Композиция и способ для иммунизации, способ продуцирования неприродного, упорядоченного и повторяющегося массива антигенов и оболочечный белок - RU2295973C2
Код документа: RU2295973C2
Чертежи




















































































































































































Описание
Предпосылки создания изобретения
Область, к которой относится изобретение
Настоящее изобретение относится к области молекулярной биологии, вирусологии, иммунологии и медицины. Настоящее изобретение относится к композиции, содержащей упорядоченный и повторяющийся массив антигенов или антигенных детерминант. Настоящее изобретение также относится к способу продуцирования антигенов или антигенных детерминант в упорядоченном и повторяющемся массиве. Упорядоченный и повторяющийся массив антигенов или антигенных детерминант может быть использован в целях продуцирования вакцин для лечения инфекционных болезней, для лечения аллергий и в качестве фармацевтической вакцины для профилактики или лечения злокачественных опухолей, а также для эффективного индуцирования специфических аутоиммунных ответов, в частности гуморальных ответов.
Предпосылки изобретения
В WO 00/3227 описаны композиции и способы продуцирования упорядоченных и повторяющихся массивов антигенов или антигенных детерминант. Эти композиции могут быть использованы в целях продуцирования вакцин для профилактики инфекционных болезней, для лечения аллергий и для лечения злокачественных опухолей. Эти композиции содержат коровую частицу, такую как вирусоподобная частица, с которой ассоциирован, по меньшей мере, один антиген или одна антигенная детерминанта посредством, по меньшей мере, одной непептидной связи, в результате чего образуется упорядоченный и повторяющийся массив антигенов.
Вирусоподобные частицы (VLP), благодаря своим структурным свойствам и неинфекционной природе, используются для продуцирования вакцин. VLP представляют собой надмолекулярные структуры, состоящие из множества симметрично расположенных молекул белка одного или нескольких типов. Они не содержат вирусного генома, а поэтому являются неинфекционными. В большинстве случаев VLP могут быть продуцированы в больших количествах посредством гетерологичной экспрессии и могут быть легко очищены.
Примерами VLP являются капсидные белки вируса гепатита В (Ulrich et al., Virus Res. 50:141-182 (1998)), вируса кори (Warnes et al., Gene 160:173-178 (1995)), вируса Синдбис, ротавируса (патент США №5071651 и патент США №5374426), вируса ящура (Twomey et al., Vaccine 13:1603-1610 (1995)), вируса Норуолк (Jiang X. et al., Science 250:1580-1583 (1990); Matsui S.M. et al., J. Clin. Invest. 87:1456-1461 (1991)), ретровирусный белок GAG (WO 96/30253), белок р1 ретротранспозона Ty, поверхностный белок вируса гепатита В (WO 92/11291) и вирус папиломы человека (WO 98/15631).
Индуцирование иммунного ответа в организме против его собственных молекул представляет определенные трудности из-за иммунологической толерантности. В частности, при стимуляции с использованием стандартных стратегий вакцинации, лимфоциты, обладающие специфичностью к собственным молекулам, обнаруживали низкую реактивность или даже ее отсутствие. Амилоидный пептид В (Аβ1-42) играет центральную роль в нейропатологии болезни Альцгеймера. Очаговое внеклеточное накопление пептида Аβ сопровождается микроглиозом, цитоскелетными изменениями, дистрофическим невритом и потерей синапсов. С этими патологическими изменениями, очевидно, связано снижение познавательной способности, которое лежит в основе данного заболевания.
Для получения мышиной модели болезни Альцгеймера были созданы трансгенные животные, у которых продуцировался Аβ1-42 (PDAPP-мышь), развивались бляшки и возникали повреждения нейронов в их головном мозге. В недавно опубликованной работе была описана иммунизация молодых PDAPP-мышей с использованием Аβ1-42, которая приводила к ингибированию образования бляшек и развития ассоциированного дистрофического неврита (Schenk D. et al., Nature 400:173-77 (1999)).
Кроме того, иммунизация более старых PDAPP-мышей с уже развившейся БА-подобной нейропатологией приводила к снижению тяжести и темпов прогрессии нейропатологий. Эти исследования проводили в соответствии со следующим протоколом иммунизации: пептид растворяли в водном буфере и смешивали 1:1 с полным адъювантом Фройнда (для первой дозы) до получения концентрации пептида 100 мкг/доза. В последующих бустер-инъекциях использовали неполный адъювант Фройнда. Мышей иммунизировали 11 раз в течение 11 месяцев. При этом достигались и поддерживались титры антител более чем 1:10000. Следовательно, иммунизация может оказывать эффективное профилактическое и терапевтическое действие против болезни Альцгеймера.
В других исследованиях периферически вводимые антитела против Аβ1-42 способны проникать через гематоэнцефалический барьер, связываться с пептидом Аβ и вызывать экскрецию уже присутствующего амилоида (Bard F. et al., Nature Medicine 6:916-19 (2000)). Это исследование проводили с использованием либо поликлональных антител против Аβ1-42, либо моноклональных антител против синтетических фрагментов, происходящих от различных областей Аβ. Таким образом, индуцирование антител может рассматриваться как возможное терапевтическое лечение болезни Альцгеймера.
Было точно установлено, что введение лишь очищенных белков обычно недостаточно для индукции сильного иммунного ответа; и выделенный антиген должен, как правило, присутствовать вместе со вспомогательными веществами, называемыми адъювантами. Вводимый антиген, находящийся в этих адъювантах, защищен от быстрой деградации, и такой адъювант обеспечивает пролонгированное высвобождение малого количества антигена.
Как уже было показано, одним из ключевых событий при развитии болезни Альцгеймера (БА) является отложение амилоида в виде нерастворимых волокнистых масс (амилоидогенез), приводящее к образованию внеклеточных невритных бляшек и их отложению вокруг кровеносных сосудов головного мозга (см. обзор Selkoe, D.J. (1999) Nature 399, A23-31). Главным компонентом невритных бляшек и конгофильной ангиопатии является амилоид β (Аβ), хотя эти отложения также включают и другие белки, такие как гликозаминогликаны и аполипопротеины. Аβ протеолитически отщепляется от более крупного гликопротеина, известного как амилоидный белок-предшественник (АРР), который содержит изоформы из 695-770 аминокислот с одной гидрофобной трансмембранной областью. Аβ образует группу пептидов длиной до 43 аминокислот, обнаруживающих значительную амино- и карбоксиконцевую гетерогенность (усечение), а также их модификаций (Roher, A.E., Palmer, K.C. Chau, V. & Ball, M.J. (1988) J. Cell. Biol. 107, 2703-2716, Roher, A.E., Palmer, K.C., Yurewicz, E.C., Ball, M.J., & Greenberg, B.D. (1993) J. Neurochem. 61, 1916-1926). Характерными изоформами являются А• 1-40 и 1-42. Эти изоформы имеют в высокой степени выраженную тенденцию к образованию β-складчатых листов, которые агрегируются в фибриллы, что, в конечном счете, приводит к образованию амилоида. Недавние исследования продемонстрировали, что снижение отложений амилоида в головном мозге, индуцированное вакцинацией, приводит к улучшению познавательной способности (Schenk D., Barbour, R. Dunn, W. Gordon, G. Grajeda, H., Guido, T. Hu, K., Huang, J. Johnson-Wood, K. Khan, K. et al., (1999) Nature, 400, 173-177).
Авторами изобретения было неожиданно обнаружено, что собственные молекулы и аутоантигены, представленные в виде высокоупорядоченного и повторяющегося массива, способны эффективно индуцировать специфические аутоиммунные ответы, а в частности гуморальные ответы. Кроме того, такие ответы могут быть даже индуцированы в отсутствие адъювантов, которые, так или иначе, неспецифически активируют ангиген-презентирующие клетки или другие иммунные клетки.
Краткое описание изобретения
Настоящее изобретение относится к композициям, которые содержат высокоупорядоченные и повторяющиеся массивы антигенов или антигенных детерминант, а также к способам их получения и использования. Таким образом, композиции настоящего изобретения могут быть использованы в целях получения вакцин для профилактики инфекционных заболеваний, для лечения аллергий и злокачественных опухолей и для эффективного индуцирования специфических аутоиммунных ответов, в частности гуморальных ответов.
В своем первом аспекте настоящее изобретение относится к новой композиции, содержащей или, альтернативно, состоящей из них, (А) неприродный молекулярный каркас и (В) антиген или антигенную детерминанту. Неприродный молекулярный каркас содержит или, альтернативно, состоит из них, (i) коровую частицу, выбранную из группы, состоящей из (1) неприродной коровой частицы и (2) природной коровой частицы; и (ii) организатор, содержащий, по меньшей мере, один первый сайт связывания, где указанный организатор связан с указанной коровой частицей, по меньшей мере, одной ковалентной связью. Антиген или антигенная детерминанта представляют собой аутоантиген или его фрагмент и имеют, по меньшей мере, один второй сайт связывания, выбранный из группы, состоящей из (i) сайта связывания, который в природе не ассоциируется с указанным антигеном или указанной антигенной детерминантой; и (ii) сайта связывания, который в природе ассоциируется с указанным антигеном или указанной антигенной детерминантой. Настоящее изобретение относится к упорядоченному и повторяющемуся массиву аутоантигенов, полученному посредством присоединения второго сайта связывания к первому сайту связывания, по меньшей мере, одной непептидной связью. Таким образом, аутоантиген или аутоантигенная детерминанта и неприродный молекулярный каркас, связанные вместе посредством указанного присоединения первого сайта связывания ко второму сайту связывания, образуют упорядоченный и повторяющийся массив антигенов.
Во втором аспекте настоящее изобретение относится к новой композиции, содержащей или, альтернативно, состоящей из них, (А) неприродный молекулярный каркас и (В) антиген или антигенную детерминанту. Неприродный молекулярный каркас содержит или, альтернативно, состоит из них, (i) коровую частицу и (ii) организатор, содержащий, по меньшей мере, один первый сайт связывания, где указанной коровой частицей является вирусоподобная частица, содержащая рекомбинантные белки или их фрагменты, или бактериофаг, и где указанный организатор связан с указанной коровой частицей, по меньшей мере, одной ковалентной связью. Антиген или антигенная детерминанта имеют, по меньшей мере, один второй сайт связывания, выбранный из группы, состоящей из (i) сайта связывания, который в природе не ассоциируется с указанным антигеном или указанной антигенной детерминантой; и (ii) сайта связывания, который в природе ассоциируется с указанным антигеном или указанной антигенной детерминантой. Настоящее изобретение относится к упорядоченному и повторяющемуся массиву антигенов, полученному путем присоединения второго сайта связывания к первому сайту связывания посредством, по меньшей мере, одной непептидной связи.
В своем третьем аспекте настоящее изобретение относится к новой композиции, содержащей или, альтернативно, состоящей из них, (А) неприродный молекулярный каркас и (В) антиген или антигенную детерминанту. Неприродный молекулярный каркас содержит или, альтернативно, состоит из них, (i) коровую частицу, выбранную из группы, состоящей из (1) неприродной коровой частицы и (2) природной коровой частицы и (ii) организатор, содержащий, по меньшей мере, один первый сайт связывания, где указанный организатор связан с указанной коровой частицей, по меньшей мере, одной ковалентной связью. Антиген или антигенная детерминанта представляют собой амилоидный бета-пептид (Аβ1-42) или его фрагмент и имеют, по меньшей мере, один второй сайт связывания, выбранный из группы, состоящей из (i) сайта связывания, который в природе не ассоциируется с указанным антигеном или указанной антигенной детерминантой, и (ii) сайта связывания, который в природе ассоциируется с указанным антигеном или указанной антигенной детерминантой. Настоящее изобретение относится к упорядоченному и повторяющемуся массиву антигенов, полученному путем присоединения второго сайта связывания к первому сайту связывания посредством, по меньшей мере, одной непептидной связи.
В своем четвертом аспекте настоящее изобретение относится к новой композиции, содержащей или, альтернативно, состоящей из них, (А) неприродный молекулярный каркас и (В) антиген или антигенную детерминанту. Неприродный молекулярный каркас содержит или, альтернативно, состоит из них, (i) коровую частицу, выбранную из группы, состоящей из (1) неприродной коровой частицы и (2) природной коровой частицы; и (ii) организатор, содержащий, по меньшей мере, один первый сайт связывания, где указанный организатор связан с указанной коровой частицей, по меньшей мере, одной ковалентной связью. Антиген или антигенная детерминанта представляют собой антиидиотипическое антитело или фрагмент антиидиотипического антитела и имеют, по меньшей мере, один второй сайт связывания, выбранный из группы, состоящей из (i) сайта связывания, который в природе не ассоциируется с указанным антигеном или указанной антигенной детерминантой, и (ii) сайта связывания, который в природе ассоциируется с указанным антигеном или указанной антигенной детерминантой. Настоящее изобретение относится к упорядоченному и повторяющемуся массиву антигенов, полученному путем присоединения второго сайта связывания к первому сайту связывания посредством, по меньшей мере, одной непептидной связи.
Другие аспекты, а также предпочтительные варианты и преимущества настоящего изобретения будут очевидны из нижеследующего описания, а в частности из подробного описания, примеров и прилагаемой формулы изобретения.
В предпочтительном варианте осуществления изобретения коровой частицей является вирусоподобная частица, содержащая рекомбинантные белки РНК-фага, предпочтительно, выбранного из группы, состоящей из а) бактериофага Qβ; b) бактериофага R17; с) бактериофага fr; d) бактериофага GA; е) бактериофага SP; f) бактериофага MS2; g) бактериофага М11; h) бактериофага МХ1; i) бактериофага NL95; k) бактериофага f2 и 1) бактериофага РР7. Наиболее предпочтительными являются бактериофаг Qβ и бактериофаг fr.
В другом предпочтительном варианте осуществления изобретения рекомбинантные белки РНК-фагов включают оболочечные белки вирусов дикого типа.
В другом предпочтительном варианте осуществления изобретения рекомбинантные белки РНК-фагов включают мутантные оболочечные белки.
В другом варианте осуществления изобретения коровая частица содержит или, альтернативно, состоит из них, один или несколько различных коровых (капсидных) белков вируса гепатита (HBcAg). В родственном варианте осуществления изобретения один или несколько цистеиновых остатков указанных HBcAg либо делетированы, либо заменены другими аминокислотными остатками (например, сериновым остатком). В конкретном варианте осуществления изобретения указанные цистеиновые остатки HBcAg, используемые для получения композиций настоящего изобретения и соответствующие аминокислотным остаткам 48 и 107 в SEQ ID NO:134, либо делетированы, либо заменены другими аминокислотными остатками (например, сериновым остатком).
Кроме того, варианты HBcAg, используемые для получения композиции настоящего изобретения, обычно представляют собой варианты, которые сохраняют способность ассоциироваться с другими HBcAg и образуют димерные и мультимерные структуры, представляющие собой упорядоченные и повторяющиеся массивы антигенов или антигенных детерминант.
В другом варианте осуществления изобретения неприродный молекулярный каркас содержит или, альтернативно, состоит из них, пили или пилеподобные структуры, которые либо были продуцированы из белков пилинов, либо выделены из бактерий. Если пили или пилеподобные структуры используются для получения композиции настоящего изобретения, то они могут быть образованы из продуктов генов пилина, которые обычно присутствуют в бактериальных клетках, но которые были модифицированы методами генной инженерии (например, посредством гомологичной рекомбинации), или генов пилина, которые были введены в эти клетки.
В родственном варианте осуществления изобретения коровая частица содержит или, альтернативно, состоит из них, пили или пилеподобные структуры, которые либо были продуцированы из белков пилинов, либо выделены из бактерий. Эти коровые частицы могут быть продуцированы из продуктов генов пилина, обычно присутствующих в бактериальных клетках.
В конкретном варианте осуществления изобретения организатор может содержать, по меньшей мере, один первый сайт связывания. Первый и второй сайты связывания являются особенно важными элементами композиций настоящего изобретения. В различных вариантах осуществления изобретения первым, и/или вторым сайтом связывания может быть антиген и антитело или фрагмент антитела; биотин и авидин; стрептавидин и биотин; рецептор и его лиганд, лиганд-связывающий белок и его лиганд; полипептиды, взаимодействующие с лейциновой молнией; аминогруппа и химическая группа, способная реагировать с этой группой; карбоксильная группа и химическая группа, способная реагировать с этой группой; сульфгидрильная группа и химическая группа, способная реагировать с этой группой; или их комбинации.
В другом предпочтительном варианте осуществления изобретения указанная композиция, кроме того, содержит аминокислотный линкер. Предпочтительно, чтобы указанный аминокислотный линкер содержал второй сайт связывания или, альтернативно, состоял из этого сайта. Указанный второй сайт связывания опосредует направленную и упорядоченную ассоциацию и связывание, соответственно, указанного антигена с указанной коровой частицей. Важной функцией аминокислотного линкера, кроме того, является обеспечение соответствующего представления и доступности второго сайта связывания, а следовательно, облегчение связывания антигена с коровой частицей, в частности посредством химического сшивания. Другим важным свойством аминокислотного линкера, кроме того, является обеспечение оптимальной доступности, а в частности, реакционной способности второго сайта связывания. Эти свойства аминокислотного линкера являются даже более важными для белковых антигенов.
В другом предпочтительном варианте осуществления изобретения указанный аминокислотный линкер выбран из группы, состоящей из (а) CGG; (b) N-концевого 1-линкера гамма; (с) N-концевого 3-линкера гамма; (d) шарнирных областей Ig; (е) N-концевых глициновых линкеров; (f) (G)kC(G)n, где n=0-12, а k=0-5; (g) N-концевых глицин-сериновых линкеров; (h) (G)kC(G)m (S)l(GGGGS)n, где n=0-3, k=0-5, m=0-10, l=0-2; (i) GGC; (k) GGC-NH2; (l) C-концевого 1-линкера гамма; (m) С-концевого 3-линкера гамма; (n) С-концевых глициновых линкеров; (o) (G)nC(G)k, где n=0-12, а k=0-5; (p) C-концевых глицин-сериновых линкеров; (q) (G)m(S)l(GGGGS)n(G)oC(G)k, где n=0-3, k=0-5, m=0-10, l=0-2 и о=0-8.
Важным свойством глициновых и глицин-сериновых линкеров является их гибкость, а в частности их структурная гибкость, обеспечивающая образование широкого ряда конформаций и укладку в затрудненные структуры, которые делают недоступным второй сайт связывания. Поскольку глициновые и глицин-сериновые линкеры содержат ограниченное количество остатков боковой цепи или вовсе не содержат их, то они имеют ограниченную способность вступать в экстенсивные взаимодействия с антигеном, а поэтому дополнительно гарантируют доступность указанного второго сайта связывания. Сериновые остатки в глицин-сериновых линкерах сообщают этим линкерам повышенную растворимость. В соответствии с этим объем настоящего изобретения включает инсерцию одной или нескольких аминокислот, расположенных тандемно или отдельно, а в частности полярных или заряженных аминокислотных остатков, в глициновый или глицин-сериновый аминокислотный линкер.
В другом предпочтительном варианте осуществления изобретения аминокислотным линкером является GGC-NH2, GGC-NMe, GGC-N(Me)2, GGC-NHET или GGC-N(Et)2, где С-конец цистеинового остатка GGC является амидированным. Эти аминокислотные линкеры являются особенно предпочтительными для пептидных антигенов, а в частности для тех вариантов осуществления изобретения, в которых антиген или антигенная детерминанта с указанным вторым сайтом связывания содержит пептиды Аβ или их фрагменты. Особенно предпочтительным является GGC-NH2. В другом варианте осуществления настоящего изобретения указанным аминокислотным линкером является шарнирная область человеческого иммуноглобулина (Ig). В объем настоящего изобретения входят также фрагменты шарнирных областей Ig, а также шарнирные области Ig, модифицированные глициновыми остатками. Предпочтительно, чтобы указанные шарнирные области Ig содержали только один цистеиновый остаток. Следует отметить, что один цистеиновый остаток указанного аминокислотного линкера шарнирной области Ig может быть локализован в нескольких положениях внутри линкерной последовательности, и каждый специалист может самостоятельно выбрать это положения исходя из описания настоящего изобретения.
В одном из своих вариантов настоящее изобретение относится к связыванию почти любого выбранного антигена с поверхностью вируса, бактериальных пилей, структуры, образованной бактериальными пилином, бактериофага, вирусоподобной частицы или вирусной капсидной частицы. В соответствии с настоящим изобретением путем введения антигена в квазикристаллическую "вирусоподобную" структуру продуцируют сильную антивирусную иммунологическую реакцию у хозяина в целях вырабатывания у него высокоэффективного иммунного ответа, то есть осуществляют вакцинацию против представленного антигена.
В еще одном варианте осуществления изобретения антиген может быть выбран из группы, состоящей из (1) белка, подходящего для индуцирования иммунного ответа против злокачественных клеток; (2) белка, подходящего для индуцирования иммунного ответа против инфекционных заболеваний; (3) белка, подходящего для индуцирования иммунного ответа против аллергенов; (4) белка, подходящего для индуцирования повышенного ответа против аутоантигенов; и (5) белка, подходящего для индуцирования иммунного ответа у сельскохозяйственных животных или домашних питомцев. В другом варианте осуществления изобретения первый сайт связывания и/или второй сайт связывания выбраны из группы, включающей (1) генетически сконструированный лизиновый остаток и (2) генетически сконструированный цистеиновый остаток, где эти два остатка могут быть химически связаны друг с другом.
В еще одном предпочтительном варианте осуществления изобретения первый сайт связывания содержит или представляет собой аминогруппу, а указанный второй сайт связывания содержит или представляет собой сульфгидрильную группу. Указанный первый сайт связывания, предпочтительно, содержит или представляет собой лизиновый остаток, а указанный второй сайт связывания, предпочтительно, содержит или представляет собой цистеиновый остаток.
Настоящее изобретение также включает варианты, в которых указанная частица-организатор имеет только один первый сайт связывания, а антиген или антигенная детерминанта имеет только один второй сайт связывания. Таким образом, при получении упорядоченного и повторяющегося массива антигенов или антигенных детерминант с использованием указанных вариантов каждый организатор может быть связан с одним антигеном или с одной антигенной детерминантой. В другом своем аспекте настоящее изобретение относится к композициям, содержащим или, альтернативно, состоящим из них, (а) неприродный молекулярный каркас, содержащий (i) коровую частицу, выбранную из группы, состоящей из неприродной коровой частицы и природной коровой частицы; и (ii) организатор, содержащий, по меньшей мере, один первый сайт связывания, где указанная коровая частица содержит или, альтернативно, состоит из них, вирусоподобную частицу, бактериальные пили, пилеподобную структуру, модифицированный HBcAg или его фрагмент; и где указанный организатор связан с указанной коровой частицей, по меньшей мере, одной ковалентной связью, и (В) антиген или антигенную детерминанту, имеющую, по меньшей мере, один второй сайт связывания, где указанный второй сайт связывания выбран из группы, состоящей из (i) сайта связывания, который в природе не ассоциируется с указанным антигеном или указанной антигенной детерминантой; и (ii) сайта связывания, который в природе ассоциируется с указанным антигеном или указанной антигенной детерминантой, и где указанный второй сайт связывания способен связываться с первым сайтом связывания посредством, по меньшей мере, одной непептидной связи, и где указанный антиген или антигенная детерминанта и указанный каркас взаимодействуют посредством связывания друг с другом, образуя упорядоченный и повторяющийся массив антигенов.
Другие варианты настоящего изобретения включают способы получения композиций настоящего изобретения и способы терапевтического лечения с использованием описанных здесь вакцинных композиций. Следует отметить, что вышеуказанное общее описание и нижеследующее подробное описание приводится лишь для иллюстрации и лучшего понимания настоящего изобретения и имеет своей целью более подробное объяснение заявленного изобретения.
В еще одном своем аспекте настоящее изобретение относится к композиции, содержащей оболочечный белок бактериофага Qβ, присоединенный ковалентной связью к белку фосфолипазы A2 или к его фрагменту. В предпочтительном варианте осуществления изобретения указанный белок фосфолипазы A2 или его фрагмент и оболочечный белок бактериофага Qβ взаимодействуют посредством ковалентной связи и образуют упорядоченный и повторяющийся массив антигенов. В другом предпочтительном варианте осуществления изобретения указанной ковалентной связью является непептидная связь. В другом предпочтительном варианте осуществления изобретения указанный белок фосфолипазы A2 включает аминокислотную последовательность, выбранную из группы, состоящей из аминокислотной последовательности SEQ ID NO:168, аминокислотной последовательности SEQ ID NO:169, аминокислотной последовательности SEQ ID NO:170, аминокислотной последовательности SEQ ID NO:171, аминокислотной последовательности SEQ ID NO:172, аминокислотной последовательности SEQ ID NO:173, аминокислотной последовательности SEQ ID NO:174 и аминокислотной последовательности SEQ ID NO:175.
Настоящее изобретение также относится к способу получения композиции, предусматривающему объединение оболочечного белка бактериофага Qβ с белком фосфолипазы A2, где указанный оболочечный белок бактериофага Qβ и указанный белок фосфолипазы A2 взаимодействуют с образованием массива антигенов.
В другом своем аспекте настоящее изобретение также относится к композиции, содержащей неприродный молекулярный каркас, включающий оболочечный белок бактериофага Qβ, и организатор, содержащий, по меньшей мере, один первый сайт связывания, где указанный организатор связан с оболочечным белком бактерифага Qβ , по меньшей мере, одной ковалентной связью; и белок фосфолипазы A2, или его фрагмент, или его вариант, имеющий, по меньшей мере, один второй сайт связывания, где указанный второй сайт связывания выбран из группы, состоящей из сайта связывания, который в природе не ассоциируется с указанным белком фосфолипазы A2 или с его фрагментом, и сайта связывания, который в природе ассоциируется с указанным белком фосфолипазы A2 или с его фрагментом, где указанный второй сайт связывания присоединен к первому сайту связывания, по меньшей мере, одной непептидной связью, и где указанный антиген или антигенная детерминанта и указанный каркас взаимодействуют посредством связывания друг с другом и образуют упорядоченный и повторяющийся массив антигенов. В предпочтительном варианте осуществления изобретения указанный белок фосфолипазы A2 включает аминокислотную последовательность, выбранную из группы, состоящей из аминокислотной последовательности SEQ ID NO:168, аминокислотной последовательности SEQ ID NO:169, аминокислотной последовательности SEQ ID NO:170, аминокислотной последовательности SEQ ID NO:171, аминокислотной последовательности SEQ ID NO:172, аминокислотной последовательности SEQ ID NO:173, аминокислотной последовательности SEQ ID NO:174 и аминокислотной последовательности SEQ ID NO:175.
Настоящее изобретение также относится к способу получения композиции, предусматривающему объединение оболочечного белка бактериофага Qβ с белком фосфолипазы A2, где указанный оболочечный белок бактериофага Qβ и указанный белок фосфолипазы A2 взаимодействуют друг с другом с образованием массива антигенов. Указанный массив антигенов является, предпочтительно, упорядоченным и/или повторяющимся.
Настоящее изобретение также относится к фармацевтической композиции, содержащей белок фосфолипаы A2 и фармацевтически приемлемый носитель. Настоящее изобретение также относится к вакцинной композиции, содержащей белок фосфолипазы A2. В предпочтительном варианте осуществления изобретения указанная вакцинная композиция по п.31, кроме того, содержит, по меньшей мере, один адъювант.
Настоящее изобретение также относится к способу лечения аллергии на пчелиный яд, предусматривающему введение фармацевтической композиции или вакцинной композиции индивидууму. В результате такого введения у индивидуума ослабляется иммунный ответ на указанный яд.
Настоящее изобретение также относится к вакцине для профилактики опосредованных прионами заболеваний путем индуцирования антител против лимфотоксина-β, лимфотоксина-α или против рецептора лимфотоксина-β. Эта вакцина содержит белок-носитель, который является чужеродным для иммунизированного человека или животного и который связывается с лимфотоксином-β или его фрагментами, с лимфотоксином-α или его фрагментами или с рецептором лимфотоксина-β или его фрагментами. Указанную вакцину инъецируют человеку или животным для вырабатывания у них антител против эндогенного лимфотоксина-β, лимфотоксина-α или рецептора лимфотоксина-β. Индуцированные таким образом антитела против эндогенного лимфотоксина-β, лимфотоксина-α или рецептора лимфотоксина-β способствуют сокращению или элиминации пула фолликулярных дендритных клеток, присутствующих в лимфоидных органах. Поскольку репликация приона в лимфоидных органах и его транспорт в центральную нервную систему нарушаются в отсутствие фолликулярных дендритных клеток, то такое лечение приводит к ингибированию прогрессирования опосредованных прионами заболеваний. Кроме того, блокирование лимфотоксина-β оказывает благоприятное действие на пациентов с аутоиммунными заболеваниями, такими как диабет типа I.
Краткое описание графического материала
Фиг.1А-1С: Модулярные эукариотические экспрессирующие векторы для экспрессии антигенов настоящего изобретения.
Фиг.2А-2С: Клонирование, экспрессия и связывание резистина с капсидным белком Qβ.
Фиг.3А-3В: Клонирование и экспрессия конструкций лимфотоксина-β для связывания с вирусоподобными частицами и пилями.
Фиг.4А-4В: Клонирование, экспрессия и связывание конструкций MIF с капсидным белком Qβ.
Фиг.4С: ELISA-анализ антител IgG, специфичных к MIF в сыворотке мышей, иммунизированных против белков MIF, связанных с капсидным белком Qβ.
Фиг.5: Связывание MIF-конструкций с капсидным белком fr и с капсидным белком HBcAg-lys-2cys-Mut, проанализированное с помощью электрофореза в ДСН-ПААГ.
Фиг.6: Клонирование и экспрессия человеческого С-RANKL.
Фиг.7: Клонирование и экспрессия белка приона.
Фиг.8А: ELISA-анализ антител IgG, специфичных к "ангио I" в сыворотке мышей, иммунизированных против пептидов ангиотензина, связанных с капсидным белком Qβ.
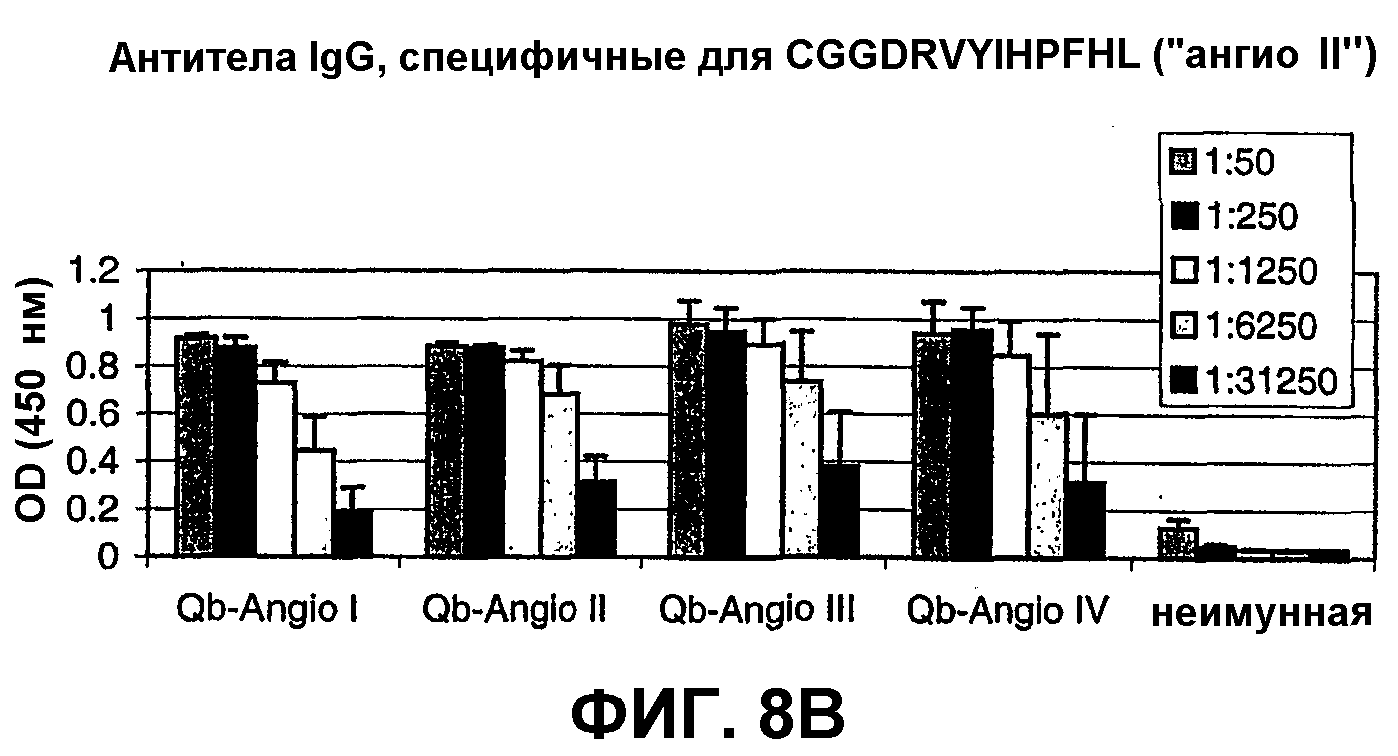
Фиг.8В: ELISA-анализ антител IgG, специфичных к "ангио II" в сыворотке мышей, иммунизированных против пептидов ангиотензина, связанных с капсидным белком Qβ.
Фиг.8С: ELISA-анализ антител IgG, специфичных к "ангио III" в сыворотке мышей, иммунизированных против пептидов ангиотензина, связанных с капсидным белком Qβ.
Фиг.8D: ELISA-анализ антител IgG, специфичных к "ангио IV" в сыворотке мышей, иммунизированных против пептидов ангиотензина, связанных с капсидным белком Qβ.
Фиг.9A: ELISA-анализ антител IgG, специфичных к "Der p I p52" в сыворотке мышей, иммунизированных против пептидов Der p I, связанных с капсидным белком Qβ.
Фиг.9D: ELISA-анализ антител IgG, специфичных к "Der p I p117" в сыворотке мышей, иммунизированных против пептидов Der p I, связанных с капсидным белком Qβ.
Фиг.10A: ELISA-анализ антител IgG, специфичных к человеческому пептиду VEGFR II в сыворотке мышей, иммунизированных против человеческого пептида VEGFR II и внеклеточного домена человеческого VEGFR II, которые оба связаны с белком пилей типа I.
Фиг.10В: ELISA-анализ антител IgG, специфичных к внеклеточному домену человеческого VEGFR II в сыворотке мышей, иммунизированных против человеческого пептида VEGFR II и внеклеточного домена человеческого VEGFR II, которые оба связаны с белком пилей типа I.
Фиг.11: ELISA-анализ антител IgG, специфичных к анти-TNFα белку в сыворотке мышей, иммунизированных против пептидов полноразмерного НВс-TNF.
Фиг.12: ELISA-анализ антител IgG, специфичных к анти-TNFα белку в сыворотке мышей, иммунизированных против 2cysLys-mut HBcAg1-149, связанного с пептидом 3'TNF II.
Фиг.13А: ДСН-ПААГ-электрофоретический анализ связывания "Аβ1-15" с капсидным белком Qβ с использованием сшивающего линкера SMPH.
Фиг.13В: ДСН-ПААГ-электрофоретический анализ связывания "Аβ33-42" с капсидным белком Qβ с использованием сшивающего линкера SMPH.
Фиг.13С: ДСН-ПААГ-электрофоретический анализ связывания "Аβ1-27" с капсидным белком Qβ с использованием сшивающего линкера SMPH.
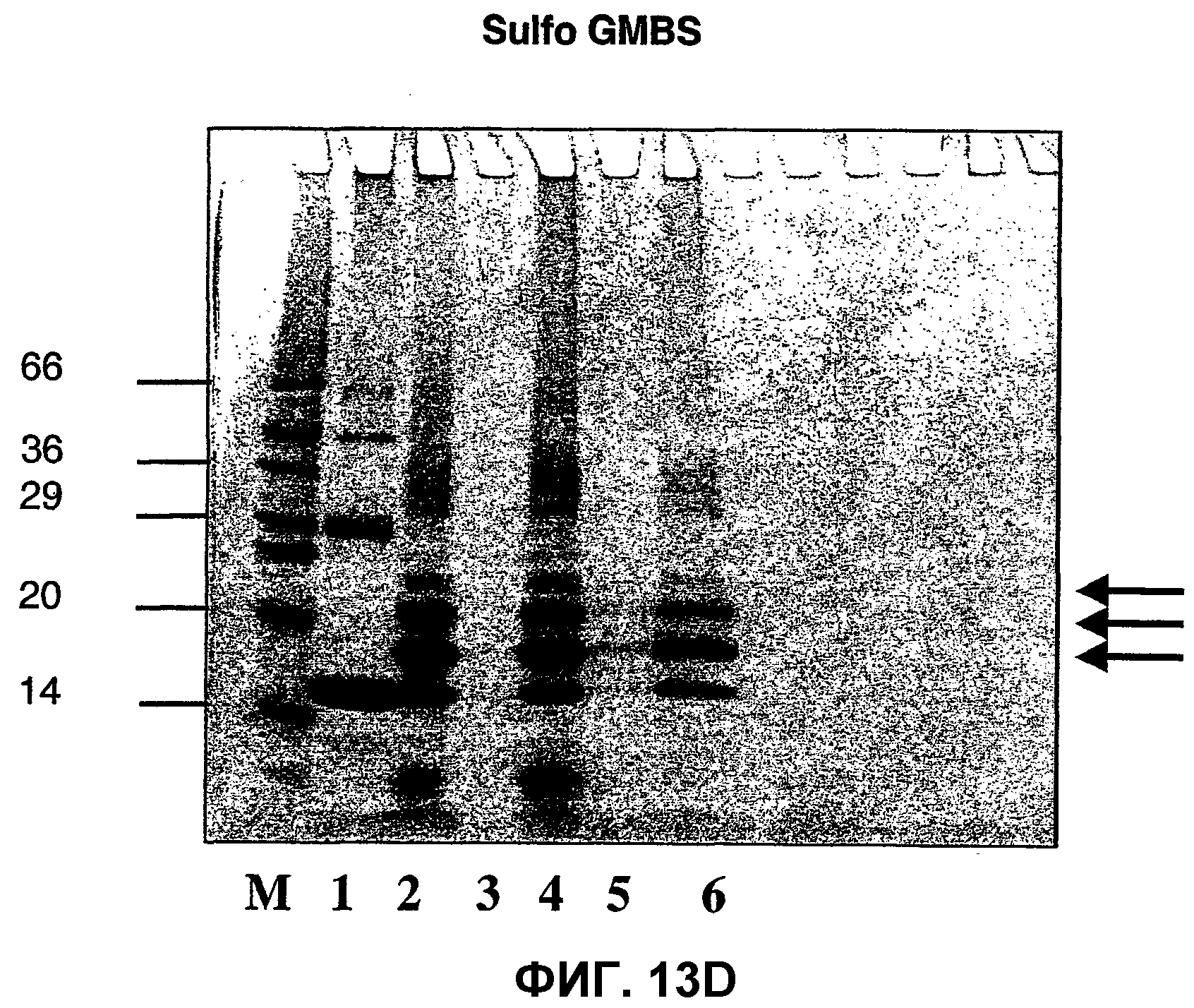
Фиг.13D: ДСН-ПААГ-электрофоретический анализ связывания "Аβ1-15" с капсидным белком Qβ с использованием сшивающего линкера сульфо-GMBS.
Фиг.13Е: ДСН-ПААГ-электрофоретический анализ связывания "Аβ1-15" с капсидным белком Qβ с использованием сшивающего линкера сульфо-MBS.
Фиг.14А: ELISA-анализ антител IgG, специфичных к "Аβ1-15" в сыворотке мышей, иммунизированных против "Аβ1-15", связанного с капсидным белком Qβ.
Фиг.14В: ELISA-анализ антител IgG, специфичных к "Аβ1-27" в сыворотке мышей, иммунизированных против "Аβ1-27", связанного с капсидным белком Qβ.
Фиг.14С: ELISA-анализ антител IgG, специфичных к "Аβ33-42" в сыворотке мышей, иммунизированных против "Аβ33-42", связанного с капсидным белком Qβ.
Фиг.15А: ДСН-ПААГ-электрофоретический анализ связывания рСС2 с капсидным белком Qβ.
Фиг.15В: ДСН-ПААГ-электрофоретический анализ связывания рСА2 с капсидным белком Qβ.
Фиг.15С: ДСН-ПААГ-электрофоретический анализ связывания рСВ2 с капсидным белком Qβ.
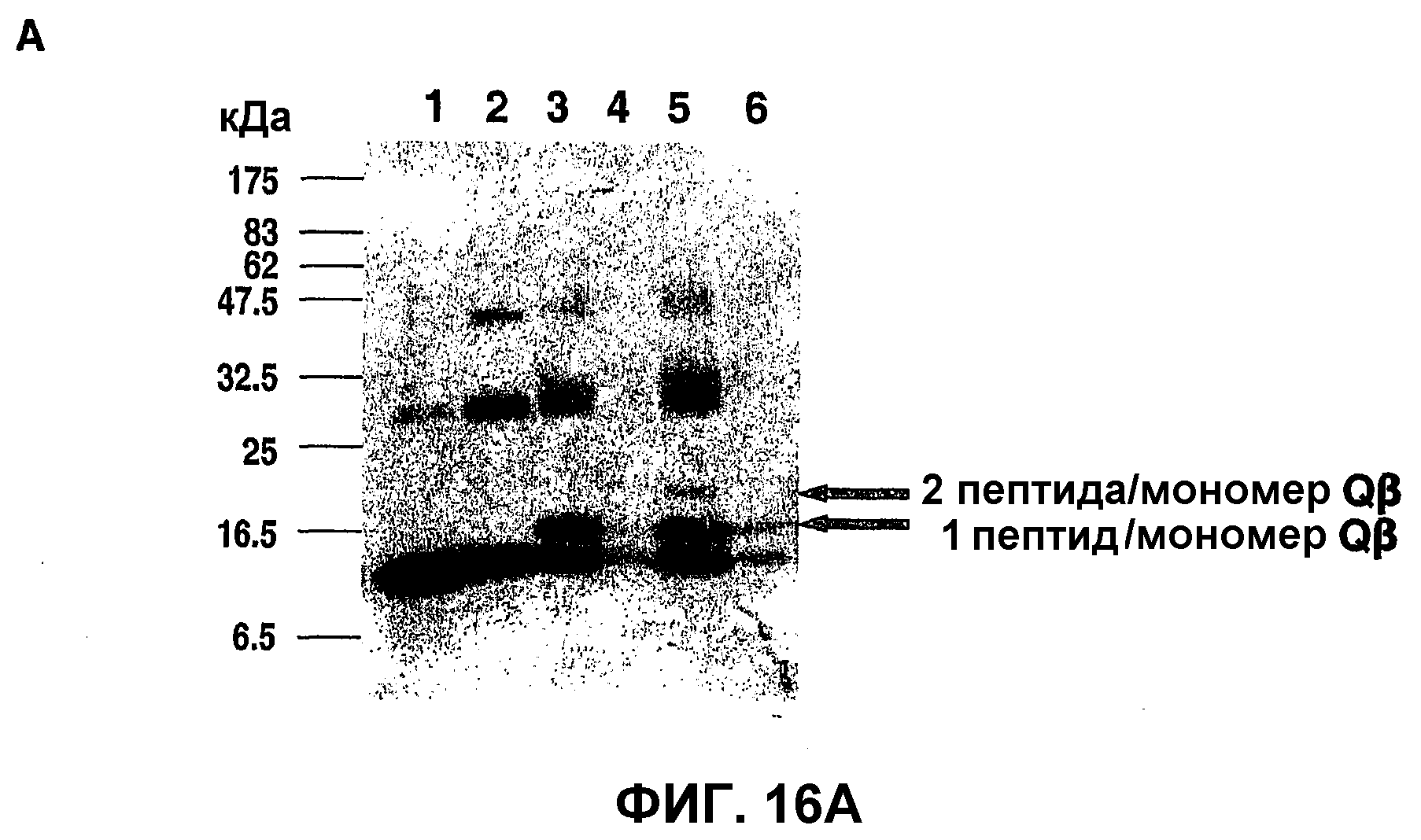
Фиг.16: Связывание пептидов приона с капсидным белком Qβ; ДСН-ПААГ-электрофоретический анализ.
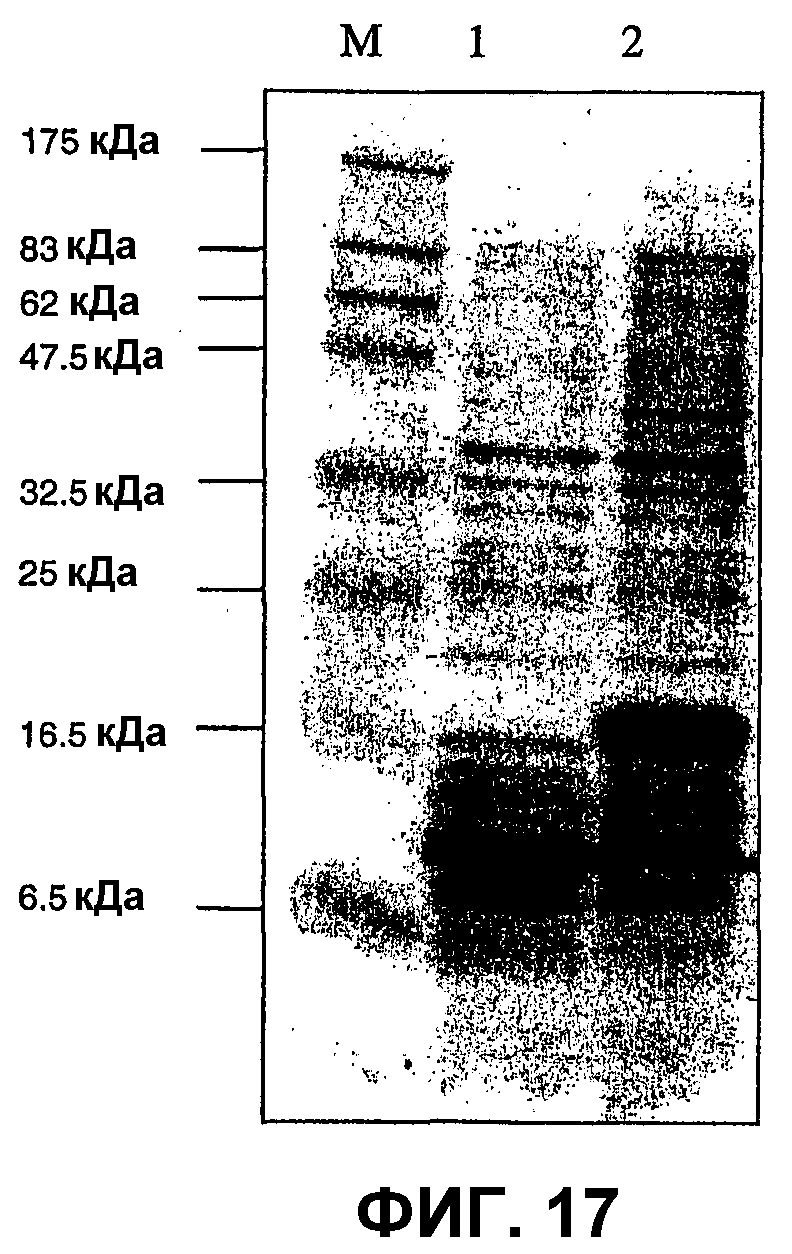
Фиг.17А: ДСН-ПААГ-электрофоретический анализ экспрессии IL-5 в бактериях.
Фиг.17В: Вестерн-блот-анализ экспрессии IL-5 и IL-13 в эукариотических клетках.
Фиг.18А: ДСН-ПААГ-электрофоретический анализ связывания мышиного пептида VEGFR-2 с пилями.
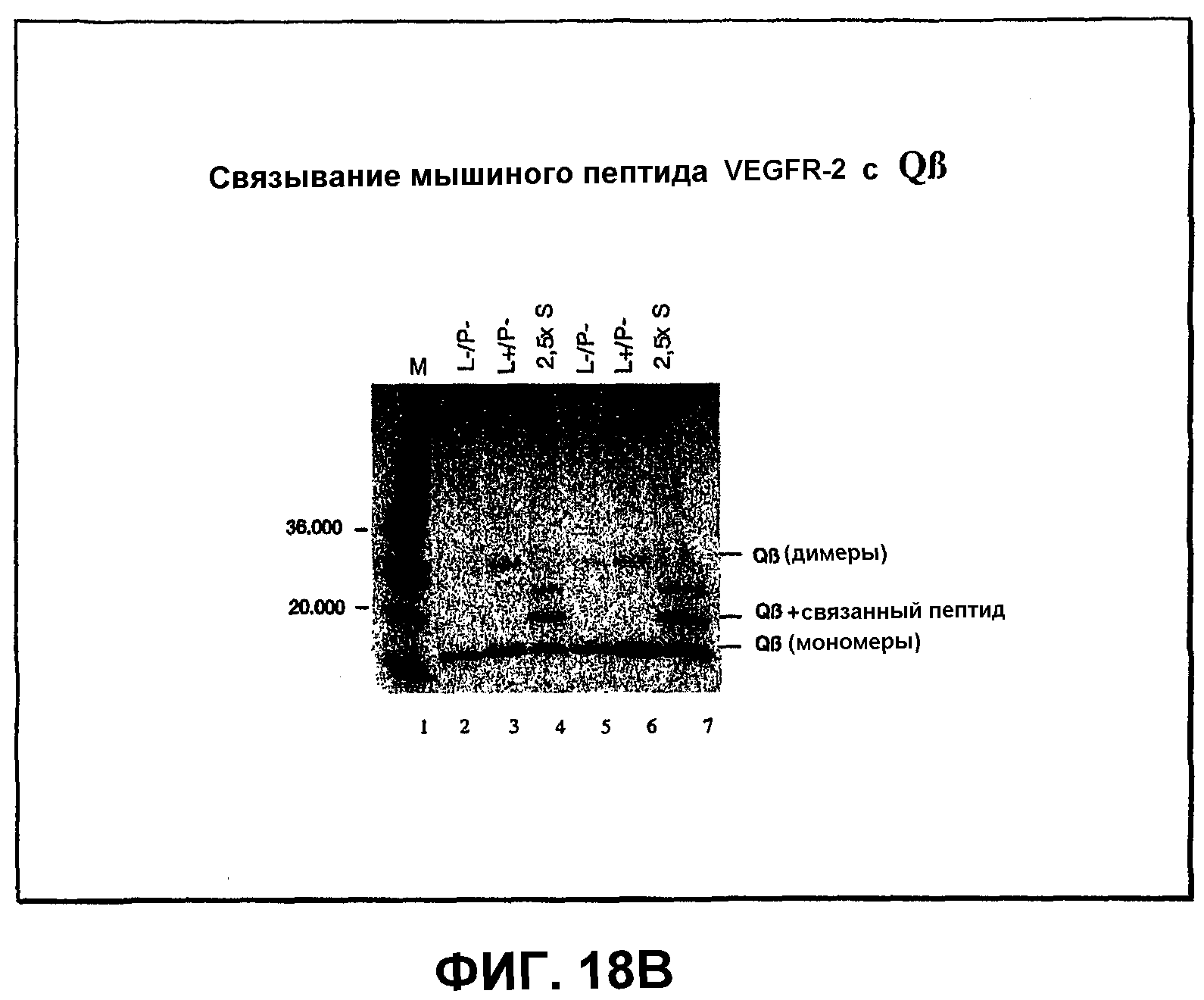
Фиг.18В: ДСН-ПААГ-электрофоретический анализ связывания мышиного пептида VEGFR-2 с капсидным белком Qβ.
Фиг.18С: ДСН-ПААГ-электрофоретический анализ связывания мышиного пептида VEGFR-2 с HBcAg-lys-2cys-Mut.
Фиг.18D: ELISA-анализ антител IgG, специфичных к мышиному пептиду VEGFR-2 в сыворотке мышей, иммунизированных против пептидов мышиного пептида VEGFR-2, связанного с пилями.
Фиг.18Е: ELISA-анализ антител IgG, специфичных к мышиному пептиду VEGFR-2 в сыворотке мышей, иммунизированных против пептидов мышиного пептида VEGFR-2, связанного с капсидным белком Qβ.
Фиг.18F: ELISA-анализ антител IgG, специфичных к мышиному пептиду VEGFR-2 в сыворотке мышей, иммунизированных против пептидов мышиного пептида VEGFR-2, связанного с HBcAg-lys-2cys-Mut.
Фиг.19А: ДСН-ПААГ-электрофоретический анализ связывания пептида Аβ1-15 с HBcAg-lys-2cys-Mut и капсидным белком fr.
Фиг.19В: ELISA-анализ антител IgG, специфичных к пептиду Аβ1-15 в сыворотке мышей, иммунизированных против пептида Аβ1-15, связанного с HBcAg-lys-2cys-Mut или с капсидным белком fr.
Фиг.20: ELISA-анализ антител IgG, специфичных к человеческому Аβ в сыворотке трансгенных мышей АРР23, иммунизированных человеческими пептидами Аβ, связанными с капсидным белком Qβ.
Фиг.21: ДСН-ПААГ-электрофоретический анализ связывания фрагмента антитела Fab с капсидным белком Qβ.
Фиг.22А: ДСН-ПААГ-электрофоретический анализ связывания пептида flag, связанного с мутантным капсидным белком Qβ с использованием сшивающего линкера сульфо-GMBS.
Фиг.22В: ДСН-ПААГ-электрофоретический анализ связывания пептида flag, связанного с мутантным капсидным белком Qβ с использованием сшивающего линкера сульфо-MBS.
Фиг.22С: ДСН-ПААГ-электрофоретический анализ связывания пептида flag, связанного с мутантным капсидным белком Qβ с использованием сшивающего линкера SMPH.
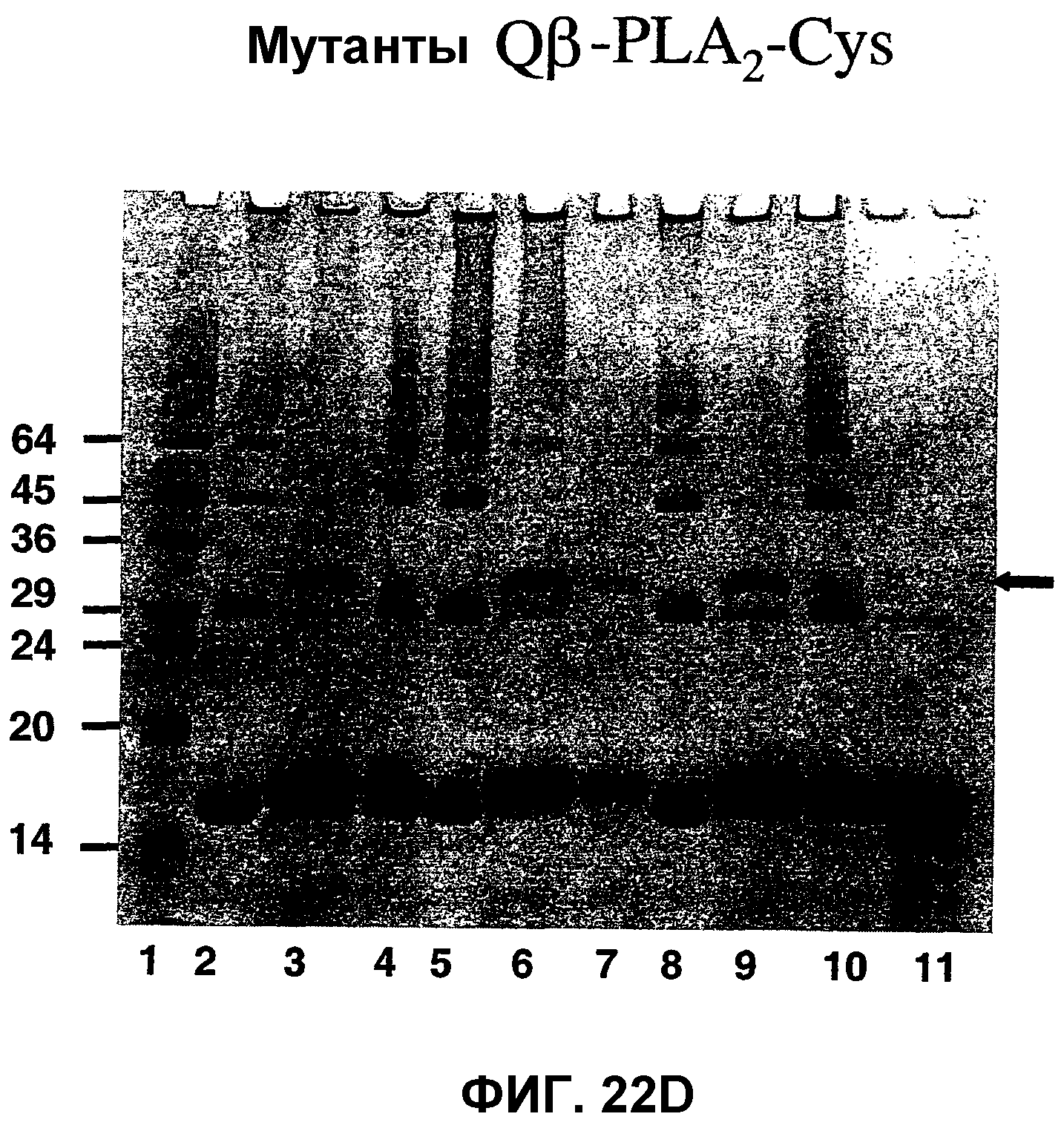
Фиг.22D: ДСН-ПААГ-электрофоретический анализ связывания белка PLA2-cys, связанного с мутантным капсидным белком Qβ с использованием сшивающего линкера SMPH.
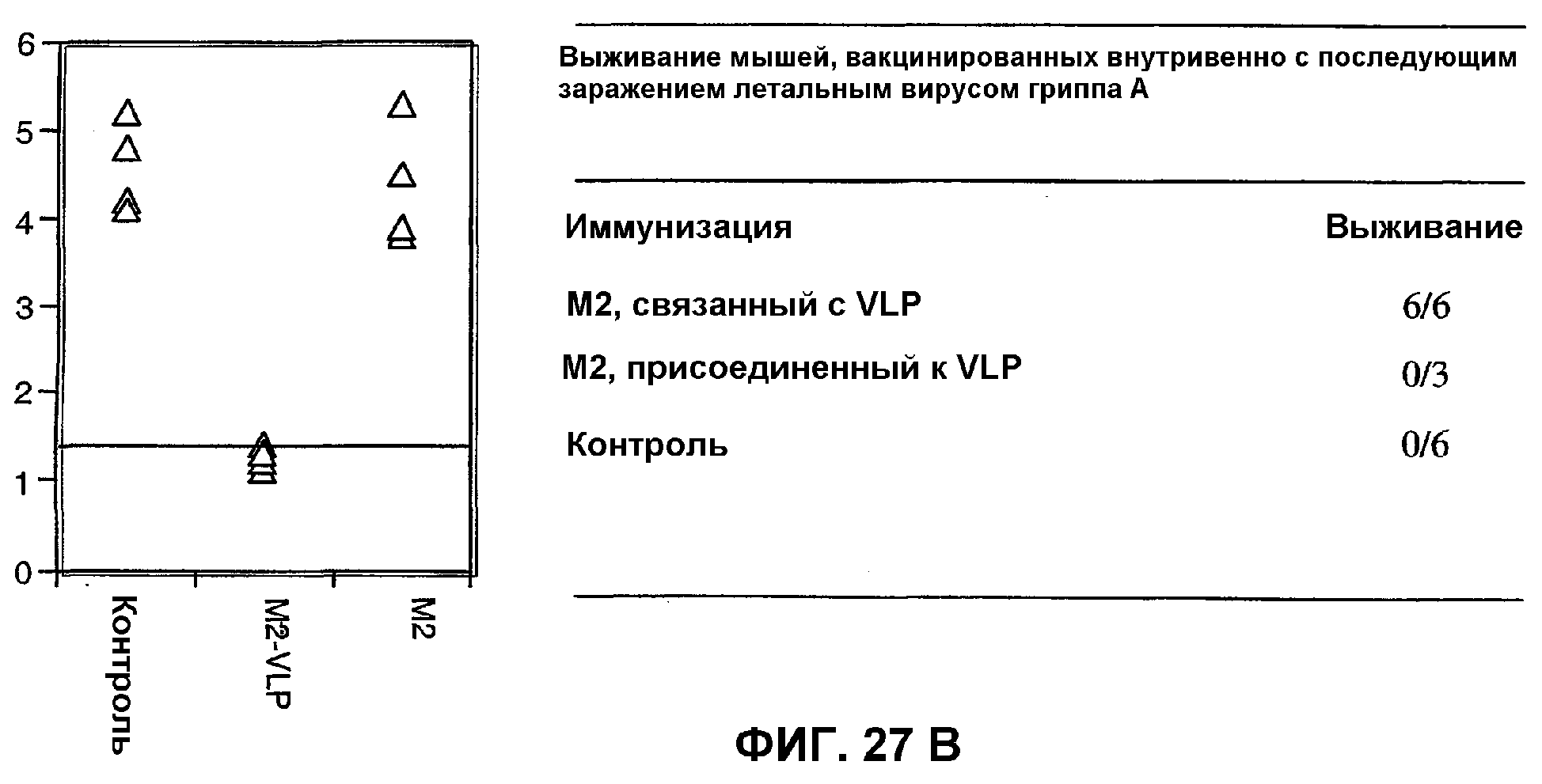
Фиг.23: ELISA-анализ иммунизации пептидом М2, связанным с мутантным капсидным белком Qβ и капсидом fr.
Фиг.24: ДСН-ПААГ-электрофоретический анализ связывания пептида DER p1,2, связанного с мутантным капсидным белком Qβ.
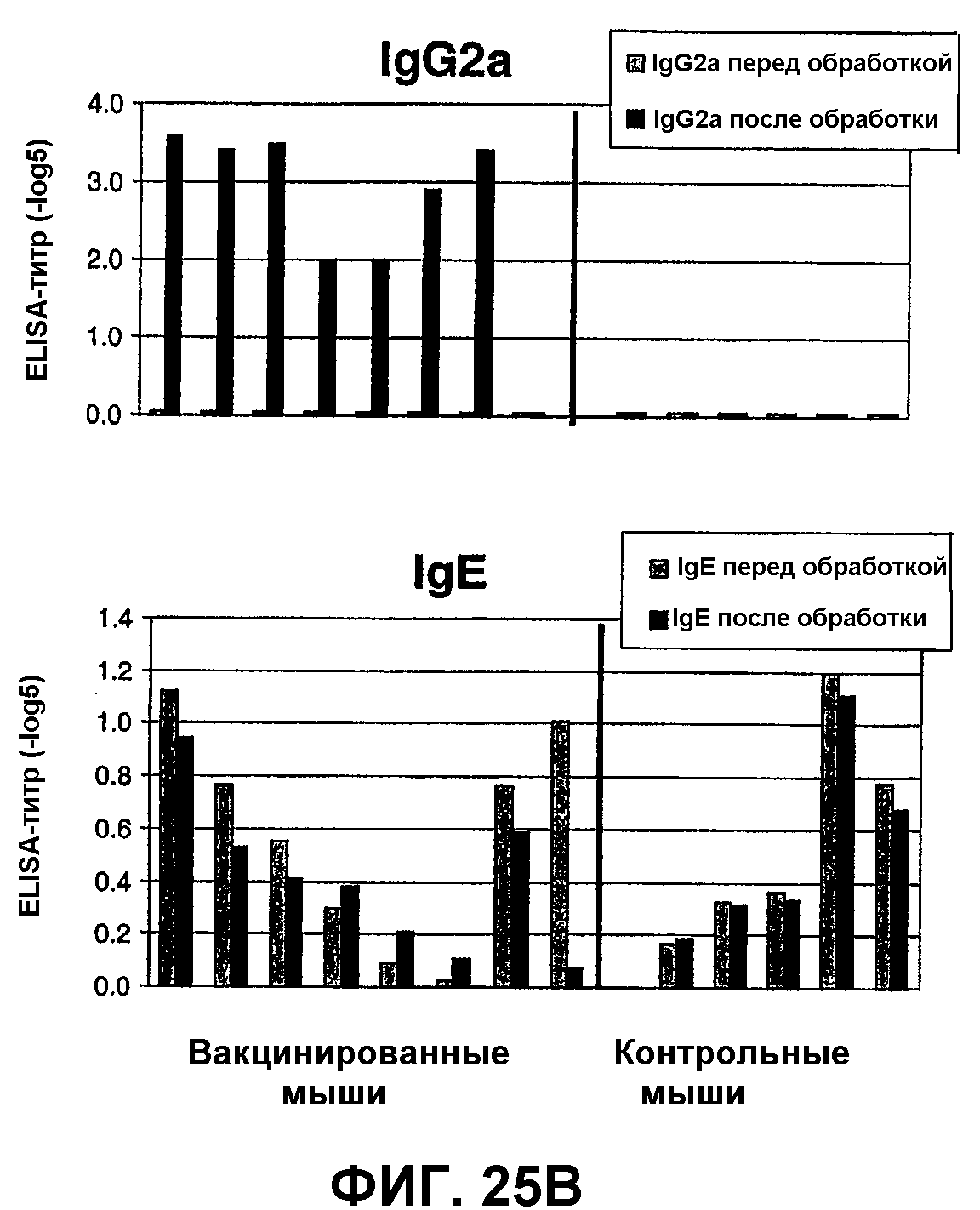
Фиг.25А: Десенсибилизация мышей, сенсибилизированных аллергеном с использованием PLA2, связанного с капсидным белком Qβ: измерения температуры.
Фиг.25В: Десенсибилизация мышей, сенсибилизированных аллергеном с использованием PLA2-cys, связанного с капсидным белком Qβ: титры IgG 2А и IgE.
Фиг.26: ДСН-ПААГ-электрофоретический анализ и Вестерн-блот-анализ связывания PLA2-cys с капсидным белком Qβ.
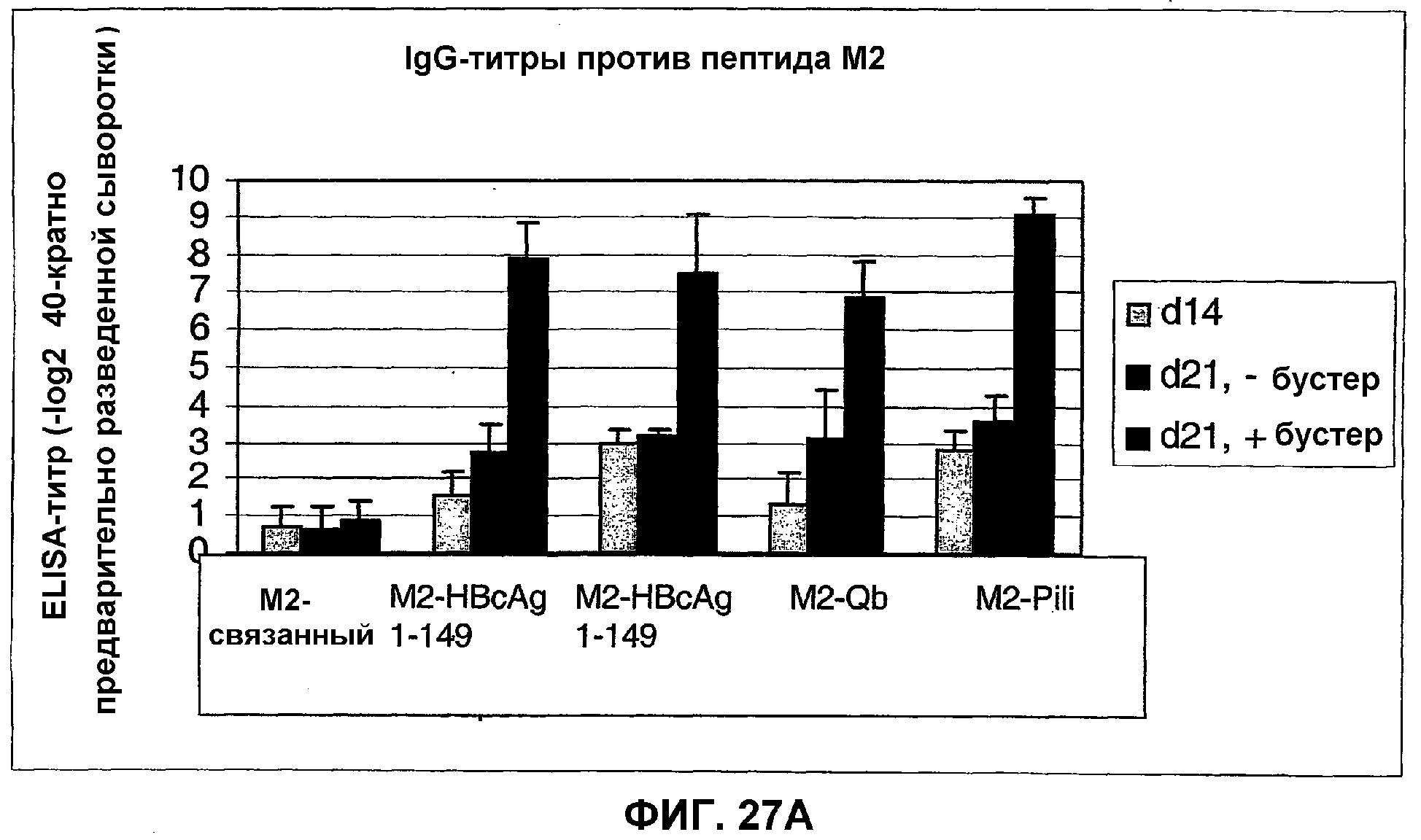
Фиг.27А: ELISA-анализ антител IgG, специфичных к пептиду М2 в сыворотке мышей, иммунизированных против пептида М2, связанного с HBcAg-lys-2cys-Mut, капсидным белком Qβ, капсидным белком fr, HBcAg-lys-1-183, и против пептида М2, связанного с HBcAg-1-183.
Фиг.28А: ДСН-ПААГ-электрофоретический анализ связывания антиидиотипического мимиоантитела IgE VAE051 с капсидным белком Qβ.
Фиг.28В: ELISA-анализ антител IgG, специфичных к антиидиотипическому антителу VAE051, и человеческого IgE в сыворотке мышей, иммунизированных против VAE051, связанного с капсидным белком Qβ.
Подробное описание изобретения
1. Определения
Альфавирус: используемый здесь термин "альфавирус" означает любые РНК-вирусы, принадлежащие к роду Alphavirus. Описание членов этого рода можно найти у Strauss & Strauss, Microbiol. Rev., 58:491-562 (1994). Примерами альфавирусов являются вирус Аура, вирус Бебару, вирус Кабассу, вирус Чикунгунья, вирус восточного энцефаломиелита лошадей, вирус Форт-Морган, вирус Гета, вирус Кизильгаха, вирус Майяро, вирус Миддельбург, вирус Мукамбо, вирус Ндуму, вирус Пиксуна, вирус Тонате, вирус Тринити, вирус Уна, вирус западного энцефаломиелита лошадей, вирус Ватароа, вирус Синдбис (SIN), вирус лесов Семлики (SFV), вирус венесуэльского энцефаломиелита лошадей (VEE) и вирус реки Росс.
Антиген: используемый здесь термин "антиген" представляет собой молекулу, способную связываться с антителом. Кроме того, антиген способен индуцировать гуморальный иммунный ответ и/или клеточный иммунный ответ, приводящий к продуцированию В- и/или Т-лимфоцитов. Антиген может иметь один или несколько эпитопов (В- и Т-эпитопы). Упомянутая выше специфическая реакция означает, что указанный антиген может взаимодействовать с высокой селективностью с его соответствующим антителом, но не с множеством других антител, которые могут вырабатываться другими антигенами.
Антигенная детерминанта: используемый здесь термин "антигенная детерминанта" означает часть антигена, которая специфически распознается В- или Т-лимфоцитами. В-лимфоциты отвечают на чужеродные антигенные детерминанты продуцированием антитела, тогда как Т-лимфоциты являются медиаторами клеточного иммунитета. Таким образом, антигенные детерминанты или эпитопы являются участками антигена, распознаваемого антителами, или, в случае МНС, Т-клеточными рецепторами.
Ассоциация: используемый здесь термин "ассоциация", если он применяется к первому и второму сайтам связывания, означает, по меньшей мере, одну непептидную связь. По своей природе такая ассоциации может быть ковалентной, ионной, гидрофобной, полярной или представлять собой любую их комбинацию.
Первый сайт связывания: используемый здесь термин "первый сайт связывания" означает элемент "организатора", который сам связан с коровой частицей нерандомизированным образом, и с которым может связываться второй сайт связывания, локализованный на антигене или антигенной детерминанте. Первым сайтом связывания может быть белок, полипептид, аминокислота, пептид, сахар, полинуклеотид природный или синтетический полимер, вторичный метаболит, или соединение (биотин, флуоресцеин, ретинол, дигоксигенин, ионы металла, фенилметилсульфонилфторид), или их комбинация, или их химически реактивная группа. На поверхности неприродного молекулярного каркаса может присутствовать множество первых сайтов связывания в повторяющейся конфигурации.
Второй сайт связывания: используемый здесь термин "второй сайт связывания" означает элемент, ассоциированный с антигеном или с антигенной детерминантой, с которой может связываться первый сайт связывания "организатора", и локализованный на поверхности неприродного молекулярного каркаса. Вторым сайтом связывания антигена или антигенной детерминанты может быть белок, полипептид, пептид, сахар, полинуклеотид, природный или синтетический полимер, вторичный метаболит или соединение (биотин, флуоресцеин, ретинол, дигоксигенин, ионы металла, фенилметилсульфонилфторид), или их комбинация, или их химически реактивная группа. На антигене или антигенной детерминанте присутствует, по меньшей мере, один второй сайт связывания. Поэтому термин "антиген или антигенная детерминанта, имеющие, по меньшей мере, один второй сайт связывания" означает антиген или антигенную конструкцию, содержащую, по меньшей мере, антиген или антигенную детерминанту и второй сайт связывания. Однако в случае второго сайта связывания, который в природе, обычно, не ассоциируется с данным антигеном или антигенной детерминантой, то этот антиген или антигенная конструкция содержит "аминокислотный линкер". Такой аминокислотный линкер или также называемый просто "линкером" в настоящем описании либо связывает антиген или антигенную детерминанту со вторым сайтом связывания, либо, что более предпочтительно, уже составляет или содержит второй сайт связывания, а обычно, но необязательно, он представляет собой аминокислотный остаток, предпочтительно, цистеиновый остаток. Однако используемый здесь термин "аминокислотный линкер" не означает, что этот аминокислотный линкер состоит исключительно из аминокислотных остатков, даже если аминокислотный линкер, состоящий из аминокислотных остатков, является предпочтительным вариантом настоящего изобретения. Аминокислотными остатками указанного аминокислотного линкера, предпочтительно, являются природные аминокислоты или неприродные аминокислоты, известные специалистам, например все L- или D-аминокислоты или их смеси. Однако аминокислотный линкер, содержащий молекулу с сульфгидрильной группой или цистеиновым остатком, также входит в объем настоящего изобретения. Такая молекула, предпочтительно, содержит С1-С6-алкильную, циклоалкильную (С5, С6), арильную или гетероарильную группу. Связывание между антигеном или антигенной детерминантой или, необязательно, вторым сайтом связывания и аминокислотным линкером, предпочтительно, осуществляется посредством, по меньшей мере, одной ковалентной связи, а более предпочтительно, по меньшей мере, одной пептидной связи.
Связанный: используемый здесь термин "связанный" относится к связыванию или присоединению, которое может быть ковалентным, например посредством химического связывания, или нековалентным, например посредством ионного взаимодействия, гидрофобного взаимодействия, водородной связи и т.п. Ковалентными связями могут быть, например, сложноэфирная, эфирная, фосфоэфирная, амидная, пептидная, имидная, углерод-серная, углерод-фосфорная связи и т.п. Термин "связанный" имеет более широкое значение и может включать понятие "сопряженный", "слитый" и "присоединенный".
Коровая частица: используемый здесь термин "коровая частица" означает жесткую структуру с присущей этой структуре повторяющейся организацией, которая является основой для присоединения "организатора". Используемой здесь коровой частицей может быть продукт синтеза или продукт биологического процесса.
Оболочечный(ые) белок(ки): используемый здесь термин "оболочечный(ые) белок(ки)" означает белок(белки) бактериофага или РНК-фага, способный включаться в капсидную структуру указанного бактериофага или РНК-фага. Однако при обозначении специфического генного продукта гена оболочечного белка РНК-фагов используется термин "СР". Так, например, специфический генный продукт гена оболочечного белка РНК-фага Qβ обозначается "Qβ СР", а термин "оболочечные белки" бактериофага Qβ включает белок "Qβ СР", а также белок А1.
Цис-действующий: используемый здесь термин "цис-действующая" последовательность относится к последовательностям нуклеиновой кислоты, с которыми связывается репликаза и катализирует РНК-зависимую репликацию РНК-молекул. Эти события репликации приводят к репликации полноразмерных и неполных РНК-молекул, а поэтому субгеномный промотор альфавируса также является "цис-действующей" последовательностью. Цис-действующие последовательности могут быть локализованы на 5'-конце, 3'-конце, или на обоих концах молекулы нуклеиновой кислоты, или вблизи от этих концов, а также внутри этой молекулы.
Слияние: используемый здесь термин "слияние" означает объединение аминокислотных последовательностей различного происхождения в полипептидной цепи путем соединения, в той же рамке считывания, кодирующих их нуклеотидных последовательностей. Очевидно, что термин "слияние" включает внутренние объединения, то есть инсерцию последовательностей различной природы в полипептидной цепи, а также присоединение к одному из их концов.
Гетерологичная последовательность: используемый здесь термин "гетерологичная последовательность" означает вторую нуклеотидную последовательность, присутствующую в векторе настоящего изобретения. Термин "гетерологичная последовательность" также означает любую аминокислотную последовательность или РНК-последовательность, кодируемую гетерологичной ДНК-последовательностью, содержащейся в векторе настоящего изобретения. Гетерологичные последовательности могут кодировать белки или РНК-молекулы, обычно экспрессируемые в клетках определенного типа, в которых они присутствуют, или молекулы, которые обычно не экспрессируются в этих клетках (например, структурные белки вируса Синдбис).
Выделенный: используемый здесь термин "выделенный" применяется к молекуле и означает, что эта молекула была удалена из ее природного окружения. Так, например, полинуклеотид или полипептид, присутствующие в своем нормальном состоянии в живом организме, не являются "выделенными", но тот же самый полинуклеотид или полипептид, отделенный от окружающего его природного материала, считается "выделенным". Кроме того, рекомбинантные ДНК-молекулы, содержащиеся в векторе, считаются выделенными в целях настоящего изобретения. Выделенными РНК-молекулами являются продукты in vivo- или in vitro-репликации РНК, полученные от ДНК- и РНК-молекул. Выделенными молекулами нуклеиновой кислоты также являются синтетически продуцированные молекулы. Кроме того, векторные молекулы, содержащиеся в рекомбинантных клетках-хозяевах, также являются выделенными. Таким образом, не все "выделенные" молекулы должны быть "очищенными".
Иммунотерапевтический: используемый здесь термин "иммунотерапевтический" относится к композиции для лечения заболеваний или расстройств. Более конкретно, это термин применяется к методу лечения аллергий или к методу лечения злокачественных опухолей.
Индивидуум: используемый здесь термин "индивидуум" означает многоклеточные организмы и включает как растения, так и животных. Предпочтительными многоклеточными организмами являются животные, более предпочтительно, позвоночные, еще более предпочтительно, млекопитающие, а наиболее предпочтительно, человек.
Низкий или необнаружимый: используемый здесь термин "низкий или необнаружимый" применяется к уровню экспрессии гена и означает, что уровень экспрессии гена является либо значительно ниже уровня максимально индуцированной экспрессии (например, ниже, по меньшей мере, в пять раз), либо этот уровень практически не детектируется методами, описанными в нижеследующих примерах.
Лектин: используемый здесь термин "лектин" означает белки, которые получают, в частности, из семян овощных растений, а также из многих других растительных источников и животных, и которые имеют специфические сайты связывания с моно- или олигосахаридами. В качестве примеров могут быть упомянуты конканавалин А и агглютинин проростков пшеницы, которые широко используются в качестве аналитических и препаративных агентов в исследовании гликопротеинов.
Мимотоп: используемый здесь термин "мимотоп" означает вещество, которое индуцирует иммунный ответ к антигену или антигенной детерминанте. Как правило, термин "мимотоп" используется применительно к конкретному антигену. Так, например, пептид, который индуцирует вырабатывание антител к фосфолипазе A2 (PLA2), представляет собой мимотоп антигенной детерминанты, с которой связывается антитело. Мимотоп может иметь, а может и не иметь, значительное структурное сходство или общие структурные свойства с антигеном или с антигенной детерминантой, против которых индуцируется иммунный ответ. Методы генерирования и идентификации мимотопов, индуцирующих иммунный ответ на конкретные антигены или антигенные детерминанты, известны специалистам и описаны в настоящей заявке.
Природное происхождение: используемый здесь термин "природное происхождение" относится к материалу и означает, что целый материал или его части не являются синтетическими и существуют или продуцируются в природе.
Неприродный: используемый здесь термин, в общих чертах, означает "не встречающийся в природе", а более конкретно, "изготовленный человеком".
Неприродного происхождения: используемый здесь термин "неприродного происхождения" в общих чертах означает синтетический или не встречающийся в природе, а более конкретно этот термин означает "изготовленный человеком".
Неприродный молекулярный каркас: используемый здесь термин "неприродный молекулярный каркас" означает любой изготавливаемый человеком продукт, который может служить для изготовления жесткого и повторяющегося массива первых сайтов связывания. В идеальном случае, но необязательно, эти первые сайты связывания могут быть расположены в геометрическом порядке. Неприродный молекулярный каркас может быть органическим или неорганическим и может быть частично или целиком синтезирован химическим или биологическим способом. Этот неприродный молекулярный каркас состоит из (а) коровой частицы природного или неприродного происхождения; и (b) организатора, который, сам по себе, содержит, по меньшей мере, один первый сайт связывания и присоединен к коровой частице посредством, по меньшей мере, одной ковалентной связи. В конкретном варианте осуществления изобретения таким неприродным молекулярным каркасом может быть вирус, вирусоподобная частица, бактериальные пили, вирусная капсидная частица, фаг, его рекомбинантная форма или синтетическая частица.
Упорядоченный и повторяющийся массив антигенов или антигенных детерминант: используемый здесь термин "упорядоченный и повторяющийся массив антигенов или антигенных детерминант" в общих чертах означает повторяющуюся конфигурацию антигенов или антигенных детерминант, характеризующуюся однородным пространственным расположением антигенов или антигенных детерминант по отношению к неприродному молекулярному каркасу. В одном из вариантов осуществления изобретения такой повторяющейся конфигурацией может быть геометрическая конфигурация. Примерами подходящих упорядоченных и повторяющихся массивов антигенов или антигенных детерминант являются массивы, которые содержат точно повторяющиеся в паракристаллическом порядке антигены или антигенные детерминанты, которые расположены друг от друга на расстоянии 5-15 нанометров.
Организатор: используемый здесь термин "организатор" означает элемент, связанный с коровой частицей нерандомизированным образом, который обеспечивает место образования ядра для создания упорядоченного и повторяющегося массива антигенов. Организатор представляет собой любой элемент, содержащий, по меньшей мере, один сайт присоединения, который связывается с коровой частицей посредством, по меньшей мере, одной ковалентной связи. Таким организатором может быть белок, полипептид, пептид, аминокислота (то есть остаток белка, полипептид или пептид), сахар, полинуклеотид, природный или синтетический полимер, вторичный метаболит или соединение (биотин, флуоресцеин, ретинол, дигоксигенин, ионы металлов, фенилметилсульфонилфторид), или их комбинация, или их химически реактивная группа. Таким образом, указанный организатор, кроме того, обеспечивает образование упорядоченного и повторяющегося массива антигенов настоящего изобретения. В типичных вариантах осуществления изобретения коровая частица является модифицированной, например методами генной инженерии или посредством химической реакции, в целях генерирования неприродного молекулярного каркаса, содержащего коровую частицу и организатор, где указанный организатор присоединен к указанной коровой частице, по меньшей мере, одной ковалентной связью. Однако в некоторых вариантах осуществления изобретения выбранным организатором является часть коровой частицы. Поэтому, в соответствии с этими вариантами осуществления изобретения, для генерирования неприродного молекулярного каркаса, содержащего коровую частицу и организатор, и для обеспечения образования упорядоченного и повторяющегося массива антигенов нет необходимости в обязательном проведении модификации коровой частицы.
Пермиссивная температура: используемый здесь термин "пермиссивная температура" означает температуры, при которых фермент имеет относительно высокие уровни каталитической активности.
Пили: используемый здесь термин "пили" (в единственном числе "пилюс") означает внеклеточные структуры бактериальных клеток, состоящих из мономеров белка (например, мономеров пилина), которые организованы в упорядоченные и повторяющиеся конфигурации. Кроме того, пили представляют собой структуры, которые участвуют в таких процессах как связывание бактериальных клеток с рецепторами поверхности клеток-хозяев, межклеточные генетические обмены и распознавание клетки другой клеткой. Примерами пилей являются пили типа 1, Р-пили, пили F1С, S-пили и 987Р-пили. Другие примеры пилей проводятся ниже.
Пилеподобная структура: используемый здесь термин "пилеподобная структура" означает структуры, имеющие свойства, аналогичные свойствам пилей, и состоящие из мономеров белка. Одним из примеров "пилеподобных структур" является структура, образованная бактериальной клеткой, экспрессирующей модицированные белки пилины, которые не образуют упорядоченные и повторяющиеся массивы, в основном, идентичные массивам природных пилей.
Полипептид: используемый здесь термин "полипептид" означает полимер, состоящий из аминокислотных остатков, обычно природных аминокислотных остатков, связанных друг с другом пептидными связями. Хотя размер полипептида не обязательно должен быть ограниченным, однако под понятием "полипептид" часто подразумевается пептид, имеющий размер примерно от десяти до 50 аминокислот.
Белок: используемый здесь термин "белок" означает полипептид, обычно имеющий размер свыше 20, а более конкретно свыше 50 аминокислотных остатков. Белки обычно имеют определенную трехмерную структуру, и хотя они необязательно должны быть уложенными, однако часто подразумевается, что они являются уложенными, в противоположность пептидам и полипептидам, которые, в большинстве случаях, не имеют определенной трехмерной структуры, но, при этом, они могут принимать большое число различных конформаций и определяются как неуложенные. Определенные трехмерные структуры белков имеют особенно важное значение для связывания между коровой частицей и антигеном, опосредуемого вторым сайтом связывания, а в частности, посредством химического сшивания между первым и вторым сайтами связывания с помощью химического сшивающего линкера. В некоторых аспектах настоящего изобретения аминокислотный линкер непосредственно зависит от структурных свойств белков.
Очищенный: используемый здесь термин "очищенный" относится к молекуле и означает, что концентрация данной очищенной молекулы превышает концентрацию молекул, ассоциированных с ней в ее природном окружении. Этими ассоциированными природными молекулами являются белки, нуклеиновые кислоты, липиды и сахара, но ими обычно не являются вода, буферы и реагенты, добавляемые для сохранения целостности или облегчения очистки очищаемых молекул. Так, например, даже если мРНК разводят водным растворителем в процессе хроматографии на колонке с oligo-dT, то молекулы мРНК могут быть очищены с помощью этой хроматографии в том случае, если ассоциированные с ней в природе нуклеиновые кислоты и другие биологические молекулы не связываются с этой колонкой и отделяются от рассматриваемых молекул мРНК.
Рецептор: используемый здесь термин "рецептор" означает белки, или гликопротеины, или их фрагменты, способные взаимодействовать с другой молекулой, называемой лигандом. Этот лиганд может принадлежать к любому классу биохимических или химических соединений. Этот рецептор необязательно должен быть мембрано-ассоциированным белком. Растворимые белки, например, такие как белок, связывающийся с мальтозой, или белок, связывающийся с ретинолом, также являются рецепторами.
Остаток: используемый здесь термин "остаток" означает конкретную аминокислоту в полипептидном остове или в боковой цепи.
Рекомбинантная клетка-хозяин: используемый здесь термин "рекомбинантная клетка-хозяин" означает клетку-хозяина, в которую были введены одна или несколько молекул нуклеиновой кислоты настоящего изобретения.
Рекомбинантный вирус: используемый здесь термин "рекомбинантный вирус" означает вирус, генетически модифицированный человеком. Этот термин охватывает любой вирус, известный специалистам. Более конкретно, этот термин означает альфавирус, генетически модифицированный человеком, а более конкретно, этот термин означает вирус Синдбис, генетически модифицированный человеком.
Рестриктивная температура: используемый здесь термин "рестриктивная температура" означает температуру, при которой фермент имеет низкие или недетектируемые уровни каталитической активности. Известно, что существуют "тепло"- и "холодо"чувствительные мутанты, а поэтому рестриктивная температура может быть выше или ниже пермиссивной температуры.
Событие РНК-зависимой репликации РНК: используемый здесь термин "событие РНК-зависимой репликации РНК" означает процессы, приводящие к образованию молекулы РНК, в которых в качестве матрицы используется молекула РНК.
РНК-зависимая РНК-полимераза: используемый здесь термин "РНК-зависимая РНК-полимераза" означает полимеразу, которая катализирует продуцирование молекулы РНК из другой молекулы РНК. Этот термин используется здесь как синоним термина "репликаза".
РНК-фаг: используемый здесь термин "РНК-фаг" означает РНК-вирусы, инфицирующие бактерии, а предпочтительно, бактерии, инфицирующие одноцепочечные РНК-вирусы с позитивным геномом.
Аутоантиген: используемый здесь термин "аутоантиген" означает белки, кодируемые ДНК хозяина, и продукты, генерируемые белками или РНК, кодируемыми указанной ДНК хозяина и определяемые как "свои". Кроме того, белки, полученные в результате комбинирования двух или нескольких собственных молекул или представляющие собой фракцию собственной молекулы, и белки, имеющие в высокой степени гомологичные две собственные молекулы, определенные выше (>95%), также могут рассматриваться как "свои".
Температурочувствительный: используемый здесь термин "температурочувствительный" относится к ферменту, который легко катализирует реакцию при одной температуре, но медленно катализирует или вообще не катализирует ту же самую реакцию при другой температуре. Примером температурочувствительного фермента является белок репликазы, кодируемый вектором pCYTts, который обладает легко детектируемой репликазной активностью при температурах ниже 34°С, либо имеет низкую или недетектируемую активность при 37°С.
Транскрипция: используемый здесь термин "транскрипция" означает продуцирование молекул РНК из ДНК-матриц, катализируемых РНК-полимеразой.
Нетранслируемая РНК: используемый здесь термин "нетранслируемая РНК" означает РНК-последовательность или молекулу, которая не кодирует открытую рамку считывания или кодирует открытую рамку считывания или ее часть, но в формате, в котором не продуцируется аминокислотная последовательность (например, в отсутствие инициирующего кодона). Примерами таких молекул являются молекулы тРНК, молекулы рРНК и рибозимы.
Вектор: используемый здесь термин "вектор" означает агент (например, плазмиду или вирус), используемый для переноса генетического материала в клетку-хозяина. Вектор может состоять из ДНК или РНК.
Вирусоподобная частица: используемый здесь термин "вирусоподобная частица" означает структуру, сходную со структурой вирусной частицы. Кроме того, в соответствии с настоящим изобретением вирусоподобная частица является нерепликативной и неинфекционной, поскольку она не содержит всего вирусного генома или его части, а в частности репликативных и инфекционных компонентов указанного вирусного генома. В соответствии с настоящим изобретением вирусоподобная частица может содержать нуклеиновую кислоту, отличающуюся от генома вирусов.
Вирусоподобная частица бактериофага: используемый здесь термин "вирусоподобная частица бактериофага" означает вирусоподобную частицу, которая по своей структуре сходна с бактериофагом, но является нерепликативной и неинфекционной, а также не содержит, по меньшей мере, одного гена или несколько генов, кодирующих репликативный комплекс бактериофага, и обычно также не содержит один или несколько генов, кодирующих белок или белки, ответственные за связывание с вирусом или проникновение в клетку-хозяина. Однако это определение должно также охватывать вирусоподобные частицы бактериофагов, в которых вышеупомянутый ген или гены еще присутствуют, но являются неактивными, что, тем самым, приводит к образованию нерепликативных и неинфекционных вирусоподобных частиц бактериофага.
Вирусная частица: используемый здесь термин "вирусная частица" означает морфологическую форму вируса. У некоторых типов вирусов она содержит геном, окруженный белковым капсидом, а другие вирусные частицы имеют дополнительные структуры (например, оболочки, хвостовые нити и т.п.)
Слово "один" и артикль "а" или "an": используемые в настоящем описании слово "один", или артикль "а" или "an" означают "по меньшей мере, один", либо "один или несколько", если это не оговорено особо.
2. Композиции упорядоченных и повторяющихся массивов антигенов или антигенных детерминант и способы их получения
Настоящее изобретение относится к композициям, содержащим упорядоченный и повторяющийся массив антигенов или антигенных детерминант. Кроме того, в соответствии с настоящим изобретением каждый специалист может сконструировать упорядоченные и повторяющиеся массивы антигенов или антигенных детерминант для различных терапевтических целей, а именно для профилактики инфекционных заболеваний, лечения аллергий и лечения злокачественных опухолей.
Композиции настоящего изобретения, в основном, содержат или, альтернативно, состоят из них, два элемента: (1) неприродный молекулярный каркас и (2) антиген или антигенную детерминанту, которые имеют, по меньшей мере, один второй сайт связывания, способный связываться с указанным первым сайтом связывания посредством, по меньшей мере, одной непептидной связи.
Композиции настоящего изобретения также содержат или, альтернативно, состоят из них, белки бактериальных пилей, с которыми непосредственно связываются антигены или антигенные детерминанты.
Неприродный молекулярный каркас содержит или, альтернативно, состоит из них, (а) коровую частицу, выбранную из группы, состоящей из (1) неприродной коровой частицы и (2) природной коровой частицы; и (b) организатор, содержащий, по меньшей мере, один первый сайт связывания, где указанный организатор связан с указанной коровой частицей, по меньшей мере, одной ковалентной связью.
Композиции настоящего изобретения также содержат или, альтернативно, состоят из них, коровые частицы, с которыми непосредственно связываются антигены или антигенные детерминанты.
Антиген или антигенная детерминанта имеют, по меньшей мере, один второй сайт связывания, выбранный из группы, состоящей из (а) сайта связывания, который в природе не ассоциируется с указанным антигеном или указанной антигенной детерминантой; и (b) сайта связывания, который в природе ассоциируется с указанным антигеном или указанной антигенной детерминантой.
Настоящее изобретение относится к упорядоченному и повторяющемуся массиву антигенов, полученному путем присоединения второго сайта связывания к первому сайту связывания посредством, по меньшей мере, одной непептидной связи. Таким образом, антиген или антигенная детерминанта и неприродный молекулярный каркас, связанные вместе посредством указанного присоединения первого сайта связывания ко второму сайту связывания, образуют упорядоченный и повторяющийся массив антигенов.
Специалист может специально сконструировать антиген или антигенную детерминанту и второй сайт связывания так, чтобы расположение всех антигенов или антигенных детерминант, связанных с неприродным молекулярным каркасом или, в некоторых вариантах осуществления изобретения, с коровыми частицами, было равномерным. Так, например, на антигене или антигенной детерминанте у карбоксильного или амино-конца может присутствовать один второй сайт связывания, в результате чего можно получить конструкцию, в которой все молекулы антигенов или антигенных детерминант, присоединенные к неприродному молекулярному каркасу, будут расположены равномерно. Таким образом, настоящее изобретение относится к удобному способу размещения любого антигена или любой антигенной детерминанты на неприродном молекулярном каркасе в определенном порядке и так, чтобы образовывалась повторяющаяся конфигурация.
Как очевидно для каждого специалиста, в некоторых вариантах осуществления изобретения предусматривается использование техники рекомбинантных ДНК, такой как клонирование, полимеразная цепная реакция, очистка ДНК и РНК, экспрессия рекомбинантных белков в прокариотических и эукариотических клетках и т.п. Такие методы хорошо известны специалистам, и их описание можно найти в опубликованных руководствах по лабораторной практике (см., например, Sambrook J. et al., (1989), Molecular Cloning: A Laboratory Manual, 2nd Edition., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, (1989); Ausubel F. et al., Current Protocols in Molecular Biology, John H. Wiley & Sons, Inc. (1997)). Фундаментальные лабораторные методы для работы с клеточными линиями тканевой культуры (Celis, J. ed., Cell Biology, Academic Press, 2nd edition, (1998)) и методы с использованием антител (Harlow, E. and Lane, D., "Antibodies: A Laboratory Manual," Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. (1988); Deutscher, M.P., "Guide to Protein Purification," Meth. Enzymol. 128, Academic Press San Diego (1990); Scopes, R.K., "Protein Purification Principles and Practice," 3rd ed., Springer-Verlag, New York (1994)) также хорошо описаны в литературе и вводятся в настоящее описание посредством ссылки.
А. Коровые частицы и неприродные молекулярные каркасы
Одним из элементов в некоторых композициях настоящего изобретения является неприродный молекулярный каркас, содержащий или, альтернативно, состоящий из них, коровую частицу и организатор. Используемый здесь термин "неприродный молекулярный каркас" относится к любому полученному человеком продукту, который может служить для изготовления жесткого и повторяющегося массива первых сайтов связывания. Более конкретно, этот неприродный молекулярный каркас включает или, альтернативно, состоит из них, (а) коровую частицу природного или неприродного происхождения, выбранную из группы, состоящей из (1) неприродной коровой частицы и (2) природной коровой частицы; и (b) организатор, содержащий, по меньшей мере, один первый сайт связывания, где указанный организатор связан с указанной коровой частицей, по меньшей мере, одной ковалентной связью.
Как очевидно для каждого специалиста, указанная коровая частица неприродного молекулярного каркаса настоящего изобретения не ограничивается какой-либо конкретной формой. Такая коровая частица может быть органической или неорганической и может быть получена методом химического синтеза или в ходе биологического процесса.
В одном из вариантов осуществления изобретения неприродная коровая частица может быть синтетическим полимером, липидной мицеллой или металлом. Такие коровые частицы известны специалистам и являются основой, на которой конструируют новый неприродный молекулярный каркас настоящего изобретения. Так, например, коровые частицы из синтетического полимера или металла описаны в патенте США №5770380, где проиллюстрировано использование каликсаренового кристаллического органического каркаса, к которому присоединяется множество пептидных петель, с образованием "антитела-миметика", а в патенте США №5334394 описаны нанокристаллические частицы, используемые в качестве вирусной ловушки, которая состоит из широкого ряда неорганических материалов, включая металлы или керамические материалы. Подходящими металлами являются хром, рубидий, железо, цинк, селен, никель, золото, серебро и платина. Подходящими керамическими материалами в этом варианте осуществления изобретения являются диоксид кремния, диоксид титана, оксид алюминия, оксид рутения и оксид олова. В этом варианте осуществления изобретения коровые частицы могут быть изготовлены из органических материалов, включая углерод (алмаз). Подходящими полимерами являются полистирол, найлон и нитроцеллюлоза. Для получения нанокристаллических частиц такого типа могут быть также использованы частицы, изготовленные из оксида олова, диоксида титана или углерода (алмаза). Липидная мицелла может быть получена любым известным методом. Так, например, мицеллы могут быть получены методом, описанным в работах: Baiselle и Miller (Biophys. Chem. 4:355-361 (1975)), или Corti et al. (Chem. Phys. Lipids 38:197-214 (1981)), или Lopez et al. (FEBS Lett. 426:314-318 (1998)), или Topchieva and Karezin (J. Colloid Interface Sci. 213:29-35 (1999)), или Morein et al., (Nature 308:451-460 (1984)), которые вводятся в настоящее описание посредством ссылки.
Коровая частица может быть также продуцирована биологическим способом, и эта частица может быть природной или неприродной. Так, например, такой вариант осуществления изобретения может включать коровую частицу, содержащую или, альтернативно, состоящую из них, вирус, вирусоподобную частицу, бактериальные пили, фаг, вирусную капсидную частицу или их рекомбинантную форму. В более конкретном варианте осуществления изобретения коровая частица может включать или, альтернативно, состоять из них, рекомбинантные белки ротавируса, рекомбинантные белки вируса Норуолк, рекомбинантные белки альфавируса, рекомбинантные белки, которые образуют бактериальные пили или пилеподобные структуры, рекомбинантные белки вируса ящура, рекомбинантные белки ретровируса, рекомбинантные белки вируса гепатита В (например, HBcAg), рекомбинантные белки вируса табачной мозаики, рекомбинантные белки вируса овечьих кошар (Flok House Virus) и рекомбинантные белки человеческого папиломавируса. Коровая частица, кроме того, может содержать или, альтернативно, состоять из них, один или несколько фрагментов указанных белков, а также варианты таких белков, которые сохраняют способность связываться друг с другом с образованием упорядоченных и повторяющихся массивов антигенов или антигенных детерминант.
Как будет объяснено ниже, варианты белков, которые сохраняют способность связываться друг с другом с образованием упорядоченных и повторяющихся массивов антигенов или антигенных детерминант, могут иметь аминокислотную последовательность, которая, например, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентична их аналогам дикого типа. Так, например, для иллюстрации, если используется HBcAg, имеющий аминокислотную последовательность, представленную в SEQ ID NO:89, то настоящее изобретение охватывает вакцинные композиции, включающие полипептиды HBcAg, содержащие или, альтернативно, состоящие из них, аминокислотные последовательности, которые, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентичны аминокислотной последовательности, представленной в SEQ ID NO:89, и формы указанного белка, который был процессирован, если это необходимо, для удаления N-концевой лидерной последовательности. Эти варианты обычно способны связываться с образованием димерных или мультимерных структур. Методами, которые могут быть использованы для того, чтобы определить, образуют ли белки указанные структуры, являются гель-фильтрация, электрофорез в агарозном геле, центрифугирование в градиенте сахарозы и электронная микроскопия (например, Koschel, M. et al., J. Virol 73:2153-2160 (1999)).
Фрагменты белков, которые сохраняют способность ассоциироваться друг с другом с образованием упорядоченных и повторяющихся массивов антигенов или антигенных детерминант, могут содержать или, альтернативно, состоять из них, полипептиды, которые имеют длину 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190 или 200 аминокислот. Примерами таких фрагментов белка являются фрагменты рассматриваемых здесь белков, которые могут быть использованы для получения коровых частиц и/или неприродных молекулярных каркасов.
Коровые частицы настоящего изобретения, независимо от того, являются ли они природными или неприродными, обычно содержат организатор, который связан с указанной природной или неприродной коровой частицей, по меньшей мере, одной ковалентной связью. Этот организатор представляет собой элемент, связанный с коровой частицей нерандомизированным образом и обеспечивающий сайт нуклеации для создания упорядоченных и повторяющихся массивов антигенов. В идеальном случае, но необязательно, этот организатор ассоциирован с коровой частицей в геометрической конфигурации. Этот организатор содержит, как минимум, первый сайт связывания.
В некоторых вариантах осуществления изобретения упорядоченный и повторяющийся массив образован посредством связывания между (1) либо коровыми частицами или неприродными молекулярными каркасами и (2) либо (а) ангигеном или антигенной детерминантой, либо (b) одним или несколькими антигенами или антигенными детерминантами. Так, например, бактериальные пили или пилеподобные структуры могут быть образованы из белков, которые организованы в упорядоченные и повторяющиеся структуры. Таким образом, во многих случаях можно получить упорядоченные массивы из антигенов или антигенных детерминант путем присоединения этих компонентов к бактериальным пилям или к пилеподобным структурами либо непосредственно, либо посредством организатора.
Как указывалось ранее, организатором может быть любой элемент, содержащий, по меньшей мере, один первый сайт связывания, который связан с коровой частицей, по меньшей мере, одной ковалентной связью. Таким организатором может быть белок, полипептид, пептид, аминокислота (то есть остаток белка, полипептида или пептида), сахар, полинуклеотид, природный или синтетический полимер, вторичный метаболит или соединение (биотин, флуоресцеин, ретинол, дигоксигенин, ионы металла, фенилметилсульфонилфторид), или их комбинации, или их химически реактивная группа. В более конкретном варианте осуществления изобретения указанный организатор может включать первый сайт связывания, содержащий антиген, антитело или фрагмент антитела, биотин, авидин, стрептавидин, рецептор, лиганд рецептора, лиганд, лиганд-связывающий белок, полипептид, взаимодействующий с лейциновой молнией, аминогруппу, химическую группу, реагирующую с аминогруппой, карбоксильную группу, химическую группу, реагирующую с карбоксильной группой, сульфгидрильную группу, химическую группу, реагирующую с сульфгидрильной группой, или их комбинации.
В одном из вариантов осуществления изобретения коровая частица неприродного молекулярного каркаса включает вирус, бактериальный пилюс, структуру, образованную бактериальным пилином, бактериофаг, вирусоподобную частицу, вирусную капсидную частицу или их рекомбинантную форму. В качестве неприродного молекулярного каркаса настоящего изобретения может быть выбран любой вирус, известный специалистам и имеющий упорядоченную и повторяющуюся структуру его оболочечного белка и/или корового белка; и примерами подходящих вирусов являются вирус Синдбис и другие альфавирусы, рабдовирусы (например, вирус везикулярного стоматита), пикорнавирусы (например, риновирус человека, вирус Эйши), тогавирусы (например, вирус краснухи), ортомиксовирусы (например, вирус Тогото, вирус Баткен, вирус чумы домашних птиц), полиомавирусы (например, полиомавирус ВК, полиомавирус JC, полиомавирус птиц BFDV), парвовирусы, ротавирусы, бактериофаг Qβ, бактериофаг R17, бактериофаг М11, бактериофаг МХ1, бактериофаг NL95, бактериофаг fr, бактериофаг GA, бактериофаг SP, бактериофаг MS2, бактериофаг f2, бактериофаг РР7, вирус Норуолк, вирус ящура, ретровирус, вирус гепатита В, вирус табачной мозаики, вирус овечьих кошар и человеческий папиломавирус (см., например, таблицу 1 в работе Bachman, M.F. & Zinkernagel, R.M., Immunol. Today 17:553-558 (1996)).
В одном из вариантов своего осуществления настоящее изобретение относится к генетическому конструированию вируса для создания гибрида между расположенным в определенном порядке и повторяющимся оболочечным белком вируса и организатором, содержащего выбранные гетерологичный белок, пептид, антигенную детерминанту или реакционноспособный аминокислотный остаток. При конструировании неприродного молекулярного каркаса могут быть проведены и другие генетические манипуляции, известные специалистам; так, например, может оказаться желательным ограничение способности рекомбинантного вируса к репликации, которое может быть осуществлено путем введения генетической мутации. Вирусный белок, выбранный для присоединения к белку (то есть к первому сайту связывания) организатора, должен иметь организованную и повторяющуюся структуру. Такой упорядоченной и повторяющейся структурой являются паракристаллические организаторы с расстоянием 5-15 нм на поверхности вируса. Создание гибридного белка такого типа позволяет получить множество упорядоченных и повторяющихся организаторов на поверхности вируса. Таким образом, полученная из этих белков упорядоченная и повторяющаяся организация первых сайтов связывания будет отражать нормальную организацию нативного вирусного белка.
Как будет более подробно обсуждаться ниже, в другом варианте осуществления изобретения неприродным молекулярным каркасом является рекомбинантный альфавирус, а более конкретно, рекомбинантный вирус Синдбис. Альфавирусы представляют собой РНК-вирусы с позитивной цепью, и их геном полностью реплицируется в цитоплазме инфицированной клетки без промежуточного ДНК-продукта (Strauss J. & Strauss, E. Microbiol. Rev. 58:491-562 (1994)). Некоторые члены семейства альфавирусов, такие как вирус Синдбис (Xiong C. et al., Science 243:1188-1191 (1989); Schlesinger S. Trends Biotechnol. 11:18-22 (1993)), вирус лесов Семлики (SFV) (Liljestrőm, P. & Garoff, H., Bio/Technology 9:1356-1361 (1991)) и другие вирусы (Davis N.L. et al., Virology 171:189-204 (1989)) заслуживают особого внимания как кандидаты на их использование в качестве экспрессирующих векторов для получения ряда различных белков (Lundstrom K. Curr. Opin. Biotechnol. 8:578-582 (1997); Liljestrőm, P., Curr. Opin. Biotechnol. 5:495-500 (1994)) и как кандидаты для получения вакцин. Недавно был выдан ряд патентов на использование альфавирусов для экспрессии гетерологичных белков и разработки вакцин (см. патенты США №№5766602, 5792462, 5739026, 5789245 и 5814482). Конструирование альфавирусного каркаса настоящего изобретения может быть осуществлено с помощью техники рекомбинантных ДНК, в общих чертах известной специалистам и описанной в вышеуказанных статьях, которые вводятся в настоящее описание посредством ссылки.
Для продуцирования коровой частицы на основе вируса для ее присоединения к антигену или антигенной детерминанте может быть использован ряд различных рекомбинантных клеток-хозяев. Так, например, известно, что альфавирусы имеют широкий круг хозяев; вирус Синдбис инфицирует культивированные клетки млекопитающих, рептилий и амфибий, а также некоторые клетки насекомых (Clark, H. J. Natl. Cancer. Inst. 51:645 (1973); Leake, C. J. Gen. Virol. 35:335 (1997); Stollar, V., The Togaviruses, R.W. Schlesinger, Ed., Academic Press, (1980); рр.583-621). Для осуществления настоящего изобретения может быть использовано множество рекомбинантных клеток-хозяев. Клетки BHK, COS, Vero, HeLa и CHO являются особенно подходящими для продуцирования гетерогенных белков, поскольку они обладают способностью гликозилировать гетерологичные белки аналогично человеческим клеткам (Watson, E. et al., Glycobiology 4:227 (1994)) и могут быть отобраны (Zang M. et al., Bio/Technology 13:389 (1995)) или генетически сконструированы (Renner, W. et al., Biotech. Bioeng. 4:476 (1995); Lee K. et al., Biotech. Bioeng. 50:336 (1996)) для культивирования в бессывороточной среде, а также в суспензии.
Введение полинуклеотидных векторов в клетки-хозяева может быть осуществлено методами, описанными в известных руководствах по лабораторной практике (см., например, Sambrook J. et al., Molecular Cloning: A Laboratory Manual, 2nd Edition., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, (1989), Chapter 9; Ausubel F. et al., Current Protocols in Molecular Biology, John H. Wiley & Sons, Inc. (1997) Chapter 16), включая такие методы как электропорация, трансфекция, опосредованная DEAE-декстраном, трансфекция, микроинжекция, трансфекция, опосредованная катионным липидом, трансдукция, загрузка путем соскабливания, введение методами биобаллистики и инфицирование. Методы введения экзогенных ДНК-последовательностей в клетки-хозяева обсуждаются в работе Felgner P. et al., в патенте США №5580859.
Для инфицирования клеток-хозяев могут быть также использованы упакованные РНК-последовательности. Эти упакованные РНК-последовательности могут быть введены в клетки-хозяева путем их добавления в культуральную среду. Так, например, получение неинфекционных альфавирусных частиц описано в ряде источников, включая "Sindbis Expression System", Version C (Invitrogen Catalog №К750-1).
В случае использования клеток млекопитающих в качестве рекомбинантных клеток-хозяев для продуцирования вирусных коровых частиц на основе вируса эти клетки обычно культивируют в тканевой культуре. Методы культивирования клеток в культуре хорошо известны специалистам (см., например, Celis, J., ed., Cell Biology, Academic Press, 2nd edition, (1998); Sambrook, J. et al, eds., Molecular Cloning, A Laboratory Manual, 2nd. edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989); Ausubel, F. et al., eds., Current Protocols in Molecular Biology, John H. Wiley & Sons, Inc. (1997); Freshney, R. Culture of Animal Cells, Alan R. Liss, Inc. (1983)).
Как очевидно для каждого специалиста, указанным первым сайтом связывания может быть или является часть любого подходящего белка, полипептида, сахара, полинуклеотида, пептида (аминокислота), природного или синтетического полимера, вторичного метаболита или их комбинации, которые могут быть использованы для специфического связывания выбранного антигена или антигенной детерминанты с неприродным молекулярным каркасом. В одном из вариантов осуществления изобретения указанным сайтом связывания является белок или пептид, который может быть выбран из белков или пептидов, известных специалистам. Так, например, первый сайт связывания может быть выбран из следующей группы молекул: лиганда, рецептора, лектина, авидина, стрептавидина, биотина, эпитопа, такого как НА или Т7-метка, Мус, Мах, доменов иммуноглобулина и любых других аминокислотных последовательностей, которые известны специалистам и которые могут быть использованы в качестве первого сайта связывания.
Следует отметить, что в соответствии с другим вариантом осуществления изобретения первый сайт связывания может быть сконструирован после получения организатора (то есть белка или полипептида), используемого при конструировании гибридного капсидного белка в той же рамке считывания. Так, например, белок может быть использован для присоединения к оболочечному белку, имеющему аминокислотную последовательность, которая, как известно, подвергается специфическому гликозилированию, а молекула сахара, затем присоединяемая посредством связывания с лектином, служащим вторым сайтом связывания антигена, может служить первым сайтом связывания вирусного каркаса. Альтернативно, последовательность организатора может быть биотинилирована in vivo, и молекула биотина может служит первым сайтом связывания настоящего изобретения, либо последовательность организатора может быть подвергнута химической модификации в различных аминокислотных остатках in vitro, где указанная модифицированная последовательность служит первым сайтом связывания.
В другом варианте осуществления изобретения первый сайт связывания выбирают так, чтобы домен белка лейциновой молнии JUN-FOS был присоединен к капсидному (коровому) белку вируса гепатита В (HBcAg) в той же самой рамке считывания. Однако для каждого специалиста очевидно, что для определения локализации первого сайта связывания в неприродном молекулярном каркасе настоящего изобретения в конструкции гибридного белка могут быть использованы и другие вирусные капсидные белки.
В другом варианте осуществления изобретения первый сайт связывания выбирают так, чтобы лизиновый или цистеиновый остаток был присоединен к HBcAg в той же самой рамке считывания. Однако для каждого специалиста очевидно, что для определения локализации первого сайта связывания в неприродном молекулярном каркасе настоящего изобретения в конструкции гибридного белка могут быть использованы и другие вирусные капсидные белки или вирусоподобные частицы.
JUN, используемый для первого сайта связывания, имеет следующую аминокислотную последовательность:
CGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNHVGC
(SEQ ID NO:59)
В этом случае предполагаемый второй сайт связывания на антигене должен представлять собой домен белка лейциновой молнии FOS и иметь следующую аминокислотную последовательность:
CGGLT·TLQAETDQVEDEKSALQTEIANLLKEKEKLEFILAAHGGC
(SEQ ID NO:60)
Эти последовательности происходят от факторов транскрипции JUN и FOS, каждый из которых фланкирован короткой последовательностью, содержащей на обоих концах цистеиновый остаток. Известно, что эти последовательности взаимодействуют друг с другом. Исходя из гипотетической структуры, предполагаемой для димера JUN-FOS, было высказано предположение, что гидрофобные боковые цепи одного мономера "переплетаются" с соответствующими боковыми цепями другого мономера подобно "молнии" (Landschulz et al., Science, 240:1759-1764 (1988)). Однако было доказано, что эта гипотеза является неправильной, и известно, что эти белки образуют α-суперспирализованную структуру (O'Shea et al., Science, 243:538-542 (1989); O'Shea et al., Cell 68:699-708 (1992); Cohen & Parry, Trends Biochem, Sci. 11:245-248 (1986)). Таким образом, термин "лейциновая молния" часто означает домены белка и скорее имеет исторические корни, чем отражает его структуру. Используемый в описании патента настоящего изобретения термин "лейциновая молния" означает последовательности, описанные выше, или последовательности, в основном, аналогичные последовательностям, описанным выше. Термины JUN и FOS используются для обозначения соответствующих доменов лейциновой молнии, но не для целых белков JUN и FOS.
Как указывалось ранее, настоящее изобретение относится к вирусным коровым частицам, которые включают или, альтернативно, состоят из них, вирус, вирусоподобную частицу, фаг, вирусную капсидную частицу или их рекомбинантную форму. Продуцирование таких коровых частиц и присоединение к ним организаторов может быть осуществлено каждым специалистом. Продуцирование вирусоподобных частиц гепатита В и капсидных частиц вируса кори в качестве коровых частиц описано в примерах 17-22 заявки WO 00/32227, которая во всей своей полноте вводится в настоящее описание посредством ссылки. В указанных вариантах осуществления настоящего изобретения домен белка лейциновой молнии JUN или домен белка лейциновой молнии FOS может быть использован в качестве организатора, а следовательно, в качестве первого сайта связывания для неприродного молекулярного каркаса настоящего изобретения.
В примерах 23-29 подробно описано продуцирование коровых частиц вируса гепатита В, несущих пептид с присоединенным внутри рамки считывания реакционноспособным лизиновым остатком, и антигенов, несущих генетически присоединенный цистеиновый остаток, в качестве первого и второго сайта связывания, соответственно.
1. В других вариантах осуществления изобретения коровые частицы, используемые в композициях настоящего изобретения, состоят из капсидного (корового) белка вируса гепатита В (HBcAg), фрагмента HBcAg или другого белка или пептида, которые могут образовывать упорядоченные массивы и которые были модифицированы так, что они либо не содержали свободных цистеиновых остатков, либо имели меньшее количество этих остатков. Zhou et al. (J. Virol. 66:5393-5398 (1992)) продемонстрировали, что HBcAg, которые были модифицированы путем удаления природных цистеиновых остатков, сохраняли свою способность к связыванию и образованию мультимерных структур. Таким образом, коровыми частицами, подходящими для использования в композициях настоящего изобретения, являются коровые частицы, содержащие модифицированные HBcAg или их фрагменты, в которых один или несколько природных цистеиновых остатков либо были делетированы, либо были заменены другим аминокислотным остатком (например, сериновым остатком).
2. HBcAg представляет собой белок, генерированный посредством процессинга корового белка-предшественника антигена вируса гепатита В. Был индентифицирован ряд изотипов HBcAg. Так, например, белок HBcAg, имеющий аминокислотную последовательность, представленную в SEQ ID NO:132, имел длину в 183 аминокислоты и был генерирован посредством процессинга корового белка-предшественника антигена вируса гепатита В длиной 212 аминокислот. Этот процессинг приводит к удалению 29 аминокислот из N-конца корового белка-предшественника антигена вируса гепатита В. Аналогичным образом, белок HBcAg, имеющий аминокислотную последовательность, представленную в SEQ ID NO:134, имеет длину в 185 аминокислоты и был генерирован посредством процессинга белка-предшественника корового антигена вируса гепатита В длиной 214 аминокислот. Аминокислотная последовательность, представленная в SEQ ID NO:134, по сравнению с аминокислотной последовательностью, представленной в SEQ ID NO:132, содержит две аминокислотные инсерции в положениях 152 и 153 в SEQ ID NO:134.
В большинстве случаев вакцинные композиции настоящего изобретения могут быть получены с использованием процессированной формы HBcAg (то есть HBcAg, из которого была удалена N-концевая лидерная последовательность (например, первые 29 аминокислотных остатков, представленные в SEQ ID NO:134) белка-предшественника корового антигена вируса гепатита В).
Кроме того, если HBcAg продуцируется в условиях, при которых процессинг обычно не происходит, то HBcAg может, в основном, экспрессироваться в "процессированной" форме. Так, например, в бактериальных системах, таких как E.coli, лидерные последовательности, также называемые "сигнальными пептидами" белков, которые нормально экспрессируются в эукариотических клетках, обычно не удаляются. Таким образом, если для продуцирования HBcAg настоящего изобретения используется экспрессирующая система E.coli, то эти белки будут, в основном, экспрессироваться так, что в них будет отсутствовать N-концевая лидерная последовательность белка-предшественника корового антигена вируса гепатита В.
В одном из вариантов осуществления изобретения модифицированный HBcAg, содержащий аминокислотную последовательность, представленную в SEQ ID NO:134, или ее подпоследовательность, используют для получения неприродных молекулярных каркасов. В частности, модифицированными HBcAg, подходящими для осуществления настоящего изобретения, являются белки, в которых один или несколько цистеиновых остатков в положениях, соответствующих положениям 48, 61, 107 и 185 белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:134, были либо делетированы, либо заменены другими аминокислотными остатками (например, сериновым остатком). При этом следует отметить, что цистеиновые остатки в аналогичных положениях вариантов HBcAg, имеющих аминокислотные последовательности, отличающиеся от последовательностей, представленных в SEQ ID NO:134, могут быть также делетированы или заменены другими аминокислотными остатками. Затем модифицированные варианты HBcAg могут быть использованы для получения вакцинных композиций настоящего изобретения.
Настоящее изобретение также относится к вариантам HBcAg, которые были модифицированы путем делеции или замены одного или нескольких дополнительных цистеиновых остатков, не обнаруживаемых в полипептидах, имеющих аминокислотную последовательность, представленную в SEQ ID NO:134. Примерами таких вариантов HBcAg являются варианты, имеющие аминокислотные последовательности, представленные в SEQ ID NO:90 и 132. Эти варианты содержат цистеиновые остатки в положении, соответствующем положению аминокислотного остатка 147 в SEQ ID NO:134. Таким образом, вакцинными композициями настоящего изобретения являются композиции, содержащие HBcAg, в которых цистеиновые остатки, не присутствующие в аминокислотной последовательности, представленной в SEQ ID NO:134, были делетированы.
В некоторых случаях (например, если для присоединения антигенов или антигенных детерминант к неприродному молекулярному каркасу используется гетеробифункциональный сшивающий реагент) присутствие свободных цистеиновых остатков в HBcAg, вероятно, приводит к ковалентному связыванию токсических компонентов к коровым частицам, а также к перекрестному сшиванию мономеров с образованием неидентифицированных молекул.
Кроме того, во многих случаях эти токсические компоненты не могут быть детектированы с помощью анализов композиций, осуществляемых в соответствии с настоящим изобретением. Именно поэтому ковалентное связывание токсических компонентов с неприродным молекулярным каркасом должно приводить к образованию популяции различных молекул, в которых токсические компоненты связаны с разными цистеиновыми остатками HBcAg или, в некоторых случаях, не имеют цистеиновых остатков. Другими словами, каждый свободный цистеиновый остаток каждого HBcAg не будет ковалентно связываться с токсическими компонентами. Кроме того, во многих случаях, ни один из цистеиновых остатков конкретного HBcAg не будет связываться с токсическими компонентами. Таким образом, присутствие этих токсических компонентов может быть трудно обнаруживаемым, поскольку они могут присутствовать в смешанной популяции молекул. Однако присоединение к отдельным молекулам HBcAg, содержащим токсические компоненты, может приводить к серьезной неблагоприятной реакции.
Хорошо известно, что свободные цистеиновые остатки могут участвовать в ряде химических побочных реакций. Такими побочными реакциями являются реакция дисульфидного обмена, реакция с химическими веществами или метаболитами, которые, например, вводятся путем инъекции или образуются при комбинированной терапии с использованием других веществ, или реакция прямого окисления и реакция с нуклеотидами при УФ-облучении. Могут быть также генерированы токсические аддукты, а особенно, если учесть, что HBcAg имеют в высокой степени выраженную тенденцию к связыванию с нуклеиновыми кислотами. Детекция таких токсических продуктов в конъюгатах "антиген-капсид" с использованием капсидов, полученных с помощью HBcAg, содержащих свободные цистеины и гетеробифункциональные сшивающие линкеры, может представлять определенные трудности, поскольку, в этом случае, должно генерироваться распределение продуктов с большим разбросом молекулярных масс. Токсичные аддукты должны быть, таким образом, распределены между множеством молекул, которые, по отдельности, могут присутствовать в низкой концентрации, но вместе они могут достигать токсических уровней.
Исходя из вышесказанного, одно из преимуществ использования HBcAg в вакцинных композициях, которые были модифицированы путем удаления природных цистеиновых остатков, заключается в том, что число сайтов, с которыми могут связываться токсичные молекулы, при присоединении антигенов или антигенных детерминант к неприродному молекулярному каркасу либо уменьшается, либо эти сайты вообще отсутствуют. Кроме того, для продуцирования в высокой степени "декорированных" частиц при беспрепятственном генерировании множества неидетифицированных перекрестно сшитых молекул HBcAg-мономеров (то есть различных смесей перекрестно сшитых мономерных HBcAg) может быть использована высокая концентрация сшивающего линкера.
Был идентифицирован ряд природных вариантов HBcAg, подходящих для осуществления настоящего изобретения. Так, например, Yuan et al. (J. Virol. 73:10122-10128 (1999)), описывают варианты, в которых изолейциновый остаток в положении, соответствующем положению 97 в SEQ ID NO:134, заменен либо лейциновым остатком, либо фенилаланиновым остатком. Аминокислотные последовательности ряда вариантов HBcAg, а также некоторых вариантов предшественников корового антигена вируса гепатита В описаны в документах Genbank AAF121240 (SEQ ID NO:89), AF121239 (SEQ ID NO:90), X85297 (SEQ ID NO:91), X02496 (SEQ ID NO:92), X85305 (SEQ ID NO:93), X85303 (SEQ ID NO:94), AF151735 (SEQ ID NO:95), X85259 (SEQ ID NO:96), X85286 (SEQ ID NO:97), X85260 (SEQ ID NO:98), X85317 (SEQ ID NO:99), X85298 (SEQ ID NO:100), AF043593 (SEQ ID NO:101), M20706 (SEQ ID NO:102), X85295 (SEQ ID NO:103), X80925 (SEQ ID NO:104), X85284 (SEQ ID NO:105), X85275 (SEQ ID NO:106), X72702 (SEQ ID NO:107), X85291 (SEQ ID NO:108), X65258 (SEQ ID NO:109), X85302 (SEQ ED NO:110), M32138 (SEQ ID NO:111), X85293 (SEQ ID NO:112), X85315 (SEQ ID NO:113), U95551 (SEQ ID NO:114), X85256 (SEQ ID NO:115), X85316 (SEQ ID NO:116), X85296 (SEQ ID NO:117), AB033559 (SEQ ID NO:118), X59795 (SEQ ID NO:119), X85299 (SEQ ID NO:120), X85307 (SEQ ID NO:121), X65257 (SEQ ID NO:122), X85311 (SEQ ID NO:123), X85301 (SEQ ID NO:124), X85314 (SEQ ID NO:125), X85287 (SEQ ID NO:126), X85272 (SEQ ID NO:127), X85319 (SEQ ID NO:128), AB010289 (SEQ ID NO:129), X85285 (SEQ ID NO:130), AB010289 (SEQ ID NO:131), AF121242 (SEQ ID NO:132), M90520 (SEQ ID N0:135), P03153 (SEQ ID NO:136), AF110999 (SEQ ID NO:137), и M95589 (SEQ ID NO:138), описание которых вводится в настоящее описание посредством ссылки. Аминокислотные последовательности этих вариантов HBcAg отличаются в ряде положений, включая аминокислотные остатки, соответствующие аминокислотным остаткам, локализованным в положениях 12, 13, 21, 22, 24, 29, 32, 33, 35, 38, 40, 42, 44, 45, 49, 51, 57, 58, 59, 64, 66, 67, 69, 74, 77, 80, 81, 87, 92, 93, 97, 98, 100, 103, 105, 106, 109, 113, 116, 121, 126, 130, 133, 135, 141, 147, 149, 157, 176, 178, 182 и 183 в SEQ ID NO:134.
HBcAg, подходящие для использования в настоящем изобретении, могут быть получены из любого организма, при условии, что они способны к связыванию с образованием упорядоченного и повторяющегося антигенного массива.
Как указывалось выше, обычно процессированные HBcAg (то есть не содержащие лидерных последовательностей) могут быть использованы в вакцинных композициях настоящего изобретения. Так, например, если для получения вакцинных композиций настоящего изобретения используется HBcAg, имеющий аминокислотную последовательность, представленную в SEQ ID NO:136, 137 или 138, то, обычно, аминокислотные остатки 30, 35-43 или 35-43 в N-конце каждого из этих белков, соответственно, могут быть исключены.
Настоящее изобретение относится к вакцинным композициям, а также к способам использования этих композиций, в которых для получения неприродных молекулярных каркасов используются вышеуказанные варианты HBcAg.
Кроме того, в объем настоящего изобретения входят дополнительные варианты HBcAg, способные связываться с образованием димерных или мультимерных структур. Таким образом, настоящее изобретение также относится к вакцинным композициям, включающим полипетиды HBcAg, содержащие или, альтернативно, состоящие из них, аминокислотные последовательности, которые, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентичны любой из аминокислотных последовательностей, представленных в SEQ ID NO:89-132 и 134-138, и формы этих белков, которые были процессированы, если это необходимо, для удаления N-концевой лидерной последовательности.
Чтобы определить, имеет ли данный полипептид аминокислотную последовательность, которая, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентична одной из аминокислотных последовательностей, представленных в SEQ ID NO:89-132 и 134-138, или их субпоследовательностей, можно использовать известные компьютерные программы, такие как программа Bestfit. Если используется программа Bestfit или любая другая программа для выравнивания последовательностей, например, для того, чтобы определить, является ли данная последовательность, например, на 95% идентичной эталонной аминокислотной последовательности настоящего изобретения, то параметры устанавливают так, чтобы можно было вычислить процент идентичности по всей длине эталонной аминокислотной последовательности, и так, чтобы "пропуски" в гомологии составляли до 5% от полного числа аминокислотных остатков в эталонной последовательности.
Варианты и предшественники HBcAg, имеющие аминокислотные последовательности, представленные в SEQ ID NO:89-132 и 134-136, имеют относительное сходство друг с другом. Так, например, выражение "аминокислотный остаток варианта HBcAg, локализованный в положении, которое соответствует конкретному положению SEQ ID NO:134", означает аминокислотный остаток, который присутствует в этом положении аминокислотной последовательности, представленной в SEQ ID NO:134. Гомология между вариантами HBcAg является, по большей части, достаточно высокой для вируса гепатита В, инфицирующего млекопитающих, а поэтому для специалиста не представляет труда проанализировать аминокислотную последовательность, представленную в SEQ ID NO:134, а также аминокислотную последовательность конкретного варианта HBcAg и идентифицировать "соответствующие" аминокислотные остатки. Так, например, аминокислотная последовательность HBcAg, показанная в SEQ ID NO:135 и представляющая собой аминокислотную последовательность HBcAg, происходящую от вируса лесного сурка, имеет достаточную гомологию с антигеном HBcAg, имеющим аминокислотную последовательность, представленную в SEQ ID NO:131, то есть легко видеть, что в SEQ ID NO:135 присутствуют три аминокислотных вставки между аминокислотными остатками 155 и 156 SEQ ID NO:134.
HBcAg вирусов гепатита В, которые инфицируют белых гусей и уток, весьма отличаются от аминокислотных последовательностей HBcAg вируса гепатита В, инфицирующего млекопитающих, а поэтому сопоставление этих форм указанного белка с аминокислотной последовательностью, представленной в SEQ ID NO:134, представляет определенные трудности. Однако настоящее изобретение относится к вакцинным композициям, которые включают варианты HBcAg вируса гепатита В, инфицирующего птиц, а также к вакцинным композициям, которые содержат фрагменты этих вариантов HBcAg. Композициями, содержащими фрагменты HBcAg, подходящие для получения вакцинных композиций настоящего изобретения, являются композиции, которые содержат полипептидные фрагменты, включающие или, альтернативно, состоящие из них, аминокислотные остатки, выбранные из группы, состоящей из 36-240, 36-269, 44-240, 44-269, 36-305 и 44-305 SEQ ID NO:137 или 36-240, 36-269, 44-240, 44-269, 36-305 и 44-305 SEQ ID NO:138. Каждому специалисту очевидно, что один, два, три или несколько цистеиновых остатков, обычно присутствующих в этих полипептидах (например, цистеиновые остатки в положении 153 SEQ ID NO:137 или в положениях 34, 43 и 196 SEQ ID NO:138), перед их включением в вакцинные композиции настоящего изобретения могут быть либо заменены другим аминокислотным остатком, либо делетированы.
В одном из вариантов осуществления настоящего изобретения цистеиновые остатки в положениях 48 и 107 белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:134, либо делетированы, либо заменены другим аминокислотным остатком, однако цистеин в положении 61 остается неизмененным. Затем этот модифицированный полипептид используют для получения вакцинных композиций настоящего изобретения.
Как показано ниже в примере 31, цистеиновые остатки в положениях 48 и 107, которые являются доступными для растворителя, могут быть удалены, например, посредством сайт-направленного мутагенеза. Кроме того, авторами настоящего изобретения было обнаружено, что мутант HBcAg с двойной заменой Cys-48-Ser, Cys-107-Ser, сконструированный, как описано в примере 31, может быть экспрессирован в E.coli.
Как обсуждалось выше, элиминация свободных цистеиновых остатков приводит к снижению числа сайтов, в которых токсические компоненты могут связываться с HBcAg, а также к удалению сайтов, в которых может происходить перекрестное сшивание лизинового и цистеинового остатков тех же самых или соседних молекул HBcAg. Цистеин в положении 61, который участвует в образовании димера и образует дисульфидный мостик с цистеином в положении 61 другого HBcAg, обычно остается интактным и служит для стабилизации димеров и мультимеров HBcAg настоящего изобретения.
Как показано в примере 32, эксперименты по перекрестному связыванию (1) с HBcAg, содержащими свободные цистеиновые остатки, и (2) с HBcAg, в которых свободные цистеиновые остатки были сделаны нереакционноспособными с помощью йодацетамида, показали, что свободные цистеиновые остатки HBcAg ответственны за перекрестное связывание между антигенами HBcAg, осуществляемое посредством реакций между лизиновыми боковыми цепями, дериватизированными гетеробифункциональным сшивающим линкером, и свободными цистеиновыми остатками. В примере 32 также показано, что сшивание субъединиц HBcAg приводит к образованию высокомолекулярных частиц неустановленного размера, которые не могут быть разделены с помощью электрофореза в полиакриламидном геле с ДСН.
При связывании антигена или антигенной детерминанты с неприродным молекулярным каркасом посредством лизинового остатка может оказаться предпочтительной замена или делеция одного или обоих природных лизиновых остатков, локализованных в положениях, соответствующих положениям 7 и 96 SEQ ID NO:134, а также других лизиновых остатков, присутствующих в вариантах HBcAg. Элиминация этих лизиновых остатков приводит к удалению сайтов связывания с антигенами или антигенными детерминантами, что может приводить к разрушению упорядоченного массива и должно приводить к улучшению качества и однородности конечной вакцинной композиции.
Во многих случаях при элиминации обоих природных лизиновых остатков в положениях, соответствующих положениям 7 и 96 SEQ ID NO:134, в HBcAg может быть введен другой лизин в качестве сайта связывания для антигена или антигенной детерминанты. Способы введения такого лизинового остатка описаны, например, ниже в примере 23. При этом часто оказывается преимущественным вводить лизиновый остаток в HBcAg, если, например, оба природных лизиновых остатка в положениях, соответствующих положениям 7 и 96 в SEQ ID NO:134, были модифицированы, при этом антиген или антигенную детерминанту следует присоединить к неприродному молекулярному каркасу с использованием гетеробифункционального сшивающего агента.
Было показано, что С-конец HBcAg обеспечивает ядерную локализацию этого белка (Eckyardt et al., J. Virol. 65:575-583 (1991)). Кроме того, эта область белка также, вероятно, сообщает HBcAg способность связываться с нуклеиновыми кислотами.
В некоторых вариантах осуществления изобретения вакцинные композиции настоящего изобретения могут содержать HBcAg, обладающие активностью связываться с нуклеиновой кислотой (например, содержащие природный домен связывания HBcAg с нуклеиновой кислотой). HBcAg, содержащие один или несколько доменов связывания с нуклеиновой кислотой, могут быть использованы для получения вакцинных композиций, обладающих повышенной активностью, стимулирующей Т-клетки. Так, например, вакцинными композициями настоящего изобретения являются композиции, которые содержат HBcAg, обладающие активностью связывания с нуклеиновой кислотой. Кроме того, рассматриваются вакцинные композиции, а также использование таких композиций в схемах вакцинации, где HBcAg связываются с нуклеиновыми кислотами. Эти HBcAg могут быть связаны с нуклеиновыми кислотами до или после их введения индивидууму.
В других вариантах осуществления изобретения вакцинные композиции настоящего изобретения могут содержать HBcAg, из которых была удалена С-концевая область (например, аминокислотные остатки 145-185 или 150-185 SEQ ID NO:134), и которые не связываются с нуклеиновыми кислотами. Таким образом, другими модифицированными HBcAg, подходящими для осуществления настоящего изобретения, являются мутанты с С-концевым усечением. Подходящими усеченными мутантами являются HBcAg, у которых из их С-конца были удалены 1, 5, 10, 15, 20, 25, 30, 34, 35, 36, 37, 38, 39, 40, 41, 42 или 48 аминокислот.
HBcAg, подходящими для осуществления настоящего изобретения, являются мутанты с N-концевым усечением. Подходящими усеченными мутантами являются HBcAg, у которых из их N-конца были удалены 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот.
Другими HBcAg, подходящими для осуществления настоящего изобретения, являются мутанты с N- и С-концевым усечением. Подходящими усеченными мутантами являются HBcAg, у которых из их N-конца были удалены 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот, а из их С-конца были удалены 1, 5, 10, 15, 20, 25, 30, 34, 35, 36, 37, 38, 39, 40, 41, 42 или 48 аминокислот.
Настоящее изобретение также относится к вакцинным композициям, содержащим полипептиды HBcAg, включающие или, альтернативно, состоящие из них, аминокислотные последовательности, которые, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентичны аминокислотным последовательностям вышеописанных усеченных мутантов.
Как обсуждалось выше, в некоторых вариантах осуществления изобретения, в полипептид, который образует неприродный молекулярный каркас, вводят лизиновый остаток в качестве первого сайта связывания. В предпочтительных вариантах осуществления изобретения вакцинные композиции получают с использованием HBcAg, содержащего или, альтернативно, состоящего из них, аминокислоты 1-144 или аминокислоты 1-149 SEQ ID NO:134, которые были модифицированы путем замены аминокислот, соответствующих положениям 79 и 80, пептидом, имеющим аминокислотную последовательность Gly-Gly-Lys-Gly-Gly (SEQ ID NO:158), и либо путем делеции цистеиновых остатков в положениях 48 и 107, либо путем их замены другими аминокислотными остатками с сохранением цистеина в положении 61. Кроме того, настоящее изобретение относится к вакцинными композициям, содержащим соответствующие фрагменты полипептидов, имеющих аминокислотные последовательности, представленные в любой из SEQ ID NO:89-132 и 135-136, которые также имеют вышеуказанные аминокислотные модификации.
Настоящее изобретение также относится к вакцинным композициям, содержащим фрагменты HBcAg, включающие или, альтернативно, состоящие из них, аминокислотную последовательность, отличающуюся от последовательности, представленной в SEQ ID NO:134, из которой был делетирован цистеиновый остаток, отсутствующий в соответствующем положении SEQ ID NO:134. Примером такого фрагмента может служить полипептид, содержащий или, альтернативно, состоящий из них, аминокислоты 1-149 SEQ ID NO:132, где цистеиновый остаток в положении 147 либо был заменен другим аминокислотным остатком, либо был делетирован.
Кроме того, настоящее изобретение относится к вакцинным композициям, содержащим полипептиды HBcAg, включающие или, альтернативно, состоящие из них, аминокислотные последовательности, которые, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентичны аминокислотам 1-144 или 1-149 SEQ ID NO:134, и соответствующие подпоследовательности полипептида, содержащего аминокислотную последовательность, представленную в любой из SEQ ID NO:89-132 или 134-136, а также аминокислоты 1-147 или 1-152 SEQ ID NO:158.
Настоящее изобретение также относится к вакцинным композициям, содержащим полипептиды HBcAg, включающие или, альтернативно, состоящие из них, аминокислотные последовательности, которые, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентичны аминокислотам 36-240, 36-269, 44-240, 44-269, 36-305 и 44-305 SEQ ID NO:137 или 36-240, 36-269, 44-240, 44-269, 36-305 и 44-305 SEQ ID NO:138.
Вакцинные композиции настоящего изобретения могут содержать смеси различных HBcAg. Таким образом, эти вакцинные композиции могут состоять из HBcAg, которые отличаются по своим аминокислотным последовательностям. Так, например, могут быть получены вакцинные композиции, содержащие HBcAg "дикого типа" и модифицированный HBcAg, в котором один или несколько аминокислотных остатков были модифицированы (например, делетированы, инсертированы или заменены). Однако в большинстве случаев может быть использован один тип HBcAg или, по меньшей мере, несколько HBcAg, имеющих, в основном, эквивалентные первые сайты связывания, поскольку вакцины, полученные с использованием таких HBcAg, будут представлять в высокой степени упорядоченные и повторяющиеся массивы антигенов или антигенных детерминант. Кроме того, предпочтительными вакцинными композициями настоящего изобретения являются композиции, которые содержат в высокой степени упорядоченные и повторяющиеся массивы антигенов или антигенных детерминант.
Настоящее изобретение также относится к вакцинными композициям, в которых неприродный молекулярный каркас получают с использованием HBcAg, присоединенного к другому белку. Как обсуждалось выше, примером такого гибридного белка является гибрид HBcAg/FOS. Другими примерами гибридных белков HBcAg, подходящих для использования в вакцинных композициях настоящего изобретения, являются гибридные белки, в которые была добавлена аминокислотная последовательность, способствующая образованию и/или стабилизации димеров и мультимеров HBcAg. Эта дополнительная аминокислотная последовательность может быть присоединена к N- или С-концу HBcAg. Одним из примеров такого гибридного белка является гибрид HBcAg со спиральной областью GСN4 Saccharomyces cerevisiae (Genbank, рег.№Р03069 (SEQ ID NO:154)).
Домен спирали белка GСN4 образует гомодимеры посредством нековалентных взаимодействий, которые могут быть использованы для получения и стабилизации димеров и мультимеров HBcAg.
В одном из вариантов своего осуществления настоящее изобретение относится к вакцинным композициям, полученным с использованием гибридных белков HBcAg, содержащих HBcAg или его фрагмент, и полипептид GСN4, имеющий последовательность из аминокислотных остатков 227-276 в SEQ ID NO:154, присоединенных к С-концу. Этот полипептид GСN4 может быть также присоединен к N-концу HBcAg.
Для получения вакцинных композиций настоящего изобретения могут быть также использованы гибридные белки с гомологичным доменом 3 (SH3) HBcAg/src. Домены SH3 представляют собой относительно небольшие домены, присутствующие в ряде белков, которые сообщают способность к взаимодействию со специфическими богатыми пролином последовательностями у партнеров по связыванию с белком (см. McPherson, Cell Signal, 11:229-238 (1999). Гибридные белки HBcAg/SH3 могут быть использованы несколькими способами. Во-первых, домен SH3 может образовывать первый сайт связывания, который взаимодействует со вторым сайтом связывания антигена или антигенной детерминанты. Аналогичным образом, богатая пролином аминокислотная последовательность может быть присоединена к HBcAg и использована в качестве первого сайта связывания для домена SH3, взаимодействующего со вторым сайтом связывания антигена или антигенной детерминанты. Во-вторых, домен SH3 может связываться с богатыми пролином областями, введенными в HBcAg. Таким образом, домены SH3 и богатые пролином сайты взаимодействия SH3 могут быть введены либо в одинаковые, либо в различные HBcAg и использованы для формирования и стабилизации димеров и мультимеров настоящего изобретения.
В других вариантах осуществления изобретения для получения вакцинных композиций настоящего изобретения используется бактериальный пилин, подпоследовательность бактериального пилина или гибридный белок, содержащий либо бактериальный пилин, либо его подпоследовательность. Примерами белков пилинов являются пилины, продуцируемые Escherichia coli, Haemophilus influenzae, Neisseria meningitidis, Neisseria gonorrhoeae, Caulobacter crescentus, Pseudomonas stutzeri и Pseudomonas aeruginosa. Аминокислотными последовательностями белка пилина, подходящими для использования в настоящем изобретении, являются последовательности, представленные в документах GenBank AJ000636 (SEQ ID NO:139), AJ132364 (SEQ ID NO:140), AF229646 (SEQ ID NO:141), AF051814 (SEQ ID NO:142), AF051815 (SEQ ID NO:143) и Х00981 (SEQ ID NO:155), полное описание которых вводится в настоящее изобретение посредством ссылки.
Бактериальные белки пилины обычно процессируются с удалением N-концевых лидерных последовательностей с последующим экспортом этих белков в бактериальную периплазму. Кроме того, для каждого специалиста очевидно, что природные бактериальные белки пилины, используемые для получения вакцинных композиций настоящего изобретения, обычно не имеют лидерной последовательности.Одним из конкретных примеров белка пилина, подходящего для использования в настоящем изобретении, является Р-пилин E.coli (документ GenBank AF237482 (SEQ ID NO:144)). Примером пилина типа 1 E.coli, подходящего для использования в настоящем изобретении, является пилин, имеющий аминокислотную последовательность, депонированную в Genbank под названием Р04128 (SEQ ID NO:146) и кодируемую нуклеиновой кислотой, имеющей нуклеотидную последовательность, депонированную в Genbank под названием М27603 (SEQ ID NO:145). Полное описание этих данных Genbank вводится в настоящее описание посредством ссылки. И в этом случае зрелая форма вышеуказанного известного белка может быть, в основном, использована для получения вакцинных композиций настоящего изобретения.
Бактериальные пилины или их подпоследовательности, подходящие для осуществления настоящего изобретения, в основном, способны связываться с образованием неприродных молекулярных каркасов.
Методы приготовления пилей и пилеподобных структур in vitro известны специалистам. Так, например, в работе Bullitt et al., Proc. Natl. Acad. Sci. USA, 93:12890-12895 (1996) описано in vitro реконструирование субъединиц Р-пилей E.coli. Кроме того, в работе Eschdat et al., J. Bacteriol. 148:308-314 (1981) описаны методы, подходящие для диссоциации пилей типа 1 E.coli, и реконструирование пилей. Вкратце, эти методы заключаются в следующем: пили подвергают диссоциации путем инкубирования при 37°С в насыщенном гидрохлориде гуанидина. Затем белки пилины очищают хроматографией, после чего получают димер пилина путем диализа против 5 мМ гидрохлорида трис(гидроксиметил)аминометана (рН 8,0). Eschdat и др. также обнаружили, что после диализа против 5 мМ трис(гидроксиметил)аминометана (рН 8,0), содержащего 5 мм MgCl2, димеры пилина подвергаются повторной сборке с образованием пилей.
Кроме того, с использованием, например, стандартной техники генной инженерии и методов модификации белков белки пилины могут быть модифицированы так, чтобы они содержали первый сайт связывания, с которым связывается антиген или антигенная детерминанта посредством второго сайта связывания. Альтернативно, антигены или антигенные детерминанты могут быть непосредственно связаны посредством второго сайта связывания с аминокислотными остатками, которые обычно присутствуют в этих белках. Затем эти модифицированные белки пилины могут быть использованы в вакцинных композициях настоящего изобретения.
Бактериальные белки пилины, используемые для получения вакцинных композиций настоящего изобретения, могут быть модифицированы тем же способом, который был описан здесь для HBcAg. Так, например, цистеиновые и лизиновые остатки могут быть либо делетировны, либо заменены другими аминокислотными остатками, и к этим белкам могут быть присоединены первые сайты связывания. Затем белки пилины могут либо экспрессированы в модифицированной форме, либо химически модифицированы после экспрессии. Аналогичным образом, интактные пили могут быть выделены из бактерий, а затем химически модифицированы.
В другом варианте осуществления изобретения пили или пилеподобные структуры выделяют из бактерий (например, E.coli) и используют для получения вакцинных композиций настоящего изобретения. Одним из примеров пилей, подходящих для получения вакцинных композиций, являются пили типа 1 E.coli, которые состоят из мономеров пилина, имеющего аминокислотную последовательность, представленную в SEQ ID NO:146.
Для сбора бактериальных пилей существует ряд методов, известных специалистам. Bullitt & Makowski (Biophys. J. 64:623-632 (1998)), например, описывают метод очистки Р-пилей, полученных из E.coli. В соответствии с этим методом пили выделяли из E.coli, несущей большое количество пилей и содержащей плазмиду Р-пилуса, и очищали путем солюбилизации и преципитации с использованием MgCl2 (1,0М). Аналогичный метод очистки описан ниже в примере 33.
После сбора пили или пилеобразные структуры могут быть модифицированы различными способами. Так, например, первый сайт связывания может быть присоединен к пилусу, к которому могут быть присоединены антигены или антигенные детерминанты через второй сайт связывания. Другими словами, бактериальные пили или пилеобразные структуры могут быть собраны и модифицированы с образованием неприродных молекулярных каркасов.
Пили или пилеобразные структуры могут быть модифицированы путем связывания антигенов или антигенных детерминант в отсутствии неприродного организатора. Так, например, антигены или антигенные детерминанты могут быть связаны с природными цистеиновыми остатками или с лизиновыми остатками. В таких случаях большое количество и повторяемость природного аминокислотного остатка должно определять связывание антигенов или антигенных детерминант с пилями или пилеобразными структурами. Так, например, пили или пилеобразные структуры могут быть связаны со вторыми сайтами связывания указанных антигенов или антигенных детерминант с использованием гетеробифункционального сшивающего агента.
Если для получения вакцинных композиций настоящего изобретения используются структуры, которые в природе синтезируются микроорганизмами (например, пили), то, в большинстве случаев, эти микроорганизмы должны быть преимущественно сконструированы так, чтобы они продуцировали структуры, имеющие нужные свойства. Так, например, если используются пили типа 1 E.coli, то E.coli, из которых собирают эти пили, могут быть модифицированы так, чтобы они продуцировали структуры, имеющие конкретные свойства. Примерами возможных модификаций белков пилинов являются введение одного или нескольких лизиновых остатков, делеция или замена одного или несколько природных лизиновых остатков и делеция или замена одного или несколько природных цистеиновых остатков (например, цистеиновых остатков в положениях 44 и 84 в SEQ ID NO:146).
Кроме того, в гены пилина могут быть внесены дополнительные модификации, которые приводят к экспрессии продуктов, содержащих первый сайт связывания, не являющийся лизиновым остатком (например, домен FOS или JUN). Само собой разумеется, что подходящие первые сайты связывания ограничены, в основном, такими сайтами, которые не предотвращают образования белками пилинами пилей или пилеподобных структур, подходящих для использования в вакцинных композициях настоящего изобретения.Гены пилины, которые обычно присутствуют в бактериальных клетках, могут быть модифицированы in vivo (например, путем гомологичной рекомбинации), либо в эти клетки могут быть введены гены пилинов с конкретными свойствами. Так, например, гены пилины могут быть введены в бактериальные клетки как компонент либо репликативного клонирующего вектора, либо вектора, который встраивается в бактериальную хромосому. Встроенные гены пилина могут быть также присоединены к экспрессирующим регуляторным последовательностям (например, к оператору lac).
В большинстве случаев пили или пилеподобные структуры, используемые в вакцинных композициях настоящего изобретения, могут состоять из субъединицы пилина одного типа. Обычно используются пили или пилеподобные структуры, состоящие из идентичных субъединиц, поскольку предполагается, что они образуют структуры, которые представляют собой в высокой степени упорядоченные и повторяющиеся массивы антигенов.
Однако композициями настоящего изобретения также являются вакцины, содержащие пили или пилеподобные структуры, образованные из гетерогенных субъединиц пилина. Эти субъединицы пилина, которые образуют эти пили или пилеподобные структуры, могут быть экспрессированы из генов, обычно присутствующих в бактериальной клетке, либо они могут быть введены в эти клетки. Если в клетке экспрессируются как природный ген пилина, так и введенный ген пилина, то в этом случае обычно будут образовываться структуры, состоящие из смеси этих белков пилинов. Кроме того, если в бактериальной клетке экспрессируются два или более генов пилинов, то относительная экспрессия каждого гена пилина обычно может служить фактором, который определяет отношение различных субъединиц пилина в пилях или пилеподобных структурах.
Если необходимо использовать пили или пилеподобные структуры, имеющие конкретный состав из смешанных субъединиц пилина, то экспрессия, по меньшей мере, одного из генов пилина может регулироваться гетерологичным индуцибельным промотором. Такие промоторы, а также другие генетические элементы могут быть использованы для регуляции относительных количеств различных субъединиц пилина, продуцируемых в бактериальной клетке, а следовательно, и регуляции состава пилей или пилеподобных структур.
Кроме того, хотя в большинстве случаев антиген или антигенная детерминанта могут связываться с бактериальными пилями или с пилеподобными структурами непептидной связью, однако бактериальные клетки, которые продуцируют пили или пилеподобные структуры, используемые в композициях настоящего изобретения, могут быть генетически сконструированы так, чтобы они генерировали белки пилины, которые присоединяются к антигену или к антигенной детерминанте. Такие гибридные белки, которые образуют пили или пилеподобные структуры, могут быть использованы в вакцинных композициях настоящего изобретения.
Как уже обсуждалось выше, вирусные капсиды могут быть использованы (1) для презентации антигена или антигенных детерминант и (2) для получения вакцинных композиций настоящего изобретения. В частности, для осуществления настоящего изобретения могут быть использованы вирусные капсидные белки, также называемые здесь "оболочечными белками", которые после экспрессии образуют капсиды или капсидоподобные структуры. Таким образом, эти капсидные белки могут образовывать коровые частицы и неприродные молекулярные каркасы. Обычно эти капсиды или капсидоподобные структуры образуют упорядоченные и повторяющиеся массивы, которые могут быть использованы для презентации антигенов или антигенных детерминант и для получения вакцинных композиций настоящего изобретения.
Один или несколько (например, один, два, три, четыре, пять и т.п.) антигенов или антигенных детерминант могут быть присоединены любыми способами к одному или нескольким (например, одному, двум, трем, четырем, пяти и т.п.) белкам, которые образуют вирусные капсиды или капсидоподобные структуры (например, оболочечные белки бактериофага), а также другие белки. Так, например, антигены или антигенные детерминанты могут быть присоединены к коровым частицам с использованием первого и второго сайтов связывания. Кроме того, один или несколько (например, один, два, три, четыре, пять и т.п.) гетеробифункциональных сшивающих линкеров могут быть использованы для присоединения антигенов или антигенных детерминант к одному или нескольким белкам, образующим вирусные капсиды или капсидоподобные структуры.
Вирусные капсидные белки или их фрагменты могут быть использованы, например, для получения коровых частиц и вакцинных композиций настоящего изобретения. Оболочечные белки бактериофага Qβ могут быть, например, рекомбинантно экспрессированы в E.coli. Кроме того, после такой экспрессии эти белки спонтанно образуют капсиды. Более того, эти капсиды образуют упорядоченные и повторяющиеся массивы антигенов или антигенных детерминант, которые могут быть использованы для презентации антигена и для получения вакцинных композиций. Как описано ниже в примере 38, оболочечные белки бактериофага Qβ могут быть использованы для получения вакцинных композиций, которые вырабатывают иммунологические ответы на антигенные детерминанты.
Конкретными примерами оболочечных белков бактериофага, которые могут быть использованы для получения композиций настоящего изобретения, являются оболочечные белки РНК-бактериофагов, таких как бактериофаг Qβ (SEQ ID NO:159; база данных PIR, регистрационный номер VCBPQβ, относящийся к Qβ СР и SEQ ID NO:217; регистрационный номер AAA16663, относящийся к белку Qβ А1), бактериофаг R17 (SEQ ID NO:160; PIR, регистрационный номер VCBPR7), бактериофаг fr (SEQ ID NO:161; PIR, регистрационный номер VCBPFR), бактериофаг GA (SEQ ID NO:162; Genbank, регистрационный номер NP-040754), бактериофаг SP (SEQ ID NO:163; Genbank, регистрационный номер САА30374, относящийся к SP СР и SEQ ID NO:254; регистрационный номер, относящийся к белку SP А1), бактериофаг MS2 (SEQ ID NO:164; PIR, регистрационный номер VCBPМ2), бактериофаг М11 (SEQ ID NO:165; Genbank, регистрационный номер ААC06250), бактериофаг МХ1 (SEQ ID NO:166; Genbank, регистрационный номер ААC14699), бактериофаг NL95 (SEQ ID NO:167; Genbank, регистрационный номер ААC14704), бактериофаг f2 (SEQ ID NO:215; Genbank, регистрационный номер Р03611), бактериофаг РР7 (SEQ ID NO:253). Каждому специалисту очевидно, что для получения вакцинных композиций настоящего изобретения может быть использован любой белок, который образует капсиды или капсидоподобные структуры. Кроме того, белок А1 бактериофага Qβ или его усеченные по С-концу формы, у которых в С-конце отсутствует до 100, 150 или 180 аминокислот, могут быть введены в капсидный комплекс оболочечных белков Qβ. Белок А1 может быть также присоединен к организатору, а следовательно, и к первому сайту связывания для присоединения антигенов, содержащих второй сайт связывания. Обычно процентное количество белка А1 по отношению к Qβ СР в капсидном комплексе, должно быть ограничено для гарантии образования капсида. Белок А1 имеет регистрационный номер ААА16663 (SEQ ID NO:217).
Было также обнаружено, что оболочечный белок Qβ, при его экспрессии в E.coli, способен к самосборке в капсиды (Kozlovska T.M. et al., GENE, 137:133-137 (1993)). Полученные капсиды или вирусоподобные частицы обнаруживают икосаэдрическую фагоподобную капсидную структуру, имеющую диаметр 25 нм и квазисимметрию Т=3. Кроме того, была установлена кристаллическая структура фага Qβ. Капсид этого фага содержит 180 копий оболочечного белка, которые соединены в ковалентные пентамеры и гексамеры дисульфидными мостиками (Golmohammadi R. et al., Structure, 4:543-5554 (1996)). Было также показано, что другие оболочечные белки РНК-фага способны к самосборке после экспрессии в бактериальном хозяине (Kastelein R.A. et al., Gene, 23:245-254 (1983), Kozlovskaya T.M. et al., Dokl. Akad. Nauk SSSR, 287: 452-455 (1986), Adhin M.R. et al., Virology, 170:238-242 (1989), Ni, CZ et al., Protein Sci. 5:2485-2493 (1996), Priano C. et al., J. Mol. Biol. 249:283-297 (1995)). Капсид фага Qβ, помимо оболочечного белка, содержит так называемый белок с "прочитанным" терминатором, А1, и белок созревания А2. А1 генерируется путем супрессии в стоп-кодоне UGA и имеет длину 329 аминокислот. Капсид фага рекомбинантного оболочечного белка фага Qβ, используемый в настоящем изобретении, не содержит белка лизина А2 и содержит РНК данного хозяина. Оболочечный белок РНК-фагов представляет собой РНК-связывающий белок и взаимодействует со структурой "стебель-петля" сайта связывания с рибосомой гена репликазы, действующего как трансляционный репрессор жизненного цикла вируса. Последовательность и структурные элементы такого взаимодействия известны специалистам (Witherell, G.W. & Uhlenbeck O.C. Biochemistry, 28:71-76 (1989); Lim F. et al. J. Biol. Chem. 271:31839-31845 (1996)). В основном, известно, что структура "стебель-петля" и РНК участвуют в сборке вируса (Golmohammadi R. et al., Structure, 4:543-5554 (1996)).
Белки или домены белка могут влиять на структуру и сборку частицы даже больше, чем короткий пептид. Так, например, "правильная" укладка антигенов, содержащих дисульфидные мостики, обычно невозможна в цитоплазме E.coli, где экспрессируются частицы Qβ. Аналогичным образом, гликозилирование обычно невозможно в прокариотических экспрессирующих системах. А поэтому преимущество рассматриваемого и описанного здесь изобретения заключается в том, что антиген присоединяют к частице, начиная с уже собранной частицы и выделенного антигена. Это позволяет осуществлять экспрессию частицы и антигена в экспрессирующем хозяине, что гарантирует "правильную" укладку антигена и правильную укладку и сборку частицы.
Преимущество настоящего изобретения заключается в том, что один или несколько антигенных молекул могут быть присоединены к одной субъединице капсидного оболочечного белка РНК-фагов. Конкретным отличительным признаком капсидного оболочечного белка РНК-фагов, а в частности капсида Qβ, является его способность связываться с несколькими антигенами на одну субъединицу. Это позволяет генерировать плотный массив антигенов. Другие вирусные капсиды, используемые для ковалентного присоединения антигенов посредством химического сшивания, таких как, например, HBcAg, модифицированные лизиновым остатком в его основной иммунодоминантной области (MIR; WO 00/32227), дают максимальную плотность связывания 0,5 антигенов на субъединицу. Такое расстояние между шипами (соответствующими MIR) HBcAg составляет 50 Е (Wynne S.A. et al., Mol. Cell. 3:771-780 (1999)), а поэтому массив антигенов с расстояниями короче чем 50 Е не может быть генерирован.
Капсиды из оболочечного белка Qβ имеют определенное число лизиновых остатков на своей поверхности с определенной топологией, где три лизиновых остатка локализованы ближе к внутренней части капсида и взаимодействуют с РНК, а четыре других лизиновых остатка расположены на поверхности этого капсида. Эти характерные свойства благоприятствуют связыванию антигенов с внешней поверхностью частицы, но не с внутренней ее частью, где лизиновые остатки взаимодействуют с РНК. Капсиды из других оболочечных белков РНК-фага также имеют определенное число лизиновых остатков на своей поверхности и определенную топологию этих лизиновых остатков. Другим преимуществом капсидов, происходящих от РНК-фагов, является их высокий уровень экспрессии в бактериях, что позволяет продуцировать большое количество материала при приемлемых метериальных затратах.
Другим отличительным признаком капсида из оболочечных белков фага Qβ является его стабильность. Субъединицы Qβ присоединены друг к другу дисульфидными мостиками, ковалентно связывающими эти субъединицы. Капсидный белок Qβ также обнаруживает необычную резистентность к органическим растворителям и к денатурирующим агентам. Авторами настоящего изобретения было неожиданно обнаружено, что ДМСО и ацетонитрил, используемые в концентрации до 30%, и гуанидиний, используемый в концентрации до 1М, не оказывают какого-либо воздействия на стабильность или способность капсида к образованию антигенных массивов. Таким образом, эти органические растворители могут быть использованы для связывания с гидрофобными пептидами. Высокая стабильность капсида, состоящего из оболочечных белков Qβ, является важным признаком, позволяющим использовать его для иммунизации и вакцинации млекопитающих и, в частности, человека. Резистентность капсида к органическому растворителю позволяет осуществлять связывание с антигенами, которые являются нерастворимыми в водных буферах.
Введение цистеинового остатка в N-концевую β-шпильку оболочечного белка РНК-фага MS-2 описано в патентной заявке США №5698424. Однако авторы настоящего изобретения отмечают, что присутствие свободных цистеиновых остатков на поверхности капсида могут приводить к олигомеризации капсидов посредством образования дисульфидного мостика. Другим типом связывания, рассматриваемым в патентной заявке США 5698424, является образование дисульфидных мостиков между антигеном и частицей Qβ. Такое связывание является лабильным для молекул, содержащих сульфгидрильную группу.
Реакция между исходным дисульфидным мостиком, образованным цис-остатком на Qβ и антигеном, содержащим свободный сульфгидрильный остаток, приводит к высвобождению сульфгидрилсодержащих молекул, не являющихся антигеном. Эти новообразованные сульфгидрилсодержащие молекулы могут снова реагировать с другими дисульфидными мостиками, присутствующими на этой частице, в результате чего устанавливается равновесие. После реакции с дисульфидным мостиком, образованным на этой частице, антиген может либо образовывать дисульфильный мостик с цис-остатком этой частицы или с цис-остатком молекулы уходящей группы, которая была образована исходным дисульфидным мостиком на этой частице. Кроме того, описанный другой метод присоединения с использованием гетеробифункционального сшивающего линкера, реагирующего с цистеином частицы Qβ на одной своей стороне и с лизиновым остатком на антигене на другой своей стороне, приводит к случайной ориентации антигенов на частице.
Авторы настоящего изобретения отмечают, что в отличие от капсида, состоящего из оболочечных белков Qβ и Fr, рекомбинантный MS-2, описанный в патентной заявке США 5698424, в основном, не содержит нуклеиновых кислот, а его РНК упакована внутри двух капсидов, упомянутых выше.
Авторы настоящего изобретения описывают новые и обладающие признаками изобретения композиции, позволяющие образовывать жесткие антигенные массивы с различной плотностью антигенов. Авторы настоящего изобретения показали, что может быть достигнута более высокая плотность эпитопов, чем плотность, которая обычно достигается с использованием других VLP. Авторы также описывают композиции с одновременным представлением нескольких антигенов, расположенных на соответствующих расстояниях друг от друга, и композиции, в которые может оказаться желательным добавление вспомогательных молекул, улучшающих растворимость, или модифицирующих капсид.
В настоящей заявке описано получение композиций капсидов, состоящих из оболочечных белков РНК-фага с высокой плотностью эпитопов. Каждому специалисту хорошо известно, что условия сборки упорядоченного и повторяющегося массива антигенов зависит от наличия подходящего участка на антигене и от выбора второго сайта связывания на этом антигене. В случае отсутствия подходящего второго сайта связывания такой второй сайт связывания с антигеном должен быть сконструирован.
Для конструирования второго сайта связывания необходимо выбрать положение, в котором он должен быть присоединен, инсертирован или генетически сконструирован. Каждому специалисту должно быть известно, как можно выбрать положение второго сайта связывания. Кристаллическая структура этого антигена может дать информацию относительно доступности С- или N-концов данной молекулы (определенной, например, исходя из их доступности для растворителя) или относительно доступности для растворителя остатков, подходящих для использования в качестве вторых сайтов связывания, таких как цистеиновые остатки. Доступные для взаимодействия дисульфидные мостики, как и в случае Fab-фрагментов, могут также служить источником для второго сайта связывания, поскольку они могут быть, в основном, превращены только в одиночные цистеиновые остатки посредством восстановления в мягких условиях. Вообще говоря, в случае, когда целью иммунизации аутоантигеном является ингибирование взаимодействия этого аутоантигена с его природными лигандами, то может быть присоединен второй сайт связывания так, чтобы он обеспечивал генерирование антител против сайта взаимодействия с природными лигандами. Таким образом, локализацию второго сайта связывания выбирают так, чтобы можно было избежать стерического затруднения, создаваемого этим вторым сайтом связывания или любым аминокислотным линкером, содержащим этот сайт. В других вариантах осуществления изобретения желательно стимулировать гуморальный ответ, направленный на сайт, отличающийся от сайта взаимодействия аутоантигена с его природным лигандом. В таких вариантах осуществления изобретения второй сайт связывания может быть выбран так, чтобы он предотвращал вырабатывание антител против сайта взаимодействия с его природными лигандами.
Другими критериями при выборе положения второго сайта связывания является состояние олигомеризации антигена, сайта олигомеризации, присутствие кофактора и доступность сайтов в антигенной структуре, определяемая по экспериментальным данным, и последовательность, в которой модификация антигена является совместимой с функцией аутоантигена или с генерированием антител, распознающих аутоантиген.
В некоторых вариантах осуществления изобретения для конструирования второго сайта связывания на антигене требуется его присоединение к аминокислотному линкеру, содержащему аминокислоту, подходящую для использования в качестве второго сайта связывания, в соответствии с описанием настоящего изобретения. В предпочтительном варианте осуществления изобретения указанной аминокислотой является цистеин. Выбор аминокислотного линкера зависит от природы аутоантигена, от его биохимических свойств, таких как pI, распределение заряда и гликозилирование. В основном, предпочтительно использовать гибкие аминокислотные линкеры. Примерами аминокислотных линкеров являются шарнирная область иммуноглобулинов, глицин-сериновые линкеры (GGGGS)n и глициновые линкеры (G)n, каждый из которых, кроме того, содержат цистеиновый остаток в качестве второго сайта связывания и необязательно дополнительные глициновые остатки. (Ниже представлены примеры указанных аминокислотных линкеров:
N-концевой гамма 1: CGDKTHTSPP
С-концевой гамма 1: DKTHTSPPCG
N-концевой гамма 3: CGGPKPSTPPGSSGGAP
С-концевой гамма 3: PKPSTPPGSSGGAPGGCG
N-концевой глициновый линкер: GCGGGG
С-концевой глициновый линкер: GGGGCG)
Было показано, что для пептидов подходящими являются линкеры GGCG у С-конца этого пептида или CGG у его N-конца. В основном, глициновые остатки встраивают между объемными аминокислотами и цистеином, используемым в качестве второго сайта связывания.
Особенно предпочтительным методом связывания антигенов с VLP, а в частности с капсидом из оболочечных белков РНК-фага, является связывание лизинового остатка на поверхности капсида из оболочечных белков РНК-фага с цистеиновым остатком на антигене. Для эффективного действия в качестве второго сайта связывания сульфгидрильная группа должна быть доступной для такого связывания. Так, например, цистеиновый остаток должен присутствовать в своем восстановленном состоянии, то есть в виде свободного цистеина или цистеинового остатка со свободной сульфгидрильной группой. В случае, когда цистеиновый остаток, предназначенный для использования в качестве второго сайта связывания, находится в окисленной форме, например, если он образует дисульфидный мостик, то необходимо восстановление этого дисульфидного мостика, например, с помощью ДТТ, ТСЕР или β-меркаптоэтанола.
Отличительной особенностью настоящего изобретения является то, что плотность эпитопов на капсиде из оболочечных белков РНК-фага может быть модулирована путем выбора сшивающего линкера и других реакционных условий. Так, например, сшивающие линкеры сульфо-GMBS и SMPH позволяют достигать более высокой плотности эпитопов, чем сшивающий линкер сульфо-MBS в одних и тех же реакционных условиях. Для регуляции числа антигенов, связанных с капсидными белками РНК-фагов, а в частности с капсидным белком Qβ, может быть осуществлена дериватизация, на которую позитивно влияет высокая концентрация реагентов и изменение реакционных условий.
Исходя из теоретических расчетов, максимально достижимое число антигенов глобулярных белков размером 17 кДа не превышает 0,5. Таким образом, несколько лизиновых остатков капсидных оболочечных белков Qβ могут быть дериватизированы сшивающей линкерной молекулой, еще не содержащей антигена. Это приводит к элиминации положительного заряда, что может оказывать негативное влияние на растворимость и стабильность этого конъюгата. Путем замены некоторых лизиновых остатков аргининами, например в случае описанного мутантного оболочечного белка Qβ, авторы настоящего изобретения смогли предотвратить чрезмерное снижение количества положительного заряда, поскольку аргининовые остатки не реагируют со сшивающим линкером.
В других вариантах осуществления изобретения авторы описывают мутантный оболочечный белок Qβ с дополнительными лизиновыми остатками, подходящий для получения массивов антигенов высокой плотности.
Была определена кристаллическая структура некоторых РНК-бактериофагов (Golmohammadi, R. et al., Structure, 4:543-5554 (1996)). С использованием такой информации каждый специалист может легко идентифицировать находящиеся на поверхности остатки и модифицировать оболочечные белки бактериофага так, чтобы можно было вводить один или несколько реакционноспособных аминокислотных остатков. Таким образом, каждый специалист может легко генерировать и идентифицировать модифицированные формы оболочечных белков бактериофага, которые могут быть использованы в настоящем изобретении. Так, например, для получения вакцинных композиций настоящего изобретения могут быть также использованы варианты белков, которые образуют капсиды или капсидоподобные структуры (например, оболочечные белки бактериофага Qβ, бактериофага R17, бактериофага fr, бактериофага GA, бактериофага SP и бактериофага MS2).
Хотя последовательность белков-вариантов, обсуждаемых выше, будет отличаться от их аналогов дикого типа, однако эти белки-варианты обычно сохраняют способность образовывать капсиды или капсидоподобные структуры. Так, например, настоящее изобретение, кроме того, относится к вакцинным композициям, которые содержат варианты белков, образующих капсиды или капсидоподобные структуры, а также к способам получения таких вакцинных композиций, к отдельным субъединицам белка, используемым для получения таких вакцинных композиций, и к молекулам нуклеиновой кислоты, кодирующим эти субъединицы белка. Таким образом, в объем настоящего изобретения также входят варианты белков дикого типа, которые образуют упорядоченные и повторяющиеся массивы антигенов (например, варианты белков, образующих капсиды или капсидоподобные структуры) и которые сохраняют способность к связыванию и образованию капсидов или капсидоподобных структур.
В соответствии с этим настоящее изобретение, кроме того, относится к вакцинным композициям, включающим или, альтернативно, состоящим из них, аминокислотные последовательности, которые, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентичны белкам дикого типа и которые образуют упорядоченные массивы. Во многих случаях эти белки могут быть процессированы для удаления сигнальных пептидов (например, гетерологичных сигнальных пептидов).
Кроме того, в объем настоящего изобретения входят молекулы нуклеиновой кислоты, кодирующие белки, используемые для получения вакцинных композиций настоящего изобретения.
В конкретных вариантах своего осуществления настоящее изобретение, кроме того, относится к вакцинным композициям, включающим или, альтернативно, состоящим из них, аминокислотные последовательности, которые, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентичны любой из аминокислотных последовательностей, представленной в SEQ ID NO:159-167, и формы этих белков, которые были процессированы, если это необходимо, для удаления N-концевой лидерной последовательности.
Белками, подходящими для осуществления настоящего изобретения, также являются мутантные белки с усечением по С-концу, которые образуют капсиды или капсидоподобные структуры, а также другие упорядоченные массивы. Конкретными примерами таких усеченных мутантов являются белки, имеющие аминокислотную последовательность, представленную в любой из аминокислотных последовательностей SEQ ID NO:159-167, где из С-конца были удалены 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот. Обычно мутанты с усечением по С-концу, используемые для осуществления настоящего изобретения, могут сохранять способность образовывать капсиды или капсидоподобные структуры.
Другими белками, подходящими для использования в настоящем изобретении, также являются мутантные белки с усечением по N-концу, которые образуют капсиды или капсидоподобные структуры, а также другие упорядоченные массивы. Конкретными примерами таких усеченных мутантов являются белки, имеющие аминокислотную последовательность, представленную в любой из аминокислотных последовательностей SEQ ID NO:159-167, где из С-конца были удалены 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот. Обычно мутанты с усечением по N-концу, используемые для осуществления настоящего изобретения, могут сохранять способность образовывать капсиды или капсидоподобные структуры.
Другими белками, подходящими для использования в настоящем изобретении, также являются мутантные белки с усечением по N-концу и по С-концу, которые образуют капсиды или капсидоподобные структуры, а также другие упорядоченные массивы. Конкретными примерами таких мутантов с усечениями являются белки, имеющие аминокислотную последовательность, представленную в любой из аминокислотных последовательностей SEQ ID NO:159-167, где из N-конца были удалены 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот, а из С-конца были удалены 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот. Обычно мутанты с усечением по N- и С-концам, используемые для осуществления настоящего изобретения, могут сохранять способность образовывать капсиды или капсидоподобные структуры.
Настоящее изобретение также относится к вакцинным композициям, содержащим белки, включающие или, альтернативно, состоящие из них, аминокислотные последовательности, которые, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентичны аминокислотным последовательностям вышеописанных усеченных мутантов.
Настоящее изобретение также относится к вакцинным композициям, полученным из белков, которые образуют упорядоченные массивы; к способам получения вакцинных композиций из отдельных субъединиц белка; к способам получения этих отдельных субъединиц белка; к молекулам нуклеиновых кислот, кодирующим указанные субъединицы, и к способам вакцинации и/или индукции иммунных ответов у индивидуумов с использованием вакцинных композиций настоящего изобретения.
В. Конструирование антигенов или антигенных детерминант со вторым сайтом связывания
Вторым элементом в композиции настоящего изобретения является антиген или антигенная детерминанта, имеющие, по меньшей мере, один второй сайт связывания, способный ассоциироваться посредством, по меньшей мере, одной непептидной связи с первым сайтом связывания неприродного молекулярного каркаса. Настоящее изобретение относится к композициям, которые могут варьироваться в зависимости от антигена или антигенной детерминанты, выбранной в целях достижения желаемого терапевтического эффекта. Другие композиции варьируются в зависимости от молекул, выбранных для второго сайта связывания.
Однако, если для получения вакцинных композиций настоящего изобретения используются бактериальные пили или пилеподобные структуры, то есть белки пилины, то антигены или антигенные детерминанты могут быть присоединены к белкам пилинам посредством экспрессии гибридных белков "пилин/антиген". Аналогичным образом, если для получения вакцинных композиций настоящего изобретения используются белки, не являющиеся пилинами (например, вирусные капсидные белки), то антигены или антигенные детерминанты могут быть присоединены к этим непилиновым белкам посредством экспрессии гибридных белков "непилиновый белок/антиген". Антигены или антигенные детерминанты могут быть также присоединены к бактериальным пилям, пилеподобным структурам, белкам пилинам и к другим белкам, которые образуют упорядоченные массивы посредством непептидных связей.
Антигены настоящего изобретения могут быть выбраны из группы, состоящей из: (а) белков, подходящих для индуцирования иммунного ответа против злокачественных клеток; (b) белков, подходящих для индуцирования иммунного ответа против инфекционных заболеваний; (с) белков, подходящих для индуцирования иммунного ответа против аллергенов; (d) белков, подходящих для индуцирования иммунного ответа у сельскохозяйственных животных, и (е) фрагментов (например, домена) любого из белков, перечисленных в (а)-(d).
В одном из конкретных вариантов осуществления изобретения антигеном или антигенной детерминантой является антиген или антигенная детерминанта, которые могут быть использованы для профилактики инфекционного заболевания. Такие антигены или антигенные детерминанты могут быть использованы для лечения инфекционных заболеваний широкого ряда, поражающих широкий круг хозяев, например человека, коров, овец, свиней, собак, кошек и млекопитающих других видов, а также видов животных, не относящихся к млекопитающим. Подвергаемые лечению инфекционные заболевания хорошо известны специалистам, и их примерами являются инфекции вирусной этиологии, такие как ВИЧ, грипп, инфекции, вызываемые герпесвирусом, вирусом гепатита, вирусом Эпштейна-Барра и полиовирусом, вирусный энцефалит, корь, оспа кур и т.п.; или инфекции бактериальной этиологии, такие как пневмония, туберкулез, сифилис и т.п.; или инфекции, вызываемые паразитами, такие как малярия, трипаносомоз, лейшманиоз, трихомониаз, амебиаз и т.п. Таким образом, антигены или антигенные детерминанты, выбранные для композиций настоящего изобретения, хорошо известны специалистам-медикам, и примерами антигенов или антигенных детерминант могут служить: антигены gp140 и gp160 вируса ВИЧ; антиген гемагглютинин вируса гриппа; белок М2 и нейраминидаза; поверхностный антиген вируса гепатита В, белок кругового спорозоита малярии.
В конкретных вариантах своего осуществления настоящее изобретение относится к вакцинным композициям, подходящим для использования в способах предупреждения и/или ослабления симптомов заболевания или состояний, которые вызываются или усиливаются продуктами "своего" гена (например, факторы некроза опухоли). Так, например, вакцинными композициями настоящего изобретения являются композиции, обеспечивающие продуцирование антител, которые способствуют предупреждению и/или ослаблению симптомов заболеваний или состояний, вызываемых или усиливаемых продуктами "своего" гена. Примерами таких заболеваний или состояний являются реакция "трансплантат против хозяина", IgE-опосредованные аллергические реакции, анафилаксия, респираторный дистресс-синдром взрослых, болезнь Крона, аллергическая астма, острый лимфолейкоз (ALL), не-ходжскинская лимфома (НХЛ), болезнь Грейвса, системная красная волчанка (СКВ), воспалительные аутоиммунные заболевания, тяжелая миастения, иммунопролиферативная лимфаденопатия (IPL), ангиоиммунопролиферативная лимфаденопатия (AIL), иммунобластная лимфаденопатия (IBL), ревматоидный артрит, диабет, рассеянный склероз, болезнь Альцгеймера и остеопороз.
В родственных конкретных вариантах осуществления изобретения, композициями настоящего изобретения являются иммунотерапевтические композиции, которые могут быть использованы для лечения аллергий или злокачественных опухолей.
Выбор антигенов или антигенных детерминант для получения композиций и их использование в методах лечения аллергий могут быть осуществлены в соответствии с известной практикой, применяемой специалистами-медиками для лечения таких заболеваний. Репрезентативными примерами таких антигенов или антигенных детерминант являются: фосфолипаза A2 пчелиного яда, Bet v I (аллерген березовой пыльцы), 5 Dol m V (аллерген яда пятнистой осы), меллитин и Der p I (аллерген клеща домашней пыли), а также фрагменты каждого из них, которые могут быть использованы для вырабатывания иммунологических ответов.
Как указывалось выше, предпочтительным антигеном или антигенной детерминантой является Der p I. Der p I представляет собой протеазу в 25 кД, обнаруженную в частицах фекалий клеща домашней пыли. Der p I представляет собой главную аллергенную молекулу клеща домашней пыли. В соответствии с этим у 80% пациентов с аллергией к этому клещу вырабатываются антитела IgE против Der p I. В частности, известно, что пептиды р52-72 и р117-133 и др. содержат эпитопы, распознаваемые антителами против нативного Der p I. Антитела IgE вырабатываются при поликлональном ответе на высокоаффинное связывание целого антигена с пептидным участком 59-94 (L.Pierson-Mullany et al., (2000) Molecular Immunology). Другие области также связываются с IgE с высокой аффинностью. Пептид р117-133 содержит свободный цистеин у своего N-конца и, предпочтительно, представляет собой второй сайт связывания настоящего изобретения. В соответствии с трехмерной моделью пептиды р52-72 и р-117-133 находятся на поверхности целого белка. Однако для настоящего изобретения подходящими и предпочтительными являются другие фрагменты белка Der p I, которые могут содержать В-клеточные эпитопы.
Выбор антигенов или антигенных детерминант для получения композиций и их использование в методах лечения злокачественных опухолей могут быть осуществлены в соответствии с известной практикой, применяемой специалистами-медиками для лечения таких заболеваний. Репрезентативными примерами таких антигенов или антигенных детерминант являются антигены и антигенные детерминанты такого типа: Her2 (рак молочной железы), GD2 (нейробластома), EGF-R (злокачественная глиобластома), СЕА (медуллярный рак щитовидной железы), CD52 (лейкоз), белок меланомы человека gp100, белок меланомы человека melan-А/MART-1, тирозиназа, белок NА17-А nt, белок MAGE-3, белок р53, белок НРV16 Е7, а также фрагменты каждого из них, которые могут быть использованы для вырабатывания иммунологических ответов. Другими предпочтительными антигенными детерминантами, которые могут быть использованы в композициях и способах лечения злокачественных опухолей, являются молекулы и антигенные детерминанты, участвующие в ангиогенезе. Ангиогенез, то есть образование новых кровеносных сосудов, играет важную роль в физиологических и патофизиологических процессах, таких как заживление ран и рост солидных опухолей, соответственно. (Folkman, J. (1995) Nat. medicine, 1, 27-31; Folkman, J., and Klagsbrun, M. (1987) Science, 235, 442-446; Martiny-Baron, G., and Marmé, D. (1995) Curr. Opin. Biotechnol. 6, 675-680; Risau, W. (1997) Nature, 386, 671-674). Быстро растущие опухоли инициируют образование новых кровеносных сосудов, и их рост зависит от образования этих сосудов, которые обеспечивают необходимое для них кровообращение. Таким образом, антиангиогенные агенты могут быть эффективными для противораковой терапии.
Среди нескольких предполагаемых ангиогенных факторов, которые были идентифицированы до настоящего времени, может быть упомянут васкулярный эндотелиальный фактор роста (VEGF), который представляет собой сильный митоген, специфичный для эндотелиальных клеток, и является главным стимулятором образования сосудов во множестве солидных опухолей. Хотя недавно полученные данные показали, что для образования функциональных сосудов действие серии ангиогенных факторов должно быть четко согласованным, однако, очевидно, что блокада даже одного фактора роста может ограничивать развитие сосудов, индуцированное данным заболеванием. Таким образом, блокада VEGF может быть основной целью для профилактики опухоль-индуцированного ангиогенеза. Для борьбы с опухолями недавно была разработана новая стратегия нацеливания на эндотелий, а не на саму опухоль. (Millauer, B., Shawver, L.K., Plate, K.H., Risau, W., and Ullrich, A. (1994) Nature, 367, 576-579; Kim, J., Li, B., Winer, J., Armanini, M., Gillett, N., Phillip, H.S., Ferrara, N, (1993) Nature, 362, 841-844). В отличие от опухолей, которые легко изменяют свои структуры-мишени, распознаваемые иммунной системой, эндотелиальные клетки обычно не выходят из-под надзора иммунной системы или других терапевтических систем.
Было описано анти-VEGFR-2 антитело (IMC-1С11) и анти-VEGF антитело (Lu, D., Kussie, P., Pytowski, B., Persaud, K., Bohlen, P., Witte, L., Zhu, Z. (2000) J. Biol. Chem. 275, 14321-14330; Presta, L.G, Chen, H., O'Connor, S.J., Chisholm, V., Meng, Y.G., Krummen, L., Winkler, M., Ferrara N. (1997) Cancer Res. 47, 4593-4599). "Первое" нейтрализующее моноклональное антитело против VEGFR-2 распознает эпитоп, который был идентифицирован как предполагаемый сайт связывания с VEGF/VEGFR-2 (Piossek, C., Schneider-Mergener, J., Schirner, M., Vakalopoulou, E., Germeroth, L., Thierauch, K.H. (1999) J. Biol. Chem. 274, 5612-5619).
Таким образом, в другом предпочтительном варианте осуществления изобретения антиген или антигенная детерминанта представляют собой пептид, происходящий от сайта взаимодействия с VEGFR-2. На основе этого были получены композиция и вакцинная композиция настоящего изобретения, которые могут обладать антиангиогенными свойствами, эффективными для лечения злокачественных опухолей. Было продемонстрировано ингибирование роста опухоли у мышей с использованием сыворотки, специфичной для VEGFR-2 (Wei, Y.Q., Wang, Q.R., Zhao, X., Yang, L., Tian, L., Lu, Y., Kang, В., Lu, C.J., Huang, M.J., Lou, Y.Y., Xiao, R., He, Q.M., Shu, J.M., Xie, X.J., Mao, Y.Q., Lei, S., Luo, F., Zhou, L.Q., Liu, C.E., Zhou, H., Jiang, Y., Peng, F., Yuan, L.P., Li, Q., Wu, Y., Liu, J.Y. (2000) Nature Medicine 6, 1160-1165). Поэтому другие предпочтительные антигенные детерминанты, подходящие для антиангиогенных вакцинных композиций, обладающих признаками изобретения, включают пептид, происходящий от человеческого VEGFR-2 и имеющий последовательность CTARTELNVGIDFNWEYPSSKHQHKK, и/или пептид, происходящий от мышиного VEGFR-2 и имеющий последовательность CTARTELNVGLDFTWHSPPSKSHHKK, и/или соответствующие внеклеточные глобулярные домены 1-3 VEGFR-2.
Поэтому в предпочтительном варианте осуществления изобретения вакцинная композиция содержит коровую частицу, выбранную из вирусоподобной частицы или бактериальных пилей и пептида, происходящего от VEGFR-2 или его фрагмента, в качестве антигена или антигенной детерминанты настоящего изобретения.
Выбор антигенов или антигенных детерминант для получения композиций и их использование в методах лечения других заболеваний и состояний, ассоциированных с аутоантигенами, могут быть осуществлены в соответствии с известной практикой, применяемой специалистами-медиками для лечения таких заболеваний. Репрезентативными примерами таких антигенов или антигенных детерминант являются, например: лимфотоксины (например, лимфотоксин α (LT· α), лимфотоксин β (LT·β)) и рецепторы лимфотоксинов, активатор рецептора лиганда ядерного фактора kB (RANKL), васкулярный эндотелиальный фактор роста (VEGF), рецептор васкулярного эндотелиального фактора роста (VEGF-R), интерлейкин-5, интерлейкин-17, интерлейкин-13, CCL21, CXCL12, SDF-1, MCP-1, эндоглин, резистин, GHRH, LHRH, TRH, MIF, эотаксин, брадикинин, BLC, фактор некроза опухоли α и амилоидный бета-пептид Аβ1-42 (SEQ ID NO:220), а также их фрагменты, каждый из которых может быть использован для вырабатывания иммунологических ответов. В предпочтительном варианте осуществления настоящего изобретения указанным антигеном является амилоидный бета-пептид (Аβ1-42) (DAEFRHDSGYEVHHQKL VFFAEDVGSNKGAIIGLMVGGVVIA (SEQ ID NO:220) или его фрагменты. Этот амилоидный бета-пептид имеет последовательность, представленную в SEQ ID NO:218. Белок-предшественник амилоидного бета-пептида имеет последовательность, представленную в SEQ ID NO:219.
В другом предпочтительном варианте осуществления настоящего изобретения антигеном или антигенной детерминантой является пептид ангиотензин или его фрагмент. Используемый здесь термин "пептид ангиотензина" означает любой пептид, содержащий последовательность ангиотензиногена, ангиотензина I или ангиотензина II или ее фрагменты. Ниже представлены их последовательности: ангиотензиноген: DRVYIHPFHLVIHN; ангиотензин I: DRVYIHPFHL; ангиотензин II: DRVYIHPF. Обычно к С- или N-концу последовательностей пептида ангиотензина могут быть добавлены одна или несколько дополнительных аминокислот. Пептидная последовательность ангиотензинов соответствует человеческой последовательности и идентична мышиной последовательности. Поэтому иммунизация человека или мыши вакцинами или композициями, содержащими, соответственно, такие пептиды ангиотензина в качестве антигенной детерминанты настоящего изобретения, предусматривает вакцинацию против аутоантигена. Такими дополнительными аминокислотами являются, в частности, аминокислоты, подходящие для направленного и упорядоченного присоединения к коровой частице.
Пептид ангиотензина, предпочтительно, имеет аминокислотную последовательность, выбранную из группы, состоящей из а) аминокислотной последовательности CGGDRVYIHPF; b) аминокислотной последовательности CGGDRVYIHPFHL; с) аминокислотной последовательности DRVYIHPFHLGGC и d) аминокислотной последовательности CDRVYIHPFH. Ангиотензин I образуется путем отщепления от ангиотензиногена (14 аминокислот) под действием почечного фермента ренина. Ангиотензин I представляет собой биологически неактивный пептид, состоящий из 10 аминокислот. Кроме того, он расщепляется у своего N-конца ангиотензин-конвертирующим ферментом (АСЕ) с образованием биологически активного ангиотензина II, состоящего из 8 аминокислот. Этот пептид связывается с рецепторами ангиотензина АТ1I и АТ2, что приводит к сужению сосудов и высвобождению альдостерона.
Вакцина настоящего изобретения содержит, по меньшей мере, один пептид ангиотензина, который может быть использован для лечения гипертензии.
В конкретном варианте осуществления изобретения антиген или антигенную детерминанту выбирают из группы, состоящей из (а) рекомбинантного белка ВИЧ, (b) рекомбинантного белка вируса гриппа (например, белка М2 вируса гриппа или его фрагмента), (с) рекомбинантного белка вируса гепатита С, (d) рекомбинантного белка Toxoplasma, (е) рекомбинантного белка Plasmodium falciparum, (f) рекомбинантного белка Plasmodium vivax, (g) рекомбинантного белка Plasmodium ovale, (h) рекомбинантного белка Plasmodium malariae, (i) рекомбинантного белка клеток рака молочной железы, (j) рекомбинантного белка клеток рака почек, (к) рекомбинантного белка клеток рака предстательной железы, (1) рекомбинантного белка клеток рака кожи, (m) рекомбинантного белка клеток рака головного мозга, (n) рекомбинантного белка клеток лейкоза, (о) рекомбинантного белка профилина, (р) рекомбинантного белка яда пчел, вызывающего аллергию, (q) рекомбинантного белка орехов, вызывающего аллергию, (r) рекомбинантных белков пищевых аллергенов, (s) рекомбинантных белков, вызывающих астму, (t) рекомбинантного белка Chlamydia и (u) фрагментов любого из белков, перечисленных в (а)-(t).
После выбора антигенов или антигенных детерминант для данной композиции он может быть присоединен, по меньшей мере, в одном втором сайте связывания к молекуле для создания конструкции упорядоченного и повторяющегося массива, ассоциированного с неприродным молекулярным каркасом настоящего изобретения. Состав подходящего второго сайта связывания может быть определен самим специалистом. Репрезентативные примеры второго сайта связывания представляют собой, без ограничения, антиген, антитело или фрагмент антитела, биотин, авидин, стрептавидин, рецептор, рецепторный лиганд, лиганд, лиганд-связывающий белок, полипептид, взаимодействующий с лейциновой молнией, аминогруппу, химическую группу, реагирующую с аминогруппой, карбоксильную группу, химическую группу, реагирующую с карбоксильной группой, сульфгидрильную группу, химическую группу, реагирующую с сульфгидрильной группой, или их комбинации.
Связывание между первым и вторым сайтом связывания может быть определено по характеристикам соответствующих выбранных молекул и содержит, по меньшей мере, одну непептидную связь. В зависимости от комбинации первого и второго сайта связывания это связывание, по своей природе, может быть ковалентным, ионным, гидрофобным, полярным или смешанным.
В одном из вариантов осуществления изобретения второй сайт связывания может представлять собой домен белка лейциновой молнии FOS или домен белка лейциновой молнии JUN.
В более конкретном варианте осуществления изобретения выбранным вторым сайтом связывания является домен белка лейциновой молнии FOS, который специфически связывается с доменом белка лейциновой молнии JUN неприродного молекулярного каркаса настоящего изобретения. Связывание доменов белка лейциновой молнии JUN и FOS дает основу для образования организованного и повторяющегося массива антигенов и антигенных детерминант на поверхности указанного каркаса. Домен белка лейциновой молнии FOS может быть присоединен к выбранному антигену или к антигенной детерминанте в той же рамке считывания либо у амино-конца, либо у карбокси-конца, либо внутри этого белка, если это необходимо.
Для иллюстрации ниже приводятся некоторые гибридные конструкции FOS: человеческий гомон роста (пример 4), фосфолипаза пчелиного яда А2 (PLA2) (пример 9), овальбумин (пример 10) и gp140 ВИЧ (пример 12).
Для облегчения получения FOS-гибридных конструкций описано несколько векторов, которые предложены на выбор для создания и конструирования антигенов или антигенных детерминант (см. пример 6). Для экспрессии FOS-гибрида в E.coli были сконструированы векторы pAV1-4, а для экспрессии FOS-гибридных белков в эукариотических клетках были сконструированы векторы pAV5 и pAV6. Свойства этих векторов вкратце описаны ниже:
1. pAV1: этот вектор был сконструирован для секреции гибридных белков, имеющих FOS на С-конце, в периплазматическое пространство E.coli. Представляющий интерес ген (п.и.г.) может быть лигирован в StuI/NotI-сайты данного вектора.
2. pAV2: этот вектор был сконструирован для секреции гибридных белков, имеющих FOS на N-конце, в периплазматическое пространство E.coli. Представляющий интерес ген (п.и.г.) может быть лигирован в NotI/EcoRV-сайты (или NotI/HindIII-сайты) данного вектора.
3. pAV3: этот вектор был сконструирован для цитоплазматического продуцирования гибридных белков, имеющих FOS на С-конце, в E.coli. Представляющий интерес ген (п.и.г.) может быть лигирован в EcoRV/NotI-сайты данного вектора.
4. pAV4: этот вектор был сконструирован для цитоплазматического продуцирования гибридных белков, имеющих FOS на N-конце, в E.coli. Представляющий интерес ген (п.и.г.) может быть лигирован в NotI/EcoRV-сайты (или NotI/HindIII-сайты) данного вектора. N-концевой метиониновый остаток был удален путем протеолитического расщепления после синтеза белка (Hirel et al., Proc. Natl. Acad. Sci. USA, 86:8247-8251 (1989)).
5. pAV5: этот вектор был сконструирован для эукариотического продуцирования гибридных белков, имеющих FOS на С-конце. Представляющий интерес ген (п.и.г.) может быть введен между последовательностями, кодирующими сигнальную последовательность hGH, и доменом FOS путем лигирования в Eco47III/NotI-сайты данного вектора. Альтернативно, ген, содержащий свою собственную сигнальную последовательность, может быть присоединен к кодирующей области FOS путем лигирования в StuI/NotI-сайты.
6. pAV6: этот вектор был сконструирован для эукариотического продуцирования гибридных белков, имеющих FOS на N-конце. Представляющий интерес ген (п.и.г.) может быть лигирован в NotI/StuI-сайты (или NotI/HindIII-сайты) данного вектора.
Следует отметить, что конструирование гибридного белка "FOS-антиген" или "FOS-антигенная детерминанта" может предусматривать добавление некоторых генетических элементов для облегчения продуцирования рекомбинантного белка. В примере 4 дано руководство по добавлению некоторых регуляторных элементов E.coli для трансляции, а в примере 7 дано руководство по добавлению эукариотической сигнальной последовательности. Другие генетические элементы могут быть выбраны в зависимости от конкретных целей, преследуемых специалистом.
Настоящее изобретение также относится к продуцированию гибридного белка "FOS-антиген" или "FOS-антигенная детерминанта" в бактериальных (пример 5) или эукариотических клетках (пример 8). Выбор типа клеток, в которых должен быть экспрессирован данный гибридный белок, может быть сделан самим специалистом в зависимости от того, имеют ли посттрансляционные модификации важное значение для конструирования данной композиции.
Как указывалось ранее, в настоящем изобретении описаны различные способы конструирования гибридного белка "FOS-антиген" или "FOS-антигенная детерминанта" с использованием векторов pAV. Помимо обеспечения экспрессии в прокариотах и эукариотах эти векторы облегчают специалисту выбор между N-концевым и С-концевым присоединением к антигену домена белка лейциновой молнии FOS. Конкретными примерами являются присоединение N- и С-концевых FOS-гибридов к PLA2 (пример 9) и к овальбумину (пример 10). В примере 11 продемонстрирована очистка гибридных белков "PLA2-FOS" и "овальбумин-FOS".
В более конкретном варианте своего осуществления настоящее изобретение относится к антигену или к антигенной детерминанте, кодируемой геномом ВИЧ. Более конкретно, антиген или антигенная детерминанта ВИЧ представляет собой gp140. Как описано в примерах 11-15, gp140 ВИЧ может быть сконструирован с использованием домена белка лейциновой молнии FOS, и синтезированный гибридный белок очищают для присоединения к неприродному молекулярному каркасу настоящего изобретения. Каждому специалисту очевидно, что для получения композиции настоящего изобретения могут быть использованы другие антигены или антигенные детерминанты ВИЧ.
В другом более конкретном варианте своего осуществления настоящее изобретение относится к вакцинным композициям, содержащим, по меньшей мере, один антиген или одну антигенную детерминанту, кодируемые нуклеиновой кислотой вируса гриппа, и к использованию таких вакцинных композиций для вырабатывания иммунных ответов. В еще более конкретном варианте осуществления изобретения антигеном или антигенной детерминантой вируса гриппа может быть белок М2 (например, белок М2, имеющий аминокислоты, представленные в SEQ ID NO:213, Genbank, регистрационный номер Р06821, или в SEQ ID NO:214, Genbank, регистрационный номер МFIV62, или его фрагмент (например, аминокислоты примерно от 2 до 24 в SEQ ID NO:213, аминокислотная последовательность в SEQ ID NO:212). Кроме того, антигены или антигенные детерминанты вируса гриппа могут быть присоединены к неприродным молекулярным каркасам или к коровым частицам посредством либо пептидной, либо непептидной связи. При присоединении антигенов или антигенных детерминант вируса гриппа к неприродным молекулярным каркасам или к коровым частицам посредством пептидных связей молекулы, которые образуют упорядоченные и повторяющиеся массивы, могут быть, в основном, получены в виде продуктов экспрессии гибридного белка. Однако в более предпочтительном варианте своего осуществления настоящее изобретение относится к композиции, в которой пептид М2 связан посредством химического сшивания с капсидным белком Qβ, капсидным белком HBcAg или белком пилей.
Части белка М2 (например, белка М2, имеющего аминокислотную последовательность, представленную в SEQ ID NO:213), а также другие белки, против которых необходимо продуцировать иммунный ответ и которые являются подходящими для использования в настоящем изобретении, могут включать или, альтернативно, состоять из них, пептиды длиной в любое число аминокислот, но обычно длиной, по меньшей мере, 6 аминокислот (например, пептиды длиной 6, 7, 8, 9, 10, 12, 15, 18, 20, 25 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или 97 аминокислот).
В другом более конкретном варианте осуществления изобретения выбранным вторым сайтом связывания является цистеиновый остаток, который специфически связывается с лизиновым остатком неприродного молекулярного каркаса или коровой частицы настоящего изобретения, либо выбранным вторым сайтом связывания является лизиновый остаток, который специфически связывается с цистеиновым остатком неприродного молекулярного каркаса или коровой частицы настоящего изобретения. Химическая связь лизинового остатка (Lys) и цистеинового остатка (Cys) дает основу для образования упорядоченных и повторяющихся массивов антигенов и антигенных детерминант на поверхности указанного каркаса или коровой частицы. Цистеиновый или лизиновый остаток могут быть введены с сохранением рамки считывания в выбранный антиген или антигенную детерминанту либо у амино-конца, либо у карбокси-конца, либо внутри этого белка, если это необходимо. Так, например, в PLA2 и gp140 ВИЧ был введен цистеиновый остаток для связывания с лизиновым остатком, присутствующим в качестве первого сайта связывания. В других конкретных вариантах своего осуществления настоящее изобретение относится к вакцинным композициям, предназначенным для профилактики и/или ослабления аллергических реакций, то есть таких аллергических реакций, которые приводят к анафилаксии. Таким образом, вакцинными композициями настоящего изобретения являются композиции, которые обеспечивают продуцирование антител, способствующих предупреждению и/или ослаблению аллергических реакций. Так, например, в некоторых вариантах осуществления изобретения вакцинными композициями настоящего изобретения являются композиции, которые вырабатывают иммунный ответ против аллергена. Примерами таких аллергенов являются фосфолипазы, такие как белки фосфолипазы A2 (PLA2), Apis mellifera (SEQ ID NO:168, Genbank, регистрационный номер 443189; SEQ ID NO:169, Genbank, регистрационный номер 229378), Apis dorsata (SEQ ID NO:170, Genbank, регистрационный номер В59055), Apis cerana (SEQ ID NO:171, Genbank, регистрационный номер А59055), Bombus pennsylvanicus (SEQ ID NO:172, Genbank, регистрационный номер В56338) и Heloderma suspectum (SEQ ID NO:173, Genbank, регистрационный номер Р80003; SEQ ID NO:174, Genbank, регистрационный номер S14764; SEQ ID NO:175, Genbank, регистрационный номер 226711).
С использованием аминокислотной последовательности белка PLA2 Apis mellifera (SEQ ID NO:168), выбранного для иллюстрации, пептиды длиной, по меньшей мере, в 60 аминокислот, которые представляют любую часть всей последовательности PLA2, могут быть также использованы в композициях для профилактики и/или ослабления аллергических реакций. Примерами таких пептидов являются пептиды, которые включают аминокислоты 1-60 SEQ ID NO:168, аминокислоты 1-70 SEQ ID NO:168, аминокислоты 10-70 SEQ ID NO:168, аминокислоты 20-80 SEQ ID NO:168, аминокислоты 30-90 SEQ ID NO:168, аминокислоты 40-100 SEQ ID NO:168, аминокислоты 47-99 SEQ ID NO:168, аминокислоты 50-110 SEQ ID NO:168, аминокислоты 60-120 SEQ ID NO:168, аминокислоты 70-130 SEQ ID NO:168 или аминокислоты 90-134 SEQ ID NO:168, а также соответствующие части других белков PLA2 (например, белка PLA2, описанных выше). Другими примерами таких пептидов являются пептиды, которые включают аминокислоты 1-10 SEQ ID NO:168, аминокислоты 5-15 SEQ ID NO:168, аминокислоты 10-20 SEQ ID NO:168, аминокислоты 20-30 SEQ ID NO:168, аминокислоты 30-40 SEQ ID NO:168, аминокислоты 40-50 SEQ ID NO:168, аминокислоты 50-60 SEQ ID NO:168, аминокислоты 60-70 SEQ ID NO:168, аминокислоты 70-80 SEQ ID NO:168, аминокислоты 80-90 SEQ ID NO:168 или аминокислоты 90-100 SEQ ID NO:168, аминокислоты 100-110 SEQ ID NO:168, аминокислоты 110-120 SEQ ID NO:168 или аминокислоты 120-130 SEQ ID NO:168, а также соответствующие части других белков PLA2 (например, белка PLA2, описанных выше).
Части белка PLA2, а также части других белков, против которых необходимо продуцировать иммунный ответ и которые являются подходящими для использования в настоящем изобретении, могут включать или, альтернативно, состоять из них, пептиды длиной в любое число аминокислот, но обычно длиной, по меньшей мере, 6 аминокислот (например, пептиды длиной 6, 7, 8, 9, 10, 12, 15, 18, 20, 25 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или 100 аминокислот).
Пептиды PLA2 (например, полноразмерные белки PLA2, обсуждаемые выше, а также каждая из их подпоследовательностей) могут быть также связаны с любым веществом, например с капсидным белком Qβ или его фрагментом), что позволяет получить упорядоченные и повторяющиеся массивы антигенов.
В другом своем аспекте настоящее изобретение относится к композициям, подходящим для ослабления и/или предупреждения симптомов заболеваний или состояний, которые вызываются или усиливаются продуктами "своего" гена.
В предпочтительном варианте осуществления изобретения указанной антигенной детерминантой является RANKL (активатор рецептора лиганда NFkb). RANKL также известен как TRANCE (цитокин, индуцированный TNF-опосредованной активацией), ODF (фактор дифференцировки остеокластов) или OPGL (лиганд остеопротегерина). Аминокислотная последовательность внеклеточной части человеческого RANKL представлена в SEQ ID NO:221 (RANKL_human: TrEMBL:О14778), а аминокислотная последовательность человеческой изоформы представлена в SEQ ID NO:222. Последовательности внеклеточной части мышиного RANKL и изоформы представлены в SEQ ID NO:223 (RANKL_mouse: TrEMBL:О35235) и в SEQ ID NO:224 (RANKL_mouse splice forms: TrEMBL:Q9JJK8 и TrEMBL:Q9JJK9), соответственно.
Было показано, что RANKL является главным фактором, ответственным за остеокластогенез. Ингибирование взаимодействия RANKL с его рецептором RANK может приводить к супрессии остеокластогенеза и, тем самым, к прекращению чрезмерной резорбции кости, наблюдаемой при остеопорозе и других состояниях. Взаимодействие RANKL/RANK ингибируется либо гибридным белком RANK-Fc, либо растворимым рецептором-"ловушкой" RANKL, называемым остеопротегерином OPG.
В иммунной системе RANKL экспрессируется на Т-клетках, а RANK обнаруживается на антиген-презентирующих клетках. Было показано, что взаимодействие RANKL-RANK имеет важное значение для CD40L-независимой активации Т-клеток-хелперов (Bachmann et al., J. Exp. Med. 7:1025 (1999)) и усиливает продолжительность жизни и адъювантные свойства дендритных клеток (Josien et al., J.Exp. Med. 191:495 (2000)).
В кости RANKL экспрессируется на стромальных клетках или на остеобластах, а RANK обнаруживается на клетках-предшественниках остеокластов. Взаимодействие RANKL и RANK имеет важное значение для развития предшественников остеокластов в зрелые остеобласты. Это взаимодействие может блокироваться остеопротегерином.
У OPG-дефицитных мышей развивается остеопороз, который может быть предотвращен инъекцией рекомбинантного OPG. Это означает, что OPG способен вызывать обратное развитие остеопороза. Таким образом, ингибирование взаимодействия RANKL-RANK путем инъекции, которая предусматривается в этом конкретном варианте осуществления изобретения, может приводить к обратному развитию остеопороза.
Кроме того, обратное развитие артериосклероза, наблюдаемого у OPG-дефицитных мышей, может быть достигнуто путем инъекции OPG (Min et al., J. Exp. Med. 4:463 (2000)). Инъекция OPG животному с моделью индуцированного адъювантом артрита может предотвращать разрежение кости и деструкцию хрящей, но не воспаление (опухание лап). Можно предположить, что активированные Т-клетки обеспечивают RANKL-опосредованное усиление остеокластогенеза и разрежение кости. OPG, присутствующий в кости мышей, ингибирует остеокластогенез, индуцированный раком предстательной железы, и предотвращает рост опухоли предстательной железы. OPG замедляет усиление болей при раке кости у мышей.
RANKL представляет собой трансмембранный белок, состоящий из 245 аминокислот и принадлежащий к надсемейству TNF. Часть внеклеточной области (178 аминокислот) может быть отщеплена TAGE-подобной протеазой (Lum et al., J. Biol. Chem. 274:13613 (1999)). Кроме того, были описаны сплайсированные варианты, у которых отсутствует трансмембранный домен (Ikeda et al., Endocrinology, 142:1419 (2001)). Такая отщепленная часть содержит домен, который в высокой степени гомологичен растворимому TNF-α. Этот внеклеточный домен RANKL образует гомодимеры, как это наблюдается для TNF-α. Очевидно, что С-конец находится в сайте контактирования с тримером. В этой области последовательности присутствует один цистеин.
Авторами настоящего изобретения была создана модель для 3-мерной структуры соответствующей области RANKL, и было обнаружено, что присутствующий в нем природный цистеин в уложенной структуре может быть недоступным для взаимодействия с первым сайтом связывания на носителе в соответствии с настоящим изобретением. Для присоединения второго сайта связывания предпочтительным является N-конец, содержащей аминокислотный линкер с дополнительным цистеиновым остатком. Конструкция человеческого RANKL с N-концевым аминокислотным линкером, содержащим цистеиновый остаток, соединенный с внеклеточной частью RANKL, является особенно предпочтительным вариантом настоящего изобретения. Однако в своих других предпочтительных вариантах настоящее изобретение относится к аминокислотному линкеру, содержащему цистеиновый остаток в качестве второго сайта связывания и присединенному к С-концу последовательности RANKL или к внеклеточной части RANKL.
Конструкции, содержащие человеческий RANKL, такие как одна из последовательностей, идентифицированная в SEQ ID NO:320, генерировали в соответствии с техникой, описанной в примере 6, и любой специалист может сравнить мышиную и человеческую последовательности RANKL в анализе выравнивания первичных последовательностей белка для идентификации части последовательности человеческого RANKL, клонируемой в векторы, описанные в примере 6. Фрагменты, содержащие аминокислоты 138-317 и соответствующие С-концевой области внеклеточного домена человеческого RANKL, являются особенно предпочтительными вариантами настоящего изобретения и могут быть модифицированы для связывания с VLP и пилями, если это необходимо в целях настоящего изобретения. Однако для экспрессии в подходящем хозяине, описанном ниже, могут быть также использованы и другие подходящие векторы. В объем настоящего изобретения также входят другие конструкции, содержащие человеческий RANKL, а в частности конструкции, содержащие часть внеклеточной области (178 аминокислот) или ее фрагменты (которые могут быть отщеплены TAGE-подобной протеазой) (Lum et al., J. Biol. Chem. 274:13613 (1999)), или содержащие последовательность, соответствующую вариантам альтернативного сплайсинга, у которых отсутствует трансмембранный домен, а также его консервативные фрагменты. Человеческие С-концевые фрагменты, включающие аминокислоты 165-317, также являются вариантами настоящего изобретения. Альтернативно, в объем настоящего изобретения также входят фрагменты, которые охватывают всю внеклеточную область (аминокислоты 71-317) и которые могут быть модифицированы для связывания с VLP и пилями, если это необходимо в целях настоящего изобретения.
RANKL был экспрессирован в различных системах (в E.coli, в клетках насекомых, в клетках млекопитающих) и оказался активным в этих системах, а поэтому для продуцирования этого антигена в данной композиции могут быть использованы несколько экспрессирующих систем. В случае, когда экспрессия белка направляется в периплазму E.coli, то сигнальный пептид RANKL или сигнальный пептид RANKL-конструкции, включающий внеклеточную часть данного белка, которые оба могут быть модифицированы так, чтобы они содержали второй сайт связывания настоящего изобретения, заменяются бактериальным сигнальным пептидом. Для экспрессии белка в цитоплазме E.coli RANKL-конструкции не должны содержать сигнального пептида.
В другом предпочтительном варианте осуществления изобретения антигенной детерминантой является MIF или его фрагмент. MIF представляет собой цитокин, который был впервые описан в 1966 г. и который действует как ингибитор миграции макрофагов. Он известен также как медленно-ранний белок 6 (DЕR6), фактор ингибирования гликозилирования или фенилпируват-таутомераза. Последнее название он получил, исходя из своей ферментативной MIF-активности, однако его эндогенный субстрат не был идентифицирован.
Было показано, что MIF ответственен за широкий ряд состояний. MIF (мРНК или белок) участвует в позитивной регуляции реакций гиперчувствительности замедленного типа (DTH), индуцированной туберкулином, и антитело против MIF ингибирует эту реакцию DTH. MIF также активирует реакцию отторжения трансплантата почки. В модели аутоиммунного заболевания глаз, экспериментального аутоиммунного увеоретинита (EAU), лечение антителом против MIF приводит к замедлению развития EAU. У пациентов с болезнью Бехчета и у пациентов, страдающих иридоциклитом, обнаруживается повышенное содержание MIF в сыворотке. Иммунизация против MIF может рассматриваться как способ лечения ревматоидного артрита.
Высокая концентрация MIF в сыворотке была обнаружена у пациентов с атопическим дерматитом. При поражениях кожи наблюдается диффузная экспрессия MIF, тогда как у контроля MIF экспрессируется в базальном клеточном слое. После лечения стероидами концентрация MIF снижается, что подтверждает роль MIF в воспалении. Было также обнаружено, что MIF играет определенную роль в развитии гломерулонефрита. У животных, обработанных антителом против MIF, наблюдалось значительное ослабление симптомов гломерулонефрита. MIF вырабатывается гипофизом, затем секретируется, например, после ЛПС-стимуляции и потенцирует эндотоксемию. В соответствии с этим анти-MIF антитело (mAb) ингибирует эндотоксемию и септический шок, тогда как рекомбинантный MIF значительно увеличивает летальность от перитонита. MIF также является индуцируемым глюкокортикоидом модулятором продуцирования цитокинов и стимулирует воспаление.
MIF продуцируется Т-клетками (Th2), поддерживает пролиферацию Т-клеток, а поэтому обработка антителами против MIF приводит к снижению уровня пролиферации Т-клеток и уровень IgG. У пациентов с рассеянным склерозом и у пациентов с нейрозаболеванием Бехчета присутствуют повышенные концентрации MIF в цереброспинальной жидкости. Высокие уровни MIF в сыворотке также обнаруживаются и у пациентов, уже давно страдающих псориазом. Высокие уровни MIF в сыворотке также обнаруживаются у пациентов с язвенным колитом, но не у пациентов с болезнью Крона.
Высокие уровни MIF в сыворотке были также обнаружены у пациентов с бронхиальной астмой. Позитивная регуляция MIF также наблюдалась в синовиальной жидкости у пациентов с ревматоидным артритом. Лечение антителами против MIF способствует эффективному ослаблению ревматоидного артрита у мышиных и крысиных моделей (Mikulowska et al., J. Immunol. 158:5514-7(1997); Leech et al., Arthritis Rheum. 41:910-7(1998), Leech et al. Arthritis Rheum. 43:827-33 (2000), Santos et al., Clin. Exp. Immunol. 123:309-14 (2001)). Таким образом, лечение, направленное на ингибирование активности MIF, с использованием композиции, содержащей MIF в качестве антигенной детерминанты, может быть эффективным для состояний, указанных выше.
Мышиный, крысиный и человеческий MIF состоит из 114 аминокислот и содержит три консервативных цистеина, как показано в SEQ ID NO:225 (MIF_rat: SwissProt), в SEQ ID NO:226 (MIF_mouse: SwissProt) и в SEQ ID NO:227 (MIF_human: SwissProt). Эти три субъединицы образуют гомотример, который не стабилизируется дисульфидными связями. Проведенный рентгеноструктурный анализ выявил три свободных цистеина (Sun et al., PNAS, 93:5191-96 (1996)), а в некоторых литературных источниках сообщается о присутствии дисульфидной связи. Тем не менее ни один их этих цистеинов не был достаточно доступным для оптимального взаимодействия с возможным первым сайтом связывания, присутствующим на носителе. Таким образом, поскольку С-конец данного белка является доступным в указанной тримерной структуре, то в предпочтительном варианте осуществления изобретения, для генерирования второго сайта связывания, желательно, чтобы аминокислотный линкер, содержащий три цистеиновых остатка, был присоединен к С-концу данного белка, как это проиллюстрировано в примере 4 для крысиного MIF.
Между мышиным и крысиным MIF имеется различие только в одной аминокислоте, и, аналогичным образом, последовательности человеческого и крысиного MIF или человеческого и мышиного MIF являются в высокой степени гомологичными (с идентичностью примерно 90%). Конструкции настоящего изобретения, содержащие человеческий MIF и мышиный MIF, описаны в настоящей заявке и могут быть получены, как описано в примере 4. Для подтверждения высокой эффективности белка MIF или его фрагментов, ассоциированных с коровой частицей настоящего изобретения, в индуцировании специфического аутоиммунного ответа мышам инъецировали конструкции, которые содержали крысиный MIF и были связаны с капсидным белком Qβ. Высокий титр антител, полученный при иммунизации мышей крысиными MIF, показал, что толерантность к иммунизации аутоантигенами значительно превышала толерантность при иммунизации MIF-конструкциями, связанными с вирусоподобными частицами, в частности с капсидным белком Qβ (пример 4). Поэтому в особенно предпочтительных вариантах своего осуществления настоящее изобретение относится к композициям, содержащим человеческий белок MIF, ассоциированный с коровой частицей, предпочтительно, с пилями или вирусоподобной частицей, более предпочтительно, с вирусоподобной частицей РНК-фага, и даже более предпочтительно, с частицей РНК-фага Qβ или fr.
Однако в другом предпочтительном варианте осуществления изобретения аминокислотный линкер, содержащий свободный цистеин, присоединен у N-конца последовательности MIF. MIF был экспрессирован в E.coli и очищен, и было обнаружено, что он является полностью функциональным (Bernhagen et al., Biochemistry 33: 14144-155 (1994). Таким образом, в предпочтительном варианте осуществления изобретения MIF может быть, предпочтительно, экспрессирован в E.coli.
Таутомеразная активность MIF ингибируется, если инициирующий метионин не отщепляется от данной конструкции. MIF-конструкции, экспрессированные в E.coli и описанные в примере 4, обнаруживали таутомеразную активность. В другом варианте осуществления изобретения мутанты MIF, где инициирующий метионин отщепляется и где пролиновый остаток, расположенный справа от этого инициирующего метионина в данной последовательности, заменен на аланин, также не обнаруживают таутомеразной активности, и эти мутанты входят в объем настоящего изобретения. В некоторых конкретных вариантах осуществления изобретения предусматривается иммунизация MIF-мутантами, которые не обладают таутомеразной активностью.
В другом предпочтительном варианте осуществления изобретения антигенной детерминантой является интерлейкин-17 (IL-17). Человеческий IL-17 имеет размер в 32 кДа и представляет собой гомодимерный белок с дисульфидными связями и с вариабельным гликозилированием (Yao Z. et al., J. Immunol. 155: 5483-5486 (1995); Fossiez F. et al., J. Exp. Med. 183:2593-2603 (1996)). Этот белок содержит 155 аминокислот и включает сигнальную последовательность, обеспечивающую N-концевую секрецию, состоящую из 19-23 остатков. Аминокислотная последовательность IL-17 имеет сходство только с белком герпесвируса (НSV13), но не имеет сходства с другими цитокинами или известными белками. Аминокислотная последовательность IL-17 представлена в SEQ ID NO:228 (регистрационный номер AAC50341), последовательность мышиного белка представлена в SEQ ID NO:229 (регистрационный номер AAA37490). Из множества оцененных тканей и клеточных линий мРНК-транскрипт, кодирующий IL-17, был обнаружен только в активированных Т-клетках и в мононуклеарных клетках периферической крови, стимулированных 12-миристат-13-ацетат/иономицином (Yao Z. et al., J. Immunol. 155: 5483-5486 (1995); Fosseiz F. et al., J. Exp. Med. 183: 2593-2603 (1996)). Как человеческая, так и мышиная последовательности содержат 6 цистеиновых остатков.
Рецептор для IL-17 широко экспрессируется во многих тканях и клеточных линиях (Yao Z. et al., Cytokine, 9:794-800 (1997)). Хотя аминокислотная последовательность человеческого рецептора IL-17 (866 аминокислот) представляет белок с одним трансмембранным доменом и одним внутриклеточным доменом длиной в 525 аминокислот, однако эта рецепторная последовательность является уникальной и не похожа ни на одну из последовательностей рецепторов, принадлежащих к семейству цитокинов/рецепторов факторов роста. Это обусловлено отсутствием сходства самого IL-17 с другими известными белками и указывает на то, что IL-17 и его рецептор могут быть частью нового семейства сигнальных белков и рецепторов. Клинические исследования показали, что IL-17 может участвовать во многих воспалительных заболеваниях. IL-17 секретируется синовиальными Т-клетками, ассоциированными с ревматоидным артритом, и стимулирует продуцирование медиаторов воспаления (Chabaud M. et al., J. Immunol. 161:409-414 (1998); Chabaud M. et al., Arthritis Rheum. 42:963-970 (1999)). Высокие уровни IL-17 были обнаружены у пациентов с ревматоидным артритом (Ziolkowska M. et al., J. Immunol. 164:2832-8 (2000)).
Было показано, что интерлейкин 17 влияет на деградацию протеогликанов в коленном суставе у мышей (Dudler J. et al., Ann. Rheum. Dis. 59:529-32 (2000)) и участвует в деструкции синовиального матрикса (Chabaud M. et al., Cytokyne, 12:1092-9 (2000). А поэтому эти мыши являются подходящей моделью артрита для проведения анализа на эффект иммунизации против MIF (Chabaud M. et al., Cytokyne, 12:1092-9 (2000). Повышенные уровни мРНК IL-17 были обнаружены в мононуклеарных клетках пациентов с рассеянным склерозом (Matusevicius, D. et al., Mult. Scler. 5:101-104 (1999). Повышенные уровни IL-17 наблюдались у пациентов, страдающих системной красной волчанкой (Wong C.K. et al., Lupus, 9:589-93 (2000)). Кроме того, отмечалось повышение уровней мРНК IL-17 в Т-клетках, выделенных из кожи, пораженной псориазом (Teunissen M.B. et al., J. Invest. Dermatol. 111:645-649 (1998)).
Было также продемонстрировано участие IL-17 в отторжении почечного трансплантата (Fosseiz F. et al., Int. Rev. Immunol. 16:541-51 (1998)). Подтверждение участия IL-17 в отторжении трансплантированного органа было также представлено в работе Antonysamy et al. (J. Immunol. 162:577-84 (1999)), где было показано, что IL-17 стимулирует функциональную дифференцировку дендритных клеток-предшественников. Полученные этими авторами данные позволяют предположить, что IL-17 играет определенную роль в пролиферации аллогенных Т-клеток, которая может быть частично опосредована индуцирующим созревание действием на DC. Кроме того, те же авторы сообщали (Tang J.L. et al., Transplantation 72:348-50 (2001), что IL-17 играет определенную роль в иммунопатогенезе острой реакции сосудистого отторжения, где антагонизм интерлейкина 17 приводит к ингибированию острой, но не хронической реакции сосудистого отторжения. Из этого следует, что IL-17 может рассматриваться как новая мишень для терапевтического лечения отторжения аллотрансплантата.
Вышеуказанные данные позволяют предположить, что IL-17 может играть главную роль в инициации или поддержании воспалительного ответа (Jovanovic D.V. et al., J. Immunol. 160:3513-3521 (1998)).
После индуцирования 50 нг/мл IL-17 моноклональное антитело против IL-17, mAb5 (Shering-Plough Research Institute) способно полностью ингибировать продуцирование IL-6 в супернатантах синовиальной жидкости, взятой от пациентов с ревматоидным артритом (РА). В этом анализе нерелевантное mAb МХ1 не давало какого-либо эффекта. mAb5 представляет собой мышиный IgG1, полученный после иммунизации мышей человеческим rIL-17 (r - рекомбинантный). Концентрация mAb5 в 1 мкг/мл способна полностью ингибировать продуцирование IL-6 в аналитической системе. (Chabaud M. et al., J. Immunol. 161:409-414 (1998)). Таким образом, иммунизация против белков IL-17 может рассматриваться как способ лечения различных состояний, описанных выше.
В другом предпочтительном варианте осуществления изобретения композиция включает линкер, содержащий второй сайт связывания и присоединенный к С-концу рекомбинантного IL-17. Однако в других предпочтительных вариантах осуществления изобретения аминокислотный линкер, содержащий свободный цистеин, присоединен к N-концу последовательности, соответствующей последовательности процессированного белка, или встроен в N-конец последовательности зрелой формы данного белка с С-конца по отношению к сигнальному пептиду. Для эукариотических экспрессирующих систем сигнальный пептид гена IL-17, как и в случае других описанных здесь аутоантигенов, может быть заменен другим сигнальным пептидом, если это необходимо. Для экспрессии в бактериях сигнальный пептид либо заменяют бактериальным сигнальным пептидом для экспрессии в периплазме, либо делетируют для экспрессии в цитоплазме. Конструкции человеческого IL-17, не содержащие сигнального пептида, предпочтительно включают остатки 24-155, 22-155, 21-155 или 20-155. Конструкции мышиного IL-17, не содержащие сигнального пептида, предпочтительно включают остатки 26-158, 25-158, 24-158 или 27-155. Человеческий IL-17 может экспрессироваться в клетках CVI/EBNA; при этом было показано, что рекомбинантный hIL-17 экспрессируется как в гликозилированной, так и в негликозилированной формах (Yao Z. et al., J. Immunol. 155:5483-5486 (1995). IL-17 может быть также экспрессирован в виде гибридного белка hIL-17/Fc с последующим отщеплением белка IL-17 от гибридного белка. IL-17 может быть также экспрессирован в дрожжах Pichia pastoris (Murphy K.P. et al., Protein Exp. Purif. 12:208-14 (1998)). Человеческий IL-17 может быть также экспрессирован в E.coli. Если экспрессия IL-17 в E.coli направлена в периплазму, то сигнальный пептид IL-17 заменяют бактериальным сигнальным пептидом. Для экспрессии белка в цитоплазме E.coli конструкции IL-17 не содержат сигнального пептида.
В другом предпочтительном варианте осуществления изобретения антигенной детерминантой является интерлейкин-13 (IL-13). IL-13 представляет собой цитокин, который секретируется посредством активированных Т-лимфоцитов и, главным образом, воздействует на моноциты, макрофаги и В-клетки. Аминокислотная последовательность предшественника человеческого IL-13 показана в SEQ ID NO:230, а аминокислотная последовательность процессированного человеческого IL-13 показана в SEQ ID NO:231. Первые 20 аминокислот этого белка-предшественника соответствуют аминокислотам сигнального пептида и отсутствуют в процессированном белке. Была также описана мышиная последовательность, и процессированная аминокислотная последовательность представлена в SEQ ID NO:232 (Brown K.D. et al., J. Immunol. 142:679-687 (1989)). В зависимости от экспрессии в хозяине конструкция IL-13 будет содержать последовательность белка-предшественника, например, для экспрессии и секреции в эукариотических хозяевах, или она будет состоять из зрелого белка, например, для цитоплазматической экспрессии в E.coli. Для экспрессии в периплазме E.coli сигнальный пептид IL-13 заменяют бактериальным сигнальным пептидом.
IL-13 представляет собой цитокин, происходящий от Т-хелпера 2 (такой как IL-4, IL-5), который, как было недавно установлено, индуцирует аллергические ответы в дыхательных путях (астма). Позитивная регуляции IL-13 и рецептора IL-13 была обнаружена в опухолях многих типов (например, в лимфоме Ходжкина). Интерлейкин 13 секретируется и клетками Ходжкина и Рида-Штернберга и стимулирует их рост (Kapp U. et al., J. Exp. Med. 189:1936-46 (1999)). Таким образом, иммунизация против IL-13 может рассматриваться как способ лечения, помимо описанных выше состояний, таких состояний как астма и лимфома Ходжкина.
Предпочтительно, указанные композиции включают аминокислотный линкер, содержащий свободный цистеиновый остаток и присоединенный к N- или С-концу последовательности зрелого IL-13 для введения второго сайта связывания в этот белок. В других предпочтительных вариантах осуществления изобретения аминокислотный линкер, содержащий свободный цистеиновый остаток, присоединяют к N-концу зрелой формы IL-13, поскольку он является легкодоступным, как было установлено для структуры IL-13 с помощью ЯМР-спектроскопии (Eisenmesser E.Z. et al., J. Mol. Biol. 310:231 (2001)). В других предпочтительных вариантах осуществления изобретения аминокислотный линкер, содержащий свободный цистеиновый остаток, присоединяют к N-концу последовательности, соответствующей последовательности процессированного белка, или вводят в N-конец последовательности зрелой формы этого белка, со стороны С-конца сигнального пептида. В других предпочтительных вариантах осуществления изобретения аминокислотный линкер IL-13, содержащий свободный цистеиновый остаток, присоединяют к С-концу последовательности белка.
IL-13 может быть экспрессирован в E.coli (Eisenmesser E.Z. et al., Protein Expr. Purif. 20:186-95 (2000)) или в клетках NS-0 (эукариотическая клеточная линия)(Cannon-Carlson S. et al., Protein Expr. Purif. 12:239-48 (1998). В примере 9 описаны конструкции и экспрессия конструкций мышиного IL-13, присоединенного к аминокислотному линкеру, содержащему цистеиновый остаток, в бактериальных и эукариотических клетках-хозяевах. Конструкции человеческого IL-13 могут быть генерированы способами, описанными в примере 9, где после экспрессии гибридных белков и расщепления фактором Ха и энтерокиназой, соответственно, продуцируются человеческие белки С-IL-13-F (SEQ ID NO:330) и С-IL-13-S (SEQ ID NO:331). В предпочтительных вариантах осуществления изобретения полученные таким образом белки могут быть присоединены к VLP и к пилям.
В еще одном варианте осуществления изобретения антигенной детерминантой является интерлейкин-5 (IL-5). IL-5 представляет собой цитокин, специфичный к линии дифференцировки и участвующий в эозинофилопоэзе, и играет важную роль в развитии заболеваний, ассоциированных с повышенным числом эозинофилов, таких как астма. Последовательность незрелого и процессированного человеческого IL-5 представлена в SEQ ID NO:233 и в SEQ ID NO:234, соответственно, а процессированная мышиная аминокислотная последовательность представлена в SEQ ID NO:235.
Биологическая функция IL-5 была выявлена в нескольких исследованиях (Coffman R.L. et al., Science, 245:308-10 (1989); Kopf et al., Immunity, 4:15-24 (1996)), в которых было установлено, что ингибирование функции IL-5 оказывает позитивное действие при заболеваниях, опосредованных эозинофилами. Таким образом, ингибирование действия IL-5 может открывать путь к лечению астмы и других заболеваний, ассоциированных с эозинофилами.
IL-5 образует димер, ковалентно связанный дисульфидным мостиком. Была описана одноцепочечная (оц) конструкция, в которой два мономера IL-5 связаны пептидным линкером.
В предпочтительных вариантах осуществления изобретения пептидный линкер, содержащий свободный цистеиновый остаток, присоединяют к N-концу последовательности процессированной формы IL-5. Присоединение линкера, содержащего свободный цистеин, также предпочтительно, может быть осуществлено у N-конца последовательности процессированной формы оцIL-5. В других предпочтительных вариантах осуществления изобретения аминокислотный линкер, содержащий свободный цистеин, присоединяют к N-концу последовательности, соответствующей последовательности процессированного белка, или вводят у N-конца последовательности зрелой формы этого белка, со стороны С-конца от сигнального пептида.
В других предпочтительных вариантах осуществления изобретения линкер, содержащий свободный цистеин, присоединяют к С-концу последовательности IL-5 или к С-концу последовательности оцIL-5.
Был описан ряд экспрессирующих систем для IL-5, и эти системы могут быть использованы для получения композиций настоящего изобретения. Бактериальная экспрессирующая система с использованием E.coli была описана Proudfoot et al., (Biochem J. 270:357-61 (1990)). В случае, если IL-5 экспрессируется в цитоплазме E.coli, то конструкция IL-5 не содержит сигнального пептида. Для создания IL-5-конструкции в целях получения композиций настоящего изобретения могут быть также использованы клетки насекомых (Pierrot C. et al., Biochem Biophys Res. Commun. 253:756-60 (1998)). Аналогичным образом, могут быть также использованы бакуловирусные экспрессирующие системы (клетки sf9; Ingley E. et al., Eur. J. Biochem. 196:623-9 (1991) and Brown P.M. et al., Protein Exp. Purif. 6:63-71 (1995)). И наконец, были также описаны экспрессирующие системы млекопитающих, которые могут быть использованы для получения указанных композиций настоящего изобретения (Kodama S. et al., J. Biochem. (Tokyo), 110:693-701 (1991)).
Для мышиного IL-5 были также описаны бакуловирусные экспрессирующие системы (Mitchell et al., Biochem. Soc. Trans. 21:332S (1993); Kunimoto D.Y. et al., Cytokine, 3:224-30 (1991)) и экспрессирующие системы клеток млекопитающих, полученные с использованием клеток СНО (Kodama S. et al., Glycobiology, 2:419-27 (1992)).
В примере 10 описана экспрессия мышиных IL-5-конструкций, где последовательность IL-5 у его N-конца присоединяют к аминокислотным линкерам, содержащим цистеиновый остаток, для связывания с VLP и пилями. В предпочтительных вариантах осуществления изобретения человеческие конструкции могут быть генерированы методами, описанными в примере 10, с получением человеческого белка С-IL-5-Е (SEQ ID NO:335), человеческого белка С-IL-5-F (SEQ ID NO:336) и человеческого белка С-IL-5-S (SEQ ID NO:337), подходящих для связывания с VLP и пилями.
В другом предпочтительном варианте осуществления изобретения антигенной детерминантой является CCL-21. CCL-21 представляет собой хемокин подсемейства СС, который также известен как небольшой индуцибельный цитокин А21, exodus-2, SLD (вторичный цитокин лимфоцитов), ТСА4 (тимусный хемотаксический агент 4) или 6С-кин.
CCL-21 ингибирует гемопоэз и стимулирует хемотаксис для тимоцитов, активированных Т-клеток и дендритных клеток, но не для В-клеток, макрофагов или нейтрофилов. Этот хемокин обнаруживает преимущественную активность по отношению к "наивным" Т-клеткам. Он также является сильным хемоаттрактантом для мезангиальных клеток. CCL-21 связывается с хемокиновыми рецепторами CСR7 и СХСR3 (в зависимости от вида). Он может инициировать быструю интегрин-зависимую остановку "катящихся" лимфоцитов в физиологических условиях и в высокой степени экспрессируется в эндотелиальных венулах.
Мышиный CCL-21 ингибирует рост опухолей и ангиогенез в мышиной модели SCID рака легких человека (Arenberg et al., Cancer Immunol. Immunother. 49:587-92 (2001)) и в мышиной модели карциномы толстой кишки человека (Vicari et al., J. Immunol. 165:1992-2000 (2001)). Ангиостатическая активность мышиного CCL-21 была также обнаружена в анализе микрокармана роговицы крыс (Soto et al., Proc. Natl. Acad. Sci. USA 95:8205-10 (1998).
Было показано, что хемокиновые рецепторы ССR7 и СХСR4 активируются в клетках рака молочной железы и что CCL-21 и СХCL12, их соответствующие лиганды, в высокой степени экспрессируются в органах, являющихся главными мишенями при метастазах рака молочной железы (Müller et al. Nature, 410:50-6 (2001)). In vitro-хемотаксис, опосредуемый CCL-21, может быть блокирован путем нейтрализации антител против CCL-21, а хемотаксис, опосредуемый СХСR4, может быть блокирован соответствующими антителами. Таким образом, иммунизация против CCL-21 является одним из способов лечения против возникновения метастазов рака, а в частности рака молочной железы.
Секретированный CCL-21 у мышей и человека состоит из 110 или 111 аминокислот, соответственно. Соответствующие последовательности представлены в SEQ ID NO:236 (Swissprot: SY21_human) и в SEQ ID NO:237 (Swissprot: SY21_mouse). В противоположность другим цитокинам СС цитокин CCL-21 содержит еще два цистеина в удлиненной области у С-конца. Предполагается, что все цистеины связаны между собой дисульфидными связями.
Ниже описаны конструкции и экспрессирующие системы для получения композиций настоящего изобретения, содержащих антигенную детерминанту CCL21. В ЯМР-структуре гомологичного белка эотаксина N- и С-концы доступны для действия растворителя. В некоторых конкретных вариантах осуществления изобретения аминокислотный линкер, содержащий свободный цистеиновый остаток в качестве второго сайта связывания, присоединен у С-конца этого белка. Был экспрессирован гибридный белок со щелочной фосфатазой (у С-конца CCL-21), и было показано, что он является функциональным, и что гибриды у С-конца CCL-21 являются подходящими для связывания с рецептором. В других конкретных вариантах осуществления изобретения аминокислотный линкер, содержащий свободный цистеин, присоединен к N-концу последовательности, соответствующей последовательности процессированного белка, или встроен у N-конца последовательности зрелой формы этого белка, у С-конца сигнального пептида.
Было описано несколько экспрессирующих систем для продуцирования CCL-21 (например, Hedrick et al., J. Immunol. 159:1589-93 (1997)). Так, например, он может быть экспрессирован в бакуловирусной системе (Nagira et al., J. Biol. Chem. 272:19518-24 (1997)).
В родственном предпочтительном варианте осуществления изобретения антигенной детерминантой является стромальный фактор 1 (SDF-1), обозначенный здесь СХCL12. СХCL12 представляет собой хемокин, продуцируемый стромальными клетками костного мозга, и был впервые идентифицирован как фактор стимуляции пре-В-клеток.
Как уже указывалось выше, было показано, что хемокиновые рецепторы ССR7 и СХСR4 позитивно регулируются в клетках рака молочной железы, и что CCL-21 и SDF-1, соответствующие лиганды, в высокой степени экспрессируются в органах, в которых, главным образом, возникают метастазы рака молочной железы (Müller et al., Nature, 410:50-6 (2001)). SDF-1/СХСR4-опосредуемый хемотаксис in vitro может быть ингибирован с помощью нейтрализующих антител против SDF-1 и против СХСR4.
В мышиной модели SCID метастазов рака молочной железы, созданной с использованием клеточной линии рака молочной железы человека MDA-МВ-231, значительное снижение метастазов в легких наблюдалось в том случае, когда мышей обрабатывали антителами против СХСR4. При дренировании лимфоузлов наблюдалось снижение метастазов в паховых и подмышечных лимфоузлах (38% вместо 100% метастазов у контроля). Таким образом, иммунизация против СХCL12 может рассматриваться как способ лечения метастазов рака, а более конкретно рака молочной железы.
Было показано, что пара хемокиновых рецепторов SDF-1/СХСR4 повышают эффективность "хоминга" более примитивных кроветворных клеток-предшественников в костный мозг. Кроме того, предполагается, что СХСR4 и SDF-1 влияют на распределение клеток хронического лимфолейкоза. Эти клетки постоянно инфильтрируют костный мозг пациентов, и было показано, что их миграция в костном мозге является СХСR4-зависимой. Клетки хронического лимфолейкоза подвергаются апоптозу, если только они не культивируются совместно со стромальными клетками. SDF-1-блокирующие антитела могут ингибировать протективный эффект стромальных клеток (Burger et al., Blood, 96:2655-63 (2000)). Таким образом, иммунизация против СХCL12 является одним из способов лечения хронического лейкоза.
Было показано, что СХCR4 является ко-рецептором, обеспечивающим внедрение ВИЧ в Т-клетки. SDF-1 ингибирует инфицирование CD4+-клеток ВИЧ-штаммами Х4 (СХСR4-зависимыми) (Oberlin et al., Nature, 382:833-5 (1996); Bleul et al., Nature, 382:829-33 (1996), Rusconi et al., Antivir. Ther. 5:199-204 (2000)). Было показано, что синтетические пептидные аналоги SDF-1 эффективно ингибируют проникновение ВИЧ-1 в клетки и инфицирование этих клеток посредством рецептора CXCR4 (WO 059928A1). Таким образом, иммунизация против СХCL12 рассматривается как способ блокирования проникновения ВИЧ в Т-клетки, а следовательно, и лечения СПИД.
Сообщалось также, что взаимодействие SDF-1 - СХСR4 играет центральную роль в аккумуляции CD4+-Т-клеток в синовиальном слое при ревматоидном артрите (Nanki et al., 2000). Таким образом, иммунизация против SDF-1 рассматривается как способ лечения ревматоидного артрита.
Известно, что человеческий и мышиный SDF-1 присутствует в двух формах, SDF-1α и SDF-1β, образующихся в результате дифференциального сплайсинга одного гена. Эти формы отличаются четырьмя С-концевыми аминокислотами, которые присутствуют в SDF-1β (74 аминокислоты) и отсутствуют в SDF-1α (70 аминокислот). Человеческая последовательность SDF-1 представлена в SEQ ID NO:238 (Swissprot: SDF1_human), а мышиная последовательность SDF-1 представлена в SEQ ID NO:239 (Swissprot: SDF1_mouse). SDF-1 содержит четыре консервативных цистеина, которые образуют две внутримолекулярных дисульфидных связи. Кристаллическая структура SDF представляет собой нековалентно связанный димер (Dealwis et al., PNAS 95:6941-46 (1998)). Структура SDF-1 также имеет длинное N-концевое удлинение.
Для идентификации (части) сайта связывания с рецептором на SDF-1 был использован аланин-сканирующий мутагенез (Ohnishi et al., J. Interferon Cytokine Res. 20:691-700 (2000)), & Elisseeva et al., (J. Biol. Chem. 275:26799-805 (2000)) & Heveker et al. (Curr. Biol. 8:369-76 (1998)) описали происходящие от SDF-1 пептиды, ингибирующие связывание с рецептором (и внедрения ВИЧ).
Ниже описаны конструкции и экспрессирующие системы, подходящие для получения композиций настоящего изобретения, относящихся к SDF-1. N- и С-концы SDF-1 доступны для действия растворителя. В конкретных вариантах осуществления изобретения аминокислотный линкер, содержащий цистеин в качестве второго сайта связывания, присоединен к С-концу последовательности белка, а в другом конкретном варианте осуществления изобретения аминокислотный линкер, содержащий цистеин в качестве второго сайта связывания, присоединен к N-концу последовательности белка. Аминокислотный линкер, содержащий свободный цистеин, присоединен к N-концу последовательности, соответствующей последовательности процессированного белка, или встроен у N-конца последовательности зрелой формы этого белка, т.е. у С-конца сигнального пептида. Гены, кодирующие эти специфические конструкции, могут быть клонированы в подходящий экспрессирующий вектор.
Была описана экспрессия SDF-1 в вирусной системе Сендай в фибробластах куриных эмбрионов (Moriya et al., FEBS Lett. 425:105-11 (1998)), а также экспрессия в E.coli (Holmes et al., Prot. Expr. Purif. 21:367-77 (2001)) и химический синтез SDF-1 (Dealwis et al., PNAS, 95:6941-46 (2001)).
В еще одном варианте осуществления изобретения антигенной детерминантой является BLC. Хемоаттрактант В-лимфоцитов (BLC, СХCL13) экспрессируется в селезенке, в пейеровых бляшках и в лимфоузлах (Gunn et al., 1998). Его экспрессия является наиболее сильной в зародышевых центрах, где В-клетки подвергаются соматической мутации и аффинному созреванию. Он принадлежит к семейству хемокинов СХС, и его наиболее близким гомологом является GROα (Gunn et al., Nature, 391:799-803 (1998)). Человеческий BLC на 64% гомологичен мышиному BLC. Его рецептором является СХСR5. BLC также имеет гомологию с IL-8. BLC обеспечивает рекрутинг В-клеток в фолликулы во вторичных лимфоидных органах, таких как селезенка и пейеровы бляшки. BLC также необходим для рекрутинга В-клеток в участок лимфоузлов, богатый фолликулярными дендритными клетками (FDC) (Ansel et al., Nature, 406:309-314 (2000)). BLC также индуцирует повышенную экспрессию лимфотоксина α1β2 (LT·α1β2) на рекрутированных В-клетках. Поскольку LT·α1β2 стимулирует экспрессию BLC, это обеспечивает образование петли с положительной обратной связью (Ansel et al., Nature, 406:309-314 (2000)). Было также показано, что BLC способен индуцировать лимфоидный неогенез (Luther et al., Immunity, 12:471-481 (2000)). Очевидно также, что FDC экспрессируют BLC. Таким образом, иммунизация против BLC может рассматриваться как способ лечения аутоиммунных заболеваний, ассоциированных с лимфоидным неогенезом, таких как ревматоидный синовит и ревматоидный артрит или диабет типа I. Была также описана конструкция BLC, несущая С-концевую his-метку, и эта конструкция является функциональной (Ansel K.M. et al., J. Exp. Med. 190:1123-1134 (1999)).
Таким образом, в предпочтительном варианте осуществления настоящего изобретения указанная композиция включает линкер, содержащий цистеиновый остаток в качестве второго сайта связывания и присоединенный у С-конца последовательности BLC.
В IL-8, который является гомологичным BLC, оба N- и С-конца являются свободными. В другом предпочтительном варианте осуществления изобретения присоединение аминокислотного линкера, содержащего цистеиновый остаток в качестве второго сайта связывания, осуществляют у N-конца BLC для получения специфической композиции настоящего изобретения.
В других предпочтительных вариантах осуществления изобретения указанная композиция включает аминокислотный линкер, содержащий свободный цистеин и присоединенный к N-концу последовательности, соответствующей последовательности процессированного белка, или встроенный у N-конца последовательности зрелой формы этого белка, у С-конца сигнального пептида. Гены, кодирующие эти специфические конструкции, могут быть клонированы в подходящий экспрессирующий вектор и экспрессированы соответствующим образом. Последовательность человеческого BLC представлена в SEQ ID NO:240 (регистрационный номер: NP_006410). Аминокислоты 1-22 этой последовательности представляют собой сигнальный пептид. Мышиная последовательность BLC представлена в SEQ ID NO:241 (регистрационный номер: NP_061354). Аминокислоты 1-21 представляют собой сигнальный пептид. Для получения композиций настоящего изобретения, содержащих BLC в качестве антигенной детерминанты, предпочтительно, используется зрелая форма этого белка.
В другом конкретном варианте осуществления изобретения указанной антигенной детерминантной является эотаксин. Эотаксин представляет собой хемокин, который является специфическим для хемокинового рецептора 3, присутствующего на эозинофилах, базофилах и Th2-клетках. Однако, очевидно, что эотаксин является в высокой степени специфичным для эозинофилов (Zimmerman et al., J. Immunol. 165:5839-46 (2000)). У мышей с дефицитом эутоксина-1 миграция эозинофилов снижается на 70%, однако при этом у них все же развивается эозинофилия (Rothenberg et al., J. Exp. Med. 185:785-90 (1997)). IL-5, очевидно, ответственен за миграцию эозинофилов из костного мозга в кровь, а эотаксин ответственен за локальную миграцию в ткань (Humbles et al., J. Exp. Med. 186:601-12 (1997)).
Человеческий геном содержит 3 гена эотаксина, эотаксинов 1-3. Они имеют 30%-ную гомологию по отношению к друг другу. До настоящего времени у мышей было обнаружено два гена эотаксина: ген эотаксина 1 и ген эотаксина 2 (Zimmerman et al., J. Immunol. 165:5839-46 (2000)). Они имеют 38%-ную гомологию по отношению к друг другу. Мышиный эотаксин-2 на 59% гомологичен эотаксину-2 человека. У мышей эотаксин-1, очевидно, экспрессируется во всех участках желудочно-кишечного тракта, а эотаксин-2, очевидно, экспрессируется преимущественно в тощей кишке (Zimmerman et al., J. Immunol. 165:5839-46 (2000)). Эотаксин-1 присутствует в бронхоальвеолярной жидкости (Teixeira et al., J. Clin. Invest. 100:1657-66 (1997)). Последовательность человеческого эотаксина-1 представлена в SEQ ID NO:242 (аминокислоты 1-23 соответствуют сигнальному пептиду), последовательность человеческого эотаксина-2 представлена в SEQ ID NO:243 (аминокислоты 1-26 соответствуют сигнальному пептиду), последовательность человеческого эотаксина-3 представлена в SEQ ID NO:244 (аминокислоты 1-23 соответствуют сигнальному пептиду), последовательность мышиного эотаксина-1 представлена в SEQ ID NO:245 (аминокислоты 1-23 соответствуют сигнальному пептиду), а последовательность мышиного эотаксина-2 представлена в SEQ ID NO:246 (аминокислоты 1-23 соответствуют сигнальному пептиду).
Эотаксин имеет м.м. 8,3 кДа. В нем сохраняется равновесие между мономерами и димерами в различных условиях широкого ряда, при этом оцененный Kd составляет 1,3 мМ при 37°С (Crump et al., J. Biol. Chem. 273:22471-9 (1998)). Однако чаще всего встречается его мономерная форма. Структура эотаксина была определена с помощью ЯМР-спектроскопии. Сайт связывания с его рецептором ССR3 находится у N-конца, а область, предшествующая первому цистеину, имеет очень важное значение (Crump et al., J. Biol. Chem. 273:22471-9 (1998)). Пептиды хемокиновых рецепторов, связанные с эотоксином, подтверждают эти результаты. Эотаксин имеет четыре цистеина, образующих два дисульфидных мостика. Поэтому в предпочтительном варианте своего осуществления настоящее изобретение относится к композиции, включающей аминокислотный линкер, содержащий цистеиновый остаток в качестве второго сайта связывания и, предпочтительно, присоединенный к С-концу последовательности эотаксина. В других предпочтительных вариантах осуществления изобретения аминокислотный линкер, содержащий свободный цистеин, присоединен к N-концу последовательности, соответствующей последовательности процессированного белка, или встроен у N-конца последовательности зрелой формы этого белка, то есть у С-конца сигнального пептида. Гены, кодирующие эти специфические конструкции, клонируют в подходящий экспрессирующий вектор.
Эотаксин может быть получен методом химического синтеза (Clark-Lewis et al., Biochemistry, 30:3128-3135 (1991)). Для эотаксина-1 была также описана экспрессия в цитоплазме E.coli (Crump et al., J. Biol. Chem. 273:22471-9 (1998)). Для эотаксина-2 была описана экспрессия в E.coli в виде телец включения с последующей повторной укладкой (Mayer et al., Biochemistry, 39:8382-95 (2000)) и экспрессия в клетках насекомых (Forssman et al., J. Exp. Med. 185:2171-6 (1997)), и указанная экспрессия также может быть использована в конкретных вариантах осуществления изобретения.
В еще одном конкретном варианте осуществления изобретения указанной антигенной детерминантой является макрофагальный колониестимулирующий фактор (М-CSF или CSF-1). М-CSF или CSF-1 является регулятором пролиферации, дифференцировки и выживания магрофагов и их предшественников в костном мозге. Рецептором для М-CSF является тирозинкиназный рецептор клеточной поверхности, кодируемый протоонкогеном cfms. Усиленная экспрессия М-CSF и его рецептора ассоциируются с плохим прогнозом при некоторых видах эпителиального рака, такого как рак молочной железы, матки и яичника. Прогрессирование опухоли исследовали на линиях мышей, полученных в результате скрещивания трансгенной мыши, восприимчивой к раку молочной железы (РуМТ), с мышью, содержащей рецессивную молчащую мутацию в гене csf-1. У этой мыши, по сравнению с мышью РуМТ, обнаруживалась аттенюированная инвазивная карцинома на поздней стадии и метастазы в легкие (Lin et al., J. Exp. Med. 193:727-739 (2001)). Это обусловлено, очевидно, отсутствием рекрутинга макрофагов в опухолевые ткани. У мышей с молчащей мутацией в гене csf-1 также наблюдался подкожный рост опухоли рака легких Льюиса. Было высказано предположение, что механизм усиления роста опухоли макрофагами может быть обусловлен ангиогенными факторами, факторами роста и протеазами, продуцированными этими макрофагами.
Имеются данные о структуре растворимой формы М-CSF (кристаллическая структура: Pandit et al., Science, 258:1358-62 (1992)), и было показано, что оба N- и С-конца указанного белка являются доступными. Однако N-конец находится возле сайта взаимодействия с рецептором. Кроме того, М-CSF присутствует как в растворимой форме, так на клеточной поверхности, где трансмембранная область находится у его С-конца. Поэтому в предпочтительном варианте осуществления изобретения композиция настоящего изобретения включает аминокислотный линкер, содержащий цистеин и, предпочтительно, присоединенный к С-концу М-CSF или его фрагментов, либо, предпочтительно, к С-концу растворимой формы М-CSF. В других предпочтительных вариантах осуществления изобретения аминокислотный линкер, содержащий свободный цистеин, присоединен к N-концу последовательности, соответствующей последовательности процессированного белка или растворимой формы белка, или встроен у N-конца последовательности зрелой формы этого белка или растворимой формы этого белка, то есть у С-конца сигнального пептида. М-CSF представляет собой димер, где два мономера связаны межцепьевым дисульфидным мостиком.
Была описана экспрессирующая система в E.coli для N-концевого (функционального) фрагмента М-CSF, состоящего из 149 аминокислот (Koths et al., Mol. Reprod. Dev. 46:31-37 (1997)). Этот фрагмент М-CSF, предпочтительно, модифицированный, как описано выше, представляет собой предпочтительную антигенную детерминанту настоящего изобретения.
Человеческая последовательность представлена в SEQ ID NO:247 (регистрационный номер: NP_000748). Другие предпочтительные антигенные детерминанты настоящего изобретения включают N-концевой фрагмент, состоящий из остатков 33-181 или 33-185 SEQ ID NO:247, соответствующих растворимой форме рецептора.
Мышиная последовательность (регистрационный номер: NP_031804) представлена в SEQ ID NO:248. Эта зрелая последовательность начинается с аминокислоты 33. Таким образом, предпочтительная антигенная детерминанта настоящего изобретения содержит аминокислоты 33-181 или 33-185.
В другом конкретном варианте осуществления изобретения указанной антигенной детерминантой является резистин (Res). Исследования по пассивной иммунизации осуществляли с использованием кроличьих поликлональных антител против гибридного белка "мышиный резистин (mRes) - GST", экспрессированного в бактериях. Эта пассивная иммунизация приводит к улучшению поглощения глюкозы у животных-моделей с ожирением/диабетом типа II (Steppan et al., Nature, 409:307-12 (2001)).
Резистин (Res) представляет собой пептидный гормон из 114 аминокислот размером 12 кД. Он содержит 11 цистеинов, из которых самый крайний N-концевой цистеин, как было показано, ответственен за димеризацию белка, а другие 10, очевидно, участвуют в образовании внутримолекулярных дисульфидных связей (Banerjee & Lazar, J. Biol. Chem. 276:25970-3 (2001)). Замена первого цистеина аланином приводит к предотвращению димеризации mRes.
Было показано, что mRes с FLAG-меткой у своего С-конца все еще сохраняет активность в животном-модели (Steppan et al., Nature, 409:307-12 (2001)), аналогично С-концевому НА-меченому (гемаглютининовой меткой) варианту резистина, который, как было показано в анализе с культивированием тканей, является активным (Kim et al., J. Biol. Chem. 276:11252-6 (2001)), что позволяет предположить, что С-конец не очень чувствителен к вводимым модификациям. Поэтому в предпочтительном варианте осуществления изобретения композиция настоящего изобретения включает аминокислотный линкер, содержащий цистеиновый остаток в качестве второго сайта связывания и присоединенный к С-концу последовательности резистина. В других предпочтительных вариантах осуществления изобретения аминокислотный линкер, содержащий свободный цистеин, присоединен к N-концу последовательности, соответствующей последовательности процессированного белка, или встроен у N-конца последовательности зрелой формы этого белка, то есть у С-конца сигнального пептида.
В предпочтительном варианте осуществления изобретения mRes или huRes могут быть также экспрессированы в виде молекул-Fc-гибридов с сайтом расщепления протеазой, введенным между резистином и Fc-частью этой конструкции, а предпочтительно, у С-конца одного или нескольких цистеиновых остатков шарнирной области Fc-части этого гибридного белка, в эукариотической экспрессирующей системе, или, более предпочтительно, так, как это описано в примере 2. Расщепление гибридного белка приводит к высвобождению резистина, который, кроме того, включает либо аминокислотный линкер, содержащий цистеиновый остаток, как описано в примере 2, либо части, либо всей шарнирной области Fc-части гибридного белка, которая включает цистеиновый остаток у его С-конца и которая может быть использована для связывания VLP или пилей. Последовательность человеческого резистина представлена в SEQ ID NO:249 (регистрационный номер AF32081). Мышиная последовательность резистина представлена в SEQ ID NO:250 (регистрационный номер AF323080). В своем предпочтительном варианте настоящее изобретение относится к человеческому белку резистину с присоединенным у его С-конца аминокислотным линкером, содержащим цистеиновый остаток. Конструкция человеческого резистина может быть получена методами, описанными в примере 2, а также путем выравнивания последовательностей белка мышиного и человеческого резистина в целях идентификации части последовательности человеческого резистина, клонируемой в векторы, описанные в примере 1 и примере 2 в соответствии с методикой, описанной в примере 2, или в другие подходящие экспрессирующие векторы. Примером конструкций человеческого резистина, подходящих для получения композиций настоящего изобретения, являются человеческий резистин-С-Ха (SEQ ID NO:325), человеческий резистин-С-ЕК (SEQ ID NO:326) и человеческий резистин-С (SEQ ID NO:327).
Полученные таким образом конструкции человеческого резистина являются предпочтительным вариантом осуществления настоящего изобретения. Таким образом, вакцинация против резистина с использованием вышеупомянутых композиций настоящего изобретения может рассматриваться как способ лечения диабета типа II и ожирения.
В другом варианте осуществления настоящего изобретения указанной антигенной детерминантой является лимфотоксин-β. Иммунизация против лимфотоксина-β может быть использована для лечения заболевания, опосредованного прионами. Репликация агента, вызывающего скрэпи (заболевание, опосредованное прионами), очевидно, происходит, главным образом, в лимфоидных тканях, и было показано, что она зависит от экспрессии белка приона фолликулярными дендритными клетками (FDC) (Brown et al., Nature Med. 11:1308-1312 (1999)). Следовательно, было показано, что у мышей, у которых отсутствуют функциональные фолликулярные дендритные клетки, наблюдается нарушение репликации прионов в селезенке и (небольшое) замедление нейроинвазии (Montrasio et al., Science, 288:1257-1259 (2000)). Это было достигнуто путем инъекции мышам растворимого гибридного белка "рецептор лимфотоксина-β-Fc" (LT·R-Fc). Эта конструкция растворимого рецептора ингибирует развитие FDC путем предотвращения играющего важную роль взаимодействия лимфотоксина-β на Т-, В- и NK-клетках с рецептором лимфотоксина-β на клетках-предшественниках FDC. Таким образом, вакцинация против лимфотоксина-β (также называемого TNFγ) может рассматриваться как способ лечения или предупреждения заболевания Крейтцфельда-Якоба (вариантная форма) или других опосредованных прионами заболеваний и предупреждения репликации и нейроинвазии прионов.
Иммунизация против лимфотоксина-β также рассматривается как способ лечения диабета. Трансгенная экспрессия растворимого гибридного белка LT·R-Fc у не склонных к ожирению мышей NOD с диабетом блокировала развитие диабета, но не инсулита. (Ettinger et al., J. Exp. Med. 193:1333-40К (2001)). Wu et al. (J. Exp. Med. 193:1327-32 (2001)) также использовали NOD-мышей для проведения исследований по вовлечению лимфотоксина-β в патологический процесс, но, вместо трансгенных животных, они использовали животных, инфицированных гибридным белком LT·R-Fc. У этих мышей наблюдалось сильное ингибирование развития диабета и ингибирование инсулита. Наиболее интересным является тот факт, что у этих мышей, после их обработки гибридным белком, даже могло наблюдаться обратное развитие уже существующего инсулита. Таким образом, образование лимфоидных фолликулярных структур в поджелудочной железе может быть обратимым. А следовательно, вакцинация против лимфотоксина-β может рассматриваться как способ лечения диабета типа I.
Последовательность внеклеточного домена человеческого лимфотоксина-β представлена в SEQ ID NO:250 (TNFC_human), а последовательность внеклеточного домена мышиного лимфотоксина-β представлена в SEQ ID NO:251 (TNFC_mouse).
В другом предпочтительном варианте осуществления изобретения композиция настоящего изобретения включает аминокислотный линкер, содержащий свободный цистеин и присоединенный к N-концу последовательности, соответствующей процессированной форме лимфотоксина-β, или встроенный между N-концом последовательности, соответствующей зрелой форме указанного белка, и сигнальным пептидом у его С-конца. В других предпочтительных вариантах осуществления изобретения внеклеточную часть лимфотоксина-β экспрессируют как гибридный белок либо с глутатион-S-трансферазой, присоединенной у N-конца к лимфотоксину-β, либо с 6-гистидиновой меткой, а затем с myc-меткой, также присоединенной у N-конца к внеклеточной части лимфотоксина-β. Аминокислотный спейсер, содержащий сайт расщепления протеазой, а также линкерную последовательность, содержащую свободный цистеин в качестве сайта связывания у С-конца по отношению к указанному сайту расщепления протеазой, присоединяют к N-концу последовательности внеклеточной части лимфотоксина-β. Внеклеточная часть лимфотоксина-β предпочтительно состоит из фрагментов, соответствующих аминокислотам 49-306 или 126-306 лимфотоксина-β. Эти специфические композиции настоящего изобретения могут быть клонированы и экспрессированы в эукариотический вектор рСЕР-Pu. В других предпочтительных вариантах осуществления изобретения композиции настоящего изобретения включают аминокислотный линкер, содержащий свободный цистеиновый остаток, подходящий в качестве второго сайта связывания и присоединенный к С-концу лимфотоксина-β или фрагментов лимфотоксина-β. В особенно предпочтительном варианте осуществления изобретения аминокислотная последовательность LACGG, содержащая аминокислотный линкер ACGG, который сам содержит цистеиновый остаток для связывания с VLPS и пилями, присоединяют к N-концу внеклеточной части лимфотоксина-β или фрагмента внеклеточной части лимфотоксина-β, с получением человеческого белка С-LT•49-306 (SEQ ID NO:346) и человеческого белка С-LT•126-306 (SEQ ID NO:347) после расщепления энтерокиназой соответствующих гибридных белков, экспрессируемых либо в векторе pCEP-SP-GST-EK, либо в векторе pCP-SP-his-myc-EK, как описано в примере 3.
В предпочтительном варианте осуществления изобретения указанным антигеном или антигенной детерминантной является белок прион, его фрагменты, а в частности пептиды указанного белка приона. В одном из вариантов осуществления изобретения указанным белком прионом является человеческий белок прион. Руководство по модификации человеческого белка приона для его связывания с частицей cpre приводится в описании настоящей заявки, а в частности, в примере 7. Описаны конструкции мышиного белка приона, и могут быть также генерированы конструкции человеческого белка приона, которые представлены, например, в последовательности SEQ ID NO:348. Дополнительные конструкции включают всю последовательность человеческого белка приона и другие фрагменты указанного человеческого белка приона, которые составляют другую композицию настоящего изобретения. Иммунизация против белка приона может быть одним из способов лечения или предупреждения болезни Крейтцфельда-Якоба (вариантной формы) или других опосредованных прионами заболеваний. Иммунизация с использованием композиций настоящего изобретения, содержащих указанный белок прион, может быть одним из способов лечения опосредуемых прионами заболеваний у других животных, и соответствующие последовательности конструкций бычьих и овечьих белков прионов представлены в SEQ ID NO:349 и в SEQ ID NO:350, соответственно. Пептиды человеческого белка приона, соответствующие мышиным пептидам, описанным в примере 8, и аминокислотная последовательность CSAMSRPIIHFGSDYEDRYYRENMHR ("человеческий длинный пептид cprplong") и CGSDYEDRYYRENMHR ("человеческий короткий пептид cprpshort"), являются предпочтительными вариантами настоящего изобретения. Эти пептиды включают N-концевой цистеиновый остаток, добавленный для связывания с VLP и пилями. Соответствующими бычьими и овечьими пептидами являются CSAMSRPLIHFGNDYEDRYYRENMHR ("бычий длинный пептид cprplong"), CGNDYEDRYYRENMHR ("бычий короткий пептид cprpshort"), CSAMSRPLIHFGNDYEDRYYRENMYR ("овечий длинный пептид cprplong") и CGNDYEDRYYRENMYR ("овечий короткий пептид cprpshort"), где все они являются вариантами настоящего изобретения.В другом предпочтительном варианте осуществления изобретения антигенная детерминанта представляет собой фактор некроза опухоли α (TNF-α) и фрагменты или пептиды TNF-α. В частности, пептиды или фрагменты TNF-α могут быть использованы для индуцирования аутоиммунного ответа, направленного против целого белка при иммунизации человека или животного вакцинами и композициями, соответственно, содержащими такие пептиды или фрагменты настоящего изобретения. В качестве коровой частицы предпочтительно использовать VLP, бактериофаги или бактериальные пили, к которым присоединяются TNF-α, пептиды или фрагменты настоящего изобретения.
Указанные ниже мышиные пептиды представляют собой мышиные гомологи человеческих пептидов, которые, как было показано, связываются с антителами, нейтрализующими активность TNF-α (Yone et al., J. Biol. Chem. 270:19509-19515), и которые, в предпочтительном варианте осуществления изобретения, модифицированы цистеиновыми остатками для их связывания с VLP, бактериофагами или бактериальными пилями.
Пептид MuTNF-α: последовательность CGG присоединена к N-концу эпитопа, состоящего из аминокислотных остатков 22-32 зрелого мышиного TNF-α: CGGVEEQLEWLSQR.
Пептид 3'TNF-α II: последовательность GGC присоединена к С-концу эпитопа, состоящего из аминокислотных остатков 4-22 зрелого мышиного TNF-α, а глутамин 21 был заменен на глицин. Этот пептид имеет последовательность: SSQNSSDKPVAHVVANHGVGGC.
Пептид 5'TNF-α II: цистеиновый остаток присоединен к N-концу эпитопа, состоящего из аминокислотных остатков 4-22 зрелого мышиного TNF-α, а глутамин 21 был заменен на глицин. Полученный пептид имеет последовательность: CSSQNSSDKPVAHVVANHGV.
Соответствующей аминокислотной последовательностью эпитопа, состоящего из остатков 4-22, является SSRTPSDKPVAHVVANPQAEGQ. Подобно мышиной последовательности, цистеин, предпочтительно, присоединен к N-концу эпитопа, либо последовательность GGC присоединена к С-концу эпитопа для ковалентного связывания с VLP, с бактериофагами или с бактериальными пилями в соответствии с настоящим изобретением. Однако в объем настоящего изобретения также входит последовательность, содержащая другой цистеиновый остаток, присоединенный к N- или С-концу эпитопов. В основном, один или два цистеина, предпочтительно, встраивают между дополнительным цистеиновым остатком и последовательностью эпитопа. Кроме того, вместо глициновых остатков могут быть также инсертированы и другие аминокислотные остатки, и эти аминокислотные остатки являются, предпочтительно, мелкими аминокислотами, такими как серин.
Человеческая последовательность, соответствующая аминокислотным остаткам 22-32, представляет собой QLQWLNRRANA. Предпочтительно, последовательность CGG присоединена к N-концу эпитопа для ковалентного связывания с VLP, с бактериофагами или с бактериальными пилями в соответствии с настоящим изобретением. Другие TNF-α-эпитопы, подходящие для использования в настоящем изобретении, описаны и определены, например, в работе Yone et al. (J. Biol. Chem. 270:19509-19515).
Настоящее изобретение также относится к композициям, содержащим мимотопы антигенов или антигенных детерминант, описанных в настоящей заявке.
Специфическая композиция настоящего изобретения содержит антитело или, предпочтительно, фрагмент антитела, представленный на вирусоподобной частице или пилях, для индуцирования иммунного ответа против указанного антитела. Для связывания с вирусоподобной частицей и иммунизации могут быть выбраны антитела или фрагменты антител, продуцируемые клетками лимфомы в целях индуцирования протективного иммунного ответа против лимфомы.
В других вариантах осуществления изобретения антитело или фрагмент антитела, имитирующие антиген, присоединены к данной частице. Антитело-миметик или фрагмент антитела могут быть генерированы путем иммунизации, и последующее выделение этого антитела-миметика или фрагмента антитела может быть осуществлено хорошо известными методами, такими как гибридомная техника (Gherardi E. et al., J. Immunol. Methods, 126:61-68 (1990)), фаговое представление (Harrison et al., Methods Enzymol. 267:83-109 (1996)), рибосомное представление (Hanes J. et al., Nat. Biotechnol. 18:1287-1292 (2000), получение двухкомпонентных дрожжевых гибридов (Visintin, M. et al., Proc. Natl. Acad. Sci. USA, 96:11273-11278 (1999)), представление на поверхности дрожжей (Boder E.T. & Wittrup, K.D. Methods. Enzym. 328:430-444 (2000)), представление на поверхности бактерий (Daugherty, P.S. et al., Protein Eng. 12:613-621 (1999)). Антитело-миметик может быть также выделено из библиотеки антител или из библиотеки наивных антител методами, известными специалистам, например методами, упомянутыми выше.
В другом варианте осуществления изобретения может быть использовано антитело, распознающее сайт связывания с другим антителом, то есть антиидиотипическое антитело, называемое далее иммунизирующим антителом. Антитело, распознаваемое этим антиидиотипическим антителом, далее будет называться нейтрализующим антителом. Таким образом, путем иммунизации против антиидиотипического антитела могут быть генерированы молекулы со специфичностью нейтрализующего антитела in situ; и эти генерированные антитела будут далее называться индуцированными антителами. В другом предпочтительном варианте осуществления изобретения иммунизирующее антитело выбирают так, что оно взаимодействовало с молекулой, являющейся лигандом для молекулы-мишени, против которой осуществляют иммунизацию. Эта лигандная молекула может представлять собой любую молекулу, взаимодействующую с указанной молекулой-мишенью, а предпочтительно, молекулу, взаимодействующую с сайтом молекулы-мишени, против которой должны вырабатываться антитела для ингибирования ее функции. Указанной лигандной молекулой может быть природный лиганд молекулы-мишени, либо она может представлять собой любой генерированный, сконструированный или выделенный лиганд, имеющий подходящие связывающие свойства.
Иммунизирующие антитела могут быть человеческими антителами, такими как антитела, выделенные из библиотеки наивных или иммунных антител человека, либо они могут быть выделены из библиотеки, генерированной из другого животного, например мыши.
Связывание антитела или фрагмента антитела с VLP или пилями достигается либо путем ограниченного восстановления экспонируемых дисульфидных мостиков (например, межцепьевых дисульфидных мостиков, находящихся между СН1 и Ck или Сλ в Fab-фрагменте), либо путем присоединения линкера, содержащего свободный цистеиновый остаток у С-конца антитела или фрагмента антитела. В другом варианте осуществления изобретения линкер, содержащий свободный цистеиновый остаток, присоединяют к N-концу антитела или фрагмента антитела для связывания с VLP или белком пилей.
Существует ряд вакцинных композиций, в которых используются мимотопы, а также известны методы генерирования и идентификации мимотопов конкретных эпитопов. Так, например, в работе Arnon et al., Immunology 101:555-562 (2000), которая во всей своей полноте вводится в настоящее описание посредством ссылки, описаны вакцины на основе пептидных мимотопов против Schistosoma mansoni. Мимотопы, используемые в этих вакцинах, были получены путем скрининга твердофазной 8-мерной рандомизированной пептидной библиотеки для идентификации мимотопов эпитопа, распознаваемого протективным моноклональным антителом против Schistosoma mansoni. Аналогичным образом, в работе Olszewska et al., Virology 272:98-105 (2000), которая во всей своей полноте вводится в настоящее описание посредством ссылки, описаны идентификация синтетических пептидов, которые имитируют эпитоп гибридного белка вируса кори, и использование этих пептидов для иммунизации мышей. Кроме того, в работе Zuercher et al., Eur. J. Immunol. 30:128-135 (2000), которая во всей своей полноте вводится в настоящее описание посредством ссылки, описаны композиции и способы пероральной иммунизации анти-IgE антителом с использованием эпитоп-представляющего фага. В частности, эпитоп-представляющие бактериофаги М13 были использованы в качестве носителей для пероральной вакцины против IgE. Тестируемые вакцинные композиции содержат мимотопы и эпитопы моноклонального анти-IgE антитела ВSW17.
Таким образом, настоящее изобретение относится к вакцинным композициям, содержащим мимотопы, которые вырабатывают иммунологические ответы против конкретных антигенов, а также к отдельным конъюгатам "мимотоп/коровая частица" и отдельным конъюгатам "мимотоп/неприродный молекулярный каркас", которые входят в состав указанных вакцинных композиций, и к использованию таких вакцинных композиций для вырабатывания иммунологических ответов против специфических антигенов или антигенных детерминант. Мимотопами могут быть также полипептиды, такие как антиидиотипические антитела. Поэтому в другом предпочтительном варианте осуществления изобретения указанным антигеном или антигенной детерминантой является антиидиотипическое антитело или фрагмент антиидиотипического антитела.
Настоящее изобретение, кроме того, относится к композициям, которые содержат описанные здесь мимотопы антигенов или антигенных детерминант.
Мимотопы конкретных антигенов могут быть генерированы и идентифицированы рядом способов, включая скрининг рандомизированных пептидных библиотек фагового представления (см., например, публикацию РСТ №WO 97/31948, полное описание которой вводится в настоящее описание посредством ссылки). В большинстве случаев скрининг таких библиотек может быть осуществлен для идентификации пептидов, которые связываются с одним или несколькими антителами, специфичными для конкретного антигена.
Мимотопами, подходящими для использования в вакцинных композициях настоящего изобретения, могут быть линейные или кольцевые пептиды. Мимотопы, которые являются линейными или кольцевыми пептидами, могут быть присоединены к неприродным молекулярным каркасам или к коровым частицам связью, которая не является пептидной связью.
Как указывалось выше, был идентифицирован ряд человеческих мимотопов и эпитопов для IgE, которые вырабатывают иммунологические ответы против человеческих молекул IgE (см., например, публикацию РСТ №WO 97/31948). Таким образом, в некоторых вариантах осуществления изобретения вакцинными композициями настоящего изобретения являются композиции, которые вырабатывают иммунологический ответ против молекул иммуноглобулина (например, молекул IgE).
Пептидами, которые могут быть использованы для вырабатывания таких иммунологических ответов, являются белки, субъединицы белков, домены молекул IgE и мимотопы, которые способны продуцировать антитела, обладающие специфичностью к молекулам IgE. В общих чертах, части молекул IgE, используемые для получения вакцинных композиций, могут быть получены из молекул IgE, происходящих от животного того же вида, что и животное, которому вводят эту композицию. Так, например, вакцинная композиция, предназначенная для введения человеку, в большинстве случаев содержит одну или несколько частей молекулы IgE человека и/или один или несколько мимотопов, способных вырабатывать иммунологические ответы против человеческих молекул IgE.
В конкретных вариантах осуществления изобретения вакцинные композиции настоящего изобретения, предназначенные для введения человеку, содержат, по меньшей мере, одну часть константной области тяжелой цепи IgE, представленной в SEQ ID NO:176; регистрационный номер ААВ59424 (SEQ ID NO:176). В более конкретных вариантах осуществления изобретения пептиды IgE, используемые для получения вакцинных композиций настоящего изобретения, включают или, альтернативно, состоят из них, пептиды, имеющие следующие аминокислотные последовательности: CGGVNLT·SRASG (SEQ ID NO:178).
В других конкретных вариантах осуществления изобретения вакцинные композиции настоящего изобретения содержат, по меньшей мере, один мимотоп, способный вырабатывать иммунный ответ, что приводит к продуцированию антител, специфичных к конкретному антигену.
Примерами мимотопов IgE, подходящих для использования в получении вакцинных композиций настоящего изобретения, являются пептиды, имеющие нижеследующие аминокислотные последовательности:
С. Получение вакцин на основе альфа-частиц
Настоящее изобретение относится к новым композициям и к способам конструирования упорядоченных и повторяющихся массивов антигенов. Каждому специалисту известно, что условия сборки упорядоченных и повторяющихся массивов антигенов в большой степени зависят от конкретного выбора первого сайта связывания неприродного молекулярного каркаса и конкретного выбора второго сайта связывания антигена или антигенной детерминанты. Таким образом, выбор, сделанный специалистом, при создании данной композиции (то есть выбор первого и второго сайтов связывания, антигена и неприродного молекулярного каркаса) будет определять конкретные условия для сборки вакцинной альфа-частицы (объединенного упорядоченного и повторяющегося массива антигенов и неприродного молекулярного каркаса). Информация, относящаяся к сборке вакцинной альфа-частицы, хорошо известна специалистам в данной области, и в помощь специалистам было опубликовано множество работ (например, Sambrook, J. et al., eds., Molecular Cloning, A Laboratory Manual, 2nd. edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989); Ausubel, F. et al., eds., Current Protocols in Molecular Biology, John H. Wiley & Sons, Inc. (1997); Celis, J., ed., Cell Biology, Academic Press, 2nd edition, (1998); Harlow, E. and Lane, D., "Antibodies: A Laboratory Manual," Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. (1988), каждая из которых вводится в настоящее описание посредством ссылки.
В конкретном варианте осуществления настоящего изобретения для первого и второго сайтов связывания настоящего изобретения, соответственно, используют домены лейциновой молнии белка JUN и FOS. При получении частиц альфавакцины антиген должен быть продуцирован и очищен в условиях, стимулирующих сборку упорядоченного и повторяющегося массива антигенов на неприродном молекулярном каркасе. В конкретном варианте домена белка "лейциновая молния" JUN/FOS, FOS-антиген или FOS-антигенная детерминанта должны быть обработаны восстановителем (например, дитиотрейтолом (DTT)) для снижения или предотвращения образования дисульфидных связей (пример 15).
Для получения неприродного молекулярного каркаса (то есть рекомбинантного вируса Синдбис) данного варианта домена белка "лейциновая молния" JUN/FOS, рекомбинантные вирусные частицы Е2-JUN должны быть концентрированы, нейтрализованы и обработаны восстановителем (см. пример 16).
Сборку упорядоченного и повторяющегося массива антигенов в варианте JUN/FOS осуществляют в присутствии окислительно-восстановительной системы. Вирусные частицы Е2-JUN объединяют с 240-кратным молярным избытком FOS-антигена или FOS-антигенных детерминант в течение 10 часов при 4°С. Затем частицу альфа-вакцины концентрируют и очищают с помощью хроматографии (пример 16).
1. В другом варианте осуществления изобретения связывание неприродного молекулярного каркаса с антигеном или антигенной детерминантой может быть осуществлено путем химического сшивания. В конкретном варианте осуществления изобретения сшивающим агентом является гетеробифункциональный сшивающий агент, такой как N-гидроксисукцинимидоэфир ∈-малеимидокапроновой кислоты (Tanimori et al., J. Pharm. Dyn. 4:812(1981); Fujiwara et al., J. Immunol. Meth. 45:195 (1981)), который содержит (1) сукцинимидную группу, реагирующую с аминогруппой, и (2) малеимидную группу, реагирующую с группами SH. Гетерологичный белок или полипептид первого сайта связывания могут быть сконструированы так, чтобы они содержали один или несколько лизиновых остатков, которые могут служить реакционноспособной группой для сукцинимидной части гетеробифункционального сшивающего агента. После химического связывания с лизиновыми остатками гетерологичного белка малеимидная группа гетеробифункционального сшивающего агента может реагировать с группой SH цистеинового остатка на антигене или антигенной детерминанте. В этом случае для получения антигена или антигенной детерминанты может потребоваться конструирование цистеинового остатка на белке или полипептиде, выбранном в качестве второго сайта связывания так, чтобы он мог реагировать со свободной малеимидной функциональной группой на сшивающем агенте, связанном с первыми сайтами связывания неприродного молекулярного каркаса. Таким образом, в этом случае гетеробифункциональный сшивающий агент связывается с первым сайтом связывания неприродного молекулярного каркаса и соединяет этот каркас со вторым сайтом связывания антигена или антигенной детерминанты.
3. Композиции, вакцины, их введение и способы лечения
Настоящее изобретение относится к вакцинным композициям, которые могут быть использованы для профилактики и/или ослабления определенных заболеваний или состояний. Кроме того, настоящее изобретение относится к способам вакцинации для профилактики и/или ослабления конкретных заболеваний или состояний у индивидуумов.
В одном из своих вариантов настоящее изобретение относится к вакцинам для профилактики инфекционных заболеваний у животных, принадлежащих к различным видам широкого ряда, а в частности у млекопитающих таких видов как человек, обезьяна, корова, собака, кошка, лошадь, свинья и т.п. Вакцины могут быть получены для лечения инфекций вирусной этиологии, таких как ВИЧ-инфекции; грипп; инфекции, вызываемые герпесвирусом, вирусным гепатитом, вирусом Эпштейна-Барр и полиовирусом; вирусный энцефалит, корь, оспа кур и т.п.; инфекции бактериальной этиологии, такие как пневмония, туберкулез, сифилис и т.п.; или инфекции, вызываемые паразитами, такие как малярия, трипаносомоз, лейшманиоз, трихомониаз, амебиаз и т.п.
В другом варианте осуществления изобретения настоящее изобретение относится к вакцинам для профилактики злокачественных опухолей у животных широкого ряда, а в частности у млекопитающих таких видов как человек, обезьяна, корова, собака, кошка, лошадь, свинья и т.п. Вакцины могут быть получены для лечения всех типов рака: лимфом, карцином, сарком, меланом и т.п.
В другом варианте осуществления изобретения композиции настоящего изобретения могут быть использованы в целях получения вакцин для лечения аллергий. Важными компонентами при аллергических реакциях являются антитела изотипа IgE. Тучные клетки связываются с антителами IgE на их поверхности и высвобождают гистамины и другие медиаторы аллергического ответа после связывания специфического антигена с молекулами IgE, связанными на поверхности тучных клеток. Поэтому ингибирование продуцирования антител IgE является перспективным методом защиты от аллергии. Это будет возможным при достижении нужного клеточного ответа с продуцированием Т-клеток-хелперов. Ответы с продуцированием Т-клеток-хелперов могут быть подразделены на ответы типа 1 (TН1) и типа 2 (TН2) (Romagnani, Immunol. Today, 18:263-266 (1997)). TН1-клетки секретируют интерферон-гамма и другие цитокины, которые стимулируют В-клетки, продуцирующие антитела IgG1-3. В противоположность этому, главным цитокином, продуцируемым TН2-клетками, является IL-4, который стимулируется В-клетками для продуцирования IgG4 и IgE. Во многих экспериментальных системах вырабатывание TН1- и TН2-ответов является взаимоисключающими событиями, поскольку TН1-клетки подавляют индуцирование TН2-клеток и наоборот. Таким образом, антигены, которые стимулируют сильный TН1-ответ, в то же самое время подавляют вырабатывание TН2-ответов, а следовательно, и продуцирование антител IgE. Интересно отметить, что фактически все вирусы индуцируют TН1-ответ у хозяина и не индуцируют вырабатывание антител IgE (Coutelier et al., J. Exp. Med. 165:64-69 (1987)). Такой характер продуцирования изотипов не ограничивается "живыми" вирусами и также наблюдается в случае инактивированных или рекомбинантных вирусных частиц (LoMan et al., Eur. J. Immunol. 28:1401-1407 (1998)). Так, например, с использованием способов настоящего изобретения (например, техника, разработанная для альфа-вакцин), вирусные частицы могут быть "декорированы" различными аллергенами и использованы для иммунизации. Благодаря получению "вирусной структуры" данного аллергена может вырабатываться TН1-ответ, то есть будут продуцироваться "протективные" антитела IgG1-3 и будет предотвращаться продуцирование антител IgE, которые вызывают аллергические реакции. Поскольку этот аллерген представлен вирусными частицами, которые распознаются другим набором Т-клеток-хелперов, отличающимся от набора Т-клеток, которые распознают сам аллерген, то очевидно, что аллерген-специфические антитела IgG1-3 будут индуцироваться даже у индивидуумов с аллергией, у которых уже имеются TН2-клетки, специфичные к данному аллергену. Присутствие высоких концентраций антител IgG может предотвращать связывание аллергенов с IgE с тучными клетками и тем самым ингибировать высвобождение гистамина. Таким образом, присутствие антител IgE может защищать от IgE-опосредованных аллергических реакций. Типичными веществами, вызывающими аллергию, являются пыльца растений, амброзии, березы или можжевельника, домашняя пыль, клещи, перхоть животных, плесень, яд насекомых или лекарственные средства (например, пенициллин). Таким образом, иммунизация субъектов несущими аллерген вирусными частицами будет выгодна не только до, но и после начала аллергии.
В конкретных вариантах своего осуществления настоящее изобретение относится к способам предупреждения и/или ослабления симптомов заболеваний или состояний, вызываемых или усиливаемых продуктами "своего" гена (например, факторами некроза опухоли), то есть используемыми здесь "аутоантигенами". В родственных вариантах своего осуществления настоящее изобретение относится к способам индуцирования иммунологических ответов у индивидуумов, что приводит к продуцированию антител, которые предупреждают и/или ослабляют заболевания или состояния, вызываемые или усиливаемые продуктами "своего" гена. Примерами таких заболеваний или состояний являются реакция "трансплантат против хозяина", IgE-опосредованные аллергические реакции, анафилаксия, респираторный дистресс-синдром взрослых, болезнь Крона, аллергическая астма, острый лимфолейкоз (ALL), не-ходжкинская лимфома (НХЛ), болезнь Грейвса, воспалительные аутоиммунные заболевания, тяжелая миастения, системная красная волчанка (СКВ), иммунопролиферативная лимфаденопатия (IPL), ангиоиммунопролиферативная лимфаденопатия (AIL), иммунобластная лимфаденопатия (IBL), ревматоидный артрит, диабет, рассеянный склероз, остеопороз и болезнь Альцгеймера.
Следует отметить, что композиции настоящего изобретения, предназначенные для введения индивидууму, могут содержать соли, буферы, адъюванты или другие вещества, которые необходимы для повышения эффективности этой композиции. Примеры материалов, подходящих для использования в получении фармацевтических композиций, указаны в различных литературных источниках, включая Remington's pharmaceuticals sciences (Osol, A. ed., Mack Publishing Co., (1990)).
Композиции настоящего изобретения называют "фармакологически приемлемыми", если индивидуум-реципиент является толерантным к их введению. Кроме того, композиции настоящего изобретения могут быть введены в "терапевтически эффективном количестве" (то есть в количестве, которое продуцирует нужный физиологический эффект).
Композиции настоящего изобретения могут быть введены различными методами, известными специалистам, но обычно их вводят путем инъекции, инфузии, ингаляции, перорально или другими подходящими физическими методами. Альтернативно, указанные композиции могут быть введены внутримышечно, внутривенно или подкожно. Компонентами композиций для введения являются стерильные водные (например, физиологические) или безводные растворы и суспензии. Примерами безводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и инъецируемые органические сложные эфиры, такие как этилолеат. Для повышения проницаемости кожи и усиления абсорбции антигена могут быть использованы носители или окклюзионные повязки.
Опосредованные прионами заболевания представляют в настоящее время все большую угрозу для общества. В частности, индуцированная прионами BSE (коровья губчатая энцефалопатия) у крупного рогатого скота представляет собой заболевание, о котором долгое время не было ничего известно и которое может поражать большое число животных по всей Европе. Кроме того, инфицирование человека после употребления мяса инфицированного прионами рогатого скота приписывается вариантной форме CJD. Хотя число инфицированных людей до настоящего времени было относительно небольшим, однако нельзя исключать, что эта болезнь не приобретет характер эпидемии. При этом отдаленный прогноз развития vCJD может представлять значительные трудности, поскольку инкубационный период между инфицированием и проявлением болезни является очень продолжительным (по оценкам специалистов, 10 лет).
Прионы представляют собой клеточные белки, присутствующие у большинства видов млекопитающих. Белки прионы существуют в двух формах, в "правильно" уложенной форме, которая обычно присутствует у здоровых индивидуумов (PrPc), и в "неправильно" уложенной форме, которая вызывает заболевание (PrpSc). В настоящее время была высказана гипотеза, что неправильно уложенный прион в форме PrpSc может катализировать повторную укладку нормального приона PrPc в прион PrpSc, вызывающий заболевание (А. Aguzzi, Haematologica, 85, 3-10 (2000)). В некоторых редких случаях такая транзиция может также происходить спонтанно и вызывать классическую CJD у человека. Некоторые мутации в PrPc ассоциируются с возрастанием этих спонтанных транзиций, что приводит к различным формам наследственной CJD. Однако PrpSc может быть также инфекционным и может передаваться путем переливания крови или через цепь питания. Эта последняя форма опосредованной прионом болезни известна под названием Куру-куру и возникает у людей-каннибалов. Однако, поскольку виды животных, которые поедают животных своего вида, являются довольно редкими, то эта форма передаваемого пероральным путем заболевания также является слишком редкой, чтобы ее можно было зарегистрировать у других видов.
Обильное кормление коров говяжьими продуктами по всей Европе изменило ситуацию, и в последнее время число коров, инфицированных трансмиссивной формой PrpSc, вызывающей BSE, резко возросло от нескольких сотен до нескольких тысяч. Такое внезапное появление огромного числа коров, пораженных BSE, вызвало большие опасения в обществе, поскольку аналогичное заболевание может быть индуцировано и у человека. Действительно, в 1996 г. был зарегистрирован первый случай обнаружения вариантной формы CJD, который быть объяснен употреблением PrpSc-инфицированной говядины. В настоящее время эта опасность еще более увеличивается, поскольку за последние годы число инфицированных людей неуклонно возрастает, а способ лечения от этой болезни пока еще не найден. Кроме того, поскольку овцы погибают от опосредованного прионами заболевания, называемого скрепи, и поскольку другие виды млекопитающих могут быть инфицированы PrpSc, то с экспериментальной точки зрения возможно, что BSE-подобные заболевания могут также возникать и у других видов животных. Был подробно исследован механизм передачи прионов. В настоящее время известно, что прионы сначала реплицируются в лимфоидных органах инфицированных мышей, а затем транспортируются в центральную нервную систему. Фолликулярные дендритные клетки (ФДК), редкая клеточная популяция в лимфоидных органах, очевидно, имеет важное значение как для репликации белков прионов в лимфоидных органах, так и для их транспорта в центральную нервную систему (S.Brandner M.A. Klein, A. Aguzzi, Transfus Clin. Biol. 6, 17-23 (1993); F.Montrasio et al., Science, 288, 1257-9 (2000)). ФДК принадлежат к плохо изученному типу клеток, однако, в настоящее время было установлено, что их развитие зависит от продуцирования лимфотоксина и/или TNF В-клетками (F.Mackay J.L. Browning, Nature, 395, 26-27 (1998)). Действительно, у мышей с отсутствием лимфотоксина не обнаруживаются ФДК (M.S. Matsumoto et al., Science, 264, 703-707 (1996)). Кроме того, эти мыши не были подвержены эффективному инфицированию прионами и не погибали от этого заболевания. Помимо ФДК, определенную роль в прогрессировании этого заболевания могут также играть и антитела (S. Brandner, M.A. Klein, A. Aguzzi, Transfus Clin Biol. 6, 17-23 (1999)).
Недавно было показано, что блокирование LTβ-пути с использованием гибридной молекулы "LTβ-рецептор Fc" не только приводит к элиминации ФДК у мышей, но также и блокируют инфекцию PrpSc (F. Montrasio et al., Science, 288, 1257-9 (2000). Таким образом, вакцина, которая индуцирует антитела, специфичные к LTβ или к его рецептору, может блокировать передачу PrpSc от одного индивидуума другому или от периферической нервной системы в центральную нервную систему.
Однако индуцировать гуморальный ответ к собственным молекулам путем обычной вакцинации достаточно трудно, а то и вообще невозможным. Одним из способов повышения эффективности вакцинации является увеличение степени повторяемости применяемого антигена, и в отличие от выделенных белков вирусы индуцируют быстрый и эффективный иммунный ответ в отсутствие какого-либо адъюванта с помощью или без помощи Т-клеток (Bachmann & Zinkemagel, Ann. Rev. Immunol: 15:235-270 (1991)). Хотя вирусы часто состоят из нескольких белков, они способны стимулировать гораздо более сильные иммунные ответы, чем их выделенные компоненты. Известно, что для продуцирования В-клеточных ответов важным фактором для иммуногенности вирусов является повторяемость и порядок расположения эпитопов на их поверхности. Многие вирусы имеют квазикристаллическую поверхность, которая отображает массив правильного ряда эпитопов, эффективно связываются с эпитоп-специфическими иммуноглобулинами на В-клетках (Bachmann & Zinkernagel, Immunol. Today, 17:553-558 (1996)). Такое перекрестное связывание иммуноглобулинов на поверхности В-клеток является сильным активирующим сигналом, который непосредственно индуцирует прохождение клеточного цикла и продуцирование антител IgM. Кроме того, указанные стимулированные В-клетки могут активировать Т-клетки-хелперы, которые, в свою очередь, индуцируют переключение продуцирования антитела IgM на продуцирование антитела IgG в В-клетках и генерирование долговременной В-клеточной памяти, которое является целью любой вакцинации (Bachmann & Zinkernagel, Ann. Rev. Immunol. 15:235-270 (1997)). Вирусная структура однозначно связана с генерированием анти-антител при аутоиммунном заболевании и является частью природного ответа на действие патогенов (см. Fehr T. et al., J. Exp. Med. 185:1785-1792 (1997)). Таким образом, антитела, презентированные высокоорганизованной вирусной поверхностью, способны индуцировать сильный гуморальный ответ.
Иммунная система, обычно, не продуцирует антитела против собственных структур. Для растворимых антигенов, присутствующих в низких концентрациях, это может быть обусловлено толерантностью на Th-клеточном уровне. В этих условиях связывание аутоантигена с носителем, способным доставлять Т-хелпер, может нарушать эту толерантность. Для растворимых антигенов, присутствующих в высоких концентрациях, или мембранных белков, присутствующих в низкой концентрации, В- и Th-клетки могут быть толерантными. Однако В-клеточная толерантность может быть обратимой (анергия) и может нарушаться путем введения антигена, который высокоорганизованным способом связан с чужеродным носителем (Bachmann & Zinkernagel, Ann. Rev. Immunol. 15:235-270 (1997). Таким образом, LTβ, LTα или рецептор LTβ, которые являются такими же высокоорганизованными, как вирус, вирусная частица, вирусоподобная частица или бактериальные пили, способны нарушать В-клеточную толерантность и индуцировать вырабатывание специфических антител против этих молекул.
Настоящее изобретение относится к области молекулярной биологии, вирусологии, иммунологии и медицины. Настоящее изобретение также относится к способу, который стимулирует индуцирование антител, специфических для эндогенного лимфотоксина (LTβ, LTα или рецептора LTβ). Настоящее изобретение, кроме того, относится к способу продуцирования антигена или антигенной детерминанты, позволяющему вырабатывать антитела, специфичные для LTβ, LTα или рецептора LTβ, которые могут быть использованы для профилактики и лечения опосредованных прионом заболеваний, таких как вариант болезни Крейтцфельда-Якоба (vCJD) или коровья губчатая энцефалопатия (BSE), а также для элиминации подобных структур в тканях лимфоидных органов при аутоиммуных заболеваниях.
Целью настоящего изобретения является получение вакцины, способной индуцировать антитела против LTβ, LTα или рецептора LTβ и тем самым удалять ФДК из лимфоидных органов. Такое лечение позволит предупреждать PrpSc-инфицирование или распространение PrpSc из периферической нервной системы в центральную нервную систему. Кроме того, это лечение блокирует генерирование подобных структур в лимфоидных органах, подверженных аутоиммунному заболеванию, и может даже растворять такие имеющиеся структуры, и тем самым, ослаблять симптомы заболевания.
LTβ, LTα или рецептор LTα или его фрагменты связываются с белком-носителем, который является чужеродным для данного хозяина. В предпочтительном варианте осуществления изобретения LTβ, LTα или рецептор LTβ или его фрагменты будут связываться с высокоорганизованной структурой и образовывать в высокой степени повторяющиеся и организованные молекулы. Такими высокоорганизованными структурами могут быть бактериальный пилус, вирусоподобная частица (VLP), генерированная рекомбинантными белками бактериофага Qβ, рекомбинантными белками ротавируса, рекомбинантными белками вируса Норуолк, рекомбинантными белками альфавируса, рекомбинантными белками вируса ящура, рекомбинантными белками ретровируса, рекомбинантными белками вируса гепатита В, рекомбинантными белками вируса табачной мозаики, рекомбинантными белками вируса овечьих кошар и рекомбинантными белками человеческого папиломавируса. Для оптимизации трехмерной организации LTβ, LTα или рецептора LTβ или его фрагментов на высокоорганизованной структуре сайт связывания, такой как химически реакционноспособная аминокислота, вводят в указанную высокоорганизованную структуру (независимо от ее природы), и этот сайт связывания, такой как химически реакционноспособная аминокислота, будет введен в LTβ, LTα или рецептор LTβ или его фрагменты (независимо от его природы). Присутствие сайта связывания на высокоорганизованной структуре и сайта связывания на LTβ, LTα или рецепторе LTβ или его фрагментах будет обеспечивать связывание этих молекул с повторяющейся структурой направленным и упорядоченным способом, что очень важно для индуцирования эффективных В-клеточных ответов.
В таком же предпочтительном варианте осуществления изобретения сайтом связывания, вводимым в повторяющуюся структуру, является биотин, специфически связывающийся со стрептавидином. Биотин может быть введен путем химической модификации. LTβ, LTα или рецептор LTβ или его фрагменты могут быть слиты или связаны со стрептавидином и присоединены к биотинилированной повторяющейся структуре.
В других вариантах своего осуществления настоящее изобретение относится к способам получения композиций настоящего изобретения и к способам терапевтического лечения с использованием этих композиций. Следует отметить, что вышеприведенное общее описание, а также нижеследующее подробное описание является иллюстративным и имеет своей целью лишь более детальное объяснение заявленного изобретения.
Помимо вакцинной технологии рассматриваются и другие варианты осуществления изобретения, в которых проиллюстрированы способы терапевтического лечения злокачественных опухолей и аллергий.
Все упомянутые здесь патенты и публикации во всей своей полноте вводятся в настоящее описание посредством ссылки.
ПРИМЕРЫ
В экспериментах были использованы следующие ферменты и реагенты: ДНК-лигаза Т4, полученная из New England Biolabs; ДНК-полимераза Taq, плазмидный набор QIAprep Spin Plasmid Kit, плазмидный набор QIAGEN Plasmid Midi Kit, набор для гель-экстракции QiaExII, набор для ПЦР-очистки QIAquick, полученный от QIAGEN; набор для микроочистки мРНК Quick Prep Micro, полученный от Pharmacia; набор для одностадийной ОТ-ПЦР Superscript, фетальная телячья сыворотка (FCS), бакто-триптон и дрожжевой экстракт, полученные от Gibco BRL; олигонуклеотиды, полученные от Microsynth (Switzerland); рестриктирующие эндонуклеазы, полученные от Boehringer Mannheim, New England Biolabs или MBI Fermentas; полимераза Pwo и dNTP, полученные от Boehringer Mannheim. Среда НР-1 была получена от Cell Culture Technologies (Glattbrugg, Switzerland). Все стандартные химические реагенты были получены от Fluka-Sigma-Aldrich, а материалы для культивирования клеток были получены из ТРР.
Манипуляции с ДНК проводили с использованием стандартной техники. ДНК получали в соответствии с инструкциями производителей либо из 2 мл бактериальной культуры с использованием набора QIAprep Spin Plasmid Kit, либо из 50 мл культуры с использованием набора QIAGEN Plasmid Midi Kit. Для гидролиза рестриктирующими ферментами ДНК инкубировали, по меньшей мере, в течение 2 часов с соответствующим рестриктирующим ферментом при концентрации 5-10 единиц (ед.) на мг ДНК в условиях, рекомендованных производителем (буфер и температура). Если реакционные условия были подходящими для всех ферментов, то гидролиз с использованием более чем одного фермента осуществляли одновременно, в противном случае этот гидролиз осуществляли последовательно. ДНК-фрагменты, выделенные для проведения последующих манипуляций, разделяли с помощью электрофореза в 0,7-1,5% агарозном геле, затем вырезали из этого геля и очищали с помощью набора для гель-экстракции QiaExII в соответствии с инструкциями производителя. Для лигирования ДНК-фрагментов 100-200 пг очищенной векторной ДНК инкубировали в течение ночи с трехкратным молярным избытком встраиваемого фрагмента при 16°С в присутствии 1 ед. ДНК-лигазы Т4 в буфере в соответствии с инструкциями производителя (полный объем: 10-20 мкл). Для трансформации XL1-Blue E.coli (Stratagene) использовали аликвоту (0,1-0,5 мкл) реакционной смеси для лигирования. Трансформацию осуществляли путем электропорации с использованием импульсного генератора Gene Pulser (BioRaD) и 0,1 см-кюветы импульсного генератора Gene Pulser Cuvettes (BioRAD) при 200 Ом, 25 мкФ, 1,7 кВ. После электропорации клетки инкубировали при встряхивании в течение 1 часа в 1 мл среды S.O.B. (Miller, 1972), а затем высевали на селективный агар S.O.B.
ПРИМЕР 1
Модулярная эукариотическая экспрессирующая система для связывания антигенов с VLP
Эта система была разработана для присоединения различных аминокислотных линкерных последовательностей, содержащих цистеиновый остаток, к антигенам для химического связывания с VLP.
А. Конструирование экспрессирующей системы, происходящей от EBNA и кодирующей цистеин-содержащий аминокислотный линкер и отщепляемую Fc-метку: рСер-Pu (Wuttke et al., J. Biol. Chem. 276:36839-48 (2001)) гидролизовали ферментами KpnI и BamHI и вводили новый сайт множественного клонирования с использованием гибридизованных олигонуклеотидов PH37 (SEQ ID NO:270) и PH38 (SEQ ID NO:271), в результате чего получали рСер-MCS.
Модулярную систему, содержащую свободный цистеин, фланкированный несколькими глицинами, сайт расщепления протеазой и константную область человеческого IgG1, генерировали следующим образом. рSec2/Hygro B (Invitrogen Cat. №V910-20) гидролизовали ферментами Bsp120I и HindIII и лигировали с гибридизованными олигонуклеотидами SU7 (SEQ ID NO:278) и SU8 (SEQ ID NO:279), в результате чего получали конструкцию pSec-B-MCS. Затем pSec-B-MCS гидролизовали ферментами NheI и HindIII и лигировали с гибридизованными олигонуклеотидами PH29 (SEQ ID NO:264) и PH30 (SEQ ID NO:265) с получением конструкции pSec 29/30. Конструкцию pSec-FL-ЕК-Fc* генерировали путем лигирования трех следующих фрагментов: первого фрагмента pSec 29/30, гидролизованного EcoRI и HindIII; гибридизованных олигонуклеотидов PH31 (SEQ ID NO:266) и PH32 (SEQ ID NO:267) и BglI/EcoRI-фрагмента плазмиды (pSP-Fc*-С1), содержащего модифицированный вариант константной области человеческого IgG1 (детали последовательности чел. IgG1 показаны в последовательности конечной конструкции рСер-Ха-Fc*, см. фиг.1А-1С). Полная последовательность рСер-Ха-Fc* представлена в SEQ ID NO:283. Полученная конструкция была обозначена pSec-FL-ЕК-Fc*. Из этой плазмиды, с помощью гидролиза ферментами NheI и PmeI, вырезали линкерную область и Fc-часть человеческого IgG1 и клонировали в рСер-MCS, гидролизованную NheI и PmeI, с получением конструкции рСер-FL-ЕК-Fc*. Таким образом был сконструирован модулярный вектор, в котором линкерная последовательность и сайт расщепления протеазой, расположенные между NheI- и HindIII-сайтами, могут быть легко заменены гибридизованными олигонуклеотидами. Для генерирования расщепляемого гибридного белка векторы рСер-FL-ЕК-Fc* гидролизовали NheI и HindIII, и сайт расщепления фактором Ха, фланкированный у N-конца аминокислотами GGGGCG, вводили с помощью гибридизованных олигонуклеотидов РН35 (SEQ ID NO:268) и РН36 (SEQ ID NO:269), а сайт энтерокиназы, фланкированный у N-конца GGGGCG, вводили с помощью гибридизованных олигонуклеотидов РН39 (SEQ ID NO:272) и РН40 (SEQ ID NO:273) с получением конструкций рСер-Ха-Fc* (см. фиг.1А) и рСер-ЕК-Fc* (см. фиг.1В), соответственно. Конструкцию рСер-SP-ЕК-Fc* (см. фиг.1С), которая, кроме того, содержала эукариотический сигнальный пептид, генерировали путем лигирования трех фрагментов: рСер-ЕК-Fc*, гидролизованного KpnI/BamHI, гибридизованных олигонуклеотидов PH41 (SEQ ID NO:274) и PH42 (SEQ ID NO:275) и гибридизованных олигонуклеотидов PH43 (SEQ ID NO:276) и PH44 (SEQ ID NO:277).
В. Крупномасштабное продуцирование гибридных белков:
Для крупномасштабного продуцирования различных гибридных белков клетки 293-EBNA (Invitrogen) трансфицировали различными экспрессирующими плазмидами рСер с использованием реагента липофектамина 2000 (Life Technologies) в соответствии с рекомендациями производителя. Через 24-36 часов после трансфекции клетки разделяли в отношении 1:3 путем отбора с использованием пуромицина (1 мкг/мл) в DMEM, в которую была добавлена 10% FCS. Затем резистентные клетки размножали в селективной среде. Для сбора гибридных белков популяцию резистентных клеток высевали на чашки, покрытые поли-L-лизином. После достижения клетками конфлюентности их два раза промывали PBS и в чашки добавляли бессывороточную среду (DMEM). Супернатант тканевой культуры собирали каждые 2-4 дня и заменяли свежей средой DMEM в течение периода времени вплоть до одного месяца. Собранные супернатанты хранили при 4°С.
С. Очистка гибридных белков:
Рекомбинантные Fc-гибридные белки очищали аффинной хроматографией с использованием связанной с белком А сефарозы CL-4В (Amersham Pharmacia Biotech AG). Вкратце, хроматографические колонки упаковывали 1-3 мл смолы, конъюгированной с белком А, и супернатанты тканевых культур, содержащие рекомбинантные белки, наносили на колонку с помощью перистальтического насоса при скорости потока 0,5-1,5 мл/мин. Затем колонку промывали 20-50 мл PBS. В зависимости от гибридного белка расщепление протеазой осуществляли на колонке либо белок элюировали, как описано ниже. Рекомбинантные гибридные белки элюировали цитратным/фосфатным буфером (рН 3,8), в который было добавлено 150 мл NaCl, и фракции, содержащие этот белок, объединяли и концентрировали с использованием фильтров для ультрацентрифугирования Ultrafree (Millipore).
D. Расщепление рекомбинантных гибридных белков протеазами (фактором Ха, энтерокиназой):
Элюированные рекомбинантные гибридные белки, содержащие сайт расщепления энтерокиназой (ЕК), расщепляли с использованием системы EKmax (Invitrogen) в соответствии с рекомендациями производителя. Отщепленную Fc-часть этого гибридного белка удаляли путем инкубирования с белком А. Затем энтерокиназу удаляли с помощью системы ЕК-Away (Invitrogen) в соответствии с рекомендациями производителя. Аналогичным образом, гибридные белки, содержащие сайт расщепления фактором Ха (Ха), гидролизовали с использованием набора для расщепления рестриктирующей протеазой, фактором Ха, и для ее удаления (Roche) в соответствии с рекомендациями производителя. Отщепленную Fc-часть удаляли путем инкубирования с белком А, а протеазу удаляли с помощью конюъгата "стрептавидин - смола", имеющегося в наборе.
Различные гибридные белки концентрировали с использованием фильтра для ультрацентрифугирования (Millipore), количественно оценивали с помощью УФ-спектрофотометрии и использовали в следующих реакциях связывания.
На фиг.1А-1С представлены неполные последовательности различных используемых эукариотических экспрессирующих векторов. Показаны только модифицированные последовательности.
Фиг.1А: рСер-Ха-Fc*: эта последовательность показана от BamHI-сайта и далее, а различные отличительные элементы показаны выше транслированной последовательности. Стрелки указывают на сайт расщепления протеазой фактора Ха.
Фиг.1В: рСер-ЕК-Fc*: эта последовательность показана от BamHI-сайта и далее, а различные отличительные элементы показаны выше транслированной последовательности. Стрелки указывают на сайт расщепления энтерокиназой. Последовательность, расположенная ниже HindIII-сайта, идентична последовательности, представленной на фиг.1А.
Фиг.1С: рСер-SP-ЕК-Fc*: эта последовательность показана, начиная от сигнального пептида и далее, а различные отличительные элементы показаны выше транслированной последовательности. Последовательность сигнального пептида, расщепляемая сигнальной пептидазой, показана жирным шрифтом. Стрелка указывает на сайт расщепления энтерокиназой. Последовательность, расположенная ниже HindIII-сайта, идентична последовательности, представленной на фиг.1А.
ПРИМЕР 2
Эукариотическая экспрессия и связывание мышиного резистина с VLP и с пилями
А. Клонирование мышиного резистина:
Полноразмерную РНК выделяли из 60 мг мышиной жировой ткани с использованием набора Qiagen RNeasy Kit в соответствии с рекомендациями производителя. РНК элюировали в 40 мкл H2O. Затем эту полноразмерную РНК использовали для обратной транскрипции с oligo-dT-праймера с использованием системы ОТ-ПЦР ThermoScriptTM (Life Technologies) в соответствии с рекомендациями производителя. Этот образец инкубировали при 50°С в течение 1 ч, нагревали до 85°С в течение 5 минут и обрабатывали РНКазой Н в течение 20 минут при 37°С.
2 мкл ОТ-содержащей реакционной смеси использовали для ПЦР-амплификации мышиного резистина. ПЦР осуществляли с использованием полимеразы Platinium TAQ (Life Technologies) в соответствии с рекомендациями производителей с помощью праймеров PH19 (SEQ ID NO:260) и PH20 (SEQ ID NO:261). Праймер РН19 (SEQ ID NO:260) соответствовал положениям 58-77, а праймер РН20 (SEQ ID NO:261) соответствовал положениям 454-435 последовательности мышиного резистина. ПЦР-смесь сначала денатурировали при 94°С в течение 2 минут, а затем осуществляли 35 циклов: 30 секунд, 94°С, 30 секунд, 56°С и 1 минута, 72°С, после чего образцы оставляли на 10 минут при 72°С. ПЦР-фрагмент очищали и субклонировали путем ТА-клонирования в вектор pGEMTeasy (Invitrogen), в результате чего получали вектор pGEMT-mRes. Для добавления соответствующих рестрикционных сайтов вторую ПЦР осуществляли на pGEMT-mRes с использованием праймеров РН21 (SEQ ID NO:262) и РН22 (SEQ ID NO:263) в соответствии с той же самой программой проведения циклов, описанной выше. Прямой праймер (РН21) (SEQ ID NO:262) содержит BamHI-сайт и нуклеотиды 81-102 последовательности мышиного резистина. Обратный праймер (РН22) (SEQ ID NO:263) содержал XbaI-сайт и нуклеотиды 426-406 последовательности мышиного резистина. Указанные положения обозначают последовательность мышиного резистина, имеющуюся в Genbank под регистрационным номером AF323080. ПЦР-продукт очищали и гидролизовали ферментами BamHI и XbaI и субклонировали в pcmv-Fc*-C1, гидролизованный ферментами BamHI и XbaI, и получали конструкции pcmv-mRes-Fc*.
Открытую рамку считывания для резистина вырезали из pcmv-Res-Fc* путем гидролиза BamHI/XbaI и клонировали в рСер-Ха-Fc* и рСер-ЕК-Fc* (см. Пример 1, раздел В), гидролизованные BamHI и NheI, с получением конструкций рСер-mRes-Xa-Fc* и рСер-mRes-EK-Fc*, соответственно.
В. Продуцирование, очистка и расщепление резистина
Конструкции рСер-mRes-Ха-Fc* и рСер-mRes-ЕК-Fc* затем использовали для трансфекции клеток 293-EBNA для продуцирования рекомбинантных белков, как описано в примере 1, в разделе В. Супернатанты тканевой культуры очищали, как описано в примере 1, в разделе С. Затем очищенные белки расщепляли, как описано в примере 1, в разделе D. Полученные рекомбинантные белки обозначали "резистин-С-Ха" или "Res-C-Ха" и "резистин-С-ЕК" или "Res-C-ЕК" в соответствии с используемым вектором (см. фиг.2А и Фиг.2В).
На фиг.2А и 2В показана последовательность рекомбинантных мышиных белков резистина, используемых для экспрессии и последующего связывания. Res-C-Ха (фиг.2А) и Res-C-ЕК (фиг.2В) показаны как транслируемые ДНК-последовательности. Сигнальная последовательность резистина, которая отщепляется после секреции белка под действием сигнальной пептидазы, показана курсивом. Аминокислотные последовательности, которые образуются под действием сигнальной пептидазы и специфической протеазы (фактора Ха или энтерокиназы), показаны жирным шрифтом. Выделенные жирным шрифтом последовательности соответствуют нужной последовательности белка, которая используется для связывания, то есть SEQ ID NO:280, SEQ ID NO:281. SEQ ID NO:282 соответствует альтернативной конструкции белка резистина, который может быть также использован для связывания с вирусоподобными частицами или с пилями в соответствии с настоящим изобретением.
С. Связывание белков "резистин-С-Ха" и "резистин-С-ЕК" с капсидным белком Qβ
Раствор 0,2 мл 2 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 5,6 мкл раствора 100 мМ SMPH (Pierce) в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С. После этого 8 мкл диализованной реакционной смеси Qβ подвергали реакции с 32 мкл раствора белка "резистин-С-Ха" (с получением конечной концентрации резистина 0,39 мг/мл), а 13 мкл реакционной смеси Qβ подвергали реакции с 27 мкл раствора белка "резистин-С-ЕК" (с получением конечной концентрации резистина 0,67 мг/мл) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ (см. фиг.2С). Дополнительная полоса размером 24 кДа присутствовала в продукте реакции связывания, но не в дериватизированном Qβ и резистине, соответственно. Размер 24 кДа соответствует ожидаемому размеру 24 кДа для данного продукта связывания (14 кДа для Qβ плюс 10 кДа для белков "резистин-С-Ха" и "резистин-С-ЕК", соответственно).
На фиг.2С показаны результаты связывания белков "резистин-С-Ха" и "резистин-С-ЕК" с Qβ. Продукты связывания анализировали с помощью электрофореза в 16% ПААГ с ДСН в восстанавливающих условиях. Дорожка 1: маркер молекулярной массы. Дорожка 2: "резистин-С-ЕК" перед связыванием. Дорожка 3: "резистин-С-ЕК-Qβ" после связывания. Дорожка 4: дериватизированный Qβ. Дорожка 5: "резистин-С-Ха" перед связыванием. Дорожка 6: "резистин-С-Ха-Qβ" после связывания. Молекулярные массы маркерных белков приведены с левого края. Связанная полоса показана стрелкой.
D. Связывание белков "резистин-С-Ха" и "резистин-С-ЕК" с капсидным белком fr
Раствор 0,2 мл 2 мг/мл капсидного белка fr в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 5,6 мкл раствора 100 мМ SMPH (Pierce) в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С. После этого, 8 мкл диализованной реакционной смеси капсидного белка fr подвергали реакции с 32 мкл раствора белка "резистин-С-Ха" (с получением конечной концентрации резистина 0,39 мг/мл), а 13 мкл реакционной смеси капсидного белка fr подвергали реакции с 27 мкл раствора белка "резистин-С-ЕК" (с получением конечной концентрации резистина 0,67 мг/мл) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ в восстанавливающих условиях.
Е. Связывание белков "резистин-С-Ха" и "резистин-С-ЕК" с HBcAg-Lys-2cys-Mut
Раствор 0,2 мл 2 мг/мл HBcAg-Lys-2cys-Mut в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 5,6 мкл раствора 100 мМ SMPH (Pierce) в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого 8 мкл диализованной реакционной смеси HBcAg-Lys-2cys-Mut подвергали реакции с 32 мкл раствора белка "резистин-С-Ха", а 13 мкл реакционной смеси HBcAg-Lys-2cys-Mut подвергали реакции с 27 мкл раствора белка "резистин-С-ЕК" в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
F. Связывание белков "резистин-С-Ха" и "резистин-С-ЕК" с пилями
Раствор 400 мкл 2,5 мг/мл пилей типа 1 E.coli в 20 мМ Hepes, рН 7,4, подвергали реакции в течение 60 минут с 50-кратным молярным избытком сшивающего линкера SMPH, разведенного из маточного раствора в ДМСО (Pierce) при комнатной температуре на ротационном шейкере. Реакционную смесь обессоливали на колонке PD-10 (Amersham-Pharmacia Biotech). Белоксодержащие фракции, элюированные с колонки, объединяли, и 8 мкл обессоленного дериватизированного белка пилей подвергали реакции с 32 мкл раствора белка "резистин-С-Ха", а 13 мкл обессоленного дериватизированного белка пилей подвергали реакции с 27 мкл раствора белка "резистин-С-ЕК" в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
ПРИМЕР 3
А. Введение cys-содержащих линкеров, экспрессия и очистка мышиного лимфотоксина-β
Внеклеточная часть мышиного лимфотоксина-• (LT-•) рекомбинантно экспрессировалась вместе с аминокислотным линкером CGG у его N-конца. Этот линкер содержал один цистеин для связывания с длинной VLP. Длинный (аминокислоты 49-306) и короткий (аминокислоты 126-306) вариант этого белка присоединяли у их N-конца либо к глутатион-S-трансферазе (GST), либо к гистидин-myc-метке для очистки. Для отщепления метки вводили сайт расщепления энтерокиназой (ЕК).
Конструирование С-LT•49-306 и С-LT•126-306
Мышиный С-LT•49-306 амплифицировали с помощью ПЦР с использованием олигонуклеотидов 5'LT• и 3'LT• из кДНК-библиотеки мышиной селезенки, встроенной в pFB-LIB. Для ПЦР-реакции использовали 0,5 мкг каждого праймера и 200 нг матричной ДНК в 50 · 1 реакционной смеси (1 единица полимеразы PFX Platinum, 0,3 мМ dNTP и 2 мМ MgSO4). Проводили следующие температурные циклы: 94°С, 2 минуты, затем 25 циклов при 94°С (15 секунд), 68°С (30 секунд), 68°С (1 минута), а затем цикл при 68°С, 10 минут. ПЦР-продукт фосфорилировали киназой Т4 и лигировали в плазмиду pEntry1A (Life technologies), которая была разрезана ферментом EcoRV и дефосфорилирована. Полученную плазмиду обозначали pEntry1A-LT•49-306.
Вторую ПЦР-реакцию осуществляли с использованием олигонуклеотидов 5'LT•long-NheI и 3'LT•stop-NotI, и соответственно, с использованием 5'LT•short-NheI и 3'LT•stop-NotI, а также с использованием pEntry1A-LT•49-306 в качестве матрицы. Олигонуклеотиды 5'LT•long-NheI и 5'LT•short-NheI имели внутренний NheI-сайт и содержали кодоны для линкера Cys-Gly-Gly, а 3'LT•stop-NotI имел внутренний NotI-сайт и содержал стоп-кодон. Для второй ПЦР-реакции использовали 0,5 мкг каждого праймера и 150 нг матричной ДНК в 50 · 1 реакционной смеси (1 единица полимеразы PFX Platinum, 0,3 мМ dNTP и 2 мМ MgSO4). Проводили следующей температурные циклы: 94°С, 2 минуты, затем 5 циклов при 94°С (15 секунд), 50°С (30 секунд), 68°С (1 минута), а затем 20 циклов при 94°С (15 секунд), 64°С (30 секунд), 68°С (1 минута), а затем цикл при 68°С, 10 минут.
ПЦР-продукты гидролизовали NheI и NotI и встраивали либо в рСЕР-SP-GST-ЕК, либо в рСЕР-SP-his-myc-ЕК (Wuttke et al., J. Biol. Chem. 276:36839-48 (2001)). Полученные плазмиды обозначали рСЕР-SP-GST-ЕК-С-LT•49-306, рСЕР-SP-GST-ЕК-С-LT•126-306, рСЕР-SP-his-myc-ЕК-С-LT•49-306, рСЕР-SP-his-myc-ЕК-С-LT•126-306, соответственно. GST означает глутатион-S-трансферазу, ЕК означает энтерокиназу, his означает гексагистидиновую метку, а myc означает антитело против эпитопа с-myc. С означает линкер CGG, содержащий дополнительный цистеин.
Все другие стадии осуществляли в соответствии со стандартными протоколами, используемыми в молекулярной биологии.
Последовательность олигонуклеотидов:
5'LT•:
5'-CTT GGT GCC GCA GGA TCA G-3' (SEQ ID NO:284)
3'LT•:
5'-CAG ATG GCT GTC ACC CCA C-3' (SEQ ID NO:285)
5'LT• long-NheI:
5'-GCC CGC TAG CCT GCG GTG GTC AGG ATC AGG GAC GTC G-3' (SEQ ID NO:286)
5'LT• short-NheI:
5'-GCC CGC TAG CCT GCG GTG GTT CTC CAG CTG CGG ATT C-3' (SEQ ID NO:287)
3'LT• stop-NotI
5'-CAA TGA CTG CGG CCG CTT ACC CCA CCA TCA CCG -3' (SEQ ID NO:288)
Экспрессия и продуцирование GST-ЕК-С-LT•49-306,
GST-ЕК-С-LT•126-306, his-myc-ЕК-С-LT•49-306 и his-myc-ЕК-С-LT•126-306
Плазмиды рСЕР-SP-GST-ЕК-С-LT•49-306, рСЕР-SP-GST-ЕК-С-LT• 126-306, рСЕР-SP-his-myc-ЕК-С-LT•49-306, рСЕР-SP-his-myc-ЕК-С-LT• 126-306 трансфицировали в клетки 293-EBNA (Invitrogen) для продуцирования белка как описано в примере 1. Полученные белки обозначали GST-ЕК-С-LT•49-306, GST-ЕК-С-LT•126-306, his-myc-ЕК-С-LT•49-306 и his-myc-ЕК-С-LT•126-306.
Последовательности гибридных белков LT• транслировались из кДНК-последовательностей:
GST-EK-C-LT•49-306: SEQ ID NO:289
GST-EK-C-LT•126-306: SEQ ID NO:290
his-myc-EK-C-LT•49.306: SEQ ID NO:291
his-myc-EK-C-LT•126-306: SEQ ID NO:292
Эти гибридные белки анализировали с помощью электрофореза в 12% ПААГ с ДСН в восстанавливающих условиях. Гели блотировали на нитроцеллюлозные мембраны. Мембраны блокировали, инкубировали с моноклональным мышиным анти-myc антителом или с анти-GST антителом. Затем блоты инкубировали с козьим антимышиным IgG, конъюгированным с пероксидазой хрена, или с кроличьим антикозьим IgG, конъюгированным с пероксидазой хрена. Результаты представлены на фиг.3. GST-ЕК-С-LT•49-306 и GST-ЕК-С-LT•126-306 могут быть детектированы с помощью анти-GST антитела при молекулярной массе 62 кДа и 48 кДа, соответственно. his-myc-ЕК-С-LT•49-306 и his-myc-ЕК-С-LT•126-306 могут быть детектированы с помощью анти-myc антитела при молекулярной массе 40-56 кДа и 33-39 кДа, соответственно.
На фиг.3А и 3В показаны результаты экспрессии гибридных белков LT•. Гибридные белки LT• анализировали с помощью электрофореза в 12% ПААГ с ДСН в восстанавливающих условиях. Гели блотировали на нитроцеллюлозные мембраны. Мембраны блокировали, инкубировали либо с моноклональным мышиным анти-myc антителом (разведение 1:2000) (фиг.3А), либо с анти-GST антителом (разведение 1:2000) (фиг.3В). Затем блоты инкубировали с козьим антимышиным IgG, конъюгированным с пероксидазой хрена (разведение 1:4000) (фиг.3А), или с кроличьим антикозьим IgG, конъюгированным с пероксидазой хрена (разведение 1:4000) (фиг.3В). А: дорожки 1 и 2: his-myc-ЕК-С-LT•126-306. Дорожки 3 и 4: his-myc-ЕК-С-LT•49-306. В: дорожки 1 и 2: GST-ЕК-С-LT•126-306. Дорожки 3 и 4: GST-ЕК-С-LT•49-306. Молекулярные массы маркерных белков приведены с левого края.
В. Очистка GST-ЕК-С-LT•49-306, GST-ЕК-С-LT•126-306 , his-myc-ЕК-С-LT•49-306 и his-myc-ЕК-С-LT•126-306
GST-ЕК-С-LT•49-306 и GST-ЕК-С-LT•126-306 очищали на колонке с глутатион-сефарозой, а his-myc-ЕК-С-LT•49-306 и his-myc-ЕК-С-LT•126-306 очищали на колонке с Ni-NTA-сефарозой в соответствии со стандартными протоколами очистки. Очищенные белки расщепляли энтерокиназой и анализировали с помощью электрофореза в 16% ПААГ с ДСН в восстанавливающих условиях.
С. Связывание С-LT•49-306 и С-LT•126-306 с капсидным белком Qβ
Раствор 120 мкМ капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 25-кратным молярным избытком SMPH (Pierce), разведенным из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого, диализованную реакционную смесь Qβ подвергали реакции с раствором С-LT•49-306 и С-LT•126-306 (конечные концентрации: 60 мкМ Qβ, 60 мкМ С-LT•49-306 и С-LT•126-306) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
D. Связывание С-LT•49-306 и С-LT•126-306 с капсидным белком fr
Раствор 120 мкМ капсида fr в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 25-кратным молярным избытком SMPH (Pierce), разведенным из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого диализованную реакционную смесь капсидного белка fr подвергали реакции с раствором С-LT•49-306 и С-LT•126-306 (конечные концентрации: 60 мкМ fr, 60 мкМ С-LT•49-306 и С-LT•126-306) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
Е. Связывание С-LT•49-306 и С-LT•126-306 с HBcAg-Lys-2cys-Mut
Раствор 120 мкМ HBcAg-Lys-2cys-Mut в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 25-кратным избытком SMPH (Pierce), разведенном из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. Диализованную реакционную смесь HBcAg-Lys-2cys-Mut подвергали реакции с раствором С-LT•49-306 и С-LT•126-306 (конечные концентрации: 60 мкМ HBcAg-Lys-2cys-Mut, 60 мкМ С-LT•49-306 и С-LT•126-306) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
F. Связывание С-LT•49-306 и С-LT•126-306 с пилями
Раствор 125 мкМ пилей типа 1 E.coli в 20 мМ Hepes, рН 7,4, подвергали реакции в течение 60 минут с 50-кратным молярным избытком сшивающего линкера SMPH, разведенного из маточного раствора в ДМСО (Pierce) при комнатной температуре на ротационном шейкере. Реакционную смесь обессоливали на колонке PD-10 (Amersham-Pharmacia Biotech). Белоксодержащие фракции, элюированные с колонки, объединяли, и обессоленный дериватизированный белок пилей подвергали реакции с раствором С-LT•49-306 и С-LT•126-306 (конечные концентрации: 60 мкМ пилей, 60 мкМ С-LT•49-306 и С-LT•126-306) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ в восстанавливающих условиях.
ПРИМЕР 4
А. Введение cys-содержащих линкеров, экспрессия, очистка и связывание крысиного ингибирующего миграцию макрофагов фактора MIF с Qβ
Крысиный фактор, ингибирующий миграцию макрофагов (rMIF), рекомбинантно экспрессировали вместе с тремя различными аминокислотными линкерами С1, С2 и С3, присоединенными у его С-концов. Каждый из этих линкеров содержал один цистеин для связывания с VLP.
Конструирование rMIF-С1, rMIF-С2 и rMIF-С3
MCS плазмиды рЕТ22b(+) (Novagen, Inc.) заменяли на GTTTAACTTTAAGAAGGAGATATACATATGGATCCGGCTAGCGCTCGAGGGTTTAAACGGCGGCCGCATGCACC путем замены исходной последовательности, простирающейся от NdeI-сайта до XhoI-сайта, гибридизованными олигонуклеотидными праймерами MCS-1F и MCS-1R (отжиг в буфере 15 мМ Трис-HCl, рН 8). Полученную плазмиду обозначали pMod00, и эта плазмида имела рестрикционные NdeI-, BamHI-, NheI-, XhoI- PmeI- и NotI-сайты в ее MCS. Гибридизованную пару олигонуклеотидов Bamhis6-ЕК-Nhe-F и Bamhis6-ЕК-Nhe-R и гибридизованную пару "oligo1F-C-глициновый линкер" и "oligo1R-C-глициновый линкер" лигировали вместе в BamHI-NotI-гидролизованную плазмиду pMod00 с получением плазмиды рМоdЕС1, которая имела N-концевую гексагистидиновую метку, сайт расщепления энтерокиназой и С-концевой аминокислотный глициновый линкер, содержащий один цистеиновый остаток. Гибридизованную пару олигонуклеотидов Bamhis6-ЕК-Nhe-F и Bamhis6-ЕК-Nhe-R и гибридизованную пару "oligo1F-C-гамма1-линкер" и "oligo1R-C-гамма1-линкер" лигировали вместе в BamHI-NotI-гидролизованную плазмиду pMod00 с получением плазмиды рМоdЕС2, которая имела N-концевую гексагистидиновую метку, сайт расщепления энтерокиназой и С-концевой • 1-линкер, происходящий от шарнирной области человеческого иммуноглобулина γ1 и содержащий один цистеиновый остаток. Гибридизованную пару олигонуклеотидов Bamhis6-ЕК-Nhe-F и Bamhis6-ЕК-NheI-R, гибридизованную пару "oligo1FA-C-гамма3-линкер" и "oligo1RА-C-гамма3-линкер" и гибридизованную пару "oligo1FB-C-гамма3-линкер" и "oligo1RB-C-гамма3-линкер" лигировали вместе в BamHI-NotI-гидролизованную плазмиду pMod00 с получением рМоdЕС3, которая имела N-концевую гексагистидиновую метку, сайт расщепления энтерокиназой и С-концевой • 3-линкер, содержащий один цистеиновый остаток и происходящий от шарнирной области мышиного иммуноглобулина • 3.
Плазмиду pBS-rMIF, содержащую крысиную кДНК MIF, амплифицировали с помощью ПЦР с использованием олигонуклеотидов rMIF-F и rMIF-Xho-R. rMIF-F имеет внутренний NdeI-сайт, а rMIF-Xho-R имеет внутренний XhoI-сайт. ПЦР-продукт гидролизовали ферментами NdeI и XhoI и лигировали в рМоdЕС1, рМоdЕС2 и рМоdЕС3, гидролизованные теми же самыми ферментами. Полученные плазмиды обозначали pMod-rMIF-С1, pMod-rMIF-С2 и pMod-rMIF-С3, соответственно.
Для ПЦР-реакции 15 пмоль каждого олигонуклеотида и 1 нг матричной ДНК использовали в 50 · 1 реакционной смеси (2 единицы полимеразы PFX, 0,3 мМ dNTP и 2 мМ MgSO4). При этом проводили следующие температурные циклы: 94°С в течение 2 минут, затем 30 циклов при 94°С (30 секунд), 60°С (30 секунд), 68°С (30 секунд), а затем цикл при 68°С в течение 2 минут.
Все другие стадии осуществляли в соответствии со стандартными протоколами, используемыми в молекулярной биологии.
Последовательность олигонуклеотидов:
праймерMCS-1F:
5'-TAT GGA TCC GGC TAG CGC TCG AGG GTT TAA ACG GCG GCC GCA T-3' (SEQ ID NO:293)
праймерMCS-1R:
5'-TCG AAT GCG GCC GCC GTT TAA ACC CTC GAG CGC TAG CCG GAT CCA-3' (SEQ ID NO:294)
Bamhis6-EK-Nhe-F:
5'-GAT CCA CAC CAC CAC CAC CAC CAC GGT TCT GGT GAG GAC GAT GAC AAA GCG CTA GCC C-3' (SEQ ID NO:295)
Bamhis6-EK-Nhe-R:5'-TCG AGG GCT AGC GCT TTG TCA TCG TCG TCA CCA GAA CCG TGG TGG TGG TGG TGG TGT G-3' (SEQ ID NO:296)
oligo1F-C-глициновый линкер:
5'-TCG AGG GTG GTG GTG GTG GTT GCG GTT AAT AAG TTT AAA CGC-3' (SEQ ID NO:297)
oligo1R-C-глициновый линкер:
5'-GGC CGC GTT TAA ACT TAT TAA CCG CAA CCA CCA CCA CCA CCC-3' (SEQ ID NO:298)
oligo1F-C-гамма1-линкер:
5'-TCG AGG ATA AAA CCC ACA CCT CTC CGC CGT GTG GTT AAT AAG TTT AAA CGC-3' (SEQ ID NO:299)
oligo1R-C-гамма1-линкер:
5'-GGC CGC GTT TAA ACT TAT TAA CCA CAC GGC GGA GAG GTG TGG GTT TTA TCC-3' (SEQ ID N0:300)
oligo1FA-C-гамма3-линкер:
5'-TCG AGC CGA AAC CGT CTA CCC CGC CGG GTT CTT CTG-3' (SEQ ID NO:301)
oligo1RA-C-гамма3-линкер:
5'-CAC CAC CAG AAG AAC CCG GCG GGG TAG ACG GTT TCG GC-3' (SEQ ID NO:302)
oligo2FB-C-гамма3-линкер:
5'-GTG GTG CTC CGG GTG GTT GCG GTT AAT AAG TTT AAA CGC-3' (SEQ ID NO:303)
oligo2RB-C-гамма3-линкер:
5'-GGC CGC GTT TAA ACT TAT TAA CCG CAA CCA CCC GGA G-3' (SEQ ID NO:304)
rMIF-F:5'-GGA ATT CCA TAT GCC TAT GTT CAT CGT GAA CAC-3' (SEQ ID NO:305)
rMIF-Xho-R:
5'-CCC GCT CGA GAG CGA AGG TGG AAC CGT TC-3' (SEQ ID NO:306)
Экспрессия и очистка rMIF-С
Компетентные клетки BL21 E.coli (DЕ3) трансформировали плазмидами pMod-rMIF-С1, pMod-rMIF-С2 и pMod-rMIF-С3. Моноколонии из планшетов с агаром, содержащим ампициллин(Amp), размножали в жидкой культуре (SB с 150 мМ MOPS, рН 7,0, 200 мкг/мл Amp, 0,5% глюкозы) и инкубировали при 30°С со встряхиванием в течение ночи при скорости 220 об/мин. Затем 1 л SB (150 мМ MOPS, pH 7,0, 200 мкг/мл Amp) инокулировали ночной культурой, 1:50 об/об, и культивировали до OD600=2,5 при 30°С. Экспрессию индуцировали 2 мМ IPTG. После культивирования в течение ночи клетки собирали и центрифугировали при 6000 об/мин. Клеточный осадок суспендировали в буфере для лизиса (10 мМ Na2HPO4, 30 мМ NaCl, 10 мМ EDTA и 0,25% Твина-20), содержащего 0,8 мг/мл лизоцима, разрушали ультразвуком и обрабатывали бензоназой. Затем 2 мл лизата пропускали через колонку 20 мл QXL- и 20 мл SPXL-колонку. Белки rMIF-C1, rMIF-C2 и rMIF-C3 находились в проходящем потоке.
Последовательности белков rMIF-C были транслированы из кДНК-последовательностей.
rMIF-C1: SEQ ID NO:307
rMIF-C2: SEQ ID NO:308
rMIF-C3: SEQ ID NO:309
Связывание rMIF-С1 с капсидным белком Q•
Раствор 1,48 мл 6 мг/мл капсидного белка Q• в 20 мМ Hepes, 150 мМ NaCl, рН 7,0, подвергали реакции в течение 30 минут с 14,8 мкл SMPH (Pierce) (из 100 мМ маточного раствора; растворенного в ДМСО) при 25°С. Затем реакционный раствор два раза диализовали в течение 3 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. Раствор 1,3 мл 3,6 мг/мл белка rMIF-C1 в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 1 часа с 9,6 мкл ТСЕР (Pierce) (из 36 мМ исходного раствора, растворенного в H2O) при 25°С. После этого 130 мкл дериватизированного и диализованного белка Q• подвергали реакции со 129 мкл восстановленного rMIF-C1 в 241 мкл 20 мМ Hepes, 150 мМ NaCl, рН 7,0, в течение ночи при 25°С.
Связывание rMIF-С2 с капсидным белком Q•
Раствор 0,9 мл 5,5 мг/мл капсидного белка Q• в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 9 мкл SMPH (Pierce) (из 100 мМ исходного раствора, растворенного в ДМСО) при 25°С. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. Раствор 850 мкл 5,80 мг/мл белка rMIF-C2 в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 1 часа с 8,5 мкл ТСЕР (Pierce) (из 36 мМ исходного раствора, растворенного в H2O) при комнатной температуре. После этого, 80 мкл дериватизированного и диализованного белка Q• подвергали реакции с 85 мкл восстановленного rMIF-C2 в 335 мкл 20 мМ Hepes, 150 мМ NaCl, рН 7,2, в течение ночи при 25°С.
Связывание rMIF-С3 с капсидным белком Q•
Раствор 1,48 мл 6 мг/мл капсидного белка Q• в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 14,8 мкл SMPH (Pierce) (из 100 мМ маточного раствора, растворенного в ДМСО) при 25°С. Затем реакционный раствор два раза диализовали в течение 3 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,0, при 4°С. Раствор 720 мкл 5,98 мг/мл белка rMIF-C3 в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 1 часа с 9,5 мкл ТСЕР (Pierce) (из 36 мМ маточного раствора, растворенного в H2O) при 25°С. После этого 130 мкл дериватизированного и диализованного белка Q• подвергали реакции с 80 мкл восстановленного rMIF-C3 в 290 мкл 20 мМ Hepes, 150 мМ NaCl, рН 7,0, в течение ночи при 25°С.
Все три продукта связывания анализировали с помощью электрофореза в 16% ПААГ с ДСН в восстанавливающих условиях. Гели либо окрашивали кумасси бриллиантовым голубым, либо блотировали на нитроцеллюлозные мембраны. Мембраны блокировали, инкубировали с поликлональной кроличьей антисывороткой против Qβ (разведение 1:2000) или с очищенным кроличьим анти-MIF антителом (Torrey Pines Biolabs, Inc.) (разведение 1:2000). Затем блоты инкубировали с козьим антикроличьм IgG, конъюгированным с пероксидазой хрена (разведение 1:2000). Результаты представлены на фиг.4А и фиг.4В. Продукты связывания были детектированы в кумасси-окрашенных гелях (фиг.4А) и помощью анти-Qβ• антисыворотки и анти-MIF антитела (фиг.4В), что явно указывало на ковалентное связывание всех трех вариантов rMIF с капсидным белком Qβ•.
На фиг.4А показано связывание конструкции MIF с Qβ . Продукты связывания анализировали с помощью электрофореза в 16% ПААГ с ДСН в восстанавливающих условиях. Гель окрашивали кумасси бриллиантовым голубым. Молекулярные массы маркерных белков приведены с левого края.
На фиг.4В показано связывание конструкции MIF-С1 с Qβ. Продукты связывания анализировали с помощью электрофореза в 16% ПААГ с ДСН в восстанавливающих условиях. Дорожка 1: MIF-С1 перед связыванием; дорожка 2: дериватизированный Qβ перед связыванием. Дорожки 3-5: Qβ-MIF-С1. Дорожки 1-3 окрашены кумасси бриллиантовым голубым. Дорожки 4-5 представляют Вестерн-блоты реакции связывания, проявленной с помощью анти-MIF антисыворотки и анти-Qβ антисыворотки, соответственно. Молекулярные массы маркерных белков приведены с левого края.
В. Иммунизация мышей MIF-С1, связанным с капсидным белком Qβ
Самок мышей Balb/С вакцинировали MIF-С1, связанным с капсидным белком Qβ, без добавления адъювантов. 25 мкл общего белка каждого образца разводили PBS до 200 мкл и подкожно инъецировали (100 мл с двух вентральных сторон) на день 0 и на день 14. На 31 день у мышей ретроорбитально брали кровь и их сыворотку анализировали с использованием MIF-специфического ELISA.
С. ELISA
ELISA-планшеты сенсибилизировали MIF-С1 при концентрации 5 мкг/мл. Эти планшеты блокировали, а затем инкубировали с серийно разведенными мышиными сыворотками. Связанные антитела детектировали с использованием ферментативно меченного антитела против мышиных IgG. В качестве контроля была также протестирована неиммунная сыворотка тех же самых мышей. Результаты представлены на фиг.4С. При этом явно наблюдалась реактивность мышиной сыворотки против MIF-С1, связанного с капсидным белком Qβ, тогда как неиммунная сыворотка не реагировала с MIF (фиг.4С, данные не приводятся). При серийном разведении антисывороткой против MIF-С1, связанного с капсидным белком Qβ, полумаксимальный титр был достигнут при разведении 1:84000.
На фиг.4С показаны ELISA-сигналы, полученные с использованием сыворотки мышей, вакцинированных MIF-С1, связанным с капсидным белком Qβ. Самок мышей Balb/с вакцинировали подкожно 25 мкг вакцины в PBS на день 0 и на день 14. На 31 день сыворотку оценивали на IgG против MIF-С1. В качестве контроля анализировали неиммунную сыворотку одной из мышей. Результаты для указанных разведений сыворотки представлены в виде оптической плотности при 450 нм. Все вакцинированные мыши имели высокие титры антител IgG. У контроля (невакцинированной мыши) MIF-специфических антител не обнаруживалось.
ПРИМЕР 5
Связывание rMIF-С1 с капсидным белком fr и с капсидным белком HBcAg-Lys-2cys-Mut
Связывание rMIF-С1 с капсидным белком fr
Раствор 100 мкл 3,1 мг/мл капсидного белка fr в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 3 мкл 100 мМ маточного раствора SMPH (Pierce), растворенного в ДМСО, при 25°С. В параллельной реакции капсидный белок fr сначала алкилировали йодацетамидом, а затем подвергали реакции с SMPH в реакционных условиях, описанных выше. Затем реакционные растворы два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. Раствор 80 мкл 5,7 мг/мл белка rMIF-C1 в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 1 часа с 1 мкл 36 мМ маточного раствора ТСЕР (Pierce), растворенного в H2O, при 25°С. 50 мкл дериватизированного и диализованного капсидного белка fr и 50 мкл дериватизированного, алкилированного и диализованного капсидного белка fr, каждого, подвергали реакции с 17 мкл восстановленного rMIF-C1 в течение двух часов при 25°С.
Продукты связывания анализировали с помощью электрофореза в 16% ПААГ с ДСН (фиг.5). Дополнительная полоса ожидаемого размера 27 кДа (rMIF-С1: кажущаяся Mw 13 кДа; капсидный белок fr: кажущаяся Mw 14 кДа) и 29 кДа (rMIF-С1: кажущаяся Mw 13 кДа; HBcAg-Lys-2cys-Mut: кажущаяся Mw 15 кДа) могла быть детектирована в реакции связывания, но не в растворах капсидного белка fr и rMIF-С1, что явно подтверждало связывание.
Связывание rMIF-С1 с капсидным белком HBcAg-Lys-2cys-Mut вируса гепатита
Раствор 100 мкл 1,2 мг/мл капсидного белка HBcAg-Lys-2cys-Mut в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 1,4 мкл SMPH (Pierce) (из 100 мМ маточного раствора, растворенного в ДМСО) при 25°С. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. Раствор 80 мкл 5,7 мг/мл белка rMIF-C1 в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 1 часа с 1 мкл ТСЕР (Pierce) (из 36 мМ маточного раствора, растворенного в H2O) при 25°С. После этого 60 мкл дериватизированного и диализованного капсидного белка HBcAg-Lys-2cys-Mut подвергали реакции с 20 мкл восстановленного rMIF-C1 в течение двух часов при 25°С.
Продукты связывания анализировали с помощью электрофореза в 16% ПААГ с ДСН (фиг.5) в восстанавливающих условиях. Дополнительная полоса ожидаемого размера 28 кДа (rMIF-С1: кажущаяся Mw 13 кДа; HBcAg-Lys-2cys-Mut: кажущаяся Mw 15 кДа) может быть детектирована в реакции связывания, но не в дериватизированном HBcAg-Lys-2cys-Mut или rMIF-С1, что явно подтверждало связывание.
На гель, фиг.5, были загружены следующие образцы: дорожка 1: маркер молекулярной массы. Дорожка 2: rMIF-С1 перед связыванием. Дорожка 3: rMIF-С1 - капсидный белок fr после связывания. Дорожка 4: дериватизированный капсидный белок fr. Дорожка 5: rMIF-С1-fr после связывания с алкилированным капсидным белком fr. Дорожка 6: алкилированный и дериватизированный капсидный белок fr. Дорожка 7: rMIF-HBcAg-Lys-2cys-Mut после связывания. Дорожки 8 и 9: дериватизированный HBcAg-Lys-2cys-Mut. Гель окрашивали кумасси бриллиантовым голубым. Молекулярные массы маркерных белков показаны с левого края.
ПРИМЕР 6
А. Введение аминокислотных линкеров, содержащих цистеиновый остаток, экспрессия и очистка мышиного RANKL
Фрагмент активатора рецептора лиганда ядерного фактора каппа b (RANKL), который также был назван фактором дифференцировки остеокластов, лигандом остеопротегерина и цитокином, индуцируемым активацией, ассоциированной с фактором некроза опухоли, рекомбинантно экспрессировали вместе с N-концевым линкером, содержащим один цистеин для связывания с VLP.
Конструирование экспрессирующей плазмиды
С-концевую кодирующую область гена RANKL подвергали ПЦР-амплификации с использованием олигонуклеотидов RANKL-UP и RANKL-DOWN. RANKL-UP имел внутренний Apal-сайт, а RANKL-DOWN имел внутренний XhoI-сайт. ПЦР-продукт гидролизовали ферментами Apal и XhoI и лигировали в pGEX-6р1 (Amersham Pharmacia). Полученную плазмиду обозначали pGEX-RANKL. Все стадии осуществляли в соответствии со стандартным протоколом, используемым в молекулярной биологии, и последовательность подтверждали. Плазмида pGEX-RANKL кодирует гибридный белок "глутатион-S-трансфераза" - сайт расщепления PreScission - цистеин-содержащий аминокислотный линкер - RANKL (GST-PS-C-RANKL). Цистеин-содержащий аминокислотный линкер имел последовательность GCGGG. Эта конструкция также содержит гексагистидиновую метку между цистеин-содержащим аминокислотным линкером и последовательностью RANKL.
Олигонуклеотиды:
RANKL-UP:5'CTGCCAGGGGCCCGGGTGCGGCGGTGGCCATCATCACCACCATCACCAGCGCTTCTCAGGAG-3' (SEQ ID NO:316)
RANKL-DOWN:
5'-CCGCTCGAGTTAGTCTATGTCCTGAACTTTGAAAG-3' (SEQ ID NO:317)
Белок GST-PS-C-RANKL (SEQ ID NO:318) и кДНК-последовательность GST-PS-C-RANKL (SEQ ID NO:319)

Экспрессия и очистка С-RANKL
Компетентные клетки BL21 E.coli (DЕ3) Gold pLys трансформировали плазмидой pGEX-RANKL. Моноколонии из планшетов с агаром, содержащим канамицин и хлорамфеникол, размножали в жидкой культуре (среда LB, 30 мкг/мл канамицина и 50 мкг/мл хлорамфеникола) и инкубировали при 30°С со встряхиванием при скорости 220 об/мин в течение ночи. Затем 1 л LB (с 30 мкг/мл канамицина) инокулировали ночной культурой, 1:100 об/об, и культивировали до OD600=1 при 24°С. Экспрессию индуцировали 0,4 мМ IPTG. Через 16 часов клетки собирали и центрифугировали при 5000 об/мин. Клеточный осадок суспендировали в буфере для лизиса (50 мМ Трис-HCl, рН=8; 25% сахарозы; 1мМ EDTA; 1% NaN3; 10 мМ DTT; 5 мМ MgCl2; 1 мг/мл лизоцима; 0,4 ед./мл ДНКазы) в течение 30 минут. Затем добавляли 2,5 объема буфера А (50 мМ Трис-HCl, рН=8; 1% тритона Х100; 100 мМ NaCl, 0,1% NaN3; 10 мМ DTT; 1 мМ PMSF) и инкубировали при 37оС в течение 15 мин. Клетки обрабатывали ультразвуком и осаждали при 9000 об/мин в течение 15 минут. Этот супернатант использовали непосредственно для GST-аффинной хроматографии.
Колонку GST-Trap FF (5 мл) (Amersham Pharmacia) уравновешивали в PBS, рН 7,3 (140 мМ NaCl, 2,7 мМ KCl, 10 мМ Na2HPO4, 1,8 мМ КН2PO4). Супернатант загружали на 5 мл-GST-Trap FF-колонку, а затем эту колонку промывали 5 колоночными объемами PBS. Белок GST-PS-C-RANKL элюировали 50 мМ Трис-HCl, рН 8,0, содержащим 10 мМ GSH.
Очищенный белок GST-PS-C-RANKL гидролизовали с использованием протеазы PreScission (Amersham Pharmacia). Гидролиз осуществляли при 37°С в течение 1 часа с использованием конъюгата GST-PS-C-RANKL - PreScission в молярном отношении 500/1.
Кроме того, реакцию гидролиза протеазой осуществляли при замене буфера с использованием обессоливающей колонки HiPrep 26/10 (Amersham Pharmacia), и фракции, содержащие эти белки, объединяли и использовали непосредственно в другой стадии GST-аффинной хроматографии в тех же самых условиях, которые были описаны выше. Очистку С-RANKL оценивали с помощью электрофореза в ДСН-ПААГ в восстанавливающих условиях, как показано на Фиг.6. На этой фигуре, молекулярные массы маркерных белков показаны с левого края геля. Гель окрашивали кумасси бриллиантовым голубым. Расщепленный С-RANKL присутствовал в проходящем потоке (несвязанная фракция), а нерасщепленный GST-PS-C-RANKL, расщепленный GST-PS и PreScission были связаны с данной колонкой. Полученный белок С-RANKL ожидаемого размера 22 кДа имел высокую степень чистоты.
На гель, фиг.6, были загружены следующие образцы: дорожка 1: маркеры низкой молекулярной массы. Дорожки 2 и 3: супернатант клеточных лизатов BL21/DE3, трансформированных "пустым" вектором pGEX6p1 и pGEX-RANKL, соответственно, после индуцирования 0,4 мМ IPTG в течение шестнадцати часов. Дорожка 4: очищенный белок GST-PS-C-RANKL после проведения очистки на GST-Trap FF-колонке. Дорожка 5: фракция, не связанная с GST-Trap FF-колонкой. Дорожка 6: очищенный белок GST-PS-C-RANKL после расщепления протеазой PreScission. Дорожка 7: фракция, не связанная с GST-Trap FF-колонкой, загруженной гидролизатом GST-RANKL, и содержащая очищенный С-RANKL. Дорожка 8: фракция, связанная с GST-Trap FF-колонкой, загруженной гидролизатом GST-PS-C-RANKL, и элюированная GSH.
В. Связывание С-RANKL с капсидным белком Qβ
Раствор 120 мкМ капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 25-кратным молярным избытком SMPH (Pierce), разведенным из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого диализованную реакционную смесь Qβ подвергали реакции с раствором С-RANKL (конечные концентрации: 60 мкМ Qβ, 60 мкМ С-RANKL) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
С. Связывание С-RANKL с капсидным белком fr
Раствор 120 мкМ капсида fr в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 25-кратным молярным избытком SMPH (Pierce), разведенным из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого диализованную реакционную смесь капсидного белка fr подвергали реакции с раствором С-RANKL (конечные концентрации: 60 мкМ капсидного белка fr, 60 мкМ С-RANKL) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
D. Связывание С-RANKL с HBcAg-Lys-2cys-Mut
Раствор 120 мкМ капсида HBcAg-Lys-2cys-Mut в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 25-кратным молярным избытком SMPH (Pierce), разведенным из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. Диализованную реакционную смесь HBcAg-Lys-2cys-Mut подвергали реакции с раствором С-RANKL (конечные концентрации: 60 мкМ HBcAg-Lys-2cys-Mut, 60 мкМ С-RANKL) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
Е. Связывание С-RANKL с пилями
Раствор 125 мкМ пилей типа 1 E.coli в 20 мМ Hepes, рН 7,4, подвергали реакции в течение 60 минут с 50-кратным молярным избытком сшивающего линкера SMPH, разведенного из маточного раствора в ДМСО при комнатной температуре на ротационном шейкере. Реакционную смесь обессоливали на колонке PD-10 (Amersham-Pharmacia Biotech). Белоксодержащие фракции, элюированные с колонки, объединяли, и обессоленный дериватизированный белок пилей подвергали реакции с раствором С-RANKL (конечные концентрации: 60 мкМ пилей, 60 мкМ С-RANKL) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
ПРИМЕР 7
А. Введение аминокислотных линкеров, содержащих цистеиновый остаток, экспрессия и очистка усеченной формы мышиного белка приона
Усеченную форму (аминокислоты 121-230) мышиного белка приона (обозначенного mPrPt) рекомбинантно экспрессировали вместе с аминокислотным линкером GGGGCG, присоединенным у его С-конца для связывания с VLP и с пилями. Этот белок присоединяли к N-концу человеческого Fc-фрагмента для очистки. Сайт расщепления энтерокиназой (ЕК) вводили позади сайта расщепления ЕК для отщепления Fc-части гибридного белка после очистки.
Конструирование mPrPt-ЕК-Fc*
Мышиный PrPt амплифицировали с помощью ПЦР с использованием праймера 5'PrP-BamHI и 3'PrP-NheI и с использованием плазмиды рВРCMVPrP-Fc в качестве матрицы. рВРCMVPrP-Fc содержала последовательность мышиного белка приона дикого типа. 5'PrP-BamHI имел внутренний BamHI-сайт и содержал ATG, а 3'PrP-NheI имел внутренний NheI-сайт.
Для ПЦР-реакции использовали 0,5 мкг каждого праймера и 200 нг матричной ДНК в 50 · 1 реакционной смеси (1 единица полимеразы PFX Platinum, 0,3 мМ dNTP и 2 мМ MgSO4). При этом проводили следующие температурные циклы: 94°С, 2 минуты, затем 5 циклов при 94°С (15 секунд), 50°С (30 секунд), 68°С (45 секунд), а затем 20 циклов при 94°С (15 секунд), 64°С (30 секунд), 68°С (45 секунд), а затем цикл при 68°С, 10 минут.
ПЦР-продукт гидролизовали BamHI и NheI и встраивали в рСер-SP-ЕК-Fc*, содержащую линкерную последовательность GGGGCG у 5'-конца последовательности расщепления ЕК. Полученную плазмиду обозначали рСер-SP-mPrPt-ЕК-Fc*.
Все другие стадии осуществляли в соответствии со стандартными протоколами, используемыми в молекулярной биологии.
Олигонуклеотиды:
Праймер 5'PrP-BamHI
5'-CGG GAT CCC ACC ATG GTG GGG GGC CTT GG-3' (SEQ ID NO:321)
Праймер 3'PrP-NheI
5'-CTA GCT AGC CTG GAT CTT CTC CCG-3' (SEQ ID NO:322)
Экспрессия и очистка mPrPt-ЕК-Fc*
Плазмиду рСер-SP-mPrPt-ЕК-Fc* трансфицировали в клетки 293-EBNA (Invitrogen) и очищали на колонке с сефарозой, связанной с белком А, как описано в примере 1.
Последовательность белка mPrPt-ЕК-Fc* идентифицирована в SEQ ID NO:323.
После отщепления mPrPt имел последовательность, идентифицированную в SEQ ID NO:324 вместе с линкером GGGGCG у его С-конца.
Этот очищенный гибридный белок mPrPt-ЕК-Fc* расщепляли энтерокиназой и анализировали с помощью электрофореза в 16% ПААГ с ДСН в восстанавливающих условиях до и после расщепления энтерокиназой. Гель окрашивали кумасси бриллиантовым голубым. Результат показан на фиг.7. На этой фигуре молекулярные массы маркерных белков показаны с левого края геля. Гибридный белок mPrPt-ЕК-Fc* мог быть детектирован в виде полосы 50 кДа. Отщепленный белок mPrPt, содержащий аминокислотный линкер GGGGCG, присоединенный к его С-концу, мог быть детектирован в виде широкой полосы размером от 18 до 25 кДа. Идентичность mPrPt подтверждали с помощью Вестерн-блоттинга (данные не приводятся). Таким образом, mPrPt с С-концевым аминокислотным линкером, содержащим цистеиновый остаток, может быть экспрессирован и очищен для связывания с VLP и с пилями.
На гель, фиг.7, были загружены следующие образцы: дорожка 1: маркеры молекулярной массы. Дорожка 2: mPrPt-ЕК-Fc* перед расщеплением. Дорожка 3: mPrPt после расщепления.
В. Связывание mPrPt с капсидным белком Qβ
Раствор 120 мкМ капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 25-кратным молярным избытком SMPH (Pierce), разведенным из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого диализованную реакционную смесь Qβ подвергали реакции с раствором mPrPt (конечные концентрации: 60 мкМ Qβ, 60 мкМ mPrPt) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
С. Связывание mPrPt с капсидным белком fr
Раствор 120 мкМ капсида fr в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 25-кратным молярным избытком SMPH (Pierce), разведенным из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого диализованную реакционную смесь капсидного белка fr подвергали реакции с раствором mPrPt (конечные концентрации: 60 мкМ fr, 60 мкМ mPrPt) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
D. Связывание mPrPt с HBcAg-Lys-2cys-Mut
Раствор 120 мкМ капсида HBcAg-Lys-2cys-Mut в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 25-кратным избытком SMPH (Pierce), разведенным из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. Диализованную реакционную смесь HBcAg-Lys-2cys-Mut подвергали реакции с раствором mPrPt (конечные концентрации: 60 мкМ HBcAg-Lys-2cys-Mut, 60 мкМ mPrPt) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
Е. Связывание mPrPt с пилями
Раствор 125 мкМ пилей типа 1 E.coli в 20 мМ Hepes, рН 7,4, подвергали реакции в течение 60 минут с 50-кратным молярным избытком сшивающего линкера SMPH (Pierce), разведенного из маточного раствора в ДМСО при комнатной температуре на ротационном шейкере. Реакционную смесь обессоливали на колонке PD-10 (Amersham-Pharmacia Biotech). Белоксодержащие фракции, элюированные с колонки, объединяли, и обессоленный дериватизированный белок пилей подвергали реакции с раствором mPrPt (конечные концентрации: 60 мкМ пилей, 60 мкМ mPrPt) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
ПРИМЕР 8
А. Связывание пептидов приона с капсидным белком Qβ: вакцины на основе белков прионов
Были химически синтезированы нижеследующие пептиды прионов: CSAMSRPMIHFGNDWEDRYYRENMYR ("cprplong") и CGNDWEDRYYRENMYR ("cprpshort"), которые содержали добавленный N-концевой цистеиновый остаток для связывания с VLP и с пилями, и эти пептиды были использованы для химического связывания с Qβ, как описано ниже.Раствор 5 мл 140 мкМ капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут со 108 мкл раствора SMPH (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 5 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С. После этого 100 мкл диализованной реакционной смеси подвергали реакции либо с 1,35 мкл 2 мМ маточного раствора (в ДМСО) пептида cprpshort (отношение пептид/капсидный белок Q•, 1:2), либо с 2,7 мкл того же самого маточного раствора (отношение пептид/Q•, 1:1). 1 мкл 10 мМ маточного раствора (в ДМСО) пептида cprplong подвергали реакции со 100 мкл диализованной реакционной смеси. Реакции связывания проводили в течение ночи при 15°С в водяной бане. Затем реакционные смеси диализовали в течение 24 часов против 2 х 5 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С.
Продукты связывания центрифугировали, и супернатанты и осадки анализировали с помощью электрофореза в 16% ПААГ с ДСН в восстанавливающих условиях. Гели окрашивали кумасси бриллиантовым голубым. Результаты представлены на фиг.16. На этой фигуре молекулярные массы маркерных белков показаны с левого края геля. Полосы с молекулярной массой от 16,5 до 25 кДа явно указывали на ковалентное связывание пептидов cprpshort и cprplong с капсидным белком Q•.
На гель, фиг.16А, были загружены следующие образцы: дорожка 1: очищенный капсидный белок Q•. Дорожка 2: дериватизированный капсидный белок Qβ перед связыванием. Дорожки 3-6: связывание "капсидный белок Qβ - cprpshort" при отношении пептид/Q·=1:2 (дорожки 3 и 4) и при отношении пептид/Q·=1:1 (дорожки 5 и 6). Также показаны растворимые фракции (дорожки 3 и 5) и нерастворимые фракции (дорожки 4 и 6).
На гель, фиг.16В, были загружены следующие образцы: дорожка 1: маркеры молекулярной массы. Дорожка 2: дериватизированный капсидный белок Qβ перед связыванием. Дорожки 3 и 4: реакции связывания "капсидный белок Qβ - cprplong". Также показана растворимая фракция (дорожка 3) и нерастворимая фракция (дорожка 4).
В. Связывание пептидов прионов с капсидным белком fr
Раствор 120 мкМ капсидного белка fr в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 10-кратным молярным избытком SMPH (Pierce), разведенным из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого диализованную реакционную смесь fr подвергали реакции с эквимолярной концентрацией пептида cprpshort или cprplong/fr в отношении 1:2 в течение ночи при 16°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
С. Связывание пептидов приона с HBcAg-Lys-2cys-Mut
Раствор 120 мкМ HBcAg-Lys-2cys-Mut в 20 мМ Hepes, 150 мМ NaCl, рН 7, 2, подвергали реакции в течение 30 минут с 10-кратным молярным избытком SMPH (Pierce), разведенным из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. Диализованную реакционную смесь HBcAg-Lys-2cys-Mut подвергали реакции с эквимолярной концентрацией пептида cprpshort или cprplong/HBcAg-Lys-2cys-Mut в отношении 1:2 в течение ночи при 16°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
D. Связывание пептидов приона с пилями
Раствор 125 мкМ пилей типа 1 E.coli в 20 мМ Hepes, рН 7,4, подвергали реакции в течение 60 минут с 50-кратным молярным избытком сшивающего линкера SMPH (Pierce), разведенного из маточного раствора в ДМСО при комнатной температуре на ротационном шейкере. Реакционную смесь обессоливали на колонке PD-10 (Amersham-Pharmacia Biotech). Белоксодержащие фракции, элюированные с колонки, объединяли, и обессоленный дериватизированный белок пилей подвергали реакции с пептидами приона в эквимолярной концентрации или в отношении 1:2 в течение ночи при 16°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
ПРИМЕР 9
Клонирование, экспрессия и очистка IL-13 для связывания с VLP и с пилями
А. Клонирование и экспрессия интерлейкина 13 (IL-13) с N-концевым аминокислотным линкером, содержащим цистеиновый остаток для связывания с VLP и пилями
а) Клонирование мышиного IL-13 (НЕК-293Т) для экспрессии в клетках млекопитающих в качестве Fc-гибридного белка
ДНК для клонирования IL-13 выделяли с помощью ОТ-ПЦР из in vitro-активированных спленоцитов, которые были получены следующим образом: CD4+-Т-клетки выделяли из клеток селезенки мышей и инкубировали в течение 3 дней в IMDM (+5% FCS + 10 нг/мл IL-4) в 6-луночных планшетах, которые были предварительно сенсибилизированы анти-CD3 и анти-CD28 антителами. РНК из этих клеток использовали для амплификации IL-13 с помощью одностадийной ОТ-ПЦР (набор для одностадийной ПЦР Qiagen). Для обратной транскрипции РНК использовали праймер XhoIL13-R, а для ПЦР-амплификации кДНК IL-13 использовали праймеры NheIL13-F (SEQ ID NO:338) и XhoIL13-R (SEQ ID NO:339). Амплифицированную кДНК IL13 лигировали в вектор pMOD с использованием рестрикционных NheI/XhoI-сайтов (с получением вектора рMODB1-IL13). рMODB1-IL13 гидролизовали BamHI/XhoI, и фрагмент, содержащий IL-13, лигировали в вектор рСер-SP-XA-Fc* (Δxho), аналог рСер-SP-XA-Fc*, из которого был удален XhoI-сайт в конце последовательности Fc и который был предварительно гидролизован BamHI/XhoI. Плазмиду, полученную в результате лигирования (pCEP-SP-IL13-Fc), секвенировали и использовали для трансфекции клеток НЕК-293Т. Полученная конструкция IL13, кодируемая этой плазмидой, имела аминокислотную последовательность ADPGCGGGGGLA, присоединенную к N-концу зрелой последовательности IL-13. Эта последовательность включает аминокислотную линкерную последовательность GCGGGGG, фланкированную дополнительными аминокислотами, введенными в процессе процедуры клонирования. IL13-Fc может быть очищен из супернатанта клеток, трансфицированных рСер-SP-IL13-Fc, с использованием смолы, связанной с белком А. Результат экспрессии представлен на фиг.17В (см. пример 10 для описания образцов). Зрелый IL-13, соединенный своим N-концом с вышеупомянутой аминокислотной последовательностью, высвобождался после расщепления гибридного белка фактором Ха, что приводило к образованию белка, называемого далее "мышиным С-IL-13-F" и имеющего последовательность, представленную в SEQ ID NO:328. Результат, представленный на фиг.17В, явно продемонстрировал экспрессию конструкции IL-13.
b) Клонирование мышиного IL-13 (НЕК-293Т) для экспрессии в клетках млекопитающих с GST (глутатион-S-трансферазой), присоединенной у его N-конца
кДНК, используемую для клонирования IL-13 с N-концевой GST, получали из кДНК ТН2-активированных Т-клеток, как описано выше (а). IL-13 амплифицировали из этой кДНК с использованием праймеров NhelinkIL13-F и IL13StopXhoNot-R. ПЦР-продукт гидролизовали NheI и XhoI и лигировали в вектор рСер-SP-GST-ЕК, предварительно гидролизованный NheI/XhoI. Плазмиду, выделенную в результате лигирования (pCEP-SP-GST-IL-13), использовали для трансфекции клеток НЕК-293Т. Полученная конструкция IL13, кодируемая этой плазмидой, имела аминокислотную последовательность LACGGGGG, присоединенную у N-конца зрелой последовательности IL-13. Эта последовательность включает аминокислотную линкерную последовательность ACGGGGG, фланкированную дополнительной аминокислотой, введенной в процессе процедуры клонирования. Супернатант клеточной культуры, трансфицированной pCEP-SP-GST-IL-13, содержал гибридный белок GST-IL-13, который мог быть очищен с помощью аффинной хроматография на глутатионе в соответствии со стандартными протоколами. Зрелый IL-13, соединенный своим N-концом с вышеупомянутой аминокислотной последовательностью, высвобождался после расщепления гибридного белка энтерокиназой, что приводило к образованию белка, называемого далее "мышиным С-IL-13-S" и имеющего последовательность, представленную в SEQ ID NO:329.
В. Связывание мышиного С-IL-13-F и мышиного С-IL-13-S с капсидным белком Qβ
Раствор 120 мкМ капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 25-кратным молярным избытком SMPH (Pierce), разведенным из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого диализованную реакционную смесь Qβ подвергали реакции с раствором мышиного С-IL-13-F или мышиного С-IL-13-S (конечные концентрации: 60 мкМ капсидного белка Qβ, 60 мкМ мышиного С-IL-13-F или мышиного С-IL-13-S) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
С. Связывание мышиного С-IL-13-F и мышиного С-IL-13-S с капсидным белком fr
Раствор 120 мкМ капсидного белка fr в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 25-кратным молярным избытком SMPH (Pierce), разведенным из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С. После этого диализованную реакционную смесь капсидного белка fr подвергали реакции с раствором мышиного С-IL-13-F или мышиного С-IL-13-S (конечные концентрации: 60 мкМ капсидного белка fr, 60 мкМ мышиного С-IL-13-F или мышиного С-IL-13-S) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
D. Связывание раствора мышиного С-IL-13-F или мышиного С-IL-13-S с HBcAg-Lys-2cys-Mut
Раствор 120 мкМ капсида HBcAg-Lys-2cys-Mut в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 25-кратным избытком SMPH (Pierce), разведенным из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. Диализованную реакционную смесь HBcAg-Lys-2cys-Mut подвергали реакции с раствором мышиного С-IL-13-F или мышиного С-IL-13-S (конечные концентрации: 60 мкМ HBcAg-Lys-2cys-Mut, 60 мкМ мышиного С-IL-13-F или мышиного С-IL-13-S) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
Е. Связывание раствора мышиного С-IL-13-F или мышиного С-IL-13-S с пилями
Раствор 125 мкМ пилей типа 1 E.coli в 20 мМ Hepes, рН 7,4, подвергали реакции в течение 60 минут с 50-кратным молярным избытком сшивающего линкера SMPH, разведенного из маточного раствора в ДМСО при комнатной температуре на ротационном шейкере. Реакционную смесь обессоливали на колонке PD-10 (Amersham-Pharmacia Biotech). Белоксодержащие фракции, элюированные с колонки, объединяли, и обессоленный дериватизированный белок пилей подвергали реакции с раствором мышиного С-IL-13-F или мышиного С-IL-13-S (конечные концентрации: 60 мкМ пилей, 60 мкМ мышиного С-IL-13-F или мышиного С-IL-13-S) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
ПРИМЕР 10
Клонирование и экспрессия интерлейкина 5 (IL-5) с N-концевым аминокислотным линкером, содержащим цистеиновый остаток для связывания с VLP и пилями
А. Клонирование IL-5 для экспрессии в качестве телец включения в E.coli
IL-5 амплифицировали из клона АТСС (pmIL5-4G; номер АТСС 37562) с помощью ПЦР с использованием следующих двух праймеров: Spelinker3-F1 (SEQ ID NO:340) и Il5StopXho-R (SEQ ID NO:342). Продукт этой ПЦР использовали в качестве матрицы для второй ПЦР, проводимой с помощью праймеров SpeNlinker3-F2 (SEQ ID NO:341) и IlStopXho-R. Вставку гидролизовали SpeI и NotI. Затем эту вставку лигировали в производное вектора рЕТ (вектор рMODEC3-8), предварительно гидролизованное NheI и NotI (не дефосфорилированное), и трансформировали в клетки TG1 E.coli. Конструкция IL-5, генерированная путем клонирования в вектор pMODEC3-8, содержит на своем N-конце гексагистидиновую метку, а затем энтерокиназный сайт, N-концевой аминокислотный линкер гамма 3, содержащий цистеиновый остаток, фланкированный у С-конца последовательностью AS, а у N-конца последовательностью ALV, и зрелую форму гена IL5. Белок, выделенный путем расщепления энтерокиназой, называли "мышиным С-IL-5-E" (SEQ ID NO:332). Плазмидную ДНК полученного клона рMODC6-IL5.2 (также обозначаемого pMODC6-IL5), последовательность которого была подтверждена путем ДНК-секвенирования, трансформировали в штамм BL21 E.coli.
Клон pMODC6-IL5/BL21 культивировали в течение ночи в 5 мл-среде LB, содержащей 1 мг/л ампициллина. 2 мл этой культуры разводили в 100 мл бульона Terrific (TB), содержащего 1 мг/л ампициллина. После того как культура достигала оптической плотности OD600=0,7, ее индуцировали добавлением 0,1 мл 1М раствора изопропил-β-D-тиогалактопиранозида (IPTG). Через каждые 2 часа брали 10 мл образца. Образцы центрифугировали в течение 10 минут при 4000 х g. Образовавшийся осадок ресуспендировали в 0,5 мл буфера для лизиса, содержащего 50 мМ трис-HCl, 2 мМ EDTA, 0,1% тритона Х-100 (рН8). После добавления 20 мкл лизоцима (40 мг/мл) и инкубирования в пробирке в течение 30 минут при 4°С клетки обрабатывали ультразвуком в течение 2 минут. Затем добавляли 100 мкл 50 мМ раствора MgCl2 и 1 мл бензоназы. После этого клетки инкубировали в течение 30 минут при комнатной температуре и центрифугировали в течение 15 минут при 13000 х g.
Супернатант отбрасывали и осадок кипятили в течение 5 минут при 98°С в 100 мкл загрузочного буфера с ДСН. 10 мкл образцов в загрузочном буфере анализировали с помощью электрофореза в ПААГ с ДСН в восстанавливающих условиях (фиг.17А). Гель, показанный на фиг.17A, явно продемонстрировал экспрессию конструкции IL-5. На гель, фиг.17А, были загружены следующие образцы: дорожка М: маркер (NEB, предварительно окрашенный маркерами широкого диапазона). Дорожка 1: клеточный экстракт 1 мл культуры перед индуцированием. Дорожка 2: клеточный экстракт 1 мл культуры через 4 ч после индуцирования.
В. Клонирование IL-5 для экспрессии в клетках млекопитающих (НЕК-293Т)
а) IL-5, который своим N-концом присоединен к аминокислотному линкеру, содержащему цистеиновый остаток, а своим С-концом присоединен к Fc-фрагменту
Матрицу, описанную в (А) (АТСС клон 37562), использовали для клонирования следующей конструкции. Плазмиду pMODВ1-IL5 (производное рЕТ) гидролизовали ферментами BamHI/XhoI с получением небольшого фрагмента, кодирующего IL-5, присоединенный к N-концевому аминокислотному линкеру, содержащему цистеин. Этот фрагмент лигировали в вектор рСЕР-SP-XA-Fc* (ΔXho), который был предварительно гидролизован BamHI и XhoI. Продукт лигирования вводили путем электропорации в штамм ТG1 E.coli, и плазмидную ДНК полученного клона pCEP-SP-IL5-Fc2, последовательность которой подтверждали с помощью ДНК-секвенирования, использовали для трансфекции клеток НЕК-293Т. Полученная конструкция IL-5, кодируемая этой плазмидой, имела аминокислотную последовательность ADPGCGGGGGLA, присоединенную у N-конца зрелой последовательности IL-5. Эта последовательность включает аминокислотную линкерную последовательность GCGGGGG, содержащую цистеин и фланкированную дополнительными аминокисотами, введенными в процессе процедуры клонирования. Белок IL-5, выделенный путем расщепления гибридного белка фактором Ха, будет далее именоваться "мышиным С-IL-5-F" (SEQ ID NO:333).
После трансфекции и отбора на пуромицине супернатант культуры анализировали с помощью Вестерн-блоттинга (фиг.17В) с использованием анти-His (мышиного) антитела и антимышиного антитела IgG, конъюгированного с пероксидазой хрена. Антимышиное антитело IgG, конъюгированное с пероксидазой хрена, также распознавало Fc-гибридные белки. Очистку этого белка осуществляли с помощью аффинной хроматографии на смоле, связанной с белком А. Результат, представленный на фиг.17В, ясно продемонстрировал экспрессию IL-5-конструкции.
На Вестерн-блот, фиг.17В, были загружены следующие образцы:
Дорожка 1: супернатант культуры клеток НЕК, экспрессирующих IL-5-Fc (20 мкл). Электрофорез в ДСН-ПААГ осуществляли в восстанавливающих условиях. Дорожка 2: супернатант культуры клеток НЕК, экспрессирующих IL-13-Fc (20 мкл). Электрофорез в ДСН-ПААГ осуществляли в восстанавливающих условиях. Дорожка 3: супернатант культуры клеток НЕК, экспрессирующих IL-5-Fc (20 мкл). Электрофорез в ДСН-ПААГ осуществляли в невосстанавливающих условиях.
b) Клонирование IL-5 с GST (глутатион-S-трансферазой) и с аминокислотным линкером, содержащим цистеиновый остаток, присоединенный у его N-концаIL-5 (АТСС 37562), амплифицировали с использованием праймеров Nhe-link1-IL13-F и IL5StopXho-R. После гидролиза NheI и XhoI вставку лигировали в вектор рСер-SP-GST-ЕК, который был предварительно гидролизован NheI и XhoI. Полученную плазмиду pCEP-SP-GST-IL5 секвенировали и использовали для трансфекции клеток НЕК-293Т. Полученная конструкция IL5, кодируемая этой плазмидой, имела аминокислотную последовательность LACGGGGG, присоединенную у N-конца зрелой последовательности IL-5. Эта последовательность включает аминокислотную линкерную последовательность ACGGGGG, содержащую цистеиновый остаток и фланкированную дополнительными аминокислотами, введенными в процессе процедуры клонирования. Белок, выделенный путем расщепления энтерокиназой, далее именуют "мышиным С-IL-5-S" (SEQ ID NO:334). Очистку полученного белка осуществляли с помощью аффинной хроматографии на смоле, обладающей сродством к глутатиону.
С. Связывание мышиного С-IL-5-F или мышиного С-IL-5-S с капсидным белком Qβ
Раствор 120 мкМ капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 25-кратным молярным избытком SMPH (Pierce), разведенным из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого диализованную реакционную смесь Qβ подвергали реакции с раствором мышиного С-IL-5-F или мышиного С-IL-5-S (конечные концентрации: 60 мкМ капсидного белка Qβ, 60 мкМ мышиного С-IL-5-F или мышиного С-IL-5-S) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
D. Связывание мышиного С-IL-5-F или мышиного С-IL-5-S с капсидным белком fr
Раствор 120 мкМ капсидного белка fr в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 25-кратным молярным избытком SMPH (Pierce), разведенным из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого диализованную реакционную смесь капсидного белка fr подвергали реакции с раствором мышиного С-IL-5-F или мышиного С-IL-5-S (конечные концентрации: 60 мкМ капсидного белка fr, 60 мкМ мышиного С-IL-5-F или мышиного С-IL-5-S) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
Е. Связывание мышиного С-IL-5-F или мышиного С-IL-5-S с HBcAg-Lys-2cys-Mut
Раствор 120 мкМ капсида HBcAg-Lys-2cys-Mut в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 25-кратным молярным избытком SMPH (Pierce), разведенным из маточного раствора в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. Диализованную реакционную смесь HBcAg-Lys-2cys-Mut подвергали реакции с раствором мышиного С-IL-5-F или мышиного С-IL-5-S (конечные концентрации: 60 мкМ HBcAg-Lys-2cys-Mut, 60 мкМ мышиного С-IL-5-F или мышиного С-IL-5-S) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
F. Связывание мышиного С-IL-5-F или мышиного С-IL-5-S с пилями
Раствор 125 мкМ пилей типа 1 E.coli в 20 мМ Hepes, рН 7,4, подвергали реакции в течение 60 минут с 50-кратным молярным избытком сшивающего линкера SMPH, разведенного из маточного раствора в ДМСО при комнатной температуре на ротационном шейкере. Реакционную смесь обессоливали на колонке PD-10 (Amersham-Pharmacia Biotech). Белоксодержащие фракции, элюированные с колонки, объединяли, и обессоленный дериватизированный белок пилей подвергали реакции с раствором мышиного С-IL-5-F или мышиного С-IL-5-S (конечные концентрации: 60 мкМ пилей, 60 мкМ мышиного С-IL-5-F или мышиного С-IL-5-S) в течение четырех часов при 25°С на ротационном шейкере. Продукты связывания анализировали с помощью электрофореза в ДСН-ПААГ.
ПРИМЕР 11
Введение аминокислотного линкера, содержащего цистеиновый остаток, экспрессия, очистка и связывание фрагмента мышиного васкулярного эндотелиального фактора роста-2 (mVEGFR2, FLK1)
Конструкцию мышиного васкулярного эндотелиального фактора роста-2 (mVEGFR-2, FLK-1), содержащую его второй и третий внеклеточные домены, рекомбинантно экспрессировали в виде Fc-гибридного белка с аминокислотным линкером, содержащим цистеиновый остаток у его С-конца для связывания с VLP и пилями. Последовательности белка mVEGFR-2(2-3) транслировали из кДНК-последовательностей мышиного FLK-1 ((Matthews et al., Proc. Natl. Acad. Sci., USA, 88:9026-9030 (1991)): регистрационный номер Х59397; Ig-подобный домен 2 С2-типа; аминокислоты 143-209; Ig-подобный домен 3 С2-типа, аминокислоты 241-306). Конструкция mVEGFR-2 (2-3) включает последовательность mVEGFR-2 от аминокислоты пролина 126 до аминокислоты лизина 329 (в нумерации белка предшественника). Эта конструкция, помимо иммуноглобулиноподобных доменов 2 и 3 С2-типа, также включает фланкирующие области, за которыми расположен домен 2 и следующий за ним домен 3 в последовательности mVEGFR-2, для присоединения аминокислотной спейсерной молекулы. Аминокислотный линкер, содержащий цистеиновый остаток, был присоединен к С-концу последовательности mVEGFR-2 путем клонирования в вектор рСЕР-SP-ЕК-Fc* (пример 1). Фрагмент mVEGFR-2, клонированный в вектор рСЕР-SP-ЕК-Fc*, кодировал следующую аминокислотную последовательность (SEQ ID NO:345):
PFIAS VSDQHGIVYI TENKNKTVVI PCRGSISNLN VSLCARYPEK RFVPDGNRIS WDSEIGFTLP SYMISYAGMV FCEAKINDET YQSIMYIVVV VGYRIYDVIL SPPHEIELSA GEKLVLNCTA RTELNVGLDF TWHSPPSKSH HKKIVNRDVK PFPGTVAKMF LSTLT·ESVT KSDQGEYTCV ASSGRMIKRN RTFVRVHTKP
Экспрессия рекомбинантного mVEGFR-2(2-3) в эукариотических клетках
Рекомбинантный mVEGFR-2(2-3) был экспрессирован в клетках EBNA 293 с использованием вектора рСЕР-SP-ЕК-Fc*. Вектор рСЕР-SP-ЕК-Fc* имеет BamHI- и NheI-сайты, кодирует аминокислотный линкер, содержащий один цистеиновый остаток, сайт расщепления энтерокиназой и С-концевую человеческую Fc-область. mVEGFR-2(2-3) амплифицировали из кДНК 7-дневного мышиного эмбриона (Marathon-Ready cDNA, Clontech) с помощью ПЦР с использованием пары праймеров BamHI-FLK1-F и NheI-FLK1-В. Для ПЦР-реакции использовали 10 пмоль каждого олигонуклеотида и 0,5 нг кДНК (кДНК 7-дневного мышиного эмбриона (Marathon-Ready cDNA, Clontech) в 50 • 1 реакционной смеси (1 • 1 полимеразной смеси Advantage 2 (50x), 0,2 мМ dNTP и буфер для ПЦР-реакции 5 • 1 10х кДНК). При этом проводили следующие температурные циклы: 5 циклов при 94°С, 1 минута, 94°С, 30 секунд, 54°С, 30 секунд, 72°С, 1 минута; 5 циклов при 94°С (30 секунд), 54°С (30 секунд), 70°С (1 минута), а затем 30 циклов при 94°С (20 секунд), 54°С (30 секунд) и 68°С (1 минута). ПЦР-продукт гидролизовали BamHI и Nhe1 и встраивали в вектор рСЕР-SP-ЕК-Fc*, гидролизованный этими же ферментами. Полученную плазмиду обозначали mVEGFR-2(2-3)-рСЕР-ЕК-Fc. Все другие стадии осуществляли в соответствии со стандартными протоколами, используемыми в молекулярной биологии.
Олигонуклеотиды:
1. Праймер BamH1-FLKl-F
5'-CGCGGATCCATTCATCGCCTCTGTC-3' (SEQ ID NO:343)
2. Праймер Nhe1-FLK1-B
5'-CTAGCTAGCTTTGTGTGAACTCGGAC-3' (SEQ ID NO:344)
Трансфекция и экспрессия рекомбинантного mVEGFR-2(2-3)
Клетки EBNA 293 трансфицировали конструкцией mVEGFR-2(2-3)-рСЕР-Ek-Fc, описанной выше, и бессывороточный супернатант клеток собирали для очистки, как описано в примере 1.
Очистка рекомбинантного mVEGFR-2(2-3)
Очистку экспрессированных белков Fc-ЕК-mVEGFR-2(2-3) осуществляли с использованием белка А, как описано в примере 1. Затем, после связывания гибридного белка с белком А, mVEGFR-2(2-3) отщепляли от связанной с белком А Fc-части с использованием энтерокиназы (EnterokinaseMax, Invitrogen). Гидролиз осуществляли в течение ночи при 37°С (2,5 единиц энтерокиназы/100 мкл гранул белка А, связанного с гибридным белком). Высвобожденный VEGFR2(2-3) отделяли от Fc-части, еще связанной с гранулами белка А, посредством непродолжительного центрифугирования на хроматографических колонках (Micro Bio Spin, Biorad). Для удаления энтерокиназы проходящий поток обрабатывали энтерокиназой Away (Invitrogen) в соответствии с инструкциями производителя.
ПРИМЕР 12
Связывание мышиного пептида VEGFR-2 с капсидным белком Qβ, с HBcAg-Lys-2cys-Mut и с пилями и иммунизация мышей вакцинами на основе VLP-пептида и пептида пилей
А. Связывание мышиных пептидов VEGFR-2 с VLP и пилями
Были химически синтезированы нижеследующие пептиды (Eurogentec, Belgium): мышиный пептид VEGFR-2 CTARTELNVGLDFTWHSPPSKSHHKK, и этот пептид использовали для химического связывания с пилями, как описано ниже.
Связывание мышиных пептидов VEGFR-2 с пилями: Раствор 1400 мкл 1 мг/мл белка пилей в 20 мМ Hepes, рН 7,4, подвергали реакции в течение 60 минут с 85 мкл 100 мМ раствора Sulfo-MBS (Pierce) (в H2O) при комнатной температуре на ротационном шейкере. Реакционную смесь обессоливали на колонке PD-10 (Amersham-Pharmacia Biotech). Белоксодержащие фракции, элюированные с колонки (содержащие приблизительно 1, 4 мг белка), объединяли и подвергали реакции с 2,5-кратным молярным избытком (конечный объем) мышиного пептида VEGFR-2, соответственно. Так, например, к 200 мкл элюата, содержащего приблизительно 0,2 мг дериватизированных пилей, добавляли 2,4 мкл 10 мМ раствора пептида (в ДМСО). Полученную смесь инкубировали в течение четырех часов при 25°С на ротационном шейкере и диализовали против 2 л 20 мМ Hepes, рН 7,2, при 4°С в течение ночи. Результаты связывания анализировали с помощью электрофореза в ДСН-ПААГ в восстанавливающих условиях, как показано на фиг.18А. На гель загружали супернатант (S) и осадок (Р) каждого образца, а также пили и пили, дериватизированные сшивающим линкером Sulfo-MBS (Pierce). На гель, фиг.18А, были загружены следующие образцы: дорожка 1: маркерные белки; дорожки 2-5: связанные образцы (мышиные пили: пили, связанные с мышиным пептидом; человеческие пили: пили, связанные с человеческим пептидом); дорожка 6: пили, дериватизованные сшивающим линкером Sulfo-MBS; дорожки 7-9: три фракции элюата колонки PD-10. Фракция 2 представляет собой фракцию пика, фракции 1 и 3 представляют собой фракции, взятые у краев пика. Полосы связывания были ясно видны на геле, что указывало на успешное связывание мышиного VEGFR-2 с пилями.
Связывание мышиного пептида VEGFR-2 с капсидным белком Qβ: Раствор 1 мл 1 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 45 минут с 20 мкл 100 мМ раствора Sulfo-MBS (Pierce) (в H2O) при комнатной температуре на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, рН 7,4, при 4°С. 1000 мкл диализованной реакционной смеси подвергали реакции с 12 мкл 10 мМ пептидного раствора (в ДМСО) в течение 4 часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение 2 часов против 2 литров 20 мМ Hepes, рН 7,4, при 4°С. Результаты связывания были проанализированы с помощью электрофореза в ДСН-ПААГ в восстанавливающих условиях и представлены на фиг.18В. На гель загружали супернатант (S) каждого образца, а также капсидный белок Qβ и капсидный белок Qβ, дериватизованный сшивающим линкером Sulfo-MBS. Связывание проводили в дубликатах. На гель были загружены следующие образцы: дорожка 1: маркерные белки; дорожки 2, 5: капсидный белок Qβ; дорожки 3, 6: капсидный белок Qβ, дериватизованный Sulfo-MBS; дорожки 4, 7: капсидный белок Qβ, связанный с мышиным пептидом VEGFR-2. Полосы связывания были ясно видны на геле, что указывало на успешное связывание мышиного VEGFR-2 с капсидным белком Qβ.
Связывание мышиного пептида VEGFR-2 с HBcAg-Lys-2cys-Mut: Раствор 3 мл 0,9 мг/мл капсидного белка HBcAg, не содержащего cys (пример 31) в PBS, рН 7,4, подвергали реакции в течение 45 минут с 37,5 мкл 100 мМ раствора Sulfo-MBS (Pierce) (в H2O) при комнатной температуре на ротационном шейкере. Затем реакционный раствор диализовали в течение ночи против 2 л 20 мМ Hepes, рН 7,4. После замены буфера реакционный раствор диализовали еще два часа против этого же буфера. Затем диализованную реакционную смесь подвергали реакции с 3 мкл 10 мМ пептидного раствора (в ДМСО) в течение 4 часов при 25°С на ротационном шейкере. После этого реакционную смесь диализовали против 2 литров 20 мМ Hepes, рН 7,4, в течение ночи при 4°С, а затем буфер заменяли и смесь снова диализовали еще в течение 2 часов против этого же буфера. Результаты связывания были проанализированы с помощью электрофореза в ДСН-ПААГ в восстанавливающих условиях и представлены на фиг.18С. На гель загружали супернатант (S) каждого образца, а также белок HBcAg-Lys-2cys-Mut и белок HBcAg-Lys-2cys-Mut, дериватизированный сшивающим линкером Sulfo-MBS. Связывание проводили в дубликатах. Реакции связывания проводили с 2,5-кратным и с 10-кратным молярным избытком пептида. На гель были загружены следующие образцы: дорожка 1: маркерные белки; дорожки 2, 4, 6, 8: супернатант (S) и осадок (Р) реакций связывания, проводимых с 10-кратным молярным избытком пептида; дорожки 3, 5, 7, 9: супернатант (S) и осадок (Р) реакций связывания, проводимых с 2,5-кратным молярным избытком пептида; дорожка 10: HBcAg-Lys-2cys-Mut, дериватизированный Sulfo-MBS; дорожка 11: HBcAg-Lys-2cys-Mut. Полосы связывания были ясно видны на геле, что указывало на успешное связывание мышиного VEGFR-2 с белком HBcAg-Lys-2cys-Mut.
В. Иммунизация мышей:
Вакцина "пили-пептид"
Самок мышей С3Н-HeJ (дефицитных по рецептору 4 типа "ловушки") и мышей С3Н-HeN (дикого типа) вакцинировали мышиным пептидом VEGFR-2, связанным с белком пилей без добавления адъювантов. Приблизительно 100 мкг общего белка от каждого образца разводили в PBS до 200 мкл и подкожно инъецировали на день 0, день 14 и день 28. У мышей брали кровь ретроорбитально на день 14, 28 и 42, и 42-дневную сыворотку анализировали с использованием ELISA, специфичного для человеческого VEGFR-2.
Вакцина "капсидный белок-Qβ-пептид"
Самок мышей Black 6 вакцинировали мышиным пептидом VEGFR-2, связанным с капсидным белком Qβ с добавлением и без добавления адъюванта (гидроксида алюминия). Приблизительно 100 мкг общего белка от каждого образца разводили в PBS до 200 мкл и подкожно инъецировали на день 0, день 14 и день 28. У мышей брали кровь ретроорбитально на день 14, 28 и 42, и 42-дневную сыворотку анализировали с использованием ELISA, специфичного для человеческого VEGFR-2.
Вакцины на основе "HBcAg-Lys-2cys-Mut"
Самок мышей Black 6 вакцинировали мышиным пептидом VEGFR-2, связанным с белком HBcAg-Lys-2cys-Mut с добавлением и без добавления адъюванта (гидроксида алюминия). Приблизительно 100 мкг общего белка от каждого образца разводили в PBS до 200 мкл и подкожно инъецировали на день 0, день 14 и день 28. У мышей брали кровь ретроорбитально на дни 14, 28 и 42, и 42-дневную сыворотку анализировали с использованием ELISA, специфичного для человеческого VEGFR-2.
С. ELISA
Сыворотки иммунизированных мышей тестировали в ELISA с иммобилизованным мышином пептидом VEGFR-2. Мышиный пептид VEGFR-2 связывали с бычьей РНКазой А с использованием химического сшивающего линкера Sulfo-SPDP. ELISA-планшеты сенсибилизировали связанной РНКазой А в концентрации 10 мкг/мл. Планшеты блокировали, а затем инкубировали с серийным разведением мышиной сыворотки. Связанные антитела детектировали ферментативно меченным антимышиным антителом IgG. В качестве контроля также тестировали неиммунную сыворотку тех же самых мышей. Контрольные ELISA-эксперименты, проводимые с использованием сыворотки мышей, иммунизированных несвязанным носителем, показали, что детектированные антитела были специфичными к соответствующему пептиду. Результаты представлены на фиг.4-6.
Вакцина "пили-пептид"
Результат ELISA приводится на фиг.18D. Результаты для указанных разведений сывороток представлены как оптическая плотность при 450 нм. Даны средние значения из трех измерений для каждой мыши (включая стандартные отклонения). Все вакцинированные мыши обнаруживали титры антител IgG против мышиного пептида VEGFR-2. Какого-либо отличия между мышами, дефицитными по рецептору 4 типа "ловушки", и мышами дикого типа не наблюдалось, что указывало на иммуногенность мышиного аутоантигена, пептида VEGFR-2, у мышей, при его связывании с пилями. Вакцины, инъецируемые мышам, были получены на основе соответствующих проанализированных сывороток.
Вакцина "капсидный белок-Qβ -пептид"
Результаты для указанных разведений сывороток представлены на фиг.18Е как оптическая плотность при 450 нм. Даны средние величины для двух мышей (включая стандартные отклонения). Все вакцинированные мыши обнаруживали титры антител IgG против мышиного пептида VEGFR-2, что свидетельствовало о том, что при связывании с капсидным белком Qβ аутоантиген, мышиный пептид VEGFR-2, становился иммуногенным у мышей. Вакцины, инъецируемые мышам, были получены на основе соответствующих проанализированных сывороток.
Вакцина на основе HBcAg-Lys-2cys-Mut
Результаты для указанных разведений сывороток представлены на фиг.18F как оптическая плотность при 450 нм. Даны средние значения из трех измерений для каждой мыши (включая стандартные отклонения). Все вакцинированные мыши обнаруживали титры антител IgG против мышиного пептида VEGFR-2, что свидетельствовало об иммуногенности мышиного аутоантигена, пептида VEGFR-2, у мышей при его связывании с капсидным белком Qβ. Вакцины, инъецируемые мышам, были получены на основе соответствующих анализированных сывороток.
ПРИМЕР 13
Связывание пептидов Аβ 1-15 с HBcAg-Lys-2cys-Mut и капсидным белком fr
Был химически синтезирован пептид Аβ (DAEFRHDSGYEVHHQGGC), который содержал аминокислотную последовательность из остатков 1-15 человеческого Аβ, присоединенных у его С-конца к последовательности GGC для связывания с VLP и с пилями.
А. а) Связывание пептида Аβ 1-15 с HBcAg-Lys-2cys-Mut с использованием сшивающего линкера SMPH
Раствор 833,3 мкл 1,2 мг/мл белка HBcAg-Lys-2cys-Mut в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 17 мкл раствора 65 мМ SMPH (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С в трубке для диализа с отсечкой молекулярной массы 10000 Да. Затем 833,3 мкл диализованной реакционной смеси подвергали реакции с 7,1 мкл 50 мМ маточного пептидного раствора (маточный исходный раствор в ДМСО) в течение двух часов при 15°С на ротационном шейкере. После этого реакционную смесь диализовали в течение ночи против 1 литра 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С. Затем образец в аликвотах замораживали в жидком азоте и хранили при -80°С вплоть до иммунизации мышей.
b) Связывание пептида Аβ 1-15 с капсидным белком fr с использованием сшивающего линкера SMPH
Раствор 500 мкл 2 мг/мл капсидного белка fr в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 23 мкл раствора 65 мМ SMPH (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, рН 7, 4, при 4°С в трубке для диализа с отсечкой молекулярной массы 10000 Да. Затем 500 мкл диализованной реакционной смеси подвергали реакции с 5,7 мкл 50 мМ маточного пептидного раствора (маточный пептидный раствор в ДМСО) в течение двух часов при 15°С на ротационном шейкере. После этого реакционную смесь диализовали в течение ночи против 1 литра 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С. Затем образец в аликвотах замораживали в жидком азоте и хранили при -80°С вплоть до иммунизации мышей. Образцы продуктов реакции связывания анализировали с помощью электрофореза в ДСН-ПААГ в восстанавливающих условиях.
Результаты экспериментов по связыванию были проанализированы с помощью электрофореза в ДСН-ПААГ и представлены на фиг.19А. На геле видны явные полосы связывания, соответствующие связыванию Аβ 1-15 либо с капсидным белком fr, либо с капсидным белком HBcAg-Lys-2cys-Mut (на фигуре эти полосы показаны стрелками), что свидетельствует об успешном связывании Аβ 1-15 с капсидным белком fr и с капсидным белком HBcAg-Lys-2cys-Mut. Для капсидного белка fr было видно множество полос связывания, а для белка HBcAg-Lys-2cys-Mut была видна, главным образом, одна полоса связывания.
На гель, фиг.19А, были загружены следующие образцы:
1: маркерный белок (kDa Marker 7708S BioLabs. Полосы маркера молекулярной массы с верхней части геля: 175, 83, 62, 47,5, 32,5, 25, 16,5, 6,5 кДа). 2: дериватизированный белок HBcAg-Lys-2cys-Mut. 3: белок HBcAg-Lys-2cys-Mut, связанный с Аβ 1-15, супернатант образца, взятый по окончании реакции связывания и центрифугированный. 4: белок HBcAg-Lys-2cys-Mut, связанный с Аβ1-15, осадок образца, взятый по окончании реакции связывания и центрифугированный. 5: дериватизированный капсидный белок fr. 6: капсидный белок fr, связанный с Аβ1-15, супернатант образца, взятый по окончании реакции связывания и центрифугированный. 4: капсидный белок fr, связанный с Аβ1-15, осадок образца, взятый по окончании реакции связывания и центрифугированный.
В. Иммунизация мышей Balb/c
Самок мышей Balb/с два раза подкожно вакцинировали, на день 0 и на день 14, либо 10 мкг капсидного белка fr, связанного с Аβ 1-15 (fr-Аβ 1-15), либо с 10 мкг HBcAg-Lys-2cys-Mut, связанного с Аβ 1-15 (НВс-Аβ 1-15), разведенными в стерильном PBS. У мышей ретроорбитально брали кровь на день 22, и сыворотку анализировали с использованием Аβ 1-15-специфического ELISA.
С. ELISA
Пептид Аβ 1-15 подвергали связыванию с бычьей РНКазой А с использованием химического сшивающего линкера Sulfo-SPDP. ELISA-планшеты сенсибилизировали конъюгатом "Аβ 1-15-РНКаза" в концентрации 10 мкг/мл. Планшеты блокировали, а затем инкубировали с образцами серийного разведения мышиной сыворотки. Связанные антитела детектировали ферментативно меченным антимышиным антителом IgG. В качестве контроля также тестировали неиммунную сыворотку тех же самых невакцинированных мышей.
На фиг.19В показаны ELISA-сигналы, полученные на день 22 с использованием сыворотки мышей, иммунизированных вакцинами fr-Аβ 1-15 и НВс-Аβ 1-15, соответственно. Также была включена контрольная сыворотка неиммунизированных мышей (неиммунная сыворотка). Результаты, полученные для различных разведений сывороток, представлены как оптическая плотность при 450 нм. Приводятся средние величины для трех вакцинированных мышей. В сыворотке все вакцинированных мышей присутствовали Аβ 1-15-специфические антитела IgG.
ПРИМЕР 14
Связывание пептидов Аβ 1-15, Аβ 1-27 и Аβ 33-42 с пилями типа I
Связывание пептидов Аβ 1-15, Аβ 1-27 и Аβ 33-42 с пилями с использованием сшивающего линкера SMPH.
Были химически синтезированы нижеследующие пептиды Аβ:
пептид DAEFRHDSGYEVHHQGGC ("Аβ 1-15"), который содержит аминокислотную последовательность от остатка 1 до остатка 15 человеческого Аβ, присоединенную у С-конца этого пептида к последовательности GGC для связывания с пилями и VLP, пептид DAEFRHDSGYEVHHQKLVFFAEDVGCNGGC ("Аβ 1-27"), который содержит аминокислотную последовательность от остатка 1 до остатка 27 человеческого Аβ, присоединенную у С-конца этого пептида к последовательности GGC для связывания с пилями и VLP, и пептид CGHGNKSGLMVGGVVIA ("Аβ 33-42"), который содержит аминокислотную последовательность от остатка 33 до остатка 42 Аβ, присоединенную у С-конца этого пептида к последовательности CGHGNKS для связывания с пилями и VLP. Все три пептида были использованы для химического связывания с пилями, как описано ниже.
Раствор 2 мл 2 мг/мл пилей в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 45 минут с 468 мкл раствора SMPH (Pierce) в H2O при 25°С в ротационном шейкере. Затем реакционный раствор загружали на колонку PD10 (Pharmacia) и элюировали 6 х 500 мкл 20 мМ Hepes, 150 мМ NaCl, рН 7,4. Фракции анализировали путем точечного нанесения на нитроцеллюлозу (Scleicher & Schuell) и окрашивали амидо-черным. Фракции 3-6 объединяли. После этого образцы в аликвотах замораживали в жидком азоте и хранили при -80°С вплоть до связывания.
Затем 200 мкл оттаянной обессоленной реакционной смеси смешивали с 200 мкл ДМСО и с 2,5 мкл каждого из соответствующих 50 мМ пептидных маточных растворов в ДМСО, в течение 3,5 часа при комнатной температуре на ротационном шейкере. После этого 400 мкл реакционной смеси три раза диализовали в течение 1 часа против 1 литра 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С в трубке для диализа с отсечкой молекулярной массы 10000 Да. Затем образцы в аликвотах замораживали в жидком азоте и хранили при -80°С.
Получение образцов для электрофореза в ДСН-ПААГ осуществляли следующим образом: 100 мкл диализованной реакционной смеси для связывания инкубировали в течение 10 минут в 10% ТСА на льду, а затем центрифугировали. Осадок ресуспендировали в 50 мкл 8,5М раствора гуанидин-HCl и инкубировали в течение 15 минут при 70°С. Затем образцы осаждали этанолом, и после проведения второй стадии центрифугирования осадок ресуспендировали в буфере для образца.
Результаты экспериментов по связыванию анализировали с помощью электрофореза в ПААГ-ДСН в восстанавливающих условиях. Явные полосы связывания были заметны для всех трех пептидов, что указывало на связывание пептидов Аβ с пилями.
ПРИМЕР 15
Вакцинация мышей АРР23 пептидами Аβ, связанными с капсидным белком Qβ
А. Иммунизация мышей АРР23
Три различных пептида Аβ (Аβ 1-27-Gly-Gly-Cys-Cys-NH2; H-Cys-Gly-His-Gly-Asn-Lys-Ser-Аβ 33-42; Аβ 1-15-Gly-Gly-Cys-NH2) были связаны с капсидным белком Qβ. Полученные вакцины обозначали "Qb-Ab 1-15", "Qb-Ab 1-27" и "Qb-Ab 33-42". Для вакцинации использовали 8-месячных самок мышей АРР23, которые имели человеческий трансген АРР (Sturchler-Pierrat et al., Proc. Natl. Acad. Sci. USA 94:13287-13292 (1997)). Мышам подкожно инъецировали 25 мкг вакцины, разведенной стерильным PBS, и через 14 дней вводили бустер-инъекцию такого же количества вакцины. Перед началом иммунизации и через 7 дней после бустер-инъекции из хвостовой вены мышей брали кровь. Сыворотку анализировали с помощью ELISA.
В. ELISA
Маточные растворы пептидов Аβ 1-40 и Аβ 1-42 приготавливали в ДМСО и перед использованием разводили в сенсибилизирующем буфере. ELISA-планшеты покрывали 0,1 мкг/лунку пептида Аβ 1-40 или Аβ 1-42. Планшеты блокировали, а затем инкубировали серийным разведением мышиной сыворотки. Связанные антитела детектировали ферментативно меченным антимышиным антителом IgG. В качестве контроля также использовали сыворотку, полученную перед вакцинацией. Вычисляли разведение сыворотки, соответствующее среднему для трех стандартных отклонений, превышающих базовый уровень, и это разведение определяли как "ELISA-титр". Все три протестированные вакцины были иммуногенными у мышей АРР23 и индуцировали высокие титры антител против пептидов Аβ 1-40 и/или Аβ 1-42. Результаты представлены на фиг.20. В неиммунной сыворотке неиммунных мышей каких-либо специфических антител не было обнаружено (данные не приводятся).
На фиг.20 показаны ELISA-сигналы, полученные на день 22 с использованием сыворотки мышей, иммунизированных вакцинами fr-Аβ 1-15 и НВс-Аβ 1-15, соответственно. Также была включена контрольная сыворотка неиммунизированных мышей (неиммунная сыворотка). Результаты, полученные для различных разведений сывороток, представлены как оптическая плотность при 450 нм. Приводятся средние величины для трех вакцинированных мышей.
Мышам А21-А30 вводили вакцину Qb-Ab 1-15, мышам А31-А40 вводили вакцину Qb-Ab 1-27, а мышам А41-А49 вводили вакцину Qb-Ab 33-42. Для каждой мыши сывороточные титры антител, специфичных к пептиду Аβ 1-40 и Аβ 1-42, определяли на день 21 с помощью ELISA. Для каждой мыши были представлены ELISA-титры, определенные как разведение сывороток, соответствующее среднему для трех стандартных отклонений выше базового значения. Мыши, вакцинированные Qb-Ab 1-15 или Qb-Ab 1-27, имели высокие титры антител против пептидов Аβ 1-40 и Аβ 1-42, тогда как мыши, вакцинированные Qb-Ab 33-42, имели высокие титры антител только против пептида Аβ 1-42.
ПРИМЕР 16
Связывание Fab-фрагментов антитела с капсидным белком Qβ
Раствор 4,0 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 2,8 мМ SMPH (Pierce) (из маточного раствора, растворенного в ДМСО) при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С.
Fab-фрагмент человеческого IgG, полученный путем гидролиза папаином человеческого IgG, закупали у Jackson Immunolab. Этот раствор (11,1 мг/мл) разводили до концентрации 2,5 мг/мл в 20 мМ Hepes, 150 мМ NaCl, рН 7,2 и оставляли для реакции с различными концентрациями (0-1000 мкМ) либо дитиотрейтола (DTT), либо трикарбоксиэтилфосфина (ТСЕР) в течение 30 минут при 25°С.
Связывание индуцировали путем смешивания дериватизированного и диализованного раствора капсидного белка Qβ с раствором невосстановленного или восстановленного Fab (конечные концентрации: 1,14 мг/мл Qβ и 1,78 мг/мл Fab) и смесь оставляли на ночь при 25°С на ротационном шейкере.
Реакционные продукты анализировали с помощью электрофореза в 16% ПААГ с ДСН в восстанавливающих условиях. Гели окрашивали кумасси бриллиантовым голубым. Результаты представлены на фиг.21. Продукт связывания примерно 40 кДа детектировался в образцах, в которых Fab был восстановлен перед связыванием с использованием 25-1000 мкМ ТСЕР и 25-100 мкМ DTT (фиг.21, показано стрелкой), но не при 10 мкМ ТСЕР, 10 мкМ DTT или 1000 мкМ DTT. Связанная полоса также реагировала с анти-Qβ антисывороткой (данные не приводятся), что явно продемонстрировало ковалентное связывание Fab-фрагмента с капсидным белком Qβ.
На гель, фиг.21, были загружены следующие образцы:
Дорожка 1: маркер молекулярной массы. Дорожки 2 и 3: дериватизированный капсидный белок Qβ перед связыванием. Дорожки 4-13: реакция связывания Qβ-Fab после восстановления Fab, где 4: реакция связывания Qβ-Fab после восстановления Fab под действием 10 мкМ ТСЕР; 5: реакция связывания Qβ-Fab после восстановления Fab под действием 25 мкМ ТСЕР; 6: реакция связывания Qβ-Fab после восстановления Fab под действием 50 мкМ ТСЕР; 7: реакция связывания Qβ-Fab после восстановления Fab под действием 100 мкМ ТСЕР; 8: реакция связывания Qβ-Fab после восстановления Fab под действием 1000 мкМ ТСЕР; 9: реакция связывания Qβ-Fab после восстановления Fab под действием 10 мкМ DTT; 10: реакция связывания Qβ-Fab после восстановления Fab под действием 25 мкМ DTT; 11: реакция связывания Qβ-Fab после восстановления Fab под действием 50 мкМ DTT; 12: реакция связывания Qβ-Fab после восстановления Fab под действием 100 мкМ DTT; 13: реакция связывания Qβ-Fab после восстановления Fab под действием 1000 мкМ DTT. Дорожка 14: Fab перед связыванием. Гель окрашивали кумасси бриллиантовым голубым. Молекулярные массы маркерных белков даны с левого края. Стрелки указывают на связанную полосу.
ПРИМЕР 17
Вакцинация мышей АРР23 пептидами Аβ, связанными с капсидным белком Qβ
А. Иммунизация мышей АРР23
Три различных пептида Аβ (Аβ 1-27-Gly-Gly-Cys-NH2; H-Cys-Gly-His-Gly-Asn-Lys-Ser-Аβ 33-42; Аβ 1-15-Gly-Gly-Cys-NH2) были связаны с капсидным белком Qβ. Полученные вакцины обозначали "Qb-Ab 1-15", "Qb-Ab 1-27" и "Qb-Ab 33-42". Для вакцинации использовали 8-месячных самок мышей АРР23, которые имели человеческий трансген АРР (Sturchler-Pierrat et al., Proc. Natl. Acad. Sci. USA 94:13287-13292 (1997)). Мышам подкожно инъецировали 25 мкг вакцины, разведенной стерильным PBS, и через 14 дней вводили бустер-инъекцию такого же количества вакцины. Перед началом иммунизации и через 7 дней после бустер-инъекции из хвостовой вены мышей брали кровь. Сыворотку анализировали с помощью ELISA.
С. ELISA
Маточные растворы пептидов Аβ 1-40 и Аβ 1-42 приготавливали в ДМСО и перед использованием разводили в сенсибилизирующем буфере. ELISA-планшеты покрывали 0,1 мкг/лунку пептида Аβ 1-40 или Аβ 1-42. Планшеты блокировали, а затем инкубировали серийным разведением мышиной сыворотки. Связанные антитела детектировали ферментативно меченным антимышиным антителом IgG. В качестве контроля также использовали сыворотку, полученную перед вакцинацией. Разведение сывороток, соответствующее среднему для трех стандартных отклонений выше базового значения, вычисляли и определяли как "ELISA-титр". Все три протестированные вакцины были иммуногенными у мышей АРР23 и индуцировали высокие титры антител против пептидов Аβ 1-40 и/или Аβ 1-42. Результаты представлены на фиг.20. В неиммунной сыворотке неиммунных мышей каких-либо специфических антител не было обнаружено (данные не приводятся).
На фиг.20 показаны ELISA-сигналы, полученные на день 22 с использованием сыворотки мышей, иммунизированных вакцинами Qb-Ab 1-15, Qb-Ab 1-27 и Qb-Ab 33-42 соответственно. Мышам А21-А30 вводили вакцину Qb-Ab 1-15, мышам А31-А40 вводили вакцину Qb-Ab 1-27, а мышам А41-А49 вводили вакцину Qb-Ab 33-42. Для каждой мыши сывороточные титры антител, специфичных к пептиду Аβ 1-40 и Аβ 1-42, определяли на день 21 с помощью ELISA. ELISA-титры определяли как разведение сывороток, соответствующее среднему для трех стандартных отклонений выше базового значения для каждой мыши. Мыши, вакцинированные Qb-Ab 1-15 или Qb-Ab 1-27, имели высокие титры антител против пептидов Аβ 1-40 и Аβ 1-42, тогда как мыши, вакцинированные Qb-Ab 33-42, имели высокие титры антител только против пептида Аβ 1-42. Очень сильные иммунные ответы, полученные с использованием человеческих пептидов Аβ у трансгенных мышей, у которых экспрессировался человеческий трансген Аβ, продемонстрировали, что при связывании пептидов Аβ с капсидным белком Qβ может быть преодолена толерантность к аутоантигену.
ПРИМЕР 18
Конструирование, экспрессия и очистка мутантных оболочечных белков Qβ
Конструирование pQβ-240
Плазмиду pQβ10 (Kozlovska T.M. et al., Gene, 137:133-137) использовали в качестве исходной плазмиды для конструирования pQβ-240. Мутацию Lys13→Arg создавали с помощью обратной ПЦР. Обратные праймеры конструировали в обратном направлении "хвост к хвосту":
5'-GGTAACATCGGTCGAGATGGAAAACAAACTCTGGTCC-3' и
5'-GGACCAGAGTTTGTTTTCCATCTCGACCGATGTTACC-3'.
Продукты первой ПЦР были использованы в качестве матриц для второй ПЦР-реакции, в которой использовали обратный праймер 5'-AGCTCGCCCGGGGATCCTCTAG-3' и прямой праймер 5'-CGATGCATTTCATCCTTAGTTATCAATACGCTGGGTTCAG-3'. Продукт второй ПЦР гидролизовали ферментами XbaI и Mph1103I и клонировали в экспрессирующий вектор pQβ10, который был расщеплен теми же самыми рестриктирующими ферментами. ПЦР-реакции осуществляли с использованием реагентов набора для ПЦР в соответствии с протоколом, представленным производителем (MBI Fermentas, Vilnius, Lithuania).
Секвенирование методом прямого включения метки подтверждало нужные мутации. Клетки E.coli, несущие pQβ-240, обеспечивали эффективный синтез 14 кД-белка, который мигрировал на ПААГ вместе с контрольным оболочечным белком Qβ, выделенным из фаговых частиц Qβ.
В результате была получена аминокислотная последовательность (SEQ ID NO:255)
AKLETVTLGNIGRDGKQTLVLNPRGVNPTNGVASLSQAGAVP
ALEKRVTVSVSQPSRNRKNYKVQVKIQNPTACTANGSCDPSVTRQ
KYADVTFSFTQYSTDEERAFVRTELAALLASPLLIDAIDQLNPAY
Конструирование pQβ-243
Плазмиду pQβ10 использовали в качестве исходной плазмиды для конструирования pQβ-243. Мутацию Asn10→Lys создавали с помощью обратной ПЦР. Обратные праймеры конструировали в обратном направлении "хвост к хвосту":
5'-GGCAAAATTAGAGACTGTTACTTTAGGTAAGATCGG-3' и
5'-CCGATCTTACCTAAAGTAACAGTCTCTAATTTTGCC-3'.
Продукты первой ПЦР были использованы в качестве матриц для второй ПЦР-реакции, в которой использовали обратный праймер 5'-AGCTCGCCCGGGGATCCTCTAG-3' и прямой праймер 5'-CGATGCATTTCATCCTTAGTTATCAATACGCTGGGTTCAG-3'. Продукт второй ПЦР гидролизовали ферментами XbaI и Mph1103I и клонировали в экспрессирующий вектор pQβ10, который был расщеплен теми же самыми рестриктирующими ферментами. ПЦР-реакции осуществляли с использованием реагентов набора для ПЦР в соответствии с протоколом, представленным производителем (MBI Fermentas, Vilnius, Lithuania).
Секвенирование методом прямого включения метки подтверждало нужные мутации. Клетки E.coli, несущие pQβ-243, обеспечивали эффективный синтез 14 кД-белка, который мигрировал на ПААГ вместе с контрольным оболочечным белком Qβ, выделенным из фаговых частиц Qβ.
В результате была получена аминокислотная последовательность (SEQ ID NO:256)
AKLETVTLGKIGKDGKQTLVLNPRGVNPTNGVASLSQAGAVP
ALEKRVTVSVSQPSRNRKNYKVQVKIQNPTACTANGSCDPSVTRQ
KYADVTFSFTQYSTDEERAFVRTELAALLASPLLIDAIDQLNPAY
Конструирование pQβ-250
Плазмиду pQβ-240 использовали в качестве исходной плазмиды для конструирования pQβ-250. Мутацию Lys2→Arg создавали с помощью сайт-направленного мутагенеза. Обратный праймер 5'-GGCCATGGCACGACTCGAGACTGTTACTTTAGG-3' и прямой праймер 5'-GATTTAGGTGACACTATAG-3' использовали для синтеза мутантного ПЦР-фрагмента, который вводили в уникальные рестрикционные NcoI- и HindIII-сайты экспрессирующего вектора pQβ-185. ПЦР-реакции осуществляли с использованием реагентов набора для ПЦР в соответствии с протоколом, представленным производителем (MBI Fermentas, Vilnius, Lithuania).
Секвенирование методом прямого включения метки подтверждало нужные мутации. Клетки E.coli, несущие pQβ-250, обеспечивали эффективный синтез 14 кД-белка, который мигрировал на ПААГ вместе с контрольным оболочечным белком Qβ, выделенным из фаговых частиц Qβ.
В результате была получена аминокислотная последовательность (SEQ ID NO:257)
ARLETVTLGNIGRDGKQTLVLNPRGVNPTNGVASLSQAGAVP
ALEKRVTVSVSQPSRNRKNYKVQVKIQNPTACTANGSCDPSVTRQ
KYADVTFSFTQYSTDEERAFVRTELAALLASPLLIDAIDQLNPAY
Конструирование pQβ-251
Плазмиду pQβ10 использовали в качестве исходной плазмиды для конструирования pQβ-251. Мутацию Lys16→Arg создавали с помощью обратной ПЦР. Обратные праймеры конструировали в обратном направлении "хвост к хвосту":
5'-GATGGACGTCAAACTCTGGTCCTCAATCCGCGTGGGG-3' и
5'-CCCCACGCGGATTGAGGACCAGAGTTTGACGTCCATC-3'.
Продукты первой ПЦР были использованы в качестве матриц для второй ПЦР-реакции, в которой использовали обратный праймер 5'-AGCTCGCCCGGGGATCCTCTAG-3' и прямой праймер 5'-CGATGCATTTCATCCTTAGTTATCAATACGCTGGGTTCAG-3'. Продукт второй ПЦР гидролизовали ферментами XbaI и Mph1103I и клонировали в экспрессирующий вектор pQβ10, который был расщеплен теми же самыми рестриктирующими ферментами. ПЦР-реакции осуществляли с использованием реагентов набора для ПЦР в соответствии с протоколом, представленным производителем (MBI Fermentas, Vilnius, Lithuania).
Секвенирование методом прямого включения метки подтверждало нужные мутации. Клетки E.coli, несущие pQβ-251, обеспечивали эффективный синтез 14 кД-белка, который мигрировал на ПААГ вместе с контрольным оболочечным белком Qβ, выделенным из фаговых частиц Qβ. Полученная аминокислотная последовательность, кодируемая этой конструкцией, представлена в SEQ ID NO:259.
Конструирование pQβ-259
Плазмиду pQβ-251 использовали в качестве исходной плазмиды для конструирования pQβ-259. Мутацию Lys2→Arg создавали с помощью сайт-направленного мутагенеза. Обратный праймер 5'-GGCCATGGCACGACTCGAGACTGTTACTTTAGG-3' и прямой праймер 5'-GATTTAGGTGACACTATAG-3' использовали для синтеза мутантного ПЦР-фрагмента, который вводили в уникальные рестрикционные NcoI- и HindIII-сайты экспрессирующего вектора pQβ-185. ПЦР-реакции осуществляли с использованием реагентов набора для ПЦР в соответствии с протоколом, представленным производителем (MBI Fermentas, Vilnius, Lithuania).
Секвенирование методом прямого включения метки подтверждало нужные мутации. Клетки E.coli, несущие pQβ-259, обеспечивали эффективный синтез 14 кД-белка, который мигрировал на ПААГ вместе с контрольным оболочечным белком Qβ, выделенным из фаговых частиц Qβ.
В результате была получена аминокислотная последовательность (SEQ ID NO:258)
AKLETVTLGNIGKDGKQTLVLNPRGVNPTNGVASLSQAGAVP
ALEKRVTVSVSQPSRNRKNYKVQVKIQNPTACTANGSCDPSVTRQ
KYADVTFSFTQYSTDEERAFVRTELAALLASPLLIDAIDQLNPAY
Общие процедуры для экспрессии и очистки Qβ и мутантов Qβ
Экспрессия
E.coli JМ109 трасформируют экспрессирующими плазмидами Q-бета. 5 мл жидкой среды LB с 20 • г/мл ампициллина инокулируют клонами, трансформированными экспрессирующими плазмидами Q-бета. Затем инкубируют при 37°С в течение 16-24 ч без встряхивания.
100-300 мл среды LB, содержащей 20 • г/мл ампициллина, 1:100, инокулируют полученным инокулятом. Затем инкубируют при 37°С в течение ночи без встряхивания. Среду в колбах, содержащую М9 + 1% казаминокислот + 0,2% глюкозы, инокулируют полученным инокулятом, 1:50, и инкубируют при 37°С в течение ночи без встряхивания.
Очистка
Растворы и буферы для процедуры очистки:
1. Буфер для лизиса LB: 50 мМ Трис-HCl, pH 8,0 с 5 мМ EDTA, 0,1% тритона Х-100 и свежеприготовленный! PMSF в концентрации до 5 микрограммов на мл. Без лизоцима и ДНКазы.
2. SAS
Насыщенный сульфат аммония в воде
3. Буфер NET
20 мМ Трис-HCl, pH 7,8 с 5 мМ EDTA и 150 мМ NaCl.
4. PEG
40% (мас./об.) полиэтиленгликоль 6000 в NET
Дизрупция и лизис
Замороженные клетки ресуспендировали в LB при концентрации клеток 2 мг/г. Смесь пять раз обрабатывали ультразвуком с 22 кГц в течение 15 секунд, с интервалами в 1 минуту для охлаждения раствора на льду. Затем лизат центрифугировали при 14000 об/мин в течение 1 ч с использованием ротора Janecki К 60. Все стадии центрифугирования, описанные ниже, осуществляли на том же самом роторе, если это не оговорено особо. Супернатант хранили при 4°С, а клеточный дебрис промывали два раза LB. После центрифугирования супернатанты лизата и промывочные фракции объединяли.
Фракционирование
К вышеуказанному объединенному лизату по каплям при перемешивании добавляли насыщенный раствор сульфата аммония. Объем SAS доводили до 1/5 от общего объема с получением 20% насыщения. Раствор оставляли на ночь и на следующий день центрифугировали при 14000 об/мин в течение 20 минут. Осадок промывали небольшим количеством 20% сульфата аммония и снова центрифугировали. Полученные супернатанты объединяли и по каплям добавляли SAS до получения 40% насыщения. Раствор оставляли на ночь и на следующий день центрифугировали при 14000 об/мин в течение 20 минут. Полученный осадок солюбилизировали в буфере NET.
Хроматография
Капсидный белок, снова солюбилизированный в буфере NET, загружали на колонку с сефарозой CL-4В. В процессе хроматографии элюировались три пика. Первый пик содержал, главным образом, мембраны и мембранные фрагменты, а поэтому его не собирали. Капсиды содержались во втором пике, а в третьем пике содержались другие белки E.coli.
Фракции пиков собирали и NaCl доводили до конечной концентрации 0,65М. Затем по каплям, при перемешивании добавляли объем раствора ПЕГ, соответствующий одной половине собранной фракции пиков. Раствор оставляли на ночь без перемешивания. Капсидный белок осаждали путем центрифугирования при 14000 об/мин в течение 20 минут. Затем его солюбилизировали в минимальном объеме NET и снова загружали на колонку с сефарозой CL-4В. Фракции пиков собирали и осаждали сульфатом аммония при 60% насыщении (мас./об.). После центрифугирования и повторной солюбилизации в буфере NET капсидный белок загружали на колонку с сефарозой CL-4В для повторного проведения хроматографии.
Диализ и сушка
Фракции пиков, полученные, как описано выше, объединяли, интенсивно диализовали против стерильной воды и лиофилизовали для хранения.
Экспрессия и очистка Qβ-240
Клетки (JM109 E.coli, трансформированные плазмидой pQβ-240) ресуспендировали в LB, пять раз обрабатывали ультразвуком в течение 15 секунд (рубашка с ледяной водой) и центрифугировали при 13000 об/мин в течение одного часа. Супернатант хранили при 4°С вплоть до последующей обработки, а дебрис два раза промывали 9 мл LB, и наконец 9 мл 0,7М мочевины в LB. Все супернатанты собирали и загружали на колонку с сефарозой CL-4В. Собранные фракции пиков осаждали сульфатом аммония и центрифугировали. Затем повторно солюбилизированный белок очищали на колонке с сефарозой 2В, а затем на колонке с сефарозой 6В. И наконец, пик капсидных белков интенсивно диализовали против воды и лиофилизовали, как описано выше. Сборка оболочечного белка в капсид была подтверждена с помощью электронной микроскопии.
Экспрессия и очистка Qβ-243
Клетки (RR1 E.coli) ресуспендировали в LB и обрабатывали, как описано в общей процедуре. Белок очищали путем проведения двух последовательных стадий гель-фильтрации на колонке с сефарозой CL-4В и, наконец, на колонке с сефарозой CL-2В. Фракции пиков собирали и лиофилизовали, как описано выше. Сборка оболочечного белка в капсид была подтверждена с помощью электронной микроскопии.
Экспрессия и очистка Qβ-250
Клетки (JM109 E.coli, трансформированные pQβ-250) ресуспендировали в LB и обрабатывали, как описано в общей процедуре. Белок очищали гель-фильтрацией на колонке с сефарозой CL-4В и, наконец, на колонке с сефарозой CL-2В и лиофилизовали, как описано выше. Сборка оболочечного белка в капсид была подтверждена с помощью электронной микроскопии.
Экспрессия и очистка Qβ -259
Клетки (JM109 E.coli, трансформированные pQβ-250) ресуспендировали в LB и обрабатывали ультразвуком. Дебрис один раз промывали 10 мл LB, а второй раз 10 мМ 0,7 М мочевины в LB. Белок очищали путем проведения двух стадий гель-фильтрации на колонке с сефарозой CL-4В. Белок диализовали и лиофилизовали, как описано выше. Сборка оболочечного белка в капсид была подтверждена с помощью электронной микроскопии.
ПРИМЕР 19
Десенсибилизация мышей с аллергией с использованием PLA2, связанного с капсидным белком Qβ
С. Десенсибилизация мышей с аллергией путем вакцинации
Самок мышей СВА/J (в возрасте 8 недель) сенсибилизировали PLA: на одну мышь 0,1 мкг PLA2 от Latoxan (France) абсорбировали на 1 мг квасцов (Imject, Pierce) в общем объеме 66 мкл при интенсивном перемешивании, а затем подкожно инъецировали. Эту процедуру повторяли через каждые 14 дней всего четыре раза. Эта обработка приводила к вырабатыванию PLA2-специфической антисыворотки IgE, но не антител IgG2а. Через 1 месяц после последней сенсибилизации мышам подкожно инъецировали 10 мкг вакцины, содержащей рекомбинантный PLA2, связанный с капсидным белком Qβ. Затем, через 1 и 2 недели, мышей снова обрабатывали тем же самым количеством вакцины. Через одну неделю после последней обработки у мышей брали кровь, а затем внутрибрюшинно инъецировали 25 мкг PLA2 (Latoxan) и измеряли ректальную температуру в течение 60 минут с использованием калиброванного цифрового термометра. В качестве контроля использовали сенсибилизированных мышей, которые были обработаны не капсидным белом Qβ, а PLA2. Все контрольные мыши обнаруживали анафилактическую реакцию, выражающуюся в резком снижении ректальной температуры после введения PLA2, тогда как вакцинированные мыши были полностью или, по меньшей мере, частично защищены. Результаты представлены на фиг.25А.
В. ELISA
ELISA-планшеты (Maxisorp, Nunc) сенсибилизировали PLA2 (Latoxan) при 5 мкг/мл. Планшеты блокировали, а затем инкубировали с серийно разведенной сывороткой. Для детекции антител IgE, сыворотку предварительно обрабатывали гранулами белка G (Pharmacia) в течение 60 минут на шейкере при комнатной температуре. Гранулы удаляли путем центрифугирования и супернатант использовали для ELISA. Антитела, связанные с PLA2, детектировали ферментативно меченными антимышиными антителами IgG2а или IgE. ELISA-титры определяли при полумаксимальной оптической плотности (OD50%) и выражали как log5 предварительно 100-кратно разведенной сыворотки для IgG2а и как -log5 предварительно 10-кратно разведенной сыворотки для IgE. Для всех мышей PLA2-сецифические IgG2а и IgE в сыворотке определяли до и в конце вакцинации. Вакцинация приводила к резкому увеличению уровня PLA2-специфического IgG2а, тогда как изменений в уровнях в IgE-титрах не наблюдалось. Эти результаты показали, что вакцинация приводила к индуцированию Th1-подобного иммунного ответа (ассоциированного с продуцированием IgG2а). Результаты представлены на фиг.25В.
Анафилактический ответ у вакцинированных и невакцинированных мышей представлен на фиг.25А. Мышей сенсибилизировали PLA2, а затем 3 раза обрабатывали путем подкожного введения 10 мкг вакцины, состоящей из PLA2, связанного с капсидным белком Qβ. Контрольных мышей сенсибилизировали, но не вакцинировали. Через одну неделю после последней вакцинации всех мышей подвергали антигенной стимуляции посредством внутрибрюшинной инъекции 25 мкг PLA2 и проводили мониторинг анафилактического ответа путем измерения ректальной температуры в течение 60 минут. Все контрольные мыши обнаруживали резкое снижение температуры тела, тогда как вакцинированные мыши были полностью или, по меньшей мере, частично защищены от анафилактической реакции.
Индуцирование PLA2-специфического IgG2а путем вакцинации показано на фиг.25В.
Мышей сенсибилизировали PLA2, а затем 3 раза обрабатывали 10 мкг вакцины, состоящей из PLA2, связанного с капсидным белком Qβ. Контрольных мышей сенсибилизировали, но не вакцинировали. У сенсибилизированных мышей брали сыворотку перед началом обработки и после завершения обработки, до антигенной стимуляции. У вакцинированных мышей (левая сторона панели) наблюдалось резкое увеличение уровня PLA2-специфического IgG2а.
ПРИМЕР 20
Экспрессия, повторная укладка, очистка и связывание PLA2-Cys (также называемого PLA2 -гибридным белком)
Экспрессия и получение телец включения
Плазмиду рЕТ11а, содержащую ген PLA2-Cys примера ххх, трансформировали в E.coli BL21De3Rill (Stratagene). Ночную культуру выращивали в среде dYT, содержащей 100 мкг/мл ампициллина и 15 мкг/мл хлорамфеникола. Культуру разводили в свежей среде dYT, содержащей ампициллин и хлорамфеникол, и культивировали при 37°С до достижения OD600нм=1. Культуру индуцировали 1 мМ IPTG и культивировали в течение еще 4 часов. Клетки собирали путем центрифугирования и ресуспендировали в буфере PBS, содержащем 0,5 мг/мл лизоцима. После инкубирования на льду клетки на льду обрабатывали ультразвуком и добавляли MgCl2 до концентрации 10 мМ. К клеточному лизату добавляли 6 мкл бензоназы (Merck) и лизат инкубировали в течение 30 минут при комнатной температуре. Затем добавляли тритон до конечной концентрации 1% и лизат инкубировали еще в течение 30 минут на льду. Осадок телец включения (IB) собирали путем центрифугирования в течение 10 минут при 13000 х g. Осадок телец включения промывали в промывочном буфере, содержащем 20 мМ Трис, 23% сахарозу, 1 мМ EDTA, рН 8,0. IB солюбилизировали в 6М гуанидиния-HCl, 20 мМ Триса, рН 8,0, содержащего 200 мМ DTT. Солюбилизированные IB центрифугировали при 50000 х g и супернатант диализовали против 6М гуанидиния-HCl, 20 мМ Триса, рН 8,0, а затем диализовали против того же буфера, содержащего 0,1 мМ DTT. Затем добавляли окисленный глутатион до конечной концентрации 50 мМ и солюбилизированные IB инкубировали в течение 1 часа при комнатной температуре. Солюбилизированные IB диализовали против 6М гуанидиния-HCl, 20 мМ Триса, рН 8,0. Концентрацию раствора IB оценивали с помощью анализа Бредфорда и электрофореза в ДСН-ПААГ.
В. Повторная укладка и очистка
Раствор IB медленно тремя порциями добавляли каждые 24 часа до конечной концентрации 3 мкМ в буфер для повторной укладки, содержащий 2 мМ EDTA, 0,2 мМ бензамидина, 0,2 мМ 6-аминокапроновой кислоты, 0,2 мМ гуанидиния-HCl, 0,4 М L-аргинина, рН 6,8, в который, перед инициацией повторной укладки при 4°С, добавляли 5 мМ восстановленного глутатиона и 0,5 мМ окисленного глутатиона. Раствор для повторной укладки концентрировали до половины его объема путем ультрафильтрации с использованием мембраны YМ10 (Millipore) и диализовали против PBS, рН 7,2, содержащего 0,1 мМ DTT. Затем белок концентрировали путем ультрафильтрации и загружали на колонку с Superdex G-75 (Pharmacia), уравновешенную в 20 мМ Hepes, 150 мМ NaCl, 0,1 мМ DTT, 4°С для очистки. рН уравновешивающего буфера доводили до 7,2 при комнатной температуре. Мономерные фракции объединяли.
С. Связывание
Раствор 1,5 мг Qβ в 0,75 мл 20 мМ Hepes, 150 мМ NaCl, рН 7,4 подвергали реакции с 0,06 мл Sulfo-SMPB (Pierce; 31 мМ маточного раствора в H2O) в течение 45 минут при комнатной температуре. Реакционную смесь диализовали в течение ночи против 20 мМ Hepes, 150 мМ NaCl, рН 7,4, и 0,75 мл этого раствора смешивали с 1,5 мл раствора PLA2-Cys в 0,1 мМ DTT (62 мкМ) и 0,43 мл 20 мМ Hepes, 150 мМ NaCl, 137 мкМ DTT, рН 7,4, скорректированного при комнатной температуре. Реакционную смесь для связывания оставляли на 4 часа при комнатной температуре и эту смесь диализовали в течение ночи против 20 мМ Hepes, 150 мМ NaCl, рН 7,4, с использованием трубки для диализа Spectra Por, с отсечкой Mw 300000 Да (Spectrum). Реакцию связывания анализировали с помощью электрофореза в ДСН-ПААГ и окрашивания кумасси, а также с помощью Вестерн-блоттинга с использованием либо кроличьей антисыворотки против пчелиного яда (разведенной 1:10000), визуализированной с помощью козьего антикроличьего антитела, конъюгированного со щелочной фосфатазой (разведенного 1:10000), или кроличьей анти-Qβ антисыворотки (разведенной 1:5000), проявленной с помощью козьего антикроличьего антитела, конъюгированного со щелочной фосфатазой (разведенного 1:10000). В обоих случаях образцы подвергали электрофорезу в восстанавливающих условиях.
Результат реакции связывания представлен на Фиг.26. Полосы, соответствующие продукту связывания капсидного белка Qβ с PLA2-Cys, отчетливо видны в окрашенном кумасси ДСН-ПААГ (левая панель), в анти-Qβ Вестерн-блоте (центральная панель) и в анти-PLA2 Вестерн-блоте (правая панель) реакций связывания между капсидным белком Qβ и PLA2-Cys и показаны стрелками. 15 мкл реакционных смесей для связывания и 50 мкл диализованных реакционных смесей для связывания загружали на гель.
Дорожка 1: маркерный белок; 2: диализованная реакционная смесь для связывания 1; 3: реакция связывания 2; 5: реакция связывания 2; 6: реакция связывания 1; 7: диализованная реакционная смесь для связывания 1; 8: маркерный белок; 9: реакция связывания 2; 10: реакция связывания 1; 11: диализованная реакционная смесь для связывания 1; 12: маркерный белок.
ПРИМЕР 21
Связывание антиидиотипического мимоантитела IgE VAE051 с Qβ, иммунизация мышей и тестирование антисыворотки
Раствор 4,0 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 10-кратным молярным избытком SMPH (Pierce) (из 100 мМ маточного раствора, растворенного в ДМСО) при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. Раствор VAE051 (2,4 мг/мл) восстанавливали с использованием эквимолярной концентрации ТСЕР в течение 60 минут при 25°С.
Затем 46 мкл диализованной реакционной смеси Qβ подвергали реакции с 340 мкл ТСЕР-обработанного раствора VAE051 (2,4 мг/мл) в полном объеме 680 мкл 50 мМ натрийацетатного буфера при 16°С в течение 2 ч на ротационном шейкере.
Реакционные продукты анализировали с помощью электрофореза в 16% ПААГ с ДСН в восстанавливающих условиях. Гели окрашивали кумасси бриллиантовым голубым. Две дополнительные полосы в реакциях связывания (которые отсутствовали в растворах VAE или Qβ) соответствовали тяжелой цепи и легкой цепи VAE051, связанного с Qβ (фиг.28А). Идентичность полос подтверждали с помощью Вестерн-блоттинга с использованием антител, специфичных к тяжелой и легкой цепям, соответственно.
Иммунизация мышей
Раствор для связывания Qβ-VAE051 диализовали против 20 мМ Hepes, 150 мМ NaCl, рН 7,2, с использованием мембраны с отсечкой 300000 Да. 50 мкг Qβ-VAE051 инъецировали внутрибрюшинно двум самкам мышей Balb/с на день 0 и на день 14. У этих мышей ретроорбитально брали кровь на день 28 и их сыворотку анализировали с использованием IgE- и VAE051-специфических ELISA-анализов.
ELISA
ELISA-планшеты сенсибилизировали человеческим IgE при концентрации 0,8 мкг/мл или 10 мкг/мл VAE051. Планшеты блокировали, а затем инкубировали с серийно разведенной мышиной сывороткой. Связанные антитела детектировали ферментативно меченным антимышиным антителом IgG (фиг.28В).
Обе мыши обнаруживали высокую реактивность к VAE051, а также к человеческому IgE. Неиммуннные сыворотки тех же самых мышей не обнаруживали какой-либо реактивности к VAE051 и IgE (фиг.28В). Это свидетельствовало о том, что были продуцированы антитела против антиидиотипического мимоантитела IgE, VAE051, которые также распознавали "родительскую" молекулу IgE.
ПРИМЕР 22
Высокий уровень связывания пептида DerpI с капсидным белком Qβ дикого типа с использованием сшивающего линкера SMPH
Пептид Derp 1,2, к N-концу которого, для связывания, был присоединен цистеин, получали методом химического синтеза, и этот пептид имел следующую последовательность: H2N-CQIYPPNANKIREALAQTHSA-COOH. Указанный пептид использовали для химического связывания с капсидным белком Qβ дикого типа, как описано ниже.
D. Связывание пептида Flag с капсидным белком Qβ
Капсидный белок Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при концентрации 2 мг/мл подвергали реакции в течение 30 минут с 5- или 20-кратным избытком сшивающего линкера SMPH (Pierce) при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. Диализованную реакционную смесь подвергали реакции с 5-кратным избытком пептида Derp 1,2 в течение 2 часов при 25°С на ротационном шейкере.
Результат реакции связывания можно видеть на фиг.24. Полосы, соответствующие 1, 2 и 3 пептидам на субъединицу, соответственно, отчетливо видны на геле и показаны стрелками. В среднем, на капсиде присутствовало два пептида на субъединицу.
На гель, фиг.24, загружали следующие образцы: дорожки 1: маркерный белок; 2: капсидный белок Qβ, дериватизированный 5-кратным избытком SMPH; 3: капсидный белок Qβ, дериватизированный 20-кратным избытком SMPH; 4: реакция связывания 5-кратно дериватизированного капсидного белка Qβ; 5: реакция связывания 20-кратно дериватизированного капсидного белка Qβ.
ПРИМЕР 23
Введение пептида, содержащего лизиновый остаток в с/е1-эпитоп HBcAg(1-149)
Эпитоп с/е1 (остатки 72-88) HBcAg расположен в концевой области на поверхности капсида вируса гепатита В (HBcAg). Часть этой области (пролин 79 и аланин 80) генетически заменяли пептидом Gly-Gly-Lys-Gly-Gly (конструкция HBcAg-Lys). Введенный лизиновый остаток, в своей боковой цепи, содержал реактивную аминогруппу, которая может быть использована для межмолекулярного химического сшивания частиц HBcAg с любым антигеном, содержащим свободную цистеиновую группу.
ДНК HBcAg-Lys, имеющую аминокислотную последовательность, представленную в SEQ ID NO:158, генерировали с помощью ПЦР. Для этого два фрагмента, кодирующие фрагменты HBcAg (аминокислотные остатки 1-78 и 81-149), отдельно амплифицировали с помощью ПЦР. Праймеры, используемые для этих ПЦР, также обеспечивали введение ДНК-последовательности, кодирующей пептид Gly-Gly-Lys-Gly-Gly. Фрагмент HBcAg (1-78) амплифицировали из рЕсо63 с использованием праймеров EcoRIHBcAg(s) и Lys-HBcAg(as). Фрагмент HBcAg (81-149) амплифицировали из рЕсо63 с использованием праймеров Lys-HBcAg(s) и HBcAg(1-149)Hind(as). Праймеры Lys-HBcAg(as) и Lys-HBcAg(s) обеспечивали введение комплементарных ДНК-последовательностей на концах двух ПЦР-продуктов, что приводило к образованию гибрида этих двух ПЦР-продуктов при последующей ПЦР-сборке. После сборки фрагменты амплифицировали с помощью ПЦР и с использованием праймеров EcoRIHBcAg(s) и HBcAg(1-149)Hind(as).
Для проведения ПЦР 100 пмоль каждого олигонуклеотида и 50 нг ДНК-матриц использовали в 50 мл реакционных смесей с 2 единицами полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. Для обеих реакций проводили следующие температурные циклы: один цикл 94°С, 2 минуты, и 30 циклов при 94°С (1 минута), 50°С (1 минута), 72°С (2 минуты).
Последовательности праймеров:EcoRIHBcAg(s):
(5'-CCGGAATTCATGGACATTGACCCTTATAAAG-3') (SEQ ID N0:79);
Lys-HBcAg(as):
(5'-CCTAGAGCCACCTTTGCCACCATCTTCTAAATTAGTACCCACCCAGGTAGC-3')
(SEQ ID NO:80);
Lys-HBcAg(s):
(5'-GAAGATGGTGGCAAAGGTGGCTCTAGGGACCTAGTAGTCAGTTATGTC-3')
(SEQ ID NO:81);
HBcAg(l-149)Hind(as):
(5'-CGCGTCCCAAGCTTCTAAACAACAGTAGTCTCCGGAAG-3')
(SEQ ID NO:82).
Для присоединения двух ПЦР-фрагментов с помощью ПЦР, 100 пмоль праймеров EcoRIHBcAg(s) и HBcAg(1-149)Hind(as) использовали вместе с 100 нг двух очищенных ПЦР-фрагментов в 50 мл реакционных смесей, содержащих 2 единицы полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. ПЦР-циклы проводили при следующих условиях: один цикл при 94°С, 2 минуты, а затем 30 циклов при 94°С (1 минута), 50°С (1 минута), 72°С (2 минуты). После сборки ПЦР-продукты анализировали с помощью электрофореза в агарозном геле, очищали и гидролизовали в течение 19 часов в соответствующем буфере рестриктирующими ферментами EcoRI и HindIII. Гидролизованный ДНК-фрагмент лигировали в EcoRI/HindIII-гидролизованный вектор рКК с получением экспрессирующего вектора рКК-HBcAg-Lys. Встраивание ПЦР-продукта в этот вектор оценивали путем EcoRI/HindIII-рестрикционного анализа и ДНК-секвенирования этой вставки.
ПРИМЕР 24
Экспрессия и неполная очистка HBcAg-Lys
Штамм E.coli XL-1 blue трансформировали вектором рКК-HBcAg-Lys. 1 мл ночной культуры бактерий использовали для инокуляции 100 мл среды LB, содержащей 100 мкг/мл ампициллина. Эту культуру выращивали в течение 4 часов при 37°С до тех пор, пока OD при 600 нм не достигала примерно 0,8. Индуцирование синтеза HBcAg-Lys осуществляли добавлением IPTG до конечной концентрации 1 мМ. После индуцирования бактерии встряхивали при 37°С в течение 16 часов. Бактерии собирали центрифугированием при 5000 х g в течение 15 минут. Осадок замораживали при -20°С. Этот осадок оттаивали и ресуспендировали в буфере для лизиса бактерий (10 мМ, Na2HPO4, рН 7,0, 30 мМ NaCl, 0,25% твина-20, 10 мМ EDTA, 10 мМ DTT), в который было добавлено 200 мкг/мл лизоцима и 10 мкл бензоназы (Merck). Клетки инкубировали в течение 30 минут при комнатной температуре и подвергали дизрупции в камере пресса Френча. К лизату добавляли тритон Х-100 до конечной концентрации 0,2% и лизат инкубировали в течение 30 минут на льду, периодически встряхивая. Клетки E.coli, содержащие экспрессирующую плазмиду рКК-HBcAg-Lys или контрольную плазмиду, использовали для IPTG-индуцирования экспрессии HBcAg-Lys. Перед добавлением IPTG образец удаляли из бактериальной культуры, несущей плазмиду рКК-HBcAg-Lys, и из культуры, несущей контрольную плазмиду. Через 16 часов после добавления IPTG образцы снова удаляли из культуры, содержащей рКК-HBcAg-Lys, и из контрольной культуры. Мониторинг экспрессии белка проводили с помощью электрофореза в ДСН-ПААГ с последующим окрашиванием кумасси.
Затем лизат центрифугировали в течение 30 минут при 12000 х g для удаления нерастворившегося клеточного дебриса. Супернатант и осадок анализировали с помощью Вестерн-блоттинга с использованием моноклонального антитела против HBcAg (YVS1841, закупленного у Accurate Chemical and Scientific Corp., Westbury, NY, USA), что указывало на то, что значительное количество белка HBcAg-Lys было растворимым. Для этого лизаты клеток E.coli, экспрессирующих HBcAg-Lys, и контрольных клеток центрифугировали при 14000 х g в течение 30 минут. Супернатант (=растворимая фракция) и осадок (=нерастворимая фракция) отделяли и разводили ДСН-буфером с образцом до получения равных объемов. Образцы анализировали с помощью электрофореза в ДСН-ПААГ, а затем посредством Вестерн-блоттинга с использованием анти-HBcAg моноклонального антитела, YVS 1841.
Осветленный клеточный лизат использовали для центрифугирования в ступенчатом градиенте сахарозы, содержащем 4 мл 65% раствора сахарозы, на который было нанесено 3 мл 15% раствора сахарозы, а затем 4 мл бактериального лизата. Образец центрифугировали в течение 3 часов при 100000 х g и при 4°С. После центрифугирования 1 мл-фракции с верхней части градиента собирали и анализировали с помощью электрофореза в ДСН-ПААГ с последующим окрашиванием кумасси. Белок HBcAg-Lys детектировали посредством окрашивания кумасси.
Количество белка HBcAg-Lys возрастало на границе между 15 и 65% сахарозы, что указывало на то, что он имел форму капсидной частицы. Большинство бактериальных белков оставалось на верхнем слое градиента, не содержащем сахарозу, а поэтому центрифугирование частиц HBcAg-Lys в ступенчатом градиенте приводило к обогащению и к неполной очистке этих частиц.
ПРИМЕР 25
Химическое связывание пептида FLAG с HBcAg-Lys с использованием гетеробифункционального сшивающего линкера SPDP
Синтетический пептид FLAG, имеющий цистеиновый остаток у своего амино-конца (аминокислотная последовательность CGGDYKDDDDK (SEQ ID NO:147)), подвергали химическому связыванию с очищенными частицами HBcAg-Lys для вырабатывания иммунного ответа против пептида FLAG. 600 мл 95% чистого раствора частиц HBcAg-Lys (2 мг/мл) инкубировали в течение 30 минут при комнатной температуре с гетеробифункциональным сшивающим линкером N-сукцинимидил-3-(2-пиридилдитио)пропионатом (SPDP), (0,5 мМ). После завершения реакции смесь диализовали в течение ночи против 1 литра 50 мМ фосфатного буфера (рН 7,2), содержащего 150 мМ NaCl для удаления свободного SPDP. Затем 500 мл дериватизированного капсида HBcAg-Lys (2 мг/мл) смешивали с 0,1 мМ пептида FLAG (содержащего аминоконцевой цистеин) в присутствии 10 мМ EDTA для профилактики окисления сульфгидрила, катализируемого металлом. Мониторинг реакции проводили по увеличению оптической плотности раствора при 343 нм, которое ассоциировалось с высвобождением пиридин-2-тиона из SPDP после реакции со свободным цистеином указанного пептида. Реакция дериватизированных остатков Lys с пептидом завершалась примерно через 30 минут.
FLAG-декорированные частицы инъецировали мышам.
ПРИМЕР 26
Конструирование pMPSV-gp140cys
Ген gp140 амплифицировали с помощью ПЦР из pCytTSgp140FOS с использованием олигонуклеотидов gp140CysEcoRI и SalIgp140. Для проведения ПЦР 100 пмоль каждого олигонуклеотида и 50 нг матричных ДНК использовали в 50 мл реакционных смесей с 2 единицами полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. Для обеих реакций осуществляли следующие температурные циклы: один цикл при 94°С, 2 минуты; 30 циклов при 94°С (0,5 минуты), 55°С (0,5 минуты), 72°С (2 минуты).
ПЦР-продукт очищали с помощью набора QiaEXII, гидролизовали ферментами SalI/EcoRI и лигировали в вектор pMPSVHE, гидролизованный теми же самыми ферментами.
Олигонуклеотидные последовательности:
Gp140CysEcoRI:
5'-GCCGAATTCCTAGCAGCTAGCACCGAATTTATCTAA-3' (SEQ ID NO:83);
SalIgpl40:
5'-GGTTAAGTCGACATGAGAGTGAAGGAGAAATAT-3' (SEQ ID NO:84).
ПРИМЕР 27
Экспрессия рМРSVgp140Cys
рМРSVgp140Cys (20 мкг) линеаризовали путем рестрикционного гидролиза. Реакцию прекращали путем экстракции смесью фенол/хлороформ, а затем линеаризованную ДНК осаждали изопропанолом. Рестрикционный гидролиз оценивали с помощью электрофореза в агарозном геле. Для трансфекции 5,4 мкг линеаризованной рМРSVgp140Cys смешивали с 0,6 мкг линеаризованного рSV2Neo в 30 мкл H2O и добавляли 30 мкл 1М раствора CaCl2. После добавления 60 мкл фосфатного буфера (50 мМ HEPES, 280 мМ NaCl, 1,5 мМ Na2НРО4, рН 7,05) раствор встряхивали в течение 5 секунд, а затем инкубировали при комнатной температуре в течение 25 секунд. Этот раствор сразу добавляли к 2 мл среды НР-1, содержащей 2% FCS (среда с 2% FCS). Затем среду с 80% конфлюентной клеточной культурой ВНК21 (6-луночный планшет) заменяли ДНК-содержащей средой. После инкубирования в течение 5 часов при 37°С в CO2-инкубаторе ДНК-содержащую среду удаляли и заменяли 2 мл среды, содержащей 15% глицерина в 2% FCS. Эту глицеринсодержащую среду удаляли после проведения 30-секундной фазы инкубирования и клетки промывали путем прополаскивания 5 мл среды НР-1, содержащей 10% FCS. И наконец, добавляли 2 мл свежей среды НР-1, содержащей 10% FCS.
Стабильно трансфецированные клетки отбирали и культивировали в селективной среде (в среде НР-1, содержащей G418) при 37°С в CO2-инкубаторе. После того как смешанная популяция достигала конфлюентности, культуру распределяли по двум чашкам, а затем культивировали в течение 12 часов при 37°С. Одну чашку с клетками доводили до 30°С для индуцирования экспрессии растворимого GP-140-FOS. Другую чашку выдерживали при 37°С.
Экспрессию растворимого GP140-Cys оценивали с помощью Вестерн-блот-анализа. Культуральную среду (0,5 мл) осаждали метанолом/хлороформом и осадок ресуспендировали в буфере для проб, используемом в электрофорезе в ДСН-ПААГ. Образцы нагревали в течение 5 минут при 95°С, а затем наносили на 15% акриламидный гель. После проведения электрофореза в ДСН-ПААГ белки переносили на нитроцеллюлозные мембраны Protan (Schleicher & Schuell, Germany), как описано Bass & Yang, Creighton, T.E. ed., Protein Function: A Practical Approach, 2nd Edn., IRL Press, Oxford (1997), pp.29-55. Мембрану блокировали 1% альбумина бычьей сыворотки (Sigma) в TBS (10xTBS на литр: 87,7 г NaCl, 66,1 г гидрохлорида Trizma (Sigma) и 9,7 г основания Trizma (Sigma), pH 7,4) в течение 1 часа при комнатной температуре, а затем инкубировали с анти-GP140 или GP-160 антителом в течение 1 часа. Блот 3 раза промывали TBS-T (TBS с 0,05% Твина-20) в течение 10 минут и инкубировали в течение 1 часа с антителом против мышиных/кроличьих/обезьяньих/человеческих IgG, конъюгированным с щелочной фосфатазой. После двухкратной промывки TBS-T в течение 10 минут и двухкратной промывки TBS в течение 10 минут проводили реакцию развития окраски с использованием реагентов для детекции щелочной фосфатазы (10 мл буфера АР (100 мМ Трис/HCl, 100 мМ NaCl, рН 9,5) с 50 мкл раствора NBT (7,7% нитротетразолиевого синего (Sigma) в 70% диметилформамида) и 37 мкл раствора Х-фосфата (5% 5-бром-4-хлор-3-индолилфосфата в диметилформамиде).
ПРИМЕР 28
Очистка gp140Cys
Анти-gp120 антитело ковалентно связывали с NHS/EDC-активированным декстраном и упаковывали на хроматографическую колонку. Супернатант, содержащий GP140Cys, загружали на колонку и после соответствующей промывки GP140Cys элюировали с использованием 0,1 М HCl. Элюат нейтрализовали непосредственно в процессе сбора с использованием 1М Триса, рН 7,2, в пробирках-сборниках.
В процессе очистки могут образовываться дисульфидные связи, а поэтому собранный образец обрабатывают 10 мМ DTT в 10 мМ Триса, рН 7,5, в течение 2 часов, при 25°С.
DTT удаляли путем последовательного диализа против 10 мМ Mes; 80 мМ NaCl, рН 6,0. Наконец, GP140Cys смешивали с альфавирусными частицами, содержащими остаток JUN в Е2, как описано в примере 16.
ПРИМЕР 29
Конструирование PLA2-Cys
Ген PLA2 амплифицировали с помощью ПЦР из pAV3PLAfos с использованием олигонуклеотидов EcoRIPLA и PLA-Cys-hind. Для проведения ПЦР 100 пмоль каждого олигонуклеотида и 50 нг матричных ДНК использовали в 50 мл реакционных смесей, содержащих 2 единицы полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. Для реакции осуществляли следующие температурные циклы: один цикл при 94°С, 2 минуты; 30 циклов при 94°С (0,5 минуты), 55°С (0,5 минуты), 72°С (2 минуты).
ПЦР-продукт очищали с использованием набора QiaEXII, гидролизовали ферментами EcoRI/HindIII и лигировали в вектор pAV3, гидролизованный теми же самыми ферментами.
Олигонуклеотидные последовательности:
EcoRIPLA:
5'-TAACCGAATTCAGGAGGTAAAAAGATATGG-3' (SEQ ID NO:85)
PLA Cys-hind:
5'-GAAGTAAAGCTTTTAACCACCGCAACCACCAGAAG-3' (SEQ ID NO:86).
ПРИМЕР 30
Экспрессия и очистка PLA2-Cys
Для цитоплазмазматического продуцирования Cys-меченых белков, штамм E.coli XL-1 blue трансформировали векторами pAV3::PLA и pPLA-Cys. Эту культуру инкубировали в обогащенной среде в присутствии ампициллина при 37°С и при встряхивании. При оптической плотности 550 нм добавляли 1 мМ IPTG и инкубирование продолжали еще 5 часов. Клетки собирали центрифугированием, ресуспендировали в соответствующем буфере (например, Трис-HCl, рН 7,2, 150 мМ NaCl), содержащем ДНКазу, РНКазу и лизоцим, и подвергали дизрупции путем пропускания через камеру пресса Френча. После центрифугирования (Sorvall RC-5C, ротор SS34, 15000 об/мин, 10 минут, 4°С) осадок ресуспендировали в 25 мл буфера для промывки телец включения (20 мМ Трис-HCl, 23% сахарозы, 0,5% тритона Х-100, 1 мМ EDTA, рН 8) при 4°С и повторно центрифугировали, как описано выше. Эту процедуру повторяли до тех пор, пока супернатант после центрифугирования не становился, в основном, чистым. Тельца включения ресуспендировали в 20 мл буфера для солюбилизации (5,5 М гидрохлорида гуанидиния, 25 мМ Трис-HCl, рН 7,5) при комнатной температуре и нерастворившийся материал удаляли центрифугированием, после чего супернатант пропускали через стерильный фильтр (0,45 мкМ). Раствор белка выдерживали при 4°С, по меньшей мере, в течение 10 часов в присутствии 10 мМ EDTA и 100 мМ DTT, а затем три раза диализовали против 10 объемов 5,5 М гидрохлорида гуанидиния, 25 мМ Трис-HCl, 10 мМ EDTA, рН 6. Раствор два раза диализовали против 5 л 2М мочевины, 4 мМ EDTA, 0,1М NH4Cl, 20 мМ бората натрия (рН 8,3) в присутствии соответствующей окислительно-восстановительной системы (окисленного глутатиона/восстановительного глутатиона; цистеин/цистеин). Затем вновь уложенный белок подвергали ионообменной хроматографии. Белок хранили в соответствующем буфере при рН, превышающем 7, в присутствии 2-10 мМ DTT для сохранения цистеиновых остатков в восстановленной форме. Перед связыванием белка с альфа-вирусными частицами DTT удаляли путем пропускания раствора этого белка через гель-фильтрационную колонку с сефадексом G-25.
ПРИМЕР 31
Конструирование HBcAg, не содержащего свободных цистеиновых остатков и содержащего встроенный лизиновый остаток
Коровый антиген вируса гепатита (HBcAg), называемого здесь HBcAg-Lys-2cys-Mut, который не содержит цистеиновых остатков в положениях, соответствующих 48 и 107 в SEQ ID NO:134, и содержит встроенный лизиновый остаток, конструировали нижеследующими методами.
Сначала были введены две мутации посредством первой отдельной амплификации трех фрагментов гена HBcAg-Lys, полученного, как описано выше в примере 23, с использованием комбинации нижеследующих ПЦР-праймеров. ПЦР-методы осуществляли, в основном, как описано в примере 1, а для получения гена HBcAg-Lys-2cys-Mut использовали стандартные методы клонирования.
Вкратце, для получения фрагмента 1 были использованы следующие праймеры:
Праймер 1: EcoRIHBcAg(s)CCGGAATTCATGGACATTGACCCTTATAAAG (SEQ ID NO:148)
Праймер 2: 48as
GTGCAGTATGGTGAGGTGAGGAATGCTCAGGAGACTC (SEQ ID NO:149)
Для получения фрагмента 2 были использованы следующие праймеры:
Праймер 3: 48as
GSGTCTCCTGAGCATTCCTCACCTCACCATACTGCAC (SEQ ID NO:150)
Праймер 4: 107as
CTTCCAAAAGTGAGGGAAGAAATGTGAAACCAC (SEQ ID NO:151)
Для получения фрагмента 3 были использованы следующие праймеры:
Праймер 5: HBcAg149hindIII-as
CGCGTCCCAAGCTTCTAAACAACAGTAGTCTCCGGAAGCGTTGATAG
(SEQ ID NO:152)
Праймер 6: 107s
GTGGTTTCACATTTCTTCCCTCACTTTTGGAAG (SEQ ID NO:153)
Затем фрагменты 1 и 2 были объединены с ПЦР-праймерами EcoRIHBcAg(s) и 107as с получением фрагмента 4. После этого фрагмент 4 и фрагмент 3 объединяли с праймерами EcoRIHBcAg(s) и HBcAg149hindIII-as для продуцирования полноразмерного гена. Затем полноразмерный ген гидролизовали ферментами EcoRI (GAATTC) и HindIII (AAGCTT) и клонировали в вектор рКК (Pharmacia), разрезанный в тех же рестрикционных сайтах.
ПРИМЕР 32
Блокирование свободных цистеиновых остатков HBcAg с последующим перекрестным связыванием
Свободные цистеиновые остатки HBcAg-Lys, полученные, как описано в примере 23, блокировали с использованием йодацетамида. Затем блокированный HBcAg-Lys сшивали с пептидом FLAG посредством гетеробифункционального сшивающего линкера м-малеимидобензоил-N-гидроксисукцинимидоэфира (Sulfo-MBS).
Для блокировния свободных цистеиновых остатков и перекрестного сшивания HBcAg-Lys были использованы следуюшие методы. HBcAg-Lys (550 мкг/мл) подвергали реакции в течение 15 минут при комнатной температуре с йодацетамидом (Fluka Chemie, Brugg, Switzerland) при концентрации 50 мМ в забуференном фосфатом физиологическом растворе (PBS)(50 мМ фосфата натрия, 150 мМ хлорида натрия), рН 7,2, в полном объеме 1 мл. Затем модифицированный таким образом HBcAg-Lys сразу подвергали реакции с Sulfo-MBS (Pierce) при концентрации 330 мкМ непосредственно в реакционной смеси, полученной в стадии 1, в течение 1 часа при комнатной температуре. Затем реакционную смесь охлаждали на льду и диализовали против 1000 объемов PBS, рН 7,2. Диализованную реакционную смесь подвергали реакции с 300 мкМ пептида FLAG (CGGDYKDDDDK, SEQ ID NO:147), содержащего N-концевой свободный остаток для связывания с активированным HBcAg-Lys, и загружали на ДСН-ПААГ для анализа.
В полученной картине полос на ДСН-ПААГ-геле обнаруживалась дополнительная полоса, которая мигрировала более медленно, чем контрольный HBcAg-Lys, дериватизированный сшивающим линкером, но эта полоса не реагировала с пептидом FLAG. Реакции, проведенные в тех же самых условиях, но без предварительной дериватизации цистеинов йодацетамидом, приводили к полному сшиванию мономеров HBcAg-Lys с молекулами более высокой молекулярной массы.
ПРИМЕР 33
Выделение и химическое сшивание пептида FLAG с пилями типа 1 Escherichia coli с использованием гетеробифункционального линкера
А. Введение
Бактериальные пили или фимбрии представляют собой нитевидные поверхностные органеллы, продуцируемые бактериями широкого ряда. Эти органеллы опосредуют связывание бактерий с поверхностными рецепторами клеток-хозяев и необходимы для развития многих бактериальных инфекций, таких как цистит, пиелонефрит, менингиты новорожденных и диарея.
Пили могут быть подразделены на различные классы по их специфичности к рецептору (агглютинация кровяных клеток различных видов), по характеру их сборки (внеклеточная нуклеация, общая секреция, путь "шаперон/проводник", другой шаперон) и по их морфологическим свойствам (толстые жесткие пили; тонкие гибкие пили; атипические структуры, включая капсулу; завитки; и т.п.). Примерами толстых жестких пилей, образующих правостороннюю спираль, сборка которых осуществляется посредством так называемого пути "шаперон/проводник" и опосредуется адгезией к гликопротеинам хозяина, являются пили типа 1, Р-пили, S-пили, F1С-пили и 987Р-пили). Наиболее типичными и наиболее охарактеризованными членами этого класса пилей являются Р-пили и пили типа 1 (обзор адгезивных структур, их сборки и ассоциированных с ними заболеваний можно найти в Soto, G.E. & Hultgren, S.J., J. Bacteriol. 181:1059-1071 (1999); Bullitt & Makowski, Biophys. J. 74:623-632 (1998); Hung, D.L. & Hultgren, S.J., J. Struct, Biol. 124:201-220 (1998)).
Пили типа 1 представляют собой длинные, нитевидные полимерные белковые структуры на поверхности E.coli. Они обладают адгезивными свойствам, которые позволяют им связываться с содержащими маннозу рецепторами, присутствующими на поверхности некоторых тканей-хозяев. Пили типа 1 могут экспрессироваться 70-80 процентами всех изолятов E.coli, и одна клетка E.coli может нести до 500 пилей. Пили этого типа обычно достигают длины 0,2-2 мкм со средним числом субъединиц белка, равным 1000, которые ассоциируются с правосторонней спиралью, содержащей 3, 125 субъединиц на виток и имеющей диаметр 6-7 нм и центральное свободное пространство 2,0-2,5 нм.
Главным компонентом пилуса типа 1, FimA, который составляет 98% от общего белка пилей, является белок в 15,8 кДа. Более мелкие компоненты FimF, FimG и FimH, находятся на кончике пилей и на одном и том же расстоянии вдоль ствола пилей (Klemm, P. & Krogleft, K.A. "Type I fimbriae of Escherichia coli", in: Fimbriae. Klemm, P. (ed), CRC Press Inc., (1994) pp.9-26). Было показано, что FimH, белок в 29,1 кДа, представляет собой связывающийся с маннозой адгезин пилей типа 1 (Krogfelt, K.A. et al., Infect. Immun. 58:1995-1998 (1990); Klemm P. et al., Mol. Microbiol. 4:553-560 (1990); Hanson M.S. & Brinton, C.C.J., Nature, 17:265-268 (1988)), и его введение, вероятно, облегчается FimG и FimF (Klemm P. & Christiansen, G. Mol. Gen. Genetics, 208:439-445 (1987); Russell P.W. & Orndorff, P.E.J. Bacteriol. 174:5923-5935 (1992)). Недавно было показано, что FimH может также образовывать тонкие концевые фибриллы на кончике пилей (Jones, C.H. et al., Proc. Natl. Acad. Sci. USA, 92:2081-2085 (1995)). Порядок расположения больших и малых компонентов в отдельных зрелых пилях почти аналогичен, что указывает на высокоупорядоченный процесс сборки (Soto, G.E. & Hultegren, S.J., J.Bacteriol. 181:1059-1071 (1999)).
Р-пили E.coli имеют очень сходную архитектуру, диаметр 6,8 нм, осевую полость диаметром 1,5 нм и 3,28 субъединиц на виток (Bullitt & Makowski, Biophys. J.74:623-632 (1998)). РарА размером 16,6 кДа является главным компонентом пилей этого типа и имеет последовательность, которая на 36% идентична и на 59% аналогична FimA (см. таблицу 1). Как и в пилях типа 1, 36,0 кДа-адгезин пилуса PapG Р и специализированные адаптерные белки составляют лишь незначительную часть общего белка пилей. Наиболее очевидным отличием от пилей типа 1 является отсутствие адгезина как интегральной части трубочки пилуса и его эксклюзивная локализация на конце фимбрии, соединенной с трубочкой пилуса посредством специализированных адаптерных белков, которые отсутствуют у пилей типа 1 (Hultgren S.J. et al., Cell. 73:887-901 (1993)).
Пили типа 1 представляют собой исключительно стабильные гетероолигомерные комплексы. ДСН-обработка, гидролиз протеазой и кипячение или добавление денатурирующих агентов не могли диссоциировать пили типа 1 на их отдельные белковые компоненты. Как было ранее обнаружено, комбинация различных методов, таких как инкубирование при 100°С, рН 1,8, приводит к деполимеризации и отделению компонентов (Eshdat, Y., et al., J. Bacteriol. 148:308-314 (1981); Brinton, C.C. J., Trans, N. Y. Acad. Sci. 27:1003-1054 (1965); Hanson, A.S., et al, J. Bacteriol., 170:3350-3358 (1988); Klemm, P. & Krogfelt, K.A., "Type I fimbriae of Escherichia coli," in: Fimbriae. Klemm, P. (ed.), CRC Press Inc., (1994) pp. 9-26). Интересно отметить, что пили типа 1, при механическом размешивании, имеют тенденцию к расщеплению в положениях, где вводится FimH, что приводит к образованию фрагментов, которые представляют адгезин FimH на своих концах. Этот факт может быть интерпретирован как бактериальный механизм укорочения пилей до эффективной длины при механическом стрессе (Klemm P. & Krogfelt, K.A. "Type I fimbriae of Esherichia coli", in: Fimbriae. Klemm, P.(ed.) CRC Press Inc., (1994) pp.9-26). Было показано, что несмотря на свою исключительную стабильность пили типа 1 частично разлагаются в присутствии 50% глицерина; причем они теряют свою спиральную структуру и образуют удлиненную и гибкую белковую цепь шириной 2 нм (Abraham S.N. et al., J. Bacteriol. 174:5145-5148 (1992)).
Р-пили и пили типа 1 кодируются кластерами из одного гена на хромосоме E.coli размером приблизительно 10 т.п.н. (Klemm, P. & Krogfelt, K.A., "Type I fimbriae of Escherichia coli," in: Fimbriae. Klemm, P. (ed.), CRC Press Inc., (1994) pp. 9-26; Orndorff, P.E. & Falkow, S., J. Bacteriol. 160:61-66 (1984)). В кластере гена пилей типа 1 обнаружено всего девять генов, в кластере Р-пилуса обнаружено 11 генов (Hultgren S.J. et al., Adv. Prot. Chem. 44:99-123 (1993)). Все кластеры имеют абсолютно одинаковую организацию.
Первые два гена fim, fimB и fimE кодируют рекомбиназы, участвующие в регуляции экспрессии пилуса (McClain, M.S. et al., J. Bacteriol. 173:5308-5314 (1991). Главный структурный белок пилуса кодируется следующим геном кластера, fimA (Klemm, P., Euro. J. Biochem. 143:395-400 (1984); Orndorff, P.E. & Falkow, S., J. Bacteriol. 160:61-66 (1984); Orndorff, P.E. & Falkow, S., J. Bacteriol. 162:454-457 (1985)). Точная роль fimI пока не выяснена. Сообщалось также, что он входит в состав пилуса (Klemm P. & Krogfelt, K.A. "Type I fimbriae of Esherichia coli", in: Fimbriae. Klemm, P. (ed.) CRC Press. Inc., (1994) pp.9-26. Смежный fimC не кодирует структурный компонент зрелого пилуса, но кодирует так называемый шаперон пилуса, который играет важную роль в сборке пилуса (Klemm P., Res. Microbiol. 143:831-838 (1992); Jones, C.H. et al., Proc. Natl. Acad. Sci. USA, 90:8397-8401 (1993)).
Платформа для сборки на внешней бактериальной мембране, в которой заякорен зрелый пилус, кодируется fimD (Klemm P. & Christiansen, G. Mol. Gen, Genetics, 220:334-338 (1990)). Три небольших компонента пилей типа 1, FimF, FimG и FimH кодируются последними тремя генами этого кластера (Klemm P. & Christiansen, G. Mol. Gen. Genetics, 208:439-445 (1987)). Кроме fimB и fimE, все гены кодируют белки-предшественники для секреции в периплазму по секреторному пути.
Сходство между различными пилями в отношении пути "шаперон/проводник" не ограничивается их морфологическими свойствами. Их гены также имеют очень сходное расположение. В основном, ген, кодирующий главную структурную субъединицу, расположен непосредственно ниже регуляторных элементов в начале генного кластера, а за этим геном следует ген, кодирующий дополнительную структурную субъединицу (fimI в случае пилей типа 1 и рарН в случае Р-пилей). Был выявлен ген РарН, и было предположено, что FimAI завершает сборку пилуса (Hultgren S.J. et al., Cell, 73: 887-901 (1993)). Эти два белка, которые регулируют процесс образования пилуса, а именно специализированного шаперона пилуса и платформы для сборки внешней мембраны, локализованы непосредственно ниже. На конце кластеров кодируется различное число мелких компонентов пилуса, включая адгезины. Сходство морфологической структуры, последовательностей (см. таблицу 1), генетической организации и регуляции указывает на близкое эволюционное родство и аналогичные процессы сборки этих клеточных органелл.
Бактерии, продуцирующие пили типа 1 обнаруживают так называемую фазовую вариацию. Эти бактерии либо полностью покрыты пилями, либо являются "голыми". Это достигается инверсией геномного ДНК-фрагмента в 314 п.н., содержащего промотор fimA, в результате чего индуцируется "включение" или "выключение" экспрессии генов пилуса (McClain, M.S. et al., J. Bacteriol. 173:5308-5314 (1991). Связь экспрессии других структурных генов пилуса с экспрессии гена fimA осуществляется по еще неизвестному механизму. Однако проведенные широкие исследования выявили механизм, который влияет на переключение этих двух фенотипов.
Первые два гена кластера пилуса типа 1, fimB и fimE кодируют рекомбиназы, распознающие 9 п.н.-ДНК-сегменты симметричной диады, которая фланкирует инвертируемый промотор fimA. FimB "включает" образование пилей, тогда как FimE изменяет ориентацию промотора, приводя его в положение "выключено". Поэтому позитивная или негативная регуляция экспрессии либо гена fimB, либо гена fimE контролирует положение так называемого "fim-переключения" (McClain, M.S. et al., J. Bacteriol. 173:5308-5314 (1991); Blomfield I.C. et al., J. Bacteriol. 173:5298-5307 (1991)).
Два регуляторных белка fimB и fimE транскрибируются с разных промоторов, и было показано, что на их транскрипцию влияет широкий ряд факторов, включая фактор интеграции в хозяине (IHF) (Blomfield I.C. et al., Mol. Microbiol. 23:705-717 (1997)) и восприимчивый к лейцину регуляторный белок (LRP) (Blomfield, I.C., et al., J. Bacteriol. 175:27-36 (1993); Gally, D.L., et al., J. Bacteriol. 175:6186-6193 (1993); Gally, D.L., et al., Microbiol. 21:725-738 (1996); Roesch, R.L. & Blomfield, I.C., Mol Microbiol, 27:751-761 (1998)). Мутации в первом факторе блокируют бактерию либо на фазе "включения", либо на фазе "выключения", тогда как мутанты LRP осуществляют переключение с меньшей частотой. Кроме того, было показано действие leuX на биогенез пилуса. Этот ген локализован на хромосоме возле fim-генов и кодирует небольшие молекулы тРНК лейцина для кодона UUG. Ген fimB содержит пять кодонов UUG, fimE содержит только два таких кодона, а усиленная транскрипция leuX более благоприятствует экспрессии FimB, чем экспрессии FimE (Burghoff, R.L., et al., Infect. Immun. 61:1293-1300 (1993); Newman, J.V., et al., FEMS Microbiol. Lett. 122:281-287 (1994); Ritter, A., et al., Mol. Microbial, 25:871-882 (1997)).
Кроме того, было показано, что на активность FimB и FimE влияют температура, состав среды и другие внешние факторы. И наконец, сообщалось о спонтанном, статистическом переключении промотора fimA. Частота такого спонтанного переключения составляет приблизительно 10-3 на генерацию (Eisenstein B.I. Science, 214:337-339 (1981); Abraham S.M. et al., Proc. Nat. Acad. Sci. USA 82:5724-5727 (1985)), но она может сильно меняться под действием вышеупомянутых факторов.
Гены fimI и fimC также транскрибируются с промотора fimA, но был также идентифицирован находящийся непосредственно ниже ДНК-сегмент fimA, который обладает сильно выраженной тенденцией к образованию вторичной структуры и который, вероятно, представляет собой терминатор частичной транскрипции (Klemm P. Euro. J. Biochem. 143:395-400 (1984)); а поэтому было высказано предположение, что он в значительной степени снижает уровень транскрипции fimI и fimC. У 3'-конца fimC дополнительный промотор регулирует транскрипцию fimD; а у 3'-конца fimD, последний известный промотор fim расположен так, что он регулирует уровни FimF, FimG и FimH. Таким образом, все небольшие белки пилей типа 1 транскрибируются в виде одной мРНК (Klemm P. & Krogfelt, K.A. "Type I fimbriae of Escherichia coli", in: Fimbriae. Klemm P. (ed), CRC Press Inc., (1994) pp.9-26). Это обеспечивает стехиометрию 1:1:1 на уровне мРНК, которая, вероятно, сохраняется и на белковом уровне.
В случае Р-пилей, при определении времени полужизни мРНК для различных генов Р-пилуса, были выявлены дополнительные регуляторные механизмы. мРНК для рарА является исключительно долгоживущей, тогда как мРНК для рарВ, гена регуляторного белка пилуса, представляет собой короткоживущую мРНК (Naureckiene S. & Uhlin, B.E. Mol. Microbiol. 21:55-68 (1996); Nilsson P. et al., J. Bacterial. 178:683-690 (1996)).
В случае пилей типа 1 ген шаперона пилуса типа 1 FimC начинается с кодона GTG вместо кодона ATG, что приводит к снижению эффективности трансляции. И наконец, анализ гена FimH выявил тенденцию мРНК FimH к образованию структуры "стебель-петля", которая может в значительной степени препятствовать трансляции. В целом, биогенез бактериального пилуса регулируется широким рядом механизмов, действующих на всех уровнях биосинтеза белков.
Периплазматические белки пилуса обычно синтезируются как предшественники, содержащие N-концевую сигнальную последовательность, которая обеспечивает транслокацию через внутреннюю мембрану посредством Sec-аппарата. После транслокации предшественники обычно расщепляются сигнальной пептидазой I. Структурные субъединицы пилуса типа 1 обычно содержат дисульфидные связи, и их образование катализируется генными продуктами DsbA, а возможно DsbC и DsbG.
Шаперон FimC пилуса типа 1 не содержит цистеиновых остатков. В противоположность этому шаперон Р-пилей, PapD, является лишь членом семейства шаперонов пилуса, который содержит дисульфидную связь, при этом была точно установлена зависимость Р-пилей от DsbA (Jacob-Dubuisson, F. et al., Proc. Natl. Acad. Sci. USA, 91:11552-11556 (1994)). PapD не аккумулируется в периплазме ΔdsbA-штамма, что указывает на то, что нарушение механизма сборки Р-пилуса вызвано отсутствием этого шаперона (Jacob-Dubuisson, F. et al., Proc. Natl. Acad. Sci. USA, 91:11552-11556 (1994)). Это соответствует имеющимся данным, указывающим на то, что сборка пилей типа 1 в ΔdsbA-штамме все же происходит, хотя и на низком уровне (Hultgren, S.J. et al., "Bacterial Adhesion and Their Assembly", in: Escherichia coli and Salmonella, Neidhardt F.C. (ed.) ASM Press, (1996) pp.2730-2756).
Пили типа 1, а также Р-пили на 98% состоят из единственной или главной структурной субъединицы, называемой FimA и РаРА, соответственно. Оба белка имеют размер ˜15,5 кДа. Дополнительные минорные компоненты, кодируемые в генных кластерах пилуса, являются почти идентичными (см. таблицу 1). Сходство последовательности и размера субъединиц, за исключением адгезинов, позволяет предположить, что все они имеют идентичный мотив укладки и отличаются только по их аффинности друг к другу. В частности, N- и С-концевые области этих белков были в высокой степени консервативными, и предполагается, что они играют важную роль во взаимодействиях шаперон/субъединица, а также во взаимодействиях субъединица/субъединица внутри пилуса (Soto, G.E. & Hultgren S.J., J. Bacteriol. 181:1059-1071 (1999)). Интересно отметить, что консервативный N-концевой сегмент может находиться в середине адгезинов пилуса, что указывает на то, что двухдоменная организация адгезинов, в которых предполагаемый С-концевой домен, начинающийся с консервативного мотива, соответствует структурной субъединице пилуса, но при этом, как было показано, N-концевой домен ответственен за распознавание рецепторов клеток-хозяев (Hultgren, S.J., et al., Proc. Natl. Acad. Sci. USA 86:4357-4361 (1989); Haslam D.B. et al., Mol. Microbiol. 14:399-409 (1994); Soto G.E. et al., EMBO J. 17:6155-6167 (1998)). Было также показано, что различные субъединицы влияют на морфологические свойства пилей. Сообщалось, что удаление нескольких генов приводит к снижению числа пилей типа 1 или Р-пилей либо к увеличению их длины (fimH, papG, papK, fimF, fimG)(Russell P.W. & Orndorff, P.E., J. Bacteriol. 174:5923-5935 (1992); Jacob-Dubuisson, R. et al., EMBO J. 12:837-847 (1993); Soto, G.E. & Hultgren, S.J., J. Bacteriol. 181:1059-1071 (1999)); причем, комбинация этих генных делеций усиливает эти эффекты или приводит к полной потере способности к образованию пилей (Jacob-Dubuisson, R. et al., EMBO J. 12:837-847 (1993)).
Нефимбриальные органеллы адгезивных клеток, также собранные с помощью систем шаперон/проводник, таких как фимбрии Myf и пили CS3, имеют отличающуюся консервативную С-концевую область. Это косвенно подтверждает важность указанных С-концевых сегментов субъединиц для взаимодействий четвертичных структур (Hultgren S.J. et al., "Bacterial Adhesion and Their Assembly", in: Escherichia coli and Salmonella, Neidhardt, F.C. (ed.) ASM Press, (1996) pp.2730-2756).
Исследования по делеции генов подтвердили, что удаление шаперонов пилуса приводит к полной потере способности к образованию Р-пилей и пилей типа 1 (Lindberg F. et al., J. Bacteriol. 171:6052-6058 (1989); Klemm P. Res. Microbiol. 143:831-838 (1992); Jones, C.H. et al., Proc. Natl. Acad. Sci. USA, 90:8397-8401 (1993)). Периплазматические экстракты ΔfimC-штамма обнаруживали аккумуляцию главной субъединицы FimA, но пили не были обнаружены (Klemm P. Res. Microbiol. 143:831-838 (1992)). Попытки осуществить сверхэкспрессию отдельных субъединиц Р-пилуса потерпели неудачу, и были обнаружены лишь протеолитически расщепленные формы при отсутствии PapD; кроме того, был очищен адгезин Р-пилуса с внутренней мембранной фракцией при отсутствии шаперона (Lindberg F. et al., J. Bacteriol. 171:6052-6058 (1989)). Однако коэкспрессия структурных белков пилуса и их шаперона позволила детектировать комплексы "шаперон/субъединица" в периплазме в случае комплекса FimC/FimH, а также в случае различных Рар-белков, включая адгезин PapG и главную субъединицу РарА (Tewari R. et al., J. Biol. Chem. 268:3009-3015 (1993); Lindberg F. et al., J. Bacteriol. 171:6052-6058 (1989)). Были также проведены in vitro исследования аффинности комплексов "шаперон/субъединица" по отношению к платформе их сборки, и было установлено, что она сильно отличается (Dodson et al., Proc.Natl. Acad. Sci., USA, 90:3670-3674 (1993)). Исходя из полученных данных, было высказано предположение, что шапероны пилуса обладают нижеследующими функциями.
Они, предположительно, разпознают неуложенные субъединицы пилуса, предупреждают их агрегацию и обеспечивают "матрицу для укладки", которая определяет образование нативной структуры.
Уложенные субъединицы, которые после укладки представляют поверхность, обеспечивающую взаимодействие между субъединицами, затем, как предполагается, обеспечивают защиту от взаимодействия с другими субъединицами и поддерживаются в мономерной и компетентной в отношении сборки форме.
И наконец, предполагается, что шапероны пилуса обеспечивают стимулируемое высвобождение субъединиц при локализации их сборки на внешней мембране и при этом с различной эффективностью влияют на состав и порядок расположения зрелых пилей (см. также ниже отдельный раздел).
После высвобождения субъединицы, присутствующей на внешней мембране, шаперон становится свободным для другого раунда связывания с субстратом и обеспечивает укладку, транспорт субъединицы через периплазму и специфическую доставку в место сборки. Поскольку в периплазме отсутствуют источники энергии, например АТР, то весь процесс сборки пилуса должен осуществляться термодинамически (Jacob-Dubuisson, F. et al., Proc. Natl. Acad. Sci. USA, 91:11552-11556 (1994)). Широкий диапазон различных функций, приписываемых шаперонам пилуса, должен обеспечивать исключительно тонко отлаженный каскад реакций.
Однако некоторые факты не так легко объяснить с помощью модели вышеописанной функции шаперона пилуса. Одним из примеров является присутствие мультимерных комплексов шаперон/субъединица (Striker R.T. et al., J. Biol. Chem. 269:12233-12239 (1994)), где один шаперон связывается с димерами или тримерами субъединиц. При этом трудно себе представить матрицу для укладки, которая может быть "дважды сброшюрованной" (double-booked). Проведенные исследования относительно деталей молекулярного взаимодействия шаперон/субъединица (см. ниже) частично подтверждают функции, перечисленные выше, но при этом также возникают новые вопросы.
Все 31 периплазматических шаперона, идентифицированные до настоящего времени с помощью генетических исследований или анализа последовательности, представляют собой белки, имеющие размер примерно в 25 кДа и очень высокие значения pI, составляющие примерно 10. Из них десять шаперонов обеспечивают сборку палочкообразных пилей, четыре шаперона участвуют в образовании тонких пилей, десять шаперонов играют важную роль в биогенезе атипично тонких структур (включая капсулоподобные структуры), а две адгезивные структуры пока еще не определены (Holmgren, A. et al., EMBO J. 11:1617-1622 (1992); Bonci A. et al., J. Mol. Evolution 44:299-309 (1997); Smyth C.J. et al., FEMS Immun. Med. Microbiol. 16:127-139 (1996); Hung D.L. & Hultgren S.J. J. Struct, Biol. 124:201-220 (1998). Идентичность между этими шаперонами и PapD, при их попарном сравнивании, варьируется от 25 до 56%, что указывает, в основном, на идентичную укладку (Hung D.L. et al., EMBO J. 15:3792-3805 (1996)).
Первые исследования, касающиеся механизма распознавания шаперон/субстрат, были основаны на обнаружении того факта, что С-концы всех известных шаперонов пилуса имеют исключительно большое сходство. Было показано, что синтетические пептиды, соответствующие С-концу белков Р-пилуса, связываются с PapD в ELISA-анализе (Kuehn M.J. et al., Science 262:1234-1241 (1993)). Более важно, что были установлены Х-структуры двух комплексов, в которых PapD совместно кристаллизован с пептидами из 19 остатков, соответствующими С-концам либо адгезина PapG, либо минорного компонента РарК (Kuehn M.J. et al., Science 262:1234-1241 (1993); Soto G.E. et al., EMBO J. 17:6155-6167 (1998)). Оба пептида, связанные в удлиненной конформации с β-цепью N-концевого домена шаперона, который ориентирован в направлении внутридоменного "кармана", обеспечивают, тем самым, удлинение β-складчатого слоя дополнительной цепью. С-концевые карбоксилатные группы этих пептидов заякорены посредством водородных связей с Arg8 и Lys112, причем эти два остатка являются инвариантными в семействе шаперонов пилуса. Исследования с помощью мутагенеза подтвердили важность этих шаперонов, поскольку проведенная в них замена на аланин приводит к аккумуляции нефункционального шаперона пилуса в периплазме (Slonim L.N. et al., EMBO J. 11:4747-4756 (1992)). Кристаллическая структура PapD указывает, что ни Arg8, ни Lys112 не обеспечивают стабилизацию шаперона и делают его полностью доступными для растворителя (Holmgren A. & Branden, C.I. Nature, 342:248-251 (1989)). При этом сообщалось, что на стороне взаимодействия с субстратом замена С-концевых остатков РарА приводит к отмене образования Р-пилуса, и аналогичные эксперименты в отношении консервативного С-концевого сегмента адгезина PapG Р-пилуса приводили к предотвращению его введения в Р-пилус (Hultgren S.J. et al., "Bacterial Adhesion and Their Assembly" in: Escherichia coli and Salmonella, Neidhardt, F.C. (ed.) ASM Press, (1996) pp.2730-2756). Поэтому все эти данные указывают на то, что распознавание субъединицы пилуса происходит через С-концевые сегменты данной субъединицы.
Более недавнее исследование эффектов С-концевой аминокислотной замены адгезина PapG Р-пилуса дало более детальную картину. Ряд аминокислотных замен в положениях -2, -4, -6 и -8 по отношению к С-концу не давали какого-либо эффекта, но влияли на стабильность пилуса (Soto, G.E. et al., EMBO J. 17:6155-6167 (1998)).
Однако при более глубокой оценке этой модели возникают определенные проблемы. Адгезивные бактериальные структуры не связывались с жесткими палочкообразными пилями, у которых отсутствуют консервативные С-концевые сегменты (Hultgren S.J. et al., "Bacterial Adhesion and Their Assembly", in: Escherichia coli and Salmonella, Neidhardt, F.C. (ed.) ASM Press, (1996) pp.2730-2756), хотя они также зависят от присутствия родственных шаперонов пилуса. Это указывает на другую общую роль С-концевых сегментов субъединиц пилуса, а именно опосредование взаимодействий четвертичных структур в зрелом пилусе. Более того, попытка определить структуру С-концевого пептида в комплексе с этим шапероном с помощью ЯМР натолкнулась на значительные трудности из-за слабого связывания этого пептида с шапероном (Walse B. et al., FEBS Lett. 412:115-120 (1997)), и это при том, что главная роль С-концевых сегментов в распознавании шаперонами заключается в обеспечении взаимодействия с относительно высокой аффинностью.
Другая проблема возникает в связи с тем, что необходимо принимать в расчет вариабельность между различными субъединицами. Хотя С-концевые сегменты являются консервативными, тем не менее был обнаружен широкий ряд консервативных замен. Так, например, у двух пептидов, кристаллизованных вместе с PapD, отличаются 15 из 19 аминокислотных остатков (Soto G.E. et al., EMBO J. 17:6155-6167 (1998)). Это можно объяснить типом взаимодействия между шапероном и субстратом, которое опосредовано, главным образом, взаимодействием их остовов, а не специфическими взаимодействиями боковых цепей. И в этом случае специфичность указанного шаперона к некоторым субстратам не поддается точному объяснению. В противоположность первому аргументу консервативные остатки были приняты за основу доказательства специфичности (Hultgren S.J. et al., "Bacterial Adhesion and Their Assembly", in: Escherichia coli and Salmonella, Neidhardt, F.C. (ed.) ASM Press, (1996) pp.2730-2756).
Платформа для сборки внешней мембраны, также называемая в литературе "проводником", образована гомоолигомерами FimD или РарС, в случае пилей типа 1 и Р-пилей, соответственно (Klemm P. & Christiansen, G. Mol. Gen. Genetics, 220:334-338 (1990); Thanassi, D.G. et al., Proc. Natl. Acad. Sci., USA, 95:3146-3151 (1998)). Исследования элонгации фимбрий типа 1, проводимые с помощью электронной микроскопии, продемонстрировали удлинение указанного пилуса от его основания (Lowe M.A. et al., J. Bacteriol. 169:157-163 (1987)). В противоположность секреции неуложенных субъединиц в периплазматическую область полностью уложенные белки должны транслоцироваться через внешнюю мембрану, возможно, в олигомерной форме (Thanassi, D.G. et al., Proc. Natl. Acad. Sci. USA, 95:3146-3151 (1998)). Для этого требуются, во-первых, присутствие достаточно широких мембранных пор для прохождения, и во-вторых, определенный термодинамически управляемый механизм транспорта (Jacob-Dubuisson, F. et al., J. Biol. Chem. 269:12447-12455 (1994)).
Было показано, что экспрессия одного FimD оказывает неблагоприятное действие на бактериальный рост, тогда как ко-экспрессия субъединиц пилуса поддерживает нормальный процесс роста (Klemm P. & Christiansen, G. Mol. Gen. Genetics, 220:334-338 (1990)). Исходя из этого, можно сделать вывод, что указанные "проводники", вероятно, образуют поры, которые полностью заполнены пилями. Электронная микроскопия на мембранных везикулах, в которые был введен PapC, подтвердила порообразующую структуру с внутренним диаметром 2 нм (Thanassi, D.G. et al., Proc. Natl. Acad. Sci. USA, 95:3146-3151 (1998)). Поскольку внутренний диаметр этой поры является слишком малым для прохождения палочки пилуса, то было высказано предположение, что спиральная структура зрелого пилуса образуется на внешней поверхности бактерии. Обнаружение того факта, что глицерин приводит к "расплетанию" пилей, которые затем образуют белковую цепь, примерно в 2 нм, хорошо согласуется с этой гипотезой, поскольку удлиненная цепь субъединиц может образовываться в этой поре на первой стадии (Abraham, S.N. et al., J. Bacteriol. 174:5145-5148 (1992); Thanassi, D.G. et al., Proc. Natl. Acad. Sci. USA, 95:3146-3151 (1998)). Образование спиральной трубочки пилуса на внешней стороне бактериальной мембраны может быть затем движущей силой, ответственной за транслокацию растущего пилуса через мембрану.
Было также продемонстрировано, что белки-"проводники" пилей типа 1 и Р-пилей образуют, с различными аффиностями, третичные комплексы с комплексами шаперон/субъединица (Dodson, K.W. et al., Proc. Natl. Acad. Sci. USA, 90:3670-3674 (1993); Saulino E.T. et al., EMBO J. 17:2177-2185 (1998)). Это может быть интерпретировано как "кинетическое распределение", которое обеспечивает определенный порядок пилусных белков в данном пилусе. Более того, было высказано предположение, что структурные белки могут иметь связывающую поверхность, совместимую только с одним пилусным белком другого типа; а для достижения в высокой степени точного порядка субъединиц в зрелом пилусе необходим другой механизм (Saulino, E.T. et al., EMBO J. 17:2177-2185 (1998)).
В. Продуцирование пилей типа 1 из Escherichia coli
Штамм E.coli W3110 высевали на планшеты со средой LB (10 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л NaCl, рН 7,5, 1% агар (мас./об)) и инкубировали в течение ночи при 37°С. Затем одиночную колонию использовали для инокуляции 5 мл исходной культуры LB (10 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л NaCl, рН 7,5). После инкубирования в течение 24 часов в условиях, благоприятных для продуцирования бактериями пилей типа 1 (37°С, без перемешивания), 5-литровые шейкерные колбы, содержащие 1 литр LB, инокулировали 1 миллилитром исходной культуры. Затем бактериальные культуры инкубировали в течение еще 48-72 часов при 37°С без перемешивания. После этого бактерии собирали центрифугированием (5000 об/мин, 4°С, 10 минут) и полученный осадок ресуспендировали в 250 миллилитрах 10 мМ Трис/HCl, рН 7,5. Пили отделяли от бактерий путем 5-минутного перемешивани в подходящем миксере при 17000 об/мин. После центрифугирования в течение 10 минут при 10000 об/мин при 4°С супернатант, содержащий пили, собирали и добавляли 1М MgCl2 до конечной концентрации 100 мМ. Раствор выдерживали при 4°С в течение 1 часа, после чего осажденные пили собирали центрифугированием (10000 об/мин, 20 минут, 4°С). Затем осадок ресуспендировали в 10 мМ HEPES, рН 7,5, и раствор пилуса осветляли путем проведения конечной стадии центрифугирования для удаления остаточного клеточного дебриса.
С. Связывание FLAG с очищенными пилями типа 1 E.coli с использованием м-малеимидобензоил-N-гидроксисукцинимидоэфира (Sulfo-MBS)
600 мкл 95% чистого раствора бактериальных пилей типа 1 (2 мг/мл) инкубировали в течение 30 минут при комнатной температуре с гетеробифункциональным сшивающим линкером Sulfo-MBS (0,5 мМ). Затем смесь диализовали в течение ночи против 1 литра 50 мМ фосфатного буфера (рН 7,2), содержащего 150 мМ NaCl, для удаления свободного Sulfo-MBS. После этого 500 мкл дериватизированных пилей (2 мг/мл) смешивали с 0,5 мМ пептида FLAG (содержащего амино-концевой цистеин) в присутствии 10 мМ EDTA для профилактики катализируемого металлом окисления сульфгидрила. Несвязанный пептид удаляли путем эксклюзионной хроматографии.
ПРИМЕР 34
Конструирование экспрессирующей плазмиды для экспрессии пилей типа 1 Escherichia coli
ДНК-последовательность, депонированная в Genbank, регистрационный номер U14003, полное описание которой вводится в настоящую заявку посредством ссылки, содержит все гены Escherichia coli, необходимые для продуцирования пилей типа 1, от нуклеотида 233947 и до нуклеотида 240543 (генный кластер fim). Эта часть последовательностей содержит последовательности генов fimA, fimI, fimC, fimD, fimF fimG и FimH. Для амплификации этой части генома E.coli и последующего клонирования в pUC19 (Genbank, регистрационные номера L09137 и Х02514) проводили три различных ПЦР, как описано ниже.
ПЦР-матрицу получали путем смешивания 10 мл глицеринсодержащего маточного раствора штамма W3110 E.coli с 90 мл воды и кипячения этой смеси в течение 10 минут при 95°С с последующим центрифугированием в течение 10 минут при 140000 об/мин в настольной центрифуге и сбором супернатанта.
Затем 10 мл супернатанта смешивали с 50 пмоль ПЦР-праймера 1 и 50 пмоль ПЦР-праймера 2, как описано ниже. После этого добавляли 5 мл 10 х ПЦР-буфера, 0,5 мл ДНК-полимеразы Taq и воду до полного объема 50 мл. Все ПЦР осуществляли в соответствии со следующей схемой: 1 цикл при 94°С, в течение 2 минут, 30 циклов, 20 секунд при 94°С, 30 секунд при 55°С, и 2 минуты при 72°С. Затем ПЦР-продукты очищали с помощью электрофореза в 1% агарозном геле.
Для амплификации последовательности от нуклеотида 233947 до нуклеотида 235863, содержащей гены fimA, fimI и fimC, использовали олигонуклеотиды, имеющие следующие последовательности:
TAGATGATTACGCCAAGCTTATAATAGAAATAGTTTTTTGAAAGGAAAGCAGCATG
(SEQ ID NO: 196)
и
GTCAAAGGCCTTGTCGACGTTATTCCATTACGCCCGTCATTTTGG
(SEQ ID NO: 197)
Эти два олигонуклеотида также содержали фланкирующие последовательности, которые позволяют осуществлять клонирование продукта амплификации в puc19 посредством рестрикционных HindIII- и SalI-сайтов. Полученную плазмиду обозначали pFIMAIC (SEQ ID NO:198).
Для амплификации последовательности от нуклеотида 235654 до нуклеотида 238666, содержащей гены fimD, использовали олигонуклеотиды, имеющие следующие последовательности:
AAGATCTTAAGCTAAGCTTGAATTCTCTGACGCTGATTAACC (SEQ ID NO: 199)
и
ACGTAAAGCATTTCTAGACCGCGGATAGTAATCGTGCTATC (SEQ ID NO:200).
Эти два олигонуклеотида также содержали фланкирующие последовательности, которые позволяют осуществлять клонирование продукта амплификации в puc19 посредством рестрикционных HindIII- и XbaI-сайтов; полученную плазмиду обозначали pFIMD (SEQ ID NO:201).
Для амплификации последовательности от нуклеотида 238575 до нуклеотида 240543, содержащей гены fimF, fimG и fimH, использовали олигонуклеотиды, имеющие следующие последовательности:
AATTACGTGAGCAAGCTTATGAGAAACAAACCTTTTTATC (SEQ ID NO:202)
и
GACTAAGGCCTTTCTAGATTATTGATAAACAAAAGTCACGC (SEQ ID NO:203).
Эти два олигонуклеотида также содержали фланкирующие последовательности, которые позволяют осуществлять клонирование продукта амплификации в puc19 посредством рестрикционных HindIII- и XbaI-сайтов. Полученную плазмиду обозначали pFIMFGH (SEQ ID NO:204).
Для генерирования плазмиды, содержащей все вышеупомянутые гены fim, осуществляли следующие процедуры клонирования:
pFIMAIC гидролизовали EcoRI и HindIII (2237-3982), pFIMD гидролизовали EcoRI и SstII (2267-5276), pFIMFGH гидролизовали SstII и HindIII (2327-2231). Затем эти фрагменты лигировали и полученную плазмиду, содержащую все гены fim, необходимые для образования пилуса, обозначали pFIMAICDFGH (SEQ ID NO:205).
ПРИМЕР 35
Конструирование экспрессирующей плазмиды для пилей типа 1 Escherichia coli, не содержащих адгезивного FimH
Плазмиду pFIMAICDFGH (SEQ ID NO:205) гидролизовали KpnI, после чего фрагмент, состоящий из нуклеотидов 8895-8509, выделяли с помощью электрофореза в 0,7% агарозном геле и подвергали рециркуляризации путем самолигирования. Полученная плазмида, которую обозначали pFIMAICDFG (SEQ ID NO:206), не содержит ген FimH и может быть использована для продуцирования пилей типа 1, не содержащих FimH.
ПРИМЕР 36
Экспрессия пилей типа 1 с использованием плазмиды pFIMAICDFGH
Штамм E.coli W3110 трансформировали плазмидой pFIMAICDFGH (SEQ ID NO:205), высевали на планшеты со средой LB (10 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л NaCl, рН 7, 5, 1% агар (мас./об)), содержащей 100 мкг/мл ампициллина, и инкубировали в течение ночи при 37°С. Затем одиночную колонию использовали для инокуляции 50 мл исходной культуры LB с глюкозой (10 г/л триптона, 5 г/л дрожжевого экстракта, 1% (мас./об) глюкозы, 5 г/л NaCl, рН 7,5, 100 мг/мл ампициллина). После инкубирования в течение 12-16 часов при 37°С и при 150 об/мин 5-литровые шейкерные колбы, содержащие 2 литра LB-глюкозы, инокулировали 20 миллилитрами исходной культуры. Затем бактериальные культуры инкубировали еще 24 часа при 37°С и при перемешивании (150 об/мин). После этого бактерии собирали центрифугированием (5000 об/мин, 4°С, 10 минут) и полученный осадок русеспендировали в 250 миллилитрах 10 мМ Трис/HCl, рН 8. Пили отделяли от бактерий путем 5-минутного перемешивания в подходящем миксере при 17000 об/мин. После 10-минутного центрифугирования при 10000 об/мин и выдерживания в течение 1 часа супернатант, содержащий пили, собирали и добавляли 1М MgCl2 до конечной концентрации 100 мМ. Раствор выдерживали при 4°С в течение 1 часа, после чего осажденные пили собирали центрифугированием (10000 об/мин, 20 минут, 4°С). Затем осадок ресуспендировали в 10 мМ HEPES, рН 7,5, в течение 30 минут при комнатной температуре и раствор пилуса осветляли путем проведения конечной стадии центрифугирования для удаления остаточного клеточного дебриса. Затем этот препарат диализовали против 20 мМ HEPES, рН 7,4.
ПРИМЕР 37
Связывание эпитопов и мимотопов для IgE с пилями типа 1 Escherichia coli
66 мкл-аликвоту 100 мкМ раствора гетеробифункционального сшивающего линкера Sulfo-MBS добавляли к 400 мкл 95% чистого раствора бактериальных пилей типа 1 (2 мг/мл, 20 мМ HEPES, рН 7,4), а затем инкубировали в течение 45 минут при комнатной температуре и при перемешивании. Затем избыток Sulfo-MBS удаляли эксклюзионной хроматографией с использованием колонки PD-10. Альтернативно, сшивающий линкер может быть удален путем диализа. После этого к 1 мл-аликвотам дериватизированных пилей (1-1,25 мг/мл, 20 мМ HEPES, рН 7,4) добавляли либо 1,3 мкл раствора, содержащего 1,1 мг/мл пептида Се3еpi (CGGVNLT·SRA SG (SEQ ID NO:207)), либо пептида Cе3Mim (CGGVNLPWSFGLE (SEQ ID NO:208). Образцы инкубировали при комнатной температуре в течение 4 часов и несвязанный пептид удаляли путем двухкратного диализа против 2 л буфера, содержащего 20 мМ HEPES (рН 7,4). Альтернативно, несвязанный пептид может быть удален эксклюзионной хроматографией.
ПРИМЕР 38
Иммунизация мышей гибридным белком фосфолипазы A2 (PLA2) пчелиного яда, связанным с капсидным белком Qβ
А. Получение альтернативного вектора для цитоплазматической экспрессии каталитически неактивного варианта гена PLA2, присоединенного к аминокислотной последовательности AAASGGCGG (SEQ ID NO:209)
Конструкцию гена PLA2 примера 9 амплифицировали с помощью ПЦР из рAV3PLAfos с использованием олигонуклеотидов ecori_Ndel_pla (последовательность представлена ниже) и PLA-Cys-hind (пример 29). Для проведения этой реакции 100 пмоль каждого олигонуклеотида и примерно 1 мкг PAV3PLAfos ДНК использовали в 50 мкл реакционных смесей с раствором, содержащим 1,2 единиц ДНК-полимеразы Pfx (Gibco), 1 мМ MgSO4, 200 мкМ dNTP и энхансер Pfx (Gibco) (с 10-кратным разведением). Для проведения реакции осуществляли нижеследующие температурные циклы: 94°С, 2 минуты, 5 циклов при 92°С (0,5 минуты), 58°С (0,5 минуты), 68°С (1 минута); 25 циклов при 92°С (0,5 минуты), 63°С (0,5 минуты), 68°С (1 минута). ПЦР-продукт очищали с помощью электрофореза в агарозном геле, фрагмент, расщепленный ферментами NdeI и HindIII, выделяли с использованием набора Qiagen Qiaquick Kit, а затем клонировали в вектор РЕТ11а (Novagen), расщепленный теми же самыми ферментами.
Олигонуклеотиды:
ecorl_Ndel_pla:
TAACCGAATTCAGGAGGTAAAAACATATGGC TATCATCTACC (SEQ ED NO:214).
Этот вектор кодировал гибридный белок, имеющий аминокислотную последовательность:
MAIIYPGTLWCGHGNKSSGPNELGRFKHTDACCRTQDMCPDVMSAG
ESKHGLT· TASHTRLSCDCDDKFYDCLKNSADTISSYFVGKMYFNLIDTK
CYKLEHPVTGCGERTEGRCLHYTVDKSKPKVYQWFDLRKYAAASGGCG
G(SEQ ID NO:210).
Связывание гибридного белка PLA2 с капсидным белком Qβ
Раствор 600 мкМ капсидного белка Qβ (2 мг/мл в 20 мМ Hepes, рН 7,4) подвергали реакции с 176 мкл Sulfo-MBS (13 мг/мл в H2O) в течение 60 минут при комнатной температуре и диализовали против 1 л 20 мМ Hepes, рН 7,4, в течение ночи (О/N) при 4°С. На следующий день 500 мкл раствора PLA2 (2,5 мг/мл), содержащего 0,1 мМ DTT, обессоливали на 5 мл-колонке Hi-Trap (Pharmacia). Восстановленный и обессоленный PLA2 (60 мкл раствора приблизительно 0,5 мг/мл) смешивали с активированным и диализованным капсидом Qβ (25 мкл 1,5 мг/мл раствора) и подвергали реакции в течение 4 часов при комнатной температуре.
Капсиды диаметром 25-30 нм были хорошо видны в изображении капсидного белка Qβ, полученного с помощью электронной микроскопии до и после связывания с PLA2.
С. Иммунизация мышей PLA2, связанным с капсидным белком Qβ
Самок мышей Balb/с иммунизировали на день 0 путем внутривенного введения 50 мкг капсида Qβ, связанного с PLA2, а затем, на день 14, вводили бустер-инъекцию того же количества антигена. На 20-й день у мышей брали кровь и сыворотку анализировали в ELISA. Титр, полученный для PLA2, составлял 1:5000.
ПРИМЕР 39
Связывание эпитопов и мимотопов для IgE с капсидным белком Qβ
Эпитопы для человеческого IgE, имеющие нижеследующие последовательности, связывали с капсидным белком Qβ с использованием N-концевого цистеинового остатка:
Се3-эпитоп: CGGVNLT·SRASG (SEQ ID NO:207)
Се3-мимотоп: CGGVNLPWSFGLE (SEQ ID NO:208).
Реакцию связывания осуществляли с использованием капсидного белка Qβ, активированного Sulfo-MBS, после чего осуществляли диализ для удаления избытка сшивающего агента. Соответствующий эпитоп и мимотоп разводили в реакционной смеси, содержащей активированный капсид Qβ и смесь оставляли на 4 часа при комнатной температуре для реакции. И наконец, реакционную смесь диализовали в течение 4 часов против PBS и инъецировали мышам.
С капсидным белком Qβ был также связан нижеследующий кольцевой мимотоп: Се4-мимотоп: GEFCINHRGYWVCGDPA (SEQ ID NO:211).
Для введения защитной сульфгидрильной группы в мимотоп его сначала подвергали реакции с химической N-сукцинимидил-S-ацетилтиоацетатной группой (SATA). Затем защитную группу удаляли путем обработки гидроксиламином и сразу подвергали реакции с активированным капсидным белком Qβ в течение 4 часов при комнатной температуре. И наконец, реакционную смесь диализовали в течение 4 часов и инъецировали мышам.
ПРИМЕР 40
Иммунизация мышей HBcAg-Lys, связанным пептидом М2
А. Связывание пептида М2 с капсидным белком HBcAg-Lys
Синтетический пептид М2, соответствующий N-концевому фрагменту белка М2 вируса гриппа, имеющему цистеиновый остаток у своего С-конца (SLLT·VETPIRNEWGCRCNGSSDGGGC (SEQ ID NO:212)), был химически присоединен к очищенным частицам HBcAg-Lys для вырабатывания иммунного ответа против пептида М2. Sulfo-MBS (232 мкл, 3 мМ) подвергали реакции с раствором 1,4 мл HBcAg-Lys (1,6 мг/мл) в PBS. Смесь диализовали в течение ночи против забуференного фосфатом физиологического раствора (PBS). Пептид М2 разводили до концентрации 24 мг/мл в ДМСО и 5 мкл этого раствора разводили в 300 мкл PBS, из которого 188 мкл добавляли к 312 мкл диализованного активированного раствора HBcAg-Lys. К реакционной смеси также добавляли EDTA (10 мкл 1М раствора), после чего смесь оставляли на 4 часа при комнатной температуре для реакции.
Иммунизация мышей HBcAg-Lys, связанным с пептидом М2
Самок мышей Balb/с иммунизировали на день 0 путем внутривенного введения 50 мкг HBcAg-Lys-М2, или только пептида М2, а через 10 дней вводили бустер-инъекцию того же самого количества антигена. Еще через 10 дней мышей интраназально инфицировали вирусом гриппа (50 б.о.е., PR/8) и проводили мониторинг выживания инфицированных мышей. Кроме того, определяли титр вируса в легких. Мыши, примированные М2-HBcAg-Lys, были полностью защищены, и у них не обнаруживался этот вирус на 7-й день.
ПРИМЕР 41
Связывание пептида М2 с пилями, с капсидным белком Qβ и с капсидным белком HBcAg, не содержащим cys, и сравнение титра антител, полученного при иммунизации мышей этими связанными пилями и капсидами, с титром, полученным при иммунизации мышей N-концевым гибридным белком пептида М2, связанного с HBcAg1-183
А. Связывание петида М2 с пилями, с капсидным белком Qβ и с капсидным белком HBcAg, не содержащим Cys
Qβ: Раствор 1 мл 1 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 93 мкл раствора 13 мг/мл Sulfo-MBS (Pierce) в H2O при комнатной температуре на ротационном шейкере. Затем реакционную смесь диализовали в течение ночи против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2. После этого диализованную реакционную смесь подвергали реакции с 58,8 мкл 25 мМ маточного раствора пептида М2 (SEQ ID NO:212) в ДМСО в течение четырех часов при комнатной температуре на ротационном шейкере. Затем реакционную смесь диализовали против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, в течение ночи при 4°С.
HBcAg, не содержащий Cys: Раствор 1,25 мл 0,8 мг/мл капсидного белка HBcAg, не содержащего cys, (пример 31) в PBS, рН 7,2, подвергали реакции в течение 30 минут с 93 мкл раствора 13 мг/мл Sulfo-MBS (Pierce) в H2O при комнатной температуре на ротационном шейкере. Затем реакционную смесь диализовали в течение ночи против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2. После этого диализованную реакционную смесь подвергали реакции с 58,8 мкл 25 мМ маточного раствора пептида М2 (SEQ ID NO:212) в ДМСО в течение четырех часов при комнатной температуре на ротационном шейкере. Затем реакционную смесь диализовали против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, в течение ночи при 4°С.
Пили: Раствор 400 мкл мл 2,5 мг/мл пилей в 20 мМ Hepes, рН 7,4, подвергали реакции в течение 45 минут с 60 мкл раствора 100 мМ Sulfo-MBS (Pierce) в H2O при комнатной температуре на ротационном шейкере. Затем реакционную смесь обессоливали на колонке PD-10 (Amersham-Pharmacia, Biotech) и вторую фракцию 500 мкл белка, элюированного с колонки (содержащую приблизительно 1 г белка), подвергали реакции с 58,8 мкл 25 мМ маточного раствора пептида М2 (SEQ ID NO:212) в ДМСО в течение четырех часов при комнатной температуре на ротационном шейкере. Затем реакционную смесь диализовали против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, в течение ночи при 4° С.
Генетическое присоединение пептида М2 к HBcAg1-183
М2 генетически присоединяли к НВс: М2 клонировали в N-конец НВс, как описано в публикации Neirynck et al., Nature Medicine, 5:1157 (1999). MD-НВс экспрессировали в E.coli и очищали с помощью гель-хроматографии. Присутствие пептида М2 на N-конце М2-НВс подтверждали секвенированием по Эдману.
Иммунизация мышей:
Самок мышей Balb/с вакцинировали пептидом М2, связанным с пилями, с белком Qβ и с белком HBcAg, не содержащим cys, и пептидом М2, генетически присоединенном к иммуногену НВс, без добавления адъювантов. Затем на день 0 и на день 14 внутрибрюшинно вводили 35 мкг белка каждого образца. У мышей брали кровь на день 27 и сыворотку анализировали с использованием ELISA, специфического к пептиду М2.
ELISA
10 мкг/мл пептида М2, связанного с РНКазой, наносили на ELISA-планшет. Планшет блокировали, а затем инкубировали с серийно разведенной мышиной сывороткой. Связанные антитела детектировали ферментативно меченным антимышиным антителом IgG. В качестве контроля также тестировали неиммунную сыворотку. Контрольные ELISA-эксперименты с использованием сыворотки мышей, иммунизированных неродственными пептидами, сшитыми с НВс или с другими носителями, показали, что эти антитела были специфичными к пептиду М2. Результаты представлены на фиг.27А и В.
ПРИМЕР 42
Связывание пептидов ангиотензина I и ангиотензина II с Qβ и иммунизация мышей вакцинами "Qβ -пептид ангиотензина"
А. Связывание пептидов ангиотензина I и ангиотензина II с капсидным белком Qβ
Были химически синтезированы следующие пептиды ангиотензина: CGGDRVYIHPF ("ангио I"), CGGDRVYIHPFHL ("ангио II"), DRVYIHPFHLGGC ("ангио III") и CDRVYIHPFHL ("ангио IV"), и эти пептиды были использованы для химического связывания с Qβ, как описано ниже.
Раствор 5 мл 2 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 507 мкл раствора 13 мг/мл Sulfo-MBS (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С. После этого 665 мл диализованной реакционной смеси подвергали реакции с 2,8 мл каждого соответствующего 100 мМ маточного раствора пептида (ДМСО) в течение двух часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С.
Иммунизация мышей:
Самок мышей Balb/с вакцинировали одним из четырех пептидов ангиотензина, связанным с капсидным белком Qβ, без добавления адъювантов. 50 мкг общего белка каждого образца разводили в PBS до 200 мл и подкожно инъецировали (100 мл с двух вентральных сторон) на день 0 и на день 14. У мышей ретроорбитально брали кровь на день 21 и сыворотку анализировали с использованием ELISA, специфичного для ангиотензина.
ELISA
Все четыре пептида ангиотензина по отдельности связывали с бычьей РНКазой А с использованием химического линкера Sulfo-SPDP. ELISA-планшеты покрывали препаратами связанной РНКазы при концентрации 10 мг/мл. Эти планшеты блокировали, а затем инкубировали с серийно разведенной мышиной сывороткой. Связанные антитела детектировали ферментативно меченным антимышиным антителом IgG. В качестве контроля также тестировали неиммунную сыворотку, взятую от тех же самых мышей. Контрольные ELISA-эксперименты с использованием сыворотки мышей, иммунизированных неродственными пептидами, сшитыми с Qβ или с другими носителями, показали, что эти детектированные антитела были специфичными к соответствующему пептиду. Результаты представлены на фиг.8А-8D.
На фиг.8А, 8В, 8С и 8D, соответственно, проиллюстрированы ELISA-анализы антител IgG, специфичных к "ангио I", "ангио II", "ангио III" и "ангио IV", соответственно, в сыворотке мышей, иммунизированных против "ангио I-IV", связанных с капсидным белком Qβ. Qβ-ангио I, Qβ-ангио II, Qβ-ангио III и Qβ-ангио IV, указанные на этих фигурах, означают вакцины, инъецированные мышам, сыворотка которых содержала пептиды, соответствующие вышеуказанному определению.
Самок мышей Balb/с подкожно вакцинировали 50 мг вакцины в PBS на день 0 и день 14. На день 21 антитела IgG в сыворотке мышей, вакцинированных Qβ-ангио I, Qβ-ангио II, Qβ-ангио III и Qβ-ангио IV, оценивали против всех четырех пептидов (связанных с РНКазой А), то есть против "ангио I" (фиг.8А), "ангио II" (фиг.8В), "ангио III" (фиг.8С) и "ангио IV" (фиг.8D). В качестве контроля также анализировали неиммунную сыворотку, взятую от тех же самых мышей. Результаты для указанных разведений сыворотки представлены как оптическая плотность при 450 нм. Приводится среднее значение из трех измерений для каждой мыши (включая стандартные отклонения). Все вакцинированные мыши продуцировали высокие титры антител IgG против всех четырех пептидов. У контролей (невакцинированных мышей) не было обнаружено каких-либо ангиотензин-специфических антител.
ПРИМЕР 43
Связывание пептидов ангиотензина I и ангиотензина II с HBcAg-149-lys-2cys-Mut, то есть с HBcAg, не содержащим cys
Были химически синтезированы следующие пептиды ангиотензина: CGGDRVYIHPF ("ангио I"), CGGDRVYIHPFHL ("ангио II"), DRVYIHPFHLGGC ("ангио III") и CDRVYIHPFHL ("ангио IV") и эти пептиды были использованы для химического связывания с HBcAg-149-Lys-2cys-Mut, то есть HBcAg, не содержащим cys.
Раствор 1,25 мл 0,8 мг/мл капсидного белка HBcAg-149-Lys-2cys-Mut (см. пример 31) в PBS, рН 7,4, подвергали реакции в течение 30 минут с 93 мкл раствора 13 мг/мл Sulfo-MBS (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение ночи против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4. После замены буфера реакционный раствор диализовали еще 2 часа, затем диализованную реакционную смесь подвергали реакции с 1,8 мкл 100 мМ маточного раствора пептида (ДМСО) в течение двух часов при 25°С на ротационном шейкере. После этого реакционную смесь два раза диализовали в течение ночи против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С, а затем буфер меняли и проводили диализ еще в течение 2 часов.
ПРИМЕР 44
Связывание пептидов ангиотензина I и ангиотензина II с пилями типа 1 E.coli
Были химически синтезированы следующие пептиды ангиотензина: CGGDRVYIHPF ("ангио I"), CGGDRVYIHPFHL ("ангио II"), DRVYIHPFHLGGC ("ангио III") и CDRVYIHPFHL ("ангио IV") и эти пептиды были использованы для химического связывания с пилями типа 1 E.coli.
Раствор 400 мкл 2,5 мг/мл пилей типа 1 E.coli в 20 мМ Hepes, рН 7,4, подвергали реакции в течение 60 минут с 60 мкл раствора 100 мМ Sulfo-MBS (Pierce) в H2O при комнатной температуре на ротационном шейкере. Затем реакционную смесь обессоливали на колонке PD-10 (Amersham-Pharmacia, Biotech). Белоксодержащие фракции элюируются с колонки (содержащие приблизительно 1 мг белка, то есть дериватизированных пилей) объединяли и подвергали реакции с трехкратным молярным избытком пептида. Так, например, к 500 мкл элюата, содержащего приблизительно 1 мг дериватизированных пилей, добавляли 2,34 мкл 100 мМ маточного пептидного раствора (ДМСО). Смесь инкубировали в течение четырех часов при 25°С на ротационном шейкере, а затем реакционную смесь диализовали против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, в течение ночи при 4°С.
ПРИМЕР 45
Связывание пептидов Der p I c Qβ и иммунизация мышей вакцинами Qβ-Der p I
Связывание пептидов Der p I с капсидным белком Qβ
Были химически синтезированы следующие пептиды, происходящие от аллергена клеща домашней пыли Der p I: CGNQSLDLAEQELVDCASQHGCH ("Der p I p52"; аминокислоты 52-72, с дополнительным цистеин-глициновым линкером у N-конца) и CQIYPPNANKIREALAQTHSA ("Der p 1 p117"; аминокислоты 117-137). Эти пептиды были использованы для химического связывания с Qβ, как описано ниже.
1 мл раствора, состоящего из 2 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут со 102 мкл раствора 13 мг/мл Sulfo-MBS (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С. После этого 440 мл диализованной реакционной смеси подвергали реакции с 1,9 мкл 100 мМ маточного раствора пептида (в ДМСО) в течение двух часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С.
Иммунизация мышей:
Самок мышей Balb/с вакцинировали одним из двух пептидов Der p I, связанным с капсидным белком Qβ, без добавления адъювантов. Были использованы две мыши на каждую вакцину. 30 мкг общего белка каждого образца разводили в PBS до 200 мл и подкожно инъецировали на день 0 и на день 14. У мышей ретроорбитально брали кровь на день 21 и сыворотку анализировали с использованием ELISA, специфичного к пептиду Der p I.
ELISA
Пептиды Der p I, "Der p I р52" и "Der p I р117" по отдельности связывали с бычьей РНКазой А с использованием химического линкера Sulfo-SPDP. ELISA-планшеты покрывали препаратами связанной РНКазы при концентрации 10 мг/мл. Эти планшеты блокировали, а затем инкубировали с серийно разведенной мышиной сывороткой. Связанные антитела детектировали ферментативно меченным антимышиным антителом IgG. В качестве контроля также тестировали неиммунную сыворотку, взятую от тех же самых мышей. Контрольные ELISA-эксперименты с использованием сыворотки мышей, иммунизированных неродственными пептидами, сшитыми с Qβ или с другими носителями, показали, что эти детектированные антитела были специфичными к соответствующему пептиду. Результаты представлены на фиг.9А-9В.
На фиг.9А и фиг.9В проиллюстрированы ELISA-анализы антител IgG, специфичных к "Der p I р52" (фиг.9А) и специфичных к "Der p I р117" (фиг.9В), в сыворотке мышей, иммунизированных против пептидов Der p I, связанных с капсидным белком Qβ. "р52" и "р117", используемые, как показано на фиг.9А и 9В, означают вакцины, инъецированные мышам, от которых были получены сыворотки.
В качестве контроля анализировали неиммунную сыворотку, взятую от тех же самых мышей (день 0). Результаты для указанных разведений сыворотки представлены как оптическая плотность при 450 нм. На день 21 все вакцинированные мыши продуцировали специфические антитела IgG против всех пептидов Der p I, которыми они были вакцинированы, но не против другого пептида Der p I. Перед вакцинацией (день 0) не было обнаружено каких-либо антител, специфичных к пептиду Der p I.
Обе вакцины, полученные на основе пептидов Der p I, были в высокой степени иммуногенными в отсутствие адъювантов. Все вакцинированнные мыши продуцировали хорошие гуморальные ответы против указанного пептида в вакцинном препарате.
ПРИМЕР 46
Связывание пептидов Der p I с HBcAg-149-Lys-2cys-Mut, то есть с HBcAg, не содержащим cys
Были химически синтезированы следующие пептиды, происходящие от аллергена клеща домашней пыли Der p I р52 (аминокислоты 52-72 с дополнительным цистеин-глициновым линкером у N-конца) CGNQSLDLAEQELVDCASQHGCH и Der p I р117 (аминокислоты 117-137) CQIYPPNANKIREALAQTHSA. Эти пептиды были использованы для химического связывания с HBcAg-149-Lys-2cys-Mut, то есть с HBcAg, не содержащим cys.
Раствор 1,25 мл 0,8 мг/мл капсидного белка HBcAg-149-Lys-2cys-Mut (см. пример 31) в PBS подвергали реакции в течение 30 минут с 93 мкл раствора 13 мг/мл Sulfo-MBS (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение ночи против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4. После замены буфера реакционный раствор диализовали еще 2 часа. Затем диализованную реакционную смесь подвергали реакции с 1,8 мкл 100 мМ маточного раствора пептида (ДМСО) в течение 2 часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение ночи против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С, а затем буфер меняли и проводили диализ еще в течение 2 часов.
ПРИМЕР 47
Связывание пептидов Der p I с пилями типа 1 E.coli
Были химически синтезированы следующие пептиды, происходящие от аллергена клеща домашней пыли Der p I: Der p I р52 (аминокислоты 52-72 с дополнительным цистеин-глициновым линкером у N-конца) CGNQSLDLAEQELVDCASQHGCH и Der p I р117 (аминокислоты 117-137) CQIYPPNANKIREALAQTHSA. Эти пептиды были использованы для химического связывания с пилями типа 1 E.coli.
Раствор 400 мкл 2,5 мг/мл пилей типа 1 E.coli в 20 мМ Hepes, рН 7,4, подвергали реакции в течение 60 минут с 60 мкл раствора 100 мМ Sulfo-MBS (Pierce) в H2O при комнатной температуре на ротационном шейкере. Затем реакционную смесь обессоливали на колонке PD-10 (Amersham-Pharmacia, Biotech). Белоксодержащие фракции, элюирующиеся с колонки (содержащие приблизительно 1 мг белка, то есть дериватизированных пилей), объединяли и подвергали реакции с трехкратным молярным избытком пептида. Так, например, к 500 мкл элюата, содержащего приблизительно 1 мг дериватизированных пилей, добавляли 2,34 мкл 100 мМ маточного раствора пептида (в ДМСО). Смесь инкубировали в течение четырех часов при 25°С на ротационном шейкере, а затем реакционную смесь диализовали против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, в течение ночи при 4°С.
ПРИМЕР 48
Связывание пептида человеческого VEGFR-2 с пилями типа 1 E.coli и иммунизация мышей вакцинами, содержащими массивы "пили типа 1-пептид человеческого VEGFR-2"
Связывание человеческого пептида VEGFR-2 с пилями типа 1 E.coli
Был химически синтезирован пептид человеческого VEGFR II с последовательностью CTARTELNVGIDFNWEYPSSKHQHKK, и этот пептид использовали для химического связывания с пилями типа 1 E.coli.
Раствор 1400 мкл 1 мг/мл белка пилей в 20 мМ Hepes, рН 7,4, подвергали реакции в течение 60 минут с 85 мкл раствора 100 мг/мл Sulfo-MBS (Pierce) в H2O при комнатной температуре на ротационном шейкере. Затем реакционную смесь обессоливали на колонке PD-10 (Amersham-Pharmacia, Biotech). Белоксодержащие фракции, элюированные с колонки (содержащие приблизительно 1,4 мг белка), объединяли и подвергали реакции с 2,5-кратным молярным избытком (конечный объем) пептида человеческого VEGFR-2. Так, например, к 200 мкл элюата, содержащего приблизительно 0,2 мг дериватизированных пилей, добавляли 2,4 мкл 10 мМ раствора пептида (в ДМСО). Смесь инкубировали в течение четырех часов при 25°С на ротационном шейкере, а затем диализовали против 2 л 20 мМ Hepes, рН 7,2, в течение ночи при 4°С.
Иммунизация мышей
Самок мышей С3Н-HeJ (мышей, дефицитных по рецептору типа "ловушки" 4 и не отвечающих на ЛПС) и мышей С3Н-HeN (дикого типа) вакцинировали пептидом человеческого VEGFR-2, связанным с белком пилей типа 1, без добавления адъювантов. Приблизительно 100 мкг общего белка каждого образца разводили в PBS до 200 мл и подкожно инъецировали на день 0, на день 14 и день 28. У мышей ретроорбитально брали кровь на дни 14, 28 и 42 и сыворотку анализировали с использованием ELISA, специфичного для человеческого VEGFR-2.
ELISA
Сыворотки иммунизированных мышей тестировали в ELISA с использованием иммобилизованного пептида человеческого VEGFR-2 и внеклеточного домена VEGFR-2 (R&D Systems GmbH, Wiesbaden).
Пептид человеческого VEGFR-2 связывали с бычьей РНКазой А с использованием химического сшивающего линкера Sulfo-SPDP. ELISA-планшеты покрывали связанной РНКазой при концентрации 10 мг/мл. Внеклеточный домен человеческого VEGFR-2 адсорбировали на планшетах при концентрации 2 мкг/мл. Эти планшеты блокировали, а затем инкубировали с серийно разведенной мышиной сывороткой. Связанные антитела детектировали ферментативно меченным антимышиным антителом IgG. В качестве контроля также тестировали неиммунную сыворотку, взятую от тех же самых мышей. Контрольные ELISA-эксперименты с использованием сыворотки мышей, иммунизированных несвязанным носителем, показали, что эти детектированные антитела были специфичными к соответствующему пептиду. Результаты связывания пептида человеческого VEGFR-2 с пилями типа 1 представлены на фиг.10. В частности, на фиг.10А и на фиг.10В представлены ELISA-анализы антител IgG, специфичных к пептиду человеческого VEGFR-2 и к внеклеточному домену человеческого VEGFR-2, соответственно, в сыворотке мышей, иммунизированных против пептида человеческого VEGFR-2 и внеклеточного домена человеческого VEGFR-2, каждый из которых был связан с белком пилей типа 1.
Самок мышей С3Н-HeJ (мышей, дефицитных по рецептору типа "ловушки" 4 и не отвечающих на ЛПС) и мышей С3Н-HeN (дикого типа) подкожно вакцинировали 100 мкг вакцины в PBS на дни 0, 14 и 28. IgG-сыворотку против этого пептида (связанного с РНКазой А) и внеклеточного домена человеческого VEGFR-2, оценивали на день 42. В качестве контроля также анализировали неиммунную сыворотку, взятую от тех же самых мышей. Результаты для указанных разведений сыворотки представлены как оптическая плотность при 450 нм. Приводится среднее значение из трех измерений для каждой мыши (включая стандартные отклонения). Все вакцинированные мыши продуцировали высокие титры антител IgG против пептида человеческого VEGFR-2, а также против внеклеточного домена человеческого VEGFR-2 (KDR), и при этом каких-либо различий между мышами, дефицитными по рецептору типа "ловушки" 4, и мышами дикого типа не наблюдалось. Это последнее наблюдение является особенно любопытным, поскольку оно продемонстрировало, что продуцирование высоких титров антитела IgG против пептида человеческого VEGFR-2, а также против внеклеточного домена человеческого VEGFR-2 не зависит от содержания эндотоксиновых примесей.
ПРИМЕР 49
Связывание пептида человеческого VEGFR-2 с капсидным белком Qβ и иммунизация мышей вакцинами, содержащими массивы "капсидный белок Qβ - пептид человеческого VEGFR-2"
Связывание человеческого пептида VEGFR-2 с капсидным белком Qβ
Был химически синтезирован пептид человеческого VEGFR-2 с последовательностью CTARTELNVGIDFNWEYPSSKHQHKK, и этот пептид использовали для химического связывания с капсидным белком Qβ.
1 мл раствора 1 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 45 минут с 20 мкл раствора 100 мМ Sulfo-MBS (Pierce) в H2O при комнатной температуре на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов в 2 л 20 мМ Hepes, рН 7,4, при 4°С. После этого 1000 мкл диализованной реакционной смеси подвергали реакции с 12 мкл 10 мМ раствора пептида человеческого VEGFR-2 (в ДМСО) в течение четырех часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, рН 7,4, при 4°С.
1 мл раствора, состоящего из 2 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 102 мкл раствора 13 мг/мл Sulfo-MBS (Pierce) в H2 O при комнатной температуре на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С. После этого 440 мкл диализованной реакционной смеси подвергали реакции с 1,9 мкл 100 мМ маточного раствора пептида (в ДМСО) в течение двух часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С.
Иммунизация мышей:
Самок мышей C57BL/6 вакцинировали пептидом человеческого VEGFR-2, связанным с белком Qβ, без добавления адъювантов. Приблизительно 50 мкг общего белка каждого образца разводили в PBS до 200 мкл и подкожно инъецировали на день 0, день 14 и на день 28. У мышей ретроорбитально брали кровь на дни 14, 28 и 42 и сыворотку, взятую на день 42, анализировали с использованием ELISA, специфичного к человеческому VEGFR-2.
ПРИМЕР 50
Связывание пептида человеческого VEGFR-2 с капсидным белком HBcAg-149-Lys-2cys-Mut, то есть с HBcAg, не содержащим cys, и иммунизация мышей вакцинами, содержащими массивы "капсидный белок HBcAg-149-Lys-2cys-Mut - пептид человеческого VEGFR-2"
Связывание человеческого пептида VEGFR-2 с капсидным белком HBcAg-149-Lys-2cys-Mut
Был химически синтезирован пептид человеческого VEGFR-2 с последовательностью CTARTELNVGIDFNWEYPSSKHQHKK, и этот пептид использовали для химического связывания с капсидным белком HBcAg-149-Lys-2cys-Mut.
3 мл раствора 0,9 мг/мл капсидного белка HBcAg, не содержащего cys (см. пример 31) в PBS, рН 7,4, подвергали реакции в течение 45 минут с 37,5 мкл 100 мМ раствора Sulfo-MBS (Pierce) в H2O при комнатной температуре на ротационном шейкере. Затем реакционный раствор диализовали в течение ночи против 2 л 20 мМ Hepes, рН 7,4. После замены буфера реакционный раствор диализовали еще 2 часа. После этого диализованную реакционную смесь подвергали реакции с 3 мкл 10 мМ раствора пептида человеческого VEGFR-2 (в ДМСО) в течение 4 часов при 25°С на ротационном шейкере. Затем реакционную смесь диализовали против 2 л 20 мМ Hepes, рН 7,4, в течение ночи при 4°С, после чего буфер меняли и проводили диализ еще в течение 2 часов.
ПРИМЕР 51
Конструирование HBcAg1-183Lys
Коровый антиген (HBcAg)1-183 вируса гепатита модифицировали, как описано в примере 23. Часть области эпитопа с/е1 (остатки 72-88) (пролин 79 и аланин 80) заменяли на генном уровне пептидом Gly-Gly-Lys-Gly-Gly (конструкция HBcAg1-183Lys). Введенный лизиновый остаток содержит в своей боковой цепи реакционноспособную аминогруппу, которая может быть использована для межмолекулярного химического сшивания частиц HBcAg с любым антигеном, содержащим свободную цистеиновую группу. Ген HBcAg1-183Lys получали методами ПЦР, описанными в общих чертах в примере 1, и стандартными методами клонирования.
Последовательность Gly-Gly-Lys-Gly-Gly вводили путем амплификации двух отдельных фрагментов гена HBcAg из рЕсо63, как описано выше в примере 23, с последующим лигированием этих двух фрагментов с помощью ПЦР для сборки полноразмерного гена. При этом были использованы следующие комбинации ПЦР-праймеров:
Фрагмент 1:
Праймер 1: EcoRIHBcAg(s) (см. пример 23)
Праймер 2: Lys-HBcAg(as) (см. пример 23)
Фрагмент 2:
Праймер 3: Lys-HBcAg(s) (см. пример 23)
Праймер 4: HBcAgwtHindIII
CGCGTCCCAAGCTTCTAACATTGAGATTCCCGAGATTG
Сборка:
Праймер 1: EcoRIHBcAg(s) (см. пример 23)
Праймер 2: HBcAgwtHindIII
Собранный полноразмерный ген гидролизовали ферментами EcoRI (GAATTC) и HindIII (AAGCTT) и клонировали в вектор рКК (Pharmacia), разрезанный в тех же самых рестрикционных сайтах.
ПРИМЕР 52
Связывание пептида muTNFa с HBcAg1-183Lys и иммунизация мышей вакцинами, содержащими массивы "HBcAg1-183Lys - пептид muTNFa"
А. Связывание пептида muTNFa с HBcAg1-183Lys
HBcAg1-183Lys в концентрации 0,6 мг/мл (29 мкМ) обрабатывали йодацетамидом, как описано в примере 32. Затем HBcAg1-183Lys подвергали реакции с 50-кратным избытком сшивающего линкера Sulfo-MBS, как описано в примере 32, и диализовали в течение ночи против 20 мМ Hepes, рН 7,2, при 4°С. Активированный (дериватизированный) HBcAg1-183Lys подвергали реакции с пятикратным молярным избытком пептида muTNFa (последовательность: CGGVEEQLEWLSQR, разведенного непосредственно в растворе HBcAg1-183Lys из 100 мМ маточного раствора в ДМСО) при комнатной температуре в течение 4 часов. Реакционную смесь для связывания (примерно 1 мл раствора) два раза диализовали против 2 литров 20 мМ Hepes, рН 7,2, при 4°С в течение 4 часов. Диализованную реакционную смесь замораживали в аликвотах в жидком азоте и хранили при -80°С вплоть до иммунизации мышей.
Иммунизация
Двух мышей (самок Balb/с) иммунизировали на день 0 и 14 путем внутривенной инъекции 100 мкг HBcAg1-183Lys, связанного с пептидом muTNFa, на животное, без адъюванта. Антитела, специфичные к пептиду muTNFa (приготовленные в виде конъюгата с рибонуклеазой А) и к нативному белку TNF-α (Sigma) в сыворотке, оценивали на день 21 с помощью ELISA.
ELISA
Мышиный белок TNF-α (Sigma) наносили в концентрации 2 мкг/мл. В качестве контроля тестировали неиммунную сыворотку от тех же самых мышей, используемых для иммунизации. На фиг.14 показан результат ELISA-эксперимента, который продемонстрировал, что при иммунизации HBcAg1-183Lys, связанным с пептидом muTNFa (полноразмерным НВс-TNF), вырабатывался иммунный ответ, специфичный к мышиному белку TNF-α. Сыворотки, полученные из крови мышей на день 0 (неиммунные) и на день 21, тестировали при трех различных разведениях. Каждый столбец соответствует среднему сигналу, полученному для сывороток, взятых от двух мышей. Таким образом, вакцинация антигеном HBcAg1-183Lys, связанным с пептидом muTNFa, индуцировала иммунный ответ против аутоантигена, поскольку аминокислотная последовательность пептида muTNFa происходит от последовательности мышиного белка TNF-α.
ПРИМЕР 53
Связывание пептида 3'TNF II с 2cysLys-mutHBcAg1-149 и иммунизация мышей вакцинами, содержащими массивы "2cysLys-mutHBcAg1-149 - пептид 3'TNF II"
Связывание пептида 3'TNF II с 2cysLys-mutHBcAg1-149
2cysLys-mutHBcAg1-149 в концентрации 2 мг/мл подвергали реакции в течение 30 минут при комнатной температуре с 50-кратным избытком сшивающего линкера в 20 мМ Hepes, 150 мМ NaCl, рН 7,2. Избыток сшивающего линкера удаляли в течение ночи путем диализа, и активированный (дериватизированный) капсидный белок 2cysLys-mutHBcAg1-149 подвергали реакции с десятикратным избытком пептида 3'TNF II (последовательность: SSQNSSDKPVAHVVANHGVGGC, разведенного из 100 мМ маточного раствора в ДМСО) при комнатной температуре в течение 4 часов. Затем реакционную смесь диализовали в течение ночи в трубке для диализа с отсечкой молекулярной массы 50000 Да, замораживали в жидком азоте и хранили при -80°С вплоть до иммунизации мышей.
Иммунизация мышей
3 самок мышей С3Н/HeN, возрастом 8 недель, вакцинировали пептидом 3'TNF II, связанным с 2cysLys-mutHBcAg1-149, без добавления адъювантов. 50 мкг всего белка разводили в PBS до 200 мл и подкожно инъецировали (100 мкл в паховые области с двух сторон) на день 0 и на день 14. У мышей ретроорбитально брали кровь на дни 0 и 21 и сыворотку анализировали с использованием ELISA, специфичного для мышиного белка TNF-α .
ELISA
Мышиный белок TNFα (Sigma) наносили в концентрации 2 мкг/мл. В качестве контроля тестировали неиммунную сыворотку тех же самых мышей, используемых для иммунизации. На фиг.15 показан результат ELISA-эксперимента, который продемонстрировал, что иммунизация антигеном 2cysLys-mutHBcAg1-149, связанным с пептидом 3'TNF II, приводила к вырабатыванию иммунного ответа, специфичного к мышиному белку TNFα. Сыворотки, полученные из крови мышей на день 0 (неиммунные) и на день 21, тестировали при трех различных разведениях. Каждый столбец соответствует среднему сигналу, полученному для сывороток, взятых от 3 мышей. Таким образом, вакцинация антигеном 2cysLys-mutHBcAg1-149, связанным с пептидом 3'TNF II, продуцировала иммунный ответ против аутоантигена, поскольку аминокислотная последовательность пептида 3'TNF II происходит от последовательности мышиного белка TNFα.
ПРИМЕР 54
Связывание пептидов Аβ 1-15, Аβ 1-27 и Аβ 33-42 с Qβ и иммунизация мышей вакцинами, содержащими массивы "Qβ - пептид Аβ"
А. Связывание пептидов Аβ 1-15 и Аβ 33-42 с капсидным белком Qβ с использованием сшивающего линкера SMPH
Были химически синтезированы следующие пептиды Аβ: пептид DAEFRHDSGYEVHHQGGC (сокращенно называемый "Аβ 1-15"), который включает аминокислотную последовательность от остатка 1 до остатка 15 человеческого Аβ, присоединенную у ее С-конца к последовательности GGC для связывания с капсидным белком Qβ, и пептид CGHGNKSGLMVGGVVIA (сокращенно называемый "Аβ 33-42"), который включает аминокислотную последовательность от остатка 33 до остатка 42 Аβ, присоединенную у ее N-конца к последовательности CGHGNKS для связывания с капсидным белком Qβ. Оба эти пептида были использованы для химического связывания с Qβ, как описано ниже.
Раствор 1,5 мл 2 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 16,6 мкл раствора 65 мМ SMPH (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С в трубке для диализа с отсечкой молекулярной массы 10000 Да. После этого 450 мкл диализованной реакционной смеси, которая содержала активированный (дериватизированный) Qβ, подвергали реакции с 6,5 мкл из каждого соответствующего 50 мМ маточного раствора пептида (в ДМСО) в течение двух часов при 15°С на ротационном шейкере. Затем 200 мкл реакционной смеси диализовали в течение ночи против 2 литров 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С, а на следующее утро, после замены буфера, диализовали в течение еще двух часов. Затем реакционную смесь в аликвотах замораживали в жидком азоте и хранили при -80°С вплоть до иммунизации мышей.
Результаты экспериментов по связыванию были проанализированы с помощью электрофореза в ДСН-ПААГ и представлены на фиг.13А и 13В. Стрелками показаны полосы, соответствующие одному и двум пептидам, связанным с одной субъединицей Qβ (фиг.13А), или одному пептиду, связанному с одной субъединицей Qβ (фиг.13В). Молекулярные массы маркерных белков показаны с левого края на фиг.13А и фиг.13В.
На гель, фиг.13А, были загружены следующие образцы: 1: дериватизированный Qβ; 2: Qβ, связанный с "Аβ 1-15"; супернатант этого образца был собран в конце реакции связывания и центрифугирован; 3: Qβ, связанный с "Аβ 1-15", осадок этого образца был собран в конце реакции связывания и центрифугирован; 4: Qβ, связанный с "Аβ 1-15"; супернатант этого образца оставляли на 24 часа при 4°С, но не диализовали, и центрифугировали; 5: Qβ, связанный с "Аβ 1-15"; осадок этого образца оставляли на 24 часа при 4°С, но не диализовали, и центрифугировали; 6: Qβ, связанный с "Аβ 1-15"; супернатант этого образца брали после диализа реакционной смеси для связывания и центрифугировали.
На гель, фиг.13В, были загружены следующие образцы: 1: дериватизированный Qβ; 2: Qβ, связанный с "Аβ 33-42"; супернатант этого образца был собран в конце реакции связывания и центрифугирован; 3: Qβ, связанный с "Аβ 33-42", осадок этого образца был собран в конце реакции связывания и центрифугирован; 4: Qβ, связанный с "Аβ 33-42"; супернатант этого образца оставляли на 24 часа при 4°С, но не диализовали, и центрифугировали; 5: Qβ, связанный с "Аβ 33-42"; осадок этого образца оставляли на 24 часа при 4°С, но не диализовали, и центрифугировали; 6: Qβ, связанный с "Аβ 33-42"; супернатант этого образца брали после диализа реакционной смеси для связывания и центрифугировали.
В. Связывание пептида "Аβ 1-27" с капсидным белком Qβ с использованием сшивающего линкера SMPH
Был химически синтезирован следующий пептид Аβ ("Аβ 1-27)": DAEFRHDSGYEVHHQKLVFFAEDVGCNGGC. Этот пептид содержит аминокислотную последовательность от остатка 1 до остатка 27 человеческого Аβ, присоединенную у ее С-конца к последовательности GGC для связывания с капсидным белком Qβ.
Первую партию "Аβ 1-27", связанного с капсидным белком Qβ, далее сокращенно называемую "партией 1 Qβ-Аβ1-27", получали следующим образом:
Раствор 1,5 мл 2 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 16,6 мкл раствора 65 мМ SMPH (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С в трубке для диализа с отсечкой молекулярной массы 10000 Да. После этого 450 мкл диализованной реакционной смеси подвергали реакции с 6,5 мкл 50 мМ маточного раствора пептида (в ДМСО) в течение двух часов при 15°С на ротационном шейкере. Затем 200 мкл образца разделяли на аликвоты, замораживали в жидком азоте и хранили при -80°С вплоть до иммунизации мышей.
Вторую партию "Аβ 1-27", связанного с капсидным белком Qβ, далее сокращенно называемую "партией 2 Qβ -Аβ1-27", получали следующим образом:
500 мкл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 11,3 мкл раствора 32,5 мМ SMPH (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С в трубке для диализа с отсечкой молекулярной массы 3500 Да (SnakeSkin, Pierce). Затем диализованную реакционную смесь подвергали реакции с 3,6 мкл 50 мМ маточного раствора пептида (в ДМСО) в течение двух часов при 15°С на ротационном шейкере. После этого реакционную смесь два раза диализовали против 1 л 20 мМ Hepes, 150 мМ NaCl, pH 7,4, в течение 1 часа и в течение ночи после последней замены буфера с использованием мембраны для диализа с отсечкой 50000 Да (Spectrapor, spectrum). После этого реакционную смесь замораживали в аликвотах в жидком азоте и хранили при -80°С вплоть до иммунизации мышей. "Партию 1 Qβ-Аβ 1-27" использовали для первой иммунизации, а "партию 2 Qβ-Аβ 1-27" использовали для бустер-иммунизации.
Результат эксперимента по связыванию анализировали с помощью электрофореза в ДСН-ПААГ и этот результат представлен на фиг.13С. Стрелками показаны полосы, соответствующие одному пептиду, связанному с одной субъединицей Qβ.
На гель, фиг.13С, были загружены следующие образцы: М: маркерный белок; 1: капсидный белок Qβ; 2: дериватизированный Qβ; супернатант этого образца был собран в конце реакции дериватизации и центрифугирован; 3: дериватизированный Qβ, осадок этого образца был собран в конце реакции связывания и центрифугирован; 4: Qβ, связанный с "Аβ 1-27"; супернатант этого образца был взят в конце реакции связывания и центрифугирован; 5: Qβ, связанный с "Аβ 1-27"; осадок этого образца был взят в конце реакции связывания и центрифугирован; 6: Qβ, связанный с "Аβ 1-27"; супернатант этого образца был взят после диализа реакционной смеси для связывания и центрифугирован; 7: Qβ, связанный с "Аβ 1-27"; осадок этого образца был взят после диализа реакционной смеси для связывания и центрифугирован.
С. Связывание пептида "Аβ 1-15" с капсидным белком Qβ с использованием сшивающего линкера Sulfo-GMBS
Раствор 500 мкл 2 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 5,5 мкл раствора 65 мМ SMPH (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С в трубке для диализа с отсечкой молекулярной массы 10000 Да. После этого 500 мкл диализованной реакционной смеси подвергали реакции с 6,5 мкл 50 мМ маточного раствора пептида (в ДМСО) в течение двух часов при 15°С на ротационном шейкере. Затем 200 мкл реакционной смеси диализовали в течение ночи против 2 литров 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С, а на следующее утро, после замены буфера, диализовали еще в течение двух часов. Затем реакционную смесь в аликвотах замораживали в жидком азоте и хранили при -80°С вплоть до иммунизации мышей.
Результаты экспериментов по связыванию были проанализированы с помощью электрофореза в ДСН-ПААГ и представлены на фиг.13D. Стрелками показаны полосы, соответствующие одному, двум и трем пептидам, соответственно, связанным с одной субъединицей Qβ.
На гель, фиг.13D, были загружены следующие образцы: М: маркерный белок; 1: дериватизированный Qβ; 2: Qβ, связанный с "Аβ 1-15", супернатант этого образца был собран в конце реакции связывания и центрифугирован; 3: Qβ, связанный с "Аβ 1-15", осадок этого образца был собран в конце реакции связывания и центрифугирован; 4: Qβ, связанный с "Аβ 1-15"; супернатант этого образца оставляли на 24 часа при 4°С, но не диализовали, и центрифугировали; 5: Qβ, связанный с "Аβ 1-15"; осадок этого образца оставляли на 24 часа при 4°С, но не диализовали, и центрифугировали; 6: Qβ, связанный с "Аβ 1-15"; супернатант этого образца брали после диализа реакционной смеси для связывания и центрифугировали.
D. Связывание пептида "Аβ 1-15" с капсидным белком Qβ с использованием сшивающего линкера Sulfo-MBS
500 мкл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 14,7 мкл раствора 100 мМ Sulfo-MBS (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С в трубке для диализа (SnakeSkin, Pierce) с отсечкой молекулярной массы 3500 Да. После этого диализованную реакционную смесь подвергали реакции с 7,2 мкл 50 мМ маточного раствора пептида (в ДМСО) в течение двух часов при 15°С на ротационном шейкере. Затем реакционную смесь три раза диализовали в течение 4 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, pH 7,4, с использованием мембраны для диализа с отсечкой 50000 Да (Spectrapor, spectrum). После этого реакционную смесь замораживали в аликвотах в жидком азоте и хранили при -80°С вплоть до иммунизации мышей.
Результат эксперимента по связыванию анализировали с помощью электрофореза в ДСН-ПААГ и этот результат представлен на фиг.13Е. Стрелками показана полоса, соответствующая одному пептиду, связанному с одной субъединицей Qβ .
На гель, фиг.13Е, были загружены следующие образцы: 1: капсидный белок Qβ; 2: дериватизированный Qβ, супернатант этого образца был собран в конце реакции дериватизации и центрифугирован; 3: дериватизированный Qβ, осадок этого образца был собран в конце реакции связывания и центрифугирован; 4: дериватизированный Qβ; супернатант этого образца был взят в конце диализа реакционной смеси для дериватизации и центрифугирован; 5: дериватизированный Qβ, осадок этого образца был взят в конце диализа реакционной смеси для дериватизации и центрифугирован; 6: Qβ, связанный с "Аβ 1-15"; супернатант этого образца был взят в конце реакции связывания и центрифугирован; 7: Qβ, связанный с "Аβ 1-15"; осадок этого образца был взят в конце реакции связывания и центрифугирован; 8: Qβ, связанный с "Аβ 1-15"; супернатант этого образца был взят после диализа реакционной смеси для связывания и центрифугирован.
Е. Иммунизация мышей
Пять групп самок мышей C57BL/6 (по три мыши на группу), возрастом 8 недель, вакцинировали каждым из пяти конъюгатов "пептид Аβ - капсидный белок Qβ", без добавления адъювантов. 25 мкг общего белка каждого образца разводили в PBS до 200 мкл и подкожно инъецировали на день 0 и на день 14. У мышей ретроорбитально брали кровь на день 0 (не вакцинированных) и на день 21 и их сыворотку анализировали с помощью ELISA. Пептид "Аβ 1-15" связывали с Qβ тремя различными сшивающими линкерами, в результате чего получали три различных вакцинных препарата ("Qb-Аβ 1-15 SMPH", "Qb-Аβ 1-15 SMBS", "Qb-Аβ 1-15 SGMBS", см. раздел "ELISA", где описаны результаты).
F. ELISA
Все три пептида Аβ по отдельности связывали с бычьей РНКазой А с использованием химического линкера SPDP следующим образом: раствор 10 мг РНКазы А в 2 мл PBS (50 мМ фосфатного буфера, 150 мМ NaCl, рН 7,2) подвергали реакции со 100 мкл 20 нМ раствора SPDP в ДМСО, при 25°С в течение 60 минут на ротационном шейкере. Избыток сшивающего линкера отделяли от активированной (дериватизированной) РНКазы путем гель-фильтрации с использованием колонки PD-10 (Pharmacia). Белоксодержащие фракции собирали и концентрировали до объема 2 мл с использованием центрифужных фильтров (5000 MwCO). Образец 333 мкл раствора дериватизированной РНКазы А подвергали реакции с 2 мкл маточного раствора пептида (50 мМ в ДМСО). Мониторинг реакции связывания осуществляли с помощью спектрофотометрии.
ELISA-планшеты покрывали РНКазой, связанной с пептидом, при концентрации 10 мкг/мл. Эти планшеты блокировали, а затем инкубировали с серийно разведенными мышиными сыворотками. Связанные антитела детектировали ферментативно меченным антимышиным антителом IgG. Неиммунная сыворотка или контрольная сыворотка, взятая от мышей, иммунизированных неродственными пептидами, конъюгированными с Qβ, показала, что эти детектированные антитела были специфичными к соответствующему пептиду. На фиг.14А, фиг.14В и фиг.14С, соответственно, показаны ELISA-анализы антител, специфичных к "Аβ 1-15", "Аβ 1-27" и "Аβ 33-42", соответственно, в сыворотке мышей, иммунизированных против "Аβ 1-15", "Аβ 1-27" и "Аβ 33-42", соответственно, и связанных с капсидным белком Qβ. Обозначения на оси абсцисс означают вакцину, инъецированную мышам, от которых была получена сыворотка, и указывают на пептид и сшивающий линкер, используемые для получения соответствующей вакцины. Все сыворотки были проанализированы против трех пептидов, связанных с РНКазой А, и результаты показали, что несмотря на то что имеется перекрестная реактивность между антителами, вырабатываемыми против пептида 1-15 и 1-27, однако против пептида 33-42, такой перекрестной реактивности не наблюдалось, что указывало на специфичность иммунного ответа. Аналогичным образом, полученные ELISA-титры, выражаемые как разведения сыворотки, дающей ELISA-сигнал с тремя стандартными отклонениями выше фонового уровня, были очень высокими и составляли в пределах от 60000 до 600000. В контрольных сыворотках (от неиммунизированных мышей) каких-либо антител, специфичных к пептиду Аβ, обнаружено не было.
ПРИМЕР 55
Введение cys-содержащих линкеров; экспрессия и очистка антиидиотипических мимоантител IgE и их связывание с капсидным белком Qβ
А. Конструирование плазмид, экспрессирующих мимоантитела, для связывания с капсидным белком Qβ
Плазмиды были получены на основе экспрессирующей плазмиды VAE051-рАSK116. Эта плазмида содержит кодирующие области тяжелой цепи и легкой цепи мимоантитела. Для введения cys-содержащих линкеров у С-конца тяжелой цепи были использованы следующие праймеры:
Праймер CA2F:
CGGCTCGAGCATCACCATCACCATCACGGTGAAGTTAAACTGCAGCTGGAGTCG
Праймер CA1R:
CATGCCATGGTTAACCACAGGTGTGGGTTTTATCACAAGATTTGGGCTCAAC
Праймер CB1R:
CATGCCATGGTTAACCACACGGCGGAGAGGTGTGGGTTTTATCACAAGATTTGGGCTCAAC
Праймер CC1R:
CCAGAAGAACCCGGCGGGGTAGACGGTTTCGGGCTAGCACAAGATTTGGGCTCAACTC
Праймер CC1F:
CGCCGGGTTCTTCTGGTGGTGCTCCGGGTGGTTGCGGTTAACCATGGAGAAAATAAAGTG
Праймер CCR2:
CTCCCGGGTAGAAGTCAC
А.1. Конструирование рСА2:
Праймеры СА2F и CA1R использовали для амплификации 741 п.н.-фрагмента, кодирующего часть тяжелой цепи, с удлиняющим сегментом, кодирующим cys-содержащую линкерную последовательность. VAE-рАSK116 служила в качестве матрицы для полимеразы Pfx (Roche) в приборе для ПЦР (Rodo), проводимой при следующих условиях: (с начальной денатурацией при 92°С); 5 циклов при 92°С, 30 с; 48°С, 30 с; 68°С, 60 с; а затем 30 циклов при 92°С; 30 с; 58°С, 30 с; 68°С, 1 мин. ПЦР-продукт соответствующего размера очищали с использованием набора для ПЦР-очистки Qiagen и гидролизовали XhoI и NcoI в соответствии с инструкциями производителя (Gibco). Этот продукт очищали на агарозном геле с помощью набора для гель-экстракции Qiagen. Параллельно, плазмиду VAE-рАSK116 расщепляли ферментами XhoI и NcoI и из агарозных гелей выделяли полосу размером 3,7 т.п.н. Соответствующие аликвоты ПЦР-продукта, гидролизованного XhoI-NcoI, и указанные плазмиды лигировали в течение ночи при 16°С с использованием ДНК-лигазы Т4 в соответствии с протоколом производителя (Gibco). Продукт лигирования трансформировали в компетентные клетки XL-1 E.coli, которые высевали на планшеты с агарозой, содержащей хлорамфеникол. Моноколонии размножали в среде LB с хлорамфениколом, получали плазмиду (набор для миниплазмид Qiagen) и тестировали ее на присутствие соответствующих XhoI-NcoI-вставок нужного размера после гидролиза соответствующими ферментами. Соответствующую позитивную плазмиду, обозначенную рСА2, использовали для секвенирования на обеих цепях, и это секвенирование подтвердило идентичность плазмиды, включающей cys-содержащий линкер.
А.2. Конструирование рСВ2:
Праймеры СА2F и СВ1R использовали для введения линкера 2 у 5'-конца последовательности, кодирующей тяжелую цепь, при тех же самых условиях, которые были описаны в разделе А1. Полученный ПЦР-продукт имел размер 750 п.н. и был клонирован в VAE051-рАSK116, как описано в разделе А.1.
А.3. Конструирование рСС2:
Плазмиду рСС2 конструировали в соответствии с двухстадийной процедурой: первый ПЦР-продукт, размером 754 п.н., амплифицировали с использованием праймеров СА2F и СС1R. Второй ПЦР-продукт, размером 560 п.н., продуцировали с использованием праймеров СС1F и СС2R. Для обеих ПЦР-реакций в качестве матрицы использовали VAE051-рАSK116 в условиях, описанных в разделе А1. Оба ПЦР-продукта выделяли из агарозных гелей, смешивали с праймерами СА2F и СС2R, и осуществляли третью ПЦР-реакцию с получением фрагмента размером 1298 п.н. Этот фрагмент выделяли и гидролизовали XhoI и NcoI. Полученный фрагмент размером 780 п.н. клонировали в VAE-рАSK100 как описано в разделе А.1.
В. Экспрессия мимоантител
Компетентные клетки E.coli W3110 трансформировали плазмидами рСА2, рСВ2 и рСС2. Моноколонии из планшетов с агарозой, содержащей хлорамфеникол, размножали в жидкой культуре (LB + 15 мкг/мл хлорамфеникола) в течение ночи при 37°С. Затем 1 л среды ТВ инокулировали ночной культурой 1:50 (об/об) и культивировали до OD600=3 при 28°С. Экспрессию индуцировали 1 мг/л безводного тетрациклина. После культивирования в течение ночи, клетки собирали и центрифугировали при 6000 об/мин. Периплазму выделяли из клеточного осадка путем инкубирования в буфере для лизиса, в который был добавлен сульфат полимиксина В, в течение 2 часов при 4°С. Сферобласты отделяли путем центрифугирования при 6000 об/мин. Полученный супернатант содержал мимоантитело, и этот супернатант диализовали против 20 мМ Трис, рН 8,0.
С. Очистка мимоантител
Введение his6-метки позволяло проводить очистку мимоантитела рСА2 и рСВ2 с помощью проточной (fast flow) хроматографии на Ni-NTA (Qiagen) в соответствии с рекомендациями производителя. Если необходимо, то может быть проведена стадия конечной очистки на проточной колонке (fast flow), связанной с белком G (Amersham Pharmacia Biotech). Мимоантитела элюировали 0,1М глицина рН 2,7, сразу нейтрализовали добавлением NaOH и диализовали против 20 мМ Hepes, 150 мМ NaCl, рН 7,2. рСС2 очищали с помощью аффинной хроматографии только на белке G. Чистоту анализировали с помощью электрофореза в ДСН-ПААГ.
Белковые последовательности мимоантител транслировались с кДНК-последовательностей. N-концевые последовательности подтверждали путем секвенирования рСА2 и рСВ2 по Эдману.
Последовательности легких цепей рСА2, рСВ2 и рСС2 являются аналогичными и представляют собой:
DIELVVTQPASVSGSPGQSITISCTGTRSDVGGYNYVSWYQQHPGKAPKL
MIYDVSNRPSGVSNRFSGSKSGNTASLT·SGLQAEDEADYYCSSYTSSSTL
GVFGGGTKLT· LGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVT
VAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLT·EQWKSHKSYSCQ
VTHEGSTVEKTVAPTECS
Последовательность тяжелой цепи рСА2 представляет собой:
EVKLQLEHHHHHHGEVKLQLESGPGLVKPSETLSLT·TVSGGSISSGGYY
WTWIRQRPGKGLEWIGYIYYSGSTSYNPSLKSRVTMSVDTSKNQFSLRLT
· SVTAADTAVYYCARERGETGLYYPYYYIDVWGTGTTVTVSSASTKGPSV
FPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALT·GVHTFPAVLQ
SSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKREPKSCDKTHTCG
Последовательность тяжелой цепи рСВ2 представляет собой:
EVKLQLEHHHHHHGEVKLQLESGPGLVKPSETLSLT·TVSGGSISSGGYY
WTWIRQRPGKGLEWIGYIYYSGSTSYNPSLKSRVTMSVDTSKNQFSLRLT·SVTAADTAVYYCARERGETGLYYPYYYIDVWGTGTTVTVSSASTKGPSV
FPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALT·GVHTFPAVLQ
SSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTS
PPCG
Последовательность тяжелой цепи рСС2 представляет собой:
EVKLQLEHHHHHHGEVKLQLESGPGLVKPSETLSLT·TVSGGSISSGGYY
WTWIRQRPGKGLEWIGYIYYSGSTSYNPSLKSRVTMSVDTSKNQFSLRLT·SVTAADTAVYYCARERGETGLYYPYYYIDVWGTGTTVTVSSASTKGPSV
FPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALT·GVHTFPAVLQ
SSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCASPKPS
TPPGSSGGAPGGC
D. Связывание мимоантител с капсидным белком Qβ
D.1. Связывание мимоантитела рСС2 с капсидным белком Qβ:
Раствор 1,25 мл 4,5 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 30 минут с 40 мкл раствора SMPH (Pierce) (из 100 мМ маточного раствора, растворенного в ДМСО) при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого 6 мкл диализованной реакционной смеси подвергали реакции с 30 мкл раствора рСС2 (2,88 мг/мл) при 25°С, в течение ночи, на ротационном шейкере.
Продукты реакции анализировали с помощью электрофореза в 16% ПААГ с ДСН в восстанавливающих условиях. Гели либо окрашивали кумасси бриллиантовым голубым, либо блотировали на нитроцеллюлозные мембраны. Затем мембраны блокировали, инкубировали с поликлональной кроличьей антисывороткой против Qβ (разведение 1:2000) или с мышиным моноклональным анти-Fab mAb (Jackson ImmunoResearch) (разведение 1:2000). После этого блоты инкубировали с козьим антикроличьим IgG, конъюгированным с пероксидазой хрена, либо с козьим антимышиным IgG, конъюгированным с пероксидазой хрена (разведение 1:7000), соответственно.
Результаты представлены на фиг.13А. Продукты связывания и выделенные продукты анализировали с помощью электрофореза в 16% ПААГ с ДСН в восстанавливающих условиях. На фиг.13А, "рСС2" обозначает мимоантитело перед связыванием. "Qβ deriv" означает дериватизированный Qβ перед связыванием. "Qβ-рСС2" означает продукт реакции связывания. Гели либо окрашивали кумасси бриллиантовым голубым, либо блотировали на нитроцеллюлозные мембраны. Затем мембраны блокировали, инкубировали с поликлональной кроличьей антисывороткой против Qβ (разведение 1:2000) или с мышиным моноклональным анти-Fab mAb (Jackson ImmunoResearch) (разведение 1:2000). После этого блоты инкубировали с козьим антикроличьим IgG, конъюгированным с пероксидазой хрена, либо с козьим антимышиным IgG, конъюгированным с пероксидазой хрена (разведение 1:7000), соответственно. Для визуализации иммунореактивных полос использовали метод усиления хемилюминесценции (набор Amersham Pharmacia ELC). Молекулярные массы маркерных белков даны с левого края.
Был детектирован продукт связывания, имеющий размер примерно 40 кДа (фиг.13А, показан стрелками). Его способность реагировать с антисывороткой против Qβ и анти-Fab антителом, распознающим мимоантитело, ясно свидетельствовало о ковалентном связывании этого мимоантитела с Qβ.
D.2. Связывание мимоантител рСА2 и рСВ2 с капсидным белком Qβ:
Раствор 1,25 мл 4,5 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 40 мкл SMPH (Pierce) (из 100 мМ маточного раствора, растворенного в ДМСО) при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. рСА2 (1,2 мг/мл) восстанавливали с использованием 20 мМ ТСЕР в течение 30 минут при 25°С, а рСВ2 (4,2 мг/мл) восстанавливали с использованием 50 мМ меркаптоэтиламина при 37°С. После этого оба мимоантитела два раза диализовали против 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. Связывание проводили путем добавления 6 мкл дериватизированного Qβ к 30 мкл мимоантитела при 25° С в течение ночи на ротационном шейкере.
Продукты реакции анализировали с помощью электрофореза в 16% ПААГ с ДСН в восстанавливающих условиях. Гели либо окрашивали кумасси бриллиантовым голубым, либо блотировали на нитроцеллюлозные мембраны. Затем мембраны блокировали, инкубировали с поликлональной кроличьей антисывороткой против Qβ (Cytos, разведение 1:2000) или мышиным моноклональным антителом (mAb) против his6 (Qiagen, разведение 1:5000). После этого блоты инкубировали с козьим антикроличьим IgG, конъюгированным с пероксидазой хрена, либо с козьим антимышиным IgG, конъюгированным с пероксидазой хрена (разведение 1:5000), соответственно.
Результаты представлены на фиг.13В и 13С. Продукты связывания и выделенные продукты анализировали с помощью электрофореза в 16% ПААГ с ДСН в восстанавливающих условиях. На фиг.15А и 15В, "рСА2" и "рСВ2" соответствуют мимоантителам перед связыванием, а "Qβ deriv" означает дериватизированный Qβ перед связыванием. "Qβ-рСА2" и "Qβ-рСВ2" означают продукты реакции связывания. Гели либо окрашивали кумасси бриллиантовым голубым, либо блотировали на нитроцеллюлозные мембраны. Затем мембраны блокировали, инкубировали с поликлональной кроличьей антисывороткой против Qβ (разведение 1:2000) или с мышиным моноклональным антителом против his6 (Qiagen) (разведение 1:5000). После этого блоты инкубировали с козьим антикроличьим IgG, конъюгированным с пероксидазой хрена, либо с козьим антимышиным IgG, конъюгированным с пероксидазой хрена (разведение 1:5000), соответственно. Для визуализации иммунореактивных полос использовали метод усиления хемилюминесценции (набор Amersham Pharmacia ELC). Молекулярные массы маркерных белков даны с левого края.
Продукты связывания размером примерно 40 кДа были детектированы для связывания рСА2 и рСВ2 (фиг.15А и фиг.15В, показаны стрелками). Их способность реагировать с анти-Qβ антисывороткой и анти-his6 антителом, распознающими тяжелую цепь мимоантитела, явно указывало на ковалентное связывание этого мимоантитела с Qβ.
ПРИМЕР 56
Связывание пептидов Flag с капсидным белком Qβ дикого типа и с мутантным Qβ с использованием сшивающего линкера Sulfo-GMBS
Был химически синтезирован пептид Flag, у которого к N-концу была присоединена последовательность CGG для связывания и который имел следующую последовательность: CGGDYKDDDDK. Этот пептид использовали для химического связывания с капсидным белком Qβ дикого типа и с мутантными капсидными белками Qβ, как описано ниже.
Е. Связывание пептида Flag с капсидным белком Qβ
Раствор 100 мкл 2 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 60 минут с 7 мкл раствора 65 мМ Sulfo-GMBS (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого 100 мкл диализованной реакционной смеси подвергали реакции с 0,58 мкл 100 мМ маточного раствора пептида Flag (в H2O) в течение двух часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение двух часов против 2 литров 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С.
В. Связывание пептида Flag с капсидным белком Qβ-240
Раствор 100 мкл 2 мг/мл капсидного белка Qβ-240 в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 60 минут с 7 мкл раствора 65 мМ Sulfo-GMBS (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого 100 мкл диализованной реакционной смеси подвергали реакции с 0,58 мкл 100 мМ маточного раствора пептида Flag (в H2O) в течение двух часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение двух часов против 2 литров 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С.
С. Связывание пептидов Flag с капсидным белком Qβ-250
Раствор 100 мкл 2 мг/мл капсидного белка Qβ-250 в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 60 минут с 7 мкл раствора 65 мМ Sulfo-GMBS (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого 100 мкл диализованной реакционной смеси подвергали реакции с 0,58 мкл 100 мМ маточного раствора пептида Flag (в H2O) в течение двух часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение двух часов против 2 литров 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С.
D. Связывание пептидов Flag с капсидным белком Qβ-259
Раствор 100 мкл 2 мг/мл капсидного белка Qβ-259 в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 60 минут с 7 мкл раствора 65 мМ Sulfo-GMBS (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С. После этого 100 мкл диализованной реакционной смеси подвергали реакции с 0,58 мкл 100 мМ маточного раствора пептида Flag (в H2O) в течение двух часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение двух часов против 2 литров 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С.
Результаты реакций связывания мутантов Qβ 240, 250 и 259 с пептидом Flag анализировали с помощью электрофореза в ДСН-ПААГ, как показано на фиг.22А. При этом осуществляли следующую загрузку:
1. Дериватизированный Qβ-240; 2. Qβ-240, связанный с пептидом Flag; 3. Дериватизированный Qβ-250; 4. Qβ-250, связанный с пептидом Flag; 5. Дериватизированный Qβ-259; 6. Qβ-259, связанный с пептидом Flag; 7. Дериватизированный Qβ дикого типа; 8. Qβ дикого типа, связанный с пептидом Flag; 9. Маркерный белок.
Сравнение продукта реакции дериватизации с продуктами реакции связывания показало, что для всех пептидов, как мутантных, так и дикого типа, полосы связывания, соответствующие пептидам 1 и 2 на субъединицу, были видимыми. Полоса, соответствующая несвязанной субъединице Qβ, была очень слабой, что указывало на то, что почти все субъединицы реагировали, по меньшей мере, с одним пептидом Flag. Для мутанта Qβ-250 и для Qβ дикого типа полоса, соответствующая трем пептидам на субъединицу, была видимой. Отношение интенсивностей полосы, соответствующей двум пептидам на субъединицу, и полосы, соответствующей 1 пептиду на субъединицу, было наиболее высоким для пептида дикого типа и составляло 1:1. Это отношение еще оставалось высоким для мутанта Qβ-250, однако оно было значительно ниже для мутанта Qβ-240 и очень низким для мутанта Qβ-259.
ПРИМЕР 57
Связывание пептида Flag с капсидным белком Qβ с использованием сшивающего линкера Sulfo-MBS
Был химически синтезирован пептид Flag, у которого к N-концу была присоединена последовательность CGG для связывания и который имел следующую последовательность: CGGDYKDDDDK. Этот пептид использовали для химического связывания с капсидным белком Qβ дикого типа и с мутантным капсидным белком Qβ, как описано ниже.
F. Связывание пептидов Flag с капсидным белком Qβ
Раствор 100 мкл 2 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 60 минут с 7 мкл раствора 65 мМ Sulfo-MBS (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого 100 мкл диализованной реакционной смеси подвергали реакции с 0,58 мкл 100 мМ маточного раствора пептида Flag (в H2O) в течение двух часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение двух часов против 2 литров 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С.
В. Связывание пептида Flag с капсидным белком Qβ-240
Раствор 100 мкл 2 мг/мл капсидного белка Qβ-240 в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 60 минут с 7 мкл раствора 65 мМ Sulfo-MBS (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого 100 мкл диализованной реакционной смеси подвергали реакции с 0,58 мкл 100 мМ маточного раствора пептида Flag (в H2O) в течение двух часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение двух часов против 2 литров 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С.
С. Связывание пептида Flag с капсидным белком Qβ-250
Раствор 100 мкл 2 мг/мл капсидного белка Qβ -250 в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 60 минут с 7 мкл раствора 65 мМ Sulfo-MBS (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого 100 мкл диализованной реакционной смеси подвергали реакции с 0,58 мкл 100 мМ маточного раствора пептида Flag (в H2O) в течение двух часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение двух часов против 2 литров 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С.
D. Связывание пептидов Flag с капсидным белком Qβ-259
Раствор 100 мкл 2 мг/мл капсидного белка Qβ-259 в 20 мМ Hepes, 150 мМ NaCl, рН 7,2, подвергали реакции в течение 60 минут с 7 мкл раствора 65 мМ Sulfo-MBS (Pierce) в H2O при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С. После этого 100 мкл диализованной реакционной смеси подвергали реакции с 0,58 мкл 100 мМ маточного раствора пептида Flag (в H2O) в течение двух часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение двух часов против 2 литров 20 мМ Hepes, 150 мМ NaCl, рН 7,2, при 4°С.
Результаты реакций связывания мутантов Qβ 240, 250 и 259 с пептидом Flag анализировали с помощью электрофореза в ДСН-ПААГ, как показано на фиг.1. При этом осуществляли следующую загрузку:
1. Маркерный белок. 2. Дериватизированный Qβ-240; 3. Qβ-240, связанный с пептидом Flag; 4. Дериватизированный Qβ -250; 5. Qβ-250, связанный с пептидом Flag; 6. Дериватизированный Qβ-259; 7. Qβ-259, связанный с пептидом Flag; 8. Дериватизированный Qβ дикого типа; 9. Qβ дикого типа, связанный с пептидом Flag.
Сравнение продукта реакции дериватизации с продуктами реакции связывания показало, что для всех пептидов, как мутантных, так и дикого типа, полоса связывания, соответствующая 1 пептиду на субъединицу, была видимой. Полосы, соответствующие 2 пептидам на субъединицу, были также видимыми для мутантного Qβ-250 и для Qβ дикого типа. Отношение интенсивностей полосы, соответствующей одному пептиду на субъединицу, и полосы, соответствующей несвязанной субъединице, соответственно, было выше для мутанта Qβ-250 и Qβ дикого типа. Слабая полоса, соответствующая двум пептидам на субъединицу, была видимой для мутанта Qβ-240.
ПРИМЕР 58
Связывание пептидов Flag с мутантами Qβ с использованием сшивающего линкера SMPH
Был химически синтезирован пептид Flag, у которого к N-концу была присоединена последовательность CGG для связывания и который имел следующую последовательность: CGGDYKDDDDK. Этот пептид использовали для химического связывания с мутантами Qβ, как описано ниже.
А. Связывание пептидов Flag с капсидным белком Qβ-240
Раствор 100 мкл 2 мг/мл капсидного белка Qβ-240 в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 2,94 мкл раствора 100 мМ SMPH (Pierce) в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С. После этого 90 мкл диализованной реакционной смеси подвергали реакции с 1,3 мкл 50 мМ маточного раствора пептида Flag (в ДМСО) в течение двух часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение двух часов против 2 литров 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С.
В. Связывание пептида Flag с капсидным белком Qβ-250
Раствор 100 мкл 2 мг/мл капсидного белка Qβ-250 в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 2,94 мкл раствора 100 мМ SMPH (Pierce) в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С. После этого 90 мкл диализованной реакционной смеси подвергали реакции с 1,3 мкл 50 мМ маточного раствора пептида Flag (в ДМСО) в течение двух часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение двух часов против 2 литров 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С.
С. Связывание пептида Flag с капсидным белком Qβ-259
Раствор 100 мкл 2 мг/мл капсидного белка Qβ-259 в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 2,94 мкл раствора 100 мМ SMPH (Pierce) в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С. После этого 90 мкл диализованной реакционной смеси подвергали реакции с 1,3 мкл 50 мМ маточного раствора пептида Flag (в ДМСО) в течение двух часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение двух часов против 2 литров 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С.
Результаты реакций связывания мутантов Qβ 240, 250 и 259 с пептидом Flag анализировали с помощью электрофореза в ДСН-ПААГ, как показано на фиг.1. При этом картина загрузки была следующей: 1. Маркерный белок. 2. Qβ-240, связанный с Flag; осадок реакции связывания; 3. Qβ-240, связанный с Flag; супернатант реакции связывания. 4. SMPH-дериватизированный Qβ-240; 5. Qβ-250, связанный с Flag; осадок реакции связывания; 6. Qβ-250, связанный с Flag; супернатант реакции связывания. 7. SMPH-дериватизированный Qβ-250; 8. Qβ-259, связанный с Flag; осадок реакции связывания; 9. Qβ-259, связанный с Flag; супернатант реакции связывания. 10. SMPH-дериватизированный Qβ-259.
Сравнение продукта реакции дериватизации с продуктами реакции связывания показало, что для всех мутантов полосы связывания, соответствующие 1 пептиду и 2 пептидам на субъединицу, соответственно, были видимыми. Полосы, соответствующие трем, и соответственно, 4 пептидам на субъединицу, были также видимыми для мутанта Qβ-250.
ПРИМЕР 59
Связывание белка PLA2-cys с мутантными капсидными белками Qβ
Лиофилизованные мутантные капсидные белки Qβ оставляли на ночь для набухания в 20 мМ Hepes, 150 мМ NaCl, рН 7,4.
А. Связывание белка PLA2-cys с капсидным белком Qβ-240
Раствор 100 мкл 2 мг/мл капсидного белка Qβ-240 в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 2,94 мкл раствора 100 мМ SMPH (Pierce) в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С. После этого 90 мкл диализованной реакционной смеси смешивали со 146 мкл 20 мМ Hepes, 150 мМ NaCl, рН 7,4, и подвергали реакции с 85,7 мкл 2,1 мг/мл маточного раствора PLA2-cys в течение четырех часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение двух часов против 2 литров 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С.
В. Связывание белка PLA2-cys с капсидным белком Qβ-250
Раствор 100 мкл 2 мг/мл капсидного белка Qβ-250 в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 2,94 мкл раствора 100 мМ SMPH (Pierce) в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С. После этого 90 мкл диализованной реакционной смеси смешивали со 146 мкл 20 мМ Hepes, 150 мМ NaCl, рН 7,4, и подвергали реакции с 85,7 мкл 2,1 мг/мл маточного раствора PLA2-cys в течение четырех часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение двух часов против 2 литров 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С.
С. Связывание белка PLA2-cys с капсидным белком Qβ-259
Раствор 100 мкл 2 мг/мл капсидного белка Qβ-259 в 20 мМ Hepes, 150 мМ NaCl, рН 7,4, подвергали реакции в течение 30 минут с 2,94 мкл раствора 100 мМ SMPH (Pierce) в ДМСО при 25°С на ротационном шейкере. Затем реакционный раствор два раза диализовали в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С. После этого 90 мкл диализованной реакционной смеси смешивали со 146 мкл 20 мМ Hepes, 150 мМ NaCl, рН 7,4, и подвергали реакции с 85,7 мкл 2,1 мг/мл маточного раствора PLA2-cys в течение четырех часов при 25°С на ротационном шейкере. Затем реакционную смесь два раза диализовали в течение двух часов против 2 литров 20 мМ Hepes, 150 мМ NaCl, рН 7,4, при 4°С.
Результаты эксперимента по связыванию анализировали с помощью электрофореза в ДСН-ПААГ, как показано на фиг.1. При этом осуществляли следующую загрузку: 1. Маркерный белок. 2. Дериватизированный Qβ-240. 3. Qβ-240, связанный с PLA2-cys; супернатант реакции связывания; 4. Qβ-240, связанный с PLA2-cys; осадок реакции связывания. 5. Дериватизированный Qβ-250. 6. Qβ-250, связанный с PLA2-cys; супернатант реакции связывания; 7. Qβ-250, связанный с PLA2-cys; осадок реакции связывания. 8. Дериватизированный Qβ-259. 9. Qβ-259, связанный с PLA2-cys; супернатант реакции связывания; 10. Qβ-259, связанный с PLA2-cys; осадок реакции связывания; 11. PLA2-cys.
Полосы связывания (показаны на фигуре стрелками) были видимыми для всех мутантов, что указывало на то, что белок PLA2-cys может связываться со всеми мутантными капсидными белками Qβ.
1. Все патенты и публикации полностью вводятся в настоящее описание посредством ссылки.
2. Все описания заявок США №09/449631 и WO 00/3227, поданных 30 ноября 1999, во всей своей полноте вводятся в настоящее описание посредством ссылки. Все вышеупомянутые публикации и патенты во всей своей полноте вводятся в настоящее описание посредством ссылки.
Реферат
Изобретение относится к области молекулярной биологии, вирусологии, иммунологии и медицине. Заявлена композиция для иммунизации. Композиция содержит неприродный молекулярный каркас и антиген или антигенную детерминанту. Неприродный молекулярный каркас содержит РНК-фаг, связанный с организатором, содержащим первый сайт связывания. Антиген или антигенная детерминанта содержит второй сайт связывания. При этом антиген или антигенная детерминанта и молекулярный каркас взаимодействуют путем связывания с образованием упорядоченного и повторяющегося массива антигенов. Предложены также фармацевтическая композиция, способ иммунизации, способ продуцирования неприродного, упорядоченного и повторяющегося массива антигенов и оболочечный белок. Изобретение может быть использовано для получения вакцин, лечения аллергий, профилактики и лечения злокачественных опухолей и для эффективного индуцирования специфических аутоиммунных ответов. 6 н. и 36 з.п. ф-лы, 62 ил., 1 табл.
Комментарии