Амплификация библиотек нуклеиновых кислот с применением кинетического исключения - RU2759690C2
Код документа: RU2759690C2
Чертежи








Описание
ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая работа претендует на полезный эффект предварительной патентной заявки US 62/442680, поданной 5 января 2017 года, а также претендует на приоритет патентной заявки Великобритании GB 1704754.9, поданной 24 марта 2017 года, которая претендует на приоритет предварительной патентной заявки US 62/442680, поданной 5 января 2017 года, содержание каждой из которых полностью включено в настоящую работу посредством ссылки.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
В современном обществе генетический анализ приобретает все большую значимость. Уже доказано, что с помощью генетических анализов можно предсказать риск развития у индивидуума некоторых заболеваний (диагностика), определить вероятность достижения терапевтического эффекта в соотношении с рисками побочных эффектов для индивидуума, который рассматривает возможность проведения определенного лечения (прогноз), а также идентифицировать пропавших без вести, преступников, жертв преступлений и жертв военных действий (судебная медицина), причем это лишь несколько возможных примеров применения генетических анализов. Однако во многих случаях подходящие генетические определения (тесты) недоступны или имеют высокий процент ошибок. Одним из источников таких проблем является то, что многие из генетических определений, применяемых в настоящее время для диагностики, прогноза и в судебной медицине, основаны на методиках, которые затрагивают лишь часть генома человека. Характерные генетические признаки человека закодированы в геноме, который содержит более 3 триллионов пар оснований, но в большинстве генетических определений исследуют мутации лишь нескольких таких пар оснований. Повышение доли исследуемого генома, в идеале вплоть до 3 триллионов пар оснований, позволило бы повысить точность генетических определений и разработать способы генетических определений для более широкого ряда диагностических и прогнозируемых ситуаций.
Одним из этапов генетического определения (генетического теста) является подготовка испытуемого генетического материала. Улавливание целого генома и поддержание его целостности является непростой задачей. Двумя способами, доступными в настоящее время для улавливания больших количеств генетического материала, являются полимеразная цепная реакция в эмульсии (англ. emulsion polymerase chain reaction, сокращенно ePCR) и кластерная амплификация (например, посредством мостиковой амплификации). Применение этих способов в клинической и диагностической практике в настоящее время ограничено.
При проведении ePCR, капли водной жидкости формируются в масляной фазе наряду с фрагментами генома и гранулами (микрошариками) носителя. Условия выбирают так, чтобы повысить вероятность того, что в каждой капле будет изолирован индивидуальный фрагмент генома и одна гранула носителя. Цель процесса состоит в том, чтобы капли образовывали микрореакторы, которые не допускают диффузии фрагментов генома между каплями и, следовательно, между различными гранулами. Затем в основном объеме эмульсии могут быть проведены несколько циклов PCR амплификации, в результате которой гранула, находящаяся в определенной капле, покрывается клональными копиями имеющегося фрагмента генома. После протекания амплификации гранулы переносят на подложку для обнаружения и исследуют в аналитическом устройстве. Одна из сложностей ePCR состоит в том, что в некоторые капли попадают некоторые гранулы, не содержащие фрагмента генома, что приводит к образованию пустых гранул. Для удаления пустых гранул до введения гранул в аналитическое устройство может быть выполнено обогащение гранул; однако этот процесс может быть сложным и неэффективным. Другая сложность ePCR состоит в том, что в некоторые капли попадает больше одного фрагмента генома, что приводит к образованию гранул, содержащих смешанные клоны. Несмотря на то, что при проведении анализа гранулы, содержащие смешанные клоны, часто могут быть идентифицированы, после чего их игнорируют, их присутствие снижает эффективность и в некоторых случаях точность анализа.
Кластерная амплификация представляет собой более рациональный подход к захвату и амплификации генетического материала. В коммерческих примерах фрагменты генома закреплены на поверхности подложки (англ. substrate), образуя "зародыши" на различных участках поверхности. После удаления избытка фрагментов генома (то есть незакрепленных фрагментов) промывкой, выполняют несколько циклов амплификации, получая клональные копии, которые образуют кластер на поверхности вокруг каждого зародыша. Преимущества кластерной амплификации по сравнению с ePCR включают отсутствие этапа обогащения гранул, отсутствие этапа переноса гранул (из эмульсии на подложку для обнаружения) и отсутствие густых и часто капризных масляных эмульсий. Однако потенциальной сложностью коммерческих методик кластерной амплификации является то, что они приводят к образованию случайного расположения кластеров на поверхности. Несмотря на то, что были разработаны протоколы регистрации изображений, позволяющие локализовать и различать неупорядоченно расположенные кластеры, такие протоколы оказывают дополнительную аналитическую нагрузку на аналитические устройства. Кроме того, заполнение поверхности неупорядоченно расположенными кластерами обычно менее эффективно по сравнению с теоретически возможным заполнением, создаваемым пространственно упорядоченным размещением кластеров.
Таким образом, существует необходимость создания усовершенствованных способов получения генетического материала для диагностических, прогностических и криминалистических исследований. Настоящее изобретение решает поставленную задачу, а также предоставляет ряд других полезных эффектов.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Один из примеров относится к способу (например, амплификации библиотек нуклеиновых кислот), который включает реакцию первого раствора и отличающегося от него второго раствора на проточной ячейке, проводимую посредством протекания первого раствора по массиву сайтов амплификации, находящихся на проточной ячейке, и затем посредством протекания второго раствора по массиву сайтов амплификации. Первый раствор включает совокупность нуклеиновых кислот-мишеней и первую смесь реагентов, которая включает нуклеозидтрифосфаты (НТФ) и один или более ферментов репликации. Находящиеся в первом растворе нуклеиновые кислоты-мишени переносятся (транспортируются) к сайтам амплификации и связываются с сайтами амплификации со скоростью переноса. Под действием первой смеси реагентов происходит амплификация нуклеиновых кислот-мишеней, которые связаны с сайтами амплификации, в результате чего образуются клональные популяции ампликонов, полученные амплификацией соответствующих нуклеиновых кислот-мишеней. Ампликоны образуются со скоростью амплификации, которая превышает скорость переноса. Второй раствор включает вторую смесь реагентов и не содержит нуклеиновых кислот-мишеней. Второй раствор предназначен для увеличения количества ампликонов в клональных популяциях, находящихся на сайтах амплификации.
В одном из примеров этого способа вторая смесь реагентов включает НТФ и один или более ферментов репликации для первой смеси реагентов.
В одном из примеров этого способа проточная ячейка включает совокупность праймеров, присоединенных к проточной ячейке в сайтах амплификации; в проточной ячейке первый раствор вступает в реакцию связывания нуклеиновых кислот-мишеней и ампликонов с первой подгруппой праймеров, и в проточной ячейке второй раствор вступает в реакцию образования дополнительных ампликонов и связывания дополнительных ампликонов с по меньшей мере некоторыми праймерами, содержащимися в подгруппе праймеров, доступных для взаимодействия и отличающихся от праймеров первой подгруппы.
Этот способ может дополнительно включать удаление первого раствора из проточной ячейки перед пропусканием второго раствора по массиву сайтов амплификации, находящихся на проточной ячейке, в результате чего единственные нуклеиновые кислоты-мишени, присутствующие в проточной ячейке во время протекания второго раствора по массиву сайтов амплификации, связаны с проточной ячейкой, и не представляют собой нуклеиновые кислоты-мишени, свободно перемещаются в первом растворе.
В одном из примеров этого способа массив сайтов амплификации расположен на поверхности проточной ячейки, и первый раствор пропускают через впускное отверстие проточной ячейки по поверхности проточной ячейки, а затем второй раствор пропускают через впускное отверстие по поверхности проточной ячейки.
В одном из примеров этого способа проточная ячейка включает совокупность праймеров, присоединенных к проточной ячейке в сайтах амплификации; по меньшей мере некоторые из праймеров связываются с нуклеиновыми кислотами-мишенями, находящимися в первом растворе, под воздействием первого раствора, протекающего по массиву сайтов амплификации, после чего протекание второго раствора, который не содержит нуклеиновых кислот-мишеней, по массиву сайтов амплификации не приводит к дополнительному связыванию праймеров с нуклеиновыми кислотами-мишенями.
В одном из примеров нуклеиновые кислоты-мишени пропускают по массиву сайтов амплификации, находящихся на проточной ячейке, только в первом растворе. В другом примере этого способа первый раствор и второй раствор пропускают в изотермических условиях по массиву сайтов амплификации.
В некоторых примерах вторая смесь реагентов включает НТФ и полимеразу, в других примерах вторая смесь реагентов включает один или более из следующих ферментов: геликазу и рекомбиназу, и в других примерах вторая смесь реагентов включает праймер, содержащий по меньшей мере одну из следующих последовательностей: последовательность праймера Р5 или последовательность праймера Р7.
В одном из примеров этого способа сайты амплификации представляют собой лунки на поверхности проточной ячейки, где лунки отделены друг от друга расположенными на поверхности промежуточными участками.
Один из примеров этого способа дополнительно включает регулирование скорости переноса нуклеиновых кислот-мишеней к сайтам амплификации с помощью одного или более следующих воздействий: регулирования концентрации нуклеиновых кислот-мишеней в первом растворе, регулирования вязкости первого раствора, регулирования среднего размера нуклеиновых кислот-мишеней и наличия или отсутствия реагента фазового исключения (также называемого «молекулярной скученностью», от англ. molecular crowding) в первом растворе.
Другой пример этого способа дополнительно включает регулирование скорости амплификации нуклеиновых кислот-мишеней с помощью одного или более следующих воздействий: регулирования концентрации НТФ в первой смеси реагентов, регулирования концентрации одного или более ферментов репликации в первой смеси реагентов и регулирования температуры на сайтах амплификации.
Следует понимать, что любые признаки способа могут быть скомбинированы друг с другом любым требуемым образом и/или в виде любой требуемой конфигурации.
Другой пример относится к жидкостной системе (например, для амплификации библиотек нуклеиновых кислот), которая включает распределитель (распределительное устройство) реагентов и контроллер. Распределитель реагентов включает по меньшей мере один клапан, находящийся в гидравлическом взаимодействии со впускным отверстием проточной ячейки, которая включает массив сайтов амплификации. Распределитель реагентов дополнительно включает совокупность каналов, соединяющих посредством гидравлического взаимодействия по меньшей мере один клапан и соответствующие резервуары для реагентов. Контроллер включает один или более процессоров. Контроллер предназначен для управления по меньшей мере одним клапаном и насосом, направляющими первый раствор через впускное отверстие по массиву сайтов амплификации, находящихся на проточной ячейке, и затем направляющими второй, отличающийся от первого раствор, через впускное отверстие по массиву сайтов амплификации, находящихся на проточной ячейке. Первый раствор включает совокупность нуклеиновых кислот-мишеней и первую смесь реагентов, которая включает нуклеозидтрифосфаты (НТФ) и один или более ферментов репликации. Количество нуклеиновых кислот-мишеней в первом растворе превышает количество сайтов амплификации в массиве. Первый раствор вступает на проточной ячейке в реакцию, приводящую к образованию на сайтах амплификации клональных популяций ампликонов соответствующих нуклеиновых кислот-мишеней. Находящиеся в первом растворе нуклеиновые кислоты-мишени переносятся к сайтам амплификации и связываются с сайтами амплификации со скоростью переноса. Под действием первой смеси реагентов происходит амплификация нуклеиновых кислот-мишеней, которые прикреплены к сайтам амплификации, что приводит к образованию ампликонов со скоростью амплификации, которая превышает скорость переноса. Второй раствор включает вторую смесь реагентов и не содержит нуклеиновых кислот-мишеней. Второй раствор вступает на проточной ячейке в реакцию, приводящую к увеличению количества ампликонов в клональных популяциях ампликонов, находящихся на сайтах амплификации.
В одном из примеров этой жидкостной системы вторая смесь реагентов имеет такой же состав, что и первая смесь реагентов.
В одном из примеров этой жидкостной системы контроллер предназначен для управления по меньшей мере одним клапаном и насосом, с помощью которых производят смешивание анализируемой матрицы, которая включает нуклеиновые кислоты-мишени, с первой смесью реагентов с образованием первого раствора, и контроллер также предназначен для управления по меньшей мере одним клапаном и насосом, с помощью которых смешиванием друг с другом второй смеси реагентов, в которую не добавляют анализируемой матрицы, получают второй раствор. В другом примере этой жидкостной системы контроллер предназначен для управления по меньшей мере одним клапаном и насосом, с помощью которых перед пропусканием второго раствора по массиву сайтов амплификации, находящихся на проточной ячейке, из проточной ячейки удаляют первый раствор, в результате чего единственные нуклеиновые кислоты-мишени, присутствующие в проточной ячейке во время протекания второго раствора по массиву сайтов амплификации, связаны с проточной ячейкой, а не свободно перемещаются в первом растворе.
Следует понимать, что любые признаки жидкостной системы могут быть скомбинированы друг с другом любым требуемым образом. Кроме того, следует понимать, что может быть применена любая комбинация признаков жидкостной системы и/или способа, и/или любые признаки любого или обоих этих аспектов могут быть скомбинированы с любыми примерами, рассмотренными в настоящей работе.
Другой пример относится к способу (например, амплификации библиотек нуклеиновых кислот), который включает смешивание в резервуаре первой смеси реагентов с количеством нуклеиновых кислот-мишеней, подходящим для получения первого раствора. Первая смесь реагентов включает нуклеозидтрифосфаты (НТФ) и один или более ферментов репликации. Способ также включает течение первого раствора из резервуара по массиву сайтов амплификации, находящихся на проточной ячейке. Находящиеся в первом растворе нуклеиновые кислоты-мишени переносятся к сайтам амплификации и связываются с сайтами амплификации со скоростью переноса. Под действием первой смеси реагентов происходит амплификация нуклеиновых кислот-мишеней, которые прикреплены к сайтам амплификации, что приводит к образованию клональных популяций ампликонов соответствующих нуклеиновых кислот-мишеней. Ампликоны образуются со скоростью амплификации, которая превышает скорость переноса. После вытекания первого раствора из резервуара, способ включает смешивание второй смеси реагентов в резервуаре без добавления в резервуар дополнительного количества нуклеиновых кислот-мишеней, что приводит к получению второго раствора. Вторая смесь реагентов включает добавочные количества свежих НТФ и один или более ферментов репликации. Способ дополнительно включает протекание второго раствора из резервуара по массиву сайтов амплификации, находящихся на проточной ячейке. Вторая смесь реагентов вступает в реакцию с ампликонами, что приводит к повышению количества ампликонов в клональных популяциях, находящихся на сайтах амплификации.
В одном из примеров этого способа как первая смесь реагентов, так и вторая смесь реагентов включает буферный компонент, причем вторая смесь реагентов включает большее количество буферного компонента, чем первая смесь реагентов, для компенсации недобавленного во второй раствор дополнительного количества нуклеиновых кислот-мишеней.
Следует понимать, что любые признаки этого примера способа могут быть скомбинированы друг с другом любым требуемым образом. Кроме того, следует понимать, что может быть применена любая комбинация признаков этого способа и/или жидкостной системы и/или другого способа, и/или что любые признаки любого или всех раскрытых аспектов могут быть скомбинированы с любыми признаками примеров, рассмотренными в настоящей работе.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Признаки примеров осуществления настоящего изобретения станут более очевидными при рассмотрении представленного ниже подробного описания с учетом фигур, в которых подобные числовые обозначения относятся к подобным, но, возможно, не идентичным компонентам. Для краткости числовые обозначения или признаки, имеющие функции, описанные ранее, могут быть описаны или могут не быть описаны при рассмотрении других фигур, на которых они изображены.
На Фиг. 1А представлено составное изображение (четыре цветных канала), полученное после проведения первого цикла секвенирования в одном из примеров структурированной проточной ячейки с применением кинетического исключения.
На Фиг. 1В представлено составное изображение (четыре цветных канала), полученное после проведения одного цикла секвенирования в примере проточной ячейки, содержащей неупорядоченно расположенные кластеры.
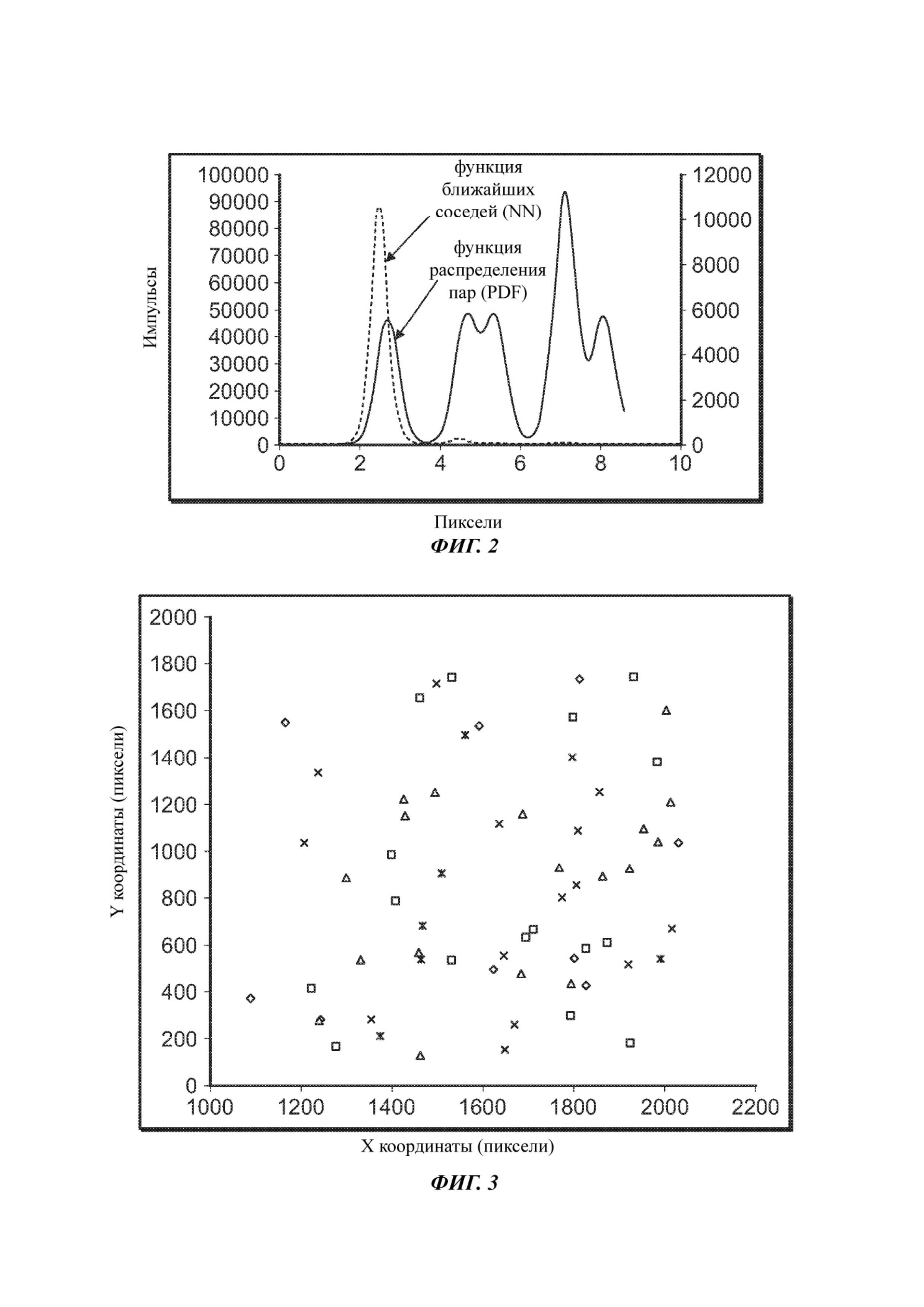
На Фиг. 2 представлена функция распределения пар (англ. pair distribution function, сокращенно PDF) и функция ближайших соседей (англ. nearest neighbor, сокращенно NN), полученные для составного изображения, полученного после первого цикла секвенирования в одном из примеров структурированной проточной ячейки с применением кинетического исключения.
На Фиг. 3 представлена полученная в одном из примеров диаграмма рассеяния пространственных положений кластеров, которые соответствуют первым 5 геномным положениям генома PhiX. Различные геномные положения обозначены крестиками, звездочками, квадратами, треугольниками и ромбами.
На Фиг. 4 (а) и (b) схематично представлены в частичном разрезе примеры архитектуры проточных ячеек для электрохимической десорбции частиц с поверхности проточной ячейки, и на Фиг. 4 (с) представлены полученные способом флуоресцентной микроскопии полного внутреннего отражения (англ. total internal reflection fluorescence, сокращенно TIRF) изображения поверхностей электродов, находящихся в проточной ячейке, имеющей конфигурацию, показанную в части (b), после применения ячейки для осаждения дезоксирибонуклеиновой кислоты (ДНК) с приложением электрического поля. Электрический потенциал может быть приложен к одной проводящей поверхности и электролиту, как показано в части (а), или к двум проводящим поверхностям, как показано в части (b). Конфигурация проточной ячейки, показанной в части (b), также может быть применена для осаждения с ДНК приложением электрического поля в режиме реального времени, что позволяет в течение нескольких секунд достичь 100× концентрации ДНК на поверхности электрода, как показано в части (с).
На Фиг. 5 представлен один из примеров блок-схемы формирвоанрия расположения биомолекул в виде схемы при приложении электрического поля, и схематические изображения в частичном разрезе примеров протекания процесса до и после приложения электрического поля.
На Фиг. 6 представлен один из примеров внесения матриц и кластерная амплификация матриц на элементах из золота размером 2 мкм (Аu), расположенных на основе из оксида индия-олова (англ. indium tin oxide, сокращенно ITO), при приложении электрического поля (а) и в отсутствие электрического поля (b). Соответствующими линии представлены профили интенсивности флуоресценции в меченых участках.
На Фиг. 7 представлен один из примеров флуоресцентных изображений на большой площади после внесения и образования кластеров при приложении электрического поля. На Фиг. 7 (а) представлена дорожка проточной ячейки, содержащая точки из золота размером 2 мкм, и на Фиг. 7 (b) представлена дорожка, содержащая точки из золота размером 200 нм. На больших площадях кластеры совмещены с микро- и наноструктурированными элементами, и пространственный порядок кластеров подтверждается соответствующими алгоритмами быстрого преобразования Фурье (англ. Fast Fourier Transform, сокращенно FFT).
На Фиг. 8 представлен пример образования кластера ДНК на сайтах из SiO2 диаметром 700 нм при приложении электрического поля. Кластеры высоко упорядочены, и в промежуточных областях наблюдает низкий флуоресцентный сигнал.
На Фиг. 9 представлены полученные в одном из примеров (а) результаты анализа гибридизации в инструментальной проточной ячейке (1) HISEQ® до прививки праймеров Р5 и Р7 с приложением электрического поля, (2) после прививки праймеров Р5 и Р7 с приложением электрического поля, (3) после прививки праймеров Р5 и Р7 с приложением электрического поля и повторной прививки праймеров Р5 и Р7, и (4) после прививки праймеров Р5 и Р7 с приложением электрического поля, повторного нанесения покрытия из акриламида, не содержащего силана (англ. silane-free acrylamide, сокращенно SFA), и повторной прививки праймеров Р5 и Р7; и (b) гистограмма, на которой показана средняя интенсивность флуоресценции (в относительных единицах, о.е.) на одну дорожку проточной ячейки после проведения каждого из этапов.
На Фиг. 10 представлены полученные согласно одному из примеров: (а) схематичное представление прямой гибридизации на диэлектрических сайтах при приложении электрического поля; (b) пространственно структурированные кластеры, полученные при приложении отталкивающего нуклеиновые кислоты электрического поля к промежуточным участкам, и (с) неупорядоченно размещенные кластеры, полученные в отсутствие отталкивающего нуклеиновые кислоты электрического поля на промежуточных участках.
На Фиг. 11 представлена блок-схема способа генерации генетических кластеров согласно одному из примеров осуществления изобретения.
На Фиг. 12 представлена гистограмма согласно одному из примеров, на которой показана интенсивность сигнала (в относительных единицах, о.е.) генетических кластеров, генерируемых на проточной ячейке различными способами генерации кластеров.
На Фиг. 13 представлена гистограмма согласно одному из примеров, на которой показано процентное прохождение фильтра (% PF) в способах генерации кластеров, показанных на Фиг. 12.
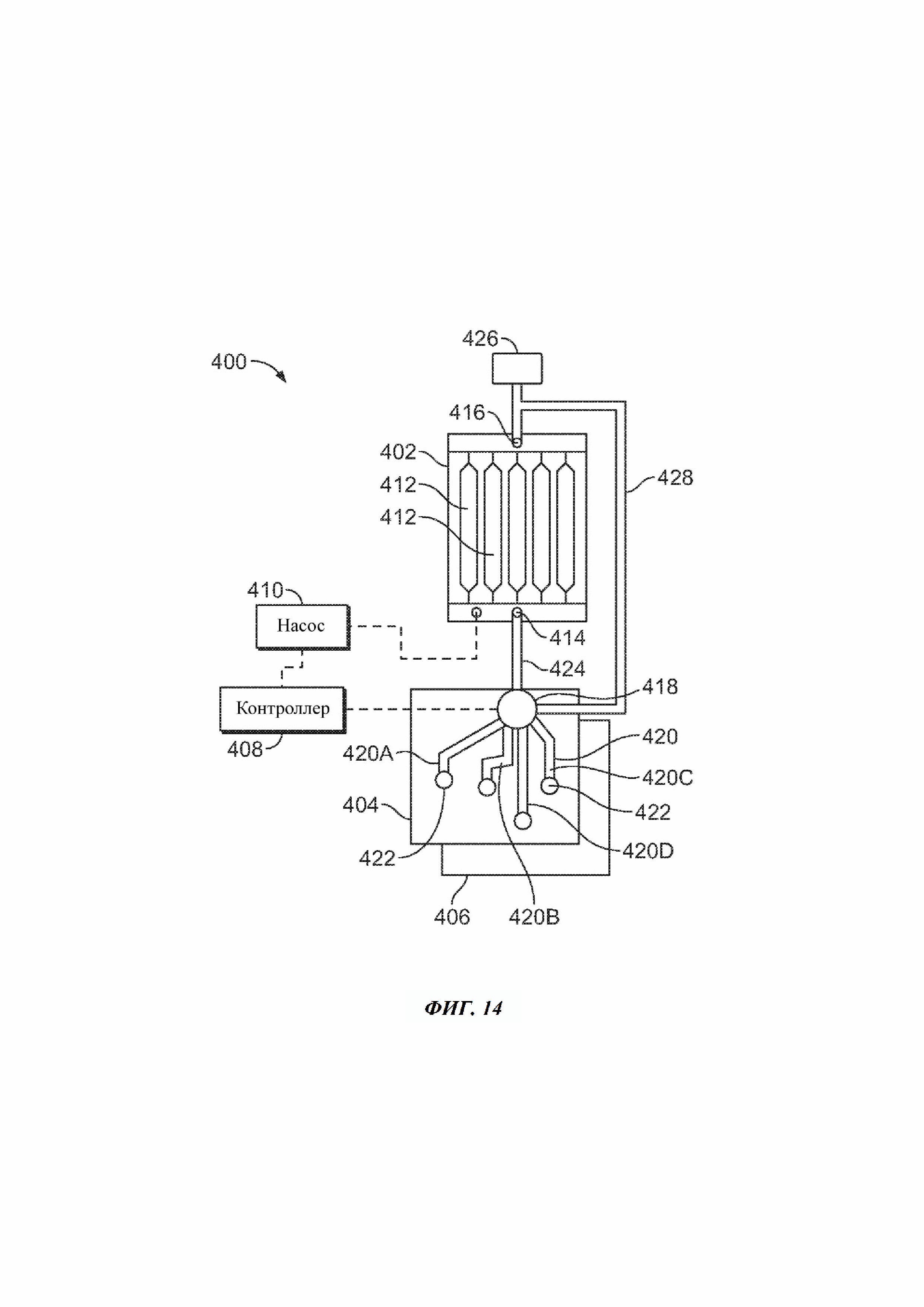
На Фиг. 14 схематически представлена жидкостная система для генерации генетических кластеров согласно одному из примеров настоящего изобретения.
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к библиотекам нуклеиновых кислот и способам получения библиотек нуклеиновых кислот. В конкретных примерах библиотека нуклеиновых кислот согласно настоящему изобретению представлена в виде массива сайтов.
Все литературные и аналогичные материалы, цитируемые в настоящей работе, включающие, без ограничений, патенты, патентные заявки, статьи, книги, руководства и интернет-страницы, независимо от формата таких литературных и аналогичных материалов, полностью включены в настоящее описание посредством ссылки. В том случае, если один или более из включенных литературных и аналогичных материалов отличается от или противоречит настоящей работе, включая, без ограничений, приведенные термины, использования терминов, рассмотренные методики или подобную информацию, преимущество имеет настоящая работа.
Массив может содержать сайты, клональные по отношению к определенным нуклеотидным последовательностям. Соответственно, каждый из индивидуальных сайтов массива может содержать множество копий одной нуклеотидной последовательности. Например, сайты могут содержать клональные копии нуклеиновой кислоты, полученной из биологического образца, такого как геном или его субфракция (например, экзом) или транскриптом (например, библиотека мРНК или библиотека кДНК) или его субфракция.
Доля клональных сайтов в массиве может превышать их долю, рассчитанную в соответствии с распределением Пуассона. Таким образом, массив, получаемый способами согласно изобретению, может иметь сверхпуассоновское распределение клональных сайтов. Сверхпуассоновское распределение может возникнуть в процессе синтеза массива, и для этого не требуется последующее проведение этапов обогащения сайтов или очистки сайта (однако при необходимости по меньшей мере некоторые примеры могут включать этапы обогащения и очистки).
В некоторых примерах сайты могут представлять собой элементы, расположенные на (или в) подложке. В таких примерах элементы могут быть клональными, доля клональных элементов в массиве может превышать долю, рассчитанную согласно распределению Пуассона, и элементы могут быть размещены в пространстве в виде повторяющегося расположения. Таким образом, сайты могут быть упорядочены в пространстве, например, в виде прямолинейной сетки, гексагональной сетки или в виде другой требуемой схемы.
Библиотека нуклеиновых кислот согласно настоящему изобретению может быть получена способом, в котором применяют кинетическое исключение. Кинетическое исключение может иметь место, если процесс протекает со скоростью, достаточно высокой для того, чтобы эффективно исключить наступление другого события или процесса. В качестве примера можно упомянуть создание массива нуклеиновых кислот, где на сайты массива неупорядоченным образом вносят из раствора нуклеиновые кислоты-мишени, и в результате амплификации генерируются копии нуклеиновой кислоты-мишени, которые полностью заполняют каждый из занятых сайтов. В соответствии со способами с применением кинетического исключения согласно настоящему изобретению, внесение и амплификация могут происходить одновременно, если скорость амплификации превышает скорость внесения. Таким образом, относительно высокая скорость образования копий на сайте, который был занят первой нуклеиновой кислотой-мишенью, будет эффективно исключать возможность внесения второй нуклеиновой кислоты на сайт для амплификации. Другие способы амплификации библиотеки нуклеиновых кислот с применением кинетического исключения рассмотрены в документе US 2016/0053310 А1, содержание которого полностью включено в настоящую работу посредством ссылки.
Для кинетического исключения может быть полезна относительно низкая скорость образования первой копии нуклеиновой кислоты-мишени по сравнению с относительно высокой скоростью образования последующих копий нуклеиновой кислоты-мишени или первой копии. В примере, приведенном в предыдущем абзаце, кинетическое исключение обусловлено относительно низкой скоростью внесения нуклеиновой кислоты-мишени (например, относительно низкой скоростью диффузии или переноса) по сравнению с относительно высокой скоростью протекания амплификации, в результате которой происходит заполнение сайта копиями вносимой нуклеиновой кислоты. В другом примере кинетическое исключение может быть обусловлено задержкой в образовании первой копии нуклеиновой кислоты-мишени, которая закреплена на сайте (например, задержкой или медленной активацией), по сравнению с относительно высокой скоростью генерации последующих копий, которые заполняют сайт. В этом примере на индивидуальный сайт могут быть внесены несколько различных нуклеиновых кислот-мишеней (например, на каждом сайте до амплификации могут присутствовать несколько нуклеиновых кислот-мишеней). Однако образование первой копии любой заданной нуклеиновой кислоты-мишени может быть активировано случайным образом, так что средняя скорость образования первой копии относительно ниже скорости генерации последующих копий. В этом случае, несмотря на то, что на индивидуальный сайт могут быть внесены несколько различных нуклеиновых кислот-мишеней, кинетическое исключение допустит амплификацию только одной из этих нуклеиновых кислот-мишеней. В частности, как только происходит приводящая к амплификации активация первой нуклеиновой кислоты-мишени, вся емкость сайта полностью заполняется ее копиями, предотвращая образование на этом сайте копий второй нуклеиновой кислоты-мишени.
Преимуществом массивов, получаемых способами согласно изобретению, является то, что клональная природа сайтов обеспечивает точность последующего анализа. Это позволяет избежать путаницы в результатах, которая возникла бы в случае исследования сайтов со смешанными популяциями.
Другое преимущество массивов согласно изобретению состоит в том, что они имеют сверхпуассоновское распределение клональных сайтов. Это повышает сложность библиотеки за счет отсутствия потерь генетического содержимого, которое происходило бы из-за изъятия в смешанных сайтах.
Другим преимуществом способов и массивов согласно изобретению является создание массива, элементы которого находятся на подложке и имеют повторяющееся расположение в пространстве. Как было указано выше, доля клональных элементов может превышать их долю, рассчитанную в соответствии с распределением Пуассона. Распределение Пуассона имеет максимум при приблизительно 37% занятости. В соответствии со способами согласно изобретению, комплемент (клональных элементов может превышать приблизительно 40%, приблизительно 50%, приблизительно 60%, приблизительно 75% или более. Массивы, получаемые способами согласно изобретению, обеспечивают более эффективное заполнение подложки по сравнению с массивами неупорядоченных кластеров. Такие массивы также легче исследовать аналитическими способами, избегая сложностей, возникающих при исследовании способами регистрации изображений, которые применяют при анализе массивов неупорядоченных кластеров.
Кроме того, преимуществом способов согласно изобретению является создание массивов на структурированных подложках, что облегчает их анализ. Например, некоторые коммерчески доступные платформы секвенирования получены на подложках, имеющих лунки, которые создают барьер для диффузии реагентов, используемых для обнаружения (например, для пирофосфата в платформах, поставляемых 454 LifeSciences (дочернее предприятие Roche, Basel Switzerland), или для протонов в платформах, поставляемых Ion Torrent (дочернее предприятие Life Technologies, Carlsbad Calif.)) в этапах определения последовательности. Преимуществом способов согласно изобретению может быть увеличение количества лунок, в большей степени заполненных клональными популяциями по сравнению со стандартными способами кластерной амплификации, которые ограничены распределением Пуассона. Способы согласно настоящему изобретению имеют преимущества по сравнению со способами, в которых используют ePCR, поскольку не требуют использования эмульсий и работы с гранулами.
Если не указано иное, то термины, употребляемые в настоящем описании, имеют значения, обычно приписываемые им в соответствующей области техники. Ниже приведены некоторые термины и их значения.
Употребляемый в настоящем описании термин "активное внесение" относится к недиффузионным силам, прилагаемым к одной или более нуклеиновым кислотам для перемещения нуклеиновой кислоты (кислот) в направлении к участку или от участка. Участком может быть сайт амплификации массива. Недиффузионные силы могут быть созданы внешним источником, таким как источник электрического или магнитного поля, или агентом, создающим в реакционном объеме фазовое исключение или химические градиенты.
Употребляемый в настоящем описании при рассмотрении нуклеиновой кислоты термин "ампликон" означает продукт копирования нуклеиновой кислоты, где продукт содержит последовательность нуклеотидов, идентичную или комплементарную по меньшей мере части нуклеотидной последовательности нуклеиновой кислоты. Ампликон может быть получен любым из множества различных способов амплификации, в которых в качестве матрицы используют нуклеиновую кислоту или ее ампликон, где способы включают, например, удлинение (достройку) с участием полимеразы, полимеразную цепную реакцию (ПЦР), амплификацию по типу катящегося кольца (англ. rolling circle amplification, сокращенно RCA), удлинение лигированием или цепную реакцию лигированием. Ампликон может представлять собой молекулу нуклеиновой кислоты, содержащую одну копию определенной нуклеотидной последовательности (например, продукт ПЦР) или множество копий нуклеотидной последовательности (например, конкатемерный продукт RCA). Первый ампликон нуклеиновой кислоты-мишени может быть комплементарной копией. Последующие ампликоны представляют собой копии, создаваемые после генерации первого ампликона из нуклеиновой кислоты-мишени или из первого ампликона. Последующий ампликон может содержать последовательность, которая по меньшей мере по существу комплементарна нуклеиновой кислоте-мишени или по меньшей мере по существу идентична нуклеиновой кислоте-мишени.
Употребляемый в настоящем описании термин "сайт амплификации" означает сайт, находящийся в или на массиве, в котором могут быть созданы один или более ампликонов. Сайту амплификации может быть дополнительно придана такая конфигурация, которая позволяет ему содержать, иметь или присоединять по меньшей мере один ампликон, созданный на сайте.
Употребляемый в настоящем описании термин "массив" относится к популяции сайтов, которые могут отличаться друг от друга относительной локализацией. Различные молекулы, находящиеся на различных сайтах массива, могут быть распознаны по локализации на сайтах массива. Индивидуальный сайт массива может включать одну или более молекул определенного типа. Например, сайт может включать одну молекулу нуклеиновой кислоты-мишени, содержащую определенную последовательность, или сайт может включать несколько молекул нуклеиновых кислот, имеющих одинаковую последовательность (и/или комплементарную ей последовательность). Сайты массива могут представлять собой различные элементы, размещенные на одной подложке. Примеры элементов включают, без ограничений, лунки в подложке, гранулы (или другие частицы) в или на подложке, выступы на подложке, ребра на подложке или каналы в подложке. Сайты массива могут представлять собой отдельные подложки, каждая из которых содержит отличную от других молекулу. Различные молекулы, присоединенные к раздельным подложкам, могут быть идентифицированы в соответствии с локализациями подложек на поверхности, с которой связаны подложки, или в соответствии с локализациями подложек в жидкости или геле. Примеры массивов, на поверхности которых расположены отдельные подложки, включают, без ограничений, массивы, содержащие гранулы в лунках.
Употребляемый в настоящем описании при рассмотрении сайта и материала нуклеиновой кислоты термин "емкость" означает максимальное количество материала нуклеиновой кислоты, которое может находиться на/в сайте. Например, этот термин может относиться к общему количеству молекул нуклеиновой кислоты, которые сайт может вмещать при определенных условиях. Могут быть использованы другие количественные характеристики, включающие, например, общую массу материала нуклеиновой кислоты или общее количество копий определенной нуклеотидной последовательности, которые сайт может вмещать при определенных условиях. Емкость сайта по отношению к нуклеиновой кислоте-мишени может быть по меньшей мере по существу эквивалентна емкости сайта по отношению к ампликонам нуклеиновой кислоты-мишени.
Употребляемый в настоящем описании термин "агент захвата" относится к материалу, химическому соединению, молекуле или ее фрагменту, который может присоединяться к, удерживаться или связываться с молекулой-мишенью (например, нуклеиновой кислотой-мишенью). Примеры агентов захвата включают, без ограничений, улавливающую (захватывающую) нуклеиновую кислоту, которая комплементарна по меньшей мере части нуклеиновой кислоты-мишени, член связывающейся пары рецептор-лиганд (например, авидин, стрептавидин, биотин, лектин, углевод, белок, связывающий нуклеиновую кислоту, эпитоп, антитело и т.д.), способный связываться с нуклеиновой кислотой-мишенью (или присоединенным к ней связующим фрагментом), или химический реагент, способный образовывать ковалентную связь с нуклеиновой кислотой-мишенью (или присоединенным к ней связующим фрагментом).
Употребляемый в настоящем описании термин "клональная популяция" относится к популяции нуклеиновых кислот, которая гомогенна по отношению к определенной нуклеотидной последовательности. Длина гомогенной последовательности может составлять по меньшей мере приблизительно 10 нуклеотидов, но она может быть длиннее и включать, например, по меньшей мере приблизительно 50, приблизительно 100, приблизительно 250, приблизительно 500 или приблизительно 1000 нуклеотидов. Клональная популяция может быть получена из одной нуклеиновой кислоты-мишени или из матричной нуклеиновой кислоты. Большинство, если не все, нуклеиновые кислоты в клональной популяции содержат одну и ту же последовательность нуклеотидов. Следует понимать, что в клональной популяции может возникать небольшое число мутаций (например, из-за артефактов амплификации), которые не оказывают существенного влияния на ее клональность.
Употребляемый в настоящем описании термин "цикл денатурации" означает одну из манипуляций при проведении реакции амплификации нуклеиновой кислоты, которая изменяет курс реакции амплификации, в результате чего происходит отделение цепочек комплементарных нуклеиновых кислот друг от друга. Неограничивающие примеры манипуляций включают введение химического реагента, который вызывает денатурацию нуклеиновых кислот, или физическое изменение реакции в результате нагревания или другой манипуляции, приводящее к денатурации нуклеиновых кислот. Циклическая реакция амплификации может включать несколько циклов денатурации. Для проведения гибридизации праймера с цепочкой нуклеиновой кислоты также может быть включен ряд других циклов, таких как циклические манипуляции. Способ согласно изобретению может не включать один или более циклов денатурации или других циклов. Таким образом, по меньшей мере в некоторых примерах настоящего изобретения реакция амплификации может быть проведена без циклических манипуляций.
Употребляемый в настоящем описании при рассмотрении нуклеиновых кислот термин "различный" означает, что нуклеиновые кислоты имеют неидентичные последовательности нуклеотидов. Две или более нуклеиновые кислоты могут иметь последовательности нуклеотидов, различающиеся по всей их длине. В альтернативном варианте две или более нуклеиновые кислоты могут иметь последовательности нуклеотидов, различающиеся на существенной части их длины. Например, две или более нуклеиновые кислоты могут иметь целевые части нуклеотидных последовательностей, отличающиеся друг от друга, но при этом иметь идентичный участок универсальной последовательности.
Употребляемый в настоящем описании при рассмотрении молекулы, находящейся в текучей среде, и сайта, находящегося в контакте с текучей средой, термин "доступ посредством текучей среды" означает способность молекулы перемещаться в или через текучую среду и контактировать с сайтом или внедряться в сайт. Термин также может означать способность молекулы отделяться от сайта или выходить из сайта в раствор. Доступ посредством текучей среды может иметь место при отсутствии барьеров, которые препятствуют внедрению молекул в сайт, контакт с сайтом, отделение от сайта и/или выход из сайта. Кроме того, следует понимать, что доступ посредством текучей среды существует, даже если диффузия замедлена, снижена или изменена, при условии, что доступ не полностью предотвращен.
Употребляемый в настоящем описании при рассмотрении молекулы нуклеиновой кислоты термин "двухцепочечный" означает, что по меньшей мере по существу все нуклеотиды в молекуле нуклеиновой кислоты связаны водородными связями с комплементарным нуклеотидом. В частично двухцепочечной нуклеиновой кислоте по меньшей мере приблизительно 10%, приблизительно 25%, приблизительно 50%, приблизительно 60%, приблизительно 70%, приблизительно 80%, приблизительно 90% или приблизительно 95% нуклеотидов могут быть связаны водородными связями с комплементарным нуклеотидом.
Употребляемый в настоящем описании при рассмотрении совокупности объектов термин "каждый" предназначен для идентификации индивидуального объекта совокупности, но не обязательно относится к каждому объекту совокупности, если из контекста не следует иное.
Употребляемый в настоящем описании при рассмотрении числовых значений термин "приблизительно" указывает на приблизительность приведенного значения, например, в пределах ±5% от приведенной величины.
Употребляемый в настоящем описании для модификации прилагательного термин "по меньшей мере по существу" означает степень, близкую или приблизительно близкую к указанному прилагательному, например, в пределах ±5% диапазона. Например, фраза "по меньшей мере по существу все нуклеиновые кислоты" может относиться ко всем (например, 100%) нуклеиновым кислотам или меньшему количеству, чем все нуклеиновые кислоты в пределах указанного диапазона, например, от 95% до 99,99% нуклеиновых кислот.
Употребляемый в настоящем описании термин "следовое количество" относится к очень низкой концентрации анализируемого вещества в смеси или растворе, такой как меньше или равной приблизительно 100 частей на миллион.
Употребляемый в настоящем описании термин "исключенный объем" относится к объему, занимаемому в пространстве определенной молекулой, в котором она не допускает присутствия других таких же молекул.
Употребляемый в настоящем описании при рассмотрении нуклеиновой кислоты, такой как праймер, термин "достраиваемый" или "состояние, подходящее для достройки" означает, что нуклеиновая кислота отвечает требованиям для добавления (присоединения) нуклеотида (например, посредством полимеразного катализа) или добавления олигонуклеотида (например, посредством лигазного катализа). "Недостраиваемая" или находящаяся в "состоянии, не подходящем для достройки" нуклеиновая кислота не отвечает в достаточной степени указанному требованию, например, из-за присутствия фрагмента, блокирующего удлинение, или отсутствия 3'-гидроксила.
Употребляемый в настоящем описании термин "промежуточный участок" относится к области в подложке или на поверхности, которая отделяет другие области подложки или поверхности. Например, промежуточный участок может отделять один элемент массива от другого элемента массива. Два участка, отделенные друг от друга, могут быть дискретными, не имеющими контакта друг с другом. В другом примере промежуточный участок может отделять первую часть элемента от второй части элемента. Отделение, обеспечиваемое промежуточным участком, может быть частичным или полным. Материал поверхности промежуточных участков может отличаться от материала поверхности элементов, находящихся на поверхности. Например, элементы массива могут содержать такое количество или концентрацию агентов захвата или праймеров, которая превышает их количество или концентрацию на промежуточных участках. В некоторых примерах агенты захвата или праймеры могут отсутствовать на промежуточных участках.
Употребляемый в настоящем описании термин "полимераза" употребляется в значении, обычно приписываемом ему в данной области техники, и означает, например, фермент, генерирующий комплементарную копию (реплику) молекулы нуклеиновой кислоты, используя нуклеиновую кислоту в качестве матричной цепочки. ДНК-полимеразы могут связываться с матричной цепочкой и затем перемещаться вдоль матричной цепочки, последовательно добавляя нуклеотиды к свободной гидроксильной группе, находящейся на 3'-конце растущей цепочки нуклеиновой кислоты. ДНК-полимеразы синтезируют комплементарные молекулы ДНК из матриц ДНК, а пол имеразы рибонуклеиновых кислот (РНК) синтезируют молекулы РНК из ДНК матриц (транскрипция). Полимеразы могут воздействовать на короткие цепочки РНК или ДНК, называемые праймерами, инициируя рост цепочки. Некоторые полимеразы могут перемещать цепочку вверх по потоку относительно сайта, где они добавляют к цепочке основания. Такие полимеразы называют замещающими цепь, поскольку они способны удалять комплементарную цепочку из матричной цепочки, прочитываемой полимеразой. Примеры полимераз, способных к замещению цепи, включают, например, крупный фрагмент полимеразы Bst (Bacillus stearothermophilus), полимераза экзофрагмента Кленова или экзополимераза секвенирования Т7. Некоторые полимеразы разрушают цепочку, находящуюся впереди них, эффективно замещая ее растущей позади них цепью (5'-экзонуклеазная активность). Некоторые полимеразы разрушают цепочку, находящуюся позади них (3'-экзонуклеазная активность). Для снижения или устранения 3'- и/или 5'-экзонуклеазной активности некоторые подходящие полимеразы модифицируют с помощью мутации или иным способом.
Используемый в настоящем описании термин "нуклеиновая кислота" употребляется в значении, обычно приписываемом ему в данной области техники, и включает природные нуклеиновые кислоты или их функциональные аналоги. Особенно подходящие функциональные аналоги способны гибридизоваться с нуклеиновой кислотой последовательность-специфичным образом или могут быть использованы в качестве матриц для репликации определенной нуклеотидной последовательности. В основных цепях встречающихся в природе нуклеиновых кислот обычно содержатся сложные фосфодиэфирные связи. Структура аналога может иметь альтернативные связи в основной цепи, которые включают любые типы связей, известных в данной области техники. Природные нуклеиновые кислоты обычно содержат дезоксирибозный сахар (например, в дезоксирибонуклеиновой кислоте (ДНК)) или рибозный сахар (например, в рибонуклеиновой кислоте (РНК)). Нуклеиновая кислота может содержать любой из множества аналогов фрагментов этих сахаров, которые известны в данной области техники. Нуклеиновая кислота может включать нативные (т.е. встречающиеся в природе) или не встречающиеся в природе основания. Так, нативная дезоксирибонуклеиновая кислота может содержать одно или более оснований, выбранных из группы, состоящей из аденина, тимина, цитозина или гуанина, а рибонуклеиновая кислота может содержать одно или более оснований, выбранных из группы, состоящей из урацила, аденина, цитозина или гуанина. Подходящие не встречающиеся в природе основания, которые могут быть включены в нуклеиновую кислоту, известны в данной области техники. Употребляемый при рассмотрении нуклеиновой кислоты термин "мишень" используется как смысловой идентификатор нуклеиновой кислоты в контексте способа или композиции согласно изобретению и не обязательно ограничивает структуру или функцию нуклеиновой кислоты до диапазона, более узкого, чем указанный.
Употребляемый в настоящем описании при рассмотрении переноса, амплификации, захвата или других химических процессов термин "скорость" имеет значение, согласующееся с его значением в химической кинетике и биохимической кинетике. Может быть произведено сравнение скоростей двух процессов по таким параметрам, как максимальная скорость (например, при насыщении), предстационарные скорости (например, до равновесия), кинетические константы скорости, или по другим параметрам, известным в данной области техники. В конкретных примерах скорость определенного процесса может быть определена по величине общего времени, требуемого для завершения процесса. Например, скорость амплификации может быть определена по времени, которое требуется для завершения амплификации. Однако скорость определенного процесса не обязательно должна быть определена по общему времени, которое требуется для завершения процесса.
Используемый в настоящем описании термин "рекомбиназа" употребляется в значении, обычно приписываемом ему в данной области техники, и включает, например, белок RecA, белок Т4 uvsX, любой гомологичный белок или белковый комплекс из любых таксонометрических групп или функциональные варианты таких белков. Эукариотические гомологи RecA обычно называются Rad51 по первому члену идентифицируемой группы. Вместо RecA могут быть применены другие негомологичные рекомбиназы, например, RecT или RecO.
Употребляемый в настоящем описании термин "одноцепочечный связывающий белок" относится к любому белку, обладающему способностью связываться с одноцепочечной нуклеиновой кислотой, например, для предотвращения преждевременной ренатурации, для защиты одноцепочечной нуклеиновой кислоты от нуклеазного расщепления, для удаления вторичной структуры нуклеиновой кислоты или для облегчения репликации нуклеиновой кислоты. Термин включает, например, белки, которые формально идентифицированы как одноцепочечные связывающие белки Комитетом по номенклатуре Международного Союза биохимии и молекулярной биологии (англ. Nomenclature Committee of the International Union of Biochemistry and Molecular Biology, сокращенно NC-IUBMB). Неограничивающие примеры одноцепочечных связывающих белков включают Е. coli SSB, Т4 gp32, Т7 ген 2.5 SSB, фаг phi 29 SSB, любой гомологичный белок или белковый комплекс из любых таксонометрических групп или функциональные варианты таких белков.
Употребляемый в настоящем описании термин "перенос" (или "транспорт") относится к перемещению молекулы через текучую среду. Термин может включать пассивный перенос, такой как перемещение молекул вдоль градиента концентрации (например, пассивную диффузию). Термин также может включать активный перенос, при котором молекулы могут перемещаться вдоль градиента их концентрации или против градиента их концентрации. Таким образом, перенос может включать подачу энергии для перемещения одной или более молекул в требуемом направлении или к требуемой локализации, такой как сайт амплификации.
Употребляемый в настоящем описании термин "универсальная последовательность" означает участок последовательности, одинаковый для двух или более молекул нуклеиновых кислот, при том, что молекулы также имеют различающиеся участки последовательности. Универсальная последовательность, присутствующая в различных членах совокупности молекул, может обеспечить захват множества различных нуклеиновых кислот с помощью популяции универсальных улавливающих нуклеиновых кислот, которые комплементарны универсальной последовательности. Аналогично, универсальная последовательность, присутствующая в различных членах совокупности молекул, позволяет проводить репликацию или амплификацию множества различных нуклеиновых кислот с помощью популяции универсальных праймеров, которые комплементарны универсальной последовательности. Таким образом, универсальная улавливающая нуклеиновая кислота или универсальный праймер включает последовательность, которая может специфично гибридизоваться с универсальной последовательностью. Молекулы нуклеиновых кислот-мишеней могут быть модифицированы для присоединения универсальных адаптеров, например, по одному или по обоим концам различных последовательностей-мишеней.
Настоящее изобретение относится к способу амплификации нуклеиновых кислот. Способ включает (а) предоставление реагента амплификации, включающего (i) массив сайтов амплификации, и (ii) раствора, содержащего совокупность различных нуклеиновых кислот-мишеней; и (b) реакцию реагента амплификации, приводящую к образованию совокупности сайтов амплификации, каждый из которых содержит клональную популяцию ампликонов индивидуальной нуклеиновой кислоты-мишени, содержавшейся в растворе, причем реакция включает одновременное протекание (i) переноса различных нуклеиновых кислот-мишеней к сайтам амплификации со средней скоростью переноса, и (ii) амплификации различных нуклеиновых кислот-мишеней на сайтах амплификации со средней скоростью амплификации, причем средняя скорость амплификации превышает среднюю скорость переноса. В конкретных примерах количество различных нуклеиновых кислот-мишеней в растворе превышает количество сайтов амплификации в массиве. Различные нуклеиновые кислоты-мишени транспортируются текучей средой к совокупности сайтов амплификации. Кроме того, каждый сайт амплификации необязательно может обладать емкостью для размещения нескольких нуклеиновых кислот совокупности различных нуклеиновых кислот.
Изобретение также относится к способу амплификации нуклеиновых кислот, который включает (а) предоставление реагента амплификации, включающего (i) массив сайтов амплификации, и (ii) раствора, содержащего совокупность различных нуклеиновых кислот-мишеней; и (b) реакцию реагента амплификации, приводящую к образованию совокупности сайтов амплификации, каждый из которых включает клональную популяцию ампликонов индивидуальной нуклеиновой кислоты-мишени, содержавшейся в растворе, причем реакция включает (i) получение первого ампликона индивидуальной нуклеиновой кислоты-мишени на каждом из сайтов амплификации, и (ii) получение последующих ампликонов индивидуальной нуклеиновой кислоты-мишени на каждом из сайтов амплификации или из первого ампликона, причем средняя скорость генерации последующих ампликонов на сайтах амплификации превышает среднюю скорость генерации первого ампликона на сайтах амплификации. В конкретных примерах количество различных нуклеиновых кислот-мишеней в растворе превышает количество сайтов амплификации в массиве. Различные нуклеиновые кислоты-мишени транспортируются текучей средой к совокупности сайтов амплификации. Кроме того, каждый из сайтов амплификации необязательно может обладать емкостью для размещения нескольких нуклеиновых кислот совокупности различных нуклеиновых кислот.
Настоящее изобретение дополнительно относится к способу амплификации нуклеиновых кислот, который включает (а) предоставление реагента амплификации, включающего (i) массив сайтов амплификации, и (ii) раствора, содержащего совокупность различных нуклеиновых кислот-мишеней; и (b) реакцию реагента амплификации, приводящую к образованию совокупности сайтов амплификации, каждый из которых содержит клональную популяцию ампликонов индивидуальной нуклеиновой кислоты-мишени, содержавшейся в растворе. Реакция включает одновременный (i) захват различных нуклеиновых кислот на сайтах амплификации, протекающий со средней скоростью захвата, и (ii) амплификацию различных нуклеиновых кислот-мишеней, захваченных на сайтах амплификации, протекающую со средней скоростью амплификации. Средняя скорость амплификации превышает среднюю скорость захвата.
Изобретение также относится к способу амплификации нуклеиновых кислот, который включает (а) предоставление реагента амплификации, включающего (i) массив сайтов амплификации, и (ii) раствора, содержащего совокупность различных нуклеиновых кислот-мишеней; и (b) реакцию реагента амплификации, приводящую к образованию совокупности сайтов амплификации, каждый из которых включает клональную популяцию ампликонов индивидуальной нуклеиновой кислоты-мишени, содержавшейся в растворе. Реакция включает (i) получение первого ампликона индивидуальной нуклеиновой кислоты-мишени, который захвачен сайтом (сайтами) амплификации, и (ii) получение последующих ампликонов индивидуальной нуклеиновой кислоты-мишени, которая захвачена каждым из сайтов амплификации, или ампликонов первого ампликона. Средняя скорость генерации последующих ампликонов на сайтах амплификации превышает среднюю скорость генерации первого ампликона на сайтах амплификации.
Изобретение дополнительно относится к способу создания структурированной поверхности из биомолекул, который включает (а) предоставление реагента, включающего (i) массив, на поверхности которого имеются не непрерывные элементы, то есть элементы разделены промежуточными участками поверхности, и (ii) раствора, содержащего совокупность различных биомолекул-мишеней; и (b) реакцию реагента, приводящую к переносу биомолекул к элементам и к присоединению индивидуальной биомолекулы к каждому из элементов. Для выталкивания биомолекул из промежуточных участков к промежуточным участкам прикладывают электрическое поле.
Массив сайтов амплификации, применяемый в способе согласно изобретению, может присутствовать в виде одной или более подложек. Примеры материалов подложки, которые могут быть применены для получения массива, включают стекло (например, модифицированное стекло, функционализированное стекло, неорганические стекла), микросферы (например, инертные и/или магнитные частицы), пластические массы, полисахариды, нейлон, нитроцеллюлозу, керамику, смолы, оксид кремния, материалы на основе оксида кремния, углерод, металлы, оптическое волокно или пучки оптических волокон, полимеры и мультилуночные планшеты (например, микротитрационные планшеты). Примеры полимеров включают акриловые полимеры, полистирол и сополимеры стирола и других материалов, полипропилен, полиэтилен, полибутилен, полиуретаны, политетрафторэтилен (например, TEFLON®, поставляемый DuPont). Примеры материалов на основе оксида кремния включают кремний и различные формы модифицированного кремния.
В конкретных примерах подложка может находиться в или составлять часть емкости, такой как лунка, трубка, канал, кювета, чашка Петри, бутылка или подобной емкости. Особенно подходящей емкостью является проточная ячейка, например, рассмотренная в опубликованной патентной заявке US 2010/0111768 А1 или в публикации Bentley с соавт., Nature 456:53-59 (2008), содержания которых полностью включены в настоящую работу посредством ссылки. Примерами проточных ячеек являются ячейки, коммерчески поставляемые Illumina, Inc. (San Diego, Calif.). Другой особенно подходящей емкостью является лунка в мультилуночном планшете или микротитрационном планшете.
В некоторых примерах сайты массива могут быть сконструированы в виде элементов, находящихся на поверхности. Элементы могут иметь любой из множества различных требуемых форматов. Например, сайты могут представлять собой лунки, углубления, каналы, ребра, приподнятые участки, выступы, столбики или подобные фигуры. Как было указано выше, сайты могут содержать гранулы. Однако в конкретных примерах сайты не обязательно содержат гранулу или частицу. Примеры сайтов включают лунки, расположенные в подложках, применяемых в коммерческих платформах секвенирования, коммерчески поставляемых 454 LifeSciences (дочернее предприятие Roche, Basel Switzerland) или Ion Torrent (дочернее предприятие Life Technologies, Carlsbad Calif.). Другие подложки, имеющие лунки, включают, например, протравленные оптические волокна и другие подложки, рассмотренные в патентах US 6266459, US 6355431, US 6770441, US 6859570, US 6210891, US 6258568, US 6274320, в опубликованной патентной заявке US 2009/0026082 А1, опубликованной патентной заявке US 2009/0127589 А1, опубликованной патентной заявке US 2010/0137143 А1, опубликованной патентной заявке US 2010/0282617 А1 или публикации международной патентной заявки WO 00/63437, содержания которых полностью включены в настоящую работу посредством ссылки. В некоторых случаях подложки, упоминаемые в приведенной выше литературе, применяют в способах, включающих размещение в лунках гранул. Подложки, содержащие лунки, могут быть применены при наличии или отсутствии в них гранул в способах или композициях согласно настоящему изобретению. В некоторых примерах лунки подложки могут содержать гелеобразный материал (при наличии или отсутствии гранул), как указано в документе US 9512422, содержание которого полностью включено в настоящую работу посредством ссылки.
Сайты массива могут представлять собой металлические элементы на неметаллической поверхности, такой как стекло, пластическая масса или другие материалы, примеры которых приведены выше. Слой металла может быть нанесен на поверхность способами, известными в данной области техники, такими как мокрое плазменное травление, сухое плазменное травление, атомно-слоевое осаждение, ионно-лучевое травление, химическое осаждение из газовой фазы, вакуумное напыление или подобные способы. Может быть применено любое из множества коммерчески доступных подходящих устройств, которые включают, например, системы FLEXAL®, OPAL™, IONFAB® 300plus или OPTOFAB® 3000 (Oxford Instruments, UK). Слой металла также может быть нанесен электроннолучевым испарением или напылением, описанным в публикации Thornton, Ann. Rev. Mater. Sci. 7:239-60 (1977), содержание которой полностью включено в настоящую работу посредством ссылки. Для создания металлических участков или областей на поверхности, методики нанесения слоя металла, например, приведенные выше, могут быть скомбинированы с методиками фотолитографии. Примеры способов, в которых комбинируют методики нанесения слоя металла и методики фотолитографии, приведены ниже в Примерах I и II и в патенте US 8778848, содержание которого полностью включено в настоящую работу посредством ссылки.
Массив элементов может выглядеть как сетка из пятен или участков. Элементы могут иметь повторяющееся расположение или неупорядоченное неповторяющееся расположение. Особенно подходящими схемами расположения являются гексагональные схемы расположения, прямолинейные схемы расположения, сетчатые схемы расположения, зеркально-симметричные схемы расположения, схемы расположения с осевой симметрией или подобные схемы. Подходящими также могут быть асимметричные схемы расположения. Шаг между различными парами ближайших соседних элементов может быть одинаковым, или шаг между различными парами ближайших соседних элементов может быть различным. В конкретных примерах каждый их элементов массива может иметь площадь, превышающую приблизительно 100 нм2, приблизительно 250 нм2, приблизительно 500 нм2, приблизительно 1 мкм2, приблизительно 2,5 мкм2, приблизительно 5 мкм2, приблизительно 10 мкм2, приблизительно 100 мкм2 или приблизительно 500 мкм2. В альтернативном варианте или дополнительно каждый из элементов массива может иметь площадь, составляющую менее приблизительно 1 мм2, приблизительно 500 мкм2, приблизительно 100 мкм2, приблизительно 25 мкм2, приблизительно 10 мкм2, приблизительно 5 мкм2, приблизительно 1 мкм2, приблизительно 500 нм2 или приблизительно 100 нм2. На практике размер участка может находиться в диапазоне между верхним и нижним пределами, выбранными из значений, примеры которых приведены выше.
В примерах, которые включают массив элементов на поверхности, элементы могут быть дискретными, разделенными промежуточными участками. Размер элементов и/или расстояние между участками может быть различным, то есть массивы могут иметь высокую плотность, среднюю плотность или пониженную плотность. Массивами с высокой плотностью считаются массивы, участки в которых разделены расстоянием менее приблизительно 15 мкм. Участки в массивах со средней плотностью разделены расстоянием, составляющим от приблизительно 15 до приблизительно 30 мкм, в то время как участки в массивах с низкой плотностью разделены расстоянием, превышающим приблизительно 30 мкм. Массив, подходящий для применения в одном или более примерах согласно изобретению, может содержать участки, разделенные расстоянием, составляющим менее приблизительно 100 мкм, приблизительно 50 мкм, приблизительно 10 мкм, приблизительно 5 мкм, приблизительно 1 мкм или приблизительно 0,5 мкм.
В конкретных примерах массив может включать совокупность гранул или других частиц. Частицы могут быть суспендированы в растворе, или они могут быть локализованы на поверхности подложки. Примерами массивов, содержащих гранулы в растворе, являются массивы, коммерчески поставляемые Luminex (Austin, Тех.). Примеры массивов, в которых гранулы расположены на поверхности, включают массивы, в которых гранулы расположены в лунках, такие как массив BEADCHIP™ (Illumina Inc., San Diego Calif.), или подложки, применяемые в платформах секвенирования, поставляемых 454 LifeSciences (дочернее предприятие Roche, Basel Switzerland) или Ion Torrent (дочернее предприятие Life Technologies, Carlsbad Calif.). Другие массивы, содержащие расположенные на их поверхности гранулы, рассмотрены в патентах US 6266459, US 6355431, US 6770441, US 6859570, US 6210891, US 6258568, US 6274320, в опубликованной патентной заявке US 2009/0026082 А1, опубликованной патентной заявке US 2009/0127589 А1, опубликованной патентной заявке US 2010/0137143 А1, опубликованной патентной заявке US 2010/0282617 А1 или патентной заявке РСТ WO 00/63437, содержания которых полностью включены в настоящую работу посредством ссылки. В некоторых из приведенных выше публикаций рассмотрены способы присоединения нуклеиновых кислот-мишеней к гранулам перед загрузкой гранул в или на подложку массива. Однако следует понимать, что в гранулы могут быть включены праймеры амплификации, и затем гранулы могут быть загружены в массив, образуя, таким образом, сайты амплификации, применяемые в способе согласно изобретению. Как было указано выше в настоящем описании, могут быть применены подложки, не содержащие гранул. Например, праймеры амплификации могут быть присоединены непосредственно к лункам или к гелеобразному материалу, находящемуся в лунках. Таким образом, в цитируемых публикациях представлены иллюстративные примеры материалов, композиций или установок, которые могут быть адаптированы для применения в способах и композициях согласно изобретению.
Сайты амплификации массива могут включать совокупность агентов захвата, способных связываться с нуклеиновыми кислотами-мишенями. Примеры агентов захвата включают рецепторы и/или лиганды, соответствующий партнер по связыванию которых присоединен к нуклеиновым кислотам-мишеням, примеры которого представлены выше в настоящем описании. Особенно подходящим агентом захвата является улавливающая нуклеиновая кислота, которая комплементарна последовательности одной или более нуклеиновых кислот-мишеней. Например, улавливающие нуклеиновые кислоты, присутствующие на сайте амплификации, могут иметь универсальную улавливающую последовательность, которая комплементарна универсальной последовательности, присутствующей в адаптерной последовательности каждой нуклеиновой кислоты-мишени. В некоторых примерах улавливающая нуклеиновая кислота также может функционировать как праймер для амплификации нуклеиновой кислоты-мишени (вне зависимости от того, содержит она или нет универсальную последовательность).
В конкретных примерах агент захвата, такой как улавливающая нуклеиновая кислота, может быть присоединен к сайту амплификации. Например, агент захвата может быть присоединен к поверхности элемента массива. Присоединение может быть осуществлено через промежуточную структуру, такую как гранула, частица или гель. Присоединение улавливающих нуклеиновых кислот к массиву через гель показано ниже в Примере I, и дополнительным примером являются проточные ячейки, коммерчески поставляемые Illumina Inc. (San Diego, Calif.) или рассмотренные в документе WO 2008/093098, содержание которого полностью включено в настоящую работу посредством ссылки. Примеры гелей, которые могут быть применены в способах и установке согласно изобретению включают, без ограничений, гели, имеющие коллоидную структуру, такие как агароза; полимерную сетчатую структуру, такие как желатин; или поперечно-сшитую полимерную структуру, такие как полиакриламид, не содержащий силана акриламид (англ. silane free acrylamide, сокращенно SFA, см., например, опубликованную патентную заявку US 2011/0059865, содержание которой полностью включено в настоящую заявку посредством ссылки) или сополимер N-(5-азидоацетамидилпентил)акриламида и акриламида (англ. poly(N-(5-azidoacetamidylpentyl)acrylamide-co-acrylamide, сокращенно PAZAM) (см., например, патент US 9012022, содержание которого полностью включено в настоящую работу посредством ссылки). Присоединение через гранулу может быть осуществлено способами, рассмотренными в настоящем описании и в приведенных выше публикациях.
В некоторых примерах элементы на поверхности подложки массива не являются непрерывными и разделены промежуточными участками поверхности. Подходящими являются промежуточные участки, имеющие существенно более низкое количество или концентрацию агентов захвата, по сравнению с элементами массива. Особенно подходящими являются промежуточные участки, не содержащие агентов захвата. Например, относительно небольшое количество или отсутствие улавливающих частиц на промежуточных участках способствует локализации нуклеиновых кислот-мишеней и образующихся впоследствии кластеров на требуемых элементах. В конкретных примерах элементы могут представлять собой находящиеся на поверхности вогнутые элементы (например, лунки), и элементы могут содержать гелеобразный материал. Гельсодержащие элементы могут быть отделены друг от друга находящимися на поверхности промежуточными участками, в которых гель по меньшей мере по существу отсутствует, или, если гель присутствует, то он по меньшей мере по существу не способен поддерживать фиксацию нуклеиновых кислот. Способы и композиции получения и применения подложек, включающих гельсодержащие элементы, такие как лунки, рассмотрены в патенте US 9512422, содержание которого полностью включено в настоящую работу посредством ссылки.
Нуклеиновые кислоты-мишени, применяемые в способе или композиции согласно настоящему изобретению, могут состоять из ДНК, РНК или их аналогов. Источником нуклеиновых кислот-мишеней может быть геномная ДНК, информационная РНК или другие нуклеиновые кислоты из природных источников. В некоторых случаях перед введением в способ или композицию нуклеиновые кислоты-мишени, полученные из таких источников, могут быть амплифицированы. Для этого может быть применена любая из множества различных известных методик амплификации, которые включают, без ограничений, полимеразную цепную реакцию (ПЦР), амплификацию по типу катящегося кольца (RCA), амплификацию с множественным вытеснением цепи (англ. multiple displacement amplification, сокращенно MDA) или случайную амплификацию с примированием (англ. random prime amplification, сокращенно RPA). Следует понимать, что амплификация нуклеиновых кислот-мишеней перед их использованием в способе или композиции согласно изобретению необязательна. Таким образом, в некоторых примерах способов и композиций согласно изобретению нуклеиновые кислоты-мишени не подвергают амплификации перед использованием. Нуклеиновые кислоты-мишени необязательно могут быть получены из синтетических библиотек. В составе синтетических нуклеиновых кислот могут быть нативные ДНК или РНК или их аналоги.
Примеры биологических образцов, из которых могут быть получены нуклеиновые кислоты-мишени, включают, например, образцы, полученные из организмов млекопитающих, таких как грызун, мышь, крыса, кролик, морская свинка, копытное животное, лошадь, овца, свинья, коза, корова, кот, собака, примат, человек или примат-нечеловек; из растения, такого как Arabidopsis thaliana, кукуруза, сорго, овес, пшеница, рис, канола, или соевые бобы; из водоросли, такой как Chlamydomonas reinhardtii; нематоды, такой как Caenorhabditis elegans; насекомого, такого как Drosophila melanogaster, комар, плодовая мушка, медоносная пчела или паук; рыбы, такой как данио; рептилии; амфибии, такой как лягушка или Xenopus laevis; dictyostelium discoideum; грибка, такого как Pneumocystis carinii, Takifugu rubripes, дрожжи, Saccharamoyces cerevisiae или Schizosaccharomyces pombe; или из Plasmodium falciparum. Нуклеиновые кислоты-мишени также могут быть получены из прокариота, такого как бактерия, Escherichia coli, стафилококк или mycoplasma pneumoniae; архебактерия; вирус, такой как вирус гепатита С или вирус иммунодефицита человека; или из вироида. Нуклеиновые кислоты-мишени могут быть получены из гомогенной культуры или их популяции перечисленных выше организмов или в альтернативном варианте из совокупности нескольких различных организмов, например, принадлежащих сообществу или экосистеме.
Нуклеиновые кислоты-мишени необязательно должны быть получены из природных источников и, напротив, могут быть синтезированы с помощью известных методик. Например, могут быть синтезированы зонды для определения экспрессии генов или зонды для генотипирования, которые в способах согласно изобретению применяют для создания массива.
В некоторых примерах нуклеиновые кислоты-мишени могут быть получены в виде фрагментов одной или более нуклеиновых кислот большей длины. Фрагментация может быть выполнена с помощью любой из множества различных методик, известных в данной области техники, которые включают, например, распыление, обработку ультразвуком, химическое расщепление, ферментативное расщепление или физическое разрезание. Фрагментация также может быть результатом применения одной из методик амплификации, согласно которой ампликоны получают копированием лишь части более крупной нуклеиновой кислоты. Например, при амплификации ПЦР получают фрагменты, размер которых определяется длиной фрагмента, находящегося между фланкирующим праймерами, используемыми для амплификации.
Популяция нуклеиновых кислот-мишеней или их ампликонов может иметь среднюю длину цепочки, которая требуется или подходит для соответствующего применения способов или композиций согласно изобретению. Например, средняя длина цепочки может составлять менее приблизительно 100000 нуклеотидов, приблизительно 50000 нуклеотидов, приблизительно 10000 нуклеотидов, приблизительно 5000 нуклеотидов, приблизительно 1000 нуклеотидов, приблизительно 500 нуклеотидов, приблизительно 100 нуклеотидов или приблизительно 50 нуклеотидов. В альтернативном варианте или дополнительно средняя длина цепочки может превышать приблизительно 10 нуклеотидов, приблизительно 50 нуклеотидов, приблизительно 100 нуклеотидов, приблизительно 500 нуклеотидов, приблизительно 1000 нуклеотидов, приблизительно 5000 нуклеотидов, приблизительно 10000 нуклеотидов, приблизительно 50000 нуклеотидов или приблизительно 100000 нуклеотидов. Средняя длина цепочки в популяции нуклеиновых кислот-мишеней или их ампликонов может находиться в диапазоне, ограниченном максимальным и минимальным значениями, указанными выше. Следует понимать, что средняя длина цепочек ампликонов, образующихся на сайте амплификации (или полученных другим образом, или используемых сайте амплификации), может находиться в диапазоне, ограниченном верхним и нижним пределами, выбранными из значений, примеры которых приведены выше.
В некоторых случаях популяция нуклеиновых кислот-мишеней может быть получена в таких условиях или иным образом модифицирована так, чтобы длина ее членов была максимальной. Например, максимальная длина ее членов, применяемых в способе согласно изобретению или присутствующих в соответствующей композиции, может составлять менее приблизительно 100000 нуклеотидов, приблизительно 50000 нуклеотидов, приблизительно 10000 нуклеотидов, приблизительно 5000 нуклеотидов, приблизительно 1000 нуклеотидов, приблизительно 500 нуклеотидов, приблизительно 100 нуклеотидов или приблизительно 50 нуклеотидов. В альтернативном варианте или дополнительно популяция нуклеиновых кислот-мишеней или их ампликонов может быть получена в таких условиях или иным образом модифицирована так, чтобы длина ее членов была минимальной. Например, минимальная длина ее членов, применяемых в способе согласно изобретению или присутствующих в соответствующей композиции, может составлять более приблизительно 10 нуклеотидов, приблизительно 50 нуклеотидов, приблизительно 100 нуклеотидов, приблизительно 500 нуклеотидов, приблизительно 1000 нуклеотидов, приблизительно 5000 нуклеотидов, приблизительно 10000 нуклеотидов, приблизительно 50000 нуклеотидов, или приблизительно 100000 нуклеотидов. Максимальная и минимальная длины цепочки в популяции нуклеиновых кислот-мишеней может находиться в диапазоне, ограниченном максимальным и минимальным значениями, указанными выше. Следует понимать, что максимальная и/или минимальная длина цепочек ампликонов, образующихся на сайте амплификации (или полученных другим образом, или используемых сайте амплификации), может находиться в диапазоне, ограниченном верхним и нижним пределами, примеры которых приведены выше.
В конкретных примерах размеры нуклеиновых кислот-мишеней выбраны в соответствии с площадью сайтов амплификации, например, для облегчения протекания кинетического исключения. Например, для проведения кинетического исключения, площадь каждого из сайтов массива может превышать диаметр исключенного объема нуклеиновых кислот-мишеней. Так, например, в тех примерах, в которых используют массив элементов, расположенных на поверхности, площадь каждого элемента может превышать диаметр исключенного объема нуклеиновых кислот-мишеней, которые переносятся к сайтам амплификации. Исключенный объем нуклеиновой кислоты-мишени и его диаметр могут быть определены, например, из длины нуклеиновой кислоты-мишени. Способы определения исключенного объема нуклеиновых кислот и диаметра исключенного объема рассмотрены, например, в патенте US 7785790 и в публикациях Rybenkov с соавт., Proc. Natl. Acad. Sci. U.S.A. 90: 5307-5311 (1993), Zimmerman с соавт., J. Mol. Biol. 222:599-620 (1991) или Sobel с соавт., Biopolymers 31:1559-1564 (1991), содержания которых полностью включены в настоящую работу посредством ссылки.
Сайты амплификации массива могут включать совокупность праймеров, которые применяют для генерации ампликонов нуклеиновой кислоты-мишени. В некоторых примерах праймеры, находящиеся на сайте амплификации, могут содержать универсальную инициирующую (примирующую) последовательность, которая комплементарна универсальной последовательности, содержащейся в адаптерной последовательности каждой нуклеиновой кислоты-мишени. В конкретных примерах к сайту амплификации может быть присоединена совокупность праймеров. Праймеры могут быть присоединены к сайту амплификации способами, описанными выше для присоединения улавливающих нуклеиновых кислот.
Как было указано выше в настоящем описании, находящиеся на поверхности подложки массива элементы могут быть не непрерывными, то есть могут быть разделены промежуточными участками поверхности. В конкретных примерах промежуточные участки содержат по существу меньшие количества или концентрации праймеров по сравнению с элементами массива. Особенно подходящими являются промежуточные участки, не содержащие праймеров. Например, относительно небольшое количество или отсутствие праймеров на промежуточных участках способствует локализации ампликонов на элементах поверхности массива. При такой конфигурации каждый элемент массива имеет границу, которая обеспечивает конечную емкость элемента в отношении ампликонов, полученных амплификацией фиксированной нуклеиновой кислоты-мишени в способах согласно изобретению.
Способ согласно настоящему изобретению может включать реакцию реагента амплификации, приводящую к образованию совокупности сайтов амплификации, каждый из которых включает клональную популяцию ампликонов индивидуальной нуклеиновой кислоты-мишени, которая была помещена на сайт. В некоторых примерах реакцию амплификации проводят до тех пор, пока не образуется количество ампликонов, достаточное для заполнения емкости соответствующего сайта амплификации. Полное заполнение емкости уже занятого сайта исключает последующее попадание на сайт других нуклеиновых кислот-мишеней и образование клональных популяций их ампликонов на сайте. Таким образом, в некоторых примерах желательно, чтобы скорость генерации ампликонов, которые заполняют емкость сайтов амплификации, соответственно превышала скорость переноса индивидуальных нуклеиновых кислот-мишеней к индивидуальным сайтам амплификации.
В некоторых примерах перед попаданием на сайт второй нуклеиновой кислоты-мишени, даже если емкость сайта амплификации не заполнена, может наблюдаться видимая клональность. В некоторых условиях амплификация первой нуклеиновой кислоты-мишени может происходить до того момента, когда образуется количество копий, достаточное для эффективно вытеснения или подавления генерации копий второй нуклеиновой кислоты-мишени, перенесенной к сайту. Так, в одном из примеров, включающих проведение мостиковой амплификации на круглых элементах, диаметр которых составляет менее приблизительно 500 нм, было обнаружено, что после проведения 14 циклов экспоненциальной амплификации первой нуклеиновой кислоты-мишени, загрязнение того же сайта второй нуклеиновой кислотой-мишенью приводит к получению такого количества загрязняющих ампликонов, которое недостаточно для негативного влияния на результаты анализа секвенирования синтезом, проводимого на платформе секвенирования Illumina.
Как было показано в приведенном выше примере, сайты амплификации массива не обязательно должны быть полностью клональными во всех примерах. Напротив, в некоторых вариантах применения, индивидуальный сайт амплификации может быть преимущественно заселен ампликонами первой нуклеиновой кислоты-мишени и также может иметь низкое содержание загрязняющих ампликонов, полученных из второй нуклеиновой кислоты-мишени. Массив может содержать один или более сайтов амплификации, имеющих низкое содержание загрязняющих ампликонов, при условии, что уровень загрязнения не оказывает неприемлемого влияния на последующее применение массива. Например, если массив применяют для обнаружений, то приемлемым уровнем загрязнения будет уровень, который не оказывает неприемлемого влияния на отношение сигнала к шуму или на разрешение методики обнаружения. Соответственно, видимая клональность обычно соответствует определенному варианту применения или использования массива, получаемого способами согласно изобретению. Примеры максимальных уровней загрязнения, которые могут быть приемлемыми для индивидуального сайта амплификации в определенных вариантах применения, включают, без ограничений, приблизительно 0,1%, приблизительно 0,5%, приблизительно 1%, приблизительно 5%, приблизительно 10% или приблизительно 25% загрязняющих ампликонов. Массив может включать один или более сайтов амплификации, имеющих приведенные уровни содержания загрязняющих ампликонов. Например, до приблизительно 5%, приблизительно 10%, приблизительно 25%, приблизительно 50%, приблизительно 75% или даже до приблизительно 100% сайтов амплификации в массиве могут содержать некоторое количество загрязняющих ампликонов.
В конкретных примерах способ согласно настоящему изобретению осуществляют для одновременного (i) переноса нуклеиновых кислот-мишеней к сайтам амплификации со средней скоростью переноса, и (ii) амплификации нуклеиновых кислот-мишеней, которые находятся на сайтах амплификации, со средней скоростью амплификации, причем средняя скорость амплификации превышает среднюю скорость переноса. Соответственно, в таких примерах кинетическое исключение может достигаться за счет относительно низкой скорости переноса. Например, для достижения требуемой средней скорости переноса может быть выбрана достаточно низкая концентрация нуклеиновых кислот-мишеней, поскольку более низкие концентрации приводят к возникновению более низких средних скоростей переноса. В альтернативном варианте или дополнительно для снижения скоростей переноса может быть применен раствор с высокой вязкостью и/или в раствор добавлены реагенты фазового исключения (далее также называемые агентами фазового исключения). Примеры подходящих реагентов фазового исключения включают, без ограничений, полиэтиленгликоль (ПЭГ), FICOLL®, декстран или поливиниловый спирт. Примеры реагентов и композиций для фазового исключения рассмотрены в патенте US 7399590, содержание которого полностью включено в настоящую работу посредством ссылки. Другим фактором, который может быть изменен для достижения требуемой скорости переноса, является средний размер нуклеиновых кислот-мишеней.
В некоторых примерах способов до начала амплификации нуклеиновые кислоты-мишени могут транспортироваться, например, в результате диффузии или другого процесса, к сайтам амплификации. В этом случае кинетическое исключение может быть достигнуто за счет относительно низкой скорости создания первого ампликона по сравнению со скоростями генерации последующих ампликонов. Например, различные скорости образования ампликонов могут быть созданы при использовании первого праймера для генерации первого ампликона, который первоначально находится в состоянии, не подходящем для достройки, в то время как другие праймеры, подходящие для генерации следующих ампликонов находятся в состоянии, подходящем для достройки посредством реакции амплификации. Таким образом, задержка при переходе первого праймера в состояние, подходящее для достройки, будет вызывать задержку в образовании первого ампликона, в то время как образование следующих ампликонов протекает без задержки. Таким образом, средняя скорость генерации последующих ампликонов на сайтах амплификации превышает среднюю скорость генерации первого ампликона на сайтах амплификации.
Ниже более подробно разобран пример кинетического исключения, обусловленного различными скоростями образования ампликонов. К сайту амплификации могут быть присоединены три субпопуляции праймеров. Первая субпопуляция праймеров служит для захвата нуклеиновой кислоты-мишени (с помощью улавливающей последовательности) и в качестве праймера для получения первого ампликона. Первая субпопуляция праймеров имеет обратную блокировку, препятствующую достройке, например, присоединенный к 3'-концу дидезоксинуклеотид. Вторая субпопуляция праймеров может включать последовательность праймера Р5, и третья популяция праймеров может включать последовательность праймера Р7. Праймеры первой и второй субпопуляций не включают дидезоксинуклеотида и, таким образом, полностью отвечают требованиям для достройки. Нуклеиновые кислоты-мишени могут быть сконструированы так, что они включают (в направлении от 5' к 3') последовательность, связывающую праймер Р7, одну из нескольких различных нуклеотидных последовательностей-мишеней, последовательность, связывающую праймер Р5, и комплемент улавливающей последовательности. Несколько различных нуклеиновых кислот-мишеней могут быть гибридизованы с первой субпопуляцией праймеров (с помощью улавливающих последовательностей). Праймеры захвата затем могут быть переведены в состояние, подходящее для достройки, например, обработкой полимеразой в условиях пирофосфоролиза (например, в присутствии избытка пирофосфата). Могут быть созданы условия, при которых в течение периода времени, требуемого для заполнения сайта амплификации образующимися последующими ампликонами, в среднем только один из праймеров захвата приобретает достраиваемую форму. Таким образом, несмотря на то, что на индивидуальном сайте амплификации могут присутствовать несколько потенциально загрязняющих нуклеиновых кислот-мишеней, кинетическое исключение приводит к образованию ампликонов только одной из нуклеиновых кислот-мишеней, в результате чего на сайте амплификации образуется клональная популяция ампликонов. С целью иллюстрации данное явление было рассмотрено на примере лишь одного сайта амплификации; однако следует понимать, что реакция может включать присоединение нуклеиновых кислот-мишеней и амплификацию на целом массиве сайтов амплификации.
В способе согласно изобретению может быть применен любой из множества различных временно недостраиваемых праймеров в комбинации с соответствующими методиками и реагентами, позволяющими переводить такие праймеры в состояние, подходящее для достройки. В приведенных выше примерах применяют дидезоксинуклеотид, который удаляют пирофосфоролизом. На праймере могут присутствовать другие недостраиваемые нуклеотиды, которые удаляют пирофосфоролизом. Кроме того, дидезоксинуклеотиды или другие недостраиваемые нуклеотиды могут быть удалены другими известными методиками, примеры которых включают экзонуклеазное воздействие полимеразы или другого подходящего фермента. В других примерах праймер может включать обратимый терминатор, такой как терминаторы, применяемые в способах секвенирования синтезом с использованием терминаторов. Примеры обратимых терминаторов и методики их удаления рассмотрены, например, в публикации Bentley с соавт., Nature 456:53-59 (2008), в патентной заявке WO 04/018497, в патентных документах US 7057026, WO 91/06678, WO 07/123744, US 7329492, US 7211414, US 7315019, US 7405281 и в опубликованной патентной заявке US 2008/0108082, содержания которых полностью включены в настоящую работу посредством ссылки.
Несмотря на то, что в приведенном выше примере, в котором нуклеиновые кислоты-мишени присутствовали на сайтах амплификации до амплификации, для создания различных скоростей генерации первого ампликона и последующих ампликонов были применены дифференциально активные праймеры, способ также может быть осуществлен в условиях переноса нуклеиновых кислот-мишеней (например, посредством диффузии) к сайтам амплификации по мере протекания амплификации. Таким образом, кинетическое исключение может быть обусловлено как относительно низкой скоростью переноса, так и относительно низкой скоростью генерации первого ампликона по сравнению с последующими ампликонами. Таким образом, реакция амплификации согласно изобретению может быть проведена в таком режиме, в котором нуклеиновые кислоты-мишени транспортируются из раствора к сайтам амплификации одновременно с (i) образованием первого ампликона, и (ii) образованием последующих ампликонов на других сайтах массива. В конкретных примерах средняя скорость генерации последующих ампликонов на сайтах амплификации может превышать среднюю скорость транспортировки нуклеиновых кислот-мишеней из раствора к сайтам амплификации. В некоторых случаях из одной нуклеиновой кислоты-мишени на индивидуальном сайте амплификации может быть получено количество ампликонов, достаточное для заполнения емкости соответствующего сайта амплификации. Скорость генерации ампликонов, с которой они заполняют емкости соответствующих сайтов амплификации, может, например, превышать скорость транспортировки индивидуальных нуклеиновых кислот-мишеней из раствора к сайтам амплификации.
Реагент амплификации, применяемый в способе согласно изобретению, предпочтительно способен быстро генерировать копии нуклеиновых кислот-мишеней на сайтах амплификации. Один или более реагентов амплификации, применяемых в способе согласно настоящему изобретению, включают полимеразу и нуклеозидтрифосфаты (НТФ). Может быть применена любая из множества различных полимераз, известных в данной области техники, но в некоторых примерах может быть желательным применение полимеразы, имеющей отрицательную активность по отношению к экзонуклеазе. В примерах получения копий ДНК НТФ могут представлять собой дезоксирибонуклеозидтрифосфаты (дНТФ). Реагент ДНК амплификации может включать четыре нативные вида дНТФ, которые включают дАТФ (дезоксрибоиаденозина трифосфат), дТТФ (дезоксириботимидина трифосфат), дГТФ (дезоксирибогуанозина трифосфат) и дЦТФ (дезоксирибоцитидина трифосфат); однако при необходимости могут быть применены их аналоги. В примерах получения копий РНК НТФ могут представлять собой рибонуклеозидтрифосфаты (рНТФ). Реагент РНК амплификации может включать четыре нативных вида рНТФ, которые включают рАТФ (рибоаденозина трифосфат), рУТФ (рибоурацила трифосфат), рГТФ (рибогуанозина трифосфат) и рЦТФ (рибоцитидина трифосфат); однако при необходимости могут быть применены их аналоги.
Реагент амплификации может включать дополнительные компоненты, способствующие образованию ампликонов и в некоторых случаях способствующие повышению скорости образования ампликонов. Одним из примеров является рекомбиназа. Рекомбиназа может способствовать образованию ампликонов за счет многократного вытеснения/удлинения. В частности, рекомбиназа может способствовать вытеснению нуклеиновой кислоты-мишени полимеразой и удлинению праймера под действием полимеразы при использовании нуклеиновой кислоты-мишени в качестве матрицы для образования ампликонов. Этот процесс может повторяться и протекать в виде цепной реакции, в которой ампликоны, получаемые в каждом цикле вытеснения/удлинения, служат матрицами для последующего цикла. Этот процесс может протекать быстрее, чем стандартная ПЦР, поскольку в нем не требуется проведения цикла денатурации (например, нагреванием или химической денатурацией). Таким образом, амплификация с применением рекомбиназы может быть проведена в изотермических условиях. Для улучшения амплификации в усиленный рекомбиназой реагент амплификации обычно желательно включить АТР или другие нуклеотиды (или в некоторых случаях их негидролизуемые аналоги). Особенно подходящей является смесь рекомбиназы и одноцепочечного связывающего (англ. single-stranded binding, сокращенно SSB) белка, поскольку SSB может дополнительно улучшать амплификацию. Примеры композиций для проведения амплификации с участием рекомбиназы включают коммерчески доступные композиции, поставляемые в наборах TWISTAMP® Компанией TwistDx (Cambridge, UK). Подходящие компоненты усиленного рекомбиназой реагента амплификации и условия реакции рассмотрены в патентах US 5223414 и US 7399590, содержания которых полностью включены в настоящую работу посредством ссылки.
Другим примером компонента, который может быть включен в реагент амплификации для улучшения образования ампликонов и в некоторых случаях для повышения скорости образования ампликонов, является геликаза. Геликаза может облегчать образование ампликонов, способствуя протеканию цепной реакции образования ампликонов. Этот процесс может протекать быстрее, чем стандартная ПЦР, поскольку в нем не требуется проведения цикла денатурации (например, нагреванием или химической денатурацией). Таким образом, амплификация с применением геликазы может быть проведена в изотермических условиях. Особенно подходящей является смесь геликазы и одноцепочечного связывающего (SSB) белка, поскольку SSB может дополнительно улучшать амплификацию. Примеры композиций для проведения амплификации с участием геликазы включают коммерчески доступные композиции, поставляемые в наборах ISOAMP® Компанией Biohelix (Beverly, Mass.). Кроме того, примеры подходящих композиций, которые включают белок геликазы, рассмотрены в патентах US 7399590 и US 7829284, содержания которых полностью включены в настоящую работу посредством ссылки.
Другим примером компонента, который может быть включен в реагент амплификации для улучшения образования ампликонов и в некоторых случаях для повышения скорости образования ампликонов, является белок, связывающий точку начала репликации.
Скорость протекания реакции амплификации может быть увеличена посредством повышения концентрации или количества одного или более активных компонентов реакции амплификации. Например, для повышения скорости амплификации может быть увеличено количество или концентрация полимеразы, нуклеозидтрифосфатов, праймеров, рекомбиназы, геликазы или SSB. В некоторых случаях один или более активных компонентов реакции амплификации, количество или концентрацию которых увеличивают (или изменяют иным образом в способе согласно изобретению), представляют собой компоненты реакции амплификации, не являющиеся нуклеиновыми кислотами.
Скорость амплификации в способе согласно изобретению также может быть повышена регулированием температуры. Например, скорость амплификации на одном или более сайтах амплификации может быть повышена при повышении температуры на сайте (сайтах) до максимальной температуры, при которой скорость реакции начинает снижаться за счет протекания денатурации или других негативных процессов. Оптимальные или желательные температуры могут быть определены на основании известных свойств применяемых компонентов амплификации или эмпирически для конкретной смеси реакции амплификации. Для повышения скорости амплификации также могут быть изменены свойства праймеров, применяемых для амплификации. Например, может быть изменена последовательность и/или длина праймеров. Изменения могут быть произведены на основании сделанных a priori прогнозов величины температуры плавления (Tm) праймера или на основании ее значений, полученных опытным путем.
Другим способом повышения скорости амплификации на сайте амплификации является повышение локальной концентрации одного или более активных компонентов реакции амплификации на сайте амплификации. Активные компоненты могут включать один или более компонентов, не являющихся нуклеиновыми кислотами. В некоторых примерах один или более активных компонентов реакции амплификации могут быть присоединены к сайту амплификации с помощью воздействия электрических полей, такого как электрофорез, изотахофорез, прямые импульсы тока или напряжения или с помощью подобного воздействия. В альтернативном варианте или дополнительно один или более компонентов амплификации могут включать аффинную метку, которая рекрутирует компонент на сайт амплификации. Аффинная метка может быть заряженной, и под воздействием электрических полей будет притягивать компонент, имеющий подходящую метку, к сайту амплификации. Также могут быть применены незаряженные аффинные метки. Например, в качестве аффинных меток для компонента реакции амплификации могут быть применены любые из множества различных лигандов или рецепторов, известных в данной области техники, такие как агенты согласно изобретению, приведенные выше в качестве примеров агентов захвата. Как и в случае агентов захвата, применяемых для захвата нуклеиновых кислот, сайт амплификации может включать партнера по связыванию аффинной метки компонента амплификации. Таким образом, локальная концентрация компонента амплификации, имеющего аффинную метку, может быть повышена в результате взаимодействия с подходящим партнером на сайте амплификации. В конкретных примерах, в которых сайт амплификации представляет собой поверхность подложки, партнер аффинной метки по связыванию может быть присоединен к поверхности.
Кроме того, для повышения локальной концентрации реагентов амплификации могут быть применены магнитные или оптические силы. В таких случаях один или более реагентов амплификации могут включать магнитную метку или оптическую метку, на которую могут воздействовать такие силы.
Скорость протекания реакции амплификации может быть увеличена посредством повышения активности одного или более реагентов амплификации. Например, в реакцию, в которой применяют полимеразу, может быть добавлен кофактор, повышающий скорость удлинения полимеразы. В некоторых примерах в полимеразную реакцию могут быть добавлены кофакторы на основе металлов, таких как магний, цинк или марганец, или в других примерах может быть добавлен бетаин.
В некоторых примерах способов согласно изобретению желательно применение популяции двухцепочечных нуклеиновых кислот-мишеней. Неожиданно было обнаружено, что образование ампликонов на массиве сайтов в условиях кинетического исключения подходит для двухцепочечных нуклеиновых кислот-мишеней. Например, в присутствии рекомбиназы и одноцепочечного связывающего белка образование совокупности сайтов амплификации, содержащих клональные популяции ампликонов, может происходить более эффективно в случае двухцепочечных нуклеиновых кислот-мишеней (чем в случае одноцепочечных нуклеиновых кислот-мишеней той же концентрации). Тем не менее, следует понимать, что в некоторых примерах способов согласно изобретению могут быть применены одноцепочечные нуклеиновые кислоты-мишени.
В способе согласно изобретению может быть применена любая из множества различных методик амплификации. Примеры методик, которые могут быть применен включают, без ограничений, полимеразную цепную реакцию (ПЦР), амплификацию по типу катящегося кольца (RCA), амплификацию с множественным вытеснением цепи (MDA) или случайную амплификацию с примированием (RPA). В некоторых примерах амплификация может быть проведена в растворе, например, если сайты амплификации могут содержать ампликоны в объеме, имеющем требуемую емкость. Предпочтительно, методику амплификации, применяемую в условиях кинетического исключения в способе согласно настоящему изобретению, осуществляют на твердой фазе. Например, один или более праймеров, применяемых для амплификации, могут быть присоединены к твердой фазе, имеющейся на сайте амплификации. В примерах проведения ПЦР один или оба праймера, применяемых для амплификации, могут быть присоединены к твердой фазе. Форматы, в которых применяют два вида праймеров, присоединенных к поверхности, часто называют мостиковой амплификацией, поскольку двухцепочечные ампликоны образуют мостиковую структуру между двумя присоединенными к поверхности праймерами, которые фланкируют копируемую матричную последовательность. Примеры реагентов и условий, которые могут быть применены для мостиковой амплификации, рассмотрены, например, в патенте US 5641658, опубликованной патентной заявке US 2002/0055100; в патенте US 7115400, опубликованной патентной заявке US 2004/0096853, опубликованной патентной заявке US 2004/0002090, опубликованной патентной заявке US 2007/0128624 и опубликованной патентной заявке US 2008/0009420, содержания которых полностью включены в настоящую работу посредством ссылки. Твердофазная ПЦР амплификация также может быть выполнена с использованием одного из праймеров амплификации, присоединенных к твердому носителю, и второго праймера, находящегося в растворе. Одним из примеров формата, в котором применяют комбинацию присоединенного к поверхности праймера и растворимого праймера, является ПЦР в эмульсии, рассмотренная, например, в публикациях Dressman с соавт., Proc. Natl. Acad. Sci. USA 100:8817-8822 (2003), WO 05/010145 или в опубликованных патентных заявках US 2005/0130173 или US 2005/0064460, содержания которых полностью включены в настоящую работу посредством ссылки. ПЦР в эмульсии представляет собой иллюстрацию указанного выше формата, и следует понимать, что для целей способов согласно изобретению применение эмульсии не является обязательным, и, действительно, в некоторых примерах эмульсию не применяют. Указанные выше методики ПЦР могут быть модифицированы для проведения нециклической амплификации (например, изотермической амплификации) посредством применения упомянутых в настоящем описании компонентов для улучшения амплификации или повышения скорости амплификации. Соответственно, указанные выше методики ПЦР могут быть применены в условиях кинетического исключения.
Методики RCA могут быть модифицированы для применения в способе согласно настоящему изобретению. Примеры компонентов, которые могут быть применены в реакции RCA, и принципы получения ампликонов в реакции RCA рассмотрены, например, в публикации Lizardi с соавт., Nat. Genet. 19:225-232 (1998) и опубликованной патентной заявке US 2007/0099208 А1, содержания которых полностью включены в настоящую работу посредством ссылки. Праймеры, применяемые в RCA, могут находиться в растворе или быть присоединенными к поверхности твердого носителя сайта амплификации. Указанные в приведенной выше литературе методики RCA могут быть модифицированы в соответствии с принципами настоящего изобретения, например, в целях повышения скорости амплификации так, чтобы она была подходящей для определенного применения. Таким образом, методики RCA могут быть применены в условиях кинетического исключения.
Методики MDA могут быть модифицированы для применения в способе согласно настоящему изобретению. Некоторые основные принципы и условия, подходящие для проведения MDA, рассмотрены, например, в публикациях Dean с соавт., Proc Natl. Acad. Sci. USA 99:5261-66 (2002); Lage с соавт., Genome Research 13:294-307 (2003), Walker с соавт., Molecular Methods for Virus Detection, Academic Press, Inc., 1995, Walker с соавт., Nucl. Acids Res. 20:1691-96 (1992), в патентах US 5455166, US 5130238 и US 6214587, содержания которых полностью включены в настоящую работу посредством ссылки. Праймеры, применяемые в MDA, могут находиться в растворе или быть присоединенными к поверхности твердого носителя сайта амплификации. Указанные в приведенной выше литературе методики MDA могут быть модифицированы в соответствии с принципами настоящего изобретения, например, в целях повышения скорости амплификации так, чтобы она была подходящей для определенного применения. Соответственно, методики MDA могут быть применены в условиях кинетического исключения.
В конкретных примерах для получения массива в условиях кинетического исключения может быть применена комбинация указанных выше методик амплификации. Например, RCA и MDA могут быть применены в комбинации, в которой RCA применяют для генерации конкатемерного ампликона в растворе (например, применяя праймеры, находящиеся в фазе раствора). Затем ампликон может быть применен в качестве матрицы для MDA, в которой участвуют праймеры, присоединенные к поверхности твердого носителя на сайте амплификации. В этом примере ампликоны, получаемые после проведения комбинации этапов RCA и MDA, присоединяются к поверхности сайта амплификации.
Как указано в некоторых приведенных выше примерах, в способе согласно настоящему изобретению применение методики циклической амплификации необязательно. Например, амплификация нуклеиновых кислот-мишеней может быть выполнена на сайтах амплификации в отсутствие цикла денатурации. Примеры циклов денатурации включают введение в реакцию амплификации денатурирующих химических веществ и/или повышение температуры реакции амплификации. Таким образом, амплификация нуклеиновых кислот-мишеней не обязательно должна включать замену раствор амплификации химическим реагентом, который вызывает денатурацию нуклеиновых кислот-мишеней и ампликонов. Аналогично, амплификация нуклеиновых кислот-мишеней не обязательно должна включать нагревание раствора до температуры, вызывающей денатурацию нуклеиновых кислот-мишеней и ампликонов. Соответственно, амплификация нуклеиновых кислот-мишеней на сайтах амплификации может быть выполнена в изотермическом режиме в течение всей продолжительности способа согласно изобретению. Действительно, способ амплификации согласно изобретению может быть проведен в отсутствие одной или более циклических манипуляций, которые выполняют при проведении некоторых методик амплификации в стандартных условиях. Кроме того, некоторые стандартные методики твердофазной амплификации после загрузки нуклеиновых кислот-мишеней на подложку и до начала амплификации включают промывку. Однако в примерах осуществления способов согласно настоящему изобретению выполнение этапа промывки между переносом нуклеиновых кислот-мишеней к реакционным сайтам и амплификацией нуклеиновых кислот-мишеней на сайтах амплификации не является обязательным. Напротив, перенос (например, посредством диффузии) и амплификация могут происходить одновременно, обеспечивая кинетическое исключение.
В некоторых примерах может быть желательным повтор цикла амплификации, проводимого в условиях кинетического исключения. Таким образом, несмотря на то, что копии нуклеиновой кислоты-мишени могут быть получены на индивидуальном сайте амплификации посредством амплификации с применением кинетического исключения и без проведения циклических манипуляций, включающих применение химических денатурирующих веществ или нагревание, массив сайтов амплификации может быть подвергнут циклической обработке для увеличения количества сайтов, которые содержат ампликоны, и/или количества ампликонов на каждом сайте после проведения каждого цикла амплификации с применением кинетического исключения. В одном или более примерах, рассмотренных в настоящем описании со ссылками на Фиг. 11-14, после выполнения первоначальной амплификации с исключением может быть проведена вторичная амплификация или стимуляция (англ. boost), включающая пропускание через сайты амплификации свежего раствора, не содержащего целевых компонентов, который включает реагенты (например, активные компоненты), но не включает нуклеиновых кислот-мишеней. Реагенты, находящиеся в растворе, не содержащем целевых компонентов, могут повышать количество ампликонов в клональных кластерах, находящихся на сайтах амплификации. Из-за отсутствия нуклеиновых кислот-мишеней в растворе, не содержащем целевых компонентов, реагент не вызывает дополнительную загрузку нуклеиновых кислот-мишеней на сайты амплификации. В конкретных примерах условия амплификации в последующем цикле могут отличаться от условий в предыдущем цикле. Например, между проведением циклов могут быть изменены одно или более из рассмотренных выше условий, приводящих к изменению скорости переноса или скорости амплификации. Таким образом, при переходе к следующему циклу может быть увеличена скорость переноса, при переходе к следующему циклу может быть снижена скорость переноса, при переходе к следующему циклу может быть увеличена скорость амплификации, или при переходе к следующему циклу может быть снижена скорость амплификации.
Способ согласно изобретению может быть адаптирован для переноса нуклеиновых кислот-мишеней к сайтам амплификации массива под действием электрического поля (е-поля). Например, каждый сайт амплификации массива может быть соединен электрическим соединением с источником питания для создания электрического заряда, который притягивает нуклеиновые кислоты-мишени. В одной из конфигураций положительный заряд на сайтах амплификации может притягивать состоящую из фосфатов сахаров отрицательно заряженную основную цепь нуклеиновых кислот. Примеры способов и установки для притяжения нуклеиновых кислот к сайтам массива под действием е-поля рассмотрены в опубликованной патентной заявке US 2009/0032401 А1, содержание которой полностью включено в настоящую работу посредством ссылки. Стимулирование е-полем может быть применено в способе согласно настоящему изобретению, например, в тех условиях, в которых совокупность различных нуклеиновых кислот-мишеней находится в растворе, то есть нуклеиновые кислоты-мишени транспортируются текучей средой к массиву сайтов амплификации в течение каждого этапа амплификации. Заряд на каждом сайте амплификации может быть отрегулирован для создания кинетического исключения. Дополнительно или в альтернативном варианте, кроме регулирования заряда, для создания кинетического исключения согласно изобретению могут быть отрегулированы другие условия, позволяющие менять скорости переноса нуклеиновых кислот-мишеней или скорости амплификации. Соответственно, заряд на сайтах амплификации массива может быть отрегулирован таким образом, чтобы он притягивал нуклеиновые кислоты-мишени, в то время как амплификация происходит одновременно на различных сайтах амплификации массива, причем средняя скорость амплификации превышает среднюю скорость переноса нуклеиновых кислот-мишеней (т.е. в условиях переноса с приложением электрического поля) к сайтам амплификации.
В конкретных примерах, в которых происходит перенос нуклеиновых кислот-мишеней к сайтам амплификации с приложением е-поля, е-поле может непрерывно воздействовать в течение проведения всей реакции амплификации. В альтернативном варианте е-поле может быть изменено (например, его величина может быть повышена или понижена) по мере протекания реакции амплификации и заполнения сайтов амплификации ампликонами. Например, при усилении е-поля может происходить увеличение количества сайтов амплификации, приобретающих нуклеиновую кислоту-мишень (которая затем подвергается амплификации с образованием клональной популяции ампликонов на каждом из сайтов). Скорость увеличения мощности е-поля и диапазон амплитуды повышения могут быть выбраны так, чтобы сбалансировать повышение скорости переноса нуклеиновой кислоты-мишени с течением времени с увеличением количества сайтов амплификации, эффективно заполняемых в течение того же периода времени. Здесь также, в зависимости от применения массивов, получаемых способом согласно изобретению, эффективное заполнение может представлять собой момент, в который емкости сайтов амплификации оказываются полностью заполненными копиями первой нуклеиновой кислоты-мишени, что предотвращает амплификацию любых вторичных нуклеиновых кислот-мишеней на том же сайте. В альтернативном варианте эффективное заполнение может представлять собой момент, в который амплификация вторичной нуклеиновой кислоты-мишени на определенном сайте амплификации будет генерировать достаточно малое количество загрязняющих ампликонов, что позволяет считать его несущественным или приемлемым для целевого использования массива.
Обычно стимулирование е-полем создает дополнительный уровень контроля за переносом нуклеиновых кислот-мишеней к одному или более сайтам амплификации массива. Несмотря на то, что применение е-поля было рассмотрено выше на примерах транспортировки нуклеиновых кислот-мишеней к массиву сайтов амплификации, протекающей одновременно с амплификацией, происходящей на различных сайтах массива, в альтернативных примерах стимулирование е-полем может быть применено для переноса нуклеиновых кислот-мишеней к сайтам амплификации до начала амплификации на сайтах. Стимулирование е-полем может быть применено в способе или композиции согласно изобретению для переноса целевых биомолекул, отличных от нуклеиновых кислот-мишеней, к интересующему сайт, такому как элемент массива.
В конкретных примерах е-поле может быть приложено ко всем сайтам амплификации массива по меньшей мере по существу равномерно. Таким образом, находящиеся в растворе нуклеиновые кислоты-мишени с равной вероятностью могут быть перенесены к любому заданному сайту амплификации. В альтернативном примере е-поле может быть приложено только к подгруппе сайтов амплификации, имеющихся в массиве. Таким образом, стимулирование е-полем может быть применено для селективного заполнения лишь некоторых сайтов. Кроме того, при необходимости, в первой подгруппе сайтов амплификации может быть создан притягивающий заряд, способствующий переносу нуклеиновых кислот-мишеней к первой подгруппе сайтов, а во второй подгруппе сайтов амплификации может быть создан отталкивающий заряд, замедляющий перенос нуклеиновых кислот-мишеней к этим сайтам или способствующий удалению (например, посредством десорбции или разрушения) нуклеиновых кислот-мишеней с сайтов второй подгруппы. Аналогично, как более подробно рассмотрено ниже и в Примере III, на промежуточных участках массива может быть создан отталкивающий заряд, замедляющий перенос нуклеиновых кислот-мишеней к промежуточным участкам или способствующий удалению (например, посредством десорбции или разрушения) нуклеиновых кислот-мишеней с промежуточных участков.
В конкретных примерах промежуточные участки массива могут быть соединены электрическим соединением с источником питания, создающим электрический заряд, который замедляет связывание нуклеиновых кислот-мишеней или других биомолекул или удаляет нуклеиновые кислоты-мишени или другие биомолекулы. В одной из конфигураций отрицательный заряд на промежуточных участках может отталкивать нуклеиновые кислоты, имеющие отрицательно заряженную основную цепь, состоящую из фосфатов сахаров. В альтернативном варианте или дополнительно заряд промежуточного участка может быть применен для создания локализованных на поверхности изменений рН, которые вызывают электрохимическое повреждение нуклеиновых кислот и биомолекул.
В способе согласно настоящему изобретению может быть применено отталкивание с помощью е-поля, например, в тех условиях, когда совокупность различных нуклеиновых кислот-мишеней находится в растворе, и нуклеиновые кислоты-мишени транспортируются текучей средой к массиву сайтов амплификации на протяжении каждого этапа амплификации. Заряд на промежуточных участках массива может быть отрегулирован для отталкивания нуклеиновых кислот (например, приводящего к удалению или ингибированию связывания) во время захвата нуклеиновых кислот на элементах массива и необязательной амплификации на элементах в условиях кинетического исключения. Дополнительно или в альтернативном варианте, кроме регулирования заряда, могут быть уточнены другие условия согласно изобретению, предназначенные для изменения скоростей переноса нуклеиновых кислот-мишеней к элементам или изменения скоростей амплификации с целью установления кинетического исключения. Соответственно, на промежуточных участках массива может быть создан такой заряд, который отталкивает нуклеиновые кислоты-мишени, в то время как на различных сайтах амплификации массива одновременно протекает амплификация, причем средняя скорость амплификации превышает среднюю скорость переноса нуклеиновых кислот-мишеней к сайтам амплификации. Соответственно, отталкивание электрическим полем на промежуточных участках может быть применено в комбинации с другими способами переноса нуклеиновых кислот (или других биомолекул) к элементам массива и установления кинетического исключения согласно изобретению.
Отталкивание, создаваемое электрическим полем на промежуточных участках в способах и установке согласно изобретению, может обеспечивать такое преимущество, как улучшенная специфическая локализация нуклеиновых кислот (или других биомолекул) на элементах, а не на промежуточных участках. Это преимущество может быть достигнуто вне зависимости от того, является ли отталкивание результатом взаимодействия зарядов, приводящим к замедлению связывания нуклеиновых кислот или других биомолекул, результатом поверхностно локализованного электрохимического повреждения нуклеиновых кислот и биомолекул или результатом иного воздействия. Отталкивание, создаваемое электрическим полем на промежуточных участках, может быть применено для улучшения специфической локализации нуклеиновых кислот или других биомолекул на интересующих элементах, особенно, если высота интересующих элементов превышает максимальный уровень поверхностно локализованного электрохимического повреждения.
В некоторых примерах перенос нуклеиновых кислот (или других биомолекул) под действием электрического поля к элементам массива может быть применен в комбинации с отталкиванием нуклеиновых кислот (или других биомолекул) под действием электрического поля от промежуточных участков массива. Притягивающее электрическое поле и отталкивающее электрическое поле могут быть приложены к массиву одновременно, или эти два поля могут быть приложены по отдельности. Например, два поля могут быть приложены по отдельности в режиме попеременного многократного включения (например, притягивающее поле может быть приложено к элементам, а отталкивающее поле в это время отключено, и затем притягивающее поле может быть отключено, а отталкивающее поле приложено к промежуточным участкам, и эта последовательность может быть повторена один или более раз).
Электрическое поле может быть приложено к участку массива и к электролиту, или оно может быть приложено к участку массива и второй поверхности. Например, на Фиг. 4(a) представлена конфигурация, в которой электрическое поле может быть приложено к промежуточному участку массива и к электролиту, и на Фиг. 4(b) представлена конфигурация, в которой электрическое поле может быть приложено к промежуточному участку массива и второй поверхности. Аналогичные конфигурации могут быть применены для приложения притягивающих полей к элементам массива. Кроме того, электрические поля, как приложенные к элементу, так и к промежуточному участку, могут генерироваться переменным током (англ. alternating current, сокращенно АС) или постоянным током (англ. direct current, сокращенно DC), подаваемым к подходящему участку массива.
Соответственно, изобретение относится к способу создания структурированной поверхности из биомолекул, который может включать: (а) предоставление реагента, включающего (i) массив, на поверхности которого имеются не непрерывные элементы, где элементы разделены промежуточными участками поверхности, и (ii) раствора, содержащего совокупность различных биомолекул-мишеней; и (b) реакцию реагента, приводящую к переносу биомолекул к элементам и к присоединению индивидуальной биомолекулы к каждому из элементов, причем к промежуточным участкам прикладывают электрическое поле для выталкивания биомолекул из промежуточных участков. Особенно подходящими для применения в способе биомолекулами являются нуклеиновые кислоты. Нуклеиновые кислоты могут быть амплифицированы на элементах способом согласно изобретению в условиях кинетического исключения, таким, как способы, рассмотренные в настоящем описании. В некоторых примерах, в которых применяют воздействие электрических полей, подложка для массива может включать слой прозрачного проводника электричества. Слой проводника электричества может служить электродом для соединения источника электрического тока, такого как батарея или генератор сигнала. При необходимости элемент массива (например, внутренние поверхности лунки в массиве лунок) может содержать открытый или изолированный проводящий слой, и к проводящим слоям может быть приложено напряжение, воздействующее на силу, действующую на нуклеиновую кислоту и/или реагенты амплификации, с целью регулирования скорости переноса к сайту, захвата на сайте, удаления с сайта и/или амплификации на сайте. В конкретных примерах электрическое поле может быть приложено к внешним поверхностям лунки таким образом, что электрическое поле, проникающее через стенки емкости, генерирует электрическую силу, воздействующую на реагенты, находящиеся в емкости, и обеспечивает, таким образом, определенную степень контроля над скоростями переноса, захвата, удаления и/или амплификации.
В массиве согласно настоящему изобретению, например, полученном способом согласно изобретению, могут быть осуществлены различные методики. Особенно подходящим применением является секвенирование нуклеиновых кислот. Одним из примеров является секвенирование синтезом (SBS). В SBS для определения последовательности нуклеотидов в матрице отслеживают достройку праймера нуклеиновой кислоты вдоль матрицы нуклеиновой кислоты (например, нуклеиновой кислоты-мишени или ее ампликона). Основополагающим химическим процессом может быть полимеризация (например, катализируемая ферментом полимеразой). В конкретном примере SBS с использованием полимеразы меченные флуоресцентной меткой нуклеотиды присоединяются к праймеру (достраивая таким образом праймер) в соответствии со структурой матрицы, и для определения последовательности матрицы может быть отслежен порядок и тип нуклеотидов, добавляемых к праймеру. Способ SBS может быть применен к совокупности различных матриц, находящихся на различных сайтах массива согласно изобретению, в условиях, в которых события, касающиеся различных матриц, можно различить по их локализации в массиве.
Проточные ячейки обеспечивают подходящий формат для вмещения массива, получаемого способами согласно настоящему изобретению, в котором выполняют SBS или применяют другую методику обнаружения, которая включает многократную подачу реагентов в течение циклов. Например, для инициирования первого цикла SBS в/через проточную ячейку, в которой находится массив матриц нуклеиновых кислот, может быть пропущен поток, содержащий один или более меченых нуклеотидов, ДНК-полимеразу и т.д. Те сайты массива, в которых при достройке праймера в него встраивается меченый нуклеотид, могут быть обнаружены. Необязательно, нуклеотиды могут дополнительно иметь свойство обратимой терминации, обеспечивающее прекращение последующей достройки праймера после добавления к праймеру нуклеотида. Например, к праймеру может быть присоединен аналог нуклеотида, содержащий фрагмент, являющийся обратимым терминатором, который препятствует последующему удлинению до тех пор, пока этот фрагмент не будет снят после добавления деблокирующего агента. Таким образом, в тех примерах, в которых применяют обратимую терминацию, в проточную ячейку может быть направлен деблокирующий реагент (до или после этапа обнаружения). Между различными этапами подачи могут быть выполнены промывки. Цикл затем может быть повторен n раз, в результате чего праймер достраивается n нуклеотидами, что приводит к определению последовательности длиной n. Примеры процедур SBS, жидкостных систем и платформ для обнаружения, которые могут быть легко адаптированы для работы с массивом, полученным способами согласно настоящему изобретению рассмотрены, например, в публикации Bentley с соавт., Nature 456:53-59 (2008), в патентных документах WO 04/018497, US 7057026, WO 91/06678, WO 07/123744, US 7329492, US 7211414, US 7315019, US 7405281 и в опубликованной патентной заявке US 2008/0108082, содержания которых полностью включены в настоящую работу посредством ссылки.
Могут быть применены другие процедуры секвенирования, в которых применяют циклические реакции, такие как пиросеквенирование. При проведении пиросеквенирования производят обнаружение высвобождения неорганического пирофосфата (PPi) по мере встраивания определенных нуклеотидов в растущую цепочку нуклеиновой кислоты (Ronaghi, с соавт., Analytical Biochemistry 242(1), 84-9 (1996); Ronaghi, Genome Res. 11(1), 3-11 (2001); Ronaghi с соавт., Science 281(5375), 363 (1998); патенты US 6210891, US 6258568 и US 6274320, содержания которых полностью включены в настоящую работу посредством ссылки). При проведении пиросеквенирования высвобожденный PPi может быть обнаружен в результате его немедленного превращения в аденозинтрифосфат (АТФ) под действием АТФ-сульфурилазы, и концентрация образующегося АТФ может быть определена по количеству фотонов, генерируемых люциферазой. Таким образом, протекание реакции секвенирования можно отслеживать с помощью системы обнаружения люминесценции. Источники излучения возбуждения, применяемые в системах обнаружения флуоресценции, для процедур пиросеквенирования не обязательны. Подходящие жидкостные системы, датчики и процедуры, которые могут быть применены для пиросеквенирования массивов согласно настоящему изобретению, рассмотрены, например, в патенте US 9096899, опубликованной патентной заявке US 2005/0191698 А1, в патентах US 7595883 и US 7244559, содержания которых полностью включены в настоящую работу посредством ссылки.
Согласно настоящему изобретению, также подходят реакции секвенирования лигированием, включающие, например, реакции, рассмотренные в публикации Shendure с соавт., Science 309:1728-1732 (2005) и в патентах US 5599675 и US 5750341, содержания которых полностью включены в настоящую работу посредством ссылки. Некоторые примеры могут включать процедуры секвенирования гибридизацией, рассмотренные, например, в публикациях Bains с соавт., Journal of Theoretical Biology 135(3), 303-7 (1988); Drmanac с соавт., Nature Biotechnology 16, 54-58 (1998); Fodor с соавт., Science 251(4995), 767-773 (1995), и в патентном документе WO 1989/10977, содержания которых полностью включены в настоящую работу посредством ссылки. Как в процедуре секвенирования лигированием, так и в процедуре секвенирования гибридизацией, проводят многократные циклы подачи и обнаружения олигонуклеотидов на нуклеиновых кислотах-мишенях (или их ампликонах), находящихся на сайтах массива. Жидкостные системы для способов SBS, согласно изобретению или рассмотренные в цитируемой в настоящем документе литературе, могут быть легко адаптированы для подачи реагентов для секвенирования лигированием или секвенирования гибридизацией. Олигонуклеотиды могут быть помечены флуоресцентной меткой и обнаружены с помощью флуоресцентных датчиков, аналогичных устройствам, рассмотренным при описании процедур SBS в настоящей работе или в литературе, цитируемой в настоящей работе.
В некоторых примерах могут быть применены способы, включающие мониторинг активности ДНК-полимеразы в режиме реального времени. Например, встраивание нуклеотидов может быть обнаружено в результате резонансного переноса энергии флуоресценции (англ. fluorescence resonance energy transfer, сокращенно FRET) между полимеразой, несущей флуорофор, и меченными γ-фосфатом нуклеотидами, или с помощью волноводов с нулевой модой (англ. zeromode waveguide, сокращенно ZMW). Методики и реагенты для секвенирования с использованием FRET рассмотрены, например, в публикациях Levene с соавт., Science 299, 682-686 (2003), Lundquist с соавт., Opt. Lett. 33, 1026-1028 (2008) и Korlach с соавт., Proc. Natl. Acad. Sci. USA 105, 1176-1181 (2008), содержания которых полностью включены в настоящую работу посредством ссылки.
Некоторые примеры SBS включают обнаружение протона, высвобождаемого при встраивании нуклеотида в продукт достройки. Например, в секвенировании с обнаружением высвобождаемых протонов может быть применен электрический датчик, и соответствующие методики, коммерчески поставляемые Ion Torrent (Guilford, Conn., дочернее предприятие Life Technologies), или способы и системы секвенирования, рассмотренные в опубликованной патентной заявке US 2009/0026082 А1, опубликованной патентной заявке US 2009/0127589 А1, опубликованной патентной заявке US 2010/0137143 А1 или опубликованной патентной заявке US 2010/0282617 А1, содержания которых полностью включены в настоящую работу посредством ссылки. Способы амплификации нуклеиновых кислот-мишеней с применением кинетического исключения согласно изобретению могут быть с успехом реализованы на подложках, применяемых для обнаружения протонов. В частности, способы согласно изобретению могут быть применены для образования клональных популяций ампликонов на сайтах массивов, которые применяют для обнаружения протонов.
Другим подходящим применением массива согласно настоящему изобретению, например, полученного способом согласно изобретению, является анализ экспрессии генов. Экспрессия генов может быть обнаружена или количественно определена с помощью методик секвенирования РНК, таких как методики, называемые цифровым секвенированием РНК. Методики секвенирования РНК могут включать способы секвенирования, известные в данной области техники, такие как методики, рассмотренные выше. Экспрессия генов также может быть обнаружена или количественно определена с помощью методики гибридизации, представляющей собой прямую гибридизацию с массивом, или с помощью мультиплексного анализа, продукты которого обнаруживают в массиве. Массив согласно настоящему изобретению, например, полученный способом согласно изобретению, также может быть применен для определения генотипов образца геномной ДНК одного или более индивидуума. Примеры способов применения массивов в анализе экспрессии и в генотипировании, в которых могут быть применены массивы согласно настоящему изобретению, рассмотрены в патентах US 7582420, US 6890741, US 6913884 или US 6355431 или в опубликованных патентных заявках US 2005/0053980 А1, US 2009/0186349 А1 или US 2005/0181440 А1, содержания которых полностью включены в настоящую работу посредством ссылки.
Преимуществом способов согласно изобретению является то, что они позволяют быстро и эффективно создавать массивы из любого множества различных библиотек нуклеиновых кислот. Соответственно, настоящее изобретение относится к интегрированным системам, которые могут создавать массив с помощью одного или более способов согласно изобретению и также могут обнаруживать в массивах нуклеиновые кислоты с помощью методик, известных в данной области техники, таких как методики, примеры которых приведены выше. Таким образом, интегрированная система согласно настоящему изобретению может включать компоненты для работы с текучими средами, которые могут подавать реагенты амплификации к массиву сайтов амплификации, такие как насосы, клапаны, резервуары, трубопроводы для текучих сред и подобные устройства. Особенно подходящим компонентом для работы с текучими средами является проточная ячейка. Проточная ячейка может быть сконструирована и/или применена в интегрированной системе для создания массива согласно настоящему изобретению и исследования массива. Примеры проточных ячеек рассмотрены, например, в опубликованной патентной заявке US 2010/0111768 А1 и в патенте US 8951781, содержания которых полностью включены в настоящую работу посредством ссылки. Как было указано при рассмотрении проточных ячеек, для осуществления способа амплификации и способа обнаружения могут быть применены один или более компонентов интегрированной системы для работы с текучими средами. В случае секвенирования нуклеиновых кислот один или более компонентов интегрированной системы для работы с текучими средами могут быть применены для осуществления способа амплификации согласно изобретению и для подачи реагентов секвенирования в способ секвенирования, такой как пример способа, приведенный выше. В альтернативном варианте интегрированная система может включать отдельные жидкостные системы для осуществления способов амплификации и осуществления способов обнаружения. Примеры интегрированных систем секвенирования, с помощью которых могут быть созданы массивы нуклеиновых кислот, а также определены последовательности нуклеиновых кислот, включают, без ограничений, инструментальную платформу MISEQ® (Illumina, Inc., San Diego, Calif.) и устройства, рассмотренные в патенте US 8951781 (см. выше). Такие устройства могут быть модифицированы для получения массивов с использованием кинетического исключения в соответствии с принципами настоящего изобретения.
Система, с помощью которой может быть осуществлен способ согласно изобретению, не обязательно должна быть интегрирована с обнаруживающим устройством. Напротив, возможно применение отдельной системы или системы, интегрированной с другими устройствами. В таких примерах могут быть применены компоненты для работы с текучими средами, аналогичные компонентам, примеры которых приведены выше при описании интегрированной системы.
Система, с помощью которой может быть осуществлен способ согласно изобретению, независимо от того, интегрирована она или нет с обнаруживающими устройствами, может включать системный контроллер, способный выполнять набор инструкций для осуществления одного или более способов, методик или процедур согласно изобретению. Например, инструкции могут регулировать выполнение этапов создания массива в условиях кинетического исключения. Необязательно, инструкции могут дополнительно регулировать выполнение этапов обнаружения нуклеиновых кислот способами, рассмотренными выше. Подходящий системный контроллер может включать любую систему на основе процессора или микропроцессора, включая системы, в которых применяют микроконтроллеры, компьютеры с сокращенным набором команд (англ. reduced instruction set computer, сокращенно RISC), специализированные интегральные схемы (англ. application specific integrated circuit, сокращенно ASIC), программируемые на месте матрицы логических элементов (англ. field programmable gate array, сокращенно FPGA), логические схемы и любые другие схемы или процессоры, способные выполнять функции, рассмотренные в настоящем описании. Набор инструкций может представлять собой программу пакета программного обеспечения. Употребляемые в настоящем описании термины "пакет программного обеспечения (программное обеспечение)" и "встроенное программное обеспечение" взаимозаменяемы и включают любую компьютерную программу, хранящуюся в памяти и предназначенную для выполнения компьютером, включающую оперативное запоминающее устройство (ОЗУ) (англ. Random Access Memory, сокращенно RAM), запоминающее устройство (ПЗУ) (англ. Read Only Memory, сокращенно ROM), память EPROM (англ. electronically programmable read only memory, рус. электрически перепрограммируемое постоянное запоминающее устройство), память EEPROM (англ. electrically erasable programmable read only memory, рус. электронно-стираемое программируемое постоянное запоминающее устройство) и энергонезависимую память RAM (NVRAM, сокр. от англ. non-volatile random access memory, рус. энергонезависимое ОЗУ). Приведенные выше типы памяти приведены только для примера и, таким образом, не ограничивают типы памяти, подходящие для хранения компьютерной программы. Программное обеспечение может иметь различные формы, такие как системное программное обеспечение или прикладное программное обеспечение. Кроме того, программное обеспечение может находиться в виде совокупности отдельных программ или программного модуля, встроенного в более крупную программу, или в виде части программного модуля. Программное обеспечение также может включать модульное программирование в виде объектно-ориентированного программирования.
Некоторые из применений массивов согласно настоящему изобретению были рассмотрены выше в контексте множественного обнаружения, при котором множество ампликонов, присутствующих на каждом сайте амплификации, обнаруживают совместно. В альтернативных примерах на каждом сайте амплификации может быть обнаружена одна нуклеиновая кислота, либо нуклеиновая кислота-мишень, либо ее ампликон. Например, сайт амплификации может быть сконструирован таким образом, чтобы он содержал одну молекулу нуклеиновой кислоты, имеющую целевую нуклеотидную последовательность, которую необходимо определить, и совокупность заполняющих нуклеиновых кислот. В этом примере заполняющие нуклеиновые кислоты служат для заполнения емкости сайта амплификации, и их необязательно следует обнаруживать. Одна определяемая молекула может быть обнаружена способом, который позволяет различать одну молекулу на фоне заполняющих нуклеиновых кислот. Могут быть применены любые из множества различных методик обнаружения одной молекулы, примеры которых включают модификации методик множественного обнаружения, рассмотренных выше, применяемые для обнаружения сайтов с повышенной интенсивностью или с использованием более чувствительных меток. Другие примеры способов обнаружения одной молекулы, которые могут быть применены, рассмотрены в опубликованной патентной заявке US 2011/0312529 А1, в патенте US 9279154 и в опубликованной патентной заявке US 2013/0085073 А1, содержания которых полностью включены в настоящую работу посредством ссылки.
Массив, подходящий для обнаружения одной молекулы нуклеиновой кислоты, может быть создан с помощью одного или более способов согласно изобретению, имеющих следующие модификации. Может быть подобрана такая совокупность различных нуклеиновых кислот-мишеней, которая включает как целевую определяемую нуклеотидную последовательность, так и одну или более заполняющих нуклеотидных последовательностей, которые амплифицируют для получения заполняющих ампликонов. Совокупность различных нуклеиновых кислот-мишеней может быть включена в реагент амплификации, такой как реагенты, рассмотренные в настоящем описании, и введена в реакцию с массивом сайтов амплификации в условиях кинетического исключения таким образом, чтобы заполняющая нуклеотидная последовательность (последовательности) заполнила сайты амплификации. Примеры конфигураций, которые могут быть применены для амплификации заполняющих последовательностей, но при этом не позволяющие протекать амплификации последовательности-мишени, включают, например, наличие в единственной молекуле-мишени первого участка, содержащего заполняющие последовательности, которые фланкированы связывающими сайтами для амплификации праймеров, присутствующих на сайте амплификации, и второго участка, содержащего последовательность-мишень, расположенную за пределами фланкированного участка. В другой конфигурации нуклеиновая кислота-мишень может включать отдельные молекулы или цепочки, несущие последовательность-мишень и заполняющую последовательность (последовательности), соответственно. Эти отдельные молекулы или цепочки могут быть присоединены к частице или сформированы в виде отростков дендримера нуклеиновой кислоты или другой разветвленной структуры.
В одном из конкретных примеров массив, содержащий сайты амплификации, каждый из которых содержит как заполняющие последовательности, так и последовательность-мишень, может быть исследован с помощью анализа с достройкой праймера или методики секвенирования синтезом. В таких случаях, в отличие от большого количества заполняющей последовательности, нуклеотидная последовательность-мишень может быть достроена определенным образом при использовании подходящим образом размещенных сайтов связывания праймера. Например, связывающие сайты для секвенирования праймеров могут быть размещены выше по ходу транскрипции относительно последовательности-мишени и могут отсутствовать в любой из заполняющих последовательностей. В альтернативном варианте или дополнительно последовательность-мишень может включать один или более не встречающихся в природе аналогов нуклеотидов, которые не способны связываться водородными связями со стандартными нуклеотидами. Не встречающийся в природе нуклеотид (нуклеотиды) может быть размещен ниже по ходу транскрипции относительно сайта связывания праймера (например, в последовательности-мишени или в участке, промежуточном между последовательностью-мишенью и сайтом связывания праймера) и, таким образом, будет препятствовать достройке или секвенированию синтезом до тех пор, пока не будет добавлен подходящий нуклеотидный партнер (т.е. нуклеотид, способный образовывать водородные связи с не встречающимся в природе аналогом (аналогами) последовательности-мишени). Особенно подходящими являются аналоги нуклеотидов изоцитозин (isoC) и изогуанин (isoG), поскольку они специфично связываются друг с другом, но не с другими стандартными нуклеотидами, применяемыми в большинстве методик достройки и секвенирования синтезом. Дополнительное преимущество размещения isoC и/или isoG в последовательности-мишени или выше по ходу транскрипции относительно последовательности-мишени состоит в предотвращении нежелательной амплификации последовательности-мишени во время этапов амплификации, если для амплификации используют смесь нуклеотидов, не содержащую соответствующего партнера.
Следует понимать, что массив согласно настоящему изобретению, например, полученный способом согласно изобретению, не обязательно должен быть применен в способе обнаружения. Напротив, массив может быть применен для хранения библиотек нуклеиновых кислот. Соответственно, массив может храниться в состоянии, которое позволяет сохранять находящиеся в нем нуклеиновые кислоты. Например, массив может храниться в сухих условиях, в замороженном состоянии (например, в жидком азоте) или в растворе, защищающем нуклеиновые кислоты. В альтернативном варианте или дополнительно массив может быть применен для репликации библиотеки нуклеиновых кислот. Например, массив может быть применен для создания реплик ампликонов, находящихся на одном или более сайтах массива.
В настоящем описании были приведены некоторые примеры, касающиеся переноса нуклеиновых кислот-мишеней к сайтам амплификации массива и создания копий захваченных нуклеиновых кислот-мишеней на сайтах амплификации. Аналогичные способы могут быть применены для молекул-мишеней, не являющихся нуклеиновыми кислотами. Таким образом, способы согласно изобретению могут быть применены для работы с другими молекулами-мишенями, а не с нуклеиновыми кислотами-мишенями, приведенными в качестве примера. Например, способ согласно настоящему изобретению может быть применен для транспортировки индивидуальных молекул-мишеней из популяции различных молекул-мишеней. Каждая молекула-мишень может быть перенесена к индивидуальному сайту массива для инициирования реакции на сайте захвата (и в некоторых случаях захвачена индивидуальным сайтом). Протекающая на каждом сайте реакция может, например, приводить к образованию копий захваченной молекулы, или реакция может изменять сайт, делая его подходящим для выделения или удаления захваченной молекулы. В любом случае в конечном итоге могут образовываться сайты массива, каждый из которых является чистым по отношению к типу присутствующей молекулы-мишени, извлеченной из популяции, содержащей различные типы молекул-мишеней.
В конкретных примерах, в которых применяют молекулы-мишени, не являющиеся нуклеиновыми кислотами, библиотека различных молекул-мишеней может быть получена способами, включающими кинетическое исключение. Например, массив молекул-мишеней может быть получен в условиях, в которых на сайты массива неупорядоченным образом вносят молекулы-мишени из раствора, после чего образуются копии молекулы-мишени, заполняющие всю емкость каждого из занятых сайтов. В соответствии со способами, выполняемыми с применением кинетического исключения согласно настоящему изобретению, внесение и копирование могут протекать одновременно, если скорость создания копий превышает скорость внесения. Таким образом, относительно высокая скорость создания копий на сайте, на который внесена первая молекула-мишень, будет эффективно исключать размещение на сайте второй молекулы-мишени. В некоторых случаях размещение молекулы-мишени будет инициировать реакцию, не представляющую собой копирование молекулы-мишени, продукты которой, тем не менее, заполняют всю емкость сайта. Например, захват молекулы-мишени на сайте может инициировать цепную реакцию, которая, в конечном итоге, делает сайт неспособным к захвату второй молекулы-мишени. Скорость цепной реакции может превышать скорость захвата молекул-мишеней, то есть процесс будет протекать в условиях кинетического исключения.
Как было показано для нуклеиновых кислот-мишеней, кинетическое исключение в случае других молекул-мишеней может быть обусловлено относительно низкой скоростью инициирования повторяющейся реакции (например, цепной реакции) на сайте массива по сравнению с относительно высокой скоростью продолжения повторяющейся реакции после ее начала. В примере, приведенном в предыдущем разделе, кинетическое исключение обусловлено относительно низкой скоростью внесения молекулы-мишени (например, относительно медленной диффузией) по сравнению с относительно высокой скоростью протекания реакции, например, заполнения сайта копиями внесенной молекулы-мишени. В другом примере кинетическое исключение может быть обусловлено задержкой образования первой копии молекулы-мишени, размещенной на сайте (например, задержкой в активации или медленной активацией), по сравнению с относительно высокой скоростью образования последующих копий, заполняющих сайт. В этом примере на индивидуальный сайт могут быть внесены несколько различных молекул-мишеней. Однако образование первой копии любой заданной молекулы-мишени может быть активировано случайным образом, так что средняя скорость образования первой копии относительно низка по сравнению со скоростью генерации последующих копий. В этом случае, несмотря на то, что на индивидуальный сайт могут быть внесены несколько различных молекул-мишеней, кинетическое исключение разрешит копирование лишь одной из этих молекул-мишеней.
Соответственно, настоящее изобретение относится к способу получения массива молекул, где способ может включать: (а) предоставление реагента, включающего (i) массив сайтов, и (ii) раствора, содержащего совокупность различных молекул-мишеней, где количество молекул-мишеней в растворе превышает количество сайтов в массиве, и различные молекулы-мишени переносятся текучей средой к совокупности сайтов, и при этом каждый из сайтов имеет емкость, достаточную для размещения нескольких молекул-мишеней, находившихся в совокупности различных молекул-мишеней; и (b) реакцию реагента, приводящую к образованию совокупности сайтов, каждый из которых содержит одну молекулу-мишень из совокупности, или приводящую к образованию совокупности сайтов, каждый из которых содержит чистую популяцию копий индивидуальных молекул-мишеней, получаемых из раствора, причем реакция включает одновременное протекание (i) переноса различных молекул к сайтам со средней скоростью переноса, и (ii) инициирования реакции, которая заполняет всю емкость сайта со средней скоростью реакции, где средняя скорость реакции превышает среднюю скорость переноса. Кроме того, в некоторых примерах этап (b) может включать реакцию реагента, приводящую к образованию совокупности сайтов, каждый из которых содержит одну молекулу-мишень из совокупности, или приводящую к образованию совокупности сайтов, каждый из которых содержит чистую популяцию копий индивидуальных молекул-мишеней, получаемых из раствора, где реакция включает (i) инициирование повторяющейся реакции (например, цепной реакции), приводящей к образованию на каждом из сайтов продукта, получаемого из молекулы-мишени, и (ii) продолжение протекания реакции на каждом из сайтов, приводящее к образованию последующих продуктов, причем средняя скорость протекания реакции на сайтах превышает среднюю скорость инициирования реакции на сайтах.
В приведенных выше примерах веществ, не являющихся нуклеиновыми кислотами, молекула-мишень может быть инициатором повторяющейся реакции, протекающей на каждом сайте массива. Например, в результате повторяющейся реакции может образовываться полимер, который препятствует заполнению сайта молекулами-мишенями. В альтернативном варианте в результате повторяющейся реакции могут образовываться один или более полимеров, которые составляют молекулярные копии молекулы-мишени, которая транспортируется к сайту.
Приведенные ниже примеры даны для иллюстрации и не ограничивают предмет настоящего изобретения.
ОПИСАНИЕ ПРИМЕРОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Пример I
Сверхпуассоновское образование массивов кластеров на проточных ячейках
В этом примере рассмотрен способ сверхпуассоновского образования массива кластеров на проточной ячейке платформы секвенирования Illumina (San Diego, Calif.). Рассмотренный способ представляет собой процесс захвата элемента библиотеки (например, фрагмента генома) на элементе массива и одновременного протекания клональной амплификации элемента библиотеки. Ключевым элементом способа в этом примере является регулирование скорости захвата относительно скорости амплификации и достижение однородности процесса. Во множестве применяемых ранее способов, разработанных для внесения высокой плотности на проточных ячейках Illumina, захват элемента библиотеки отделен от процесса клональной амплификации. В этом примере событие захвата инициирует событие клональной амплификации на элементе.
Структурированную проточную ячейку получают следующим образом. На стеклянные проточные ячейки (Illumina, Inc., San Diego, Calif.) с помощью взрывной литографии наносят покрытие из золотых пластинок. Вкратце, слой фоторезиста равномерно наносят на поверхность стеклянной проточной ячейки, и участки фоторезиста удаляют фотолитографией, открывая участки стеклянной поверхности. Затем на поверхность наносят слой золота, который образует непрерывную тонкую пленку на участках фоторезиста и участках стекла. Золото может быть нанесено электронно-лучевым испарением или напылением, как это рассмотрено в публикации Thornton, Ann. Rev. Mater. Sci. 7:239-60 (1977), содержание которой полностью включено в настоящую работу посредством ссылки. Затем слой фоторезиста удаляют ацетоном, в результате чего остаются участки золота круглой формы диаметром менее 1 микрона, окруженные промежуточными участками стеклянной поверхности. Затем на проточную ячейку, имеющую структурированное покрытие из золота, наносят покрытие из не содержащего силана акриламида (SFA), рассмотренного в патентном документе WO 2008/093098 (включенном в настоящее описание). К полимеризованному SFA через нитробензильный фрагмент, расщепляемый под действием УФ излучения, прививают праймеры Р5 и Р7 (Glenn Research, Sterling, Va.). Проточную ячейку располагают на источнике УФ света (302 нм) таким образом, чтобы участки золота создавали маску (экран) для праймеров, присоединенных на участках золота, в то время как праймеры, присоединенные к промежуточным участкам, отщепляются под действием UV света. Праймеры Р5 и Р7, оставшиеся на участках золота, могут поддерживать клональную амплификацию библиотек (Р5/Р7).
Элементы библиотеки получают следующим образом. Библиотеку геномных ДНК (гДНК) фрагментируют, и раздвоенные адаптеры, имеющие сайты связывания праймера, которые комплементарны праймерам Р5 и Р7 лигируют с фрагментами гДНК в соответствии с коммерчески доступными протоколами получения образцов, предоставляемыми Illumina.
Сверхпуассоновское образование массива кластеров выполняют следующим образом. Подготавливают раствор, содержащий элементы библиотеки (в двухцепочечной форме) и реагент TWISTAMP® Basic (TwistDx, Cambridge UK). Реагент TWISTAMP® Basic содержит смесь ферментов, которая может поддерживать зависимую от матрицы амплификацию на поверхности (ДНК-полимераза, одноцепочечный связывающий белок и рекомбиназа). Концентрацию элементов библиотеки в растворе регулируют таким образом, чтобы скорость захвата с гибридизацией элемента библиотеки любым элементом массива была намного ниже, чем скорость клональной амплификации, и таким образом, чтобы на элементе массива оставалось достаточно мало олигонуклеотидов, подходящих для захвата другого элемента библиотеки.
Оптимальная или в ином случае требуемая концентрация элементов библиотеки в растворе может быть определена эмпирически титрованием в соответствии с приведенным выше протоколом сверхпуассоновского образования массива кластеров с последующим проведением секвенирования на секвенаторе Illumina (например, на устройствах GENOMEANALYZER®, HISEQ® или MISEQ®).
Пример II
Характеристика структурированных массивов кластеров, полученных в условиях кинетического исключения
В этом примере показано сверхпуассоновское размещение моноклональных кластеров на структурированных элементах в условиях кинетического исключения.
Структурированную проточную ячейку получали следующим образом. На стеклянные проточные ячейки (Illumina, Inc., San Diego, Calif.) с помощью взрывной литографии наносили покрытие из золотых пластинок, как рассмотрено в патенте US 8778848, содержание которого полностью включено в настоящую работу посредством ссылки. Вкратце, слой фоторезиста равномерно наносили на поверхность стеклянной проточной ячейки, и участки фоторезиста удаляли фотолитографией, открывая участки стеклянной поверхности. Затем на поверхность наносили слой золота, который образовывал непрерывную тонкую пленку на участках фоторезиста и участках стекла. Золото наносили электронно-лучевым испарением, как рассмотрено в публикации Thornton, Ann. Rev. Mater. Sci. 7:239-60 (1977), содержание которой полностью включено в настоящую работу посредством ссылки. Затем слой фоторезиста удаляли ацетоном, в результате чего получали гексагональную схему расположения золотых пластинок, где каждая золотая пластинка имела круглую форму диаметром 500 нм и была окружена промежуточными участками стеклянной поверхности. Затем на проточную ячейку, имеющую структурированное покрытие из золота, наносили покрытие из не содержащего силана акриламида (SFA), рассмотренного в патентном документе WO 2008/093098 (включенном в настоящее описание). К полимеризованному SFA через нитробензильный фрагмент, расщепляемый под действием УФ излучения, прививали праймеры (Glenn Research, Sterling, Va.). Проточную ячейку располагали на источнике УФ света (302 нм) таким образом, чтобы пластинки золота создавали маску (экран) для праймеров, присоединенных к пластинкам, в то время как праймеры, присоединенные к промежуточным участкам, отщеплялись под действием UV света. Отщепленные праймеры удаляли промывкой, в результате которой на месте оставались праймеры, присоединенные к золотым пластинкам.
Кластеры выращивали на золотых пластинках, применяя набор TWISTAMP® Basic (TwistDx, Cambridge UK), следующим образом. Двухцепочечную библиотеку PhiX ДНК смешивали в различных концентрациях с буфером для регидратации TWISTAMP® Basic и ацетатом магния. Концентрации испытуемых PhiX ДНК составляли приблизительно 72 пМ (пикоМоль), приблизительно 144 пМ, приблизительно 432 пМ и приблизительно 864 пМ. Эти концентрации соответствуют избытку по сравнению с типичным диапазоном, составляющим приблизительно 9-10 пМ ДНК, применяемым при стандартном внесении в проточные ячейки Illumina. Кроме того, PhiX ДНК представляет собой двухцепочечную молекулу в отличие от стандартного внесения в проточные ячейки Illumina, при котором матричная ДНК находится в одноцепочечной форме. Смеси, содержащие PhiX ДНК, применяли для регидратации лиофилизированных гранул TWISTAMP® Basic и затем направляли в соответствующие дорожки структурированной проточной ячейки при приблизительно 38°С. Инкубацию выполняли в течение приблизительно 1 часа при приблизительно 38°С, затем промывали промывочным буфером НТ2 (Illumina, Inc., San Diego Calif.) и окрашивали кластеры Зеленым SyBr. Затем для секвенирования кластеры подвергали обработке LMX1 в течение приблизительно 30 минут для придания ДНК в кластерах линейной формы, приблизительно 0,1н NaOH для денатурации и гибридизации праймера секвенирования. Затем на устройстве Illumina HISEQ® 2000 производили секвенирование проточной ячейки, включающее 26 циклов.
Визуальный осмотр изображений проточной ячейки показал, что кластеры пространственно упорядочены в виде схемы, соответствующей схеме расположения на поверхности золотых пластинок. На Фиг. 1(a) представлено составное изображение всех четырех цветных каналов, полученное после проведения первого цикла секвенирования в проточной ячейке, полученной способами с применением кинетического исключения, рассмотренными выше. Для сравнения на Фиг. 1(b) представлено составное изображение, полученное после проведения одного цикла секвенирования в стандартной проточной ячейке, содержащей неупорядоченно расположенные кластеры.
Анализ функции распределения пар (англ. pair distribution function, сокращенно PDF) и функции ближайших соседей (англ. nearest neighbor, сокращенно NN) в составном изображении проточной ячейки также указывает на высокую степень упорядоченности. Рассчитанная групповая плотность кластеров составила приблизительно 640000 кластеров на квадратный миллиметр изображения. Функцию NN применяли для определения среднего расстояния между ближайшими соседними кластерами на изображении. Как показано на Фиг. 2, функция NN имеет преимущественно один пик при приблизительно 2,3 пикселях. Это согласуется с ожидаемым шагом схемы расположения пластинок, составляющим 1 микрон, то есть подтверждает высокую упорядоченность массива кластеров. Напротив, при неупорядоченном образовании кластеров образуется гораздо более широкий пик, с меньшими значениями, приближающимися к порогу обнаружения кластеров в соответствии с корреляционным алгоритмом (1,2 пикселя). Функция PDF на Фиг. 2 согласуется с ожидаемой структурой упорядоченного гексагонального массива. В этом примере функция PDF имеет ожидаемый первичный пик при приблизительно 2,66 пикселя, и пики более высокого порядка, соответствующие соседним элементам, расположенными за пределами ближайших, ясно видны и имеют ожидаемые соотношения пиков. Наблюдался лишь слабый сдвиг локализации пика при переходе между функциями NN и PDF. Такой низкий уровень сдвига указывает на то, что отклонение от теоретически идеальных положений было достаточно низким и в пределах приемлемых значений.
Визуальный осмотр композитных четырехцветных изображений также показывает отсутствие нежелательного проскока подложки. Проскоком подложки называется процесс, при котором на нескольких соседних участках происходит амплификация одной и той же матричной последовательности. На четырехцветном изображении проскок подложки визуально характеризуется непрерывными участками кластеров одного цвета. Отсутствие участков одного цвета в проточных ячейках, полученных в условиях кинетического исключения согласно этому примеру, указывает на то, что нежелательные уровни проскока подложки не наблюдаются. На Фиг. 3 представлено более количественно выраженное изображение цвета и пространственного положения кластера, указывающее на то, что проскок подложки не представляет собой проблему. В частности, на Фиг. 3 представлена диаграмма рассеяния пространственных положений кластеров, которые выровнены по первым 5 геномным положениям генома PhiX. Различные геномные положения показаны крестиками, звездочками, квадратами, треугольниками и ромбами. Обозначения 5 положения неупорядоченным образом распределены по диаграмме и не собраны в группу, что указывает на то, что проскок подложки не является проблемой.
В проточных ячейках, полученных в условиях кинетического исключения, проводили анализ последовательности из 26 циклов. Результаты показывают, что приблизительно 64% участков (пластинок) были заняты, и приблизительно 56% участков содержали клональные кластеры. Таким образом, предлагаемые способы обеспечивают почти 2-кратное увеличение количества клональных кластеров по сравнению с ожидаемым при пуассоновском заполнении, согласно которому по оценкам приблизительно 36% участков будут клональными, если приблизительно 64% из них будут заняты. Полученные результаты ясно показывают наличие сверхпуассоновского заполнения.
Пример III
Активная десорбция под действием электрического поля и структурированное расположение биомолекул
В этом примере рассмотрен способ пространственного расположения биомолекул под действием электрических полей. Способы, рассмотренные в этом примере, позволяют быстро располагать ДНК на целевых сайтах и электрохимическими способами выталкивать биомолекулы с промежуточных участков, что приводит к получению хорошо структурированных, адресуемых массивов, содержащих кластеры ДНК. Представленные результаты подтверждают образование проточных ячеек с упорядоченным расположением кластеров моноклональных нуклеиновых кислот.
В способе, рассмотренном в этом примере, для проведения активной десорбции молекул, присоединенных либо в результате физической адсорбции, либо в результате химического связывания, от одной или обеих находящихся под действием электрического поля поверхностей применяют электрический потенциал, приложенный к одной проводящей поверхности и электролиту или к двум проводящим поверхностям. Такой способ активной десорбции не требует обработки поверхности химическим веществами/модификации поверхности, позволяет производить очень быструю десорбцию молекул (менее 5 минут) и менее чувствителен условиям проведения, чем способы пассивной десорбции. Проводящие поверхности могут быть металлическими (например, из титана, оксида индия-олова) или полупроводниковыми, и прикладываемый ток может быть переменным или постоянным и приводить к электрохимической реакции на границе раздела электрод/электролит. Приложение электрического поля повышает соотношение сигнал (на интересующих сайтах)/шум (на промежуточных участках) на порядок величины. Способ, рассмотренный в этом примере, также может быть применен к плоским электродам для проведения селективной десорбции, селективной рефункционализации электродов и электрохимического упорядоченного расположения частиц.
Архитектура проточной ячейки
Два вида архитектуры, рассмотренные выше для электрохимической десорбции биомолекул, показаны на Фиг. 4(a) и 4(b). В частности, в качестве проводящего прозрачного материала электрода применяли оксид индия-олова (ITO). ITO осаждали на поверхность D263 радиочастотным напылением. На Фиг. 4(a) представлено приложение электрического потенциала к проводящему слою ITO и электролиту. На Фиг. 4(b) представлено приложение электрического потенциала к двум параллельным проводящим пластинам из ITO, разделенным жидкой средой. Оба типа архитектуры могут быть применены для электрической десорбции частиц с поверхности ITO. Сайты с наноструктурой из золота (Аu) подходят для целенаправленного захвата тиолированных биомолекул (например, тиолированного авидина). Для предотвращения протекания электрохимической реакции на золоте, сайты с Au отделены от нижележащего ITO диэлектрическим разделителем (например, из SiO2, SiN, алмазоподобного углерода).
Представленная на Фиг. 4(b) архитектура также может быть применена для одновременной быстрой концентрации ДНК (например, в 100 раз) на поверхности проточной ячейки под действием электрических полей, как показано на Фиг. 4(c) на изображениях, полученных замедленной съемкой. В этих экспериментах потенциал (V), составляющий приблизительно 2В, приложен к зазору, составляющему приблизительно 100 мкм, который разделяет две поверхности ITO. Повышение флуоресценции с течением времени, наблюдаемое определением флюоресценции полного внутреннего отражения (англ. total internal reflection fluorescence, сокращенно TIRF), показанное на Фиг. 4(c), обусловлено сильным увеличением поверхностной концентрации контрольной PhiX ДНК (меченной красителем YOYO) после приложения электрического поля к верхней поверхности проточной ячейки. Таким образом, описанная в настоящем документе методика может быть применена для одновременной электрохимической десорбции биомолекул с промежуточных участков и проведения быстрого внесения.
Протокол эксперимента
Протокол экспериментов активной десорбции представлен на Фиг. 5. Способ включает нанесение покрытия из авидина на поверхность проточной ячейки, последующее нанесение покрытия из не содержащего силана акриламида (SFA) и прививку праймеров к SFA. Нанесение покрытия из SFA и прививку праймеров Р5 и Р7 выполняют как указано в документе WO 2008/093098 (содержание которого полностью включено в настоящую работу посредством ссылки). Однако в способах согласно изобретению авидин наносят электрохимическим способом в виде структуры на Аu или на диэлектрические сайты (разделенные промежуточными участками из ITO), имеющиеся на поверхности проточной ячейки, выполняя этап электрической десорбции перед нанесением покрытия из SFA. Кроме того, после прививки праймеров Р5 и Р7, прикладывают электрическое поле для быстрого размещения ДНК на Au или диэлектрических сайтах и для электрохимической десорбции биомолекул (ДНК, авидин, праймеры) с промежуточных участков из ITO. В одном из примеров для эффективной десорбции молекул прикладывают напряжение, составляющее приблизительно 2В. Продолжительность приложения поля, составляющая приблизительно 5 минут, достаточна для эффективной десорбции большей части молекул с промежуточных участков. Кроме того, результаты показывают, что концентрация праймеров на промежуточных участках также снижается после выполнения этапа приложения электрического поля. Затем выполняют кластерную амплификацию способом, рассмотренным в публикации Bentley с соавт., Nature 456:53-59 (2008), и затем выполняют окрашивание кластеров интеркалирующим красителем для двухцепочечной ДНК, после чего получают изображения с помощью микроскопа. Затем с помощью секвенатора ДНК HISEQ® 2000 (Illumina, Inc. San Diego) в проточной ячейке выполняют секвенирование для определения клональности кластеров. Результаты внесения с приложением электрического поля и электрохимической десорбции схематически представлены на Фиг. 5.
Результаты экспериментов
На Фиг. 6 представлены результаты, достигнутые при использовании архитектуры проточной ячейки, показанной на Фиг. 4(b), с приложением электрического поля (Фиг. 6(a)) и в отсутствие электрического поля (Фиг. 6(b)). При приложении электрического поля, кластеры плотно локализованы на сайтах из Аu размером приблизительно 2 мкм, и в промежуточных областях наблюдали очень низкую флуоресценцию. На сайтах размером приблизительно 2 мкм кластеры имеют высокую поликлональность из-за большого размера пластинки из Аu. Степень поликлональности может быть понижена посредством уменьшения размера пластинки, поскольку в этом случае внесение множества матриц предотвращается за счет стерического исключения, или поликлональность может быть понижена в условиях кинетического исключения. Также следует отметить, что пиксельная интенсивность в промежуточных участках близка к 0 (профиль линии на Фиг. 6(a)). Напротив, в отсутствие электрического поля кластеры присутствуют как на участках из Аu, так и на промежуточных поверхностях из ITO. Периодическое изменение профиля, наблюдаемое на Фиг. 6(a), отсутствует на Фиг. 6(b), подтверждая, что локализация кластеров является результатом воздействия электрического поля.
Методика с приложением электрического поля может быть применена к кластерам, упорядоченно расположенным как на сайтах микронного размера, так и на сайтах, расположенных в виде наноструктур на больших площадях. Изображения больших площадей упорядоченно расположенных в виде структуры кластеров, введенных на сайты из Au диаметром приблизительно 2 мкм и на сайты из Au диаметром приблизительно 200 нм, показаны на Фиг. 7(a) и 7(b), соответственно, в сопровождении соответствующих схем, полученных с помощью быстрого преобразования Фурье (англ. Fast Fourier Transform, сокращенно FFT). Кластеры явно выражены и высоко упорядочены, так что в промежуточных областях ITO наблюдается очень слабое неспецифическое связывание. Это дополнительно подтверждается четко выраженными пятнами на FFT диаграмме, что указывает на наличие упорядоченной или структурированной стеки. Занятость кластерами расположенных в виде наноструктуры элементов, показанных на Фиг. 7(b), составляет приблизительно 40-50%, но она может быть дополнительно повышена посредством добавления более высоких концентраций авидина или изменения формы колебаний напряжения. Такая же химическая обработка/способ может быть применена к размещению кластеров с высокой пространственной точностью на диэлектрических сайтах. Упорядоченные кластеры, находящиеся на сайтах из SiO2 диаметром приблизительно 700 нм, показаны на Фиг. 8.
Механизм
Полученные данные показывают, что пространственному расположению кластеров способствует приложение электрического поля. Это, вероятно, объясняется электрохимическим удалением биомолекул (например, ДНК, белков и праймеров) с промежуточных участков. Как показано в анализе гибридизации с использованием зондов, меченных красителем Texas Red (TR), интенсивность сигнала привитых праймеров снижается при приложении электрического поля. На Фиг. 9(a) представлены результаты сканирования устройством Typhoon, которые показывают интенсивность флуоресценции TR (показана количественно на Фиг. 9(b)), полученную в анализе гибридизации в проточной ячейке до и после приложения электрического поля. После приложения электрического поля интенсивность флуоресценции снижается более чем в два раза, подтверждая удаление праймеров с SFA. Для повышения интенсивности сигналов кластеров, на проточную ячейку повторно наносят покрытие из SFA и повторно прививают праймеры Р5, Р7. Это приводит к существенному повышению интенсивности сигнала TR. Таким образом, для достижения высокой интенсивности сигналов пространственно упорядоченных кластеров, вероятно, необходимо произвести внесение ДНК, электрохимическое удаление неспецифично связанных молекул с промежуточных участков, повторное нанесение SFA и повторную прививку праймеров.
Непосредственная гибридизация ДНК
Упорядоченное пространственное расположение кластеров также наблюдали в экспериментах, включающих непосредственную гибридизацию одноцепочечной ДНК phiX с газоном праймеров Р5, Р7. Схематически этот процесс показан на Фиг. 10(a). Эксперименты проводили на сайтах Sift размером приблизительно 2 мкм, расположенных на ITO. Тот же способ может быть применен к сайтам, расположенным в виде наноструктуры и полученным из различных диэлектрических материалов. В этих экспериментах не требуется применение биотинилированной ДНК или авидина, что, таким образом, сокращает количество этапов химической обработки, но сохраняет специфичность расположения кластеров на сайтах. Специфичность, вероятно, является результатом электрохимической десорбции праймеров с промежуточных участков. На Фиг. 10(b) представлены кластеры, образованные на сайтах Sift размерами приблизительно 2 мкм при приложении электрического поля (приблизительно 2В, приблизительно 0,1 Гц) способом непосредственной гибридизации. На фигуре показаны кластеры, расположенные в виде структуры так, что в промежуточных участках находится очень малое их количество. На Фиг. 10(c) показаны результаты того же эксперимента в отсутствие электрического поля, откуда очевидно, что кластеры неупорядоченным образом ориентированы на SFA и на промежуточных участках из ITO, и что в отсутствие электрического поля упорядоченность не наблюдается. Полученные результаты показывают, что электрическое поле может быть применено для пространственного упорядочивания образования кластеров нуклеиновой кислоты.
На Фиг. 11 представлена блок-схема способа 100 генерации генетических кластеров на проточной ячейке согласно одному из примеров. Способ 100 осуществляют в соответствии с принципами настоящего изобретения. В этапе 102 первую смесь реагентов смешивают с количеством нуклеиновых кислот-мишеней, получая первый раствор. Первый раствор здесь и далее также называется целевым раствором, поскольку первый раствор включает существенное, не следовое количество нуклеиновых кислот-мишеней.
Нуклеиновые кислоты-мишени могут представлять собой ДНК подвергаемой секвенированию генетической библиотеки. Нуклеиновые кислоты-мишени получают в виде цепочек с адаптерами на концах, которые предназначены для связывания соответствующих праймеров, находящихся на сайтах амплификации проточной ячейки. Первая смесь реагентов состоит из различных реакционноспособных компонентов, которые включают, без ограничений, НТФ и один или более ферментов репликации. Один или более ферментов репликации может включать полимеразу, рекомбиназу, геликазу и/или подобный фермент. Кроме НТФ и одного или более ферментов репликации смесь реагентов может включать дополнительные реакционноспособные компоненты, такие как праймеры, одноцепочечные связывающие белки, буферы для жидкостного транспорта (например, воду, поверхностно-активное вещество и т.д.), агент фазового исключения, магний и/или подобные вещества. В одном из примеров первую смесь реагентов смешивают с нуклеиновыми кислотами-мишенями в смесительном резервуаре или резервуаре-хранилище с помощью распределителя реагентов, который соединен посредством гидравлического взаимодействия с проточной ячейкой. Нуклеиновые кислоты-мишени свободно перемещаются в целевом растворе.
При выполнении этапа 104 целевой раствор пропускают по массиву сайтов амплификации, находящихся на проточной ячейке, для образования клональных популяций ампликонов на проточной ячейке. Пропускание целевого раствора по сайтам амплификации приводит к образованию клональных популяций ампликонов на сайтах амплификации в результате амплификации с применением кинетического исключения, рассмотренной в настоящем описании. В настоящем описании клональные популяции ампликонов также называются клональными кластерами и генетическими кластерами. В одном из примеров целевой раствор пропускают из резервуара для смешивания распределителя реагентов в проточную ячейку, и втекающий через впускное отверстие проточной ячейки раствор вступает в контакт с массивом сайтов амплификации.
Сайты амплификации могут быть локализованы на структурных элементах поверхности проточной ячейки. В одном из примеров структурные элементы представляют собой вогнутые лунки на поверхности, но в других примерах они могут представлять собой другие элементы, такие как гранулы. Лунки отделены друг от друга промежуточными участками поверхности проточной ячейки. В одном из примеров перед пропусканием целевого раствора по сайтам амплификации проточную ячейку подготавливают, присоединяя праймеры Р5 и Р7 у поверхности проточной ячейки, находящейся на сайтах амплификации. В одном из примеров праймеры могут быть присоединены к проточной ячейке посредством нанесения на структурированную проточную ячейку покрытия из не содержащего силана акриламида (SFA) и последующей прививкой праймеров к полимеризованному SFA через расщепляемый под действием УФ света нитробензильный фрагмент. Праймеры необязательно могут быть направлены к сайтам амплификации и от промежуточных участков с помощью переноса с приложением е-поля. В альтернативном варианте праймеры могут быть привиты как к лункам, так и к промежуточным участкам, но праймеры на промежуточных участках удаляются под действием УФ облучения, шлифовки или другого способа, позволяющего удалять праймеры с промежуточных участков, не удаляя праймеры, находящиеся в лунках. Подготовленная проточная ячейка включает газон праймеров на каждом из сайтов амплификации (например, в каждой лунке) и содержит лишь небольшое количество праймеров, если они все же имеются, на промежуточных участках.
Целевой раствор может быть пропущен через сайты амплификации с помощью одного или более насосов, которые перекачивают целевой раствор к проточной ячейке, и для этого требуется источник энергии. В другом примере целевой раствор может самопроизвольно течь через сайты амплификации, и после открытия одного или более клапанов целевой раствор под действием силы тяжести и/или диффузии перемещается через сайты амплификации. Нуклеиновые кислоты-мишени свободно перемещаются в целевом растворе по мере его протекания через сайты амплификации. Совокупность нуклеиновых кислот-мишеней в целевом растворе превышает количество расположенных в проточной ячейке сайтов амплификации, таких как лунки.
В этапе 106 целевой раствор инкубируют на проточной ячейке. Во время инкубации целевого раствора, раствор контактирует с массивом сайтов амплификации, находящихся на проточной ячейке, в течение заданного периода времени и в заданных условиях (например, при заданных температуре, давлении, влажности и т.д.). Например, инкубация целевого раствора на проточной ячейке может происходить при температуре, составляющей от приблизительно 20 градусов Цельсия до приблизительно 50 градусов Цельсия, например, от приблизительно 30 до приблизительно 42 градусов Цельсия, в течение времени, составляющего приблизительно 30 минут до приблизительно 90 минут, например, приблизительно 40 минут или приблизительно 60 минут. В альтернативных примерах условия инкубации могут быть другими.
При проведении инкубации целевой раствор реагирует в проточной ячейке с образованием клональных популяций ампликонов на сайтах амплификации. Клональные популяции ампликонов образуются из соответствующих нуклеиновых кислот-мишеней. Например, нуклеиновые кислоты-мишени связываются с праймерами, присоединенными к сайтам амплификации, посредством гибридизации между концами адаптера и праймерами, то приводит к присоединению нуклеиновых кислот-мишеней к проточной ячейке. Нуклеиновые кислоты-мишени транспортируются к сайтам амплификации и связываются с праймерами со скоростью переноса. Скоростью переноса называют скорость внесения нуклеиновой кислоты-мишени на сайт амплификации, такой как лунка. Скорость переноса может зависеть от концентрации нуклеиновых кислот-мишеней в целевом растворе, который пропускают через сайты амплификации. Например, более высокая концентрация нуклеиновых кислот-мишеней в целевом растворе может повысить скорость переноса по сравнению со скоростью переноса, наблюдаемой при более низкой концентрации нуклеиновых кислот-мишеней в целевом растворе. Скорость переноса можно регулировать, регулируя концентрацию нуклеиновых кислот-мишеней в целевом растворе. Скорость переноса также можно регулировать, регулируя вязкость целевого раствора, регулируя средний размер нуклеиновых кислот-мишеней и/или добавлением или отсутствием реагента фазового исключения в целевом растворе. Например, скорость переноса может быть снижена при повышении вязкости целевого раствора, повышении среднего размера или длины цепочек нуклеиновых кислот-мишеней и/или при добавлении реагента фазового исключения (который затрудняет свободно перемещающимся нуклеиновым кислотам возможность перемещаться к праймерам и связываться с праймерами, находящимися на сайтах амплификации). Скорость переноса может быть увеличена за счет снижения вязкости целевого раствора, уменьшения среднего размера нуклеиновых кислот-мишеней и/или в отсутствие реагента фазового исключения.
Присутствие смеси реагентов в целевом растворе позволяет производить одновременную транспортировку нуклеиновых кислот-мишеней к сайтам амплификации и амплификацию нуклеиновых кислот-мишеней, уже связанных с сайтами амплификации. Например, на основе нуклеотидов НТФ ферменты производят ампликоны, которые являются копиями нуклеиновых кислот-мишеней. Ампликоны связываются с сайтами амплификации. Ампликоны образуются со скоростью амплификации, которую можно регулировать в зависимости от концентраций реакционноспособных компонентов в первой смеси реагентов, например, концентраций НТФ и одного или более ферментов репликации. Например, увеличение концентраций полимеразы и рекомбиназы может привести к повышению скорости амплификации по сравнению со скоростью амплификации при более низких концентрациях полимеразы и рекомбиназы. На скорость амплификации также может влиять изменение температуры сайтов амплификации и/или изменение свойств праймеров, находящихся на сайтах амплификации. Например, для изменения скорости амплификации может быть модифицирована или подходящим образом выбрана последовательность, длина и/или тип праймеров.
Для проведения амплификации с применением кинетического исключения скорость амплификации регулируют таким образом, чтобы она превышала скорость переноса. Например, скорость амплификации может значительно превышать скорость переноса, и после связывания первой нуклеиновой кислоты-мишени с одним праймером, находящимся на сайте амплификации, на других праймерах, находящихся на сайте амплификации, множество ампликонов первой нуклеиновой кислоты-мишени образуется еще до того, как другие нуклеиновые кислоты-мишени связываются с праймерами того же сайта амплификации. В результате на сайте амплификации образуется клональная популяция (или кластер) ампликонов первой нуклеиновой кислоты-мишени. На различных сайтах амплификации находятся клональные популяции ампликонов, полученных из разных нуклеиновых кислот-мишеней, то есть клональная популяция ампликонов на втором сайте амплификации получена из второй нуклеиновой кислоты-мишени, отличающейся от первой нуклеиновой кислоты-мишени. Таким образом, одна лунка проточной ячейки может быть заполнена ампликонами первой нуклеиновой кислоты-мишени, а вторая лунка может быть заполнена ампликонами второй нуклеиновой кислоты-мишени, находившейся в целевом растворе. Скорость амплификации превышает скорость переноса на определенную величину или значение, поэтому последующие нуклеиновые кислоты-мишени, подходящие к определенному сайту амплификации после попадания на сайт первой нуклеиновой кислоты-мишени, подвергаются кинетическому исключению и не закрепляются на этом сайте.
Пространственно разделенные клональные кластеры ампликонов применяют для генетического секвенирования. Например, при секвенировании синтезом цепочки ампликонов могут быть применены в качестве матриц, к которым присоединяются нуклеотиды, меченные флуоресцентной меткой. При возбуждении нуклеотиды, меченные флуоресцентной меткой, испускают характеристический сигнал, который используют для определения последовательности нуклеиновых кислот-мишеней.
В этапе 108 целевой раствор удаляют из проточной ячейки. Например, целевой раствор удаляют из проточной ячейки таким образом, что на поверхности проточной ячейки не остается свободно перемещающихся нуклеиновых кислот-мишеней, не связанных с поверхностью. Таким образом, нуклеиновые кислоты-мишени, находящиеся в целевом растворе, которые не были гибридизованы с одним из праймеров, находящихся на поверхности проточной ячейки, удаляют промывкой из проточной ячейки. Целевой раствор может быть удален откачиванием из проточной ячейки, промывкой проточной ячейки нейтральным раствором для удаления целевого раствора из проточной ячейки, сливом целевого раствора из проточной ячейки под действием силы тяжести и/или подобным способом. Клональные кластеры ампликонов и нуклеиновые кислоты-мишени, связанные с сайтами амплификации проточной ячейки остаются на проточной ячейке после удаления целевого раствора.
В одном из примеров для повышения количества ампликонов в клональных кластерах, находящихся на сайтах амплификации в проточной ячейке, этап вторичной амплификации с исключением выполняют после этапа первоначальной амплификации с исключением. Большее количество ампликонов в кластерах на сайтах амплификации обеспечивает сигналы большей интенсивности и улучшает соотношение сигнал/шум при проведении секвенирования синтезом, чем в случае кластеров, содержащих меньшее количество ампликонов. Например, интенсивность сигнала может повышаться при увеличении количества ампликонов в кластерах, но при этом фоновый шум остается постоянным, что приводит к более высокому соотношению сигнал/шум. Более высокое соотношение сигнал/шум позволяет производить более точные определения и повышает эффективность секвенирования, снижая вероятность ошибок на базовом уровне.
В этапе 110 смешивают вторую смесь реагентов, получая второй раствор, не содержащий целевых компонентов, в котором отсутствуют добавочные нуклеиновые кислоты-мишени. В одном из примеров раствор, не содержащий целевых компонентов, не содержит нуклеиновых кислот-мишеней. Например, вторая смесь реагентов может быть смешана во втором смесительном резервуаре, который отличается от смесительного резервуара, применяемого для смешивания первой смеси реагентов с нуклеиновыми кислотами-мишенями с образованием целевого раствора. Несмотря на то, что этот этап 110 смешивания показан в способе 100 после удаления целевого раствора из проточной ячейки, вторая смесь реагентов необязательно может быть смешана до удаления целевого раствора из проточной ячейки, например, она может быть смешана одновременно со смешиванием первой смеси реагентов и нуклеиновых кислот-мишеней с образованием целевого раствора, во время инкубации целевого раствора на проточной ячейке или в подобный момент времени. В одном из примеров раствор, не содержащий целевых компонентов, представляет собой свежий раствор, который не включает рециркулируемой части целевого раствора, которую уже пропускали через сайты амплификации.
В альтернативном примере раствор, не содержащий целевых компонентов, может включать ненулевое следовое количество нуклеиновых кислот-мишеней. Например, если первый целевой раствор направляют из смесительного резервуара в проточную ячейку, остаточное количество целевого раствора может оставаться в смесительном резервуаре и не поступать в проточную ячейку. Кроме того, вторая смесь реагентов затем может быть смешана в том же смесительном резервуаре, в результате чего остаточное количество целевого раствора смешивается со второй смесью реагентов с образованием раствора, не содержащего целевых компонентов. Полученный раствор, не содержащий целевых компонентов, может содержать следовые количества нуклеиновых кислот-мишеней, поступающих из остаточного количества целевого раствора, находившегося в смесительном резервуаре. Концентрация нуклеиновых кислот-мишеней в растворе, не содержащем целевых компонентов, составляет менее 100 частей на миллион, что может быть достаточно мало, чтобы реакция раствора, не содержащего целевых компонентов, с сайтами амплификации, не приводила к образованию дополнительных ампликонов.
Вторая смесь реагентов в растворе, не содержащем целевых компонентов, может включать те же самые реакционноспособные компоненты, что и первая смесь реагентов в целевом растворе, или другие реакционноспособные компоненты. Например, вторая смесь реагентов может включать НТФ тех же типов и ферменты репликации тех же типов, что и первая смесь реагентов. Следует отметить, что несмотря на то, что по меньшей мере некоторые из одних и тех же реакционноспособных компонентов могут быть добавлены в оба раствора, вторая смесь реагентов включает свежие реакционноспособные компоненты. Например, после протекания первого целевого раствора через проточную ячейку, его не рециркулируют, даже частично, в смесительный резервуар для получения раствора, не содержащего целевых компонентов.
Реакционноспособные компоненты, находящиеся во второй смеси реагентов, могут включать один или более из следующих реагентов: НТФ, полимеразу, рекомбиназу, геликазу, одноцепочечный связывающий белок, агенты фазового исключения, буферы и/или подобные вещества. Вторая смесь реагентов может включать количества и/или концентрации реакционноспособных компонентов, отличающиеся от их количеств в первой смеси реагентов, содержащейся в целевом растворе. Например, для повышения скорости репликации, с которой происходит образование ампликонов на сайтах амплификации, вторая смесь реагентов может включать большую концентрацию ферментов репликации, таких как полимераза, рекомбиназа и/или геликаза, по сравнению с первой смесью реагентов. В другом примере вторая смесь реагентов может включать большее количество буферного компонента (например, воды, поверхностно-активного вещества или смеси воды и поверхностно-активного вещества) по сравнению с первой смесью реагентов, что компенсирует отсутствие нуклеиновых кислот-мишеней, добавляемых во вторую смесь реагентов. Вторая смесь реагентов необязательно может включать один или более типов реакционноспособных компонентов, отличающихся от компонентов первой смеси реагентов, таких как различные белки, праймеры или подобные соединения. Например, вторая смесь реагентов может включать праймеры, которые отсутствуют в первой смеси реагентов. Праймеры могут представлять собой праймеры Р5, содержащие последовательность Р5 праймера, и/или праймеры, содержащие последовательность Р7 праймера.
В этапе 112 раствор, не содержащий целевых компонентов, пропускают по массиву сайтов амплификации, находящихся на проточной ячейке. Раствор, не содержащий целевых компонентов, может быть перекачан или выпущен из соответствующего резервуара, содержащего раствор, не включающий целевых компонентов, в проточную ячейку. Как было указано выше, в растворе, не содержащем целевых компонентов, могут полностью отсутствовать нуклеиновые кислоты-мишени, или в альтернативном варианте он может включать только следовые количества нуклеиновых кислот-мишеней, обусловленные наличием остаточного количества целевого раствора в смесительном резервуаре, применяемом для приготовления раствора, не содержащего целевых компонентов. Поскольку раствор, не содержащий целевых компонентов, не включает количества нуклеиновых кислот-мишеней, превышающие следовые, пропускание раствора, не содержащего целевых компонентов, через сайты амплификации не приводит к дополнительному внесению существенного количества нуклеиновых кислот-мишеней на сайты амплификации. В частности, с праймерами, находящимися на сайтах амплификации, не связываются дополнительные нуклеиновые кислоты-мишени. Нуклеиновые кислоты-мишени генетической библиотеки связываются с праймерами на сайтах амплификации обычно только при пропускании через проточную ячейку целевого раствора.
В этапе 114 раствор, не содержащий целевых компонентов, инкубируют на проточной ячейке. Раствор, не содержащий целевых компонентов, может быть оставлен контактировать с массивом сайтов амплификации, находящихся на проточной ячейке, в течение заданного периода времени в заданных условиях (например, при заданных температуре, влажности, давлении и т.д.). Период времени и/или условия могут быть аналогичными или идентичными периоду времени и/или условиям инкубации целевого раствора. В одном из примеров раствор, не содержащий целевых компонентов, может быть подвергнут инкубации на проточной ячейке при температуре, составляющей приблизительно 38 градусов Цельсия, в течение времени, составляющего от приблизительно 20 до приблизительно 60 минут, хотя в других примерах условия инкубации могут быть другими.
Инкубация раствора, не содержащего целевых компонентов, на проточной ячейке повышает количество ампликонов в клональных популяциях или кластерах на сайтах амплификации, обеспечивая более полное образование кластеров амплификации. Например, в течение первоначальной реакции амплификации с исключением нуклеиновые кислоты-мишени и полученные ампликоны могут связываться с некоторыми, но не со всеми праймерами на каждом сайте амплификации. Таким образом, на сайтах амплификации могут находиться праймеры, не связанные ни с нуклеиновой кислотой-мишенью, ни с ампликоном, которые готовы к связыванию. Такие праймеры образуют на каждом сайте амплификации подгруппу доступных праймеров. Смесь реагентов в растворе, не содержащем целевых компонентов, образует дополнительные ампликоны на несвязанных праймерах, что приводит к снижению количества праймеров в подгруппе доступных праймеров. Например, вторая смесь реагентов реагирует с существующими ампликонами клонального кластера, образуя новые ампликоны, которые связываются с доступными праймерами.
Вместо или наряду с образованием ампликонов на уже существующих доступных праймерах, находящихся на сайтах амплификации, вторая смесь реагентов может включать дополнительные праймеры. В одном из примеров вторая смесь реагентов включает праймер, такой как праймер Р5. Перед пропусканием раствора, не содержащего целевых компонентов, по проточной ячейке, уже существующие на сайтах амплификации праймеры Р5 (включающие цепочки Р5 ампликонов в кластерах) подвергают линеаризации и удаляют, в результате чего в кластерах остаются в основном цепочки Р7 ампликонов и оставшиеся доступные (например, неиспользованные) связанные с поверхностью праймеры Р7. Поскольку раствор, не содержащий целевых компонентов, включает праймеры Р5 из второй смеси реагентов, то пропускание раствора, не содержащего целевых компонентов, по проточной ячейке вызывает дополнительную амплификацию кластеров на доступных связанных с поверхностью праймерах Р7. Некоторые из поверхностных праймеров Р7 становятся доступными после протекания первоначальной амплификации с исключением, поскольку цепочки кластеров ДНК должны образовать мостик, так чтобы оба их конца оставались связанными с поверхностью. Стерические затруднения могут ограничивать доступ ко всем доступным имеющимся на поверхности праймерам. После удаления одного из праймеров (например, праймера Р5) с поверхности после проведения этапа первоначальной амплификации с исключением с использованием первого целевого раствора, повышается доступность другого праймера (например, праймера Р7), остающегося на поверхности. Кроме того, один конец цепочки кластера ампликонов освобождается и, не будучи связан с поверхностью, пригоден для протекания амплификации.
В другом примере вторая смесь реагентов, не содержащая дополнительных праймеров, может вызывать вторичную амплификацию. Например, связанные с поверхностью праймеры Р5 могут быть линеаризованы и удалены с сайтов амплификации перед пропусканием раствора, не содержащего целевых компонентов, но при этом не будет происходить денатурации кластеров ампликонов, то есть цепочки Р5 кластеров остаются связанными с поверхностью. Раствор, не содержащий целевых компонентов, пропускаемый через сайты амплификации в таком состоянии, может вызывать образование большего числа копий цепочек Р5 вследствие вытеснения цепи с участием неиспользованных праймеров Р7 в соответствии с механизмом линейной амплификации. В этом примере вторичная амплификация может протекать за счет того, что неиспользованные праймеры Р7 становятся более доступными в результате линеаризации одного из концов кластеров.
Необязательно, вторая смесь реагентов включает праймеры, которые присоединяются к поверхности проточной ячейки образом, сходным со вторым этапом прививки, в особенности, в стабильных условиях, подходящих для прививки. Например, связанные с BCN (сокр. от "bicylononyne", рус. бициклононин) Р5/Р7 могут быть привиты к PAZAM в водной среде или в буфере с низким содержанием солей. Второй этап прививки может быть выполнен после пропускания первого целевого раствора и перед пропусканием второго раствора, не содержащего целевых компонентов. В альтернативном варианте праймеры для второго этапа прививки могут быть смешаны со второй смесью реагентов, при условии, что прививка совместима со второй смесью реагентов. Праймеры, находящиеся в растворе, не содержащем целевых компонентов, повышают общее число праймеров на сайтах амплификации и обеспечивают новые участки, на которых могут быть образованы дополнительные ампликоны, что увеличивает количество ампликонов в кластерах.
Поскольку при получении раствора, не содержащего целевых компонентов, не добавляют дополнительных нуклеиновых кислот-мишеней, количество ампликонов в кластерах может быть повышено без добавления дополнительного генетического материала, получение которого может быть относительно сложным и/или дорогостоящим по сравнению с реакционноспособными компонентами.
В этапе 116 второй раствор, не содержащий целевых компонентов, удаляют из проточной ячейки перед выполнением последующих этапов секвенирования в проточной ячейке для секвенирования генетической библиотеки. Например, раствор, не содержащий целевых компонентов, может быть откачан из проточной ячейки, вымыт из проточной ячейки нейтральным раствором или выпущен из проточной ячейки.
Необязательно, и целевой раствор, и растворы, не содержащие целевых компонентов, пропускают через сайты амплификации в изотермических условиях, при которых ни растворы, ни проточная ячейка не подвергаются активному нагреванию. Кроме того, клональные кластеры образуются без применения химических денатурирующих веществ, способных вызывать денатурацию нуклеиновых кислот-мишеней и ампликонов на проточной ячейке. Таким образом, в отличие от некоторых других методик амплификации, таких как мостиковая амплификация, методики RCA и методики MDA, в способе 100 повышение выхода и/или качества нуклеотидных цепочек в клональных кластерах, находящихся на проточной ячейке, возможно без применения нагревания или химических денатурирующих веществ.
В альтернативном примере способа 100 генерации клональных кластеров перед введением в проточную ячейку второго раствора, не содержащего целевых компонентов, первый целевой раствор не удаляют из проточной ячейки. Таким образом, в способе 100 может отсутствовать этап 108, направленный на удаление целевого раствора из проточной ячейки. Напротив, после инкубации целевого раствора в проточной ячейке в течение первого периода времени в этапе 106, при выполнении этапа 112 раствор, не содержащий целевых компонентов, пропускают по массиву сайтов амплификации, находящихся на проточной ячейке, пока целевой раствор еще находится в проточной ячейке. Например, целевой раствор может быть подвергнут инкубации в проточной ячейке в течение приблизительно 20 минут, приблизительно 40 минут, приблизительно 60 минут или аналогичного периода перед введением в проточную ячейку раствора, не содержащего целевых компонентов. Как только при выполнении этапа 112 раствор, не содержащий целевых компонентов, пропускают по массиву сайтов амплификации, раствор, не содержащий целевых компонентов, смешивается с целевым раствором, находящимся в проточной ячейке. Вторая смесь реагентов, содержащаяся растворе, не содержащем целевых компонентов, повышает общую концентрацию реагентов в проточной ячейке, что может приводить к повышению скорости амплификации, т.е. скорости образования ампликонов. В этапе 114 раствор, не содержащий целевых компонентов, инкубируют на проточной ячейке в течение второго периода времени в присутствии целевого раствора. Таким образом, целевой раствор инкубируют в проточной ячейке в течение времени, которое в целом представляет собой сумму первого периода времени этапа 106 и второго периода времени этапа 114. В этапе 116 объединенный раствор, состоящий из первого целевого раствора и второго раствора, не содержащего целевых компонентов, удаляют из проточной ячейки, например, откачивая, промывая нейтральным раствором, сливая или аналогичным способом, и затем в проточной ячейке выполняют последующие этапы секвенирования.
На Фиг. 12 представлена гистограмма 200, на которой показана полученная в одном из примеров интенсивность сигнала генетических кластеров, зарегистрированного на проточной ячейке в различных вариантах генерации кластеров. На гистограмме 200 представлено сравнение интенсивностей сигнала в различных вариантах генерации кластеров в аналогичных условиях окружающей среды. Сравнение представлено на гистограмме 200 в относительных единицах и не отражает реальных результатов эксперимента. По оси 202 у представлена интенсивность сигнала кластеров, измеренная при проведении секвенирования синтезом после возбуждения кластеров, содержащих меченные флуоресцентными метками нуклеотиды, в диапазоне от 200 до 550. Произведено сравнение трех различных способов генерации кластеров, которые представлены на гистограмме 200 соответствующими столбиками. Столбиком 204 представлена амплификация с применением кинетического исключения (ExAmp) без проведения дополнительных этапов амплификации с целью повышения количества ампликонов в кластерах. Столбиком 206 представлена амплификация с применением кинетического исключения с последующей традиционной мостиковой амплификацией. Столбиком 208 представлена амплификация с применением кинетического исключения согласно примерам, рассмотренным в настоящем описании, в которых за первым этапом амплификации с применением кинетического исключения следует вторичный этап амплификации с применением кинетического исключения в присутствии раствора, не содержащего целевых компонентов, который не содержит нуклеиновых кислот-мишеней.
Как показано на Фиг. 12, амплификация с исключением с последующим проведением вторичного этапа амплификации с исключением (например, столбик 208) обладает наибольшей интенсивностью, меньшую интенсивность имеет мостиковая амплификация (например, столбик 206), и наименьшую интенсивность имеет амплификация с исключением без проведения вторичной амплификации (например, столбик 204). Например, если в одном этапе амплификации с применением кинетического исключения, иллюстрируемом столбиком 204, образуются генетические кластеры, интенсивность сигнала которых составляет приблизительно 285, то амплификация с применением кинетического исключения с последующим проведением мостиковой амплификации, как показано столбиком 206, приводит к образованию генетических кластеров, интенсивность сигнала которых составляет приблизительно 320, и амплификация с применением кинетического исключения с последующим проведением второго этапа амплификации с исключением, как показано столбиком 208, приводит к образованию генетических кластеров, интенсивность сигнала которых составляет приблизительно 420. Более высокая интенсивность сигнала указывает на то, что амплификация с исключением с последующим проведением вторичного этапа амплификации с исключением обеспечивает более высокое отношение сигнал/шум, чем другие два способа. Более высокое отношение сигнал/шум может повышать точность и/или эффективность секвенирования и анализа генетической библиотеки по сравнению с другими двумя способами.
На Фиг. 13 представлена гистограмма 300, на которой показаны полученные в одном из примеров значения процентного прохождения фильтра (% PF) в способах генерации кластеров, которые применяли для получения гистограммы 200. На гистограмме 300 представлено сравнение значений % PF, получаемых в различных вариантах генерации кластеров, которые выполняют в аналогичных условиях окружающей среды. Сравнение представлено на гистограмме 300 в относительных единицах и не отражает реальных результатов эксперимента. По оси 302 у представлена процентная доля кластеров, полученных каждым из названных трех способов, которые соответствуют установленным условиям чистоты фильтра, указывая на то, что эти кластеры имеют качество, достаточное для применения в секвенировании; диапазон полученных значений соответствует 60%-78%. Столбиком 304 представлена амплификация с применением кинетического исключения без проведения вторичного этапа амплификации, столбиком 306 представлена амплификация с применением кинетического исключения с последующим проведением мостиковой амплификации, и столбиком 308 представлена амплификация с применением кинетического исключения с последующим проведением вторичного этапа амплификации с применением кинетического исключения. Как показано на Фиг. 13, если в одном этапе амплификации с применением кинетического исключения, как показано столбиком 304, образуются генетические кластеры, приблизительно 65% которых проходят через фильтр, то в амплификации с применением кинетического исключения с последующим проведением мостиковой амплификации, показанной столбиком 306, образуются генетические кластеры, приблизительно 71% которых проходят через фильтр, и в амплификации с применением кинетического исключения с последующим проведением второго этапа амплификации с исключением, показанной столбиком 308, образуются генетические кластеры, приблизительно 73% которых проходят через фильтр. Таким образом, амплификация с исключением с последующим проведением вторичного этапа амплификации с исключением с использованием раствора, не содержащего целевых веществ, может представлять собой вариант, обеспечивающий наибольший выход качественных кластеров по сравнению с другими вариантами образования кластеров в аналогичных условиях окружающей среды.
На Фиг. 14 представлена схема жидкостной системы 400 для генерации клональных кластеров согласно одному из примеров осуществления изобретения. Жидкостная система 400 может быть применена для осуществления способа 100. Жидкостная система 400 может быть аналогична жидкостным системам, рассмотренным в патенте US 9410977, содержание которого полностью включено в настоящую работу посредством ссылки. В показанном примере жидкостная система 400 включает структурированную проточную ячейку 402, распределитель 404 реагентов, лоток или картридж 406 для реагентов, контроллер 408 и насос 410. В альтернативном варианте жидкостная система 400 может включать дополнительные компоненты, меньшее число компонентов и/или по меньшей мере один компонент, отличающийся от показанных в примере.
Проточная ячейка 402 включает совокупность дорожек 412, пролегающих между впускным отверстием 414 и выпускным отверстием 416 и соединенных посредством гидравлического взаимодействия с впускным отверстием 414 и выпускным отверстием 416 проточной ячейки 402. На поверхностях дорожек 412 могут быть расположены лунки или другие элементы, ограничивающие сайты амплификации, на которых происходит генерация клональных популяций или кластеров ампликонов. Распределитель 404 реагентов включает клапан 418, находящийся в гидравлическом взаимодействии (например, соединенный посредством гидравлического взаимодействия) с впускным отверстием 414 проточной ячейки 402. Клапан 418 может включать или быть соединенным с по меньшей мере одним смесительным резервуаром (также называемым резервуаром-хранилищем), который не показан на Фиг. 14. Распределитель 404 реагентов дополнительно включает совокупность каналов 420 и отверстия 422. Каналы 420 проходят между отверстиями 422 и клапаном 418 и соединяют посредством гидравлического взаимодействия отверстия 422 с клапаном 418 (и смесительные резервуары, связанные с клапаном 418). В других примерах распределитель 404 реагентов может включать совокупность клапанов 418. Несмотря на то, что это не показано, распределитель 404 реагентов может включать шприцы сиппера, расположенные в соответствующих отверстиях 422.
Лоток 406 для реагентов включает совокупность резервуаров для реагентов (не показаны), которые содержат различные реагенты. Распределитель 404 реагентов устанавливают над лотком 406 для реагентов, совмещая шприцы сиппера с резервуарами для реагентов таким образом, чтобы шприцы сиппера входили в соответствующие резервуары для реагентов и контактировали с реагентами, находящимися внутри резервуаров. С помощью шприцев сиппера реагенты извлекают из резервуаров для реагентов лотка 406, откуда реагенты направляют в проточную ячейку 402 через каналы 420 и клапан 418.
Контроллер 408 включает один или более процессоров (не показаны) или другие логические устройства, которые выполняют операции на основании инструкций, хранящихся на подходящем для компьютерного считывания материальном носителе информации или памяти для долговременного хранения информации (не показаны). Дополнительно или в альтернативном варианте контроллер 408 может включать одно или более проводных (жестко смонтированных) устройств, выполняющих операции, основываясь на жесткой логике устройств. Контроллер 408 может представлять собой технические средства, работающие на программном обеспечении или жестких инструкциях, программном обеспечении, которое дает команды техническим средствам выполнять определенные операции, или на комбинации таких программ. Контроллер 408 соединен с возможностью связи (например, через одно или более проводных или беспроводных соединений) с насосом 410 и клапаном 418. Например, контроллер 408 может управлять работой насоса 410 и клапана 418, передавая сигналы управления по проводным или беспроводным соединениям к соответствующим устройствам. Контроллер 408 может управлять клапаном 418, положения которого позволяют текучим средам течь через клапан 418 в каналы 420 и в проточную ячейку 402. Контроллер 408 управляет насосом 410, регулируя разность давлений, создаваемую насосом 410 для перекачивания текучих сред по жидкостной системе 400. В показанном примере насос 410 соединен рабочим соединением с проточной ячейкой 402; однако, в альтернативном варианте насос 410 может быть соединен с распределителем 404 реагентов.
В одном из примеров контроллер 408 управляет насосом 410 и клапаном 418, с помощью которых производят смешивание первого целевого раствора и его перемещение через впускное отверстие 414 по массиву сайтов амплификации, находящихся на проточной ячейке 402. Целевой раствор включает нуклеиновые кислоты-мишени и первую смесь реагентов. Нуклеиновые кислоты-мишени могут быть смешаны с первой смесью реагентов в распределителе 404 реагентов, например, в одном из смесительных резервуаров (не показаны), связанных с клапаном 418. Например, первый канал 420А совокупности каналов 420 может содержать матрицу генетического образца, который включает нуклеиновые кислоты-мишени, находящиеся в буфере, таком как вода или другой растворитель. Анализируемая матрица может быть извлечена из лотка 406 для реагентов шприцем сиппера, или в альтернативном варианте она может поступать в канал 420А из другого источника, не являющегося лотком 406. Второй канал 420В совокупности каналов 420 может содержать один или более реакционноспособных компонентов первой смеси реагентов, такой как НТФ, полимераза, одноцепочечный связывающий белок, и один или более буферов (например, воду, поверхностно-активные вещества или подобные вещества). Третий канал 420С и четвертый канал 420D могут содержать другие реакционноспособные компоненты первой смеси реагентов, например, рекомбиназа может находиться в третьем канале 420С, и магний и агент фазового исключения могут находиться в четвертом канале 420D. Несмотря на то, что это не четко указано, все реакционноспособные компоненты могут быть растворены в растворителе, таком как вода или другой буфер. Раздельное хранение реакционноспособных компонентов до полной готовности целевого раствора к использованию может повышать срок годности смеси реагентов, поскольку некоторые из реакционноспособных компонентов могут оказаться нестабильными при смешивании с другими реакционноспособными компонентами.
В одном из примеров контроллер 408 управляет насосом 410 и клапаном 418, с помощью которых производят смешивание определенных требуемых количеств анализируемой матрицы с реакционноспособными компонентами, содержащимися в различных каналах 420A-D, в результате чего в распределителе 404 образуется целевой раствор, который затем пропускают через впускное отверстие 414 проточной ячейки 402. Например, компоненты целевого раствора могут быть смешаны в одном из смесительных резервуаров. В альтернативном варианте компоненты целевого раствора могут находиться отдельно друг от друга до тех пор, пока они не смешиваются во впускном трубопроводе (например, трубе, трубке или канале) 424, который соединяет клапан 418 со впускным отверстием 414, или в проточной ячейке 402.
Как было указано выше, целевой раствор реагирует на проточной ячейке 402 с образованием на сайтах амплификации клональных популяций ампликонов, получаемых из соответствующих нуклеиновых кислот-мишеней. При проведении амплификации с исключением нуклеиновые кислоты-мишени транспортируются к сайтам амплификации одновременно с протеканием амплификации нуклеиновых кислот-мишеней, уже связанных с сайтами амплификации, в процессе которой образуются ампликоны. Контроллер 408 может управлять насосом 410 и/или клапаном 418, регулируя концентрацию нуклеиновых кислот-мишеней и концентрацию первой смеси реагентов в целевом растворе таким образом, чтобы скорость амплификации превышала скорость переноса. Например, контроллер 408 может снижать концентрацию анализируемой матрицы и/или повышать концентрацию одного или более компонентов первой смеси реагентов для того, чтобы создаваемая скорость амплификации значительно превышала скорость переноса, что обеспечивает кинетическое исключение.
Контроллер 408 также может быть предназначен для регулирования условий инкубации целевого раствора на проточной ячейке 402, таких как температура, давление, влажность и подобные условия, создаваемые в проточной ячейке 402. По истечении заданного периода инкубации, контроллер 408 может подавать насосу 410 команду для удаления целевого раствора из проточной ячейки 402. Целевой раствор может вытекать через выпускное отверстие 416 в резервуар 426 для отходов. Необязательно, часть целевого раствора может быть подана рециклом через трубопровод 428 в распределитель 404 реагентов и/или лоток 406 для добавления свежей загрузки или подобного действия. Целевой раствор удаляют из проточной ячейки 402 таким образом, чтобы после удаления целевого раствора в проточной ячейке 402 отсутствовали свободно перемещающиеся нуклеиновые кислоты-мишени. Например, после удаления целевого раствора все нуклеиновые кислоты-мишени, присутствующие на сайтах амплификации, присоединены к праймерам, содержащимся в клональных кластерах.
В одном из примеров, после пропускания целевого раствора через проточную ячейку 402, контроллер 408 подает насосу 410 и/или клапану 418 команду, вызывающую смешивание и подачу раствора, не содержащего целевых компонентов, через впускное отверстие 414 по массиву сайтов амплификации, находящихся на проточной ячейке 402. Раствор, не содержащий целевых компонентов, включает вторую смесь реагентов и не содержит (например, полностью лишен) дополнительных нуклеиновых кислот-мишеней. Например, контроллер 408 может управлять смешиванием реакционноспособных компонентов во втором, третьем и четвертом каналах 420B-D, находящихся в смесительном резервуаре (не показан) распределителя 404, в результате чего образуется раствор, не содержащий целевых компонентов. Контроллер 408 управляет клапаном 418, который не допускает добавление анализируемой матрицы, находящейся в первом канале 420А, в смесительный резервуар, чтобы полученный раствор, не содержащий целевых компонентов, не включал дополнительных нуклеиновых кислот-мишеней. В одном из примеров вторую смесь реагентов смешивают в другом смесительном резервуаре, а не в смесительном резервуаре, применяемом для смешивания первой смеси реагентов с анализируемой матрицей для образования целевого раствора. В альтернативном примере вторую смесь реагентов смешивают в том же смесительном резервуаре, который ранее содержал целевой раствор, в результате чего в растворе, не содержащем целевых компонентов, может присутствовать остаточное количество целевого раствора (включающее следовые количества нуклеиновых кислот-мишеней). Таким образом, второй раствор, не содержащий целевых компонентов, либо не содержит нуклеиновых кислот-мишеней, либо включает следовые количества нуклеиновых кислот-мишеней, которые не вносятся дополнительно на сайты амплификации проточной ячейки 402.
Вторая смесь реагентов может отличаться от первой смеси реагентов типами, количествами и/или концентрациями реакционноспособных компонентов. Например, и первая смесь реагентов, и вторая смесь реагентов могут включать буферный компонент, такой как вода, поверхностно-активное вещество или раствор поверхностно-активного вещества в воде. Поскольку раствор, не содержащий целевых компонентов, не содержит анализируемой матрицы, контроллер 408 необязательно может включать в раствор, не содержащий целевых компонентов, большее количество буферного компонента по сравнению с количеством того же буферного компонента в первой смеси реагентов. Дополнительное количество буфера компенсирует снижение объема, обусловленное отсутствием анализируемой матрицы. В результате увеличения объема буфера объем раствора, не содержащего целевых компонентов, может быть близок или идентичен объему целевого раствора. Кроме того, регулирование количества буфера также влияет на концентрации реакционноспособных компонентов во второй смеси реагентов, и, таким образом, могут быть созданы концентрации, аналогичные или идентичные концентрациям реакционноспособных компонентов в целевом растворе. Необязательно, контроллер 408 может изменять по меньшей мере некоторые из следующих параметров: тип (например, различных ферментов, белков, олигонуклеотидов или подобных веществ), количества и/или концентрации реакционноспособных компонентов второй смеси реагентов по сравнению с первой смесью реагентов, что позволяет изменять скорость амплификации.
Контроллер 408 дает насосу 410 и/или клапану 418 команду подавать раствор, не содержащий целевых компонентов, в проточную ячейку 402. Раствор, не содержащий целевых компонентов, инкубируют на проточной ячейке 402, в которой раствор реагирует с ампликонами, находящимися на сайтах амплификации проточной ячейки 402, образуя дополнительные ампликоны в клональных кластерах. Как было указано выше, вторичный этап амплификации с исключением с использованием раствора, не содержащего целевых компонентов, приводит к достижению большего выхода кластеров удовлетворительного качества по сравнению со способом, не включающим пропускание раствора, не содержащего целевых компонентов, через проточную ячейку 402. Повышенный выход качественных кластеров может повышать точность и/или эффективность секвенирования, поскольку в этом случае в кластерах содержатся дополнительные нуклеотидные цепочки, генерирующие флуоресцентные сигналы в способе секвенирования синтезом. Кроме того, вторичный этап амплификации с исключением проводят без использования дополнительного количества анализируемой матрицы, которая включает нуклеиновые кислоты-мишени.
Дополнительные замечания
В настоящей работе упоминаются различные публикации, патенты и патентные заявки. Содержания этих публикаций полностью включены в настоящую работу для более полного описания текущего состояния области техники, к которой принадлежит предмет настоящего изобретения.
Термины "включают", "содержат" и т.д., а также их варианты, которые упоминаются в описании и пунктах формулы настоящего изобретения, имеют неисключающий характер и включают не только перечисленные элементы, но также включают любые дополнительные элементы. Упоминаемые в настоящей работе термины "один пример", "другой пример", "один из примеров" и т.д. означают, что определенный элемент (например, элемент, структура и/или характеристика), рассмотренный при описании примера, включен в по меньшей мере один пример, рассмотренный в настоящем описании, и может присутствовать или может отсутствовать в других примерах. Кроме того, следует понимать, что, если из контекста не следует иное, элементы, рассмотренные в любом из примеров, могут быть скомбинированы любым подходящим образом в различных примерах.
Следует понимать, что все комбинации приведенных выше признаков и дополнительных признаков, более подробно раскрытых ниже (при условии, что эти признаки не являются взаимоисключающими) составляют часть предмета изобретения, рассмотренного в настоящем описании. В частности, все комбинации раскрытых предметов изобретения, очевидные после прочтения предлагаемого описания, составляют часть предмета изобретения, рассмотренного в настоящем описании. Также следует понимать, что применяемая в настоящем описании терминология, которая также может употребляться в любом документе, включенном в настоящее описание посредством ссылки, должна иметь значение, в наибольшей мере соответствующее конкретным признакам, рассмотренным в настоящем описании.
Следует понимать, что приведенные в настоящем описании диапазоны включают указанные диапазоны и любые величины или поддиапазоны, находящиеся в пределах указанного диапазона. Например, следует понимать, что диапазон, составляющий от приблизительно 20 градусов Цельсия до приблизительно 50 градусов Цельсия, включает не только явным образом упомянутые пределы от приблизительно 20 градусов Цельсия до приблизительно 50 градусов Цельсия, но также включает индивидуальные величины, такие как приблизительно 28 градусов Цельсия, приблизительно 35 градусов Цельсия, приблизительно 46,5 градусов Цельсия и т.д., и поддиапазоны, такие как от приблизительно 25 градусов Цельсия до приблизительно 49 градусов Цельсия, от приблизительно 30 градусов Цельсия до приблизительно 40 градусов Цельсия и т.д. Кроме того, если для описания величины используют модификаторы "приблизительно" и/или "по существу", то они составляют небольшие вариации (до +/-10%) от означенной величины.
Несмотря на то, что некоторые примеры были описаны подробно, следует понимать, что рассмотренные примеры могут быть модифицированы. Таким образом, приведенное выше описание не является ограничивающим. Кроме того, несмотря на то, что предмет изобретения был рассмотрен с помощью приведенных выше примеров, следует понимать, что в примеры могут быть внесены различные изменения, не выходящие за пределы объема предмета изобретения. Соответственно, объем предмета изобретения ограничен только прилагаемыми пунктами формулы изобретения.
Реферат
Группа изобретений относится к способам и жидкостной системе для увеличения количества ампликонов в клональных популяциях ампликонов на сайтах амплификации. Проводят реакцию первого раствора и отличающегося от него второго раствора на проточной ячейке в жидкостной системе посредством протекания первого раствора по сайтам амплификации, находящимся на проточной ячейке, и затем посредством протекания второго раствора по сайтам амплификации. Первый раствор включает нуклеиновые кислоты-мишени и первую смесь реагентов, которая включает нуклеозидтрифосфаты и ферменты репликации. Находящиеся в первом растворе нуклеиновые кислоты-мишени переносятся к сайтам амплификации и связываются с ними со скоростью переноса. Под действием первой смеси реагентов происходит амплификация и образуются ампликоны со скоростью амплификации, которая превышает скорость переноса. Второй раствор включает вторую смесь реагентов и не содержит нуклеиновых кислот-мишеней. Второй раствор предназначен для увеличения количества ампликонов. Дополнительные ампликоны образуются посредством амплификации исключения со скоростью амплификации, которая превышает скорость переноса. Изобретения повышают количество ампликонов в клональных популяциях или кластерах на сайтах амплификации, обеспечивая более полное образование кластеров амплификации, а также обеспечивают результат со значительно большей интенсивностью и более высоким отношением сигнал/шум, повышая точность и/или эффективность секвенирования и анализа генетической библиотеки. 3 н. и 13 з.п. ф-лы, 14 ил., 3 пр.
Формула
Документы, цитированные в отчёте о поиске
Прямой захват, амплификация и секвенирование днк-мишени с использованием иммобилизированных праймеров
Комментарии