Способ получения капсидного белка цирковируса свиней 2 типа и содержащая его фармацевтическая композиция - RU2701816C1
Код документа: RU2701816C1
Чертежи

Описание
УРОВЕНЬ ТЕХНИКИ
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[1] Настоящее изобретение относится к способу получения капсидного белка PCV2, конкретно к способу получения капсидного белка PCV2, с использованием экспрессирующей системы на основе прокариотической клетки.
ОПИСАНИЕ УРОВНЯ ТЕХНИКИ
[2] Цирковирус свиней 2 типа (PCV2) представляет собой патоген вирусной природы, который оказывает значительное влияние на свиноводческую промышленность во всем мире свинья. PCV2 в основном вызывает мультисистемный синдром истощения после отъема (PMWS), симптомами которого являются лихорадка, лимфаденопатия, потеря массы или слабость, затрудненное дыхание, диарея, бледность тела, в редких случаях желтуха и т.д. Он также может вызывать синдром дерматита и нефропатии поросят (PDNS), инфекционное врожденное дрожание (ICT) и нарушения со стороны репродуктивной системы. Кроме того, инфекция PCV2 в сочетании с другими патогенами вирусной или бактериальной природы вызывает комплекс респираторных заболеваний у свиней (PRDC). Заболевание, вызываемое инфекцией PCV2 у свиней, приводит к снижению коэффициента выживаемости и коэффициента конверсии корма, что приводит к тяжелым экономическим потерям для свиноводов.
[3] Предложены двадцать пунктов по кормлению и ведению для профилактики и борьбы с PCV2 в полевых условиях, таких как система "пусто-занято" (AIAO), хорошее ведение гигиены, уничтожение или изоляция свиней с тяжелым заболеванием и вакцинация. Среди перечисленного вакцинация может эффективно снижать скорость распространения инфекции PCV2 и дополнительно повышать коэффициент выживаемости. Вакцины против PCV2 в данной области разделяют на три категории, включая инактивированные вакцины против PCV2, инактивированные субъединичные вакцины на основе бакуловируса и инактивированные вакцины на основе химерного вируса PCV1-PCV2 (Beach and Meng, 2012; Chanhee, 2012).
[4] Инактивированную вакцину на основе PCV2 получают путем инфицирования линии клеток почки свиньи PK-15 PCV2, сбора вируса, инактивации вируса и смешивания вируса с адъювантом. Для инактивированных субъединичных вакцин на основе бакуловируса, клетки насекомых трансфицируют бакуловирусом, несущим ген ORF2, кодирующий PCV2 капсидный белок для экспрессии антигена ORF2. Если антиген экспрессируется в клетке, вакцина получают посредством разрушения с помощью ультразвука среды для культивирования, содержащей клетки, инактивации вируса и смешивания вируса с адъювантом. Если антиген секретируется во внеклеточное окружение, вакцину получают посредством сбора супернатанта культуры клеток, инактивацией вируса и смешивания вируса с адъювантом. Вакцины на основе инактивированного химерного вируса PCV1-PCV2 получают заменой ORF2 PCV1 на ORF2 PCV2, инфицированием клеток, сбором вируса, инактивацией вируса и смешиванием вируса с адъювантом.
[5] Принимая во внимание тот факт, что все существующие способы получения вакцины на основе PCV2 основаны на способе культивации вирусов, эти способы имеют недостатки, которые заключаются в длительном времени подготовки и высокой стоимости производства. Для снижения стоимости вакцин против PCV2 исследователи в данной области пытались использовать рекомбинантную E. coli с небольшой стоимостью культивирования для получения вакцинного антигена ORF2. Однако способ имеет ограничения, которые заключаются в низкой продукции ORF2, неспособности получать вирусоподобные частицы рекомбинантного ORF2, сложные процессы или низкий иммунитет.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[6] Таким образом, цель настоящего изобретения является разработка способа получения капсидного белка PCV2 для уменьшения времени его продукции и стоимости вакцины на основе PCV2.
[7] Другой целью настоящего изобретения является получение композиции для профилактики инфекции PCV2. В композиции используют капсидный белок PCV2 в качестве активного компонента, и она содержит подходящий адъювант для улучшения средства профилактики инфекции PCV2 для промышленности.
[8] Другой целью настоящего изобретения является разработка способа получения интерферона свиньи для уменьшения времени продукции и стоимости интерферона свиньи и для облегчения применения интерферона свиньи в композиции для профилактики инфекции PCV2.
[9] Для достижения указанных выше целей настоящее изобретение относится к способу экспрессии белка, включающему: (a) получение индуцируемого арабинозой экспрессирующего вектора, где индуцируемый арабинозой экспрессирующий вектор содержит экспрессирующий элемент и нуклеотидную последовательность, кодирующую целевой белок; где экспрессирующий элемент содержит: промотор; усиливающий трансляцию элемент фага T7, содержащий SEQ ID NO:01; и участок связывания рибосомы, содержащий SEQ ID NO:02; (b) трансформацию индуцируемого арабинозой экспрессирующий вектор в E. coli-хозяина и индукцию экспрессии целевого белка; где целевой белок представляет собой капсидный белок PCV2 или интерферон свиньи.
[10] Предпочтительно участок -16 промотора содержит SEQ ID NO:03.
[11] Предпочтительно экспрессирующий элемент содержит SEQ ID NO:04.
[12] Предпочтительно индуцируемый арабинозой экспрессирующий вектор дополнительно содержит нуклеотидную последовательность, кодирующую партнер по слиянию, и/или нуклеотидную последовательность, кодирующую молекула-маркер. Предпочтительно партнер по слиянию представляет собой MsyB E. coli, YjgD E. coli, D-белок фага лямбда, белок SUMO пекарских дрожжей или их сочетание. Предпочтительно молекула-маркер представляет собой: метку His, метку Strep II, метку FLAG или их сочетание.
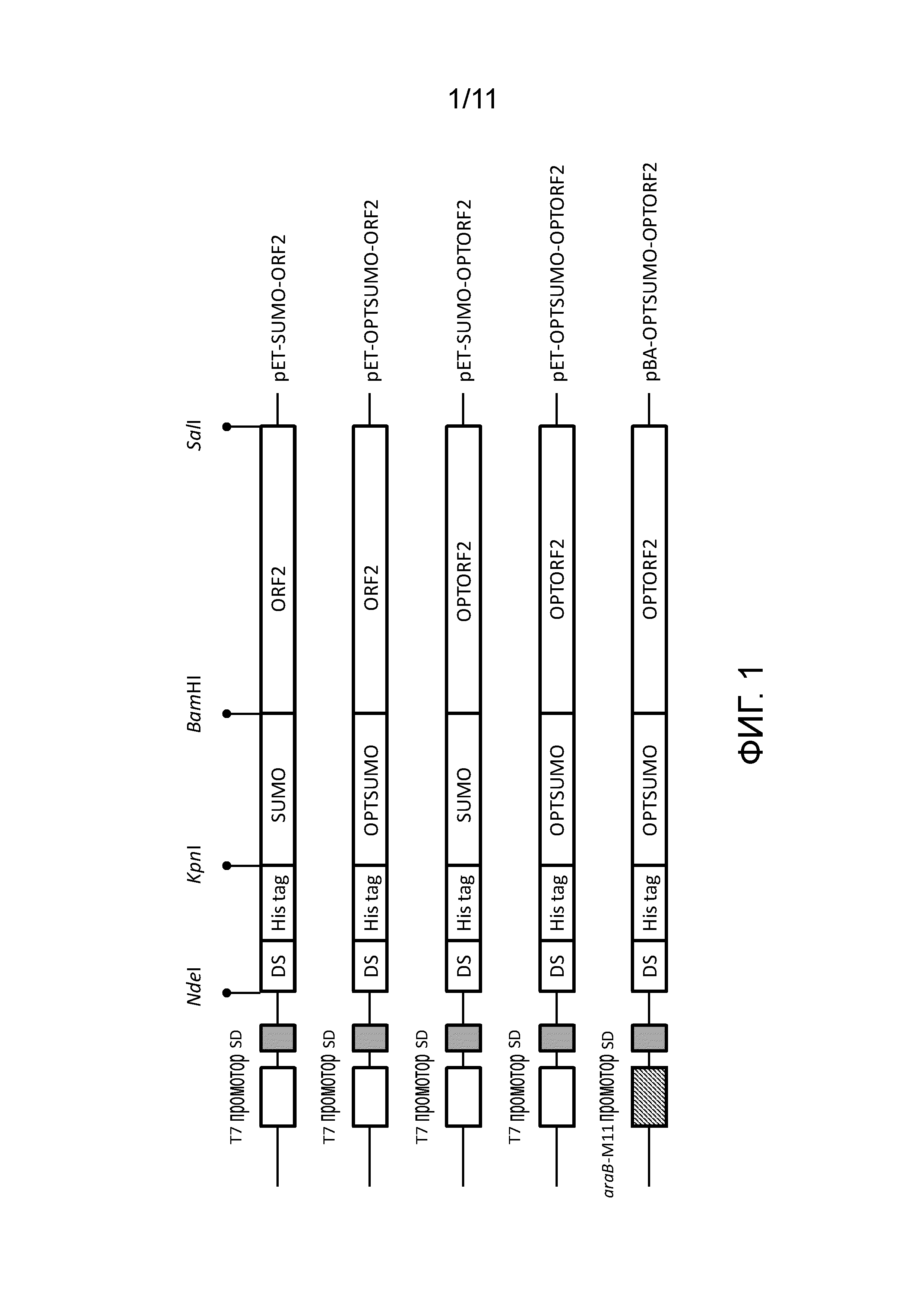
[13] Предпочтительно целевой белок представляет собой капсидный белок PCV2, кодируемый SEQ ID NO:09 или SEQ ID NO:24. Предпочтительно индуцируемый арабинозой экспрессирующий вектор содержит SEQ ID NO:46.
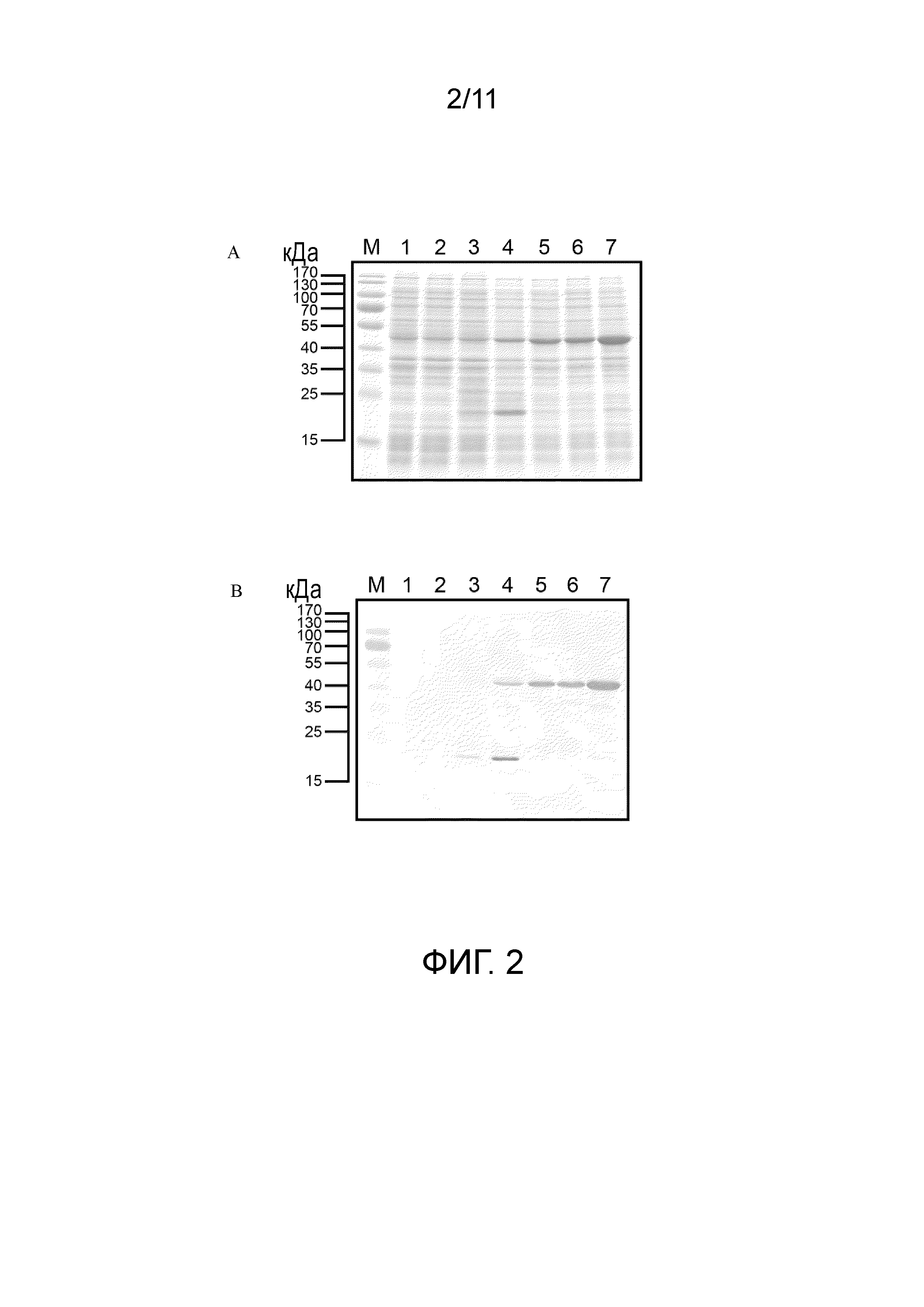
[14] Предпочтительно интерферон свиньи представляет собой интерферон α свиньи или интерферон γ свиньи. Предпочтительно целевой белок представляет собой интерферон свиньи и интерферон свиньи, кодируемый SEQ ID NO:64 или SEQ ID NO:76. Предпочтительно индуцируемый арабинозой экспрессирующий вектор содержит SEQ ID NO:80, SEQ ID NO:87 или SEQ ID NO:95. Предпочтительно способ не включает этап рефолдинга интерферона свиньи.
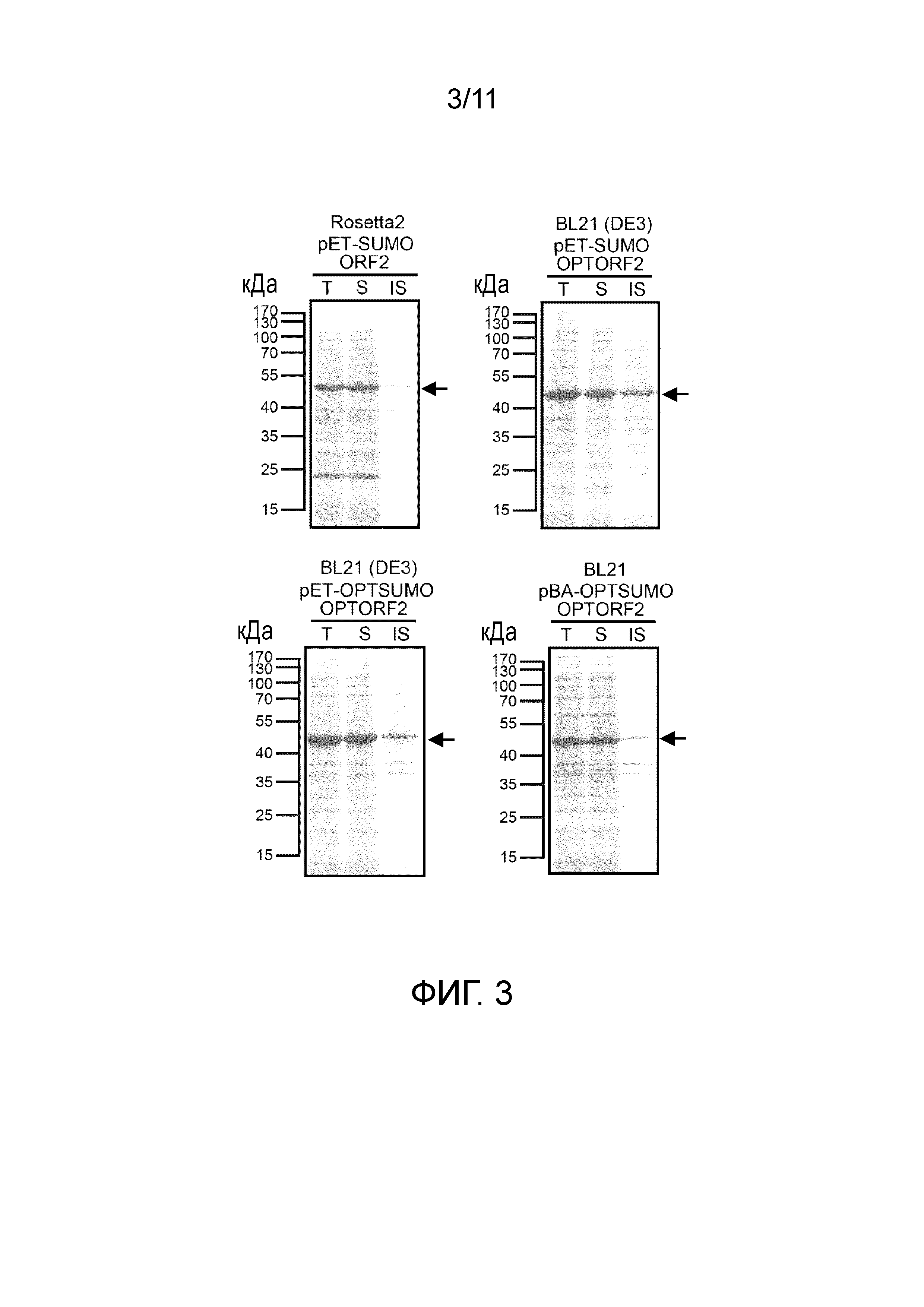
[15] Предпочтительно способ дополнительно включает стадию (c) после стадии (b): очистки целевого белка. Предпочтительно способ дополнительно включает стадию (d) после стадии (c): обработки целевого белка протеазой SUMO. Предпочтительно на стадии (d) массовое отношение целевого белка к протеазе SUMO составляет от 4 до 20.
[16] Настоящее изобретение дополнительно относится к композиции для профилактики инфекции PCV2, содержащей: от 2,5 до 250 мкг/мл капсидного белка PCV2; от 2,5 до 25 мкг/мл интерферона α свиньи; от 2,5 до 25 мкг/мл интерферона γ свиньи; и фармацевтически приемлемый носитель.
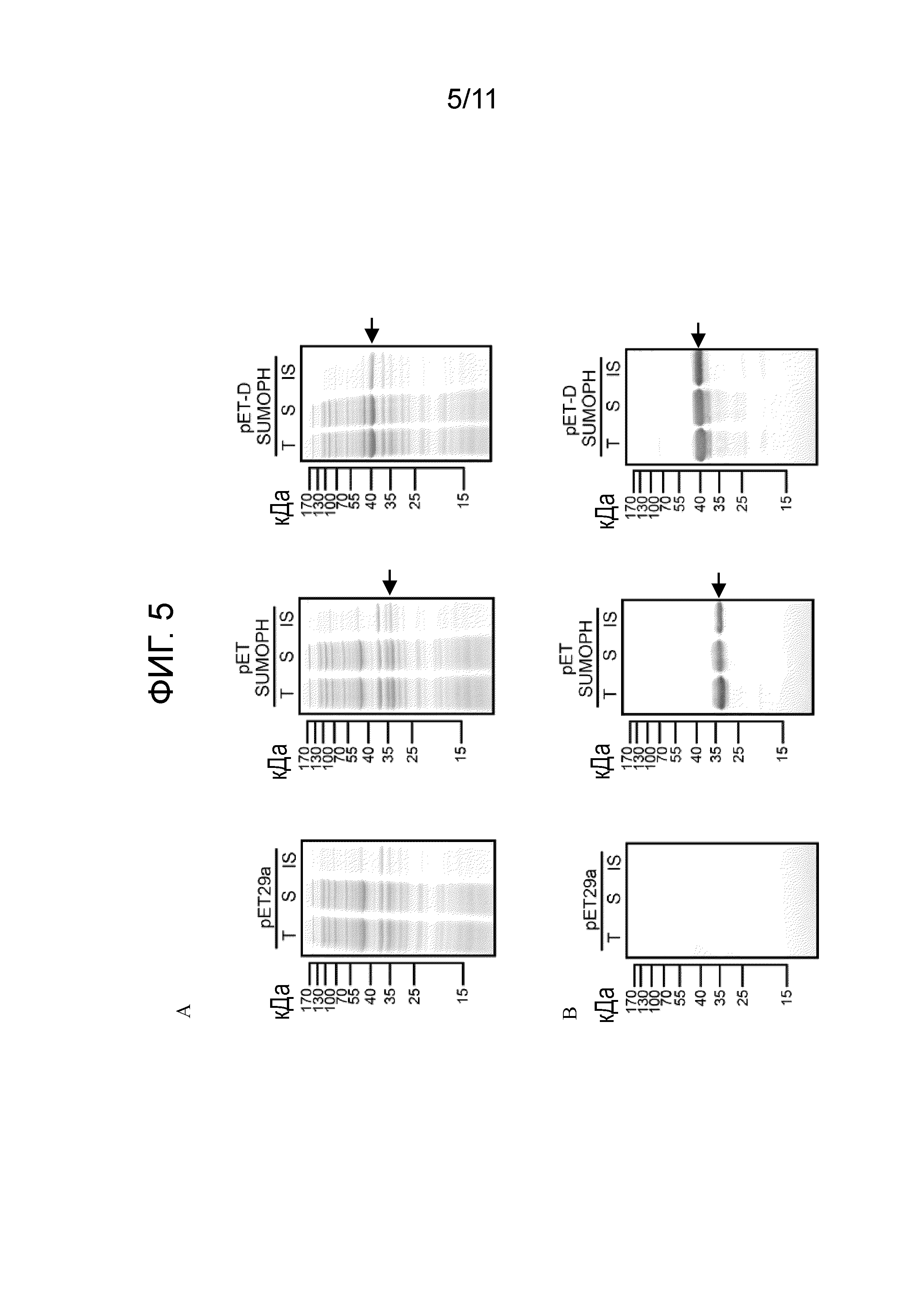
[17] Предпочтительно композиция дополнительно содержит фармацевтически приемлемый адъювант. Предпочтительно фармацевтически приемлемый адъювант представляет собой: адъювант MONTANIDE™ ISA 536 VG, адъювант MONTANIDE™ GEL 01, полный или неполный адъювант Фрейнда, гель на основе алюминия, поверхностно-активное вещество, полианионные полимеры, пептиды, масляные эмульсии или их сочетание.
[18] Предпочтительно композиция содержит: от 3,5 до 170 мкг/мл капсидного белка PCV2; от 5 до 20 мкгинтерферона α свиньи; от 5 до 20 мкгинтерферона γ свиньи и фармацевтически приемлемый носитель.
[19] В итоге, настоящее изобретение преимущественно относится к способу экспрессии белка путем использования индуцируемого арабинозой экспрессирующего вектора. Способ по настоящему изобретению эффективно облегчает синтез капсидного белка PCV2 и интерферона свиньи, используемого в качестве адъюванта в вакцине. С другой стороны, фармацевтическая композиция по настоящему изобретению объединяет капсидный белок и другие предпочтительные компоненты в соответствующем отношении для получения высокого индуцирующего иммуногенность действия. Таким образом, раскрытие настоящего изобретения характеризуется значительными положительными эффектами в отношении профилактики и лечения PCV2 в данной области.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[20] Фигура 1 представляет собой схематическую диаграмму пяти экспрессирующих векторов капсидных белков PCV2, получаемых в примере 1.
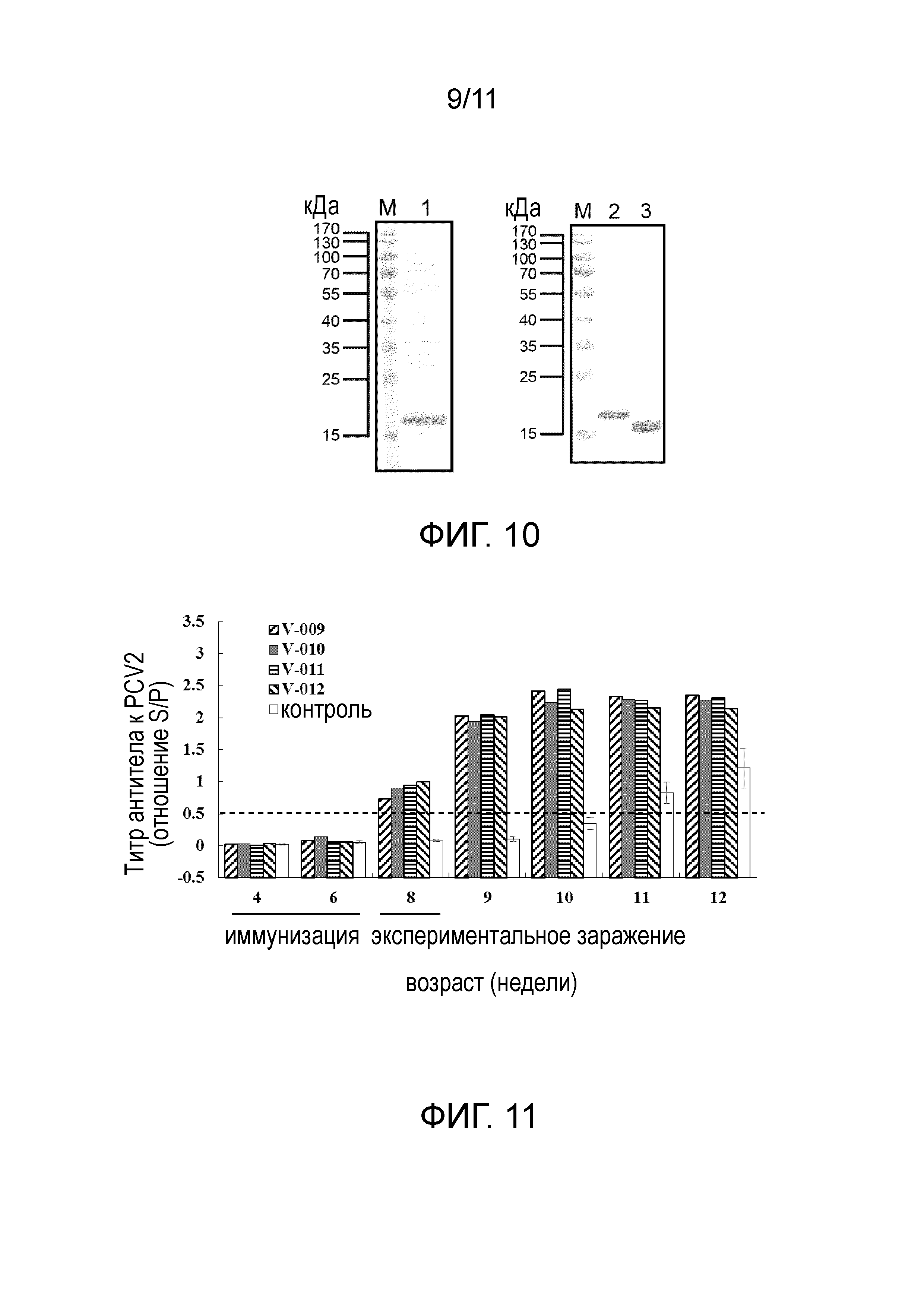
[21] На фигуре 2 представлены результаты экспрессии белка пяти экспрессирующих векторов, получаемых в примере 1 в E. coli-хозяевах после их трансформации с использованием электрофореза белков и вестерн-блоттинга. (A) Результаты электрофореза белка. (B) Результаты вестерн-блоттинга; дорожка 1: BL21(DE3)/pET29a; дорожка 2: BL21(DE3)/pET-SUMO-ORF2; дорожка 3: BL21(DE3)/pET-OPTSUMO-ORF2; дорожка 4: Rosetta2/pET-SUMO-ORF2; дорожка 5: BL21(DE3)/pET-SUMO-OPTORF2; дорожка 6: BL21(DE3)/pET-OPTSUMO-OPTORF2; дорожка 7: BL21/pBA-OPTSUMO-OPTORF2.
[22] На фигуре 3 представлена растворимость слитых белков, экспрессируемых четырьмя экспрессирующими векторами, получаемыми в примере 1, в E. coli-хозяевах после их трансформации с использованием электрофорез белков. T: общий клеточный лизат; S: фракция растворимых белков; IS: фракция нерастворимых белков. Стрелками указаны целевые белки.
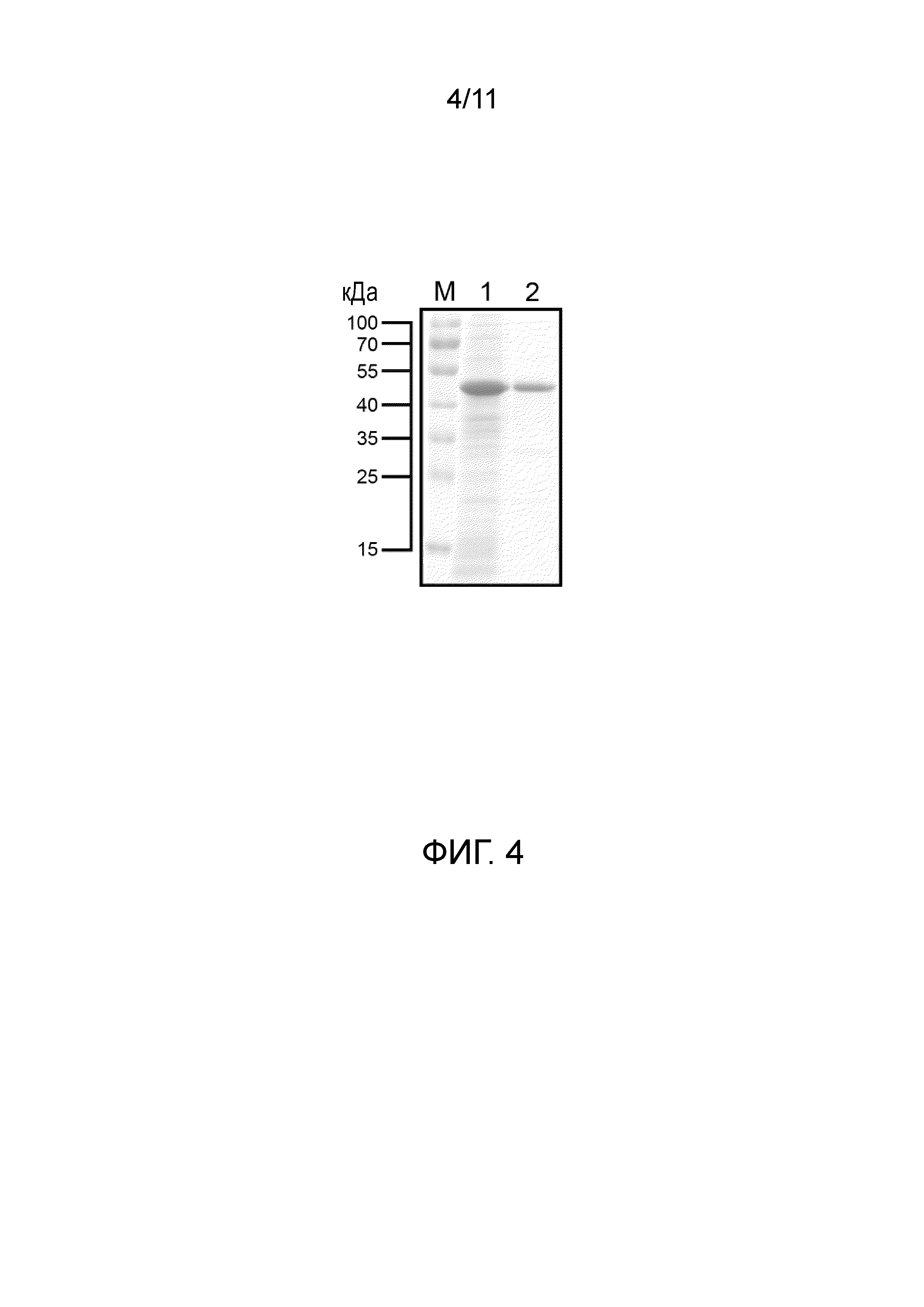
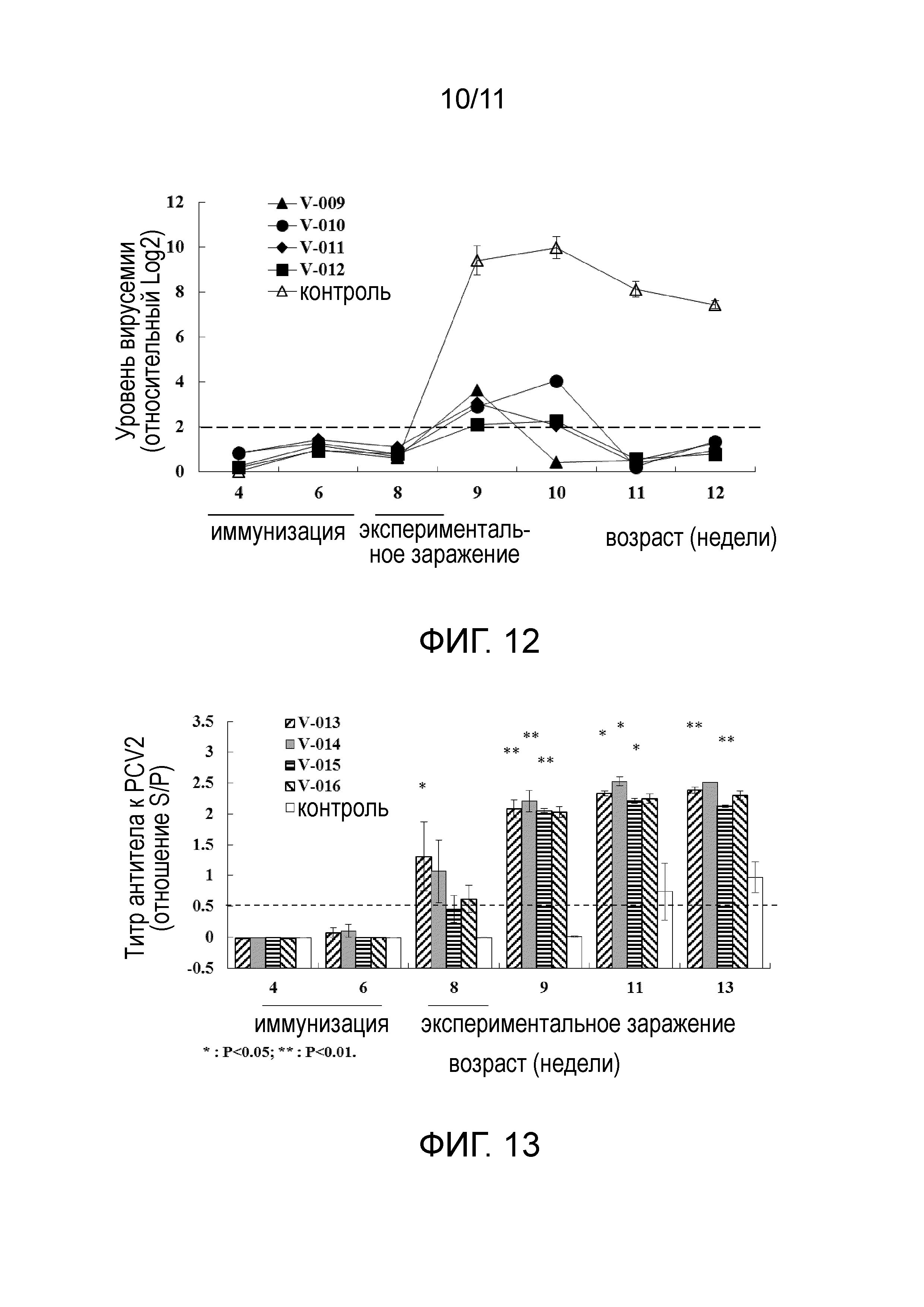
[23] На фигуре 4 представлена электрофореграмма белков, экспрессируемых E. coli BL21(DE3), несущих плазмиду pBA-OPTSUMO-OPTORF2, и очистка белка аффинной хроматографией на иммобилизованных ионах металла. Дорожка 1: общий клеточный лизат E. coli BL21 (pBA-OPTSUMO-OPTORF2); дорожка 2: очищенный слитый белок.
[24] На фигуре 5 представлены результаты экспрессии белка рекомбинантной протеазы SUMO (SUMOPH) и рекомбинантной протеазы D-SUMO (DSUMOPH) в клетках-хозяевах [E. coli BL21 (DE3)] в примере 2 с использованием электрофореза белков и вестерн-блоттинга. (A) Результаты электрофореза белка. (B) Результаты вестерн-блоттинга. T: общий клеточный лизат; S: фракция растворимых белков; IS: фракция нерастворимых белков. Стрелками указаны целевые белки.
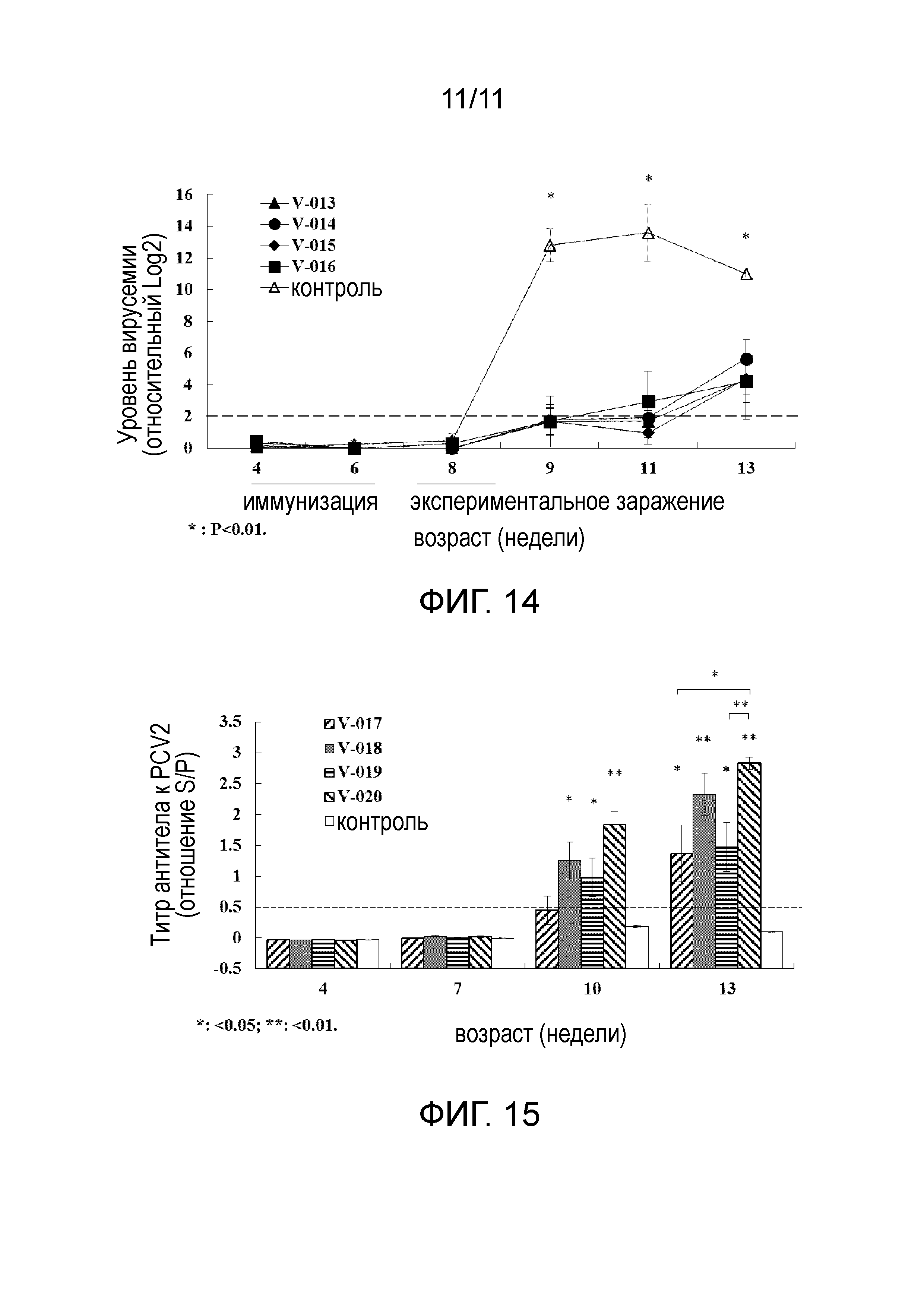
[25] На фигуре 6 представлена электрофореграмма белков, экспрессируемых E. coli BL21 (DE3), несущими плазмиды pET-SUMOPH и pET-D-SUMOPH, и очистка белка аффинной хроматографией на иммобилизованных ионах металла. Дорожка 1: очищенная протеаза SUMO (SUMOPH); дорожка 2: очищенная протеаза D-SUMO (DSUMOPH).
[26] На фигуре 7 представлена электрофореграмма очищенного SUMO-ORF2, расщепленного SUMO-ORF2 и очищенной ORF2. Дорожка 1: очищенный слитый белок SUMO-ORF2. Дорожка 2: смесь для расщепления SUMO-ORF2 с протеазой D-SUMO. Дорожка 3: очищенная ORF2 (смесь для расщепления, отфильтрованная сна мембране с порогом пропускания молекулярной массы 100 кДа).
[27] На фигуре 8 представлены изображения трансмиссионной электронной микроскопии вирусоподобных частиц слитого белка SUMO-ORF2 (A), расщепленный SUMO-ORF2 (слитый белок SUMO-ORF2, расщепленный протеазой) (B), очищенная ORF2 (C).
[28] На фигуре 9 представлены электрофореграммы экспрессии белка рекомбинантного интерферона свиньи в примере 3; T: общий клеточный лизат; S: фракция растворимых белков. (A) pET-OPTPIFNAH/E. coli Shuffle; (B) pBA-OPTPIFNAH/E. coli Shuffle; (C) pET-SUMO-OPTPIFNAH/E. coli Shuffle; (D) pET-OPTSUMO-OPTPIFNAH/E. coli Shuffle; (E) pBA-OPTSUMO-OPTPIFNAH/E. coli Shuffle; (F) pET-OPTPIFNRH/E. coli BL21 (DE3); (G) pET-SUMO-OPTPIFNRH/E. coli BL21 (DE3); (H) pET-OPTSUMO-OPTPIFNRH/E. coli BL21(DE3); (I) pBA-OPTSUMO-OPTPIFNRH/E. coli BL21(DE3). Стрелками указаны целевые белки.
[29] На фигуре 10 представлены электрофореграммы очищенного рекомбинантного интерферона свиньи, экспрессируемого в примере 3. Дорожка 1: очищенный слитый белок, экспрессируемый E. coli Shuffle (pET-OPTPIFNAH); дорожка 2: слитый белок, экспрессируемый E. coli Shuffle (pBA-OPTSUMO-OPTPIFNAH) после расщепления протеазой D-SUMO [pET-D-SUMOP/E. coli BL21(DE3) разрушение клеток] и очищенный; дорожка 3: слитый белок, экспрессируемый E. coli BL21(DE3) (pET-OPTSUMO-OPTPIFNRH) после расщепления протеазой D-SUMO [pET-D-SUMOP/E. coli BL21(DE3) разрушение клеток] и очищенный.
[30] На фигуре 11 представлены результаты ELISA титров антитела к PCV2 у свиней, индуцированных образцами эксперимента 3 в примере 4.
[31] На фигуре 12 представлен уровень вирусемии у свиней, понижаемый образцами эксперимента 3 в примере 4.
[32] На фигуре 13 представлены результаты ELISA титров антител к PCV2 у свиней, которые индуцировали образцы эксперимента 4 в примере 4.
[33] На фигуре 14 представлен уровень вирусемии у свиней, понижаемый образцами эксперимента 4 в примере 4.
[34] На фигуре 15 представлены результаты ELISA титров антител к PCV2 у свиней, индуцированных образцами эксперимента 5 в примере 4.
ПОДРОБНОЕ ОПИСАНИЕ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
[35] Как описано выше, несмотря на предпринимаемые в данной области попытки получить капсидный белок PCV2 посредством экспрессирующей системы на основе E. coli, недостатки, заключающиеся в низких выходах продукции, на момент настоящего изобретения еще не преодолели. Таким образом, существуют препятствия на пути достижения успеха в профилактики эпидемии PCV2.
[36] В способе по настоящему изобретению для получения капсидных белков PCV2 используют индуцируемый арабинозой экспрессирующий элемент, раскрываемый авторами настоящего изобретения в патентной заявке Тайваня № 103146225 (дата подачи: 30 декабря 2014 года). Полное содержание патентной заявки Тайваня № 103146225 включено в настоящее изобретение посредством ссылок.
[37] В рамках изобретения, "целевой белок" относится к белку, который должен экспрессироваться прокариотической экспрессирующей системой. В настоящем изобретении указанный выше целевой белок представляет собой капсидный белок PCV2, интерферон α свиньи, или интерферон γ свиньи.
[38] В рамках изобретения, "нуклеотидная последовательность, кодирующая целевой белок," или другое аналогичное описание относится к нуклеотидной последовательности, которая может образовывать указанный выше целевой белок посредством механизмов транскрипции/трансляции in vivo или in vitro. Таким образом, "нуклеотидная последовательность, кодирующая капсидный белок PCV2," или "нуклеотидная последовательность, кодирующая интерферон свиньи," по настоящему изобретению имеет такое же определение, как указано выше. Подобным образом, "нуклеотидная последовательность, кодирующая партнер по слиянию," или "нуклеотидная последовательность, кодирующая молекулу-маркер," по настоящему изобретению имеет такое же определение, как указано выше.
[39] В рамках изобретения, "партнер по слиянию" относится к молекуле, которую используют для увеличения растворимости указанного выше синтезированного целевого белка. Для указанно выше цели нуклеотидную последовательность, кодирующую партнер по слиянию, и нуклеотидную последовательность, кодирующую указанный выше целевой белок, конструируют указанный выше экспрессирующий вектор способом генетической инженерии, таким образом, что указанный выше целевой белок синтезируют с указанным выше партнером по слиянию в виде слитого белка. Указанный выше партнер по слиянию, например, представляет собой, но не ограничивается ими, MsyB E. coli, YjgD E. coli, белок D фага лямбда, белок SUMO пекарских дрожжей или их сочетание.
[40] В рамках изобретения, "молекула-маркер" относится к молекуле, которая облегчает наблюдение за синтезом указанного выше целевого белка или облегчает очистку указанного выше целевого белка. Для указанной выше цели нуклеотидную последовательность молекулы-маркера и нуклеотидную последовательность указанного выше целевого белка конструируют в одном и том же экспрессирующем векторе способом генетической инженерии, таким образом, что указанный выше целевой белок синтезируют с указанной выше молекулой-маркером в виде слитого белка. Указанная выше молекула-маркер, например, представляет собой, но не ограничивается ими, метка His, метка Strep II, метка FLAG или их сочетание.
[41] Первый аспект настоящего изобретения относится к способу получения капсидного белка PCV2, интерферона α свиньи или интерферона γ свиньи. Указанный выше способ включает (a) получение индуцируемого арабинозой экспрессирующего вектора, где индуцируемый арабинозой экспрессирующий вектор содержит экспрессирующий элемент и нуклеотидную последовательность, кодирующую целевой белок; и (b) трансформацию индуцируемого арабинозой экспрессирующего вектора в E. coli-хозяина для индукции экспрессии целевого белка.
[42] В альтернативном варианте осуществления указанный выше целевой белок представляет собой капсидный белок PCV2. В альтернативном варианте осуществления указанный выше целевой белок представляет собой интерферон α свиньи или интерферон γ свиньи.
[43] В предпочтительном варианте осуществления указанные выше экспрессирующие элементы являются такими, как описано в патентной заявке Тайваня № 103146225 (дата подачи: 30 декабря 2014 года) авторами настоящего изобретения. Конкретно, указанный выше экспрессирующий элемент содержит: промотор; усиливающий трансляцию элемент фага T7 и участок связывания рибосомы. Например, указанный выше обеспечивающий рабочие характеристики элемент представляет собой экспрессирующий элемент araB-M11, описанный в патентной заявке Тайваня № 103146225.
[44] В предпочтительном варианте осуществления указанный выше усиливающий трансляцию элемент фага T7 содержит SEQ ID NO:01. В предпочтительном варианте осуществления указанный выше участок связывания рибосомы содержит SEQ ID NO:02. В предпочтительном варианте осуществления участок -16 указанного выше промотора содержит SEQ ID NO:03. В предпочтительном варианте осуществления указанный выше экспрессирующий элемент содержит SEQ ID NO:04.
[45] В альтернативном варианте осуществления после указанной выше стадии (b) дополнительно следует стадия (c) очистки указанного выше целевого белка. В случае, когда в способе по настоящему изобретению в качестве молекулы-маркера используют метку His, целевой белок можно очищать аффинной хроматографией на иммобилизованных ионах металла.
[46] В альтернативном варианте осуществления в случае, когда в способе по настоящему изобретению белок SUMO используют как указанный выше партнер по слиянию, дополнительно вводят стадию (d) после указанной выше стадии (c): целевой белок обрабатывают протеазой SUMO. Указанное выше "обрабатывают" относится к тому, что партнер по слиянию SUMO разрезают протеазой SUMO, таким образом, целевой белок отделяют от белка SUMO.
[47] В альтернативном варианте осуществления протеазу SUMO получают посредством экспрессирующего вектора T7. В предпочтительном варианте осуществления в указанной выше обработке массовое отношение целевого белка к протеазе SUMO составляет от 4 до 20.
[48] В предпочтительном варианте осуществления указанный выше способ не включает стадию рефолдинга интерферона свиньи. Специалист в данной области может понимать, что "стадия рефолдинга" в экспрессирующей системе на основе прокариотической клетки означает процесс формирования третичной структуры или четвертичной структуры полипептида в результате растворения тельце включения с использованием мочевины или гидрохлорида гуанидина, а затем рефолдинг получаемого полипептида посредством диализа и другие стадии. Таким образом, специалисты в данной области могут понимать, что "не включает стадию рефолдинга интерферона свиньи" по настоящему изобретению означает, что полипептиды, получаемые способом по настоящему изобретению, могут сами сворачиваться в желаемый белок без необходимости использования мочевины или гидрохлорида гуанидина и диализа.
[49] В альтернативном варианте осуществления хозяин представляет собой E. coli. Предпочтительно E. coli представляет собой BL21, BL21 (DE3), Rosetta 2 или Shuffle.
[50] Второй аспект настоящего изобретения представляет собой композицию для профилактики инфекции PCV2, содержащую капсидный белок PCV2, интерферон α свиньи, интерферон γ свиньи и фармацевтически приемлемый носитель.
[51] В предпочтительном варианте осуществления композиция для профилактики инфекции PCV2 содержит от 2,5 до 250 мкг/мл капсидного белка PCV2; от 2,5 до 25 мкг/мл интерферона α свиньи; от 2,5 до 25 мкг/мл интерферона γ свиньи и фармацевтически приемлемый носитель. В другом предпочтительном варианте осуществления композиция для профилактики инфекции PCV2 содержит от 3,5 до 170 мкг/мл капсидного белка PCV2; от 5 до 20 мкг/мл интерферона α свиньи; от 5 до 20 мкг/мл интерферона γ свиньи и фармацевтически приемлемый носитель.
[52] В предпочтительном варианте осуществления капсидный белок PCV2 получают способом по настоящему изобретению. В предпочтительном варианте осуществления интерферон α свиньи и/или интерферон γ свиньи представляют собой интерфероны, получаемые способами по настоящему изобретению.
[53] "Фармацевтически приемлемый носитель" по настоящему изобретению относится к веществу, которое не оказывает отрицательного влияния на цели профилактики инфекции PCV2 капсидным белком PCV2, интерфероном α свиньи и/или интерфероном γ свиньи в композиции с точки зрения медицинских/фармацевтических аспектов. В альтернативном варианте осуществления фармацевтически приемлемый носитель, например, представляет собой, но не ограничивается ими, воду, фосфатно-солевой буфер, спирт, глицерин, хитин, альгинат, хондроитин, витамин E, минералы или их сочетания.
[54] В предпочтительном варианте осуществления композиция дополнительно содержит фармацевтически приемлемый адъювант. "Фармацевтически приемлемый адъювант" по настоящему изобретению относится к веществу, которое облегчает цель профилактики инфекции PCV2 капсидным белком PCV2, интерфероном α свиньи и/или интерфероном γ свиньи в композиции и повышает иммунитет с точки зрения медицинских/фармацевтических аспектов. В альтернативном варианте осуществления фармацевтически приемлемый адъювант, например, представляет собой, но не ограничивается ими, адъювант MONTANIDE™ ISA 536 VG, адъювант MONTANIDE™ GEL 01, полный или неполный адъювант Фрейнда, гель на основе алюминия, поверхностно-активное вещество, полианионные полимеры, пептиды, масляные эмульсии или их сочетания. В предпочтительном варианте осуществления фармацевтически приемлемый адъювант представляет собой адъювант MONTANIDE™ ISA 536 VG, адъювант MONTANIDE™ GEL 01 или их сочетание.
[55] Процесс исследования по настоящему изобретению дополнительно подробно описан в следующих ниже примерах. Однако следующее ниже содержание только иллюстрирует признаки настоящего изобретения для лучшего понимания. Специалисты в данной области могут изменять следующее ниже содержание, не выходя за рамки сущности настоящего изобретения и изменять его на основании общих знаний в данной области, а все еще оставаясь в рамках объема настоящего изобретения.
Пример 1: Конструкция вектора для экспрессии капсидного белка PCV2 (PCV2 ORF2)
Выделение и секвенирование вируса PCV2
[56] Лимфоидные органы, такие как селезенка и лимфоузлы, больных свиней получали из свиноводческих ферм с инфекцией PCV2 (Yunlin, Taiwan). После вырезания стерильными ножницами лимфоидные органы измельчали с использованием стерильного пестика для растирания и палочки для растирания и добавляли соответствующее количество раствора стерильного фосфатного буфера и перемешивали с получением эмульсии. Эмульсию центрифугировали (6000×g, 20 минут) для сбора супернатанта, а затем фильтровали супернатант через фильтр для удаления остатков ткани. Выделение ДНК проводили с использованием набора для очистки ДНК (DNeasy Blood&Tissue kit; Qiagen, USA). Сто (100) мкл супернатанта в виде эмульсии добавляли к 180 мкл буфера ATL и 20 мкл протеиназы K (10 мг/мл) и инкубировали при 56°C в течение 2 часов. После этого добавляли 200 мкл абсолютного спирта и хорошо перемешивали. Все растворы пипетировали на центрифужную колонку, которую помещали в пробирку для сбора элюата и центрифугировали при 6000×g в течение 1 минуты. Центрифужную колонку помещали в новую пробирку для сбора, 500 мкл буфера AW1 добавляли в пробирку и центрифугировали пробирку при 6000×g в течение 1 минуты. Помещали центрифужную колонку в новую пробирку для сбора, добавляли 500 мкл буфера AW2 в центрифужную колонку и центрифугировали центрифужную колонку при 20630×g в течение 5 минут. Центрифужную колонку помещали в стерильную пробирку Eppendorf и добавляли соответствующее количество стерильной деионизированной воды для элюирования ДНК.
[57] Конструировали праймеры PCVF (5'-ACCAGCGCACTTCGGCAGC-3'; SEQ ID NO:05) и PCVR (5'-AATACTTACAGCGCACTTCTTTCGTTTTC-3'; SEQ ID NO:06) и амплифицировали геномную ДНК PCV2 полимеразной цепной реакцией (ПЦР). Объем реакционной смеси для ПЦР составлял 100 мкл, включая 10 мкл ДНК, экстрагированной из лимфоидных органов, 10 мкл буфера 10× Taq, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации и 2,5 U ДНК-полимеразы DreamTaq. (Thermo, USA). Условия реакции ПЦР являлись следующими 94°C в течение 5 минут (1 цикл); 94°C в течение 30 секунд, 59°C в течение 30 секунд, 72°C в течение 1 минуты и 30 секунд (35 циклов); 72°C в течение 7 минут (1 цикл). Для подтверждения присутствия фрагмента ДНК с теоретически рассчитанным размером использовали электрофорез ДНК.
[58] Продукт ПЦР восстанавливали с использованием набора PCR-M™ Clean Up (GMbiolab, Taiwan) и подвергали клонированию TA с использованием набора yT&A Cloning Vector (Yeastern, Taiwan). Экспериментальную процедуру проводили на основе руководства производителя для набора yT&A Cloning Vector. Пять (5) мкл восстановленного и очищенного продукта ПЦР хорошо смешивали с 2 мкл вектора yT&A, 1 мкл буфера для лигирования A, 1 мкл буфера для лигирования B и 1 мкл ДНК-лигазы T4 (2 ед./мкл). Смесь инкубировали при 22°C в течение 30 минут. Один (1) мкл лигирующей смеси трансформировали в E. coli ECOS 9-5 (Yeastern, Taiwan). Трансформированные клетки добавляли к 1 мл среды для восстановления SOC и встряхивали при 37°C, 250 об./мин. в течение 60 минут. После этого подходящее количество раствора бактерий наносили на твердую среду, содержащую ампициллин (конечная концентрация 100 мкг/мл) и культивировали при 37°C в течение 16 часов.
[59] Затем отбирали трансформантов посредством полимеразной цепной реакции для отбора колоний. Способ полимеразной цепной реакции для отбора колоний описан ниже. Сначаал в пробирку Eppendorf добавляли 50 мкл 2× Premix Reaction Buffer (GMbiolab, Taiwan), 0,5 мкл 100 мМ праймера PCVF, 0,5 мкл праймера 100 мМ PCVR и 49 мкл стерильной воды и хорошо перемешивали. Раствор для реакции ПЦР дозировали в пробирки для ПЦР (10 мкл/пробирку). После того как колонию помещали в пробирку для ПЦР с помощью зубной палочки проводили ПЦР. Условия реакции ПЦР являлись следующими 95°C в течение 5 минут (1 цикл); 95°C в течение 30 секунд, 59°C в течение 30 секунд, 72°C в течение 1 минуты и 30 секунд (25 циклов); 72°C в течение 7 минут (1 цикл). Для подтверждения присутствия фрагмента ДНК с теоретически рассчитанным размером использовали электрофорез ДНК. После подтверждения того, что рекомбинантные плазмиды в трансформантах несли вставку ДНК, экстрагировали плазмиды в трансформантах и проводили секвенирование ДНК (Tri-I Biotech, Inc.). Плазмиду, содержащую ДНК PCV2 называли pTA-PCV2.
Амплификация и оптимизация кодона гена ORF2 (Т.е. гена, кодирующего капсидный белок)
(1) Амплификация гена ORF2
[60] Испльзование pTA-PCV2 в качестве матрица и проведение амплификации гена ORF2 с использованием набора праймеров ORF2F/ORF2R (ORF2F; 5'-CAATATGGATCCATGACGTATCCAAGGAGGCGTTTC-3'; SEQ ID NO:07 и ORF2R; 5'-GATATAGTCGACTTAGGGTTTAAGTGGGGGGTCTTTAAGATTAA-3'; SEQ ID NO:08). 50 мкл реакционной смеси для ПЦР содержало 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 100 нг pTA-PCV2 и 1 ед. GDP-HiFi ДНК-полимеразы. Условие реакции ПЦР являлось следующим: 96°C в течение 2 минут (1 цикл); 94°C в течение 30 секунд, 60°C в течение 30 секунд, 68°C в течение 30 секунд (35 циклов); 68°C в течение 5 минут (1 цикл). Для подтверждения содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем восстанавливали продукт ПЦР с использованием набора PCR-M™ Clean Up. На основании результатов секвенирования последовательность гена ORF2 представлена как SEQ ID NO:09.
(2) Синтез гена ORF2 с оптимирозованным кодоном (OPTORF2)
[61] Аминокислотную последовательность ORF2 восстанавливали до нуклеотидной последовательности на основании предпочтительных кодонов E. coli. Праймеры конструировали на основании указанных выше нуклеотидных последовательностей: OPTORF2-T1, OPTORF2-T2, OPTORF2-T3, OPTORF2-T4, OPTORF2-T5, OPTORF2-T6, OPTORF2-T7, OPTORF2-T8, OPTORF2-T9, OPTORF2-T10, OPTORF2-T11, OPTORF2-T12, OPTORF2F и OPTORF2R. Последовательности праймеров приведены в таблице 1.
Таблица 1: Праймеры, используемые для синтеза гена ORF2 (OPTORF2) с оптимизированным кодоном.
[62] В качестве матрицы для праймеров использовали OPTORF2-T1-OPTORF2-T12 и OPTORF2 и OPTORF2R использовали в качестве праймеров для амплификации. Полимеразную цепную реакцию (OEPCR) с перекрывающимися участками достройки использовали для масштабной амплификации ген ORF2 с оптимизацией кодона. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ каждого праймера и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 2 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 30 секунд (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем восстанавливали продукт ПЦР с использованием набора PCR-M™ Clean Up system. На основании результатов секвенирования последовательность гена ORF2 с оптимизацией кодона представлена как SEQ ID NO:24.
Амплификация и оптимизация кодона генов SUMO
(1) Амплификация гена SUMO
[63] Пекарские дрожжи (Saccharomyces cerevisiae), выделенные из быстрорастворимых дрожжей DIY Sun Right Food Co. инокулировали в среду YPD (20% пептон, 10% дрожжевой экстракт, 20% глюкоза; pH 6,5) и культивировали при встряхивании при 30°C, 200 об./мин. в течение 16 часов. После культивации проводили выделение генома of дрожжей с использованием набора YeaStar™ Genomic DNA (Zymo Research, USA). 1,5 мл культируемого в течение ночи бульона добавляли в пробирка Eppendorf, собирали бактериальные фракции центрифугированием (2000×g, 5 минут, комнатная температура) и тщательно смешивали с 120 мкл буфера для расщепления YD и 5 мкл R-Zymolase, и инкубировали при 37°C в течение 1 часа. Затем добавляли в смесь 120 мкл лизирующего буфера YD и аккуратно перемешивали несколько раз. Добавляли к смеси двести пятьдесят (250) мкл хлороформа и встряхивали в течение 1 минуты. Супернатант собирали центрифугированием (10000×g, 2 минуты, комнатная температура). Центрифужную колонку помещали в пробирку для сбора и добавляли супернатант в центрифужную колонку. После центрифугирования (10000×g, 1 минута, комнатная температура) удаляли фильтрат. В центрифужную колонку добавляли 300 мкл буфера для промывания ДНК, центрифугировали центрифужную колонку (10000×g, 1 минута, комнатная температура), удаляли фильтрат и повторяли эту процедуру один раз. Центрифужную колонка помещали в стерильную пробирку Eppendorf, добавляли соответствующее количество элюирующего раствора в центрифужную колонку и центрифугировали центрифужную колонку и пробирку Eppendorf (10000×g, 2 минуты, комнатная температура) для элюирования геномной ДНК.
[64] Ген SUMO амплифицировали с использованием геномной ДНК Saccharomyces cerevisiae, получаемой, как указано в предшествующем абзаце, в качестве матрице и с использованием SUMOF (5'-GATATAGGTACCATGTCGGACTCAGAAGTCAATCAAG-3'; SEQ ID NO:25)/SUMOR (5'-CAATATGGATCCACCACCAATCTG TTCTCTGTGAGC-3; SEQ ID NO:26) в качестве набора праймеров. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 200 нг геномной ДНК Saccharomyces cerevisiae и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 5 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 30 секунд (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером использовали электрофорез в агарозном геле. Затем восстанавливали продукт ПЦР с использованием набора PCR-M™ Clean Up system.
(2) Синтез гена SUMO (OPTSUMO)с оптимизированным кодоном
[65] Аминокислотную последовательность SUMO преобразовывали в нуклеотидную последовательность на основании от предпочтительных кодонов E. coli. Праймеры конструировали на основании указанных выше нуклеотидных последовательностей: OPTSUMO-T1, OPTSUMO-T2, OPTSUMO-T3, OPTSUMO-T4, OPTSUMO-T5, OPTSUMO-T6, OPTSUMO-T7, OPTSUMO-T8, OPTSUMOF и OPTSUMOR. Последовательности приведены в таблице 2.
Таблица 2: Праймеры, используемые для синтеза гена SUMO (OPTSUMO) с оптимизированным кодоном.
[66] OPTSUMO-T1-OPTSUMO-T8 использовали в качестве праймеров для матрицы и OPTSUMOF и OPTSUMOR использовали в качестве праймеров для амплификации. Для масштабной амплификации гена SUMO с оптимизированным кодоном использовали полимеразную цепную реакцию с перекрывающимися участками достройки. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ каждого праймера и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 2 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 30 секунд (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции использовали электрофорез в агарозном геле для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером. Затем восстанавливали продукт ПЦР с использованием набора PCR-M™ Clean Up. На основании результатов секвенирования последовательность гена SUMO с оптимизированным кодоном представлена как SEQ ID NO:37.
Конструкция экспрессирующего вектора слитого белка ORF2
(1) Конструкция pET-DRAHIS
[67] Реакцию ПЦР проводили с использованием pET29a в качестве матрицы и DRAF (5'- GATATACATATGAAAAAAAAATTCGTATCGCATCACCATCACCATCACAGCGGTGGTGGTACCCCAGATCTGGGTACCCTGG-3'; SEQ ID NO:38)/терминатора T7 (GCTAGTTATTGCTCAGCGG; SEQ ID NO:39) в качестве набора праймеров. 50 мкл реакционной смеси для ПЦР содержали 1× буфер Ex Taq™, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 100 нг pET29a и 1,25 ед. ДНК-полимеразы TakaRa Ex Taq™ (Takara, Japan). Условие реакции ПЦР являлось следующим: 94°C в течение 5 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 72°C в течение 50 секунд (35 циклов); 72°C в течение 7 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем восстанавливали продукт ПЦР с использованием набора PCR-M™ Clean Up system.
[68] После того, как продукт ПЦР разрезали NdeI и SalI, фрагменты ДНК лигировали в pET29a, который разрезали с теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированные продукты трансформировали в E. coli XL1-blue (Protech, Taiwan). Трансформанты случайным образом отбирали для подтверждения последовательностей ДНК. Плазмиду с правильной последовательностью ДНК называли pET-DRAHIS. Эта плазмида содержит старт-кодон с последующей нижележащей последовательностью (DS) AAAAAAAAATTCGTATCG (SEQ ID NO:40) и последовательностью ДНК с меткой His CATCACCATCACCATCAC (SEQ ID NO:41).
(2) Конструкция экспрессирующего вектора pET-SUMO-ORF2
[69] После того, как ген SUMO амплифицировали из генома Saccharomyces cerevisiae и разрезали KpnI и BamHI, фрагмент ДНК лигировали в pET-DRAHIS, разрезанную теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения того, что рекомбинантные плазмиды в трансформантах не несли встраиваемую ДНК, посредством электрофорез ДНК, плазмиды в трансформантах экстрагировали и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называли pET-SUMO.
[70] После того, как ген ORF2, амплифицированный из генома вируса PCV2 Yunlin, разрезали BamHI и SalI, фрагмент ДНК встраивали в pET-SUMO, разрезанную теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения того, что рекомбинантные плазмиды в трансформантах не несли встраиваемую ДНК, посредством электрофореза ДНК, выделяли плазмиды в трансформанте и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называют pET-SUMO-ORF2, которая содержит последовательность SEQ ID NO:42.
(3) Конструкция экспрессирующего вектора pET-OPTSUMO-ORF2
[71] После того, как синтетический ген OPTSUMO разрезали KpnI и BamHI, фрагмент ДНК лигировали в pET-DRAHIS, разрезанную теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения того, что рекомбинантная плазмида в трансформантах не несла встраиваемую ДНК, посредством электрофореза ДНК, выделяли плазмиды в трансформантах и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называли pET-OPTSUMO.
[72] После того, как ген ORF2, амплифицированный из генома вируса PCV2 Yunlin, разрезали BamHI и SalI, фрагмент ДНК лигировали в pET-OPTSUMO, разрезанную теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения того, что рекомбинантная плазмида в трансформантах не несла встраиваемую ДНК, посредством электрофореза ДНК, выделяли плазмиды в трансформантах и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называют pET-OPTSUMO-ORF2, которая содержит SEQ ID NO:43.
(4) Конструкция экспрессирующего вектора pET-SUMO-OPTORF2:
[73] После того, как синтетический ген OPTORF2 разрезали BamHI и SalI, фрагмент ДНК лигировали в pET-SUMO, разрезанную теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения того, что рекомбинантная плазмида в трансформантах не несла встраиваемую ДНК, посредством электрофореза ДНК, выделяли плазмиды в трансформанте и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называют pET-SUMO-OPTORF2, которая содержит SEQ ID NO:44.
(5) Конструкция экспрессирующего вектора pET-OPTSUMO-OPTORF2
[74] После того, как синтетический ген OPTORF2 разрезали BamHI и SalI, фрагмент ДНК встраивали в pET-OPTSUMO, разрезанную теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения посредством электрофореза ДНК того, что рекомбинантная плазмида в трансформанте не несла встраиваемую ДНК, выделяли плазмиды в трансформантах и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называют pET-OPTSUMO-OPTORF2, которая содержит SEQ ID NO:45.
(6) Конструкция экспрессирующого вектора pBA-OPTSUMO-OPTORF2
[75] pBA-OPTSUMO-OPTORF2, сконструированный в этом эксперименте, получали встраиванием фрагмента ДНК OPTSUMO-OPTORF2 в новый индуцируемый арабинозой экспрессирующий вектор pBCM-araM11. pBCM-araM11 конструировали с индуцируемым арабинозой экспрессирующим элементом и pBRCMMCS (SEQ ID NO:100), раскрытым в патентной заявке Тайваня № 103146225 (дата подачи: 30 декабря 2014 года) и № 103142753 (дата подачи: 9 декабря 2014 года) авторами настоящего изобретения. Способ конструирования экспрессирующего вектора описан ниже.
[76] После того, как pARABM11-GFPT разрезали EcoRI и NdeI, фрагмент ДНК, содержащий экспрессирующие элементы araC и araB-M11, восстанавливали с использованием набора Gel-M™ gel extraction system (GMbiolab, Taiwan). Экспрессирующие элементы araC и araB-M11 лигировали в pBRCMMCS, разрезанную теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний и выделяли плазмиды для подтверждения последовательности ДНК. Плазмиду с правильной последовательностью называют pBCM-araM11, которая содержит SEQ ID NO:98.
[77] После того, как pET-OPTSUMO-OPTORF2 разрезали NdeI и SalI, фрагмент ДНК, содержащий OPTSUMO-OPTORF2, восстанавливали с использованием набора Gel-M™ gel extraction system. OPTSUMO-OPTORF2 лигировали в pBCM-araM11, разрезанную теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний и выделяли плазмиды для подтверждения секвенирования ДНК. Плазмиду с правильной последовательностью называют pBA-OPTSUMO-OPTORF2, которая содержит SEQ ID NO:46.
[78] Фрагмент ДНК, содержащий экспрессирующий элемент araB-M11, представляет собой индуцируемый арабинозой экспрессирующий элемент по настоящему изобретению, который содержит промотор (часть -16 представлена как SEQ ID NO:03) и усиливающий трансляцию элемент фага T7 (SEQ ID NO:01), и участок связывания рибосомы (SEQ ID NO:02). Индуцируемый арабинозой экспрессирующий элемент является таким, как приведено в патентной заявке Тайваня № 103146225 (дата подачи: 30 декабря 2014 года), который содержит SEQ ID NO:04.
Выводы
[79] В итоге, в этом примере получали пять экспрессирующих векторов капсидного белка PCV2, а именно: pET-SUMO-ORF2 (SEQ ID NO:42) и pET-OPTSUMO-ORF2 (SEQ ID. NO: 43), pET-SUMO-OPTORF2 (SEQ ID NO:44), pET-OPTSUMO-OPTORF2 (SEQ ID NO:45) и pBA-OPTSUMO-OPTORF2 (SEQ ID NO:46). См. фигуру 1.
Пример 2 Получение капсидных белков PCV2 по настоящему изобретению.
[80] Как описано выше, каждый из векторов, получаемых в примере 1 (SEQ ID NO: 42-46), содержит ДНК капсидного белка ORF2, и его можно использовать для получения капсидных белков. Кроме того, для очистки и характеристик растворимости целевые белки сливали с белком SUMO и меткой His. Такой слитый белок обозначают в настоящем описании как слитый белок SUMO-ORF2, и тот факт, что слитый белок содержит метку His, больше не будет упоминаться. В этом примере для получения слитого белка SUMO-ORF2 по настоящему изобретению будут использовать экспрессирующий вектор, описанный в пример 1.
Трансформация E. coli и индуцируемая экспрессия рекомбинантного слитого белка SUMO-ORF2
(1) Экспериментальный способ
[81] Экспрессирующие векторы, такие как pET-SUMO-ORF2, pET-OPTSUMO-ORF2, pET-SUMO-OPTORF2 и pET-OPTSUMO-OPTORF2, трансформировали в E. coli BL21 (DE3) (Yeastern, Taiwan). pET-SUMO-ORF2 трансформировали в E. coli Rosetta2 (EMD Millipore, USA). pBA-OPTSUMO-OPTORF2 трансформировали в E. coli BL21 (New England Biolabs, USA). После способа трансформации следовала последовательность действий, предлагаемая производителями.
[82] Трансформант E. coli BL21(DE3) инокулировали в среду LB, содержащую канамицин (конечная концентрация: 30 мкг/мл), и культивировали при встряхивании при 37°C и 180 об./мин. После инкубации в течение ночи бактериальный раствор инокулировали в отношении 1:100 в среде LB, содержащей канамицин (конечная концентрация 30 мкг/мл). Культивирование при встряхивании проводили при 37°C и 180 об./мин. Бактерии культивировали до концентрации приблизительно от 0,4 до 0,6 OD600, измеряемой спектрофотометром, и добавляли 0,1 мМ изопропил-β-D-тиогалактозида (IPTG) для индукции экспрессии белка. Через 4 часа индукции собирали бактериальные фракции центрифугированием (8000×g, 30 минут, 4°C) и наблюдали экспрессию слитого белка SUMO-ORF2 посредством электрофореза белков и Вестерн-блоттинга. Первичные и вторичные антитела, используемые в способах Вестерн-блоттинга, представляли собой антитело кролика к поликлональным антителам с 6×His (Protech, Taiwan) и конъюгированным с щелочной фосфатазой антителам козы к IgG кролика (H+L), соответственно. Используемый краситель представлял собой NBT/BCIP (Thermo, USA). Также различали растворимые и нерастворимые белки бактерий и наблюдали растворимость слитого белка SUMO-ORF2 посредством электрофореза белков.
[83] Трансформант E. coli Rosetta2 инокулировали в среду LB, содержащую хлорамфеникол (конечная концентрация 34 мкг/мл) и канамицин (конечная концентрация 30 мкг/мл). Культивирование при встряхивании проводили при 37°C и 180 об./мин. После инкубации в течение ночи бактериальный раствор инокулировали в отношении 1:100 в среде LB, содержащей хлорамфеникол (конечная концентрация 34 мкг/мл) и канамицин (конечная концентрация 30 мкг/мл). Культивирование при встряхивании проводили при 37°C и 180 об./мин. Бактерии культивировали до концентрации приблизительно от 0,4 до 0,6 OD600, измеряемой спектрофотометром, и добавляли 0,1 мМ IPTG для индукции экспрессии белка. Через 4 часа индукции собирали бактериальные фракции центрифугированием (8000×g, 30 минут, 4°C) и наблюдали экспрессию слитого белка SUMO-ORF2 посредством электрофореза белков и Вестерн-блоттинга. Также различали растворимые и нерастворимые белки бактерий и наблюдали растворимость слитого белка SUMO-ORF2 посредством электрофореза белков.
[84] Трансформант E. coli BL21 инокулировали в среду LB, содержащую хлорамфеникол (25 мкг/мл). Культивирование при встряхивании проводили при 37°C и 180 об./мин. После инкубации в течение ночи бактериальный раствор инокулировали в отношении 1:100 в среде LB, содержащей хлорамфеникол (25 мкг/мл). Культивирование при встряхивании проводили при 37°C и 180 об./мин. Бактерии культивировали до концентрации приблизительно от 0,4 до 0,6 OD600, измеряемой спектрофотометром, и добавляли 0,2% арабиноза для индукции экспрессии белка. Через 4 часа индукции собирали бактериальные фракции центрифугированием (8000×g, 30 минут, 4°C) и наблюдали экспрессию слитого белка SUMO-ORF2 посредством электрофореза белков и Вестерн-блоттинга. Также различали растворимые и нерастворимые белки бактерий и наблюдали растворимость слитого белка SUMO-ORF2 посредством электрофореза белков.
[85] После того, как сканировали электрофоретическую пленку с белком, определяли процент экспрессии рекомбинантного слитого белка SUMO-ORF2 с использованием программного обеспечения Image Quant TL 7.0 (GE Healthcare Life Sciences, USA), и дополнительно рассчитывали выход слитого белка.
(2) Результаты эксперимента:
[86] Результаты демонстрировали, что в группе, трансформированной pET-SUMO-ORF2 и pET-OPTSUMO-ORF2 и индуцированной в E. coli BL21 (DE3), рекомбинантный слитый белок SUMO-ORF2 полностью отсутствовал (фигура 2). В группе, в которой трансформировали pET-SUMO-ORF2 и индуцировали в E. coli Rosetta2, которая способна продуцировать соответствующую тРНК с редким кодоном, рекомбинантный слитый белок SUMO-ORF2 может экспрессироваться (фигура 2), и большая его часть являлась растворимой (фигура 3), согласно результатам. Выход растворимого рекомбинантного слитого белка SUMO-ORF2 составляет 46,81 мг/л. Тот факт, что указанный выше ген ORF2 не может экспрессироваться в E. coli BL21 (DE3), свидетельствует о том, что кодоны, которые нес ORF2, в значительной степени влияют на характеристики слитого белка SUMO-ORF2 в E. coli.
[87] pET-SUMO-OPTORF2 с геном ORF2 с оптимизацией кодона трансформировали в E. coli BL21(DE3) и индуцировали. Результаты демонстрировали, что рекомбинантный слитый белок SUMO-ORF2 эффективно экспрессировался (фигура 2) и являлся в основном растворимым белком (фигура 3); выход растворимого рекомбинантного слитого белка SUMO-ORF2 составлял 54,62 мг/л. Этот результат демонстрирует, что после оптимизации кодона ORF2, характеристики слитого белка SUMO-ORF2 в E. coli BL21 (DE3) можно улучшать.
[88] Экспрессирующий вектор pET-OPTSUMO-OPTORF2, несущий полноразмерный ген ORF2 с оптимизацией кодона и ген SUMO с оптимизацией кодона, трансформировали в E. coli BL21 (DE3) и индуцировали. Результаты демонстрировали, что рекомбинантный слитый белок SUMO-ORF2 может эффективно экспрессироваться (фигура 2), и является в основном растворимым белком (фигура 3); выход растворимого рекомбинантного слитого белка SUMO-ORF2 составляет 81,66 мг/л. Этот результат демонстрирует, что после того, как частоту использования кодона гена партнера по слиянию оптимизируют, характеристики слитого белка ORF2 в E. coli можно дополнительно улучшать. Проводимые ранее исследования никогда не демонстрировали, что оптимизация кодонов гена SUMO может увеличивать экспрессию слитого белка. Авторы настоящего изобретения подтвердили, что оптимизация кодона гена SUMO может увеличивать продукцию слитого белка SUMO-ORF2.
[89] Для продукции рекомбинантного слитого белка SUMO-ORF2 фрагмент ДНК, несущий 5'-3' последовательность-метка His ДНК-ген SUMO с оптимизацией кодона-ген ORF2 с оптимизацией кодона встраивали в индуцируемый арабинозой экспрессирующий вектор pBCM-araM11 и трансформировали в E. coli BL21. Результаты демонстрируют, что рекомбинантный слитый белок SUMO-ORF2 (фигура 2) также можно получать с использованием индуцируемых арабинозой экспрессирующих систем, и он является в основном растворимым белком (фигура 3). Путем использования такого экспрессирующего вектора для продукции слитого белка SUMO-ORF2, можно получать наибольший выход продукта (103,04 мг/л). По сравнению с наибольшим выходом продукта (81,66 мг/л) экспрессирующей системы на основе T7, выход продукта можно увеличивать приблизительно в 1,27 раза. Каждый экспрессирующий вектор примера 1 по настоящему изобретению дает выход растворимого слитого белка SUMO-ORF2 в этом эксперименте такой, как обобщенно в таблице 3 ниже.
Таблица 3: Выход растворимого слитого белка SUMO-ORF2
Очистка рекомбинантного слитого белка SUMO-ORF2 с использованием аффинной хроматографии на иммобилизованных ионах металла
[90] Белок очищали аффинной хроматографией на иммобилизованных ионах металла, используя преимущество свойства рекомбинантного слитого белка SUMO-ORF2, заключающегося в том, что он содержит метку His на N-конце, которая может образовывать ковалентную связь с ионом никеля или кобальта. Очистку проводили с использованием системы жидкостной хроматографии для очистки белка ÄKTA prime plus (GE Healthcare, Sweden) с колонкой 5 мл HiTrap™ Ni excel (GE Healthcare, Sweden).
[91] Осадки суспендировали в лизирующем буфере (50 мМ Tris-HCl, 500 мМ NaCl, pH 8,0) и разрушали ультразвуковым дезинтегратором. Супернатант собирали центрифугированием (8000×g, 15 минут). После уравновешивания колонки 25 мл лизирующим буфером супернатант после обработки дезинтегратором инъецировали в колонку HiTrap™ Ni excel. После завершения инъекции образца неспецифически связанные белки отмывали 100 мл промывающего буфера (50 мМ Tris-HCl, 500 мМ NaCl, 30 мМ имидазол, pH 8,0). В заключении рекомбинантный белок на смоле элюировали 150 мл элюирующего буфера (50 мМ Tris-HCl, 500 мМ NaCl, 250 мМ имидазол, pH 8,0), который конкурировал с рекомбинантным белком за связывание с участком связывания смолы, с помощью высокой концентрации имидазола, что приводило к элюированию слитого белка рекомбинации SUMO-ORF2 из смолы. Для наблюдения очистки рекомбинантного слитого белка SUMO-ORF2 использовали электрофорез белков. Результаты эксперимента приведены на фигуре 4.
Слитый белок SUMO-ORF2 по настоящему изобретению расщепляется протеазой SUMO
[92] В этом эксперименте использовали протеазу SUMO для расщепления слитого белка ORF2, получаемого из экспрессирующей системы на основе E. coli. После расщепления SUMO можно получать фрагменты партнера по слиянию с меткой His и фрагменты капсидного белка. В этом эксперименте протеазу SUMO получали посредством экспрессирующей системы на основе E. coli и применяли в указанных выше способах применения. Специалисты в данной области также могут выполнять эту стадию с использованием протеазы SUMO, получаемой другими путями.
(1) Конструкция рекомбинантного экспрессирующего вектора протеазы SUMO pET-SUMOPH
[93] Ген протеазы SUMO амплифицировали с использованием генома Saccharomyces cerevisiae в качестве матрицы и SUMOPF (5'-CAATATGGATCCCTTGTTCCTGAATTAAATGAAAAAGACG-3'; SEQ ID NO:47)/SUMOPENZHISR (5'-GATATACTCGAGTTAGTGATGGTGATGGTGATGACCACTGCCGCTACCTTTTAAAGCGTCGGTTAAAATCAAATG-3; SEQ ID NO:48) в качестве набора праймеров. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 200 нг геномной ДНК Saccharomyces cerevisiae и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 5 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 30 секунд (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем восстанавливали продукт ПЦР с использованием набора PCR-M™ Clean Up system.
[96] After the D белок ген amplified from Lambda phage ДНК разрезали KpnI и BamHI, фрагмент ДНК лигировали в pET29a, разрезанную теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения посредством электрофореза ДНК того, что рекомбинантная плазмида в трансформанте не несла встраиваемую ДНК, выделяли плазмиды в трансформанте и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называют pET-D, которая содержит SEQ ID NO:99.
[97] После того, как амплифицированный из генома дрожжей ген протеазы SUMO разрезали BamHI и XhoI, фрагмент ДНК лигировали в pET-D, разрезанную BamHI и SalI, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения посредством электрофореза ДНК того, что рекомбинантная плазмида в трансформанте не несла встраиваемую ДНК, выделяли плазмиды в трансформанте и секвенировали ДНК. Плазмида с правильной ДНК называют pET-D-SUMOPH, которая содержит SEQ ID NO:52.
(3) Индуцированная экспрессия и очистка рекомбинантной протеазы
[98] Экспрессирующие векторы pET-SUMOPH и pET-D-SUMOPH трансформировали в E. coli BL21(DE3), соответственно. Трансформант E. coli BL21(DE3) инокулировали в среду LB, содержащую канамицин (конечная концентрация: 30 мкг/мл) и культивировали при встряхивании при 37°C и 180 об./мин. После инкубации в течение ночи бактериальный раствор инокулировали в отношении 1:100 в среде LB, содержащей канамицин (конечная концентрация 30 мкг/мл). Культивирование при встряхивании проводили при 37°C и 180 об./мин. Бактерии культивировали до концентрации приблизительно от 0,4 до 0,6 OD600, измеряемой спектрофотометром, и добавляли 0,1 мМ IPTG для индукции экспрессии белка. Через 4 часа индукции собирали бактериальные фракции центрифугированием (8000×g, 30 минут, 4°C) для дифференцировки растворимых и нерастворимых белков. Растворимость рекомбинантной протеазы наблюдали посредством электрофореза белков и Вестерн-блоттинга. Первичные и вторичные антитела, используемые в способах Вестерн-блоттинга, представляли собой поликлональные антитела кролика к метке His и конъюгированные с щелочной фосфатазой антитела козы к антитела кролика, соответственно. Используемый краситель представлял собой NBT/BCIP. Способ очистки рекомбинантной протеазы является таким же, как способ очистки рекомбинантного слитого белка ORF2.
[99] Результаты демонстрировали, что протеаза SUMO и протеаза D-SUMO можно экспрессировать в E. coli BL21 (DE3) (фигура 5), с выходом продукта 20,55 мг/л и 46,94 мг/л, соответственно, где выход протеазы D-SUMO является выше. Молярное число являлось приблизительно в 2,2 раза выше, чем молярное число протеазы SUMO. Этот результат демонстрирует, что стратегия экспрессии слияния может увеличивать уровень экспрессии протеазы SUMO в E. coli.
[0100] Затем белок очищали аффинной хроматографией на иммобилизованных ионах металла, используя преимущество того, что рекомбинантная протеаза содержит His-метка на C-конце. Результаты демонстрировали, что растворимая рекомбинантная протеаза SUMO и протеаза D-SUMO можно очищать с использованием аффинной колонки на иммобилизованных ионах металла (фигура 6), где выход очищенного продукта протеазы D-SUMO являлся выше. Из 1 литра среды можно очищать двадцать один целый, пять десятых (21,50) мг белка, что является приблизительно в 1,4 раза больше по сравнению с выходом очищенного продукта протеазы SUMO (15,33 мг).
(4) Расщепление рекомбинантного слитого белка SUMO-ORF2 и наблюдение за образованием вирусоподобных частиц
[0101] Очищенные рекомбинантный слитый белок SUMO-ORF2 смешивали с рекомбинантной протеазой (протеазой SUMO или протеазой D-SUMO) в отношении 1:0,05 по массе (например, 1 мг рекомбинантного слитого белка ORF2 и 0,05 мг рекомбинантной протеазы) и инкубировали смесь при 4°C в течение 16 часов. Раствор с расщепленным белком помещали в центрифужную колонку Amicon ultra-15 ultracel-100K (Merck Millipore, USA) и центрифугировали при 2600×g при 4°C до соответствующего объема. После этого расщепленный белок фильтровали с использованием фильтра из регенерированной целлюлозы 100 кДа. Результаты демонстрировали, что использование мембраны 100 кДа может эффективно удалять партнеров по слиянию, устраняя необходимость использовать колоночную хроматографию для разделения ORF2 от его партнера по слиянию, что значительно снижает затраты на получение антигена (фигура 7).
[0102] Затем слитый белок SUMO-ORF2, расщепленный протеазой слитый белок SUMO-ORF2 и слитый белок ORF2, получаемый расщеплением протеазой и фильтрованием, помещали соответственно на медную сетку и оставляли при комнатной температуре в течение 3 минут. Затем удаляли избыток воды фильтровальной бумагой и добавляли краску уранил ацетат для отрицательного окрашивания. Время окрашивания составляло приблизительно от 40 секунд до 1 минуты. Затем удаляли избыток краски фильтровальной бумагой и наблюдали вирусоподобные частицы с помощью автоэмиссионного трансмиссионного электронного микроскопа JEM-2100F (JEOL, Japan).
[0103] Результаты демонстрировали, что слитый белок SUMO-ORF2 не мог образовывать вирусоподобные частицы, а рекомбинантный слитый белок SUMO-ORF2, расщепленный протеазой, и слитый белок ORF2, получаемый расщеплением протеазой и прошедший фильтрацию могли образовывать вирусоподобные частицы (фигура 8). Средний размер вирусоподобных частиц, рассчитываемый посредством электронная микрофотография в проходящем свете, составлял приблизительно 19 нм.
Пример 3: Получение интерферона свиньи
[0104] Настоящее изобретение раскрывает тот факт, что интерферон свиньи можно использовать в качестве адъюванта, который является особенно подходящим для субъединичной вакцины PCV2. Таким образом, интерферон α свиньи и интерферон γ свиньи, необходимые для субъединичной вакцины по настоящему изобретению, получают в клетках-хозяевах E coli в этом примере.
Синтез генов рекомбинантного интерферона α свиньи (IFN-α) и γ (IFN-γ)
(1) Синтез гена IFN-α
[0105] Аминокислотную последовательность зрелого интерферона α-6 свиньи обратно преобразовывали как нуклеотидную последовательность на основании предпочтительного кодона для E. coli. Праймеры конструировали на основании нуклеотидной последовательности: OPTIFNA-T1, OPTIFNA-T2, OPTIFNA-T3, OPTIFNA-T4, OPTIFNA-T5, OPTIFNA-T6, OPTIFNA-T7, OPTIFNA-T8, OPTIFNAF и OPTIFNAR. Последовательности приведены в таблице 4.
Таблица 4: Праймеры для синтеза гена интерферона α-6 свиньи с оптимизацией кодона
[0106] В качестве праймеров для матрицы использовали OPTIFNA-T1-OPTIFNA-T8 и OPTIFNAF и OPTIFNAR использовали в качестве праймеров для амплификации. Полимеразную цепную реакцию с перекрывающимися участками достройки использовали для масштабной амплификации гена IFN-α с оптимизацией кодона. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ каждого праймера и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 2 минут (1 цикл); 94°C в течение 30 секунд, 58°C в течение 30 секунд, 68°C в течение 30 секунд (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем восстанавливали продукт ПЦР с использованием набора PCR-M™ Clean Up system.
[0107] Клонирование гена проводили с использованием набора CloneJET ПЦР Cloning (Thermo, USA) и смесь лигирования трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения посредством электрофореза ДНК того, что рекомбинантная плазмида в трансформанте не несла встраиваемую ДНК, выделяли плазмиды в трансформанте и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называют pJET-IFNA-6, которая содержит SEQ ID NO:63. После проверки последовательности ген IFN-α с оптимизацией кодона содержит SEQ ID NO:64.
(2) Синтез IFN-γ
[0108] Аминокислотная последовательность зрелого интерферона γ свиньи обратно преобразовывали до нуклеотидной последовательности на основании предпочтительного кодона для E. coli. Праймеры конструировали на основании указанных выше нуклеотидных последовательностей: OPTIFNR-T1, OPTIFNR-T2, OPTIFNR-T3, OPTIFNR-T4, OPTIFNR-T5, OPTIFNR-T6, OPTIFNR-T7, OPTIFNR-T8, OPTIFNRF и OPTIFNRR. Последовательности приведены в таблице 5.
Таблица 5: Праймеры для синтеза гена интерферона γ свиньи с оптимизированным кодоном
[0109] В качестве праймеров для матрицы использовали OPTIFNR-T1-OPTIFNR-T8 и OPTIFNRF и OPTIFNRR использовали в качестве праймеров для амплификации. Полимеразную цепную реакцию с перекрывающимися участками достройки использовали для масштабной амплификации гена IFN-γ с оптимизацией кодона. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ каждого праймера и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 2 минут (1 цикл); 94°C в течение 30 секунд, 57°C в течение 30 секунд, 68°C в течение 30 секунд (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем восстанавливали продукт ПЦР с использованием набора PCR-M™ Clean Up system.
[0110] Клонирование гена проводили с использованием набора CloneJET PCR Cloning и смесь лигирования трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения посредством электрофореза ДНК того, что рекомбинантная плазмида в трансформанте не несла встраиваемую ДНК, выделяли плазмиды в трансформанте и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называют pJET-IFNR, которая содержит SEQ ID NO:75. После проверки последовательности ген IFN-γ с оптимизацией кодона содержит SEQ ID NO:76.
Конструкция экспрессирующих векторов для интерферона α и γ свиньи
[0111] Ген IFN-α амплифицировали с использованием плазмиды pJET-IFNA-6 в качестве матрицы и PIFNANDEIF (5'-CAATATCATATGTGCGATCTGCCGCAAACC-3'; SEQ ID NO:77)/PIFNAHISSalIR (5'-GATATAGTCGACTTATTAGTGATGGTG ATGGTGATGTTCCTTTTTACGCAGGCGGTC-3'; SEQ ID NO:78) в качестве набора праймеров. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 100 нг pJET-IFNA-6 и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 2 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 30 секунд (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем восстанавливали продукт ПЦР с использованием набора PCR-M™ Clean Up system.
[0112] После того, как продукт ПЦР разрезали NdeI и SalI, фрагмент ДНК лигировали в pET29a, разрезанный теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения посредством электрофореза ДНК того, что рекомбинантная плазмида в трансформанте не несла встраиваемую ДНК, выделяли плазмиды в трансформанте и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называют pET-OPTPIFNAH, которая содержит SEQ ID NO:79.
(2) Конструкция экспрессирующего вектора pBA-OPTPIFNAH
[0113] После того, как амплифицированный ПЦР ген IFN-α разрезали NdeI и SalI, фрагменты ДНК лигировали соответственно в pBCM-araM11, разрезанный теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения посредством электрофореза ДНК того, что рекомбинантная плазмида в трансформанте не несла встраиваемую ДНК, выделяли плазмиды в трансформанте и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называют pBA-OPTPIFNAH, которая содержит SEQ ID NO:80.
(3) Конструкция экспрессирующего вектора pET-SUMO-OPTPIFNAH
[0114] The ген SUMO амплифицировали с использованием генома Saccharomyces cerevisiae в качестве матрицы и SUMOF (SEQ ID NO:25)/SUMOR2 (5'-ACCACCAATCTGTTCTCTGTGAGC-3'; SEQ ID NO:81) в качестве набора праймеров. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 200 нг геномной ДНК Saccharomyces cerevisiae и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 5 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 30 секунд (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем продукт ПЦР восстанавливали с использованием набора Gel-M™ gel extraction system.
[0115] Ген IFN-α амплифицировали с использованием плазмиды pJET-IFNA-6 в качестве матрицы и SUMOIFNAF (5'-GCTCACAGAGAACAGATTGGTGGTTGCGATCTGCCGCAAACC-3'; SEQ ID NO:82)/PIFNAHISSalIR (SEQ ID NO:78) в качестве набора праймеров. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 100 нг плазмиды pJET-IFNA-6 и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 5 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 30 секунд (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем продукт ПЦР восстанавливали с использованием набора Gel-M™ gel extraction system.
[0116] Слитый ген SUMO-IFN-αполучали полимеразной цепной реакцией с использованием двух продуктов ПЦР, получаемых выше, в качестве матрицы и SUMOF (SEQ ID NO:25)/PIFNAHISSalIR (SEQ ID NO:78) в качестве набора праймеров. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 100 нг продукта ПЦР SUMO, 100 нг продукта ПЦР IFN-α и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 2 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 1 минуты (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем восстанавливали продукт ПЦР с использованием набора PCR-M™ Clean Up system.
[0117] После того, как продукт ПЦР разрезали KpnI и SalI, фрагмент ДНК лигировали в pET29a, разрезанный теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения посредством электрофореза ДНК того, что рекомбинантная плазмида в трансформанте не несла встраиваемую ДНК, выделяли плазмиды в трансформанте и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называют pET-SUMO-OPTPIFNAH, которая содержит SEQ ID NO:83.
(4) Конструкция экспрессирующего вектора pET-OPTSUMO-OPTPIFNAH
[0118] Ген OPTSUMO амплифицировали с использованием pET-OPTSUMO-ORF2 (SEQ ID NO:43) в качестве матрицы и OPTSUMOF (SEQ ID NO:35)/OPTSUMOR2 (5'-GCCGCCGATTTGTTCACGG-3'; SEQ ID NO:84) в качестве набора праймеров.
[0119] 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 100 нг pET-OPTSUMO-ORF2 и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 2 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 30 секунд (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем продукт ПЦР восстанавливали с использованием набора Gel-M™ gel extraction system.
[0120] Ген IFN-α амплифицировали с использованием плазмиды pJET-IFNA-6 (SEQ ID NO:63) в качестве матрицы и OPTSUMOIFNAF (CCGTGAACAAATCGGCGGCTGCGATCTGCCGCAAACC; SEQ ID NO:85)/PIFNAHISSalIR (SEQ ID NO:78) в качестве набора праймеров. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 100 нг pJET-IFNA-6 и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 2 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 30 секунд (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем продукт ПЦР восстанавливали с использованием набора Gel-M™ gel extraction system.
[0121] Слитый ген OPTSUMO-IFN-α получали полимеразной цепной реакцией с использованием указанных выше двух продуктов ПЦР в качестве матрицы и OPTSUMOF (SEQ ID NO:35)/PIFNAHISSalIR (SEQ ID NO:78) в качестве набора праймеров. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 100 нг продукта ПЦР OPTSUMO, 100 нг продукта ПЦР IFN-α и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 2 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 1 минуты (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем восстанавливали продукт ПЦР с использованием набора PCR-M™ Clean Up system.
[0122] После того, как продукт ПЦР разрезали KpnI и SalI, фрагмент ДНК лигировали в pET29a, разрезанный теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения того, что рекомбинантные плазмиды в трансформанте не несли встраиваемую ДНК, посредством электрофореза ДНК, выделяли плазмиды в трансформанте и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называют pET-OPTSUMO-OPTPIFNAH, которая содержит SEQ ID NO:86.
(5) Конструкция экспрессирующего вектора pBA-OPTSUMO-OPTPIFNAH
[0123] После того, как pET-OPTSUMO-OPTPIFNAH разрезали NdeI и SalI, фрагмент ДНК, содержащий слитый ген OPTSUMO-IFN-α восстанавливали с использованием набора Gel-M™ gel extraction system. Фрагмент ДНК лигировали в pBCM-araM11, разрезанный теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения того, что рекомбинантные плазмиды в трансформанте не несли встраиваемую ДНК, посредством электрофореза ДНК, выделяли плазмиды в трансформанте и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называют pBA-OPTSUMO-OPTPIFNAH, которая содержит SEQ ID NO:87.
(6) Конструкция экспрессирующего вектора pET-OPTPIFNRH
[0124] Ген IFN-γ амплифицировали с использованием плазмиды pJET-IFNR в качестве матрицы, PIFNRNDEIF (5'- CAATATCATATGCAAGCCCCGTTTTTCAAAGAA-3'; SEQ ID NO:88)/PIFNRHISSalIR (5'- GATATAGTCGACTTATTAGTGATG GTGATGGTGATGTTTGCTGGCACGCTGACC-3'; SEQ ID NO:89) в качестве набора праймеров. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 100 нг плазмиды pJET-IFNR и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 2 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 30 секунд (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем восстанавливали продукт ПЦР с использованием набора PCR-M™ Clean Up system.
[0125] После того, как продукт ПЦР разрезали NdeI и SalI, фрагмент ДНК лигировали в pET29a, разрезанный теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения того, что рекомбинантные плазмиды в трансформанте не несли встраиваемую ДНК, посредством электрофореза ДНК, выделяли плазмиды в трансформанте и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называют pET-OPTPIFNRH, которая содержит SEQ ID NO:90.
(7) Конструкция экспрессирующего вектора pET-SUMO-OPTPIFNRH
[0126] Ген SUMO амплифицировали с использованием гена Saccharomyces cerevisiae в качестве матрицы и SUMOF (SEQ ID NO:25)/SUMOR2 (SEQ ID NO:81) в качестве набора праймеров. Условия амплификации и способы восстановления продукта ПЦР являются такими, как описано ранее.
[0127] Ген IFN-γ амплифицировали с использованием плазмиды pJET-IFNR (SEQ ID NO:75) в качестве матрицы и SUMOIFNRF (5'- GCTCACAGAGAACAGATTGGTGGTCAAGCCCCGTTTTTCAAAGAA-3'; SEQ ID NO:91)/PIFNRHISSalIR (SEQ ID NO:89) в качестве набора праймеров. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 100 нг плазмиды pJET-IFNR и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 2 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 30 секунд (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем продукт ПЦР восстанавливали с использованием набора Gel-M™ gel extraction system.
[0128] Слитый ген SUMO-IFN-γ получали полимеразной цепной реакцией с использованием двух продуктов ПЦР, описанных выше, в качестве матрицы и с использованием SUMOF (SEQ ID NO:25)/PIFNRHISSalIR (SEQ ID NO:89) в качестве набора праймеров. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 100 нг продукта ПЦР SUMO, 100 нг продукта ПЦР IFN-γ и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 2 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 1 минуты (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем восстанавливали продукт ПЦР с использованием набора PCR-M™ Clean Up system.
[0129] После того, как продукт ПЦР разрезали KpnI и SalI, фрагмент ДНК лигировали в pET29a, разрезанный таким же рестрикционным ферментом, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения того, что рекомбинантные плазмиды в трансформанте не несли встраиваемую ДНК, посредством электрофореза ДНК, выделяли плазмиды в трансформанте и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называют pET-SUMO-OPTPIFNRH, которая содержит SEQ ID NO:92.
(8) Конструкция экспрессирующего вектора pET-OPTSUMO-OPTPIFNRH
[0130] Ген OPTSUMO амплифицировали с использованием pET-OPTSUMO-ORF2 (SEQ ID NO:43) в качестве матрицы и OPTSUMOF (SEQ ID NO:35)/OPTSUMOR2 (SEQ ID NO:84) в качестве набора праймеров. Условия амплификации и способы восстановления продукта ПЦР являются такими, как описано ранее.
[0131] Ген интерферона γ свиньи амплифицировали с использованием плазмиды pJET-IFNR (SEQ ID NO:75) в качестве матрицы и OPTSUMOIFNRF (5'- CCGTGAACAAATCGGCGGCCAAGCCCCGTTTTTCAAAGAAATC-3'; SEQ ID NO:93)/PIFNRHISSalIR (SEQ ID NO:89) в качестве набора праймеров. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 100 нг плазмиды pJET-IFNR и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 2 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 30 секунд (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем продукт ПЦР восстанавливали с использованием набора Gel-M™ gel extraction system.
[0132] Слитый ген OPTSUMO-IFN-γ получали полимеразной цепной реакцией с использованием указанных выше двух продуктов ПЦР в качестве матриц и OPTSUMOF (SEQ ID NO:35)/PIFNRHISSalIR (SEQ ID NO:89) в качестве набора праймеров. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 100 нг продукта ПЦР OPTSUMO, 100 нг продукта ПЦР IFN-γ свиньи и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 2 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 1 минуты (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем восстанавливали продукт ПЦР с использованием набора PCR-M™ Clean Up system.
[0133] После того, как продукт ПЦР разрезали KpnI и SalI, фрагмент ДНК лигировали в pET29a, разрезанный таким же рестрикционным ферментом, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения того, что рекомбинантные плазмиды в трансформанте не несли встраиваемую ДНК, посредством электрофореза ДНК, выделяли плазмиды в трансформанте и секвенировали ДНК. Плазмида с правильной ДНК плазмиды называют pET-OPTSUMO-OPTPIFNRH, которая содержит SEQ ID NO:94.
(9) Конструкция экспрессирующего вектора pBA-OPTSUMO-OPTPIFNRH
[0134] После того, как pET-OPTSUMO-OPTPIFNRH разрезали NdeI и SalI, фрагмент ДНК, содержащий OPTSUMO-IFR-γ слитый ген, восстанавливали с использованием набора Gel-M™ gel extraction system. Фрагмент ДНК встраивали в pBCM-araM11, разрезанный теми же рестрикционными ферментами, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения того, что рекомбинантные плазмиды в трансформанте не несли встраиваемую ДНК, посредством электрофореза ДНК, выделяли плазмиды в трансформанте и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называют pBA-OPTSUMO-OPTPIFNRH, которая содержит SEQ ID NO:95.
Экспрессия и очистка рекомбинантного интерферона свиньи
(1) Экспрессия рекомбинантного интерферона свиньи
[0135] pET-OPTPIFNAH (SEQ ID NO:79), pBA-OPTPIFNAH (SEQ ID NO:80), pET-SUMO-OPTPIFNAH (SEQ ID NO:83), pET-OPTSUMO-OPTPIFNAH (SEQ ID NO:86) и pBA-OPTSUMO-OPTPIFNAH (SEQ ID NO:87) трансформировали в E. coli Shuffle (NEB, USA), соответственно. pET-OPTPIFNRH (SEQ ID NO:90), pET-SUMO-OPTPIFNHR (SEQ ID NO:92), pET-OPTSUMO-OPTPIFNRH (SEQ ID NO:94) и pBA-OPTSUMO-OPTPIFNRH (SEQ ID NO:95) трансформировали в E. coli BL21(DE3), соответственно. Трансформанты инокулировали в среду LB, содержащую канамицин (конечная концентрация: 30 мкг/мл), и культивирование при встряхивании проводили при 37°C и 180 об./мин. После инкубации в течение ночи бактериальный раствор инокулировали в отношении 1:100 к среде LB, содержащей конечную концентрацию 30 мкг/мл кономицина. Культивирование при встряхивании проводили при 37°C и 180 об./мин. Бактерии культивировали до концентрации приблизительно от 0,4 до 0,6 OD600, измеряемой спектрофотометром, и добавляли 0,1 мМ IPTG для индукции экспрессии белка при 25°C и 180 об./мин. Через 4 часа индукции собирали бактериальные фракции центрифугированием (8000×g, 30 минут, 4°C) и наблюдали экспрессию рекомбинантного интерферона свиньи посредством электрофореза белков. Кроме того, также различали растворимые и нерастворимые белки бактерий и наблюдали растворимость рекомбинантного интерферона свиньи посредством электрофореза белков.
[0136] См. результаты экспериментов на фигуре 9 (A-E). Результаты демонстрируют, что настоящее изобретение может эффективно продуцировать растворимый рекомбинантный IFN-α свиньи и слитый белок SUMO-IFN-α с использованием хозяина E. coli Shuffle, и можно исключать стадию рефолдинга, таким образом, чтобы избегать проблемы, заключающейся в том, что на биологическую активность может оказывать влияние слабая эффективность рефолдинга. Для экспрессии слитого белка SUMO-IFN-α продукцию слитого белка SUMO-IFN-α можно повышать оптимизацией кодона гена SUMO. Эффект различных экспрессирующих систем на экспрессию слитого белка SUMO-IFN-α указывало на то, что продукция SUMO-IFN-α являлась выше при использовании мутантной индуцируемый арабинозой экспрессирующей системы (155,07 мг/л; фигура 9(E)). См. результаты эксперимента на фигуре 9 (F to I). Результаты демонстрируют, что стратегия присутствия слитого белка SUMO может увеличивать растворимость рекомбинантного IFN-γ свиньи. После оптимизации кодонов гена SUMO может повышаться продукция слитых белков SUMO-IFN-γ. Эффект различных экспрессирующих систем на экспрессию слитого белка SUMO-IFN-γ свидетельствовал о том, что продукция SUMO-IFN-γ с использованием индуцируемой T7 экспрессионной системы и мутантной индуцируемой арабинозой экспрессионной системы являлась полностью удовлетворительной.
(2) Конструкция и экспрессия экспрессирующего вектора pET-D-SUMOP рекомбинантной протеазы SUMO
[0137] Для расщепления интерферона свиньи, экспрессируемого в экспрессирующей системе E. coli, описанного в предшествующих абзацах, для получения интерферона свиньи без белкового фрагмента SUMO, в этом эксперименте получали протеазу SUMO посредством экспрессирующей системы E. coli. Специалисты в данной области также могут проводить эту стадию с использованием протеазы SUMO, получаемой другими путями.
[0138] Ген протеазы SUMO амплифицировали с использованием генома Saccharomyces cerevisiae в качестве матрицы и SUMPOF (SEQ ID NO:47)/SUMOPENZR (5'-GATATACTCGAGTTATTTTAAAGCGTCGGT TAAAATCAAATG-3; SEQ ID NO:96) в качестве набора праймеров. 50 мкл реакционной смеси для ПЦР содержали 1× буфер B GDP-HiFi PCR, 200 мкМ dATP, dTTP, dGTP и dCTP, 1 мкМ праймеров для амплификации, 200 нг генома Saccharomyces cerevisiae и 1 ед. ДНК-полимеразы GDP-HiFi. Условие реакции ПЦР являлось следующим: 96°C в течение 5 минут (1 цикл); 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 30 секунд (35 циклов); 68°C в течение 5 минут (1 цикл). После реакции для подтверждения, содержал ли продукт ПЦР фрагменты ДНК с теоретически рассчитанным размером, использовали электрофорез в агарозном геле. Затем восстанавливали продукт ПЦР с использованием набора PCR-M™ Clean Up system.
[0139] После того, как амплифицированный из генома дрожжей ген протеазы SUMO разрезали BamHI и XhoI, фрагмент ДНК встраивали в pET-D, разрезанный BamHI и SalI, с использованием ДНК-лигазы T4. Лигированный продукт трансформировали в E. coli ECOS 9-5. Трансформанты отбирали посредством полимеразной цепной реакции для отбора колоний. После подтверждения того, что рекомбинантные плазмиды в трансформанте не несли встраиваемую ДНК, посредством электрофореза ДНК, выделяли плазмиды в трансформанте и секвенировали ДНК. Плазмиду с правильной последовательностью ДНК называют pET-D-SUMOP, которая содержит SEQ ID NO:97.
[0140] pET-D-SUMOP (SEQ ID NO:97) трансформировали в E. coli BL21 (DE3). E. coli BL21(DE3) инокулировали в среду LB, содержащую канамицин (конечная концентрация: 30 мкг/мл) и культивировали при встряхивании при 37°C и 180 об./мин. После инкубации в течение ночи бактериальный раствор инокулировали в отношении 1:100 в среде LB, содержащей канамицин (конечная концентрация 30 мкг/мл). Культивирование при встряхивании проводили при 37°C и 180 об./мин. Бактерии культивировали до концентрации приблизительно от 0,4 до 0,6 OD600, измеряемой спектрофотометром, и добавляли 0,1 мМ IPTG для индукции экспрессии белка при 28°C и 180 об./мин. Через 4 часа после индукции собирали бактериальные фракция центрифугированием (8000×g, 30 минут, 4°C).
(3) Расщепление и очистка рекомбинантного интерферона свиньи
[0141] После индуцированной экспрессии трансформантов, несущих экспрессирующий вектор слитого белка SUMO-интерферон свиньи и экспрессирующий вектор протеазы SUMO, собирали бактериальные фракции центрифугированием (8000×g, 30 минут, 4°C). Собранные бактерии суспендировали в соответствующем количестве лизирующего буфера (20 мМ фосфата натрия, 500 мМ NaCl, pH 7,4) с получением оптической плотности 50 при 600 нм. После разрушения бактерий с использованием ультразвукового гомогенизатора супернатант собирали центрифугированием (8000×g, 15 минут, 4°C). Очищенный слитый белок рекомбинантной SUMO-интерферона свиньи и рекомбинантную протеазу (протеазу SUMO) смешивали в массовом отношении 4 и инкубировали при 4°C в течение 16 часов. Во время этого периода слитый белок SUMO-интерферон свиньи расщеплялся протеазой SUMO до белка SUMO и интерферона свиньи с His-меткой на C-конце.
[0142] Затем очищали белок с использованием аффинной хроматография на иммобилизованных ионах металл. Очистку проводили с использованием системы жидкостной хроматографии для очистки белка ÄKTA prime plus с 5 мл колонкой HiTrap™ Ni excel. После уравновешивания колонки 25 мл лизирующего буфера инъецировали раствор для расщепления слитого белка в колонку HiTrap™ Ni excel. После завершения инъекции образца отмывали неспецифически связанные белки 100 мл промывающего буфера (20 мМ фосфата натрия, 500 мМ NaCl, 30 мМ имидазола, pH 7,4). В заключении элюировали рекомбинантный интерферон свиньи на смоле 150 мл элюирующего буфера (20 мМ фосфата натрия, 500 мМ NaCl, 250 мМ имидазола, pH 7,4) и наблюдали очистку посредством электрофореза белков (как показано на фигуре 10).
Пример 4: Получение и применение композиции по настоящему изобретению для профилактики инфекции PCV2
[0143] В этом примере кодирующий белок ORF2, слитый белок SUMO-ORF2 и интерферон свиньи, получаемые в указанном выше примере 2 и примере 3, использовали для получения композиции для профилактики и лечения инфекции PCV2. Во многих образцах, используемых в настоящем описании, композиция дополнительно содержит адъювант MONTANIDE™ ISA 563 VG (SEPPIC, France) и/или адъювант MONTANIDE™ GEL 01 (SEPPIC, France). Компоненты смешивают на основании следующей ниже схемы эксперимента, а затем инокулируют поросятам для наблюдения индуцируемого иммунного ответа, или присутствуют ли неблагоприятные воздействия (такие как рвота, тремор, депрессия, затрудненное дыхание и отек в месте нанесения; если частота встречаемости по меньшей мере трех неблагоприятных воздействий является выше 50%, то безопасность композиции является низкой.)
(1) Эксперимент 1: Влияние содержания интерферона свиньи на безопасность настоящей композиции
[0144] Четырнадцать полевых поросят в возрасте трех недель отбирали и распределяли по группам случайным образом. Их разделяли на 7 групп A-G, и число поросят в каждой группе составляло два. В каждой группе вводили одну внутримышечную инъекцию, и доза иммунизации составляла 2 мл. Компоненты каждой вакцины приведены в таблице 6 ниже. Наблюдения проводили на сутки вакцинации и на следующие сутки после нее, и регистрировали относительное число клинических неблагоприятных воздействий.
Таблица 6: Схема эксперимента 1
[0145] Результаты эксперимента (таблица 7) демонстрировали, что у свиней, вакцинированных V-001, демонстрировали клинические симптомы депрессии, но не других каких-либо неблагоприятных воздействий. У свиньей, вакцинированных V-002, демонстрировали симптомы рвоты и тремора. Кроме того, показатели безопасности образца V-003 являлись в большей степени скептическими, и неблагоприятные воздействия V-004, V-005 или V-006 на свиней, которым проводили инокуляцию, являлись более умеренными. На основании этих результатов в последующих экспериментах содержание интерферона свиньи можно сохранять на уровне 25 мкг на дозу (2 мл).
Таблица 7: Результаты эксперимента 1
(2) Эксперимент 2: Влияние содержания адъюванта на показатели безопасности настоящей композиции
[0146] Семьдесят три полевых поросенка в возрасте трех недель отбирали и распределяли по группам A и B случайным образом. Количество поросят в группе A составляло 38, и число поросят в группе B составляло 35. В каждой группе проводили одну внутримышечную инъекцию, и доза иммунизации составляла 2 мл. Композиция каждой вакцины приведена в таблице 8 ниже. Наблюдения проводили на сутки вакцинации и на следующие сутки после вакцинации, и регистрировали относительное число клинических неблагоприятных воздействий. Результаты эксперимента демонстрируют (таблица 9), что параметры безопасности образца V-009 являются высокими, но параметры безопасности образца V-008 также являются приемлемыми.
Таблица 8: Схема эксперимента 2
Таблица 9: Результаты эксперимента 2
(3) Эксперимент 3: Влияние различных адъювантов на индукцию иммунного ответа настоящей композиции
[0147] Этот эксперимент проводили на животное в животноводстве генетически модифицированных организмов (GMO) в отделе инспекции лекарственных средств для животных Научно-исследовательского института здоровья животных (AHRI). Одиннадцать поросят в возрасте 4 недель без инфекции какими-либо патогенами, случайным образом группировали и распределяли по 5 группам A-E. Группы A-D являлись экспериментальными группами, где число поросят в каждой группе составляет 2, и группа E являлась контрольной группой, в который было 3 поросенка. Поросят в группах A-D иммунизировали внутримышечно в возрасте 4 и 6 недель соответственно, и доза иммунизации составляла 2 мл. В группе E иммунизацию не проводили. Компоненты каждой вакцины приведены в таблице 10 ниже.
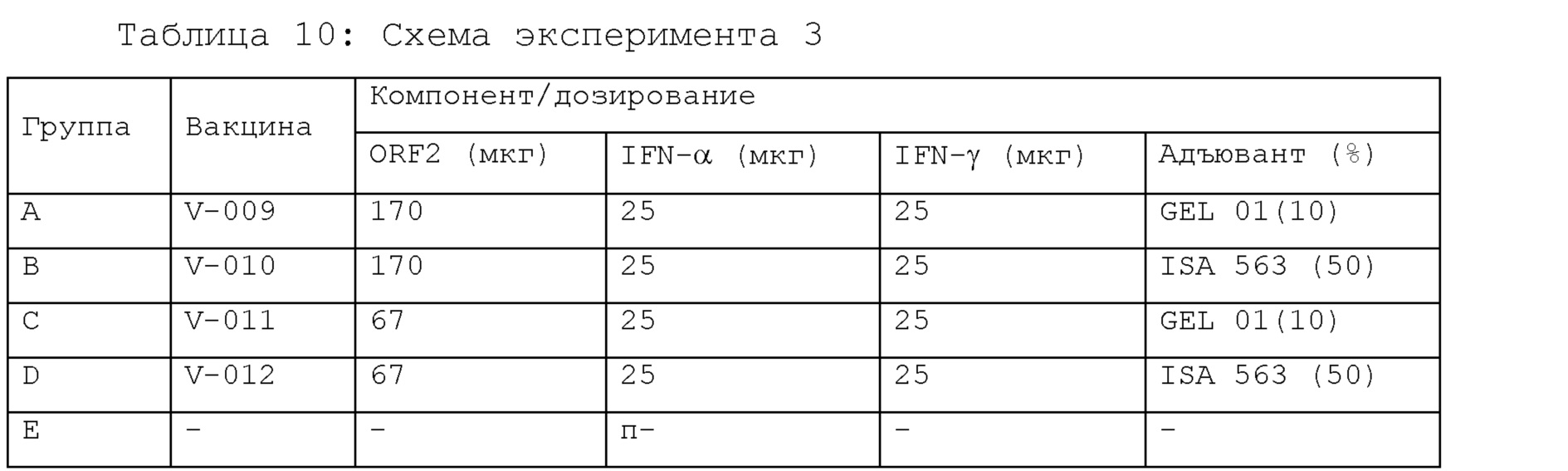
[0148] Поросят в каждой группе экспериментально заражали PCV2 в возрасте 8 недель и подвергали всех животных аутопсии через четыре недели после экспериментального заражения. Образцы сыворотки и плазмы у свиней собирали перед иммунизацией (в возрасте 4 недель), после иммунизации (в возрасте 6 и 8 недель) и после экспериментального заражения (в возрасте 9, 10, 11 и 12 недель). Титр антитела к PCV2 в сыворотке определяли с использованием коммерчески доступного набора ELISA (BioCheck, Netherlands). Количество вируса в плазме определяли с использованием количественной полимеразной цепной реакции в режиме реального времени.
[0149] Результаты эксперимента демонстрировали, что все V-009, V-010, V-011 и V-012 индуцировали антитело к ORF2 (фигура 11) и снижали вирусемию у свиней в эксперименте (фигура 12). На основании результатов эксперимента также продемонстрировано, что каждая доза (2 мл), которая содержит 67 мкг ORF2, может вызывать достаточный уровень иммунных ответов (V-011 и V-012).
(4) Эксперимент 4: Влияние слитого белка SUMO-ORF2 и OFR2 на индукцию иммунного ответа настоящей композиции
[0150] Этот эксперимент проводили в животноводстве генетически модифицированных организмов (GMO) в отделе инспекции лекарственных средств для животных Научно-исследовательского института здоровья животных (AHRI). Шестнадцать поросят в возрасте 4 недель без инфекции какими-либо конкретными патогенами, случайным образом группировали и распределяли по 5 группам A-E. Группы A-D являлись экспериментальными группами, в которых число поросят каждой группы составляло 3, и группа E являлась контрольной группой, в которой было 4 поросенка. Свиней в группах A-D иммунизировали внутримышечно в возрасте 4 и 6 недель соответственно, и доза иммунизации составляла 2 мл. В группе E иммунизацию е проводили. Компоненты каждой вакцины приведены в таблице 11 ниже.
Таблица 11: Схема эксперимента 4
[0151] Свиней в каждой группе экспериментально заражали PCV2 в возрасте 8 недель и подвергали всех животных аутопсии через 5 недель после экспериментального заражения. Образцы сыворотка и плазма собирали в конкретные моменты времени. Титр антитела к PCV2 в сыворотке определяли с использованием коммерчески доступного набора ELISA. Количество вируса в плазме определяли с использованием количественной полимеразной цепной реакции в режиме реального времени.
[0152] Результаты демонстрировали, что каждый образец мог индуцировать у свиней образование антител к PCV2, и лучшие результаты получали с образцами V-013 (содержащими 27 мкг ORF2) (фигура 13). Кроме того, все образцы снижали вирусемию у свиней (фигура 14).
(5) Эксперимент 5: Влияние интерферона α свиней и интерферона γ свиней на индукцию иммунного ответа настоящей композиции
[0153] Этот эксперимент проводили на свиноводческой форме с низкими уровнями загрязнения патогенами и без инфекции PCV2. Двадцать поросят SPF в возрасте 4 недель без инфекции PCV2 выбирали и распределяли случайным образом по 5 группам A-E, в каждой группе было 4 поросенка. Группы A-D являлись экспериментальными группами, и группа E являлась контрольной группой. Свиней в группах A-D иммунизировали внутримышечно в возрасте 4 и 7 недель соответственно, и доза иммунизации составляла 2 мл. В группа E иммунизацию не проводили. Компоненты каждой вакцины приведены в таблице 12 ниже. Образцы сыворотка собирали в конкретные моменты времени. Титр антитела к PCV2 в сыворотке определяли с использованием коммерчески доступного набора ELISA.
Таблица 12: Схема эксперимента 5
[0154] Результаты эксперимента демонстрируют, что добавление IFN-α (V-018) или IFN-γ (V-019) по отдельности в композиции по настоящему изобретению характеризовалось повышенным влиянием на индукцию иммунных ответов. Эффект от добавления IFN-α является более выраженным, чем эффект от добавления IFN-γ. С другой стороны, добавление IFN-α и IFN-γ (V-020) в композиции по настоящему изобретению индуцировало лучший иммунный ответ. Результаты указывают на то, что в композициях по настоящему изобретению IFN-α и IFN-γ обладают синергическим действием на индукцию иммунного ответа (фигура 15).
Реферат
Изобретение относится к области биотехнологии. Изобретение представляет собой способ получения капсидного белка PCV2 и фармацевтической композиции, содержащей указанный капсидный белок. В способе по настоящему изобретению используют новый индуцируемый арабинозой экспрессирующий вектор и таким образом повышают эффективность синтеза указанного капсидного белка PCV2. С другой стороны, настоящая фармацевтическая композиция сочетает в себе указанный капсидный белок и другие благоприятные компоненты в подходящем отношении, таким образом, чтобы достигать превосходного индуцирующего иммунный ответ действия. 15 з.п. ф-лы, 15 ил., 11 табл., 4 пр.
Комментарии