Анальгетическое средство на основе плазмидной днк, кодирующей hnp-1, либо hnp-2, либо hnp-3 (варианты) - RU2597789C2
Код документа: RU2597789C2
Чертежи








Описание
Изобретение (варианты) относится к области медицины, фармакологии, биотехнологии, молекулярной биологии, генной инженерии и может быть использована для анальгезии.
Важно признать, что боль является не просто индикатором основного заболевания или процесса повреждения, а самостоятельной проблемой, наносящей большой урон отдельным людям и обществу в целом. Для улучшения качества жизни облегчение боли как таковой должно стать терапевтической целью. Перспективный подход к лечению боли, к которому переходят в настоящее время, - выявление механизма и осуществление таргетной (нацеленной на конкретные молекулы) фармакологической терапии [WHO Normative Guidelines on Pain Management, Geneva June 2007].
Известно использование пептидных амидов [US4459225 (А)], фенилгидразоновых производных, в качестве противовоспалительных или анальгетических агентов [ЕР 0952159 (В1)], синтетических пептидных амидов для профилактики и лечения боли [RU 2500685 С2]. Известно использование семейства пептидов, обладающих анальгетической активностью, общей формулы A-B-Tyr-Pro (DPro, dHPro, DdHPro, DLdhPro, Hyp)-C-X [WO 2008020778 (A1)], производного пептида, вводимого подкожно или орально, имеющего анальгетическую активность, либо анти-ноцицепивную [US 2008009448 (A1)].
Недостатком химически синтезированных пептидов и их производных является возможность получения смеси искомых веществ с энантиомерами, что может привести к непредсказуемым результатам. Понятие энантиомерии играет важную роль в фармацевтике, поскольку разные энантиомеры лекарственных веществ, как правило, имеют различную биологическую активность. Использование живых систем для получения пептидных молекул позволяет получить молекулы природной структуры, гомогенную смесь.
Известна рекомбинантная плазмида, для бактериальной экспрессии, содержащая гомологичный регулон или гетерологичную ДНК, последняя может кодировать полипептид с анальгетическим эффектом [SK 278170 (В6)]. Известно моно соединение индола с новой структурой, содержащее глубоководный макрогеномный кластер генов, синтезируемое в клетках Escherichia coli [CN 102477436 (А)]. Известен полипептид из актинии Heteractis crispa, обладающий выраженным анальгетическим действием [RU 2368621 C1, RU 2404245 C1], и способ его получения [RU 2415866 C1]. Известны пептиды с большим числом мостиковых связей, выделенных их ACTINOMADURA NAMIBIENSIS, для лечения невропатической боли, вызванной воспалением [RU 2498995 (С2)]. Однако в связи с тем, что такие молекулы происходят от организмов, далеких от человека, их действие должно быть изучено особо тщательно перед применением в человеке.
Известен гибридный белок на основе конотоксина (яда улитки) MVó А и Trx, который можно использовать в подготовке обезболивающего для последних стадий рака и СПИДа, пост-операционной боли, ожогов и т.д. [CN 1487085 (А)]. Известен способ получения конотоксина с использованием бакуловируса и клеток насекомых, либо насекомых [CN 102876683 (А)]. Известны способы анальгезии и улучшения опиатной анальгезии введением омега-конопептидов TVIA (SNX-185) или MVIIA (SNX-111) [ЕР 0625162 (В1)], способы анальгезии с использованием альфа-конотоксинов [WO 2014023129 (A1), CN 102286079 (В), CN 103374066 (А)], конотоксина Lt7b [CN 102628048 (А)]. Известно использование омега-конопептида для производства медикамента для ингибирования прогресса нейропатической боли [ЕР 1336409 (В1)]. Известен полипептид скорпиона, обладающий анальгетическим и противоопухолевым действием [US 7592309 (В2), CN 101591668 (А)], и способ его применения [US 7592309 (В2)] и получения [CN 101591668 (А)], сколопендры Scolopendra subspinipes mutilans - mu-SLPTX-Ssm6a, обладающий анальгетическим эффектом [CN 102977201 (А)]. Известны пептиды яда паука Grammostola spatulata, обладающие анальгетическим действием [US 5776896 (А)], пептид HWAP-I китайского паука-птицеееда [US 6670329 (В2)]. Известны пептидные молекулы из яда змеи и их производные, гомологи, аналоги и миметики, способные индуцировать анальгезию или облегчение боли отдельно или в комбинации с другими анальгетическими молекулами [US 6613745 (В1), US 7902152 (В2)], а также укороченный нейротоксин морской змеи Lapemis hardwickii для анальгезии [US 7294697 (В2)].
Недостатком данной группы изобретений является то, что использование токсинов, даже фармацевтически чистых, все же является риском - при передозировке вплоть до смерти.
Известно использование нового полипептида - субъединицы 9.01 [CN 1345751 (А)], субъединицы 12.65 [WO 0212310 (A1)], субъединицы 9.46 [WO 0204504 (A1)], субъединицы 11.44 [CN 1352195 (А)] G-белка человека для анальгезии. Известны полипептиды на основе рецептора G-белка HFIAO41 [US 2001016336 (A1)], HLYAZ61 [ЕР 0837128 (А2)], HUVCT36 [US 5912335 (А)], Н7ТВА62 [US 5955309 (A)], org10 [US 2003162945 (A1)], org11 [WO 0200725 (A2)] а также относящиеся к семейству IGS4 [AU 779993 (В2), US 6998255 (В1)]. Известны варианты сплайсированного рецептора G-белка, индуцированного вирусом Эпштейна-Барр EBI 3 [US 5874252 (А)].
Известны способы лечения боли, обусловленной раком кости, остеоартритом, послеоперационной боли путем введения антагониста фактора роста нервов (антитела) [RU 2389509 С2, RU 2429013 С2, RU 2338555 С2], известны партнеры специфического связывания с данной молекулой [RU 2406728 С2]. Показано, что молекулы, способные ингибировать связывание между NGF и рецептором TrkA, можно использовать в качестве анальгетиков с пролонгированным эффектом [RU 2427387 С2]. Известны и антитела против фактора роста нервов, обладающие повышенной стабильностью in vivo [RU 2011149263 А], высокой аффинностью к NGF [RU 2473564 С2], и лекарственная форма препарата гуманизированного антитела против фактора роста нервной ткани [RU 2427387 С2].
Известны способы лечения:
- боли и воспаления, с использованием пептида [RU 2011105280 А], разделением взаимодействия CRMP-2 и CaV2.2, в результате чего не происходит активация кальциевого канала N-типа (CaV2.2) [WO 2012009075 (A1)], а разделением взаимодействия LI-CAM, анкирина и потенциалзависимых кальциевых каналов лечат повреждение аксонов, ингибируют выброс нейромедиатора и передачу боли и блокируют кальциевую помпу в нейронах [US 7737250 (В2)], известен аналог пептидного токсина натриевых каналов Navl.7 для анальгезии [US2013296247 (A1)].
- боли и воспаления в нейронной ткани с применением антагонистов IL-31, гуманизированного моноклонального антитела или химерного антитела [RU 2440130 С2], простатической кислой фосфатазы ("РАР"), ее активного варианта, фрагмента или производного [WO 2009064497 (A1)],
- невропатической боли с использованием антител против CCR2 [RU 2011110169 А], нейропатической боли с применением пептидов, являющихся производными просапонина [RU 2007119313 А], пептида, содержащего аминокислотную последовательность Thr-R1-Lue-Ile-Asp-Asn-Asn-Ala-Thr-Glu-Glu-Ile-Leu-Tyr, где R1 является D-аланином, оказывающего воздействие на нейродегенеративное нарушение или нарушение миелинизации [RU 2266129 С2], белков с доменом цинковые пальцы, связанным с регуляторным доменом, которые способны активировать, либо подавлять экспрессию целевого гена, связанного с нейропатической болью (VR1, NaV1.8, и TrkA) [US8466267 (В2)].
- невропатической или центральной сенсибилизационной боли, с использованием последовательности выделенного гена, который регулируется в спинном мозге млекопитающего в ответ на механистически различные первую и вторую модели невропатической или центральной сенсибилизационной боли [US 2003108906 (A1), US 2003134301 (A1), US2003138803 (A1), US2004058326 (A1)],
- хронической боли с использованием полипептидов TORC [RU 98118091 А], гистогранина и его химически стабильного аналога [СА2219437 (A1)], линейных, циклических гистограниновых пептидов и псевдопептидов на их основе [US6566327 (В1)], гибридного белка, содержащего IL-10 и IL-4 [WO 2013070076 (A1)], полипептидов на основе IL-10 [US 7261882 (В2)],
- острой и хронической боли (различных форм боли) с использованием IL-13-связывающих белков [RU 2472807 С2], 1b-12/р40-связывающих белков [RU 2012124438 А], IL-12/р41-связывающих белков [RU 2008103312 А], IL-17-связывающих белков [RU 2011140335 А], простогландин Е2-связывающих белков [RU 2011104228 А], иммуноглобулинов с двойным вариабельным доменом [RU 2515108 С2], иммуноглобулинов с двойным вариабельным доменом против простогландина Е2 [RU 2011104348 А], борьбы с хроническими или острыми приступами боли с использованием ЕЕ3-семейства белков [RU 2004114989 А],
- висцеральной боли с использованием полипептида, повышающего гуанилатциклазную активность рецепторов [RU 2433133 (С2)],
- невропатической боли, остеоартритической боли, воспалительной боли с использованием IL-1-связывающих белков [RU 2012154210 А], воспалительных процессов артрита и других TNF-a опосредованных нарушений или патофизиологических механизмов, в частности различных форм боли, с применением стабильных и растворимых антител - ингибиторов TNF-a [RU 2415151 С2], боли при артрите, с применением гуманизированного антитела-антагониста CGRP [RU 2467765 С2], боли, индуцированной по меньшей мере одним противораковым агентом, с применением как минимум одного ботулинического токсина или пептида на его основе [RU 2483747 С2], в том числе совместно с опиатным производным [RU 2434637 С2], боли с применением РАМ, ассоциированного с Мус белком [RU 2345785 С2], с применением подобного рецептору галанина GPCR полипептида [US 2004053244 (A1), US 2003073115 (A1), WO 0168843 (A1)], контулакина-G и его дегликозилированной формы [US 6696408 (В1), US 6525021 (В1)], с применением пептидных ингибиторов протеинкиназы С [US 7939493 (В2)], висцеральной боли путем введения антител-антагонистов, направленных против пептида, связанного с геном кальцитонина [RU 2012106468 А],
- боли в животе, с применением пептидов предпочтительно из Lactobacillus rhamnosus [RU 2011136454 А].
Известны способы снижения болевой чувствительности при физиологических и патофизиологических условиях (например, при аллодинии и гипералгезии), особенно восприятия боли, которая связана или опосредована механической чувствительностью через TRPA1, с использованием антагонистических антител TRPA1, предпочтительно моноклональных антител [RU 2430750 С2]. Известны противоболевые/ противовоспалительные агенты, выбранные из следующих классов антагонистов рецепторов, агонистов рецепторов и ингибиторов ферментов, каждый класс, действующий по различным молекулярным механизмам действия на боль и подавления воспаления: (4) антагонисты брадикининового рецептора (пептиды), антагонисты кальцитонин-ген опосредованного пептидного (CGRP) рецептора (альфа-CGRP-(8-37), (8) антагонисты интерлейкинового рецептора (Lys-D-Pro-Thr) [RU 2180852 С2], белка бета-целлюлина (ВТС) [JP 2000198744 (А)]. Известны способы уменьшения боли при модулируемом CRF2R расстройстве с использованием агонистов рецептора кортикотропин-рилизинг фактора (CRF 2R) [RU 2385878 С2], лечения боли с использованием пептида, обладающего свойствами амилина [RU 2385878 С2], пептидного агониста NOP [RU 2012106820 А], пептидов, содержащих два и более аминокислотных остатка фрагмента 11-28 вазоактивного интестинального пептида (VIP), доставляемых в ЦНС [WO 9104041 (A1)], производного пептида на основе кальцитонина [US 5446026A], полипептидов человека CGRP-RCF [US 5710024 (A), WO 9803534 (A1)], полипептидов - гуманизированных рецепторов CGRP, в том числе для лечения головной боли, хронической головной боли от напряжения, сильной приступообразной головной боли с периодическими рецидивами, профилактики мигреней [US 7193070 (В2)], полипептидов - новых сплайсированных вариантов 11cb человека [US 6033872 (А)], агонистов и антагонистов последнего белка [WO 9928492 (A1)], рецепторов с семью трансмембранными доменами [US 5824504 (A), US 6277960 (В1), US 6277977 (В1), US 5955308 (A), US 6221627 (B1), US 2002106766 (A1), US 5994098 (A), US 6174994 (B1), US 6037146 (A), US 5874243 (А)], пептида, имеющего фосфодиэстеразную активность [WO 0166716 (A1)], полипептидов - человеческих протеин-киназ hYAK3 и HOACF72 [ЕР 0870825 (В1) и US 5972606 (А), соответственно].
Известен гибридный белок на основе нейротрофического фактора головного мозга, получаемый с использованием прокариотической системы экспрессии растворимого белка [CN 1978466 (В), WO 9942480 (A1)]. Известны полипептиды на основе рецептора нейротензина типа 2 для лечения боли [US 6008050 (А)]. Известен конъюгат нейротензина или его аналогов с терапевтическим пептидом, который может быть использован для индукции гипотермии или анальгезии [WO 2010063122 (A1)]. Известны конформационно оптимизированные аналоги альфа-меланотропина, оказывающие специфическое воздействие на ЦНС [US 4649191 (А)]. Известен синтетический пептид, модулирующий выброс нейротрансмиттера, сходный со SNARE [KR 20090041066 (А)].
Известны пептиды, обладающие множеством функций, в том числе анальгезией, такие как пептиды с большим количеством мостиковых связей, выделяемые из Actinomadura namibiensis [RU 2010144778 А], пептид структуры DGSVVVNKVSELPAGHGLNVNTLSYGDLAAD [RU 2508296 С2], пептидный модулятор пуринергических рецепторов РТ1 [RU 2422459 C1], дипептиды, содержащие на N-концевом аминокислотном остатке 2-тиоацильную группу, в качестве ингибиторов вазопептидазы [RU 2298559 С2], тимус-специфический белок [RU 2398776 С2], полипептид, улучшающий метаболизм кальция [JPH03178993 (А)], а также пролекарственные композиции с высокой степенью проникновения на основе пептидов и родственных пептидам соединений [RU 2011149796 А].
Известен комбинированный подход к анальгезии - с использованием синергического сочетания селективного ингибитора нейронального транспортера норадреналина и анальгезирующего средства в терапевтическом лечении позвоночных животных, включая человека, для обезболивания или для профилактики или облегчения боли [US 8188048 В2]. Известны пептиды-аналоги динорфина, которые при совместном введении с наркотическими или анальгетическими средствами усиливают действие последних, в том числе энкефалинов и аналогов бета-эндорфина [ЕР 0096592 В1].
Известны способы увеличения фармакологической активности веществ при оральном и парентеральном введении композиций, с использованием пептида Tyr-Gly-Gly-Phe-Met, связанного с группой-переносчиком, формирующего таким образом предшественник лекарства, используемый в том числе для лечения или облегчения боли [AU 2002228260 (В2)].
Использование одного высокоэффективного агента для анальгезии, простого и дешевого в производстве, является более выгодным вариантом.
Известно использование пептида дефенсина Нnр-1, одного или в комбинации с нейропептидом М, в качестве терапевтического агента, для профилактики и/или лечения в том числе периферических сосудистых заболеваний, включая анестезию, распространяющуюся боль, каузалгию (жгучая боль) [WO 2009/043461 А1].
Использование молекул белковой природы, которые не задумывались в качестве индукторов иммунного ответа, в том числе антительного, может быть осложнено именно этими последствиями, что может привести к побочным эффектам, например, связанным с формированием аутоантител.
Известно использование малой интерферирующей РНК, а также короткой шпилечной РНК для генного нокдауна субъединицы NR1 подкожного рецептора N-метил-D-аспартата для снижения воспалительной боли или невыносимой хронической боли, в особенности клинической хронической боли и боли при ожогах [US8372817 (В2), US8575330 (В2), US 8361985 (В2)]. Известен РНК-интерферирующий агент для лечения хронической боли [WO 2013126963 (A1)]. Однако препараты РНК довольно быстро разрушаются вне организма, также требуются специальные условия хранения, что ставит промышленную применимость препаратов на основе РНК под вопрос.
Известен способ длительной анальгезии, по которому пациенту вводят миогенные клетки, в которых с соответствующей ДНК синтезируется пептид, активирующий опиоидный рецептор, либо опосредующий связывание субстанции Ρ с рецептором [US 7166279 (В2)]. Однако осуществление данного способа довольно сложное и связано со многими рисками.
Известны методы лечения депрессии и боли, агентом может быть белок, РНК или ДНК [WO 2013177484 (A1)]. Известны лентивирусные векторы для лечения боли, содержащие G-белок вируса бешенства [ЕР 1425403 (В1)], а также аденовирус, кодирующий IL-24 [CN 101518655 (А)], в том числе для облегчения боли при раке. Известен онкостатин М, либо его гомолог, синтезирующиеся с вектора для переноса генов: лентивируса, ретровируса, вируса Сендай, аденовируса и адено-ассоциированного вируса, в том числе для лечения боли [JP 4803789 (В2)]. Известно лечение аллодинии, гипералгезии, спонтанных болей и фантомных болей с использованием кометина - полипептида, который может быть доставлен как полипептид, либо введением экспрессионного вектора для экспрессии кометина, клеточная линия, трансформированная, либо трансфецированная данным белком и капсула, содержащая указанные клетки [WO 2013034157 (A1)]. Вирусные векторы имеют ряд недостатков, в их числе то, что они дорогостоящи и могут вызывать воспалительную реакцию, что исключает повторное введение вектора. Также вирусные вектора обладают способностью реплицироваться, что снижает степень контроля над экспрессией таргетного белка, что, в свою очередь, не всегда желательно и применимо, в особенности в отношении анальгезии.
Известна рекомбинантная ДНК-вакцина на основе вектора pVAX1, нацеливающегося на опухоль, в том числе для лечения боли при онкологии, содержащего ген, кодирующий циклооксигеназу-2 (СОХ-2), мышиный убиквитин Mubi, а также ISS иммуностимулирующие последовательности ДНК (pVAX1-mUbi-ISS-COX-2-ISS) [CN 101648011 (А)]. Однако данная конструкция нацелена вызвать иммунный ответ на нее, цитотоксический, для чего в нее введены элементы, обуславливающие и усиливающие иммуногенность. Известна в том числе фармацевтическая композиция, в которой содержится вектор, в том числе плазмидная ДНК, и фармацевтически приемлемый эксципиент, либо адъювант, для облегчения, профилактики или лечения боли, вектор содержит нуклеотидную последовательность, содержащую хотя бы одну область, модулирующую экспрессию рецептора VR1 [US 2006154886 (A1)]. В данном случае анальгезия осуществляется через блокирование рецептора, связанного с ионным каналом - неспецифическим проводником катионов. Использование плазмидной ДНК, несущей ген, с которого в клетках млекопитающих синтезируется дефенсин, в качестве средства, индуцирующего анальгезию, позволит также минимизировать возможность развития патогенов в организме: как правило, при болях любой этиологии ухудшается самочувствие и уменьшается способность организма противостоять инфекциям, в результате, после, например, оперативного вмешательства, могут назначать антибиотики, а использование природного противомикробного пептида позволит объединить и анальгезию, и защитный эффект, в результате, можно отказаться от приема антибиотиков.
Близкими аналогами предлагаемого изобретения являются изобретения, описанные в патенте CN 101648011 (А) и заявке на изобретение US 2006154886 (A1), в которых раскрыты анальгетики в форме плазмидной ДНК.
Показано, что при внутримышечном введении экспрессионного ДНК вектора он поглощается мышечными клетками, и происходит экспрессия белка, закодированного в вводимом векторе [J.A. Wolff et al., Science 247, 1465 (1990); G. Ascadi et al., Nature 352, 815 (1991)]. Показано, что плазмиды поддерживаются в виде эписомы и не реплицируются. Постоянная экспрессия была продемонстрирована после внутримышечной инъекции также в скелетные мышцы крыс, рыб и приматов, а также сердечную мышцу крыс [H. Lin et al., Circulation 82, 2217 (1990); R.N. Kitsis et al., Proc. Nati Acad. Sci. (USA) 88, 4138 (1991); E. Hansen et. al., FEBS Lett. 290, 73 (1991); S. Jiao et al., Hum. Gene Therapy 3, 21 (1992); J.A. Wolff et al., Human Mol. Genet. 1,363 (1992)]. Возможно и использование иных технологий доставки плазмидной ДНК в клетки различных тканей, например, безыгольного шприца, позволяющего осуществить доставку в клетки различных тканей живых животных [Furth et al., Analytical Biochemistry, 205, 365-368, (1992)].
Показано, что в норме мышечные волокна не экспрессируют антигены МНС, однако при воспалении, связанном с инфекцией, или в присутствии интерферона гамма мышечные волокна способны продуцировать антигены в составе комплекса с МНС первого класса, либо даже второго класса [MECKERT, Р.С, HONTEBEYRIE-JOSKOWICZ, М., CHAMBO, J., LEVIN, M., and LAGUENS, R.P. (1991). Trypanosoma cruzi: Aberrant expression of class II major histocompatibility complex molecules in skeletal and heart muscle cells of chronically infected mice. Exp. Parasitol. 72, 8-14; Hohlfeld and ENGEL, A.G. (1984) The immunobiology of muscle. Immunol. Today 15, 269-274.; MANTEGAZZA, R., and BERNASCONI, P. (1994). Cellular aspects of myositis. Curr. Opin. Rheumatol. 6, 568-574, Hartikka J, Sawdey M, Cornefert-Jensen F, Margalith M, Barnhart K, Nolasco M, Vahlsing HL, Meek J, Marquet M, Hobart P, Norman J, Manthorpe M. An improved plasmid DNA expression vector for direct injection into skeletal muscle. Hum Gene Ther. 1996 Jun 20; 7(10):1205-17], в связи с чем предпочтительным является введение плазмидной ДНК, при котором травмирование мышечной ткани минимизировано.
Показано, что даже при иммуногенности в испытаниях на лабораторных животных у человека препараты на основе плазмидной ДНК не являются иммуногенными [Saade F, Petrovsky N. Technologies for enhanced efficacy of DNA vaccines. Expert Rev Vaccines. 2012 Feb; 11 (2): 189-209. doi: 10.1586/erv. 11.188]. Исследователям также не удалось найти доказательства патологических изменений после повторных инъекций ДНК у мышей или кроликов [Parker SE, Borellini F, Wenk ML, et al. Plasmid DNA malaria vaccine: tissue distribution and safety studies in mice and rabbits. Hum. Gene Ther. 1999; 10(5):741-758.]. В 2007 году в руководстве FDA США по ДНК-вакцинам пришли к выводу, что доклинические исследования не требуются для оценки эффекта в отношении аутоиммунных заболеваний.
Плазмидная ДНК должна содержать существенные для организмов ее поддержания и использования элементы, вкупе с соответствующими регуляторными последовательностями. Регуляторные последовательности - нуклеотидные последовательности, способные повлиять на экспрессию гена на уровне транскрипции и/или трансляции, а также на механизмы, обеспечивающие существование и поддержание функционирования плазмидной ДНК.
Существенными для прокариотической системы являются ориджин репликации и репортерный ген. Бактериальные элементы плазмидной ДНК не должны отрицательно влиять на экспрессию в клетках млекопитающих и обуславливать побочный эффект от применения плазмидной ДНК. В литературе имеются данные о том, что использование гена устойчивости к ампициллину в качестве репортерного гена может быть нежелательным в связи с развитием реакции у пациентов на ампициллин, однако авторы считают такие последствия связанными с низким качеством очистки плазмидной ДНК, но не самим элементом.
Существенными элементами плазмид для использования у млекопитающих являются промотор, лидерная последовательность мРНК, терминирующая последовательность.
Промотор является важным компонентом плазмиды, который запускает экспрессию интересующего гена. Классические промоторы для плазмидных ДНК-компонентов препаратов - это CMV человека/ немедленно-ранний или CMV-chicken-β actin (CAGG) промотор. Промоторы CMV используется для большинства ДНК-вакцин, так как они опосредуют высокие уровни конститутивной экспрессии в широком диапазоне тканей млекопитающих [Manthorpe M, Cornefert-Jensen F, Hartikka J, et al. Gene therapy by intramuscular injection of plasmid DNA: studies on firefly luciferase gene expression in mice. Hum. Gene Ther. 1993; 4(4):419-431] и не подавляют пропитывание downstream. Увеличение уровня экспрессии наблюдают при изменении CMV промотора, например, включением HTLV-1R-U5 downstream от промотора цитомегаловируса или при использовании химерного SV40-CMV промотора [Williams JA, Carnes АЕ, Hodgson СР. Plasmid DNA vaccine vector design: impact on efficacy, safety and upstream production. Biotechnol. Adv. 2009; 27(4):353-370]. Альтернативой CMV промоторам служат тканеспецифические промоторы хозяина, которые позволяют избежать конститутивной экспрессии антигенов в неподходящих тканях, что в целом приводит к снижению иммуногенности [Cazeaux Ν, Bennasser Y, Vidal PL, Li Z, Paulin D, Bahraoui E. Comparative study of immune responses induced after immunization with plasmids encoding the HIV-1 Nef protein under the control of the CMV-IE or the muscle-specific desmin promoter. Vaccine. 2002; 20(27-28):3322-3331].
Лидерная последовательность мРНК также играет большую роль. Последовательность Козака непосредственно перед стартовым кодоном ATG позволяет увеличить экспрессию [Kozak M. Recognition of AUG and alternative initiator codons is augmented by G in position +4 but is not generally affected by the nucleotides in positions +5 and +6. EMBO J. 1997; 16(9):2482-2492]. Наличие интрона в плазмиде downstream от промотора может повысить стабильность мРНК и увеличить экспрессию гена.
Использование видоспецифичных кодонов позволяет увеличить экспрессию гена [Frelin L, Ahlen G, Alheim M, et al. Codon optimization and mRNA amplification effectively enhances the immunogenicity of the hepatitis С virus nonstructural 3/4A gene. Gene Ther. 2004; 11(6):522-533].
На экспрессию генов можно повлиять путем изменения терминирующей последовательности, которая необходима для сохранения стабильности мРНК, надлежащего прекращения транскрипции и экспорта мРНК из ядра, в том числе ее укорачиванием. Во многих современных ДНК-вакцинах используют последовательность терминатора транскрипции бычьего гормона роста [Montgomery DL, Shiver JW, Leander KR, et al. Heterologous and homologous protection against influenza A by DNA vaccination: optimization of DNA vectors. DNA Cell Biol. 1993; 12(9):777-783]. Полиаденилирование (полиА) необходимо для стабилизации транскрипта. Изменение последовательности полиА может привести к увеличению уровня экспрессии гена [Norman JA, Hobart Ρ, Manthorpe Μ, Feigner Ρ, Wheeler С.Development of improved vectors for DNA-based immunization and other gene therapy applications. Vaccine. 1997;15(8):801-803]. В плазмиде pVAXl (Invitrogen, Carlsbad, С А) область терминатора бычьего гормона роста содержит область гомопурина, которая чувствительна к нуклеазе. Показано, что альтернативная полиА последовательность может значительно улучшить стабильность плазмиды к нуклеазе [Azzoni AR, Ribeiro SC, Monteiro GA, Prazeres DMF. The impact of polyadenylation signals on plasmid nuclease-resistance and transgene expression. J Gene Med. 2007; 9:392-402]. Введение двух стоп-кодонов позволяет увеличить эффективность терминатора транскрипции.
Оптимальная конструкция плазмиды для осуществления назначения должна объединять «бактериальные» и «эукариотические» элементы, с соответствующими регуляторными последовательностями, чтобы обеспечить высокую копийность в процессе производства и высокий уровень экспрессии у млекопитающих [Saade F, Petrovsky Ν. Technologies for enhanced efficacy of DNA vaccines. Expert Rev Vaccines. 2012 Feb; 11 (2): 189-209. doi: 10.1586/erv.l 1.188].
Для создания анальгетического средства по настоящему изобретению можно использовать плазмидную ДНК, как минимум, стандартно применяющуюся для доставки генов и их экспрессии в организме млекопитающего, в том числе человека, а также оптимизированную по вышеперечисленным параметрам, в том числе подробно описанным в статье Williams et al., 2009 [Williams JA, Carnes AE, Hodgson CP. Plasmid DNA vaccine vector design: impact on efficacy, safety and upstream production. Biotechnol. Adv. 2009; 27(4):353-370]. В руководстве FDA (2007) заявлено, что исследования биораспределения вещества после его введения в организм могут быть отменены для ДНК-вакцин, производимых клонированием нового гена в плазмидный вектор, в отношении которого ранее документально установлено приемлемые биораспределение и профиль интеграции. В руководстве ВОЗ (2007) заявлено, что исследования биологического распределения и сохранения требуются, если еще не имеется значительный опыт работы с почти идентичным или аналогичным продуктом. В руководстве ЕМЕА (2006) заявлено, что опыт работы с векторной системой позволит оптимизировать и сфокусироваться на доклинических исследованиях. Исследования по безопасности с использованием ДНК-векторов с различными клонированными генами продемонстрировали аналогичное биораспределение [Sheets RL, Stein J, Manetz TS, Duffy С, Nason M, Andrews C, Kong WP, Nabel GJ, Gomez PL. Biodistribution of DNA plasmid vaccines against HIV-1, Ebola, Severe Acute Respiratory Syndrome, or West Nile virus is similar, without integration, despite differing plasmid backbones or gene inserts. Sheets RL, Stein J, Manetz TS, Duffy C, Nason M, Andrews C, Kong WP, Nabel GJ, Gomez PL. Toxicol Sci. 2006 Jun; 91(2):610-9. Epub 2006 Mar 28.] и токсикологию [Sheets RL, Stein J, Manetz TS, Andrews C, Bailer R, Rathmann J, Gomez PL. Toxicological safety evaluation of DNA plasmid vaccines against HIV-1, Ebola, Severe Acute Respiratory Syndrome, or West Nile virus is similar despite differing plasmid backbones or gene-inserts. Toxicol Sci. 2006 Jun; 91(2):620-30. Epub 2006 Mar 28]. Для плазмидной ДНК для применения у млекопитающих, кроме человека, требования менее строгие, в связи с чем возможно использование более широкого спектра плазмид.
Для ДНК, предназначенной для использования человеком, может быть полезно иметь конечный продукт ДНК в фармацевтически приемлемом носителе или буферном растворе. Фармацевтически приемлемые носители или буферные растворы известны из уровня техники и включают те, которые описаны в различных текстах, таких как, например, Remington's Pharmaceutical Sciences.
Прототипом вариантов изобретения является изобретение, раскрытое в заявке на изобретение WO 2009/043461 A1: применение дефенсина HNP-1 для лечения в том числе боли.
Дефенсины составляют большое семейство низкомолекулярных (3-5-кДа) цистеин-богатых катионных пептидов, стабилизированных несколькими (как правило, тремя) дисульфидными связями [Ganz, Т. 2002. Immunology: Versatile defensins. Science 298: 977-979, Lehrer, R.I. and Ganz, T. 2002. Defensins of vertebrate animals. Curr. Opin. Immunol. 14: 96-102.], которые способны к киллингу широкого спектра патогенов, включая разнообразные бактерии, грибы, а также оболочечные вирусы. У человека это семейство представлено α-субсемейством (HNP) и β-субсемейством (hBD) дефенсинов. Альфа-дефенсины наиболее представлены в нейтрофилах и клетках Панета. HNP-1 и HNP-3 содержат в своем составе всего 30 аминокислотных остатков, эти пептиды идентичны друг другу за исключением замены аланина на аспарагин в положении 1. HNP-2 - протеолитический продукт HNP-1 и HNP-3, содержит 29 аминокислотных остатков (отсутствует первая аминокислота с N-конца). Наличие дисульфидных связей обеспечивает сохранение устойчивости молекул дефенсинов к многочисленным лейкоцитарным и микробным протеиназам и сохранение антибиотических свойств в очаге воспаления и тканевой деструкции [Levy О. Antimicrobial proteins and peptides: anti-infective molecules of mammalian leukocytes. J. of Leukocyte Biology.2004; 76: 909-926]. Получение данных молекул правильной пространственной структуры является довольно сложной задачей.
Гены, кодирующие дефенсины, образуют кластер в локусе р22-23 на 8 хромосоме. HNP 1-3 кодируются двумя генами HDEFA1 и HDEFA3. Каждый ген дефенсинов содержит несколько экзонов, которые кодируют препропептид.
Вначале дефенсины синтезируются в виде предшественников, как препропептиды, длина которых составляет 94 аминокислотных остатка. Препропептиды содержат сигнальный участок (в среднем 19 аминокислотных остатков), анионный участок (в среднем 45 аминокислотных остатков) и собственно «зрелый» пептид. В результате протеолитического отщепления в эндоплазматическом ретикулуме от препропептида происходит удаление сигнального участка и образование продефенсина (в среднем 75 аминокислотных остатков). Последующее созревание (отщепление 45 аминокислотных) остатков происходит в зрелых гранулах нейтрофилов. α-Дефенсины обнаружены также в NK клетках, В-лимфоцитах, γδ Т-лимфоцитах, моноцитах/макрофагах и эпителиальных клетках [А.С. Будихина, Б.В. Пинегин. Дефенсины - мультифункциональные катионные пептиды человека. Иммунопатология, аллергология, инфектология. 2008, №2:31-40]. Авторами настоящего изобретения впервые продемонстрировано, что введение плазмидной ДНК, несущей ген, кодирующий ΗΝΡ-1/ ΗΝΡ-2/ ΗΝΡ-3, приводит к анальгезии, причем при этом наблюдали увеличение уровня бета-эндорфина в сыворотке.
Недостатки аналогов и прототипа приведены непосредственно после их описания. Заявленное изобретение (варианты) свободно от этих недостатков.
Технический результат от использования вариантов изобретения выражается, во-первых, в расширении спектра анальгетических средств. При плохой переносимости или непереносимости аналогов представитель предложенных вариантов анальгетика позволит осуществить анальгезию, за счет чего пациент получит возможность осуществить действия, которые не мог, либо не хотел осуществить без анальгезии, например, решиться на требуемую процедуру, а также улучшит качество жизни, например, получит меньше неприятных ощущений, чем без использования анальгетика, либо сможет облегчить острую, либо хроническую боль. Указанный технический результат достигается тем, что используют анальгетик по настоящему изобретению (варианты).
Кроме того, технический результат заключается в увеличении длительности анальгезии и достигается тем, что используют плазмидную ДНК, с которой после введения в организм синтезируется альфа дефенсин человека HNP-1/HNP-2/HNP-3; а также тем, что нуклеотидная последовательность, кодирующая дефенсин человека, содержит элементы, обуславливающие стабильность мРНК и, соответственно, увеличивающие время полужизни мРНК, в результате синтез белка с одной молекулы мРНК осуществляется большее количество раз, а также в результате увеличивается количество синтезируемого белка; а также тем, что нуклеотидная последовательность, в которой закодирован непосредственно препробелок альфа дефенсин человека, кодонно оптимизирована для экспрессии в млекопитающих, в результате синтез белка идет интенсивнее. При внедрении в практику это позволит существенно снизить количество вводимой плазмидной ДНК (в 10-50 раз), по сравнению с дозами, используемыми в настоящее время в отечественной и мировой практике при генной терапии.
Помимо этого, технический результат заключается в увеличении безопасности анальгезии. Указанный технический результат достигается тем, что в качестве действующего вещества используется не белок, на который могут образоваться антитела, что при применении молекулы, используемой в организме ее происхождения, может вызвать серьезные побочные эффекты, а плазмидная ДНК, которая существует в виде эписомы и не интегрирует в геном, с которой синтезируется и затем секретируется из клетки белок человека, в результате осуществляется неиммуногенное, безопасное использование дефенсина человека для анальгезии. Указанный технический результат также достигается тем, что синтезируемый в организме с плазмидной ДНК белок подвергается естественному посттрансляционному процессингу, а также обеспечивается правильный фолдинг белка за счет клеточных шаперонов. Данные модификации труднодостижимы при производстве белков, что может драматически сказаться на ряде их функций. Указанный технический результат также достигается за счет того, что используются природные механизмы метаболизма и катаболизма действующего вещества, без образования токсичных продуктов, благодаря естественной природе плазмидной конструкции и кодируемого ею белка, а также идентичности образуемого белка эндогенному аналогу. Указанный технический результат также достигается тем, что плазмидная ДНК не реплицируется после введения в организм млекопитающего, что позволяет осуществлять контроль над количеством синтезируемого белка, и, соответственно, над анальгезией. Указанный технический результат достигается и за счет наличия в плазмидной ДНК таких регуляторных последовательностей, как сайленсер и/или инсулятор, в одном из вариантов изобретения, благодаря чему также осуществляется контроль над количеством синтезируемого белка, и, в принципе, синтезом белка, как таковым, и, соответственно, над анальгезией: при необходимости есть возможность в быстрый срок остановить, либо уменьшить экспрессию гена. В последнем варианте возможно и осуществление тканеспецифической экспрессии при необходимости.
Технический результат также заключается в упрощении и удешевлении производства анальгетика за счет избегания сложностей производства и процессов очистки белковых препаратов in vitro, благодаря тому, что синтез белка происходит in vivo. Производство, очистка и хранение ДНК препаратов экономически выгоднее, чем белковых, так как первые более стабильны, их можно нарабатывать в больших количествах и с меньшими затратами.
Показано, что изменение концентрации бета-эндорфина в плазме крови находится в прямой зависимости от вида болевого синдрома, его интенсивности и эффективности анальгезии, что может служить критерием оценки эффективности обезболивания [Бета-эндорфин - маркер эффективности обезболивания при острой боли и хроническом болевом синдроме у онкологических больных / З.В. Павлова [и др.] // Проблемы клинической медицины. - 2007. - N1. - С. 36-40. - ISSN 1817-8359]. При введении анальгетика по настоящему изобретению наблюдали увеличение уровня бета-эндорфина в сыворотке (Фиг. 3).
По временным характеристикам можно выделить два типа боли:
- острую боль - новую, недавнюю боль, неразрывно связанную с вызвавшим ее повреждением и, как правило, являющуюся симптомом какого-либо заболевания, исчезает при устранении повреждения [Eddy N.B., Leimbach D. J. // Pharmacol Exp Ther. - Mar; 107(3):385-93. - 1953] (в том числе пред- и послеоперационная, посттравматическая, при ожогах, острая боль во время рождения ребенка, боль, вызванная травмой спинного мозга, острая головная боль, боль при ВИЧ/СПИДе, кризисе серповидных клеток, при невралгии тройничного нерва, панкреатите и других болях в ЖКТ, при инфаркте миокарда и других крупных сердечных событиях, интервенционная боль (при диагностических и терапевтических процедурах), острая при хронической боли [WHO Normative Guidelines on Pain Management, Geneva June 2007], которая длится до 2-3 месяцев, причем может иррадиировать, и
- хроническую боль - продолжающуюся длительный период времени (свыше 2-3 месяцев) даже после устранения причины, ее вызвавшей, часто приобретает статус самостоятельной болезни, например, воспалительного процесса [Eddy N.B., Leimbach D.J. // Pharmacol Exp Ther. - Mar; 107(3):385-93. - 1953], причем наблюдается снижение эффективности анальгетиков. К хронической боли можно отнести хроническую боль при злокачественной болезни (включая боль у больных раком, ВИЧ/СПИД, при боковом амиотрофическом склерозе (ALS), рассеянном склерозе, при конечной стадии отказа органа, расширенной хронической обструктивной болезни легких, расширенной застойной сердечной недостаточности, паркинсонизме) и хроническую боль, не связанную со злокачественной болезнью (хронические скелетно-мышечные боли, такие как спинная боль или боли в пояснице, при хроническом дегенеративном артрите, остеоартрите, ревматоидном артрите, миофасциальная и ревматическая боли, хроническая головная боль, мигрень, боли в костях; невропатические боли (в том числе боли при нервных сжатиях, травмах, боли после повреждения нерва и ампутации), при диабетической невропатии, сложные региональные болевые синдромы (тип I и тип II), при спазмах скелетных мышц, при постгерпетической невралгии, хронические боли после хирургических вмешательств; висцеральная боль (при растяжении полости внутренностей и коликах); и хроническая боль при серповидно-клеточной анемии [WHO Normative Guidelines on Pain Management, Geneva June 2007].
С использованием настоящего анальгетика возможно облегчить и нивелировать оба описанных типа боли. Как видно из уровня техники, плазмидный анальгетик, с которого в клетках млекопитающих синтезируется альфа дефенсин человека, неизвестен. Множественность и характер элементов плазмидной ДНК требуют уточняющих характеристик. Произведенная характеристика изобретения, в части «сущность изобретения», в части «подробное описание изобретение», поясняет «формулу изобретения» и включает подробное описание существенных элементов плазмидной ДНК, что позволяет осуществить изобретение во всех представленных вариантах, за счет чего достигается описанный технический результат.
Сущность изобретения
Предложена плазмидная ДНК для транзиентной экспрессии в клетках млекопитающих, представленная остовом, содержащим прокариотические элементы, ориджин репликации и репортерный ген, и эукариотические элементы, сильный промотор, лидерную последовательность мРНК, а также регуляторные последовательности для указанных элементов, от одного сайта для клонирования гена интереса и от одного сайта для посадки от одного праймера для анализа состава плазмидной ДНК, и полинуклеотидом, представленным секреторной последовательностью, фрагментом, кодирующим альфа дефенсин человека HNP-1, либо HNP-2, либо HNP-3, кодонно оптимизированными для экспрессии в клетках млекопитающих, и терминирующей последовательностью. В одном из вариантов изобретения препробелок представлен SEQ ID NO: 1, либо SEQ ID NO: 2, либо SEQ ID NO: 7, либо SEQ ID NO: 8. В одном из вариантов изобретения полинуклеотид представлен SEQ ID NO: 3, либо SEQ ID NO: 4, либо SEQ ID NO: 5, либо SEQ ID NO: 6, либо SEQ ID NO: 9, либо SEQ ID NO: 10, либо SEQ ID NO: 11, либо SEQ ID NO: 12.
Также предложены продуцент такой плазмидной ДНК на основе бактериальной клетки (варианты) и анальгетическое средство на основе такой плазмидной ДНК в эффективном количестве, также содержащее фармацевтически приемлемый эксципиент, для применения у млекопитающих, в частности, человека (варианты).
Подробное описание изобретения
Плазмидную ДНК экономически наиболее выгодно нарабатывать в прокариотических клетках, преимущественно бактериальных клетках. В связи с этим плазмидная ДНК по настоящему изобретению содержит элементы для поддержания и амплификации, преимущественно в больших количествах, в клетках бактерий. Такими существенными элементами являются бактериальные ориджин репликации, для поддержания в клетке со средней, предпочтительно высокой, копийностью, и репортерный ген для возможности селекции штамма-продуцента. Подходящий ориджин репликации представлен pM1 (der.), ColE1 (der.) и F1, pUC и F1, но не ограничивается ими. Подходящий репортерный ген представлен геном устойчивости к антибиотику, например, ампициллином, премущественно канамицином, но не ограничивается ими.
Плазмидная ДНК по настоящему изобретению содержит элементы для эффективного функционирования в клетках млекопитающих.
В одном из вариантов изобретения препробелок альфа дефенсин человека HNP-1, либо HNP-2, либо HNP-3, содержит гетерологичную секреторную последовательность вместо нативной, и, в частности, охарактеризован SEQ ID NO: 7, SEQ ID NO: 8, в результате нуклеотидная последовательность гена альфа дефенсин человека охарактеризована SEQ ID NO: 9, SEQ ID NO: 10, либо SEQ ID NO: 11, SEQ ID NO: 12, соответственно.
Подробное описание изобретения
Настоящее изобретение представляет собой анальгетическое средство на основе плазмидной ДНК для транзиентной экспрессии в организме млекопитающего, в том числе человека, с отсутствующей способностью к репликации в клетках такого организма, содержащей нуклеотидную последовательность, с которой синтезируется альфа дефенсин человека HNP-1, либо HNP-2, либо HNP-3, препробелок альфа дефенсин человека HNP-1, либо HNP-2 охарактеризован SEQ ID NO: 1, препробелок альфа дефенсин человека HNP-3, либо HNP-2 охарактеризован SEQ ID NO: 2. Анальгетическое средство также содержит фармацевтически приемлемый эксципиент.
Плазмидная ДНК содержит остов, представленный «прокариотическими» и «эукариотическими» элементами.
Плазмидную ДНК экономически наиболее выгодно нарабатывать в прокариотических клетках, преимущественно бактериальных клетках. В связи с этим плазмидная ДНК по настоящему изобретению содержит элементы для поддержания и амплификации, преимущественно в больших количествах, в клетках бактерий. Такими существенными элементами являются бактериальные ориджин репликации, для поддержания в клетке со средней, предпочтительно высокой, копийностью, и репортерный ген для возможности селекции штамма-продуцента. Подходящий ориджин репликации представлен pM1 (der.), ColE1 (der.) и F1, pUC и F1, но не ограничивается ими. Подходящий репортерный ген представлен геном устойчивости к антибиотику, например, ампициллином, премущественно канамицином, но не ограничивается ими. Для уменьшения вероятности возникновения мутаций благодаря метилированию плазмидной ДНК использовали предпочтительно штамм микроорганизма, не содержащий метилазу в геноме.
Плазмидная ДНК по настоящему изобретению содержит элементы для эффективного функционирования в клетках млекопитающих.
Плазмидная ДНК по настоящему изобретению содержит и такой важный элемент, как лидерную последовательность мРНК, содержащую последовательность Козака непосредственно перед стартовым кодоном ATG.
Плазмидная ДНК также содержит сайт, преимущественно сайты, разные, для клонирования таргетного гена, для осуществления правильной ориентации таргетного гена в плазмидной ДНК, и сайт, преимущественно сайты, для посадки праймеров для его секвенирования.
Плазмидная ДНК также содержит терминирующую последовательность, содержащую последовательно стоп-кодон, 3′ нетранслируемую область с сигналом и сайтом полиаденилирования, стоп-кодон, за счет которой сохраняется стабильность мРНК, и осуществляется надлежащее прекращение траскрипции и экспорт мРНК из ядра. Терминирующая последовательность представлена нативной, т.е. присущей таргетному гену, либо иной, более сильной, которая представлена, например, терминирующей последовательностью бычьего гормона роста (BGH), но ею не ограничивается, и во втором варианте может содержать дополнительный стоп-кодон перед 3′ нетранслируемой областью. Опираясь на существующий уровень техники, на известные и очевидные варианты такого элемента, плазмидная ДНК по настоящему изобретению может содержать любую отвечающую вышеуказанным условиям терминирующую последовательность, при которой с плазмидной ДНК осуществляется синтез альфа дефенсина HNP-1, либо HNP-2, либо HNP-3 в клетках млекопитающих.
Плазмидная ДНК также содержит нативную, либо гетерологичную секреторную последовательность, кодонно оптимизированную для млекопитающих. В одном варианте изобретения содержит, например, секреторную последовательность ТРА (tissue-type plasminogen activator isoform 1 preproprotein [Homo sapiens], NCBI Reference Sequence: NP 000921.1), но ею не ограничивается. Преимущество использования секреторной последовательности ТРА - в обширном предшествующем клиническом опыте, а также в том, что показана ее высокая производительность в отношении экспрессии секретируемого белка с различных генов-мишеней.
Плазмидная ДНК содержит и фрагмент, кодирующий альфа дефенсин человека HNP-1, либо HNP-2, либо HNP-3, кодонно оптимизированный для экспрессии в клетках млекопитающих.
Плазмидная ДНК также содержит сайт, преимущественно сайты, разные, для клонирования таргентного гена, для осуществления правильной ориентации таргетного гена в плазмидной ДНК, и сайты для посадки праймеров для его секвенирования.
Плазмидная ДНК также содержит терминирующую последовательность, содержащую последовательно стоп-кодон, 3` нетранслируемую область с сигналом и сайтом полиаденилирования, стоп-кодон, за счет которой сохраняется стабильность мРНК, и осуществляется надлежащее прекращение траскрипции и экспорт мРНК из ядра. Терминирующая последовательность представлена нативной, т.е. присущей таргетному гену, либо иной, более сильной, которая представлена, например, терминирующей последовательностью бычьего гормона роста (BGH), но ею не ограничивается, и во втором варианте может содержать дополнительный стоп-кодон перед 3` нетранслируемой областью. Опираясь на существующий уровень техники, на известные и очевидные варианты такого элемента, плазмидная ДНК по настоящему изобретению может содержать любую отвечающую вышеуказанным условиям терминирующую последовательность, при которой с плазмидной ДНК осуществляется синтез альфа дефенсина HNP-1, либо HNP-2, либо HNP-3 в клетках млекопитающих.
Плазмидная ДНК содержит и нуклеотидную последовательность, в которой закодирован непосредственно препробелок указанного альфа дефенсина человека HNP-1, либо HNP-2, либо HNP-3, охарактеризованный SEQ ID NO: 1, SEQ ID NO: 2, соответственно, в одном варианте изобретения содержащий гетерологичную секреторную последовательность вместо нативной, например, ТРА (tissue-type plasminogen activator isoform 1 preproprotein [Homo sapiens], NCBI Reference Sequence: NP 000921.1), и охарактеризованный SEQ ID NO: 7, SEQ ID NO: 8, содержащую стоп-кодон, предпочтительно два стоп-кодона, и охарактеризованную SEQ ID NO: 5, SEQ ID NO: 6, либо SEQ ID NO: 11, SEQ ID NO: 12, соответственно. Преимущество использования секреторной последовательности ТРА - в обширном предшествующем клиническом опыте, а также в том, что показана ее высокая производительность в отношении экспрессии секретируемого белка с различных генов-мишеней. Плазмидная ДНК также может дополнительно содержать исходные для кДНК указанных генов элементы, обуславливающие стабильность данной мРНК (SEQ ID NO: 3, SEQ ID NO: 4), такие как терминирующую последовательность (3`нетранслируемая область, содержащая сигнал и сайт полиаденилирования, а также сигнал терминации транскрипции - стоп-кодон), в данном случае нуклеотидная последовательности гена альфа дефенсина человека охарактеризована SEQ ID NO: 3, SEQ ID NO: 4, либо SEQ ID NO: 10, SEQ ID NO. 11, соответственно, и в остове плазмидной ДНК отсутствуют, либо не функционируют аналогичные элементы. В вариантах и с нативной, и гетерологичной сигнальными последовательностями производили кодонную оптимизацию последовательности, кодирующей непосредственно препробелок дефенсин альфа человека HNP-1, либо HNP-2, либо HNP-3, для экспрессии в клетках млекопитающих для увеличения экспрессии таргетного гена за счет увеличения эффективности считывания информации с данной мРНК на рибосомах.
Плазмидная ДНК для доставки гена, обуславливающего анальгезию, сочетает в себе такие свойства ДНК-вакцины на основе плазмидной ДНК, как высокий уровень экспрессии гена интереса в клетках млекопитающих, и такие свойства вектора для генной терапии, как отсутствие иммуногенности и длительность экспрессии гена, однако, например, за счет увеличения стабильности мРНК. Такая ДНК не встраивается в геном и не реплицируется в клетках млекопитающих.
Подходящие векторы для экспрессии в клетках млекопитающих представлены известными среднему специалисту в данной области и описанными в литературе [Hartikka J, Sawdey M, Cornefert-Jensen F, Margalith M, Barnhart K, Nolasco M, Vahlsing HL, Meek J, Marquer M, Hobart P, Norman J, Manthorpe M. An improved plasmid DNA expression vector for direct injection into skeletal muscle. Hum Gene Ther. 1996 Jun 20; 7(10):1205-17 и др.], a также плазмидами, которые могут быть созданы средним специалистом в данной области с использованием рекомендаций по элементам векторов ["Cloning Vectors", ed. Pouwls et al., Elsevier, Amsterdam- New York-Oxford, 1985, ISBN 0 444 904018, Williams JA, Carnes AE, Hodgson СР. Plasmid DNA vaccine vector design: impact on efficacy, safety and upstream production. Biotechnol Adv. 2009 Jul-Aug; 27(4):353-70. doi: 10.1016/j.biotechadv.2009.02.003. Epub 2009 Feb 20. Review и др.]. Предпочтительными плазмидными ДНК для использования у человека являются векторы, проверенные на людях, содержащие описанные выше элементы с соответствующими регуляторными последовательностями, возможно, модифицированные для соответствия заявленным критериям, что позволяет уменьшить количество требуемых исследований для регистрации средства. Однако возможно и использование иных плазмидных ДНК, содержащих требуемые описанные элементы.
Последовательность расположения описанных элементов в плазмидной ДНК понятна среднему специалисту в данной области.
Нуклеотидные последовательности, кодирующие ген альфа дефенсина человека HNP-1, либо HNP-2, либо HNP-3 согласно изобретению, клонируют в вектор, создают прокариотический, преимущественно, бактериальный (на основе клеток, преимущественно, Escherichia coli, Streptomyces, Bacillus, Pseudomonas, но не ограничиваясь ими), штамм-продуцент плазмидной ДНК - действующего агента анальгетического средства, - стандартными методами, например, трансфекцией, электропорацией или пушкой с частицами, осуществляют ее выделение и очистку и вводят полученную плазмидную ДНК в организм, описанная нуклеотидная последовательность затем экспрессируется в таком организме in vivo. При использовании сайленсера, либо инсулятора в составе конструкции возможно регулировать экспрессию таргентного гена, гена описанного альфа дефенсина человека HNP-1, либо HNP-2, либо HNP-3 (вариантов).
Среднему специалисту в данной области понятно, что используя удовлетворяющие приведенным условиям элементы для создания плазмидной ДНК и удовлетворяющий приведенным условиям прокариотический продуцент можно получить активный агент анальгезирующего средства по изобретению.
Охарактеризованную плазмидную ДНК (варианты) используют для получения фармацевтической композиции для лечения, облегчения и/или профилактики боли, в частности острой или хронической боли, нарушений чувствительности. "Больными" в контексте настоящего изобретения являются, помимо человека, также животные, млекопитающие, в частности, грызуны, например, мышь, крыса, морская свинка, кролик, и домашние или стадные животные, например, курица, гусь, утка, коза, овца, свинья, крупный рогатый скот, лошади, собаки и кошки, но ими не ограничиваются.
Авторами настоящего изобретения проведены лабораторные исследования, подтверждающие возможность реализации охарактеризованного изобретения (вариантов). Полученные результаты исследований проиллюстрированы примерами (1-3) и Фиг. 1-3.
Краткое описание чертежей.
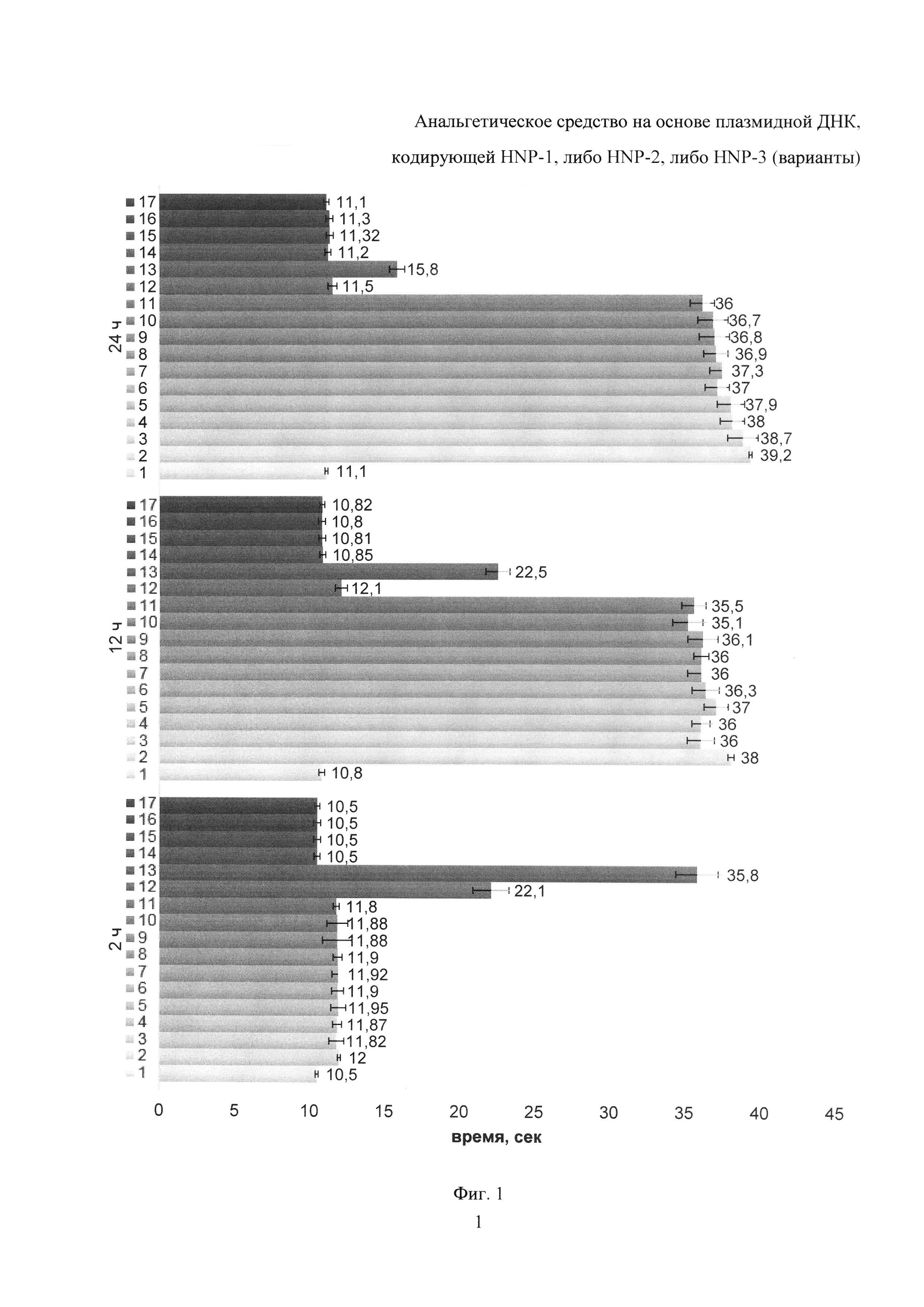
Фиг. 1. Результаты теста «горячая пластина». Ось абсцисс - латентное время облизывания передних и задних лап, ось ординат - время после введения исследуемого вещества. Легенда: 1 - отрицательный контроль (вводили физ. раствор), 2 - pVAXlseq3, 3 - pVR1012seq4, 4 - pVR1012seq5, 5 - pVAXlseq6, 6 - pVR1012seq9, 7 - pVAXlseq10, 8 - pVAXlseq11, 9 - pVR1012seq12, 10 - pcDNA3.1+ seq11, 11 - pcDNA3.1+ (var)seq12, 12 - анальгин (50 мг/кг), 13 - морфин гидрохлорид (10 мг/кг), 14 - pVAX1, 15 - pVR1012, 16 - pcDNA3.1+, 17 - pcDNA3.1+ (var). Плазмидные ДНК вводили в количестве 5 мг/кг.
Фиг. 2. Результаты исследования различных доз плазмидной ДНК pVAXlseq3 в тесте «горячая пластина». Ось абсцисс - латентное время облизывания передних и задних лап, ось ординат - время после введения исследуемого вещества. Легенда: 1 -отрицательный контроль (вводили физ. раствор), 2 - pVAXlseq3 1 мг/кг, 3 - pVAXlseq3 5 мг/кг, 4 - pVAXlseq3 10 мг/кг, 6 - анальгин (50 мг/кг), 7 - морфин гидрохлорид (10 мг/кг).
Фиг. 3. Результаты исследования изменения концентрации бета-эндорфина в сыворотке мышей. Ось абсцисс - время после введения исследуемого вещества, ось ординат - концентрация бета-эндорфина в сыворотке. Легенда: 1 - отрицательный контроль (вводили физ. раствор), 2 - pVR1012, 3 - pVR1012seq4. Плазмидные ДНК вводили в количестве 5 мг/кг.
Пример 1. Получение плазмидных ДНК, кодирующих альфа дефенсин HNP-1, либо HNP-2, либо HNP-3.
1.1. Осуществляли химический синтез ДНК, охарактеризованной SEQ ID NO: 3 - SEQ ID NO: 6 и SEQ ID NO: 9 - SEQ ID NO: 12, дополнительно фланкированной сайтами рестрикции, с добавлением последовательности Козака. Для SEQ ID NO: 3 и SEQ ID NO: 10 использовали сайты рестрикции NheI (5), BamHI (3`), для SEQ ID NO: 4 и SEQ ID NO: 9 использовали сайты рестрикции SalI (5`), BamHΙ (3`), для SEQ ID NO: 5 и SEQ ID NO: 12 использовали сайты рестрикции SalI (5`), BamHI (3`), для SEQ ID NO: 6 и SEQ ID NO: 11 использовали сайты рестрикции NheI (5`), Apal (3`).
1.2. Осуществляли клонирование синтезированных генов в плазмиды pVAXl (Invitrogen) и pVR1012 (Vical). Также осуществили клонирование последовательности SEQ ID NO: 11 в вектор pcDNA3.1+ (Invitrogen) по сайтам рестрикции NheI (5`), ApaI (3`). Также получили вектор pcDNA3.1+, неспособный к экспрессии неомицина, за счет рестрикции данного вектора рестриктазой NsiI, в области SV40 промотора (-71 п.о.). В полученный вектор клонировали SEQ ID NO: 12 по сайтам рестрикции NheI (5), ApaI (3).
1.3. На реакцию лигирования брали 3 мкл раствора синтезированной ДНК, 1 мкл раствора готового вектора, 5 мкл буфера для лигирования ×2 и 1 мкл Т4-лигазы. Реакцию проводили при +20°С в течение 2 часов.
После этого смесь прогревали при +95°С в течение 10 мин и очищали от солей диализом на нитроцелюлозных фильтрах с диаметром пор 0,025 мкм (Millipore, США). Диализ проводили против раствора, содержащего 0,5 мМ ЭДТА в 10% глицерине, в течение 10 мин.
Получили следующие плазмидные ДНК: pVAXlseq3, pVR1012seq4, pVR1012seq5, pVAXlseq6, pVR1012seq9, pVAXlseq10, pVAXlseq11, pVR1012seq12, pcDNA3.1+seq11, pcDNA3.1+ (var)seq12.
1.4. Затем трансформировали клетки E.coli штамма DH10B/R (F-mcrA, Δ(mrr-hsdRMS-mcrBC), φ80dlacZAM 15, ΔlacX74, deoR, recA1, endA1, araD139, Δ(ara,leu)769, galU, galKλ-, rpsL, nupG) полученными плазмидными ДНК методом электропорации с использованием электропоратора MicroPulser (BioRad). Данный штамм не содержит метилазу, что позволяет минимизировать возможность возникновения мутаций в ДНК, в том числе в клонированном в плазмиде, поддерживаемой в данном штамме, гене. К 12 мкл компетентных клеток добавляли 1 мкл диализованной лигазной смеси, помещали между электродами порационной ячейки и обрабатывали импульсом тока.
После трансформации клетки помещали в 1 мл SOC-среды (2% бакто-триптон, 0.5% дрожжевой экстракт, 10 мМ NaCl, 2.5 мМ КСl, 10 мМ MgCl2, 10 мМ MgSO4, 20 мМ глюкоза) и инкубировали в течение 40 мин при +37°С.
1.5. Проводили выявление клонов клеток E.coli, содержащих полученную плазмидную ДНК, на селективной среде, содержащей LB-arap, 50 мкг/мл канамицина, либо ампициллина (для плазмидных ДНК на основе pcDNA3.1+).
Из выросших клонов выделяли плазмидную ДНК. Выделение плазмидной ДНК проводили с использованием набора Wizard Minipreps DNA Purification System (Promega, США). Очищенную рекомбинантную плазмидную ДНК проверяли с помощью секвенирования.
1.6. Секвенирование клонированных фрагментов проводили по методу Сэнджера с использованием набора Applied Biosystems BigDye® Terminator (BDT) v3.1 Cycle Sequencing Kit (Applied Biosystems, США) по прилагающейся к нему инструкции. Для мечения продуктов реакции использовали меченные флуоресцентным красителем ddNTP, причем каждому ddNTP соответствовал свой краситель. Для секвенирования использовали немеченные специфические для плазмид праймеры. Проводили ПЦР-реакцию, затем реакционную смесь очищали от свободных меченых ddNTP по инструкции к набору BigDye X-Terminator Purification Kit (Applied Biosystems, США) и разделяли продукты реакции секвенирования с использованием капиллярного секвенатора Applied Biosystems 3500/3500xL Genetic Analyzer (Applied Biosystems, США) и реактива 3500/3500xL Genetic Analyzer Polymer "POP-6™" (Applied Biosystems, США).
Результаты разделения продуктов реакции секвенирования регистрировались путем сканирования лазером и детекции четырех флуоресцентных красителей, включенных во все типы ddNTP. 1.7. Компьютерный анализ последовательностей ДНК проводили с помощью
персонального компьютера с использованием программ Chromas и BioEdit. Нуклеотидные последовательности исследованных фрагментов ДНК были выровнены относительно рассчитанных, была продемонстрирована идентичность синтезированных фрагментов рассчитанным. В результате были отобраны клоны клеток E.coli, содержащие полноразмерные последовательности таргетных генов в составе плазмид последовательности ДНК, кодирующие альфа дефенсин HNP-1, либо HNP-2, либо HNP-3.
Пример 2. Наработка плазмидных ДНК, кодирующих альфа дефенсин HNP-1, либо HNP-2, либо HNP-3, для проведения исследования на лабораторных животных.
Отдельную колонию E.coli, выращенную на LB-arape в чашке Петри с добавлением канамицина (либо ампициллина), помещали в 10 мл селективной среды. Клетки растили в течение ночи при +37°С в условиях постоянного перемешивания (250 об/мин.). Полученные клетки собирали центрифугированием при 4000g. Дальнейшее выделение и очистку плазмидной ДНК осуществляли с использованием набора EndoFree Plasmid Mega Kit (Qiagen), позволяющего получить апирогенную ДНК. Выделенную плазмидную ДНК анализировали электрофорезом в 0,8%-ном агарозном геле, измеряли ее концентрацию с помощью флуориметрии.
В качестве контрольного раствора использовали воду без добавления тестируемого препарата. В ячейку для измерения оптической плотности объемом 2 мл вносили 1,950 мл воды и 0,05 мл тестируемого раствора (концентрация 1 мг/мл), перемешивали и измеряли оптическую плотность при длине волны 260 нм. Определение концентрации ДНК проводили по формуле:
С(мкг/мл)=40А260К,
где А260-оптическая плотность препарата, измеренная при длине волны 260 нм; К(мкг/мл)- для ДНК 50 мкг/мл (50 мкг/мл двухцепочечной ДНК в воде); 40 - разведение тестируемого препарата.
В итоге определили, что получили плазмидную ДНК pVAXlseq3 с концентрацией 3,4 мг/мл, pVR1012seq4 - 3,7 мг/мл, pVR1012seq5 - 4 мг/мл, pVAXlseq6 - 3,8 мг/мл, pVR1012seq9 - 4,1 мг/мл, pVAXlseq10 - 4 мг/мл, pVAXlseq11 - 4,3 мг/мл, pVR1012seq12 - 4,5 мг/мл, pcDNA3.1+seq11 - 4,4 мг/мл, pcDNA3.1+ (var)seq12 - 4,3 мг/мл. Выход плазмидной ДНК составил от 3,4 мг до 4,5 мг из 1 л питательной среды.
О чистоте полученного препарата плазмидной ДНК судили по отношению оптической плотности препарата, измеренной при длине волны 260 нм, к оптической плотности препарата, измеренной при длине волны 280 нм (А260/А280) и отношению оптической плотности препарата, измеренной при длине волны 260 нм к оптической плотности препарата, измеренной при длине волны 230 нм (А260/А230). Измерения проводили в водном растворе, в качестве контрольного раствора использовали воду без добавления тестируемого препарата.
Для чистых препаратов ДНК характерно А260/А280>1,80 и А260/А230>1,80. Определенные в эксперименте значения соответствовали значениям отношений А260/А280 и А260/А230 для чистых препаратов, для всех полученных препаратов плазмидной ДНК.
Также проводили количественное определение примесей белка в полученных препаратах плазмидных ДНК pVAXlseq3, pVR1012seq4, pVR1012seq5, pVAXlseq6, pVR1012seq9, pVAXlseq10, pVAXlseq11, pVR1012seq12, pcDNA3.1+seq11, pcDNA3.1+ (var)seq12 с помощью microBCA assay [Smith, Р.К., et all, Measurement of protein using bicinchoninic acid. Analyt. Biochem. 150, 76-85 (1985)], измеряя оптическую плотность образующихся окрашенных белковых комплексов с медью и бицинхониновой кислотой при длине волны 562 нм. Чувствительность метода microBCA assay составляет 0.5-20 мкг/мл белка. Концентрация тотального белка ни в одном из исследуемых препаратов плазмидной ДНК не превышала норму (от 0,5 до 12 мкг/мг плазмидной ДНК).
Определяли и содержание бактериального липополисахарида в препаратах плазмидных ДНК, с использованием гель-тромб варианта ЛАЛ-теста, с чувствительностью >0,25 EU/мл (ToxinSensor, GenScript, США). ЛАЛ-реагентом служил лизат амебоцитов подковообразного краба Limulus polyphemus. ЛАЛ-реактив специфически реагирует с бактериальными эндотоксинами, в результате ферментативной реакции происходит изменение реакционной смеси, пропорциональное концентрации эндотоксина. Результаты оценивали по наличию или отсутствию плотного тромба на дне пробирки путем переворачивания пробирки. Гель-тромб не образовался при исследовании образца, разведенного в 10 раз, для препаратов плазмидных ДНК pVAXlseq3, pVR1012seq4, pVR1012seq5, pcDNA3.1+ (var)seq12 и в 5 раз для препаратов плазмидных ДНК pVAXlseq6, pVR1012seq9, pVAXlseq10, pVAXlseq11, pVR1012seq12, pcDNA3.1+seq11 т.е. при чувствительности метода 2,5 EU/мл и 1,25 EU/мл, соответственно, что, учитывая концентрацию плазмидной ДНК в образце, говорит о допустимом показателе очистки от эндотоксинов.
Пример 3. Определение анальгетического эффекта плазмидной ДНК, кодирующей альфа дефенсин HNP-1, либо HNP-2, либо HNP-3.
3.1. Тест «Горячая пластина»
Проводили исследование анальгетической активности плазмидных ДНК, кодирующих альфа дефенсин HNP-1, либо HNP-2, либо HNP-3, с использованием теста «Горячая пластина» (hot plate)
Тест «Горячая пластина» (Hot plate) проводили для измерения порога острой болевой чувствительности и потенциального анальгезирующего эффекта изучаемых препаратов плазмидной ДНК [Вальдман А.В., Игнатов Ю.Д. Центральные механизмы боли. - Л.: Наука. - 1976]. Тест является базисным для исследования анальгетической активности, его используют для выявления анальгетически активных соединений.
При помещении на горячую поверхность, с достижением порога болевой чувствительности со стороны животного наблюдаются двигательные реакции беспокойства: одергивание лап, облизывание подушечек лап и подпрыгивание. В данном тесте учитывали латентное время с момента помещения животного на горячую поверхность до первого облизывания лап. Данная методика позволяет определять показатели: анальгетическая активность тестируемого объекта, пиковое время анальгезии, длительность анальгезии.
В исследовании использовали мышей линии BALB/C, самок, массой 15-22 грамма, возраста 18 недель. В опыте было сформировано 17 групп животных, включая контрольные группы, в каждой группе по 3 мыши:
1 - отрицательный контроль (вводили физраствор)
2 - pVAXlseq3
3 - pVR1012seq4
4 - pVR1012seq5
5 - pVAXlseq6
6 - pVR1012seq9
7 - pVAXlseq10
8 - pVAXlseq11
9 - pVR1012seq12
10 - pcDNA3.1+seq11,
11 - pcDNA3.1+(var)seq12
12 - анальгин (50 мг/кг)
13 - морфин гидрохлорид (10 мг/кг)
14 - pVAX1
15 - pVR1012
16 - pcDNA3.1+
17 - pcDNA3.1+(var)
Вводили по 5 мг/кг плазмидных ДНК.
Использовали прибор Hot plate-метр (Hotplate Analgesia Meter, Columbus Instruments, USA).
До начала испытания в течение 10 минут давали тест-системам акклиматизироваться в комнате для проведения исследования. Животных использовали однократно, так как повторное помещение животного на термостатируемую пластину вызывает незамедлительную реакцию на касание поверхности. Устанавливали температуру термостата 55°С. После инъекции исследуемого вещества животное аккуратно помещали на нагревательную пластину и в тот же момент нажимали кнопку «старт» на панели прибора. Отмечали латентное время облизывания передних и задних лап (с момента помещения животного на поверхность прибора до первого облизывания). После этого нажимали на кнопку «стоп» и убирали животное с горячей поверхности. Иные поведенческие реакции игнорировали. Для уменьшения вероятности теплового повреждения подушечек лап максимальное время эксперимента не превышало 60 сек. Для определения пикового времени анальгетического действия препарата измеряли латентное время облизывания передних и задних лап у контрольной группы (физраствор) (точка 0) и через 2 ч, 12 ч, 24 ч после введения препарата у тестируемых групп. Поверхность прибора протирали салфеткой, смоченной дезинфектантом (0,5% хлоргексидин-биглюконат на 70% этаноле), перед помещением на нее очередного животного [Методические рекомендации для обучаемых. Фармацевтический факультет ГОУ ВПО ММА им. И.М. Сеченова Росздрава. - Москва. - 2006].
Проводили анализ данных определения анальгетической активности препарата. Для плазмидной ДНК, показавшей наилучший результат, определяли ED50, т.е. дозу, необходимую для проявления 50%-й анальгетической активности препарата.
Результаты опыта представлены на Фиг. 1. Препараты всех исследуемых плазмидных ДНК, кодирующих дефенсин HNP-1, либо HNP-2, либо HNP-3, проявляют выраженное анальгезирущее действие, не уступающее по эффективности морфину и превышающее анальгетическую активность анальгина. Наблюдали небольшую степень анальгезии через 2 ч после введения плазмид, затем анальгезия усиливалась, и через 12 и 24 ч наблюдали одинаковый высокий уровень анальгезии, что говорит о поддержании эффекта.
В эксперименте с применением различных доз плазмидной ДНК, показавшей наилучший результат в предыдущем опыте (pVAXlseq3), продемонстрирован дозозависимый характер анальгезии в интервале от 1 до 10 мг/кг (Фиг. 2). При этом по прошествии 12 часов показатели при использовании плазмидной ДНК pVAXlseq3 в минимальной концентрации 1 мг/кг были практически идентичны показателям в группе использования морфина гидрохлорида 10 мг/кг. Через 24 ч показатель в группе плазмидной ДНК в минимальной концентрации превысил таковой группы использования морфина гидрохлорида 10 мг/кг, практически в 2 раза. Действие анальгина наблюдали только по прошествии 2 ч после введения анальгетика, ответ через 12 ч и 24 ч был аналогичен таковому в контрольной группе. При использовании плазмидной ДНК pVAXlseq3 в концентрации 5 мг/кг и 10 мг/кг наблюдали значительное увеличение (до 15%) латентного периода, по сравнению с группой с минимальной концентрацией плазмидной ДНК, при этом различие между этими двумя группами было небольшое.
3.2. Исследование концентрации бета-эндорфина в сыворотке мышей.
Поскольку изменение концентрации бета-эндорфина в плазме крови может служить критерием оценки эффективности обезболивания [Бета-эндорфин - маркер эффективности обезболивания при острой боли и хроническом болевом синдроме у онкологических больных / З.В. Павлова [и др.] // Проблемы клинической медицины. - 2007. - N1. - С. 36-40. - ISSN 1817-8359], проводили исследование концентрации бета-эндорфина в сыворотке исследуемых мышей после введения плазмидной ДНК, на примере введения плазмидной ДНК, кодирующей HNP-2, либо HNP-3 (pVR1012seq4), с использованием набора ELISA Kit for Beta-Endorphin (bEP) Mus musculus (Mouse) CEA806Mu (Life science Inc.).
Результаты исследования приведены на Фиг. 3. Видно, что уровень бета-эндорфина значительно поднялся при введении плазмидной ДНК pVR1012seq4, при том, что при введении плазмидной ДНК без вставки таргетного гена и в контрольной группе значительное увеличение уровня бета-эндорфина не наблюдали, на протяжении всего эксперимента.
Таким образом, продемонстрирована возможность создания различных вариантов плазмидной ДНК, кодирующей HNP-1, либо HNP-2, либо HNP-3, а также анальгетическое действие такой плазмидной ДНК, в любом из вариантов, соответствующих указанным критериям, причем даже в минимальной концентрации 1 мг/кг (меньше в 10 раз, чем морфина гидрохлорида), а также продемонстрирована безопасность применения такой плазмидной ДНК: животные не погибли, побочные эффекты не наблюдали.
Реферат
Изобретения относятся к области медицины и биотехнологии и могут быть использованы для анальгезии. Плазмидная ДНК для транзиентной экспрессии в клетках млекопитающих представлена остовом, содержащим прокариотические элементы, ориджин репликации и репортерный ген, и эукариотические элементы, сильный промотор, лидерную последовательность мРНК, а также регуляторные последовательности для указанных элементов, от одного сайта для клонирования гена интереса и от одного сайта для посадки от одного праймера для анализа состава плазмидной ДНК, и полинуклеотидом, представленным секреторной последовательностью, фрагментом, кодирующим альфа дефенсин человека HNP-1, либо HNP-2, либо HNP-3, кодонно оптимизированными для экспрессии в клетках млекопитающих, и терминирующей последовательностью. Также предложены продуцент плазмидной ДНК и анальгетическое средство на его основе. Использование изобретений позволяет расширить спектр анальгетических средств, увеличить длительность и безопасность анальгезии. 3 н. и 2 з.п. ф-лы, 3 ил., 3 пр.
Формула
Документы, цитированные в отчёте о поиске
Слитые конструкции лекарственного средства и конъюгаты
Комментарии