Аденовирусный/альфа-вирусный гибридный вектор для эффективного введения и экспрессии терапевтических генов в опухолевые клетки - RU2394104C2
Код документа: RU2394104C2
Чертежи














Описание
Область техники, к которой относится изобретение
Настоящее изобретение относится к векторам для экспрессии гена, полученным из аденовирусов, для получения терапевтических продуктов.
Уровень техники
Одной из основных проблем, с которыми сталкивается обычная противоопухолевая терапия, является отсутствие специфичности к опухолям, ситуация, которая часто приводит к проявлению тяжелых побочных эффектов и ограничивает применяемую терапевтическую дозу. Несмотря на то, что генная терапия остается многообещающей для лечения злокачественных опухолей, для нее также характерна проблема специфичности: целенаправленная трансгенная экспрессия в области опухоли. На основании результатов ряда исследований можно предположить, что когда вирусные векторы вводят в область поражения, несмотря на то, что трансгенная экспрессия в основном ограничена областью, смежной с ходом иглы, такая экспрессия также может иметь место в других тканях. Следовательно, важной целью генной терапии по отношению к злокачественным опухолям является направление экспрессии терапевтического гена к опухолям посредством специфического введения в ткань новообразований («тканевая цель») и/или специфической активации («транскрипционная цель») в ткани новообразований без влияния на здоровые клетки. «Тканевой цели» можно достичь созданием направленного вектора посредством модификаций взаимодействия рецептор-лиганд, что позволяет инфицировать клетки, которые экспрессируют специфический рецептор. «Транскрипционной цели» можно достичь при использовании опухолеспецифического промотора для контроля трансгенной экспрессии. В предшествующих исследованиях использовались различные опухолеспецифические промоторы. Однако таковые представляют серьезное ограничение в том плане, что они не приводят к экспрессии на высоком уровне, в результате чего противоопухолевая активность ограничивается.
Обзор самых последних достижений в разработке вирусных векторов для генной терапии можно найти у Lundstrom K. «Latest development in viral vectors for gene therapy»; Trends in Biotechnology, 2003, 21, 118-122.
Используемые в настоящее время вирусные векторы включают альфавирусы. Альфавирусы представляют вирусы с оболочкой, которые содержат простую «плюс-цепь» РНК в качестве генома. Были сконструированы и разработаны экспрессирующие векторы, полученные из альфавирусов, вируса Синдбай (SIN), вируса леса Семлики (SFV) и вируса венесуэльского энцефалита лошадей (VEE). Альфавирусные векторы основаны на применении самореплицирующихся молекул РНК, полученной из геномов альфавирусов, в которой сохраняются 5'- и 3'-последовательности, необходимые для репликации, и ген репликазы (Rep), в то время как гены, кодирующие вирусные структурные белки, подвергаются делеции и замещаются трансгеном. После трансфекции клеток данными векторами Rep транслируется, и РНК-вектор копируется в «минус-цепь» РНК, которая будет использоваться в качестве матрицы для амплификации РНК-вектора. Rep также может узнавать субгеномный промотор в «минус-цепи» РНК, из которой синтезируется меньший субгеномный сегмент РНК, который, в свою очередь, может транслироваться с продукцией гетерологичных белков на высоком уровне. Альфавирусные векторы можно непосредственно использовать в качестве РНК при транскрипции в условиях in vitro от прокариотического промотора, такого как SP6 или Е7, или в качестве ДНК, когда последовательность репликона расположена ниже эукариотического промотора, такого как CMV. РНК-вектор может быть упакован в вирусные частицы посредством котрансфекции в клетки вместе с одной или более «хелперной» РНК, которая кодирует вирусные структурные белки. Альфавирусные векторы обладают рядом свойств, которые делают их привлекательными для генной терапии: очень высокий тропизм, низкая иммуногенность и экспрессия гетерологичных белков на высоком уровне. Однако такая экспрессия является временной за счет индукции апоптоза клеток, в которых происходит репликация. В статье Rayner J.O., Dryga S.A., Kamrud K.I. «Alphavirus vectors and vaccination»; Rev. Med. Virol. 2002; 12, 279-296, описывается разработка экспрессирующих векторов на основе альфавирусов для применения в области вакцин.
Другие группы вирусных векторов основаны на аденовирусах. Имеется большое количество литературы по применению аденовирусов, которые были разработаны для преодоления некоторых недостатков генной терапии, и в качестве источника для создания экспрессирующих векторов. Статья о самых последних достижениях в области аденовирусных векторов опубликована Volpers C., Kochanek S. «Adenoviral vectors for gene transfer and therapy»; J. Gene. Med. 2004; 6: S164-S171. Аденовирусы обладают преимуществом достижения высокой эффективности трансдукции и способностью сохраняться в эписомальной форме. Однако экспрессия аденовирусных белков вызывает сильные иммунные ответные реакции, которые ограничивают продолжительность трансгенной экспрессии и вызывают токсичность в клетках, инфицированных вектором. Для решения данных проблем были получены аденовирусы «с усеченным геномом» («gutless adenoviruses»). Данные аденовирусы «с усеченным геномом» лишены всех соответствующих аденовирусных генов (сохраняются только последовательности, представляющие собой последовательности двух инвертированных концевых повторов и сигналы для упаковки), следовательно, трансдуцированные клетки не экспрессируют аденовирусного продукта и не индуцируют иммунного ответа на вектор. Кратко, элиминация всех аденовирусных генов оставляет достаточное пространство для размещения крупных экспрессирующих кассет и по этой причине аденовирусы «с усеченным геномом» также называют аденовирусными векторами с высокой емкостью. Опубликована статья, описывающая конкретные аспекты аденовирусных векторов, относящаяся к делеции всех последовательностей, кодирующих вирусные белки, Morsy M.A. et al. «An adenoviral vector deleted for all viral coding sequences results in enhanced safety and extended expression of a leptin transgene»; Proc. Natl. Acad. Sci. USA 1998, 95: 7866-7871. В статье, опубликованной Schiedner G. et al., «Variables affecting in vivo performance of high-capacity adenovirus vectors»; J. Virol. 2002, 76: 1600-1609, описывается применение «лишней» ДНК в экспрессирующих векторах на основе аденовирусов «с усеченным геномом», показывая, что наличие такой «лишней» ДНК важно для достижения значительного усиления экспрессии гена и что, в основном, конструирование векторов на основе аденовируса («с усеченным геномом») с высокой емкостью может существенно модифицировать степень и продолжительность экспрессии гена.
С другой стороны, в патенте США № 5981225 описывается вектор для переноса гена на основе аденовирусов, который включает последовательности инвертированного концевого повтора (ITR), по меньшей мере, одну последовательность сигнала для упаковки и аденовирусный ген VAI и/или аденовирусный ген VAII; и включает ген, являющийся чужеродным для аденовируса, функционально связанный с функциональным промотором в клетках-мишенях для аденовируса.
В патенте США № 5985846 описывается вектор для переноса гена, включающий последовательности инвертированного концевого повтора (ITR) аденовируса и рекомбинантные частицы аденовируса, содержащие данные последовательности.
В патенте США № 6566093 описываются векторы кДНК, полученные из альфавирусов, которые состоят из ДНК, комплементарной, по меньшей мере, в отношении части РНК альфавируса, необходимой для репликации альфавируса, и гетерологичную кДНК, например кДНК, кодирующую желаемое соединение. Последнее может представлять собой биологически активный белок или полипептид, а также иммуногенный или антигенный белок или полипептид, терапевтически активный белок или полипептид, или терапевтически активную РНК.
Целью настоящего изобретения является повышение трансгенной экспрессии и индукции апоптоза опухолевых клеток, опосредуемое гибридными векторами в условиях in vitro и in vivo. Дополнительной целью является повышение эффективности терапии опухолей на моделях на животных с помощью гибридных векторов.
Дополнительной целью также является разработка способа генной терапии, в частности, для лечения злокачественных опухолей с помощью применения гибридных векторов.
Цели настоящего изобретения достигаются объединением следующего в одном векторе:
- высокой инфицирующей способности путем использования системы высвобождения аденовируса,
- высокой трансгенной экспрессии и индукции апоптоза путем использования вектора, полученного из альфавируса, такого как SFV, и
- специфичности к опухолям путем использования опухолеспецифического промотора.
Сущность изобретения
Настоящее изобретение, во-первых, относится к аденовирусному гибридному вектору для экспрессии гена, отличающемуся тем, что он содержит, по меньшей мере, следующие элементы, ориентированные в направлении от 5' к 3':
i) первую цепь из аденовируса, содержащую последовательность первого инвертированного концевого повтора (ITR) и сигнальную последовательность для упаковки аденовируса;
ii) первую некодирующую «лишнюю» последовательность;
iii) последовательность, соответствующую тканеспецифическому промотору;
iv) цепь кДНК, полученную из альфавируса, последовательность которой частично комплементарна РНК альфавируса, включающую, по меньшей мере, последовательность, кодирующую, по меньшей мере, один интересующий экзогенный ген;
v) последовательность полиаденилирования; и
vi) последовательность второго инвертированного концевого повтора аденовируса (ITR).
Конкретнее, настоящее изобретение относится к конструкции аденовирусного гибридного вектора, содержащей в качестве элемента iv цепь кДНК, полученной из альфавируса, соответствующую последовательности рекомбинантного репликона SFV под контролем транскрипции опухолеспецифического промотора (элемента iii), который представляет собой промотор альфа-фетопротеина (AFP). В данной конструкции трансген можно вставить в репликон SFV, управляемый субгеномным промотором SFV. После инфицирования опухолевых клеток данным гибридным вектором мРНК репликона SFV транскрибируется от опухолеспецифического промотора, и неструктурные белки - nsP, которые составляют ген репликазы SFV, транслируются из указанной мРНК репликона SFV. Данные белки nsP вирусной репликазы инициируют репликацию мРНК репликона SFV с получением субгеномной РНК SFV. Следовательно, трансген можно экспрессировать на высоком уровне из субгеномной РНК SFV. Данный общий процесс репликации вируса будет приводить к апоптозу инфицированных клеток. В том случае, когда данный гибрид заражает неопухолевые клетки, мРНК SFV не будет транскрибироваться под действием опухолеспецифического промотора, который не будет активным в данных клетках. Таким образом, трансген не будет экспрессироваться, и апоптоз будет отсутствовать в нормальных клетках, инфицированных гибридным вектором.
Настоящее изобретение дополнительно относится к способу получения указанного аденовирусного гибридного вектора, предусматривающему сборку элементов i-vi аденовирусного гибридного вектора, определенного выше, с помощью методов генной инженерии.
Также настоящее изобретение относится к применению указанного гибридного вектора для переноса генетического материала в клетку, и конкретнее, для введения и экспрессии чужеродных генов в эукариотических клетках, которые могут представлять собой клетки-мишени для аденовируса.
Перенос генетического материала предпочтительно приводит к индукции иммунного ответа против чужеродных антигенов в указанной клетке.
Настоящее изобретение также относится к фармацевтической композиции, содержащей указанный аденовирусный/альфавирусный гибридный вектор, и ее применению в терапевтическом лечении злокачественной опухоли, предусматривающему введение указанной фармацевтической композиции субъекту.
Настоящее изобретение также относится к способу лечения злокачественной опухоли с помощью применения определенного выше гибридного вектора, предусматривающему введение указанного гибридного вектора субъекту.
Настоящее изобретение относится аденовирусному гибридному вектору для экспрессии гена, отличающемуся тем, что он содержит, по меньшей мере, следующие элементы, ориентированные в направлении от 5' к 3':
i) первую цепь из аденовируса, содержащую последовательность первого инвертированного концевого повтора (ITR) и сигнальную последовательность для упаковки аденовируса;
ii) первую некодирующую «лишнюю» последовательность;
iii) последовательность, соответствующую тканеспецифическому промотору;
iv) цепь кДНК, полученную из альфавируса, последовательность которой частично комплементарна РНК альфавируса, включающую, по меньшей мере, последовательность, кодирующую, по меньшей мере, один интересующий экзогенный ген;
v) последовательность полиаденилирования; и
vi) последовательность второго инвертированного концевого повтора аденовируса (ITR).
Происхождение элемента i в альфавирусном/аденовирусном гибридном векторе по настоящему изобретению, т.е. происхождение цепи из аденовируса, содержащей последовательность первого инвертированного концевого повтора (ITR) и сигнальную последовательность для упаковки (ψ) аденовируса, не является критическим аспектом для настоящего изобретения, и он может происходить из аденовируса любого серотипа. Указанные серотипы хорошо известны в данной области и включают, например, Ad12 (подрод А), Ad3 и Ad7 (подрод В), Ad2 и Ad5 (подрод С), Ad8 (подрод D), Ad4 (подрод Е), Ad40 (подрод F) и другие известные не относящиеся к человеческим аденовирусы, которые могут происходить от таких видов, как свиньи, овцы, коровы и птицы. Следовательно, данная последовательность первого инвертированного концевого повтора, которая может содержать примерно от 100 до 500 п.н. в длину, может варьировать в зависимости от используемого серотипа аденовируса. Аналогично сигнальная последовательность для упаковки аденовируса может варьировать в зависимости от используемого серотипа аденовируса.
По предпочтительному конкретному варианту осуществления изобретения указанный аденовирусный вектор для экспрессии гена содержит элемент i, имеющий последовательность SEQ ID № 1 или какую-либо другую последовательность, обладающую достаточной гомологией с SEQ ID № 1, для выполнения такой же функции.
Происхождение элемента ii в аденовирусном гибридном векторе по настоящему изобретению не является его критическим аспектом. Указанный элемент ii, функция которого заключается в увеличении общего размера конструкции, может представлять любую некодирующую «лишнюю» последовательность. Указанная последовательность предпочтительно является некодирующей последовательностью человека. Даже более предпочтительно, когда данная некодирующая «лишняя» последовательность представляет собой область интрона человеческой геномной гипоксантинфосфорибозилтрансферазы (HPRT).
Предпочтительно определенный аденовирусный гибридный вектор также содержит элемент vii, который представляет вторую некодирующую «лишнюю» последовательность, расположенную между элементом v и элементом vi, определенными выше.
Происхождение элемента iii в аденовирусном гибридном векторе по настоящему изобретению не является его критическим аспектом. Тканеспецифический промотор iii предпочтительно представляет опухолеспецифический промотор. Примеры опухолеспецифических промоторов включают промоторы AFP, теломеразы TERT, РАР (связанный с поджелудочной железой протеин), E2F и HIF.
По предпочтительному конкретному варианту осуществления изобретения опухолеспецифический промотор имеет последовательность SEQ ID №7, соответствующую промотору/энхансеру AFP (AFP р+е), или последовательность SEQ ID № 15, соответствующую теломеразе TERT, или любую другую последовательность, обладающую достаточной гомологией соответственно с последовательностью SEQ ID №7 или с последовательностью SEQ ID № 15, для выполнения такой же функции.
Происхождение элемента iv в аденовирусном гибридном векторе по настоящему изобретению не является его критическим аспектом. Альфавирусные последовательности элемента iv предпочтительно происходят из вируса леса Семлики (SFV). Однако возможно использовать другие альфавирусные последовательности, полученные из любых видов, относящихся к семейству Togaviridae, например, SIN, RRV и VEE.
Указанная цепь iv кДНК, полученная из альфавируса, последовательность которой частично комплементарна РНК альфавируса, предпочтительно содержит (в дополнении к последовательности, кодирующей, по меньшей мере, один интересующий экзогенный ген):
а) 5'-последовательность, необходимую для репликации альфавируса,
b) последовательность, кодирующую неструктурные белки, необходимые для репликации РНК альфавируса,
с) по меньшей мере, один субгеномный промотор альфавируса, и
d) 3'-последовательность, необходимую для репликации альфавируса.
Элемент iv предпочтительно образует репликон, функционально контролируемый промотором iii, и где, в свою очередь, альфавирусный субгеномный промотор, входящий в состав iv с), функционально контролирует экспрессию интересующего экзогенного гена.
По предпочтительному конкретному варианту осуществления изобретения последовательности а) - с) элемента iv в целом имеют последовательность, выбранную из SEQ ID № 3 (SFV 5'-rep-Psg), или любую другую последовательность, обладающую достаточной гомологией с SEQ ID № 3 для выполнения такой же функции, и последовательность SEQ ID № 4 (SFV 5'-rep-Psg-enh) или любую другую последовательность, обладающую достаточной гомологией с SEQ ID № 4, для выполнения такой же функции.
По особо предпочтительному конкретному варианту осуществления изобретения элемент iv d) имеет последовательность SEQ ID № 5 (SFV3') или любую другую последовательность, обладающую достаточной гомологией с SEQ ID № 5, для выполнения такой же функции.
В элементе iv альфавирусного-аденовирусного гибридного вектора по настоящему изобретению интересующий экзогенный ген предпочтительно представляет собой терапевтический ген или ген-репортер или комбинацию обоих. Без рассмотрения в качестве ограничения, терапевтический ген предпочтительно выбран из генов интерлейкина млекопитающих IL-12, колониестимулирующего фактора GMCSF, альфа-интерферона и тимидинкиназы вируса герпеса простого.
Интересующий экзогенный ген в элементе iv дополнительно может быть геном-репортером. Без рассмотрения в качестве ограничения, ген-репортер может быть выбран из генов LacZ, люциферазы, tk и GFP.
Терапевтический ген особо предпочтительно представляет ген интерлейкина IL-12 и даже более предпочтительно терапевтический ген является геном человеческого интерлейкина hIL-12.
Аденовирусный гибридный вектор для экспрессии гена может включать в элементе iv одну или нескольких подгрупп (субгеномный промотор + интересующий экзогенный ген).
Происхождение элемента v в аденовирусном гибридном векторе по настоящему изобретению не является его критическим аспектом. Элемент v предпочтительно представляет последовательность полиаденилирования SV40. Указанная последовательность полиаденилирования SV40 особо предпочтительно является последовательностью SEQ ID № 6 или любой другой последовательностью, обладающей достаточной гомологией с SEQ ID № 6, для выполнения такой же функции.
Происхождение элемента vi в аденовирусном гибридном векторе по настоящему изобретению не является его критическим аспектом. По предпочтительному варианту осуществления указанный аденовирусный вектор для экспрессии гена содержит последовательность инвертированного концевого повтора (ITR) в качестве элемента vi, который имеет последовательность SEQ ID № 2 или любую другую последовательность, обладающую достаточной гомологией с SEQ ID № 2, способную выполнять такую же функцию.
Происхождение элемента vii в аденовирусном гибридном векторе по настоящему изобретению не является его критическим аспектом. Вторая некодирующая «лишняя» последовательность может представлять любую такую последовательность. Предпочтительно это некодирующая последовательность человека и особо предпочтительно это последовательность из человеческой космиды С346.
Аденовирусный гибридный вектор для экспрессии гена по настоящему изобретению может иметь различную длину, и предпочтительно его длина находится в пределах от 27 до 38 т.п.н.
По предпочтительному конкретному варианту осуществления изобретения аденовирусный гибридный вектор содержит ITR 5' в качестве последовательности первого инвертированного концевого повтора; HPRT, область интрона человеческой геномной гипоксантинфосфорибозилтрансферазы в качестве первой «лишней» последовательности; AFP (p+e), опухолеспецифический промотор; последовательность репликона SFV, содержащую mIL-12, мышиный интерлейкин-12; полиА SV40, последовательность полиаденилирования SV40; С346, человеческую геномную космиду С346 в качестве второй «лишней» последовательности и ITR 3' в качестве последовательности второго инвертированного концевого повтора.
По дополнительному предпочтительному конкретному варианту осуществления изобретения аденовирусный гибридный вектор содержит ITR 5' в качестве последовательности первого инвертированного концевого повтора; HPRT, область интрона человеческой геномной гипоксантинфосфорибозилтрансферазы в качестве первой «лишней» последовательности; AFP (p+e), опухолеспецифический промотор; последовательность репликона SFV, содержащую LacZ; полиА SV40, последовательность полиаденилирования SV40; С346, человеческую геномную космиду С346 в качестве второй «лишней» последовательности и ITR 3' в качестве последовательности второго инвертированного концевого повтора.
По дополнительному предпочтительному конкретному варианту осуществления изобретения аденовирусный гибридный вектор содержит ITR 5' в качестве последовательности первого инвертированного концевого повтора; HPRT, область интрона человеческой геномной гипоксантинфосфорибозилтрансферазы в качестве первой «лишней» последовательности; AFP (p+e), опухолеспецифический промотор; последовательность репликона SFV, содержащую hIL-12, человеческий интерлейкин-12; полиА SV40, последовательность полиаденилирования SV40; С346, человеческую космиду С346 в качестве второй «лишней» последовательности и ITR 3' в качестве последовательности второго инвертированного концевого повтора.
По предпочтительному конкретному варианту осуществления изобретения аденовирусный гибридный вектор для экспрессии гена имеет последовательность SEQ ID № 8 или любую другую последовательность, обладающую достаточной гомологией с SEQ ID № 8, для выполнения такой же функции.
По дополнительному предпочтительному конкретному варианту осуществления изобретения аденовирусный гибридный вектор для экспрессии гена имеет последовательность SEQ ID № 9 или любую другую последовательность, обладающую достаточной гомологией с SEQ ID № 9, для выполнения такой же функции.
По дополнительному предпочтительному конкретному варианту осуществления изобретения аденовирусный гибридный вектор для экспрессии гена имеет последовательность SEQ ID № 10 или любую другую последовательность, обладающую достаточной гомологией с SEQ ID № 10, для выполнения такой же функции.
Настоящее изобретение дополнительно относится к способу получения указанного аденовирусного гибридного вектора, включающему сборку элементов i-vi или i-vii аденовирусного гибридного вектора, определенного выше, с помощью методов генной инженерии.
Также настоящее изобретение относится к применению указанного гибридного вектора для переноса генетического материала в клетку, и конкретнее, для введения и экспрессии чужеродных генов в эукариотических клетках, которые могут представлять собой клетки-мишени для аденовирусов. Указанное применение предусматривает введение указанного гибридного вектора субъекту.
Инфицирование опухолевых клеток аденовирусным гибридным вектором по изобретению приводит к транскрипции мРНК репликона альфавируса SFV от опухолеспецифического промотора, следовательно, ген Rep будет транслироваться, и РНК SFV будет амплифицироваться. Rep также продуцирует субгеномную РНК SFV, из которой будут экспрессироваться на высоких уровнях терапевтический или ген-репортер. Продукт терапевтического гена, секретируемый инфицированными клетками, будет активировать иммуноциты в области заражения. Репликация SFV будет дополнительно индуцировать апоптоз инфицированных клеток, приводя к высвобождению опухолевых антигенов из клеток, подвергшихся апоптозу; которые могут захватываться антигенпредставляющими клетками (APC), тем самым активируя иммунный ответ, направленный против опухоли. Однако если данный гибридный вектор инфицирует неопухолевые клетки, то мРНК репликона SFV не будет транскрибироваться, следовательно, трансгенная экспрессия или апоптоз будут отсутствовать.
Опухолевые клетки предпочтительно инфицируются аденовирусным гибридным вектором по изобретению таким образом, что мРНК репликона SFV транскрибируется от опухолеспецифического промотора, следовательно, ген Rep будет транслироваться, и РНК SFV будет амплифицироваться. Rep также будет продуцировать субгеномную РНК SFV, из которой будут экспрессироваться на высоких уровнях mIL-12 или hIL-12. Секретируемые инфицированными клетками mIL-12 или hIL-12 будут активировать иммуноциты в области заражения. Репликация SFV будет дополнительно индуцировать апоптоз инфицированных клеток, приводя к высвобождению опухолевых антигенов из клеток, подвергшихся апоптозу; которые могут захватываться антигенпредставляющими клетками (APC), тем самым активируя иммунный ответ, направленный против опухоли. Однако если данный гибридный вектор инфицирует неопухолевые клетки, то мРНК репликона SFV не будет транскрибироваться, следовательно, трансгенная экспрессия или апоптоз будут отсутствовать.
Дополнительным предметом настоящего изобретения является применение аденовирусного гибридного вектора, определенного ранее, в способе переноса генетического материала в клетку, предпочтительно опухолевую клетку, который включает введение указанного гибридного вектора субъекту. Еще более предпочтительно указанная клетка представляет собой опухолевую клетку, которая экспрессирует AFP.
Дополнительным предметом настоящего изобретения является применение определенного аденовирусного гибридного вектора для получения эффективного лекарственного препарата при лечении опухолей, и его применение для индукции иммунного ответа против чужеродных антигенов. Указанное применение предусматривает введение указанного лекарственного препарата субъекту.
Дополнительным предметом настоящего изобретения является фармацевтическая композиция, содержащая, по меньшей мере, аденовирусный гибридный вектор, определенный по настоящему изобретению, и ее применение в способе лечения опухолей или индукции иммунного ответа против чужеродных антигенов.
Указанная фармацевтическая композиция предпочтительно содержит аденовирусный гибридный вектор по настоящему изобретению, в котором интересующий экзогенный ген представляет ген интерлейкина млекопитающих, IL-12, предпочтительно человеческого интерлейкина hIL-12. Указанное применение предусматривает введение фармацевтической композиции, содержащей указанный гибридный вектор субъекту.
Настоящее изобретение также относится к способу лечения злокачественной опухоли с помощью применения гибридного вектора по настоящему изобретению, где указанный способ включает введение указанного гибридного вектора субъекту.
Следовательно, по предпочтительным вариантам осуществления настоящего изобретения AFP (p+e) выбран в качестве опухолеспецифического промотора, было сконструировано два аденовирусных гибридных вектора, в которых репликон SFV контролируется промотором AFP, и ген-репортер LacZ и терапевтический ген IL-12 вставлены под контролем субгеномного промотора SFV, соответственно - Ad/AFP-SFV-LacZ и Ad/AFP-SFV-mIL-12 и два аденовирусных вектора «с усеченным геномом», несущих LacZ и IL-12 от мышей, непосредственно контролируемых промотором AFP - Ad/AFP-LacZ и Ad/AFP-mIL-12 в качестве контрольных векторов. Было показано, что гибридный вектор по настоящему изобретению функционирует более эффективно по сравнению с контрольными векторами, использованными до настоящего времени.
По настоящему изобретению было показано, что вектор Ad/AFP-SFV-mIL-12 может быть пригодным вектором в лечении опухолей НСС (гепатокарцином), экспрессирующих AFP.
Также было показано, что применение других опухолевых промоторов, таких как промотор теломеразы, TERT, который находится в высоко активном состоянии в большинстве злокачественных опухолей, для контроля SFV, может превратить применение гибридного вектора, такого как по настоящему изобретению, в общую стратегию лечения всех злокачественных опухолей.
Дополнительно и преимущественно, гибридный вектор по настоящему изобретению избирательно функционирует с опухолевыми клетками и разрушает опухолевые клетки без необходимости включения терапевтического гена. Кроме того, было показано, что гибридный вектор по настоящему изобретению преимущественно индуцирует высокую противоопухолевую активность, когда он включает терапевтический ген, такой как ген IL-12.
Краткое описание фигур
Фиг. 1 (А): представлена схема гибридного вектора по варианту осуществления изобретения, Ad/SFV, содержащего аденовирусную последовательность «с усеченным геномом», в которой репликон SFV вставлен под контролем промотора/энхансера AFP (AFP (р+е)), и содержащего гетерологичный ген mIL-12, который помещен под контролем субгеномного промотора SFV (Pr sg). На фиг. 1 (В) показана противоопухолевая активность гибридного вектора по настоящему изобретению: после инфицирования опухолевых клеток НСС данным гибридным вектором (справа), благодаря наличию в гибридном векторе репликона SFV, включающего mIL-12, имеет место экспрессия mIL-12 на высоком уровне, который будет активировать иммуноциты в области заражения. Затем репликация SFV будет индуцировать апоптоз инфицированных клеток. Однако если данный гибридный вектор инфицирует неопухолевые клетки, то мРНК репликона SFV не будет транскрибироваться и, следовательно, будут отсутствовать трансгенная экспрессия или апоптоз. На данной фигуре:
- ITR, последовательности аденовирусного инвертированного концевого повтора;
- ψ, аденовирусный сигнал упаковки;
- HPRT и С346, последовательности «лишней» ДНК соответственно из области интрона человеческой геномной гипоксантин-фосфори-бозил-трансферазы или из человеческой космиды С346;
- ПолиА, сигнал полиаденилирования (например, из SV40);
- APC, антигенпрезентующие клетки.
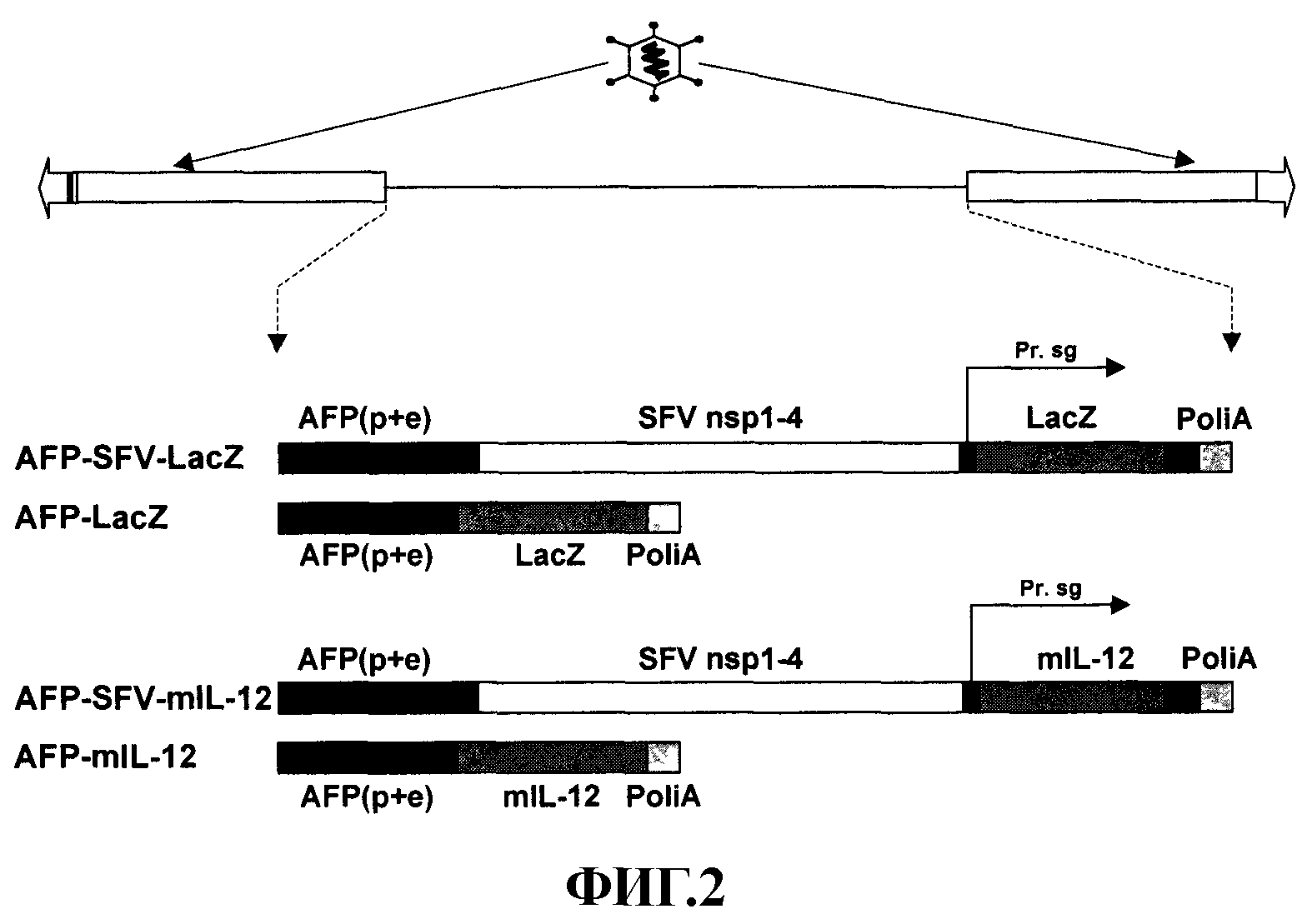
На фиг. 2 представлено строение аденовирусных гибридных векторов «с усеченным геномом» и аденовирусных векторов «с усеченным геномом». AFP-SFV-LacZ и AFP-SFV-mIL-12 являются гибридными аденовирусными векторами, в которых последовательность репликона SFV находится под контролем промотора/энхансера AFP (AFP (р+е)), и гетерологичные гены LacZ или mIL-12 клонированы соответственно под контролем субгеномного промотора SFV (Pr sg). AFP-lacZ и AFP-mIL-12 представляют аденовирусные векторы, содержащие LacZ или mIL-12, непосредственно контролируемые AFP (р+е). SFV nsp1-4, неструктурные белки SFV.
На фиг. 3 показана специфическая экспрессия mIL-12 в условиях in vitro в клетках гепатокарциномы, экспрессирующих AFP, НСС (А), и в клетках, полученных не из НСС (В), после заражения гибридными векторами Ad-SFV: AFP-mIL-12 (AFP-12), AFP-SFV-mIL-12 (AFP-SFV-12) или контрольным вектором AdCMVmIL-12 (CMV-12). Тестировали различные множественности заражения «moi» (10, 100 и 1000). Hep3B, Huh-7, HepG2 и PLC/PRF/5: клеточные линии НСС; Hela, A549, MHC1, SK-Hep-1 и Clone 9: линии, полученные не из НСС. На фиг. 3В показана только экспрессия, соответствующая moi, равной 1000.
На фиг. 4 показана специфическая экспрессия β-gal в 4 клеточных линиях НСС: (Hep3B, Huh-7, HepG2 и PLC/PRF/5) после инфицирования в условиях in vitro гибридным вектором AFP-SFV-lacZ или контрольным вектором AFP-LacZ при различных значениях «moi» (10, 100 и 1000).
На фиг. 5 представлен анализ экспрессии β-gal в клеточных линиях НСС, инфицированных AFP-LacZ и AFP-SFV-LacZ. Микрофотографии клеток, инфицированных AFP-LacZ (А-С) или AFP-SFV-LacZ (D-F) и окрашенных X-Gal. А и D, Hep3B; В и Е, Huh-7; С и F, HepG2.
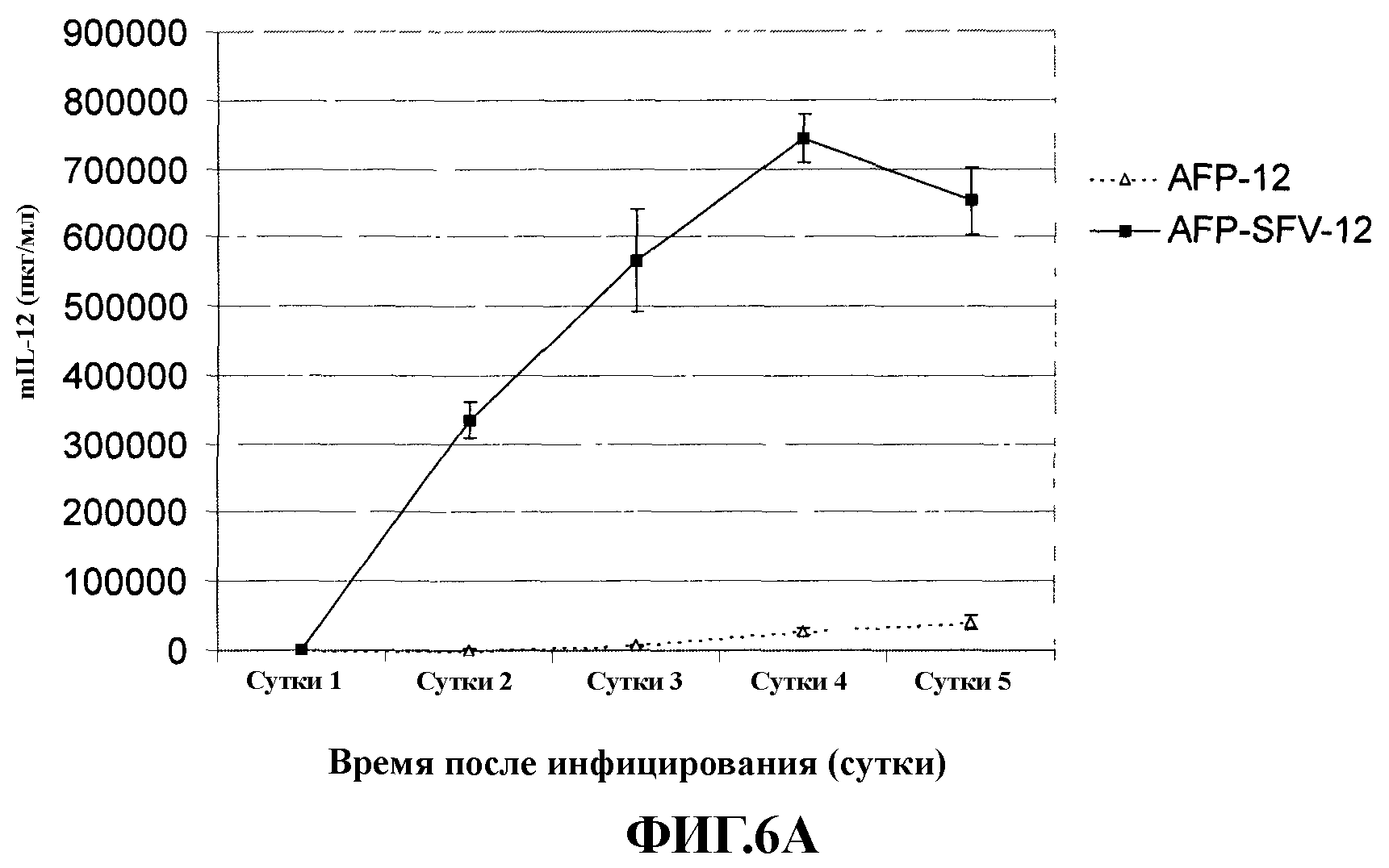
На фиг. 6 представлена кинетика экспрессии IL-12 в клеточных линиях НСС Hep3B (А) и Huh-7 (В), инфицированных в условиях in vitro аденовирусными векторами AFP-mIL-12 (AFP-12) или AFP-SFV-mIL-12 (AFP-SFV-12) при значении «moi», равном 1000.
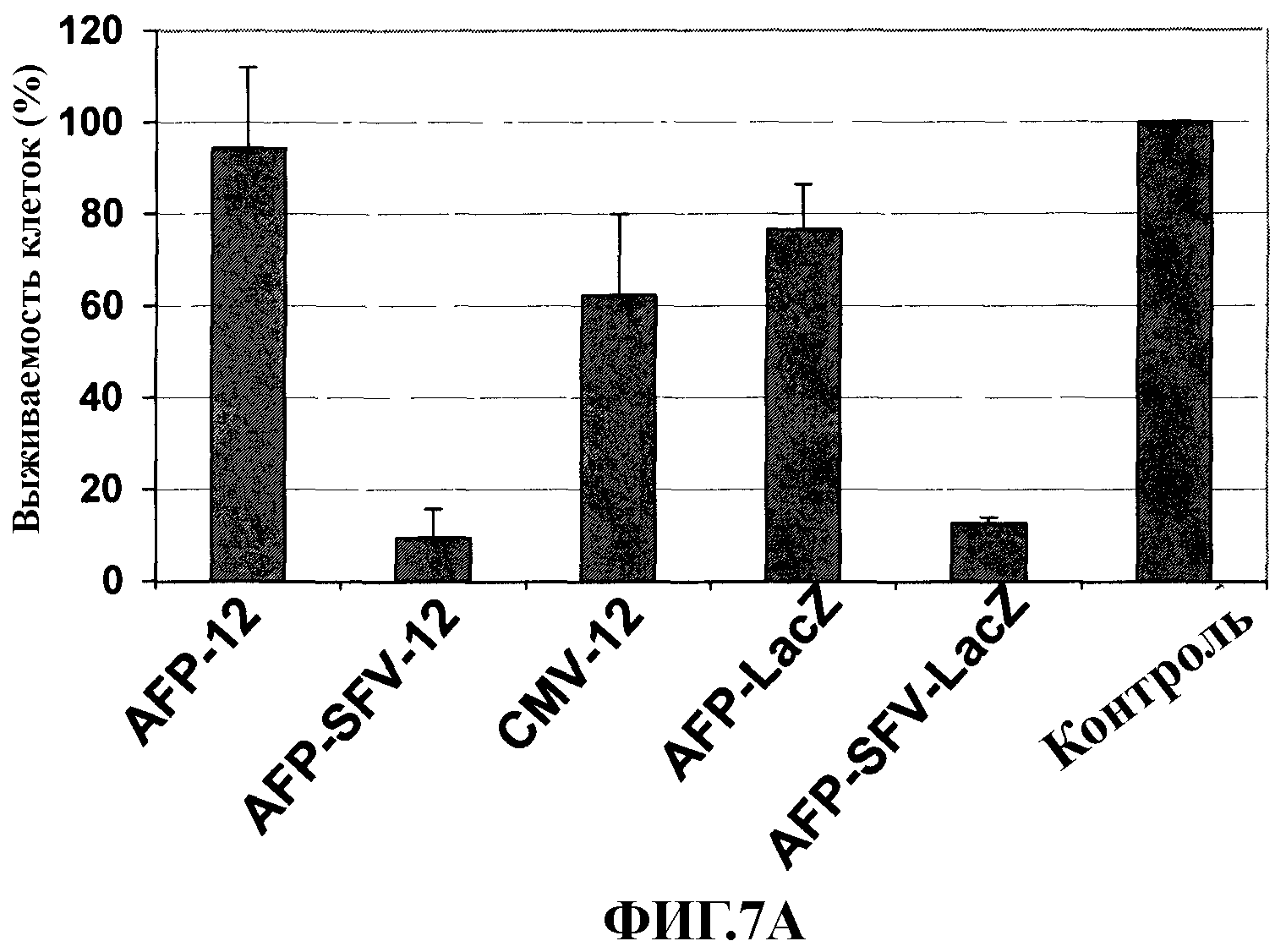
На фиг. 7 показана индукция гибели клеток после инфицирования в условиях in vitro клеточных линий НСС - Hep3B (А) и McA-RH7777 (B)- векторами AFP-IL-12 (AFP-12), AFP-SFV-IL-12 (AFP-SFV-12), AFP-LacZ, AFP-SFV-LacZ или контрольным вектором Ad/CMVmIL-12 (CMV-12). Выживаемость клеток представлена в виде процента живых клеток в зараженных лунках по сравнению с живыми клетками в незараженных контрольных лунках.
На фиг. 8 представлена экспрессия Rep SFV в клетках НСС -Hep3B (А-D) и Huh-7 (E и F) - после инфицирования векторами AFP-mIL-12 (А и В) или AFP-SFV-mIL-12 (С и F) при значении «moi», равном 1000. Через 2 суток после инфицирования клетки фиксировали и анализировали иммунофлуоресценцией со специфическим антителом против Rep. Клетки, экспрессирующие Rep, визуализировали под флуоресцентным микроскопом с фильтром FITC (А, С и Е), в то время как ядра, окрашенные DAPI во всех клетках, визуализировали с использованием Уф фильтра (В, D и F).
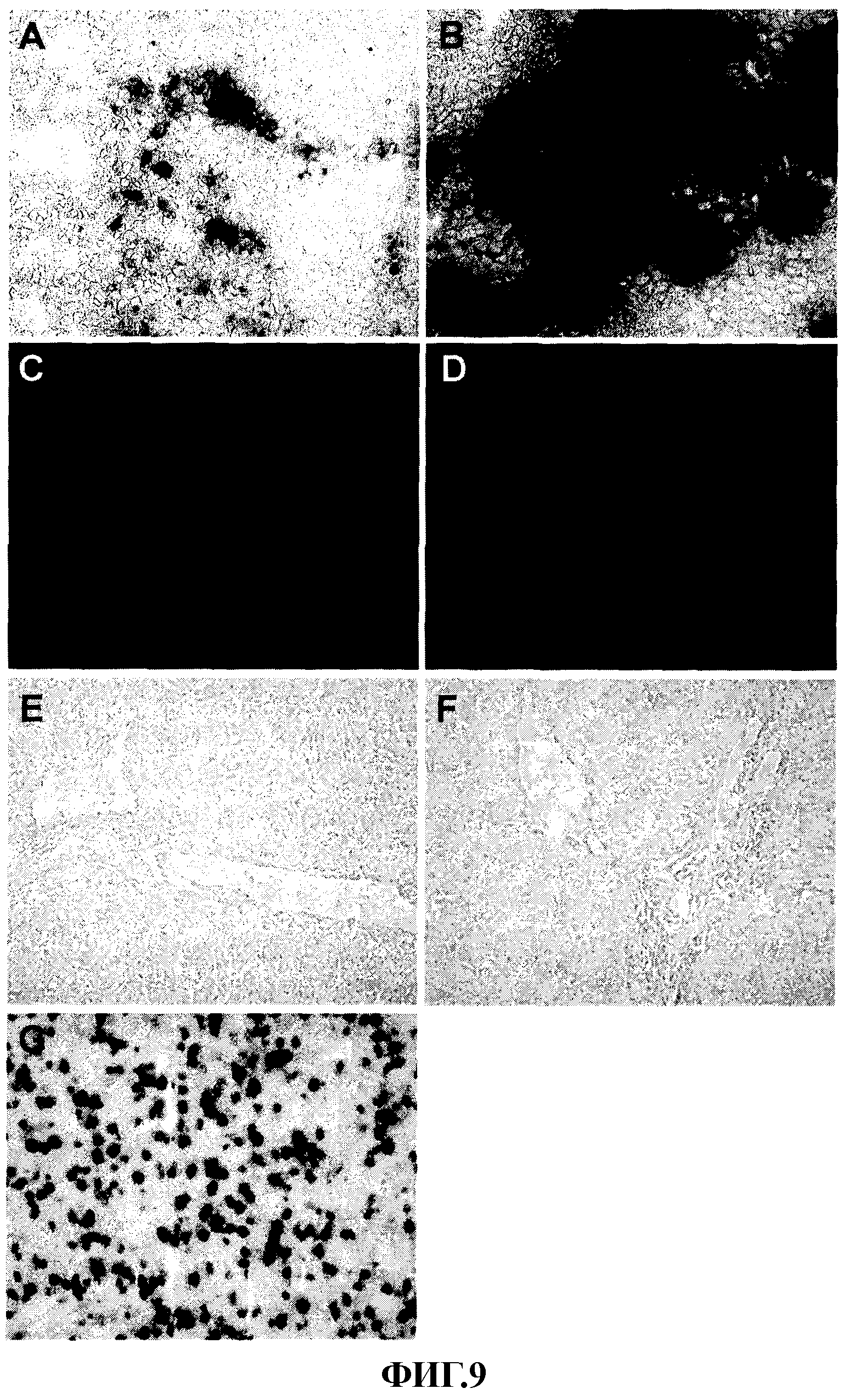
На фиг. 9 представлен перенос гена с помощью гибридных векторов «с усеченным геномом» в условиях in vivo, (A-D), эффективность переноса гена и индукция апоптоза в опухолях Huh-7. Человеческие опухоли Huh-7, прижившиеся у голых мышей с иммунодефицитом, обрабатывали инъекцией в опухоль векторов AFP-LacZ (n=4) или AFP-SFV-LacZ (n=4) из расчета 1×1010 вирусных частиц/животное. Через 3 суток после введения вируса мышей умерщвляли и анализировали срезы опухоли для оценки экспрессии трансгена окрашиванием X-Gal (А-В) или для оценки индукции апоптоза с помощью TUNEL (C-D). А, С; Микрофотографии опухолей, получивших AFP-LacZ. В, D. Микрофотографии опухолей, получивших AFP-SFV-LacZ. (E-G). Специфичность экспрессии гена с гибридными векторами «с усеченным геномом». Здоровым мышам Balb/с вводили внутривенно 1×1010 вирусных частиц векторов AFP-LacZ (Е), AFP-SFV-LacZ (F) или Ad/CMV-LacZ (G). Представлены микрофотографии срезов ткани печени, отобранной через 3 суток после заражения, и окрашенной X-Gal.
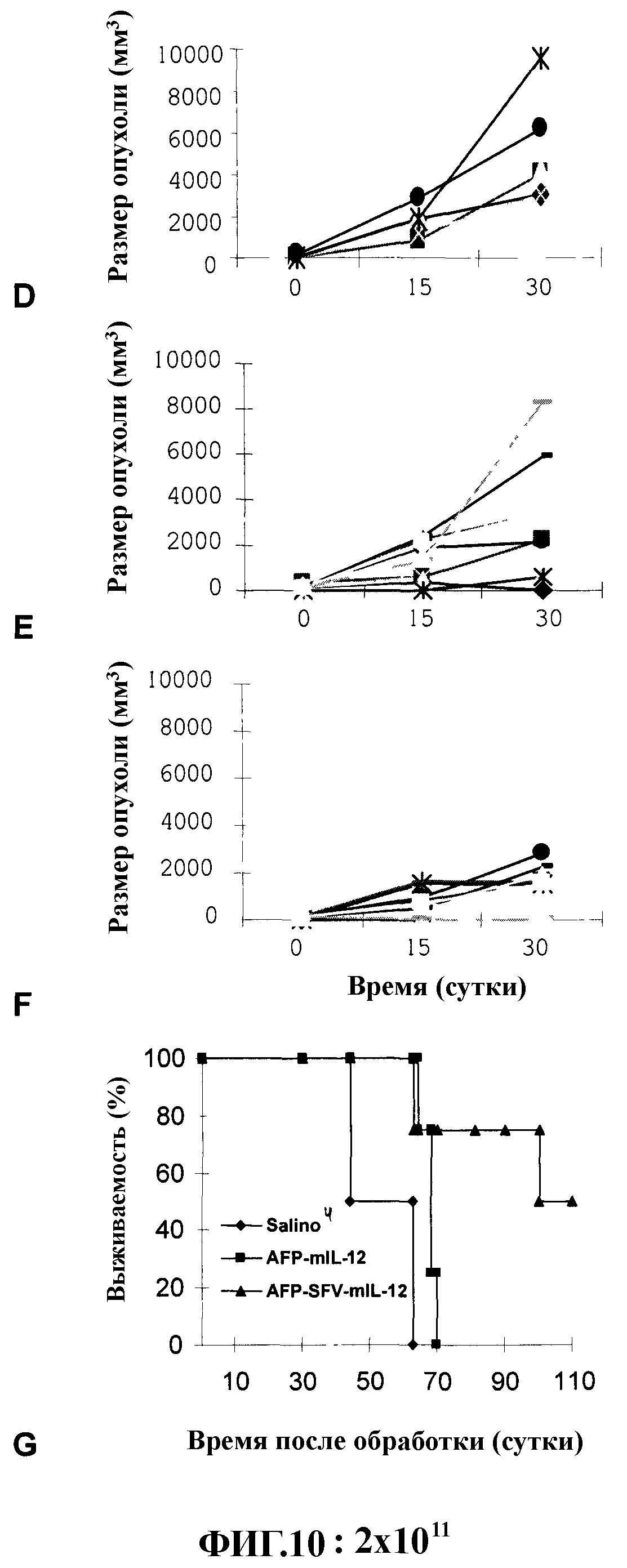
На фиг. 10 показана обработка опухолей НСС гибридными векторами. Ортотопические опухоли НСС вводили имплантацией клеток McH-RH7777 в печень крыс. Когда опухоли достигали размера 7-10 мм в диаметре, животных обрабатывали 1011 (А-С) или 2×1011 (D-G) вирусных частиц AFP-mIL-12, AFP-SFV-mIL-12 или физиологическим раствором в качестве контроля. Размер опухолей определяли на 15 и 30 сутки после введения физиологического раствора (А и Е), AFP-mIL-12 (B и F) или AFP-SFV-mIL-12 (С и G). G; выживаемость животных.
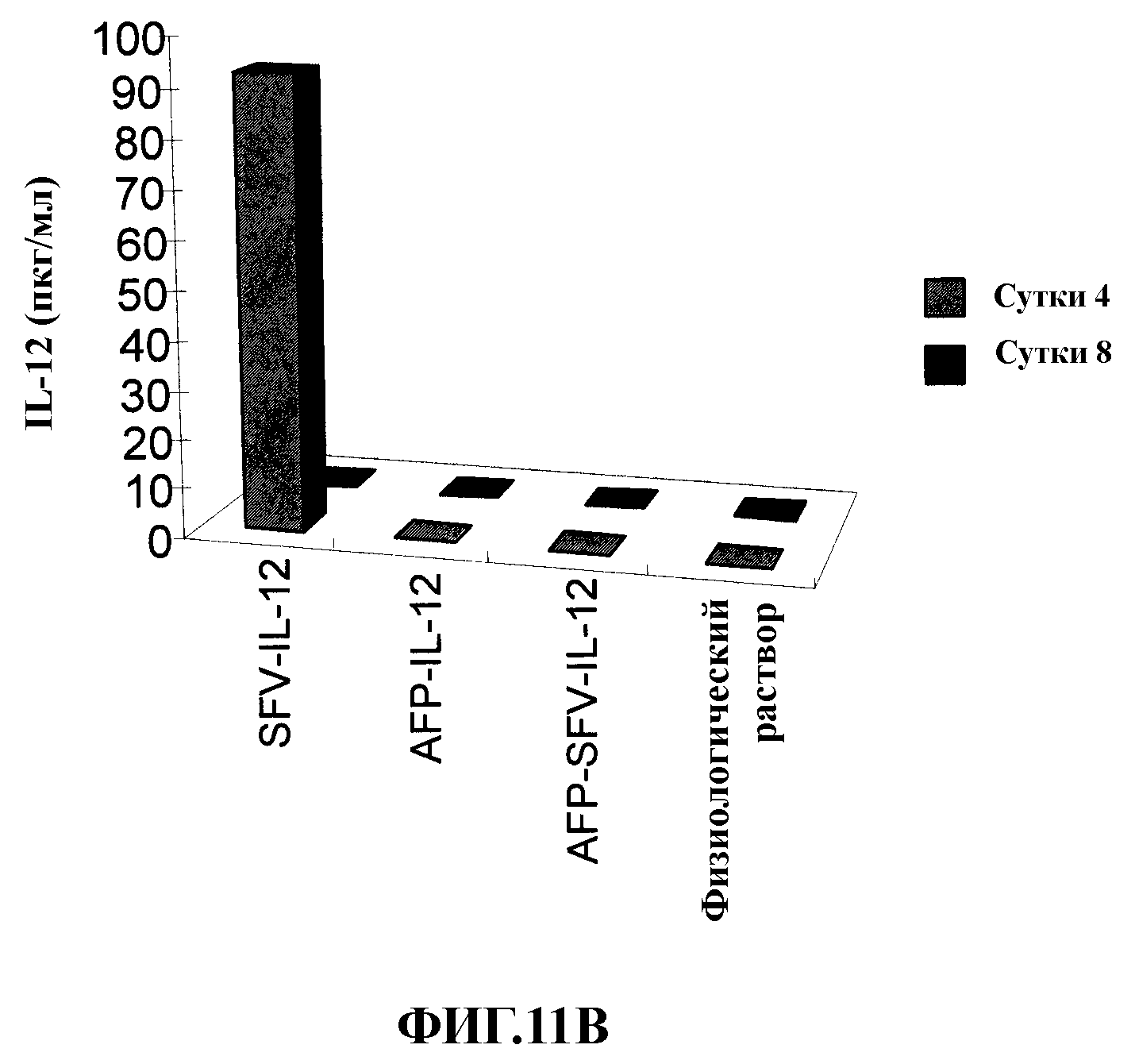
На фиг. 11 представлены результаты токсикологического исследования на крысах, которым вводили векторы, экспрессирующие IL-12. Определяли уровень трансаминаз (GPT, GOLT и GGTL) (А) или IL-12 (В) в сыворотке крови крыс с опухолями НСС в печени, которым вводили в опухоль аденовирусные векторы AFP-SFV-IL-12, AFP-SFV-mIL-12, альфавирусный вектор SFV-IL-12 или физиологический раствор. Определение проводили на 4 и 8 сутки после обработки.
На фиг. 12 показаны результаты окрашивания гематоксилин/эозином срезов печени крыс, обработанных аденовирусными гибридными векторами. Крыс с опухолями НСС обрабатывали инъекцией в опухоль физиологическим раствором (А), аденовирусными векторами AFP-IL-12 (В), AFP-SFV-IL-12 (С) или вирусными частицами SFV-IL-12 (D). Через 3 суток после обработки животных умерщвляли, печени удаляли и фиксировали в формалине, делали срезы и окрашивали гематоксилин/эозином. Черные стрелки указывают зоны с эозинофильными гепатоцитами.
На фиг. 13А и 13В представлены рестрикационные карты соответственно плазмид pGL3/AFP и pBS/mIL-12.
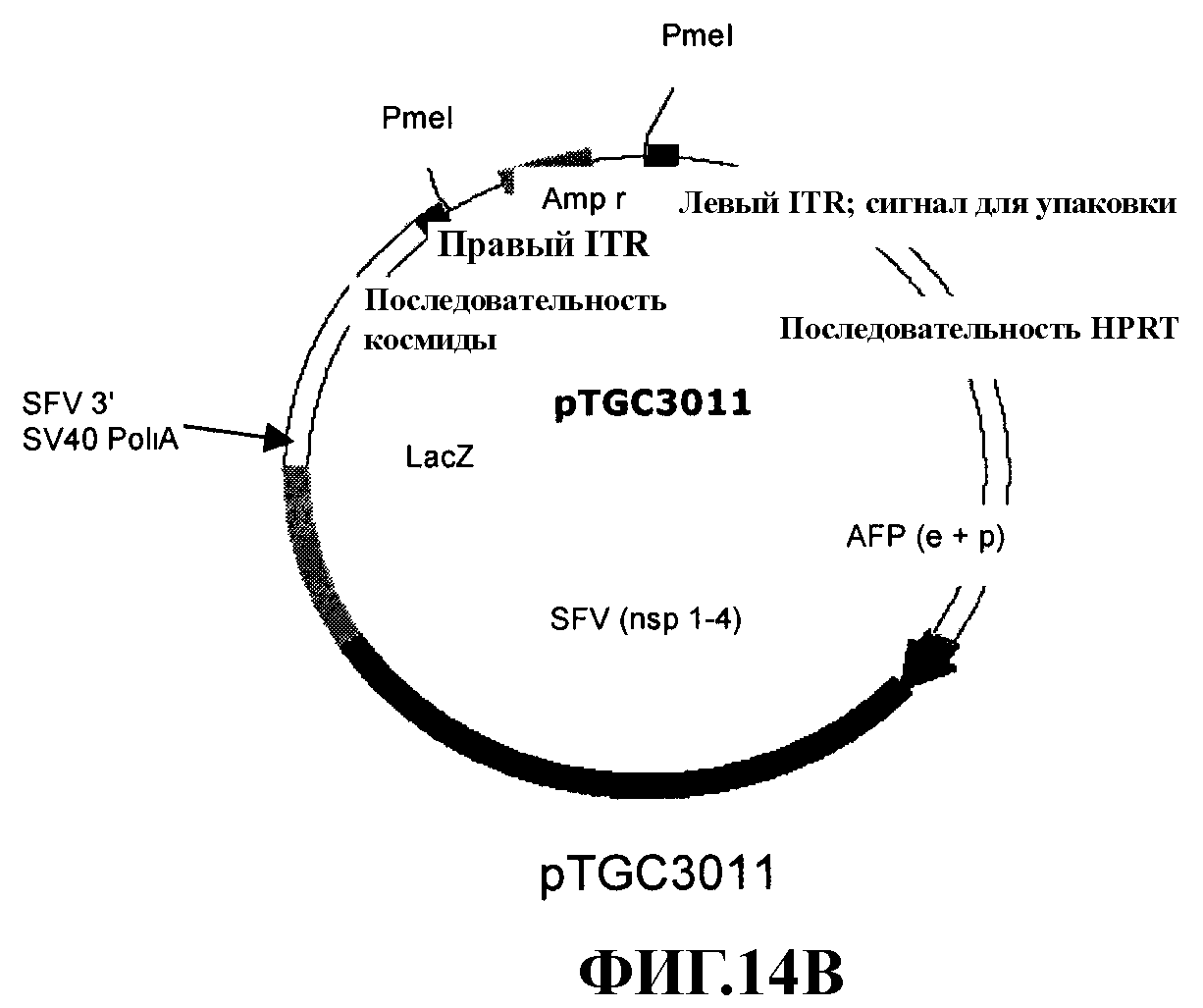
На фиг. 14А и 14В представлены рестрикационные карты соответственно плазмид pTGC3001 и pTGC3011.
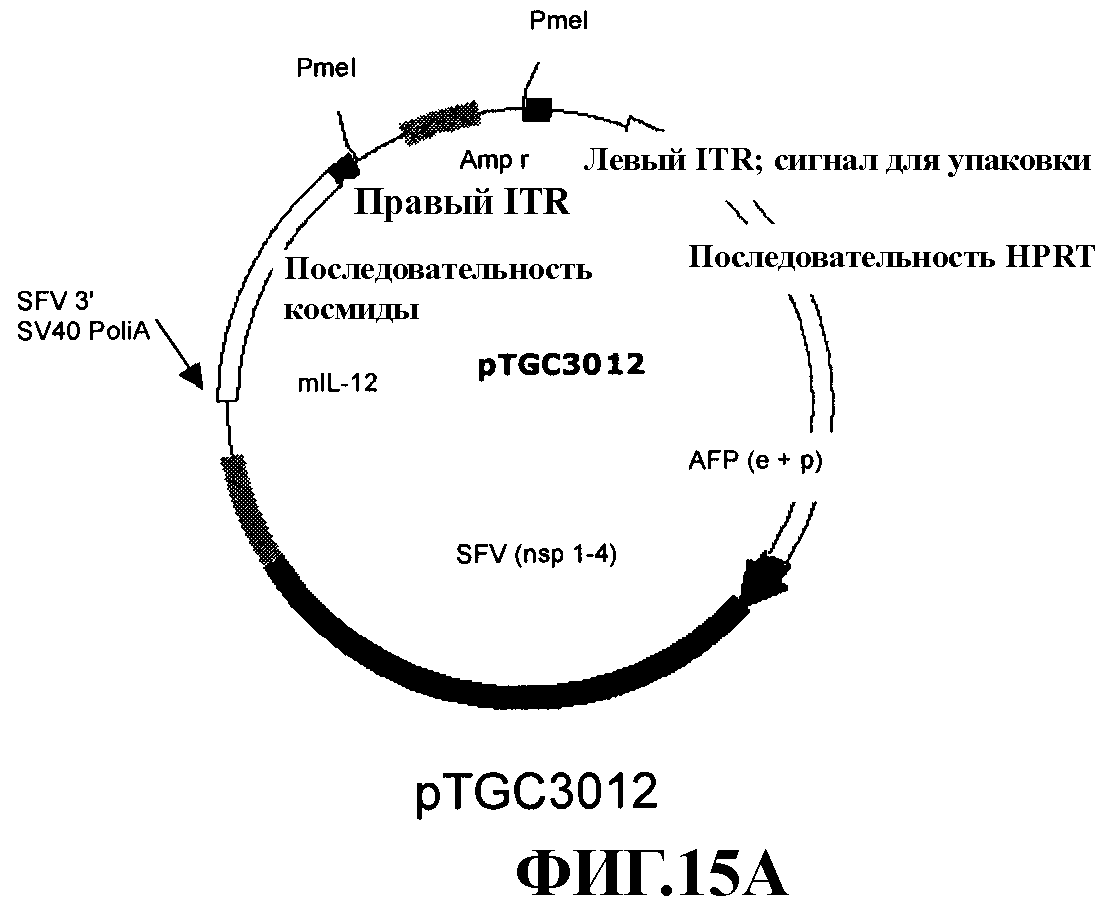
На фиг. 15А и 15В представлены рестрикационные карты соответственно плазмид pTGC3012 и pTGC3013.
На фиг. 16 представлена рестрикационная карта плазмиды pTGC3014.
Описание предпочтительных вариантов осуществления
Плазмиды
pGEM-T «легкую» и pCMVb получали от Promega, USA, и pBS-SK+ от Stratagene, USA. pSTK120 была предоставлена Dr. Kochanek (University of Ulm, Германия). pBK-SFV-1 и pBK-SFV-3 были описаны Berglund P. et al. «Enhancing immune responses using suicidal DNA vaccines». Nature Biotechnology 1998, 16: 562-565. pGL3/AFP и pBS/mIL-12 (Yonglian Sun, Cheng Qian, Dacheng Peng and Jesus Prieto. 2000. Gene transfer to liver cancer cells of B7-1 in addition to IL-12 changes immunoeffector mechanisms and supresses Th1 cytokine production induced by IL-12 alone. Human Gene Therapy 11: 127-138) получали в лаборатории заявителей.
Для конструирования pGL3/AFP получали области промотора/энхансера AFP (р+е) амплификацией ПЦР человеческой геномной ДНК. Праймеры, использованные для амплификации промотора AFP (AFP pro), представляли собой CTCTAGATTTTCTGCCCCAAAGAGCTC и CGGGATCCTGTTATTGGCAGTGGTGGAA. Праймеры, использованные для амплификации энхансера AFP, представляли собой CGGAATTCGCCTGTCATACAGCTAATAA и CTCTAGACTGTCAAATAAGTGGCCTGG. Последовательности промотора (217 пар нуклеотидов) и энхансера (785 пар нуклеотидов) клонировали в плазмиды pGEM-T. Затем проводили подтверждение амплифицированных фрагментов секвенированием. Промотор AFP вырезали из плазмиды pGEM-T/AFP-р рестрикцией XbaI/BamHI и вставляли лигированием по тупым концам в pGL3-основную плазмиду, расщепленную Sma I. Таким образом, получали плазмиду pGEM/AFP-р. Энхансер AFP вырезали из плазмиды pGEM-T/AFP-е рестрикцией Xba I/Eco RI и вставляли лигированием по тупым концам в плазмиду pGL3/ AFP-р, расщепленную Nhe I, с получением в конце плазмиды pGL3/AFP.
Клеточные линии и культуры тканей
Человеческие клеточные линии НСС Hep3B, PLC/PRF/5, HepG2 и SK-Hep-1, клеточную линию человеческой эпителиальной аденокарциномы шейки матки HeLa, клеточную линию человеческой карциномы легких А549, клеточную линию эмбриональных почечных клеток 293, крысиные клетки НСС McA-RH7777, MHC1 и Clone 9 нормальных крысиных гепатоцитов и клеточную линию крысиных НСС Нера1-6 получали из АТСС. Клетки 293, экспрессирующие рекомбиназу Cre (293Cre4) получали от Merck Research Laboratories. Клетки Hep3B, PLC/PRF/5, HeLa, SK-Hep-1, Clone 9, Huh-7 и Hepa1-6 культивировали в среде DMEM с добавлением 10% фетальной бычьей сыворотки (FBS), инактивированной тепловой обработкой, и пенициллина/стрептомицина. Клетки HepG2 и А549 культивировали в среде RPMI 1640 с добавлением 10% фетальной бычьей сыворотки (FBS), инактивированной тепловой обработкой, и пенициллина/стрептомицина. Клетки МсН-RH7777 и МНС1 культивировали в среде DMEM с добавлением 20% лошадиной сыворотки и 5% FBS. Клетки 293Cre4 культивировали в среде DMEM с добавлением 10% FBS и 0,4 мг/мл G418.
Животные
Голых мышей-самок с иммунодефицитом BALB/с в возрасте 7 недель получали из Charles Rivers laboratories (Barcelona, Испания). Крыс-самцов Буффало в возрасте 4-6 недель получали из CIFA (Animal Installations of the University of Navarra). Мышей и крыс содержали в обычных условиях в CIFA. Голые мыши получали обработанный облучением корм с автоклавированной питьевой водой. Манипуляции с голыми мышами всегда проводили в ламинарной камере. Все процедуры с животными проводили согласно стандартным рекомендациям и протоколам по уходу и использованию лабораторных животных.
Конструирование векторов
Конструирование экспрессирующих кассет AFP-SFV
5'-концевую последовательность SFV (1-292 нт) амплифицировали ПЦР с использованием плазмиды pBK-SFV-1 (содержащей полноразмерную последовательность репликона SFV) в качестве матрицы. Праймер 1 содержал сайт рестрикции Spe I в 5'-конце (подчеркнут), за которым следовали 50 нт последовательности промотора AFP и первые 20 нт последовательности SFV (курсивом): 5'-ACT AGTTAA CAG GCA TTG CCT GAA AAG AGT ATA AAA GAA TTTCAG CAT GAT TTT CCA TGG CGG ATG GAC ATA C-3'. Праймер 2 содержал сайт рестрикции Xho I (подчеркнут), за которым следовали 19 нт последовательности SFV (курсивом): 5'-CTC GAG GAT ATC CAA GAT GAG TGT GT-3'. Фрагмент ДНК размером 342 п.н. получали ПЦР и непосредственно клонировали в плазмиду pGEM-T-«легкую» с получением плазмиды pGEM-Te-SFV-1. Отсутствие ошибок ПЦР в данной плазмиде подтверждали секвенированием. Фрагмент размером 342 п.н. вырезали из плазмиды pGEM-Te-SFV-1 расщеплением Spe I и Xho I и клонировали в плазмиду pGL3/AFP, расщепленную теми же ферментами, с получением плазмиды pGL3/AFP-SFV-1, которая содержит полный промотор AFP (217 п.н.) и энхансер (785 п.н.), за которыми следует 5'-концевая последовательность SFV (SFV, включающая 292 п.н.). Кассету AFP-SFV-1 (1342 п.н.) получали из pGL3/AFP-SFV-1 расщеплением Mlu I/Xho I, обрабатывали фрагментом Кленова и клонировали в pBS-SK+, расщепленную EcoR V, с получением pBS/AFP-SFV-1. Позднюю полиА SV40 (262 п.н.) вырезали из pGL3/AFP расщеплением Xho I/BamH I, затупляли фрагментом Кленова и вставляли в сайт Sal I в pGL3/AFP-SFV-1, также затупляли фрагментом Кленова с получением таким образом pBS /AFP-SFV-1-рА. Полилинкер, содержащий оба уникальных сайта Apa I и Nru I, вставляли между сайтами BamH I и Xma I в pBS /AFP-SFV-1-рА. 3'-концевую последовательность SFV, включающую 7985 п.н., вырезали из pBK-SFV-1 расщеплением Spe I/EcoR V, затупляли фрагментом Кленова и вставляли в положение EcoR V в pBS /AFP-SFV-1-рА с получением pBS /AFP-SFV-рА.
Ген-репортер LacZ получали из pCMVb расщеплением not I, обрабатывали фрагментом Кленова и вставляли в сайт BamH I в pBS /AFP-SFV-рА, обработанной фрагментом Кленова, с получением pBS /AFP-SFV-LacZ-рА. Кассету mIL-12, содержащую гены, кодирующие субъединицы р35 и р40, связанные с внутренним сайтом входа рибосомы (IRES) отделяли от pBS/mIL-12 расщеплением Spe I/Xho I, обрабатывали фрагментом Кленова и вставляли в положение BamH I в pBS/AFP-SFV-mIL-12-рА, также затупляли фрагментом Кленова, получали плазмиду pBS/AFP-SFV-mIL-12-рА.
Конструирование гибридных аденовирусных векторов «с усеченным геномом»
Конструировали четыре аденовирусных вектора «с усеченным геномом», представленных на фиг. 2. AFP-SFV-lacZ и AFP-SFV-mIL-12 включали последовательность рекомбинантного репликона SFV, контролируемого промотором и энхансером AFP. В данных векторах ген-репортер LacZ или терапевтический ген mIL-12 клонировали соответственно под контролем субгеномного промотора SFV. AFP-lacZ и AFP-mIL-12 также представляют аденовирусные векторы «с усеченным геномом», которые включают гены LacZ и mIL-12, непосредственно контролируемые соответственно промотором/энхансером AFP. Способ конструирования данных векторов описан ниже. Для получения аденовирусного вектора с достаточной клонирующей емкостью для расположения экспрессирующей кассеты AFP-SFV-IL-12 модифицировали плазмиду pSTK120, содержащую последовательность аденовируса «с усеченным геномом». В конце элиминировали фрагмент размером 9 т.п.н. из плазмиды pSTK120 расщеплением Apa I. Кроме того, полилинкер, содержащий сайты Asc I и Sbf I, вставляли в данную новую плазмиду, получая pTGC3001. Данная плазмида содержала левый ITR, сигнал упаковки, «лишнюю» ДНК из HPRT и С346, и правый ITR. Кассету AFP-SFV-LacZ вырезали расщеплением Apa I из pBS/AFP-SFV-lacZ-рА и вставляли в сайт Apa I в pTGC3001 с получением pTGC3011. Аналогично кассету AFP-SFV-mIL-12 вырезали из pBS/AFP-SFV-mIL-12-рА расщеплением BssH II, обрабатывали фрагментом Кленова и вставляли в сайт Asc I в pTGC3001, также обрабатывали фрагментом Кленова с получением pTGC3012.
Конструирование контрольных векторов
Конструирование аденовирусного вектора «с усеченным геномом» AFP-LacZ
Последовательность энхансера/промотора AFP (AFP р+е) вырезали из pGL3/AFP расщеплением Mlu I/Xho I, обрабатывали фрагментом Кленова и вставляли в pCMVb, которую ранее расщепляли EcoR I/Xho I и обрабатывали фрагментом Кленова. При этом сильный ранний промотор CMV вырезали из pCMVb и замещали на AFP р+е с получением pAFPb. Затем кассету AFP-LacZ (5077 т.п.н.) вырезали из pAFPb расщеплением Xba I/Nar I, обрабатывали фрагментом Кленова и вставляли в сайт Swa I в pSTK120, также затупляли фрагментом Кленова с получением плазмиды pTGC3013.
Конструирование аденовирусного вектора «с усеченным геномом» AFP-mIL-12
Кассету mIL-12 вырезали из pBS/mIL-12 расщеплением Xho I/Spe I и вставляли в pGL3/AFP, которую ранее расщепляли Xho I/Xba I, в результате чего ген люциферазы удалялся из данной последней плазмиды и получали pAFP-mIL-12. Кассету AFP-mIL-12 (3760 т.п.н.) вырезали из pAFP-mIL-12 расщеплением Bam I/Sca I, обрабатывали фрагментом Кленова и вставляли в pSTK120, расщепленной Swa I, и аналогично обрабатывали фрагментом Кленова с получением pTGC3014.
«Спасение» аденовирусных векторов «с усеченным геномом»
После расщепления Pme I, экстракции фенолом/хлороформом и осаждения этанолом 2 мкг ДНК pTGC3011, pTGC3012, pTGC3013 и pTGC3014 трансфектировали соответственно в клетки 293Cre4. После трансфекции клетки инфицировали вирусом-помощником AdLC8cluc. Последующую крупномасштабную амплификацию и получение проводили, как описано ранее (Philip Ng., Robin J. Parks and Frank L. Graham. Preparation of helper-dependent adenoviral vectors. Methods in Molecular Medicine, Vol. 69, Gene Therapy Protocols, 2nd. Ed. 69, 371-88, 2002; H. Zhou, L. Pastore, A.L. Beaudet. Helper-dependent adenoviral vectors. Methods in Enzymology, Vol. 346, 177-198, 2002; Hillgenberg M., et al. System for efficient helper-dependent minimal adenovirus constructions and rescue. Hum. Gene Ther., 12, 643-657, 2001). Все векторные препараты дважды очищали центрифугированием в градиенте CsCl. Очищенные ДНК-векторы анализировали расщеплением рестриктазами, и не было обнаружено реаранжировки последовательностей. Титрование загрязнения аденовируса «с усеченным геномом» и вируса-помощника оценивали с использованием количественной ПЦР. Соотношение вирусных частиц к инфекционным единицам (iu) составляло 20:1. Загрязнение частицами вируса-помощника равнялось примерно 0,5-1%.
Количественная ПЦР
Для определения степени загрязнения вирусом-помощника конструировали зонд и праймеры для постановки количественной ПЦР области Ad5 E4 с использованием программы TaqMan (TaqMan Probe #2) и синтезировали в Sigma-Genosys Ltd. (праймер) и Applied Biosystems (зонд). Для определения титра аденовирусов «с усеченным геномом» конструировали зонды и праймеры для количественной ПЦР последовательностей LacZ и mIL-12 от мышей с использованием программы TaqMan (TaqMan Probe #2) и синтезировали в Sigma-Genosys Ltd. (праймер) и Applied Biosystems (зонд). Для определения загрязнения диким типом Ad конструировали зонды и праймеры для постановки количественной ПЦР области Ad5 E1 с использованием программы TaqMan (TaqMan Probe #2) и синтезировали в Sigma-Genosys Ltd. (праймер) и Applied Biosystems (зонд).
Опыты в условиях in vitro
Трансгенная экспрессия в клетках, инфицированных аденовирусными векторами «с усеченным геномом»
Клеточные линии, производные НСС (Hep3B, Huh-7, HepG2 и PLC/PRF/5) и клеточные линии, не относящиеся к НСС (A549, HeLa, MHC1 и Clone 9), или клеточные линии, производные НСС, но не экспрессирующие AFP (SK-Hep-1), заражали каждым из четырех аденовирусных векторов «с усеченным геномом» (AFP-LacZ, AFP-SFV-LacZ, AFP-mIL-12 или AFP-SFV-mIL-12) при значении «moi», соответственно равном 1000, 100 или 10 (частиц/клетку). Три первых поколения аденовирусов (Ad/CMV-mIL-12, Ad/CMV-LacZ, Ad/AFP-LacZ) использовали в качестве контроля. Собирали супернатанты из клеток, инфицированных векторами mIL-12, и лизаты из клеток, инфицированных векторами LacZ, в двойных параллелях из лунок для определения концентрации соответственно mIL-12 и β-галактозидазы (β-gal). Клетки, инфицированные векторами LacZ, также окрашивали Х-gal. Концентрацию mIL-12 (р70) определяли с помощью набора для постановки ELISA (Pharmingen, San Diego, CF). Уровень β-gal определяли с использованием набора для ELISA (Roche, Швейцария). Продолжительность экспрессии mIL-12 определяли на клетках НСС (Hep3B, Huh-7) после их инфицирования AFP-mIL-12, AFP-SFV-mIL-12 или контрольным вектором Ad/CMV-mIL-12 при значении «moi», равном 1000. Ежедневно собирали супернатанты до 5 суток после заражения.
Анализ специфичности трансгенной экспрессии с использованием гибридных векторв Ad-SFV в условиях in vitro
Для оценки специфичности трансгенной экспрессии с использованием ранее описанных рекомбинантных векторов, четыре человеческие клеточные линии НСС (Hep3B, HepG2, Huh-7 и PLC/PRF/5) и две человеческие клеточные линии, не относящиеся к НСС (HeLa и A549), или которые являются производными НСС, но не экспрессируют AFP (SK-Hep-1), инфицировали AFP-mIL-12, AFP-SFV-mIL-12 или Ad-CMV-mIL-12 в качестве положительного контроля при различных значениях «moi» (10, 100 или 1000). Через 2 суток после заражения собирали супернатанты и определяли содержание в них mIL-12. Результаты приведены на фиг. 3 (А) и (В). Не было отмечено экспрессии mIL-12 в человеческих клетках НСС после инфицирования AFP-mIL-12 при значениях «moi», равных 10 или 100, и только при «moi», равном 1000, обнаруживали небольшую концентрацию mIL-12 в некоторых клеточных линиях (фиг. 3А). В противоположность инфицирование данных клеток AFP-SFV-mIL-12 при «moi», равном 10, 100 или 1000, приводило к экспрессии mIL-12 в зависимости от дозы (фиг. 3А). Уровень экспрессии mIL-12 в клетках, инфицированных AFP-SFV-mIL-12, при значении «moi», равном 10, был сравним с уровнем, полученным в клетках, зараженных AFP-mIL-12, при «moi», равном 1000. Кроме того, концентрация mIL-12 в клетках НСС, инфицированных AFP-SFV-mIL-12 при различных значениях «moi», была сравнима с таковой, полученной с контрольным вектором Ad-CMV-mIL-12. Однако заражение клеток AFP-mIL-12 или AFP-SFV-mIL-12, которые не экспрессируют AFP, не приводило к детектируемым уровням mIL-12 даже при использовании наиболее высокого значения «moi» (1000) (фиг. 3В). В данных клетках только контрольный вектор Ad-CMV-mIL-12 был способен приводить к экспрессии mIL-12 на высоком уровне.
С другой стороны, четыре клеточные линии НСС (Hep3B, Huh-7, HepG2 и PLC/PRF/5) инфицировали гибридными векторами LacZ-AFP-lacZ или AFP-SFV-lacZ при различных значениях «moi» (10, 100 или 1000) и обнаруживали специфическую экспрессию β-gal. В данном случае были получены аналогичные данные, и эти результаты приведены на фиг. 4.
На фиг. 5 представлены микрофотографии клеток НСС, инфицированных аденовирусными векторами «с усеченным геномом» AFP-lacZ и AFP-SFV-lacZ, с последующим окрашиванием Х-gal. Заражение клеток НСС AFP-lacZ приводило к низкой экспрессии в инфицированных клетках, которые окрашивались очень слабо. В противоположность, заражение клеток НСС AFP-SFV-lacZ давало экспрессию β-gal на высоком уровне, о чем судили по интенсивному окрашиванию с помощью Х-gal. Эти данные указывают на то, что гибридный вектор Ad-SFV, содержащий репликон SFV под контролем промотора AFP, может привести к высокому уровню интенсивной трансгенной экспрессии в опухолевых клетках, экспрессирующих AFP.
Продолжительность экспрессии mIL-12 в клетках НСС в условиях in vitro
Для оценки продукции mIL-12 в различные периоды времени после инфицирования гибридными векторами Ad-SFV две клеточные линии НСС (Hep3B и Huh-7) инфицировали AFP-mIL-12 или AFP-SFV-mIL-12, и ежедневно собирали супернатанты до 5 суток после заражения. На фиг. 6 приведены результаты оценки трансгенной экспрессии, полученные после заражения указанных клеток. Указанные результаты свидетельствуют о постоянном увеличении экспрессии mIL-12 с 1 по 4 сутки после инфицирования в клетках, зараженных AFP-SFV-mIL-12 (фиг. 6). Однако на 5 сутки после заражения концентрация mIL-12 несколько снижалась. В клетках, инфицированных AFP-mIL-12, уровень экспрессии был очень низким, и только незначительное повышение продукции mIL-12 отмечали с течением времени.
Тестирование цитотоксичности - оценка пролиферации клеток по включению МТТ
Клеточные линии (Hep3B, Huh-7, MCH-RH7777, Hep1-6) инфицировали AFP-LacZ, AFP-SFV-LacZ, AFP-mIL-12, AFP-SFV-mIL-12 или Ad-CMV-mIL-12 при значении «moi», равном 1000. Через 5 суток после заражения определяли выживаемость клеток с помощью теста МТТ (3-(4,5-диметилтиазолил)-2,5-дифенилтетразолий бромид) Mosmann, T. (1983) J. Immunol. Meth. 65, 55-63; Tada, H. et al. (1986) J. Immunol. Meth. 93, 157-65. Кратко, клетки промывали один раз PBS и в лунку вносили 200 мкл свежеприготовленного раствора краски МТТ (в 48-луночных планшетах). Затем клетки культивировали в течение 3-4 ч с последующим добавлением 500 мкл буфера для солюбилизации. Отбирали 100 мкл каждой пробы для определения поглощения на спектрофотометре при длине волны 570 нм.
Индукция гибели клеток после инфицирования клеток НСС гибридными векторами Ad-SFV в условиях in vitro
Как уже указывалось, репликация векторов SFV индуцирует гибель клеток, опосредованную апоптозом, в большинстве клеток позвоночных. Для оценки того, насколько это также имеет место в случае клеток НСС, инфицированных гибридными векторами Ad-SFV, клетки Hep3B и Huh-7 заражали данными векторами и определяли выживаемость клеток на 5 сутки после заражения. Как следует из данных фиг. 7, выживаемость на данной временной точке после заражения составляла менее 20% для клеток, инфицированных AFP-SFV-mIL-12 или AFP-SFV-lacZ. Однако инфицирование этих же клеток AFP-mIL-12 или AFP-lacZ или контрольным вектором Ad-CMV-mIL-12 не оказывало влияния на выживаемость клеток. Данные результаты указывают на то, что SFV реплицируется в клетках, инфицированных векторами AFP-SFV.
Детектирование Rep SFV иммунофлуоресценцией
Клетки НСС (Hep3B, Huh-7, MCH-RH7777) высевали на стеклянные покровные стекла в 6-луночных планшетах (1×105 клеток/лунку) и инфицировали AFP-mIL-12, AFP-SFV-mIL-12 или Ad-CMV-mIL-12 при значении «moi», равном 1000. Через 2 суток после заражения покровные стекла дважды промывали PBS и клетки фиксировали в метаноле при -20°С в течение 6 мин. Планшеты вновь промывали три раза PBS и инкубировали при комнатной температуре (RT) в течение 30 мин с PBS, содержащим 0,5% желатина и 0,25% BSA, для блокирования неспецифического связывания. Затем блокирующий буфер заменяли на первичные антитела (антирепликазные MAb) в разведении 1:10 в блокирующем буфере и инкубировали при комнатной температуре в течение 30 мин. Клетки вновь промывали три раза PBS-0,25% BSA и инкубировали в течение 30 мин при комнатной температуре со вторичными антителами (антимышиная кроличья сыворотка, конъюгированная с FITC, Sigma) в разведении 1:250 в блокирующем буфере. Наконец, клетки промывали три раза PBS-0,25% BSA, один раз водой и помещали на предметные стекла с использованием щитка Vecta с Dapi для окрашивания клеточных ядер.
Экспрессия Rep SFV в клетках НСС, инфицированных гибридными векторами Ad-SFV в условиях in vitro
Экспрессию Rep SFV оценивали в клетках НСС, инфицированных гибридными векторами Ad-SFV, иммунофлуоресценцией с использованием специфических моноклональных антител к данному белку. На фиг. 8 показано, что цитоплазма клеток НСС, инфицированных AFP-SFV-mIL-12 или AFP-SFV-lacZ, была положительной на Rep. В противоположность, клетки, инфицированные AFP-mIL-12 или AFP-lacZ, не были окрашены.
Опыты в условиях in vivo
Индукция ксенотрансплантатов НСС и исследование в условиях in vivo эффективности и специфичности переноса гена
Клетки Huh-7 собирали и дважды промывали не содержащей сыворотку средой.
2×106 клеток ресуспендировали в 100 мкл физиологического раствора и вводили подкожно (п/к) в правый бок голых мышей BALB/с. Через 4 недели после перевивки клеток и когда размер опухолевых узелков достигал 6-8 мм в диаметре, 1×1010 вирусных частиц AFP-LacZ (n=4) или AFP-SFV-LacZ (n=4), разведенные в 80 мкл физиологического раствора, вводили в опухоль. Контрольным животным (n=3) вводили в опухоль 80 мкл физиологического раствора. Мышей умерщвляли на 3 или 6 сутки после заражения. В это время у каждого животного извлекали опухоли и печень, заливали в О.С.Т. (Sakura, Голландия) и замораживали при -80°С. Делали срезы замороженных тканей и помещали на предметные стекла для окрашивания X-gal или анализа с помощью TUNEL. Для исследования специфичности инфицирования гибридным вектором в условиях in vivo нормальным мышам BALB/с вводили внутривенно AFP-LacZ (n=4), AFP-SFV-LacZ (n=4) или Ad/CMV-LacZ в дозе 1010 вирусных частиц/мышь. Животных умерщвляли на 3 сутки после заражения и отбирали основные органы для анализа экспрессии LacZ с помощью окрашивания X-gal.
Эффективность переноса гена гибридных векторов Ad-SFV в человеческие ксенотрансплантаты НСС у голых мышей с иммунодефицитом
Для оценки эффективности трансдукции гибридных векторов «с усеченным геномом» в условиях in vivo использовали человеческую модель НСС на основе клеток Huh-7, способных экспрессировать AFP. Клетки Huh-7 вводили подкожно голым мышам BALB/с иммунодефицитом и после получения опухолевых узелков через 30 суток животным вводили в опухоли 1×1010 вирусных частиц AFP-SFV-LacZ или AFP-LacZ в качестве контроля. Мышей умерщвляли на 3 или 6 сутки после введения вируса, извлекали опухоли и печень и исследовали окрашиванием X-gal. Как следует из данных, представленных на фиг. 9А, имела место слабая трансгенная экспрессия в срезах опухолей у животных, которым вводили AFP-LacZ. В противоположность, имела место интенсивная экспрессия LacZ в срезах опухолей у животных, которые получали AFP-SFV-LacZ (фиг. 9В). Трансгенная экспрессия отсутствовала в срезах печени животных, которым вводили AFP-LacZ или AFP-SFV-LacZ, данный факт указывает на то, что расположение векторов, вероятно, было ограничено местом введения (данные не представлены). С целью исследования того, насколько гибридные векторы Ad-SFV индуцируют апоптоз в инфицированных опухолевых клетках, срезы обработанных опухолевых клеток анализировали методом TUNEL. В пробах от мышей, которых заражали AFP-LacZ, апоптоз не наблюдали (фиг. 9С). Однако большое количество подвергшихся апоптозу клеток отмечали в опухолях мышей, которым вводили AFP-SFV-LacZ (фиг. 9D). Эти данные указывают на то, что гибридные векторы Ad-SFV не только индуцируют экспрессию гена избирательно в опухолях, но также они избирательно индуцируют гибель клеток в результате апоптоза в этих же клетках.
Специфичность гибридных векторов Ad-SFV в условиях in vivo
Для демонстрации специфичности гибридных векторов 1010 вирусных частиц AFP-LacZ, AFP-SFV-LacZ или контрольного вектора Ad/CMV-LacZ вводили внутривенно мышам BALB/с. Через 3 суток после введения векторов анализировали экспрессию β-галактозидазы в печени. Как представлено на фиг. 9 (E-F) ни AFP-LacZ, ни AFP-SFV-LacZ не были способны индуцировать детектируемую экспрессию трансгена в печени. Однако большое количество положительных на β-галактозидазу клеток в срезах ткани печени обнаруживали у тех животных, которым вводили Ad/CMV-LacZ (фиг. 9G). Данные факты подтверждают то, что экспрессия, опосредуемая гибридными векторами, является специфической для опухолевых клеток.
Индукция ортотопических НСС и генная терапия в условиях in vivo
5×105 клеток McA-RH7777 вводили в левую долю печени крыс Буффало. Через 10 суток после перевивки опухолевых клеток у каждого животного отмечали появление одного опухолевого узелка размером 7-10 мм в диаметре. Опухоли обрабатывали 1011 или 2×1011вирусных частиц AFP-mIL-12, AFP-SFV-mIL-12 или физиологическим раствором в качестве контроля. Через 2 и 4 недели после обработки животных подвергали анестезии и проводили лапаротомию для оценки роста опухоли. Также анализировали выживаемость животных. Размер опухолей оценивали по определению длины и ширины каждого узелка и данные обрабатывали по формуле: объем опухоли=(длина в мм)×(ширина в мм)×0,5236 (Janic et al., 1975).
Эффективность обработки ортотопических НСС у крыс Буффало
Для оценки противоопухолевой эффективности гибридных векторов Ad-SFV, содержащих IL-12, ортотопические опухоли НСС получали у крыс имплантацией крысиных клеток McA-RH7777 в печень. Данную модель выбрали ввиду того, что на ней была показана экспрессия AFP клетками McA-RH7777. В первом опыте животных обрабатывали одной инъекцией в опухоль 1×1011 вирусных частиц AFP-mIL-12, AFP-SFV-mIL-12 или физиологическим раствором в качестве контроля (фиг. 10А-С). У животных, получивших AFP-mIL-12, наблюдали уменьшенные размеры опухолей по сравнению с контрольными животными, у которых имело место постоянное увеличение размера опухолей в течение опыта (фиг. 10А-В). Однако обработка AFP-SFV-mIL-12 приводила к полной регрессии опухоли у 1 из 4 крыс, подвергшихся обработке, стабилизации заболевания у 2 крыс и отсутствию ответной реакции у 1 животного (фиг. 10С). Для исследования того, насколько более высокие дозы гибридного вектора могли повысить противоопухолевый эффект, проводили второй опыт, в котором животных обрабатывали инъекцией в опухоль 2×1011 вирусных частиц AFP-mIL-12, AFP-SFV-mIL-12 или физиологического раствора в качестве контроля (фиг. 10D-G). Как и в предыдущем опыте, у животных, которые получали вектор AFP/IL-12, наблюдали только слабую противоопухолевую ответную реакцию, которая выражалась только в одной полной ремиссии, у 4 животных рост опухолей замедлялся по сравнению с контролем и у 7 животных ответная реакция отсутствовала в целом из 12 обработанных животных (фиг. 10Е). Однако обработка вектором AFP-SFV-IL-12 имела более высокий эффект, приводя к полной регрессии опухолей у 4 животных (33%), частичной регрессии у 6 животных (50%), замедленному росту опухолей у 2 животных (16%), и ответная реакция отсутствовала у 2 животных (16%) в целом из 12, подвергшихся обработке животных (фиг. 10F).В данном втором опыте вектор AFP-SFV-IL-12 позволил достичь выживаемости 50% обработанных животных против 0% выживаемости у животных, обработанных AFP-IL-12 или физиологическим раствором (фиг. 10G).
Опыт по определению токсичности в условиях in vivo: определение уровней сывороточных трансаминаз и IL-12, и гистологическое исследование печени
Отбирали пробы крови у крыс, обработанных инъекцией в опухоль аденовирусными векторами AFP-SFV-IL-12 или AFP-IL-12 в дозе 2×1011 частиц или физиологическим раствором, через 4 и 8 суток после заражения. В данном опыте также участвовали крысы, которым вводили 108 частиц альфавируса SFV-IL-12. Сыворотку отделяли от крови центрифугированием при 2000 об/мин в течение 15 мин. Уровень трансаминаз определяли на автоматическом анализаторе Hitachi 911 (Boehringer Mannheim, Германия). Концентрацию IL-12 определяли с помощью ELISA. Гистологическое исследование проводили при извлечении печени у зараженных животных через 3 суток после обработки. Орган фиксировали в формалине, заливали в парафин и на микротоме делали срезы с толщиной 6 микрон. Затем данные срезы окрашивали гематоксилин/эозином.
Определение токсичности гибридных векторов Ad-SFV у крыс
Для оценки токсичности, связанной с введением гибридного вектора AFP-SFV-IL-12 определяли уровень трансаминаз (GOT, GPT и GGTL) в сыворотке крови крыс, которым вводили в опухоль 2×1011 вирусных частиц различных векторов (см. предыдущий раздел). Данное исследование также включало группу крыс, которым в опухоль вводили 108 вирусных частиц альфавирусного вектора SFV-IL-12 (фиг. 11А). У крыс, получивших аденовирусные векторы AFP-SFV-IL-12 или AFP-IL-12, определяли очень низкие уровни трансаминаз, значения которой были очень близки уровням у контрольных животных, которым вводили физиологический раствор. Однако уровни трансаминаз у животных, обработанных частицами SFV-IL-12, была достоверно выше по сравнению с другими группами (р<0,05). В данном опыте также проводили определение уровня IL-12 в сыворотке крови животных в те же временные точки. IL-12 не определяли в сыворотке крови животных, которым вводили Ad векторы AFP-SFV-IL-12 или AFP-IL-12 или физиологический раствор (фиг. 11В), указывая на то, что экспрессия трансгена в данных векторах ограничена опухолями, на основании чего можно предположить, что токсичность гибридных векторов Ad-SFV является очень низкой. Однако обработка вирусными частицами SFV-IL-12 повышала уровни IL-12 в сыворотке крови после короткого периода времени создавалась ситуация, которая могла вызвать токсичность для печени. Наконец, опыт по токсичности завершали гистологическим исследованием срезов печени, окрашенных гематоксилин/эозином, от крыс, обработанных введением в опухоль тех же векторов и в тех же дозах, которые уже описаны (фиг. 12). В данном исследовании не было выявлено гистологических различий у крыс, получивших физиологический раствор и аденовирусные векторы AFP-SFV-IL-12 или AFP-IL-12. Однако в срезах печени от крыс, обработанных вирусными частицами SFV-IL-12, наблюдали зоны с эозинофильными гепатоцитами, а также слияние данных гепатоцитов, что указывает на некоторую степень токсичности (черные стрелки, фиг. 12D).






































































Реферат
Изобретение относится к области биотехнологии, генной инженерии и медицины. Аденовирусный гибридный вектор содержит первую цепь из аденовируса, содержащую последовательность первого инвертированного концевого повтора (ITR) и сигнальную последовательность для упаковки аденовируса; первую некодирующую последовательность наполнителя; последовательность, соответствующую тканеспецифическому промотору; цепь кДНК, полученную из альфавируса, последовательность которой частично комплементарна РНК альфавируса, включающую, последовательность, кодирующую, один интересующий экзогенный ген; последовательность полиаденилирования; и последовательность второго инвертированного концевого повтора (ITR). Раскрыты также фармацевтические композиции, содержащие такой аденовирусный гибридный вектор, и их применение. Заявляемая группа изобретений может быть использована в способе переноса генетического материала в клетку, в частности, опухолевую клетку, которая предпочтительно экспрессирует альфа-фетопротеин (AFP), и к его применению для индукции иммунного ответа против чужеродных антигенов. 8 н. и 30 з.п. ф-лы, 16 ил.
Формула
i. первую цепь из аденовируса, содержащую последовательность первого инвертированного концевого повтора (ITR) и сигнальную последовательность для упаковки аденовируса;
ii. первую некодирующую последовательность наполнителя;
iii. последовательность, соответствующую тканеспецифическому промотору;
iv. цепь кДНК, полученную из альфа-вируса, последовательность которой частично комплементарна РНК альфа-вируса, включающую, по меньшей мере, последовательность, кодирующую, по меньшей мере, один интересующий экзогенный ген;
v. последовательность полиаденилирования и
vi. последовательность второго аденовирусного инвертированного концевого повтора (ITR).
а) 5'-последовательность, необходимую для репликации альфа-вируса,
b) последовательность, кодирующую неструктурные белки, необходимые для репликации РНК альфа-вируса,
c) по меньшей мере, один субгеномный промотор альфа-вируса и
d) 3'-последовательность, необходимую для репликации альфа-вируса.
i. первую цепь из аденовируса, содержащую последовательность первого инвертированного концевого повтора (ITR) и сигнальную последовательность для упаковки аденовируса;
ii. первую некодирующую последовательность наполнителя, которая представляет собой область интрона человеческой геномной гипоксантинфосфорибозилтрансферазы(НРRТ);
iii. последовательность, соответствующую тканеспецифическому промотору, который представляет собой промотор AFP;
iv. цепь кДНК, полученную из альфа-вируса, последовательность которой частично комплементарна РНК альфа-вируса, полученной из вируса SFV, которая включает последовательность, кодирующую интересующий экзогенный ген, который представляет собой hIL-12;
v. последовательность полиаденилирования из SV40;
vi. последовательность второго инвертированного концевого повтора (ITR) и
vii. вторую некодирующую последовательность наполнителя, которая представляет собой человеческую геномную С346, расположенную между элементом v и элементом vi.
i. первую цепь из аденовируса, содержащую последовательность первого инвертированного концевого повтора (ITR) и сигнальную последовательность для упаковки аденовируса;
ii. первую некодирующую последовательность наполнителя, которая представляет собой область интрона человеческой геномной гипоксантинфосфорибозилтрансферазы (HPRT);
iii. последовательность, соответствующую тканеспецифическому промотору, который представляет собой промотор AFP;
iv. цепь кДНК, полученную из альфа-вируса, последовательность которой частично комплементарна РНК альфа-вируса, полученной из вируса SFV, которая включает последовательность, кодирующую интересующий экзогенный ген, выбранный из mIL-12 и LacZ;
v. последовательность полиаденилирования из SV40;
vi. последовательность второго инвертированного концевого повтора (ITR) и
vii. вторую некодирующую последовательность наполнителя, которая
представляет собой человеческую геномную С346, расположенную между элементом v и элементом vi.
Комментарии