Продуцирующие антитела млекопитающие, не являющиеся человеком - RU2559524C2
Код документа: RU2559524C2
Чертежи






































































Описание
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Изобретение относится к получению и применению не являющихся человеком животных, способных продуцировать антитела и их производные, которые экспрессируются по меньшей мере частично, из экзогенных нуклеиновых кислот (трансгенов). Описаны трансгены для продуцирования и способы получения таких гетерологичных антител; также описаны способы и векторы для получения таких трансгенных животных.
УРОВЕНЬ ТЕХНИКИ
Гуморальный иммунитет опосредуется B-клетками путем продуцирования специфических антител. Основной структурной субъединицей антитела (Ab) является молекула иммуноглобулина (Ig). Молекулы Ig состоят из комплекса двух идентичных тяжелых (H) и двух идентичных легких (L) полипептидных цепей. На аминном конце каждой H-цепи и L-цепи находится участок с изменяющейся аминокислотной последовательностью, называемый вариабельной (V) областью. Остальная часть H- и L-цепей сравнительно постоянна по своей аминокислотной последовательности, и называется константной (C) областью. В молекуле Ig V-области H- и L-цепей (VH и VL) накладываются друг на друга с образованием потенциального участка связывания антигена. Гены, которые кодируют V-области H- и L-цепей, собираются соматически из сегментов ДНК зародышевой линии во время дифференциации предшественника B-клетки (pre-B): сегменты гена V, D и J для H-цепи и сегменты гена V и J для L-цепи. Среди V-участков Ig имеется три участка с наибольшей изменчивостью аминокислотной последовательности, которые взаимодействуют с образованием участка распознавания антигена, и поэтому называются гипервариабельными участками, отвечающими за комплементарное взаимодействие с антигеном (CDR).
Сегмент гена V кодирует основной объем домена V-участка, включая CDR1 и CDR2. Вариабельность в CDR1 и CDR2 происходит из гетерогенности последовательности для множества различных закодированных в зародышевой линии сегментов V. CDR3 кодируется последовательностями, которые образуются путем соединения сегментов гена V, D и J H-цепи и сегментов гена V и J L-цепи и посредством механизмов, которые создают гетерогенность нуклеотидной последовательности при соединении данных сегментов. Дополнительная вариабельность может происходить из соединения различных V-областей H и L-цепей. Вместе данные процессы производят иммунный репертуар антител, кодируемых сегментами генов зародышевой линии, и экспрессируемых вновь образовавшимися B-клетками.
Дополнительный источник вариабельности антител налагается поверх вариабельности, сгенерированной рекомбинацией сегментов генов Ig. B-клетки имеют возможность внесения мутаций в V-области антитела, которое они экспрессируют, посредством процесса, называемого соматической сверхмутацией. Соответственно, когда животному впервые встречается антиген, антиген связывается со специфической B-клеткой, которая оказалась носителем антител, имеющих V-область, связывающую антиген. Такой первичный ответ может активировать данную B-клетку для продолжения секретирования родственного антитела. Данные активированные B-клетки также могут направлять процесс соматической мутации для своих перестроенных сегментов генов антитела, что позволяет осуществлять продуцирование дочерних клеток, которые создают варианты антител первичного ответа. В процессе выбора распространяются те вариантные потомки B-клетки, которые создают антитело с повышенной аффинностью антигена. В B-клетках соматические сверхмутации нацелены на ограниченный участок генома, содержащий перестроенные гена для VH и VL. Таким образом, соматическая мутация позволяет осуществлять созревание аффинности - продуцирование и выбор высокоаффинных антител. Следовательно, соматическая мутация является важной для генерации высокоаффинных антител.
Сильная специфичность и высокая аффинность антител и изобретение технологии гибридом, позволяющей генерировать моноклональные антитела (mAb), дало большие надежды на их использование в качестве адресного терапевтического средства для болезней человека. MAb идентичны, поскольку они продуцируются единичной B-клеткой и ее потомством. MAb создаются путем слияния клеток селезенки мыши, которая была иммунизирована необходимым антигеном, с клетками миеломы с целью создания иммортализованных гибридом. Одним из главных препятствий для разработки in vivo приложений для mAb у человека является иммуногенность, свойственная нечеловеческим Ig. Пациенты отвечают на терапевтические дозы mAb мыши посредством создания антител против последовательностей Ig мыши (антимышиные антитела человека; HAMA), вызывающих острую токсичность, изменения их биораспределения и ускорения выведения, что в результате снижает эффективность последовательных введений (Mirick, et al., (2004) Q. Nucl. Med. Mol. Imaging 48, 251-257).
С той целью, чтобы обойти создание HAMA, были разработаны методы гуманизации антител в попытке продуцирования mAb с пониженной иммуногенностью при применении у человека. Результатом таких попыток стали различные подходы, основанные на рекомбинантных ДНК и направленные на повышение содержания аминокислотных последовательностей человека в mAb при условии сохранения специфичности и аффинности родительского антитела, не принадлежащего человеку. Гуманизация началась с создания химерных антител мыши-человека (Morrison, S. L., et al., (1984). Proc. Natl. Acad. Sci. USA., 81, 6851-5), в которых С-области Ig в mAb мыши были заменены С-областями человека. Химерные mAb содержат 60-70% аминокислотных последовательностей человека и имеют значительно меньшую иммуногенность, чем их аналоги мыши, при инъецировании человеку, хотя ответ человека против химерного антитела все еще наблюдался (Hwang, W. Y., et al. (2005). Methods, 36, 3-10).
В попытках дальнейшей гуманизации mAb мыши была разработана прививка CDR. При прививке CDR антитела мыши гуманизируются путем прививки их CDR в VL- и VH-каркасы молекул Ig человека при сохранении остатков каркаса мыши, считающихся важными для специфичности и аффинности (Jones, P.T., et al., (1986). Nature, 321, 522). В целом, антитела с привитыми CDR состоят более чем на 80% из аминокислотных последовательностей человека (Queen, C. et al. (1989) Proc. Natl. Acad. Sci. U. S. A. 86, 10029; Carter, P. et al. (1992) Proc. Natl. Acad. Sci. U. S. A. 89, 4285). Несмотря на эти попытки, было показано, что гуманизированные антитела все еще вызывали образование антител против привитой V-области (Hwang, W. Y., et al. (2005). Methods, 36, 3).
После прививки CDR были разработаны методы гуманизации, основанные на различных парадигмах, таких как ремоделирование (Padlan, E. A., et al., (1991). Mol. Immunol., 28, 489), сверхгуманизация (Tan, P., D. A., et al., (2002) J. Immunol., 169, 1119), оптимизация содержания последовательностей человека (Lazar, G. A., et al., (2007). Mol. Immunol., 44, 1986) и гуманиринг (humaneering), в попытке дальнейшего снижения содержания последовательностей, не принадлежащих человеку, в терапевтических mAb (Almagro, J. C, et al., (2008). Frontiers in Bioscience 13, 1619). Так же как и подходы, основанные на прививке CDR, данные методы опираются на анализ структуры антитела и сравнение последовательности нечеловеческих и человеческих mAb с целью оценки влияния процесса гуманизации на иммуногенность конечного продукта. При сравнении иммуногенности химерных и гуманизированных антител оказывается, что гуманизация вариабельных областей еще больше снижает иммуногенность (Hwang, W. Y., et al. (2005). Methods, 36, 3-10).
Еще одним подходом, разработанным для снижения иммуногенности химерных антител или антител мыши, является деиммунизация. Она включает идентификацию линейных эпитопов T-клеток в исследуемом антителе, с применением биоинформатики, и их последующую замену посредством сайт-направленного мутагенеза на последовательности человека или неиммуногенные последовательности (WO09852976A1). Несмотря на то, что деиммунизированные антитела продемонстрировали сниженную иммуногенность в приматах, по сравнению с их химерными аналогами, наблюдалась некоторая потеря связывающей способности (Jain, M., et al., (2007). Trends in Biotechnol. 25, 307).
Разработка технологии фагового дисплея дополнила и расширила подходы к иммунизации в попытке получения менее иммуногенных mAb для терапии людей. В фаговом дисплее большие наборы («библиотеки») VH- и VL-областей антител человека экспрессируются на поверхности нитеобразных частиц бактериофага. Из этих библиотек выбираются редкие фаги с помощью связывающего взаимодействия с антигеном; растворимые фрагменты антител экспрессируются зараженными бактериями, и связывающая способность выбранных антител улучшается посредством мутации (Winter, G., et al. (1994). Annu. Rev. Immunol. 12, 433). Данный процесс имитирует иммунный отбор, и с использованием данного подхода были выделены антитела со многими различными специфичностями связывания (Hoogenboom, H. R., et al. (2005). Nat. Biotechnol., 23, 1105). Различные источники V-области H- и L-цепей были использованы для построения фагового дисплея, включая выделение из неиммунных или иммунных доноров. Кроме того, библиотеки фагового дисплея были сформированы из V-областей, которые содержали искусственно рандомизированные синтетические CDR-области, с целью создания дополнительного разнообразия. Часто антитела, полученные из библиотек фагового дисплея, подвергаются созреванию аффинности in vitro с целью получения высокоаффинных антител (Hoogenboom, H. R., et al. (2005). Nat. Biotechnol., 23, 1105).
Создание трансгенных линий мыши, продуцирующих антитела человека при отсутствии антител мыши, обеспечило создание еще одной технологической платформы для создания специфических и высокоаффинных mAb мыши для применения в людях. У таких трансгенных животных эндогенный механизм продуцирования антител инактивирован и заменен локусами Ig человека с целью, по существу, воспроизведения гуморальной иммунной системы человека в мышах (Jakobovits, A., et al. (2007). Nat. Biotechnol. 25, 1134. Lonberg, N. (2005). Nat. Biotechnol. 23, 1117). Развитие B-клеток, а также диверсификация Ig посредством рекомбинации сегментов генов корректно воспроизводится в этих мышах, в результате чего получается широкий репертуар B-клеток мыши, экспрессирующих Ig человека. Посредством иммунизации этих мышей антигенами было дополнительно продемонстрировано, что данные трансгенные животные накопили соматические мутации в V-областях как тяжелой, так и легкой цепей, достаточные для продукции широкого разнообразия высокоаффинных mAb человека (Lonberg, N. (2005). Nat. Biotechnol. 23, 1117).
На вопрос, являются ли «полностью человеческие» mAb, такие как полученные из библиотек фагового дисплея или трансгенных мышей, менее иммуногенными, чем гуманизированные mAb, пока еще нет ответа, поскольку полные данные по иммуногенности доступны только для двух mAb человека. mAb против фактора некроза опухоли, разработанное на основе библиотек фагового дисплея человека, вызывает иммунный ответ у 12% пациентов - на верхней границе уровня противоантительных ответов гуманизированных антител (Hwang, W. Y., et al. (2005). Methods, 36, 3-10).
Оценка иммуногенности первого зарегистрированного mAb человека, созданного посредством трансгенного подхода, показала, что обработка с помощью mAb привела к образованию антител примерно у 5,5% подвергавшихся обработке раковых пациентов (Jakobovits, A., et al. (2007). Nat. Biotechnol. 25, 1134., Lofgren, J. A., et al. (2007). J. Immunol. 178, 7467).
Следовательно, по-прежнему остается необходимость в способе и средствах получения антител, являющихся специфичными для своих мишеней, но менее иммуногенных. В соответствии с изобретением, снижение иммуногенности по меньшей мере частично достигается путем получения не являющегося человеком трансгенного млекопитающего, содержащего по меньшей мере в клеточной линии своих B-клеток, нуклеиновую кислоту, кодирующую по меньшей мере легкую цепь и тяжелую цепь иммуноглобулина, при этом кодирующая тяжелую или легкую цепь последовательность предоставлена вместе со средствами, которые делают ее устойчивой к перестройкам ДНК и/или к соматическим гипермутациям, при этом предпочтительное не являющееся человеком млекопитающее является грызуном, конкретнее, мышью. Нуклеиновая кислота предпочтительно кодирует цепь человеческого, подобного человеческому или гуманизированного иммуноглобулина.
Остальное описание относится к примерам, в которых обычно в качестве не являющихся человеком млекопитающих используются мыши. Трансгенные не являющиеся человеком млекопитающие-хозяева способны индуцировать иммунный ответ на антиген, при этом ответ заключается в продукции антител, имеющих вариабельные области примата, в частности, человека. Могут быть использованы различные трансгенные хозяева, в частности, мыши и крысы, зайцеобразные, овечьи, жвачные, свиноподобные, лошадиные, псовые, кошачьи и т.п. Мыши были использованы для продукции B-лимфоцитов для иммортализации в целях продуцирования антитела. Поскольку с мышами легко работать, они могут быть разведены в большом количестве, и о них известно, что они имеют широкий иммунный репертуар, то мыши будут обычно являться выбираемыми животными. Следовательно, в приведенном ниже обсуждении будут рассматриваться мыши, но следует понимать, что другие животные, в частности, не являющиеся приматами млекопитающие, могут быть с легкостью использованы вместо мышей при следовании тем же процедурам.
Причина предотвращения перестроек и гипермутации состоит в том, что таким образом неиммуногенный полипептид может быть выбран заблаговременно, поскольку будет известно, что его полипептидная цепь останется неиммуногенной. По меньшей мере одна из цепей полученного в результате иммуноглобулина будет, таким образом, менее иммуногенной. Полученное в результате антитело должно (обычно) иметь и легкую, и тяжелую цепь. Следовательно, неиммуногенная цепь должна быть способна к соединению с другой цепью. Другая цепь может являться эндогенной цепью, экзогенной цепью или гибридом обеих. Для терапии человека неиммуногенная цепь должна быть близка к цепи человека насколько это возможно.
Средства получения гена, кодирующего цепь (или цепи) иммуноглобулина, устойчивую(-ые) к перестройке и/или мутации ДНК, состоят, очевидно, в удалении всех генетических элементов, ответственных за вышеупомянутую перестройку и/или мутацию. Их недостаток заключается в том, что устраняется вариабельность двух цепей, тогда как в изобретении предпочтительно сохранение вариабельности в одной из цепей (предпочтительно, в тяжелой цепи), и подавляется и/или предотвращается перестройка-мутация другой цепи (предпочтительно, легкой цепи).
Элементы для перестройки и/или гипермутации, охарактеризованные ранее, размещаются в пределах локусов для иммуноглобулинов. Следовательно, средство получения кодирующей иммуноглобулин последовательности, устойчивой к перестройке и/или мутации ДНК, состоит во вставке гена в локус вне локусов иммуноглобулина.
Таким образом, изобретение относится к трансгенному не являющемуся человеком млекопитающему, у которого последовательность, кодирующая легкую/тяжелую цепь, интегрирована в геном не являющегося человеком млекопитающего в локусе вне локусов иммуноглобулина. Предпочтительно, вставка производится в локусе, устойчивом к сайленсингу генов. В соответствии с изобретением, интеграция происходит в локусе Rosa или сопоставимом локусе.
Предпочтительным является экспрессионная кассета, которая может быть вставлена в локус Rosa со средствами, допускающими экспрессию цепи(-ей) иммуноглобулина, в основном ограниченную клетками линии B-клеток, предпочтительно, со средствами, которые допускают экспрессию легкой цепи, кодирующей нуклеиновую кислоту, в течение определенной стадии развития B-клетки. Термин «по существу ограниченная экспрессия» означает, что экспрессия происходит преимущественно в клетках клеточной линии B-клеток, но при этом допускается возможность более низких уровней экспрессии, по сравнению с экспрессией в B-клетках, в других клетках. В предпочтительном варианте осуществления термин «по существу ограниченная экспрессия» означает, что экспрессия имеет место исключительно в клетках из клеточной линии B-клеток. Такие средства обычно и предпочтительно включают специфические промоторы B-клетки (стадия развития), такие как CD19, CD20, μHC (все V-гены), VpreB1, VpreB2, VpreB3, λ5, Igα, Igβ, κLC (все гены), λLC (все гены), BSAP (Pax5). Несмотря на то, что имеется хорошая возможность управления экспрессией цепи, устойчивой к перестройкам и/или мутациям ДНК, посредством таких промоторов, они являются относительно слабыми. Сильный промотор обычно будет требоваться для обеспечения адекватной поверхностной экспрессии рецептора B-клетки (сформированного из прикрепленных к мембране H и L цепей Ig) и для конкурирования с экспрессией и соединением эндогенных цепей (при наличии) посредством исключения аллеля. Однако такой промотор обычно не является тканеспецифичным. Для обеспечения тканеспецифичности предпочтительной является непрямая система, использующая Cre/lox и т.п. Нужная цепь помещается под управление сильного промотора, ингибируемого элементом, который может быть удален посредством воздействия Cre-белка, что приведет к активации нужного гена, кодирующего иммуноглобулин. Данная система подробно описана в работе Wunderlich F. T. (2004), "Generation of inducible Cre systems for conditional gene inactivation in mice", Inauguraldissertation zur Erlangung des Doktorgrades der Mathematisch- Naturwissenschaftlichen Fakultat der Universitat zu KoIn; http://deposit.ddb.de/cgi-bin/dokserv?idn= 97557230x&dok_var=dl&dok_ext=pdf&filename=97557230x.pdf.
Предпочтительно, цепь иммуноглобулина, продуцируемая устойчивой к перестройкам и гипермутации, является легкой цепью, способной к соединению с различными тяжелыми цепями, кодируемыми не являющимся человеком млекопитающим. Таким образом, легкая цепь будет одной и той же (и менее иммуногенной) во всех антителах, но разнообразие специфичности сохраняется посредством перестроек и гипермутаций в тяжелых цепях. В этом случае может быть предпочтительным сайленсинг по меньшей мере одного из эндогенных локусов, кодирующего легкую цепь, хотя исключение аллеля может привести к отсутствию такой необходимости.
В соответствии с данным вариантом осуществления, предпочтительным является функциональный сайленсинг эндогенной легкой цепи каппа (κ).
Если сайленсинг выполнен для локуса эндогенной легкой цепи κ, но также и по другим причинам, предпочтительно, чтобы устойчивая легкая цепь представляла собой легкую цепь κ, предпочтительно, легкую цепь, имеющую последовательность, подобную последовательности зародышевой линии. В соответствии с изобретением, такая легкая цепь приводила бы к наличию антитела со сниженной иммуногенностью. Предпочтительная последовательность зародышевой линии, основывающая на IGKV1-39 человека (O12) в качестве такой легкой цепи, очень часто наблюдается в репертуаре человека (de Wildt et al. 1999. J. Mol. Biol. 285(3):895) и имеет исключительную термодинамическую стабильность, выход и растворимость (Ewert et al. 2003. J. Mol. Biol. 325(3):531).
Ниже приведены более конкретные варианты осуществления экспрессионной кассеты, с помощью которой может быть получено не являющееся человеком животное по изобретению. Несмотря на то, что это обычно является благоприятным для иммуноглобулинов, также предполагаются и другие представляющие интерес гены.
Таким образом, изобретение в конкретном варианте осуществления относится к трансгенному не являющемуся человеком млекопитающему, у которого легкая цепь, кодирующая нуклеиновую кислоту, содержит в направлении 5'-3': специфичный для B-клетки промотор, лидерную последовательность, перестроенный V-ген человека, необязательно, энхансер MoEκi, константную область (κ) и, необязательно, (сокращенный) энхансер MoEκ3'. Нюбергером был обнаружен и исследован новый специфичный для B-клеток энхансер, расположенный по ходу транскрипции относительно константной области каппа (EP004690251). Было показано, что данный энхансер играет ключевую роль в экспрессии каппа-генов, поскольку удаление состоящего из 808 п.н. энхансера сильно снижает экспрессию. Удаление 3'-каппа энхансера также сильно снижает уровень соматических гипермутаций (SHM). При изучении трансгенной и клеточной экспрессии было обнаружено, что сокращение, мутирование или удаление 3'-каппа энхансеров не только снижает уровни экспрессии, но также снижает уровень соматических гипермутаций. В настоящее время нельзя определить, вовлечен ли 3'-каппа энхансер в процессы SHM, регуляцию экспрессии, или в оба процесса (обзор Odegard, V. H., et al.(2006). Nat. Rev. Immunol. 6, 573; Inlay, M., et al. (2002). Nat. Immunol. 3, 463).
Всесторонние исследования экспрессии с использованием вариантов 3'-каппа энхансера, созданных способами генной инженерии показали, что 50-нуклеотидный участок является достаточным для контроля экспрессии. Однако для надлежащей экспрессии предпочтительна сокращенная последовательность из 145 нуклеотидов (EP04690251; Meyer, K. B., et al. (1990) Nucleic Acids Res. 18(19):5609-15)
Таким образом, изобретение в одном из аспектов относится к нуклеиновой кислоте для встраивания в геном не являющегося человеком животного, представляющую собой экспрессионную кассету для экспрессии необходимой белковой молекулы в клетках, развивающихся в зрелые B-клетки, в течение определенной стадии развития, при этом вышеупомянутая кассета содержит средства для предотвращения сайленсинга экспрессии необходимой белковой молекулы после введения в клетку хозяина и средства для согласования времени экспрессии необходимой белковой молекулы с необходимой стадией развития клетки-хозяина.
Экспрессионная кассета определена как нуклеиновая кислота, которая была введена средствами внедрения в геном клетки хозяина, такими как последовательности, которые допускают гомологичную рекомбинацию с определенным участком в геноме. Как правило, нуклеиновой кислотой является ДНК, обычно двухцепочечная. Обычно экспрессионную кассету вводят в клетку с помощью вектора, из которого она переносится в геном клетки. Экспрессионная кассета также содержит все элементы, необходимые для экспрессии гена в клетке-хозяине, хотя в определенных вариантах осуществления некоторые из таких элементов могут присутствовать во второй нуклеиновой кислоте, предназначенной для внесения, посредством чего данные элементы действуют в процессе передачи. Элементы, необходимые для экспрессии в клетке-хозяине, включают промоторы, энхансеры и другие регуляторные элементы. Необходимыми являются только те элементы, которые не предоставляются клеткой-хозяином.
В соответствии с изобретением важно, чтобы экспрессия представляющего интерес гена не подвергалась сайленсингу в геноме клетки-хозяина, особенно на стадии развития, на которой требуется экспрессия. Эта цель может быть достигнута различными средствами, такими как вставка в эндогенный локус, или посредством предоставления кассеты с элементами нуклеиновой кислоты, которые предотвращают сайленсинг (Kwaks et al. (2006) Trends Biotechnol. 24(3), p.137-142; включенная в настоящий документ посредством ссылки). Предпочтительно, чтобы экспрессионная кассета вставлялась в локус, который не подвергался сайленсингу в клетках-хозяевах (EP 01439234; включенная в настоящий документ посредством ссылки).
Вышеупомянутые средства для предотвращения сайленсинга включают стабилизирующие последовательности против репрессии (STAR®-последовательности) и участки прикрепления к матриксу (MAR). STAR-последовательность представляет собой последовательность нуклеиновой кислоты, которая имеет способность влияния на транскрипцию генов вcis-положении. Как правило, хотя и не обязательно, STAR-последовательность сама не кодирует функциональный белковый элемент. В одном из вариантов осуществления используется один STAR-элемент. Однако предпочтительно использовать более одного STAR-элемента. В особенно предпочтительном варианте осуществления экспрессионная кассета по изобретению имеет две STAR-последовательности; одна STAR-последовательность на 5'-стороне кодирующей последовательности гена иммуноглобулина и одна STAR-последовательность на 3'-стороне кодирующей последовательности гена иммуноглобулина. MAR представляют собой последовательности ДНК, которые вовлечены в фиксирование ДНК/хроматина на ядерном матриксе; они были описаны для млекопитающих и растений. MAR обладают рядом характеристик, которые способствуют открытию и поддержке эухроматина. MAR могут повышать экспрессию трансгенов и ограничивать позиционные эффекты.
В соответствии с изобретением важно, чтобы экспрессия из кассеты имела место только в определенный период развития клетки, в частности, развития B-клетки, конкретнее, B-клетки в трансгенном не являющемся человеком животном, в частности, в мыши. В данном конкретном случае период развития выбирается таким образом, чтобы экспрессия гена из кассеты (полипептида, подобного легкой или тяжелой цепи) не являлась значительной помехой для нормальной дифференциации и/или созревания клетки и, при необходимости, допускала соединение продуцированной полипептидной цепи с ее копией.
В соответствии с изобретением, это может быть достигнуто, в одном из вариантов осуществления, путем получения нуклеиновой кислоты по изобретению, при этом вышеупомянутые средства для согласования времени экспрессии представляют собой промотор, активность которого в основном ограничена определенной стадией развития. В процессе развития B-клетка, которая, например, созревает и/или дифференцируется после иммунизации, экспрессия представляющего интерес гена, когда он представляет собой одну из полипептидных цепей иммуноглобулина, не должна являться (значительной) помехой для вышеупомянутого созревания и/или дифференциации, и она должна быть согласована по времени таким образом, чтобы результирующий полипептид мог соединяться со своими аналогами. Следовательно, изобретение относится к нуклеиновой кислоте по изобретению, при этом вышеупомянутая определенная стадия начинается сразу после, или совпадает с началом, экспрессии молекул легкой цепи вышеупомянутыми клетками на определенной стадии развития в зрелые B-клетки.
Этого можно достигнуть посредством выбора промотора, который является активным только в течение вышеупомянутого подходящего периода. Такой промотор может представлять собой промотор CD19, промотор Ig-α, промотор Ig-β, промотор μhc (все гены), промотор Vk или их аналоги и гомологи.
В конкретном варианте осуществления настоящего изобретения промотор, как описано выше, не контролирует экспрессию представляющего интерес гена напрямую. Вместо этого он контролирует экспрессию гена, продукт которого активирует в транс-положении экспрессию представляющего интерес гена. Таким активируемым геном может быть ген, кодирующий так называемую Cre-рекомбиназу или Cre-подобный белок. Экспрессионная кассета для представляющего интерес гена может, например, вводиться с последовательностью, которая подавляет экспрессию представляющего интерес гена. Вышеупомянутая последовательность может быть удалена посредством воздействия Cre-рекомбиназы, находящейся под управлением желаемого промотора (активного во время соответствующей стадии развития). В данном варианте осуществления требуется множество экспрессионных кассет.
Таким образом, изобретение относится к множеству нуклеиновых кислот, представляющих собой экспрессионные кассеты, при этом одна нуклеиновая кислота содержит экспрессионную кассету, кодирующую Cre-подобный белок под управлением промотора, активного во время желаемой стадии развития клетки-хозяина, и вторая нуклеиновая кислота содержит последовательность, кодирующую желаемую белковую молекулу под управлением конститутивного промотора, который может быть активирован посредством воздействия Cre-подобного белка. Вышеупомянутая активация предпочтительно достигается путем удаления стоп-последовательности, фланкированной сайтами loxP. Система Cre/lox подробно описана в работе Rajewsky et al. (1996) J. Clin. Invest. 98, p.600-603, которая включена в настоящий документ посредством ссылки. Обзор таких систем приведен в работе Wunderlich F. T. (2004), "Generation of inducible Cre systems for conditional gene inactivation in mice", Inauguraldissertation zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen Fakultat der Universitat zu KoIn; http://deposit.ddb.de/cgi-bin/dokserv?idn=97557230x&dok_var=dl&dok_ext=pdf&filename=9755723ox.pd, которая включена в настоящий документ посредством ссылки.
Изобретение также относится к трансгенному не являющемуся человеком животному, в которое была введена экспрессионная кассета по изобретению, при этом желаемая белковая молекула является полипептидной цепью иммуноглобулина. Предпочтительной полипептидной цепью является легкая цепь. Более предпочтительным полипептидом является легкая цепь зародышевой линии или подобная ей. Наиболее предпочтительным полипептидом является O12, предпочтительно, перестроенная каппа-легкая цепь зародышевой линии IGKV1-39*01/IGKJ1*01 (номенклатура в соответствии с базой данных IMGT, http://www.imgt.org).
Также предпочтительно, чтобы полипептидная цепь приводилась в состояние неспособности к перестройке и/или исключения любой модификации последовательности, такой как обычно происходящая с Ig в течение процесса созревания аффинности B-клетки. Таким образом, изобретение относится к трансгенному не являющемуся человеком животному, в которое была введена экспрессионная кассета по изобретению, при этом вышеупомянутая перестройка и/или модификация последовательности предотвращается посредством отсутствия элементов по меньшей мере частично ответственных за соматическую гипермутацию, таких как, например, энхансер MoEκi.
Предпочтительная экспрессионная кассета по изобретению содержит средства для предотвращения сайленсинга. В одном из вариантов осуществления, вышеупомянутые средства для предотвращения сайленсинга представляют собой средства для вставки в локус генома клетки-хозяина, который является устойчивым к сайленсингу. Вышеупомянутые средства для вставки предпочтительно представляют собой средства для гомологичной рекомбинации в вышеупомянутом сайте, устойчивом к сайленсингу. Предпочтительным локусом в случае, когда не являющееся человеком животное представляет собой мышь, является rosa-локус.
Еще одна предпочтительная экспрессионная кассета по изобретению содержит в направлении 5'- 3': VK-промотор, лидерную последовательность мыши, V-ген человека, необязательно, энхансер MoEκi, константную область (Сκ) крысы и, необязательно, (сокращенный) энхансер MoEκ3'.
Еще одна предпочтительная экспрессионная кассета по изобретению содержит в направлении 5'- 3': VK-промотор, лидерную последовательность человека, V-ген человека, необязательно, энхансер MoEκi, константную область (Сκ) крысы и, необязательно, (сокращенный) энхансер MoEκ3'.
Естественно, конечная цель изобретения состоит в получении антител для использования в терапии человека. Таким образом, изобретение относится к способу получения желаемого антитела, включающему воздействие на не являющегося человеком млекопитающего по изобретению антигеном таким образом, что индуцируется выработка антител, и выделение антител, специфичных для антигена.
В альтернативном варианте осуществления, изобретение относится к способу получения желаемого антитела, включающему воздействие на не являющегося человеком млекопитающего по изобретению антигеном таким образом, что индуцируется выработка антител, и выделение клеток, продуцирующих такие антитела, выращивание и, необязательно, иммортализацию вышеупомянутых клеток и сбор вышеупомянутых антител.
В еще одном варианте осуществления, изобретение относится к способу получения желаемого антитела, включающему воздействие на не являющегося человеком млекопитающего по изобретению антигеном таким образом, что индуцируется выработка антител, и выделение нуклеиновой кислоты, кодирующей по меньшей мере часть такого антитела, вставку вышеупомянутой нуклеиновой кислоты, или ее копии или производной, в экспрессионную кассету и экспрессирование вышеупомянутого антитела в клетке-хозяине.
Способы получения антител из трансгенных мышей известны специалистам в данной области техники. Особенно предпочтительными являются способы получения смеси антител из одной клетки, посредством чего, по изобретению, из мышей выделяются нуклеиновые кислоты, кодирующие данные антитела.
Такое так называемое олигоклонирование изложено в документах WO04106375 и WO05068622, которые включены в настоящий документ посредством ссылки.
Настоящее изобретение относится к трансгенным не являющимся человеком млекопитающим, предпочтительно, мышам, способным к созданию специфичных и высокоаффинных гибридных антител мыши-человека, предпочтительно, с вариабельными областями (VL) легкой цепи иммуноглобулина человека в конфигурации, близкой к зародышевой линии, и, предпочтительно, с вариабельными областями (VH) тяжелой цепи иммуноглобулина мыши, которые могут иметь накопленные соматические мутации во время процесса, контролируемого антигеном созревания аффинности. Предусматривается, что VH-области гибридных антител мыши могут подвергаться процедурам гуманизации с целью получения mAb, имеющих пониженную иммуногенность при применении в организме человека, на основании VL-областей зародышевой линии, или близких к ней, и VH-областей мыши, подвергшихся гуманизации.
В частности, в настоящем изобретении было показано, что трансгенные мыши, которые содержат конструкцию экспрессии ДНК, кодирующую перестроенную VL-область человека под контролем действующих в cis-положении генетических элементов, которые обеспечивают своевременную и регулируемую экспрессию трасгена в значительной части B-клеток в процессе развития B-клеток, и не имеют элементов, управляющих механизмом соматической гипермутации трансгена, способны создавать специфичные и высокоаффинные гибридные антитела мыши-человека в основном, с немутировавшими L-цепями. Было показано, что перестроенный трансген человека может соединяться с множеством эндогенных H-цепей иммуноглобулинов мыши с образованием гибридных иммуноглобулинов мыши-человека, экспрессируемых на поверхности B-клетки, и может в достаточной степени способствовать развитию B-клеток мыши для получения многочисленных и разнообразных ячеек периферийных B-клеток.
В предпочтительном варианте осуществления, конструкция для экспрессии трансгена содержит кодирующие последовательности перестроенного V-участка L-цепи человека под управлением VL-промотора человека с целью управления специфичной экспрессией B-клеток. Кроме того, конструкция содержит последовательность 3'Ck-энхансера мыши для специфичной и индуцируемой экспрессии трансгена на высоком уровне в B-клетке. Кроме того, конструкция задумана как не имеющая регуляторных элементов, которые способствуют привлечению механизма соматической гипермутации для трансгена, таких как энхансер интрона и 3'C-каппа энхансер.
В связанном варианте осуществления, перестроенный VL-ген человека вставляется в локус Rosa26 мыши посредством сайт-специфичного встраивания. Локус Rosa26 полезен в контексте подхода «направленного трансгенеза» для эффективного создания трансгенных организмов (таких как мыши) с предсказуемым профилем экспрессии трансгена.
В предпочтительном варианте осуществления перестроенная VL-область человека выбирается по ее способности к соединению с множеством различных VH-генов мыши с тем, чтобы обеспечить создание популяции B-клеток с разнообразным репертуаром VH-генов. Способ получения таких VL-областей включает амплификацию репертуара перестроенных VH-генов из B-клеток мышей и репертуара перестроенных VL-областей зародышевой линии человека из B-клеток человека, а также их клонирование в векторы фагмидного дисплея для приготовления разнообразных библиотек гибридных иммуноглобулинов в бактериях. Посредством анализа нуклеотидных последовательностей не выбранных и выбранных антигеном пар VH/VL, идентифицируются VL-гены зародышевой линии человека, которые соединяются с множеством различных VH-генов мыши. Описана коллекция VL-генов зародышевой линии человека с такой способностью.
В одном из вариантов осуществления, было показано, что после иммунизации антигеном B-клетки способны устанавливать иммунный ответ, приводящий к образованию B-клеток, которые секретируют гибридные антитела с высокой специфичностью и аффинностью. V-области, кодирующие данные антитела, характеризуются трансгенной легкой цепью человека, которая содержит очень мало мутаций, или не содержит их вообще, и тяжелой цепью мыши, которая содержит переменное количество мутаций, внесенных посредством механизма соматической гипермутации.
В связанном варианте осуществления, предусматриваются стратегии получения высокоаффинных гибридных моноклональных антител из трансгенных мышей посредством технологий гибридом и дисплея, а также процедура гуманизации VH-областей мыши с целью получении менее иммуногенных антител для применения в организме человека.
В одном из вариантов осуществления, изобретение относится к конструкции из трансгенной L-цепи иммуноглобулина, содержащей последовательности ДНК, которые кодируют VL-область иммуноглобулина человека вместе с константной областью (CL) легкой цепи иммуноглобулина животного, при этом данные последовательности функционально соединены с последовательностями регуляции транскрипции таким образом, что при интеграции в не являющееся человеком трансгенное животное продуцируется Ig-полипептид VL-CL с VL-областью человека, который не подвержен или незначительно подвержен соматической гипермутации. VL Ig имеет способность соединения с перестроенными полипептидами VH-CH, которые образуются в процессе развития B-клетки в не являющемся человеком трансгенном животном, при этом вышеупомянутые полипептиды VH-CH сохраняют способность подвергаться соматической гипермутации при стимуляции. CL-область может принадлежать животному произвольного вида, и обычно может соединяться с CH-областями не являющегося человеком трансгенного животного.
Изобретение также относится к применению вышеуказанной трансгенной конструкции для получения трансгенного не являющегося человеком животного, способного продуцировать гибридные антитела, состоящие из VL-CL полипептидов и VH-CH полипептидов, в которых VL-область имеет человеческое происхождение и CL, VH и CH могут принадлежать животным произвольного вида, включая человека. После иммунизации данные трансгенные животные могут создавать высокоаффинные антитела, кодируемые подвергавшимися соматической гипермутации VH-генами, и в основном немутированные VL-гены, кодируемые трансгеном.
В другом аспекте, изобретение относится к способу получения трансгенного не являющегося человеком животного, способного к продукции гибридных антител в ответ на антигенный стимул, включающий функциональное прерывание эндогенного локуса легкой цепи иммуноглобулина и вставку в геном животного трансгенной конструкции изобретения.
Изобретение относится к применению животных, получаемых посредством данного процесса, для получения B-клеток, которые продуцируют иммуноглобулин, имеющий VL легкой цепи человека. В другом аспекте изобретение относится к процессу получения B-клеток, которые продуцируют иммуноглобулин, имеющий VL человека и связывающийся с выбранным антигеном, включающий стимулирование животного, получаемого посредством вышеописанного процесса, вышеупомянутым антигеном и отбор B-клеток вышеупомянутого животного, которые связывают вышеупомянутый антиген. Изобретение также относится к B-клеткам, получаемым с помощью данного процесса, и гибридомы, получаемые с помощью иммортализации таких B-клеток, например, гибридомы, получаемые путем слияния B-клеток, указанных выше, с клетками миеломы. Изобретение также относится к способу получения моноклонального антитела, включающему культивирование такой гибридомы. В еще одном аспекте изобретение относится к применению указанной выше B-клетки для получения гибридомы или соответствующего моноклонального антитела.
Еще в одном аспекте изобретение относится к способу получения иммуноглобулина, имеющего VL-цепь человека и связывающегося с выбранным антигеном, включающему стимулирование животного, полученного в соответствии с описанным выше, вышеупомянутым антигеном, и получение из него иммуноглобулина.
В одной из стратегий, в качестве отдельного этапа, перестроенная VL-область, кодируемая сегментами генов V и J зародышевой линии человека, и константная область легкой цепи животного произвольного вида, но, предпочтительно, константная область мыши, вводятся в зародышевую линию мыши. Трансгенная ДНК может быть внесена в пронуклеус оплодотворенных ооцитов или стволовые клетки эмбриона. Интеграция может быть случайной или гомологичной в зависимости от конкретной применяемой стратегии. Например, VL-трансген может быть внесен посредством случайной вставки, в результате чего мыши будут нести одну или множество копий трансгена в геноме. В качестве альтернативы, VL-трансген человека может быть нацелен на конкретный локус генома с использованием сайт-специфичной рекомбинации, в соответствии с описанным в технике.
В одном из предпочтительных вариантов осуществления, VL-трансген нацелен на локус ROSA26 мыши, который является подходящим местом интеграции, допускающим сильную и предсказуемую экспрессию вставленных трансгенов (EP 1439234). Целевой вектор допускает вставку одной копии экспрессионной кассеты гена, посредством чего предотвращается модуляция экспрессии трансгена путем расположения множества копий. За счет выбор аутосомного локуса Rosa26 в качестве места вставки, профиль экспрессии вставленного трансгена в не являющемся человеком животном является предсказуемым. Кроме того, можно предотвратить эффекты случайной X-инактивации и/или модуляции по позиции в хромосоме. Это также устраняет потребность в создании и анализе множества трансгенных штаммов для произвольного заданного трансгена. В конечном итоге, целевой вектор Rosa26 для сайт-специфичной интеграции может быть использован для множества экспрессионных кассет генов. Таким образом, можно предусмотреть, чтобы 2 или более различных перестроенных VL-области зародышевой линии человека вставлялись в локус Rosa26 с целью дальнейшего увеличения разнообразия репертуара гибридных антител или антител человека.
В другом варианте осуществления, перестроенная VL-область человека может быть нацелена на локус каппа- или лямбда-легкой цепи Ig мыши, с тем чтобы функционально инактивировать эндогенный локус, или мыши, содержащие перестроенную VL-область человека, могут скрещиваться с мышами, не имеющими функциональных каппа- или лямбда локусов Ig, или может быть сделано и то, и другое. Таким образом, путем применения трансформации, с использованием повторяющихся шагов или в комбинации со скрещиванием, могут быть получены трансгенные животные, способные к продуцированию антител, содержащих VL-трансген человека при отсутствии, в основном, эндогенных легких цепей иммуноглобулина хозяина.
В одном из вариантов осуществления, VL-трансген человека выбирают по его способности к соединению со значительной частью VH-областей мыши с образованием разнообразного репертуара функциональных гибридных антител мыши-человека, экспрессируемых на поверхности B-клетки. Под значительной частью VH-областей мыши понимается, что VL человека соединяется по меньшей мере с 0,1% VH-областей мыши, созданных в процессе развития B-клетки, более предпочтительно, по меньшей мере с 1%, и, наиболее предпочтительно, по меньшей мере с 10%. Способы идентификации VL-генов человека с данной характеристикой включают парное соединение репертуара VL-областей человека с репертуаром VH-областей мыши, коэкспрессию VH и VL-областей в соответствующих эукариотических и прокариотических экспрессионных векторах и отбор VL-областей человека, которые соединяются со значительной частью VH-областей мыши. В одном из вариантов осуществления, фагмидные вектора могут быть использованы для контроля экспрессии фрагментов антитела мыши-человека в бактериальных клетках или на поверхности нитеобразного фага и анализа связывающей способности фрагментов антител с помощью методов, известных в технике.
В другом варианте осуществления, VL-трансген человека отбирается по его способности к соединению со значительной частью VH-областей человека с образованием разнообразного репертуара антител человека, экспрессируемых на поверхности B-клетки. Под значительной частью VH-областей человека понимается, что VL человека соединяется по меньшей мере с 0,1% VH-областей человека, созданных в процессе развития B-клетки, более предпочтительно, по меньшей мере с 1%, и, наиболее предпочтительно, по меньшей мере с 10%.
В последнем варианте осуществления, трансгенные мыши с VL человека скрещиваются с мышами, содержащими функционально перестроенные или неперестроенные локусы H-цепи иммуноглобулина человека и функционально инактивированные эндогенные локусы H-цепи Ig, в соответствии с описанным в технике. Функциональная инактивация двух копий из каждых трех локусов Ig хозяина (тяжелая цепь, каппа- и лямбда-легкая цепь), где хозяин содержит IgH человека и перестроенный VL-трансген человека, позволила бы продуцировать чисто молекулы антител человека без продуцирования антител хозяина или химерных антител хозяина-человека. Такой штамм-хозяин, за счет иммунизации специфическими антигенами, отвечал бы продуцированием B-клеток мыши, продуцирующих специфические антитела человека, при этом B-клетки позднее сливаются с клетками миеломы мыши или иммортализируются любым другим способом в целях непрерывного стабильного продуцирования моноклональных антител человека. В качестве альтернативы, вышеупомянутая популяция B-клеток используется в качестве источника VH-областей, которые могут быть получены путем создания библиотек кДНК или ПЦР-амплификации с использованием праймеров для VH-областей человека, в соответствии с известным в технике.
Перестроенный VL-ген человека реконструируется в подходящем эукариотическом или прокариотическом микроорганизме, и полученные фрагменты ДНК могут быть внесены в пронуклеус оплодотворенных ооцитов или стволовые клетки эмбриона. Различные конструкции, направляющие специфичную для B-клеток экспрессию VL-трансгенов, были описаны в технике и имеют следующий общий формат: лидерная последовательность и соответствующие последовательности, расположенные против хода транскрипции, для управления специфичной для B-клеток экспрессией трансгена, кодирующая последовательность VL-трансгена человека, энхансерная последовательность, которая направляет специфичную для B-клеток и высокоуровневую экспрессию трансгена, и ген константной области мыши. В предпочтительном варианте, энхансер представляет собой энхансер C-каппа 3', поскольку он управляет высокоуровневой экспрессией в клеточной линии B-клеток, но не мобилизует соматическую гипермутацию при использовании в трансгенных конструкциях.
В одном из вариантов осуществления, животные, предпочтительно, мыши, содержащие одну или множество копий трансгена в своем геноме, изолируются и анализируются на предмет устойчивой экспрессии. Выбирают животных, которые демонстрируют устойчивую экспрессию трансгена в течение более длительных периодов времени, предпочтительно, в B-клетках. При необходимости, различные линии животных, содержащие независимые вставки одной копии или множества копий трансгена, предпочтительно, в различных хромосомах, скрещиваются с целью получения животных с различными вставками одной копии или множества копий трансгена с целью повышения уровня экспрессии у животных, предпочтительно, в B-клетках.
Изобретение также относится к потомству трансгенных не являющихся человеком животных по изобретению, при этом потомство содержит по меньшей мере в своей клеточной линии B-клеток, последовательность, кодирующую тяжелую или легкую цепь, вместе со средствами, которые обеспечивают устойчивость последовательности к перестройкам и/или соматическим гипермутациям ДНК.
Изобретение также относится к потомству трансгенных не являющихся человеком животных по изобретению, при этом потомство содержит экспрессионную кассету для экспрессии желаемой белковой молекулы в клетках в течение определенной стадии развития для клеток, развивающихся в зрелые B-клетки.
Кроме того, изобретение относится к клетке, которая выделена из трансгенного не являющегося человеком животного по изобретению, при этом клетка содержит последовательность, кодирующую тяжелую или легкую цепь, вместе со средствами, которые обеспечивают устойчивость последовательности к перестройкам и/или соматическим гипермутациям ДНК. Кроме того, изобретение относится к клетке, которая выделена из трансгенного не являющегося человеком животного по изобретению, при этом клетка содержит экспрессионную кассету для экспрессии желаемой белковой молекулы в клетках в течение определенной стадии развития для клеток, развивающихся в зрелые B-клетки. Клетка по изобретению, предпочтительно, продуцирующая антитело B-клетка или клетка, способная к дифференциации или созреванию в продуцирующее антитело B-клетку, может быть использована для продукции антител in vitro, как известно специалисту, например, из работы Gascan et al. 1991. J. Exp. Med. 173: 747-750. Способы иммортализации клетки по изобретению известны в технике и включают создание гибридом, например, путем слияния с клеткой миеломы, трансформацию с помощью вируса Эпштейна-Барр; экспрессию трансдуктора сигналов активации и транскрипции (STAT), активацию через передачу сигналов по рецепторам CD40 и IL4 и/или экспрессию Bcl6 (Shvarts et al. 2002. Genes Dev 16: 681-686).
На отдельном этапе эндогенные локусы каппа- и лямбда-легких цепей мыши делаются, в основном, нефункциональными таким образом, что по меньшей мере большинство B-клеток в трансгенных мышах имеют Ig-рецепторы, которые содержат трансгенную VL-область человека. Инактивация эндогенных локусов иммуноглобулина мыши достигается путем направленной дезорганизации соответствующих локусов посредством гомологичной рекомбинации в стволовых клетках эмбриона мыши. Вышеупомянутая направленная дезорганизация включает изменение геномной последовательности таким образом, что, в основном, не осуществляется продуцирование функциональных эндогенных каппа- и/или лямбда-легких цепей иммуноглобулина мыши. Термин «в основном, отсутствие функциональных эндогенных иммуноглобулинов мыши» означает, что эндогенные локусы каппа- и/или лямбда-легких цепей подверглись функциональному сайленсингу, в результате чего уровень экспрессии функциональных белков эндогенных локусов каппа- и/или лямбда-легких цепей, предпочтительно, локуса каппа-легкой цепи, снижается до около 20% от уровня экспрессии у мыши-контроля, более предпочтительно, до около 10%, более предпочтительно, до около 5%, более предпочтительно, до около 2% и, более предпочтительно, до около 1%. В наиболее предпочтительном варианте осуществления, уровень экспрессии функциональных белков локусов каппа- и/или лямбда-легких цепей сокращается до 0%. Уровень экспрессии функциональных белков может быть определен с помощью средств, известных специалисту, включая вестерн-блоттинг и парное соединение с тяжелой цепью мыши. Вышеупомянутая мышь-контроль представляет собой мышь, в которой эндогенные локусы каппа- и/или лямбда-легких цепей не подвергались дезорганизации. Вышеупомянутое изменение включает мутацию и/или удаление последовательностей генов, которые требуются для функциональной экспрессии генов эндогенных иммуноглобулинов. В качестве альтернативы, вышеупомянутое изменение включает вставку нуклеиновой кислоты в эндогенные локусы каппа- и/или лямбда-легких цепей мыши с тем, чтобы снизить функциональную экспрессию генов эндогенных иммуноглобулинов. В одном из вариантов осуществления, вышеупомянутая нуклеиновая кислота содержит элемент сайленсинга, что приводит к транскрипционного сайленсингу гена эндогенного иммуноглобулина. В еще одном варианте осуществления, или в дополнение, вышеупомянутая нуклеиновая кислота содержит последовательность, которая нарушает сплайсинг и/или трансляцию гена эндогенного иммуноглобулина, например, путем введения экзона, который вызывает сдвиг рамки считывания в кодирующей последовательности, или который содержит преждевременный стоп-кодон. В каждом случае создаются химерные животные, которые частично получены из модифицированных эмбриональных стволовых клеток, и способны передавать генетические модификации через зародышевую линию. Спаривание штаммов мыши с локусами иммуноглобулинов человека со штаммами с инактивированными локусами мыши дает животных, которые продуцируют антитела, содержащие, в основном, только легкие цепи человека.
Конструкцию для гомологичной рекомбинации можно получить средствами, известными из уровня техники, и все нежелательные последовательности, например, прокариотические последовательности, удаляются. Может быть использована любая удобная методика введения конструкции для гомологичной рекомбинации в клетку-мишень. Данные методики включают слияние сферобластов, липофекцию, электропорацию, опосредованный фосфатом кальция перенос ДНК или прямую микроинъекцию. После трансформации или трансфекции клеток-мишеней, клетки-мишени выбираются с помощью положительных и/или отрицательных маркеров, например, устойчивости к неомицину и/или устойчивости к ацикловиру и/или ганцикловиру. Клетки, демонстрирующие желаемый фенотип, могут затем подвергаться дополнительному анализу посредством рестрикционного анализа, электрофореза, саузерн-анализа, ПЦР и т.п. Путем идентификации фрагментов, которые демонстрируют наличие поражения(-ий) в целевом локусе, идентифицируются клетки, в которых оказалось, что гомологичная рекомбинация инактивировала копию целевого локуса.
Кроме того, показано, что после иммунизации VH-области мыши и человека у вышеупомянутых трансгенных мышей, но не VL-области, способны подвергаться соматическим гипермутациям для создания высокоаффинных антител. Является благоприятным, что было предсказано, что данные антитела, кодируемые VL-областями зародышевой линии, способствуют более низкой иммуногенности при применении для людей, и в результате получаются более стабильные антитела, которые меньше подвержены агрегации и, следовательно, являются более безопасными для терапевтического применения на людях.
Все MAb, полученные из вышеупомянутых не являющихся человеком трансгенных животных или клеток, имеют одни и те же идентичные VL-области человека. Было описано, что mAb, которые имеют одни и те же идентичные VL-области, могут коэкспрессироваться в одной клональной клетке с целью продуцирования смеси рекомбинантных антител с функциональными сайтами связывания (см. WO04106375 и WO05068622). Соответственно, изобретение относится к платформе для создания специфичных и высокоаффинных mAb, которые составляют основу для смесей mAb, продуцируемых клональными клетками.
Предпочтительно, чтобы MAb, полученные из вышеупомянутых не являющихся человеком трансгенных животных или клеток, были направлены против клеточных мишеней. Предпочтительными мишенями являются экспрессируемые на поверхности или растворимые белки человека или молекулы углеводов. Более предпочтительными мишенями являются экспрессируемые на поверхности или растворимые белки или молекулы углеводов, которые экспрессируются на поверхности бактерий, вирусов, других патогенных микроорганизмов, особенно человека.
Конкретно, предпочтительные мишени включают цитокины и хемокины, включая, но не ограничиваясь перечисленным, интерлейкин 1бета (IL1бета), IL2, IL4, IL5, IL7, IL8, IL12, IL13, IL15, IL18, IL21, IL23 и хемокины, такие как, например, CXC-хемокины, CC-хемокины, C-хемокины (или γ-хемокины), такие как XCL1 (лимфотактин-α) и XCL2 (лимфотактин-β) и CX3C-хемокины. Также включаются в число предпочтительных мишеней молекулы рецепторов цитокинов и хемокинов, включая цитокиновые рецепторы типа I, такие как, например, рецептор IL-2, цитокиновые рецепторы типа II, такие как, например, рецепторы интерферона, рецепторы суперсемейства иммуноглобулинов (Ig), семейство рецепторов фактора некроза опухоли, включая рецепторы для CD40, CD27 и CD30, рецепторы серин/треониновой протеинкиназы, такие как рецепторы TGF-бета, соединенные рецепторы G-белка, такие как CXCR1-CXCR7, и рецепторы тирозинкиназы, такие как члены семейства рецепторов фактора роста фибробластов (FGFR), члены семейства EGF-рецептора, включая erbB1 (EGF-R; HER1), erbB2, (HER2), erbB3 (HER3) и erbB4 (HER4), члены семейства инсулиновых рецепторов, члены семейства рецепторов фактора роста гепатоцитов, включая c-Met (HGF-R), члены семейства рецепторов Trk, члены семейства рецепторов AXL, члены семейства рецепторов LTK, члены семейства рецепторов TIE, члены семейства рецепторов ROR, члены семейства рецепторов DDR, члены семейства рецепторов KLG, члены семейства рецепторов RYK, члены семейства рецепторов MuSK и члены семейства рецепторов фактора роста эндотелия сосудов (VEGFR).
Кроме того, предпочтительными мишенями являются мишени, которые сверхэкспрессированы или избирательно экспрессированы в опухолях, такие как, например, VEGF, CD20, CD38, CD33, CEA, EpCAM, PSMA, CD54, Lewis Y, CD52, CD40, CD22, CD51/CD61, CD74, MUC-1, CD38, CD19, CD262 (TRAIL-R2), RANKL, CTLA4, и CD30; мишени, которые вовлечены в хроническое воспаление, такие как, например, CD25, CD11a, TNF, CD4, CD80, CD23, CD3, CD14, IFNгамма, CD40L, CD50, CD 122, TGFбета и TGFальфа.
Предпочтительные экспрессируемые на поверхности белки или молекулы углеводов, которые экспрессируются на поверхности бактерий, вирусов, других патогенных паразитных микроорганизмов, особенно человека, включают маркеры поверхности вирусов гриппа A и B, такие как гемагглютинин (HA) и нейраминидаза (NA), филовирусы, такие как вирусы Эбола, бешенства, кори, краснухи, инфекционного паротита, флавивирусы, такие как вирусы Денге типа 1-4, вирус клещевого энцефалита, вирус Западного Нила, вирус японского энцефалита и вирус желтой лихорадки, парамиксовирусы, включая парамиксовирус, такой как парагрипп 1, 3, рубулавирус, такой как вирус инфекционного паротита и парагриппа 2, 4, вирус кори и пневмовирус, такие как респираторный синцитальный вирус, коровья оспа, натуральная оспа, коронавирусы, включая вирус тяжелого острого респираторного синдрома (SARS), вирус гепатита A, B и C, вирус иммунодефицита человека, вирусы герпеса, включая цитомегаловирус, вирус Эпштейна-Барр, вирус простого герпеса и вирус ветряной оспы, парвовирусы, такие как, например, B19; Legionella pneumophila; Listeria monocytogenes; Campylobacter jejuni; Staphylococcus aureus; E. coli O157:H7; Borrelia burgdorferi; Helicobacter pylori; Ehrlichia chaffeensis; Clostridium difficile; Vibrio cholera; Salmonella enterica Serotype Typhimurium; Bartonella henselae; Streptococcus pyogenes (стрептококк группы A); Streptococcus agalactiae (стрептококк группы B); устойчивый к множеству лекарств S. aureus (например, MRSA); Chlamydia pneumoniae; Clostridium botulinum; Vibrio vulnificus; Parachlamydia pneumonia; Corynebacterium amycolatum; Klebsiella pneumonia; устойчивые к линезолиду энтерококки (E. faecalis и E. faecium); и устойчивый к множеству лекарств Acinetobacter baumannii.
Наиболее предпочтительными мишенями являются IL-6 и его рецептор, IL-6Rальфа, гликопротеин-определенный gp130, RSV, особенно поверхностные белки F, G и SH и неструктурные белки, такие как N и M, и рецепторы тирозинкиназ, в частности, erbB1 (EGF-R; HER1), erbB2, (HER2), erbB3 (HER3), erbB4 (HER4), IGF-R1 и IGF-RII, c-Met (HGF-R).
Таким образом, изобретение относится к основе создания специфичных и высокоаффинных mAb против вышеупомянутых мишеней, которые составляют основу для смесей mAb, продуцируемых клональными клетками. В предпочтительном варианте осуществления вышеупомянутые специфичные и высокоаффинные mAb включают mAb, которые направлены против различных эпитопов по меньшей мере на одной из мишеней. В еще одном предпочтительном варианте осуществления, вышеупомянутые специфичные и высокоаффинные mAb включают mAb, которые направлены против различных мишеней, таких как, например, один или более членов семейства EGF-рецепторов, включая erbB1 (EGF-R; HER1), erbB2, (HER2), erbB3 (HER3) и erbB4 (HER4).
Если не определено другое, то научные и технические термины, используемые в связи с настоящим изобретением, должны иметь значения, которые хорошо известны и понятны специалистам в данной области техники. Кроме того, если другое не предусматривается контекстом, термины в единственном числе должны включать множественное число, и термины во множественном числе должны включать единственное. Как правило, номенклатуры, а также методики, используемые в связи с выращиванием клеток и тканей, молекулярной биологией, химией белков и олиго- или полинуклеотидов и гибридизацией, описанные в настоящем документе, представляют собой номенклатуры и методики, хорошо известные и обычно используемые в технике. Стандартные методики используются для рекомбинантных ДНК, синтеза олигонуклеотидов и выращивания и трансформации тканей (например, электропорация, липофекция). Ферментативные реакции и методики очистки осуществляются в соответствии со спецификациями производителя, в соответствии с общепринятым в технике или в соответствии с описанным в настоящем документе. Вышеупомянутые методики и процедуры обычно выполняются в соответствии с обычными методами, хорошо известными из уровня техники, и в соответствии с описанным в различных общих и более конкретных ссылках, которые цитируются и обсуждаются в настоящей спецификации. См., например, Sambrook et al. Molecular Cloning: A Laboratory Manual (3rd edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001)), которая включена в настоящий документ посредством ссылки. Номенклатуры, а также лабораторные процедуры и методики, используемые в связи с аналитической химией, синтетической органической химией и медицинской и фармацевтической химией, описанные в настоящем документе, представляют собой номенклатуры, процедуры и методики, хорошо известные и обычно используемые в технике. Стандартные методики используются для химического синтеза, химических анализов, приготовления фармацевтических препаратов, составления рецептур, доставки препаратов и лечения пациентов.
Описание чертежей
Фигура 1
Схема топологии мест отжига специфичных для мыши VH-праймеров и положение требуемых сайтов рестрикции, которые вводятся в виде последовательностей с висящими краями на 3'-конце праймеров.
Фигура 2
Этапы ПЦР-амплификации (амплификация, промежуточный этап и введение сайтов). Положение и название праймеров для амплификации VH мыши (и смесей праймеров) указаны для каждого этапа.
Фигура 3
Топология вектора MV1043. Данный вектор используется для клонирования VH-фрагментов мыши и человека. O12 (IGKV1-39) указан как VL-ген. Продукты данного вектора, совместно со вспомогательными фагами в клетках E. coli, позволяют получать фаги, которые отображают Fab-фрагменты на поверхности фаговых частиц в форме слитого с белком g3 продукта и наличие вектора в фаге в форме генетического содержания (F1 ORI).
Фигура 4
Топология C-каппа локуса мыши по ходу транскрипции относительно J-сегментов. Указаны оба энхансера и область C-каппа. Стрелка внизу указывает участок, который удаляется с целью выполнения сайленсинга локуса.
Фигура 5
Топология C-лямбда локуса мыши. Указаны все три активных V-области (Igl-V1, V2 и V3), J-сегменты (Igl-J1, Igl-J2, Igl-J3, Igl-J4 и псевдосегмент Igl-J3p) и константные области (Igl-C1, Igl-C2, Igl-C3 и Igl-C4). Области, которые удаляются с целью выполнения сайленсинга локуса, отмечены маркерами удаления. Данные удаления включают все активные V-гены (1, 2 и 3) и межгенный сегмент между V2 и V3.
Фигура 6
Топология конструкции IGKV1-39/J-Ck с интроном, расположенным в лидерной последовательности открытой рамки считывания (ORF).
Фигура 7
Топология конструкции IGLV2-14/J-Ck с интроном, расположенным в лидерной последовательности открытой рамки считывания (ORF).
Фигура 8
Топология конструкции VkP-IGKV1-39/J-Ck (VkP-O12). Промотор происходит из гена IGKV1-39 и помещается непосредственно перед необходимыми элементами для эффективной транскрипции и трансляции. Межгенные последовательности (включая энхансеры) извлечены из мышей и получены из BAC-клонов. Последовательность C-каппа кодирует каппа-константную область крысы.
Фигура 9
Топология конструкции VkP-IGLV2-14/J-Ck (VkP-2a2). Промотор происходит из гена IGKV1-39 и помещается непосредственно перед необходимыми элементами для эффективной транскрипции и трансляции. Межгенные последовательности (включая энхансеры) извлечены из мышей и получены из BAC-клонов. Последовательность C-каппа кодирует каппа-константную область крысы.
Фигура 10
Топология конструкции VkP-IGKV1-39/J-Ck-Δ1 (VkP-O12-del1) идентична VkP-IGKV1-39/J-Ck с Фигуры 9, за исключением того, что энхансерная область интрона удалена.
Фигура 11
Топология конструкции VkP-IGKV1-39/J-Ck-Δ2 (VkP-O12-del2) идентична VkP-IGKV1-39/J-Ck-Δ1 с Фигуры 10, за исключением того, что удалена большая часть межгенного участка между геном Ck 3'-энхансером. Кроме того, 3'-энхансер сокращен в размер с 809 п.н. до 125 п.н.
Фигура 12
Общие сведения о последовательностях, используемых в настоящей заявке, или на которые имеются ссылки в настоящей заявке.
Фигура 13
Создание аллеля Rosa26-IgVk1-39 KI. (A) Схематическое изображение целевого вектора pCAGGS-IgVK1-39. (B) Нуклеотидная последовательность целевого вектора pCAGGS-IgVK1-39. (C) Стратегия нацеливания.
Фигура 14
(A) Анализ саузерн-блоттингом геномной ДНК ES-клонов, содержащих вставку целевого вектора pCAGGS-IgVK1-39. Геномная ДНК из 4 независимых клонов была расщеплена с AseI и зондирована с 5e1, указывающим 5'-границу целевого вектора. Все клоны содержат правильную вставку целевого вектора на 5'-конце.
(B) Анализ саузерн-блоттингом геномной ДНК ES-клонов, содержащих вставку целевого вектора pCAGGS-IgVK1-39. Геномная ДНК из 4 независимых клонов была расщеплена с MscI и зондирована с 3e1, указывающим 3'-границу целевого вектора. Все клоны содержат правильную вставку целевого вектора на 3'-конце.
(C) Анализ саузерн-блоттингом геномной ДНК ES-клонов, содержащих вставку целевого вектора pCAGGS-IgVK1-39. Геномная ДНК из 4 независимых клонов была расщеплена с BamHI и зондирована с внутренним зондом Neo, указывающим 5'-границу целевого вектора. Все клоны содержат правильную единичную вставку целевого вектора.
Фигура 15
Создание аллеля Rosa26-IgV12-14 KI. (A) Схематическое изображение целевого вектора pCAGGS-IgVL2-14. (B) Нуклеотидная последовательность целевого вектора pCAGGS-IgVL2-14, содержащего экспрессионную вставку CAGGS, основанную на перестроенной V-лябмда области IGLV2-14/J зародышевой линии (IGLV2-14/J-Ck). (C) Стратегия нацеливания.
Фигура 16
Epibase®-профиль остатков 1-107 IGKV1-39. Подфигура A показывает связывающую способность для аллотипов DRB1, тогда как C показывает связывающую способность для аллотипов DRB3/4/5, DQ и DP. Значения на Фигуре представляют собой константы диссоциации (Kd) и изображены на логарифмической шкале в диапазоне 0,01 мкМ-0,1 мкМ (очень сильные связи могли выйти за пределы графика). Для пептидов со средним связыванием показаны только качественные значения, пептиды со слабым связыванием и несвязывающиеся не показаны. Значения показаны на первом остатке пептида в целевой последовательности (сам пептид продолжается другими 9-ю остатками). Важно, что показан только рецептор с наибольшей связывающей способностью для каждого пептида: имеющие перекрестную реакцию аллотипы с более низкой аффинностью не видны на данном графике. Рецептор с наибольшей связывающей способностью указан под своим серотипическим наименованием. Наконец, любые профильтрованные по зародышевой линии пептиды изображены более светлым цветом на карте эпитопов (в данном случае не было обнаружено несобственных эпитопов). Подфигура B показывает разнородность связывания HLA для каждого декамерного пептида (Y-ось: число аллотипов HLA, распознающих критические эпитопы в каждом пептиде, начиная с указанного остатка, показанного на X-оси). Разнородность измерялась как количество аллотипов из суммарных 47, для которых пептид является критическим связывающим агентом. Белые столбцы относятся к аутопептидам, а черные столбцы (здесь отсутствуют) - к несобственным пептидам.
Фигура 17
Карта эпитопов IGKV1-39, показывающая наличие связывающих пептидов, предсказанных в последовательности IGKV1-39 по серотипу в формате 15-меров. Каждый 15-мер пронумерован в соответствии с указанным вверху фигуры. Полная последовательности соответствующего 15-мера приведена в Таблице 7. Черные прямоугольники показывают наличие одного или более критических аутоэпитопов в 15-мере для серотипа, показанного слева. Критические эпитопы функционально определяются как имеющие сильное или среднее связывание с DRB1 и сильное связывание с DRB3/4/5 или DP или DQ.
Фигура 18
Конститутивный нокаут (KO) каппа-локуса Ig. (A) Стратегия нацеливания. (B) Схематическое изображение целевого вектора pIgКаппа.
Фигура 19
Конститутивный KO лямбда-локуса Ig. (A) Первый этап стратегии нацеливания. (B) Второй этап стратегии нацеливания.
Фигура 20
Схематическое изображение целевых векторов. (A) pVkP-O12 (VkP-IGKV1-39/J-Ck); (B) pVkP-O12-del1 (VkP-IGKV1-39/J-Ck-Δ1); (C) pVkP-O12-del2 (VkP-IGKV1-39/J-Ck-Δ2).
Фигура 21
Стратегии нацеливания для вставки трансгена в локус Rosa26 с помощью нацеленного трансгенеза с использованием RMCE. (A) VkP-O12 (VkP-IGKV1-39/J-Ck); (B) VkP-O12-del1 (VkP-IGKV1-39/J-Ck-Δ1); (C) VkP-O12-del2 (VkP-IGKV1-39/J-Ck-Δ2).
Фигура 22
Топология вектора MV1057. Замена указанного «лишнего» фрагмента на VH-фрагмент приводит к тому, что экспрессионный вектор может быть трансфицирован в эукариотические клетки для продукции антител IgG1 с легкими цепями, содержащими VL-ген O12 (IGKV1-39).
Фигура 23
Недостаточная экспрессия трансгенного Vk1 легкой цепи человека в популяциях клеток селезенки, не являющихся B-клетками.
Фигура 24
Легкая цепь трансгенного Vk1 человека экспрессируется во всех популяциях B-клеток селезенки.
Фигура 25
Легкая цепь трансгенного Vk1 человека экспрессируется B1-клетках брюшной полости.
Фигура 26
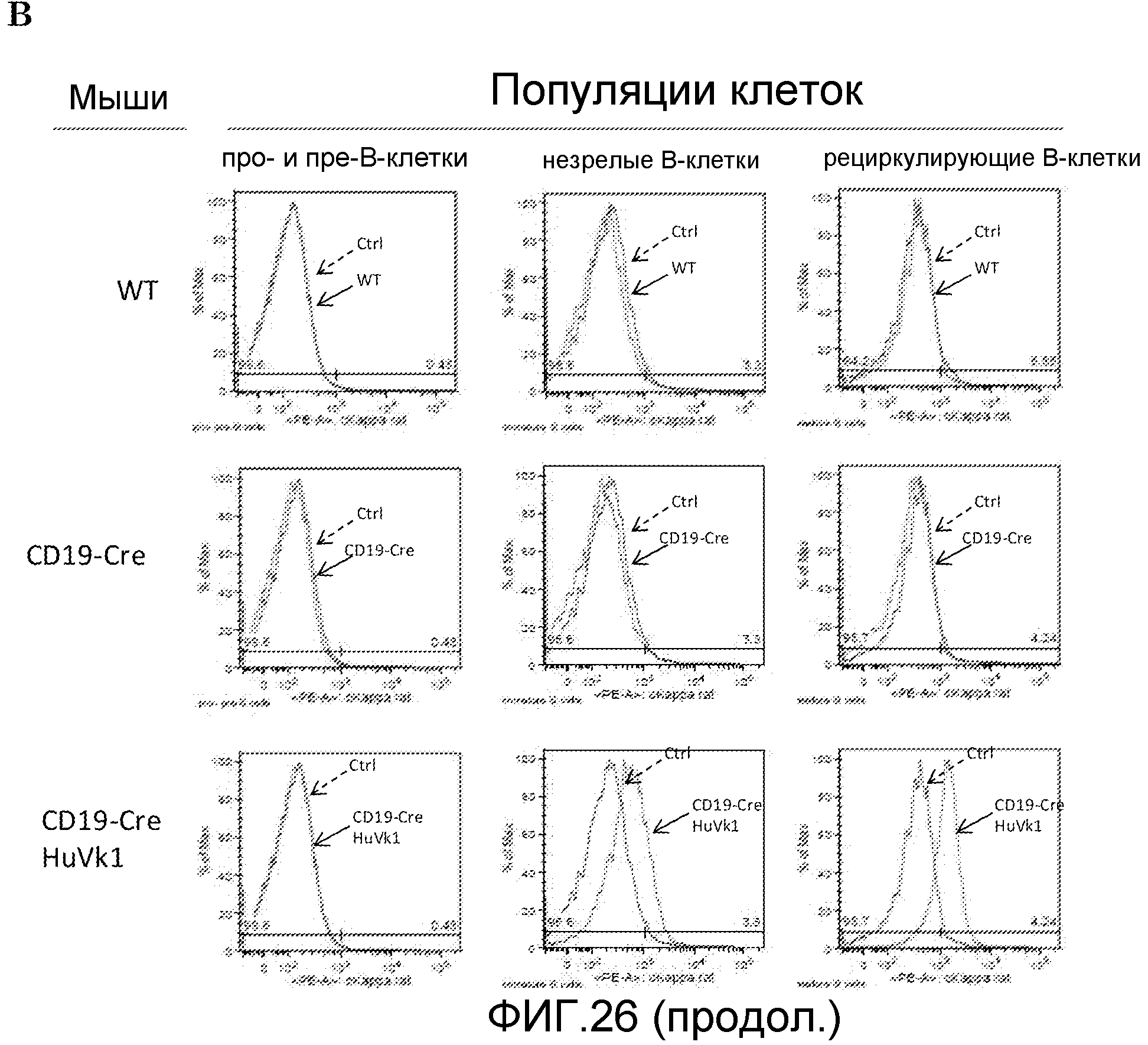
Легкая цепь трансгенного Vk1 человека не экспрессируется в про- и пре-B-клетках, но экспрессируется в незрелых и рециркуляционных популяциях B-клеток костного мозга. (A) Открытие мембранного канала клеток костного мозга. (B) Гистограммы экспрессии трансгена с наложением одного контроля для дикого типа.
Фигура 27
Легкая цепь трансгенного Vk1 человека напрямую связана с экспрессией эндогенной легкой цепи и IgM в циркулирующих в крови B-клетках.
ПРИМЕРЫ
Пример 1.
Клоны V-гена легкой цепи человека
В данном примере описывается логическое обоснование выбора двух V-генов легкой цепи человека, одного гена каппа-типа и одного гена лямбда-типа, которые используются в качестве доказательства концепции для трансгенных мышей, экспрессирующих легкую цепь. de Wildt et al. 1999 (de Wildt et al. (1999) J. Mol. Biol. 285(3):895) проанализировали экспрессию легких цепей человека в периферийных IgG-положительных B-клетках. На основании этих данных, IGKV1-39 (O12) и IGLV2-14 (2a2) были выбраны в качестве легких цепей, поскольку они очень хорошо представлены в репертуаре B-клеток. Последовательность J-сегмента легких цепей была выбрана на основании последовательностей, представленных под кодом GenBank ABA26122 для IGKV1-39 (Rabquer,B.J., Smithson,S.L., Shriner,A.K. and Westerink,M.A.J.) и кодом GenBank AAF20450 для IGLV2-14 (Ignatovich,O., Tomlinson,I.M., Popov,A.V., Bruggemann,M. and Winter,G. J. Mol. Biol. 294 (2), 457-465 (1999)).
Все каркасные сегменты конвертируются в аминокислотные последовательности зародышевой линии с целью обеспечения наименьшей возможной иммуногенности в потенциальных клинических применениях.
Пример 2.
Получение V-генов тяжелой цепи мыши, которые образуют пары с генным сегментом IGKV1-39 человека с образованием функциональных сайтов связывания антитела
В данном примере описывается идентификация V-генов тяжелой цепи мыши, которые способны образовывать пары с единичной перестроенной областью IGKV1-39/J зародышевой линии человека. VH-репертуар селезенки для мышей, которые были иммунизированы столбнячным токсином, был клонирован в Fab-векторе фагового дисплея с единственной каппа-легкой цепью IGKV1-39-C человека и подвергнут пэннингу через столбнячный токсин. Клоны, полученные после первого цикла пэннинга, анализировались на предмет их специфичности связывания. VH-гены мыши, кодирующие специфичные для столбнячного токсина Fab-фрагменты, подвергались анализу последовательности с целью идентификации уникальных клонов и назначения использования VH, DH и JH.
Многие протоколы, описанные здесь, представляют собой стандартные протоколы для создания библиотек фагового дисплея и пэннинга фагов для связывания с представляющим интерес антигеном, и описаны в работе Antibody Phage Display: Methods and Protocols (editor(s): Philippa M. O'Brien, Robert Aitken).
Иммунизации
BALB/c мыши получали одну иммунизацию столбнячным токсином и подвергались стимуляции столбнячным токсином после 6 недель.
Выделение спленоцитов
Приготовление взвеси клеток селезенки. После препарирования селезенка промывалась PBS и переносилась на 60 мм чашку Петри с 20 мл PBS. Шприц, заполненный 20 мл PBS и с иглой G20, был использован для многократного промывания селезенки. После промывания промытых клеток PBS, клетки аккуратно помещались в суспензию с использованием 20 мл PBS и оставлялись на рабочей площадке на 5 минут для отделения спленоцитов от дебриса и клеточных кластеров. Суспензия спленоцитов переносилась на заполненную фиколл-пак плюс (Ficoll-Paque™ PLUS) тубу и обрабатывалась в соответствии с процедурами производителя с целью выделения лимфоцитов (Amersham Biosciences).
Выделение РНК и синтез кДНК
После выделения и осаждения лимфоцитов, клетки суспендировались в реагенте TRIzol LS (Invitrogen) для выделения тотальной РНК в соответствии с прилагаемым производителем протоколом и подвергались реакции обратной транскрипции с использованием 1 микрограмма РНК, Superscript® III RT вместе с dT20, в соответствии с процедурами производителя (Invitrogen).
ПЦР-амплификация кДНК
кДНК амплифицировалась в ПЦР-реакции с использованием комбинаций праймеров, которые позволяли амплифицировать приблизительно 110 различных V-генов мыши, принадлежащих к 15 VH-семействам (таблица 1; идентификатор последовательности NG_005838; Thiebe et al., 1999. European Journal of Immunology 29: 2072 - 2081). В первом цикле были использованы комбинации праймеров, которые связывались с 5'-концом V-генов и 3'-концом J-областей. Во втором цикле ПЦР-продукты, которые были синтезированы с праймером MJH-Rev2, подвергались амплификации с целью внесения модификаций в 3'-область для того, чтобы можно было осуществлять эффективное клонирование продуктов. В последнем цикле амплификации все ПЦР-продукты были амплифицированы с использованием праймеров, которые вводят сайт рестрикции SfiI на 5'-конце и сайт рестрикции BstEII на 3'-конце (см. фигуры 1 и 2 и таблицу 1).
Условия реакции для первого цикла ПЦР: 4 различных реакции, объединяющие все 25 прямых праймеров (MVH1-MVH25, таблица 1 и Фигура 2) и 1 обратный праймер в каждой реакции (MJH-Rev1, MJH-Rev2, MJH-Rev3 или MJH-Rev4; см. таблицу 1 и Фигуру 2). 50-микролитровые емкости ПЦР состояли из 2 микролитров кДНК (из RT-реакций), 10 микролитров полимеразного HF-буфера 5* Phusion, 40 нМ каждого из 25 прямых праймеров (суммарная концентрация - 1 микромоль), 1-микромолярного обратного праймера, 1 микролитра 10 мМ исходного раствора dNTP, 1,25 единиц полимеразы Phusion и стерильной MQ-воды. Программа термоциклера состояла из программы по схеме «touchdown»: 1 цикл при 98°C в течение 30 секунд, 30 циклов при 98°C в течение 10 секунд, снижение до 58°C с шагом 0,2°C на 10-секундный цикл, 72°C в течение 20 секунд и 1 цикл при 72°C в течение 3 минут. Второй цикл программы ПЦР устанавливался только для продуктов первой ПЦР, которые содержали праймер MJH-Rev2: 2 различных реакции, объединяющие праймеры ExtMVH-1 или ExtMVH-2 (таблица 1 и Фигура 2) вместе с обратным праймером ExtMJH-Rev2int (таблица 1 и Фигура 2). 50-микролитровые емкости ПЦР состояли из 50 нг ПЦР-продукта (из первого цикла ПЦР), 10 микролитров полимеразного HF-буфера 5* Phusion, 500 нМ каждого прямого праймера, 1-микромолярного обратного праймера, 1 микролитра 10 мМ исходного раствора dNTP, 1,25 единиц полимеразы Phusion и стерильной MQ-воды. Программа термоциклера состояла из программы по схеме «touchdown», после которой проводился этап обычной амплификации: 1 цикл при 98°C в течение 30 секунд, 10 циклов при 98°C в течение 10 секунд, снижение до 65°C с шагом 1,5°C на 10-секундный цикл, 72°C в течение 20 секунд, 10 циклов при 98°C в течение 10 секунд, 55°C в течение 10 секунд, 72°C в течение 20 секунд и 1 цикл при 72°C в течение 3 минут. Третий цикл программы ПЦР устанавливался в соответствии с описанным на Фигуре 2. 50-микролитровые емкости ПЦР состояли из 50 нг ПЦР-продукта (из предыдущих циклов ПЦР, Фигура 2), 10 микролитров полимеразного HF-буфера 5* Phusion, 1-микромолярного прямого праймера (таблица 1 и Фигура 2), 1-микромолярного обратного праймера, 1 микролитра 10 мМ исходного раствора dNTP, 1,25 единиц полимеразы Phusion и стерильной MQ-воды. Программа термоциклера состояла из программы по схеме «touchdown», после которой проводился этап обычной амплификации: 1 цикл при 98°C в течение 30 секунд, 10 циклов при 98°C в течение 10 секунд, снижение до 65°C с шагом 1,5°C на 10-секундный цикл, 72°C в течение 20 секунд, 10 циклов при 98°C в течение 10 секунд, 55°C в течение 10 секунд, 72°C в течение 20 секунд и 1 цикл при 72°C в течение 3 минут. После ПЦР-амплификаций все ПЦР-продукты очищались в геле с использованием Qiaex II в соответствии с протоколами производителя.
Расщепление рестрикционными ферментами
Очищенные продукты были расщеплены BstEII и SfiI в два этапа. Сначала 1 микрограмм ДНК расщеплялся в 100 микролитровых реакциях, состоящих из 10 микролитров 10* NEB буфера 3 (New England Biolabs), 1 микролитра 100* BSA, 12,5 единиц BstEII и стерильной воды в течение 6 часов при 60°C в сушильной камере. Продукт были очищены с использованием набора для ПЦР-очистки Qiaquick ПЦР от Qiagen в соответствии с инструкциями пользователя, элюированы в 40 микролитрах воды. Затем все продукты подвергались дальнейшему расщеплению с SfiI в 100 микролитровых реакциях, состоящих из 10 микролитров 10* NEB буфера 2 (New England Biolabs), 1 микролитра 100* BSA, 12,5 единиц SfiI и стерильной воды в течение 12 часов при 50°C в сушильной камере. Расщепленные фрагменты были очищены с помощью набора для извлечения из геля Qiaquick, после разделения геля на 20 см 1,5% агарозной TBE плюс гель бромида этидия на 80 В. 100 микрограмм вектора-акцептора (MV1043, фигуры 3 и 12) были расщеплены с помощью 50 единиц Eco91I в 600 микролитрах при стандартных условиях (буфер Tango) и затем очищены на 0,9% агарозном геле. После второго этапа расщепления при заданных условиях с 400 единицами SfiI в 500 микролитрах в течение 12 часов, было добавлено 100 единиц BsrGI на 3 часа при 50°C.
Сшивание
Каждый ПЦР-продукт подвергался отдельному сшиванию в соответствии со следующей схемой: 70 нг расщепленных ПЦР-продуктов, 300 нг расщепленного вектора-акцептора, 100 единиц лигазы T4 (NEB), 1* лиганзный буфер в 30 микролитрах в течение 16 часов при 12°C. Реакции сшивания подвергались очистке с помощью экстракций фенола/хлороформа/изоамилового спирта, после чего следовало осаждение гликогеном (Sigma-Aldrich #G1767) в соответствии с протоколом производителя, и в конце выполнялось растворение в 25 микролитрах стерильной воды.
Трансформации и хранение в библиотеке
Очищенные продукты сшивания были трансформированы посредством электропорации с использованием 1200 микролитров электрокомпетентной бактерии TG1 (Stratagene #200123) на партию сшитого продукта и высеяны на LB-карбенициллиновые чашки, содержащие 4% глюкозу. Библиотеки были собраны посредством соскабливания бактерий в 50 мл LB-карбенициллине. После центрифугирования на 2000g в течение 20 минут при 4°C, бактериальный осадок был аккуратно ресуспендирован в 2 мл ледяного 2*TY/30% глицерола в ледяной воде и заморожен на сухом льде/этаноле до хранения при -80°C.
Амплификация библиотеки
Библиотеки были выращены и собраны в соответствии с процедурами, описанными Kramer et al. 2003 (Kramer et al. 2003. Nucleic Acids Res. 31(11): e59) с использованием VCSM13 (Stratagene) в качестве вспомогательного штамма фага.
Выбор фагов на покрытых иммунотубах (immunotubes)
Столбнячный токсин был растворен в PBS в концентрации 2 мкг/мл и нанесен слоем на Nunc-иммунотубы MaxiSorp (Nunc 444474) и оставлен в течение ночи при 4°C. После исключения покрывающего раствора, тубы были блокированы 2% обезжиренным молоком (ELK) в PBS (блокирующий буфер) в течение 1 часа при комнатной температуре. Параллельно, 0,5 мл фаговой библиотеки было смешано с 1 мл блокирующего буфера и инкубировалось в течение 20 минут при комнатной температуре. После блокирования фагов фаговый раствор был добавлен к покрытым столбнячным токсином тубам и инкубировался в течение 2 часов при комнатной температуре на медленно вращающейся платформе с тем, чтобы допустить связывание. Затем тубы промывались 10 раз PBS/0,05% Tween-20, после чего проводилось элюирование фагов посредством инкубации с 1 мл 50 мМ глицина-HCl pH 2,2 в течение 10 минут при комнатной температуре на вращающемся колесе, после чего сразу проводилась нейтрализация собранного элюента с помощью 0,5 мл 1 M трис-HCl pH 7,5.
Сбор фаговых клонов
5 мл культуры XL1-Blue MRF (Stratagene) при O.D. 0,4 было добавлено к собранному раствору фагов, и была проведена инкубация в течение 30 минут при 37°C без перемешивания с целью допущения инфицирования фагов. Бактерии высевались на чашки с карбенициллином/тетрациклином и 4% глюкозой 2*TY и выращивались в течение ночи при 37°C.
Продуцирование фагов
Фаги были выращены и обработаны в соответствии с описанным Kramer et al. 2003 (Kramer et al. 2003. Nucleic Acids Res. 31(11): e59) с использованием VCSM13 в качестве вспомогательного штамма фага.
Иммуноферментный анализ фагов (ELISA)
Чашки для иммуноферментного анализа (ELISA) были покрыты 100 микролитрами столбнячного токсина на 1 лунку при концентрации 2 микрограмма/мл в PBS, и оставлены на ночь при 4°C. Чашки, покрытые 100 микролитрами тироглобулина при концентрации 2 микрограмма/мл в PBS были использованы в качестве отрицательного контроля. Лунки были опустошены, высушены посредством выпуска жидкости на бумажном полотенце, заполнены полностью PBS-4% обезжиренным молоком (ELK) и инкубировались в течение 1 часа при комнатной температуре для блокирования лунок. После исключения блокирующего раствора, фаговые миникомпозиции, предварительно смешанные с 50 мкл блокирующего раствора, были добавлены и инкубировались в течение 1 часа при комнатной температуре. Затем посредством проведения 5 этапов промывания PBS-0,05% Tween-20 были удалены несвязанные фаги. Связанные фаги были обнаружены путем инкубирования лунок в 100 микролитрах анти-M13-HRP конъюгата антитела (разведенного 1/5000 в блокирующем буфере) в течение 1 часа при комнатной температуре. Свободные антитела были удалены посредством повторения этапов промывания, описанных выше, после чего проводилось инкубирование с помощью субстрата TMB до момента, когда формирование цвета стало заметным. Реакция была остановлена путем добавления 100 микролитров 2M H2SO4 на лунку, после чего был проведен анализ на аппарате для чтения ELISA при длине волны излучения 450 нм (Таблица 2). Большие числа указывают более сильные сигналы, и, следовательно, более высокую частоту специфичного связывания комплекса фаг-Fab.
Секвенирование
Клоны, которые давали сигналы по меньшей мере в 3 раза превышающие фоновый сигнал (Таблица 2) были размножены, использованы для miniprep-процедур ДНК (см. процедуры из руководства Qiagen miniPrep) и подвергнуты анализу нуклеотидной последовательности. Секвенирование выполнялось в соответствии с инструкцией, прилагаемой к набору Big Dye 1.1 (Applied Biosystems) с использованием обратного праймера (CH1_Rev1, таблица 1), распознающего 5'-последовательность CH1-области тяжелой цепи IgG1 человека (присутствует в векторе дисплея Fab MV1043, фигуры 3 и 12). Последовательности VH мыши для 28 связывающих столбнячный токсин клонов приведены в Таблице 3. Результаты показывают, что выбранные VH-гены мыши принадлежат к различным семействам генов, и различные отдельные представители этих семейств генов могут попарно соединяться с перестроенной VH-областью IGKV1-39/J человека с образованием функциональных сайтов связывания антитела, специфичных для столбнячного токсина. На основании анализа последовательностей был сделан вывод о том, что VH-области мыши используют разнообразие сегментов генов DH и JH.
Пример 3
Сайленсинг локуса каппа-легкой цепи мыши
В данном примере описывается сайленсинг эндогенного локуса каппа-легкой цепи мыши. Эндогенный локус каппа-легкой цепи модифицируется посредством гомологичной рекомбинации в ES-клетках, после чего в эмбрионы мыши вводятся генетически модифицированные ES-клетки с целью получения генетически приспособленного потомства.
Вектор, который содержит собранную нуклеотидную последовательность, состоящую из части, содержащей J-область до 338 п.н. по ходу транскрипции от сегмента гена J5, слитой с последовательностью, заканчивающейся на 3'-конце энхансером 3'-CK, используется для гомологичной рекомбинации в ES-клетках. Собранная последовательность используется для удаления фрагмента геномной ДНК, проходящей от 3'-конца JK-области точно до 3'-конца 3'-CK энхансера. В результате этой процедуры удаляются постоянный ген CK, 3'-энхансер и некоторые межгенные участки (см. фигуры 4 и 18).
Конструирование целевого вектора
Вектор, получивший 4,5-8 т.п.н. фланкирующие «плечи» на 3'- и 5'-конце, слитый с сегментом удаления, был использован для нацеленной гомологичной рекомбинации в клеточной линии ES. Оба «плеча» были получены средствами ПЦР, обеспечивающими максимальную гомологию. Стратегия нацеливания допускает создание конститутивного KO-аллеля. Геномная последовательность мыши, охватывающая интронный энхансер Igk, константную область Igk 3'-энхансер Igk, была заменена кассетой PuroR, которая была фланкирована F3-сайтами и вставлена по ходу транскрипции от Jk-элементов. Flp-опосредованное удаление маркера выбора привело к образованию конститутивного KO-аллеля. Замена Igk области генома MiEk-Igk C-Igk 3'E (приблизительно, 10 т.п.н.) на кассету F3-Puro (приблизительно 3 т.п.н.), по-видимому, понизила эффективность гомологичной рекомбинации. Следовательно, «плечи» гомологии были соответствующим образом расширены и было проанализировано большее количество клеточных колоний ES после трансфекции с целью идентификации гомологичных рекомбинантных клонов.
Получение ES-клеток, несущих удаленный каппа-фрагмент
Получение генетически модифицированных ES-клеток было выполнено в основном в соответствии с описанным (Seibler et al. Nucleic Acids Res. 2003 Feb 15;31(4):el2). См. также подробное описание в примере 14.
Получение ES-мышей посредством комплементации тетраплоидного зародыша.
Получение мышей посредством комплементации тетраплоидного зародыша с использованием модифицированных ES-клеток было выполнено в основном в соответствии с описанным (Eggan et al., PNAS 98, 6209-6214; Seibler J, et al. Nucleic Acids Res. 2003 Feb 15;31(4):el2; Hogan et al., (Summary of mouse development. Manipulating the Mouse Embryo, (1994) Cold Spring Harbor Laboratory Press, Cold Spring Harbor NY.), pp. 253-289.)).
Пример 4
Сайленсинг локуса лямбда-легкой цепи мыши
В данном примере описывается сайленсинг эндогенного локуса лямбда-легкой цепи мыши. Эндогенный локус лямбда-легкой цепи мыши модифицируется посредством гомологичной рекомбинации в ES-клетках, после чего в эмбрионы мыши вводятся генетически модифицированные ES-клетки с целью получения генетически приспособленного потомства.
Удалению подвергаются две области лямбда-локуса мыши, которые вместе содержат все функциональные лямбда-V-области.
Первая область, намеченная для основанного на гомологичной рекомбинации удаления, представляет собой область, находящуюся на 408 п.н. против хода транскрипции от точки начала сегмента гена IGLV2 и заканчивающуюся на 215 п.н. по ходу транскрипции от сегмента гена IGLV3, включая межгенную последовательность, находящуюся между сегментами генов IGLV. Вторая область, подвергающаяся удалению, включает сегмент IGLVL-гена, состоящий из фрагмента, охватывающего область от 392 п.н. против хода транскрипции до 171 п.н. по ходу транскрипции относительно сегмента IGLVL-гена. В результате двух этапов удаления, удаляются все функциональные сегменты генов V-лямбда, что делает локус функционально неактивным (фигуры 5 и 19).
Конструирование целевых векторов
Вектор, получивший 3-9,6 т.п.н. фланкирующие «плечи» на 3'- и 5'-конце, слитый с сегментом удаления, был использован для нацеленной гомологичной рекомбинации в клеточной линии ES. Оба «плеча» были получены средствами ПЦР, обеспечивающими максимальную гомологию. На первом этапе геномная последовательность мыши, охватывающая области Igl V2-V3, была заменена кассетой PuroR, которая была фланкирована F3-сайтами, что приводит к образованию конститутивного KO-аллеля после Flp-опосредованного удаления маркера выбора (см. Фигуру 19A). На втором этапе геномная последовательность мыши, охватывающая VL-область Igl, была заменена кассетой Neo в клонах ES-клеток, в которых уже было проведено удаление областей Igl V2-V3 (см. Фигуру 19B). Маркер выбора (NeoR) был фланкирован FRT-сайтами. Конститутивный KO-аллель был получен после Flp-опосредованного удаления маркеров выбора.
Получение ES-клеток, несущих удаленный лямбда-фрагмент
Получение генетически модифицированных ES-клеток было выполнено в основном в соответствии с описанным (Seibler J, Zevnik B, Kuter-Luks B, Andreas S, Kern H, Hennek T, Rode A, Heimann C, Faust N, Kauselmann G, Schoor M, Jaenisch R, Rajewsky K, Kiihn R, Schwenk F. Nucleic Acids Res. 2003 Feb 15;31(4):el2). См. также подробное описание в примере 14. С целью демонстрации того, что оба нацеливания имели место на одной и той же хромосоме, клоны с двойным нацеливанием были выбраны для удаления in vitro с помощью pCMV C31дельтаCpG. Клоны были размножены под давлением антибиотиков на митотически инактивированном фидерном "питающем" слое, содержащем фибробласты эмбрионов мыши в DMEM-среде с высоким содержанием глюкозы, содержащей 20% FCS (PAN) и 1200 единиц/мл фактора подавления лейкемии (Millipore ESG 1107). 1×107 клеток из каждого клона были подвергнуты электропорации с 20 мкг циркулярного pCMV C31дельтаCpG при 240 В и 500 мкФ, и нанесены на четыре 10 см чашки для каждого клона. Через 2-3 дня после электропорации клетки были собраны и проанализированы посредством ПЦР. Использовались следующие праймеры:
2005_5: CCCTTTCCAATCTTTATGGG
2005_7: AGGTGGATTGGTGTCTTTTTCTC
2005_9: GTCATGTCGGCGACCCTACGCC
Реакции ПЦР выполнялись в смесях, содержащих 5 мкл ПЦР-буфера 10x (Invitrogen), 2 мкл MgCl2 (50 мМ), 1 мкл dNTP (10 мМ), 1 мкл первого праймера (5 мкМ), 1 мкл второго праймера (5 мкМ), 0,4 мкл Taq (5 единиц/мкл, Invitrogen), 37,6 мкл H2O и 2 мкл ДНК. Использовалась программа: 95°C, 5 минут; после чего проводилось 35 циклов при 95°C в течение 30 секунд; 60°C в течение 30 секунд; 72°C в течение 2 минуты; затем - 72°C в течение 10 секунд.
Получение ES-мышей посредством комплементации тетраплоидного зародыша.
Получение мышей посредством комплементации тетраплоидного зародыша с использованием модифицированных ES-клеток было в выполнено в основном в соответствии с описанным (Eggan et al., PNAS 98, 6209-6214; Seibler J, Zevnik B, Kuter-Luks B, Andreas S, Kern H, Hennek T, Rode A, HeimannC, Faust N, Kauselmann G, Schoor M, Jaenisch R, Rajewsky K, Kiihn R, Schwenk F. Nucleic Acids Res. 2003 Feb 15;31(4):el2; Hogan et al., (Cold Spring Harbor Laboratory Press, Cold Spring Harbor NY.), pp. 253-289).
Пример 5
Конструирование экспрессионной вставки CAGGS на основании перестроенного гена IGKV1-39/J-Ck человека (IGKV1-39/J-Ck) зародышевой линии
В данном примере описывается конструирование экспрессионной кассеты CAGGS, содержащей перестроенную область IGKV1-39/J зародышевой линии человека. Данная вставляемая экспрессионная кассета охватывает сайты клонирования, последовательность Козака, лидерную последовательность, содержащую интрон, открытую рамку считывания перестроенной области IGKV1-39, константную CK-область крысы из аллеля «a» и последовательность завершения трансляции (IGKV1-39/J-Ck; Фигура 6). Первичная конструкция состоит из встречающихся в природе последовательностей, и была проанализирована и оптимизирована путем удаления нежелательных действующих в cis-положении элементов, таких как внутренние TATA-боксы, сигналы полиаденилирования, chi-сайты, сайты исходного положения рибосомы, AT-богатые или GC-богатые фрагменты последовательности, элементы последовательности ARE-, INS- и CRS, последовательности повторов, вторичные структуры РНК, (латентные) сайты донора и акцептора сплайсированного фрагмента и точки разветвления сплайсинга (GeneArt GmbH). Кроме того, частота использования кодонов в участках открытой рамки считывания оптимизируется для экспрессии в мышах. Последовательность интрона не изменяется и, следовательно, представляет собой последовательность, идентичную кодирующей части лидерного интрона IGKV1-39 человека.
На 5'-конце экспрессионной кассеты был внесен сайт NotI, и на 3'-конце - сайт NheI. Оба сайта используются для клонирования в модуле экспрессии CAGGS. После сборки гена в соответствии с методами, используемыми GeneArt, вырезается вставка с NotI-NheI и клонируется в экспрессионный модуль, содержащий CAGGS-промотор, стопперную последовательность, фланкированную сайтами LoxP ('флоксированную'), последовательность сигнала полиаденилирования и, на 5'- и 3'-концах, последовательности, способствующие гомологичной рекомбинации в локус Rosa26 клеточных линий ES мыши. Промотор и/или фрагменты кДНК были амплифицированы посредством ПЦР, подтверждены секвенированием и/или клонированием напрямую из доставленных плазмид в вектор обмена RMCE, обладающий указанными характеристиками. Схематическое изображение и подтвержденная последовательность конечного целевого вектора pCAGGS-IgVK1-39 показаны на Фигурах 13A и 13B. Стратегия нацеливания изображена на Фигуре 13C.
Пример 6
Экспрессионная вставка CAGGS, основанная на перестроенной V-лямбда-области зародышевой линии IGLV2-14/J (IGLV2-14/J-Ck)
В данном примере описывается последовательность и вставка экспрессионной кассеты, содержащей перестроенную V-лямбда-область зародышевой линии IGLV2-14/J. Данная вставка охватывает сайты клонирования, последовательность Козака, лидерную последовательность, содержащую интрон, открытую рамку считывания перестроенной области IGLV2-14/J, константную CK-область крысы из аллеля «a» и последовательность завершения трансляции (IGLV2-14/J-Ck; Фигура 7). Первичная конструкция состоит из встречающихся в природе последовательностей, и была проанализирована и оптимизирована путем удаления нежелательных действующих в cis-положении элементов, таких как внутренние TATA-боксы, сигналы полиаденилирования, chi-сайты, сайты исходного положения рибосомы, AT-богатые или GC-богатые фрагменты последовательности, элементы последовательности ARE-, INS- и CRS, последовательности повторов, вторичные структуры РНК, (латентные) сайты донора и акцептора сплайсированного фрагмента и точки разветвления сплайсинга (GeneArt GmbH). Кроме того, частота использования кодонов в участках открытой рамки считывания оптимизируется для экспрессии в мышах. Последовательность интрона не изменяется и, следовательно, представляет собой последовательность, идентичную кодирующей части лидерного интрона IGKV1-39 человека.
На 5'-конце экспрессионной кассеты был внесен сайт NotI, и на 3'-конце - сайт NheI. Оба сайта используются для клонирования в модуле экспрессии CAGGS, как описано Taconic Artemis. После сборки гена в соответствии с методами, используемыми GeneArt, вырезается вставка с NotI-NheI и клонируется в экспрессионный модуль, содержащий CAGGS-промотор, стопперную последовательность, фланкированную сайтами LoxP ('флоксированную'), последовательность сигнала полиаденилирования и, на 5'- и 3'-концах, последовательности, способствующие гомологичной рекомбинации в локус Rosa26 клеточных линий ES мыши. Для создания конечного нацеленного на ROSA26 RMCE вектора, промотор и/или фрагменты кДНК были амплифицированы посредством ПЦР. Амплифицированные продукты были подтверждены секвенированием и/или клонированием напрямую из доставленных плазмид в вектор обмена RMCE, обладающий указанными характеристиками. Схематическое изображение и подтвержденная последовательность конечного целевого вектора pCAGGS-IgVL2-14 показана на Фигурах 15A и 15B. Стратегия нацеливания изображена на Фигуре 15C.
Пример 7
Экспрессия IGKV1-39/J-Ck в клеточных линиях HEK293 (pSELECT-IGKV1-39/J-Ck)
В данном примере описывается способ проверки того, что конструкции IGKV1-39/J-Ck, описанные в примере 5, дают возможность экспрессии и обнаружения L-цепи IGKV1-39/J-Ck в клетках HEK293. Вставка IGKV1-39/J (Фигура 6) была модифицирована на 5'-конце путем замены сайта NotI на сайт SalI. Данная замена требуется для клонирования продукта в плазмиду экспрессионной кассеты pSELECT-hygro (InvivoGen). CAGGS-экспрессионная вставка IGKV1-39/J-Ck и pSELECT-hygro были расщеплены с SalI и NheI, сшиты и использованы для трансформации компетентных XL1-Blue клеток с использованием стандартных методик. Колонии были собраны и подвергнуты очистке ДНК с использованием колонок Qiagen Midi-prep в соответствии с процедурами производителя. Результирующий экспрессионный вектор легкой цепи (LC), названный 0817676_pSELECT_0815426, был использован для трансфекции клеток HEK293 препаратом Fugene6 (Roche) в соответствии с протоколами производителя. Был проведен скрининг супернатантов на предмет наличия легких цепей IGKV1-39/J-Ck посредством ELISA и вестерн-блоттинга с использованием антитела против Ck крысы (Beckton Dickinson #550336 и 553871) и протоколов, используемых в технике.
VH IgG MG1494 против столбнячного токсина (TT) была клонирована в экспрессионный вектор IgG MV1056 с использованием сайтов рестрикции SfiI и BstEII. У полученного в результате клона была проверена последовательность. Клетки HEK293T были трансфицированы пятью различными комбинациями векторов, как показано в Таблице 4 (см. пример 8 для подробностей о векторе 0817678_pSELECT_0815427). Супернатанты были собраны и были определены концентрации IgG (см. Таблицу 4). IgG не были обнаружены для супернатантов A и B, содержащих только легкую цепь, как и ожидалось (детекторное антитело распознавало Fc-часть IgG). Концентрация IgG в супернатантах C и D была сравнимой с концентрацией для супернатанта положительного контроля E, что указывает на правильную экспрессию конструкций легкой цепи.
Связывание с TT было проанализировано посредством ELISA для проверки функциональности продуцированных антител, с использованием гемоглобина в качества антигена отрицательного контроля. TT-специфическое связывание не могло быть обнаружено для супернатантов A и B, содержащих только легкую цепь, как и ожидалось. TT-специфическое связывание для супернатантов C и D было по меньшей мере настолько же хорошим, как и для супернатанта положительного контроля E, что подтверждает правильность экспрессии конструкций легкой цепи и функциональность их сборки с тяжелой цепью. Антитела были обнаружены не только с использованием вторичного антитела против IgG человека, но также и с использованием вторичного антитела против легкой цепи Скаппа крысы. Результаты подтверждают, что антитело против Cкаппа крысы (BD Pharmingen #553871, клон MRK-I) распознает легкую цепь, экспрессируемую векторами pSELECT.
Супернатанты были проанализированы с помощью нередуцирующего SDS-PAGE и вестерн-блоттинга (не показано). Детектирование с использованием антитела против тяжелой цепи IgG человека не показало полосы для супернатантов A и B, содержащих только легкую цепь, как и ожидалось. Результаты для супернатантов C и D были сравнимы с супернатантом положительного контроля E, при этом имелась полоса вблизи 170 кДа-маркера, как и ожидалось для интактного IgG. Дополнительные полосы для низких молекулярных масс также наблюдались для супернатантов C, D и E, которые могут соответствовать продуктам распада, фрагментам IgG, получающимся из-за (частичного) редуцирования и/или полосам для не относящихся к делу белков из-за неспецифического связывания детекторного антитела.
Детектирование с использованием антитела против легкой цепи Скаппа крысы показало полосу, близкую к 26 кДа-маркеру для супернатантов A и B, как и ожидалось только для легкой цепи. Данная полоса была намного более выраженной для A по сравнению с B, что указывает на то, что свободная легкая цепь IGKV1-39 может быть лучше экспрессирована и/или более стабильна, чем свободная легкая цепь IGLV2-14. Как и ожидалось, для супернатанта положительного контроля E не было обнаружено полос, поскольку экспрессированный IgG содержит Cкаппа-легкую цепь человека. Для супернатантов C и D наблюдались ожидаемые полосы вблизи 170 кДа-маркера; полосы для низких молекулярных масс также наблюдались, но в меньшей степени, чем описывалось выше для антитела против IgG человека.
В заключение необходимо отметить, что трансфекция экспрессионных конструкций легкой цепи, объединенных с тяжелой цепью противостолбнячного токсина (TT) IgG MG 1494, привела к продуцированию IgG, сравнимому с конструкцией положительного контроля, для конструкций pSELECT для обеих легких цепей каппа- и лямбда. Продуцирование IgG в обоих случаях дало сигналы ELISA в TT ELISA, которые были лучше чем, или сравнимы с контрольным IgG. SDS-PAGE и вестерн-блоттинг анализ подтвердили наличие интактного IgG. Протестированное антитело против крысиной Cкаппа работало эффективно и в ELISA, и на вестерн-блоте. Культура супернатантов из клеток, трансфицированных конструкциями только с легкой цепью, не привело ни к детектируемому продуцированию IgG, ни к детектируемому TT-специфическому связыванию, тогда как свободная легкая цепь детектировалась на вестерн-блоте.
Пример 8
Экспрессия IGLV2-14/J-Ck в клеточных линиях HEK293 (pSELECT-IGLV2-14/J-Ck)
В данном примере описывается способ проверки того, что конструкции IGLV2-14/J, описанные в примере 6, дают возможность экспрессии и обнаружения L-цепи IGLV2-14/J-Ck в клетках HEK293. Вставка IGLV2-14/J-Ck (Фигура 7) была модифицирована на 5'-конце путем замены сайта NotI на сайт SalI. Данная замена требуется для клонирования продукта в плазмиду экспрессионной кассеты pSELECT-hygro (InvivoGen). CAGGS-экспрессионная вставка IGLV2-14/J-Ck и были расщеплены с SalI и NheI, сшиты и использованы для трансформации компетентных XL1-Blue клеток с использованием стандартных методик. Колонии были собраны и подвергнуты очистке ДНК с использованием колонок Qiagen Midi-prep в соответствии с процедурами производителя. Результирующий экспрессионный вектор легкой цепи (LC), названный 0817678_pSELECT_0815427, был использован для трансфекции клеток HEK293 препаратом Fugene6 (Roche) в соответствии с протоколами производителя. Был проведен скрининг супернатантов на предмет наличия легких цепей IGLV2-14/J-Ck посредством ELISA и вестерн-блоттинга с использованием антитела против Ck крысы (Becton Dickinson #550336 и 553871) и протоколов, используемых в технике. Подробности и результаты см. в примере 7.
Пример 9
Конструирование управляемой промотором экспрессионной конструкции VK, содержащей вставку IGKV1-39/J и множество энхансерных элементов, полученных из CK-локуса мыши (VkP-IGKV1-39/J-Ck; VkP-O12)
В данном примере описывается конструкция экспрессионной кассеты, которая содержит необходимые элементы для обеспечения специфичной для B-клеток и фазы развития/дифференциации экспрессии перестроенной VK-области IGKV1-39 человека, основанная на области VK-промотора IGKV1-39, лидерной последовательности, содержащей интрон, V-гене зародышевой линии, CDR3, сегменте IGKJ, межгенной области мыши, расположенной между Jk и CK, открытой рамке считывания Ck-аллеля «a» крысы и межгенном фрагменте мыши от 3'-конца CK-гена мыши, заканчивающемся точно перед 3'-концом 3'CK-энхансера.
Оптимизированные открытые рамки считывания лидерной последовательности, перестроенный ген IGKV1-39, и ген крысиного CK-аллеля «a», в соответствии с описанным в примере 5, были использованы для конструирования экспрессионной кассеты. Область VK-промотора была получена с помощью процедур синтеза генов (GeneArt, GmbH), и является практически идентичной последовательности области IGKV1-39 человека между -500 п.н. и ATG (точка начала) гена. Единственное отклонение от природной последовательности состоит во внесении последовательности Козака GCCACCATGG в точку сайта ATG (стартовую) с целью способствования трансляции. Фрагмент генома из BAC-клона мыши (TaconicArtemis) использовался в качестве основы для внесения отдельных элементов. Данный фрагмент является идентичным последовательности VK-локуса мыши, начиная с донорного сайта интрона, расположенного точно у 3'-конца области JK5, и заканчивая точно перед 3'-концом 3' CK- энхансера, и покрывает приблизительно 12,5 т.п.н.
Конечная конструкция содержит от 5'- до 3'-конца следующие элементы: геномный промотор IGKV1-39 человека (500 п.н.), последовательность Козака, лидерная последовательность IGKV1-39 человека, часть 1 (оптимизированная), лидерный интрон IGKV1-39 человека, лидерная последовательность IGKV1-39 человека, часть 2 (оптимизированная), ген зародышевой линии IGKV1-39 человека (оптимизированный), J-область человека (оптимизирована), межгенная область мыши, содержащая интронный энхансерный элемент, константная каппа-область крысы (Rattus norvegicus) (оптимизирована) и межгенная область мыши, содержащая 3'каппа-энхансер. Элементы данной экспрессионной кассеты показаны на Фигуре 8 и названы VkP-IGKV1-39/J-Ck (VkP-O12). Схема вектора pVkP-O12 и стратегия нацеливания изображены на Фигурах 20A и 21A. Вектор был внесен в ES-клетки с использованием стандартных процедур (см. пример 14).
Пример 10
Конструирование управляемой промотором экспрессионной конструкции VK, содержащей клон IGLV2-14/J и множество энхансерных элементов, полученных из CK-локуса мыши (VkP- IGLVL2-14/J-Ck; VkP-2a2).
В данном примере описывается та же конструкция, что была описана в примере 9, за исключением того, что ген IGKV1-39 и J-область заменены оптимизированным геном IGLV2-14 зародышевой линии человека, содержащим область V-J (VkP-IGLV2-14/J-Ck; VkP-2a2; Фигура 9).
Пример 11
Конструирование управляемой промотором экспрессионной конструкции VK, содержащей клон IGKV1-39, не имеющий CK-интронного энхансерного элемента (VkP-IGKV1-39/J-Ck-Δ1; VkP-O12-del1)
Конструкция, описанная в примере 9, была модифицирована путем удаления CK-интронного энхансерного элемента, расположенного в межгенной области между J-областью человека и CK-областью крысы, посредством стандартных методологий ПЦР-модификации и клонирования ДНК (GeneArt, GmBH). Результирующая экспрессионная кассета показана на Фигуре 10 и названа VkP-IGKV1-39/J-Ck-Δ1 (VkP-O12-del1).
Схема вектора pVkP-O12-del1 и стратегия нацеливания изображены на Фигурах 20B и 21B. Вектор был внесен в ES-клетки с использованием стандартных процедур (см. пример 14).
Пример 12
Конструирование управляемой промотором экспрессионной конструкции VK, содержащей клон IGKV1-39, не имеющий CK-интронного энхансерного элемента и имеющий укороченный 3' CK-энхансерный элемент (VkP-IGKV1-39/J-Ck-Δ2; VkP-O12-del2)
Конструкция, описанная в примере 11, была модифицирована путем укорачивания 3'CK-энхансерного элемента и удаления части межгенной 3'-области для Ck-гена крысы, с целью удаления потенциальных ингибирующих элементов. Это было достигнуто путем удаления межгенной последовательности между сайтом EcoRV (расположенном на 3'-конце Ck-гена крысы) и сайтом NcoI, присутствующем в 3'-энхансере (5993 п.н.) и дальнейшим удалением последовательности между сайтом BstXI 3'-энхансера и сайтом BstXI-3' 3'-энхансера (474 н.п.) с использованием стандартных методов. Результирующая экспрессионная кассета показана на Фигуре 11 и названа VkP-IGKV1-39/J-Ck-Δ2 (VkP-O12-del2).
Схема вектора pVkP-O12-del2 и стратегия нацеливания изображены на Фигурах 20C и 21C. Вектор был внесен в ES-клетки с использованием стандартных процедур (см. пример 14).
Пример 13
Экспрессия Vk-конструкций в клеточных линиях
Конструкции, описанные в примерах 9-12, проверяются на предмет их способности к продуцированию белков легкой цепи в клеточных линиях миеломы MPC11 (ATCC CCL167), B-клетках лимфомы WEHI231 (ATCC CRL-1702), T-клетках лимфомы EL4 (ATCC TIB-39) и в HEK293 (ATCC CRL1573). Энхансерный и промоторный элементы в конструкции позволяют осуществлять экспрессию в линиях B-клеток, но не в клеточных линиях, полученных из других тканей. После трансфекции клеточных линий с использованием очищенной линеаризованной ДНК и Fugene6 (Roche), клетки культивируются для транзиентной экспрессии. Клетки и супернатант собираются и подвергаются анализу SDS-PAGE, после чего проводится вестерн-блоттинг с использованием специфического антитела против C-каппа крысы. Супернатанты анализируются с помощью ELISA на предмет секретирования L-цепей с использованием антитела против CK крысы (Beckton Dickinson #550336).
Пример 14
Получение трансгенных ES-линий
Все конструкции, описанные в примерах 3, 4, 5, 6, 9, 10, 11 и 12, были использованы для создания отдельных стабильных трансгенных ES-линий посредством гомологичной рекомбинации. Методы получения трансгенных ES-линий через гомологичную рекомбинацию известны в данной области техники (например, Eggan et al., PNAS 98, 6209-6214; Seibler J, Zevnik B, Kuter-Luks B, Andreas S, Kern H, Hennek T, Rode A, HeimannC, Faust N, Kauselmann G, Schoor M, Jaenisch R, Rajewsky K, Kiihn R, Schwenk F. Nucleic Acids Res. 2003 Feb 15;31(4):el2; Hogan et al. (Cold Spring Harbor Laboratory Press, Cold Spring Harbor NY.), pp. 253-289.).
Для всех конструкций, описанных в примерах 5-6 и примерах 9-12, клеточная линия RMCE ES (полученная из штамма мыши 129S6B6F1-Gt(ROSA)26Sortm10Arte) была выращена на митотически инактивированном фидерном "питающем" слое, содержащем фибробласты эмбриона мыши (MEF) в DMEM-среде с высоким содержанием глюкозы, содержащей 15% FBS (PAN 1302-P220821). Фактор ингибирования лейкемии (Chemicon ESG 1107) был добавлен в среду в концентрации 900 единиц/мл. Для проведения манипуляций, 2×105 ES-клеток были высеяны на 3,5 см чашки в 2 мл среды. Непосредственно перед трансфекцией к клеткам было добавлено 2 мл свежей среды. 3 мкл реагента Fugene6 (Roche; каталожный номер 1 814 443) было смешано со 100 мкл бессывороточой среды (OptiMEM I с Glutamax I; Invitrogen; каталожный номер 51985-035) и инкубировалось в течение 5 минут. 100 мкл раствора Fugene/OptiMEM было добавлено к 2 мкг циркулярного вектора и 2 мкг CAGGS-Flp, после чего проводилась инкубация в течение 20 минут. Данный трансфекционный комплекс добавлялся к клеткам по каплям и перемешивался. Свежая среда была добавлена к клеткам на следующий день. Начиная с дня 2, среда заменялась ежедневно на среду, содержащую 250 мкг/мл G418 (Geneticin; Invitrogen; каталожный номер 10131-019). Через семь дней после трансфекции единичные клоны были выделены, было проведено их наращивание и проведен молекулярный анализ посредством саузерн-блоттинга в соответствии со стандартными процедурами.
Для каждой конструкции был проведен анализ множества клонов путем расщепления рестрикционными ферментами геномной ДНК единичных клонов, после чего проводилась гибридизация с 5'-зондами, 3'-зондами и внутренними зондами, в результате чего были получены клоны, которые содержали правильную единичную вставку в правильной позиции локуса Rosa26. Пример представлен на Фигуре 14.
Пример 15
Получение трансгенных штаммов мыши
Все линии ES-клеток, которые были получены и проверены на предмет их модификации в соответствии с описанным в примере 14, были использованы для получения стабильных трансгенных мышей посредством тетраплоидной рекомбинации. Методы известны в данной области техники. Как правило, после введения гормонов сверховулированные Balb/c самки скрещивались с Balb/c самцами. Бластоцисты были выделены из матки при dpc 3,5. Для микроинъекции бластоцисты были помещены в каплю DMEM с 15% FCS под минеральным маслом. Пьезоактивируемая пипетка для микроинъекций с плоским наконечником с внутренним диаметром 12-15 микрометров была использована для инъецирования 10-15 целевых ES-клеток C57BL/6 N.tac в каждую бластоцисту. После выделения, инъецированные бластоцисты были перенесены в каждый рог матки через 2,5 дня после спаривания самкам NMRI с ложной беременностью. Химеризм измерялся в химерах (G0) посредством вклада ES-клеток в цвет меха Balb/c-хозяина (черный/белый). Мыши с высоким химеризмом скрещивались с самками штамма C57BL/6. В зависимости от требований проекта, партнерши для спаривания C57BL/6 являются немутантными (W) или мутантными относительно наличия гена рекомбиназы (Flp-удалитель, или Cre-удалитель, или индуцируемый CreER удалитель, или комбинация Flp-удалитель/CreER). Передача зародышевой линии идентифицировалась по наличию потомства штамма C57BL/6 с черным мехом (G1).
Например, клон ESC IgVK1-39 2683 8 (см. примеры 5 и 14) был суммарно инъецирован в 62 бластоцисты в 3 независимых экспериментах. Было получено 3 помета, суммарно содержащих 6 детенышей. Все детеныши были химерными. Было получено 3 гетерозиготных потомка, которые были использованы для дальнейшего скрещивания.
ESC-клон каппа 2692 A-C10 (см. примеры 3 и 14) был инъецирован суммарно в 54 бластоцисты в 3 независимых экспериментах. Было получено 3 помета, суммарно содержащих 11 детенышей, 10 из которых были химерными. Было получено 8 гетерозиготных потомков, которые были использованы для дальнейшего скрещивания.
ESC-клон каппа 2692 B-C1 (см. примеры 3 и 14) был инъецирован суммарно в 51 бластоцисту в 3 независимых экспериментах. Было получено 2 помета, суммарно содержащих 6 детенышей, 4 из которых были химерными. Было получено 3 гетерозиготных потомка, которые были использованы для дальнейшего скрещивания.
Пример 16
Селекция
В данном примере описывается селекция с целью получения мышей, которые содержат трансгенные экспрессионные кассеты, в соответствии с описанным в примере 14, и нокаутные мыши, в которых был выполнен сайленсинг лямбда- и каппа-локусов. Локализация V-лямбда на хромосоме 16 и CD19 на хромосоме 7 позволяет применять стандартные процедуры селекции. Селекция совместно локализованных Vk-локуса и Rosa26-локуса на хромосоме 6 с расстоянием около 24 cM требует специального внимания в процессе скрининга, поскольку только часть потомства демонстрирует такой кроссинговер, что обе модификации сводятся вместе на одной хромосоме.
Все четыре локуса должны быть объединены в одном штамме мыши, который является гомо- или гетерозиготным по CD19-cre (не описан) и модифицированному трансгену Rosa26 и гомозиготным по другим локусам. Селекция выполняется соответствующим образом посредством стандартных методик селекции и скрининга в соответствии с предлагаемым компаниями коммерческой селекции (например, TaconicArtemis).
Пример 17
Иммунизация мыши
Первичная и вторичная иммунизация мышей выполняется с использованием стандартных протоколов.
В целях подтверждения трансгенной экспрессии перестроенного Vк O12 человека (IGKV1-39) - Cк-легких цепей крысы (см. примеры 5, 14-16) в B-клетки из мышей CD19-HuVκ1 и для оценки ее влияния на размер VH-репертуара, разнообразие использования VH-семейства и рекомбинацию V(D)J после иммунизации, трансгенные мыши CD19-HuVκ1 были иммунизированы вакциной столбнячного токсина (TT-вакциной), и разнообразие последовательностей VH из случайно выбранных клонов мышей CD19-HuVκ1 сравнивалось с TT-иммунизированными мышами дикого типа и CD19-Cre HuVk1 отрицательными детенышами одного помета. Были получены данные по частоте SHM для трансгена Vк O12 человека в иммунизированных мышах. Разнообразная коллекция по меньшей мере из 40 TT-специфических, не связанных клонально mAb, содержащих Vк O12 человека, была получена из мышей CD19-HuVκ1 посредством фагового дисплея.
Для этого три взрослых мыши CD19-HuVκ1 вакцинировались TT-вакциной TT с применением стандартных процедур иммунизации. После иммунизации проводилось измерение титров сыворотки с использованием TT-специфической ELISA (TT: Statens Serum Institute, номер артикула 2674), и суспензии селезенки подвергались клеточному сортингу посредством FACS-процедуры после окрашивания с Cк-специфическим моноклональным антителом крысы с целью выделения трансгенных B-клеток (клон RG7/9.1; BD Pharmingen# 553901, Lot# 06548). Были извлечены РНК из Cκ-положительной B-клетки крысы, и полученный в результате кДНК-материал был использован для построения библиотек и SHM-анализа.
Стандартное моноклональное антитело мыши против CK крысы (клон RG 7/9.1; BD Pharmingen# 553901, Lot# 06548) использовалось в FACS-анализе экспрессирующих трансген B-клеток (Meyer et al., 1996, Int. Immunol., 8: 1561). Антитело клона RG7/9.1 реагирует с монотипичной (общей) детерминантой каппа-цепи. Данное антитело против Cк крысы (клон RG7/9.1 (BD Pharmingen# 553901, Lot# 06548) помечено R-фикоэритрином (PE) с использованием набора для быстрой конъюгации LYNX в соответствии с инструкциями производителя для FACS-анализа и сортинга. Помеченное антитело сначала тестируется проточной цитометрией на предмет связывания с функциональными Cκ-содержащими белками легкой цепи крысы, продуцированными в транзиентно трансфицированных клетках HEK-293T; неконъюгированное антитело служит в качестве положительного контроля. Два других антитела, связывание которых с Cк крысы было показано посредством ELISA и вестерн-блоттинга (см. пример 7), были также протестированы с помощью проточной цитометрии.
Построение Fab-библиотеки фагового дисплея проводилось со множеством оптимизированных вырожденных ПЦР-праймеров, разработанных для амплификации VH-генов C57BL/6; минимальный размер библиотеки составил 106 клонов, и минимальная частота вставки составляет 80%. Используемый вектор MV1043 (фигуры 3 и 12) содержит Vк O12 человека, слитый с Cк-областью человека. Таким образом, Cк крысы заменяется на аналог человека в процессе создания библиотеки.
До отбора выполняется VH-секвенирование 96 случайно выбранных клонов с целью проверки разнообразия VH-репертуара, которое сравнивается с разнообразием из невыбранной библиотеки, ранее созданной с использованием тех же процедур из мышей BALB/c, иммунизированных TT. Библиотека из мышей дикого типа C57B1/6, которые иммунизировались таким же образом, позволяет провести сравнение разнообразия между двумя предварительно отобранными библиотеками, имеющими одну и ту же вакцину и один и тот же генетический фон.
Выполняется несколько независимых отборов на TT-покрытых иммунотубах. Изменения, которые могут иметь место, включают выбор с использованием биотинилированных антигенов в растворе или выбор на захваченных TT. На основании количества и разнообразия положительных клонов, согласно анализу ELISA, в первых выборках, принимаются решения о дополнительных этапах выборки. Клоны считаются положительными, когда они более чем в три раза превосходят клоны отрицательного контроля. Положительные клоны анализируются посредством ELISA относительно панели антигенов отрицательного контроля с целью проверки антигенной специфичности. Цель состоит в идентификации по меньшей мере 40 уникальных VH-областей как основанных на уникальных CDR3-последовательностях и VHDJH-перестройках.
Амплификация кДНК-материала крысиных Cκ-положительных отсортированных B-клеток выполняется с прямым ПЦР-праймером, специфическим для лидерной последовательности человека и обратным ПЦР-праймером, специфическим для Cк-последовательности крысы, в области, не избыточной по Cк-последовательности мыши, как сообщалось в недавнем исследовании (Brady et al., 2006, JIM, 315: 61). Комбинации праймеров и температуры отжига вначале тестируются на кДНК из клеток HEK-293T, трансфицированных вектором 0817676_pSELECT_0815426=pSELECT с ДНК-кассетой IGKV1-39 (см. пример 7).
Продукты амплификации клонируются в вектор pJET-1 и, после трансформации XL1-blue, 96 колоний секвенируется для оценки частоты SHM для VL путем прямого сравнения с зародышевой последовательностью VK O12 (IGKV1-39). Метод R/S, в соответствии с описанным в нашем исследовании на TT-специфических антителах человека (de Kruif et al., 2009, J. Mol. Biol., 387: 548), позволяет разделять случайные мутации и направляемые антигеном мутации, которые появляются в VL-последовательностях.
Пример 18
Иммунофлуоресцентный анализ популяций B-клеток в трансгенных линиях мыши.
В данном примере описывается использование антител и проточной цитометрии для анализа популяций B-клеток в первичных (костный мозг) и вторичных (селезенка, брюшина) лимфоидных органах и крови. Методы и реагенты описаны в работах Middendorp et al. (2002) J. Immunol. 168:2695 и Middendorp et al. (2004) J. Immunol. 172:1371. В целях анализа раннего развития B-клеток в костном мозге, клетки подвергались поверхностному окрашиванию комбинациями антител (Becton Dickinson), специфичными для B220, CD19, CD25, IgM, IgD, Cкаппа мыши, Cлямбда мыши и Cкаппа крысы для обнаружения популяций про-B-клеток, пре-B-клеток, больших пре-B-клеток, ранних и поздних незрелых B-клеток и рециркулирующих B-клеток, экспрессирующих трансген на своей поверхности. DAPI-окрашивание (Invitrogen) было применено для исключения мертвых клеток из анализа, и FC-блок (Becton Dickinson) применялся для подавления взаимодействия антитела с Fc-рецепторами на миелоидных клетках. В целях анализа экспрессии трансгена на поверхности популяций B-клеток в периферийных лимфоидных органах и крови, клетки были окрашены комбинациями антител (Becton Dickinson), специфичными для B220, CD5, CD19, CD21, CD23, IgM, IgD, Cкаппа мыши, Cлямбда мыши и Cкаппа крысы. DAPI-окрашивание было применено для исключения мертвых клеток из анализа, и FC-блок применялся для подавления взаимодействия антитела с Fc-рецепторами на миелоидных клетках. Кроме того, были включены комбинации антител (Becton Dickinson), специфичные для CD3, CD4, CD11b, CD11c и NK1.1, с целью определения, имеет ли место экспрессия трансгена в клеточных типах вне ячейки B-клеток.
Были проанализированы три мыши, гетерозиготные по трансгену IGKV1-39 человека/Cкаппа крысы и гетерозиготные по трансгену CD19-Cre на фоне C57BL6 (HuVkl/CD19-Cre). В качестве контроля для FACS-анализа были включены три детеныша того же помета дикого типа для трансгена IGKV1-39 человека/Cкаппа крысы и гетерозиготные по трансгену CD19-Cre на фоне C57BL6 (CD19-Cre) и две мыши C57BL6/NTac (дикий тип). Все животные акклиматизировались в виварии в течение 1 недели до начала анализа, и все мыши были самцами в возрасте 6 недель. Лимфоциты были выделены из бедра, селезенки, брюшинной полости и крови мышей с использованием обычных методик, в соответствии с описанным ранее (Middendorp et al. (2002) J. Immunol. 168:2695 и Middendorp et al. (2004) J. Immunol. 172:1371). Антитела были предварительно объединены в соответствии с показанным в Таблице 10, и было проведено окрашивание в 96-луночных планшетах. Инкубация с PE-конъюгированным противокрысиным C каппа (описано выше) была проведена до окрашивания анти-мышиными антителами крысы с целью предотвращения неспецифичного связывания. После завершения клеточного окрашивания, помеченные клетки были проанализированы на FACS-машине Becton Dickinson LSR II, и полученные данные были проанализированы с помощью программного обеспечения FlowJo (v6.4.7).
Трансгенные мыши были сходны с мышами дикого типа по весу, внешнему виду и активности. Во время сбора тканей не наблюдалось сильных анатомических изменений. Между трансгенными мышами и мышами дикого типа не наблюдалось разницы в количестве B-клеток в костном мозге (BM) и селезенке (Таблица 11) или в количестве B-клеток, T-клеток и миелоидных клеток в периферических органах. Кроме того, частота или доля клеток в различных каналах развития лимфоцитов не изменялась в трансгенных мышах по сравнению с мышами дикого типа. Таким образом, у двойных трансгенных (HuVk I/CD19-Cre) и трансгенных (CD19-Cre) мышей развитие лимфоидных, и, самое важное, B-клеток было неотличимым от мышей дикого типа.
В периферических лимфоидных органах окрашивание специфичным для трансгена антителом (анти-крысиное Cкаппа-PE) наблюдалось только в популяциях B-клеток. Популяции T-клеток, миелоидных клеток и NK-клеток были отрицательными по поверхностной экспрессии трансгена в селезенке (Фигура 23). В отличие от этого, в клетках, окрашенных pan-маркерами B-клеток B220 и CD19, все клетки были сдвинуты направо на FACS-плоте, что указывает на экспрессию трансгена на поверхности клетки (Фигура 24). Аналогичное трансген-специфическое окрашивание измерялось в клетках CD5+ B1 брюшной полости (отличающейся по развитию клеточной популяции B-клеток) (Фигура 25).
Дифференциация B-клеток из мультилинейных предшественников в зрелые B-клетки происходит в костном мозге. В проанализированных лимфоцитах из костного мозга, внеклеточная экспрессия и экспрессия трансгена были недетектируемыми, в самых ранних предках B-клеток, про- и пре-B-клетках, экспрессия была согласована с профилем нормальной экспрессии легкой цепи (Фигура 26). Экспрессия трансгена впервые становится детектируемой в незрелых B-клетках на стадии развития, в которых легкая цепь зародышевой линии мыши претерпевает перестройку и экспрессируются на клеточной поверхности в контексте предварительно выбранной тяжелой цепи (Фигура 26). Экспрессия трансгенной легкой цепи также обнаружена в зрелых рециркулирующих B-клетках (Фигура 26), что согласуется с окрашиванием в селезенке. Таким образом, управляемая CD19-Cre экспрессия трансгена согласуется с нормальным профилем экспрессии легкой цепи. Окрашивание специфичным для эндогенной легкой цепи антителом более интенсивно, чем для антитела, специфического для легкой цепи трансгена. Это может указывать на более высокий уровень экспрессии эндогенной легкой цепи, большую чувствительность окрашивания специфичным для эндогенной легкой цепи антителом, или на комбинацию и того, и другого. Важно, что интенсивность поверхностной экспрессии трансгенной легкой цепи коррелирует с поверхностной экспрессией как эндогенной легкой цепи, так и IgM, что наблюдалось при окрашивании циркулирующих B-клеток в крови (Фигура 27).
Таким образом, в общем данный анализ демонстрирует, что экспрессия трансгена IGKV1-39/Cкаппа человека ограничена ячейкой B-клеток и временная регуляция его экспрессии аналогична эндогенным каппа- и лямбда-легким цепям, в результате чего происходит нормальное развитие популяций B-клеток. Явно более низкий уровень экспрессии трансгена может объясняться силой промотора по сравнению с промотором и энхансерами, присутствующими для генов эндогенной легкой цепи, или задержкой в экспрессии трансгена, которая дает эндогенным легким цепям конкурентное преимущество при спаривании с перестроенной тяжелой цепью. Это согласуется с наблюдением, что при созревании B-клеток относительная интенсивность окрашивания трансгена возрастает по сравнению с эндогенными легкими цепями. Кроме того, наблюдение, что количество B-клеток является нормальным, и что каждая поверхностная Ig+ B-клетка коэкспрессирует эндогенную и трансгенную легкую цепь, подтверждает вывод о том, что вариабельная область IGKV1-39 может соединяться с нормальным репертуаром различных вариабельных областей тяжелой цепи мыши. Из этого анализа заявители сделали вывод, что вставка трансгена IGKV1-39/Cкаппа крысы, управляемая активируемым CD19-Cre CAGGS-промотором в локусе Rosa, способствует своевременной и специфичной для B-клеток экспрессии трансгена, и что трансген может соединяться с нормальным репертуаром тяжелых цепей мыши.
Пример 19
Epibase®-профиль T-клеток для IGKV1-39.
Белковая последовательность IGKV1-39 (Фигура 12, зародышевая линия человека, белок IGKV1-39/J) сканировалась на предмет наличия предполагаемых ограниченных эпитопов HLA класса II, также известных как TH-эпитопы. Для этого, платформа Algonomics' Epibase® применялась для IGKV1-39. Коротко, эта платформа анализирует специфичности связывания HLA для всех возможных 10-мерных пептидов из целевой последовательности (Desmet et al. Nature 1992, 356:539-542; Desmet et al. FASEB J. 1997, 11:164-172; Desmet et al. Proteins 2002, 48:31-43; Desmet et al. Proteins 2005, 58:53-69). Анализ профиля выполняется на уровне аллотипа для 20 DRB1, 7 DRB3/4/5, 13 DQ и 7 DP, то есть, суммарно для 47 HLA-рецепторов класса II (см. Таблицу 5). Epibase® рассчитывает количественную оценку свободной энергии связывания ΔGbind пептида для каждого из 47 HLA-рецепторов класса II. Эти данные подвергались дальнейшей обработке следующим образом:
- Свободные энергии преобразовывались в Kd-значения по формуле ΔGbind=RT ln(Kd).
- Пептиды классифицировались как имеющие сильное (S), среднее (M), слабое связывание или как несвязывающие (N). Использовались следующие граничные значения:
S: сильное связывание: Kd<0,1 мкМ.
M: среднее связывание: 0,1 мкМ≤Kd<0,8 мкМ.
N: слабое связывание и отсутствие связывания: 0,8 мкМ ≤ Kd.
- Пептиды, соответствующие собственным пептидам, обрабатывались отдельно. Список собственных пептидов был получен из 293 последовательностей антител зародышевой линии. Они обозначаются как пептиды, 'фильтрованные по зародышевой линии'.
S- и M-пептиды отображаются на целевую последовательность в так называемых картах эпитопов; S-аффинности изображаются количественно; M-значения показываются качественно. В качестве общего обзора результатов, в Таблице 6 представлено количество белков с сильной и средней связывающей способностью среди анализируемых белков, для групп HLA-рецепторов класса II, соответствующих генам DRB1, DQ, DP и DRB3/4/5. Расчет выполнялся отдельно для связей с сильной и средней аффинностью. Пептиды, связывающиеся с множеством аллотипов одной и той же группы, считались как один. Значения в скобках относятся к пептидам, фильтрованным по зародышевой линии. В Таблице 7 показана последовательность в формате, подходящем для экспериментальной работы. Последовательность разделена на последовательные 15-меры, перекрывающиеся на 12 остатков. Для каждого l5-мера приведена разнородность (количество аллотипов из суммарных 47, для которых 15-мер содержит критический связыватель), а также предполагаемые серотипы. Epibase®-профиль и карты эпитопов показаны на Фигурах 16 и 17.
Был сделан вывод о том, что IGKV1-39 не содержит сильных несобственных связывателей DRB1. Обычно для генов DRB1 находилось значительно больше связывателей, чем для других HLA-генов. Это согласуется с экспериментальными свидетельствами того, что аллотипы, принадлежащие к группе DRB1, являются более сильными связывателями пептидов. Ожидается, что эпитопы средней силы для аллотипов DRB1 вносят вклад в ответ популяции, и не могут не рассматриваться. Снова, несобственных связывателей DRB1 не было обнаружено в IGKV1-39.
В гуморальном ответе, возникающем против антигена, наблюдаемая активация/пролиферация TH-клеток обычно интерпретируется в отношении DRB1-специфичности. Однако нельзя игнорировать возможный вклад генов DRB3/4/5, DQ и DP. Учитывая, что данные гены имеют более низкие уровни экспрессии по сравнению с DRB1, большее внимание уделялось классу сильных эпитопов для DRB3/4/5, DQ и DP. 'Критичные эпитопы' - это эпитопы, имеющие сильную связывающую способность для любого аллотипа DRB1, DRB3/4/5, DQ или DP, или имеющие среднюю связывающую способность для DRB1. IGKV1-39 не содержит сильных или средних несобственных связывателей для DRB3/4/5, DQ или DP.
Ряд пептидов также присутствует в последовательностях зародышевой линии (значения в скобках в Таблице 6). Такие пептиды могут очень хорошо связываться с HLA, но предполагается, что они являются собственными и, следовательно, неиммуногенными. Суммарно в IGKV1-39 было обнаружено 6 сильных и 16 средних фильтрованных по зародышевой линии связывателей. Участок от каркасной области 1 до каркасной области 3 имеет точное соответствие с V-сегментом зародышевой линии VKI 2-1-(1) O12 (VBase), также известным как IGKV1-39*O1 (IMGT). Каркасная область 4 имеет точное соответствие с J-сегментом зародышевой линии JK1 (V-base) также известным как IGKJ1*O1(IMGT). Не является неожиданным, что данные сегменты не содержат ни одного несобственного эпитопа.
Пример 20
Характеристики продуцирования IGKV1-39
Существует большая потребность в платформах обнаружения антител, которые выдавали бы терапевтические антитела, являющиеся термодинамически стабильными и обеспечивали бы хорошую экспрессию. Эти характеристики являются важными для обеспечения стабильности лекарственного вещества во время продуцирования и после инъецирования фармацевтического продукта пациенту. Кроме того, хороший выход продуктов экспрессии напрямую влияет на стоимость производства лекарства и, следовательно, на его цену, доступность для пациентов и прибыльность. Практически все терапевтические антитела, находящиеся в данный момент в клиническом использовании, состоят из IgG1 человека и каппа-константных областей человека, но используют различные вариабельные области тяжелой и легкой цепи, что обеспечивает специфичность. Вариабельные домены тяжелой и легкой цепи человека могут быть разделены на семейства, имеющие расхождение в последовательностях более 80%. Когда перестроенные примеры данных семейств в конфигурации зародышевой линии объединяются и сравниваются по стабильности и выходу, становится ясным, что семейства генов не являются равнозначными по своим биофизическим свойствам. В частности, VH3, VH1 и VH5 обладают хорошей стабильностью для тяжелых цепей, и Vk1 и Vk3 имеют наилучшую стабильность и выход для легких цепей. Кроме того, при внесении мутаций в рамках процесса соматической гипермутации, они могут вызывать помехи для спаривания VH/VL. Для оценки влияния, которое различные гены легкой цепи с различными уровнями мутаций имеют на характеристики продуцирования фиксированной VH-цепи, была построена библиотека Fab-фагового дисплея для легких цепей (каппа- и лямбда) шести ранее не подвергавшихся экспериментам доноров, объединенная с панелью из 44 TT-связывающих тяжелых цепей из иммунизированных доноров. После одного цикла селективного TT-связывания Fab-клоны были изолированы. Некоторые из них имели такой же VH-ген, что и TT-клон PG1433, в комбинации с различными легким цепями. Fab-фрагменты легкой цепи были повторно клонированы в каппа-экспрессионном векторе и трансфицированы вместе с ДНК, кодирующей тяжелую цепь PG1433, в 293 клетки, и специфическое продуцирование IgG измерялось посредством ELISA. Как показано в таблице 8, выбранные клоны, содержащие PG1433 VH , объединялись с различными легкими цепями, имеющими от 5 до 10 раз более низкую экспрессию белка PG1433 VH, объединенного с IGKV1-39. Стоит отметить, что все легкие цепи содержали в своих кодирующих областях аминокислотные мутации, которые могли помешать спариванию с VH и могли понизить стабильность продуцирования. Таким образом, ожидается, что помимо снижения вероятности нежелательной иммуногенности, использование легкой цепи IGKV1-39 без мутации также вносит вклад в улучшение стабильности продуцирования и повышение выхода для различных вносящих вклад в специфичность VH-генов. Действительно, все стабильные клоны, созданные посредством трансфекции различных VH-генов, спаренных с IGKV1-39, могли подвергаться активному пассированию и при этом сохранять устойчивые характеристики продуцирования, как показано в таблице 9.
Пример 21
Получение мышей, экспрессирующих VH и VL-области полностью человека.
Трансгенные мыши по изобретению скрещиваются с мышами, уже содержащими локус VH человека. Примеры подходящих мышей, содержащих локус VH человека, описаны в работах (Taylor et al. (1992). Nucleic Acids Res 20: 6287-95; Lonberg et al. (1994). Nature 368: 856-9; Green et al. (1994). Nat Genet 7: 13-21; Dechiara et al. (2009). Methods Mol Biol 530: 311-24.).
После скрещивания и отбора мышей, которые были по меньшей мере гетерозиготными по трансгену IGKV1-39 и локусу VH человека, выбранные мыши иммунизировались целевым веществом. VH-гены собирались в соответствии с описанным выше в настоящем документе. Данный способ имеет преимущество в том, что VH-гены уже являются полностью человеческими и, следовательно, не требуют гуманизации.
Пример 22
Выделение, характеризация и олигоклональное форматирование и продуцирование антител, нацеленных на IL6 человека, для лечения заболеваний хронического воспаления, таких как ревматоидный артрит
Репертуар VH селезенки из трансгенных мышей, иммунизированных рекомбинантным IL6 человека, клонировался в Fab-векторе фагового дисплея с одной IGKV1-39-C каппа легкой цепью человека (идентичной трансгену мыши) и подвергался пэннингу с иммуногенным IL6 человека. Клоны, которые были получены после от двух до четырех циклов пэннинга, анализировались на предмет их специфичности связывания. VH-гены, кодирующие IL6-специфические Fab-фрагменты, подвергались анализу последовательности с целью идентификации уникальных клонов и назначения использования VH, DH и JH. Fab-фрагменты реформатировались как молекулы IgG1 и транзиентно экспрессировались. Уникальные клоны затем группировались на основании отсутствия конкурентности при анализе связывания, и подвергались анализу аффинности и функциональности. Наиболее сильные анти-IL6 IgG1 mAb последовательно экспрессировались как комбинации двух, трех, четырех или пяти тяжелых цепей, содержащих различные VH-области в формате олигоклонирования, вместе с одной основанной на IGKV1-39-C каппа-легкой цепью, и тестировались in vitro на предмет образования комплекса с IL-6. Олигоклонирование также тестировалось in vivo на предмет выведения IL-6 человека из мышей. Было выбрано олигоклонирование с наиболее сильной выводящей активностью, и VH-гены мыши гуманизировались в соответствии с обычными методами. Гуманизированный IgG1 трансфицировался в клеточную линию млекопитающего для создания стабильного клона. Выбирался оптимальный субклон для создания банка базовых клеток и получения материала для клинических исследований.
Многие протоколы, описанные здесь, являются стандартными протоколами для конструирования библиотек фагового дисплея и пэннинга фагов для связывания с представляющим интерес антигеном, и описаны, например, в Antibody Phage Display: Methods and Protocols. 2002. Editor(s): Philippa M. O'Brien, Robert Aitken. Humana Press, Totowa, New Jersey, USA.
Иммунизации
Трансгенные мыши получали три иммунизации IL6 человека каждые две недели с использованием адъюванта Sigma titerMax в соответствии с инструкциями производителя.
Выделение РНК и синтез кДНК
Через три дня после последней иммунизации, у мышей удалялись селезенки и лимфоузлы и проводились через 70-микронный фильтр в тубу, содержащую PBS, pH 7,4, для получения одноклеточной суспензии. После промывания и осаждения лимфоцитов, клетки суспендировались в реагенте TRIzol LS (Invitrogen) для выделения тотальной РНК, в соответствии с протоколом производителя, и подвергались реакции обратной транскрипции с использованием 1 микрограмма РНК Superscript® III RT, в комбинации с dT20, в соответствии с процедурами производителя (Invitrogen).
Создание Fab-библиотек фагового дисплея проводилось в соответствии с описанным в Примере 2.
Отбор фагов на покрытых иммунотубах
Рекомбинатный IL6 человека был растворен в PBS в концентрации 5 мкг/мл и нанесен слоем на Nunc-иммунотубы MaxiSorp (Nunc 444474) и оставлен в течение ночи при 4°C. После исключения покрывающего раствора, тубы были блокированы 2% обезжиренным молоком (ELK) в PBS (блокирующий буфер) в течение 1 часа при комнатной температуре (RT). Параллельно, 0,5 мл фаговой библиотеки было смешано с 1 мл блокирующего буфера и инкубировалось в течение 20 минут при комнатной температуре. После блокирования фагов фаговый раствор был добавлен к покрытым IL6 тубам и инкубировался в течение 2 часов при комнатной температуре на медленно вращающейся платформе с тем, чтобы допустить связывание. Затем тубы промывались 10 раз PBS/0,05% Tween-20, после чего проводилось элюирование фагов посредством инкубации с 1 мл 50 мМ глицина-HCl pH 2,2 в течение 10 минут при комнатной температуре на вращающемся колесе, после чего сразу проводилась нейтрализации собранного элюента с помощью 0,5 мл 1 M трис-HCl pH 7,5.
Сбор фаговых клонов
5 мл культуры XL1-Blue MRF (Stratagene) при O.D. 0,4 было добавлено к собранному раствору фагов, и была проведена инкубация в течение 30 минут при 37°C без перемешивания с целью допущения инфицирования фагов. Бактерии высевались на чашки с карбенициллином/тетрациклином и 4% глюкозой 2*TY и выращивались в течение ночи при 37°C.
Получение фагов
Фаги были выращены и обработаны в соответствии с описанным Крамером и соавт. 2003 (Kramer et al. 2003. Nucleic Acids Res. 31(11): e59) с использованием VCSM13 в качестве вспомогательного штамма фага.
Иммуноферментный анализ фагов (ELISA)
Чашки для иммуноферментного анализа (ELISA) были покрыты 100 микролитрами рекомбинантного IL6 человека на 1 лунку при концентрации 2,5 микрограмма/мл в PBS, и оставлены на ночь при 4°C. Чашки, покрытые 100 микролитрами тироглобулина при концентрации 2 микрограмма/мл в PBS были использованы в качестве отрицательного контроля. Лунки были опустошены, высушены посредством выпуска жидкости на бумажном полотенце, заполнены полностью PBS-4% обезжиренным молоком (ELK) и инкубировались в течение 1 часа при комнатной температуре для блокирования лунок. После исключения блокирующего раствора, фаговые миникомпозиции (miniprep), предварительно смешанные с 50 мкл блокирующего раствора, были добавлены и инкубировались в течение 1 часа при комнатной температуре. Затем посредством проведения 5 этапов промывания PBS-0,05% Tween-20 были удалены несвязанные фаги. Связанные фаги были обнаружены путем инкубирования лунок в 100 микролитрах анти-M13-HRP конъюгата антитела (разведенного 1/5000 в блокирующем буфере) в течение 1 часа при комнатной температуре. Свободные антитела были удалены посредством повторения этапов промывания, описанных выше, после чего проводилось инкубирование с помощью субстрата TMB до момента, когда формирование цвета стало заметным. Реакция была остановлена путем добавления 100 микролитров 2M H2SO4 на лунку, после чего был проведен анализ на аппарате для чтения ELISA при длине волны излучения 450 нм.
Секвенирование
Клоны, которые давали сигналы по меньшей мере в 3 раза превышающие фоновый сигнал (Таблица 2) были размножены, использованы для miniprep-процедур ДНК (см. процедуры из руководства Qiagen miniPrep) и подвергнуты анализу нуклеотидной последовательности. Секвенирование выполнялось в соответствии с инструкцией, прилагаемой к набору Big Dye 1.1 (Applied Biosystems) с использованием обратного праймера (CH1_Rev1, таблица 1), распознающего 5'-последовательность CH1-области тяжелой цепи IgG1 человека (присутствует в векторе дисплея Fab MV1043, фигуры 3 и 12). Последовательности VH-областей мыши анализировались на предмет разнообразия сегментов генов DH и JH.
Конструирование и экспрессия химерного IgG1
Вектор MV1057 (фигуры 12 и 22) был получен путем клонирования фрагмента L-цепи трансгена (IGKV1-39) в производную вектора pcDNA3000Neo (Crucell, Leiden, Нидерланды), которая содержала константные области IgG1 человека и каппа-константные области человека. VH-области клонировались в MV1057, и нуклеотидные последовательности для всех конструкций проверялись в соответствии со стандартными методиками. Полученные в результате конструкции транзиентно экспрессировались в клетках HEK293T, и супернатанты, содержащие химерный IgG1, были получены и очищены с использованием стандартных процедур, как было описано ранее (Throsby, M. 2006. J Virol 80: 6982-92).
Анализ связывания и конкуренции IgG1
Антитела IgG1 титровались в ELISA с использованием покрытых IL6 планшетов, в соответствии с описанным выше, и конъюгата пероксидазы против IgG человека. Конкурентные иммуноферментные анализы для группировки антител на основании распознавания эпитопов были выполнены посредством инкубирования Fab-фагов вместе с IgG1 или с коммерческими антителами против IL6 (например Abeam, каталожный номер ab9324) на покрытых IL6 планшетах, после чего проводилось детектирование связанных Fab-фагов с использованием конъюгата анти-M13 пероксидазы.
Измерения аффинности IgG1
Аффинности антител против IL6 определялись в соответствии с протоколом количественной кинетики с использованием Octet (ForteBio). Антитела захватывались захватывающим биосенсором против Fc IgG человека и подвергались воздействию свободного IL6, после чего анализировались с помощью специализированного программного обеспечения для вычисления Kd для каждого антитела.
Функциональная активность антител IL6
Для тестирования способности отобранных антител к ингибированию связывания между IL6 и рецептором IL6 (IL6R) было использовано исследование на основе ELISA. Различные концентрации антитела смешивались с фиксированной концентрацией (10 нг/мл) биотинилированного IL6, как описано в работе Naoko et al. 2007, Can. Res. 67: 817-875. Иммунный комплекс IL6-антитела добавлялся для иммобилизации IL6R. Связывание биотинилированного IL6 с IL6R определялось с помощью стрептавидина, конъюгированного с пероксидазой хрена. Снижение сигнала ELISA являлось мерой ингибирования. В качестве положительного контроля для ингибирования связывания между IL6 и IL6R использовалось анти-IL6R антитело (Abeam, каталожный номер ab34351; клон B-R6) или анти-IL6 антитело (Abeam, каталожный номер ab9324).
Блокирующая активность in vitro отобранных антител против IL6 измерялась в пролиферационном анализе с использованием антитела IL6-зависимой клеточной линии 7TD1. Кратко, клетки инкубировались с различными концентрациями IL6 человека при наличии или в отсутствие анти-IL6 антитела. Доступное количество IL6 определяет степень пролиферации. Таким образом, если добавленное антитело блокирует IL6, то регистрируемая пролиферация снижается по сравнению с контролем для несвязывающего антитела. Пролиферация измерялась путем включения 5-бромо-2'-дезокси-уридина (BrdU) в ДНК с использованием набора для пролиферации с BrdU (Roche, каталожный номер 11444611001) в соответствии с инструкциями производителя.
Получение олигоклонов анти-IL6
Наиболее сильные анти-IL6 антитела выбирались для каждой группы эпитопов. Экспрессионные конструкции, экспрессирующие данные антитела, трансфицировались в клетки HEK293T в неконкурирующих группах по три в различных соотношениях (1:1:1; 3:1:1; 1:3:1; 1:1:3; 3:3:1; 1:3:3; 3:1:3; 10:1:1; 1:10:1; 1:1:10; 10:10:1; 1:10:10; 10:1:10; 3:10:1; 10:3:1; 1:10:3; 3:1:10; 10:1:3; 1:3:10). Антитела, содержащие супернатанты, собирались, очищались и характеризовались в соответствии с описанным выше.
Образование комплекса и in vivo выведение олигоклонов анти-IL6
Для измерения способности олигоклонов анти-IL6 образовывать иммунные комплексы и для анализа этих комплексов использовалась эксклюзионная хроматография (SEC) в соответствии с подходом, изложенным в работе Min-Soo Kim et al. (2007) JMB 374: 1374-1388, в целях характеризации иммунных комплексов, образованных с различными антителами к TNFα. Различные молярные концентрации олигоклонов анти-IL6 смешивались с IL6 человека и инкубировались в течение 20 часов при 4°C или 25°C. Смесь анализировалась на HPLC-системе, оборудованной колонкой для эксклюзии по размерам; различные временные интервалы элюции были связаны с молекулярным весом с использованием стандартов молекулярных весов.
Способность антител образовывать комплексы с IL6 коррелирует с их способностью к быстрому выведению цитокина из циркуляции in vivo. Это подтверждается измерением выведения помеченного радиоактивной меткой IL6 из мышей. Коротко, были получены самки мышей Balb/c в возрасте от 6 до 8 недель, и за 18 часов до эксперимента животные подвергались внутривенной (IV) инъекции через латеральную хвостовую вену различных доз очищенных олигоклонов анти-IL6. В день 0 мышам внутривенно инъецировалось 50 микролитров помеченного радиоактивной меткой IL-6 (1×10E7 cpm/мл) при тех же условиях. Образцы крови (приблизительно, 50 микролитров) собирались в различные временные интервалы и хранились при 4°C. Образцы подвергались центрифугированию в течение 5 минут на 4000 xg, и измерялась радиоактивность сыворотки. Все фармакокинетические эксперименты выполнялись одновременно для трех животных для каждой обработки.
Получение стабильных олигоклонов анти-IL6 и доклиническое развитие
Доминирующий олигоклон анти-IL6 выбирался на основании способности in vitro и in vivo, в соответствии с определенным выше. Гены VH мыши гуманизировались в соответствии со стандартными методами и объединялись с полностью легкой цепью IGKV1-39 человека в экспрессионном векторе, в соответствии с описанным выше. Примеры методов гуманизации включают методы, основанные на таких парадигмах, как ремоделирование (Padlan, E. A., et al., (1991). Mol. Immunol., 28, 489), сверхгуманизация (Tan, P., D. A., et al., (2002) J. Immunol., 169, 1119) и оптимизация содержания последовательностей человека (Lazar, G. A., et al., (2007). Mol. Immunol., 44, 1986). Три конструкции трансфицировались в клетки PER.C6 в заранее определенном оптимальном соотношении (описано выше) под селективным давлением G418 в соответствии со стандартными методами. Отбирался имеющий стабильно высокое продуцирование клон олигоклонов анти-IL6, и создавался банк квалифицированных базовых клеток.






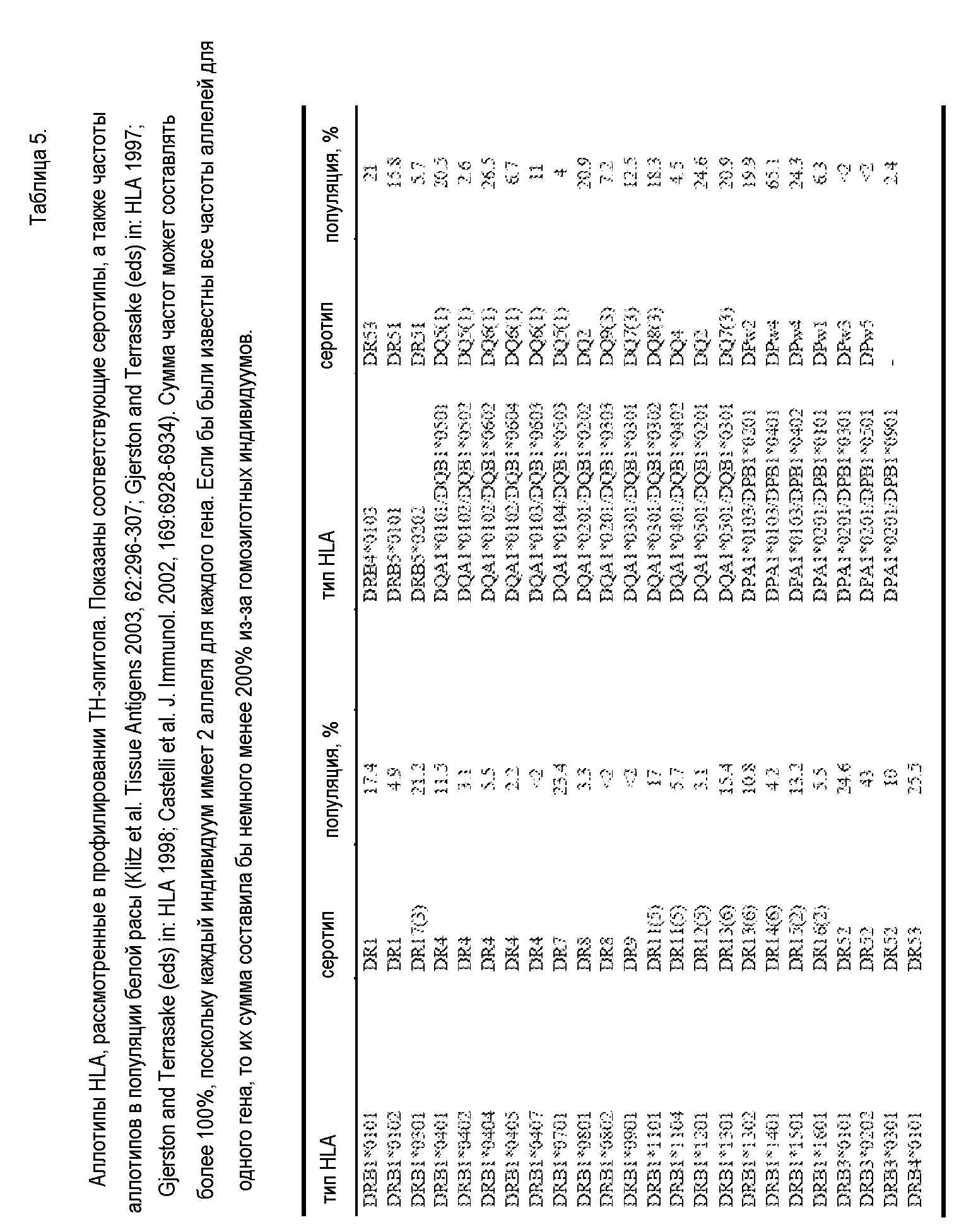



Реферат
Изобретение относится к области биотехнологии. Предложено трансгенное животное, представляющее собой мышь или крысу, в геном которого встроена нуклеиновая кислота, кодирующая перестроенную вариабельную область легкой цепи иммуноглобулина человека, функционально связанную с константной областью иммуноглобулина указанного животного и функционально связанную с промотором, обеспечивающим экспрессию трансгенной легкой цепи в В-клетках млекопитающего, таким образом, что она образует пары с различными эндогенными тяжелыми цепями, обеспечивая получение антител. Также рассмотрены способы получения антител с использованием трансгенного животного, В-клетка трансгенного животного, применение животного и его В-клетки для получения антител и способ получения трансгенного животного по изобретению. Данное изобретение позволяет продуцировать антитела, содержащие легкую цепь с вариабельной областью иммуноглобулина человека в паре с различными тяжелыми цепями иммуноглобулинов животного. 8 н. и 16 з.п. ф-лы, 27 ил., 11 табл., 22 пр.
Комментарии