Растворимые антитела, содержащие только тяжелые цепи - RU2528737C2
Код документа: RU2528737C2
Чертежи

















































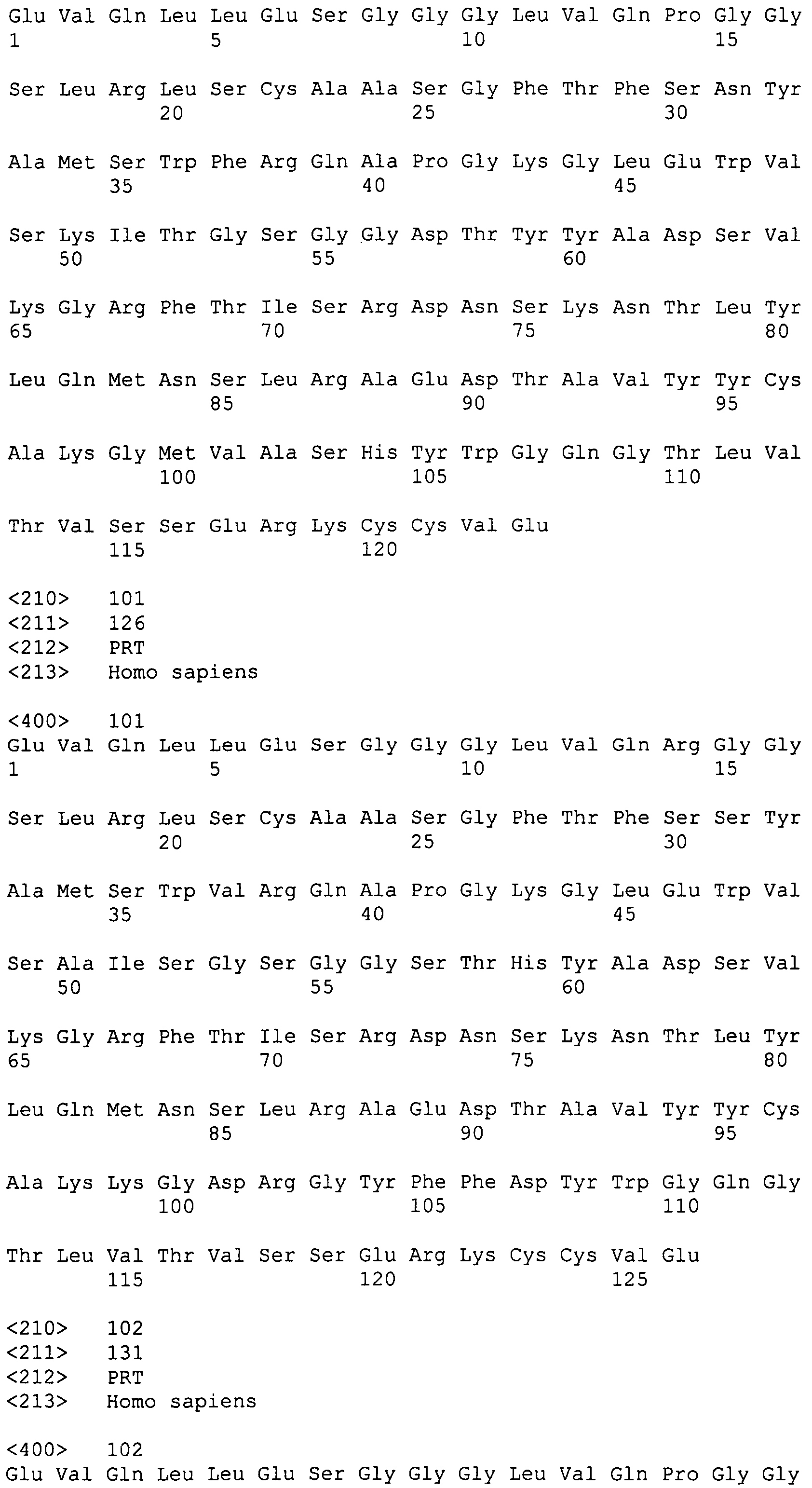
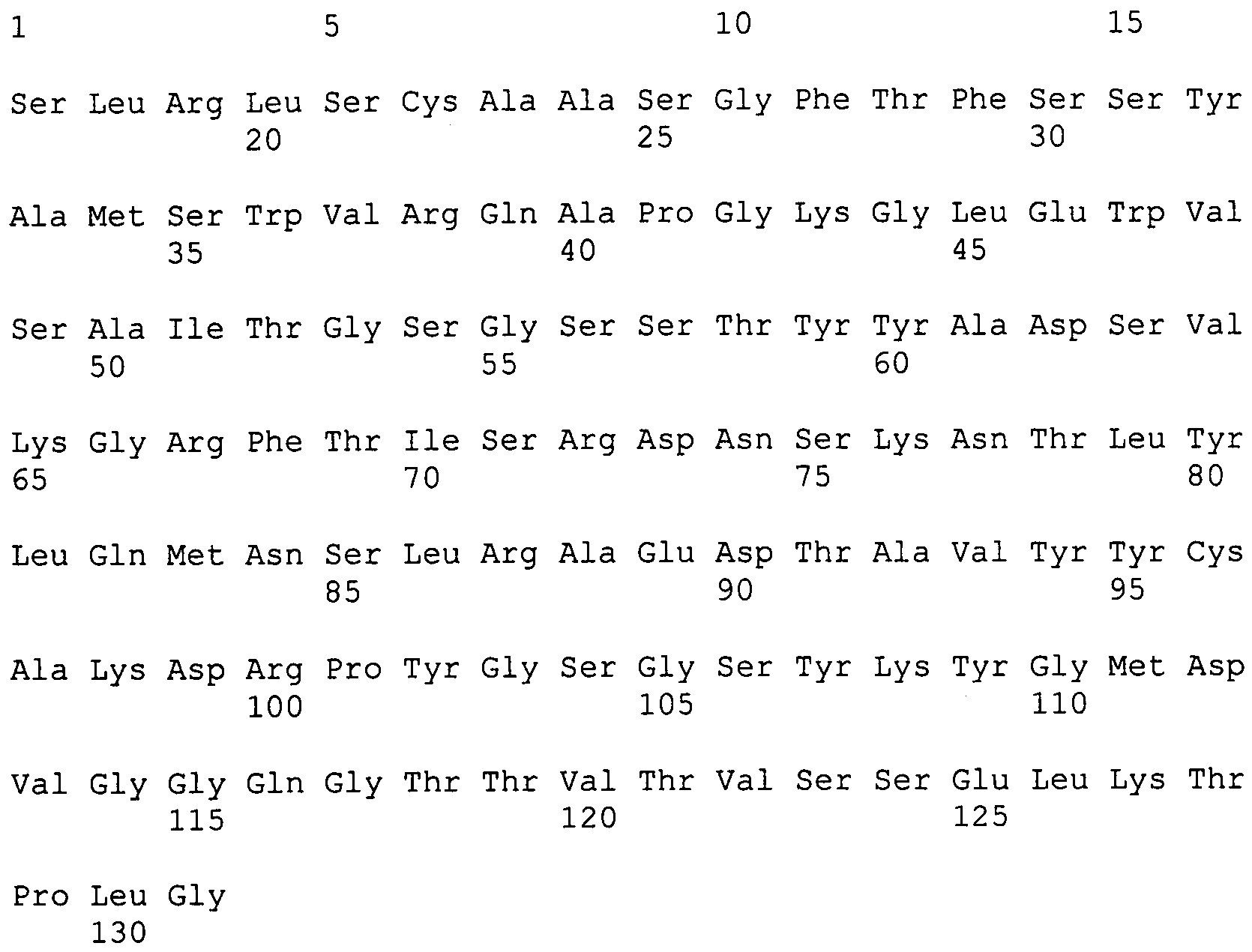
Описание
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к усовершенствованным способам выделения из трансгенных млекопитающих, не относящихся к человеку, широкого репертуара функциональных растворимых антител, содержащих только тяжелые цепи, и растворимых доменов VH, полученных из них, которые структурно отличны от доменов VH, полученных из антител, содержащих тяжелые и легкие цепи. Предпочтительно, антитела, содержащие только тяжелые цепи, и растворимые домены VH получают из долгоживущих плазматических клеток или В-клеток памяти.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Моноклональные антитела или их варианты составляют большую часть новых лекарственных средств, выпущенных в XXI веке. Терапия моноклональным антителами уже принята в качестве предпочтительного пути лечения ревматоидного артрита и болезни Крона, а также имеет место впечатляющий прогресс в лечении злокачественных опухолей. Также в разработке находятся продукты на основе антител для лечения сердечнососудистых и инфекционных заболеваний. Большинство представленных на рынке продуктов моноклональных антител распознают и связывают один четко определенный эпитоп на целевом лиганде (например, TNFα).
Структура антител хорошо известна в данной области. Большинство природных антител являются тетрамерами и содержат две тяжелых цепи и две легких цепи. Тяжелые цепи соединены друг с другом через дисульфидные связи между шарнирными доменами, расположенными приблизительно посередине каждой тяжелой цепи. Легкая цепь связана с каждой тяжелой цепью на N-концевой стороне шарнирного домена. Каждая легкая цепь обычно связана с соответствующей ей тяжелой цепью дисульфидной связью рядом с шарнирным доменом.
Когда молекула антитела уложена правильно, каждая цепь уложена в определенное число отдельных глобулярных доменов, соединенных более линейной полипептидной последовательностью. Например, легкая цепь уложена в вариабельный (VL) и константный (CL) домены. Тяжелая цепь имеет один вариабельный домен (VH) рядом с вариабельным доменом (VL) легкой цепи, первый константный домен (СН1), шарнирный домен и еще два или три константных домена. Взаимодействие вариабельных доменов тяжелой (VH) и легкой (VL) цепей приводит к образованию антигенсвязывающей области (Fv).
Взаимодействие между тяжелыми и легкими цепями опосредовано первым константным доменом тяжелой цепи (СН1) и константным доменом (CL) легкой цепи. Однако также существует поверхность контакта между вариабельным доменом тяжелой цепи (VH) и вариабельным доменом легкой цепи (VL), которая участвует во взаимодействии между тяжелой цепью и легкой цепью.
Поскольку вариабельные домены имеют различные аминокислотные последовательности, разработана система нумерации аминокислотных остатков в этих доменах. Эта система нумерации описана авторами Kabat et al. ((1991) US Public Health Services, NIH publication 91-3242) и использована в настоящем описании. В вариабельном домене тяжелой цепи каркасная область 1 (FR1) содержит остатки с 1 до 30, каркасная область 2 (FR2) содержит остатки с 36 до 49, каркасная область 3 (FR3) содержит остатки с 66 до 94, а каркасная область 4 (FR4) содержит остатки с 103 до 113. Также в вариабельном домене тяжелой цепи определяющие комплементарность области (CDR) содержат остатки с 30 до 35 (CDR1), остатки с 50 до 65 (CDR2) и с 95 до 102 (CDR3).
Нормальные В-клетки человека содержат один локус тяжелой цепи на хромосоме 14, из которого посредством перестройки возникает ген, кодирующий тяжелую цепь. У мышей локус тяжелой цепи расположен на хромосоме 12. Нормальный локус тяжелой цепи содержит несколько генных сегментов V, несколько генных сегментов D и несколько генных сегментов J. Наибольшую часть домена VH кодирует генный сегмент V, но С-конец каждого домена VH кодирует генный сегмент D и генный сегмент J. Антигенсвязывающую специфичность каждого домена VH обеспечивает перестройка VDJ в В-клетках и последующее созревание аффинности. Анализ последовательностей нормальных тетрамерных антител демонстрирует, что источником разнообразия в основном является сочетание перестройки VDJ и соматической гипермутации (Xu and Davies, (2000) Immunity, 13, 37-45). Существует более 50 генных сегментов V человека, присутствующих в геноме человека, из которых только 39 функциональны. Среди нормальных диплоидных антителопродуцирующих В-клеток каждая клетка продуцирует тетрамерное антитело из одного набора локусов тяжелой и легкой цепей антитела. Другой набор локусов используется, но не продуктивно, в результате процесса, называемого аллельным исключением (см. Singh et al., (2003) J. Exp. Med., 197, 743-750 и Immunology 5th edition, R.Goldsby, T.Kindt, B.Osborne, J.Kuby (2003) W.H.Freeman and Company NY, NY и цитируемые в них источники).
Способность «помнить» встречу с патогеном является определяющим признаком иммунной системы высших позвоночных. По-видимому, вклад В-клеток в «память» является функцией двух отдельных популяций В-клеток, долгоживущих плазматических клеток и В-клеток памяти (обзор см. в Tangye and Tarlinton (2009) Eur. J. Immunol., (2009) 39, 2065-2075). Их образование происходит в результате начального первичного иммунного ответа (антигенный стимул) наивных В-клеток. Одна популяция представлена долгоживущими плазматическими клетками, В-клетками, которые продолжают секретировать высокие уровни нейтрализующих антител в течение длительных периодов (месяцев) после элиминации антигена. Плазматические клетки прошли окончательную дифференциацию и содержат перестроенные локусы иммуноглобулинов, прошедшие созревание аффинности, которые кодируют антиген-специфические антитела с высокой аффинностью. Они отличаются от короткоживущих плазматических клеток со сроком жизни в несколько дней, которые погибают в результате стресса, вызванного интенсивным синтезом антиген-специфического антитела (см. Radbruch et al. (2006) Nature Reviews Immunology, 6, 741-750). В отличие от этого, вторая популяция антиген-специфических В-клеток памяти представляет популяцию В-клеток, также содержащих перестроенные локусы иммуноглобулинов, прошедших созревание аффинности, которые кодируют антиген-специфические антитела с высокой аффинностью, но которые не секретируют антитела в больших количествах. В-клетки памяти обладают способностью к быстрому делению и дифференциации в секретирующие антитела плазматические клетки после повторного воздействия начального иммунизирующего антигена. Это ведет к быстрому обогащению пула антиген-специфических иммуноглобулинов, доступных для ответа на данный антигенный стимул. Работа по вакцинации против натуральной оспы демонстрирует ответ В-клеток памяти у иммунизированных индивидуумов в течение периода в 50 лет (Grotty et al. (2003) J. Immunology, 171, 4969-4973).
У человека В-клетки памяти персистируют во вторичных лимфоидных органах, содержат перестроенные соматически мутировавшие гены иммуноглобулинов и опосредуют вторичный иммунный ответ на повторную сенсибилизацию (см. Bernasconi et al. (2002) Science, 298, 2199-2202). По-видимому, селезенка человека является одним из основных мест локализации клеток памяти (обзор см. в Tangye and Tarlinton (2009) Eur. J. Immunol., (2009) 39, 2065-2075).
Долгоживущие секретирующие антитела плазматические клетки персистируют во вторичных лимфоидных органах и костном мозге в течение месяцев. Например, антигензависимые секретирующие иммуноглобулины клетки персистируют в селезенке и костном мозге человека (Ellyard et al. (2004) Blood, 103, 3805-3812).
По-видимому, у других млекопитающих, таких как мыши, первичный длительный ответ с образованием антител является функцией долгоживущих плазматических клеток. Таким образом, у мышей окончательно дифференцированные долгоживущие плазматические клетки, присутствующие в костном мозге и селезенке, персистируют и продолжают секретировать антитела в течение длительных периодов времени, превышающих один год (см. Slifka et al. (1998) Immunity, 8, 363-372; Maruyama et al. (2000) Nature, 407, 636-641). Хотя у различных млекопитающих еще нужно достоверно установить точное взаимоотношение между клетками памяти и окончательно дифференцированными долгоживущими плазматическими клетками, у млекопитающих существует два пула долгоживущих В-клеток, каждый содержит перестроенные локусы иммуноглобулинов, прошедшие созревание аффинности, которые кодируют антиген-специфические антитела с высокой аффинностью. Один обладает способностью к быстрому делению. Другой не делится, но сохраняет способность секретировать антитело в течение длительных периодов.
Антиген-специфические тетрамерные антитела человека, продуцируемые пациентами, являются потенциальным источником клинически и коммерчески значимых антиген-специфических антител. Также их можно получить непосредственно из трансформированных EBV В-клеток человека, предпочтительно из трансформированных EBV В-клеток памяти человека, необязательно трансформированных в присутствие поликлональных активаторов В-клеток (PCT/IB04/01071).
До сих пор клетки памяти и долгоживущие плазматические клетки не были использованы в качестве источника кодируемых трансгенами антител с высокой аффинностью, в частности, антител человека или гибридных антител, содержащих VH и VL домены человека.
Создание доменов VН и антител, содержащих только тяжелые цепи
Способность антител, содержащих только тяжелые цепи и лишенных легких цепей, связывать антиген, установлена в 1960-х (см. Jaton et al. (1968) Biochemistry, 7, 4185-4195). Тяжелые цепи иммуноглобулина, физически отделенные от легких цепей, сохраняют 8 0% антигенсвязывающей активности по отношению к тетрамерному антителу. Кроме того, активность связывания связывали с N-концевым фрагментом (Fd) тяжелой цепи. Пока в этих экспериментах использовали точно охарактеризованное антиген-специфическое поликлональное антитело кролика, они были репрезентативны для любой поликлональной популяции, которая содержит тетрамерное антитело, полученное у млекопитающего, включая человека. В димера тяжелых цепей сохранен связывающий домен СН1 легкой цепи.
Во многих публикациях 1980-х годов описаны манипуляции с генами тяжелых цепей in vitro с целью создания новых антител. Большинство этих работ основано на перестроенном гене µ антитела (IgM) мыши, который кодирует антитело, индуцированное против точно охарактеризованного антигена. Особенность этого антитела состоит в том, что известно, что антигенсвязывающую специфичность определяет домен VH, поскольку сборка и секреция с использованием несоответствующей легкой цепи показала сохранение связывания антигена (см. Neuberger and Williams (1986) Phil. Trans. R. Soc. Lond., A317, 425-432). Используя эту систему, было показано, что антиген-специфический связывающий домен VH мыши можно использовать для получения нового антитела, которое содержит константную область ε человека, слитую с антиген-специфическим доменом VH мыши. Полученное химерное IgE сохраняло антигенную специфичность и проявляло эффекторную активность, ожидаемую от IgE (см. Neuberger et al. (1985) Nature, 314, 268-270).
Другие опубликованные примеры создания тяжелой цепи включают получение химерного антитела мыши-человека, которое содержит VH мыши, слитый с константными областями IgA или IgG человека (см. Morrison et al. (1984) PNAS, 81, 6851-6855; Sun et al. (1987) PNAS, 84, 214-218); и Heinrich et al. (1989) J. Immunol, 143, 3589-97). Таким образом, к концу 19080-х годов концепция создания тяжелых цепей была успешно сформулирована. Любой охарактеризованный связывающий домен VH можно слить с любой константной областью антитела с сохранением антиген-специфической активности связывания. Эффекторную функцию определяет выбранная константная область тяжелой цепи (например, IgGl, IgG2, IgG3, IgG4, IgA, IgE и т.д.). Итоговое экспрессируемое химерное антитело во всех случаях содержит тетрамерное антитело.
Для создания целой тяжелой цепи человека необходим источник доменов VH человека с установленной связывающей специфичностью. Эта проблема частично решена посредством разработки библиотек последовательностей связывающих доменов VH человека, полученных с использованием сочетаний сегментов V, D и J наивной зародышевой линии человека или прошедших созревание аффинности доменов VH, полученных из тетрамерных антител, источником которых служат лимфоциты человека. Затем можно выделить антиген-специфические домены VH с аффинностями связывания в диапазоне от 20 нМ до 100 нМ из библиотек дисплеев связывающих доменов VH, полученных из мРНК антителопродуцирующих В-клеток периферической крови человека (см. Ward et al. (1989) Nature, 341, 544-546) или из случайных последовательностей VDJ, полученных из ДНК наивной зародышевой линии человека (см. ЕР-А-0368684). Эти подходы предоставляют источник антиген-специфических доменов VH млекопитающих и, в частности, доменов VH человека.
Предполагают, что:
«Выделенные вариабельные (VH) домены могут представлять собой альтернативу моноклональным антителам и выполняют ключевую функцию в связывании антител человека с высокой аффинностью»;
«Соединяя вариабельные домены с константными доменами легких или подходящих тяжелых цепей и экспрессируя собранные гены в клетках млекопитающих, можно получить целые антитела, которые будут выполнять естественные эффекторные функции, такие как лизис комплимента»; и
«Этот подход может подтвердить ценность создания терапевтически значимых антител человека» (см. Ward et al. (1989) Nature, 341, 544-546 и EP-A-0368684).
При использовании альтернативного подхода Sitia et al. ((1990) Cell, 60, 781-790) продемонстрировано, что удаление домена СН1 из перестроенного гена µ мыши приводит к синтезу и секреции антитела, содержащего только тяжелые цепи и лишенного легких цепей, клетками млекопитающих в культуре. В этом случае секретируемая тяжелая цепь, содержащая связывающий домен VH и константную область, состоящую из гибкого шарнирного домена и константных доменов СН2, СН3 и СН4 из IgM, обеспечивает димеризацию и эффекторную функцию тяжелых цепей.
Таким образом, к 1990 году стали доступны основные инструменты для создания гомодимеров антитела человека, содержащих только тяжелые цепи, in vitro (иногда их обозначают как Dabs-Fc) с установленными специфичностями связывания VH и для выбора эффекторных областей тяжелых цепей, лишенных СН1.
С этим подходом связано две проблемы:
домены VH, выбранные посредством массовых подходов, обладали относительно низкой антигенсвязывающей аффинностью (20-100 нМ); и
домены VH в отсутствие легких цепей были «липкими», нерастворимыми и склонными к агрегации и поэтому не приспособленными для фармацевтического применения.
Склонность выделенных доменов VH к агрегации является следствием отсутствия легкой цепи. Когда легкая цепь отсутствует, обнажены определенные гидрофобные боковые цепи, которые обычно скрыты в поверхности контакта тяжелой/легкой цепи. Кроме того, нарушена термодинамическая стабильность, поскольку стабильность доменов VH зависит от контактов на поверхности контакта легкой цепи (см. Worn and Pluckthun (1999) Biochemistry, 38, 8739-8750).
Приведенное авторами Hamers-Casterman описание встречающегося в природе гомодимерного антитела IgG, содержащего только тяжелые цепи и лишенного легких цепей, которое циркулирует у верблюдовых, в дополнение к нормальным тетрамерным антителам верблюдовых (см. Hamers-Casterman et al. (1993) Nature, 363, 446-448) проливает свет на проблему растворимости. В соответствии с молекулой, созданной авторами Sitia et al. ((1990) Cell, 60, 781-790), функциональность CHI домена отсутствовала в антителах верблюдовых, содержащих только тяжелые цепи, в связи с сайтом альтернативного сплайсинга, который устраняет СН1 из процессированной мРНК тяжелой цепи антитела.
Домены VH антител верблюдовых, содержащих только тяжелые цепи (которые в соответствии с широким использованием далее в настоящем документе обозначают как домены VHH), полученные посредством сенсибилизации антигеном, проходили созревание аффинности в В-клетках in vivo, обладают аффинностями связывания в нижнем наномолярном диапазоне, растворимы и не склонны к агрегации (см. Ewert et al. (2002) Biochemistry, 41, 3628-3636). Как правило, домены VHH верблюдовых также проявляют повышенную тепловую стабильность по отношению к доменам VH, полученным из зародышевой линии и выделенным посредством подходов с использованием дисплеев, и в некоторой степени сохраняют активность связывания в присутствие денатурирующих средств, таких как детергенты и отбеливающие средства, а также после тепловой обработки (см. Dumoulin et al. (2002) Protein Science, 11, 500-511 и Dolk et al. (2005) Applied Environmental Microbiology, 71, 442-450).
Антиген-специфические антитела, содержащие только тяжелые цепи, можно получить, используя мРНК В-клеток верблюдовых в стандартных способах клонирования или способами фаговых или других дисплеев (см. Riechmann and Muyldermans (1999) J. Immunol. Methods, 231, 25-38). Антитела, содержащие только тяжелые цепи, полученные из верблюдовых, обладают высокой аффинностью. Анализ последовательностей мРНК, кодирующей антитело, содержащее только тяжелые цепи, демонстрирует, что источником разнообразия в первую очередь является сочетание перестройки VHHDJ и соматической гипермутации, что также наблюдают при образовании нормальных тетрамерных антител. Однако остается необходимым установить, образуют ли нормальные В-клетки верблюдовых антитела, содержащие только тяжелые цепи, в противоположность тетрамерным антителам. В действительности известно немногое о развитии пре-В клеток у верблюдовых (см. WO 2004/049794 и Zou et al. (2005) J. Immunology, 175, 3769-3779). Кроме того, остается необходимым установить, участвуют ли антитела верблюдовых, содержащие только тяжелые цепи, подобно тетрамерам иммуноглобулинов, содержащим тяжелые и легкие цепи, в длительном ответе с образованием антител посредством клеток памяти или посредством долгоживущих плазматических клеток.
Специфическим признаком антител верблюдовых, содержащих только тяжелые цепи, является то, что в домене VHH мутации подвергаются некоторые аминокислотные остатки, которые в нормальном домене VH взаимодействуют с VL доменом. Таким образом, лейцин в положении 45 в 98% VH последовательностей человека и мыши подвергается мутации. Предполагают, что эти мутации у верблюдовых, консервативные в зародышевой линии, согласуются с поддержанием растворимости VHH при отсутствии легкой цепи. Также описаны другие характерные мутации аминокислот в доменах VHH верблюдовых по сравнению с нормальными доменами VH. Среди них самые распространенные и важные изменения происходят в четырех положениях (тетрада VHH) во второй каркасной области, которые отвечают за снижение гидрофобности упомянутой поверхности контакта с легкой цепью. Как правило, выявляют Glu-44 и Arg-45 на месте менее гидрофильных Gly-44 и Leu-45, представленных в нормальных доменах VH, тогда как повышение гидрофильности в положении 4 7 происходит за счет замены Тyr на остатки меньшего размера. Четвертое изменение состоит в замене Val-37 на Phe или Тyr. Структурный анализ показывает, что это формирует малое гидрофобное ядро, типично включающее Tyr-91, Trp-103, Arg-45 и гидрофобные остатки, присутствующие в петле CDR3 домена VHH. Эти изменения повышают гидрофильность упомянутой поверхности контакта с легкой цепью посредством прямой замены на более гидрофильные боковые цепи или посредством извлечения гидрофобных боковых цепей из растворителя посредством взаимодействия между CDR3 и каркасными остатками (см. Bond et al. (2003) J. Mol. Biol., 332, 643-655).
В домене VHH петля CDR3 увеличена по отношению к той, что присутствует в тетрамерных антителах верблюдовых и других точно охарактеризованных тетрамерных антителах человека и мыши (см. Riechmann and Muyldermans (1999) J. Immunol. Methods, 231, 25-38). Роль ключевых гидрофобных остатков, присутствующих в петле CDR3 VHH, например, в домене VHH ламы, демонстрирует структурную роль CDR3 в стабильности и растворимости доменов VHH (см. Bond et al. (2003) J. Mol. Biol, 332, 643-655). В совокупности структурные исследования доменов VHH демонстрируют, что стабильность и растворимость этих доменов зависят, в дополнение к каркасным остаткам, от CDR3, а также что требования к ключевым гидрофобным остаткам накладывает значительные ограничения на тип петли CDR3, совместимой с благоприятными биофизическими характеристиками доменов VHH по отношению к доменам VH (см. Barthelemy et al. (2008) J. Biol. Chem., 283, 3639-3654).
Однако несмотря на высокую степень консервативности между последовательностями верблюдовых и человека, домены VHH верблюдовых сохраняют родство с верблюдовыми и поэтому являются потенциально антигенными для человека. Гуманизация in vitro может снизить этот риск, но ценой возможного снижения аффинности и растворимости в результате использования способов создания in vitro.
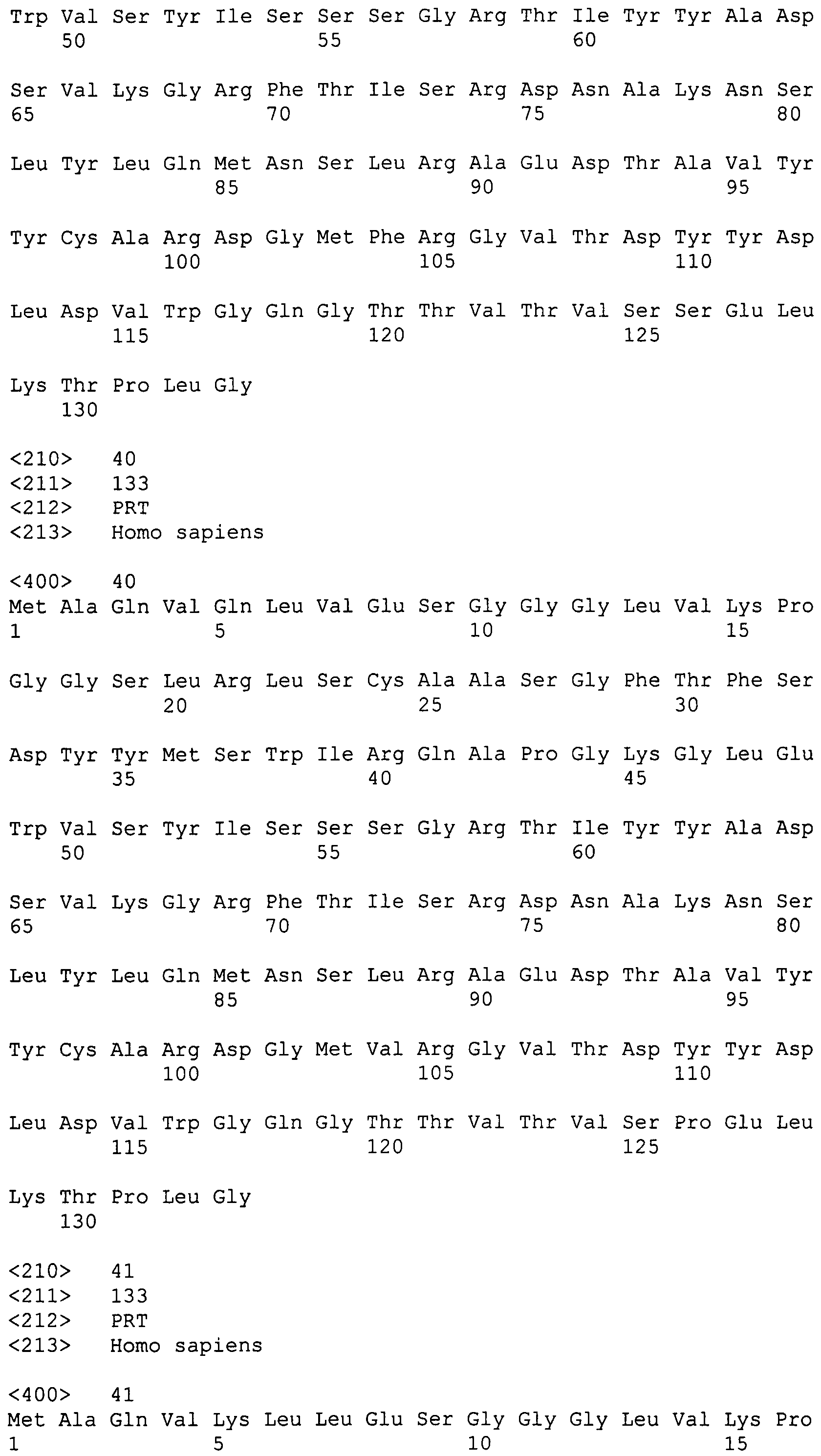
Связывающие домены УН человека и проблема растворимости
Плохие биофизические характеристики доменов VH человека и других млекопитающих, полученных из последовательностей зародышевой линии или из нормальных тетрамерных антител, по сравнению с доменами VHH верблюдовых, в частности, склонность к агрегации, в настоящее время ограничивают их полезность в качестве реактивов, диагностических и лекарственных средств (см. Rosenberg (2006) The AAPS Journal, 8(3), Article 59 E501-E507 и Fahrner et al. (2001) Biotechnol. Gen. Eng. Rev., 18, 301-327).
Первые попытки решить проблему растворимости были основаны на введении камелизирующих аминокислотных последовательностей в выбранные связывающие VH области человека на поверхности контакта VH/VL. Доказана несостоятельность этого подхода, поскольку на стабильность и растворимость нельзя повлиять посредством «камелизации» в положении 45 поверхности контакта VH/VL в отдельности (см. Domantis submissions, датированные 18-м мая 2007 года и 5-м июня 2007 года, в противовес ЕР-В-0656946). Действительно, введения направленных «камелизирующих» мутаций не достаточно для того, чтобы решить проблему «слипания» у выделенных доменов VH человека (см. Riechmann and Muyldermans (1999) J. Immunol. Methods, 231, 25-38). Введение камелизирующих мутаций ведет к деформации β-складчатого слоя каркасной области, которая возможно отвечает за пониженную стабильность белка, наблюдаемую в результате этих экспериментов (см. Riechmann (1996) J. Mol. Biol., 259, 957-969). Кроме того, роль CDR3, содержащей ключевые гидрофобные остатки, в поддержании структурной стабильности и растворимости доменов VHH в отсутствие легкой цепи, добавляет сложности, что потенциально ограничивает разнообразие последовательностей CDR3 теми, что совместимы с каркасной областью верблюдовых (см. Barthelemy et al. (2008) J. Biol. Chem., 283, 3639-3654).
Таким образом, домены VH млекопитающих, включая полученные у человека, которые получены из прошедших созревание аффинности природных тетрамерных антител или из последовательностей наивных сегментов VDJ зародышевой линии, в отсутствие легких цепей, относительно нерастворимым и подвержены агрегации. У таких доменов VH отсутствуют необходимые биофизические характеристики растворимого домена VH, подходящего для использования в качестве лекарственного средства.
Антитела, содержащие только тяжелые цепи, и растворимые домены УН человека, полученные трансгенезом in vivo.
Растворимые связывающие домены VH для фармацевтического применения предпочтительно получены у человека и обладают признаками, такими как высокая антигенсвязывающая аффинность и растворимость при физиологических условиях при отсутствии агрегации. Такие растворимые связывающие домены VH также можно использовать в качестве диагностических средств и реактивов и они получат широкое распространение в промышленном и сельскохозяйственном применениях.
Альтернативный подход к получению антиген-специфических антител, содержащих только тяжелые цепи, состоит в использовании трансгенных мышей, которые содержат локусы тяжелых цепей, лишенных функциональности СН1 (см. Janssens et al. (2006) PNAS, 103:15130-5 и WO 02/085944).
Вначале не было известно, будут ли нормальные В-клетки мыши активироваться и секретировать антитела, содержащие только тяжелые цепи, в результате сенсибилизации антигеном или такие антитела будут обладать растворимостью. Кроме того, не было известно, необходимы ли IgM или, как у верблюдовых, ограничена ли экспрессия классами антител Igγ2 и Igγ3, или нормальные В-клетки мыши могут секретировать даже потенциально нерастворимые домены VH.
Авторы Janssens et al. ((2006) PNAS, 103:15130-5) описали локус гена химерной тяжелой цепи, который содержит два VHH генных сегмента ламы, все генные сегменты D и J человека и сочетания генных сегментов, кодирующих константные области Сµ и Сγ2 и Сγ3 человека, у которых отсутствует функциональность СН1. Выбор генных сегментов VHH ламы обеспечил то, что мутации каркасной области зародышевой линии, сохраняемые у верблюдовых на поверхности контакта VH/VL, также присутствуют в экспрессируемом камелизованном локусе человека с тем, чтобы увеличить возможность того, что полученный «камелизованный» домен VH может сохранять характеристики растворимости и стабильности, проявляемые VHH верблюдовых, таким образом повышая вероятность функционального ответа на сенсибилизацию антигеном даже в отсутствие петли CDR3, полученной у верблюдовых.
В результате было показано, что экспрессия трансгена антитела, содержащего только тяжелую цепь, происходит специфичным для В-клеток образом, что происходит размножение В-клеток и что в отсутствие функциональности СН1 присутствие любого функционального гена легкой цепи нецелесообразно. Трансгены, содержащие только тяжелые цепи, подвергаются перестройке VDJ, за которой следует активация В-клеток и созревание аффинности в ответ на сенсибилизацию антигеном. Также происходит переключение класса между IgM и IgG, содержащими только тяжелые цепи, равно как и переключение изотипа Сγ. В отсутствие Сµ в локусе, использование Сγ происходит с равным успехом. Специфичность антител в значительной степени определяется перестройкой VDJ (CDR3), тогда как анализ соматических мутаций показывает, что это происходит на всем протяжении области VH, предпочтительно в областях CDR1 и CDR2 экспрессируемого генного сегмента VHH ламы. Среди ограниченного числа охарактеризованных камелизованных антител человека, содержащих только тяжелые цепи, для всех подтвердили растворимость, будь они получены из В-клеток селезенки посредством фагового дисплея доменов VH или гибридомной селекцией, несмотря на тот факт, что петля CDR3, которую получали полностью из последовательностей человека, меньше тех, что найдены в доменах VHH верблюдовых.
Лучшие гибридные антитела ламы/человека, содержащие только тяжелые цепи, из этой ограниченной серии обладали аффинностями в диапазоне 1-3 нМ. Организм мыши также использует трансген в качестве другого локуса тяжелой цепи в этом размножении В-клеток, а селезенка имеет по существу нормальную структуру даже в отсутствие перестройки легкой цепи (см. Janssens et al. (2006) PNAS, 103:15130-5). Кроме того, механизмы аллельного исключения определяют продуктивную экспрессию эндогенного локуса тяжелой цепи мыши или трансгена тяжелой цепи, у которой отсутствует функциональность СН1 (см. WO 2007/09677).
К удивлению, наблюдения Janssens et al. ((2006) PNAS, 103:15130-5) указывают на то, что стабильные растворимые камелизованные антитела, содержащие только тяжелые цепи, можно получить с использованием каркасной области верблюдовых даже в отсутствие последовательностей верблюдовых, содержащих петлю CDR3.
В качестве расширения этих экспериментов (см. WO 2006/008548) созданы дополнительные линии трансгенных мышей, которые содержат полностью человеческие локусы генов только тяжелых цепей, у которых, таким образом, отсутствует последовательность, кодирующая каркасную область VHH верблюдовых, и которые, кроме того, содержат петли CDR3, полученные не из последовательностей верблюдовых.
В этом подходе вводили четыре естественных генных сегмента VH зародышевой линии человека (естественный локус V4), таким образом, полагаясь полностью на естественную селекцию посредством созревания аффинности для образования антигенной специфичности и структурной стабильности полученных растворимых доменов VH человека. Оба локуса содержат все генные сегменты D и J человека, генные сегменты Сγ2 и Сγ3 константной области человека, у каждого отсутствует СН1, энхансерные регуляторные элементы иммуноглобулина человека и, предпочтительно, LCR тяжелой цепи антитела.
Локус V4 человека функционален и антитела человека, содержащие только тяжелые цепи, циркулируют в плазме животных, которых не сенсибилизировали антигеном. После сенсибилизации антигеном, определяют антиген-специфические антитела человека, используя стандартный анализ ELISA. Локусы человека, содержащие сконструированные генные сегменты V (локус V17) (также см. WO 2008/035216), также функциональны (также см. Antibody Engineering and Antibody Therapeutics Meeting. F. Grosveld presentation, San Diego Antibody Meeting, December 3-4, 2007).
Кроме того, к удивлению сенсибилизация антигеном трансгенных мышей, содержащих локусы 44, позволяет выделить и охарактеризовать растворимые антиген-специфические домены VH человека с высокой аффинностью в отсутствие характерных мутаций каркасной области зародышевой линии верблюдовых и петли CDR3, похожей на петлю CDR3 верблюдовых, типичной для доменов VHH верблюдовых. Кроме того, домены VH, полученные из антиген-специфических антител человека, содержащих только тяжелые цепи, растворимы (см. WO 2006/008548). После сенсибилизации антигеном, перестройки VDJ и последующего размножения В-клеток, структура селезенки у мышей, содержащих локус V4 человека, по существу схожа с гуморальным ответом мышей дикого типа на сенсибилизацию антигеном. Световая микроскопия выявляет расщепление кластеризации Т-клеток в периартериолярном слое лимфоцитов (PALS), окруженном областями, богатыми В-клетками, включая фолликулы и краевые зоны, присутствующие на наружных границах белой пульпы. Также существуют структуры, схожие с терминальным центрами в В-клеточных фолликулах вторичных лимфоидных тканей, сопоставимые с мышами дикого типа в ходе Т-клеточно-зависимого гуморального ответа
Альтернативный подход к идентификации и конструированию синтетических доменов VH человека с использованием дисплея, который не зависит от прогностического анализа естественных последовательностей доменов VHH верблюдовых, также демонстрирует, что автономные домены VH со структурными свойствами, не входящие в спектр природных каркасных областей, можно получить, используя неприродные мутации, которые отличаются от тех, что выявлены у верблюдовых (см. Barthelemy et al. (2008) J. Biol. Chem., 283, 3639-3654 и патентную заявку США №11/102,502). Однако данные ничего не говорят о том, сохраняют ли такие домены VH биофизические характеристики растворимости при физиологических условиях при отсутствии агрегации в сочетании с аффинностями связывания в нижнем наномолярном диапазоне.
Несмотря на элегантность, подходы к идентификации и конструированию синтетических доменов VH человека посредством дисплея являются трудоемкими и ограничены объемом доступности доменов VH человека и трехмерной структурной информации о VHH верблюдовых. Тонкие различия возникают в зависимости от используемых сочетаний VDJ, которые ограничивают разработку библиотек VH на основе синтетических последовательностей с использованием разнообразий CDR3, не ограниченных структурными требованиями, которые обеспечивают связывание антигена с высокой аффинностью. В то время как результаты показывают, что в отличие от доменов VHH верблюдовых, растворимые стабильные конформации доменов VH человека не зависят от взаимодействий между петлей CDR3 и характерными заменами аминокислот верблюдовых в упомянутой поверхности контакта с легкой цепью, данные ограничены биофизическими, характеристиками. Например, в контексте этих изменений не учитывали сохранение связывания антигена с высокой аффинностью.
Изобретение
В настоящем изобретении описаны новые растворимые антитела с высокой аффинностью, содержащие только тяжелые цепи, и растворимые домены VH, структурно отличные от доменов VH, получаемых из нормальных тетрамерных антител или последовательностей зародышевой линии, подходящие для использования в качестве лекарственных средств, и предпочтительные пути для выделения таких новых антител, содержащих только тяжелые цепи, и растворимых доменов VH из трансгенных животных после сенсибилизации антигеном.
Таким образом, первый аспект настоящего изобретения относится к новому антиген-специфическому растворимому антителу с высокой аффинностью, содержащему только тяжелые цепи, которое:
не содержит характерные замены аминокислот верблюдовых и содержит замены FR2, которые обнаружены в нормальных тетрамерных антителах;
обладает повышенной суммарной гидрофобностью в CDR1 и содержит увеличенное число заряженных аминокислот, присутствующих в CDR3; и
содержит одну или более замен аминокислот в β-складчатом слое каркасной области, которые ведут к повышению суммарной гидрофобности в FR1 и увеличению числа заряженных аминокислот, представленных в FR3.
Антитело, содержащее только тяжелые цепи, предпочтительно представляет собой антитело человека. Антитело человека, содержащее только тяжелые цепи, не содержит характерные для верблюдовых замены аминокислот и содержит замены FR2, которые не найдены в нормальных тетрамерных антителах человека.
Антитело, содержащее только тяжелые цепи, предпочтительно имеет аффинность связывания 10 нМ или лучше и ведет себя как растворимый мономер при гель-фильтрации при физиологических буферных условиях.
Настоящее изобретение также относится к антиген-специфическому растворимому домену VH с высокой аффинностью, который:
не содержит характерные для верблюдовых замены аминокислот и содержит замены FR2, которые не найдены в нормальных тетрамерных антителах;
обладает повышенной суммарной гидрофобностью в CDR1 и содержит увеличенное число заряженных аминокислот, присутствующих в CDR3; и
содержит одну или более замен аминокислот в β-складчатом слое каркасной области, которые выдут к повышению гидрофобности в FR1 и увеличению числа заряженных аминокислот, присутствующих в FR3.
Растворимый домен VH предпочтительно получен у человека. Растворимый домен VH человека не содержит характерные для верблюдовых замены аминокислот и содержит замены FR2, которые не найдены в нормальных тетрамерных антителах человека.
Растворимый домен VH предпочтительно имеет аффинность связывания 10 нМ или лучше и ведет себя как растворимый мономер при гель-фильтрации при физиологических буферных условиях.
Настоящее изобретение дополнительно относится к генному сегменту V, который после рекомбинации с генным сегментом D и генным сегментом J и последующего созревания аффинности, кодирует растворимый домен VH в соответствии с изобретением.
Необязательно, последовательность генного сегмента V можно сконструировать посредством по меньшей мере одной замены, вставки или делеции или их сочетания по отношению к естественной последовательности зародышевой линии генного сегмента V с тем, чтобы он содержал кодон(ы), кодирующие аминокислотные остатки или аминокислотные последовательности, которые отличны от последовательности зародышевой линии животного-хозяина, не представляют собой характерные для верблюдовых замены аминокислот, и могут представлять собой замены FR2, которые, как правило, не обнаруживают в нормальных тетрамерных антителах. Такие изменения могут содержать замены, вставки или делеции или их сочетание в областях CDR1 и/или CDR2 генного сегмента V с тем, чтобы увеличить разнообразие аминокислот, доступных для связывания антигена, по сравнению с последовательностями зародышевой линии.
Настоящее изобретение дополнительно относится к генному сегменту D, который после рекомбинации с генным сегментом V в соответствии с изобретением и генным сегментом J. кодирует домен VH в соответствии с изобретением.
Необязательно последовательность генного сегмента D можно сконструировать посредством по меньшей мере одной замены, вставки или делеции или их сочетания по отношению к естественной последовательности генного сегмента D зародышевой линии. Такая замена, вставка или делеция, или их сочетание, такова, что генный сегмент D может содержать кодоны, которые кодируют аминокислотные остатки, для которых установлено, что они содействуют сохранению растворимости и стабильности растворимого домена VH, включая последовательности, которые кодирует генный сегмент D, или альтернативно могут увеличивать разнообразие генного сегмента D в любой получаемой петле CDR3 по сравнению с последовательностями зародышевой линии.
Также настоящее изобретение дополнительно относится к генному сегменту J, который после рекомбинации с генным сегментом V в соответствии с изобретением и генным сегментом D, кодирует растворимый домен VH в соответствии с изобретением.
Необязательно последовательность генного сегмента J можно сконструировать посредством по меньшей мере одной замены, вставки или делеции, или их сочетания, по отношению к естественной последовательности генного сегмента J зародышевой линии. Такая замена, вставка или делеция, или их сочетание, такова, что генный сегмент J может содержать кодоны, которые кодируют аминокислотные остатки, для которых установлено, что они содействуют сохранению растворимости и стабильности домена VH, включая последовательность, кодируемую генным сегментом J, или альтернативно могут увеличивать разнообразие генного сегмента J в любой получаемой петле CDR3 по сравнению с последовательностями зародышевой линии.
Настоящее изобретение предпочтительно относится к гетерологичному локусу, содержащему только тяжелую цепь, который содержит один или более генных сегментов V в соответствии с изобретением, один или более генных сегментов D в соответствии с изобретением, один или более генных сегментов J в соответствии с изобретением и один или более генных сегментов константных эффекторных областей, каждая из которых кодирует константную эффекторную область антитела, у которой отсутствует функциональность СН1, где генные сегменты расположены так, что генные сегменты V, D и J и генный сегмент константной области можно рекомбинировать для того, чтобы получить перестроенный ген, кодирующий антиген-специфическое растворимое антитело с высокой аффинностью, содержащее только тяжелые цепи, в соответствии с изобретением.
Локус, содержащий только тяжелую цепь, содержит константную эффекторную область(и) тяжелой цепи позвоночного, которая предпочтительно получена у млекопитающего, более предпочтительно, получена от человека, мыши, крысы или верблюдового.
Предпочтительно локус тяжелой цепи устроен так, чтобы получить антитело, содержащее только тяжелые цепи, которое содержит растворимый домен VH человека и константную эффекторную область, полученную у млекопитающего.
Также изобретение дополнительно относится к трансгенному млекопитающему, не относящемуся к человеку, которое содержит гетерологичный локус тяжелой цепи в соответствии с изобретением. Предпочтительно млекопитающим является грызун, наиболее предпочтительно мышь или крыса. Предпочтительно, функциональность эндогенного локуса тяжелой цепи трансгенного млекопитающего, не относящегося к человеку, ослабляют любым известным в данной области способом или вводят делецию, которая переводит ген в молчащее состояние (см. US 5591669; US 6162963; US 5877397; Kitamura and Rajewsky (1992) Nature, Mar 12; 356(6365):154-6). Альтернативно можно удалить В-клетки, которые, в противоположность трансгенам, экспрессируют эндогенные гены иммуноглобулинов. Эндогенные гены легких цепей k и λ могут сохранять функциональность или необязательно можно сделать нефункционирующими или удалить (ЕР 139959). Менее предпочтительно трансген может представлять собой локус нормальной тяжелой цепи антитела, который содержит функциональность СН1 при том, что в генотипе организма-хозяина отсутствует функциональность генов эндогенной тяжелой или легкой цепей (см. ЕР 139959 и Zou et al. J. Exp. Med., 204, 3271-3283).
Трансген иммуноглобулина, содержащего только тяжелые цепи, также может содержать локусы гена эндогенной тяжелой цепи иммуноглобулина, которые сконструированы для того, чтоб устранить функциональность СН1, и необязательно, кроме того, сконструированы для того, чтобы ввести или заменить один или более эндогенных генных сегментов V, D и J на альтернативные генные сегменты V, D и J, предпочтительно полученные у человека.
В другом аспекте настоящее изобретение относится к способу получения антиген-специфического растворимого антитела с высокой аффинностью, содержащего только тяжелые цепи, который включает стимуляцию антигеном трансгена млекопитающего, не относящегося к человеку, в соответствии с изобретением. Предпочтительно, растворимое антитело, содержащее только тяжелые цепи, относится к человеку.
Предпочтительно, способ включает стадию иммортализации антителопродуцирующих клеток млекопитающего. Клетки можно иммортализовать посредством слияния с миеломной клеткой или можно иммортализовать посредством трансформации вирусом, таким как EBV.
Предпочтительно, способ дополнительно содержит стадию выделения или растворимого домена VH или нуклеиновой кислоты, которая кодирует растворимый домен VH, из В-клетки или иммортализованной клеточной линии, которая продуцирует антитело, содержащее только тяжелые цепи, с желаемой антигенной специфичностью.
Альтернативно, способ может дополнительно содержать стадию выделения или растворимого домена VH или нуклеиновой кислоты, которая кодирует растворимый домен VH, из клетки, продуцирующей антитело, содержащее только тяжелые цепи, полученной у млекопитающего или из иммортализованной клетки, продуцирующей антитело, содержащее только тяжелые цепи, полученной у млекопитающего или подхода с использованием дисплея.
Настоящее изобретение также относится к использованию растворимого домена VH или нуклеиновой кислоты, кодирующей растворимый домен VH, полученный способом по изобретению, в получении антитела, содержащего только тяжелые цепи, VH интраантитела, VH полипротеина, комплекса с доменом VH или слитого VH.
Настоящее изобретение дополнительно относится к антителу, содержащему только тяжелые цепи, VH интраантителу, VH полипротеину, комплексу с доменом VH или слитому VH, которые содержат по меньшей мере один растворимый домен VH в соответствии с изобретением.
Также настоящее изобретение дополнительно относится к антителу, содержащему только тяжелые цепи, VH интраантителу, VH полипротеину, комплексу с доменом VH или слитому VH, которые содержат по меньшей мере один растворимый домен VH в соответствии с изобретением для использования в терапии.
Настоящее изобретение также относится к использованию антитела, содержащего только тяжелые цепи, VH интраантитела, VH полипротеина, комплекса с доменом VH или слитого VH, которые содержат по меньшей мере один растворимый домен VH в соответствии с изобретением, в получении лекарственного средства для использования в лечении заболеваний или нарушений, включая в качестве неограничивающих примеров: заживление ран, нарушения клеточной пролиферации, включая новообразования, меланому, остеосаркому легких, ободочной и прямой кишки, саркому прямой кишки, яичника, солидные опухоли шейки матки, пищевода, груди, поджелудочной железы, мочевого пузыря, головы и шеи и другие; миелопролиферативые нарушения, такие как лейкемия, неходжкинская лимфома, лейкопения, тромбоцитопения, нарушение ангиогенеза, саркома Капоши; аутоиммунные/воспалительные нарушения, включая аллергию, воспалительное заболевание кишечника, артрит, псориаз и воспаление дыхательных путей, астму, иммунные нарушения и отторжение трансплантата органа; сердечнососудистые и сосудистые нарушения, включая гипертензию, отек, стенокардию, атеросклероз, тромбоз, сепсис, шок, реперфузионное повреждение и ишемию; неврологические нарушения, включая заболевания центральной нервной системы, болезнь Альцгеймера, травму головного мозга, амиотрофический боковой склероз и боль; нарушения развития; нарушения метаболизма, включая сахарный диабет, остеопороз и ожирение, СПИД и почечную недостаточность; инфекции, включая вирусную инфекцию, бактериальную инфекцию, грибковую инфекцию и паразитарную инфекцию, патологические состояния, связанные с плацентой и другие патологические состояния, а также для использования в иммунотерапии.
Также настоящее изобретение дополнительно относится к кассете для получения растворимого домена VH в соответствии с изобретением, которая содержит последовательности генов, которые кодируют FR1, FR2 и FR3, где последовательности генов, кодирующих FR1 и FR2 и FR2 и FR3, разделены последовательностями, которые позволяют вводить подходящие последовательности, кодирующие CDR1 и CDR2, а на 3'-конце последовательности, кодирующей FR3, она содержит последовательность, которая позволяет вводить последовательность, которая кодирует подходящие последовательности, кодирующие CDR3.
В изобретении используют способность естественных механизмов В-клеток млекопитающего в трансгенном млекопитающем, не относящемся к человеку, предпочтительно в мыши или крысе, генерировать новые растворимые антитела с высокой аффинностью, содержащие только тяжелые цепи, в ответ на сенсибилизацию антигеном. Растворимые антитела по изобретению, содержащие только тяжелые цепи, обладают аффинностями связывания 10 нм или лучше.
Анализ последовательностей растворимых связывающих доменов VH человека, полученных из антиген-специфических антител человека с высокой аффинностью, содержащих только тяжелые цепи и лишенных легких цепей, к удивлению, показывает, что отсутствуют все характерные консенсусные аминокислотные последовательности, типичные для доменов VHH верблюдовых. Действительно, естественная селекция не компенсирует селекцию последовательностей, характерных для верблюдовых, в результате созревания аффинности. Таким образом, когда в зародышевой линии отсутствуют характерные для верблюдовых последовательности, различные механизмы, действующие in vivo, компенсируют отсутствие легкой цепи с тем, чтобы сохранить структурную стабильность и растворимость домена VH человека.
Наиболее удивительное отличие состоит не только в том, что растворимы антиген-специфические антитела человека с высокой аффинностью, содержащие только тяжелые цепи и лишенные легких цепей, но и в том, что антигенсвязывающие области (CDR) должны принимать участие в структурной стабильности и растворимости в дополнение к связыванию антигена, и что характерные консенсусные замены аминокислот в каркасной области, кодируемые VHH зародышевой линии верблюдовых, которые компенсируют отсутствие легкой цепи на упомянутой поверхности контакта с легкой цепью, не только отсутствуют в трансгенных локусах, содержащих только тяжелые цепи, но также не вставлены в результате созревания аффинности для того, чтобы компенсировать растворимость, во время процесса естественной селекции. В кодируемых трансгенами растворимых доменах VH человека после созревания аффинности альтернативные замены аминокислот могут иметь место во всех трех каркасных областях. Мутации каркасных областей могут находиться преимущественно, но не исключительно, в β-складчатом слое на упомянутой поверхности контакта с легкой цепью, таким образом компенсируя отсутствие легкой цепи.
Сравнительный анализ суммарных заряда и гидрофобности каркасной области и областей CDR антигенсвязывающих доменов VH человека, полученных в присутствие и отсутствие легкой цепи, выявляет значительные изменения в отсутствие легких цепей. В отсутствие легких цепей повышена суммарная гидрофобность аминокислот, присутствующих в CDR1, и повышено число заряженных аминокислот, присутствующих в CDR3. Замены аминокислот в β-складчатых слоях каркасной области ведут к снижению общей гидрофобности и повышению числа заряженных аминокислот, присутствующих в каркасной области 3. В каркасной области 1 увеличено число заряженных аминокислот и повышена гидрофобность, а в каркасной области 2 немного снижена гидрофобность. Это несколько отличается от доменов VHH верблюдовых, таких как лама, где сравнительный анализ выявляет признаки, общие с доменом VH человека в отсутствие легкой цепи, а также отличия. У верблюдовых, таких как лама, в каркасной области 2 значительно снижена суммарная гидрофобность, сопоставимая с повышенной гидрофильностью, введенной характерными заменам аминокислот в упомянутой поверхностью контакта с легкой цепью, и повышенным суммарным зарядом в CDR3.
Таким образом, после сенсибилизации антигеном, В-клетки компенсируют отсутствие легкой цепи, даже когда в зародышевой линии отсутствуют необходимые замены аминокислот, которые найдены у верблюдовых, таких как ламы и родственные виды.
В отсутствие последовательностей зародышевой линии, которые компенсируют отсутствие легкой цепи, найдены различные решения in vivo, все они ведут к образованию стабильных растворимых антител с высокой аффинностью, содержащих только тяжелые цепи, и растворимых доменов VH в отсутствие легкой цепи.
Эти наблюдения согласуются с основной функцией антигенсвязывающих областей (CDR), которая, в дополнение к связыванию антигена, состоит в стабилизации домена VH. Дополнением этому служат последующие редкие или новые замены аминокислот в каркасной область, в частности, в β-складчатом слое на упомянутой поверхности контакта с легкой цепью. В растворимых доменах VH, полученных не у верблюдовых, а посредством трансгенеза in vivo, отсутствуют характерные для верблюдовых замены аминокислот, которые играют важную для растворимости и стабильности VHH роль.
Антиген-специфические антитела не верблюдовых, содержащие только тяжелые цепи и эти признаки, растворимы и не подвержены агрегации при нормальных физиологических условиях. Кроме того, растворимые домены VH, выделенные из этих, сохраняют благоприятные признаки в отношении стабильности и растворимости домена VH. После сенсибилизации антигеном эти антитела, содержащие только тяжелые цепи, и полученные из них растворимые домены VH развиваются посредством естественных механизмов селекции специфическим для В-клеток образом. Антитела с высокой аффинностью (10 нм или лучше), получаемые посредством той же перестройки VDJ, как правило, содержат больше замен каркасной области, чем наблюдают у антител с более низкой аффинностью, содержащих только тяжелые цепи.
Таким образом, антигенсвязывающие домены (CDR) и β-складчатый слой каркасной области вносят вклад в общую стабильность, растворимость и аффинность связывания этих антител, содержащих только тяжелые цепи и лишенных легких цепей, или полученных из них растворимых доменов VH.
Анализ последовательностей стабильных антиген-специфических растворимых связывающих доменов VH с высокой аффинностью показывает, что:
селекция и созревание аффинности в антигенсвязывающих областях (CDR) соответствует общему увеличению гидрофобности и заряда, что ведет к специфичности и аффинности связывания антигена и структурной стабильности;
мутации, присутствующие в каркасных областях 2 и 3 вокруг упомянутой поверхности контакта с легкой цепью, соответствуют усовершенствованным растворимости и стабилизации домена VH и, таким образом, усовершенствованной аффинности связывания антигена;
мутации с каркасными областями 1 и 3 соответствуют компенсаторным структурным мутациям, которые повышают общую стабильность домена VH.
Таким образом, в отсутствие легкой цепи антитела, В-клетка млекопитающего может компенсировать свойственную нестабильность и нерастворимость связывающего домена VH тяжелой цепи посредством созревания аффинности и селекции, при этом сохраняя аффинность связывания антигена.
Анализа авторов настоящего изобретения показывает, что естественные механизмы, по-видимому, действуют независимо от используемых генных сегментов V. Таким образом, массовое секвенирование доменов VH человека, генерируемых в селезенке мыши, содержащей трансген V4 человека, в отсутствие легкой цепи позволяет провести сравнительный анализ частоты мутаций. Самую высокую мутационную нагрузку наблюдали в CDR1 и CDR2. Однако участки с повышенной мутационной нагрузкой можно различить в каркасной области 1, каркасной области 2 и по всей каркасной области 3. Возникающий общий паттерн для конкретного использованного генного сегмента V не носит специфического характера, поскольку в этой серии экспериментов антитела с высокой аффинностью получали, используя V3-23, V3-11 и V1-46 со схожими возникающими паттернами мутационной нагрузки.
Аналогичным образом, в CDR3 преобладает J4, но использовали не только этот сегмент.
По-видимому, В-клеточные механизмы созревания аффинности имеют множество функций в процессинге и секреции растворимых доменов VH в отсутствие легких цепей. Гипермутация в CDR в первую очередь отражает аффинность связывания антигена, а также, по-видимому, играет роль в структурной стабильности. Гипермутация в каркасных областях компенсирует отсутствие легкой цепи, но также, по-видимому, повышает стабильность и, таким образом, растворимость домена VH, а также посредством этого максимизирует аффинность связывания антигена. Авторы настоящего изобретения исходят из того, что растворимость и стабильность антител, содержащих только тяжелые цепи и не содержащих легкие цепи, являются условием для эффективного внутриклеточного транспорта к клеточной поверхности, презентации и последующего размножения В-клеток.
Благоприятные мутации каркасной области также можно встроить в новые локусы антител, содержащих только тяжелые цепи, относящиеся к зародышевой линии, которые содержат сконструированные сегменты V человека, каждый из которых содержит предпочтительные замены аминокислот.
Аналогичным образом, предпочтительные замены аминокислот можно встроить в генные сегменты J и D для того, чтобы дополнительно повысить растворимость и стабильность доменов VH после перестройки VDJ.
Таким образом, предоставлены растворимые антитела человека с высокой аффинностью, содержащие только тяжелые цепи, в которых отсутствуют характерные для верблюдовых замены аминокислот и которые имеют повышенную гидрофобность в CDR1 и увеличенное число заряженных аминокислот в CDR3 по отношению к доменам VH, найденным в нормальных тетрамерных антителах человека. Растворимые антитела человека с высокой аффинностью, содержащие только тяжелые цепи, также содержат одну или более замен аминокислот в β-складчатом слое каркасной области, которые ведут к повышению общей гидрофобности в каркасной области 1, снижению гидрофобности в каркасной области 3 и увеличению числа заряженных аминокислот, присутствующих в каркасной области 3 по отношению к нормальным тетрамерным антителам человека.
Подход позволяет осуществлять естественную селекцию новых антител, содержащих только тяжелые цепи. Растворимые домены VH, полученные из таких антител, содержат мутации, которые редко или вообще не встречаются среди наблюдаемых в естественных тетрамерных антителах, где растворимость VH зависит' от взаимодействия VL доменом легких цепей к или А иммуноглобулина. В отсутствие легких цепей наблюдаемые замены аминокислот придают предпочтительные биофизические характеристики получаемым антиген-специфическим антителам, содержащим только тяжелые цепи, и растворимым доменам VH. В частности, антитела, содержащие только тяжелые цепи, и растворимые домены VH, полученные этим способом, не имеют склонности к «липкости», относительной нерастворимости и агрегации, которой обладают естественные домены VH, полученные из последовательностей зародышевой линии или естественных тетрамерных антител. Эти антитела, содержащие только тяжелые цепи, и растворимые домены VH по изобретению обладают высокой аффинностью (10 нМ или лучше), растворимы при отсутствии агрегации и подходят, в частности, для использования в качестве лекарственных средств
Наложение замен аминокислот, найденных в растворимых VH антигенсвязывающих доменах человека с высокой аффинностью на известную трехмерную структуру домена VH человека, присутствующую в нормальном тетрамерном антителе, проливает свет на положение и функциональное влияние определяемых замен аминокислот. Например:
Наблюдали замены в каркасной области 1 с 13-й до 15-й аминокислоты, преимущественно в положении 13, которые ведут к общему повышению гидрофобности. Изменения в положении 13, расположенные в малой петле, ведут к локальному повышению гидрофильности;
В β-складчатом слое на упомянутой поверхности контакта с легкой цепью могут происходить замены в каркасной области 2, сосредоточенные вокруг положений 35 и 50; и
По-видимому, в каркасной области 3 замены преобладают в участках контакта каркасной области/CDR, например, в конце CDR2 (положения 56 и 57) и в начале CDR3 (положение 93).
Изобретение необязательно относится ко вставкам или заменам аминокислот в предпочтительных новых каркасных областях, которые встроены в природные генные сегменты V для использования в оптимизированных трансгенных локусах антител, содержащих только тяжелые цепи, для получения антител с высокой аффинностью, содержащих только тяжелые цепи, и доменов VH с предпочтительными биофизическими характеристиками, включая растворимость при отсутствии агрегации.
Таким образом, предоставлены выбранные растворимые домены VH человека с предпочтительными биофизическими характеристиками, такими как растворимость при отсутствии агрегации. Это предоставляет источник природных каркасных областей тяжелых цепей, которые, после объединения с одним или более CDR, предусматривают дальнейшее образование библиотек связывающих белков VH, источник новых антигенсвязывающих белков (см. патентные заявки США №№11/102502 и 11/745644).
Также авторы настоящего изобретения с удивлением обнаружили, что экспрессировать кодируемые трансгенами антитела, содержащие только тяжелые цепи, также могут долгоживущие плазматические клетки мышей или В-клетки памяти, выделенные из вторичных лимфатических желез, костного мозга и селезенки трансгенных животных, сенсибилизированных антигеном, которые содержат локусы генов только тяжелых цепей. Кроме того, разработаны значительно более высокоэффективные способы, для которых не нужны гибридомы или классические подходы с использованием дисплеев для получения антиген-специфических антител, содержащих только тяжелые цепи, или растворимых доменов VH.
Предпочтительно клонированную кДНК, кодирующую антитело, содержащее только тяжелые цепи, или растворимые домены VH, полученные из выделенной из популяций плазматических клеток или клеток памяти сенсибилизированных антигеном трансгенных животных, может экспрессировать любая предпочитаемая клеточная линия, которая способна экспрессировать, накапливать, секретировать и представлять слитый белок домена VH или антитело, содержащее только тяжелые цепи, на клеточной поверхности. Предпочтительно выбирают линии бактериальных клеток или линии клеток млекопитающих, подходящие для производства веществ для клинического применения.
Таким образом, согласно второму аспекту по изобретению, антитела, содержащие только тяжелые цепи, и растворимые домены VH по первому аспекту изобретения получают из В-клеток памяти или долгоживущих плазматических клеток организма-хозяина. Второй аспект изобретения относится к способу получения антитела, содержащего только тяжелые цепи, которое специфично связывает антиген, способ содержит:
(a) иммунизация трансгенного млекопитающего, не относящегося к человеку, антигеном, где млекопитающее экспрессирует антитела, содержащие только тяжелые цепи, у которых отсутствует функциональность СН1 в транскрибируемой и процессируемой мРНК тяжелой цепи;
(b) выделение долгоживущих плазматических клеток или В-клеток памяти из иммунизированного млекопитающего;
(c) выделение популяции мРНК из клеток, полученных на стадии (b);
(d1) клонирование популяции кДНК, полученных из мРНК, выделенных на стадии (с), в экспрессионный вектор и экспрессия кДНК в предпочитаемой клеточной линии;
(e1) выбор по меньшей мере одной клеточной линии, которая продуцирует антитело, содержащее только тяжелые цепи, которое специфично связывает антиген;
или
(d2) клонирование популяции кДНК, содержащих домены VH, полученные из мРНК, выделенных на стадии (с), в предпочитаемый экспрессионный вектор так, чтобы слитый белок VH, который может содержать эффекторную область тяжелой цепи, экспрессировать в предпочитаемой клеточной линии;
(е2) выбор по меньшей мере одной клеточной линии, которая продуцирует слитый белок VH, который специфично связывается с антигеном.
Предпочитаемая клеточная линия предпочтительно получена у млекопитающего, но может быть получена и из дрожжей или любого организма, способного накапливать или секретировать антитела, содержащие только тяжелые цепи, и слитые белки домена VH или данные указанные антитела, содержащие только тяжелые цепи, и слитые белки домен VH на клеточной поверхности.
Способ подходит для выделения антител, содержащих только тяжелые цепи, которые получены или естественным путем в организме-хозяине, таком как лама, или в результате экспрессии трансгенного локуса в организме-хозяине, не относящемся к человеку.
Предпочтительно, предпочитаемая клеточная линия млекопитающего, например, клетки СНО, подходит для получения вещества для клинического применения.
Способ прост, высокоэффективен и хорошо подходит для автоматизации, поскольку антитела, содержащие только тяжелые цепи, являются гомодимерами, а не комплексными тетрамерами. Нет необходимости в поиске родственной легкой цепи, что представляет собой проблему, которая значительно усложняет получение антиген-специфических тетрамерных антител из популяций клеток памяти и плазматических клеток (см. Meijer et al. (2009) Therapeutic Antibodies: Methods and Protocols, 525, 261-277).
Популяции В-клеток, обогащенные долгоживущими плазматическими клетками или клетками памяти, можно выделить из организма-хозяина многими способами, которые хорошо отлажены в данной области. Предпочтительным организмом-хозяином является мышь. У мышей популяции В-клеток подробно охарактеризованы, а для того, чтобы различать различные субпопуляции, можно использовать поверхностные клеточные маркеры. Например, плазматические клетки мыши экспрессируют CD138 (Syndecan-1) на клеточной поверхности. Экспрессия CD138 на клетках линии дифференцировки В-клеток соответствует стадии развития, локализации и адгезии (см. Sanderson et al. (1989) Cell Regulation, 1, 27-35; Kim et al (1994) Mol. Biol. Cell., 5, 797-805). Плазматические клетки CD138+выявлены преимущественно в селезенке, лимфатических железах и костном мозге. Плазматические клетки дополнительно можно отличить от популяций наивных В-клеток, В-клеток памяти и не В-клеток мыши, поскольку они положительны по CD138, слабо положительны/отрицательны по CD4 5R (В220) и слабо положительны/отрицательны по CD19.
Доступность антител, распознающих поверхностные клеточные маркеры, позволяет отделять плазматические клетки от других популяций В-клеток посредством сортировки флуоресцентно-активированных клеток или хорошо отлаженными способами разделения клеток, которые широко доступны в наборах на основе, например, процедур разделения на магнитах, колонках или в центрифуге с использованием антител, конъюгированных с твердой подложкой, что позволяет отделить связыванием одну клеточную популяцию от другой. Таким образом, например магнитные бусы, содержащие антитела против CD45R (когда-то известные как В220), будут устранять большинство не плазматических клеток из смешанной популяции В-клеток. Последующая инкубация с магнитными бусами, содержащими антитела против CD138, будут захватывать высокообогащенную популяцию плазматических клеток (см. набор для магнитной сортировки для выделения CD138+ плазматических клеток мыши компании Miltenyi Biotec). Также можно использовать другие поверхностные клеточные маркеры для выделения этих клеток (например, Anderson et al. (2007) J. Exp. Med., 204, 2103-14). Также исходную клеточную популяцию можно получать из лимфатических желез и/или Пейеровых бляшек.
Когда не доступны смеси антител, которые позволяют дифференцировать данные линии дифференцировки В-клеток хозяина, тогда можно использовать комбинацию выделения В-клеток из костного мозга и лимфатических узлов с последующей очисткой В-клеток с использованием антитела против общего поверхностного маркера В-клеток, что позволяет получить высокообогащенную популяцию плазматических клеток.
В частности, это уместно для таких видов, как корова, кролик, овца и т.п., и даже человек, где поверхностные клеточные маркеры, такие как CD138, могут быть недоступны. Для других видов доступны маркеры для выбора конкретных популяций В-клеток (например, http://www.miltenvibiotec.com/en/PG_91_625_CD138_Plasma_Cell_Iso lation_Kit.aspx; Klein et al., Human immunoglobulin (Ig)M+IgD+peripheral blood B-cells expressing the CD27 cell surface antigen carry somatically mutated variable region genes: CD27 as a general marker for somatically mutated (memory) В cells, (1998) J. Exp. Med., 188, (9), 1679-89; Denham et al., Monoclonal antibodies putatively identifying porcine В cells, (1998) Vet Immunol Immunopathol. 30;60(3-4):317-28; Boersma et al., Summary of workshop findings for porcine B-cell markers, (2001) Vet. Immunol Immunopathol., 80, (1-2), 63-78; Naessens, Surface Ig on В lymphocytes from cattle and sheep, (1991) Int. Immunol., 9, (3), 349-54; Sehgal et al., Distinct clonal Ig diversification patterns in young appendix compared to antigen-specific splenic clones, (2002) J. Immunol., 168, (11), 5424-33).
После выделения мРНК из выделенных популяций плазматических клеток или клеток памяти, кДНК (которая представляет или антитело, содержащее только тяжелые цепи, или отдельно домены VH) клонируют в предпочитаемый экспрессионный вектор, и вектором трансфицируют популяцию клеток-мишеней для экспрессии антител, содержащих только тяжелые цепи, или слитые белки VH. Клетки, экспрессирующие антиген-специфические антитела, содержащие только тяжелые цепи, можно отобрать, например, стандартным ELISA в 96-луночном формате или в формате миркочипа или сортировки флуоресцентно-активированных клеток (FACS) для получения клеточных линий.
Предпочтительно для того, чтобы максимизировать эффективность процесса, экспрессионным вектором сначала трансфицируют бактерии, а колонии, содержащие клонируемую кДНК в экспрессионном векторе, собирают в 96-луночном формате (или используют в другом высокопроизводительном автоматизированном способе), растят в 96-луночном формате и выделяют ДНК. Плазмидной ДНК впоследствии трансфицируют в том же формате экспрессирующие клетки (например, клетки НЕК или клетки дрожжей и т.п.), а трансформанты растят в том же 96-луночном формате. Затем супернатант клеток тестируют в том же формате (или в другом, предпочтительно высокоплотном, формате, таком как микрочип, использующем меньшее количество антигена) на связывание антигена. Затем можно дополнительно размножать антиген-положительные клетки, экспрессирующие антитела, содержащие только тяжелые цепи, и положительные кДНК, непосредственно доступные из исходного 96-луночного формата, в котором ДНК получали для дальнейшей характеризации, такой как анализ последовательностей, или соответствующий домен VH можно непосредственно клонировать на другом остове, например, в другой константной области от того же вида или от другого вида. ДНК также можно использовать непосредственно для трансфекции клеток других типов для создания других клеточных линий.
Затем можно размножать клеточные линии, экспрессирующие антитела с высокой аффинностью, создавать банки клеток и использовать их в качестве источника целевых антител, содержащих только тяжелые цепи, для исследовательских целей или, возможно, для доклинических и клинических исследований. Таким образом, способ дополнительно содержит стадию поддержания выбранной клеточной линии, которая продуцирует антитело, содержащее только тяжелые цепи, или слитый белок VH, который специфично связывает антиген мишени.
Этот способ направленного клонирования также будет полезен для нормальных тетрамерных антител, полученных посредством трансгенов, где в каждой лунке используют отдельные CD138+ клетки. (Избыточную) мРНК, которая кодирует тяжелые и легкие цепи, раздельно амплифицируют посредством ПЦР с использованием праймеров со специфичностью к цепи и затем клонируют в экспрессионный вектор. Совместная трансфекция двумя векторами приведет к экспрессии тетрамерного антитела, которое можно идентифицировать таким же образом, как описано для антитела, содержащего только тяжелые цепи.
Предпочитаемые экспрессионные векторы известны специалисты в данной области и разработаны для осуществления эффективной экспрессии антитела, содержащего только тяжелые цепи, или слитого белка VH в предпочитаемом типе клеток. Когда предусмотрена секреция, вектор будет содержать сигнальный пептид на N-конце. Когда предусмотрена экспрессия на клеточной поверхности, тогда также будет иметь место гидрофобная пептидная последовательность на С-конце. Предпочтительно, но не по существу, такие N- и С-концевые пептиды можно получить из тяжелых цепей иммуноглобулинов млекопитающего. Векторы конструируют в формате кассеты так, чтобы растворимые домены VH, клонированные из антигенспецифических антител, содержащие только тяжелые цепи, можно было получать в виде слитых белков. Получаемый слитый белок может представлять собой антитело, содержащее только тяжелые цепи, и содержит растворимый домен VH и предпочитаемый эффекторный домен тяжелой цепи. Если растворимый домен VH и предпочитаемый эффекторный домен получены у человека, то полученный слитый белок представляет собой антитело человека, содержащее только тяжелые цепи, которое принадлежит к классу (и субтипу), определяемому посредством выбора эффекторной области (например, IgM, IgG, IgA, IgE или IgD). В любом антителе, содержащем только тяжелые цепи, растворимый домен VH и константные эффекторные области связаны шарнирной областью. Предпочтительно, но не по существу, она представляет собой естественную шарнирную область тяжелой цепи иммуноглобулина, полученную из соответствующего класса или субтипа.
Преимущество указанного выше способа относительно других способов, таких как фаговый дисплей или гибридомы с использованием веществ, полученных из цельной селезенки, состоит в том, что способ позволяет отбирать антитела с высокой аффинностью, содержащие только тяжелые цепи, (10 нм или лучше), продуцируемые плазматическими клетками или клетками памяти после все еще не известного процесса селекции in vivo. Такие популяции В-клеток продуцируют антитела, содержащие только тяжелые цепи, которые растворимы и не накапливаются во внутриклеточных компартментах (в ином случае такое накопление будет вести к апоптозу). Такие антитела, содержащие только тяжелые цепи, можно эффективно экспрессировать с использованием стандартных экспрессирующих систем, в частности, в созданных линиях клеток млекопитающих, таких как НЕК или СНО. Затем трансфицированные клетки можно упорядочить, используя налаженную технологию, типично автоматизированный микролуночный формат (например, 96 лунок). Затем те клетки, которые экспрессируют антигенсвязывающие антитела с высокой аффинностью, содержащие только тяжелые цепи, или альтернативно клетки, которые экспрессируют антитела, содержащие только тяжелые цепи, которые связывают антиген в определенном диапазоне аффинностей, можно отобрать для дальнейшего анализа. Альтернативно связывающие средства с высокой аффинностью можно отобрать посредством анализа FACS. Использование клеток памяти или плазматических клеток, полученных из одного или более сенсибилизированных антигеном трансгенных животных, не относящихся к человеку, например, из мыши, которая экспрессирует антиген-специфические антитела, содержащие только тяжелые цепи, в качестве источника мРНК антитела, содержащего только тяжелые цепи, предоставляет чрезвычайно эффективный путь для селекции антител, содержащих только тяжелые цепи, который позволяет осуществлять селекцию потенциально миллионов клеток, продуцирующих антитела, содержащие только тяжелые цепи, для последующего анализа. Затем прошедшие селекцию клетки, экспрессирующие антигенсвязывающие антитела с высокой аффинностью, содержащие только тяжелые цепи, можно размножать в культуре, обеспечивая источник клеток, которые продуцируют антитела, содержащие только тяжелые цепи, в отсутствие дополнительных стадий клонирования.
Необязательно, выделенные долгоживущие плазматические клетки или В-клетки памяти можно иммортализовать перед выделением мРНК и клонированием антиген-специфических антител, содержащих только тяжелые цепи, или доменов VH.
Настоящее изобретение также относится к трансгенному млекопитающему, не относящемуся к человеку, которое содержит трансгенный локус тяжелой цепи или локус, содержащий только тяжелую цепь (отсутствует функциональность СН1), который содержит доминантный селекционный маркерный ген. Также предусмотрено введение более чем одного селективного маркера, каждый маркер связан с отдельным локусом или группой локусов, что допускает предпочтительный отбор гибридом, экспрессирующих один в отличие от другого локуса или группы локусов, способных экспрессировать антитела, содержащие только тяжелые цепи.
Использование нескольких селективных маркеров в различных локусах основано на обнаружении того, что когда трансгенное млекопитающее, не относящееся к человеку, содержит несколько локусов, содержащих только тяжелые цепи, эти локусы подвержены аллельному исключению (PCT/IB2007/001491). Следовательно, после сенсибилизации антигеном происходит случайный выбор только одного локуса и успешная рекомбинация, которая ведет к образованию антитела, содержащего только тяжелые цепи. Следовательно можно использовать несколько VH локусов, содержащих только тяжелые цепи, в одном и том же трансгенном млекопитающем, не относящемся к человеку, чтобы максимизировать репертуар и разнообразие антител, которые можно получить у млекопитающего. После сенсибилизации антигеном, трансгенное млекопитающее, не относящееся к человеку, будет осуществлять случайные рекомбинации в локусе за локусом, не отдавая предпочтения какому-либо данному локусу, пока одна из рекомбинаций не будет «защитной», т.е. не приведет к образованию антитела. Происходит остановка дальнейшей рекомбинации и тем самым «исключение» других аллелей (локусов). Клетки, которые продуцируют антитело с высокой аффинностью, предпочтительно размножают.
Типично доминантные селективные маркерные гены получают у прокариотов и выбирают из группы, которая или придает устойчивость к токсическим лекарственным средствам, таким как пуромицин, гигромицин и G418, или содержит гены, которые устраняют определенные требования к питательной среде, так что их экспрессия превращает токсичный субстрат в незаменимую аминокислоту, например, превращает индол в триптофан или превращает гистидинол в гистидин. Необходимое требование состоит в том, что если используют доминантный селективный маркер, то происходит его совместная экспрессия с желаемым аллелем иммуноглобулина, содержащим только тяжелые цепи, таким образом гарантируя В-клеточную экспрессию и интеграцию сайт-независимой экспрессии трансгенного локуса. Альтернативно, ген устойчивости к лекарственному средству можно встроить в эндогенный или экзогенный (трансгенный) локус иммуноглобулина с использованием гомологичной рекомбинации в сочетании с клетками ES или подходов, основанных на ядерной передаче.
Таким образом, предоставлен способ получения моноклонального антитела, который требует осуществления селекции линии гибридомы или трансформированных В-клеток, предпочтительно полученных из клеток памяти или плазматических клеток, экспрессирующих определенный локус иммуноглобулина, содержащего только тяжелые цепи, содержащих один из нескольких локусов генов, содержащих только тяжелые цепи, которые присутствуют в трансгенном млекопитающем, не относящемся к человеку, указанный локус содержит совместно экспрессируемый доминантный селективный маркерный ген, вставленный в указанный локус.
Предпочтительно, этот локус представляет собой локус, содержащий только тяжелую цепь, доминантный селективный маркерный ген, этот локус экспрессирует антитело, содержащее только тяжелые цепи. Предпочтительно, антитело, содержащее только тяжелые цепи, получают из эндогенного локуса, содержащего только тяжелую цепь, который сконструирован без функциональности СН1, или из введенного трансгена тяжелой цепи, у которого отсутствует функциональность СН1, необязательно в отсутствие экспрессии гена легкой цепи иммуноглобулина. Указанный локус также может содержать доминантный селективный маркер и локус тяжелой цепи в отсутствие локусов легких цепей, указанный ген тяжелой цепи, во время экспрессии, спонтанно повышает уровень мРНК транскрипта, у которого отсутствует функциональность СН1, что ведет к экспрессии антитела, содержащего только тяжелые цепи, зависимым от В-клеток образом.
Согласно третьему аспекту по изобретению, когда для иммортализации плазматических клеток или клеток памяти предусмотрены подходы с использованием гибридом или трансформации В-клеток, тогда включение функционального доминантного селективного маркерного гена в каждый локус тяжелой цепи позволяет осуществлять последовательную селекцию и стабильное поддержание гибридомы или трансформированных линий В-клеток, экспрессирующих локус, представляющий интерес, после сенсибилизации антигеном, и при этом теряются все гибридомные клетки или трансформированные В-клетки, которые не экспрессируют этот локус, представляющий интерес. Для цели по изобретению можно использовать любой доминантный селективный маркерный ген при условии, что экспрессия гена обеспечивает гибридомы селективным преимуществом в присутствие селективного стимула, например, токсического стимула (см. Vara et al. (1986) NAR5 14, 4617-4624; Santerre et al. (1984) Gene, 30, 147-156; Colbere-Garapin et al. (1981) 150, 1-14; Hartmann and Mulligan (1988) PNAS, 85, 8047-8051).
Гетерологичный локус, содержащий только тяжелую цепь
В контексте настоящего изобретения термин «гетерологичный» обозначает нуклеотидную последовательность или локус, как описано в настоящем документе, который не является эндогенным для млекопитающего, в котором он расположен, или эндогенный локус, модифицированный посредством замены или удаления эндогенной последовательности.
«Локус, содержащий только тяжелую цепь» в контексте настоящего изобретения относится к локусу, который кодирует домен VH, содержащий один или более генных сегментов V, один или более генных сегментов D и один или более генных сегментов J, которые функционально связаны с одной или более эффекторными областями тяжелых цепей (у каждой отсутствует функциональность СН1 домена).
Сложность генного сегмента V можно повысить путем увеличения числа генных сегментов V, присутствующих в локусе, или путем использования различных локусов, каждый из которых содержит различные генные сегменты V.
Предпочтительно локус, содержащий только тяжелую цепь, содержит от пяти до двадцати различных генных сегментов V, полученных у любого вида позвоночных.
Предпочтительно генные сегменты V получены у человека, необязательно прошли селекцию или получены конструированием для повышения растворимости.
Предпочтительно локус, содержащий только тяжелую цепь, содержит от двух до сорока (2, 3, 4, 5, 6, 7, 8, 9, 10, 12, 14, 16, 18, 20, 30 или 40) или более генных сегментов D. Генные сегменты D можно получить из любого вида позвоночных, но наиболее предпочтительно генные сегменты D представляют собой генные сегменты D человека (в норме 25 функциональных генных сегментов D).
Предпочтительно, локус, содержащий только тяжелую цепь, содержит от двух до двадцати (2, 3, 4, 5, 6, 7, 8, 9, 10, 12, 14, 16, 18 или 20) или более генных сегментов J. Генные сегменты J можно получить из любого вида позвоночных, наиболее предпочтительно генные сегменты J представляют собой генные сегменты J человека (в норме 6 генных сегментов J).
Предпочтительно локус, содержащий только тяжелую цепь, содержит два или более генных сегмента V, двадцать пять функциональных генных сегментов D человека и 6 генных сегментов J человека.
Термин «генный сегмент V» охватывает встречающийся в природе генный сегмент V, полученный у позвоночного, включая верблюдовых и человека, который необязательно прошел селекцию, созревание или получен конструированием для повышения характеристик, таких как растворимость. Генные сегменты V также обнаружены у других видов позвоночных, таких как акула (см. Kokubu et al. (1988) EMBO J., 7, 3413-3422), или эволюционировали для того, чтобы предоставить разнообразные VH-подобные семейства связывающих белков, примером чего служит, например, эволюция репертуара VL легких цепей антител или репертуара VH Т-клеточного рецептора.
Генный сегмент V должен быть способен к рекомбинации с генным сегментом D, генным сегментом J и константной (эффекторной) областью тяжелой цепи (которая может содержать несколько экзонов, но не содержит экзон СН1) в соответствии с настоящим изобретением для создания антитела, содержащего только тяжелые цепи, когда экспрессируют нуклеиновую кислоту.
Объем генного сегмента V согласно настоящему изобретению также включает любую последовательность гена, кодирующую гомолог, производное или фрагмент белка, которая способна к рекомбинации с генным сегментом D, генным сегментом J и константной эффекторной областью тяжелой цепи (содержит один или более экзонов, но не функциональный экзон СН1) в соответствии с настоящим изобретением для создания антитела, содержащего только тяжелые цепи, как определено в настоящем документе. Константная эффекторная область тяжелой цепи может содержать СН1 домен в условиях, где локус тяжелой цепи иммуноглобулина экспрессируют у животного-хозяина с генотипом, лишенным экспрессии гена легкой цепи иммуноглобулина.
Таким образом, VH кодирующие последовательности можно получить из встречающегося в природе источника или их можно сконструировать или синтезировать, используя способы, известные специалистам в данной области.
«Растворимый домен VH» в контексте настоящего изобретения относится к продукту экспрессии генного сегмента V после рекомбинации с генным сегментом D и генным сегментом J, как определено выше. После естественной селекции и созревания аффинности родительского антитела, содержащего только тяжелые цепи, растворимый домен VH, как применяют в настоящем документе, после экспрессии и выделения остается в растворе при отсутствии агрегации и сохраняет активность в физиологической среде без необходимости в любом другом факторе для поддержания растворимости. Также можно сконструировать отдельно растворимый домен VH с использованием различных белковых доменов, чтобы получить слитые белки для целей таргетной терапии и диагностики, например с токсинами, ферментами или средствами для визуализации или с белками крови, такими как альбумин, чтобы фармакокинетикой управлять растворимого домена VH и распределением в тканях in vivo (см. US 5843440 и Smith et al. (2001) Bioconjugate Chem., 12, 750-756).
В контексте настоящего изобретения термины «генный сегмент D» и «генный сегмент J» содержат встречающиеся в природе последовательности генных сегментов D и J. Предпочтительно генные сегменты D и J получают у того же позвоночного, у которого получают генный сегмент V. Например, если генные сегмент V получают у человека и затем повышают его растворимость или используют в конструировании, то генные сегменты D и J предпочтительно также получают у человека. Альтернативно можно получить генные сегменты V, например, у крысы или мыши, а генные сегменты D и J у верблюда или человека.
Также объем терминов «генный сегмент D» и «генный сегмент J» включает их производные, гомологи и фрагменты при условии, что получаемый сегмент можно рекомбинировать с остальными компонентами локуса тяжелой цепи антитело, как описано в настоящем документе, для создания антитела, содержащего только тяжелые цепи, как описано в настоящем документе. Генные сегменты D и J можно получить из встречающихся в природе источников или их можно синтезировать, используя способы, известные специалистам в данной области и описанные в настоящем документе. Генные сегменты D и J могут содержать определенные дополнительные аминокислотные остатки или определенные замены аминокислот или делеции для того, чтобы повысить разнообразие CDR3.
Генные сегменты V, D и J способны к рекомбинации и предпочтительно подвержены соматическим мутациям.
Генные сегменты V, D и J предпочтительно получают у одного вида позвоночных. Он может представлять собой любой вид позвоночных, но предпочтительно он представляет собой человека.
Характерные последовательности представляют собой четыре точно определенные замены в аминокислотной последовательности, кодируемые зародышевой линией (тетрада VHH), представленные в доменах VHH верблюдовых, но не в растворимых доменах VH. Они все присутствуют во второй каркасной области и выполняют функцию снижения гидрофобности упомянутой поверхность контакта с легкой цепью. Glu-44 и Arg-45 обнаруживают вместо менее гидрофильных Gly-4 4 и Leu-4 5, которые присутствуют в доменах VH, тогда как гидрофильность в положении 4 7 повышают замены Тгр на остатки меньших размеров. Четвертый заряд требует замещения Val-37 на Phe или Туr.
Константная область тяжелой цепи
Функционально константную область тяжелой цепи кодирует встречающийся в природе или сконструированный генный сегмент, который способен к рекомбинации с генным сегментом V, генным сегментом D и генным сегментом J в В-клетке. Предпочтительно константную область тяжелой цепи получают из локуса антитела.
Каждая константная область тяжелой цепи по существу содержит по меньшей мере один ген константной области тяжелой цепи, который экспрессируется без функционального СН1 домена с тем, чтобы могло произойти образование антитела, содержащего только тяжелые цепи. Когда отсутствует экспрессия эндогенного гена легкой цепи иммуноглобулина, константная область тяжелой цепи может содержать функциональность СН1, которая может с низкой частотой спонтанно исчезать во время экспрессии гена тяжелой цепи. Каждая константная область тяжелой цепи также может содержать одну или более дополнительных экзонов константной области тяжелой цепи, которые выбирают из группы, состоящей из Сδ, Сγ1-4, Сµ, Сε и Сα1-2. Генные сегменты константной области тяжелой цепи выбирают в зависимости от предпочтительного класса или необходимой смеси классов антител. Необязательно, в гетерологичном локусе тяжелой цепи не достает Сµ и Сδ.
Например, экспрессия всего или части локуса Сγ гетерологичной тяжелой цепи, лишенной СН1, будет создавать необязательно некоторые или все изотипы IgG, в зависимости от изотипов IgG1, IgG2, IgG3 и IgG4, присутствующих в гетерологичном локусе IgG.
Альтернативно можно получить выбранные смеси антител. Например, когда константная область тяжелой цепи содержит ген Сα и Сµ, можно получить IgA и IgM.
Константная область тяжелой цепи, присутствующая в трансгенном локусе, может быть получена у человека, но может быть такой же как или близкородственной к таковой млекопитающего-хозяина с тем, чтобы максимизировать ответ на антигены in vivo. Таким образом, для получения антител, содержащих только тяжелые цепи, или растворимых доменов VH для применения в терапии людей, гены VDJ, присутствующее в локусе, получают из необязательно модифицированных последовательностей зародышевой линии человека, а константные области можно получать у грызуна, если животным-хозяином является грызун, например, мышь. Затем прошедшие селекцию антитела, содержащие только тяжелые цепи, которые содержат растворимые связывающие домены VH человека и константные эффекторные области грызуна, можно клонировать и заменить эффекторные области грызуна предпочитаемыми константными эффекторными областями человека или растворимыми доменом VH, используемым в полученных альтернативных слитых белках VH, комплексах домена VH антитела и т.п.
Когда антитела, содержащие только тяжелые цепи, подлежат использованию в ветеринарных или альтернативных целях, генные сегменты V, D и J предпочтительно получают у позвоночного или млекопитающего, наилучшим образом подходящего для предполагаемой цели. Например, генные сегменты V и генные сегменты D и J можно получить у других млекопитающих (например, у мыши, крысы, свиньи, коровы, козы, овцы, верблюда и т.п.) в зависимости от предполагаемого использования, например, в ветеринарном, промышленном, сельскохозяйственном, диагностическом применении или применении в качестве реактива.
«Экзон константной области тяжелой цепи» («экзон СН»), как определено в настоящем документе, содержит последовательности встречающихся в природе экзонов СН позвоночных и, в особенности, млекопитающих. Они отличаются в зависимости от класса. Например, природные IgG и IgA лишены СН4 домена. Объем термина «экзон СН» также включает его производные, гомологи и фрагменты при условии, что экзон СН способен к формированию функционального антитела, содержащего только тяжелые цепи, как определено в настоящем документе, когда он является компонентом константной области тяжелой цепи.
Млекопитающие
Трансгенное млекопитающее, используемое в способах по изобретению, не является человеком. Трансгенными млекопитающими предпочтительно являются грызуны, такие как кролик, морская свинка, крыса или мышь. Мыши и крысы особенно предпочтительны. Также можно использовать альтернативных млекопитающих, таких как козы, свиньи, рогатый скот, овцы, кошки, собаки или другие животные.
Предпочтительно, трансгенных животных, содержащих гетерологичные локусы антител, содержащих только тяжелые цепи, интегрированные в зародышевую линию, создают с использованием налаженной технологии инъекции в ооцит и, если налажено, технологии клеток ES, технологии клеток iPS или технологии ядерной передачи (клонирования). Альтернативно локусы можно вводить в указанные выше клетки или непосредственно в оплодотворенные яйцеклетки с использованием нуклеаз, содержащих цинковые пальцы, (Reray et al. (2009) Transgenic Res., 2009 Sep 26. [электронная публикация до выхода в печать], Kandavelou et al. (2009) Biochem. Biophys. Res. Commun., 388, (1), 56-61) или технологии рекомбинантных ферментов (например, Sakurai et al. (2010) Nucleic Acids Res., Jan 13 [электронная публикация до выхода в печать] и цитируемые в них источники), такие как Сrе и т.д.
Стандартную гомологичную рекомбинацию в клетках ES также можно использовать для устранения функциональности СН1 из эндогенных генов тяжелых цепей и для замены элементов или последовательностей в эндогенных сегментах генов V или генных сегментах D и J, что ведет к образованию антител, содержащих только тяжелые цепи, из эндогенных локусов тяжелых цепей.
Предпочтительно усиливают трансгенную экспрессию гетерологичных локусов генов, содержащих только тяжелые цепи, где локусы тяжелых цепей антитела, эндогенные для млекопитающего, удаляют или переводят их молчащее состояние или они имеют сниженную способность к образованию эндогенных антител. Когда в эндогенных локусах тяжелых цепей организма-хозяина путем конструирования устраняют функциональность CH1, а VDJ и необязательно константные эффекторные области заменяют последовательностями других видов, предпочтительно человека, тогда добавление дополнительных трансгенных локусов тяжелых цепей не является необходимым.
Этот подход к созданию антител, содержащих только тяжелые цепи, и растворимые домены VH, как описано выше, может иметь особое значение при создании растворимых антител и комплексов антител, не образующих агрегаты, для терапевтического использования у человека. Следовательно, в дополнительном аспекте настоящее изобретение относится к трансгенному млекопитающему, которое экспрессирует гетерологичный локус тяжелой цепи в соответствии с настоящим изобретением в ответ на сенсибилизацию антигеном.
Антителопродуцирующие клетки можно получить из трансгенных животных в соответствии с настоящим изобретением и использовать, например, в предпочтительном варианте осуществления, для получения клеток памяти или долгоживущих плазматических клеток, а также для получения антител, содержащих только тяжелые цепи, от гибридом или посредством массовых подходов, как определено в настоящем документе. Кроме того или альтернативно, последовательности нуклеиновых кислот, кодирующие антитела, содержащие только тяжелые цепи, и/или растворимые домены VH, можно выделять из В-клеток, в особенности, из клеток памяти или долгоживущих плазматических клеток трансгенных млекопитающих в соответствии с настоящим изобретением после сенсибилизации антигеном и использовать для получения растворимых антител с доменом VH, содержащих только тяжелые цепи, растворимых биспецифических/бифункциональных комплексов с доменом VH или растворимых полипротеинов доменов VH (см. WO 99/23221), используя способы рекомбинантной ДНК, которые известны специалистам в данной области.
Альтернативно или кроме того, поликлональные антиген-специфические антитела, содержащие только тяжелые цепи, можно создавать посредством иммунизации трансгенного животного в соответствии с настоящим изобретением.
Таким образом, в дополнительном аспекте настоящее изобретение относится к способу получения антител с высокой аффинностью, содержащих только тяжелые цепи, посредством иммунизации трансгенного млекопитающего в соответствии с настоящим изобретением, с использованием антигена.
В предпочтительном варианте осуществления этого аспекта по изобретению млекопитающим является грызун, предпочтительно мышь или крыса.
Получение антител, содержащих только тяжелые цепи, и слитых белков растворимых доменов УН
Системы для получения антител, содержащих только тяжелые цепи, и слитых белков растворимых доменов VH включают клетки млекопитающих в культуре (например, клетки СНО), растения (например, маис), трансгенных коз, кроликов, рогатый скот, овец, кур и личинки насекомых, подходящие для технологий массового размножения. Другие системы для получения, включая вирусную инфекцию (например, бакуловирус в личинках насекомых и клеточных линиях), являются альтернативой для подходов с использованием клеточных культур и зародышевой линии. Другие способы получения также известны специалистам в данной области. Когда необходим сборка содержащих только тяжелые цепи IgA или IgM, совместная экспрессия «цепи J» может быть полезной. Подходящие способы получения антител, содержащих только тяжелые цепи, или растворимых связывающих доменов VH в отдельности или комплексов связывающих доменов VH известны в данной области. Например, в бактериальных системах получали связывающие домены VH и комплексы связывающих доменов VH, а в гибридомах и трансфицированных клетках млекопитающих получали гомодимеры, содержащие только тяжелые цепи, (см. Reichmann and Muyldermans, (1999) J. Immunol. Methods, 231, 25-38).
Также хорошо отлажены способы для экспрессии сконструированных связывающих доменов VH человека, полученных с использованием технологии фагового дисплея (Tanha et al. (2001) J. Biol. Chem., 276, 24774-24789 и цитируемые в них источники).
Настоящее изобретение также относится к полинуклеотидной последовательности, состоящей из гетерологичного локуса тяжелой цепи, выделенного полинуклеотида, кодирующего антитело по изобретению, содержащее только тяжелые цепи, и к вектору, содержащему гетерологичный локус тяжелой цепи или его фрагмент или выделенный полинуклеотид, который кодирует антитело, содержащее только тяжелые цепи, в соответствии с настоящим изобретением. Изобретение также относится к полинуклеотидным последовательностям, которые кодируют растворимые домены VH и слитые белки доменов VH, и к вектору, содержащему такие последовательности.
Настоящее изобретение также относится к клетке-хозяину, которую трансформировали гетерологичным локусом, содержащим только тяжелую цепь, или его фрагментом или выделенным полинуклеотидом, который кодирует антитело, содержащее только тяжелые цепи, растворимый домен VH или слитый белок домена VH в соответствии с настоящим изобретением.
Использование антител, содержащих только тяжелые цепи, растворимых доменов УН и полипептидных комплексов и полипротеинов доменов VН по изобретению
Растворимые антитела с высокой аффинностью, содержащие только тяжелые цепи, слитые белки доменов VH, полипептидные комплексы доменов VH и растворимые домены VH по настоящему изобретению, в частности, подходят для использования как в качестве парентеральных, так и в качестве непарентеральных лекарственных средств. В контексте изобретения предпочтительно их полностью получают у человека, но необязательно и менее предпочтительно они могут содержать генные сегменты V, D или J других видов.
Антитела, содержащие только тяжелые цепи, содержащие растворимые домены VH, выделенные растворимые домены VH и связывающие комплексы растворимых доменов VH и полипротеинов с повышенной растворимостью по изобретению особенно подходят для использования в качестве реактивов, а также, в дополнение к терапии, в качестве диагностических средств.
Все они подходят для фармацевтического использования у людей и, таким образом, изобретение относится к фармацевтической композиции, которая содержит антитело, содержащее только тяжелые цепи, содержащие растворимые домены VH, выделенные растворимые домены VH, связывающие комплексы растворимых доменов VH или полипротеины растворимых доменов VH. Изобретение также относится к использованию антител, содержащих только тяжелые цепи, содержащих растворимые домены VH, выделенных растворимых доменов VH, связывающих комплексов растворимых доменов VH или полипротеинов растворимых домен VH по настоящему изобретению в получении лекарственного средства для профилактики и/или лечения заболевания.
Перед введением пациентам фармацевтические композиции и лекарственные средства имеют типичный состав.
Например, антитело, содержащее только тяжелые цепи, содержащие растворимые домены VH, выделенные растворимые домены VH, связывающие комплексы растворимых доменов VH или полипротеины растворимых доменов VH можно смешать со стабилизаторами, в частности, если они подлежат лиофилизации. Чтобы придать стабильность во время лиофилизации, типично добавляют сахара (например, маннит, сахарозу или трегалозу), а предпочтительным стабилизатором является маннит. В качестве стабилизатора также можно добавлять сывороточный альбумин человека (предпочтительно, рекомбинантный). Также можно использовать смеси сахаров, например, сахарозу и маннит, трегалозу и маннит и т.д.
В композицию можно добавлять буфер например, буфер Tris, гистидиновый буфер, глициновый буфер или предпочтительно фосфатный буфер (например, содержащий дигидрофосфат натрия и гидрофосфат динатрия). Предпочтительно добавляют буфер, чтобы получить pH между 7,2 и 7,8 и, в частности, pH приблизительно 7,5.
Для восстановления после лиофилизации можно использовать стерильную воду для инъекций. Также можно восстанавливать лиофилизат с использованием водных композиций, содержащих сывороточный альбумин человека (предпочтительно, рекомбинантный).
Как правило, антитела, содержащие только тяжелые цепи, содержащие растворимые домены VH, выделенные растворимые домены VH, связывающие комплексы растворимых доменов VH или полипротеины растворимых доменов VH используют в очищенной форме вместе с фармакологически приемлемыми носителями.
Таким образом, изобретение относится к способу лечения пациента, который содержит введение фармацевтической композиции по изобретению пациенту. Пациентом предпочтительно является человек и, возможно, ребенок (например, ребенок ясельного возраста или младенец), подросток или взрослый, но, как правило, взрослый.
Изобретение также относится к антителу, содержащему только тяжелые цепи, содержащие растворимые домены VH, выделенным растворимым доменам VH, связывающим комплексам растворимых доменов VH и к полипротеинам растворимых доменов VH по изобретению для применения в качестве лекарственного средства.
Изобретение также относится к использованию антител, содержащих только тяжелые цепи, содержащих растворимые домены VH, выделенных растворимых доменов VH, связывающих комплексов растворимых доменов VH и полипротеинов растворимых доменов VH по изобретению в производстве лекарственного средства для лечения пациента.
Эти применения, способы и лекарственные средства предпочтительно предназначены для лечения заболеваний или нарушений, которые включают в качестве неограничивающих примеров: заживление ран, нарушения клеточной пролиферации, включая новообразования, меланому, остеосаркому легких, ободочной и прямой кишки, саркому прямой кишки, яичника, солидные опухоли шейки матки, пищевода, груди, поджелудочной железы, мочевого пузыря, головы и шеи и другие; миелопролиферативые нарушения, такие как лейкемия, неходжкинская лимфома, лейкопения, тромбоцитопения, нарушение ангиогенеза, саркома Капоши; аутоиммунные/воспалительные нарушения, включая аллергию, воспалительное заболевание кишечника, артрит, псориаз и воспаление дыхательных путей, астму, иммунные нарушения и отторжение трансплантата органа; сердечнососудистые и сосудистые нарушения, включая гипертензию, отек, стенокардию, атеросклероз, тромбоз, сепсис, шок, реперфузионное повреждение и ишемию; неврологические нарушения, включая заболевания центральной нервной системы, болезнь Альцгеймера, травму головного мозга, амиотрофический боковой склероз и боль; нарушения развития; нарушения метаболизма, включая сахарный диабет, остеопороз и ожирение, СПИД и почечную недостаточность; инфекции, включая вирусную инфекцию, бактериальную инфекцию, грибковую инфекцию и паразитарную инфекцию, патологические состояния, связанные с плацентой и другие патологические состояния, а также для использования в иммунотерапии.
В еще одном дополнительном аспекте настоящее изобретение относится к использованию антитела, содержащего только тяжелые цепи, содержащих растворимые домены VH, выделенных растворимых доменов VH, связывающих комплексов растворимых доменов VH или полипротеинов растворимых доменов VH по настоящему изобретению в качестве диагностического, прогностического средства или терапевтического средства для визуализации по отдельности или в сочетании с подходящими эффекторными средствами.
Настоящее изобретение относится к использованию антитела, содержащего только тяжелые цепи, содержащие растворимые домены VH, выделенные растворимые домены VH, связывающие комплексы растворимых доменов VH или полипротеинов растворимых доменов VH, как описано в настоящем документе, в качестве внутриклеточного связывающего реактива или абзима. Предпочтительны антигенспецифические растворимые связывающие домены VH, связывающие комплексы растворимых доменов VH и полипротеины растворимых доменов VH.
Настоящее изобретение также относится к использованию антигенспецифического антитела, содержащего только тяжелые цепи, или растворимого связывающего домен VH в соответствии с настоящим изобретением в качестве ингибитора фермента или блокатора рецептора. Предпочтительными фрагментами антител, содержащими только тяжелые цепи, являются антигенспецифические растворимые связывающие домены VH.
Настоящее изобретение также относится к использованию одного или более растворимых доменов VH, слитых с эффекторной молекулой, для применения в качестве терапевтического, диагностического средства, средства для визуализации, абзима или реактива.
Далее, только в качестве примера, приведено следующее подробное описание изобретения, относящееся к следующим фигурам.
ФИГУРЫ
Фиг.1. Два локуса иммуноглобулинов человека, содержащих только тяжелые цепи, один несет 4 генных сегмента VH, другой несет 18 генных сегментов VH. Локусы, содержащее генные сегменты 18VH, содержат константные эффекторные области мыши (-СН1), которые покрывают более чем 90% используемых генных сегментов VH в нормальных тетравалентных антителах человека.
Фиг.2. Протокол иммунизации и выделения для домена VH человека посредством фагового дисплея или антител человека, содержащих только тяжелые цепи, посредством стандартной гибридомной технологии.
Фиг.3. ELISA (вверху) и вестерн-блоттинг (внизу) иммунизированной сыворотки Luk-sPV (V4 в сравнении с нормальными мышами дикого типа) и вестерн-блоттинг отдельно выделенных доменов VH с использованием электрофореза белка Lulc-sPV в геле, его блоттинга и определения с использованием различных положительных клонов из скрининга фагового дисплея.
Фиг.4. Пример двух последовательностей домена VH после иммунизации с использованием Luks-PV, который показывает, что получены как IgG2, так и IgG3 (переключение класса), и что одна и та же перестройка VDJ ведет к различным мутациям.
Фиг.5. Последовательности домена VH, полученные с использованием генного сегмента V3-23 после иммунизации с использованием CR1 человека, которые показывают, что получены как IgG2, так и IgG3 (переключение класса), и что та же перестройка VDJ ведет к различным мутациям, когда сравнивают с последовательностью сегмента V3-23 зародышевой линии, который представлен вверху. Части последовательностей получали посредством гибридом, а остальные посредством библиотек дисплеев.
Фиг.6. Последовательности домена VH, полученные с использованием генного сегмента V3-11 после иммунизации с использованием CR1 человека, которые показывают, что получены как IgG2, так и IgG3 (переключение класса), и что та же перестройка VDJ ведет к различным мутациям, когда сравнивают с последовательностью сегмента V3-11 зародышевой линии, представленной вверху. Части последовательностей получали посредством гибридом (стрелки), а остальные посредством библиотек дисплеев.
Фиг.7. Пример электрофореза в геле (слева) доменов VH (αCR1) после очистки и экспрессии в системе гибридного белка SUMO (InVitrogen). В примере показаны домены VH трех различных длин с правильной молекулярной массой. Дорожка с маркером расположена слева. В средней части представлен профиль элюции доменов VH на системе Sephadex 75 Smart. Правая часть содержит аналогичную информацию, но для полного IgG, содержащего только тяжелые цепи, на Sephadex 200. В нижней части представлены профили элюции двух других доменов VH, сконцентрированных до >4 мг/мл.
Фиг.8. А. График светорассеяния и В. Тепловая стабильность доменов VH.
Фиг.9. Измерения аффинности нескольких антител против αCR1, содержащих только тяжелые цепи, (слева) и профили связывании и диссоциации одного из антител на системе Octet, чтобы получить аффинность связывания.
Фиг.10. Растворимость нескольких доменов VH против CR1.
Фиг.11. Массовое параллельное секвенирование для определения предпочтительных мутаций.
Фиг.12. Анализ коробчатой диаграммы для каркасной области 1 (FR1) и области CDR1 антител человека, содержащих только тяжелые цепи, в сравнении с теми же областями тяжелых цепей нормальных тетрамерных антител верблюда, ламы и человека, которые содержат тяжелые/легкие цепи. Суммарный заряд определяли по уравнению Хендерсона-Хассельбаха при pH=7,4; суммарную гидрофобность определяли по индексу Коана-Уиттакера при pH=7,5. Числа, расположенные ниже различных антител, представляют собой число уникальных входных последовательностей.
Фиг.13. Анализ коробчатой диаграммы каркасной области 2 (FR2) и области CDR2 антител человека, содержащих только тяжелые цепи, в сравнении с теми же областями тяжелых цепей нормальных тетрамерных антител верблюда, ламы и человека, которые содержат тяжелые/легкие цепи. Суммарный заряд определяли по уравнению Хендерсона-Хассельбаха при pH=7,4; суммарную гидрофобность определяли по индексу Коана-Уиттакера при pH=7,5. Числа, расположенные ниже различных антител, представляют собой число уникальных входных последовательностей.
Фиг.14. Анализ коробчатой диаграммы каркасной области 3 (FR3) и области CDR3 антител человека, содержащих только тяжелые цепи, в сравнении с теми же областями тяжелых цепей нормальных тетрамерных антител верблюда, ламы и человека, которые содержат тяжелые/легкие цепи. Суммарный заряд определяли по уравнению Хендерсона-Хассельбаха при pH=7,4; суммарную гидрофобность определяли по индексу Коана-Уиттакера при pH=7,5. Числа, расположенные ниже различных антител, представляют собой число уникальных входных последовательностей
Фиг.15. Трехмерная структура доменов VH, связывающих или CR1 человека или Luk-sPV стафилококка. Антигенсвязывающий участок приведен в верхнем правом положении, а упомянутый VL домен взаимодействия обращен к читателю. Изменения аминокислот в трехмерной структуре показаны цветом.
Фиг.16. Различия в средней гидрофобности в каждом положении домена VH (за исключением CDR3), которые указывают на сдвиг в сторону повышения гидрофобности антител, содержащих только тяжелые цепи, в частности в N-конце домена VH. Множество горячих точек в отношении различий гидрофобности видны в положениях 4 9, 62 и 72 по сравнению с усредненным тетрамерным VH человека.
Фиг.17. Протокол иммунизации и выделения антитела человека, содержащего только тяжелые цепи, посредством сортировки CD138 положительных клеток и непосредственного экспрессионного клонирования в НЕК клетки.
Фиг.18. Вектор для экспрессии антитела, содержащего только тяжелые цепи, в клетке млекопитающего. Вектор имеет стандартный остов pUC (не показан) с геном устойчивости к ампициллину и геном устойчивости млекопитающего (гигромицин). Соответствующая часть содержит стандартный энхансер CMV и промотор β-актина курицы (InVitrogen), за которым идет участок связывания рибосомы Козака, за которым следует сигнальная последовательность VH3-23. Задний конец кассеты состоит из константной области IgG2 или IgG3, которая начинается с шарнирного и СН2 доменов. После амплификации с использованием праймеров, показанных стрелками, кДНК VH клонируют между этими двумя областями.
Фиг.19. Пример и краткое изложение непосредственного экспрессионного клонирования антитела, содержащего только тяжелые цепи, в клетки НЕК. Слева вверху представлен стандартный ELISA для определения той иммунизированной мыши, которая положительно отвечала на иммунизацию антигеном НА (мышей, обозначенных красным, использовали для экспрессионного клонирования в клетках НЕК). Справа вверху представлены два примера ELISA, который осуществляли для трансфицированных клеток НЕК в 96-луночном формате.
Фиг.20. Анализ последовательностей ELISA-положительного антитела, содержащего только тяжелые цепи, полученные из последовательности VH 3-11 зародышевой линии.
Фиг.21. Анализ последовательностей ELISA-положительного антитела, содержащего только тяжелые цепи, полученные из последовательности VH 3-23 зародышевой линии.
Фиг.22. Профиль элюции полных IgG, содержащих только тяжелые цепи, на колонках Sephadex 200 Smart.
Фиг.23. Измерения аффинности двух антител против НА, содержащих только тяжелые цепи, на Octet.
Фиг.24. Аминокислоты по всей VH области антител против НА, содержащих только тяжелые цепи, в сравнении с тем, что наблюдают в антителах человека против CR1, которые содержат только тяжелые цепи, (см. выше), тетрамерных антителах человека и ламы, содержащих только тяжелые цепи.
Фиг.25. Протокол иммунизации и выделения антител человека, содержащих только тяжелые цепи, после одноклеточной сортировки и непосредственного экспрессионного клонирования в клетки НЕК.
Фиг.26. Протокол иммунизации и выделения нормальных тетрамерных антител человека, содержащих тяжелые и легкие цепи, после одноклеточной сортировки и непосредственного экспрессионного клонирования в клетки НЕК.
Пример 1. Создание антител против Luk-sFV
Осуществляли иммунизацию трансгенных мышей, содержащих локус тяжелой цепи антитела, полностью полученный у человека, белком Luks-PV из Staph А, как описано в WO 2006/008548. Эти мыши несли по 4 VH сегмента человека, все из областей D тяжелой цепи человека, все из областей JH человека и областей Сγ2 и Сγ3 человека (фиг.1).
СН1 домены удаляли из константных областей Су человека для того, чтобы сделать возможным получение антител человека, содержащих только тяжелые цепи, как описано (Janssens et al. (2006) PNAS, 103, 15130-15135). Мышей иммунизировали, используя стандартные протоколы, и выделяли антитела, как показано на схеме на фиг.2.
Мышей иммунизировали четыре раза, используя стандартные протоколы, и проверяли сыворотку на положительный ответ посредством стандартного ELISA (фиг.3).
После иммунизации собирали клетки селезенки и стандартными способами осуществляли обратную транскрипцию и амплификацию мРНК в кДНК, используя специфичные к доменам VH праймеры (см., например, Janssens et al. (2006) PNAS, 103, 15130-15135). Полученные фрагменты VH клонировали в вектор фагового дисплея. После 3 раундов скрининга и пэннинга по стандартным протоколам для способов с использованием дисплеев, домены VH (VH-D-J) клонировали в бактериальный экспрессионный вектор. Положительные клоны подтверждали вестерн-блоттингом, а клонированную ДНК затем секвенировали стандартными способами, в результате чего определяли аминокислотную последовательность области домена VH, которая показана фиг.4.
Альтернативно, антитела можно получать с помощью стандартной гибридомной технологии. Домены VH, полученные посредством технологии фагового дисплея, можно встроить обратно в предпочитаемую константную эффекторную область тяжелой цепи (лишенную функциональности СН1) стандартными способами клонирования, чтобы получить предпочитаемое целое антитело, содержащее только тяжелые цепи, в отношении константной области (например, Сα, а не Сγ). Аналогичным образом, антитела, содержащие только тяжелые цепи, полученные посредством гибридомной технологии, можно клонировать посредством стандартных технологий кДНК и ПЦР. Это позволяет отдельно выделять домены VH. Таким образом, оба подхода можно использовать для получения антиген-специфических доменов VH или антиген-специфических антител, содержащих только тяжелые цепи.
Пример 2. Создание антител против CR1 человека
Пример 2 очень схож с примером 1 за тем исключением, что иммунизацию осуществляли, используя в качестве антигена белок CR1, который в норме экспрессируют на своей поверхности красные клетки крови человека. Иммунизацию также осуществляли введением красных клеток крови мыши, которые экспрессируют CR1 человека на своей клеточной поверхности. Результат других иммунизации очень схож. Осуществляли иммунизацию трансгенных мышей, содержащих локус тяжелой цепи антитела, полностью полученный у человека, как описано в WO 2006/008548. Эти мыши несли по 4 VH сегмента человека, все из областей D тяжелой цепи человека, все из областей JH человека и областей Сγ2 и Сγ3 человека (фиг.1).
Области СН1 удаляли из константных областей Сγ человека для того, чтобы сделать возможным получение антител человека, содержащих только тяжелые цепи, как описано (Janssens et al. (2006) PNAS, 103, 15130-15135). Мышей иммунизировали, используя стандартные протоколы, и выделяли антитела, как показано на схеме на фиг.2.
После иммунизации собирали клетки селезенки и стандартными способами осуществляли обратную транскрипцию и амплификацию мРНК в кДНК, используя специфичные к доменам VH праймеры (см., например, Janssens et al (2006) PNAS, 103, 15130-15135). Полученные фрагменты VH клонировали в вектор фагового дисплея. После 3 раундов скрининга и пэннинга по стандартным протоколам для способов с использованием дисплеев, домены VH (VH-D-J) клонировали в бактериальный экспрессионный вектор. Затем клонированную последовательность секвенировали стандартными способами, в результате чего получали область домена VH, представленную на фиг.5 и 6. Альтернативно, антитела можно получать с помощью стандартной гибридомной технологии (фигуры 5 и 6). Домены VH, полученные посредством технологии фагового дисплея, можно встроить обратно в предпочитаемую константную эффекторную область тяжелой цепи (лишенную функциональности СН1) стандартными способами клонирования, чтобы получить целое антитело, содержащее только тяжелые цепи, в отношении константной области (например, Сα, а не Сγ). Аналогичным образом, антитела, содержащие только тяжелые цепи, полученные посредством гибридомной технологии, можно клонировать посредством стандартных технологий кДНК и ПЦР, что позволяет отдельно выделять домены VH. Таким образом, оба подхода можно использовать для получения антиген-специфических доменов VH или антиген-специфических антител, содержащих только тяжелые цепи.
Пример 3. Экспрессия доменов VН и антител, содержащих только тяжелые цепи, аффинность и растворимость
Домены VH из примеров 1 и 2 можно экспрессировать в системе бактериальной экспрессии для того, чтобы получать большие количества очищенных доменов VH. Например, домен VH можно экспрессировать в бактериях в виде гибридного белка, используя систему гибридной экспрессии SUMO (InVitrogen). После очистки, в денатурирующем электрофорезе в геле он идет как один фрагмент (фиг.7). После прохождения через хроматографическую систему Smart Sephadex при неденатурирующих условиях эти домены VH преимущественно идут в виде одного основного пика правильной молекулярной массы (фиг.7), с небольшими признаками растворимых агрегатов.
Антитело, содержащее только тяжелые цепи, полученное из супернатанта гибриды, также преимущественно идет в виде одного основного пика, с признаками небольшой доли димеров антитела на переднем фронте пика.
Один пик доменов VH указывает на то, что они представляют собой растворимые мономеры, что подтверждали графиком рассеяния при прохождении (фиг.8А), поскольку пик светорассеяния совпадает с пиком поглощения белка во время прохождения. Наконец, при нагревании домены VH стабильны при температуре до 55°С, как измеряли посредством стандартного анализа кругового дихроизма (фиг.8В).
Наконец, растворимые домены VH и антитела, содержащие только тяжелые цепи, тестировали на их аффинность связывания, используя системы BiaCore и Octet. Полученные аффинности находились в нижнем наномолярном диапазоне или лучше (фиг.9).
Также концентрировали несколько доменов VH, полученных из антител, содержащих только тяжелые цепи, посредством вакуумного центрифугирования и удаляли коллоид посредством высокоскоростного центрифугирования. Получали значения растворимости, которые представляют собой минимальные оценки, поскольку малые количества образцов препятствовали дальнейшему концентрированию. Все тестируемые домены VH обладали растворимостью более 4 мг/мл (фиг.10).
Пример 4. Массовое секвенирование для определения паттернов мутаций
После проверки всех последовательностей, полученных из антител, связывающих Luk-sPV, CR1 и другие антигены, становится ясно, что рекомбинация VDJ и созревание (гипермутация) растворимых доменов VH человека, полученных из трансгенов антител, содержащих только тяжелые цепи, экспрессируемых в В-клетках мыши, значительно отличалась от наблюдаемой в доменах VHH верблюда и ламы или доменах VH человека, полученных из нормальных тетрамерных антител, содержащих тяжелые и легкие цепи. Поэтому осуществляли массовое параллельное секвенирование (секвенатор Roche 454), повторяя процесс от рекомбинации до гипермутации. Это использовали для того, чтобы подтвердить, что процесс действительно отличается, и понять, какие параметры и часть(и) антитела важны для создания растворимого антитела человека с высокой аффинностью, содержащего только тяжелые цепи, после начального выбора конкретной рекомбинации VDJ. Результаты дают общее представление о том, как протекает процесс (фиг.11).
Этот анализ показывает, что организм мыши может вводить множество разнообразных замен аминокислот по всему домену VH экспрессируемого трансгена. Большинство расположено в положении CDR1 и CDR2. Однако замены также видны в FR1 и FR2 и распределены вдоль FR3. Области контакта CDR/FR выглядели как горячие точки для замен. Из этого анализа ясно, что процесс мутации в отношении положений и замен аминокислот сильно отличается от наблюдаемого в доменах VHH верблюдовых и доменах VH, присутствующих в нормальных тетрамерных антителах человека.
Пример 5. Сравнительный анализ отдельной каркасной области и областей CDR, присутствующих в растворимых доменах VН, доменах VHH и доменах VН, полученных из тетрамерных антител
Все мутации растворимых антигенсвязывающих доменов VH человека вносили в таблицы и анализировали различные области (FR1, CDR1, FR2, CDR2, FR3, CDR3), сравнивая с доменами VHH из антител верблюдовых, содержащих только тяжелые цепи, антител лам, содержащих только тяжелые цепи, и с доменами VH человека, полученными из нормальных тетрамерных антител человека.
Это сравнение включало 10 известных последовательностей доменов VHH верблюда, 58 известных последовательностей доменов VHH ламы и более чем 4000 известных последовательностей доменов VH человека, полученных из нормальных тетрамерных антител, все получены из литературы и публичных баз данных об антителах.
Область FR1 растворимых доменов VH человека (фиг.12) показывает увеличение суммарного заряда по сравнению с верблюдом и ламой и повышение гидрофобности по сравнению с верблюдом, ламой и доменами VH, полученными из нормальных тетрамерных антител человека, несмотря на увеличение числа заряженных аминокислот.
Область CDR1 (фиг.12) имеет очень маленькое изменение суммарного заряда (незначительно снижен по сравнению с ламой) и повышение гидрофобности по отношению к эквивалентному домену VH из тетрамерных антител человека, доменов VHH ламы и верблюда. Число заряженных аминокислот имеет небольшое значимое отличие.
Область FR2 растворимого домена VH человека (фиг.13) не имеет отличий суммарного заряда по сравнению с доменами VHH верблюда и ламы и имеет существенное повышение гидрофобности по сравнению с доменами VHH верблюда и ламы, несмотря на то, что число заряженных аминокислот снижено по сравнению с доменами VHH верблюда и ламы. В обоих аспектах это похоже на домены VH человека, полученные из тетрамерных антител. Область CDR2 (фиг.13) имеет суммарный заряд, схожий с ламой, который выше, чем у тетрамерных антител верблюда и человека, и гидрофобность, очень схожую с другими, несмотря на то, что число заряженных аминокислот схоже с доменами VH человека, полученными из тетрамерных антител, но несколько ниже, чем у других.
Область FR3 растворимого домена VH человека (фиг.14) показывает увеличение суммарного заряда по сравнению с доменами VH человека, полученными из тетрамерных антител, и снижение гидрофобности по сравнению с доменами VH человека, полученными из тетрамерных антител, несмотря на то, что число заряженных аминокислот повышено по сравнению с доменами VH человека, полученными из тетрамерных антител. Во всех аспектах это схоже с доменами VHH верблюда и ламы. Область CDR3 растворимых доменов VH человека (фиг.14.) имеет сниженный суммарный заряд по отношению к доменам VHH ламы и верблюдовых и доменам VH человека, полученным из тетрамерных антител, хотя число заряженных аминокислот повышено по сравнению со всеми остальными. Область CDR3 растворимых доменов VH человека имеет очень схожую гидрофобность с VHH ламы, но ниже, чем наблюдаемая в доменах VH, присутствующих в нормальных тетрамерных антителах человека и нормальных тетравалентных антителах верблюда.
Результаты показывают, что среди начальной рекомбинации VDJ происходит селекция событий рекомбинации с более высоким числом заряженных аминокислот по сравнению с тетрамерными (CDR3) антителами человека. За этим следует созревание аффинности в областях CDR1 и CDR2 и во всех трех каркасных областях (FR1, FR2, FR3). Это ведет к образованию антител человека, содержащих только тяжелые цепи, с растворимыми доменами VH, которые имеют повышенную гидрофобность в FR1, но пониженную в FR3 по сравнению с доменами VH, полученными из тетрамерных антител человека. Имеет место очень небольшое различие в областях FR2 или CDR1 и CDR2. Следовательно, общий паттерн показывает, что гидрофобность распространена по-разному на протяжении растворимого домена VH (сдвинута в сторону FR1 и от FR3 и CDR3) по отношению к доменам VH, полученным из тетрамерных антител, которые менее растворимы и обладают склонностью к агрегации. Этот анализ не показывает, есть ли конкретные мутации в конкретных положениях или областях домена VH (см. пример 6), но показывает, что есть признаки, связанные с распределением заряда и гидрофобностью по всему растворимому домену VH человека, которые отличают его от доменов VHH ламы и верблюдовых и доменов VH человека, полученных из тетрамерных антител.
Пример 6. Трехмерные структуры
Анализ из примера 4 показывает, что изменения в каркасных областях (FR1, FR2, FR3) растворимого домена VH человека после начальной перестройки VDJ в значительной мере сфокусированы в конкретных положениях, когда их размещают на трехмерной структуре домена VH человека, полученного из тетрамерного антитела (фиг.15).
В растворимом домене VH человека мутации FR1 наблюдали в аминокислотах 13-18, преимущественно изменения в положении 13, ведущие к повышению гидрофобности, даже если повышено число заряженных аминокислот, но не суммарный заряд. Важно, что изменения вокруг положения 13 расположены в малой петле, у которой повышена гидрофильность, которая предположительно повышает растворимость.
В FR2 растворимых доменов VH человека имеет место четкий фокус вокруг положения 35 и еще один вокруг положения 50, без общего изменения гидрофобности по сравнению с доменами VH, полученными из тетрамерных антител человека.
Изменения в FR3 растворимых доменов VH по отношению к доменам VH, полученным из тетрамерных антител человека, совпадают с концом области CDR2 и области рядом с концом FR3 (непосредственно после CDR3).
Когда эти изменения, наблюдаемые в растворимых доменах VH человека, полученных из антител, содержащих только тяжелые цепи, накладывают на трехмерную структуру домена VH, сразу становится ясно, что мутации FR2 происходят в β-складчатых слоях, которые в норме образуют поверхность взаимодействия VH с VL доменом в тетрамерных антителах. В FR1 изменения также приводят к повышению гидрофобности домена. Результаты указывают на то, что N-концевая половина домена VH имеет повышенную гидрофобность для повышения стабильности/растворимости домена VH и компенсации отсутствия легкой цепи. В отличие от этого, С-концевая половина менее гидрофобна. Наиболее важными изменениями являются «перераспределение» паттерна гидрофобности (N-конец более гидрофобный, С-конец менее гидрофобный, фиг.16) и мутации каркасной области, которые влияют на упомянутую поверхность контакта с легкой цепью.
Общий сдвиг гидрофобности и изменения в специфических мотивах, которые осуществляет иммунная система мыши, позволяют создавать растворимые антитела с высокой аффинностью, содержащие только тяжелые цепи, и растворимые домены VH со структурными признаками, которые отличают их от доменов VH, склонных к агрегации, которые получены из тетрамерных антител человека.
Пример 7. Создание и экспрессионное клонирование антител человека против НА
В примере 7 использовали процедуры иммунизации, как описано в примерах 1 и 2, за исключением того, что иммунизацию осуществляли, используя НА гриппа (H3N2) в качестве антигена. Иммунизацию осуществляли посредством инъекции белка трансгенным мышам, содержащим локус тяжелой цепи антитела, полностью полученный у человека, как описано в Janssens et al. (2006) PNAS, 103, 15130-15135. Эти мыши несли по 4 VH сегмента человека, все из областей D тяжелой цепи человека, все из областей JH человека и областей Сγ2 и Сγ3 человека (фиг.1).
Области СН1 удаляли из константных областей Сγ человека для того, чтобы сделать возможным получение антител человека, содержащих только тяжелые цепи, как описано (Janssens et al. (2006) PNAS, 103, 15130-15135). Мышей иммунизировали, используя стандартные протоколы, и выделяли антитела, как показано на схеме на фиг.17.
После иммунизации собирали плазматические клетки и сортировали (FACS) по наличию антигена CD138 и отсутствию маркеров линии дифференцировки стандартными способами (см. Sanderson et al. (1989) Cell Regulation 1, 27-35; Kim et al. (1994) Mol.Biol. Cell, 5, 797-805 и см. набор для магнитной сортировки для выделения CD138+ плазматических клеток мыши компании Miltenyi Biotec). Клетки собирали и осуществляли обратную транскрипцию и амплификацию мРНК в кДНК с использованием специфичных к доменам VH праймеров стандартными способами (например, Janssens et al (2006) PNAS, 103, 15130-15135). На 5'-конце использовали праймеры, которые сконструированы для введения сайта PvuII для целей клонирования (фиг.18). На 31-конце использовали праймеры из шарнирной области СН2 IgG2 или IgG3, которые сконструированы с сайтом BstEII. Если это невозможно, праймер для IgG3 содержал сайт Sad для целей клонирования (фиг.18). Получаемые фрагменты VH надлежащим образом клонировали в два экспрессионных вектора млекопитающих, содержащие энхансер CMV и промотор β-актина курицы, за которыми следует последовательность Козака и сигнальная последовательность из VH3-23, граничащая с ее сайтом PvuII в VH сегменте. В первом векторе (фиг.18) за этим следовал IgG2 человека, начиная с шарнирной области (с сайта BstEli) СН2 с СН2, СН3, стоп-кодоном и полиадениловой последовательностью. Во втором векторе все последовательности IgG2 заменяли эквивалентными последовательностями IgG3 человека. Последнее позволяет клонировать антитела, содержащие только тяжелые цепи, которые не содержат сайт BstEII, используя сайт Sad в качестве альтернативы. Остальная часть плазмиды содержала стандартный селективный маркер для клеток млекопитающих, такой как кассета устойчивости к гигромицину в плазмиде на основе pUC. Очевидно, что IgG2 или 3 человека можно заменить любой другой СН1-дефицитной константной областью, полученной у человека или любого другого вида. В равной мере очевидно, что аналогичное справедливо для VH части вектора. Затем кДНК клонировали между сайтами PvuII и BstEII (или PvuII и Sad) и вектором трансфицировали Е.coli стандартными способами. Колонии собирали в 96-луночные (или 384-луночные) микротитровальные планшеты, содержащие бактериальную среду для выращивания. Планшеты (типично >10) инкубировали так, чтобы позволить бактериям расти, и в каждой лунке выделяли ДНК. Затем ДНК из каждой лунки, т.е. из каждого целого планшета, добавляли в новый 96-луночный планшет, содержащий клетки НЕК млекопитающего (или другие клетки для экспрессионных целей), для трансформации с помощью стандартных способов и реактивов для трансфекции. Трансформированные клетки растили в селекционных условиях на основе присутствия селекционного маркера на экспрессионной плазмиде (например, гигромицина). Затем среду, содержащую экспрессируемое антитело, анализировали на присутствие антигенспецифического антитела, содержащего только тяжелые цепи, посредством стандартных анализов ELISA. Их можно осуществлять в том же 96-луночном формате, но также можно осуществлять в других форматах, например, посредством микрочипов, чтобы уменьшить расходуемое количество антигена. Положительные лунки непосредственно указывают на плазмиды, которые содержат антитела человека, содержащие только тяжелые цепи, которые кодируют антиген-специфические антитела, содержащие только тяжелые цепи. На фиг.19 показан ELISA для сывороток мышей, которых иммунизировали, и примеры ELISA для трансфицированных клеток НЕК в 96-луночном формате. На ней также обобщен эксперимент по клонированию НА, менее одного миллиона кДНК использовали в эксперименте с использованием в целом 960 лунок. Тем самым получили 28 различных антигенспецифических антител, отнесенных к 13 группам посредством анализа последовательностей, полученных из двух областей VH - VH3-11 (фиг.21) и VH3-23 (фиг.22). Использовали как IgG2 (SSERKCCVE), так и IgG3 (SSELKTPLG), области VH подвергались гипермутации и, подобно человеку, область J4 использовали наиболее часто, хотя другие также использовали. Секретируемые антитела, содержащие только тяжелые цепи, растворимы и представляют собой мономеры, как определяли по их профилям элюции на стандартной колонке SMART (фиг.23). Аффинности антител находились в диапазоне от наномолярных до субнаномолярных значений, как определяли по стандартным профилям связывания и диссоциации в анализе на Octet (фиг.24) или BiaCore. Анализ мутаций и сравнение с другими последовательностями иммуноглобулинов (фиг.25) выявил такие же свойства различных областей, которые показаны на фиг.11-13.
Как описано выше, растворимые домены VH можно выделить из растворимых антител, содержащих только тяжелые цепи, полученных из плазматических клеток.
Пример 8. Создание и экспрессионное клонирование антител человека, содержащих только тяжелые цепи, из отдельных клеток
Вариант схемы, описанной выше посредством клонирования в клетки НЕК, состоит в сортировке CD138+ клеток негативной линии дифференцировки и получении кДНК из отдельных клеток, которые можно клонировать непосредственно в клетки НЕК или другие клетки млекопитающих (фиг.26). Это можно анализировать очень похожими способами, как описано выше.
Пример 9. Создание и экспрессионное клонирование нормальных (тетрамерных) антител человека, содержащих тяжелые и легкие цепи, из отдельных клеток
Вариант способа, описанного в примере 8, также можно использовать для клонирования трансгенных экспрессируемых нормальных тетрамерных антител непосредственно посредством экспрессионного клонирования в клетки НЕК или другие клетки млекопитающих. В этом варианте отдельные клетки снова выделяли сортировкой. РНК выделяли из отдельных клеток и VH и VL домены клонировали посредством ПЦР амплификации в экспрессионный вектор тяжелой и легкой цепи, аналогичный тому, что описано выше. Индивидуальным сочетанием клонируемых экспрессионных плазмид тяжелых и легких цепей совместно трансфицировали клетки НЕК или другие клетки млекопитающих. Форматы, скрининг и анализ полностью аналогичны описанному выше для 96-луночного формата для антител, содержащих только тяжелые цепи, (примеры 8 и 9).
Реферат
Настоящее изобретение относится к области биотехнологии и иммунологии. Раскрыт антиген-специфическое растворимое антитело с высокой аффинностью, содержащее только тяжелые цепи. Антитело не содержит характерные для верблюдовых замены аминокислот и содержит замены FR2, которые не найдены в антителах, которые содержат тяжелую и легкую цепи. Антитело обладает повышенной суммарной гидрофобностью в CDR1 и содержит увеличенное число заряженных аминокислот, присутствующих в CDR3, и содержит одну или более замен аминокислот в β-складчатом слое каркасной области, которые ведут к повышению суммарной гидрофобности в FR1 и к увеличению числа заряженных аминокислот, присутствующих в FR3. Также предоставлены домены VH, которые обладают теми же свойствами, генные сегменты для их получения, способы их получения и использование антитела доменов VH в терапии. 3 н. и 11 з.п. ф-лы, 26 ил., 9 пр.
Формула
(a) иммунизацию трансгенного млекопитающего, не относящегося к человеку, антигеном, где млекопитающее экспрессирует антитела, содержащие только тяжелые цепи, у которых отсутствует функциональность СН1 в транскрибируемой и процессируемой мРНК тяжелой цепи;
(b) выделение долгоживущих плазматических клеток или В-клеток памяти из иммунизированного млекопитающего;
(c) выделение популяции мРНК из клеток, полученных на стадии (b);
(d) клонирование популяции кДНК, полученных из мРНК, выделенных на стадии (с), в экспрессионный вектор и экспрессирование указанного экспрессионного вектора в клеточной линии; и
(e) отбор, по меньшей мере, одной клеточной линии, которая продуцирует антитело, содержащее только тяжелые цепи, которое специфично связывает антиген.
(a) иммунизацию трансгенного млекопитающего, не относящегося к человеку, антигеном, где млекопитающее экспрессирует антитела, содержащие только тяжелые цепи, у которых отсутствует функциональность СH1 в транскрибируемой и процессируемой мРНК тяжелой цепи;
(b) выделение долгоживущих плазматических клеток или В-клеток памяти из иммунизированного млекопитающего;
(c) выделение популяции мРНК из клеток, полученных на стадии (b);
(d) клонирование популяции кДНК, содержащей кДНК, которые кодируют домены VH, полученные из мРНК на стадии (c), в экспрессионный вектор так, чтобы в клеточной линии экспрессировался домен VH или домен VH, слитый с эффекторной областью тяжелой цепи;
(e) отбор, по меньшей мере, одной клеточной линии, которая продуцирует домен VH или домен VH, слитый с эффекторной областью тяжелой цепи, которые специфично связываются с антигеном.
Комментарии