Генетически модифицированные мыши, экспрессирующие химерные молекулы главного комплекса гистосовместимости - RU2660564C2
Код документа: RU2660564C2
Чертежи















Описание
Ссылка на родственную заявку
По настоящей заявке испрашивается приоритет в соответствии с предварительной заявкой на выдачу патента США №61/552584, поданной 28 октября 2011 г., которая полностью включена в настоящий документ посредством ссылки.
Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится к не являющемуся человеком животному, например, грызуну (например, мыши или крысе), которое генетически сконструировано для экспрессии гуманизированного белка главного комплекса гистосовместимости (МНС) II класса, а также к зародышам, тканям и клеткам, его экспрессирующим. Настоящее изобретение дополнительно относится к способам получения генетически модифицированного не являющегося человеком животного, которое экспрессирует гуманизированный белок МНС II. Также предусматриваются способы применения не яляющихся человеком животных, клеток и тканей, которые экспрессируют гуманизированный белок МНС II класса, для идентификации пептидов, которые активируют лимфоциты и привлекают Т-клетки, и для разработки вакцин и других терапевтических средств для человека.
Предшествующий уровень техники настоящего изобретения
В приобретенном иммунном ответе чужеродные антигены распознаются рецепторными молекулами на В-лимфоцитах (например, иммуноглобулинами) и Т-лимфоцитах (например, Т-клеточным рецептором, или TCR). Эти чужеродные антигены презентируются на поверхности клеток в виде пептидных фрагментов специализированными белками, имеющими общее название молекулы главного комплекса гистосовместимости (МНС). Молекулы МНС кодируются множественными локусами, которые встречаются в виде соединенного кластера генов, который охватывает приблизительно 4 м.п.н. У мышей гены МНС находятся на хромосоме 17, и исторически они называются гены гистосовместимости 2 (Н-2). У людей гены находятся на хромосоме 6 и называются гены антигена лейкоцита человека (HLA). Локусы у мышей и людей являются полигенными; они включают в себя три высоко полиморфных класса генов МНС (I, II и III класс), которые проявляют сходную организацию в геноме человека и мыши (смотрите фиг. 2 и фиг. 3 соответственно).
Локусы МНС проявляют самый высокий полиморфизм в геноме; некоторые гены представлены >300 аллелями (например, HLA-DR-β человека и HLA-B человека). Все гены МНС I и II класса могут презентировать пептидные фрагменты, но каждый ген экспрессирует белок с различными характеристиками связывания, отражая полиморфизмы и аллельные варианты. Любой рассматриваемый индивидуум содержит уникальный спектр пептидных фрагментов, которые могут быть презентированы на клеточной поверхности В- и Т-клеткам в ходе иммунного ответа.
Как люди, так и мыши содержат гены МНС II класса (смотрите фиг. 2 и 3). У людей классические гены МНС II называются HLA-DP, HLA-DQ и HLA-DR, тогда как у мышей они представляют собой Н-2А и Н-2Е (часто сокращенно I-Α и 1-Е соответственно). Дополнительные белки, кодируемые генами в локусе МНС II, HLA-DM и HLA-DO у людей и Н-2М и Н-20 у мышей, не встречаются на клеточной поверхности, но локализуются в компартменте эндоцитоза и обеспечивают правильную загрузку молекул МНС II пептидами. Молекулы II класса состоят из двух полипептидных цепей: α цепи и β цепи. Внеклеточная часть цепи α содержит два внеклеточных домена, α1 и а2; и внеклеточная часть цепи β также содержит два внеклеточных домена, β1 и β2 (смотрите фиг. 1). Цепи α и β нековалентно соединены друг с другом.
Молекулы МНС II класса экспрессируются на антигенпрезентирующих клетках (АПК), например, В-клетках, макрофагах, дендритных клетках, эндотелиальных клетках в ходе воспаления и т.д. Молекулы МНС II, экспрессирующиеся на поверхности АПК, как правило, презентируют образованные во внутриклеточных везикулах антигены CD4+ Т-клеткам. Чтобы принимать участие в CD4+Т-клеточной активации, комплекс МНС класса II с представляющим интерес антигеном должен быть достаточно стабильным, чтобы длительно существовать для привлечения CD4+ Т-клетки. Когда CD4+ Т-хелперная клетка привлекается комплексом чужеродный пептид/МНС II на поверхности АПК, Т-клетка активируется для высвобождения цитокинов, которые принимают участие в иммунном ответе на возбудителя.
Не все антигены будут вызывать активацию Т-клеток вследствие механизмов толерантности. Тем не менее, при некоторых заболеваниях (например, злокачественной опухоли, аутоиммунных заболеваниях) пептиды, происходящие из собственных белков, становятся мишенью клеточного компонента иммунной системы, что приводит к разрушению клеток, презентирующих такие пептиды. Произошел значительный прогресс в распознавании антигенов, являющихся клинически значимыми (например, антигенов, связанных с различными типами злокачественных опухолей). Тем не менее, для улучшения идентификации и выбора пептидов, которые будут вызывать подходящий ответ в Т-клетке человека, в частности пептидов клинически значимых антигенов, остается потребность в системах in vivo и in vitro, которые имитируют аспекты иммунной системы человека. Таким образом, существует необходимость в биологических системах (например, генетически модифицированных не относящихся к человеку животных и клетках), которые могут проявлять компоненты иммунной системы человека.
Краткое раскрытие настоящего изобретения
Предусматривается биологическая система для получения или идентификации пептидов, которые ассоциируются с белками МНС II класса человека и их химерами и связываются с CD4+ Т-клетками. Предусматриваются не являющиеся человеком животные, содержащие не являющиеся человеческими клетки, которые экспрессируют гуманизированные молекулы, функционирующие в клеточном иммунном ответе. Также предусматриваются гуманизированные локусы грызуна, которые кодируют гуманизированные белки МНС II. Также предусматриваются гуманизированные клетки грызуна, которые экспрессируют гуманизированные молекулы МНС. Предусматриваются системы in vivo и in vitro, которые содержат гуманизированные клетки грызуна, причем клетки грызуна экспрессируют одну или несколько гуманизированных молекул иммунной системы.
В настоящем документе предусматривается не являющееся человеком животное, например, грызун (например, мышь или крыса), содержащее в своем геноме нуклеотидную последовательность, кодирующую гуманизированный комплекс МНС II, причем человеческая часть гуманизированного комплекса МНС II содержит внеклеточный домен комплекса МНС II человека, например, гуманизированный внеклеточный домен α МНС II и гуманизированный внеклеточный домен β МНС II.
Согласно одному аспекту в настоящем документе предусматривается не являющееся человеком животное, содержащее на эндогенном локусе гена α МНС II нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид α МНС II. Согласно одному варианту осуществления человеческая часть такого химерного человеческого/не относящегося к человеку полипептида α МНС II содержит внеклеточный домен α МНС II человека. Согласно одному варианту осуществления не являющееся человеком животное экспрессирует функциональный комплекс МНС II на поверхности клетки животного. Согласно одному варианту осуществления внеклеточный домен α МНС II человека у животного содержит домены α1 и α2 МНС II человека; согласно одному варианту осуществления не относящаяся к человеку часть химерного человеческого/не относящегося к человеку полипептида α МНС II содержит трансмембранный и цитоплазматический домены эндогенного не относящегося к человеку полипептида α МНС II. Согласно одному варианту осуществления нуклеотидная последовательность, кодирующая химерный человеческий/не относящийся к человеку полипептид α МНС II, экспрессируется под регуляторным контролем эндогенных не относящихся к человеку α промоторных и регуляторных элементов МНС II. Согласно одному варианту осуществления человеческая часть химерного полипептида происходит из белка HLAII класса человека, выбранного из группы, состоящей из HLA-DR, HLA-DQ и HLA-DP, например, человеческая часть происходит из белка HLA-DR4. Не являющееся человеком животное может представлять собой грызуна, например, мышь. Согласно одному аспекту не являющееся человеком животное, содержащее на эндогенном локусе гена α МНС II нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид α МНС II, дополнительно содержит на эндогенном локусе гена β МНС II нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид β МНС II. Также в настоящем документе предусматривается способ получения генетически модифицированного не являющегося человеком животного, содержащего на эндогенном локусе гена α МНС II нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид α МНС II. Такой способ может предусматривать замещение на эндогенном локусе гена α МНС II нуклеотидной последовательности, кодирующей эндогенный не относящийся к человеку полипептид α МНС II, на нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид α МНС II.
Также в настоящем документе предусматривается не являющееся человеком животное, содержащее на эндогенном локусе гена β МНС II нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид β МНС II. Согласно одному варианту осуществления человеческая часть такого химерного человеческого/не относящегося к человеку полипептида β МНС II содержит внеклеточный домен β МНС II человека. Согласно одному варианту осуществления не являющееся человеком животное экспрессирует функциональный комплекс МНС II на поверхности клетки животного. Согласно одному варианту осуществления человеческий внеклеточный домен β МНС II у животного содержит домены β1 и β2 МНС II человека; согласно одному варианту осуществления не относящаяся к человеку часть химерного человеческого/не относящегося к человеку полипептида β МНС II содержит трансмембранный и цитоплазматический домены эндогенного не относящегося к человеку полипептида β МНС II. Согласно одному варианту осуществления нуклеотидная последовательность, кодирующая химерный человеческий/не относящийся к человеку полипептид β МНС II, экспрессируется под регуляторным контролем эндогенных не относящихся к человеку β промоторных и регуляторных элементов МНС II. Согласно одному варианту осуществления человеческая часть химерного полипептида происходит из белка HLAII класса человека, выбранного из группы, состоящей из HLA-DR, HLA-DQ и HLA-DP, например, человеческая часть происходит из белка HLA-DR4. Не являющееся человеком животное может представлять собой грызуна, например, мышь. Согласно одному аспекту не являющееся человеком животное, содержащее на эндогенном локусе гена β МНС II нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид β МНС II, дополнительно содержит на эндогенном локусе гена α МНС II нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид α МНС II. Также в настоящем документе предусматривается способ получения генетически модифицированного не являющегося человеком животного, содержащего на эндогенном локусе гена β МНС II нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид β МНС II. Такой способ может предусматривать замещение на эндогенном локусе гена β МНС II нуклеотидной последовательности, кодирующей эндогенный не относящийся к человеку полипептид β МНС II, на нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид β МНС II.
Согласно одному аспекту предусматривается не являющееся человеком животное, содержащее на эндогенном локусе гена МНС II первую нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид α МНС II, и вторую нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид β МНС II, причем человеческая часть химерного человеческого/не относящегося к человеку полипептида α МНС II содержит человеческий внеклеточный домен α МНС II, и человеческая часть химерного человеческого/не относящегося к человеку полипептида β МНС II содержит человеческий внеклеточный домен β МНС II. Согласно одному варианту осуществления химерные человеческие/не относящиеся к человеку полипептиды α и β МНС II образуют функциональный химерный комплекс МНС II (например, человеческий/не относящийся к человеку комплекс МНС II) на поверхности клетки. Согласно одному варианту осуществления человеческий внеклеточный домен α МНС II содержит домены α1 и α2 человека МНС II человека. Согласно одному варианту осуществления человеческий внеклеточный домен β МНС II содержит домены β1 и β2 человека МНС II человека. Согласно различным аспектам первая нуклеотидная последовательность экспрессируется под регуляторным контролем эндогенных не относящихся к человеку α промоторных и регуляторных элементов МНС II. Согласно различным аспектам вторая нуклеотидная последовательность экспрессируется под регуляторным контролем эндогенных не относящихся к человеку β промоторных и регуляторных элементов МНС II. Согласно некоторым вариантам осуществления не относящаяся к человеку часть химерного человеческого/не относящегося к человеку полипептида α МНС II содержит трансмембранный и цитоплазматический домены эндогенного не относящегося к человеку полипептида α МНС II. Согласно некоторым вариантам осуществления не относящаяся к человеку часть химерного человеческого/не относящегося к человеку полипептида β МНС II содержит трансмембранный и цитоплазматический домены эндогенного не относящегося к человеку полипептида β МНС II.
Согласно различным вариантам осуществления не являющееся человеком животное представляет собой грызуна, и человеческие части химерных человеческих/относящихся к грызуну полипептидов α и β МНС II содержат последовательности человека, происходящие из белка HLA II класса, выбранного из группы, состоящей из HLA-DR, HLA-DQ и HLA-DP. Согласно некоторым вариантам осуществления настоящего изобретения человеческие части химерных человеческих/относящихся к грызуну последовательностей α и β МНС II происходят из последовательности HLA-DR4 человека; таким образом, нуклеотидная последовательность, кодирующая внеклеточный домен α МНС II, происходит из последовательности гена HLA-DRα*01, и нуклеотидная последовательность, кодирующая внеклеточный домен β МНС II, происходит из последовательности, кодирующей ген ΗLΑ-DRβ1*04.
Согласно различным вариантам осуществления настоящего изобретения первая и вторая нуклеотидные последовательности расположены на одной хромосоме. Согласно некоторым аспектам животное содержит две копии локуса МНС II, содержащего первую и вторую нуклеотидные последовательности, тогда как согласно другим аспектам животное содержит одну копию локуса МНС II, содержащего первую и вторую нуклеотидные последовательности. Таким образом, животное может являться гомозиготным или гетерозиготным в отношении локуса МНС II, содержащего первую и вторую нуклеотидные последовательности.
Согласно некоторым аспектам химерный полипептид α МНС II и/или химерный полипептид β МНС II функционально связан с не относящейся к человеку лидерной последовательностью.
Согласно одному аспекту генетически сконструированное не являющееся человеком животное представляет собой грызуна. Согласно одному варианту осуществления грызуна выбирают из группы, состоящей из мыши и крысы. Таким образом, согласно некоторым вариантам осуществления не относящиеся к человеку последовательности химерных генов α и β МНС II происходят из нуклеотидных последовательностей, кодирующих белок МНС II мыши, например, белок Н-2Е мыши. Согласно одному варианту осуществления грызун (например, мышь или крыса) согласно настоящему изобретению не экспрессирует функциональные эндогенные полипептиды МНС II из их эндогенных локусов. Согласно одному варианту осуществления, в которых грызун представляет собой мышь, мышь не экспрессирует функциональные эндогенные полипептиды Н-2Е и Н-2А из их эндогенных локусов.
Таким образом, согласно некоторым вариантам осуществления предусматривается мышь, содержащая на эндогенном локусе МНС II мыши первую нуклеотидную последовательность, кодирующую химерный человеческий/мышиный полипептид α МНС II, и вторую нуклеотидную последовательность, кодирующую химерный человеческий/мышиный полипептид β МНС II, причем человеческая часть химерного полипептида α МНС II содержит внеклеточный домен, происходящий из полипептида α белка HLA-DR4 человека, и человеческая часть химерного человеческого/мышиного полипептида β МНС II содержит внеклеточный домен, происходящий из полипептида β белка HLA-DR4 человека, причем мишиная часть химерного полипептида α МНС II содержит трансмембранный и цитоплазматический домены цепи α Н-2Е мыши, и мышиная часть химерного полипептида β МНС II содержит трансмембранный и цитоплазматический домены цепи β Н-2Е мыши, и при этом мышь экспрессирует функциональный химерный HLA-DR4/H-2E комплекс МНС II. Согласно некоторым аспектам внеклеточный домен химерного полипептида α МНС II содержит домены α1 и а2 человека; согласно некоторым аспектам внеклеточный домен химерного полипептида β МНС II содержит домены β1 и β2 человека. Согласно некоторым вариантам осуществления первая нуклеотидная последовательность экспрессируется под регуляторным контролем эндогенных α промоторных и регуляторных элементов МНС II мыши, и вторая нуклеотидная последовательность экспрессируется под регуляторным контролем эндогенных β промоторных и регуляторных элементов МНС II мыши. Согласно различным вариантам осуществления мышь не экспрессирует функциональные эндогенные полипептиды МНС II, например, полипептиды Н-2Е и Н-2А, из их эндогенных локусов. Согласно некоторым аспектам мышь содержит две копии локуса МНС II, содержащего первую и вторую нуклеотидные последовательности, тогда как согласно другим аспектам мышь содержит одну копию локуса МНС II, содержащего первую и вторую нуклеотидные последовательности.
Также предусматриваются способы получения генетически сконструированных не являющихся человеком животных (например, грызунов, например, мышей или крыс), описанных в настоящем документе. Согласно различным вариантам осуществления не являющихся человеком животных (например, грызунов, например, мышей или крыс) по настоящему изобретению получают путем замещения эндогенных последовательностей МНС II нуклеотидными последовательностями, кодирующими химерные человеческие/не относящиеся к человеку (например, человеческие/мышиные) полипептиды α и β МНС II. Согласно одному варианту осуществления настоящее изобретение относится к способу модификации локуса МНС II грызуна (например, мыши или крысы) для экспрессии химерного человеческого/относящегося к грызуну комплекса МНС II, предусматривающему замещение на эндогенном локусе МНС II мыши нуклеотидной последовательности, кодирующей комплекс МНС II грызуна, на нуклеотидную последовательность, кодирующую химерный человеческий/относящийся к грызуну комплекс МНС II. Согласно одному аспекту способа нуклеотидная последовательность, кодирующая химерный человеческий/относящийся к грызуну комплекс МНС II, содержит первую нуклеотидную последовательность, кодирующую внеклеточный домен цепи α МНС II человека и трансмембранный и цитоплазматический домены цепи α МНС II грызуна, и вторую нуклеотидную последовательность, кодирующую внеклеточный домен цепи β МНС II человека и трансмембранный и цитоплазматический домены цепи β МНС II грызуна. Согласно некоторым аспектам относящаяся к грызуну часть химерного комплекса МНС II происходит из белка Н-2Е мыши, и человеческая часть происходит из белка HLA-DR4 человека. Согласно некоторым вариантам осуществления замещение описанных в настоящем документе эндогенных локусов МНС II проводят в одной ES (эмбриональной стволовой) клетке, и одну ES клетку вводят в зародыш грызуна (например, мыши или крысы) для получения генетически модифицированного грызуна (например, мыши или крысы).
Также в настоящем документе предусматриваются клетки, например, выделенные антигенпрезентирующие клетки, полученные от описанных в настоящем документе не являющихся человеком животных (например, грызуны, например, мыши или крысы). Также предусматриваются ткани и зародыши, полученные от описанных в настоящем документе не являющихся человеком животных.
Любой из описанных в настоящем документе вариантов осуществления и аспектов может использоваться совместно друг с другом, если иное не указано или не очевидно из контекста. Другие варианты осуществления станут очевидными специалистам в настоящей области техники из обзора последующего подробного раскрытия. Последующее подробное раскрытие включает в себя иллюстративные представления различных вариантов осуществления настоящего изобретения, которые не ограничивают заявленное настоящее изобретение. Прилагаемые фигуры составляют часть настоящего описания изобретения и вместе с описанием служат исключительно для иллюстрации вариантов осуществления, а не для ограничения настоящего изобретения.
Краткое описание графических материалов
На фиг. 1 представлено схематическое изображение молекулы МНС II класса, экспрессирующейся на поверхности антигенпрезентирующей клетки (АПК), содержащей четыре домена: α1, α2, β1 и β2. Серые кружки представляют пептид, связанный на пептидсвязывающей бороздке.
На фиг. 2 представлено схематическое изображение (без соблюдения масштаба) относительной геномной структуры HLA человека, показывающее гены I, II и III класса.
На фиг. 3 представлено схематическое изображение (без соблюдения масштаба) относительной геномной структуры МНС мыши, показывающее гены I, II и III класса.
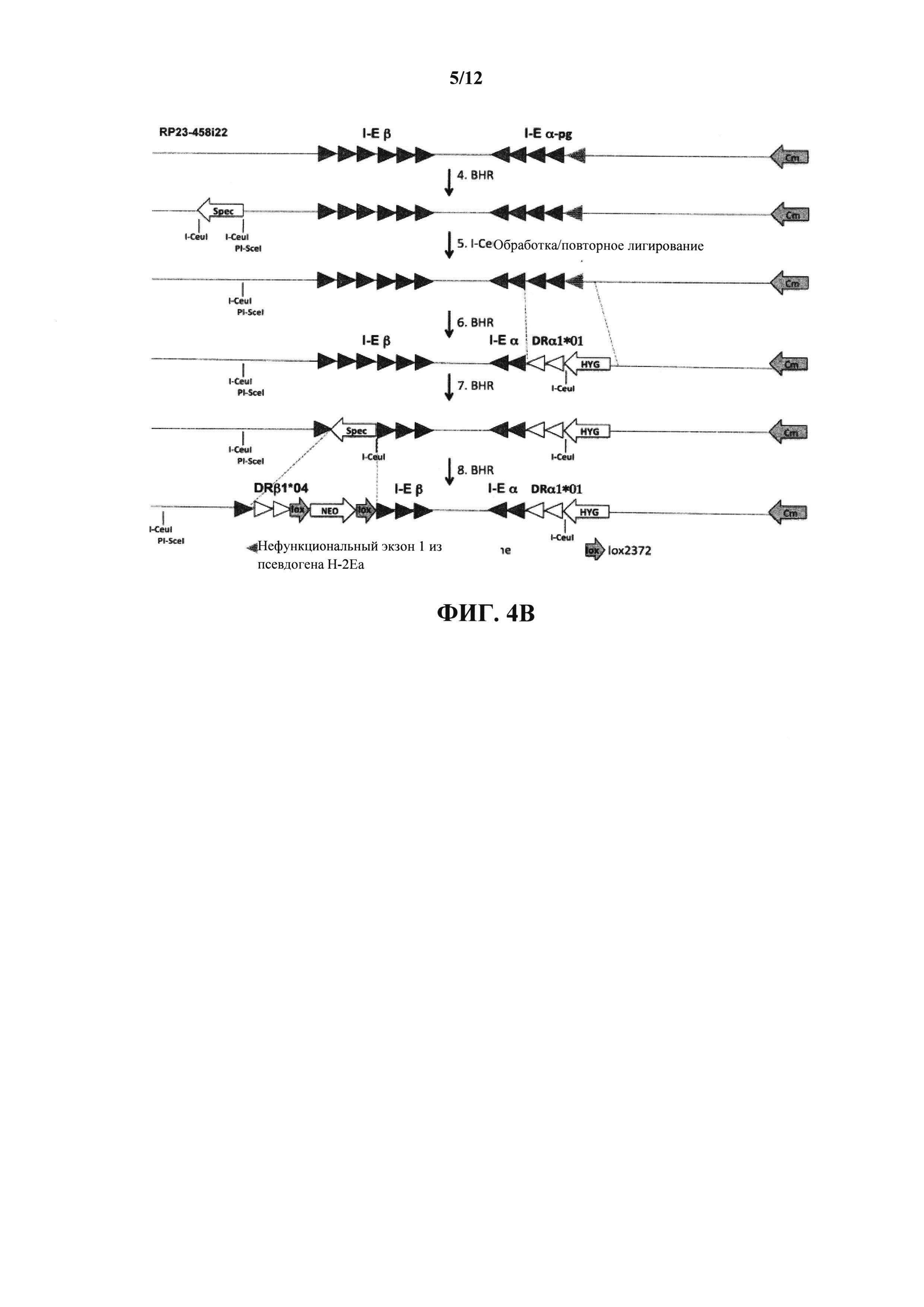
На фиг. 4 (A-D) схематически проиллюстрирована (без соблюдения масштаба) стратегия создания нацеливающего вектора, содержащего гуманизированные I-Е β и I-Е α (т.е. химеру Η-2Εβ/ΗLΑ-DRβ1*04 и H-2Eα/HLA-DRα*01 соответственно). На фиг. 4С конечную гуманизированную последовательность МНС II из фиг. 4В лигируют между сайтами рестрикции PI-SceI и I-CeuI конечной конструкции из фиг. 4А для создания конструкции, содержащей гуманизированный МНС II и экзон 1 I-Eα из BALB/c. Pg=псевдоген; BHR=бактериальная гомологичная рекомбинация; СМ=хлорамфеникол; spec=спектиномицин; hyg=гигромицин; neo=неомицин; ЕР=электропорация. Треугольники представляют экзоны, закрашенные треугольники представляют экзоны мыши от мыши C57BL/6 (за исключением заштрихованных треугольников, которые представляют экзон 1 I-Eα от мыши BALB/c) и незакрашенные треугольники представляют экзоны человека.
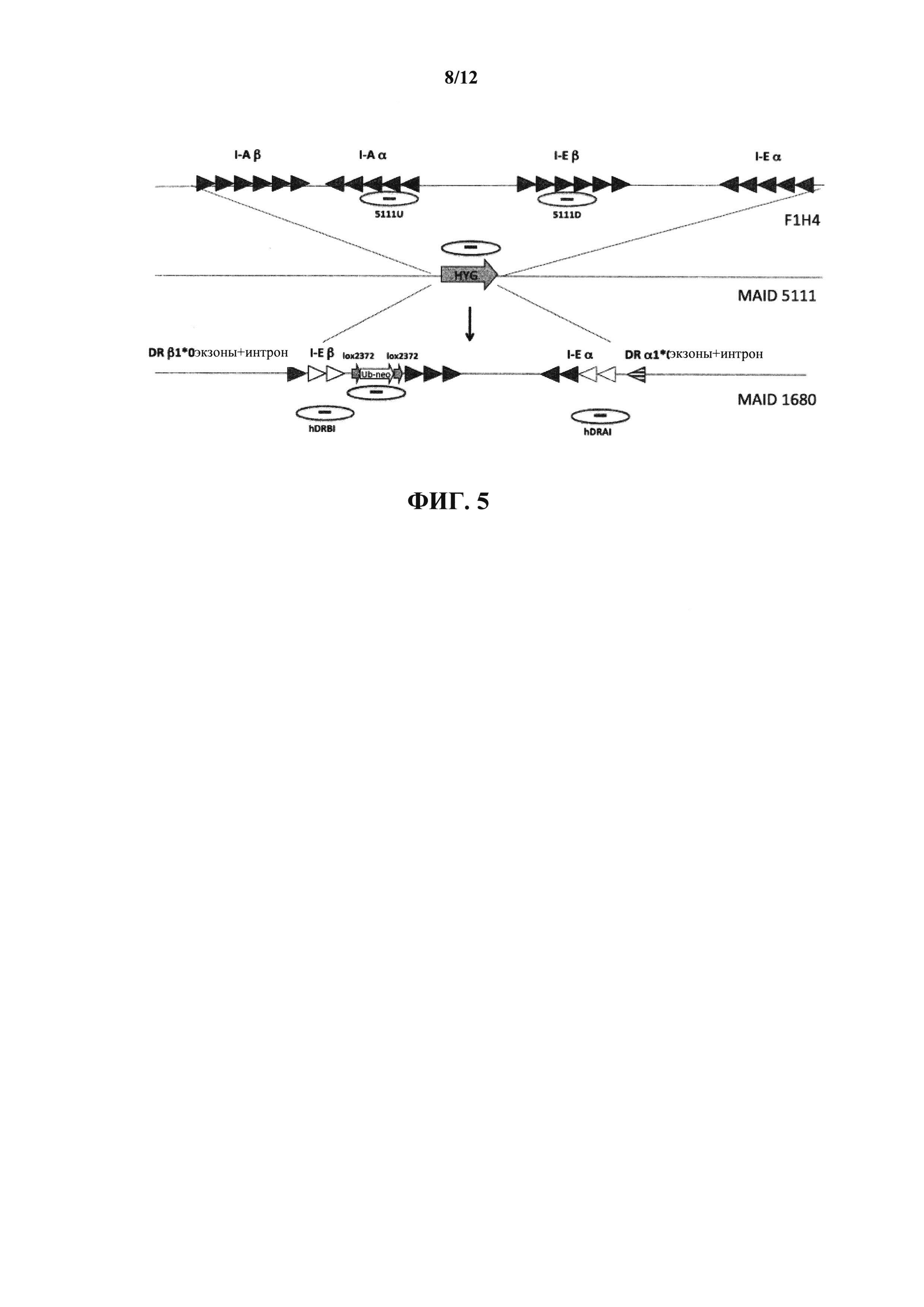
На фиг. 5 показано схематическое изображение, без соблюдения масштаба, генов ΙΕ и I-Α МНС II класса, показывающее нокаут мышиного локуса с использованием кассеты гигромицина с последующим введением вектора, содержащего гуманизированные I-Е β и I-Е α (т.е. химеру Η-2Εβ/ΗLΑ-DRβ1*04 и H-2Eα/HLA-DRα*01 соответственно). Незакрашенные треугольники представляют экзоны человека; закрашенные треугольники представляют экзоны мыши. Используемые для генотипирования зонды обведены кружком.
На фиг. 6 показано схематическое изображение, без соблюдения масштаба, Cre-опосредованного удаления кассеты неомицина из фиг. 5. Незакрашенные треугольники представляют экзоны человека; закрашенные треугольники представляют экзоны мыши. Две верхние цепи представляют локусы МНС II у гетерозиготной мыши с гуманизированным МНС II, несущей селективную кассету неомицина, и две нижние цепи представляют локусы МНС II у гетерозиготной мыши с гуманизированным МНС II с удаленной кассетой неомицина.
На фиг. 7 показано схематическое сравнительное изображение, без соблюдения масштаба, локусов II класса мыши и человека. Гены II класса представлены прямоугольниками, а пустые прямоугольники представляют псевдогены. Представлены относительные размеры (т.п.н.) различных фрагментов нуклеиновой кислоты.
На фиг. 8, на левой панели, представлено схематическое изображение (без соблюдения масштаба) стратегии гуманизации для цепи α МНС II; в частности на фигуре показано замещение доменов α1 и α2, кодируемых экзонами 2 и 3 гена α МНС II, при сохранении трансмембранной и цитоплазматической хвостовых последовательностей мыши. В гуманизированном локусе лидерная последовательность α МНС II получена от штамма BALB/c мыши. На правой панели показана гуманизация цепи β МНС II; в частности на фигуре показано замещение доменов β1 и β2, кодируемых экзонами 2 и 3 гена β МНС II, при сохранении мышиной лидерной и трансмембранной и цитоплазматической хвостовых последовательностей мыши. В верхнем ряду представлены все последовательности человека; в среднем ряду представлены все последовательности мыши; в нижнем ряду представлены все гуманизированные последовательности, с экзонами 2 и 3, происходящими из генов HLA-DR человека.
На фиг. 9 показан анализ FACS с антителом к HLA-DR В-клеток от мыши, гетерозиготной в отношении химерного HLA-DR4 (кассета neo удалена) в присутствии (1681НЕТ + поли(I:С) или при отсутствии (1681НЕТ ) поли(I:С), и от мыши дикого типа (мыши WT).
Подробное раскрытие настоящего изобретения
Определения
Настоящее изобретение относится к генетически модифицированным не являющимся человеком животным (например, мышам, крысам, кроликам и т.д.), которые экспрессируют относящийся к человеку или гуманизированный полипептид МНС II; к зародышам, клеткам и тканям, его содержащим; способам их получения; а также способам их применения. Если не указано иное, все используемые в настоящем документе термины и фразы включают значения, которые подразумеваются под терминами и фразами в настоящей области техники, если противоположное ясно не указано или ясно не следует из контекста, в котором используется термин или фраза.
Термин "консервативная", используемый для описания консервативной аминокислотной замены, включает замену аминокислотного остатка другим аминокислотным остатком с R-группой боковой цепи со сходными химическими свойствами (например, зарядом или гидрофобностью). Консервативные аминокислотные замены могут быть достигнуты путем модификации нуклеотидной последовательности так, чтобы ввести изменение нуклеотида, которое будет кодировать консервативную замену. Как правило, консервативная аминокислотная замена не будет существенно изменять представляющие интерес функциональные свойства белка, например, способность МНС II к презентации представляющего интерес пептида. Примеры групп аминокислот, которые содержат боковые цепи со сходными химическими свойствами, включают такие алифатические боковые цепи, как глицин, аланин, валин, лейцин и изолейцин; такие алифатические гидроксильные боковые цепи, как серии и треонин; такие амидсодержащие боковые цепи, как аспарагин и глутамин; такие ароматические боковые цепи, как фенилаланин, тирозин и триптофан; такие основные боковые цепи, как лизин, аргинин и гистидин; такие кислотные боковые цепи, как аспарагиновая кислота и глутаминовая кислота; и такие серосодержащие боковые цепи, как цистеин и метионин. Группы консервативных аминокислотных замен включают, например, валин/лейцин/изолейцин, фенилаланин/тирозин, лизин/аргинин, аланин/валин, глутамат/аспартат и аспарагин/глутамин. Согласно некоторым вариантам осуществления консервативная аминокислотная замена может представлять собой замену любого нативного остатка в белке на аланин, что используется, например, в сканирующем аланином мутагенезе. Согласно некоторым вариантам осуществления проводят консервативную замену, которая характеризуется положительным значением в матрице логарифмического правдоподобия РАМ250, раскрытой в Gonnet et al. ((1992) Exhaustive Matching of the Entire Protein Sequence Database, Science 256:1443-45), включенной в настоящий документ посредством ссылки. Согласно некоторым вариантам осуществления замена представляет собой умеренно консервативную замену, причем замена характеризуется неотрицательным значением в матрице логарифмического правдоподобия РАМ250.
Таким образом, в настоящем изобретении также предусматривается генетически модифицированное не являющееся человеком животное, геном которого содержит нуклеотидную последовательность, кодирующую человеческий или гуманизированный полипептид МНС II, причем полипептид содержит консервативные аминокислотные замены в описанной в настоящем документе аминокислотной последовательности.
Специалисту в настоящей области техники понятно, что в дополнение к остаткам нуклеиновой кислоты, кодирующим описанный в настоящем документе человеческий или гуманизированный полипептид МНС II, вследствие вырожденности генетического кода, другие нуклеиновые кислоты могут кодировать полипептид по настоящему изобретению. Следовательно, в дополнение к генетически модифицированному не являющемуся человеком животному, которое содержит в своем геноме нуклеотидную последовательность, кодирующую полипептид МНС II с консервативными аминокислотными заменами, также предусматривается не являющееся человеком животное, геном которого содержит нуклеотидную последовательность, которая отличается от описанной в настоящем документе нуклеотидной последовательности вследствие вырожденности генетического кода.
Термин "идентичность" при использовании в связи с последовательностью включает идентичность, определяемую с помощью набора различных известных в настоящей области техники алгоритмов, которые могут использоваться для измерения идентичности нуклеотидной и/или аминокислотной последовательности. Согласно некоторым описанным в настоящем документе вариантам осуществления, идентичности определяют с использованием ClustalW v. 1.83 (медленного) выравнивания с использованием штрафа за открытие делеции, составляющего 10,0, штрафа за продление делеции, составляющего 0,1, и с использованием матрицы сравнения согласно Gonnet (MacVector™ 10.0.2, MacVector Inc., 2008). Длина последовательностей, сравниваемых в отношении идентичности последовательностей, будет зависеть от конкретных последовательностей. Согласно различным вариантам осуществления идентичность определяют путем сравнения последовательности зрелого белка в направлении от его N-конца к его С-концу. Согласно различным вариантам осуществления при сравнении химерной человеческой/не относящейся к человеку последовательности с последовательностью человека, человеческая часть химерной человеческой/не относящейся к человеку последовательности (но не часть, не относящаяся к человеку) используется в осуществления сравнения с целью выяснения уровня идентичности между последовательностью человека и человеческой частью химерной человеческий/не относящейся к человеку последовательности (например, сравнивая эктодомен человека химерного человеческого/мыши белка с эктодоменом белка человека).
Термины "гомология" или "гомологичный" в отношении последовательностей, например, нуклеотидных или аминокислотных последовательностей, означает, что две последовательности при оптимальном выравнивании и сравнении являются идентичными по меньшей мере приблизительно в 75% нуклеотидов или аминокислот, по меньшей мере приблизительно в 80% нуклеотидов или аминокислот, по меньшей мере приблизительно в 90-95% нуклеотидов или аминокислот, например, больше чем 97% нуклеотидов или аминокислот. Специалисту в настоящей области техники будет понятно, что для оптимального нацеленного воздействия на ген нацеливающий конструкт должен содержать плечи, гомологичные эндогенным последовательностям ДНК (т.е. "плечи гомологии"); таким образом, гомологичная рекомбинация может происходить между нацеливающей конструкцией и нацеленной эндогенной последовательностью.
Термин "функционально связанный" относится к смежному положению, причем описанные таким образом компоненты находятся во взаимодействии, позволяющем им функционировать предусмотренным для них образом. В связи с этим, кодирующая белок последовательность нуклеиновой кислоты может быть функционально связана с регуляторными последовательностями (например, последовательностью промотора, энхансера, сайленсера и т.д.) так, чтобы сохранять надлежащую транскрипционную регуляцию. Кроме того, различные части химерного или гуманизированного белка по настоящему изобретению могут быть функционально связаны для сохранения надлежащей укладки, процессинга, нацеленного воздействия, экспрессии и других функциональных свойств белка в клетке. Если не указано иное, различные домены химерного или гуманизированного белка по настоящему изобретению функционально связаны друг с другом.
Используемые в настоящем документе термины " комплекс МНС II", "белок МНС II" или подобное включают комплекс между полипептидом α МНС II и полипептидом β МНС II. Используемый в настоящем документе термин "полипептид α МНС II" или "полипептид β МНС II" (или подобное) включает полипептид α МНС I отдельно или полипептид β МНС II отдельно соответственно. Аналогично, термины "комплекс HLA-DR4", "белок HLA-DR4", "комплекс Н-2Е", "белок Н-2Е" или подобное относятся к комплексу между полипептидами α и β. Как правило, термины "МНС человека" и "HLA" используются взаимозаменяемо.
Термин "замещение" в отношении замещения гена относится к размещению экзогенного генетического материала на эндогенном генетическом локусе, тем самым замещая весь эндогенный ген или его часть ортологичной или гомологичной последовательностью нуклеиновой кислоты. Как показано в примерах ниже, последовательность нуклеиновой кислоты эндогенного локуса МНС II замещали нуклеотидной последовательностью, содержащей последовательности, кодирующие части полипептидов α и β МНС II человека; в частности, кодирующие внеклеточные части полипептидов α и β МНС II.
Используемый в настоящем документе термин "функциональный", например, по отношению к функциональному полипептиду, относится к полипептиду, который сохраняет по меньшей мере одну биологическую активность, в норме связанную с нативным белком. Например, согласно некоторым вариантам осуществления настоящего изобретения замещение на эндогенном локус (например, замещение на эндогенном не относящемся к человеку локусе МНС II) дает в результате локус, который не способен экспрессировать функциональный эндогенный полипептид.
Генетически модифицированные в отношении МНС II животные
Согласно различным аспектам настоящее изобретение в основном относится к генетически модифицированным не являющимся человеком животным, которые содержат в своем геноме нуклеотидную последовательность, кодирующую человеческий или гуманизированный комплекс МНС II; таким образом, животные экспрессируют человеческий или гуманизированный комплекс МНС II (например, полипептиды α и β МНС II).
Гены МНС относятся к трем классам: I классу, II классу и III классу, причем все указанные классы кодируются либо на хромосоме 6 человека, либо на хромосоме 17 мыши. Схематическая иллюстрация относительной организации классов МНС человека и мыши представлена на фиг. 2 и 3 соответственно. Большинство генов МНС являются полиморфными, фактически, они представляют собой наиболее полиморфные гены геномов мыши и человека. Предполагают, что полиморфизмы МНС являются важными в обеспечении эволюционного преимущества; изменения в последовательности могут привести к различиям в связывании пептида, что обеспечивает возможность лучшей презентации антигена. Одно исключение составляет цепь HLA-DRα человека и ее мышиный гомолог, Еα (т.е. Н-2Еа), которые являются мономорфными.
Комплекс МНС II класса содержит два нековалентно связанных домена: цепь α и цепь β, также в настоящем документе имеющие название полипептид α и полипептид β (фиг. 1). Белок охватывает плазматическую мембрану; таким образом, он содержит внеклеточный домен, трансмембранный домен и цитоплазматический домен. Внеклеточная часть цепи α включает в себя домены α1 и α2, и внеклеточная часть цепи β включает в себя домены β1 и β2. Домены α1 и β1 образуют пептидсвязывающую бороздку на клеточной поверхности. Вследствие трехмерной конформации пептидсвязывающей бороздки комплекса МНС II теоретически не существует верхнего предела в отношении длины связанного антигена, но, как правило, презентируемые с помощью МНС II пептиды составляют от 13 до 17 аминокислот в длину.
В дополнение к своему взаимодействию с антигенными пептидами пептидсвязывающая бороздка молекулы МНС II взаимодействует с инвариантной цепью (Ii) в ходе процессов образования комплекса МНС II и захвата пептида. Димеры α/β МНС II собираются в эндоплазматическом ретикулуме и соединяются с цепью Ii, которая отвечает за контроль связывания пептида и направление МНС II в эндоцитозный путь. В эндосоме И подвергается протеолизу, и небольшой фрагмент К, пептид связанной с II классом инвариантной цепи (CLIP), остается на пептидсвязывающей бороздке. В эндосоме под контролем HLA-DM (у людей) CLIP обменивается на антигенные пептиды.
МНС II взаимодействует с Т-клеточным корецептором CD4 в гидрофобном кармане на стыке между доменами а2 и β2. Wang and Reinherz (2002) Structural Basis of Τ Cell Recognition of Peptides Bound to MHC Molecules, Molecular Immunology, 38:1039-49. Когда CD4 и Т-клеточный рецептор связываются с одной и той же молекулой МНС II в комплексе с пептидом, чувствительность Т-клетки к антигену увеличивается, и для активации требуется в 100 раз меньше антигена. Смотрите, Janeway's Immunobiology, 7th Ed., Murphy et al. eds., Garland Science, 2008, включенную в настоящий документ посредством ссылки.
В отношении трансмембранного и цитоплазматического доменов МНС II предполагали разнообразные функции. В случае цитоплазматического домена, было показано, что он является важным для внутриклеточной передачи сигналов, транспорта к плазматической мембране и, в конце концов, презентации антигена. Например, было показано, что Т-клеточные гибридомы слабо отвечают на антигенпрезентирующие клетки (АПК), трансфицированные цепями β МНС II, процессированные на цитоплазматическом домене, а также затрудняется индукция В-клеточной дифференцировки. Смотрите, например, Smiley et al. (1996) Truncation of the class II β-chain cytoplasmic domain influences the level of class Il/invariant chain-derived peptide complexes, Proc. Natl. Acad. Sci. USA, 93:241-44. Вероятно, процессирование молекул II класса нарушает продукцию сАМР. Предполагается, что делеция цитоплазматического хвоста МНС II влияет на внутриклеточный транспорт, таким образом предотвращая контакт комплекса с соответствующими антигенами в эндоцитозном пути. Smiley с соавт. (ранее) показали, что процессирование молекул II класса на цитоплазматическом домене снижает количество комплексов CLIP/II класс, позволяя предположить, что это влияет на способность CLIP эффективно регулировать презентацию антигена.
Выдвинули гипотезу о том, что поскольку кластеризация МНС II важна для запуска Т-клеточного рецептора (TCR), если предотвращалось связывание процессированных на цитоплазматическом домене молекул МНС II с цитоскелетом и, таким образом, предотвращалась агрегация, то нарушалась презентация антигена Т-клеткам. Ostrand-Rosenberg et al. (1991) Abrogation of Tumorigenicity by МНС Class II Antigen Expression Requires the Cytoplasmic Domain of the Class II Molecule, J. Immunol. 147:2419-22. Действительно, недавно было показано, что процесированный на цитоплазматическом домене HLA-DR не мог связаться с цитоскелетом после олигомеризации. El Fakhy et al. (2004) Delineation of the HLA-DR Region and the Residues Involved in the Association with the Cytoskeleton, J. Biol. Chem. 279:18472-80. Важно, что актиновый цитоскелет представляет собой место действия локализованной передачи сигнала, что может оказывать влияние на презентацию антигена. Кроме ассоциации с цитоскелетом недавние исследования также показали, что до 20% всех молекул HLA-DR постоянно находятся в липидных рафтах АПК, которые представляют собой микродомены, обогащенные холестерином и гликосфинголипидами, и что такая локализация важна для презентации антигена, формирования иммунного синапса и опосредованной МНС II передачи сигнала. Смотрите, например, Dolan et al. (2004) Invariant Chain and the МНС II Cytoplasmic Domains Regulate Localization of МНС Class II Molecules to Lipid Rafts in Tumor Cell-Based Vaccines, J. Immunol. 172:907-14. Dolan с соавт.предположили, что процессирование цитоплазматического домена МНС II снижает постоянную локализацию МНС II в липидных рафтах.
Кроме того, цитоплазматический домен МНС II, в частности цепь β, содержит остаток лейцина, который подвергается убиквитинированию с помощью убиквитинлигизы, мембраноассоциированной RING-CH I (MARCH I), которая контролирует эндоцитозный транспорт, интернализацию и деградацию МНС II; и было показано, что опосредованное MARCH убиквитинирование прекращается при созревании дендритных клеток, приводя к повышенному содержанию МНС II на плазматической мембране. Shin et al. (2006) Surface expression of МНС class II in dendritic cells is controlled by regulated ubiquitination, Nature 444:115-18; De Gassart et al. (2008) МНС class II stabilization at the surface of human dendritic cells is the result of maturation-dependent MARCH I down-regulation, Proc. Natl. Acad. Sci. USA 105:3491-96.
Трансмембранные домены цепей α и β МНС II взаимодействуют друг с другом, и указанное взаимодействие является важным для правильной сборки комплекса МНС II класса. Cosson and Bonifacino (1992) Role of Transmembrane Domain Interactions in the Assembly of Class IIMHC Molecules, Nature 258:659-62. Действительно, молекулы МНС II, в которых трансмембранные домены цепей α и β замещены цепью α рецептора IL-2, сохранялись в эндоплазматическом ретикулуме и с трудом обнаруживались на клеточной поверхности (там же). Посредством исследований с помощью мутагенеза было обнаружено, что консервативные остатки Gly на трансмембранных доменах α и β отвечают за сборку МНС II на клеточной поверхности (там же). Таким образом, как трансмембранные, так и цитоплазматические домены являются критически важными для правильного функционирования комплекса МНС II
Согласно различным вариантам осуществления настоящее изобретение относится к генетически модифицированному не являющемуся человеком животному (например, мыши, крысе, кролику и т.д.), которое содержит в своем геноме нуклеотидную последовательность, кодирующую человеческий или гуманизированный комплекс МНС II, например, челевеческий(е) или гуманизированный(е) полипептид(ы) α и/или β МНС II. Не являющееся человеком животное может содержать в своем геноме нуклеотидную последовательность, которая кодирует комплекс МНС II, который частично является человеческим и частично не относящимся к человеку, например, не являющееся человеком животное, которое экспрессирует химерный человеческий/не относящийся к человеку комплекс МНС II (например, не являющееся человеком животное, которое экспрессирует химерные человеческие/не относящиеся к человеку полипептиды α и β МНС II). Согласно одному аспекту не являющееся человеком животное экспрессирует только человеческий или гуманизированный комплекс МНС II, например, химерный человеческий/не относящийся к человеку комплекс МНС II, и не экспрессирует эндогенный не относящийся к человеку комплекс МНС II из эндогенного локуса МНС II. Согласно некоторым вариантам осуществления животное не способно экспрессировать какой-либо эндогенный не относящийся к человеку комплекс МНС II из эндогенного локуса МНС II, но экспрессирует только человеческий или гуманизированный комплекс МНС II. Согласно различным вариантам осуществления генетически модифицированное не являющееся человеком животное (например, мышь, крыса, кролик и т.д.) содержит в своей зародышевой линии нуклеотидную последовательность, кодирующую человеческий или гуманизированный комплекс МНС II, например, человеческий(е) или гуманизированный(е) полипептид(ы) α и/или β МНС II.
Согласно одному аспекту предусматривается химерный человеческий/не относящийся к человеку комплекс МНС II. Согласно одному варианту осуществления химерный человеческий/не относящийся к человеку комплекс МНС II содержит химерный человеческий/не относящийся к человеку полипептид α МНС II и химерный человеческий/не относящийся к человеку полипептид β МНС II. Согласно одному аспекту человеческая часть химерного полипептида α МНС II и/или человеческая часть химерного полипептида β МНС II содержит пептидсвязывающий домен полипептида α МНС II человека и/или полипептида β МНС II человека соответственно. Согласно одному аспекту человеческая часть химерного полипептида α и/или β МНС II содержит внеклеточный домен полипептида α и/или β МНС II человека соответственно. Согласно одному варианту осуществления человеческая часть химерного полипептида α МНС II содержит домен α1 полипептида α МНС II человека; согласно другому варианту осуществления человеческая часть химерного полипептида α МНС II содержит домены α1 и а2 полипептида α МНС II человека. Согласно дополнительному варианту осуществления человеческая часть химерного полипептида β МНС II содержит домен β1 полипептида β МНС II человека; согласно другому варианту осуществления человеческая часть химерного полипептида β МНС II содержит домены β1 и β2 полипептида β МНС II человека.
Человеческая часть описанных в настоящем документе полипептидов α и β МНС II может кодироваться любым из локусов HLA-DP, -DQ, и -DR. Перечень широко используемых антигенов и аллелей HLA представлен в Shankarkumar et al. ((2004) The Human Leukocyte Antigen (HLA) System, Int. J. Hum. Genet. 4(2):91-103), включенной в настоящий документ посредством ссылки. Shankarkumar с соавт. также представляют краткое объяснение используемой в настоящей области техники номенклатуры HLA. Дополнительную информацию относительно номенклатуры HLA и различных аллелей HLA можно найти в Holdsworth et al. (2009) The HLA dictionary 2008: a summary of HLA-A, -B, -C, -DRB1/3/4/5, and DQB1 alleles and their association with serologically defined HLA-A, -B, -C, -DR, and -DQ antigens, Tissue Antigens 73:95-170, и недавно обновленной редакции Marsh et al. (2010) Nomenclature for factors of the HLA system, 2010, Tissue Antigens 75:291-455, включенных в настоящий документ посредством ссылки. Таким образом, человеческий или гуманизированный полипептид МНС II может происходить из любых описанных в настоящем документе функциональных молекул HLA человека.
Согласно одному конкретному аспекту человеческие части описанного в настоящем документе гуманизированного комплекса МНС II происходят из HLA-DR человека, например, HLA-DR4. Как правило, цепи α HLA-DR являются мономорфными, например, цепь α комплекса HLA-DR кодируется геном HLA-DRA (например, геном HLA-DRα*01). С другой стороны, цепь β HLA-DR является полиморфной. Таким образом, HLA-DR4 содержит цепь а, кодируемую геном HLA-DRA, и цепь β, кодируемую геном HLA-DRB1 (например, геном HLA-DRβ1*04). Как описано в настоящем документе ниже, известно, что HLA-DR4 связан с возникновением ряда аутоиммунных заболеваний, например, ревматоидного артрита, диабета I типа, рассеянного склероза и т.д. Согласно одному варианту осуществления настоящего изобретения аллель HLA-DRA представляет собой аллель HLA-DRα*01, например, HLA-DRα*01:01:01:01. Согласно другому варианту осуществления аллель HLA-DRB представляет собой HLA-DRβ1*04, например, HLA-DRβ1*04:01:01. Несмотря на то что настоящие примеры описывают указанные конкретные последовательности HLA, в настоящем документе предусматриваются любые подходящие последовательности HLA-DR, например, полиморфные варианты, проявляющиеся в человеческой популяции, последовательности с одной или несколькими консервативными или неконсервативными аминокислотными модификациями, последовательности нуклеиновой кислоты, отличающиеся от описанных в настоящем документе последовательностей вследствие вырожденности генетического кода и т.д.
Человеческие части гуманизированного комплекса МНС II могут кодироваться нуклеотидными последовательностями аллелей HLA, которые, как известно, связаны с распространенными заболеваниями человека. Такие аллели HLA включают без ограничения HLA-DRB1*0401, -DRB1*0301, -DQA1*0501, -DQB1*0201, -DRB1*1501, -DRB1*1502, -DQB1*0602, -DQA1*0102, -DQA1*0201, -DQB1*0202, -DQA1*0501 и их комбинации. Обобщенное изложение взаимосвязей аллелей HLA и заболеваний можно найти в Bakker et al. (2006) A high-resolution HLA and SNP haplotype map for disease association studies in the extended human MHC, Nature Genetics 38:1166-72 и Supplementary Information, включенных в настоящий документ посредством ссылки.
Согласно одному аспекту не относящаяся к человеку часть химерного человеческого/не относящегося к человеку комплекса МНС II содержит трансмембранный и/или цитоплазматический домены эндогенного не относящегося к человеку (например, относящегося к грызуну, например, мыши, крысе и т.д.) комплекса МНС II. Таким образом, не относящаяся к человеку часть химерного человеческого/не относящегося к человеку полипептида α МНС II может содержать трансмембранный и/или цитоплазматический домены эндогенного не относящегося к человеку полипептида α МНС II. Не относящаяся к человеку часть химерного человеческого/не относящегося к человеку полипептида β МНС II может содержать трансмембранный и/или цитоплазматический домены эндогенного не относящегося к человеку полипептида β МНС II. Согласно одному аспекту животное представляет собой мышь, и не относящиеся к человеку части химерных полипептидов α и β происходят из белка Н-2Е мыши. Таким образом, не относящиеся к человеку части химерных полипептидов α и β могут содержать трансмембранный и цитоплазматический домены, происходящие из белка Н-2Е мыши. Хотя в примерах рассматриваются конкретные последовательности Н-2Е, в настоящем документе предусматриваются любые подходящие последовательности, например, полиморфные варианты, консервативные/неконсервативные аминокислотные замены и т.д.
Согласно различным аспектам настоящего изобретения последовательность(и), кодирующая(ие) химерный человеческий/не относящийся к человеку комплекс МНС II, расположены на эндогенном не относящимся к человеку локусе МНС II (например, локусе Н-2А и/или Н-2Е мыши). Согласно одному варианту осуществления это приводит к замещению эндогенного(ых) гена(ов) МНС II или его(их) части нуклеотидной(ыми) последовательностью(ями), кодирующей(ими) человеческий или гуманизированный белок МНС II, например, химерным геном, кодирующим химерный человеческий/не относящийся к человеку белок МНС II, описанный в настоящем документе. Поскольку нуклеотидные последовательности, кодирующие полипептиды α и β МНС II, расположены на хромосоме в непосредственной близости друг от друга, замещение можно разработать для нацеленного воздействия на два гена либо независимо, либо вместе; причем обе эти возможности предусматриваются в настоящем документе. Согласно одному варианту осуществления замещение предусматривает замещение эндогенной нуклеотидной последовательности, кодирующей полипептиды α и β МНС II, нуклеотидной последовательностью, кодирующей химерный человеческий/не относящийся к человеку полипептид α МНС и химерный человеческий/не относящийся к человеку полипептид β МНС. Согласно одному аспекту замещение предусматривает замещение нуклеотидных последовательностей, представляющих один или несколько (например, два) эндогенных гена МНС II. Таким образом, не являющееся человеком животное содержит химерную человеческую/не относящуюся к человеку нуклеотидную последовательность на эндогенном локусе МНС II, и экспрессирует химерный человеческий/не относящийся к человеку белок МНС II из эндогенного не относящегося к человеку локуса.
Таким образом, в настоящем документе предусматривается не являющееся человеком животное, содержащее на эндогенном генном локусе МНС II первую нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид α МНС II, и вторую нуклеотидную последовательность, кодирующую химерный человеческий/не относящийся к человеку полипептид β МНС II, причем человеческая часть химерного человеческого/не относящегося к человеку полипептида α МНС II содержит внеклеточный домен α МНС II человека, и человеческая часть химерного человеческого/не относящегося к человеку полипептида β МНС II содержит внеклеточный домен β МНС II человека, и при этом химерные человеческие/не относящиеся к человеку полипептиды α МНС II и полипептиды β МНС II образуют функциональный комплекс МНС II на поверхности клетки.
Химерный человеческий/не относящийся к человеку полипептид может быть таким, что он содержит человеческую или не относящуюся к человеку лидерную (сигнальную) последовательность. Согласно одному варианту осуществления химерный полипептид α МНС II содержит не относящуюся к человеку лидерную последовательность эндогенного полипептида α МНС II. Согласно одному варианту осуществления химерный полипептид β МНС II содержит не относящуюся к человеку лидерную последовательность эндогенного полипептида β МНС II. Согласно альтернативному варианту осуществления химерный полипептид α МНС II и/или полипептид β МНС II содержит не относящуюся к человеку лидерную последовательность полипептида α МНС II и/или полипептида β МНС II соответственно от другого не являющегося человеком животного, например, другого штамма грызуна или другого штамма мыши. Таким образом, нуклеотидная последовательность, кодирующая химерный полипептид α МНС II и/или полипептид β МНС II, может быть функционально связана с нуклеотидной последовательностью, кодирующей не относящуюся к человеку лидерную последовательность α МНС II и/или лидерную последовательность β МНС II соответственно. Согласно другому варианту осуществления химерный полипептид α МНС II и/или полипептид β МНС II содержит лидерную последовательность человека полипептида α МНС II человека и/или полипептида β МНС II человека соответственно (например, лидерную последовательность HLA-DRA человека и/или HLA-DRβ1*04 человека соответственно).
Химерный человеческий/не относящийся к человеку полипептид α МНС II и/или полипептид β МНС II может содержать в своей человеческой части полный или по существу полный внеклеточный домен полипептида α МНС II человека и/или полипептида β МНС II человека соответственно. Таким образом, человеческая часть может содержать по меньшей мере 80%, предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90%, например, 95% или больше аминокислот, кодирующих внеклеточный домен полипептида α МНС II человека и/или полипептида β МНС II человека (например, HLA-DRA человека и/или HLA-DRβ1*04 человека). Согласно одному примеру по существу полный внеклеточный домен полипептида α МНС II человека и/или полипептида β МНС II человека не содержит лидерной последовательности человека. Согласно другому примеру химерный человеческий/не относящийся к человеку полипептид α МНС II и/или химерный человеческий/не относящийся к человеку полипептид β МНС II содержит лидерную последовательность человека.
Более того, химерный полипептид α МНС II и/или полипептид β МНС II может экспрессироваться под контролем эндогенных не относящихся к человеку промоторных и регуляторных элементов, например, α регуляторных элементов МНС II и/или β регуляторных элементов МНС II мыши соответственно. Такое расположение будет облегчать надлежащую экспрессию химерных полипептидов МНС II у не являющегося человеком животного, например, в ходе иммунного ответа у не являющегося человеком животного.
Генетически модифицированное не являющееся человеком животное может быть выбрано из группы, состоящей из мыши, крысы, кролика, свиньи, крупного рогатого скота (например, коровы, быка, буйвола), оленя, овцы, козы, курицы, кошки, собаки, хорька, примата (например, игрунки, макака-резуса). Для не являющихся человеком животных, у которых подходящие генетически модифицируемые ES клетки не являются общедоступными, используются другие способы для получения не являющегося человеком животного, содержащего генетическую модификацию. Такие способы включают, например, модификацию генома не относящихся к ES клеток (например, фибробласта или индуцированной плюрипотентной клетки) и использование ядерного транспорта для переноса модифицированного генома в подходящую клетку, например, ооцит, и гестацию модифицированной клетки (например, модифицированного ооцита) в не являющемся человеком животном при подходящих условиях для образования зародыша.
Согласно одному аспекту не являющееся человеком животное представляет собой млекопитающее. Согласно одному аспекту не являющееся человеком животное представляет собой небольшое млекопитающее, например, из надсемейства Dipodoidea или Muroidea. Согласно одному варианту осуществления генетически модифицированное животное представляет собой грызуна. Согласно одному варианту осуществления грызуна выбирают из мыши, крысы и хомяка. Согласно одному варианту осуществления грызуна выбирают из надсемейства Muroidea. Согласно одному варианту осуществления генетически модифицированное животное происходит из семейства, выбранного из Calomyscidae (например, мышеподобные хомяки), Cricetidae (например, хомяк, крысы и мыши Нового Света, полевки), Muridae (настоящие мыши и крысы, карликовые песчанки, иглистые мыши, косматые хомяки), Nesomyidae (рипидомисы, скалистые хомячки, белохвостые крысы, мадагаскарские крысы и мыши), Platacanthomyidae (например, колючие соневидные хомяки), и Spalacidae (например, скальные крысы, бамбуковые крысы и цокоры). Согласно конкретному варианту осуществления генетически модифицированного грызуна выбирают из настоящей мыши или крысы (семейство Muridae), карликовой песчанки, иглистой мыши и косматого хомяка. Согласно одному варианту осуществления генетически модифицированную мышь получают из представителя семейства Muridae. Согласно одному варианту осуществления животное представляет собой грызуна. Согласно конкретному варианту осуществления грызуна выбирают из мыши и крысы. Согласно одному варианту осуществления не являющееся человеком животное представляет собой мышь.
Согласно конкретному варианту осуществления не являющееся человеком животное представляет собой грызуна, который представляет собой мышь штамма C57BL, выбранного из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/01a. Согласно другому варианту осуществления мышь представляет собой штамм 129, выбранный из группы, состоящей из штамма, который представляет собой 129Р1, 129Р2, 129РЗ, 129X1, 129S1 (например, 129S1/SV, 129Sl/SvIm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129Т1, 129Т2 (смотрите, например, Festing et al. (1999) Revised nomenclature for strain 129 mice, Mammalian Genome 10:836, смотрите также Auerbach et al (2000) Establishment and Chimera Analysis of 129/SvEv- and C57BL/6-Derived Mouse Embryonic Stem Cell Lines). Согласно конкретному варианту осуществления генетически модифицированная мышь представляет собой смесь вышеупомянутого штамма 129 и вышеупомянутого штамма C57BL/6. Согласно другому конкретному варианту осуществления мышь представляет собой смесь вышеупомянутых штаммов 129 или смесь вышеупомянутых штаммов BL/6. Согласно конкретному варианту осуществления штамм 129 смеси представляет собой штамм 129S6 (129/SvEvTac). Согласно другому варианту осуществления мышь представляет собой штамм BALB, например, штамм BALB/c. Согласно другому варианту осуществления мышь представляет собой смесь штамма BALB и другого вышеупомянутого штамма.
Согласно одному варианту осуществления не являющееся человеком животное представляет собой крысу. Согласно одному варианту осуществления крысу выбирают из крысы штамма Вистар, штамма LEA, штамма Спрага-Доули, штамма Фишера, F344, F6 и Dark Agouti. Согласно одному варианту осуществления штамм крысы представляет собой смесь двух или больше штаммов, выбранных из группы, состоящей из штамма Вистар, LEA, Спрага-Доули, Фишера, F344, F6 и Dark Agouti.
Таким образом, согласно одному варианту осуществления настоящее изобретение относится к генетически модифицированной мыши, которая содержит в своем геноме нуклеотидную последовательность, кодирующую химерный человеческий/мышиный комплекс МНС II, например, химерные человеческие/мышиные полипептиды α и β МНС II. Согласно одному варианту осуществления человеческая часть химерного человеческого/мышиного полипептида α МНС II содержит связывающий или внеклеточный домен пептида α МНС II человека, и человеческая часть химерного человеческого/мышиного полипептида β МНС II содержит связывающий или внеклеточный домен пептида β МНС II человека. Согласно некоторым вариантам осуществления мышь не экспрессирует пептидсвязывающий или внеклеточный домен эндогенного мышиного полипептида α и/или β из эндогенного локуса мыши (например, локуса Н-2А и/или Н-2Е). Согласно некоторым вариантам осуществления мышь содержит геном, который не содержит ген, кодирующий функциональную молекулу II класса МНС, содержащую Н-2Ab1, Н-2Аа, Н-2Eb1, Н-2Eb2, Н-2Еа и их комбинацию. Пептидсвязывающий домен полипептида α МНС II человека может содержать домен α1, и пептидсвязывающий домен полипептида β МНС II человека может содержать домен β1; таким образом, пептидсвязывающий домен химерного комплекса МНС II может содержать домены α1 и β1 человека. Внеклеточный домен полипептида α МНС II человека может содержать домены α1 и а2, и внеклеточный домен полипептида β МНС II человека может содержать домены β1 и β2; таким образом, внеклеточный домен химерного комплекса МНС II может содержать домены α1, α2, β1 и β2 человека. Согласно одному варианту осуществления мышиная часть химерного комплекса МНС II содержит трансмембранный и цитозольный домены МНС II мыши, например Н-2Е мыши (например, трансмембранный и цитозольный домены цепей α и β Н-2Е мыши).
Следовательно, согласно одному варианту осуществления предусматривается генетически модифицированная мышь, причем мышь содержит на эндогенном локусе МНС II мыши первую нуклеотидную последовательность, кодирующую химерный человеческий/мышиный полипептид α МНС II, и вторую нуклеотидную последовательность, кодирующую химерный человеческий/мышиный полипептид β МНС II, причем человеческая часть химерного полипептида α МНС II содержит внеклеточный домен, происходящий из полипептида α белка HLA-DR4 человека, и человеческая часть химерного полипептида β МНС II содержит внеклеточный домен, происходящий из полипептида β белка HLA-DR4 человека, причем мышиная часть химерного полипептида α МНС II содержит трансмембранный и цитоплазматический домены цепи α Н-2Е мыши, и мышиная часть химерного полипептида β МНС II содержит трансмембранный и цитоплазматический домены цепи β Н-2Е мыши, и при этом мышь экспрессирует функциональный химерный комплекс МНС II HLA-DR4/H-2E. Согласно одному варианту осуществления химерный комплекс МНС II HLA-DR4/H-2E содержит цепь α МНС II, которая включает внеклеточные домены (например, домены α1 и а2), происходящие из белка HLA-DR4 (домены α1 и а2 HLA-DRA) и трансмембранный и цитоплазматический домены из цепи α Н-2Е мыши, а также цепь β МНС II, которая включает внеклеточные домены (например, домены β1 и β2), происходящие из HLA-DR4 (домены β1 и β2 HLA-DRβ1*04) и трансмембранные и цитоплазматические домены из цепи β Н-2Е мыши. Согласно одному аспекту мышь не экспрессирует функциональные эндогенные полипептиды Н-2А и Н-2Е из их эндогенных локусов мыши (например, мышь не экспрессирует полипептиды Н-2Ab1, Н-2Аа, Н-2Eb1, Н-2Eb2 и Н-2Еа). Согласно различным вариантам осуществления экспрессия первой и второй нуклеотидных последовательностей находится под контролем соответствующих эндогенных промоторов и регуляторных элементов мыши. Согласно различным вариантам осуществления настоящего изобретения первая и вторая нуклеотидные последовательности расположены на одной хромосоме. Согласно некоторым аспектам мышь содержит две копии химерного локуса МНС II, содержащего первую и вторую нуклеотидные последовательности, тогда как согласно другим аспектам мышь содержит одну копию локуса МНС II, содержащего первую и вторую нуклеотидные последовательности. Таким образом, мышь может являться гомозиготной или гетерозиготной в отношении химерного локуса МНС II, содержащего первую и вторую нуклеотидные последовательности. Согласно различным вариантам осуществления первая и вторая нуклеотидные последовательности содержатся в зародышевой линии мыши.
Согласно некоторым описанным в настоящем документе вариантам осуществления предусматривается мышь, которая содержит химерный локус МНС II на эндогенном локусе МНС II мыши, например, посредством замещения эндогенных генов Н-2А и Н-2Е мыши. Согласно некоторым аспектам химерный локус содержит нуклеотидную последовательность, которая кодирует внеклеточный домен HLA-DRA человека и трансмембранный и цитоплазматический домены цепи α Н-2Е мыши, а также внеклеточный домен HLA-DRβ1*04 человека и трансмембранный и цитоплазматический домены цепи β Н-2Е мыши. Различные домены химерного локуса соединены таким образом, чтобы локус экспрессировал функциональный химерный человеческий/мышиный комплекс МНС II.
Согласно различным вариантам осуществления не являющееся человеком животное (например, грызун, например, мышь или крыса), которое экспрессирует функциональный химерный белок МНС II из описанного в настоящем документе химерного локуса МНС II, проявляет химерный белок на клеточной поверхности. Согласно одному варианту осуществления не являющееся человеком животное экспрессирует химерный белок МНС II на клеточной поверхности в клеточной локализации, которая аналогична наблюдаемой у человека. Согласно одному аспекту клетка проявляет пептидный фрагмент (фрагмент антигена), связанный с внеклеточной частью (например, внеклеточной частью HLA-DR4 человека) химерного белка МНС II.
Согласно различным вариантам осуществления клетка, проявляющая химерный белок МНС II, например, белок HLA-DR4/H-2E, представляет собой антигенпрезентирующую клетку (АПК), например, макрофаг, дендритную клетку или В-клетку. Согласно некоторым вариантам осуществления пептидный фрагмент, презентируемый химерным белком, происходит из опухоли. Согласно другим вариантам осуществления пептидный фрагмент, презентируемый химерным белком МНС II, происходит из патогена, например, бактерии, вируса или паразита.
Описанный в настоящем документе химерный белок МНС II может взаимодействовать с другими белками на поверхности той же клетки или второй клетки. Согласно некоторым вариантам осуществления химерный белок МНС II взаимодействует с эндогенными не относящимися к человеку белками на поверхности указанной клетки. Химерный белок МНС II также может взаимодействовать с человеческими или гуманизированными белками на поверхности той же клетки или второй клетки. Согласно некоторым вариантам осуществления вторая клетка представляет собой Т-клетку, и химерный белок МНС II взаимодействует с Т-клеточным рецептором (TCR) и его корецептором CD4. Согласно некоторым вариантам осуществления Т-клетка представляет собой эндогенную Т-клетку мыши. Согласно другим вариантам осуществления Т-клетка представляет собой Т-клетку человека. Согласно некоторым вариантам осуществления TCR представляет собой человеческий или гуманизированный TCR. Согласно дополнительным вариантам осуществления CD4 представляет собой человеческий или гуманизированный CD4. Согласно другому варианту осуществления или один из TCR и CD4, или оба не относятся к человеку, например, относятся к мыши или крысе.
Согласно одному варианту осуществления предусматривается описанное в настоящем документе генетически модифицированное не являющееся человеком животное, у которого опухоли не развиваются с большей скоростью, чем у животного дикого типа, которое не содержит химерный ген МНС II. Согласно некоторым вариантам осуществления у животного не развивается гематологические злокачественные опухоли, например, различные Т- и В-клеточные лимфомы, лейкозы, композитные лимфомы (например, лимфома Ходжкина), с большей скоростью, чем у животного дикого типа.
В дополнение к генетически сконструированному не являющемуся человеком животному, также предусматривается не являющийся человеческим зародыш (например, зародыш грызуна, например, мыши или крысы), причем зародыш содержит донорную ES клетку, которая получена от описанного в настоящем документе не являющегося человеком животного (например, грызуна, например, мыши или крысы). Согласно одному аспекту зародыш содержит ES донорную клетку, которая содержит химерный ген МНС II и клетки зародыша-хозяина.
Также предусматривается ткань, причем ткань получена от описанного в настоящем документе не являющегося человеком животного (например, грызуна, например, мыши или крысы) и экспрессирует химерный белок МНС II (например, белок HLA-DR4/H-2E).
Кроме того, предусматривается не являющаяся человеческой клетка, выделенная из описанного в настоящем документе не являющегося человеком животного. Согласно одному варианту осуществления клетка представляет собой ES клетку. Согласно одному варианту осуществления клетка представляет собой антигенпрезентирующую клетку, например, дендритную клетку, макрофаг, В-клетку. Согласно одному варианту осуществления клетка представляет собой иммунную клетку. Согласно одному варианту осуществления иммунная клетка представляет собой лимфоцит.
Также предусматривается не являющаяся человеческой клетка, содержащая хромосому или ее фрагмент описанного в настоящем документе не являющегося человеком животного. Согласно одному варианту осуществления не являющаяся человеческой клетка содержит ядро описанного в настоящем документе не являющегося человеком животного. Согласно одному варианту осуществления не являющаяся человеческой клетка содержит хромосому или ее фрагмент как результат ядерного транспорта.
Согласно одному аспекту предусматривается не являющаяся человеческой индуцированная плюрипотентная клетка, содержащая ген, кодирующий описанный в настоящем документе химерный белок МНС II (например, белок HLA-DR4/H-2E). Согласно одному варианту осуществления индуцированная плюрипотентная клетка получена из описанного в настоящем документе не являющегося человеком животного.
Согласно одному аспекту предусматривается гибридома или квадрома, происходящая из клетки описанного в настоящем документе не являющегося человеком животного. Согласно одному варианту осуществления не являющееся человеком животное представляет собой мышь или крысу.
Согласно одному аспекту предусматривается препарат in vitro, который содержит первую клетку, несущую химерный человеческий/относящийся к грызуну поверхностный белок МНС II, который содержит связанный пептид для образования химерного человеческого/относящегося к грызуну комплекса МНС II/пептид, и вторую клетку, которая связывает химерный человеческий/относящийся к грызуну комплекс МНС II/пептид. Согласно одному варианту осуществления вторая клетка содержит человеческий или гуманизированный Т-клеточный рецептор, и согласно одному варианту осуществления дополнительно содержит человеческий или гуманизированный CD4. Согласно одному варианту осуществления вторая клетка представляет собой клетку грызуна (например, мыши или крысы), содержащую человеческий или гуманизированный Т-клеточный рецептор и человеческий или гуманизированный белок CD4. Согласно одному варианту осуществления вторая клетка представляет собой клетку человека.
Также предусматривается способ получения описанного в настоящем документе генетически сконструированного не являющегося человеком животного (например, генетически сконструированного грызуна, например, мыши или крысы). Способ получения генетически сконструированного не являющегося человеком животного дает в результате животного, геном которого содержит нуклеотидную последовательность, кодирующую химерный белок МНС II (например, химерные полипептиды α и β МНС II). Согласно одному варианту осуществления способ дает в результате генетически сконструированную мышь, геном которой содержит на эндогенном локусе МНС II нуклеотидную последовательность, кодирующую химерный человеческий/мышиный белок МНС II, причем человеческая часть химерного белка МНС II содержит внеклеточный домен HLA-DR4 человека, и мышиная часть содержит трансмембранные и цитоплазматические домены Н-2Е мыши. Согласно некоторым вариантам осуществления в способе используется нацеливание конструкции, полученной с использованием технологии VELOCIGENE®, введение конструкции в ES клетки и введение нацеленных ES клеточный клонов к мышиный зародыш с использованием технологии VELOCIMOUSE®, описанной в примерах. Согласно одному варианту осуществления ES клетки представляют собой смесь штаммов мышей 129 и C57BL/6; согласно одному варианту осуществления ES клетки представляют собой смесь штаммов мышей BALB/c и 129.
Также предусматривается нуклеотидная конструкция, используемая для создания генетически сконструированных описанных в настоящем документе не являющихся человеком животных. Согласно одному аспекту нуклеотидная конструкция содержит: 5' и 3' не относящиеся к человеку плечи гомологии, фрагмент ДНК, содержащий последовательности цепей α и β HLA-DR человека, и кассету селекции, фланкированную сайтами рекомбинации. Согласно одному варианту осуществления последовательности цепей α и β HLA-DR человека представляют собой геномные последовательности, которые содержат интроны и экзоны генов цепей α и β HLA-DR человека. Согласно одному варианту осуществления не относящиеся к человеку плечи гомологии являются гомологичными по отношению к не относящейся к человеку геномной последовательности МНС II.
Согласно одному варианту осуществления последовательность цепи α HLA-DR человека содержит кодирующую домен α1 и α2 последовательность. Согласно конкретному варианту осуществления она содержит, в направлении 5'-3': экзон α1 (экзон 2), интрон α1/α2 (интрон 2), и экзон а2 (экзон 3). Согласно одному варианту осуществления последовательность цепи β HLA-DR человека содержит кодирующую домен β1 и β2 последовательность. Согласно конкретному варианту осуществления она содержит, в направлении 5'-3': экзон β1 (экзон 2), интрон β1/β2 (интрон 2) и экзон β2 (экзон 3).
Кассета селекции представляет собой нуклеотидную последовательность, вставленную в нацеливающую конструкцию для облегчения селекции клеток (например, ES клеток), которые интегрировали представляющей интерес конструкцией. Ряд подходящих кассет селекции известен в настоящей области техники. Как правило, кассета селекции обеспечивает положительную селекцию в присутствии конкретного антибиотика (например, Neo, Hyg, Pur, CM, SPEC и т.д.). Кроме того, кассета селекции может быть фланкирована сайтами рекомбинации, которые обеспечивают делецию кассеты селекции при обработке ферментами рекомбиназами. Широко используемые сайты рекомбинации представляют собой lοxΡ и Frt, распознаваемые ферментами Cre и Flp соответственно, но в настоящей области техники известны и другие. Кассета селекции может быть расположена где-либо в конструкции вне кодирующей области. Согласно одному варианту осуществления кассета селекции расположена в интроне цепи β, например, интроне β2/трансмембранный домен (интрон 3).
Согласно одному варианту осуществления 5' и 3' плечи гомологии содержат геномные последовательность в положениях 5' и 3' эндогенного не относящегося к человеку локуса МНС II. Согласно одному варианту осуществления 5' плечо гомологии содержит геномную последовательность против хода транскрипции по отношению к гену Н-2Ab1 мыши, и 3' плечо гомологии содержит геномную последовательность по ходу транскрипции по отношению к гену Н-2Еа мыши. Согласно настоящему варианту осуществления конструкт обеспечивает замещение как генов Н-2Е, так и Н-2А мыши.
Таким образом, согласно одному аспекту предусматривается нуклеотидная конструкция, содержащая в направлении 5'-3':5' плечо гомологии, содержащее мышиную геномную последовательность против хода транскрипции по отношению к гену Н-2Ab1 мыши, первую нуклеотидную последовательность, содержащую последовательность, кодирующую химерную человеческую/мышиную цепь β МНС II, вторую нуклеотидную последовательность, содержащую последовательность, кодирующую химерную человеческую/мышиную цепь α МНС II, и 3' плечо гомологии, содержащее мышиную геномную последовательность по ходу транскрипции по отношению к гену Н-2Еа мыши. Согласно конкретному варианту осуществления первая нуклеотидная последовательность, содержащая последовательность, кодирующую химерную человеческую/мышиную цепь β МНС II, содержит человеческий экзон β1, интрон β1/β2, экзон β2, кассету селекции, фланкированную сайтами рекомбинации, вставленными в интронную область между последовательностью экзона β2 человека и последовательностью экзона трансмембранного домена мыши. Согласно конкретному варианту осуществления вторая нуклеотидная последовательность, содержащая последовательность, кодирующую химерную человеческую/мышиную цепь α МНС II, содержит человеческий экзон α1, интрон α1/α2 и экзон α2 человека. Иллюстративная конструкция по настоящему изобретению изображена на фиг. 5 (MAID 1680).
При завершении нацеленного воздействия на ген ES клетки или генетически модифицированных не являющихся человеком животных подвергают скринингу для подтверждения успешного встраивания представляющей интерес экзогенной нуклеотидной последовательности или экспрессии экзогенного полипептида. Различные техники известны специалистам в настоящей области техники и включают в себя (без ограничения) саузерн-блоттинг, ПЦР длинных фрагментов, количественную ПЦР (например, ПЦР в реальном времени с использованием TAQMAN®), флуоресцентную гибридизацию in situ, нозерн-блоттинг, проточную цитометрию, вестерн-блоттинг, иммуноцитохимию, иммуногистохимию и т.д. Согласно одному примеру не являющиеся человеком животные (например, мыши), несущие представляющую интерес генетическую модификацию, могут быть идентифицированы путем скрининга в отношении потери аллеля мыши и/или приобретения аллеля человека с использованием модификации аллельного анализа, описанного в Valenzuela et al. (2003) High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nature Biotech. 21(6):652-659. Другие анализы, которые идентифицируют конкретную нуклеотидную или аминокислотную последовательность у генетически модифицированных животных, известны специалистам в настоящей области техники.
Настоящее раскрытие также относится к способу модификации локуса МНС II не являющегося человеком животного для экспрессии описанного в настоящем документе химерного человеческого/не относящегося к человеку комплекса МНС II. Согласно одному варианту осуществления настоящее изобретение относится к способу модификации локуса МНС II мыши для экспрессии химерного человеческого/мышиного комплекса МНС II, предусматривающему замещение на эндогенном локусе МНС II мыши нуклеотидной последовательности, кодирующей комплекс МНС II мыши, нуклеотидной последовательностью, кодирующей химерный человеческий/мышиный комплекс МНС II. Согласно конкретному аспекту нуклеотидная последовательность, кодирующая химерный человеческий/мышиный комплекс МНС II, содержит первую нуклеотидную последовательность, кодирующую внеклеточный домен цепи α МНС II человека (например, цепи α HLA-DR4) и трансмембранный и цитоплазматический домены цепи α МНС II мыши (например, цепи α Н-2Е), и вторую нуклеотидную последовательность, кодирующую внеклеточный домен цепи β МНС II человека (например, цепи β HLA-DR4) и трансмембранный и цитоплазматический домены цепи β МНС II мыши (например, цепи β Н-2Е, например, цепи Н-2Eb1). Согласно некоторым вариантам осуществления модифицированный локус МНС II мыши экспрессирует химерный белок HLA-DR4/H-2E.
Согласно одному аспекту предусматривается способ получения химерной человеческой HLA II класса /не относящейся к человеку МНС II класса молекулы, предусматривающий экспрессию в одной клетке химерного белка HLA-DR4/H-2E из описанной в настоящем документе нуклеотидной конструкции. Согласно одному варианту осуществления нуклеотидная конструкция представляет собой вирусный вектор; согласно конкретному варианту осуществления вирусный вектор представляет собой лентивирусный вектор. Согласно одному варианту осуществления клетку выбирают из СНО, COS, 293, HeLa и ретинальной клетки, экспрессирующей вирусную последовательность нуклеиновой кислоты (например, клетка PERC.6™).
Согласно одному аспекту предусматривается клетка, которая экспрессирует химерный белок HLA-DR4/H-2E. Согласно одному варианту осуществления клетка содержит вектор экспрессии, содержащий описанную в настоящем документе химерную последовательность МНС II класса. Согласно одному варианту осуществления клетку выбирают из СНО, COS, 293, HeLa и ретинальной клетки, экспрессирующей вирусную последовательность нуклеиновой кислоты (например, клетка PERC.6™).
Также предусматривается химерная молекула МНС II класса, полученная с помощью описанного в настоящем документе не являющегося человеком животного, причем химерная молекула МНС II класса содержит домены α1, α2, β1 и β2 из белка МНС II человека, например, белка HLA-DR4, и трансмембранный и цитоплазматический домены из не относящегося к человеку белка МНС II, например, белка Н-2Е мыши. Химерный комплекс МНС II, содержащий внеклеточный домен HLA-DR4, описанного в настоящем документе, может быть обнаружен с помощью антител к HLA-DR. Таким образом, клетка, проявляющая химерный человеческий/не относящийся к человеку полипептид МНС II, может быть обнаружена и/или выбрана с использованием антитела к HLA-DR.
Несмотря на то что представленные ниже примеры описывают генетически сконструированное животное, геном которого содержит замещение нуклеотидной последовательности, кодирующей белки Н-2А и Н-2Е мыши, нуклеотидной последовательностью, кодирующей химерный человеческий/мышиный белок HLA-DR4/H-2E, специалисту в настоящей области техники будет понятно, что аналогичная стратегия может использоваться для введения химер, содержащих другие человеческие гены МНС II (HLA-DP и HLA-DQ). Таким образом, дополнительный вариант осуществления настоящего изобретения направлен на генетически сконструированное животное, геном которого содержит нуклеотидную последовательность, кодирующую химерный белок HLA-DQ/H-2A. Согласно одному варианту осуществления нуклеотидная последовательность кодирует химерный белок HLA-DQ2.5/H-2A. Согласно другому варианту осуществления нуклеотидная последовательность кодирует химерный белок HLA-DQ8/H-2A. Кроме того, также предусматривается введение множественных гуманизированных молекул МНС II (например, химерных HLA-DR/H-2E и HLA-DQ/H-2А).
Применение генетически модифицированных животных
Согласно различным вариантам осуществления описанные в настоящем документе генетически модифицированные не являющиеся человеком животные производят АПК с человеческими или гуманизированным МНС II на клеточной поверхности и, в результате этого, презентируют пептиды, происходящие из цитозольных белков в качестве эпитопов Т-клеткам характерным для человека образом, поскольку по существу все компоненты комплекса являются человеческими или гуманизированными. Генетически модифицированные не являющиеся человеком животные по настоящему изобретению могут использоваться для исследования функции иммунной системы человека у гуманизированного животного; для идентификации антигенов и антигенных эпитопов, которые вызывают иммунный ответ (например, Т-клеточных эпитопов, например, уникальных эпитопов злокачественных опухолей человека), например, для применения в разработке вакцин; для оценки кандидатных вакцин и других стратегий разработки вакцин; для исследования аутоиммунитета человека; для исследования инфекционных заболеваний человека; и в других случаях для разработки лучших терапевтических стратегий на основании экспрессии МНС человека.
Комплекс МНС II связывает пептиды, происходящие из внеклеточных белков, например, внеклеточной бактерии, соседних клеток или полипептидов, связанных В-клеточными рецепторами и интернализованных в В-клетку. Как только внеклеточные белки попадают в эндоцитозный путь, они распадаются на пептиды, и пептиды связываются и презентируются с помощью МНС II. Как только пептид, презентируемый МНС II, распознается CD4+ Т-клетками, Т-клетки активируются, пролиферируют, дифференцируются до различных подтипов Т-хелперов (например, TH1, TH2), и приводят к ряду событий, включающих активацию опосредованного макрофагами лизиса патогена, В-клеточную пролиферацию и продукцию антител. Вследствие роли МНС II в иммунном ответе, понимание презентации пептида МНС II важна в разработке способов лечения патологий у человека. Тем не менее, презентация антигенов в сочетании с МНС II мыши лишь в некоторой степени имеет отношение к заболеванию человека, поскольку комплексы МНС человека и мыши распознают антигены различным образом, например, МНС II мыши может не распознавать одинаковые антигены или может презентировать другие эпитопы, в отличие от МНС II человека. Таким образом, наиболее релевантные данные для патологий человека получают посредством исследования презентации антигенных эпитопов МНС II человека.
Таким образом, согласно различным вариантам осуществления генетически сконструированные животные по настоящему изобретению являются применимыми, среди прочего, для оценки способности антигена инициировать иммунный ответ у человека, и для создания разнообразных антигенов и идентификации специфического антигена, который может использоваться для разработки вакцины для человека.
Согласно одному аспекту предусматривается способ определения у человека антигенности пептидной последовательности, предусматривающий воздействие на описанное в настоящем документе генетически модифицированное не являющееся человеком животное молекулой, содержащей пептидную последовательность, предоставление не являющемуся человеком животному возможности развить иммунный ответ и обнаружение у не являющегося человеком животного клетки, которая связывает последовательность пептида, презентируемого описанным в настоящем документе гуманизированным комплексом МНС II.
Согласно одному аспекту предусматривается способ определения того, будет ли пептид вызывать иммунный ответ у человека, предусматривающий воздействие на описанное в настоящем документе генетически модифицированное не являющееся человеком животное пептидом, предоставление не являющемуся человеком животному возможности развить иммунный ответ и обнаружение у не являющегося человеком животного клетки, которая связывает последовательность пептида описанной в настоящем документе химерной человеческой/не относящейся к человеку молекулой МНС II класса. Согласно одному варианту осуществления не являющееся человеком животное после воздействия содержит рестриктированную по МНС II класса CD4+ Т-клетку, которая связывает пептид.
Согласно одному аспекту предусматривается способ идентификации CD4+ Т-клеточного эпитопа человека, предусматривающий воздействие на описанное в настоящем документе не являющееся человеком животное антигена, содержащего предполагаемый Т-клеточный эпитоп, предоставление не являющемуся человеком животному возможности развить иммунный ответ и идентификацию эпитопа, связанного рестриктированной по МНС II класса CD4+ Т-клеткой.
Согласно одному аспекту предусматривается способ идентификации антигена, который вызывает CD4+ Т-клеточный ответ у человека, предусматривающий воздействие предполагаемого антигена на описанную в настоящем документе мышь, предоставление мыши возможности развития иммунного ответа, обнаружение CD4+ Т-клеточного ответа, который является специфическим для антигена в сочетании с молекулой МНС II человека (например, молекулой HLA-DR), и идентификацию антигена, связанного рестриктированной по МНС II молекулой человека (например, рестриктированной по HLA-DR молекулой человека).
Согласно одному варианту осуществления антиген содержит бактериальный белок. Согласно одному варианту осуществления антиген содержит антиген опухолевой клетки человека. Согласно одному варианту осуществления антиген содержит предполагаемую вакцину для применения у человека, или другое биофармацевтическое средство. Согласно одному варианту осуществления антиген содержит эпитоп человека, который вызывает образование антител у человека. Согласно другому варианту осуществления антиген содержит антиген дрожжевой или грибковой клетки. Согласно другому варианту осуществления антиген происходит из паразита человека.
Согласно одному аспекту предусматривается способ определения, содержит ли предполагаемый антиген эпитоп, который при воздействии на иммунную систему человека будет вызывать рестриктированный по HLA-DR иммунный ответ (например, рестриктированньш по HLA-DR4 ответ), предусматривающий воздействие на описанную в настоящем документе мышь предполагаемого антигена и измерение антигенспецифического рестриктированного по HLA-DR (например, рестриктированного по HLA-DR4) иммунного ответа у мыши. Согласно другому аспекту предусматривается способ определения, причем предполагаемый антиген содержит эпитоп, который при воздействии на иммунную систему человека, будет вызывать рестриктированньш по HLA-DQ ответ.
Также предусматривается способ получения антител к антигену, например, антигену, происходящему из бактерии, паразита и т.д., презентированному в сочетании с комплексом МНС II человека, предусматривающий воздействие на описанную в настоящем документе мышь антигеном, предоставлении мыши возможности развить иммунный ответ, причем иммунный ответ включает продукцию антител, и выделение антитела, которой распознает антиген, презентированный в сочетании с комплексом МНС II человека. Согласно одному варианту осуществления для получения антител к комплексу пептид-МНС II гуманизированную в отношении МНС II мышь иммунизируют с помощью иммуногена пептид-МНС II.
Согласно одному аспекту предусматривается способ идентификации вариабельного домена Т-клеточного рецептора, который распознает антиген, презентированный в сочетании с МНС II (например, опухолевый антиген человека, вакцина и т.д.), предусматривающий воздействие на мышь, содержащую описанный в настоящем документе гуманизированный комплекс МНС II, антигеном, предоставлении мыши возможности развить иммунный ответ и выделение из мыши последовательности нуклеиновой кислоты, кодирующей вариабельный домен Т-клеточного рецептора, который связывает рестриктированный по МНС II антиген. Согласно одному варианту осуществления антиген презентируется в сочетании с гуманизированным МНС II (например, эктодоменом HLA II человек/трансмембранным и/или цитоплазматическим доменом МНС II мыши).
Последствие взаимодействия между Т-клеткой и АПК, представляющей пептид в сочетании с МНС II (например, эктодоменом HLA II человека/трансмембранным и/или цитоплазматическим доменом МНС II мыши), может быть измерено с помощью ряда методов, известных в настоящей области техники, например, с помощью анализов Т-клеточной пролиферации, анализов высвобождения цитокинов и т.д.
В дополнение к способности идентифицировать антигены и их Т-клеточные эпитопы из патогенов или новообразований, генетически модифицированные животные по настоящему изобретению могут использоваться для идентификации аутоантигенов, имеющих отношение к аутоиммунному заболеванию человека, и для других исследований прогрессирования аутоиммунного заболевания человека. Известно, что полиморфизмы в пределах локусов HLA играют роль в предрасположенности к аутоиммунному заболеванию человека. Действительно установили, что специфические полиморфизмы в локусах HLA-DR и HLA-DQ коррелируют с развитием ревматоидного артрита, диабета I типа, тиреоидита Хашимото, рассеянного склероза, тяжелой миастении, диффузного токсического зоба, системной красной волчанки, глютеновой болезни, болезни Крона, язвенного колита и других аутоиммунных нарушений. Смотрите, например, Wong and Wen (2004) What can the HLA transgenic mouse tell us about autoimmune diabetes?, Diabetologia 47:1476-87; Taneja and David (1998) HLA Transgenic Mice as Humanized Mouse Models of Disease and Immunity, J. Clin. Invest. 101:921-26; Bakker et al. (2006), ранее; и International МНС and Autoimmunity Genetics Network (2009) Mapping of multiple susceptibility variants within the MHC region for 7 immune-mediated diseases, Proc. Natl. Acad. Sci. USA 106:18680-85.
Таким образом, способы получения описанных в настоящем документе гуманизированных в отношении комплекса МНС II животных могут использоваться для введения молекул МНС II, которые, как полагают, связаны с конкретными аутоиммунными заболеваниями человека, и может изучаться прогрессирование аутоиммунного заболевания человека. Кроме того, описанные в настоящем документе не являющиеся человеком животные могут использоваться для разработки животных моделей аутоиммунного заболевания человека. Мыши согласно настоящему изобретению, несущие описанные в настоящем документе гуманизированные белки МНС II, могут использоваться для идентификации потенциальных аутоантигенов, для картирования эпитопов, вовлеченных в прогрессирование заболевания, и для разработки стратегий для модулирования аутоиммунных заболеваний.
Кроме того, описанные в настоящем документе генетически модифицированные животные могут использоваться в изучении аллергического ответа человека. Поскольку аллергические ответы, вероятно, связаны с аллелями МНС II, описанные в настоящем документе генетически модифицированные животные могут использоваться для определения рестрикции по HLA специфического в отношении аллергена Т-клеточного ответа и для разработки стратегий борьбы с аллергическим ответом.
ПРИМЕРЫ
Настоящее изобретение будет дополнительно проиллюстрированы представленными ниже не ограничивающими примерами. Эти примеры представлены для содействия в понимании настоящего изобретения, но не предусмотрены для ограничения и не должны рассматриваться как ограничивающие его объем каким-либо образом. Примеры не включают подробные описания общепринятых методов, которые будут хорошо известны специалистам в настоящей области техники (методы молекулярного клонирования и т.д.). Если не указано иное, части представляют собой части по весу, молекулярный вес представляет собой средний молекулярный вес, температура указана в градусах Цельсия и давление равно или близко атмосферному.
Пример 1. Делеция эндогенных локусов Н-2А и Н-2Е МНС II класса
Нацеливающий вектор для введения делеции эндогенных генов МНС II класса Н-2Ab1, Н-2Аа, Н-2Eb1, Н-2Eb2 и Н-2Еа получали с использованием технологии генной инженерии VELOCIGENE® (смотрите, например, патент США №6586251 и Valenzuela et al., ранее). Бактериальную искусственную хромосомную (ВАС) RP23-458Í22 (Invitrogen) ДНК модифицировали для удаления эндогенных генов МНС II класса Н-2Ab1, Н-2Аа, Н-2Eb1, Н-2Eb2 и Н-2Еа.
Кратко, плечи гомологии против хода транскрипции и по ходу транскрипции получали с помощью ПЦР ВАС ДНК мыши из положений 5' гена Н-2Ab1 и 3' гена Н-2Еа соответственно. Как показано на фиг. 5, эти плечи гомологии использовали для получения кассеты, которая удаляла ~79 т.п.н. RP23-458i22, содержащей гены Н-2Ab1, Н-2Аа, Н-2Eb1, Н-2Eb2 и Н-2Еа локуса МНС II класса с помощью бактериальной гомологичной рекомбинации (BHR). Эту область замещали кассетой гигромицина, фланкированной сайтами lox66 и lox71. Конечный нацеливающий вектор в направлении 5'-3' включал в себя плечо гомологии длиной 34 т.п.н., содержащее геномную последовательность мыши 5' в отношении гена Н-2Ab1 эндогенного локуса МНС II класса, 5' сайт lox66, кассету гигромицина, 3' сайт lox71 и плечо гомологии длиной 63 т.п.н., содержащее геномную последовательность мыши 3' по отношению к гену Н-2Еа эндогенного локуса МНС II класса (MAID 5111, смотрите фиг. 5).
Нацеливающий вектор ВАС ДНК (описанный выше) использовали для электропорации ES клеток мыши для получения модифицированных ES клеток, содержащих делецию эндогенного локуса МНС II класса. Положительные ES клетки, содержащие подвергнутый делеции эндогенный локус МНС II класса идентифицировали с помощью анализа количественной ПЦР с использованием зондов TAQMAN™ (Lie and Petropoulos (1998) Curr. Opin. Biotechnology 9:43-48). Область против хода транскрипции подвергнутого делеции локуса подтверждали с помощью ПЦР с использованием праймеров 5111U F (CAGAACGCCAGGCTGTAAC; SEQ ID NO:1) и 5111U R (GGAGAGCAGGGTCAGTCAAC; SEQ ID NO:2) и зонда 5111U Ρ (CACCGCCACTCACAGCTCCTTACA; SEQ ID NO:3), тогда как область по ходу транскрипции подвергнутого делеции локуса подтверждали с использованием праймеров 5111D F (GTGGGCACCATCTTCATCATTC; SEQ ID NO:4) и 5111D R (CTTCCTTTCCAGGGTGTGACTC; SEQ ID NO:5) и зонда 5111D Ρ (AGGCCTGCGATCAGGTGGCACCT; SEQ ID NO:6). Присутствие кассеты гигромицина из нацеливающего вектора подтверждали с использованием праймеров HYGF (TGCGGCCGATCTTAGCC; SEQ ID NO:7) и HYGR (TTGACCGATTCCTTGCGG; SEQ ID NO:8) и зонда HYGP (ACGAGCGGGTTCGGCCCATTC; SEQ ID NO:9). Нуклеотидная последовательность, охватывающая точку делеции против хода транскрипции (SEQ ID NO:10), включала в себя следующее, что показывает эндогенную мышиную последовательность против хода транскрипции по отношению к точке делеции (которая находится в скобках ниже), соединенную смежным образом с последовательностью кассеты, присутствующей в точке делеции: (TTTGTAAACA AAGTCTACCC AGAGACAGAT GACAGACTTC AGCTCCAATG CTGATTGGTT CCTCACTTGG GACCAACCCT) CTCGAGTACC GTTCGTATAA TGTATGCTAT ACGAAGTTAT ATGCATCCGG GTAGGGGAGG. Нуклеотидная последовательность, охватывающая точку делеции по ходу транскрипции (SEQ ID NO: 11), включала в себя следующее, что показывает последовательность кассеты, смежную с эндогенной мышиной последовательностью по ходу транскрипции по отношению к точке делеции (которая содержится в скобках ниже): CCTCGACCTG CAGCCCTAGG ATAACTTCGT ATAATGTATG CTATACGAAC GGTAGAGCTC (CACAGGCATT TGGGTGGGCA GGGATGGACG GTGACTGGGA CAATCGGGAT GGAAGAGCAT AGAATGGGAG TTAGGGAAGA). Положительные ES клеточные клоны затем использовали для имплантации самкам мышей с использованием способа VELOCIMOUSE® (описанного ниже) для получения помета детенышей, содержащих делецию эндогенного локуса МНС II класса.
Описанные выше нацеленные ES клетки использовали в качестве донорных ES клеток и вводили в зародыш мыши на стадии 8 клеток с помощью способа VELOCIMOUSE® (смотрите, например, патент США №7294754 и Poueymirou et al. (2007) F0 generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses, Nature Biotech. 25(1):91-99). Мышей, несущих делецию генов Н-2Ab1, Н-2Аа, Н-2Eb1, Н-2Eb2 и Н-2Еа в эндогенном локусе МНС II класса, идентифицировали путем генотипирования с использованием модификации аллельного анализа (Valenzuela et al, ранее), который обнаруживал присутствие кассеты гигромицина и подтверждал отсутствие эндогенных последовательностей МНС II класса.
Мышей, несущих делецию генов Н-2Ab1, Н-2Аа, Н-2Eb1, Н-2Eb2 и Н-2Еа в эндогенном локусе МНС II класса, могут скрещивать с Cre-детекторным штаммом мышей (смотрите, например, публикацию международной патентной заявки № WO 2009/114400) для удаления какой-либо фланкированной сайтами lox кассеты гигромицина, введенной с помощью нацеливающего вектора, которая не удалена, например, на стадии ES клеток или у зародыша. Необязательно, у мышей сохраняют кассету гигромицина.
Пример 2. Получение большого нацеливающего вектора (LTVEC), содержащего гуманизированные гены Н-2Eb1 и Н-2Еа
Нацеливающий вектор для введения гуманизированных последовательностей МНС II разрабатывали, как показано на фиг. 4. с использованием технологии генной инженерии VELOCIGENE®, ДНК бактериальной искусственной хромосомы (ВАС) RP23-458i22 модифицировали на различных стадиях для: (1) создания вектора, содержащего функциональный I-Е α экзон 1 из гена BALB/c Н-2Еа (ФИГ. 4А); (2) создания вектора, содержащего замещение экзонов 2 и 3 гена β I-Е мыши экзонами из DRβ1*04 человека и замещение экзонов 2 и 3 α I-Е мыши экзонами DRα1*01 человека (фиг. 4В); (3) создания вектора, несущего экзоны 2 и 3 DRβ1*04 человека среди остальных I-Е β экзонов мыши, и экзоны 2 и 3 DRα1*01 человека среди остальных I-Е α экзонов мыши, включая в себя функциональный I-Е α экзон 1 из BALB/c мыши (стадия (1) (фиг. 4С); и (4) удаления скрытого сайта сплайсинга в векторе, созданном в (3) (фиг. 4D).
В частности, поскольку у мышей С57В1/6 I-Е α ген представляет собой псевдоген вследствие присутствия нефункционального экзона 1, вначале создавали вектор, содержащий функциональный I-Е α экзон 1 из гена BALB/c Н-2Еа (фиг. 4А). RP23-458i22 ВАС модифицировали с помощью бактериальной гомологичной рекомбинации (1.BHR) для замещения гена устойчивости к хлорамфениколу на ген устойчивости к спектромицину. Полученный вектор дополнительно модифицировали с помощью BHR для замещения полной I-Α и I-Е кодирующей области на кассету неомицина, фланкированную сайтами рекомбинации (2.BHR). Другой цикл BHR (3. BHR) с конструкцией, содержащей экзон, кодирующий BALB/c Ι-Εα лидерную последовательность (экзон 1) и ген устойчивости к хлорамфениколу, фланкированный сайтами рестрикции PI-SceI и I-CeuI, давал в результате вектор, содержащий функциональный экзон 1 BALB/c Н-2Еа.
Независимо, для создания вектора, содержащего замещение экзонов 2 и 3 I-Е β гена мыши экзонами DRβ1*04 человека и замещение экзонов 2 и 3 I-Е α мыши экзонами DRα1*01 человека, RP23-458i22 ВАС модифицировали посредством нескольких стадий гомологичной рекомбинации, 4. BHR - 8. BHR (фиг. 4В). Полученную последовательность нуклеиновой кислоты фланкировали сайтами рестрикции PI-SceI/I-CeuI для обеспечения лигирования в конструкцию, несущую экзон 1 BALB/c I-Ea, упомянутый выше (фиг. 4С).
Последовательность конечной конструкции, изображенной на фиг. 4С, содержала скрытый сайт сплайсинга на 3'-конце интрона BALB/c. Несколько стадий BHR (11. BHR - 12. BHR) с последующей стадией делеции проводили для получения конечного нацеливающего вектора (MAID 1680), который использовали для электропорации в ES клетки (фиг. 4D).
Подробно, конечный нацеливающий вектор (MAID 1680), в направлении 5'-3', состоял из 5' мышиного плеча гомологии, состоящего из ~26 т.п.н. мышиной геномной последовательности, заканчивающейся сразу выше против хода транскрипции по отношению к гену Н-2Ab1 эндогенного локуса МНС II класса; вставки ~59 т.п.н., содержащей гуманизированный ген МНС II β цепи (гуманизированный ген Н-2Eb1) и гуманизированный ген МНС II α цепи (гуманизированный ген Н-2Еа) и фланкированную flox кассету неомицина; и 3' мышиного плеча гомологии, состоящей из ~57 т.п.н. мышиной геномной последовательности, начинающейся сразу ниже по ходу транскрипции по отношению к гену Н-2Еа эндогенного локуса МНС II класса. Нуклеотидная последовательность, охватывающая соединение между 5' плечом и вставкой (SEQ ID NO:12), включала в себя следующее: (TGCTGATTGG ТТССТСАСТТ GGGACCAACC С) TAAGCTTTA TCTATGTCGG GTGCGGAGAA AGAGGTAATG AAATGGCACA AGGAGATCAC АСАСССАААС CAAACTCGCC, где последовательность, выделенная курсивом, представляет собой уникальный сайт PI-SceI, и мышиная геномная последовательность в 5' плече гомологии представлена в скобках. Нуклеотидная последовательность, охватывающая соединение между вставкой и 3' плечом (SEQ ID NO:13), включала в себя следующее: CACATCAGTG AGGCTAGAAT АААТТААААТ CGCTAATATG AAAATGGGG (ATTTGTACCT CTGAGTGTGA AGGCTGGGAA GACTGCTTTC AAGGGAC), где мышиная геномная последовательность в 3' плече гомологии представлена в скобках.
В пределах вставки ~59 т.п.н. ген Н-2Eb1 модифицировали следующим образом: 5136 п. н. область Н-2Eb1, включающую в себя последние 153 п.н. интрона 1, экзон 2, интрон 2, экзон 3 и первые 22 п.н. интрона 3, замещали 3111 п.н. гомологичной областью HLA-DRB1*04 человека, включающей в себя последние 148 п.н. интрона 1, экзон 2, интрон 2, экзон 3 и первые 132 п.н. интрона 3. На соединении между последовательностями человека и мыши интрона 3, вводили кассету, состоящую из сайта 5' lox2372, промотора UbC, гена устойчивости к неомицину и сайта 3' lox2372. Полученный ген кодировал химерный белок HLA-DRB1*04/H-2Eb1, состоящий из Н-2Eb1 лидерной последовательности мыши, β1 и β2 доменов человека из DRB1*04 и мышиного трансмембранного домена и цитоплазматического хвоста. Нуклеотидная последовательность, охватывающая соединение последовательностей мыши/человека в интроне 1 (SEQ ID NO:14), включала в себя следующее: (ТССАТСАСТТ CACTGGGTAG CACAGCTGTA ACTGTCCAGC CTG) GGTACCGAGC TCGGATCCAC TAGTAACGGC CGCCAGTGTG CTGGAATTC GCCCTTGATC GAGCTCCCTG GGCTGCAGGT GGTGGGCGTT GCGGGTGGGG CCGGTTAA, где последовательность, выделенная курсивом, представляет собой сайт множественного клонирования, введенный в ходе стадий клонирования, и последовательности интрона 1 мыши представлены в скобках. Нуклеотидная последовательность, охватывающая соединение между интроном 3 человека и кассетой неомицина (SEQ ID NO:15), включала в себя следующее: (АТСТССАТСА GAAGGGCACC GGT) АТААСТТ CGTATAAGGT ATCCTATACG AAGTTATATG CATGGCCTCC GCGCCGGGTT, где сайт 5' lox2372 представлен курсивом, и последовательность интрона 3 человека представлена в скобках. Нуклеотидная последовательность, охватывающая соединение между кассетой неомицина и интроном 3 мыши (SEQ ID NO: 16), включала в себя следующее: ATAACTTCGT ATAAGGTATC CTATACGAAG TTATCTCGAG (TGGCTTACAG GTAGGTGCGT GAAGCTTCTA CAAGCACAGT TGCCCCCTGG), где сайт 3' lox2372 представлен курсивом, и последовательность интрона 3 мыши представлена в скобках.
Также в пределах вставки длиной ~59 т.п.н. ген Н-2Еа модифицировали следующим образом: 1185 п. н. область Н-2Еа, включающую в себя последние 101 п.н. интрона 1, экзон 2, интрон 2, экзон 3 и первые 66 п.н. интрона 3, замещали 1189 п.н. гомологичной областью HLA-DRA1*01 человека, включающей в себя последние 104 п.н. интрона 1, экзон 2, интрон 2, экзон 3 и первые 66 п.н. интрона 3. Как описано выше, поскольку экзон 1 аллеля C57BL/6 Н-2Еа содержит делецию, которая делает ген нефункциональным, экзон 1 Н-2Еа и оставшуюся часть интрона 1 замещали эквивалентной 2616 п.н. областью из аллеля BALB/c Н-2Еа, который является функциональным. Полученный ген кодировал химерный белок H-2Ea/HLA-DRA1*01, состоящий из Н-2Еа лидерной последовательности мыши из BALB/c, α1 и α2 доменов человека из DRA1*01 и мышиного трансмембранного домена и цитоплазматического хвоста. Нуклеотидная последовательность, охватывающая соединение последовательностей мыши/человека в интроне 1 (SEQ ID NO: 17), включала в себя следующее: (CTGTTTCTTC ССТААСТССС ATTCTATGCT СТТССАТССС GA) CCGCGGCCCA АТСТСТСТСС АСТАСТТССТ GCCTACATGT ATGTAGGT, где последовательность, выделенная курсивом, представляет собой сайт фермента рестрикции, введенный в ходе стадий клонирования, и последовательности интрона 1 BALB/c представлены в скобках. Нуклеотидная последовательность, охватывающая соединение последовательностей человека/мыши в интроне 3 (SEQ ID NO:18), включала в себя следующее: CAAGGTTTCC TCCTATGATG CTTGTGTGAA ACTCGGGGCC GGCC (AGCATTTAAC AGTACAGGGA TGGGAGCACA GCTCAC), где последовательность, выделенная курсивом, представляет собой сайт фермента рестрикции, введенный в ходе стадий клонирования, и последовательности интрона 3 мыши представлены в скобках. Нуклеотидная последовательность, охватывающая соединение C57BL/6-BALB/C 5' от экзона 1 (SEQ ID NO:19), включала в себя следующее: (GAAAGCAGTC TTCCCAGCCT TCACACTCAG AGGTACAAAT) ССССАТТТТС ATATTAGCGA ТТТТААТТТА TTCTAGCCTC, где C57BL/6-специфические последовательности представлены в скобках. Нуклеотидная последовательность, охватывающая соединение BALB/C-C57BL/6 3' от экзона 1 (SEQ ID NO:20), включала в себя следующее: ТСТТСССТАА СТСССАТТСТ ATGCTCTTCC ATCCCGA CCG CGG (СССААТС ТСТСТССАСТ ACTTCCTGCC TACATGTATG), где сайт рекстрикции SacII представлен курсивом, а последовательности C57BL/6 представлены в скобках.
Пример 3. Получение гуманизированных в отношении МНС II мышей
Упрощенные диаграммы стратегии получения гуманизированных в отношении МНС II мышей с использованием вектора из примера 2 представлены на фиг. 5 и 8.
В частности, ВАС ДНК MAID1680 (описанную выше) использовали для электропорации MAID5111 ES клеток для создания модифицированных ES клеток, содержащих замещение эндогенных локусов I-Α и I-Е мыши геномным фрагментом, содержащим химерный локус DR4 человека/ I-Е мыши. Положительные ES клетки, содержащие подвергнутые делеции эндогенные локусы I-Α и I-Е, замещенные геномным фрагментом, содержащим химерный локус DR4 человека/ I-Е мыши, идентифицировали с помощью анализа количественной ПЦР с использованием зондов TAQMAN™ (Lie and Petropoulos, ранее). Вставку последовательностей DRa человека подтверждали с помощью ПЦР с использованием праймеров hDRA1F (CTGGCGGCTTGAAGAATTTGG; SEQ ID NO:21), hDRA1R (CATGATTTCCAGGTTGGCTTTGTC; SEQ ID NO:22) и зонда hDRA1P (CGATTTGCCAGCTTTGAGGCTCAAGG; SEQ ID NO:23). Вставку последовательностей DRP человека подтверждали с помощью ПЦР с использованием праймеров hDRB1F (AGGCTTGGGTGCTCCACTTG; SEQ ID NO:24), hDRB1R (GACCCTGGTGATGCTGGAAAC; SEQ ID NO:25) и зонда hDRB1P (CAGGTGTAAACCTCTCCACTCCGAGGA; SEQ ID NO:26). Потерю кассеты гигромицина из нацеливающего вектора подтверждали с помощью праймеров HYGF (TGCGGCCGATCTTAGCC; SEQ ID NO:7) и HYGR (TTGACCGATTCCTTGCGG; SEQ ID NO:8) и зонда HYGP (ACGAGCGGGTTCGGCCCATTC; SEQ ID NO:9).
Положительные ES клеточные клоны затем использовали для имплантации самкам мышей с использованием способа VELOCIMOUSE® (ранее) для получения помета детенышей, содержащих замещение эндогенных локусов I-Α и I-Е химерным локусом DR4 человека/ I-Е мыши. Описанные выше нацеленные ES клетки использовали в качестве донорных ES клеток и вводили в зародыш мыши на стадии 8 клеток с помощью способа VELOCIMOUSE®. Мышей, несущих химерный локус DR4 человека/ I-Е мыши, идентифицировали путем генотипирования с использованием модификации аллельного анализа (Valenzuela et al., ранее), который обнаруживает присутствие химерного локуса DR4 человека/ I-Е мыши.
Мышей, несущих химерный локус DR4 человека/ I-Е мыши, могут скрестить с Cre-детекторным штаммом мышей (смотрите, например, публикацию международной патентной заявки № WO 2009/114400) для удаления какой-либо фланкированной lox кассеты неомицина, введенной с помощью нацеливающего вектора, которая не удалена, например, на стадии ES клеток или у зародыша (смотрите фиг. 6).
Пример 4. Экспрессия химерного HLA-DR4 у генетически модифицированных мышей
Селезенки от мышей дикого типа (WT) или гетерозиготных гуманизированных HLA-DR4 мышей ("1681 НЕТ") перфузировали Collagenase D (Roche Bioscience) и эритроциты лизировали с помощью лизирующего буфера АСК. Спленоциты культивировали в течение двух дней с помощью 25 мкг/мл поли(I:С) для стимуляции экспрессии генов МНС-II Экспрессию HLA-DR4 человека на клеточной поверхности анализировали с помощью FACS с использованием конъюгированного с флуорохромом антитела к CD3 (17А2), антитела к CD19 (1D3), антитела к CD11c (N418), антитела к F480 (ВМ8), антитела к I-A/I-E (M15) и антитела к HLADR (L243). Проточную цитометрию проводили с использованием BD-LSRII. Экспрессия HLA-DR4 человека была ясно обнаруживаемой на поверхности CD19+ В-клеток и в значительной степени положительно регулировалась при стимуляции агонистом toll-подобного рецептора поли(I:С) (смотрите фиг. 9).
Эквиваленты
Специалистам в настоящей области техники будут понятны, или с использованием не более чем рутинного экспериментирования они смогут установить многие эквиваленты описанных в настоящем документе конкретных вариантов осуществления настоящего изобретения. Предусматривается, что такие эквиваленты включены в представленную ниже формулу изобретения.
Полное содержание всех процитированных в настоящей заявке непатентных документов, патентных заявок и патентов полностью включено посредством ссылки в настоящем документе.
Реферат
Изобретение относится к области биохимии, в частности к грызуну, который экспрессирует химерный человеческий/относящийся к грызуну белковый комплекс МНС II, а также к его клетке и ткане. Также раскрыт способ модификации локуса МНС II мыши для экспрессии химерного человеческого/мышиного комплекса МНС II, а также способ идентификации рестриктированного по HLA класса II пептида, презентация которого клеткой человека и связывание лимфоцитом человека будет приводить к цитотоксичности несущей пептид клетки. Изобретение также относится к химерному человеческому/относящимуся к грызуну белковому комплексу МНС II для идентификации и отбора пептидов, которые будут обеспечивать подходящий ответ в Т-клетке человека. Изобретение позволяет эффективно получать грызуна, экспрессирующего химерный человеческий/относящийся к грызуну белковый комплекс МНС II. 7 н. и 15 з.п. ф-лы, 9 ил., 4 пр.
Комментарии