Мутантные полипептиды суаа и производные полипептидов, подходящие для доставки иммуногенных молекул в клетку - RU2585216C2
Код документа: RU2585216C2
Чертежи


























































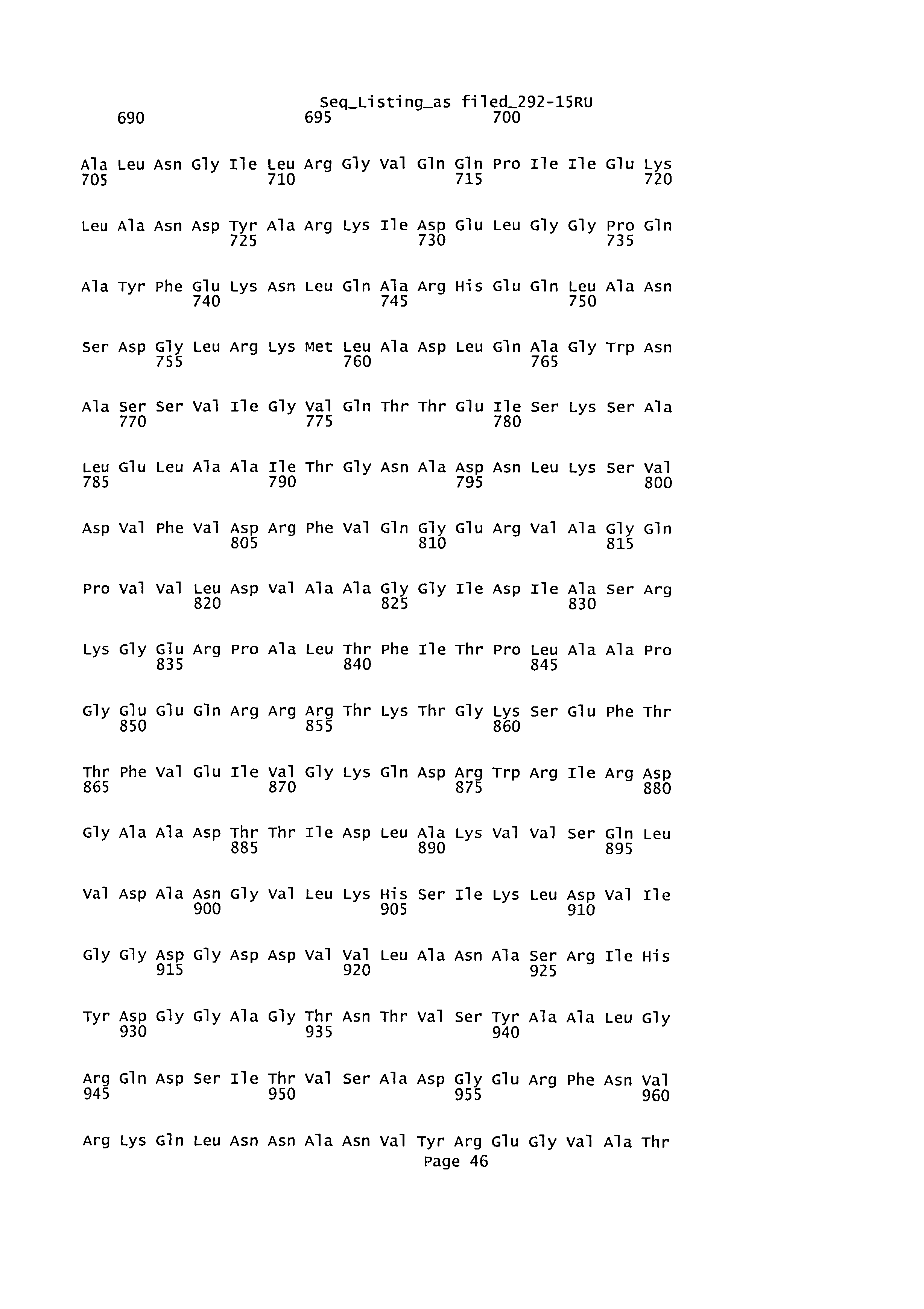


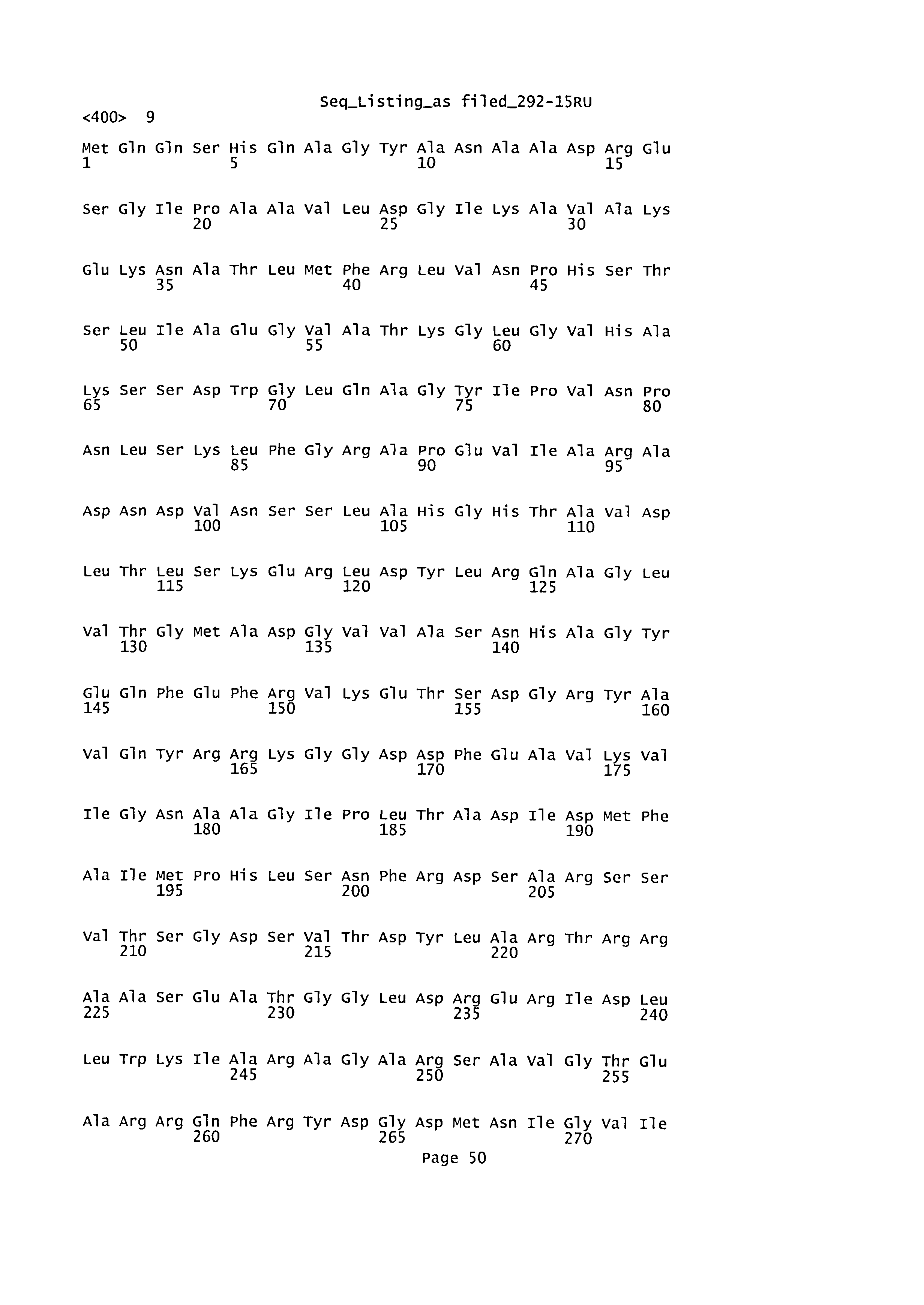






Описание
Настоящее изобретение относится к полипептидам, которые можно применять для доставки одной или более молекулы в клетку.
В частности, настоящее изобретение относится к полипептидам, которые можно применять для доставки одной или более молекулы, способной вызывать иммунный ответ у хозяина, главным образом за счет направленного воздействия на клетки, экспрессирующие рецепторы CD11b/CD18 (которые также упоминаются в настоящей заявке как "клетки, экспрессирующие CD11b").
В частности, настоящее изобретение направлено на полипептиды, полученные на основе белка аденилатциклазы (СуаА), при этом указанные полипептиды применяют либо в форме токсина, либо в форме обезвреженного белка, либо в форме анатоксина, который представляет собой мутантный вариант полипептида. Указанные мутантные полипептиды способны сохранять активность связывания с клетками-мишенями, характерную для природной формы СуаА, и также, предпочтительно, сохраняют способность к транслокации в клетки-мишени с помощью N-концевого домена, характерную для природной формы СуаА, и, кроме того, их порообразующая способность уменьшена или подавлена, по сравнению с природным вариантом токсина СуаА.
Настоящее изобретение относится, в частности, к применению указанных полипептидов в качестве белковых векторов. Соответственно, указанные мутантные варианты полипептидов дополнительно комбинируют с отличными от СуаА молекулами, и таким образом получают производные полипептидов, при этом указанные молекулы важны для превентивной вакцинации и/или терапии при введении хозяину.
Полипептиды согласно настоящему изобретению можно применять в качестве белковых векторов для доставки молекулы, в частности полипептидной молекулы, которая имеет аминокислотную последовательность, включающую один или более эпитоп, в частности антигенов, в клетку, в частности, в клетку, экспрессирующую CD11b.
Таким образом, настоящее изобретение также относится к полипептидному производному (производному мутантного полипептида согласно настоящему изобретению), которое содержит или состоит из мутантного полипептида согласно настоящему изобретению, объединенного посредством рекомбинации с одной или более молекулами, в частности, с одной или более молекулами, которые способны вызывать иммунный ответ, и таким образом представляет собой рекомбинантный полипептид или слитый полипептид. Изобретение также относится к производным полипептидов, полученным путем химического переноса указанных молекул на мутантные полипептиды.
Согласно одному варианту реализации изобретения, производные полипептидов в соответствии с настоящим изобретением можно применять для профилактического воздействия, в частности, при вакцинации и при терапии, в том числе при иммунотерапии, в частности, для индукции иммунного ответа у пациента.
Природная СуаА, применимая в контексте настоящего изобретения для конструирования полипептидов согласно настоящему изобретению, представляет собой аденилатциклазу, продуцируемую главным образом организмами Bordetella, и в частности Bordetella pertussis, и которая имеет характеристики и свойства, описанные в настоящей заявке с целью охарактеризовать указанный белок в контексте настоящего изобретения.
Бифункциональный токсин-гемолизин RTX семейства аденилатциклаз (обозначаемый в настоящей заявке как токсин аденилатциклазы (СуаА, ACT или AC-Hly) является одним из ключевых факторов вирулентности Bordetella pertussis, которая является возбудителем коклюша (1). Данный полипептид длиной 1706 остатков представляет собой продукт слияния N-концевого домена фермента аденилатциклазы (АЦ, англ. АС) или его части (~400 остатков) с порообразующим гемолизином RTX (Repeat in ToXin cytolysin) размером ~1306 остатков, составляющим С-конец или домен (2). Последний несет сайты активации ргоСуаА с образованием СуаА путем ковалентного посттрансляционного пальмитоилирования ε-аминогруппы Lys860 и Lys983, a также многочисленные повторы RTX, формирующие ~40 кальций-связывающих сайтов, загрузка которых необходима для цитотоксической активности СуаА (3, 4). Белок СуаА синтезируются в виде неактивного протоксина, который преобразуется в активный токсин в результате посттрансляционного пальмитоилирования двух внутренних остатков лизина (лизины 860 и 983). Для такой посттрансляционной модификации наряду с экспрессией гена суаА необходима экспрессия вспомогательного гена, а именно суаС, который расположен рядом с суаА на хромосоме B. pertussis.
Указанный токсин, в первую очередь, целенаправленно воздействует на миелоидные фагоциты, экспрессирующие рецептор интегрина αMβ2, известный также как CD11b/CD18, CR3 или Мас-1 (5). Указанный токсин, в частности, связывается с рецептором CD11b/CD18 клеток, экспрессирующих CD11b/CD18, через сайт связывания с рецептором, расположенный в его C-концевой части. Такие клетки, соответственно, представляют собой клетки-мишени для нативного токсина, а также для полипептидов согласно настоящему изобретению. СуаА встраивается в цитоплазматическую мембрану клеток и транслоцирует ферментативный домен АЦ в цитозоль указанной клетки-мишени (6, 7). Внутри клетки АЦ активируется кальмодулином и катализирует неконтролируемое превращение клеточного АТФ в цАМФ, молекулу, представляющую собой ключевой вторичный посредник, в результате чего нарушается бактерицидная функция фагоцитов (1). При высоких дозах (>100 нг/мл) катализируемое СуаА превращение АТФ в цАМФ оказывается цитотоксическим и приводит к апоптозу или даже к быстрой некротической гибели и лизису CD11b+ моноцитов (8, 9).
Недавно авторы настоящего изобретения показали, что СуаА связывается с N-связанными олигосахаридами на рецепторе CD11b/CD18 (10). Это факт позволяет предположить, что низкая специфичность взаимодействия с гликанами убиквитарных белков или гликолипидов клеточной поверхности может объяснять сниженную примерно на два порядка, но легко детектируемую способность СуаА проникать также в клетки, не несущие CD11b/CD18. Действительно, как было показано, вследствие чрезвычайно высокой удельной каталитической активности домена АЦ, СуаА существенно увеличивает уровень цАМФ также и в эритроцитах млекопитающих и птиц, в лимфоцитах, в клетках лимфомы, нейробластомы, СНО или в трахеальных эпителиальных клетках (1, 11).
Ранее в данной области техники был предложен обезвреженный токсин, также называемый анатоксином, у которого снижена, в частности, в существенной степени, активность аденилатциклазы. Такой анатоксин СуаА/АС можно применять для получения полипептидов согласно настоящему изобретению.
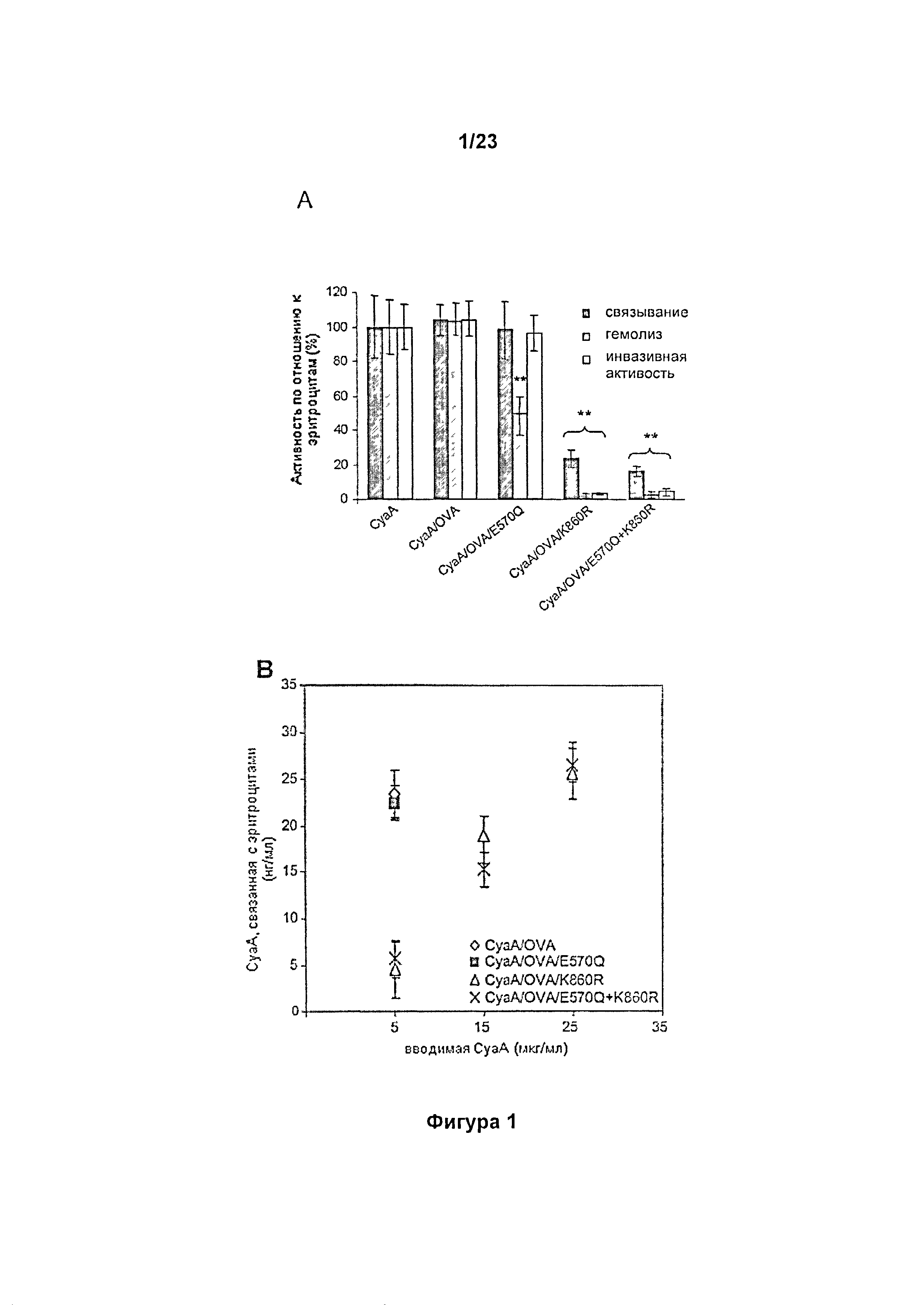
Помимо увеличения уровня цАМФ, токсин также демонстрирует умеренную гемолитическую активность по отношению к эритроцитам млекопитающих и птиц. Такая активность обусловлена способностью к образованию небольших катион-селективных пор, диаметром, по оценкам, от 0,6 до 0,8 нм, которые пермеабилизируют клеточную мембрану и в конечном итоге приводят к коллоидно-осмотическому лизису клеток (12). Недавно авторами настоящего изобретения и другими исследователями был показан синергетический эффект между порообразующей активностью СуаА и клеточно-инвазивной ферментативной активностью АЦ этого белка, что вносит вклад в общую цитолитическую активность СуаА по отношению к CD11b+ клеткам (13, 14). Благодаря интактной порообразующей (гемолитической) активности, в отсутствие осмопротекторов, таких как сыворотка, ферментативно неактивный анатоксин СуаА/АС- (15) сохраняет полную гемолитическую активностью по отношению к эритроцитам и остаточную, сниженную примерно в десять раз, цитолитическую активность по отношению к моноцитам, экспрессирующим CD11b (8), что накладывает ограничения на его применение в терапии.
Гемолитическая (порообразующая) активность и способность АЦ к транслокации через мембрану (клеточно-инвазивная активность) СуаА, как было показано ранее, могут быть разобщены при низкой концентрации кальция, низкой температуре (16), а также при определенной степени и характере ацилирования СуаА (4, 12, 17). Более того, эти две активности существенно различаются по чувствительности к заменам глутамата, которые изменяют заряд или являются нейтральными, в положениях 509, 516, 570 и 581 в гидрофобной области (8, 13, 18). Таким образом, было высказано предположение, что клеточно-инвазивная и порообразующая активность СуаА являются независимыми друг от друга и осуществляются параллельно в мембране клеток-мишеней. Модель, показанная на Фигуре 5А, предполагает, что два различных конформера СуаА встраиваются в мембрану клетки-мишени параллельно, при этом один из них является транслоцируемым предшественником, обуславливающим доставку домена АЦ через клеточную мембрану, что сопровождается притоком ионов кальция в клетку, другой является порообразующим предшественником, который определяет формирование олигомерных пор (13, 18, 19).
Авторы настоящего изобретения проверили данную модель и улучшили ее, показав, что порообразующая активность не вовлечена в транслокацию АЦ домена в клетки-мишени.
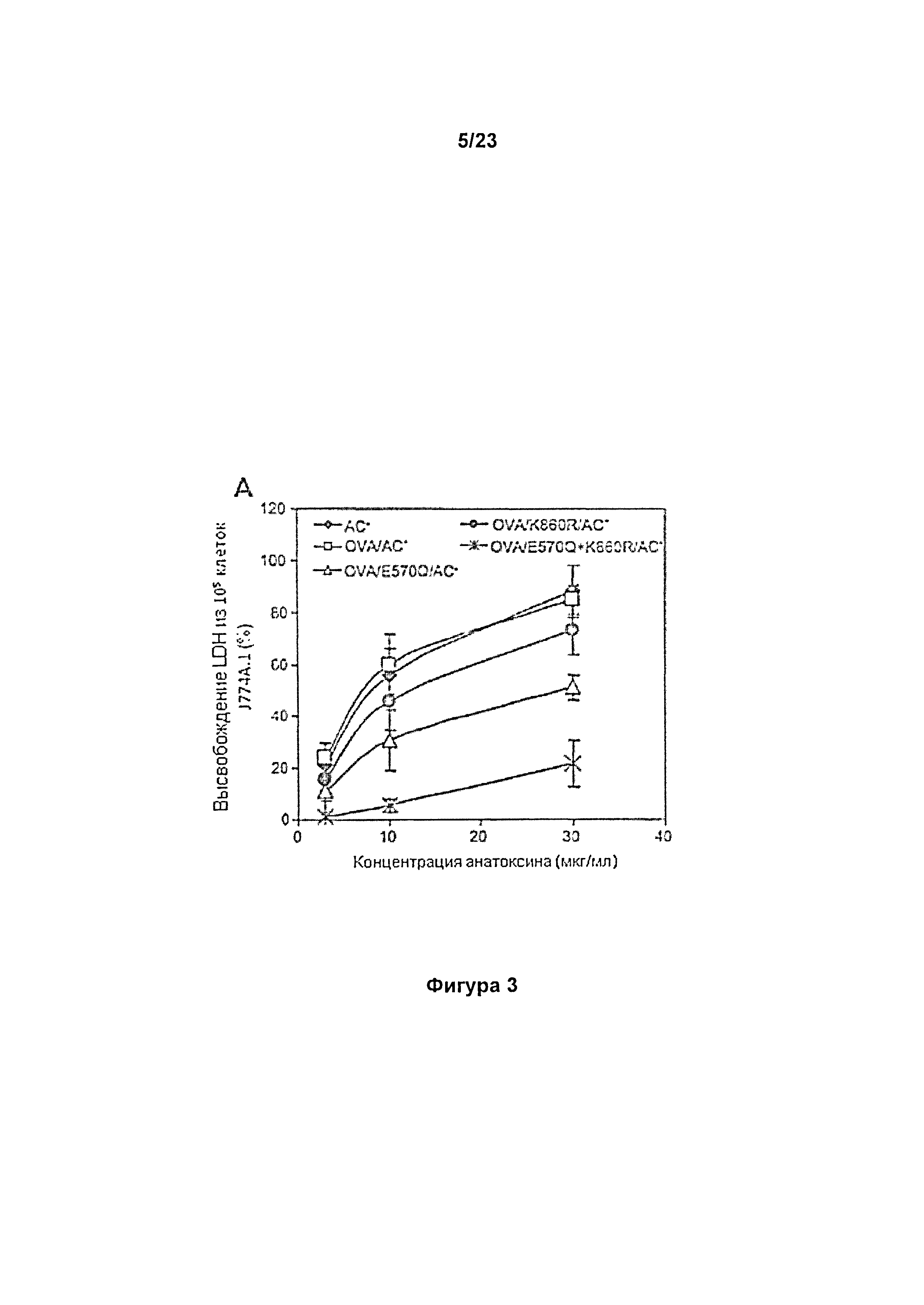
В настоящем изобретении авторы первоначально разработали мутантные варианты полипептидов СуаА на основе, в частности, аденилатциклазы из Bordetella pertussis, либо в виде токсина, либо анатоксина, несущие комбинации замен в порообразующем домене (E570Q), либо в домене, содержащем сайт ацилирования (K860R), и показали, что данная конкретная комбинация замен специфичным образом устраняет пермеабилизирующую клетки активность СуаА, что приводит к исчезновениюостаточной цитолитической активности анатоксинов СуаА/АС- в отношении клеток CD11b+. В то же время, конструкция E570Q+K860R сохраняет в полной мере способность к транслокации домена АЦ в цитозоль, приводящей к увеличению уровня цАМФ в клетке, и анатоксин на его основе в полной мере способен осуществлять доставку содержащих эпитопы молекул, встроенных в указанную конструкцию, в цитозоль дендритных клеток для опосредованной белками ГКГС I класса презентации и индукции специфических цитотоксических Т-клеточных ответов in vivo.
Мутантный вариант CyaA/233OVA/E570Q+K860R, разработанный авторами настоящего изобретения, со встроенным антигенным пептидом OVA, как описано в примерах, представляет собой первую конструкцию, которая иллюстрирует способность мутантного варианта СуаА обеспечивать значимое снижение способности пермеабилизировать клетки с сохранением при этом в полной мере способности транслоцировать домен АЦ через клеточную мембрану.
Авторы настоящего изобретения разработали конкретные конструкции, например, такие как анатоксин CyaA/E570Q+K860R/AC-, и показали, что, несмотря на значительное снижение активности по пермеабилизации клеток (цитолитической активности), такой анатоксин в полной мере сохраняет способность доставлять антиген в антигенпрезентирующие клетки (АПК) CD11b+. Кроме того, авторы настоящего изобретения показали, что общая цитолитическая активность такого анатоксина, как например, CyaA/E570Q+K860R/AC- является очень низкой. Таким образом, он лишен остаточной токсичности по отношению к клеткам животного или человека, и поэтому его легко можно применять в терапии.
Таким образом, настоящее изобретение обеспечивает новые полипептиды, представляющие собой анатоксины, которые имеют повышенный профиль безопасности и которые можно применять в качестве белковых векторов для доставки представляющих интерес молекул, в частности иммуногенных пептидных последовательностей, в клетки нуждающегося в лечении пациента, и, в частности, в клетки, экспрессирующие CD11b.
На основании экспериментов, проведенных авторами настоящего изобретение, таким образом, стало возможным выявление и получение полипептида, который является мутантным вариантом белка аденилатциклазы (мутантный полипептид), и аминокислотная последовательность которого представляет собой или включает одну из следующих последовательностей:
a) аминокислотную последовательность аденилатциклазы (СуаА) из Bordetella pertussis, Bordetella parapertussis или Bordetella hinzii, в которую внесены следующие мутации:
(i) замена остатка глутаминовой кислоты в положении 570 на остаток глутамина (E570Q) или консервативный аминокислотный остаток, а также
(ii) замена остатка лизина в положении 860 на остаток аргинина (K860R) или консервативный аминокислотный остаток, или;
b) аминокислотную последовательность фрагмента аденилатциклазы из Bordetella pertussis, Bordetella parapertussis или Bordetella hinzii, причем указанный фрагмент обладает способностью белка СуаА из Bordetella pertussis связываться с клеткой-мишенью и способностью транслоцировать N-концевой домен фермента аденилатциклазы или его часть в указанную клетку, при этом указанный фрагмент также содержит следующие измененные аминокислотные остатки, расположенные в положениях 570 и 860 в указанной аденилатциклазы: E570Q и K860R, или
c) аминокислотную последовательность, которая отличается от аминокислотной последовательности, определенной в пункте а) или b), заменой и/или вставкой одной или более аминокислоты, и которая обладает способностью белка СуаА из Bordetella pertussis связываться с клеткой-мишенью и способностью транслоцировать N-концевой домен фермента аденилатциклазы или его часть в указанную клетку, при этом указанная аминокислотная последовательность также содержит следующие измененные аминокислотные остатки, расположенные в положениях 570 и 860 в указанной аденилатциклазе: E570Q и K860R, или
d) аминокислотную последовательность аденилатциклазы (СуаА) из Bordetella parapertussis, в которую внесены следующие мутации:
(i) замена остатка глутаминовой кислоты в положении 569 на остаток глутамина (E569Q) или консервативный аминокислотный остаток, а также
(ii) замена остатка лизина в положении 859 на остаток аргинина (K859R) или консервативный аминокислотный остаток, или;
e) аминокислотную последовательность фрагмента аденилатциклазы из Bordetella bronchisepticai, причем указанный фрагмент обладает способностью белка СуаА из Bordetella pertussis связываться с клеткой-мишенью и способностью транслоцировать N-концевой домен фермента аденилатциклазы или его часть в указанную клетку, при этом указанный фрагмент также содержит следующие измененные аминокислотные остатки, расположенные в положениях 569 и 859 в указанной аденилатциклазе: E569Q и K859R, или
f) аминокислотную последовательность, которая отличается от аминокислотной последовательности, определенной в пункте d) или е), аменой и/или вставкой одной или более аминокислоты, и которая обладает способностью белка СуаА из Bordetella pertussis связываться с клеткой-мишенью и способностью транслоцировать N-концевой домен фермента аденилатциклазы или его часть в указанную клетку, при этом указанная аминокислотная последовательность также содержит следующие измененные аминокислотные остатки, расположенные в положениях 569 и 859 в указанной аденилатциклазе: E569Q и K859R.
Для целей настоящего изобретения, N-концевой домен описанного фрагмента представляет собой аминокислотную последовательность фрагмента, которая включает в себя последовательно расположенные аминокислотные остатки N-концевой части природного белка СуаА, например, N-концевая часть фрагмента представляет собой полностью или частично последовательные остатки, составляющие последовательность из 400 аминокислотных остатков N-концевого домена белка СуаА Bordetella pertussis.
В настоящей заявке "E570Q" охватывает замену остатка глутаминовой кислоты в положении 570 нативной СуаА из Bordetella pertussis, Bordetella parapertussis или Bordetella hinzii на остаток глутамина или на другой консервативный остаток, в частности, остаток, размер боковой цепи которого и гидрофильные свойства близки таковым у глутаминовой кислоты. Остаток глутаминовой кислоты в положении 570 предпочтительно замещен аминокислотным остатком, выбранным из Gln, Asn, Met, Thr, Ser, Gly, Arg, Lys, Val, Leu, Cys, Ile, Asp.
В настоящей заявке "K860R" охватывает замену остатка лизина в положении 860 нативной СуаА из Bordetella pertussis, Bordetella parapertussis или Bordetella hinzii на остаток аргинина или на другой консервативный остаток, в частности, остаток, размер боковой цепи которого и гидрофильные свойства близки таковым у лизина. Указанный остаток лизина в положении 860 предпочтительно замещен аминокислотным остатком, выбранным из Arg, Asn, Gln, Met, Thr, Ser, Gly, Val, Leu, Cys, Ile.
В настоящей заявке "E569Q" охватывает замену остатка глутаминовой кислоты в положении 569 нативной СуаА из Bordetella bronchiseptica на остаток глутамина или другой консервативный остаток, в частности, остаток, размер боковой цепи которого и гидрофильные свойства близки к таковым у глутаминовой кислоты. Указанный остаток глутаминовой кислоты в положении 569 предпочтительно замещен аминокислотным остатком, выбранным из Gln, Asn, Met, Thr, Ser, Gly, Arg, Lys, Val, Leu, Cys, Ile, Asp.
В настоящей заявке "K859R" охватывает замену остатка лизина в положении 859 нативной СуаА из Bordetella bronchiseptica на остаток аргинина или другой консервативный остаток, в частности, остаток, размер боковой цепи которого и гидрофильные свойства близки к таковым у лизина. Указанный остаток лизина в положении 859 предпочтительно замещен аминокислотным остатком, выбранным из Arg, Asn, Gln, Met, Thr, Ser, Gly, Val, Leu, Cys, Ile.
В вариантах реализации, описанных ниже, мутантные белки СуаА Bordetella pertussis или фрагменты таких белков, несущие замены "E570Q" и "K860R", могут быть заменены мутантными белками СуаА Bordetella parapertussis или Bordetella hinzii или фрагментами таких белков, несущими замены, эквивалентные "E570Q" и "K860R", или мутантными белками СуаА Bordetella bronchiseptica или фрагментами таких белков, несущих замены, эквивалентные "E569Q" и "K859R".
Последовательность аминокислот и нуклеотидная последовательность нативной СуаА из Bordetella pertussis также была описана в работе Glaser, P. et al., 1988, Molecular Microbiology 2(1), 19-30. Данная последовательность обозначена как SEQ ID №1, как показано на фигуре 6. Соответственно, при упоминании аминокислотных остатков или последовательностей или нуклеотидов или нуклеотидных последовательностей белка СуаА В. pertussis в настоящем изобретении, их положения указаны относительно последовательностей, описанных в упомянутой публикации Glaser et al. 1988.
В одном из вариантов реализации настоящего изобретения аминокислотная последовательность аденилатциклазы Bordetella pertussis представляет собой последовательность, представленную в SEQ ID №1.
Следует подчеркнуть, что при упоминании в настоящей заявке SEQ ID №1 или SEQ ID №2, если это уместно с технической точки зрения, описываемые характеристики аналогичным образом относятся к последовательности, модифицированной в результате вставки остатков в SEQ ID №1 и SEQ ID №2 с целью детоксикации (обезвреживания) белка СуаА. В таком случае, нумерацию аминокислотных остатков необходимо откорректировать (в частности, когда речь идет о положениях 570 и 860 нативной последовательности).
Предпочтительно, белок СуаА или его фрагмент представляет собой белок или его фрагмент, который является результатом совместной экспрессии в клетке, в частности, в рекомбинантной клетке, обоих генов - суаА и суаС. Действительно, было показано, что для приобретения инвазивных свойств по отношению к клеткам-мишеням белок СуаА должен пройти посттрансляционные модификации, для осуществления которых необходима экспрессия как суаА, так и суаС гена (WO 93/21324).
В конкретном варианте реализации изобретения белок СуаА представляет собой бактериальный белок. В предпочтительном варианте реализации изобретения белок СуаА получен из вида Bordetella.
Одним из видов Bordetella, представляющих интерес в соответствии с настоящим изобретением, является Bordetella pertussis. Другими штаммами Bordetella, представляющими интерес, являются Bordetella parapertussis, Bordetella hinzii или Bordetella bronchiseptica. Последовательность белка СуаА из В. parapertussis, в частности, описана под номером доступа NC 002928.3 (в виде последовательности из 1740 аминокислот) и в работе Parkhill J. et al. (Nat. Genet. DOI, 10 (2003)), последовательность из В. hinzii описана в работе Donate G.M. et al. (J. Bacteriol. 2005 Nov. 187(22):7579-88), a последовательность из В. bronchiseptica - в работе Betsou F. et al (Gene 1995, August 30; 162(1):165-6). Последовательность из Bordetella parapertussis описана под номером доступа САВ76450 и обозначена в настоящей заявке как SEQ ID №7, как показано на Фигуре 14. Последовательность из Bordetella hinzii описана под номером доступа AAY57201 и обозначена в настоящей заявке как SEQ ID №8, как показано на Фигуре 15. Последовательность из Bordetella bronchiseptica описана под номером доступа САА85481 и обозначена в настоящей заявке как SEQ ID №9, как показано на Фигуре 16. Соответственно, при упоминании в настоящем изобретении аминокислотных остатков или последовательностей белка СуаА из Bordetella parapertussis, Bordetella hinzii или Bordetella bronchiseptica, их положения указаны относительно последовательностей, представленных в SEQ ID №7, 8 и 9, соответственно.
Выражение "мутантный пептид белка аденилатциклазы" не охватывает нативную аденилатциклазу, экспрессируемую Bordetella. Как указано выше, упомянутый мутантный полипептид характеризуется основным отличием от природного белка, которое заключается в комбинированной замене двух конкретных аминокислотных остатков. Такой мутантный полипептид может быть дополнительно модифицирован, по сравнению с указанным нативным белком, и, в частности, может представлять собой фрагмент такого мутантного белка, например, укороченный вариант указанного мутантного белка, в котором удалены остатки на одном или на обоих концах. В частности, могут быть удалены остатки на С-конце, при условии, что такая делеция не затрагивает сайт узнавания и сайт связывания клеточного рецептора CD11b/CD18. В качестве альтернативы или дополнения, могут быть удалены остатки на N-конце, при условии, что такая делеция не влияет на транслокационную способность полученного мутантного полипептида Также фрагмент может быть получен после удаления одного или более внутренних остатков мутантного варианта нативного белка СуаА.
Если изобретение относится к мутантному полипептиду, который представляет собой фрагмент, как указано в настоящем описании, указанный фрагмент, который обязательно включает в себя измененные остатки E570Q и K860R (при отсылке на аминокислотную последовательность белка СуаА из Bordetella pertussis), также сохраняет способность мутантного полноразмерного белка СуаА связываться с клетками и транслоцировать N-концевой домен в цитозоль клеток-мишеней, в частности, клеток, экспрессирующих CD11b/CD18.
Изобретение обеспечивает, таким образом, мутантные полипептиды, которые можно применять для разработки средств доставки одной или более молекул в клетки, в частности, в клетки-мишени, экспрессирующие рецептор CD11b/CD18.
В частности, изобретение обеспечивает мутантные полипептиды на основе белка СуаА, где указанный белок либо получен на основании токсина СуаА или предпочтительно получен на основании его анатоксина, в частности, анатоксина СуаА/АС-. Мутантные полипептиды, способные связываться с клеткой, в частности, с клеткой-мишенью, в частности, с клеткой-мишенью, экспрессирующей рецептор CD11b/CD18, способны траснлоцировать свой N-концевой домен или молекулы, встроенные в указанный домен, или пересаженные на него, в клетку, и при этом порообразующая активность таких мутантных полипептидов полностью или частично подавлена, по сравнению с токсином или анатоксином СуаА.
Способность мутантного полипептида целенаправленно воздействовать на клетки CD11b/CD18 можно проанализировать, в частности, с помощью методики, описанной в ЕР 03291486.3, в работе EI-Azami-EI-Idrissi M. et al., J. Biol. Chem., 278 (40) 38514-21 или в международной публикации WO 02/22169. Кроме того, способность мутантного полипептида транслоцировать эпитоп (эпитопы) или полипептид (полипептиды), содержащий указанный эпитоп (эпитопы), в цитозоль клетки-мишени может быть проанализирована с помощью методики, описанной в WO 02/22169.
Полное или частичное подавление порообразующей активности или клеточно-пермеабилизирующей способности токсина или анатоксина СуаА следует понимать как полное или частичное подавление способности образовывать поры, в частности, катион-селективные поры диаметром по оценкам от 0,6 до 0,8 нм, которые пермеабилизируют клеточную мембрану и в конечном итоге вызывают коллоидно-осмотический лизис клеток. Порообразующую активность можно измерить с помощью исследования методом локальной фиксации потенциала ("пэтч-кламп") всей клетки, как описано в примерах.
Порообразующая активность токсина СуаА вносит свой вклад в общую цитолитическую или гемолитическую активность по отношению к клеткам. Действительно, в контексте настоящего изобретения, под общей цитолитической или гемолитической активностью СуаА (или его "общей цитотоксической активностью") следует понимать результирующую активность, включающую, по меньшей мере, аденилатциклазную и порообразующую активность токсина СуаА. Таким образом, полное или частичное подавление порообразующей активности токсина СуаА обеспечивает, по меньшей мере, частичное подавление его цитолитической активности.
В предпочтительном варианте реализации общая цитолитическая активность полипептида согласно настоящему изобретению, в частности, по отношению к клеткам, которые экспрессируют рецепторы CD11b/CD18, полностью или частично подавлена, по сравнению с токсином СуаА из Bordetella pertussis. Цитолитичесую активность полипептида согласно настоящему изобретению можно определить путем измерения количества гемоглобина (для эритроцитов) или лактатдегидрогеназы (для моноцитов), высвобождаемых клетками при инкубации с тестируемым полипептидом, как описано в примерах.
В предпочтительном варианте реализации общая цитолитическая активность полипептида согласно настоящему изобретению по отношению к клеткам, которые экспрессируют рецептор CD11b/CD18, снижена по меньшей мере на 75%, желательно по меньшей мере на 80%, 85%, 90% или 95%снижена, по сравнению с активностью токсина СуаА из Bordetella pertussis или белка СуаА Bordetella pertussis, аденилатциклазная активность которого частично или полностью подавлена (или "анатоксин СуаА"). В предпочтительном варианте реализации общая цитолитическая активность полипептида согласно настоящему изобретению по отношению к клеткам, которые экспрессируют рецептор CD11b/CD18, снижена по меньшей мере на 75%, желательно по меньшей мере на 80% или 85% по сравнению с активностью анатоксина СуаА из Bordetella pertussis, аминокислотная последовательность которого представлена на фигуре 2 (SEQ ID №2).
В предпочтительном варианте реализации настоящее изобретение относится к полипептиду, представляющему собой мутантный вариант аденилатциклазы, аминокислотная последовательность которого представляет собой или состоит из аминокислотной последовательности (последовательностей), измененной (мутированной), по сравнению с аминокислотной последовательностью, представленной в SEQ ID N°1 таким образом, что аминокислотная последовательность указанного мутантного варианта содержит замены E570Q и K860R, при этом указанный полипептид способен связываться с клеткой-мишенью и способен транслоцировать свой N-концевой домен в клетку.
В конкретном варианте осуществления настоящего изобретения фрагмент, который содержит замену остатка глутаминовой кислоты в положении 570 из SEQ ID №1 на остаток глутамина (упоминаемую как "E570Q"), и замену остатка лизина в положении 860 из SEQ ID №1 на остаток аргинина (упоминаемую как "K860R"), включает, по меньшей мере, аминокислотную последовательность белка СуаА, которая начинается с первого N-концевого остатка или с одного из аминокислотных остатков, находящихся между положениями 1 и 400, предпочтительно, положениями от 1 до 380, и продолжается до остатков, формирующих сайт узнавания и сайт связывания клеточного рецептора CD11b/CD18, и при этом указанный фрагмент содержит остатки, соответствующие измененным остаткам E570Q и K860R, или состоит из указанной аминокислотной последовательности. В конкретном варианте реализации фрагмент, содержащий замены E570Q и K860R, не содержит последовательность аминокислот, начиная от аминокислоты в положении 1 из SEQ ID №1 до аминокислоты в положении 372 из SEQ ID №1.
В предпочтительном варианте реализации изобретения фрагмент, сконструированный таким образом, по существу лишен ферментативной активности аденилатциклазы (активности АЦ).
В предпочтительном варианте реализации мутантный полипептид согласно настоящему изобретению получен путем совместной экспрессии в рекомбинантных клетках измененного (мутантного) гена, кодирующего аминокислотную последовательность мутантной СуаА с заменами E570Q и R860R, и гена суаС с последующим выделением выбранных экспрессируемых фрагментов мутантной СуаА.
Предпочтительно мутантный полипептид согласно настоящему изобретению содержит остаток лизина, соответствующий остатку лизина в положении 983 аминокислотной последовательности СуаА, представленной в SEQ ID №1, который ацилирован, в частности, пальмитоилирован или пальмитолеилирован.
Предпочтительно мутантный полипептид соласно настоящему изобретению содержит остаток лизина, соответствующий остатку лизина в положении 983 из аминокислотной последовательности СуаА, представленной в SEQ ID №1, который не ацилирован.
В конкретном варианте реализации мутантный полипептид согласно изобретению имеет аминокислотную последовательность, полученную на основе аминокислотной последовательности СуаА, представленной в SEQ ID №1, в результате мутации остатков приводящей к заменам E570Q и K860R, и имеет аминокислотную последовательность, которая по меньшей мере на 50%, предпочтительно по меньшей мере на 60%, 70%, 75%, 80%, 85%, 90%, 95% или 99% идентична последовательности, представленной в SEQ ID №1.
В другом конкретном варианте реализации мутантный полипептид согласно настоящему изобретению имеет аминокислотную последовательность, отличающуюся от аминокислотной последовательности СуаА, соответствующей SEQ ID №1, в результате мутации остатков приводящей к заменам E570Q и K860R, и в результате дополнительных мутаций, приводящим к от 1 до 500, в частности, к от 1 до 400, от 1 до 300, от 1 до 200, от 1 до 100, от 1 до 50, от 1 до 40, от 1 до 30, от 1 до 25, от 1 до 20, от 1 до 15, от 1 до 10 или от 1 до 5 заменам, делециям и/или вставкам аминокислотных остатков, включая замены E570Q K860R.
В конкретном варианте реализации мутантный полипептид согласно настоящему изобретению не несет никаких замен, делеций и/или вставок остатков аминокислот, по сравнению с аминокислотной последовательностью СуаА из Bordetella pertussis, кроме замен E570Q и K860R. В конкретном варианте реализации мутантный полипептид имеет последовательность аминокислот, представленную в SEQ ID №2, как показано на Фигуре 7. В другом конкретном варианте настоящего изобретения единственные дополнительные замены, делеций и/или вставки аминокислот, по сравнению с аминокислотной последовательностью SEQ ID №2, включают замены, делеций и/или вставки аминокислот, которые полностью или частично подавляют ферментативную активность аденилатциклазы белка СуаА, например, в частности, вставка дипептида, например, дипептида "LQ", или "GS" между аминокислотами в положениях 188 и 189.
В конкретном варианте реализации мутантный полипептид согласно настоящему изобретению отличается от аминокислотной последовательности СуаА, представленной в SEQ ID №1, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 заменами, делециями и/или вставками аминокислотных остатков в дополнение к заменам E570Q и K860R.
В конкретном варианте реализации в дополнение к заменам E570Q и K860R остаток лейцина в положении 247 нативного белка СуаА из Bordetella pertussis замещен на остаток глутамина (L247Q) или другой аминокислотный остаток, в частности, консервативный аминокислотный остаток.
Мутантный полипептид согласно настоящему изобретению, который представляет собой фрагмент аминокислотной последовательности, представленной в SEQ ID №1, как описано в настоящей заявке, следует понимать как последовательность, включающую один или более фрагмент, имеющий по меньшей мере от примерно 350 аминокислотных остатков до примерно 1705 аминокислотных остатков из аминокислотной последовательности SEQ ID №1, в частности, фрагмент, включающий участок по меньшей мере из 400, 500, 600, 700, 800, 900, 1000, 1100, 1200, 1300, 1400, 1500, 1600 аминокислотных остатков из SEQ ID №1, содержащий остатки E570Q и K860R. Мутантный полипептид согласно настоящему изобретению можно также определить как фрагмент аминокислотной последовательности, представленной в SEQ ID №2, который включает один или более фрагмент, имеющий по меньшей мере от примерно 350 аминокислотных остатков до примерное 1705 аминокислотных остатков из аминокислотной последовательности SEQ ID №2, в частности, фрагмент, включающий участок из по меньшей мере 400, 500, 600, 700, 800, 900, 1000, 1100, 1200, 1300, 1400, 1500, 1600 аминокислотных остатков из SEQ ID №2, содержащий остатки 570 и 860. Указанные фрагменты предпочтительно сохраняют способность связываться с рецептором CD11b/CD18 клеток и способностью транслоцировать свой N-концевой домен в клетки-мишени. Предпочтительно мутантный полипептид согласно настоящему изобретению, представляющий собой такой фрагмент, содержит участок из аминокислот, включающий аминокислотные остатки с 570, в виде E570Q по 860 в виде K860R, или с 1 по 860, или со 2 по 860 из SEQ ID №1, при условии, что мутации E570Q и K860R содержатся в исходной последовательности SEQ ID №1.
В предпочтительном варианте реализации настоящего изобретения указанный фрагмент дополнительно включает аминокислотные остатки с 1166 по 1281 или аминокислотные остатки с 1208 по 1243 из аминокислотной последовательности СуаА, представленной в SEQ ID №1 белка СуаА, для взаимодействия с клетками-мишенями CD11b/CD18.
Конкретный фрагмент, таким образом, включает весь С-концевой участок, или его часть, нативного белка, при этом указанный участок отвечает за связывание полипептида согласно настоящему изобретению с мембраной клетки-мишени и/или рецептором CD11b/CD18, а также за последующую доставку N-концевого домена полипептида в цитозоль клетки. Конкретный полипептид согласно настоящему изобретению представляет собой фрагмент белка СуаА, содержащий аминокислотные остатки с 373 до 1706 из белка СуаА, в частности, SEQ ID №1, при этом остатки 570 и 860 изменены как E570Q и K860R.
В другом предпочтительном варианте реализации мутантный полипептид, который представляет собой такой фрагмент, включает:
а) первую аминокислотную последовательность, которая соответствует участку по меньшей мере из 100 последовательных аминокислотных остатков из SEQ ID №1, включающему аминокислотный остаток 570 в виде E570Q, и дополнительно включающий 0, 1, 2, 3, 4 или 5 делеции, замены или вставки, по сравнению с SEQ ID №1 и
б) вторую аминокислотную последовательность, которая соответствует участку по меньшей мере из 100 последовательных аминокислотных остатков из SEQ ID №1, включающему аминокислотный остаток 860 в виде K860R, и дополнительно включающий 0, 1, 2, 3, 4 или 5 делеции, замены или вставки, по сравнению с SEQ ID №1 и
в) третью аминокислотную последовательность, включающую аминокислотные остатки с 1166 по 1281 или аминокислотные остатки с 1208 по 1243 из аминокислотной последовательности СуаА, представленной в SEQ ID №1 белка СуаА, для взаимодействия с клетками-мишенями CD11D/CD18.
Другой конкретный полипептид согласно настоящему изобретению представляет собой фрагмент, который соответствует мутантному белку СуаА с заменами E570Q и K860R, в котором удалены аминокислотные остатки с 225 по 234, в результате чего получен фрагмент, содержащий остатки с 1 по 224 и с 235 по 1706 из последовательности мутантного белка.
В особенно предпочтительном варианте реализации полипептидный фрагмент согласно настоящему изобретению связывается с клеткой, экспрессирующей рецепторы CD11D/CD18, посредством специфичного связывания с указанным рецептором.
В предпочтительном варианте реализации аденилатциклазная активность указанного полипептида в клетке полностью или частично подавлена, по сравнению с токсином СуаА из Bordetella pertussis. Как указано выше, термин "белок СуаА "относится к соответствующему белку либо в форме токсина, либо в форме анатоксина. Соответственно, каждый вариант реализации изобретения, относящийся к полипептиду, представляющему собой мутантный белок СуаА, относится к соответствующему белку либо в форме токсина, либо в форме анатоксина.
Полное или частичное подавление аденилатциклазной или ферментативной активности СуаА следует понимать как полное, или частичное подавление способности преобразовывать АТФ в цАМФ в клеточной среде, по сравнению с токсином СуаА, продуцируемым в результате совместной экспрессии генов суаА и суаС в клетке. Способность преобразования АТФ в цАМФ можно оценить с помощью измерения уровня внутриклеточного цАМФ, как описано в примерах.
Такое полное или частичное подавление может быть получено как в результате генетической инактивации, например, путем введения короткой аминокислотной последовательности, содержащей, например от одной до десяти аминокислот, в частности дипептида, в сайт аминокислотной последовательности СуаА, который является частью каталитического центра, то есть в участок, расположенный в пределах первых 400 аминокислот (АС домен) SEQ ID №1, или путем делеции или замены части аминокислотной последовательности СуаА, изложенной в SEQ ID №1, которая важна для ферментативной активности. В предпочтительном варианте реализации полного или частичного подавления ферментативной активности СуаА достигают путем вставки дипептида, например, "LQ" или дипептида "GS" между аминокислотами в положениях 188 и 189 последовательности СуаА, представленной в SEQ ID №1. Это, в свою очередь, может быть достигнуто путем вставки олигонуклеотида, такого как "CTGCAG" или "CGATCC", по сайту рестрикции EcoRV в положении 564 в кодирующей области гена суаА. См. публикацию Ladant et al., 1992. Кроме того, полное или частичное подавление ферментативной активности также может быть осуществлено с помощью направленного мутагенеза, например, путем замены остатка лизина в положении 58 или 65 нативного белка СуаА из Bordetella pertussis (Glaser et al., 1989) на остаток Gln.
Полипептид согласно настоящему изобретению также можно определить как полипептид, который может быть получен на основе полипептида СуаА, имеющего аминокислотную последовательность согласно SEQ ID №1, 7, 8 или 9, в результате:
a) замены остатка глутаминовой кислоты в положении 570 из SEQ ID №1, 7, 8 или в положении 569 из SEQ ID №9 на остаток глутамина или консервативный аминокислотный остаток;
b) замены остатка лизина в положении 860 из SEQ ID №1, 7, 8 или в положении 859 из SEQ ID №9 на остаток аргинина или консервативный аминокислотный остаток, а также
c) при необходимости, осуществления одной или более замены, вставки и/или делеции остатков аминокислот в положениях, отличающихся от положений, указанных в пунктах (а) и (b), при условии, что полученный таким образом полипептид обладает способностью белка СуаА от Bordetella pertussis связываться с клеткой-мишенью и транслоцировать N-концевой домен фермента аденилатциклазы или его часть в указанную клетку.
Предпочтительно на этапе (а) остаток глутаминовой кислоты заменяют на аминокислотный остаток, выбранный из Gln, Asn, Met, Thr, Ser, Gly, Arg, Lys, Val, Leu, Cys, Ile, Asp, наиболее предпочтительно на Gln. Предпочтительно на этапе (b) остаток лизина заменяют на аминокислотный остаток, выбранный из Arg, Asn, Gln, Met, Thr, Ser, Gly, Val, Leu, Cys, Ile, наиболее предпочтительно на Arg.
В конкретном варианте реализации на этапе (с) не осуществляют дополнительных замен, вставок и/или делеций остатков аминокислот.
В частности, этап (с) может включать укорочение одного или обоих концевых участков молекулы. В частности, могут быть удалены остатки на С-конце, при условии, что такая делеция не затрагивает сайт узнавания и сайт связывания клеточного рецептора CD11b/CD18. В качестве альтернативы или дополнения, могут быть удалены остатки на N-конце, при условии, что такая делеция не влияет на транслокационную способность полученного мутантного полипептида. Также могут быть осуществлены внутренние делеций одного или более остатков нативного белка СуаА, локализованных в положениях, отличающихся от положений аминокислот, указанных в пунктах (а) и (b). В конкретном варианте реализации этап (с) включает делецию до 380 или до 400 аминокислот в N-терминальной аминокислотной последовательности полипептида СуаА, предпочтительно делецию участка аминокислотной последовательности, начиная с аминокислоты в положении 1 из SEQ ID №1, 7, 8 или 9 до аминокислоты в положении 372 из SEQ ID №1, 7, 8 или 9.
Предпочтительно, после осуществления этапа (с) полученный полипептид включает весь C-концевой участок, или его часть, нативного белка, при этом указанный участок отвечает за связывание полипептида согласно настоящему изобретению с мембраной клетки-мишени и/или рецептором CD11b/CD18, а также за последующую доставку N-концевого домена полипептида в цитозоль клетки. В конкретном варианте реализации на стадии (с) аминокислотные остатки с 373 по 1706 из SEQ ID №1, 7 или 8, или аминокислотные остатки с 373 по 1705 из SEQ ID №9, остаются неделетированными. В конкретном варианте реализации на стадии (с) аминокислотные остатки с 1208 по 1243 из SEQ ID №1, 7 или 8, или аминокислотные остатки с 1207 по 1242 из SEQ ID №9, остаются неделетированными.
В предпочтительном варианте реализации этап (с) включает аминокислотные замены, делеции и/или вставки, которые полностью или частично подавляют аденилатциклазную ферментативную активность белка СуаА. Такое полное или частичное подавление ферментативной активности аденилатциклазы может быть осуществлено путем встраивания коротких последовательностей аминокислот, содержащих, например от одной до десяти аминокислот, в частности, дипептида, в участок, расположенный в пределах первых 400 аминокислот (домен АЦ) SEQ ID №1, 7, 8 или 9, или делеции или замены части аминокислотной последовательности СуаА, изложенной в SEQ ID №1, 7, 8 или 9, которая существенна для ферментативной активности. В предпочтительном варианте реализации полного или частичного подавления ферментативной активности СуаА достигают путем вставки дипептида, например, "LQ" или дипептида "GS" между аминокислотами в положениях 188 и 189 последовательности СуаА, представленной в SEQ ID №1, 7, 8 или 9. Кроме того, полного или частичного подавления ферментативной активности также можно достичь с помощью направленного мутагенеза, например, путем замены остатка лизина в положении 58 или 65 нативного белка СуаА из Bordetella pertussis (Glaser et al., 1989) на остаток Gln.
Предпочтительно, на стадии (с) остаток лизина в положении 983 аминокислотной последовательности СуаА, представленной в SEQ ID №1, 7, 8, или в положении 982 в SEQ ID N° 9 не замещают и не удаляют. В одном из вариантов реализации указанный остаток лизина ацилируют, в частности пальмитоилируют или пальмитолеилируют. В качестве альтернативы, указанный остаток лизина не ацилирован.
Предпочтительно, после осуществления этапа (с) полученный полипептид имеет аминокислотную последовательность, имеющую по меньшей мере 50%, предпочтительно по меньшей мере 60%, 70%, 75%, 80%, 85%, 90%, 95% или 99% идентичности с последовательностью, представленной в SEQ ID №1, 7, 8 или 9.
Кроме того, предпочтительно после осуществления этапа (с) полученный полипептид имеет аминокислотную последовательность, которая отличается от последовательности, изложенной в SEQ ID №1, 7, 8 или 9, аминокислотными заменами, произведенными на стадиях (а) и (b), и от 1 до 500, в частности, от 1 до 400, от 1 до 300, от 1 до 200, от 1 до 100, от 1 до 50, от 1 до 40, от 1 до 30, от 1 до 25, от 1 до 20, от 1 до 15, от 1 до 10 или от 1 до 5 дополнительными заменами, делециями и/или вставками аминокислот. В конкретном варианте реализации после осуществления этапа (с) полученный полипептид отличается от аминокислотной последовательности СуаА, представленной в SEQ ID №1, 7, 8 или 9, 1,2, 3, 4, 5, 6, 7, 8, 9 или 10 заменами, делециями и/или вставками аминокислотных остатков в дополнение к заменам, осуществленным на этапах (а) и (b).
Изобретение также относится к полипептидному производному, включающему или состоящему из мутантного полипептида согласно настоящему изобретению, который дополнительно объединен с одной или более представляющей интерес молекулой. В предпочтительном варианте реализации представляющая интерес молекула представляет собой биологически активную молекулу, взятую в отдельности или объединенную с полипептидом настоящего изобретения. Указанные молекулы могут, в частности, иметь профилактическую или терапевтическую ценность, т.е. могут обладать профилактической или терапевтической активностью, или могут усиливать профилактическую или терапевтическую активность.
В конкретных вариантах реализации представляющие интерес молекулы выбраны из группы, включающей: пептиды, гликопептиды, липопептиды, полисахариды, олигосахариды, нуклеиновые кислоты, липиды и химические вещества.
В конкретном варианте реализации одна или более представляющая интерес молекула представляет собой молекулу полипептида или содержит молекулу полипептида. Аминокислотная последовательность такой молекулы может содержать от 2 до 1000, предпочтительно 5-800, от 5 до 500, от 5 до 200, от 5 до 100, от 8 до 50, 5 до 25, от 5 до 20 или 8 до 16, или 300-600, 400-500 аминокислотных остатков.
В предпочтительном варианте реализации одна или более представляющая интерес молекула представляет собой гетерологичную аминокислотную последовательность, способную индуцировать иммунный ответ (также упоминаемая как "гетерологичный антиген"), в частности, аминокислотную последовательность, содержащую или состоящую из эпитопа, включающего антиген. Используемый в настоящей заявке термин "гетерологичный" относится к антигену, отличному от мутантного полипептида, который используется в самом векторе. Используемый в настоящей заявке термин "эпитоп" относится к гетерологичной молекуле, и в частности, к гетерологичному пептиду, который может вызвать иммунный ответ при его презентации иммунной системе хозяина. В частности, такой эпитоп может включать или состоять из участка из 8, 9, 10, 11, 12, 13, 14, 15 или 16 аминокислотных остатков. Как альтернатива, такой эпитоп может включать полноразмерный антиген или включать фрагмент (фрагменты) антигена (антигенов).
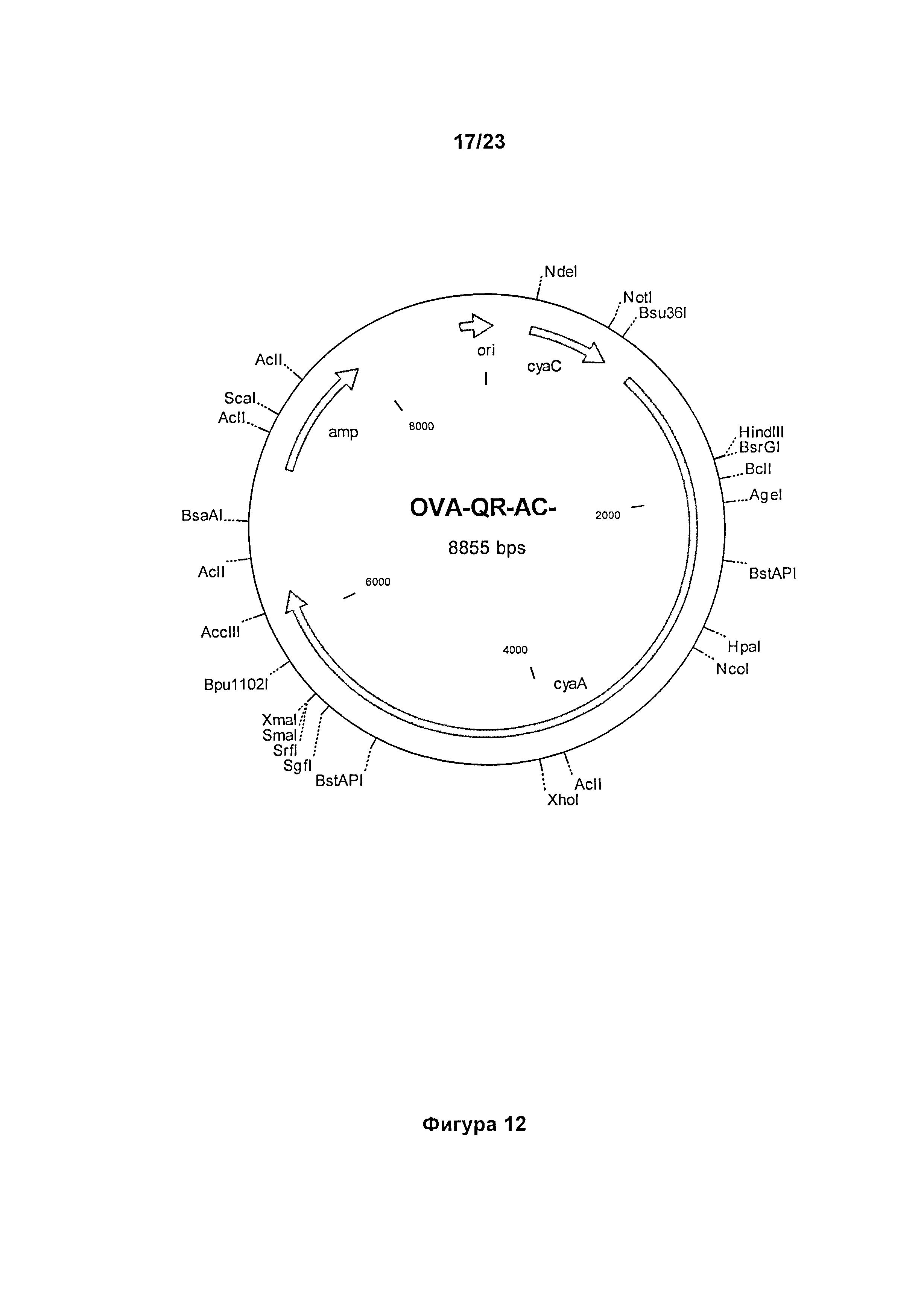
В конкретном варианте реализации производное полипептида в соответствии с настоящим изобретением может кодироваться плазмидой, соответствующей плазмиде OVA-QR-AC", депонированной под номером доступа CNCM I-4137 (фиг.12), в которой последовательность ДНК, кодирующая антигенную последовательность "OVA", замещена последовательностью ДНК, кодирующей антигенную последовательность, включающую один или более эпитоп.
Полипептидная молекула, которая подходит для индукции иммунного ответа, в частности, представляет собой молекулу, вызывающую Т-клеточный иммунный ответ, в том числе, например, ответ ЦТЛ (цитотоксических Т-лимфоцитов). Полипептидная молекула, которая подходит для индукции иммунного ответа, также может представлять собой молекулу, вызывающую В-клеточный иммунный ответ.
В конкретных вариантах реализации гетерологичный антиген выбран из группы, включающей антиген бактериального патогена, антиген опухолевой клетки, вирусный антиген, ретровирусный антиген, грибковый антиген или антиген клеточного паразита.
Представляющая интерес молекула может представлять собой, в частности, антиген, выбранный из группы, включающей антиген хламидии, антиген микоплазмы, антиген вируса гепатита, антиген вируса полиомиелита, антиген ВИЧ, антиген вируса гриппа, антиген вируса хориоменингита, опухолевый антиген или часть любого из указанных антигенов, который содержит по меньшей мере один эпитоп.
В предпочтительном варианте реализации производного полипептида согласно настоящему изобретению, аминокислотная последовательность каждой из указанных молекул, подходящей для запуска иммунного ответа, включает в себя или состоит из аминокислотной последовательности антигена хламидии, антигена микоплазмы, антигена вируса гепатита, антигена вируса полиомиелита, антигена ВИЧ, антигена вируса гриппа, антигена вируса хориоменингита, опухолевого антигена или состоит из или включает в себя часть любого из указанных антигенов, которая содержит по меньшей мере один эпитоп.
В особенно предпочтительном варианте реализации представляющая интерес молекула представляет собой антиген, ассоциированный с опухолью (ТАА, tumor associated antigen). Ассоциированные с опухолями антигены характерны для ряда опухолей, таких как, например: меланома, в частности метастатическая меланома, рак легких, рак головы и шеи, рак шейки матки, рак пищевода, рак мочевого пузыря, в частности, инфильтрирующий рак мочевого пузыря, карцинома предстательной железы; рак молочной железы, колоректальный рак, рак почки, саркома, лейкоз, миелома. Для таких различных гистологических типов рака было показано, что антигенные пептиды специфично экспрессируются в опухолевом образце и узнаются Т-клетками, в частности CD8+ Т-клетками или CD4+ Т-клетками.
Обзор пептидов, выявленных в качестве ассоциированных с опухолями антигенов в упомянутых типах опухолей сделан в работе Van der Bruggen P. et al. (Immunological Reviews, 2002, vol 188:51-64). В частности, пептиды, указанные в таблице 3 упомянутого обзора, приведены в настоящей заявке как примеры таких ассоциированных с опухолями антигенов, и указанная таблица 3 включена в настоящую заявку посредством ссылки.
Следующие антигены также приведены в качестве примеров ассоциированных с опухолями антигенов, узнаваемых Т-клетками, согласно публикации Kawakami Y. et al. (Cancer Sci, October 2004, vol.95, no. 10, p784-791), в которой также представлены способы скрининга таких антигенов или одного из указанных: антигены, общие для различных видов рака, в том числе MAGE (в частности, в меланоме), NY-ESO-1, Her2/neu, WT1, сурвивин, hTERT, CEA, AFP, SART3, GnT-V, антигены, специфичные для некоторых конкретных форм рака, таких как β-катенин, CDK4, МАРТ-2, MUM3, gp100, MART-1, тирозиназы для меланомы, BCR-ABL, TEL-AML1 для лейкемии, PSA, PAP, PSM, PSMA для рака предстательной железы; протеиназа 3 для миелоидной лейкемии, MUC-1 для рака молочной железы, яичников или поджелудочной железы; EBV-EBNA, HTLV-1 tax для лимфомы, ATL для рака шейки матки; мутантная форма HLA-A2 для почечноклеточного рак; НА1 для лейкоза/лимфомы. Также описаны опухолевые антигены у животных, такие как циклин D1 и циклин D2 в опухолях, поражающих кошек или собак.
Опухолевые антигены, узнаваемые Т-клетками, также описаны в работе Novellino L. et al. (Immunol Immunother 2004, 54:187-207).
В целом, ассоциированные с опухолями антигены (ТАА), представляющие интерес согласно настоящему изобретению, представляют собой антигены, соответствующие мутантным формам антигенов, или антигены, сверхэкспрессированные на поверхности опухолевых клеток, общие для различных опухолей антигены, тканеспецифичные дифференцированные антигены или вирусные антигены.
В конкретном варианте реализации изобретения указанный антиген, ассоциированный с опухолью, представляет собой антиген вируса папилломы человека (ВПЧ) или представляет собой тирозиназу.
В соответствии с другим конкретным вариантом реализации изобретения аминокислотные последовательности полипептидных молекул, включающих эпитоп или состоящих из эпитопа, модифицированы относительно их природной аминокислотной последовательности, например, с целью уменьшения числа отрицательно заряженных аминокислотных остатков в последовательности. Такая модификация может быть получена путем удаления некоторых из них отрицательно заряженных аминокислотных остатков, или также путем добавления некоторые положительно заряженных аминокислотных остатков, в частности, в виде фланкирующих остатков эпитопов. Полипептиды, которые в результате содержат меньше отрицательно заряженных остатков, могут с большим предпочтением транслоцировать каталитический домен производного полипептида согласно настоящему изобретению в цитозоль клеток-мишеней.
Аминокислотные последовательности полипептидных молекул, которые включают или состоят из эпитопа или антигена также можно сконструировать таким образом, что они "развернуты" при встраивании в производное полипептида согласно настоящему изобретению, что повышает эффективность интернализации представляющей интерес молекулы (молекул) согласно настоящему изобретению клетками-мишенями. Такие "развернутые" полипептиды, которые претерпевают укладку согласно их аминокислотному составу, могут быть получены, например, путем удаления или замены остатков цистеина с целью избежать образования дисульфидных связей, которые могут обуславливать укладку полипептидов. В некоторых случаях можно предотвратить сворачивание (укладку) полипептидов, получая их в присутствии восстановителей, что позволяет избежать рефолдинга in vivo.
В конкретном варианте реализации аминокислотная последовательность, в частности антигена, может состоять из или включать криптические эпитопы.
Авторами настоящего изобретения установлено, что конструкции полипептидных производных, которые включают (i) полипептид согласно настоящему изобретению, представляющий собой мутантный вариант белка СуаА (мутантный полипептид) в соответствии с определениями, принятыми в настоящей заявке, и (ii) полипептидную молекулу, имеющую аминокислотную последовательность, которая содержит один или несколько антигенных фрагментов одного или нескольких антигенов, позволяют криптическим эпитопам указанных антигенов становиться иммуногенными в результате их презентации в таких конструкциях. В частности, указанные конструкции, включающие мутантные полипептиды, как это определено в настоящем изобретении, включают полипептидную молекулу (молекулы), полученную из представляющих интерес антигенов, в частности для профилактических или терапевтических целей, в том числе иммунотерапевтической вакцинации, претерпевают процессинг в клетках-мишенях, при этом полипептидная молекула (молекулы) способна интернализоваться в результате транслокации N-концевого домена мутантного полипептида. Такой процессинг делает возможной презентацию эпитопов посредством молекул ГКГС I класса клеток-мишеней, и указанные эпитопы могут включать криптические эпитопы антигена, что позволяет им становиться иммуногенными и, в частности, вызывать Т-клеточный ответ у хозяина, в частности, ответ ЦТЛ.
Таким образом, настоящее изобретение также относится к полипептидному производному, в частности, рекомбинантному белку, включающему одну или несколько полипептидных молекул, имеющих аминокислотную последовательность, которая содержит или несет один или несколько эпитопов одного или нескольких антигенов, указанная аминокислотную последовательность (последовательности) указанного антигена (антигенов) указанной полипептидной молекулы (молекул), встроены в один и тот же или в различные сайты, в различные пермиссивные сайты мутантного полипептида согласно настоящему изобретению, при этом указанный рекомбинантный белок сохраняет свойство токсина СуаА специфично взаимодействовать с антигенпрезентирующими клетками (АПК), при этом по меньшей мере один из указанных эпитопов представляет собой субдоминантный криптический Т-клеточный эпитоп, и при этом указанное производное полипептида, в частности указанный рекомбинантный белок, способен вызывать антиген-специфичный ответ против указанной полипептидной молекулы (молекул).
В конкретном варианте реализации производного полипептида согласно настоящему изобретению одна или более аминокислотная последовательность встроена в один или более сайт, в частности, в пермиссивный сайт.
В контексте настоящего изобретения "пермиссивный сайт" представляет собой сайт последовательности белка СуаА, в который может быть встроен полипептид без существенного изменения желаемых функциональных свойств белка СуаА, в частности, без существенного изменения способности целенаправленно воздействовать на клетки, в частности, целенаправленно воздействовать на антигенпрезентирующие клетки (АПК), в том числе без существенного влияния на специфичное связывание с рецептором CD11b/CD18 и преимущественно без существенного изменения доменов белка, вовлеченных в процесс транслокации N-концевого домена СуаА в клетки-мишени.
Способы выбора пермиссивного сайта представлены, например, в публикации WO93/21324, в работе Ladant et al., 1992, и в работе Osicka et al., 2000 (Infection and Immunity, 2000, 68(1):247-256). В частности, методология с применением двойной селекции (устойчивость к антибиотику и колориметрический тест на чашках с помощью α-комплементации) позволяет легко определить вставки олигонуклеотидов (сохраняющие рамку считывания) в область гена, кодирующую N-концевой каталитический домен токсина. Функциональные последствия таких мутаций на каталитическую активность токсина можно легко проанализировать, как с помощью генетических подходов (функциональная комплементация штамма Е.coli суа-) и биохимических подходов (характеристика стабильности модифицированной аденилатциклазы, ее ферментативной активности или ее взаимодействия с саМ и т.д.). Такая методология позволила выявить большое количество мутаций с целью выявления участков, которые потенциально предпочтительны для вставки антигенных детерминант.
Пермиссивные сайты аденилатциклазы Bordetella pertussis, которые способны к транслокации каталитического домена СуаА и, следовательно, транслокации аминокислотных последовательностей, включенных в такие пермиссивные сайты включают, но не ограничены перечисленными, остатки 137-138 (Val-Ala), остатки 224-225 (Arg-Ala), остатки 228-229 (Glu-Ala), остатки 235-236 (Arg-Glu), и остатки 317-318 (Ser-Ala) (Sebo et al., 1995). Следующие дополнительные пермиссивные сайты также включены в варианты осуществления изобретения: остатки 107-108 (Gly-His), остатки 132-133 (Met-Ala), остатки 232-233 (Gly-Leu), 335-336 (Gly-Gln) и 336-337. Кроме того, в настоящем изобретении можно применять другие пермиссивные сайты, которые могут быть идентифицированы, например, с помощью методологии, указанной выше, в частности, сайты между остатками 400 и 1700 из белка СуаА.
Для других видов Bordetella соответствующие пермиссивные сайты могут быть определены путем сравнения последовательностей и определения соответствующих остатков.
Согласно другому варианту реализации одна или более аминокислотная последовательность полипептида так или иначе может быть встроена по одному и/или другому концевому участку (концу) полипептида настоящего изобретения, предпочтительно по N-концу мутантного полипептида СуаА, лишенного целиком или частично N-концевого каталитического домена белка СуаА Bordetella pertussis, в частности, лишенного остатков 1-373.
Согласно конкретному варианту реализации одна или несколько аминокислотных последовательностей, способные вызывать иммунный ответ, присоединены к аминокислотному остатку указанного полипептида.
В соответствии с настоящим изобретением "объединение/комбинирование" (или вставка, встраивание) аминокислотной последовательности с мутантным полипептидом СуаА с получением так называемого производного полипептида, также упоминаемого как "рекомбинантный белок" или "гибридный белок", охватывает генетические вставки, в частности, с помощью доступных технологий рекомбинантной ДНК. Кроме того, "комбинирование" также охватывает негенетическое объединение, включающее химическое встраивание, например, ковалентное присоединение, которое осуществляется, в частности, по одному концу аминокислотной последовательности, или нековалентное присоединение. Негенетическое встраивание, в частности, может быть предпочтительно, если встраиваемая аминокислотная последовательность является синтетической или полусинтетической. Способы присоединения лекарственного средства к полипептиду хорошо известны в данной области и включают, например, образование дисульфидной связи с помощью N-пиридилсульфонил-активированного сульфгидрила.
В частности, можно присоединить молекулы к полипептидам настоящего изобретения с помощью химического связывания или генетического встраивания для целенаправленной доставки in vivo в клетки-мишени СуаА, такие как АПК, например CD11b/CD18-положительные клетки, в частности в цитозоль указанных клеток. Действительно, при связывании молекулы, соответствующей данному CD8+ Т-клеточному эпитопу, с каталитическим доменом обезвреженного СуаА, либо с помощью дисульфидных связей или генетического встраивания, было показано, что сконструированная молекула может вызвать in vivo специфичный ответ ЦТЛ, таким образом, демонстрируя, что указанный CD8+ Т-клеточный эпитоп транслоцируется в цитозоль CD11b-экспрессирующих клеток.
В предпочтительном варианте изобретения мутантный полипептид СуаА применяют в производстве белковых векторов или в получении композиции, специально разработанной для примирования ответа CD8+цитотоксических Т-клеток (ЦТЛ), при этом указанный ответ сопровождается направленным воздействием мутантного полипептида СуаА, модифицированного (в частности, рекомбинированного или конъюгированного) с представляющей интерес молекулой, на CD11b-экспрессирующие клетки, с последующей траснлокацией представляющей интерес молекулы в цитозоль указанных CD11b-экспрессирующих клеток, и в частности, миелоидных дендритных клеток. В этом контексте представляющая интерес молекула предпочтительно представляет собой или включает эпитоп или антиген.
В другом предпочтительном варианте реализации настоящего изобретения мутантный полипептид СуаА применяют в производстве белковых векторов или в получении композиции, специально разработанной для примирования ответа CD4+ цитотоксических Т-клеток (ЦТЛ), при этом указанный ответ сопровождается направленным воздействием модифицированной аденилатциклазы (в частности, рекомбинированной или конъюгированной) с представляющей интерес молекулой, на CD11b-экспрессирующие клетки, в частности, миелоидные дендритные клетки. В этом контексте, представляющая интерес молекула предпочтительно представляет собой или включает эпитоп или антиген.
Мутантные полипептиды также можно применять в производстве белковых векторов для адресной доставки соединения профилактического или терапевтического значения в CD11b-экспрессирующие клетки. В этом контексте, в одном конкретном варианте реализации изобретения, так называемая представляющая интерес молекула имеет профилактическое или терапевтическое значение, и в частности, представляет собой лекарственное средство. Указанное соединение профилактического или терапевтического значения, и в частности, указанное лекарственное средство, может быть соединено химическим или генетическим способом с мутантным полипептидом. Способы присоединения соединения к полипептиду хорошо известны в данной области и включают, например, образование дисульфидной связи с помощью N-пиридилсульфонил-активированного сульфгидрила. В одном варианте реализации представляющая интерес молекула представляет собой противовоспалительное соединение, которое, будучи соединенным с мутантным полипептидом, специфичным образом доставляется к поверхности клеток, участвующих в воспалительной реакции, например, дендритных клеток или нейтрофилов.
В частности, презентация антигена для селективной активации CD8+ цитотоксических клеток в основном осуществляется миелоидными дендритными клетками.
Соответственно, в конкретном варианте реализации мутантный полипептид СуаА, применяемый для производства белковых векторов, представляет собой генетически модифицированную аденилатциклазу, содержащую одну или более молекулу (молекулы), химически соединенную посредством дисульфидных связей с генетически встроенным остатком цистеина, расположенным в пределах каталитического домена мутантного полипептида СуаА. Действительно, несколько молекул могут быть химически соединены с мутантным полипептидом СуаА с помощью дисульфидных связей с различными остатками цистеина, расположенными в разных пермиссивных сайтах в пределах каталитического домена.
Мутантные полипептиды или полипептидные производные согласно настоящему изобретению подходят для применения в терапии или с целью профилактики.
Под терапией или терапевтическим эффектом подразумевают любой эффект, который оказывает благотворное действие на состояние пациента, будь то лечебное воздействие или достаточное для ограничения симптомов или последствий патологического состояния, в том числе ограничение прогрессирования патологического состояния. Терапия или терапевтический эффект также охватывает предотвращение развития патологического состояния.
Мутантные полипептиды или полипептидные производные в соответствии с настоящим изобретением, в частности, применимы для запуска клеточного иммунного ответа, такого как Т-клеточный иммунный ответ, или В-клеточный иммунный ответ в организме хозяина, нуждающегося в этом. Такой ответ включает ответ ЦТЛ, в частности ответ Th1, в том числе CD4+ Т-клеточный ответ и/или CD8+ Т-клеточный ответ.
Способность полипептида, полученного на основе белка СуаА, запускать клеточный иммунный ответ может быть достаточной для предотвращения опухолевого роста in vivo или может даже обуславливать регрессию опухоли у животного. Такая способность может быть усилена за счет активации врожденного компонента иммунного ответа посредством активации TLR и подавления регуляторных компонентов иммунного ответа с помощью химиотерапевтических агентов. Настоящее изобретение обеспечивает средства, которые позволят достичь таких результатов в условиях более высокой безопасности в результате выбранных комбинированных мутаций E570Q и K860R.
Настоящее изобретение, таким образом, также относится к терапевтическим способам, включающим введение животному или пациенту-человеку мутантного полипептида или полипептидного производного в соответствии с настоящим изобретением, с целью запуска у пациента Т-клеточного иммунного ответа или В-клеточного иммунного ответа в организме хозяина, нуждающемуся в этом.
Мутантные полипептиды или полипептидные производные в соответствии с настоящим изобретением можно, в частности, применять для профилактики или лечения заболеваний, выбранных из неоплазии, рака и инфекционных заболеваний, выбранных из заболеваний, вызываемых вирусом, ретровирусом, бактерией или x грибком. В частности, полипептидные производные можно применять для лечения ВИЧ-инфекции у пациента.
В частности, предусмотрено, что в конкретном варианте реализации изобретения мутантный полипептид СуаА или производное полипептида, применимое для лечения инфильтрирующих или васкуляризированных опухолей, в отличие от поверхностных опухолей, или для лечения метастатических опухолей, в отличие от первичных опухолей, в соответствии с принятыми клиническими критериями классификации опухолей.
В частности, солидные опухоли представляют собой мишень для лечения с помощью полипептидных производных настоящего изобретения.
К опухолям, которые подходят для лечения с помощью производного полипептида согласно настоящему изобретению, относятся следующие примеры опухолей, для которых охарактеризованы ассоциированные с опухолью антигены:
меланома, в частности метастатическая меланома, рак легких, рак головы и шеи, рак шейки матки, рак пищевода, рак мочевого пузыря, в частности, инфильтрирующий рак мочевого пузыря, карцинома предстательной железы; рак молочной железы, колоректальный рак, рак почки, саркома, лейкоз, миелома. Для таких различных гистологических типов рака было показано, что антигенные пептиды специфично экспрессируются в опухолевом образце и узнаются Т-клетками, в частности CD8+ Т-клетками или CD4+ Т-клетками.
Изобретение также относится к применению производного полипептида согласно настоящему изобретению для получения терапевтической композиции для лечения заболевания, выбранного из неоплазии, рака и инфекционного заболевания, выбранного из заболеваний, вызванных вирусами или ретровирусами.
В предпочтительном варианте полипептид или производное полипептида согласно настоящему изобретению можно вводить пациенту в сочетании с адъювантном и/или в сочетании с другой терапевтически активной молекулой или агентом.
В контексте настоящего изобретения указанная "другая терапевтически активная молекула или агент" представляет собой молекулу или агент, которая (который) может оказать благоприятный эффект на патологическое состояние пациента, которому вводят такую молекулу или агент. Такая молекула или агент представляет собой, в частности, действующий агент, подходящий для применения в производстве лекарственного средства. Такая молекула или агент может представлять собой соединение, подходящее либо для усиления действия или изменения эффекта терапевтически активного действующего агента.
Мутантный полипептид СуаА или производное полипептида можно вводить с терапевтически активной молекулой или агентом, в частности, молекулой или агентом, способным индуцировать иммунный ответ у пациента.
В частности, мутантный полипептид СуаА или его производное можно вводить с терапевтически активным агентом, применимым для модулирования клеточного ответа у пациента, в частности за счет снижения или подавления иммуносупрессорной активности регуляторных Т-клеток.
В соответствии с конкретным вариантом реализации изобретения, такой эффект на ответ регуляторных клеток может быть достигнут с помощью агента, модулирующего супрессорный ответ регуляторной Т-клетки и/или другой клетки, например, как супрессорный ответ миелоидной клетки, при этом указанный агент целенаправленно воздействует на указанные регуляторные клетки, в частности, Т-клетки, за счет элиминации или инактивации таких клеток (например, с помощью CD25-специфичного антитела, или циклофосфамида), изменяя миграцию указанных клеток, в частности, регуляторных Т-клеток (например, с помощью CCL22-специфичных антител) или изменяя дифференцировку и сигналинг указанных клеток (например, путем блокирования сигнала FOXP3 (forkhead box Р3)).
В соответствии с конкретным вариантом реализации изобретения, агент, модулирующий ответ регуляторной клетки, целенаправленно воздействует на супрессорные молекулы, в частности, молекулы, присутствующие на АПК (например, В7-Н1, В7-Н4, IDO (индоламин 2,3-диоксигеназа) или аргиназа) или на Т-клетки (например, CTLA4 (антиген, ассоциированный с цитотоксическими Т-лимфоцитами 4) или PD1 (программированная гибель клеток 1)), или целенаправленно воздействует на растворимые иммуносупрессорные молекулы (такие как TGF бета (трансформирующий фактор роста), IL-10, VEGF (фактор роста сосудов эндотелия), СОХ2 (циклооксигеназа 2)).
В качестве примеров агентов, оказывающих влияние на ответ регуляторных клеток, предложены цитостатические агенты, которые могут убить регуляторные Т-клетки или другие иммуносупрессорные клетки, или которые могут блокировать их активность и/или развитие и/или накопление.
В конкретном варианте реализации изобретения, агент, модулирующий ответ регуляторных клеток, в частности, ответ регуляторных Т-клеток, представляет собой химиотерапевтический агент. В частности, такой агент выбран из химиотерапевтических агентов, известных как противораковые агенты и применяемых в химиотерапии. Такие агенты включают агенты, способствующие уменьшению опухолевой массы, усиливающие чувствительность опухолевых клеток к лечению, или агенты, способные убивать или инактивировать иммунные регуляторные клетки. Химиотерапевтические агенты, применяемые в рамках изобретения, таким образом, усиливают противоопухолевый иммунитет.
В конкретном варианте реализации изобретения химиотерапевтический агент представляет собой алкилирующий агент. В частности, такой агент представляет собой циклофосфамид (СТХ) (Sigma, Steinheim, Germany). Циклофосфамид способен элиминировать или инактивировать регуляторные Т-клетки.
В конкретном варианте реализации изобретения химиотерапевтический агент представляет собой интеркалирующий агент.
В конкретном варианте реализации химиотерапевтический агент представляет собой доксорубицин (DOX) (Calbiochem, La Jolla, CA, USA).
Химиотерапевтический агент преимущественно вводят в низких дозах.
Мутантный полипептид СуаА или производное полипептида можно также вводить совместно с адъювантом, подходящим для активации врожденного иммунного ответа, примированного опухолью у пациента.
В конкретном варианте реализации изобретения адъювант выбран из группы компонентов, включающей нуклеиновые кислоты, пептидогликаны, углеводы, пептиды, цитокины, гормоны и низкомолекулярные соединения, при этом указанный адъювант способен запускать сигнальный каскад через паттерн-распознающие рецепторы (ПРР, англ. pattern-recognition receptors).
Известно, что ПРР опосредуют врожденный иммунный ответ на патогены, а также опухоли, узнавая так называемые эволюционно консервативные паттерны патогенов (патоген-ассоциированные молекулярные паттерны, ПАМП). ПРР присутствуют на поверхности различных иммунных клеток, включая дендритные клетки, натуральные киллеры, В-клетки, а также на некоторых не иммунных клетках, таких как эпителиальные клетки и эндотелиальные клетки. ПРР и их участие во врожденном иммунном ответе описаны в работе Pashine A. et al. (Nature medicine supplement volume 11, N°4, April 2005).
В частности, адъювант для активации врожденного иммунного ответа может направленно воздействовать на ПРР и, таким образом, активировать сигналинг через ПРР, при этом указанные ПРР охватывают Toll-подобные рецепторы или нуклеотид-связывающий домен олигомеризации (NOD) или лектины С-типа.
В конкретном варианте реализации изобретения указанный адъювант представляет собой агонист Toll-подобного рецептора (Toll-like receptor, TLR). Агонист Toll-подобного рецептора, в частности, добавляют в композицию для эффективной активации иммунной системы пациента. Указанный агонист TLR способен связываться с TLR, т.е. представляет собой лиганд TLR, и более того, способен усиливать иммунный ответ, запущенный с участием указанного TLR.
Для примера, агонисты TLR выбраны из группы агонистов TLR-9, TLR-8, TLR-3 и TLR-7. Однако для осуществления изобретения можно применять агонисты других рецепторов TLR, например, агонисты рецепторов TLR2, TLR4, TLR5.
Указанный агонист TLR, применяемый в настоящем изобретении, может представлять собой природный или синтетический агониста. Агонист может представлять собой сочетание различных агонистов одного и того же или различных toll-подобных рецепторов.
В соответствии с конкретным вариантом реализации настоящего изобретения, агонист TLR представляет собой иммуностимулирующую нуклеотидную последовательность, в частности, стабилизированную нуклеотидную последовательность, например, стабилизированную в результате структурной модификации, такой как фосфоротиоатная модификация. Нуклеотидная последовательность также может быть защищена от деградации благодаря конкретному составу композиции. В частности, липосомная композиция, например, липосомная суспензия, может быть предпочтительна для эффективного введения иммуностимулирующей нуклеотидной последовательности.
В конкретном варианте реализации изобретения иммуностимулирующая нуклеотидная последовательность представляет собой одноцепочечную РНК.
В конкретном варианте реализации изобретения иммуностимулирующая нуклеотидная последовательность включает CpG-мотив, и в частности, представляет собой олигонуклеотид CpG (CpG ODN). В качестве примера подходящих олигонуклеотидов CpG изобретение обеспечивает лиганды TLR-9, такие как CpG ODN А-типа, а именно CpG 2216, имеющий нуклеотидную последовательность 5'-GGGGGACGATCGTCGGGGGG-3', или CpG ODN В-типа, а именно CpG 1826, имеющий нуклеотидную последовательность 5'-TCCATGACGTTCCTGACGTT-3'.
Олигонуклеотид CpG можно применять после образования комплекса с DOTAP (Roche Manheim, Germany), с целью его защиты от деградации и для облегчения его усвоения.
В конкретном варианте реализации изобретения агонист TLR представляет собой низкомолекулярное соединение. Низкомолекулярное соединение, применимое в качестве агониста TLR, представляет собой, например, производное имидазохинолинамина, например, известное как R848 (резихимод), т.е. 4-амино-2-этоксиметил-а,а,диметил-1-Н-имидазо[4,5с]хинолин-1-этанол, доступный от Invivogen, например, лиганд TLR-7 или известный как R837 (имихимод), доступный от Aldara как агонист TLR-7.
Другие молекулы, которые можно применять в качестве агонистов TLR, представляют собой полиуридин (ПУ) как лиганд TLR-3, или полицитидиловую кислоту (ПЦК) как лиганд TLR-7.
Такие молекулы могут входить в состав препарата для облегчения его поглощения и/или для защиты его от деградации. Такие молекулы также могут быть получены в виде липосомной композиции, в частности, в виде липосомной суспензии, для введения пациенту.
В соответствии с другим конкретным вариантом реализации изобретения адъювантный компонент может представлять собой адъювантный компонент на основе клетки. Примером такого являются дендритные клетки, которые, как известно, способны примировать ответ лимфоцитов, при это такие дендритные клетки можно кондиционировать ех vivo перед их введением с целью повышения их активности к стимуляции ответа Т-клеток. Дендритные клетки могут, таким образом, могут стимулироваться адъювантами, взаимодействующими с ПРР, в том числе лигандами или агонистами TLR (Pashine A. et al. Nature Medicine Supplement Volume 11, №4, April 2005 p S63-S68).
Кроме того, полипептид или производное полипептида согласно настоящему изобретению можно вводить пациенту без адъюванта.
Действительно, авторы настоящего изобретения ранее показали, что ЦТЛ, специфичный для антигена в векторе, может быть примирован in vivo после однократной внутривенной инъекции рекомбинантного токсина, в частности, без необходимости гетерологичного адъюванта. Такие результаты, а также, в частности, специфичное целенаправленное воздействие эпитопа на миелоидные дендритные клетки, позволяют обходиться без помощи адъюванта и CD4+ Т-клеток.
Таким образом, изобретение также относится к применению мутантного полипептида СуаА, рекомбинированного с молекулой и, в частности, пептидом, представляющего интерес для получения композиции, предназначенной для внутривенного введения, и обуславливающего иммунный ответ CD8+ Т-клеток in vivo, при этом указанная композиция не содержит гетерологичный адъювант. Изобретение также относится к указанной композиции как таковой.
Настоящее изобретение дополнительно направлено на терапевтические способы, включающие введение мутантного полипептида или производного полипептида в соответствии с изобретением животному или человеку, страдающего от заболевания, выбранного из неоплазии, рака и инфекционных заболеваний, выбранных из вирусных, ретровирусных, бактериальных или грибковых заболеваний.
Мутантный полипептид или производное полипептида можно, в частности, вводить с терапевтически активной молекулой и/или адъювантом.
Мутантный полипептид СуаА или его производное полипептида, терапевтически активную молекулу и/или адъювант можно вводить вместе в составе фармацевтической композиции, которая дополнительно включает фармацевтически приемлемый носитель или эксципиент (эксципиенты).
Кроме того, различные типы молекул, описанные в настоящей заявке, применяемые для реализации настоящего изобретения, можно вводить по отдельности либо единовременно (в частности, для мутантного полипептида СуаА или его производного полипептида и адъюванта) либо в разные моменты времени (в частности, для мутантного полипептида СуаА).
Введение терапевтически активной молекулы можно, как альтернатива, осуществлять до и после введения мутантного полипептида СуаА или производного полипептида и/или адъюванта. Введение можно также осуществлять последовательно во времени.
Конкретный режим терапии, который можно применять, включает протокол с повторным введением, в частности, протокол, включающий два или более раунда введения по меньшей мере одного из соединений, выбранных из мутантного полипептида СуаА или его производного полипептида, терапевтически активного агента и/или адъюванта.
Изобретение также относится к фармацевтической композиции, которая включает мутантный полипептид СуаА или его производное полипептида в соответствии с настоящим изобретением, фармацевтически приемлемый носитель или эксципиент (эксципиенты), и, при необходимости, адъювант и/или другую терапевтически активную молекулу.
Изобретение также относится к набору компонентов, включающих мутантный полипептид СуаА или его производное полипептида, терапевтически активную молекулу и/или адъювант.
Соединения из набора компонентов или композицию согласно настоящему изобретению, в частности, можно давать пациенту посредством внутривенного введения, внутриопухолевого введения или подкожного введения.
Набор компонентов или композиция обладает способностью (I) направленно воздействовать на адаптивный иммунный ответ посредством мутантного полипептида СуаА или его производного полипептида, описанного в настоящей заявке, (II) подавлять регуляторный иммунный ответ посредством терапевтически активного агента и адъюванта, (при его наличии), (III) направленно воздействовать на компоненты врожденного иммунного ответа путем активации указанного врожденного ответа посредством адъюванта.
Изобретение также относится к способу лечения пациента, нуждающегося в таком лечении, пациента-человека или животного, включающему стадию введения компонентов набора или композиции, описанных в настоящей заявке.
Изобретение, в частности, также относится к новой иммуногенной композиции, разработанной для введения, в частности, внутривенного введения, в организм животного или человека, отличающейся тем, что она содержит производное рекомбинантного полипептида СуаА, в состав которого входит антиген, встроенный в каталитический домен.
Изобретение также относится к фармацевтической композиции для введения человеку или животному, предназначенной для целенаправленной доставки представляющей интерес молекулы в клетки, экспрессирующие CD11b, отличающейся тем, что указанная представляющая интерес молекула соединена с мутантным полипептидом СуаА, как описано в настоящей заявке.
В другом конкретном варианте реализации фармацевтическая или иммуногенная композиция включает конструкцию нуклеиновой кислоты, кодирующей рекомбинантное производное полипептида СуаА, включающее мутантный полипептид СуаА, как определено в настоящей заявке, соединенный с представляющей интерес молекулой.
Кроме того, изобретение также относится к применению иммуногенной композиции, как определено в настоящей заявке, для получения вакцины или иммунотерапевтической композиции для введения в организм животного или человека.
Применяемый в настоящей заявке термин "иммунотерапевтическая композиция" относится к композиции, которая приводит к иммунологической реакции и которая ассоциирована с терапевтическим лечением, таким как лечение неоплазии, рака, вирусных инфекций, грибковых инфекций, паразитических инфекции или бактериальных инфекций.
Изобретение, кроме того, относится к способу иммунизации организма животного или человека, где указанный способ включает следующие стадии:
a) получение иммуногенной композиции, как определено выше;
b) введение указанной иммуногенной композиции, предпочтительно посредством внутривенного введения, указанному организму-хозяину с целью индукции иммунного ответа.
В частности, иммуногенные композиции согласно настоящему изобретению способны индуцировать или стимулировать in vivo или in vitro иммунный ответ, в частности, с участием дендритных клеток. Иммуногенные композиции согласно изобретению можно, в частности, применять для профилактики или терапевтической вакцинации пациента.
Как следствие, в конкретном варианте реализации иммуногенная или фармацевтическая композиция, преимущественно, не содержит примирующих адъювантов, обычно применяемых в данной области, например, гидроксида алюминия.
Изобретение относится к способу получения белкового вектора, подходящего для доставки представляющей интерес молекулы в клетку, включающему связывание представляющей интерес молекулы с мутантным полипептидом СуаА, как определено в настоящей заявке.
Изобретение также относится к молекулам нуклеиновых кислот, в частности, ДНК или РНК, которые кодируют полипептид или производное полипептида, как определено выше.
Изобретение также относится к эукариотическим или прокариотическим клеткам, которые включают молекулы нуклеиновой кислоты, как указано выше.
Изобретение также относится к эукариотическим клеткам, предпочтительно клеткам млекопитающих, которые включают мутантный полипептид СуаА или его производное полипептида, как определено выше. В предпочтительном варианте реализации клетки представляют собой клетки человека.
Изобретение также относится к эукариотическим клеткам, предпочтительно клеткам млекопитающих, трансформированным с помощью белкового вектора, как определено выше.
Фигуры
Фиг.1. Замены в порообразующем домене и домене ацилирования взаимно усиливают действие друг друга в снижении специфичной гемолитической активности СуаА. (А) Эритроциты барана (5×108/мл) в буфере TNC инкубировали с 5 µг/мл ферментативно активными белками СуаА при температуре 37°С. Через 30 мин аликвоты клеточной суспензии промывали несколько раз для удаления несвязавшихся молекул СуаА и использовали для определения количества клеточно-ассоциированной и клеточно-инвазивной активности АЦ. Гемолитическую активность измеряли через 5 часов инкубации по количеству высвобождаемого гемоглобина с помощью фотометрического анализа (A54i). Активность интактной СуаА принимали за 100%. (В) Эритроциты инкубировали, как указано выше, с указанными концентрациями ферментативно активных белков на основе СуаА, в течение 30 мин, промывали, и определяли количество клеточно-ассоциированной активности фермента АЦ. (С) Снижение способности связываться с клетками для белков с заменой K860R было компенсировано за счет увеличения их концентрации от 5 мкг/мл до 25 мкг/мл. Активность CyaA/233OVA (CyaA/OVA) принимали за 100%. Результаты представляют собой средние значения по меньшей мере из трех независимых экспериментов, которые проводили в двух повторностях. Звездочками обозначены статистически значимые различия (**, р<0,001), по сравнению с активностью СуаА (фиг.1А) или CyaA/OVA (фиг.1C).
Фиг.2. CyaA/OVA/E570Q+K860R связывается с моноцитами CD11b+ и транслоцируется в них. (А) Клетки J774A.1 (106/мл) инкубировали в D-МЕМ в течение 30 мин при 4°C с 2,5 мкг/мл СуаА, промывали несколько раз, и определяли количество клеточно-ассоциированной активности АЦ в клеточных лизатах. Для блокирования рецептора CD11b/CD18 клетки инкубировали в течение 30 мин с CD11b-специфичными антителами М1/70 (Exbio, Чехия) в конечной концентрации 10 µ г/мл до добавления СуаА (**, p<0,001). (В) Оценивали транслокационную способность домена АЦ конструкций как способность проникать в клетки и преобразовывать цитозольный АТФ в цАМФ. Клетки J774A.1 инкубировали с конструкциями СуаА в течение 30 минут при температуре 37°С, и определяли количество цАМФ, накопленного в клеточных лизатах (41). В качестве контроля рецепторы CD11b/CD18 блокировали с помощью антитела к CD11b M1/70, как указано выше. Проникновение через мембрану CD11b/CD18-связанного и претерпевшего эндоцитоз токсина проверяли с помощью варианта с двойной мутацией СуаА/E570K+Е581Р, который не изменен в отношении связывания с рецептором, но не способен перемещать АЦ-домен через клеточную мембрану и повышать концентрацию цАМФ в цитозоле (Vojtova-Vodolanova et al., 2009). (С) Клетки J774A.1 инкубировали с K+-чувствительным флуоресцентным зондом PBFI/AM при конечной концентрации 9,5 µ М при 25°С в течение 45 мин в присутствии Pluronic F-127 [0,05% (в./в.)]. Клетки промывали в HBSS перед добавлением 3 µ г мл-1 указанного токсина. Соотношение интенсивности флуоресценции PBFI (длина волны возбуждения 340 нм, длина волны излучения 450 и 510 нм), отражающее концентрацию внутриклеточного K+, регистрировали каждые 15 сек. На правой шкале показаны значения концентрации внутриклеточного [K+], полученные в калибровочных экспериментах (см. Процедуры эксперимента). За время исследования не наблюдали лизиса клеток, оцениваемого по высвобождению лактатдегидрогеназы (LDH). Представлены результаты, типичные для трех независимых экспериментов, которые проводили в двух повторностях.
Фиг.3. Анатоксин E570Q+K860R не пермеабилизирует клетки J774A.1. (А) Лизис клеток J774A.1 определяли по количеству лактатдегидрогеназы (LDH), высвобожденной из 105 клеток при инкубации в течение 3 ч с 3, 10 и 30 мкг мл-1 указанного белка в 37% в DMEM без сыворотки. Результаты представляют собой средние значения, полученные в двух независимых экспериментах, которые проводили в двух повторностях. (В) Пэтч-кламп анализ всей клетки проводили при комнатной температуре на отдельных клетках J774A.1, обработанных 1 или 10 мкг/мл белков CyaA/233OVA/AC- или CyaA/233OVA/E570Q+K860R/AC-, как описано в Материалах Методах. Показаны кривые, типичные для шести измерений в 3 независимых экспериментах.
Фиг.4. Анатоксин с заменами E570Q+K860R обеспечивает доставку Т-клеточного эпитопа OVA для презентации молекулами ГКГС I класса и индукции CD8+ ЦТЛ. (A) BMDC (3×105 клеток/лунку), используемые в качестве АПК, инкубировали в присутствии указанных концентраций (от 0 до 60 нМ) анатоксинов, несущих эпитоп OVA или с имитатором СуаА/АС-. После кокультивации в течение 24 часов с клетками B3Z Т (1×105 клеток на лунку) оценивали секрецию IL-2 стимулированными клетками B3Z при помощи анализа пролиферации CTLL. Результаты представлены как число Δ имп/мин включившегося [3H]-тимидина (имп/мин в присутствии анатоксина - имп/мин в отсутствие анатоксина) ± СО, и являются типичными для пяти независимых экспериментов. (В) Анализ индукции OVA (SIINFEKL)-специфичного ответа ЦТЛ с помощью анализа киллинга in vivo. На день 0 мыши в.в. получали 50 мкг имитатора АС- или анатоксина OVA/AC-, а на 7 день им вводили в.в. смесь (1:1) меченных OVA (SIINFEKL) CFSEвысок и немеченных CFSEнизк спленоцитов. Число CFSE-положительных клеток, оставшихся в селезенке после 20 ч, определяли с помощью флуоресцентной сортировки клеток (FACS-анализ), как представлено для одного типичного анализа киллинга in vivo на верхней панели графиков, на которых указан процент учтенных клеток. На нижней панели собраны результаты по анализу киллинга in vivo, полученные в трех независимых экспериментах. Статистическую значимость определяли по t-критерию Стьюдента (р=0,75 для OVA/AC- в сравнении с OVA/E570Q+K860R/AC-).
Фиг.5. Модель действия СуаА на мембрану. (А) модель предсказывает существование равновесия между двумя конформерами СуаА в растворе, при этом каждый из них встраивается в клеточную мембрану в различной конформации. На выходе ожидается мономерный транслоцируемый предшественник СуаА, доставку домена АЦ в цитозоль и сопутствующий приток ионов кальция в клетки. Конформер, как ожидается, встраивается в виде порообразующего предшественника, олигомеризующегося с образованием поры СуаА. (В) ожидается, что синергетический эффект замен E570Q и K860R выборочно блокирует способность порообразующего предшественника СуаА олигомеризоваться с образованием поры, тогда как способность транслоцируемых предшественников к переносу домена АЦ через мембрану будет оставаться неизменной.
Фиг.6. Аминокислотная последовательность токсина СуаА от Bordetella pertussis (SEQ ID №1)
Фиг.7. Аминокислотная последовательность мутантного варианта CyaA/E570Q+K860R от Bordetella pertussis (SEQ ID №2)
Фиг.8. Аминокислотная последовательность мутантного варианта CyaA/E570Q+K860R/AC- от Bordetella pertussis (SEQ ID №3)
Фиг.9. Аминокислотная последовательность мутантного варианта CyaA/233OVA/E570Q+K860R/AC- от Bordetella pertussis (SEQ ID №4)
Фиг.10. Плазмида, кодирующая мутантный вариант CyaA/E570Q+K860R/AC- (QR-AC-).
Фиг.11. Последовательность ДНК плазмиды QR-AC-, кодирующей мутантный вариант CyaA/E570Q+K860R/AC- (SEQ ID №5)
Фиг.12. Плазмида, кодирующая мутантный вариант CyaA/233OVA/E570Q+K860R/AC- (OVA-QR-AC-).
Фиг.13. Последовательность ДНК плазмиды OVA-QR-AC-, кодирующей мутантный вариант CyaA/233OVA/E570Q+K860R/AC- (SEQ ID №6)
Фиг.14. Аминокислотная последовательность токсина СуаА от Bordetella parapertussis (номер доступа САВ76450, SEQ ID №7)
Фиг.15. Аминокислотная последовательность токсина СуаА от Bordetella hinzii (номер доступа AAY57201, SEQ ID №8)
Фиг.16. Аминокислотная последовательность токсина СуаА от Bordetella bronchiseptica (номер доступа САА85481, SEQ ID №9)
Примеры
Токсин аденилатциклаза транслоцируется через мембрану клетки-мишени без формирования пор
Материалы и методы
Конструирование, получение и очистка белков СуаА.
Модификации для получения конструкций CyaA/AC-, CyaA/233OVA, CyaA/E570Q и CyaA/K860R, описаны ранее (13, 20, 21), и их ввели в CyaA/233OVA/AC- по отдельности или в комбинации. Белки-производные СуаА получали в Е.coli XL-1 Blue и очищали до состояния близкого к однородности, как описано выше (29). При проведении гидрофобной хроматографии смолу со связанным токсином многократно промывали 60% изопропанолом (30) с целью уменьшения содержания эндотоксинов в образцах СуаА до уровня 100 МЕ/мг белка, которое определяли с помощью ЛАМ-анализа (лизат амебоцитов мечехвоста) и QCL-1000 (Cambrex).
Штамм Escherichia coli XLI-Blue (Stratagene), содержащий плазмиду QR-AC- (Фигура 10), которая кодирует мутантный CyaA/E570Q+K860R/AC-, был депонирован на хранение 18 марта 2009 года в CNCM (Collection Nationale de Cultures de Microorganismes, Франция) под номером доступа CNCM 1-4136 (Фигура 10). Последовательность ДНК плазмиды QR-AC- (SEQ ID №5) показана на Фигуре 11.
Штамм Escherichia coli XLI-Blue (Stratagene), содержащий плазмиду OVA-QR-AC- (Фигура 12), которая кодирует мутантный СуаА CyaA/233OVA/E570Q+K860R/AC-, был депонирован на хранение 18 марта 2009 года в CNCM (Collection Nationale de Cultures de Microorganismes, Франция) под номером доступа CNCM I-4137. ДНК плазмиды OVA-QR-AC- (SEQ ID №6) показана на Фигуре 13.
Связывание с клетками и гемолитическая активность на модели эритроцитов барана. Отмытые эритроциты барана в количестве 5×108 клеток в растворе, содержащем 50 мМ Трис с рН 7,4, 150 мМ NaCl и 2 мМ CaCl2 (TNC-буфер), инкубировали при 37°С в присутствии 5 мкг/мл белков СуаА и затем определяли их активность связывания с клетками, клеточно-инвазивную активность АЦ и гемолитическую активность способами, детально описанными ранее (13). Достоверность различий значений активности оценивали с использованием одностороннего дисперсионного анализа (ANOVA) с применение критерия Бонферрони (SigmaStat v. 3.11, Systat, San Jose, CA).
Способность СуаА связываться с макрофагами, увеличивать уровень цАМФ и вызывать лизис клеток. Мышиные моноциты/макрофаги J774.AI (номер в АТСС - TIB-67) культивировали при 37°С в атмосфере увлажненной смеси воздух/CO2 (19:1) в среде RPMI с добавлением 10% (об/об) инактивированной путем нагревания эмбриональной бычьей сыворотки, пенициллина (100 МЕ/мл), стрептомицина (100 мкг/мл) и амфотерицина В (250 нг/мл). Перед проведением анализа RPMI заменяли на среду Игла в модификации Дульбекко (D-MEM) (1,9 мМ Са2+) без добавления ФТС и клетки оставляли в D-MEM в течение 1 ч при 37°С в увлажненной атмосфере с 5% CO2 (8). Макрофаги J774A.1 (106) инкубировали в D-MEM, содержащей варианты СуаА в концентрации 2,5 мкг/мл, в течение 30 мин при 4°С, после чего удаляли несвязавшийся токсин путем трехкратной промывки клеток в D-MEM. Для определения активности АЦ, связавшейся с клетками, клетки лизировали в 0,1% Тритон Х-100. Для анализа внутриклеточного образования цАМФ 105 клеток инкубировали с СуаА в течение 30 минут в среде D-MEM, содержащей 100 мкМ ИБМК (3-изобутил-1-метилксантин), реакцию останавливали путем добавления 0,2% Tween-20 в 100 мМ HCl, образцы кипятили в течение 15 минут при 100°С, смесь нейтрализовали путем добавления 150 мМ имидазола в отсутствии буфера, количество цАМФ измеряли, как описано в литературе (29). С целью блокирования рецепторов CD11b/CD18 перед добавлением СуаА клетки предварительно инкубировали в течение 30 минут на льду с блокирующими антителами MAb M1/70 (Exbio, Чехия), специфичными к CD11b, при их конечной концентрации 10 мкг/мл. Лизис клеток J774A.1, индуцированный токсином, оценивали с помощью набора CytoTox 96 (Promega) по количеству высвобожденной лактатдегидрогеназы из 105 клеток за 3 часа инкубации, при концентрации соответствующего белка 10 мкг/мл, при 37°С, в D-MEM, как описано в литературе (8). Достоверность различий значений активности анализировали, как описано выше.
Анализ методом локальной фиксации потенциала ("пэтч-кламп").
Пэтч-кламп анализ целых клеток осуществляли на клетках линии J774A.1, находящихся в среде HBSS (140 мМ NaCl, 5 мМ KCl, 2 мМ CaCl2, 3 мМ MgCl2, 10 мМ HEPES-Na, 50 мМ глюкозы, рН 7,4).Оплавленные на огне стеклянные микропипетки с внешним диаметром около 3 мкм заполняли раствором, содержащим 125 мМ калия глюконата, 15 мМ KCl, 0,5 мМ CaCl2, 1 мМ MgCl2, 5 мМ EGTA, 10 мМ HEPES-KOH рН 7,2. Результирующее сопротивление микроэлектродов составляло от 3 до 5 МОм. Разность потенциалов оценивали при токе -40 мВ, регистрируемый сигнал фильтровали при частоте 1 кГц и оцифровывали при частоте 2 кГц при помощи усилителя Axopatch 200A, цифрового преобразователя DIGIDATA 1320A и пакета программного обеспечения PClamp-9 (Axon Instruments, Foster City, CA).
Определение снижения концентрации ионов K+ в цитозоле. Клетки, выращенные на покровных стеклах, промывали средой HBSS и погружали в 9,5 мкМ раствор эфира PBFI и ацетоксиметила (PBFI/AM, Molecular Probes, Eugene, OR, USA) на 30 минут при 25°C с 0,05% (в/в) Pluronic F-127 (Sigma, St.Louis, МО) в темноте. Логометрические измерения проводили при 25°C с использованием спектрофлуориметра FluoroMax-3 (Jobin Yvon Horriba, Франция) и прилагаемого программного обеспечения Datamax. Интенсивность флуоресценции PBFI (длина волны возбуждения 340, длины волн излучения 450 и 510 нм) регистрировали в течение 15 секунд, с временем интеграции для каждой длины волны 3 секунды. Отношение длины волн 450/510 нм показано на рисунке. Исследованная область покровного стекла, размещенного в 1 см кювете, составила приблизительно 10 мм2, что соответствует количеству примерно 104клеток. В калибровочных экспериментах использовали клетки, которые предварительно пермеабилизовали валиномицином или нигерицином в течение 30 минут, с использованием 50 мМ раствора HEPES, рН 7, содержащего различные концентрации ацетата калия (10, 30, 60 или 140 мМ) и ацетата натрия (135, 115, 85 или 5 мМ), соответственно. Конечное отношение интенсивности (450/510 нм) показано на правой вертикальной оси диаграммы.
Мыши и клеточные линии. Самок мышей линии C57BL/6, полученных из Charles River Laboratories, содержали в специальных условиях в отсутствии патогенных микроорганизмов и ухаживали за ними в соответствии с правилами, принятыми в институте. Клетки линии CTLL-2 получены из АТСС. Клетки B3Z, CD8+-специфичная Т-клеточная гибридома, Kb-рестриктированный эпитоп OVA (SIINFEKL) получены от Н. Шастри (University of California, Berkeley), и их поддерживали в полной питательной среде RPMI 1640 (Invitrogen Life Technologies), содержащей 10% ФТС, инактивированной нагреванием, 100 ед/мл пенициллина, 100 мкг/мл стрептомицина, и 5-кратный 10-5 М раствор 2-МЕ, в присутствии 1 мг/мл G418 и 400 мкг/мл гигромицина В.
Исследование презентации антигена. Дендритные клетки из костного мозга (ДККМ, 3×105 клеток на лунку), используемые в качестве АПК (антигенпрезентирующих клеток), инкубировали в присутствии различных концентраций рекомбинантных CyaA/OVA/AC- (от 0 до 60 нМ), несущих эпитоп OVA (SIINFEKL) или его имитацию, и культивировали в течение 24 часов совместно с Т-клетками B3Z (1×105 на лунку), селективно распознающими комплексы белков ГКГС I класса с OVA SIINFEKL/H-2Kb. После 18 ч культивирования супернатанты замораживали и выдерживали в течение по меньшей мере 2 ч при -80°С. Затем определяли количество IL-2, продуцируемого стимулированными клетками B3Z, на основании пролиферации CTLL. Кратко: клетки IL-2-зависимой клеточной линии CTLL в количестве 104 на лунку культивировали с добавлением 100 мкл супернатанта до конечного объема 200 мкл. Спустя двадцать четыре часа к клеткам добавляли [3Н]-тимидин (50 мкКи/лунка) и спустя 6 часов клетки собирали с использованием автоматизированного сборщика клеток. Включенный [3Н]-тимидин оценивали путем сцинтилляционного измерения. Каждый вариант оценивали дважды, а эксперимент повторяли пять раз. Результаты представлены как Δимп/мин для включившегося [3H]-тимидина (имп/мин в присутствии анатоксина - имп/мин в отсутствие анатоксина).
Анализ киллинга in vivo. Для примирования ЦТЛ мышей иммунизировали путем внутривенного введения 50 мкг рекомбинантного CyaA/OVA/AC-, несущего эпитоп OVA (SIINFEKL) или его имитацию СуаА/АС-. Через семь дней после иммунизации нативные сингенные спленоциты, сенсибилизировали с помощью пептида OVA (SIINFEKL) (10 мкг/мл, 30 минут при 37°С), тщательно промывали и проводили мечение, обрабатывая карбоксифлуоресцеина сукцинимидил эфиром (CFSE; Molecular Probes, Eugene, OR) в высокой концентрации (1,25 мкМ). Популяцию не сенсибилизированных контрольных клеток метили раствором CFSE низкой концентрации (0,125 мкМ). Клетки, меченные растворами высокой (CFSEвысок) и низкой (CFSEнизк) концентрации, смешивали в соотношении 1:1 (5×106клеток каждой популяции) и вводили внутривенно мышам. Клетки селезенки собирали спустя 20 часов, промывали и ресуспендировали в FACS-буфере (PBS с добавлением 1% БСА и 0,1% NaN3). Число CFSE-положительных клеток, оставшихся в селезенке после 20 часов, определяли с помощью флуоресцентной сортировки клеток (FACS). Процент специфичного лизиса рассчитывали следующим образом: процент специфичного лизиса=100-[100×(% CFSEвысок иммунизированные мыши/% CFSEнизк иммунизированные мыши)/(% CFSEвысок контрольные мыши/% CFSEнизк контрольные мыши)].
Статистический анализ: Достоверность различий значений оценивали с использованием одностороннего дисперсионного анализа (ANOVA) с применение критерия Бонферрони (SigmaStat v. 3.11, Systat, San Jose, CA).
Результаты
Совместное удаление отрицательно заряженного глутамата 570 и ацилированного лизина 860 снижает клеточно-пермеабилизирующую активность СуаА. Рабочая модель действия СуаА предполагает, что СуаА может быть модифицирован таким образом, что порообразующая (гемолитическая) активность будет потеряна при сохранении способности транслоцировать домен АЦ в цитозоль клеток-мишеней. Для проверки данной гипотезы авторы изобретения попытались получить конструкции СуаА, демонстрирующие наиболее низкую из возможных гемолитическую и цитолитическую активности, опираясь на предыдущие наблюдения, которые показывают, что способность анатоксинов СуаА/АС- лизировать клетки может быть изменена, как в сторону увеличения, так и в сторону уменьшения, путем введения замен в порообразующий домен (8, 12-14, 18). С целью сделать возможным оценку проницаемости клеток-мишеней, в том числе и для анатоксинов СуаА/АС-, авторы изобретения получали такие мутанты на основе токсина CyaA/233OVA, которые ранее были маркированы путем вставки пептида SIINFEKL из овальбумина (OVA). Данный вариант СуаА был выбран ввиду того, что вставка репортера - Kb-рестриктированного эпитопа CD8+ Т-клеток по остатку 233 не влияет на активность АЦ и позволяет количественно оценить транслокацию фермента OVA/AC в клетки по увеличению концентрации цитозольного цАМФ. Что еще более важно, наличие эпитопа OVA позволяет также оценить способность ферментативно неактивных анатоксинов CyaA/233OVA/AC- доставлять домен OVA/AC- в цитозоль CD11b+ антиген презентирующих клеток (АПК), поскольку это делает возможным протеасомный процессинг и обеспечивает презентацию на поверхности клеток эпитопа OVA с гликопротеинами ГКГС I класса, которые можно детектировать с помощью OVA-специфичных CD8+ Т-клеток, как in vitro, так и in vivo (20).
Для создания анатоксинов СуаА/АС-, по возможности не обладающих цитолитической активностью, авторы изобретения совместили замены E570Q и K860R, при наличии которых ранее было показано снижение специфической гемолитической активности СуаА на эритроцитах барана, кроме того, для замены E570Q также была показано уменьшение цитолитической активности СуаА/АС- на модели CD11b+ моноцитов J774A.1 (8, 13). Данные замены вводили в CyaA/233OVA/AC по отдельности и совместно, и специфичную гемолитическую и цитолитическую активности полученных анатоксинов сравнивали параллельно с помощью эритроцитов барана как модельных CD11b- мишеней, так и с помощью клеток линии J774A.1 как модельных CD11b+ мишеней (Таблица I). В соответствии с ранее полученными результатами, при изучении анатоксинов без включенного эпитопа OVA (4, 8, 13, 21) в условиях данного эксперимента анатоксины OVA/AC-, несущие единичные замены E570Q или K860R, демонстрировали двукратное (55±8) и нулевое (1±1), соответственно, снижение гемолитической активности на эритроцитах, и относительная цитолитическая активность анатоксина с заменой E570Q по отношению к CD11b-экспрессирующим клеткам J774A.1 была также снижена (37±10), по сравнению с OVA/AC-. В свою очередь, как и ожидалось при анализе ферментативно активной конструкции с заменой K860R, несмотря на низкую гемолитическую активность на CD11b- эритроцитах, анатоксин с заменой K860R показал лишь незначительное снижение относительной цитолитической активности на CD11b+ клетках J774A.1 (72±22%), подтверждая тем самым, что структурный дефект, вызванный заменой K860R, был восстановлен при взаимодействии с рецепторами CD11b/CD18 (4). Тем не менее, в сочетании с заменой E570Q, замена K860R демонстрировала явный синергетический эффект, выраженный в снижении относительной цитолитической активности конструкции с заменами E570Q и K860R по отношению к клеткам J774A.1 до 14±7%.
Для возможности количественной оценки способности конструкции с заменами E570Q и K860R траснлоцировать домен АЦ в цитозоль клетки в ферментативно активную конструкцию, полученную на основе CyaA/233OVA (CyaA/OVA), вводили замены E570Q и K860R. Данные конструкции получали и очищали так же, как это делали для анатоксинов АС- (данные не представлены), и анализировали их связывание с клетками, их гемолитическую активность и их способность к транслокации АЦ на модели эритроцитов барана. Как показано на Фигуре 1А и как ожидалось по результатам исследования с токсинами без эпитопа OVA (4, 13, 21), замена E570Q никак не повлияла на связывание эритроцитов или способность CyaA/OVA транслоцировать домен АЦ в цитозоль эритроцитов, и лишь селективно снизила его относительную гемолитической активность. Также как и ожидалось (4), замена K860R значительно снизила способность CyaA/OVA связываться с и проникать внутрь эритроцитов, что приводило к резкому снижению относительной гемолитической и клеточно-инвазивной активностей АЦ мутантов с заменами E570Q и E570Q+K860R по отношению к эритроцитам.
Следует отметить, что гемолитическая активность СуаА в высокой степени зависит от количества СуаА, ассоциированной с клетками (коэффициент Хилла >3), что позволяет предположить, что необходимым условием для образования пор является олигомеризация СуаА (22).
Таким образом, при оценке воздействия совместных замен E570Q и K860R на гемолитическую активность, потеря способности конструкции с заменой K860R связываться с эритроцитами должна быть компенсирована за счет увеличения ее концентрации до 25 мкг/мл (по сравнению без воздействия токсина 5 мкг/мл) для достижения эквивалентного количества всех белков, связывающихся с эритроцитами, как показано на Фигуре 1C.
В этих условиях совместное присутствие замен E570Q и K860R демонстрирует явный синергетический эффект, выражающийся в сокращении в два раза уже нарушенной гемолитической активности конструкций, несущей только замену E570Q (~50%) или замену K860R (~30%), как показано на фигуре 2С. Это свидетельствует о влиянии комбинации этих двух замен на клеточно-пермеабилизирующую способность СуаА.
Порообразующая активность СуаА не является необходимой для транслокации через мембрану домена АЦ. В отличие от влияния замены K860R на токсичность по отношению к эритроцитам, ранее было обнаружено, что замены E570Q и K860R не влияют на способность СуаА связываться и проникать внутрь моноцитов J774A.1, экспрессирующих рецептор CD11b/CD18 (4, 8). Более того, как показано на Фигуре 2, при комбинировании двух замен в одной молекуле токсина конструкция CyaA/OVA/E570Q+K860R демонстрировала эквивалентную способность связываться с клетками J774A.1 (Фигура 2А) и доставлять домен АЦ в цитозоль с увеличением концентрации цитозольного цАМФ (Фигура 2В), что и исходный СуаА. Однако в то же время, анатоксин с двойной заменой E570Q+K860R демонстрировал сниженную в 7 раз (14±7%) относительную цитолитическую активность по отношению к клеткам (см. Таблицу I).
Как показано на Фигуре 2С, по сравнению с исходным СуаА, у конструкции с одиночной мутацией E570Q и с двойной мутацией E570Q+K860R была значительно нарушена их способность вызывать снижение внутриклеточной концентрации ионов калия ([K+]ц) при обработке клеток J774A.1 токсином. Хотя не было выявлено лизиса клеток в течение более 20 минут при проведении теста на высвобождение лактатдегидрогеназы, [K+]ц в клетках J774A.1, обработанных исходным СуаА в концентрации 3 мкг/мл, уменьшилась от 140 мМ до менее, чем 30 мМ уже через 10 минут после добавления токсина. В свою очередь, при использовании конструкции с заменами ER570Q и K860R (3 мкг/мл), уровень [K+]ц в клетках не опускался ниже 100 мМ (Фигура 2С). Действительно, ранее было показано, что отток калия из клеток является отличительной чертой внедрения СуаА - перового предшественника в клеточную мембране (32). Это позволило предположить, что сочетание замен E570Q и K860R селективно нарушает только способность анатоксина пермеабилизовать клетки J774A.1, но не его способность транслоцировать домен АЦ через клеточную мембрану.
Данное заключение также подтверждается фактом значительного снижения относительной цитолитической активности соответствующих анатоксинов с заменой E570Q/AC и заменами E570Q и K860R/AC, что показано выше (Таблица I) и показано на Фигуре 3А. Анатоксин с двойной мутацией E570Q+K860R при 3 мкг/мл по сути был не в состоянии вызвать заметного высвобождения лактатдегидрогеназы из клеток J774A.1 за 3 часа инкубации, в то время как 20% клеток подвергалось лизису в присутствии эквивалентного количества исходного анатоксина.
Для проверки данного факта авторы изобретения проанализировали клеточно-пермеабилизирующую способность конструкции с заменами E570Q и K860R в исследованиях методом пэтч-кламп всей клетки. Для этого было необходимо использовать АС- анатоксины с целью избежать массового раффлинга клеток J774A., провоцируемого цАМФ, возникающего под действием токсина (23). Как показано на Фигуре 3А, согласно репрезентативной записи ионных токов через мембрану единичной клетки J774A.1 (пэтч-кламп анализ), подвергшейся воздействию CyaA/OVA/AC- в концентрации 1 мкг/мл, при инициальном лаг-периоде в 3 минуты, клетки J774A.1 были постепенно и массово пермеабилизованы под воздействием CyaA/OVA/AC-, и ток через клеточные мембраны достигал - 3000 пА в течение 10 минут. В отличие от этого, как показано на Фигуре 3 В, воздействие CyaA/OVA/E570Q+K860R/AC- воспроизводимо вызывало только кратковременную и минимальную пермеабилизацию клеток на начальном периоде, с токами через клеточные мембраны не превышающем -200 пА и возвращающемся к близкому к нулю значению в течение 10 минут после добавления анатоксина. Совершенно аналогичная картина наблюдалась при увеличении концентрации анатоксина до 10 мкг/мл, концентрации, которую использовали при сравнении относительной цитолитической активности анатоксинов, что приведено в Таблице I. 10-кратное увеличение концентрации от 1 до 10 мкг/мл привело к двукратному увеличение токов через клеточную мембрану при воздействии OVA/AC-, в то время как практически не происходило усиление клеточной пермеабилизации даже при повышенной концентрации OVA/E570Q+K860R/AC- (следует обратить внимание на то, что для отображения данных при использовании концентрации 10 мкг/мл, y-ось была увеличена). Представленные записи являются репрезентативными по меньшей мере для шести измерений трех независимых экспериментов, и результаты показывают, что комбинация замен E570Q и K860R оказала значительное влияние на способность анатоксина пермеабилизовать мембраны клеток J774A.1 клеток. Учитывая, что ферментативно активный вариант той же самой конструкции является способным транслоцировать домен АЦ в клетки J774A.1 (см. Фигуру 2В), эти результаты убедительно свидетельствуют, что клеточно-пермеабилизирующая (порообразующая) активность СуаА не является необходимой для транслокации домена АЦ через клеточную мембрану. Мембранно-пермеабилизующая активность СуаА не является необходимой для доставки переносимых антигенов в цитозоль по пути ГКГС I класса. Поскольку определение цитозольного цАМФ нельзя использовать для оценки способности АС- анатоксинов проникать в клетки, для оценки их способности доставлять репортерный эпитоп OVA к сайту процессинга презентации антигена ГКГС I класса использовали альтернативный подход (7, 24). С этой целью авторы изобретения определяли способность дендритных клеток из костного мозга мышей C57BL/6 (ДККМ) стимулировать высвобождение IL-2 Т-клетками B3Z, которые селективно распознают комплекс Kb ГКГС I класса с пептидом SIINFEKL (OVA) на АПК. Действительно, ранее было показано, что способность СуаА/АС-анатоксинов к транслокации домена АЦ через цитоплазматическую мембрану в цитозоль ДККМ играет важную роль в способности анатоксинов стимулировать презентацию доставленного эпитопа OVA в комплексе с молекулами Н-2Kb ГКГС I класса. Это, в свою очередь, важно для специфичной стимуляции цитотоксических Т-клеток in vivo (29). Тем не менее, было важно подтвердить, что доставка эпитопа OVA для протеасомного процессинга и последующей презентации связана с транслокацией домена АЦ через цитоплазматическую мембрану в цитозоль ДККМ, а не с поступлением добавленного антигена в клетку в ходе пассивного транспорта или эндоцитоза. С этой целью использовали не способный к переносу, содержащий двойную замену вариант анатоксина OVA/E570K+E581P/АС-, который дополнительно содержал меняющие заряд и нарушающие структуры спирали замены аминокислоты глутамата в трансмембранном домене в положениях 570 и 581 (33). Ранее было обнаружено, что эта конструкция демонстрирует полную способность связываться с клетками, экспрессирующими CD11b/CD18 (см. Фигуру 2А), в то время как ее способность транслоцировать домен АЦ через мембрану клеток-мишеней полностью нарушена за счет комбинации замен E570K и Е581Р (см. Фигура 2В).
Как показано на Фигуре 4А, гибридомные клетки B3Z были эффективно стимулированы при совместной инкубации с ДККМ с добавлением анатоксинов OVAE570Q/AC- и OVA/E570Q+K860R/AC-. С другой стороны, стимуляцию клеток B3Z не наблюдали при совместной инкубации их с ДККМ с добавлением анатоксина OVA/E570K+E581P/АС-, который дефектен по транслокации домена АЦ через клеточную мембрану. Более того, анатоксины OVA/E570Q/AC- и OVA/E570Q+K860R/AC- индуцировали стимуляцию лимфоцитов B3Z in vitro в присутствии АПК, так же эффективно, как и исходный анатоксин OVA/AC-. Эти результаты подтверждают, что двойной мутант с заменами E570Q и K860R полностью способен транслоцировать домен АЦ в цитозоль ДККМ для процессинга и презентации эпитопа OVA с молекулами Kb ГКГС I класса, будучи по существу не способным пермеабилизовать клетки J774A.1. Эти результаты позволяют предположить, что клеточно-пермеабилизирующая (порообразующая) активность СуаА не является необходимой для транслокации домена АЦ через клеточную мембрану и не играет никакой роли в способности СуаА доставлять переносимый эпитоп в цитозоль АПК.
Для подтвердждения наблюдаемой способности нецитолитических анатоксинов осуществлять доставку антигена in vitro, авторы изобретения оценили их способность примировать OVA-специфичные цитотоксические CD8+ Т-лимфоциты (ЦТЛ) in vivo. 50 мкг различных OVA-анатоксинов внутривенно вводили мышам линии C57BL/6 и через неделю оценивали OVA-специфичный ответ ЦТЛ иммунизированных животных in vivo путем анализа киллинга. Мыши линии C57BL/6 получали внутривенную инъекцию смеси (1:1) меченных OVA (SIINFEKL) CFSEвысок и немеченных CFSEнизк спленоцитов, и затем, через день, проводили анализ FACS CFSE-меченых клеток. Как показано на Фигуре 4В, иммунизация мышей имитатором анатоксина не индуцировала какую-либо SIINFEKL-специфичную активность ЦТЛ in vivo. В свою очередь, иммунизация анатоксином с заменами E570Q и K860R индуцировала такой же OVA-специфичный ответ киллинга ЦТЛ in vivo, как и при применении исходного анатоксина, который использовали как положительный контроль, с небольшой разницей в средних значениях оценки ответа между исходным анатоксином и анатоксином с двойной мутацией, которая не была статистически достоверной (р=0,065).
Эти результаты показывают, что клеточно-пермеабилизирующая активность СуаА не является необходимой для способности анатоксинов CyaA/233OVA/AC- доставлять встроенный в АЦ переносимый антиген в цитозоль АПК in vivo.
Обсуждение
Авторами настоящего изобретения показано, что перенос домена АЦ СуаА через мембрану миелоидных клеток-мишеней, которые экспрессируют рецепторы CD11b/CD18, не зависит от способности токсина формировать поры и пермеабилизовать клеточную мембрану.
Как резюмировано в модели, предложенной на Фигуре 5, авторы настоящего изобретения ранее сообщали о том, что баланс между двумя активностями СуаА может быть сдвинут за счет мутаций или альтернативного ацилирования СуаА. Действительно, усиление порообразующей (гемолитической) активности за счет способности транслоцировать АЦ внутрь клеток наблюдалось при замене на лизин глутаматов 509, 516 и 581 (13, 18) или при блокировании транслокации АЦ с помощью моноклональных антител (МАт) 3D1 (25). В свою очередь, сдвиг в противоположном направлении наблюдался у рекомбинанта r-Ес-СуаА, ацилированного в E.coli остатками пальмитолеила (С16:1), в отличие от нативного пальмитоилированного (С16:0) Вр-СуаА, продуцируемого B. pertussis. Было обнаружено, что r-Ес-СуаА демонстрирует гемолитическую активность, уменьшенную в четыре раза и примерно в десять раз уменьшенную порообразующую активность в планарных липидных бислоях по сравнению с Вр-СуаА (12); в то же время обе формы СуаА проявляли одинаковую активность при проникновении через клеточную мембрану и транслокации домена АЦ в эритроциты (17, 26). Более того, недавно было показано, что конструкция CyaA/E570Q демонстрирует полную способность доставлять АЦ и в эритроциты, и в макрофаги J774A.1, проявляя в то же время уменьшенную гемолитическую активность и сниженную способность формировать специфические поры в планарных липидных бислоях, по сравнению с интактным СуаА, при этом анатоксин CyaA/E570Q/AC- демонстрировал уменьшенную в два раза цитолитическую активность на клетках J774A.1 (8, 13).
Несмотря на вышесказанное и на большое количество мутантов, которые охарактеризовали авторы настоящего изобретения, остается нерешенным вопрос о том, требуется ли формирование поры СуаА в мембране для переноса домена АЦ через мембрану клеток, экспрессирующих CD11b. Стоит упомянуть, что, если основываться на предыдущих сравнениях гемолитической способности у интактного r-Ес-СуаА с таковой у нативного Вр-СуаА, выделенного из В. pertussis, специфичная гемолитическая активность описанного в настоящей заявке анатоксина r-Ес-CyaA/OVA/E570Q+K860R/AC- по отношению к эритроцитам барана должна быть уменьшена примерно на три порядка по величине. Остаточная специфичная цитолитическая активность r-Ec-CyaA/OVA/E570Q+K860R/AC- на клетках, экспрессирующих CD11b, по оценкам будет примерно в пятьдесят раз меньше, чем таковая у Bp-СуаА, в то время как специфическая способность обоих белков доставлять домен АС в эти клетки будет одинаковой (если принять интактные r-Ес-СуаА как стандарт 100% инвазивной активности АС для сравнения). Описанный в настоящей заявке мутант CyaA/233OVA/E570Q+K860R является первой конструкцией со значительно уменьшенной способностью проникать через клетки, которая остается полностью способной транслоцировать домен АЦ через клеточную мембрану. Это показывает, что на своем пути в цитозоль переносящийся домен АС может обходить катион-селективную пору, формируемую СуаА.
Способ и путь транслокации домена АЦ через клеточную мембрану, однако, все еще необходимо изучить более детально. Если принять во внимание различающиеся эффекты замен глутаматов 509, 516, 570 и 581 на порообразующую активность и активность доставки АС у СуаА (8, 13, 18), при которых баланс между двумя активностями может быть изменен на практически обратный в обоих направлениях за счет конкретных замен, амфипатические спирали, содержащие указанные глутаматные остатки, оказываются вовлеченными в обе активности СуаА альтернативным образом. В пользу этого говорит эффект комбинированной замены E509K+E516K, результатом которой является гипергемолитический СуаА, неспособный доставлять домен АЦ в клетки (8, 18), в то время как результатом описанной в настоящей заявке комбинации замен E570Q+K860R является, напротив, в высшей степени нецитолитический СуаА, который полностью подходит для транслокации домена АЦ в клетки J774A.1 (CD11b+).
Более того, эти наблюдения подтверждают предложенную модель, согласно которой две мембранные активности СуаА зависят от двух различных конформеров, встраивающихся в мембрану, из которых один обеспечивает транслокацию домена АЦ мономерами токсина, а другой обеспечивает формирование олигомерных пор (13, 18). Было предложено, что трансмембранные сегменты, содержащие ключевые остатки глутамата 509, 516, 570 и 581, могут участвовать в формировании олигомерной катион-селективной поры только в том случае, если у встроенного в мембрану конформера-предшественника поры домен АЦ локализован вне клетки. В транслоцируемом АЦ конформере те же самые трансмембранные сегменты принимают другую конформацию в мембране, будучи, вероятно, выдвинутыми, и не имея возможности вводить олигомеры СуаА с помощью полипептида, связывающего С-конец домена АЦ с трансмембранными сегментами. Подтверждение такой интерпретации может быть выведено из результатов, полученных Греем и соавторами (25). Эти авторы показали, что делеция домена АЦ вместе с сегментом, связывающим его с пороформирующим доменом (до остатка 489), или связывание с антителами 3D1, которые блокируют перенос через мембрану терминального сегмента домена АЦ между остатками 373 и 399, значительно усиливают порообразующую (гемолитическую) активность токсина. Вероятно, это происходит из-за принятия трансмембранными сегментами СуаА конформации, которая способствует формированию олигомерных пор. Нужно все еще определить, какие сегменты СуаА снаружи от порообразующего домена вовлечены в перенос домена АЦ через мембрану. Большой домен повторов RTX (остатки с 1006 по 1706), вероятно, принимает участие в переносе АС в клетки, если принять во внимание необходимость его структурной целостности (27). Он должен быть достаточного размера (700 остатков) для того, чтобы формировать гидрофильную поверхность переноса внутри клеточной мембраны, которая может позволить пройти несвернутому домену АС через мембрану без сопутствующего формирования настоящей поры, пермеабилизирующей клетку. В качестве альтернативы, СуаА может способствовать формированию обращенных неламеллярных (обращенная гексагональная фаза) липидных структур (28), которые, вероятно, могут входить в состав изолированной белково-липидной границы, через которую домен АС может проскользнуть в цитозоль.
Действительно, было предварительно показано, что СуаА способствует формированию обращенных неламеллярных (обращенная гексагональная фаза) липидных структур (28). Они, вероятно, принимают участие в формировании изолированной белково-липидной границы, делая возможным, таким образом, транслокацию домена АЦ через мембрану без пермеабилизации клеток. Для формирования неламеллярных липидных структур благоприятными являются обогащенные холестеролом липидные рафты, и, действительно, было недавно показано, что СуаА переносится в рафты в комплексе со своим рецептором CD11b/CD18. Более того, авторы настоящего изобретения недавно показали, что перенос домена АС через мембрану завершается только после переноса СуаА в рафты (L., Bumba, J., Masin, R., Fiser, и Р., Sebo, принято к печати). Интересно, что было ранее обнаружено, что перенос каталитической субъединицы дифтерийного токсина (ДТ) через клеточную цитоплазматическую мембран происходит без выявляемой пермеабилизации клетки, когда ДТ проникал в клетки за счет низкого рН после связывания с укороченным рецептором ДТ с якорем GPI(34). Авторы не исследовали, локализован ли рецептор ДТ с якорем GPI в липидных рафтах, но это весьма вероятно. Это, таким образом, является попыткой предположить, что специфический липидный состав мембраны в рафте может способствовать переносу различных белковых токсинов в клетки-мишени, не требуя при этом формирования настоящей пермеабилизующей клетки поры, которая пропускает белки.
Последнее по порядку, но не по значению практическое открытие, описанное в настоящей заявке, заключается в том, что анатоксин CyaA/E570Q+K860R/AC- с сильно уменьшенной пермеабилизирующей клетки (цитолитической) активностью, сохраняет полную активность транслокации антигена в АПК CD11b+. Это имеет большое значение в свете его потенциального применения как профильного инструмента с усиленной безопасностью для доставки опухоль-специфичных антигенов во втором поколении вакцин-производных СуаА/АС- для иммунотерапии рака.
Источники информации
1. Vojtova J, Kamanova J, Sebo P (2006) Bordetella adenylate cyclase toxin: a swift saboteur of host defense. Curr Opin Microbiol 9:69-75.
2. Glaser P, Sakamoto H, Bellalou J, Ullmann A, Danchin A (1988) Secretion of cyclolysin, the calmodulin-sensitive adenylate cyclase-haemolysin bifunctional protein of Bordetella pertussis. Embo J 7:3997-4004.
3. Rose Т, Sebo P, Bellalou J, Ladant D (1995) Interaction of calcium with Bordetella pertussis adenylate cyclase toxin. Characterization of multiple calcium-binding sites and calcium-induced conformational changes. J Biol Chem 270:26370-26376.
4. Masin J, et al. (2005) Acylation of lysine 860 allows tight binding and cytotoxicity of Bordetella adenylate cyclase on CD11b-expressing cells. Biochemistry 44:12759-12766.
5. Guermonprez P, et al. (2001) The adenylate cyclase toxin of Bordetella pertussis binds to target cells via the alpha(M)beta(2) integrin (CD11b/CD18). J Fxp Med 193:1035-1044.
6. Gordon VM, Leppla SH, Hewlett EL (1988) Inhibitors of receptor-mediated endocytosis block the entry of Bacillus anthracis adenylate cyclase toxin but not that of Bordetella pertussis adenylate cyclase toxin. Infect Immun 56:1066-1069.
7. Schlecht G, Loucka J, Najar H, Sebo P, Leclerc С (2004) Antigen targeting to CD11b allows efficient presentation of CD4+ and CD8+ Т cell epitopes and in vivo Th1-polarized Т cell priming. J Immunol 173:6089-6097.
8. Basler M, Masin J, Osicka R, Sebo P (2006) Pore-forming and enzymatic activities of Bordetella pertussis adenylate cyclase toxin synergize in promoting lysis of monocytes. Infect Immun 74:2207-2214.
9. Khelef N, Zychlinsky A, Guiso N (1993) Bordetella pertussis induces apoptosis in macrophages: role of adenylate cyclase-hemolysin. Infect Immun 61:4064-4071.
10. Morova J, Osicka R, Masin J, Sebo P (2008) RTX cytotoxins recognize {beta}2 integrin receptors through N-linked oligosaccharides. Proc Natl Acad Sci USA.
11. Paccani SR, et al. (2008) Suppression of T-lymphocyte activation and chemotaxis by the adenylate cyclase toxin of Bordetella pertussis. Infect Immun 76:2822-2832.
12. Benz R, Maier E, Ladant D, Ullmann A, Sebo P (1994) Adenylate cyclase toxin (CyaA) of Bordetella pertussis. Evidence for the formation of small ion-permeable channels and comparison with HlyA of Escherichia coli. J Biol Chem 269:27231-27239.
13. Basler M, et al. (2007) Segments crucial for membrane translocation and pore-forming activity of Bordetella adenylate cyclase toxin. J Biol Chem 282:12419-12429.
14. Hewlett EL, Donate GM, Gray MC (2006) Macrophage cytotoxicity produced by adenylate cyclase toxin from Bordetella pertussis: more than just making cyclic AMP! Mol Microbiol 59:447-459.
15. Fayolle С, Sebo P, Ladant D, Ullmann A, Leclerc С (1996) In vivo induction of CTL responses by recombinant adenylate cyclase of Bordetella pertussis carrying viral CD8+ Т cell epitopes. J Immunol 156:4697-4706.
16. Rogel A, Hanski E (1992) Distinct steps in the penetration of adenylate cyclase toxin of Bordetella pertussis into sheep erythrocytes. Translocation of the toxin across the membrane. J Biol Chem 267:22599-22605.
17. Havlicek V, et al. (2001) Mass spectrometric analysis of recombinant adenylate cyclase toxin from Bordetella pertussis strain 18323/pHSP9. J Mass Spectrom 36:384-391.
18. Osickova A, Osicka R, Maier E, Benz R, Sebo P (1999) An amphipathic alpha-helix including glutamates 509 and 516 is crucial for membrane translocation of adenylate cyclase toxin and modulates formation and cation selectivity of its membrane channels. J Biol Chem 274:37644-37650.
19. Fiser R, et al. (2007) Third activity of Bordetella adenylate cyclase (AC) toxin-hemolysin. Membrane translocation of AC domain polypeptide promotes calcium influx into CD11b+ monocytes independently of the catalytic and hemolytic activities. J Biol Chem 282:2808-2820.
20. Osicka R, et al. (2000) Delivery of CD8(+) T-cell epitopes into major histocompatibility complex class I antigen presentation pathway by Bordetella pertussis adenylate cyclase: delineation of cell invasive structures and permissive insertion sites. Infect Immun 68:247-256.
21. Basar Т, et al. (1999) The conserved lysine 860 in the additional fatty-acylation site of Bordetella pertussis adenylate cyclase is crucial for toxin function independently of its acylation status. J Biol Chem 274:10777-10783.
22. Szabo G, Gray MC, Hewlett EL (1994) Adenylate cyclase toxin from Bordetella pertussis produces ion conductance across artificial lipid bilayers in a calcium- and polarity-dependent manner. J Biol Chem 269:22496-22499.
23. Kamanova J, et al. (2008) Adenylate cyclase toxin subverts phagocyte function by RhoA inhibition and unproductive ruffling. J Immunol 181:5587-5597.
24. Guermonprez P, Ladant D, Karimova G, Ullmann A, Leclerc С (1999) Direct delivery of the Bordetella pertussis adenylate cyclase toxin to the MHC class I antigen presentation pathway. J Immunol 162:1910-1916.
25. Gray MC, et al. (2001) Translocation-specific conformation of adenylate cyclase toxin from Bordetella pertussis inhibits toxin-mediated hemolysis. J Bacteriol 183:5904-5910.
26. Hackett M, et al. (1995) Hemolytic, but not cell-invasive activity, of adenylate cyclase toxin is selectively affected by differential fatty-acylation in Escherichia coli. J Biol Chem 270:20250-20253.
27. Iwaki M, Ullmann A, Sebo P (1995) Identification by in vitro complementation of regions required for cell-invasive activity of Bordetella pertussis adenylate cyclase toxin. Mol Microbiol 17:1015-1024.
28. Martin С, et al. (2004) Membrane restructuring by Bordetella pertussis adenylate cyclase toxin, a member of the RTX toxin family. J Bacteriol 186:3760-3765.
29. Karimova G, Pidoux J, Ullmann A, Ladant D (1998) A bacterial two-hybrid system based on a reconstituted signal transduction pathway. Proc Natl Acad Sci USA 95:5752-5756.
30. Franken KL, er al. (2000) Purification of his-tagged proteins by immobilized chelate affinity chromatography: the benefits from the use of organic solvent. Protein Expr Purif 18:95-99.
31. Bellalou J, Sakamoto H, Ladant D, Geoffroy C, Ullmann A (1990) Deletions affecting hemolytic and toxin activities of Bordetella pertussis adenylate cyclase. Infect Immun 58:3242-3247.
32. Gray M.C., et al. (1998) Distinct mechanisms for K+ efflux, intoxication, and hemolysis by Bordetella pertussis AC toxin. J Biol CHem 273:18260-18267
33. Vojvova-Vodolanova J., et al. (2009) Oligomerization is involved in pore formation by Bordetella pertussis adenylate cyclase toxin. FASEB J 23:2331-2343
34. Lanzrein M., et al. (1996) GPI anchored diphteria toxin receptor allows membrane translocation of the toxin without detectable ion channel activity. EMBO J. 15:725-734
Реферат
Группа изобретений относится к области биотехнологии. Предложен мутантный полипептид, содержащий или представляющий собой фрагмент аденилатциклазы, соответствующей SEQ ID NO:1, 7, 8 или 9. Также предложен рекомбинантный полипептид, представляющий собой вышеуказанный полипептид, который дополнительно объединен с одной или более представляющей интерес молекулой, способной запускать иммунный ответ. Предложена фармацевтическая композиция, содержащая один из вышеуказанных полипептидов. Группа изобретений позволяет вызывать Т-клеточный и/или В-клеточный иммунный ответ в организме хозяина для предотвращения или лечения заболевания, выбранного из неоплазии, рака и инфекционных заболеваний, вызываемых вирусом, ретровирусом, бактерией, паразитом или грибком. 9 н. и 12 з.п. ф-лы, 16 ил., 1 табл., 1 пр.
Формула
(1) фрагмента из по меньшей мере 350 остатков аминокислот аденилатциклазы, соответствующей SEQ ID NO: 1, 7 или 8, в которой: (а) остаток глутаминовой кислоты, соответствующий положению 570 в последовательности нативной СуаА из Bordetella pertussis, Bordetella parapertussis или Bordetella hinzii, определенной в SEQ ID NO: 1, 7 или 8 соответственно, заменен остатком глутамина, или консервативным остатком, выбранным из группы, которая состоит из: Asn, Met, Thr, Ser, Gly, Arg, Lys, Val, Leu, Cys, Ile и Asp, (b) остаток лизина, соответствующий положению 860 в последовательности нативной СуаА из Bordetella pertussis, Bordetella parapertussis или Bordetella hinzii, определенной в SEQ ID NO: 1, 7 или 8 соответственно, заменен остатком аргинина, или консервативным остатком, выбранным из группы, которая состоит из: Asn, Gln, Met, Thr, Ser, Gly, Val, Leu, Cys и Ile, и (с) осуществлено от 0 до 50 замен, делеций и/или вставок, и
(2) фрагмента из по меньшей мере 350 остатков аминокислот последовательности аденилатциклазы, соответствующей SEQ ID NO: 9, в которой (а) остаток глутаминовой кислоты, соответствующий положению 569 в последовательности нативной СуаА из Bordetella bronchiseptica, определенной в SEQ ID NO: 9, заменен остатком глутамина, или консервативным остатком, выбранным из группы, которая состоит из: Asn, Met, Thr, Ser, Gly, Arg, Lys, Val, Leu, Cys, Ile и Asp, (b) остаток лизина, соответствующий положению 859 в последовательности нативной СуаА из Bordetella bronchiseptica, определенной в SEQ ID NO: 9, заменен остатком аргинина, или консервативным остатком, выбранным из группы, которая состоит из: Asn, Gln, Met, Thr, Ser, Gly, Val, Leu, Cys и Ile, и (с) осуществлено от 0 до 50 замен, делеций и/или вставок;
при этом указанный полипептид обладает способностью белка СуаА из Bordetella pertussis, Bordetella parapertussis, Bordetella hinzii или Bordetella bronchiseptica связывать рецептор CD11b/CD18 клеток, которые его экспрессируют, и обладает способностью транслоцировать свой N-концевой домен фермента аденилатциклазы или его часть в указанные клетки, и порообразующая способность указанного пептида уменьшена или подавлена по сравнению с нативным токсином СуаА.
(i) остаток глутаминовой кислоты, соответствующий положению 570 в последовательности SEQ ID NO: 1, 7 или 8 или соответствующий положению 569 в последовательности SEQ ID NO: 9, заменен остатком глутамина; и
(ii) остаток лизина, соответствующий положению 860 в последовательности SEQ ID NO: 1, 7, или 8 или соответствующий положению 859 в последовательности SEQ ID NO: 9 заменен остатком аргинина.
(1) последовательности SEQ ID NO: 1, 7 или 8, в которой осуществили следующие мутации: (а) замену остатка глутаминовой кислоты, соответствующего положению 570 в последовательности нативной СуаА из Bordetella pertussis, Bordetella parapertussis или Bordetella hinzii, определенной в SEQ ID NO: 1, 7 или 8 соответственно, остатком глутамина, или консервативным остатком, выбранным из группы, которая состоит из: Asn, Met, Thr, Ser, Gly, Arg, Lys, Val, Leu, Cys, Ile и Asp, и (b) замену остатка лизина, соответствующего положению 860 в последовательности нативной СуаА из Bordetella pertussis, Bordetella parapertussis или Bordetella hinzii, определенной в SEQ ID NO: 1, 7 или 8 соответственно, остатком аргинина, или консервативным остатком, выбранным из группы, которая состоит из: Asn, Gln, Met, Thr, Ser, Gly, Val, Leu, Cys и Ile; или
(2) последовательности, определенной в SEQ ID NO: 9, в которой осуществили следующие мутации: (а) замену остатка глутаминовой кислоты, соответствующего положению 569 в последовательности нативной СуаА из Bordetella bronchiseptica, определенной в SEQ ID NO: 9, остатком глутамина или консервативным остатком, выбранным из группы, которая состоит из: Asn, Met, Thr, Ser, Gly, Arg, Lys, Val, Leu, Cys, Ile и Asp, и (b) замену остатка лизина, соответствующего положению 859 в последовательности нативной СуаА из Bordetella bronchiseptica, определенной в SEQ ID NO: 9, остатком аргинина или консервативным остатком, выбранным из группы, которая состоит из: Asn, Gln, Met, Thr, Ser, Gly, Val, Leu, Cys и Ile.
(1) последовательности SEQ ID NO: 1, 7 или 8, в которой осуществили следующие мутации: (а) замену остатка глутаминовой кислоты, соответствующего положению 570 в последовательности нативной СуаА из Bordetella pertussis, Bordetella parapertussis или Bordetella hinzii, определенной в SEQ ID NO: 1, 7 или 8 соответственно, остатком глутамина, и (b) замену остатка лизина, соответствующего положению 860 в последовательности нативной СуаА из Bordetella pertussis, Bordetella parapertussis или Bordetella hinzii, определенной в SEQ ID NO: 1, 7 или 8 соответственно, остатком аргинина; или
(2) последовательности SEQ ID NO: 9, в которой осуществили следующие мутации: (а) замену остатка глутаминовой кислоты, соответствующего положению 569 в последовательности нативной СуаА из Bordetella bronchiseptica, определенной в SEQ ID NO: 9, остатком глутамина, и (b) замену остатка лизина, соответствующего положению 859 в последовательности нативной СуаА из Bordetella bronchiseptica, определенной в SEQ ID NO: 9, остатком аргинина.
Комментарии