Не являющиеся человеком животные с модифицированными последовательностями тяжелых цепей иммуноглобулинов - RU2618886C2
Код документа: RU2618886C2
Чертежи





































































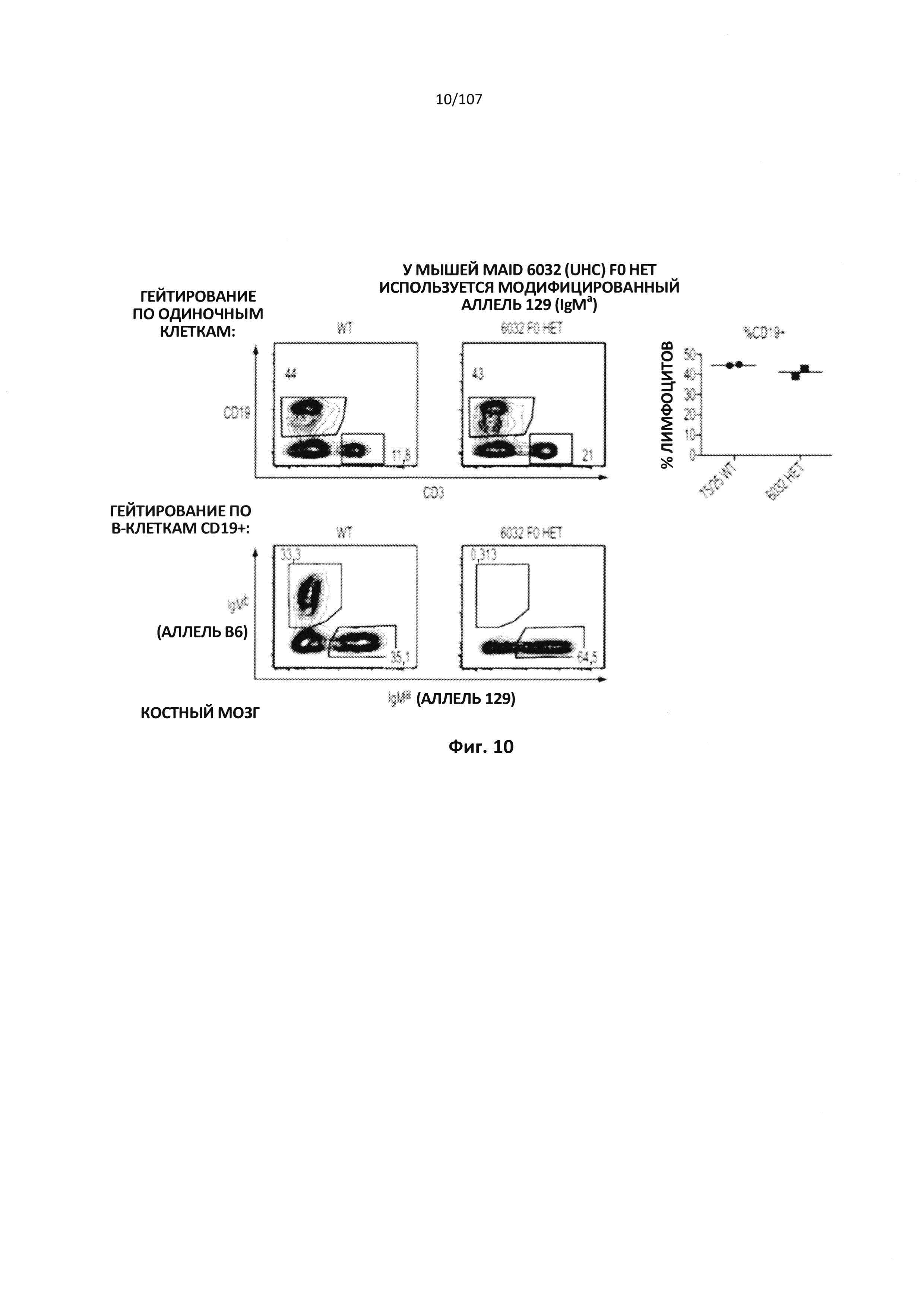


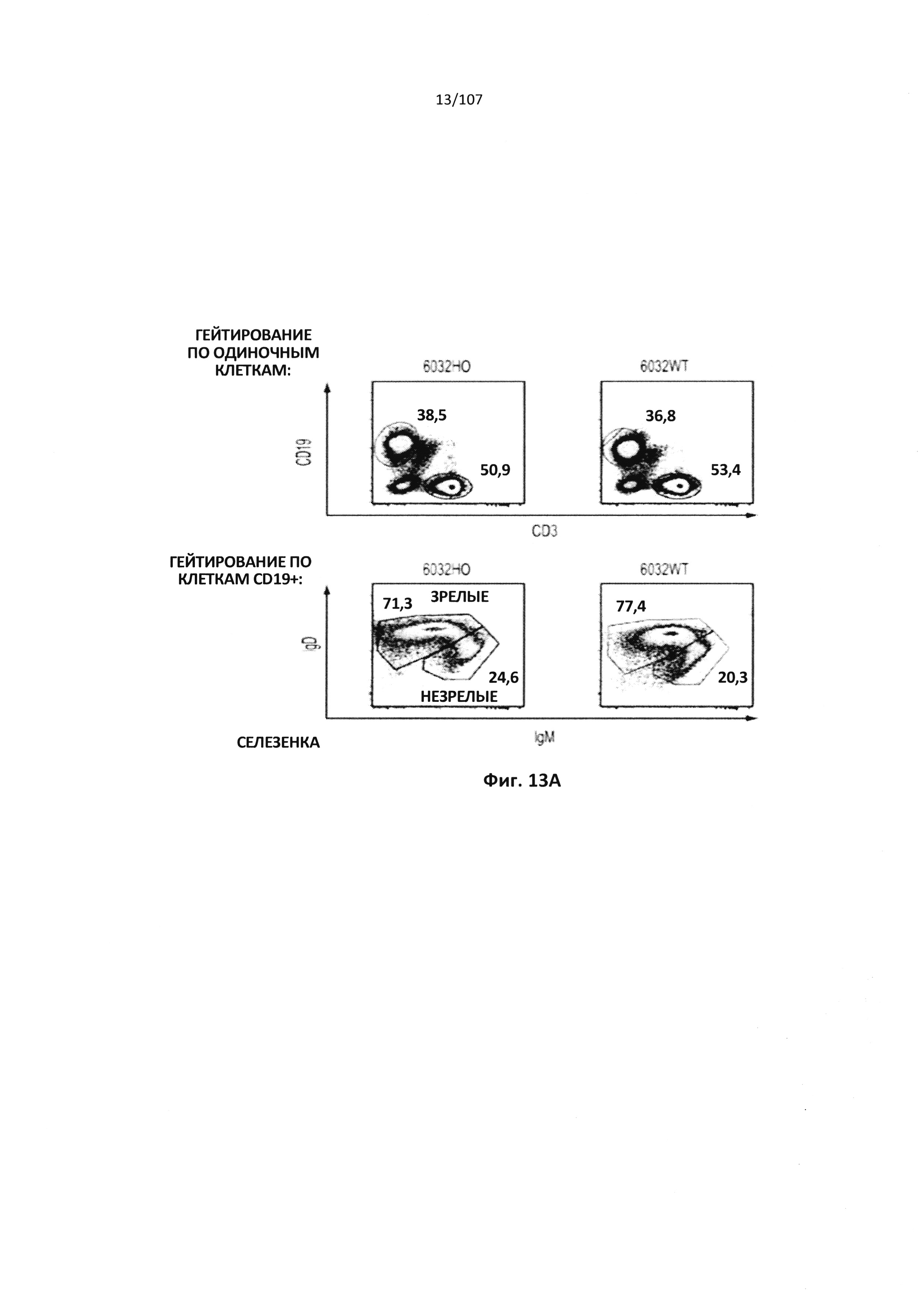

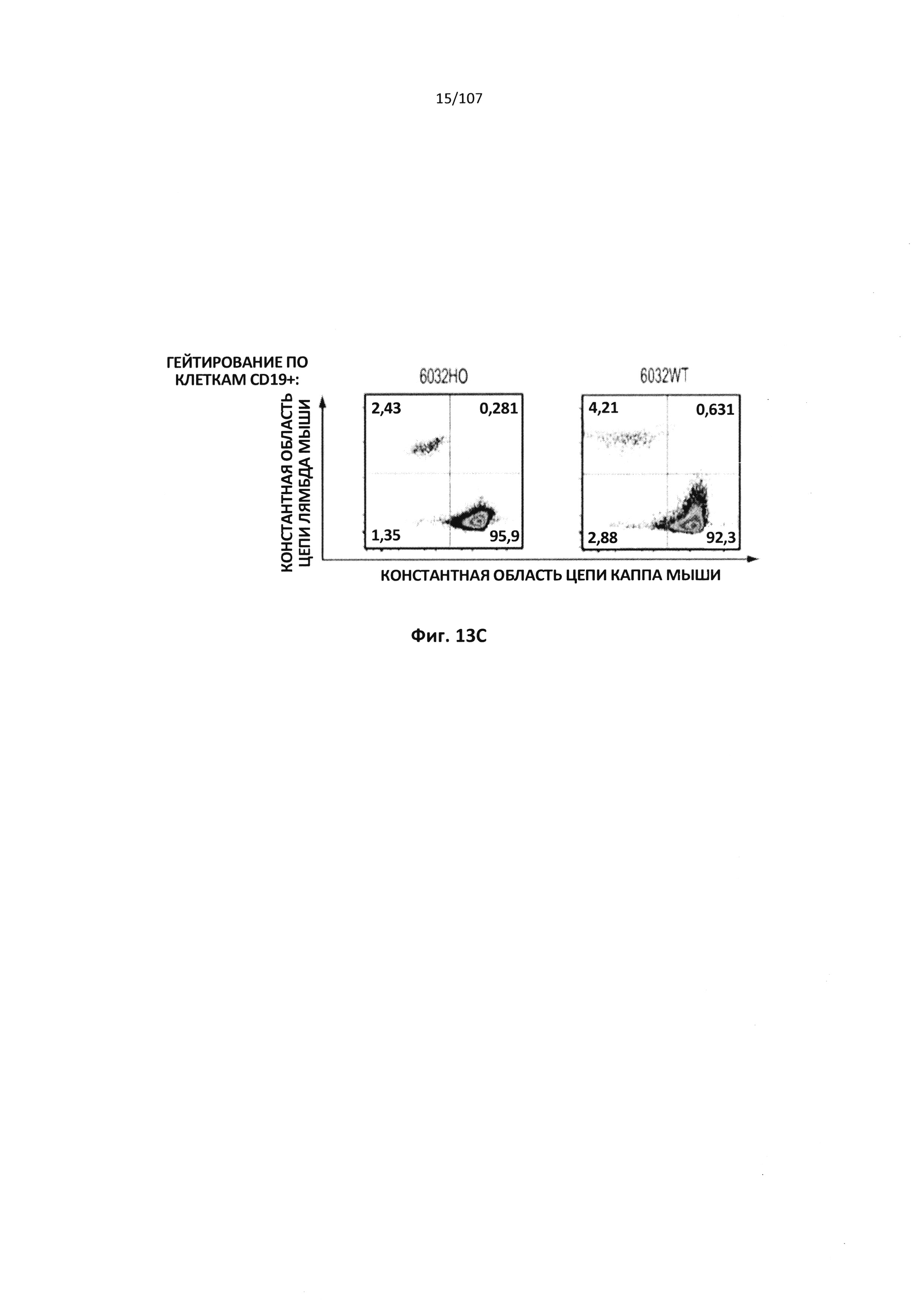
































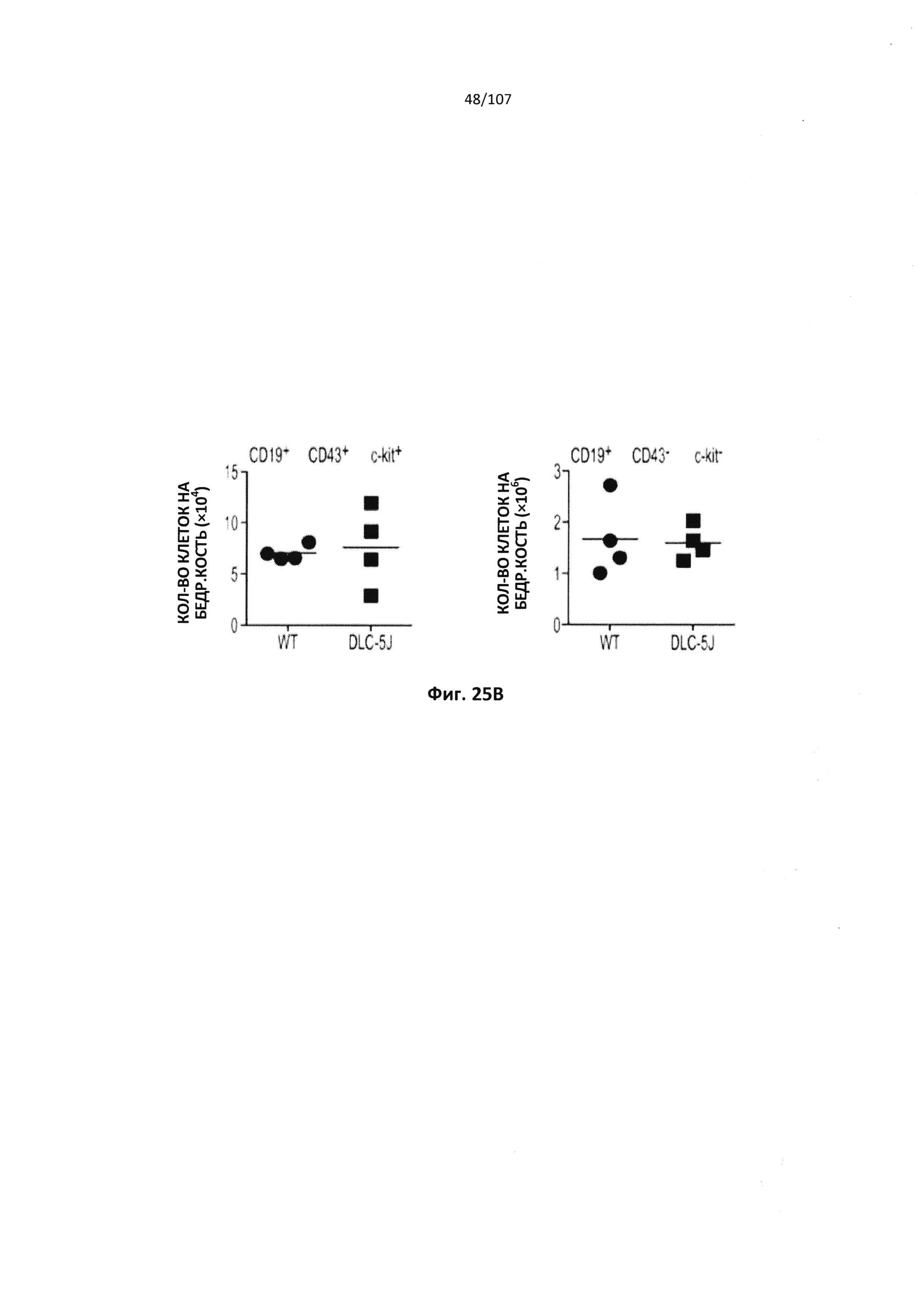


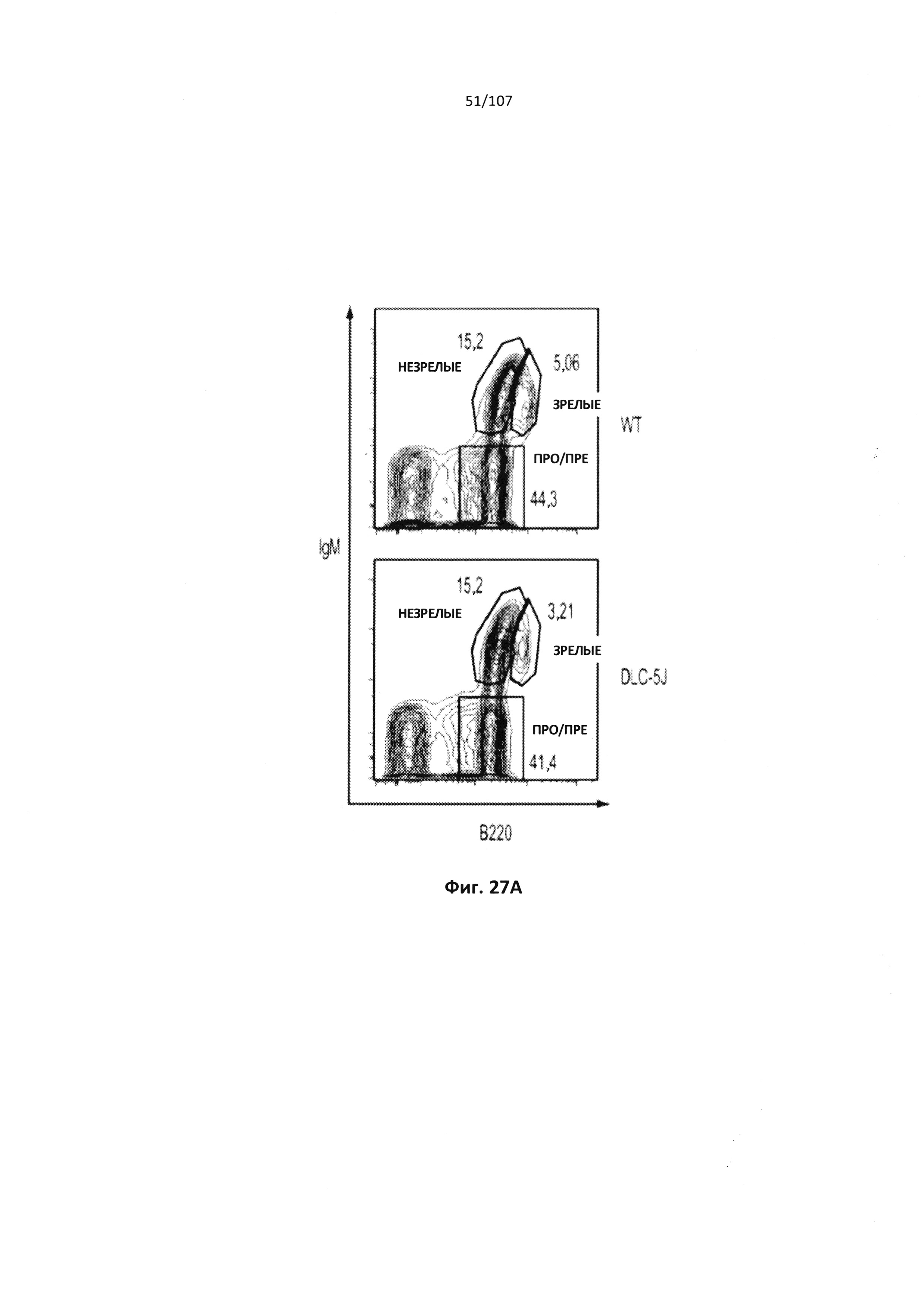

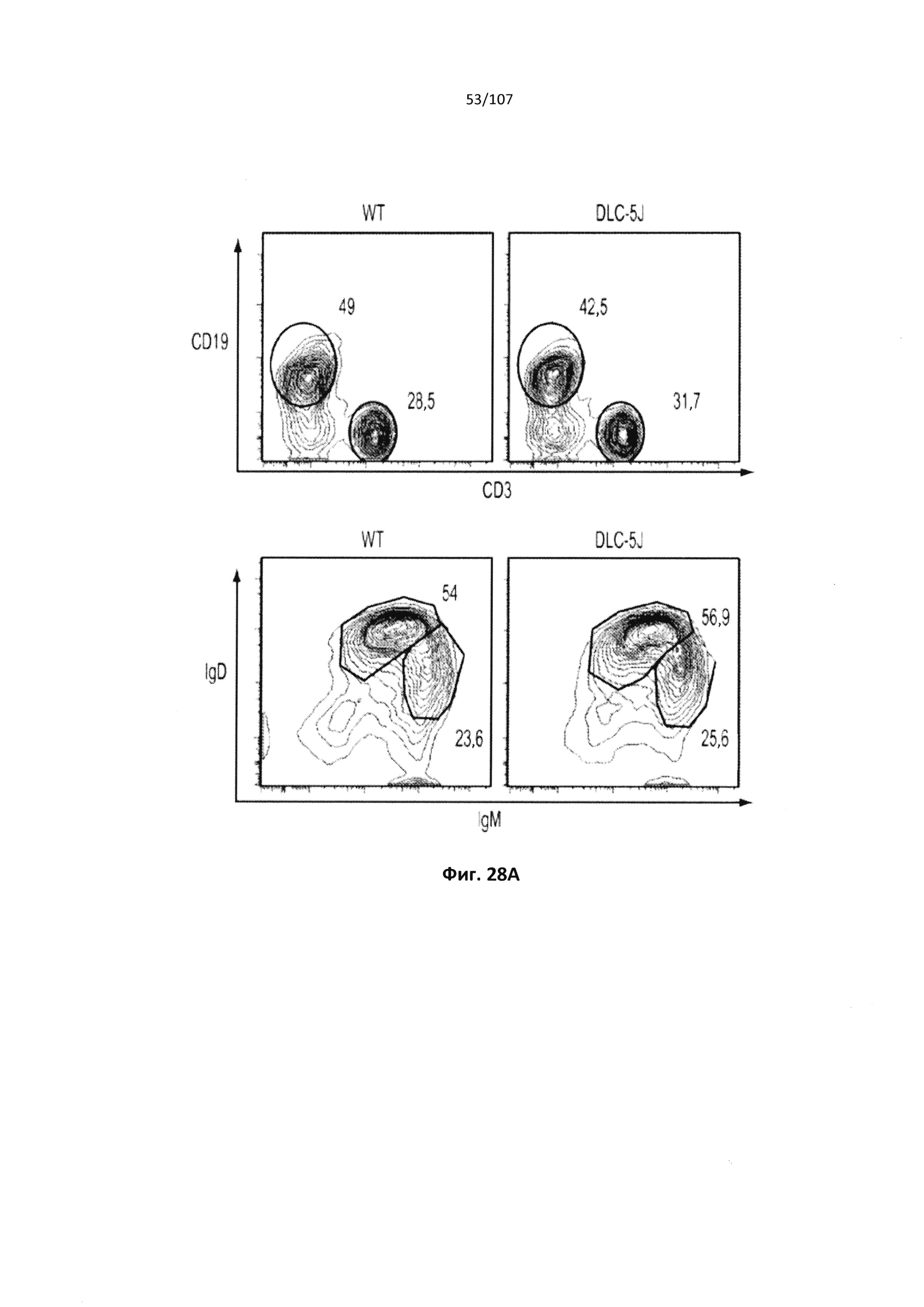



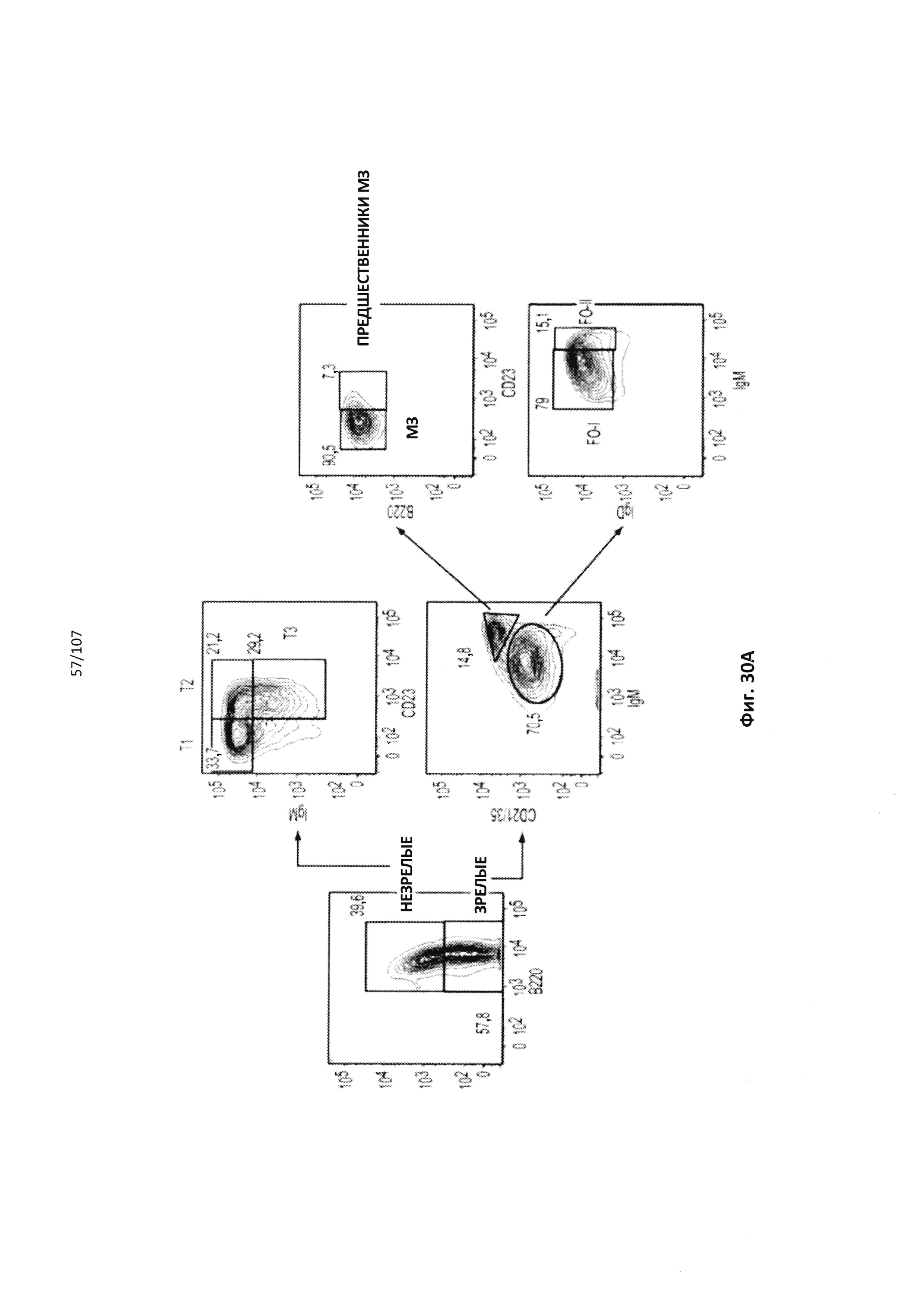

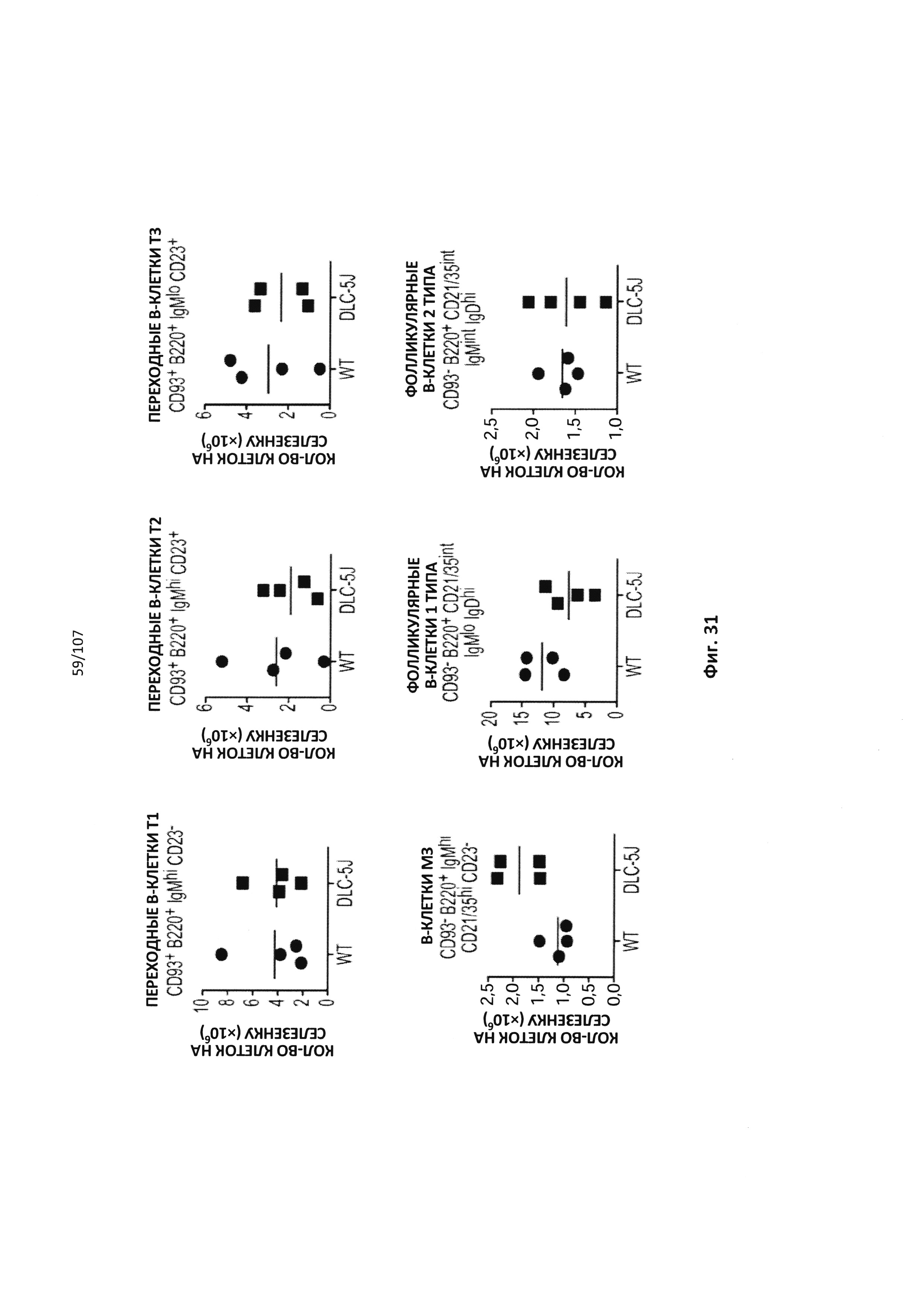










































Описание
С ПЕРЕКРЕСТНЫМИ ССЫЛКАМИ НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка испрашивает приоритет по предварительной заявке на патент США №61/766765, поданной 20 февраля 2013 г., и предварительной заявке на патент США №61/879338, поданной 18 сентября 2013 г., которые полностью включены в настоящий документ посредством ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Перечень последовательностей в виде текстового файла (озаглавленного «1270WO_SL.txt», созданный 20 февраля 2014 г. и имеющий размер 87000 байт, полностью включен в настоящий документ посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
[0003] Генетически модифицированные не являющиеся человеком животные, например, грызуны, такие как мыши и крысы, у которых имеется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи человека (т.е. реаранжированная последовательность VDJ тяжелой цепи), функционально связанная с нуклеотидной последовательностью константной области. Согласно некоторым вариантам реализации указанные животные генетически сконструированы таким образом, что у них имеется иммуноглобулиновый локус, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи (последовательность VDJ), функционально связанную с последовательностью гена константной области иммуноглобулина, при этом последовательность VDJ представляет собой последовательность VDJ человека, а последовательность гена константной области является человеческой или имеет нечеловеческое происхождение. Согласно некоторым вариантам реализации указанные не являющиеся человеком животные, у которых имеется генетически модифицированный иммуноглобулиновый локус, содержат: (1) первую нуклеотидную последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. первая нуклеотидная последовательность представляет собой реаранжированную нуклеотидную последовательность вариабельной области тяжелой цепи иммуноглобулина человека), при этом указанная первая нуклеотидная последовательность функционально связана с последовательностью гена константной области легкой цепи (например, легкой цепи κ или λ); и (2) вторую нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи человеческого происхождения или нечеловеческого происхождения (например, легкой цепи κ или λ) (т.е. вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области легкой цепи иммуноглобулина человека), при этом указанная вторая нуклеотидная последовательность функционально связана с последовательностью гена константной области тяжелой цепи. Согласно некоторым вариантам реализации указанные не являющиеся человеком животные содержат генетически модифицированный локус тяжелой цепи иммуноглобулина, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, при этом указанный реаранжированный вариабельный домен тяжелой цепи содержит последовательность сегмента V гена тяжелой цепи (VH), функционально связанную через спейсер с последовательностью сегмента J гена тяжелой цепи (JH), и при этом указанный спейсер содержит по меньшей мере один аминокислотный остаток. Предложены генетически модифицированные не являющиеся человеком животные, например, грызуны, такие как мыши и крысы, в геноме которых содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи человека, функционально связанная с нуклеотидной последовательностью константной области; и (ii) локус легкой цепи иммуноглобулина, содержащий один или большее количество, но меньшее, чем у дикого типа, сегментов гена вариабельной области легкой цепи человека. Предложены генетически модифицированные не являющиеся человеком животные, в геноме которых содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи человека, функционально связанная с нуклеотидной последовательностью константной области; и (ii) локус легкой цепи иммуноглобулина, содержащий один или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека. Согласно некоторым вариантам реализации по меньшей мере один из сегментов гена вариабельной области кодирует один или большее количество остатков гистидина, не кодируемых соответствующим сегментом гена вариабельной области легкой цепи зародышевой линии человека. Предложены способы получения описанных в настоящем документе генетически модифицированных не являющихся человеком животных. Предложены способы получения последовательностей вариабельной области легких цепей иммуноглобулина (например, легкой цепи κ или λ), которые способны связывать антиген в отсутствие тяжелой цепи, и/или могут быть связаны с реаранжированным вариабельным доменом тяжелой цепи, и/или проявляют рН-зависимые антигенсвязывающие свойства, подходящие для получения биспецифических антител.
УРОВЕНЬ ТЕХНИКИ
[0004] Биспецифические антитела представляют собой многофункциональные антитела, содержащие антигенсвязывающие сайты, которые могут связывать две отдельные антигенные детерминанты, возникшие в качестве одного из основных биологических терапевтических средств для лечения многих заболеваний, в том числе раковых. Несмотря на то, что за последнее время были разработаны разнообразные биспецифические антитела с двойными антигенсвязывающими свойствами, специфичностью и аффинностью вариабельных доменов легких цепей или тяжелых цепей иммуноглобулина в обычных биспецифических антителах приходится в определенной степени жертвовать, поскольку в обычных биспецифических антителах в связывание с каждой антигенной детерминантой вносит вклад вариабельный домен либо только тяжелой цепи, либо только легкой цепи, тогда как в случае обычных антител в связывании одной и той же антигенной детерминанты могут участвовать вариабельные области как легких, так и тяжелых цепей. Кроме того, для достижения требуемого уровня эффективности терапевтических антител, например, биспецифических терапевтических антител, часто требуются высокие или множественные дозы антител ввиду их ограниченной циркуляцией in vivo.
[0005] Большинство разработанных на сегодняшний день антигенсвязывающих белков, нацеленных на два антигена или эпитопа, содержат два антигенсвязывающих фрагмента: (i) первый антигенсвязывающий фрагмент, содержащий пару вариабельный домен тяжелой цепи-вариабельный домен легкой цепи иммуноглобулина, участвующую в связывании с первым антигеном или эпитопом; и (ii) второй антигенсвязывающий фрагмент, содержащий вторую пару вариабельный домен тяжелой цепи-вариабельный домен легкой цепи, участвующую в связывании со вторым антигеном или эпитопом. Указанные антигенсвязывающие белки, несмотря на то, что они являются биспецифическими в контексте полного антигенсвязывающего белка, не обязательно являются биспецифическими в каждом антигенсвязывающем фрагменте, что ограничивает применение указанных антигенсвязывающих белков в мультиспецифическом формате, например, триспецифических антигенсвязывающих белков. Согласно настоящему описанию не являющееся человеком животное, у которого экспрессируется универсальный вариабельный домен тяжелой цепи, может быть использовано в качестве общего инструмента для получения антигенсвязывающих белков для применения в антигенсвязывающих белках разнообразных форматов.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0006] В данной области техники существует потребность в получении последовательностей вариабельных доменов легких цепей иммуноглобулинов, отличающихся тем, что специфичность и аффинность в отношении антигена происходит полностью или в основном, и/или основана полностью или в основном на разнообразии вариабельных доменов легких цепей иммуноглобулинов. Такие последовательности были бы чрезвычайно полезны при разработке антигенсвязывающих белков, например, биспецифических антител, в которых каждый вариабельный домен по отдельности отвечает за специфическое связывание отдельного антигена. Различные аспекты и варианты реализации, описанные в настоящем документе, основаны отчасти на неожиданном открытии того, что генетически модифицированные не являющиеся человеком животные, у которых имеются вариабельные домены тяжелых цепей иммуноглобулинов, кодируемые последовательностью гена реаранжированной вариабельной области тяжелой цепи (например, реаранжированной последовательностью VDJ тяжелой цепи), могут удовлетворять указанную потребность. У не являющихся человеком животных, кодирующих реаранжированный вариабельный домен тяжелой цепи иммуноглобулина (т.е. универсальный вариабельный домен тяжелой цепи) механизмы обеспечения разнообразия антител основаны в основном на нереаранжированных (т.е. диверсифицируемых) вариабельных доменах легких цепей антител. Не являющиеся человеком животные включают, например, млекопитающих и, согласно конкретным вариантам реализации, грызунов (например, мышей, крыс или хомяков).
[0007] Предложены генетически модифицированные не являющиеся человеком животные, которые при стимуляции представляющим интерес антигеном продуцируют антитела, обладающие антигенсвязывающей специфичностью, основанной полностью или в основном на вариабельных доменах легких цепей антител. Могут быть идентифицированы аминокислоты вариабельных доменов легких цепей антител и соответствующие нуклеотидные последовательности антител, продуцируемых такими генетически модифицированными животными, и указанные последовательности могут использоваться в рекомбинантных антителах или других антигенсвязывающих белках для разработки вариабельных доменов легких цепей, связывающих антигенную детерминанту независимо (и при этом с достаточной специфичностью и аффинностью) от вариабельных доменов тяжелых цепей. Кроме того, полезные свойства генетически модифицированных животных с реаранжированным вариабельным доменом тяжелой цепи (т.е. содержащих последовательность гена реаранжированного вариабельного домена тяжелой цепи) могут применяться путем введения нуклеотидной последовательности, кодирующей реаранжированный вариабельный домен тяжелой цепи в разнообразные геномные контексты, например, в различные иммуноглобулиновые локусы. Последовательности генов реаранжированных вариабельных доменов тяжелых цепей могут быть нацелены в локус тяжелой цепи или локус легкой цепи таким образом, что реаранжированные последовательности вариабельных доменов тяжелых цепей могут быть функционально связаны с последовательностью константной области тяжелой или легкой цепи, человеческого происхождения или нечеловеческого происхождения. Последовательности генов реаранжированных вариабельных доменов тяжелых цепей могут быть размещены в любом месте генома, при этом они функционально связаны человеческими, имеющими нечеловеческое происхождение или смешанными человеческими/имеющими нечеловеческое происхождение последовательностями константных областей иммуноглобулинов. Кроме того, не являющиеся человеком животные, у которых имеется нуклеотидная последовательность, кодирующая реаранжированный вариабельный домен тяжелой цепи могут быть скомбинированы с дополнительными генетическими модификациями иммуноглобулиновых локусов (например, могут скрещиваться с животными, у которых имеется дополнительные генетические модификации иммуноглобулиновых локусов). Например, направленное разнообразие, обеспечиваемое реаранжированной последовательностью гена вариабельного домена тяжелой цепи, нацеленного на локус легкой цепи, может сочетаться с последовательностью гена вариабельного домена легкой цепи, встроенного в локус тяжелой цепи, с получением таким образом животных, у которых в полной мере используются временные характеристики и разнообразие выбранного геномного контекста (например, механизмы, обеспечивающие разнообразие локуса тяжелой цепи) для увеличения разнообразия последовательности выбранного гена вариабельной области антитела (например, последовательностей гена вариабельной области легкой цепи антитела). Кроме того, за счет использования мышей, имеющих ограниченный (лимитированный) репертуар сегментов гена вариабельной области легкой цепи (например, ограниченное число последовательностей гена вариабельной области легкой цепи, которые содержат один или более, но меньше, чем у дикого типа, сегментов гена VL человека в комбинации с единственной реаранжированной последовательностью тяжелой цепи, описанной выше), можно получить вариабельный домен легкой цепи иммуноглобулина, способный более эффективно сочетаться с вариабельным доменом тяжелой цепи иммуноглобулина.
[0008] Соответственно, предложены генетически модифицированные не являющиеся человеком животные (например, грызуны такие как мыши, крысы или хомяки), у которых имеется иммуноглобулиновый локус, содержащий реаранжированную вариабельную область тяжелой цепи иммуноглобулина человека (т.е. нуклеотидную последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи; т.е. реаранжированная последовательность VDJ тяжелой цепи).
[0009] Согласно различным аспектам единственная геномная нуклеотидная последовательность, кодирующая вариабельный домен тяжелой цепи, экспрессируемые генетически модифицированными не являющимися человеком животными, представлена реаранжированный вариабельным доменом тяжелой цепи. Соответственно, разнообразие вариабельных доменов тяжелых цепей антител, продуцированных генетически модифицированными не являющимися человеком животными, крайне ограничено.
[00010] Согласно некоторым вариантам реализации предложены генетически модифицированные не являющиеся человеком животные, в геноме которых имеется иммуноглобулиновый локус, который был генетически модифицирован таким образом, что последовательности его вариабельных областей состоят по существу из единственной реаранжированной вариабельной области тяжелой цепи человека. Понятно, что разные клетки таких генетически модифицированных не являющихся человеком животных не обязательно всегда содержат полностью идентичные последовательности в указанной единственной реаранжированной вариабельной области тяжелой цепи человека (например, за счет ошибок репликации, соматических гипермутаций или других механизмов), но в любом случае у таких генетически модифицированных не являющихся человеком животных наблюдается резкое ограничение разнообразия вариабельных доменов тяжелых цепей антител по сравнению с животными, у которых имеются нереаранжированные вариабельные последовательности тяжелых цепей, и/или животными, в геномах которых содержится несколько сегментов гена вариабельной области тяжелых цепей (например, несколько сегментов V, D и/или J, в частности, нереаранжированных).
[00011] Согласно различным аспектам предложен генетически модифицированный локус тяжелой цепи иммуноглобулина, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (т.е. содержащий нуклеотидную последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи). Согласно различным аспектам нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека и реаранжированный вариабельный домен тяжелой цепи, который она кодирует, происходят из сегмента V, D и J гена человека. Согласно различным аспектам нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека и реаранжированный вариабельный домен тяжелой цепи, который она кодирует, происходят из гена VH человека и сегмента JH человека. Согласно различным аспектам нуклеотидная последовательность реаранжированной вариабельной области тяжелой. цепи иммуноглобулина человека функционально связана с последовательностью гена константной области тяжелой цепи. Согласно различным аспектам нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области легкой цепи. Согласно различным аспектам генетически модифицированный иммуноглобулиновый локус присутствует в зародышевой линии не являющегося человеком животного. Согласно различным аспектам у указанных генетически модифицированных не являющихся человеком животных имеется полный набор сегментов гена нереаранжированной вариабельной области легкой цепи, способных к реаранжировке с образованием гена легкой цепи, функционально связанного с последовательностью гена константной области легкой цепи. Согласно другим аспектам у указанных генетически модифицированных не являющихся человеком животных имеется совокупность, но не полный набор (т.е. меньшее количество, чем у животных дикого типа) сегментов гена нереаранжированной вариабельной области легкой цепи. Согласно различным аспектам сегменты гена неаранжированной вариабельной области легкой цепи функционально связаны с последовательностью гена константной области тяжелой цепи. Согласно конкретным вариантам реализации указанное не являющееся человеком животное представляет собой грызуна, например, мышь, крысу или хомяка. Согласно другому аспекту предложена содержащая нуклеиновые кислоты конструкция, содержащая реаранжированную вариабельную область тяжелой цепи иммуноглобулина человека (т.е. содержащая нуклеотидную последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи; т.е. предварительно реаранжированную последовательность VDJ тяжелой цепи) согласно описанию в настоящем документе.
[00012] В настоящем документы описаны разнообразные варианты генетически модифицированных не являющихся человеком животных, у которых имеется иммуноглобулиновый локус, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (т.е. иммуноглобулиновый локус, содержащий нуклеотидную последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи). Каждый вариант способен обеспечивать механизмы разнообразия антител за счет нуклеотидных последовательностей вариабельных областей легких цепей иммуноглобулина.
[00013] Согласно различным аспектам нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. вариабельный домен тяжелой цепи, кодируемый нуклеотидной последовательностью реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), функционально связана с человеческой или имеющей нечеловеческое происхождение последовательностью гена константной области тяжелой цепи (например, последовательность гена константной области тяжелой цепи, которая кодирует изотип иммуноглобулина, выбранный из IgM, IgD, IgA, IgE, IgG и их комбинаций). Например, предложены генетически модифицированные не являющиеся человеком животные, у которых имеются иммуноглобулиновые локусы, в которых: (а) первая нуклеотидная последовательность кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. первая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), при этом указанная первая нуклеотидная последовательность функционально связана с человеческой или имеющей нечеловеческое происхождение последовательностью (или комбинированной последовательностью человека/не человека) гена константной области тяжелой цепи; и (b) вторая нуклеотидная последовательность кодирует вариабельный домен легкой цепи (т.е. вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека), при этом указанная вторая нуклеотидная последовательность функционально связана с человеческой или имеющей нечеловеческое происхождение последовательностью гена константной области легкой цепи.
[00014] Согласно другому аспекту предложены модифицированные не являющиеся человеком животные, отличающиеся тем, что у них имеется реаранжированная нуклеотидная последовательность, которая кодирует вариабельный домен тяжелой цепи, при этом указанный вариабельный домен тяжелой цепи содержит последовательность вариабельной области тяжелой цепи (VH), функционально связанную через спейсер с последовательностью сегмента J тяжелой цепи (JH), причем указанный спейсер кодирует по меньшей мере один аминокислотный остаток.
[00015] Согласно другому аспекту предложено не являющееся человеком животное, у которого имеется генетически модифицированный иммуноглобулиновый локус, который содержит нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (т.е. имеется нуклеотидная последовательность, кодирующая реаранжированный вариабельный домен тяжелой цепи; т.е. реаранжированную последовательность VDJ тяжелой цепи), при этом указанный генетически модифицированный иммуноглобулиновый локус присутствует в зародышевой линии не являющегося человеком животного. Согласно некоторым вариантам реализации указанный генетически модифицированный иммуноглобулиновый локус представляет собой локус тяжелой цепи. Согласно некоторым вариантам реализации указанный генетически модифицированный иммуноглобулиновый локус представляет собой локус легкой цепи
[00016] Согласно другому аспекту предложены генетически модифицированные не являющиеся человеком животные (например, грызуны, такие как мыши, крысы или хомяки), у которых имеется генетически модифицированный иммуноглобулиновый геномный локус с нуклеотидной последовательностью реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, при этом указанная нуклеотидная последовательность функционально связана с человеческой или имеющей нечеловеческое происхождение последовательностью гена константной области легкой цепи (например, легкой цепи κ или λ).
[00017] Согласно другому аспекту предложено не являющееся человеком животное, у которого имеется генетически модифицированный иммуноглобулиновый локус, содержащий: (а) нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанную с последовательностью гена константной области легкой цепи; и (b) нереаранжированную человеческую или имеющую нечеловеческое происхождение нуклеотидную последовательность вариабельной области легкой цепи (например, легкой цепи κ или λ), функционально связанную с человеческой или имеющей нечеловеческое происхождение последовательностью гена константной области тяжелой цепи (например, последовательностью гена константной области тяжелой цепи, которая кодирует изотип иммуноглобулина, выбранный из IgM, IgD, IgA, IgE, IgG и их комбинации).
[00018] Согласно другому аспекту предложено генетически модифицированное не являющееся человеком животное с иммуноглобулиновым локусом, содержащим:
(а) первый аллель, содержащий:
(i) первую нуклеотидную последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. первая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), функционально связанную с последовательностью гена константной области тяжелой цепи, и
(ii) вторую нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи (т.е. вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека), функционально связанную с последовательностью гена константной области легкой цепи; и
(b) второй аллель, содержащий
(i) третью нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи (т.е. третья нуклеотидная последовательность представляет собой нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека), функционально связанную с последовательностью гена константной области тяжелой цепи, и
(ii) четвертую нуклеотидную последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. четвертая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), функционально связанную с последовательностью гена константной области легкой цепи.
[00019] Согласно различным аспектам предложены генетически модифицированные не являющиеся человеком животные с локусами или последовательностями гена неаранжированной вариабельной области тяжелой цепи. Согласно некоторым вариантам реализации у указанных генетически модифицированных не являющихся человеком животных имеется соответствующее дикому типу число сегментов (т.е. все или по существу все сегменты) гена вариабельной области легкой цепи иммуноглобулина человека (т.е. последовательностей). Согласно другим аспектам у не являющихся человеком животных, описанных в настоящем документе, имеется ограниченный набор сегментов гена вариабельной области легкой цепи, например, (i) один, два или более, но меньше, чем у дикого типа, сегментов гена VL человека; и (ii) один или большее количество сегментов гена JL человека, функционально связанных с имеющей нечеловеческое происхождение нуклеотидной последовательностью константной области легкой цепи. Нуклеотидная последовательность тяжелой цепи и/или сегменты легких цепей могут содержаться, например, в трансгене или в эндогенном иммуноглобулиновом локусе.
[00020] Согласно различным аспектам предложены генетически модифицированные не являющиеся человеком животные, отличающиеся тем, что все вариабельные домены тяжелых цепей иммуноглобулина у указанного животного происходят из одной и той же последовательности гена реаранжированной вариабельной области тяжелой цепи, и тем, что указанные вариабельные домены экспрессируются совместно с вариабельным доменом легкой цепи, происходящим из одного из по меньшей мере одного, двух, трех или большего количества сегментов гена VL и по меньшей мере одного, двух, трех или большего количества сегментов гена JL. Дополнительно предложены генетически модифицированные не являющиеся человеком животные (например, грызуны, такие как мыши и крысы), в геноме которых содержатся: (i) локус тяжелой цепи иммуноглобулина, который содержит нуклеотидную последовательности реаранжированной вариабельной области тяжелой цепи человека, функционально связанную с человеческой или имеющей нечеловеческое происхождение нуклеотидной последовательностью константной области тяжелой цепи; и (ii) локус легкой цепи иммуноглобулина, содержащий один или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека (например, два сегмента гена Vκ человека и один или большее количество сегментов гена Jκ человека), функционально связанных с человеческой или имеющей нечеловеческое происхождение нуклеотидной последовательностью константной области легкой цепи. Согласно некоторым вариантам реализации указанная константная область легкой цепи представляет собой константную область крысы или мыши, например, константную область Сκ крысы или мыши.
[00021] Согласно другому аспекту генетически модифицированные не являющиеся человеком животные согласно описанию в настоящем документе при стимуляции представляющим интерес антигеном экспрессируют антигенсвязывающий белок, содержащий последовательность аминокислот тяжелой цепи и легкой цепи иммуноглобулина, при этом указанная последовательность аминокислот тяжелой цепи получена из генетически модифицированного локуса тяжелой цепи, содержащего нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи человека, функционально связанную с нуклеотидной последовательностью константной области тяжелой цепи. Согласно определенным аспектам последовательность аминокислот легкой цепи получена из генетически модифицированного локуса легкой цепи иммуноглобулина, содержащего один или более, но меньше, чем у дикого типа, сегментов гена VL человека и (ii) два или большее количество сегментов гена JL человека, функционально связанных с имеющей нечеловеческое происхождение нуклеотидной последовательностью константной области легкой цепи.
[00022] Предложены генетически модифицированные не являющиеся человеком животные, в геноме которых имеется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, которая содержит последовательность сегмента V гена тяжелой цепи (VH), функционально связанную через спейсер с последовательностью сегмента J гена тяжелой цепи (JH), причем указанный спейсер кодирует по меньшей мере одну аминокислоту (например, 2 аминокислоты, 3 аминокислоты или 4 аминокислоты), и/или модифицированный сегмент D гена. Согласно различным вариантам реализации нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи функционально связана с человеческой или имеющей нечеловеческое происхождение нуклеотидной последовательностью константной области тяжелой цепи. Согласно различным вариантам реализации в геномах указанных не являющихся человеком животных дополнительно содержится генетически модифицированный локус легкой цепи иммуноглобулина, содержащий один или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека, например, два сегмента гена Vκ человека и один или большее количество сегментов гена Jκ человека, функционально связанных с человеческой или имеющей нечеловеческое происхождение нуклеотидной последовательностью константной области легкой цепи.
[00023] Также предложены способы получения и применения генетически модифицированных не являющихся человеком животных, описанных в настоящем документе. Предложены способы обеспечения функциональной связи нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи человека с нуклеотидной последовательностью константной области тяжелой или легкой цепи иммуноглобулина в геноме не являющегося человеком животного.
[00024] Согласно другому аспекту предложены способы получения последовательности аминокислот вариабельной области легкой цепи иммуноглобулина (VL), способных к связыванию антигена независимо от последовательности аминокислот вариабельной области тяжелой цепи.
[00025] Согласно другому аспекту предложен генетически модифицированный иммуноглобулиновый локус, который может быть получен с помощью любого из способов согласно описанию в настоящем документе.
[00026] Согласно различным аспектам предложены антигенсвязывающие белки (например, антитела), продуцированные генетически модифицированными не являющимися человеком животными или происходящие из генетически модифицированных не являющихся человеком животных, описанных в настоящем документе. Также предложены способы получения антигенсвязывающих белков, в том числе мультиспецифических (например, биспецифических или триспецифических) антигенсвязывающих белков. Также предложены способы получения эффекторных вариабельных доменов легкой цепи иммуноглобулина.
[00027] Согласно другому аспекту предложены плюрипотентная клетка, индуцированные плюрипотентные или тотипотентные стволовые клетки, происходящие из не являющегося человеком животного, у которого имеются различные геномные модификации согласно настоящему описанию. Также предложены клетки, имеющие ядро, содержащее генетическую модификацию согласно описанию в настоящем документе, например, модификацию, введенную в клетку посредством пронуклеарной инъекции.
[00028] Согласно различным аспектам предложен эмбрион не являющегося человеком животного, содержащий клетку, геном которой содержит локус тяжелой цепи иммуноглобулина, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи человека, функционально связанную с нуклеотидной последовательностью константной области. Согласно определенным аспектам указанный эмбрион не являющегося человеком животного также содержит локус легкой цепи иммуноглобулина, содержащий два или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека, функционально связанных с нуклеотидной последовательностью константной области легкой цепи.
[00029] Также предложены способы получения нуклеотидных последовательностей, которые кодируют последовательность аминокислот вариабельной области легкой цепи иммуноглобулина (VL), способную к связыванию антигена или его эпитопа независимо от вариабельного домена тяжелой цепи, включающие: (а) иммунизацию не являющегося человеком животного представляющим интерес антигеном или его иммуногеном, при этом в геноме указанного не являющегося человеком животного содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области, и (ii) нуклеотидная последовательность реаранжированной вариабельной области легкой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью константной области легкой цепи; (b) обеспечение возможности развития иммунного ответа у указанного не являющегося человеком животного; (с) выделение из указанного иммунизированного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, способный связывать антиген; и (d) получение из указанной клетки нуклеотидной последовательности, которая кодирует указанный вариабельный домен легкой цепи (домен VL), способный связывать антиген. Согласно различным вариантам реализации указанная клетка представляет собой лимфоцит, в том числе, но не ограничиваясь перечисленными, естественные киллерные клетки, Т-клетки и В-клетки. Согласно различным вариантам реализации указанный способ дополнительно включает (с)' слияния указанного лимфоцита с раковыми клетками, например, клеткой миеломы.
[00030] Также предложены способы получения нуклеотидных последовательностей, которые кодируют последовательность аминокислот вариабельной области легкой цепи иммуноглобулина (VL), способную к связыванию антигена или его эпитопа независимо от вариабельного домена тяжелой цепи, включающие: (а) иммунизацию не являющегося человеком животного представляющим интерес антигеном или его иммуногеном, при этом в геноме указанного не являющегося человеком животного содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области, и (ii) два или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL), функционально связанных с нуклеотидной последовательностью константной области легкой цепи; (с) выделение из указанного иммунизированного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, способный связывать антиген; и (d) получение из указанной клетки нуклеотидной последовательности, которая кодирует указанный вариабельный домен легкой цепи (VL домен), способный связывать антиген. Согласно различным вариантам реализации указанная клетка представляет собой лимфоцит, в том числе, но не ограничиваясь перечисленными, естественные киллерные клетки, Т-клетки и В-клетки. Согласно различным вариантам реализации указанный способ дополнительно включает (с)' слияния указанного лимфоцита с раковыми клетками, например, клеткой миеломы.
[00031] Также предложены способы получения нуклеотидных последовательностей, которые кодируют последовательность аминокислот вариабельной области легкой цепи (VL) иммуноглобулина, способную к связыванию антигена или его эпитопа независимо от вариабельного домена тяжелой цепи, включающие: (а) иммунизацию не являющегося человеком животного представляющим интерес антигеном или его иммуногеном, при этом в геноме указанного не являющегося человеком животного содержатся: (i) нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью константной области легкой цепи, и (ii) сегменты гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL), функционально связанные с нуклеотидной последовательностью тяжелой цепи константной области; (с) выделение из указанного иммунизированного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, способный связывать антиген; и (d) получение из указанной клетки нуклеотидной последовательности, которая кодирует указанный вариабельный домен легкой цепи (VL домен), способный связывать антиген. Согласно различным вариантам реализации указанная клетка представляет собой лимфоцит, в том числе, но не ограничиваясь перечисленными, естественные киллерные клетки, Т-клетки и В-клетки. Согласно различным вариантам реализации указанный способ дополнительно включает (с)' слияния указанного лимфоцита с раковыми клетками, например, клеткой миеломы.
[00032] Также предложены способы получения антигенсвязывающих белков, включающие:
(a) иммунизацию не являющегося человеком животного первым антигеном, который содержит первый эпитоп или его иммуногенную часть, при этом в геноме указанного не являющегося человеком животного содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области, и (ii) нуклеотидная последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью константной области легкой цепи;
(b) обеспечение возможности развития иммунного ответа на первый эпитоп или его иммуногенную часть у указанного не являющегося человеком животного;
(c) выделение из указанного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(d) получение из указанной клетки по (с) нуклеотидной последовательности, которая кодирует вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(e) применение нуклеотидной последовательности по (с) в экспрессионной конструкции, соединенной с нуклеотидной последовательностью константной области иммуноглобулина человека; и
(f) экспрессию нуклеотидной последовательности по (с) в продукционной клеточной линии, экспрессирующей тяжелую цепь иммуноглобулина человека, которая специфически связывает второй антиген или его эпитоп, для получения легкой цепи антигенсвязывающего белка, кодируемой нуклеиновой кислотой по (с) и связывающей первый эпитоп или его иммуногенную часть независимо от тяжелой цепи, тяжелая цепь которого специфически связывает указанный второй антиген или эпитоп.
[00033] Также предложены способы получения антигенсвязывающих белков, включающие:
(a) иммунизацию не являющегося человеком животного первым антигеном, который содержит первый эпитоп или его иммуногенную часть, при этом в геноме указанного не являющегося человеком животного содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области, и (ii) два или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL), функционально связанных с нуклеотидной последовательностью константной области легкой цепи;
(b) обеспечение возможности развития иммунного ответа на первый эпитоп или его иммуногенную часть у указанного не являющегося человеком животного;
(c) выделение из указанного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(d) получение из указанной клетки по (с) нуклеотидной последовательности, которая кодирует вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(e) применение нуклеотидной последовательности по (с) в экспрессионной конструкции, соединенной с нуклеотидной последовательностью константной области иммуноглобулина человека; и
(f) экспрессию нуклеотидной последовательности по (с) в продукционной клеточной линии, экспрессирующей тяжелую цепь иммуноглобулина человека, которая специфически связывает второй антиген или его эпитоп для получения легкой цепи антигенсвязывающего белка, кодируемую нуклеиновой кислотой по (с) и связывающую первый эпитоп или его иммуногенную часть независимо от тяжелой цепи, тяжелая цепь которого специфически связывает указанный второй антиген или эпитоп.
[00034] Также предложены способы получения антигенсвязывающих белков, включающие:
(a) иммунизацию не являющегося человеком животного первым антигеном, который содержит первый эпитоп или его иммуногенную часть, при этом в геноме указанного не являющегося человеком животного содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью константной области легкой цепи, и (ii) сегменты гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL), функционально связанные с нуклеотидной последовательностью тяжелой цепи константной области;
(b) обеспечение возможности развития иммунного ответа на первый эпитоп или его иммуногенную часть у указанного не являющегося человеком животного;
(c) выделение из указанного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(d) получение из указанной клетки по (с) нуклеотидной последовательности, которая кодирует вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(e) применение нуклеотидной последовательности по (с) в экспрессионной конструкции, соединенной с нуклеотидной последовательностью константной области иммуноглобулина человека; и
(f) экспрессию нуклеотидной последовательности по (с) в продукционной клеточной линии, экспрессирующей тяжелую цепь иммуноглобулина человека, которая специфически связывает второй антиген или его эпитоп, для получения легкой цепи антигенсвязывающего белка, кодируемой нуклеиновой кислотой по (с) и связывающей первый эпитоп или его иммуногенную часть независимо от тяжелой цепи, тяжелая цепь которого специфически связывает указанный второй антиген или эпитоп.
[00035] Согласно различным аспектам предложено не являющееся человеком животное, в геноме зародышевой линии которого содержится локус тяжелой цепи иммуноглобулина, который содержит нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека. Согласно некоторым вариантам реализации указанное не являющееся человеком животное представляет собой млекопитающее. Согласно некоторым вариантам реализации указанное млекопитающее представляет собой грызуна. Согласно некоторым вариантам реализации указанный грызун выбран из группы, состоящей из мыши, крысы и хомяка. Согласно некоторым вариантам реализации указанное не являющееся человеком животное является гомозиготным по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с имеющей нечеловеческое происхождение последовательностью гена константной области тяжелой цепи. Согласно определенным вариантам реализации указанная имеющая нечеловеческое происхождение последовательность гена константной области тяжелой цепи кодирует Fc. Согласно конкретным вариантам реализации указанная имеющая нечеловеческое происхождение последовательность гена константной области тяжелой цепи представляет собой последовательность гена константной области тяжелой цепи мыши или крысы. Согласно некоторым вариантам реализации указанное не являющееся человеком животное представляет собой грызуна, и указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области тяжелой цепи человека. Согласно конкретным вариантам реализации указанную последовательность гена константной области тяжелой цепи выбирают из CH1, шарнира, CH2, CH3 и их комбинации. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека получена из сегмента VH гена тяжелой цепи человека, сегмента D гена тяжелой цепи человека и сегмента JH гена тяжелой цепи человека. Согласно определенным вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека получена из сегмента VH тяжелой цепи зародышевой линии человека, сегмента D тяжелой цепи зародышевой линии человека и сегмента JH тяжелой цепи зародышевой линии человека. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека кодирует последовательность VH3-23/GY/JH4-4 человека. Согласно некоторым вариантам реализации по существу все эндогенные функциональные сегменты генов VH, D и JH удаляют из локуса тяжелой цепи иммуноглобулина не являющегося человеком животного, или устраняют их функциональность. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного имеется модификация, которая обеспечивает удаление или устранение функциональности эндогенных функциональных сегментов гена VH, D и JH; и у указанного не являющегося человеком животного имеется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, при этом указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека присутствует эктопически. Согласно некоторым вариантам реализации вариабельный домен тяжелой цепи иммуноглобулина, кодируемый указанной нуклеотидной последовательностью реаранжированной вариабельной области тяжелой цепи, не является иммуногенным для указанного не являющегося человеком животного. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного имеется ген Adam6a, ген Adam6b или оба указанных гена. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного также имеется нуклеотидная последовательность, кодирующая сегмент гена (VL) нереаранжированной легкой цепи иммуноглобулина человека и сегмент J гена нереаранжированной легкой цепи иммуноглобулина человека. Согласно определенным вариантам реализации нуклеотидная последовательность, кодирующая сегмент V гена нереаранжированной легкой цепи (VL) и сегмент гена (JL) нереаранжированной легкой цепи, функционально связана с последовательностью гена константной области легкой цепи иммуноглобулина. Согласно конкретным вариантам реализации последовательность гена константной области легкой цепи выбирают из последовательности гена константной области грызуна и человека. Согласно более конкретным вариантам реализации грызуна выбирают из мыши, крысы и хомяка. Согласно определенным вариантам реализации сегмент гена (VL) нереаранжированной легкой цепи иммуноглобулина человека и сегмент гена (JL) нереаранжированного иммуноглобулина человека функционально связаны в эндогенном локусе грызуна с последовательностью гена константной области с иммуноглобулина грызуна. Согласно некоторым вариантам реализации локус тяжелой цепи иммуноглобулина содержит несколько копий нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека.
[00036] Согласно дополнительным аспектам предложен локус тяжелой цепи имеющего нечеловеческое происхождение иммуноглобулина в геноме имеющей нечеловеческое происхождение зародышевой клетки, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанную с последовательностью гена константной области тяжелой цепи, при этом указанная последовательность гена константной области содержит имеющую нечеловеческое происхождение последовательность, человеческую последовательность или их комбинацию. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с эндогенной последовательностью гена константной области имеющего нечеловеческое происхождение иммуноглобулина. Согласно определенным вариантам реализации указанная эндогенная последовательность гена константной области имеющего нечеловеческое происхождение иммуноглобулина представляет собой последовательность гена константной области тяжелой цепи мыши или крысы.
[00037] Согласно дополнительным аспектам предложены способы получения не являющегося человеком животного, включающие:
(a) модификацию генома не являющегося человеком животного с удалением или устранением функциональности эндогенных функциональных сегментов VH, D и JH гена тяжелых цепей иммуноглобулина; и
(b) введение в указанный геном нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека.
Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области тяжелой цепи имеющего нечеловеческое происхождение иммуноглобулина. Согласно определенным вариантам реализации указанная последовательность гена константной области тяжелой цепи имеющего нечеловеческое происхождение иммуноглобулина представляет собой последовательность гена константной области тяжелой цепи иммуноглобулина мыши или крысы. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека расположена в эндогенном локусе тяжелой цепи иммуноглобулина в геноме. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека находится в геноме зародышевой линии не являющегося человеком животного. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека находится в эктопическом локусе в геноме. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного имеется несколько копий нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного имеется ген Adam6a, ген Adam6b или оба указанных гена. Согласно некоторым вариантам реализации указанное не являющееся человеком животное представляет собой грызуна, выбранного из группы, состоящей из мыши, крысы или хомяка. Согласно некоторым вариантам реализации предложено не являющееся человеком животное, гетерозиготное по локусу тяжелой цепи иммуноглобулина согласно описанию в настоящем документе, отличающееся тем, что у указанного не являющегося человеком животного экспрессируется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека преимущественно из локуса тяжелой цепи иммуноглобулина.
[00038] Согласно дополнительным аспектам предложены не являющиеся человеком животные, у которых имеется генетически модифицированный иммуноглобулиновый локус, содержащий:
(a) нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанную с последовательностью гена константной области легкой цепи; и
(b) нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека, функционально связанную с последовательностью гена константной области тяжелой цепи.
Согласно некоторым вариантам реализации указанное не являющееся человеком животное представляет собой млекопитающее. Согласно конкретным вариантам реализации указанное млекопитающее выбирают из группы, состоящей из мыши, крысы и хомяка. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области легкой цепи κ. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области легкой цепи λ. Согласно некоторым вариантам реализации указанная последовательность гена константной области легкой цепи представляет собой последовательность гена константной области легкой цепи мыши или крысы. Согласно некоторым вариантам реализации указанная последовательность гена константной области легкой цепи представляет собой последовательность гена константной области легкой цепи человека. Согласно некоторым вариантам реализации указанная последовательность гена константной области тяжелой цепи представляет собой последовательность гена константной области тяжелой цепи мыши или крысы. Согласно некоторым вариантам реализации указанная последовательность гена константной области тяжелой цепи представляет собой последовательность гена константной области тяжелой цепи человека. Согласно некоторым вариантам реализации указанная последовательность гена константной области тяжелой цепи кодирует последовательность, выбранную из СН1, шарнира, СН2, СН3 и их комбинации. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека содержит последовательность гена к вариабельного домена легкой цепи человека. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека содержит последовательность гена вариабельного домена легкой цепи λ человека. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека получена из сегмента VH гена тяжелой цепи человека, сегмента D гена тяжелой цепи человека и сегмента JH гена тяжелой цепи человека. Согласно определенным вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека получена из сегмента VH тяжелой цепи зародышевой линии человека, сегмента D тяжелой цепи зародышевой линии человека и сегмента JH тяжелой цепи зародышевой линии человека. Согласно определенным вариантам реализации указанный сегмент гена VH человека выбирают из группы, состоящей из VH1-2, VH1-3, VH1-8, VH1-18, VH1-24, VH1-45, VH1-46, VH1-58, VH1-69, VH2-5, VH2-26, VH2-70, VH3-7, VH3-9, VH3-11, VH3-13, VH3-15, VH3-16, VH3-20, VH3-21, VH3-23, VH3-30, VH3-30-3, VH3-30-5, VH3-33, VH3-35, VH3-38, VH3-43, VH3-48, VH3-49, VH3-53, VH3-64, VH3-66, VH3-72, VH3-73, VH3-74, VH4-4, VH4-28, VH4-30-1, VH4-30-2, VH4-30-4, VH4-31, VH4-34, VH4-39, VH4-59, VH4-61, VH5-51, VH6-1, VH7-4-1, VH7-81 и их полиморфных вариантов. Согласно определенным вариантам реализации указанный сегмент D гена человека выбирают из группы, состоящей из D1-1, D1-7, D1-14, D1-20, D1-26, D2-2, D2-8, D2-15, D2-21, D3-3, D3-9, D3-10, D3-16, D3-22, D4-4, D4-11, D4-17, D4-23, D5-12, D5-5, D5-18, D5-24, D6-6, D6-13, D6-19, D6-25, D7-27 и их полиморфных вариантов. Согласно определенным вариантам реализации указанный сегмент гена JH человека выбирают из группы, состоящей из JH1, JH2, JH3, JH4, JH5, JH6 и их полиморфных вариантов. Согласно некоторым вариантам реализации, при этом указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека кодирует последовательность человека VH3-23/GY/JH4-4 (SEQ ID NO: 137). Согласно некоторым вариантам реализации указанный генетически модифицированный иммуноглобулиновый локус присутствует в зародышевой линии не являющегося человеком животного. Согласно некоторым вариантам реализации по существу все эндогенные функциональные сегменты генов VH, D и JH удаляют из локуса тяжелой цепи иммуноглобулина не являющегося человеком животного, или устраняют их функциональность. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного имеется модификация, которая обеспечивает удаление или устранение функциональности эндогенных функциональных сегментов гена VH, D и JH; и у указанного не являющегося человеком животного имеется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с последовательностью гена константной области легкой цепи, при этом указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека находится в эктопическом локусе в геноме. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного имеется ген Adam6a, ген Adam6b или оба указанных гена. Согласно некоторым вариантам реализации вариабельный домен тяжелой цепи, кодируемый нуклеотидной последовательностью реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, не является иммуногенным для указанного не являющегося человеком животного. Согласно некоторым вариантам реализации указанный генетически модифицированный иммуноглобулиновый локус содержит несколько копий нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанную с последовательностью гена константной области легкой цепи.
[00039] Согласно дополнительным аспектам предложен иммуноглобулиновый локус в геноме зародышевой линии не являющегося человеком животного, содержащий:
(1) нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанную с последовательностью гена константной области легкой цепи, и
(2) нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека, функционально связанную с последовательностью гена константной области тяжелой цепи.
Согласно некоторым вариантам реализации указанная последовательность гена константной области легкой цепи представляет собой последовательность гена константной области легкой цепи κ. Согласно некоторым вариантам реализации указанная последовательность гена константной области легкой цепи представляет собой последовательность гена константной области легкой цепи λ. Согласно некоторым вариантам реализации указанная последовательность гена константной области легкой цепи представляет собой последовательность гена константной области легкой цепи мыши или крысы.
[00040] Согласно дополнительным аспектам предложены способы получения не являющегося человеком животного, у которого имеется модифицированный иммуноглобулиновый локус, включающие:
(a) модификацию генома не являющегося человеком животного с удалением или устранением функциональности:
(i) эндогенных функциональных сегментов V, D и J гена тяжелых цепей иммуноглобулина, и
(ii) эндогенных функциональных сегментов V и J генов легкой цепи иммуноглобулина; и
(b) введение в указанный геном:
(i) нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанной с последовательностью гена константной области легкой цепи, и
(ii) нуклеотидной последовательности нереаранжированной вариабельной области легкой цепи иммуноглобулина человека, функционально связанной с последовательностью гена константной области тяжелой цепи.
Согласно некоторым вариантам реализации нуклеотидная последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека кодирует вариабельный домен легкой цепи κ. Согласно некоторым вариантам реализации нуклеотидная последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека кодирует вариабельный домен легкой цепи λ. Согласно некоторым вариантам реализации последовательность гена константной области тяжелой цепи представляет собой последовательность гена константной области тяжелой цепи имеющего нечеловеческое происхождение иммуноглобулина. Согласно определенным вариантам реализации последовательность гена константной области тяжелой цепи имеющего нечеловеческое происхождение иммуноглобулина представляет собой последовательность гена константной области тяжелой цепи мыши или крысы. Согласно конкретным вариантам реализации последовательность гена константной области тяжелой цепи кодирует последовательность, выбранную из СН1, шарнира, СН2, СН3 и их комбинации. Согласно некоторым вариантам реализации не являющееся человеком животное представляет собой грызуна, выбранного и'; группы, состоящей из мыши, крысы или хомяка. Согласно некоторым вариантам реализации модифицированный иммуноглобулиновый локус находится в геноме зародышевой линии не являющегося человеком животного. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного имеется ген Adam6a, ген Adam6b или оба указанных гена.
[00041] Также предложены не являющиеся человеком животные, у которых имеется модифицированный локус тяжелой цепи иммуноглобулина, который содержит нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, содержащую последовательность сегмента V тяжелой цепи (VH), функционально связанную через спейсер с последовательностью сегмента J тяжелой цепи (JH), причем указанный спейсер содержит по меньшей мере один аминокислотный остаток. Согласно некоторым вариантам реализации указанное не являющееся человеком животное представляет собой грызуна. Согласно некоторым вариантам реализации грызуна выбирают из группы, состоящей из мыши, крысы и хомяка. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с имеющей нечеловеческое происхождение последовательностью гена константной области тяжелой цепи. Согласно некоторым вариантам реализации указанная имеющая нечеловеческое происхождение последовательность гена константной области тяжелой цепи представляет собой последовательность гена константной области мыши или крысы. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области тяжелой цепи человека. Согласно определенным вариантам реализации указанная последовательность гена константной области тяжелой цепи кодирует последовательность, выбранную из СН1, шарнира, СН2, СН3 и их комбинации. Согласно некоторым вариантам реализации указанные последовательность VH и последовательность JH происходят из сегмента гена VH человека и сегмента гена JH человека. Согласно определенным вариантам реализации сегмент гена VH человека выбирают из группы, состоящей из VH1-2, VH1-3, VH1-8, VH1-18, VH1-24, VH1-45, VH1-46, VH1-58, VH1-69, VH2-5, VH2-26, VH2-70, VH3-7, VH3-9, VH3-11, VH3-13, VH3-15, VH3-16, VH3-20, VH3-21, VH3-23, VH3-30, VH3-30-3, VH3-30-5, VH3-33, VH3-35, VH3-38, VH3-43, VH3-48, VH3-49, VH3-53, VH3-64, VH3-66, VH3-72, VH3-73, VH3-74, VH4-4, VH4-28, VH4-30-1, VH4-30-2, VH4-30-4, VH4-31, VH4-34, VH4-39, VH4-59, VH4-61, VH5-51, VH6-1, VH7-4-1, VH7-81 и их полиморфных вариантов. Согласно конкретным вариантам реализации указанный сегмент гена VH человека представляет собой VH3-23 или его полиморфный вариант. Согласно некоторым вариантам реализации указанный спейсер кодирует последовательность, происходящую из сегмента D гена человека. Согласно конкретным вариантам реализации указанный сегмент D гена человека выбирают из группы, состоящей из D1-1, D1-7, D1-14, D1-20, D1-26, D2-2, D2-8, D2-15, D2-21, D3-3, D3-9, D3-10, D3-16, D3-22, D4-4, D4-11, D4-17, D4-23, D5-12, D5-5, D5-18, D5-24, D6-6, D6-13, D6-19, D6-25, D7-27 и их полиморфных вариантов. Согласно конкретным вариантам реализации указанный спейсер кодирует последовательность D4-4 или его полиморфного варианта. Согласно определенным вариантам реализации указанный сегмент JH гена человека выбирают из группы, состоящей из JH1, JH2, JH3, JH4, JH5, JH6 и их полиморфных вариантов. Согласно определенным вариантам реализации указанный сегмент JH человека представляет собой JH4-4 или его полиморфный вариант. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина кодирует последовательность человека VH3-23/GY/JH4-4 (SEQ ID NO: 137). Согласно некоторым вариантам реализации по существу все эндогенные функциональные сегменты генов VH, D и JH удаляют из локуса вариабельной области тяжелой цепи иммуноглобулина не являющегося человеком животного, или устраняют их функциональность. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного имеется модификация, которая обеспечивает удаление или устранение функциональности эндогенных функциональных сегментов гена VH, D и JH; и у указанного не являющегося человеком животного имеется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека в эктопическом локусе генома. Согласно некоторым вариантам реализации вариабельный домен тяжелой цепи, кодируемый нуклеотидной последовательностью реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, не является иммуногенным для указанного не являющегося человеком животного. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного имеется ген Adam6a, ген Adam6b или оба указанных гена.
[00042] Согласно дополнительным аспектам предложен локус тяжелой цепи иммуноглобулина в геноме зародышевой линии не являющегося человеком животного, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, содержащую сегмент вариабельной области тяжелой гена цепи (VH), который функционально связан через спейсер с сегментом J гена тяжелой цепи (JH), причем указанный спейсер кодирует по меньшей мере один аминокислотный остаток. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с имеющей нечеловеческое происхождение последовательностью гена константной области тяжелой цепи. Согласно определенным вариантам реализации указанная имеющая нечеловеческое происхождение последовательность гена константной области тяжелой цепи представляет собой последовательность гена константной области тяжелой цепи мыши или крысы. Согласно некоторым вариантам реализации указанный иммуноглобулиновый локус содержит несколько копий реаранжированной нуклеотидной последовательности вариабельной области тяжелой цепи.
[00043] Согласно дополнительным аспектам предложены способы получения не являющегося человеком животного, у которого имеется модифицированный локус тяжелой цепи иммуноглобулина, включающие:
(a) модификацию генома не являющегося человеком животного с удалением, или устранением функциональности эндогенных функциональных сегментов гена VH, D и JH тяжелых цепей иммуноглобулина; и
(b) введение в указанный геном нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, содержащей сегмент гена вариабельной области тяжелой цепи (VH), который функционально связан через спейсер с сегментом гена J тяжелой цепи (JH), причем указанный спейсер содержит по меньшей мере один аминокислотный остаток.
Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реанжированной тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области тяжелой цепи имеющего нечеловеческое происхождение иммуноглобулина. Согласно определенным вариантам реализации указанная последовательность гена константной области тяжелой цепи имеющего нечеловеческое происхождение иммуноглобулина представляет собой последовательность гена константной области тяжелой цепи мыши или крысы. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека расположена в эндогенном локусе тяжелой цепи иммуноглобулина в геноме. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека находится в эктопическом локусе в геноме. Согласно некоторым вариантам реализации модифицированный локус тяжелой цепи содержит несколько копий нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека. Согласно некоторым вариантам реализации указанное не являющееся человеком животное представляет собой грызуна, выбранного из группы, состоящей из мыши, крысы или хомяка. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного имеется ген Adam6a, ген Adam6b или оба указанных гена. Согласно некоторым вариантам реализации модифицированный локус тяжелой цепи иммуноглобулина находится в геноме зародышевой линии не являющегося человеком животного.
[00044] Согласно дополнительным аспектам предложено модифицированное не являющееся человеком животное, в геноме которого содержатся:
(a) локус тяжелой цепи иммуноглобулина, который содержит нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи человека, функционально связанную с нуклеотидной последовательностью тяжелой цепи константной области; и
(b) локус легкой цепи иммуноглобулина, который содержит один или более, но меньше, чем у дикого типа, сегментов VL и JL гена легкой цепи иммуноглобулина человека, функционально связанных с нуклеотидной последовательностью константной области легкой цепи.
Согласно некоторым вариантам реализации указанное не являющееся человеком животное представляет собой млекопитающее. Согласно определенным вариантам реализации указанное млекопитающее представляет собой грызуна. Согласно конкретным вариантам реализации грызуна выбирают из группы, состоящей из мыши, крысы и хомяка. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи представляет собой последовательность константной области грызуна, которая кодирует иммуноглобулин изотипа, выбранного из IgM, IgD, IgG, IgE, IgA и их комбинации. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области легкой цепи представляет собой нуклеотидная последовательность константной области κ или λ грызуна. Согласно некоторым вариантам реализации указанную нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи человека выбирают из сегмента VH зародышевой линии человека, сегмента D зародышевой линии человека и сегмента JH зародышевой линии человека. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи человека функционально связана с нуклеотидной последовательностью константной области в эндогенном локусе. Согласно некоторым вариантам реализации один или более, но меньше, чем у дикого типа, сегментов VL и JL гена легкой цепи иммуноглобулина человека функционально связаны с нуклеотидной последовательностью константной области легкой цепи в эндогенном локусе. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного имеется не более чем два нереаранжированных сегмента гена VL иммуноглобулина человека, и один, два, три или большее количество человека нереаранжированных сегментов гена JL. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи получена из сегмента гена VH зародышевой линии человека, выбранного из группы, состоящей из VH1-2, VH1-3, VH1-8, VH1-18. VH1-24, VH1-45, VH1-46, VH1-58, VH1-69, VH2-5, VH2-26, VH2-70, VH3-7, VH3-9, VH3-11, VH3-13, VH3-15, VH3-16, VH3-20, VH3-21, VH3-23, VH3-30, VH3-30-3, VH3-30-5, VH3-33, VH3-35, VH3-38, VH3-43, VH3-48, VH3-49, VH3-53, VH3-64, VH3-66, VH3-72, VH3-73, VH3-74, VH4-4, VH4-28, VH4-30-1, VH4-30-2, VH4-30-4, VH4-31, VH4-34, VH4-39, VH4-59, VH4-61, VH5-51, VH6-1, VH7-4-l, VH7-81 и их полиморфных вариантов. Согласно определенным вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи получена из сегмента гена VH3-23 зародышевой линии человека. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи кодирует последовательность человека VH3-23/GY/JH4-4 (SEQ ID NO: 137). Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи кодирует последовательность VH3-23/X1X2/J, где X1 представляет собой любую аминокислоту, и Х2 представляет собой любую аминокислоту. Согласно определенным вариантам реализации X1 представляет собой Gly и Х2 представляет собой Tyr. Согласно некоторым вариантам реализации указанный локус тяжелой цепи иммуноглобулина содержит функциональный ген Adam6a, ген Adam6b или оба указанных гена. Согласно определенным вариантам реализации указанные ген Adam6a, ген Adam6b или оба указанных гена представляют собой эндогенные гены Adam6. Согласно некоторым вариантам реализации указанное генетически модифицированное не являющееся человеком животное содержит ген Adam6a, ген Adam6b или оба указанных гена в эктопическом локусе генома. Согласно некоторым вариантам реализации указанные сегменты генов вариабельной области VL и JL человека способны к реаранжировке и кодированию вариабельного домена легкой цепи иммуноглобулина человека. Согласно некоторым вариантам реализации указанный локус легкой цепи иммуноглобулина содержит два сегмента гена VL, Vκ1-39 и Vκ3-20 человека. Согласно определенным вариантам реализации указанные сегменты генов представляют собой сегменты генов зародышевой линии. Согласно определенным вариантам реализации у указанного не являющегося человеком животного имеются сегменты генов Jκ1, Jκ2, Jκ3, Jκ4, и Jκ5. Согласно некоторым вариантам реализации два или большее количество, три или большее количество, четыре или большее количество, или пять или большее количество сегментов гена VL человека и два или большее количество сегментов гена JL человека присутствуют в эндогенном локусе легкой цепи. Согласно некоторым вариантам реализации по меньшей мере один из сегментов гена легкой цепи VL или JL человека кодируют один или большее количество гистидиновых кодонов, не кодируемых соответствующим сегментом гена вариабельной области легкой цепи зародышевой линии человека. Согласно некоторым вариантам реализации по меньшей мере один из сегментов гена VL содержит добавление гистидинового кодона или замену гистидиновым кодоном по меньшей мере одного негистидинового кодона, кодируемого соответствующей последовательностью сегмента VL зародышевой линии человека гистидиновым кодоном. Согласно определенным вариантам реализации указанный добавленный или заменяющий гистидиновый кодон находится в CDR3. Согласно некоторым вариантам реализации указанные сегменты генов VL человека представляют собой сегменты генов Vκ1-39 и Vκ3-20 человека, и каждый из указанных сегментов гена Vκ1-39 и Vκ3-20 человека содержит замену по меньшей мере одного негистидинового кодона, кодируемого соответствующим сегментом гена VL зародышевой линии человека, гистидиновым кодоном. Согласно определенным вариантам реализации указанная замена представляет собой замену трех негистидиновых кодонов сегмента гена Vκ1-39 человека, при этом указанная замена предназначена для обеспечения экспрессии гистидинов в положениях 106, 108 и 111. Согласно конкретным вариантам реализации указанная замена представляет собой замену четырех негистидиновых кодонов сегмента гена Vκ1-39 человека, и указанная замена предназначена для обеспечения экспрессии гистидинов в положениях 105, 106, 108 и 111. Согласно конкретным вариантам реализации указанная замена представляет собой замену трех негистидиновых кодонов сегмента гена Vκ3-20 человека, и указанная замена предназначена для обеспечения экспрессии гистидинов в положениях 105, 106 и 109. Согласно конкретным вариантам реализации указанная замена представляет собой замену четырех негистидиновых кодонов сегмента гена Vκ3-20 человека, и указанная замена предназначена для обеспечения экспрессии гистидинов в положениях 105, 106, 107 и 109. Согласно некоторым вариантам реализации не являющееся человеком животное по п. 107, отличающееся тем, что у указанного не являющегося человеком животного при стимуляции представляющим интерес антигеном, экспрессируется антигенсвязывающий белок, который специфически связывает указанный антиген, при этом указанный антигенсвязывающий белок содержит последовательность аминокислот, происходящую из сегментов гена VL и JL человека, и указанный антигенсвязывающий белок содержит по меньшей мере один гистидиновый остаток в положении аминокислоты, кодируемом сегментом гена VL человека. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного экспрессируется популяция антигенсвязывающих белков в ответ на антиген, при этом все антигенсвязывающие белки в указанной популяции содержат: (а) тяжелые цепи иммуноглобулина, содержащие вариабельные домены тяжелых цепей человека, происходящие из нуклеиновой последовательности реаранжированной вариабельной области человека; и (b) легкие цепи иммуноглобулина, содержащие вариабельные домены легких цепей иммуноглобулина, полученные в результате реаранжировки сегментов гена VL и сегментов гена JL человека в геноме не являющегося человеком животного, и при этом по меньшей мере один из сегментов гена VL человека кодирует один или большее количество гистидиновых кодонов, не кодируемых соответствующим сегментом гена VL зародышевой линии человека.
[00045] Согласно дополнительным аспектам предложены способы получения не являющегося человеком животного, у которого имеется модифицированный иммуноглобулиновый локус, включающие:
(а) модификацию генома не являющегося человеком животного с удалением, или устранением функциональности:
(i) эндогенных функциональных сегментов гена VH, D и/или JH тяжелой цепи иммуноглобулина, и
(ii) эндогенных функциональных сегментов гена легкой цепи VL и JL иммуноглобулина; и
(b) введение в модифицированный геном не являющегося человеком животного:
(i) нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанной с нуклеотидной последовательностью константной области тяжелой цепи иммуноглобулина; и
(ii) один или более, но меньше, чем у дикого типа, сегментов VL и JL гена легкой цепи иммуноглобулина человека, функционально связанных с нуклеотидной последовательностью константной области легкой цепи иммуноглобулина.
Согласно некоторым вариантам реализации указанное не являющееся человеком животное представляет собой грызуна. Согласно определенным вариантам реализации указанный грызун представляет собой мышь, крысу или хомяка. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области мыши или крысы тяжелой цепи, выбранной из СН1, шарнира, СН2, СН3 и их комбинации. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека и сегменты генов VL и JL легкой цепи иммуноглобулина человека вводят в соответствующую нуклеотидную последовательность или возле соответствующей нуклеотидной последовательности не являющегося человеком животного дикого типа. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека и сегменты генов VL и JL легкой цепи иммуноглобулина человека вводят в эктопический локус в геноме. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного имеется локус тяжелой цепи иммуноглобулина, содержащий эндогенный ген Adam6a, ген Adam6b или оба указанных гена. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного имеется ген Adam6a, ген Adam6b или оба указанных гена в эктопическом локусе генома. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека кодирует последовательность человека VH3-23/GY/JH4-4 (SEQ ID NO: 137). Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области легкой цепи иммуноглобулина представляет собой нуклеотидная последовательность константной области Сκ крысы или мыши. Согласно некоторым вариантам реализации указанные сегменты генов легкой цепи VL и JL иммуноглобулина человека способны к реаранжировке и кодированию вариабельного домена легкой цепи иммуноглобулина человека. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного имеется локус легкой цепи иммуноглобулина, содержащий два сегменты генов VL человека, Vκ1-39 и Vκ3-20. Согласно определенным вариантам реализации у указанного не являющегося человеком животного имеются сегменты генов Jκ1, Jκ2, Jκ3, Jκ4 и Jκ5. Согласно некоторым вариантам реализации в эндогенном локусе легкой цепи присутствуют два или большее количество, три или большее количество, четыре или большее количество, или пять или большее количество сегментов гена VL человека, и два или большее количество сегментов гена JL человека. Согласно некоторым вариантам реализации по меньшей мере один из сегментов гена VL или JL легкой цепи иммуноглобулина человека кодирует один или большее количество гистидиновых кодонов, не кодируемых соответствующим сегментом гена вариабельной области легкой цепи зародышевой линии человека. Согласно некоторым вариантам реализации по меньшей мере один из сегментов гена VL содержит добавление гистидинового кодона или замену на гистидиновый кодон по меньшей мере одного негистидинового кодона, кодируемого соответствующей последовательностью сегмента VL зародышевой линии человека. Согласно определенным вариантам реализации указанный добавленный или заменяющий гистидиновый кодон находится в CDR3. Согласно определенным вариантам реализации указанные сегменты генов VL человека представляют собой сегменты генов Vκ1-39 и Vκ3-20 человека, и каждый из указанных сегментов гена Vκ1-39 и Vκ3-20 человека содержит замену по меньшей мере одного негистидинового кодона, кодируемого соответствующим сегментом гена VL зародышевой линии человека, на гистидиновый кодон. Согласно конкретным вариантам реализации указанная замена представляет собой замену трех негистидиновых кодонов сегмента гена Vκ1-39 человека, и при этом указанная замена предназначена для обеспечения экспрессии гистидинов в положениях 106, 108 и 111. Согласно конкретным вариантам реализации указанная замена представляет собой замену четырех негистидиновых кодонов сегмента гена Vκ1-39 человека, при этом указанная замена предназначена для обеспечения экспрессии гистидинов в положениях 105, 106, 108 и 111. Согласно конкретным вариантам реализации указанная замена представляет собой замену трех негистидиновых кодонов сегмента гена Vκ3-20 человека, при этом указанная замена предназначена для обеспечения экспрессии гистидинов в положениях 105, 106 и 109. Согласно конкретным вариантам реализации указанная замена представляет собой замену четырех негистидиновых кодонов сегмента гена Vκ3-20 человека, и указанная замена предназначена для обеспечения экспрессии гистидинов в положениях 105, 106, 107 и 109.
[00046] Согласно дополнительным аспектам предложены способы получения нуклеотидной последовательности, которая кодирует вариабельный домен легкой цепи иммуноглобулина (VL) способный к связыванию антигена независимо от вариабельного домена тяжелой цепи, включающие:
(a) иммунизацию не являющегося человеком животного представляющим интерес антигеном или его иммуногеном, при этом в геноме указанного не являющегося человеком животного содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области; и (ii) два или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL), функционально связанных с нуклеотидной последовательностью константной области легкой цепи;
(b) обеспечение возможности развития иммунного ответа у указанного не являющегося человеком животного;
(c) выделение из указанного иммунизированного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, способный связывать антиген; и
(d) получение из указанной клетки нуклеотидной последовательности, которая кодирует указанный вариабельный домен легкой цепи (VL домен), способный связывать антиген.
Согласно некоторым вариантам реализации указанный этап выделения (с) проводят путем сортировки клеток с активированной флуоресценцией (FACS) или проточной цитометрии. Согласно некоторым вариантам реализации указанная клетка, содержащая нуклеотидную последовательность, которая кодирует указанный вариабельный домен легкой цепи, связывающий антиген, представляет собой лимфоцит. Согласно определенным вариантам реализации указанный лимфоцит включает естественные киллерные клетки, Т-клетки или В-клетки. Согласно некоторым вариантам реализации указанный способ дополнительно включает этап по (с)' слияния указанного лимфоцита с раковой клеткой. Согласно определенным вариантам реализации указанная раковая клетка представляет собой клетку миеломы. Согласно некоторым вариантам реализации нуклеотидная последовательность по (d) соединяют с нуклеотидной последовательностью, кодирующей нуклеотидную последовательность константной области иммуноглобулина. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области легкой цепи представляет собой каппа-последовательность человека или лямбда-последовательность человека. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи представляет собой последовательность человека, выбранную из СН1, шарнира, СН2, СН3 и их комбинации. Согласно некоторым вариантам реализации нуклеотидная последовательность по (d) содержит одну или большее количество замен или вставок гистидиновых кодонов, происходящих из нереаранжированного сегмента гена VL в геноме указанного животного.
[00047] Согласно дополнительным аспектам предложены способы получения антигенсвязывающего белка, который содержит вариабельный домен легкой цепи иммуноглобулина, способный связывать антиген независимо от вариабельного домена тяжелой цепи, включающие:
(a) иммунизацию генетически модифицированного не являющегося человеком животного первым антигеном, который содержит первый эпитоп или его иммуногенную часть, при этом в геноме указанного не являющегося человеком животного содержится:
(i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области; и
(ii) два или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL), функционально связанных с нуклеотидной последовательностью константной области легкой цепи иммуноглобулина;
(b) обеспечение возможности развития иммунного ответа на первый эпитоп или его иммуногенную часть у указанного не являющегося человеком животного;
(c) выделение из указанного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(d) получение из указанной клетки по (с) нуклеотидной последовательности, которая кодирует указанный вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(e) применение нуклеотидной последовательности по (d) в экспрессионной конструкции, соединенной с нуклеотидной последовательностью константной области иммуноглобулина человека; и
(f) экспрессию нуклеотидной последовательности по (d) в продукционной клеточной линии, экспрессирующей тяжелую цепь иммуноглобулина человека, которая специфически связывает второй антиген или эпитоп, для получения легкой цепи антигенсвязывающего белка, кодируемой нуклеиновой кислотой по (d) и связывающей первый эпитоп или его иммуногенную часть независимо от тяжелой цепи, тяжелая цепь которого специфически связывает указанный второй антиген или эпитоп.
Согласно некоторым вариантам реализации по меньшей мере один из сегментов гена легкой цепи VL или JL человека кодирует один или большее количество гистидиновых кодонов, не кодируемых соответствующим сегментом гена вариабельной области легкой цепи зародышевой линии человека. Согласно некоторым вариантам реализации указанный первый эпитоп получен из рецептора клеточной поверхности. Согласно конкретным вариантам реализации указанный рецептор клеточной поверхности представляет собой Fc-рецептор. Согласно более конкретным вариантам реализации указанный Fc-рецептор представляет собой FcRn. Согласно некоторым вариантам реализации указанный второй антиген или эпитоп получен из растворимого антигена. Согласно некоторым вариантам реализации указанный второй антиген или эпитоп получен из рецептора клеточной поверхности. Согласно некоторым вариантам реализации первый антиген представляет собой Fc-рецептор, второй антиген представляет собой растворимый белок, а антигенсвязывающий белок содержит одну или большее количество замен и вставок гистидина, происходящих из сегмента гена VL в геноме не являющегося человеком животного.
[00048] Согласно дополнительным аспектам предложены способы получения нуклеотидной последовательности, которая кодирует вариабельный домен легкой цепи иммуноглобулина (VL), способный к связыванию антигена независимо от вариабельного домена тяжелой цепи; включающие:
(a) иммунизацию не являющегося человеком животного первым антигеном, который содержит первый эпитоп или его иммуногенную часть, при этом в геноме указанного не являющегося человеком животного содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области, и (ii) нуклеотидная последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью константной области легкой цепи;
(b) обеспечение возможности развития иммунного ответа у указанного не являющегося человеком животного;
(c) выделение из указанного иммунизированного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, способный связывать антиген; и
(d) получение из указанной клетки нуклеотидной последовательности, которая кодирует указанный вариабельный домен легкой цепи (VL домен), способный связывать антиген.
Согласно некоторым вариантам реализации указанный этап выделения (с) проводят путем сортировки клеток с активированной флуоресценцией (FACS) или проточной цитометрии. Согласно некоторым вариантам реализации указанная клетка, содержащая нуклеотидную последовательность, которая кодирует указанный вариабельный домен легкой цепи, связывающий антиген, представляет собой лимфоцит. Согласно определенным вариантам реализации указанный лимфоцит включает естественные киллерные клетки, Т-клетки или В-клетки. Согласно некоторым вариантам реализации указанный способ дополнительно включает: (с)' слияние указанного лимфоцита с раковой клеткой. Согласно определенным вариантам реализации указанная раковая клетка представляет собой клетку миеломы. Согласно некоторым вариантам реализации нуклеотидная последовательность по (d) соединяют с нуклеотидной последовательностью, кодирующей нуклеотидную последовательность константной области иммуноглобулина. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области легкой цепи представляет собой каппа-последовательность человека или лямбда-последовательность человека. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи представляет собой последовательность человека, выбранную из СН1, шарнира, СН2, СН3 и их комбинации. Согласно некоторым вариантам реализации нуклеотидная последовательность по (d) содержит одну или большее количество замен или вставок гистидиновых кодонов, происходящих из нереаранжированного сегмента гена VL в геноме указанного животного.
[00049] Согласно дополнительному аспекту предложены способы получения нуклеотидной последовательности, которая кодирует вариабельный домен легкой цепи иммуноглобулина (VL), способный к связыванию антигена независимо от вариабельного домена тяжелой цепи, включающие:
(a) иммунизацию не являющегося человеком животного первым антигеном, который содержит первый эпитоп или его иммуногенную часть, при этом в геноме указанного не являющегося человеком животного содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью константной области легкой цепи, и (ii) сегменты гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL), функционально связанные с нуклеотидной последовательностью тяжелой цепи константной области;
(b) обеспечение возможности развития иммунного ответа у указанного не являющегося человеком животного;
(c) выделение из указанного иммунизированного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, способный связывать антиген; и
(d) получение из указанной клетки нуклеотидной последовательности, которая кодирует указанный вариабельный домен легкой цепи (VL домен), способный связывать антиген.
Согласно некоторым вариантам реализации указанный этап выделения (с) проводят путем сортировки клеток с активированной флуоресценцией (FACS) или проточной цитометрии. Согласно некоторым вариантам реализации указанная клетка, содержащая нуклеотидную последовательность, которая кодирует указанный вариабельный домен легкой цепи, связывающий антиген, представляет собой лимфоцит. Согласно определенным вариантам реализации указанный лимфоцит включает естественные киллерные клетки, Т-клетки или В-клетки. Согласно некоторым вариантам реализации указанный способ дополнительно включает: (с)' слияния указанного лимфоцита с раковой клеткой. Согласно определенным вариантам реализации указанная раковая клетка представляет собой клетку миеломы. Согласно некоторым вариантам реализации нуклеотидную последовательность по (d) соединяют с нуклеотидной последовательностью, кодирующей нуклеотидную последовательность константной области иммуноглобулина. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области легкой цепи представляет собой каппа-последовательность человека или лямбда-последовательность человека. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи представляет собой последовательность человека, выбранную из СН1, шарнира, СН2, СН3 и их комбинации. Согласно некоторым вариантам реализации нуклеотидная последовательность по (d) содержит одну или большее количество замен или вставок гистидиновых кодонов, происходящих из нереаранжированного сегмента гена VL в геноме указанного животного.
[00050] Согласно дополнительному аспекту предложены способы получения антигенсвязывающего белка, который содержит вариабельный домен легкой цепи иммуноглобулина, способный связывать антиген независимо от вариабельного домена тяжелой цепи, включающие:
(a) иммунизацию генетически модифицированного не являющегося человеком животного первым антигеном, который содержит первый эпитоп или его иммуногенную часть, при этом в геноме указанного не являющегося человеком животного содержатся:
(i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области; и
(ii) нереаранжированные сегменты гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL), функционально связанные с нуклеотидной последовательностью константной области легкой цепи;
(b) обеспечение возможности развития иммунного ответа на первый эпитоп или его иммуногенную часть у указанного не являющегося человеком животного;
(c) выделение из указанного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(d) получение из указанной клетки по (с) нуклеотидной последовательности, которая кодирует указанный вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(e) применение нуклеотидной последовательности по (d) в экспрессионной конструкции, соединенной с нуклеотидной последовательностью константной области иммуноглобулина человека; и
(f) экспрессию нуклеотидной последовательности по (d) в продукционной клеточной линии, экспрессирующей тяжелую цепь иммуноглобулина человека, которая специфически связывает второй антиген или эпитоп, для получения легкой цепи антигенсвязывающего белка, кодируемой нуклеиновой кислотой по (d) и связывающей первый эпитоп или его иммуногенную часть независимо от тяжелой цепи, тяжелая цепь которого специфически связывает указанный второй антиген или эпитоп.
Согласно некоторым вариантам реализации по меньшей мере один из сегментов гена легкой цепи VL или JL человека кодирует один или большее количество гистидиновых кодонов, не кодируемых соответствующим сегментом гена вариабельной области легкой цепи зародышевой линии человека. Согласно некоторым вариантам реализации указанный первый эпитоп получен из рецептора клеточной поверхности. Согласно определенным вариантам реализации указанный рецептор клеточной поверхности представляет собой Fc-рецептор. Согласно конкретным вариантам реализации указанный Fc-рецептор представляет собой FcRn. Согласно некоторым вариантам реализации указанный второй антиген или эпитоп получен из растворимого антигена. Согласно некоторым вариантам реализации указанный второй антиген или эпитоп получен из рецептора клеточной поверхности. Согласно некоторым вариантам реализации первый антиген представляет собой Fc-рецептор, второй антиген представляет собой растворимый белок, а антигенсвязывающий белок содержит одну или большее количество замен и вставок гистидина, происходящих из сегмента гена VL в геноме не являющегося человеком животного.
[00051] Согласно дополнительным аспектам предложены способы получения антигенсвязывающего белка, который содержит вариабельный домен легкой цепи иммуноглобулина, способный связывать антиген независимо от вариабельного домена тяжелой цепи, включающие:
(а) иммунизацию генетически модифицированного не являющегося человеком животного первым антигеном, который содержит первый эпитоп или его иммуногенную часть, при этом в геноме указанного не являющегося человеком животного содержится:
(i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи человека, функционально связанная с нуклеотидной последовательностью константной области легкой цепи; и
(ii) нереаранжированные сегменты гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL), функционально связанные с нуклеотидной последовательностью тяжелой цепи константной области;
(b) обеспечение возможности развития иммунного ответа на первый эпитоп или его иммуногенную часть у указанного не являющегося человеком животного;
(c) выделение из указанного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(d) получение из указанной клетки по (с) нуклеотидной последовательности, которая кодирует указанный вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(e) применение нуклеотидной последовательности по (d) в экспрессионной конструкции, соединенной с нуклеотидной последовательностью константной области иммуноглобулина человека; и
(f) экспрессию нуклеотидной последовательности по (d) в продукционной клеточной линии, экспрессирующей тяжелую цепь иммуноглобулина человека, которая специфически связывает второй антиген или эпитоп, для получения легкой цепи антигенсвязывающего белка, кодируемой нуклеиновой кислотой по (d) и связывающей первый эпитоп или его иммуногенную часть независимо от тяжелой цепи, тяжелая цепь которого специфически связывает указанный второй антиген или эпитоп.
Согласно некоторым вариантам реализации по меньшей мере один из сегментов гена легкой цепи VL или JL человека кодирует один или большее количество гистидиновых кодонов, не кодируемых соответствующим сегментом гена вариабельной области легкой цепи зародышевой линии человека. Согласно некоторым вариантам реализации указанный первый эпитоп получек из рецептора клеточной поверхности. Согласно определенным вариантам реализации указанный рецептор клеточной поверхности представляет собой Fc-рецептор. Согласно конкретным вариантам реализации указанный Fc-рецептор представляет собой FcRn. Согласно некоторым вариантам реализации указанный второй антиген или эпитоп получен из растворимого антигена. Согласно некоторым вариантам реализации указанный второй антиген или эпитоп получен из рецептора клеточной поверхности. Согласно некоторым вариантам реализации первый антиген представляет собой Fc-рецептор, второй антиген представляет собой растворимый белок, а антигенсвязывающий белок содержит одну или большее количество замен и вставок гистидина, происходящих из сегмента гена VL в геноме не являющегося человеком животного.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[00052] На фиг. 1 приведены схемы конструирования мини-локуса реаранжированного вариабельного домена тяжелой цепи («мини-локус UHC»), содержащего реаранжированную нуклеотидную последовательность вариабельной области иммуноглобулина человека (VH3-23/D/JH4; SEQ ID NO: 136) и интрон JH4 (SEQ ID NO: 140), которые функционально связаны с человеческим промотором VH3-23 (SEQ ID NO: 139). Мини-локус UHC был фланкирован с 5' и 3'-стороны гомологичными фрагментами мыши. На этапе 1 (лигирование I-Ceul/Spel (Amp + Spec)) вводили спектиномициновую селекционную кассету в 5'-направлении от промотора между сайтами I-CeuI и SpeI для получения pJSh0038 (мини-локус UHC).
[00053] На фиг. 2 приведены схемы для (А) нацеливания гигромициновой селекционной кассеты (EM7-HYG) на 5'-конец клона MAID 1115 ВАС (2. BHR (Hyg+Kan)); и (В) нацеливания на мини-локус UHC (pJSh0038) в 5'-направлении от локуса IgM в клоне VI432 ВАС (3. BHR (Spec+Hyg)).
[00054] На фиг. 3 приведены схемы (А) нацеливания конструкции pDBa0049, содержащей хлорамфеникольную кассету, на З'-конец клона VI421, который содержит, от 5' к 3', ген Adam6a (расположен в направлении 3'-5'); неомициновую кассету (расположена в направлении 3'-5') фланкированную FRT-сайтами; ген Adam6b (расположен в направлении 3'-5'); межгенную контрольную область 1 (IGCR1; ключевую регулирующую рекомбинацию область V(D)J); и спектиномициновую кассету (расположена в направлении 3') (4. BHR (Cm+Kan)); и (В) нацеливания на геномный локус клона VI444 ВАС, содержащего гены Adam6a и 6b в 5'-направлении от геномного локуса универсальной тяжелой цепи (UHC) клона VI443 ВАС между сайтами I-Ceul и AscI путем рестрикционного расщепления и лигирования (5. Лигирование I-CeuI/AscI (Hyg+Kan)).
[00055] На фиг. 4 приведены схемы для (А) нацеливания готовой конструкции (ДНК MAID6031 ВАС) в ЭС-клетки, выделенные из гетерозиготной мыши 1661; и представлена (В) геномная локализация зондов и праймеров, которые используются в скрининговых анализах.
[00056] На фиг. 5 приведен перечень антител из базы данных ASAP Regeneron Pharmaceuticals, содержащих последовательности CDR3, аналогичные последовательности UHC CDR3 (AKDYSNYYFDY; SEQ ID NO: 143).
[00057] Фиг. 6 иллюстрирует геномную организацию ДНК бактериальной искусственной хромосомы (ВАС) 6031 и гетерозиготных ЭС-клеток 6031, и геномную локализацию праймеров и зондов, которые используются в скрининговых анализах.
[00058] На фиг. 7 приведен перечень праймеров и зондов, которые используются для подтверждения утраты аллеля (LOA), появления аллеля (GOA) или родительского аллеля (PARENTAL) в скрининговых анализах.
[00059] На фиг. 8 приведены последовательности праймеров и зондов, которые используются в скрининговых анализах.
[00060] Фиг. 9 иллюстрирует геномную структуру локуса тяжелой цепи иммуноглобулина генетически модифицированных мышей F0, который содержит одну копию целевого аллеля (в том числе генов Adam6a/6b и нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (hVH3-23(D)JH4;). (A) MAID 6031 het: гетерозиготная мышь F0, у которой имеется генетически модифицированный локус тяжелой цепи иммуноглобулина с селекционной кассетой; (В) MAID 6032 het: гетерозиготная мышь F0, у которой имеется генетически модифицированный локус тяжелой цепи иммуноглобулина без селекционной кассеты.
[00061] На фиг. 10 показан результат анализа с сортировкой клеток с активированной флуоресценцией (FACS) клеток костного мозга, выделенных из мыши дикого типа или гетерозиготной мыши 6032. Верхняя панель: Клетки костного мозга, выделенные из мыши дикого типа или гетерозиготной мыши F0 6032, гейтировали по одиночным клеткам и сортировали на основании экспрессии CD19 (маркер В-клеток) и экспрессии CD3 (маркер Т-клеток). Нижняя панель: гейтированные по CD19+ В-клетки сортировали на основании присутствия антител IgMb (антител, полученных из аллеля дикого типа; аллель В6) или антител IgMa (антител, полученных из генетически модифицированного аллеля (аллель 129), кодирующих реаранжированный вариабельный домен тяжелой цепи (hVH3-23(D)JH4).
[00062] На фиг. 11 показан результат FACS-анализа клеток селезенки, выделенных из гетерозиготной мыши дикого типа или 6032. Верхняя панель: Клетки селезенки, выделенные из гетерозиготной мыши дикого типа или F0 6032, гейтировали по одиночным клеткам и сортировали на основании экспрессии CD19 (маркер В-клеток) и экспрессии CD3 (маркер Т-клеток). Нижняя панель: гейтированные CD19+ В-клетки сортировали на основании присутствия антител IgMb (антител, полученных из аллеля дикого типа; аллеля В6) или антител IgMa (антител, полученных из генетически модифицированного аллеля (аллель 129), кодирующего реаранжированный вариабельный домен тяжелой цепи (hVH3-23(D)JH4).
[00063] На фиг. 12 показан результат FACS-анализа клеток крови, выделенных из гетерозиготной мыши дикого типа или 6032. Верхняя панель: Клетки крови, выделенные из гетерозиготной мыши дикого типа или F0 6032, гейтировали по одиночным клеткам и сортировали на основании экспрессии CD19 (маркер В-клеток) и экспрессии CD3 (маркер Т-клеток). Нижняя панель: гейтированные CD19+ В-клетки сортировали на основании присутствия антител IgMb (антител, полученных из аллеля дикого типа; аллеля В6) или антител IgMa (антител, полученных из генетически модифицированного аллеля (аллеля 129), кодирующего реаранжированный вариабельный домен тяжелой цепи (hVH3-23(D)JH4).
[00064] На фиг. 13А представлены результаты FACS-анализа на общее количество CD19+ В-клеток, незрелых В-клеток (CD19+IgDintIgMhi) и зрелых В-клеток (CD19+IgMloIgDhi) в собранных селезенках мышей дикого типа (WT) и мышей, гомозиготных (6032НО) по реаранжированной нуклеотидной последовательности вариабельной области иммуноглобулина человека (VH3-23/D/JH4). Верхняя панель: Клетки селезенки, выделенные из гомозиготной мыши дикого типа или F2 6032, гейтировали по одиночным клеткам и сортировали на основании экспрессии CD19 (маркер В-клеток) и экспрессии CD3 (маркер Т-клеток). На нижней панели представлены репрезентативные контурные диаграммы для спленоцитов, гейтированных по CD19+ и окрашенных на иммуноглобулин D (IgD) и иммуноглобулин М (IgM), от мыши дикого типа (WT) и мыши, гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (VH3-23/D/JH4). Приведен процент клеток в каждой гейтированной области.
[00065] На фиг. 13В показано общее количество В-клеток (CD 19+), зрелых В-клеток (CD19+IgDhi IgMlo) и незрелых В-клеток (CD19+IgDintIgMhi) в собранных селезенках от мышей дикого типа (WT) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (VH3-23/D/JH4).
[00066] На фиг. 13С представлены репрезентативные контурные диаграммы Igλ+ и Igκ+ спленоцитов, тестированных по CD19+, от мыши дикого типа (WT) и мыши (6032НО), гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (VH3-23/D/JH4).
[00067] На фиг. 13D приведено общее количество В-клеток (CD19+), Igκ+ В-клеток (CD19+Ig каппа+) и Igλ+ В-клеток (CD19+Ig лямбда+) в собранных селезенках мышей дикого типа (WT) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (VH3-23/D/JH4).
[00068] На фиг. 13Е показано развитие периферических В-клеток у мыши дикого типа и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (VH3-23/D/JH4). В первом столбце (слева) на контурных диаграммах показаны спленоциты CD93+ и В220+, гейтированные по CD19+, указывающие на незрелые и зрелые В-клетки. Во втором столбце (в середине) на контурной диаграмме показана экспрессия IgM+ и CD23+ в незрелых В-клетках, указывающая на популяции В-клеток T1 (IgD-IgM+CD2lloCD23-), Т2 (IgDhiIgMhiCD21midCD23+) и Т3. В третьем столбце (справа) контурных диаграмм показана экспрессия CD21+ (CD35+) и IgM+ в зрелых В-клетках, что указывает на меньшую первую популяцию, дающую начало В-клеткам маргинальной зоны, и вторую популяцию, дающую начало фолликулярным (FO) В-клеткам. Приведен процент клеток в каждой гейтированной области.
[00069] На фиг. 14А представлены репрезентативные контурные диаграммы для костного мозга, окрашенного на В-клетки и Т-клетки (CD19+ и CD3+, соответственно), мышей дикого типа (WT) и мыши, гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (VH3-23/D/JH4).
[00070] На фиг. 14В приведено абсолютное количество клеток (слева), общее количество клеток (в середине) и общее количество В-клеток (CD19+) (справа) в костном мозге, выделенном из бедренных костей мышей дикого типа (WT) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (VH3-23/D/JH4).
[00071] На фиг. 14С представлены репрезентативные контурные диаграммы для костного мозга, гейтированного по одиночным клеткам, окрашенным на иммуноглобулин М (IgM) и В220, от мышей дикого типа (WT) и мыши, гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (VH3-23/D/JH4). Незрелые, зрелые и про/пре-В-клетки отмечены на каждой из контурных диаграмм.
[00072] На фиг. 14D показано общее количество В-клеток, количество зрелых В-клеток (B220hiIgM+) и незрелых В-клеток (B220intIgM+) в костном мозге, выделенном из бедренных костей мышей дикого типа (WT) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (VH3-23/D/JH4).
[00073] На фиг. 14Е представлены репрезентативные контурные диаграммы для костного мозга, гейтированного по одиночным клеткам, окрашенным на иммуноглобулин М (IgM) и В220, от мыши дикого типа (WT) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (VH3-23/D/JH4). Незрелые, зрелые и про/пре-В-клетки отмечены на каждой из контурных диаграмм.
[00074] На фиг. 14F представлены репрезентативные контурные диаграммы для костного мозга, гейтированного по незрелым (B220intIgM+) и зрелым (B220hiIgM+) В-клеткам, окрашенным для определения экспрессии Igκ и Igλ, выделенным из бедренных костей мыши дикого типа (WT) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (VH3-23/D/JH4).
[00075] На фиг. 15 приведены уровни антигенспецифических mIgG в сыворотке мышей (дикого типа или 6031 НЕТ, F0 и F1) на 15 день и 24 день после иммунизации через подушечки лап.
[00076] На фиг. 16 представлена кодон-оптимизированная нуклеотидная последовательность и выведенная последовательность аминокислот hVH3-23(D4-4, рамка считывания 3)JH6 (SEQ ID NO: 145).
[00077] На фиг. 17 представлена кодон-оптимизированная нуклеотидная последовательность и выведенная последовательность аминокислот hVH3-23(D4-4, рамка считывания 2)JH6 (SEQ ID NO: 146).
[00078] На фиг. 18 представлена кодон-оптимизированная нуклеотидная последовательность и выведенная последовательность аминокислот hVH3-23(D4-4, рамка считывания 3)JH4 (SEQ ID NO: 147).
[00079] На фиг. 19 представлена кодон-оптимизированная нуклеотидная последовательность и выведенная последовательность аминокислот hVH3-23(D4-4, рамка считывания 2)JH4 (SEQ ID NO: 148).
[00080] На фиг. 20 представлены примеры двух генетически модифицированных локусов двойных легких цепей (DLC). Локус в верхней части (DLC-5J) содержит сконструированный фрагмент ДНК человека, содержащий два сегмента гена Vκ человека и пять сегментов гена Jκ человека. Локус в нижней части (DLC-1J) содержит сконструированный фрагмент ДНК человека, содержащий два сегмента гена Vκ человека и один сегмент гена Jκ человека. Каждый локус способен к реаранжировке с образованием области Vκ человека, функционально связанной с эндогенной константной областью легкой цепи (например, Сκ). Показаны промоторы иммуноглобулинов (Р, незакрашенная стрелка над локусом), лидерные экзоны (L, короткие незакрашенные стрелки) и два сегмента гена Vκ человека (длинные незакрашенные стрелки) фланкированы с 5'-стороны неомициновой кассетой, содержащей сайты Frt-рекомбинации. Рекомбинационные сигнальные последовательности, встроенные в каждый из сегментов гена (Vκ и J Jκ) человека, обозначены незакрашенными эллипсами рядом с каждым сегментом гена. Согласно большинству вариантов реализации, если не указано иное, закрашенными фигурами и сплошными линиями обозначены последовательности мыши, а незакрашенными фигурами и двойными линиями обозначены последовательности человека. Диаграммы представлены без соблюдения масштаба.
[00081] На фиг. 21А-21С представлена общая стратегия конструирования направленного вектора для получения локуса цепи каппа иммуноглобулина, содержащего два сегмента Vκ человека (hVκ1-39 и hVκ3-20) и один сегмент Jκ человека (Jκ5), а также энхансеры и фрагмент IgκC мыши. На фиг. 21D показано введение указанного направленного вектора в ЭС-клетки и получение гетерозиготных мышей с указанным вектором; а на фиг. 21Е показано удаление селекционной кассеты в ЭС-клетках с применением фермента FLP. Согласно большинству вариантов реализации, если не указано иное, закрашенными фигурами и сплошными линиями обозначены последовательности мыши, а незакрашенными фигурами и двойными линиями обозначены последовательности человека. Диаграммы представлены без соблюдения масштаба.
[00082] На фиг. 22A-22D приведена нуклеотидная последовательность (SEQ ID NO: 82) сконструированной части локуса к иммуноглобулина, содержащей два сегмента Vκ человека (hVκ1-39 и hVκ3-20) и один сегмент Jκ человека; указанная нуклеотидная последовательность захватывает сконструированную последовательность человека и содержащую 100 пар оснований эндогенную последовательность мыши с обоих концов, 5' и 3'. Внизу на фиг. 22D перечислены разные шрифты, использованные для обозначения различных последовательностей.
[00083] На фиг. 23А-23В показана общая стратегия конструирования направленного вектора для получения локуса цепи каппа иммуноглобулина, содержащего два сегмента Vκ человека (hVκ1-39 и hVκ3-20) и пять сегментов Jκ человека, а также энхансеры и фрагмент IgκC мыши. На фиг. 23С показано введение указанного направленного вектора в ЭС-клетки и получение гетерозиготных мышей с указанным вектором; а на фиг. 23D показано удаление селекционной кассеты в ЭС-клетках с применением фермента FLP. Согласно большинству вариантов реализации, если не указано иное, закрашенными фигурами и сплошными линиями обозначены последовательности мыши, а незакрашенными фигурами и двойными линиями обозначены последовательности человека. Диаграммы представлены без соблюдения масштаба.
[00084] На фиг. 24A-24D показана нуклеотидная последовательность (SEQ ID NO: 83) сконструированного локуса к иммуноглобулина, содержащего два сегмента Vκ человека (hVκ1-39 и hVκ3-20) и пять сегментов Jκ человека; нуклеотидная последовательность захватывает указанную сконструированную последовательность и 100 пар оснований эндогенной последовательности мыши и с 5', и с 3'-конца. Снизу на фиг. 24D объясняется, какие шрифты использованы для обозначения различных последовательностей.
[00085] На фиг. 25А, на верхней панели, представлены репрезентативные контурные диаграммы для костного мозга, окрашенного на В-клетки и Т-клетки (CD19+ и CD3+, соответственно), от мышей дикого типа (WT) и мыши, гомозиготной по двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека (DLC-5J). На нижней панели представлены репрезентативные контурные диаграммы для костного мозга, гейтированного по CD19+ и окрашенного на ckit+ и CD43+, от мышей дикого типа (WT) и мыши, гомозиготной по двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека (DLC-5J). На контурных диаграммах нижней панели отмечены про- и пре-В-клетки.
[00086] На фиг. 25В приведено число про-(CD19+CD43+ckit+) и пре-(CD19+CD43-ckit-) В-клеток в костном мозге, полученном из бедренных костей мышей дикого типа (WT) и мышей, гомозиготных по двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека (DLC-5J).
[00087] На фиг. 26А представлены репрезентативные контурные диаграммы для костного мозга, гейтированного по одиночным клеткам, окрашенным на иммуноглобулин М (IgM) и В220, от мышей дикого типа (WT) и мыши, гомозиготной по двум человека Vκ и пяти сегментам генов Jκ человека (DLC-5J). Незрелые, зрелые и про/пре В-клетки отмечены на каждой из контурных диаграмм.
[00088) На фиг. 26В показано общее количество В-клеток (CD19+), незрелых В-клеток (B220intIgM+) и зрелых В-клеток (B220hiIgM+) в костном мозге, выделенном из бедренных костей мышей дикого типа (WT) и мышей, гомозиготных по двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека (DLC-5J).
[00089] На фиг. 27А представлены репрезентативные контурные диаграммы для костного мозга, гейтированного по одиночным клеткам, окрашенным на иммуноглобулин М (IgM) и В220, от мышей дикого типа (WT) и мыши, гомозиготной по двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека (DLC-5J). Незрелые, зрелые и про/пре В-клетки отмечены на каждой из контурных диаграмм.
[00090] На фиг. 27В представлены репрезентативные контурные диаграммы для костного мозга, гейтированного по незрелым (B220intIgM+) и зрелым (B220hiIgM+) В-клеткам, окрашенным на экспрессию Igκ и Igλ, выделенным из бедренных костей мыши дикого типа (WT) и мыши, гомозиготной по двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека (DLC-5J).
[00091] На фиг. 28А, на верхней панели, представлены репрезентативные контурные диаграммы для спленоцитов, гейтированных по одиночным клеткам и окрашенных на В-клетки и Т-клетки (CD19+ и CD3+, соответственно), от мышей дикого типа (WT) и мыши, гомозиготной по двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека (DLC-5J). На нижней панели представлены репрезентативные контурные диаграммы для спленоцитов, гейтированных по CD19+ и окрашенных на иммуноглобулин D (IgD) и иммуноглобулин М (IgM), от мышей дикого типа (WT) и мыши, гомозиготной по двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека (DLC-5J). Зрелые (54 для WT, 56,9 для DLC-5J) и переходные (23,6 для WT, 25,6 для DLC-5J) В-клетки отмечены на каждой из контурных диаграмм.
[00092] На фиг. 28В показано общее количество CD19+ В-клеток, переходных В-клеток (CD19+IgMhiIgDlo) и зрелых В-клеток (CD19+IgMloIgDhi) в собранных селезенках мышей дикого типа (WT) и мышей, гомозиготных по двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека (DLC-5J).
[00093] На фиг. 29А представлены репрезентативные контурные диаграммы спленоцитов Igλ+ и Igκ+, гейтированных по CD19+, от мышей дикого типа (WT) и мыши, гомозиготной по двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека (DLC-5J).
[00094] На фиг. 29В показано общее количество В-клеток (CD 19+), IgK+ В-клеток (CD19+Igκ+) и Igλ+ В-клеток (CD19+Igλ+) в собранных селезенках мышей дикого типа (WT) и мышей, гомозиготных по двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека (DLC-5J).
[00095] На фиг. 30А показано развитие периферических В-клеток у мышей, гомозиготных по двум сегментам Vκ и пяти сегментам Jκ генов человека. На первой (крайней слева) контурной диаграмме представлены спленоциты CD93+ и В220+, гейтированные по CD19+, указывающие на незрелые (39,6) и зрелые (57,8) В-клетки. На второй (верхней средней) контурной диаграмме представлена экспрессия IgM+ и CD23+ в незрелых В-клетках, указывающая на популяции В-клеток Т1 (33,7; IgD-IgM+CD21loCD23-), Т2 (21,2; IgDhiIgMhiCD21midCD23+) и Т3 (29,1). На третьей (нижней средней) контурной диаграмме представлена экспрессия CD21+ (CD35+) и IgM+ зрелых В-клеток, указывающая на небольшую популяцию (14,8), дающую начало В-клеткам маргинальной зоны, и вторую популяцию (70,5), дающую начало фолликулярным (FO) В-клеткам. На четвертой (верхней правой) контурной диаграмме представлена экспрессия В220+ и CD23+ в зрелых В-клетках, указывающая на популяции В-клеток маргинальной зоны (90,5; МЗ) и предшественников маргинальной зоны (7,3; IgMhiIgDhiCD21hiCD23+). На пятой (снизу справа) контурной диаграмме представлена экспрессия IgD+ и IgM+ в зрелых В-клетках, указывающая на популяции В-клеток FO-I (79,0; IgDhiIgMloCD21midCD23+) и FO-II (15,1; IgDhiIgMhiCD21midCD23+). Приведен процент клеток в каждой гейтированной области.
[00096] На фиг. 30В показано развитие периферических В-клеток у мышей дикого типа. На первой (крайней слева) контурной диаграмме представлены спленоциты CD93+и В220+гейтированные по CD19+, указывающие на незрелые (31,1) и зрелые (64,4) В-клетки. На второй (верхней средней) контурной диаграмме представлена экспрессия IgM+ и CD23+ в незрелых В-клетках, указывающая на популяции В-клеток Т1 (28,5; IgD-IgM+CD2lloCD23-), Т2 (28,7; IgDhiIgMhiCD21midCD23+) и Т3 (30,7). На третьей (нижней средней) контурной диаграмме представлена экспрессия CD21+ (CD35+) и IgM+ в зрелых В-клетках, указывающая на небольшую популяцию (7,69), дающую начало В-клеткам маргинальной зоны, и вторую популяцию (78,5), дающую начало фолликулярным (FO) В-клеткам. На четвертой (верхней правой) контурной диаграмме представлена экспрессия В220+ и CD23+ в зрелых В-клетках, указывающая на популяции В-клеток маргинальной зоны (79,9; МЗ) и предшественников маргинальной зоны (19,4; IgMhiIgDhiCD21hiCD23+). На пятой (снизу справа) контурной диаграмме представлена экспрессий IgD+ и IgM+ в зрелых В-клетках, указывающая на популяции В-клеток FO-I (83,6; IgDhiIgMloCD21midCD23+) и FO-II (13,1; IgDhiIgMhiCD21midCD23+). Приведен процент клеток в каждой гейтированной области.
[00097] На фиг. 31 показано общее количество популяций переходных В-клеток, В-клеток маргинальной зоны и фолликулярных В-клеток в собранных селезенках мышей дикого типа (WT) и мышей, гомозиготных по двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека (DLC-5J).
[00098] На фиг. 32 показана относительная экспрессия мРНК в костном мозге (ось у) происходящих из Vκ3-20 и происходящих из Vκ1-39 легких цепей в количественном ПЦР-анализе с применением зондов, специфических в отношении сегменты генов Vκ3-20 или Vκ1-39 у мышей, гомозиготных по замене указанных эндогенных сегментов гена Vκ и Jκ на сегменты генов Vκ и Jκ человека (Нκ) (легкая цепь человека у мыши VELOCIMMUNE®), мышей дикого типа. (WT), мышей, гомозиготных по двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека (DLC-5J), и мышей, гомозиготных по двум сегментам генов Vκ человека и одному сегменту гена Jκ человека (DLC-1J). Сигналы нормированы по экспрессии Сκ мыши. Н/д: не детектировано.
[00099] На фиг. 33 показана относительная экспрессия в цельных селезенках мРНК (ось у), происходящей из легких цепей Vκ3-20 и Vκ1-39, при количественном ПЦР-анализе с применением зондов, специфических в отношении сегментов гена Vκ3-20 или Vκ1-39 у мышей, гомозиготных по замене указанных эндогенных сегментов гена Vκ и Jκ на сегменты генов Vκ и Jκ человека (Нк) (легкая цепь человека у мыши VELOCIMMUNE®), мышей дикого типа (WT), мышей, гомозиготных по двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека (DLC-5J), и мышей, гомозиготных по двум сегментам генов Vκ человека и одному сегменту гена Jκ человека (DLC-1J). Сигналы нормированы по экспрессии Сκ мыши. Н/Д: не детектируется.
[000100] На фиг. 34 приведена последовательность и свойства (% содержание GC, N, % несовпадений, Tm) выбранных праймеров для мутагенеза, используемых для встраивания четырех остатков гистидина в области CDR3 последовательности легких цепей Vκ1-39 и Vκ3-20 человека. Последовательности SEQ ID NO указанных праймеров, использованных в Перечне последовательностей, включены в таблицу ниже. F = прямой праймер, R = обратный праймер.
[000101] На фиг. 35А показано введение направленного вектора, содержащего два сегмента легкой цепи Vκ человека, каждый из которых замещен четырьмя остатками гистидина, и пять Jκ человека, в ЭС-клетки и получение гетерозиготных мышей с указанными вектором; а на фиг. 35В показано удаление селекционной кассеты в ЭС-клетках с применением фермента FLPo. Согласно большинству вариантов реализации, если не указано иное, закрашенными фигурами и сплошными линиями обозначены последовательности мыши, а незакрашенными фигурами и двойными линиями обозначены последовательности человека. Диаграммы представлены без соблюдения масштаба.
[000102] На фиг. 36 приведена последовательность и указаны свойства (процентное содержание GC, N, % несовпадений, Tm) выбранных праймеров для мутагенеза, использованных для встраивания трех остатков гистидина в последовательность областей CDR3 легких цепей Vκ1-39 и Vκ3-20 человека. Последовательности (SEQ ID NO) указанных праймеров, приведенные в Перечне последовательностей, включены в таблица ниже. Р = прямой праймер, R = обратный праймер.
[000103] На фиг. 37А показано введение направленного вектора, содержащего два сегмента Vκ легкой цепи человека, каждый из которых замещен тремя остатками гистидина, и пять сегментов Jκ человека, в ЭС-клетки и получение гетерозиготных мышей с указанными вектором; а на фиг. 37В показано удаление селекционной кассеты в ЭС-клетках с применением фермента FLPo. Согласно большинству вариантов реализации, если не указано иное, закрашенными фигурами, и сплошными линиями обозначены последовательности мыши, а незакрашенными фигурами и двойными линиями обозначены последовательности человека. Диаграммы представлены без соблюдения масштаба.
[000104] На фиг. 38А представлено выравнивание последовательности аминокислот, кодируемой последовательностью Vκ3-20 зародышевой линии человека (нижняя последовательность), с примером трансляции в аминокислоты последовательности вариабельной области легкой цепи каппа IgM, экспрессируемой у мыши, у которой имеется два сегмента V каппа (Vκ3-20 и Vκ1-39), каждый из которых замещен на 3 остатка гистидина в последовательности CDR3 (верхняя последовательность); на выравнивании показана вариабельная последовательность цепи каппа IgM, экспрессируемой у мыши, сохраняющей все три замены гистидина, введенные в последовательность зародышевой линии. На фиг. 38 В представлено выравнивание последовательности аминокислот, кодируемых последовательностью Vκ1-39 зародышевой линии человека (нижняя последовательность в каждом выравнивании) с примером трансляции аминокислот вариабельной последовательности легкой цепи каппа IgM, экспрессируемой у мыши, у которой имеется два сегмента V каппа (Vκ3-20 и Vκ1-39), каждый из которых замещен на 3 остатка гистидина в последовательности CDR3 (верхняя последовательность в каждом выравнивании); на верхнем выравнивании показана вариабельная последовательность цепи каппа IgM, экспрессируемой у мыши, сохраняющей все три гистидиновые модификации, введенные в последовательность зародышевой линии, на нижнем выравнивании показана последовательность вариабельной цепи каппа IgM, экспрессируемой у мыши, сохраняющей две из трех гистидиновых модификаций, введенных в последовательность зародышевой линии. Согласно некоторым вариантам реализации гистидин, введенный в последнее положение Vκ, может быть утрачен во время V-J-реаранжировки.
[000105] Фиг. 39 иллюстрирует геномную структуру генетически модифицированных мышей F2, у которых имеется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи в локусах тяжелой цепи (MAID6032; «мышь UHC») и имеются также генетически сконструированные локусы легкой цепи, содержащие два сегмента гена Vκ человека (например, сегмент гена VκI-39 человека и Vκ3-20 человека) и пять сегментов гена Jκ человека (hJκ1-5; DLC-5J) (MAID 1912НО).
[000106] На фиг. 40А, на верхней панели, представлены репрезентативные контурные диаграммы для спленоцитов, гейтированных по одиночным клеткам и окрашенных на В-клетки и Т-клетки (CD19+ и CD3+, соответственно), от генетически модифицированных контрольных мышей (VI3; 1293НО 1460НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусах тяжелой цепи, и двум сегментам Vκ человека и пяти сегментам генов Jκ человека в локусах легкой цепи (MAID 1912НО 6032 НО; DLC × UHC). На нижней панели представлены репрезентативные контурные диаграммы для спленоцитов, гейтированных по CD 19+ и окрашенных на иммуноглобулин D (IgD) и иммуноглобулин М (IgM), от генетически модифицированных контрольных мышей (VI3; 1293НО 1460НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусах тяжелой цепи, и двум сегментам генов Vκ человека и пяти сегментам Jκ человека в локусах легкой цепи (MAID 1912НО 6032 НО; DLC × UHC). Зрелые и незрелые В-клетки отмечены на каждой из контурных диаграмм.
[000107] На фиг. 40В показано общее количество CD19+ В-клеток, зрелых В-клеток (CD19+IgMloIgDhi) и незрелых В-клеток (CD19+IgMhiIgDint) в собранных селезенках генетически модифицированных контрольных мышей (VI3; 1293НО 1460НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусах тяжелой цепи, и двум сегментам Vκ человека и пяти сегментам генов Jκ человека в локусах легкой цепи (MAID 1912НО 6032 НО; DLC × UHC).
[000108] На фиг. 41А представлены репрезентативные контурные диаграммы для Igλ+ и Igκ+ спленоцитов, гейтированных по CD19+, от генетически модифицированных контрольных мышей (VI3; 1293НО 1460НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусах тяжелой цепи, и двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека в локусах легкой цепи (MAID 1912НО 6032 НО; DLC × UHC).
[000109] На фиг. 41В приведено общее количество В-клеток (CD19+), Igκ+ В-клеток (CD19+Igκ+) и Igλ+ В-клеток (CD19+Igλ+) в собранных селезенках от генетически модифицированных контрольных мышей (VI3; 1293НО 1460НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусах тяжелой цепи, и двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека в локусах легкой цепи (MAID 1912НО 6032 НО; DLC × UHC).
[000110] На фиг. 42 представлен проточно-цитометрический анализ поверхностной экспрессии IgM на В-клетках в собранных селезенках генетически модифицированных контрольных мышей (VI3; 1293НО 1460НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусах тяжелой цепи, и двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека в локусах легкой цепи (MAID 1912НО 60.32 НО; DLC × UHC). Клетки окрашивали флуоресцентным (ФЭ-Су7-конъюгированным) антителом против IgM.
[000111] На фиг. 43А показано развитие периферических В-клеток у генетически модифицированных контрольных мышей (VI3; 1293НО 1460НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусах тяжелой цепи и двух сегментам генов Vκ человека и пяти сегментам генов Jκ человека в локусах легкой цепи (MAID 1912НО 6032 НО; DLC × UHC). На первой (крайней слева) контурной диаграмме представлены CD93+ и В220+ спленоциты, гейтированные по CD19+, что указывает на незрелые и зрелые В-клетки. На второй (в середине) контурной диаграмме представлена экспрессия IgM+ и CD23+ незрелых В-клеток, указывающая на популяции В-клеток T1 (IgD-IgM+CD21loCD23-), Т2 (IgDhiIgMhiCD21midCD23+) и Т3. На третьей (справа) контурной диаграмме представлена экспрессия CD21+ (CD35+) и IgM+ зрелых В-клеток, указывающая на первые меньшие популяции, дающие начало В-клеткам маргинальной зоны, и вторые большие популяции, дающие начало фолликулярным (FO) В-клеткам. Приведен процент клеток в каждой гейтированной области.
[000112] На фиг. 43В показано развитие периферических В-клеток у генетически модифицированных контрольных мышей (VI3; 1293НО 1460НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусах тяжелой цепи, и двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека в локусах легкой цепи (MAID 1912НО 6032 НО; DLC × UHC). На первой (слева) контурной диаграмме представлена экспрессия CD21+ (CD35+) и IgM+ зрелых В-клеток, указывающая на небольшую популяцию, дающую начало В-клеткам маргинальной зоны, и вторую популяцию, дающую начало фолликулярным (FO) В-клеткам. На второй (в середине) контурной диаграмме представлена экспрессия В220+ и CD23+ в зрелых В-клетках, указывающая на популяции В-клеток маргинальной зоны (МЗ) и предшественников маргинальной зоны (IgMhiIgDhiCD21hiCD23+). На третьей (справа) контурной диаграмме представлена экспрессия IgD+ и IgM+ в зрелых В-клетках, указывающая на популяции В-клеток FO-I (IgDhiIgMintCD21intCD23+) и FO-II (IgDhiIgMintCD21intCD23+). Приведен процент клеток в каждой гейтированной области.
[000113] На фиг. 44А представлены репрезентативные контурные диаграммы для костного мозга, окрашенного на В-клетки и Т-клетки (CD19+ и CD3+, соответственно), генетически модифицированной контрольной мыши (VI3; 1293НО 1460НО) и мыши, гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусах тяжелой цепи, и двум сегментам генов человека Vκ и пяти сегментам генов Jκ человека в локусах легкой цепи (MAID 1912НО 6032 НО; DLC × UHC).
[000114] На фиг. 44В приведен процент лимфоцитов, общее количество клеток на бедренную кость и число CD19+ В-клеток в костном мозге, полученном из бедренных костей генетически модифицированных контрольных мышей (VI3; 1293НО 1460НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусах тяжелой цепи, и двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека в локусах легкой цепи (MAID 1912НО 6032 НО; DLC × UHC).
[000115] На фиг. 45А представлены репрезентативные контурные диаграммы для костного мозга, гейтированного по CD19+ и окрашенного на ckit+ и CD43+, от генетически модифицированной контрольной мыши (VI3; 1293НО 1460НО) и мыши, гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусах тяжелой цепи, и двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека в локусах легких цепей (MAID 1912НО 6032 НО; DLC × UHC). На контурных диаграммах отмечены про- и пре-В-клетки.
[000116] На фиг. 45В приведено число пре-(CD19+CD43-ckit-) и про-(CD19+CD43+ckit+) В-клеток в костном мозге, полученном из бедренных костей генетически модифицированных контрольных мышей (VI3; 1293НО 1460НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусах тяжелой цепи и двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека в локусах легких цепей (MAID 1912НО 6032НО; DLC × UHC).
[000117] На фиг. 46А представлены репрезентативные контурные диаграммы для костного мозга, гейтированного по одиночным клеткам, окрашенным на иммуноглобулин М (IgM) и В220, от генетически модифицированной контрольной мыши (VI3; 1293НО 1460НО) и мыши, гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусах тяжелой цепи и двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека в локусах легких цепей (MAID 1912НО 6032 НО; DLC × UHC). Незрелые, зрелые и про/пре В-клетки отмечены на каждой из контурных диаграмм.
[000118] На фиг. 46В показано общее количество клеток на бедренную кость, незрелых В-клеток (B220intIgM+) и зрелых В-клеток (B220hiIgM+) в костном мозге, выделенном из бедренных костей генетически модифицированных контрольных мышей (VI3; 1293НО 1460НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусах тяжелой цепи и двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека в локусах легких цепей (MAID 1912НО 6032 НО; DLC × UHC).
[000119] На фиг. 47 представлены репрезентативные контурные диаграммы для костного мозга, гейтированного по незрелым (B220intIgM+) и зрелым (B220hiIgM+) В-клетки окрашенного на Igλ и Igκ экспрессия выделенные из бедренных костей генетически модифицированной контрольной мыши (VI3; 1293НО 1460НО) и мыши, гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусах тяжелой цепи и двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека в локусах легких цепей (MAID 1912НО 6032 НО; DLC × UHC).
[000120] На фиг. 48 представлены уровни антигенспецифических mIgG в сыворотке мышей (дикого типа или 1912НО 6031 НЕТ (гомозиготный DLC × гетерозиготный UHC)) до иммунизации через подушечки лап, через 23 дня после первого раунда иммунизации через подушечки лап, через 5 недель после первого раунда иммунизации через подушечки лап, и после второго раунда иммунизации через подушечки лап.
[000121] Фиг. 49 иллюстрирует геномную структуру генетически модифицированных мышей F1, у которых имеются нуклеотидные последовательности реаранжированной вариабельной области тяжелой цепи в локусах легкой цепи каппа (т.е. реаранжированная последовательность VDJ тяжелой цепи, функционально связанная с нуклеотидной последовательностью константной области легкой цепи каппа).
[000122] На фиг. 50А, на верхней панели, представлены репрезентативные контурные диаграммы для костного мозга, окрашенного на В-клетки и Т-клетки (CD19+ и CD3+, соответственно), от мышей дикого типа (WT) и мыши, гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи (hVH3-23/D/JH4) в локусе легкой цепи каппа. На нижней панели представлены репрезентативные контурные диаграммы для костного мозга, гейтированного по CD19+ и окрашенного на ckit+ и CD43+, от мышей дикого типа (WT) и мыши, гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи (hVH3-23/D/JH4) в локусе легкой цепи каппа. На контурных диаграммах нижней панели отмечены про- и пре-В-клетки.
[000123] На фиг. 50В приведено число про-(CD19+CD43+ckit+) и пре-(CD19+CD43-ckit-) В-клеток в костном мозге, полученном из бедренных костей мышей дикого типа (WT) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи (hVH3-23/D/JH4) в локусе легкой цепи каппа.
[000124] На фиг. 51А представлены репрезентативные контурные диаграммы для костного мозга, гейтированного по одиночным клеткам, окрашенным на иммуноглобулин М (IgM) и В220, от мышей дикого типа (WT) и мыши, гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи (hVH3-23/D/JH4) в локусе легкой цепи каппа. Незрелые, зрелые и про/пре-В-клетки отмечены на каждой из контурных диаграмм.
[000125] На фиг. 51В показано общее количество В-(CD19+) и про/пре-В-клеток (IgM-B220+) в костном мозге, выделенном из бедренных костей мышей дикого типа (WT) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи (hVH3-23/D/JH4) в локусе легкой цепи каппа.
[000126] На фиг. 51С приведено число незрелых В-клеток (B220intIgM+) и зрелых В-клеток (B220hiIgM+) в костном мозге, выделенном из бедренных костей мышей дикого типа (WT) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи (hVH3-23/D/JH4) в локусе легкой цепи каппа.
[000127] На фиг. 52 представлены репрезентативные контурные диаграммы для костного мозга, гейтированного по незрелым (B220intIgM+) и зрелым (B220hiIgM+) В-клеткам, окрашенным на экспрессию Igλ, и Igκ, выделенным из бедренных костей мышей дикого типа (WT) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи (hVH3-23/D/JH4) в локусе легкой цепи каппа.
[000128] На фиг. 53А, на верхней панели, представлены репрезентативные контурные диаграммы для спленоцитов, гейтированных по одиночным клеткам и окрашенных на В- и Т-клетки (CD19+ и CD3+, соответственно), от мышей дикого типа (WT) и мыши, гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи (hVH3-23/D/JH4) в локусе легкой цепи каппа. На нижней панели представлены репрезентативные контурные диаграммы для спленоцитов, гейтированных по CD19+ и окрашенных на иммуноглобулин D (IgD) и иммуноглобулин М (IgM), от мышей дикого типа (WT) и мыши, гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи (hVH3-23/D/JH4) в локусе легкой цепи каппа. Зрелые (56,9 для WT, 43 для hVH3-23/D/JH4 на каппа) и переходные (26,8 для WT, 34 для hVH3-23/D/JH4 на каппа) В-клетки отмечены на каждой из контурных диаграмм.
[000129] На фиг. 53В показано общее количество CD19+ В-клеток, зрелых В-клеток (CD19+IgMloIgDhi) и переходных В-клеток (CD19+IgMhiIgDint) в собранных селезенках мышей дикого типа (WT) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи (hVH3-23/D/JH4) в локусе легкой цепи каппа.
[000130] На фиг. 54А представлены репрезентативные контурные диаграммы спленоцитов Igλ+ и Igκ+, гейтированных по CD19+ от мыши дикого типа (WT) и нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи (hVH3-23/D/JH4) в локусе легкой цепи каппа.
[000131] На фиг. 54В показано общее количество В-клеток (CD19+), Igκ+ В-клеток (CD19+Igκ+) и Igλ+ В-клеток (CD19+Igλ+) в собранных селезенках мышей дикого типа (WT) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи (hVH3-23/D/JH4) в локусе легкой цепи каппа.
[000132] На фиг. 55 показано развитие периферических В-клеток в компартменте селезенки мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи (hVH3-23/D/JH4) в локусе легкой цепи каппа, по сравнению с мышами дикого типа. На первой (слева) контурной диаграмме представлены CD93+ и В220+ спленоциты, гейтированные по CD19+, указывающие на незрелые и зрелые В-клетки. На второй (в середине) контурной диаграмме представлена экспрессия IgM+ и CD23+ в незрелых В-клетках, указывающая на популяции В-клеток T1, Т2 и Т3. На третьей (справа) контурной диаграмме представлена экспрессия CD21+ (CD35+) и IgM+ зрелых В-клеток, указывающая на первую меньшую популяцию, дающую начало В-клеткам маргинальной зоны, и вторую большую популяцию, дающую начало фолликулярным (FO) В-клеткам. Приведен процент клеток в каждой гейтированной области.
[000133] Фиг. 56 иллюстрирует структуру генома генетически модифицированных мышей F2, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусе легкой цепи каппа (MAID 6079НО; гомозиготная «мышь UHC-на- каппа») и гомозиготных по нуклеотидной последовательности вариабельной области легкой цепи каппа в локусе тяжелой цепи (MAID 1994НО; мышь «каппа на тяжелой цепи» («КоН»)).
[000134] На фиг. 57А на верхней панели, представлены репрезентативные контурные диаграммы для костного мозга, окрашенного на В-клетки и Т-клетки (CD19+ и CD3+, соответственно) от мыши VELOCIMMUNE® (VI3) и мыши, гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусе легкой цепи каппа и гомозиготной по нуклеотидной последовательности вариабельной области легкой цепи каппа в локусе тяжелой цепи. На нижней панели представлены репрезентативные контурные диаграммы для костного мозга, гейтированного по CD19+ и окрашенного на ckit+ и CD43+, от VELOCIMMUNE® (VI3) и мыши, гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусе легкой цепи каппа и гомозиготной по нуклеотидной последовательности вариабельной области легкой цепи каппа в локусе тяжелой цепи. На контурных диаграммах нижней панели отмечены про- и пре-В-клетки.
[000135] На фиг. 57В показано общее количество В-клеток (CD19+) и количества про-(CD19+CD43+ckit+)B-клеток и пре-(CD19+CD43-ckit-)B-клеток в костном мозге, полученных из бедренных костей мышей VELOCIMMUNE® (1242НО 1640НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусе легкой цепи каппа и гомозиготных по нуклеотидной последовательности вариабельной области легкой цепи каппа в локусе тяжелой цепи (1994НО 6079НО). Количества приведены в виде абсолютного количества клеток на бедренную кость и процента клеток.
[000136] На фиг. 58А представлены репрезентативные контурные диаграммы для костного мозга, гейтированного по одиночным клеткам, окрашенным на иммуноглобулин М (IgM) и В220, от мыши VELOCIMMUNE® (1242НО 1640НО) и мыши, гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусе легкой цепи каппа и гомозиготной по нуклеотидной последовательности вариабельной области легкой цепи каппа в локусе тяжелой цепи (1994НО 6079НО). На каждой из контурных диаграмм отмечены незрелые, зрелые и про/пре В-клетки.
[000137] На фиг. 58В приведено число незрелых В (B220intIgM+) и зрелых В-клеток (B220hiIgM+) в костном мозге, выделенном из бедренных костей мышей VELOCIMMUNE® (1242НО 1640НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусе легкой цепи каппа и гомозиготных по нуклеотидной последовательности вариабельной области легкой цепи каппа в локусе тяжелой цепи (1994НО 6079НО). Количества приведены в виде абсолютного количества клеток на бедренную кость и процента клеток.
[000138] На фиг. 59 представлены репрезентативные контурные диаграммы для костного мозга, гейтированного по незрелым (B220intIgM+) и зрелым (B220hiIgM+) В-клеткам, окрашенным на экспрессию Igλ, и Igκ, выделенным из бедренных костей мышей VELOCIMMUNE® (1242НО 1640НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусе легкой цепи каппа и гомозиготных по нуклеотидной последовательности вариабельной области легкой цепи каппа в локусе тяжелой цепи (1994НО 6079НО).
[000139] На фиг. 60А представлены репрезентативные контурные диаграммы для спленоцитов, гейтированных по CD19+ и окрашенных на иммуноглобулин D (IgD) и иммуноглобулин М (IgM), от мыши VELOCIMMUNE® (VI3; 1242НО 1640НО) и мыши, гомозиготной по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусе легкой цепи каппа и гомозиготной по нуклеотидной последовательности вариабельной области легкой цепи каппа в локусе тяжелой цепи (1994НО 6079НО). Зрелые и переходные/незрелые В-клетки отмечены на каждой из контурных диаграмм.
[000140] На фиг. 60В показано общее количество CD19+ В-клеток, зрелых В-клеток (CD19+IgMloIgDhi) и переходных В-клеток (CD19+IgMhiIgDlo) в собранных селезенках мышей VELOCIMMUNE® (1242НО 1640НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусе легкой цепи каппа и гомозиготных по нуклеотидной последовательности вариабельной области легкой цепи каппа в локусе тяжелой цепи (1994НО 6079НО).
[000141] На фиг. 61 показано общее количество В-клеток (CD19+), Igκ+ В-клеток (CD19+Igκ+) и Igλ+ В-клеток (CD19+Igλ+) в собранных селезенках мышей VELOCIMMUNE® (1242НО 1640НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусе легкой цепи каппа и гомозиготных по нуклеотидной последовательности вариабельной области легкой цепи каппа в локусе тяжелой цепи (1994НО 6079НО). Количества приведены в виде абсолютного количества клеток и как процент лимфоцитов от числа клеток.
[000142] На фиг. 62 показано развитие периферических В-клеток в компартменте селезенки мышей VELOCIMMUNE® (1242НО 1640НО) и мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусе легкой цепи каппа и гомозиготных по нуклеотидной последовательности вариабельной области легкой цепи каппа в локусе тяжелой цепи (1994НО 6079НО). На верхней контурной диаграмме представлены CD93+ и В220+ спленоциты, гейтированные по CD19+, что указывает на незрелые и зрелые В-клетки. На нижней контурной диаграмме представлена экспрессия IgM+ и CD23+ в незрелых В-клетках, указывающая на популяции В-клеток T1, Т2 и Т3. Приведен процент клеток в каждой гейтированной области.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[000143] Указанное изобретение не ограничивается конкретными описанными способами и экспериментальными условиями, поскольку такие способы и условия могут отличаться. Следует также понимать, что использованная в настоящем документе терминология предназначена исключительно для описания частных вариантов реализации, и не предназначена для ограничения, поскольку объем настоящего изобретения определен формулой изобретения.
[000144] Если не указано иное, все используемые термины и выражения в настоящем документе включают значения, которые указанные термины и выражения в данной области техники, если иное явным образом не указано или не очевидно из контекста, в котором используется указанный(ое) термин или выражение. Хотя при осуществлении или тестировании настоящего изобретения могут использоваться любые способы и материалы, аналогичные или эквивалентные описанным в настоящем документе, ниже будут описаны конкретные способы и материалы. Все упоминаемые публикации включены в настоящий документ посредством ссылки.
[000145] Термин «антитело» в настоящем документе включает молекулы иммуноглобулина, содержащие четыре полипептидных цепи, две тяжелых (Н) цепи и две легких (L) цепи, соединенные между собой дисульфидными связями. Каждая тяжелая цепь содержит вариабельный домен тяжелой цепи и константную область тяжелой цепи (CH). Константная область тяжелой цепи содержит три домена, CH1, CH2 и CH3. Каждая легкая цепь содержит вариабельный домен легкой цепи и константную область легкой цепи (CL). Вариабельные домены тяжелых цепей и легких цепей могут дополнительно подразделяться на области гипервариабельности, называемые определяющими комплементарность областями (CDR), которые перемежаются с более консервативными областями, называемыми каркасными областями (FR). Каждый вариабельный домен тяжелой и легкой цепи содержит три CDR и четыре FR, расположенные, от аминоконца к карбоксильному концу, в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4 (области CDR тяжелых цепей могут обозначаться как HCDR1, HCDR2 и HCDR3; области CDR легких цепей могут обозначаться как LCDR1, LCDR2 и LCDR3). Термин «высокоаффинное» антитело относится к антителу, KD которого в отношении целевого эпитопа составляет приблизительно 10-9 М или менее (например, приблизительно 1×10-9 М, 1×10-10 М, 1×10-11 М или приблизительно 1×10-12 М). Согласно одному варианту реализации KD измеряют с помощью поверхностного плазмонного резонанса, например, BIACORE™; согласно другому варианту реализации KD измеряют с применением ELISA.
[000146] Выражение «биспецифические антитело» включает антитело, способное избирательно связывать два или большее количество эпитопов. Биспецифические антитела обычно содержат две неидентичных тяжелых цепи, при этом каждый тяжелая цепь специфически связывает отличный эпитоп - либо на двух разных молекулах (например, разные эпитопы на двух разных иммуногенах) или на одной молекуле (например, разные эпитопы на одном иммуногене). Если биспецифическое антитело способно избирательно связывать два разные эпитопа (первый эпитоп и второй эпитоп), аффинность первой тяжелой цепи в отношении первого эпитопа обычно меньше по меньшей мере на один, два, три или четыре, или большее количество порядков, чем аффинность первой тяжелой цепи в отношении второго эпитопа, и наоборот. Эпитопы, специфически связываемые биспецифическим антителом, могут быть расположены на одной или разных мишенях (например, на одном или на разных белках). Примеры биспецифических антител включают антитела, содержащие первую тяжелую цепь, специфическую в отношении опухолевого антигена, и вторую тяжелую цепь, специфическую в отношении цитотоксического маркера, например, Fc-рецептора (например, FcγRI, FcγRII, FcγRIII и т.п.) или маркера Т-клеток (например, CD3, CD28 и т.п.). Кроме того, второй вариабельный домен тяжелой цепи может быть заменен на вариабельный домен тяжелой цепи, обладающий другой требуемой специфичностью. Например, биспецифические антитело с первой тяжелой цепью, специфической в отношении опухолевого антигена, и второй тяжелой цепью, специфической в отношении токсина, может быть подобран таки образом, чтобы доставлять токсин (например, сапорин, алкалоид барвинка и т.п.) в опухолевую клетку. Другие примеры биспецифических антител включают биспецифические антитела с первой тяжелой цепью, специфической в отношении активирующего рецептора (например, В-клеточный рецептор, FcγRI, FcγRIIA, FcγRIIIA, FcαRI, Т-клеточный рецептор и т.п.). и второй тяжелой цепью, специфической в отношении ингибирующего рецептора (например, FcγRIIB, CD5, CD22, CD72, CD300a и т.п.). Такие биспецифические антитела могут быть сконструированы для терапевтических состояний, связанных с активацией клеток (как, например, аллергия и астма). Биспецифические антитела могут быть получены, например, путем комбинирования тяжелых цепей, распознающих разные эпитопы одного и того же иммуногена. Например, нуклеотидные последовательности, кодирующие вариабельные последовательности тяжелых цепей, распознающие разные эпитопы одного иммуногена могут быть соединены с нуклеотидными последовательностями, кодирующими одни и те же или разные константные области тяжелой цепи, и такие последовательности могут экспрессироваться в клетке, которая экспрессирует легкую цепь иммуноглобулина. Типичное биспецифическое антитело содержит две тяжелых цепи, каждая из который содержит три области CDR тяжелых цепей, за которыми следуют (от N-конца к С-концу) домен CH1, шарнир, домен CH2 и домен CH3, и легкую цепь иммуноглобулина, которая либо не придает эпитоп-связывающую специфичность, но способна связываться с каждой тяжелой цепью, либо способна связываться с каждой тяжелой цепью л может связывать один или большее количество эпитопов, связываемых эпитоп-связывающими областями тяжелых цепей, либо способна связываться с каждой тяжелой цепью и обеспечивать связывание одной или обоих тяжелых цепей с одним или обоими эпитопами. Аналогичным образом, термин «триспецифическое антитело» включает антитело, способное избирательно связывать три или большее количество эпитопов.
[000147] Термин «клетка» включает любую клетку, подходящую для экспрессии рекомбинантной нуклеотидной последовательности. Клетки включают клетки прокариот и эукариот (одноклеточные или многоклеточные), бактериальные клетки (например, штаммы Е. coli, Bacillus spp., Streptomyces spp. и т.п.), клетки микобактерий, клетки грибов, дрожжевые клетки (например, S. cerevisiae, S. pombe, P. pastor is, P. methanolica и т.п.), растительные клетки, клетки насекомых (например, SF-9, SF-21, инфицированные бакуловирусом клетки насекомых, Trichoplusia ni и т.п.), клетки не являющихся человеком животных, клетки человека или слитые клетки, такие как, например, гибридомы или квадромы. Согласно некоторым вариантам реализации указанная клетка представляет собой клетку человека, обезьяны, примата, хомяка, крысы или мыши. Согласно некоторым вариантам реализации указанная клетка является эукариотической и выбрана из следующих клеток: СНО (например, СНО K1, DXB-11 СНО, Veggie-CHO), COS (например, COS-7), ретинальной клетки, Vero, CV1, клеток почки (например, НЕК293, 293 EBNA, MSR 293, MDCK, НаК, ВНК), HeLa, HepG2, WI38, MRC 5, Colo205, НВ 8065, HL-60, (например, ВНК21), Jurkat, Daudi, А431 (эпидермальной), CV-1, U937, 3Т3, L-клетки, клетки С127, SP2/0, NS-0, ММТ 060562, клетки Сертоли, клетки BRL 3А, клетки НТ1080, клетки миеломы, опухолевой клетки и клеточной линии, происходящей из вышеупомянутых клеток. Согласно некоторым вариантам реализации указанная клетка содержит один или большее количество вирусных генов, например, ретинальная клетка, которая экспрессирует вирусный ген (например, клетка PER.C6™).
[000148] Выражение «определяющая комплементарность область», или термин «CDR», включает последовательность аминокислот, кодируемую нуклеотидной последовательностью иммуноглобулиновых генов организма, которая в норме (т.е. у животного дикого типа) присутствует между двумя каркасными областями вариабельной области легкой либо тяжелой цепи молекулы иммуноглобулина (например, антитела или Т-клеточного рецептора). Кодировать область CDR может, например, последовательность зародышевой линии, либо реаранжированная или нереаранжированная последовательность, например, необученной или зрелой В-клетки или Т-клетки. Область CDR может быть соматически мутированной (например, отличаться от последовательности, кодируемой в зародышевой линии животного), гуманизированной и/или модифицированной в результате замен, добавлений или удалений аминокислот. В некоторых обстоятельствах (например, в случае CDR3), области CDR могут кодировать две или большее количество последовательностей (например, последовательностей зародышевой линии), не являющихся непрерывными (например, в нереаранжированной нуклеотидной последовательности), но непрерывных в нуклеотидной последовательности В-клетки, например, в результате сплайсинга или соединения последовательностей (например, V-D-J-рекомбинации с образованием CDR3 тяжелой цепи).
[000149] Термин «консервативный» при использовании для описания консервативной замены аминокислоты, включает замену аминокислотного остатка другим аминокислотным остатком, содержащим R-группу боковой цепи с аналогичными химическими свойствами (например, зарядом или гидрофобностью). В общем случае, консервативная замена аминокислоты по существу не меняет функциональных свойств представляющего интерес белка, например, способность вариабельной области специфически связывать целевой эпитоп с требуемым сродством. Примеры групп аминокислот, содержащих боковые цепи с аналогичными химическими свойствами, включают алифатические боковые цепи, такие как глицин, аланин, валин, лейцин и изолейцин; алифатические гидроксильные боковые цепи, такие как серии и треонин; амидсодержащие боковые цепи, такие как аспарагин и глутамин; ароматические боковые цепи, такие как фенилаланин, тирозин, и триптофан; основные боковые цепи, такие как лизин, аргинин и гистидин; кислые боковые цепи, такие как аспарагиновая кислота и глутаминовая кислота; и серосодержащие боковые цепи, такие как цистеин и метионин. Группы консервативной замены аминокислот включают, например, валин/лейцин/изолейцин, фенилаланин/тирозин, лизин/аргинин, аланин/валин, глутамат/аспартат и аспарагин/глутамин. Согласно некоторым вариантам реализации консервативная замена аминокислоты может представлять собой замену любого нативного остатка белка на аланин, что используется, например, для аланин-сканирующего мутагенеза. Согласно некоторым вариантам реализации осуществляют консервативную замену, имеющую положительное значение в матрице логарифмического правдоподобия РАМ250, описанной в источнике: Gonnet et al. (1992) Exhaustive Matching of the Entire Protein Sequence Database, Science 256:1443-45, включенном в настоящий документ посредством ссылки. Согласно некоторым вариантам реализации указанная замена представляет собой умеренно консервативную замену, имеющую неотрицательное значение в матрице логарифмического правдоподобия РАМ250.
[000150] Согласно некоторым вариантам реализации положения остатков в легкой цепи или тяжелой цепи иммуноглобулина отличаются за счет одной или большего количества консервативных замен аминокислот. Согласно некоторым вариантам реализации положения остатков в легкой цепи иммуноглобулина или ее функциональном фрагменте (например, фрагменте, обеспечивающем экспрессию и секрецию, например, В-клеткой), не идентичны с легкой цепью, последовательность аминокислот которой приведена в настоящем документе, но отличается одной или большим количеством консервативных замен аминокислот.
[000151] Выражение «эпитоп-связывающий белок» включает белок, содержащий по меньшей мере одну область CDR и способный избирательно распознавать эпитоп, например, способный связывать эпитоп с KD, составляющей приблизительно 1 мкМ или менее (например, KD, составляющей приблизительно 1×10-6 М, 1×10-7 М, 1×10-9 М, 1×10-9 М, 1×10-10 М, 1×10-11 М или приблизительно 1×10-12 М). Для терапевтических эпитоп-связывающих белков (например, терапевтических антител) часто требуется KD в наномолярном или пикомолярном диапазоне.
[000152] Выражение «функциональный фрагмент» включает фрагменты эпитоп-связывающих белков, которые могут экспрессироваться, секретироваться и специфически связываться с эпитопом с KD в микромолярном, наномолярном или пикомолярном диапазоне. Специфическое распознавание включает наличие KD по меньшей мере в микромолярном диапазоне, наномолярном диапазоне или пикомолярном диапазоне.
[000153] Термин «зародышевая линия» в отношении нуклеотидной последовательности иммуноглобулинов включает нуклеотидную последовательность, которая может быть переданы потомству.
[000154] Выражение «тяжелая цепь» или «тяжелая цепь иммуноглобулина» включает последовательность тяжелой цепи иммуноглобулина, в том числе последовательность константной области тяжелой цепи иммуноглобулина любого организма. Вариабельные домены тяжелых цепей включают три области CDR тяжелых цепей и четыре области FR, если не указало иное. Фрагменты тяжелых цепей включают области CDR, CDR и FR, и их комбинации. Типичная тяжелая цепь содержит, за вариабельным доменом (от N-конца к С-концу), домен CH1, шарнир, домен CH2 и домен CH3. Функциональный фрагмент тяжелой цепи включает фрагмент, способный специфически распознавать эпитоп (например, распознавать эпитоп с KD в микромолярном, наномолярном, или пикомолярном диапазоне), который может быть экспрессирован и секретирован клеткой, и который содержит по меньшей мере одну область CDR. Вариабельный домен тяжелой цепи кодирует последовательность гена вариабельной области, которая обычно содержит сегменты VH, DH, и JH, происходящие из репертуара сегментов VH, DH, и JH, имеющихся в зародышевой линии. Последовательности, локализация и номенклатура сегментов V, D и J тяжелой цепи различных организмов можно найти в базе данных IMGT, www.imgt.org.
[000155] Термин «идентичность» в отношении последовательности включает идентичность согласно определению с применением ряда различных алгоритмов, известных в данной области техники, которые могут использоваться для оценки идентичности последовательности нуклеотидов и/или аминокислот. Согласно некоторым вариантам реализации, описанным в настоящем документе, идентичность определяют с применением выравнивания ClustalW v. 1.83 (медленное), со штрафом за внесение пропуска 10,0, штрафом за продолжение пропуска 0,1 и с применением матрицы сходства по Gonnet (MACVECTOR™ 10.0.2, Mac Vector Inc., 2008). Длина сравниваемых в отношении идентичности последовательностей зависит от конкретных последовательностей, но в случае константного домена легкой цепи последовательность должна быть достаточной длины для формирования константного домена легкой цепи, способной к самоассоциации с образованием канонического константного домена легкой цепи, например, способной к образованию двух бета-слоев, содержащих бета-тяжи, и способной взаимодействовать по меньшей мере с одним доменом CH1 человека или мыши. В случае домена CH1, длина последовательности должна быть достаточной для того, чтобы она формировала домен CH1, способный образовывать два бета-слоя, содержащие бета-тяжи, и способный взаимодействовать с по меньшей мере одним константным доменом легкой цепи мыши или человека.
[000156] Выражение «молекула иммуноглобулина» включает две тяжелые цепи иммуноглобулина и две легкие цепи иммуноглобулина. Тяжелые цепи могут быть идентичными или разными, и легкие цепи могут быть идентичными или разными.
[000157] Выражение «легкая цепь» включает последовательность легкой цепи иммуноглобулина любого организма, и, если не указано иное, включает легкие цепи каппа и лямбда человека и VpreB, а также суррогатные легкие цепи. Вариабельные домены легких цепей, как правило, включают три области CDR легких цепей и четыре каркасные (FR) области, если не указано иное. В общем случае, полноразмерная легкая цепь включает, от аминоконца к карбоксильному концу, вариабельный домен, который включает FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, и константную область легкой цепи. Вариабельный домен легкой цепи кодирует последовательность генов вариабельной области легкой цепи, которая обычно содержит сегменты VL и JL, происходящие из репертуара сегментов V и J, присутствующих в зародышевой линии. Последовательности, локализацию и номенклатуру сегментов V и J легких цепей различных организмов можно найти в базе данных IMGT, www.imgt.org. Легкие цепи включают, например, такие легкие цепи, которые не связывают избирательно либо первый, либо второй эпитоп, избирательно связываемый эпитоп-связывающим белком, в состав которого они входят. Легкие цепи также включают связывающие и распознающие один или большее количество эпитопов, или способствующие связыванию и распознаванию тяжелой цепью одного или большего количества эпитопов, избирательно связываемых эпитоп-связывающим белком, в состав которого они входят. Общие или универсальные легкие цепи включают происходящие из гена Vκ1-39Jκ5 человека или гена Vκ3-20Jκ1 человека, и включают их соматически мутированные (например, с созревшей аффинностью) варианты. Двойные легкие цепи (DLC) включают происходящие из локуса легкой цепи, содержащие не более чем два сегмента Vκ человека, например, сегмент гена Vκ1-39 человека и сегмент гена Vκ3-20 человека, и включают их соматически мутированные (например, с созревшей аффинностью) варианты.
[000158] Выражение «соматически гипермутированный» включает нуклеотидную последовательность В-клетки, в которой произошло переключение классов, при этом указанная нуклеотидная последовательность вариабельной области иммуноглобулина (например, кодирующая вариабельный домен тяжелой цепи или включающая последовательность CDR или FR тяжелой цепи) в В-клетке, в которой произошло переключение классов, не идентична нуклеотидной последовательности в В-клетке до переключения классов, например, например, различаются нуклеотидные последовательности участков CDR или каркасных участков В-клетки, в которой не произошло переключение классов, и В-клетки, в которой произошло переключение класса. «Соматически мутированные» последовательности включает нуклеотидные последовательности из В-клеток с созревшей аффинностью, не идентичные соответствующим последовательностям вариабельных областей иммуноглобулина в В-клетках с несозревшей аффинностью (т.е. последовательностям в геноме клеток зародышевой линии). Выражение «соматически мутированный» также включает нуклеотидную последовательность вариабельной области иммуноглобулина из В-клетки после воздействия на указанную В-клетку представляющим интерес эпитопом, при этом указанная нуклеотидная последовательность отличается от соответствующей нуклеотидные последовательности до воздействия на указанную В-клетку указанным представляющим интерес эпитопом. Выражение «соматически мутированный» относится к последовательностям антител, которые образовались у животного, например, мыши, у которой имеются нуклеотидные последовательности вариабельных областей иммуноглобулина человека, в ответ на стимуляцию иммуногеном, и образующиеся в результате врожденных процессов селекции, происходящих у такого животного.
[000159] Термин «нереаранжированная» в отношении нуклеотидные последовательности включает нуклеотидные последовательности, существующие в зародышевой линии животной клетки.
[000160] Выражение «вариабельный домен» включает последовательность аминокислот легкой или тяжелой цепи иммуноглобулина (модифицированную требуемым образом), которая содержит следующие аминокислотные области, от N-конца к С-концу (если не указано иное): FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4.
[000161] Термин «функционально связанный» относится к такой взаимосвязи компонентов, когда указанные функционально связанные компоненты функционируют в соответствии с предполагаемым назначением. В одном случае нуклеотидная последовательность, кодирующая белок, может быть функционально связана с регуляторными последовательностями (например, последовательностью промотора, энхансера, сайленсера и т.п.) таким образом, чтобы сохранялась надлежащая транскрипционная регуляция. В одном случае нуклеотидная последовательность вариабельной области иммуноглобулина (или сегменты V(D)J) может быть функционально связана с нуклеотидной последовательностью константной области иммуноглобулина таким образом, чтобы обеспечивать надлежащую рекомбинацию последовательностей с получением последовательности тяжелой или легкой цепи иммуноглобулина.
[000162] «Функциональный» в настоящем документе, например, в отношении функционального полипептида, включает полипептид, сохраняющий по меньшей мере один тип биологической активности, в норме связанный с нативным белком. В другом случае функциональный сегмент гена иммуноглобулина может включать сегмент гена вариабельной области, способный к продуктивной реаранжировки для получения реаранжированной последовательности гена иммуноглобулина.
[000163] «Нейтральные значения рН» включают значения рН от приблизительно 7,0 до приблизительно 8,0, например, значения рН от приблизительно 7,0 до приблизительно 7,4, например, от приблизительно 7,2 до приблизительно 7,4, например, физиологические значения рН. «Кислотные значения рН» включают значения рН, составляющие 6,0 или менее, например, значения рН от приблизительно 5,0 до приблизительно 6,0, значения рН от приблизительно 5,75 до приблизительно 6,0, например, значения рН в эндосомальном или лизосомальном компартменте.
[000164] Термин «полиморфный вариант» в настоящем документе включает последовательность, в которой один или большее количество нуклеотидов или аминокислот были заменены другими нуклеотидами или аминокислотами по сравнению с заданной последовательностью. Полиморфные аллели сегментов гена вариабельной области тяжелой цепи (генов VH) иммуноглобулина человека в значительной степени представляли собой результат вставки/удаления сегментов гена и однонуклеотидных отличий в кодирующих областях; и первое, и второе потенциально может иметь функциональные последствия для молекулы иммуноглобулина. Примеры распространенных полиморфных аллелей генов VH иммуноглобулинов человека хорошо известны в данной области техники (см., например, US 13/653456, полностью включенный в настоящий документ посредством ссылки).
[000165] Термин «существенный» или «по существу все» по отношению к количеству сегментов гена (например, «по существу все» сегменты генов V, D, или J) включает и функциональные, и нефункциональные сегменты генов и включает, согласно различным вариантам реализации, например, 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более, 98% или более, или 99% или более всех сегментов гена V, D или J. Согласно различным вариантам реализации «по существу все» сегменты генов включают, например, по меньшей мере 95%, 96%, 97%, 98%, или 99% функциональных (т.е. не псевдогенов) сегментов гена).
Не являющиеся человеком животные, у которых имеется реаранжированная последовательность гена вариабельной области тяжелой цепи и, необязательно, ограниченный репертуар нереаранжированных сегментов гена вариабельной области легкой цепи
[000166] Несмотря на то, что были разработаны разнообразные биспецифические антитела с двойными антигенсвязывающими свойствами, специфичностью и аффинностью вариабельных областей легкой цепи или тяжелой цепи в обычных биспецифических антителах приходится в определенной степени жертвовать, поскольку в обычных биспецифических антителах вклад в связывание каждой отдельной антигенной детерминанты вносит вариабельная область только либо тяжелой цепи, либо легкой цепи, тогда как в случае обычных антител в связывании одной и той же антигенной детерминанты могут участвовать вариабельные области как легких, так и тяжелых цепей.
[000167] Таким образом, получение вариабельных областей легких цепей, обладающих способностью связывать антиген независимо от вариабельной области тяжелой цепи, может быть полезным для получения вариабельных доменов легких цепей (VL) для применения в антигенсвязывающих молекулах (например, биспецифических связывающих молекулах, которые содержат константную область тяжелой цепи (например, выбранную из СН1, шарнира, СН2, СН3 и их комбинации), соединенную с VL), в частности, не содержащих вариабельного домена тяжелой цепи, в том числе гетеродимеров, содержащих такую же или аналогичную константную область тяжелой цепи, но имеющих домены VL с другой специфичностью и/или аффинностью.
[000168] Один из подходов к получению таких вариабельных доменов легких цепей, которые могут связываться с антигеном независимо от вариабельной области тяжелой цепи, заключается р использовании селективного давления на нуклеотидные последовательности, которые кодируют вариабельную область или домен легкой цепи (VL) для получения областей CDR3 легкой цепи с более разнообразным антигенсвязывающим репертуаром. Согласно настоящему описанию это может достигаться путем получения генетически модифицированного не являющегося человеком животного, в геноме у которого имеется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека. Поскольку у указанных животных последовательность тяжелой цепи ограничена общими или универсальными (т.е. идентичными или в значительной степени сходными) последовательностями, нуклеотидные последовательности вариабельных областей легких цепей (т.е. гены) вынужденно будут обеспечивать образование областей CDR3 легких цепей с более разнообразными и эффективными антигенсвязывающими свойствами, способными связывать антигенную детерминанту независимо от вариабельных областей тяжелых цепей. Кроме того, согласно настоящему описанию точная замена сегментов гена вариабельной области зародышевой линии (например, путем опосредованного гомологичной рекомбинацией генного таргетинга) обеспечивает получение животных (например, мышей), у которых имеются частично человеческие иммуноглобулиновые локусы. Поскольку в указанных частично человеческих иммуноглобулиновых локусах происходят нормальные реаранжировка, гипермутация и соматическая мутация (например, переключение классов), указанные частично человеческие иммуноглобулиновые локусы продуцируют у указанного животного антитела, которые содержат вариабельные области человека. Гуморальная иммунная система указанных животных по существу аналогична системе животных дикого типа, и содержит нормальные клеточные популяции и нормальные структуры лимфоидных органов - даже если у указанных животных отсутствует полный репертуар сегментов гена вариабельных областей человека. Иммунизация указанных животных (например, мышей) приводит к мощному гуморальному ответу, при котором наблюдается использование большого разнообразия сегментов гена вариабельных областей. Нуклеотидные последовательности, которые кодируют вариабельные области, могут быть идентифицированы и клонированы, затем соединены (например, в системе in vitro) с любыми выбранными последовательностями, например, иммуноглобулином любого изотипа, подходящего для конкретного применения, что приводит к получению антитела или антигенсвязывающего белка, происходящего полностью из последовательностей человека.
[000169] Кроме того, за счет использования животных (например, мышей или крыс), имеющих ограниченный (лимитированный) репертуар сегментов гена вариабельной области легкой цепи, например, ограниченный репертуар сегментов вариабельных областей легких цепей, содержащий один или более, но меньше, чем у дикого типа, сегментов гена VL человека (например, двойной легкой цепи, или «DLC», см. опубликованную заявку на патент США №2013/0198880, полностью включенную в настоящий документ посредством ссылки), в комбинации с нуклеотидной последовательностью реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека согласно описанию выше, может быть получен вариабельный домен легкой цепи иммуноглобулина, способный более эффективно сочетаться с вариабельным доменом тяжелой цепи иммуноглобулина. Кроме того, за счет введения гистидиновых кодонов, например, путем добавления одного или большего количества гистидиновых кодонов или замены одного или большего количества негистидиновых кодонов на гистидиновые кодоны в лимитированных сегментах генов вариабельной области легкой цепи в геноме описанных в настоящем документе не являющихся человеком животных, могут быть получены последовательности аминокислот вариабельных областей легких цепей, которые могут придавать антигенсвязывающим белкам улучшенную способность к рН-зависимой рециклизации (например, биспецифические или триспецифические антитела).
[000170] Согласно некоторым вариантам реализации генетически модифицированные не являющиеся человеком животные согласно описанию в настоящем документе обеспечивают получение большего количества антител при их ограниченном разнообразии, с повышением таким образом вероятности успешного сочетания легких цепей с тяжелыми цепями, образующимися у не являющегося человеком животного, у которого имеется единственная реаранжированная вариабельная область легкой цепи (например, мыши, у которой имеется универсальная легкая цепь (мышь «ULC»); см., например, опубликованную через 18 месяцев с даты приоритета патентную заявку США 2013/0185821, включенную в настоящий документ посредством ссылки). Согласно некоторым вариантам реализации указанные легкие цепи могут сами по себе проявлять антигенсвязывающие свойства. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного может быть индуцирован синтез антигенсвязывающих белков, проявляющих антигенспецифичность, обеспечиваемую их легкими цепями (например, путем ограничения репертуара тяжелых цепей иммуноглобулина мыши или крысы; например, путем замены локуса тяжелой цепи мыши или крысы на локус, содержащий единственную нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека). Согласно некоторым вариантам реализации антигенсвязывающие белки (например, антитела), продуцированные такими животными, являются специфическими в отношении конкретного первого эпитопа (например, эффекторных антигенов, цитотоксических молекул, Fc-рецепторов, токсинов, активирующих или ингибирующих рецепторов, маркеров Т-клеток, транспортеров иммуноглобулинов и т.п.) за счет связывания их легкими цепями. Такие эпитоп-специфические легкие цепи человека, происходящие из указанных не являющихся человеком животных, могут коэкспрессироваться вместе с тяжелыми цепями человека, полученными из мыши с лимитированным репертуаром легких цепей, например, мыши или крысы ULC, при этом указанную тяжелую цепь выбирают на основании способности связывать второй эпитоп (например, второй эпитоп на другом антигене).
[000171] Согласно различным аспектам предложено не являющееся человеком животное, геном зародышевой линии которого содвержит локус тяжелой цепи иммуноглобулина, который содержит нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (т.е. реаранжированную последовательность VDJ тяжелой цепи). Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с человеческой или имеющей нечеловеческое происхождение последовательность константной области тяжелой цепи. Согласно некоторым вариантам реализации вариабельный домен тяжелой цепи иммуноглобулина, кодируемый указанной нуклеотидной последовательностью реаранжированной вариабельной области тяжелой цепи, не является иммуногенным для указанного не являющегося человеком животного. Согласно некоторым вариантам реализации указанное не являющееся человеком животное модифицируют таким образом, чтобы у него имелась нуклеотидная последовательность, которая кодирует две копии, три копии, четыре или более копии реаранжированного вариабельного домена тяжелой цепи, функционально связанного с константным доменом тяжелой цепи. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность кодирует несколько копий нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека. Например, указанная нуклеотидная последовательность может кодировать по меньшей мере одну, два, три, четыре, пять копий нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность кодирует 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 копий нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека. Согласно некоторым вариантам реализации указанный локус содержит несколько копий нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанной с последовательностью гена константного домена тяжелой цепи.
[000172] Согласно другим аспектам предложено не являющееся человеком животное, генетически сконструированное таким образом, что у него имеется локус легкой цепи иммуноглобулина, кодирующий реаранжированный вариабельный домен тяжелой цепи (т.е. локус легкой цепи, который содержит нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), функционально связанную с человеческой или имеющей нечеловеческое происхождение последовательностью гена константной области легкой цепи. Например, согласно некоторым вариантам реализации нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (т.е. предварительно сконструированная область VDJ; т.е. общая или универсальная последовательность тяжелой цепи) может быть функционально связана с последовательностью гена константной области легкой цепи путем нацеливания реаранжированной последовательности тяжелой цепи в локусы легких цепей мыши или крысы, либо каппа, либо лямбда. Соответственно, согласно некоторым вариантам реализации указанная нуклеотидная последовательность, кодирующая реаранжированный вариабельный домен тяжелой цепи, присутствует в геноме зародышевой линии не являющегося человеком животного. Согласно некоторым вариантам реализации указанный реаранжированный вариабельный домен тяжелой цепи, экспрессируемый генетически модифицированным не являющимся человеком животным, не является иммуногенным для указанного не являющегося человеком животного. Согласно некоторым вариантам реализации указанное не являющееся человеком животное модифицируют таким образом, чтобы у него имелась нуклеотидная последовательность, которая кодирует две копии, три копии, четыре или более копии нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанной о константным доменом легкой цепи. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность может кодировать несколько копий нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека. Например, указанная нуклеотидная последовательность кодирует по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 копий нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека. Согласно некоторым вариантам реализации указанный локус содержит несколько копий нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связанный с константным доменом легкой цепи последовательность гена.
[000173] Согласно различным аспектам локус легкой цепи иммуноглобулина у не являющихся человеком животных, описанных в настоящем документе, содержит ограниченный репертуар сегментов гена вариабельной области легкой цепи, например, один или более, но меньше, чем у дикого типа, сегментов гена VL человека; и один или большее количество сегменты генов JL человека, функционально связанных с имеющей нечеловеческое происхождение нуклеотидной последовательностью константной области легкой цепи. Соответственно, предложены генетически модифицированные не являющиеся человеком животные, в геноме которых содержатся: (i) локус тяжелой цепи иммуноглобулина, который содержит нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи человека, функционально связанную с человеческой или имеющей нечеловеческое происхождение нуклеотидной последовательностью константной области тяжелой цепи; и (ii) локус легкой цепи иммуноглобулина, содержащий два или более, но меньше, чем у дикого типа, сегментов гена VL и JL вариабельной области легкой цепи иммуноглобулина человека, функционально связанных с нуклеотидной последовательностью константной области легкой цепи. Согласно некоторым вариантам реализации указанная константная область легкой цепи представляет собой константную область крысы или мыши, например, константную область Сκ крысы или мыши. Согласно некоторым вариантам реализации указанные сегменты генов вариабельной области человека способны к реаранжировке и кодированию вариабельных доменов человека в антителе, и у указанного не являющегося человеком животного не содержится эндогенный сегмент гена VL. Согласно некоторым вариантам реализации у указанного не являющегося человеком животного имеется пять сегментов гена Jκ человека, например, сегменты генов Jκ1, Jκ2, Jκ3, Jκ4 и Jκ5. Согласно некоторым вариантам реализации указанный локус легкой цепи иммуноглобулина содержит два сегмента гена VL человека, Vκ1-39 и Vκ3-20. Согласно некоторым вариантам реализации один или большее количество (например, 2, 3, 4 или 5) сегментов гена VL человека, и два или большее количество сегментов гена JL человека присутствуют в эндогенном локусе легкой цепи, например, в эндогенном локусе легкой цепи каппа. Согласно некоторым вариантам реализации у указанной мыши имеется функциональный локус легкой цепи λ. Согласно некоторым вариантам реализации у указанной мыши имеется нефункциональный локус легкой цепи λ. Согласно некоторым вариантам реализации указанные один или большее количество сегментов гена VH человека, один или большее количество сегментов гена DH человека, и один или большее количество сегментов гена JH человека функционально связаны с последовательностью константной области тяжелой цепи мыши или крысы.
[000174] Согласно некоторым вариантам реализации генетически модифицированные мыши, в геноме которых имеются (i) локус тяжелой цепи иммуноглобулина, который содержит нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи человека, функционально связанную с человеческой или имеющей нечеловеческое происхождение нуклеотидной последовательностью константной области тяжелой цепи, и (ii) локус легкой цепи иммуноглобулина, содержащий два или более, но меньше, чем у дикого типа, сегментов гена VL и JL вариабельной области легкой цепи иммуноглобулина человека, функционально связанных с нуклеотидной последовательностью константной области легкой цепи, наблюдались количества CD19+ В-клеток и зрелых В-клеток, по существу совпадающие с количествами, наблюдаемыми у мышей дикого типа или мышей, у которых имелись другие модификации иммуноглобулиновых локусов (т.е. генетически модифицированных контрольных мышей; например, мышей VELOCIMMUNE®, гуморальная иммунная система которых функционирует так же, как и у мыши дикого типа). Согласно некоторым вариантам реализации у таких мышей наблюдалось увеличение количества незрелых В-клеток в селезенке по сравнению с генетически модифицированными контрольными мышами. Согласно конкретным вариантам реализации у таких мышей наблюдалось приблизительно 2-кратное, приблизительно 3-кратное, приблизительно 4-кратное или приблизительно 5-кратное или более увеличение количества незрелых В-клеток в селезенке по сравнению с генетически модифицированными контрольными мышами. Согласно некоторым вариантам реализации такие мыши также по существу аналогичны мышам дикого типа или генетически модифицированным контрольным мышам в отношении частоты использования легкой цепи каппа и гамма в В-клетках селезенки. Согласно некоторым вариантам реализации у таких мышей наблюдалось повышенные уровни поверхностных IgM на В-клетках селезенки (т.е. более высокие уровни поверхностной экспрессии IgM на клетку) по сравнению с генетически модифицированными контрольными мышами. Согласно некоторым вариантам реализации у таких мышей наблюдалось измененное развитие периферических В-клеток на различных стадиях развития В-клеток в компартменте селезенки по сравнению с генетически модифицированными контрольными мышами, например, увеличение количества незрелых В-клеток, В-клеток Т1 и/или В-клеток маргинальной зоны. Согласно некоторым вариантам реализации в компартменте костного мозга у таких мышей наблюдались количества CD19+ В-клеток, по существу аналогичные количествам, наблюдаемым у генетически модифицированных контрольных мышей. Согласно некоторым вариантам реализации у таких мышей наблюдалось меньшее количество про-В-клеток в костном мозге по сравнению с генетически модифицированными контрольными мышами. Согласно конкретным вариантам реализации количества про-В-клеток в компартменте костного мозга снижаются приблизительно в 2 раза, приблизительно в 5 раз, приблизительно в 10 раз, приблизительно в 15 раз, приблизительно в 20 раз, приблизительно в 25 раз или более по сравнению с генетически модифицированными контрольными мышами. Согласно некоторым вариантам реализации у таких мышей наблюдалось приблизительно в 2 раза, приблизительно в 3 раза, приблизительно в 4 раза, приблизительно в 5 раз и т.п. меньше незрелых и/или зрелых В клеток в костном мозге по сравнению с генетически модифицированными контрольными мышами. Согласно некоторым вариантам реализации у таких мышей наблюдается незначительное повышение (например, 2-кратное увеличение) частоты использования генов легкой цепи лямбда в компартменте костного мозга по сравнению с генетически модифицированными контрольными мышами.
[000175] Согласно другому аспекту предложено не являющееся человеком животное, у которого имеется генетически модифицированный иммуноглобулиновый локус, содержащий: (а) первую нуклеотидную последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи {т.е. первая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), при этом указанная первая нуклеотидная последовательность функционально связана с последовательностью гена константной области легкой цепи; и (b) вторая нуклеотидная последовательность, которая кодирует вариабельный домен легкой цепи человека (т.е. вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека), при этом указанная вторая нуклеотидная последовательность функционально связана с последовательностью гена константной области тяжелой цепи. Например, согласно некоторым вариантам реализации реаранжированная тяжелая цепь предварительно сконструированной области VDJ (т.е. нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека; т.е. общая или универсальная последовательность тяжелой цепи) может быть функционально связана с последовательностью гена константной области легкой цепи путем нацеливания реаранжированной последовательности тяжелой цепи в локусы легких цепей мыши, либо каппа, либо лямбда. Соответственно, как и в других вариантах реализации, указанный генетически сконструированный иммуноглобулиновый локус может присутствовать в геноме зародышевой линии не являющегося человеком животного. Генетически модифицированные не являющиеся человеком животные, у которых имеются нуклеотидные последовательности вариабельных областей легких цепей иммуноглобулина человека, функционально связанные с последовательностями генов константных областей тяжелых цепей, описаны в опубликованной через 18 месяцев с даты приоритета патентной заявке США №2012/0096572, включенной в настоящий документ посредством ссылки. Согласно некоторым вариантам реализации указанная первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи, функционально связана с последовательностью гена константной области легкой цепи κ. Согласно некоторым вариантам реализации указанная первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи, функционально связана с последовательностью гена константной области мыши или крысы легкой цепи κ. Согласно некоторым вариантам реализации указанная первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи, функционально связана с последовательностью гена константной области легкой цепи человека κ. Согласно некоторым вариантам реализации указанная первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи, функционально связана с последовательностью гена константной области легкой цепи λ. Согласно некоторым вариантам реализации указанная первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи, функционально связана с последовательностью гена константной области мыши или крысы легкой цепи λ. Согласно некоторым вариантам реализации указанная первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи, функционально связана с последовательностью гена константной области легкой цепи λ человека.
[000176] Согласно некоторым вариантам реализации у генетически модифицированной мыши, у которой имеется локус легкой цепи иммуноглобулина, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, и локус тяжелой цепи иммуноглобулина, содержащий нереаранжированные последовательности вариабельного домена легкой цепи иммуноглобулина человека (например, гены легкой цепи каппа), частота встречаемости CD19+ и пре-В-клеток в костном мозге изменена по сравнению с мышью дикого типа или генетически модифицированной мышью с другими модификациями в иммуноглобулиновом локусе (т.е. генетически модифицированными контрольными мышами; например, мышами VELOCIMMUNE®, гуморальная иммунная система которых функционирует так же, как и у мыши дикого типа). Согласно конкретным вариантам реализации количества CD19+ В-клеток и пре-В-клеток в костном мозге в 2 раза ниже, в 3 раза ниже, в 4 раза ниже или в 5 раз ниже по сравнению с мышью дикого типа или контрольной мышью с генетически модифицированным иммуноглобулиновым локусом. Согласно конкретным вариантам реализации количество незрелых В-клеток в костном мозге в 2 раза меньше, в 3 раза меньше, в 4 раза меньше или в 5 раз меньше, чем у мыши дикого типа или контрольной мыши с генетически модифицированным иммуноглобулиновым локусом. Согласно некоторым вариантам реализации у генетически модифицированной мыши, у которой имеется локус легкой цепи иммуноглобулина, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, и локус тяжелой цепи иммуноглобулина, содержащий нереаранжированные последовательности вариабельного домена легкой цепи иммуноглобулина человека (например, гены легкой цепи каппа), в клетках костного мозга не экспрессируются или по существу не экспрессируются гены легких цепей лямбде-. Согласно некоторым вариантам реализации у генетически модифицированной мыши, у которой имеется локус легкой цепи иммуноглобулина, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, и локус тяжелой цепи иммуноглобулина, содержащий нереаранжированные последовательности вариабельного домена легкой цепи иммуноглобулина человека (например, гены легкой цепи каппа), понижены уровни В-клеток селезенки по сравнению с мышью дикого типа или контрольной мышью с генетически модифицированным иммуноглобулиновым локусом. Согласно конкретным вариантам реализации уровни В-клеток селезенки и зрелых В-клеток в 2 раза ниже, в 3 раза ниже, в 4 раза ниже или в 5 раз ниже, чем у мыши дикого типа или контрольной мыши с генетически модифицированным иммуноглобулиновым локусом. Согласно некоторым вариантам реализации у генетически модифицированной мыши, у которой имеется локус легкой цепи иммуноглобулина, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, и локус тяжелой цепи иммуноглобулина, содержащий нереаранжированные последовательности вариабельного домена легкой цепи иммуноглобулина человека (например, гены легкой цепи каппа), в В-клетках селезенки не экспрессируются или по существу не экспрессируются гены легкой цепи лямбда. Согласно конкретным вариантам реализации генетически модифицированная мышь, у которой имеется локус легкой цепи иммуноглобулина, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, и локус тяжелой цепи иммуноглобулина, содержащий нереаранжированные последовательности вариабельного домена легкой цепи иммуноглобулина человека (например, гены легкой цепи каппа), чаще встречаются клетки в фазе Т1 в селезенке по сравнению с мышью дикого типа или контрольной мышью с генетически модифицированным иммуноглобулиновым локусом.
[000177] Согласно некоторым вариантам реализации указанное не являющееся человеком животное представляет собой млекопитающее. Хотя в настоящем документе подробно обсуждаются варианты реализации, задействующие реаранжированный вариабельный домен тяжелой цепи человека у мыши (т.е. у мыши имеется иммуноглобулиновый локус, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), также предложены другие не являющиеся человеком животные, у которых имеется генетически модифицированный иммуноглобулиновый локус, кодирующий реаранжированный вариабельный домен тяжелой цепи человека. Такие не являющиеся человеком животные включают любых животных, которые могут быть генетически модифицированы для экспрессии нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека согласно настоящему описанию, в том числе, например, млекопитающих, например, мышь, крысу, кролика, свинью, крупный рогатый скот (например. корову, бык, буйвол), оленя, овцу, козу, курицу, кошку, собаку, хорька, примата (например. игрунку, макака-резус) и т.п. Например, для не являющихся человеком животные, подходящие генетически модифицируемые ЭС-клетки которых не являются легкодоступными, используются другие способы получения не являющегося человеком животного, у которого имеется генетическая модификация. Такие способы включают, например, модификацию генома не являющейся ЭС клетки (например, фибробласта или индуцированной плюрипотентной клетки) н использование переноса ядра соматической клетки (SCNT) для переноса указанного генетически модифицированного генома в подходящую клетку, например, лишенный ядра ооцит, и гестацию модифицированной клетки (например, модифицированного ооцита) у не являющегося человеком животного в подходящих условиях для формирования эмбриона. Способы модификации генома не являющегося человеком животного (например, генома свиньи, коровы, грызуна, курицы и т.п.) включают, например, использование цинк-пальцевой нуклеазы (ZFN) или подобной транскрипционному активатору эффекторной нуклеазы (TALEN) для модификации генома с включением в него нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека.
[000178] Согласно некоторым вариантам реализации указанное не являющееся человеком животное представляет собой небольшое млекопитающее, например, из надсемейства Dipodoidea или Muroidea. Согласно некоторым вариантам реализации указанное генетически модифицированное животное представляет собой грызуна. Согласно некоторым вариантам реализации грызуна выбирают из мыши, крысы и хомяка. Согласно некоторым вариантам реализации грызуна выбирают из надсемейства Muroidea. Согласно некоторым вариантам реализации указанное генетически модифицированное животное относится к семейству, выбранному из Calomyscidae (например, мышевидные хомяки), Cricetidae (например, хомяк, крысы и мыши Нового Света, полевки), Muridae (настоящие мыши и крысы, песчанки, иглистые мыши, косматые хомяки), Nesomyidae (лазающие мыши, скальные хомячки, белохвостые крысы, малагасийские крысы (воалаво) и мыши), Platacanthomyidae (например, колючие соневидные хомяки) и Spalacidae (например, слепыши, бамбуковые крысы и цокоры). Согласно конкретному варианту реализации указанного генетически модифицированного грызуна выбирают из настоящих мышей или крыс (семейство Muridae), песчанки, иглистой мыши и косматого хомяка. Согласно некоторым вариантам реализации указанная генетически модифицированная мышь является представителем семейства Muridae. Согласно некоторым вариантам реализации указанное животное представляет собой грызуна. Согласно конкретным вариантам реализации грызуна выбирают из мыши и крысы. Согласно некоторым вариантам реализации указанное не являющееся человеком животное представляет собой мышь.
[000179] Согласно некоторым вариантам реализации указанное не являющееся человеком животное представляет собой грызуна, который представляет собой мышь линии C57BL, выбранную из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6N, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/01a. Согласно другому варианту реализации указанная мышь относится к линии 129. Согласно некоторым вариантам реализации указанную линию 129 выбирают из группы, состоящей из 129Р1, 129Р2, 129РЗ, 129X1, 129S1 (например, 129S1/SV, 129S1/SvIm), 129S2, 129S4, 129S5, 129S9/SvEVH, 129S6 (129/SvEvTac), 129S7, 129S8, 129Т1, 129Т2 (см., например, Festing etal. (1999) Revised nomenclature for strain 129 mice, Mammalian Genome 10:836, см. также, Auerbach et al. (2000) Establishment and Chimera Analysis of 129/SvEv- and C57BL/6-Derived Mouse Embryonic Stem Cell Lines). Согласно некоторым вариантам реализации указанная генетически модифицированная мышь представляет собой гибрид вышеупомянутой линии 129 и вышеупомянутой линии C57BL (например, линию C57BL/6). Согласно другому варианту реализации указанная мышь представляет собой гибрид вышеупомянутых линий 129, или гибрид вышеупомянутых линий C57BL/6. Согласно некоторым вариантам реализации указанная линия 129 указанной смеси относится к линии 129S6 (129/SvEvTac). Согласно другому варианту реализации указанная мышь представляет собой гибрид линий, происходящих из 129/SvEv и C57BL/6. Согласно конкретному варианту реализации указанная мышь представляет собой гибрид линий, происходящих из 129/SvEv и C57BL/6, согласно описанию у Auerbach et al. 2000 BioTechniques 29:1024-1032. Согласно другому варианту реализации указанная мышь относится к линии BALB, например, линии BALB/c. Согласно другому варианту реализации указанная мышь представляет собой гибрид линии BALB (например, BALB/c линия) и другой вышеупомянутой линии.
[000180] Согласно некоторым вариантам реализации указанное не являющееся человеком животное представляет собой крысу. Согласно некоторым вариантам реализации указанную крысу выбирают из крысы линии Wistar, линии LEA, линии Sprague Dawley, линии Fischer, F344, F6, ACI и Dark Agouti (DA). Согласно некоторым вариантам реализации указанная крыса линия представляет собой гибрид двух или более линий, выбранных из группы, состоящей из Wistar, LEA, Sprague Dawley, Fischer, F344, F6, ACI и Dark Agouti (DA).
[000181] Согласно некоторым вариантам реализации генетически модифицированная мышь, в геноме зародышевой линии которой имеется локус тяжелой цепи иммуноглобулина, который содержит нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, производит популяции зрелых и незрелых В-клеток селезенки, по существу нормальных при сравнении с мышью дикого типа. Согласно некоторым вариантам реализации в В-клетках селезенки такой генетически модифицированной мыши наблюдается незначительное снижение использования последовательностей генов легкой цепи лямбда по сравнению с диким типом. Согласно конкретным вариантам реализации в В-клетках селезенки такой генетически модифицированной мыши последовательности генов легкой цепи лямбда используются в 2 раза, 3 раза, 4 раза или 5 раз реже, чем у дикого типа. Согласно некоторым вариантам реализации у такой генетически модифицированной мыши наблюдается незначительное уменьшение популяции В-клеток Т1 селезенки и увеличение популяции В-клеток маргинальной зоны селезенки по сравнению с диким типом. Согласно некоторым вариантам реализации у такой генетически модифицированной мыши имеются практически нормальные популяции В-клеток в костном мозге. Согласно некоторым вариантам реализации у такой генетически модифицированной мыши последовательности генов лямбда используются с частотой в два или более чем в два раза меньшей, чем последовательности генов лямбда у дикого типа.
[000182] Согласно различным вариантам реализации, описанным в настоящем документе, реаранжированный вариабельный домен тяжелой цепи (например, нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека) получен из последовательности гена или сегмента V, D и J человека. Согласно некоторым вариантам реализации указанный реаранжированный вариабельный домен тяжелой цепи получен из сегмента V зародышевой линии человека, сегмента D зародышевой линии человека и сегмента J зародышевой линии человека. Согласно некоторым вариантам реализации указанный сегмент VH человека соответствует наблюдаемым в популяции человека вариантам.
[000183] Согласно различным вариантам реализации, описанным в настоящем документе, сегмент V гена человека выбирают из группы, состоящей из VH1-2, VH1-3, VH1-8, VH11-18, VH1-24, VH1-45, VH1-46, VH1-58, VH1-69, VH2-5, VH2-26, VH2-70, VH3-7, VH3-9, VH3-11, VH3-13, VH3-15, VH3-16, VH3-20, VH3-21, VH3-23, VH3-30, VH3-30-3, VH 3-30-5, VH3-33, VH3-35, VH3-38, VH3-43, VH3-48, VH3-49, VH3-53, VH3-64, VH3-66, VH3-72, VH3-73, VH3-74, VH4-4, VH4-28, VH4-30-1, VH4-30-2, VH4-30-4, VH4-31, VH4-34, VH4-39, VH4-59, VH4-61, VH5-51, VH6-1, VH7-4-l, VH7-81 и их полиморфных вариантов. Согласно некоторым вариантам реализации указанный сегмент V человека представляет собой VH3-23 или его полиморфный вариант. Согласно различным вариантам реализации, описанным в настоящем документе, сегмент О гена человека выбирают из группы, состоящей из D1-1, D1-7, D1-14, D1-20, D1-26, D2-2, D2-8, D2-15, D2-21, D3-3, D3-9, D3-10, D3-16, D3-22, D4-4, D4-11, D4-17, D4-23, D5-12, D5-5, D5-18, D5-24, D6-6, D6-13, D6-19, D6-25, D7-27 и их полиморфных вариантов. Согласно некоторым вариантам реализации последовательность константной области тяжелой цепи человека или не являющегося человеком животного включает последовательность, выбранную из СН1, шарнира, СН2, СН3 и их комбинации. Согласно конкретным вариантам реализации указанная последовательность константной области включает CH1, шарнир, CH2 и CH3. Согласно различным вариантам реализации, описанным в настоящем документе, сегмент J гена человека выбирают из группы, состоящей из JH1, JH2, JH3, JH4, JH5, JH6 и их полиморфных вариантов. Согласно некоторым вариантам реализации нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека кодирует последовательность человека VH3-23/GY/JH4-4 (SEQ ID NO: 137). Согласно некоторым вариантам реализации реаранжированный вариабельный домен тяжелой цепи, кодируемый и экспрессируемый из указанной нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина, человека, содержит последовательность человека VH3-23/X1X2/J (где X1 представляет собой любую аминокислоту, и Х2 представляет собой любую аминокислоту). Согласно некоторым вариантам реализации X1 представляет собой Gly и Х2 представляет собой Tyr. Согласно некоторым вариантам реализации указанный реаранжированный вариабельный домен тяжелой цепи содержит последовательность человека VH3-23/X1X2/JH4-4 (где X1 представляет собой любую аминокислоту, и Х2 представляет собой любую аминокислоту). Согласно некоторым вариантам реализации Х2 представляет собой аминокислоту, содержащую фенильную группу. Согласно конкретным вариантам реализации Х2 выбирают из Tyr и Phe.
[000184] Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека содержит сегмент D человека, который не является аутореактивным (является неиммуногенным) для указанного животного. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность содержит способный экспрессироваться сегмент D человека в вариабельной последовательности тяжелой цепи зрелой В-клетки мыши. Согласно некоторым вариантам реализации указанный D сегмент представляет собой сегмент, который был экспрессирован у мыши, у которой имеется гуманизированный иммуноглобулиновый локус содержащий сегмент VH человека, сегмент D человека и сегмент JH человека.
[000185] В различных вариантах реализации используются или содержатся признаки или информация о последовательностях, происходящих из гуманизированных мышей VELOCIMMUNE®. У гуманизированных мышей VELOCIMMUNE® в зародышевой линий происходит точная масштабная замена вариабельных областей тяжелых цепей иммуноглобулина (IgH) и легких цепей иммуноглобулина (например, легкой цепи к, IgK) мыши на соответствующие вариабельные области иммуноглобулина человека, в указанных эндогенных локусах (см., например, US 6596541 и US 8502018, содержание которых полностью включено в настоящий документ посредством ссылки). Всего приблизительно шесть мегабаз локусов мыши заменяют на приблизительно 1,5 мегабаз геномной последовательности человека. Указанная точная замена приводит к получению мыши с гибридными иммуноглобулиновыми локусами, продуцирующей тяжелые и легкие цепи с вариабельными областями человека и константной областью мыши. При точной замене сегментов VH-D-JH и Vκ-Jκ мыши фланкирующие последовательности мыши в гибридных иммуноглобулиновых локусах остаются интактными и функциональными. Гуморальная иммунная система указанной мыши функционирует так же, как и у мыши дикого типа. Развитие В-клеток происходит беспрепятственно во всех значимых отношениях, и при стимуляции антигеном у мыши образуется значительное разнообразие вариабельных областей человека. Кроме того, у гуманизированных мышей VELOCIMMUNE® наблюдается по существу нормальный ответ дикого типа на иммунизацию, отличающийся только в одном значимом аспекте от ответа у мышей дикого типа - вариабельные области, синтезируемые в ответ на иммунизацию, являются полностью человеческими. Получение гуманизированных мышей VELOCIMMUNE® возможно благодаря тому, что реаранжировка сегментов гена тяжелых и легких цепей к иммуноглобулинов у человека и у мыши происходит аналогичным образом. Хотя указанные локусы неидентичны, они достаточно сходны для того, чтобы гуманизация генного локуса вариабельной области тяжелой цепи могла осуществляться путем замены приблизительно трех миллионов пар оснований непрерывной последовательности мыши, которая содержит все сегменты генов VH, D и JH, приблизительно 1 миллионом оснований непрерывной геномной последовательности человека, захватывающей по существу эквивалентную последовательность кз локуса иммуноглобулина человека. Например, согласно некоторым вариантам реализации сегмент D получен из тяжелой цепи, экспрессируемой в зрелой В-клетке гуманизированной мыши VELOCIMMUNE®, иммунизированной антигеном, при этом указанный сегмент D вносит вклад не более чем в две аминокислоты в последовательность CDR3 тяжелой цепи.
[000186] Согласно конкретным вариантам реализации предложена мышь VELOCIMMUNE®, у которой имеется локус тяжелой цепи иммуноглобулина, кодирующий реаранжированный вариабельный домен тяжелой цепи (т.е. имеется локус тяжелой цепи иммуноглобулина, который содержит нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека). У модифицированной таким образом мыши VELOCIMMUNE® происходит замена сегментов гена вариабельной области тяжелой цепи иммуноглобулина мыши на нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (т.е. универсальную последовательность тяжелой цепи в эндогенном локусе тяжелой цепи) и замена сегментов гена к вариабельной области легкой цепи иммуноглобулина мыши по меньшей мере на 40 сегментов гена Vκ человека и пять сегментов гена Jκ человека. Согласно некоторым вариантам реализации указанные сегменты генов человека Vκ выбирают из группы, состоящей из Vκ1-5, Vκ1-6, Vκ1-8, Vκ1-9, Vκ1-12, Vκ1-13, Vκ1-16, Vκ1-17, Vκ1-22, Vκ1-27, Vκ1-32, Vκ1-33, Vκ1-35, Vκ1-37, Vκ1-39, Vκ1D-8, Vκ1D-12, Vκ1D-13, Vκ1D-16, Vκ1D-17, Vκ1D-22, Vκ1D-27, Vκ1D-32, Vκ1D-33, Vκ1D-35, Vκ1D-37, Vκ1D-39, Vκ1D-42, Vκ1D-43, Vκ1-NL1, Vκ2-4, Vκ2-10, Vκ2-14, Vκ2-18, Vκ2-19, Vκ2-23, Vκ2-24, Vκ2-26, Vκ2-28, Vκ2-29, Vκ2-30, Vκ2-36, Vκ2-38, Vκ2-40, Vκ2D-10, Vκ2D-14, Vκ2D-18, Vκ2D-19, Vκ2D-23, Vκ2D-24. Vκ2D-26, Vκ2D-28, Vκ2D-29, Vκ2D-30, Vκ2D-36, Vκ2D-38, Vκ2D-40, Vκ3-7, Vκ3-11, Vκ3-15, Vκ3-20, Vκ3-25, Vκ3-31, Vκ3-34, Vκ3D-7, Vκ3D-7, Vκ3D-11, Vκ3D-15, Vκ3D-15, Vκ3D-20, Vic3D-25, Vκ3D-31, Vκ3D-34, Vκ3-NL1, Vκ3-NL2, Vκ3-NL3, Vκ3-NL4, Vκ3-NL5, Vκ4-1, Vκ5-2, Vκ6-21, Vκ6D-21, Vκ6D-41 и Vκ7-3. Согласно некоторым вариантам реализации указанные сегменты генов человека Vκ содержат Vκ4-1, Vκ5-2, Vκ7-3, Vκ2-4, Vκ1-5 и Vκ1-6. Согласно одному варианту реализации сегменты генов Vκ содержат Vκ3-7, Vκ1-8, VκI-9, Vκ2-10, Vκ3-1 1, Vκ1-12, VκI-13, Vκ2-14, Vκ3-15 и Vκ1-16. Согласно некоторым вариантам реализации указанный человека Vκ сегменты генов содержат Vκ1-17, Vκ2-18, Vκ2-19, Vκ3-20, Vκ6-21, Vκ1-22, Vκ1-23, Vκ2-24, Vκ3-25, Vκ2-26, VκI-27, Vκ2-28, Vκ2-29 и Vκ2-30. Согласно некоторым вариантам реализации указанный человека Vκ сегменты генов содержат Vκ3-31, VκI-32, Vκ1-33, Vκ3-34, VκI-35, Vκ2-36, Vκ1-37, Vκ2-38, Vκ1-39 и Vκ2-40. Согласно конкретным вариантам реализации указанные сегменты генов Vκ содержат непрерывные сегменты генов к иммуноглобулина человека, захватывающие локус легкой цепи к иммуноглобулина человека от Vκ4-1 до Vκ2-40 включительно, а сегменты генов Jκ содержат непрерывные сегменты генов, захватывающие локус легкой цепи к иммуноглобулина человека от Jκ1 до Jκ5 включительно. Согласно некоторым вариантам реализации указанная реаранжированная нуклеотидная последовательность вариабельного домена тяжелой цепи человека функционально связана с последовательностью константной области тяжелой цепи мыши. Мышь VELOCIMMUNE®, у которой имеется локус тяжелой цепи иммуноглобулина, кодирующий реаранжированный вариабельный домен тяжелой цепи (т.е. у которой имеется локус тяжелой цепи иммуноглобулина, который содержит нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), может использоваться в любых описанных в настоящем документе аспектах, вариантам реализации, способах и т.п.
[000187] Согласно различным вариантам реализации нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана человеческой или мышиной последовательностью гена константной области тяжелой цепи (например, последовательность гена константной области тяжелой цепи, которая кодирует изотип иммуноглобулина, выбранный из IgM, IgD, IgA, IgE, IgG, и их комбинаций). Например, предложены генетически модифицированные не являющиеся человеком животные, у которых имеются иммуноглобулиновые локусы, в которых: (а) первая нуклеотидная последовательность кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. первая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), при этом указанная первая нуклеотидная последовательность функционально связана с человеческой или имеющей нечеловеческое происхождение последовательностью гена константной области тяжелой цепи; и (b) вторая нуклеотидная последовательность кодирует вариабельный домен легкой цепи (т.е. вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека), при этом указанная вторая нуклеотидная последовательность функционально связана с человеческой или имеющей нечеловеческое происхождение последовательностью гена константной области легкой цепи. Согласно некоторым вариантам реализации указанную последовательность гена константной области тяжелой цепи человека выбирают из CH1, шарнир, CH2, CH3 и их комбинаций. Согласно некоторым вариантам реализации последовательность гена константной области тяжелой цепи мыши выбирают из CH1, шарнира, CH2, CH3 и их комбинаций. Согласно некоторым вариантам реализации дальнейшая замена определенных последовательностей генов константных областей не являющегося человеком животного на последовательности генов человека (например, замена последовательности CH1 мыши на последовательность CH1 человека, и замена последовательности CL мыши на последовательность CL человека) приводит к получению генетически модифицированных не являющихся человеком животных с гибридными иммуноглобулиновыми локусами, продуцирующих антитела, содержащие вариабельные области человека и частично человеческие константные области, подходящие для, например, получения полностью человеческих фрагментов антител, например, полностью человеческих фрагментов Fab. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области тяжелой цепи крысы. Согласно некоторым вариантам реализации указанную последовательность гена константной области тяжелой цепи крысы выбирают из CH1, шарнира, CH2, CH3 и их комбинаций. Согласно различным вариантам реализации указанный генетически модифицированный локус тяжелой цепи иммуноглобулина не являющегося человеком животного содержит две копии, три копии, четыре или более копии нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связанный с последовательностью гена константного домена тяжелой цепи. Согласно конкретным вариантам реализации указанный локус содержит несколько копий нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанной с последовательностью гена константного домена тяжелой цепи.
[000188] Согласно различным вариантам реализации нуклеотидная последовательность константной области тяжелой цепи содержит модификацию в CH2 или CH3, при этом указанная модификация повышает аффинность последовательности аминокислот константной области тяжелой цепи в отношении FcRn в кислой среде (например, в эндосоме, где значение рН варьирует от приблизительно 5,5 до приблизительно 6,0). Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи кодирует последовательность аминокислот константной области тяжелой цепи человека, содержащую модификацию в положении 250 согласно нумерации EU (263 согласно нумерации Kabat) (например, Е или Q); 250 согласно нумерации EU (263 согласно нумерации Kabat) и 428 согласно нумерации EU (459 согласно нумерации Kabat) (например, L или F); 252 согласно нумерации EU (265 согласно нумерации Kabat) (например, L/Y/F/W или Т), 254 согласно нумерации EU (267 согласно нумерации Kabat) (например, S или Т), и 256 согласно нумерации EU (269 согласно нумерации Kabat)(например, S/R/Q/E/D или Т); или модификацию в положении 428 согласно нумерации EU (459 согласно нумерации Kabat) и/или 433 согласно нумерации EU (464 согласно нумерации Kabat) (например, L/R/S/P/Q или К) и/или 434 согласно нумерации EU (465 согласно нумерации Kabat) (например, H/F или Y); или модификацию в положении 250 согласно нумерации EU (263 согласно нумерации Kabat) и/или 428 согласно нумерации EU (459 согласно нумерации Kabat); или модификацию в положении 307 согласно нумерации EU (326 согласно нумерации Kabat) или 308 согласно нумерации EU (327 согласно нумерации Kabat) (например, 308F, V308F), и 434 согласно нумерации EU (465 согласно нумерации Kabat). Согласно одному варианту реализации указанная модификация включает модификацию 428L (например, M428L) и 434S (например, N434S) согласно нумерации EU (459, например, M459L, и модификацию 465S (например, N465S) согласно нумерации Kabat); модификацию 428L, 2591 (например, V259I), и 308F (например, V308F) согласно нумерации EU (459L, 2721 (например, V272I), и модификацию 327F (например, V327F) согласно нумерации Kabat; а 433К (например, Н433К) и модификацию 434 (например, 434Y) согласно нумерации EU (модификацию 464К (например, Н464К) и 465 (например, 465Y) согласно нумерации Kabat; модификацию 252, 254, и 256 (например, 252Y, 254Т, и 256Е) согласно нумерации EU (модификацию 265, 267, 269 (например, 265Y, 267Т, и 269Е) согласно нумерации Kabat; модификацию 250Q и 428L (например, T250Q и M428L) согласно нумерации EU (модификацию 263Q и 459L, например, T263Q и M459L, согласно нумерации Kabat); и модификацию 307 и/или 308 (например, 307F или 308Р) согласно нумерации EU (326 и/или модификацию 327, например, 326F или 308Р, согласно нумерации Kabat), при этом указанная модификация повышает сродство последовательности аминокислот константной области тяжелой цепи к FcRn в кислой среде (например, в эндосоме, где значение рН варьирует от приблизительно 5,5 до приблизительно 6,0). Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи кодирует последовательность CH2 человека аминокислот, содержащую по меньшей мере одну модификацию между аминокислотными остатками в положениях 252 и 257 согласно нумерации EU (т.е. по меньшей мере одну модификацию между аминокислотными остатками в положениях 265 и 270 согласно нумерации Kabat), при этом указанная модификация повышает сродство последовательности аминокислот CH2 человека к FcRn в кислой среде (например, в эндосоме, где значение рН варьирует от приблизительно 5,5 до приблизительно 6,0). Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи кодирует последовательность аминокислот CH2 человека, содержащую по меньшей мере одну модификацию между аминокислотными остатками в положениях 307 и 311 (т.е. по меньшей мере одну модификацию между аминокислотными остатками в положениях 326 и 330 согласно нумерации Kabat), при этом указанная модификация повышает сродство последовательности аминокислот CH2 к FcRn в кислой среде (например, в эндосоме, где значение рН варьирует от приблизительно 5,5 до приблизительно 6,0). Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи кодирует последовательность аминокислот CH3 человека, при этом указанная CH3 последовательность аминокислот содержит по меньшей мере одну модификация между аминокислотными остатками в положениях 433 и 436 согласно нумерации EU (т.е. по меньшей мере одну модификацию между аминокислотными остатками в положениях 464 и 467 согласно нумерации Kabat), при этом указанная модификация повышает сродство последовательности аминокислот CH3 к FcRn в кислой среде (например, в эндосоме, где значение рН варьирует от приблизительно 5,5 до приблизительно 6,0). Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи кодирует последовательность аминокислот константной области тяжелой цепи человека, содержащую мутацию, выбранную из группы, состоящей из M428L согласно нумерации EU (459 согласно нумерации Kabat), N434S согласно нумерации EU (465 согласно нумерации Kabat) и их комбинации. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи кодирует последовательность аминокислот константной области тяжелой цепи человека, содержащую мутацию, выбранную из группы, состоящей из M428L согласно нумерации EU (M459L согласно нумерации Kabat), V259I согласно нумерации EU (V272I согласно нумерации Kabat), V308F согласно нумерации EU (V327 согласно нумерации Kabat) и их комбинации. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи кодирует последовательность аминокислот константной области тяжелой цепи человека, содержащую мутацию N434A согласно нумерации EU (мутацию N465A согласно нумерации Kabat). Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи кодирует последовательность аминокислот константной области тяжелой цепи человека, содержащую мутацию, выбранную из группы, состоящей из M252Y согласно нумерации EU (M265Y согласно нумерации Kabat), S254T согласно нумерации EU (S267T согласно нумерации Kabat), Т256Е согласно нумерации EU (Т269Е согласно нумерации Kabat) и их комбинации. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи кодирует последовательность аминокислот константной области тяжелой цепи человека, содержащую мутацию, выбранную из группы, состоящей из T250Q согласно нумерации EU (T263Q согласно нумерации Kabat), M428L согласно нумерации EU (M459L согласно нумерации Kabat), или обе мутации. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи кодирует последовательность аминокислот константной области тяжелой цепи человека, содержащую мутацию, выбранную из группы, состоящей из Н433К согласно нумерации EU (Н464К согласно нумерации Kabat),N434Y согласно нумерации EU (N465Y согласно нумерации Kabat), или обе мутации.
[000189] Согласно некоторым вариантам реализации генетически модифицированный иммуноглобулиновый локус содержит: (1) первый аллель, при этом указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека согласно описанию в настоящем документе функционально связана с первой нуклеотидной последовательностью константной области тяжелой цепи, кодирующей первую последовательность аминокислот СН3 IgG человека, выбранного из IgG1, IgG2, IgG4 и их комбинации; и (2) второй аллель, при этом указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека согласно описанию в настоящем документе функционально связана с второй нуклеотидной последовательностью константной области тяжелой цепи, кодирующей вторую последовательность аминокислот CH3 IgG человека, выбранный из IgG1, IgG2, IgG4, и их комбинации, и при этом вторая последовательность аминокислот СН3 содержит модификацию, которая снижает или элиминирует связывание второй последовательности аминокислот СН3 с белком А (см., например, патент США №8586713, полностью включенный посредством ссылки 6 настоящий документ). Согласно некоторым вариантам реализации указанная вторая последовательность аминокислот СН3 содержит модификацию H95R (согласно нумерации экзонов IMGT; H435R согласно нумерации EU). Согласно одному варианту реализации вторая последовательность аминокислот СН3 также содержит модификацию Y96F (согласно нумерации экзонов IMGT; H436F по EU). Согласно другому варианту реализации вторая последовательность аминокислот СН3 содержит обе модификации, модификацию H95R (согласно нумерации экзонов IMGT; H435R согласно нумерации EU) и модификацию Y96F (согласно нумерации экзонов IMGT; H436F по EU). Согласно некоторым вариантам реализации указанная вторая последовательность аминокислот СН3 происходит из модифицированного IgG1 человека и также содержит мутацию, выбранную из группы, состоящей из D16E, L18M, N44S, K52N, V57M и V82I (IMGT; D356E, L38M, N384S, K392N, V397M и V422I по EU). Согласно некоторым вариантам реализации указанная второй СН3 последовательность аминокислот происходит из модифицированного IgG2 человека и также содержит мутацию, выбранную из группы, состоящей из N44S, K52N и V82I (IMGT: N384S, K392N, и V422I по EU). Согласно некоторым вариантам реализации указанная вторая последовательность аминокислот СН3 происходит из модифицированного IgG4 человека и также содержит мутацию, выбранную из группы, состоящей из Q15R, N44S, K52N, V57M, R69K, E79Q и V82I (IMGT: Q355R, N384S, K392N, V397M, R409K, E419Q и V422I по EU). Согласно некоторым вариантам реализации указанная последовательность аминокислот константной области тяжелой цепи представляет собой имеющую нечеловеческое происхождение последовательность аминокислот константной области, и указанная последовательность аминокислот константной области тяжелой цепи содержит одну или большее количество любых типов модификаций, описанных выше.
[000190] Согласно различным вариантам реализации Fc-домены модифицируют таким образом, чтобы изменить связывание с Fc-рецепторами, что, в свою очередь, влияет на эффекторную функцию. Согласно некоторым вариантам реализации сконструированная константная область тяжелой цепи (CH), которая включает Fc-домен, является гибридной. Таким образом, в гибридной области CH сочетаются домены CH, происходящие из более чем одного изотипа иммуноглобулина. Например, гибридная область CH содержит часть домена CH2 или полный домен CH2, происходящий из молекулы IgGl человека, IgG2 человека или IgG4 человека, скомбинированный с частью домена Сн3 или полным доменом Сн3, происходящим из молекулы IgG1 человека, IgG2 человека или IgG4 человека. Согласно некоторым вариантам реализации гибридная область CH содержит гибридную шарнирную область. Например, гибридный шарнир может содержать последовательность аминокислот «верхнего шарнира» (аминокислотные остатки в положениях 216-227 в соответствии с нумерацией EU; аминокислотные остатки в положениях 226-240 в соответствии с нумерацией Kabat), происходящую из шарнирной области IgG1 человека, IgG2 человека или IgG4 человека, скомбинированную с последовательностью «нижнего шарнира» (аминокислотные остатки в положениях 228-236 в соответствии с нумерацией EU; аминокислоты в положениях 241 -249 в соответствии с нумерацией Kabat), происходящей из шарнирной области IgG1 человека, IgG2 человека или IgG4 человека. Согласно некоторым вариантам реализации указанная гибридная шарнирная область содержит аминокислотные остатки, происходящие из верхнего шарнира IgG1 человека или IgG4 человека, и аминокислотные остатки, происходящие из нижнего шарнира IgG2 человека.
[000191] Согласно некоторым вариантам реализации может быть сконструирован Fc-домен для активации всех, некоторых или ни одного из нормальных эффекторных функций Fc, без влияния на требуемые фармакокинетические свойства Fc-содержащего белка (например, антитела). Примеры белков, содержащих гибридные области CH и обладающих измененными эффекторными функциями, см. предварительную заявку на патент США 61/759578, поданную 1 февраля 2013 г., полностью включенную в настоящий документ посредством ссылки.
[000192] Согласно различным аспектам геном не являющихся человеком животных модифицируют (i) с удалением или устранением функциональности (например, путем вставки нуклеотидной последовательности (например, экзогенной нуклеотидной последовательности)) в иммуноглобулиновый локус или путем нефункциональной реаранжировки или инверсии всех или по существу всех эндогенных функциональных сегментов VH, D, JH иммуноглобулина; и (ii) для включения нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, при этом указанная нуклеотидная последовательность присутствует в эндогенном локусе (т.е. там, где указанная нуклеотидная последовательность расположена у не являющегося человеком животного дикого типа). Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека располагается в эктопическом локусе в геноме (например, в локусе, отличном от эндогенного локуса цепи иммуноглобулина в геноме, или в эндогенном локусе, например, в составе локуса вариабельной области иммуноглобулина, при этом указанный эндогенный локус расположен или перемещен в другую локализацию в геноме). Согласно некоторым вариантам реализации например, приблизительно 80% или более, приблизительно 85% или более, приблизительно 90% или более, приблизительно 95% или более, приблизительно 96% или более, приблизительно 97% или более, приблизительно 98% или более, или приблизительно 99% или более из всех эндогенных функциональных сегментов гена тяжелых цепей V, D или J удаляют или устраняют их функциональность. Согласно некоторым вариантам реализации например, по меньшей мере 95%, 96%, 97%, 98% или 99% эндогенных функциональных сегментов гена тяжелых цепей V, D или J удаляют или устраняют их функциональность. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с человеческой или имеющей нечеловеческое происхождение последовательностью гена константной области тяжелой цепи. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с человеческой или имеющей нечеловеческое происхождение последовательностью гена константной области легкой цепи, либо каппа, либо лямбда.
[000193] Согласно некоторым вариантам реализации у указанного генетически модифицированного не являющегося человеком животного имеется модификация, которая обеспечивает удаление или устранение функциональности эндогенных функциональных сегментов гена VH, D и JH вариабельных областей тяжелой цепи и эндогенных функциональных сегментов гена вариабельной области легкой цепи VL и JL; и имеется (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека и (ii) нуклеотидная последовательность, кодирующая нереаранжированные сегменты генов V легкой цепи (VL) иммуноглобулина человека и нереаранжированные сегменты генов J легкой цепи (JL) иммуноглобулина человека (т.е. там, где указанная нуклеотидная последовательность расположена у не являющегося человеком животного дикого типа) или в эктопической локализации (например, в локусе, отличном от эндогенного локуса цепи иммуноглобулина в геноме, или в эндогенном локусе, например, в локусе вариабельной области иммуноглобулина, причем указанный эндогенный локус расположен или перемещен в другую локализацию в геноме). Согласно некоторым вариантам реализации у указанного генетически модифицированного не являющегося человеком животного имеется модификация, которая обеспечивает удаление или устранение функциональности эндогенных функциональных сегментов гена VH, D и JH вариабельных областей тяжелой цепи и эндогенных функциональных сегментов гена вариабельной области легкой цепи VL и JL; и имеется (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека и (ii) один или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL) в эндогенной локализации (т.е. там, где указанная нуклеотидная последовательность расположена у не являющегося человеком животного дикого типа) или в эктопической локализации (например, в локусе, отличном от эндогенного локуса цепи иммуноглобулина в геноме, или в эндогенном локусе, например, в локусе вариабельной области иммуноглобулина, причем указанный эндогенный локус расположен или перемещен в другую локализацию в геноме). Согласно некоторым вариантам реализации например, приблизительно 80% или более, приблизительно 85% или более, приблизительно 90% или более, приблизительно 95% или более, приблизительно 96% или более, приблизительно 97% или более, приблизительно 98% или более, или приблизительно 99% или более из всех эндогенных функциональных сегментов гена тяжелых цепей V, D или J удаляют или устраняют их функциональность. Согласно некоторым вариантам реализации, например, по меньшей мере 95%, 96%, 97%, 98%, или 99% эндогенных функциональных сегментов гена тяжелых цепей V, D или J удаляют или устраняют их функциональность. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с человеческой или имеющей нечеловеческое происхождение последовательностью гена константной области тяжелой цепи. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с человеческой или имеющей нечеловеческое происхождение последовательностью гена константной области легкой цепи, либо каппа, либо лямбда.
[000194] Различные варианты реализации охватывают вариабельные домены легких цепей. Нуклеотидные последовательности, кодирующие вариабельные домены легких цепей, могут использоваться при получении генетически модифицированных не являющихся человеком животных, описанных в настоящем документе, могут экспрессироваться такими животными, и/или могут кодировать аминокислоты, присутствующие в антителах, продуцированных такими животными (или происходящих из последовательностей, диверсифицированных такими животными). Согласно некоторым вариантам реализации вариабельный домен легкой цепи представляет собой вариабельный домен легкой цепи к человека. Согласно некоторым вариантам реализации указанный вариабельный домен легкой цепи представляет собой вариабельный домен легкой цепи к мыши. Согласно некоторым вариантам реализации указанный вариабельный домен легкой цепи представляет собой вариабельный домен легкой цепи к крысы. Согласно некоторым вариантам реализации указанный вариабельный домен легкой цепи представляет собой вариабельный домен легкой цепи λ человека. Согласно некоторым вариантам реализации указанный вариабельный домен легкой цепи представляет собой вариабельный домен легкой цепи λ мыши. Согласно некоторым вариантам реализации указанный вариабельный домен легкой цепи представляет собой вариабельный домен легкой цепи λ крысы.
[000195] Согласно различным вариантам реализации вариабельные домены легких цепей, продуцированные генетически модифицированными не являющимися человеком животными, описанными в настоящем документе, кодируют один или большее количество сегментов гена вариабельной области легкой цепи к иммуноглобулина мыши или человека. Согласно некоторым вариантам реализации указанные один или большее количество сегментов гена вариабельной области легкой цепи к иммуноглобулина мыши содержат приблизительно три мегабазы локуса легкой цепи к иммуноглобулина мыши. Согласно некоторым вариантам реализации указанные один или большее количество сегментов гена вариабельной области легкой цепи к иммуноглобулина мыши содержат по меньшей мере 137 сегментов гена Vκ, по меньшей мере пять сегментов гена Jκ или их комбинации, локуса легкой цепи к иммуноглобулина мыши. Согласно некоторым вариантам реализации указанные один или большее количество сегментов гена вариабельной области легкой цепи к иммуноглобулина человека содержат приблизительно 1/2 мегабазы локуса легкой цепи к иммуноглобулина человека. Согласно конкретным вариантам реализации указанные один или большее количество сегментов гена вариабельной области легкой цепи к иммуноглобулина человека содержат проксимальный повтор (в отношении константной области цепи к иммуноглобулина) локуса легкой цепи к иммуноглобулина человека. Согласно некоторым вариантам реализации указанные один или большее количество сегментов гена вариабельной области легкой цепи к иммуноглобулина человека содержат по меньшей мере 40 сегментов гена Vκ, по меньшей мере пять сегментов гена Jκ или их комбинацию, из локуса легкой цепи к иммуноглобулина человека.
[000196] Согласно конкретным вариантам реализации у указанных генетически модифицированных не являющихся человеком животных дополнительно содержится нуклеотидная последовательность, кодирующая сегмент гена нереаранжированной легкой цепи иммуноглобулина человека (VL) и сегмент гена нереаранжированной легкой цепи иммуноглобулина человека (JL). Согласно некоторым вариантам реализации указанная нуклеотидная последовательность, кодирующая сегмент V гена нереаранжированной легкой цепи и сегмент J гена нереаранжированной легкой цепи, функционально связана с последовательностью гена константной области легкой цепи иммуноглобулина. Согласно другим вариантам реализации нуклеотидная последовательность, кодирующая сегмент V гена нереаранжированной легкой цепи и сегмент J гена нереаранжированной легкой цепи, функционально связана с последовательностью гена константной области иммуноглобулина тяжелой цепи. Согласно некоторым вариантам реализации сегмент V гена нереаранжированной легкой цепи иммуноглобулина человека (VL) и нереаранжированный сегмент J гена (JL) иммуноглобулина человека функционально связаны в эндогенном локусе грызуна с геном константной области легкой цепи иммуноглобулина грызуна; например, геном константной области легкой цепи κ или λ.
[000197] Согласно различным вариантам реализации сегменты гена неаранжированной вариабельной области человека (например, сегменты генов Vκ человека) способны к реаранжировке и кодированию человеческих вариабельных доменов антитела. Согласно некоторым вариантам реализации указанное не являющееся человеком животное не содержит эндогенного сегмента гена VL. Согласно некоторым вариантам реализации указанные сегменты генов Vκ человека, экспрессируемые не являющимися человеком животными, выбирают из группы, состоящей из Vκ1-5, Vκ1-6, Vκ1-8, Vκ1-9, Vκ1-12, Vκ1-13, Vκ1-16, Vκ1-17, Vκ1-22, Vκ1-27, Vκ1-32, Vκ1-33, Vκ1-35, Vκ1-37, Vκ1-39, Vκ1D-8, Vκ1D-12, Vκ1D-13, Vκ1D-16, Vκ1D-171 Vκ1D-22, Vκ1D-27, Vκ1D-32, Vκ1D-33, Vκ1D-35, Vκ1D-37, Vκ1D-39, Vκ1D-42, Vκ1D-43, Vκ1-NL1, Vκ2-4, Vκ2-10, Vκ2-14, Vκ2-18, Vκ2-19, Vκ2-23, Vκ2-24, Vκ2-26, Vκ2-28, Vκ2-29, Vκ2-30, Vκ2-36, Vκ2-38, Vκ2-40, Vκ2D-10, Vκ2D-14, Vκ2D-18, Vκ2D-19, Vκ2D-23, Vκ2D-24, Vκ2D-26, Vκ2D-28, Vκ2D-29, Vκ2D-30, Vκ2D-36, Vκ2D-38, Vκ2D-40, Vκ3-7, Vκ3-11, Vκ3-15, Vκ3-20, Vκ3-25, Vκ3-31, Vκ3-34, Vκ3D-7, Vκ3D-7, Vκ3D-11, Vκ3D-15, Vκ3D-15, Vκ3D-20, Vκ3D-25, Vκ3D-31, Vκ3D-34, Vκ3-NL1, Vκ3-NL2, Vκ3-NL3, Vκ3-NL4, Vκ3-NL5, Vκ4-1, Vκ5-2, Vκ6-21, Vκ6D-21, Vκ6D-41 и Vκ7-3. Согласно некоторым вариантам реализации у указанных генетически модифицированных не являющихся человеком животных, описанных в настоящем документе экспрессируются все функциональные гены Vκ человека. Согласно некоторым вариантам реализации указанные сегменты генов Vκ человека содержат Vκ4-1, Vκ5-2, Vκ7-3, Vκ2-4, Vκ1-5 и Vκ1-6. Согласно некоторым вариантам реализации указанные сегменты генов Vκ содержат Vκ3-7, Vκ1-8, Vκ1-9, Vκ2-10, Vκ3-11, Vκ1-12, Vκ1-13, Vκ2-14, Vκ3-15 и Vκ1-16. Согласно некоторым вариантам реализации указанные сегменты генов Vκ человека содержат Vκ1-17, Vκ2-18, Vκ2-19, Vκ3-20, Vκ6-21, Vκ1-22, Vκ1-23, Vκ2-24, Vκ3-25, Vκ2-26, Vκ1-27, Vκ2-28, Vκ2-29 и Vκ2-30. Согласно некоторым вариантам реализации указанные сегменты генов Vκ человека содержат Vκ3-31, Vκ1-32, Vκ1-33, Vκ3-34, Vκ1-35, Vκ2-36, Vκ1-37, Vκ2-38, Vκ1-39 и Vκ2-40. Согласно различным вариантам реализации у указанного не являющегося человеком животного имеется пять сегментов гена Jκ человека, например, сегменты генов Jκ1, Jκ2, Jκ3, Jκ4 и Jκ5. Согласии конкретным вариантам реализации сегменты генов Vκ включают непрерывные сегменты генов к иммуноглобулина человека, захватывающие локус легкой цепи к иммуноглобулина человека от Vκ4-1 до Vκ2-40 включительно, и сегменты генов Jκ включают непрерывные сегменты генов, захватывающие локус легкой цепи к иммуноглобулина человека от JκI до Jκ5 включительно. Согласно некоторым вариантам реализации указанный локус легкой цепи иммуноглобулина содержит два сегмента гена VL человека, Vκ1-39 и Vκ3-20. Согласно некоторым вариантам реализации один или большее количество (например, 2, 3, 4 или 5) сегментов гена VL человека и два или большее количество сегментов гена JL человека присутствует в эндогенном локусе легкой цепи, например, в эндогенном локусе легкой цепи каппа. Согласно некоторым вариантам реализации указанная генетически модифицированное не являющееся человеком животное представляет собой мышь, у которой имеется функциональный λ локус легкой цепи. Согласно другим вариантам реализации у указанной мыши имеется нефункциональный локус легкой цепи λ. Согласно некоторым вариантам реализации указанные один или большее количество сегментов гена VH человека, один или большее количество сегментов гена DH человека и один или большее количество сегментов гена JH человека функционально связаны с последовательностью константной области тяжелой цепи мыши или крысы (т.е. один или большее количество сегментов гена VL человека и два или большее количество сегментов гена JL человека присутствуют в эндогенном локусе тяжелой цепи).
[000198] Согласно некоторым вариантам реализации у генетически модифицированного не являющегося человеком животного (например, мыши или крысы) согласно описанию в настоящем документе экспрессируется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (т.е. продуцируется антигенсвязывающий белок, содержащий реаранжированный вариабельный домен тяжелой цепи) и один или большее количество, два или большее количество, три или большее количество, четыре или большее количество, пять или большее количество и т.п. вариабельных доменов легких цепей, кодируемых генами Vκ, выбранными из группы, состоящей из Vκ1-5, Vκ1-6, Vκ1-8, Vκ1-9, Vκ1-12, Vκ1-13, Vκ1-16, Vκ1-17, Vκ1-22, Vκ1-27, Vκ1-32, Vκ1-33, Vκ1-35, Vκ1-37, Vκ1-39, Vκ1D-8, Vκ1D-12, Vκ1D-13, Vκ1D-16, Vκ1D-17, Vκ1D-22, Vκ1D-27, Vκ1D-32, Vκ1D-33, Vκ1D-35, Vκ1D-37, Vκ1D-39, Vκ1D-42, Vκ1D-43, Vκ1-NLI, Vκ2-4, Vκ2-10, Vκ2-14, Vκ2-18, Vκ2-19, Vκ2-23, Vκ2-24, Vκ2-26, Vκ2-28, Vκ2-29, Vκ2-30, Vκ2-36, Vκ2-38, Vκ2-40, Vκ2D-10, Vκ2D-14, Vκ2D-18, Vκ2D-19, Vκ2D-23, Vκ2D-24, Vκ2D-26, Vκ2D-28, Vκ2D-29, Vκ2D-30, Vκ2D-36, Vκ2D-38, Vκ2D-40, Vκ3-7, Vκ3-11, Vκ3-15, Vκ3-20, Vκ3-25, Vκ3-31, Vκ3-34, Vκ3D-7, Vκ3D-7, Vκ3D-11, Vκ3D-15, Vκ3D-15, Vκ3D-20, Vκ3D-25, Vκ3D-31, Vκ3D-34, Vκ3-NL1, Vκ3-NL2, Vκ3-NL3, Vκ3-NL4, Vκ3-NL5, Vκ4-1, Vκ5-2, Vκ6-21, Vκ6D-21, Vκ6D-41 и Vκ7-3.
[000199] Согласно различным вариантам реализации по меньшей мере один из сегментов гена вариабельной области легкой цепи (например, сегментов гена вариабельной области легкой цепи человека) кодирует один или большее количество гистидиновых кодонов, не кодируемых соответствующим сегментом гена вариабельной области легкой цепи зародышевой линии человека. Согласно некоторым вариантам реализации при кислотных значениях рН по сравнению с нейтральными значениями рН наблюдается уменьшение периода полураспада (t1/2) вариабельного домена легкой цепи согласно описанию в настоящем документе по меньшей мере приблизительно в 2 раза, по меньшей мере приблизительно в 3 раза, по меньшей мере приблизительно в 4 раза, по меньшей мере приблизительно в 5 раз, по меньшей мере приблизительно в 10 раз, по меньшей мере приблизительно в 15 раз, по меньшей мере приблизительно в 20 раз, по меньшей мере приблизительно в 25 раз или по меньшей мере приблизительно в 30 раз. Согласно некоторым вариантам реализации при кислотных значениях рН происходит уменьшение t1/2 приблизительно в 30 раз или более по сравнению с нейтральными значениями рН. Согласно некоторым вариантам реализации по меньшей мере один из сегментов гена VL содержит замену по меньшей мере одного негистидинового кодона, кодируемого соответствующей последовательностью сегмента VL зародышевой линии человека, на гистидиновый кодон. Согласно некоторым вариантам реализации указанная замена представляет собой замену одного, двух, трех или четырех кодонов (например, трех или четырех кодонов). Согласно некоторым вариантам реализации указанная замена происходит в кодоне(ах) области CDR3. Согласно некоторым вариантам реализации указанные сегменты генов VL человека представляют собой сегменты генов Vκ1-39 и Vκ3-20 человека, и каждый из сегментов гена Vκ1-39 и Vκ3-20 человека содержит замену по меньшей мере одного негистидинового кодона, кодируемого соответствующим сегментом гена VL зародышевой линии человека, на гистидиновый кодон. Согласно некоторым вариантам реализации каждый из сегментов гена Vκ1-39 и Vκ3-20 человека содержит замену трех или четырех гистидиновых кодонов. Согласно некоторым вариантам реализации указанные три или четыре замены происходят в области CDR3. Согласно некоторым вариантам реализации указанная замена представляет собой замену трех негистидиновых кодонов сегмента гена Vκ1-39 человека, при этом указанная замена предназначена для экспрессирования гистидинов в положениях 106, 108 и 111. Согласно некоторым вариантам реализации указанная замена представляет собой замену четырех негистидиновых кодонов сегмента гена Vκ1-39 человека, и указанная замена предназначена для обеспечения экспрессии гистидинов в положениях 105, 106, 108 и 111 (см., например, US 2013/0247234А1 и WO 2013/138680, включенные в настоящий документ посредством ссылки). Согласно некоторым вариантам реализации указанная замена представляет собой замену трех негистидиновых кодонов сегмента гена Vκ3-20 человека, и указанная замена предназначена для обеспечения экспрессии гистидинов в положениях 105, 106 и 109. Согласно дополнительным вариантам реализации указанная замена представляет собой замену четырех негистидиновых кодонов сегмента гена Vκ3-20 человека, и указанная замена предназначена для обеспечения экспрессии гистидинов в положениях 105, 106, 107 и 109. Согласно некоторым вариантам реализации указанный локус легкой цепи иммуноглобулина содержит один или более, но меньше, чем у дикого типа, сегментов гена человека VL и один или большее количество, например, два или большее количество, сегментов гена JL человека, при этом каждый из указанных сегментов гена VL человека содержит по меньшей мере один гистидиновый кодон, не кодируемый соответствующим сегментом гена VL зародышевой линии человека. Согласно различным вариантам реализации у указанного не являющегося человеком животного, у которого имеются генетически модифицированные иммуноглобулиновые локусы согласно описанию в настоящем документе, при стимуляции представляющим интерес антигеном экспрессируется антигенсвязывающий белок, содержащий последовательность аминокислот, происходящую из сегментов гена VL человека, при этом указанный антигенсвязывающий белок сохраняет по меньшей мере один гистидиновый остаток в положении аминокислоты, кодируемой по меньшей мере одним гистидиновым кодоном, введенным в сегмент гена VL человека. Согласно некоторым вариантам реализации у указанного животного экспрессируется популяция антигенсвязывающих белков в ответ на антиген, при этом все антигенсвязывающие белки в указанной популяции содержат (а) вариабельные домены легких цепей иммуноглобулина, полученные в результате реаранжировки сегментов гена VL человека и сегментов гена JL, при этом по меньшей мере один из сегментов гена VL человека кодирует один или большее количество гистидиновых кодонов, не кодируемых соответствующим сегментом гена VL человека зародышевой линии, и (b) тяжелые цепи иммуноглобулина, содержащие вариабельные домены тяжелых цепей человека, кодируемые нуклеотидной последовательностью реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека.
[000200] Различные варианты реализации охватывают последовательности константной области легкой цепи. Согласно некоторым вариантам реализации например, первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. первая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), функционально связана с последовательностью гена константной области тяжелой цепи, и вторая нуклеотидная последовательность, которая кодирует вариабельный домен легкой цепи человека (т.е. вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека), функционально связана с последовательностью гена константной области легкой цепи. Согласно некоторым вариантам реализации первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. первая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), функционально связана с последовательностью гена константной области легкой цепи, и вторая нуклеотидная последовательность, которая кодирует вариабельный домен легкой цепи человека (т.е. вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека), функционально связана с последовательностью гена константной области тяжелой цепи. Согласно различным вариантам реализации указанная последовательность константной области легкой цепи, функционально связанная с нуклеотидной последовательностью реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека представляет собой последовательность константной области легкой цепи к человека. Согласно некоторым вариантам реализации указанная последовательность константной области легкой цепи, функционально связанная с реаранжированным вариабельным доменом тяжелой цепи, представляет собой последовательность константной области легкой цепи к мыши. Согласно некоторым вариантам реализации указанная последовательность константной области легкой цепи, функционально связанная с реаранжированным вариабельным доменом тяжелой цепи, представляет собой последовательность константной области легкой цепи к крысы. Согласно некоторым вариантам реализации указанная последовательность константной области легкой цепи, функционально связанная с реаранжированным вариабельным доменом тяжелой цепи, представляет собой человека легкая цепь λ константная область последовательность. Согласно некоторым вариантам реализации указанная последовательность константной области легкой цепи, функционально связанная с реаранжированным вариабельным доменом тяжелой цепи, представляет собой последовательность константной области легкой цепи λ мыши. Согласно некоторым вариантам реализации указанная последовательность константной области легкой цепи, функционально связанная с реаранжированным вариабельным доменом тяжелой цепи, представляет собой последовательность константной области легкой цепи λ крысы.
[000201] Согласно различным аспектам предложены не являющиеся человеком животные, у которых имеется генетически модифицированный иммуноглобулиновый локус, кодирующий реаранжированный вариабельный домен тяжелой цепи (т.е. иммуноглобулиновый локус содержит нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), при этом указанный реаранжированный вариабельный домен тяжелой цепи содержит последовательность вариабельной области тяжелой цепи (VH), функционально связанную через спейсер с последовательностью сегмента тяжелой цепи J (JH), причем указанный спейсер содержит по меньшей мере один аминокислотный остаток. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с имеющей нечеловеческое происхождение последовательностью гена константной области тяжелой цепи. Согласно некоторым вариантам реализации указанная имеющая нечеловеческое происхождение последовательность гена константной области тяжелой цепи представляет собой последовательность гена константной области мыши или крысы. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области тяжелой цепи человека. Согласно некоторым вариантам реализации указанная константная область тяжелой цепи содержит последовательность, выбранную из СН1, шарнира, СН2, СН3 и их комбинации. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с имеющей нечеловеческое происхождение последовательностью гена константной области легкой цепи. Согласно некоторым вариантам реализации указанная имеющая нечеловеческое происхождение последовательность гена константной области легкой цепи представляет собой последовательность гена константной области мыши или крысы. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области легкой цепи человека. Согласно некоторым вариантам реализации указанная спейсер представляет собой единственный аминокислотный остаток. Согласно некоторым вариантам реализации указанные спейсеры представлены двумя аминокислотными остатками. Согласно некоторым вариантам реализации указанные спейсеры представлены тремя аминокислотными остатками. Согласно некоторым вариантам реализации указанные спейсеры представлены четырьмя аминокислотными остатками. Согласно некоторым вариантам реализации указанные спейсеры представлены пятью аминокислотными остатками. Согласно некоторым вариантам реализации указанные спейсеры представлены шестью аминокислотными остатками.
[000202] Согласно другому аспекту предложены генетически модифицированные не являющиеся человеком животные и способы получения указанных животных, отличающиеся тем, что у указанных животных имеется локус функциональной универсальной легкой цепи («ULC») иммуноглобулина. Согласно некоторым вариантам реализации у таких животных дополнительно содержится реаранжированный локус вариабельного домена тяжелой цепи (т.е. иммуноглобулиновый локус вариабельного домен тяжелой цепи, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека). ULC представляет собой общую легкую цепь, которая может использоваться в биспецифическом формате, включающем функцию, например, модификацию, которая влияет на связывание FcRn, для увеличения периода полужизни, например, биспецифическое антитело, которое содержит тяжелую цепь, связывающую антиген, и легкую цепь, связывающую FcRn. Например, генетически модифицированных мышей согласно описанию в настоящем документе иммунизируют FcRN для получения антител, которые связывают FcRN исключительно за счет легких цепей. Указанные легкие цепи, продуцированные указанным генетически модифицированным не являющимся человеком животным, используют в качестве ULC, помогающих биспецифическому антителу связываться с FcRn, способствуя таким образом увеличению периода полужизни. Остальную часть антитела (например, либо вторую, отличную легкую цепь, либо тяжелую цепь, связывающую антиген, отличный от FcRn) выбирают для осуществления второй функции. Согласно описанным в настоящем документе вариантам реализации ULC может также использоваться для получения последовательностей вариабельных цепей антител, разнообразие которых обусловлено в основном процессами соматических мутаций (например, гипермутацией), с выявлением таким образом последовательностей вариабельных цепей антител, антигенсвязывающая способность которых улучшается в результате постгеномных событий.
[000203] Различные аспекты включают генетически модифицированных не являющихся человеком животных, в геномах которых имеется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи человека, а также нуклеиновая кислота, кодирующая вариабельный домен легкой цепи согласно описанию в настоящем документе, клонированная на нуклеотидную последовательность константной области, выбранную из константной области каппа, константной области лямбда, константной области тяжелой цепи (например, выбранной из группы, состоящей из СН1, шарнира, СН2, СН3 и их комбинации). Согласно некоторым вариантам реализации нуклеотидную последовательность вариабельней области легкой цепи клонируют на первую нуклеотидную последовательность константной области тяжелой цепи человека, и второй вариабельный домен легкой цепи клонируют на вторую нуклеотидную последовательность константной области тяжелой цепи человека; при этом указанные первая и вторая нуклеотидные последовательности константной области тяжелой цепи человека одинаковы, первый вариабельный домен легкой цепи специфически связывает первый антиген, и второй вариабельный домен легкой цепи специфически связывает второй антиген. Согласно указанным вариантам реализации образуется димер двух полипептидов, отличающийся тем, что каждый из вариабельных доменов легких цепей, соединенных с константной областью тяжелой цепи, проявляет отличную антигенсвязывающую специфичность.
[000204] Согласно другому аспекту предложено генетически модифицированное не являющееся человеком животное (например, мышь), способное продуцировать легкую цепь, связывающую рецептор или другой фрагмент, проникающий через гематоэнцефалический барьер, например, трансферриновый рецептор. Предыдущие исследования показали, что низкоаффинные антитела против трансферринового рецептора проникают через гематоэнцефалический барьер и высвобождаются благодаря низкой аффинности. Соответственно, согласно некоторым вариантам реализации описанных в настоящем документе генетически модифицированных животных (например, мышей) используют для получения низкоаффинного антитела к фрагменту, способному проникать через гематоэнцефалический барьер (например, к трансферриновому рецептору), при этом указанное низкоаффинное антитело является биспецифическим, и обладает второй связывающей специфичностью в отношении нужной мишени (т.е. указанное антитело связывает проникающий фрагмент, а также другую мишень, отличную от проникающего фрагмента).
[000205] Предложены способы получения и применения описанных в настоящем документе генетически модифицированных не являющихся человеком животных. Предложены способы функционального связывания нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи человека с нуклеотидной последовательностью константной области тяжелой или легкой цепи иммуноглобулина в геноме не являющегося человеком животного. Согласно различным вариантам реализации нуклеотидная последовательность константной области является человеческой или имеет нечеловеческое происхождение, и указанное не являющееся человеком животное представляет собой грызуна. Согласно различным вариантам реализации указанные способы включают получение не являющегося человеком животного, у которого также имеется локус легкой цепи иммуноглобулина, содержащий один или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи человека, например, два сегмента гена Vκ человека и один или большее количество сегментов гена Jκ человека, функционально связанных с человеческой или имеющей нечеловеческое происхождение нуклеотидной последовательностью константной области легкой цепи. Согласно различном аспектам указанные способы включают введение вышеупомянутых последовательностей в зародышевую линию не являющегося человеком животного, например, грызуна, с применением, например, трансгенной технологии, в том числе, например, с применением модифицированных плюрипотентных или тотипотентных донорских клеток (например, ЭС-клеток или ИПСК) и эмбрионов-хозяев, клеток зародышевой линии (например, ооцитов) и т.п. Соответственно, варианты реализации включают локус тяжелой цепи имеющего нечеловеческое происхождение иммуноглобулина в геноме имеющей нечеловеческое происхождение зародышевой клетки, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанную с последовательностью гена константной области тяжелой цепи, при этом указанная последовательность гена константной области содержит имеющую нечеловеческое происхождение последовательность, человеческую последовательность или их комбинацию. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с эндогенной последовательностью гена константной области имеющего нечеловеческое происхождение иммуноглобулина. Согласно некоторым вариантам реализации указанная эндогенная последовательность гена константной области имеющего нечеловеческое происхождение иммуноглобулина представляет собой последовательность гена константной области тяжелой цепи мыши или крысы.
[000206] Согласно различным аспектам предложен способ получения не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус, при этом указанный способ включает: (а) модификацию генома не являющегося человеком животного с удалением или устранением функциональности эндогенных функциональных сегментов гена V, D и J тяжелых цепей иммуноглобулина; и (b) введение в геном нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека. Согласно одному такому аспекту предложен способ получения не являющегося человеком животного, у которого экспрессируется единственная тяжелая цепь иммуноглобулина из реаранжированной последовательности гена тяжелой цепи в зародышевой линии не являющегося человеком животного, включающий этап генетической модификации не являющегося человеком животного таким образом, что вся имеющаяся у него популяция экспрессирующих антитела зрелых В-клеток экспрессирует тяжелую цепь, происходящую из (i) единственного сегмента гена VH; (ii) аминокислотного спейсера, включающего одну, две, три, четыре, пять, или шесть аминокислот; и (iii) единственного сегмента гена JH. Согласно некоторым аспектам указанный способ включает инактивацию или замену эндогенного локуса вариабельной области тяжелой цепи иммуноглобулина единственным реаранжированным геном тяжелой цепи согласно описанию в настоящем документе.
[000207] Согласно другому аспекту предложены способы получения не являющегося человеком животного, у которого имеется генетически модифицированный локус тяжелой цепи иммуноглобулина, включающие: (а) модификацию генома не являющегося человеком животного с удалением или устранением функциональности эндогенных функциональных сегментов гена V, D и J тяжелых цепей иммуноглобулина; и (b) введение в указанный геном нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека. Согласно некоторым вариантам реализации по существу все эндогенные функциональные сегменты генов VH, D и JH удаляют из локуса тяжелой цепи иммуноглобулина не являющегося человеком животного, или устраняют их функциональность {например, путем вставки нуклеотидной последовательности {например, экзогенной нуклеотидной последовательности в иммуноглобулиновый локус, или путем нефункциональной реаранжировки или инверсии эндогенных сегментов VH, D, JH). Согласно некоторым вариантам реализации указанный способ включает встраивание нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека {т.е. нуклеотидной последовательности, которая кодирует реаранжированный вариабельный домен тяжелой цепи) в эндогенную локализацию (т.е. нацеливание на локализацию указанной нуклеотидной последовательности у не являющегося человеком животного дикого типа). Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека присутствует эктопически (например, в локусе, отличном от эндогенного локуса цепи иммуноглобулина в геноме, или з эндогенном локусе, например, в локусе вариабельной области иммуноглобулина, при этом указанный эндогенный локус расположен или перемещен в другую локализацию в геноме). Согласно некоторым вариантам реализации, например, приблизительно 80% или более, приблизительно 85% или более, приблизительно 90% или более, приблизительно 95% или более, приблизительно 96% или более, приблизительно 97% или более, приблизительно 98% или более, или приблизительно 99% или более из всех эндогенных функциональных сегментов гена V, D, или J удаляют или устраняют их функциональность. Согласно некоторым вариантам реализации, например, по меньшей мере 95%, 96%, 97%, 98%, или 99% эндогенных функциональных сегментов гена тяжелых цепей V, D или J удаляют или устраняют их функциональность.
[000208] Согласно другому аспекту предложены способы получения не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус, включающие: (а) модификацию генома не являющегося человеком животного с удалением или устранением функциональности эндогенных функциональных сегментов V и J генов легкой цепи иммуноглобулина; и (b) введение в эндогенный локус легкой цепи иммуноглобулина нуклеотидной последовательности реаранжированной вариабельной области тяжелой Цепи иммуноглобулина человека (т.е. нуклеотидной последовательности, которая кодирует реаранжированный вариабельный домен тяжелой цепи), при этом указанная нуклеотидная последовательность функционально связана с последовательностью гена константной области легкой цепи. Согласно некоторым вариантам реализации указанный генетически сконструированный иммуноглобулиновый локус присутствует в геноме зародышевой линии не являющегося человеком животного. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области легкой цепи κ. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области мыши или крысы легкой цепи κ. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области легкой цепи человека κ. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области легкой цепи λ. Согласно некоторым вариантам реализации нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области мыши или крысы легкой цепи λ. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области легкой цепи λ человека.
[000209] Согласно другому аспекту предложены способы получения не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус, включающие: (а) модификацию генома не являющегося человеком животного с удалением или устранением функциональности: (i) эндогенных функциональных сегментов гена V, D и J тяжелых цепей иммуноглобулина, и (ii) эндогенных функциональных сегментов V и J генов легкой цепи иммуноглобулина; и (b) введение в указанный геном: (i) первой нуклеотидной последовательности, которая кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. первая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), при этом указанная первая нуклеотидная последовательность функционально связана с последовательностью гена константной области легкой цепи, и (ii) второй нуклеотидной последовательности, которая кодирует вариабельный домен легкой цепи иммуноглобулина человека (т.е. вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека), при этом указанная вторая нуклеотидная последовательность функционально связана с последовательностью гена константной области тяжелой цепи. Согласно некоторым вариантам реализации указанный генетически сконструированный иммуноглобулиновый локус присутствует в геноме зародышевой линии не являющегося человеком животного. Согласно некоторым вариантам реализации указанная первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи, функционально связана с последовательностью гена константной области легкой цепи κ. Согласно некоторым вариантам реализации указанная первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи, функционально связана с последовательностью гена константной области мыши или крысы легкой цепи κ. Согласно некоторым вариантам реализации указанная первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи, функционально связана с последовательностью гена константной области легкой цепи к человека. Согласно некоторым вариантам реализации первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи, функционально связана с последовательностью гена константной области легкой цепи λ. Согласно некоторым вариантам реализации первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи, функционально связана с последовательностью гена константной области легкой цепи λ мыши или крысы. Согласно некоторым вариантам реализации первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи, функционально связана с последовательностью гена константной области легкой цепи λ человека. Согласно некоторым вариантам реализации вариабельный домен легкой цепи иммуноглобулина человека представляет собой вариабельный домен легкой цепи κ. Соответственно, согласно некоторым вариантам реализации вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность вариабельной области легкой цепи каппа человека. Согласно некоторым вариантам реализации вариабельный домен легкой цепи иммуноглобулина человека представляет собой вариабельный домен легкой цепи λ. Соответственно, согласно некоторым вариантам реализации вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность вариабельной области легкой цепи лямбда человека. Согласно некоторым вариантам реализации последовательность гена константной области тяжелой цепи представляет собой последовательность гена константной области тяжелой цепи имеющего нечеловеческое происхождение иммуноглобулина. Согласно некоторым вариантам реализации последовательность гена константной области тяжелой цепи имеющего нечеловеческое происхождение иммуноглобулина представляет собой последовательность гена константной области тяжелой цепи мыши или крысы.
[000210] Согласно другому аспекту предложены способы получения не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус, включающие: (а) модификацию генома не являющегося человеком животного с удалением или устранением функциональности: (i) эндогенных функциональных сегментов гена V, D и J тяжелых цепей иммуноглобулина, и (ii) эндогенных функциональных сегментов V и J генов легкой цепи иммуноглобулина; и (b) введение в указанный геном: (i) первой нуклеотидной последовательности, которая кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. первая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), при этом указанная первая нуклеотидная последовательность функционально связана с последовательностью гена константной области тяжелой цепи, и (ii) второй нуклеотидной последовательности, которая кодирует вариабельный домен легкой цепи иммуноглобулина человека (т.е. вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека), при этом указанная вторая нуклеотидная последовательность функционально связана с последовательностью гена константной области легкой цепи. Согласно некоторым вариантам реализации указанная последовательность гена константной области легкой цепи представляет собой последовательность гена константной области легкой цепи κ. Согласно некоторым вариантам реализации указанная последовательность гена константной области легкой цепи представляет собой последовательность гена константной области легкой цепи к мыши или крысы. Согласно некоторым вариантам реализации указанная последовательность гена. константной области легкой цепи представляет собой последовательность гена константной области легкой цепи к человека. Согласно некоторым вариантам реализации указанная последовательность гена константной области легкой цепи представляет собой последовательность гена константной области легкой цепи λ. Согласно некоторым вариантам реализации указанная последовательность гена константной области легкой цепи представляет собой последовательность гена константной области легкой цепи λ мыши или крысы. Согласно некоторым вариантам реализации указанная последовательность гена константной области легкой цепи представляет собой последовательность гена константной области легкой цепи λ человека. Согласно некоторым вариантам реализации вариабельный домен легкой цепи иммуноглобулина человека представляет собой вариабельный домен легкой цепи κ. Согласно некоторым вариантам реализации вариабельный домен легкой цепи иммуноглобулина человека представляет собой вариабельный домен легкой цепи λ. Согласно некоторым вариантам реализации последовательность гена константной области тяжелой цепи представляет собой последовательность гена константной области тяжелой цепи имеющего нечеловеческое происхождение иммуноглобулина. Согласно некоторым вариантам реализации; последовательность гена константной области тяжелой цепи имеющего нечеловеческое происхождение иммуноглобулина представляет собой последовательность гена константной области тяжелой цепи мыши или крысы.
[000211] Согласно другому аспекту предложен способ получения не являющегося человеком животного, у которого имеется генетически модифицированный локус тяжелой цепи иммуноглобулина, включающий: (а) модификацию генома не являющегося человеком животного с удалением или устранением функциональности эндогенных функциональных сегментов гена V, D и J тяжелых цепей иммуноглобулина; и (b) введение в указанный геном нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, при этом указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека содержит последовательность гена V сегмента тяжелой цепи (VH), функционально связанную через спейсер с последовательностью гена J сегмента тяжелой цепи (JH), причем указанный спейсер содержит по меньшей мере один аминокислотный остаток. Согласно некоторым вариантам реализации нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области тяжелой цепи имеющего нечеловеческое происхождение иммуноглобулина. Согласно некоторым вариантам реализации последовательность гена константной области тяжелой цепи имеющего нечеловеческое происхождение иммуноглобулина представляет собой последовательность гена константной области тяжелой цепи иммуноглобулина мыши или крысы. Согласно некоторым вариантам реализации нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области легкой цепи имеющего нечеловеческое происхождение иммуноглобулина. Согласно некоторым вариантам реализации последовательность гена константной области легкой цепи имеющего нечеловеческое происхождение иммуноглобулина представляет собой последовательность гена константной области легкой цепи иммуноглобулина мыши или крысы. Согласно некоторым вариантам реализации нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека присутствует в эндогенной локализации (т.е. там, где указанная нуклеотидная последовательность расположена у не являющегося человеком животного дикого типа). Согласно некоторым вариантам реализации нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека присутствует эктопически (например, в локусе, отличном от эндогенного локуса цепи иммуноглобулина в геноме, или в эндогенном локусе, например, в локусе вариабельной области иммуноглобулина, при этом указанный эндогенный локус расположен или перемещен в другую локализацию в геноме). Согласно некоторым вариантам реализации указанные спейсеры представлены единственным аминокислотным остатком. Согласно некоторым вариантам реализации указанные спейсеры представлены двумя аминокислотными остатками. Согласно некоторым вариантам реализации указанные спейсеры представлены тремя аминокислотными остатками. Согласно некоторым вариантам реализации указанные спейсеры представлены четырьмя аминокислотными остатками. Согласно некоторым вариантам реализации указанные спейсеры представлены пятью аминокислотными остатками. Согласно некоторым вариантам реализации указанные спейсеры представлены шестью аминокислотными остатками. Согласно некоторым вариантам реализации указанные нуклеотидные последовательности кодируют две копии, три копии, четыре копии, или более нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанной с последовательностью гена константного домена тяжелой цепи. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность кодирует несколько копий нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанной с последовательностью гена константного домена тяжелой цепи.
[000212] Предложены способы получения не являющегося человеком животного, включающие: (а) модификацию генома не являющегося человеком животного с удалением или устранением функциональности (i) эндогенных функциональных сегментов гена тяжелой цепи иммуноглобулина VH, D и и/или JH, и (ii) эндогенных функциональных сегментов V и J генов легкой цепи иммуноглобулина; и (b) введение (i) нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локус тяжелой цепи, при этом указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи содержит последовательность гена V сегмента тяжелой цепи (VH), функционально связанную через спейсер с последовательностью гена J сегмента тяжелой цепи (JH), причем указанный спейсер содержит по меньшей мере один аминокислотный остаток; и (ii) одного или большего количества, но меньшего, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека (например, двух сегментов гена Vκ человека и по меньшей мере одного сегмента гена Jκ человека), функционально связанных с человеческой или имеющей нечеловеческое происхождение нуклеотидной последовательностью константной области легкой цепи. Согласно некоторым вариантам реализации по меньшей мере один из сегментов гена вариабельной области легкой цепи кодирует один или большее количество гистидиновых кодонов, не кодируемых соответствующим сегментом гена вариабельной области легкой цепи зародышевой линии человека.
[000213] Согласно некоторым аспектам предложен способ получения не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус, включающий:
(a) модификацию генома не являющегося человеком животного с удалением или устранением функциональности эндогенных функциональных сегментов V и J генов легкой цепи иммуноглобулина; и
(b) введение в указанный геном не являющегося человеком животного реаранжированной нуклеотидной последовательности вариабельной области тяжелой цепи человека, функционально связанной с нуклеотидной последовательностью константной области легкой цепи.
Согласно различным вариантам реализации указанное не являющееся человеком животное представляет собой грызуна, например, мышь, крысу или хомяка. Согласно некоторым вариантам реализации указанный грызун представляет собой мышь. Согласно некоторым вариантам реализации указанная константная область легкой цепи представляет собой константную область крысы или мыши, например, константную область Cκ крысы или мыши.
[000214] Согласно другому аспекту предложен способ получения не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус, включающий:
(a) модификацию генома не являющегося человеком животного с удалением или устранением функциональности:
(i) эндогенных функциональных сегментов V, D и/или J генов тяжелых цепей иммуноглобулина, и
(ii) эндогенных функциональных сегментов V и J генов легкой цепи иммуноглобулина; и
(b) введение в указанный геном не являющегося человеком животного:
(i) первой нуклеотидной последовательности, которая кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. первая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), при этом указанная первая нуклеотидная последовательность функционально связана с последовательностью гена константной области легкой цепи, и
(ii) второй нуклеотидной последовательности, которая кодирует человеческий или имеющий нечеловеческое происхождение вариабельный домен легкой цепи (т.е. вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека), при этом указанная вторая нуклеотидная последовательность функционально связана с последовательностью гена константной области тяжелой цепи.
Согласно различным вариантам реализации указанное не являющееся человеком животное представляет собой грызуна, например, мышь, крысу или хомяка. Согласно некоторым вариантам реализации указанный грызун представляет собой мышь. Согласно некоторым вариантам реализации указанная константная область легкой цепи представляет собой константную область крысы или мыши, например, константную область Cκ крысы или мыши. Согласно некоторым вариантам реализации указанная вторая нуклеотидная последовательность функционально связана с последовательностью гена константной области тяжелой цепи мыши или крысы, выбранной из СН1, шарнира, СН2, СН3 и их комбинации. Согласно некоторым вариантам реализации указанная вторая нуклеотидная последовательность функционально связана с последовательностью гена константной области тяжелой цепи человека, выбранной из СН1, шарнира, СН2, СН3 и их комбинации.
[000215] Согласно другому аспекту предложен способ получения не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус, включающие:
(a) модификацию генома не являющегося человеком животного с удалением пли устранением функциональности:
(i) эндогенных функциональных сегментов V, D и/или J генов тяжелых цепей иммуноглобулина, и
(ii) эндогенных функциональных сегментов V и J генов легкой цепи иммуноглобулина; и
(b) введение в указанный геном не являющегося человеком животного:
(i) первой нуклеотидной последовательности, которая кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. первая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), при этом указанная первая нуклеотидная последовательность функционально связана с последовательностью гена константной области тяжелой цепи; и
(ii) второй нуклеотидной последовательности, которая кодирует вариабельный домен легкой цепи (т.е. вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека), при этом указанная вторая нуклеотидная последовательность функционально связана с последовательностью гена константной области легкой цепи.
[000216] Согласно другому аспекту предложен способ получения не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый лэкус, включающие:
(a) модификацию генома не являющегося человеком животного с удалением или устранением функциональности:
(i) эндогенных функциональных сегментов V, D и/или J генов тяжелых цепей иммуноглобулина, и
(ii) эндогенных функциональных сегментов V и J генов легкой цепи иммуноглобулина; и
(b) введение в указанный геном не являющегося человеком животного:
(i) первого аллеля, содержащего:
(1) первую нуклеотидную последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. первая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), функционально связанную с последовательностью гена константной области тяжелой цепи, и
(2) вторую нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи (т.е. вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека), функционально связанную с последовательностью гена константной области легкой цепи; и
(ii) второго аллеля, содержащего
(1) третью нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи (т.е. третья нуклеотидная последовательность представляет собой нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека), функционально связанную с последовательностью гена константной области тяжелой цепи, и
(2) четвертую нуклеотидную последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. четвертая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), функционально связанную с последовательностью гена константной области легкой цепи.
[000217] Согласно другому аспекту предложен способ получения не являющегося человеком животного, у которого имеется генетически модифицированный локус тяжелой цепи иммуноглобулина, включающий: (а) модификацию генома не являющегося человеком животного с удалением или устранением функциональности эндогенных функциональных сегментов V, D и/или J генов тяжелых цепей иммуноглобулина; и (b) введение в указанный геном нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, при этом указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека содержит последовательность гена V сегмента тяжелой цепи (VH), функционально связанную через спейсер с последовательностью гена J сегмента тяжелой цепи (JH), причем указанный спейсер содержит по меньшей мере один аминокислотный остаток.
[000218] Согласно другому аспекту предложен способ получения не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус, включающий:
(а) модификацию генома не являющегося человеком животного с удалением или устранением функциональности:
(i) эндогенных функциональных сегментов V, D и/или J генов тяжелых цепей иммуноглобулина, и
(ii) эндогенных функциональных сегментов V и J генов легкой цепи иммуноглобулина; и
(b) введение в указанный геном не являющегося человеком животного:
(i) нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанной с нуклеотидной последовательностью константной области тяжелой цепи; и
(ii) одного или большего количества, но меньшего, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека, функционально связанных с нуклеотидной последовательностью константной области легкой цепи.
Согласно различным вариантам реализации указанное не являющееся человеком животное представляет собой грызуна, например, мышь, крысу или хомяка. Согласно некоторым вариантам реализации указанный грызун представляет собой мышь. Согласно некоторым вариантам реализации указанная константная область легкой цепи представляет собой константную область крысы или мыши, например, константную область Cκ крысы или мыши. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи человека функционально связана с последовательностью гена константной области тяжелой цепи мыши или крысы, выбранной из СН1, шарнира, СН2, СН3 и их комбинации. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи функционально связана с последовательностью гена константной области тяжелой цепи человека, выбранной из СН1, шарнира, СН2, СН3 и их комбинации.
[000219] Согласно другому аспекту предложен способ получения не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус, включающий:
(а) модификацию генома не являющегося человеком животного с удалением или устранением функциональности:
(i) эндогенных функциональных сегментов V, D и/или J генов тяжелых цепей иммуноглобулина, и
(ii) эндогенных функциональных сегментов V и J генов легкой цепи иммуноглобулина; и
(b) введение в указанный геном не являющегося человеком животного:
(i) нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанной с нуклеотидной последовательностью константной области легкой цепи; и
(ii) одного или большего количества, но меньшего, чем у дикого типа, сегментов гена VL и JL вариабельной области легкой цепи иммуноглобулина человека, функционально связанных с нуклеотидной последовательностью константной области тяжелой цепи.
Согласно различным вариантам реализации указанное не являющееся человеком животное представляет собой грызуна, например, мышь, крысу или хомяка. Согласно некоторым вариантам реализации указанный грызун представляет собой мышь. Согласно некоторым вариантам реализации указанная константная область легкой цепи представляет собой константную область крысы или мыши, например, константную область Cκ крысы или мыши.
[000220] Согласно другому аспекту предложены нуклеотидные последовательности, кодирующие реаранжированный вариабельный домен тяжелой цепи (т.е. нуклеотидные последовательности, представляющие собой реаранжированные нуклеотидные последовательности вариабельной области тяжелой цепи иммуноглобулина человека; т.е. предварительно реаранжированная нуклеотидная последовательность вариабельной области VDJ тяжелой цепи). Согласно некоторым вариантам реализации указанная нуклеотидная последовательность получена из последовательности гена или сегмента V, D и J человека. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность получена из сегмента V зародышевой линии человека, сегмента D зародышевой линии человека и сегмента J зародышевой линии человека. Согласно некоторым вариантам реализации указанный сегмент VH человека соответствует наблюдаемым в популяции человека вариантам. Согласно различным вариантам реализации указанная нуклеотидная последовательность содержит ген V человека, выбранный из группы, состоящей из VH1-2, VH1-3, VH1-8, VH1-18, VH1-24, VH1-45, VH1-46, VH1-58; VH1-69, VH2-5, VH2-26, VH2-70, VH3-7, VH3-9, VH3-11, VH3-13, VH3-15, VH3-16, VH3-20, VH3-21, VH3-23, VH3-30, VH3-30-3, VH3-30-5, VH3-33, VH3-35, VH3-38, VH3-43, VH3-48, VH3-49, VH3-53, VH3-64, VH3-66, VH3-72, VH3-73, VH3-74, VH4-4, VH4-28, VH4-30-1, VH4-30-2, VH4-30-4, VH4-31, VH4-34, VH4-39, VH4-59, VH4-61, VH5-51, VH6-1, VH7-4-l, VH7-81 и их полиморфный вариант. Согласно некоторым вариантам реализации указанный сегмент V человека представляет собой VH3-23 или его полиморфный вариант. Согласно различным вариантам реализации указанная нуклеотидная последовательность содержит сегмент D гена человека, выбранный из' группы, состоящей из D1-1, D1-7, D1-14, D1-20, D1-26, D2-2, D2-8, D2-15, D2-21, D3-3, D3-9, D3-10, D3-16, D3-22, D4-4, D4-11, D4-17, D4-23, D5-12, D5-5, D5-18, D5-24, D6-6, D6-13, D6-19, D6-25, D7-27 и их полиморфных вариантов. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность содержит сегмент D человека, который не является аутореактивным (является неиммуногенным) для указанного животного. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность содержит способный экспрессироваться сегмент D человека в вариабельной последовательности тяжелой цепи зрелой В-клетки мыши. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность также содержит последовательность гена константной области тяжелой цепи человека или не являющегося человеком животного, выбранную из СН1, шарнира, СН2, СН3 и их комбинации. Согласно конкретным вариантам реализации указанная нуклеиновая кислота включает последовательность гена константной области, содержащую CH1, шарнир, CH2 и CH3. Согласно различным вариантам реализации указанная нуклеотидная последовательность содержит сегмент J гена человека выбранный из группы, состоящей из JHI, JH2, JH3, JH4, JH5, JH6 и их полиморфных вариантов Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека кодирует последовательность человека VH3-23/GY/JH4-4 (SEQ ID NO: 137). Согласно некоторым вариантам реализации указанная нуклеотидная последовательность кодирует реаранжированный вариабельный домен тяжелой цепи, содержащий последовательность человека VH3-23/X1X2/J (где X1 представляет собой любую аминокислоту, и Х2 представляет собой любую аминокислоту). Согласно некоторым вариантам реализации X1 представляет собой Gly и Х2 представляет собой Туг. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность кодирует реаранжированный вариабельный домен тяжелой цепи, содержащий последовательность человека VH3-23/X1X2/JH4-4 (где X1 представляет собой любую аминокислоту, и Х2 представляет собой любую аминокислоту). Согласно некоторым вариантам реализации Х2 представляет собой аминокислоту, содержащую фенильную группу. Согласно конкретным вариантам реализации Х2 выбирают из Tyr и Phe. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность также содержит последовательность гена константной области легкой цепи человека или не являющегося человеком животного.
[000221] Согласно другому аспекту предложена содержащая нуклеиновые кислоты конструкция, содержащая нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (т.е. предварительно реаранжированную последовательность VDJ тяжелой цепи) согласно описанию в настоящем документе. Согласно некоторым вариантам реализации указанная содержащая нуклеиновые кислоты конструкция сконструирована таким образом, что нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с последовательностью гена константной области тяжелой цепи человека или не являющегося человеком животного. Согласно некоторым вариантам реализации указанная содержащая нуклеиновые кислоты конструкция содержит две копии, три копии, четыре копии, или более нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанной с последовательностью гена константной области тяжелой цепи. Согласно некоторым вариантам реализации указанная содержащая нуклеиновые кислоты конструкция представляет собой направленный вектор. Согласно некоторым вариантам реализации указанный направленный вектор содержит ген Adam6a, ген Adam6b или оба указанных гена, для предотвращения проблем с фертильностью, связанных с удалением генов Adam6a/6b (см., например, источник US 2012-0322108А1, полностью включенный посредством ссылки). Согласно некоторым вариантам реализации гены Adam6a и Adam6b вводят в направлении 5' от транскрипционной единицы универсальной последовательности тяжелой цепи. Согласно некоторым вариантам реализации указанный направленный вектор содержит селекционную кассету, фланкированную сайтами рекомбинации. Согласно некоторым вариантам реализации указанный направленный вектор содержит один или большее количество сайтов специфической рекомбинации (например, сайт 1охР или FRT).
[000222] Согласно другому аспекту предложены способы получения последовательности аминокислот вариабельной области легкой цепи (VL), способной к связыванию антигена независимо от последовательности аминокислот вариабельной области тяжелой цепи, включающие: (а) иммунизацию генетически модифицированного не являющегося человеком животного согласно описанию в настоящем документе (например, генетически модифицированного животного, у которого имеется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи человека, функционально связанная с нуклеотидной последовательностью константной области легкой или тяжелой цепи) представляющим интерес антигеном, при этом у указанного не являющегося человеком животного развивается иммунный ответ на указанный антиген; и (b) получение нуклеотидные последовательности реаранжированной легкой цепи (VJ) из вариабельного домена легкой цепи, специфически связывающего антиген, из клетки (например, зрелой В-клетки) указанного генетически модифицированного не являющегося человеком животного. Согласно различным вариантам реализации предложены вариабельные области легких цепей, продуцированные с применением таких способов.
[000223] Согласно некоторым аспектам предложены способы получения нуклеотидные последовательности, которая кодирует домен вариабельной области легкой цепи (VL) иммуноглобулина, способный к связыванию антигена независимо от вариабельной области тяжелой цепи, включающие: (а) иммунизацию не являющегося человеком животного представляющим интерес антигеном или его иммуногеном, при этом в геноме указанного не являющегося человеком животного содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области, (b) обеспечение возможности развития иммунного ответа у указанного не являющегося человеком животного, (с) выделение из указанного иммунизированного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, способный связывать антиген, и (d) получение из указанной клетки нуклеотидные последовательности, которая кодирует указанный вариабельный домен легкой цепи (VL домен), способный связывать антиген. Согласно некоторым вариантам реализации указанная последовательность гена константной области тяжелой цепи представляет собой последовательность гена константной области тяжелой цепи мыши или крысы. Согласно некоторым вариантам реализации указанная последовательность гена константной области тяжелой цепи представляет собой последовательность гена константной области тяжелой цепи человека. Согласно некоторым вариантам реализации указанный реаранжированный вариабельный домен тяжелой цепи, экспрессируемый указанным генетически модифицированным локусом, не является аутореактивным, т.е. является неиммуногенным для указанного не являющегося человеком животного. Согласно некоторым вариантам реализации в геноме указанного не являющегося человеком животного также имеется два или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL). Согласно некоторым вариантам реализации указанные сегменты гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL) функционально связаны с нуклеотидной последовательностью константной области легкой цепи. Согласно некоторым вариантам реализации указанный этап выделения (с) проводят путем сортировки клеток с активированной флуоресценцией (FACS) или проточной цитометрии. Согласно некоторым вариантам реализации указанная клетка, содержащая нуклеотидную последовательность, которая кодирует указанный вариабельный домен легкой цепи, связывающий антиген, представляет собой лимфоцит. Согласно некоторым вариантам реализации указанный лимфоцит включает естественные киллерные клетки, Т-клетки или В-клетки. Согласно некоторым вариантам реализации указанный способ дополнительно включает этап по (с)' слияния указанного лимфоцита с раковой клеткой. Согласно определенным вариантам реализации указанная раковая клетка представляет собой клетку миеломы.
[000224] Соответственно, согласно различным аспектам предложены способы получения нуклеотидные последовательности, которая кодирует вариабельный домен легкой цепи иммуноглобулина (VL), способный к связыванию антигена независимо от вариабельного домена тяжелой цепи, включающие:
(a) иммунизацию не являющегося человеком животного представляющим интерес антигеном или его иммуногеном, при этом в геноме указанного не являющегося человеком животного содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области; и (ii) нереаранжированные сегменты гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL), функционально связанные с нуклеотидной последовательностью константной области легкой цепи;
(b) обеспечение возможности развития иммунного ответа у указанного не являющегося человеком животного;
(c) выделение из указанного иммунизированного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, способный связывать антиген; и
(d) получение из указанной клетки нуклеотидные последовательности, которая кодирует указанный вариабельный домен легкой цепи (VL домен), способный связывать антиген.
Согласно некоторым вариантам реализации указанный этап выделения (с) проводят путем сортировки клеток с активированной флуоресценцией (FACS) или проточной цитометрии. Согласно некоторым вариантам реализации указанная клетка, содержащая нуклеотидную последовательность, которая кодирует указанный вариабельный домен легкой цепи, связывающий антиген, представляет собой лимфоцит. Согласно конкретным вариантам реализации указанный лимфоцит включает естественные киллерные клетки, Т-клетки или В-клетки. Согласно некоторым вариантам реализации указанные способы дополнительно включают этап по (с)' слияния указанного лимфоцита с раковой клеткой. Согласно конкретным вариантам реализации указанная раковая клетка представляет собой клетку миеломы. Согласно некоторым вариантам реализации нуклеотидную последовательность по (d) соединяют с нуклеотидной последовательностью, кодирующей нуклеотидную последовательность константной области иммуноглобулина. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области легкой цепи представляет собой каппа-последовательность человека или лямбда-последовательность человека. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области легкой цепи представляет собой каппа-последовательность мыши или лямбда-последовательность мыши. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области легкой цепь представляет собой каппа-последовательность крысы или лямбда-последовательность крысы. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи представляет собой последовательность человека, выбранную из СН1, шарнира, СН2, СН3 и их комбинации. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи представляет собой последовательность мыши или крысы, выбранную из СН1, шарнира, СН2, СН3 и их комбинации. Согласно некоторым вариантам реализации нуклеотидная последовательность по (d) содержит одну или большее количество замен или вставок гистидиновых кодонов, происходящих из нереаранжированного сегмента гена VL в геноме указанного животного.
[000225] Согласно различным аспектам предложены способы получения нуклеотидные последовательности, которая кодирует вариабельный домен легкой цепи иммуноглобулина (VL), способный к связыванию антигена независимо от вариабельного домена тяжелой цепи, включающие:
(а) иммунизацию не являющегося человеком животного представляющим интерес антигеном или его иммуногеном, при этом в геноме указанного не являющегося человеком животного содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью константной области легкой цепи; и (ii) нереаранжированные сегменты гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL), функционально связанные с нуклеотидной последовательностью тяжелой цепи константной области;
(b) обеспечение возможности развития иммунного ответа у указанного не являющегося человеком животного;
(c) выделение из указанного иммунизированного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, способный связывать антиген; и
(d) получение из указанной клетки нуклеотидные последовательности, которая кодирует указанный вариабельный домен легкой цепи (домен VL), способный связывать антиген.
Согласно некоторым вариантам реализации указанный этап выделения (с) проводят путем сортировки клеток с активированной флуоресценцией (FACS) или проточной цитометрии. Согласно некоторым вариантам реализации указанная клетка, содержащая нуклеотидную последовательность, которая кодирует указанный вариабельный домен легкой цепи, связывающий антиген, представляет собой лимфоцит. Согласно конкретным вариантам реализации указанный лимфоцит включает естественные киллерные клетки, Т-клетки или В-клетки. Согласно некоторым вариантам реализации указанные способы дополнительно включают этап по (с)' слияния указанного лимфоцита с раковой клеткой. Согласно конкретным вариантам реализации указанная раковая клетка представляет собой клетку миеломы. Согласно некоторым вариантам реализации нуклеотидную последовательность по (d) соединяют с нуклеотидной последовательностью, кодирующей нуклеотидную последовательность константной области иммуноглобулина. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области легкой цепи представляет собой каппа-последовательность человека или лямбда-последовательность человека. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области легкой цепи представляет собой каппа-последовательность мыши или лямбда-последовательность мыши. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области легкой цепи представляет собой каппа-последовательность крысы или лямбда-последовательность крысы. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи представляет собой последовательность человека, выбранную из СН1, шарнира, СН2, СН3 и их комбинации. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи представляет собой последовательность мыши или крысы, выбранную из СН1, шарнира, СН2, СН3 и их комбинации. Согласно некоторым вариантам реализации нуклеотидная последовательность по (d) содержит одну или большее количество замен или вставок гистидиновых кодонов, происходящих из нереаранжированного сегмента гена VL в геноме указанного животного.
[000226] Согласно некоторым аспектам предложены способы получения нуклеотидные последовательности, которая кодирует вариабельный домен легкой цепи иммуноглобулина (VL), способный к связыванию антигена независимо от вариабельного домена тяжелой цепи, включающие:
(a) иммунизацию не являющегося человеком животного представляющим интерес антигеном или его иммуногеном, при этом в геноме указанного не являющегося человеком животного содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанную с нуклеотидной последовательностью тяжелой цепи константной области; и (ii) два или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL), функционально связанных с нуклеотидной последовательностью константной области легкой цепи;
(b) обеспечение возможности развития иммунного ответа у указанного не являющегося человеком животного;
(c) выделение из указанного иммунизированного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, способный связывать антиген; и
(d) получение из указанной клетки нуклеотидные последовательности, которая кодирует указанный вариабельный домен легкой цепи (домен VL), способный связывать антиген.
Согласно некоторым вариантам реализации этап выделения (с) проводят путем сортировки клеток с активированной флуоресценцией (FACS) или проточной цитометрии. Согласно некоторым вариантам реализации указанная клетка, содержащая нуклеотидную последовательность, которая кодирует указанный вариабельный домен легкой цепи, связывающий антиген, представляет собой лимфоцит. Согласно конкретным вариантам реализации указанный лимфоцит включает естественные киллерные клетки, Т-клетки или В-клетки. Согласно некоторым вариантам реализации указанные способы дополнительно включают этап по (с)' слияния указанного лимфоцита с раковой клеткой. Согласно конкретным вариантам реализации указанная раковая клетка представляет собой клетку миеломы. Согласно некоторым вариантам реализации нуклеотидную последовательность по (d) соединяют с нуклеотидной последовательностью, кодирующей нуклеотидную последовательность константной области иммуноглобулина. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области легкой цепи представляет собой каппа-последовательность человека или лямбда-последовательность человека. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области легкой цепи представляет собой каппа-последовательность мыши или лямбда-последовательность мыши. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области легкой цепи представляет собой каппа-последовательность крысы или лямбда-последовательность крысы. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи представляет собой последовательность человека, выбранную из СН1, шарнира, СН2, СН3 и их комбинации. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность константной области тяжелой цепи представляет собой последовательности мыши или крысы, выбранную из СН1, шарнира, СН2, СН3 и их комбинации. Согласно некоторым вариантам реализации нуклеотидная последовательность по (d) содержит одну или большее количество замен или вставок гистидиновых кодонов, происходящих из нереаранжированного сегмента гена VL в геноме указанного животного.
[000227] Согласно некоторым аспектам предложены способы получения нуклеотидные последовательности, которая кодирует вариабельный домен легкой цепи иммуноглобулина (VL), способный к связыванию антигена независимо от вариабельного домена тяжелой цепи, включающие:
(a) иммунизацию не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус согласно описанию в настоящем документе, представляющим интерес антигеном, при этом в геноме указанного не являющегося человеком животного содержится нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области;
(b) обеспечение возможности развития иммунного ответа у указанного не являющегося человеком животного;
(c) получение лимфоцита {например, В-клетки) из указанного иммунизированного не являющегося человеком животного;
(d) слияние указанного лимфоцита с клеткой миеломы для образования клетки гибридомы; и
е) получение из клетки гибридомы нуклеотидные последовательности, которая кодирует вариабельный домен легкой цепи (VL домен), способный связывать антиген.
[000228] Согласно другому аспекту предложены способы получения нуклеотидные последовательности, которая кодирует нуклеотидную последовательность вариабельного домена легкой цепи иммуноглобулина (VL), из легкой цепи иммуноглобулина, способной к связыванию антигена независимо от вариабельной области тяжелой цепи, включающие:
(a) иммунизацию не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус согласно описанию в настоящем документе, представляющим интерес антигеном, при этом в геноме указанного не являющегося человеком животного содержится нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области;
(b) обеспечение возможности развития иммунного ответа у указанного не являющегося человеком животного;
(c) идентификацию лимфоцита (например, В-клетки) из указанного иммунизированного не являющегося человеком животного, которая экспрессирует последовательность аминокислот VL, связывающую указанный антиген независимо от вариабельной области тяжелой цепи; и
(d) клонирование нуклеотидные последовательности, кодирующей последовательность аминокислот VL по (с) из лимфоцита по (с).
[000229] Согласно другому аспекту предложены способы получения последовательности аминокислот вариабельной области легкой цепи иммуноглобулина (VL), способной к связыванию антигена независимо от вариабельной области тяжелой цепи, включающие:
(a) иммунизацию не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус согласно описанию в настоящем документе, представляющим интерес антигеном, при этом в геноме указанного не являющегося человеком животного содержатся: (i) первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. первая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), при этом указанная первая нуклеотидная последовательность функционально связана с последовательностью гена константной области легкой цепи; и (ii) вторая нуклеотидная последовательность, которая кодирует человеческий или имеющий нечеловеческое происхождение вариабельный домен легкой цепи (т.е. вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека), при этом указанная вторая нуклеотидная последовательность функционально связана с последовательностью гена константной области тяжелой цепи;
(b) обеспечение возможности развития иммунного ответа у указанного не являющегося человеком животного;
(c) получение лимфоцита (например, В-клетки) из указанного иммунизированного не являющегося человеком животного;
(d) слияние указанного лимфоцита с клеткой миеломы для образования клетки гибридомы; и
е) получение из клетки гибридомы нуклеотидные последовательности, которая кодирует вариабельный домен легкой цепи (домен VL), способный связывать антиген.
[000230] Согласно другому аспекту предложены способы получения нуклеотидные последовательности, которая кодирует вариабельный домен легкой цепи иммуноглобулина (VL), способный к связыванию антигена независимо от вариабельного домена тяжелой цепи, включающие:
(a) иммунизацию не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус согласно описанию в настоящем документе, представляющим интерес антигеном, при этом в геноме указанного не являющегося человеком животного содержатся: (i) первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. первая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), при этом указанная первая нуклеотидная последовательность функционально связана с последовательностью гена константной области легкой цепи; и (ii) вторая нуклеотидная последовательность, которая кодирует человеческую или имеющую нечеловеческое происхождение вариабельный домен легкой цепи (т.е. вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека), при этом указанная вторая нуклеотидная последовательность функционально связана с последовательностью гена константной области тяжелой цепи;
(b) обеспечение возможности развития иммунного ответа у указанного не являющегося человеком животного;
(c) идентификацию лимфоцита (например, В-клетки) из указанного иммунизированного не являющегося человеком животного, которая экспрессирует последовательность аминокислот VL, связывающую указанный антиген независимо от вариабельной области тяжелой цепи; и,
(d) клонирование нуклеотидные последовательности, кодирующей последовательность аминокислот VL по (с) из лимфоцита по (с).
[000231] Согласно другому аспекту предложены способы получения последовательности аминокислот вариабельной области легкой цепи иммуноглобулина (VL), способной к связыванию антигена независимо от вариабельной области тяжелой цепи, включающие:
(a) иммунизацию не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус согласно описанию в настоящем документе, представляющим интерес антигеном, при этом в геноме указанного не являющегося человеком животного содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области; и (ii) два или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL);
(b) обеспечение возможности развития иммунного ответа у указанного не являющегося человеком животного;
(c) получение лимфоцита (например, В-клетки) из указанного иммунизированного не являющегося человеком животного;
(d) слияние указанного лимфоцита с клеткой миеломы для образования клетки гибридомы; и
е) получение из клетки гибридомы нуклеотидные последовательности, которая кодирует вариабельный домен легкой цепи (VL домен), способный связывать антиген.
[000232] Согласно другому аспекту предложены способы получения нуклеотидные последовательности вариабельной области легкой цепи иммуноглобулина (VL), из легкой цепи иммуноглобулина, способной к связыванию антигена независимо от вариабельной области тяжелой цепи, включающие:
(а) иммунизацию не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус согласно описанию в настоящем документе, представляющим интерес антигеном, при этом в геноме указанного не являющегося человеком животного содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области; и (ii) два или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL);
(b) обеспечение возможности развития иммунного ответа у указанного не являющегося человеком животного;
(c) идентификацию лимфоцита (например, В-клетки) из указанного иммунизированного не являющегося человеком животного, которая экспрессирует последовательность аминокислот VL, связывающую указанный антиген независимо от вариабельной области тяжелой цепи; и,
(d) клонирование нуклеотидные последовательности, кодирующей последовательность аминокислот VL по (с) из лимфоцита по (с).
[000233] Согласно различным вариантам реализации описанный в настоящем документе вариабельный домен легкой цепи представляет собой эффекторный вариабельный домен легкой цепи. Согласно некоторым вариантам реализации указанный эффекторный вариабельный домен легкой цепи специфически связывает FcRn, увеличивая период полужизни мультиспецифических антител. Например, биспецифическое антитело содержит вариабельный домен тяжелой цепи, связывающий антиген, и вариабельный домен легкой цепи, связывающий FcRn. Согласно некоторым вариантам реализации генетически модифицированных мышей согласно описанию в настоящем документе иммунизируют FcRN для получения антител, которые связывают FcRN исключительно за счет легких цепей. Указанные легкие цепи, продуцированные указанным генетически модифицированным не являющимся человеком животным, используют в качестве универсальных или общих легких цепей, помогающих указанному биспецифическому антителу связываться с FcRn, способствуя таким образом увеличению периода полужизни. Остальную часть антитела (например, либо вторую, отличную легкую цепь, либо тяжелую цепь, связывающую антиген, отличный от FcRn) выбирают для осуществления второй функции.
[000234] Согласно дополнительным аспектам предложен генетически модифицированный иммуноглобулиновый локус, который может быть получен любыми способами согласно описанию в настоящем документе. Согласно различным вариантам реализации также предложены вариабельные области легких цепей, полученные с помощью способов согласно описанию в настоящем документе, и нуклеотидная последовательность, кодирующая такие вариабельные области легких цепей.
[000235] Согласно некоторым аспектам предложен иммуноглобулиновый локус в геноме зародышевой линии не являющегося человеком животного, содержащий (1) нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанную с последовательностью гена константной области тяжелой цепи, и (2) нуклеотидную последовательность нереаранжированной вариабельной области лег кой цепи иммуноглобулина человека, функционально связанную с последовательностью гена константной области легкой цепи. Согласно некоторым аспектам предложен иммуноглобулиновый локус в геноме зародышевой линии не являющегося человеком животного, содержащий (1) нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанную с последовательностью гена константной области легкой цепи, и (2) нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека, функционально связанную с последовательностью гена константной области тяжелой цепи. Согласно некоторым аспектам предложен иммуноглобулиновый локус в геноме зародышевой линии не являющегося человеком животного, содержащий (1) нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанную с последовательностью гена константной области тяжелой цепи, и (2) нуклеотидную последовательность, которая кодирует два или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL). Согласно некоторым вариантам реализации указанная последовательность гена константной области легкой цепи представляет собой последовательность гена константной области легкой цепи κ. Согласую некоторым вариантам реализации указанная последовательность гена константной области легкой цепи представляет собой последовательность гена константной области легкой цепи λ. Согласно некоторым вариантам реализации указанная последовательность гена константной области легкой цепи представляет собой последовательность гена константной области легкой цепи мыши или крысы. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность вариабельной области легкой цепи представляет собой последовательность гена вариабельной области легкой цепи κ. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность вариабельной области легкой цепи представляет собой последовательность гена вариабельной области легкой цепи λ. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность вариабельной области легкой цепи представляет собой последовательность гена вариабельной области легкой цепи мыши или крысы.
[000236] Дополнительные аспекты включают антигенсвязывающие белки (например, антитела), продуцированные генетически модифицированными не являющимися человеком животными, описанными в настоящем документе. Также предложены антигенсвязывающие белки {например, рекомбинантной антител) с последовательностями вариабельной области легкой цепи (VL), происходящими из или продуцированными (т.е. экспрессируемыми из сегментов гена нереаранжированной вариабельной области легкой цепи иммуноглобулина человека) генетически модифицированными не являющимися человеком животными, описанные в настоящем документе. Согласно некоторым вариантам реализации антигенсвязывающие белки, полученные с применением способов, описанных в настоящем документе, содержат тяжелую цепь и легкую цепь, при этом указанная тяжелая цепь не влияет на связывание легкой цепи с антигеном, и/или тяжелая цепь не связывает антиген в отсутствие легкой цепи. Согласно некоторым вариантам реализации указанный вариабельный домен легкой цепи связывает представляющий интерес антиген с KD не более чем на один порядок выше в отсутствие тяжелой цепи, чем в присутствии тяжелой цепи {например, KD ~ 10-10 в присутствии тяжелой цепи или KD ~10-9 в отсутствие тяжелой цепи). Согласно некоторым вариантам реализации антигенсвязывающие белки согласно описанию в настоящем документе включают легкую цепь иммуноглобулина, которая может специфически связывать представляющий интерес антиген со сродством (KD) менее 10-6, 10-7, 10-8, 10-9 или 10-10. Согласно некоторым вариантам реализации указанная легкая цепь иммуноглобулина, полученная с помощью указанных способов, способна специфически связывать представляющий интерес антиген в отсутствие вариабельной области тяжелой цепи со сродством (KD) менее 10-6, 10-7, 10-8, 10-9 или 10-10.
[000237] Согласно различным вариантам реализации вариабельные домены легких цепей, полученные согласно описанию в настоящем документе, специфически связывают целевую молекулу («Т»). Целевая молекула представляет собой любой белок, полипептид или другую макромолекулу, активность или внеклеточную концентрацию которой требуется ослабить, снизить или элиминировать. Во многих случаях целевая молекула, с которой связывается вариабельная область легкой цепи, представляет собой белок или полипептид (т.е. «целевой белок»); при этом также предложены варианты реализации, отличающиеся тем, что целевая молекула («Т») представляет собой углевод, гликопротеин, липид, липопротеин, липополисахарид или другой небелковый полимер или молекулу, с которой связывается вариабельная область легкой цепи. Согласно различным вариантам реализации Т может представлять собой экспрессируемый на клеточной поверхности целевой белок или растворимый целевой белок. Связывание мишени антигенсвязывающей молекулой может происходить во внеклеточной среде или на клеточной поверхности. Однако согласно определенным вариантам реализации антигенсвязывающая молекула связывает целевую молекулу внутри клетки, например, во внутриклеточном компоненте, таком как эндоплазматический ретикулум, аппарат Гольджи, эндосома, лизосома и т.п.Примеры экспрессируемых на клеточной поверхности целевых молекул включают экспрессируемые на клеточной поверхности рецепторы, мембраносвязанные лиганды, ионные каналы и любой другой мономерный или мультимерный полипептидный компонент с внеклеточной частью, присоединенной или связанной с клеточной мембраной. Неограничивающие примеры экспрессируемых на клеточной поверхности целевых молекул, на которые могут быть нацелены мультиспецифические антигенсвязывающие молекулы, предложенные в настоящем документе, включают, например, цитокиновые рецепторы (например, рецепторы IL-1, IL-4, IL-6, IL-13, IL-22, IL-25, IL-33 и т.п.), а также мишени на клеточной поверхности, в том числе другие трансмембранные рецепторы 1 типа, такие как PRLR, сопряженные с G-белками рецепторы, такие как GCGR, ионные каналы, такие как NaV1.7, ASIC1 или ASIC2, нерецепторные поверхностные белки, такие как MHC-I (например, HLA-B*27) и т.п. Согласно вариантам реализации, в которых Т представляет собой экспрессируемый на клеточной поверхности целевой белок, компонент D1 мультиспецифической антигенсвязывающей молекулы может представлять собой, например, антитело или антигенсвязывающие фрагмент антитела, которое специфически связывает Т, или лиганд или часть лиганда, который специфически взаимодействует с указанным экспрессируемым на клеточной поверхности целевым белком. Например, если Т представляет собой IL-4R, компонент D1 может содержать или состоять из IL-4 или его связывающей рецептор части. Примеры растворимых целевых молекул включают цитокины, факторы роста и другие лиганды и сигнальные белки. Неограниченные примерь; растворимых целевых белков, на которые могут быть нацелены мультиспецифические антигенсвязывающие молекулы, предложенные в настоящем документе, включают, например, IL-1, IL-4, IL-6, IL-13, IL-22, IL-25, IL-33, SOST, DKK1 и т.п.Растворимые целевые молекулы также включают, например, не человеческие целевые молекулы, такие как аллергены (например, Fel D1, BetVl, CryJ1), патогены (например, Candida albicans, S. aureus и т.п.) и патогенные молекулы (например, липополисахарид (LPS), липотейхоевая кислота (LTA), белок А, токсины и т.п.). Согласно вариантам реализации, в которых Т представляет собой растворимую целевую молекулу, компонент D1 мультиспецифической антигенсвязывающей молекулы может представлять собой, например, антитело или антигенсвязывающий фрагмент антитела, которое специфически связывает Т, или рецептор или часть рецептора, который специфически взаимодействует с указанной растворимой целевой молекулой. Например, если Т представляет собой IL-4, компонент D1 может содержать или состоять из IL-4R или его лиганд-связывающей части. Целевые молекулы также включают опухолеассоциированные антигены.
[000238] Согласно другому аспекту антигенсвязывающие белки (например, биспецифические или триспецифические антитела) могут быть получены с применением антигенспецифических вариабельных доменов легких цепей, происходящих от не являющегося человеком животного (т.е. содержащих продуцированные указанным животным последовательности вариабельных областей легких цепей (VL) человека), у которого имеется иммуноглобулиновый локус с реаранжированной нуклеотидной последовательностью вариабельной области тяжелой цепи человека (т.е. животного, у которого имеется предварительно сконструированная реаранжированная последовательность VDJ тяжелой цепи). Такие антигенспецифические обратно-гибридные (например, содержащие вариабельную область человека/константную область мыши) легкие цепи могут быть использованы для получения последовательностей антигенспецифических вариабельных областей легких цепей, которые могут быть клонированы внутрь рамки в экспрессионный вектор с подходящей последовательностью константной области легкой цепи человека. Антигенспецифическая(ие) вариабельная(ые) область(и) тяжелой цепи человека (специфическая(ие) в отношении эпитопа на том же или другом антигене, отличного от эпитопа антигенспецифической легкой цепи) животного, у которого имеется иммуноглобулиновый локус с реаранжированной нуклеотидной последовательностью вариабельной области тяжелой цепи человека (т.е. мыши, у которой имеется предварительно сконструированная реаранжированная последовательность VDJ тяжелой цепи), может(гут) быть клонирована(ы) внутри рамки в экспрессионный вектор, содержащий последовательность константной области тяжелой цепи человека, и антигенспецифические легкие и тяжелые цепи человека могут коэкспрессироваться в подходящей клетке для получения антигенсвязывающего белка (например, биспецифического или триспецифического антитела человека). Как вариант, ранее выбранная антигенспецифические тяжелая цепь, например, тяжелая цепь из антитела, которое содержит легкую цепь, происходящую из того же сегмента гена вариабельной области, что и используемый в нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи человека, может быть клонирована внутри рамки в экспрессионный вектор, содержащий последовательность константной области тяжелой цепи человека, и антигенспецифические легкие и тяжелые цепи человека могут коэкспрессироваться в подходящей клетке для получения антигенсвязывающего белка (например, биспецифического или триспецифического антитела человека). Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с имеющей нечеловеческое происхождение последовательностью гена константной области тяжелой цепи (например, каппа или лямбда мыши или крысы). Согласно некоторым вариантам реализации указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека функционально связана с имеющей нечеловеческое происхождение последовательностью гена константной области легкой цепи (например, каппа или лямбда мыши или крысы). Согласно некоторым вариантам реализации последовательности вариабельной области легкой цепи (VL) человека представляют собой последовательности генов каппа.
[000239] Согласно другому аспекту предложен способ получения мультиспецифического антигенсвязывающего белка, включающий:
(a) иммунизацию представляющим интерес антигеном первого не являющегося человеком животного, у которого имеется первый генетически модифицированный иммуноглобулиновый локус согласно описанию в настоящем документе, при этом в геноме указанного первого не являющегося человеком животного содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области;
(b) обеспечение возможности развития иммунного ответа у первого не являющегося человеком животного;
(c) получение первого лимфоцита (например, В-клетки) из иммунизированного первого не являющегося человеком животного, при этом указанный первая лимфоцит экспрессирует антитела с созревшей аффинностью, при этом указанные антитела с созревшей аффинностью содержат вариабельный домен человека, соединенный с не человеческим константным доменом;
(d) идентификация нуклеотидные последовательности, которая кодирует человеческий вариабельный домен легкой цепи антител с созревшей аффинностью;
(e) клонирование нуклеотидные последовательности по (d) в первую экспрессионную конструкцию внутри рамки с подходящей нуклеотидной последовательностью константной области человека (например, последовательностью лямбда или каппа человека) с образованием первого гена полипептида;
(f) иммунизацию второго не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус согласно описанию в настоящем документе, вторым представляющим интерес антигеном, при этом в геноме указанного второго не являющегося человеком животного содержатся: (i) нереаранжированные сегменты генов человека V, D и J, связанные с имеющей нечеловеческое происхождение нуклеотидной последовательностью константной области тяжелой цепи; и (ii) единственная реаранжированная последовательность вариабельной области легкой цепи человека;
(g) обеспечение возможности развития иммунного ответа у второго не являющегося человеком животного;
(h) получение второго лимфоцита от иммунизированного второго не являющегося человеком животного, при этом указанный второй лимфоцит экспрессирует антитела с созревшей аффинностью, при этом указанные антитела с созревшей аффинностью содержат вариабельный домен тяжелой цепи человека, соединенный с не человеческим константным доменом;
(i) идентификация нуклеотидные последовательности, которая кодирует вариабельный домен тяжелой цепи человека специфически связывающих второй антиген антител с созревшей аффинностью;
(j) клонирование нуклеотидные последовательности по (i) во второй экспрессионной конструкции внутри рамки с подходящей нуклеотидной последовательностью константной области человека (например, последовательностью константной области IgG1 человека) с образованием второго гена полипептида; и
(k) введение первой экспрессионной конструкции и второй экспрессионной конструкции в клетку, подходящую для экспрессирования первого гена полипептида и второго гена полипептида для получения антигенсвязывающего белка, содержащего димер второго полипептида, отличающегося тем, что каждый мономер второго полипептида связан с мономером первого полипептида.
Согласно различным вариантам реализации указанные первая экспрессионная конструкция и вторая экспрессионная конструкция располагаются на отдельных векторах. Согласно различным вариантам реализации указанные первая экспрессионная конструкция и вторая экспрессионная конструкция располагаются на одном векторе. Согласно различным вариантам реализации указанные первый антиген и второй антиген отличаются. Согласно одному варианту реализации указанные первый антиген и второй антиген являются одинаковыми. Согласно различным вариантам реализации первый антиген представляет собой рецептор клеточной поверхности, и второй антиген выбирают из растворимого антигена и антигена, связанного с клеточной поверхностью. Согласно конкретным вариантам реализации первый антиген представляет собой Fc-рецептор {например, FcRN), второй антиген представляет собой растворимый белок, а антигенсвязывающий белок содержит одну или большее количество замен и вставок гистидина, происходящих из сегмента гена VL в геноме не являющегося человеком животного.
[000240] Согласно другому аспекту предложен способ получения мультиспецифического антигенсвязывающего белка, включающий:
(а) иммунизацию первого не являющегося человеком животного, у которого имеется первый генетически модифицированный иммуноглобулиновый локус согласно описанию в настоящем документе, представляющим интерес антигеном, при этом в геноме указанного первого не являющегося человеком животного имеется (i) первая нуклеотидная последовательность, которая кодирует реаранжированный вариабельный домен тяжелой цепи (т.е. первая нуклеотидная последовательность представляет собой нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека), при этом указанная первая нуклеотидная последовательность функционально связана с последовательностью гена константной области легкой цепи; и (ii) вторая нуклеотидная последовательность, которая кодирует человеческий или имеющий нечеловеческое происхождение вариабельный домен легкой цепи (т.е. вторая нуклеотидная последовательность представляет собой нуклеотидную последовательность нереаранжированной вариабельной области легкой цепи иммуноглобулина человека), при этом указанная вторая нуклеотидная последовательность функционально связана с последовательностью гена константной области тяжелой цепи;
(b) обеспечение возможности развития иммунного ответа у первого не являющегося человеком животного;
(c) получение первого лимфоцита (например, В-клетки) из иммунизированного первого не являющегося человеком животного, при этом указанный первый лимфоцит экспрессирует антитела с созревшей аффинностью, при этом указанные антитела с созревшей аффинностью содержат вариабельный домен человека, соединенный с не человеческим константным доменом;
(d) идентификация нуклеотидные последовательности, которая кодирует человеческий вариабельный домен легкой цепи антител с созревшей аффинностью;
(e) клонирование нуклеотидные последовательности по (d) в первой экспрессионной конструкции внутри рамки с подходящей нуклеотидной последовательностью константной области человека (например, последовательностью лямбда или каппа человека) с образованием первого гена полипептида;
(f) иммунизацию второго не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус согласно описанию в настоящем документе, вторым представляющим интерес антигеном, при этом в геноме указанного второго не являющегося человеком животного содержатся: (i) нереаранжированные сегменты генов человека V, D и J, связанные с имеющей нечеловеческое происхождение нуклеотидной последовательностью константной области тяжелой цепи; и (ii) единственная реаранжированная последовательность вариабельной области легкой цепи человека;
(g) обеспечение у второго не являющегося человеком животного развития иммунного ответа;
(h) получение второго лимфоцита от иммунизированного второго не являющееся человеком животного, при этом указанный второй лимфоцит экспрессирует антитела с созревшей аффинностью, при этом указанные антитела с созревшей аффинностью содержат вариабельный домен тяжелой цепи человека, соединенный с не человеческим константным доменом;
(i) идентификация нуклеотидные последовательности, которая кодирует вариабельный домен тяжелой цепи человека антител с созревшей аффинностью, которая специфически связывает второй антиген;
(j) клонирование нуклеотидные последовательности по (i) во второй экспрессионной конструкции внутри рамки с подходящей нуклеотидной последовательностью константной области человека (например, последовательностью константной области IgG1 человека) с образованием второго гена полипептида; и
(к) введение первой экспрессионной конструкции и второй экспрессионной конструкции в клетку, подходящую для экспрессирования первого гена полипептида и второго гена полипептида, для получения антигенсвязывающего белка, содержащего димер второго полипептида, отличающегося тем, что каждый мономер второго полипептида связан с мономером первого полипептида.
Согласно различным вариантам реализации указанные первая экспрессионная конструкция и вторая экспрессионная конструкция располагаются на отдельных векторах. Согласно различным вариантам реализации указанные первая экспрессионная конструкция и вторая экспрессионная конструкция располагаются на одном векторе. Согласно некоторым вариантам реализации первый антиген и второй антиген отличаются. Согласно некоторым вариантам реализации первый антиген и второй антиген являются одинаковыми. Согласно различным вариантам реализации первый антиген представляет собой рецептор клеточной поверхности, и второй антиген выбирают из растворимого антигена и антигена, связанного с клеточной поверхностью. Согласно конкретным вариантам реализации первый антиген представляет собой Fc-рецептор (например, FcRN), второй антиген представляет собой растворимый белок, а антигенсвязывающий белок содержит одну или большее количество замен и вставок гистидина, происходящих из сегмента гена VL в геноме не являющегося человеком животного.
[000241] Согласно другому аспекту предложен способ получения мультиспецифического антигенсвязывающего белка, включающий:
(a) иммунизацию первого не являющегося человеком животного, у которого имеется первый генетически модифицированный иммуноглобулиновый локус согласно описанию в настоящем документе, представляющим интерес антигеном, при этом в геноме указанного первого не являющегося человеком животного имеется (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области; и (ii) два или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL);
(b) обеспечение возможности развития иммунного ответа у первого не являющегося человеком животного;
(c) получение первого лимфоцита (например, В-клетки) из иммунизированного первого не являющегося человеком животного, при этом указанный первый лимфоцит экспрессирует антитела с созревшей аффинностью, при этом указанные антитела с созревшей аффинностью содержат вариабельный домен человека, соединенный с не человеческим константным доменом;
(d) идентификация нуклеотидные последовательности, которая кодирует указанный человеческий вариабельный домен легкой цепи антител с созревшей аффинностью;
(e) клонирование нуклеотидные последовательности по (d) в первой экспрессионной конструкции внутри рамки с подходящей нуклеотидной последовательностью константной области человека (например, последовательностью лямбда или каппа человека) с образованием первого гена полипептида;
(f) иммунизацию второго не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус согласно описанию в настоящем документе, вторым представляющим интерес антигеном, при этом в геноме указанного второго не являющегося человеком животного содержатся: (i) нереаранжированные сегменты генов человека V, D и J, связанные с имеющей нечеловеческое происхождение нуклеотидной последовательностью константной области тяжелой цепи; и (ii) единственная реаранжированная последовательность вариабельной области легкой цепи человека, при этом указанная единственная реаранжированная последовательность вариабельной области легкой цепи человека получена из того же сегмента гена VL, что и сегмент гена VL, кодирующий вариабельный домен легкой цепи этапа (с);
(g) обеспечение у второго не являющегося человеком животного развития иммунного ответа;
(h) получение второго лимфоцита от иммунизированного второго не являющееся человеком животного, при этом указанный второй лимфоцит экспрессирует антитела с созревшей аффинностью, при этом указанные антитела с созревшей аффинностью содержат вариабельный домен тяжелой цепи человека, соединенный с не человеческим константным доменом;
(i) идентификация нуклеотидные последовательности, которая кодирует вариабельный домен тяжелой цепи человека антител с созревшей аффинностью, который специфически связывает второй антиген;
(j) клонирование нуклеотидные последовательности по (i) во второй экспрессионной конструкции внутри рамки с подходящей нуклеотидной последовательностью константной области человека (например, последовательностью константной области IgG1 человека) с образованием второго гена полипептида; и
(k) введение первой экспрессионной конструкции и второй экспрессионной конструкции в клетку, подходящую для экспрессирования первого гена полипептида и второго гена полипептида, для получения антигенсвязывающего белка, содержащего димер второго полипептида, отличающегося тем, что каждый мономер второго полипептида связан с мономером первого полипептида.
Согласно различным вариантам реализации указанные первая экспрессионная конструкция и вторая экспрессионная конструкция располагаются на отдельных векторах. Согласно различным вариантам реализации указанные первая экспрессионная конструкция и вторая экспрессионная конструкция располагаются на одном векторе. Согласно различным вариантам реализации указанные первый антиген и второй антиген отличаются. Согласно различным вариантам реализации указанные первый антиген и второй антиген являются одинаковыми. Согласно различным вариантам реализации первый антиген представляет собой рецептор клеточной поверхности, и второй антиген выбирают из растворимого антигена и антигена, связанного с клеточной поверхностью. Согласно различным вариантам реализации первый антиген представляет собой Fc-рецептор (например, FcRN), второй антиген представляет собой растворимый белок, а антигенсвязывающий белок содержит одну или большее количество замен и вставок гистидина, происходящих из сегмента гена VL в геноме не являющегося человеком животного.
[000242] Согласно другому аспекту предложен способ получения мультиспецифического антигенсвязывающего белка, включающий:
(a) иммунизацию не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус согласно описанию в настоящем документе, первым антигеном, при этом в геноме указанного не являющегося человеком животного содержатся: (i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области; и (ii) один или большее количество сегментов гена VL и JL иммуноглобулина человека;
(b) обеспечение возможности развития иммунного ответа у указанного не являющегося человеком животного;
(c) получение лимфоцита (например, В-клетки) из указанного иммунизированного не являющегося человеком животного, при этом указанный лимфоцит экспрессирует антитела с созревшей аффинностью, содержащие вариабельный домен легкой цепи иммуноглобулина человека, соединенный с константным доменом иммуноглобулина мыши;
(d) идентификация первой нуклеотидные последовательности, которая кодирует указанный человеческий вариабельный домен легкой цепи антител с созревшей аффинностью;
(e) клонирование первой нуклеотидные последовательности по (d) в первый экспрессионный вектор внутри рамки с человеческой нуклеотидной последовательностью константной области легкой цепи;
(f) введение в клетку-хозяина: (i) первого экспрессионного вектора, содержащего первую нуклеотидную последовательность внутри рамки с нуклеотидной последовательностью константной области легкой цепи человека; и (ii) второго экспрессионного вектора, содержащего вторую нуклеотидную последовательность, которая кодирует первый антигенспецифический вариабельный домен тяжелой цепи, соединенный с константной областью тяжелой цепи человека;
(g) культивирование клетки-хозяина для обеспечения экспрессии мультиспецифических антител; и
(h) выделение мультиспецифических антител, отличающихся тем, что указанные мультиспецифические антитела содержат первую антигенспецифическую тяжелую цепь и вариабельный домен легкой цепи, при этом вариабельный домен тяжелой цепи указанных мультиспецифических антител проявляет антигенсвязывающую специфичность, отличную от вариабельного домена легкой цепи.
Согласно различным вариантам реализации указанные мультиспецифические антител представляют собой биспецифические антитела. Согласно некоторым вариантам реализации указанные мультиспецифические антитела представляют собой триспецифические антитела, и этап (f) также включает введение третьего экспрессионного вектора, содержащего третью нуклеотидную последовательность, которая кодирует второй антигенспецифический вариабельный домен тяжелой цепи, соединенный с последовательностью константной области тяжелой цепи человека.
[000243] Согласно другому аспекту предложен способ получения мультиспецифического антигенсвязывающего белка, включающий:
(a) иммунизацию не являющегося человеком животного, у которого имеется генетически модифицированный иммуноглобулиновый локус согласно описанию в настоящем документе первым антигеном, отличающееся тем, что в геноме указанного не являющегося человеком животного содержатся: (i) реаранжированный вариабельный домен тяжелой цепи, функционально связанный с нуклеотидной последовательностью тяжелой цепи константной области; и (ii) один или более, но меньше, чем у дикого типа, сегментов гена VL и JL иммуноглобулина человека;
(b) обеспечение возможности развития иммунного ответа у указанного не являющегося человеком животного;
(c) получение лимфоцита (например, В-клетки) из указанного иммунизированного не являющегося человеком животного, при этом указанный лимфоцит экспрессирует антитела с созревшей аффинностью, содержащие вариабельный домен легкой цепи иммуноглобулина человека, соединенный с константным доменом иммуноглобулина мыши;
(d) идентификация первой нуклеотидные последовательности, которая кодирует человеческий вариабельный домен легкой цепи антител с созревшей аффинностью;
(e) клонирование первой нуклеотидные последовательности по (d) в первый экспрессионный вектор внутри рамки с человеческой нуклеотидной последовательностью константной области легкой цепи;
(f) введение в клетку-хозяина: (i) первого экспрессионного вектора, содержащего первую нуклеотидную последовательность внутри рамки с нуклеотидной последовательностью константной области легкой цепи человека; и (ii) второго экспрессионного вектора, содержащего вторую нуклеотидную последовательность, которая кодирует первый антигенспецифический вариабельный домен тяжелой цепи, соединенный с константной областью тяжелой цепи человека;
(g) культивирование клетки-хозяина для обеспечения экспрессии мультиспецифических антител; и
(h) выделение мультиспецифических антител, отличающихся тем, что указанные мультиспецифические антитела содержат первую антигенспецифическую тяжелую цепь и вариабельный домен легкой цепи, при этом вариабельный домен тяжелой цепи указанных мультиспецифических антител проявляет антигенсвязывающую специфичность, отличную от вариабельного домена легкой цепи.
Согласно различным вариантам реализации указанные мультиспецифические антитела представляют собой биспецифические антитела. Согласно некоторым вариантам реализации указанные мультиспецифические антитела представляют собой триспецифические антитела, и этап (f) также включает введение третьего экспрессионного вектора, содержащего третью нуклеотидную последовательность, которая кодирует второй антигенспецифический вариабельный домен тяжелой цепи, соединенный с последовательностью константной области тяжелой цепи человека.
[000244] Согласно другому аспекту предложены способы получения антигенсвязывающего белка, который содержит вариабельный домен легкой цепи иммуноглобулина, способный связывать антиген независимо от вариабельного домена тяжелой цепи. Такие способы включают
(a) иммунизацию генетически модифицированного не являющегося человеком животного первым антигеном, который содержит первый эпитоп или его иммуногенную часть, при этом в геноме указанного не являющегося человеком животного содержится:
(i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области; и
(ii) сегменты гена вариабельной области нереаранжированной легкой цепи иммуноглобулина человека (VL и JL), функционально связанные с нуклеотидной последовательностью константной области легкой цепи иммуноглобулина;
(b) обеспечение возможности развития иммунного ответа на первый эпитоп или его иммуногенную часть у указанного не являющегося человеком животного;
(c) выделение из указанного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(d) получение из указанной клетки по (с) нуклеотидной последовательности, которая кодирует указанный вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(e) применение нуклеотидной последовательности по (d) в экспрессионной конструкции, соединенной с нуклеотидной последовательностью константной области иммуноглобулина человека; и
(f) экспрессию нуклеотидной последовательности по (d) в продукционной клеточной линии, экспрессирующей тяжелую цепь иммуноглобулина человека, которая специфически связывает второй антиген или эпитоп, для получения легкой цепи антигенсвязывающего белка, кодируемой нуклеиновой кислотой по (d) и связывающей первый эпитоп или его иммуногенную часть независимо от тяжелой цепи, тяжелая цепь которого специфически связывает указанный второй антиген или эпитоп.
Согласно некоторым вариантам реализации по меньшей мере один из нереаранжированных сегментов гена легкой цепи VL или JL человека кодирует один или большее количество гистидиновых кодонов, не кодируемых соответствующим сегментом гена вариабельной области легкой цепи зародышевой линии человека. Согласно некоторым вариантам реализации указанный первый эпитоп получен из рецептора клеточной поверхности. Согласно некоторым вариантам реализации указанный рецептор клеточной поверхности представляет собой Fc-рецептор. Согласно конкретным вариантам реализации указанный Fc-рецептор представляет собой FcRn. Согласно некоторым вариантам реализации указанный второй антиген или эпитоп получен из растворимого антигена. Согласно некоторым вариантам реализации указанный второй антиген или эпитоп получен из рецептора клеточной поверхности. Согласно некоторым вариантам реализации первый антиген представляет собой Fc-рецептор, второй антиген представляет собой растворимый белок, а антигенсвязывающий белок содержит одну или большее количество замен и вставок гистидина, происходящих из сегмента гена VL в геноме не являющегося человеком животного.
[000245] Согласно другому аспекту предложены способы получения антигенсвязывающего белка, который содержит вариабельный домен легкой цепи иммуноглобулина, способный связывать антиген независимо от вариабельного домена тяжелой цепи. Такие способы включают
(a) иммунизацию генетически модифицированного не являющегося человеком животного первым антигеном, который содержит первый эпитоп или его иммуногенную часть, при этом в геноме указанного не являющегося человеком животного содержится:
(i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи человека, функционально связанная с нуклеотидной последовательностью константной области легкой цепи; и
(ii) нереаранжированные сегменты гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL), функционально связанные с нуклеотидной последовательностью константной области тяжелой цепи иммуноглобулина;
(b) обеспечение возможности развития иммунного ответа на первый эпитоп или его иммуногенную часть у указанного не являющегося человеком животного;
(c) выделение из указанного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(d) получение из указанной клетки по (с) нуклеотидной последовательности, которая кодирует указанный вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(e) применение нуклеотидной последовательности по (d) в экспрессионной конструкции, соединенной с нуклеотидной последовательностью константной области иммуноглобулина человека; и
(f) экспрессию нуклеотидной последовательности по (d) в продукционной клеточной линии, экспрессирующей тяжелую цепь иммуноглобулина человека, которая специфически связывает второй антиген или эпитоп, для получения легкой цепи антигенсвязывающего белка, кодируемой нуклеиновой кислотой по (d) и связывающей первый эпитоп или его иммуногенную часть независимо от тяжелой цепи, тяжелая цепь которого специфически связывает указанный второй антиген или эпитоп.
Согласно некоторым вариантам реализации по меньшей мере один из нереаранжированных сегментов гена VL или JL легкой цепи человека кодирует один или большее количество гистидиновых кодонов, не кодируемых соответствующим сегментом гена вариабельной области легкой цепи зародышевой линии человека. Согласно некоторым вариантам реализации указанный первый эпитоп получен из рецептора клеточной поверхности. Согласно некоторым вариантам реализации указанный рецептор клеточной поверхности представляет собой Fc-рецептор. Согласно конкретным вариантам реализации указанный Fc-рецептор представляет собой FcRn. Согласно некоторым вариантам реализации указанный второй антиген или эпитоп получен из растворимого антигена. Согласно некоторым вариантам реализации указанный второй антиген или эпитоп получен из рецептора клеточной поверхности. Согласно некоторым вариантам реализации первый антиген представляет собой Fc-рецептор, второй антиген представляет собой растворимый белок, а антигенсвязывающий белок содержит одну или большее количество замен и вставок гистидина, происходящих из сегмента гена VL в геноме не являющегося человеком животного.
[000246] Согласно другому аспекту предложены способы получения антигенсвязывающего белка, который содержит вариабельный домен легкой цепи иммуноглобулина, способный связывать антиген независимо от вариабельного домена тяжелой цепи. Такие способы включают
(a) иммунизацию генетически модифицированного не являющегося человеком животного первым антигеном, который содержит первый эпитоп или его иммуногенную часть, при этом в геноме указанного не являющегося человеком животного содержатся:
(i) нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи человека, функционально связанная с нуклеотидной последовательностью тяжелой цепи константной области; и
(ii) два или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека (VL и JL), функционально связанных с нуклеотидной последовательностью константной области легкой цепи иммуноглобулина;
(b) обеспечение возможности развития иммунного ответа на первый эпитоп или его иммуногенную часть у указанного не являющегося человеком животного;
(c) выделение из указанного не являющегося человеком животного клетки, содержащей нуклеотидную последовательность, которая кодирует вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(d) получение из указанной клетки по (с) нуклеотидной последовательности, которая кодирует указанный вариабельный домен легкой цепи, специфически связывающий указанный первый эпитоп или его иммуногенную часть;
(e) применение нуклеотидной последовательности по (d) в экспрессионной конструкции, соединенной с нуклеотидной последовательностью константной области иммуноглобулина человека; и
(f) экспрессию нуклеотидной последовательности по (d) в продукционной клеточной линии, экспрессирующей тяжелую цепь иммуноглобулина человека, которая специфически связывает второй антиген или эпитоп, для получения легкой цепи антигенсвязывающего белка, кодируемой нуклеиновой кислотой по (d) и связывающей первый эпитоп или его иммуногенную часть независимо от тяжелой цепи, тяжелая цепь которого специфически связывает указанный второй антиген или эпитоп.
Согласно некоторым вариантам реализации по меньшей мере один из сегментов гена легкой цепи VL или JL человека кодирует один или большее количество гистидиновых кодонов, не кодируемых соответствующим сегментом гена вариабельной области легкой цепи зародышевой линии человека. Согласно некоторым вариантам реализации указанный первый эпитоп получен из рецептора клеточной поверхности. Согласно некоторым вариантам реализации указанный рецептор клеточной поверхности представляет собой Fc-рецептор. Согласно конкретным вариантам реализации указанный Fc-рецептор представляет собой FcRn. Согласно некоторым вариантам реализации указанный второй антиген или эпитоп получен из растворимого антигена. Согласно некоторым вариантам реализации указанный второй антиген или эпитоп получен из рецептора клеточной поверхности. Согласно некоторым вариантам реализации первый антиген представляет собой Fc-рецептор, второй антиген представляет собой растворимый белок, а антигенсвязывающий белок содержит одну или большее количество замен и вставок гистидина, происходящих из сегмента гена VL в геноме не являющегося человеком животного.
[000247] Согласно другому аспекту для обеспечения простого разделения антигенсвязывающих белков, описанных в настоящем документе, одну из тяжелых цепей модифицируют, чтобы устранить связывающую белок А детерминанту, что обуславливает различное сродство к связыванию белка А гомодимерного связывающего белка и гетеродимерного связывающего белка. Композиции и способы для решения указанной проблемы описаны в патенте США №8586713, выданном 19 ноября 2013 г. и озаглавленном «Readily Isolated Bispecific Antibodies with Native Immunoglobulin Format)) («Простое выделение биспецифических антител в нативном формате иммуноглобулинов»), включенном в настоящий документ посредством ссылки. После выбора вида, содержащего гетеродимерную тяжелую цепь с идентичной легкой цепью, может быть проведен скрининг указанного биспецифического антигенсвязывающего белка для подтверждения сохранения рН-зависимых антигенсвязывающих свойств.
[000248] Согласно различным аспектам предложены плюрипотентная клетка, индуцированные плюрипотентные или тотипотентные стволовые клетки, происходящие из не являющегося человеком животного, у которого имеются различные геномные модификации согласно настоящему описанию. Согласно некоторым вариантам реализации указанная плюрипотентная или тотипотентная клетка получена из не являющегося человеком животного. Согласно некоторым вариантам реализации указанное не являющееся человеком животное представляет собой грызуна, например, мышь, крысу или хомяка. Согласно некоторым вариантам реализации указанный грызун представляет собой мышь. Согласно конкретным вариантам реализации указанная плюрипотентная клетка представляет собой эмбриональную стволовую (ЭС) клетку. Согласно некоторым вариантам реализации в геноме указанной плюрипотентной клетки имеется: (i) локус тяжелой цепи иммуноглобулина, который содержит нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи человека, функционально связанную с нуклеотидной последовательностью тяжелой цепи константной области; и (ii) локус легкой цени иммуноглобулина, содержащий один или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи VL и JL иммуноглобулина человека, функционально связанных с нуклеотидной последовательностью константной области легкой цепи. Согласно конкретным вариантам реализации указанные плюрипотентные, индуцированные плюрипотентные или тотипотентные стволовые клетки представляют собой эмбриональные стволовые (ЭС) клетки мыши или крысы. Согласно некоторым вариантам реализации указанная плюрипотентные, индуцированные плюрипотентные или тотипотентные стволовые клетки имеют кариотип XX или кариотип XY.
[000249] Также предложены клетки, которые содержат ядро с генетической модификацией согласно описанию в настоящем документе, например, модификацией, введенной в клетку посредством пронуклеарной инъекции. Согласно другому аспекту предложена гибридома или квадрома, происходящая из клетки не являющегося человеком животного согласно описанию в настоящем документе. Согласно некоторым вариантам реализации указанное не являющееся человеком животное представляет собой грызуна, такого как мышь, крыса или хомяк.
[000250] Согласно другому аспекту предложен лимфоцит, выделенный из генетически модифицированного не являющегося человеком животного согласно описанию в настоящем документе. Согласно некоторым вариантам реализации указанный лимфоцит представляет собой В-клетку, при этом указанная В-клетка содержит иммуноглобулиновый геномный локус, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, функционально связанную с человеческой или не являющемуся человеком животному (например, мыши или крысе) последовательностью гена константной области тяжелой цепи или легкой цепи. Согласно некоторым вариантам реализации указанная В-клетка способна продуцировать антитела, отличающиеся тем, что реаранжированный вариабельный домен тяжелой цепи согласно описанию в настоящем документе функционально связан с константным доменом тяжелой цепи или легкой цепи.
[000251] Согласно другому аспекту предложен эмбрион не являющегося человеком животного, содержащий клетку, в геноме которой имеется: (i) локус тяжелой цепи иммуноглобулина, содержащий нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи человека, функционально связанную с нуклеотидной последовательностью константной области; и (ii) локус легкой цепи иммуноглобулина, содержащий два или более, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека, функционально связанных с нуклеотидной последовательностью константной области легкой цепи.
[000252] Согласно различным вариантам реализации у указанных генетически модифицированных не являющихся человеком животных экспрессируется репертуар антител (например, репертуар IgG), полученных из нуклеотидной последовательности, которая кодирует реаранжированный вариабельный домен тяжелой цепи, и совокупность сегментов V легких цепей (и совокупность сегментов J легких цепей). Согласно некоторым вариантам реализации указанный генетически модифицированный локус продуцирует популяцию антител, которая содержит легкую цепь иммуноглобулина, способную специфически связывать представляющий интерес антиген со сродством (KD) менее 10-6, 10-7, 10-8, 10-9 или 10-10. Согласно некоторым вариантам реализации указанная легкая цепь иммуноглобулина, экспрессируемая указанным генетически модифицированным локусом, способна специфически связывать представляющий интерес антиген в отсутствие вариабельной области тяжелой цепи со сродством (KD) менее 10-6, 10-7, 10-8, 10-9 или 10-10.
[000253] Согласно различным вариантам реализации генетические модификации, описанные 6 настоящем документе, не влияют на фертильность не являющегося человеком животного (см., например, US 2012-0322108 А1, полностью включенный посредством ссылки). Согласно некоторым вариантам реализации указанный локус тяжелой цепи содержит эндогенный ген Adam6a, ген Adam6b или оба указанных гена, и указанная генетическая модификация не влияет на экспрессию и/или функцию указанного эндогенного гена Adam6a, гена Adam6b или обоих указанных генов. Согласно некоторым вариантам реализации геном указанного генетически модифицированного не являющегося человеком животного содержит расположенный эктопически ген Adam6a, ген Adam6b или оба указанных гена. Согласно некоторым вариантам реализации ген Adam6a и/или Adam6b расположен в направлении 5' от транскрипционной единицы нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи. Согласно некоторым вариантам реализации указанный ген Adam6a и/или ген Adam6b расположен в направлении 3' от транскрипционной единицы нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи.
[000254] Согласно некоторым вариантам реализации указанный генетически модифицированный локус тяжелой цепи не содержит нуклеотидную последовательность межгенной контрольной области 1 (IGCR1). Согласно некоторым вариантам реализации указанный генетически модифицированный локус тяжелой цепи содержит последовательность IGCR1 в направлении 3' от нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи. Согласно некоторым вариантам реализации указанная нуклеотидная последовательность IGCR1 расположена между нуклеотидной последовательностью реаранжированной вариабельной области тяжелой цепи и расположенным ближе всего к сегменту V сегментом гена DH.
[000255] Согласно некоторым аспектам, как отмечалось ранее, локус легкой цепи иммуноглобулина описанных в настоящем документе не являющихся человеком животных содержит ограниченный репертуар сегментов гена вариабельной области легкой цепи, например, (i) один, два или более, но меньше, чем у дикого типа, сегментов гена VL человека. Согласно некоторым вариантам реализации указанное не являющееся человеком животное представляет собой мышь; и вариабельный домен легкой цепи иммуноглобулина получают в результате реаранжировки одного из двух сегментов гена Vκ человека и одного из 1, 2, 3, 4 или 5 сегментов гена Jκ человека. Согласно некоторым вариантам реализации у указанной мыши наблюдается отношение (а) В-клеток в костном мозге, экспрессирующих иммуноглобулин, содержащий легкую цепь λ, и (b) В-клеток в костном мозге, экспрессирующих иммуноглобулин, содержащий легкую цепь к, составляющее от приблизительно 1 до приблизительно 15. Согласно некоторым вариантам реализации реаранжировка включает сегмент гена Vκ1-39 человека. Согласно некоторым вариантам реализации указанная реаранжировка включает сегмент гена Vκ3-20 человека. Согласно некоторым вариантам реализации указанные два сегмента гена Vκ человека расположены в эндогенном иммуноглобулиновом локусе Vκ, и, согласно некоторым вариантам реализации, указанные два сегмента гена Vκ человека заменяют все или по существу все сегменты генов Vκ иммуноглобулина мыши. Согласно некоторым вариантам реализации указанные два сегмента гена человека Vκ расположены в эндогенном иммуноглобулиновом локусе Vκ, и, согласно некоторым вариантам реализации, указанные два сегмента гена Vκ человека заменяют все или по существу все сегменты генов Vκ и Jκ иммуноглобулина мыши. Согласно некоторым вариантам реализации указанные два сегмента гена Vκ человека функционально связаны с двумя или большим количеством (например, 2, 3, 4, 5) сегментов гена Jκ человека. Согласно некоторым другим вариантам реализации вариабельный домен легкой цепи мыши образуется путем реаранжировки сегмента гена Vκ1-39 человека или сегмента гена Vκ3-20 человека, и одного из двух или большего количества (например, 2, 3, 4 или 5) сегментов гена Jκ человека. Согласно некоторым таким вариантам реализации отношение незрелых В-клеток в костном мозге, экспрессирующих иммуноглобулин, содержащий легкую цепь X, и незрелых В-клеток, экспрессирующих иммуноглобулин, содержащий легкую цепь к, составляет от приблизительно 1 до приблизительно 13. Согласно некоторым другим вариантам реализации вариабельный домен легкой цепи мыши образуется путем реаранжировки сегмента гена Vκ1-39 человека или сегмента гена Vκ3-20 человека, и одного из двух или большего количества (например, 2, 3, 4 или 5) сегментов гена Jκ человека, и отношение зрелых В-клеток в костном мозге, экспрессирующих иммуноглобулин, содержащий легкую цепь X, и незрелых В-клеток, экспрессирующих иммуноглобулин, содержащий легкую цепь к, составляет от приблизительно 1 до приблизительно 7.
[000256] Согласно конкретным вариантам реализации вариабельный домен легкой цепи генетически модифицированной мыши согласно описанию в настоящем документе образуется путем реаранжировки сегмента гена Vκ1-39 человека или сегмента гена Vκ3-20 человека, и одного из двух или большего количества (например, 2, 3, 4 или 5) сегментов гена Jκ человека, и имеется популяция про-В-клеток в костном мозге численностью в диапазоне от приблизительно 2,5×104 до приблизительно 1,5×105 клеток включительно, например, приблизительно 2,5×104, 3,0×104, 3,5×104, 4,0×104, 4,5×104, 5,0×104, 5,5×104, 6,0×104, 6,5×104, 7,0×104, 7,5×104, 8,0×104, 8,5×104, 9,0×104, 9,5×104, 1,0×105 или 1,5×105 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция про-В-клеток в костном мозге численностью приблизительно 2,88×104 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция про-В-клеток в костном мозге численностью приблизительно 6,42×104 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция про-В-клеток в костном мозге численностью приблизительно 9,16×104 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция про-В-клеток в костном мозге численностью приблизительно 1,19×105 клеток. Примеры про-В-клеток в костном мозге генетически модифицированных грызунов (например, мыши) согласно описанию в настоящем документе характеризуются экспрессией CD19, CD43, c-kit и/или их комбинации (например, CD19+, CD43+, c-kit+). Согласно некоторым вариантам реализации у грызуна (например, мышь) согласно описанию в настоящем документе экспрессируется легкая цепь, образованная путем реаранжировки сегмента гена Vκ1-39 человека или сегмента гена Vκ3-20 человека, и одного из двух или большего количества (например, 2, 3, 4 или 5) сегментов гена Jκ человека, и имеется популяция пре-В-клеток в костном мозге численностью в диапазоне от приблизительно 1×106 до приблизительно 2×106 клеток включительно, например, приблизительно 1,0×106, 1,1×106, 1,2×106, 1,3×106, 1,4×106, 1,5×106, 1,6×106, 1,7×106, 1,8×106, 1,9×106, или 2,0×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция пре-В-клеток в костном мозге численностью приблизительно 1,25×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция пре-В-клеток в костном мозге численностью приблизительно 1,46×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция пре-В-клеток в костном мозге численностью приблизительно 1,64×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция пре-В-клеток в костном мозге численностью приблизительно 2,03×106 клеток. Примеры пре-В-клеток в костном мозге генетически модифицированных грызунов (например, мыши) согласно описанию в настоящем документе характеризуются экспрессией CD19, CD43, с-kit и/или их комбинации (например, CD19+, CD43-, c-kit-).
[000257] Согласно различным вариантам реализации у генетически модифицированной мыши согласно описанию в настоящем документе экспрессируется легкая цепь, образованная путем реаранжировки сегмента гена Vκ1-39 человека или сегмента гена Vκ3-20 человека, и одного из двух или большего количества (например, 2, 3, 4 или 5) сегментов гена Jκ человека, и имеется популяция незрелых В-клеток в костном мозге, количество которых находится в диапазоне от приблизительно 5×105 до приблизительно 7×105 клеток включительно, например, приблизительно 5,0×105, 5,1×105, 5,2×105, 5,3×105, 5,4×105, 5,5×105, 5,6×105, 5,7×105, 5,8×105, 5,9×105, 6,0×105, 6,1×105, 6,2×105, 6,3×105, 6,4×105, 6,5×105, 6,6×105, 6,7×105, 6,8×105, 6,9×105 или 7,0×105 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция незрелых В-клеток в костном мозге численностью приблизительно 5,33×105 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция незрелых В-клеток в костном мозге численностью приблизительно 5,80×105 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция незрелых В-клеток в костном мозге численностью приблизительно 5,92×105 клеток; согласно некоторым вариантам реализации у указанного грызуна (например, мыши) имеется популяция незрелых В-клеток в костном мозге численностью приблизительно 6,67×105 клеток. Примеры незрелых В-клеток в костном мозге генетически модифицированных грызунов (например, мышей) согласно описанию * в настоящем документе характеризуются экспрессией IgM, В220 и/или их комбинации (например, IgM+, B220int).
[000258] Согласно различным вариантам реализации у генетически модифицированной мыши согласно описанию в настоящем документе экспрессируется легкая цепь, образованная путем реаранжировки сегмента гена Vκ1-39 человека или сегмента гена Vκ3-20 человека и одного из двух или большего количества (например, 2, 3, 4 или 5) сегментов гена Jκ человека, и имеется популяция зрелых В-клеток в костном мозге, количество которых находится в диапазоне от приблизительно 3×104 до приблизительно 1,5×105 клеток включительно, например, приблизительно 3,0×104, 3,5×104, 4,0×104, 4,5×104, 5,0×104, 5,5×104, 6,0×104, 6,5×104, 7,0×104, 7,5×104, 8,0×104, 8,5×104, 9,0×104, 9,5×104, 1,0×105, или 1,5×105 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция зрелых В-клеток в костном мозге численностью приблизительно 3,11×104 клеток; согласно некоторым вариантам реализации у описанного в настоящем документе модифицированного грызуна (например, мыши) имеется популяция зрелых В-клеток в костном мозге численностью приблизительно 1,09×105 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция зрелых В-клеток в костном мозге численностью приблизительно 1,16×105 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция зрелых В-клеток в костном мозге численностью приблизительно 1,44×105 клеток. Примеры зрелых В-клеток в костном мозге генетически модифицированных грызунов (например, мышей) согласно описанию в настоящем документе характеризуются экспрессией IgM, В220 и/или их комбинации (например, IgM+, B220hi).
[000259] Согласно различным вариантам реализации у генетически модифицированного грызуна (например, мыши) согласно описанию в настоящем документе экспрессируется легкая цепь, образованная путем реаранжировки сегмента гена Vκ1-39 человека или сегмента гена Vκ3-20 человека и одного из двух или большего количества (например, 2, 3, 4 или 5) сегментов гена Jκ человека, и имеется популяция тотальных В-клеток в костном мозге численностью в диапазоне от приблизительно 1×106 до приблизительно 3×106 клеток включительно, например, приблизительно 1,0×106, 1,1×106, 1,2×106, 1,3×106, 1,4×106, 1,5×106, 1,6×106, 1,7×106, 1,8×106, 1,9×106, 2,0×106, 2,1×106, 2,2×106, 2,3×106, 2,4×106, 2,5×106, 2,6×106, 2,7×106, 2,8×106, 2,9×106 или 2,0×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция тотальных В-клеток в костном мозге численностью приблизительно 1,59×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция тотальных В-клеток в костном мозге численностью приблизительно 1,75×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция тотальных В-клеток в костном мозге численностью приблизительно 2,13×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция тотальных В-клеток в костном мозге численностью приблизительно 2,55×106 клеток. Пример тотальных В-клеток в костном мозге генетически модифицированных грызунов (например, мыши) согласно описанию в настоящем документе характеризуется экспрессией CD19, CD20 и/или их комбинации (например, CD19+).
[000260] Согласно различным вариантам реализации у генетически модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется локус легкой цепи к иммуноглобулина, который содержит два нереаранжированных сегмента гена Vκ иммуноглобулина человека и два или большее количество (например, 2, 3, 4 или 5) нереаранжированных сегментов гена Jκ человека, при этом у указанного грызуна (например, мыши) имеется популяция периферических В-клеток селезенки, содержащая популяции переходных (например, T1, Т2 и Т3) В-клеток, приблизительно такие же, что и у грызуна (например, мыши), у которого имеется набор дикого типа, представленный сегментами генов V и J легких цепей к иммуноглобулинов. Примеры популяции переходных В-клеток (например, T1, Т2 и Т3) в селезенке генетически модифицированного грызуна (например, мыши) согласно описанию в настоящем документе характеризуются экспрессией IgM, CD23, CD93, В220 и/или их комбинации.
[000261] Согласно различным вариантам реализации у генетически модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция В-клеток Т1 в селезенке (например, CD93+, В220+, IgMhi, CD23-) численностью в диапазоне от приблизительно 2×106 до приблизительно 7×106 клеток включительно, например, приблизительно 2,0×106, 2,5×106, 3,0×106, 3,5×106, 4,0×106, 4,5×106, 5,0×106, 5,5×106, 6,0×106, 6,5×106 или 7,0×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция В-клеток Т1 в селезенке численностью приблизительно 2,16×106 клеток; согласно некоторым вариантам реализации у грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция В-клеток Т1 в селезенке численностью приблизительно 3,63×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция В-клеток Т1 в селезенке численностью приблизительно 3,91×106; согласно некоторым вариантам реализации у модифицированного грызуна (например), мыши) согласно описанию в настоящем документе имеется популяция В-клеток Т1 в селезенке численностью приблизительно 6,83×106 клеток.
[000262] Согласно различным вариантам реализации у генетически модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция Т2 В-клеток в селезенке (например, CD93+, В220+, IgMhi, CD23+) в диапазоне от приблизительно 1×106 до приблизительно 7×106 клеток включительно, например, приблизительно 1,0×106, 1,5×106, 2,0×106, 2,5×106, 3,0×106, 3,5×106, 4,0×106, 4,5×106, 5,0×106, 5,5×106, 6,0×106, 6,5×106 или 7,0×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция Т2 В-клеток в селезенке численностью приблизительно 1,30×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция Т2 В-клеток в селезенке численностью приблизительно 2,46×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция Т2 В-клеток в селезенке численностью приблизительно 3,24×106; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция Т2 В-клеток в селезенке численностью приблизительно 6,52×106 клеток.
[000263] Согласно различным вариантам реализации у генетически модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция В-клеток Т3 в селезенке (например, CD93+, В220+, IgMlo, CD23+) в диапазоне от приблизительно 1×106 до приблизительно 4×106 клеток включительно, например, приблизительно 1,0×106, 1,5×106, 2,0×106, 2,5×106, 3,0×106, 3,5×106 или 4,0×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция В-клеток Т3 в селезенке численностью приблизительно 1,08×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция В-клеток Т3 в селезенке численностью приблизительно 1,35×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция В-клеток Т3 в селезенке численностью приблизительно 3,37×106; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция В-клеток Т1 в селезенке численностью приблизительно 3,63×106 клеток.
[000264] В-клетки маргинальной зоны представляют собой нециркулирующие зрелые В-клетки, анатомически обособленные в маргинальной зоне (МЗ) селезенки. У грызунов В-клетки МЗ являются сессильными и располагаются во внешней белой пульпе селезенки между маргинальным синусом и красной пульпой. Указанная область содержит множество подтипов макрофагов, дендритных клеток и В-клеток МЗ; она не полностью сформирована до 2-3 недель после рождения у грызунов и до 1-2 лет у человека. В-клетки МЗ в указанной области, как правило, экспрессируют высокие уровни sIgM, CD21, CD1, CD9 наряду с уровнями sIgD, CD23, CD5, и CD11b от низких до пренебрежимо малых, что помогает фенотипически отличить их от фолликулярных (FO) В-клеток и В-клеток В1. Аналогично В-клеткам В1, В-клетки МЗ могут быть быстро рекрутированы в ходе раннего адаптивного иммунного ответа независимым от Т-клеток образом. Расположение В-клеток М, в частности, хорошо подходит для обеспечения первой линии защиты против системных антигенов в крови, поступающих в кровоток и удерживаемых в селезенке. Считается, что они, в частности, являются реактивными в отношении компонентов бактериальной клеточной стенки и представляют собой важный источник липид-специфических антител. В-клетки МЗ также демонстрируют более низкий порог активации по сравнению с эквивалентными В-клетками FO, при повышенной склонности к дифференцировке в клетки плазмы, что вносит дополнительный вклад в ускоренный ответ первичных антител.
[000265] Согласно различным вариантам реализации генетически модифицированный грызун (например, мышь) согласно описанию в настоящем документе, у которого имеется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (например, VH3-23/D/JH4), содержатся повышенные уровни В-клеток маргинальной зоны по сравнению с уровнями у грызунов дикого типа (например, мышей дикого типа). Согласно некоторым вариантам реализации уровни В-клеток маргинальной зоны у генетически модифицированного грызуна (например, мыши), у которого имеется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека, повышены на 10%, 20%, 30%, 40%, 50% или более по сравнению с уровнями у грызунов дикого типа (например, мышей дикого типа).
[000266] Согласно различным вариантам реализации у генетически модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется локус легкой цепи к иммуноглобулина, который содержит два нереаранжированных сегмента гена Vκ иммуноглобулина человека, и 1, 2, 3, 4 или 5 нереаранжированных сегментов гена иммуноглобулина Jκ человека, и при этом у указанного грызуна (например, мыши) имеется популяция периферических В-клеток селезенки, включающая популяции В-клеток маргинальной зоны и предшественников В-клеток маргинальной зоны, приблизительно такие же, что и у грызуна (например, мыши), у которого имеется набор сегменты генов Vκ и Jκ иммуноглобулина дикого типа. Примеры популяций В-клеток маргинальной зоны в селезенке генетически модифицированного грызуна (например, мыши) согласно описанию в настоящем документе характеризуются экспрессией IgM, CD21/35, CD23, CD93, В220 и/или их комбинации.
[000267] Согласно различным вариантам реализации у генетически модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция В-клеток маргинальной зоны в селезенке (например, CD93-, В220+, IgMhi, CD21/35hi, CD23-) численностью в диапазоне от приблизительно 1×106 до приблизительно 3×106 клеток включительно, например, приблизительно 1,0×106, 1,5×106, 2,0×106, 2,5×106 или 3,0×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция В-клеток маргинальной зоны в селезенке численностью приблизительно 1,47×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция В-клеток маргинальной зоны в селезенке численностью приблизительно 1,49×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция В-клеток маргинальной зоны в селезенке численностью приблизительно 2,26×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция В-клеток маргинальной зоны в селезенке численностью приблизительно 2,33×106 клеток.
[000268] Согласно различным вариантам реализации предложен генетически модифицированный грызун (например, мышь), отличающийся тем, что у указанного грызуна (например, мыши) имеется локус легкой цепи к иммуноглобулина, который содержит два нереаранжированных сегмента гена Vκ иммуноглобулина человека и 1, 2, 3, 4 или 5 нереаранжированных сегментов гена Jκ иммуноглобулина человека, и при этом у указанного грызуна (например, мыши) имеется популяция периферических В-клеток селезенки, содержащая популяцию(и) фолликулярных (например, FO-I и FO-II) В-клеток, приблизительно такую же, как у грызуна (например, мыши), у которого имеется набор сегментов гена Vκ и Jκ иммуноглобулина дикого типа. Примеры популяций фолликулярных В-клеток (например, FO-I и FO-II) в селезенке генетически модифицированного грызуна (например, мышь) согласно описанию в настоящем документе характеризуются экспрессией IgM, IgD, CD21/35, CD93, В220 и/или их комбинации.
[000269] Согласно различным вариантам реализации у генетически модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция фолликулярных В-клеток 1 типа в селезенке (например, CD93-, В220+, CD21/35int, IgMlo, IgDhi) численностью в диапазоне от приблизительно 3×106 до приблизительно 1,5×107 клеток включительно, например, приблизительно 3,0×106, 3,5×106, 4,0×106, 4,5×106, 5,0×106, 5,5×106, 6,0×106, 6,5×106, 7,0×106, 7,5×106, 8,0×106, 8,5×106, 9,0×106, 9,5×106, 1,0×107 или 1,5×107 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция фолликулярных В-клеток 1 типа в селезенке численностью приблизительно 3,57×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция фолликулярных В-клеток 1 типа в селезенке численностью приблизительно 6,31×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция фолликулярных В-клеток 1 типа в селезенке численностью приблизительно 9,42×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция фолликулярных В-клеток 1 типа в селезенке численностью приблизительно 1,14×107 клеток.
[000270] Согласно различным вариантам реализации у генетически модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция фолликулярных В-клеток 2 типа в селезенке (например, CD93-, В220+, CD21/35int, IgMint, IgDhi) численностью в диапазоне от приблизительно 1×106 до приблизительно 2×106 клеток включительно, например, 1,0×106, 1,25×106, 1,5×106, 1,75×106 или 2,0×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция фолликулярных В-клеток 2 типа в селезенке численностью приблизительно 1,14×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция фолликулярных В-клеток 2 типа в селезенке численностью приблизительно 1,45×106 клеток; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция фолликулярных В-клеток 2 типа в селезенке численностью приблизительно 1,80×106; согласно некоторым вариантам реализации у модифицированного грызуна (например, мыши) согласно описанию в настоящем документе имеется популяция фолликулярных В-клеток 2 типа в селезенке численностью приблизительно 2,06×106 клеток.
[000271] Способность генетически модифицированных не являющихся человеком животных, описанных в настоящем документе, обеспечивать селективное давление на гены или полинуклеотиды, кодирующие вариабельные области или домены легких цепей (например,' CDR3 легких цепей) может использоваться для получения разнообразных последовательностей генов вариабельных областей легких цепей. Другими словами, реаранжированные последовательности генов вариабельных доменов тяжелых цепей, раскрытые в настоящем документе, могут сочетаться с одной или большим количеством генетических модификаций локуса легкой цепи и/или вставок нуклеотидных последовательностей, кодирующих вариабельные домены легких цепей, в локус тяжелой цепи. Это может осуществляться путем, например, спаривания (а именно, кроссбридинга или скрещивания животных с единственной модификацией) не являющихся человеком животных, описанных в настоящем документе (ограниченных общим или универсальным вариабельным доменом тяжелой цепи) с не являющимися человеком животными, у которых имеются генетические модификации в одном или нескольких кодирующих легкие цепи локусах. Генетически модифицированные не являющиеся человеком животные, у которых имеются иммуноглобулиновые локусы, содержащие модификации как реаранжированного вариабельного домена тяжелой цепи, так и одной или большего числа легких цепей, могут также быть получены путем направленной замены нескольких генных локусов, одновременной или последовательной (например, путем последовательной рекомбинации в эмбриональных стволовых клетках). Ни тип, ни способ модификации в локусах легких цепей не ограничивают варианты реализации, описанные в настоящем документе, если специально не указано иное. Напротив, селективное давление с помощью вариантов реализации, описанных в настоящем документе, могут применяться практически для любой полинуклеотидной последовательности, которая может экспрессироваться и функционировать в качестве антигенсвязывающей последовательности легкой цепи, запуская таким образом эволюцию наиболее подходящих вариабельных областей антител.
[000272] Например, согласно описанию в настоящем документе, генетически модифицированные не являющиеся человеком животные, у которых имеется иммуноглобулиновый локус с последовательностью гена реаранжированного вариабельного домена тяжелой цепи, может дополнительно иметься (например, путем кроссбридинга или различных стратегий нацеливания на гены) одну или большее количество модификаций согласно описанию в WO 2011/072204, WO 2011/163311, WO 2011/163314, WO 2012/018764, WO 2012/141798, U.S. 2013/0185821, WO 2013/022782, WO 2013/096142, WO 2013/116609; указанные публикации полностью включены в настоящий документ посредством ссылки. Согласно конкретным вариантам реализации генетически модифицированную мышь, у которой имеется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи в локусе легкой цепи (т.е. реаранжированная последовательность гена вариабельного домена тяжелой цепи, функционально связанная с человеческой или имеющей нечеловеческое происхождение последовательностью гена константной области легкой цепи к) скрещивают с генетически модифицированной мышью, у которой имеется локус тяжелой цепи иммуноглобулина, содержащий сегменты генов вариабельных областей легких цепей человека (например, 40 генов Vκ человека и все гены Jκ человека, встроенные в локус тяжелой цепи мыши; см., например, опубликованную через 18 месяцев с даты приоритета патентную заявку США 2012/0096572, включенную в настоящий документ посредством ссылки). Согласно конкретным вариантам реализации генетически модифицированную мышь, у которой имеется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи в локусе легкой цепи (т.е. реаранжированная последовательность гена вариабельного домена тяжелой цепи, функционально связанная с человеческой или имеющей нечеловеческое происхождение последовательностью гена константной области легкой цепи к) скрещивают с генетически модифицированной мышью, у которой имеется локус тяжелой цепи иммуноглобулина, содержащий один или большее количество (например, два), но меньше, чем у чем дикого типа, сегментов гена вариабельной области легкой цепи человека. Полученные мыши способны продуцировать каппа+ В-клетки с вариабельными областями тяжелых цепей, происходящими из геномной последовательности вариабельных областей легкой цепи, с облегчением таким образом идентификации последовательностей VJ каппа, связывающихся со специфическими мишенями, которые затем могут быть преобразованы обратно в легкую цепь и скомбинированы с различными тяжелыми цепями с получением биспецифических или триспецифических антител.
ПРИМЕРЫ
[000273] Приведенные ниже примеры предназначены для того, чтобы обеспечить для специалистов в данной области техники полное раскрытие и описание способов получения и применения не являющихся человеком животных согласно описанию в настоящем документе, и не предназначены для ограничения объема заявленного авторами изобретения, а также не означают, что ниже представлены все или единственные проведенные эксперименты. Были приложены усилия, чтобы обеспечить точность приводимых численных значений (например, количеств, температуры и т.п.), однако необходимо учитывать возможные экспериментальные погрешности и отклонения. Если не указано иное, части представлены массовыми частями, молекулярная масса представлена средней молекулярной массой, температуры приведены в градусах Цельсия, и давление соответствует или приближено к атмосферному давлению.
Пример 1. Анализ клонирования и экспрессии кандидатных универсальных последовательностей тяжелых цепей
[000274] Предыдущие исследования показали, что hVH3-23 представляет собой термостабильный сегмент гена вариабельной области тяжелой цепи человека, а также она представляет собой один из наиболее часто используемых сегментов вариабельной области репертуара человека. Соответственно, для конструирования реаранжированной последовательности вариабельной области тяжелой цепи (далее «универсальная тяжелая цепь», или «UHC») отбирали кодон-оптимизированные сегменты генов человека VH3-23, D4-4 (рамка считывания 2 или 3) и JH4 (или JH6).
[000275] Вкратце, следующие четыре кандидатные реаранжированные последовательности VDJ синтезировали de novo (в IDT) и клонировали в экспрессионные ЦМВ-векторы (например, pRG1301 или pRG1368): (1) hVH3-23(D4-4_RF2)JH4 (SEQ ID NO: 148); (2) hVH3-23(D4-4_RF2)JH6 (SEQ ID NO: 146); (3) hVH3-23(D4-4_RF3)JH4 (SEQ ID NO: 147); (4) hVH3-23(D4-4_RF3)JH6 (SEQ ID NO: 145). Все указанные конструкции конструировали таким образом, чтобы синтезированные гены UHC могли быть лигированы в векторы pRG1301 (hIgG1) или pRG1368 (mIgG1) после расщепления с применением Xho I/Sap I. Для анализа экспрессии четыре гена UHC (1-4) в pIDTSMART субклонировали в сайты Xho VSap I вектора pRG1301 (hlgG1) или pRG1368 (mIgG1), и каждую экспрессионную конструкцию трансфицировали по отдельности в клетки СНО. После трансфекции все четыре кандидатные последовательности VDJ экспрессировались на достаточном уровне, и экспрессируемые пептиды были способны сочетаться с разными легкими цепями κ и λ.
[000276] Чтобы избежать появления потенциально аутореактивных антител, что может приводить к истощению В-клеток у генетически модифицированных мышей, в базе данных антител ASAP Regeneron Pharmaceuticals, основанной на антителах, продуцированных гуманизированными мышами VELOCIMMUNE®, проводили поиск антител, содержащих последовательность аминокислот, аналогичную hVH3-23(D4-4)JH4 (Фиг. 5). В частности, критерии, использованные для идентификации неаутореактивных антител, включали последовательность аминокислот по DYSNY (SEQ ID NO: 144) или последовательности, аналогичные DYSNY (SEQ ID NO: 144). Исследования экспрессии в клетках СНО, однако, показали, что последовательности UHC, содержащие DYSNY (SEQ ID NO: 144), не экспрессируются удовлетворительным образом в клетках млекопитающих. Таким образом, вместо них выбирали другую последовательность, где отсутствует D, но содержится последовательность YSNY (т.е. антитело Н1Н2002 В; AKGYYFDY (SEQ ID NO: 143); где АК происходит из 3-23; GY получен путем добавления D или N, или добавлений N и Р; и YFDY представляет собой JH4) и тестировали ее экспрессию в клетках СНО. Модифицированная последовательность UHC экспрессировалась в клетках СНО на достаточном уровне. Указанные результаты предполагают, что необходимо наличие некоторых аминокислотных остатков (т.е. спейсера) между последовательностью, кодируемой сегментом гена V тяжелой цепи, и последовательностью, кодируемой сегментом гена J тяжелой цепи, для надлежащей экспрессии реаранжированной последовательности VDJ в клетках млекопитающих.
[000277] Кроме того, уровни экспрессии пептида AKGYYFDY, происходящего из реаранжированной последовательности VDJ (VH3-23/GY/JH4; HIH2002B) в клетках СНО сравнивали с пептидом, происходящим из VH3-23/D4-4 (Рамка считывания 2)/JH4 (SEQ ID NO: 148), в отношении экспрессии реаранжированных последовательностей VDJ с пятью цепями κ человека, тремя цепями λ человека и других реаранжированных последовательностей VDJ (т.е. VH3-20 и VH1-39). Для выбранной реаранжированной последовательности VDJ (VH3-23/GY/JH4) наблюдалась экспрессия на уровнях, эквивалентных уровням у контроля.
[000278] На основании указанных данных в качестве реаранжированной последовательности вариабельного домена тяжелой цепи для получения генетически модифицированной мыши выбирали VH3-23/GY/JH4 (SEQ ID NO: 137; HIH2002B). Подробные стратегии нацеливания для получения мыши, у которой имеется генетически модифицированный иммуноглобулиновый локус, кодирующий реаранжированный вариабельный домен тяжелой цепи (т.е. мыши, у которой имеется иммуноглобулиновый локус, содержащий реаранжированную вариабельную область тяжелой цепи иммуноглобулина человека), представлены на фиг. 1-9 и описаны ниже.
Пример 2. Конструирование локусов тяжелых цепей иммуноглобулина, содержащих реаранжированную последовательность VDJ
[000279] Конструирование локусов тяжелой цепи иммуноглобулина, содержащие а реаранжированный последовательность VDJ человека осуществляли посредством серии реакций гомологичной рекомбинации в бактериальных клетках (BHR) с применением ДНК бактериальной искусственной хромосомой (ВАС). Получали несколько направленных конструкций для получения генетически сконструированной мыши, у которой экспрессируется реаранжированный вариабельный домен тяжелой цепи, с применением генноинженерной технологии VELOCIGENE® (см., например, патент США №6586251 и Valenzuela, D.М. et al. (2003), High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nature Biotechnology 21(6):652-659, полностью включенные в настоящий документ посредством ссылки)
[000280] Вкратце, конструировали направленные векторы для введения нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека (т.е. hVH3-23(D)JH4; SEQ ID NO: 136) в генетически модифицированную мышь, у которой были удалены все или по существу все эндогенные функциональные сегменты генов V, D, J тяжелых цепей иммуноглобулинов. Кроме того, направленные векторы включали геномную область, содержащую гены Adam6a и Adam6b, для предотвращения проблем с фертильностью, связанных с удалением у мышей геномной области, содержащей гены Adam6a/6b (см., например, US 2012-0322108А1, полностью включенную посредством ссылки в настоящий документ).
[000281] Сначала конструировали донор BHR для модификации клона ВАС мыши, содержащий лидерную последовательность (направляющую тяжелую цепь через эндоплазматический ретикулум), нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи (VH3-23(D)JH4; SEQ ID NO: 136) и интрон hJH4 (SEQ ID NO: 140), которые функционально связаны с промотором VH3-23 2239 длиной п.о. (SEQ ID NO: 139). Дополнительно геномный локус фланкировали с 5'- и 3'-стороны гомологичными фрагментами IgH мыши для гомологичной рекомбинации с клоном ВАС MAID1115 (Фиг. 1).
[000282] Кроме того, осуществляли следующие пять модификаций для получения направленной конструкции, содержащей нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека.
[000283] Во-первых, вводили спектиномициновую селекционную кассета в направлении 5' от промотора VH3-23 (между сайтами I-CeuI и SpeI) для получения pJSh0038 (мини-локус UHC: SEQ ID NO: 142) (фиг. 1, 1. Лигирование I-CeuI/SpeI (Amp + Spec)). Мини-локус UHC содержит: (1) спектиномициновую (Spec) кассету с сайтами I-CeuI/AscI для лигирования; (2) Промотор hVH3-23 длиной п.о. (SEQ ID NO: 139); (3) реаранжированную нуклеотидную последовательность hVH3-23(D)JH4 (SEQ ID NO: 136); (4) интрон hJH4 (SEQ ID NO: 140); и (5) гомологичные фрагменты мыши для BHR (MAID 1115).
[000284] Во-вторых, направленно вводили гигромициновую селекционную кассету (EM7-HYG) в 5'-конец геномной области клона ВАС MAID 1115, которая содержит фланкированную loxP неомициновую кассету (Pgk-Neo) в направлении 5' от геномной области IgM. Вставка гигромициновой кассеты удаляла сайт loxP, расположенный на 5'-конце клона ВАС MAID 1115. Бактериальные клетки, содержащие генетически модифицированный клон ВАС (VI432), отбирали путем селекции по устойчивости к гигромицину/канамицину (фиг. 2, 2. BHR (Hyg+Kan)).
[000285] В-третьих, мини-локус UHC, который конструировали на этапе 1, направленно вводили в направлении 5' от локуса IgM клона ВАС VI432. При введении мини-локуса UHC происходила замена фланкированной 1охР неомициновой селекционной кассеты новой спектиномициновой кассетой (VI443). Бактериальные клетки, содержащие генетически модифицированные клоны ВАС (VI443), отбирали путем селекции по устойчивости к спектиномицину и гигромицину (фиг. 2; 3. BHR (Spec+Hyg)).
[000286] В-четвертых, на клон ВАС VI421, который содержит, от 5' к 3', (1) ген Adam6a (расположен в направлении 5'); (2) неомициновую кассету (расположена в направлении 5') фланкированную FRT-сайтами; (3) ген Adam6b (расположен в направлении 5'); (4) межгенную контрольную область 1 (IGCR1; т.е. ключевая регулирующая рекомбинацию V(D)J область); и (5) спектиномициновую кассету (расположена в направлении 3'), нацеливали конструкцию pDBa0049, которая содержит: хлорамфеникольную (Cm) кассету; сайт рестрикции AscI в направлении 5' от гена хлорамфеникола; и гомологичные фрагменты с 5'- и 3'-стороны. При направленном введении конструкции pDBa0049 происходило удаление IGCR1 и спектиномициновой кассеты из клона VI421; и введение нового сайта рестрикции AscI и хлорамфеникольной кассеты в направлении 3' от гена Adam6b. Бактериальные клетки, содержащие успешно таргетированный клон (VI444), отбирали путем селекции по устойчивости к хлорамфениколу и канамицину (Фиг. 3; 4. BHR (Cm+Kan)).
[000287] В-пятых, вводили геномную область клона ВАС VI444, содержащую гены Adam6a и/или 6b, в 5'-направлении от геномного локуса универсальной тяжелой цепи в клон ВАС VI443 между сайтами I-CeuI и AscI посредством рестрикционного расщепления и лигирования (фиг. 3). При указанной модификации в клон вводятся гены Adam6a и/или 6b и происходит замена спектиномициновой кассеты на неомициновую кассету, с получением готовой направленной конструкции (MAID6031; VI445). Бактериальные клетки (BHR), содержащие готовую направленную конструкцию (MAID 6031; VI445), отбирали на основании устойчивости к гигромицину и канамицину (Фиг. 3, 5. Лигирование I-CeuI/AscI (Hyg+Kan)).
[000288] Готовая направленная конструкция (MAID6031) для получения геномного локуса, содержащего реаранжированную последовательность вариабельного домена тяжелой цепи человека, содержит, от 5' к 3', (1) гомологичный 5'-фрагмент, содержащий приблизительно 20000 п.о. геномной последовательности мыши в направлении 5' от указанного эндогенного локуса тяжелой цепи Ig; (2) ген Adam6a; (3) 5' FRT-сайт; (4) неомициновую кассету; (5) 3' FRT-сайт, (6) ген Adam6b; (7) 2239 п. о. промотора hVH3-23 (SEQ ID NO: 139); (8) реаранжированную нуклеотидную последовательность тяжелой цепи иммуноглобулина человека (hVH3-23(D)JH4; SEQ ID NO: 136); (9) интрон hJH4 (SEQ ID NO: 140); и (10) гомологичный 3'-фрагмент, содержащий приблизительно 7400 п.о. геномной последовательности мыши в направлении 3' от сегментов гена JH мыши.
[000289] Готовую направленную конструкцию, MAID6031 ВАС DNA, линеаризировали и вводили путем электропорации в ЭС-клетки, выделенные из гетерозиготной мыши 1661 (фиг. 4.), у которой имеются геномные локусы VDJ тяжелых цепей Ig дикого типа и мутированные геномные локусы VDJ, в которых все VH, D, JH гены были удалены. Проводили скрининг успешно таргетированных ЭС-клеток мыши с применением праймеров и зондов, представленных на фиг. 6-8. Успешно таргетированные ЭС-клетки мыши вводили хозяевам - эмбрионам мышей с применением технологии VELOCIMOUSE® для получения генетически модифицированной гетерозиготной мыши F0. Для получения мыши (MAID 6032 het) без селекционной кассеты (т.е. FRT-Ub-Neo-FRT), в успешно таргетированные ЭС-клетки вводили путем электропорации плазмиду, которая экспрессирует рекомбиназу Flp, до введения эмбрионам-хозяевам. Как вариант, MAID6031 гетерозиготных самцов мышей, несущих селекционную кассету, скрещивали с самками мышей, экспрессирующих рекомбиназу Flp, для удаления кассеты. Гетерозиготных мышей, несущих указанную модификацию, скрещивали друг с другом для получения гомозигот (MAID 6032 НО), способных продуцировать тяжелые цепи иммуноглобулина только из генетически модифицированного локуса.
Пример 3. Характеризация генетически модифицированных мышей, экспрессирующих реаранжированный вариабельный домен тяжелой цепи
[000290] Всех мышей размещали и разводили в специальных беспатогенных условиях в Regeneron Pharmaceuticals. Умерщвляли три однопометных контрольных мыши (возраст 16 недель, самцы, n=2; Происхождение: 75% C57/BL6 и 25% 129) дикого типа (WT) и 2-4 мыши MAID 6032 НЕТ F0 (фиг. 9; возраст 9 недель, самцы, n=2; Происхождение: 50% C57/BL6 и 50% 129), и собирали кровь, селезенки и костный мозг животных. Дополнительно умерщвляли четыре однопометных контрольных мыши (возраст 10 недель; 2 самца и 2 самки) дикого типа (WT) и четыре гомозиготных («НО») мыши F2 MAID 6032 (возраст 10 недель; 3 самца; 1 самка), и собирали кровь, селезенки и костный мозг животных. Кровь собирали в пробирки BD Microtainer с ЭДТК (кат. №365973). Костный мозг собирали из бедренных костей, промывая полной средой RPMI (среда RPMI с добавлением фетальной телячьей сыворотки, пирувата натрия, Hepes, 2-меркаптоэтанола, заменимых аминокислот и гентамицина). Препараты эритроцитов периферической крови, селезенки и костного мозга лизировали лизирующим буфером АСК и промывали полной средой RPMI.
Проточная цитометрия
[000291] Для исследования способности описанных в настоящем документе генетически модифицированных гетерозиготных мышей F0 (MAID 6032 НЕТ) продуцировать антитела, происходящие из генетически модифицированного аллеля (т.е. из аллеля, который содержит единственную копию реаранжированного VH3-23/D/JH4), проводили анализ методом сортировки клеток с активированной флуоресценцией (FACS) с применением клеток крови, селезенки и костного мозга, выделенных из мыши дикого типа или гетерозиготной мыши 6032.
[000292] Вкратце, 1×106 клеток инкубировали с антителами мыши против CD16/CD32 (2.4G2, BD) на льду на протяжении 10 минут, с последующим мечением следующим коктейлем антител на протяжении 30 минут на льду: конъюгированное с ФИТЦ антитело против IgMa мыши (DS-1, BD Biosciences), конъюгированное с Pacific blue против CD3 мыши (17А2, BicLegend), конъюгированное с АРС-Н7 против CD 19 мыши (1D3, BD Biosciences) и конъюгированное с ФЭ антитело против IgMb мыши (AF6-78, BioLegend). Окрашенные клетки промывали и фиксировали в 2% формальдегиде. Данные регистрировали на проточном цитометре BD LSRFortessa и анализировали с применением FlowJo. Клетки костного мозга, клетки селезенки и клетки крови, выделенные из гетерозиготной мыши дикого типа или F0 6032, гейтировали по одиночным клеткам и сортировали на основании экспрессии CD19 (маркер В-клеток) или экспрессии CD3 (маркер Т-клеток). Кроме того, гейтированные CD 19+В-клетки сортировали на основании присутствия антител IgMb (IgM-антител, полученных из аллеля дикого типа (В6 аллель)) или антител IgMa (антител, полученных из генетически модифицированного аллеля (аллель 129), содержащих нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи (hVH3-23(D)JH4;). Результаты FACS-анализа (фиг. 11-12) предполагают, что мыши, гетерозиготные по целевому аллелю (т.е. содержащие одну копию реаранжированной последовательности вариабельной области тяжелой цепи; MAID 6032 het) были способны продуцировать антитела IgM, в основном происходящие из генетически модифицированного аллеля 129 (IgMa).
[000293] Для исследования способности генетически модифицированных гомозиготных мышей F2 (MAID 6032 НО), описанных в настоящем документе, продуцировать антитела, происходящие из генетически модифицированного аллеля (т.е. из аллеля, который содержит единственную копию реаранжированного VH3-23/D/JH4), проводили анализ методом сортировки клеток с активированной флуоресценцией (FACS) согласно описанию выше, с применением клеток селезенки и костного мозга, выделенных из мыши дикого типа или гомозиготной мыши 6032.
[000294] Только зрелые В-лимфоциты могут входить в лимфоидные фолликулы селезенки и лимфатические узлы, и, соответственно, эффективно участвовать в иммунном ответе. Зрелые долгоживущие В-лимфоциты происходят из короткоживущих предшественников, образующихся в костном мозге. Отбор в пул зрелых клеток представляет собой активный процесс, который происходит в селезенке. Две популяции В-клеток селезенки были идентифицированы как предшественники зрелых В-клеток. Переходные В-клетки 1 типа (Т1) представляют собой недавно мигрировавшие из костного мозга клетки. Они развиваются в переходные В-клетки 2 типа (Т2) которые находятся в активном клеточном цикле и обнаруживаются исключительно в первичных фолликулах селезенки. Зрелые В-клетки могут быть получены из В-клеток Т1 или Т2. Loder, F. et al., J. Exp. Med., 190(1): 75-89, 1999.
[000295] Результаты FACS-анализа (фиг. 13A и 13В) предполагают, что мыши, гомозиготные по целевому аллелю (т.е. содержащие две копии реаранжированной последовательности вариабельной области тяжелой цепи: MAID 6032 НО) были способны продуцировать нормальные популяции зрелых и незрелых В-клеток селезенки, хотя и с незначительным снижением уровня последовательностей лямбда по сравнению с мышами дикого типа (фиг. 13С и 13D). Также в селезенке мышей MAID 6032НО наблюдалось незначительное уменьшение популяции В-клеток Т1 и увеличение популяции В-клеток маргинальной зоны (Фиг. 13Е).
[000296] В костном мозге мышей MAID6032 НО формировались практически нормальные популяции В-клеток (фиг. 14А-14Е) с частотой использования последовательностей лямбда, составляющей1/2 от частоты для дикого типа (Фиг. 14F).
Исследования иммунизации
[000297] Пять мышей WT (происхождение: 75% C57BL6 / 25% 129) и 3-4 мышей MAID 6032 НЕТ иммунизировали через подушечки лап введением 0,025 мл смеси, содержащей 2,35 мкг антигена X, 10 мкг CpG-олигонуклеотида (ODN 1826, InvivoGen, кат. № tlrl-1826) и 25 мкг адъюванта - геля алюминия фосфата (Brenntag кат. №7784-30-7). Мыши получали бустерную иммунизацию шестикратно в той же дозировке. На 15 и 24 день после первичной иммунизации у анестезированных мышей забирали кровь через ретроорбитальный синус в пробирки для отделения сыворотки BD (BD, кат. №365956), и получали сыворотку согласно указаниям производителя.
[000298] Для измерения уровней антигенспецифических антител IgG и обратного скрининга метки mmh (myc-myc-his), планшеты для ИФА ELISA (Nunc) покрывали либо 1 мкг/мл антигена X, инкубированного в течение ночи при 4°С. Избыток антигена отмывали перед блокированием ФСБ + 1% БСА в течение 1 ч при комнатной температуре. Получали серийные разведения сыворотки, и планшеты инкубировали в течение 1 ч при комнатной температуре до промывания. Планшеты инкубировали с конъюгированным с пероксидазой хрена (HRP) антителом против IgG (кат. №1030-05, Southern Biotech) в течение 1 ч при комнатной температуре. После промывания планшеты инкубировали с ТМВ-субстратом (кат. №555214, BD). Реакции останавливали 1Н серной кислотой и регистрировали оптическую плотность при 450 нм, с применением ридера Victor Х5 Perkin Elmer Reader. Анализировали данные с помощью GraphPad Prism для расчета разведения сыворотки, дающего значения в 2 раза ниже исходных. Все эксперименты на животных были одобрены Институциональным комитетом по содержанию и использованию животных (IACUC) и Regeneron Pharmaceuticals.
[000299] Как видно из фиг. 15, генетически модифицированные мыши F0 и F1 (MAID 6032 het), гетерозиготные по целевому аллелю (т.е. содержащие одну копию реаранжированной нуклеотидной последовательности VH3-23/D/JH4), были способны продуцировать антигенспецифические антитела IgG на уровнях, сравнимых с уровнями, продуцируемыми мышами дикого типа как на 15, так и на 24 день после первичной иммунизации.
Пример 3. Получение и анализ мышей, у которых имеется два сегмента V легкой цепи человека
Пример 3.1: Конструирование направленного вектора для получения мышей, у которых имеется два сегмента V легкой цепи человека
[000300] Было создано два сконструированных локуса легких цепей, содержащих два сегмента гена Vκ человека (например, сегмент гена Vκ1-39 человека и Vκ3-20 человека; т.е. двойную легкую цепь («DLC»)) (фиг. 20). Один сконструированный локус легкой цепи содержал два сегменты генов Vκ человека и пять сегментов гена Jκ человека в нереаранжированной конфигурации (DLC-5J). Второй сконструированный локус легкой цепи содержал два сегмента гена Vκ человека и один сегмент гена Jκ человека в нереаранжированной конфигурации (DLC-1J). В каждом из двух дополнительных сконструированных локусов легких цепей сегменты генов человека были фланкированы со стороны 3' рекомбинационными сигнальными последовательностями для обеспечения реаранжировки in vivo сегментов гена человека в В-клетках.
[000301] Конструирование и получение мышей DLC-1J. Этапы конструирования, приводящие к получению локуса легкой цепи, содержащего два сегмента гена Vκ человека (Vκ1-39 и Vκ3-20) и один сегмент гена Jκ человека (Jκ5), называемый также DLC-1J, представлены на фиг. 21. В частности, последовательности Vκ1-39 и Vκ3-20 человека амплифицировали посредством ПЦР с матриц ВАС (Invitrogen), и вместе с амплифицированной последовательностью, содержащей рекомбинационную сигнальную последовательность (rss) и сегмент Jκ5 человека, клонировали посредством крестообразного лигирования в плазмиду, содержащую UB-гигромициновую селекционную кассету (Фиг. 21А). 5' и 3' фрагменты присоединяли согласно фиг. 21В и 21С.
[000302] Полученная направленная конструкция изображена на фиг. 21С (нижняя диаграмма; DLC-1J), рекомбинационные сигнальные последовательности (RSS) обозначены незакрашенными эллипсами. Модифицированный клон ДНК ВАС сконструированного локуса DLC-1J легкой цепи функционально связан с последовательностями мыши (т.е. в направлении 5' и 3' от последовательностей указанного эндогенного локуса легкой цепи к иммуноглобулина) подтверждали посредством ПЦР с применением праймеров, расположенных в последовательностях в составе сконструированного локуса легкой цепи, содержащего два сегмента гена Vκ человека, с последующим электропорацией в ЭС-клетки, в которых был удален локус вариабельной области IgK мыши (содержащий сегменты генов к вариабельной и соединительной областей) (Фиг. 21D) для получения мыши, у которой экспрессируется любой из двух сегментов гена Vκ человека. Позитивные клоны ЭС-клеток, которые содержали сконструированный локус DLC-1J легкой цепи, подтверждали посредством скрининга и кариотипирования Taqman™ с применением зондов, специфических в отношении сконструированного локуса DLC-1J легкой цепи. Последовательности праймеров и зондов, используемые для скрининга ЭС-клеток DLC-1J, представлены в таблице 8 ниже и включены в Перечень последовательностей.


[000304] Подтвержденные клоны ЭС-клеток затем использовались для имплантации самкам мышей для получения помета детенышей, у которых имеется DLC-1J локус легкой цепи и экспрессируется вариабельный домен легкой цепи человека, соединенный с доменом Cκ мыши. Последовательности праймеров и зондов, используемых для генотипирования детенышей, перечислены в таблице 8 выше. Последовательность, включающая сконструированный локус DLC-1J, в том числе 100 нуклеотидов последовательности мыши в 5' и в 3' направлении от встроенной сконструированной последовательности, представлена на фиг. 22A-22D и приведена в последовательности SEQ ID NO: 82.
[000305] ЭС-клетки, несущие сконструированный локус легкой цепи, могут быть трансфицированы конструкцией, которая экспрессирует FLP, для удаления неомициновой кассеты FRTed, введенной направленной конструкцией (см. фиг. 21Е). Необязательно неомициновую кассету удаляют путем скрещивания с мышами, экспрессирующими рекомбиназу FLP (например, US 6774279). Необязательно неомициновую кассету у мышей сохраняют.
[000306] Конструирование и получение мышей DLC-5J. Для получения локуса легкой цепи, содержащего два сегмента гена Vκ человека (Vκ1-39 и Vκ3-20) и пять сегментов гена Jκ человека (JκI, Jκ2, Jκ3, Jκ4 и Jκ5), называемого также DLC-5J, амплифицированную последовательность длиной 2000 п.о., содержащую все 5 сегментов Jκ человека, лигировали в вектор, содержащий два сегмента гена Vκ человека и один Jκ человека, представленные на фиг. 21В (в середине) (см. фиг. 23А). Последующие этапы конструирования включали присоединение 3'- и 5'-фрагментов, показанных на фиг. 23В.
[000307] Полученная направленная конструкция представлена на фиг. 23В (нижняя диаграмма, DLC-5J), рекомбинационные сигнальные последовательности (RSS) выделены незакрашенными эллипсами. Модифицированный клон ДНК ВАС сконструированного локуса легкой цепи DLC-5J, функционально связанный с последовательностями мыши (т.е. в направлении 5' и в 3' от последовательностей указанного эндогенного локуса легкой цепи к иммуноглобулина), подтверждали посредством ПЦР с применением праймеров, локализованных в последовательностях в составе сконструированного локуса легкой цепи, содержащего два сегмента гена Vκ человека, с последующей электропорацией в ЭС-клетки, в которых был удален локус вариабельной области IgK мыши (содержащий сегменты генов к вариабельной и соединительной областей) (фиг. 23С) для получения мыши, у которой экспрессируется любой из двух сегментов гена Vκ человека. Позитивные клоны ЭС-клеток, которые содержали сконструированный локус легкой цепи DLC-5J, подтверждали посредством скрининга и кариотипирования Taqman™ с применением зондов, специфических в отношении сконструированного локуса легкой цепи DLC-5J. Последовательности праймеров и зондов, использованные для скрининга ЭС-клеток DLC-5J, представлены в таблице 9 ниже и включены в Перечень последовательностей.



[000308] Подтвержденные клоны ЭС-клеток затем использовали для имплантации самкам мышей для получения помета детенышей, у которых имелся, локус DLC-5J легкой цепи и экспрессировался вариабельный домен легкой цепи человека, соединенный с доменом Cκ мыши. Последовательности праймеров и зондов, использованных для генотипирования детенышей, приведены в таблице 9 выше. Последовательность, включающая сконструированный локус DLC-5J, в том числе 100 нуклеотидов последовательности мыши в направлении 5' и в направлении 3' от встроенной сконструированной последовательности, представлена на фиг. 24A-24D и приведена в последовательности SEQ ID NO: 83.
[000309] ЭС-клетки, несущие сконструированный локус легкой цепи, могут быть трансфицированы конструкцией, которая экспрессирует FLP, для удаления неомициновой кассеты FRTed, введенной направленной конструкцией (см. фиг. 23D). Необязательно неомициновую кассету удаляют путем скрещивания с мышами, экспрессирующими рекомбиназу FLP (например, US 6774279). Необязательно неомициновую кассета у мышей сохраняют.
Пример 3.2: Характеризация мышей, у которых имеется два сегмента V человека
[000310] Проточная цитометрия. Популяции В-клеток и развитие В-клеток у мышей DLC подтверждали путем анализа препаратов спленоцитов и костного мозга с применением проточной цитометрии. Суспензии клеток от мышей, гомозиготных по двум сегментам генов Vκ человека и пяти сегментам генов Jκ человека (n=4), мышей, гомозиготных по двум сегментам генов Vκ человека и одному сегменту человека Jκ гена (n=4), и мышей дикого типа (n=4) получали с применением стандартных способов и окрашивали флуоресцентно мечеными антителами.
[000311] Вкратце, 1×106 клеток инкубировали с CD16/CD32 против мыши (клон 2.4G2, BD Pharmigen) на льду на протяжении 10 минут, с последующим мечением следующим коктейлем антител на протяжении 30 минут на льду: конъюгированным с АРС-Н7 против CD 19 мыши (клон 1D3, BD Pharmigen), конъюгированным с Pacific Blue против CD3 мыши (клон 17А2, BioLegend), конъюгированным с ФИТЦ против IgK мыши (клон 187.1, BD Pharmigen) или против CD43 мыши (клон 1 В11, BioLegend), конъюгированным с ФЭ против Ig λ мыши (клон RML-42, BioLegend) или против c-kit мыши (клон 2 В8, BioLegend), конъюгированным с РегСР-Су5.5 против IgD мыши (BioLegend), конъюгированным с ФЭ-Су7 против IgM мыши (клон П/41, eBioscience), конъюгированным с АРС против В220 мыши (клон RA3-6B2, eBioscience). После окрашивания клетки промывали и фиксировали в 2% формальдегиде. Данные регистрировали на проточном цитометре LSRII и анализировали с применением FlowJo (Tree Star, Inc.). Гейтирование: общее количество В-клеток (CD19+CD3-), Igκ+ В-клеток (Igκ+Igλ-CD19+CD3-), Igλ+ В-клеток (Igκ-Igλ+CD19+CD3-). Результаты для компартмента костного мозга представлены на фиг. 25А-27В. Результаты для компартмента селезенки представлены на фиг. 28А-фиг. 31.
[000312] Как видно из указанного примера, у мышей DLC-5J наблюдались нормальные популяции В-клеток в компартментах селезенки и костного мозга (фиг. 25А-31). У мышей DLC-5J наблюдались популяции незрелых, зрелых и пре/про-В-клеток в компартменте костного мозга, по существу совпадающие с наблюдаемыми у однопометных животных дикого типа. Фактически, локус DLC-5J был способен конкурировать с указанным эндогенным локусом легкой цепи лямбда с получением соотношения каппа : лямбда, по существу совпадающим с наблюдаемым у мышей дикого типа (фиг. 27В). Также у мышей DLC-5J наблюдалось нормальное развитие периферических В-клеток по мере прохождения В-клетками различных стадий в компартменте селезенки (например, незрелые, зрелые, T1, Т2 Т3, предшественники маргинальной зоны, клетки маргинальной зоны, фолликулярные-I, фолликулярные-II и т.п.), которое происходит по существу совпадающим с наблюдаемым у мышей дикого типа образом (фиг. 30А-31). Напротив, у мышей DLC-1J наблюдалось меньшее общее количество В-клеток и повышенная частота использования легких цепей лямбда по сравнению со сконструированной легкой цепью каппа (данные не показаны).
[000313] Экспрессия двойных легких цепей. Экспрессию обоих сегментов гена Vκ человека анализировали у гомозиготных мышей с применением количественного ПЦР-анализа. Вкратце, CD19+ В-клетки очищали из костного мозга и цельных селезенок мышей дикого типа, мышей, гомозиготных по замене локусов вариабельных областей тяжелой цепи и легкой цепи к мыши на соответствующие локусы вариабельных областей тяжелой цепи и легкой цепи κ человека (Нκ), a также мышей, гомозиготных по сконструированным локусам легких цепей κ, содержащим два сегмента гена Vκ человека, и либо пять сегментов гена Jκ человека (DLC-5J), либо один сегмент гена Jκ человека (DLC-1J). Относительную экспрессию нормировали по экспрессии области Cκ мыши (n=3-5 мышей на группу). Результаты представлены на фиг. 32 и фиг. 33.
[000314] Экспрессию легких цепей, содержащих реаранжированный сегмент гена Vκ3-20 или Vκ1-39 человека, детектировали как в костном мозге, так и в селезенке мышей DLC-5J и DLC-1J (фиг. 32 и фиг. 33). В компартменте костного мозга экспрессия обеих происходящих из Vκ3-20 человека и происходящих из Vκ1-39 человека легких цепей в обеих линиях мышей DLC была значительно выше по сравнению с мышами, у которых была произведена замена сегмента гена Vκ и Jκ мыши на соответствующие сегменты генов Vκ и Jκ человека (Нκ; фиг. 32). Наблюдаемая экспрессия происходящих из Vκ3-20 человека легких цепей была приблизительно в 6 (DLC-5J) - 15 раз (DLC-1J) выше, чем у мышей Нκ. В компартменте костного мозга у мышей DLC-1J наблюдался приблизительно в два раза более высокий уровень экспрессии происходящих из Vκ3-20 человека легких цепей по сравнению с мышами DLC-5J. Наблюдаемая экспрессия происходящих из Vκ1-39 человека легких цепей была приблизительно в 6 (DLC-5J) - 13 раз (DLC-1J) выше по сравнению с мышами Нκ. У DLC-1J мышей в компартменте костного мозга наблюдался приблизительно в два раза более высокий уровень экспрессии происходящих из Vκ1-39 человека легких цепей по сравнению с мышами DLC-5J.
[000315] В компартменте селезенки экспрессия обеих происходящих из Vκ3-20 человека и происходящих из Vκ1-39 человека легких цепей в обеих линиях мышей DLC была значительно выше по сравнению с мышами Нκ (фиг. 33). Наблюдаемая экспрессия происходящих из Vκ3-20-человека легких цепей была приблизительно в 4 раза (DLC-5J) и 8 раз (DLC-1J) выше, чем у мышей Нκ. У мышей DLC-1J в компартменте селезенки наблюдался приблизительно в два раза более высокий уровень экспрессии происходящих из Vκ3-20 человека легких цепей по сравнению с мышами DLC-5J. Наблюдаемая экспрессия происходящих из Vκ1-39 человека легких цепей была приблизительно в 4 (DLC-5J) - 5 раз (DLC-1J) выше, чем у мышей Нκ. У мышей DLC-1J наблюдалась сходная с мышами DLC-5J экспрессия происходящих из Vκ1-39 человека легких цепей в компартменте селезенки.
[000316] Частота использования Vκ/Jκ человека у мышей DLC-5J. у мышей, гомозиготных по двум нереаранжированным сегментам генов Vκ человека и пяти нереаранжированным сегментам генов Jκ человека (DLC-5J) анализировали частоту использования сегмента гена Vκ/Jκ человека в В-клетках селезенки с применением полимеразной цепной реакции с обратной транскриптазой (ОТ-ПЦР).
[000317] Вкратце, селезенки гомозиготных мышей DLC-5J (n=3) и мышей дикого типа (n=2) собирали и измельчали в 10 мл RPMI 1640 (Sigma), содержащей 10% термоинактивированной фетальной бычьей сыворотки, с применением матовых предметных стекол для получения суспензии отдельных клеток. Спленоциты осаждали центрифугированием (1200 об/мин в течение пяти минут), и эритроциты лизировали в 5 мл лизирующего буфера АСК (GIBCO) на протяжении трех минут. Спленоциты разводили ФСБ (Irvine Scientific), фильтровали через клеточный фильтре размером пор 0,7 мкм и центрифугировали повторно для осаждения клеток, с последующим ресуспендированием в 1 мл ФСБ.
[000318] РНК выделяли из осажденных спленоцитов с применением мини-набора AllPrep DNA/RNA (Qiagen) в соответствии с инструкциями производителя. ОТ-ПЦР выполняли на РНК спленоцитах с применением системы 5' RACE (быстрой амплификации концов кДНК, «Rapid Amplification of cDNA ends») с праймерами, специфическими в отношении гена Cκ мыши в соответствии с инструкциями производителя (Invitrogen). Использовались праймеры, специфические в отношении гена Cκ мыши: 3' mIgκC RACE1 (AAGAAGCACA CGACTGAGGC AC; SEQ ID NO: 90) и mIgκC3'-1 (CTCACTGGAT GGTGGGAAGA TGGA; SEQ ID NO: 91). Продукты ПЦР очищали гель-фильтрацией и клонировали в вектор pCR®2.1-ТОРО® (набор ТОРО® ТА Cloning® Kit, Invitrogen) и секвенировали с прямым (М13 Forward, GTAAAACGAC GGCCAG; SEQ ID NO: 92) и обратным (М13 Reverse, CAGGAAACAG CTATGAC; SEQ ID NO: 93) праймерами, присутствующими в составе указанного вектора в положениях, фланкирующих сайт клонирования. Секвенировали 10 клонов из каждого образца селезенки. Последовательности сравнивали с наборами иммуноглобулинов мыши и человека из наборов референсного каталога IMGT/V-QUEST для определения частоты использования Vκ/Jκ. В таблице 10 приведены комбинации Vκ/Jκ для выбранных клонов, наблюдаемые у клонов ОТ-ПЦР из каждого образца спленоцитов. В таблице 11 приведена последовательность аминокислот соединительных областей Vκ человека / Jκ человека и Jκ человека / Cκ мыши, выбранных клонов ОТ-ПЦР от гомозиготных мышей DLC-5J. Строчными буквами обозначены мутации в последовательности аминокислот вариабельной области или нешаблонные добавления, обусловленные добавлениями N и/или Р при рекомбинации.
[000319] Как видно из указанного примера, мыши, гомозиготные по двум нереаранжированным сегментам генов Vκ человека и пяти нереаранжированным сегментам генов Jκ человека (DLC-5J), функционально связанным с геном Cκ мыши, способны к продуктивной рекомбинации как сегментов гена Vκ человека с различными сегментами генов Jκ человека с образованием ограниченного репертуара легких цепей иммуноглобулина. Среди реаранжировок у гомозиготных мышей DLC-5J, приведенных в таблице 10, уникальные реаранжировки Vκ/Jκ человека наблюдались для Vκ1-39/Jκ2 (1), Vκ1-39/Jκ3 (1), Vκ3-20/Jκ1 (7), Vκ3-20/Jκ2 (4) и Vκ3-20/Jκ3 (1). Кроме того, при таких уникальных реаранжировках наблюдалось разнообразие J-сегментов за счет присутствия уникальных аминокислот в области CDR3 легкой цепи (таблица 11), обусловленного или мутацией, и/или рекомбинацией сегментов гена Vκ и Jκ человека в ходе развития. Для всех указанных реаранжировок наблюдалась функциональная сквозная транскрипция в Cκ мыши (таблица 11).
[000320] В совокупности эти данные показывают, что у мышей, сконструированных для обеспечения вариантов выбора из не более чем двух сегментов гена Vκ человека, из которых оба способны к реаранжировке (например, с одним или большим количеством сегментов гена и, согласно некоторым вариантам реализации, до пяти сегментов гена Jκ человека) и кодированию домена VL легкой цепи иммуноглобулина человека, количества и развитие В-клеток почти соответствует дикому типу во всех аспектах. Такие мыши продуцируют набор антител, содержащий легкие цепи иммуноглобулина, включающие один из двух возможных сегментов гена VL человека, присутствующих в наборе. Мышь продуцирует указанный набор антител в ответ на стимуляцию антигеном, и указанный набор антител связан с разнообразием обратно-гибридных (с вариабельными областями человека/константными областями мыши) тяжелых цепей.




Пример 4: Получение и характеризация мышей, у которых имеются две легкие гистидин-замещенные цепи человека
Пример 4.1: Конструирование и получение мышей, у которых имеется два сегмента V каппа, каждый из которых содержит четыре замены гистидина
[00235] Замены гистидина вводили в локус двойной легкой цепи согласно описанию выше для Vκ1-39 и Vκ3-20 ULC мыши. Вкратце, последовательность DLC, представленную на фиг. 23А, (снизу) подвергали сайт-специфическому мутагенезу, с модификацией сначала последовательности Vκ1-39 и последующей модификацией последовательности Vκ3-20, с применением праймеров, представленных на фиг. 34. Полученная последовательность двойной легкой цепи содержала сегмент Vκ1-39 с гистидинами, введенными в последовательность зародышевой линии в положениях 105, 106, 108 и 111, сегмент Vκ3-20 с гистидинами, введенными в последовательность зародышевой линии в положениях 105, 106, 107 и 109, а также все пять сегментов Jκ (Jκ1, Jκ2, Jκ3, Jκ4, и Jκ5). Следующий этап конструирования включал присоединение 5'-фрагмента, несущего кассету FRT-UB-NEO-FRT, и 3'-фрагмент, несущий энхансеры Igκ и константную область мышь. Указанный направленный вектор вводили путем электропорации е ЭС-клетки, в которых был удален локус вариабельной области Igκ мыши (содержащие сегменты генов к вариабельной и соединительной областей), согласно представленному на фиг. 35А (рекомбинационные сигнальные последовательности, RSS, не показаны на указанном чертеже). Проводили скрининг таргетированных ЭС-клеток путем анализа модификации аллеля согласно описанию выше, с применением праймеров и зондов для детекции областей, описанных выше в таблицах 1, 5, 8 и 9 (в частности, 1633h2, 1635h2, neo, Jxn 1-39/3-20, mIgKD2 и mIgKp15), а также два дополнительных набора праймеров и зондов, перечисленных в таблице 12 ниже. Последовательности указанных двух дополнительных наборов праймеров и зондов включены в Перечень последовательностей.


[00236] Подтвержденный клон ЭС-клеток затем используют для имплантации самкам мышей для получения помета детенышей, у которых имеется локус DLC-5J легкой цепи с четырьмя гистидиновыми модификациями в каждой из двух присутствующих последовательностей сегментов VL, и экспрессирующих вариабельный домен легкой цепи человека, соединенный с доменом Cκ мышь. Некоторые из последовательностей, используемых для скрининга ЭС-клеток, также используют для генотипирования детенышей.
[00237] ЭС-клетки, несущие сконструированный локус легкой цепи, могут быт» трансфицированы конструкцией, которая экспрессирует FLP (например, FLPo) для удаления неомициновой кассеты FRTed, введенной направленной конструкцией (см. фиг. 35В, RSS не показаны на указанном чертеже). Необязательно неомициновую кассету удаляют путем скрещивания с мышами, экспрессирующими рекомбиназу FLP (например, US 6774279). Необязательно неомициновую кассету у мышей сохраняют.
Пример 4.2: Конструирование и получение мышей, у которых имеется два сегмента V каппа, каждый из которых содержит три замены гистидина
[00238] Три замены гистидина вводили в каждый Vκ1-39 и Vκ3-20 двойной легкой цепи мыши. Вкратце, последовательность DLC, представленную на фиг. 23А (внизу), подвергать сайт-специфическому мутагенезу, с модификацией сначала последовательности Vκ1-39, и последующей модификацией последовательности Vκ3-20, с применением праймеров, представленных на фиг. 36. Полученная последовательность двойной легкой цепи содержала сегмент Vκ1-39 с гистидинами, введенными в последовательность зародышевой линии в положениях 106, 108 и 111, Vκ3-20 сегмент с гистидинами, введенными в последовательность зародышевой линии в положениях 105, 106 и 109, а также все пять Jκ сегментов (Jκ1, Jκ2, Jκ3. Jκ4 и Jκ5). Последующий этап конструирования включал присоединение 5'-фрагмента, несущего кассету FRT-UB-NEO-FRT, и 3'-фрагмента, несущего энхансеры и константную область IgK мыши. Указанный направленный вектор вводили путем электропорации в ЭС-клетки, в которых был удален локус вариабельной области IgK мыши (содержащий сегменты генов вариабельной и соединительной областей κ), как показано на фиг. 37А (RSS не показаны на указанном чертеже). Проводили скрининг таргетированных ЭС-клеток путем анализа модификации аллеля согласно описанию выше, с применением праймеров и зондов для детекции областей, описанных выше в таблицах 1, 5, 8, и 9 (в частности, 1633h2, 1635h2, neo, Jxn 1-39/3-20, mIgKD2 и mIgKp15), а также два дополнительных набора праймеров и зондов, перечисленные в таблице 13 ниже. Последовательности указанных двух дополнительных наборов праймеров и зондов включены в Перечень последовательностей.

[00239] Подтвержденный клон ЭС-клеток затем используют для имплантации самкам мышей для получения помета детенышей, у которых имеется локус легкой цепи DLC-5J с четырьмя гистидиновыми модификациями в каждой из двух присутствующих последовательностей сегментов VL, и экспрессируется вариабельный домен легкой цепи человека, соединенный с доменом Cκ мыши. Некоторые из последовательностей, используемых для скрининга ЭС-клеток, также используют для генотипирования детенышей.
[00240] ЭС-клетки, несущие сконструированный локус легкой цепи, могут быть трансфицированы конструкцией, которая экспрессирует FLP (например, FLPo) для удаления неомициновой кассеты FRTed, введенной направленной конструкцией (см. фиг. 37 В, RSS не показаны на указанном чертеже). Необязательно неомициновую кассету удаляют путем скрещивания с мышами, экспрессирующими рекомбиназу FLP (например, US 6774279). Необязательно неомициновая кассета у мышей сохраняется.
Пример 4.3: Разведение мышей, у которых имеются гистидин-замещенные двойные легкие цепи человека
[00241] Мышей, несущих сконструированный локус гистидин-замещенных двойных легких цепей человека, скрещивают с мышами, у которых удален эндогенный локус легкой цепи X, для получения потомства, у которого экспрессируются в качестве единственных легких цепей сконструированные гистидин-замещенные легкие цепи, происходящие из локуса двойной легкой цепи.
[00242] Мышей, несущих сконструированный локус гистидин-замещенных двойных легких цепей человека, скрещивают с мышами, у которых эндогенный локус вариабельной области тяжелой цепи мыши заменен на локус вариабельной области тяжелой цепи человека (см. US 6596541 и US 8502018; мышь VELOCIMMUNE®, Regeneron Pharmaceuticals, Inc.).
Пример 4.4: Детекция гистидиновых модификаций в легких цепях иммуноглобулина, полученных от мыши, у которой имеется два сегмента v каппа, каждый из которых содержит три замены гистидина
[00243] Ампликоны V каппа из мРНК В-клеток селезенки получали с применением ПЦР с обратной транскриптазой (ОТ-ПЦР) и высокопроизводительного скрининга.
[00244] Вкратце, селезенки пяти гетерозиготных мышей, у которых имеется два сегмента V каппа (Vκ1-39 и Vκ3-20), каждый из которых содержит три замены гистидина (мыши, каппа-локус которых изображен на фиг. 35) и эндогенные тяжелые цепи мыши, собирали и гомогенизировали в 1× ФСБ (Gibco) с применением предметных стекол. Клетки осаждали центрифугированием (500×g в течение 5 минут), и эритроциты лизировали в лизирующем буфере АСК (Gibco) в течение 3 минут. Клеток промывали 1×фСБ и фильтровали с применением клеточного фильтра с размером пор 0,7 мкм. В-клеток выделяли из клеток селезенки с применением сортировки позитивной селекцией с использованием MACS на CD 19 (Miltenyi Biotec). Тотальную РНК выделяли из осажденных В-клеток с применением набора RNeasy Plus (Qiagen). ПолиА+ мРНК выделяли из тотальной РНК с применением мини-набора для выделения мРНК Oligotex Direct mRNA Mini Kit (Qiagen).
[00245] Двуцепочечную кДНК получали из В-клеток селезенки мРНК посредством 5' RACE с применением набора для синтеза SMARTer Pico cDNA Synthesis Kit (Clontech). Обратную транскриптазу и дНТФ от Clontech заменяли на Superscript II и дНТФ от Invitrogen. Наборы легких цепей иммуноглобулина выделяли из кДНК с применением праймера, специфического в отношении константной области IgK и праймера SMARTer 5' RACE (Таблица 14). Продукты ПЦР очищали с применением набора для очищения QIAquick PCR Purification Kit (Qiagen). Второй раунд ПЦР проводили с применением того же праймера для 5' RACE и вложенного 3'-праймера, специфического в отношении константной области IgK (Таблица 15). Продукты второго раунда ПЦР очищали с применением системы SizeSelect E-gel (Invitrogen). Третью ПЦР проводили с использованием праймеров, добавлявших 454-адаптеры и штрихкоды. Продукты третьего раунда ПЦР очищали с применением гранул Agencourt AMPure ХР Beads. Очищенные продукты ПЦР количественно определяли с помощью SYBR-qPCR с применением набора КАРА Library Quantification Kit (КАРА Biosystems). Объединенные библиотеки подвергали эмульсионной ПЦР (emPCR) с применением набора 454 GS Junior Titanium Series Lib-A emPCR Kit (Roche Diagnostics) и двунаправленному секвенированию с применением инструмента Roche 454 GS Junior в соответствии с протоколами производителя.


[00246] При биоинформационном анализе риды последовательностей 454-секвенирования сортировали на основании точного совпадения штрихкода образца и удаляли низкокачественные основания. Последовательности аннотировали на основании выравнивания реаранжированных последовательностей Ig на сегменты V и J зародышевой линии человека из базы данных с применением локально установленного IgBLAST (NCBI, v2.2.25+). Последовательность помечали как неоднозначную и удаляли из анализа, если обнаруживалось несколько лучших совпадений с идентичным показателем. Набор скриптов Perl был разработан для анализа результатов и хранения данных в базе данных MySQL. Область CDR3 легкой цепи каппа определяли как последовательность между консервативным С-кодоном и мотивом FGXG.
[00247] На фиг. 38 представлены выравнивания последовательности аминокислот, кодируемой последовательностью IGKV3-20 (Фиг. 38А) или IGKV1-39 (Фиг. 38В) зародышевой линии человека, с трансляцией в аминокислоты примеров последовательностей Vκ, полученных из продуктивно реаранжированных антител, продуцированных мышами, у которых имеется гистидин-модифицированный DLC-5J (у которых имеется локус вариабельной области легкой цепи, содержащий сегменты генов Vκ1-39 и Vκ3-20, при этом каждый сегмент включает три гистидиновых модификации согласно описанию выше). Риды последовательностей показали, что в большинстве продуктивно реаранжированных легких цепей сохраняется по меньшей мере один гистидин, введенный в CDR3 зародышевой линии. В некоторых случаях в большинстве продуктивно реаранжированных легких цепей человека, содержащих последовательность Vκ3-20, в которой сохраняется по меньшей мере один гистидиновый остаток, сохраняются все три гистидиновые модификации, введенные в CDR3 зародышевой линии (см. фиг. 38А). В некоторых случаях в продуктивно реаранжированных легких цепях человека, содержащих последовательность Vκ1-39, в которой сохраняется по меньшей мере один гистидиновый остаток, приблизительно в 50% легких цепей сохраняются все три гистидина, введенные в CDR3 зародышевой линии (см. верхнее выравнивание на фиг. 38В), и приблизительно в 50% легких цепей сохраняются два из трех гистидинов, введенных в CDR3 зародышевой линии (см. нижнее выравнивание на фиг. 38В). В некоторых случаях гистидины в последнем положении последовательности сегмента V могут утрачиваться в результате V-J-реаранжировки.
Пример 5. Получение и анализ мышей, у которых имеется единственная нуклеотидная последовательность реанжированной тяжелой цепи иммуноглобулина человека и два сегмента гена V каппа
[000321] Мышей, у которых имеется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи в локусе тяжелой цепи (MAID6031; «мышь UHC»), получали согласно описанию выше. Вкратце, у мыши UHC удаляли все эндогенные функциональные сегменты генов вариабельных областей тяжелой цепи и заменяли на одну нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи, которая кодирует hVH3-23/D/JH4, функционально связанную с эндогенной нуклеотидной последовательностью константной области тяжелой цепи.
[000322] Мышей, у которых имеются генетически сконструированные локусы легких цепей; содержащие два сегмента гена Vκ человека (например, сегмент гена Vκ1-39 человека и сегмент гена Vκ3-20 человека) и любой из сегментов Jκ человека (Jκ5; DLC-1J) или пяти сегментов гена Jκ человека (hJκ1-5; DLC-5J), получали согласно описанию выше. Вкратце, один сконструированный локус легкой цепи содержит два сегмента гена Vκ человека и пять сегментов гена Jκ человека (Jκ1-5) в нереаранжированной конфигурации, и функционально связан с эндогенной последовательностью к константной области мыши (MAID 1911 (DLC-5J); фиг. 19Е). Другой сконструированный локус легкой цепи содержит два сегмента гена Vκ человека и один сегмент гена Jκ человека (Jκ1) в нереаранжированной конфигурации, и функционально связан с эндогенной последовательностью константной области к мыши (MAID 1913(DLC-1J); фиг. 21D). В каждом из двух дополнительных сконструированных локусов легких цепей сегменты генов человека были фланкированы со стороны 3' рекомбинационными сигнальными последовательностями для обеспечения реаранжировки in vivo сегментов гена человека в В-клетках.
[000323] Описанных выше гомозиготных мышей UHC (MAID6031) скрещивали с гомозиготными мышами DLC-5J (MAID1911) для получения мыши, гетерозиготной по аллелю UHC и аллелю DLC-5J. Аналогичным образом гомозиготных мышей UHC (MAID6031) скрещивали с гомозиготными мышами DLC-1J (MAID1913) для получения мыши, гетерозиготной по аллелю UHC и аллелю DLC-1J. Гетерозиготных мышей F1, полученных в результате указанных скрещиваний, скрещивали друг с другом для получения мышей, гомозиготных по каждому аллелю. Присутствие генетически модифицированных аллелей в локусах тяжелых цепей и легких цепей иммуноглобулина подтверждали посредством скрининга и кариотипирования TAQMAN™ с применением специфических зондов и праймеров, описанных выше.
[000324] Мышей, гетерозиготных по аллелю UHC, и DLC-5J скрещивали друг с другом для получения гомозигот (MAID 1912НО 6032НО; «DLC × UHC») экспрессирующих «легкие» цепи иммуноглобулина в основном из генетически модифицированного локуса. У мышей MAID 1912НО 6032НО (гомозиготных DLC × UHC) имеется вставка описанной в настоящем документе универсальной тяжелой цепи (например, hVH3-23/hD/hJH4) в локусе тяжелой цепи мыши, в котором были удалены все эндогенные гены VDJ вариабельных областей тяжелых цепей, и локусе легкой цепи каппа (κ) мыши DLC-5J (hVκ1-39 hVκ3-20 hJκ1-5), в котором были удалены все гены Vκ и Jκ мыши.
[000325] Всех мышей размещали и разводили в специальных беспатогенных условиях в компании Regeneron Pharmaceuticals. Трех мышей F5 VELOCIMMUNE® (MAID 12930 1640НО ("VI3"); см. патент США №8502018, включенный в настоящий документ посредством ссылки) (возраст 14 недель, самцы; Происхождение: 26,5% C57/BL6, 22,75% 129 и 50,75% Balb/c) и трех мышей F2 MAID 1912НО 6032НО (фиг. 39; возраст 7-8 недель, самки; Происхождение: 18,75 C57/BL6, 18,75% 129 и 62,5% Balb/c) умерщвляли, и собирали селезенки и костный мозг животных. Костный мозг собирали из бедренных костей, промывая полной средой RPMI (среда RPMI с добавлением фетальной телячьей сыворотки, пирувата натрия, Hepes, 2-меркаптоэтанола, заменимых аминокислот и гентамицина). Препараты эритроцитов селезенки и костного мозга лизировали лизирующим буфером АСК и промывали полной средой RPMI.
Проточная цитометрии
[000326] Для исследования способности описанных в настоящем документе генетически модифицированных гомозиготных мышей «DLC × UHC» (MAID 1912НО 6032НО) продуцировать антитела, происходящие из генетически модифицированных аллелей (например, из аллеля который содержит единственную копию реаранжированного VH3-23/D/JH4 в локусе тяжелой цепи и аллеля, который содержит два гена Vκ человека и пять генов Jκ человека в локусе легкой цепи), проводили анализ с сортировкой клеток с активированной флуоресценцией (FACS) согласно примеру 3.
[000327] В компартменте селезенки у мышей MAID 1912НО 6032НО наблюдаются количества CD19+ В-клеток и зрелых В-клеток, по существу совпадающие с количествами, наблюдаемыми у мышей VELOCIMMUNE® (VI3) (фиг. 40А-40В), которых используют в качестве контрольных, для определения специфических эффектов, наблюдаемых у мышей MAID 1912НО 6032НО по сравнению с мышами с другими генетическими модификациями в иммуноглобулиновых локусах; также и гуморальная иммунная система мышей VELOCIMMUNE® функционирует как у мышей дикого типа (выше). У мышей MAID 1912НО 6032НО наблюдалось 2-кратное увеличение количества незрелых В-клеток в селезенке по сравнению с мышами VI3 (фиг. 40А-40 В). Мыши MAID 1912НО 6032НО также были по существу аналогичны мышам VI3 в отношении использования легких цепей каппа и гамма (фиг. 41А-41 В). У мышей MAID 1912НО 6032НО (DLC × UHC) также наблюдались повышенные уровни поверхностного IgM на В-клетки селезенки (т.е. повышенная поверхностная экспрессия IgM на клетку) по сравнению с мышами VI3 (фиг. 42).
[000328] Также у мышей MAID 1912НО 6032НО (DLC × UHC) наблюдалось измененное развитие периферических В-клеток в ходе прохождения В-клеток через различные стадии в компартменте селезенки (например, незрелые, зрелые, T1, Т2 Т3, клетки-предшественники маргинальной зоны, клетки маргинальной зоны, фолликулярные клетки типа I, фолликулярные клетки типа II и т.п.), которое происходило отличным от наблюдаемого у мышей VI3 образом (фиг. 43А). В частности, у мышей MAID 1912НО 6032НО (DLC × UHC) наблюдалось больше незрелых В-клеток, В-клеток Т1 и В-клеток маргинальной зоны (МЗ) в компартменте селезенки По сравнению с мышами VI3. Количества фолликулярных клеток I и фолликулярных клеток II у мышей MAID 1912НО 6032НО (DLC × UHC) по существу совпадали с наблюдаемыми у мышей VI3 (фиг. 43 В).
[000329] В компартменте костного мозга у мышей MAID 1912НО 6032НО (DLC × UHC) наблюдались количества CD19+ В-клеток, аналогичные количествам у контрольных мышей VI3 (фиг. 44А-44В). Однако у мышей MAID 1912НО 6032НО (DLC × UHC) наблюдалось приблизительно в 25 раз меньше про-В-клеток в костном мозге по сравнению с мышами VI3 (фиг. 45А-45В). У мышей MAID 1912НО 6032НО (DLC × UHC) также наблюдались приблизительно в 2 раза меньше незрелых В-клеток и в 2 раза меньше зрелых В-клеток в костном мозге по сравнению с мышами VI3 (фиг. 46А-46В). Также у мышей MAID 1912НО 6032НО (DLC × UHC) наблюдалась предпочтительная экспрессия (2-кратное увеличение экспрессии) лямбда по сравнению с мышами VI3 (фиг. 47).
Исследования иммунизации
[000330] Пять мышей WT (происхождение: 75% C57BL6/25% 129) и семь мышей F2 MAID1912HO 6031 НЕТ (гомозиготная DLC × гетерозиготная UHC) иммунизировали через подушечки лап введением 0,025 мл смеси, содержащей 2,35 мкг антигена X, 10 мкг CpG-олигонуклеотида (ODN 1826, InvivoGen, кат. № tlrl-1826) и 25 мкг адъюванта - геля алюминия фосфата (Brenntag кат. №7784-30-7). Мыши получали бустерную иммунизацию шестикратно в той же дозировке. На 0, 15 и 23 день после первичной иммунизации собирали кровь оп: анестезированных мышей через ретроорбитальный синус в пробирки для отделения сыворотки BD (BD, кат. №365956), и получали сыворотку согласно указаниям производителя. Второй раунд иммунизации проводили согласно описанию выше через пять недель после первого раунда иммунизации.
[000331] Для измерения уровней антигенспецифических антител IgG, проводили анализ ИФА ELISA согласно примеру 3. Как видно из фиг. 48, генетически модифицированный мыши, гетерозиготные по целевому аллелю, содержащему реаранжированную нуклеотидную последовательность VH3-23/D/JH4, и гомозиготные по целевому аллелю, содержащему DLC-5J, были способны продуцировать антигенспецифические IgG-антитела на уровнях, сравнимых с уровнями, продуцируемыми мышами дикого типа, как на 23 день, так и через 5 недель после первичной иммунизации. Мыши MAID1912HO 6031НЕТ (гомозиготные DLC × гетерозиготные UHC) также были способны продуцировать антигенспецифические IgG-антитела на уровнях, сравнимых с уровнями, продуцируемыми мышами дикого типа после второго раунда иммунизации.
Соответственно, указанные мыши продуцируют антитела, содержащие обратно-гибридную легкую цепь (вариабельный домен легкой цепи человека и Cκ мыши), полученную в результате реаранжировки одного из двух сегментов гена VL человека (сегментов гена Vκ1-39 или Vκ3-20) и сегментов Jκ человека, и обратно-гибридную тяжелую цепь (вариабельный домен тяжелой цепи человека и CH мыши), происходящую из единственного реаранжированного сегмента гена вариабельной области тяжелой цепи человека. После иммунизации представляющим интерес антигеном получают обратно-гибридные антитела (т.е. антитела, состоящие из указанных обратно-гибридных цепей).
Пример 6. Получение и анализ мышей, у которых имеется единственная нуклеотидная последовательность реанжированной тяжелой цепи иммуноглобулина человека и два сегмента гена v каппа, содержащие три замены гистидина
[000332] Аналогичным образом, мышей, несущих сконструированный локус легкой цепи человека, содержащий гистидин-модифицированную двойную легкую цепь (например, мыши, у которых имеется два сегмента гена человека VL с гистидиновыми модификациями, описанными выше в настоящем документе), скрещивают с мышами, у которых эндогенный локус вариабельной области тяжелой цепи мыши заменен на локус универсальной тяжелой цепи человека (локус, содержащий единственный реаранжированный вариабельный домен тяжелой цепи человека согласно описанию выше в настоящем документе). Соответственно, указанные мыши продуцируют антитела, содержащие обратно-гибридную легкую цепь (вариабельный домен легкой цепи человека и Cκ мыши), полученную в результате реаранжировки одного из двух гистидин-модифицированных сегментов гена VL человека (сегментов гена Vκ1-39 или Vκ3-20) и сегментов Jκ человека, и обратно-гибридную тяжелую цепь (вариабельный домен тяжелой цепи человека и CH мыши), происходящую из единственного реаранжированного вариабельного домена тяжелой цепи человека. Обратно-гибридные антитела получают после иммунизации представляющим интерес антигеном. рН-зависимые антитела человека, продуцированные такими мышами, идентифицируют с применением способов выделения и скрининга антител, известных в данной области техники или описанных выше.
[000333] Идентифицируют нуклеотидные последовательности вариабельных областей легких и тяжелых цепей в В-клетках, экспрессирующих указанные антитела, и получают полностью человеческие легкие и тяжелые цепи путем соединения нуклеотидных последовательностей вариабельных областей легких и тяжелых цепей с нуклеотидными последовательностями CL и CH человека, соответственно. Представляющие интерес легкие цепи, например, легкие цепи, которые связываются с представляющим интерес антигеном (например, легкие цепи антител, которые также проявляют рН-зависимые антигенные свойства при использовании разнообразных известных в данной области техники анализов, например, анализа BIACORE™), коэкспрессируют в подходящей экспрессионной системе с тяжелыми цепями, происходящими из других антител, например, тяжелыми цепями, происходящими из антител, которые содержат легкие цепи, происходящие из того же сегмента гена VL, что и представляющая интерес легкая цепь (например, Vκ1-39 или Vκ3-20), и тестируют способность восстановленного антитела сохранять антигенсвязывающие и рН-зависимые антигенсвязывающие свойства.
Пример 7. Конструирование мышей, у которых имеется локус легкой цепи иммуноглобулина, содержащий реаранжированную последовательность VDJ тяжелой цепи
[000334] Мышей, у которых имеется нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи в локусе легкой цепи каппа (MAID6079; «мышь UHC-на-каппа») получали способами, аналогичными описанным выше для нацеливания на локус тяжелой цепи. Вкратце, у мыши «UHC-на-каппа» удаляли все эндогенные функциональные сегменты генов легкой цепи каппа вариабельной области Vκ и Jκ и заменяли на одну нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи, которая кодирует hVH3-23/D/JH4, функционально связанный с эндогенной нуклеотидной последовательностью константной области легкой цепи. Готовая направленная конструкция для получения геномного локуса, содержащего реаранжированную последовательность вариабельного домена тяжелой цепи человека, содержит, от 5'- к 3'-концу, (1) гомологичный 5'-фрагмент, содержащий приблизительно 22500 п.о. геномной последовательности мыши в направлении 5' от указанного эндогенного локуса легкой цепи Ig; (2) 5' FRT-сайт; (3) неомициновую кассету; (4) 3' FRT-сайт, (5) 2239 п.о. промотора hVH3-23 (SEQ ID NO: 139); (6) реаранжированную нуклеотидную последовательность тяжелой цепи иммуноглобулина человека (hVH3-23/D/JH4; SEQ ID NO: 136); (7) интрон hJH4 (SEQ ID NO: 140); и (8) гомологичный 3'-фрагмент, содержащий приблизительно 75000 п. о. геномной последовательности мыши в направлении 3' от сегментов гена JL мыши. Гетерозиготных мышей, несущих указанную модификацию, скрещивали друг с другом для получения гомозигот (MAID 6079НО), способных продуцировать «легкие» цепи иммуноглобулина, происходящие исключительно из генетически модифицированного локуса. У мышей MAID 6079НО (гомозиготных UHC-на-каппа) имеется вставка универсальной тяжелой цепи, описанной в настоящем документе (например, hVH3-23/hD/hJH4) в локусе легкой цепи каппа (к) мыши, в котором были удалены все гены Vκ и Jκ мыши.
[000335] Всех мышей размещали и разводили в специальных беспатогенных условиях в компании Regeneron Pharmaceuticals. Четыре мыши MAID 6079НО F1 (фиг. 49; возраст 7-12,5 недель, самцы и самки) и четыре однопометных контрольных мыши MAID 6079 FI дикого типа (возраст 7-12,5 недель, самцы и самки) умерщвляли, и собирали селезенки и костный мозг животных. Костный мозг собирали из бедренных костей, промывая полной средой RPMI (среда RPMI с добавлением фетальной телячьей сыворотки, пирувата натрия, Hepes, 2-меркаптоэтанола, заменимых аминокислот и гентамицина). Препараты эритроцитов из селезенки и костного мозга лизировали лизирующим буфером АСК и промывали полной средой RPMI.
Проточная цитометрии
[000336] Для исследования способности описанных в настоящем документе генетически модифицированных гомозиготных мышей «UHC-на-каппа» (MAID 6079НО) продуцировать антитела, происходящие из генетически модифицированного аллеля (например, из аллеля, который содержит единственную копию реаранжированного VH3-23/D/JH4 в локусе легкой цепи каппа), проводили анализ методом сортировки клеток с активированной флуоресценцией (FACS) согласно примеру 3.
[000337] У мышей MAID 6079НО в компартменте костного мозга наблюдались количества про- и пре-В-клеток, по существу совпадающие с наблюдаемыми у однопометных животных дикого типа (фиг. 50А-50В). С другой стороны, у них наблюдались меньшие количества незрелых и зрелых В-клеток в компартменте костного мозга по сравнению с однопометными животными дикого типа (фиг. 51А-51С). Фактически, у указанных мышей имелось в 2 раза меньше незрелых В-клеток и почти в 4 раза меньше зрелых В-клеток. У мышей MAID 6079НО использовались почти исключительно последовательности легких цепей лямбда в незрелых и зрелых В-клетках в костном мозге (Фиг. 52).
[000338] В компартменте селезенки у мышей MAID 6079НО наблюдалось меньше зрелых В-клеток по сравнению с однопометными животными дикого типа (фиг. 53А-53В). Аналогично наблюдаемому для костного мозга, у мышей MAID 6079НО использовались почти исключительно последовательности легких цепей лямбда в компартменте селезенки (фиг. 54А-54В). Также у них наблюдалось меньше незрелых клеток, увеличение количества В-клеток маргинальной зоны и уменьшение количества фолликулярных В-клеток по сравнению с однопометными животными дикого типа (фиг. 55).
Пример 8. Получение и анализ мышей, у которых имеется локус легкой цепи иммуноглобулина, содержащий реаранжированную последовательность VDJ тяжелой цепи и локус тяжелой цепи иммуноглобулина, содержащий последовательность вариабельного домена легкой цепи человека
[000339] Мышей, гомозиготных по нуклеотидной последовательности реаранжированной вариабельной области тяжелой цепи в локусе легкой цепи (MAID 6079НО; гомозиготная «мышь с UHC на цепи каппа»), получали согласно описанию выше. Указанных мышей скрещивали с мышами, гомозиготными (MAID 1994НО) по нуклеотидной последовательности вариабельной области легкой цепи каппа в локусе тяжелой цепи (мышь «каппа на тяжелой цепи» («КоН»)). У гомозиготных КоН мышей MAID 1994 имеется 40 генов Vκ человека и все гены Jκ человека, с длинной IGCR и с ADAM6 мыши, встроенными в локус константной области тяжелой цепи Ig мыши (т.е. удален локус тяжелой цепи Ig мыши)). КоН были описаны ранее; см., например, опубликованную через 18 месяцев с даты приоритета патентную заявку США 2012/0096572, включенную в настоящий документ посредством ссылки.
[000340] Всех мышей размещали и разводили в специальных беспатогенных условиях в компании Regeneron Pharmaceuticals. Две мыши VELOCIMMUNE® (MAID 1242НО 1640НО («VI3»); см. патент США №8502018, включенный в настоящий документ посредством ссылки) (возрастом 15 недель, самки, n=2; Происхождение: 28% C57/BL6, 13% 129 и 59% Balb/c), четыре мыши MAID 1994НО 6079НО F2 (фиг. 56; возраст 13-14 недель, самцы, n=2; Происхождение: 25% C57/BL6, 25% 129, и 50% Balb/c), и однопометных контрольных мышей MAID 6079 дикого типа умерщвляли, и собирали селезенки и костный мозг животных. Костный мозг собирали из бедренных костей, промывая полной средой RPMI (среда RPMI с добавлением фетальной телячьей сыворотки, пирувата натрия, Hepes, 2-меркаптоэтанола, заменимых аминокислот и гентамицина). Эритроциты селезенки и препараты костного мозга лизировали лизирующим буфером АСК и промывали полной средой RPMI.
Проточная цитометрии
[000341] Для исследования способности описанных в настоящем документе генетически модифицированных гомозиготных мышей «КоН × UHC на каппа» (MAID 1994НО 6079НО) продуцировать антитела, происходящие из генетически модифицированных аллелей (например, из аллеля, который содержит единственную копию реаранжированного VH3-23/D/JH4, и аллеля, который содержит нуклеотидную последовательность вариабельной области легкой цепи каппа в локусе тяжелой цепи), выполняли анализ с применением сортировки клеток с активированной флуоресценцией (FACS) согласно примеру 3.
[000342] У мышей MAID 1994НО 6079НО наблюдалась более низкая частота CD19+ и пре-В-клеток в компартменте костного мозга по сравнению с мышами VI3 (фиг. 57А). В частности, у мышей MAID 19940 6079НО наблюдались приблизительно в 2 раза меньшие количества CD19+ и пре-В-клеток в костном мозге по сравнению с мышами VI3 (фиг. 57В). Кроме того, у мышей MAID 1994НО 6079НО наблюдалось приблизительно в 3 раза меньше незрелых В-клеток в компартменте костного мозга по сравнению с мышами VI3 (фиг. 58А и 58В). Также было обнаружено, что в В-клетках мышей MAID 1994НО 6079НО по существу отсутствует экспрессия легкой цепи лямбда в костном мозге (фиг. 59).
[000343] У мышей MAID1994HO 6079НО наблюдалось меньшая частота В-клеток в компартменте селезенки. В частности, у мышей MAID 1994НО 6079НО имеется меньшее количество В-клеток селезенки (приблизительно в 2 раза) и зрелых В-клеток (приблизительно в 3 раза) по сравнению с мышами VI3 (фиг. 60А-60В. У них также наблюдается отсутствие экспрессии легкой цепь лямбда по сравнению с мышами VI3 (Фиг. 61).
[000344] В отношении развития периферических В-клеток в селезенке FACS-анализ показал, что в селезенке мышей MAID1994HO 6079НО чаще встречаются клетки в фазе Т1, чем у мышей VI3 (Фиг. 62).
[00248] Специалистам в данной области техники известны, или могут быть выявлены с применением не более чем рутинных экспериментов, многие эквиваленты конкретных вариантов реализации настоящего изобретения, описанных в настоящем документе. Такие эквиваленты охвачены приведенной ниже формулой изобретения.
[00249] Содержание всех непатентных документов, патентных заявок и патентов, цитируемых в тексте настоящей заявки, полностью включено посредством ссылки в настоящий документ.
[000345] Хотя настоящее изобретение было описано со ссылками на конкретные аспекты и варианты реализации, специалистам в данной области техники будет понятно, что могут быть реализованы различные изменения и эквиваленты без отступления от действительных существа и объема настоящего изобретения. Кроме того, различные модификации могут быть внесены в объективные объем и существо описанного изобретения для адаптации к конкретной ситуации, материалу, составу вещества, способу, этапу или этапам способа. Все такие модификации включены в объем прилагаемой формулы изобретения.
Реферат
Предложены не являющиеся человеком животные, например млекопитающие, например мыши или крысы, содержащие локус тяжелой цепи иммуноглобулина, который содержит нуклеотидную последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека. Указанная нуклеотидная последовательность реаранжированной вариабельной области тяжелой цепи иммуноглобулина человека может быть функционально связана с нуклеотидной последовательностью константной области легкой или тяжелой цепи. Также описаны генетически модифицированные не являющиеся человеком животные, у которых имеется локус легкой цепи иммуноглобулина, содержащий один или большее количество, но меньше, чем у дикого типа, сегментов гена вариабельной области легкой цепи иммуноглобулина человека, которые могут быть функционально связаны с нуклеотидной последовательностью константной области легкой цепи. Также предложены способы получения нуклеотидных последовательностей, которые кодируют вариабельные домены легких цепей иммуноглобулина, способные к связыванию антигена в отсутствие тяжелой цепи. 3 н. и 24 з.п. ф-лы, 107 ил., 8 табл., 8 пр.
Формула
Документы, цитированные в отчёте о поиске
Человеческое моноклональное антитело против ailim, костимулирующей молекулы передачи сигнала, и его фармацевтическое применение
Комментарии