Химерные вакцинные антигены против вируса классической чумы свиней - RU2406534C2
Код документа: RU2406534C2
Чертежи






Описание
ОБЛАСТЬ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к ветеринарии, в частности к новым химерным антигенам, включающим вирусные субъединицы вируса классической чумы свиней (ВКЧС), соединенные с белками, способными стимулировать клеточную и гуморальную иммунную систему, вызывая сильный и ранний иммунный ответ на такие вирусы у свиней.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Полагают, что классическая чума свиней (КЧС), также известная как холера свиней за свой высокоинфекционный характер, является наиболее важным заболеванием у свиней, и она включена в официальный перечень заболеваний Всемирной ветеринарной организации. Этиологическим агентом этого заболевания, КЧС, является вирус рода Pestvirus из семейства Flaviviridae. Известно, что этот вирус имеет липидную оболочку, диаметр от 40 до 60 нм и гексагональную симметрию, с одноцепочечной рибонуклеиновой кислотой (РНК) в качестве генетического материала (Kummerer et al. (2000). The genetic basis of cytopathogenicity of pestviruses. Vet. Microbiol. 77:117-128; Moenning et al. (2003)).
КЧС представляет собой высококонтагиозное заболевание, в своей острой форме представленное лихорадкой, разрушением капиллярных сосудов, некрозом внутренних органов и смертью. Первые клинические симптомы появляются после инкубационного периода длительностью от 2 до 6 дней, вызывая гипертермию, снижение подвижности и потерю аппетита, с ухудшением в последующие дни, и температура может достигать 42°С. Также развивается лейкопения с количествами белых кровяных клеток в крови менее 8000/мм3. У свиней также развиваются конъюнктивиты, запоры с последующей диареей, рвота, потеря координации движений, судороги и парезы мышц в терминальной фазе. Это проявляется красным цветом кожи, распространяющимся по всему брюху, морде, ушам и внутренней части конечностей. В большинстве фатальных случаев при гистопатологическом исследовании мозга выявляется негнойный энцефалит с высокой васкуляризацией (Moenning et al. (2002) Clinical Signs and Epidemiology of Classical Swine Fever. A review of knowledge. Vet. Journal 161:1-10).
ВКЧС действует как иммуносупрессор во время протекания инфекции (Susa et al. (1992) Pathogenesis of Classical Swine Fever: B-lymphocyte deficiemcy caused by Hog Cholera virus. J. Virol. 66:1171-1175), и обнаружение нейтрализующих антител начинается на 2 и 3 неделе после инфицирования (Laevents et al. (1998)) An experimental infection with a classical swine fever virus introduction in natural outbreaks. Vet. Q. 20: 46-49). Терминальная стадия инфекции ассоциирована со значительным снижением уровня В-лимфоцитов в циркулирующей крови так же, как и в лимфоидной ткани (Susa et al. (1992) Pathogenesis of Classical Swine Fever: B-lymphocyte deficiency caused hog cholera virus. J. Virol. 66: 1171-1175). Большинство свиней, которые заболели, умирали в интервале времени от 10-го до 20-го дня после инфицирования, с уровнем смертности более 95%. Особенностью поражений при КЧС на аутопсии являлся геморрагический диатез с петехиями в большинстве систем органов. Наиболее постоянно эти изменения встречаются в почках, мочевом пузыре и лимфатических ганглиях, хотя они могут появляться также в селезенке, гортани, на слизистых и серозных оболочках (Mouwen et al. (1983) Atlas of Veterinary Pathology, Bunge, Utrecht The Netherlands).
Трансплацентарное инфицирование представляет собой другую клиническую форму КЧС; в этом случае вирус способен проникать через плаценту беременных свиноматок, инфицируя плоды. Последствиями этого инфицирования может являться выкидыш, рождение мертвого потомства, мумификации, мальформации, рождение слабых свиней и проблемы, связанные с дифференцировкой органов. В зависимости от срока беременности, на котором произошло инфицирование, в результате заражения от свиноматок (вертикальной передачи) может рождаться обладающее иммунитетом потомство. Поросята остаются инфицированными и виремическими до смерти, создавая стабильный очаг распространения ВКЧС в стаде (Moenning et al., (2003) Clinical Signs and Epidemiology of Classical Swine Fever: a rewiew of new knowledge. Vet. Journal 165: 11-20). Смертность, ассоциированная с КЧС, представляет собой экономическую проблему для затронутых ею стран, негативно влияющую на экономическую и социальную ситуацию развивающихся государств. По этой причине в странах, имеющих высокую плотность свиней и большое распространение вируса, становится необходимостью применение контролирующих программ, основанных на вакцинации. В высокоразвитых странах, в которых разведение свиней в основном субсидируется правительствами, таких как Европа, Соединенные Штаты и Канада, применяется способ эрадикации посредством полного санитарного убоя. Однако стоимость такого способа является очень высокой, и такие страны являются все еще чувствительными к возможным эпизоотиям.
Европейский Союз (ЕС) рассматривается как высокая зона риска повторных экстренных случаев новых ВКЧС-эпизоотий вследствие высокой плотности популяции свиней, своей политикой невакцинирования и своей географической близостью к восточно-европейским странам, в которых ВКЧС является энзоотическим заболеванием. Одной из проблем, связанной с угрозой возникновения новых эпизоотий в этом регионе, является наличие диких кабанов с эндемическим инфицированием КЧС (Laddomada (2000) Incidence and control of CSF in wild boars in Europe. Vet. Microb, 73:(121-30)). Эти появления новых эпизоотий происходили вопреки жестким программам контроля, которые были реализованы в Европейском Союзе, которые включали санитарное уничтожение всей контагиозной популяции и ограничение экспорта свинины из пораженных районов в районы, свободные от заболевания (van Oirschot (2003) Vaccinology of Classical Swine Fever: from lab to field. Vet Microbiol, 96:367-384). Таким образом, неотложной является потребность в разработке вакцин, которые вызывают иммунный ответ, ранний и надежный, гарантирующий защиту от инфицирования и передачи вируса.
Были разработаны вакцины против ВКЧС на основе интактного вируса: вакцины с вирусом, инактивированным кристаллическим фиолетовым или формалином (Biront et al. (1998) Classical swine fever related infections. Liess B.M. Ed. Martinus Nijhoff Publishing, Boston: 181-200), вакцины с вирусом, аттенуированным посредством пассирования в кролике, например штамма Sinlak (Baibikov et al. RU 2182495) и штамма Lapinizied Chinese (Dahle et al. (1995) Assessment of safety and protective value of a cell culture modified strain C vaccine of hog cholera/classical swine fever virus. Berl-Munch. Tieraztl.Wsch, 108:20-25), или вакцины с вирусом, аттенуированным в культурах тканей, полученных из кролика, морской свинки и свиньи (Kachiku et al. JP 73001484; Terpstra et al. (1990) Development and properties of a cell culture produced for hog cholera based on Chinese strain. Ditsh. Tierarztl.Wsch. 97: 77-79). Применение этих типов вакцин представляют собой риск вследствие вероятности содержания фракций активного вируса, который, привитый восприимчивым животным, будет вызывать новые вспышки КЧС. Кроме того, в некоторых случаях для получения защитного иммунологического ответа необходимы повторяющиеся иммунизации, поскольку инактивация влияет на иммуногенетические свойства вируса.
В отдельных случаях применения живой вакцины с аттенуированной вирулентностью, они имеют высокий риск того, что произойдет частичная аттенуация или восстановление вирулентности. В некоторых случаях они будут формировать патогенные вирусные частицы, которые, будучи введенными восприимчивым животным, сделают возможным инфицирование, клиническое заболевание и распространение КЧС в стадах. Эти проблемы обуславливают еще больший риск для беременных свиноматок, поскольку вирус может инфицировать плоды, которые являются высоковосприимчивыми, и инфицированное потомство будет распространять заболевание.
Существуют вакцины на основе штаммов ВКЧС, которые были аттенуированы, такие как штамм C Chinese, штамм PAV 250, штамм Thierval и штамм IFFA/A-49 (Björlund, H.JV. et al. (1998) Molecular characterization of the 3'noncoding region of classical swine fever virus vaccine strains. Virus Genes 16: 307-312, Launais et al. (1978) Hog Cholera Virus: Active immunization of piglets with the Thiverval strain in the presence and absence of calostral passive immunity. Vet. Microbiology 3:31-43). Эти штаммы применяются только в странах, где заболевание является энзоотическим, поскольку они имеют такое неудобство, что они не позволяют отличить вакцинированное животное и животное, инфицированное нативным вирусом. Животные, вакцинированные этими штаммами, показывают такие же ответы в серологических тестах, как и инфицированные животные. Специфические антитела анти-ВКЧС, которые вырабатываются при применении вакцин на основе аттенуированного вируса, мешают диагностике инфицирования КЧС. Диагностика осуществляется посредством иммунологического определения инфицирующего вируса в небных миндалинах и размножения вакцинного вирусного штамма, происходящего в небных миндалинах. По этой причине аттенуированные штаммы являются непригодными для применения в программах эрадикации. Вакцинация штаммом LK-VNIIVVM и дополнительная гипериммунизация очищенным штаммом Shi-Myng, составленным с адъювантом Фрейнда, является другим примером. Но иммунизация в 40-45 местах является невозможной при кампании вакцинации, в которой сотни животных должны быть вакцинированы ежедневно (Balashova et al. RU2183972).
Иммунизация этими вакцинами, содержащими целый вирус, препятствует также дифференциальной диагностике инфекций, вызванных ВКЧС, и инфекций, вызванных другими членами рода Pestvirus, которые могут инфицировать свиней, такими как вирус вирусной диареи коров (сокращенно ВВДК) и вирус пограничной болезни овец (сокращенно ВПБО), (Dahle et al. (1991) Clinical Post Mortem and Virologocal Findings after Simultaneous Inoculation of Pig with Hog Cholera and BVD Virus. J. Med. Vet. 38: 764-772).
Для избежания неудобств вакцин на основе целого вируса получены пригодные для применения вакцины, полностью безопасные, такие как варианты на основе субъединиц или вирусных белков, полученных рекомбинантным путем. Эти варианты должны защищать стада от повторного внедрения штаммов вируса, а также позволяют различать вакцинированных и инфицированных животных простыми серологическими способами. С этой целью были разработаны вакцины на основе вирусных субъединиц. Вакцины, содержащие вирусные белки, такие как гликопротеин Е2 оболочки вируса (Bourna et al. (2000) Duration of the onset of the herd immunity induced by E2 subunit vaccine classical Swine Fever virus 18: 1374-1381), являются безопасными, поскольку их применение не содержит риска возврата вирулентности и не препятствует диагностике. Эти вакцины позволяют дифференцировать инфицированных животных и вакцинированных животных, поскольку антитела, которые вырабатываются, реагируют только против вирусного сегмента. Таким образом, они целесообразны для программы эрадикации КЧС.
Были разработаны несколько рекомбинантных вакцин на основе экспрессии белка Е2 в прокариотах, и вакцины на основе синтетических пептидов этого белка (Chen et al. WO 200232453). В этих случаях белок является негликозилированным, так что страдают его иммуногенность и защитные свойства. Другие варианты вакцин используют вирусные векторы для экспрессии гетерологичного гена Е2 в эукариотических клетках подобно вирусу псевдобешенства свиней (Peeterrs et al. (1997). Biologically safe, non-transmissible pseudorabies virus vector vaccine protect pigs against both Aujeszky's disease and classical swine fever. J. Gen. Virol. 78: 3311-3315), вирусу натуральной оспы свиней (Gibbs et al. US62117882) и аденовирусу свиней (Nagy et al. WO 200183737). В этих случаях инфицирование вирусом дикого типа вызывает выработку нейтрализующих антител против вирусного вектора такого же серотипа. Таким образом, страдает вызывание иммунного ответа против ВКЧС. Также вирусные векторы на основе вируса псевдобешенства свиней и вируса натуральной оспы свиней не могут применяться в странах, признанными свободными от этих вирусов, вследствие проблем законодательства. Также был применен в качестве вектора вирус осповакцины, но постановления Всемирной организации здравоохранения затрудняют его применение (Meyeers et al. EP 1087014).
Вакцины на основе лишенной оболочки дезоксирибонуклеиновой кислоты (ДНК) для экспрессии белка Е2 в миоцитах и остеоцитах имеют неудобство, связанное с тем, что для того, чтобы вызвать иммунный ответ, требуются более высокие концентрации ДНК, поскольку трансфекция с применением лишенной оболочки ДНК является очень неэффективной. Эта вакцина подвергается строгому законодательному контролю, который затрудняет ее применение (Audonnet et al. WO 20152888).
Получение Е2-ВКЧС в качестве антигена в экспрессионных системах клеток насекомых, опосредованное бакуловирусом, стало возможной альтернативой (Van-Rjin et al. (1990). An experimental marker vaccine and accompanying serological diagnostic test both based on enveloped glycoprotein E2 of classical swine fever virus. Vaccine, 17: 433-440; Kretzdom et al. US 20040028701). В этой системе рекомбинантный Е2 получают в виде гликопротеина, повышая иммуногенность относительно негликозилированной изоформы. Бакуловирус дополнительно инактивируется, не оказывая патогенных эффектов на свиней. Однако эффективная защита против инфицирования возникает через три недели после вакцинации, и защита против внутриматочного инфицирования является неполной. Следовательно, важной проблемой в предотвращении ВЧС является отсутствие рекомбинантных вакцин на основе субъединиц, делающих возможной проведение дифференциальной диагностики между вакцинированными и инфицированными животными, и способных к формированию ранней защиты после вакцинации, ликвидируя трансплацентарную передачу от беременных свиноматок своему потомству.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение решает вышеописанную проблему. Новая вакцина содержит химерные антигены, содержащие вирусные субъединицы в комбинации с молекулами, стимулирующими иммунную систему, которые делают возможным развитие раннего иммунного ответа, который защищает свиней от инфицирования ВКЧС. Другим преимуществом предложенного решения является то, что оно прекращает передачу вируса от инфицированных беременных свиноматок потомству вследствие эффекта активации иммунитета стимулирующими молекулами, которые объединены с вирусными белками в химерных антигенах.
В частности, изобретение относится к химерным антигенам против КЧС, которые имеют в качестве главного компонента гликопротеин Е2 из оболочки ВКЧС. Внеклеточный сегмент гликопротеина Е2 применяется в качестве иммуногена, объединенного с белком, стимулирующим иммунную систему (называемым в контексте данного изобретения «молекулярный адъювант»), для усиления и стимуляции раннего клеточного иммунного ответа, и вызывания выработки более высоких титров нейтрализующих антител к ВКЧС.
В конкретном варианте осуществления изобретения белок, стимулирующий иммунную систему, представляет собой интерферон альфа или внеклеточный сегмент молекулы CD154. В предпочтительном варианте осуществления интерферон альфа или внеклеточный сегмент молекулы CD154 может быть получен из любого млекопитающего.
Вакцинные антигены настоящего изобретения на основе химерных белков гарантируют защиту вакцинированных свиней с первой недели после иммунизации, когда производится их контрольное заражение 105 DL50 (доза вируса, которая вызывает смерть 50% животных, инфицированных ВКЧС). Такая защита опосредуется сильным клеточным иммунным ответом против ВКЧС, который непосредственно связан с комбинацией элементов, которые сочетаются в химерном антигене. Также наблюдается уменьшение времени индукции нейтрализующих антител, которые появлялись на второй неделе, следующей после вакцинации. Это способствует повышению защиты против ВКЧС вакцинированных свиней. Иммунизированные животные не имеют симптомов клинического заболевания, и выделения ВКЧС из биологических жидкостей не могло быть получено ни в один день после контрольного заражения таким вирусом.
Химерные антигены с молекулярным адъювантом Е2 предотвращают вертикальную передачу ВКЧС от свиноматок к плодам. Эти белки вызывают раннюю защиту у беременных свиноматок, которая задерживает развитие клинического заболевания и делает невозможным размножение вируса, не только у матерей, но также и у плодов, после контрольного заражения 105 DL50 ВКЧС.
В предпочтительном варианте осуществления химерный вакцинный антиген характеризуется содержанием главным образом аминокислотной последовательности внеклеточного сегмента (или домена) гликопротеина Е2 ВКЧС, который представлен в Списке последовательностей как SEQ ID NO: 1; и внеклеточного сегмента молекулы CD154 свиней, который представлен как SEQ ID NO: 2. Химерный вакцинный антиген фактически содержит такие аминокислотные последовательности, но он может также включать в себя внеклеточный сегмент Е2 любого изолированного ВКЧС.
Другим аспектом настоящего изобретения является то, что химерный вакцинный антиген может быть получен рекомбинантным синтетическим путем или посредством химической конъюгации. В конкретном варианте осуществления изобретения вариант на основе химерного белка, содержащего Е2his (внеклеточный сегмент Е2, слитый с хвостом 6 гистидинов), и молекулярного адъюванта был получен в виде слитого белка. С этой целью спейсерный пептид, состоящий из 4 повторяющихся единиц Gly4Ser (4G4S), и молекула, стимулирующая иммунную систему, были добавлены на С-конец Е2his. Включение в состав пептида 4G4S допускает определенную степень релаксации полипептидной цепи. Это гарантирует правильную третичную структурную укладку структуры белка для получения слитых белков с такой же третичной структурой, как и нативная. Один из вакцинных антигенов, объект изобретения, имеет внеклеточный домен молекулы CD154 свиней, слитый с его С-концом, в качестве молекулярного адъюванта (Е2his-СD154).
До настоящего времени не было изучено получение рекомбинантных вакцин-кандидатов против ВКЧС, опосредованное экспрессионными системами животных в качестве биореакторов. Несмотря на это, способность грудной железы экспрессировать гликозилированные рекомбинантные белки с правильной укладкой их третичной структуры, делает ее адекватной экспрессионной системой для получения гликопротеина Е2 с высокой иммуногенностью и защитными свойствами. Временная экспрессионная система в грудной железе жвачного животного, опосредованная аденовирусными векторами, представляет собой средство для получения высоких уровней экспрессии рекомбинантных белков быстрым и простым путем (Toledo et al., WO 2004/034780). Этот способ является очень полезным для получения рекомбинантного Е2 для целей применения в программах вакцинации, направленных на эрадикацию КЧС.
В одном варианте осуществления изобретения вакцинные антигены, объект настоящего изобретения, экспрессируются в эпителиальных клетках молочной железы генетически модифицированных млекопитающих во время процесса лактации и секретируются в молоко. Рекомбинантные химерные молекулы получают в молоке трансгенных млекопитающих или посредством прямой трансформации эпителия молочных желез нетрансгенных млекопитающих с применением аденовирусных векторов. В другом варианте осуществления изобретения химерные вакцинные антигены получают в генетически модифицированных дрожжевых клетках. Такие антигены вырабатываются в культуральной среде дрожжей, трансформированных с применением химерного гена и регуляторных последовательностей, делающих возможными экспрессию и секрецию рекомбинантного белка в культуральную среду.
Белок Е2 нативного ВКЧС экспонируется на вирусной оболочке в виде гомодимера, стабилизированного при помощи межцепочечных дисульфидных связей. Это определяет то, что нейтрализация и защитные антитела вырабатываются против конформационных эпитопов, представленных на гомодимерах. Вакцинные антигены, разработанные в настоящем изобретении, получают в экспрессионных системах, которые делают возможным правильное свертывание этих рекомбинантных белков. Конфигурация генетических конструкций гарантирует отсутствие изменений третичной структуры слитых белков. Рекомбинантные вакцинные антигены легко очищают с применением простой процедуры аффинной хроматографии на ионах металлов.
Конфигурация генетических конструкций, применение экспрессионных систем и относительная простота процедуры очистки гарантирует, что вакцинные антигены против ВКЧС, описанные в настоящем изобретении, имеют такие же, как у вирусного белка Е2, антигенные и иммуногенетические свойства. Иммунизация химерными молекулами, полученными в таких экспрессионных системах, как Pichia pastoris или молочная железа козы, приводит к сильному и раннему иммунному ответу. Внеклеточный домен Е2 образует гомодимеры, которые обеспечивают конформационные эпитопы для образования нейтрализующих и защитных антител. Сегмент из CD154 действует в качестве молекулярного адъюванта, который стимулирует иммунную систему вакцинированных свиней, вызывает клеточный иммунный ответ, который защищает животных от ВКЧС с первой следующей после вакцинации недели. Комбинация обеих молекул в химерном белке, который содержит спейсерный пептид, гарантирует правильное свертывание каждой молекулы. Примененные экспрессионные системы позволили экспрессировать рекомбинантные белки в гликозилированной изоформе. Это также помогает получать молекулы с правильной третичной структурой.
Другим аспектом настоящего изобретения являются вакцинные композиции со способностью вызывать защищающий иммунный ответ против ВКЧС, которые характеризуются содержанием химерных антигенов, описанных ранее, содержащих внеклеточный домен гликопротеина Е2 и молекулярный адъювант. Такие вакцинные композиции могут вводиться животным системным путем или через слизистые оболочки для предотвращения КЧС и избежания материальных и экономических потерь, которые вызывает инфицирование ВКЧС свиных стад.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг.1. Анализ с помощью SDS-электрофореза в ПААГ, в восстанавливающих условиях экспрессии Е2his в клетках РК-15, трансдукцированными аденовирусными векторами Ad-E2his-sec. (А) SDS-электрофорез в ПААГ, дорожка 1: культуральная среда от трансдуцированных клеток, дорожка 2: культуральная среда от необработанных клеток, MWM: маркер молекулярной массы. (В) Иммуноидентификация Е2his при помощи иммуноблоттинга с применением моноклональных антител, направленных на гистидиновый хвост, дорожка 1: культуральная среда трансдуцированных клеток, дорожка 2: культуральная среда от необработанных клеток, дорожка 3: положительный контроль для гистидинового хвоста, MWM: маркер молекулярной массы. (С) Иммуноидентификация Е2his при помощи иммуноблоттинга с применением поликлональной сыворотки, полученной из инфицированных ВКЧС свиней, дорожка 1: культуральная среда от необработанных клеток, дорожка 2: культуральная среда от трансдуцированных Ad-E2his клеток, MWM: маркер молекулярной массы.
Фиг.2. Анализ условий экспрессии Е2his и E2his-CD154 в клетках РК-15, трансдукцированных аденовирусными векторами Ad-E2his-sec и Ad-E2hisCD154-sec. Белки в культуральной среде сепарируют с применением SDS-электрофореза в ПААГ в невосстанавливающих условиях. Иммуноидентификация интересующих молекул осуществлялась при помощи иммуноблоттинга с применением моноклональных антител (Mab) против белка Е2 ВКЧС (Mab-1G6). Дорожка 1: культуральная среда от трансдуцированных вектором Ad-E2his-sec клеток, дорожка 2: культуральная среда от трансдуцированных вектором Ad-E2hisCD154-sec клеток, MWM: маркер молекулярной массы.
Фиг.3. Кинетика экспрессии Е2his в молоко коз, трансдукцированных вектором Ad-E2his-sec. Белки из образцов молочной сыворотки, соответствующих каждому дню доения, разделяют с применением SDS-электрофореза в ПААГ в невосстанавливающих условиях. Иммуноидентификация интересующих молекул исследовалась при помощи иммуноблоттинга с применением Mab-1G6. Дорожка РК: положительный контроль Е2his, экспрессированного в культуральную среду клеток РК-15, трансдукцированных вектором Ad-E2his-sec, дорожка С-: образцы молочной сыворотки от коз, не подвергавшихся обработке, дорожки 1-8: образцы молочной сыворотки от коз, трансдукцированных вектором Ad-E2his-sec, соответствующие каждому из 8 дней доения, следующих за аденовирусной трансдукцией.
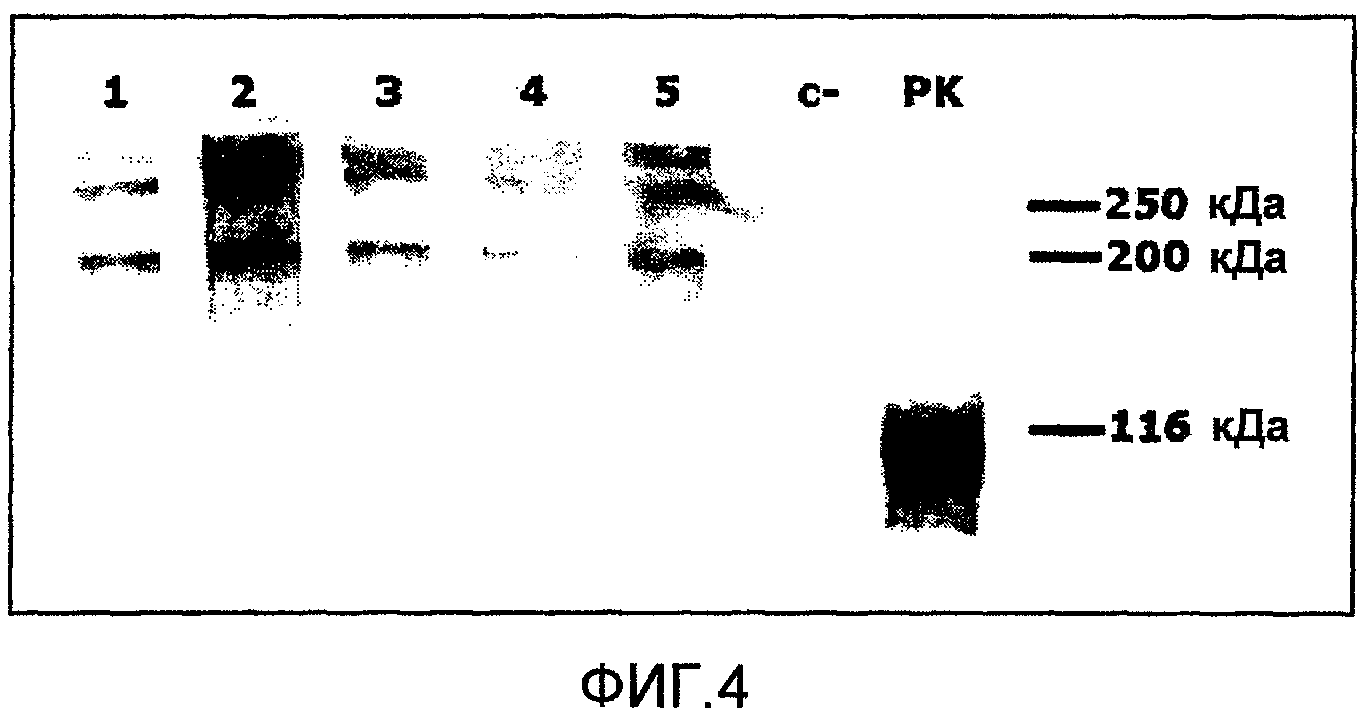
Фиг.4. Кинетика экспрессии Е2his-CD154 в молоко коз, трансдукцированных вектором Ad-E2hisCD154-sec. Белки из образцов молочной сыворотки, соответствующих каждому дню доения, разделяют с применением SDS-электрофореза в ПААГ в невосстанавливающих условиях. Иммуноидентификация интересующих молекул исследовалась при помощи иммуноблоттинга с применением Mab-1G6. Дорожки 1-5: образцы молочной сыворотки от коз, трансдукцированных вектором Ad-E2hisCD154-sec, соответствующие каждому из 5 дней доения, следующих за аденовирусной трансдукцией, дорожка С-: образцы молочной сыворотки от коз, не подвергавшихся обработке, дорожка РК: положительный контроль Е2his-CD154, экспрессированного в культуральную среду клеток РК-15, трансдукцированных вектором Ad-E2hisCD154-sec.
Фиг.5. Анализ чистоты и идентификация Е2his, подвергнутого SDS-электрофорезу в ПААГ в невосстанавливающих условиях. Белок экспрессировался в молоко коз, трансдукцированных вектором Ad-E2his-sec, и очистка осуществлялась при помощи аффинной хроматографии с ионами металлов. (А) SDS-электрофорез в ПААГ различных стадий очистки. (В) Иммуноидентификация при помощи иммуноблоттинга с применением Mab-1G6. Дорожка 1: положительный контроль Е2his, экспрессированного в культуральную среду клеток РК-15, трансдукцированных вектором Ad-E2his-sec, дорожка 2: образцы молочной сыворотки от коз, не подвергавшихся обработке, дорожка 3: образцы молочной сыворотки от коз, трансдукцированных вектором Ad-E2his-sec, взятые в качестве исходного материала для хроматографии, дорожка 4: материал, не связавшийся с матриксом, дорожка 5: промывание 20 мМ имидазолом, дорожка 6: промывание 50 мМ имидазолом, дорожка 7: элюция 200 мМ имидазолом.
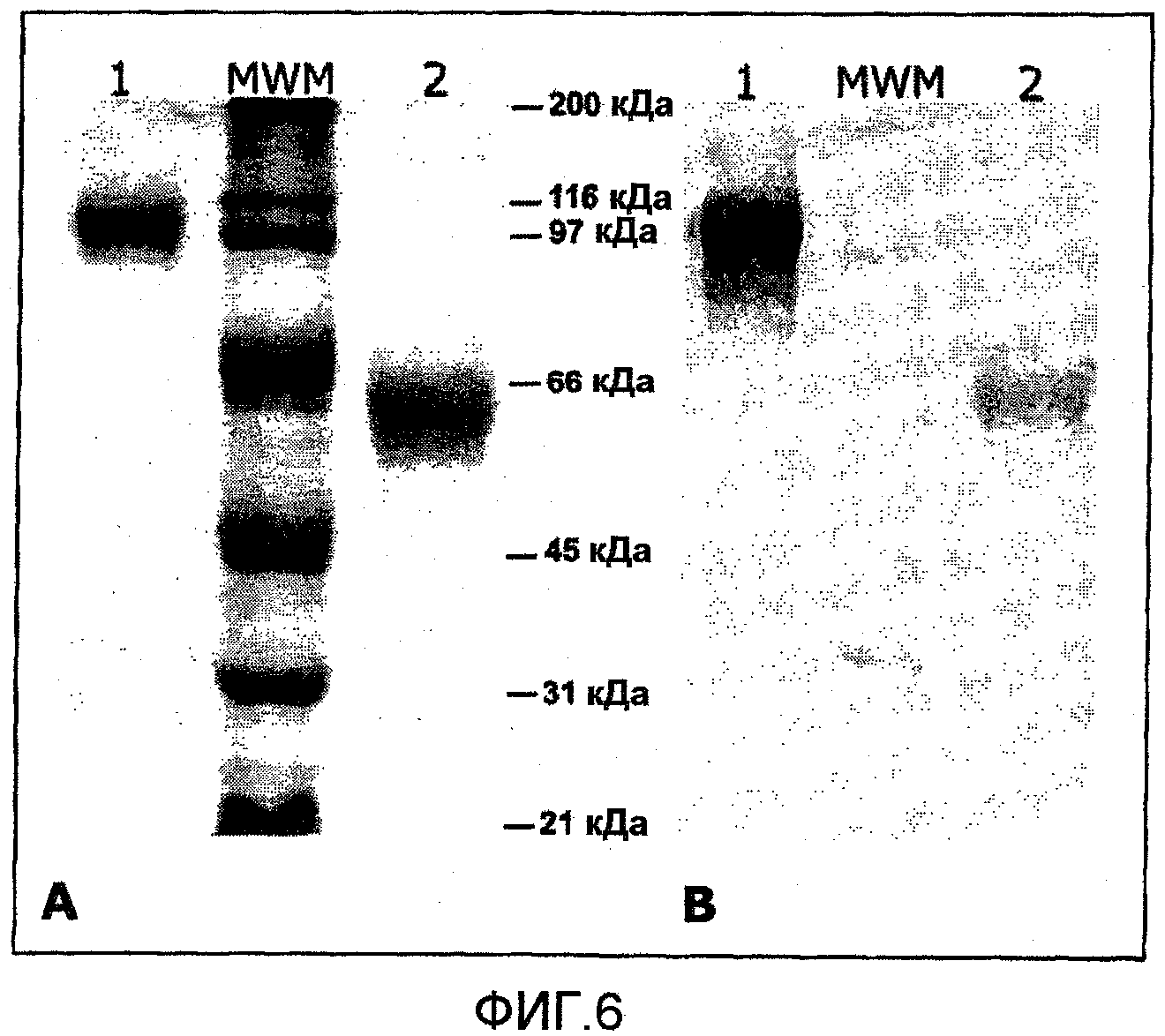
Фиг.6. Сравнение антигенного распознавания двух изоформ вакцинного антигена E2his антителами, присутствующими в сыворотке свиней, инфицированных вирулентным штаммом ВКЧС. E2his, очищенный из молока коз, трасформированных аденовирусным вектором Ad-E2his-sec, исследовали при помощи электрофореза и иммуноблоттинга в восстанавливающих условиях (мономер) и в невосстанавливающих условиях (гомодимер). (А) SDS-электрофорез в ПААГ. (В) иммуноблоттинг с применением поликлональной сыворотки от инфицированных КЧС свиней, дорожка 1: E2his, подвергнутый электрофорезу в невосстанавливающих условиях, дорожка 2: E2his, повергнутый электрофорезу в восстанавливающих условиях.
Фиг.7. Кинетика нейтрализующих антител, полученных в двух группах свиней, вакцинированных одной дозой вакцинного антигена E2his, титры антител определялись при помощи теста на нейтрализацию, основанного на пероксидазе. Группа А была привита дозой 30 мкг на каждое животное и группа В - дозой 50 мкг на каждое животное. Обе группы подвергали контрольному заражению через три недели после вакцинации дозой вируса ВКЧС 105 DL50. Результаты показаны в виде среднего геометрического соответствующих титров.
Фиг.8. Лимфопролиферативный анализ с применением лимфоцитов, выделенных от свиней на 5 день после вакцинации, с антигенами Е2-CD154 (группы D и Е) и Е2his (группа F). Результаты выражены в виде индекса стимуляции (SI), определенного как отношение между значениями счета в минуту (cpm) стимулированной культуры и уровнями cpm контрольной культуры, не подвергавшейся стимуляции. Лимфопролиферативный ответ, для которого определялось SI≥2, рассматривался как положительный. Была оценена пролиферация в культурах, обработанных ВКЧС так же, как и ингибирование пролиферации в культурах, обработанных ВКЧС и Mab против домена CD4 свиней.
Фиг.9. Исследование антивирусной активности клеток РК-15 с применением сыворотки, полученной от свиней, вакцинированных антигенами Е2-CD154 (группы D и Е) и Е2his (группа F). Результаты выражены в виде среднего геометрического соответствующих титров.
Фиг.10. Кинетика нейтрализующих антител, полученных в двух группах свиней, вакцинированных антигенами Е2-CD154 (группа Н) и Е2his (группа I), с применением дозы, равной 50 мкг на животное. Титры антител определялись посредством теста на нейтрализацию на основе пероксидазы. Результаты показаны в виде среднего геометрического соответствующих титров.
Фиг.11. Анализ условий экспрессии E2his и E2his-CD154 в клетках GMGE, трансдуцированных лентивирусными векторами Lv-E2his и Lv-E2his-CD154. Белки в культуральной среде разделяли методом SDS-электрофореза в ПААГ в невосстанавливающих 4 условиях. Иммуноидентификацию интересующих молекул проводили вестерн-блот анализом, используя Mab против E2 ВКЧС (αE2-1G6). Дорожка 1: культуральная среда из клеток, трансдуцированных вектором E2his-CD154; дорожка 2: культуральная среда из клеток, трансдуцированных вектором E2his; дорожка 3: культуральная среда из необработанных клеток; дорожка 4: положительный контроль, E2his, экспрессированные в культуральной среде клеток РК-15, трансдуцированных вектором Ad-E2his-sec.
ПРИМЕРЫ
Пример 1: Амплификация участков гена, кодирующих внеклеточные домены ВКЧС E2 и CD154 свиней и клонирование плазмиды pMOS-E2his-CD154
Участок гена, кодирующий внеклеточный домен Е2 из 363 аминокислот, амплифицируют обратной транскрипцией и полимеразной цепной реакцией (RT-PCR) с вирусного генома кубинского варианта ВКЧС, штамма «Маргарита», номер доступа AJ704817 в базе данных Национального Центра Биотехнологической Информации (National Center for Biotechnology Information, NCBI). В 3'-олигонуклеотид включают последовательность, кодирующую «хвост» из 6 остатков гистидина, для обеспечения возможности легкой очистки антигена.
Кодирующую последовательность для внеклеточного домена CD154 свиньи из 210 аминокислот получают химическим синтезом с применением в качестве образца гена CD154 свиньи Sus scrofa (номер доступа NCBI AB040443). В 5'-конец кодирующей последовательности этой молекулы включают участок, кодирующий пептид из четырех повторяющихся единиц Gly-Gly-Gly-Gly-Ser (4G4S). При субклонировании в плазмиду pMOS-BLUE (Amersham, USA) синтезированная последовательность (4G4S-CD154) вставляется непосредственно после «хвоста» из 6 гистидинов в кодирующей последовательности Е2his. Получают плазмиду pMOS-2Ehis-CD154.
Пример 2: Генетические конструкции молекул E2his и E2his-CD154 с сигналами секреции для клеток млекопитающих
Последовательность, соответствующую E2his и полученную RT-PCR, вставляют по сайтам Bgl II-EcoR V в плазмиду pAEC-SPT (Herrera et al. (2000) Biochem. Biophys. Res. Commun. 279:548-551). Тем самым получают вектор pE2his-sec, содержащий кодирующую последовательность для E2his, которой предшествует сигнал секреции тканевого активатора плазминогена человека (htPA) и которая находится под транскрипционным контролем предраннего промотора цитомегаловируса человека (PCMV).
Последовательность, соответствующую E2his-CD154 и субклонированную в вектор pMOS-BLUE, вставляют по сайтам для эндонуклеаз рестрикции Bgl II-Sal I в плазмиду pAEC-SPT. Тем самым получают вектор pE2his СD154-sec, содержащий кодирующую последовательность для E2his-СD154, которой предшествует сигнал секреции htPA и которая находится под транскрипционным контролем PCMV.
Пример 3: Формирование рекомбинантных аденовирусных векторов, содержащих кодирующие последовательности для E2his и E2his-CD154 с сигналами секреции для клеток млекопитающих
Дефектные по репликации аденовирусные векторы (Ad-ΔE1, ΔE3) получают как описано в инструкции к системе AdEasy (AdEasytm-Vector System Quantum Biotechnology, EE.UU). Плазмиду pAdTrack-CMV используют как переносящий вектор. Кодирующую последовательность для E2his с сигналом секреции htPA (E2his-sec) вырезают из плазмиды рE2his-sec энзиматическим перевариванием эндонуклеазами рестрикции Nco I и EcoR V и вставляют по сайту рестрикции EcoR V в вектор pAdTrack. Получают рекомбинантный pAdT-E2his-sec с секретируемым вариантом E2his под транскрипционным контролем PCMV.
Кодирующую последовательность для секретируемого E2his-CD154 вырезают из плазмиды рE2his-CD154-sec энзиматическим перевариванием эндонуклеазами рестрикции Nco I и Sal I и вставляют по сайтам рестрикции Kpn I-Xho I в вектор pAdTrack. Получают рекомбинантный pAdT-E2hisCD154-sec с E2his-CD154sec под транскрипционным контролем PCMV.
Переносящие аденовирусные векторы pAdT-E2his-sec и pAdT-E2hisCD154-sec линеаризуют энзиматическим перевариванием эндонуклеазой рестрикции Pme I для формирования рекомбинантных аденовирусных геномов.
Каждый из линейных векторов вводят электропорацией вместе с вектором pAdEASY-1 в штамм Escherichia coli BJ5183. Рекомбинантные геномы векторов pAd-E2his-sec и pAd-E2hisCD154-sec получают рекомбинацией гомологов. Один из них содержит кодирующую последовательность для молекулы E2his-sec, а другой содержит кодирующую последовательность для молекулы E2his-CD154-sec. В обоих случаях они остаются под транскрипционным контролем PCMV.
Рекомбинантные аденовирусные геномы затем переваривают эндонуклеазой Pac I и трансфецируют в комплементарные клетки линии НЕК-293А и получают инфекционные вирионы. Формируются два аденовирусных вектора: Ad-E2his-sec и Ad-E2his-CD154-sec. Векторы независимо размножаются в клетках линии НЕК-293А до достижения титра 1×1012 колониеобразующих единиц/мл (CFU/мл) и очищаются двукратным центрифугированием в градиенте CsCl. Векторы затем переводят диализом против буфера для хранения (10 мМ Трис, рН 8,0, 2 мМ MgCl2, 4% сахароза) и хранят при -70°С. Способность аденовирусных векторов Ad-E2his-sec и Ad-E2hisCD154-sec трансформировать клетки млекопитающих и опосредовать экспрессию и секрецию молекул E2his и E2his-CD154 в среду культивирования была подтверждена тестами на инфекционность на клетках свиньи линии РК15. Образцы белков, присутствовавших в среде культивирования инфицированных клеток, разделялись электрофорезом в полиакриламидном геле в присутствии додецилсульфата натрия в невосстанавливающих условиях и проанализированы иммуноблоттингом с моноклональными антителами против Е2 ВКЧС (αE2-1G6) (фиг.1 и 2).
Анализ молекулярных масс гликопротеинов E2his и E2his-CD154 подтвердил, что они соответствовали димерным и тримерным изоформам. На дорожке 1 фиг.2 видны две полосы, соответствующие димерным (180 кДа) и тримерным изоформам E2-CD154 (270 кДа).
Пример 4: In situ трансдукция эпителия молочной железы козы для получения E2his и E2his-CD154 в молоке
Для трансформации эпителия молочной железы экспрессионными кассетами E2his и E2his-CD154 применяются рекомбинантные аденовирусные векторы Ad-E2his-sec и Ad-E2hisCD154-sec. В обоих случаях векторы вводятся в молочную железу коз, производящих молоко, прямым вливанием в вымя через канал соска. Аденовирусные векторы инфицируют секреторные эпителиальные клетки, которые составляют эпителий молочной железы, разрешая экспрессию рекомбинантных белков.
Используются козы на втором месяце лактации, с производительностью в среднем 1 литр в день. Для введения аденовирусных векторов самки сначала подвергаются доению до удаления молока из вымени, затем изоосмотический солевой раствор вливается в цистерны непосредственно через канал соска при мягком массировании вымени для гарантии полного промывания молочной железы. Солевой раствор удаляется путем тщательного доения вымени, и процесс повторяют дважды. Впоследствии вводится аденовирусный инокулят с титром 109 CFU/мл в солевом растворе, содержащем 36 мМ ЭГТА. Объем вливаемого препарата варьирует в зависимости от емкости вымени и гарантированно его заполняет. После вливания вымя массируют для облегчения гомогенного распределения инокулята в железе и достижения секреторных эпителиальных клеток альвеол. Через 24 часа аденовирусный инокулят удаляется посредством доения. С целью удаления остаточных аденовирусных векторов из цистерны или молочных протоков молочные железы промывают снова путем вливания солевого раствора.
Через двадцать четыре часа после этого путем ручного доения начинают собирать молоко от трансформированных животных. Ежедневно производят два доения с 12-часовым интервалом. Собранное молоко сохраняют при -70°С. Кинетику экспрессии рекомбинантных белков E2his и E2his-CD154 в молоке анализируют для каждого надаиваемого образца (фиг.3 и 4). Доказано, что молекулярные размеры рекомбинантных белков соответствуют димерным и тримерным изоформам. Для E2his была получена средняя экспрессия 1,03 г/л на 2-8 дни после инокуляции, со средним выходом 5,22 г из одного животного. Для рекомбинантной молекулы E2his-CD154 была получена средняя экспрессия 0,73 г/л, со средним выходом 3,04 г из одного животного.
Пример 5: Выделение антигенов E2his и E2his-CD154 из молока коз
Образцы от каждого дня доения, содержавшие рекомбинантные вакцинные антигены E2his и E2his-CD154, соответственно, смешивают и центрифугируют при 15000g в течение 30 минут при 4°С. Растворимую фазу (сыворотку молока) сепарируют и жировую фазу отбрасывают. Собранную сыворотку разводят буфером для сепарирования молока (10 мМ Трис-HCl, 10 мМ СаСl2, pH 8,0) в соотношении 1:4. Смесь охлаждают на льду в течение 30 минут и центрифугируют при 15000g в течение 30 минут при 4°С. Супернатанты и осадки анализируют SDS-электрофорезом в ПААГ и иммуноблоттингом. Было определено, что наибольшее количество таких рекомбинантных белков присутствует в растворимой фазе, а осадки содержат казеины.
Фракции сыворотки, содержащие интересующие рекомбинантные антигены (E2his и E2his-CD154), осветляют последовательным фильтрованием через мембраны с порами 0,8 мкм и 0,4 мкм (Millipore) и затем наносят на ХК16 колонку (Amersham, USA), заполненную Ni-NTA-агарозной смолой (Qiagen, USA). Ее промывают в две стадии, сначала 100 мМ фосфатным буфером с 20 мМ имидазола, рН 7,2, затем 100 мМ фосфатом с 50 мМ имидазолом, рН 7,2. После промывания интересующий белок элюируют 100 мМ фосфатным буфером с 200 мМ имидазола, рН 7,2. Пик, соответствующий чистой фракции, переводят диализом в 10 мМ фосфатного буфера, рН 7,2 (фиг.5).
Процедура очистки E2his и E2his-CD154 из козьего молока одинакова для обоих вакцинных антигенов. Два белка получают с уровнем чистоты, превышающим 90%. Выход E2his составляет 70%, для E2his-CD154 получен выход 58%. Очищенные белки анализируют SDS-электрофорезом в ПААГ и иммуноблоттингом с целью выявления образования белковых агрегатов. Может быть определено, что димерные изоформы (гомодимеры) E2his, полученные в молоке, эффективно узнаются поликлональной сывороткой из свиней, инфицированных ВКЧС, что указывает на то, что эта специфическая конформация повышает молекулярную антигенность (фиг.6).
Пример 6. Конструирование экспрессионных векторов в метилотрофных дрожжах Pichia pastoris
Вектор pPS10 для экспрессии в P. pastoris переваривают эндонуклеазой рестрикции Nae I для того, чтобы вставить интересующую кодирующую последовательность на 3'-конец сигнала секреции инвертазы сахарозы Saccharomyces cerevisiae (Suc2). Кодирующую Е2 последовательность, амплифицированную с помощью ПЦР, вставляют по сайту рестрикции Nae I в плазмиду pPS10. Последовательность, кодирующую E2his-CD154, вырезают из плазмиды pMOS-E2his-CD154 энзиматическим перевариванием эндонуклеазами рестрикции Sma I-EcoR V и вставляют по сайту рестрикции Nae I в плазмиду pPS10. Таким образом, получают плазмиды pPS-E2his и pPS-E2his-CD154. Кодирующие последовательности для обеих молекул соединяются с сигналом секреции Suc2 из S. cerevisiae и остаются под транскрипционным контролем промотора фермента алкоголь-оксидазы (AOX1) из дрожжей P. pastoris.
Рекомбинантные плазмиды линеаризуют эндонуклеазой рестрикции Pvu II и вставляют электропорацией в электрокомпетентные клетки P. pastoris, штамм MP36. Таким образом, получают несколько клонов P. pastoris, штамм MP36, стабильно трансформированных плазмидами pPS-E2his и pPS-E2his-CD154. Этот штамм является мутантом, ауксотрофным по гистидину, поэтому рекомбинантные дрожжи приобретают His+ фенотип, что позволяет его ауксотрофную селекцию.
Рекомбинантные дрожжи, сначала идентифицированные в дот-блоттинге, анализируют также блоттингом по Саузерну для определения характера интегрирования, которое происходит при замене гена AOX1 P. pastoris с получением Muts-His фенотипа (низкая концентрация метанола). Генетическая замена AOX1 происходит путем рекомбинации между 5'-районом промотора AOX1 и 3'-районом AOX1, присутствующим в геноме дрожжей, и другим, присутствующим в плазмиде, что приводит к уничтожению участка, кодирующего ген AOX1. Рекомбинантные дрожжи с Mutsфенотипом поддерживают производство фермента алкоголь-оксидазы с гена AOX2, но скорость их роста в метаноле низка. Интегрирование фенотипа Mut+-His также может быть получено заменой.
Последовательности, кодирующие варианты E2his и E2his-CD154, остаются под регулирующим контролем промотора AOX1, индуцируемым метанолом. P. pastoris секретирует белки в низкой концентрации, и среда для его культивирования не нуждается в дополнительных белках, поэтому можно ожидать, что секретируемый гетерологический белок составляет большую часть суммарных белков в среде (до более чем 80%). Производство рекомбинантного белка осуществляют в 5-литровых ферментерах. Индукцию экспрессии проводят добавлением метанола в культуру в течение 5 дней, и рекомбинантные белки получают в культуральной среде ферментирования. E2his секретируется в среду культивирования рекомбинантных дрожжей на уровне 0,143 мг/мл. В случае E2his-CD154 получен уровень 0,122 мг/мл.
Пример 7: Выделение антигенов E2his и E2his-CD154 из среды культивирования Pichia pastoris
Продукт ферментирования центрифугируют при 10000g в течение 30 минут при 4°С для отделения биомассы от жидкой фазы. Среду культивирования фильтруют через мембраны с порами 0,8 мкм и 0,2 мкм (Millipore) и наносят на колонку XK16 (Amersham, USA), заполненную Ni-NTA агарозным матриксом (Qiagen, USA). Проводят промывание 100 мМ фосфатным буфером с 30 мМ имидазолом, рН 7,2, и интересующий белок элюируют 100 мМ фосфатным буфером с 200 мМ имидазолом, рН 7,2. Чистую фракцию диализом переводят в 10 мМ фосфатный буфер. Процедура выделения E2his и E2-CD154 из супернатанта ферментации генетически трансформированных P. pastoris одинакова для обоих вакцинных антигенов. Два белка получали с уровнем чистоты выше 95%. Выход E2his составлял 83%, в случае E2his-CD154 выход составлял 78%.
Пример 8: Испытание иммунитета у свиней, вакцинированных секретируемым вариантом E2his
В этом тесте используются двадцать четыре здоровых свиньи весом около 20 кг, с негативной в отношении ВКЧС серологией, принадлежащие не вакцинированному и свободному от КЧС стаду. Свиней распределяют по группам, по 8 животных в каждой, и помещают в три отдельные экспериментальные комнаты (А, В и С) с водой и кормом в неограниченном объеме.
Животных из группы А и группы В иммунизируют композицией вакцины, содержащей антиген E2his, в виде отдельной дозы 30 мкг (группа А) и 50 мкг (группа В) на животное, группу С иммунизируют плацебо. Антиген готовят как эмульсию воды в масле, и 2 мл вводят внутримышечно в шею. Плацебо составляют из адъюванта и фосфатного солевого раствора в соотношении 1:1 (объем/объем) и вводят при таких же условиях. На третьей неделе после иммунизации проводят контрольное заражение всех животных 105DL50 гомологичного штамма ВКЧС, «Маргарита», путем внутримышечной инъекции.
Введение препарата вакцины E2his не вызывает побочных реакций, судя по тому, что не наблюдается изменений нормальных клинических параметров. Титры нейтрализующих антител, превышающие 1:50 (считается иммунитетом) получают у вакцинированных групп со второй недели иммунизации. После третьей недели титры возрастают до 1:1600-1:6400 (фиг.7), но между вакцинированными группами (А и В) разницы в иммунном ответе не наблюдается. У вакцинированных свиней не развиваются пирексия или клинические симптомы заболевания, и из лимфоцитов не выделяются вирусы в последующие после контрольного заражения дни. Однако в группе плацебо развиваются все клинические симптомы заболевания, включая повышение температуры, кровоизлияния и негнойный энцефалит. В этой группе вирусные изоляты получают с четвертого дня после контрольного заражения до дня забоя. Здесь продемонстрировано, что отнятые от груди свиньи, вакцинированные препаратом E2his, принятым в дозе 30 мкг по предлагаемой схеме вакцинации, остаются защищенными от клинических симптомов и инфекции ВКЧС.
Пример 9: Испытание наследственного иммунитета у беременных свиноматок, вакцинированных секретируемым антигеном E2his
Берут десять свиноматок, серологически негативных относительно ВКЧС, из свободного от ВКЧС и не имеющего истории вакцинации (в течение 3 предыдущих лет) стада. После отнятия от груди индуцируют эстральный цикл гормональной обработкой, и через три дня все свиноматки оказываются оплодотворенными. Оплодотворение осуществляют одновременно с иммунизацией. Берут группу из 5 свиноматок и вводят им внутримышечной инъекцией в шею по 2 мл препарата вакцины, упомянутой в примере 7 (группа В). Остальные 5 свиноматок берут как отрицательный контроль и инъецируют их 2 мл плацебо, составленным из адъюванта и солевого раствора в пропорции 1:1 (объем/объем). Через 21 день вакцинированную группу ревакцинируют. Беременных свиноматок обследуют путем измерения клинической триады (температуры, частоты сердечных сокращений и частоты дыхания) и еженедельного забора крови для гематологии и детектирования нейтрализующих антител против ВКЧС. Двумя месяцами позже производят контрольное заражение беременных свиноматок 105 DL50 гомологичного штамма ВКЧС «Маргарита» путем внутримышечной инъекции. На 3 и 5 день после контрольного заражения производят выделение вируса из лимфоцитов периферической крови с целью обнаружения присутствия ВКЧС. Через две недели после контрольного заражения свиноматок забивают и извлекают плоды для морфологического и патологоанатомического анализа и теста на выделение вируса. В процессе эксперимента свиноматки имеют неограниченный доступ к воде и корму.
Вакцина оказалась безвредной для всех беременных свиноматок, случаев выкидышей или клинических нарушений после иммунизации не было. Вакцинированные животные вырабатывали специфические нейтрализующие антитела против ВКЧС с титрами от 1:50 до 1:51200. Свиноматки из вакцинированной группы оставались полностью здоровыми после контрольного заражения. Ни одно из этих животных не показало повышения температуры, лейкопении, тромбоцитоза или любого другого клинического симптома КЧС.
Морфометрический и патологоанатомический анализ позволяет определить, что плоды вакцинированных свиноматок имеют нормальный размер, и в них не присутствуют гистопатологические нарушения. ВКЧС не был изолирован ни из лейкоцитов в образцах крови маток, взятых после контрольного заражения, ни из крови или органов забитых плодов.
Свиноматки из группы плацебо после контрольного заражения имели повышенную температуру и лейкопению. Одна из свиноматок перенесла выкидыш на 8 день после контрольного заражения и была забита на 9 день после контрольного заражения. Патологические признаки, такие как малый размер и мумифицирование, увеличение селезенки, несколько точечных кровоизлияний в почках и мочевом пузыре и негнойный энцефалит, наблюдались у плодов от свиноматок, забитых на 2 неделе после контрольного заражения и у абортированных плодов. ВКЧС был изолирован из крови и из всех органов от плодов этой группы. Вакцинация свиней препаратом вакцины E2his предотвращала передачу ВКЧС от свиноматок потомству.
Пример 10: Испытание раннего иммунитета у свиней, вакцинированных препаратом вакцины E2his-CD154
Берут четыре группы по 6 свиней в каждой (в тех же условиях, что в примере 8) и применяют к ним композицию вакцины со следующими количествами антигена: 50 мкг E2his-CD154 (группа D), 80 мкг E2his-CD154 (группа E), 50 мкг E2his (группа F). Группу G берут как плацебо. Антигены готовят в виде эмульсии «вода-в-масле» и по 2 мл прививают путем внутримышечной инъекции, группу плацебо прививают адъювантом без белков. Вакцины применяют в виде однократной дозы. На 8 день после иммунизации проводят контрольное заражение животных 105 DL50 штаммом ВКЧС «Маргарита» посредством внутримышечного введения. Клинические признаки регистрируют ежедневно в течение эксперимента и еженедельно берут кровь для гематологического анализа и определения нейтрализующих антител. Кроме того, образцы крови, взятые на 1, 3, 5 и 7 день после вакцинации, используют для оценки клеточного иммунологического ответа по пролиферации лимфоцитов и в тесте на «антивирусную активность сыворотки».
После вакцинации наблюдались нормальные клинические признаки, и не наблюдалось побочных реакций в месте прививки. Было детектировано повышенное количество лимфоцитов в культурах лимфоцитов от животных, вакцинированных антигеном E2-CD154 (группы D и Е) и усиленное действие митогена фитогемагглютинина в тесте на пролиферацию лимфоцитов. Этот ответ блокировался моноклональными антителами против домена CD4, что указывает на то, что иммунный ответ был опосредован Т-хелперными лимфоцитами. Лимфоциты из образцов от животных из групп F и G (плацебо) не отвечали в этом тесте на стимуляцию митогеном или ВКЧС (фиг.8).
На 3, 5 и 7 день после вакцинации антигеном E2-CD154 в образцах из групп D и Е наблюдался высокий титр интерферона альфа. Однако в течение эксперимента интерферон не был детектирован у животных, вакцинированных антигеном E2his (группа F) и в группе плацебо (G). Тест на «антивирусную активность» против вируса трансмиссивного гастроэнтерита проводят на клетках РК-15. В группах D и Е был получен титр антивирусной активности 1:512; однако антивирусная защита не была детектирована в образцах от свиней, иммунизированных E2his, или группы плацебо (фиг.9). Эти эксперименты определили, что антиген Е2, связанный с молекулой CD154, усиливает клеточный иммунный ответ на ВКЧС, который имеет отношение к иммуностимулирующей активности CD154.
Пример 11: Сравнение кинетики нейтрализующих антител у свиней, вакцинированных однократной дозой композиций вакцин, содержавших E2his и E2his-CD154
Берут три группы по 6 свиней, весом приблизительно по 20 кг, серологически негативных в отношении ВКЧС, из стада, в котором в предыдущие 3 года не было случаев заболеваний КЧС или истории вакцинации. Животных обеспечивают водой и ежедневным кормом в неограниченном объеме.
Каждое животное вакцинируют 50 мкг E2his-CD154 в группе Н, 50 мкг E2his в группе I, и группа J берется как плацебо. Антигены готовят в виде эмульсии «вода-в-масле» и 2 мл прививают путем внутримышечной инъекции, группу плацебо прививают адъювантом без белков. Вводят единственную дозу, и в течение 5 недель после иммунизации измеряют уровень нейтрализующих антител с помощью теста на основе нейтрализации перекиси водорода (NPLA).
Нейтрализующие антитела детектируются начиная со второй недели иммунизации, с титром, превышающим 1:50 (рассматривается как иммунитет) в группах, вакцинированных E2-CD154 и E2his (H и I). В течение испытаний не было обнаружено антител у животных группы плацебо. На второй неделе после иммунизации титры нейтрализующих антител у животных из группы Н (антиген E2-CD154) были выше, чем титры у группы, иммунизированной антигеном E2his. Эти результаты дают основание предположить более высокую стимуляцию гуморального ответа у животных группы Н (фиг.10).
Был сделан вывод о том, что композиция вакцины E2his-CD154 в дозе на уровне 50 мкг является безопасной, иммуногенной и индуцирует ранний гуморальный ответ у вакцинированных свиней, по сравнению с композицией вакцины E2his.
Пример 12: Испытание наследственного иммунитета у беременных свиноматок, вакцинированных композицией вакцины E2his-CD154
Отбирают десять свиноматок такого же состояния здоровья и происхождения, что были использованы в примере 8. После отнятия от груди гормональной обработкой индуцируют эстральный цикл, и через три дня все свиноматки оказываются оплодотворенными. Одновременно группу из 5 свиней иммунизируют 2 мл композиции вакцины E2his-CD154 (80 мкг/животное; композиция, использованная в примере 10 для группы Е), с помощью внутримышечной иммунизации позади уха, на шее. Группу из 5 оставшихся свиней иммунизируют адъювантом как плацебо.
Беременных свиноматок обследуют, измеряя клиническую триаду (температуру, частоту сердечных сокращений и частоту дыхания), еженедельно берут кровь для гематологии и детектирования нейтрализующих антител против ВКЧС. На 2 месяце беременности проводят контрольное заражение животных 105 DL50 штамма ВКЧС «Маргарита». Наличие вируса тестируют в крови, взятой на 3 и 5 день после контрольного заражения. Через две недели свиноматок забивают, и плоды извлекают для вирусологического, морфологического и патологического анализа. На протяжении эксперимента животные имеют неограниченный доступ к воде и корму.
После иммунизации не наблюдалось случаев выкидышей или других клинических проявлений КЧС. Таким образом, композиция вакцины E2his-CD154 при однократной иммунизации оказалась безвредной для беременных животных. Вакцинированные животные вырабатывали специфические нейтрализующие антитела против ВКЧС в титрах от 1:50 до 1:16000.
После контрольного заражения в группе вакцинированных свиноматок не наблюдалось повышения температуры, лейкопении или тромбоцитоза. В этой группе плоды имели нормальный размер, и не было обнаружено никаких гистопатологических изменений, судя по результатам морфометрии и патологоанатомического анализа. ВКЧС не был обнаружен в лейкоцитах ни в образцах крови, взятых у вакцинированных маток после контрольного заражения, ни в органах или крови их плодов.
Свиноматки из группы плацебо имели повышенную температуру, лейкопению и снижение аппетита после контрольного заражения. Плоды из этой группы имели малый размер и демонстрировали гистопатологические изменения, совместимые с КЧС, такие как увеличение селезенки, точечные кровоизлияния в почках и мочевом пузыре, некроз кишечника, несколько кровоизлияний во внутренних органах и негнойный энцефалит. ВКЧС был изолирован из всех органов и крови плодов этой группы. Вакцинация беременных свиноматок препаратом вакцины E2his-CD154, проведенная по апробированной схеме, препятствовала передаче ВКЧС от свиноматок потомству.
Пример 13: Генетические конструкции pL-E2his и pL-E2his-CD154 лентивирусных плазмид
Последовательность, соответствующую E2his, полученную способом ПЦР в реальном времени, субклонировали на участках Bd1 II-EcoR V плазмиды pAEC-SPT (Herrera et al. (2000) Biochem. Byophys Res. Commun. 279:548-551) с получением вектора E2his-sec. Затем этот вектор расщепляли рестрикционным ферментом Nco I-Sma I с выделением генного сегмента Е2, которому предшествует сигнал секреции htPA. Полученную полосу обрабатывали фрагментом I полимеразы Кленова для получения тупого конца из 5' выступа, образованного эндонуклеазой Nco I. Экстрагированную полосу встраивали в сайт EcoR V вектора pBSM-αSIc под транскрипционным контролем αSl-казеинового промотора буйвола (αSlcP), специфичного для молочных желез. Получали плазмиду pBSM-αSlc-E2his.
Для создания вектора pL-E2his, из pLenti6/V5-GW/lacZ (ViraPowerTM Lentiviral Expression System) удаляли кассету экспрессии lacZ путем расщепления Cla I и Kpn I, и в это место (вектор pL) встраивали синтетический фрагмент ДНК, содержащий центральный полипуриновый участок, фланкированный двумя мультиклонирующими сайтами. В сайт ВаmН I в направлении 3' полипуринового участка (вектор pLW) дополнительно встраивали пострегуляторный элемент лесного сурка (woodchuck post-regulatory element, WPRE). Наконец, фрагмент, содержащий промотор αS1с и ген E2his, амплифицировали методом ПЦР из вектора pBSM-αS1c-E2his и клонировали в сайтах Sal I-Xho I между полипуриновым участком и последовательностью WPRE, получая векгор pL-E2his.
Синтезированную последовательность (4G4S-CD154) встраивали сразу за хвостовым участком, состоящим из 6 гистидинов, в кодирующую последовательность E2his вектора pL-E2his. Получали вектор pL-E2his-CD154, содержащий кодирующую последовательность для E2his-CD154, которой предшествовал сигнал секреции htPA, и когорая находилась под транскрипционным контролем тканеспецифичного промотора αSlc.
Пример 14: Создание рекомбинантного лентивирусного вектора Lv-E2his и Lv-E2his-CDl54 с помощью трансфекции с использованием полиэтиленимина в клетках 293Т
Ленгивирусные частицы получали с помощью котрансфекции плазмидных векторов (pL-E2his и pL-E2his-CD154, независимо) и вспомогательных средств для упаковки лентивирусной плазмиды (pLI, pL2, pLP-VSVG, ViraPower™ Lentiviral Expression System, Invitrogen-Life Technologies, USA) в клетки 293Т, используя полиэтилениминовый (РЕI) способ. Вирусный супернатант собирали после инкубации, проводимой в течение 48 часов при температуре 37°С в 5% СO2. Этап очистки включал центрифугирование (2000 об/мин, 10 мин, 4°С) и фильтрование (0,45 мкм фильтры, покрытые ПВДФ (поливинилиденфторид)). Концентрацию лентивирусных частиц увеличивали ультрацентрифугированием (25000 об/мин, 90 мин, 4°С) супернатанта. Вирусные частицы в пеллетах суспендировали в среде DMEM. Титрование осуществляли методом ПЦР в реальном времени, собрав примерно 108 единиц трансдукции (ТU)/мл.
Способность лентивирусных векторов Lv-E2his и Lv-E2his-CD154 к трансформации клеток млекопитающих и способность обосредовать экспрессию и секрецию молекул E2his и E2his-CDl54 подтверждали оценкой трансдукции в линии эпителиальных клеток молочной железы коз (GMGE). Эти клетки высевали с плотностью примерно 2,5×105клеток/см2 в среде DMEM, дополненной 10% фетальной бычьей сывороткой, эпидермальным фактором роста (10 нг/мл) и инсулином (10 мкг/мл). Образцы белков, находящиеся в культуральной среде инфицированных клеток, разделяли методом SDS-электрофореза в ПААГ в нередуцирующих условиях и анализировали вестерн-блот анализом, используя Mab против Е2 ВКЧС (αE2-lG6) (Фиг.11).
Пример 15: Создание трансгенных коров для получения E2his и E2his-CDl54 в молоке, и очистка антигенов
Трансгенные животные были получены путем субзональной инъекции лентивирусных векторов (Lv-E2his и Lv-E2his-CD154, независимо) в перивителлиновое пространство in vitro зрелых ооцитов с последующей in vitro фертилизацией, культивированием эмбрионов и переносом эмбрионов в синхронизированные реципиенты. Коровьи ооцит-кумулюсные комплексы собирали аспирацией овариальных фолликул, полученных от забитых животных, и доводили до созревания in vitro в течение 22 часов при температуре 39°С в 5% СO2. Для инъекции ооцитов кумулюсные клетки полностью удаляли встряхиванием в присутствии гиалуронидазы перед проведением субзональной вирусной инъекции и 18 часовой in vitro фертилизации. Эмбрионы культивировали в модифицированной синтетической жидкости яйцевода, дополненной 10% (v/v) эстральной коровьей сывороткой при температуре 39°С во влажной атмосфере, состоящей из 5% CO2, 5% O2 и 90% N2.
В качестве реципиентов использовали телок с нормальным половым циклом в возрасте от 22 до 36 месяцев на стадии диэструс. Через семь дней были пересажены бластоциты нехирургическим способом. Реципиентов исследовали на 28 день после переноса эмбрионов с помощью ультразвука на наличие оплодотворенного яйца и ректальным пальпированием на 42, 60 и 90 дни беременности. Телок-основателей (F0) идентифицировали с помощью анализа геномной ДНК, полученной из крови и ткани. Образцы молока трансгенных самок получали в результате естественной и индуцированной (ручным способом или гормонально) лактации. Затем из молока трансгенных коров проводили очистку антигенов E2his и E2his-CD154, используя такую же процедуру, как описано в примере 5. Получали два белка с уровнем очистки до 90%.
Пример 16: Эксперимент по защите свиней, перорально инокулированных рекомбинантными аденовирусами Ad-E2his-sec и Ad-E2hisCD154-sec
В исследовании использовали двадцать четыре здоровые свиньи весом примерно 20 кг с негативной серологией к ВКЧС, которые принадлежали к стаду невакцинированных и свободных от КЧС животных. Свиней распределяли в группы по 8 животных в каждой и содержали в трех отдельных экспериментальных комнатах (К, L и М) со свободным доступом к воде и пище.
Животных из группы К иммунизировали антигеном Ad-E2his-sec, из группы L иммунизировали антигеном Ad-E2hisCD154-sec, а из группы М иммунизировали плацебо. Каждая свинья получала единичную пероральную дозу в количестве 2 мл, содержащую титр вируса, равный 5х106 бляшкообразующих единиц (p.f.u.)/мл. В таких же условиях вводили плацебо, состоящее из раствора фосфата. Вводили единичную дозу и измеряли уровни нейтрализующих антител в сыворотке с помощью анализа нейтрализации связанной пероксидазы (NPLA) в течение 5 недель после иммунизации. Через три недели после иммунизации все животные были спровоцированы внутримышечной инъекцией штамма "Маргарита" гомологичного ВКЧС 105 DL50 дозой.
Инокуляция рекомбинантного аденовируса не спровоцировала негативной реакции, поскольку не наблюдалось каких-либо изменений нормальных клинических параметров. Титры нейтрализующих антител, превышающие 1/50 (считающиеся защитными), наблюдали у вакцинированных групп после второй недели иммунизации. После третьей недели титры увеличились до 1/800-1/2500 (таблица 1). У животных из группы плацебо во время эксперимента не было зарегистрировано никаких антител. На второй неделе после иммунизации титры нейтрализующих антител у животных группы L (AdE2-CD154) были выше титров группы, иммунизированной AdE2his. Эти результаты подтверждают более высокий уровень стимуляции гуморального ответа у животных группы L. У иммунизированных свиней не развивалась лихорадка или клинические симптомы заболевания и не были получены вирусные изоляты из лимфоцитов в последующие дни после провокации. Однако у группы плацебо появились все клинические симптомы заболевания, включая лихорадку, геморрагию и негнойный энцефалит. Животные этой группы умерли через неделю после провокации, а образцы крови были непригодными для проведения теста NPLA.


Реферат
Настоящее изобретение относится к области ветеринарии и описывает химерный вакцинный антиген против вируса классической чумы свиней (ВКЧС) характеризующийся тем, что он состоит из внеклеточного сегмента гликопротеина Е2 вирусной оболочки ВКЧС на N-конце химерного антигена и внеклеточного домена молекулы CD154 свиньи, идентифицированной как SEQ ID NO: 2, на С-конце химерного антигена. Химерные антигены могут быть получены в экспрессионных системах, которые гарантируют правильное свертывание третичной структуры химерных молекул. Вакцинные композиции, которые содержат такие химерные антигены, вызывают ранний и сильный иммунный ответ у вакцинированных свиней и создают полную защиту против ВКЧС. Более того, полученные вакцинные композиции предотвращают передачу вируса от свиноматок их потомству. Химерные антигены так же, как и полученные вакцинные композиции, могут применяться в ветеринарии в качестве вакцин для превентивного использования у свиней. 2 н. и 8 з.п. ф-лы, 11 ил., 1 табл.
Комментарии