Способ получения трансгенной овцы - RU2085587C1
Код документа: RU2085587C1
Чертежи











Описание
Изобретение относится к способу получения слитого полипептида. Более конкретно, изобретение относится к получению белка и к получению биологического материала, чье образование катализируется ферментивными белками.
Для получения важных в промышленном отношении биологических продуктов в последние десять лет во все больших масштабах используются методы биотехнологии. В этой связи осуществлено клонирование ДНК-последовательностей, кодирующих важные с точки зрения медицины белки человека. В число таких белков входят инсулин, активатор плазминогена, альфа1 -антитрипсин и факторы коагуляции VIII и IX. В настоящее время, даже при постоянно появляющихся новых методах биотехнологии, такие белки, как правило, выделяемые из крови и тканей с использованием дорогостоящих и требующих больших затрат времени способов, могут привести к риску занесения инфекционных агентов, например, таких, которые вызывают СПИД или гепатит.
Несмотря на то, что экспрессия ДНК-последовательностей в бактериях с целью получения целевых важных в медицинском отношении белков выглядит весьма привлекательно, на практике бактерии часто оказываются неудовлетворительными в роли хозяев, поскольку в бактериальной клетке посторонние белки неустойчивы и не процессируются соответствующим образом.
С пониманием данной проблемы связаны попытки экспрессии клонированных генов в культуре тканей млекопитающих, которые в некоторых случаях доказали свою жизнеспособность. Однако широкомасштабная ферментация клеток животного является дорогостоящим и требующим широкого технологического обеспечения процессом.
Таким образом, существует необходимость в высокопродуктивном дешевом способе получения биологических веществ, таких как необходимым образом модифицированные эукариотные полипептиды. Отсутствие инфекционных для человека агентов будет служить преимуществом такого способа.
Согласно первому аспекту изобретения предложен способ получения трансгенной овцы, предусматривающий конструирование рекомбинантной плазмидной ДНК, кодирующей слитый белок, выделение фрагмента SalI XbaI плазмиды PSST tg XS или PSSI tg XS FIX, кодирующего овечий бета лактоглобулин, слитый с фактором IX или антитрипсин, введение полученного фрагмента ДНК в яйцо или эмбрион овцы в результате микроинъекции, трансплантацию яйца или эмбриона в рецинистных овцематок с последующим отбором трансгенных овцематок.
α Антитрипсин или фактор IX может быть полипептидом или модифицированным полипептидом (например, гликозилированным), или же полипептид может быть сшит путем посттрансляционной модификации полипептида.
Изобретение может быть использовано, к примеру, для получения a1 - антитрипсина или фактора IX.
Получаемые настоящим изобретением ферменты могут оказаться способны действовать на продуцирующие их субстраты in vitro в молочной железе, т.е. на получение вещества, являющегося продуктом реакции фермента. Продукт реакции, как правило, может быть выделен из молока взрослой женской особи млекопитающего.
ДНК-последовательность, кодирующую α1 -антитрипсин фактор IX ("представляющая интерес ДНК-последовательность") вводят in vitro в ген белка сыворотки молока, способный экспрессироваться в молочной железе взрослой женской особи млекопитающего с образованием гена слияния, который вводят в зародышевую линию путем инъекции в оплодотворенное яйцо млекопитающего, после чего оплодотворенное инъецированное яйцо трансплантируют во взрослую женскую особь млекопитающего. Следует ожидать, что не все оплодотворенные инъецированные яйца разовьются во взрослые женские особи, экспрессирующие представляющую интерес ДНК-последовательность. Помимо всего прочего примерно половина животных будет мужского пола, от которых женские особи могут быть получены в следующих поколениях.
Приемлемые
методики, в которых чужеродные ДНК-последовательности вводят в зародышевую линию млекопитающего, апробированы на мышах. В
настоящее время наиболее эффективный подход заключается в прямом
микроинъецировании нескольких сот линейных молекул ДНК в проядро оплодотворенного одноклеточного яйца, и такой путь является
предпочтительным методом настоящего изобретения. Микроинъецированные яйца
могут быть затем перенесены в яйцеводы псевдобеременных приемных матерей для дальнейшего развития. В работе сообщается, что
около 25% развивающихся мышей наследуют одну или несколько копий
микроинъектированной ДНК [1] Известен способ получения трансгенной овцы, предусматривающий конструирование рекомбинантной плазмидной
ДНК, выделение слитого гена из полученной ДНК, инъекцию слитого
гена в яйцо эмбрион овцы, пересадку эмбрионов реципиентным овцематкам с последующим отбором трансгенной овцы [2]
После
внедрения в зародышевую линию чужеродная ДНК может быть экспрессирована
на высоком уровне в выбранной ткани с образованием функционального белка, который может быть извлечен из животного. Для этого
ДНК-последовательность, кодирующая представляющий интерес пептид, как
правило, сливается с ДНК-последовательностью или последовательностями, обеспечивающими ее экспрессию в соответствующей ткани.
Хотя отдельные начальные опыты на мышах показывают, что специфическая
экспрессия внедренной ДНК в тканях до некоторой степени изменчива [3] существующее в настоящее время общее мнение заключается в
том, что нужная специфическая экспрессия большинства трансфекированных
генов достижима [4-7] Для необходимой тканевой специфичности, видимо, важно удалить все прокариотные векторные
последовательности, используемые в клонировании представляющей интерес
ДНК-последовательности, перед микроинъецированием.
Несмотря на то, что приведенные публикации и указывают на некоторые различные ткани, которые могут быть выбраны в качестве мишеней для создания трансгенного животного, ни в одной из них не предполагается возможность использования какой-то конкретной ткани для промышленного получения содержащих полипептид веществ. В работах также не предполагается возможность использования конкретного гена, экспрессируемого в данной ткани, и который мог бы служить для направленной экспрессии внедренной ДНК.
В связи со слитым полипептидом и тканью, в которой он будет экспрессироваться, следует учесть целый ряд факторов. Многие белки для проявления ими полной биологической активности требуют интенсивной посттрансляционной модификации. Например, фактор IX для проявления биологической активности требует гамма-карбоксилирования специфических участков остатков глутаминовой кислоты. Печень, являющаяся местом синтеза фактора IX в природе, избирательно выдает именно эту модификацию. Фибробласты также способны осуществлять гамма-карбоксилирование, хотя и не столь эффективно [10] Однако некоторые белки могут быть нужным образом модифицированы, только если их синтез осуществляется в специфических тканях, и в некоторых случаях может оказаться необходимым связать место экспрессии с требованиями, предъявляемыми к получаемому белку. Полагают, что использование молочной железы в качестве ткани для экспрессии позволяет преодолеть либо полностью, либо до удовлетворительного предела эти затруднения.
Предпочтителен сбор целевого продукта из биологических жидкостей, а не твердых тканей, поскольку в этом случае этот источник в большей степени возобновляем, и кроме того, большая часть представляющих биомедицинскую ценность белков сами по себе секретируются в биологические жидкости. Возможным путем является секреция в кровеносный поток из печени или B-лимфоцитов, однако коагулирующие свойства крови и присутствие в ней биологически активных пептидов, также как и антигенных молекул может создать затруднения при последующем нисходящем процессинге.
В соответствии с настоящим изобретением эти трудности могут быть преодолены с использованием в качестве ткани для экспрессии молочной железы. Молоко легко собирать, образуется оно в больших количествах и хорошо характеризуется биохимически. Кроме того, большая часть молочных белков присутствует в молоке в высоких концентрациях (от примерно 1 до 15 г/л).
Для овец интервал молочной продуктивности составляет 1-3 л в день в зависимости от породы. Следует отметить, что промышленностью освоен выпуск специального оборудования для дойки овец и, разумеется, для дойки крупного рогатого скота. Таким образом, выбранными животными стали овцы, в частности выращиваемые на фермах овцы, такие как Восточно-Фризляндской породы. В качестве вполне возможной альтернативы может рассматриваться линия, полученная скрещиванием подходящей высокопродуктивной молочной породы овец с черноголовыми овцами.
Лактирующие молочные железы представляют собой высоко специализированный орган, состоящий из разветвленной системы каналов, осушающих сложные дольки секреторных клеток. Молочные клетки самым разным образом приспособлены к высокому уровню секреции. Например, эти клетки обладают специализированным транспортным механизмом, обеспечивающим эффективный прием предшественников из крови, и интенсивной системой межклеточных мембран (грубая эндоплазмическая сетка, аппарат Гольджи и т.п.), позволяющих эффективно проводить синтез белка, его посттрансляционное модифицирование и вынос из клеток.
Молочные железы зависят от гормонов на всем протяжении их роста и функционирования. Действующие на молочные железы гормоны вызывают в них в период беременности и лактации поразительные изменения. Например, у овцематок их действие приводит к почти к двукратному увеличению общего числа клеток, а также к изменению соотношения между различными типами клеток и к конечному дифференцированию секреторных клеток.
Молочная железа секретирует в молоко ряд различных белков. Существуют качественные и количественные различия состава молока различных видов животных, хотя общее отличие может быть сделано между казеинами и растворимыми /сыворотка/ белками [13] У основных жвачных животных к белкам сыворотки, которые синтезируются молочной железой, принадлежат альфа-лактаальбумин и бета-лактоглобулин.
Существует три основных типа казеина. Это альфа-казеин, бета-казеин и каппа-казеин, и они присутствуют в молоке большей части рассматриваемых видов животных. Их роль в молоке заключается в изоляции кальция, с которым они агрегируются в виде мицелл. Роль бета-лактоглобулина основного белка сыворотки жвачных неизвестна, хотя он, видимо, взаимодействует с каппа-казеином. Альфа-лактоальбумин является существенным кофактором превращения глюкозы и галактозы в лактозу. В табл. 1 приведен белковый состав молока различных животных.
По большей части считают, что различные молочные белки кодируются одной копией [16] Гены казеина, видимо, связаны с мышами [18] и коровами. Поскольку гены казеина, видимо, экспрессируются в молоке на гораздо более высоком уровне по сравнению с генами белка сыворотки, вполне естественно выбрать гены казеина для введения чужеродной ДНК с целью ее экспрессии. Однако в настоящем изобретении реализуется введение ДНК-последовательности, кодирующей представляющий интерес пептид, в ген, кодирующий белок сыворотки.
Ген альфа-лактоальбумина крыс состоит из четырех экзонов и находится в хромосомной ДНК размером 2,5 кb [19] Ген коровьего альфа-лактоальбумина, видимо, организован аналогично. В настоящее время установлено, что ген овечьего бета-лактоглобулина наиболее вероятно имеет одну копию, состоит из семи экзонов в пределах транскрипционного звена в 4,9 kb, и именно этот ген рекомендуется использовать в настоящем изобретении.
Изменения в уровнях молочного белка (мРНК) в ходе развития молочной
железы у крыс [20] у кроликов [21] и мышей [22] выявлены с использованием как трансляции
in vitro, так и гибридизации кДНК. Молочная железа аккумулирует где-то около 80000-100000 молекул мРНК,
специфичных для белка молока, образовавшихся в результате действия согласованного механизма,
включающего как синтез на высоком уровне молочного белка мРНК, так и эффективную стабилизацию этих молекул.
Существуют, однако, и исключения. Для овцематок бета-казеин составляет до 45% всего
молочного белка, но только 17% клонов кДНК, выделенных из библиотеки кДНК, соответствуют этому гену [25] У кроличьих
количество альфа-лактоальбумина, собранного в микросомных пузырьках, значительно
меньше, чем можно было бы ожидать на основании скорости синтеза белка в отсутствие микросомных пузырьков, возможно
вследствие того, что сигнальный пептид не обладает оптимальной конфигурацией [26]
Первичные продукты трансляции многих генов молочного белка подвергаются интенсивной модификации. Например,
альфа-лактоальбумин N-гликозилируется, каппа-казеин O-гликозилируется, альфа- и бета-казеин
O-фосфорилируется. Кроме того, свою роль в посттрансляционной модификации выполняет и комплекс Гольджи,
осуществляя конденсацию и упаковку казеинов в мицеллы. Существует вероятность того, что казеины
и белки сыворотки выводятся из клетки различными путями. В этом смысле интересно отметить, что как при
между-, так и при внутриспецифическом сравнении сигнальный пептид казеина (в сравнении с большей
частью молекулы) в высшей степени консервативен и, следовательно, этот пептид может играть роль в
выделении предшественника пептида в правильном направлении [24]
Как указано выше, гены
молочного белка в изобилии экспрессируются в лактирующей молочной железе. Для овцематки, например, мРНК
альфаSI-казеина составляет примерно 30% и мДНК бета-лактоглобулина составляет
примерно 5% полиА+РНК [25] Допустим, что эти транскрипты происходят из единичных копий генов, приведенные
концентрации в этом случае указывают на высокую скорость транскрипции и эффективную
стабилизацию мРНК [27] Специфическая стабилизация мРНК может быть передана через последовательности, присутствующие
в зрелой мРНК. В рекомендуемых вариантах настоящего изобретения могут быть
достигнуты одинаковые уровни экспрессии ДНК-последовательности, вводимой в зародышевую линию овцы. Это достигается ее
лигированием с ДНК-последовательностями, связанными с геном белка молочной
сыворотки, придающего высокие значения специфичной для тканей экспрессии и стабилизации мРНК, для этих целей рекомендуется
ген бета-лактоглобулина.
Перенос новых генов, осуществляемый на мышах, является в настоящее время обычным способом. У трансгенных мышей, показывающих высокие значения специфичной для тканей экспрессии чужеродного гена, экзогенная ДНК включает не только структурный ген, но также 5'- и 3'-фланкирующие последовательности. Несмотря на то, что существуют многочисленные факты, подтверждающие, что многие важные регулирующие элементы расположены в интервале с 5' до сар сайта мРНК [28-31] а также очевидно, что важные регулирующие последовательности, в частности последовательности, определяющие специфичную для тканей экспрессию, могут быть расположены в пределах структурного гена или даже в интервале с 3' до этого гена [32-33] Более того, экзонные последовательности (т. е. последовательности зрелой мРНК) могут содержать последовательности, определяющие стабильность мРНК. В табл. 2 показаны некоторые детали специфичной для ткани экспрессии чужеродного гена у мышей.
Именно по той причине (о чем сказано выше), вышеуказанные гены слияния будут использованы для направления экспрессии ДНК-последовательности, кодирующей представляющий интерес пептид, эти гены включают один или более промотр, старт-сайт для транскрипции, одну или более /предположительно/ дистальных 5' регулирующих последовательностей гена молочного белка, 3' последовательности, фланкирующие ген молочного белка. Наиболее рекомендуемый ген слияния включает последовательности всех указанных типов.
Выбранный ген слияния состоит из кДНК, кодирующей представляющий интерес пептид, внедренный в первый экзон гена белка молочной сыворотки. Рекомендуется, чтобы несколько кb 5' фланкирующих последовательностей гена молочного белка были включены в такой ген слияния. В таких конструкциях секреция представляющего интерес пептида предпочтительно будет обеспечиваться его собственным сигнальным пептидом, вследствие чего рекомендуется, чтобы ген слияния включал сигнальный пептид для представляющего интерес пептида. Однако специфичный по отношению к ткани сигнальный пептид может оказаться важным и для направления предшественника пептида на правильный секреторный путь (о чем речь шла выше). Таким образом, особенно рекомендуется, чтобы кодирующие сигнальный пептид белка сыворотки ДНК-последовательности были точнейшим образом сцеплены с ДНК-последовательностями вставки, кодирующими N-концевые аминокислоты зрелого белка. 3'-конец места вставки предпочтительно обрывается после его стоп-кодона, но перед его собственным сайтом расщепления при трансляции и полиаденилирования. В нисходящем направлении от сайта вставки остальная часть структурного гена, как правило, сохраняется, так же как и отдельные 3' фланкирующие последовательности.
У трансгенных животных такая конструкция будет максимально увеличивать уровень специфичной для молочной железы экспрессии. Необходимо, чтобы первичный транскрипт был правильно полиаденилирован и подвергнут сплайсингу в сохранившихся сайтах гена белка молочной сыворотки. Необходимо, чтобы зрелая мРНК имела последовательности для эффективной стабилизации. Необходимо, чтобы в конструкциях, использующих сигнальный пептид гена молока, зрелая мРНК была транслирована с образованием предшественника полипептида слияния, в котором сигнальный пептид, происходящий из гена молочного белка, эффективно направляет секрецию зрелого пептида, закодированного внедренной кДНК.
Как было указано выше, бета-лактоглобулин овцы является выбранным геном молочной сыворотки, который "нагружают" кДНК, кодирующей представляющий интерес пептид. У овец бета-лактоглобулин является наиболее изобильно экспрессируемым в молочной железе белком сыворотки, и его мРНК составляет примерно 8% всей полиА+РНК. Этот ген в достаточной степени охарактеризован и был использован для создания конструкций с кДНК-последовательностями, кодирующими фактор IX человека и альфаI-антитрипсин человека.
На фиг. 1-4 схематично показаны основные стадии создания гена сцепления бета-лактоглобулина способом настоящего изобретения; на фиг.5 - блоттинг-анализ по Саузерну ДНК овцы; на фиг.6 блоттинг-анализ по Саузерну ДНК и наследуемость гена сцепления путем реальной генерации; на фиг.7 анализ белка сыворотки молока мышей и овец с помощью НДС-ПАГЭ; на фиг.8 - Вестерн-блоттинг белков мышей и овец; на фиг.9 P и А молока, полученного от трансгенной овцематки, несущей описанный ген сцепления бета-лактоглобулина.
Пример 1.
А. Методика с
рекомбинантной ДНК
Если нет специальных указаний, использованы методики с рекомбинантной ДНК [34-35] Если не указано особо, то все химикаты фирмы были BDH
Chemical Ltd, Пул, Дорсет, Англия
или фирмы Sigma Chemical Company, Пул, Дорсет, Англия.
Конструкция гена сцепления бета-лактоглобулина овцы
Получение ДНК из селезенки овцы
Из только что убитой овцы
черноголовой суффольской породы получают ткани селезенки, из которых выделяют ядра [36] Ядерные шарики лизируют в 0,3 М растворе NaCl, 10мМ трис-HCl, 10 мМ ЭДТК, 1% НДС
/pH 7,4/ и 400 мкг/мл
протеиназы К /Sigma Co. Fancy Road, Пул, Дорсет ВН17 711Н/ и инкубируют 2 ч при 37oC. Проводят непрерывную экстракцию смесью фенола с хлороформом до полной
депротеинизации препарата. ДНК
осаждают этанолом и сматывают с помощью стеклянной палочки, промывают 70% EtОН-30% ТЭ /ТЭ 10 мМ трис-HCl, 1 мМ ЭДТК, pH 8/, высушивают на воздухе и вновь суспендируют в
ТЭ до концентрации 1 мг/мл.
Конструкция лямбда-гена сцепления ДНК селезенки овцы
Для построения геномной библиотеки используют лямбда-фаг ЕМВ13 30 мкг ДНК бактериофага
/обрабатывают 5-кратным избытком
ферментов рестрикции EсоRI и Bam HI /представлены Amersham и Intgernational pIc, Линкольн Плайс, Грийн Энд, Олесбери, Букингемпшир, Англия/ в условиях рекомендованных
изготовителем. После обработки
добавляют хлоргидрат спермина до концентрации 5мМ с целью осаждения лямбда-ДНК. После инкубирования на льду в течение часа ДНК превращают в шарики при 10,000g в течение
15 мин с помощью ступенчатой
микроцентрифуги, промывают в 70% EtOH, 300 мМ NaAc, 100 mM MgCl2, снова превращают в шарики и, наконец, вновь суспендируют в ТЭ до концентрации 1 мг/мл.
ДНК овцы частично обрабатывают с помощью фермента рестрикции Sau3A /Amersham/. Аликвоты по 100 мкг ДНК овцы гидролизуют различными количествами Sau3A /от 5-40 единиц/ в течение 20 мин при 37oC. Реакции прекращают добавлением ЭДТК до концентрации 15 мМ. Степень рестрикции выявляют с помощью электрофореза на 0,6% агарозном геле. Надлежащим образом гидролизованные образцы объединяют и наносят на 38 мл 10-40% градиента сахарозы, приготовленного в IM NaCl, 20 mM трис-HCl, 5 мМ ЭДТК при pH 8. Такие градиенты центрифугируют 24 ч при скорости 26000 об/мин на роторе Beckman SW28. Градиенты сахарозы фракционируют сверху с отбором фракций по 1 мл. Распределение молекул ДНК по размерам в каждой фракции выявляют с помощью электрофореза на агарозном геле, фракции, содержащие молекулы ДНК размером в 14-21 кb, объединяют. После двукратного разбавления в ТЭ добавляют 2 объема EtOH и в течение суток при -20oC осаждают ДНК. После этого ДНК вновь суспендируют в ТЭ до концентрации 300 мкг/мл.
Обработанную BamHI-ECORI фракцию ЕМВ13 в 7,5 мкг и 2,5 мкг ДНК из селезенки овцы, частично обработанные Sau3A, смешивают в 50 мкл раствора, содержащего 60 мМ трис-HCl, 6 мM MgCl2, 10 мМ ДДТ, 0,01% желатина, 0,25 мМ рАТФ и 25 единиц Т4ДНК-лигазы /Boehringer Company, Boehringer Mannheim Haise Белл Лейн, Лийвес, Ист Сассекс/ и инкубируют при 14oC в течение суток.
После лигации аликвоты по 1 мкг ДНК упаковывают с помощью прибора фирмы Amersham согласно методике, рекомендованной изготовителем. Упакованную библиотеку титруют на штамме E.coli ED 8654. Установленный размер библиотеки составляет 5,7х106 образующих бляшки единиц /о.б.е./. Сразу же после титрования аликвоты неамплифицированной библиотеки наносят на чашечки Петри размером 10х22 см2 /мегапластинки/ при использовании штамма E.coli ED 8654 при плотности примерно 50000 о.б.е. на пластинку.
Отбор геномномной лямбды-библиотеки.
На мембраны из нитроцеллюлозы в 20 см2 осуществляют перенос с мегапластин вышеполученные бляшки. Клон кДНК бета-лактоглобулина /р931/ никтранслируют с помощью32P дЦТФ до удельной активности > 108 dpm/мкг [39] кДНК бета-лактоглобулина была клонирована [40] Заключительную промывку проводят 1хSET/SET 0,15 мМ NaCl, 2 мМ ЭДТК, 0,03 М трис-HCl с pH 8/. Фильтры в сухом виде метят32P с целью их ориентации перед экспозицией на рентгеновскую пленку. Области, содержащие положительно гибридизированные бляшки, располагают на мегапластинах со ссылкой на меченные32P пятна, собранные с использованием стерильного тупого конца пипетки Пастера. Первоначально перенесенные бляшки титруют на штамме E.coli ED 8654 и помещают в чашечки Петри диаметром 15 см при плотности бляшек примерно 500 на чашку. Эти чашки подвергают повторному отбору по вышеприведенной методике, в результате чего отбирают отдельные положительно гибридизованные бляшки с использованием зубочистки в 1 мл фагового буфера/фаговый буфер 10 мМ трис-HCl, 10 мМ MgCl2, 0,01% желатина, pH 7, 4/.
Получение клонированной ДНК бета-лактоглобулина.
К E. coli ED 8654 добавляют 0,4 мл раствора фага [41] и помещают в чашечки Петри диаметром 9 см с целью получения зоны лизиса бактерий. Получают зону лизиса, с которых в 10 мл фагового буфера снимают верхний слой агара и инкубируют сутки с несколькими каплями хлороформа. Бактериальные лизаты превращают в гранулы с помощью центрифугирования со скоростью 5000 об./мин, а фаговое сырье хранят при 4oC. Полученное сырье титруют на E.coli ED 8654 для определения значения о.б.е./мл.
На 7х109 клетках E.coli в 10 мл мМ MgSO4 при 37oC адсорбируют 8х107 о.б. е. Спустя 15 мин в колбе на один литр к 100 мл 1-бульона-10 мМ MgSO4 добавляют аликвоты по 2,5 мл. Бактериальную суспензию несколько часов интенсивно встряхивают и каждый час проверяют значение ОД540. Лизис, как определено по падению значения ОД540 , происходит через несколько часов. После завершения лизиса к каждым 100 мл культуры добавляют 0,2 мл хлороформа и культуру оставляют при 4oC на сутки.
Бактериальные лизаты удаляют центрифугированием в течение 15 мин при скорости 10000 об. /мин. К центрифугату добавляют 10 мкг/мл РНКазы А и 10 мкг/мл ДНКазы 1, после чего инкубируют один час при 37oC. После инкубирования добавляют NaCl до концентрации 40 г/л и полиэтиленгликоль /ПЭГ/ до концентрации 10% Препарат охлаждают до 4oC и оставляют по меньшей мере на два часа для осаждения фага. Фаговые бляшки превращают в шарики при 10000 об/мин в течение 15 мин и вновь суспендируют в 16 мл фагового буфера. 8 мл полученной суспензии наносят в виде слоя на ступенчатый градиент, содержащий 1, 5 мл 56% CsCl, 1,6 мл 45% CsCl и 2,5 мл 31% CsCI /в растворе фагового буфера/, помещенного в ультрацентрифужную пробирку на 14 мл. Ступенчатые градиенты центрифугируют 1,5 ч при скорости 35000 об./мин в роторе с колебательно-вращательным движением и 20oC. С помощью иглы и шприца удаляют полосу фага и для завершения очистки частиц фага проводят центрифугирование второго ступенчатого градиента.
Очищенные частицы фага подвергают диализу в 0,1 М NaCl, 10 мМ трис-HCl, 1 мМ ЭДТК /pH 8/, после чего депротонизируют последовательной экстракцией фенолом и хлороформом. Добавляют NaCl до окончательной концентрации 0,3 М, после чего добавлением 2 объемов Et ОН осаждают ДНК фага. Центрифугированием при 10000 в течение 15 мин ДНК превращают в шарики, промывают 70% EtOH, 30% ТЭ, сушат и вновь суспендируют в ТЭ до конечной концентрации 200-400 мкг/мл.
Характеристика рекомбинантных клонов бета-глобулина.
Аликвоты по 0, 5 мкг вышеописанных препаратов ДНК подвергают рестрикции различными рестрикционными ферментами и продукты единичного и двойного расщепления анализируют с помощью электрофореза на 0,6% и 1% агарозном геле. ДНК с таких гелей переносят на фильтры из нитроцеллюлозы или мембраны Hybond (Amersham Intenational, Литтл Чалфонт, Бакс) [42] и гибридизуют с клоном p931, меченным32Р. Использованная процедура по существу аналогична вышеприведенной, и гибридизованные фильтры анализируют с помощью афторадиографии. С использованием различных рестрикционных ферментов и специфических зондов из 5'- и 3'-концов p931 построена карта рестрикции, на основании которой определен размер и ориентация гена/ов/ бета-лактоглобулина /фиг.1/.
Идентичность клонов
бета-лактоглобулина и точное положение 5'- и
3'-концов гена непосредственно подтверждено определением ДНК-последовательности. С использованием подходящих фрагменты субклонируют в рестрикционные сайты
плазмидных векторов и векторов М13.
Установление последовательности проводят с помощью дидезокси-метода [43]
Обработка генов сцепления бета-глобулина.
Подход, используемый для обработки генов сцепления, включающих бета-глобулин и представляющих интерес генов, экспрессируемых в молочной железе, отражен на фиг. 1-4. В таком подходе используют последовательности, образованные лямбда-клоном, характеристика которых и выделение описано выше. Такой подход включает внедрение представляющей интерес ДНК-последовательности в область ДНК, соответствующую 5' нетрансляционной области мРНК бета-лактоглобулина. Белки, транслируемые с транскриптов мРНК такого гена, будут содержать секреторный пептид целевого белка.
Субклон pss-Itg SE
конструируют так, как показано на фиг.
1, путем лигирования фрагмента SphI-EcoRI 1,4 кb лямбда-фага SS-1 с векторной плазмидой pPOIy, которую также разрезают с помощью SphI + EcoRI и с последующей
трансформацией штамма E.coli D ДН1 для
придания ему компетентности [44] Выделяют устойчивые к амплицилину клоны и из них получают ДНК [45]
На фиг. 1 верхняя стрелка определяет ориентацию и
протяженность /примерно 4,9
к.о./ транскрипционного звена бета-лактоглобулина, присутствующего в пределах SS 1, тот же масштаб сохранен повсюду. В более общем смысле следует отметить, что на фиг. 1-4
показаны только относящиеся
к делу рестрикционные сайты. Большие открытые прямоугольники представляют ветви лямбда-EMBL3, узкие открытые прямоугольники представляют pPoIy, узкий закрашенный
прямоугольник представляет
экспрессируемую целевую последовательность, линии представляют последовательности для овцы, соответствующие гену бета-лактоглобулина и его фланкированным
последовательностям.
Субклон PSS-ItgSE линеаризуют путем гидролиза с рестрикционной эндонуклеазой PVU11 /фиг.2/, которая разрезает плазмиду в уникальном сайте той области ДНК, которая соответствует 5' нетранслированных мРНК-последовательностей бета-лактоглобулина. 5 мкг полностью гидролизованной плазмиды растворяют в 0,5 М трис-HCl, 10 мМ MgCl2, 1 мM MnCl2, 10 мM спермидина /pH 9/ и обрабатывают 0,01-0,04 единицами кишечной фосфатазы теленка /Boehringer/ в течение 30 мин при 56oC. Кишечную фосфатазу теленка дезактивируют в 0,5 НДС, а ДНК выделяют экстракгированием фенолом-хлороформом и осаждением EtOH.
Клон ДНК фактора 1Х p5'G3'CV1 получен от Др.G.Brownlee из школы Паталогии cэра Вильяма Дунна, Оксфордского
Университета, Оксфорд. Такой клон
имеет вставку в 1579 по плазмиде, полученной из PAT153 [46] Она простирается от TagI-сайта у-7 от предполагаемого старт-сайта мРНК до +1572 и содержит
полную кодирующую
последовательность
фактора 1Х человека [47]
Фрагмент Nhel-Hind II, включающий 1553 по последовательность фактора IХ, отрезают из векторных последовательностей и очищают
нижеописанными способами. Hind III и Nhel
концы фрагмента затупляют с помощью полимеразы Кленова [48] и фрагмент лигируют в PVU II сайт, обработанной рестриктазой и фосфитазой плазмиды pSSItg SE
/см.выше/ с образованием pSSItg SE-фактор IХ
после трансформирования штамма Е.coli H-I на устойчивость к амплициллину.
Последовательность кДНК фактора IХ, далее обозначаемая как TARG, что можно видеть на фиг.2.
Плазмидную ДНК получают, как указано выше, и проверяют путем обработки соответствующим рестрикционным ферментом. Плазмидную ДНК обрабатывают с помощью Sphl
+ ECORI и подвергают электрофорезу на 1%
агарозном геле, содержащем 0,5 мкг/мл этидий-бромида /Sigma/. Нужный Sphl ECORI фрагмент локализуют освещением УФ-лампой. Кусок диализной мембраны вставляют в
переднюю часть полосы и затем ДНК
подвергают электрофорезу на мембране. ДНК вымывают с диализной мембраны и изолируют с использованием "EIU-tip" [49]
Плазмиду pSSItgXS /фиг.3/ конструируют
путем лигирования фрагмента
XbaI-SaII лямбда-SSI и XbaI SaII и фрагмента плазмиды pPoIy. Клоны изолируют и плазмидную ДНК получают, как описано ранее. Из этой плазмиды выделяют независимо два
фрагмента ДНК или более мелкие
субклоны, образовавшиеся из плазмиды (фрагмент 10,5 к.в. SphI) частичный Hind II и фрагмент 1,2 к.в. EcoRI-Hind III /фиг. 4/. Эти фрагменты выделяют электрофорезом на
геле, как описано выше.
Фрагменты SphI-EcoRi, SphI-HindII и EcoRI-HindIII лигируют совместно в примерно равных отношениях и ДНК, используемой для трансформации ДН-1. Плазмидную ДНК /pSSItgXS-TARC/ получают, как описано выше, и гидролизуют с помощью Xbal и Sall с отщеплением от вектора гена слияния бета-лактоглобулина. Такой фрагмент очищают электрофорезом на геле с последующим использованием "Е1u-tip". Осажденную этанолом ДНК превращают в шарики, вновь суспендируют в ТЭ, экстрагируют фенол-хлороформом и снова суспендируют. Наконец ДНК вновь суспендируют в ТЭ и непосредственно применяют для микроинъекций.
В. Создание трансгенных животных
Отбор оплодотворенных яиц
Использованные методики аналогичны.
Эмбрионы извлекают из овцематок, у которых во время течки, регулируемой действием прогестагена, вызвана суперовуляция. Зрелые овцематки с доказанной оплодотворяемостью обрабатывают 12-16 дн внутривагинально тампоном, пропитанным 60 мг медрогпроксиэстеронацетата. Фолликулстимулирующий гормон лошади вводят в виде 2 равных внутримышечных инъекций /3,5-4,3 мг в водном растворе на овцематки/ за 28 ч до конца обработки прогестагеном и во время удаления тампона. Во время течки, происходящей через 20-72 после удаления тампона, овцематок несколько раз спаривают. Овцематки ежедневно в 8.00, 12.00, 16.00 и 20.00 осматривают на предмет начала повышения температуры. Через 36-72 ч после начала течки хирургическим путем извлекают эмбрионы на 1-4 клеточной стадии развития.
Анастезию вызывают внутривенной инъекцией тиопентона натрия /Intraval, Mayano Baker/ и поддерживают смесью кислорода с закисью азота в полузамкнутой циркуляционной системе. Извлечение эмбрионов осуществляют [49] Репродуктивный тракт обнажают через средне-центральный надрез и через фимбрию в яйцевод вводят найлоновый катетер. Через затупленную иглу 18 размера в просвет матки вводят среду, которую пропускают через соединение матки с фаллопиевыми трубами и яйцевод. Эмбрионы извлекают в фосфатном буферном солевом растворе, содержащем источники энергии и белок. Во время хранения и микроинъекции яиц такую среду дополняют 20% плодной сыворотки теленка.
Инъецирование ДНК
ДНК инъецируют в количестве 1-2 мкг/мл в одно проядро единичных одноклеточных яиц в одно или более ядер 2 и 4
клеточных яиц. Обработку яиц
проводят в камере, заполненной культуральной средой яиц. Камера состоит из силиконизированного слайда микроскопа со стеклянными держателями /25 мм х 2 мм х 3 мм/,
параллельными длинной стороне слайда.
Верхняя часть держателей накрыта лентой, места соединения герметизируют силиконовой смазкой. Открытый конец камеры заполнен 50 мл жидкости Dow-Corning 200 (ВДН
Chemicals).
Предназначенные для инъецирования яйца удерживают путем засасывания в тупую стеклянную пипетку. Проядра или ядра осматривают с использованием инвертируемого микроскопа Nihon Diaphet (Nikoon) /Великобритания/Ltd. Нейбрук, Нэйсфилд 9, Тэлфорд, Шропшир/. ДНК инъецируют с помощью микропипетки, вытянутой из капиллярной трубочки/боросиликатное стекло, внутренний диаметр 1 мм, тонкостенная с нитью Clarc Electromedical Instrument, PO Box 8, Пэнгбурн, Рийдинг, R68 7HU/, в проядро или ядро на микроэлектродном выталкивателе /Campdem Instruments 186 Кэмпден Нил Роуд, Лондон/. Положение двух микроинструментов регулируют с применением микроманипуляторов /leitz Mechanical Micromanipulators, E. Leitz (Instruments) Ltd, 48 Парк Стрит, Лутон, Англия/. Микропипетку с инъецируемой ДНК соединяют воздухонепроницаемой трубочкой со стеклянным шприцем на 100 мл. Инъецирование осуществляют созданием с помощью шприца давления. Успешное инъецирование видно по набуханию проядра или ядра. Инъецированные яйца инкубируют минимум 30 мин при комнатной температуре, чтобы выявить поврежденные яйца по их вырождению.
Эмбрионы, которые считают выжившими после инъецирования, пересаживают неспаривавшимся рецепиентным овцематкам, у которых цикл течки синхронизирован с циклом доноров введением прогестагена /Veromix, Upjohn Ltd/. Эмбрионы пересаживают в яйцевод с применением мелко вытянутой пипетки для рта. Каждой овцематки пересаживают до 4 эмбрионов. Эмбрионы распределяют между яйцеводами.
Стенку тела овцематки зашивают растворимой нитью /Descon, Davisand Creek/, а шкуру затягивают зажимами Мишеля. Каждой овце дают антибиотики /Duphapen L. A. Duphar, Amersham/ во время хирургической операции. Через 10-20 дн после операции зажимы снимают.
Развитие и рост
После того как овцы очнутся от анестезии, их возвращают в загон, где они остаются на весь срок беременности. По мере необходимости овцы
получают дополнительное сено,
турнепс и концентраты. В ходе третьего месяца беременности ультразвуковым сканированием определяют число плодов [50] Уход за беременными овцами после этого устанавливают
в зависимости от числа плодов.
По мере приближения ожидаемой даты родов овец помещают в помещение, облегчающее их осмотр и помощь во время ягнения.
Анализ трансгенных ягнят
По меньшей мере через 2 нед после
рождения у ягнят из вены с помощью медицинского шприца отбирают по 10 мл образца крови, которую собирают в гепаринизированную пробирку. Из образцов крови
приготовляют ДНК следующим образом: к 10 мл
образца крови добавляют 30 мл раствора лизиса /155 мМ NH4Cl, 10 мM KHCO3, 1 мM ЭДТК/ и смесь инкубируют 15 мин на льду. Белые
кровяные тельца центрифугируют при 1500 g в течение
10 мин при 40oC и вновь суспендируют в 10 мл SE /75 мM NaCl, 2 мM ЭДТК/ и затем один раз промывают SE/. К 100 мкг/мл добавляют
протеиназу К, затем 1 мл 20% SDS и препарат инкубируют 4 ч.
Проводят непрерывную экстракцию фенолхлороформом до полной депротонизации препарата. Для осаждения ДНК к водной фазе прибавляют 1/30 об. 0,
3 M NaAc-1 об. изобропанола, ДНК подцепляют крючком,
промывают 70% Et OH и вновь суспендируют в SE.
Аликвоты ДНК по 10 мкг обрабатывают приемлемым рестрикционным ферментом /напр. EcoRI/ и подвергают электрофорезу на 0,8% гелях. Такие гели анализируют блоттингом по Саузерну, гибридизацию проводят по существу так, как было описано.
Положительно гибридизированные животные, т.е. животные, имеющие ген слияния /предположительно расположенный в хромосомной области/, оставляют расти до зрелого возраста. Женские особи спаривают и после начала лактации их молоко анализируют на содержание представляющего интерес вещества /см. пример 7/. Положительно гибридизированные особи спаривают и родившихся от них дочерей отбирают на предмет присутствия экзогенных ДНК-последовательностей, а их молоко затем анализируют на присутствие представляющего интерес вещества.
Пример 2
Воспроизводят методику примера 1 за тем исключением, что
ДНК-последовательность, кодирующая представляющий интерес
полипептид /TARG-последовательность/, кодирует альфа1 антитрипсин /получена от R. Cortese EMБL, Майерхофштрассе 1, D-6900
Гейдельберг, ФРГ/. Фрагмент TagI-bstIII 1294 п.о. [51] отрезают,
выделяют из таких клонов и клонируют в Pvu II сайт p SSItgSE способами, описанными в примере 1.
Пример 3
Воспроизведена методика примера 1 за исключением того, что вместо
плазмиды pPoIy используют плазмиду pVC18 /Pharmacia Ltd. Pharmacia House, Мидсаммер Бульвар, Милтон Кейнес, Англия/.
Пример 4
Воспроизведена методика примера 1 за исключением
того, что вместо плазмиды pPoIy используют плазмиду pVC19 /Pharmacia Ltd. Pharmacia House, Мидсаммер Бульвар, Милтон Кейнс,
Англия/.
Пример 5
Создание трансгенной овцы
Фрагмент Sal_ XbaI, вырезанный из плазмиды pSSItgXS /обозначаемая также как pSSItgXS, см. пример 1/, инъецируют в яйца
овцы. Инъецируют примерно 200 копий на оплодотворенное яйцо. Из 252
одноклеточных яиц, инъецированных и пересаженных овцам-реципиентам, родилось 52 живых ягненка. На основе анализа ДНК,
приготовленной из образца крови, установлено, что четыре из числа этих животных
являлись носителями экзогенных последовательностей бета-лактоглобулина-фактора IX /табл.3/. Все яйца инъецируют на
стадии проядра и не подвергают центрифугированию.
Число копий определяют денситометрией с количественным сканированием.
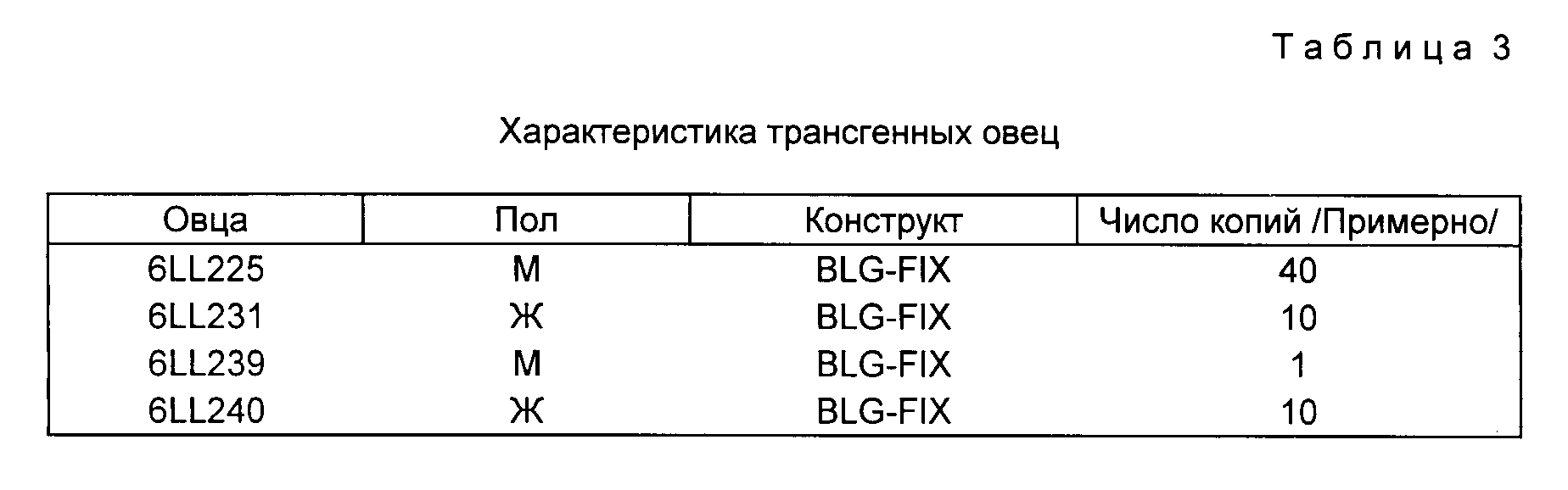
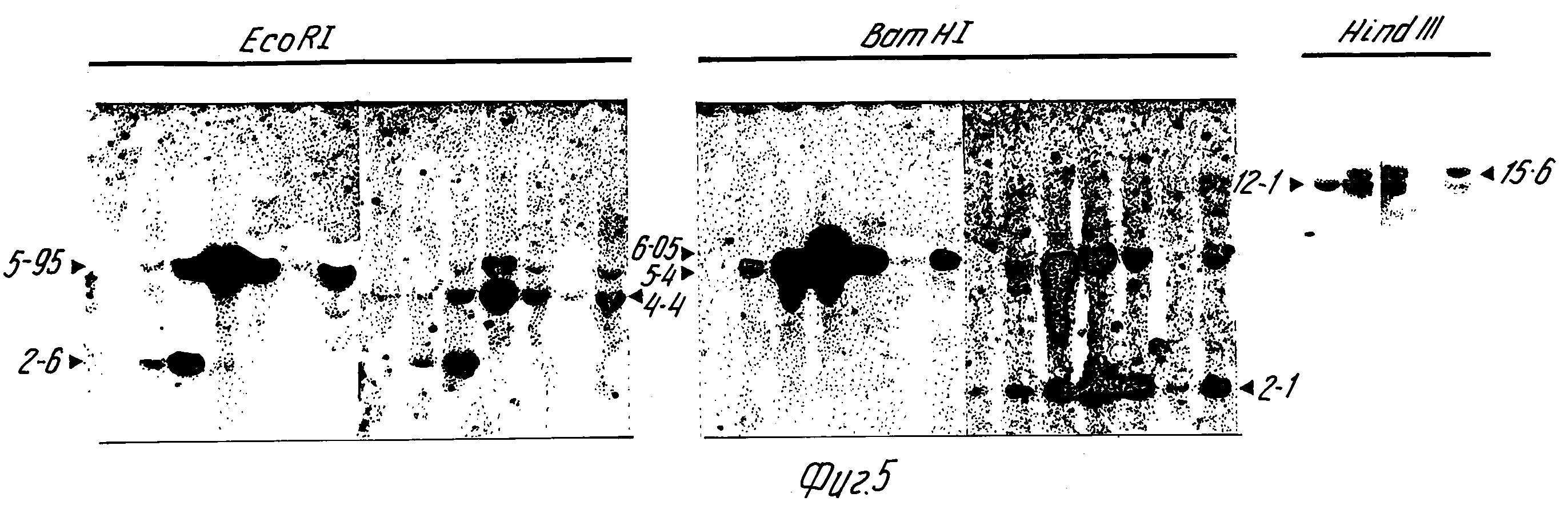
На фиг. 5 приведены результаты блоттинг-анализа по Саузерну. ДНК овцы получают, как описано, обрабатывают указанным рестрикционным ферментом, подвергают электрофорезу на 0,8% агарозном геле и переносят на мембраны Hybond /Amersham International, Литтл Чалфонт, Бакс, Великобритания/. Фильтр зондируют меченной32P плазмидой, pSSItgXS-FIX /полосы 1-7/ и затем /после отделения от мембраны/ /плазмидой p931 /полосы 1' 7'/. Контроль копий pSSItgXS-FIX проведен на геле там, где указано. Полоса 1 контрольные ДНК овцы /нетрансгенной/; полосы 2 и 3 контрольная ДНК плюс 1 и 5 копия эквивалентов p5 C3 C VI: Полосы 4-7 ДНК трансгенных овец 6LL 225, 6LL 231, 6LL 239 и 6LL 240. Каждая трансгенная овца дает полосы 5,95 к.b /EcoRI/ и 6,05 к.b. /BamHI/, идентичные по размерам полосам, образовавшимся из pSSItgXS-FIX, что указывает на интактность 5'-концов трансгенов. Значительной гибридизиции в случае фактора IX овцы не наблюдалось. Гибридизация с p931 выявила фрагменты 4,4 к.b /EcoRI/ и 2,1 к.b /BamHI/. Хотя гены бета-лактоглобулина эндогенной овцы вносят вклад в гибридизацию этих полос, повышенная интенсивность в образцах от 6LL 225, 6LL 231, 6LL 240 указывает на происхождение этих полос, в первую очередь, из инъецированного гена слияния бета-глобулин-фактор IX, что подтверждает неприкосновенность 3'-концов конструкта. ДНК, расщепленная HindIII-полоса 1, 1, 12,1 к, b, плазмиды pSSItgX-FIX; полосы 2-5 ДНК от овец 6LL 225, 6LL 231, 6LL 239 и 6LL 240. Зондом служит p5'G3'GYI. Гибридизованный фрагмент HindIII в 12,1 к. b. /6LL 225, 6LL 231 и 6LL 240/, идентичный по размерам инъецированному фрагменту, указывает на расположение голова к хвосту, фрагмент в 15,6 к. b. общий для той же овцы, указывает на присутствие также повторов с расположением голова к голове. Эти данные указывают, что у овец 6LL 225, 6LL 231 и 6LL 240 фрагмент pSSItgX-FIX, образовавшийся из pSSItgX-FIX /ген слияния бета-лактоглобулин-фактор IX/, интегрирован без обнаруживаемой перегруппировки расположения тандема. Для овцы 6II239 данные сопоставимы с интегрированием единичной неперегруппированной копии этого фрагмента.
Пример 6
Передача последовательностей фактора IX следующим поколениям.
Трансгенного барана 6LL 225 примера 5 /несущего примерно 40 копий фрагмента SaII XbaI, приготовленного из pSSItgXS-FIX/ успешно спаривают с рядом овцематок Финн-Дорсетской и Восточно Фризояндской пород. ДНК из образцов крови его потомства подвергают блоттинг-анализу по Саузарену с применением плазмидного зонда p5'G3'GYI фактора IX человека /см. выше/. Результаты для отдельных представителей этого потомства приведены на фиг. 6. Полосы под NN 1, 3, 4, 5, 6, 7, 9, 11, 43, 45, 48 и 49 ДНК от потомства барана 6LL 225 /обозначены как 7R1, 7R3 7R49/, полоса с 211 ДНК от контрольной /нетрансгенной/ овцы 6LL 211, полосы 5-211 и 1-211 ДНК от овцы 6LL 211 плюс 5 и 1 копии эквивалентов pSSIIgXS-FIX. Полученные данные показывают, что 6 и 12 представителей потомства трансгенного барана 6LL 225 унаследовали последовательности фактора IX, т.е. то, что эти последовательности оказались включенными в зародышевую линию.
Пример 7
Экспрессия
гена, кодирующего бета-лактоглобулин овцы, в трансгенных мышах.
Выращивают трансгенных мышей [52] Получено несколько трансгенных мышей, несущих фрагмент SaII клона лямбда I /фиг. 3/. Одна из этих мышей /самка B-Lac 7/, как было показано, является носительницей 15-20 копий фрагмента SaII. Мышь B-Lac 7 многократно спаривалась и произвела многочисленное потомство, унаследовавшее SS-I последовательности.
Через 8-12 дн после рождения приплода у мыши забирают молоко. Для этого проводят внутрибрюшинную инъекцию 0,31 Е окситоцина /Sigma/ и 7 мкл/г зверька Гипнорм-Гипновеля [53] после предварительного удаления мышат на 1 ч, выжидания в течение 20 мин и затем миссирования вручную каждой отдельной молочной железы. Молоко отбирают в капиллярную трубочку на 50 мкл.
Мышиное молоко разбавляют дистиллированной водой /1:5/, центрифугируют в течение короткого промежутка на ступенчатой центрифуге для обезжиривания, казеины осаждают добавлением I н HCl до окончательного pH 4,6. После центрифугирования на ступенчатой центрифуге удаляют белки сыворотки, осаждают 5% трихлоруксусной кислотой и анализируют с помощью электрофореза на полиакриламидном геле согласно LaemmIi [54] На фиг. 7 приведен НДС ПАГЭ анализ белков сыворотки молока мыши и овцы. Полоса 1 белки-метки; 2 - нормальный белок мыши; 3 белок овцы; 4 белок нормальной мыши; 5 белок мыши B-Lac7; 6 белок B-Lac7 /2,5 х 5/. Антисыворотка, выработанная у кроликов к бета-лактоглобулину овцы, использована для обнаружения бета-лактоглобулина овцы вестерн-блоттингом [55] образцов, разделенных электрофорезом на геле. На фиг. 8 приведены результаты вестерн-блоттинг анализа. Пятна подвергают реакции с анти-бета-лактоглобулин сывороткой кролика и анти-I -пероксидаза сывороткой кролика. /Полоса 1 белки-метки; 2 белок овцы; 3 белок мыши B; 4 белок нормальной мыши; 5 очищенный бета-лактоглобулин; 6 окрашенный красителем Кумасси белок овцы /параллельный опыт/.
Результаты этого анализа показывают, что бета-лактоглобулин в больших количествах секретируется в молоко мыши, указывая на высокую степень экспрессии SS-I у мыши B-Lac7. Такой клон предположительно содержит все необходимые последовательности, гарантирующие высокий уровень экспрессии в молочной железе трансгенной мыши, и таким образом можно ожидать, что этот клон будет действовать столь же эффективно, если не еще лучше, в гомологичных видах, т.е. в трансгенной овце. Следовательно, образовавшийся из такого клона ген слияния будет экспрессироваться /успешно/ в молочной железе овцы.
Пример 8
Экспрессия фактора IX человека в молоке
трансгенной овцематки
Две овцематки /6LL 231 и 6LL 240/, каждая является носителем примерно 10
копий фрагмента SaII-XbaI, полученного из pSSItg XS-FIX/, успешно спаривают с баранами Восточно
Фриэляндской породы. После рождения оставляют сосать естественным путем примерно на две недели с целью
стимулирования лактации. В стерильные пластиковые сосуды вручную собирают молоко /примерно по 25
мл от каждой овцы/. Образцы молока замораживают до -20oC и отсылают в Национальную службу
переливания крови Шотландии при Королевской Лечебнице, где проводят радиоиммунологические испытания
/РИИ/ на фактор IX человека.
Образцы трансгенного и контрольного молока диализуют по дистиллированной воде при 4oC в течение суток и затем сушат вымораживанием. Высушенный вымораживанием образец вновь суспендируют в дистиллированной воде и затем центрифугируют и исследуют на присутствие фактора IX человека /РИИ/ следующим образом. С использованием смешанной плазмы нормального человека, разбавленной РИИ-буфером /50 мм Трис-HCl, 0,25% желатина, 1% Твин-20, 10 мм HCl, pH 7,2/, получают стандартную кривую при разбавлениях 1/10-1/1280 /фиг.9/. Для того чтобы выявить влияние на испытание молока, одновременно нормальную смешанную плазму разбавляют в контрольном молоке и получают стандартную кривую. Испытаниям подвергают разбавленные образцы высушенного вымораживанием молока овец 6LL 231 и 6LL 240. Содержимое каждой пробирки для РИИ следующее: 50 мкл РИИ-буфера, 50 мкл разбавленного образца, 50 мкл поликлональных антител анти-фактора IX кролика с разбавлением 1/30000 и 50 мкл1125меченного фактора IX. Был установлен контроль для максимального связывания, содержащий 100 мкл РИИ-буфера, 50 мкл антител и 50 мкл1125фактора IX. Установлен контроль для неспецифичевкого связывания, содержащий 150 мкл РИИ-буфера, 50 мкл антител,1125фактор IX-следы.
После инкубирования в течение суток 50 мл Сефарозы-SI000, сопряженные со вспомогательным анти-IgG кролика, смешивают в пробирках и сепарацией с сахарозой получают шарики. РИИ чувствителен в пределах примерно 0,125 международных единиц (iu)/dI.
В РИИ откладывают связывания по отношению к обратному фактору разбавления для
следующих образцов:
1. Нормальная смешанная плазма
2. Нормальная смешанная
плазма плюс молоко
3. Высушенные вымораживанием образцы молока овцы 6LL 231 (TI)
4.
Высушенные вымораживанием образцы молока овцы 6LL 240 (T2).
Полученные результаты приведены на фиг. 9.
В молоке обеих овец 6LL 231 (TI) и 6II240 (T2) обнаруживаются уровни фактора IX соответственно при 2,5 iw/dI и 8 iu/dI. Никакой активности не обнаруживается в контрольном молоке на уровне чувствительности испытания. Полученные данные показывают, что трансгенные овцематки, являющиеся носителем гена слияния бета-лактоглобулин-фактор IX (особенно фрагмент SaII XbaI, образовавшийся из pSSItgXS-FIX (также часто обозначаемый как pSSItgXS -TARG), экспрессируют этот ген и секретируют белок человека в молоко. Тем самым создается основа для получения подобными способами белка человека.
СПИСОК ЛИТЕРАТУРЫ
1. Brinster et al. Proc. Acad.
Sci, 82/1985/4438-4442.
2. Nature, vol. 315, 20.06.85, R. E. Hammer et. al. "Production of trasgenic rablits, sheep. and pigs by microinfection" pp. 680-683 (прототип).
3. Laceу et. al. Cell, 34 (1983), 343-346.
4. Brinster et. al. Cell, 27 (1981), 223-231.
5. Swift et. al. Cell, 38 (1984), 639-646.
6. Shani, Nature, 314 (1985), 283-286.
7. Magram et. al. Nature, 315 (1985), 338-340.
8. Krumlauf. et. al. mol. Cell Biol. 5 (1985), 1639-1698.
9. De Slipio and Davie, Biochemistry 18 (1979), 899-904.
10. de le Jalle et. al. Nature 316 (1985) 268-270.
11. Topper and Treeman, Physiol Rev. (1980), 1049-1106.
12. Биохимия лектации, под ред. Mepham, Elsevier (1982), 309-349.
13. I. Daicy Sci 64 (1981), 1038-1054.
14. Brew, Nature, 223 (1969), 671-672.
15. Mercier and Gaye "Биохимия лактации"./ Под ред. Mepham, Elsevier (1983), 177-225.
16. Rosen et. al. Biochem. Soc. Trans. 9 (1982), 112.
17. Gross Claude, Proc. 16th, 2n tl. Cons, Animal. Blood Groups Biohem. Polymorphy, 1 (1979), 54-59.
18. Qasba u Jafaya, Nature, 311 (1984), 377-580.
19. Narhasi and Qasba, I, Biol. Chem. 254 (1979 6016-6025).
20. Shuster et al. Eur. I. Biochem. 71 (1976), 193-199.
21. Panley et al. Nature, 275 (1978), 455-457.
22. Merciev and Cuyelle et. al. Cell 17 (1979) 1013 1073.
23. Mercies et al. Biochimie 67, (1985), 959-971.
24. Gaye et al. Biochimie 64 (1982), 173-184.
25. Teyss ot and Houde fine, Eur. I, Biochem, 110 (1980) 263-272.
26. Mc Knight and Kinsgs bury, Science 217 (1982) 316-324.
27. Payvar et al. Cell 35 (1983), 381-392.
28. Renka witz et al. Cell, 37 (1984), 503-510.
29. Korin et Nature, 308 (1984), 513-578.
30. Charuay et al. Cell 38 (1984) 251-263.
31.Gillies et al. Cell 33 (1983), 717-718.
32.Молекулярное клонирование Гоулд Сиринг Нарбор (1982).
33. Методы энзимологии, т. 68, 100 и 101 (Wr. под ред. Crossman Moldave), Academic Press.
34. Burch and Weintraub, Cell 33, (1983) 65.
35. Trischauf et al. J Hcol. Biol. 170, (1983), 827.
36. Science, 196, 180 (1977).
37. Rigby et. al. J Hcol. 113 (1977), 237.
38. Mescier et. al. Biochimie 67 (1985), 959, 971.
39. Borg and et al. Mol. Con. Genetics, 146 (1976), 199-207.
40. Southern J. Mol. Biol. 98/1975, 503.
41. Sanger et al. PNAS, 74 (1977), 5463.
42. Hanahan and Mesel Son Gene 10 (1980), 63.
43. Birnboim and Doty, Huc. Acid. Res. 7 (1979), 1513.
44. Twig et al. Hature 283 (1980), 216-218.
45. Anson et al. Embo J. 3 (1984), 1053-1060.
46. Maniatis et. al. Молекулярное клонирование, 1982, Колд.
47. Sch leicher anol Schull, Pestfach, 4, Д-335 4, Дассель, ДЕ.
48. Hunter et. al. J Agric Sci, 46/1955,143-149.
49. White et al. Vet Rec. 115 (1986), 140-143.
50. Ciliberto et. al. Cell 41 (1985), 531-540.
51. Gordon and Ruddle. Методы энзимологии, т. 101, 1983, под. ред. Grossman and Moldave, Academic Press с. 411-432.
52. Flecrnoil. Vet. Rec. 10.12.83, p. 574.
53. Nature 277 (1970), 680-684.
54. Burnett Anal. Biochem. 112 (1981), 195-203.
Реферат
Использование: биотехнология, медицина. Сущность изобретения: конструируют рекомбинантную плазмиду psstgxs-Targ, содержащую последовательность ДНК, которая кодирует овечий β - лактоглобулин, слитый либо с фактором IX, либо с a1 - антитрипсином, из полученной плазмиды изолируют фрагмент, включающий по крайней мере регуляторную и секреторную сигнальную последовательности гена β\ -лактоглобулина, слитую с последовательностью гена целевого белка, вводят его в оплодотворенную яйцеклетку или эмбрион овцы, трансплантируют полученную яйцеклетку или эмбрион в реципиентных овцематок и затем отбирают трансгенных животных. 9 ил., 3 табл.

Комментарии