Высокогликозилированный слитый белок на основе фактора свертывания крови человека viii, способ его получения и его применение - RU2722374C1
Код документа: RU2722374C1
Чертежи







Описание
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ НАСТОЯЩЕЕ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к области слитых белков и, более конкретно, к слитому белку на основе фактора свертывания крови человека VIII (FVIII), способу его получения и путям его применения, особенно к путям применения при лечении различных заболеваний, связанных со свертыванием крови.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Фактор свертывания крови VIII (FVIII), также известный как антигемофильный фактор, играет крайне важную роль в эндогенной системе свертывания крови. На основании результатов большого количества исследований по молекулярной генетике FVIII, отсутствие генов FVIII, сцепленных с половой Х-хромосомой, будет приводить к гемофилии типа А. Согласно статистическим данным гемофилия А характеризуется показателем распространенности 1/5000 среди мужского населения, которое составляет более 80% от общего числа пациентов с гемофилией. В настоящее время общепринятым лечением гемофилии А является заместительная терапия, то есть восполнение фактора свертывания VIII, который отсутствует у пациентов с гемофилией.
FVIII представляет собой многодоменный макромолекулярный гликопротеин, который разделен на шесть доменов: три домена A (A1, A2, A3), один богатый углеводами и не играющий важной роли центральный домен B (B) и два домена C (C1, C2). Зрелый белок состоит из легкой цепи и тяжелой цепи и имеет молекулярную массу приблизительно 280 кДа. Легкая цепь характеризуется молекулярной массой приблизительно 80 кДа и содержит домены A3, C1 и C2 со схемой связывания A3-C1-C2. Тяжелая цепь характеризуется молекулярной массой приблизительно 90-200 кДа и содержит домены А1, А2 и В со схемой связывания А1-А2-В. Связь между тяжелой цепью и легкой цепью является зависимой от ионов металлов. В плазме крови димер, образованный тяжелой цепью и легкой цепью, защищен от преждевременного разрушения за счет высокоаффинного связывания с фактором Виллебранда (vWF). Период полужизни неактивированного FVIII, связанного с vWF в плазме крови, составляет приблизительно 12 часов. FVIII активируется активированным фактором FX (FXa) и тромбином посредством протеолитического расщепления по аминокислотам Arg 372 и Arg 740 в тяжелой цепи и по Arg 1689 в легкой цепи, что приводит к высвобождению фактора vWF и образованию активированного димера FVIII (FVIIIa). В присутствии Ca2+ FVIIIa образует тесный комплекс с активированным фактором свертывания крови FIX (FIXa) и FX на фосфолипидной поверхности. Затем FIXa активирует FX, и активированный FX (FXa) диссоциируется из комплекса. FXa превращает протромбин в тромбин, который превращает фибриноген непосредственно в фибрин. В качестве кофактора системы коагуляции FVIII может на несколько порядков повышать эффективность FIXa в отношении активации FX.
На сегодняшний день молекула FVIII является одним из наиболее длинных клонированных генных фрагментов и является наиболее высокомолекулярным белковым лекарственным средством, применяемым в клинической практике. Для рекомбинантных белков с высокой молекулярной массой и высокой степенью гликозилирования оптимальной системой экспрессии являются клетки млекопитающих. Однако выход при in vitro экспрессии рекомбинантного FVIII значительно ниже, чем в случае других генов схожей природы. Например, уровень экспрессии FVIII составляет всего лишь 1% от FIX. Низкий уровень экспрессии FVIII может быть отражением потребности организма в FVIII, но это, несомненно, является основным препятствием для in vitro экспрессии рекомбинантного FVIII. Кроме того, поскольку период полужизни FVIII в крови является коротким, всего лишь 8-12 ч, получающим профилактическое лечение пациентам с тяжелой формой гемофилии А должны вводить внутривенные (i.v.) инъекции приблизительно 3 раза в неделю.
Для пролонгирования функционального периода полужизни FVIII in vivo современные методики должны связывать FVIII с фрагментами, пролонгирующими период полужизни, такими как ПЭГ, человеческий сывороточный альбумин (HSA), трансферрин или Fc IgG. Например, фармацевтическими компаниями Novo Nordisk, Bayer и Baxter были разработаны пегилированные продукты FVIII длительного действия под названиями N8-GP, BAY94-9027 и BAX 855 соответственно, которые вошли в стадию клинических исследований. Однако дополнительная стадия химического конъюгирования ПЭГ с FVIII в способе получения белка привела к снижению конечного выхода и увеличению стоимости получения. Фармакокинетические данные демонстрируют, что пегилированный FVIII не достигает значимо более длительного периода полужизни. Например, у пациентов с гемофилией A период полужизни N8-GP в кровотоке составлял приблизительно 18 ч (Tiede A et al., J Thromb Haemost, 2013, 11:670-678). Клиническое исследование I фазы в отношении BAY94-9027 продемонстрировало, что период его полужизни у здоровых людей составлял приблизительно 18,2 ч, что приблизительно в 1,4 раза больше, чем у FVIII дикого типа (Coyle T et al., Haemophilia, 2012, 18 (Suppl 3):22). Период полужизни Bax 855 составлял приблизительно 18 ч (Turecek PL et al., Hamostaseologie, 2012, 32 Suppl 1:S29-38).
Слитый белок rFVIIIFc, Eloctate®, представляющий собой мономер-димерный гибрид, разработанный компанией Biogen Idec, США, был одобрен Управлением по контролю за продуктами и лекарствами США в июне 2014 года. Клинические данные продемонстрировали, что Eloctate® характеризовался увеличением периода полужизни у людей в 1,5-1,7 раза (Dumont J A et al., Blood, 2012, 119:3024-3030; Powell JS et al., Blood, 2012, 119:3031-3037), при этом его необходимо было вводить инъекцией раз в 3-5 дней. Сообщалось, что компания Biogen сконструировала двойной вектор экспрессии, содержащий гены как rFVIIIFc, так и Fc. После трансфекции вектора в клетки HEK-293 среди экспрессированных продуктов не выявляли ожидаемое слияние rFVIIIFc в гомодимерной форме, а выявляли только слитый белок rFVIIIFc, представляющий собой мономер-димерный гибрид, и димер Fc. Исследователи из компании предположили следующее. Поскольку молекулярный размер гомодимерной формы был слишком велик для системы экспрессии, клетки-хозяева не могли секретировать гомодимерный белок rFVIIIFc с молекулярной массой приблизительно 400 кДа, или мономер rFVIIIFc не агрегировался из-за эффекта стерического затруднения (Peters RT et al., J Thromb Haemost, 2013, 11(1):132-141). Следовательно, экспрессия гомодимерной формы слитого белка на основе FVIII была довольно затруднительной.
Карбокси-терминальный пептид (в дальнейшем называемый CTP) бета-цепи хорионического гонадотропина человека (hCG) обладает эффектом пролонгирования периода полужизни in vivo у некоторых белков. Таким образом, фрагменты, пролонгирующие периоды полужизни, можно выбрать из Fc-фрагмента иммуноглобулина, HSA, CTP или других, которые встречаются в слитых белках, раскрытых в некоторых патентных документах. Кроме того, CTP также можно применять в качестве линкера, главным образом, применять для связывания различных субъединиц одного и того же белка. Например, CTP применяют в качестве линкера для связывания бета- и альфа-субъединиц фолликулостимулирующего гормона, как раскрыто в китайских патентных заявках №№ CN 103539860A, CN 103539861 A, CN 103539868 A и CN 103539869 A. В качестве другого примера, CTP применяют в качестве линкера для связывания бета- и альфа-субъединиц гликопротеинового гормона, который раскрыт в патентном документе WO 2005058953 A2.
Авторы настоящего изобретения не применяют CTP в качестве линкера или в качестве пролонгирующего период полужизни фрагмента, как было предложено в предшествующем уровне техники, а вместо этого соединяют его с гибким пептидным линкером (например, (GGGGS)n) с образованием новой линкерной последовательности. Новый линкер расположен между FVIII и пролонгирующим период полужизни фрагментом (например, Fc-фрагментом иммуноглобулина, который не содержит CTP, как было предложено в предшествующем уровне техники), с образованием нового слитого белка на основе FVIII, дополнительно пролонгируя период полужизни и сохраняя биологическую активность и функциональность FVIII.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В настоящем изобретении представлен высокогликозилированный слитый белок, состоящий из Fc и фактора свертывания крови VIII, в гомодимерной форме. Слитый белок обладает пролонгированным активным периодом полужизни in vivo и биологической активностью, подобной рекомбинантному FVIII. Кроме того, в настоящем изобретении представлен способ эффективной и стабильной экспрессии слитого белка. Слитый белок, экспрессируемый с помощью такого способа, обладает преимуществами высокого выхода, хорошей стабильности в способах получения и при хранении и биологической активностью, подобной таковой у представленных на рынке рекомбинантных факторов FVIII.
В первом аспекте настоящего изобретения представлен высокогликозилированный слитый белок на основе FVIII (в дальнейшем сокращенно обозначаемый как слитый белок), содержащий, последовательно от N- к С-концу, фактор свертывания крови человека VIII (hFVIII), гибкий пептидный линкер (L), по меньшей мере одно жесткое карбокси-терминальное пептидное звено бета-субъединицы хорионического гонадотропина человека (в дальнейшем сокращенно обозначаемое как жесткое CTP-звено, изображаемое как (CTP)n, при этом n предпочтительно равно 1, 2, 3, 4 или 5) и пролонгирующий период полужизни фрагмент (например, Fc-фрагмент иммуноглобулина, альбумин, трансферрин или ПЭГ, предпочтительно вариант Fc IgG человека (изображаемый как vFc)). В некоторых предпочтительных вариантах осуществления настоящего изобретения слитый белок экспрессируется в виде hFVIII-L-CTPn-vFc.
При этом hFVIII представляет собой белок дикого типа или его мутант; кроме того, hFVIII дикого типа имеет аминокислотную последовательность, представленную под SEQ ID NO: 1; предпочтительно, мутант hFVIII характеризуется по меньшей мере 85% идентичностью с аминокислотной последовательностью, представленной под SEQ ID NO: 1; более предпочтительно, мутант hFVIII характеризуется по меньшей мере 90% идентичностью с аминокислотной последовательностью, представленной под SEQ ID NO: 1; и, наиболее предпочтительно, мутант hFVIII характеризуется по меньшей мере 95% идентичностью с аминокислотной последовательностью, представленной под SEQ ID NO: 1.
При этом предпочтительно, гибкий пептидный линкер является неиммуногенным и может создавать достаточное пространственное расстояние между hFVIII и Fc, чтобы минимизировать стерическое затруднение между ними. Предпочтительно, гибкий пептидный линкер состоит из двух или более аминокислотных остатков, выбранных из следующих нескольких аминокислот: Gly (G), Ser (S), Ala (A) и Thr (T).
Более предпочтительно, гибкий пептидный линкер содержит остатки G и S. Длина пептидного линкера важна для активности слитого белка. В настоящем изобретении пептидный линкер предпочтительно может характеризоваться общей формулой аминокислотной последовательности, образованной объединением повторяющегося(повторяющихся) звена(звеньев), (GS)a(GGS)b(GGGS)c(GGGGS)d, где a, b, c и d представляют собой целые числа, превышающие или равные 0, и a + b + c + d ≥ 1.
В частности, в вариантах осуществления настоящего изобретения пептидный линкер предпочтительно может содержать следующие последовательности:
(i) L1: GSGGGSGGGGSGGGGS (представленную под SEQ ID NO: 2);
(ii) L2: GSGGGGSGGGGSGGGGSGGGGSGGGGS (представленную под SEQ ID NO: 3);
(iii) L3: GGGGSGGGGSGGGGSGGGGS (представленную под SEQ ID NO: 4);
(iv) L4: GSGGGGSGGGGSGGGGSGGGGSGGGGSGGGGSGGGGS (представленную под SEQ ID NO: 5);
(v) L5: GGGSGGGSGGGSGGGSGGGS (представленную под SEQ ID NO: 6);
При этом жесткое CTP-звено выбрано из полноразмерной последовательности, состоящей из карбокси-терминальных аминокислот 113-145 β-субъединицы хорионического гонадотропина человека или ее фрагмента. В частности, жесткое CTP-звено содержит аминокислотную последовательность, представленную под SEQ ID NO: 7, или ее укороченную последовательность. Во-первых, CTP-пептид, который в естественных условиях встречается в организме человека и содержит несколько сайтов гликозилирования, является неиммуногенным. Во-вторых, по сравнению со случайной спиралью гибкого пептидного линкера жесткий CTP-пептидный линкер, содержащий несколько сайтов гликозилирования, может образовывать стабильную стерическую конформацию, которая позволяет сегментам FVIII и Fc независимо укладываться в правильные трехмерные конформации, при этом они не воздействуют на показатели биологической активности друг друга. Более того, гликозильные боковые цепи CTP обладают защитным эффектом, который может снижать чувствительность пептидного линкера к протеазам.
Предпочтительно, жесткое CTP-звено содержит по меньшей мере 2 сайта гликозилирования. Например, в предпочтительном варианте осуществления настоящего изобретения жесткое CTP-звено содержит 2 сайта гликозилирования. В качестве иллюстрации, жесткое CTP-звено содержит 10 N-терминальных аминокислот из SEQ ID NO: 7, т.е. SSSS*KAPPPS*, или жесткое CTP-звено содержит 14 C-терминальных аминокислот из SEQ ID NO: 7, т.е. S*RLPGPS*DTPILPQ. В качестве другого примера, в другом варианте осуществления жесткое CTP-звено содержит 3 сайта гликозилирования. В качестве иллюстрации, жесткое CTP-звено содержит 16 N-терминальных аминокислот из SEQ ID NO: 7, т.е. SSSS*KAPPPS*LPSPS*R. В качестве еще одного примера, в других вариантах осуществления жесткое CTP-звено содержит 4 сайта гликозилирования. В качестве иллюстрации, жесткое CTP-звено содержит 28, 29, 30, 31, 32 или 33 аминокислоты, которые начинаются с положения 113, 114, 115, 116, 117 или 118 и заканчиваются положением 145 бета-субъединицы хорионического гонадотропина человека. В частности, жесткое CTP-звено содержит 28 N-терминальных аминокислот из SEQ ID NO: 7, т.е. SSSS*KAPPPS*LPSPS*RLPGPS*DTPILPQ. В данном контексте * представляет сайт гликозилирования. Каждый возможный вариант представляет отдельный вариант осуществления настоящего изобретения.
В других вариантах осуществления жесткие CTP-звенья, представленные в настоящем изобретении, характеризуются по меньшей мере 70% идентичностью с нативной аминокислотной последовательностью CTP. В других вариантах осуществления жесткие CTP-звенья, представленные в настоящем изобретении, характеризуются по меньшей мере 80% идентичностью с нативной аминокислотной последовательностью CTP. В других вариантах осуществления жесткие CTP-звенья, представленные в настоящем изобретении, характеризуются по меньшей мере 90% идентичностью с нативной аминокислотной последовательностью CTP. В других вариантах осуществления жесткие CTP-звенья, представленные в настоящем изобретении, характеризуются по меньшей мере 95% идентичностью с нативной аминокислотной последовательностью CTP.
Предпочтительно, жесткие CTP-звенья, описываемые в конкретных вариантах осуществления настоящего изобретения, могут содержать следующие последовательности:
(i) CTP1: PRFQDSSSSKAPPPSLPSPSRLPGPSDTPILPQ (представленную под SEQ ID NO: 7);
(ii) CTP2: SSSSKAPPPSLPSPSRLPGPSDTPILPQ (представленную под SEQ ID NO: 8);
(iii) CTP3: SSSSKAPPPS (представленную под SEQ ID NO: 9);
(iv) CTP4: SRLPGPSDTPILPQ (представленную под SEQ ID NO: 10).
В некоторых вариантах осуществления настоящего изобретения слитый белок содержит одно из описанных выше жестких CTP-звеньев.
В других вариантах осуществления настоящего изобретения слитый белок содержит более одного из описанных выше жестких CTP-звеньев, предпочтительно содержит 2, 3, 4 или 5 описанных выше жестких CTP-звеньев. Например, в одном варианте осуществления настоящего изобретения слитый белок содержит два жестких CTP3-звена: SSSSKAPPPSSSSSKAPPPS (CTP3-CTP3 или изображаемые как (CTP3)2).
При этом, предпочтительно, пролонгирующий период полужизни фрагмент выбран из Fc-фрагмента иммуноглобулина IgG, IgM или IgA и, более предпочтительно, из Fc-фрагмента IgG1, IgG2, IgG3 или IgG4 человека и их вариантов. Кроме того, варианты Fc IgG человека содержат по меньшей мере одну аминокислотную модификацию относительно Fc IgG человека дикого типа и обладают сниженными эффекторными функциями (эффекты ADCC и/или CDC) и/или повышенной аффинностью связывания с неонатальным рецептором FcRn. Кроме того, варианты Fc IgG человека можно выбрать из следующей группы:
(i) vFcγl: шарнирная область, области CH2 и CH3 IgG1 человека, содержащие мутации Leu234Val, Leu235Ala и Pro331Ser и имеющие аминокислотную последовательность, представленную под SEQ ID NO: 11;
(ii) vFcγ2-l: шарнирная область, области CH2 и CH3 IgG2 человека, содержащие мутацию Pro331Ser и имеющие аминокислотную последовательность, представленную под SEQ ID NO: 12;
(iii) vFcγ2-2: шарнирная область, области CH2 и CH3 IgG2 человека, содержащие мутации Thr250Gln и Met428Leu и имеющие аминокислотную последовательность, представленную под SEQ ID NO: 13;
(iv) vFcγ2-3: шарнирная область, области CH2 и CH3 IgG2 человека, содержащие мутации Pro331Ser, Thr250Gln и Met428Leu и имеющие аминокислотную последовательность, представленную под SEQ ID NO: 14;
(v) vFcγ4: шарнирная область, области CH2 и CH3 IgG4 человека, содержащие мутации Ser228Pro и Leu235Ala и имеющие аминокислотную последовательность, представленную под SEQ ID NO: 15.
Варианты Fc IgG, представленные в настоящем изобретении, включают без ограничения пять вариантов, описанных в (i) - (v), а также могут представлять собой варианты, полученные путем объединения или добавления сайтов мутаций двух функциональных вариантов IgG одного и того же подтипа. Например, вариант, описанный выше в (iv), является новым комбинационным вариантом Fc IgG2, полученным за счет добавления сайтов мутаций в (ii) и (iii).
Варианты Fc (vFc) в слитых белках по настоящему изобретению содержат шарнирную область, области CH2 и CH3 IgG человека, например, IgG1, IgG2 и IgG4 человека. Область CH2 содержит аминокислотные мутации в положениях 228, 234, 235 и 331, определяемые по системе нумерации EU, и полагают, что эти аминокислотные мутации могут снижать эффекторные функции, опосредуемые областью Fc. IgG2 человека не связывается с FcγR и характеризуется очень слабой активацией комплемента. Варианты Fcγ2 с мутацией Pro331Ser характеризуются более низкой активацией комплемента, чем нативный Fcγ2, и при этом не связываются с FcγR. Fc IgG4 не способен активировать каскад комплемента, а его аффинность связывания с FcγR приблизительно на порядок ниже, чем у Fc IgGl. Варианты Fcγ4 с мутацией Leu235Ala проявляют минимальные эффекторные функции в сравнении с нативным Fcγ4. Варианты Fcγ1 с мутациями Leu234Val, Leu235Ala и Pro331Ser также проявляют сниженные эффекторные функции в сравнении с нативным Fcγ1. Вышеупомянутые варианты Fc более пригодны для получения слитых белков на основе FVIII, чем нативные Fc IgG человека. Аминокислотные мутации в положениях 250 и 428, определяемые по системе нумерации EU, увеличивают аффинность связывания области Fc с неонатальным рецептором FcRn, тем самым дополнительно пролонгируя период полужизни (Paul R et al., J Biol Chem, 2004, 279:6213-6216). Два вышеупомянутых типа функциональных вариантов объединяют или добавляют друг к другу с получением новых комбинированных вариантов для снижения эффекторных функций, при этом пролонгируя период полужизни. Варианты Fc по настоящему изобретению содержат мутации, которые не ограничиваются вышеописанными сайтами, но также могут вводить замены в другие сайты, вследствие чего Fc характеризуются сниженными эффекторными функциями и/или повышенным связыванием с FcRn. В то же самое время, эти мутации не приводят к снижению функции/активности вариантов Fc или к нежелательным конформационным изменениям. Характерные сайты мутаций можно найти в Shields RL et al., J Biol Chem, 2001, 276(9):6591-604.
В предпочтительном варианте осуществления настоящего изобретения аминокислотная последовательность слитого белка представлена под SEQ ID NO: 16.
Во втором аспекте настоящего изобретения представлена молекула ДНК, кодирующая любой из вышеописанных слитых белков.
В предпочтительном варианте осуществления настоящего изобретения последовательность ДНК слитого белка представлена под SEQ ID NO: 17.
В еще одном аспекте настоящего изобретения представлен вектор, содержащий вышеописанную молекулу ДНК.
В еще одном аспекте настоящего изобретения представлена клетка-хозяин, содержащая вышеописанный вектор или трансфицированная им.
В конкретном варианте осуществления настоящего изобретения клетка-хозяин представляет собой клеточный штамм DG44, производный от СНО.
В пятом аспекте настоящего изобретение представлена фармацевтическая композиция. Фармацевтическая композиция содержит фармацевтически приемлемый носитель, наполнитель и/или разбавитель и эффективное количество вышеописанного слитого белка.
В другом аспекте настоящего изобретения представлен способ получения или производства слитого белка за счет линии клеток млекопитающих, например клеточной линии, производной от СНО, который предусматривает стадии:
(а) введения молекулы ДНК, кодирующей слитый белок, в линию клеток млекопитающих с получением клеточной линии, производной от СНО;
(b) скрининга клеточных штаммов из стадии (а) с получением высокопродуктивного клеточного штамма, приводящего к образованию более 1 МЕ/106 клеток за 24 ч в своей ростовой среде;
(c) культивирования клеточного штамма, полученного на стадии (b), для экспрессии слитого белка;
(d) сбора ферментационного бульона из стадии (с) и выделения и очистки слитого белка.
Кроме того, клеточной линией, производной от CHO, на стадии (a) является DG44.
Кроме того, культивирование клеток на стадии (с) можно осуществлять с помощью способа периодической культуры, перфузионной культуры или способа подпитываемой культуры.
Кроме того, на стадии (d) слитый белок очищают с помощью четырехстадийной процедуры хроматографии, т.е. аффинной хроматографии, гидрофобной хроматографии, анионообменной хроматографии и хроматографии на молекулярных ситах. В примере 5 настоящего изобретения дополнительно приведены предпочтительные условия очистки.
В предпочтительном варианте осуществления настоящего изобретения слитый белок, полученный вышеуказанным способом, характеризуется активностью >6000 МЕ/мг.
В шестом аспекте настоящего изобретения представлено применение слитого белка при изготовлении лекарственного средства для предупреждения или лечения геморрагической болезни или эпизода, вызванных дефицитом или функциональным дефектом FVIII.
Кроме того, данное заболевание предусматривает гемофилию типа A. Функция слитых белков по настоящему изобретению заключается в контроле или предупреждении возникновения кровотечения при спонтанных эпизодах кровотечения, хирургической профилактике, ведении периоперационного периода или хирургическом вмешательстве у пациентов с гемофилией А.
Авторами настоящего изобретения было обнаружено, что преимущества как слитых белков, так и способов их получения, описанных и/или раскрытых в настоящем изобретении, могут быть подытожены следующим образом.
1. Слитый белок на основе FVIII, сконструированный в соответствии с настоящим изобретением, является нелитическим, то есть за счет мутирования области связывания комплемента и рецептора у Fc-сегмента и корректировки аффинности связывания Fc с соответствующим рецептором эффекты ADCC и CDC снижаются или устраняются, при этом сохраняется только функция Fc-сегмента в пролонгировании периода полужизни in vivo активного белка, не вызывая цитотоксичность. Fc-сегмент слитого белка на основе FVIII, разработанного компанией Biogen, происходит из природного источника. Вполне предсказуемо, что неблагоприятные эффекторные функции, опосредованные Fc, будут увеличивать риски лечения для пациентов.
2. В настоящем изобретении предусмотрено применение клеток СНО для экспрессии слитых белков. Среди продуктов экспрессии встречается только гомодимерный слитый белок на основе Fc и FVIII, и стадия очистки является простой и эффективной. Для экспрессии (мономерного) слитого белка на основе FVIII, представляющего собой мономер-димерный гибрид, компания Biogen сконструировала двойной вектор экспрессии, который экспрессировал как rFVIIIFc, так и Fc, который необходимо было трансфицировать в клетки HEK-293 (публикация США № US 20130274194 A1). Ожидали, что в ферментационном бульоне будут присутствовать три формы белковых продуктов экспрессии: гомодимерный (димерный) слитый белок FVIII-Fc:FVIII-Fc, (мономерный) слитый белок FVIII-Fc:Fc, представляющий собой мономер-димерный гибрид, и димер Fc:Fc. В процессе экспрессии слитого белка клетки-хозяева должны были одновременно экспрессировать обе одноцепочечные молекулы FVIII-Fc и Fc, а затем эти молекулы связывались друг с другом с образованием трех вышеуказанных продуктов, так что конечная эффективность экспрессии целевого продукта значительно снижалась. В дополнение, в процессе очистки также следовало удалять остальные две формы, являющиеся побочными продуктами. Следовательно, процесс очистки был более сложным, а эффективность производства была ниже, вследствие чего значительно увеличивались затраты при производстве. По сравнению со способом получения мономерного слитого белка rFVIIIFc, разработанного компанией Biogen, способ получения по настоящему изобретению обладает определенными техническими и ценовыми преимуществами. Процесс экспрессии и очистки по настоящему изобретению проще и эффективнее, а затраты при производстве ниже.
3. Мышам HemA вводили слитый белок на основе FVIII, FP-B, по настоящему изобретению в дозах 30 МЕ/кг, 90 МЕ/кг и 270 МЕ/кг соответственно. В группах средней и высокой дозы острая геморрагия у мышей HemA могла эффективно контролироваться, а выживаемость мышей в группах, которым давали каждую дозу FP-B, была выше, чем в группе, которой давали рекомбинантный FVIII под названием Xyntha (Pfizer), что свидетельствует о том, что слитый белок FP-B характеризовался более длительным фармакодинамическим эффектом по сравнению с Xyntha. В то же самое время, результаты, связанные как с продолжительностью, так и с объемом кровотечения в группах высокой и низкой дозы FP-B, показали дозозависимые связи.
4. Можно ожидать, что по сравнению с рекомбинантным FVIII, Xyntha, слитый белок по настоящему изобретению будет характеризоваться сниженной иммуногенностью и сниженной выработкой нейтрализующих антител у пациентов.
5. Слитые белки, представленные в настоящем изобретении, характеризуются высокими показателями биологической активности. Активность каждой партии очищенных слитых белков находится в диапазоне 6000-10000 МЕ/мг. Если выразить ее в виде удельной молярной активности, то она составляет приблизительно 2340-3900 МЕ/нМ слитого белка, что соответствует 1170-1950 МЕ/нМ FVIII, поскольку каждая молекула слитого белка содержит две молекулы FVIII. В некоторых партиях активность очищенного слитого белка составляет даже более 12000 МЕ/мг, выраженная в виде удельной молярной активности как приблизительно 4680 МЕ/нМ слитого белка, что соответствует 2340 МЕ/нМ FVIII. Таким образом, активность слитого белка, представленного в настоящем изобретении, сравнима или даже выше активности слитого белка rFVIIIFc, представляющего собой мономер-димерный гибрид (1660-1770 МЕ/нМ), разработанного компанией Biogen (J. McCue et al., Biologicals, 2015,43:213-219), и активности представленного на рынке рекомбинантного FVIII ReFacto (1521-2287 МЕ/нМ) (публикация США № US 20130274194 A1). Это свидетельствует о том, что Fc-сегмент, слитый с С-концом слитого белка по настоящему изобретению, производит незначительный эффект на активность FVIII.
6. Слитый белок, представленный в настоящем изобретении, содержит жесткий CTP-полипептид с несколькими гликозильными боковыми цепями, который может образовывать стабильную стерическую конформацию, в сравнении со случайной спиралью гибкого линкера, такого как (GGGGS)n. Данный эффект «разделения» обусловливает то, что сегменты FVIII и Fc независимо укладываются в правильные трехмерные конформации, при этом они не воздействуют на показатели биологической активности друг друга. На CTP располагаются гликозильные группы. Высокосиалированный, отрицательно заряженный CTP может выдерживать клиренс почками, дополнительно пролонгируя период полужизни слитого белка. Более того, защитный эффект боковых цепей CTP может снижать чувствительность пептидного линкера к протеазам, вследствие чего слитый белок с трудом расщепляется в области связывания.
7. Слитый белок по настоящему изобретению характеризуется хорошей стабильностью в процессах ферментации, очистки и хранения.
8. Способ получения слитого белка, представленный в настоящем изобретении, характеризуется преимуществом высокого выхода. После культивирования клеток в 300 мл встряхиваемой колбе в течение 14 дней совокупный выход может достигать по меньшей мере 150 мг/л, причем его можно увеличивать в масштабе для реализации крупномасштабного промышленного производства.
Следует понимать, что в пределах объема настоящего изобретения вышеописанные технические характеристики настоящего изобретения и характеристики, конкретно описанные далее (например, в примерах), можно объединять друг с другом для получения нового или предпочтительного технического решения.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Карбокси-терминальный пептид (CTP) hCG-β
CTP представляет собой короткий пептид на карбокси-конце бета-субъединицы хорионического гонадотропина человека (hCG). Четыре вида связанных с репродукцией полипептидных гормонов, фолликулостимулирующий гормон (FSH), лютеинизирующий гормон (LH), тиреотропный гормон (TSH) и хорионический гонадотропин человека (hCG), содержат одинаковую альфа-субъединицу и свои соответствующие специфические бета-субъединицы. По сравнению с остальными тремя гормонами, hCG характеризуется значительно более пролонгированным in vivo периодом полужизни, что в основном обусловлено специфическим карбокси-терминальным пептидом (CTP) на β-субъединице hCG (Fares FA et al., Proc Natl Acad Sci USA, 1992, 89 (10):4304-4308). Природный CTP содержит 37 аминокислотных остатков и имеет четыре сайта O-гликозилирования. На конце находятся остатки сиаловой кислоты. Высокосиалированный, отрицательно заряженный CTP может выдерживать клиренс почками, тем самым пролонгируя период полужизни in vivo у белка (Fares FA et al., Proc Natl Acad Sci USA, 1992, 89(10):4304-4308). Авторы настоящего изобретения творчески связывают по меньшей мере один CTP-пептид с гибким пептидным линкером соответствующей длины с составлением нового пептидного линкера, который связывает FVIII с пролонгирующим период полужизни фрагментом, например, с Fc-фрагментом иммуноглобулина.
Авторами настоящего изобретения было обнаружено, что добавление CTP-пептида между FVIII и вариантом Fc равноценно добавлению жесткого пептидного линкера. С одной стороны, добавление CTP-пептида обеспечивает то, что слитый на N-конце FVIII не воздействует на сайт связывания варианта Fc с FcRn, таким образом не производя эффект на период полужизни. Кроме того, сайт связывания Fc с белком A важен для стадии очистки. Добавление CTP-пептида обеспечивает то, что слитый на N-конце FVIII не будет «блокировать» этот сайт связывания с белком A, что позволит выбрать более дешевые и более подходящие гранулы для очистки слитого белка и уменьшить затраты при очистке. С другой стороны, добавление CTP-пептида не допускает, чтобы Fc-сегмент размером приблизительно 25 кДа препятствовал правильной укладке слитого на N-конце FVIII, что приводит к отсутствию потери или снижения биологической активности/функции FVIII. Жесткий CTP-пептид, содержащий несколько гликозильных боковых цепей, может образовывать стабильную стерическую конформацию, в сравнении со случайной спиралью гибкого линкера, такого как (GGGGS)n. Данный эффект «разделения» обусловливает то, что сегменты FVIII и Fc независимо укладываются в правильные трехмерные конформации, при этом они не воздействуют на показатели биологической активности друг друга. Более того, защитный эффект гликозильных боковых цепей CTP снижает чувствительность пептидного линкера к протеазам, вследствие чего слитый белок с трудом расщепляется в области связывания.
Варианты Fc IgG
Нелитические варианты Fc
Элемент Fc происходит от Fc-фрагмента константной области иммуноглобулина IgG, и он играет важную роль в уничтожении патогенов при иммунной защите. Эффекторные функции IgG, опосредованные Fc, осуществляются посредством двух механизмов. (1) После связывания с рецепторами Fc (FcγR) на клеточной поверхности патоген расщепляется за счет фагоцитоза или лизиса или с помощью клетки-киллера посредством пути антителозависимой клеточноопосредованной цитотоксичности (ADCC). В качестве альтернативы (2) после связывания с C1q, компонентом первой системы комплемента C1, запускается путь комплементзависимой цитотоксичности (CDC), и, тем самым, патоген подвергается лизису. Из четырех подтипов IgG человека IgG1 и IgG3 способны эффективно связываться с FcγR, а IgG4 характеризуется более низкой аффинностью связывания с FcγR. Связывание IgG2 с FcγR слишком низкое, чтобы его можно было измерить, поэтому IgG2 человека оказывает незначительные эффекты ADCC. Кроме того, IgG1 и IgG3 человека также могут эффективно связываться с C1q, активируя каскад системы комплемента. IgG2 человека слабо связывается с C1q, а IgG4 не связывается с C1q (Jefferis R et al., Immunol Rev, 1998, 163: 59-76), поэтому эффект CDC IgG2 человека также является слабым. Очевидно, что ни один из нативных подтипов IgG не подходит в достаточной степени для конструирования слитых белков FVIII-Fc. Наиболее эффективным способом получения нелитических вариантов Fc без эффекторных функций является мутирование областей связывания комплемента и рецептора у Fc-сегмента и корректировка аффинности связывания Fc с его родственными рецепторами для снижения или устранения эффектов ADCC и CDC. В результате сохраняются биологическая активность функционального белка и длительный период полужизни in vivo у FVIII, при этом не вызывается цитотоксичность. Больше сайтов мутаций, содержащихся в нелитических вариантах Fc, можно найти в публикации Shields RL et al., J Biol Chem, 2001, 276(9):6591-604 или в патенте Китая № CN 201280031137.2.
Варианты Fc с повышенной аффинностью к неонатальному рецептору FcRn
Период полужизни IgG в плазме зависит от его связывания с FcRn. Как правило, IgG связывается с FcRn при рН 6,0 и диссоциирует от FcRn при рН 7,4 (рН плазмы крови). По результатам исследования обоих сайтов связывания модифицируют сайты IgG, которые связываются с FcRn, для увеличения аффинности связывания при рН 6,0. Было показано, что мутации некоторых остатков в домене Fcγ, которые важны для связывания IgG с FcRn, могут увеличивать период полужизни IgG в плазме. Ранее сообщалось, что мутации остатков T250, M252, S254, T256, V308, E380, M428 и N434 увеличивают или уменьшают аффинность связывания с FcRn (Roopenian et al., Nat. Review Immunology 7:715-725, 2007). Варианты трастузумаба (герцептин, Genentech), раскрытые в патенте Кореи № KR 10-1027427, характеризуются повышенной аффинностью связывания с FcRn, и эти варианты содержат одну или несколько аминокислотных модификаций, выбранных из 257C, 257M, 257L, 257N, 257Y, 279Q, 279Y, 308F и 308Y. Варианты бевацизумаба (авастин, Genentech), представленные в патенте Кореи № KR 2010-0099179, характеризуются пролонгированным периодом полужизни in vivo в связи с тем, что они содержат аминокислотные модификации N434S, M252Y/M428L, M252Y/N434S и M428L/N434S. Кроме того, Hinton и соавт. также обнаружили, что два варианта T250Q и M428L увеличивали аффинность связывания с FcRn в 3 и 7 раз соответственно. Если два сайта мутировали одновременно, аффинность связывания увеличивалась в 28 раз. У макака-резуса вариант M428L или T250Q/M428L демонстрирует 2-кратное увеличение периода полужизни в плазме крови (Paul R. Hinton et al., J Immunol, 2006, 176:346-356). Больше сайтов мутаций, содержащихся в вариантах Fc с повышенной аффинностью связывания с FcRn, можно найти в патенте Китая № CN 201280066663.2. Кроме того, исследования демонстрируют, что мутации T250Q/M428L в областях Fc пяти гуманизированных антител улучшают взаимодействие между доменом Fc и FcRn. Более того, в последующих in vivo фармакокинетических тестах при сравнении с антителами дикого типа антитела с мутантными Fc демонстрируют улучшенные фармакокинетические параметры, такие как повышенное время воздействия in vivo, сниженный клиренс и повышенная подкожная биодоступность, при введении с помощью подкожной инъекции (Datta-Mannan A et al., MAbs. Taylor & Francis, 2012, 4(2): 267-273.).
Слитые белки и способы их получения
Ген слитого белка по настоящему изобретению синтезируют искусственно после оптимизации кодонов. На основании нуклеотидной последовательности по настоящему изобретению специалист в данной области успешно может получать кодирующую молекулу нуклеиновой кислоты различными известными способами. Эти способы не ограничены искусственным синтезом и традиционным субклонированием и т.д., и конкретные способы можно найти в Molecular Cloning: A Laboratory Manual by J. Sambrook. В одном варианте осуществления настоящего изобретения кодирующую последовательность нуклеиновой кислоты по настоящему изобретению конструируют путем синтеза сегментов нуклеотидных последовательностей с последующим субклонированием.
В настоящем изобретении также представлен вектор экспрессии для клетки млекопитающего, содержащий последовательность, кодирующую слитый белок по настоящему изобретению, и регулирующую экспрессию последовательность, которая функционально связана с ней. Термин «функционально связанный» относится к состоянию, при котором определенные части линейной последовательности ДНК могут регулировать или управлять активностью других частей той же линейной последовательности ДНК. Например, если промотор управляет транскрипцией последовательности ДНК, то он функционально связан с кодирующей последовательностью.
Векторы экспрессии в клетках млекопитающих могут быть коммерчески доступны, например, включая без ограничения pcDNA3, pIRES, pDR, pBK, pSPORT и т.д. Эти векторы можно применять для экспрессии в системе экспрессии в эукариотических клетках. Специалист в данной области также может подобрать подходящий вектор экспрессии на основании клетки-хозяина.
На основании рестрикционной карты известного пустого вектора экспрессии специалисты в данной области могут сконструировать рекомбинантный вектор экспрессии по настоящему изобретению, применяя стандартный способ, т.е. вставку кодирующей последовательности слитого белка по настоящему изобретению, в подходящие сайты рестрикции посредством расщепления рестрикционными ферментами и лигирования.
В настоящем изобретении также представлена клетка-хозяин для экспрессии слитого белка по настоящему изобретению, содержащая кодирующую последовательность слитого белка по настоящему изобретению. Предпочтительно клетки-хозяева представляют собой эукариотические клетки, такие как без ограничения клетки СНО, клетки COS, клетки 293, клетки RSF и т.д. В предпочтительном варианте осуществления настоящего изобретения клетки представляют собой клетки СНО, которые способны лучше экспрессировать слитый белок по настоящему изобретению и продуцировать слитый белок с хорошей активностью и стабильностью.
В настоящем изобретении также представлен способ получения слитого белка по настоящему изобретению с помощью технологии рекомбинантной ДНК, который предусматривает стадии:
(1) получения последовательности нуклеиновой кислоты, кодирующей слитый белок;
(2) вставки последовательности нуклеиновой кислоты из стадии (1) в подходящий вектор экспрессии для конструирования рекомбинантного вектора экспрессии;
(3) введения рекомбинантного вектора экспрессии (2) в подходящую клетку-хозяина;
(4) культивирования трансфицированных клеток-хозяев в подходящих для экспрессии условиях;
(5) сбора надосадочной жидкости и очистки полученного слитого белка.
Введение кодирующей последовательности в клетку-хозяина можно производить различными методиками, известными в данной области техники, такими как без ограничения осаждение фосфатом кальция, опосредованная липосомами трансфекция, электропорация, микроинъекция, способ вирусной инфекции и способ с применением ионов щелочных металлов.
Информацию о культивировании клеток-хозяев и экспрессии белка можно найти в Olander RM et al., Dev Biol Stand 1996, 86:338. Клетки и клеточный дебрис в суспензии можно удалять за счет центрифугирования для сбора надосадочной жидкости.
Слитый белок, подготовленный и полученный так, как описано выше, можно очистить, как указано дальше, до практически однородной формы, которая, например, демонстрирует одну или специфические полосы на геле после SDS-PAGE. Сначала концентрируют надосадочную жидкость, а концентрат можно дополнительно очистить с помощью гель-фильтрационной хроматографии или ионообменной хроматографии, такой как анионообменная или катионообменная хроматография. Гелевая матрица может представлять собой агаровую, глюкановую, полиамидную и другие среды, обычно применяемые при очистке белка. Q- или SP-группа является идеальной группой для ионного обмена. Очищенный продукт можно дополнительно очищать с помощью адсорбционной хроматографии на гидроксиапатите, металл-хелатной хроматографии, хроматографии гидрофобного взаимодействия и высокоэффективной жидкостной хроматографии с обращенной фазой и т.д. Все вышеперечисленные стадии очистки можно проводить в различных комбинациях, чтобы в конечном итоге достичь высокой степени чистоты и однородности белка. Экспрессированный слитый белок также можно очистить с применением колонки для аффинной хроматографии, содержащей антитело, рецептор или лиганд, специфические к слитому белку. В зависимости от свойств применяемой колонки для аффинной хроматографии слитый белок, связавшийся на колонке для аффинной хроматографии, можно элюировать с применением традиционных способов, таких как применение буфера с высоким содержанием солей, изменение рН и т.д.
Фармацевтическая композиция
В настоящем изобретении также представлена фармацевтическая композиция, содержащая эффективную дозу (предпочтительно приблизительно 2-10 мкг/кг) слитого белка по настоящему изобретению и фармацевтически приемлемый носитель. В большинстве случаев эффективное количество слитого белка по настоящему изобретению можно составить в нетоксичной, инертной и фармацевтически приемлемой водной среде-носителе, рН которой обычно составляет приблизительно 5-8, предпочтительно приблизительно 6-8. Термин «эффективное количество» или «эффективная доза» относится к количеству, которое оказывает функциональные или активные эффекты на людей и/или животных и является приемлемым для людей и/или животных. «Фармацевтически приемлемыми» ингредиентами являются ингредиенты, которые подходят для применения на людях и/или млекопитающих без чрезмерных неблагоприятных побочных эффектов (например, токсичности, раздражения и аллергии), т.е. вещества с разумным соотношением польза/риск. Термин «фармацевтически приемлемый носитель» относится к носителю для доставки терапевтического средства, и такой носитель включает различные наполнители и разбавители.
Фармацевтически приемлемые носители включают без ограничения солевой раствор, буфер, глюкозу, воду, глицерин, этанол и их комбинации. В большинстве случаев фармацевтический состав должен быть совместим со способом введения. Фармацевтические композиции по настоящему изобретению можно приготовить в форме инъекций, например, приготовленных традиционными способами с применением физиологического солевого раствора или водных растворов, содержащих глюкозу и другие вспомогательные вещества. Описанные выше фармацевтические композиции предпочтительно изготавливают в асептических условиях. Количество вводимого активного ингредиента представляет собой терапевтически эффективное количество. Фармацевтический состав по настоящему изобретению также можно приготовить в форме с замедленным высвобождением.
Эффективное количество слитого белка по настоящему изобретению может варьироваться в зависимости от способа введения и тяжести подлежащего лечению заболевания. Предпочтительное эффективное количество может определить специалист в данной области на основании различных факторов, например, с помощью клинических испытаний. К таким факторам относятся без ограничения фармакокинетические параметры слитого белка, такие как биодоступность, метаболизм, период полужизни и т.д., тяжесть подлежащего лечению заболевания у пациента, вес пациента, иммунный статус пациента, путь введения и т.д.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1. Нуклеотидная последовательность FP-B во фрагменте SpeI-EcoRI (сайты рестрикции подчеркнуты) в векторе экспрессии pcDNA3 и производная аминокислотная последовательность в соответствии с примером по настоящему изобретению. FVIII человека состоит из сигнального пептида (аминокислоты 1-19, подчеркнуты с помощью «……») и зрелого белка FVIII (аминокислоты 20-1457). Зрелый слитый белок содержит hFVIII (аминокислоты 20-1457), гибкий пептидный линкер (аминокислоты 1458-1484, подчеркнуты с помощью «___»), жесткое CTP-звено (аминокислоты 1485-1512, подчеркнуты с помощью

Фиг. 2. Результаты SEC-HPLC-хроматографии очищенного белка FP-B.
Фиг. 3. Результаты SDS-PAGE-анализа очищенного белка FP-B.
Фиг. 4. Объем кровотечения (мкл) для каждой мыши после перерезания хвоста. Примечание: * p <0,05, ** p <0,01.
Фиг. 5. Продолжительность кровотечения (с) для каждой мыши после перерезания хвоста. Примечание: * p <0,05, ** p <0,01.
Примеры
Пример 1. Конструирование плазмиды экспрессии, кодирующей слитый белок на основе FVIII
Генные последовательности, кодирующие сигнальный пептид FVIII, зрелый белок, гибкий пептидный линкер, жесткое CTP-звено и вариант vFc IgG человека, подвергали искусственной оптимизации кодонов для экспрессии в клетках CHO и синтезировали искусственно. Синтезированный полноразмерный фрагмент ДНК слитого белка имел сайт рестрикции SpeI на 5'-конце и сайт рестрикции BamHI на 3'-конце. Полноразмерный фрагмент ДНК вставляли в соответствующие сайты рестрикции вектора для переноса pUC57 и подтверждали с помощью секвенирования ДНК.
Полноразмерный генный фрагмент слитого белка, полученный выше, клонировали из промежуточного вектора в соответствующие сайты рестрикции плазмиды экспрессии PTY1A1 с конструированием плазмиды для высокого уровня экспрессии слитого белка. Плазмида PTY1A1 была получена из pcDNA3.1 посредством модификации. Плазмида PTY1A1 содержала без ограничения следующие важные элементы экспрессии: 1) ранний промотор цитомегаловируса человека и энхансер, необходимые для высокого уровня экзогенной экспрессии в клетках млекопитающих; 2) двойные маркеры скрининга с устойчивостью к канамицину у бактерий и устойчивостью к G418 у клеток млекопитающих; 3) кассету экспрессии гена дигидрофолатредуктазы (DHFR) мыши. Если клетки-хозяева относились к типу с дефицитом по гену DHFR, метотрексат (MTX) приводить к совместной амплификации слитого гена с геном DHFR (см. патент США №4399216). Плазмиду экспрессии слитого белка трансфицировали в линию клеток-хозяев млекопитающих. Чтобы обеспечить достижение стабильного и высокого уровня экспрессии, предпочтительной линией клеток-хозяев была линия клеток CHO с дефицитом по ферменту DHFR (см. патент США №4818679). Через два дня после трансфекции среду заменяли на среду для скрининга, содержащую 0,6 мг/мл G418. Клетки высевали в 96-луночный планшет при определенной концентрации (5000-10000 жизнеспособных клеток/лунка) и культивировали в течение 10-14 дней до появления больших отдельных клеточных клонов. Устойчивых к выбранному антибиотику трансфектантов подвергали скринингу с помощью анализа ELISA. Лунки, в которых слитый белок продуцировался на высоком уровне, субклонировали путем предельного разведения в 96-луночном культуральном планшете.
Как показано в таблице 1, в настоящем изобретении была сконструирована серия слитых белков на основе hFVIII, которые содержали линкеры различной длины, жесткие CTP-звенья различного состава и элементы вариантов Fc (vFc) IgG нескольких разных подтипов. Для подтверждения того, что по меньшей мере одно жесткое CTP-звено различной длины могло в значительной степени улучшать активность слитого белка, авторы настоящего изобретения сконструировали слитые белки: FP-A, FP-B, FP-C, FP-D и FP-E. Аминокислоты и кодирующие нуклеотиды FP-B показаны на фиг. 1. Для подтверждения важности жесткого CTP-звена для активности слитого белка авторы настоящего изобретения также сконструировали слитые белки с Fc, FP-G и FP-H, без жесткого CTP-звена. Плазмиды экспрессии конструировали, как описано выше. В дополнение, для подтверждения важности положения жесткого CTP-звена авторы настоящего изобретения также сконструировали FP-F, который имел жесткое CTP-звено на C-конце Fc. Подробности см. в таблице 1. Аминокислотные последовательности каждого компонента были показаны в перечнях последовательностей.
Таблица 1. Составы различных слитых белков на основе FVIII

Пример 2. Временная экспрессия и определение активности различных слитых белков
Восемь плазмид экспрессии, полученных в примере 1, соответственно трансфицировали в 3×107 клеток CHO-K1 с применением реагента DNAFect LT (ATGCell) в 30-мл встряхиваемой колбе, и трансфицированные клетки культивировали в бессывороточной ростовой среде, содержащей 1000 нг/мл витамина K1, в течение 5 дней. Измеряли концентрацию слитого белка в надосадочной жидкости и определяли его активность с помощью способа, описанного в примере 6 или 7. Результаты ELISA показали, что в этих условиях уровни временной экспрессии белка для восьми плазмид были аналогичными, но для этих слитых белков показаны большие различия в показателях активности свертывания крови.
Авторы настоящего изобретения определили удельную молярную активность FP-A как равную 100%. Слитый белок FP-G, секретированный в надосадочную жидкость культуры клеток, преимущественно находился в форме неактивных агрегатов. Плазмиды FP-F и FP-H экспрессировали малоактивные слитые белки, при этом их показатели их активности составляли приблизительно 20,5% и 15,2% от активности FP-A соответственно. Подобно FP-G большая часть слитых белков FP-F и FP-H находилась в форме агрегатов. Более того, слитые белки FP-F, FP-G и FP-H были склонны к разложению, что свидетельствует о неудовлетворительной стабильности. Сообщалось, что липид-связывающая область FVIII (аминокислоты 2303-2332) была крайне важна для его функции, и небольшие конформационные изменения в этой области вызывали агрегацию белка и приводили к утрате активности (Gilbert GE et al., Biochemistry, 1933,32(37): 9577-9585). Следовательно, авторы настоящего изобретения сделали предположение, что конформации липид-связывающих областей в слитых белках на основе FVIII, FP-F, FP-G и FP-H, менялись под влиянием C-терминальных Fc-лигандов, что приводило к агрегации белков и значительному снижению показателей активности. Показатели активности FP-B, FP-C, FP-D и FP-E, содержавших CTP, составляли 113,4%, 96,0%, 87,4% и 93,7% от активности FP-A соответственно.
На основании различий в активности у FP-B, FP-F и FP-H, можно было понять, что, только увеличением длины пептидного линкера нельзя было ни эффективно повысить активность слитого белка, ни решить проблему того, что слитый белок склонен к агрегации и разложению. Добавление CTP-звена приводило к значимому увеличению активности слитого белка FP-B. Авторы настоящего изобретения предположили, что причины были следующими. Слишком длинные гибкие пептидные линкеры придавали FVIII большую гибкость, так что FVIII мог свободно вращаться относительно домена Fc. Как результат, трехмерная структура FVIII располагалась близко к домену Fc. С одной стороны, добавление жесткого CTP-звена между FVIII и Fc эквивалентно добавлению жесткого пептидного линкера, обеспечивающего расположение доменов FVIII и Fc на расстоянии друг от друга. Что еще более важно, в сравнении со случайной спиралью гибкого пептидного линкера, жесткий CTP-пептид, содержащий несколько гликозильных боковых цепей, мог образовывать стабильную стерическую конформацию и эффективно отделять различные функциональные области слитого белка. Таким образом, FVIII- и Fc-части могли независимо складываться в правильные трехмерные конформации с сохранением высоких показателей активности. Авторы настоящего изобретения подтверждали правильность этой гипотезы путем сравнения показателей активности FP-B и FP-F. Активность FP-F составляла менее 20% от активности FP-B. В FP-F жесткое CTP-звено было размещено на C-конце Fc, тогда как в FP-B жесткое CTP-звено было размещено на N-конце Fc. Приведенные выше результаты продемонстрировали, что жесткое CTP-звено было крайне важным для активности слитого белка, и размещение жесткого CTP-звена на N-конце Fc могло эффективно повышать активность слитого белка.
Пример 3. Скрининг на стабильно трансфицированные линии клеток с высокой экспрессией слитых белков
Плазмиды экспрессии FP-A, FP-B, FP-C, FP-D и FP-E трансфицировали в линии клеток-хозяев млекопитающих для экспрессии слитых белков на основе FVIII. Для поддержания стабильного высокого уровня экспрессии предпочтительной клеткой-хозяином была клетка CHO с дефицитом по DHFR (патент США №4818679). Одним предпочтительным способом трансфекции была электропорация, но при этом можно было применять и другие способы, включая совместное осаждение с фосфатом кальция, липосомную трансфекцию, микроинъекцию и т.д. В случае способа электропорации применяли электропоратор Gene Pulser (Bio-Rad Laboratories), установленный на напряжение 300В и емкость 1050 мкФд. 50 мкг линеаризованной плазмиды экспрессии PvuI добавляли к 2-3×107 клеток, размещенных в кювете. После электропорации клетки переносили во встряхиваемую колбу, содержащую 30 мл ростовой среды. Через два дня после трансфекции среду заменяли на среду для скрининга, содержащую 0,6 мг/мл G418. Клетки высевали в 96-луночный планшет при определенной концентрации (5000-10000 жизнеспособных клеток/лунка) и культивировали в течение 10-12 дней до появления больших отдельных клеточных клонов. Для скрининга трансфектантов, которые были устойчивы к выбранному лекарственному средству, использовали анализ ELISA с антителами к Fc IgG человека. Количественное определение экспрессии слитого белка также можно было осуществлять с применением анализа ELISA с антителами к FVIII. Затем лунки, в которых слитые белки продуцировались на высоком уровне, субклонировали путем предельного разведения.
Для достижения более высокого уровня экспрессии слитого белка предпочтительным было осуществление совместной амплификации с использованием гена DHFR, который мог ингибироваться лекарственным средством MTX. В ростовой среде, содержащей повышенные концентрации МТХ, трансфицированный ген слитого белка амплифицировался совместно с геном DHFR. Положительные по DHFR субклоны подвергали предельному разведению, и за счет возрастающего давления скрининга отбирали трансфектантов, которые были способны расти в среде, содержащей до 6 мкМ MTX. Определяли значения эффективности их секреции и посредством скрининга отбирали клеточные линии с высоким уровнем экспрессии экзогенных белков. Линии клеток, эффективность секреции которых составляла более чем приблизительно 1 (предпочтительно приблизительно 3) МЕ на 106 клеток за 24 часа, адаптировали к суспензионному культивированию с применением бессывороточной среды, а затем из кондиционированной среды очищали слитый белок.
В приведенных ниже примерах FP-B брали в качестве примера для иллюстрации способа ферментации и очистки слитого белка. Способы ферментации и очистки FP-A, FP-C, FP-D и FP-E были такими же, как и для FP-B, и еще раз описываться в данном документе не будут.
Пример 4. Получение слитого белка
Клеточный штамм с высоким уровнем экспрессии, полученный в примере 3, сначала акклиматизировали к бессывороточной среде в чашке Петри, а затем переносили во встряхиваемую колбу для получения суспензионной культуры. После адаптации клеток к данным условиям культивирования клетки культивировали методом периодических культур с подпиткой в 300-мл встряхиваемой колбе или культивировали путем ежедневной замены среды для имитации перфузионной системы. Клеточный штамм, производный от CHO, который экспрессирует слитый белок FP-B, полученный в примере 3, культивировали методом периодических культур с подпиткой в 300-мл колбе в течение 14 дней, и совокупный выход экспрессированного рекомбинантного слитого белка достигал 200 мг/л, в то время как наиболее высокая плотность жизнеспособных клеток могла достигать 15×106 клеток/мл. Для получения большего количества слитого белка можно было применять 1000-мл встряхиваемые колбы. При другом способе культивирования указанный выше клеточный штамм, производный от СНО, культивировали в 100-мл встряхиваемой колбе с ежедневной заменой среды. Совокупный выход экспрессированного рекомбинантного слитого белка достигал приблизительно 20 мг/л в день. Наиболее высокая плотность жизнеспособных клеток во встряхиваемой колбе составляла до 30×106 клеток/мл. Показатели биологической активности рекомбинантных слитых белков, полученных двумя вышеупомянутыми способами, были эквивалентны.
Пример 5. Очистка и качественный анализ слитого белка
В настоящем изобретением в основном предусмотрено применение четырехстадийной процедуры хроматографии для очистки слитого белка FP-B, т.е. аффинной хроматографии, гидрофобной хроматографии, анионообменной хроматографии и хроматографии на молекулярных ситах. В данном примере устройством, применяемым для очистки белка, была система AKTA pure 25 M (GE Healthcare, США). Все реагенты, которые применяли в данном примере, приобретали у компании Sinopharm Chemical Reagent Co., и они имели степень чистоты «чистый для анализа».
Стадия 1. Аффинная хроматография: захват образца, концентрирование и удаление части примесей осуществляли с применением устойчивой к щелочной среде смолы Diamond с белком A (Bestchrom, Шанхай) или других коммерчески доступных смол для аффинной хроматографии с рекомбинантным белком A. Другие смолы включали, например, MabSelect (GE Healthcare), MabSelect SuRe (GE Healthcare), Toyopearl AF-рекомбинантный белок A-650F (Tosoh Bioscience), гранулы с рекомбинантным белком A (Smart-Lifesciences, Чанчжоу, Китай), MabPurix (Sepax Technologies) и керамический сорбент с белком A HyperD (Pall Life Sciences). Колонку уравновешивали при линейной скорости потока 50-100 см/ч с помощью 3-5 объемов колонки (CV) равновесного буфера: 20 мM His-HCl, 150 мM NaCl, 5 мM CaCl2, 0,02% Tween 80, pH 6,8-7,2. Подвергнутую центрифугированию надосадочную жидкость после ферментации загружали на колонку из расчета не более 50000 МЕ белка/мл смолы при линейной скорости потока 50-100 см/ч. После загрузки колонку уравновешивали с помощью 3-5 CV равновесного буфера при линейной скорости потока 50-100 см/ч для отмывки несвязавшихся материалов. Затем колонку промывали с помощью 3-5 CV деконтаминационного буфера 1: 20 мM His-HCl, 2M NaCl, 4M мочевины, 5 мM CaCl2, 0,02% Tween 80, pH 6,8-7,2, при линейной скорости потока 50-100 см/ч для удаления части примесей. Колонку уравновешивали с помощью 3-5 CV равновесного буфера при линейной скорости потока 50-100 см/ч. Колонку дополнительно промывали с помощью 3-5 CV деконтаминационного буфера 2: 20 мM His-HCl, 5 мM EDTA, 150 мM NaCl, 30% этиленгликоля, 5 мM CaCl2, 0,02% Tween 80, pH 6,8-7,2, при линейной скорости потока 50-100 см/ч для удаления части примесей. Колонку уравновешивали с помощью 3-5 CV равновесного буфера при линейной скорости потока 50-100 см/ч. Затем целевой продукт элюировали и собирали с помощью элюирующего буфера: 20 мM His-HCl, 5 мM CaCl2, 0,02% Tween 80, 50% этиленгликоля, pH 5,0, при линейной скорости потока, не превышающей 50 см/ч. В элюат добавляли Tris, pH 9,0, для доведения pH до нейтрального значения (7,0-8,0).
Стадия 2. Гидрофобная хроматография: на промежуточной стадии очистки для снижения количества агрегатов применяли смолу бутил-бестароза HP (Bestchrom, Шанхай) или другие коммерчески доступные смолы для гидрофобной хроматографии. Другие смолы включали бутил-сефарозу HP (GE Healthcare), Toyopearl бутил-650 (Tosoh Bioscience), гранулы бутил-4FF (Smart-Lifesciences, Чанчжоу, Китай), Generic MC 30-HIC-бутил (Sepax Technologies) и Fractogel EMD-пропил (Merck). Элюат аффинной хроматографии на первой стадии все еще содержал определенную долю агрегатов. Агрегаты образовывались вследствие целого ряда причин. Некоторые агрегаты содержали белки, все еще находящиеся в нативной конформации, в то время как другие содержали белки, конформация которых была изменена. Для агрегатов в различных конформационных формах показаны значимые отличия в биологической активности, которые приводили к сильным искажениям при анализе активности. Таким образом, после завершения захвата белка на первой стадии очистки затем необходимо удалить агрегаты. После агрегации целевого белка неагрегированные и агрегированные формы проявляют отличающиеся свойства, включая характеристики заряда и гидрофобность. Различие в гидрофобности применяли для разделения этих двух форм. Поскольку последней стадией очистки была хроматография на молекулярных ситах, слитый белок, захваченный в ходе аффинной хроматографии на первой стадии, подвергали дополнительной очистке с помощью колонки с бутил HP для осуществления второй стадии очистки с целью частичного удаления агрегатов, вследствие чего содержание агрегатов составляло менее 10%. Сначала колонку уравновешивали с помощью 3-5 CV равновесного буфера: 20 мM His-HCl, 1,5 M NaCl, 5 мM CaCl2, 0,02% Tween 80, pH 6,8-7,2, при линейной скорости потока 50-100 см/ч. Подвергнутый аффинному захвату образец разводили в два раза равновесным буфером для снижения содержания органического растворителя, а затем к образцу добавляли равный объем концентрированного буфера: 20 мM His-HCl, 3M NaCl, 5 мM CaCl2, 0,02% Tween 80, pH 6,8-7,2. Затем образец загружали на колонку из расчета менее 20000 МЕ белка/мл смолы. После загрузки колонку промывали с помощью 3-5 CV равновесного буфера при линейной скорости потока 50-100 см/ч и промывали с помощью 3-5 CV промывочного буфера: 20 мM His-HCl, 1,5 M NaCl, 5M CaCl2, 0,02% Tween 80, 20% этиленгликоля, pH 6,8-7,2, для удаления некоторой части агрегатов. Наконец, целевой белок элюировали с помощью элюирующего буфера: 20 мM His-HCl, 5 мM CaCl2, 0,02% Tween 80, 50% этиленгликоля, pH 6,8-7,2, при линейной скорости потока, не превышающей 60 см/ч, и элюированные фракции собирали и анализировали с помощью SEC-HPLC. Целевые фракции с процентным содержанием неагрегированных форм более 90% объединяли и подвергали следующей стадии очистки.
Стадия 3. Анионообменная хроматография: на промежуточной стадии очистки применяли смолу Q-HP (Bestchrom, Шанхай) или другие коммерчески доступные смолы для анионообменной хроматографии с целью разделения структурных вариантов и дополнительного удаления примесей, таких как HCP, ДНК и т.д. Другие смолы включали Q HP (GE Healthcare), Toyopearl GigaCap Q-650 (Tosoh Bioscience), DEAE Beads 6FF (Smart-Lifesciences, Чанчжоу, Китай), Generik MC-Q (Sepax Technologies), Fractogel EMD TMAE (Merck) и Q Ceramic HyperD F (Pall Life Sciences). Сначала колонку промывали с помощью 3-5 CV равновесного буфера: 20 мM His-HCl, 200 мM NaCl, 5 мM CaCl2, 0,02% Tween 80, pH 6,8-7,2, при линейной скорости потока 50-100 см/ч. Целевой белок, выделенный с помощью гидрофобной хроматографии на второй стадии, разводили в два раза для снижения содержания органического растворителя и загружали на колонку из расчета менее 5000-10000 МЕ белка/мл смолы. После загрузки колонку промывали с помощью 3-5 CV равновесного буфера при линейной скорости потока 50-100 см/ч, с последующим элюированием при линейном градиенте концентрации солей с применением элюирующего буфера: 20 мM His-HCl, 1M NaCl, 5 мM CaCl2, 0,02% Tween 80, pH 6,8-7,2. Условия элюирования представляли собой градиент от 0 до 100% элюирующего буфера на протяжении 2 ч при линейной скорости потока, не превышающей 50 см/ч. Элюированные фракции собирали и анализировали на содержание белка, SEC-HPLC, активность и содержание HCP. После определения концентрации и активности белка рассчитывали удельную активность белка, которая составила приблизительно 10000 МЕ/мг.
Стадия 4. Хроматография с молекулярными ситами: для разделения применяли смолу для препаративного анализа Chromdex 200 (Bestchrom) или другие коммерчески доступные смолы с молекулярными ситами (например, Superdex 200 от GE Healthcare) с целью снижения содержания агрегатов до <5% и дополнительного снижения содержания ключевых примесей. Колонку промывали с помощью 2 CV равновесного буфера: 20 мM His-HCl, 200 мM NaCl, 5 мM CaCl2, 0,02% Tween 80, pH 6,8-7,2, при линейной скорости потока 20-40 см/ч. Объем загружаемого образца не превышал 3% объема колонки. Образец белка элюировали при линейной скорости потока 20 см/ч, и элюированные фракции собирали и подвергали анализу SEC-HPLC с последующим объединением.
Результаты анализов чистоты с помощью SEC-HPLC и SDS-PAGE-электрофореза очищенного FP-B показаны на фиг. 2 и 3 соответственно. Результаты SEC-HPLC показали, что чистота главного пика очищенного слитого белка FP-B превышала 97%. Паттерны полосок при SDS-PAGE-электрофорезе выглядели так, как и ожидалось. Дорожка электрофореза в невосстанавливающих условиях содержала полоску непроцессированного слитого белка FP-B (390 кДа), полоску HC-LC-L-CTP-Fc:LC-L-CTP-Fc (300 кДа), у которого один фрагмент тяжелой цепи FVIII был утерян во время электрофореза, полоску димера (LC-L-CTP-Fc)2 (210 кДа), у которого два фрагмента тяжелой цепи были утеряны во время электрофореза, и полоску фрагмента тяжелой цепи HC (90 кДа). Дорожка электрофореза в восстанавливающих условиях содержала полоску HC-LC-L-CTP-Fc (190 кДа), полоску LC-L-CTP-Fc (105 кДа) и полоску одноцепочечной HC (90 кДа).
Пример 6. Непрямое определение in vitro активности слитого белка с помощью анализа с хромогенным субстратом
Активность слитого белка на основе FVIII можно было определять с помощью анализа с хромогенным субстратом. В данном примере применяли набор Chromogenix Coatest SP FVIII (Chromogenix, № по кат. K824086), и принцип такого анализа был следующим. При активации под действием тромбина FVIIIa связывался с FIXa в присутствии фосфолипида и ионов кальция с образованием ферментного комплекса, который, в свою очередь, переводил фактор X в его активную форму Xa. Затем активированный фактор Xa разлагал свой специфический хромогенный субстрат (Chromogenix S-2765), высвобождая хромофор pNA. Количество полученного pNA измеряли при 405 нм и, таким образом, определяли уровень активности FXa, который был прямо пропорционален количеству pNA. Поскольку количество фактора IXa и фактора X в системе анализа было избыточным и постоянным, активность FXa была напрямую связана только с количеством FVIIIa. В данном анализе определили, что показатели удельной активности слитых белков на основе FVIII составляли приблизительно 6000-10000 МЕ/мг.
Пример 7. Прямое определение биологической активности слитого белка с помощью анализа коагуляции
Анализ коагуляции для определения биологической активности FVIII был основан на свойстве FVIII корректировать пролонгированное время свертывания плазмы крови с дефицитом FVIII. За счет применения набора плазмы с дефицитом фактора свертывания крови VIII (№ по кат. OTXW17) от немецкой компании Siemens осуществляли следующий способ определения активности FVIII. Вначале стандарт FVIII с известным значением активности, полученный от Национального института по контролю за продуктами и лекарственными средствами (Китай), разводили до 10 МЕ/мл с помощью 5% плазмы с дефицитом FVIII, а затем его дополнительно разводили в 10, 20, 40 раз и 80 раз соответственно. Активированное частичное тромбопластиновое время (APTT) определяли с помощью автоматического анализатора гемагглютинации (CA500, Sysmex). Стандартную кривую строили с помощью стандарта FVIII путем откладывания на графике линейной регрессии логарифма значений активности (МЕ/мл) растворов стандарта FVIII относительно логарифма их соответствующих значений времени коагуляции (с). Затем тестируемый образец надлежащим образом разводили и смешивали с субстратной плазмой с дефицитом FVIII для проведения анализа APTT. Значение активности тестируемого образца FVIII (МЕ/мл) можно было рассчитать путем подстановки времени коагуляции в уравнение стандартной кривой. Таким образом можно было рассчитать удельную активность тестируемого образца VIII в единицах МЕ/мг. В данном анализе определили, что показатели удельной активности слитых белков на основе FVIII составляли приблизительно 6000-10000 МЕ/мг.
Пример 8. Кровоостанавливающий эффект слитых белков при острой геморрагии у мышей с гемофилией A
Авторы настоящего изобретения оценивали кровоостанавливающую активность слитого белка FP-B, полученного в примере 5, на модели кровотечения с отсечением хвоста у гомозиготных мышей HemA с нокаутом гена фактора VIII. Самцов мышей HemA (возрастом 8-12 недель, Shanghai Model Organisms Center, Inc.) для адаптации кормили в течение одной недели, а затем случайным образом делили на 6 групп. Кроме того, одну группу мышей HemA брали в качестве отрицательного контроля, а другую группу нормальных мышей С57 брали в качестве положительного контроля. 8 группам вводили различные активные дозы слитого белка FP-B или контрольного лекарственного средства Xyntha (Pfizer) в виде однократной инъекции в хвостовую вену. В таблице 2 показана схема эксперимента и распределение животных по группам.
Таблица 2. Распределение животных по группам в исследовании кровоостанавливающего эффекта слитых белков у мышей HemA
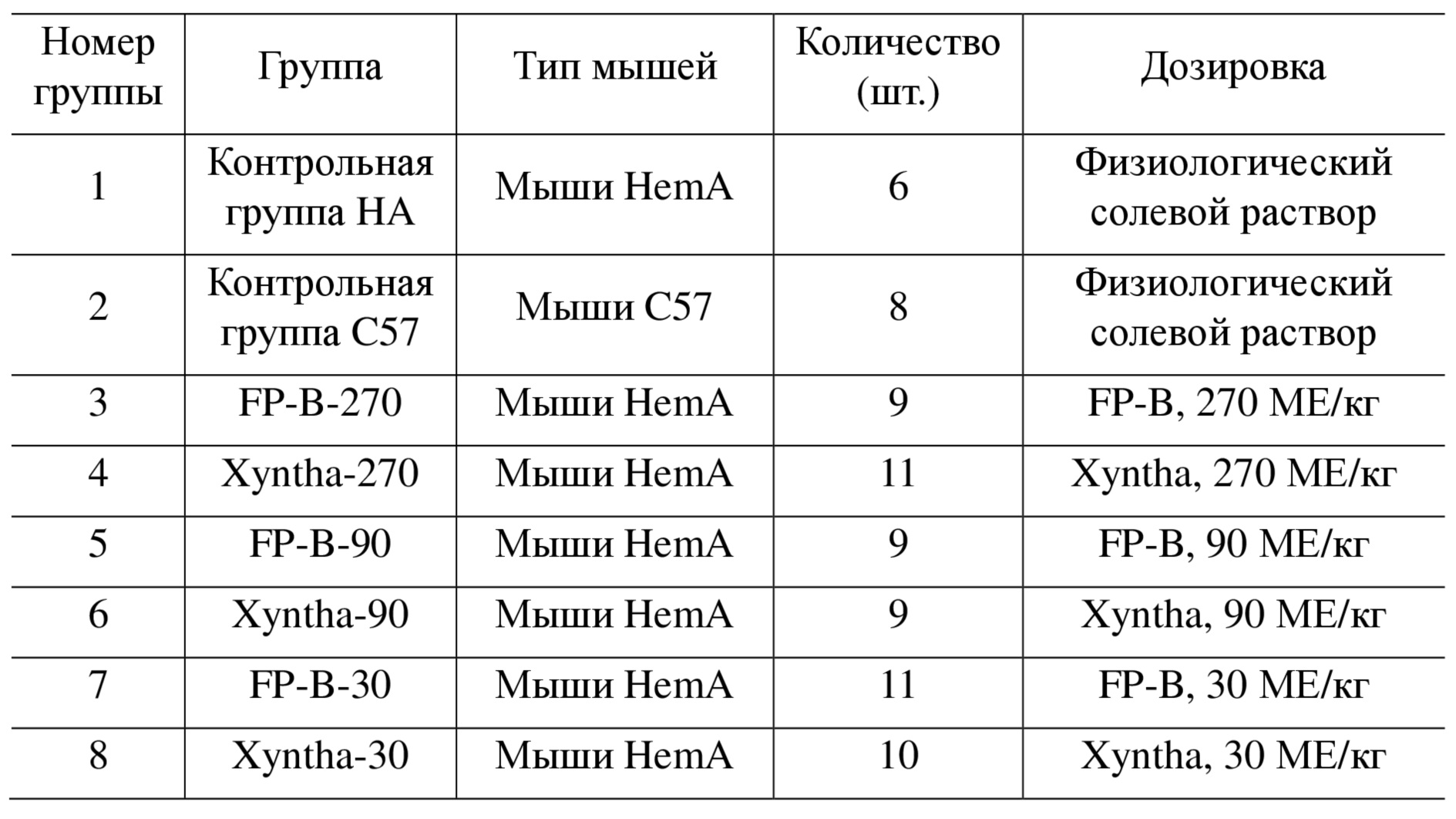
Перед осуществлением введения каждую мышь анестезировали путем внутрибрюшинной инъекции 1,0% пентобарбитала натрия (Sigma) в дозе 0,1мл/10г, а затем ее помещали на грелку-подушку с температурой 37°C для поддержания температуры тела. Хвост мыши погружали в теплую воду с температурой 37°С на 10 минут для расширения хвостовой вены, а затем вводили соответствующую дозу, которая указана в таблице 2. Через 10 минут после введения хвост отрезали на расстоянии 1,5 см от кончика хвоста и хвост быстро погружали в приблизительно 13 мл предварительно нагретого солевого раствора, содержащегося в центрифужной пробирке. Начинали отсчет времени. Если кровотечение останавливалось менее, чем за 30 минут, регистрировали время и объем кровотечения. Если время кровотечения превышало 30 минут, его регистрировали как 30 минут. Объем кровотечения (мл) = (масса центрифужной пробирки после сбора крови (г) - масса центрифужной пробирки до сбора крови (г)) / 1,05. Через 30 минут хвост убирали из пробирки, содержащей солевой раствор. На протяжении 24 ч через каждые 10 минут наблюдали и регистрировали рецидивирующее кровотечение и регистрировали количество выживших мышей. Все данные выражали как среднее ± стандартная ошибка (± SEM). Для сравнения экспериментальных групп применяли t-критерий. Программным обеспечением для анализа было GraphPad Prism 5.0. P<0,05 считали статистически значимым.
На фиг. 4 и 5 показаны результаты статистических анализов времени и объема кровотечения для каждой группы животных. Через 10 минут после введения мышам HemA 270 МЕ/кг FP-B время и объем кровотечения в группе FP-B-270 были близки к таковым у контрольной группы C57. Прокоагулянтный эффект FP-B был очевидным, что свидетельствовало о том, что FP-B можно применять в качестве эффективного средства свертывая крови при острой геморрагии у пациентов с дефицитами фактора свертывания крови, такими как гемофилия. Когда мышам вводили 90 МЕ/кг FP-B, время и объем кровотечения в группе FP-B-90 также были близки к таковым у контрольной группы C57. Значимое отличие в объеме кровотечения между мышами HemA, которым вводили одинаковое количество активной дозы FP-B и Xyntha, отсутствовало, но время кровотечения в группе FP-B при каждой дозе было немного меньше, чем в группе Xyntha, что свидетельствовало о том, что FP-B может иметь определенное преимущество в эффективности по сравнению с Xyntha. По сравнению с группой 30 МЕ/кг FP-B для группы 90 МЕ/кг FP-B показано значимо более короткое время кровотечения (p<0,05), а для группы 270 МЕ/кг FP-B показано значимо более короткое время кровотечения (p<0,05), а также значимо сниженный объем кровотечения (p<0,05). Это свидетельствовало о том, что слитый белок FP-B оказывал дозозависимый эффект на гемостаз при острой геморрагии у мышей HemA (подробности см. в таблице 3).
Согласно данным послеоперационного восстановления, при введении одинакового количества активной дозы FP-B и Xyntha группа FP-B при каждой дозе характеризовалась более высокой выживаемостью мышей, чем группа Xyntha при такой же активной дозе, что свидетельствовало о том, что слитый белок FP-B оказывал более длительный эффект, чем Xyntha (см. таблицу 3).
Таблица 3. Статистические данные по времени, объему кровотечения, рецидивному кровотечению и выживаемости для каждой группы мышей HemA после отсечения хвоста

Примечание:aПоскольку были мыши, которые погибли в диапазоне 24-48 ч, представлена выживаемость через 48 ч.
bЕсли время кровотечения превышало 30 минут, его регистрировали как 1800 с.
Все документы, упомянутые в настоящем изобретении, настоящим включены посредством ссылки в том же объеме, как если бы каждый из этих документов был отдельно изложен по ссылке. Следует понимать, что после прочтения вышеизложенных идей настоящего изобретения, которые также подпадают под объем прилагаемой к ним формулы изобретения, специалистами в данной области могут быть сделаны различные изменения и модификации.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> АМПСОРС БИОФАРМА ИНК.
ФУЖЭНЬ ФАРМАСЬЮТИКАЛ ГРУП КО., ЛТД.
ФАРМАБ, ИНК.
КАЙФЫН ФАРМАСЬЮТИКАЛ (ГРУП) КО., ЛТД.
<120> ВЫСОКОГЛИКОЗИЛИРОВАННЫЙ СЛИТЫЙ БЕЛОК НА ОСНОВЕ ФАКТОРА
СВЕРТЫВАНИЯ КРОВИ ЧЕЛОВЕКА VIII, СПОСОБ ЕГО ПОЛУЧЕНИЯ
И ЕГО ПРИМЕНЕНИЕ
<130> 2016
<160> 17
<170> PatentIn версия 3.3
<210> 1
<211> 1438
<212> БЕЛОК
<213> аминокислотная последовательность зрелого белка FVIII
<400> 1
Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser Trp Asp Tyr
1 5 10 15
Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg Phe Pro Pro
20 25 30
Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val Tyr Lys Lys
35 40 45
Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile Ala Lys Pro
50 55 60
Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln Ala Glu Val
65 70 75 80
Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser His Pro Val
85 90 95
Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser Glu Gly Ala
100 105 110
Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp Asp Lys Val
115 120 125
Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu Lys Glu Asn
130 135 140
Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser Tyr Leu Ser
145 150 155 160
His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile Gly Ala Leu
165 170 175
Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr Gln Thr Leu
180 185 190
His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly Lys Ser Trp
195 200 205
His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp Ala Ala Ser
210 215 220
Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr Val Asn Arg
225 230 235 240
Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val Tyr Trp His
245 250 255
Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile Phe Leu Glu
260 265 270
Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser Leu Glu Ile
275 280 285
Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met Asp Leu Gly
290 295 300
Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His Asp Gly Met
305 310 315 320
Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro Gln Leu Arg
325 330 335
Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu Thr Asp
340 345 350
Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro Ser Phe
355 360 365
Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr Trp Val His
370 375 380
Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro Leu Val Leu
385 390 395 400
Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn Asn Gly Pro
405 410 415
Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met Ala Tyr Thr
420 425 430
Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu Ser Gly Ile
435 440 445
Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu Leu Ile Ile
450 455 460
Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro His Gly Ile
465 470 475 480
Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys Gly Val Lys
485 490 495
His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe Lys Tyr Lys
500 505 510
Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp Pro Arg Cys
515 520 525
Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg Asp Leu Ala
530 535 540
Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu Ser Val Asp
545 550 555 560
Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val Ile Leu Phe
565 570 575
Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu Asn Ile Gln
580 585 590
Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp Pro Glu Phe
595 600 605
Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val Phe Asp Ser
610 615 620
Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp Tyr Ile Leu
625 630 635 640
Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe Ser Gly Tyr
645 650 655
Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr Leu Phe Pro
660 665 670
Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro Gly Leu Trp
675 680 685
Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly Met Thr Ala
690 695 700
Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp Tyr Tyr Glu
705 710 715 720
Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala
725 730 735
Ile Glu Pro Arg Ser Phe Ser Gln Asn Pro Pro Val Leu Lys Arg His
740 745 750
Gln Arg Glu Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile
755 760 765
Asp Tyr Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp
770 775 780
Ile Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys
785 790 795 800
Thr Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr Gly
805 810 815
Met Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser Gly Ser
820 825 830
Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr Asp Gly Ser
835 840 845
Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu His Leu Gly Leu
850 855 860
Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp Asn Ile Met Val Thr
865 870 875 880
Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser Phe Tyr Ser Ser Leu Ile
885 890 895
Ser Tyr Glu Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg Lys Asn Phe
900 905 910
Val Lys Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys Val Gln His His
915 920 925
Met Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys Ala Trp Ala Tyr Phe
930 935 940
Ser Asp Val Asp Leu Glu Lys Asp Val His Ser Gly Leu Ile Gly Pro
945 950 955 960
Leu Leu Val Cys His Thr Asn Thr Leu Asn Pro Ala His Gly Arg Gln
965 970 975
Val Thr Val Gln Glu Phe Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr
980 985 990
Lys Ser Trp Tyr Phe Thr Glu Asn Met Glu Arg Asn Cys Arg Ala Pro
995 1000 1005
Cys Asn Ile Gln Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr Arg
1010 1015 1020
Phe His Ala Ile Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly Leu
1025 1030 1035
Val Met Ala Gln Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser Met
1040 1045 1050
Gly Ser Asn Glu Asn Ile His Ser Ile His Phe Ser Gly His Val
1055 1060 1065
Phe Thr Val Arg Lys Lys Glu Glu Tyr Lys Met Ala Leu Tyr Asn
1070 1075 1080
Leu Tyr Pro Gly Val Phe Glu Thr Val Glu Met Leu Pro Ser Lys
1085 1090 1095
Ala Gly Ile Trp Arg Val Glu Cys Leu Ile Gly Glu His Leu His
1100 1105 1110
Ala Gly Met Ser Thr Leu Phe Leu Val Tyr Ser Asn Lys Cys Gln
1115 1120 1125
Thr Pro Leu Gly Met Ala Ser Gly His Ile Arg Asp Phe Gln Ile
1130 1135 1140
Thr Ala Ser Gly Gln Tyr Gly Gln Trp Ala Pro Lys Leu Ala Arg
1145 1150 1155
Leu His Tyr Ser Gly Ser Ile Asn Ala Trp Ser Thr Lys Glu Pro
1160 1165 1170
Phe Ser Trp Ile Lys Val Asp Leu Leu Ala Pro Met Ile Ile His
1175 1180 1185
Gly Ile Lys Thr Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu Tyr
1190 1195 1200
Ile Ser Gln Phe Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys Trp
1205 1210 1215
Gln Thr Tyr Arg Gly Asn Ser Thr Gly Thr Leu Met Val Phe Phe
1220 1225 1230
Gly Asn Val Asp Ser Ser Gly Ile Lys His Asn Ile Phe Asn Pro
1235 1240 1245
Pro Ile Ile Ala Arg Tyr Ile Arg Leu His Pro Thr His Tyr Ser
1250 1255 1260
Ile Arg Ser Thr Leu Arg Met Glu Leu Met Gly Cys Asp Leu Asn
1265 1270 1275
Ser Cys Ser Met Pro Leu Gly Met Glu Ser Lys Ala Ile Ser Asp
1280 1285 1290
Ala Gln Ile Thr Ala Ser Ser Tyr Phe Thr Asn Met Phe Ala Thr
1295 1300 1305
Trp Ser Pro Ser Lys Ala Arg Leu His Leu Gln Gly Arg Ser Asn
1310 1315 1320
Ala Trp Arg Pro Gln Val Asn Asn Pro Lys Glu Trp Leu Gln Val
1325 1330 1335
Asp Phe Gln Lys Thr Met Lys Val Thr Gly Val Thr Thr Gln Gly
1340 1345 1350
Val Lys Ser Leu Leu Thr Ser Met Tyr Val Lys Glu Phe Leu Ile
1355 1360 1365
Ser Ser Ser Gln Asp Gly His Gln Trp Thr Leu Phe Phe Gln Asn
1370 1375 1380
Gly Lys Val Lys Val Phe Gln Gly Asn Gln Asp Ser Phe Thr Pro
1385 1390 1395
Val Val Asn Ser Leu Asp Pro Pro Leu Leu Thr Arg Tyr Leu Arg
1400 1405 1410
Ile His Pro Gln Ser Trp Val His Gln Ile Ala Leu Arg Met Glu
1415 1420 1425
Val Leu Gly Cys Glu Ala Gln Asp Leu Tyr
1430 1435
<210> 2
<211> 16
<212> БЕЛОК
<213> аминокислотная последовательность L1
<400> 2
Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 3
<211> 27
<212> БЕЛОК
<213> аминокислотная последовательность L2
<400> 3
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
1 5 10 15
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25
<210> 4
<211> 20
<212> БЕЛОК
<213> аминокислотная последовательность L3
<400> 4
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 5
<211> 37
<212> БЕЛОК
<213> аминокислотная последовательность L4
<400> 5
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
1 5 10 15
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25 30
Gly Gly Gly Gly Ser
35
<210> 6
<211> 20
<212> БЕЛОК
<213> аминокислотная последовательность L5
<400> 6
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser
1 5 10 15
Gly Gly Gly Ser
20
<210> 7
<211> 33
<212> БЕЛОК
<213> аминокислотная последовательность CTP1
<400> 7
Pro Arg Phe Gln Asp Ser Ser Ser Ser Lys Ala Pro Pro Pro Ser Leu
1 5 10 15
Pro Ser Pro Ser Arg Leu Pro Gly Pro Ser Asp Thr Pro Ile Leu Pro
20 25 30
Gln
<210> 8
<211> 28
<212> БЕЛОК
<213> аминокислотная последовательность CTP2
<400> 8
Ser Ser Ser Ser Lys Ala Pro Pro Pro Ser Leu Pro Ser Pro Ser Arg
1 5 10 15
Leu Pro Gly Pro Ser Asp Thr Pro Ile Leu Pro Gln
20 25
<210> 9
<211> 10
<212> БЕЛОК
<213> аминокислотная последовательность CTP3
<400> 9
Ser Ser Ser Ser Lys Ala Pro Pro Pro Ser
1 5 10
<210> 10
<211> 14
<212> БЕЛОК
<213> аминокислотная последовательность CTP4
<400> 10
Ser Arg Leu Pro Gly Pro Ser Asp Thr Pro Ile Leu Pro Gln
1 5 10
<210> 11
<211> 227
<212> БЕЛОК
<213> аминокислотная последовательность vFc?1
<400> 11
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Val Ala Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Ser Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 12
<211> 223
<212> БЕЛОК
<213> аминокислотная последовательность vFc?2-1
<400> 12
Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
1 5 10 15
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
20 25 30
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
35 40 45
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
50 55 60
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser
65 70 75 80
Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
85 90 95
Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Ser Ile Glu Lys Thr Ile
100 105 110
Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
115 120 125
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
130 135 140
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
145 150 155 160
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser
165 170 175
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
180 185 190
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
195 200 205
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215 220
<210> 13
<211> 223
<212> БЕЛОК
<213> аминокислотная последовательность vFc?2-2
<400> 13
Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
1 5 10 15
Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg Thr
20 25 30
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
35 40 45
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
50 55 60
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser
65 70 75 80
Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
85 90 95
Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile
100 105 110
Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
115 120 125
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
130 135 140
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
145 150 155 160
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser
165 170 175
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
180 185 190
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu
195 200 205
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215 220
<210> 14
<211> 223
<212> БЕЛОК
<213> аминокислотная последовательность vFc?2-3
<400> 14
Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
1 5 10 15
Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg Thr
20 25 30
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
35 40 45
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
50 55 60
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser
65 70 75 80
Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
85 90 95
Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Ser Ile Glu Lys Thr Ile
100 105 110
Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
115 120 125
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
130 135 140
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
145 150 155 160
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser
165 170 175
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
180 185 190
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu
195 200 205
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215 220
<210> 15
<211> 229
<212> БЕЛОК
<213> аминокислотная последовательность vFc?4
<400> 15
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
1 5 10 15
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 16
<211> 1716
<212> БЕЛОК
<213> аминокислотная последовательность FP-B
<400> 16
Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser Trp Asp Tyr
1 5 10 15
Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg Phe Pro Pro
20 25 30
Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val Tyr Lys Lys
35 40 45
Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile Ala Lys Pro
50 55 60
Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln Ala Glu Val
65 70 75 80
Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser His Pro Val
85 90 95
Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser Glu Gly Ala
100 105 110
Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp Asp Lys Val
115 120 125
Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu Lys Glu Asn
130 135 140
Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser Tyr Leu Ser
145 150 155 160
His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile Gly Ala Leu
165 170 175
Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr Gln Thr Leu
180 185 190
His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly Lys Ser Trp
195 200 205
His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp Ala Ala Ser
210 215 220
Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr Val Asn Arg
225 230 235 240
Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val Tyr Trp His
245 250 255
Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile Phe Leu Glu
260 265 270
Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser Leu Glu Ile
275 280 285
Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met Asp Leu Gly
290 295 300
Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His Asp Gly Met
305 310 315 320
Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro Gln Leu Arg
325 330 335
Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu Thr Asp
340 345 350
Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro Ser Phe
355 360 365
Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr Trp Val His
370 375 380
Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro Leu Val Leu
385 390 395 400
Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn Asn Gly Pro
405 410 415
Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met Ala Tyr Thr
420 425 430
Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu Ser Gly Ile
435 440 445
Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu Leu Ile Ile
450 455 460
Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro His Gly Ile
465 470 475 480
Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys Gly Val Lys
485 490 495
His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe Lys Tyr Lys
500 505 510
Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp Pro Arg Cys
515 520 525
Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg Asp Leu Ala
530 535 540
Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu Ser Val Asp
545 550 555 560
Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val Ile Leu Phe
565 570 575
Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu Asn Ile Gln
580 585 590
Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp Pro Glu Phe
595 600 605
Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val Phe Asp Ser
610 615 620
Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp Tyr Ile Leu
625 630 635 640
Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe Ser Gly Tyr
645 650 655
Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr Leu Phe Pro
660 665 670
Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro Gly Leu Trp
675 680 685
Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly Met Thr Ala
690 695 700
Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp Tyr Tyr Glu
705 710 715 720
Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala
725 730 735
Ile Glu Pro Arg Ser Phe Ser Gln Asn Pro Pro Val Leu Lys Arg His
740 745 750
Gln Arg Glu Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile
755 760 765
Asp Tyr Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp
770 775 780
Ile Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys
785 790 795 800
Thr Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr Gly
805 810 815
Met Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser Gly Ser
820 825 830
Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr Asp Gly Ser
835 840 845
Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu His Leu Gly Leu
850 855 860
Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp Asn Ile Met Val Thr
865 870 875 880
Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser Phe Tyr Ser Ser Leu Ile
885 890 895
Ser Tyr Glu Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg Lys Asn Phe
900 905 910
Val Lys Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys Val Gln His His
915 920 925
Met Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys Ala Trp Ala Tyr Phe
930 935 940
Ser Asp Val Asp Leu Glu Lys Asp Val His Ser Gly Leu Ile Gly Pro
945 950 955 960
Leu Leu Val Cys His Thr Asn Thr Leu Asn Pro Ala His Gly Arg Gln
965 970 975
Val Thr Val Gln Glu Phe Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr
980 985 990
Lys Ser Trp Tyr Phe Thr Glu Asn Met Glu Arg Asn Cys Arg Ala Pro
995 1000 1005
Cys Asn Ile Gln Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr Arg
1010 1015 1020
Phe His Ala Ile Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly Leu
1025 1030 1035
Val Met Ala Gln Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser Met
1040 1045 1050
Gly Ser Asn Glu Asn Ile His Ser Ile His Phe Ser Gly His Val
1055 1060 1065
Phe Thr Val Arg Lys Lys Glu Glu Tyr Lys Met Ala Leu Tyr Asn
1070 1075 1080
Leu Tyr Pro Gly Val Phe Glu Thr Val Glu Met Leu Pro Ser Lys
1085 1090 1095
Ala Gly Ile Trp Arg Val Glu Cys Leu Ile Gly Glu His Leu His
1100 1105 1110
Ala Gly Met Ser Thr Leu Phe Leu Val Tyr Ser Asn Lys Cys Gln
1115 1120 1125
Thr Pro Leu Gly Met Ala Ser Gly His Ile Arg Asp Phe Gln Ile
1130 1135 1140
Thr Ala Ser Gly Gln Tyr Gly Gln Trp Ala Pro Lys Leu Ala Arg
1145 1150 1155
Leu His Tyr Ser Gly Ser Ile Asn Ala Trp Ser Thr Lys Glu Pro
1160 1165 1170
Phe Ser Trp Ile Lys Val Asp Leu Leu Ala Pro Met Ile Ile His
1175 1180 1185
Gly Ile Lys Thr Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu Tyr
1190 1195 1200
Ile Ser Gln Phe Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys Trp
1205 1210 1215
Gln Thr Tyr Arg Gly Asn Ser Thr Gly Thr Leu Met Val Phe Phe
1220 1225 1230
Gly Asn Val Asp Ser Ser Gly Ile Lys His Asn Ile Phe Asn Pro
1235 1240 1245
Pro Ile Ile Ala Arg Tyr Ile Arg Leu His Pro Thr His Tyr Ser
1250 1255 1260
Ile Arg Ser Thr Leu Arg Met Glu Leu Met Gly Cys Asp Leu Asn
1265 1270 1275
Ser Cys Ser Met Pro Leu Gly Met Glu Ser Lys Ala Ile Ser Asp
1280 1285 1290
Ala Gln Ile Thr Ala Ser Ser Tyr Phe Thr Asn Met Phe Ala Thr
1295 1300 1305
Trp Ser Pro Ser Lys Ala Arg Leu His Leu Gln Gly Arg Ser Asn
1310 1315 1320
Ala Trp Arg Pro Gln Val Asn Asn Pro Lys Glu Trp Leu Gln Val
1325 1330 1335
Asp Phe Gln Lys Thr Met Lys Val Thr Gly Val Thr Thr Gln Gly
1340 1345 1350
Val Lys Ser Leu Leu Thr Ser Met Tyr Val Lys Glu Phe Leu Ile
1355 1360 1365
Ser Ser Ser Gln Asp Gly His Gln Trp Thr Leu Phe Phe Gln Asn
1370 1375 1380
Gly Lys Val Lys Val Phe Gln Gly Asn Gln Asp Ser Phe Thr Pro
1385 1390 1395
Val Val Asn Ser Leu Asp Pro Pro Leu Leu Thr Arg Tyr Leu Arg
1400 1405 1410
Ile His Pro Gln Ser Trp Val His Gln Ile Ala Leu Arg Met Glu
1415 1420 1425
Val Leu Gly Cys Glu Ala Gln Asp Leu Tyr Gly Ser Gly Gly Gly
1430 1435 1440
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
1445 1450 1455
Gly Ser Gly Gly Gly Gly Ser Ser Ser Ser Ser Lys Ala Pro Pro
1460 1465 1470
Pro Ser Leu Pro Ser Pro Ser Arg Leu Pro Gly Pro Ser Asp Thr
1475 1480 1485
Pro Ile Leu Pro Gln Val Glu Cys Pro Pro Cys Pro Ala Pro Pro
1490 1495 1500
Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
1505 1510 1515
Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
1520 1525 1530
Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val
1535 1540 1545
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
1550 1555 1560
Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val
1565 1570 1575
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
1580 1585 1590
Asn Lys Gly Leu Pro Ala Ser Ile Glu Lys Thr Ile Ser Lys Thr
1595 1600 1605
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
1610 1615 1620
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
1625 1630 1635
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
1640 1645 1650
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp
1655 1660 1665
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
1670 1675 1680
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Leu His
1685 1690 1695
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
1700 1705 1710
Pro Gly Lys
1715
<210> 17
<211> 5208
<212> ДНК
<213> нуклеотидная последовательность FP-B
<400> 17
atgcagatcg aactgtcaac ttgtttcttc ctgtgcctgc tgagattttg cttttccgcc 60
actcgtcgtt actacctagg agccgtggaa ctgagctggg attacatgca gtctgacctg 120
ggagagctgc cagtggacgc tagatttccc cctcgcgtgc ctaagagttt ccccttcaac 180
acctcagtgg tctataagaa aacactgttc gtggagttta ctgatcacct gttcaacatc 240
gccaagccaa gaccaccctg gatgggactg ctgggaccta caatccaggc tgaggtgtac 300
gacactgtgg tcattaccct gaaaaacatg gcaagtcacc cagtgtcact gcatgccgtc 360
ggggtgtcat actggaaggc ttccgaaggt gcagagtatg acgatcagac ctctcagcgc 420
gaaaaagagg acgataaggt gtttcccggc ggaagccata catacgtctg gcaggtgctg 480
aaggagaatg gccccatggc cagcgaccct ctgtgcctga cctactcata tctgtcccac 540
gtggacctgg tgaaggatct gaacagcggg ctgatcggtg cactgctggt gtgtagagaa 600
ggctctctgg ccaaggagaa aactcagacc ctgcataagt tcattctgct gttcgccgtg 660
tttgacgaag gaaaaagctg gcactctgag actaagaact ccctgatgca ggacagggat 720
gcagcaagcg cacgagcttg gcccaaaatg cataccgtca acggctacgt gaatcgaagt 780
ctgcctggcc tgatcggatg ccaccgtaag tccgtctatt ggcatgtgat cgggatgggc 840
accacacccg aagtccacag cattttcctg gagggtcata cctttctggt gagaaaccac 900
cgccaggcat ccctggagat cagccctatt actttcctga ccgcccagac actgctgatg 960
gatctgggcc agttcctgct gttttgccac atctccagcc accagcatga tggaatggag 1020
gcatacgtca aagtggactc ttgtcctgag gaaccacaac tgaggatgaa gaacaatgag 1080
gaagccgaag actatgacga tgacctgaca gactccgaga tggatgtggt ccgcttcgat 1140
gacgataact cccctagctt tatccagatt cgaagcgtcg ccaagaaaca cccaaagact 1200
tgggtgcatt acatcgcagc cgaggaagag gactgggatt atgctccact ggtgctggca 1260
cccgatgatc ggagttacaa atcacagtat ctgaacaatg ggcctcagcg aattggtcgt 1320
aagtacaaga aagtgcgatt catggcctat actgatgaaa cctttaagac acgtgaagct 1380
atccagcacg agtctgggat tctgggtcca ctgctgtacg gcgaagtggg agacacactg 1440
ctgatcattt ttaagaacca ggcaagcaga ccttacaata tctatccaca tggaattact 1500
gatgtccggc ctctgtactc taggcggctg ccaaaggggg tgaaacacct gaaggacttc 1560
cccatcctgc ctggtgaaat ttttaagtac aagtggacag tcactgtgga ggatgggcca 1620
acaaagtctg accctcgatg cctgactcgt tactattcta gtttcgtgaa tatggaaaga 1680
gacctggcct ccgggctgat cggtcctctg ctgatttgtt acaaagagtc tgtggatcag 1740
aggggcaacc agatcatgag tgacaagcgg aatgtcattc tgttcagcgt gtttgacgaa 1800
aacaggtctt ggtatctgac cgagaacatc cagcggttcc tgccaaatcc cgcaggcgtg 1860
cagcttgaag atccagagtt tcaggccagc aacatcatgc attctattaa tggatacgtg 1920
ttcgactctc tgcagttgag tgtctgtctg cacgaggtgg cctactggta tatcctgtct 1980
attggcgctc agactgattt cctgtcagtg ttcttttccg gatacacctt taagcataaa 2040
atggtgtatg aggacaccct gacactgttc ccctttagtg gcgaaaccgt gtttatgtca 2100
atggagaatc ctggcctgtg gattctggga tgccacaact ccgatttcag aaatcgcggg 2160
atgaccgctc tgctgaaagt gtcatcctgt gacaagaaca ctggtgacta ctatgaagat 2220
agttacgagg acatctcagc ttatctgctg tccaaaaaca atgcaattga accacgatct 2280
tttagtcaga atcctccagt gctgaagagg caccagcggg agatcacaag gactaccctg 2340
cagagtgatc aggaagagat cgactacgac gatactattt ccgtggaaat gaagaaagag 2400
gacttcgaca tctatgacga agatgagaac cagtccccca ggagcttcca gaagaaaacc 2460
cgtcattact ttattgctgc agtggagcgc ctgtgggatt atggcatgag ctctagtcca 2520
cacgtcctgc gaaatcgtgc ccagtcaggc tccgtgcccc agttcaagaa agtggtcttc 2580
caggagttta cagacggctc ctttactcag ccactgtaca gaggagaact gaacgagcat 2640
ctgggcctgc tgggacccta tatccgcgcc gaagtcgagg ataacattat ggtgaccttc 2700
agaaatcagg ccagccgccc ctactctttt tattcatccc tgatcagcta cgaagaggac 2760
cagagacagg gcgctgaacc ccgcaaaaac ttcgtgaagc ctaatgagac taaaacctac 2820
ttttggaagg tgcagcacca catggcacct acaaaagacg agttcgattg caaggcatgg 2880
gcctattttt cagacgtcga tctggagaag gacgtgcatt ctgggctgat cggtcccctg 2940
ctggtgtgtc atacaaacac tctgaatcct gctcacggca ggcaggtcac cgtgcaggaa 3000
tttgcactgt tctttaccat ctttgatgag acaaagtctt ggtactttac agaaaacatg 3060
gagagaaatt gccgcgctcc ttgtaatatt cagatggaag acccaacttt caaggagaac 3120
tatcggtttc atgcaatcaa tggctatatt atggataccc tgcctggact ggtcatggcc 3180
caggaccaga ggattcggtg gtatctgctg tctatgggga gtaacgagaa tatccacagt 3240
attcatttct caggtcacgt ctttaccgtg aggaagaaag aagagtataa aatggccctg 3300
tacaacctgt atccaggcgt cttcgaaaca gtggagatgc tgccctccaa ggctggaatc 3360
tggcgggtgg aatgcctgat tggggagcac ctgcatgcag gcatgtccac actgtttctg 3420
gtgtacagca ataagtgtca gactccactg gggatggcca gcggtcatat ccgggatttc 3480
cagattaccg cttctggcca gtacggacag tgggctccca agctggctag actgcactat 3540
agcggctcta tcaacgcctg gtccactaaa gagcccttct cctggattaa ggtggacctg 3600
ctggctccca tgatcattca tgggatcaaa acccagggtg cacgccagaa gttcagctct 3660
ctgtacatct ctcagtttat catcatgtac agtctggatg gaaagaaatg gcagacctac 3720
cgaggcaatt ccaccggaac actgatggtc ttctttggca acgtggacag ttcaggaatc 3780
aagcacaaca ttttcaatcc ccctatcatt gctcgataca tccgtctgca ccctacccat 3840
tattcaatta ggtccacact gcggatggaa ctgatggggt gcgatctgaa cagttgttca 3900
atgccactgg gtatggagtc caaggcaatc agcgacgccc agattaccgc ttccagctac 3960
ttcactaata tgtttgccac ctggtccccc agcaaagcta ggctgcatct gcagggccga 4020
agcaacgcct ggcgtccaca ggtcaacaat cccaaggagt ggctgcaggt ggattttcag 4080
aaaacaatga aggtcactgg cgtgacaact cagggagtca aatctctgct gacaagtatg 4140
tacgtgaagg agttcctgat ctctagttca caggacggac accagtggac tctgttcttt 4200
cagaacggga aggtcaaagt gttccagggt aatcaggatt ccttcacccc tgtggtcaac 4260
tctctagacc cacccctgct gaccaggtat ctgcgaatcc acccacagag ctgggtccat 4320
cagattgctc tgagaatgga agtgctgggg tgcgaggcac aggatctgta tggatccggt 4380
ggcggtggct ccggtggagg cggaagcggc ggtggaggat caggcggtgg aggtagcggc 4440
ggaggcggta gctccagctc tagtaaagct ccccctcctt ccctgccctc accctcaaga 4500
ctgcctggac cttccgacac tcccatcctg ccacaggtgg agtgccctcc atgtccagca 4560
ccccctgtcg caggtccatc tgtgttcctg tttccaccca agcctaaaga ccagctgatg 4620
atctcccgca ccccagaagt cacctgtgtg gtcgtggatg tgagccatga agaccccgag 4680
gtccagttca attggtacgt ggatggcgtc gaggtgcaca acgctaagac aaaacctaga 4740
gaagagcagt tcaactctac ctttcgcgtc gtgagtgtgc tgacagtcgt gcaccaggac 4800
tggctgaatg gcaaggagta taagtgcaaa gtgagcaaca aaggactgcc tgcctcaatc 4860
gaaaagacta tttccaagac caaaggacag ccaagagagc cccaggtgta caccctgcct 4920
ccaagccgcg aagagatgac taaaaatcag gtctctctga cctgtctggt gaaggggttt 4980
tatcctagtg atatcgccgt ggaatgggag tcaaacggtc agccagagaa caattacaag 5040
accacacccc ctatgctgga cagcgatggg tctttctttc tgtatagcaa actgacagtg 5100
gacaagtctc ggtggcagca gggtaacgtc ttctcttgca gtgtgctgca cgaagcactg 5160
cacaatcatt acacccagaa gtcactgtca ctgagcccag gaaaatga 5208
<---
Реферат
Изобретение относится к области биотехнологии, конкретно к получению высокогликозилированных слитых белков на основе фактора свертывания крови человека VIII (FVIII), и может быть использовано в медицине для предупреждения или лечения геморрагических болезней у пациентов с дефицитом FVIII. Предложенный гомодимерный слитый белок состоит из, последовательно от N- к C-концу, фактора свертывания крови человека VIII, гибкого пептидного линкера, по меньшей мере одного жесткого карбокси-терминального пептидного звена бета-субъединицы хорионического гонадотропина человека и Fc-фрагмента иммуноглобулина человека. Изобретение позволяет получить белок с уровнем биологической активности, аналогичным уровню рекомбинантного FVIII, и продленным периодом полужизни in vivo, т.е. лекарственное средство на основе FVIII с улучшенной фармакокинетикой и эффективностью. 7 н. и 4 з.п. ф-лы, 5 ил., 3 табл.
Формула
Документы, цитированные в отчёте о поиске
Физиологически активный полипептидный конъюгат, обладающий пролонгированным периодом полувыведения in vivo
Комментарии