоящего изобретения, относятся, но не ограничиваясь ими, крупы, масло, или разрушенные, или целые зерна, или семена растения, или любой пищевой продукт, включая любые крупы, масло, или разрушенные, или целые зерна рекомбинантного растения или семени, содержащего одну или более последовательностей настоящего изобретения. Детекция одной или более последовательностей настоящего изобретения в одном или более сырье или сырьевых продуктах, рассмотренных здесь, является явным доказательством того, что сырьё или сырьевой продукт составлен из трансгенного растения, разработанного для экспрессии одной или более нуклеотидных последовательностей настоящего изобретения с целью контроля поражения насекомым, с помощью способов дцРНК-опосредованной супрессии генов.
D. Получение нуклеиновых кислот
Настоящее изобретение предоставляет способ получения нуклеиновой кислоты, включающей нуклеотидную последовательность для продукции дцРНК или siРНК. В одном варианте осуществления такой способ включает: (a) исследование библиотеки кДНК или gДНК с зондом гибридизации, включающим всю или часть нуклеотидной последовательности или ее гомолога от заданного насекомого; (b) идентификация клона ДНК, который гибридизирует с зондом гибридизации; (c) изоляция клона ДНК, идентифицированного в шаге (b); и (d) секвенирование кДНК или фрагмента gДНК, который включает клон, выделенный в шаге (c), в котором секвенированная молекула нуклеиновой кислоты транскрибирует всю или основную часть последовательности нуклеиновой кислоты РНК или ее гомолога.
В другом варианте осуществления способ настоящего изобретения для получения фрагмента нуклеиновой кислоты, включающей нуклеотидную последовательность для продукции основной части дцРНК или siРНК, включает: (a) синтез первого и второго олигонуклеотидных праймеров, соответствующих части одной из нуклеотидных последовательностей у заданного насекомого; и (b) амплификация матрицы кДНК или gДНК в вектор клонирования, применяя первый и второй олигонуклеотидные праймеры шага (a), при этом амплифицированная молекула нуклеиновой кислоты транскрибирует основную часть дцРНК или siРНК настоящего изобретения.
При осуществлении настоящего изобретения ген-мишень может быть получен из кукурузного жука (CRW), такого как WCR или SCR, или любых видов насекомых, которые вызывает повреждение хлебных злаков и последующие потери урожая. Предполагается, что при выборе предпочтительных генов-мишеней можно использовать несколько критериев. Интересующим является ген, белковый продукт которого подвергается быстрому метаболизму так, что ингибирование дцРНК может привести к быстрому уменьшению содержания белка. В некоторых вариантах осуществления эффективно выбрать ген, для которого малое снижение уровня экспрессии приводит к отрицательным воздействиям на насекомого. Если желательно воздействовать на широкий диапазон видов насекомых, выбирают ген, который является эволюционно высоко стабильным среди этих видов. Наоборот, с целью придания специфичности в некоторых вариантах осуществления изобретения выбирают ген, который содержит области, которые имеют плохую эволюционную стабильность среди отдельных видов насекомых, или среди насекомых и других организмов. В некоторых вариантах осуществления может быть желательным выбор гена, который не имеет известных гомологов в других организмах.
Применяемый здесь термин "полученный из" относится к точно установленной нуклеотидной последовательности, которая может быть получена из конкретного установленного источника или видов, хотя не обязательно непосредственно из этого установленного источника или видов.
В одном варианте осуществления выбирают ген, который экспрессируется в кишечнике насекомого. Направленное воздействие на гены, экспрессируемые в кишечнике, позволяет избежать потребности в распространении дцРНК в насекомом. Гены-мишени для применения в настоящем изобретении могут включать, например, гены, в которых сочетаются основные гомологии к нуклеотидным последовательностям известных экспрессируемых кишечником генов, которые кодируют белковые компоненты протонной V-АТФазы вакуоли и плазматической мембраны (Dow и др., 1997; Dow, 1999). Этот белковый комплекс является единственным активатором эпителиального ионного транспорта и ответственен за ощелачивание полости средней кишки. V-АТФаза также экспрессируется в Мальпигиевых сосудах, отростках задней кишки насекомого, функция которых состоит в регуляции жидкостного баланса и детоксикации чужеродных соединений по аналогии с почками млекопитающих. В другом варианте осуществления V-АТФаза может быть Vha68-2, или ее гомологом, или ортологом (например, как указано в SEQ ID NO:821).
В другом варианте осуществления выбирают ген, который в основном задействован в росте, развитии и репродукции насекомого. К примерам генов относятся, но не ограничиваясь ими, ген CHD3, ген β-тубулина и ген, кодирующий белок, который, как предполагается, задействован в транспорте. Ген CHD3 у Drosophila melanogaster кодирует белок с активностью АТФ-зависимой ДНК геликазы, которая задействована в сборке/разборке хроматина в ядре. Подобные последовательности были найдены в различных организмах, таких как Arabidopsis thaliana, Caenorhabditis elegans и Saccharomyces cerevisiae. Семейство генов бета-тубулина кодирует белки, связанные с микроканальцами, которые являются компонентом клеточного цитоскелета. Родственные последовательности обнаружены в таких различных организмах, как C. elegans и Manduca sexta. Белки, которые по расчетам являются субъединицами комплекса эндосомального распознавания, необходимого для транспорта (ESCRT) - III (Babst и др., 2002), например Dv49, найдены в различных организмах, включая млекопитающих, дрожжей и насекомых, таких как D. virgifera. Другим связанным с транспортом белком является β'-коатомерный белок, сокращенно β'Cop, который кодирует продукт, задействованный в ретроградном (от Golgi до ER) транспорте. Подобные или предлагаемые последовательности были идентифицированы у C. elegans и/или, virgifera, например, Dv248 (SEQ ID NO:.
К другим генам-мишеням для применения в настоящем изобретении могут относиться, например, гены, которые играют важные роли в жизнеспособности, росте, развитии, репродукции и инфекционности. Эти гены-мишени могут относиться к генам поддержания жилища, факторам транскрипции и генам, специфичным для насекомого, или смертельным нокаут мутациям у Drosophila. Гены-мишени для применения в настоящем изобретении могут также быть генами от других организмов, например от нематод (например, C. elegans). Дополнительно, нуклеотидные последовательности для применения в настоящем изобретении также могут быть получены из растительных, вирусных, бактериальных или грибковых генов, функции которых были установлены из литературы и нуклеотидные последовательности которых в основном подобны генам-мишеням в геноме насекомого. Согласно одному аспекту настоящего изобретения для контроля WCR, заданные последовательности могут в основном быть получены от заданного насекомого WCR. Некоторые из примеров заданных последовательностей из библиотеки кДНК от WCR, которые кодируют белки или их фрагменты у D. virgifera, которые являются гомологами известных белков, могут быть найдены в Списке последовательностей. Известны молекулы нуклеиновых кислот от D. virgifera, кодирующие гомологи известных белков (Andersen и др., заявка на патент США Сер. № 10/205189).
С целью настоящего изобретения молекулы дцРНК или siРНК могут быть получены от CRW с помощью полимеразной цепной (ПЦР™) амплификации заданных генных последовательностей CRW, полученных из gДНК кукурузного жука или библиотеки кДНК, или их частей. Личинки WCR могут быть получены с помощью способов, известных специалисту в области техники, а ДНК/РНК могут быть экстрагированы. В целях настоящего изобретения для экстракции ДНК/РНК могут применяться личинки различных размеров в пределах от 1 возрастных стадий до полностью выращенных CRWs. Для ПЦР™ амплификации для получения дцРНК или siРНК могут применяться геномные библиотеки ДНК или кДНК, созданные от WCR.
Гены-мишени можно затем амплифицировать с помощью ПЦР™ и упорядочить с помощью способов, легко доступных в области техники. Специалист в области техники способен модифицировать условия ПЦР™ для гарантии оптимального формирования продукта ПЦР™. Подтвержденный продукт ПЦР™ может применяться в качестве матрицы для транскрипции in vitro, для создания смысловой и антисмысловой РНК с включенными минимальными промоторами.
Настоящие изобретатели предполагают, что для контроля WCR и других заданных насекомых в настоящем изобретении могут применяться последовательности нуклеиновых кислот, идентифицированные и выделенные от любых видов насекомых в царстве насекомых. В одном аспекте настоящего изобретения нуклеиновая кислота может быть получена от вида жесткокрылых. В частности, нуклеиновая кислота может быть получена от жуков листоедов, принадлежащих к роду Diabrotica (жесткокрылые, Chrysomelidae), и а именно молекулы нуклеиновой кислоты настоящего изобретения могут быть получены из видов группы virgifera. Точнее, молекулы нуклеиновых кислот настоящего изобретения могут быть получены из Diabrotica virgifera virgifera LeConte, который обычно упоминается как WCR. Выделенные нуклеиновые кислоты могут быть эффективными, например, в идентификации гена-мишени и в создании рекомбинантного вектора, который продуцирует стабилизированные дцРНК или siРНКs настоящего изобретения, для защиты растений от поражений насекомыми WCR.
Таким образом, в одном варианте осуществления настоящее изобретение включает выделенные и очищенные нуклеотидные последовательности от WCR или Lygus, которые можно применять в качестве агентов контроля насекомых. Выделенные и очищенные нуклеотидные последовательности могут включать последовательности, приведенные в списке последовательностей.
Нуклеиновые кислоты от WCR или других насекомых, которые можно применять в настоящем изобретении, могут также включать выделенные и в основном очищенные Unigenes и молекулы нуклеиновых кислот EST или молекулы фрагментов этих нуклеиновых кислот. Молекулы нуклеиновых кислот EST могут кодировать значительные части или фактически большие части полипептидов. В альтернативном варианте фрагменты могут включать меньшие олигонуклеотиды от приблизительно 15 приблизительно до 250 нуклеотидных остатков, и более предпочтительно от приблизительно 15 до приблизительно 30 нуклеотидных остатков. В альтернативном варианте молекулы нуклеиновых кислот для применения в настоящем изобретении могут быть получены из библиотек кДНК от WCR или от любых других видов жесткокрылых паразитов.
Молекулы нуклеиновых кислот и их фрагменты от WCR или других видов жесткокрылых паразитов можно применять для получения других молекул нуклеиновых кислот от других видов для применения в настоящем изобретении, для продукции желательных молекул дцРНК и siРНК. Такие молекулы нуклеиновых кислот включают молекулы нуклеиновых кислот, которые кодируют всю последовательность, кодирующую белок, и промоторы и фланкирующие последовательности таких молекул. Кроме того, такие молекулы нуклеиновых кислот содержат молекулы нуклеиновых кислот, которые кодируют члены семейства генов. Такие молекулы могут быть быстро получены при использовании вышеописанных молекул нуклеиновой кислоты или их фрагментов для скрининга, например, библиотеки кДНК или gДНК получены от D. v. virgifera или других жесткокрылых, или от Lygus hesperus. Способы формирования таких библиотек известны в области техники.
Применяемые здесь фразы "кодирующая последовательность", "структурная нуклеотидная последовательность" или "структурная молекула нуклеиновой кислоты" относятся к нуклеотидной последовательности, которая транслируется в полипептид, обычно посредством мРНК, при помещении под контролем подходящих регуляторных последовательностей. Границы кодирующей последовательности определяют с помощью трансляции стартового кодона на 5'-конце и терминирующего трансляцию кодона на 3'-конце. Кодирующая последовательность может включать, но не ограничиваясь ей, геномную ДНК, кДНК, EST и рекомбинантные нуклеотидные последовательности.
Термины "рекомбинантная ДНК" или "рекомбинантная нуклеотидная последовательность" относятся к ДНК, которая содержит модификацию, созданную методами генной инженерии посредством манипуляции с помощью мутагенеза, рестрикционных энзимов и т.п.
Для многих насекомых, которые являются потенциальными мишенями для контроля настоящим изобретением, может быть ограничена информация относительно последовательностей большинства генов или фенотипа, вследствие мутации специфических генов. Поэтому настоящие изобретатели полагают, что выбор подходящих генов от насекомых паразитов для применения в настоящем изобретении может быть осуществлен с помощью информации, доступной от исследования соответствующих генов в организме-модели, таком как Drosophila, в некоторых других видах насекомых, или даже в видах нематод, в видах грибов, в видах растений, в которых были характеризованы гены. В некоторых случаях будет возможно получить последовательность соответствующего гена от насекомого-мишени с помощью поиска баз данных, таких как GenBank, применяя либо название гена, либо последовательность от, например, Drosophila, другого насекомого, нематоды, гриба или растения, от которого был клонирован ген. После получения последовательности можно применить ПЦР™ для амплификации соответственно выбранного сегмента гена в насекомом для применения в настоящем изобретении.
Для получения сегмента ДНК от соответствующего гена вида насекомого на основании последовательности, выявленной в WCR или других насекомых, от которых был клонирован ген, могут быть разработаны праймеры ПЦР™. Праймеры разрабатывают для амплификации сегмента ДНК достаточной длины для применения в настоящем изобретении. ДНК (или геномную ДНК или кДНК) получают от видов насекомых, а праймеры ПЦР™ применяют для амплификации сегмента ДНК. Условия амплификации выбирают так, чтобы амплификация происходила даже если праймеры не будут точно соответствовать заданной последовательности. В альтернативном варианте ген (или его часть) может быть клонирован от библиотеки gДНК или кДНК, полученной от видов насекомых паразитов, применяя ген WCR или другой известный ген насекомого в качестве зонда. Технологии осуществления ПЦР™ и клонирования от библиотек известны. Дальнейшие детали процесса, с помощью которого могут быть изолированы сегменты ДНК от заданных видов насекомых паразитов, основанные на последовательности генов, заранее клонированных от WCR или других видов насекомых, предоставлены в Примерах. Специалисту в области техники ясно, что для выделения сегментов гена от видов насекомых паразитов, которые соответствуют генам, предварительно выделенным от других видов, можно применять множество технологий.
Если паразитом-мишенью для настоящего изобретения является насекомое, к таким паразитам относятся, но не ограничиваясь ими: насекомые из отряда Чешуекрылых, например,
Acleris spp., Adoxophyes spp., Aegeria spp., Agrotis spp., Alabama argillaceae, Amylois spp., Anticarsia gemmatalis, Archips spp, Argyrotaenia spp., Autographa spp., Busseola fusca, Cadra cautella, Carposina nipponensis, Chilo spp., Choristoneura spp., Clysia ambiguella, Cnaphalocrocis spp., Cnephasia spp., Cochylis spp., Coleophora spp., Crocidolomia binotalis, Cryptophlebia leucotreta, Cydia spp., Diatraea spp., Diparopsis castanea, Earias spp., Ephestia spp., Eucosma spp., Eupoecilia ambiguella, Euproctis spp., Euxoa spp., Grapholita spp., Hedya nubiferana, Heliothis spp., Hellula undalis, Hyphantria cunea, Keiferia lycopersicella, Leucoptera scitella, Lithocollethis spp., Lobesia botrana, Lymantria spp., Lyonetia spp., Malacosoma spp., Mamestra brassicae, Manduca sexta, Operophtera spp., Ostrinia Nubilalis, Pammene spp., Pandemis spp., Panolis flammea, Pectinophora gossypiella, Phthorimaea operculella, Pieris rapae, Pieris spp., Plutella xylostella, Prays spp., Scirpophaga spp., Sesamia spp., Sparganothis spp., Spodoptera spp., Synanthedon spp., Thanmetopoea spp., Tortrix spp., Trichoplusia ni и Yponomeuta spp.;
насекомые из отряда Жесткокрылых, например,
Agriotes spp., Anthonomus spp., Atomaria linearis, Chaetocnema tibialis, Cosmopolites spp., Curculio spp., Dermestes spp., Diabrotica spp., Epilachna spp., Eremnus spp., Leptinotarsa decemlineata, Lissorhoptrus spp., Melolontha spp., Orycaephilus spp., Otiorhynchus spp., Phlyctinus spp., Popillia spp., Psylliodes spp., Rhizopertha spp., Scarabeidae, Sitophilus spp., Sitotroga spp., Tenebrio spp., Tribolium spp. и Trogoderma spp.;
из отряда Прямокрылых, например,
Blatta spp., Blattella spp., Gryllotalpa spp., Leucophaea maderae, Locusta spp., Periplaneta ssp. и Schistocerca spp.;
из отряда Термитов, например,
Reticulitemes ssp;
из отряда Сеноедов, например,
Liposcelis spp.;
из отряда Вшей, например,
Haematopinus spp., Linognathus spp., Pediculus spp., Pemphigus spp. и Phylloxera spp.;
из отряда Пухоедов, например,
Damalinea spp. и Trichodectes spp.;
из отряда Трипсов, например,
Franklinella spp., Hercinothrips spp., Taeniothrips spp., Thrips palmi, Thrips tabaci и Scirtothrips aurantii;
из отряда Полужесткокрылых, например,
Cimex spp., Distantiella theobroma, Dysdercus spp., Euchistus spp., Eurygaster spp., Leptocorisa spp., Nezara spp., Piesma spp., Rhodnius spp., Sahlbergella singularis, Scotinophara spp., Triatoma spp., Miridae family spp., такие как Lygus hesperus и Lygus lineoloris, Lygaeidae family spp., такие как Blissus leucopterus и Pentatomidae family spp.;
из отряда Равнокрылых, например,
Aleurothrixus floccosus, Aleyrodes brassicae, Aonidiella spp., Aphididae, Aphis spp., Aspidiotus spp., Bemisia tabaci, Ceroplaster spp., Chrysomphalus aonidium, Chrysomphalus dictyospermi, Coccus hesperidum, Empoasca spp., Eriosoma larigerum, Erythroneura spp., Gascardia spp., Laodelphax spp., Lacanium corni, Lepidosaphes spp., Macrosiphus spp., Myzus spp., Nehotettix spp., Nilaparvata spp., Paratoria spp., Pemphigus spp., Planococcus spp., Pseudaulacaspis spp., Pseudococcus spp., Psylla ssp., Pulvinaria, aethiopica, Quadraspidiotus spp., Rhopalosiphum spp., Saissetia spp., Scaphoideus spp., Schizaphis spp., Sitobion spp., Trialeurodes vaporariorum, Trioza erytreae и Unaspis citri;
из отряда Перепончатокрылых, например,
Acromyrmex, Atta spp., Cephus spp., Diprion spp., Diprionidae, Gilpinia polytoma, Hoplocampa spp., Lasius sppp., Monomorium pharaonis, Neodiprion spp, Solenopsis spp. и Vespa ssp.,
из отряда Двукрылых, например,
Aedes spp., Antherigona soccata, Bibio hortulanus, Calliphora erythrocephala, Ceratitis spp., Chrysomyia spp., Culex spp., Cuterebra spp., Dacus spp., Drosophila melanogaster, Fannia spp., Gastrophilus spp., Glossina spp., Hypoderma spp., Hyppobosca spp., Liriomysa spp., Lucilia spp., Melanagromyza spp., Musca ssp., Oestrus spp., Orseolia spp., Oscinella frit, Pegomyia hyoscyami, Phorbia spp., Rhagoletis pomonella, Sciara spp., Stomoxys spp., Tabanus spp., Tannia spp. и Tipula spp.,
из отряда блох, например,
Ceratophyllus spp. и Xenopsylla cheopis и
из отряда Щетинохвосток, например,
Lepisma saccharina.
Установлено, что настоящее изобретение особенно эффективно, когда насекомым паразитом является Diabrotica spp., и особенно когда паразитом является Diabrotica virgifera virgifera (Западный кукурузный жук, WCR), Diabrotica barberi (Северный кукурузный жук, NCR), Diabrotica virgifera zea (Мексиканский кукурузный жук, MCR), Diabrotica balteata (Бразильский кукурузный жук (BZR) или комплекс Бразильского кукурузного жука (BCR), состоящий из Diabrotica viridula и Diabrotica speciosa), или Diabrotica undecimpunctata howardi (Южный кукурузный жук, SCR).
Примеры
Изобретатели идентифицировали здесь средства для контроля поражения жесткокрылыми паразитами с помощью внесения двухцепочечных молекул рибонуклеиновой кислоты в пищевой рацион паразитов. К удивлению, изобретатели обнаружили двухцепочечные молекулы рибонуклеиновой кислоты, которые после приема их внутрь паразитом, функционируют ингибируя биологическую функцию у паразита, приводя к одному или более следующих признаков: уменьшение пищевой активности паразита, уменьшение жизнеспособности паразита, гибель паразита, ингибирование дифференцировки и развития паразита, отсутствие или уменьшение способности паразита к половому размножению, образование мышц, ювенильное образование гормонов, ювенильная гормональная регуляция, ионная регуляция и транспорт, поддержание мембранного потенциала клетки, биосинтез аминокислот, распад аминокислот, образование спермы, синтеза феромонов, чувствительность к феромонам, образование щупалец, образование крыльев, образование конечностей, развитие и дифференцировку, образование яиц, созревания личинок, образование пищеварительных ферментов, синтез гемолимфы, поддержание гемолимфы, передачу нервных импульсов, деление клетки, энергетический метаболизм, дыхание, апоптоз и любой компонент структуры цитоскелета эукариотических клеток, такой как, например, актин и тубулин. Любой или любая комбинация этих признаков могут привести к эффективному ингибированию поражения паразитами, и в случае паразита растений - ингибированию поражения растения. Например, при применении пищевой композиции, содержащей достаточное для ингибирования паразита количество одной или более двухцепочечных молекул рибонуклеиновой кислоты, наносимой местно на растение в виде обработки семян, в виде нанесения на почву вокруг растения, или при продукции растением от рекомбинантной молекулы ДНК, присутствующей в клетках растения, поражение паразитами растения неожиданно резко уменьшается. Примеры, приведенные ниже, являются иллюстративными для изобретения в отношении отдельного паразита. Однако, специалисту ясно, что способы, формулы и идеи, представленные в Примерах, не предназначены для ограничения и применимы ко всем видам жесткокрылых паразитов, которые могут потреблять источники пищи, которые могут быть сформулированы таким образом, чтобы содержать достаточное количество агента, ингибирующего паразита, состоящего по крайней мере из одной или более двухцепочечных молекул РНК, представленных здесь для супрессии некоторых жизненно важных свойств или функций паразита.
Пример 1
Идентификация заданных нуклеотидных последовательностей для получения дцРНК, полезной для контроля кукурузных жуков
Из целых личинок, куколок и из препарированных участков средней кишки создали библиотеки кДНК кукурузного жука (LIB 149, LIB 150, LIB3027, LIB3373), получили информацию о нуклеотидной последовательности (см. Andersen и др., Заявка на патент США Сер. № 10/205,189, поданная 24 июля 2002, специально приведенная здесь в качестве ссылки в полном объеме). Кроме того, библиотеки кДНК создали из целых личинок в различных стадиях развития и в разное время в пределах каждой стадии развития для максимального увеличения числа различных последовательностей EST от видов Diabrotica. Библиотеки LIB5444 и LIB5462 создали соответственно из пулов мРНК, полученных от личинок Западного кукурузного жука первой (1 грамм) и третьей (2,9 грамм) возрастной стадии. Собранных насекомых быстро заморозили с помощью помещения в жидкий азот. Насекомых растерли в ступке с пестиком, удерживая при -20°C или ниже с помощью охлаждения на сухом льду и/или добавления жидкого азота в ступку, пока ткань не измельчили до мелкодисперсного порошка. РНК экстрагировали с помощью реагента TRIzol® (Invitrogen) согласно инструкциям изготовителя. Поли А+РНК изолировали от всей РНК, полученной с помощью DYNABEADS Oligo dT (Invitrogen), следуя инструкциям изготовителя. Библиотеку кДНК создали из Поли А+РНК, применяя плазмидную систему Superscript™ (Invitrogen). кДНК фракционировали на участки меньшего размера с помощью хроматографии. Четвертые и пятые фракции забрали и лигировали в вектор pSPORTl (Life Technologies Inc, Gaithersburg MD) между участками распознавания рестрикционных эндонуклеаз Sal1 и Notl, и трансформировали в электро-компетентные клетки E. coli DH10B с помощью электропорации. Библиотека личинок первой возрастной стадии включала приблизительно 420000 колониеобразующих единиц. Библиотека личинок третьей возрастной стадии включала приблизительно 2,78×106 колониеобразующих единиц. Колонии от LIB 149, LIB 150 промыли от чашек, смешали до однородности с помощью кратковременного интенсивного перемешивания и объединили с буфером Трис-EDTA. Половину промывки поместили в 10% глицерин, расфасовали аликвотами по криопробиркам и хранили при -70°C. Другую половину применяли для продукции плазмидной ДНК, применяя колонку очистки Quiagen midi-prep, или ее эквивалент. Очищенную плазмидную ДНК расфасовали аликвотами по микроцентрифужным пробиркам и хранили при -20°C.
Колонии от библиотек кДНК Diabrotica virgifera LIB5444 и LIB5462 амплифицировали по отдельности на среде высокой вязкости. Приблизительно 200000 колониеобразующих единиц от LIB5444 и 600000 колониеобразующих единиц от LIB5462 смешали на пластине для смешивания отдельно в среде 500 мл LB, содержащей 0,3% SeaPrep agarose® и 50 мг/л карбенициллина при 37°C, и затем быстро охладили в воде/ванне со льдом в течение 1 часа, позволяя формироваться однородной суспензии бактериальных колоний. Затем инокулированные библиотеки выращивали при 30°C в течение 42 часов. После инкубации клетки перемешивали в течение 5 минут на пластине для смешивания. Затем среду переместили в два центрифужных флакона объемом 250 мл. Бактериальные клетки осадили центрифугированием при 10000×g в течение 10 минут. Носитель удалили из флаконов, и клетки повторно суспендировали в общей сложности в 20 мл среды LB с 50 мг/л карбенициллина. Диметилсульфоксид добавили до 10% для поддержания клеток в сохранности при заморозке. Обе библиотеки амплифицировали до результирующего титра 108 колониеобразующих единиц на миллилитр. Образцы библиотеки кДНК Diabrotica virgifera LIB5444 и LIB5462 объединили и довели до концентрации ДНК приблизительно 1,25 микрограмм на микролитр в стерильной дистиллированной и деионизированной воде и расфасовали аликвотами в двадцать пять криопробирок, чтобы каждая криопробирка содержала приблизительно 8,75 микрограмм ДНК. Эти образцы были внесены на хранение заявителем(ями)/изобретателями в American Type Culture Collection (ATCC), расположенную по адресу 10801 University Boulevard, Manassas, Virginia, USA ZIP 20110-2209 10 июня 2004 и назвали LIB5444/62. ATCC предоставила Заявителю охранную расписку, назначив ATCC депозитный номер PTA-6072.
Библиотеки кДНК высокой молекулярной массы от кукурузного жука, то есть LIB5496 и LIB5498, изготовили в основном, как описано выше, для продукции библиотеки кДНК кукурузного жука. Библиотеки LIB5496 и LIB5498 создали соответственно из пулов мРНК, полученных от первой (1 грамм), и второй, и третьей (1 грамм) возрастной стадии личинок Западного кукурузного жука. Вкратце, насекомых быстро заморозили в жидком азоте. Замороженных насекомых измельчили до тонкодисперсного порошка с помощью перемалывания в ступке с пестиком. РНК экстрагировали, применяя реагент TRIzol® (Invitrogen), следуя инструкциям изготовителя. Поли А+РНК изолировали от всей полученной РНК с помощью DYNABEADS Oligo dT (Invitrogen). Библиотеку кДНК высокой молекулярной массы изготовили из 20 микрограмм Поли А+РНК, применяя Плазмидную систему Superscript™ (Invitrogen). кДНК фракционировали на фрагменты меньшего размера, на 1% агарозном геле в TAЕ, и кДНК в диапазоне 1 Kb до 10 Kb собрали и лигировали в вектор pSPORTl между участками рестрикции Sal1 и Not1 и трансформировали в электро-компетентные клетки E. coli DH10B с помощью электропорации. LIB5496 достигла общего титра приблизительно 3,5×106 колониеобразующих единиц. LIB5498 достигла общего титра приблизительно 1,0×106 колониеобразующих единиц. Колонии от библиотек LIB5496 и LIB5498 кДНК высокой молекулярной массы от кукурузного жука амплифицировали по отдельности в среде высокой вязкости. Приблизительно 600000 колониеобразующих единиц от LIB5496 и LDB5498 смешали на пластине для смешивания раздельно в 500 мл среды LB, содержащей 0,3% SeaPrep agarose® и 50 мг/л карбенициллина при 37°C, и затем быстро охладили в воде/ванне со льдом в течение 1 часа, позволяя образоваться однородной суспензии бактериальных колоний. Затем библиотеки выращивались при 30°C в течение 42 часов. После инкубации клетки перемешивали в течение 5 минут на пластине для смешивания. Затем носитель переместили в два центрифужных флакона по 250 мл. Бактериальные клетки осадили центрифугированием при 10000×g в течение 10 минут. Среду удалили из флаконов, и клетки повторно суспендировали в общей сложности в 20 мл среды LB с 50 мг/л карбенициллина. Добавили диметилсульфоксид до 10% для поддержания клеток в сохранности при заморозке. Обе библиотеки амплифицировали до результирующего титра 108 колониеобразующих единиц на миллилитр. Информацию о вставленной последовательности кДНК получили из библиотек видоспецифичных плазмид кукурузного жука.
Библиотеки жуков Andersen и др. вместе с дополнительными последовательностями из библиотек LIB5444 и LIB5462 первоначально продуцировали приблизительно 18415 одиночных последовательностей EST, состоящих из приблизительно 1,0×107 нуклеотидных остатков. Средняя длина последовательности EST составляла приблизительно 586 нуклеотидных остатков. Эти последовательности EST были подвергнуты биоинформатическим алгоритмам, которые привели к сборке последовательностей, упомянутых здесь как последовательности UNIGENE, и индивидуальных последовательностей EST, которые не могли быть собраны с помощью перекрытия идентичности с другими последовательностями EST, упомянутыми здесь как синглтоны. Затем библиотеки LIB5444 и LIB5462 упорядочили намного глубже, что привело к дополнительным одиночным последовательностям EST. Последовательности EST, полученные от библиотек, то есть LIB149, LIB150, LIB3027, LIB3373, LIB5444, LIB5462, LIB5496 и LIB5503, выбрали для дальнейшего исследования в биологических анализах пищевой активности, как сформулировано ниже, и соответствующие последовательности приведены в списке последовательностей.
Последовательности EST, выделенные от библиотеки кДНК CRW, собрали, по возможности, в наборы UNIGENE, и эти собранные последовательности Unigene включены в список последовательностей. UNIGENE является ген-ориентированным кластером, сформированным из перекрытия одиночных последовательностей EST в областях идентичности последовательности, для формирования большей последовательности (Pontius и др., 2003). Каждую нуклеотидную последовательность, указанную в списке последовательностей, проанализировали для идентификации наличия открытых рамок считывания. Информацию аминокислотной последовательности, полученную из открытых рамок считывания, сравнили с известной информацией аминокислотной последовательности, доступной в общественных базах данных, чтобы установить степень идентичности или подобия аминокислотной последовательности известным аминокислотным последовательностям. Биологическая функция, если она имеется, связана с известными аминокислотными последовательностями в общественных базах данных, аннотируется к аминокислотным последовательностям, полученным из информации о нуклеотидной последовательности из библиотеки кДНК. Комментарии предоставляют информацию, которая была суггестивной для функции белка, который может экспрессироваться от конкретного гена, который давал начало конкретной последовательности кДНК, но не был определяющим для результата. На основании суггестивной аннотирующей информации некоторые последовательности кДНК были охарактеризованы как кодирующие белок, который вероятно задействован в некоторой биологической функции в клетках кукурузного жука, которая либо необходима для жизни или которая необходима для гарантии здоровья и жизнеспособности клетки, или, вероятно, участвует в поддержании целостности клетки, поддержании жизнедеятельности клетки, репродуктивной способности и т.п.
Последовательности, выбранные для дальнейшего исследования, применяли в создании двухцепочечных молекул РНК для инкорпорации в диету CRW. На основе исходных последовательностей кДНК и EST разработали пары праймеров тепловой амплификации для получения последовательностей, применяемых в анализах пищевой активности. Либо пары праймеров были созданы в виде пар нуклеотидных последовательностей, в которой каждый элемент пары праймера имел прекрасную комплементарность либо к смысловой, либо к антисмысловой последовательности. Некоторые последовательности пары праймеров создали так, чтобы каждый элемент пары имел последовательность, Т7 фаг промотор РНК полимеразы на 5' конце. Предпочтительно первую реакцию амплификации более высокой точности осуществляли, применяя первую пару праймеров, не содержащую промотора T7, для создания первого ампликона, применяя геномную ДНК CRW в качестве матрицы. Предпочтительно, в качестве матрицы для синтеза молекулы дцРНК для применения в настоящем изобретении применяли последовательность кДНК или мРНК, потому что в области техники установлено, что последовательности эукариотического генома содержат последовательности, которые не присутствуют в зрелой молекуле РНК. Образец первого ампликона, полученного от первой реакции амплификации более высокой точности, затем применяли в качестве матрицы во второй реакции тепловой амплификации со второй парой праймеров, включающих промоторную последовательность T7, для продукции второго ампликона, который содержал промотор T7 на 5' конце или встроенный в него в каждой цепи второго ампликона. Полную нуклеотидную последовательность второго ампликона получили в обоих направлениях и сравнили с нуклеотидной последовательностью, указанной для кДНК, и указали несоответствия между этими двумя последовательностями, если таковые имелись. В основном, последовательности, полученные, применяя геномную ДНК в качестве матрицы, были несовместимы с дальнейшим применением в виде молекул дцРНК для применения в достижении значимых уровней супрессии вследствие вариаций в геномных последовательностях, которые не присутствовали в пределах последовательностей мРНК или кДНК.
Реакция транскрипции in vitro обычно включает от приблизительно 1 до приблизительно 2 микрограмм матрицы линеаризованной ДНК, буфера полимеразной реакции T7 от 10X концентрата, рибонуклеотиды, ATP, CTP, GTP и UTP в конечной концентрации от 50 до 100 мМ каждый, и 1 единицу фермента T7 РНК полимеразы. РНК полимеразную реакцию инкубировали при приблизительно 37°C, в зависимости от температуры, оптимальной для РНК полимеразы, применяемой согласно инструкциям изготовителя, на период времени в пределах от нескольких минут до нескольких часов. В основном, реакции осуществляли от приблизительно 2 до приблизительно 6 часов для транскрипции матричных последовательностей до приблизительно 400 нуклеотидов в длину и в течение 20 часов для транскрипции матричных последовательностей, более приблизительно 400 нуклеотидов в длину. Нагревание реакции до 65°C в течение пятнадцати минут приводит к терминации транскрипции РНК. Продукты транскрипции РНК подвергли преципитации в этаноле, промыли, высушили воздухом и повторно суспендировали в воде, не содержащей РНК-азу, до концентрации, приблизительно, 1 микрограмм на микролитр. Большинство транскриптов, обладающих преимуществами стратегии противоположного промотора T7, выделенной выше, продуцировали двухцепочечную РНК в реакции транскрипции in vitro, однако, более высокий выход продукта двухцепочечной РНК был получен с помощью нагревания очищенной РНК до 65°C и затем медленного охлаждения до комнатной температуры для гарантии надлежащей ренатурации смыслового и антисмыслового сегментов РНК. Затем двухцепочечные продукты РНК инкубировали с ДНК-азой I и РНК-азой при 37°C в течение одного часа для удаления любой ДНК или одноцепочечной РНК, присутствующей в смеси. Двухцепочечные продукты РНК очистили на колонке согласно инструкциям изготовителя (AMBION MEGAscript® РНКi KIT) и повторно суспендировали в буфере 10 мМ Трис-HCl (pH 7,5) или в воде, не содержащей РНК-азу, до концентрации от 0,1 до 1,0 микрограмм на микролитр.
Следующие нуклеотидные последовательности были получены первоначально в виде последовательностей кДНК, идентифицированных в библиотеке кДНК средней кишки кукурузного жука (Andersen и др., там же), и были приспособлены к применению в построении двухцепочечных молекул РНК для применения в тестировании эффективности ингибирования биологической функции у паразита с помощью подкрепления двухцепочечными молекулами РНК в пищевой рацион паразита.
A. Последовательности, гомологичные Chd3
Гены CHD идентифицировали у многочисленных эукариот, и соответствующие белки были предложены для функционирования в качестве хроматин-ремоделирующих факторов. Термин CHD получен из трех доменов гомологии последовательностей, обнаруженной в белках CHD: хромо (модификатор организации хроматина) домен, домен SNF2-связанной геликазы/АТФ-азы и ДНК-связанный домен, каждый из которых, как полагают, придает различную связанную с хроматином активность. Белки CHD разделены на две категории на основании наличия или отсутствия другого домена гомологии последовательностей, домена цинковых пальцев PHD, обычно ассоциированного с связанной с хроматином активностью. CHD3-связанные белки обладают доменом цинковых пальцев PHD, но CHD1-связанные белки не имеют его. Экспериментальные наблюдения позволяют предположить, что белки CHD3 играют определенную роль в репрессии транскрипции и показано, что у некоторых видов они являются компонентом комплекса, который содержит гистон деацетилазы в качестве субъединицы. Деацетилирование гистонов коррелирует с инактивацией транскрипции, и, таким образом, белки CHD3 задействованы в функционировании в качестве репрессоров транскрипции вследствие того, что они являются компонентом комплекса гистона деацетилазы (Ogas и др., 1999). Таким образом, супрессия синтеза белка CHD3 может быть эффективной мишенью для опосредованного двухцепочечной РНК ингибирования жесткокрылых паразитов.
B. Последовательности, гомологичные бета-тубулину
Белки тубулина являются важными структурными компонентами многих клеточных структур во всех эукариотических клетках и преимущественно в формировании микротрубочек. Ингибирование формирования микротрубочек в клетках приводит к катастрофическим эффектам, включая интерференцию с формированием митотических шпинделей, блокирование деления клетки и т.п. Поэтому супрессия формирования белка тубулина может быть эффективной мишенью для опосредованного двухцепочечной РНК ингибирования.
C. Последовательности, гомологичные 40 кДа V-АТФ-азе
Энергетический метаболизм в субклеточных органеллах в эукариотических системах является жизненно важной функцией. Вакуолярные ATP синтазы задействованы в поддержании достаточных уровней ATP в вакуолях. Поэтому вакуолярные ATP синтазы могут быть эффективной мишенью для ингибирования, опосредованного двухцепочечной РНК.
D. Последовательности, гомологичные EF1α
Факторы элонгации транскрипции и терминации транскрипции являются жизненно важными для метаболизма и могут быть эффективными мишенями для ингибирования, опосредованного двухцепочечной РНК.
E. Последовательности, гомологичные субъединице p28 протеасомы 26S
Протеасома 26S является большой, ATP-зависимой мультисубъединичной протеазой, которая в большом количестве содержится во всех эукариотах. Ее основной функцией является селективное удаление различных короткоживущих белков, которые первоначально связываются ковалентной связью с убиквитином, а затем последовательно распадаются под воздействием комплекса протеасомы 26S. Путь убиквитина играет важную роль в контроле клеточного цикла посредством специфичного разложения множества регуляторных белков, включая митотические циклины и ингибиторы циклин-зависимых киназ, таких как p27 в клетках млекопитающих. Таким образом, супрессия синтеза протеасомы 26S и супрессия синтеза ее составляющих субъединиц может быть предпочтительной мишенью для ингибирования, опосредованного двухцепочечной РНК (Smith и др., 1997).
F. Последовательности, гомологичные ювенильному гормону эпоксидгидролазе
Ювенильный гормон насекомых контролирует и регулирует множество необходимых биологических процессов в ходе жизненного цикла насекомого, включая, но не ограничиваясь ими, метаморфозы, репродукцию и диапаузу. Концентрации ювенильного гормона (JH) дожны достигать пика в определенное время в гемолимфе личиночной формы насекомого паразита, в частности в личинках чешуекрылых и жесткокрылых, а затем должны распадаться для терминации эффектов гормональной реакции. Ферменты, задействованные в уменьшении концентрации ювенильного гормона, действуют через два основных пути метаболического разложения. Один путь включает ювенильную гормональную эстеразу, которая гидролизует метиловый эфир, предоставляя соответствующую кислоту. Второй путь включает эпоксидгидролазу ювенильного гормона (JHEH) для достижения гидролиза эпоксида, приводя к образованию диола. Вклад JHE в разложение JH хорошо изучен и, как установлено, является неизменным среди видов жесткокрылых и чешуекрылых. Ингибирование эстеразы JH было связано с тяжелыми морфологическими изменениями, включая, но не ограничиваясь ими, миграцию личинок, отсроченное окукливание и развитие уродливых интермедиатов. Напротив, вклад JHEH в метаболизм JH менее ясен и, как показано, варьирует между видами, но недавние исследования указывают на признак, который позволяет предположить, что JHEH может быть основным путем метаболизма JH (Brandon J. Fetterolf, Doctoral Dissertation, North Carolina State University (Feb. 10, 2002) Synthesis and Analysis of Mechanism Based Inhibitors of Juvenile Hormone Epoxide Hydrolase from Insect Trichoplusia ni). В любом случае, нарушение любого пути разложения JH с помощью технологии генной супрессии, может быть эффективной мишенью для ингибирования паразита, опосредованного двухцепочечной РНК.
G. Последовательности, гомологичные белку хлорных каналов, зависимых от набухания
Установлено, что белки хлорных каналов, зависимых от набухания, играют критическую роль в осморегуляции в системах эукариотических животных клеток. Поэтому, нуклеотидная последовательность, обладающая способностью экспрессировать аминокислотную последовательность, которая обладает гомологией к предварительно идентифицированным белкам хлорных каналов, зависимых от набухания, может быть эффективной мишенью для ингибирования РНК у паразита.
H. Последовательности, гомологичные белку глюкозо-6-фосфат 1-дегидрогеназы
Белок глюкозо-6-фосфат 1-дегидрогеназы (G6PD) катализирует окисление глюкозо-6-фосфата до 6-фосфоглюконата, одновременно восстанавливая окисленную форму никотинамид аденин динуклеотид фосфата (NADP+) до NADPH. NADPH известен в области техники как необходимый кофактор во многих эукариотических реакциях биосинтеза, и известно, что он поддерживает глутатион в его восстановленной форме. Восстановленный глутатион действует как нейтрализатор опасных окислительных метаболитов в эукариотических клетках, и с помощью фермента глутатитон пероксидазы преобразует опасную перекись водорода в воду (Beutler и др., 1991). Поэтому G6PD может быть предпочтительной мишенью для ингибирования, опосредованного двухцепочечной РНК, у жесткокрылого паразита.
I. Последовательности, гомологичные белку Act42A
Актин является повсеместно распространенным и высоко консервативным эукариотическим белком, необходимым для подвижности клетки и локомоции (Lovato и др., 2001). Идентифицировано множество последовательностей кДНК у CRW, которые, как предполагается, кодируют актин или белки, имеющие структуру аминокислотной последовательности, относящуюся к актиновым белкам. Поэтому, гены, кодирующие гомологи актина в клетке паразита, могут быть эффективными мишенями для ингибирования, опосредованного двухцепочечной РНК.
J. Последовательности, гомологичные фактору ADP-рибозилирования 1
Показано, что факторы рибозилирования ADP являются жизненно важными для функции клетки, так как они играют существенные роли в процессах восстановления повреждения ДНК, канцерогенезе, гибели клетки и геномной стабильности. Таким образом, была бы эффективной возможность селективного прерывания транскрипции факторов ADP-рибозилирования у видов жесткокрылых паразитов, применяя ингибирование, опосредованное двухцепочечной РНК.
K. Последовательности, гомологичные белку фактора транскрипции IIB
Факторы элонгации транскрипции и терминации транскрипции, как указано выше, являются жизненно важными для метаболизма и могут быть благоприятными мишенями для ингибирования, опосредованного двухцепочечной РНК, для контроля или устранения поражения жесткокрылыми паразитами.
L. Последовательности, гомологичные Chitinase
Хитин является β(1→4)гомополимером N-ацетилглюкозамина и обнаружен в экзоскелете насекомых. Хитин образуется из UDP-N-ацетилглюкозамина в реакции, катализируемой хитинсинтазой. Хитин является структурным гомополимерным полисахаридом и в конструкции этой высоко разветвленной и поперечно-сшитой структуры задействовано много ферментных шагов. Хитин придает форму, ригидность и поддержку насекомым и обеспечивает опору, к которой прикрепляются внутренние органы, такие как мышцы. Хитин должен также распадаться до некоторой степени, чтобы опосредовать шаги, задействованные в процессе линьки насекомого. Поэтому полагают, что ингибирование белков, опосредованное двухцепочечной РНК, в этих путях будет эффективно в качестве средства контроля поражения жесткокрылыми паразитами.
M. Последовательности, гомологичные убиквитин конъюгирующему ферменту
Убиквитиновый путь играет важную роль в контроле клеточного цикла посредством специфичного разложения множества регуляторных белков, включая митотические циклины и ингибиторы циклин-зависимых киназ, таких как p27 в клетках млекопитающих. Таким образом, гены, кодирующие убиквитин, и сопутствующие компоненты могут быть предпочтительной мишенью для ингибирования, опосредованного двухцепочечной РНК (Smith и др., 1997). Убиквитин-зависимый протеолитический путь является одним из главных путей, посредством которого внутриклеточные белки селективно разрушаются у эукариот. Конъюгация убиквитина с белками субстрата опосредуется необычайно разнообразным рядом ферментов. Протеолитическое наведение может также подвергаться регуляции в шагах между убиквитинацией субстрата и его разложением до пептидов посредством мультисубъединичной протеазы 26S. Сложность убиквитиновой системы позволяет предположить о центральной роли круговорота белков в регуляции эукариотических клеток и участии других белков в пути, включая убиквитин-активирующий фермент, убиквитин-конъюгирующий фермент, убиквитин-протеин лигазу и компоненты субъединицы протеасомы 26S. Таким образом, полагают, что ингибирование белков, опосредованное двухцепочечной РНК, в этом пути будет эффективным в качестве средства контроля поражения жесткокрылыми паразитами.
N. Последовательности, гомологичные глицеральдегид-3-фосфат дегидрогеназе
Гликолитический путь является жизненно важным путем в большинстве организмов и задействован в продукции метаболической энергии в результате расщепления глюкозы. Одним важным ферментом во второй ступени гликолитического пути является глицеральдегид-3-фосфат дегидрогеназа (G3PDH), которая в присутствии NAD+ и неорганического фосфата катализирует окисление 3-фосфо-глицеральдегида до 3-фосфоглицерол-фосфата с одновременным образованием NADH. Важным компонентом этой реакции является сохранение энергии посредством формирования NADH. Гены, кодирующие ферменты, связанные с гликолитическим путем, и в частности гены, кодирующие ферменты, задействованные в шагах, эффективных в формировании резервов энергии, могут быть особенно эффективными мишенями для ингибирования, опосредованного двухцепочечной РНК, у видов жесткокрылых паразитов.
O. Последовательности, гомологичные убиквитину B
Как описано выше, убиквитиновый путь разложения белков играет важную роль в контроле клеточного цикла посредством специфичного разложения множества регуляторных белков, включая митотические циклины и ингибиторы циклин-зависимых киназ, такие как p27 в клетках млекопитающих. Таким образом, ген, кодирующий убиквитин, и сопутствующие компоненты могут быть предпочтительными мишенями для ингибирования, опосредованного двухцепочечной РНК (Smith и др., 1997).
P. Гомологи эстеразы ювенильных гормонов
Как указано выше, ювенильный гормон насекомых контролирует и регулирует множество необходимых биологических процессов в жизненном цикле насекомых, включая, но не обязательно ограничиваясь ими, метаморфоз, репродукцию и диапаузу. Нарушение синтеза JH или путей разложения с помощью технологии генной супрессии может быть эффективной мишенью для ингибирования паразитов, опосредованного двухцепочечной РНК.
Q. Последовательности, гомологичные альфа-тубулину
Структурные элементы цитоскелета эукариотических клеток, которые являются важными, в основном используются не только в качестве механической опоры, но также и для поддержания формы клетки. Полугибкие микрофиламенты делают клетки мобильными, помогают им делиться при митозе (цитокинез) и у позвоночных и беспозвоночных животных ответственны за мышечное сокращение. Относительно жесткие микротрубочки, которые состоят из белков альфа и бета тубулина, играют важную роль в активности, как своеобразная магистраль для транспорта везикул и органелл, и в разделении хромосом во время митоза (кариокинез). Гибкие промежуточные филаменты обеспечивают по крайней мере дополнительную прочность всей клеточной структуре. Также известно, что цитоскелет задействован в передаче сигналов через цитоплазму клетки. Принимая во внимание эти функции, полагают, что любое разрушение цитоскелета или даже малые изменения его целостности могут привести к патологическим последствиям для клетки.
R. Последовательности, связанные с транспортом
Как указано выше, сортировка и транспорт различных молекул в пределах клетки, включая транспорт до соответствующих органелл, а также их секреция являются важными физиологическими функциями. Такие пути сортировки могут включать пути, которые основаны на комплексе эндосомальной сортировки, необходимом для транспорта (ESCRT), комплексах I-III и др. Таким образом, функции, связанные с транспортом полипептидов и других молекул, могут также быть предпочтительной мишенью для дцРНК-опосредованного ингибирования.
Пример 2
Биологические анализы пищевого рациона насекомых
Образцы двухцепочечной РНК (дцРНК) подвергли биологическому анализу с выбранным числом паразитов-мишеней. дцРНК получили от последовательностей, идентифицированных согласно Примеру 1, применяя либо полную непрерывную последовательность, в случае SEQ ID Nos:1-6, либо последовательность, амплифицированную от собранной непрерывной последовательности, применяя пары праймеров, как указано в списке последовательностей. Варьирующие дозы дцРНК применяли в качестве добавки к искусственному пищевому субстрату для кукурузного жука согласно следующей процедуре. Яйца Diabrotica virgifera virgifera (WCR) получили от Crop Characteristics, Inc., Farmington, Minnesota. Яйца WCR не в диапаузе инкубировали в почве в течение приблизительно 13 дней при 24°C, относительной влажности 60%, в полной темноте. На 13 день почву, содержащую яйца WCR, поместили в сита от #30 до #60, и яйца вымыли из почвы, применяя садовый шланг под высоким давлением. Яйца поверхностно продезинфицировали с помощью пропитывания в LYSOL в течение трех минут, прополоскали три раза в стерильной воде, однократно промыли 10% раствором формалина и затем три дополнительных раза прополоскали в стерильной воде. Яйца, обработанные таким образом, распределили на стерильных кофейных фильтрах и инкубировали в течение ночи при 27°C, относительной влажности 60%, в полной темноте.
Для получения дцРНК ампликоны выбранных последовательностей клонировали в плазмидный вектор, способный к репликации в E. coli, и достаточные количества плазмидной ДНК восстановили для возможности транскрипции in vitro T7 РНК полимеразы от встроенных конвергентных промоторов T7 с обоих концов клонированного фрагмента. Двухцепочечную РНК продуцировали и подвергли биологическому анализу; один сегмент РНК включал последовательность, как указано в списке последовательностей, другой сегмент РНК являлся в основном обратным комплементом нуклеотидной последовательности, соответственно на месте тимидиновых остатков располагались уридиновые. Образец двухцепочечной РНК (дцРНК) обработали DICER или РНКаза III для получения достаточных количеств малых интерферирующих РНК (siРНК). Образцы, содержащие siРНК или дцРНК, были добавлены в биологический анализ пищевого рациона CRW, как описано выше, и личинкам позволили питаться, как указано ниже.
Образец двухцепочечной РНК либо непосредственно добавили в каждую лунку, содержащую пищевой субстрат насекомого, как указано выше, либо модифицировали до добавления к пищевому субстрату насекомого. Модификацию двухцепочечной РНК проводили следуя инструкциям для РНКазы III (AMBION CORPORATION, Austin, Texas) или DICER (STRATAGENE, La Jolla, California), предоставленных изготовителями. Ферментирование РНКазой III двухцепочечной РНК привело к образованию двадцати одного и двадцати двух нуклеотидных дуплексов, содержащих 5' фосфорилированные концы, и 3' гидроксильные концы с 2-3 основными липкими концами, подобно ~21-26 основной паре дуплексных коротких интерферирующих фрагментов РНК (siРНК), продуцированных посредством режущего фермента в эукариотическом пути, идентифицированном Hamilton et. al. (1999) и Elbashir et. al. (200la). Эту коллекцию коротких интерферирующих дуплексов РНК далее очистили и образец охарактеризовали с помощью электрофореза в полиакриломидном геле для определения целостности и эффективности формирования дуплексов. Чистоту и величину образца затем определили с помощью спектрофотометрии с длиной волны 250 нанометров, а неиспользованный образец сохранили для дальнейшего применения, удерживая при -20°C.
Пищевой субстрат для насекомых изготовили в основном согласно Pleau и др. (2002), со следующими модификациями. 9,4 грамм агара SERVA распределили в 540 миллилитрах очищенной воды и перемешивали до тех пор, пока агар не был полностью распределен. Смесь воды/агара нагрели до кипения для полного растворения агара, а затем влили в измельчитель WARING. Измельчитель поддерживали на низкой скорости, одновременно добавляя в горячую смесь агара 62,7 грамма смеси BIO-SERV DIET (F9757), 3,75 грамма лиофилизированного корня кукурузы, 1,25 миллилитра зеленого пищевого красителя и 0,6 миллилитров формалина. Затем pH смеси отрегулировали до 9,0 с помощью добавления 10% концентрированного раствора гидроксида калия. Жидкий пищевой субстрат в объеме приблизительно 600 миллилитров непрерывно перемешивали с высокой скоростью и удерживали при температуре от приблизительно 48°C до приблизительно 60°C, используя стерильный NALGENE, накрытый магнитный мешальник на нагревательной пластине магнитной мешалки, при распределении аликвотами по 200 микролитров в каждую лунку 96-луночных титрационных микропланшетов с круглым основанием FALCON. Пищевому субстрату в планшетах позволили затвердеть и высушивали воздухом в стерильном вытяжном шкафу для биологических образцов в течение приблизительно десяти минут.
Тестовые образцы в объеме тридцать (30) микролитров, содержащие либо контрольные реагенты, либо двухцепочечную РНК в варьирующих количествах, добавили на поверхность пищевого субстрата для насекомых в каждой лунке, применяя микропипеттор повторитель. Пищевой субстрат для насекомых оставили в стерильном вытяжном шкафу для биологических образцов в течение до одного получаса после нанесения контрольных образцов для возможности диффузии реагентов в пищевой субстрат и возможности высыхания поверхности пищевого субстрата. По одной новорожденной личинке WCR поместили в каждую лунку с помощью тонкой кисти. Затем планшеты герметично закрыли с MYLAR и создали обмен воздуха с помощью энтомологической булавки. 12-72 личинки насекомых тестировали на дозу в зависимости от исполнения анализа. Планшеты для биологического анализа инкубировали при 27°C, относительной влажности 60% в полной темноте в течение 12-14 дней. Число выживших личинок на дозу регистрировали в момент времени 12-14 дней. Личиночную массу определяли с помощью подходящих микровесов для каждой выжившей личинки. Данные проанализировали с помощью статистического программного обеспечения JMP®4 (SAS Institute, 1995) и для обнаружения эффектов обработки в сравнении с контролем без обработки (P<0,05) провели полный факториальный ANOVA с тестом Dunnet's. Затем провели тест Tukey-Kramer для сравнения всех пар (P<0,05) обработки. Результаты анализов пищевой активности личинок CRW показали значительное ингибирование роста и смертность в сравнении с контрольными группами, как объяснено ниже.
Пример 3
Результаты биологических анализов пищевой активности насекомых
Искусственный пищевой субстрат, подходящий для выведения личинок кукурузного жука, изготовили с помощью нанесения образцов последовательностей двухцепочечной РНК, идентифицированных, как описано в Примере 1, применяя биологические анализы, проводимые, как описано в Примере 2. Личинкам кукурузного жука обычно позволяют поедать пищевой субстрат в течение двенадцати дней и проводят мониторинг смертности и остановки роста в сравнении с жуками, которым позволяют питаться только негативным и позитивным контрольными пищевыми субстратами. Результаты исследований подтверждают существенные уровни (p<0,05) остановки роста и/или смертности личинок, применяя дцРНК, содержащие части последовательности, гомологичные множеству различных классов генов. Последовательности и векторы, вызывающие существенную остановку роста и/или смертность и соответствующие SEQ ID NO для последовательности, экспрессируемой в виде дцРНК, приведены ниже в Таблицах 1-5.
Пример 4
Трансформация и биологические анализы трансгенных растений
Вкратце последовательность, кодирующая конструкт дцРНК, как описано выше, связана на 5' конце с последовательностью, состоящей из промотора 35S, операбельно связанного с интроном кукурузы hsp70, и на 3' конце с последовательностью терминации транскрипции и полиаденилирования Nos3'. Эта кассета экспрессии расположена ниже кассеты селекции глифозата. Затем эти связанные кассеты помещают в функциональный вектор трансформации растения Agrobacterium tumefaciens, применяемый для трансформации ткани кукурузы для толерантности к глифозату, и объектам, выбранным и помещенным в почву. Корни растения R0 скармливают личинкам Западного кукурузного жука (WCR, Diabrotica virgifera). Корни трансгенной кукурузы помещают в чашки Петри со средой MS0D, содержащей антибиотики и глифозат для селекции in vitro. Двумя личинками WCR заражают корень в каждой чашке с помощью кисти с тонким наконечником. Чашки герметично закрывают парафильмом для предотвращения уползания личинок. Образцы помещают в термостат Percival при 27°C, 60% RH в полной темноте. Проводят мониторинг контаминации и качества личинок. После шести дней питания тканью корня личинок перемещают в пищевой субстрат WCR в 96-луночный планшет. Личинкам позволяют поедать пищевой субстрат в течение восьми дней, проводя полный анализ в течение четырнадцати дней. Личиночную массу и выживаемость регистрируют для анализа. Односторонний анализ ANOVA и тест Dunnett's выполняют по данным личиночной массы для определения статистической значимости в сравнении с нетрансформированным отрицательным контролем. Остановку роста личинок WCR определяют после поедания двух объектов и в сравнении с ростом личинок, питавшихся растениями отрицательного контроля.
Созданные трансгенные растения (R0) высаживают в 10-дюймовые резервуары, содержащие почву Metromix, после достижения подходящего размера. Когда растения достигают фазы роста V4, приблизительно 1000 яиц Западного кукурузного жука (WCR, Diabrotica virgifera} помещают в зону корня. Нетрансгенную кукурузу того же генотипа заражают в подобной фазе роста для отрицательного контроля. Яйца предварительно инкубируют так, что выход личинок из яиц происходит в течение 24 часов после заражения. Личинкам позволяют поедать корневые системы в течение 3 недель. Растения извлекают из почвы и промывают так, чтобы можно было оценить поедание корней личинками. Повреждение корней оценивают, применяя Шкалу Повреждения Узла (NIS), для определения уровня повреждения, где 0 указывает на отсутствие повреждения, 1 указывает, что один узел корней подрезан в пределах 1,5 дюймов, 2 указывает, что 2 узла подрезаны, тогда как 3 указывает, что 3 узла подрезаны. Поскольку растения, применяемые для оценки, находятся непосредственно вне культуры тканей после трансформации и поскольку объекты трансформации уникальны, только одно растение оценивают на объект в это время. Растения в анализе, которые имеют признаки или симптомы поедания личинками, указывают, что достигнуто успешное заражение. Корни растений отрицательного контроля усредненно имеют повреждения от средней до сильной степени, тогда как корни трансгенных растений обеспечивают существенный контроль поедания личинками с показателем приблизительно 0,2 или меньше по Шкале Повреждения Узла.
Пример 5
Обеспечение супрессии генов насекомых-паразитов с помощью ta-siРНК опосредованного сайленсинга
Способ
В альтернативном способе сайленсинга генов у вредителя растений применяют обнаруженный в последнее время класс действующих в трансположении малых интерферирующих РНК (ta-siРНК) (Dalmay и др., 2000; Mourrain и др., 2000; Peragine и др., 2004; Vazquez и др., 2004). ta-siРНК получают из транскриптов одноцепочечных РНК, которые направляют с помощью miРНК природного происхождения в пределах клетки. Способы применения микроРНК для инициирования ta-siРНК для сайленсинга генов в растениях, описанные в предварительной заявке на патент США Сер. № 60/643,136 (Carrington и др. 2004), приведены здесь в качестве ссылки в полном объеме. Идентифицирована по крайней мере одна паразит-специфичная miРНК, экспрессируемая в эпителиоцитах кишечника личинок кукурузного жука. Эту паразит-специфичную miРНК затем применяют для идентификации по крайней мере одной последовательности заданного транскрипта РНК, комплементарной miРНК, которая экспрессируется в клетке. Соответствующая заданная последовательность является короткой последовательностью из не более чем 21 смежного нуклеотида, которая, когда является частью транскрипта РНК и контактирует его соответствующей miРНК в типе клеток с функциональным путем РНКi, приводит к опосредованному отклонением расщеплению указанного транскрипта. После идентификации заданных последовательностей miРНК по крайней мере одна заданная последовательность miРНК сливается со второй последовательностью, которая соответствует части гена паразита, который должен быть заглушен с помощью этого способа. Например, заданная последовательность(и) miРНК сливается с любой с SEQ ID NO:1 по SEQ ID NO:906 или ее фрагментом, например с последовательностью гена вакуолярной АТФазы кукурузного жука (V-ATPase). Заданная последовательность miРНК может быть помещена на 5' конец, 3' конец или встроена в середине заданной последовательности. Может быть предпочтительно применение множества заданных последовательностей miРНК, соответствующих множеству генов miРНК, или применение той же заданной последовательности miРНК множество раз в химере заданной последовательности miРНК и последовательности гена-мишени. Последовательность гена-мишени может иметь любую длину с минимумом 21 bp.
Химера заданной последовательности(ей) miРНК и последовательность гена-мишени экспрессируются в растительных клетках, применяя любой из множества подходящих промоторов и других регулирующих транскрипцию элементов, пока транскрипция происходит в типах клеток, предназначенных для обеспечения в пищевом рационе паразита, например в корнях кукурузы для контроля кукурузного жука.
Этот способ может иметь дополнительное преимущество в доставке более длинных молекул РНК к заданному паразиту. Как правило, дцРНК, продуцируемые в растениях, быстро перерабатываются с помощью Dicer в короткие РНК, которые могут быть неэффективны при экзогенном скармливании некоторым паразитам. В этом способе одноцепочечный транскрипт продуцируется в растительной клетке, поедаемой паразитом, и преобразуется в дцРНК в клетке паразита, в которой она затем преобразуется в ta-siРНК, способную к посттранскрипционному сайленсингу одного или более генов в одном или более паразитах-мишенях.
Пример 6
Способы предоставления последовательности ДНК для ДцРНК-опосредованного саленсинга генов
В этом примере показан способ предоставления последовательности ДНК для дцРНК-опосредованного сайленсинга генов. В частности, в этом примере описана селекция улучшенной ДНК, эффективной в дцРНК-опосредованном сайленсинге генов с помощью (a) селекции из гена-мишени начальной последовательности ДНК, включающей более 21 смежного нуклеотида; (b) идентификации по крайней мере одной более короткой последовательности ДНК, полученной из областей начальной последовательности ДНК, состоящей из областей, которые как предполагается не генерируют нежелательные полипептиды и не обладают идентичностью известным последовательностям, как гомологи/ортологи, и (c), выбора последовательности ДНК для дцРНК-опосредованного саленсинга генов, которая включает по крайней мере одну более короткую последовательность ДНК. Нежелательные полипептиды включают, но не ограничены, полипептиды, гомологичные аллергенным полипептидам и полипептидам, гомологичным известным полипептидным токсинам.
Показано, что V-АТФаза WCR функционирует в анализах пищевой активности кукурузного жука для тестирования дцРНК опосредованного сайленсинга в качестве средства контроля личиночного роста. Последовательность кДНК от гена-мишени, такая как ген вакуолярной АТФазы (V-ATPase) от Западного кукурузного жука (WCR) (Diabrotica virgifera virgifera LeConte), выбрана для применения в качестве начальной последовательности ДНК. Эта начальная последовательность ДНК может быть подвергнута скринингу для идентификации областей, в пределах которых каждый смежный фрагмент, включающий по крайней мере 21 нуклеотид, соответствует менее чем 21 из 21 смежного нуклеотида известных последовательностей позвоночных. Идентифицированы сегменты последовательности, состоящие более чем из приблизительно 100 смежных нуклеотидов, не содержащих таких участков 21/21. Таким образом, критерии, включая длину сегмента, содержание GC, последовательность, предсказанную функцию, основанную на последовательности или функции соответствующего гена в организме-модели, и предсказанную вторичную структуру (например, Elbashir и др., 2001b) можно применять для выбора и конструкции последовательности(ей) для применения. Различные комбинации этих сегментов последовательности объединяют для создания химерных последовательностей ДНК для экспрессии в виде дцРНК и применения в анализах пищевой активности насекомых, как описано выше.
Пример 7
Дополнительные результаты анализа пищевой активности насекомого с последовательностями, выбранными из EST
База данных
Этот пример иллюстрирует дополнительные последовательности, которые, как установлено, являются эффективными в вызывании остановки личиночного роста и/или гибели при их поедании личинками жуков в виде двухцепочечных последовательностей РНК. Способы разведения личинок кукурузного жука, применения дцРНК и биологических анализов насекомых соответствуют описанным в Примерах 1-3. Результаты исследований подтверждают значительные уровни (p<0,05) остановки роста личинок и/или гибели, применяя дцРНК, содержащие части последовательностей, гомологичные множеству различных классов генов. Последовательности и векторы, приводящие к существенной остановке роста и/или смертности, и соответствующие SEQ ID NO для последовательности, экспрессируемой в виде дцРНК, приведены ниже в Таблице 6. pMON98503 - пример бинарного вектора, применяемого для трансформации кукурузы, содержит следующие элементы между правой и левой границами T-ДНК для перемещения в растительную клетку: e35S - HSP70 - DV49 (антисмысловая ориентация) - универсальный спейсер - DV49 (смысловая ориентация) - hsp17; ACT (промотор и интрон) - сигнал транзита CTP2 - CP4 - NOS.: pMON98504, другой пример бинарного вектора, применяемого при трансформации кукурузы, включает следующие элементы между правой и левой границами: e35S - HSP70 - C1 (антисмысловая ориентация) - универсальный спейсер - C1 (смысловая ориентация) - hsp17; ACT (промотор и интрон) - сигнал транзита CTP2 - CP4 - NOS.
Тесты эффективности проводили следующим образом, используя потомство зерновых растений, трансформированное с выбранными конструктами контроля насекомых:
1. Семь дней после посадки: инкубируют 10000 яиц WCR на объект (10 растений на объект) при 25°C, 60% RH в полной темноте в течение семи дней.
2. Четырнадцать дней после поражения: растения трансплантируют из торфоперегнойных горшочков 4'' в горшочки 8''; образцы кончиков корней v4 можно забирать для исследований экспрессии генов.
3. Четырнадцать дней после посадки: вымывают яйца WCR из почвы. Помещают яйца и почву в сито с 60 ячейками и помещают сито с 30 ячейками на вершину сита с 60 ячейками для защиты яиц от потока воды. Полностью прополаскивают теплой водой, применяя распылитель до удаления почвы.
4. Суспендируют яйца в 2% (вес./об.) растворе Difco агара, 25 мл раствора на 1 мл яиц. Яйцами заражают почву в количестве приблизительно 3 или 4 аликвоты, применяя, например, пипетку-повторитель Eppendorf, приблизительно 1000 яиц на растение. С помощью лопатки в почве делают отверстия перед заражением и закрывают после заражения.
5. Через двадцать восемь дней после посадки v8 образцы кончиков корней можно взять для исследований экспрессии генов.
6. Тридцать пять дней после посева; проводят оценку анализа. Растения срезают, применяя режущие ножницы, оставляя приблизительно 6" ножку. В частях растения, содержащих информацию об объекте, продавливают отверстия, и крепко привязывают к ножке. Как можно больше почвы удаляют из корневой системы. Остаточную почву вымывают, поливая из поливного шланга.
7. Корни исследуют и дают оценку повреждения корня с помощью шкалы Olsen (0-3) NTS для повреждения личинками WCR.
На Фиг. 1 и Фиг. 2 показаны результаты контроля насекомых, полученные после проверочного заражения WCR зерновых растений F1 (полученных из растений, трансформированных либо с pMON98503, либо с pMON98504). Тесты эффективности в камере роста выполняли в основном, как описано выше, у потомства объектов, полученных с помощью трансформации с pMON98503 или pMON98504, были отмечены показатели NIS на уровне или ниже порога повреждения.
Последовательности ДНК EST полной длины были собраны для выбранных генов, описанных в Примерах 3 и 7, отображающих существенную активность в отношении Западного кукурузного жука. Эти последовательности EST внесены в список в Таблице 7.
Пример 8
Результаты создания и эффективности последовательностей Dv49 и Dv248
Части собранной EST и смежных последовательностей Dv49 (SEQ ID NO:818) и Dv248 (SEQ ID NO:819) отобрали для дальнейших анализов биоактивности, основанных на критериях, включающих предполагаемую функцию, фенотип нокаут-мутантов или соответствующих кодирующих областей в других организмах, длину сегмента, содержание GC, подобие известным последовательностям и предполагаемую вторичную структуру (например, Elbashir и др., 2001b). Соответствующие последовательности Dv49 и Dv248 синтезировали in vitro на основании их предполагаемой активности в отношении WCR каждого в отдельности, а также группировали, как показано на Фиг. 3-4, и в Таблице 8, и нанесли на личинки WCR.
Фрагменты F1-F3 соответствуют частям транскрипта Dv49 полной длины. Фрагменты F4-F6 соответствуют частям транскрипта Dv248 или фланкирующей области. Фрагменты F7-F13 являются конкатемерами двух или более фрагментов F1-F6, как показано на рисунке 4. Фрагмент F13 (SEQ ID NO:834) представляет конкатемер C38 (Dv49-Dv248).
Данные эффекта дозы для F1-F13 показаны на Фиг. 5-7. Как показано на Фиг. 5, активность (% смертности личинок) фрагментов F4-F6, включая последовательности, производные от Dv248 (Фиг. 4), была значительно лучше, чем в контроле при скармливании личинкам WCR в дозе 0,1 ppm. Фрагмент F6 также показал существенную активность при скармливании в дозе 0,02 ppm. Активность фрагмента F5 в наименьшей дозе похожа на артефакт, т.к. у выживших личинок не наблюдалось никакой остановки роста.
Как показано на Фиг. 6, каждый из фрагментов F7-F10 обладает статистически значимой активностью при скармливании личинкам WCR в дозе 0,1 ppm. Фрагменты F9 и F10 также обладают статистически значимой активностью при скармливании личинкам WCR в дозе 0,02 ppm, а фрагмент F10 также обладает статистически значимой активностью при скармливании личинкам WCR в дозе 0,01 ppm. Активность F8 в дозе 0,01 ppm может быть артефактом.
Как показано на Фиг. 7, фрагменты F11-F13 обладают статистически значимой активностью при скармливании личинкам WCR в дозе 0,02 ppm или выше. Дополнительно, наибольшие фрагменты (F12 и F13) обладают активностью в дозе 0,005 ppm и выше.
Пример 9
Дополнительные последовательности активного конкатемера, полученные от конкатемера C6
Как показано в Таблице 9, части активного конкатемера C6 (SEQ ID NO:806), полученного из pMON98368, были идентифицированы в биологических анализах дополнения пищевого рациона (выполненных, как описано, например, в Примере 2), как ингибирующие рост и/или выживание кукурузного жука (WCR). Конкатемер полной длины C6 содержит 7 заданных субфрагментов, каждый по 70 bp, как указано в Таблице 6 и Таблице 9.
Конкатемер полной длины C7 содержит 8 заданных субфрагментов, каждый по 70 bp, как указано в Таблице 6. Подобным образом, конкатемер полной длины C12 содержит 7 заданных суб-фрагментов от Dv23-Dv13-Dv26-Dv18-Dv49-Dv22-Dv20, варьирующих по длине между 53-80 bp, как указано в Таблице 6 и Таблице 10.
В таблице 11 перечислены дополнительные последовательности для применения в направленном воздействии на других жесткокрылых паразитов, включая Diabrotica sp. и др. Coccinellidae и Chrysomelidae.
* * *
Все композиции и способы, раскрытые и испрошенные здесь, могут быть созданы и выполнены без неправомерного экспериментирования в свете настоящего раскрытия. В то время как композиции и способы этого изобретения были описаны в контексте предшествующих иллюстративных вариантов осуществления, специалистам в области техники ясно, что к композиции, способам и в этапах или в последовательности этапов описанных здесь способов могут быть применены вариации, изменения, модификации и альтерации, не отступая от истинной концепции, сущности и объема изобретения. В частности, ясно, что некоторые агенты, которые являются родственными как химически, так и физиологически, можно заменить на агенты, описанные здесь, при том, что могут быть достигнуты такие же или подобные результаты. Считается, что все такие подобные замещения и модификации, известные специалистам в области техники, находятся в пределах сущности, объема и концепции изобретения, как определено в соответствии с приложенной формулой изобретения.
Библиография
Следующие ссылки, при условии, что они предоставляют примеры процедур или других деталей, дополняющих указанные здесь, специфически приведены здесь в качестве ссылки.
Патент США 4245432; Патент США 4272417; Патент США 4339456; Патент США 4372080; Патент США 4383391; Патент США 4465017 ; Патент США 4536475; Патент США 4634587; Патент США 4693977; Патент США 4735015; Патент США 4759945; Патент США 4876197; Патент США 4880734; Патент США 4886937; Патент США 4959317; Патент США 5080925; Патент США 5107065; Патент США 5107787; Патент США 5231020; Патент США 5283184; Патент США 5300127; Патент США 5302523; Патент США 5328942; Патент США 5384253; Патент США 5389399; Патент США 5464765; Патент США 5501967; Патент США 5527695; Патент США 5538877; Патент США 5538880; Патент США 5550318; Патент США 5554445; Патент США 5563055; Патент США 5580544; Патент США 5591616; Патент США 5593 874; Патент США 5622003; Патент США 5633435; Патент США 5661103; Патент США 5698425; Патент США 5712135; Патент США 5759829; Патент США 5780708; Патент США 5789214; Патент США 5791084; Патент США 5804693; Патент США 5834447; Патент США 5837848; Патент США 5849320; Патент США 5876739; Патент США 5882713; Патент США 5891246; Патент США 5918413; Патент США 5939356; Патент США 6118047; Патент США 6326193; Патент США 6489542; Патент США 6506559;
Заявка на патент США Сер. 10/205189; Заявка на патент США Сер. 10/465800; Заявка на патент США Сер. 60/643136.
Публикация патента США 2002/0048814; Публикация патента США 2002/0048814; Публикация патента США 2003/0018993; Публикация патента США 2003/0061626; Публикация патента США 2003/0150017; Публикация патента США 2003/0175965; Публикация патента США 2004/0029283;
Altschul et al., J. Mol. Biol, 215:403-410, 1990.
Ausubel et al., In: Current Protocols in Molecular Biology, John, Wiley & Sons, Inc, New York, 1998.
Babst et al., Dev. Cell, 3:271-282, 2002.
Beutler et al., N. Engl. J. Med., 324(19): 1370, 1991.
Bevan et al., Nature, 304:184-187, 1983.
Botstein et al., Gene, 8(1):17-24, 1979.
Brake et al., Proc. Natl. Acad. Sci. USA, 81(15):4642-4646, 1984.
Brutlag et al., Computers and Chemistry, 17:203-207, 1993.
Current Protocols in Molecular Biology, Ausubel et al. (Eds.), Greene Publishing and Wiley-Interscience, NY, 1989.
Dalmay et al., Cell, 101:543-553, 2000.
Dellaporta et al., In: Chromosome Structure and Function: Impact of New Concepts, 18th Stadler Genetics Symposium, 11:263-282, 1988.
Dow et al., J. Exp. Biol., 200(Pt 2):237-245, 1997.
Dow, J. Bioenerg. Biomembr., 31(1):75-83, 1999.
Elbashir et al., Genes Dev., 5(2): 188-200, 2001a.
Elbashir et al., EMBO J. 20: 6877-6888, 2001b.
Elbashir et al., Methods, 26:199-213, 2002.
Европейская заявка 0 120 516.
Европейская заявка 0 122 791.
Европейская заявка 0 127 328.
Hamilton and Baulcombe, Science, 286(5441):950-952, 1999.
Hannon et al., J. Mol. Neurosci., 18(1-2): 15-27, 2002.
Haymes et al., In: Nucleic acid hybridization, a practical approach, IRL Press, Washington, DC, 1985.
Herrera-Estrella et al., Nature, 303:209-213, 1983.
Ikatu et al., Bio/Technol., 8:241-242, 1990.
Jefferson et al., EMBO J., 6:3901-3907, 1987a.
Jefferson et al., Plant Mol. Biol., Rep., 5:387-405, 1987b.
Kaeppler et al., Plant Cell Rep., 8:415-418, 1990.
Katz et al., J. Gen. Microbiol., 129:2703-2714, 1983.
Kennerdell and Carthew, Cell, 95:1017-1026, 1998.
Kennerdell and Carthew, Nat. Biotechnol., 18:896-898, 2000.
Klee et al., Bio-Technology, 3(7):637-642, 1985.
Lovato et al., Insect. Mol. Biol, 10(4):333-340, 2001.
Lu et al., Phytochemistry, 53(8):941-946, 2000.
Mourrain et al., Cell, 101:533, 2000.
Myanohara et al., Proc. Natl. Acad. Sci. USA, 80(1): 1-5, 1983.
Odell et al., Nature, 313:810-812, 1985.
Ogas et al., Proc. Natl. Acad. Sci. USA, 96(24): 13839-13844, 1999.
Omirulleh et al., Plant Mol. Biol., 21 (3):415-428, 1993.
Orr-Weaver et al., Proc. Natl. Acad. Sci. USA, 80(14):4417-4421, 1983.
Заявка PCT WO 97/32016.
Заявка PCT WO 99/53050.
Заявка PCT WO 99/49029.
Заявка PCT WO 94/01550.
Заявка PCT WO 98/05770.
Peragine et al., Genes Dev., 18(19):2368-2379, 2004.
Pleau et al., Entomol. Exp. Appl. 105:1-11, 2002.
Pontius et al., In: UniGene: A unified view of the transcriptome. NCBI handbook. National Center for Biotechnology Information, Bethesda, MD, 2003.
Potrykus et al., Mol. Gen. Genet, 199(2): 169-177, 1985.
Rajagopal et. al., J. Biol. Chem., 277:46849-46851, 2002.
Rine et al., Proc. Natl. Acad. Sci. USA, 80(22):6750-6754. 1983.
Sambrook et al., In: Molecular cloning: a laboratory manual, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989.
Schnepf et al., Microbiol. Mol. Biol. Rev., 62(3):775-806, 1998.
Smith et al., Cell Dev. Biol., 33:75-79; 1997.
Stinchcomb et al., J. Mol. Biol., 158(2):157-190, 1982.
Sutcliffe et al., Proc. Natl. Acad. Sci. USA, 75:3737-3741, 1978.
Tabara et al., Science, 282(5388):430-431, 1998.
Tabashnik, Appl. Environ. Microbiol., 58(10):3343-3346, 1992.
Van Heeke and Schuster, J. Biol. Chem., 264(33): 19475-19477, 1989.
Vazquez et al., Mol. Cell, 16(1):69-79, 2004.
Yadav et al., Molec. Biochem. Parasitol, doi:10.1016/ j.molbiopara.2006.03.013; в прессе, 2006.
Zukowsky et al., Proc. Natl. Acad. Sci. USA, 80:1101-1105, 1983.
Реферат
Изобретение относится к области биохимии. Представлен выделенный полинуклеотид для ингибирования экспрессии одного или более генов-мишеней у жесткокрылого паразита Diabrotica spp., выбранный из группы, состоящей из полинуклеотида, включающего последовательность нуклеиновой кислоты с SEQ ID NO:179,818,822-824, фрагментов из, по крайней мере, 21 смежного нуклеотида из последовательности нуклеиновой кислоты с SEQ ID NO:179,818,822-824, а также полинуклеотидов, по меньшей мере, на 95% идентичных указанным последовательностям нуклеиновых кислот. Также представлены вектор и клетка, содержащие указанный выделенный полинуклеотид. Раскрыты растение и его семя, содержащие указанный выделенный полинуклеотид. Представлен способ контроля жесткокрылых вредителей Diabrotica spp. с использованием данного выделенного полинуклеотида. Также раскрыт способ улучшения выхода урожая, полученного от сельскохозяйственной культуры, подвергнутой поражению жесткокрылым вредителем Diabrotica spp., а также способ понижения потери устойчивости к засухе вследствие поражения жесткокрылым вредителем Diabrotica spp. урожая, полученного от сельскохозяйственной культуры, подверженной поражению насекомыми-вредителями при включении указанного выделенного полинуклеотида в сельскохозяйственную культуру. Изобретение позволяет создавать растения, устойчивые к поражению жестокрылыми паразитами Diabrotica spp. 12 н. и 15 з.п. ф-лы, 7 ил., 11 табл., 9 пр.
Формула
(a) полинуклеотида, включающего последовательность нуклеиновой кислоты с SEQ ID NO:179,818,822-824;
(b) фрагмента из, по крайней мере, 21 смежного нуклеотида из последовательности нуклеиновой кислоты с SEQ ID NO:179, 818, 822-824, где прием внутрь жесткокрылым вредителем растений двухцепочечной рибонуклеотидной последовательности, включающей, по крайней мере, одну цепь, которая является комплементарной указанному фрагменту, ингибирует рост указанного вредителя; и
(c) полинуклеотида, включающего последовательность, которая по меньшей мере на 95% идентична последовательности нуклеиновой кислоты с SEQ ID NO:179, 818, 822-824; и
(d) комплемента последовательности (а) или (b).
a) включения полинуклеотида по п.1 в указанную сельскохозяйственную культуру,
b) культивирования сельскохозяйственной культуры для возможности экспрессии указанного полинуклеотида, где экспрессия полинуклеотида ингибирует поедание насекомыми-вредителями и снижение выхода урожая вследствие его поражения вредителями.
a) включения полинуклеотида по п.1 в указанную сельскохозяйственную культуру,
b) культивирования сельскохозяйственной культуры для возможности экспрессии указанного полинуклеотида, где экспрессия полинуклеотида ингибирует поедание насекомыми-вредителями и потерю устойчивости к засухе вследствие поражения вредителями.
Документы, цитированные в отчёте о поиске
Способ секвенирования полинуклеотида и устройство для его осуществления
Комментарии