Генетически модифицированные в отношении т-клеточного рецептора мыши - RU2661106C2
Код документа: RU2661106C2
Чертежи






































Описание
Ссылка на родственные заявки
Согласно настоящей заявке испрашивается приоритет в соответствии с предварительной заявкой на выдачу патента США №61/552582, поданной 28 октября 2011 года, предварительной заявкой на выдачу патента США №61/621198, поданной 6 апреля 2012 года, и предварительной заявкой на выдачу патента США №61/700908, поданной 14 сентября 2012 года, полное содержание которых включено в настоящий документ посредством ссылки.
Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится к генетически модифицированному не относящемуся к человеку животному, например, грызуну (например, мыши или крысе), которое содержит в своем геноме относящиеся к человеку или гуманизированные вариабельные генные локусы Т-клеточного рецептора (TCR) (например, вариабельные генные локусы TCR α и TCRβ и/или вариабельные генные локусы TCRδ и TCRγ), и экспрессирует относящиеся к человеку или гуманизированные полипептиды TCR (например, полипептиды TCRα и TCRβ и/или полипептиды TCRδ и TCRγ) из относящихся к человеку или гуманизированных вариабельных генных локусов TCR. Не относящееся к человеку животное с относящимися к человеку или гуманизированными вариабельными генными локусами TCR по настоящему изобретению содержит нереаранжированные генные сегменты вариабельной области TCR человека (например, сегменты V, D и/или J) на эндогенных не относящихся к человеку генных локусах TCR. Настоящее изобретение также относится к зародышам, тканям и клеткам (например, Т-клеткам), которые содержат относящиеся к человеку или гуманизированные вариабельные генные локусы TCR и экспрессируют относящиеся к человеку или гуманизированные полипептиды TCR. Также предусмотрены способы получения генетически модифицированного не относящегося к человеку животного, содержащего относящиеся к человеку или гуманизированные вариабельные генные локусы TCR; и способы применения не относящихся к человеку животных, зародышей, тканей и клеток, которые содержат относящиеся к человеку или гуманизированные вариабельные генные локусы TCR и экспрессируют относящиеся к человеку или гуманизированные полипептиды TCR из их локусов.
Предшествующий уровень техники настоящего изобретения
В приобретенном иммунном ответе чужеродные антигены распознаются рецепторными молекулами на В-лимфоцитах (например, иммуноглобулинами) и Т-лимфоцитах (например, Т-клеточным рецептором, или TCR). В то время как патогены в крови и внеклеточном пространстве распознаются антителами в ходе гуморального иммунного ответа, разрушение патогенов внутри клеток опосредуется Т-клетками в ходе клеточного иммунного ответа.
Т-клетки распознают и атакуют антигены, презентированные им в контексте главного комплекса гистосовместимости (МНС) на клеточной поверхности. Распознавание антигена опосредуется TCR, экспрессированными на поверхности Т-клеток. Два основных класса Т-клеток выполняют эту функцию: цитотоксические Т-клетки, которые экспрессируют белок клеточной поверхности CD8, и хелперные Т-клетки, которые экспрессируют белок клеточной поверхности CD4. Цитотоксические Т-клетки активируют каскады передачи сигнала, которые приводят к непосредственному разрушению клетки, презентирующей антиген (в контексте МНС I), тогда как хелперные Т-клетки дифференцируются в несколько классов, и их активация (примированная распознаванием антигена, презентированного в контексте МНС II) приводит к опосредованному макрофагами разрушению патогена и стимуляции продукции антител В-клетками.
Вследствие антигенной специфичности антител в настоящее время проводят обширные исследования антител в отношении их терапевтического потенциала против многочисленных нарушений у людей. Для создания антител, способных нейтрализовать мишени у людей, одновременно избегая активации иммунных ответов против таких антител, ученые сконцентрировали свои усилия на получении относящихся к человеку или гуманизированных иммуноглобулинов. Одним из путей получения гуманизированных антител in vivo является использование мыши VELOCIMMUNE®, гуманизированной мыши, содержащей (1) репертуар нереаранжированных сегментов V, D и J иммуноглобулина человека, функционально связанных друг с другом, и константную область мыши на эндогенном локусе тяжелой цепи иммуноглобулина мыши и (2) репертуар нереаранжированных сегментов Vκ и Jκ человека, функционально связанных друг с другом, и константную κ область мыши на эндогенном локусе κ легкой цепи иммуноглобулина мыши. В связи с этим, мыши VELOCIMMUNE® обеспечивают богатый источник реаранжированных вариабельных доменов антитела с высокой степенью разнообразия для применения в конструировании антител человека.
Аналогично антителу Т-клеточный рецептор содержит вариабельную область, кодируемую переаранжированными локусами (α и β локусами, или δ и γ локусами), содержащими сегменты V(D)J вариабельной области, и эта вариабельная область обеспечивает Т-клетке ее антигенсвязывающую специфичность. Также аналогично антителу специфичность TCR к его антигену может использоваться для разработки новых терапевтических средств. Таким образом, в настоящей области техники существует потребность в не относящихся к человеку животных (например, грызунах, например, крысах или мышах), которые содержат нереаранжированные генные сегменты вариабельной области Т-клетки человека, способные к реаранжировке для образования генов, которые кодируют вариабельные домены Т-клеточного рецептора человека, включающие в себя домены, которые являются родственными друг для друга, и включающие в себя домены, которые специфически связывают представляющий интерес антиген. Также существует потребность в не относящихся к человеку животных, которые содержат локусы вариабельной области Т-клетки, которые содержат консервативные гуманизированные сегменты, включающих в себя не относящихся к человеку животных, которые содержат нереаранжированные генные сегменты человека, которые могут реаранжироваться для образования генов вариабельной области Т-клеточного рецептора, которые связаны с не относящимися к человеку (эндогенными) генными последовательностями константной области Т-клеточного рецептора. Остается необходимость в не относящихся к человеку животных, которые способные создавать разнообразный репертуар вариабельных последовательностей Т-клеточного рецептора человека. Существует потребность в не относящихся к человеку животных, которые способны к реаранжировке большинства или всех функциональных сегментов вариабельной области Т-клеточного рецептора, в ответ на представляющий интерес антиген, для образования полипептидов Т-клеточного рецептора, которые содержат полностью человеческие вариабельные домены.
Краткое раскрытие настоящего изобретения
Предусмотрены не относящиеся к человеку животные, например, грызуны, содержащие не относящиеся к человеку клетки, которые экспрессируют гуманизированные молекулы, выполняющие функции в клеточном иммунном ответе. Также предусмотрены не относящиеся к человеку животные, которые содержат нереаранжированные вариабельные генные локусы TCR. Предусмотрены системы in vivo и in vitro, которые содержат гуманизированные клетки грызуна, причем клетки грызуна экспрессируют одну или несколько гуманизированных молекул иммунной системы. Также предусмотрены нереаранжированные гуманизированные локусы TCR грызуна, которые кодируют гуманизированные белки TCR.
Согласно одному аспекту в настоящем документе предусмотрено генетически модифицированное не относящееся к человеку животное (например, грызун, например, мышь или крыса), которое содержит в своем геноме (а) нереаранжированный вариабельный генный локус TCRα, содержащий по меньшей мере один сегмент Vα человека и по меньшей мере один сегмент Jα человека, функционально связанный с не относящейся к человеку (например, относящейся к грызуну, например, мыши или крысе) константной генной последовательностью TCRα, и/или (b) нереаранжированный вариабельный генный локус TCRβ, содержащий по меньшей мере один сегмент Vβ человека, по меньшей мере один сегмент Dβ человека и по меньшей мере один сегмент Ιβ человека, функционально связанный с не относящейся к человеку (например, относящейся к грызуну, например, мыши или крысе) константной генной последовательностью ΤCRβ.
Согласно одному варианту осуществления нереаранжированный вариабельный генный локус TCRα замещает эндогенный не относящийся к человеку (например, относящийся к грызуну) вариабельный генный локус TCRα на эндогенном вариабельном генном локусе TCRα. Согласно одному варианту осуществления нереаранжированный вариабельный генный локус ΤCRβ замещает эндогенный не относящийся к человеку (например, относящийся к грызуну) вариабельный генный локус ΤCRβ на эндогенном вариабельном генном локусе ΤCRβ. Согласно одному варианту осуществления эндогенные не относящиеся к человеку (например, относящиеся к грызуну) сегменты Vα и Jα являются неспособными к реаранжировке для образования реаранжированной последовательности Vα/Jα. Согласно одному варианту осуществления эндогенные не относящиеся к человеку (например, относящиеся к грызуну) сегменты Vβ, Dβ и Jβ являются неспособными к реаранжировке для образования реаранжированной последовательности Vβ/Dβ/Jβ. Согласно одному варианту осуществления не относящееся к человеку животное содержит такую делецию, что геном животного не содержит функциональный сегмент Vα и функциональный сегмент Jα. Согласно одному варианту осуществления не относящееся к человеку животное содержит такую делецию, что геном животного не содержит функциональный эндогенный сегмент Vβ, функциональный эндогенный сегмент Dβ и функциональный эндогенный сегмент Jβ. Согласно одному варианту осуществления животное содержит делецию всех функциональных эндогенных сегментов Vα и Jα. Согласно одному варианту осуществления грызун содержит делению всех функциональных эндогенных сегментов Vβ, Dβ и Jβ. Согласно некоторым вариантам осуществления сегменты Vα и Jα человека реаранжируются для образования реаранжированной последовательности Vα/Jα. Согласно некоторым вариантам осуществления сегменты Vβ, Dβ и Jβ человека реаранжируются для образования реаранжированной последовательности Vβ/Dβ/Jβ. Таким образом, согласно различным вариантам осуществления не относящееся к человеку животное (например, грызун) экспрессирует Т-клеточный рецептор, содержащий вариабельную область человека и не относящуюся к человеку (например, относящуюся к грызуну) константную область на поверхности Т-клетки.
Согласно некоторым аспектам Т-клетки не относящегося к человеку животного проходят развитие Т-клеток в тимусе для образования одинарных позитивных в отношении CD4 и CD8 Т-клеток. Согласно некоторым аспектам не относящееся к человеку животное содержит нормальное соотношение CD3+ Т-клеток селезенки к общему количеству спленоцитов. Согласно различным вариантам осуществления не относящееся к человеку животное производит популяцию центральных и эффекторных Т-клеток памяти в периферических органах.
Согласно одному варианту осуществления нереаранжированный вариабельный генный локус TCRα у описанного в настоящем документе не относящегося к человеку животного содержит 61 сегмент Jα человека и 8 сегментов Vα человека. Согласно другому варианту осуществления нереаранжированный вариабельный генный локус TCRα у не относящегося к человеку животного содержит полный репертуар сегментов Jα человека и полный репертуар сегментов Vα человека.
Согласно одному варианту осуществления нереаранжированный вариабельный генный локус ΤCRβ у описанного в настоящем документе не относящегося к человеку животного содержит 14 сегментов Jβ человека, 2 сегмента Dβ человека и 14 Vβ сегментов человека. Согласно другому варианту осуществления нереаранжированный вариабельный генный локус ΤCRβ у не относящегося к человеку животного содержит полный репертуар сегментов Jβ человека, полный репертуар сегментов Dβ человека и полный репертуар сегментов Vβ человека.
Согласно дополнительному варианту осуществления описанное в настоящем документе не относящееся к человеку животное (например, грызун) дополнительно содержит нуклеотидные последовательности вариабельных сегментов TCRα человека на гуманизированном локусе TCRα. Согласно одному варианту осуществления не относящееся к человеку животное (например, грызун) дополнительно содержит по меньшей мере один из сегментов Vδ, Dδ и Jδ человека, например, полный репертуар сегментов Vδ, Dδ и Jδ человека на гуманизированном локусе TCRα.
Согласно одному варианту осуществления не относящееся к человеку животное сохраняет эндогенный не относящийся к человеку локус TCRα и/или TCRβ, причем локус представляет собой нефункциональный локус.
Согласно одному варианту осуществления не относящееся к человеку животное представляет собой грызуна. Согласно одному варианту осуществления грызун выбран из мыши и крысы. Согласно одному варианту осуществления грызун представляет собой мышь.
Согласно одному аспекту настоящее изобретение относится к генетически модифицированной мыши, содержащей в своем геноме (а) нереаранжированный вариабельный генный локус TCRα, содержащий репертуар сегментов Jα человека и репертуар сегментов Vα человека, функционально связанный с не относящейся к человеку (например, относящейся к мыши или крысе) константной генной последовательностью TCRα, и/или (b) нереаранжированный вариабельный генный локус ΤCRβ, содержащий репертуар сегментов Jβ человека, репертуар сегментов Dβ человека и репертуар сегментов Vβ человека, функционально связанный с не относящейся к человеку (например, относящейся к мыши или крысе) константной генной последовательностью TCRβ. Согласно одному варианту осуществления мышь содержит полный репертуар сегментов Vα человека. Согласно одному варианту осуществления мышь содержит полный репертуар сегментов Vβ человека. Согласно одному варианту осуществления мышь содержит полный репертуар сегментов Vα человека и сегментов Jα человека. Согласно одному варианту осуществления мышь содержит полный репертуар сегментов Vα человека и сегментов Vβ человека. Согласно одному варианту осуществления мышь содержит полный репертуар сегментов Vα человека, сегментов Jα человека, сегментов Vβ человека, сегментов Dβ человека и сегментов Jβ человека.
Согласно одному варианту осуществления мышь содержит по меньшей мере один эндогенный сегмент Vα мыши и по меньшей мере один эндогенный сегмент Jα мыши, причем эндогенные сегменты являются неспособными к реаранжировке для образования реаранжированной последовательности Vα/Jα, а также содержит по меньшей мере один эндогенный сегмент Vβ мыши, по меньшей мере один эндогенный сегмент Dβ мыши и по меньшей мере один эндогенный сегмент Jβ мыши, причем эндогенные сегменты являются неспособными к реаранжировке для образования реаранжированной последовательности Vβ/Dβ/Jβ.
Согласно одному варианту осуществления нереаранжированный вариабельный генный локус TCRα, который содержит сегменты вариабельной области TCRα человека замещает вариабельный гены TCRα мыши на эндогенном вариабельном локусе TCRα мыши, и нереаранжированный вариабельный генный локус TCRβ, который содержит сегменты вариабельной области ΤCRβ человека замещает вариабельные гены TCRβ мыши на эндогенном вариабельном локусе TCRβ мыши.
Согласно одному варианту осуществления сегменты Vα и Jα человека реаранжируются для образования реаранжированной последовательности Vα/Jα человека, и сегменты Vβ, Dβ и Jβ человека реаранжируются для образования реаранжированной последовательности Vβ/Dβ/Jβ человека. Согласно одному варианту осуществления реаранжированная последовательность Vα/Jα человека функционально связана с последовательностью константной области TCRα мыши. Согласно одному варианту осуществления реаранжированная последовательность Vβ/Dβ/Jβ человека функционально связана с последовательностью константной области ΤCRβ мыши. Таким образом, согласно различным вариантам осуществления мышь экспрессирует Т-клеточный рецептор на поверхности Т-клетки, причем Т-клеточный рецептор содержит вариабельную область человека и константную область мыши.
Согласно одному варианту осуществления мышь дополнительно содержит репертуар сегментов вариабельной области TCRδ человека (например, сегменты Vδ, Jδ и Dδ человека) на гуманизированном локусе TCRα. Согласно одному варианту осуществления репертуар сегментов вариабельной области TCRδ человека представляет собой полный репертуар сегментов вариабельной области TCRδ человека. Согласно одному варианту осуществления сегменты вариабельной области TCRδ человека находятся на эндогенном локусе TCRα. Согласно одному варианту осуществления сегменты вариабельной области TCRδ человека замещают эндогенные сегменты вариабельной области TCRδ мыши.
Согласно одному варианту осуществления генетически модифицированная мышь экспрессирует Т-клеточный рецептор, содержащий вариабельную область человека и константную область мыши на поверхности Т-клетки. Согласно одному аспекту Т-клетки мыши проходят развитие Т-клеток в тимусе для образования одинарных позитивных в отношении CD4 и CD8 Т-клеток. Согласно одному аспекту мышь содержит нормальное соотношение CD3+ Т-клеток селезенки к общему количеству спленоцитов; согласно одному аспекту мышь производит популяцию центральных и эффекторных Т-клеток памяти к представляющему интерес антигену.
Также предусмотрены способы получения описанных в настоящем документе генетически модифицированных не относящихся к человеку животных (например, грызунов, например, мышей или крыс).
Согласно одному аспекту предусмотрен способ получения гуманизированного грызуна (например, мыши или крысы), предусматривающий замещение сегментов вариабельной области TCRα и ΤCRβ грызуна, но не константных генов грызуна, переаранжированными сегментами вариабельной области TCRα и ΤCRβ человека на эндогенных локусах TCR грызуна. Согласно одному варианту осуществления способ предусматривает замещение сегментов вариабельной области TCRα грызуна (Vα и/или Jα) сегментами вариабельной области TCRα человека (Vα и/или Jα), причем сегменты вариабельной области TCRα функционально связаны с не относящимся к человеку геном константной области TCR для образования гуманизированного локуса TCRα; и замещение сегменты вариабельной области ΤCRβ грызуна (Vβ и/или Dβ и/или Jβ) сегментам вариабельной области ΤCRβ человека (Vβ и/или Dβ и/или Jβ), причем сегменты вариабельной области ΤCRβ функционально связаны с не относящимся к геном константной области TCR человека для образования гуманизированного локуса TCRβ. Согласно одному варианту осуществления гуманизированный грызун представляет собой мышь, и зародышевая линия мыши содержит сегменты вариабельной области TCRα человека, функционально связанные с эндогенной константной последовательностью TCRα мыши на эндогенном локусе TCRα; и зародышевая линия мыши содержит сегменты вариабельной области ΤCRβ человека, функционально связанные с эндогенной константной последовательностью ΤCRβ мыши на эндогенном локусе TCRβ.
Согласно одному варианту осуществления в настоящем документе предусмотрен способ получения генетически модифицированного не относящегося к человеку животного (например, грызуна, например, мыши или крысы), которое экспрессирует Т-клеточный рецептор, содержащий относящуюся к человеку или гуманизированную вариабельную область и не относящуюся к человеку (например, относящуюся к грызуну) константную область на поверхности Т-клетки, предусматривающий: замещение у первого не относящегося к человеку животного эндогенного не относящегося к человеку вариабельного генного локуса TCRα нереаранжированным гуманизированным вариабельным генным локусом TCRα, содержащим по меньшей мере один сегмент Vα человека и по меньшей мере один сегмент Jα человека, причем гуманизированный вариабельный генный локус TCRα функционально связан с эндогенной не относящейся к человеку константной областью TCRα; замещение у второго не относящегося к человеку животного эндогенного не относящегося к человеку вариабельного генного локуса ΤCRβ нереаранжированным гуманизированным вариабельным генным локусом ΤCRβ, содержащим по меньшей мере один сегмент Vβ человека, по меньшей мере один сегмент Dβ человека и по меньшей мере один сегмент Jβ человека, причем гуманизированный вариабельный генный локус TCRβ функционально связан с эндогенной константной областью ΤCRβ; и скрещивание первого и второго не относящегося к человеку животного для получения не относящегося к человеку животного, которое экспрессирует Т-клеточный рецептор, содержащий относящуюся к человеку или гуманизированную вариабельную область и не относящуюся к человеку константную область.
Согласно одному варианту осуществления способа эндогенные не относящиеся к человеку (например, относящиеся к грызуну) сегменты Vα и Jα являются неспособными к реаранжировке для образования реаранжированной последовательности Vα/Jα, и эндогенные не относящиеся к человеку (например, относящиеся к грызуну) сегменты Vβ, Dβ и Jβ являются неспособными к реаранжировке для образования реаранжированной последовательности Vβ/Dβ/Jβ. Согласно одному варианту осуществления способа сегменты Vα и Jα человека реаранжируются для образования реаранжированной последовательности Vα/Jα, и сегменты Vβ, Dβ и Jβ человека реаранжируются для образования реаранжированной последовательности Vβ/Dβ/Jβ. Согласно одному варианту осуществления способа нереаранжированный гуманизированный вариабельный генный локус TCRα содержит 61 сегмент Jα человека и 8 сегментов Vα человека, и нереаранжированный гуманизированный вариабельный генный локус TCRβ содержит 14 сегментов Vβ человека, 2 сегмента Dβ человека и 14 сегментов Jβ человека. Согласно другому варианту осуществления способа нереаранжированный гуманизированный вариабельный генный локус TCRα содержит полный репертуар сегментов Jα человека и полный репертуар сегментов Vα человека, и нереаранжированный гуманизированный вариабельный генный локус TCRβ содержит полный репертуар сегментов Vβ человека, полный репертуар сегментов Dβ человека и полный репертуар сегментов Jβ человека.
Согласно одному аспекту способа Т-клетки не относящегося к человеку животного (например, грызуна) проходят развитие Т-клеток в тимусе для образования одинарных позитивных в отношении CD4 и CD8 Т-клеток. Согласно одному аспекту не относящееся к человеку животное (например, грызун) содержит нормальное соотношение CD3+ Т-клеток селезенки к общему количеству спленоцитов. Согласно одному аспекту не относящееся к человеку животное (например, грызун) производит популяцию центральных и эффекторных Т-клеток памяти к представляющему интерес антигену.
Согласно некоторым вариантам осуществления замещение описанного в настоящем документе эндогенного не относящегося к человеку вариабельного генного локуса TCRα проводят в единственной ES клетке и единственную ES клетку вводят в зародыш не относящегося к человеку (например, относящегося к грызуну, например, мыши или крысе) для получения генетически модифицированного не относящегося к человеку животного (т.е. первого не относящегося к человеку животного, например, первого грызуна); и замещение описанного в настоящем документе эндогенного не относящегося к человеку вариабельного генного локуса TCRβ проводят в единственной ES клетке и единственную ES клетку вводят в зародыш не относящегося к человеку (например, относящегося к грызуну, например, мыши или крысе) для получения генетически модифицированного не относящегося к человеку животного (т.е. второго не относящегося к человеку животного, например, второго грызуна). Согласно одному варианту осуществления первого грызуна и второго грызуна скрещивают для получения потомства, причем потомство содержит в своей зародышевой линии гуманизированный вариабельный локус TCRα и гуманизированный вариабельный локус TCRβ.
Согласно одному варианту осуществления способа не относящееся к человеку животное представляет собой грызуна, например, мышь. Таким образом, настоящее изобретение также относится к способу получения генетически модифицированной мыши.
Также в настоящем документе предусмотрены клетки, например, выделенные Т-клетки (например, цитотоксические Т-клетки, Т-хелперы, Т-клетки памяти и т.д.), полученные от описанных в настоящем документе не относящихся к человеку животных (например, грызунов, например, мышей или крыс). Также предусмотрены ткани и зародыши, полученные от описанных в настоящем документе не относящихся к человеку животных.
Согласно одному аспекту предусмотрен способ получения вариабельного домена TCR человека, предусматривающий генетическую модификацию грызуна, описанного в настоящем документе, для введения гуманизированного локуса TCRα и/или гуманизированного локуса ΤCRβ, содержание грызуна в условиях, достаточных для образования Т-клетки, причем Т-клетка экспрессирует вариабельный домен TCRα человека /или вариабельный домен ΤCRβ человека.
Согласно одному аспекту предусмотрен способ получения последовательности нуклеиновой кислоты, кодирующей вариабельный домен TCR человека, который связывает представляющий интерес эпитоп, предусматривающий воздействие на описанное в настоящем документе не относящееся к человеку животное представляющего интерес эпитопа, содержание не относящегося к человеку животного в условиях, достаточных для того, чтобы животное презентировало представляющий интерес эпитоп гуманизированному TCR животного, и идентификацию нуклеиновой кислоты животного, которая кодирует полипептид вариабельного домена TCR человека, который связывает представляющий интерес эпитоп.
Согласно одному аспекту предусмотрено применение описанного в настоящем документе не относящегося к человеку животного для получения гуманизированного рецептора TCR. Согласно одному аспекту предусмотрено применение описанного в настоящем документе не относящегося к человеку животного для получения вариабельного домена TCR человека. Согласно одному аспекту предусмотрено применение описанного в настоящем документе не относящегося к человеку животного для получения последовательности нуклеиновой кислоты, кодирующей вариабельный домен TCR человека.
Согласно одному аспекту предусмотрено применение последовательности нуклеиновой кислоты, кодирующей вариабельный домен TCR человека или его фрагмент, для получения антигенсвязывающего белка. Согласно одному варианту осуществления антигенсвязывающий белок содержит вариабельный домен TCR, содержащий вариабельный домен TCRα человека и/или вариабельный домен TCRβ человека, который связывает представляющий интерес антиген.
Согласно одному аспекту предусмотрено применение описанного в настоящем документе не относящегося к человеку животного для получения не относящейся к человеку клетки, которая экспрессирует на своей поверхности гуманизированный Т-клеточный рецептор.
Согласно одному аспекту предусмотрен гуманизированный Т-клеточный рецептор от описанного в настоящем документе не относящегося к человеку животного.
Согласно одному аспекту предусмотрена последовательность нуклеиновой кислоты, кодирующая вариабельный домен TCR человека или его фрагмент, полученная у описанного в настоящем документе не относящегося к человеку животного.
Любой из описанных в настоящем документе вариантов осуществления и аспектов может использоваться совместно друг с другом, если иное не указано или не очевидно из контекста. Другие варианты осуществления станут очевидными специалистам в настоящей области техники из обзора последующего подробного раскрытия. Последующее подробное раскрытие включает в себя иллюстративные представления различных вариантов осуществления настоящего изобретения, которые не ограничивают заявленное настоящее изобретение. Прилагаемые фигуры составляют часть настоящего описания изобретения и вместе с описанием служат исключительно для иллюстрации вариантов осуществления, а не для ограничения настоящего изобретения.
Краткое описание графических материалов
На фиг. 1 показано взаимодействие у мыши между молекулой TCR и молекулой МНС: на левой панели показана Т-клетка (сверху) мыши от гуманизированной в отношении TCR, содержащая Т-клеточный рецептор с вариабельными доменами TCR человека и константными доменами TCR мыши, которая распознает антиген (серый кружок), презентированный посредством МНС I класса антигенпрезентирующей клеткой (внизу); на правой панели показано то же для МНС II класса. Комплексы МНС I и МНС II показаны вместе со своими соответствующими корецепторами, CD8 и CD4. Относящиеся к мыши области показаны черным цветом и относящиеся к человеку области показаны белым цветом.
На фиг. 2 показана (без соблюдения масштаба) общая организация локуса TCRα мыши (верхняя панель, первый локус) и человека (верхняя панель, второй локус). На нижней панели показана стратегия замещения сегментов вариабельной области TCRα у мыши (закрашенные символы) сегментами вариабельной области TCRα человека (незакрашенные символы) на эндогенном локусе мыши на 14 хромосоме; гуманизированный локус TCRα с сегментами Vα и Jα человека показан с константной областью мыши и энхансером мыши; в показанном варианте осуществления локус TCRδ удален в ходе гуманизации.
На фиг. 3 показана (без соблюдения масштаба) поэтапная стратегия гуманизации локуса TCRα мыши, причем генные сегменты вариабельной области TCRα последовательно добавляют выше против хода транскрипции от исходного участка гуманизации подвергнутого делеции локуса мыши (MAID 1540). Последовательность мыши показана закрашенными символами; последовательность человека показана незакрашенными символами. MAID относится к идентификационному номеру (ID) модифицированного аллеля. TRAV = сегмент Vα TCR, TRAJ = сегмент Jα TCR (hTRAJ = TRAJ человека), TRAC = домен Cα TCR, TCRD = TCRδ.
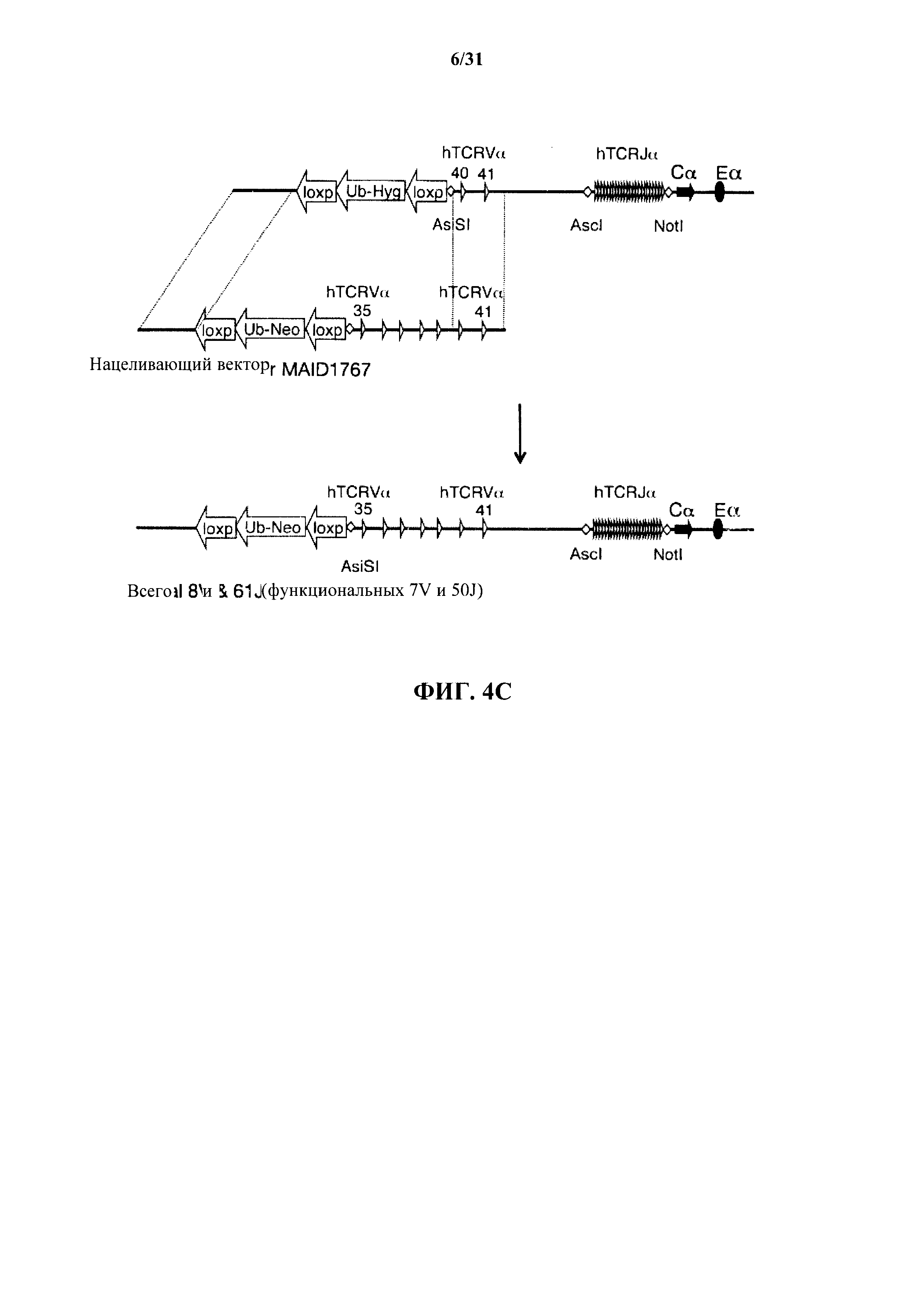
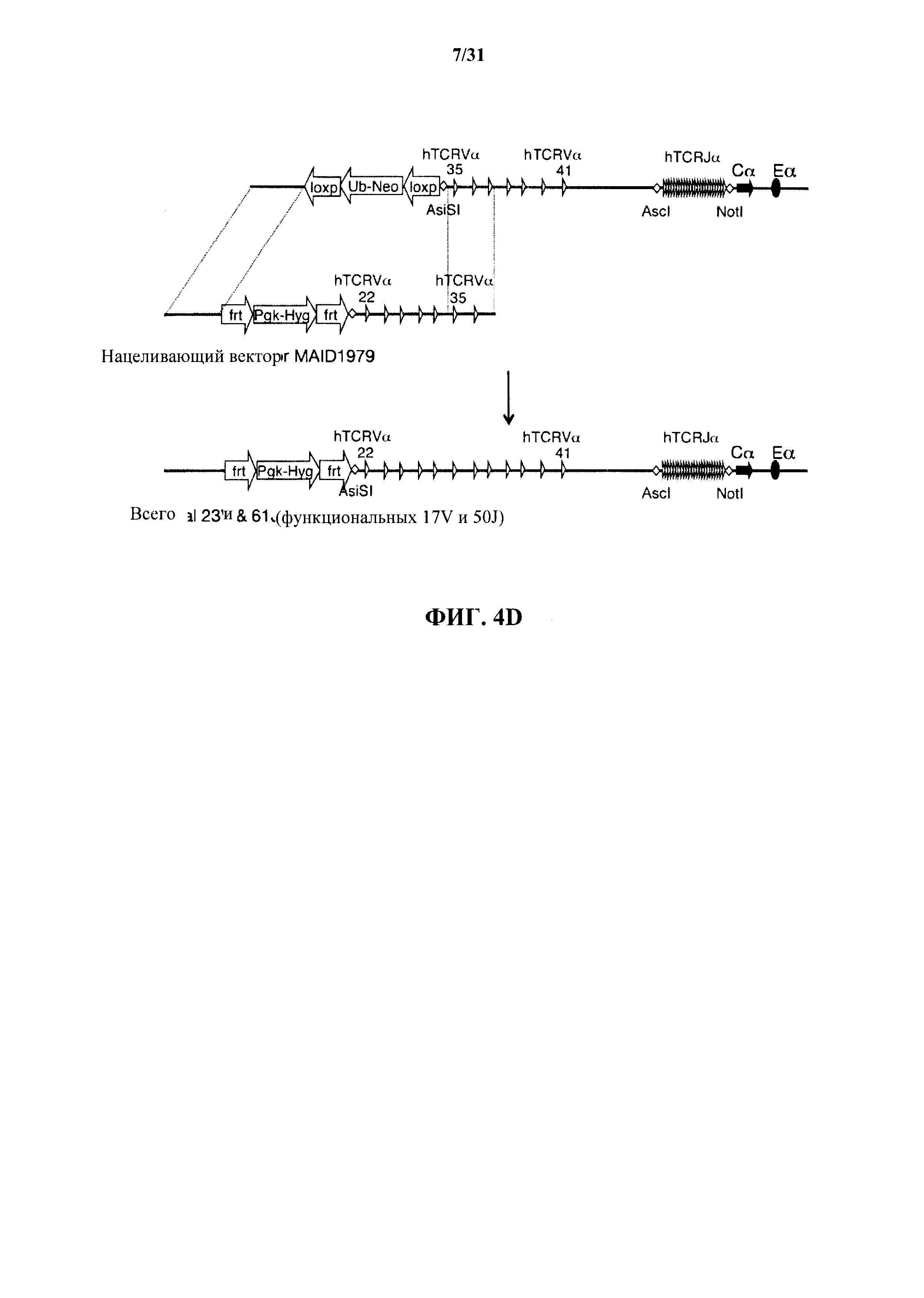
На фиг. 4 представляет собой подробное описание (без соблюдения масштаба) поэтапной стратегии гуманизации на локусе TCRα. На фиг. 4А показана делеция сегментов V и J TCRα мыши; на фиг. 4В показана стратегия вставки 2 сегментов V и 61 сегмента J человека в подвергнутый делеции локус TCRα мыши; на фиг. 4С показана стратегия вставки дополнительных сегментов V человека, дающей в результате в общем 8 сегментов V и 61 сегмент J человека; на фиг. 4D показана стратегия вставки дополнительных сегментов V человека, дающей в результате в общем 23 V и 61 J сегмент человека; фиг. 4Е показана стратегия вставки дополнительных сегментов V человека, дающая в результате 35 V и 61 J сегмент человека; фиг. 4F показана стратегия вставки дополнительных сегментов человека, дающая в результате 48 V и 61 J сегмент человека; и фиг. 4G показана стратегия вставки дополнительных сегментов человека, дающая в результате 54 V и 61 J сегмент человека. MAID относится к идентификационному номеру модифицированного аллеля.
На фиг. 5 показан (без соблюдения масштаба) один вариант осуществления стратегии гуманизации локуса TCRα мыши, при которой последовательности TCR□δ человека (TCRδ Vs, TCRδ Ds, TCRδ Js, TCRδ enh (энхансер) и константную последовательность TCRδ (С)) также замещают на гуманизированном локусе TCRα. Последовательность мыши показана закрашенными символами; последовательность человека показана незакрашенными символами. LTVEC относится к большому нацеливающему вектору; hTRD = TCRδ человека.
На фиг. 6 показана (без соблюдения масштаба) общая организация локусов TCRβ мыши (верхняя панель, первый локус; на 6 хромосоме мыши) и человека (верхняя панель, второй локус; на 7 хромосоме человека). На нижней панели показана стратегия замещения сегментов вариабельной области TCRβ у мыши (закрашенные символы) сегментами вариабельной области ΤCRβ человека (незакрашенные символы) на эндогенном локусе мыши на 6 хромосоме мыши. Гуманизированный локус ΤCRβ, содержащий сегменты Vβ, Dβ и Jβ человека, показан с константными областями мыши и энхансером мыши; согласно показанному варианту осуществления гуманизированный локус сохраняет гены трипсиногена мыши (сплошные прямоугольники); и согласно показанному конкретному варианту осуществления единственный сегмент V мыши сохраняют против хода транскрипции 5' от генов трипсиногена мыши.
На фиг. 7 показана (без соблюдения масштаба) поэтапная стратегия гуманизации локуса ΤCRβ мыши, причем генные сегменты вариабельной области TCRβ последовательно добавляют к подвергнутому делеции вариабельному локусу TCRβ мыши. Последовательность мыши показана закрашенными символами; последовательность человека показана незакрашенными символами. MAID относится к идентификационному номеру модифицированного аллеля. TRBV или TCRBV = сегмент V TCRβ.
На фиг. 8 показано подробное описание поэтапной стратегии гуманизации на локусе TCRβ. На фиг. 8А показана стратегия делеции сегментов V ΤCRβ мыши; на фиг. 8В показана стратегия вставки 14 сегментов V сегментов в подвергнутый делеции локус ΤCRβ; на фиг. 8С показана стратегия вставки 2 D и 14 J сегментов в локус ΤCRβ (i), с последующей делецией сайта loxP (ii), дающей в результате 14 V, 2 D и 14 J сегментов человека; на фиг. 8D показана стратегия вставки дополнительных сегментов V человека, дающая в результате 40 V, 2 D и 14 J сегментов человека; и на фиг. 8Е показана стратегия вставки дополнительных сегментов V человека, дающей в результате 66 V, 2 D и 14 J сегментов человека; на фиг. 8F показано замещение сегмента V мыши ниже по ходу транскрипции от энхансера мыши, дающее в результате 67 V, 2 D и 14 J сегментов человека. Согласно этому конкретному варианту осуществления один сегмент V мыши сохраняют 5' от генов трипсиногена мыши.
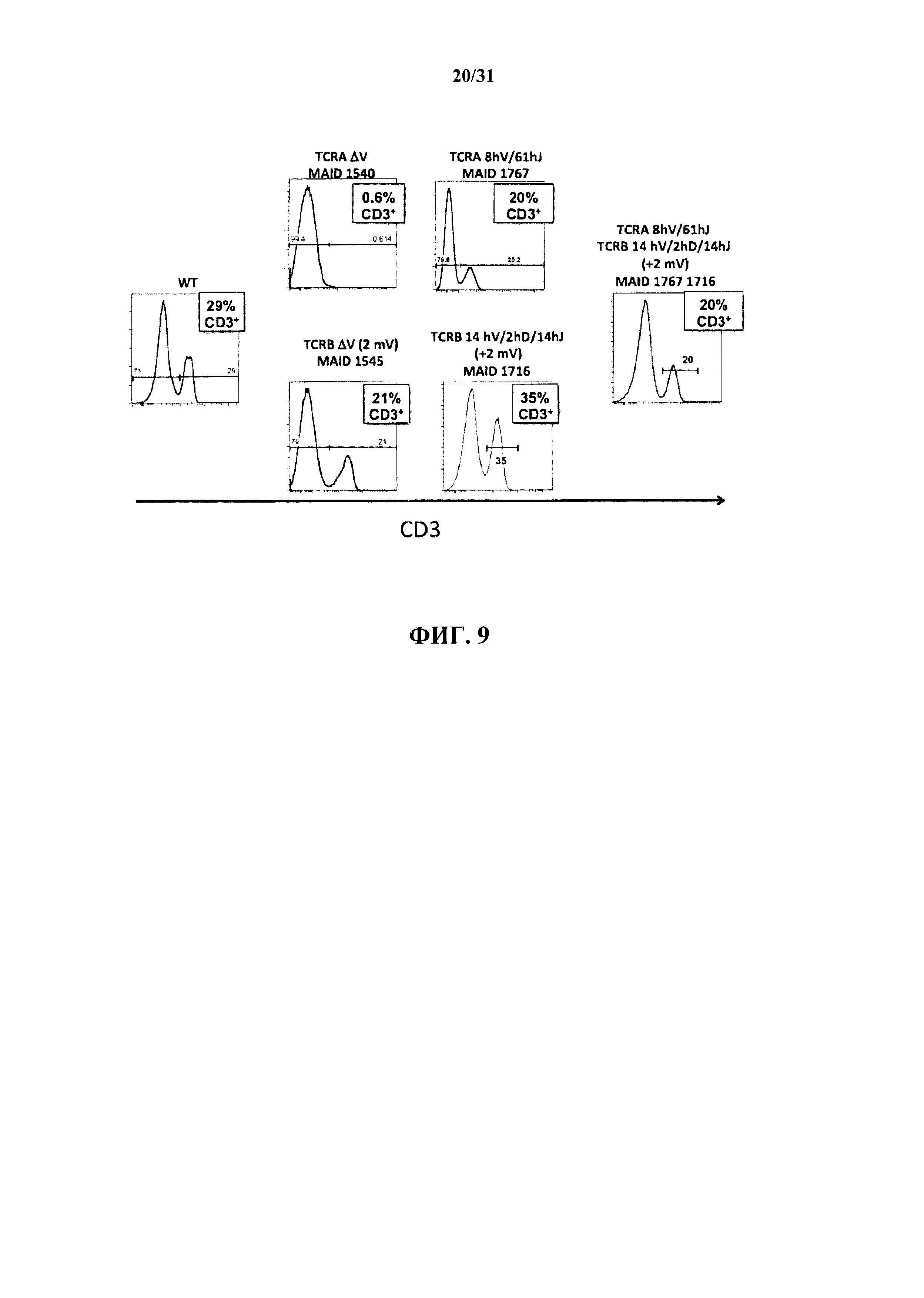
На фиг. 9 показана репрезентативные гистограммы анализа FACS для процентного содержания клеток селезенки (где ось Y представляет собой количество клеток, ось X представляет собой среднюю интенсивность флуоресценции, и гейт показывает частоту встречаемости CD3+ Т-клеток в пределах отдельной популяции лимфоцитов), окрашенных антителом к CD3 у мыши дикого типа(WT); мыши, гомозиготной в отношении подвергнутого делении локуса TCRα (первая верхняя панель; MAID 1540 из фиг. 3); мыши, гомозиготной в отношении подвергнутого делеции локуса TCRα и содержащей 8 сегментов Vα человека и 61 сегмент Jα человека (вторая верхняя панель; MAID 1767 из фиг. 3 или гуманизированной в отношении TCRα мыши); мыши, гомозиготной в отношении подвергнутого делеции локуса ΤCRβ за исключением одного вышележащего и одного нижележащего сегментов Vβ мыши (первая нижняя панель; MAID 1545 из фиг. 7); мыши, гомозиготной в отношении подвергнутого делеции локуса ΤCRβ с одним вышележащим и одним нижележащим сегментами Vβ мыши и содержащей 14 сегментов Vβ человека, 2 сегмента Dβ человека и 14 сегментов Jβ человека (вторая нижняя панель; MAID 1716 из фиг. 7 или гуманизированной в отношении TCRβ мыши); и мыши, гомозиготной в отношении делеций как локусов TCRα, так и локусов ΤCRβ (за исключением указанных двух сегментов Vβ мыши) и содержащей 8 сегментов Vα человека и 61 сегментов Jα человека на эндогенном локусе TCRα, а также 14 сегментов Vβ человека, 2 сегмента Dβ человека и 14 сегментов Jβ человека на эндогенных локусах ΤCRβ (MAID 1767/1716 или гуманизированной в отношении TCRα/β мыши).
На фиг. 10 представляет собой репрезентативный контурный график FACS клеток тимуса мыши от WT, гомозиготной гуманизированной TCRα (1767 НО; hTCRα); гомозиготной гуманизированной ΤCRβ (1716 НО; hΤCRβ); и гомозиготной гуманизированной TCRα/β мыши (1716 НО 1767 НО; hTCRα/β), окрашенных антителом к CD4 (ось Υ) и антителом к CD8 (ось X) (верхняя панель), и антителом к CD44 (ось Υ) и антителом к CD25 (ось X) (нижняя панель). График FACS на верхней панели позволяет различить двойные негативные (DN), двойные позитивные (DP), CD4 одинарные позитивные (CD4 SP), и CD8 одинарные позитивные (SP CD8) Т-клетки. График FACS на нижней панели позволяет различить различные стадии двойных негативных Т-клеток в ходе развития Т-клеток (DN1, DN2, DN3 и DN4). 1716 и 1767 относятся к номерам MAID, как показано на фигурах 3 и 7.
На фиг. 11 показана или частота встречаемости (верхняя панель), или абсолютное количество (нижняя панель) DN, DP, CD4 SP и CD SP Т-клеток в тимусе или мышей WT, или hTCRα (1767 НО); hTCRβ (1716 НО); или hTCRα/β (1716 НО 1767 НО) мышей (n=4).
На фиг. 12 представляет собой репрезентативный анализ FACS клеток селезенки мыши WT, hTCRα (1767 НО); hTCRβ (1716 НО); или hTCRα/β (1716 НО 1767 НО) мышей: на левой панели представлен анализ одинарных Т-клеток на основании окрашивания антителом к CD 19 (ось Y; окрашивание в отношении В-лимфоцитов) или антителом к CD3 (ось X; окрашивание в отношении Т-лимфоцитов); на средней панели представлен анализ CD3+ клеток на основании окрашивания антителом к CD4 (ось Y) или антителом к CD8 (ось X); и на правой панели представлен анализ или CD4+, или CD8+ клеток на основании окрашивания антителом к CD44 (ось Y) или антителом к CD62L (ось X), окрашивания позволяют различить различные типы Т-клеток в периферических органах (наивные Т-клетки в отличие от центральных Т-клеток памяти (Tcm) в отличие от эффекторных Т-клеток или эффекторных Т-клеток памяти (Teff/Tem)).
На фиг. 13 показано количество CD4+ (левая панель) или CD8+ (правая панель) Т-клеток на селезенку (оси Y) мышей WT, hTCRα (1767 НО); hTCRβ (1716 НО); или hTCRα/β (1716 НО 1767 НО) мышей (n=4).
На фиг. 14 показано количество наивных Т-клеток, Tcm и Teff/em-клеток на селезенку (оси Y) CD4+ (верхняя панель) или CD8+ (нижняя панель) Т-клеток мышей WT, hTCRα (1767 НО); hΙCRβ (1716 НО); или hTCRα/β (1716 НО 1767 НО) мышей (n=4).
На фиг. 15 представлены таблицы, обобщающие данные относительно экспрессии (определенной с помощью анализа FACS с использованием специфических к вариабельному сегменту антител) различных V сегментов ΤCRβ человека в CD8+ Т-клетках селезенки (фиг. 15А) или CD4+ Т-клетках селезенки (фиг. 15В) у мышей WT, hTCRβ (1716 НО) или hTCRα/β (1716 НО 1767 НО) мышей. Данные представлены как среднее ± SD (среднеквадратичное отклонение) (n=4 мыши на группу).
На фиг. 16 показана экспрессия мРНК (оси Υ) различных V сегментов ΤCRβ человека, присутствующих у мышей WT, hTCRα (1767 НО); hTCRβ (1716 НО); или hTCRα/β (1716 НО 1767 НО) мышей в Т-клетках тимуса или селезенки; на фиг. 16А представлен анализ экспрессии мРНК вариабельных сегментов ΤCRβ человека (hTRBV) 18, 19, 20 и 24; и на фиг. 16В представлен анализ экспрессии мРНК hTRBV 25, 27, 28 и 29.
На фиг. 17 показаны репрезентативные гистограммы FACS клеток селезенки (где ось Y представляет собой количество клеток, ось X представляет собой среднюю интенсивность флуоресценции, и гейт показывает частоту встречаемости CD3+ Т-клеток в пределах отдельной популяции лимфоцитов), окрашенных антителом к CD3 у мыши WT, мыши, гомозиготной в отношении подвергнутого делеции локуса TCRα (TCRA AV), мыши, гомозиготной в отношении подвергнутого делеции локуса TCRα с 2 сегментами V человека и 61 сегментом J человека (TCRA 2 hV; MAID 1626 из фиг. 3), мыши, гомозиготной в отношении подвергнутого делеции локуса TCRα с 8 сегментами V человека и 61 сегментом J человека (TCRA 8 hV; MAID 1767 из фиг. 3), и мыши, гомозиготной в отношении подвергнутого делеции локуса TCRα с 23 сегментами V человека и 61 сегментом J человека (TCRA 23 hV; MAID 1979 из фиг. 3).
На фиг. 18, на левой верхней панели, представлен репрезентативный анализ FACS CD3+ Т-клеток тимуса, полученных от либо мыши WT, либо гомозиготной hTCRα мыши с 23 сегментами V человека и 61 сегментом J человека (1979 НО), окрашенных либо с помощью антитела к CD4 (ось Y), либо с помощью антитела к CD8 (ось X); на левой нижней панели, представлен анализ FACS DN Т-клеток либо от мыши WT, либо от 1979 мыши, окрашенных либо с помощью антитела к CD44 (ось Y), либо с помощью антитела к CD25 (ось X); на правой панели представлены графики количества тимоцитов в % (ось Y), которые представляют собой DN, DP, CD4 SP или CD8 SP либо у мышей WT, либо у 1979 НО мышей (n=4).
На фиг. 19, на левой панели, представлен репрезентативный анализ FACS лимфоцитов селезенки от мыши WT или мыши 1979 НО, окрашенных либо антителом к CD 19, либо антителом к CD3; на правой панели, представлены графики количества спленоцитов в % (ось Y), полученных от мышей WT и мышей 1979 НО (n=4), которые являются CD3+.
Подробное раскрытие настоящего изобретения
Определения
Настоящее изобретение относится к генетически модифицированным не относящимся к человеку животным, например, грызунам, например, мышам или крысам, которые экспрессируют гуманизированные Т-клеточные рецепторы. Настоящее изобретение также относится к генетически модифицированным не относящимся к человеку животным, которые содержат в своей зародышевой линии нереаранжированные вариабельные генные локусы Т-клеточного рецептора. Также предусмотрены содержащие их зародыши, клетки и ткани; способы их получения; а также способы их применения. Если не указано иное, все используемые в настоящем документе термины и фразы включают в себя значения, которые подразумеваются под терминами и фразами в настоящей области техники, если противоположное ясно не указано или ясно не следует из контекста, в котором используется термин или фраза.
Термин "консервативная", используемый для описания консервативной аминокислотной замены, включает в себя замену аминокислотного остатка другим аминокислотным остатком с R-группой боковой цепи со сходными химическими свойствами (например, зарядом или гидрофобностью). Консервативные аминокислотные замены могут быть достигнуты путем модификации нуклеотидной последовательности так, чтобы ввести изменение нуклеотида, которое будет кодировать консервативную замену. Как правило, консервативная аминокислотная замена не будет существенно изменять представляющие интерес функциональные свойства белка, например, способность Т-клетки распознавать пептид, презентируемый молекулой МНС. Примеры групп аминокислот, которые содержат боковые цепи со сходными химическими свойствами, включают в себя такие алифатические боковые цепи глицин, аланин, валин, лейцин и изолейцин; такие алифатические гидроксильные боковые цепи серии и треонин; такие амидсодержащие боковые цепи, как аспарагин и глутамин; такие ароматические боковые цепи, как фенилаланин, тирозин и триптофан; такие основные боковые цепи, как лизин, аргинин и гистидин; такие кислотные боковые цепи, как аспарагиновая кислота и глутаминовая кислота; и такие серосодержащие боковые цепи, как цистеин и метионин. Группы консервативных аминокислотных замен включают в себя, например, валин/лейцин/изолейцин, фенилаланин/тирозин, лизин/аргинин, аланин/валин, глутамат/аспартат, и аспарагин/глутамин. Согласно некоторым вариантам осуществления консервативная аминокислотная замена может представлять собой замену любого нативного остатка в белке на аланин, что используется, например, в сканирующем аланином мутагенезе. Согласно некоторым вариантам осуществления проводят консервативную замену, которая характеризуется положительным значением в матрице логарифмического правдоподобия РАМ250, раскрытой в Gonnet et al. ((1992) Exhaustive Matching of the Entire Protein Sequence Database, Science 256: 1443-45), включенной в настоящий документ посредством ссылки. Согласно некоторым вариантам осуществления представляет собой умеренно консервативную замену, причем замена характеризуется неотрицательным значением в матрице логарифмического правдоподобия РАМ250.
Таким образом, настоящим изобретением предусмотрено генетически модифицированное не относящееся к человеку животное, экспрессирующее гуманизированные полипептиды TCRα и TCRβ (и/или гуманизированные полипептиды TCRδ и TCRγ), содержащие консервативные аминокислотные замены в описанной в настоящем документе аминокислотной последовательности.
Специалисту в настоящей области техники понятно, что в дополнение к остаткам нуклеиновой кислоты, кодирующим описанные в настоящем документе гуманизированные полипептиды TCRα и TCRβ, другие нуклеиновые кислоты, вследствие вырожденности генетического кода, могут кодировать полипептиды по настоящему изобретению. Следовательно, в дополнение к генетически модифицированному не относящемуся к человеку животному, которое содержит в своем геноме нуклеотидные последовательности, кодирующие описанные в настоящем документе гуманизированные полипептиды TCR, также предусмотрено не относящееся к человеку животное, которое содержит в своем геноме нуклеотидные последовательности, которые отличаются от описанных в настоящем документе нуклеотидных последовательностей вследствие вырожденности генетического кода.
Термин "идентичность" при использовании в связи с последовательностью включает в себя идентичность, определяемую с помощью набора различных известных в настоящей области техники алгоритмов, которые могут использоваться для измерения идентичности нуклеотидной и/или аминокислотной последовательности. Согласно некоторым описанным в настоящем документе вариантам осуществления идентичности определяют с использованием ClustalW v. 1.83 (медленного) выравнивания с использованием штрафа за открытие делеции, составляющего 10,0, штрафа за продление делеции, составляющего 0,1, и с использованием матрицы сравнения согласно Gonnet (MacVector™ 10.0.2, Mac Vector Inc., 2008). Длина последовательностей, сравниваемых в отношении идентичности последовательностей, будет зависеть от конкретных последовательностей. Согласно различным вариантам осуществления идентичность определяют путем сравнения последовательности зрелого белка в направлении от его N-конца к его С-концу. Согласно различным вариантам осуществления при сравнении химерной относящейся к человеку/не относящейся к человеку последовательности с последовательностью человека, относящаяся к человеку часть химерной относящейся к человеку/не относящейся к человеку последовательности (но не часть, не относящаяся к человеку) используется в осуществления сравнения с целью выяснения уровня идентичности между последовательностью человека и относящейся к человеку части химерной относящейся к человеку/не относящейся к человеку последовательности (например, сравнивая эктодомен человека химерного относящегося к человеку/мыши белка с относящимся к человеку эктодоменом белка человека).
Термины "гомология" или "гомологичный" в отношении последовательностей, например, нуклеотидных или аминокислотных последовательностей, означает две последовательности, которые при оптимальном выравнивании и сравнении являются идентичными по меньшей мере приблизительно в 75% нуклеотидов или аминокислот, по меньшей мере приблизительно в 80% нуклеотидов или аминокислот, по меньшей мере приблизительно в 90-95% нуклеотидов или аминокислот, например, больше чем в 97% нуклеотидов или аминокислот. Специалисту в настоящей области техники будет понятно, что для оптимального нацеленного воздействия на ген нацеливающая конструкция должна содержать плечи, гомологичные эндогенным последовательностям ДНК (т.е. "плечи гомологии"); таким образом, может происходить гомологичная рекомбинация между нацеливающей конструкцией и нацеленной эндогенной последовательностью.
Термин "функционально связанный" относится к смежному положению, причем описанные таким образом компоненты находятся во взаимодействии, позволяющем им функционировать предусмотренным для них образом. В связи с этим, кодирующая белок последовательность нуклеиновой кислоты может быть функционально связана с регуляторными последовательностями (например, последовательностью промотора, энхансера, сайленсера и т.д.) так, чтобы сохранять надлежащую транскрипционную регуляцию. Кроме того, различные части гуманизированного белка по настоящему изобретению могут быть функционально связаны для сохранения надлежащей укладки, процессинга, нацеленного воздействия, экспрессии и других функциональных свойств белка в клетке. Если не указано иное, различные домены гуманизированного белка по настоящему изобретению функционально связаны друг с другом.
Термин "замещение" в отношении замещения гена относится к размещению экзогенного генетического материала на эндогенном генетическом локусе, тем самым замещая весь эндогенный ген или его часть ортологичной или гомологичной последовательностью нуклеиновой кислоты. В одном случае эндогенный не относящийся к человеку ген или его фрагмент замещают соответствующим геном человека или его фрагментом. Соответствующий ген человека или его фрагмент представляет собой ген человека или фрагмент, которые представляет собой ортолог, гомолог или является по существу идентичным или одинаковым по структуре и/или функции по отношению к эндогенным не относящимся к человеку геном или его фрагментом, который замещают. Как показано в примерах ниже, нуклеотидные последовательности эндогенных не относящихся к человеку вариабельных генных локусов TCRα и β замещают нуклеотидными последовательностями, соответствующими вариабельным генным локусам TCRα и β человека.
Используемый в настоящем документе термин "функциональный", например, по отношению к функциональному белку, относится к белку, который сохраняет по меньшей мере одну биологическую активность, в норме связанную с нативным белком. Например, согласно некоторым вариантам осуществления настоящего изобретения замещение на эндогенном локусе (например, замещение на эндогенных не относящихся к человеку вариабельных генных локусах TCRα, TCRβ, TCRδ и/или TCRγ) дает в результате локус, который не способен экспрессировать функциональный эндогенный белок.
Используемые в настоящем документе локус TCR или генный локус TCR (например, локус TCRα или локус ΤCRβ) относятся к геномной ДНК, содержащей кодирующую область TCR, включающую в себя полную кодирующую область TCR, включающую в себя нереаранжированные последовательности V(D)J, энхансерную последовательность, константную(ые) последователыюсть(и) и любые вышележащие или нижележащие (UTR, регуляторные области и т.д.) или прерывающую последовательность ДНК (интроны и т.д.). Вариабельный локус TCR или вариабельный генный локус TCR (например, вариабельный генный локус TCRα или вариабельный генный локус ΤCRβ) относится к геномной ДНК, содержащей область, которая включает в себя сегменты вариабельной области TCR (области V(D)J), но не включает в себя константные последовательности TCR и, согласно различным вариантам осуществления, энхансерные последовательности. Другие последовательности могут быть включены в вариабельный генный локус TCR для целей генетической манипуляции (например, кассеты селекции, сайты рестрикции и т.д.), и они предусмотрены в настоящем документе.
Генетически модифицированные в отношении TCR животные
Согласно различным вариантам осуществления настоящее изобретение в основном относится к генетически модифицированным не относящимся к человеку животным, причем не относящиеся к человеку животные содержат в геноме нереаранжированные гуманизированные вариабельные генные локусы TCR.
Т-клетки связывают эпитопы на малых антигенных детерминантах на поверхности антигенпрезентирующих клеток, которые ассоциированы с главным комплексом гистосовместимости (МНС; у мышей) или комплексом лейкоцитарного антигена человека (HLA; у людей). Т-клетки связывают эти эпитопы через комплекс Т-клеточного рецептора (TCR) на поверхности Т-клетки. Т-клеточные рецепторы представляют собой гетеродимерные структуры, состоящие из двух типов цепей: α (альфа) и β (бета) цепи, или γ (гамма) и δ (дельта) цепи, α цепь кодируется последовательностью нуклеиновой кислоты, расположенной в пределах α локуса (на 14 хромосоме человека или мыши), которая также охватывает полный δ локус, и β цепь кодируется последовательностью нуклеиновой кислоты, расположенной в пределах β локуса (на 6 хромосоме мыши или 7 хромосоме человека). Большинство Т-клеток содержат αβ TCR; тогда как небольшое количество Т-клеток несут γδ TCR. Взаимодействия TCR с молекулами МНС I класса (осуществляющих презентацию CD8+ Т-клеткам) и молекулами МНС II класса (осуществляющих презентацию CD4+ Т-клеткам) показано на фиг. 1 (закрашенные символы представляют не относящиеся к человеку последовательности; незакрашенные символы представляют последовательности человека, демонстрируя один конкретный вариант осуществления белка TCR по настоящему изобретению).
α и β полипептиды Т-клеточного рецептора (и аналогично γ и δ полипептиды) связаны друг с другом посредством дисульфидной связи. Каждый из двух полипептидов, которые образуют TCR, содержит внеклеточный домен, содержащий константную и вариабельную области, трансмембранный домен и цитоплазматический хвост (трансмембранный домен и цитоплазматический хвост также являются частью константной области). Вариабельная область TCR определяет его антигенную специфичность, и аналогично иммуноглобулинам, содержит 3 определяющих комплементарность области (CDR). Также аналогично генам иммуноглобулина, вариабельные генные локусы Т-клеточного рецептора (например, локусы TCRα и ΤCRβ) содержат ряд нереаранжированных сегментов V(D)J (вариабельные (V), соединительные (J) сегменты, и в TCR β и δ, дополнительные (D) сегменты). В ходе развития Т-клеток в тимусе вариабельный генный локус TCRα подвергается реаранжировке, так что полученная α цепь TCR кодируется специфической комбинацией сегментов VJ (последовательностью Vα/Jα); и вариабельный генный локус ΤCRβ подвергается реаранжировке, так что полученная β цепь TCR кодируется специфической комбинацией сегментов VDJ сегментов (последовательностью Vβ/Dβ/Jβ).
Взаимодействия со стромой тимуса запускают прохождение тимоцитов через несколько стадий развития, характеризующихся экспрессией различных маркеров клеточной поверхности. Обобщение характерных маркеров клеточной поверхности на различных стадиях развития в тимусе представлена в таблице 1. Реаранжировка на вариабельном генном локусе ΤCRβ начинается на стадии DN2 и заканчивается в течение стадии DN4, тогда как реаранжировка вариабельного генного локуса TCRα происходит на стадии DP. После завершения реаранжировки локуса TCRβ клетки экспрессируют цепь ΤCRβ на клеточной поверхности вместе с α цепью-имитатором, рТα. Смотрите, Janeway's Immunobiology, Chapter 7, ранее.

Наивные CD4+ и CD8+ Т-клетки покидают тимус и проникают в периферические лимфоидные органы (например, селезенку), где на них воздействуют антигены, и они активируются для клонального размножения и дифференцировки в ряд эффекторных Т-клеток (Teff), например, цитотоксические Т-клетки, TREG клетки, ТН17 клетки, ТН1 клетки, ТН2 клетки и т.д. После инфекции ряд Т-клеток персистирует в качестве Т-клеток памяти, и их классифицируют либо как центральные Т-клетки памяти (Tcm), либо как эффекторные Т-клетки памяти (Tem). Sallusto et al. (1999) Two subsets of memory Т lymphocytes with distinet homing potentials and effector funetions, Nature 401: 708-12 и Commentary в Mackay (1999) Dual personality of memory Т cells, Nature 401: 659-60. Sallusto и коллеги предположили, что после начальной инфекции, Tem-клетки представляют легко доступный пул примированных антигеном Т-клеток памяти в периферических тканях с эффекторными функциями, тогда как Tcm-клетки представляют примированные антигеном Т-клетки памяти в периферических лимфоидных органах, которые при вторичной сенсибилизации могут становиться новыми эффекторными Т-клетками. Наряду с тем, что все Т-клетки памяти экспрессируют изоформу CD45RO CD45 (наивные Т-клетки экспрессируют изоформу CD45RA), Тст характеризуются экспрессией L-селектина (также известного как CD62L) и CCR7+, которые важны для связывания и передачи сигнала в периферических лимфоидных органах и лимфатических узлах (там же). Таким образом, все обнаруженные в периферических лимфоидных органах Т-клетки (например, наивные Т-клетки, Tcm-клетки и т.д.) экспрессируют CD62L. Известно, что кроме в дополнение к CD45RO, все Т-клетки памяти экспрессируют ряд различных маркеров клеточной поверхности, например, CD44. Для обобщения различных маркеров клеточной поверхности на Т-клетках, смотрите Janeway's Immunobiology, Chapter 10, ранее.
Наряду с тем, что вариабельный домен TCR функционирует в первую очередь в распознавании антигена, внеклеточная часть константного домена, а также трансмембранный и цитоплазматический домены TCR также выполняют важные функции. Для полного рецепторного комплекса TCR необходимы не только α и β или γ и δ полипептиды; необходимые дополнительные молекулы включают в себя CD3γ, CD3δ, и CD3ε, а также гомодимер ζ цепи (ζζ). При завершении реаранжировки TCRβ, когда клетки экспрессируют TCRβ/ρΤα, этот пре-TCR комплекс существует вместе с CD3 на клеточной поверхности. TCRα (или рТα) на клеточной поверхности содержит в своем трансмембранном домене два основных остатка, один из которых обеспечивает рекрутинг гетеродимера CD3γε, а другой обеспечивает рекрутинг ζζ через их соответствующие кислотные остатки. TCRβ в своем трансмембранном домене содержит дополнительный основный остаток, который, как полагают, обеспечивает рекрутинг гетеродимера CD3δε. Смотрите, например, Kuhns et al. (2006) Deconstructing the Form and Function of the TCR/CD3 Complex, Immunity 24:133-39; Wucherpfennig et al. (2009) Structural Biology of the T-cell Receptor: Insights into Receptor Assembly, Ligand Recognition, and Initiation of Signaling, Cold Spring Harb. Perspect. Biol. 2:a005140. Собранный комплекс, содержащей гетеродимер TCRαβ, CD3γε, CD3δε и ζζ, экспрессируется на Т-клеточной поверхности. Предполагали, что полярные остатки в трансмембранном домене служат в качестве контроля качества для выхода из эндоплазматического ретикулума; было показано, что при отсутствии субъединиц CD3, цепи TCR остаются в эндоплазматическом ретикулуме и направляются на разрушение. Смотрите, например, Call and Wucherpfennig (2005) The Τ Cell Receptor: Critical Role of the Membrane Environment in Receptor Assembly and Function, Annu. Rev. Immunol. 23: 101-25.
CD3 и ζ цепи собранного комплекса обеспечивают компоненты для передачи сигнала TCR, поскольку сам гетеродимер TCRαβ (или гетеродимер TCRγδ) не обладает активностью передачи сигнала. Каждая из CD3 цепей содержит один иммунорецепторный тирозиновый активирующий мотив (ITAM), тогда как ζ цепь содержит три тандемных ΙΤΑΜ. ΙΤΑΜ содержат тирозиновые остатки, способные фосфорилироваться ассоциированными киназами. Таким образом, собранный комплекс TCR-CD3 содержит 10 мотивов ΙΤΑΜ. Смотрите, например, Love and Hayes (2010) ITAM-Mediated Signaling by the T-Cell Antigen Receptor, Cold Spring Harb. Perspect. Biol. 2:e002485. После вовлечения TCR мотивы ΙΤΑΜ фосфорилируются тирозинкиназами Src семейства, Lck и Fyn, которые инициируют каскад передачи сигнала, приводящий к активации Ras, мобилизации кальция, реаранжировкам актинового цитоскелета и активации факторов транскрипции, что все в итоге приводит к Т-клеточной дифференциации, пролиферации и эффекторным действиям (там же), смотрите также, Janeway's Immunobiology, 7th Ed., Murphy et al. eds., Garland Science, 2008; обе включенные в настоящий документ посредством ссылки.
Кроме того, полагают, что трансмембранный и цитоплазматический домены TCRβ играют роль в митохондриальном нацеливании и индукции апоптоза; действительно, встречающиеся в природе процессированные по N-концу молекулы TCRβ существуют в тимоцитах. Shani et al. (2009) Incomplete T-cell receptor-β peptides target the mitochondrion and induce apoptosis, Blood 113: 3530-41. Таким образом, некоторые важные функции осуществляются константной областью TCR (которая, согласно различным вариантам осуществления, содержит часть внеклеточного домена, а также трансмембранный и цитоплазматический домены); и согласно различным вариантам осуществления структура этой области должна приниматься во внимание при конструировании гуманизированных TCR или генетически модифицированных не относящихся к человеку животных, их экспрессирующих.
В настоящей области техники известны мыши, трансгенные в отношении реаранжированных последовательностей Т-клеточного рецептора. Настоящее изобретение относится к генетически модифицированным не относящимся к человеку животным (например, грызунам, например, крысам, мышам), которые содержат нереаранжированные относящиеся к человеку или гуманизированные Т-клеточные вариабельные генные локусы, которые способны к реаранжировке для образования последовательностей нуклеиновой кислоты, которые кодируют вариабельные домены Т-клеточного рецептора человека, включающих в себя животных, которые содержат Т-клетки, которые содержат реаранжированные вариабельные домены человека и не относящиеся к человеку (например, относящиеся к мыши или крысе) константные области. Настоящее изобретение также относится к не относящимся к человеку животным (например, грызунам, например, крысам, мышам), которые способны создавать разнообразный репертуар последовательностей вариабельной области Т-клеточного рецептора человека; таким образом, настоящее изобретение относится к не относящимся к человеку животным, которые экспрессируют TCR с полностью человеческими вариабельными доменами в ответ на представляющий интерес антиген и которые связывают эпитоп представляющего интерес антигена. Согласно некоторым вариантам осуществления предусмотрены не относящиеся к человеку животные, которые создают разнообразный репертуар Т-клеточного рецептора, способный реагировать с различными антигенами, включающими в себя без ограничения презентируемые АПК антигены.
Согласно одному варианту осуществления настоящее изобретение относится к генетически модифицированным не относящимся к человеку животным (например, грызунам, например, крысам, мышам), которые содержат в своем геноме нереаранжированные сегменты вариабельной области TCR человека (сегменты V(D)J), причем нереаранжированные сегменты вариабельной области TCR человека замещают на эндогенном не относящемся к человеку (например, относящемся к грызуну) вариабельном генном локусе TCR (например, вариабельном генном локусе TCRα, β, δ, и/или γ), эндогенные не относящиеся к человеку сегменты вариабельной области TCR. Согласно одному варианту осуществления нереаранжированный вариабельный генный локус TCR человека замещает эндогенный не относящийся к человеку вариабельный генный локус TCR.
Согласно другому варианту осуществления настоящее изобретение относится к генетически модифицированным не относящимся к человеку животным (например, грызунам, например, крысам, мышам), которые содержат в своем геноме переаранжированные сегменты вариабельной области TCR человека (сегменты V(D)J), причем нереаранжированные сегменты вариабельной области TCR человека функционально связаны с не относящейся к человеку генной последовательностью константной области TCR, давая в результате гуманизированный локус TCR, причем гуманизированный локус TCR находится в положении в геноме, отличном от эндогенного не относящегося к человеку локуса TCR. Таким образом, согласно одному варианту осуществления также предусмотрено не относящееся к человеку животное (например, грызун, например, мышь, крыса), содержащее трансген, который содержит нереаранжированные сегменты вариабельной области TCR человека, функционально связанные с не относящейся к человеку последовательностью константной области TCR.
Согласно одному аспекту генетически модифицированные не относящиеся к человеку животные по настоящему изобретению содержат в своем геноме сегменты вариабельной области TCR человека при сохранении не относящихся к человеку (например, относящихся к грызуну, например, мыши, крысе) константных генных сегментов TCR. Согласно различным вариантам осуществления константные области включают в себя трансмембранный домен и цитоплазматический хвост TCR. Таким образом, согласно различным вариантам осуществления настоящего изобретения генетически модифицированные не относящиеся к человеку животные сохраняют эндогенный не относящийся к человеку трансмембранный домен TCR и цитоплазматический хвост. Согласно другим вариантам осуществления не относящиеся к человеку животные содержат не относящиеся к человеку не являющиеся эндогенными константные генные последовательности TCR, например, не относящийся к человеку не являющийся эндогенным трансмембранный домен и цитоплазматический хвост TCR. Как указано выше, константная область TCR принимает участие в каскаде передачи сигнала, который инициируется в ходе примированной антигеном активации Т-клетки; таким образом, эндогенная константная область TCR взаимодействует с разнообразными не относящимися к человеку якорными и сигнальными белками в Т-клетке. Таким образом, согласно одному аспекту генетически модифицированные не относящиеся к человеку животные по настоящему изобретению экспрессируют гуманизированные Т-клеточные рецепторы, которые сохраняют способность обеспечивать рекрутинг разнообразных эндогенных не относящихся к человеку якорных или сигнальных молекул, например, CD3 молекул (например, CD3γ, CD3δ, CD3ε), ζ цепи, Lck, Fyn, ZAP-70 и т.д. He ограничивающий перечень молекул, которые подвергаются рекрутингу в комплекс TCR, представлен в Janeway's Immunobiology, ранее. Кроме того, аналогично мышам VELOCIMMUNE®, которые проявляют нормальное развитие В-клеток и нормальные процессы клональной селекции, как полагают, вследствие по меньшей мере частичного замещения вариабельных областей на эндогенных локусах мыши и сохранения константных доменов мыши, согласно одному аспекту не относящиеся к человеку животные по настоящему изобретению проявляют нормальные процессы развития Т-клеток и Т-клеточной дифференциации.
Согласно некоторым вариантам осуществления предусмотрено не относящееся к человеку животное, которое содержит в своем геноме переаранжированные сегменты вариабельной области TCRα человека, причем переаранжированные сегменты вариабельной области TCRα человека функционально связаны с не относящейся к человеку генной последовательностью константной области TCRα, давая в результате гуманизированный локус TCRα. Согласно одному варианту осуществления гуманизированный локус TCRα находится в положении в геноме, отличном от эндогенного не относящегося к человеку локуса TCRα. Согласно другому варианту осуществления нереаранжированные сегменты вариабельной области TCRα человека замещают эндогенные не относящиеся к человеку сегменты вариабельной области TCRα при сохранении эндогенной не относящейся к человеку константной области TCRα. Согласно одному варианту осуществления нереаранжированный вариабельный генный локус TCRα человека замещает эндогенный не относящийся к человеку вариабельный генный локус TCRα. Согласно некоторым вариантам осуществления животное сохраняет эндогенные не относящиеся к человеку генные последовательности вариабельной области и константной области TCRβ. Таким образом, животное экспрессирует TCR, который содержит химерную относящуюся к человеку/не относящуюся к человеку (т.е. гуманизированную) цепь TCRα и не относящуюся к человеку цепь TCRβ.
Согласно другим вариантам осуществления предусмотрено не относящееся к человеку животное, которое содержит в своем геноме нереаранжированные сегменты вариабельной области TCRβ человека, причем нереаранжированные сегменты вариабельной области ΤCRβ человека функционально связаны с не относящейся к человеку генной последовательностью константной области TCRβ, давая в результате гуманизированный локус TCR-β. Согласно одному варианту осуществления гуманизированный локус ΤCRβ находится в положении в геноме, отличном от эндогенного не относящегося к человеку локуса TCRβ.Согласно другому варианту осуществления нереаранжированные сегменты вариабельной области ΤCRβ человека замещают эндогенные не относящиеся к человеку сегменты вариабельной области TCRβ при сохранении эндогенной не относящейся к человеку ΤCRβ константной области. Согласно одному варианту осуществления нереаранжированный вариабельный генный локус ΤCRβ человека замещает эндогенный не относящийся к человеку вариабельный генный локус TCRβ. Согласно некоторым вариантам осуществления животное сохраняет эндогенные не относящиеся к человеку генные последовательности вариабельной области и константной области TCRα. Таким образом, животное экспрессирует TCR, который содержит химерную относящуюся к человеку/не относящуюся к человеку (т.е. гуманизированную) цепь TCRβ и не относящуюся к человеку цепь TCRα.
Согласно некоторым конкретным вариантам осуществления настоящее изобретение относится к генетически модифицированному не относящемуся к человеку животному (например, грызуну, например, мыши или крысе), которое содержит в своем геноме (а) нереаранжированный вариабельный генный локус Т-клеточного рецептора (TCR) α, содержащий по меньшей мере один сегмент Vα человека и по меньшей мере один сегмент Jα человека, функционально связанный с эндогенными не относящимися к человеку (например, относящимися к грызуну, например, мыши или крысе) константными генными последовательностями TCRα, и/или (b) нереаранжированный вариабельный генный локус ΤCRβ, содержащий по меньшей мере один сегмент Vβ человека, по меньшей мере один сегмент Dβ человека и по меньшей мере один сегмент Jβ человека, функционально связанный с эндогенной не относящейся к человеку (например, относящейся к грызуну, например, мыши или крысе) константной генной последовательностью TCRβ.
Согласно различным вариантам осуществления настоящего изобретения нереаранжированные относящийся к человеку или гуманизированный вариабельный генный локус TCR (например, вариабельный генный локус TCRα и/или вариабельный генный локус TCRβ) содержится в зародышевой линии не относящегося к человеку животного (например, грызуна, например, мыши или крысы). Согласно различным вариантам осуществления замещения сегментов V(D)J TCR переаранжированными сегментами V(D)J TCR человека (например, сегментами Vα и Jα и/или Vβ и Dβ и Jβ) происходят на эндогенном не относящийся к человеку вариабельном локусе TCR (или локусах), причем переаранжированные сегменты V и J и/или V и D и J человека функционально связаны с не относящимися к человеку генами константной области TCR.
Согласно некоторым вариантам осуществления настоящего изобретения не относящееся к человеку животное содержит две копии нереаранжированного относящегося к человеку или гуманизированного вариабельного генного локуса TCRα и/или две копии нереаранжированного относящегося к человеку или гуманизированного вариабельного генного локуса TCRβ. Таким образом, не относящееся к человеку животное является гомозиготным в отношении одного или обоих нереаранжированных относящихся к человеку или гуманизированных вариабельных генных локусов TCRα и TCRβ. Согласно некоторым вариантам осуществления настоящего изобретения не относящееся к человеку животное содержит одну копию нереаранжированного относящегося к человеку или гуманизированного вариабельного генного локуса TCRα и/или одну копию нереаранжированного относящегося к человеку или гуманизированного вариабельного генного локуса TCRβ. Таким образом, не относящееся к человеку животное является гетерозиготным в отношении одного или обоих нереаранжированных относящихся к человеку или гуманизированных вариабельных генных локусов TCRα и TCRβ.
Согласно одному варианту осуществления нереаранжированный вариабельный генный локус TCRα, содержащий сегменты вариабельной области человека (например, сегменты Vα и Jα человека), расположен в не относящемся к человеку геноме так, что сегменты вариабельной области человека замещают соответствующие не относящиеся к человеку сегменты вариабельной области. Согласно одному варианту осуществления нереаранжированный вариабельный генный локус TCRα, содержащий сегменты вариабельной области человека, замещает эндогенный вариабельный генный локус TCRα. Согласно одному аспекту эндогенные не относящиеся к человеку сегменты Vα и Jα являются неспособными к реаранжировке для образования реаранжированной последовательности Vα/Jα. Таким образом, согласно одному аспекту сегменты Vα и Jα человека в нереаранжированном вариабельном генном локусе TCRα способны к реаранжировке для образования реаранжированной последовательности Vα/Jα человека.
Аналогично, согласно одному варианту осуществления нереаранжированный вариабельный генный локус TCRβ, содержащий сегменты вариабельной области человека (например, сегменты Vβ, Dβ и Jβ человека), расположен в не относящемся к человеку геноме так, что сегменты вариабельной области человека замещают соответствующие не относящиеся к человеку сегменты вариабельной области. Согласно одному варианту осуществления нереаранжированный вариабельный генный локус ΤCRβ, содержащий сегменты вариабельной области человека, замещает эндогенный вариабельный генный локус TCR-β. Согласно одному аспекту эндогенные не относящиеся к человеку сегменты Vβ, Dβ и Jβ являются неспособными к реаранжировке для образования реаранжированной последовательности Vβ/Dβ/Jβ. Таким образом, согласно одному аспекту сегменты Vβ, Dβ и Jβ человека в нереаранжированном вариабельном генном локусе ΤCRβ способны к реаранжировке для образования реаранжированной последовательности Vαβ/Dβ/Jβ человека.
Согласно другому варианту осуществления оба нереаранжированных вариабельных генных локуса TCRα и β, содержащие сегменты вариабельной области человека, замещают соответствующие эндогенные вариабельные генные локусы TCRα и β. Согласно одному аспекту эндогенные не относящиеся к человеку сегменты Vα и Jα являются неспособными к реаранжировке для образования реаранжированной последовательности Vα/Jα, и эндогенные не относящиеся к человеку сегменты Vβ, Dβ и Jβ являются неспособными к реаранжировке для образования реаранжированной последовательности Vβ/Dβ/Jβ. Таким образом, согласно одному аспекту сегменты Vα и Jα человека в нереаранжированном вариабельном генном локусе TCRα способны к реаранжировке для образования реаранжированной последовательности Vα/Jα человека, и сегменты Vβ, Οβ и Jβ человека в нереаранжированном вариабельном генном локусе TCRβ способны к реаранжировке для образования реаранжированной последовательности Vαβ/Dβ/Jβ человека.
Согласно некоторым аспектам настоящего изобретения не относящееся к человеку животное, содержащее гуманизированный генный локус TCRα и/или TCRβ (содержащий нереаранжированный вариабельный генный локус TCRα и/или ΤCRβ), сохраняет эндогенный не относящийся к человеку вариабельный генный локус TCRα и/или TCRβ. Согласно одному варианту осуществления эндогенный не относящийся к человеку TCRα и/или вариабельный генный локус TCRβ представляет собой нефункциональный локус. Согласно одному варианту осуществления нефункциональный локус представляет собой инактивированный локус, например, инвертированный локус (например, кодирующая последовательность нуклеиновой кислоты вариабельного генного локуса находится в инвертированной ориентации по отношению к последовательности константной области так, что никакие успешные реаранжировки не способны использовать сегменты вариабельной области из инвертированного локуса). Согласно одному варианту осуществления гуманизированный вариабельный генный локус TCRα и/или ΤCRβ расположен между эндогенным не относящимся к человеку вариабельным генным локусом TCRα и/или TCRβ и эндогенным не относящимся к человеку константным генным локусом TCRα и/или TCRβ.
Количества, номенклатура, положение, а также другие аспекты сегментов V и J и/или V, D и J локусов TCR человека и мыши могут быть установлены с использованием базы данных IMGT, доступной на www.imgt.org. Вариабельный локус TCRα мыши составляет приблизительно 1,5 м.п.н. и содержит в общем 110 Vα и 60 Jα сегментов (фиг. 2). Вариабельный локус TCRα человека составляет приблизительно 1 м.п.н. и содержит в общем 54 Vα и 61 Jα сегментов, причем полагают, что 45 Vα и 50 Jα сегментов являются функциональными. Если не указано иное, количества сегментов V(D)J человека, которые указаны в настоящем описании, относятся к общему количеству сегментов V(D).T. Согласно одному варианту осуществления настоящего изобретения генетически модифицированное не относящееся к человеку животное (например, грызун, например, мышь или крыса) содержит по меньшей мере один сегмент Vα человека и по меньшей мере один сегмент Jα человека. Согласно одному варианту осуществления не относящееся к человеку животное содержит гуманизированный локус TCRα, который содержит 1, 2, 3, 4, 5, 6, 7, 8. 9, 10, 15, 20, 23, 25, 30, 35, 40, 45, 48, 50 или до 54 сегментов Vα человека. Согласно некоторым вариантам осуществления гуманизированный локус TCRα содержит 2, 8, 23, 35, 48 или 54 сегмента Vα человека. Таким образом, согласно некоторым вариантам осуществления гуманизированный локус TCRα у не относящегося к человеку животного может содержать 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% 99% или 100% сегментов Vα человека; согласно некоторым вариантам осуществления он может содержать приблизительно 2%, приблизительно 3%, приблизительно 15%, приблизительно 65%, приблизительно 90% или 100% сегментов Vα человека.
Согласно одному варианту осуществления не относящееся к человеку животное содержит гуманизированный локус TCRα, который содержит фрагмент ДНК, содержащий смежную последовательность человека сегмента Vα40 - Vα41 человека (сегмент Vα также называется "TRAV" или "TCRAV"), и фрагмент ДНК, содержащий смежную последовательность человека 61 сегмента Jα человека (сегмент Jα также называется "TRAJ" или "TCRAJ"). Согласно одному варианту осуществления не относящееся к человеку животное содержит гуманизированный локус TCRα, который содержит фрагмент ДНК, содержащий смежную последовательность человека сегментов TRAV35 - TRAV41 человека, и фрагмент ДНК, содержащий смежную последовательность человека 61 сегмента TRAJ человека. Согласно одному варианту осуществления не относящееся к человеку животное содержит гуманизированный локус TCRα, который содержит фрагмент ДНК, содержащий смежную последовательность человека сегментов TRAV22 - TRAV41 человека, и фрагмент ДНК, содержащий смежную последовательность человека 61 сегмента TRAJ человека. Согласно одному варианту осуществления не относящееся к человеку животное содержит гуманизированный локус TCRα, который содержит фрагмент ДНК, содержащий смежную последовательность человека сегментов TRAV13-2 - TRAV41 человека, и фрагмент ДНК, содержащий смежную последовательность человека 61 сегмента TRAJ человека. Согласно одному варианту осуществления не относящееся к человеку животное содержит гуманизированный локус TCRα, который содержит фрагмент ДНК, содержащий смежную последовательность человека сегментов TRAV6 - TRAV41 человека и 61 сегмента TRAJ человека. Согласно одному варианту осуществления не относящееся к человеку животное содержит гуманизированный локус TCRα, который содержит фрагмент ДНК, содержащий смежную последовательность человека сегментов TRAV1-1 - TRAV 41 человека и 61 сегмента TRAJ человека. Согласно различным вариантам осуществления фрагменты ДНК, содержащие смежные последовательности человека сегментов вариабельной области TCRα человека, также содержат сайты фермента рестрикции, кассеты селекции, сайты эндонуклеаз или другие сайты, введенные для облегчения клонирования и селекции в течение процесса гуманизации локуса. Согласно различным вариантам осуществления эти дополнительные сайты не влияют на надлежащее функционирование (например, реаранжировку, сплайсинг и т.д.) различных генов на локусе TCRα.
Согласно одному варианту осуществления гуманизированный локус TCRα содержит 61 сегмент Jα человека или 100% сегментов Jα человека. Согласно конкретному варианту осуществления гуманизированный локус TCRα содержит 8 сегментов Vα человека и 61 сегмент Jα человека; согласно другому конкретному варианту осуществления гуманизированный локус TCRα содержит 23 сегмента Vα человека и 61 сегмент Jα человека. Согласно другому конкретному варианту осуществления гуманизированный TCRα локус содержит полный репертуар сегментов Vα и Jα человека, т.е. все генные сегменты вариабельной α области человека, кодируемые α локусом, или 54 сегмента Vα человека и 61 сегмент Jα человека. Согласно различным вариантам осуществления не относящееся к человеку животное не содержит любые эндогенные не относящиеся к человеку сегменты Vα или Jα на локусе TCRα.
Вариабельный локус TCRβ мыши составляет приблизительно 0,6 м.п.н. и содержит в общем 33 Vβ, 2 Dβ и 14 Jβ сегментов (фиг. 6). Вариабельный локус ΤCRβ человека составляет приблизительно 0,6 м.п.н. и содержит в общем 67 Vβ, 2 Dβ и 14 Jβ сегментов. Согласно одному варианту осуществления настоящего изобретения генетически модифицированное не относящееся к человеку животное (например, грызун, например, мышь или крыса) содержит по меньшей мере один сегмент Vβ человека, по меньшей мере один сегмент Dβ человека и по меньшей мере один сегмент Jα человека. Согласно одному варианту осуществления не относящееся к человеку животное содержит гуманизированный локус ΊΧΡVβ, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 23, 25, 30, 35, 40, 45, 48, 50, 55, 60 или до 67 сегментов Vβ человека. Согласно некоторым вариантам осуществления гуманизированный локус TCRβ содержит 8, 14, 40, 66 или 67 сегментов Vβ человека. Таким образом, согласно некоторым вариантам осуществления гуманизированный локус TCRβ у не относящегося к человеку животного может содержать 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% 99% или 100% Vβ человека; согласно некоторым вариантам осуществления он может содержать приблизительно 20%, приблизительно 60%), приблизительно 15%, приблизительно 98% или 100% Vβ человека.
Согласно одному варианту осуществления не относящееся к человеку животное содержит гуманизированный локус ΤΟΡVβ, который содержит фрагмент ДНК, содержащий смежную последовательность человека сегментов Vβ18 - Vβ29-1 человека (сегмент Vβ также называется "TRBV" или "TCRBV"). Согласно одному варианту осуществления не относящееся к человеку животное содержит гуманизированный локус ΤΟΡVβ, который содержит фрагмент ДНК, содержащий смежную последовательность человека сегментов TRBV18 - TRBV29-1 человека, отдельный фрагмент ДНК, содержащий смежную последовательность человека сегментов Dβ1-Jβ1 человека (т.е. сегментов Dβ1-Jβ1-1-Jβ1-6 человека), и отдельный фрагмент ДНК, содержащий смежную последовательность человека сегментов Dβ2-Jβ2 человека (т.е. сегментов Dβ2-Jβ2-1-Jβ2-7 человека). Согласно одному варианту осуществления не относящееся к человеку животное содержит гуманизированный локус ΤCRβ, который содержит фрагмент ДНК, содержащий смежную последовательность человека сегментов TRBV6-5 - TRBV29-1 человека, отдельный фрагмент ДНК, содержащий смежную последовательность человека сегментов ΟβΜβΙ человека (т.е. сегментов Dβ1-Jβ1-1-Jβ1-6 человека) и отдельный фрагмент ДНК, содержащий смежную последовательность человека сегментов Dβ2-Jβ2 человека (т.е. сегментов Dβ2-Jβ2-1-Jβ2-7 человека). Согласно одному варианту осуществления не относящееся к человеку животное содержит гуманизированный локус TCRβ, который содержит фрагмент ДНК, содержащий смежную последовательность человека сегментов TRBV1 - TRBV29-1 человека, отдельный фрагмент ДНК, содержащий смежную последовательность человека сегментов Dβ1-Jβ1 человека и отдельный фрагмент ДНК, содержащий смежную последовательность человека сегментов Dβ2-Jβ2 человека.
Согласно одному варианту осуществления не относящееся к человеку животное содержит гуманизированный локус TCRβ, который содержит фрагмент ДНК, содержащий смежную последовательность человека сегментов TRBV1 - TRBV29-1 человека, отдельный фрагмент ДНК, содержащий смежную последовательность человека сегментов Dβ1-Jβ1 человека, отдельный фрагмент ДНК, содержащий смежную последовательность человека сегментов Dβ2-Jβ2 человека, и отдельный фрагмент ДНК, содержащий последовательность сегмента TRBV30 человека. Согласно различным вариантам осуществления фрагменты ДНК, содержащие смежные последовательности человека сегментов вариабельной области ΤCRβ человека, также содержат сайты фермента рестрикции, кассеты селекции, сайты эндонуклеаз или другие сайты, введенные для облегчения клонирования и селекции в ходе процесса гуманизации локуса. Согласно различным вариантам осуществления эти дополнительные сайты не влияют на надлежащее функционирование (например, реаранжировку, сплайсинг и т.д.) различных генов на локусе TCRβ.
Согласно одному варианту осуществления гуманизированный локус TCRβ содержит 14 сегментов Jβ человека или 100% сегментов Jβ человека и 2 сегмента Dβ человека или 100% сегментов Jβ человека. Согласно другому варианту осуществления гуманизированный локус ΤCRβ содержит по меньшей мере один сегмент Vβ человека, например, 14 сегментов Vβ человека и все сегменты Dβ и Jβ мыши. Согласно конкретному варианту осуществления гуманизированный локус ΤCRβ содержит 14 сегментов Vβ человека, 2 сегмента Dβ человека и 14 сегментов Jβ человека. Согласно другому конкретному варианту осуществления гуманизированный локус ΤCRβ содержит полный репертуар сегментов Vβ, Dβ и Jβ человека, т.е. все генные сегменты вариабельной β области человека, кодируемые β локусом или 67 сегментов Vβ человека, 2 сегмента Dβ человека и 14 сегментов Jβ человека. Согласно одному варианту осуществления не относящееся к человеку животное содержит один (например, 5') не относящийся к человеку сегмент Vβ на гуманизированном локусе TCRβ. Согласно различным вариантам осуществления не относящееся к человеку животное не содержит любые эндогенные не относящиеся к человеку сегменты Vβ, Dβ или Jβ на локусе TCRβ.
Согласно различным вариантам осуществления, в которых не относящееся к человеку животное (например, грызун) содержит репертуар сегментов вариабельной области (например, полный репертуар сегментов вариабельной области) TCRα и ΤCRβ человека (и необязательно TCRδ и TCRγ человека), репертуар различных сегментов (например, полный репертуар различных сегментов) используется животным для создания разнообразного репертуара молекул TCR к различным антигенам.
Согласно различным аспектам не относящиеся к человеку животные содержат смежные части геномных вариабельных локусов TCR человека, которые содержат сегменты V, D и J, или D и J, или V и J, или V, расположенные, как в нереаранжированном геномном вариабельном локусе человека, например, включая в себя промоторные последовательности, лидерные последовательности, межгенные последовательности, регуляторные последовательности и т.д., расположенные, как в геномном вариабельном локусе TCR человека. Согласно другим аспектам различные сегменты расположены, как в нереаранжированном не относящемся к человеку геномном вариабельном локусе TCR. Согласно различным вариантам осуществления гуманизированного локуса TCRα и/или β, гуманизированный локус может содержать два или больше соединенных геномных сегментов человека, которые не наблюдаются в геноме человека, например, фрагмент сегментов V человека локуса V, расположенный в геноме человека вблизи константной области, соединенный с фрагментом сегментов V человека локуса V, расположенным в геноме человека на вышележащем конце локуса V человека.
Как у мыши, так и у человека генные сегменты TCRδ расположены вместе с локусом TCRα (смотрите фигуры 2 и 5). Сегменты J и D TCRδ расположены между сегментами Vα и Jα, тогда как сегменты V TCRδ рассеяны по локусу TCRα, причем большинство расположено среди различных сегментов Vα. Количество и положения различных сегментов TCRδ можно определить из базы данных IMGT. Вследствие геномного расположения генных сегментов TCRδ в пределах локуса TCRα, успешная реаранжировка на локусе TCRα, как правило, удаляет генные сегменты TCRδ.
Согласно некоторым вариантам осуществления настоящего изобретения не относящееся к человеку животное, содержащее нереаранжированный вариабельный генный локус TCRα человека, также содержит по меньшей мере один сегмент Vδ человека, например, вплоть до полного репертуара сегментов Vδ человека. Таким образом, согласно некоторым вариантам осуществления замещение эндогенного вариабельного генного локуса TCRα приводит к замещению по меньшей мере одного не относящегося к человеку сегмента Vδ сегментом Vδ человека. Согласно другим вариантам осуществления не относящееся к человеку животное по настоящему изобретению содержит полный репертуар сегментов Vδ, Dδ и Jδ человека на нереаранжированном гуманизированном локусе TCRα; согласно другим вариантам осуществления не относящееся к человеку животное содержит полный нереаранжированный локус TCRδ человека на нереаранжированном гуманизированном локусе TCRα (т.е. локус TCRδ, включающий в себя сегменты вариабельной области человека, а также энхансер и константную область человека). Иллюстративный вариант осуществления конструирования переаранжированного гуманизированного локуса TCRα, содержащего полный нереаранжированный локус TCRδ, показан на фиг. 5.
Согласно другому варианту осуществления не относящееся к человеку животное по настоящему изобретению дополнительно содержит нереаранжированный гуманизированный локус TCRγ, например, локус TCRγ, содержащий по меньшей мере один сегмент Vγ человека и по меньшей мере один сегмент Jγ человека (например, полный репертуар сегментов Vγ человека и Jγ человека вариабельной области). Локус TCRγ человека находится на 7 хромосоме человека, тогда как локус TCRγ мыши находится на 13 хромосоме мыши. Смотрите базу данных IMGT для подробной информации о локусе TCRγ.
Согласно одному аспекту не относящееся к человеку животное (например, грызун, например, мышь или крыса), содержащее описанные в настоящем документе гуманизированные вариабельные генные локусы TCRα и β (и необязательно гуманизированные вариабельные генные локусы TCRδ/γ), экспрессирует гуманизированный Т-клеточный рецептор, содержащий вариабельную область человека и не относящуюся к человеку (например, относящуюся к грызуну, например, мыши или крысе) константную область на поверхности Т-клетки, Согласно некоторым аспектам не относящееся к человеку животное способно экспрессировать или экспрессирует разнообразный репертуар гуманизированных Т-клеточных рецепторов, которые распознают разнообразные презентированные антигены.
Согласно различным вариантам осуществления настоящего изобретения, описанные в настоящем документе гуманизированные полипептиды Т-клеточного рецептора содержат лидерные последовательности человека. Согласно альтернативным вариантам осуществления гуманизированные TCR-рецепторные последовательности нуклеиновой кислоты сконструированы так, что гуманизированные полипептиды TCR содержат не относящиеся к человеку лидерные последовательности.
Описанные в настоящем документе гуманизированные полипептиды TCR могут эксирессироваться под контролем эндогенных не относящихся к человеку регуляторных элементов (например, регуляторных элементов грызуна), например, промотора, сайленсера, энхансера и т.д. Описанные в настоящем документе гуманизированные полипептиды TCR могут альтернативно экспрессироваться под контролем регуляторных элементов человека. Согласно различным вариантам осуществления описанные в настоящем документе не относящиеся к человеку животные дополнительно содержат все регуляторные и другие последовательности, встречающиеся в норме in situ в геноме человека.
Согласно различным вариантам осуществления вариабельная область человека гуманизированного белка TCR способна взаимодействовать с различными белками на поверхности той же клетки или другой клетки. Согласно одному варианту осуществления вариабельная область человека гуманизированного TCR взаимодействует с белками МНС (например, белками МНС I или II класса), презентирующими антигены на поверхности второй клетки, например, антигенпрезентирующей клетки (АПК). Согласно некоторым вариантам осуществления белок МНС I или II представляет собой не относящийся к человеку (например, относящийся к грызуну, например, мыши или крысе) белок. Согласно другим вариантам осуществления белок МНС I или II представляет собой белок человека. Согласно одному аспекту вторая клетка, например, АПК, представляет собой эндогенную не относящуюся к человеку клетку, экспрессирующую относящуюся к человеку или гуманизированную молекулу МНС. Согласно другому варианту осуществления вторая клетка представляет собой клетку человека, экспрессирующую молекулу МНС человека.
Согласно одному аспекту не относящееся к человеку животное экспрессирует гуманизированный Т-клеточный рецептор с не относящейся к человеку константной областью на поверхности Т-клетки, причем рецептор способен взаимодействовать с не относящимися к человеку молекулами, например, якорными или сигнальными молекулами, экспрессируемыми в Т-клетке (например, CD3 молекулами, ζ цепью или другими белками, прикрепленными к TCR посредством CD3 молекул или ζ цепи).
Таким образом, согласно одному аспекту предусмотрен клеточный комплекс, содержащий не относящуюся к человеку Т-клетку, которая экспрессирует TCR, который содержит описанную в настоящем документе гуманизированную цепь TCRα и описанную в настоящем документе гуманизированную цепь TCRβ, и не относящуюся к человеку антигенпрезентирующую клетку, содержащую антиген, связанный с МНС I или МНС II. Согласно одному варианту осуществления не относящиеся к человеку константные цепи TCRα и ΤCRβ образуют комплекс с не относящимися к человеку гомодимером дзета (ζ) цепей и гетеродимерами CD3. Согласно одному варианту осуществления клеточный комплекс представляет собой клеточный комплекс in vivo. Согласно одному варианту осуществления клеточный комплекс представляет собой клеточный комплекс in vitro.
Генетически модифицированное не относящееся к человеку животное может быть выбрано из группы, состоящей из мыши, крысы, кролика, свиньи, крупного рогатого скота (например, коровы, быка, буйвола), оленя, овцы, козы, курицы, кошки, собаки, хорька, примата (например, игрунки, макака-резуса). Для не относящихся к человеку животных, у которых подходящие генетически модифицируемые ES клетки не являются общедоступными, используются другие способы для получения не относящегося к человеку животного, содержащего генетическую модификацию. Такие способы включают в себя, например, модификацию генома не относящихся к ES клеток (например, фибробласта или индуцированной плюрипотентной клетки) и использование ядерного транспорта для переноса модифицированного генома в подходящую клетку, например, ооцит, и гестацию модифицированной клетки (например, модифицированного ооцита) в не относящемся к человеку животном при подходящих условиях для образования зародыша.
Согласно одному аспекту не относящееся к человеку животное представляет собой млекопитающее. Согласно одному аспекту не относящееся к человеку животное представляет собой небольшое млекопитающее, например, из надсемейства Dipodoidea или Muroidea. Согласно одному варианту осуществления генетически модифицированное животное представляет собой грызуна. Согласно одному варианту осуществления грызуна выбирают из мыши, крысы и хомяка. Согласно одному варианту осуществления грызуна выбирают из надсемейства Muroidea. Согласно одному варианту осуществления генетически модифицированное животное происходит из семейства, выбранного из Calomyscidae (например, мышевидные хомячки), Cricetidae (например, хомячок, крысы и мыши Нового Света, полевки), Muridae (настоящие мыши и крысы, песчанки, иглистые мыши, косматые хомяки), Nesomyidae (рипидомисы, скалистые хомячки, белохвостые крысы, магадаскарские крысы и мыши), Platacanthomyidae (например, колючие соневидные хомяки) и Spalacidae (например, слепыши, бамбуковые крысы и цокоры). Согласно конкретному варианту осуществления генетически модифицированного грызуна выбирают из настоящей мыши или крысы (семейство Muridae), карликовой песчанки, иглистой мыши и косматого хомяка. Согласно одному варианту осуществления генетически модифицированную мышь получают из представителя семейства Muridae. Согласно одному варианту осуществления животное представляет собой грызуна. Согласно конкретному варианту осуществления грызуна выбирают из мыши и крысы. Согласно одному варианту осуществления не относящееся к человеку животное представляет собой мышь.
Согласно конкретному варианту осуществления не относящееся к человеку животное представляет собой грызуна, который представляет собой мышь линии C57BL, выбранной из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/O1a. Согласно другому варианту осуществления мышь представляет собой мышь линии 129, выбранной из группы, состоящей из линии 129Р1, 129Р2, 129Р3, 129X1, 129S1 (например, 129S1/SV, 129S1/SvIm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129Т1, 129Т2 (смотрите, например, Festing et al. (1999) Revised nomenclature for strain 129 mice, Mammalian Genome 10:836, смотрите также, Auerbach et al (2000) Establishment and Chimera Analysis of 129/SvEv- and C57BL/6-Derived Mouse Embryonic Stem Cell Lines). Согласно конкретному варианту осуществления генетически модифицированная мышь представляет собой сочетание вышеупомянутой линии 129 и вышеупомянутой линии C57BL/6. Согласно другому конкретному варианту осуществления мышь представляет собой сочетание вышеупомянутых линий 129 или сочетание вышеупомянутых линий BL/6. Согласно конкретному варианту осуществления линия 129 сочетания представляет собой линию 129S6 (129/SvEvTac). Согласно другому варианту осуществления мышь представляет собой мышь линии BALB, например, линии BALB/c. Согласно другому варианту осуществления мышь представляет собой сочетание линии BALB и другой вышеупомянутой линии.
Согласно одному варианту осуществления не относящееся к человеку животное представляет собой крысу. Согласно одному варианту осуществления крысу выбирают из крысы линии Wistar, линии LEA, линии Sprague Dawley, линии Fischer, F344, F6 и Dark Agouti. Согласно одному варианту осуществления линия крысы представляет собой сочетание двух или больше линий, выбранных из группы, состоящей из Wistar, LEA, Sprague Dawley, Fischer, F344, F6 и Dark Agouti.
Таким образом, согласно одному варианту осуществления настоящее изобретение относится к генетически модифицированной мыши, содержащей в своем геноме нереаранжированный относящийся к человеку или гуманизированный вариабельный генный локус TCR, например, вариабельный генный локус TCRα, TCRβ, TCRδ и/или TCRγ. Согласно некоторым вариантам осуществления нереаранжированный относящийся к человеку или гуманизированный вариабельный генный локус TCR замещает эндогенный вариабельный генный локус TCR мыши. Согласно другим вариантам осуществления нереаранжированный относящийся к человеку или гуманизированный вариабельный генный локус TCR находится в положении в геноме, отличном от соответствующего эндогенного локуса TCR мыши. Согласно некоторым вариантам осуществления относящийся к человеку или гуманизированный нереаранжированный вариабельный генный локус TCR функционально связан с константной областью TCR мыши.
Согласно одному варианту осуществления предусмотрена генетически модифицированная мышь, причем мышь содержит в своем геноме нереаранжированный вариабельный генный локус Т-клеточного рецептора (TCR) а, содержащий по меньшей мере один сегмент Jα человека и по меньшей мере один сегмент Vα человека, функционально связанные с константной генной последовательностью TCRα мыши, и нереаранжированный вариабельный генный локус ΤCRβ, содержащий по меньшей мере один сегмент Jβ человека, по меньшей мере один сегмент Dβ человека и по меньшей мере один сегмент Vβ человека, функционально связанные с константной генной последовательностью ΤCRβ мыши. Согласно одному конкретному варианту осуществления мышь содержит в своем геноме нереаранжированный вариабельный генный локус TCRα, содержащий полный репертуар сегментов Jα человека и полный репертуар сегментов Vα человека, функционально связанных с константной генной последовательностью TCRα мыши, и нереаранжированный вариабельный генный локус TCRβ, содержащий полный репертуар сегментов Jβ человека, полный репертуар сегментов Dβ человека и полный репертуар сегментов Vβ человека, функционально связанных с константной генной последовательностью TCRβ мыши.
Согласно некоторым вариантам осуществления нереаранжированный вариабельный генный локус TCRα, содержащий TCRα сегменты вариабельной области человека, замещает эндогенный вариабельный генный локус TCRα мыши, и нереаранжированный вариабельный генный локус ΤCRβ, содержащий сегменты вариабельной области TCRβ человека, замещает эндогенный вариабельный генный локус TCRβ мыши. Согласно некоторым вариантам осуществления эндогенные сегменты Vα и Jα мыши являются неспособными к реаранжировке для образования реаранжированной последовательности Vα/Jα, и эндогенный сегменты Vβ, DP и Jβ мыши являются неспособными к реаранжировке для образования реаранжированной последовательности Vβ/Dβ/Jβ. Согласно некоторым вариантам осуществления сегменты Vα и Jα человека реаранжируются для образования реаранжированной последовательности Vα/Jα человека, и сегменты Vβ, Dβ и Jβ человека реаранжируются для образования реаранжированной последовательности Vβ/Dβ/Jβ человека.
Согласно различным вариантам осуществления описанные в настоящем документе не относящиеся к человеку животные (например, грызуны, например, мыши или крысы) производят Т-клетки, которые способны проходить развитие в тимусе, развиваясь от DN1 до DN2 до DN3 до DN4 до DP и до CD4 или CD8 SP Т-клеток. Такие Т-клетки не относящегося к человеку животного по настоящему изобретению экспрессируют молекулы клеточной поверхности, как правило, производимые Т-клеткой в течение конкретной стадии развития в тимусе (например, CD25, CD44, Kit, CD3, ρΤα и т.д.). Таким образом, описанные в настоящем документе не относящиеся к человеку животные экспрессируют ρΤα в комплексе с TCRβ на стадии DN3 развития в тимусе. Описанные в настоящем документе не относящиеся к человеку животные экспрессируют Т-клетки, способные проходить развитие в тимусе для получения CD4+ и CD8+ Т-клеток. В норме в тимусе физиологическое соотношение CD4+ к CD8+ Т-клеткам составляет приблизительно 2:1-3:1. Смотрите, например, Ge and Stanley (2008) The O-fucose glycan in the ligand-binding domain of Notch 1 regulates embryogenesis and Τ cell development, Proc. Natl. Acad. Sci. USA 105: 1539-44. Таким образом, согласно одному варианту осуществления описанные в настоящем документе не относящиеся к человеку животные производят CD4+ и CD8+ Т-клетки в тимусе в соотношении от приблизительно 2:1 до 3:1 (CD4+:CD8+).
Согласно различным вариантам осуществления описанные в настоящем документе не относящиеся к человеку животные производят Т-клетки, которые способны проходить нормальную Т-клеточную дифференциацию в периферических органах. Согласно некоторым вариантам осуществления описанные в настоящем документе не относящиеся к человеку животные способны производить нормальный репертуар эффекторных Т-клеток, например, CTL, (цитотоксические Т-лимфоциты), ТН1, ТН2, TREG, ТН17 и т.д. Таким образом, согласно этим вариантам осуществления описанные в настоящем документе не относящиеся к человеку животные производят эффекторные Т-клетки, которые выполняют различные функции, типичные для конкретного типа Т-клеток, например, распознают, связывают и отвечают на чужеродные антигены. Согласно различным вариантам осуществления описанные в настоящем документе не относящиеся к человеку животные производят эффекторные Т-клетки, которые лизируют клетки, проявляющие пептидные фрагменты цитозольных патогенов, экспрессированные в контексте молекул МНС I; распознают пептиды, происходящие из антигенов, разрушенных во внутриклеточных везикулах и презентированных молекулами МНС II на поверхности макрофагов и индуцируют макрофаги для лизиса микроорганизмов; производят цитокины, которые управляют В-клеточной дифференциацией; активируют В-клетки для получения опсонизирующих антител; индуцируют эпителиальные клетки для получения хемокинов, которые производят рекрутинг нейтрофилов в места инфекции; и т.д.
Согласно дополнительным вариантам осуществления описанные в настоящем документе не относящиеся к человеку животные содержат нормальное количество CD3+ Т-клеток в периферических органах, например, в селезенке. Согласно некоторым вариантам осуществления процентное отношение периферических CD3+ Т-клеток у описанных в настоящем документе не относящихся к человеку животных сопоставим с их процентным отношением у животных дикого типа (т.е. животных, содержащих все эндогенные сегменты вариабельной области TCR). Согласно одному варианту осуществления описанные в настоящем документе не относящиеся к человеку животные содержат нормальное соотношение CD3+ Т-клеток селезенки к общему количеству спленоцитов.
Согласно другим аспектам описанные в настоящем документе не относящиеся к человеку животные способны производить популяцию Т-клеток памяти в ответ на представляющий интерес антиген. Например, не относящиеся к человеку животные производят как центральные Т-клетки памяти (Tcm), так и эффекторные Т-клетки памяти (Tem) на антиген, например, представляющий интерес антиген (например, антиген, исследуемый для разработки вакцины и т.д.).
Клетки DN1 и DN2, которые не получают достаточных сигналов (например, сигналов Notch), могут развиваться в В-клетки, миелоидные клетки (например, дендритные клетки), тучные клетки и NK-клетки. Смотрите, например. Yashiro-Ohtani et al. (2010) Notch regulation of early thymocyte development, Seminars in Immunology 22: 261-69. Согласно некоторым вариантам осуществления у описанных в настоящем документе не относящихся к человеку животных развиваются нормальные количества В-клеток, миелоидных клеток (например, дендритных клеток), тучных клеток и NK-клеток. Согласно некоторым вариантам осуществления у описанных в настоящем документе не относящихся к человеку животных в тимусе развивается популяция нормальных дендритных клеток.
Преобладающий тип Т-клеточных рецепторов, экспрессируемых на поверхности Т-клеток, представляет собой TCRα/β, при этом меньшая часть клеток экспрессирует TCRδ/γ. Согласно некоторым вариантам осуществления настоящего изобретения Т-клетки не относящихся к человеку животных, содержащих гуманизированные локусы TCRα и/или β, проявляют нормальное использование локусов TCRα/β и TCRδ/γ, например, использование локусов TCRα/β и TCRδ/γ, которое аналогично использованию у животного дикого типа (например, Т-клеткам описанных в настоящем документе не относящихся к человеку животных экспрессируют белки TCRα/β и TCRδ/γ в сопоставимом соотношении к белкам, экспрессированным животными дикого типа). Таким образом, согласно некоторым вариантам осуществления не относящиеся к человеку животные содержащее гуманизированные TCRα/β и эндогенные не относящиеся к человеку локусы TCR5/γ, проявляют нормальное использование всех локусов.
В дополнение к генетически сконструированным описанным в настоящем документе не относящимся к человеку животным, также предусмотрен не относящийся к человеку зародыш (например, зародыш грызуна, например, зародыш мыши или крысы), причем зародыш содержит донорную ES клетку, которая получена от описанного в настоящем документе не относящегося к человеку животного (например, грызуна, например, мыши или крысы). Согласно одному аспекту зародыш содержит ES донорную клетку, которая содержит нереаранжированный гуманизированный локус TCR, и клетки зародыша-хозяина.
Также предусмотрена ткань, причем ткань получена от описанного в настоящем документе не относящегося к человеку животного (например, мыши или крысы), и экспрессирует гуманизированный полипептид TCR (например, полипептид TCRα и/или TCRβ, или TCRδ, и/или TCRγ).
Кроме того, предусмотрена не относящаяся к человеку клетка, выделенная из описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления клетка представляет собой ES клетку. Согласно одному варианту осуществления клетка представляет собой Т-клетку. Согласно одному варианту осуществления Т-клетка представляет собой CD4+ Т-клетку. Согласно другому варианту осуществления Т-клетка представляет собой CD8+ Т-клетку.
Также предусмотрена не относящаяся к человеку клетка, содержащая хромосому или ее фрагмент описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления не относящаяся к человеку клетка содержит ядро описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления не относящаяся к человеку клетка содержит хромосому или ее фрагмент как результат ядерного транспорта.
Также предусмотрена не относящаяся к человеку клетка, которая экспрессирует белок TCR, содержащий вариабельную область человека и не относящуюся к человеку константную область. Белок TCR может содержать TCRα, TCRβ или их комбинацию. Согласно одному варианту осуществления клетка представляет собой Т-клетку, например, CD4+ или CD8+ Т-клетку.
Согласно одному аспекту предусмотрена не относящаяся к человеку индуцированная плюрипотентная клетка, содержащая нереаранжированный гуманизированный локус TCR, кодирующий гуманизированный полипептид TCR, описанный в настоящем документе. Согласно одному варианту осуществления индуцированная плюрипотентная клетка получена от описанного в настоящем документе не относящегося к человеку животного.
Согласно одному аспекту предусмотрена гибридома или квадрома, происходящая из клетки описанного в настоящем документе не относящегося к человеку животного. Согласно одному варианту осуществления не относящееся к человеку животное представляет собой грызуна, например, мышь или крысу.
Также предусмотрен способ получения описанного в настоящем документе генетически модифицированного не относящегося к человеку животного (например, грызуна, например, мыши или крысы). Способ получения генетически модифицированного не относящегося к человеку животного дает в результате животное, чей геном содержит гуманизированный нереаранжированный локус TCR (например, гуманизированный нереаранжированный локус TCRα, TCRβ, TCRδ и/или TCRγ). Согласно одному варианту осуществления предусмотрен способ получения генетически модифицированного не относящегося к человеку животного (например, грызуна, например, мыши или крысы), которое экспрессирует Т-клеточный рецептор, содержащий вариабельную область человека и не относящуюся к человеку (например, относящуюся к грызуну, например, мыши или крысе) константную область на поверхности Т-клетки, причем способ предусматривает замещение у первого не относящегося к человеку животного эндогенного не относящегося к человеку вариабельного генного локуса TCRα нереаранжированный гуманизированным вариабельным генным локусом TCRα, содержащим по меньшей мере один сегмент Vα человека и по меньшей мере один сегмент Jα человека, причем гуманизированный вариабельный генный локус TCRα функционально связан с эндогенной константной областью TCRα; замещение у второго не относящегося к человеку животного эндогенного не относящегося к человеку вариабельного генного локуса TCRβ нереаранжированный гуманизированным вариабельным генным локусом TCRβ, содержащим по меньшей мере один сегмент Vβ человека, один сегмент Dβ человека и один сегмент Jβ человека, причем гуманизированный вариабельный генный локус ΤCRβ функционально связан с эндогенной константной областью ΤCRβ; и скрещивание первого и второго не относящегося к человеку животного для получения не относящегося к человеку животного, которое экспрессирует Т-клеточный рецептор, содержащий вариабельную область человека и не относящуюся к человеку константную область. Согласно другим вариантам осуществления настоящее изобретение относится к способам получения генетически модифицированного не относящегося к человеку животного, чей геном содержит гуманизированный нереаранжированный локус TCRα, или не относящегося к человеку животного, чей геном содержит гуманизированный нереаранжированный локус ΤCRβ, полученный согласно описанным в настоящем документе способам. Согласно различным вариантам осуществления замещения проводят на эндогенных локусах. Согласно некоторым вариантам осуществления в способе используют одну или несколько нацеливающих конструкций, полученных с использованием технологии VELOCIGENE®, введение конструкции(ий) в ES клетки и введение нацеленных ES клеточных клонов в зародыш мыши с использованием технология VELOCIMOUSE®, описанной в примерах. Согласно некоторым вариантам осуществления ES клетки получают от мыши, которая представляет собой сочетание линий 129 и C57BL/6. Согласно различным вариантам осуществления способ предусматривает поэтапную стратегию гуманизации, причем конструкцию, содержащую дополнительные сегменты вариабельной области, вводят в ES клетки на каждой последующей стадии гуманизации, в итоге получая мышь, содержащую полный репертуар сегментов вариабельной области человека (смотрите, например, фигуры 3 и 7).
Таким образом, также предусмотрены нуклеотидные конструкции, используемые для получения генетически сконструированных описанных в настоящем документе не относящихся к человеку животных. Согласно одному аспекту нуклеотидная конструкция содержит: 5' и 3' плечи гомологии, фрагмент ДНК человека, содержащий генный(е) сегмент(ы) вариабельной области TCR человека, и кассету селекции, фланкированную сайтами рекомбинации. Согласно одному варианту осуществления фрагмент ДНК человека представляет собой генный фрагмент TCRα и он содержит по меньшей мере один сегмент вариабельной области TCRα человека. Согласно другому варианту осуществления фрагмент ДНК человека представляет собой фрагмент ΤCRβ и он содержит по меньшей мере один генный сегмент вариабельной области TCRβ человека. Согласно одному аспекту по меньшей мере одно плечо гомологии представляет собой не относящееся к человеку плечо гомологии и оно гомологично не относящемуся к человеку локусу TCR (например, не относящемуся к человеку локусу TCRα или ΤCRβ).
Кассета селекции представляет собой нуклеотидную последовательность, вставленную в нацеливающую конструкцию для облегчения селекции клеток (например, ES клеток), которые интегрировали представляющую интерес конструкцию. В настоящей области техники известен ряд подходящих кассет селекции. Как правило, кассета селекции обеспечивает положительную селекцию в присутствии конкретного антибиотика (например, Neo, Hyg, Pur, CM, Spec и т.д.). Кроме того, кассета селекции может быть фланкирована сайтами рекомбинации, которые обеспечивают делецию кассеты селекции при обработке ферментами рекомбиназами. Широко используемые сайты рекомбинации представляют собой lοxΡ и Frt, распознаваемые ферментами Cre и Flp, соответственно, но в настоящей области техники известны и другие.
Согласно одному варианту осуществления кассета селекции расположена на 5'-конце фрагмента ДНК человека. Согласно другому варианту осуществления кассета селекции расположена на 3'-конце фрагмента ДНК человека. Согласно другому варианту осуществления кассета селекции расположена в пределах фрагмента ДНК человека, например, в пределах интрона человека. Согласно другому варианту осуществления кассета селекции расположена на стыке фрагмента ДНК человека и мыши.
Различные иллюстративные варианты осуществления стратегии нацеливания для получения генетически сконструированных не относящихся к человеку животных, конструкций и нацеливающих векторов, используемых для нее, представлены на фигурах 3, 4, 5, 7 и 8.
После завершения нацеленного воздействия на ген ES клетки или генетически модифицированных не относящихся к человеку животных подвергают скринингу для подтверждения успешного встраивания представляющей интерес экзогенной нуклеотидной последовательности или экспрессии экзогенного полипептида (например, сегментов вариабельной области TCR человека). Специалистам в настоящей области техники известны различные техники, и они включают в себя (без ограничения) саузерн-блоттинг, ПЦР длинных фрагментов, количественную ПЦР (например, ПЦР в реальном времени с использованием TAQMAN®), флуоресцентную гибридизацию in situ, нозерн-блоттинг, проточную цитометрию, вестерн-блоттинг, иммуноцитохимию, иммуногистохимию и т.д. Согласно одному примеру не относящиеся к человеку животные (например, мыши), несущие представляющую интерес генетическую модификацию, могут быть идентифицированы путем скрининга в отношении потери аллеля мыши и/или приобретения аллеля человека с использованием модификации аллельного анализа, описанного в Vαlenzuela et al. (2003) High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nature Biotech. 21(6): 652-659. Специалистам в настоящей области техники известны другие анализы, которые идентифицируют конкретную нуклеотидную или аминокислотную последовательность у генетически модифицированных животных.
Настоящее раскрытие также относится к способу модификации вариабельного генного локуса TCR (например, генного локуса TCRα, TCRβ, TCRδ и/или TCRγ) не относящегося к человеку животного для экспрессии описанного в настоящем документе гуманизированного белка TCR. Согласно одному варианту осуществления настоящее изобретение относится к способу модификации вариабельного генного локуса TCR для экспрессии гуманизированного белка TCR на поверхности Т-клетки, причем способ предусматривает замещение у не относящегося к человеку животного эндогенного не относящегося к человеку вариабельного генного локуса TCR нереаранжированным гуманизированным вариабельным генным локусом TCR. Согласно одному варианту осуществления, в котором вариабельный генный локус TCR представляет собой вариабельный генный локус TCRα, нереаранжированный гуманизированный вариабельный генный локус TCR содержит по меньшей мере один сегмент Vα человека и по меньшей мере один сегмент Jα человека. Согласно одному варианту осуществления, в котором вариабельный генный локус TCR представляет собой вариабельный генный локус ΤCRβ, нереаранжированный гуманизированный вариабельный генный локус TCR содержит по меньшей мере один сегмент Vβ человека, по меньшей мере один сегмент Dβ человека и по меньшей мере один сегмент Jβ человека. Согласно различным аспектам нереаранжированный гуманизированный вариабельный генный локус TCR функционально связан с соответствующей эндогенной не относящейся к человеку константной областью TCR.
Также предусмотрен гуманизированный белок TCR, производимый описанным в настоящем документе не относящимся к человеку животным (например, грызуном, например, мышью или крысой), причем гуманизированный белок TCR содержит вариабельную область человека и не относящуюся к человеку константную область. Таким образом, гуманизированный белок TCR содержит определяющие комплементарность области человека (т.е. CDR1, 2 и 3 человека) в своем вариабельном домене и не относящуюся к человеку константную область.
Несмотря на то, что в представленных ниже примерах описано генетически сконструированное не относящееся к человеку животное, чей геном содержит гуманизированный вариабельный генный локус TCRα и/или гуманизированный вариабельный генный локус TCRβ, специалисту в настоящей области техники будет понятно, что аналогичная стратегия может использоваться для получения генетически сконструированных животных, чей геном содержит гуманизированный вариабельный генный локус TCRδ и/или TCRγ. Также предусмотрено генетически сконструированное не относящееся к человеку животное, у которого все четыре вариабельных генных локуса TCR гуманизированы.
Применение генетически модифицированных в отношении TCR животных
Согласно различным вариантам осуществления генетически модифицированные не относящиеся к человеку животные по настоящему изобретению производят Т-клетки с гуманизированными молекулами TCR на своей поверхности и в результате этого будут распознавать пептиды, презентированные им комплексами МНС, характерным для человека образом. Описанные в настоящем документе генетически модифицированные не относящиеся к человеку животные могут использоваться для исследования развития и функции Т-клеток человека и процессов иммунологической толерантности; для испытания кандидатных вакцин для человека; для получения TCR с определенными специфичностями для генной терапии на основе TCR; для создания библиотек TCR для ассоциированных с заболеваниями антигенов (например, ассоциированных с опухолью антигенов (ТАА); и т.д.
В настоящей области техники существует растущий интерес к Т-клеточной терапии, поскольку Т-клетки (например, цитотоксические Т-клетки) могут быть направлены, чтобы атаковать и привести к разрушению представляющего интерес антигена, например, вирусного антигена, бактериального антигена, опухолевого антигена и т.д., или клеток, которые его презентируют. Первые исследования Т-клеточной терапии злокачественных опухолей были направлены на выделение проникающих в опухолевые ткани лимфоцитов (TIL; популяций лимфоцитов в массе опухоли, которые предположительно содержат Т-клетки, реактивные по отношению к опухолевым антигенам) из клеточной массы опухоли, их размножение in vitro с использованием факторов роста Т-клеток и перенос их обратно пациенту в процессе, имеющем название адоптивный перенос Т-клеток. Смотрите, например, Restiro et al. (2012) Adoptive immunotherapy for cancer: harnessing the Τ cell response, Nature Reviews 12:269-81; Linnermann et al. (2011) T-Cell Receptor Gene Therapy: Critical Parameters for Clinical Success, J. Invest. Dermatol. 131: 1806-16. Тем не менее, успех в этих способах терапии на сегодняшний день ограничен меланомой и почечно-клеточной карциномой; и адоптивный перенос TIL не направлен специфически на определенные ассоциированные с опухолью антигены (ТАА). Linnermann et al., ранее.
Были предприняты попытки инициировать генную терапию на основе TCR, где Т-клетки либо выбирали, либо программировали для нацеленного воздействия на представляющий интерес антиген, например, ТАА. Современная генная терапия на основе TCR основана на идентификации последовательностей TCR, которые направлены на конкретные антигены, например, ассоциированные с опухолью антигены. Например, Rosenberg и коллеги опубликовали несколько исследований, в которых они трансдуцировали лимфоциты периферической крови, полученные от пациента с меланомой, с генами, кодирующими цепи TCRα и β, специфические для эпитопов ассоциированного с меланомой антигена MART-1, и использовали полученные размноженные лимфоциты для адоптивной Т-клеточной терапии. Johnson et al. (2009) Gene therapy with human and mouse T-cell receptors mediates cancer regression and targets normal tissues expressing cognate antigen, Blood 114: 535-46; Morgan et al. (2006) Cancer Regression in Patients After Transfer of Genetically Engineered Lymphocytes, Science 314: 126-29. Специфические для MART-1 TCR выделяли из пациентов, у которых наблюдалось развитие опухоли после терапии на основе TIL. Тем не менее, идентификация таких TCR, в частности высокоавидных TCR (которые наиболее вероятно являются терапевтически применимыми), осложняется тем фактом, что большинство опухолевых антигенов представляют собой собственные антигены, и TCR, нацеленно воздействующие на эти антигены, часто либо подвергаются удалению, либо обладают субоптимальной аффинностью, главным образом, вследствие иммунологической толерантности.
Согласно различным вариантам осуществления настоящее изобретение решает эту проблему путем обеспечения генетически сконструированных не относящихся к человеку животных, содержащих в своем геноме нереаранжированный вариабельный генный локус TCR человека. Описанное в настоящем документе не относящееся к человеку животное способно производить Т-клетки с разнообразным репертуаром гуманизированных Т-клеточных рецепторов. Таким образом, описанные в настоящем документе не относящиеся к человеку животные могут представлять собой источник разнообразного репертуара гуманизированных Т-клеточных рецепторов, например, высокоавидных гуманизированных Т-клеточных рецепторов для применения в адоптивном переносе Т-клеток.
Таким образом, согласно одному варианту осуществления настоящее изобретение относится к способу создания Т-клеточного рецептора в ответ на антиген человека, предусматривающему иммунизацию описанного в настоящем документе не относящегося к человеку животного (например, грызуна, например, мыши или крысы) представляющим интерес антигеном, предоставление животному возможности развивать иммунный ответ, выделение из животного активированной Т-клетки со специфичностью к представляющему интерес антигену и определение последовательности нуклеиновой кислоты Т-клеточного рецептора, экспрессированной антигенспецифической Т-клеткой.
Согласно одному варианту осуществления настоящее изобретение относится к способу получения Т-клеточного рецептора человека, специфического к представляющему интерес антигену (например, ассоциированному с заболеванием антигену), предусматривающему иммунизацию описанного в настоящем документе не относящегося к человеку животного представляющим интерес антигеном; предоставление животному возможности развивать иммунный ответ; выделение из животного Т-клетки, реактивной по отношению к представляющему интерес антигену; определение последовательности нуклеиновой кислоты вариабельной области TCR человека, экспрессированной Т-клеткой; клонирование вариабельной области TCR человека в нуклеотидную конструкцию, содержащую последовательность нуклеиновой кислоты константной области TCR человека так, чтобы вариабельная область TCR человека была функционально связана с константной областью TCR человека; и экспрессию из конструкции Т-клеточного рецептора человека, специфического по отношению к представляющему интерес антигену. Согласно одному варианту осуществления стадии выделения Т-клетки, определения последовательности нуклеиновой кислоты вариабельной области TCR человека, экспрессированной Т-клеткой, клонирования вариабельной области TCR человека в нуклеотидную конструкцию, содержащую последовательность нуклеиновой кислоты константной области TCR человека, и экспрессии Т-клеточного рецептора человека проводят с использованием стандартных методов, известных специалистам в настоящей области техники.
Согласно одному варианту осуществления в клетке экспрессируется нуклеотидная последовательность, кодирующая Т-клеточный рецептор, специфический по отношению к представляющему интерес антигену. Согласно одному варианту осуществления клетка, экспрессирующая TCR, выбрана из клетки СНО, COS, 293, HeLa, PERC.6™ и т.д.
Представляющий интерес антиген может представлять собой любой антиген, который, как известно, вызывает заболевание или состояние или ассоциирован с заболеванием или состоянием, например, ассоциированный с опухолью антиген; антиген вирусного, бактериального или другого патогенного происхождения и т.д. В настоящей области техники известны многие ассоциированные с опухолью антигены. Отбор ассоциированных с опухолью антигенов представлена в Cancer Immunity (A Journal of the Cancer Research Institute) Peptide Database (archive.cancerimmunity.org/peptidedatabase/Tcellepitopes.htm). Согласно некоторым вариантам осуществления настоящего изобретения представляющий интерес антиген представляет собой антиген человека, например, ассоциированный с опухолью антиген человека. Согласно некоторым вариантам осуществления антиген представляет собой специфический для клеточного типа внутриклеточный антиген, и Т-клеточный рецептор используется для цитолиза клетки, экспрессирующей антиген.
Согласно одному варианту осуществления в настоящем документе предусмотрен способ идентификации Т-клетки со специфичностью против представляющего интерес антигена, например, ассоциированного с опухолью антигена, предусматривающий иммунизацию описанного в настоящем документе не относящегося к человеку животного представляющим интерес антигеном, предоставление животному возможности развивать иммунный ответ и выделение из не относящегося к человеку животного Т-клетки со специфичностью к антигену.
Настоящее изобретение относится к новым способам адоптивной Т-клеточной терапии. Таким образом, в настоящем документе предусмотрен способ лечения или улучшения заболевания или состояния (например, злокачественной опухоли) у субъекта (например, субъекта-млекопитающего, например, субъекта-человека), предусматривающим иммунизацию описанного в настоящем документе не относящегося к человеку животного антигеном, ассоциированным с заболеванием или состоянием, предоставление животному возможности развивать иммунный ответ, выделение из животного популяции антигенспецифических Т-клеток и инфузию выделенных антигенспецифических Т-клеток субъекту. Согласно одному варианту осуществления настоящее изобретение относится к способу лечения или улучшения заболевания или состояния у субъекта-человека, предусматривающему иммунизацию описанного в настоящем документе не относящегося к человеку животного представляющим интерес антигеном (например, ассоциированным с заболеванием или состоянием антигеном, например, ассоциированным с опухолью антигеном), предоставление животному возможности развивать иммунный ответ, выделение из животного популяции антигенспецифических Т-клеток, определение последовательности нуклеиновой кислоты Т-клеточного рецептора, экспрессированного антигенспецифическими Т-клетками, клонирование последовательности нуклеиновой кислоты Т-клеточного рецептора в вектор экспрессии (например, ретровирусный вектор), введение векторы в Т-клетки, полученные от субъекта так, чтобы Т-клетки экспрессировали антигенспецифический Т-клеточный рецептор, и инфузию Т-клеток субъекту. Согласно одному варианту осуществления последовательность нуклеиновой кислоты Т-клеточного рецептора дополнительно гуманизируют перед введением в Т-клетки, полученные от субъекта, например, последовательность, кодирующую не относящуюся к человеку константную область, модифицируют для дополнительного сходства с константной областью TCR человека (например, не относящуюся к человеку константную область замещают константной областью человека). Согласно некоторым вариантам осуществления заболевание или состояние представляет собой злокачественную опухоль. Согласно некоторым вариантам осуществления популяцию антигенспецифических Т-клеток увеличивают перед инфузией субъекту. Согласно некоторым вариантам осуществления проводят иммунное истощение популяции иммунных клеток субъекта перед инфузией антигенспецифических Т-клеток. Согласно некоторым вариантам осуществления антигенспецифический TCR представляет собой высокоавидный TCR, например, высокоавидный TCR по отношению к ассоциированному с опухолью антигену. Согласно некоторым вариантам осуществления Т-клетка представляет собой цитотоксическую Т-клетку. Согласно другим вариантам осуществления заболевание или состояние вызвано вирусом или бактерией.
Согласно другому варианту осуществления заболевание или состояние представляет собой аутоиммунное заболевание. TREG клетки представляют собой субпопуляцию Т-клеток, которые поддерживают толерантность к собственным антигенам и предотвращают патологическую аутореактивность. Таким образом, в настоящем документе также предусмотрены способы лечения аутоиммунного заболевания, которые основаны на создании антигенспецифических TREG клеток у описанного в настоящем документе не относящегося к человеку животного по настоящему изобретению.
Также в настоящем документе предусмотрен способ лечения или улучшения заболевания или состояния (например, злокачественной опухоли) у субъекта, предусматривающий введение клеток, пораженных заболеванием или состоянием (например, злокачественных клеток) от субъекта, не относящемуся к человеку животному, предоставление животному возможности развивать иммунный ответ на клетки, выделение из животного популяции Т-клеток, реактивных по отношению к клеткам, определение последовательности нуклеиновой кислоты Т-клеточного рецептора, экспрессированного Т-клеткам, клонирование последовательности Т-клеточного рецептора в вектор, введение вектора в Т-клетки, полученные от субъекта, и инфузию субъекту Т-клеток субъекта, несущих Т-клеточный рецептор.
Также в настоящем документе предусмотрено применение описанного в настоящем документе не относящегося к человеку животного для получения последовательностей нуклеиновой кислоты, кодирующих вариабельные домены TCR человека (например, вариабельные домены TCR α и/или β). Согласно одному варианту осуществления предусмотрен способ получения последовательности нуклеиновой кислоты, кодирующей вариабельный домен TCR человека, предусматривающий иммунизацию описанного в настоящем документе не относящегося к человеку животного представляющим интерес антигеном, предоставление не относящемуся к человеку животному возможности развивать иммунный ответ на представляющий интерес антиген и получение от него последовательности нуклеиновой кислоты, кодирующей вариабельный домен TCR человека, который связывает представляющий интерес антиген. Согласно одному варианту осуществления способ дополнительно предусматривает получение последовательности нуклеиновой кислоты, кодирующей вариабельный домен TCR человека, который функционально связан с не относящейся к человеку константной областью TCR, предусматривая выделение Т-клетки из описанного в настоящем документе не относящегося к человеку животного и получение из него последовательности нуклеиновой кислоты, кодирующей вариабельный домен TCR, связанный с константной областью TCR.
Также в настоящем документе предусмотрено применение описанного в настоящем документе не относящегося к человеку животного для получения терапевтического средства для человека, предусматривающее иммунизацию не относящегося к человеку животного представляющим интерес антигеном (например, ассоциированным с опухолью антигеном), предоставление не относящемуся к человеку животному возможности развивать иммунный ответ, получение от животного Т-клеток, реактивных по отношению к представляющему интерес антигену, получение из Т-клеток последовательности(ей) нуклеиновой кислоты, кодирующей(их) гуманизированный белок TCR, которые связывает представляющий интерес антиген, и использование последовательности(ей) нуклеиновой кислоты, кодирующей(их) гуманизированный белок TCR в терапевтическом средстве для человека.
Таким образом, также предусмотрен способ получения терапевтического средства для человека, предусматривающий иммунизацию описанного в настоящем документе не относящегося к человеку животного представляющим интерес антигеном, предоставление не относящемуся к человеку животному возможности развивать иммунный ответ, получение от животного Т-клеток, реактивных по отношению к представляющему интерес антигену, получение из Т-клеток последовательности(ей) нуклеиновой кислоты, кодирующей(их) гуманизированный Т-клеточный рецептор, которые связывает представляющий интерес антиген, и использование гуманизированного Т-клеточного рецептора в терапевтическом средстве для человека.
Согласно одному варианту осуществления терапевтическое средство для человека представляет собой Т-клетку (например, Т-клетку человека, например, Т-клетку, полученную от субъекта-человека), содержащую представляющую интерес последовательность нуклеиновой кислоты (например, трансфектированную или трансдуцированную с представляющей интерес нуклеиновой кислотой или содержащую иным образом введенную представляющую интерес нуклеиновую кислоту) так, чтобы Т-клетка экспрессировала гуманизированный белок TCR с аффинностью к представляющему интерес антигену. Согласно одному аспекту субъект, по отношению к которому используют терапевтическое средство, нуждается в терапии конкретного заболевания или состояния, и антиген ассоциирован с заболеванием или состоянием. Согласно одному аспекту Т-клетка представляет собой цитотоксическую Т-клетку, антиген представляет собой ассоциированный с опухолью антиген, и заболевание или состояние представляет собой злокачественную опухоль. Согласно одному аспекту Т-клетка получена от субъекта.
Согласно другому варианту осуществления терапевтическое средство для человека представляет собой Т-клеточный рецептор. Согласно одному варианту осуществления терапевтический рецептор представляет собой растворимый Т-клеточный рецептор. Много усилий было направлено на создание растворимых Т-клеточных рецепторов или вариабельных областей TCR для применения в качестве терапевтических средств. Создание растворимых Т-клеточных рецепторов зависит от получения реаранжированных вариабельных областей TCR. Один подход заключается в разработке одноцепочечных TCR, содержащих TCRα и ΤCRβ, и, аналогично формату scFv иммуноглобулина, слиянии их вместе посредством линкера (смотрите, например, международную патентную заявку № WO 2011/044186). Полученный scTv, если он аналогичен scFv, будет обеспечивать термически стабильную и растворимую форму TCRα/β связывающего белка. Альтернативные подходы включали в себя разработку растворимого TCR с константными доменами ΤCRβ (смотрите, например, Chung et al., (1994) Functional three-domain single-chain T-cell receptors, Proc. Natl. Acad. Sci. USA. 91: 12654-58); а также конструирование негативной дисульфидной связи на границе между константными доменами TCR (смотрите обзор в Boulter and Jakobsen (2005) Stable, soluble, high-affinity, engineered Τ cell receptors: novel antibody-like proteins for specific targeting of peptide antigens, Clinical and Experimental Immunology 142: 454-60; смотрите также, патент США №7569664). Были описаны другие форматы растворимых Т-клеточных рецепторов. Описанные в настоящем документе не относящиеся к человеку животные могут использоваться для определения последовательности Т-клеточного рецептора, который с высокой аффинностью связывается с представляющим интерес антигеном, и последующей разработки растворимого Т-клеточного рецептора на основе последовательности.
Растворимый Т-клеточный рецептор, полученный из последовательности TCR рецептора, экспрессированной не относящимся к человеку животным, может использоваться для блокирования функции представляющего интерес белка, например, вирусного, бактериального или ассоциированного с опухолью белка. Альтернативно, растворимый Т-клеточный рецептор может быть слит с фрагментом, который может лизировать инфицированную или злокачественную клетку, например, цитотоксическими молекулами (например, химиотерапевтическим средством), токсином, радионуклидом, пролекарственным средством, антителом и т.д. Растворимый Т-клеточный рецептор также может быть слит с иммуномодулирующей молекулой, например, цитокином, хемокином и т.д. Растворимый Т-клеточный рецептор также может быть слит с иммунной интибирующей молекулой, например, молекулой, которая ингибирует цитолиз Т-клеткой других клеток, содержащих антиген, распознанный Т-клеткой. Такие растворимые Т-клеточные рецепторы, слитые с иммунными ингибирующими молекулами, могут использоваться, например, в блокировании аутоиммунитета. Обзор различных иллюстративных иммунных ингибирующих молекул, которые могут быть слиты с растворимым Т-клеточным рецептором, представлен в Ravetch and Lanier (2000) Immune Inhibitory Receptors, Science 290: 84-8, включенной в настоящий документ посредством ссылки.
Настоящее изобретение также относится к способам исследования иммунологического ответа у контексте TCR человека, включая в себя реаранжировку TCR человека, развитие Т-клеток, активацию Т-клеток, иммунологическую толерантность и т.д.
Также предусмотрены способы испытания кандидатных вакцин. Согласно одному варианту осуществления в настоящем документе предусмотрен способ определения того, будет ли вакцина активировать иммунологический ответ (например, Т-клеточную пролиферацию, высвобождение цитокинов и т.д.) и приводить к созданию эффекторных Т-клеток, а также Т-клеток памяти (например, центральных и эффекторных Т-клеток памяти).
ПРИМЕРЫ
Настоящее изобретение будет дополнительно проиллюстрировано представленными ниже не ограничивающими примерами. Эти примеры представлены для содействия в понимании настоящего изобретения, но не предусмотрены для ограничения и не должны рассматриваться как ограничивающие его объем каким-либо образом. Примеры не включают в себя подробные описания общепринятых способов, которые будут хорошо известны специалистам в настоящей области техники (техники молекулярного клонирования и т.д.). Если не указано иное, части представляют собой части по весу, молекулярный вес представляет собой средний молекулярный вес, температура указана в градусах Цельсия и давление равно или близко атмосферному.
Пример 1. Получение мышей с гуманизированными вариабельными генными локусами TCR
Мышей, содержащих делецию эндогенных вариабельных локусов TCR (α или β) и замещение эндогенных сегментов V и J или V, D и J получали с использованием технологии генной инженерии VELOCIGENE® (смотрите, например, патент США №6586251 и Valenzuela, D.M., et al. (2003) High-throughput engineering of the mouse genome coupled with high-resolution expression analysis. Nat. Biotech. 21(6): 652-659), причем последовательности человека, полученные из библиотек ВАС с использованием бактериальной гомологичной рекомбинации, используются для получения больших нацеливающих векторов (LTVEC), содержащих геномные фрагменты вариабельных локусов TCR человека, фланкированные нацеливающими плечами для нацеливания LTVEC на эндогенные вариабельные локусы TCR мыши в ES клетках мыши. LTVEC релианизировали и подвергали электропорации в ES клеточную линию мыши согласно Valenzuela с соавт. ES клетки выбирали по устойчивости к гигромицину или неомицину и подвергали скринингу в отношении потери аллеля мыши или приобретения аллеля человека.
Нацеленные ES клеточные клоны вводили зародышам мыши на стадии 8 клеток (или на более ранней стадии) способом VELOCIMOUSE® (Poueymirou, W.T. et al. (2007). F0 generation mice fully derived from gene-targeted embryonic stem cells allowing immediate phenotypic analyses. Nat. Biotech. 25: 91-99.). VELOCIMICE® (F0 мышей, полностью полученных из донорной ES клетки), несущих гуманизированные локусы TCR идентифицируют с помощью скрининга в отношении потери эндогенного вариабельного аллеля TCR и приобретения аллеля человека с использованием модификации аллельного анализа (Valenzuela et al). F0 детенышей генотипируют и скрещивают до гомозиготности. Мышей, гомозиготных в отношении гуманизированных вариабельных локусов TCRα и/или ΤCRβ (например, содержащих подгруппу вариабельных сегментов TCRα и/или ΤCRβ человек), получали и фенотипировали, как описано в настоящем документе.
Всех мышей содержали и скрещивали в специальном свободном от патогенов виварии в Regeneran Pharmaceuticals. Все эксперименты с использованием животных были одобрены IACUC и Regeneran Pharmaceuticals.
Пример 2: Поэтапная гуманизация вариабельного локуса TCRα
1,5 м.п.н. ДНК на локусе TCRα мыши, соответствующей 110 V и 60 J сегментам мыши, замещали 1 м.п.н. ДНК, соответствующей 54 сегментам V и 61 сегменту J TCRα человека с использованием поэтапной стратегии гуманизации, обобщенно представленной на фигурах 2 и 3. Пограничные последовательности нуклеиновой кислоты различных нацеливающих векторов, используемых для поэтапной стратегии гуманизации локуса TCRα, обобщенно представлены в таблице 2, и включены в список последовательностей.


Сегменты вариабельной области человека TCRα нумеруют согласно базе данных IMGT. По меньшей мере 100 п.н. на каждом стыке (приблизительно 50 п.н. с каждого конца) включены в список последовательностей.
Конкретно, как показано на фиг. 4А, ДНК от BAC клона RP23-6А14 мыши (Invitrogen) модифицировали с помощью гомологичной рекомбинации и использовали в качестве нацеливающего вектора (MAID 1539) для замещения области TCRAJ1-TCRAJ28 эндогенного локуса TCRα мыши кассетой Ub-гигромицина, за которой следует сайт loxP ДНК от ВАС клона RP23-117i19 мыши (Invitrogen) модифировали с помощью гомологичной рекомбинации и использовали в качестве нацеливающего вектора (MAID 1535) для замещения ~15 т.п.н. области, окружающей (и включающей в себя) TCKAV1 эндогенного локуса ТСКα и δ мыши, кассетой PGK- неомицина, за которой следует сайт loxP. Е8 клетки, несущие дважды нацеленную хромосому (т.е. один эндогенный локус TCRα мыши, нацеленный с помощью обоих нацеливающих векторов) подтверждали с помощью известные в настоящей области техники способов кариотипирования и скрининга (например, TAQMAN™). Модифицированные ES клетки обрабатывали с помощью рекомбиназы CRE, тем самым опосредуя делецию области между двумя сайтами loxP (т.е. области, состоящей из эндогенного локуса TCRα мыши от TCRAV1 до TCRAJ1) и оставляя только один сайт loxP, кассету неомицина и константную и энхансерную области мыши. Эта стратегия приводила к созданию подвергнутого делеции локуса TCR α/δ мыши (MAID 1540).
Первый нацеливающий вектор человека для TCRα содержал 191660 п.н. ДНК человека из клонов CTD2216p1 и CTD2285m07 ВАС (Invitrogen), которые содержали первых два следующие друг за другом генных сегмента TCRαV человека (TKAV40 и 41) и 61 TCRαJ (50 функциональных) генный сегмент. Эту ВАС модифировали с помощью гомологичной рекомбинации, чтобы она содержала сайт Notl 403 п.н. ниже (3') от генного сегмента TCRαJ для лигирования 3' плеча гомологии мыши и 5' сайт AsiSI для лигирования 5' плеча гомологии мыши. Два различных плеча гомологии использовали для лигирования с этим фрагментом человека: 3' плечо гомологии содержало эндогенные последовательности TCRα мыши из клона RP23-6A14 ВАС, и 5' плечо гомологии содержало эндогенную последовательность TCRα 5' от TCRαV мыши из ВАС клона RP23-117i19 мыши. Эту относящуюся к мыши-человеку химерную ВАС использовали в качестве нацеливающего вектора (MAID 1626) для получения начальной вставки генных сегментов TCRα человека вместе с вышележащей кассетой loxp-ub-гигромицин-loxp на локусах TCRα мыши (фиг. 4В). Пограничные последовательности нуклеиновой кислоты (SEQ ID NO: 1-3) для нацеливающего вектора MAID 1626 описаны в таблице 2.
Впоследствии, получали серию нацеливающих векторов человека, в которых использовались такое же 5' плечо мыши, которое содержало эндогенную последовательность TCRα 5' от TCRαV мыши из ВАС клона RP23-117i19 мыши с чередующимися кассетами селекции loxP-неомицин-loxP и loxP-гигромицин-loxP (или frt-гигромицин-frt для MAID 1979).
Для создания минилокуса TCRα человека, содержащего в общем 8 генных сегментов TCRαV (7 функциональных) человека и 61 генный сегмент TCRαJ (50 функциональных) человека, ДНК из ВАС клона RP11-349р11 человека (Invitrogen) модифировали с помощью гомологичной рекомбинации и использовали в качестве нацеливающего вектора (MAID 1767) (фиг. 4С). Он добавлял 104846 п.н. ДНК человека, содержащую следующие 6 (5 функциональных) последовательных генных сегментов TCRαV человека (TRAV35 - TRAV39) и 5' кассету loxP-ub-неомицин-loxP. Полученный локус TCRα содержал 5' кассету loxp-ub-неомицин-loxP вместе с 8 генными сегментами TCRαV (7 функциональных) человека и 61 генным сегментом TCRαJ человека, функционально связанными с константными генами и энхансерами TCRα мыши. Пограничные последовательности нуклеиновой кислоты (SEQ ID NO: 4 и 5) для нацеливающего вектора MAID 1767 описаны в таблице 2.
Для создания минилокуса TCRα человека, содержащего в общем 23 генных сегмента TCRαV (17 функциональных) человека и 61 генный сегмент TCRαJ человека, ДНК из ВАС клона мыши, содержащую в направлении 5'-3': уникальный сайт I-Ceul, 20 т.п.н. плечо TCRA мыши 5' от локуса TCRA мыши для использования в гомологичной рекомбинации в ES клетки, и кассету loxP-Ub-Hyg-loxP в обратной ориентации, модифировали с помощью бактериальной гомологичной рекомбинации, чтобы она содержала в направлении 5'-3': уникальный сайт I-Ceul, 20 т.п.н. плечо TCRA мыши 5' от локуса TCRA мыши, кассету frt-pgk-Hyg-frt и уникальный сайт AsiSI. ДНК из ВАС клона RP11-622o20 человека (Invitrogen), содержащую TCRαV22-V34 человека, модифировали с помощью гомологичной рекомбинации, чтобы она содержала кассету Spec, фланкированную уникальными сайтами I-CeuI и AsiSI. Впоследствии, кассету Spec в модифицированном ВАС клоне человека замещали последовательностью, содержащейся между сайтами I-Ceul и AsiSI в модифицированном ВАС клоне мыши с помощью стандартных техник обработки рестриктазами/лигирования. Полученный нацеливающий вектор (MAID 1979; фиг. 4D) добавлял 136557 п.н. ДНК человека, которая содержала следующие 15 (10 функциональных) последовательных генных сегментов TCRαJ человека□ (TRAV22 - TRAV34) и 5' кассету frt-pgk-Uyg-frt. Полученный локус TCRα содержал 5' кассету frt-pgk-Uyg-frt вместе с 23 генными сегментами TCRαV человека (17 функциональными) и 61 генным сегментом TCRαJ человека, функционально связанными с константными генами и энхансерами TCRα мыши. Пограничные последовательности нуклеиновой кислоты (SEQ ID NO: 6 и 7) для нацеливающего вектора MAID 1979 описаны в таблице 2.
Для создания минилокуса TCRα человека, содержащего в общем 35 генных сегментов TCRαV человека (28 функциональных) и 61 генный сегмент TCRαJ человека, ДНК из ВАС клона CTD2501-k5 человека (Invitrogen) модифировали с помощью гомологичной рекомбинации и использовали в качестве нацеливающего вектора (MAID (фиг. 4Е). Он добавлял 124118 п.н. ДНК человека, которая содержала следующие 12 (11 функциональных) следующих друг за другом генных сегментов TCRαV человека (TRAV13-2 - TRAV21) и 5' кассету loxp-ub-неомицин-loxP. Полученный локус TCRα содержал 5' кассету loxp-ub-неомицин-loxP вместе с 35 генными сегментами TCRαV (28 функциональными) человека и 61 генным сегментом TCRαJ человека, функционально связанными с константными генами и энхансерами TCRα мыши. Пограничные последовательности нуклеиновой кислоты (SEQ ID NO: 8 и 9) для нацеливающего вектора MAID 1769 описаны в таблице 2.
Для создания минилокуса TCRα человека, содержащего в общем 48 генных сегментов TCRαV (39 функциональных) человека и 61 генный сегмент TCRαJ человека, ДНК из ВАС клона RP11-92F11 человека (Invitrogen) модифировали с помощью гомологичной рекомбинации и использовали в качестве нацеливающего вектора (MAID (фиг. 4F). Он добавлял 145505 п.н. ДНК человека, которая содержала следующие 13 (11 функциональных) следующих друг за другом генных сегментов TCRαV □ человека (TRAV6 - TRAV8.5) и 5' кассету loxp-ub-гигромицин-loxP. Полученный локус TCRα содержит 5'' кассету loxp-ub-гигромицин-loxP вместе с 48 генными сегментами TCRαV (39 функциональными) человека и 61 генным сегментом TCRαJ человека, □ функционально связанными с константными генами и энхансерами TCRα мыши. Пограничные последовательности нуклеиновой кислоты (SEQ ID NO: 10 и 11) для нацеливающего вектора MAID 1770 описаны в таблице 2.
Для создания минилокуса TCRα человека, содержащего в общем 54 генных сегмента ТСКаУ (45 функциональных) человека и 61 генный сегмент ТСКαJ человека, ДНК из ВАС клона RP11-780М2 человека (Invitrogen) модифировали с помощью гомологичной рекомбинации и использовали в качестве нацеливающего вектора (MAID 1771) (ФИГ. 40). Он добавлял 148496 п.н. ДНК человека, которая содержала следующие 6 (6 функциональных) следующих друг за другом генных сегментов ТСКαV человека (TRAV1-1 - TRAV5) и 5' кассету loxp-ub-неомицин-loxP. Полученный локус ТСКа содержит 5' кассету loxp-ub-неомицин-loxP вместе с 54 генными сегментами ТСКаУ (45 функциональными) человека и 61 □ генным сегментом ТСКαJ человека, функционально связанными с константными генами и энхансерами ТСКα мыши. Пограничные последовательности нуклеиновой кислоты (SEQ ID NO: 12 и 13) для нацеливающего вектора MAID 1771 описаны в таблице 2.
В любой из описанных выше стадий кассеты селекции удаляют путем делеции с помощью рекомбиназы Cre или Flp. Кроме того, могут вводить локус TCRδ человека, как показано на фиг. 5.
Пример 3: Поэтапная гуманизация вариабельного локуса TCRβ
0,6 м.п.н. ДНК на локусе TCRβ мыши, соответствующей 33 V, 2 D и 14 J сегментам мыши, замещали 0,6 м.п.н. ДНК, соответствующей 67 V, 20 и 14 I сегментам TCRβ человека с использованием поэтапной стратегии гуманизации, обобщенно представленной на фигурах 6 и 7. Пограничные последовательности нуклеиновой кислоты различных нацеливающих векторов, используемых для поэтапной стратегии гуманизации локуса TCRβ, обобщенно представлены в таблице 3 и включены в список последовательностей.


Сегменты вариабельной области ΤCRβ человека нумеруют, как в базе данных IMGT. По меньшей мере 100 п.н. на каждом стыке (приблизительно 50 п.н. с каждого конца) включены в список последовательностей.
Конкретно, ДНК из ВАС клона RP23-153p19 мыши (Invitrogen) модифировали с помощью гомологичной рекомбинации и использовали в качестве нацеливающего вектора (MAID 1544) для замещения 17 т.п.н. области (включающей в себя TCRBV30) сразу выше против хода транскрипции от 3' генного кластера трипсиногена в эндогенном локусе ΤCRβ мыши кассетой PGK-neo, за которой следует сайт loxP (фиг. 8А). ДНК из ВАС клона RP23-461M5 мыши (Invitrogen) модифировали с помощью гомологичной рекомбинации и использовали в качестве нацеливающего вектора (MAID 1542) для замещения 8355 п.н. области (включающей в себя TCRBV2 и TCRBV3) ниже по ходу транскрипции от 5' генного кластера трипсиноген в эндогенном локусе ΤCRβ мыши кассетой Ub-гигромицин, за которой следует сайт loxP. ES клетки, несущие дважды нацеленную хромосому (т.е. один эндогенный локус TCRβ мыши, нацеленный обоими нацеливающими векторами), подтверждали с помощью известных в настоящей области техники способами кариотипирования и скрининга (например, TAQMAN™). Модифицированные ES клетки обрабатывали с помощью рекомбиназы CRE, опосредуя делецию области между сайтами 5' и 3' loxP (состоящую из эндогенного локуса ΤCRβ мыши от TCRBV2 до TCRBV30) и оставляя только один сайт loxP, гигромициновую кассету и TCRBD, TCRBJ, константную и энхансерную последовательности мыши. Один TCRβvp мыши оставляли выше против хода транскрипции от 5' кластера генов трипсиногена, и один TCRBP мыши оставляли ниже по ходу транскрипции от Ер мыши, как показано на фиг. 8А.
Первый нацеливающий вектор человека для TCRβ содержал 125781 п.н. ДНК человека из ВАС клона CTD2559j2 (Invitrogen), которая содержала первые 14 следующих друг за другом генных сегментов TCRβV человека (TRBV18-TRBV29-1). Эту ВАС модифировали с помощью гомологичной рекомбинации, чтобы она содержала 5' сайт AsiSI и 3' сайт AscI для лигирования 5' и 3' плечей гомологии мыши. Два различных плеча гомологии использовали для лигирования с фрагментом человека: один набор плечей гомологии содержал эндогенную последовательность TCRβ, окружающую нижележащие гены трипсиногена мыши из ВАС клона RP23-153p19, и другой набор содержал эндогенную последовательность TCRβ, окружающую вышележащие гены трипсиногена мыши из ВАС клона RP23-461h15 мыши. Эту относящуюся к мыши человеку химерную ВАС использовали в качестве нацеливающего вектора (MAID 1625) для получения начальной вставки генных сегментов TCRβ человека вместе с вышележащей кассетой frt-ub-неомицин-frt на локусе TCRβ мыши, и это давало в результате минилокус TCRβ человека, содержащий 14 TCRβV (8 функциональных) человека (фиг. 8В). Пограничные последовательности нуклеиновой кислоты (SEQ ID NO: 14-16) для нацеливающего вектора MAID 1625 описаны в таблице 3.
Для замещения D и J сегментов ΤCRβ мыши D и J сегментами TCRβ человека, ДНК из ВАС клона RP23-302p18 мыши (Invitrogen) и из ВАС клона RP11-701D14 человека (Invitrogen) модифировали с помощью гомологичной рекомбинации и использовали в качестве нацеливающего вектора (MAID 1715) в ES клетках, которые содержали описанный выше минилокус TCRβV (т.е. MAID 1625). Эта модификация замещала -18540 п.н. область (от 100 п.н. ниже по ходу транскрипции от polyA 3' генов трипсиногена до 100 п.н. ниже против хода транскрипции от J сегментов в D2 кластере, который включал в себя TCRBD1-J1 мыши, константную область 1 мыши к TCRBD2-J2 мыши) в эндогенном локусе TCRβ мыши на ~25425 п.н. последовательности, содержащей TCRBD1-J1 человека, кассету loxP Ub-гигромицин-loxP, константную область 1 мыши, TCRBD2-J2 человека (фиг. 8C(i)). ES клетки, несущие дважды нацеленную хромосому (т.е. один эндогенный локус ΤCRβ мыши, нацеленный обоими нацеливающими векторами), подтверждали известными в настоящей области техники способами кариотипирования и скрининга (например, TAQMAN™). Модифицированные ES клетки обрабатывали с помощью рекомбиназы CRE, тем самым опосредуя делецию гигромициновой кассеты, оставляя только один сайт loxP, ниже по ходу транскрипции от J сегментов человека в кластере D1J (фиг. 8C(ii)). Пограничные последовательности нуклеиновой кислоты (SEQ ID NO: 17-21) для нацеливающего вектора MAID 1715 описаны в таблице 3.
Впоследствии, получали серию нацеливающих векторов человека, в которых использовались такое же 5' плечо мыши, которое содержало эндогенную последовательность TCRβ, окружающую вышележащие гены трипсиногена мыши из ВАС клона RP23-461h15 мыши с чередующейся кассетой селекции.
Для создания минилокуса TCRβ человека, содержащего в общем 40 сегментов TCRβV (30 функциональных) человека и D и J сегменты TCRβ человека, ДНК из ВАС клонов RP11-134h14 и RP11-785k24 человека (Invitrogen) модифировали с помощью гомологичной рекомбинации и комбинировали в нацеливающий вектор (MAID 1791) с использованием стандартных техник бактериальной гомологичной рекомбинации, обработки рестриктазой/лигирования и других техник клонирования. Введение нацеливающего вектора MAID 1791 приводило к добавлению 198172 п.н. ДНК человека, которая содержала следующие 26 (22 функциональных) следующих друг за другом генных сегментов TCRβV человека (TRBV6-5 - TRBV17) и 5' кассету frt-ub-гигромицин-frt. Полученный локус ΤCRβ содержал 5' кассету frt-ub-гигромицин-frt вместе с 40 генными сегментами ΤCRβV (30 функциональными) человека и D и J генными сегментами ΤCRβ человека, функционально связанными с константными генами и энхансерами TCRβ мыши (фиг. 8D). Пограничные последовательности нуклеиновой кислоты (SEQ ID NO: 22 и 23) для нацеливающего вектора MAID 1791 описаны в таблице 3.
Для создания минилокуса ΤCRβ человека, содержащего в общем 66 сегментов ΤCRβV (47 функциональных) человека и D и J сегменты TCRβ человека, ДНК из ВАС клона RP И-902 В 7 человека (Invitrogen) модифировали с помощью гомологичной рекомбинации и использовали в качестве нацеливающего вектора (MAID 1792). Это приводило к добавлению 159742 п.н. ДНК человека, которая содержала следующие 26 (17 функциональных) следующих друг за другом генных сегментов ΤCRβV человека (TRBV1 - TRBV12-2) и кассету 5' frt-ub-неомицин-frt. Полученный локус ΤCRβ содержал 5' кассету frt-ub-неомицин-frt вместе с 66 генными сегментами TCRβV (47 функциональными) человека и D и J генными сегментами ΤCRβ человека, функционально связанными с константными генами и энхансерами TCRβ мыши. (фиг. 8Е). Пограничные последовательности нуклеиновой кислоты (SEQ ID NO: 24 и 25) для нацеливающего вектора MAID 1792 описаны в таблице 3.
На любой из описанной выше стадий кассеты селекции удаляют путем делеции с помощью рекомбиназы Cre или Flp. Например, как показано на фиг. 7, MAID 1716 соответствует MAID 1715 с делецией гигромициновой кассеты.
В итоге, создавали минилокус TCRβ человека, содержащий в общем 67 TCRβV (48 функциональных) человека и D и J сегменты TCRβ человека. TCRBV31 мыши расположен ~9.4 т.п.н. 3' от TCRBC2 (вторая последовательность константной области TCRB) и находится к противоположной ориентации по отношению к другим сегментам TCRBV. Эквивалентный V сегмент человека представляет собой TCRBV30, который расположен в аналогичном положении в локусе TCRB человека.
Для гуманизации TCRBV31 ВАС клон мыши, содержащий TCRBV31 мыши, модифировали путем бактериальной гомологичной рекомбинации для получения LTVEC MAID 6192 (фиг. 8F). Полную кодирующую область, начинающуюся на старт-кодоне в экзоне 1, интрон, 3' UTR и сигнальные последовательности рекомбинации (RSS) TCRBV31 замещали гомологичными последовательностями TCRBV30 человека. 5' UTR сохраняли в качестве последовательности мыши. Для селекции самоудаляющуюся кассету (lox2372-убиквитиновый промотор-Hyg-PGKpolyA-протаминовый промотор-Cre-SV40polyA-lox2372) вводили в интрон (72 п.н. 3' от экзона 1289 п.н. 5' от экзона 2). Для простоты на фигурах 7 и 8 показана кассета селекции 3' от hTCRBV30, тогда как ее сконструировали, чтобы она располагалась в интроне между экзоном 1 и экзоном 2 гена hTCRBV30. Управляемая промотором протамина экспрессия Cre транскрибируется исключительно в постмейотических сперматидах, поэтому кассета "самостоятельно удаляется" в F1 поколении мышей.
Пограничные последовательности нуклеиновой кислоты (SEQ ID NO: 26 и 27) для нацеливающего вектора MAID 6192 описаны в таблице 3. ДНК MAID 6192 подвергают электропорации в ES клетки MAID 1792. ES клеточные клоны подвергают селекции в отношении устойчивости к гигромицину и скринингу в отношении потери аллеля TCRB31 мыши и приобретения аллеля TCRB30 человека.
Аналогичную стратегию конструирования используют для необязательной делеции оставшегося 5' V сегмента ΤCRβ мышь.
Пример 4: Создание мышей с TCRα/TCRβ
На каждой стадии поэтапной гуманизации локусов TCRα и TCRβ мышей, гомозиготных в отношении гуманизированного вариабельного локуса TCRα, могут скрещивать с мышами, гомозиготными в отношении гуманизированного вариабельного локуса TCRβ, для получения потомства, содержащего гуманизированные вариабельный локусы TCRα и TCRβ. Потомство скрещивают до гомозиготности по отношению к гуманизированным локусам TCRα и TCRβ.
Согласно одному варианту осуществления мышей, гомозиготных в отношении гуманизированного вариабельного локуса TCRα, содержащего 8 Vα человека и 61 Jα человека (MAID 1767; "1767 НО") скрещивали с мышами, гомозиготными в отношении гуманизированного вариабельного локуса ΤCRβ, содержащего 14 Vβ человека, 2 Dβ человека и 14 Jβ человека (MAID 1716; "1716 НО"). Потомство скрещивали до гомозиготности по отношению к обоим гуманизированным локусам.
Пример 5: Продукция Т-клеток селезенки у мышей, гомозиготных в отношении гуманизированного локуса TCRα и/или TCRβ
Селезенки от мышей дикого типа (WT); мышей с подвергнутым делеции локусом TCRα мыши ("MAID1540", смотрите фиг. 3); мышей, гомозиготных в отношении локус TCRα человека ("MAID 1767", смотрите фиг. 3); мышей с подвергнутыми делеции V сегментами TCRβ за исключением двух оставшихся V сегментов мыши ("MAID 1545", смотрите фиг. 7); мышей, гомозиготных в отношении локуса ΤCRβ человека, также содержащим два оставшихся V сегмента мыши ("MAID 1716", смотрите фиг. 7); и мышей, гомозиготных в отношении обоих локусов TCRα и TCRβ человека, с локусом TCRβ, также содержащим два оставшихся V сегмента мыши ("MAID 1767 1716"), перфузировали с коллагеназой D (Roche Bioscience) и эритроциты лизировали с помощью лизирующего буфера АСК с последующей отмывкой в среде RPMI.
Спленоциты от одного WT, MAID 1540, 1767, 1545, 1716, и 1716 1767 репрезентативного животного оценивали с помощью проточной цитометрии. Кратко, клеточные суспензии получали с использованием стандартных способов. 1×106 клеток инкубировали с антителом к CD16/CD32 мыши (2.4G2, BD) на льду в течение 10 минут, окрашивали с помощью соответствующего коктейля антител в течение 30 минут на льду. После окрашивания клетки отмывали и затем фиксировали в 2% формальдегида. Регистрацию данных проводили на проточном цитометре LSRII/CantoII/LSRFortessa и анализировали с помощью FlowJo.
Для окрашивания спленоцитов использовали антитела к FITC-CD3 мыши (17А2, BD). Как показано на фиг. 9, мыши с сегментами TCR человека были способны производить существенные количества CD3+ Т-клеток, тогда как мыши с делецией локуса TCRα мыши не могли этого делать. Мыши с делецией локуса TCRβ также производили CD3+ Т-клетки, предположительно вследствие использования оставшегося 3' V сегмента мыши (смотрите ниже).
Пример 6: Развитие Т-клеток в тимусе у мышей, гомозиготных в отношении гуманизированного локуса TCRα и/или TCRβ
Для определения того, проявляются ли мыши, гомозиготные в отношении гуманизированного локуса TCRα и/или TCRβ, нормальное развитие Т-клеток в тимусе, спленоциты от четырех из каждой группы совпадающих по возрасту (7-10 недель) животных WT, 1767 НО, 1716 НО, и 1716 НО 1767 НО использовали в проточной цитометрии для оценки продукции Т-клеток на различных стадиях развития, а также для оценки частоты встречаемости и абсолютного количества каждой из DN, DP, CD4 SP и CD8 SP Т-клеток.
Определения клеточного типа получали на основании присутствия маркеров клеточной поверхности CD4, CD8, CD44 и CD25, как обобщенно представлено в таблице 1. Корреляция между обозначением клеточного типа и экспрессией маркеров клеточной поверхности в тимусе является следующей: двойные негативные (DN) клетки (CD4- CD8), двойные позитивные (DP) клетки (CD4+ CD8+), CD4 одинарные позитивные клетки (CD4+ CD8-), CD8 одинарные позитивные клетки (CD4- CD8+), двойные негативные 1/DN1 клетки (CD4- CD8-, CD25- CD44+), двойные негативные 2/DN2 клетки (CD4- CD8-, CD25+CD44+), двойные негативные 3/DN3 клетки (CD4- CD8-, CD25+ CD44-), двойные негативные 4/DN4 клетки (CD4- CD8-, CD25- CD44-).
Тимоциты оценивали с помощью проточной цитометрии. Кратко, клеточные суспензии получали с использованием стандартных способов. Проточную цитометрию проводили, как описано в примере 5. Используемые антитела представляли собой: антимышиные антитела PE-CD44 (IM7, BioLegend), PeCy7-CD25 (РС61, BioLegend), АРС-H7-CD8a (53-6.7, BD) и APC-CD4 (GK1.5, eBioscience).
Как показано на фигурах 10 и 11, мыши, гомозиготные в отношении гуманизированных TCRα, ΤCRβ, и обоих TCRα и TCRβ, были способны производить DN1, DN2, DN3, DN4, DP, CD4 SP и CD8 SP Т-клетки, указывая на то, что Т-клетки, полученные из гуманизированных локусов, способны проходить развитие Т-клеток в тимусе.
Пример 7: Дифференциация Т-клеток селезенки у мышей, гомозиготных в отношении гуманизированного локуса TCRα и/или TCRβ
Для определения того, проявляются ли мыши, гомозиготные в отношении гуманизированного локуса TCRα и/или TCRβ, нормальную Т-клеточную дифференциацию в периферических органах (например, селезенке), четыре из каждой группы совпадающих по возрасту (7-10 недель) животных WT, 1767 НО, 1716 НО, и 1716 НО 1767 НО использовали в проточной цитометрии для оценки продукции различных типов Т-клеток в селезенке (CD3+, CD4+, CD8+, Τ наивные, Tcm и Teff/em), а также для оценки абсолютного количества каждого типа Т-клеток в селезенке.
Определения клеточного типа получали на основании присутствия маркеров клеточной поверхности: CD 19 (В-клеточного маркера), CD3 (Т-клеточного маркера), CD4, CD8, CD44, и CD62L (L-селектина). Корреляция между обозначением клеточного типа и экспрессией маркеров клеточной поверхности в селезенке является следующей: Т-клетки (CD3+), CD4 Т-клетки (CD3+ CD4+ CD8-), CD8 Т-клетки (CD3+ CD4- CD8+), CD4 эффекторные/эффекторные Т-клетки памяти (CD3+ CD4+ CD8- CD62L- CD44+), CD4 центральные Т-клетки памяти (CD3+ CD4+ CD8- CD62L+ CD44+), CD4 наивные Т-клетки (CD3+ CD4+ CD8- CD62L+ CD44-), CD8 эффекторные/эффекторные Т-клетки памяти (CD3+ CD4- CD8+ CD62L- CD44+), CD8 центральные Т-клетки памяти (CD3+ CD4- CD8+ CD62L+ CD44+), CD8 наивные Т-клетки (CD3+ CD4- CD8+ CD62L+ CD44-).
Спленоциты оценивали с помощью проточной цитометрии. Кратко, клеточные суспензии получали с использованием стандартных способов. Проточную цитометрию проводили, как описано в примере 5. Используемые антитела представляли собой: антимышиные антитела FITC-CD3 (17А2, BD), PE-CD44 (IM7, BioLegend), PerCP-Cy5.5-CD62L (Mel-14, BioLegend), APC-H7-CD8a (53-6.7, BD), APC-CD4 (GK1.5, eBioscience) и V450-CD19(1D3,BD).
Как показано на фигурах 12-14, Т-клетки в селезенке мышей, гомозиготных в отношении гуманизированного TCRα, ΤCRβ и обоих TCRα и TCRβ, были способны проходить Т-клеточную дифференциацию и присутствовали как CD4+, так и CD8+ Т-клетки. Кроме того, в селезенках исследуемых мышей обнаруживали Т-клетки памяти.
Пример 8: Использование V сегментов человека у гуманизированных в отношении TCR мышей
Экспрессию V сегментов ΤCRβ человека оценивали на уровне белка и РНК с использованием проточной цитометрии и ПЦР в реальном времени TAQMAN™, соответственно, у мышей, гомозиготных в отношении гуманизированного локуса ΤCRβ (1716 НО), и мышей, гомозиготных в отношении обоих гуманизированных локусов ΤCRβ и TCRα (1716 НО 1767 НО).
Для проточной цитометрии получали Т-клетки селезенки и анализ проводили, как описано в примере 5. Для проточной цитометрии использовали набор репертуара TCRβ (IOTEST® Beta Mark, Beckman Coulter). Набор содержит антитела к человеку, специфические к ряду TCRBV человека, например, hTRBV-18, -19, -20, -25, -27, -28 и -29.
Результаты обобщают на фиг. 15. На таблицах, представленных на фиг. 15А (покрытие CD8 Т-клеток) и фиг. 15В (покрытие CD4 Т-клеток), показано, что Т-клетки селезенки как у 1716 НО, так и у 1716 НО 1767 НО мышей использовали ряд V сегментов ΤCRβ человека. Мышей дикого типа использовали в качестве отрицательного контроля.
Для ПЦР в реальном времени общую РНК очищали из селезенки и тимуса с использованием набора для выделения общей РНК MAGMAX™-96 для микрочипов (Ambion by Life Technologies) согласно инструкциями производителя. Геномную ДНК удаляли с использованием буфера ДНазы MAGMAX™TURBO™ и TURBO ДНазы из приведенного выше набора MAGMAX (Ambion by Life Technologies). мРНК (до 2,5 мкг) обратно транскрибировали в сДНК с использованием мастер-микса SUPERSCRIPT® VILO™ (Invitrogen by Life Technologies). кДНК разбавляли до 2-5 нг/мкл и 10-25 нг кДНК амплифицировали с мастер-миксом для генной экспрессии TAQMAN® (Applied Biosystems by Life Technologies) с использованием системы обнаружения последовательности ABI 7900НТ (Applied Biosystems), с использованием праймеров и зондов Taqman MGB (Applied Biosystems) или зондов BHQ1/BHQ-Plus (Biosearcgh Technologies), показанных в таблице 4, согласно инструкциям производителя. Относительную экспрессию каждого гена нормировали к контролю константной области 1 TCR-бета (TRBC1) мыши.

Как показано на фигурах 16А-В, мыши, гомозиготные в отношении гуманизированного локуса TCRβ (1716 НО), и мыши, гомозиготные в отношении обоих гуманизированных локусов TCRβ и TCRα (1716 НО 1767 НО), проявляли экспрессию РНК различных сегментов TCRβ человека как в тимусе, так и в селезенке. Мыши также проявляли экспрессию РНК сегментов TRBV-1 и TRBV-31 мыши (данные не показаны), но ни один белок TRBV-1 мыши не обнаружили с помощью проточной цитометрии (данные не показаны).
Сегмент TRBV-31 мыши замещают сегментом TRBV-30 человека, как показано на фиг. 8F, и мышей получают из ES клеток MAID 6192, как описано в настоящем документе. Селезенки и тимусы полученных гомозиготных животных исследуют в отношении использования сегментов VP человека, включающих в себя TRBV-30, с помощью проточной цитометрии и/или ПЦР в реальном времени, как описано в настоящем документе, сегмент mTRBV-1 также могут подвергать делеции.
Пример 9: Развитие Т-клеток у мышей, гомозиготных в отношении 23 сегментов Vα TCR человека
Гомозиготные гуманизированные в отношении TCRα мыши, характеристики которых определили в предыдущих примерах, содержали 8 сегментов Vα человека и 61 сегмент Jα человека (1767 НО, смотрите фиг. 3). Гомозиготных гуманизированных в отношении TCRα мышей, содержащих 23 сегмента Vα человека и 61 сегмент Jα человека (1979 НО, смотрите фиг. 3), исследовали в отношении их способности производить CD3+ Т-клетки селезенки и проходить развитие Т-клеток в тимусе.
Экспериментальные данные получали с использованием проточной цитометрии с использованием соответствующих антител, описанных в предыдущих примерах. Как показано на фиг. 17, мышь, гомозиготная в отношении 23 сегментов Vα человека и 61 сегмента Jα человека, производила значительное количество CD3+ Т-клеток селезенки, и процент периферических CD3+ Т-клеток был сопоставим с процентом от животных дикого типа (фиг. 19).
Более того, тимоциты у 1979 НО мышей были способны проходить развитие Т-клеток и содержали Т-клетки на стадиях DN1, DN2, DN3, DN4, DP, CD4 SP и CD8 SP (фиг. 18).
Пример 10: Развитие Т-клеток и дифференциация у мышей, гомозиготных в отношении полного репертуара сегменты вариабельной области как TCRα, так и TCRβ человека
Мышей, гомозиготных в отношении полного репертуара сегментов вариабельной области TCRα человека (т.е. 54 Vα человека и 61 Jα человека) и гомозиготных в отношении полного репертуара сегментов вариабельной области ΤCRβ человека (67 Vβ человека, 2 Dβ человека и 14 Jβ человека), "1771 НО 6192 НО" (смотрите фигуры 3 и 7) исследовали в отношении их способности производить тимоциты, которые проходят нормальное развитие Т-клеток, производить Т-клетки, которые проходят нормальную Т-клеточную дифференциацию в периферических органах, и использовать полный репертуар их Vα и Vβ сегментов человека.
Проточную цитометрию проводят для определения присутствия DN1, DN2, DN3, DN4, DP, CD4 SP и CD8 SP Т-клеток в тимусе с использованием антимышиных антител CD4, CD8, CD25 и CD44, как описано выше в примерах 5 и 6. Проточную цитометрию также проводят для определения количества CD3+ Т-клеток в периферических органах, а также для оценки Т-клеточной дифференциации в периферических органах (например, присутствия эффекторных Т-клеток и Т-клеток памяти в периферических органах). Эксперимент проводят с использованием антимышиных антител CD3, CD 19, CD4, CD8, CD44 и CD62L, описанных выше в примерах 5 и 7.
В итоге, проточную цитометрию и/или ПЦР в реальном времени проводят для определения того, используют ли Т-клетки у 1771 НО 6192 НО мышей полный репертуар V сегментов TCRB и TCRA. Для экспрессии белка с использованием проточной цитометрии используют набор репертуара ΤCRβ (IOTEST® Beta Mark, Beckman Coulter), содержащий антитела, специфические к hTCRBV человека (смотрите пример 8). Для экспрессии РНК с использованием ПЦР в реальном времени кДНК из селезенок или тимусов амплифицируют с использованием праймеров TCR-V человека и зондов Taqman согласно инструкциям производителя и как описано выше в примере 8.
Эквиваленты
Специалистам в настоящей области техники будут понятны, или с использованием не более чем рутинного экспериментирования они смогут установить многие эквиваленты описанных в настоящем документе конкретных вариантов осуществления настоящего изобретения. Предусматривается, что такие эквиваленты включены в представленную ниже формулу изобретения.
Полное содержание всех процитированных в настоящей заявке непатентных документов, патентных заявок и патентов полностью включено посредством ссылки в настоящем документе.
Реферат
Изобретение относится к области биохимии, в частности к генетически модифицированному грызуну, экспрессирующему гуманизированный Т-клеточный рецептор (TCR) и содержащему в геноме своей зародышевой линии: нереаранжированную последовательность вариабельной области TCRα, и/или нереаранжированную последовательность вариабельной области TCRβ, к способу его получения, а также к его выделенной эмбриональной клетке и ткани. Также раскрыт способ получения Т-клеточного рецептора человека к представляющему интерес антигену, а также способ получения Т-клетки со специфичностью против представляющего интерес антигена с использование вышеуказанного грызуна. Изобретение также относится к нуклеиновой кислоте, кодирующей гуманизированный TCRα полипептид, а также к нуклеиновой кислоте, кодирующей гуманизированный TCRβ полипептид. Изобретение позволяет эффективно получать гуманизированный Т-клеточный рецептор. 8 н. и 28 з.п. ф-лы, 19 ил., 4 табл., 10 пр.
Комментарии