Применение расар в качестве молекулярного адъюванта для вакцин - RU2580294C2
Код документа: RU2580294C2
Чертежи


Описание
Область техники
Настоящее изобретение относится к области медицины, молекулярной биологии, иммунологии, в частности к разработке вакцин и адъюванта для вакцин. В частности, в настоящем изобретении раскрыто применение питуитарного пептида, активирующего аденилатциклазу (PACAP), в качестве молекулярного адъюванта для вакцин, которые используются в стратегиях иммунизации.
Предпосылки изобретения
Основная цель вакцинации состоит в том, чтобы индуцировать специфический и эффективный иммунный ответ против патогена, который также вызывает защиту от инфекции и/или заболевания и приводит к ее ликвидации. Концепция о том, что иммунный ответ против специфического антигена может быть усилен путем добавления определенных соединений в состав вакцины, была показана около 100 лет назад, когда соли алюминия вводили в составы и были названы «адъювантом» (G. Leroux-Roels (2010) Vaccine 28S C25-C36).
Традиционные вакцины состоят из инактивированных или ослабленных патогенов или токсинов, полученных из этих микроорганизмов. Несмотря на то что применение инактивированных или ослабленных возбудителей обладает высокой иммуногенностью, в настоящее время оно является непривлекательным в области вакцинологии из-за высокой токсичности этих препаратов.
Следовательно, растет интерес к адъювантам в исследованиях, связанных с разработкой новых поколений вакцин на основе рекомбинантных белковых субъединиц, синтетических пептидов и плазмид дезоксирибонуклеиновой кислоты (ДНК). Эти новые вакцинные варианты, несмотря на меньшую токсичность, в основном являются менее иммуногенными при введении без иммуностимулирующего адъюванта. По этой причине в последние годы возросла потребность в более безопасных и более эффективных адъювантах (Saenz et al. (2010) Vaccine 28 (47): 7556-7562).
Хотя применение адъювантов хорошо известно, механизм их действия остается менее понятными. Считается, что, как правило, они повышают эффективность вакцин посредством нескольких механизмов, в том числе: 1) усиление процессинга антигена и представления дендритными клетками, 2) индукция сигнала «опасности» через сигнальные пути, опосредованные рецепторами, которые узнают структуры, связанные с патогенами, такими как «Toll»-рецепторы, и 3) путем активации костимуляторных сигналов, которые активируют лимфоциты. Эти механизмы запускаются с помощью индукции цитокинов и регуляции экспрессии соответствующих костимуляторных сигналов (Secombes (2010) Fish & Shellfish Immunology, 409-416).
Несколько эндогенных молекул и белков (Yin and Kwang (2000) Fish & Shellfish Immunology 10, 375-378; Lingnau et al. (2007) Expert Rev Vaccines 6 (5): 741-6; Zhang et al. (2010) Vaccine 28: 5114-5127) были изучены в качестве адъювантов. Кроме того, известно, что некоторые иммуностимулирующие пептиды могут выступать в качестве адъюванта in vivo, стимулируя иммунный ответ белковых или пептидных антигенов (Saenz et al. (2010) Vaccine 28 (47): 7556-7562).
PACAP принадлежит к суперсемейству секретин/глюкагон/вазоактивный кишечный пептид (Miyata et al (1989) Biochem Biophys Res Commun. 164: 567-574). Он представляет собой многофункциональный нейропептид, который играет важные роли в качестве нейротрофического фактора и тропного гормона гипофиза, в качестве нейромедиатора, нейромодулятора и сосудорасширяющей молекулы у млекопитающих (Arimura A. (1998) Japanese Journal of Physiology 48: 301-31). Показана его роль в делении и дифференцировке клеток, а также в гибели клеток (Sherwood et al. (2000) Endocrine Review 21: 619-670). Этот пептид существует в двух молекулярных формах в 38 (PACAP38) и 27 (PACAP27) аминокислот (Miyata et al. (1990) Biochemical and Biophysical Research Communications 170: 643-8). Биологические действия PACAP проявляются посредством семейства трех VIP/PACAP рецепторов, которые принадлежат к секретин G-белок связанному рецептору: рецептору I типа, который высоко специфичен для PACAP и называется PAC-1; и рецепторам II типа, которые проявляют такую же активность к PACAP, что и к вазоактивному кишечному пептиду (VIP), которые известны как VPAC-1 и VPAC-2 (Vaudry et al. (2000) Pharmacol. Rev 52: 269-324).
PACAP широко распространен в различных тканях, в том числе связанных с иммунной системой, хотя присутствие этого пептида и его рецепторов в клетках иммунной системы млекопитающих лишь частично выявлено (Gaytan et al. (1994) Cell Tissue Res 276: 223-7, Abad et al. (2002) Neuroimmunomodulation 10: 177-86). PACAP модулирует воспалительную реакцию посредством регулирования интерлейкина-6 (ИЛ-6) и интерлейкина-10 (ИЛ-10) (Martinez et al. (1996) J Immunol. 156 30(11): 4128-36; Martinez et al. (1998) J Neuroimmunol. 85(2): 155-67, Martinez et al. (1998) J Leukoc Biol. 63 (5): 591-601).
В активированных макрофагах PACAP ингибирует выработку провоспалительных цитокинов и стимулирует выработку противовоспалительных цитокинов, обеспечивая, таким образом, гомеостаз иммунной системы. Кроме того, известно, что PACAP снижает экспрессию костимуляторных молекул B7.2/B7.1 и последующую активацию Т-хелперов (Th). С другой стороны, PACAP ингибирует в активированных макрофагах продукцию IL-6 через его PAC-1 рецептор, подавляя воспаление (Martinez et al. (1998) J Neuroimmunol. 85(2): 155-67, Martinez et al. (1998) J Leukoc Biol. 1998, 63(5): 591-601). Ингибирующее действие PACAP на транскрипцию IL-6 в ответ на интенсивные воспалительные стимулы помогает защите тканей и иммунной системы гомеостаза (Martinez et al. (1998) J Neuroimmunol. 85(2): 155-67; Martinez et al. (1998) J Leukoc Biol. 1998 года; 63(5): 591-601). В отличие от этого, PACAP индуцируют экспрессию B7.2 и способствует клеточной дифференциации в Th2 в нестимулированных макрофагах (Delgado and Ganea (2001) Arch Immunol. Ther Exp. (Warsz) 49(2): 101-10). Сообщалось о наличии PACAP в лимфоидных органах уток (Squillacioti et al. (2005) Anatomia, Histologia, Embryologia. Vol.34, Issue Supplements 1, p.49). Было выдвинуто предположение о применении PACAP у млекопитающих в качестве терапевтического агента для лечения аутоиммунных заболеваний, таких как септический шок, ревматоидный артрит и болезнь Крона (Gomariz et al. (2006) Ann. NY Acad. Sci 1070: 51-74). Известно, что при аутоиммунных заболеваниях происходит бесконтрольный иммунный ответ против веществ и тканей организма. В связи с этим PACAP предотвращает воспалительные процессы в животных моделях аутоиммунных заболеваний через правильный баланс цитокинов и хемокинов и их рецепторов, посредством привлечения иммунных клеток и путем регулирования образования и активации Th1-клеток и цитокинов, которые эти клетки секретируют.
Кроме того, известно, что молекулы, участвующие в гомеостазе иммунной системы, такие как «Toll»-рецепторы, необходимы для активации врожденного иммунного ответа, который повышает адаптивный иммунный ответ. Однако эта реакция может привести к патогенезу острого и/или хронического воспаления, аутоиммунного заболевания и рака (Gomariz et al. (2010) Current Pharmaceutical. Design 16: 1063-1080). В связи с этим известно, что система VIP-PACAP участвует в регуляции экспрессии генов, кодирующих эти рецепторы (Gomariz et al. (2006) Ann. NY Acad. Sci 1070: 51-74). Таким образом, оба пептида могут вызывать нарушения в регуляции путей продуцирования этих рецепторов.
PACAP уменьшает уровни циркулирующих цитокинов, опосредованных эндогенной молекулой HMGB1 (high mobility group box 1) (Tang et al. (2008) International Immunopharmacology 8 (12): 1646-1651) и ингибирует их высвобождение. Известно, что молекула HMGB1 и производные от нее пептиды способны выступать в качестве адъюванта, усиливая иммунный ответ против пептидного антигена и белка (Saenz et al. (2010) Vaccine 28 (47): 7556-7562).
Знания о функции PACAP в модуляции иммунного ответа у рыб ограничены исследованиями, выполненными нашей исследовательской группой. Мы показали, что рекомбинантный PACAP Clarias gariepinus, введенный путем иммерсионных ванн или путем инъекции, не только способствует росту, но также стимулирует врожденные иммунные параметры (лизоцим, метаболиты-производные оксида азота и антиоксидантные защиты) и приобретенный иммунитет (IgM) у личинок и молоди рыб (Carpio et al. (2008) Fish and Shellfish Immunology 25: 439-45; Lugo et al. (2010) Fish and Shellfish Immunology 29: 513-520). Эти свойства PACAP были описаны в международной патентной заявке WO 2007/059714, «Neuropeptides para el cultivo de organismos acuaticos».
В настоящее время в сфере медицины и ветеринарии по-прежнему существует интерес к идентификации соединений, которые могут быть использованы в качестве безопасных и более эффективных адъювантов, которые могут быть включены в существующие вакцины и в те, которые находятся в стадии разработки.
Подробное описание изобретения
Настоящее изобретение разрешает описанные выше проблемы, предоставляя новую альтернативу адъюванта, способного значительно усиливать иммунный ответ на антиген, который вводится совместно, и, следовательно, она представляет собой весьма эффективный адъювант. Применение «питуитарного активирующего аденилатциклазу пептида» (PACAP) в качестве молекулярного адъюванта для антигенов, которые присутствуют в вакцинах, используемых в различных стратегиях иммунизации, является предметом настоящего изобретения. В одном варианте осуществления изобретения указанные вакцины, содержащие PACAP в качестве адъюванта, предназначены для иммунизации против инфекционных агентов. Такие инфекционные агенты могут представлять собой, среди прочих, вирусы, бактерии и эктопаразиты. В конкретном варианте осуществления эти инфекционные агенты влияют на млекопитающих, птиц и рыб. Термин «питуитарного активирующего аденилатциклазу пептида (PACAP)», используемый в настоящем изобретении, включает эту молекулу в любом варианте (PACAP27 или PACAP38), либо изолированную из своего естественного источника, произведенную путем химического синтеза или произведенную с помощью технологии рекомбинантной ДНК, а также его применение в качестве пептида или в виде нуклеиновых кислот.
В контексте этого изобретения термин «молекулярный адъювант» относится к любой молекуле белковой природы, способной модулировать иммунный ответ вакцины против антигена, увеличивая его.
До сих пор ни в одном исследовании не было показано использование PACAP в качестве молекулярного адъюванта. Как указано выше, было предложено использовать этот пептид млекопитающих в качестве терапевтического агента для лечения аутоиммунных заболеваний, таких как септический шок, ревматоидный артрит и болезнь Крона (Gomariz et al. (2006) Ann. NY Acad. Sci. 1070: 51-74). В связи с этим PACAP предотвращает воспалительные процессы в животных моделях аутоиммунных заболеваний с помощью надлежащего баланса цитокинов и хемокинов и их рецепторов, путем привлечения иммунокомпетентных клеток, а также путем регулирования образования и активации Th1-клеток и цитокинов, которые эти клетки секретируют. Учитывая эти результаты, неожиданным является эффект PACAP, обнаруженный как часть наших исследований, показывающий, что его введение в сочетании с антигеном повышает иммунный ответ против указанного антигена через стимулирование простимулирующих цитокинов и других элементов гуморального иммунного ответа.
Кроме того, известно, что система VIP-PACAP участвует в регуляции экспрессии генов, кодирующих рецепторы «Toll» (Gomariz et al. (2006) Ann. NY Acad. Sci 1070: 51-74). В этом смысле оба пептида могут вызывать нарушения в путях регуляции производства этих рецепторов, что делает неочевидным их использование в качестве вакцинного адъюванта. Еще один элемент, контрастирующий с новым использованием PACAP, раскрытый в данном изобретении, является демонстрацией того, что этот пептид ослабляет циркулирующие уровни и ингибирует высвобождение цитокинов, опосредуемых эндогенной молекулой HMGB1 (Tang et al. (2008) International Immunopharmacology 8 (12): 1646-1651).
В настоящем изобретении впервые показано, что усиление иммунного ответа, специфического к вакцинному антигену, совместно вводимому с PACAP, наблюдается на уровне как гуморального, так и клеточного иммунного ответа.
Другой объект настоящего изобретения представляет собой вакцинную композицию, содержащую PACAP в качестве адъюванта, по меньшей мере вакцинного антигена, и фармацевтически приемлемые носители или разбавители. Эта вакцинная композиция, содержащая PACAP в качестве адъюванта и по меньшей мере один представляющий интерес антиген, вызывает иммунный ответ против антигена или антигенов, который является либо местным, или системным. Вакцины и вакцинные композиции по настоящему изобретению могут быть основаны, например, на рекомбинантных белках, аттенюированных микроорганизмах или нуклеиновых кислотах. В одном варианте осуществления изобретения упомянутые вакцинные композиции вводят перорально, при помощи инъекции или при помощи иммерсионных ванн.
В контексте настоящего изобретения в таких вакцинных композициях PACAP вводят в качестве пептида, изолированного из своего естественного источника, полученного путем химического синтеза или с помощью технологии рекомбинантной ДНК. PACAP может также использоваться в качестве нуклеиновых кислот.
В различных вариантах осуществления настоящего изобретения вакцины, содержащие PACAP в комбинации с несколькими антигенами, вводят млекопитающим, птицам или рыбам, впервые демонстрируя адъювантный эффект для PACAP. Неожиданно было отмечено, что введение нейропептида PACAP в комбинации с антигеном повышало уровень специфических антител против указанного антигена у млекопитающих, птиц или рыб. Кроме того, впервые было показано in vitro, что PACAP значительно увеличивает транскрипты интерлейкина-1 бета (IL-1β) и интерлейкина 15 (IL-15) в лейкоцитах рыб.
В разных вариантах осуществления настоящего изобретения, которые раскрывают вакцинные композиции или комбинации, включающие PACAP в качестве адъюванта, используются разнообразные антигены, такие как овальбумин (ОВА), E2-гликопротеин вируса классической чумы свиней (CSFV), гемагглютинин (НА) вируса птичьего гриппа, полипептид my32 Caligus rogercresseyi, инактивированные клетки Aeromonas hydrophila, 106 кДа белок (NH2-VP2-VP4VP3-COOH) вируса инфекционного некроза поджелудочной железы (IPNV), гликопротеин вируса вирусной геморрагической септицемии (VHSV) и реснитчатый паразитарный Ichthyophthirius multifiliis.
В специфическом варианте осуществления изобретения в вакцинных композициях PACAP применяют в качестве комбикорма, в концентрации 50-750 мкг/кг корма. Во втором варианте осуществления изобретения PACAP применяется путем инъекции, в концентрации 0,1-10 мкг на грамм массы тела. В другом варианте осуществления изобретения, PACAP применяется для водных организмов способом погружения в ванну, в концентрации 50-1000 мкг на литр воды.
В другом варианте осуществления изобретения антиген и адъювант PACAP может быть частью вакцинной комбинации, элементы которой вводят одновременно, по отдельности или последовательно во время одного и того же протокола иммунизации. В определенном варианте осуществления изобретения комбинация используется для профилактики вирусных, бактериальных и паразитарных заболеваний. Для этой вакцинной комбинации PACAP и антиген или антигены вводят перорально, путем инъекции или способом погружения в ванну. В определенном варианте осуществления PACAP применяется как комбикорма в концентрации 50-750 мкг/кг корма. Во втором варианте осуществления PACAP-5 применяется путем инъекции в концентрации 0,1-10 мкг пептида на грамм массы тела. В другом варианте осуществления изобретения PACAP применяется для рыб способом погружения в ванну в концентрации 50-1000 мкг PACAP на литр воды.
Другой аспект этого изобретения представляет собой способ повышения иммунного ответа против антигена, в котором используется вакцинная композиция, содержащая «питуитарный активирующий аденилатциклазу пептид» (PACAP) в качестве вспомогательного средства для указанного антигена. В соответствии со способом, раскрытым в изобретении, PACAP в качестве адъюванта и антиген можно вводить одновременно, раздельно или последовательно в течение одного и того же графика иммунизации. PACAP используется в качестве вспомогательного способа изобретения как пептид, который был получен из своего естественного источника, путем химического синтеза или с помощью технологии рекомбинантной ДНК. Также изобретение включает в себя способ, в котором PACAP, используемый как адъювант, вводят в виде нуклеиновых кислот. В соответствии со способом по изобретению увеличение иммунного ответа, специфического для вакцинного антигена, вводимого одновременно с PACAP, имеет место как при гуморальном, так и при клеточном специфическом иммунном ответе.
Как продемонстрировано в нескольких вариантах осуществления изобретения, повышение или увеличение иммунного ответа против представляющего интерес антигена выражается в виде более высокого уровня защиты против различных инфекционных агентов, в том числе вирусных, бактериальных и эктопаразитарных сущностей.
Краткое описание чертежей
Фиг.1. Титры общего иммуноглобулина G (IgG) (A), IgG1 (B) и IgG2a (C), индуцированные у мышей путем иммунизации OVA при совместном введении с нейропептидом PACAP38. Использовали две экспериментальные группы по 6 животных в каждой. Группа отрицательного контроля (буферизованный фосфатом солевой раствор (PBS)/OVA) была инокулирована внутрибрюшинно в день 0 и 7 в дозе 6 мкг OVA в 0,2 мл PBS. Группа, которая также получила PACAP38 (PBS/OVA+пептид), была инокулирована внутрибрюшинно в день 0 и 7 в дозе 6 мкг OVA + 0,5 мкг PACAP38 в 0,2 мл PBS. Разные буквы указывают на значительные различия.
Фиг.2. Оценка клеточного иммунного ответа у свиней, вакцинированных вакцинной композицией на основе E2-PACAP. Клеточный иммунный ответ измеряли в свиных лимфоцитах, выделенных на 5 день после вакцинации E2 (1-я группа), E2+PACAP (2-я группа) и плацебо (группа 3). (A) анализ лимфопролиферации: результаты выражаются в виде индекса стимуляции (SI), определяемого как отношение импульсов в минуту (cpm) стимулированной культуры (cpm) к cpm необработанной контрольной группы. Лимфопролиферативный ответ с SI>2 считался положительным (B). Определение IFN-γ секреции методом ПЦР в реальном времени. Значения выражены как среднеарифметическое от 2-∆∆ct. (**)p<0,01.
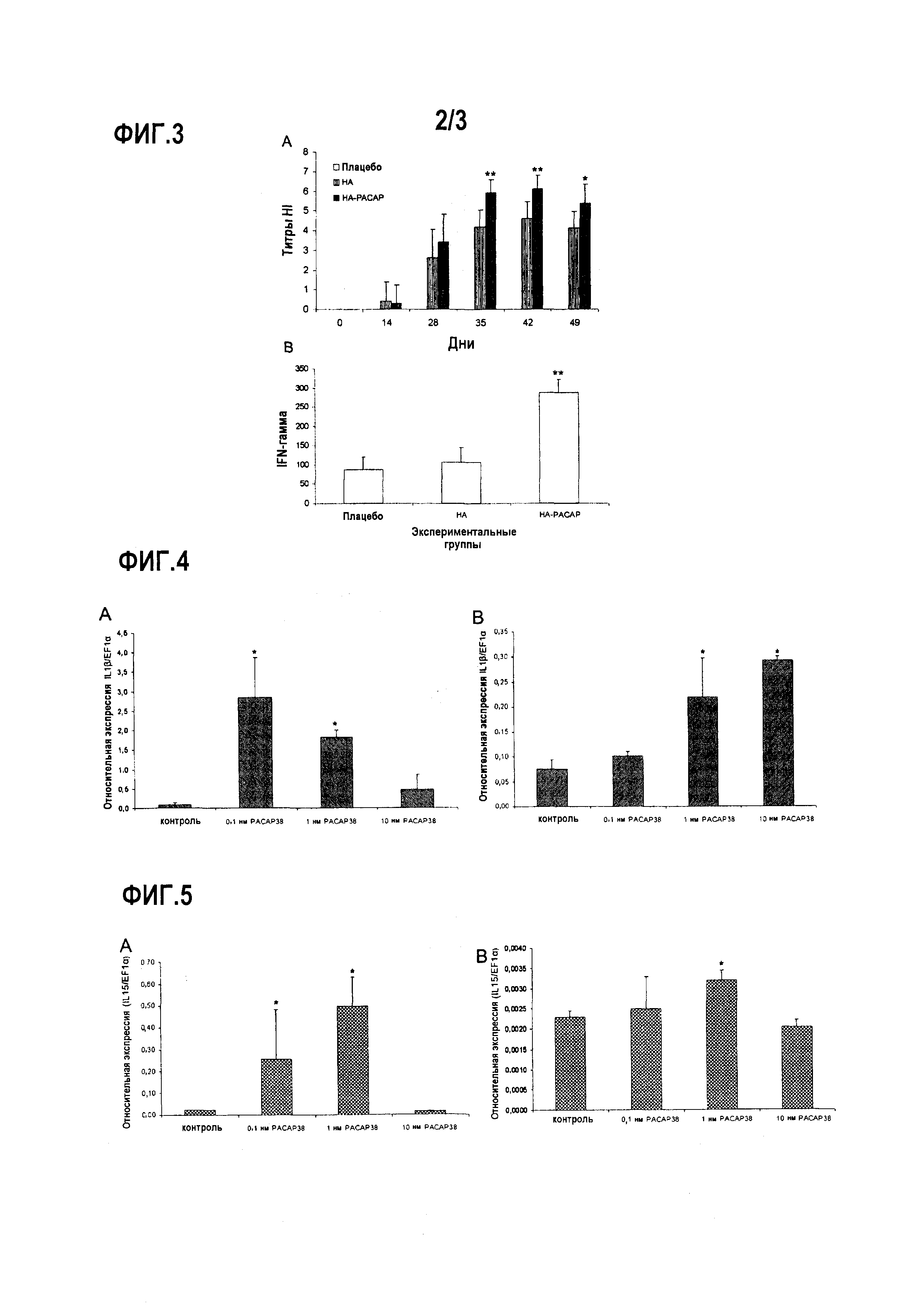
Фиг.3. Титры ингибирования гемагглютинации у кур, иммунизированных НА или НА-PACAP (А). Среднее арифметическое титров антител было выражено как log2 реципрокного значения наибольшего разбавления сыворотки, которая производила ингибирование гемагглютинации. (B) Клеточный иммунный ответ у вакцинированных кур. Секрецию IFN-γ определяли с помощью ПЦР в режиме реального времени. Значения выражены как среднеарифметическое 2-∆∆ct. (**)p<0,01.
Фиг.4. Анализ экспрессии ПЦР в реальном времени эффекта PACAP38 in vitro на транскрипцию IL-1β в лейкоцитах периферической крови (A) и головных почек (B) у наивной радужной форели. Эффект введения PACAP в концентрации 10-10 М, 10-9 М и 10-8 М оценивали через 48 ч после лечения. Эксперимент был повторен 4 раза. Культуру лейкоцитов обрабатывали в повторностях, и ПЦР проводились в трех повторностях. Данные представлены в виде среднего относительной экспрессии IL-1β относительно эндогенного фактора элонгации EF1α ± стандартное отклонение (SD). (*) p<0,05). * (p<0,05) указывает на то, что относительная экспрессия представляющего интерес гена была статистически значимо выше, чем его относительная экспрессия в необработанной культуре лейкоцитов (отрицательный контроль)
Фиг.5. Анализ экспрессии путем ПЦР в реальном времени эффекта PACAP38 in vitro на транскрипцию IL-15 в лейкоцитах периферической крови (A) и головных почек (B) у наивной радужной форели. Последствия PACAP введения в концентрации 10-10 М, 10-9 М и 10-8 М оценивалась через 48 ч после лечения. Эксперимент был повторен 4 раза. Культуру лейкоцитов обрабатывали в повторностях, и ПЦР проводились в трех повторностях. Данные представлены в виде среднего относительной экспрессии IL-15 относительно эндогенного фактора элонгации EF1α ± стандартное отклонение (SD). (*) p<0,05.
Фиг.6. Титры агглютинирующих антител против A. hydrophila у канального карпа. Значения на оси Y представляют собой среднее арифметическое титров антител ± стандартная ошибка. (*) p<0,05) (**) p<0,01. Группа 1: инъецирована PBS, группа 2: инъецирована инактивированными формалином клетками A. hydrophila, группа 3: инъецирована инактивированными формалином клетками A. hydrophila и PACAP (1 мкг/рыбу).
Фиг.7. Эффект совместного введения при помощи внутримышечной инъекции ДНК-вакцины (pP-IPNV) в дозе 1 мкг/рыба с плазмидной ДНК, содержащей кДНК PACAP Clarias gariepinus, под контролем быстрого раннего промотора цитомегаловируса человека (pCMV-PACAP). В день 30 после вакцинации рыбы подверглись воздействию вируса путем внутрибрюшинного введения 100 мкл IPNV (1×107 (TCID50) мл-1 на рыбу). В 7 день после заражения 10 рыб из группы были забиты для того, чтобы оценить вирусную нагрузку в головных почках с помощью ПЦР в реальном времени. Значения представляют среднее ± стандартное отклонение. Разные надстрочные буквы представляют собой статистически значимые различия между группами. Группа 1: инъецирована PBS, группа 2: инъецирована pP-IPNV в дозе 1 мкг/рыбу, группа 3: совместно инъецирована pP-IPNV (1 мкг/рыбу) и pCMV-PACAP (0,5 мкг/рыбу), группа 4: совместно инъецирована pP-IPNV (1 мкг/рыбу) и pCMV (0,5 мкг/рыбу), группа 5: совместно инъецирована pP-IPNV (1 мкг/рыбу) и 20 pCMV-PACAP (0,05 мкг/рыбу), группа 6: совместно инъецирована pP-IPNV (1 мкг/рыбу) и pCMV (0,05 мкг/рыбу).
Фиг.8. Суммарная смертность радужной форели, иммунизированной внутримышечно ДНК-вакциной на основе гена, кодирующего гликопротеин G VHSV (pG-VHSV) и пептид PACAP. Через 4 недели после вакцинации рыб подвергали воздействию вируса (1×105 TCID50 мл-1 на рыбу) способом погружения в ванну. Суммарную смертность оценивали в течение 4 недель после заражения. Группа 1: инъецирована PBS. Группа 2: инъецирована pG-VHSV (0,01 мкг/рыбу), группа 3: инъецирована pG-VHSV (0,01 мкг/рыбу) и PACAP (0,1 мкг/рыбу), группа 4: инъецирована pG-VHSV (0,01 мкг/рыбу) и PACAP (0,5 мкг/рыбу).
Примеры
Пример 1
Эффект совместной иммунизации с OVA и PACAP на гуморальный иммунный ответ у мышей
Мыши BALB/c (n=12) с массой тела 20 г были разделены на две экспериментальные группы по 6 животных в каждой. Группе отрицательного контроля (PBS/OVA) внутрибрюшинно инъецировали на 0 и 7 день дозой 6 мкг OVA в 0,2 мл PBS. Группе, получавшей PACAP38 (PBS/OVA+пептид), внутрибрюшинно вводили на 0-й и 7-й день дозу 6 мкг OVA + 0,5 мкг PACAP38 в 0,2 мл PBS. На 15-й день протокола иммунизации кровь из каждой рыбы была взята с целью оценки общих титров IgG, IgG1 и IgG2a.
Фиг.1А, В и С показывают общие титры IgG, lgG1 и lgG2a, соответственно, вызванные иммунизацией мышей OVA при совместном введении с PACAP38. Животные в группе PBS/OVA+пептид показали специфический общий титр IgG против OVA, статистически более высокий по сравнению с контрольной группой (фиг.1А). Аналогичным образом, заметили, что титры IgG1 и IgG2a, специфических против OVA, в группе иммунизации OVA+PACAP были значительно выше, чем те, которые наблюдались в группе иммунизации только OVA (фиг.1B и C).
Пример 2
Оценка клеточного иммунного ответа на вакцинацию свиней в состав вакцины на основе E2-PACAP
Антиген E2 является основным гликопротеином оболочки CSFV. В целях оценки клеточного иммунного ответа у свиней, вакцинированных вакцинной композицией на основе E2-PACAP, и сравнения этого ответа с ответом, произведенным E2-антиген, выбрали 18 здоровых свиней со средней массой 20 кг и серологически отрицательных по CSFV, на ферме без истории этого заболевания и не вакцинированных против CSFV в 3 предыдущих года. Свиньи были распределены в 3 различные группы по 6 животных в каждой, с водой и кормом ad libitum. Каждую вакцинную композицию применяли как однократную иммунизацию следующим образом: 25 мкг рекомбинантного E2 (группа 1) и 25 мкг Е2 совместно вводили с аналогичным количеством PACAP38 (группа 2). 3 группу использовали в качестве плацебо-группы. Иммуногены составляли в виде эмульсии масляного адъюванта и инокулировали путем внутримышечной инъекции в конечном объеме 2 мл. Животные были заражены на 8-й день после иммунизации путем внутримышечной инъекции CSFV (105 LD50 изоляции «Маргарита»). Проводили ежедневный анализ клинических признаков, и образцы крови были взяты в дни 3 и 5 для оценки пролиферации лимфоцитов и экспрессии IFN-γ в качестве индикаторов клеточного иммунного ответа. У животных, иммунизированных Е2-PACAP, было обнаружено увеличение в лимфоцитном ответе (фиг.2А) и высочайшие уровни IFN-γ на 5-й день, по сравнению с другими группами (фиг.2B). Эти результаты показывают, что одновременное введение E2-антигена с PACAP производит клеточный иммунный ответ против CSFV у свиней.
Пример 3
Эффект совместной иммунизации НА вируса птичьего гриппа (вирус A/VietNam1203/2004) и PACAP на гуморальный и клеточный иммунный ответ у кур
Для иммунизации были использованы куры белый леггорн 3-недельного возраста. Были использованы три экспериментальные группы по десять цыплят.
Группа 1: инъецированы 20 мкг НА.
Группа 2: инъецированы 20 мкг НА + 0,5 мкг из PACAP27.
Группа 3: инъецированы 20 мкг бычьего сывороточного альбумина.
HA был получен путем рекомбинантной экспрессии в клетках млекопитающих. Вакцины составляли с Montanide 888 и вводили в окончательном объеме 0,5 мл способом подкожно. Животных повторно иммунизировали в день 28 после иммунизации. Образцы крови были взяты в дни 0, 14, 28, 35, 42 и 49 для того, чтобы оценить титры защитных антител с помощью теста ингибирования гемагглютинации, а также для изолирования лейкоцитов для оценки клеточного иммунного ответа, произведенного вакцинами in vitro.
Куры, иммунизированные НА-PACAP, показали превосходную реакцию антител в сравнении с группой, иммунизированной НА, от дня 35 повторной иммунизации (фиг.3А). Группа, получавшая НА-PACAP, показала рост в клеточном иммунном ответе, измеренном как стимуляция IFN-γ, в сравнении с группами, иммунизированными НА или контрольной группой (фиг.3B).
Пример 4
Эффект PACAP на экспрессию IL1-β и IL-15 в периферической крови и лейкоцитов головных почек наивной радужной форели (Oncorhynchus mykiss)
Использовали мальков радужной форели (O. mykiss) приблизительно по 50 г, не зараженных VHSV и IPNV. Рыбы (n=5) были анестезированы солью метансульфоновой кислоты (Sigma, USA), и асептически отбирали периферическую кровь из хвостовой вены от каждой рыбы, а затем собирали головную почку.
Периферическая кровь и лейкоциты головной почки выделяли, следуя способу, описанному Graham и Secombes (Graham and Secombes (1998) Immunology 65: 293-7).
Клетки ресуспендировали в L-15 с 5% FCS в концентрации 5×106 5 клеток/мл и раскапывали в 24-луночные планшеты по 1 мл на лунку. Лейкоциты обрабатывали 3 дозами Clarias gariepinus PACAP38 (10-10 М, 10-9 М и 10-8 М), полученным путем химического синтеза, в двух повторностях. Необработанные лейкоциты, раскапанные в двух повторностях, были использованы в качестве отрицательного контроля.
Уровни IL-1β и IL-15 оценивали через 48 часов после обработки. Для достижения этой цели тотальную РНК очищали от культуры лейкоцитов, обработанных PACAP при помощи способа, описанного Chomczynski и Sacchi (Chomczynski and Sacchi (1987) Anal. Biochem. 162: 156-9).
Из-за того, что спроектированные праймеры для амплификации IL-1β и IL-15 не различают кДНК и геномной ДНК, суммарную РНК, очищенную из различных тканей, обрабатывали ДНК-нуклеазой, в частности свободной от РНКаз ДНКазой RQ1 (Promega).
Для кДНК-синтеза применяли коммерческий набор, в котором используется обратная транскриптаза SuperScript III (Invitrogen). Наконец, для количественного PCR (qPCR) была использована коммерческая смесь для ПЦР: Power SYBR Green PCR Master Mix (Applied 20 Biosystems). Результаты qPCR нормализовали против эндогенного гена с конститутивной экспрессией, в частности против элонгации фактора 1α (EF 1а), и выполняли в трех повторностях. Результаты выражали как 2-∆Ct, где ∆Ct равно остатку значения Ct гена-мишени минус значение Ct нормированного гена EF 1α.
Уровни белка IL-1β увеличивались через 48 после обработки лейкоцитов периферической крови, обработанной 10-10 М PACAP38. В дозе 10-9 М также есть стимулирующий эффект PACAP38 на транскрипцию IL-1β, по сравнению с группой отрицательного контроля, но уровни экспрессии, обнаруженные в данном случае, были ниже тех, полученных при дозе 10-10 М. Эти результаты показывают положительный эффект PACAP38 на транскрипцию IL-1β при низких концентрациях (10-9 М и 10-10 М) (фиг.4А). В лейкоцитах головных почек эффект PACAP 38 на транскрипцию IL-1β был умеренным, по сравнению с таковым, полученным в лейкоцитах периферической крови, показывая стимулирующий эффект только в дозе 10-9 (фиг.4В).
Кроме того, наблюдался стимулирующий эффект PACAP38 на экспрессию IL-15 в лейкоцитах периферической крови, стимулированных PACAP при дозах (10-10 М и 10-9 М). Наиболее высокие значения были получены в дозе 10-10 М (фиг.5A). В лейкоцитах головных почек, как раньше наблюдалось с транскрипцией IL-1β, эффект был умеренным. Уровни экспрессии IL-15 в группе, получавшей PACAP, были статистически выше, чем в группе отрицательного контроля в дозе 10-9 М (фиг.5В).
Пример 5
Контролируемый тест на заражение Caligus rogercresseyi у Salmo salar, ранее иммунизированных внутрибрюшинно антигеном my32, и my32, вводимым совместно с PACAP
Белок my32 был получен в рекомбинантной форме, в осадке разрушенных клеток BL21(DE3) E. coli, трансформированных pET28a-my32. Известно, что этот белок, введенный в брюшную полость (IP), производил 57% случаев ингибирования инфекции во втором поколении паразитов в тесте заражения C. rogercresseyi у S. salar (Carpio et al. (2011) Vaccine. 29 (15): 2810-20).
Чтобы продемонстрировать адъювантный эффект PACAP на этот белок, разработали эксперимент по вакцинации-заражению у S. salar в контролируемых условиях. В этом эксперименте были использованы шесть экспериментальных групп по 25 животных (средняя масса 80 г) в каждой из них:
Группа 1: Инъецированная IP с PBS.
Группа 2: Инъецированная IP с PACAP38 в дозе 1 мкг/рыбу.
Группа 3: Инъецированная IP с my32 в дозе 3 мкг/г массы тела (гмт).
Группа 4: Совместное введение с помощью IP-инъекции my32 3 мкг/гмт и PACAP в дозе 1 мкг/рыбу.
Группа 5: Рыба, инъецированная IP с my32 в дозе 3 мкг/гмт и получавшая питание 250 мкг из PACAP/кг корма в день, в течение одной недели до и после иммунизации my32. В ходе эксперимента рыб кормили два раза в день коммерческой формулой без PACAP в количестве 1% от массы их тела, за исключением группы 5, которой предоставлялась в течение одной недели до и после иммунизации та же коммерческая формула с PACAP, которая описана в Adelmann et al. ((2008) Vaccine 26, 837-844) для перорального введения ослабленного штамма вируса VHSV.
После 500 произвольных термических емкостей рыбы адаптировались к морской воде в течение двух недель. Впоследствии они были заражены 2000±200 веслоногих на каждую емкость. Рыб держали без потока воды в темноте и в условиях температуры, солености и кислорода, предложенных в Stone et al. (Dis Aquat Organ, 2000, 41: 141-149) в течение 24 дней. Оборот воды и фильтрацию выполняли вручную каждые 48 часов. Через 24 дня рыбы были анестезированы и убиты для подсчета паразитов под стереомикроскопом. Результаты в таблице 1 показывают снижение уровней заражения в группах, получавших PACAP, my32 и PACAP + my32. Наибольшее сокращение произошло в группах 4 и 5, что показывает адъювантный эффект PACAP на антиген.

Пример 6
Адъювантный эффект PACAP на гуморальный иммунный ответ у карпа (Cyprinus carpio) в сравнении с Aeromonas hydrophila
Эксперимент проводился с карпом (C. carpio) 40±10 г. Эти аквариумы поддерживали с 600 л при температуре 28±2°C. Использовали три экспериментальные группы по 10 карпов в каждой, и им вводили внутрибрюшинно следующие иммуногены:
Группа 1: PBS.
Группа 2: Инактивированные клетки A. hydrophila.
Группа 3: Инактивированные клетки A. hydrophila плюс 1 мкг на рыбу PACAP38.
Рыб инъецировали в день 0 и 14, и кровь собирали из хвостовой вены в дни 0 и 21. Результаты показали, что титры агглютинирующих антител были значительно выше в группе, иммунизированной бактерией плюс PACAP, по сравнению с группой, иммунизированной только бактерией (фиг.6). Эти результаты демонстрируют эффект PACAP в качестве молекулярного адъюванта. Препарат клеток A. hydrophila и измерения титров антител были сделаны в соответствии с Yin et al. ((1996) Fish & Shellfish Immunology 6, 57-69).
Пример 7
Контролируемый тест на заражение А. hydrophila у карпа (C. carpio), ранее иммунизированного внутрибрюшинно инактивированными бактериями и инактивированными бактериями, введенными совместно с PACAP
Эксперимент проводился с карпом (C. carpio) 30±5 г. Рыб поддерживали в 250 л аквариумах при температуре 30±2°С. Использовали три экспериментальные группы по 20 карпов в каждой, которых инъецировали внутрибрюшинно. Группа 1: PBS.
Группа 2: Инактивированные клетки A. hydrophila.
Группа 3: Инактивированные клетки A. hydrophila плюс 1 мкг PACAP38 на рыбу. Рыб инъецировали в день 0 и 14. В 21-й день заражение было выполнено путем инъекции IP LD50 бактерий и смертность была зафиксирована в течение 7 дней. Рассчитали относительный процент выживания (RPS) как:
RPS (%)=(% контролей на смертность - % смертности у подвергнутых лечению)/(% контролей на смертность)×100.
Результат составил 65% в группе 2 и 95% в группе 3, что свидетельствует о том, что введение PACAP увеличивает степень устойчивости к патогену вакцинированных и зараженных рыб.
Пример 8
Эффект совместного введения PACAP с ДНК-вакциной на основе гена, кодирующего 106 кДа полипротеин (VP2-VP4VP3-NH2-COOH) IPNV, у радужной форели (Oncorhynchus mykiss), экспериментально зараженной этим вирусом
Был проведен эксперимент по оценке эффекта внутримышечного совместного введения PACAP с ДНК-вакциной на основе гена, кодирующего 106 кДа полипротеин (VP2-VP4VP3-NH2-COOH) IPNV, у радужной форели, экспериментально зараженной этим вирусом. Было составлено шесть экспериментальных групп по 15 рыб в каждой из них (12±1 г), и рыб держали в воде при 10-12°C:
Группа 1: Рыбы, инъецированные PBS.
Группа 2: Рыбы, инъецированные ДНК-вакциной (pP-IPNV) в дозе 1 мкг/рыбу.
Группа 3: Рыбы, совместно инъецированные ДНК-вакциной (pP-IPNV) в дозе 1 мкг/рыбу и плазмидой, содержащей последовательность кДНК PACAP C. gariepinus под контролем быстродействующего раннего промотора цитомегаловируса (pCMV-PACAP) человека в дозе 0,5 мкг pCMV-PACAP/рыбу.
Группа 4: Рыбы, совместно инъецированные вакциной pP-IPNV (1 мкг/рыбу) с плазмидой негативного контроля в дозе 0,5 мкг pCMV/рыбу.
Группа 5: Рыбы, совместно инъецированные вакциной pP-IPNV (1 мкг/рыбу) с плазмидой, обладающей последовательностью кДНК PACAP C. gariepinus (pCMV-PACAP) в дозе 0,05 мкг pCMV-PACAP/рыбу.
Группа 6: Рыбы, совместно инъецированные вакциной pP-IPNV (1 мкг/рыбу) с плазмидой негативного контроля в дозе 0,05 мкг pCMV/рыбу.
Через 30 дней после вакцинации рыбы подверглись воздействию вируса путем внутрибрюшинного введения 100 мкл IPNV (1×107 TCID50 мл-1/рыбу).
Через 7 дней после заражения 10 рыб в группе забивали и вирусную нагрузку оценивали в головных почках.
Для определения вирусной нагрузки РНК выделяли из отдельных образцов, и был проведен RT-PCR с 1 мкг РНК. Детекцию экспрессии гена VP1 оценивали способом ПЦР в реальном времени. Результаты показаны на фиг.7.
Совместное введение pP-IPNV и pCMV-PACAP значительно снижало вирусную нагрузку, по сравнению с группой, которая была совместно инъецирована pP-IPNV и пустым вектором. Не было никаких различий между проверенными дозами pCMV-PACAP.
Пример 9
Эффект совместного введения PACAP с ДНК-вакциной на основе гена, кодирующего G гликопротеин VHSV, у радужной форели (O. mykiss), экспериментально зараженной указанным вирусом
Был проведен эксперимент по оценке влияния совместного введения с помощью внутримышечной инъекции PACAP C. gariepinus с ДНК-вакцины на основе гена, кодирующего G гликопротеин VHSV (pG-VHSV) у радужной форели, экспериментально зараженной этим вирусом. Четыре экспериментальные группы были сформированы, по 20 рыб в каждой (10±2 г) и держали в воде при 10-12°C:
Группа 1: Рыбы, инъецированные PBS.
Группа 2: Рыбы, инъецированные ДНК-вакциной pG-VHSV (0,01 мкг/рыбу).
Группа 3: Рыбы, совместно инъецированные ДНК-вакциной pG-VHSV в дозе 0,01 мкг/рыбу и PACAP C. gariepinus в дозе 0,1 мкг PACAP/рыбу.
Группа 4: Рыбы, совместно инъецированные ДНК-вакциной pG-VHSV в дозе 0,01 мкг/рыбу и PACAP C. gariepinus в дозе 0,5 мкг PACAP/рыбу.
Через 4 недели после вакцинации рыб подвергали воздействию вируса методом погружения в ванну. Введение осуществляли в течение 2 ч в воде, содержащей инфицирующую дозу VHSV (1×105 TCID50 на рыбу мл-1). Суммарная смертность была оценена через 4 недели после заражения и результаты показаны на фиг.8.
Совместное введение пептида с ДНК-вакциной снижало смертность на 15% и 29% по сравнению с введением только ДНК-вакцины, в дозах 0,1 и 0,5 мкг PACAP/рыбу, соответственно.
Пример 10
Эффект введения PACAP на выживание канального сома (Ictalurus punctatus) после иммунизации теронтами реснитчатого паразита Ichthyophthirius multifiliis
Был проведен эксперимент по оценке эффекта введения PACAP, после иммунизации теронтами реснитчатого паразита I. multifiliis, на выживание канального сома (I. punctatus). Вакцинацию и процедуру заражения выполняли, как предложено Wang and Dickerson ((2002) Clinical and Diagnostic Laboratory Immunology 9 (1), 176-181). Четыре экспериментальные группы по 25 рыб (12±5 г) были созданы:
Группа 1: Рыбы, иммунизированные в дни 1 и 35 PBS.
Группа 2: Рыбы, иммунизированные в 1 день 8000 живыми теронтами I. multifiliis и в день 35 с 10000 живых теронтов паразита.
Группа 3: Рыба, иммунизированные в 1 день 8000 живыми теронтами I. multifiliis и в день 35 10000 живых теронтов паразита. Эти рыбы также получали методом погружения в ванну нейропептид PACAP в дозе 100 мкг PACAP/л воды. Лечение проводили в течение 1 часа, через день, в течение двух недель, предшествующих заражению (шесть иммерсионных ванн).
Группа 4: Рыбы, иммунизированные в 1 день 8000 живых теронтов I. multifiliis и в день 35 10000 живых теронтов паразита. Эти рыбы были также подвергнуты погружению в ванну в течение 1 часа без РАСА, с помощью процедуры, похожей на таковую у группы 3. Перед иммунизацией рыбы были обработаны формалином, чтобы удалить существующих эктопаразитов. Рыбы поддерживались при 23±2°С и в непрерывном потоке воды. В день 84 рыбы столкнулись с 15000 теронтов. Данные о смертности были получены в течение 30 дней после заражения. Результаты приведены в таблице 2. Эти результаты показывают увеличение на 27% выживаемости иммунизированных рыб, которые одновременно получали PACAP.
Реферат
Группа изобретений относится к применению питуитарного активирующего аденилатциклазу пептида (PACAP) в качестве молекулярного адъюванта для вакцин для профилактики заболеваний, вызываемых инфекционными антигенами у рыб, к соответствующим вакцинной композиции, вакцинной комбинации, обе того же назначения, и к способу повышения иммунного ответа на вакцинный антиген при профилактике инфекционных заболеваний у рыб от инфекционных агентов, таких как вирусы, бактерии и эктопаразиты. Технический результат состоит в повышении иммунного ответа рыб против этого инфекционного агента. 4 н. и 8 з.п. ф-лы, 8 ил., 2 табл.
Комментарии