Способы лечения рака с использованием антагонистов, связывающих с осью pd-1, и ингибиторов tigit - RU2702108C2
Код документа: RU2702108C2
Чертежи
























































Описание
Перекрестные ссылки на родственные заявки
Настоящая заявка заявляет приоритет предварительной заявки США № 61/846941, поданной 16 июля 2013, предварительной заявки США № 61/865582, поданной 13 августа 2013, предварительной заявки США № 61/950754, поданной 10 марта 2014, предварительной заявки США № 61/985884, поданной 29 апреля 2014, и предварительной заявки США 61/992109, поданной 12 мая 2014 года, каждая из которых включена сюда во всей ее полноте в качестве ссылки.
Представление перечня последовательностей в текстовом файле ASCII
Содержание следующего представления в формате текстового файла ASCII включено в данное описание посредством ссылки во всей своей полноте: машиночитаемая форма (CRF) перечня последовательностей (название файла: 146392025940SEQLISTING.TXT, зарегистрированная дата: 16 июля 2014, размер: 25 kb).
Уровень техники
Поступление двух различных сигналов к Т-клеткам является широко распространенной моделью активации покоящихся Т-лимфоцитов антигенпредставляющими клетками (АРС). Lafferty et al., Aust. J. Exp. Biol. Med. ScL, 53:27-42 (1975). Эта модель также устанавливает различие между аутотолерантностью и иммунологической толерантностью. Bretscher et al., Science, 169: 1042-1049 (1970); Bretscher, P.A., P.N.A.S. USA, 96:185-190 (1999); Jenkins et al., J. Exp. Med., 165:302-319 (1987). Первичный сигнал или антиген-специфический сигнал передается через Т-клеточный рецептор (TCR) после распознавания антигена-пептида, представленного в главном комплексе гистосовместимости (MHC). Вторичный или костимулирующий сигнал передается Т-клеткам костимулирующими молекулами, которые экспрессируются на антигенпрезентирующих клетках (АРС), и индуцирует Т-клетки на клональную экспансию, секрецию цитокинов и эффекторную функцию. Lenschow et al., Ann. Rev. Immunol., 14:233 (1996). При отсутствии костимуляции Т-клетки могут стать невосприимчивыми к антигенной стимуляции, что приводит к толерогенному ответу на любой чужеродный или эндогенный антиген.
В двухсигнальной модели Т-клетки получают положительные костимулирующие и отрицательные коингибирующие сигналы. Регуляция таких положительных и отрицательных сигналов является критической для максимального повышения защитных иммунных ответов хозяина, одновременно сохраняя иммунологическую толерантность и предупреждая развитие аутоиммунитета. Вероятно, отрицательные сигналы необходимы для индукции толерантности Т-клеток, в то время как положительные сигналы способствуют активации Т-клеток.
Как костимулирующие, так и коингибирующие сигналы поступают на подвергшиеся воздействию антигена Т-клетки, и необходимо взаимодействие костимулирующих и коингибирующих сигналов для контроля степени иммунного ответа. Кроме того, сигналы, поступающие на Т-клетки, изменяются по мере того, как инфекция или иммунная провокация разрешаются, ухудшаются или сохраняются, и эти изменения оказывают сильное влияние на реагирующие Т-клетки и видоизменяют иммунный ответ.
Механизм костимуляции представляет интерес с точки зрения терапии, поскольку было показано, что манипулирование костимулирующими сигналами обеспечивает средства для усиления или подавления клеточного иммунного ответа. Недавно было установлено, что дисфункция или анергия Т-клеток происходит одновременно с индуцированной и устойчивой экспрессией ингибирующего рецептора, полипептида 1 программируемой гибели (PD-1). В результате направленное терапевтическое воздействие на PD-1 и другие молекулы, которые передают сигналы посредством взаимодействия с PD-1, такие как лиганд 1 программируемой гибели (PD-L1) и лиганд 2 запрограммированной гибели (PD-L2), находится в области повышенного интереса.
PD-L1 сверхэкспрессируется во многих видах злокачественных опухолей и часто ассоциируется с неблагоприятным прогнозом (Okazaki T et al., Intern. Immun., 2007 19(7):813; Thompson RH et al., Cancer Res, 2006, 66(7):3381). Интересно отметить, что большинство инфильтрирующих опухоль Т-лимфоцитов преимущественно экспрессируют PD-1 в противоположность Т-лимфоцитам в нормальных тканях и Т-лимфоцитам периферической крови, указывая, что положительная регуляция PD-1 на опухолереактивных Т-клетках может способствовать подавлению противоопухолевых иммунных реакций (Blood, 2009, 114(8):1537). Данное явление может возникнуть за счет использования сигнального пути с участием PD-L1, опосредованного взаимодействием PD-L1-экспрессирующих опухолевых клеток с PD-1-экспрессирующими Т-клетками, приводя к ослаблению активации Т-клеток и ускользанию от иммунологического контроля (Sharpe et al., Nat. Rev., 2002; Keir M.E. et al., 2008, Annu. Rev. Immunol., 26:677). Таким образом, ингибирование взаимодействия PD-L1/PD-1 может усилить опосредованную CD8+ Т-клетками элиминацию опухолей.
Ингибирование оси сигнального пути PD-1 через его непосредственные лиганды (например, PD-LL, PD-L2) было предложено в качестве средства усиления Т-клеточного иммунитета для лечения рака (например, противоопухолевого иммунитета). Кроме того, подобное усиление T-клеточного иммунитета было отмечено в результате ингибирования связывания PD-L1 с партнером по связыванию В7-1. Кроме того, комбинируя ингибирование сигнального пути PD-1 с другими сигнальными путями, которые дисрегулированы в опухолевых клетках, можно дополнительно повысить эффективность лечения. Существует потребность в такой оптимальной терапии для лечения, стабилизации, предупреждения и/или замедления развития различных видов рака.
Все ссылки, публикации и патентные заявки, приведенные в настоящем документе, включены здесь в полном объеме в качестве ссылки.
Сущность изобретения
Настоящее изобретение относится к комбинированному лечению, включающему антагонист, связывающийся с осью PD-1, и агент, который снижает или ингибирует экспрессию и/или активность TIGIT.
Обеспечиваются способы лечения или замедления прогрессирования рака у индивидуума, включающие введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT.
Также обеспечиваются способы снижения или ингибирования рецидива или прогрессирования рака у индивидуума, включающие введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT.
Также обеспечиваются способы лечения или замедления прогрессирования связанного с иммунитетом заболевания у индивидуума, включающие введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT.
Также обеспечиваются способы снижения или ингибирования прогрессирования связанного с иммунитетом заболевания у индивидуума, включающие введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT.
В некоторых вариантах осуществления связанное с иммунитетом заболевание ассоциировано с Т-клеточным дисфункциональным расстройством. В некоторых вариантах осуществления Т-клеточное дисфункциональное расстройство характеризуется снижением реактивности на антигенную стимуляцию. В некоторых вариантах осуществления Т-клеточное дисфункциональное расстройство характеризуется анергией Т-клеток или пониженной способностью к секреции цитокинов, пролиферации или осуществлению цитолитической активности. В некоторых вариантах осуществления Т-клеточное дисфункциональное расстройство характеризуется истощением Т-клеток. В некоторых вариантах осуществления Т-клетки представляют собой CD4+ и CD8+ Т-клетки. В некоторых вариантах осуществления Т-клеточное дисфункциональное расстройство выбрано из группы, состоящей из неразрешенной острой инфекции, хронической инфекции и иммунитета к опухоли.
Также обеспечиваются способы повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума введением индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT.
Также обеспечиваются способы лечения или замедления прогрессирования рака у индивидуума, включающие введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226.
Также обеспечиваются способы снижения или ингибирования рецидива и прогрессирования рака у индивидуума, включающие введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226.
Также обеспечиваются способы лечения или замедления прогрессирования связанного с иммунитетом заболевания у индивидуума, включающие введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226.
Также обеспечиваются способы снижения или ингибирования прогрессирования связанного с иммунитетом заболевания у индивидуума, включающие введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226.
В некоторых вариантах осуществления связанное с иммунитетом заболевание ассоциировано с Т-клеточным дисфункциональным расстройством. В некоторых вариантах осуществления Т-клеточное дисфункциональное расстройство характеризуется снижением реактивности на антигенную стимуляцию. В некоторых вариантах осуществления Т-клеточное дисфункциональное расстройство характеризуется анергией Т-клеток или пониженной способностью к секреции цитокинов, пролиферации или осуществлению цитолитической активности. В некоторых вариантах осуществления Т-клеточное дисфункциональное расстройство характеризуется истощением Т-клеток. В некоторых вариантах осуществления Т-клетки представляют собой CD4+ и CD8+ Т-клетки. В некоторых вариантах осуществления Т-клеточное дисфункциональное расстройство выбрано из группы, состоящей из неразрешенной острой инфекции, хронической инфекции и иммунитета к опухоли.
Также обеспечиваются способы повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума введением индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226.
В некоторых вариантах осуществления агент, который модулирует экспрессию и/или активность CD226, способен повышать или стимулировать экспрессию и/или активность CD226.
В некоторых вариантах осуществления агент, который модулирует экспрессию и/или активность CD226, выбран из агента, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, антагониста экспрессии и/или активности TIGIT, антагониста экспрессии и/или активности PVR, агента, который ингибирует и/или блокирует взаимодействия TIGIT с PVR, агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR.
В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, представляет анти-TIGIT-антитело или его антигенсвязывающий фрагмент.
В некоторых вариантах осуществления антагонист экспрессии и/или активности TIGIT представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления антагонист экспрессии и/или активности TIGIT представляет анти-TIGIT-антитело или его антигенсвязывающий фрагмент.
В некоторых вариантах осуществления антагонист экспрессии и/или активности PVR представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид.
В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVR, представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид.
В некоторых вариантах осуществления агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR, представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид.
Настоящее изобретение также относится к комбинированному лечению, включающему агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и агент, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов.
Обеспечиваются способы повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума введением индивидууму эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов.
В некоторых вариантах осуществления один или более дополнительных иммунных коингибирующих рецепторов выбран из группы, состоящей из PD-1, CTLA-4, LAG3, TIM3, BTLA и VISTA. В некоторых вариантах осуществления один или более дополнительных иммунных коингибирующих рецепторов выбран из группы PD-1, CTLA-4, LAG3 и TIM3.
Настоящее изобретение также относится к комбинированному лечению, включающему агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и агент, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов.
Обеспечиваются способы повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума введением индивидууму эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов.
В некоторых вариантах осуществления один или более дополнительных иммунных костимулирующих рецепторов выбран из группы, состоящей из CD226, ОХ-40, CD28, CD27, CD137, HVEM и GITR. В некоторых вариантах осуществления один или более дополнительных иммунных костимулирующих рецепторов выбран из группы, состоящей CD226, OX40, CD27, CD137, HVEM и GITR. В некоторых вариантах осуществления один или более дополнительных иммунных костимулирующих рецепторов выбран из группы, состоящей из ОХ-40 и CD27.
В некоторых вариантах осуществления любой из вышеописанных способов дополнительно включает введение, по меньшей мере, одного химиотерапевтического агента.
В некоторых вариантах у индивидуума в любом из вышеописанных способов имеется рак. В некоторых вариантах осуществления индивидуумом в любом из вышеописанных способов является человек.
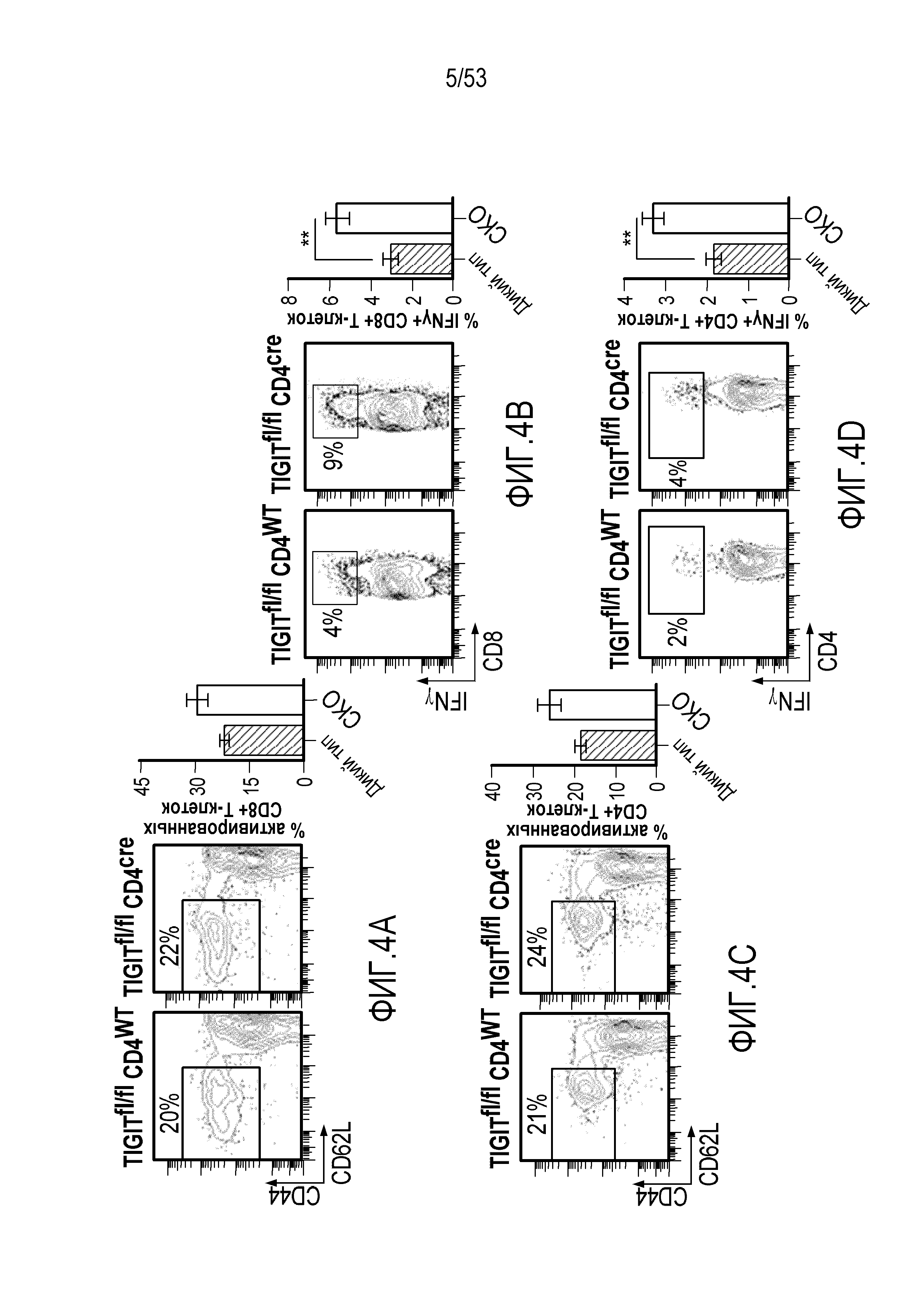
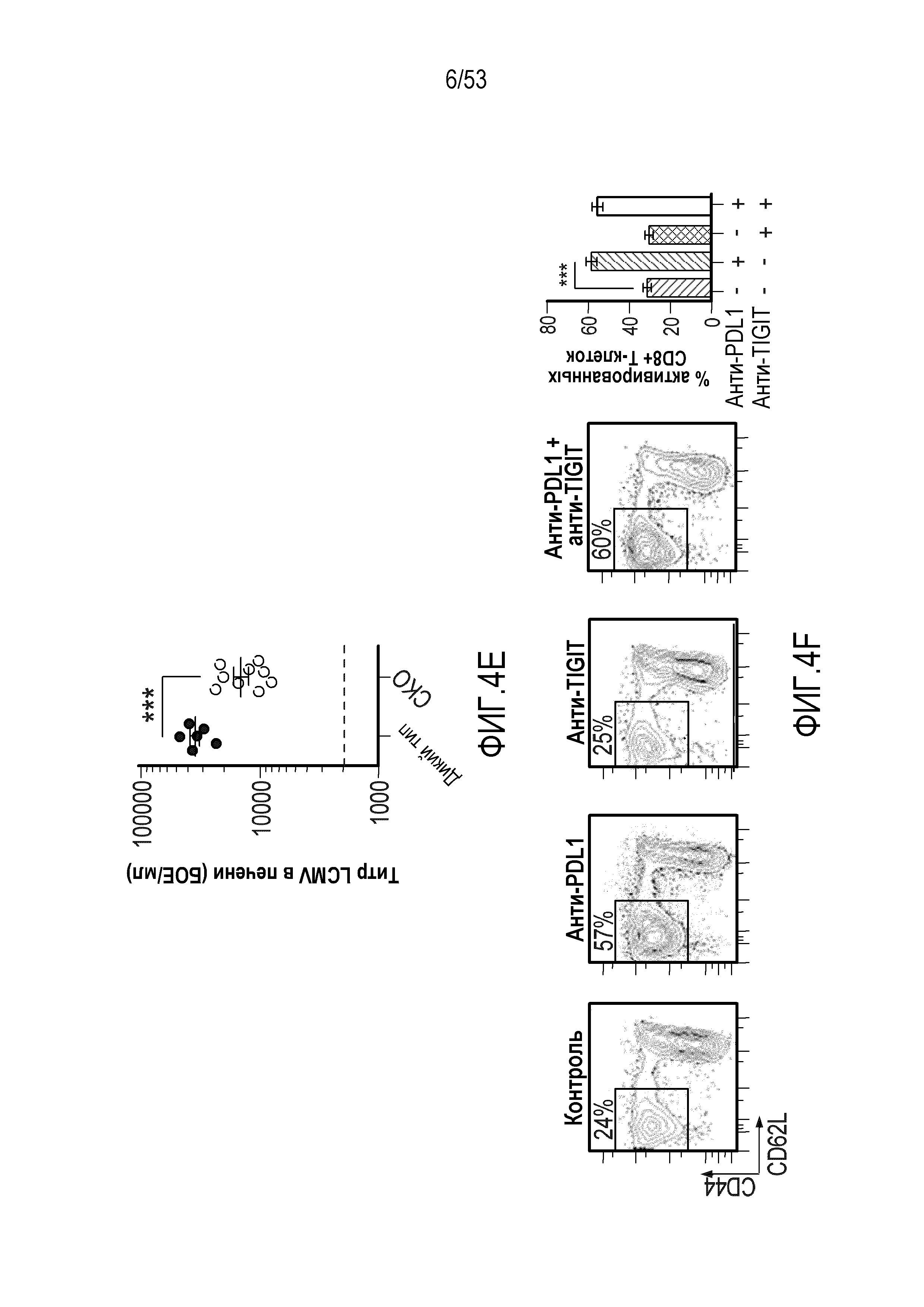
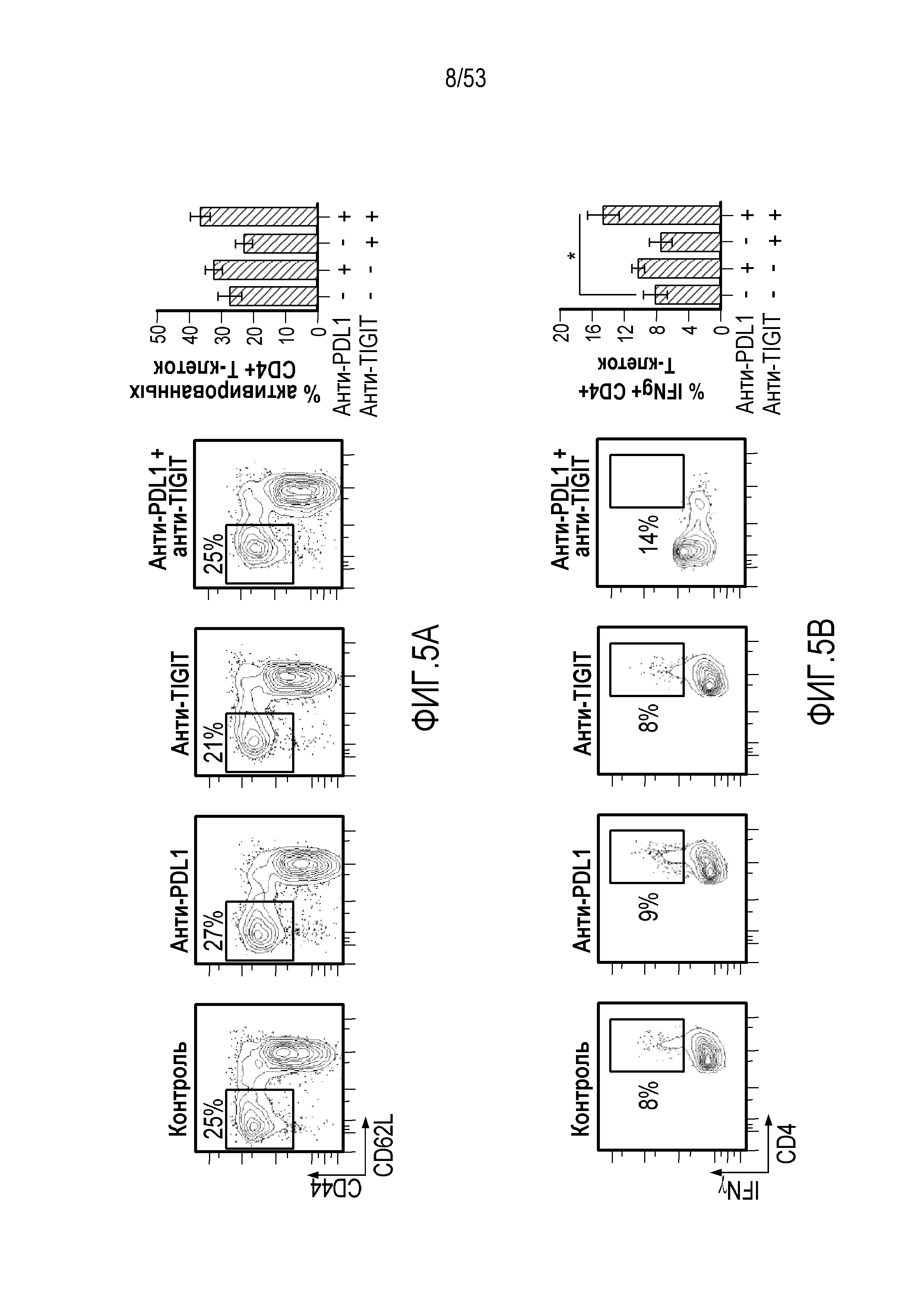
В некоторых вариантах осуществления CD4 и/или CD8 Т-клетки у индивидуума имеют повышенное или усиленное праймирование, активацию, пролиферацию, высвобождение цитокинов и/или цитолитическую активность по сравнению с периодом до введения комбинации.
В некоторых вариантах осуществления количество CD4 и/или CD8 Т-клеток повышено по сравнению с периодом до введения комбинации. В некоторых вариантах осуществления количество активированных CD4 и/или CD8 Т-клеток повышено по сравнению с периодом до введения комбинации. В некоторых вариантах осуществления активированные CD4 и/или CD8 Т-клетки характеризуются CD4 и/или CD8 Т-клетками, продуцирующими γ-IFN+, и/или повышенной цитолитической активностью по сравнению с периодом до введения комбинации. В некоторых вариантах осуществления CD4 и/или CD8 Т-клетки проявляют повышенное высвобождение цитокинов, выбранных из группы, состоящей из IFN-γ, TNF-α и интерлейкинов.
В некоторых вариантах осуществления CD4 и/или CD8 Т-клетка является эффекторной Т-клеткой памяти. В некоторых вариантах осуществления CD4 и/или CD8 эффекторная Т-клетка памяти характеризуется CD4 и/или CD8 Т-клетками, продуцирующими γ-IFN+, и/или повышенной цитолитической активностью. В некоторых вариантах осуществления CD4 и/или CD8 эффекторная Т-клетка памяти характеризуется наличием экспрессии CD44высокий CD62Lнизкий.
В некоторых вариантах осуществления злокачественная опухоль в любом из вышеуказанных способов имеет повышенные уровни Т-клеточной инфильтрации.
В некоторых вариантах осуществления агент, который снижает или ингибирует экспрессию и/или активность TIGIT, выбран из группы, состоящей из антагониста экспрессии и/или активности TIGIT, антагониста экспрессии и/или активности PVR, и агента, который ингибирует взаимодействие и/или внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR.
В некоторых вариантах осуществления антагонист экспрессии и/или активности TIGIT выбран из группы, состоящей низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида.
В некоторых вариантах осуществления антагонист экспрессии и/или активности PVR выбран из группы, состоящей низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида.
В некоторых вариантах осуществления агент, который ингибирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR, выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида.
В некоторых вариантах осуществления антагонист экспрессии и/или активности TIGIT представляет собой анти-TIGIT-антитело или его антигенсвязывающий фрагмент.
В некоторых вариантах осуществления изобретения анти-TIGIT-антитело или его антигенсвязывающий фрагмент содержит, по меньшей мере, один HVR, включающий аминокислотную последовательность, выбранную из аминокислотных последовательностей KSSQSLYYSGVKENLLA (SEQ ID NO:1), ASIRFT (SEQ ID NO:2), QQGINNPLT (SEQ ID NO:3), GFTFSSFTMH (SEQ ID NO:4), FIRSGSGIVFYADAVRG (SEQ ID NO:5) и RPLGHNTFDS (SEQ ID NO:6) или RSSQSLVNSYGNTFLS (SEQ ID NO:7), GISNRFS (SEQ ID NO:8), LQGTHQPPT (SEQ ID NO:9), GYSFTGHLMN (SEQ ID NO:10), LIIPYNGGTSYNQKFKG (SEQ ID NO:11) и GLRGFYAMDY (SEQ ID NO:12).
В некоторых вариантах осуществления анти-TIGIT-антитело или его антигенсвязывающий фрагмент содержит легкую цепь, включающую аминокислотную последовательность, представленную в DIVMTQSPSSLAVSPGEKVTMTCKSSQSLYYSGVKENLLAWYQQKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAEDMGQYFCQQGINNPLTFGDGTKLEIKR (SEQ ID NO:13) или DVVLTQTPLSLSVSFGDQVSISCRSSQSLVNSYGNTFLSWYLHKPGQSPQLLIFGISNRFSGVPRFSGSGSGTDFTLKISTIKPEDLGMYYCLQGTHQPPTFGPGTKLEVK (SEQ ID NO:14).
В некоторых вариантах осуществления анти-TIGIT-антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, включающую аминокислотную последовательность, представленную в EVQLVESGGGLTQPGKSLKLSCEASGFTFSSFTMHWVRQSPGKGLEWVAFIRSGSGIVFYADAVGRFTISRDNAKNLLFLQMNDLKSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS (SEQ ID NO:15) или EVQLQQSGPELVKPGTSMKISCKASGYSFTGHLMNWVKQSHGKNLEWIGLIIPYNGGTSYNQKFGKATLTVDKSSSTAYMELLSLTSDDSAVYFCSRGLRGFYAMDYWGQGTSVTVSS (SEQ ID NO:16).
В некоторых вариантах осуществления анти-TIGIT-антитело или его антигенсвязывающий фрагмент содержит легкую цепь, включающую аминокислотную последовательность, представленную в DIVMTQSPSSLAVSPGEKVTMTCKSSQSLYYSGVKENLLAWYQQKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAEDMGQYFCQQGINNPLTFGDGTKLEIKR (SEQ ID NO:13) или DVVLTQTPLSLSVSFGDQVSISCRSSQSLVNSYGNTFLSWYLHKPGQSPQLLIFGISNRFSGVPDRFSGSGSGTDFTLKISTIKPEDLGMYYCLQGTHQPPTFGPGTKLEVK (SEQ ID NO:14), и тяжелую цепь антитела, включающую аминокислотную последовательность, представленную в EVQLVESGGGLTQPGKSLKLSCEASGFTFSSFTMHWVRQSPGKGLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDLKSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS (SEQ ID NO:15) или EVQLQQSGPELVKPGTSMKISCKASGYSFTGHLMNWVKQSHGKNLEWIGLIIPYNGGTSYNQKFKGKATLTVDKSSSTAYMELLSLTSDDSAVYFCSRGLRGFYAMDYWGQGTSVTVSS (SEQ ID NO:16).
В некоторых вариантах осуществления обеспечивается анти-TIGIT-антитело или его антигенсвязывающий фрагмент, где антитело выбрано из гуманизированного антитела, химерного антитела, биспецифического антитела, гетероконъюгатного антитела и иммунотоксина.
В некоторых вариантах осуществления анти-TIGIT-антитело или его антигенсвязывающий фрагмент содержит, по меньшей мере, один HVR, по меньшей мере, на 90% идентичный HVR, представленному в любой из последовательностей KSSQSLYYSGVKENLLA (SEQ ID NO:1), ASIRFT (SEQ ID NO:2), QQGINNPLT (SEQ ID NO:3), GFTFSSFTMH (SEQ ID NO:4), FIRSGSGIVFYADAVRG (SEQ ID NO:5) и RPLGHNTFDS (SEQ ID NO:6) или RSSQSLVNSYGNTFLS (SEQ ID NO:7), GISNRFS (SEQ ID NO:8), LQGTHQPPT (SEQ ID NO:9), GYSFTGHLMN (SEQ ID NO:10), LIIPYNGGTSYNQKFKG (SEQ ID NO:11) и GLRGFYAMDY (SEQ ID NO:12).
В некоторых вариантах осуществления анти-TIGIT-антитело или его антигенсвязывающий фрагмент содержит легкую цепь и/или тяжелую цепь, включающие аминокислотные последовательности, по меньшей мере, на 90% идентичные аминокислотным последовательностям, представленным в DIVMTQSPSSLAVSPGEKVTMTCKSSQSLYYSGVKENLLAWYQQKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAEDMGQYFCQQGINNPLTFGDGTKLEIKR (SEQ ID NO:13) или DVVLTQTPLSLSVSFGDQVSISCRSSQSLVNSYGNTFLSWYLHKPGQSPQLLIFGISNRFSGVPDRFSGSGSGTDFTLKISTIKPEDLGMYYCLQGTHQPPTFGPGTKLEVK (SEQ ID NO: 14) или EVQLVESGGGLTQPGKSLKLSCEASGFTFSSFTMHWVRQSPGKGLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDLKSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS (SEQ ID NO:15) или EVQLQQSGPELVKPGTSMKISCKASGYSFTGHLMNWVKQSHGKNLEWIGLIIPYNGGTSYNQKFKGKATLTVDKSSSTAYMELLSLTSDDSAVYFCSRGLRGFYAMDYWGQGTSVTVSS (SEQ ID NO:16) соответственно.
В некоторых вариантах осуществления антагонист, связывающийся с осью PD-1, выбран из группы, состоящей из PD-1-связывающего антагониста, PD-L1-связывающего антагониста и PD-L2-связывающего антагониста.
В некоторых вариантах осуществления антагонист, связывающийся с осью PD-1, представляет PD-1-связывающий антагонист. В некоторых вариантах осуществления PD-1-связывающий антагонист ингибирует связывание PD-1 с лигандом его партнеров по связыванию. В некоторых вариантах осуществления PD-1-связывающий антагонист ингибирует связывание PD-1 с PD-L1. В некоторых вариантах осуществления PD-1-связывающий антагонист ингибирует связывание PD-1 с PD-L2. В некоторых вариантах осуществления PD-1-связывающий антагонист ингибирует связывание PD-1 с PD-L1 и PD-L2. В некоторых вариантах осуществления PD-1-связывающий антагонист представляет собой антитело. В некоторых вариантах осуществления PD-1-связывающий антагонист представляет антитело MDX-1106 (ниволумаб). В некоторых вариантах осуществления PD-1-связывающий антагонист представляет антитело Merck 3475 (ламбролизумаб). В некоторых вариантах осуществления PD-1-связывающий антагонист представляет антитело CT-011 (пидилизумаб). В некоторых вариантах осуществления PD-1-связывающий антагонист представляет антитело AMP-224.
В некоторых вариантах осуществления антагонист, связывающийся с осью PD-1, представляет PD-L1-связывающий антагонист. В некоторых вариантах осуществления PD-L1-связывающий антагонист ингибирует связывание PD-L1 с PD-1. В некоторых вариантах осуществления PD-L1-связывающий антагонист ингибирует связывание PD-L1 с В7-1. В некоторых вариантах осуществления PD-L1-связывающий антагонист ингибирует связывание PD-L1 с обоими PD-1 и B7-1. В некоторых вариантах осуществления PD-L1-связывающий антагонист представляет собой антитело.
В некоторых вариантах осуществления PD-L1-связывающий антагонист выбран из группы, состоящей из YW243.55.S70, MPDL3280A, MDX-1105 и 4736 MEDI.
В некоторых вариантах осуществления анти-PD-L1-антитело содержит тяжелую цепь, включающую последовательность HVR-H1 GFTFSDSWIH (SEQ ID NO:17), последовательность HVR-H2 AWISPYGGSTYYADSVKG (SEQ ID NO:18) и последовательность HVR-Н3 RHWPGGFDY (SEQ ID NO:19); и легкую цепь, включающую последовательность HVR-L1 RASQDVSTAVA (SEQ ID NO:20), последовательность HVR-L2 SASFLYS (SEQ ID NO:21) и последовательность HVR-L3 QQYLYHPAT (SEQ ID NO:22).
В некоторых вариантах осуществления анти-PD-L1-антитело содержит вариабельную область тяжелой цепи, включающую аминокислотную последовательность EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSA (SEQ ID NO:23), и вариабельную область легкой цепи, включающую аминокислотную последовательность DIQMTQSPSSLSASVGDRVTITCRASQDVSTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYLYHPATFGQGTKVEIKR (SEQ ID NO:24).
В некоторых вариантах осуществления антагонист, связывающийся с осью PD-1, представляет PD-L2-связывающий антагонист. В некоторых вариантах осуществления PD-L2-связывающий антагонист представляет собой антитело. В некоторых вариантах осуществления PD-L2-связывающий антагонист представляет иммуноадгезин.
В некоторых вариантах осуществления рак, подлежащий лечению, выбран из группы, состоящей из немелкоклеточного рака легких, мелкоклеточного рака легких, почечно-клеточного рака, колоректального рака, рака яичников, рака молочной железы, рака поджелудочной железы, карциномы желудка, рака мочевого пузыря, рака пищевода, мезотелиомы, меланомы, злокачественных опухолей головы и шеи, рака щитовидной железы, саркомы, рака предстательной железы, глиобластомы, рака шейки матки, карциномы тимуса, лейкоза, лимфомы, миеломы, грибовидного микоза, рака из клеток Меркеля и других злокачественных заболеваний крови.
В некоторых вариантах осуществления агент, который снижает или ингибирует экспрессию и/или активность TIGIT, вводят непрерывно. В некоторых вариантах осуществления агент, который снижает или ингибирует экспрессию и/или активность TIGIT, вводят периодически. В некоторых вариантах осуществления агент, который снижает или ингибирует экспрессию и/или активность TIGIT, вводят до антагониста, связывающегося с осью PD-1. В некоторых вариантах осуществления агент, который снижает или ингибирует экспрессию и/или активность TIGIT, вводят одновременно с антагонистом, связывающимся с осью PD-1. В некоторых вариантах осуществления, агент, который снижает или ингибирует экспрессию и/или активность TIGIT, вводят после антагониста, связывающегося с осью PD-1.
Также обеспечиваются наборы, содержащие антагонист, связывающийся с осью PD-1, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT, для лечения или замедления прогрессирования рака у индивидуума.
Также обеспечиваются наборы, содержащие антагонист, связывающийся с осью PD-1, и агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT, для лечения или замедления прогрессирования рака у индивидуума.
Также обеспечиваются наборы, содержащие агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в комбинации с антагонистом, связывающимся с осью PD-1, для лечения или замедления прогрессирования рака у индивидуума.
Также обеспечиваются наборы, содержащие антагонист, связывающийся с осью PD-1, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT, для усиления иммунной функции индивидуума, имеющего рак.
Также обеспечиваются наборы, содержащие антагонист, связывающийся с осью PD-1, и агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT, для усиления иммунной функции индивидуума, имеющего рак.
Также обеспечиваются наборы, содержащие агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в комбинации с антагонистом, связывающимся с осью PD-1, для усиления иммунной функции индивидуума, имеющего рак.
Также обеспечиваются наборы, содержащие антагонист, связывающийся с осью PD-1, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, в комбинации с агентом, который модулирует экспрессию и/или активность CD226, для лечения или замедления прогрессирования рака у индивидуума.
Также обеспечиваются наборы, содержащие антагонист, связывающийся с осью PD-1, и агент, который модулирует экспрессию и/или активность CD226, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226, для лечения или замедления прогрессирования рака у индивидуума.
Также обеспечиваются наборы, содержащие агент, который модулирует экспрессию и/или активность CD226, и вкладыш в упаковке, содержащий инструкции по применению агента, который модулирует экспрессию и/или активность CD226, в комбинации с антагонистом, связывающимся с осью PD-1, для лечения или замедления прогрессирования рака у индивидуума.
Также обеспечиваются наборы, содержащие антагонист, связывающийся с осью PD-1, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, в комбинации с агентом, который модулирует экспрессию и/или активность CD226, для усиления иммунной функции у индивидуума, имеющего с рак.
Также обеспечиваются наборы, содержащие антагонист, связывающийся с осью PD-1, и агент, который модулирует экспрессию и/или активность CD226, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226, для усиления иммунной функции у индивидуума, имеющего рак.
Также обеспечиваются наборы, содержащие агент, который модулирует экспрессию и/или активность CD226, и вкладыш в упаковке, содержащий инструкции по применению агента, который модулирует экспрессию и/или активность CD226, в комбинации с антагонистом, связывающимся с осью PD-1, для усиления иммунной функции у индивидуума, имеющего рак.
В некоторых вариантах осуществления наборы содержат антагонист, связывающийся с осью PD-1, который представляет собой анти-PD-L1-антитело. В некоторых вариантах осуществления наборы содержат антагонист, связывающийся с осью PD-1, который представляет собой анти-PD-1-антитело. В некоторых вариантах осуществления наборы содержат агент, который снижает или ингибирует экспрессию и/или активность TIGIT, выбранный из группы, состоящей из антагониста экспрессии и/или активности TIGIT, антагониста экспрессии и/или активности PVR, и агента, который ингибирует взаимодействие и/или внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR. В некоторых вариантах осуществления наборы содержат антагонист экспрессии и/или активности TIGIT, представляющий собой анти-TIGIT-антитело или его антигенсвязывающий фрагмент.
В некоторых вариантах осуществления наборы содержат агент, который модулирует экспрессию и/или активность CD226, который способен повышать и/или стимулировать экспрессию и/или активность CD226. В некоторых вариантах осуществления наборы содержат агент, который модулирует экспрессию и/или активность CD226, выбранный из агента, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, антагониста экспрессии и/или активности TIGIT, антагониста экспрессии и/или активности PVR, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVR, агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR. В некоторых вариантах осуществления наборы содержат агент, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, и/или антагонист экспрессии и/или активности TIGIT, представляющий собой анти-TIGIT-антитело или его антигенсвязывающий фрагмент.
В некоторых аспектах настоящее изобретение относится к способу лечения или замедления прогрессирования рака у индивидуума, включающему введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT. В других аспектах настоящее изобретение относится к применению эффективного количества антагониста, связывающегося с осью PD-1, в производстве лекарственного средства для лечения или замедления прогрессирования рака у индивидуума, где антагонист, связывающийся с осью PD-1, используется в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к применению эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в производстве лекарственного средства для лечения или замедления прогрессирования рака у индивидуума, где агент, который снижает или ингибирует экспрессию и/или активность TIGIT, используется в комбинации с антагонистом, связывающимся с осью PD-1. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей антагонист, связывающийся с осью PD-1, для применения в лечении или замедлении прогрессирования рака в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей агент, который снижает или ингибирует экспрессию и/или активность TIGIT, для применения в лечении или замедлении прогрессирования рака в комбинации с антагонистом, связывающимся с осью PD-1.
В еще одних аспектах настоящее изобретение относится к способу снижения или ингибирования рецидива или прогрессирования рака у индивидуума, включающему введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к применению эффективного количества антагониста, связывающегося с осью PD-1, в производстве лекарственного средства для снижения или ингибирования рецидива или прогрессирования рака у индивидуума, где антагонист, связывающийся с осью PD-1, применяется в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к применению эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в производстве лекарственного средства для снижения или ингибирования рецидива или прогрессирования рака у индивидуума, где агент, который снижает или ингибирует экспрессию и/или активность TIGIT, применяется в комбинации с антагонистом, связывающимся с осью PD-1. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей антагонист, связывающийся с осью PD-1, для применения в снижении или ингибировании рецидива или прогрессирования рака в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей агент, который снижает или ингибирует экспрессию и/или активность TIGIT, для применения в снижении или ингибировании рецидива или прогрессирования рака в комбинации с антагонистом, связывающимся с осью PD-1.
В еще одних аспектах настоящее изобретение относится к способу лечения или замедления прогрессирования связанного с иммунитетом заболевания у индивидуума, включающему введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к применению эффективного количества антагониста, связывающегося с осью PD-1, в производстве лекарственного средства для лечения или замедления прогрессирования связанного с иммунитетом заболевания у индивидуума, где антагонист, связывающийся с осью PD-1, применяется в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к применению эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в производстве лекарственного средства для лечения или замедления прогрессирования связанного с иммунитетом заболевания у индивидуума, где агент, который снижает или ингибирует экспрессию и/или активность TIGIT, применяется в комбинации с антагонистом, связывающимся с осью PD-1. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей антагонист, связывающийся с осью PD-1, для применения в лечении или замедлении прогрессирования связанного с иммунитетом заболевания, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей агент, который снижает или ингибирует экспрессию и/или активность TIGIT, для применения в лечении или замедлении прогрессирования связанного с иммунитетом заболевания, в комбинации с антагонистом, связывающимся с осью PD-1.
В еще одних аспектах настоящее изобретение относится к комбинации, содержащей эффективное количество антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT.
В еще одних аспектах настоящее изобретение относится к способу снижения или ингибирования прогрессирования связанного с иммунитетом заболевания у индивидуума, включающему введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к применению эффективного количества антагониста, связывающегося с осью PD-1, в производстве лекарственного средства для снижения или ингибирования прогрессирования связанного с иммунитетом заболевания у индивидуума, где антагонист, связывающийся с осью PD-1, применяется в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к применению эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в производстве лекарственного средства для снижения или ингибирования прогрессирования связанного с иммунитетом заболевания у индивидуума, где агент, который снижает или ингибирует экспрессию и/или активность TIGIT, применяется в комбинации с антагонистом, связывающимся с осью PD-1. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей антагонист, связывающийся с осью PD-1, для применения в снижении или ингибирования прогрессирования связанного с иммунитетом заболевания в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей агент, который снижает или ингибирует экспрессию и/или активность TIGIT, для применения в снижении или ингибирования прогрессирования связанного с иммунитетом заболевания в комбинации с антагонистом, связывающимся с осью PD-1.
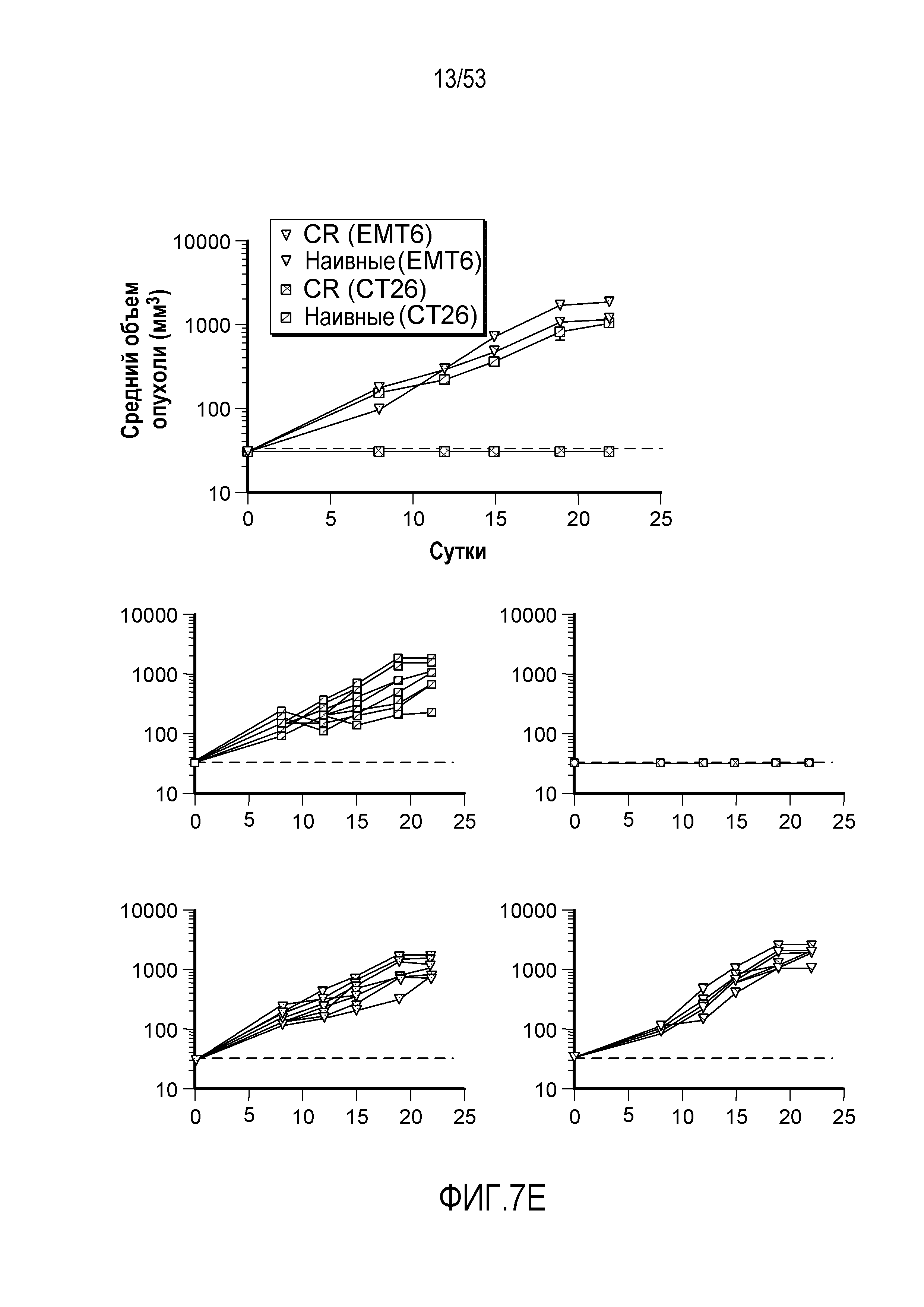
В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, связанное с иммунитетом заболевание ассоциировано с Т-клеточным дисфункциональным расстройством. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, связанное с иммунитетом заболевание представляет вирусную инфекцию. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, вирусная инфекция представляет собой хроническую вирусную инфекцию. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, Т-клеточное дисфункциональное расстройство характеризуется снижением реактивности на антигенную стимуляцию. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, Т-клеточное дисфункциональное расстройство характеризуется анергией Т-клеток или пониженной способностью к секреции цитокинов, пролиферации или осуществлению цитолитической активности. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, Т-клеточное дисфункциональное расстройство характеризуется истощением Т-клеток. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, Т-клетки представляют собой CD4+ и CD8+ Т-клетки. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, связанное с иммунитетом заболевание выбрано из группы, состоящей из неразрешенной острой инфекции, хронической инфекции и иммунитета к опухоли.
В еще одних аспектах настоящее изобретение относится к способу повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, включающему введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к применению эффективного количества антагониста, связывающегося с осью PD-1, в производстве лекарственного средства для усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, где антагонист, связывающийся с осью PD-1, применяется в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к применению эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в производстве лекарственного средства для усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, где агент, который снижает или ингибирует экспрессию и/или активность TIGIT, применяется в комбинации с антагонистом, связывающимся с осью PD-1. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей антагонист, связывающийся с осью PD-1, для применения в усилении или стимуляции иммунного ответа или иммунной функции у индивидуума в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей агент, который снижает или ингибирует экспрессию и/или активность TIGIT, для применения в усилении или стимуляции иммунного ответа или иммунной функции у индивидуума в комбинации с антагонистом, связывающимся с осью PD-1. В еще одних аспектах настоящее изобретение относится к комбинации, содержащей эффективное количество антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT.
В еще одних аспектах настоящее изобретение относится к способу лечения или замедления прогрессирования рака у индивидуума, включающему введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226. В еще одних аспектах настоящее изобретение относится к применению эффективного количества антагониста, связывающегося с осью PD-1, в производстве лекарственного средства для лечения или замедления прогрессирования рака у индивидуума, где антагонист, связывающийся с осью PD-1, применяется в комбинации с агентом, который модулирует экспрессию и/или активность CD226. В еще одних аспектах настоящее изобретение относится к применению эффективного количества агента, который модулирует экспрессию и/или активность CD226, в производстве лекарственного средства для лечения или замедления прогрессирования рака у индивидуума, где агент, который модулирует экспрессию и/или активность CD226, применяется в комбинации с антагонистом, связывающимся с осью PD-1. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей антагонист, связывающийся с осью PD-1, для применения в лечении или замедлении прогрессирования рака в комбинации с агентом, который модулирует экспрессию и/или активность CD226. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей агент, который модулирует экспрессию и/или активность CD226, для применения в лечении или замедлении прогрессирования рака в комбинации с антагонистом, связывающимся с осью PD-1.
В еще одних аспектах настоящее изобретение относится к способу снижения или ингибирования рецидива или прогрессирования рака у индивидуума, включающему введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226. В еще одних аспектах настоящее изобретение относится к применению эффективного количества антагониста, связывающегося с осью PD-1, в производстве лекарственного средства для снижения или ингибирования рецидива или прогрессирования рака у индивидуума, где антагонист, связывающийся с осью PD-1, применяется в комбинации с агентом, который модулирует экспрессию и/или активность CD226. В еще одних аспектах настоящее изобретение относится к применению эффективного количества агента, который модулирует экспрессию и/или активность CD226, в производстве лекарственного средства для снижения или ингибирования рецидива или прогрессирования рака у индивидуума, где агент, который модулирует экспрессию и/или активность CD226, применяется в комбинации с антагонистом, связывающимся с осью PD-1. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей антагонист, связывающийся с осью PD-1, для применения в снижении или ингибировании рецидива или прогрессирования рака в комбинации с агентом, который модулирует экспрессию и/или активность CD226. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей агент, который модулирует экспрессию и/или активность CD226, для применения в снижении или ингибировании рецидива или прогрессирования рака в комбинации с антагонистом, связывающимся с осью PD-1.
В еще одних аспектах настоящее изобретение относится к способу лечения или замедления прогрессирования связанного с иммунитетом заболевания у индивидуума, включающему введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226. В еще одних аспектах настоящее изобретение относится к применению эффективного количества антагониста, связывающегося с осью PD-1, в производстве лекарственного средства для лечения или замедления прогрессирования связанного с иммунитетом заболевания у индивидуума, где антагонист, связывающийся с осью PD-1, применяется в комбинации с агентом, который модулирует экспрессию и/или активность CD226. В еще одних аспектах настоящее изобретение относится к применению эффективного количества агента, который модулирует экспрессию и/или активность CD226, в производстве лекарственного средства для лечения или замедления прогрессирования связанного с иммунитетом заболевания у индивидуума, где агент, который модулирует экспрессию и/или активность CD226, применяется в комбинации с антагонистом, связывающимся с осью PD-1. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей антагонист, связывающийся с осью PD-1, для применения в лечении или замедлении прогрессирования связанного с иммунитетом заболевания в комбинации с агентом, который модулирует экспрессию и/или активность CD226. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей агент, который модулирует экспрессию и/или активность CD226, для применения в лечении или замедлении прогрессирования связанного с иммунитетом заболевания в комбинации с антагонистом, связывающимся с осью PD-1.
В еще одних аспектах настоящее изобретение относится к комбинации, содержащей эффективное количество антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226.
В еще одних аспектах настоящее изобретение относится к способу снижения или ингибирования прогрессирования связанного с иммунитетом заболевания у индивидуума, включающему введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226. В еще одних аспектах настоящее изобретение относится к применению эффективного количества антагониста, связывающегося с осью PD-1, в производстве лекарственного средства для снижения или ингибирования прогрессирования связанного с иммунитетом заболевания у индивидуума, где антагонист, связывающийся с осью PD-1, применяется в комбинации с агентом, который модулирует экспрессию и/или активность CD226. В еще одних аспектах настоящее изобретение относится к применению эффективного количества агента, который модулирует экспрессию и/или активность CD226, в производстве лекарственного средства для снижения или ингибирования прогрессирования связанного с иммунитетом заболевания у индивидуума, где агент, который модулирует экспрессию и/или активность CD226, применяется в комбинации с антагонистом, связывающимся с осью PD-1. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей антагонист, связывающийся с осью PD-1, для применения в снижении или ингибировании прогрессирования связанного с иммунитетом заболевания в комбинации с агентом, который модулирует экспрессию и/или активность CD226. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей агент, который модулирует экспрессию и/или активность CD226, для применения в снижении или ингибировании прогрессирования связанного с иммунитетом заболевания в комбинации с антагонистом, связывающимся с осью PD-1.
В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, связанное с иммунитетом заболевание ассоциировано с Т-клеточным дисфункциональным расстройством. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, связанное с иммунитетом заболевание представляет вирусную инфекцию. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, вирусная инфекция представляет собой хроническую вирусную инфекцию. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, Т-клеточное дисфункциональное расстройство характеризуется снижением реактивности на антигенную стимуляцию. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, Т-клеточное дисфункциональное расстройство характеризуется анергией Т-клеток или пониженной способностью к секреции цитокинов, пролиферации или осуществлению цитолитической активности. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, Т-клеточное дисфункциональное расстройство характеризуется истощением Т-клеток. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, Т-клетки представляют собой CD4+ и CD8+ Т-клетки. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, связанное с иммунитетом заболевание выбрано из группы, состоящей из неразрешенной острой инфекции, хронической инфекции и иммунитета к опухоли.
В еще одних аспектах настоящее изобретение относится к способу повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, включающему введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226. В еще одних аспектах настоящее изобретение относится к применению эффективного количества антагониста, связывающегося с осью PD-1, в производстве лекарственного средства для усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, где антагонист, связывающийся с осью PD-1, применяется в комбинации с агентом, который модулирует экспрессию и/или активность CD226. В еще одних аспектах настоящее изобретение относится к применению эффективного количества агента, который модулирует экспрессию и/или активность CD226, в производстве лекарственного средства для усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, где агент, который модулирует экспрессию и/или активность CD226, применяется в комбинации с антагонистом, связывающимся с осью PD-1. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей антагонист, связывающийся с осью PD-1, для применения в усилении или стимуляции иммунного ответа или иммунной функции в комбинации с агентом, который модулирует экспрессию и/или активность CD226. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей агент, который модулирует экспрессию и/или активность CD226, для применения в усилении или стимуляции иммунного ответа или иммунной функции в комбинации с антагонистом, связывающимся с осью PD-1. В еще одних аспектах настоящее изобретение относится к комбинации, содержащей эффективное количество антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226.
В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который модулирует экспрессию и/или активность CD226, представляет агент, который повышает и/или стимулирует экспрессию и/или активность CD226. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который модулирует экспрессию и/или активность CD226, представляет агент, который повышает и/или стимулирует взаимодействие CD226 с PVR. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который модулирует экспрессию и/или активность CD226, представляет агент, который повышает и/или стимулирует внутриклеточную передачу сигналов, опосредованную связыванием CD226 с PVR. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который модулирует экспрессию и/или активность CD226, выбран из группы, состоящей из агента, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, антагониста экспрессии и/или активности TIGIT, антагониста экспрессии и/или активности PVR, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVR, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL2, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL3, агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR, агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL2, агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL3, и их комбинаций. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который модулирует экспрессию и/или активность CD226, представляет агент, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, представляет анти-TIGIT-антитело или его антигенсвязывающий фрагмент. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, представляет ингибирующую нуклеиновую кислоту, выбранную из группы, состоящей из антисмыслового полинуклеотида, интерферирующей РНК, каталитической РНК и химеры РНК-ДНК. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антисмысловой полинуклеотид направлен на TIGIT. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, интерферирующая РНК направлена на TIGIT. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, каталитическая РНК направлена на TIGIT. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, химера РНК-ДНК направлена на TIGIT. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который модулирует экспрессию и/или активность CD226, представляет антагонист экспрессии и/или активности TIGIT. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист экспрессии и/или активности TIGIT представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист экспрессии и/или активности TIGIT представляет анти-TIGIT-антитело или его антигенсвязывающий фрагмент. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист экспрессии и/или активности TIGIT представляет ингибирующую нуклеиновую кислоту, выбранную из группы, состоящей из антисмыслового полинуклеотида, интерферирующей РНК, каталитической РНК и химеры РНК-ДНК. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист экспрессии и/или активности PVR, выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVR, выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL2, выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/ или блокирует взаимодействие TIGIT с PVRL3, выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR, выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL2, выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL3, выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида.
В еще одних аспектах настоящее изобретение относится к способу повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, включающему введение индивидууму эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который снижает или ингибирует один или более дополнительных иммунных ко-ингибирующих рецепторов. В еще одних аспектах настоящее изобретение относится к применению эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT в производстве лекарственного средства для усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, где агент, который снижает или ингибирует экспрессию и/или активность TIGIT, применяется в комбинации с агентом, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов. В еще одних аспектах настоящее изобретение относится к применению эффективного количества агента, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов, в производстве лекарственного средства для усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, где агент, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов, применяют в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей агент, который снижает или ингибирует экспрессию и/или активность TIGIT, для применения в усиления или стимуляции иммунного ответа или иммунной функции в комбинации с агентом, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей агент, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов, для применения в усилении или стимуляции иммунного ответа или иммунной функции в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к комбинации, содержащей эффективное количество агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, один или более дополнительных иммунных коингибирующих рецепторов выбран из группы, состоящей из PD-1, CTLA-4, LAG3, TIM3, BTLA, VISTA, B7H4 и CD96. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, один или более дополнительных иммунных коингибирующих рецепторов выбран из группы, состоящей из PD-1, CTLA-4, LAG3 и TIM3.
В еще одних аспектах настоящее изобретение относится к способу повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, включающему введение индивидууму эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов. В еще одних аспектах настоящее изобретение относится к применению эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в производстве лекарственного средства для усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, где агент, который снижает или ингибирует экспрессию и/или активность TIGIT, применяется в комбинации с агентом, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов. В еще одних аспектах настоящее изобретение относится к применению эффективного количества агента, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, в производстве лекарственного средства для усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, где агент, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, применяется в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей агент, который снижает или ингибирует экспрессию и/или активность TIGIT, для применения в усилении или стимуляции иммунного ответа или иммунной функции в комбинации с агентом, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов. В еще одних аспектах настоящее изобретение относится к фармацевтической композиции, содержащей агент, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, для применения в усилении или стимуляции иммунного ответа или иммунной функции в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. В еще одних аспектах настоящее изобретение относится к комбинации, содержащей эффективное количество агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, один или более дополнительных иммунных костимулирующих рецепторов выбран из группы, состоящей из CD226, ОХ-40, CD28, CD27, CD137, HVEM, GITR, MICA, ICOS, NKG2D и 2В4. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, один или более дополнительных иммунных костимулирующих рецепторов выбран из группы, состоящей из CD226, ОХ-40, CD27, CD137, HVEM и GITR. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, один или более дополнительных иммунных костимулирующих рецепторов выбран из группы, состоящей из ОХ-40 и CD27.
В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, способ дополнительно включает введение, по меньшей мере, одного химиотерапевтического агента. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, у индивидуума имеется рак. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, индивидуум является человеком. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, CD4 и/или CD8 Т-клетки у индивидуума имеют повышенное или усиленное праймирование, активацию, пролиферацию, высвобождение цитокинов и/или цитолитическую активность по сравнению с периодом до введения комбинации. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, количество CD4 и/или CD8 Т-клеток повышено по сравнению с периодом до введения комбинации. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, количество активированных CD4 и/или CD8 Т-клеток повышено по сравнению с периодом до введения комбинации. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, активированные CD4 и/или CD8 Т-клетки характеризуются CD4 и/или CD8 Т-клетками, продуцирующими γ-IFN+, и/или повышенной цитолитической активностью по сравнению с периодом до введения комбинации. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, CD4 и/или CD8 Т-клетки проявляют повышенное высвобождение цитокинов, выбранных из группы, состоящей из IFN-γ, TNF-α и интерлейкинов. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, CD4 и/или CD8 Т-клетки представляют эффекторные Т-клетки памяти. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, CD4 и/или CD8 эффекторные Т-клетки памяти характеризуются CD4 и/или CD8 Т-клетками, продуцирующими γ-IFN+, и/или повышенной цитолитической активностью. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, CD4 и/или CD8 эффекторные Т-клетки памяти характеризуется наличием экспрессии CD44высокий CD62Lнизкий. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, злокачественная опухоль имеет повышенные уровни Т-клеточной инфильтрации. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который снижает или ингибирует экспрессию и/или активность TIGIT, выбран из группы, состоящей из антагониста экспрессии и/или активности TIGIT, антагониста экспрессии и/или активности PVR, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVR, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL2, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL3, агента, который ингибирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR, агента, который ингибирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL2, агента, который ингибирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL3, и их комбинаций. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист экспрессии и/или активности TIGIT выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист экспрессии и/или активности PVR выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVR, выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL2, выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL3, выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR, выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL2, выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL3, выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист экспрессии и/или активности TIGIT представляет ингибирующую нуклеиновую кислоту, выбранную из группы, состоящей из антисмыслового полинуклеотида, интерферирующей РНК, каталитической РНК и химеры РНК-ДНК. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антисмысловой полинуклеотид направлен на TIGIT.
В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, интерферирующая РНК направлена на TIGIT. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, каталитическая РНК направлена на TIGIT. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, химера РНК-ДНК направлена на TIGIT. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист экспрессии и/или активности TIGIT представляет анти-TIGIT-антитело или его антигенсвязывающий фрагмент. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, анти-TIGIT-антитело или его антигенсвязывающий фрагмент содержит, по меньшей мере, один HVR, включающий аминокислотную последовательность, выбранную из аминокислотных последовательностей (1) KSSQSLYYSGVKENLLA (SEQ ID NO:1), ASIRFT (SEQ ID NO:2), QQGINNPLT (SEQ ID NO:3), GFTFSSFTMH (SEQ ID NO:4), FIRSGSGIVFYADAVRG (SEQ ID NO:5) и RPLGHNTFDS (SEQ ID NO: 6); или (2) RSSQSLVNSYGNTFLS (SEQ ID NO:7), GISNRFS (SEQ ID NO:8), LQGTHQPPT (SEQ ID NO:9), GYSFTGHLMN (SEQ ID NO:10), LIIPYNGGTSYNQKFKG (SEQ ID NO:11) и GLRGFYAMDY (SEQ ID NO:12). В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, обеспечивается анти-TIGIT-антитело или его антигенсвязывающий, фрагмент, где легкая цепь антитела содержит аминокислотную последовательность, представленную в DIVMTQSPSSLAVSPGEKVTMTCKSSQSLYYSGVKENLLAWYQQKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAEDMGQYFCQQGINNPLTFGDGTKLEIKR (SEQ ID NO:13) или DVVLTQTPLSLSVSFGDQVSISCRSSQSLVNSYGNTFLSWYLHKPGQSPQLLIFGISNRFSGVPDRFSGSGSGTDFTLKISTIKPEDLGMYYCLQGTHQPPTFGPGTKLEVK (SEQ ID NO:14). В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, обеспечивается анти-TIGIT-антитело или его антигенсвязывающий фрагмент, где тяжелая цепь антитела содержит аминокислотную последовательность, представленную в EVQLVESGGGLTQPGKSLKLSCEASGFTFSSFTMHWVRQSPGKGLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDLKSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS (SEQ ID NO:15) или VQLQQSGPELVKPGTSMKISCKASGYSFTGHLMNWVKQSHGKNLEWIGLIIPYNGGTSYNQKFKGKATLTVDKSSSTAYMELLSLTSDDSAVYFCSRGLRGFYAMDYWGQGTSVTVSS (SEQ ID NO:16). В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, обеспечивается анти-TIGIT-антитело или его антигенсвязывающий фрагмент, где легкая цепь антитела содержит аминокислотную последовательность, представленную в DIVMTQSPSSLAVSPGEKVTMTCKSSQSLYYSGVKENLLAWYQQKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAEDMGQYFCQQGINNPLTFGDGTKLEIKR (SEQ ID NO:13) или DVVLTQTPLSLSVSFGDQVSISCRSSQSLVNSYGNTFLSWYLHKPGQSPQLLIFGISNRFSGVPDRFSGSGSGTDFTLKISTIKPEDLGMYYCLQGTHQPPTFGPGTKLEVK (SEQ ID NO:14), и тяжелая цепь антитела содержит аминокислотную последовательность, представленную в EVQLVESGGGLTQPGKSLKLSCEASGFTFSSFTMHWVRQSPGKGLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDLKSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS (SEQ ID NO:15) или EVQLQQSGPELVKPGTSMKISCKASGYSFTGHLMNWVKQSHGKNLEWIGLIIPYNGGTSYNQKFKGKATLTVDKSSSTAYMELLSLTSDDSAVYFCSRGLRGFYAMDYWGQGTSVTVSS SEQ ID NO:16). В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, анти-TIGIT-антитело или его антигенсвязывающий фрагмент, где антитело выбрано из группы, состоящей из гуманизированного антитела, химерного антитела, биспецифического антитела, гетероконъюгатного антитела и иммунотоксина. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, анти-TIGIT-антитело или его антигенсвязывающий фрагмент содержит, по меньшей мере, один HVR, который, по меньшей мере, на 90% идентичен HVR, представленному в любой из последовательностей (1) KSSQSLYYSGVKENLLA (SEQ ID NO:1), ASIRFT (SEQ ID NO:2), QQGINNPLT (SEQ ID NO:3), GFTFSSFTMH (SEQ ID NO:4), FIRSGSGIVFYADAVRG (SEQ ID NO:5) и RPLGHNTFDS (SEQ ID NO:6); или (2) RSSQSLVNSYGNTFLS (SEQ ID NO:7), GISNRFS (SEQ ID NO:8), LQGTHQPPT (SEQ ID NO:9), GYSFTGHLMN (SEQ ID NO:10), LIIPYNGGTSYNQKFKG (SEQ ID NO:11) и GLRGFYAMDY (SEQ ID NO:12). В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, анти-TIGIT-антитело или его антигенсвязывающий фрагмент содержит легкую цепь, включающую аминокислотные последовательности, по меньшей мере, на 90% идентичные аминокислотным последовательностям, представленным в DIVMTQSPSSLAVSPGEKVTMTCKSSQSLYYSGVKENLLAWYQQKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAEDMGQYFCQQGINNPLTFGDGTKLEIKR (SEQ ID NO:13) или DVVLTQTPLSLSVSFGDQVSISCRSSQSLVNSYGNTFLSWYLHKPGQSPQLLIFGISNRFSGVPDRFSGSGSGTDFTLKISTIKPEDLGMYYCLQGTHQPPTFGPGTKLEVK (SEQ ID NO:14); и/или тяжелую цепь, включающую аминокислотные последовательности, по меньшей мере, на 90% идентичные аминокислотным последовательностям, представленным в EVQLVESGGGLTQPGKSLKLSCEASGFTFSSFTMHWVRQSPGKGLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDLKSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS (SEQ ID NO:15) или EVQLQQSGPELVKPGTSMKISCKASGYSFTGHLMNWVKQSHGKNLEWIGLIIPYNGGTSYNQKFKGKATLTVDKSSSTAYMELLSLTSDDSAVYFCSRGLRGFYAMDYWGQGTSVTVSS (SEQ ID NO:16). В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист, связывающийся с осью PD-1, выбран из группы, состоящей из PD-1-связывающего антагониста, PD-L1-связывающего антагониста и PD-L2-связывающего антагониста. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист, связывающийся с осью PD-1, представляет PD-1-связывающий антагонист. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-1-связывающий антагонист ингибирует связывание PD-1 с лигандами его партнеров по связыванию. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-1-связывающий антагонист ингибирует связывание PD-1 с PD-L1. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-1-связывающий антагонист ингибирует связывание PD-1 с PD-L2. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-1-связывающий антагонист ингибирует связывание PD-1 с обоими PD-L1 и PD-L2. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-1-связывающий антагонист представляет собой антитело. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-1-связывающий антагонист представляет антитело MDX-1106. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-1-связывающий антагонист представляет антитело МК-3475. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-1-связывающий антагонист представляет антитело СТ-011. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-1-связывающий антагонист представляет антитело AMP-224. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист, связывающийся с осью PD-1, представляет PD-L1-связывающий антагонист. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-L1-связывающий антагонист ингибирует связывание PD-L1 с PD-1. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-L1-связывающий антагонист ингибирует связывание PD-L1 с B7-1. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-L1-связывающий антагонист ингибирует связывание PD-L1 с обоими PD-1 и B7-1. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-L1-связывающий антагонист представляет анти-PD-L1-антитело. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-L1-связывающий антагонист выбран из группы, состоящей из YW243.55.S70, MPDL3280A, MDX-1105 и MEDI4736. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, анти-PD-L1-антитело содержит тяжелую цепь, включающую последовательность HVR-H1 GFTFSDSWIH (SEQ ID NO:17), последовательность HVR-H2 AWISPYGGSTYYADSVKG (SEQ ID NO:18) и последовательность HVR-H3 RHWPGGFDY (SEQ ID NO:19); и легкую цепь, включающую последовательность HVR-L1 RASQDVSTAVA (SEQ ID NO:20), последовательность HVR-L2 SASFLYS (SEQ ID NO:21) и последовательность HVR-L3 QQYLYHPAT (SEQ ID NO:22). В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, анти-PD-L1-антитело содержит вариабельную область тяжелой цепи, включающую аминокислотную последовательность EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSA (SEQ ID NO:23), EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSSASTK (SEQ ID NO:40) или EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS (SEQ ID NO:41), и вариабельную область легкой цепи, включающую аминокислотную последовательность DIQMTQSPSSLSASVGDRVTITCRASQDVSTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYLYHPATFGQGTKVEIKR (SEQ ID NO:24). В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист, связывающийся с осью PD-1, представляет PD-L2-связывающий антагонист. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-L2-связывающий антагонист представляет собой антитело. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-L2-связывающий антагонист представляет собой иммуноадгезин. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, рак выбран из группы, состоящей из немелкоклеточного рака легких, мелкоклеточного рака легких, почечно-клеточного рака, колоректального рака, рака яичников, рака молочной железы, рака поджелудочной железы, карциномы желудка, рака мочевого пузыря, рака пищевода, мезотелиомы, меланомы, злокачественных опухолей головы и шеи, рака щитовидной железы, саркомы, рака предстательной железы, глиобластомы, рака шейки матки, карциномы тимуса, лейкоза, лимфомы, миеломы, грибовидного микоза, рака из клеток Меркеля и других злокачественных заболеваний крови. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который снижает или ингибирует экспрессию и/или активность TIGIT, вводят непрерывно. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который снижает или ингибирует экспрессию и/или активность TIGIT, вводят периодически. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который снижает или ингибирует экспрессию и/или активность TIGIT, вводят до введения антагониста, связывающегося с осью PD-1. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который снижает или ингибирует экспрессию и/или активность TIGIT, вводят одновременно с антагонистом, связывающимся с осью PD-1. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который снижает или ингибирует экспрессию и/или активность TIGIT вводят после введения антагониста, связывающегося с осью PD-1. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист, связывающийся с осью PD-1, вводят до агента, который модулирует экспрессию и/или активность CD226. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист, связывающийся с осью PD-1, вводят одновременно с агентом, который модулирует экспрессию и/или активность CD226. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист, связывающийся с осью PD-1, вводят после агента, который модулирует экспрессию и/или активность CD226. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который снижает или ингибирует экспрессию и/или активность TIGIT, вводят до агента, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который снижает или ингибирует экспрессию и/или активность TIGIT, вводят одновременно с агентом, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который снижает или ингибирует экспрессию и/или активность TIGIT, вводят после агента, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который снижает или ингибирует экспрессию и/или активность TIGIT, вводят до агента, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который снижает или ингибирует экспрессию и/или активность TIGIT, вводят одновременно с агентом, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который снижает или ингибирует экспрессию и/или активность TIGIT, вводят после агента, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов.
В еще одних аспектах настоящее изобретение относится к набору, содержащему антагонист, связывающийся с осью PD-1, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT, для лечения или замедления прогрессирования рака у индивидуума.
В еще одних аспектах настоящее изобретение относится к набору, содержащему антагонист, связывающийся с осью PD-1, и агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT, для лечения или замедления прогрессирования рака у индивидуума.
В еще одних аспектах настоящее изобретение относится к набору, содержащему агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в комбинации с антагонистом, связывающимся с осью PD-1, для лечения или замедления прогрессирования рака у индивидуума.
В еще одних аспектах настоящее изобретение относится к набору, содержащему антагонист, связывающийся с осью PD-1, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT для усиления иммунной функции индивидуума, имеющего рак.
В еще одних аспектах настоящее изобретение относится к набору, содержащему антагонист, связывающийся с осью PD-1, и агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT для усиления иммунной функции у индивидуума, имеющего рак.
В еще одних аспектах настоящее изобретение относится к набору, содержащий агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в комбинации с антагонистом, связывающимся с осью PD-1, для усиления иммунной функции у индивидуума, имеющего рак.
В еще одних аспектах настоящее изобретение относится к набору, содержащему антагонист, связывающийся с осью PD-1, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, в комбинации с агентом, который модулирует экспрессию и/или активность CD226, для лечения или замедления прогрессирования рака у индивидуума.
В еще одних аспектах настоящее изобретение относится к набору, содержащему антагонист, связывающийся с осью PD-1, и агент, который модулирует экспрессию и/или активность CD226, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226, для лечения или замедления прогрессирования рака у индивидуума.
В еще одних аспектах настоящее изобретение относится к набору, содержащему агент, который модулирует экспрессию и/или активность CD226, и вкладыш в упаковке, содержащий инструкции по применению агент, который модулирует экспрессию и/или активность CD226, в комбинации с антагонистом, связывающимся с осью PD-1, для лечения или замедления прогрессирования рака у индивидуума.
В еще одних аспектах настоящее изобретение относится к набору, содержащему антагонист, связывающийся с осью PD-1, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, в комбинации с агентом, который модулирует экспрессию и/или активность CD226, для усиления иммунной функции у индивидуума, имеющего рак.
В еще одних аспектах настоящее изобретение относится к набору, содержащему антагонист, связывающийся с осью PD-1, и агент, который модулирует экспрессию и/или активность CD226, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226, для усиления иммунной функции у индивидуума, имеющего рак.
В еще одних аспектах настоящее изобретение относится к набору, содержащему агент, который модулирует экспрессию и/или активность CD226, и вкладыш в упаковке, содержащий инструкции по применению агента, который модулирует экспрессию и/или активность CD226, в комбинации с антагонистом, связывающимся с осью PD-1, для усиления иммунной функции индивидуума, имеющего рак.
В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист, связывающийся с осью PD-1, представляет собой анти-PD-L1-антитело. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, анти-PD-L1-антитело выбрано из группы, состоящей из YW243.55.S70, MPDL3280A, MDX-1105 и MEDI4736. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, анти-PD-L1-антитело содержит тяжелую цепь, включающую последовательность HVR-H1 GFTFSDSWIH (SEQ ID NO:17), последовательность HVR-H2 AWISPYGGSTYYADSVKG (SEQ ID NO:18) и последовательность HVR-H3 RHWPGGFDY (SEQ ID NO:19); и легкую цепь, включающую последовательность HVR-L1 RASQDVSTAVA (SEQ ID NO:20), последовательность HVR-L2 SASFLYS (SEQ ID NO:21) и последовательность HVR-L3 QQYLYHPAT (SEQ ID NO:22). В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, анти-PD-L1-антитело содержит вариабельную область тяжелой цепи, включающую аминокислотную последовательность EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSA (SEQ ID NO:23), EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSSASTK (SEQ ID NO:40) или EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS (SEQ ID NO:41), и вариабельную область легкой цепи, включающую аминокислотную последовательность DIQMTQSPSSLSASVGDRVTITCRASQDVSTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYLYHPATFGQGTKVEIKR (SEQ ID NO:24). В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист, связывающийся с осью PD-1, представляет анти-PD-1-антитело. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, анти-PD-1-антитело представляет MDX-1106, МK-3475 или СТ-011. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист, связывающийся с осью PD-1, представляет антитело AMP-224. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист, связывающийся с осью PD-1, представляет PD-L2-связывающий антагонист. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-L2-связывающий антагонист представляет собой антитело. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, PD-L2-связывающий антагонист, представляет собой иммуноадгезин.
В еще одних аспектах настоящее изобретение относится к набору, содержащему агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в комбинации с агентом, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов для лечения или замедления прогрессирования рака у индивидуума. В еще одних аспектах настоящее изобретение относится к набору, содержащему агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и агент, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов, и вкладыш в упаковке, содержащую инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов, для лечения или замедления прогрессирования рака у индивидуума. В еще одних аспектах настоящее изобретение относится к набору, содержащему агент, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT, для лечения или замедления прогрессирования рака у индивидуума. В еще одних аспектах настоящее изобретение относится к набору, содержащему агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в комбинации с агентом, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов, для усиления иммунной функции у индивидуума, имеющего рак. В еще одних аспектах настоящее изобретение относится к набору, содержащему агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и агент, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов, для усиления иммунной функции у индивидуума, имеющего рак. В еще одних аспектах настоящее изобретение относится к набору, содержащему агент, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT, для усиления иммунной функции у индивидуума, имеющего рак. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, один или более дополнительных иммунных коингибирующих рецепторов выбран из группы, состоящей из PD-1, CTLA-4, LAG3, TIM3, BTLA, VISTA, B7H4 и CD96. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, один или более дополнительных иммунных коингибирующих рецепторов выбран из группы, состоящей из PD-1, CTLA-4, LAG3 и TIM3.
В еще одних аспектах настоящее изобретение относится к набору, содержащему агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в комбинации с агентом, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, для лечения или замедления прогрессирования рака у индивидуума. В еще одних аспектах настоящее изобретение относится к набору, содержащему агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и агент, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, для лечения или замедления прогрессирования рака у индивидуума. В еще одних аспектах настоящее изобретение относится к набору, содержащему агент, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, и вкладыш в упаковке, содержащий инструкции по применению агента, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT, для лечения или замедления прогрессирования рака у индивидуума. В еще одних аспектах настоящее изобретение относится к набору, содержащему агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в комбинации с агентом, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, для усиления иммунной функции у индивидуума, имеющего рак. В еще одних аспектах настоящее изобретение относится к набору, содержащему агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и агент, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, для усиления иммунной функции у индивидуума, имеющего рак. В еще одних аспектах настоящее изобретение относится к набору, содержащему агент, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, и вкладыш в упаковке, содержащий инструкции по применению агента, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT, для усиления иммунной функции у индивидуума, имеющего рак. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, один или более дополнительных иммунных костимулирующих рецепторов выбран из группы, состоящей из CD226, ОХ-40, CD28, CD27, CD137, HVEM, GITR, MICA, ICOS, NKG2D и 2В4. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, один или более дополнительных иммунных костимулирующих рецепторов выбран из группы, состоящей из CD226, ОХ-40, CD27, CD137, HVEM и GITR. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, один или более дополнительных иммунных костимулирующих рецепторов выбран из группы, состоящей из ОХ-40 и CD27.
В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, индивидуум является человеком. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который снижает или ингибирует экспрессию и/или активность TIGIT, выбран из группы, состоящей из антагониста экспрессии и/или активности TIGIT, антагониста экспрессии и/или активности PVR, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVR, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL2, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL3, агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR, агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVL2, и агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVL3. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, антагонист экспрессии и/или активности TIGIT представляет собой анти-TIGIT-антитело или его антигенсвязывающий фрагмент. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который модулирует экспрессию и/или активность CD226, представляет агент, который повышает и/или стимулирует экспрессию и/или активность CD226. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который модулирует экспрессию и/или активность CD226, представляет агент, который повышает и/или стимулирует взаимодействие CD226 с PVR. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который модулирует экспрессию и/или активность CD226, представляет агент, который повышает и/или стимулирует внутриклеточную передачу сигналов, опосредованную связыванием CD226 с PVR. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который модулирует экспрессию и/или активность CD226, выбран из группы, состоящей из агента, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, антагониста экспрессии и/или активности TIGIT, антагониста экспрессии и/или активности PVR, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVR, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL2, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL3, агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR, агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL2, и агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL3. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который модулирует экспрессию и/или активность CD226, представляет агент, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, агент, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, представляет анти-TIGIT-антитело или его антигенсвязывающий фрагмент. В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, анти-TIGIT-антитело или его антигенсвязывающий фрагмент содержит, по меньшей мере, один HVR, включающий аминокислотную последовательность, выбранную из аминокислотных последовательностей (1) KSSQSLYYSGVKENLLA (SEQ ID NO:1), ASIRFT (SEQ ID NO:2), QQGINNPLT (SEQ ID NO:3), GFTFSSFTMH (SEQ ID NO:4), FIRSGSGIVFYADAVRG (SEQ ID NO:5) и RPLGHNTFDS (SEQ ID NO:6); или (2) RSSQSLVNSYGNTFLS (SEQ ID NO:7), GISNRFS (SEQ ID NO:8), LQGTHQPPT (SEQ ID NO:9), GYSFTGHLMN (SEQ ID NO:10), LIIPYNGGTSYNQKFKG (SEQ ID NO:11) и GLRGFYAMDY (SEQ ID NO:12). В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, обеспечивается анти-TIGIT-антитело или его антигенсвязывающий фрагмент, где легкая цепь антитела включает аминокислотную последовательность, представленную в DIVMTQSPSSLAVSPGEKVTMTCKSSQSLYYSGVKENLLAWYQQKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAEDMGQYFCQQGINNPLTFGDGTKLEIKR (SEQ ID NO:13) или DVVLTQTPLSLSVSFGDQVSISCRSSQSLVNSYGNTFLSWYLHKPGQSPQLLIFGISNRFSGVPDRFSGSGSGTDFTLKISTIKPEDLGMYYCLQGTHQPPTFGPGTKLEVK (SEQ ID NO:14). В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, обеспечивается анти-TIGIT-антитело или его антигенсвязывающий фрагмент, где тяжелая цепь антитела включает аминокислотную последовательность, представленную в EVQLVESGGGLTQPGKSLKLSCEASGFTFSSFTMHWVRQSPGKGLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDLKSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS (SEQ ID NO:15) или EVQLQQSGPELVKPGTSMKISCKASGYSFTGHLMNWVKQSHGKNLEWIGLIIPYNGGTSYNQKFKGKATLTVDKSSSTAYMELLSLTSDDSAVYFCSRGLRGFYAMDYWGQGTSVTVSS (SEQ ID NO:16). В некоторых вариантах осуществления, которые могут быть объединены с любым из предшествующих вариантов осуществления, обеспечивается анти-TIGIT-антитело или его антигенсвязывающий фрагмент, где легкая цепь антитела включает аминокислотную последовательность, представленную в DIVMTQSPSSLAVSPGEKVTMTCKSSQSLYYSGVKENLLAWYQQKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAEDMGQYFCQQGINNPLTFGDGTKLEIKR (SEQ ID NO:13) или DVVLTQTPLSLSVSFGDQVSISCRSSQSLVNSYGNTFLSWYLHKPGQSPQLLIFGISNRFSGVPDRFSGSGSGTDFTLKISTIKPEDLGMYYCLQGTHQPPTFGPGTKLEVK (SEQ ID NO:14), и тяжелая цепь антитела включает аминокислотную последовательность, представленную в EVQLVESGGGLTQPGKSLKLSCEASGFTFSSFTMHWVRQSPGKGLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDLKSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS (SEQ ID NO:15) или EVQLQQSGPELVKPGTSMKISCKASGYSFTGHLMNWVKQSHGKNLEWIGLIIPYNGGTSYNQKFKGKATLTVDKSSSTAYMELLSLTSDDSAVYFCSRGLRGFYAMDYWGQGTSVTVSS (SEQ ID NO:16).
Краткое описание фигур
На фиг. 1 показано, что TIGIT экспрессируется на высоком уровне на истощенных CD8+ и CD4+ Т-клетках. На фиг. 1A показаны CD8+ Т-клетки селезенки мышей C57BL6/J, обогащенные с использованием MACS, которые стимулировали фиксированными на планшетах анти-CD3- и анти-CD28-антителами в течение 24-48 ч in vitro. Гистограммы проточной цитометрии представляют экспрессию TIGIT (красный цвет) по сравнению с изотипным окрашиванием (серый цвет). Также представлены результаты количественного определения TIGIT MFI. ***Р <0,001. Данные представляют результаты 2 независимых опытов; n = 3. На фиг. 1B-1C, мышей C57BL6/J заражали штаммом Armstrong LCMV и спленоциты анализировали через 7 суток после заражения. Данные представляют результаты двух независимых опытов; n = 5. На фиг. 1В представлена гистограмма проточной цитометрии, показывающая экспрессию TIGIT наивными (CD44низкий CD62Lвысокий) и эффекторными CD4+ и CD8+ Т-клетками памяти (CD44высокий CD62Lнизкий). Также представлены результаты количественного определения TIGIT MFI. ***Р<0,001. На фиг. 1С представлена гистограмма проточной цитометрии, показывающая экспрессию TIGIT PD-1высокий и PD-1низкий эффекторными CD8+ Т-клетками. Также представлены результаты количественного определения TIGIT MFI. ***Р<0,001. На фиг. 1D показано, что у мышей C57BL6/J быстро истощали CD4+ Т-клетки и животных заражали клоном 13 штамма LCMV. Спленоциты анализировали через 42 суток после заражения. Гистограмма проточной цитометрии показывает экспрессию TIGIT наивными CD8+ Т-клетками (CD44низкий CD62Lвысокий), CD8+ Т-клетками центральной памяти (CD44высокий CD62Lвысокий) и эффекторными (CD44высокий CD62Lнизкий) CD8+ Т-клетками памяти. Также представлены результаты количественного определения TIGIT MFI. ***Р <0,001. Данные представляют результаты двух независимых опыта; n = 5. Вертикальные линии показывают стандартную ошибку среднего.
На фиг. 2 показано получение мыши TIGITloxP/loxP. Экзон 1 TIGIT фланкировали сайтами loxP с использованием стандартных методик.
На фиг. 3 показано, что клетки TIGIT-дефицитные CD8+ и CD4+ Т-клетки нормально отвечают на острую вирусную инфекцию. Мышей TIGITfl/fl CD4cre (СКО) и мышей из этого же помета TIGITfl/fl (WT) заражали штаммом Armstrong LCMV. Спленоциты анализировали через 7 суток после заражения. Данные представляют результаты двух независимых опытов; n = 5. На фиг. 3А показаны репрезентативные диаграммы FACS, гейтированные на CD8+ Т-клетках, где активированные (CD44высокий) клетки находятся в рамке. Результаты количественного определения активированных CD8+ Т-клеток в процентах от общего числа CD8+ Т-клеток. На фиг. 3B показаны репрезентативные диаграммы FACS, гейтированные на CD8+ Т-клетках, после стимуляции in vitro, где IFNγ-продуцирующие клетки находятся в рамке. Результаты количественного определения IFNγ-продуцирующих клеток в процентах от общего числа CD8+ Т-клеток. На фиг. 3C показаны репрезентативные диаграммы FACS, гейтированные на CD4+ Т-клетках, где активированные (CD44высокий) клетки находятся в рамке. Результаты количественного определения активированных CD4+ Т-клеток в процентах от общего числа CD4+ Т-клеток. На фиг. 3D показаны репрезентативные диаграммы FACS, гейтированные на CD4+ Т-клетках, после стимуляции in vitro, где IFNγ-продуцирующие клетки находятся в рамке. Результаты количественного определения IFNγ-продуцирующих клеток в процентах от общего числа CD4+ Т-клеток. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 4 показано, что TIGIT и PD-1 синергически регулируют эффекторную функцию истощенных Т-клеток in vivo. На фиг. 4A-4E, у мышей TIGITfl/flCD4-cre- (WT) и TIGITfl/fl CD4-cre+ (СКО) быстро истощали CD4+ Т-клетки и животных заражали клоном 13 штамма LCMV. Титры вируса в спленоцитах и печени определяли через 42 суток после заражения. Данные представляют результаты двух независимых опытов, и n=9 в группе. На фиг. 4A показаны репрезентативные диаграммы FACS, гейтированные на CD8+ Т-клетках, где активированные (CD44высокий CD62Lнизкий) клетки находятся в рамке. Результаты количественного определения активированных CD8+ Т-клеток в процентах от общего числа CD8+ Т-клеток. На фиг. 4В показаны репрезентативные диаграммы FACS, гейтированные на CD8+ Т-клетках, после стимуляции in vitro, где IFNγ-продуцирующие клетки находятся в рамке. Результаты количественного определения активированных IFNγ-продуцирующих клеток в процентах от общего числа CD8+ Т-клеток. На фиг. 4C показаны репрезентативные диаграммы FACS, гейтированные на CD4+ Т-клетках, где активированные (CD44высокий CD62Lнизкий) клетки находятся в рамке. Результаты количественного определения активированных CD4+ Т-клеток в процентах от общего числа CD4+ Т-клеток. На фиг. 4D показаны репрезентативные диаграммы FACS, гейтированные на CD4+ Т-клетках, после стимуляции in vitro, где IFNγ-продуцирующие клетки находятся в рамке. Результаты количественного определения IFNγ-продуцирующих клеток в процентах от общего числа CD4+ Т-клеток. На фиг. 4E показаны результаты количественного определения титров LCMV в печени. ***Р <0,0001. На фиг. 4F-4H, у мышей C57BL6/J быстро истощали CD4+ Т-клетки и животных заражали клоном 13 штамма LCMV. Мышей обрабатывали изотипически сходными контрольными антителами, анти-PD-L1-, анти-TIGIT- или анти-PD-L1-+ анти-TIGIT-антителами, начиная с 28 суток после заражения. Титры вируса в спленоцитах и печени определяли через 42 суток после заражения. Данные представляют результаты двух независимых опытов; n=10 в группе. На фиг. 4F показаны репрезентативные диаграммы FACS, гейтированные на CD8+ Т-клетках, где активированные (CD44высокий CD62Lнизкий) CD8+ Т-клетки находятся в рамке. Результаты количественного определения активированных CD8+ Т-клеток в процентах от общего числа CD8+ Т-клеток. ***Р <0,0001. На фиг. 4G показаны репрезентативные диаграммы FACS, гейтированные на активированных CD8+ Т-клетках, после стимуляции in vitro, где IFNγ-продуцирующие клетки находятся в рамке. Результаты количественного определения IFNγ-продуцирующих клеток в процентах от общего числа CD8+ Т-клеток. *Р = 0,0352. **P = 0,0047. На фиг. 4H показаны результаты количественного определения титров LCMV в печени. *P = 0,0106. **P = 0,0047. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 5 показано, что совместное блокирование TIGIT/PD-L1 усиливало эффекторную функцию CD4+ Т-клеток во время хронической вирусной инфекции. У мышей C57BL6/J истощали CD8+ Т-клетки и животных заражали клоном 13 штамма LCMV. Мышей обрабатывали изотипически сходными контрольными антителами, анти-PD-L1-, анти-TIGIT- или анти-PD-L1-+ анти-TIGIT-антителами, начиная с 28 суток после заражения. Титры вируса в спленоцитах и печени определяли через 42 суток после заражения. Данные представляют результаты двух независимых опытов; n=10 в группе. На фиг. 5A показаны репрезентативные диаграммы FACS, гейтированные на CD4+ Т-клетках, где активированные (CD44высокий CD62Lнизкий) клетки находятся в рамке. Результаты количественного определения активированных CD4+ Т-клеток в процентах от общего числа CD4+ Т-клеток. На фиг. 5B показаны репрезентативные диаграммы FACS, гейтированные на CD4+ Т-клетках, после стимуляции in vitro, где IFNγ-продуцирующие клетки находятся в рамке. *P = 0,019. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 6 показано, что экспрессия TIGIT повышена при раке молочной железы человека и коррелирует с экспрессией CD8 и ингибирующих корецепторов. Анализировали данные по экспрессии генов злокачественной опухоли молочной железы с использованием микрочипового анализа, полученные в проекте Атлас ракового генома. Данные по экспрессии генов нормализовали и выражали в виде относительных значений (log2). На фиг. 6A показана экспрессия TIGIT в нормальных образцах и образцах злокачественной опухоли молочной железы (слева) и в подтипах злокачественной опухоли молочной железы (справа). ***Р = 6×10-12. Показаны рамочные и коробчатые диаграммы. На фиг. 6В показана корреляция экспрессии TIGIT и CD3ε. R2 = 0,61. На фиг. 6C показана корреляция TIGIT и CD8α (слева, R2 = 0,80) или CD4 (справа, R2 = 0,42). На фиг. 6D показана корреляция TIGIT и PD-1 (слева, R2 = 0,87), LAG3 (в центре, R2 = 0,80), и CTLA4 (справа, R2 = 0,76).
На фиг. 7 показано, что TIGIT и PD-1 ингибируют противоопухолевые Т-клеточные ответы. На фиг. 7A-7В, мышам линии BALB/с прививали клетки колоректальной карциномы СТ26. Спленоциты и инфильтрирующие опухоль лимфоциты (TIL) анализировали через 14 суток после прививки, когда опухоли достигали размера примерно 200 мм3. Данные представляют результаты одного опыта; n=6. На фиг. 7A представлена гистограмма проточной цитометрии, показывающая экспрессию TIGIT CD8+ Т-клетками селезенки и инфильтрирующими опухоль CD8+ Т-клетками. Также представлены результаты количественного определения TIGIT MFI. **P = 0,0023. На фиг. 7В представлена гистограмма проточной цитометрии, показывающая экспрессию TIGIT CD4+ Т-клетками селезенки и инфильтрирующими опухоль CD4+ Т-клетками. Также представлены результаты количественного определения TIGIT MFI. ***P = 0,0002. На фиг. 7C-7E, мышам BALB/с прививали клетки колоректальной карциномы СТ26. Когда опухоли достигали размера примерно 200 мм3, мышей обрабатывали изотипически сходными контрольными антителами, анти-PD-L1-, анти-TIGIT- или анти-PD-L1-+ анти-TIGIT-антителами в течение трех недель. Данные представляют результаты двух независимых опытов; n=10-20 (фиг. 7С-7D) или 7-10 (фиг. 7E). На фиг. 7C показаны средние объемы опухолей СТ26 во времени. На фиг. 7D показана выживаемость мышей. На фиг. 7E показано, что примерно через 60 суток после первоначальной прививки мышам с состоянием полной ремиссии (CR), которые ранее получали анти-TIGIT-+ анти-PD-L1-антитела, а также нативным мышам линии BALB/с, прививали клетки СТ26 с левой стороны грудной клетки и прививали клетки карциномы молочной железы EMT6 в жировую подушку молочной железы. Показаны средние (слева) и индивидуальные (справа) объемы опухоли CT26 (квадраты) и EMT6 (треугольники) у мышей с CR (фиолетовый и зеленый цвет) и нативных мышей (черный и оранжевый цвет). На фиг. 7F показано, что мышам прививали опухоли СТ26 и обрабатывали, как показано на фиг. 7C. Анализировали проточной цитометрией инфильтрирующие опухоль Т-клетки и резидентные Т-клетки дренирующих опухоль лимфатических узлов. Репрезентативные диаграммы FACS CD8+ TIL после стимуляции в in vitro, где IFNγ-продуцирующие клетки находятся в рамке. Результаты количественного определения IFNγ-продуцирующих CD8+ TIL в процентах от общего числа CD8+ TIL. ***Р = 0,0003. Данные представляют результаты двух независимых опытов; n=5. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 8 показано, что экспрессия TIGIT инфильтрирующими опухоль CT26 лимфоцитами коррелирует с экспрессией Tim-3. Мышам линии BALB/с прививали клетки колоректальной карциномы СТ26. Спленоциты и инфильтрирующие опухоль лимфоциты (TIL) анализировали примерно через 14 суток после прививки, когда опухоли достигали размера примерно 200 мм3. Данные представляют результаты одного опыта; n=6. На фиг. 8A приведена репрезентативная гистограмма экспрессии TIGIT CD8+ Т-клетками селезенки и инфильтрирующими опухоль CD8+ Т-клетками. Результаты количественного определения TIGIT MFI. **P = 0,0026. На фиг. 8B приведена репрезентативная гистограмма экспрессии TIGIT CD4+ Т-клетками селезенки и инфильтрирующими опухоль CD4+ Т-клетками. Результаты количественного определения TIGIT MFI. ***Р <0,0001. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 9 показано, что экспрессия TIGIT инфильтрирующими опухоль МС38 лимфоцитами коррелирует с экспрессией PD-1 и Tim-3. Мышам C57BL6/J прививали клетки колоректальной карциномы МС38. Спленоциты и инфильтрирующие опухоль лимфоциты (TIL) анализировали примерно через 14 суток после прививки, когда опухоли достигали размера примерно 200 мм3. Данные представляют результаты одного опыта; n=5. На фиг. 9А приведена репрезентативная гистограмма экспрессии TIGIT CD8+ Т-клетками селезенки и инфильтрирующими опухоль CD8+ Т-клетками. Результаты количественного определения TIGIT MFI. ***Р <0,0001. На фиг. 9В приведена репрезентативная гистограмма экспрессии TIGIT CD4+ Т-клетками селезенки и инфильтрирующими опухоль CD4+ Т-клетками. Результаты количественного определения TIGIT MFI. *P = 0,0136. **P = 0,0029. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 10 показан рост опухоли СТ26 у мышей, обработанных анти-PD-L1- и/или анти-TIGIT-антителами. Нативным мышам BALB/с прививали опухолевые клетки СТ26 и обрабатывали анти-PD-L1- и/или анти-TIGIT-антителами или изотипически сходными контрольными антителами, как показано на фиг. 4D-4F. Показаны объемы опухолей во времени для отдельных мышей в каждой опытной группе согласно обработке. Данные представляют результаты двух независимых опытов.
На фиг. 11 показан анализ проточной цитометрией CD4+ TIL и Т-клеток дренирующих опухоль лимфатических узлов. Мышам BALB/с прививали клетки колоректальной карциномы СТ26. Когда опухоли достигали размера примерно 200 мм3, мышей обрабатывали изотипически сходными контрольными антителами, анти-PD-L1-, анти-TIGIT-, или анти-PD-L1-+ анти-TIGIT-антителами в течение 7 суток. Отбирали опухоли и дренирующие опухоль лимфатические узлы. Данные представляют результаты двух независимых опытов; n=5. Показаны репрезентативные гистограммы FACS, гейтированные на CD8+ Т-клетках дренирующих опухоль лимфатических узлов, после стимуляции in vitro, где IFNγ-продуцирующие клетки находятся в рамке. Результаты количественного определения IFNγ-продуцирующих клеток в процентах от общего числа CD8+ Т-клеток. ***Р <0,001. Результаты количественного определения CD8+ Т-клеток в процентах от общего числа TIL. **Р = 0,0065. Результаты количественного определения активированных (CD44высокий CD62Lнизкий) CD8+ Т-клеток в процентах от общего числа CD8+ TIL. *Р = 0,012. Результаты количественного определения CD8+ Т-клеток в процентах от общего числа клеток в дренирующих опухоль лимфатических узлах. Результаты количественного определения активированных CD8+ Т-клеток в процентах от общего числа CD8+ Т-клеток в дренирующих опухоль лимфатических узлах. *Р <0,05. На фиг. 11C показаны результаты количественного определения CD4+ Т-клеток в процентах от общего числа TIL. *P = 0,016. На фиг. 11D показаны результаты количественного определения активированных CD4+ Т-клеток в процентах от общего числа CD4+ TIL. На фиг. 11E показаны результаты количественного определения CD4+ Т-клеток в процентах от общего числа клеток в дренирующих опухоль лимфатических узлах. На фиг. 11F показаны результаты количественного определения активированных CD4+ Т-клеток в процентах от общего числа CD4+ Т-клеток в дренирующих опухоль лимфатических узлах. На фиг. 11А показаны результаты количественного определения IFNγ-продуцирующих клеток в процентах от общего числа CD4+ TIL после стимуляции in vitro. На фиг. 11B показаны результаты количественного определения IFNγ-продуцирующих клеток в процентах от общего числа CD4+ TIL в дренирующих опухоль лимфатических узлах после стимуляции in vitro. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 12 показан дополнительный анализ проточной цитометрией CD8+ TIL. Мышам BALB/с прививали клетки колоректальной карциномы СТ26 и обрабатывали изотипически сходными контрольными антителами, анти-PD-L1-, анти-TIGIT- или анти-PD-L1-+ анти-TIGIT-антителами, как показано на фиг. 4. Опухоли отбирали через 7 суток после обработки и анализировали проточной цитометрией. Данные представляют результаты двух независимых опытов; n=5. На фиг. 12А показаны результаты количественного определения TNFα+ клеток в процентах от общего числа CD8+ TIL. **Р <0,01. На фиг. 12В показаны результаты количественного определения CD8+ TIL в процентах от общего числа TIL. **Р <0,01. На фиг. 12C показаны результаты количественного определения активированных (CD44высокий CD62Lнизкий) CD8+ TIL в процентах от общего числа CD8+ TIL. *Р <0,05. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 13 показан анализ проточной цитометрией резидентных CD8+ Т-клеток в дренирующих опухоль лимфатических узлах. Мышам BALB/с прививали клетки колоректальной карциномы СТ26 и обрабатывали изотипически сходными контрольными антителами, анти-PD-L1-, анти-TIGIT- или анти-PD-L1-+ анти-TIGIT-антителами как показано на фиг. 4. Отбирали дренирующие опухоль лимфатические узлы через 7 суток после обработки и анализировали проточной цитометрией. Данные представляют результаты двух независимых опытов; n=5. На фиг. 13А представлены репрезентативные гистограммы FACS, гейтированные на резидентных CD8+ Т-клеток в дренирующих опухоль лимфатических узлах после стимуляции в in vitro, где IFNγ-продуцирующие клетки находятся в рамке. Результаты количественного определения IFNγ-продуцирующих+ клеток в процентах от общего числа CD8+ Т-клеток. ***Р <0,001. На фиг. 13B представлены результаты количественного определения CD8+ Т-клеток в процентах от общего числа клеток в дренирующих опухоль лимфатических узлах. На фиг. 13C представлены результаты количественного определения активированных (CD44высокий CD62Lнизкий) CD8+ Т-клеток в процентах от общего объема CD8+ Т-клеток. *Р <0,05. Вертикальные линии представляют стандартное отклонение от среднего. На фиг. 13D представлены результаты количественного определения TNFα-продуцирующих+ клеток в процентах от общего числа CD8+ Т-клеток в дренирующих опухоль лимфатических узлах.
На фиг. 14 показана коэкспрессия CD226 и TIGIT инфильтрирующими опухоль CD8+ Т-клетками. Мышам C57BL6/J прививали клетки колоректальной карциномы МС38. Спленоциты и инфильтрирующие опухоль лимфоциты (TIL) анализировали примерно 14 суток после прививки, когда опухоли достигли размера примерно 200 мм3. Репрезентативные гистограммы экспрессии CD226 В-клетками селезенки (серый цвет), CD8+ Т-клетками (синий цвет) и TIGT+ инфильтрирующими опухоль CD8+ Т-клетками (красный цвет). Данные представляют результаты двух независимых опытов; n=5.
На фиг. 15 показано, что CD226 и TIGIT коиммунопреципитируют (со-IP) на трансфектированных клетках. Клетки COS7 котрансфектировали экспрессионными плазмидами, содержащими кДНК меченных белков TIGIT-HA (5 нг) или CD226-Flag (10 нг), или контрольной плазмидой (pRK). После трансфекции клетки промывали и центрифугировали, и клеточный осадок лизировали. Полученный супернатант предварительно очищали и центрифугировали, и затем разделяли поровну на две пробирки и иммунопреципитировали с анти-НА- или анти-Flag-антителами с использованием стандартных процедур. Иммуннопреципитированные белки подвергали SDS-PAGE и вестерн-блоттингу. Вестерн-блоты зондировали анти-FLAG-HRP- или анти-НА-HRP-антителами.
На фиг. 16 показано, что TIGIT и CD226 взаимодействуют на первичных CD8+ Т-клетках. CD8+ Т-клетки селезенки мышей C57BL6/J, обогащенные с использованием MACS, стимулировали фиксированными на планшетах анти-CD3- и анти-CD28-антителами и рекомбинантным IL-2 в течение 48 ч и лизировали. Клеточные лизаты подвергали иммунопреципитации с анти-TIGIT-антителом и зондировали анти-CD226-антителом. Дорожки: маркер молекулярной массы (1), ввод (2), проточная коиммунопреципитация (3) и коиммунопреципитат. Стрелка показывает предполагаемую молекулярную массу CD226.
На фиг. 17 показано детектирование взаимодействия TIGIT/CD226 с использованием методологии TR-FRET. На фиг. 17А показана диссоциация гомодимеров Flag-SТ-CD226 под действием HA-TIGIT. Отношение FRET между Flag-ST-CD226, измеренное на клетках COS-7, экспрессирующих постоянное количество Flag-ST-CD226 и возрастающие концентрации HA-TIGIT. На фиг. 17B показано отношение FRET между Flag-ST-CD226, зарегистрированное после 15-минутной инкубации с PBS (белый столбик) или анти-TIGIT-антителом (черный столбик). На фиг. 17C показана ассоциация Flag-ST-CD226 с HA-TIGIT. Интенсивность FRET между Flag-ST-CD226 и HA-TIGIT по экспрессии Flag-ST-CD226, как определено анти-Flag ELISA на той же партии трансфектированных клеток COS-7. На фиг. 17D показано изменение FRET между Flag-ST-CD226 и HA-TIGIT после 15-мин инкубации с PBS (белый столбик) или анти-TIGIT-антителом (черный столбик). Данные на фиг. А и С представляют результаты четырех независимых опытов, каждый из которых проводили в трех повторностях. Данные на фиг. В и D представляют результаты двух независимых опытов, каждый из которых проводили в трех повторностях.
На фиг. 18 показана экспрессия на клеточной поверхности Flag-ST-CD226 и HA-TIGIT. Анти-Flag и анти-HA ELISA на интактных клетках COS-7, экспрессирующих указанные меченные конструкции. Данные представляют трех независимых опыта, каждый из которых проводили в трех повторностях.
На фиг. 19 показано, что блокирование CD226 отменяет повышенный противовирусный ответ Т-клеток, индуцированный совместным блокированием TIGIT/PD-L1. На фиг. 19A-19D, у мышей C57BL6/J быстро истощали CD4+ Т-клетки и животных заражали клоном 13 штамма LCMV. Мышей обрабатывали изотипически сходными контрольными антителами, анти-CD226-, анти-PD-L1-+ анти-TIGIT- или анти-PD-L1-+ анти-TIGIT-+ анти-CD226-антителами, начиная через 28 суток после заражения. Титры вируса в спленоцитах и печени определяли через 42 суток после заражения. На фиг. 19А представлены результаты количественного определения CD8+ Т-клеток в процентах от спленоцитов. На фиг. 19B представлены результаты количественного определения CD8+ Т-клеток в процентах от общего числа CD8+ Т-клеток. ***Р <0,001. На фиг. 19C представлены результаты количественного определения IFNg-продуцирующих клеток в процентах от общего числа активированных CD8+ Т-клеток. ***Р <0,001. На фиг. 19D представлены результаты количественного определения титров LCMV в печени. *** Р <0,001. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 20 показано, что экспрессия TIGIT повышена при раке человека и высоко коррелирует с экспрессией CD8 и PD-1. Анализ экспрессии генов при различных типах рака человека проводили, как описано в примере 11. Диаграммы рассеивания показывают данные расчета на ген, нормализованные к размеру библиотеки. Рамочные и коробчатые диаграммы показывают стабилизированную дисперсию соотношение экспрессии TIGIT и CD3e. На фиг. 20А показана корреляция экспрессии РНК TIGIT и CD3e в LUSC (серый цвет) и нормальных легких (черный цвет). ρ = 0,86. Также представлены результаты количественного определения соотношений экспрессии TIGIT/CD3e. Повышение соотношения в LUSC = 372%. ***Р = 1,46×10-46. На фиг. 20В показана корреляция экспрессии РНК TIGIT и CD3e в COAD (серый цвет) и нормальной ободочной кишке (черный цвет). ρ = 0,83. Также представлены результаты количественного определения соотношений экспрессии TIGIT/CD3e. Повышение соотношения в COAD = 116%. ***Р = 3,66×10-6. На фиг. 20C показана корреляция экспрессии РНК TIGIT и CD3e в UCEC (серый цвет) и эндометрии нормальной матки (черный цвет). ρ = 0,87. Также представлены результаты количественного определения соотношений экспрессии TIGIT/CD3e. Повышение соотношения в UCEC = 419%. ***Р = 7,41×10-5. На фиг. 20D показана корреляция экспрессии РНК TIGIT и CD3e в BRCA (серый цвет) и нормальной молочной железе (черный цвет). ρ = 0,82. Также представлены результаты количественного определения соотношений экспрессии TIGIT/CD3e. Повышение соотношения в BRCA = 313%. ***Р = 4,6×10-44. На фиг. 20E показана корреляция экспрессии РНК TIGIT и CD3e в светлой почечноклеточной карциноме (серый цвет) и нормальных почках (черный цвет). ρ = 0,94. Также представлены результаты количественного определения соотношений экспрессии TIGIT/CD3e. На фиг. 20F представлена корреляция TIGIT и CD8A (слева) или TIGIT и CD4 (справа) в плоскоклеточной карциноме легких (серый цвет) и нормальных легких (черный цвет). ρ = 0,77 и 0,48 соответственно. На фиг. 20G показана корреляция TIGIT и PD-1 (Pdcd1) в плоскоклеточной карциноме легких (серый цвет) и нормальных легких (черный цвет). ρ = 0,82. На фиг. 20H показана корреляция TIGIT и CD226 в плоскоклеточной карциноме легких (красный цвет) и нормальных легких (черный цвет). ρ= 0,64.
На фиг. 21 представлен анализ экспрессии генов, ассоциированных с Т-клетками, в плоскоклеточной карциноме легких (LUSC). Экспрессию генов в образцах LUSC и нормальной ткани анализировали, как описано в примере 11, и готовили тепловую карту генов, наилучшим образом коррелирующую с сигнатурой генов в LUSC. Гены и образцы кластеризовали с помощью иерархического группирования, используя метод Варда на матрице евклидовых расстояний для центрированных и масштабных данных по экспрессии.
На фиг. 22 показано, что TIGIT и PD-1 координировано экспрессируются инфильтрирующими опухоль лимфоцитами человека и мыши. На фиг. 22А-22С показан анализ лимфоцитов из только что удаленной опухоли NSCLC человека, соответствующей опухоли периферической крови и периферической крови здорового донора. Данные представляют результаты по двум независимо анализированным опухолям. На фиг. 22А представлены репрезентативные гистограммы FACS экспрессии TIGIT периферическими и инфильтрирующими опухоль CD8+ Т-клетками, где TIGIT+ клетки находятся в рамке. На фиг. 22В представлены репрезентативные гистограммы FACS экспрессии TIGIT периферическими и инфильтрирующими опухоль CD4+ Т-клетками, где TIGIT+ клетки находятся в рамке. На фиг. 22С представлена гистограмма проточной цитометрии, показывающая экспрессию TIGIT PD-1высокий (красный цвет) и PD-1низкий (синий цвет) NSCLC-инфильтрирующими CD8+ (слева) и CD4+ (справа) Т-клетками. На фиг. 22D-22G, мышам BALB/C прививали сингенные клетки колоректальной карциномы СТ26. Спленоциты и инфильтрирующие опухоль лимфоциты (TIL) анализировали через 14 суток после прививки, когда опухоли достигали размера примерно 200 мм3. Данные представляют результаты двух независимых опытов; n=5-6. На фиг. 22D представлены репрезентативные гистограммы FACS экспрессии TIGIT инфильтрирующими опухоль CD8+ Т-клетками, где TIGIT+ клетки находятся в рамке. На фиг. 22E представлены репрезентативные гистограммы FACS экспрессии TIGIT инфильтрирующими опухоль CD4+ Т-клетками, где TIGIT+ клетки находятся в рамке. Также представлены результаты количественного определения частоты встречаемости TIGIT+ Т-клеток в процентах от общего количества Т-клеток. *P = 0,0134. ***Р <0,0001. На фиг. 22F представлена гистограмма проточной цитометрии, показывающая экспрессию TIGIT PD-1высокий и PD-1низкий инфильтрирующими опухоль CD8+ Т-клетками и CD8+ Т-клетками селезенки. Также представлены результаты количественного определения TIGIT MFI. **P = 0,0023. На фиг. 22G представлена гистограмма проточной цитометрии, показывающая экспрессию TIGIT PD-1высокий и PD-1низкий инфильтрирующими опухоль CD4+ Т-клетками и CD4+ Т-клетками селезенки. Также представлены результаты количественного определения TIGIT MFI. ***P = 0,0002. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 23 представлена характеристика экспрессии TIGIT человеческими инфильтрирующими опухоль Т-клетками. На фиг. 23A-23B представлены гистограммы FACS, показывающие экспрессию TIGIT NSCLC-инфильтрирующими CD8+ и CD4+ Т-клетками (фиг. 23A) и РВМС CD8+ и CD4+ Т-клетками от совпадающих доноров (фиг. 23В), где TIGIT+ клетки находятся в рамке. На фиг. 23C-23D представлены гистограммы FACS, показывающие экспрессию TIGIT CRC-инфильтрирующими CD8+ и CD4+ Т-клетками (фиг. 23С) и РВМС CD8+ и CD4+ Т-клетками от совпадающих доноров (фиг. 23D), где TIGIT+ клетки находятся в рамке.
На фиг. 24 показано, что взаимодействие TIGIT:CD226 не регулируется взаимодействиями PVR TIGIT:CD226 и TIGIT Q56R:CD226, что было детектировано с использованием методологии TR-FRET, и отношение FRET между Flag-ST-CD226 и HA-TIGIT или HA-TIGIT Q56R показывает, что WT и Q56R TIGIT связываются с CD226 с одинаковой эффективностью. Данные представляют результаты трех независимых опытов, проведенных в трех повторностях.
На фиг. 25 показана эффективность совместного блокирования TIGIT/PD-L1 антител у мышей, несущих опухоли МС38. На фиг. 25A-25C, получали мышей, несущих опухоли МС38, как описано выше, и обрабатывали блокирующими антителами против PD-L1 (красный цвет), TIGIT (синий цвет), TIGIT и PD-L1 (фиолетовый цвет) или изотипически сходными контрольными антителами (черный цвет) в течение трех недель. N = 10 (контроль, одни анти-PD-L1-антитела, одни анти-TIGIT-антитела) или 20 (анти-TIGIT-антитела+ анти-PD-L1-антитела). На фиг. 25А показы средние (слева) и индивидуальные (справа) объемы опухолей МС38 во времени. На фиг. 25B показаны объемы опухолей МС38 через 14 суток обработки антителами. ***Р = 0,0005. **P = 0,0093. *P = 0,0433. На фиг. 25C приведены данные по выживаемости мышей во времени. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 26 показана дополнительная характеристика экспрессии TIGIT мышиными инфильтрирующими опухоль Т-клетками. На фиг. 26А показано, что CD8+ Т-клетки селезенки мышей C57BL6/J обогащали с использованием MACS и культивировали с фиксированными на планшетах агонистическими анти-CD3- и анти-CD28-антителами. Репрезентативные гистограммы окрашивания TIGIT (красный цвет) и изотипически сходными контрольными антителами (сплошная серая линия) во времени. Результаты количественного определения TIGIT MFI. ***Р <0,001. Стимулированные клетки индуцибельно экспрессировали PD-1 и конститутивно экспрессировали CD226 (данные не представлены). Данные представляют результаты двух независимых опытов; n=5. На фиг. 26В-26Е, мышам C57BL6/J дикого типа подкожно прививали сингенные клетки колоректальной карциномы МС38. Опухолям давали расти без вмешательства, пока они не достигали размера 150-200 мм3. Данные представляют результаты двух независимых опытов; n=5. На фиг. 26В представлена репрезентативная гистограмма FACS инфильтрирующих опухоль CD8+ Т-клеток, где TIGIT+ клетки находятся в рамке. Результаты количественного определения частоты встречаемости TIGIT+ клеток в процентах от общего числа инфильтрирующих опухоль CD8+ Т-клеток или CD8+ Т-клеток селезенки. ***Р <0,0001. На фиг. 26C представлена репрезентативная гистограмма FACS инфильтрирующих опухоли CD4+ Т-клеток, где TIGIT+ клетки находятся в рамке. Результаты количественного определения частоты встречаемости TIGIT+ клеток в процентах от общего числа инфильтрирующих опухоль CD4+ Т-клеток или CD4+ Т-клеток селезенки. ***Р <0,0001. На фиг. 26D представлена репрезентативная гистограмма экспрессии TIGIT PD-1высокий и PD-1низкий инфильтрирующими опухоли CD8+ Т-клетками (красный и синий цвета соответственно) и CD8+ Т-клетками селезенки (серый цвет). Результаты количественного определения TIGIT MFI. ***Р <0,0001. На фиг. 26E представлена репрезентативная гистограмма экспрессии TIGIT PD-1высокий и PD-1низкий инфильтрирующими опухоль CD4+ Т-клетками (красный и синий цвета соответственно) и CD4+ Т-клетками селезенки (серый цвет). Результаты количественного определения TIGIT MFI. *P = 0,0136. **P = 0,0029. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 27 показано, что инфильтрирующие опухоль CD8+ и CD4+ Т-клетки поддерживают высокий уровень экспрессии CD226. Мышам BALB/с дикого типа прививали опухолевые клетки СТ26, как здесь описано. После того, как опухоли достигали размера примерно 150-200 мм3, опухоли и селезенки анализировали проточной цитометрией. На фиг. 27A приведены результаты количественного определения CD226+ CD8+ Т-клеток, CD4+ Т-клеток и клеток, отличных от Т-клеток, в процентах от общего числа CD8+Т-клеток, CD4+ Т-клеток и клеток, отличных от Т-клеток, соответственно. На фиг. 27В представлены репрезентативные гистограммы экспрессии CD226 в опухоли и селезенки. Данные представляют результаты двух независимых опытов; n=5. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 28 показано, что TIGIT супрессия CD8+ Т-клеточных ответов зависит от CD226. Мышам BALB/с подкожно прививали клетки колоректальной карциномы СТ26 с правой стороны грудной клетки. Когда опухоли достигали размера примерно 200 мм3, мышей обрабатывали изотипически сходными контрольными антителами (черный цвет), анти-CD226-антителом (оранжевый цвет), анти-PD-L1-антителом (красный цвет), анти-TIGIT-+ анти-PD-L1-антителами (фиолетовый цвет) или анти- TIGIT-+ анти-PD-L1-+ анти-CD226-антителами (зеленый цвет) в течение трех недель. Данные представляют результаты одного опыта; n=10 (А-В) или 5 (С-F). На фиг. 28А представлены средние (слева) и индивидуальные (справа) объемы опухолей CT26 во времени. На фиг. 28B представлена выживаемость мышей во времени. На фиг. 28С-28F, через 7 суток обработки резидентные лимфоциты в дренирующих опухоль лимфатических узлах анализировали проточной цитометрией. На фиг. 28С представлены результаты количественного определения IFNγ-продуцирующих CD8+ TIL в процентах от общего числа CD8+ TIL после стимуляции in vitro. **Р <0,01. На фиг. 28D представлены результаты количественного определения IFNγ-продуцирующих клеток в процентах от общего числа CD8+ Т-клеток после стимуляции in vitro. *Р <0,05. На фиг. 28E представлены результаты количественного определения CD8+ TIL в процентах от общего числа TIL. **Р <0,01. На фиг. 28F представлены результаты количественного определения CD8+ Т-клеток в процентах от общего количества резидентных лимфоцитов в дренирующих опухоль лимфатических узлах. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 29 показано, что TIGIT подавляет функцию CD226, непосредственно нарушая гомодимеризацию CD226. На фиг. 29A показано, что CD8+ Т-клетки от TIGITfl/fl CD4cre (WT) и TIGITfl/fl CD4wt мышей из одного помета обогащали с использованием MACS и стимулировали в присутствии анти-CD226-антител или изотипически сходных контрольных антител, как показано. Поглощение Н3-тимидина показано как отношение клеток, культивированных с анти-CD3-антителом+ PVR-Fc, к клеткам, культивированным с одним анти-СD3-антителом. **P = 0,0061. ***Р <0,0001. Данные представляют результаты двух независимых опытов; n=5. На фиг. 29B показано, что CD8+ Т-клетки у мышей C57BL6/J дикого типа обогащали с использованием MACS и стимулировали в присутствии анти-TIGIT-антител, анти-CD226-антител или изотипически сходных контрольных антител, как указано. Поглощение Н3-тимидина показано как отношение клеток, культивированных с анти-CD3-антителом+ PVR-Fc, к клеткам, культивированным с одним анти-СD3-антителом. ***Р <0,001 в парных t-тестах. На фиг. 29C показано, что CD8+ Т-клетки обогащали с использованием MACS и стимулировали с субоптимальными концентрациями фиксированных на планшете анти-CD3-антител в присутствии или отсутствии человеческого рекомбинантного PVR-Fc. Добавляли анти-TIGIT-антитела или изотипически сходные контрольные антитела, как указано. Результаты количественного определения поглощения3Н-тимидина. **P = 0,0071 и 0,0014 соответственно. На фиг. 29D показано, что клетки СНО транзиентно трансфектировали возрастающими концентрациями акцептора и донора FLAG-ST-CD226, как указано. Результаты количественной оценки FRET интенсивности относительно эмиссии донора. Данные представляют результаты трех независимых опытов; n=3. На фиг. 29E-29F, клетки СНО транзиентно трансфектировали FLAG-ST-CD226 и возрастающими концентрациями HA-TIGIT, как указано. Данные представляют результаты двух или более независимых опытов; n=4. Данные нормализовали к максимальному сигналу. На фиг. 29E представлены результаты количественного определения FRET отношения TIGIT:CD226 (FRET отношение 1). На фиг. 29F представлены результаты количественного определения FRET отношения TIGIT:CD226 (FRET отношение 2). На фиг. 29G представлены иммуноблоты анти-Flag (слева) и анти-HA (справа), полученные на анти-FLAG или анти-НА иммунопреципитатах, приготовленных из клеток COS-7, трансфектированных пустым вектором pRK или комбинацией Flag-CD226 и HA-TIGIT. Данные представляют результаты двух независимых опытов. На фиг. 29H представлены результаты количественного определения FRET отношения TIGIT:CD226 после инкубации с PBS (белый цвет) или анти-TIGIT-антителом (красный цвет). ***Р <0,001. Данные представляют результаты четырех независимых опытов; n=3. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 30 показано, что первичные Т-клетки человека обогащали из крови с использованием MACS и стимулировали анти-CD3- и анти-CD28-антителами. Клетки TIGIT+ и TIGIT- были подвергнуты сортингу, выдерживали, повторно стимулировали и метили FRET указанными антителами. Данные представляют результаты двух независимых опытов. ***Р <0,001. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 31 показано, что совместное блокирование TIGIT и PD-1 не восстанавливало эффекторную функцию истощенных CD4+ Т-клеток при хронической вирусной инфекции. На фиг. 31A показаны результаты количественного определения CD8+ Т-клеток в процентах от общего числа спленоцитов. На фиг. 31B показаны результаты количественного определения gp33 Pentamer+ клеток в процентах от общего числа CD8+ Т-клеток селезенки. **P = 0,0040. На фиг. 31C представлены репрезентативные гистограммы FACS, гейтированные на gp33 Pentamer+ CD8+ Т-клетках, после стимуляции in vitro, где IFNγ+ клетки находятся в рамке. Результаты количественного определения IFNγ-продуцирующих клеток в процентах от общего числа gp33 Pentamer+CD8+ клеток. *Р = 0,0319. **P = 0,0030. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 32 показано, что эффективность совместного блокирования TIGIT/PD-L1 зависит от CD8+ Т-клеток. На фиг. 32A-32B, мышам дикого типа BALB/с прививали опухоли СТ26, как показано на фиг. 7. Когда опухоли достигали размера 100-150 мм3, у мышей транзиентно истощали CD8+ Т-клетки и обрабатывали анти-TIGIT-+ анти-PD-L1-антителами. Данные представляют результаты одного опыта; n=10/группу. На фиг. 32А показаны средние (слева) и индивидуальные (справа) объемы опухолей CT26 во времени. На фиг. 32B представлены результаты количественного определения объемов опухолей СТ26 через 17 суток после начала обработки. ***Р = 0,0004. На фиг. 32C, мышам BALB/c дикого типа прививали опухоли СТ26 и обрабатывали анти-TIGIT-+ анти-PD-L1-антителами, и затем повторно прививали опухоли СТ26 с транзиентным истощением CD8+ Т-клеток на момент повторной прививки. Данные представляют результаты двух независимых опытов; n=5. На фиг. 32C показаны средние (слева) и индивидуальные (справа) объемы опухолей CT26 во времени. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 33 показано, что экспрессия PVR на опухолевых клетках не является обязательной для проявления эффективности совместного блокирования TIGIT/PD-L1. Мышам BALB/с дикого типа прививали опухоли дикого типа или PVR-дефицитные опухоли (PVR.KO), как описано. Когда опухоли достигали размера 150-200 мм3, мышей обрабатывали анти-TIGIT-+ анти-PD-L1-антителами или изотипически сходных контрольных антител. Данные представляют результаты одного опыта; n=10/группу. На фиг. 33 показаны средние (слева) и индивидуальные (справа) объемы опухолей CT26 во времени.
На фиг. 34 показана эффективность совместного блокирования TIGIT/PD-L1 антитела у мышей, несущих опухоли EMT6. Мышей, несущих опухоли EMT6, получали, как описано выше, и обрабатывали блокирующими антителами против PD-L1 (красный цвет), TIGIT (синий цвет), TIGIT и PD-L1 (фиолетовый цвет) или изотипически сходными контрольными антителами (черный цвет) в течение трех недель. N = 10 (контроль, одно анти-PD-L1-антитело, одно анти-TIGIT-антитело) или 20 (анти-TIGIT-+ анти-PD-L1-антитела). На фиг. 34 показаны средние (слева) и индивидуальные (справа) объемы опухолей EMT26 во времени.
На фиг. 35 показано, что TIGIT регулирует эффекторную функцию инфильтрирующих опухоли CD8+ Т-клеток. Мышам BALB/с подкожно прививали клетки колоректальной карциномы СТ26 с правой стороны грудной клетки и обрабатывали анти-PD-L1-, анти-TIGIT- или анти-PD-L1-+ анти-TIGIT-антителами, как показано на фиг. 7. Резидентные Т-клетки в дренирующих опухоль лимфатических узлах (dLN) и инфильтрирующие опухоль Т-клетки анализировали проточной цитометрией через 7 суток после начала обработки. Данные представляют результаты двух независимых опытов; n=5. На фиг. 35А представлены результаты количественного определения CD8+ и CD4+ Т-клеток двойных продуцентов IFNγ/TNF в процентах от общего числа резидентных в dLN CD8+ и CD4+ Т-клеток соответственно. Также показана продукция двух цитокинов нестимулированными Т-клетками. **P = 0,002, 0,003, 0,001 соответственно. На фиг. 35В представлены результаты количественного определения инфильтрирующих опухоль CD8+ и CD4+ Т-клеток двойных продуцентов IFNγ/TNF в процентах от общего числа инфильтрирующих опухоль CD8+ и CD4+ Т-клеток соответственно. Также показана продукция двух цитокинов нестимулированными Т-клетками. ***Р <0,0001. Вертикальные линии представляют стандартное отклонение от среднего.
На фиг. 36 показан анализ лимфоцитов из подвергшихся резекции опухолей NSCLC человека, совпадающей с опухолью периферической крови и периферической крови здоровых доноров. Объединяли данные по трем независимым коллекциям образцов. На фиг. 36A представлены результаты количественного определения TIGIT+ клеток в процентах от общего числа CD8+ Т-клеток. *Р <0,05. На фиг. 36B представлены результаты количественного определения TIGIT+ клеток в процентах от общего числа CD4+ Т-клеток.
На фиг. 37 представлена характеристика экспрессии TIGIT в опухолях человека. На фиг. 37A представлены репрезентативные гистограммы проточной цитометрии экспрессии TIGIT резидентными лимфоцитами в NSCLC (красный цвет, CD45+ FSCнизкий), миелоидными клетками (синий цвет, CD45+ FSCвысокий) и негематопоэтическими клетками (зеленый цвет, CD45-) по отношению к окрашиванию изотипически сходных субпопуляций (серый цвет). На фиг. 37B показана стратегия гейтирования для PD-1высокий и PD-1низкий инфильтрирующих NSCLC-опухоль CD8+ и CD4+ Т-клеток.
Подробное описание изобретения
I. Общие методы
Методики и процедуры, описанные или на которые сделана ссылка в настоящем описании изобретения, как правило, хорошо известны и обычно применяются специалистами в данной области с использованием обычных методологий, описанных, например, в руководствах Sambrook et al., Molecular Cloning: A Laboratory Manual, 3d edition (2001), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.; Current Protocols in Molecular Biology (F.M. Ausubel, et al., eds., (2003)); изданиях Methods in Enzymology (Academic Press, Inc.): PCR 2: A Practical Approach (M.J. MacPherson, B.D. Hames and G.R. Taylor, eds. (1995)), Harlow and Lane, eds. (1988) Antibodies, A Laboratory Manual, и Animal Cell Culture (R.I. Freshney, ed. (1987)); Oligonucleotide Synthesis (M.J. Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J.E. Cellis, ed., 1998) Academic Press; Animal Cell Culture (R.I. Freshney), ed., 1987); Introduction to Cell and Tissue Culture (J.P. Mather and P.E. Roberts, 1998) Plenum Press; Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J.B. Griffiths, and D.G. Newell, eds., 1993-8) J. Wiley and Sons; Handbook of Experimental Immunology (D.M. Weir and C.C. Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J.M. Miller and M.P. Calos, eds., 1987); PCR: The Polymerase Chain Reaction (Mullis et al., eds., 1994); Current Protocols in Immunology (J.E. Coligan et al., eds., 1991); Short Protocols in Molecular Biology (Wiley and Sons, 1999); Immunobiology (C.A. Janeway and P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: A Practical Approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal Antibodies: A Practical Approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using Antibodies: A Laboratory Manual (E. Harlow and D. Lane (Cold Spring Harbor Laboratory Press, 1999); The Antibodies (M. Zanetti and J. D. Capra, eds., Harwood Academic Publishers, 1995); и Cancer: Principles and Practice of Oncology (V.T. DeVita et al., eds., J.B. Lippincott Company, 1993).
II. Определения
Термин «антагонист, связывающийся с осью PD-1» представляет собой молекулу, которая ингибирует взаимодействие партнера по связыванию оси PD-1 с одним или более его партнерами по связыванию, с целью устранения Т-клеточной дисфункции, возникшей в результате передачи сигналов по сигнальной оси PD-1, что позволяет восстановить или усилить функцию Т-клеток (например, пролиферацию, продукцию цитокинов, индуцированную гибель клеток-мишеней). В том смысле, в котором здесь используется термин «антагонист, связывающийся с осью PD-1», он включает PD-1-связывающий антагонист, PD-L1-связывающий антагонист и PD-L2-связывающий антагонист.
Термин «PD-1-связывающие антагонисты» представляют молекулу, которая снижает, блокирует, ингибирует, отменяет или препятствует передаче сигналов, возникающих в результате взаимодействия PD-1 с одним или более его партнерами по связыванию, такими как PD-L1, PD-L2. В некоторых вариантах осуществления PD-1-связывающий антагонист представляет собой молекулу, которая ингибирует связывание PD-1 с его партнерами по связыванию. В конкретном аспекте PD-1-связывающий антагонист ингибирует связывание PD-1 с PD-L1 и/или PD-L2. Например, PD-1-связывающие антагонисты включают анти-PD-1-антитела, их антигенсвязывающие фрагменты, иммуноадгезины, гибридные белки, олигопептиды и другие молекулы, которые снижают, блокируют, ингибируют, отменяют или препятствуют передаче сигналов, возникающих в результате взаимодействия PD-1 с PD-L1 и/или PD-L2. В одном варианте осуществления PD-1-связывающий антагонист снижает отрицательный костимулирующий сигнал, опосредованный или вызываемый поверхностными клеточными белками, экспрессированными на Т-лимфоцитах, которые опосредуют передачу сигналов через PD-1 таким образом, чтобы восстановить дисфункциональные Т-клетки в менее дисфункциональные (например, усиление эффекторных ответов на распознавание антигена). В некоторых вариантах осуществления PD-1-связывающий антагонист представляет собой анти-PD-1-антитело. В конкретном аспекте PD-1-связывающий антагонист представляет антитело MDX-1106, описанное здесь. В еще одном конкретном аспекте PD-1-связывающий антагонист представляет антитело Merck 3745, описанное здесь. В еще одном конкретном аспекте PD-1-связывающий антагонист представляет антитело СТ-011, описанное здесь. В еще одном конкретном аспекте PD-1-связывающий антагонист представляет антитело AMP-224, описанное здесь.
Термин «PD-L1-связывающие антагонисты» представляют молекулу, которая снижает, блокирует, ингибирует, отменяет или препятствует передаче сигналов, возникающих в результате взаимодействием PD-L1 с одним или более его партнерами по связыванию, такими как PD-1, B71. В некоторых вариантах осуществления PD-L1-связывающий антагонист представляет собой молекулу, которая ингибирует связывание PD-L1 с его партнерами по связыванию. В конкретном аспекте PD-L1-связывающий антагонист ингибирует связывание PD-L1 с PD-1 и/или B71. В некоторых вариантах осуществления PD-L1-связывающие антагонисты включают анти-PD-1-антитела, их антигенсвязывающие фрагменты, иммуноадгезины, гибридные белки, олигопептиды и другие молекулы, которые снижают, блокируют, ингибируют, отменяют или препятствуют передаче сигналов, возникающих в результате взаимодействия PD-L1 с одним или более его партнерами по связыванию, такими как PD-1, В7-1. В одном варианте осуществления PD-L1-связывающий антагонист снижает отрицательный костимулирующий сигнал, опосредованный или вызываемый поверхностными клеточными белками, экспрессированными на Т-лимфоцитах, которые опосредуют передачу сигналов через PD-L1 таким образом, чтобы восстановить дисфункциональные Т-клетки в менее дисфункциональные (например, усиление эффекторных ответов на распознавание антигена). В некоторых вариантах осуществления PD-L1-связывающий антагонист представляет собой анти-PD-L1-антитело. В конкретном аспекте анти-PD-L1-антитело представляет антитело YW243.55.S70, описанное здесь. В другом конкретном аспекте анти-PD-L1-антитело представляет антитело MDX-1105, описанное здесь. В еще одном конкретном аспекте анти-PD-L1-антитело представляет антитело MPDL3280A, описанное здесь. В еще одном конкретном аспекте анти-PD-L1-антитело представляет антитело MEDI 4736, описанное здесь.
Термин «PD-L2-связывающие антагонисты» представляют молекулу, которая снижает, блокирует, ингибирует, отменяет или препятствует передаче сигналов, возникающих в результате взаимодействием PD-L2 с одним или более его партнерами по связыванию, такими как PD-1. В некоторых вариантах осуществления PD-L2-связывающий антагонист представляет собой молекулу, которая ингибирует связывание PD-L2 с его партнерами по связыванию. В конкретном аспекте PD-L2-связывающий антагонист ингибирует связывание PD-L2 с PD-1. В некоторых вариантах осуществления PD-L2-связывающие антагонисты включают анти-PD-L2-антитела, их антигенсвязывающие фрагменты, иммуноадгезины, гибридные белки, олигопептиды и другие молекулы, которые снижают, блокируют, ингибируют, отменяют или препятствуют передаче сигналов, возникающих в результате взаимодействия PD-L2 с одним или более его партнерами по связыванию, такими как PD-1. В одном варианте осуществления PD-L2-связывающий антагонист снижает отрицательный костимулирующий сигнал, опосредованный или вызываемый поверхностными клеточными белками, экспрессированными на Т-лимфоцитах, которые опосредуют передачу сигналов через PD-L2 таким образом, чтобы восстановить дисфункциональные Т-клетки в менее дисфункциональные (например, усиление эффекторных ответов на распознавание антигена). В некоторых вариантах осуществления PD-L2-связывающий антагонист представляет собой иммуноадгезин.
Термин «аптамер» относится к молекуле нуклеиновой кислоты, которая способна связываться с молекулой-мишенью, такой как полипептид. Например, аптамер по изобретению может специфически связываться с полипептидом TIGIT, или с молекулой в сигнальном пути, который модулирует экспрессию TIGIT. Получение и терапевтическое применение аптамеров хорошо известно в данной области. Смотри, например, патент США № 5475096, и информацию по терапевтической эффективности Macugen® (Eyetech, Нью-Йорк) для лечения возрастной макулярной дегенерации.
Термин «антагонист» используется в самом широком смысле и включает любую молекулу, которая частично или полностью блокирует, ингибирует или нейтрализует биологическую активность нативного полипептида, раскрытого здесь. Аналогичным образом, термин «агонист» используется в самом широком смысле и включает любую молекулу, которая имитирует биологическую активность нативного полипептида, раскрытого здесь. Подходящие агонистические или антагонистические молекулы, в частности, включают агонистические или антагонистические антитела или фрагменты антител, фрагменты или варианты аминокислотных последовательностей нативных полипептидов, пептиды, антисмысловые олигонуклеотиды, небольшие органические молекулы и т.д. Способы идентификации агонистов или антагонистов полипептида могут включать контактирование полипептида с агонистической или антагонистической молекулой-кандидатом и определение детектируемого изменения одной или более биологических активностей, обычно присущих полипептиду.
Термины «антагонист TIGIT» и «антагонист активности TIGIT или экспрессии TIGIT» используются взаимозаменяемо и относятся к соединению, которое нарушает нормальное функционирование TIGIT, либо в результате снижения транскрипции или трансляции нуклеиновой кислоты, кодирующей TIGIT, либо в результате ингибирования или блокирования активности полипептида TIGIT, или обоих событий. Примеры антагонистов TIGIT включают, не ограничиваясь этим, антисмысловые полинуклеотиды, интерферирующие РНК, каталитические РНК, химеры РНК-ДНК, TIGIT-специфические аптамеры, анти-TIGIT-антитела, TIGIT-связывающие фрагменты анти-TIGIT-антител, небольшие молекулы, связывающиеся с TIGIT, пептиды, связывающие с TIGIT, и другие полипептиды, которые специфически связываются с TIGIT (включая, не ограничиваясь этим, TIGIT-связывающиеся фрагменты одного или более лигандов TIGIT, необязательно слитые с одним или более дополнительными доменами), таким образом, что взаимодействие между антагонистом TIGIT и TIGIT приводит к снижению или прекращению активности или экспрессии TIGIT. Специалистам с обычной квалификацией в данной области должно быть понятно, что в некоторых случаях антагонист TIGIT может оказывать антагонистическое действие в отношении одной активности TIGIT, не оказывая влияния на другую активность TIGIT. Например, желательным антагонистом TIGIT для применения в некоторых способах, описанных здесь, является антагонист TIGIT, который оказывает антагонистическое действие активности TIGIT в отношении только взаимодействия с PVR, взаимодействия с PVRL3 или взаимодействия с PVRL2, например, не оказывая влияния или оказывая минимальное влияние на другие взаимодействия TIGIT.
Термины «антагонист PVR» и «антагонист активности PVR или экспрессии PVR» используются взаимозаменяемо и относятся к соединению, которое нарушает нормальное функционирование PVR, либо в результате снижения транскрипции или трансляции нуклеиновой кислоты, кодирующей PVR, либо в результате ингибирования или блокирования активности полипептида PVR, или обоих событий. Примеры антагонистов PVR включают, не ограничиваясь этим, антисмысловые полинуклеотиды, интерферирующие РНК, каталитические РНК, химеры РНК-ДНК, PVR-специфические аптамеры, анти-PVR-антитела, PVR-связывающие фрагменты анти-PVR-антител, небольшие молекулы, связывающиеся с PVR, пептиды, связывающие с PVR, и другие полипептиды, которые специфически связываются с PVR (включая, не ограничиваясь этим, PVR-связывающиеся фрагменты одного или более лигандов PVR, необязательно слитые с одним или более дополнительными доменами), таким образом, что взаимодействие между антагонистом PVR и PVR приводит к снижению или прекращению активности или экспрессии PVR. Специалистам с обычной квалификацией в данной области должно быть понятно, что в некоторых случаях антагонист PVR может оказывать антагонистическое действие в отношении только одной активности PVR, не оказывая влияния на другую активность PVR. Например, желательным антагонистом PVR для применения в некоторых способах, описанных здесь, является антагонист PVR, который оказывает антагонистическое действие активности PVR в отношении только взаимодействия с TIGIT, не оказывая влияния на взаимодействия PVR-CD96 и/или PVR-CD226.
Термин «дисфункция» в контексте иммунной дисфункции относится к состоянию пониженной иммунной реактивности на антигенную стимуляцию. Термин включает общие элементы истощения и/или анергии, при которых может происходить распознавание антигена, но последующий иммунный ответ является неэффективным для контроля инфекции или роста опухоли.
В том смысле, в котором здесь используется термин «дисфункциональный», он включает рефракторное или безответное распознавание антигена, в частности, нарушенную способность транслировать распознавание антигена ниже на эффекторные функции Т-клеток, такие как пролиферация, продукция цитокинов (например, IL-2) и/или индукция гибели клеток-мишеней.
Термин «анергия» относится к состоянию отсутствия реакции на антигенную стимуляцию в результате неполных или недостаточных сигналов, подаваемых через Т-клеточный рецептор (например, повышение уровня внутриклеточного Ca+2 в отсутствии ras-активации). Т-клеточная анергия также может возникнуть в результате стимуляции антигеном в отсутствие костимуляции, в результате чего клетка становится невосприимчивой к последующей активации антигеном даже в случае костимуляции. Состояние отсутствия реакции часто может быть преодолено в присутствии интерлейкина-2. Анергические Т-клетки не подвергаются клональной экспансии и/или не приобретают эффекторные функции.
Термин «истощение» относится к истощению Т-клеток в виде состояния Т-клеточной дисфункции, возникающей в результате непрерывной передачи сигналов TCR, которая имеет место при многих хронических инфекций и при раке. Оно отличается от анергии тем, что возникает не в результате неполной или недостаточной передачи сигналов, а в результате продолжительной передачи сигналов. Оно определяется слабой эффекторной функцией, устойчивой экспрессией ингибирующих рецепторов и состоянием транскрипции, отличным от такового у функциональных эффекторных Т-клеток или Т-клеток памяти. Истощение препятствует оптимальному контролю инфекции и опухоли. Истощение может быть вызвано внешними отрицательными регуляторными путями (например, иммунорегуляторными цитокинами), а также внутренними отрицательными регуляторными (костимулирующими) путями клетки (PD-1, В7-Н3, B4-H7 и т.д.).
«Усиление функции Т-клеток» означает индукцию, обеспечение или стимуляцию Т-клеток на наличие продолжительной или усиленной биологической функции, или восстановление или реактивацию истощенных или неактивных Т-клеток. Примеры усиления функции Т-клеток включают: повышенную секрецию γ-интерферона CD8+ Т-клетками, повышенную пролиферацию, повышенную реактивность на антигены (например, элиминация вируса, патогена или опухоли) по сравнению с такими уровнями до вмешательства. В одном варианте осуществления степень усиления составляет, по меньшей мере, 50%, альтернативно, 60%, 70%, 80%, 90%, 100%, 120%, 150%, 200%. Специалисту с обычной квалификацией в данной области техники известны методы измерения данного усиления.
«Т-клеточное дисфункциональное расстройство» представляет расстройство или состояние Т-клеток, характеризующееся пониженной реактивностью на антигенную стимуляцию. В конкретном варианте осуществления Т-клеточное дисфункциональное расстройство представляет расстройство, которое специфически связано с неадекватной повышенной передачей сигналов с участием PD-1. В еще одном варианте осуществления Т-клеточное дисфункциональное расстройство представляет расстройство, при котором Т-клетки являются анергичными или обладают пониженной способностью к секреции цитокинов, пролиферации или утрачивается цитолитическая активность. В конкретном аспекте пониженная реактивность приводит к неэффективному контролю патогена или опухоли, экспрессирующих иммуноген. Примеры Т-клеточных дисфункциональных расстройств, характеризующихся дисфункцией Т-клеток, включают неразрешенную острую инфекцию, хроническую инфекцию и иммунитет к опухоли.
«Иммунитет к опухоли» относится к процессу, при котором опухоли уклоняются от иммунологического распознавания и элиминации. Таким образом, с терапевтической точки зрения иммунитет к опухоли «лечится», когда такое уклонение ослабляется, и опухоли распознаются и атакуются иммунной системой. Примеры распознавания опухолей включают связывание опухолей, уменьшение размера опухолей и элиминацию опухолей.
«Иммуногенность» относится к способности конкретного вещества индуцировать иммунный ответ. Опухоли являются иммуногенными, и усиление иммуногенности опухолей направлено на элиминацию опухолевых клеток под действием иммунного ответа. Примеры усиления иммуногенности опухолей включают, но не ограничиваются этим, обработку антагонистом, связывающимся с осью PD-1 (например, анти-PD-L1-антителом и ингибитором TIGIT (например, анти-TIGIT-антителом).
«Продолжительная реакция» относится к продолжительному эффекту на снижение роста опухоли после прекращения лечения. Например, размер опухоли может оставаться таким же или стать меньше по сравнению с ее размером в начале фазы лечения. В некоторых вариантах осуществления продолжительная реакция имеет продолжительность, по меньшей мере, такую же, как продолжительность лечения, по меньшей мере, в 1,5 раза, 2,0 раза, 2,5 раза или 3,0 раза превышающую продолжительность лечения.
Термин «антитело» включает моноклональные антитела (включая полноразмерные антитела, которые содержат Fc-область иммуноглобулина), композиции антител с полиэпитопной специфичностью, мультиспецифические антитела (например, биспецифические антитела, димеры и одноцепочечные молекулы, а также фрагменты антитела (например, Fab, F(аb')2и Fv). Термин «иммуноглобулин» (Ig) используется взаимозаменяемо с «антителом».
Основная четырехцепочечная структурная единица антитела представляет собой гетеротетрамерный гликопротеин, состоящий из двух идентичных легких (L) цепей и двух идентичных тяжелых (Н) цепей. IgM антитело состоит из 5 основных гетеротетрамерных единиц, а также дополнительного полипептида, называемого J-цепью, и содержит 10 антигенсвязывающих сайтов, в то время, как IgA антитела содержат 2-5 основных 4-цепочечных структурных единиц, которые могут полимеризоваться с образованием поливалентных ансамблей в комбинации с J-цепью). В случае IgG 4-цепочечная структурная единица обычно имеет молекулярную массу примерно 150000 дальтон. Каждая L-цепь связана с Н-цепью одной ковалентной дисульфидной связью, тогда как две Н-цепи связаны друг с другом одной или более дисульфидными связями в зависимости от изотипа Н-цепи. Каждая Н- и L-цепь имеет также регулярно расположенные внутрицепочечные дисульфидные мостики. Каждая Н-цепь имеет на N-конце вариабельную область (VH), за которой следуют три константных домена (CH), для каждой из α и γ цепей, и четыре CH домена для μ и ε изотипов. Каждая L-цепь имеет на N-конце вариабельную область (VL), за которой следует константный домен (CL) на другом конце. VL выравнен с VH, a CL выравнен с первым константным доменом тяжелой цепи (CH1). Полагают, что определенные аминокислотные остатки образуют поверхность раздела между вариабельными областями легкой цепи и тяжелой цепи. В результате спаривания VH и VL образуется один антигенсвязывающий сайт. О структуре и свойствах различных классов антител смотри, например, публикации Basic and Clinical Immunology, 8th edition, Daniel P. Stites, Abba I. Terr and Tristram G. Parslow (eds.), Appleton & Lange, Norwalk, CT, 1994, page 71 and Chapter 6. L-цепь различных видов позвоночных можно отнести к одному из двух четко различающихся типов, называемых каппа и лямбда, на основании аминокислотных последовательностей их константных доменов. В зависимости от аминокислотной последовательности константного домена их тяжелых цепей (CH) иммуноглобулины можно отнести к различным классам или изотипам. Существует пять классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, имеющих тяжелые цепи, обозначаемые α, σ, γ, ε и μ соответственно. Классы γ и α дополнительно подразделяются на подклассы на основе сравнительно незначительных различий в последовательности CH и функции, например, в организме человека экспрессируются следующие подклассы: IgG1, IgG2А, IgG2В, IgG3, IgG4, IgA1 и IgA2.
Термин «вариабельная область» или «вариабельный домен» антитела относится к аминоконцевым областям тяжелой или легкой цепи антитела. Вариабельные области тяжелой цепи и легкой цепи могут обозначаться соответственно «VH» и «VL». Как правило, эти области являются наиболее вариабельными частями антитела (по сравнению с другими антителами того же класса) и содержат антигенсвязывающие участки.
Термин «вариабельный» относится к тому факту, что некоторые сегменты вариабельных областей антител очень значительно различаются в последовательности некоторых антител. V домен опосредует связывание антигена и определяет специфичность конкретного антитела к его конкретному антигену. Однако вариабельность распределяется неравномерно на полном участке вариабельных областей. Напротив, она концентрируется в трех сегментах, называемых гипервариабельными областями (HVR), в вариабельных областях как легкой, так и тяжелой цепи. Более высококонсервативные участки вариабельных областей называются каркасными областями (FR). Каждая из вариабельных областей нативной тяжелой и легкой цепей содержит четыре области FR, преимущественно имеющие конфигурацию бета-складок, связанных тремя HVR, которые образуют петли, связывающие бета-складчатые структуры, и в некоторых случаях образующие их часть. HVR в каждой цепи удерживаются вместе в тесной близости посредством FR областей и, с HVR из другой цепи, вносят свой вклад в образование антигенсвязывающего сайта антител (смотри Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, National Institute of Health, Bethesda, MD (1991)). Константные области непосредственно не участвуют в связывании антитела с антигеном, но проявляют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной токсичности.
В том смысле, в котором здесь используется термин «моноклональное антитело», он относится к антителу, полученному из популяции по существу гомогенных антител, т.е. индивидуальные антитела, составляющие популяцию, являются идентичными за исключением возможных природных мутаций и/или посттрансляционных модификаций (например, изомеризации, амидирования), которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифичными, направленными против одного антигенного сайта. В отличие от препаратов поликлональных антител, которые обычно включают различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигене. В дополнение к их специфичности моноклональные антитела обладают тем преимуществом, что они синтезируются гибридомной культурой, не загрязненной другими иммуноглобулинами. Определение «моноклональное» указывает на характер антитела, полученного из практически гомогенной популяции антител, и не должно быть истолковано как требующее получения антитела каким-либо конкретным способом. Например, моноклональные антитела, которые будут использоваться согласно настоящему изобретению, могут быть получены различными методами, в том числе, например, методом гибридом (например, Kohler and Milstein., Nature, 256:495-97 (1975); Hongo et al., Hybridoma, 14 (3):253-260 (1995), Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988); Hammerling et al., in: Monoclonal Antibodies and T-Cell Hybridomas, 563-681 (Elsevier, N.Y., 1981)), методами рекомбинантной ДНК (смотри, например, патент США № 4816567), технологиями фагового дисплея (смотри, например, Clackson et al., Nature, 352:624-628 (1991); Marks et al., J. Mol. Biol., 222:581-597 (1992); Sidhu et al., J. Mol. Biol., 338(2):299-310 (2004); Lee et al., J. Mol. Biol., 340(5):1073-1093 (2004); Fellouse, Proc. Natl. Acad. Sci. USA, 101(34): 12467-12472 (2004); и Lee et al., J. Immunol. Methods, 284(1-2): 119-132 (2004), и технологиями получения человеческих или подобных человеческим антител у животных, которые имеют фрагменты или все человеческие иммуноглобулиновые локусы или гены, кодирующие человеческие иммуноглобулиновые последовательности (смотри, например, WO 1998/24893; WO 1996/34096; WO 1996/33735; WO 1991/10741; Jakobovits et al., Proc. Natl. Acad. Sci. USA, 90:2551 (1993); Jakobovits et al., Nature, 362:255-258 (1993); Bruggemann et al., Year in Immunol., 7:33 (1993); патенты США № 5545807; 5545806; 5569825; 5625126; 5633425 и 5661016; Marks et al., Bio/Technology, 10:779-783 (1992); Lonberg et al., Nature, 368:856-859 (1994); Morrison, Nature, 368:812-813 (1994); Fishwild et al., Nature Biotechnol., 14: 845-851 (1996); Neuberger, Nature Biotechnol., 14: 826 (1996) и Lonberg and Huszar, Intern. Rev. Immunol., 13: 65-93 (1995).
Термин «голое антитело» относится к антителу, которое не конъюгировано с цитотоксическим фрагментом или радиоактивной меткой.
Термины «полноразмерное антитело», «интактное антитело» или «целое антитело» используются взаимозаменяемо по отношению к антителу в его по существу интактной форме, в отличие от фрагмента антитела. В частности, целые антитела включают антитела с тяжелой и легкой цепями, включая Fc-область. Константные области могут представлять константные области с нативной последовательностью (например, константные области с нативной последовательностью человека) или варианты их аминокислотной последовательности. В некоторых случаях интактное антитело может обладать одной или более эффекторными функциями.
«Фрагмент антитела» включает фрагмент интактного антитела, предпочтительно антигенсвязывающий участок и/или вариабельную область интактного антитела. Примеры фрагментов антитела включают фрагменты Fab, Fab', F(аb')2 и Fv; диатела; линейные антитела (смотри патент США № 5641870, пример 2; Zapata et al., Protein Eng., 8(10):1057-1062 [1995]); молекулы одноцепочечного антитела и мультиспецифические антитела, полученные из фрагментов антител. Расщепление антител папаином дает два идентичных антигенсвязывающих фрагмента, называемых «Fab»-фрагментами, и остаточный «Fc»-фрагмент, его обозначение отражает способность легко кристаллизоваться. Fab-фрагмент состоит из целой L-цепи, а также вариабельной области Н-цепи (VH) и первого константного домена одной тяжелой цепи (CH1). Обработка антитела пепсином дает один большой фрагмент F(ab')2, который в грубом приближении соответствует двум связанным дисульфидной связью Fab-фрагментам, обладающим различной антигенсвязывающей активностью и все еще способным к перекрестному связыванию антигена. Fab'-фрагменты отличаются от Fab-фрагментов наличием нескольких дополнительных остатков на карбокси-конце CH1 домена, включая один или более цистеиновых остатков из шарнирной области антитела. Fab'-SH обозначает в данном описании Fab', в котором цистеиновый остаток(и) константной области несет свободную тиоловую группу. Первоначально фрагменты F(ab')2 антитела были получены в виде пары Fab'-фрагментов, которые имеют между ними шарнирные цистеиновые остатки. Известны также другие примеры связывания фрагментов антитела химической связью.
Fc-фрагмент содержит карбокси-концевые участки обеих Н-цепей, связанные дисульфидной связью. Эффекторные функции антител определяются последовательностями в Fc-области, области, которая распознается Fc-рецепторами (FcR), имеющимися на некоторых типах клетках.
«Fv» представляет минимальный фрагмент антитела, который содержит полный антигенраспознающий и антигенсвязывающий сайт. Данный фрагмент состоит из димера, состоящего из одной вариабельной области тяжелой цепи и одной вариабельной области легкой цепи, прочно связанных нековалентной связью. В результате укладки данных двух доменов образуется шесть гипервариабельных петель (по 3 петли из каждой Н- и L-цепи), которые входят в аминокислотные остатки, участвующие в связывании антигена, и придают антителу антигенсвязывающую специфичность. Однако даже одна вариабельная область (или половина Fv, содержащая только три HVR, специфичная для антигена) способна распознавать и связывать антиген, хотя, с более низкой аффинностью, чем целый сайт связывания.
«Одноцепочечный Fv», также сокращенно обозначаемый, как «sFv» или «scFv», представляет собой фрагмент антитела, который содержит VH- и VL-области антитела, соединенные в единую полипептидную цепь. Предпочтительно полипептид sFv дополнительно содержит полипептидный линкер между VH- и VL-областями, который способствует образованию фрагментом sFv структуры, необходимой для связывания антигена. Обзор по sFv смотри у Pluckthun in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp.269-315 (1994).
«Функциональные фрагменты» антител по изобретению содержат фрагмент интактного антитела, обычно включающий антигенсвязывающий участок или вариабельную область интактного антитела или Fc-область антитела, которое сохраняет или имеет модифицированную способность связываться с FcR. Примеры фрагментов антитела включают линейное антитело, молекулы одноцепочечных антител и мультиспецифические антитела, полученные из фрагментов антител.
Термин «диатела» относится к небольшим фрагментам антител, полученным при конструировании sFv фрагментов (смотри предыдущий абзац), с короткими линкерами (примерно, 5-10 остатков) между VH- и VL-доменами так, чтобы достигалось межцепочечное, а не внутрицепочечное спаривание V доменов, с получением в результате бивалентного фрагмента, т.е. фрагмента, имеющего два антигенсвязывающих сайта. Биспецифические диатела представляют собой гетеродимеры двух «перекрестных» sFv-фрагментов, в которых VH- и VL-домены двух антител находятся в различных полипептидных цепях. Более подробно диатела описаны, например, в ЕР 404097; WO 93/11161; и в публикации Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993).
Моноклональные антитела по настоящему изобретению, в частности, включают «химерные» антитела (иммуноглобулины), в которых фрагмент тяжелой и/или легкой цепи идентичен или гомологичен соответствующим последовательностям в антителах, полученных от конкретных видов или относящихся к конкретному классу или подклассу антител, в то время, как остальная часть цепи(ей) является идентичной или гомологичной соответствующим последовательностям в антителах, полученных от другого вида, или относящихся к другому классу или подклассу антител, а также фрагменты таких антител, при условии, что они проявляют желаемую биологическую активность (патент США № 4816567; Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855 (1984)). Представляющие интерес для настоящего изобретения химерные антитела включают приматизированные антитела (PRIMATIZED®), где антигенсвязывающий участок антитела происходит из антитела, полученного, например, иммунизацией макак представляющим интерес антигеном. В том смысле, в котором здесь используется термин «гуманизированное антитело», он включает субпопуляцию «химерных антител».
«Гуманизированные» формы антител, отличных от человеческих антител (например, мышиные), представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из антитела, отличного от человеческого антитела. В одном варианте осуществления гуманизированное антитело представляет собой человеческий иммуноглобулин (реципиентное антитело), в котором остатки из HVR (определяются ниже) реципиента заменены на остатки из HVR от вида, отличного от человека (донорное антитело), такого как мышь, крыса, кролик или примат, отличный от человека, имеющие требуемую специфичность, аффинность и/или активность. В некоторых случаях остатки каркасной области (FR) человеческого иммуноглобулина заменены на соответствующие остатки нечеловеческого иммуноглобулина. Кроме того, гуманизированные антитела могут содержать остатки, которые не обнаруживаются в реципиентном антителе или в донорном антителе. Эти модификации можно получить для того, чтобы дополнительно усовершенствовать характеристики антитела, такие как аффинность связывания. Как правило, «гуманизированное» антитело содержит практически все или, по меньшей мере, один, обычно два, вариабельных домена, в которых все или практически все гипервариабельные петли соответствуют гипервариабельным петлям последовательности иммуноглобулина, отличного от человеческого, и все или практически все FR области представляют собой FR области последовательности человеческого иммуноглобулин, хотя, FR области могут включать одну или более замен отдельных остатков FR, которые улучшают характеристики антитела, такие как аффинность связывания, изомеризация, иммуногенность и т.д. Как правило, количество таких аминокислотных замен в FR составляет не более 6 в Н-цепи и не более 3 в L-цепи. Гуманизированное антитело, необязательно, содержит, по меньшей мере, фрагмент константной области иммуноглобулина (Fc), как правило, человеческого иммуноглобулина. Более подробные сведения смотри, например, в публикациях Jones et al., Nature, 321:522-525 (1986); Riechmaim et al., Nature 332: 323-329 (1988); и Presta, Curr. Op. Struct. Biol., 2: 593-596 (1992). Также смотри, например, Vaswani and Hamilton, Ann. Allergy, Asthma & Immunol., 1:105-115 (1998); Harris, Biochem. Soc. Transactions, 23:1035-1038 (1995); Hurle and Gross, Curr. Op. Biotech., 5:428-433 (1994) и патенты США № 6982321 и 7087409.
«Человеческое антитело» представляет антитело, которое обладает аминокислотной последовательностью, соответствующей последовательности антитела, продуцированного человеком, и/или было получено с использованием любого из методов получения человеческих антител, описанных в настоящем документе. Это определение человеческого антитела специфически исключает гуманизированное антитело, содержащее антигенсвязывающие остатки, отличные от человеческих. Человеческие антитела могут быть получены с использованием различных методов, известных в данной области техники, включая библиотеки фагового дисплея. Hoogenboom and Winter, J. Mol. Biol., 227:381 (1991); Marks et al., J. Mol. Biol., 222:581 (1991). Также для получения человеческих моноклональных антител имеются методы, описанные Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p. 77 (1985); Boerner et al., J. Immunol., 147(1):86-95 (1991). Также смотри публикацию van Dijk and van de Winkel, Curr. Opin. Pharmacol., 5: 368-74 (2001). Также смотри van Dijk and van de Winkel, Curr. Opin. Pharmacol., 5: 368-74 (2001). Человеческие антитела могут быть получены введением антигена трансгенному животному, которое было модифицировано с целью продукции таких антител в ответ на антигенную стимуляцию, но эндогенные локусы которых были заблокированы, например, иммунизированные ксеномыши (смотри, например, патенты США № 6075181 и 6150584, касающиеся технологии XENOMOUSE™). Смотри также, например, публикацию Li et al., Proc. Natl. Acad. Sci. USA, 103:3557-3562 (2006) о человеческих антителах, полученных с помощью технологи гибридом на основе человеческих В-клеток.
Используемый здесь термин «гипервариабельная область», «HVR» или «HV» относится к участкам вариабельной области антитела, которые являются гипервариабельными по последовательности и/или образуют структурно определенные петли. Как правило, антитела содержат шесть HVR; три в VH (H1, H2, H3) и три в VL (L1, L2, L3). В нативных антителах Н3 и L3 проявляют наибольшее разнообразие шести HVR, и как полагают, H3, в частности, играет уникальную роль в придании тонкой специфичности антител. Смотри, например, Xu et al., Immunity, 13:37-45 (2000); Johnson and Wu, in Methods in Molecular Biology, 248:1-25 (Lo, ed., Human Press, Totowa, NJ, 2003). Действительно, встречающиеся в природе антитела верблюда, состоящие только из тяжелой цепи, являются функциональными и стабильными в отсутствие легкой цепи. Смотри, например, Hamers-Casterman et al., Nature, 363:446-448 (1993); Sheriff et al., Nature Struct. Biol., 3:733-736 (1996).
Ряд описаний HVR используется и охватывается в данном документе. Области, определяющие комплементарность по Kabat (CDR) основаны на вариабельности последовательностей и обычно являются наиболее широко используемыми (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Напротив, Chothia определяет гипервариабельные участки на основе локализации структурных петель (Chothia and Lesk, J. Mol. Biol., 196:901-917 (1987)). AbM HVR представляют компромисс между HVR Kabat и структурными петлями Chothia и используются Оксфордской программой молекулярного моделирования AbM антител (Oxford Molecular's antibody AbM modeling software). «Контактные» HVR основаны на анализе имеющихся сложных кристаллических структур. Остатки из каждого из этих HVR отмечены ниже.
Области HVR могут включать «удлиненные HVR», например: 24-36 или 24-34 (L1), 46-56 или 50-56 (L2) и 89-97 или 89-96 (L3) в VL и 26-35 (H1), 50-65 или 49-65 (Н2) и 93-102, 94-102 или 95-102 (Н3) в VH. Остатки в вариабельных доменах нумеруются по Kabat et al., выше, для каждого из этих определений.
Выражения «нумерация остатков вариабельных доменов по Kabat» или «нумерация аминокислотных положений по Kabat», и их варианты относятся к системе нумерации, используемой для вариабельных областей тяжелой цепи или вариабельных областей легкой цепи при составлении антител в приведенной выше публикации Kabat et al. При использовании этой системы нумерации действительная линейная аминокислотная последовательность может содержать меньшее или дополнительное число аминокислот, соответствующее укорочению или инсерции в FR или HVR вариабельной области. Например, вариабельная область тяжелой цепи может включать инсерцию одной аминокислоты (остаток 52а по Kabat) после остатка 52 участка Н2 и инсерции остатков (например, 82а, 82b и 82с и т.д. по Kabat) после остатка 82 FR тяжелой цепи. Нумерацию остатков по Kabat можно определить для данного антитела выравниванием гомологичных областей последовательности антитела со «стандартной» последовательностью с нумерацией по Kabat.
Остатки «каркасной области» или «FR» представляют остатки вариабельных областей, отличные от остатков HVR, определенных здесь.
«Человеческая консенсусная каркасная область» или «акцепторная человеческая каркасная область» представляет каркасную область, которая содержит наиболее часто встречающиеся аминокислотные остатки при выборе каркасных последовательностей VL или VH человеческого иммуноглобулина. Обычно выбор VL или VH последовательностей человеческого иммуноглобулина проводят из подгруппы последовательностей вариабельных областей. Как правило, подгруппа последовательностей представляет подгруппу по Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991). Примеры подгруппы для VL могут включать подгруппу каппа I, каппа II, каппа III или IV каппа, как приведено в публикации Kabat et al., выше. Кроме того, подгруппа для VH может включать подгруппу I, подгруппу II или подгруппу III, как приведено в публикации Kabat et al., выше. Альтернативно человеческую консенсусную последовательность можно получить из вышеприведенной последовательности, в которой определенные остатки, например, остатки человеческой каркасной области, выбирают на основании их гомологии с донорной каркасной областью, выравниванием последовательности донорной каркасной области с рядом различных последовательностей человеческой каркасной области. Человеческая акцепторная каркасная область «происходящая из» каркасной области человеческого иммуноглобулина или человеческой консенсусной каркасной области может содержать такую же аминокислотную последовательность, или она может содержать уже существующие изменения аминокислотных последовательностей. В некоторых вариантах осуществления количество уже существующих аминокислотных изменений составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее, или 2, или менее.
«Консенсусная каркасная область VH подгруппы III» включает консенсусную последовательность, полученную из аминокислотных последовательностей в вариабельной области тяжелой цепи подгруппы III по Kabat et al., выше. В одном варианте осуществления консенсусная аминокислотная последовательность каркасной области VH подгруппы III включает, по меньшей мере, фрагмент или всю каждую из следующих последовательностей: EVQLVESGGGLVQPGGSLRLSCAAS (HC-FR1) (SEQ ID NO:25), WVRQAPGKGLEWV (HC-FR2) (SEQ не ID NO:26), RFTISADTSKNTAYLQMNSLRAEDTAVYYCAR (HC-FR3, SEQ ID NO:27), WGQGTLVTVSA (HC-FR4), (SEQ ID NO:28).
«Консенсусная каркасная последовательность VL подгруппы I» включает консенсусную последовательность, полученную из аминокислотных последовательностей в вариабельной области легкой цепи каппа подгруппы I по Kabat et al. В одном варианте осуществления консенсусная аминокислотная последовательность каркасной области VL подгруппы I включает, по меньшей мере, фрагмент или всю каждую из следующих последовательностей: DIQMTQSPSSLSASVGDRVTITC (LC-FR1) (SEQ ID NO:29), WYQQKPGKAPKLLIY (LC-FR2) (SEQ ID NO:30), GVPSRFSGSGSGTDFTLTISSLQPEDFATYYC (LC-FR3) (SEQ ID NO:31), FGQGTKVEIKR (LC-FR4) (SEQ ID NO:32).
Термин «модификация аминокислоты» в указанном положении, например, Fc-области, относится к замене или делеции указанного остатка или инсерции, по меньшей мере, одного аминокислотного остатка, смежного с указанным остатком. Инсерция «смежная» с указанным остатком означает инсерцию одного-двух остатков. Инсерция может быть N-концевой или С-концевой по отношению к указанному остатку. Предпочтительной аминокислотной модификацией здесь является замена.
Термин «аффинно зрелое» антитело представляет собой антитело с одним или более изменениями в одном или более HVR, что приводит к повышению аффинности антитела к антигену по сравнению с исходным антителом, которое не обладает таким изменением(ями). В одном варианте осуществления аффинно зрелое антитело обладает аффинностью на уровне наномолей или даже пикомолей к антигену-мишени. Аффинно зрелые антитела получают с использованием процедур, известных в данной области. Например, Marks et al., Bio/Technology, 10:779-783 (1992) описывает созревание аффинности посредством перестановки VH- и VL-доменов. Случайный мутагенез остатков HVR и/или каркасных областей описан, например, Barbas et al. Proc. Nat. Acad. Sci. USA, 91:3809-3813 (1994); Schier et al., Gene, 169:147-155 (1995); Yelton et al., J. Immunol., 155:1994-2004 (1995); Jackson et al., J. Immunol., 154(7):3310-9 (1995) и Hawkins et al., J. Mol. Biol., 226:889-896 (1992).
В том смысле, в котором здесь используется термин «специфически связывается с» или «специфичный к», он относится к определяемому и воспроизводимому взаимодействию, такому как связывание мишени и антитела, которое определяется присутствием мишени в присутствии гетерогенной популяции молекул, включая биологические молекулы. Например, антитело, которое специфически связывается с мишенью (которая может быть эпитопом), является антителом, которое связывается с данной мишенью с большей аффинностью, авидностью, легче и/или с большей продолжительностью, чем оно связывается с другими мишенями. В одном варианте осуществления степень связывания антитела с неродственной мишенью составляет менее чем примерно 10% от связывания антитела с родственной мишенью, как определено, например, с помощью радиоиммуноанализа (RIA). В некоторых вариантах осуществления антитело, которое специфически связывается с мишенью, имеет константу диссоциации (Kd), равную ≤1 мкМ, ≤100 нМ, ≤10 нм, ≤1 нМ или ≤0,1 нм. В некоторых вариантах осуществления антитело специфически связывается с эпитопом на белке, который является консервативным белком среди различных видов. В другом варианте осуществления специфическое связывание может включать, но не обязательно, связывание, исключающее связывание с другими мишенями.
В том смысле, в котором здесь используется термин «иммуноадгезин», он обозначает антитело-подобные молекулы, которые сочетают специфичность связывания гетерологичного белка («адгезина») с эффекторными функциями константных областей иммуноглобулина. В структурном отношении иммуноадгезины представляют гибрид аминокислотной последовательности с требуемой специфичностью связывания, которая, является иной, чем сайт распознавания антигена и связывания антитела (т.е. является «гетерологичной»), и последовательности константной области иммуноглобулина. Адгезиновый фрагмент молекулы иммуноадгезина обычно представляет собой смежную аминокислотную последовательность, содержащую, по меньшей мере, сайт связывания рецептора или лиганда. Последовательность константной области иммуноглобулина в иммуноадгезине может быть получена из любого иммуноглобулина, такого как подтипы IgG1, IgG2 (включая IgG2a и IgG2b), IgG3 или IgG4 IgA, (включая IgA1 и IgA2), IgE, IgD или IgM. Гибридные Ig предпочтительно включают замену, по меньшей мере, одной вариабельной области внутри молекулы Ig доменом полипептида или антитела по настоящему изобретению. В особенно предпочтительном варианте осуществления гибридный иммуноглобулин включает шарнирную область, СН2 и СН3, или шарнирную область, CH1, CH2 и CH3 молекулы IgG1. Информацию по получению гибридов иммуноглобулина смотри также в патенте США № 5428130, выданном 27 июня 1995 года. Например, полезные иммуноадгезины, используемые в качестве вторых лекарственных средств, пригодных здесь для комбинированной терапии, включают полипептиды, которые содержат внеклеточный или связывающийся с PD-1 фрагменты PD-L1 или PD L2, или внеклеточный или связывающийся с PD-L1 или PD-L2 фрагменты PD-1, слитые с константной областью последовательности иммуноглобулина, такие как PD-L1 ECD-Fc, PD-L2 ECD-Fc и PD-1 ECD-Fс соответственно. В некоторых случаях иммуноадгезиновые комбинации Ig Fc и ECD рецепторов клеточной поверхности иногда называют растворимыми рецепторами.
«Гибридный белок» и «гибридный полипептид» относятся к полипептиду, имеющему два фрагмента, ковалентно связанных друг с другом, где каждый из фрагментов представляет собой полипептид, обладающий различными свойствами. Свойство может представлять биологическое свойство, такое как активность in vitro или in vivo. Свойство также может быть простым химическим или физическим свойством, таким как связывание с молекулой-мишенью, катализ реакции и т.д. Эти два фрагмента могут быть связаны непосредственно простой пептидной связью или через пептидный линкер, но оба находятся в рамке считывания друг с другом.
«Олигопептид PD-1», «олигопептид PD-L1», или «олигопептид PD-L2» представляют олигопептид, который связывается, предпочтительно специфически с отрицательным костимулирующим полипептидом PD-1, PD-L1 или PD-L2 соответственно, включая рецептор, лиганд или компонент сигнального пути соответственно, как здесь описано. Такие олигопептиды можно химически синтезировать с использованием известной методологии синтеза олигопептидов или можно получить и очистить с использованием рекомбинантной технологии. Такие олигопептиды обычно имеют длину, по меньшей мере, примерно из 5 аминокислот, альтернативно, по меньшей мере, примерно из 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22 , 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72 , 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100 аминокислот или более. Такие олигопептиды можно идентифицировать с использованием хорошо известных методов. В этой связи следует отметить, что в данной области хорошо известны методы скрининга олигопептидных библиотек с целью выявления олигопептидов, которые способны специфически связываться с полипептидом-мишенью (смотри, например, патенты США № 5556762, 5750373, 4708871, 4833092, 5223409, 5403484, 5571689, 5663143; публикации РСТ WO 84/03506 и WO84/03564; публикации Geysen et al., Proc. Natl. Acad. Sci. U.S.A., 81:3998-4002 (1984); Geysen et al., Proc. Natl. Acad. Sci. U.S.A., 82:178-182 (1985); Geysen et al., in Synthetic Peptides as Antigens, 130-149 (1986); Geysen et al., J. Immunol. Meth., 102:259-274 (1987); Schoofs et al., J. Immunol., 140:611-616 (1988), Cwirla, S. E. et al. Proc. Natl. Acad. Sci. USA, 87:6378 (1990); Lowman, H.B. et al. Biochemistry, 30:10832 (1991); Clackson, T. et al. Nature, 352: 624 (1991); Marks, J. D. et al., J. Mol. Biol., 222:581 (1991); Kang, A.S. et al. Proc. Natl. Acad. Sci. USA, 88:8363 (1991) и Smith, G. P., Current Opin. Biotechnol., 2:668 (1991).
«Блокирующее» антитело или «антагонистическое» антитело представляет собой антитело, которое ингибирует или снижает биологическую активность антигена, с которым оно связывается. В некоторых вариантах осуществления, блокирующие антитела или антагонистические антитела в основном или полностью ингибируют биологическую активность антигена. Анти-PD-L1-антитела по изобретению блокируют передачу сигналов через PD-1, с восстановлением функционального ответа Т-клеток (например, пролиферацию, продукцию цитокинов, индукцию гибели клеток-мишеней) из дисфункционального состояния на антигенную стимуляцию.
Термин «агонист» или активирующее антитело представляет собой антитело, которое усиливает или инициирует передачу сигналов антигеном, с которым оно связывается. В некоторых вариантах осуществления антитела-агонисты вызывают или активируют передачу сигналов без присутствия природного лиганда.
Термин «Fc-область» используется здесь для определения С-концевой области тяжелой цепи иммуноглобулина, включая Fc-области с нативной последовательностью и вариантные Fc-области. Несмотря на то, что границы Fc-области тяжелой цепи иммуноглобулина могут изменяться, Fc-область тяжелой цепи человеческого IgG обычно определяется как простирающаяся от аминокислотного остатка в положении Cys226 или от Pro230 его карбоксильного конца. C-концевой лизин (остаток 447 согласно системе нумерации EU) Fc-области может быть удален, например, при получении или очистке антитела, или рекомбинантным методом можно сконструировать нуклеиновую кислоту, кодирующую тяжелую цепь антитела. Следовательно, композиция интактных антител может включать популяции антител со всеми удаленными остатками K447, популяции антител без удаленных остатков K447 и популяции антител, содержащих смесь популяций антител с удаленными и неудаленными остатками K447. Подходящие Fc-области с нативной последовательностью для использования в антителах по изобретению включают человеческий IgG1, IgG2 (IgG2a, IgG2b), IgG3 и IgG4.
«Fc-рецептор» или «FcR» описывает рецептор, который связывается с Fc-областью антитела. Предпочтительным FcR является FcR с нативной последовательностью человека. Кроме того, предпочтительным FcR является рецептор, который связывается с IgG антителом (гамма-рецептор) и он включает рецепторы подклассов FcγRI, FcγRII и FcγRIII, в том числе, аллельные варианты и альтернативно сплайсированные формы этих рецепторов, FcγRII рецепторы включают FcγRIIA («активирующий рецептор») и FcγRIIB («ингибирующий рецептор»), которые имеют сходные аминокислотные последовательности, которые отличаются главным образом своими цитоплазматическими доменами. Активирующий рецептор FcγRIIA содержит мотив активации иммунорецептора на основе тирозина (ITAM) в его цитоплазматическом домене. Ингибирующий рецептор FcγRIIB содержит мотив ингибирования иммунорецептора на основе тирозина (ITAM) в его цитоплазматическом домене (смотри, например, M. Daëron, Annu. Rev. Immunol., 15:203-234 (1997). Обзор по FcR представлен Ravetch and Kinet, Annu. Rev. Immunol., 9:457-92 (1991); Capel et al., Immunomethods, 4:25-34 (1994); и de Haas et al., J. Lab. Clin. Med., 126:330-41 (1995). Другие FcR, в том числе те, которые будут идентифицированы в будущем, охватываются термином «FcR» в данном документе.
Термин «Fc-рецептор» или «FcR» также включает неонатальный рецептор FcRn, который ответственен за перенос материнских IgG антител плоду. Guyer et al., J. Immunol., 117:587 (1976) и Kim et al., J. Immunol., 24:249 (1994). Известны методы количественного определения связывания с FcRn (смотри, например, Ghetie and Ward, Immunol. Today, 18:(12): 592-8 (1997); Ghetie et al., Nature Biotechnology, 15(7):637-40 (1997); Hinton et al., J. Biol. Chem., 279(8):6213-6 (2004); WO 2004/92219 (Hinton et al.)). Связывание с FcRn in vivo и период полураспада в сыворотке крови человека высокоаффинных полипептидов, связывающихся с FcRn, можно определить, например, на трансгенных мышах или в трансфектированных клеточных линиях человека, экспрессирующих человеческий FcRn, или на приматах, которым вводят полипептиды, содержащие вариантную Fc-область. В WO 2004/42072 (Presta) описаны варианты антител, которые обладают улучшенным или пониженным связыванием с FcR. Смотри также, например, Shields et al., J. Biol. Chem., 9(2):6591-6604 (2001).
В том смысле, в котором здесь используется выражение «существенно уменьшена» или «существенно отличается», оно обозначает достаточно высокую степень различия между двумя числовыми значениями (обычно одно, относящееся к определяемой молекуле, и другое, относящееся к стандарту/молекуле сравнения), что позволяет рассматривать специалисту в данной области различие между двумя значениями на уровне статистически значимости в контексте биологической характеристики, определяемой указанными значениями (например, значения Kd). Различие между указанными двумя значениями, например, составляет более чем примерно 10%, более чем примерно 20%, более чем примерно 30%, более чем примерно 40% и/или более чем примерно 50% по сравнению со значением для стандарта/молекулы сравнения.
В том смысле, в котором здесь используется выражение «по существу аналогичный» или «по существу одинаковый», оно обозначает достаточно высокую степень сходства двух числовых значений (обычно одно, относящееся к определяемой молекуле, и другое, относящееся к стандарту/молекуле сравнения), что позволяет специалисту в данной области рассматривать различие между двумя значениями как незначительное или не имеющее биологической и/или статистической значимости в контексте биологической характеристики, определяемой указанными значениями (например, значения Kd). Различие между указанными двумя значениями, например, составляет менее чем примерно 50%, менее чем примерно 40%, менее чем примерно 30%, менее чем примерно 20% и/или менее чем примерно 10% по сравнению со значением для стандарта/молекулы сравнения.
В том смысле, в котором здесь используется термин «носители», он включает фармацевтически приемлемые носители, эксципиенты или стабилизаторы, которые являются нетоксичными для клетки или млекопитающего, подвергающихся их воздействию, в используемых дозах и концентрациях. Часто физиологически приемлемый носитель представляет собой водный раствор с забуференным рН. Примеры физиологически приемлемых носителей включают буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту; низкомолекулярный полипептид (менее чем примерно 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахарные спирты, такие как маннит или сорбит; солеобразующие противоионы, такие как натрий; и/или неионногенные поверхностно-активные вещества, такие как Твин™, полиэтиленгликоль (ПЭГ) и PLURONICS™.
«Вкладыш в упаковке» относится к инструкциям, которые обычно входят в коммерческие упаковки лекарственных средств, которые содержат информацию о показаниях, применении, дозировке, введении, противопоказаниях, других лекарственных средствах, которые применяются совместно с упакованным продуктом, и/или предупреждения, касающиеся применения таких лекарственных средств и т.д.
В том смысле, в котором здесь используется термин «лечение», он относится к клиническому вмешательству, направленному на изменение естественного процесса в организме индивидуума или клетке, подвергаемых лечению в случае клинической патологии. Желательные эффекты лечения включают снижение скорости прогрессирования заболевания, облегчение или ослабление болезненного состояния, и ремиссию или более благоприятный прогноз. Например, индивидуум успешно «лечится», если один или более симптомов, связанных с раком, ослабляются или устраняются, включая, не ограничиваясь этим, снижение пролиферации (или гибель) раковых клеток, ослабление симптомов, возникших в результате заболевания, улучшение качества жизни индивидуумов, страдающих этим заболеванием, снижение дозы других препаратов, необходимых для лечения заболевания, замедление прогрессирования заболевания и/или повышение выживаемости индивидуумов.
В том смысле, в котором здесь используется термин «замедление прогрессирования заболевания», он означает торможение, предупреждение, замедление, отмену, стабилизацию и/или отсрочку развития заболевания (например, рака). Такое замедление может иметь различную продолжительность времени, в зависимости от истории болезни и/или индивидуума, который подвергается лечению. Как очевидно специалисту в данной области, достаточное или значительное замедление может, в действительности, включать предупреждение заболевания, в том числе ситуацию, когда болезнь у субъекта не разовьется. Например, поздняя стадия рака, такая как развитие метастазов, может быть задержана.
В том смысле, в котором здесь используется термин «снижение или ингибирование рецидива рака», он означает снижение или ингибирование рецидива опухоли или рака или прогрессирования опухоли или рака.
В том смысле, в котором здесь используются термины «рак» и «раковый», они относятся или описывают физиологическое состояние у млекопитающих, которое обычно характеризуется неконтролируемым ростом клеток. В это определение входят доброкачественные и злокачественные опухоли, а также «дремлющие» опухоли или микрометастазы. Примеры рака включают, не ограничиваясь этим, карциному, лимфому, бластому, саркому и лейкоз. Более конкретные примеры таких раковых заболеваний включают плоскоклеточный рак, рак легких (в том числе мелкоклеточный рак легких, немелкоклеточный рак легких, аденокарциному легких и плоскоклеточный рак легких), рак брюшной полости, гепатоцеллюлярный рак, рак желудка или карциному желудка (в том числе рак органов желудочно-кишечного тракта), рак поджелудочной железы, глиобластому, рак шейки матки, рак яичников, рак печени, рак мочевого пузыря, гепатому, рак молочной железы, рак ободочной кишки, рак прямой кишки, рак эндометрия или карциному матки, карциному слюнной железы, рак почек или почечно-клеточный рак, рак печени, рак предстательной железы, рак вульвы, рак щитовидной железы, карциному печени и различные злокачественные опухоли головы и шеи, а также B-клеточную лимфому (включая низкодифференцированную/фолликулярную неходжкинскую лимфому (НХЛ); мелкоклеточную лимфоцитарную (SL) НХЛ; среднедифференцированную/фолликулярную НХЛ; среднедифференцированную диффузную НХЛ; высокодифференцированную иммунобластную НХЛ; высокодифференцированную лимфобластную НХЛ; высокодифференцированную мелкоклеточную с нерасщепленными ядрами НХЛ; массивное НХЛ поражение; мантийно-клеточную лимфому, связанную со СПИДом, и макроглобулинемию Вальденстрема), хронический лимфоцитарный лейкоз (CLL); острый лимфобластный лейкоз (ALL); волосатоклеточный лейкоз; хронический миелобластный лейкоз; и лимфопролиферативное расстройство после трансплантации (PTLD), а также аномальную пролиферацию сосудов, связанную с факоматозами, отек (такой как связанный с опухолями головного мозга) и синдром Мейгса.
В том смысле, в котором здесь используется термин «метастазы», он означает распространение рака из места его первичной локализации в другие места в организме. Раковые клетки могут оторваться от первичной опухоли, проникнуть в лимфатические и кровеносные сосуды, циркулировать в кровотоке, и расти в отдаленном месте (метастазировать) в нормальных тканях где угодно в организме. Метастазы могут быть локальными или отдаленными. Метастазирование это последовательный процесс, зависящий от опухолевых клеток, отрывающихся от первичной опухоли, «путешествуя» через кровоток, и останавливаясь в отдаленном месте. На новом месте клетки устанавливают кровоснабжение и могут расти с образованием угрожающей жизни массы. Оба стимулирующие и ингибирующие молекулярные пути в опухолевой клетке регулируют этот процесс, и также имеют значение взаимодействия между опухолевой клеткой и клеткой-хозяином в отдаленном месте.
Термин «эффективное количество» означает, по меньшей мере, минимальную концентрацию, необходимую для достижения определяемого улучшения или предупреждения конкретного заболевания. Эффективное количество может варьировать в зависимости от таких факторов, как состояние болезни, возраст, пол и масса тела пациента, а также от способности антитела вызывать желаемый ответ у индивидуума. Эффективное количество также представляет количество, при котором любые токсические или отрицательные эффекты лечения перевешиваются терапевтически благоприятными эффектами. Для профилактического применения благоприятные или желаемые результаты включают такие результаты, как устранение или снижение риска, ослабление тяжести или задержку начала развития болезни, включая биохимические, гистологические и/или поведенческие симптомы заболевания, его осложнения и промежуточные патологические фенотипы, возникающие во время развития заболевания. Для терапевтического применения благоприятные или желаемые результаты включают клинические результаты, такие как снижение одного или более симптомов, возникших в результате заболевания, улучшение качества жизни индивидуума, страдающего данным заболеванием, снижение дозы других лекарственных средств, необходимых для лечения заболевания, усиление действия другого препарата, например, благодаря направленному действию, замедление прогрессирования заболевания и/или повышение выживаемости. В случае рака или опухоли эффективное количество лекарственного средства может иметь эффект, проявляющийся в снижении количества раковых клеток; уменьшении размера опухоли; ингибировании (т.е. замедлении до некоторой степени или желательно остановки) инфильтрации раковых клеток в периферические органы; ингибировании (т.е. замедлении до некоторой степени и желательно остановки) метастазирования опухоли; ингибирования до некоторой степени роста опухоли; и/или облегчение до некоторой степени одного или более симптомов, связанных с расстройством. Эффективное количество можно вводить в один или несколько приемов. Для целей настоящего изобретения эффективное количество лекарственного средства, соединения или фармацевтической композиции представляет количество, достаточное для достижения профилактического или терапевтического лечения прямо или опосредованно. Как известно в клинической практике, эффективное количество лекарственного средства, соединения или фармацевтической композиции может или может не быть достигнуто в комбинации с другим лекарственным средством, соединением или фармацевтической композицией. Таким образом, «эффективное количество» может рассматриваться с учетом введения одного или более терапевтических агентов, и один агент может считаться введенным в эффективном количестве, если в комбинации с одним или более другими агентами, может быть или достигается желаемый результат.
В том смысле, в котором здесь используется выражение «в комбинации с», оно относится к введению одного лекарственного средства в комбинации с другим лекарственным средством. Таким образом, выражение «в комбинации с» относится к введению одного лекарственного средства до, во время или после введения другого лекарственного средства индивидууму.
В том смысле, в котором здесь используется термин «субъект», он означает млекопитающее, включая, не ограничиваясь этим, человека или млекопитающих, отличных от человека, таких как представители семейств бычьих, лошадиных, собачьих, овцы или представители семейства кошачьих. Предпочтительно субъект является человеком. Пациенты также являются субъектами в данном документе.
В том смысле, в котором здесь используется термин «полный ответ или «CR», он относится к исчезновению всех патологических очагов-мишеней; «частичный ответ» или «PR» относится, по меньшей мере, к 30% уменьшению суммы наибольших диаметров (SLD) патологических очагов-мишеней, взяв в качестве стандарта исходное значение SLD; и «стабильное заболевание» или «SD» относится к состоянию, при котором не происходит достаточного уменьшения патологических очагов-мишеней, чтобы квалифицировать это как PR, ни достаточного их увеличения, чтобы квалифицировать их как PD, взяв в качестве стандарта наименьшее значение SLD с момента начала лечения.
В том смысле, в котором здесь используется термин «прогрессирующее заболевание» или «PD», он относится, по меньшей мере, к увеличению на 20% SLD патологических очагов-мишеней, взяв в качестве стандарта наименьшее значение SLD с момента начала лечения, или образованию одного или более новых очагов.
В том смысле, в котором здесь используется термин «выживаемость без прогрессирования» (PFS), он относится к продолжительности периода во время и после лечения, в течение которого заболевание, которое подвергается лечению (например, рак), не ухудшается. Выживаемость без прогрессирования может включать период времени, когда у пациентов имеется полный ответ или частичный ответ, а также период времени, когда у пациентов имеется стабильное заболевание.
В том смысле, в котором здесь используется термин «общая частота ответов» (ORR), он относится к общей частоте полных ответов (CR) и частоте частичных ответов (PR).
В том смысле, в котором здесь используется термин «общая выживаемость», он относится к проценту индивидуумов в группе, которые, вероятно, останутся живы по истечении определенного периода времени.
«Химиотерапевтическое средство» представляет химическое соединение, пригодное для лечения опухолей. Примеры химиотерапевтических средств включают алкилирующие агенты, такие как тиотепа и циклофосфамид (CYTOXAN®); алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, включая алтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилолмеламин; ацетогенины (в частности, буллатацин и буллатацинон); дельта-9-тетрагидроканнабинол (дронабинол, MARINOL®); бета-лапахон; лапахол; колхицины; бетулиновую кислоту; камптотецин (включая синтетический аналог топотекан (HYCAMTIN®); CPT-11 (иринотекан, CAMTOSAR®), ацетилкамптотецин, скололектин и 9-аминокамптотеуин); бриостатин; пеметрексед; каллистатин; СС-1065 (включая его синтетические аналоги адозелезин, карзелезин и бизелезин); подофиллотоксин; подофиллиновую кислоту; тенипозид; криптофицины (в частности, криптофицин 1 и криптофицин 8); доластатин; дуокармуцин (включая синтетические аналоги KW-2189 и СВ1-ТМ1); элеутеробин; панкратистатин; TLK-286; CDP323, ингибитор альфа-4-интегрина для перорального введения; саркодиктин; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, холофосфамид, эстрамустин, ифосфамид, мехлоретамин, гидрохлорид мехлоретаминоксида, мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урациловый иприт; нитрозомочевины, такие как кармустин, хлорзотоцин, фотемустин, ломустин, нимустин и ранимустин; антибиотики, такие как энедииновые антибиотики (например, калихеамицин, в частности, калихеамицин гамма 1I и калихеамицин омега I1 (смотри, например, публикацию Nicolaou et al., Angew. Chem. Intl. Ed. Engl., 33: 183-186 (1994)); динемицин, включая динемицин А; эсперамицин; а также неокарциностатиновый хромофор и родственные хромофоры хромопротеиновых энедииновых антибиотиков), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карцинофиллин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин (включая ADRIAMYCIN®, морфолинодоксорубицин, цианоморфолинодоксорубицин, 2-пирролинодоксорубицин и липосомальный дезоксидоксорубицин HCl для инъекций (DOXIL® и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марселломицин, митомицины, такие как митомицин С, микофеноловую кислоту, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат, гемцитабин (GEMZAR®), тегафур (UFTORAL®), капецитабин (XELODA®), эпотилон и 5-фторурацил (5-ФУ); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметотрексат; аналоги пуринов, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидинов, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин и иматиниб (производное 2-фениламинопиримидина), а также другие игибиторы c-Kit; антиадреналиновые средства, такие как аминоглютетимид, митотан, трилостан; восполнитель фолиевой кислоты, такой как фролиновая кислота; ацеглатон; альдофосфамидгликозид; аминолевулиновую кислоту; энилурацил; амсакрин; бестрабуцил; бисантрен; эдатраксат; дефофамин; демекольцин; диазиквон; элфорнитин; эллиптиния ацетат; этоглуцид; галлия нитрат; гидроксимочевину; лентинан; лонидаинин; майтансиноиды, такие как майтанзин и анзамитоцины; митогуазон; митоксантрон; мопиданмол; нитраэрин; пентостатин; фенамет; пирарубицин; лозоксантрон; 2-этилгидразид; прокарбазин; PSK® полисахаридный комплекс (JHS Natural Produсts, Eugene, OR); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновую кислоту; триазиквон; 2,2',2''-трихлортриэтиламин; трихотецены (в частности, токсин Т-2, верракурин А, роридин А и ангуидин); уретан; виндесин (ELDISINE®, FILDESIN®); дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид («Ara-C»); тиотепа; таксоиды, например паклитаксел TAXOL®, препарат паклитаксела в виде наночастиц на основе альбумина (ABRAXANE™) и доксетаксел TAXOTERE®; хлоранбуцил; 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин (VELBAN®); платину; этопозид (VP-16); ифосфамид; митоксантрон; винкристин (ONCOVIN®); оксалиплатин; лейкововин; винорелбин (NAVELBINE®); новантрон; эдатрексат; дауномицин; аминоптерин; ибандронат; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота; фармацевтически приемлемые соли, кислоты или производные любого из приведенных выше лекарственных средств; а также комбинации двух или более вышеуказанных лекарственных средств, такие как CHOP, сокращенное название комбинированной терапии циклофосфамидом, доксорубицином, винкристином и преднизолоном, и FOLFOX, сокращенное название для схемы лечения оксалиплатином (ELOXATIN™) в сочетании с 5-FU и лейковорином.
В данное определение также входят антигормональные агенты, которые регулируют, снижают, блокируют или ингибируют действие гормонов, которые могут способствовать росту опухоли, и часто применяются в виде системного или лечения всего организма. Они сами могут представлять гормоны. Примеры включают антиэстрогены и селективные модуляторы рецепторов эстрогенов (SERM), в том числе, например, тамоксифен (включая тамоксифен NOLVADEX®), ралоксифен (EVISTA®), дролоксифен, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и торемифен (FARESTON®); антипрогестероны; отрицательные регуляторы рецепторов эстрогенов (ERD); агенты, которые действуют, подавляя или блокируя функцию яичников, например агонисты лютеинизирующего гормона-высвобождающего гормона (LHRH), такие как лейпролида ацетат (LUPRON® и ELIGARD®), гозерелина ацетат, бузерелина ацетат и триптерелин; антиандрогены, такие как флутамид, нилутамид и бикалутамид; и ингибиторы ароматазы, которые ингибируют фермент ароматазу, который регулирует выработку эстрогена в надпочечниках, такие как, например, 4(5)-имидазолы, аминоглутетимид, мегестрола ацетат (MEGASE®), экземестан (AROMASIN®), форместан, фадрозол, ворозол (RIVISOR®), летрозол (FEMARA®) и анастрозол (ARIMIDEX®). Кроме того, в определение химиотерапевтических средств входят бифосфонаты, такие как клодронат (например, BONEFOS® или OSTAC®), этидронат (DIDROCAL®), NE-58095, золедроновая кислота/золедронат (ZOMETA®), алендронат (FOSAMAX®), памидронат (AREDIA®), тилудронат (SKELID®) или ризедронат (ACTONEL®); а также троксацитабин (аналог нуклеозида цитозина на основе 1,3-диоксолана); антисмысловые олигонуклеотиды, в частности, олигонуклеотиды, которые ингибируют экспрессию генов сигнальных путей, вовлеченных в нарушенную клеточную пролиферацию, такие как, например, PKC-альфа, Raf, H-Ras, и рецептор эпидермального фактора роста (EGF-R); вакцины, такие как вакцина THERATORE® и вакцины для генной терапии, например вакцина ALLOVECTIN®, вакцина LEUVECTIN® и вакцина VAXID®; ингибитор топоизомеразы 1 (например, LUPTOTECAN®); антиэстроген, такой как фулвестрант; ингибитор Kit, такой как иматиниб или EXEL-0862 (ингибитор тирозинкиназы); ингибитор EGFR, такой как эрлотиниб или цетуксимаб; ингибитор VEGF, такой как бевацизумаб; аринотекан; rmRH (например, ABARELIX®); лапатиниб и лапатиниб дитозилат (низкомолекулярный двойной ингибитор тирозинкиназы ErbB-2 и EGFR, также известный как GW572016); 17AAG (производное гелданамицина, которое является ядом для белка теплового шока (Hsp)90) и фармацевтически приемлемые соли, кислоты или производные любого из вышеупомянутых лекарственных средств.
В том смысле, в котором здесь используется термин «цитокин», в общем, он относится к белкам, высвобождаемым одной популяцией клеток, которые воздействуют на другую клетку в качестве межклеточных медиаторов или оказывают аутокринное действие на клетки, продуцирующие белки. Примерами таких цитокинов являются лимфокины, монокины; интерлейкины («IL»), такие как IL-1, IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-11, IL-12, IL-13, IL-15, IL-17А-F, IL-18-IL-29 (такие как IL-23), IL-31, включая rIL-2 PROLEUKIN®; фактор некроза опухолей, такой как TNF-α или TNF-β, TNF-β1-3; и другие полипептидные факторы, включая лейкоз-ингибирующий фактор («LIF»), цилиарный нейротрофический фактор («CNTF»), CNTF-подобный цитокин («CLC»), кардиотрофин («CT») и лиганд kit (KL).
В том смысле, в котором здесь используется термин «хемокин», он относится к растворимых факторам (например, цитокинам), которые обладают способностью избирательно индуцировать хемотаксис и активацию лейкоцитов. Они также «запускают» процессы ангиогенеза, воспаления, заживления ран и онкогенеза. Примерные хемокины включают IL-8, человеческий гомолог мышиного хемоаттрактанта кератиноцитов (KC).
В настоящем описании и в прилагаемой формуле изобретения, формы единственного числа «а», «или» и «the» включают ссылки на множественное число, если из контекста явно не следует иное.
Определение значения или параметра термином «примерно» в описании, включает (и описывает) вариации, которые относятся к этому значению или параметру как таковым. Например, описание, относящееся к «примерно X» включает описание «Х».
В том смысле, в котором здесь используется выражение «фармацевтически приемлемая соль», оно относится к фармацевтически приемлемым органическим или неорганическим солям соединения по настоящему изобретению. Примеры солей включают, не ограничиваясь этим, сульфат, цитрат, ацетат, оксалат, хлорид, бромид, иодид, нитрат, бисульфат, фосфат, кислый фосфат, изоникотинат, лактат, салицилат, кислый цитрат, тартрат, олеат, таннат, пантотенат, битартрат, аскорбат, сукцинат, малеат, гентизинат, фумарат, глюконат, глюкуронат, сахарат, формиат, бензоат, глутамат, метансульфонат «мезилат», этансульфонат, бензолсульфонат, п-толуолсульфонат, памоат (т.е. 1,1'-метилен-бис(2-гидрокси-3-нафтоат)), соли щелочных металлов (например, соли натрия и калия), соли щелочноземельных металлов (например, соли магния) и соли аммония. Фармацевтически приемлемая соль может содержать включение другой молекулы, такой как ион ацетата, ион сукцината или другого противоиона. Противоион может представлять любую органическую или неорганическую группу, которая стабилизирует заряд на исходном соединении. Кроме того, его фармацевтически приемлемая соль может иметь более одного заряженного атома в ее структуре. В тех случаях, когда несколько заряженных атомов являются частью фармацевтически приемлемой соли, то соединение может иметь несколько противоионов. Следовательно, его фармацевтически приемлемая соль может иметь один или более заряженных атомов и/или один или более противоионов.
Если соединение по изобретению представляет собой основание, то требуемая фармацевтически приемлемая соль может быть получена любым подходящим способом, имеющимся в данной области, например, обработкой свободного основания неорганической кислотой, такой как соляная кислота, бромистоводородная кислота, серная кислота, азотная кислота, метансульфоновая кислота, фосфорная кислота и тому подобное, или органической кислотой, такой как уксусная кислота, малеиновая кислота, янтарная кислота, миндальная кислота, фумаровая кислота, малоновая кислота, пировиноградная кислота, щавелевая кислота, гликолевая кислота, салициловая кислота, пиранозидиловая кислота, такая как глюкуроновая кислота или галактуроновая кислота, альфа-оксикислота, такая как лимонная кислота или винная кислота, аминокислота, такая как аспарагиновая кислота или глутаминовая кислота, ароматическая кислота, такая как бензойная кислота или коричная кислота, сульфоновая кислота, такая как п-толуолсульфоновая кислота или этансульфоновая кислота, или тому подобное.
Если соединение по изобретению представляет собой кислоту, то требуемая фармацевтически приемлемая соль может быть получена любым подходящим способом, например, обработкой свободной кислоты неорганическим или органическим основанием, таким как амин (первичный, вторичный или третичный), гидроксид щелочного металла или гидроксид щелочноземельного металла или тому подобное. Иллюстративные примеры подходящих солей включают, не ограничиваясь этим, органические соли, полученные из аминокислот, таких как глицин и аргинин, аммиака, первичных, вторичных и третичных аминов, и циклических аминов, таких как пиперидин, морфолин и пиперазин, и неорганические соли, полученные из натрия, кальция, калия, магния, марганца, железа, меди, цинка, алюминия и лития.
Выражение «фармацевтически приемлемый» означает, что соединение или композиция должны быть совместимы химически и/или токсикологически с другими ингредиентами препарата, входящими в состав препарата, и/или млекопитающим, подлежащим лечению ими.
Следует понимать, что аспекты и варианты осуществления изобретения, описанные здесь, включают «состоящие из» и/или «состоящие по существу из» аспектов и вариантов.
III. Способы
В одном аспекте обеспечивается способ лечения или замедления прогрессирования рака у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT.
В другом аспекте обеспечивается способ снижения или ингибирования рецидива или прогрессирования рака у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. Как здесь раскрывается, рецидив и/или прогрессирование рака включает, не ограничиваясь этим, метастазирование рака.
В еще одном аспекте обеспечивается способ лечения или замедления прогрессирования связанного с иммунитетом заболевания у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT.
В еще одном аспекте обеспечивается способ снижения или ингибирования прогрессирования связанного с иммунитетом заболевания у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT.
В некоторых вариантах осуществления связанное с иммунитетом заболевание ассоциировано с Т-клеточным дисфункциональным расстройством. В некоторых вариантах осуществления связанное с иммунитетом заболевание представляет вирусную инфекцию. В некоторых вариантах осуществления вирусная инфекция представляет хроническую вирусную инфекцию. В некоторых вариантах осуществления Т-клеточное дисфункциональное расстройство характеризуется снижением реактивности на антигенную стимуляцию. В некоторых вариантах осуществления Т-клеточное дисфункциональное расстройство характеризуется анергией Т-клеток или пониженной способностью к секреции цитокинов, пролиферации или осуществлению цитолитической активности. В некоторых вариантах осуществления Т-клеточное дисфункциональное расстройство характеризуется истощением Т-клеток. В некоторых вариантах осуществления Т-клетки представляют собой CD4+ и CD8+ Т-клетки. В некоторых вариантах осуществления Т-клеточное дисфункциональное расстройство выбрано из группы, состоящей из неразрешенной острой инфекции, хронической инфекции и иммунитета к опухоли.
В еще одном аспекте обеспечивается способ повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT.
В еще одном аспекте обеспечивается способ лечения или замедления прогрессирования рака у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226.
В еще одном аспекте обеспечивается способ снижения или ингибирования рецидива или прогрессирования рака у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226.
В еще одном аспекте обеспечивается способ лечения или замедления прогрессирования связанного с иммунитетом заболевания у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226.
В еще одном аспекте обеспечивается способ снижения или ингибирования прогрессирования связанного с иммунитетом заболевания у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226.
В некоторых вариантах осуществления связанное с иммунитетом заболевание ассоциировано с Т-клеточным дисфункциональным расстройством. В некоторых вариантах осуществления связанное с иммунитетом заболевание представляет вирусную инфекцию. В некоторых вариантах осуществления вирусная инфекция представляет хроническую вирусную инфекцию. В некоторых вариантах осуществления Т-клеточное дисфункциональное расстройство характеризуется снижением реактивности на антигенную стимуляцию. В некоторых вариантах осуществления Т-клеточное дисфункциональное расстройство характеризуется анергией Т-клеток или пониженной способностью к секреции цитокинов, пролиферации или осуществлению цитолитической активности. В некоторых вариантах осуществления Т-клеточное дисфункциональное расстройство характеризуется истощением Т-клеток. В некоторых вариантах осуществления Т-клетки представляют собой CD4+ и CD8+ Т-клетки. В некоторых вариантах осуществления Т-клеточное дисфункциональное расстройство выбрано из группы, состоящей из неразрешенной острой инфекции, хронической инфекции и иммунитета к опухоли.
В еще одном аспекте обеспечивается способ повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума введением индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226.
В некоторых вариантах осуществления, агент, который модулирует экспрессию и/или активность CD226, способен повышать и/или стимулировать экспрессию и/или активность CD226; повышать и/или стимулировать взаимодействие CD226 с PVR, PVRL2 и/или PVRL3; и повышать и/или стимулировать внутриклеточную передачу сигналов, опосредованную связыванием CD226 с PVR, PVRL2 и/или PVRL3. В том смысле, в котором здесь используется термин «агент, который способен повышать и/или стимулировать экспрессию и/или активность CD226», он включает, не ограничиваясь этим, агенты, которые повышают и/или стимулируют экспрессию и/или активность CD226. В том смысле, в котором здесь используется термин «агент, который способен повышать и/или стимулировать взаимодействие CD226 с PVR, PVRL2, и/или PVRL3», он включает, не ограничиваясь этим, агенты, которые повышают и/или стимулируют взаимодействие CD226 с PVR, PVRL2 и/или PVRL3. В том смысле, в котором здесь используется термин «агент, который способен повышать и/или стимулировать внутриклеточную передачу сигналов, опосредованную связыванием CD226 с PVR, PVRL2 и/или PVRL3», он включает, не ограничиваясь этим, агенты, которые повышают и/или стимулируют внутриклеточную передачу сигналов, опосредованную связыванием CD226 с PVR, PVRL2 и/или PVRL3.
В некоторых вариантах осуществления агент, который модулирует экспрессию и/или активность CD226, выбран из агента, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, антагониста экспрессии и/или активности TIGIT, антагониста экспрессии и/или активности PVR, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVR, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL2, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL3, агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR, агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL2, агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL3, и их комбинаций.
В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, представляет анти-TIGIT-антитело или его антигенсвязывающий фрагмент. В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, представляет ингибирующую нуклеиновую кислоту, выбранную из антисмыслового полинуклеотида, интерферирующей РНК, каталитической РНК и химеры РНК-ДНК.
В некоторых вариантах осуществления антагонист экспрессии и/или активности TIGIT представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления антагонист экспрессии и/или активности TIGIT представляет анти-TIGIT-антитело или его антигенсвязывающий фрагмент. В некоторых вариантах осуществления антагонист экспрессии и/или активности TIGIT представляет ингибирующую нуклеиновую кислоту, выбранную из антисмыслового полинуклеотида, интерферирующей РНК, каталитической РНК и химеры РНК-ДНК.
В некоторых вариантах осуществления антагонист экспрессии и/или активности PVR представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления антагонист экспрессии и/или активности PVR выбран из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида.
В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVR, представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVR, выбран из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида.
В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL2, выбран из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида.
В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL3, выбран из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида.
В некоторых вариантах осуществления агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR, представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR, выбран из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида.
В некоторых вариантах осуществления агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL2, выбран из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида.
В некоторых вариантах осуществления агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL3, выбран из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида.
В еще одном аспекте обеспечивается способ повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума введением индивидууму эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов. В некоторых вариантах осуществления один или более дополнительных иммунных коингибирующих рецепторов выбран из PD-1, CTLA-4, LAG3, TIM3, BTLA, VISTA, B7H4 и CD96. В некоторых вариантах осуществления один или более дополнительных иммунных коингибирующих рецепторов выбран из PD-1, CTLA-4, LAG3 и TIM3.
В еще одном аспекте обеспечивается способ повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума введением индивидууму эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который повышает или активирует экспрессию и/или активность одного или более дополнительных иммунных костимулирующих рецепторов. В некоторых вариантах осуществления один или более дополнительных иммунных костимулирующих рецепторов выбран из CD226, ОХ-40, CD28, CD27, CD137, HVEM, GITR, MICA, ICOS, NKG2D и 2B4. В некоторых вариантах осуществления один или более дополнительных иммунных костимулирующих рецепторов выбран из CD226, OX40, CD28, CD27, CD137, HVEM и GITR. В некоторых вариантах осуществления один или более дополнительных иммунных костимулирующих рецепторов выбран из группы, состоящей из ОХ-40 и CD27.
Способы по настоящему изобретению могут найти применение при лечении состояний, где требуется повышенная иммуногенность, такая как повышенная иммуногенность опухоли, для лечения рака или Т-клеточных дисфункциональных расстройств.
Можно лечить различные виды рака или можно замедлить их прогрессирование.
В некоторых вариантах осуществления у индивидуума имеется немелкоклеточный рак легких. Немелкоклеточный рак легких может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется мелкоклеточный рак легких. Мелкоклеточный рак легких может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется почечно-клеточный рак. Почечно-клеточный рак может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется колоректальный рак. Колоректальный рак может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется рак яичников. Рак яичников может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется рак молочной железы. Рак молочной железы может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется рак поджелудочной железы. Рак поджелудочной железы может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется карцинома желудка. Карцинома желудка может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется рак мочевого пузыря. Рак мочевого пузыря может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется рак пищевода. Рак пищевода может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется мезотелиома. Мезотелиома может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется меланома. Меланома может быть на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеются злокачественные опухоли головы и шеи. Злокачественные опухоли головы и шеи могут находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется рак щитовидной железы. Рак щитовидной железы может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется саркома. Саркома может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется рак предстательной железы. Рак предстательной железы может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется глиобластома. Глиобластома может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется рак шейки матки. Рак шейки матки может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется карцинома тимуса. Карцинома тимуса может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется лейкоз. Лейкоз может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется лимфома. Лимфома может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется миелома. Миелома может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется грибовидный микоз. Грибовидный микоз может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеется рак из клеток Меркеля. Рак из клеток Меркеля может находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления у индивидуума имеются злокачественные заболевания крови. Злокачественные заболевания крови могут находиться на ранней стадии или на поздней стадии. В некоторых вариантах осуществления индивидуумом является человек.
В некоторых вариантах осуществления способов по настоящему изобретению CD4 и/или CD8 Т-клетки у индивидуума имеют повышенное или усиленное праймирование, активацию, пролиферацию, высвобождение цитокинов и/или цитолитическую активность по сравнению с периодом до введения комбинации.
В некоторых вариантах осуществления способов по настоящему изобретению количество CD4 и/или CD8 Т-клеток повышено по сравнению с периодом до введения комбинации. В некоторых вариантах осуществления способов по настоящему изобретению количество активированных CD4 и/или CD8 Т-клеток повышено по сравнению с периодом до введения комбинации.
В некоторых вариантах осуществления способов по настоящему изобретению активированные CD4 и/или CD8 Т-клетки характеризуются CD4 и/или CD8 Т-клетками, продуцирующими γ-IFN+, и/или повышенной цитолитической активностью по сравнению с периодом до введения комбинации.
В некоторых вариантах осуществления способов по настоящему изобретению CD4 и/или CD8 Т-клетки проявляют повышенное высвобождение цитокинов, выбранных из группы, состоящей из IFN-γ, TNF-α и интерлейкинов.
В некоторых вариантах осуществления способов по настоящему изобретению CD4 и/или CD8 Т-клетка является эффекторной Т-клеткой памяти. В некоторых вариантах осуществления способов по настоящему изобретению CD4 и/или CD8 эффекторная Т-клетка памяти характеризуется CD4 и/или CD8 Т-клетками, продуцирующими γ-IFN+, и/или повышенной цитолитической активностью. В некоторых вариантах осуществления способов по настоящему изобретению CD4 и/или CD8 эффекторная Т-клетка памяти характеризуется наличием экспрессии CD44высокий CD62Lнизкий.
В некоторых вариантах осуществления способов по настоящему изобретению злокачественная опухоль имеет повышенные уровни Т-клеточной инфильтрации.
В некоторых вариантах осуществления способы по настоящему изобретению могут дополнительно включать применение дополнительной терапии. Дополнительная терапия может представлять лучевую терапию, хирургическое вмешательство, химиотерапию, генную терапию, терапию ДНК, вирусную терапию, терапию РНК, иммунотерапию, трансплантацию костного мозга, нанотерапию, терапию моноклональными антителами или комбинацию вышеперечисленного. Дополнительная терапия может быть в виде адъювантной или неоадъювантной терапии. В некоторых вариантах осуществления дополнительная терапия представляет введение агентов, снижающих побочные эффекты (например, агентов, предназначенных для уменьшения развития и/или тяжести побочных эффектов, возникших в результате лечения, таких как противорвотные средства и т.д.). В некоторых вариантах осуществления дополнительная терапия представляет лучевую терапию. В некоторых вариантах осуществления дополнительная терапия является хирургическим вмешательством. В некоторых вариантах осуществления дополнительная терапия может представлять один или более химиотерапевтических агентов, представленных выше.
Любой из антагонистов, связывающихся с осью PD-1, и агентов, которые снижают или ингибируют экспрессию и/или активность TIGIT, описанных ниже, может использоваться в способах по настоящему изобретению.
В некоторых вариантах осуществления любая из мишеней, описанных здесь (например, PD-1, PD-L1, PD-L2, CTLA-4, LAG3, TIM3, BTLA, VISTA, B7H4, CD96, В7-1, TIGIT, CD226, ОХ-40, CD28, CD27, CD137, HVEM, GITR, MICA, ICOS, NKG2D, 2В4 и т.д.) представляет человеческий белок.
Антагонисты, связывающиеся с осью PD-1
Обеспечивается способ лечения или замедления прогрессирования рака у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. Обеспечивается способ снижения или ингибирования рецидива или прогрессирования рака у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. Обеспечивается способ лечения или замедления прогрессирования связанного с иммунитетом заболевания у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. Обеспечивается способ снижения или ингибирования прогрессирования связанного с иммунитетом заболевания у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. Обеспечивается способ повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT.
Например, антагонист, связывающийся с осью PD-1, включает PD-1-связывающий антагонист, PD-L1-связывающий антагонист и PD-L2-связывающий антагонист.
В некоторых вариантах осуществления PD-1-связывающий антагонист представляет молекулу, которая ингибирует связывание PD-1 с лигандом его партнеров по связыванию. В конкретном аспекте лиганд партнеров по связыванию PD-1 представляет PD-L1 и/или PD-L2. В еще одном варианте осуществления PD-L1-связывающий антагонист представляет молекулу, которая ингибирует связывание PD-L1 с его партнерами по связыванию. В конкретном аспекте партнеры по связыванию с PD-L1 представляют PD-1 и/или В7-1. В еще одном варианте осуществления PD-L2-связывающий антагонист представляет молекулу, которая ингибирует связывание PD-L2 с его партнерами по связыванию. В конкретном аспекте партнером по связыванию PD-L2 является PD-1. Антагонист может представлять собой антитело, его антигенсвязывающий фрагмент, иммуноадгезин, слитый белок или олигопептид.
В некоторых вариантах осуществления PD-1-связывающий антагонист выбран из MDX-1106 (ниволумаб), Merck 3475 (ламбролизумаб), CT-011 (пидилизумаб) и AMP-224. В некоторых вариантах осуществления PD-L1-связывающий антагонист выбран из группы, состоящей из YW243.55.S70, MPDL3280A, MDX-1105 и MEDI 4736. В некоторых вариантах осуществления PD-L2-связывающий антагонист представляет антитело АМР-224. В некоторых вариантах осуществления PD-1-связывающий антагонист представляет антитело АМР-224. Антитело MDX-1105, также известное как BMS-936559, представляет анти-PD-L1-антитело, описанное в WO2007/005874. Антитело YW243.55.S70 (SEQ ID NO:20) представляет анти-PD-L1-антитело, описанное в WO 2010/077634 А1 и патенте США № 8217149, которые включены здесь в качестве ссылки. Антитело MDX-1106, также известное как MDX-1106-04, ONO-4538, BMS-936558 или ниволумаб, представляет анти-PD-1-антитело, описанное в WO2006/121168. Антитело Merck 3745, также известное как МК 3475, МК-3475, SCH-900475 или ламбролизумаб, представляет анти-PD-1-антитело, описанное в WO2009/114335. Антитело СТ-011, также известное как hBAT, hBAT-1 или пидилизумаб, представляет анти-PD-1-антитело, описанное в WO2009/101611. Антитело AMP-224, также известное как B7-DCIg, представляет слитый растворимый рецептор PD-L2-Fc, описанный в WO2010/027827 и WO2011/066342.
Примеры анти-PD-L1-антител, пригодных для способов по настоящему изобретению, и способы их получения описаны в заявке на патент PCT WO 2010/077634 А1 и патенте США № 8217149, которые включены здесь в качестве ссылки.
В некоторых вариантах осуществления антагонист, связывающийся с осью PD-1, представляет собой анти-PD-L1-антитело. В некоторых вариантах осуществления изобретения анти-PD-L1-антитело способно ингибировать связывание PD-L1 и PD-1 и/или PD-L1 и В7-1. В некоторых вариантах осуществления анти-PD-L1-антитело представляет собой моноклональное антитело. В некоторых вариантах осуществления анти-PD-L1-антитело представляет фрагмент антитела, выбранный из группы, состоящей из фрагментов Fab, Fab'-SH, Fv, ScFv и (Fab')2. В некоторых вариантах осуществления изобретения анти-PD-L1-антитело представляет гуманизированное антитело. В некоторых вариантах осуществления изобретения анти-PD-L1-антитело представляет человеческое антитело.
Анти-PD-L1-антитела, пригодные в данном изобретении, включая композиции, содержащие такие антитела, например, описанные в WO 2010/077634 А1 и патенте США № 8217149, можно использовать в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT, с или без дополнительной терапии (например, химиотерапии), для лечения рака или связанного с иммунитетом заболевания (например, Т-клеточного дисфункционального расстройства, вирусной инфекции, хронической вирусной инфекции и т.д.).
В одном варианте осуществления анти-PD-L1-антитело содержит полипептид вариабельной области тяжелой цепи, включающий последовательность HVR-H1, HVR-H2 и HVR-H3, где:
(а) последовательность HVR-H1 представляет GFTFSX1SWIH (SEQ ID NO:33);
(b) последовательность HVR-H2 представляет AWIX2PYGGSX3YYADSVKG (SEQ ID NO:34);
(с) последовательность HVR-H3 представляет RHWPGGFDY (SEQ ID NO:19);
где далее: X1 представляет D или G, Х2 представляет S или L; X3 представляет Т или S.
В одном конкретном аспекте X1 представляет D; Х2 представляет S, и Х3 представляет Т. В еще одном аспекте полипептид дополнительно содержит последовательности каркасной области вариабельной области тяжелой цепи, находящиеся между HVR, согласно формуле: (HC-FR1)-(HVR-H1)-(HC-FR2)-(HVR-H2)-(HC-FR3)-(HVR-H3)-(HC-FR4). В еще одном аспекте последовательности каркасной области получены из консенсусных последовательностей человеческой каркасной области. В дополнительном аспекте последовательности каркасной области представляют консенсусную каркасную область VH подгруппы III. В еще одном аспекте, по меньшей мере, одна из последовательностей каркасной области представляет следующее:
HC-FR1 представляет EVQLVESGGGLVQPGGSLRLSCAAS (SEQ ID NO:25)
HC-FR2 представляет WVRQAPGKGLEWV (SEQ ID NO:26)
HC-FR3 представляет RFTISADTSKNTAYLQMNSLRAEDTAVYYCAR (SEQ ID NO:27)
HC-FR4 представляет WGQGTLVTVSA (SEQ ID NO:28).
В еще одном аспекте полипептид тяжелой цепи дополнительно объединен с вариабельной областью легкой цепи, содержащей HVR-L1, HVR-L2 и HVR-L3, где:
(а) последовательность HVR-L1 представляет RASQX4X5X6TX7X8A (SEQ ID NO:35);
(b) последовательность HVR-L2 представляет SASX9LX10S, (SEQ ID NO:36);
(с) последовательность HVR-L3 представляет QQX11X12X13X14PX15T (SEQ ID NO:37);
где далее: Х4 представляет D или V; Х5 представляет V или I; Х6 представляет S или N; X7 представляет А или F; X8 представляет V или L; X9 представляет F или Т; X10 представляет Y или А; X11 представляет Y, G, F или S; X12 представляет L, Y, F или W; X13 представляет Y, N, A, T, G, F или I; X14 представляет Н, V, Р, Т или I; X15 представляет A, W, R, P или Т.
В еще одном дополнительном аспекте Х4 представляет D; Х5 представляет V; X6 представляет S; X7 представляет А; X8 представляет V; X9 представляет F; X10 представляет Y; X11 представляет Y; Х12 представляет L; X13 представляет Y; X14 представляет Н; X15 представляет А. В еще одном дополнительном аспекте легкая цепь дополнительно содержит последовательности каркасной области вариабельной области легкой цепи, находящиеся между HVR, согласно формуле: (LC-FR1)-(HVR-L1)-(LC-FR2)-(HVR-L2)-(LC-FR3)-(HVR-L3)- (LC-FR4). В еще одном аспекте последовательности каркасной области получены из консенсусных последовательностей человеческой каркасной области. В дополнительном аспекте последовательности каркасной области представляют консенсусную каркасную область VL каппа I. В еще одном аспекте, по меньшей мере, одна из последовательностей каркасной области представляет следующее:
LC-FR1 представляет DIQMTQSPSSLSASVGDRVTITC (SEQ ID NO:29)
LC-FR2 представляет WYQQKPGKAPKLLIY (SEQ ID NO:30)
LC-FR3 представляет GVPSRFSGSGSGTDFTLTISSLQPEDFATYYC (SEQ ID NO:31)
LC-FR4 представляет FGQGTKVEIKR (SEQ ID NO:32).
В еще одном варианте осуществления обеспечивается выделенное анти-PD-L1-антитело или его антигенсвязывающий фрагмент, содержащее последовательность вариабельной области тяжелой цепи и легкой цепи, где:
(а) тяжелая цепь содержит HVR-H1, HVR-H2, и HVR-H3, где далее:
(i) последовательность HVR-H1 представляет GFTFSX1SWIH; (SEQ ID NO:33)
(ii) последовательность HVR-H2 представляет AWIX2PYGGSX3YYADSVKG (SEQ ID NO:34)
(iii) последовательность HVR-H3 представляет RHWPGGFDY и (SEQ ID NO:19)
(b) легкая цепь содержит HVR-L1, HVR-L2 и HVR-L3, где далее:
(i) последовательность HVR-L1 представляет RASQX4X5X6TX7X8A (SEQ ID NO:35)
(ii) последовательность HVR-L2 представляет SASX9LX10S; и (SEQ ID NO:36)
(iii) последовательность HVR-L3 представляет QQX11X12X13X14PX15T; (SEQ ID NO:37)
где далее: X1 представляет D или G, Х2 представляет S или L; X3 представляет Т или S; Х4 представляет D или V; Х5 представляет V или I; Х6 представляет S или N; X7 представляет А или F; X8 представляет V или L; X9 представляет F или Т; X10 представляет Y или А; X11 представляет Y, G, F или S; X12 представляет L, Y, F или W; X13 представляет Y, N, A, T, G, F или I; X14 представляет Н, V, Р, Т или I; X15 представляет A, W, R, P или Т.
В конкретном аспекте X1 представляет D; Х2 представляет S, и Х3 представляет T. В еще одном аспекте Х4 и представляет D; Х5 представляет V; X6 представляет S; X7 представляет А; X8 представляет V; X9 представляет F; X10 представляет Y; X11 представляет Y; Х12 представляет L; X13 представляет Y; X14 представляет Н; X15 представляет А. В еще одном аспекте X1 представляет D; Х2 представляет S; и X3 представляет Т; Х4 представляет D; Х5 представляет V; X6 представляет S; X7 представляет А; X8 представляет V; X9 представляет F; X10 представляет Y; X11 представляет Y; Х12 представляет L; X13 представляет Y; X14 представляет Н, и X15 представляет А.
В еще одном дополнительном аспекте вариабельная область тяжелой цепи содержит одну или более последовательностей каркасной области, находящихся между HVR, согласно формуле: (HC-FR1)-(HVR-H1)-(HC-FR2)-(HVR-H2)-(HC-FR3)-(HVR-H3)-(HC-FR4), и вариабельные области легкой цепи содержат одну или более последовательностей каркасной области, находящихся между HVR, согласно формуле: (LC-FR1)-(HVR-L1)-(LC- FR2)-(HVR-L2)-(LC-FR3)-(HVR-L3)-(LC-FR4). В еще одном аспекте последовательности каркасной области получены из консенсусных последовательностей человеческой каркасной области. В еще одном дополнительном аспекте последовательности каркасной области тяжелой цепи получены из последовательности подгруппы I, II, III по Kabat. В еще одном дополнительном аспекте последовательность каркасной области тяжелой цепи представляет консенсусную каркасную область VH подгруппы III. В еще одном дополнительном аспекте одна или более последовательностей каркасной области тяжелой цепи представляют следующее:
HC-FR1 EVQLVESGGGLVQPGGSLRLSCAAS (SEQ ID NO:25)
HC-FR2 WVRQAPGKGLEWV (SEQ ID NO:26)
HC-FR3 RFTISADTSKNTAYLQMNSLRAEDTAVYYCAR (SEQ ID NO:27)
HC-FR4 WGQGTLVTVSA (SEQ ID NO:28).
В еще одном дополнительном аспекте последовательности каркасной области легкой цепи получены из последовательности подгруппы I, II, III или IV каппа по Kabat. В еще одном дополнительном аспекте последовательности каркасной области легкой цепи представляют консенсусную каркасную область VL каппа подгруппы I. В еще одном аспекте одна или более последовательностей каркасной области легкой цепи представляют следующее:
LC-FR1 DIQMTQSPSSLSASVGDRVTITC (SEQ ID NO:29)
LC-FR2 WYQQKPGKAPKLLIY (SEQ ID NO:30)
LC-FR3 GVPSRFSGSGSGTDFTLTISSLQPEDFATYYC (SEQ ID NO:31)
LC-FR4 FGQGTKVEIKR (SEQ ID NO:32).
В еще одном дополнительном конкретном аспекте антитело дополнительно содержит человеческую или мышиную константную область. В еще одном дополнительном аспекте человеческая константная область выбрана из группы, состоящей из IgG1, IgG2, IgG2, IgG3, IgG4. В еще одном дополнительном конкретном аспекте человеческая константная область представляет IgG1. В еще одном дополнительном аспекте мышиная константная область выбрана из группы, состоящей из IgG1, IgG2a, IgG2b, IgG3. В еще одном дополнительном аспекте мышиная константная область представляет IgG2a. В еще одном дополнительном конкретном аспекте антитело обладает пониженной или минимальной эффекторной функцией. В еще одном дополнительном конкретном аспекте минимальная эффекторная функция является результатом «мутации со снижением эффекторной функции Fc-области» или агликозилирования. В еще одном дополнительном варианте осуществления мутация со снижением эффекторной функции Fc-области представляет N297A или замену D265A/N297A в константной области.
В еще одном варианте осуществления обеспечивается анти-PD-L1-антитело, содержащее последовательность вариабельной области тяжелой цепи и легкой цепи, где:
(а) тяжелая цепь дополнительно содержит последовательность HVR-H1, HVR-H2 и HVR-H3, обладающую, по меньшей мере, 85% идентичностью последовательности с GFTFSDSWIH (SEQ ID NO:17), AWISPYGGSTYYADSVKG (SEQ ID NO:18) и RHWPGGFDY (SEQ ID NO:19) соответственно, или
(b) легкая цепь дополнительно содержит последовательность HVR-L1, HVR-L2 и HVR-L3, обладающую, по меньшей мере, 85% идентичностью последовательности с RASQDVSTAVA (SEQ ID NO:20), SASFLYS (SEQ ID NO:21) и QQYLYHPAT (SEQ ID NO:22) соответственно,
(c) в конкретном аспекте идентичность последовательности составляет 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%. В еще одном аспекте вариабельная область тяжелой цепи содержит одну или более последовательностей каркасной области, расположенных между HVR, согласно формуле: (HC-FR1)-(HVR-H1)-(HC-FR2)-(HVR-H2)-(HC-FR3)-(HVR-H3)-(HC-FR4), и вариабельные области легкой цепи содержат одну или более последовательностей каркасной области, расположенных между HVR, согласно формуле: (LC-FR1)-(HVR-L1)-(LC-FR2)-(HVR-L2)-(LC-FR3)-(HVR-L3)-(LC-FR4). В еще одном аспекте последовательности каркасной области получены из консенсусных последовательностей человеческой каркасной области. В еще одном дополнительном аспекте последовательности каркасной области тяжелой цепи получены из последовательности подгруппы I, II или III по Kabat. В еще одном дополнительном аспекте последовательность каркасной обалсти тяжелой цепи представляет консенсусную каркасную область VH подгруппы III. В еще одном аспекте одна или более последовательностей каркасной области тяжелой цепи представляют следующее:
HC-FR1 EVQLVESGGGLVQPGGSLRLSCAAS (SEQ ID NO:25)
HC-FR2 WVRQAPGKGLEWV (SEQ ID NO:26)
HC-FR3 RFTISADTSKNTAYLQMNSLRAEDTAVYYCAR (SEQ ID NO:27)
HC-FR4 WGQGTLVTVSA (SEQ ID NO:28).
В еще одном дополнительном аспекте последовательности каркасной области легкой цепи получены из последовательности подгруппы I, II, III или IV каппа по Kabat. В еще одном дополнительном аспекте последовательности каркасной области легкой цепи представляют консенсусную каркасную область VL каппа подгруппы I. В еще одном аспекте одна или более последовательностей каркасной области легкой цепи представляют следующее:
LC-FR1 DIQMTQSPSSLSASVGDRVTITC (SEQ ID NO:29)
LC-FR2 WYQQKPGKAPKLLIY (SEQ ID NO:30)
LC-FR3 GVPSRFSGSGSGTDFTLTISSLQPEDFATYYC (SEQ ID NO:31)
LC-FR4 FGQGTKVEIKR (SEQ ID NO:32).
В еще одном дополнительном конкретном аспекте антитело дополнительно содержит человеческую или мышиную константную область. В еще одном дополнительном аспекте человеческая константная область выбрана из группы, состоящей из IgG1, IgG2, IgG2, IgG3, IgG4. В еще одном дополнительном конкретном аспекте человеческая константная область представляет IgG1. В еще одном аспекте мышиная константная область выбрана из группы, состоящей из IgG1, IgG2a, IgG2b, IgG3. В еще одном дополнительном аспекте мышиная константная область представляет IgG2a. В еще одном дополнительном конкретном аспекте антитело обладает пониженной или минимальной эффекторной функцией. В еще одном дополнительном конкретном аспекте минимальная эффекторная функция является результатом «мутации со снижением эффекторной функции Fc-области» или агликозилирования. В еще одном дополнительном варианте осуществления мутация со снижением эффекторной функции Fc-области представляет N297A или замену D265A/N297A в константной области.
В еще одном варианте осуществления обеспечивается анти-PD-L1-антитело, содержащее последовательность вариабельной области тяжелой цепь и легкой цепи, где:
(а) последовательность тяжелой цепи обладает, по меньшей мере, 85% идентичностью последовательности с последовательностью тяжелой цепи:
EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSA (SEQ ID NO:23), EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSSASTK (SEQ ID NO:40) или EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS (SEQ ID NO:41), или
(b) последовательность легкой цепи обладает, по меньшей мере, 85% идентичностью последовательности с последовательностью легкой цепи:
DIQMTQSPSSLSASVGDRVTITCRASQDVSTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYLYHPATFGQGTKVEIKR (SEQ ID NO:24).
В конкретном аспекте идентичность последовательностей составляет 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%. В еще одном аспекте вариабельная область тяжелой цепи содержит одну или более каркасных последовательностей, расположенных между HVR, согласно формуле: (HC-FR1)-(HVR-H1)-(HC-FR2)-(HVR-H2)-(HC-FR3)-(HVR-H3)-(HC-FR4), и вариабельные области легкой цепи содержат одну или более последовательностей каркасной области, расположенных между HVR, согласно формуле: (LC-FR1)-(HVR-L1)-(LC-FR2)-(HVR-L2)-(LC-FR3)-(HVR-L3)-(LC-FR4). В еще одном аспекте последовательности каркасной области получены из консенсусных последовательностей человеческой каркасной области. В дополнительном аспекте последовательности каркасной области тяжелой цепи получены из последовательности подгруппы I, II или III по Kabat. В еще одном дополнительном аспекте последовательность каркасной области тяжелой цепи представляет консенсусную каркасную область VH подгруппы III. В еще одном дополнительном аспекте одна или более последовательностей каркасной области тяжелой цепи представляют следующее:
HC-FR1 EVQLVESGGGLVQPGGSLRLSCAAS (SEQ ID NO:25)
HC-FR2 WVRQAPGKGLEWV (SEQ ID NO:26)
HC-FR3 RFTISADTSKNTAYLQMNSLRAEDTAVYYCAR (SEQ ID NO:27)
HC-FR4 WGQGTLVTVSA (SEQ ID NO:28).
В еще одном дополнительном аспекте последовательности каркасной области легкой цепи получены из последовательности подгруппы I, II, III или IV каппа по Kabat. В еще одном дополнительном аспекте последовательности каркасной области легкой цепи представляют консенсусную каркасную область VL каппа подгруппы I. В еще одном дополнительном аспекте одна или более последовательностей каркасной области легкой цепи представляют следующее:
LC-FR1 DIQMTQSPSSLSASVGDRVTITC (SEQ ID NO:29)
LC-FR2 WYQQKPGKAPKLLIY (SEQ ID NO:30)
LC-FR3 GVPSRFSGSGSGTDFTLTISSLQPEDFATYYC (SEQ ID NO:31)
LC-FR4 FGQGTKVEIKR (SEQ ID NO:32).
В еще одном дополнительном конкретном аспекте антитело дополнительно содержит человеческую или мышиную константную область. В еще одном дополнительном аспекте человеческая константная область выбрана из группы, состоящей из IgG1, IgG2, IgG2, IgG3, IgG4. В еще одном дополнительном конкретном аспекте человеческая константная область представляет IgG1. В еще одном аспекте мышиная константная область выбрана из группы, состоящей из IgG1, IgG2a, IgG2b, IgG3. В еще одном дополнительном аспекте мышиная константная область представляет IgG2a. В еще одном дополнительном конкретном аспекте антитело обладает пониженной или минимальной эффекторной функцией. В еще одном дополнительном конкретном аспекте минимальная эффекторная функция является результатом продукции в прокариотических клетках. В еще одном дополнительном конкретном аспекте минимальная эффекторная функция является результатом «мутации со снижением эффекторной функции Fc-области» или агликозилирования. В еще одном дополнительном варианте осуществления мутация со снижением эффекторной функции Fc-области представляет N297A или замену D265A/N297A в константной области.
В еще одном дополнительном варианте осуществления настоящее изобретение относится к композициям, содержащим любое из описанных выше анти-PD-L1-антител, в комбинации, по меньшей мере, с одним фармацевтически приемлемым носителем.
В еще одном дополнительном варианте осуществления обеспечивается выделенная нуклеиновая кислота, кодирующая вариабельную область легкой цепи или тяжелой цепи анти-PD-L1-антитела, где:
(а) тяжелая цепь дополнительно содержит последовательность HVR-H1, HVR-H2 и HVR-H3, обладающую, по меньшей мере, 85% идентичностью последовательности с GFTFSDSWIH (SEQ ID NO:17), AWISPYGGSTYYADSVKG (SEQ ID NO:18) и RHWPGGFDY (SEQ ID NO:19) соответственно, и
(b) легкая цепь дополнительно содержит последовательность HVR-L1, HVR-L2 и HVR-L3, обладающую, по меньшей мере, 85% идентичностью последовательности с RASQDVSTAVA (SEQ ID NO:20), SASFLYS (SEQ ID NO:21) и QQYLYHPAT (SEQ ID NO:22) соответственно.
В конкретном варианте осуществления идентичность последовательностей составляет 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%. В еще одном аспекте вариабельная область тяжелой цепи содержит одну или более последовательностей каркасной области, расположенных между HVR, согласно формуле: (HC-FR1)-(HVR-H1)-(HC-FR2)-(HVR-H2)-(HC-FR3)-(HVR-H3)-(HC-FR4), и вариабельные области легкой цепи содержат одну или более последовательностей каркасной области, расположенных между HVR, согласно формуле: (LC-FR1)-(HVR-L1)-(LC-FR2)-(HVR-L2)-(LC-FR3)-(HVR-L3)-(LC-FR4). В еще одном аспекте последовательности каркасной области получены из консенсусных последовательностей человеческой каркасной области. В еще одном дополнительном аспекте последовательности каркасной области тяжелой цепи получены из последовательности подгруппы I, II или III по Kabat. В еще одном дополнительном аспекте последовательность каркасной области тяжелой цепи представляет консенсусную каркасную область VH подгруппы III. В еще одном дополнительном аспекте одна или более последовательностей каркасной области тяжелой цепи представляют следующее:
HC-FR1 EVQLVESGGGLVQPGGSLRLSCAAS (SEQ ID NO:25)
HC-FR2 WVRQAPGKGLEWV (SEQ ID NO:26)
HC-FR3 RFTISADTSKNTAYLQMNSLRAEDTAVYYCAR (SEQ ID NO:27)
HC-FR4 WGQGTLVTVSA (SEQ ID NO:28).
В еще одном дополнительном аспекте последовательности каркасной области легкой цепи получены из последовательности подгруппы I, II, III или IV каппа по Kabat. В еще одном дополнительном аспекте последовательности каркасной области легкой цепи представляют консенсусную каркасную область VL каппа подгруппы I. В еще одном аспекте одна или более последовательностей каркасной области легкой цепи представляют следующее:
LC-FR1 DIQMTQSPSSLSASVGDRVTITC (SEQ ID NO:29)
LC-FR2 WYQQKPGKAPKLLIY (SEQ ID NO:30)
LC-FR3 GVPSRFSGSGSGTDFTLTISSLQPEDFATYYC (SEQ ID NO:31)
LC-FR4 FGQGTKVEIKR (SEQ ID NO:32).
В еще одном дополнительном конкретном аспекте антитело дополнительно содержит человеческую или мышиную константную область. В еще одном дополнительном аспекте человеческая константная область выбрана из группы, состоящей из IgG1, IgG2, IgG2, IgG3, IgG4. В еще одном дополнительном конкретном аспекте человеческая константная область представляет IgG1. В еще одном аспекте мышиная константная область выбрана из группы, состоящей из IgG1, IgG2a, IgG2b, IgG3. В еще одном дополнительном аспекте мышиная константная область представляет IgG2a. В еще одном дополнительном конкретном аспекте антитело обладает пониженной или минимальной эффекторной функцией. В еще одном дополнительном конкретном аспекте минимальная эффекторная функция является результатом продукции в прокариотических клетках. В еще одном дополнительном конкретном аспекте минимальная эффекторная функция является результатом «мутации со снижением эффекторной функции Fc-области» или агликозилирования. В еще одном дополнительном варианте осуществления мутация со снижением эффекторной функции Fc-области представляет N297A или замену D265A/N297A в константной области.
В еще одном дополнительном варианте осуществления обеспечивается анти-PD-L1-антитело, содержащее последовательность вариабельной области тяжелой цепи и легкой цепи, где:
(а) последовательность тяжелой цепи обладает, по меньшей мере, 85% идентичностью последовательности с последовательностью тяжелой цепи:
EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS (SEQ ID NO:41), или
(b) последовательность легкой цепи обладает, по меньшей мере, 85% идентичностью последовательности с последовательностью легкой цепи:
DIQMTQSPSSLSASVGDRVTITCRASQDVSTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYLYHPATFGQGTKVEIKR (SEQ ID NO:24).
В конкретном аспекте идентичность последовательностей со-ставляет 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%. В еще одном аспекте вариабельная область тяжелой цепи содержит одну или более последовательностей каркасной области, расположенных между HVR, согласно формуле: (HC-FR1)-(HVR-H1)-(HC-FR2)-(HVR-H2)-(HC-FR3)-(HVR-H3)-(HC-FR4), и вариабельные области легкой цепи содержат одну или более последовательностей каркасной области, расположенных между HVR, согласно формуле: (LC-FR1)-(HVR-L1)-(LC-FR2)-(HVR-L2)-(LC-FR3)-(HVR-L3)-(LC-FR4). В еще одном аспекте последовательности каркасной области получены из консенсусных последовательностей человеческой каркасной области. В дополнительном аспекте последовательности каркасной области тяжелой цепи получены из последовательности подгруппы I, II или III по Kabat. В еще одном дополнительном аспекте последовательность каркасной области тяжелой цепи представляет консенсусную каркасную область VH подгруппы III. В еще одном дополнительном аспекте одна или более последовательностей каркасной области тяжелой цепи представляют следующее:
HC-FR1 EVQLVESGGGLVQPGGSLRLSCAAS (SEQ ID NO:25)
HC-FR2 WVRQAPGKGLEWV (SEQ ID NO:26)
HC-FR3 RFTISADTSKNTAYLQMNSLRAEDTAVYYCAR (SEQ ID NO:27)
HC-FR4 WGQGTLVTVSS (SEQ ID NO:42).
В еще одном дополнительном аспекте последовательности каркасной области легкой цепи получены из последовательности подгруппы I, II, III или IV каппа по Kabat. В еще одном дополнительном аспекте последовательности каркасной области легкой цепи представляют консенсусную каркасную область VL каппа подгруппы I. В еще одном дополнительном аспекте одна или более последовательностей каркасной области легкой цепи представляют следующее:
LC-FR1 DIQMTQSPSSLSASVGDRVTITC (SEQ ID NO:29)
LC-FR2 WYQQKPGKAPKLLIY (SEQ ID NO:30)
LC-FR3 GVPSRFSGSGSGTDFTLTISSLQPEDFATYYC (SEQ ID NO:31)
LC-FR4 FGQGTKVEIKR (SEQ ID NO:32).
В еще одном дополнительном конкретном аспекте антитело дополнительно содержит человеческую или мышиную константную область. В еще одном дополнительном аспекте человеческая константная область выбрана из группы, состоящей из IgG1, IgG2, IgG2, IgG3, IgG4. В еще одном дополнительном конкретном аспекте человеческая константная область представляет IgG1. В еще одном аспекте мышиная константная область выбрана из группы, состоящей из IgG1, IgG2a, IgG2b, IgG3. В еще одном дополнительном аспекте мышиная константная область представляет IgG2a. В еще одном дополнительном конкретном аспекте антитело обладает пониженной или минимальной эффекторной функцией. В еще одном дополнительном конкретном аспекте минимальная эффекторная функция является результатом продукции в прокариотических клетках. В еще одном дополнительном конкретном аспекте минимальная эффекторная функция является результатом «мутации со снижением эффекторной функции Fc-области» или агликозилирования. В еще одном дополнительном варианте осуществления мутация со снижением эффекторной функции Fc-области представляет N297A или замену D265A/N297A в константной области.
В дополнительном аспекте вариабельная область тяжелой цепи содержит одну или более последовательностей каркасной области, расположенных между HVR, согласно формуле: (HC-FR1)-(HVR-H1)-(HC-FR2)-(HVR-H2)-(HC-FR3)-(HVR-H3)-(HC-FR4), и вариабельные области легкой цепи содержат одну или более последовательностей каркасной области, расположенных между HVR, согласно формуле: (LC-FR1)-(HVR-L1)-(LC-FR2)-(HVR-L2)-(LC-FR3)-(HVR-L3)-(LC-FR4). В еще одном дополнительном аспекте последовательности каркасной области получены из консенсусных последовательностей человеческой каркасной области. В дополнительном аспекте последовательности каркасной области тяжелой цепи получены из последовательности подгруппы I, II или III по Kabat. В еще одном дополнительном аспекте последовательность каркасной области тяжелой цепи представляет консенсусную каркасную область VH подгруппы III. В еще одном дополнительном аспекте одна или более последовательностей каркасной области тяжелой цепи представляют следующее:
HC-FR1 EVQLVESGGGLVQPGGSLRLSCAASGFTFS (SEQ ID NO:43)
HC-FR2 WVRQAPGKGLEWVA (SEQ ID NO:44)
HC-FR3 RFTISADTSKNTAYLQMNSLRAEDTAVYYCAR (SEQ ID NO:27)
HC-FR4 WGQGTLVTVSS (SEQ ID NO:45).
В еще одном дополнительном аспекте последовательности каркасной области легкой цепи получены из последовательности подгруппы I, II, III или IV каппа по Kabat. В еще одном дополнительном аспекте последовательности каркасной области легкой цепи представляют консенсусную каркасную область VL каппа подгруппы I. В еще одном дополнительном аспекте одна или более последовательностей каркасной области легкой цепи представляют следующее:
LC-FR1 DIQMTQSPSSLSASVGDRVTITC (SEQ ID NO:29)
LC-FR2 WYQQKPGKAPKLLIY (SEQ ID NO:30)
LC-FR3 GVPSRFSGSGSGTDFTLTISSLQPEDFATYYC (SEQ ID NO:31)
LC-FR4 FGQGTKVEIK (SEQ ID NO: 46).
В еще одном дополнительном конкретном аспекте антитело дополнительно содержит человеческую или мышиную константную область. В еще одном дополнительном аспекте человеческая константная область выбрана из группы, состоящей из IgG1, IgG2, IgG2, IgG3, IgG4. В еще одном дополнительном конкретном аспекте человеческая константная область представляет IgG1. В еще одном аспекте мышиная константная область выбрана из группы, состоящей из IgG1, IgG2a, IgG2b, IgG3. В еще одном дополнительном аспекте мышиная константная область представляет IgG2a. В еще одном дополнительном конкретном аспекте антитело обладает пониженной или минимальной эффекторной функцией. В еще одном дополнительном конкретном аспекте минимальная эффекторная функция является результатом «мутации со снижением эффекторной функции Fc-области» или агликозилирования. В еще одном дополнительном варианте осуществления мутация со снижением эффекторной функции Fc-области представляет N297A или замену D265A/N297A в константной области.
В еще одном варианте осуществления обеспечивается анти-PD-L1-антитело, содержащее последовательность вариабельной области тяжелой цепи и легкой цепи, где:
(d) тяжелая цепь дополнительно содержит последовательность HVR-H1, HVR-H2 и HVR-H3, обладающую, по меньшей мере, 85% идентичностью последовательности с GFTFSDSWIH (SEQ ID NO:17), AWISPYGGSTYYADSVKG (SEQ ID NO:18) и RHWPGGFDY (SEQ ID NO:19) соответственно, и
(e) легкая цепь дополнительно содержит последовательность HVR-L1, HVR-L2 и HVR-L3, обладающую, по меньшей мере, 85% идентичностью последовательности с RASQDVSTAVA (SEQ ID NO:20), SASFLYS (SEQ ID NO:21) и QQYLYHPAT (SEQ ID NO:22) соответственно.
В конкретном аспекте идентичность последовательностей со-ставляет 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%. В еще одном аспекте вариабельная область тяжелой цепи содержит одну или более последовательностей каркасной области, расположенных между HVR, согласно формуле: (HC-FR1)-(HVR-H1)-(HC-FR2)-(HVR-H2)-(HC-FR3)-(HVR-H3)-(HC-FR4), и вариабельные области легкой цепи содержат одну или более последовательностей каркасной области, расположенных между HVR, согласно формуле: (LC-FR1)-(HVR-L1)-(LC-FR2)-(HVR-L2)-(LC-FR3)-(HVR-L3)-(LC-FR4). В еще одном аспекте последовательности каркасной области получены из консенсусных последовательностей человеческой каркасной области. В еще одном дополнительном аспекте последовательности каркасной области тяжелой цепи получены из последовательности подгруппы I, II или III по Kabat. В еще одном дополнительном аспекте последовательность каркасной области тяжелой цепи представляет консенсусную каркасную область VH подгруппы III. В еще одном дополнительном аспекте одна или более последовательностей каркасной области тяжелой цепи представляют следующее:
HC-FR1 EVQLVESGGGLVQPGGSLRLSCAAS (SEQ ID NO:25)
HC-FR2 WVRQAPGKGLEWV (SEQ ID NO:26)
HC-FR3 RFTISADTSKNTAYLQMNSLRAEDTAVYYCAR (SEQ ID NO:27)
HC-FR4 WGQGTLVTVSSASTK (SEQ ID NO: 47).
В еще одном дополнительном аспекте последовательности каркасной области легкой цепи получены из последовательности подгруппы I, II, III или IV каппа по Kabat. В еще одном дополнительном аспекте последовательности каркасной области легкой цепи представляют консенсусную каркасную область VL каппа подгруппы I. В еще одном дополнительном аспекте одна или более последовательностей каркасной области легкой цепи представляют следующее:
LC-FR1 DIQMTQSPSSLSASVGDRVTITC (SEQ ID NO:29)
LC-FR2 WYQQKPGKAPKLLIY (SEQ ID NO:30)
LC-FR3 GVPSRFSGSGSGTDFTLTISSLQPEDFATYYC (SEQ ID NO:31)
LC-FR4 FGQGTKVEIKR (SEQ ID NO:32).
В еще одном дополнительном конкретном аспекте антитело дополнительно содержит человеческую или мышиную константную область. В еще одном дополнительном аспекте человеческая константная область выбрана из группы, состоящей из IgG1, IgG2, IgG2, IgG3, IgG4. В еще одном дополнительном конкретном аспекте человеческая константная область представляет IgG1. В еще одном аспекте мышиная константная область выбрана из группы, состоящей из IgG1, IgG2a, IgG2b, IgG3. В еще одном дополнительном аспекте мышиная константная область представляет IgG2a. В еще одном дополнительном конкретном аспекте антитело обладает пониженной или минимальной эффекторной функцией. В еще одном дополнительном конкретном аспекте минимальная эффекторная функция является результатом «мутации со снижением эффекторной функции Fc-области» или агликозилирования. В еще одном дополнительном варианте осуществления мутация со снижением эффекторной функции Fc-области представляет N297A или замену D265A/N297A в константной области.
В еще одном дополнительном варианте осуществления обеспечивается анти-PD-L1-антитело, содержащее последовательность вариабельной области тяжелой цепь и легкой цепи, где:
(а) последовательность тяжелой цепи обладает, по меньшей мере, 85% идентичностью последовательности с последовательностью тяжелой цепи:
EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSSASTK (SEQ ID NO:40), или
(b) последовательность легкой цепи обладает, по меньшей мере, 85% идентичностью последовательности с последовательностью легкой цепи:
DIQMTQSPSSLSASVGDRVTITCRASQDVSTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYLYHPATFGQGTKVEIKR (SEQ ID NO:24).
В некоторых вариантах осуществления обеспечивается выделенное анти-PDL1-антитело, содержащее последовательность вариабельной области тяжелой цепи и легкой цепи, где последовательность вариабельной области легкой цепи обладает, по меньшей мере, 85%, по меньшей мере, 86%, по меньшей мере, 87%, по меньшей мере, 88%, по меньшей мере, 89%, по меньшей мере, 90%, по меньшей мере, 91%, по меньшей мере, 92%, по меньшей мере, 93%, по меньшей мере, 94%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98%, или, по меньшей мере, 99% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:24. В некоторых вариантах осуществления обеспечивается выделенное анти-PDL1-антитело, содержащее последовательность вариабельной области тяжелой цепи и легкой цепи, где последовательность вариабельной области тяжелой цепи обладает, по меньшей мере, 85%, по меньшей мере, 86%, по меньшей мере, 87%, по меньшей мере, 88%, по меньшей мере, 89%, по меньшей мере, 90%, по меньшей мере, 91%, по меньшей мере, 92%, по меньшей мере, 93%, по меньшей мере, 94%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:40. В некоторых вариантах осуществления обеспечивается выделенное анти-PDL1-антитело, содержащее последовательность вариабельной области тяжелой цепи и легкой цепи, где последовательность вариабельной области легкой цепи обладает, по меньшей мере, 85%, по меньшей мере, 86%, по меньшей мере, 87%, по меньшей мере, 88%, по меньшей мере, 89%, по меньшей мере, 90%, по меньшей мере, 91%, по меньшей мере, 92%, по меньшей мере, 93%, по меньшей мере, 94%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:24, и последовательность вариабельной области тяжелой цепи обладает, по меньшей мере, 85%, по меньшей мере, 86%, по меньшей мере, 87%, по меньшей мере, 88%, по меньшей мере, 89%, по меньшей мере, 90%, по меньшей мере, 91%, по меньшей мере, 92%, по меньшей мере, 93%, по меньшей мере, 94%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:40.
В еще одном дополнительном варианте осуществления обеспечивается анти-PDL1-антитело, содержащее последовательность тяжелой цепь и легкой цепи, где:
(а) последовательность тяжелой цепи обладает, по меньшей мере, 85% идентичностью последовательности с последовательностью тяжелой цепи:
EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYASTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG (SEQ ID NO:48), или
(b) последовательность легкой цепи обладает, по меньшей мере, 85% идентичностью последовательности с последовательностью легкой цепи:
DIQMTQSPSSLSASVGDRVTITCRASQDVSTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYLYHPATFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO:49).
В некоторых вариантах осуществления обеспечивается выделенное анти-PDL1-антитело, содержащее последовательность тяжелой цепи и легкой цепи, где последовательность легкой цепи обладает, по меньшей мере, 85%, по меньшей мере, 86%, по меньшей мере, 87%, по меньшей мере, 88%, по меньшей мере, 89%, по меньшей мере, 90%, по меньшей мере, 91%, по меньшей мере, 92%, по меньшей мере, 93%, по меньшей мере, 94%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98%, или, по меньшей мере, 99% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:49. В некоторых вариантах осуществления обеспечивается выделенное анти-PDL1-антитело, содержащее последовательность тяжелой цепи и легкой цепи, где последовательность тяжелой цепи обладает, по меньшей мере, 85%, по меньшей мере, 86%, по меньшей мере, 87%, по меньшей мере, 88%, по меньшей мере, 89%, по меньшей мере, 90%, по меньшей мере, 91%, по меньшей мере, 92%, по меньшей мере, 93%, по меньшей мере, 94%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:48. В некоторых вариантах осуществления обеспечивается выделенное анти-PDL1-антитело, содержащее последовательность тяжелой цепи и легкой цепи, где последовательность легкой цепи обладает, по меньшей мере, 85%, по меньшей мере, 86%, по меньшей мере, 87%, по меньшей мере, 88%, по меньшей мере, 89%, по меньшей мере, 90%, по меньшей мере, 91%, по меньшей мере, 92%, по меньшей мере, 93%, по меньшей мере, 94%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:49, и последовательность тяжелой цепи обладает, по меньшей мере, 85%, по меньшей мере, 86%, по меньшей мере, 87%, по меньшей мере, 88%, по меньшей мере, 89%, по меньшей мере, 90%, по меньшей мере, 91%, по меньшей мере, 92%, по меньшей мере, 93%, по меньшей мере, 94%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:48.
В еще одном дополнительном аспекте нуклеиновая кислота дополнительно включает вектор, подходящий для экспрессии нуклеиновой кислоты, кодирующей любое из вышеописанных анти-PD-L1-антител. В еще одном дополнительном конкретном аспекте вектор дополнительно включает клетку-хозяин, подходящую для экспрессии нуклеиновой кислоты. В еще одном дополнительном конкретном аспекте клетка-хозяин представляет эукариотическую клетку или прокариотическую клетку. В еще одном дополнительном конкретном аспекте эукариотическая клетка представляет собой клетку млекопитающего, такую как клетка яичника китайского хомячка (СНО).
Анти-PD-L1-антитело или его антигенсвязывающий фрагмент можно получить с использованием способов, известных в данной области, например, способом, включающим культивирование клетки-хозяина, содержащей нуклеиновую кислоту, кодирующую любое из вышеописанных анти-PD-L1-антител или их антигенсвязывающий фрагмент, в форме, подходящей для экспрессии, в условиях, подходящих для получения такого антитела или фрагмента, и выделение антитела или фрагмента.
В еще одном дополнительном варианте осуществления настоящее изобретение относится к композиции, содержащей анти-PD-L1-антитело или его антигенсвязывающий фрагмент, описанные здесь, и, по меньшей мере, один фармацевтически приемлемый носитель.
Агенты, которые снижают или ингибируют экспрессию и/или активность TIGIT
Обеспечивается способ лечения или замедления прогрессирования рака у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. Также обеспечивается способ снижения или ингибирования рецидива или прогрессирования рака у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. Также обеспечивается способ лечения или замедления прогрессирования связанного с иммунитетом заболевания у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. Также обеспечивается способ снижения или ингибирования прогрессирования связанного с иммунитетом заболевания у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. Также обеспечивается способ повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, включающий введением индивидууму эффективного количества антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT. Также обеспечивается способ повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, включающий введение индивидууму эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов. Также обеспечивается способ повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума, включающий введение индивидууму эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов. Например, агент, который снижает или ингибирует экспрессию и/или активность TIGIT включает антагонист экспрессии и/или активности TIGIT, антагонист экспрессии и/или активности PVR, агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVR, агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL2, агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL3, агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR, агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL2, агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL3, и их комбинации.
В некоторых вариантах осуществления антагонист экспрессии и/или активности TIGIT включает низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид.
В некоторых вариантах осуществления антагонист экспрессии и/или активности PVR включает низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид.
В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVR, включает низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид.
В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL2, включает низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид.
В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL3, включает низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид.
В некоторых вариантах осуществления агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR, включает низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид.
В некоторых вариантах осуществления агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL2, включает низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид.
В некоторых вариантах осуществления агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL3, включает низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид.
В некоторых вариантах осуществления антагонист экспрессии и/или активности TIGIT представляет ингибирующую нуклеиновую кислоту, выбранную из группы, состоящей из антисмыслового полинуклеотида, интерферирующей РНК, каталитической РНК и химеры РНК-ДНК.
В некоторых вариантах осуществления антагонист экспрессии и/или активности TIGIT представляет анти-TIGIT-антитело или его антигенсвязывающий фрагмент.
Анти-TIGIT-антитела, пригодные в данном изобретении, включая композиции, содержащие такие антитела, например, описанные в WO 2009/126688, могут быть использованы в комбинации с антагонистами, связывающимися с осью PD-1.
Анти-TIGIT-антитела
Настоящее изобретение относится к анти-TIGIT-антителам. Примерные антитела включают поликлональные, моноклональные, гуманизированные, биспецифические и гетероконъюгатные антитела. Специалисту обычной квалификации в данной области должно быть понятно, что изобретение также относится к антителам к другим полипептидам (т.е. анти-PVR-антителам), и что любое из приведенного здесь описания относится к способу создания, получения, вариететам, применению или другим аспектам анти-TIGIT-антител, будет применимо к антителам, специфичным к другим полипептидам, не относящимся к TIGIT.
Поликлональные антитела
Анти-TIGIT-антитела могут включать поликлональные антитела. Способы получения поликлональных антител известны специалистам в данной области. Выработку поликлональных антител можно индуцировать у млекопитающего, например, одной или несколькими инъекциями иммунизирующего агента и, если желательно, адъюванта. Как правило, иммунизирующий агент и/или адъювант вводят млекопитающему несколькими подкожными или внутрибрюшинными инъекциями. Иммунизирующий агент может включать полипептид TIGIT или его гибридный белок. Может быть полезным конъюгировать иммунизирующий агент с белком, о котором известно, что он является иммуногенным у млекопитающего, которое иммунизируют. Примеры таких иммуногенных белков включают, не ограничиваясь этим, гемоцианин лимфы улитки, сывороточный альбумин, бычий тиреоглобулин и соевый ингибитор трипсина. Примеры адъювантов, которые могут быть использованы, включают полный адъювант Фрейнда и адъювант MPL-TDM (монофосфориллипид А, синтетический дикориномиколат трегалозы). Протокол иммунизации может выбрать специалист в данной области без излишнего экспериментирования.
Моноклональные антитела
Анти-TIGIT-антитела могут, альтернативно, представлять моноклональные антитела. Моноклональные антитела могут быть получены с использованием гибридомных методов, таких как описаны Kohler and Milstein, Nature, 256: 495 (1975). В гибридомном методе мышь, хомяка или другое подходящее животное-хозяин, как правило, иммунизируют иммунизирующим агентом для индукции выработки лимфоцитов, которые продуцируют или способны продуцировать антитела, которые будут специфически связываться с иммунизирующим агентом. Альтернативно лимфоциты могут быть иммунизированы in vitro.
Иммунизирующий агент, как правило, включает полипептид TIGIT или его гибридный белок. Как правило, используются лимфоциты периферической крови («PBL»), если желательны клетки человеческого происхождения, или используются клетки селезенки или клетки лимфатических узлов, если в качестве источника желательны млекопитающие, отличные от человека. Затем лимфоциты сливают с иммортализированной клеточной линии с использованием подходящего агента для слияния, такого как полиэтиленгликоль, с образованием гибридомной клетки [Goding, Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York, (1987) pp. 51-63]. Иммортализированные клеточные линии, как правило, представляют трансформированные клетки млекопитающих, в частности, миеломные клетки грызуна, быка и человека. Как правило, используются крысиные или мышиные миеломные клеточные линии. Клетки гибридомы можно культивировать в подходящей культуральной среде, которая предпочтительно содержит одно или более веществ, которые ингибируют рост или выживаемость неслитых, иммортализованных клеток. Например, если в родительских клетках отсутствует фермент гипоксантингуанинфосфорибозилтрансфераза (HGPRT или HPRT), то культуральная среда для гибридом обычно будет включать гипоксантин, аминоптерин и тимидин («среда НАТ»), где эти вещества предотвращают рост HGPRT-дефицитных клеток.
Предпочтительными иммортализированными клеточными линиями являются те, которые эффективно сливаются, поддерживают стабильный высокий уровень экспрессии антитела выбранными антителопродуцирующими клетками и чувствительны к среде, такой как среда НАТ. Более предпочтительными иммортализированными клеточными линиями являются мышиные миеломные линии, которые могут быть получены, например, из Центра клеточных культур в Сан Диего (Salk Institute Cell Distribution Center, San Diego, California) и Американской коллекции типовых культур, Manassas, Virginia. Также были описаны человеческие миеломные и мышиные-человеческие гетеромиеломные клеточные линии для получения моноклональных антител человека [Kozbor, J. Immunol, 133:. 3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York, (1987) pp. 51-63].
Затем культуральную среду, в которой культивируют гибридомные клетки, анализируют на присутствие моноклональных антител к полипептиду. Предпочтительно специфичность связывания моноклональных антител, продуцированных клетками гибридомы, определяют иммунопреципитацией или анализом на связывание in vitro, таким как радиоиммуноанализ (РИА) или твердофазный иммуноферментный анализ (ELISA). Такие методики и анализы известны в данной области. Аффинность связывания моноклонального антитела можно, например, определить анализом по Скэтчарду, описанным Munson and Pollard, Anal. Biochem., 107:220 (1980).
После идентификации желаемых гибридомных клеток клоны можно субклонировать процедурами ограничивающего разведения и культивировать стандартными методами [Goding, выше]. Подходящие культуральные среды для этой цели включают, например, среду Игла, модифицированную по способу Дульбекко, и среду RPMI-1640. Кроме того, гибридомные клетки можно культивировать in vivo в асцитной жидкости у млекопитающего.
Моноклональные антитела, секретируемые субклонами, могут быть выделены или очищены из культуральной среды или асцитной жидкости общепринятыми процедурами очистки иммуноглобулинов, такими как, например, хроматография на белок А-сефарозе, хроматография на гидроксилапатите, гель-электрофорез, диализ или аффинная хроматография.
Моноклональные антитела также можно получить методами рекомбинантной ДНК, такими как методы, описанные в патенте США № 4816567. ДНК, кодирующую моноклональные антитела по изобретению, можно легко выделить и секвенировать, используя обычные процедуры (например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелые и легкие цепи мышиных антител). Гибридомные клетки по изобретению служат в качестве предпочтительного источника такой ДНК. После выделения ДНК можно поместить в экспрессионные векторы, которые затем трансфектируют в клетки-хозяева, такие как клетки обезьяны COS, клетки яичника китайского хомячка (СНО) или миеломные клетки, которые иначе не продуцируют белок иммуноглобулина, с обеспечением синтеза моноклональных антител в рекомбинантных клетках-хозяевах. ДНК также можно модифицировать, например, заменой последовательности, кодирующей константные области тяжелой и легкой цепи, гомологичными мышиными последовательностями [патент США № 4816567; Morrison et al., выше] или ковалентным присоединением к кодирующей последовательности иммуноглобулина всей или части кодирующей последовательности для полипептида, отличного от иммуноглобулина. Такой полипептид, отличный от иммуноглобулина, можно заменить константной областью антитела по изобретению, или можно заменить вариабельной областью одного антигенсвязывающего сайта антитела по изобретению с получением химерного двухвалентного антитела.
Антитела могут представлять моновалентные антитела. Способы получения моновалентных антител хорошо известны в данной области техники. Например, один метод включает рекомбинантную экспрессию легкой цепи иммуноглобулина и модифицированной тяжелой цепи. Обычно тяжелая цепь усекается в любой точке Fc-области для предупреждения поперечного сшивания тяжелой цепи. Альтернативно соответствующие остатки цистеина замещают другим аминокислотным остатком или подвергаются делеции таким образом, чтобы предупредить поперечное сшивание.
Также пригодны способы получения моновалентных антител in vitro. Расщепление антител для получения их фрагментов, в частности, Fab-фрагментов, можно провести с использованием обычных методов, известных в данной области.
Человеческие и гуманизированные антитела
Анти-TIGIT-антитела по изобретению могут дополнительно включать гуманизированные антитела или человеческие антитела. Гуманизированные формы нечеловеческих (например, мышиных) антител представляют собой химерные иммуноглобулины, иммуноглобулиновые цепи или фрагменты (например, Fv, Fab, Fab', F(аb')2 или другие антигенсвязывающие субпоследовательности антител), которых содержат минимальную последовательность, полученную из иммуноглобулина, отличного от человеческого. Гуманизированные антитела включают человеческие иммуноглобулины (реципиентное антитело), в которых остатки из гипервариабельной области (CDR) реципиента замещены остатками из CDR из вида, отличного от человека (донорное антитело), такого как мышь, крыса или кролик, имеющих требуемую специфичность, аффинность и активность. В некоторых случаях остатки каркасной области Fv иммуноглобулина человека замещены соответствующими не принадлежащими человеку остатками. Гуманизированные антитела также могут содержать остатки, которые не обнаруживаются в реципиентном антителе или в импортируемых CDR или каркасных последовательностях. В общем, гуманизированное антитело будет по существу содержать все, по меньшей мере, одно, и обычно два вариабельных домена, в которых все или по существу все области CDR соответствуют таковым нечеловеческого иммуноглобулина, и все или по существу все области FR из консенсусной последовательности человеческого иммуноглобулина. Гуманизированное антитело оптимально также содержит, по меньшей мере, часть константной области иммуноглобулина (Fc), обычно человеческого иммуноглобулина [Jones et al., Nature, 321:522-525 (1986); Riechmann et al., Nature, 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol., 2:593-596 (1992)].
Способы гуманизации нечеловеческих антител хорошо известны в данной области техники. Как правило, гуманизированное антитело содержит один или более аминокислотных остатков, введенных в него из источника, который не является человеком. Такие «не-человеческие» аминокислотные остатки часто называют «импортными» остатками, которые обычно берут из «импортной» вариабельной области. Гуманизация в основном может быть выполнена, следуя способу Winter и соавторов [Jones et al., Nature, 321:522-525 (1986); Riechmann et al., Nature, 332:323-327 (1988); Verhoeyen et al., Science, 239:1534-1536 (1988)], замещением CDR грызунов или последовательностей CDR на соответствующие последовательности человеческого антитела. Соответственно, такие «гуманизированные» антитела представляют собой химерные антитела (патент США № 4816567), в которых по существу менее чем интактный человеческий вариабельный домен был замещен соответствующей последовательностью от вида, отличного от человека. На практике гуманизированные антитела, как правило, представляют человеческие антитела, в которых некоторые остатки CDR и, возможно, некоторые остатки FR замещены остатками из аналогичных сайтов антител грызунов.
Человеческие антитела также могут быть получены с использованием различных методов, известных в данной области техники, включая библиотеки фагового дисплея [Hoogenboom and Winter, J. Mol. Biol., 227:381 (1991); Marks et al., J. Mol. Biol., 222:581 (1991)]. Также доступны методы Cole et al. и Boerner et al. для получения человеческих моноклональных антител (Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p. 77 (1985) and Boerner et al., J. Immunol., 147(1):86-95 (1991)]. Аналогично человеческие антитела можно получить введением локусов человеческого иммуноглобулина трансгенным животным, например, мышам, у которых гены эндогенных иммуноглобулинов были частично или полностью инактивированы. При стимуляции наблюдается продукция человеческого антитела, что близко напоминает то, что имеет место у людей во всех отношениях, включая перестановку генов, сборку и репертуар антител. Такой подход описан, например, в патентах США № 5545807; 5545806; 5569825; 5625126; 5633425; 5661016 и в следующих научных публикациях: Marks et al., Bio/Technology 10, 779-783 (1992); Lonberg et al., Nature 368, 856-859 (1994); Morrison, Nature 368, 812-13 (1994); Fishwild et al., Nature Biotechnology, 14, 845-51 (1996); Neuberger, Nature Biotechnology, 14, 826 (1996); Lonberg and Huszar, Intern. Rev. Immunol. 13, 65-93 (1995).
Такие антитела также можно сделать аффинно зрелыми с использованием известных методов селекции и/или мутагенеза, как описано выше. Предпочтительные аффинно зрелые антитела обладают аффинностью, которая в 5 раз, более предпочтительно в 10 раз, еще более предпочтительно 20 или 30 раз выше, чем аффинность исходного антитела (как правило, мышиного гуманизированного или человеческого), из которого получено зрелое антитело.
Биспецифические антитела
Биспецифические антитела являются моноклональными, предпочтительно человеческими или гуманизированными антителами, которые обладают специфичностью связывания, по меньшей мере, для двух различных антигенов. В данном случае одна из специфичностей связывания представляет специфичность для TIGIT, другая для любого другого антигена, и предпочтительно для белка или рецептора или субъединицы рецептора клеточной поверхности.
В данной области известны методы получения биспецифических антител. Обычно рекомбинантная продукция биспецифических антител основана на коэкспрессии двух пар иммуноглобулиновых тяжелой цепи/легкой цепи, где две тяжелые цепи обладают различными специфичностями [Milstein and Cuello, Nature, 305:537-539 (1983)]. В результате случайного набора тяжелых и легких цепей иммуноглобулина, такие гибридомы (квадромы) продуцируют потенциальную смесь из десяти различных молекул антител, из которых только одна имеет правильную биспецифическую структуру. Очистку правильной молекулы обычно проводят стадиями аффинной хроматографии. Аналогичные процедуры раскрыты в WO 93/08829, опубликованной 13 мая 1993 года, и в публикации Traunecker et al., EMBO J., 10:3655-3659 (1991).
Вариабельные области антител с желаемой специфичностью связывания (сайты связывания антитело-антиген) могут быть слиты с последовательностями константных областей иммуноглобулинов. Слияние предпочтительно проводят с константной областью тяжелой цепи иммуноглобулина, содержащей, по меньшей мере, часть шарнирной области, СН2 и СН3. Предпочтительно иметь первую константную область тяжелой цепи (CH1), содержащую сайт, необходимый для связывания легкой цепи, присутствующий, по меньшей мере, в одной из слитых конструкций. ДНК, кодирующие слитые конструкции тяжелой цепи иммуноглобулина и, при желании, легкой цепи иммуноглобулина, вставляют в отдельные экспрессионные векторы и котрансфектируют в подходящий организм-хозяин. Более подробную информацию о получении биспецифических антител смотри, например, в публикации Suresh et al., Methods in Enzymology, 121:210 (1986).
Согласно другому подходу, описанному в WO 96/27011, границу раздела между парой молекул антител можно сконструировать таким образом, чтобы максимизировать процент гетеродимеров, которые выделяют из культуры рекомбинантных клеток. Предпочтительная граница раздела включает, по меньшей мере, часть CH3 домена константной области антитела. В данном методе одну или более небольших аминокислотных боковых цепей на границе раздела первой молекулы антитела заменяют более крупными боковыми цепями (например, тирозина или триптофана). Компенсаторные «полости» идентичного или близкого размера относительно крупной боковой цепи(ей), создают на поверхности раздела второй молекулы антитела заменой крупных аминокислотных боковых цепей более мелкими (например, аланина или треонина). Это обеспечивает механизм повышения выхода гетеродимера относительно нежелательных конечных продуктов, таких как гомодимеры.
Биспецифические антитела можно получить в виде полноразмерных антител или фрагментов антител (например, F(ab')2 биспецифические антитела). В литературе описаны способы получения биспецифических антител из фрагментов антител. Например, биспецифические антитела могут быть получены с использованием химического связывания. Brennan et al., Science, 229:81 (1985) описывают процедуру, в которой интактные антитела подвергают протеолитическому расщеплению с получением фрагментов F(ab')2. Эти фрагменты восстанавливают в присутствии арсенита натрия, образующего комплекс с дитиолами, для стабилизации вицинальных дитиоловых групп и предупреждения образования межмолекулярных дисульфидных связей. Образовавшиеся при этом Fab'-фрагменты затем превращают в производные тионитробензоата (TNB). Одно из таких производных, Fab'-TNB, затем вновь превращают в Fab'-тиол восстановлением меркаптоэтиламином и смешивают с эквимолярным количеством другого Fab'-TNB-производного с образованием биспецифического антитела. Полученные биспецифические антитела можно использовать в качестве агентов для селективной иммобилизации ферментов.
Fab'-фрагменты также можно непосредственно выделить из E. coli и химически связать с получением биспецифических антител. Shalaby et al., J. Exp. Med.. 175: 217-225 (1992) описывают получение молекулы полностью гуманизированного биспецифического антитела F(ab')2. Каждый Fab'-фрагмент отдельно секретируют из E. coli и подвергают непосредственному химическому связыванию in vitro с образованием биспецифического антитела. Полученное таким образом биспецифическое антитело способно связываться с клетками, сверхэкспрессирующими рецептор ErbB2 и нормальными человеческими Т-клетками, а также «запускают» литическую активность человеческих цитотоксических лимфоцитов против злокачественной опухоли-мишени молочной железы человека.
В литературе описаны различные методики получения и выделения фрагментов биспецифических антител непосредственно из культуры рекомбинантных клеток. Например, биспецифические антитела получали с использованием лейциновых застежек-молний (Kostelny et al., J. Immunol., 148(5):1547-1553 (1992)). Лейциновые застежки-молнии из Fos и Jun белков связывают с Fab'-фрагментами двух различных антител слиянием генов. Гомодимеры антител восстанавливают в шарнирной области с образованием мономеров и затем повторно окисляют с получением гетеродимеров антител. Этот метод может также использоваться для получения гомодимеров антител. Технология «диател», описанная Hollinger et al., Proc. Natl Acad. Sci. USA, 90:6444-6448 (1993), представляет альтернативный механизм получения биспецифических фрагментов антител. Фрагменты включают вариабельную область тяжелой цепи (VH), соединенную с вариабельной областью легкой цепи (VL) через линкер, который является слишком коротким для того, чтобы происходило спаривание между двумя доменами одной и той же цепи. Соответственно области VH и VL одного фрагмента вынуждены спариваться с комплементарными областями VL и VH другого фрагмента, тем самым образуя два антигенсвязывающих сайта. Также была описана еще одна стратегия создания биспецифических фрагментов антител за счет использования димеров одноцепочечных Fv (sFv) (смотри Gruber et al, J. Immunol, 152:5368 (1994)).
Также предусматриваются антитела, обладающие более чем двумя валентностями. В качестве одного неограничивающего примера, могут быть получены триспецифические антитела (Tuft et al. J. Immunol. 147: 60 (1991)).
Примерные биспецифические антитела могут связываться с двумя различными эпитопами на полипептиде TIGIT по настоящему изобретению. Альтернативно плечо анти-полипептид TIGIT можно объединить с плечом, которое связывается с триггерной молекулой на лейкоците, такой как молекула рецептора Т-клеток (например, CD2, CD3, CD28 или В7), или Fc-рецепторы для IgG (FcγR), такие как FcγRI (CD64), FcγRII (CD32) и FcγRIII (CD16), таким образом, чтобы сфокусировать защитные механизмы клетки в направлении клетки, экспрессирующей конкретный полипептид TIGIT. Биспецифические антитела также можно использовать для доставки цитотоксических агентов к клеткам, которые экспрессируют конкретный полипептид TIGIT. Такие антитела имеют TIGIT-связывающее плечо и плечо, которое связывает цитотоксический агент или хелатор радионуклидов, такой как EOTUBE, DPTA, DOTA или TETA. Другое представляющее интерес биспецифическое антитело связывает полипептид TIGIT и, кроме того, связывает тканевой фактор (TF).
Гетероконъюгатные антитела
Гетероконъюгатные антитела также находятся в объеме настоящего изобретения. Гетероконъюгатные антитела состоят из двух ковалентно связанных антител. Такие антитела, например, были предложены для нацеливания клеток иммунной системы на нежелательные клетки [патент США № 4676980] и для лечения ВИЧ-инфекции [WO 91/00360; WO 92/200373; ЕР 03089]. Предполагается, что антитела можно получить in vitro с использованием методов, известных в химии синтетических белков, включая методы с участием поперечно-сшивающих агентов. Например, можно сконструировать иммунотоксины с использованием реакции дисульфидного обмена или образования тиоэфирной связи. Примеры подходящих реагентов для этой цели включают иминотиолат и метил-4-меркаптобутиримидат и реагенты, описанные, например, в патенте США № 4676980.
Конструирование эффекторной функции
Может быть желательным модифицировать антитело по настоящему изобретению в отношении его эффекторной функции для того, чтобы повысить эффективность антитела в лечении рака. Например, один или более цистеиновых остатков можно ввести в Fc-область, что будет способствовать образованию в этом месте межцепочечной дисульфидной связи. Полученное таким образом гомодимерное антитело может обладать повышенной способностью к интернализации и/или повышенной способностью к лизису клеток, опосредованному комплементом, и повышенной антителозависимой клеточной цитотоксичностью (ADCC) (смотри Caron et al., J. Exp Med., 176:1191-1195 (1992); и Shopes, B. J. Immunol., 148:2918-2922 (1992)). Также могут быть получены гомодимерные антитела с повышенной противоопухолевой активностью при использовании гетеробифункциональных поперечно-сшивающих линкеров, описанных в публикации Wolff et al. Cancer Research, 53:2560-2565 (1993)). Альтернативно может быть сконструировано антитело таким образом, что оно будет иметь двойные Fc-области и за счет этого будет обладать повышенной способностью к комплемент-зависимому лизису и ADCC (смотри Stevenson et al., Anti-Cancer Drug Design, 3:219-230 (1989)).
В некоторых вариантах осуществления получены анти-TIGIT-антитела, которые представляют хомяк-анти-мышь-антитела. Два антитела, 10А7 и 1F4, также специфически связывали с человеческим TIGIT. Были определены аминокислотные последовательности легкой и тяжелой цепей антитела 10А7 с использованием стандартных методик. Последовательность легкой цепи антитела представляет: DIVMTQSPSSLAVSPGEKVTMTCKSSQSLYYSGVKENLLAWYQQKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAEDMGQYFCQQGINNPLTFGDGTKLEIKR (SEQ ID NO:13) и последовательность тяжелой цепи этого антитела представляет: EVQLVESGGGLTQPGKSLKLSCEASGFTFSSFTMHWVRQSPGKGLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDLKSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS (SEQ ID NO:15), где гипервариабельные участки (CDR) каждой цепи выделены жирным шрифтом. Таким образом, CDR1 легкой цепи антитела 10А7 имеет последовательность KSSQSLYYSGVKENLLA (SEQ ID NO:1), CDR2 легкой цепи антитела 10А7 имеет последовательность ASIRFT (SEQ ID NO:2), и CDR3 легкой цепи антитела 10А7 имеет последовательность QQGINNPLT (SEQ ID NO:3). CDR1 тяжелой цепи антитела 10А7 имеет последовательность GFTFSSFTMH (SEQ ID NO:4), CDR2 тяжелой цепи антитела 10А7 имеет последовательность FIRSGSGIVFYADAVRG (SEQ ID NO:5), и CDR3 тяжелой цепи антитела 10А7 имеет RPLGHNTFDS последовательность (SEQ ID NO:6).
Также были определены аминокислотные последовательности легкой и тяжелой цепей антитела 1F4. Последовательность легкой цепи данного антитела представляет: DVVLTQTPLSLSVSFGDQVSISCRSSQSLVNSYGNTFLSWYLHKPGQSPQLLIFGISNRFSGVPDRFSGSGSGTDFTLKISTIKPEDLGMYYCLQGTHQPPTFGPGTKLEVK (SEQ ID NO:14) и последовательность тяжелой цепи этого антитела представляет: EVQLQQSGPELVKPGTSMKISCKASGYSFTGHLMNWVKQSHGKNLEWIGLIIPYNGGTSYNQKFKGKATLTVDKSSSTAYMELLSLTSDDSAVYFCSRGLRGFYAMDYWGQGTSVTVSS (SEQ ID NO:16), где гипервариабельные участки (CDR) из каждой цепи выделены жирным шрифтом. Таким образом, CDR1 легкой цепи антитела 1F4 имеет последовательность RSSQSLVNSYGNTFLS (SEQ ID NO:7), CDR2 легкой цепи антитела 1F4 имеет последовательность GISNRFS (SEQ ID NO:8), и CDR3 легкой цепи антитела 1F4 имеет последовательность LQGTHQPPT (SEQ ID NO:9). CDR1 тяжелой цепи антитела 1F4 имеет последовательность GYSFTGHLMN (SEQ ID NO:10), CDR2 тяжелой цепи антитела 1F4 имеет последовательность LIIPYNGGTSYNQKFKG (SEQ ID NO:11), и CDR3 тяжелой цепи антитела 1F4 имеет последовательность GLRGFYAMDY (SEQ ID NO:12).
Была определена нуклеотидная последовательность, кодирующая легкую цепь антитела 1F4: GATGTTGTGTTGACTCAAACTCCACTCTCCCTGTCTGTCAGCTTTGGAGATCAAGTTTCTATCTCTTGCAGGTCTAGTCAGAGTCTTGTAAACAGTTATGGGAACACCTTTTTGTCTTGGTACCTGCACAAGCCTGGCCAGTCTCCACAGCTCCTCATCTTTGGGATTTCCAACAGATTTTCTGGGGTGCCAGACAGGTTCAGTGGCAGTGGTTCAGGGACAGATTTCACACTCAAGATCAGCACAATAAAGCCTGAGGACTTGGGAATGTATTACTGCTTACAAGGTACGCATCAGCCTCCCACGTTCGGTCCTGGGACCAAGCTGGAGGTGAAA (SEQ ID NO:38) и нуклеотидная последовательность, кодирующая тяжелую цепь антитела 1F4: GAGGTCCAGCTGCAACAGTCTGGACCTGAGCTGGTGAAGCCTGGAACTTCAATGAAGATATCCTGCAAGGCTTCTGGTTACTCATTCACTGGCCATCTTATGAACTGGGTGAAGCAGAGCCATGGAAAGAACCTTGAGTGGATTGGACTTATTATTCCTTACAATGGTGGTACAAGCTATAACCAGAAGTTCAAGGGCAAGGCCACATTGACTGTAGACAAGTCATCCAGCACAGCCTACATGGAGCTCCTCAGTCTGACTTCTGATGACTCTGCAGTCTATTTCTGTTCAAGAGGCCTTAGGGGCTTCTATGCTATGGACTACTGGGGTCAAGGAACCTCAGTCACCGTCTCCTCA (SEQ ID NO:39).
В некоторых вариантах осуществления анти-TIGIT-антитело или его антигенсвязывающий фрагмент содержит, по меньшей мере, один HVR, включающий аминокислотную последовательность, выбранную из аминокислотных последовательностей, представленных в (1) KSSQSLYYSGVKENLLA (SEQ ID NO:1), ASIRFT (SEQ ID NO:2), QQGINNPLT (SEQ ID NO:3), GFTFSSFTMH (SEQ ID NO:4), FIRSGSGIVFYADAVRG (SEQ ID NO:5), и RPLGHNTFDS (SEQ ID NO:6), или (2) RSSQSLVNSYGNTFLS (SEQ ID NO:7), GISNRFS (SEQ ID NO:8), LQGTHQPPT (SEQ ID NO:9), GYSFTGHLMN (SEQ ID NO:10), LIIPYNGGTSYNQKFKG (SEQ ID NO:11) и GLRGFYAMDY (SEQ ID NO:12).
В некоторых вариантах осуществления обеспечивается анти-TIGIT-антитело или его антигенсвязывающий фрагмент, где легкая цепь антитела включает аминокислотную последовательность, представленную в DIVMTQSPSSLAVSPGEKVTMTCKSSQSLYYSGVKENLLAWYQQKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAEDMGQYFCQQGINNPLTFGDGTKLEIKR (SEQ ID NO:13) или DVVLTQTPLSLSVSFGDQVSISCRSSQSLVNSYGNTFLSWYLHKPGQSPQLLIFGISNRFSGVPDRFSGSGSGTDFTLKISTIKPEDLGMYYCLQGTHQPPTFGPGTKLEVK (SEQ ID NO:14).
В некоторых вариантах осуществления обеспечивается анти-TIGIT-антитело или его антигенсвязывающий фрагмент, где тяжелая цепь антитела включает аминокислотную последовательность, представленную в EVQLVESGGGLTQPGKSLKLSCEASGFTFSSFTMHWVRQSPGKGLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDLKSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS (SEQ ID NO:15) или EVQLQQSGPELVKPGTSMKISCKASGYSFTGHLMNWVKQSHGKNLEWIGLIIPYNGGTSYNQKFKGKATLTVDKSSSTAYMELLSLTSDDSAVYFCSRGLRGFYAMDYWGQGTSVTVSS (SEQ ID NO:16).
В некоторых вариантах осуществления обеспечивается анти-TIGIT-антитело или его антигенсвязывающий фрагмент, где легкая цепь антитела включает аминокислотную последовательность, представленную в DIVMTQSPSSLAVSPGEKVTMTCKSSQSLYYSGVKENLLAWYQQKPGQS PKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAEDMGQYFCQQGINNPLTFGDGTKLEIKR (SEQ ID NO:13) или DVVLTQTPLSLSVSFGDQVSISCRSSQSLVNSYGNTFLSWYLHKPGQSPQLLIFGISNRFSGVPDRFSGSGSGTDFTLKISTIKPEDLGMYYCLQGTHQPPTFGPGTKLEVK (SEQ ID NO:14), и тяжелая цепь антитела включает аминокислотную последовательность, представленную в EVQLVESGGGLTQPGKSLKLSCEASGFTFSSFTMHWVRQSPGKGLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDLKSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS (SEQ ID NO:15) или EVQLQQSGPELVKPGTSMKISCKASGYSFTGHLMNWVKQSHGKNLEWIGLIIPYNGGTSYNQKFKGKATLTVDKSSSTAYMELLSLTSDDSAVYFCSRGLRGFYAMDYWGQGTSVTVSS (SEQ ID NO:16).
В некоторых вариантах осуществления обеспечивается анти-TIGIT-антитело или его антигенсвязывающий фрагмент, где антитело выбрано из гуманизированного антитела, химерного антитела, биспецифического антитела, гетероконъюгатного антитела и иммунотоксина.
В некоторых вариантах осуществления изобретения анти-TIGIT-антитело или его антигенсвязывающий фрагмент содержит, по меньшей мере, один HVR, который, по меньшей мере, на 90% идентичен HVR, представленному в любой из последовательностей (1) KSSQSLYYSGVKENLLA (SEQ ID NO:1), ASIRFT (SEQ ID NO:2), QQGINNPLT (SEQ ID NO:3), GFTFSSFTMH (SEQ ID NO:4), FIRSGSGIVFYADAVRG (SEQ ID NO:5) и RPLGHNTFDS (SEQ ID NO:6), или (2) RSSQSLVNSYGNTFLS (SEQ ID NO:7), GISNRFS (SEQ ID NO:8), LQGTHQPPT (SEQ ID NO:9), GYSFTGHLMN (SEQ ID NO:10), LIIPYNGGTSYNQKFKG (SEQ ID NO:11) и GLRGFYAMDY (SEQ ID NO:12).
В некоторых вариантах осуществления анти-TIGIT-антитело или его антигенсвязывающий фрагмент содержит легкую цепь и/или тяжелую цепь, включающую аминокислотные последовательности, по меньшей мере, на 90% идентичные аминокислотным последовательностям, представленным в DIVMTQSPSSLAVSPGEKVTMTCKSSQSLYYSGVKENLLAWYQQKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAEDMGQYFCQQGINNPLTFGDGTKLEIKR (SEQ ID NO:13) или DVVLTQTPLSLSVSFGDQVSISCRSSQSLVNSYGNTFLSWYLHKPGQSPQLLIFGISNRFSGVPDRFSGSGSGTDFTLKISTIKPEDLGMYYCLQGTHQPPTFGPGTKLEVK (SEQ ID NO:14), или EVQLVESGGGLTQPGKSLKLSCEASGFTFSSFTMHWVRQSPGKGLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDLKSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS (SEQ ID NO:15) или EVQLQQSGPELVKPGTSMKISCKASGYSFTGHLMNWVKQSHGKNLEWIGLIIPYNGGTSYNQKFKGKATLTVDKSSSTAYMELLSLTSDDSAVYFCSRGLRGFYAMDYWGQGTSVTVSS (SEQ ID NO:16) соответственно.
Агенты, которые модулируют экспрессию и/или активность CD226
Обеспечивается способ лечения или замедления прогрессирования рака у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226. Также обеспечивается способ снижения или ингибирования рецидива или прогрессирования рака у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226. Также обеспечивается способ лечения или замедления прогрессирования связанного с иммунитетом заболевания у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226. Также обеспечивается способ снижения или ингибирования прогрессирования связанного с иммунитетом заболевания у индивидуума, включающий введение индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226. Также обеспечивается способ повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума введением индивидууму эффективного количества антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226.
Например, агенты, которые модулируют экспрессию и/или активность CD226, представляют агенты, способные повышать или стимулировать экспрессию и/или активность CD226, повышать и/или стимулировать взаимодействие CD226 с PVR, PVRL2 и/или PVRL3, и повышать и/или стимулировать внутриклеточную передачу сигналов, опосредованную связыванием CD226 с PVR, PVRL2 и/или PVRL3. В некоторых вариантах осуществления, агенты, способные повышать и/или стимулировать экспрессию и/или активность CD226, представляют агенты, которые повышают и/или стимулируют экспрессию и/или активность CD226. В некоторых вариантах осуществления, агенты, способные повышать и/или стимулировать взаимодействие CD226 с PVR, PVRL2 и/или PVRL3, представляют агенты, которые повышают и/или стимулируют взаимодействие CD226 с PVR, PVRL2 и/или PVRL3. В некоторых вариантах осуществления, агенты, способные повышать и/или стимулировать внутриклеточную передачу сигналов, опосредованную связыванием CD226 с PVR, PVRL2 и/или PVRL3, представляют агенты, которые повышают и/или стимулируют внутриклеточную передачу сигналов, опосредованную связыванием CD226 с PVR, PVRL2 и/или PVRL3.
В некоторых вариантах осуществления агент, который модулирует экспрессию и/или активность CD226, выбран из агента, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, антагониста экспрессии и/или активности TIGIT, антагониста экспрессии и/или активности PVR, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVR, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL2, агента, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL3, агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR, агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL2, агента, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL3, и их комбинаций. В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, выбран из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида. В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, представляет анти-TIGIT-антитело или его антигенсвязывающий фрагмент. В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие CD226 с TIGIT, представляет ингибирующую нуклеиновую кислоту, выбранную из антисмыслового полинуклеотида, интерферирующей РНК, каталитической РНК и химеры РНК-ДНК.
В некоторых вариантах осуществления антагонист экспрессии и/или активности TIGIT представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления антагонист экспрессии и/или активности TIGIT представляет анти-TIGIT-антитело или его антигенсвязывающий фрагмент. В некоторых вариантах осуществления антагонист экспрессии и/или активности TIGIT представляет ингибирующую нуклеиновую кислоту, выбранную из антисмыслового полинуклеотида, интерферирующей РНК, каталитической РНК и химеры РНК-ДНК. В некоторых вариантах осуществления антагонист экспрессии и/или активности PVR представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVR, представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL2, представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления агент, который ингибирует и/или блокирует взаимодействие TIGIT с PVRL3, представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR, представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL2, представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVRL3, представляет низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид.
В некоторых вариантах осуществления антагонист экспрессии и/или активности TIGIT включает низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления антагонист экспрессии и/или активности PVR включает низкомолекулярный ингибитор, ингибирующее антитело или его антигенсвязывающий фрагмент, аптамер, ингибирующую нуклеиновую кислоту и ингибирующий полипептид. В некоторых вариантах осуществления агент, который ингибирует и/или блокирует внутриклеточную передачу сигналов, опосредованную связыванием TIGIT с PVR, выбран из группы, состоящей из низкомолекулярного ингибитора, ингибирующего антитела или его антигенсвязывающего фрагмента, аптамера, ингибирующей нуклеиновой кислоты и ингибирующего полипептида. В некоторых вариантах осуществления антагонист экспрессии и/или активности TIGIT, представляет анти-TIGIT-антитело или его антигенсвязывающий фрагмент. В некоторых вариантах осуществления антагонист экспрессии и/или активности TIGIT представляет ингибирующую нуклеиновую кислоту, выбранную из антисмыслового полинуклеотида, интерферирующей РНК, каталитической РНК и химеры РНК-ДНК.
Комбинации Т-клеточных мишеней для терапии иммунорегуляторными антителами
В дополнение к специфическому распознаванию антигена через TCR активация Т-клеток регулируется посредством баланса положительных и отрицательных сигналов от костимулирующих рецепторов. Эти поверхностные белки, как правило, являются членами суперсемейств рецепторов TNF или В7. Активирующие костимулирующие рецепторы включают CD226, CD28, OX40, GITR, CD137, CD27, HVEM, MICA, ICOS, NKG2D и 2В4. Ингибирующие костимулирующие рецепторы включают CTLA-4, PD-1, TIM-3, BTLA, VISTA, LAG-3, B7H4 и CD96. Агонистические антитела, направленные против активирующих костимулирующих молекул, и блокирующие антитела против отрицательных костимулирующих молекул, могут усилить стимуляцию Т-клеток, что способствует распаду опухолей.
Обеспечивается способ повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума введением индивидууму эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов. В некоторых вариантах осуществления один или более дополнительных иммунных коингибирующих рецепторов выбран из PD-1, CTLA-4, LAG3, TIM3, BTLA, VISTA, B7H4 и CD96. В некоторых вариантах осуществления один или более дополнительных иммунных коингибирующих рецепторов выбран из PD-1, CTLA-4, LAG3 и TIM3.
Также обеспечивается способ повышения, усиления или стимуляции иммунного ответа или иммунной функции у индивидуума введением индивидууму эффективного количества агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов. В некоторых вариантах осуществления один или более дополнительных иммунных костимулирующих рецепторов выбран из CD226, ОХ-40, CD28, CD27, CD137, HVEM, GITR, MICA, ICOS, NKG2D и 2В4. В некоторых вариантах осуществления один или более дополнительных иммунных костимулирующих рецепторов выбран из CD226, ОХ-40, CD27, CD137, HVEM и GITR. В некоторых вариантах осуществления один или более дополнительных иммунных костимулирующих рецепторов выбран из ОХ-40 и CD27.
IV Наборы
В еще одном аспекте обеспечивается набор, содержащий антагонист, связывающийся с осью PD-1, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT, для лечения или замедления прогрессирования рака у индивидуума или для усиления иммунной функции у индивидуума, имеющего рак. Любой из антагонистов, связывающихся с осью PD-1, и/или агентов, которые снижают или ингибируют экспрессию и/или активность TIGIT, описанных здесь, может быть включен в набор.
В еще одном аспекте обеспечивается набор, содержащий антагонист, связывающийся с осью PD-1, и агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, и агента, который снижает или ингибирует экспрессию и/или активность TIGIT, для лечения или замедления прогрессирования рака у индивидуума или для усиления иммунной функции у индивидуума, имеющего рак. Любой из антагонистов, связывающихся с осью PD-1, и/или агентов, которые снижают или ингибируют экспрессию и/или активность TIGIT, описанных здесь, может быть включен в набор.
В еще одном аспекте обеспечивается набор, содержащий агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в комбинации с антагонистом, связывающимся с осью PD-1, для лечения или замедления прогрессирования рака у индивидуума или для усиления иммунной функции у индивидуума, имеющего рак. Любой из антагонистов, связывающихся с осью PD-1, и/или агентов, которые снижают или ингибируют экспрессию и/или активность TIGIT, описанных здесь, может быть включен в набор.
В еще одном аспекте обеспечивается набор, содержащий антагонист, связывающийся с осью PD-1, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, в комбинации с агентом, который модулирует экспрессию и/или активность CD226, для лечения или замедления прогрессирования рака у индивидуума. Любой из антагонистов, связывающихся с осью PD-1, и/или агентов, которые модулируют экспрессию и/или активность CD226, описанных здесь, может быть включен в набор.
В еще одном аспекте обеспечивается набор, содержащий антагонист, связывающийся с осью PD-1, и агент, который модулирует экспрессию и/или активность CD226, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226, для лечения или замедления прогрессирования рака у индивидуума. Любой из антагонистов, связывающихся с осью PD-1, и/или агентов, которые модулируют экспрессию и/или активность CD226, описанных здесь, может быть включен в набор.
В еще одном аспекте обеспечивается набор, содержащий агент, который модулирует экспрессию и/или активность CD226, и вкладыш в упаковке, содержащий инструкции по применению агента, который модулирует экспрессию и/или активность CD226, в комбинации с антагонистом, связывающимся с осью PD-1, для лечения или замедления прогрессирования рака у индивидуума. Любой из антагонистов, связывающихся с осью PD-1, и/или агентов, которые модулируют экспрессию и/или активность CD226, описанных здесь, может быть включен в набор.
В еще одном аспекте обеспечивается набор, содержащий антагонист, связывающийся с осью PD-1, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, в комбинации с агентом, который модулирует экспрессию и/или активность CD226, для усиления иммунной функции у индивидуума, имеющего рак. Любой из антагонистов, связывающихся с осью PD-1, и/или агентов, которые модулируют экспрессию и/или активность CD226, описанных здесь, может быть включен в набор.
В еще одном аспекте обеспечивается набор, содержащий антагонист, связывающийся с осью PD-1, и агент, который модулирует экспрессию и/или активность CD226, и вкладыш в упаковке, содержащий инструкции по применению антагониста, связывающегося с осью PD-1, и агента, который модулирует экспрессию и/или активность CD226, для усиления иммунной функции у индивидуума, имеющего рак. Любой из антагонистов, связывающихся с осью PD-1, и/или агентов, которые модулируют экспрессию и/или активность CD226, описанных здесь, может быть включен в набор.
В еще одном аспекте обеспечивается набор, содержащий агент, который модулирует экспрессию и/или активность CD226, и вкладыш в упаковке, содержащий инструкции по применению агента, который модулирует экспрессию и/или активность CD226, в комбинации с антагонистом, связывающимся с осью PD-1, для усиления иммунной функции у индивидуума, имеющего рак. Любой из антагонистов, связывающихся с осью PD-1, и/или агентов, которые модулируют экспрессию и/или активность CD226, описанных здесь, может быть включен в набор.
В еще одном аспекте обеспечивается набор, содержащий агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в комбинации с агентом, который снижает или ингибирует один или более иммунных коингибирующих рецепторов, для лечения или замедления прогрессирования рака у индивидуума или усиления иммунной функции у индивидуума, имеющего рак. Любой из агентов, которые снижают или ингибируют экспрессию и/или активность TIGIT, и/или агентов, которые снижают или ингибируют один или более иммунных коингибирующих рецепторов, описанных здесь, может быть включен в набор.
В еще одном аспекте обеспечивается набор, содержащий агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и агент, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов, для лечения или замедления прогрессирования рака у индивидуума или усиления иммунной функции у индивидуума, имеющего рак. Любой из агентов, которые снижают или ингибируют экспрессию и/или активность TIGIT, и/или агентов, которые снижают или ингибируют один или более иммунных коингибирующих рецепторов, описанных здесь, может быть включен в набор.
В еще одном аспекте обеспечивается набор, содержащий агент, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует один или более дополнительных иммунных коингибирующих рецепторов, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT, для лечения или замедления прогрессирования рака у индивидуума или усиления иммунной функции у индивидуума, имеющего рак. Любой из агентов, которые снижают или ингибируют экспрессию и/или активность TIGIT, и/или агентов, которые снижают или ингибируют один или более иммунных коингибирующих рецепторов, описанных здесь, может быть включен в набор.
В еще одном аспекте обеспечивается набор, содержащий агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, в комбинации с агентом, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, для лечения или замедления прогрессирования рака у индивидуума или усиления иммунной функции у индивидуума, имеющего рак. Любой из агентов, которые снижают или ингибируют экспрессию и/или активность TIGIT, и/или агентов, которые повышают или активируют один или более иммунных костимулирующих рецепторов, описанных здесь, может быть включен в набор.
В еще одном аспекте обеспечивается набор, содержащий агент, который снижает или ингибирует экспрессию и/или активность TIGIT, и агент, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, и вкладыш в упаковке, содержащий инструкции по применению агента, который снижает или ингибирует экспрессию и/или активность TIGIT, и агента, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, для лечения или замедления прогрессирования рака у индивидуума или усиления иммунной функции у индивидуума, имеющего рак. Любой из агентов, которые снижают или ингибируют экспрессию и/или активность TIGIT, и/или агентов, которые повышают или активируют один или более иммунных костимулирующих рецепторов, описанных здесь, может быть включен в набор.
В еще одном аспекте обеспечивается набор, содержащий агент, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, и вкладыш в упаковке, содержащий инструкции по применению агента, который повышает или активирует один или более дополнительных иммунных костимулирующих рецепторов, в комбинации с агентом, который снижает или ингибирует экспрессию и/или активность TIGIT, для лечения или замедления прогрессирования рака у индивидуума или усиления иммунной функции у индивидуума, имеющего рак. Любой из агентов, которые снижают или ингибируют экспрессию и/или активность TIGIT, и/или агентов, которые повышают или активируют один или более иммунных костимулирующих рецепторов, описанных здесь, может быть включен в набор.
В некоторых вариантах осуществления набор содержит контейнер, включающий один или более антагонистов, связывающихся с осью PD-1, и агенты, которые снижают или ингибируют экспрессию и/или активность TIGIT, описанные здесь. В некоторых вариантах осуществления набор содержит контейнер, включающий один или более антагонистов, связывающихся с осью PD-1, и агенты, которые модулируют экспрессию и/или активность CD226, описанные здесь. В некоторых вариантах осуществления набор содержит контейнер, включающий один или более агентов, которые снижают или ингибируют экспрессию и/или активность TIGIT, и агенты, которые снижают или ингибируют один или более дополнительных иммунных ко-ингибирующих рецепторов, описанных здесь. В некоторых вариантах осуществления набор содержит контейнер, включающий агенты, которые снижают или ингибируют экспрессию и/или активность TIGIT, и агенты, которые повышают или активируют один или более дополнительных иммунных костимулирующих рецепторов, описанных здесь. Подходящие контейнеры включают, например, бутыли, флаконы (например, двухкамерные флаконы), шприцы (например, однокамерные или двухкамерные шприцы) и пробирки. Контейнер может быть изготовлен из различных материалов, таких как стекло или пластик. В некоторых вариантах осуществления набор может иметь этикетку (например, на или связанную с контейнером) или вкладыш в упаковке. На этикетке или вкладыше в упаковке может быть указано, что соединение, находящееся в нем, может быть пригодно и предназначено для лечения или замедления прогрессирования рака у индивидуума, или для усиления иммунной функции у индивидуума, имеющего рак. Набор может дополнительно включать другие материалы, желательные с коммерческой точки зрения и с точки зрения пользователя, включая другие буферы, разбавители, фильтры, иглы и шприцы.
Примеры
Настоящее изобретение может быть лучше понято при обращении к следующим примерам, которые приведены в качестве иллюстрации и не предназначены для ограничения объема изобретения.
Пример 1: TIGIT экспрессируется на высоком уровне на истощенных CD8+ и CD4+ Т-клетках, и его экспрессия коррелирует с экспрессией PD-1
Для подтверждения того, что CD8+ Т-клетки являются компетентными для экспрессии TIGIT после стимуляции in vitro CD8+ Т-клетки селезенки мышей C57BL6/J, обогащенные с использованием MACS, стимулировали фиксированными на планшетах анти-CD3- и анти-CD28-антителами в течение 24-48 ч in vitro. Использовали проточную цитометрию для определения экспрессии TIGIT. Согласуясь с ранее полученными данными по экспрессии TIGIT СD4+ Т-клетками (Yu, X. et al., The surface protein TIGIT suppresses T cell activation by promoting the generation of mature immunoregulatory dendritic cells. Nature immunology, 10, 48-57 (2009)), мышиные CD8+ Т-клетки экспрессировали TIGIT в течение 48 ч стимуляции in vitro (фиг. 1А).
Для оценки экспрессии TIGIT активированными CD8+ Т-клетками in vivo мышей C57BL6/J заражали штаммом Armstrong вируса лимфобластного хориоменингита (LCMV), и спленоциты анализировали через 7 суток после заражения. Вкратце, для воспроизведения острой инфекции мышей заражали внутривенно 2×106 бляшкообразующих единиц (БОЕ) штамма Armstrong LCMV. Использовали проточную цитометрию для определения экспрессии TIGIT наивными (CD44низкий CD62Lвысокий) и CD8+ и CD4+ эффекторными Т-клетками памяти (CD44высокий CD62Lнизкий). На пике ответа Т-клеток на LCMV субпопуляция CD4+ эффекторных Т-клеток памяти (ТЕА) и почти все CD8+ TEM клетки экспрессировали TIGIT на высоком уровне (фиг. 1B). Использовали проточную цитометрию для определения экспрессии TIGIT PD-1высокий и PD-1низкий эффекторными CD8+Т-клетками памяти. Интересно отметить, что экспрессия TIGIT практически идеально коррелировала с экспрессией PD-1 (фиг. 1С).
Поскольку PD-1 ассоциирован с истощением Т-клеток, то определяли экспрессию TIGIT на хронически стимулированных Т-клетках. Вкратце, для воспроизведения хронической инфекции мышей заражали внутривенно 2×106 БОЕ клона 13 LCMV и обрабатывали 500 мкг и 250 мкг истощающих анти-CD4-антител (клон GK1.5) в течение 3 суток до и в течение 4 суток после заражения соответственно. Там, где это было показано, мышам, зараженным клоном 13 штамма LCMV, внутрибрюшинно вводили 200 мкг антител контрольного изотипа, 200 мкг анти-PD-L1-антител и/или 500 мкг анти-TIGIT-антител 3 раза в неделю, начиная с суток 28 по 42 сутки после заражения. Спленоциты анализировали через 42 суток после заражения. Использовали проточную цитометрию для определения экспрессии TIGIT наивными (CD44низкий CD62Lвысокий), CD8+ и CD4+ Т-клетками центральной памяти (CD44высокий CD62Lнизкий) и эффекторными CD8+ и CD4+ Т-клетками памяти (CD44высокий CD62Lнизкий). Фактически у мышей, хронически зараженных клоном 13 штамма LCMV, TIGIT экспрессировался на высоком уровне преимущественно на PD-1высокий Т-клетках, но не на наивных PD-1низкий ТЕМ клетках, или Т-клетках центральный памяти (фиг. 1D).
Пример 2: роль TIGIT в истощении Т-клеток у мышей с дефицитом TIGIT
Для характеристики роли TIGIT в истощении Т-клеток, получали мышей, у которых TIGIT был условно делецирован в Т-клетках, (TIGITfl/fl CD4-cre+ (СКО), фиг. 2). Вкратце, мышей CD4-cre и мышей TIGITLoxP/LoxP получали на основе мышей C57BL/6J с использованием стандартных методов и перекрестных скрещиваний. Клеточную линию ES с проверенным качеством (Art B6/3.6 (генетическая основа: C57BL/6 NTAC) культивировали на митотически инактивированном фидерном слое, состоящем из мышиных эмбриональных фибробластов, в клеточной культуральной среде ES, содержащей лейкоз-ингибирующий фактор и фетальную бычью сыворотку. Клетки подвергали электропорации с линеаризованным направленным ДНК вектором согласно Стандартным операционным процедурам Taconic Artemis'. Селективную среду, содержащую G418 и ганцикловир, использовали в качестве механизмов для обогащения гомологично рекомбинированных клонов. Резистентные клеточные колонии ES (ES клоны) с различимой морфологией выделяли на 8 сутки после трансфекции и анализировали саузерн-блоттингом и/или ПЦР в первичном скрининге. Гомологично рекомбинантные клеточные клоны ES размножали и замораживали в жидком азоте после тщательного молекулярного валидирования. Кассету neo удаляли с использованием рекомбиназы flpE до микроинъекции мышам-самкам альбиносам BL/6, которых использовали в качестве доноров. Получали химерное потомство и материал из хвоста исследовали с помощью ПЦР в отношении передачи зародышевой линии. Экспрессия TIGIT была отменена с 96% эффективностью из Т-клеток у мышей TIGITfl/FL CD4cre.
У мышей с отсутствием на Т-клетках экспрессии TIGIT индуцировали ответ CD4+ и CD8+ Т-клеток на острое заражение штаммом Armstrong LCMV, аналогично заражали мышей дикого типа (фиг. 3).
Для оценки эффекта в формате хронической инфекции у мышей TIGITfl/fl CD4-cre-(WT) и TIGITfl/fl CD4-cre+ (CKO) быстро истощали CD4+ Т-клетки и животных заражали клоном 13 штамма LCMV. Титры вируса в спленоцитах и печени определяли через 42 суток после заражения. При хронической инфекции, вызванной заражением клоном 13 штамма LCMV, достоверно большее количество CD8+ и CD4+ Т-клеток у мышей TIGITfl/fl CD4-cre+ (CKO) было компетентным для продукции интерферона-гамма (IFN-γ) по сравнению с Т-клетками от мышей дикого типа из того же помета (TIGITfl/fl CD4-cre- (WT)) (повышение на 82-86%, P<0,01, фиг. 4A-4D). Кроме того, вирусная нагрузка достоверно снижалась у хронически инфицированных мышей TIGITfl/fl CD4-cre+(СКО) (снижение на 68%, Р <0,0001, фиг. 4E).
Полученные результаты дают основание предположить, что TIGIT играет важную роль в регуляции активности и ответа Т-клеток при хронических иммунных реакциях, например, при хронической вирусной инфекции, и что TIGIT может регулировать эффекторную функцию, в частности, компетентность в отношении продукции эффекторных цитокинов, таких как IFNγ и TNFα, хронически стимулированными или истощенными CD8+ и CD4+ Т-клетками.
Пример 3: TIGIT и PD-1 синергически регулируют эффекторную функцию истощенных Т-клеток in vivo
Поскольку экспрессия TIGIT тесно коррелировала с экспрессией PD-1, в частности, на CD8+ Т-клетках при острой и хронической вирусной инфекции (фиг. 1), то блокирование TIGIT и PD-1 в комбинации может восстановить эффекторную функцию Т-клеток на более высоком уровне, чем можно получить путем блокирования каждого корецептора по отдельности.
Для проверки этой гипотезы у мышей C57BL6/J быстро истощали CD4+ Т-клетки и животных заражали клоном 13 штамма LCMV. Для воспроизведения хронической инфекции мышей заражали внутривенно 2×106 БОЕ клона 13 LCMV и обрабатывали 500 мкг и 250 мкг истощающих анти-CD4-антител (клон GK1.5) в течение 3 суток до и в течение 4 суток после заражения соответственно. Там, где это было показано, мышам, зараженным клоном 13 штамма LCMV, внутрибрюшинно вводили 200 мкг изотипически сходных контрольных антител, 200 мкг анти-PD-L1-антител и/или 500 мкг анти-TIGIT-антител 3 раза в неделю, начиная с суток 28 по 42 сутки после заражения. Обработку начинали на 28 сутки после заражения, поскольку Т-клеточный ответ преимущественно был истощен на данной временной точке на данной модели хронической вирусной инфекции ((Wherry et al., Molecular Signature of CD8+ T cell Exhaustion During Chronic Viral Infection, Immunity, 2007 Oct;27(4):670-84). Титры вируса в спленоцитах и печени определяли через 42 суток после заражения.
У этих мышей обработка анти-PD-L1-антителом индуцировала более сильную активацию CD8+ Т-клеток по сравнению с обработкой изотипически сходными контрольными антителами (повышение на 88%, P <0,0001, фиг. 4F), как сообщалось ранее (Barber, D.L., et al., Restoring function in exhausted CD8 T cells during chronic viral infection. Nature, 439, 682-687 (2006)). Обработка одними анти-TIGIT-антителами или в комбинации с анти-PD-L1-антителами не оказала заметного влияния на активацию CD8+ Т-клеток (фиг. 4F). Аналогично блокирование одного PD-1 приводила к умеренному повышению компетенции CD8+ Т-клеток к продукции цитокинов, в то время как блокирование одного TIGIT не оказывала никакого эффекта (фиг. 4G). Однако, частота встречаемости IFNγ-продуцирующих CD8+ Т-клеток резко возрастала у мышей, обработанных обоими анти-TIGIT-антителами и анти-PD-L1-антителами, и в достоверно большей степени, чем это наблюдали у мышей, обработанных только анти-PD-L1-антителами (фиг. 4G, повышение на 93%, P = 0,0050). Аналогичный эффект наблюдали для CD4+ Т-клеток (фиг. 5). Как также показано на фигуре 31, одновременное блокирование TIGIT/PD-L1 достоверно усиливало эффекторную функцию CD8+ Т-клеток, но не эффекторную функцию CD4+ Т-клеток у мышей по сравнению с мышами, обработанными только одним анти-PD-L1-антителом. Аналогичные эффекты также наблюдали в отношении экспансии Т-клеток и эффекторной функции LCMV gp33 антиген-специфических Т-клеток (фиг. 31). Полученные результаты свидетельствуют о наличии выраженной синергии между PD-1 и TIGIT на истощенных CD8+ Т-клетках, и указывают на то, что TIGIT специфически регулирует компетентность CD8+ Т-клеток в отношении продукции цитокинов и эффекторной функции.
Согласуясь с этими результатами, вирусная нагрузка LCMV умеренно снижалась у мышей, обработанных одними анти-PD-L1-антителами, не уменьшалась у мышей, обработанных только одним анти-TIGIT-антителом, и существенно снижалась у мышей, обработанных обоими анти-TIGIT- и анти-PD-L1-антителами (снижение титра вируса на 68% при обработке анти-PD-L1-антителом, P = 0,0004; снижение титра вируса на 92% при обработке анти-TIGIT-антителом+ анти-PD-L1-антителом, P <0,0001, фиг. 4H). Полученные данные демонстрируют наличие высокой синергии между ингибирующими эффектами PD-1 и TIGIT, и на их основании можно предположить, что в отличие от PD-1, TIGIT не является ингибитором широкого действия в отношении активации эффекторных Т-клеток, а играет незначительную роль в ограничении компетентности Т-клеток в отношении продукции цитокинов и эффекторной функции.
Пример 4: экспрессия TIGIT повышена при раке молочной железы человека, и его экспрессия коррелирует с экспрессией CD8 и ингибирующих корецепторов
Истощение Т-клеток также является основным иммунологическим признаком рака с наличием большого количества инфильтрирующих опухоль лимфоцитов (TIL), с высоким уровнем ингибирующих корецепторов и отсутствием способности продуцировать эффекторные цитокины (Wherry E.J., T cell exhaustion. Nature immunology, 12, 492-499 (2011); Rabinovich G.A., Gabrilovich D. & Sotomayor E.M., Immunosuppressive strategies that are mediated by tumor cells, Annual review of immunology, 25, 267-296 (2007)).
Для определения того, насколько TIGIT ингибирует эффекторную функцию TIL, анализировали данные по экспрессии генов злокачественной опухоли молочной железы с использованием микрочипового анализа, полученные в проекте Атлас ракового генома (CGAN) (Network, C.G.A. Comprehensive molecular portraits of human breast tumours, Nature, 490, 61-70 (2012)).
Экспрессия TIGIT была достоверно повышена в злокачественных опухолях молочной железы в целом (повышение на 135% по сравнению с нормальными образцами молочной железы, Р = 6×10-12, фиг. 6А) и в четырех основных молекулярных субтипах рака молочной железы (фиг. 6А) (Perou C.M., et al., Molecular portraits of human breast tumours, Nature, 406, 747-752 (2000); Sorlie T., et al., Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications, Proceedings of the National Academy of Sciences of the United States of America, 98, 10869-10874 (2001)). Экспрессия TIGIT высоко коррелировала с экспрессией CD3ε, соответственно с экспрессией TIL (R2 = 0,61, фиг. 6B). Интересно отметить, что экспрессия TIGIT высоко коррелировала с CD8α, но не с CD4, или только умеренно коррелировала с CD4, на основании чего можно предположить, что в первую очередь TIGIT регулирует функцию CD8+ TIL (CD8α, R2 = 0,80; CD4, R2 = 0,42; фиг. 6C).
С учетом коэкспрессии TIGIT и PD-1 при хронической вирусной инфекции, также оценивали корреляцию PD-1 и других ингибирующих корецепторов с TIGIT при раке молочной железы. Корреляция между TIGIT и PD-1, CTLA4 и LAG3 была очень высокой (PD-1, R2 = 0,87; CTLA4, R2 = 0,76; LAG3, R2 = 0,80; фиг. 6D). В совокупности полученные данные свидетельствуют о том, что TIGIT экспрессировался TIL, в частности, CD8+ Т-клетками, и что он может подавлять их функцию.
Пример 5: TIGIT и PD-1 ингибируют противоопухолевые ответы Т-клеток
Для того чтобы лучше охарактеризовать TIGIT из клеток TIL у мышей, мышам BALB/с прививали клетки колоректальной карциномы СТ26. Вкратце, мышам BALB/с подкожно прививали 1×105 клеток карциномы ободочной кишки СТ26, суспендированных в матригеле (BD Biosciences) с правой стороны грудной клетки. Через две недели мышей, несущих опухоли размером примерно 200 мм3, произвольно разделяли на опытные группы согласно обработкам, которым внутрибрюшинно вводили 35 мг/кг изотипически сходных контрольных антител, анти-PD-L1-антител и/или анти-TIGIT-антител 3 раза в неделю в течение 3 недель. Размер опухолей определяли 2 раза в неделю с использованием штангенциркуля. Животные, у которых опухоли изъязвлялись/некротизировались, или их размер превышал 2000 мм3, подвергались эвтаназии. Спленоциты и инфильтрирующие опухоль лимфоциты (TIL) анализировали через 14 суток после прививки, когда опухоли достигали размера примерно 200 мм3.
Согласуясь с данными по экспрессии TIGIT в опухолях человека (фиг. 6), CD8+ и CD4+ CT26 TIL экспрессировали TIGIT на высоком уровне (фиг. 7А-7В). Кроме того, согласуясь с данными опытов с хронической вирусной инфекции, экспрессия TIGIT клетками TIL тесно коррелировала с экспрессией других ингибирующих корецепторов, включая PD-1 (фиг. 7A-7B) и Tim-3 (фиг. 8). Подобный паттерн экспрессии TIGIT был обнаружен в карциноме ободочной кишки МС38 (фиг. 9).
Для тестирования физиологического значения экспрессии TIGIT в контексте противоопухолевого иммунного ответа, мышей BALB/с с развившимися опухолями СТ26 (размером примерно 200 мм3) обрабатывали 200 мкг изотипически сходных контрольных антител, 200 мкг анти-PD-L1-антител, 500 мкг анти-TIGIT-антител или 200 мкг анти-PD1-антител+ 500 анти-TIGIT-антител в течение 3 недель.
Рост опухолей CT26 незначительно замедлялся при обработке анти-TIGIT-антителами или анти-PD-L1-антителами по отдельности, оба которых показали незначительное повышение медианы выживаемости на 3 сутки (фиг. 7С-7D). Однако комбинированная терапия обоими анти-PD-L1-антителом и анти-TIGIT-антителом приводила к резкому снижению роста опухолей (снижение на 75% среднего объема опухолей на 16 сутки, P<0,0001, фиг. 7С и фиг. 10). Кроме того, у 70% мышей, получавших комбинацию анти-TIGIT-антитела и анти-PD-L1-антитела, отмечали полную и продолжительную ремиссию опухолей, и они оставались живыми в течение всего опытного периода, даже в отсутствие дополнительной обработки антителами (фиг. 7С-7D). Эти эффекты также наблюдали у мышей с опухолями, обработанными комбинацией блокирующих антител против TIGIT и PD-1.
Для исследования иммунитета к опухолевым клеткам СТ26 у этих выживших мышей, примерно через 60 суток после первоначальной прививки, мышам с состоянием полной ремиссии (CR), которые получали анти-TIGIT-антитела+ анти-PD-L1-антитела, а также нативным мышам BALB/с, повторно прививали клетки СТ26 с левой стороны грудной клетки (куда ранее прививка не проводилась). Этим мышам также прививали 1×105 клеток карциномы молочной железы EMT6 в матригеле в жировую подушку молочной железы. Размер опухолей определяли 2 раза в неделю. Животные, у которых опухоли изъязвлялись/некротизировались, или их размер превышал 2000 мм3, подвергались эвтаназии.
Как показано на фигуре 7E, оба вида опухолей интенсивно росли у нативных контрольных мышей, но опухоли EMT6 росли только у мышей, которые ранее освободились от опухолей CT26. Эти результаты показывают, что совместное блокирование TIGIT и PD-1 во время онкогенеза обеспечивало состояние специфического иммунитета к опухолевым клеткам СТ26.
Для определения того, опосредована ли эффективность совместного блокирования TIGIT/PD-L1 CD8+ Т-клетками, мышей с опухолями СТ26 подвергали истощению CD8+ Т-клеток с использованием истощающих антител в начале обработки анти-TIGIT-антителом и анти-PDL1-антителом. У мышей, обработанных анти-TIGIT-антителом и анти-PD-L1-антителом, отсутствовало отторжение клеток опухоли СТ26, когда имело место истощение CD8+ Т-клеток в начале обработки (повышение среднего объема опухолей на 1532% через 17 суток после обработки, Р = 0,0004, фиг. 32А-32В). Кроме того, истощение CD8+ Т-клеток нарушало способность ранее обработанных мышей с CR контролировать повторную прививку опухоли CT26 (фиг. 32С). В совокупности полученные результаты показывают, что анти-TIGIT-антитело и анти-PD-L1-антитело функционировали через CD8+ Т-клетки с индукцией эффективных первичных и вторичных противоопухолевых иммунных ответов.
Для определения того, насколько экспрессия PVR опухолевыми клетками не является обязательной для эффективности совместного блокирования TIGIT/PD-L1, мышам BALB/с дикого типа прививали опухоли СТ26 дикого типа (которые экспрессируют PVR) или PVR-дефицитные опухоли СТ26. Вкратце, опухолевые клетки СТ26 дикого типа транзиентно трансфектировали нуклеиновой кислотой, которая снижала экспрессию PVR. Примерно через две недели после трансфекции клетки СТ26 субклонировали на основе потери экспрессии PVR, с использованием проточной цитометрии и количественной ПЦР. Когда опухоли достигали размера 150-200 мм3, мышей обрабатывали анти-TIGIT-антителом и анти-PD-L1-антителом, или изотипически сходных контрольных антител. Мыши, обработанные анти-TIGIT-антителом и анти-PD-L1-антителом, были способны к отторжению опухолей дикого типа и PVR-дефицитных опухолей, в отличие от мышей с привитыми опухолями, которых обработали контрольными антителами (фиг. 33). Полученные результаты показали, что анти-TIGIT-антитело и анти-PD-L1-антитело функционируют независимо от PVR, экспрессированного в опухоли.
Также была тестирована и подтверждена эффективность совместного блокирования TIGIT/PD-L1 на модели опухоли МС38. Мышам C57BL6/J дикого типа подкожно прививали сингенные клетки колоректальной карциномы МС38 и обрабатывали развившиеся опухоли комбинацией блокирующих антител против TIGIT и PD-L1, как описано выше. В отличие от модели CT26, обработка одним анти-PD-L1-антителом была достаточной для индукции у некоторых мышей полного ответа (фиг. 25). Однако, как и на модели CT26, обработка мышей с опухолями МС38 обоими анти-TIGIT-антителом и анти-PD-L1-антителом синергически предупреждала рост опухолей и индуцировала элиминацию опухолей у большинства мышей (фиг. 26). Эти эффекты наблюдали также у мышей, которым прививали сингенные клетки карциномы молочной железы EMT6 (фиг. 34).
Полученные результаты показали, что совместное блокирование TIGIT и PD-1 могло индуцировать стойкий и антиген-специфический противоопухолевый иммунный ответ. На основании этих результатов также можно предположить, что адаптивные противоопухолевые ответы были полностью функциональными и реактивировались у обработанных мышей.
Для оценки функциональных эффектов блокирования TIGIT и PD-1 на сами инфильтрирующие опухоль лимфоциты, мышам прививали опухолевые клетки СТ26 и обрабатывали анти-TIGIT-антителом и/или анти-PD-L1-антителом, как описано выше. Через семь суток после начала обработки отбирали опухоли и дренирующие опухоль лимфатические узлы для анализа проточной цитометрией.
Инфильтрирующие опухоль и резидентные CD4+ Т-клетки в дренирующих опухоль лимфатических узлах продуцировали IFNγ на низком уровне, и не продуцировали этот цитокин при блокировании TIGIT/PD-1 (фиг. 11). Однако, инфильтрирующие опухоль CD8+ Т-клетки от мышей, обработанных как анти-TIGIT-антителом, так и анти-PD-L1-антителом, но не клетки от мышей, обработанных по отдельности анти-TIGIT-антителом или анти-PD-L1-антителом, были достоверно более компетентными в отношении продукции IFNγ при стимуляции in vitro (повышение на 174% по сравнению с контролем, Р = 0,0001, фиг. 13D). Аналогичные результаты были получены для CD8+ TIL в отношении продукции TNFγ (фиг. 12).
Интересно отметить, что у мышей, обработанных только анти-TIGIT-антителом или анти-PD-L1-антителом, или обоими антителами, повышалась компетентность в отношении продукции цитокинов резидентными CD8+ Т-клетками в дренирующих опухоль лимфатических узлах (повышение на 75-113%, P<0,001, фиг. 13), на основании чего можно предположить, что резидентные CD8+ Т-клетки в дренирующих опухоль лимфатических узлах подвергались ингибированию в меньшей степени, чем их инфильтрующие опухоль аналоги. Накопление и фенотипическая активации инфильтрирующих опухоль и резидентных CD8+ Т-клеток и CD4+ Т-клеток в дренирующих опухоль лимфатических узлах не изменялись и незначительно повышались после обработки одним антителом и обработки двумя антителами соответственно (фиг. 12-13). Частота встречаемости CD8+ Т-клеток, продуцирующих одновременно IFNγ/TNFα, в опухолях и дренирующих опухоль лимфатических узлах, имела аналогичные паттерны (фиг. 35А-35В).
Следовательно, в то время как блокирование одним TIGIT или одним PD-L1 было достаточным для усиления эффекторной функции CD8+ Т-клеток в дренирующих опухоль лимфатических узлах, блокирование обоих рецепторов необходимо для восстановления функции истощенных CD8+ Т-клеток в самой опухоли, что согласуется с представлением о том, что опухолевая микросреда является высоко иммуносупрессивной.
Пример 6: коэкспрессия TIGIT с CD226 CD8+ Т-клетками, инфильтрирующими опухоль
TIGIT конкурирует с костимулирующим рецептором CD226 за связывание с рецептором полиовируса (PVR) (Yu, X., et al. The surface protein TIGIT suppresses T cell activation by promoting the generation of mature immunoregulatory dendritic cells, Nature immunology, 10, 48-57 (2009)). С учетом того, что дефицит CD226 может усилить истощение Т-клеток при хронической вирусной инфекции (Cella, M., et al. Loss of DNAM-1 contributes to CD8+ T-cell exhaustion in chronic HIV-1 infection, European Journal of Immunology 40(4), 949-954 (2010); Welch, M., et al., CD8 T cell defect of TNA-a and IL-2 in DNAM-1 deficient mice delays clearance in vivo of a persistent virus infection, Virology, 429(2), 163-170 (2012)), то возможно, что TIGIT может ингибировать Т-клеточные ответы в отношении влияния на активность CD226.
Для оценки того, существует ли взаимосвязь между CD226 и TIGIT в ингибировании Т-клеточных ответов, определяли экспрессию TIGIT и CD226 на инфильтрирующих опухоль CD8+ Т-клетках.
Как показано на фигуре 14, мышам C57BL6/J прививали клетки колоректальной карциномы МС38. Спленоциты и инфильтрирующие опухоль лимфоциты (TIL) анализировали с использованием анализа FACS примерно через 14 суток после прививки, когда опухоли достигали размера примерно 200 мм3. Репрезентативная гистограмма экспрессии CD226 В-клетками селезенки (серый цвет), CD8+ Т-клетками селезенки (синий цвет) и TIGIT+ CD8+ Т-клетками, инфильтрирующими опухоль (красный цвет). Данные представляют результаты двух независимых опытов; n = 5. На фигуре 14 показано, что CD8+ Т-клетки селезенки экспрессировали CD226 на высоком уровне и TIGIT+ CD8+ Т-клетки, инфильтрирующие опухоль, также экспрессировали CD226 на высоком уровне. Данные показывают, что TIGIT и CD226 координировано экспрессируются на мышиных CD8+ Т-клетках, инфильтрирующих опухоль, и могут регулировать функцию друг друга на CD8+ Т-клетках. Это наблюдение аналогично тому, что имеет место в активированных CD8+ Т-клетках и NK-клетках, которые также коэкспрессируют TIGIT и CD226.
Пример 7: коиммунопреципитация TIGIT с CD226 на трансфектированных клетках
Для определения того, насколько TIGIT взаимодействует с CD226 на поверхности клеток, клетки котрансфектировали с человеческим TIGIT и человеческим CD226, и подвергали иммунопреципитации. Вкратце, клетки COS 7 на чашках размером 15 см котрансфектировали экспрессионными плазмидами, содержащими кДНК меченных белков TIGIT-HA (5 нг) или CD226-Flag (10 нг), или контрольной плазмидой (pRK). Через 23 ч после трансфекции клетки промывали PBS и собирали в 4 мл охлажденного на льду PBS, и центрифугировали при 300×g в течение 5 мин и клеточные осадки ресуспендировали в 2 мл лизирующего буфера при 4°С. Клетки лизировали в течение 50 мин с перемешиванием на вортексе каждые 15 мин и затем центрифугировали при 10000×g в течение 15 мин при 4°С. Полученный супернатант предварительно осветляли с использованием 160 мкл суспензии сефарозы CL6B при вращении в течение 30 мин при 4°С и центрифугировали в течение 2 мин при 3000×g. Супернатант разделяли поровну на две пробирки и иммунопреципитировали с анти-НА или анти-flag с использованием стандартных процедур. Иммуннопреципитированные белки подвергали SDS-PAGE и вестерн-блоттингу. Вестерн-блоты зондировали анти-Flag-HRP или анти-НА-HRP.
Как показано на фигуре 15, анти-TIGIT-антитело преципитировало CD226 и анти-CD226-антитело преципитировало TIGIT, демонстрируя, что TIGIT и CD226 находятся в физическом контакте на поверхности клеток.
Пример 8: TIGIT и CD226 взаимодействуют на CD8+ Т-клетках
В дополнение к демонстрации способности CD226 и TIGIT взаимодействовать в трансфектированных клетках, также оценивали взаимодействие CD226 и TIGIT в первичных CD8+ Т-клетках. Вкратце, CD8+ Т-клетки селезенки мышей C57BL6/J, обогащенные с использованием MACS, стимулировали фиксированными на планшетах анти-CD3- и анти-CD28-антителами и рекомбинантным IL-2 в течение 48 ч и лизировали. Клеточные лизаты подвергали иммунопреципитации с анти-TIGIT-антителом и зондировали анти-CD226-антителом. На фигуре 16 показано, что TIGIT и CD226 взаимодействуют в активированных первичных CD8+ клетках, поскольку оба были обнаружены в коиммунопреципитате. Полученные данные показывают, что CD226 и TIGIT также взаимодействуют друг с другом на первичных клетках.
Пример 9: взаимодействие TIGIT/CD226 на трансфектированных клетках с использованием для анализа TR-FRET (времяразрешенного флуоресцентного индуктивно-резонансного переноса энергии)
Для оценки того, существует ли молекулярное взаимодействие между TIGIT и CD226, использовали методологию TR-FRET. FRET (флуоресцентный индуктивно-резонансный перенос энергии) основан на передаче энергии между двумя флуорофорами, донором и акцептором, когда они находятся в непосредственной близости друг от друга. Молекулярные взаимодействия между биомолекулами можно оценить связыванием каждого партнера с флуоресцентной меткой и определением уровня передачи энергии. Когда два соединения подходят достаточно близко друг к другу, то возбуждение донора источником энергии «запускает» перенос энергии к акцептору, который в свою очередь испускает специфическую флуоресценции при определенной длине волны. За счет этих спектральных свойств донорно-акцепторный комплекс может быть детектирован без необходимости в физическом разделении от несвязанных партнеров. Комбинация времяразрешенного измерения FRET (TR) позволяет очистить сигнал от фоновой флуоресценции. Обычно это осуществляется введением временной задержки между возбуждением системы и измерением флуоресценции, чтобы сигнал очистился от всех неспецифических короткоживущих эмиссий.
С использованием TR-FRET было показано, что TIGIT и CD226 индуцировали FRET, когда экспрессировались в одной той же клетке, что свидетельствует о молекулярном взаимодействии этих двух молекул. Вкратце, клетки COS-7 трансфектировали SNAP-меченным (ST) CD226 и HA-TIGIT с использованием липофектамина 2000 (Life Technologies) и высевали в белый 96-луночный планшет (Costar) из расчета 100000 клеток на лунку. Через 24 ч клетки метили 100 нМ конъюгированного с донором бензилгуанин SNAP-Lumi-4Tb (Cisbio) и 1 мкМ конъюгированного с донором бензилгуанин SNAP-А647 (New England Biolabs), разведенных в DMEM с 10% FCS, в течение 1 ч при 37°С, в атмосфере 5% СО2. После трех промываний PBS сигнал FRET регистрировали при 665 нм в течение 400 мкс после задержки 60 мкс после лазерного возбуждения при 343 нм с использованием ридера для планшетов Safire2 (Tecan). Когда тестировали анти-TIGIT-антитело, то сигнал FRET также регистрировали через 15 мин инкубации. Отношение FRET рассчитывали в виде интенсивности FRET, деленной на эмиссию донора при 620 нм. Интенсивность FRET представляла: (сигнал при 665 нм от клеток, меченных SNAP-донором и акцептором) - (сигнал при 665 нм от той же партии трансфектированных клеток, меченных только SNAP-донором).
Как показано на фигуре 17 TIGIT был способен непосредственно разрушать и вызывать диссоциацию гомодимеров CD226. Как показано на фигуре 17А, диссоциацию гомодимеров Flag-SТ-CD226 наблюдали с увеличением концентраций HA-TIGIT, что показано снижением соотношения Flag-SТ-CD226, измеренного на клетках COS-7, экспрессирующих постоянное количество Flag-SТ-CD226 и возрастающие концентрации HA-TIGIT. Однако, как показано на фигуре 17b, когда анти-TIGIT-антитело вносили в культуру клеток, то это блокировало способность TIGIT и CD226 к ассоциации. Данный факт иллюстрируется отсутствием снижения интенсивности гомодимеров Flag-SТ-CD226. Это показывает, что CD226 и TIGIT связаны в виде комплексов, но что анти-TIGIT-антитела могут нарушить эти взаимодействия (фиг. 17A и 17B).
С использованием методологии TR-FRET также была показана способность TIGIT к ассоциации с CD226, и это показано на фиг. 17C и 17D. Вкратце, после мечения SNAP-меткой с использованием 1 мкМ конъюгированного с донором Flag-SТ-А647 (смотри выше), клетки промывали три раза PBS и инкубировали с 2 нМ анти-НА конъюгированного с донором Lumi-4Tb (Cisbio), разведенного в PBS+ 0,2% BSA, в течение 2 ч при комнатной температуре. Регистрировали сигнал FRET, интенсивность FRET представляла: (сигнал при 665 нм от клеток, меченных SNAP-акцептором и анти-НА донором) - (сигнал при 665 нм от той же партии трансфектированных клеток, меченных только SNAP-акцептором и анти-НА донором).
Как показано на фигуре 17C, наблюдали ассоциацию Flag-SТ-CD226 с HA-TIGIT, о чем свидетельствует возрастающая интенсивность FRET между Flag-SТ-CD226 и HA-TIGIT, измеренная на клетках COS-7, экспрессирующих постоянное количество Flag-SТ-CD226 и возрастающие концентрации HA-TIGIT. При добавлении анти-TIGIT-антитела интенсивность FRET между Flag-SТ-CD226 и HA-TIGIT снижалась, как показано на фигуре 17D, на основании чего можно предположить, что взаимодействие TIGIT с CD226 может быть блокировано блокирующим анти-TIGIT-антителом.
Для подтверждения экспрессии на клеточной поверхности Flag-SТ-CD226 и HA-TIGIT в опытах с использованием методологии FRET, ставили ELISA с анти-Flag-антителом и анти-HA-антителом на интактных клетках COS-7, экспрессирующих указанные меченные конструкции. Вкратце, клетки COS7 фиксировали в 4% параформальдегиде, дважды промывали и блокировали забуференным фосфатом физиологическим раствором+1% фетальной телячьей сыворотки (FCS). Затем клетки инкубировали с моноклональным анти-HA-антителом (клон 3F10, Roche Applied Science) или моноклональным анти-Flag-М2-антителом (Sigma), и конъюгировали с пероксидазой хрена. После промывки клетки инкубировали с субстратом ELISA SuperSignal (Pierce) и измеряли хемилюминесценцию на ридере для планшетов Safire2 (Tecan). Специфический сигнал рассчитывали вычитанием сигнала, зарегистрированного для контрольных трансфектированных клеток. Как показано на фигуре 18, была подтверждена экспрессия CD226 и TIGIT на клеточной поверхности в анализе ELISA.
Для подтверждения того, что взаимодействие TIGIT:CD226 не было обусловлено связыванием с PVR, получали Flag-SТ-CD226 и HA-TIGIT (WT) или HA-TIGIT Q56R, как описано в публикации Stengel et al., (2012) PNAS, 109(14):5399-5904, и определяли отношения FRET, как описано. Как показано на фигуре 24, WT TIGIT и Q56R TIGIT связывали CD226 с одинаковой эффективностью.
Полученные данные не только показывают, что CD226 и TIGIT ассоциированы в виде комплексов, но и что анти-TIGIT-антитело может нарушить эти взаимодействия и что взаимодействие TIGIT:CD226 не обусловлено связыванием с PVR. Данные подтверждают роль TIGIT в ограничении CD226-опосредованной активации Т-клеток, и свидетельствуют о том, что влияние на активность CD226 может быть важным механизмом действия, посредством которого TIGIT ингибирует Т-клеточные ответы и активность.
Пример 10: блокирование CD226 отменяет эффекты блокирования TIGIT/PD-L1 in vivo
Для исследования физиологического значения взаимодействия CD226 и TIGIT у мышей воспроизводили хроническую инфекцию заражением клоном 13 LCMV и затем животных обрабатывали анти-TIGIT-+ анти-PD-L1-антителами в отсутствие или в присутствии блокирующих анти-CD226-антител. Вкратце, у мышей C57BL6/J быстро истощали CD4+ Т-клетки и заражали клоном 13 штамма LCMV. Для воспроизведения хронической инфекции мышей заражали внутривенно 2×106 БОЕ клона 13 LCMV и обрабатывали 500 мкг и 250 мкг истощающих анти-CD4-антител (клон GK1.5) в течение 3 суток до и в течение 4 суток после заражения соответственно. Там, где это было показано, мышам, зараженным клоном 13 штамма LCMV, внутрибрюшинно вводили 200 мкг изотипически сходных контрольных антител, 500 мкг анти-CD226-антитела, 200 мкг анти-PD-L1-антитела+500 мкг анти-TIGIT-антитела, или 500 мкг анти-CD226-антитела+200 мкг анти-PD-L1-антитела+500 мкг анти-TIGIT-антитела 3 раза в неделю, начиная с 28 по 42 сутки после заражения. Обработку начинали на 28 сутки после заражения, поскольку Т-клеточный ответ преимущественно был истощен на данной временной точке на данной модели хронической вирусной инфекции ((Wherry et al., Molecular Signature of CD8+ T cell Exhaustion During Chronic Viral Infection, Immunity, 2007 Oct;27(4):670-84). Титры вируса в спленоцитах и печени определяли через 42 суток после заражения.
У этих мышей обработка только анти-CD226-антителом оказывала ограниченное влияние на частоту встречаемости, активацию или компетентность в отношении продукции цитокинов CD8+ Т-клеток (фиг. 19А-19С). Однако обработка анти-CD226-антителом существенно отменяла усиление активации CD8+ Т-клеток и продукции IFNg, наблюдаемые у мышей, получавших анти-PD-L1-антитела+анти-TIGIT-антитела (снижение соответственно на 59% и 58%, P <0,001; фиг. 19B-19D).
Согласуясь с этими результатами, вирусная нагрузка LCMV была достоверно выше у мышей, получавших анти-CD226-антитело+ анти-PD-L1-антитело+ анти-TIGIT-антитело, чем у мышей, обработанных только анти-PD-L1-антителом+анти-TIGIT-антителом (повышение на 272%, Р <0,001, фиг. 19D).
На основании полученных данных можно предположить, что основной механизм, посредством которого TIGIT ограничивает хронические Т-клеточные ответы, это интерференция с CD226-опосредованной костимуляцией. Данные показывают ранее неизвестную роль TIGIT во взаимодействии с CD226 и нарушении димеризации CD226, приводя к снижению или потере ключевого костимулирующего сигнала на CD8+ Т-клетках. Данные показывают, что интерференция с CD226-опосредованной Т-клеточной костимуляцией может быть основным механизмом, посредством которого TIGIT ограничивает хронические Т-клеточные ответы, какие имеются при раке или хронической вирусной инфекции. Полученные данные также определяют основной параметр для анти-TIGIT-антител, предназначенных для восстановления эффекторной функции хронически стимулированных или истощенных CD8+илиCD4+ Т-клеток посредством влияния на способность TIGIT к взаимодействию с CD226 и/или способность TIGIT к нарушению димеризации CD226.
Материалы и методы
Мыши. Мышей C57BL/6J и BALB/с получали из питомников Jackson Laboratory и Charles River Laboratories. Мышей CD4cre и мышей TIGITloxP/LoxP получали на основе мышей C57BL/6J с использованием стандартных методов и перекрестных скрещиваний. Экспрессия TIGIT Т-клетками у мышей TIGITloxP/loxPCD4cre была отменена с эффективностью 96%.
Проточная цитометрия. Получали суспензии отдельных клеток селезенки, лимфатических узлов и опухоли осторожным механическим разрушением. Поверхностное окрашивание проводили с использованием коммерчески доступных антител к CD4, CD8, CD44, CD62L, PD-1 (eBiosciences) и CD226 (BioLegend). Антитела к TIGIT получали в Genentech, как описано ранее (Yu, X. et al. The surface protein TIGIT suppresses T cell activation by promoting the generation of mature immunoregulatory dendritic cells, Nature immunology 10, 48-57 (2009))) и конъюгировали с Alexa Fluor 647 согласно инструкциям изготовителя (Molecular Probes).
Для внутриклеточного окрашивания цитокинов (ICS) клетки стимулировали в течение 4 ч с 20 нг/мл форбол 12-миристат-13-ацетата (РМА, Sigma) и 1 мкМ иономицина (Sigma) в присутствии 3 мкг/мл брефелдина А (eBiosciences). После стимуляции клетки окрашивали на поверхностные маркеры, как описано, и фиксировали и пермеабилизовали буфером для фиксации FoxP3 (eBioscience), согласно указаниям изготовителя. Фиксированные клетки окрашивали антителами к TNFγ и IFNα (eBiosciences).
Блокирующие антитела. Блокирующее моноклональное IgG2a анти-TIGIT-антитело (клон 10А7, реагирующее на мышиный и человеческий TIGIT) получали, как описано ранее, и клонировали в мышиный IgG2a остов. Блокирующее моноклональное IgG2a анти-PD-L1-антитело (клон 25A1) получали иммунизацией мышей PDL1-/- гибридным белком PD-1-Fc и клонировали в мышиный IgG2a остов. Клон 25A1 модифицировали ранее описанными мутациями, отменяющими связывание с Fcγ-рецепторами. Блокирующее моноклональное IgG2a анти-CD226-антитело (клон 37F6) получали иммунизацией хомяков рекомбинантным мышиным CD226 и клонировали в мышиный IgG2a остов. Эти антитела также использовали в тестах, описанных в других примерах, приведенных здесь.
Вирусные инфекции. Для воспроизведения острой инфекции мышей заражали внутривенно 2×106 бляшкообразующих единиц (БОЕ) штамма Armstrong LCMV. Для воспроизведения хронической инфекции мышей внутривенно заражали 2×106 БОЕ клона 13 штамма LCMV и обрабатывали 500 мкг и 250 мкг истощающих анти-CD4-антител (клон GK1.5) в течение 3 суток до и 4 суток после заражения соответственно. Там, где это было показано, мышам, зараженным клоном 13 штамма LCMV, внутрибрюшинно вводили 200 мкг изотипически сходных контрольных антител, 200 мкг анти-PD-L1-антител и/или 500 мкг анти-TIGIT-антител 3 раза в неделю в период с 28 по 42 сутки после заражения.
Анализ титра вируса. Монослои клеток MC57 культивировали с нанесением поверх 1% метилцеллюлозы и инфицировали серийными разведениями гомогената печени от зараженных LCMV мышей. Через 72 ч после заражения клетки фиксировали в 4% параформальдегиде и пермеабилизовали 0,5% Triton-X. Вирусные бляшки окрашивали анти- LCMV NP (клон VL-4) и HRP-конъюгированным анти-крысиным IgG и визуализировали о-фенилендиамином (OPD, Sigma).
Биоинформатика. Данные по экспрессии генов злокачественной опухоли молочной железы с использованием микрочипового анализа получали из проекта Атлас ракового генома (CGAN) (Network, C.G.A. Comprehensive molecular portraits of human breast tumours, Nature, 490, 61-70 (2012)). Обработку и нормализацию данных микрочипов проводили с использованием R языка программирования (http://r-project.org) и пакета limma Bioconductor (http://bioconductor.org). Значения интенсивности биологических микрочипов из каждого канала предварительно обрабатывали с использованием метода поправок на основе нормального+ экспоненциального распределения, как описано ранее в ссылке 22. Затем скорректированные значения интенсивности нормализовали с использованием квантильной нормализации, как описано ранее в ссылке 23. Нормализованные log-значения рассчитывали вычитанием референтного канала из опытного канала для каждого микрочипа. Данные были дополнительно профильтрованы с использованием неспецифического фильтра, как описано ранее в ссылке 24, удалением зондов, которые не картируют известные гены, и уменьшением базы данных до зависимости «один зонд на ген». Для дифференциального анализа экспрессии модерированные t-статистики рассчитывали с использованием пакета limma, как описано ранее (Smyth G.K., Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Statistical applications in genetics and molecular biology, 3, Article3 (2004)). Для оценки корреляции использовали коэффициенты корреляции Пирсона.
Карцинома ободочной кишки CT26. Мышам BALB/с подкожно прививали 1×105клеток карциномы ободочной кишки СТ26, суспендированных в матригеле (BD Biosciences) с правой стороны грудной клетки. Через две недели мышей, несущих опухоли размером примерно 200 мм3, произвольно распределяли на опытные группы согласно обработкам, которым внутрибрюшинно вводили 35 мг/кг изотипически сходных контрольных антител, анти-PD-L1-антител и/или анти-TIGIT-антител 3 раза в неделю в течение 3 недель. Размер опухолей определяли 2 раза в неделю с использованием штангенциркуля. Животные, у которых опухоли изъязвлялись/некротизировались, или их размер превышал 2000 мм3, подвергались эвтаназии.
Карцинома молочной железы EMT6. Мышам BALB/с подкожно прививали в жировую подушку четвертой молочной железы 1×105 сингенных клеток карциномы молочной железы EMT6 в матригеле (BD Biosciences). Через две недели мышей, несущих опухоли размером 150-200 мм3, произвольно распределяли на опытные группы согласно обработкам, которым внутрибрюшинно вводили 35 мг/кг изотипически сходных контрольных антител, анти-PD-L1-антител и/или анти-TIGIT-антител 3 раза в неделю в течение 3 недель. Размер опухолей определяли 2 раза в неделю с использованием штангенциркуля, и объем опухолей рассчитывали с помощью модифицированной формулы для эллисоидов, 1/2 × (длина × ширина2). Животные, у которых размер опухолей уменьшался до 32 мм3 или менее, рассматривались в качестве животных с полным ответом (CR). Животные, у которых размер опухолей увеличивался более чем до 2000 мм3, рассматривались в качестве животных, у которых заболевание прогрессировало, и их подвергали эвтаназии. Животные, у которых опухоли изъязвлялись/некротизировались до прогрессирования заболевания или полного ответа, подвергались эвтаназии и исключались из опыта.
Повторная прививка CT26. Там, где это было показано, мышам, BALB/с мышей, которым ранее прививали клетки карциномы ободочной кишки СТ26, как описано выше, повторно прививали клетки СТ26 с левой стороны грудной клетки (куда ранее прививку не проводили). Этим мышам также прививали 1×105 клеток карциномы молочной железы EMT6 в матригеле в жировую подушку четвертой молочной железы. Размер опухолей измеряли 2 раза в неделю. Животные, у которых опухоли изъязвлялись/некротизировались, или их размер превышал 2000 мм3, подвергались эвтаназии.
Статистика. Статистические тесты проводили с использованием непарного (парного, где указано) двустороннего теста Стьюдента. Вертикальные линии показывают стандартное отклонение от среднего.
Контроль за проведением исследований на животных. Все исследования на животных были одобрены Институциональным комитетом по контролю за уходом за животными и их использованию Genentech's.
Пример 11: экспрессия TIGIT повышена при раке человека и, его экспрессия коррелирует с экспрессией CD8 и PD-1 и CD8+Т-клеточной инфильтрацией
Материалы и методы
Биоинформатика. Обработку и анализ данных по секвенированию РНК проводили с использованием R языка программирования (http://www.r-project.org) вместе с несколькими пакетами из проекта Bioconductor (http://www.bioconductor.org). Данные секвенирования РНК для злокачественных опухолей и соответствующих нормальных образцов тканей получали из TCGA по пяти различным показаниям: рак молочной железы (Network, C.G.A. Comprehensive molecular portraits of human breast tumours, Nature, 490, 61-70 (2012)), аденокарцинома ободочной кишки ((Network, T.C.G.A. Comprehensive molecular characterization of human colon and rectal cancer, Nature, 487, 330-337 (2012)), светлоклеточная карцинома почек (Network, C.G.A. Comprehensive molecular characterization of clear cell renal cell carcinoma, Nature, 499, 43-49 (2013)), плоскоклеточная карцинома легких (Network, T.C.G.A. Comprehensive genomic characterization of squamous cell lung cancers, Nature, 489, 519-525 (2012)) и карцинома эндометрия (Network, T.C.G.A. Integrated genomic characterization of endometrial carcinoma, Nature, 497, 67-73 (2012)).
Необработаные данные по РНК-seq обрабатывали с использованием пакета HTSeqGenie Bioconductor. Вкратце, данные выравнивали с геномом человека (NCBI build 37), используя алгоритм GSNAP (Wu T.D. & Nacu S, Fast and SNP-tolerant detection of complex variants and splicing in short reads. Bioinformatics (Oxford, England) 26, 873-881 (2010)). Уникально выравненные пары данных, которые попадали в экзоны, рассчитывали для оценки уровня экспрессии генов для отдельных генов. Использовали значение размера библиотек из пакета edgeR (Robinson M.D., McCarthy D.J. & Smyth G.K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics (Oxford, England) 26, 139-140 (2010)) для нормализации различных образцов по отношению к их соответствующим глубинам секвенирования.
Для получения генной сигнатуры, специфической для Т-клеток, вручную курировали гены Т-клеток, идентифицированные в рамках проекта IRIS, удалением генов, ассоциированных с процессами клеточного цикла, генов с высокой экспрессией в других тканях и известных коактивирующих и коингибирующих рецепторов. Это давало сигнатуру из 15 генов, специфичных для Т-клеток. Для расчета сигнатурного балла экспрессии генов в T-клетках в данных по плоскоклеточной карциноме легких, заявители впервые провели преобразование, стабилизирующее дисперсию, на необработанных расчетных данных с использованием voom функции из пакета limma Bioconductor. Затем рассчитывали первый собственный вектор центрированных и масштабных данных со стабилизированной дисперсией по 15-генной сигнатуре Т-клеток. Такой подход обеспечивает надежную оценку относительной представленности в Т-клетках. Затем для каждого гена была подобрана линейная модель, включающая сигнатурный балл Т-клеток, вновь с использованием пакета limma. Затем гены ранжировали их соотношением с Т-клеточной сигнатурой в полученной линейной модели, выбирая только гены, которые положительно коррелировали с Т-клеточной сигнатурой. Для визуализации генов, ассоциированных с Т-клетками, в виде тепловой карты, центрировали и масштабировали данные со стабилизированной дисперсией к единичной дисперсии, что позволило сравнить гены с разными средними уровнями экспрессии.
Для определения корреляции между экспрессией TIGIT и других генов заявители нормализовали расчетные данные по секвенированию РНК с учетом различий в размере библиотеки, используя метод из edgeR пакета Bioconductor (Robinson M.D., McCarthy D.J. & Smyth G.K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data, Bioinformatics (Oxford, England) 26, 139-140 (2010)). Затем рассчитали коэффициент ранговой корреляции Спирмена по нормализованным значениям. Считали, что rpo >0,75 свидетельствует о наличии высокой корреляции, Ро ≤ 0,75, но >0,5 указывает на наличие умеренной корреляции и rpo ≤ 0,5, но >0,25 свидетельствует о наличии низкой корреляции.
Для расчета отношений TIGIT/CD3ε по каждому показанию вначале рассчитывали данные со стабилизированной дисперсией для каждого ряда данных по секвенирования РНК. Затем рассчитывали отношение log2 данных со стабилизированной дисперсией для TIGIT и CD3ε. Для расчета разницы между опухолью и нормальными образцами провели стандартный линейной анализ с использованием стандартных R-функций. Р-значение <0,01 принимали в качестве доказательства наличия статистически значимого различия между опухолью и нормальной тканью.
Для идентификации генов, ассоциированных с инфльтрирующими опухоль Т-клетками, использовали подход, основанный на сигнатуре генов, с запросом данных по экспрессии генов из проекта Атлас ракового генома (TCGA) по коллекции плоскоклеточной карциномы (LUSC) (Network, T.C.G.A. Comprehensive genomic characterization of squamous cell lung cancers, Nature 489, 519-525 (2012)). С использованием ряда генов, специфичных для иммунных клеток, определенных в проекте Immune Response In Silico (Abbas, A.R. et al. Immune response in silico (IRIS): immune-specific genes identified from a compendium of microarray expression data. Genes and immunity 6, 319-331 (2005))), а также методов, описанных выше, разработали высокоспецифичную сигнатуру из 15 генов. При исследовании генов, наиболее ассоциированных с Т-клеточной сигнатурой, идентифицировали несколько коингибирующих рецепторов, ранее ассоциированных с Т-клеточной дисфункцией при опухолях, в частности, PD-1 (фиг. 21). В LUSC экспрессия TIGIT и CD3ε высоко коррелировала с коэффициентом ранговой корреляции Спирмена (ρ равен 0,82 (фиг. 20А)). Фактически экспрессия TIGIT и CD3ε также высоко коррелировала во многих дополнительных базах данных по экспрессии генов опухолей TCGA, включая аденокарциному ободочной кишки (COAD), эндометриоидную карциному тела матки (UCEC), карциному молочной железы (BRCA) и светлоклеточную карциному почек (KIRC) со значением ρ в диапазоне от 0,83 до 0,94 (фиг. 20B-20E). Кроме того, экспрессия TIGIT была выше по сравнению с экспрессией CD3ε во многих образцах опухолей, с повышенным отношением TIGIT/CD3ε в образцах опухолей LUSC, COAC, UCEC и BRCA по сравнению с соответствующими нормальными тканями (повышение на 116%-419%, фиг. 20A-20D). Отношение экспрессии TIGIT к CD3ε в образцах KIRC не изменялось, хотя, экспрессия TIGIT и CD3ε была существенно выше в образцах KIRC, чем в образцах нормальных ткани (фиг. 20Е). Эти данные указывают, что экспрессия TIGIT была повышена в лимфоцитах, инфильтрирующих опухоль (TIL) в широком ряду солидных опухолей.
TIGIT был ранее описан в качестве ингибитора праймирования CD4+ Т-клеток, его функция для CD8+ Т-клеток оставалась не известной. Однако экспрессия TIGIT в образцах LUSC высоко коррелировала с CD8A и слабо коррелировала с CD4 (ρ = 0,77 и 0,48 соответственно; фиг. 20F). Также экспрессия TIGIT коррелировала с экспрессией его дополнительного костимулирующего рецептора, CD226, а также с экспрессией PD-1, ключевого медиатора супрессии Т-клеток в опухолях и при других хронических иммунных ответах (ρ = 0,64 и 0,82 соответственно; фиг. 20G-20H). Хотя, эти гены имеются в некоторых клетках, отличных от лимфоцитов, полученные данные убедительно показывают, что Т-клетки, инфильтрирующие опухоли, в частности, «истощенные» CD8+ Т-клетки, экспрессировали TIGIT на высоких уровнях.
Пример 12: TIGIT и PD-1 координировано экспрессируются человеческими и мышиными лимфоцитами, инфильтрирующими опухоли
Материалы и методы
Образцы опухоли человека и PBMC. Соответствующую цельную кровь и только что хирургически удаленные опухолевые ткани получали из Conversant Biosciences или Foundation Bio. Все образцы получали с письменного информированного согласия и отбирали, используя протокол, одобренный Наблюдательным советом госпиталя Хартфорд (IRB) (пациент 1 с NSCLC, данные по нему представлены на фигуре 22) или Западным институциональным наблюдательным советом (пациент 2 с NSCLC и пациент 1 с CRC, данные по ним представлены на фигуре 23 и фигуре 37). Цельную кровь здорового взрослого человека получали от здорового добровольца. РВМС выделяли из цельной крови центрифугированием в градиенте Ficoll. Опухолевые ткани разрезали на небольшие кусочки и инкубировали с коллагеназой и ДНКазой (Roche) и клетки разделяли с помощью аппарата GentleMACS Disassociator (Miltenyi).
Проточная цитометрия. Получали суспензии мышиных отдельных клеток селезенки, лимфатических узлов и опухоли осторожным механическим разрушением. Поверхностное окрашивание проводили с использованием коммерчески доступных антител к CD4, CD8, CD44, CD62L, PD-1 (eBiosciences) и CD226 (BioLegend). Антитела к TIGIT получали в Genentech и конъюгировали с Alexa Fluor 647 согласно инструкциям изготовителя (Molecular Probes).
Для внутриклеточного окрашивания цитокинов (ICS) клетки стимулировали в течение 4 ч с 20 нг/мл форбол 12-миристат-13-ацетата (РМА, Sigma) и 1 мкМ иономицина (Sigma) в присутствии 3 мкг/мл брефелдина А (eBiosciences). После стимуляции клетки окрашивали на поверхностные маркеры, как описано, и фиксировали и пермеабилизовали буфером для фиксации FoxP3 (eBioscience), согласно указаниям изготовителя. Фиксированные клетки окрашивали антителами к TNFγ и IFNα (eBiosciences).
Образцы опухолей человека и РВМС готовили, как описано выше. Поверхностное окрашивание проводили с помощью витального красителя (Molecular Probes), коммерческих антител к CD45 (eBiosciences), CD3, CD4, CD8, PD-1 (BD Biosciences), и анти- TIGIT-антител, полученных, как описано выше.
Все образцы исследовали на приборах LSR-II или LSR-Fortessa (BD Biosciences) и анализировали с использованием программного обеспечения FlowJo (Treestar).
Для подтверждения положительной регуляции TIGIT в Т-клетках, инфильтрирующих опухоли, оценивали экспрессию белка TIGIT на Т-клетках, инфильтрирующих немелкоклеточную карциному легких человека, соответствующих периферических Т-клетках и периферических Т-клетках от здорового донора. TIGIT на клеточной поверхности экспрессировался субпопуляциями CD8+ и CD4+ Т-клеток, инфильтрирующих NSCLC (51% и 39% соответственно, фиг. 22А-22В). На фигуре 36 также показано, что TIGIT клеточной поверхности экспрессировался большим процентом CD8+ и CD4+ Т-клеток, инфильтрирующих NSCLC (количество TIGIT+клеток соответственно 58% и 28%, фиг. 36A-36B). Интересно отметить, что периферические CD8+ и CD4+ Т-клетки от донора с опухолью NSCLC также экспрессировали TIGIT на более высоких уровнях по сравнению с клетками от здоровых доноров (фиг. 22A-22В и фиг. 36A-36B). Аналогичные результаты были получены со второй коллекцией соответствующих образцов NSCLC и РВМС и коллекцией соответствующих образцов колоректальной карциномы (CRC) и РВМС (фиг. 23 и фиг. 37). Почти все Т-клетки, инфильтрирующие опухоль, экспрессирующие высокие уровни TIGIT, коэкспрессировали PD-1, что согласуется с корреляцией между экспрессией TIGIT и PD-1, показанной на фигуре 1 (фиг. 22С).
Для расширения наблюдений на доклинических моделях рака человека охарактеризовали экспрессию TIGIT Т-клетками, инфильтрирующими подкожные колоректальные опухоли CT26 и МС38, у мышей BALB/с дикого типа и мышей C57BL6/J соответственно. Через две недели после прививки, когда опухоли CT26 и МС38 развились и достигли размера 150-200 мм3, TIGIT экспрессировался примерно 50% CD8+ Т-клетками, инфильтрующими опухоль, и 25% CD4+ Т-клетками, инфильтрующими опухоль, на уровнях, аналогичных таковым для первичных CD8+ Т-клеток, стимулированных in vitro (фиг. 22D-22E и фиг. 26). На обоих видах мышиных CD8+ и CD4+ TIL CD226 экспрессировался конститутивно, и экспрессия TIGIT и PD-1 вновь тесно коррелировала (фиг. 22F-22G).
Полученные результаты подтвердили, что TIGIT экспрессировался на высоком уровне Т-клетками, инфильтрирующими опухоль, и что экспрессия TIGIT происходила параллельно с экспрессией других коингибирующих рецепторов, в первую очередь PD-1.
Пример 13: супрессия под действием TIGIT CD8+ Т-клеточных ответов зависит от CD226
В отличие от PD-1 или CTLA-4 отсутствует прямое биохимическое доказательство наличия Т-клеточного ингибирующего сигнального каскада, инициированного TIGIT in cis. Однако коингибирующие рецепторы также могут функционировать посредством снижения активности комплементарного костимулирующего рецептора, например, подавлением сигнального пути CD28 под действием CTLA-4. Установив, что TIGIT является отрицательным регулятором инфильтрирующих опухоль и противовирусных CD8+ Т-клеток, заявители задались вопросом, индуцирует ли TIGIT истощение Т-клеток опосредованно через подавление комплементарного костимулирующего рецептора, CD226, который экспрессируется на высоком уровне периферическими и инфильтрирующими опухоль CD8+ T-клетками (фиг. 27).
Мышей BALB/с дикого типа, несущих опухоли СТ26 размером 150-200 мм3, обрабатывали комбинацией анти-PD-L1-антитела и анти-TIGIT-антитела в присутствии или отсутствие блокирующего анти-CD226-антитела, или одним анти-CD226-антителом. Обработка одним анти-CD226-антителом несколько ускоряла рост опухолей по сравнению с контрольными мышами, приводя к снижению медианы выживаемости на 2 суток (одно анти-CD226-антитело по сравнению с контролем, P = 0,0118, фиг. 28A-28B). Удивительно, но дополнительное введение блокирующих анти-CD226-антител мышам, обработанным совместным блокированием анти-TIGIT-антителом и анти-PD-L1-антителом, приводила к существенно усилению роста опухолей и полностью восстанавливала эффективность совместного блокирования TIGIT/PD-L1 в отношении регрессии опухоли и выживаемости (фиг. 28A-28B). Аналогичный эффект наблюдали на титрах LCMV у мышей с хронической вирусной инфекцией, получавших анти-TIGIT-антитело, анти-PD-L1-антитело и/или анти-CD226-антитело (фиг. 19D). Полученные данные указывали на то, что CD226 способствовал проявлению противоопухолевого и других хронических ответов Т-клеток, и что TIGIT подавлял эти ответы, по меньшей мере, частично супрессией CD226.
Для более полного понимания того, каким образом активность TIGIT и CD226 влияет на противоопухолевые Т-клеточные ответы, тестировали как один CD226, так и в комбинации с TIGIT, влияет на активацию, инфильтрацию опухоли и эффекторной функцию Т-клеток. Анализировали опухоли и дренирующие опухоль лимфатические узлы от мышей с опухолями СТ26, обработанных как описано выше, в течение семи суток. Как было установлено ранее, совместное блокирование PD-L1 и TIGIT повышало продукцию IFNγ как инфильтрирующими опухоль CD8+ T-клетками, так и CD8+ T-клетками дренирующих опухоль лимфатических узлов (повышение на 130% и 99% соответственно, P <0,001, фиг. 28C-28D). Блокирование одного CD226 не оказывало никакого эффекта на продукцию IFNγ инфильтрирующими опухоль CD8+ T-клетками и резидентными CD8+ T-клетками дренирующих опухоль лимфатических узлов, на основании чего можно предположить, что эффекты костимуляции CD226 уже ингибированы в истощенных Т-клетках (фиг. 28C-28E). Однако блокирование CD226 снижало частоту встречаемости и эффекторную функцию инфильтрирующих опухоль CD8+ T-клеток у мышей, обработанных комбинацией анти-TIGIT-антитела и анти-PD-L1-антитела (снижение на 57%, Р = 0,0015, фиг. 28D). Обработка анти-CD226-антителом не оказывала такого эффекта на CD8+ T-клетки, находящиеся в дренирующих опухоль лимфатических узлах, в то время как одно анти-PD-L1-антитело повышало эффекторную функцию CD8+ T-клеток, на основании чего можно предположить, что блокирование PD-L1 было достаточным для повышения эффекторной функции CD8+ T-клеток даже в отсутствие CD226. Блокирование CD226 также приводила к снижению часоты встречаемости инфильтрирующих опухоль CD8+ T-клеток (снижение на 53%, Р = 0,0044, фиг. 28Е-28F). В совокупности полученные данные свидетельствуют о том, что CD226 функционирует для поддержания как накопления, так и эффекторной функции инфильтрирующих опухоль CD8+ T-клеток, и что TIGIT препятствует последнему.
Пример 14: TIGIT ослабляет функцию CD226, непосредственно нарушая гомодимеризацию CD226
Для тестирования того, насколько TIGIT может подавлять активность CD226 in cis, исследовали влияние TIGIT на костимуляцию CD226 in vitro. TIGIT-дефицитные CD8+ T-клетки, стимулированные субоптимальными концентрациями анти-CD3-антитела, сильнее отвечали на костимуляцию PVR, чем аналогичные CD8+ T-клетки дикого типа, и такая усиленная реакция зависела от CD226 (повышение пролиферации на 46%, P = 0,0061 фиг. 29A). Согласуясь с этими данными, CD8+ T-клетки дикого типа, стимулированные субоптимальными концентрациями анти-CD3-антитела и PVR, пролиферировали в большей степени в присутствии анти-TIGIT-антител, чем это имело место в присутствии изопипически сходных контрольных антител, и данный эффект зависел от CD226 (повышение пролиферации на 105%, P = 0,0010, фиг. 29B).
Для того чтобы исследовать значение TIGIT для первичных CD8+ T-клеток человека, выделяли CD8+ T-клетки из крови здоровых доноров и стимулировали их субоптимальными концентрациями фиксированного на планшете анти-CD3-антитела и рекомбинантным гибридным человеческим белком PVR-Fc. В присутствии изопипически сходных контрольных антител костимуляция PVR умеренно повышала стимуляцию и пролиферацию Т-клеток. Кроме того, добавление блокирующих анти-TIGIT-антител достоверно повышало эффекты костимуляции PVR, что согласуется с эффектами TIGIT на первичные мышиные CD8+ T-клетки (повышение пролиферации на 69%, P = 0,0071, фиг. 29C). Полученные данные продемонстрировали внутриклеточную роль в ингибировании под действием TIGIT функции CD226 на первичных мышиных и человеческих CD8+ T-клетках.
Использовали технологию TR-FRET (времяразрешенный флуоресцентный индуктивно-резонансный перенос энергии) для определения молекулярного механизма, посредством которого TIGIT нарушал активность CD226. Вначале экспрессировали и метили человеческий ST-CD226 непостоянными донорным и акцепторным флуорофорами. Эти клетки давали сильный сигнал FRET, что подтверждает способность CD226 к гомодимеризации (фиг. 29D). Для исследования взаимодействий CD226 и TIGIT на поверхности клеток ST-CD226 экспрессировали в отсутствии или в присутствии человеческого HA-TIGIT, меченного субстратом SNAP-tag и анти-НА-антителом соответственно. Удивительно, но коэкспрессия возрастающих количеств TIGIT (контролировали ELISA) ослабляла сигнал FRET CD226/CD226, указывая на то, что TIGIT мог нарушить гомодимеризацию CD226 (фиг. 29Е). Действительно, акцептор CD226 и донор TIGIT также приводили к воспроизведению высокого сигнала FRET, что указывает на прямое взаимодействие между этими двумя белками (фиг. 29F). Это взаимодействие дополнительно подтверждалось коиммунопреципитацией (фиг. 29G). Полученные данные показали, что TIGIT и CD226 непосредственно взаимодействуют на клеточной поверхности, и это взаимодействие может нарушить гомодимеризацию CD226.
Для исследования эффектов блокирования антителами TIGIT взаимодействия TIGIT-CD226, вновь коэкспрессировали человеческие ST-CD226 и HA-TIGIT, на этот раз в присутствии или отсутствии блокирующих антител к человеческому TIGIT. Внесение анти-TIGIT-антител в клеточные культуры достоверно снижало способность TIGIT и CD226 к ассоциации (рис. 29Н). Полученные данные дают основание предположить, что обработка анти-TIGIT-антителом может ограничить взаимодействие TIGIT с CD226, и этот факт согласуется с наблюдением о том, что подавление активности CD226 является ключевым механизмом действия, посредством которого TIGIT приводит к истощению CD8+ T-клеток. Это также согласуется со способностью анти-TIGIT-антител усиливать костимуляцию CD226.
Затем подтверждали способность эндогенных TIGIT и CD226 к взаимодействию (фиг. 30). Первичные человеческие Т-клетки стимулировали in vitro анти-CD3- и анти-CD28-антителами, сортированными на основе экспрессии TIGIT, выдерживали, повторно стимулировали и метили антителами против эндогенных TIGIT и CD226, которые были конъюгированы с флуорофорами, совместимыми с TR-FRET. Т-клетки, экспрессирующие TIGIT, меченные конъюгированными с донором анти-TIGIT-антителами и конъюгированными с акцептором анти-CD226-антителами, давали сильный сигнал FRET (фиг. 30). В противоположность детектировали только низкий сигнал FRET на Т-клетках, которые не экспрессировали TIGIT или которые были мечены конъюгированными с донором анти-TIGIT-антителами и конъюгированными с акцептором анти-HVEM-антителами (фиг. 30), что подтверждает специфический характер обнаруженного взаимодействия между эндогенными TIGIT и CD226.
Полученные результаты показывают, что эндогенные TIGIT и CD226 могут непосредственно взаимодействовать на клеточной поверхности, и что это взаимодействие нарушает гомодимеризацию CD226. С учетом роли CD226 в качестве костимулятора Т-клеточных ответов in vivo, и, не желая связываться с теорией, полагается, что супрессия CD226 может быть ключевым механизмом действия, посредством которого TIGIT вызывает истощение CD8+ T-клеток при хронической вирусной инфекции и раке.
Материалы и методы
Времяразрешенный флуоресцентный индуктивно-резонансный перенос энергии с трансфектированными клеточными линиями. СНО-клетки трансфектировали CD226 N-концевой SNAP-tag (ST) и N-концевым HA-TIGIT с помощью липофектамина 2000 (Life Technologies) и высевали в белый 96-луночный планшет (Costar) из расчета 100000 клеток на лунку. Через 24 ч клетки метили для измерения TR-FRET между SNAP-донор/SNAP-акцептор или между SNAP-акцептор/анти-HA-донор. 1) Мечение SNAP-донор/SNAP-акцептор: клетки инкубировали с 100 нМ конъюгированного с донором бензилгуанина SNAP-Lumi-4Tb (Cisbio) и 1 мкМ конъюгированного с акцептором бензилгуанин SNAP-A647 (New England Biolabs), разведенного в DMEM с 10% FCS, в течение 1 ч при 37°С, в атмосфере 5% СО2. Затем клетки промывали три раза PBS перед измерением FRET сигнала. 2) SNAP-акцептор/анти-НА-донор: клетки инкубировали с 1 мкМ конъюгированного с акцептором бензилгуанин SNAP-A647, разведенного в DMEM с 10% FCS в течение 1 ч при 37°С, в атмосфере 5% СО2. После трех промываний PBS клетки инкубировали в течение 2 ч с 2 нМ анти-HA Lumi-4Tb (Cisbio) в PBS+ 0,2% BSA при комнатной температуре. Сигнал FRET регистрировали при 665 нм в течение 400 мкс после задержки 60 мкс после лазерного возбуждения при 343 нм с использованием ридера для планшетов Safire2 (Tecan). Когда анти-TIGIT-антитело тестировали в конечной концентрации 10 мкг/мл, то сигнал FRET также регистрировали после 15 мин инкубации. При исследовании взаимодействия Flag-ST-CD226/Flag-ST-CD226 рассчитывали отношение FRET как интенсивность FRET деленную на эмиссию донора при 620 нм, которое пропорционально экспрессии CD226. Интенсивность FRET представляла: (сигнал при 665 нм от клеток, меченных SNAP-донором и акцептором) - (сигнал при 665 нм от той же партии трансфектированных клеток, меченных только SNAP-донором). При исследовании взаимодействия Flag-ST-CD226/HA-TIGIT отношение FRET представляет интенсивность FRET деленную на экспрессию Flag-ST-CD226, что определяли ELISA с анти-Flag-антителом. В этом случае интенсивность FRET представляла: (сигнал при 665 нм от клеток, меченных SNAP-донор и анти-НА-донор) - (сигнал при 665 нм от контрольных трансфектированных клеток, меченных SNAP-акцептор и анти-НА-донор).
Времяразрешенный флуоресцентный индуктивно-резонансный перенос энергии с человеческими Т-клетками. Человеческие анти-TIGIT-антитела (Genentech клон 1F4), анти-CD226-антитела (Santa Cruz Biotechnology) и анти-HVEM-антитела (eBioscience) конъюгировали с флуорофорами, совместимыми с TR-FRET (Cisbio). Первичные Т-клетки человека, выделенные из крови с использованием MACS, стимулировали in vitro фиксированными на планшетах анти-CD3- и анти-CD28-антителами в течение 72 ч. Затем экспрессирующие и не-экспрессирующие TIGIT Т-клетки (все экспрессирующие CD226) подвергали сортингу, выдерживали без стимуляции в течение 72 ч и повторно стимулировали в течение 48 ч. Затем каждую популяцию один раз промывали Трис-буфером Кребса (20 мМ Трис, рН 7,4, 118 мМ NaCl, 5,6 мМ глюкозы, 1,2 мМ KH2PO4, 1,2 мМ MgSO4, 4,7 мМ KCl, 1,8 мМ CaCl2) и культивировали в следующих условиях, в трех повторностях: 1) анти-TIGIT Ab-Lumi4-Tb (5 мкг/мл), 2) анти-TIGIT Ab-Lumi4-Tb (5 мкг/мл)+ анти-HVEM-d2 (10 мкг/мл), 3) анти- TIGIT Ab-Lumi4-Tb (5 мкг/мл)+ анти-CD226 (10 мкг/мл), 4) анти-TIGIT Ab-Lumi4-Tb (5 мкг/мл)+ анти-CD226 (10 мкг/мл)+ немеченные анти-TIGIT Ab (клон 1F4) (50 мкг/мл). Указанные концентрации были оптимизированы для гарантии получения наиболее высокого сигнал FRET. Клетки инкубировали в течение 2 ч при комнатной температуре на ротаторе и затем промывали 3 раза Трис-буфером Кребса. Затем Т-клетки высевали из расчета 400000 клеток/лунку в белом 96-луночном планшете (Costar) и сигнал TR-FRET регистрировали при 665 нм в течение 400 мкс после 60 мкс задержки после лазерного возбуждения при 343 нм с использованием ридера для планшетов PHERAstar (BMG Labtech). FRET интенсивность выражали в виде сигнала при 665 нм от клеток, меченных Аб-Lumi4-Tb+ Ab-d2 минус сигнал при 665 нм от той же партии клеток, меченных только Ab-Lumi4-Tb. Неспецифический сигнал FRET давали Т-клетки, инкубированные с Lumi4Tb+ d2+ избыток немеченных Ab.
Коиммунопреципитация. Вкратце, клетки COS 7 на чашках размером 15 см котрансфектировали экспрессионными плазмидами, содержащими кДНК для меченных белков TIGIT-HA (5 нг) или CD226-Flag (10 нг), или контрольной плазмидой (pRK). Через 23 ч после трансфекции клетки промывали PBS с и собирали в 4 мл охлажденного на льду PBS, и центрифугировали при 300×g в течение 5 мин и клеточные осадки ресуспендировали в 2 мл лизирующего буфера при 4°С. Клетки лизировали в течение 50 мин с перемешиванием на вортексе каждые 15 мин и затем центрифугировали при 10000×g в течение 15 мин при 4°С. Полученный супернатант предварительно осветляли с использованием 160 мкл суспензии сефарозы CL6B при вращении в течение 30 мин при 4°С и центрифугировали в течение 2 мин при 3000×g. Супернатант разделяли поровну на две пробирки и иммунопреципитировали с анти-НА или анти-flag с использованием стандартных процедур. Иммуннопреципитированные белки подвергали SDS-PAGE и вестерн-блоттингу. Вестерн-блоты зондировали анти-Flag-HRP или анти-НА-HRP.
Все патенты, заявки на патент, документы и статьи, цитированные в настоящем описании, включены здесь во всей их полноте в качестве ссылки.
Реферат
Группа изобретений относится к медицине и может быть использована для лечения или замедления прогрессирования рака у индивидуума. Для этого индивидууму вводят эффективное количество антагониста анти-PD-L1-антитела, или его антигенсвязывающего фрагмента и антагониста анти-TIGIT-антитела, или его антигенсвязывающего фрагмента. Также предложен способ снижения или ингибирования рецидива рака у индивидуума. Группа изобретений обеспечивает снижение роста опухоли и индукцию элиминации опухоли за счет синергетического действия антагонистов анти-PD-L1-антитела и анти-TIGIT-антитела. 2 н. и 14 з.п. ф-лы, 37 ил., 14 пр.
Комментарии