Онколитический hsv-вектор - RU2719190C2
Код документа: RU2719190C2
Чертежи









Описание
Перекрестная ссылка на родственные заявки
В настоящей патентной заявке испрашивается приоритет предварительной заявки на патент США 61/896497, поданной 28 октября, 2013, и содержание этой заявки во всей своей полноте вводится в настоящее описание посредством ссылки.
Указание на спонсирование научных исследований и разработок Федеральными органами
Настоящее изобретение было разработано при поддержке Правительства в соответствии с грантами NN CA119298, CA163205, CA175052, NS040923 и DK044935, выданными Национальным Институтом Здравоохранения. Правительство имеет определенные права на данное изобретение.
Предшествующий уровень техники
Мультиформная глиобластома (МГБ), несмотря на существующие в настоящее время методы комбинированной терапии, все еще остается смертельным заболеванием. Преклинические исследования дают основание предположить, что компетентные по репликации вирусы, включая онколитические HSV-векторы («oHSV»), представляют собой перспективную альтернативу существующим методам лечения, но как показали испытания с участием пациентов, терапевтическая эффективность этих вирусов ограничена. Безопасность вектора может быть достигнута путем снижения числа мутаций вектора, которые могут также оказывать негативное воздействие на литическую репликацию в опухолевых клетках.
Краткое описание сущности изобретения
Настоящее изобретение относится к oHSV, способному к опухолеспецифической репликации вектора без его аттенюирования, где указанный oHSV получают путем перенацеливания вектора на рецепторы опухолеассоциированной клеточной поверхности в комбинации с ингибированием репликации вектора под действием клеточной микроРНК («miR»), которая в высокой степени экспрессируется в здоровом головном мозге, но фактически отсутствует в опухолевых клетках. miR-реактиные элементы предотвращают патогенез вектора в головном мозге «голых» мышей, и, при этом, не оказывают негативного влияния на репликацию литического вектора в первичных опухолевых клетках in vitro или у модели с ксеногенной опухолью головного мозга. Такая конструкция нового вектора может быть положена в основу обеспечения безопасности и более высокой эффективности вектора и может быть адаптирована в целях его применения для лечения опухолей у пациентов.
Краткое описание иллюстративного материала нескольких видов
На фигуре 1 представлены данные результатов экспериментов, проводимых для оценки эффективности и специфичности элемента T124. Плазмиды, экспрессирующие люциферазу светляка (fLuc) и содержащие T124 (pfLuc-T124) или регуляторную последовательность (pfLuc-Ctrl) в 3'UTR, были котрансфецированы с внутренней регуляторной плазмидой, экспрессирующей люциферазу морской маргаритки (renilla) (prLuc), в клетки HEK293AD, которые за 24 часа до этого были трансфецированы синтетической пре-miR-124 или пре-miR-21. Люциферазную активность определяли через 48 часов. Результаты представлены как среднее ± стнадратное отклонение по трем измерениям активности fLuc, нормализованной к активности rLuc. Статистически значимые различия между парами указаны в скобках ниже соответствующих величин Р (непарный t-критерий).
На фигуре 2 представлены данные результатов экспериментов по репликации вирусов в клетках глиомы. (A) Векторные диаграммы. Родительская BAC KOS-37 содержит loxP-фланкированную BAC, последовательность резистентности к хлорамфениколу и последовательность lacZ («BAC»), расположенные между вирусными генами UL37 и UL38 (Gierasch et al., 2006). Модификации, вводимые для получения KGBAC и KG4:T124BAC, представлены ниже: gB:NT, двойная мутация в гене gB, повышающая способность проникновения вируса; gC-eGFP, гибрид полной ОРС gC и GFP, связанные пептидной последовательностью 2A; ΔJoint, делеция полноразмерной области внутренних повторов, включающей одну копию гена ICP4; ICP4:T124, инсерция T124 в 3'UTR остальной части гена ICP4. UL, уникальный длинный сегмент вирусного генома; US, уникальный короткий сегмент. (B) Влияние T124 на репликацию вируса в клеточной культуре, взятой у пациента с глиомой. Клетки Gli68 и GBM30 инфицировали вирусами KG или KG4:T124 с тремя повторностями при множественности заражения (MOI) 0,01. Через определенные периоды времени после инфицирования, клеточные лизаты и супернатанты собирали и титровали на клетках U2OS. Величины представлены как среднее ± стандартное отклонение. (C) Экспрессия MiR-124 в LV124-инфицированных клетках Gli68. Клетки инфицировали до достижения плотности 5 к.о.е./клетки, отбирали на следующий день и в течение последующих 3 дней в пуромицин-содержащей среде, а затем собирали для экстракции всех РНК. Контрольные РНК были получены из неинфицированных клеток Gli68 и U2OS. Уровни miR-124 определяли с тремя повторностями с помощью кол.ОТ-ПЦР и нормализовали к уровням RNU43. На этой фигуре показано кратное увеличение ± стандартное отклонение по отношению к клеткам U2OS. P<0,05 для всех пар (непарный t-критерий). (D) Репликация вируса с модификациями KG и KG4:T124 в miR-124-трансдуцированных и контрольных клетках GBM30 и Gli68. Клетки инфицировали LV124 или LV137R до достижения плотности 5 к.о.е./клетку, отбирали с использованием пуромицина в течение 3 дней и подвергали сверхинфицированию вирусом KG или KG4:T124 при MOI=0,01. Инфекционный HSV в объединенных клеточных лизатах и в супернатантах собирали через 72 и 96 часов, а затем тировали на клетках U2OS. Результаты представлены как средние величины ± стандартное отклонение по результатам HSV-инфицирования с тремя повторностями. В скобках указаны значимые различия пар с указанием соответствующих величин P (непарный t-критерий).
На фигуре 3 представлены данные результатов экспериментов по репликации вируса KG4:T124 и оценки токсичности в головном мозге «голых» мышей. 4,8×109 геномных копий KG или KG4:T124 интракраниально инъецировали каждой из 4 «голых» мышей BALB/c (n=4/группу). (A) Масса животных в течение определенного периода времени после инъекции вектора. Слева: KG-инъецированные животные; X, погибшие животные. Справа: KG4:T124-инъецированные мыши; заштрихованные кружки - умерщвленные животные. (B) Все копии вирусного генома, взятые в течение определенного периода времени в головном мозге мышей после инъекции вектора. Головной мозг, взятый от каждой из KG4:T124-инъецированной мышей, умерщвленных через 5, 14, 21 и 33 дня после инъекции вектора, и последнего выжившего животного из KG-инъецированной группы, умерщвленного путем эвтаназии на день 5 и имеющего тяжелые симптомы заболевания, собирали, а затем выделяли ДНК и определяли общее число геномов вирусного вектора на головной мозг определяли с помощью кол.ПЦР. (C) Кривая выживаемости Каплана-Мейера для животных, используемых в этом эксперименте. Стрелками показаны дни умерщвления отдельных животных из KG4:T124-инъецированной группы. P=0,0058, log-ранговый критерий.
На фигуре 4 представлены данные результатов экспериментов по обработке «голых» мышей с моделью человеческой глиобластомы miR-124-чувствительным HSV-вектором, перенацеленным на EGFR. Измельченные клетки GBM30 интракраниально имплантировали мышам, и через 5 дней этим мышам инъецировали PBS или 1,8×108 геномных копий (г.к.) вируса KGE или KGE-4:T124 в тех же самых координатах. (A) Кривая выживаемости Каплана-Мейера. Log-ранговый статистический критерий: KGE по сравнению с PBS, P=0,0188; KGE-4:T124 по сравнению с PBS, P=0,0009; KGE по сравнению с KGE-4:T124, P=0,8327. (B) Массы тела животных после имплантации опухолевых клеток. X - погибшее животное или животное, умерщвленное путем эвтаназии.
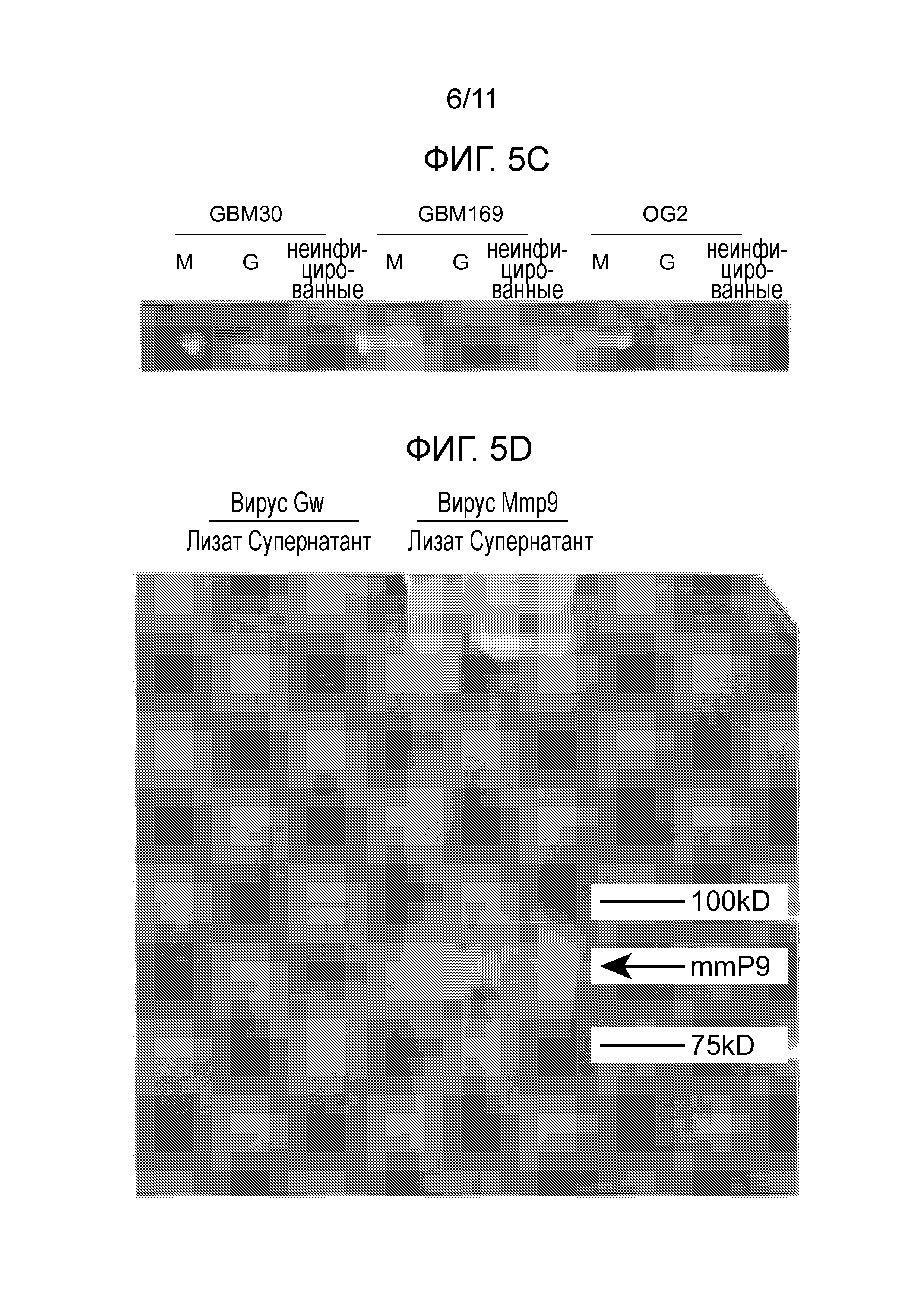
На фигуре 5 представлены данные, которые показали, что KMMP9 опосредует сверхэкспрессию ферментативно активного MMP9. (A) Структура KMMP9 и KGw. (B) Вестерн-блот-анализ лизатов клеток Vero, инфицированных KMMP9, KGw или KG (MOI=0,1). β-тубулин и гликопротеин В HSV визуализировали в виде клеточного и вирусного загрузочного контроля, соответственно. Первичные клеточные линии GBM (C) или клетки Vero (D) инфицировали KGw или KMMP9 при MOI=1. Через 24 часа после инфицирования, клеточные лизаты и супернатант собирали и объединяли (C), либо загружали по отдельности (D) на 10% полиакриламидный гель/0,1% желатиновый гель. После электрофореза, гель инкубировали в течение ночи при 37°C, окрашивали 0,05% кумасси синим и обесцвечивали, а затем получали изобрежение. Сокращения: M, KMMP9; G, KGw; KG, контрольный вирус; un., неинфицированные; gB, гликопротеин B; Sup., супернатант.
На фигуре 6 представлены данные, которые показали, что KMMP9 и KGw имеют сравнимые паттерны связывания с клеткой и размножения в клетке. (A) Клетки, перечисленные на панели слева, инфицировали вирусами, указанными сверху, при множественности заражения, выраженной в гономных копиях на клетку (г.к./клетку). Через 6 часов, клетки фиксировали и подвергали иммунологическому окрашиванию на ICP4. Клетки (B) GBM30 и (C) GBM169 разрушали и инфицировали KMMP9 или KGw при 200 г.к./клетку. Клеточные лизаты собирали через 1, 2, 4 и 6 дней после инфицирования, и титры вирусных геномных копий определяли с помощью кол.ПЦР. Какого-либо значимого различия между двумя вирусами во всех клеточных линиях хозяевах не наблюдалось (GBM30: P=0,20; GBM169: P=0,11).
На фигуре 7 представлены данные, которые показали, что KMMP9уничтожает опухолевые клетки на таком же уровне, как и KGw in vitro, или на более высоком уровне. Клетки U87, SNB19 или GBM30 инфицировали при множественности заражения 10 или 100 г.к./клетку в течение 3 или 7 дней. Процент выживаемости инфицированных клеток по отношению к выживаемости неинфицированных клеток определяли с помощью анализа, проводимого с использованием MTT (n=3; звездочка: P<0,05, непарный t-критерий Стьюдента).
На фигуре 8 представлены данные, которые показали, что KMMP9повышает инфекционность oHSV в сфероидах. Клетки GBM30 культивировали в суспензии и инфицировали 1×103 б.о.е. KMMP9 или KGw. Флуоресценцию в зеленом диапазоне спектра, излучаемую gC-T2a-eGFP в обоих векторах, визуализировали ежедневно через 2-6 дней после инфицирования в сфероидах с четким контуром. (A) Репрезентативные изображения, полученные через 3 и 5 дней после инфицирования. (B) Усредненная количественная оценка eGFP-сигнала в 6 сфероидах на вектор указывала приблизительно на 2-кратное превышение инфекционности KMMP9 по сравнению с KGw (P=0,006). (C-E) Две группы сфероидов GBM30 инфицировали геномными копиями 4×107KMMP9 и KGw на сфероид. Сфероиды фиксировали, окрашивали DAPI, и конфокальные изображения Z- секций регистрировали с интервалами в 5 мкм. На панели (C) показаны 2 репрезентативных сфероида от каждой из групп KMMP9 и KGw после 3D-реконструирования от 0 мкм до 150 мкм. Синий, DAPI; зеленый, eGFP. На панели (D) показаны Z-секции двух сфероидов от каждой группы при Z=100 мкм. (E) Каждый сфероид разделяли на 5 сегментов по глубине оси Z (снизу вверх: 0-20 мкм, 25-50 мкм, 55-80 мкм, 85-100 мкм, 105-120 мкм и 125-140 мкм). Относительную интенсивность сигнала в каждом сегменте сфероида вычисляли путем усреднения eGFP-сигнала, деленного на DAPI-сигнал. n=7; звездочка: P<0,05.
На фигуре 9 представлены данныеKMMP9-обработки «голых» мышей с моделью МГБ. Клетки GBM30 интракраниально имплантировали, и через 5 дней инъекцировали KMMP9, KGw или PBS в тех же самых пространственных координатах (на расстоянии 0,5 мм до брегмы, на 2 мм сбоку от брегмы (справа) и на 3 мм вглубь брегмы). Животных ежедневно осматривали, и при появлении признаков болезненного состояния умерщвляли. Данные представлены в виде кривой выживаемости Каплана-Мейера. Животные, обработанные KMMP9 или KGw, имели большую продолжительность жизни, чем животные, обработанные PBS (P<0,01). Какого-либо значимого различия между KMMP9 и KGw не обнаруживалось (n=4; P=0,61, log-ранговый критерий).
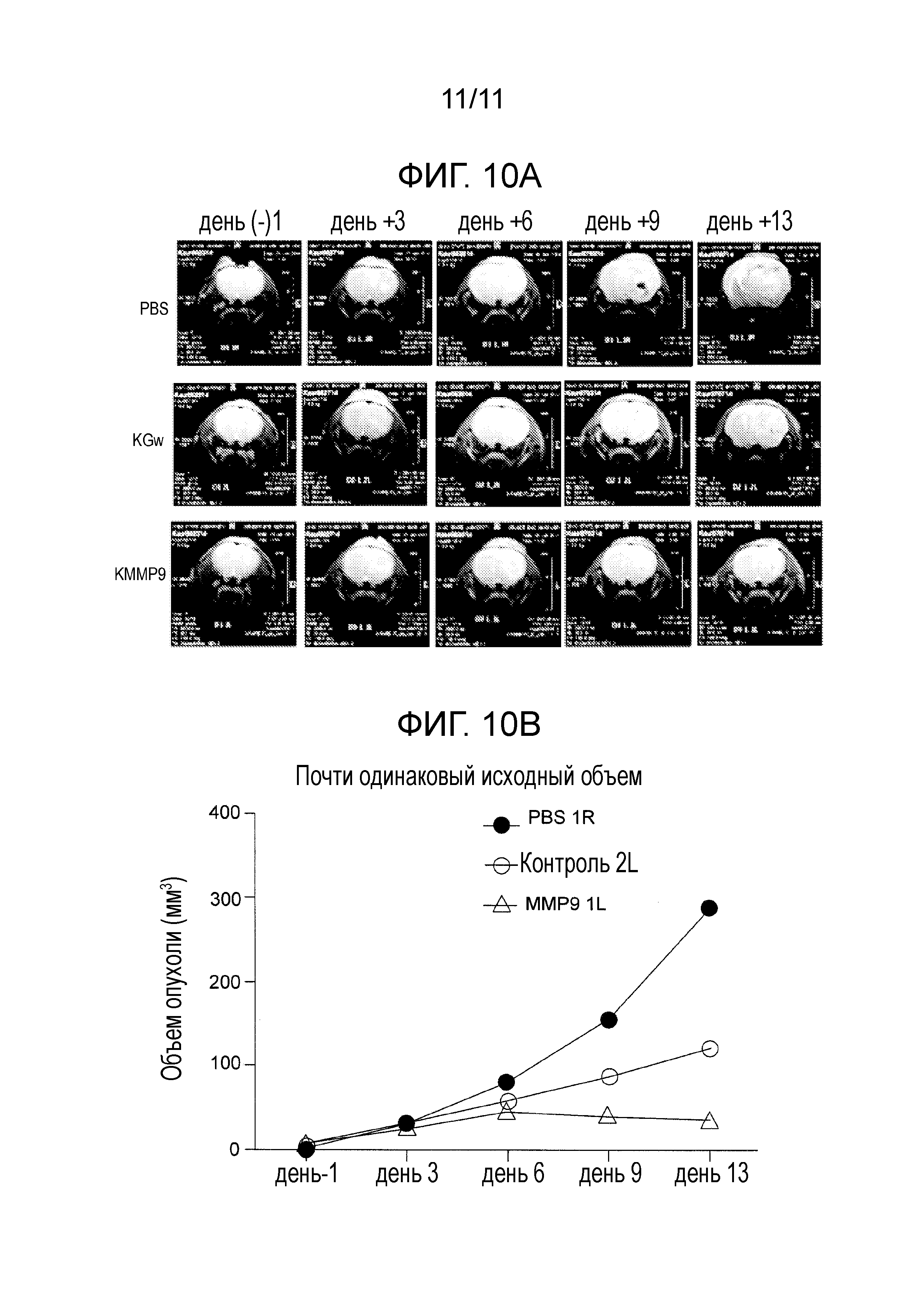
На фигуре 10 представлены T2-взвешенные МРТ-изображения головного мозга одного животного с GBM30 в группе обработки вирусом или животного с GBM30 контрольной группы, обработанной PBS. (A) Обработку проводили через 10 дней после имплантации GBM30, и изображение получали за 1 день до обработки (на день: -1) и через 3, 6, 9 и 13 дней после обработки. (B) Объемы опухолей вычисляли в те же самые дни.
Подробное описание изобретения
Настоящее изобретение относится к рекомбинантному oHSV, содержащему не-HSV-лиганд, специфичный к молекуле (белковой, липидной или углеводной детерминанте), присутствующей на поверхности клетки (такой как раковая клетка), и одну или более копий одной или более последовательностей-мишеней микроРНК, встроенных в один или более локусов гена HSV, а предпочтительно, в один или более генов HSV, необходимых для репликации HSV в нормальных (то есть, не-раковых) клетках. Настоящее изобретение также относится к штаммам oHSV и фармацевтическим композициям, содержащим oHSV согласно изобретению, и к способам уничтожения опухолевых клеток с использованием oHSV согласно изобретению.
Не-HSV-лиганд oHSV согласно изобретению вводят в гликопротеин, находящийся на поверхности oHSV, такой как gD или gC, для облегчения нацеливания данного лиганда на нужную клетку. Так, например, лиганд может быть введен в положения между остатками 1 и 25 gD. Предпочтительными лигандами для нацеливания на клетки МГБ и другие раковые клетки являются лиганды, связывающиеся с EGFR и EGFRvIII, CD133, CXCR4, карциноэмбриональным антигеном (CEA), ClC-3/аннексином-2/MMP-2, человеческим рецептором трансферрина и EpCAM, и этот лиганд может быть нацелен на такой рецептор или на молекулу клеточной поверхности, то есть, этот лиганд может специфически связываться с таким рецептором или с такой молекулой клеточной поверхности. EGFR- и EGFRvIII-специфические лиганды, такие как антитела, scFv (одноцепочечные антитела) и VHH (однодоменные антитела) описаны в литературе (Kuan et al., Int. J. Cancer, 88, 962-69 (2000); Wickstrand et al., Cancer Res., 55(14):3140-8 (1995); Omidfar et al., Tumor Biology, 25:296-305 (2004); см. также Uchida et al., Molecular Therapy, 21:561-9 (2013); см. также Braidwood et al., Gene Ther., 15, 1579-92 (2008)).
Кроме того, или альтернативно, oHSV может быть доставлен путем введения лигандов в другие молекулы клеточной поверхности или в рецепторы, которые необязательно ассоциируются с раковой опухолью. Так, например, лиганды могут включать связывающие домены природных лигандов (например, факторов роста (таких как EGF, который может связываться с EGFR; NGF, который может связываться с trkA, и т.п.)), пептидные или непептидные гормоны, пептиды, специфически связывающеся с молекулой-мишенью (например, сконструированые белки анкириновых повторов (DARPins)) и т.п. oHSV согласно изобретению может также включать мутантную форму gB и/или gH, которая способствует проникновению вектора посредством неканонических рецепторов (а также, предпочтительно, имеет такие мутации в одном или в обоих этих генах в геноме oHSV).
Предпочтительной последовательностью-мишенью микроРНК для ее включения в вектор согласно изобретению (предпочтительно, в виде множества ее копий в тандеме друг с другом) является miR-124, которая, в частности, может быть использована для лечения нервных расстройств (например, для защиты не-раковых нейронов при использовании oHSV согласно изобретению для лечения опухолей нервной системы, таких как МГБ). Альтернативно, для защиты других типов ткани могут быть использованы и другие последовательности-мишени микроРНК, и специалист в данной области может выбрать последовательность-мишень микроРНК, подходящую для защиты ткани или клетки нужного типа. Так, например, miR-122 и miR-199 экспрессируются в нормальных клетках печени, но не в первичных раковых клетках печени, и таким образом, одна из последовательностей-мишеней микроРНК miR-122 и/или miR-199 или их комбинации могут быть применены в одном из вариантов осуществления изобретения, относящихся к лечению рака печени с использованием oHSV согласно изобретению. Аналогичнным образом, последовательности-мишени микроРНК miR-128 и/или miR-137 могут быть введены в oHSV для защиты здорового головного мозга. Репрезентативной последовательностью-мишенью микроРНК может быть обратный комплемент микроРНК.
Последовательность(и)-мишень(и) микроРНК предпочтительно вводят в 3'-нетранслируемую область («UTR») гена HSV для сайленсинга этого гена в присутствии микроРНК. При этом предпочтительно, чтобы множество копий (например, две копии, три копии, четыре копии, пять копий, шесть копий или более) последовательности-мишени микроРНК были встроены в тандеме. Также предпочтительно, чтобы множество копий последовательности-мишени микроРНК были отделены друг от друга спейсерами длиной в четыре или более нуклеотидов (а более предпочтительно, длиной в восемь или более нуклеотидов). Не ограничиваясь какой-либо конкретной теорией, авторы лишь отмечают, что чем больше расстояние (например, приблизительно более, чем 8 нуклеотидов), тем выше стабильность.
Более предпочтительно, для защиты не-раковых клеток от литического действия HSV-инфекции, множество копий последовательности-мишени микроРНК встраивают в 3'UTR-область гена HSV, которая, как известно специалистам в данной области, играет важную роль в репликации не-раковых клеток. Предпочтительно, указанным сайтом является 3'UTR гена со встроенной микроРНК в его нормальном (или природном) локусе в геноме HSV. Предпочтительный oHSV согласно изобретению включает множество копий последовательности-мишени микроРНК, встроенных в 3'UTR гена ICP4, таких как одна или две копии гена ICP4 в векторах, которые имеют обе нативные копии гена ICP4.
Геном HSV-вектора согласно изобретению может дополнительно содержать один или более экзогенных экспрессионных кластеров (то есть, кластеров, содержащих кодирующие последовательности, функционально присоединенные к промоторам, энхансерам и другим подходящим регуляторным элементам), таких как кластеры, кодирующие репортерный белок (такой как белок, флуоресцирующий в зеленом диапазоне спектра), онколитический фактор или агент, усиливающий активность, направленную на уничтожение опухолей (например, фактор некроза опухоли («TNF») или TNF-родственный апоптоз-индуцирующий лиганд («TRAIL»), или другой терапевтически ценный генный продукт (например, пептиды, ферменты, активирующие лекарственные средства, антитела, терапевтические РНК и т.п.). Предпочтительный экзогенный экспрессионный кластер кодирует металлопротеиназу матрикса, такую как металлопротеиназа матрикса 9 («MMP9»), которая разлагает коллаген типа IV, то есть, главный компонент внеклеточного матрикса (ECM) и базальные мембраны глиобластом (Mammato et al., Am. J. Pathol., 183(4): 1293-1305 (2013), doi: 10.1016/j.ajpath.2013.06.026. Epub 2013 Aug 5), что приводит к повышению уровня инфицирования опухолевых клеток вектором согласно изобретению благодаря латеральному распределению и повышению активности, направленной на уничтожение опухолей. Предпочтительными также являются экспрессионные кластеры, содержащие другие гены, повышающие уровень латерального распределения HSV согласно изобретению.
Другими предпочтительными экзогенными экспрессионными кластерами являются кластеры, которые кодируют белки или полипептиды, индуцирующие у пациента иммунные ответы против рака или опухоли, которые могут быть подвергнуты лечению с использованием HSV согласно изобретению. Так, например, такие экспрессионные кластеры могут включать одну или более нуклеиновых кислот, кодирующих такие факторы, как цитокины (например, IL-2 и IFN B), антитело, направленное против белка 4, ассоциированного с цитотоксическими Т-лимфоцитами («CTLA-4») (Hodi et al., N. Engl. J. Med., 363(8): 711-23 (2010)), антитело, направленное против лиганда белка программируемой клеточной гибели 1 («PD1») или самого рецептора (Topalian et al., N. Engl. J. Med., 366(26): 2443-54 (2012)), и адгезивная молекула эпителиальных клеток («EpCAM») (Patriarca et al., Cancer Treatment Rev., 38: 68-75 (2012)). Как упоминалось выше, EpCAM может также служить в качестве таргетирующего маркера, распознаваемого вектором согласно изобретению. Кроме того, если раковым заболеванием, подвергаемым лечению, не является рак ЦНС, а в частности, глиома или глиобластома, то может быть использован другой трансген, который кодирует гранулоцитарный-макрофагальный колониестимулирующий фактор («GM-CSF»).
Другие предпочтительные экспрессионные кластеры кодируют белки или полипептиды, которые катализируют превращение пролекарств в активные агенты. Так, например, такие экспрессионные кластеры могут кодировать ферменты, например, цитозиндезаминазу, которая может превращать 5-фторцитозин («5-FC») в 5-фторурацил («5-FU») в определенном участке опухоли или раковых клеток, инфицированных вектором согласно изобретению (см., например, Akimoto et al., J. Ophthalmol., 86(5): 581-86 (2002)), в результате чего, 5-FU может действовать на этом участке клеток или опухолей и минимизировать системное воздействие этого 5-FU. Аналогичнным образом, такой экспрессионный кластер может кодировать тимидинкиназу (tk) (например, функционально присоединенную к предраннему промотору или к сильному конститутивному промотору HSV), которая может активировать ганцикловир или пурин-нуклеозид-фосфорилазу (PNP), которая, в свою очередь, может блокировать или снижать активность рибонуклеотид-редуктазы. В некоторых вариантах осуществления изобретения, векторы согласно изобретению могут также содержать функциональный нативный ген tk HSV.
В векторах согласно изобретению, кодирующие последовательности в экзогенных экспрессионных кластерах могут быть функционально присоединены к любой нужной генной регуляторной последовательности, такой как конститутивный промотор, индуцибельный промотор или тканеспецифический промотор, многие из которых известны специалистам. Так, например, широко используемым конститутивным промотором является промотор человеческого цитомегаловируса (hCMV), но могут быть также использованы и другие промоторы, например, ранний энхансер CMV/промотор гена куриного бета-актина (CAG) и предранний промотор HSV (например, промотор ICP4) и т.п.
Кроме того, в некоторых вариантах осуществления изобретения, геном вектора согласно изобретению имеет делецию области внутренних повторов (стыка), содержащей одну копию каждого из диплоидных генов ICP0, ICP34.5, LAT и ICP4 вместе с промотором гена ICP47. В других вариантах осуществления изобретения, вместо делеции стыка, экспрессия генов в области стыка, а в частности, ICP0 и/или ICP47, может быть ингибирована путем делеции этих генов, либо какими-либо другими мнтодами лимитирующего мутагенеза.
Вектор согласно изобретению может быть продуцирован стандартными методами, известными специалистам в области HSV-вирусологии. Однако, настоящее изобретение также относится к нуклеиновой кислоте, кодирующей вектор согласно изобретению и облегчающей модификацию генома HSV и продуцирование вектора согласно изобретению. Предпочтительной нуклеиновой кислотой является бактериальная искусственная хромосома («BAC»), кодирующая вектор согласно изобретению, который облегчает модификацию HSV в бактериальной системе.
При этом, следует отметить, что oHSV согласно изобретению может быть использован для нацеливания на раковые клетки и их уничтожения in vivo или in vitro. Вектор согласно изобретению, предпочтительно, применяется в терапии, а в частности, для лечения пациентов и/или для уничтожения человеческих опухолей/клеток (которые могут представлять собой ксенотрансплантаты у млекопитающих различных видов). Однако, этот способ может быть также применен для лечения животных, таких как животные-компаньоны (например, кошки и собаки), сельскохозяйственные животные (например, крупный рогатый скот, овцы, лошади и т.п.) или животные, содержащиеся в зоопарках. Репрезентативными опухолевыми/раковыми клетками, которые могут быть подвергнуты лечению с использованием векторов согласно изобретению, являются раковые клетки центральной нервной системы, а в частности, мультиформной глиобластомы.
В общих чертах, oHSV-вектор согласно изобретению является особенно ценным, если в клеточную популяцию доставляется достаточное количество вируса для гарантии того, что эти клетки будут атакованы соответствующим числом вирусов. Таким образом, настоящее изобретение относится к штамму, предпочтительно, гомогенному штамму, включающему oHSV-вектор согласно изобретению. Способы получения и анализа штаммов HSV хорошо известны специалистам. Так, например, вирусный штамм может быть получен в роллер-флаконах, содержащих клетки, трансдуцированные oHSV-вектором. Затем вирусный штамм может быть очищен в непрерывном градиенте плотности порошка Nycodenz, разделен на аликвоты и помещен на хранение до его использования. Титры вирусных штаммов могут значительно варьироваться в зависимости, главным образом, от генотипа вируса, от способа его получения и от типа клеточных линий, используемых для его получения. При этом, предпочтительно, чтобы такой штамм имел титр вируса, составляющий по меньшей мере приблизительно 105 бляшкообразующих единиц (б.о.е.), например, по меньшей мере приблизительно 106 б.о.е. или еще более предпочтительно, по меньшей мере приблизительно 107б.о.е. В еще более предпочтительных вариантах осуществления изобретения, титр может составлять по меньшей мере приблизительно 108 б.о.е. или по меньшей мере приблизительно 109 б.о.е., при этом, наиболее предпочтительными являются штаммы с высоким тиром, составляющим по меньшей мере приблизительно 1010 б.о.е. или по меньшей мере приблизительно 1011 б.о.е. Такие титры могут быть получены с использованием клеток, экспрессирующих рецептор, на который нацелен, например, указанный вектор.
Настоящее изобретение также относится к композиции, содержащей oHSV-вектор согласно изобретению и носитель, а предпочтительно, физиологически приемлемый носитель. Носителем в данной композиции может быть любой носитель, подходящий для этого вектора. Таким носителем обычно является жидкость, но может быть также и твердое вещество или комбинация жидких и твердых компонентов. При этом, желательно, чтобы такой носитель был фармацевтически приемлемым (например, физиологически или фармакологически приемлемым) носителем (например, наполнителем или разбавителем). Фармацевтически приемлемые носители хорошо известны специалистам и являются легко доступными. Выбор конкретного носителя зависит, по меньшей мере частично, от конкретного вектора и от конкретного способа введения композиции. Композиция может также содержать любые другие подходящие компоненты, а в частности, компоненты, повышающие стабильность композиции и/или конечного продукта. В соответствии с этим, может быть получен широкий ряд препаратов на основе композиции согласно изобретению. Представленные ниже композиции и способы приводятся лишь в иллюстративных целях и не ограничивают объем изобретения.
Препаратами, подходящими для парентерального введения, являются водные и безводные изотонические стерильные растворы для инъекций, которые могут содержать антиоксиданты, буферы, бактериостаты и растворенные вещества, сообщающие препарату изотоничность в крови данного реципиента, и водные и безводные стерильные суспензии, которые могут включать суспендирующие агенты, солюбилизаторы, загустители, стабилизаторы и консерванты. Препараты могут быть упакованы в герметично закрытые контейнеры для одноразового введения или многократного введения, такие как ампулы и флаконы, которые могут храниться в замороженном (лиофилизованном) состоянии, и в эти препараты, непосредственно перед их использованием, необходимо лишь добавить стерильный жидкий наполнитель, например, воду для инъекций. Растворы и суспензии в виде инъекций для немедленного приема могут быть приготовлены из стерильных порошков, гранул и таблеток различных видов, описанных ранее.
Кроме того, композиция может содержать дополнительные терапевтические средства или биологически активные агенты. Так, например, эти композиции могут включать терапевтические средства, подходящие для лечения конкретного заболевания. В состав композиции могут входить средства, снижающие воспаление, такие как ибупрофен или стероиды, которые снижают степень опухания и воспаления, ассоциированного с введением вектора in vivo, и ослабляют симптомы физиологического дистресса. В данную композицию могут быть введены супрессоры иммунной системы для снижения иммунного ответа на любую иммунную реакцию, ассоциированную с самим вектором или данным расстройством. Альтернативно, в композицию могут быть включены усилители иммунного ответа для индуцирования природных защитных свойств организма против заболевания, а в частности, против рака или опухоли, лечение которых осуществляют с использованием вектора согласно изобретению. В эту композицию могут быть включены антибиотики, то есть, микробициды и фунгициды для снижения риска инфицирования, ассоциированного с переносом генов, и риска развития других расстройств.
Пример 1
Цель: Мультиформная глиобластома (МГБ) представляет собой агрессивную опухоль головного мозга, которая не поддается эффективному лечению. Для лечения животных с моделью человеческой МГБ были разработаны oHSV-векторы, однако, испытания на их эффективность для пациентов не дали желаемых результатов. Авторами настоящего изобретения были предприняты попытки разработать новую oHSV-конструкцию, которая позволила бы осуществлять в высокой степени селективный лизис опухоли без аттюниирования вектора.
Программа эксперимента: Авторами настоящего изобретения был сконструирован oHSV для селективного инфицирования опухолевых клеток и для его селективной репликации в опухолевых клетках посредством полного перенацеливания инфекции посредством EGFR и блокирования репликации вектора в нормальных нейронах посредством введения множества копий последовательности, распознаваемой нейрон-специфичной miR-124, в 3'UTR основного предраннего гена ICP4 HSV. Область miR-124 была выбрана потому, что она в высокой степени экспрессируется в нейронах, но почти не детектируется в МГБ. Вектор тестировали в экспериментах на эффективность лечения ксеногенной опухоли головного мозга.
Результаты: Интракраниальная инокуляция «голым» мышам высокой дозы miR-124-чувствительного вируса не приводила к какому-либо патогенезу или репликации вируса, что обусловлено блокадой репликации вируса в нормальном головном мозге в результате взаимодействия miR-124 с мРНК ICP4. Лечение «голых» мышей с ортотопической моделью первичной человеческой МГБ путем введения EGFR-перенацеленного miR124-чувствительно HSV продемонстрировало повышение выживаемости (≥50%) по сравнению с выживаемостью при лечении родительским EGFR-перенацеленным вирусом, что указывало на то, что элементы распознавания miR-124 не оказывают негативного влияния на эффективность лечения.
Выводы: Авторами настоящего изобретениябыл сделан вывод, что специфичность неаттенюированного oHSV может быть максимизирована путем нацеливания векторной инфекции на опухоль в комбинации с элиминацией нежелательной репликации вектора посредством клеточных микроРНК, которые отсутствуют в опухолях, но в высокой степени экспрессируются в нормальной ткани.
Введение
МГБ представляет собой одну из самых злокачественных форм рака, для которого пока не разработан способ эффективного лечения. В медицинской практике обычно используются такие методы лечения, как хирургическая операция, лучевая терапия и химиотерапия, но эти методы дают лишь неполный и недолговременный клинический эффект. Онколитические векторы, включая векторы, происходящие от вируса простого герпеса типа-1 (oHSV-1), находятся на стадии разработки в нескольких лабораториях как возможная альтернатива существующим терапевтическим методам (1). Было обнаружено, что oHSV-векторы являются перспективными с точки зрения их использования для лечения животных с моделью МГБ, но несмотря на хороший профиль безопасности, результаты, полученные на ранних стадиях клинических испытаний, не указывали на какое-либо эффективное уничтожение опухоли или на значительное увеличение продолжительности жизни пациента (2), (3).
Самым распространенным методом достижения аттенюирования HSV является функциональная делеция не-основных генов, которые «ускользают» от природного «надзора» иммунной системы хозяина в ответ на инфицирование; получение нуклеотидных пулов для репликации в неделящихся клетках, таких как нейроны; и предотвращение апоптоза клеток (2). Репликация вирусов в раковых клетках облегчается благодаря утрате некоторых природных иммунных ответов (4), а также благодаря быстрому делению клеток и неактивным путям апоптоза (2). Однако, эти свойства не являются в одинаковой степени достаточными для интенсивной репликации oHSV, присутствующих в опухолях.
В качестве первой стадии повышения эффективности вектора, авторами настоящего изобретения были предварительно разработаны способы полного ретагретинга HSV для перенацеливания инфекционного канонического вируса HSV с рецепторов, с которыми он обычно связывается, на рецепторы, которые в высокой степени экспрессируются на поверхности опухолевых клеток (например, EGFR и EGFRvIII) (5). Перенацеленный oHSV обнаруживает надежную онколитическую активность и высокую степень специфичности к человеческим клеткам МГБ, что приводит к высокому уровню деструкции человеческой опухоли у мышей с ортотопической моделью этой опухоли. Кроме того, такой терапевтический вектор обеспечивает более высокий уровень выживаемости у большинства обработанных животных без какой-либо токсичности, ассоциированной с этим вектором. Однако, большинство в высокой степени экспрессирующихся маркеров поверхности опухолевых клеток имеют некоторое сходство с маркерами нормальных клеток, а поэтому, авторами настоящего изобретения было проведено исследование по повышению безопасности нацеленного на опухоль неаттенюированного вектора по независимому механизму блокирования репликации вируса в нормальном головном мозге без снижения степени репликации этого вируса в опухоли.
В основу недавно проведенных исследований было положено различие профилей экспрессии микроРНК (миРНК) в нормальных и в раковых клетках для разработки способа, который может служить альтернативой нацеливанию на опухоль (6). Было идентифицировано по меньшей мере 30 миРНК, которые обнаруживают дифференциальную экспрессию в глиобластомах, в нейронах и в клетках-предшественниках нейнонов (NPC) (7), (8), что дает основание предположить, что эти различия могут быть использованы для ограничения репликации вируса в нормальных клетках головного мозга, но, при этом, неограниченной репликации в опухолевых клетках. Авторами настоящего изобретения было продемонстрировано, что введение элементов распознавания miR-124 в главный ген ICP4, в основном, вируса дикого типа будет предотвращать репликацию HSV в нормальной ткани головного мозга, где наблюдается высокий уровень экспрессии miR-124. Кроме того, авторами настоящего изобретения было обнаружено, что элементы распознавания miR-124 не снижают онколитическую активность EGFR-ретаргетированного вектора. Важно отметить, что поскольку фенотип опухоли зависит от продолжительности отсутствия miR-124, то потенциальная активация miR-124 по клеточному механизму «ускользания» от репликации литического вируса будет ограничивать неконтролируемую пролиферативную активность клеток, но, при этом, не будет снижать эффективность вектора. Продуцирование вектора осуществляли в клетках, не содержащих miR-124, что, тем самым, позволяло отключить давление отбора при продуцировании miR-124-резистентных вирусных мутантов в процессе получения штамма. В целом, эти свойства обеспечивают безопасность вектора и его селективность к опухоли, что дает основание предположить, что эта общая стратегия конструирования онколитического вектора может быть применена для лечения опухолей широкого ряда.
Результаты
Подтверждение присутствия элемента распознавания miR-124. Среди множества миРНК, которые экспрессируются в нейронах на более высоких уровнях, чем в клетках МГБ, наиболее распространенной является miR-124 с самой низкой экспрессией в МГБ (6). Авторами настоящего изобретения был сконструирован элемент распознавания miR-124 (T124), состоящий из 4 тандемных копий обратного комплемента зрелой miR-124, разделенных различными спейсерами длиной в 8 нуклеотидов (нк). Для оценки функциональности этой последовательности, авторами настоящего изобретения была введена данная последовательность в 3'UTR экспрессионной плазмиды, экспрессирующей люциферазу светляка (fLuc) и были осуществлены эксперименты по котрансфекции предшественников специфичесой (пре-miR-124) или неспецифической (пре-miR-21) миРНК на клетки остеосаркомы U2OS, которые, вероятно, экспрессируют небольшое количество или вообще не экспрессируют miR-124 (9); причем, для нормализации была использована плазмида, экспрессирующая люциферазу Renilla (rLuc). Эти результаты (pfLuc-T124, фиг. 1) показали значительное снижение активности fLuc через 24 часа в клетках, котрансфецированных пре-miR-124 по сравнению с клетками, котрансфецированными контролем, или с клетками, котрансфецированными пре-miR-21. В противоположность этому, небольшое различие в уровнях экспрессии fLuc наблюдалось между клетками, трансфецированными контрольной плазмидой fLuc, содержащей 4 копии последовательности miR-21 в обратной ориентации (pfLuc-Ctrl, контроль), и клетками, трансфецированными pfLuc-Ctrl либо с пре-miR-21 либо с пре-miR-124 (фиг.1). Эти результаты продемонстрировали функциональность элемента T124 как эффективной и специфичной мишени для miR-124-опосредуемой рестрикции экспрессии гена.
Репликативная чувствительность T124-модифицированного HSV к экспрессии miR-124.Авторами настоящего изобретения была использована двойная рекомбинация Red в E.coli (10) для введения серии модификаций в BAC KOS-37, то есть, в полноразмерный геномный клон штамма KOS HSV-1 на бактериальной искусственной хромосоме (BAC) (11). Этот продукт, KGBAC (фиг. 2A), был делетирован по области внутренних повторов (стыка), содержащей одну копию каждой из диплоидных генов ICP0, ICP34.5, LAT и ICP4 вместе с промотором для гена ICP47. Эта делеция облегчает модификацию остальных копий 4 делетированных генов, создает большое пространство для потенциального включения трансгенов, которые усиливают онколитическую активность вируса и повышают специфичность к опухоли благодаря снижению уровня экспрессии нейровирулентного фактора ICP34.5 (12); где элиминация экспрессии ICP47 улучшает иммунное распознавание инфицированных раковых клеток вирус-специфическими Т-клетками (4). KGBAC также содержит открытую рамку считывания (ОРС) GFP, связанную с ОРС гликопротеина С (gC) посредством пептидной последовательности 2A (13), (14), что позволяет осуществлять мониторинг экспрессии позднего вирусного гена (после репликации). И наконец, KGBAC содержит пару мутаций в гене gB, введенную авторами для повышения способности HSV связываться посредством неканонических рецепторов (15), (16). Последовательность T124 была помещена авторами настоящего изобретения в 3'UTR оставшегося гена ICP4 KGBAC для генерирования KG4:T124BAC (фиг. 2A). Обе BAC-конструкции были преобразованы в вирусные частицы с одновременным удалением последовательностей BAC, расположенных между сайтами loxP, посредством трансфекции клеток U2OS-Cre. После очистки бляшек приготавливали вирусные штаммы KG и KG4:T124, а затем эти штаммы титровали на клетках U2OS.
Сначала, авторами настоящего изобретения было определено, может ли включение 4 тандемных сайтов-мишеней miR-124 в 3'UTR ICP4 влиять на репликацию вируса в культурах человеческих клеток МГБ. Эти результаты (фиг. 2B) показали, что KG4:T124 реплицировался с такой же кинетикой, как и KG в сфероидах двух первичных глиобластомных линий, Gli68 и GBM30, и в любой момент времени, выходы этих 2 вирусов значительно не отличались. Затем авторами было определено, может ли репликация и выход вируса зависеть от трансдуции этих линий человеческим лентивирусом, экспрессирующим miR-124 (LV124). На фиг. 2C показаны относительные уровни miR-124 в клетках U2OS, Gli68 и Gli68-LV124, измеренные с помощью кол.ПЦР в реальном времени для обратно транскрибируемых небольших РНК и стандартизированные по эндогенным уровням RNU43. KG размножался одинаково хорошо и с одинаковыми титрами на клетках Gli68-LV124 и Gli68, трандуцированных лентивирусной конструкцией, экспрессирующей обратный комплемент человеческого miR-137 (LV137R) (фиг. 2D). В противоположность этому, KG4:T124 размножался на первых клетках хуже, чем на последних, и аналогичные результаты были получены как для LV124-трансдуцированных клеток GBM30, так и для LV137R-трансдуцированных клеток GBM30 (фиг. 2D). В целом, эти наблюдения со всей очевидностью показали, что (i) элемент T124 в гене ICP4 является эффективным как средство для ограничения репликации HSV по miR-124-зависимому механизму и (ii) уровни эндогенной miR-124 в 2 линиях МГБ были достаточно низкими для минимизации этого эффекта. Кроме того, данные кол.ПЦР подтвердили, что клетки U2OS являются более подходящими для нормального размножения и титрования KG4:T124, а не KG.
KG4:T124 не реплицируется в мышином головном мозге или не вызывает заболевания. После того, как было показано, что экспрессия экзогенной miR-124 в культуре первичных клeток глиомы является в высокой степени эффективной для предупреждения репликации вектора KG4:T124, авторами настоящего изобретения был проведен анализ для того, чтобы определить, являются ли эндогенные уровни miR-124 в мышином головном мозге достаточными для предупреждения репликации вектора и типичного нейропатогенеза, ассоциированного с вирусом дикого типа, и было замечено, что последовательности зрелой человеческой и мышиной miR-124 являются идентичными (17). В этих экспериментах были использованы «голые» мыши для ограничения антивирусного ответа у хозяина и, тем самым, для облегчения идентификации прямых эффектов инсерции T124 в вирус. Для этого были выбраны мыши BALB/cnu/nu, поскольку эти животные являются в высокой степени чувствительными к репликации и патогенезу HSV (18), (19), (20) и были использованы ранее в экспериментах по оценке эффективности лечения опухоли, а именно, элиминации человеческих опухолевых клеток (21), (12, 22), (5). Авторами настоящего изобретения было проведено сравнение контрольного вектора KG и miR-124-чувствительного тестируемого вектора KG4:T124 на их способность реплицироваться в головном мозге «голых» мышей и вызывать летальную инфекцию после интракраниальной инокуляции равного количества геномных копий (г.к.)(4,8×109 г.к.) в правое полушарие головного мозга. Полученные результаты показали, что инъекция контрольного вектора приводила к быстрой гибели животного через 5 дней (фиг. 3A, C) с двухкратным увеличением общего числа геномных копий в инфицированном головном мозге (фиг. 3B). В противоположность этому, какого-либо заметного изменения состояния у мышей, которым инъецировали KG4:T124, не наблюдалось в течение 33-дневного периода проведения эксперимента, на что указывает нормальное увеличение массы тела до их умерщвления (фиг. 3A), и устойчивое снижение содержания геномных копий вируса в течение всего этого периода времени приблизительно до 0,4% от исходного уровня (фиг. 3B). Различия в выживаемости животных, которым вводили контрольный вектор, и животных, которым вводили тестируемый вектор (фиг. 3С) были в высокой степени значимыми (P=0,0058, log-ранговый критерий), что указывает на то, что 4 копии последовательности распознавания miR-124, встроенные в 3'UTR гена ICP4, обладали способностью блокировать репликацию летального вектора в головном мозге у «голых» мышей, которые являются в высокой степени чувствительными к HSV. Таким образом, эти последовательности, взятые отдельно, были достаточными для предупреждения токсичности вектора в головном мозге.
Для подтверждения предположения, сделанного исходя из этих результатов о том, что потеря или мутационная инактивация сайтов-мишеней miR-124 в процессе приготовления вирусного штамма является в лучшем случае редкой, из вирусного штамма KG4:T124 выделяли ДНК, и эту ДНК подвергали ПЦР-анализу по всему сайту инсерции T124 в 3'UTR ICP4. Анализ продуктов, проведенный с помощью гель-электрофореза и секвенирования ДНК, указывал на отсутствие ПЦР-продукта аномальных размеров или нуклеотидной вариабельности (данные не приводятся). Аналогичным образом, ПЦР-анализы и секвенирующие анализы общей ДНК головного мозга, выделенной через 3 часа или через 21 день после интракраниальной инокуляции вируса KG4:T124 (1,5×1010г.к.) нормальным мышам BALB/c, не выявили каких-либо аномальных изменений по всей области T124 (данные не приводятся). Это позволяет решить проблемы, связанные с потенциальным отбором miR-124-нечувствительных вариантов в процессе размножения вируса KG4:T124 или in vivo.
miR-124-чувствительные элементы не оказывают негативного влияния на активность EGFR-таргетированного онколитического HSV. Авторами настоящего изобретения была предпринята попытка определить, могут ли протективные элементы распознавания miR-124 негативно влиять на способность вируса уничтожать опухоль у «голых» мышей с моделью человеческой МГБ. Поскольку KG является в высокой степени токсичным при его инокуляции в головной мозг этих животных (фиг. 3C), то использование этого вируса в качестве терапевтического контроля в экспериментах на выживаемость мышей с опухолью может приводить к гибели животных, вызванной вирусом, но не опухолью, а поэтому, этот вирус не является привлекательным для такого применения. Вместо этого, авторами настоящего изобретения было введено 4 копии miR-124-связывающего сайта в полностью EGFR-ретаргетированное производное KG, поскольку опубликованные авторами наблюдения показали, что полностью EGFR-ретаргетированный HSV-1 KOS дикого типа является нетоксичным для головного мозга «голых» мышей, но эффективным для лечения «голых» мышей с ортотопической человеческой МГБ (5). Таким образом, сравнение EGFR-ретаргетированных вариантов KG и KG4:T124, обозначаемых здесь KGE и KGE-4:T124, соответственно, должно выявить любое ограничивающее действие сайтов miR-124 на вирусную онколитическую активность. Авторами были использованы взятые от пациента сферообразующие клетки GBM30 для создания агрессивных интракраниальных опухолей у «голых» мышей (5). Животных ежедневно наблюдали, а при появлении признаков болезненного состояния, умерщвляли. В соответствии с опубликованными авторами результатами, мыши, которым инъецировали PBS через 5 дней после инокуляции опухолевых клеток в тех же самых стереотактических координатах, погибали через несколько недель после имплантации опухолевых клеток (в среднем через 21,5 дней; фиг. 4A, B). В противоположность этому, лечение опухоли с использованием EGFR-ретаргетированного контрольного вируса, KGE, или T124-содержащего ретаргетированного вектора, KGE-4:T124, предупреждало развитие заболевания у половины животных на протежении всего эксперимента (90 дней), а медианы продолжительности жизни для этих двух групп были сравнимыми (79,5 и 85,5 дней, соответственно; P=0,83, log-ранговый критерий). Эти результаты показали, что сайты miR-124 в гене ICP4 KGE-4:T124 не оказывали негативного влияния на эффективность лечения опухоли GBM30.
Обсуждение
Целью настоящего изобретения является конструирование онколитического вектора HSV, который способен экспрессировать полный набор вирусных функций, но который может инфицировать только клетки, экспрессирующие МГБ-ассоциированный рецептор, и реплицироваться с высокой эффективностью только в опухоли, но не в нормальных клетках головного мозга. Опухоль-специфическая инфекция и размножение литического вируса основаны на комбинации полного перенацеливания связывания вируса (5) и клеточной miRNA-опосредуемой рестрикции репликации вируса в нормальной ткани головного мозга. Эта комбинация трансдукционного и посттранскрипционного нацеливания на опухоль является перспективной с точки зрения обеспечения высокой безопасности и эффективности oHSV, поскольку для осуществления литической инфекции необходимо наличие двух отдельных свойств клетки-мишени, которые играют важную роль в сохранении опухолевого фенотипа, таргетированного рецептора и профиля экспрессии опухолеспецифической миРНК. Эта общая стратегия может широко применяться для лечения различных раковых заболеваний с использованием таргетирующих и миРНК-распознающих элементов, и такое применение может быть оптимизировано для индивидуальной терапии после выявления потенциальных различий в уровнях экспрессии специфических антигенов и миРНК между отдельными опухолями одного и того же типа.
При МГБ, экспрессия модифицированного гена означает значительное ингибирование множества миРНК по сравнению с уровнем миРНК в нормальных тканях головного мозга (23-25), что указывает на то, что имеется несколько миРНК, которые могут быть использованы преимущественно для аттенюирования репликации сконструированного вируса в нормальном головном мозге. Поскольку miR-124 распознается как сильный индуктор дифференцировки нейронов (26) и обладает самой высокой активностью ингибирования миРНК в МГБ (6), то авторы сосредоточили свое внимание на том, что эта миРНК может служить в качестве средства для блокирования репликации oHSV в нормальной ткани головного мозга. Сайты распознавания повторов для miR-124 (T124) были введены в 3'UTR вирусного гена ICP4, продукт которого абсолютно необходим для запуска литического цикла HSV. Авторами настоящего изобретения было обнаружено, что в глиомных клетках, T124+-вирус может реплицироваться на таком же высоком уровне, как и контрольный вирус, не содержащий T124, тогда как лентивирусная экспрессия miR-124 селективно блокирует такую репликацию. Кроме того, элемент T124 является достаточным для полной защиты «голых» мышей от очень высокой интракраниальной дозы вектора (4,8×109 частиц), тогда как введение контрольного вектора приводит к гибели всех животных через пять дней. Определение общего числа геномных копий вируса в головном мозге этих животных не выявило какой-либо репликации T124+-вектора, но указывало лишь на постепенное снижение содержания вирусного генома в течение всего периода времени. Последовательность T124 была стабильной, на что указывал анализ размера и последовательности 3'UTR ICP4, амплифицированной на очищенной ДНК, выделенной из вирусных штаммов и инфицированных животных, и эти данные подтверждают полное отсутствие нейропатогенеза у животных без опухоли или высокую продолжительность жизни животных, выживших после их лечения способами, описанными в эксперименте по лечению опухоли согласно изобретению. И наконец, авторами настоящего изобретения был использован ретаргетированный вирус, который не инфицирует мышиные клетки, для того, чтобы продемонстрировать, что элемент T124 не снижает онколитическую эффективность этого вируса у «голых» мышей с моделью человеческой МГБ.
Комбинация нацеливания вируса на опухолевые рецепторы и миРНК-опосредуемого блокирования репликации вируса в нормальных клетках способствует повышению специфичности литического вируса к мишени благодаря блокированию продуктивного инфицирования нормальных клеток, которые могут иметь с опухолью общий таргетированный рецептор (например, EGFR). Хотя полученные авторами результаты показали, что инсерция четырех копий последовательности-мишени для miR-124 в 3'UTR гена ICP4 полностью блокирует нейропатогенез у «голых» мышей, индуцированный очень высокой дозой вируса, однако, было обнаружено, что не все клетки головного мозга экспрессируют miR-124. Так, например, предполагается, что последовательности-мишени miR-124 не обеспечивают защиту клеток-предшественников нейронов (NPC), находящихся в гиппокампе и в субвентрикулярной области (SVZ), поскольку эти клетки имеют профиль экспрессии миРНК, аналогичный профилю экспрессии miR-124 в клетках МГБ, включая минимальный уровень экспрессии miR-124 (27). Однако, несколько миРНК экспрессируются в NPC на уровне, в 100 раз превышающем уровни миРНК в глиомах (27), (28), (29), (30), что позволяет предположить, что сайты-мишени для дополнительных миРНК, сконструированные в одном и том же или в других главных генах того же самого вируса, будут блокировать репликацию более широкого диапазона клеток головного мозга и, при этом не будут негативно влиять на репликацию опухолеспецифического вируса.
Хотя проведенное авторами исследование позволяет предположить, что комбинация нацеливания вируса на опухолевый антиген и миРНК-рестриктированной репликации в нормальных тканях представляют собой привлекательную стратегию для проведения эффкктивной и в высокой степени специфической противоопухолевой вирусной терапии, однако, очевидно, что отдельные опухоли будут иметь различные восприимчивость к лечению, что обусловлено вариабельностью уровней опухолевых антигенов и, вероятно, содержания миРНК. Так, например, имеются значимые различия между опухолями, классифицированными как МГБ, и даже в МГБ некоторых подтипов с хорошо определенной молекулярной структурой сохраняется гетерогенность профилей экспрессии генов (31). Таким образом, отдельный ретаргетированный вирус не является эффективным для уничтожения всех клеток МГБ или всех клеток МГБ одного и того же подтипа. Кроме того, можно предположить, что популяции резистентных клеток могут появляться в очень oHSV-чувствительных опухолях в результате уже существующей вариабельности клеток в опухоли или вариабельности, индуцированной лечением. Исследования, проводимые в последние несколько лет, позволяют предположить, что небольшая популяция самообновляющихся раковых стволовых клеток, резистентных к химиотерапии и к лучевой терапии (CSC) и идентифицированных в опухолях многих различных типов, представляет собой наиболее подходящие мишени для терапии (32). Хотя сравнение отдельных CSC, происходящих от данной опухоли, является проблематичным, однако, очевидно, что их вариабельность в опухоли ограничена по сравнению с вариабельностью всей популяции опухолевых клеток. В литературе имеются описание различных маркеров глиомных стволовых клеток (GSC) (33), и для оценки роли каждого из них в развитии и поддержании человеческой МГБ у «голых» мышей могут быть использованы ретаргетированные онколитические вирусы. Авторами было высказано предположение, что опухоли, продуцирующие частичный ответ на отдельные ретаргетированные векторы, могут быть подвергнуты более эффективному лечению с использованием комбинаций векторов, перенацеленных на другие маркеры-кандидаты для распознавания GSC. Поскольку каждый из этих векторов может быть также нацелен на некоторые нормальные клетки, то, по аналогии с EGFR-ретаргетированными вирусами согласно изобретению, миРНК-опосредуемая блокада репликации вируса в этих нормальных клетках приобретает еще более важное значение. Кроме того, еще большая специфичность может быть достигнута с использованием промоторов, специфичных к клеткам определенного типа или к стадии их развития, для регуляции экспрессии ключевых признаков репликации вируса, как было впервые обнаружено Камбарой (Kambara) и сотрудниками, занимающимися исследованиями в области oHSV (34). Эти признаки могут сообщать смесям онколитических векторов высокую активность и специфичность, причем, интересно отметить, что такие векторы, как KGE-4:T124 имеют большое пространство для размещения в них трансгенов, которые могут повышать терапевтическую эффективность, таких как гены, кодирующие иммуномодуляторы, ингибиторы миграции опухолевых клеток или протеолитические ферменты, разлагающие внеклеточный матрикс опухоли, и тем самым облегчать распространение вируса внутри опухоли.
В целом, вектор KGE-4:T124, описанный в этом примере, представляет собой oHSV нового типа, который содержит полный набор репликативных функций вируса, но вирусная специфичность к опухоли обеспечивается комбинацией перенацеливания вирусной оболочки на опухолеассоциированные рецепторы и репликативной чувствительности к миРНК, которые экспрессируются в нормальной ткани, но не в опухоли. Эта комбинация регуляторных систем может быть применена и к опухолям других типов, однако, ранее не было каких-либо сообщений о применении этой комбинации для онколитических векторов. Важные преимущества стратегии согласно изобретению заключаются в том, что (i) этот вектор не содержит каких-либо дефектных генов, что обеспечивает максимальную репликацию вируса в опухолях и позволяет разработать оптимальную вирусную терапию на основе онколитического вируса, и что (ii) для репликации этого вектора необходимы экспрессия важных опухолеассоциированных маркеров клеточной поверхности и опухолеспецифический профиль экспрессии миРНК, который, в основном, отличается от профиля экспрессии миРНК в нормальной ткани. Веским аргументом в пользу стратегии согласно изобретению является то, что миРНК, выбранные для регуляции репликации вектора в нормальном головном мозге, не могут активироваться в глиобластоме и при этом не изменять фенотип опухоли (7, 25,35); причем, утрата таргетированного рецептора, такого как опухолеспецифический вариант EGFRvIII, распознаваемый вектором согласно изобретению, может давать аналогичный эффект. Таким образом, хотя в большинстве способов противораковой терапии, опухоль приобретает способность «ускользать» от лечения, однако, этот эффект будет менее вероятным при использовании миРНК-регулируемых вирусов, специфичных к опухолевому антигену. В целом, эти аргументы подтвердили предположения авторов, что такой способ согласно изобретению представляет собой в высокой степени селективную, безопасную и эффективную стратегию применения систем онколитического вектора HSV для лечения МГБ и других раковых заболеваний.
Материалы и методы
Клеточная культура. Клетки U2OS, HEK293T и HEK293AD получали из ATCC (Manassas, VA) и культивировали в 5% CO2-инкубаторе при 37°C в ATCC-рекомендованной среде, в которую была добавлена 5-10% (об./об.) фетальная бычья сыворотка (FBS; Sigma, St. Louis, MO). Клеточную линию U2OS, стабильно экспрессирующую рекомбиназу Cre (U2OS-Cre). получали путем ретровирусной трансдукции (Y.M. и J.C.G., неопубликованные результаты). Первичные глиомные сфероидные линии GMB30 и Gli68, взятые у пациента и любезно предоставленные E.A. Chiocca (Harvard Medical School, MA), были культивированы в нейробазальной среде (Gibco/Invitrogen/Life Technologies, Carlsbad, CA), содержащей 2% (об./об.) B27 без витамина A, 2 мг/мл амфотерицина B (Lonza, Walkersville, MD), 100 мкг/мл гентамицина (Lonza), 2 мМ L-глутамина (Cellgro, Manassas, VA), а также 10 нг/мл рекомбинантного человеческого эпидермального фактора роста (rhEGF) и 10 нг/мл рекомбинантного человеческого базального фактора роста фибробластов (bFGF) (оба эти фактора были получены от Shenandoah Biotechnology, Warwick, PA).
Плазмиды. pfLuc-T124 содержала четыре тандемных повтора обратного комплемента последовательности hsa-miR-124, разделенных 8 нуклеотидами, а pfLuc-Ctrl содержала четыре тандемных повтора обратной последовательности hsa-miR-21, разделенных 8 нуклеотидами. Обе плазмиды были сконструированы путем встраивания гибридизованных комплементарных олигонуклеотидов в 3'UTR гена люциферазы в pMIR-REPORT™ (миРНК Expression Reporter Vector System; Ambion, Austin, TX). Этими олигонуклеотидами являются T124-F, T124-R, TconF и TconR (таблица 1). Гибридизованные олигонуклеотиды гидролизовали ферментами SpeI и SacI и лигировали с SpeI-SacI-гидролизованным вектором pMIR-REPORT™.
Конструирование генома HSV. Хромосома BAC KOS-37 (34), содержащая полноразмерный геном штамма HSV-1 KOS в бактериальном искусственном геноме (BAC), была любезно предоставлена David Leib (Dartmouth Medical School, NH). Уникальная короткая область (US) HSV в этой хромосоме BAC находится в обратной ориентации по отношению к известной последовательности (в положениях 132, 275-145, 608) HSV-1 KOS (GenBank рег.№ JQ673480). Модификации, подробно описанные ниже, вводили посредством двойной рекомбинации Red, в основном, как описано Tischer et al. (10). Плазмиды pEPkan-S и pBAD-I-sceI (10) были любезно предоставлены Nikolaus Osterrieder (Free University of Berlin, Germany). Изменения были подтверждены посредством ПЦР-анализа, FIGE-анализа гидролизатов рестриктирующих ферментов и локального секвенирования ДНК.
Векторы, используемые в этом исследовании, были получены следующим образом. KGBAC получали из BAC KOS-37 путем делеции полноразмерной области внутренних повторов HSV или «стыка» (IRL, IRS), присоединения открытой рамки считывания (ОРС) белка, флуоресцирующего в зеленом диапазоне спектра (GFP), к ОРС гликопротеина С (gC) посредством последовательности терминации/реинициации трансляции вируса 2A (T2A) Thosea asigna (13) (37), и введения двух миссенс-мутаций в кодирующую последовательность gB (gB:N/T); (15). KG4:T124BAC получали из KGBACпутем инсерции элемента Т124 pfLuc-T124 в 3'UTR гена ICP4. Ретаргетированный вектор KGEBACполучали из KGBACпутем замены амино-концевой области гена gD соответствующей областью gD-scEGFR, содержащей последовательность, кодирующую человеческое EGFR-специфичное одноцепочечное антитело, между положениями gD 1 и 25, и миссенс-мутацию в кодоне 38 (5). KGE-4:T124BAC представляет собой комбинацию модификаций KG4:T124BAC и KGEBAC.
Размножение и очистка вируса. ДНКBAC была преобразована в инфекционный вирус путем трнасфекции клеток U2OS-Cre с использованием реагента липофектамина (Lipofectamine™ LTX) (Invitrogen); причем, рекомбиназа Cre, экспрессируемая в этих клетках, позволяла удалять элементы BAC, ингибирующие размножение вируса, и смежный ген lacZ, расположенный в BAC KOS-37 и его производные, между сигналами рекомбинации loxP (11). Отдельные бляшки выделяли путем лимитирующего разведения и тестировали на элиминацию гена lacZ по окрашиванию ферментом X-gal (38). Бесцветные блюшки подвергали двум дополнительным раундам лимитирующего разведения, и полное удаление области BAC/lacZ подтверждали путем локального секвенирования очищенной ДНК вириона. Биологические титры вирусных штаммов (б.о.е./мл) были стабилизированы на клетках U2OS, а физические титры в геномных копиях (г.к.)/мл определяли с помощью количественной ПЦР (кол.ПЦР) в реальном времени для вирусного гена gD как описано ниже.
Анализ с использованием люциферазы. Клетки HEK293AD трансфецировали плазмидой, экспрессирующей люциферазу Renilla prLuc, вместе с комбинациями других плазмид, экспрессирующих люциферазу светляка, и предшественников миРНК пре-miR™ (Ambion) с использованием липофектамина 2000 (Invitrogen). На следующий день, клетки подвергали лизису и определяли отношение экспрессии люциферазы светляка и люциферазы Renilla на люминометре Berthold LB-953 AutoLumat luminometer (Berthold Technologies USA, Oak Ridge, TN).
Экспрессия миРНК лентивирусами. Геномную ДНК, происходящую от клеток человеческой глиобластомы U-87, использовали в качестве матрицы для ПЦР-амплификации человеческой последовательности pri-miR-124 гена hsa-miR-124-3 с использованием GC-богатой ДНК-полимеразы High Fidelity Accuprime (Invitrogen) и пар праймеров miR-124, перечисленных в таблице 1. 320 п.н.-продукт гидролизовали ферментами BamHI и NheI, клонировали между соответствующими сайтами в интроне вектора pEP-miR miRNASelect (Cell Biolabs, San Diego, CA), и последовательность подтверждали. Затем область «промотор-интрон-pri-miR-124» переносили в pCDH-CMV-MCS-EF1-Puro (System Biosciences, Mountain View, CA) путем замены резидентного промотора EF1 с получением лентивирусной экспрессионной плазмиды pCDH-miR-124. Те же самые процедуры осуществляли для конструирования контрольной лентивирусной плазмиды (pCDH-miR-137R), содержащей последовательность pri-miR-137 в обратной ориентации, где ПЦР-праймеры, используемые для клонирования pri-miR-137, перечислены в таблице 1. Лентивирусы LV124 и LV137R получали путем котрансфекции pCDH-miR-124 или pCDH-miR-137R, соответственно, вместе с упаковывающими плазмидами pLP1, pLP2, pLP-VSVG (Invitrogen) в клетки HEK293T. Через 72 часа собирали супернатанты, а затем их пропускали через 0,45 мкм-фильтр (Millipore, Billerica, MA) и концентрировали путем центрифугирования в течение 16 часов при 4°C и 6800×g. Осадок ресуспендировали в DMEM и титровали как резистентные к пуромицину колониеобразующие единицы (к.о.е.) на мл клеток HEK293T.
2×105 измельченных клеток Gli68 или GBM30 инфицировали в суспензии либо вирусом LV124, либо вирусом LV137R до плотности 5 к.о.е./клетку в присутствии 8 мкг/мл полибрена в течение 90 минут и высевали. На следующий день, эти клетки подпитывали свежей средой, содержащей 30 мкг/мл пуромицина и через 72 часа подвергали суперинфицированию вирусом KG или KG4:T124 при MOI 0,01 б.о.е./клетку. Через 72 и 96 часов после HSV-инфицирования, инфекционные вирусные частицы собирали из клеток и супернатантов и титровали на клетках U2OS. РНК выделяли из параллельных культур LV124-инфицированных клеток Gli68 после 72-часового отбора на пуромицин для определения уровней miR-124 с помощью кол.ОТ-ПЦР как описано ниже.
Выделение РНК и кол.ПЦР с обратной транскриптазой. Всю РНК экстрагировали из клеток U2OS, Gli68 и LV124-инфицированных клеток Gli68 cells с использованием реагента TRIzol (Invitrogen) в соответствии с инструкциями производителей. Образцы РНК обрабатывали ДНКазой I (Invitrogen), количественно оценивали на спектрофотометре NanoDrop 2000c (Thermo-Fisher, Pittsburgh, PA) и визуализировали на MOPS-формальдегидном геле для оценки качества. Уровни зрелой hsa-miR-124 определяли по отношению к RNU43 в соответствии с протоколом проведения анализов небольших РНК TaqMan (Applied Biosystems/Life Technologies, Carlsbad, CA). Праймеры и зонды TaqMan были получены от Applied Biosystems. Все ПЦР-реакции TaqMan осуществляли с тремя повторностями.
Животные. 3-4-недельных бестимусных nu/nu-мышей BALB/c закупали в лаборатории Charles River Laboratory (Wilmington, MA) и содержали в клетках для животных BSL2. Все эксперименты на животных осуществляли в соответствии с требованиями и рекомендациями, утвержденными специалистами по уходу и содержанию лабораторных животных (Institute for Laboratory Animal Research, 1985) и одобренными Комитетом по уходу и содержанию лабораторных животных при Университете Питсбурга (IACUC).
Интракраниальная токсичность. Интаркраниальную инокуляцию вируса осуществляли, как описано в литературе (5). Мышам вводили 4,8×109 г.к. вируса KG или KG4:T124 (n=4/группу). Животных ежедневно наблюдали на признаки болезненного состояния и взвешивали каждый день. Все мыши KG-группы погибали на день 5, а одну мышь другой группы умерщвляли в тот же день. Остальных животных KG4:T124-группы умерщвляли на дни 14, 21 и 33. У мышей, подвергнутых эвтаназии, брали головной мозг целиком для экстрации всех ДНК, а затем проводили кол.ПЦР-анализ на вирусные геномы как описано ниже.
Кол.ПЦР на вирусные геномы. ДНК экстрагировали из мышиного головного мозга или из вирусных штаммов с использованием набора DNeasy Blood & Tissue (Qiagen, Valencia, CA) в соответствии с инструкциями производителей. Затем строили стандартную кривую по данным кол.ПЦР для ДНК, происходящей от плазмиды pENTR1A (Invitrogen), содержащей полноразмерную последовательность, кодирующую gD HSV-1 (штамм KOS) (pE-gD18) в соответствии с протоколом, описанным в руководстве по проведению ПЦР-анализа в реальном времени Applied Biosystems StepOne™ и StepOnePlus™. Последовательности праймеров и зондов перечислены в таблице 1.
Животные с моделью опухоли и их обработка. Интаркраниальную имплантацию человеческих клеток GBM30 «голым» мышам осуществляли как описано в литературе (5). Через 5 дней мышам вводили вирусы (1,8x108 г.к. вируса KGE или KGE-4:T124, n=8/группу) или PBS (n=2) в тех же самых координатах как описано в (5). Состояние здоровья и поведение животных оценивали как описано выше в разделе «Интаркраниальная токсичность». При появлении признаков болезненного состояния, животных подвергали эвтаназии.
Статистический анализ. Для данного анализа применяли непарный t-критерий с поправкой Велча с использованием программы GraphPad Prism для Windows версии 6.01 (GraphPad Software, La Jolla, CA; www. graphpad.com). По данным выживаемости животных строили кривые Каплана-Мейера, и эти кривые сравнивали с применением log-рангового критерия Мантеля-Кокса с использованием той же самой программы.
Библиография для примера 1
1. Parker JN, Bauer DF, Cody JJ, Markert JM. Oncolytic viral therapy of malignant glioma. Neurotherapeutics. 2009; 6:558-69.
2. Grandi P, Peruzzi P, Reinhart B, Cohen JB, Chiocca EA, Glorioso JC. Design and application of oncolytic HSV vectors for glioblastoma therapy. Expert Rev Neurother. 2009; 9:505-17.
3. Markert JM, Medlock MD, Rabkin SD, Gillespie GY, Todo T, Hunter WD, et al. Conditionally replicating herpes simplex virus mutant, G207 for the treatment of malignant glioma: results of a phase I trial. Gene therapy. 2000; 7:867-74.
4. Todo T. Oncolytic virus therapy using genetically engineered herpes simplex viruses. Frontiers in bioscience: a journal and virtual library. 2008; 13:2060-4.
5. Uchida H, Marzulli M, Nakano K, Goins WF, Chan J, Hong CS, et al. Effective treatment of an orthotopic xenograft model of human glioblastoma using an EGFR-retargeted oncolytic herpes simplex virus. Molecular therapy: the journal of the American Society of Gene Therapy. 2013; 21:561-9.
6. Gaur A, Jewell DA, Liang Y, Ridzon D, Moore JH, Chen C, et al. Characterization of микроРНК expression levels and their biological correlates in human cancer cell lines. Cancer research. 2007; 67:2456-68.
7. Karsy M, Arslan E, Moy F. Current Progress on Understanding MicroRNAs in Glioblastoma Multiforme. Genes & cancer. 2012; 3:3-15.
8. Riddick G, Fine HA. Integration and analysis of genome-scale data from gliomas. Nature reviews Neurology. 2011; 7:439-50.
9. Kumar MS, Lu J, Mercer KL, Golub TR, Jacks T. Impaired микроРНК processing enhances cellular transformation and tumorigenesis. Nature genetics. 2007; 39:673-7.
10. Tischer BK, von Einem J, Kaufer B, Osterrieder N. Two-step red-mediated recombination for versatile high-efficiency markerless DNA manipulation in Escherichia coli. Biotechniques. 2006; 40:191-7.
11. Gierasch WW, Zimmerman DL, Ward SL, Vanheyningen TK, Romine JD, Leib DA. Construction and characterization of bacterial artificial chromosomes containing HSV-1 strains 17 and KOS. J Virol Methods. 2006; 135:197-206.
12. Bennett JJ, Delman KA, Burt BM, Mariotti A, Malhotra S, Zager J, et al. Comparison of safety, delivery, and efficacy of two oncolytic herpes viruses (G207 and NV1020) for peritoneal cancer. Cancer gene therapy. 2002; 9:935-45.
13. Szymczak AL, Vignali DA. Development of 2A peptide-based strategies in the design of multicistronic vectors. Expert Opin Biol Ther. 2005; 5:627-38.
14. Doronina VA, Wu C, de Felipe P, Sachs MS, Ryan MD, Brown JD. Site-specific release of nascent chains from ribosomes at a sense codon. Mol Cell Biol. 2008; 28:4227-39.
15. Uchida H, Chan J, Goins WF, Grandi P, Kumagai I, Cohen JB, et al. A double mutation in glycoprotein gB compensates for ineffective gD-dependent initiation of herpes simplex virus type 1 infection. Journal of virology. 2010; 84:12200-9.
16. Uchida H, Chan J, Shrivastava I, Reinhart B, Grandi P, Glorioso JC, et al. Novel Mutations in gB and gH Circumvent the Requirement for Known gD Receptors in Herpes Simplex Virus 1 Entry and Cell-to-Cell Spread. Journal of virology. 2013; 87:1430-42.
17. Cao X, Pfaff SL, Gage FH. A functional study of miR-124 in the developing neural tube. Genes & development. 2007; 21:531-6.
18. Fujioka N, Akazawa R, Ohashi K, Fujii M, Ikeda M, Kurimoto M. Interleukin-18 protects mice against acute herpes simplex virus type 1 infection. Journal of virology. 1999; 73:2401-9.
19. Manickan E, Rouse RJ, Yu Z, Wire WS, Rouse BT. Genetic immunization against herpes simplex virus. Protection is mediated by CD4+ T lymphocytes. Journal of immunology. 1995; 155:259-65.
20. Sethi KK, Omata Y, Schneweis KE. Protection of mice from fatal herpes simplex virus type 1 infection by adoptive transfer of cloned virus-specific and H-2-restricted cytotoxic T lymphocytes. The Journal of general virology. 1983; 64 (Pt 2):443-7.
21. Currier MA, Gillespie RA, Sawtell NM, Mahller YY, Stroup G, Collins MH, et al. Efficacy and safety of the oncolytic herpes simplex virus rRp450 alone and combined with cyclophosphamide. Molecular therapy: the journal of the American Society of Gene Therapy. 2008; 16:879-85.
22. Hong CS, Fellows W, Niranjan A, Alber S, Watkins S, Cohen JB, et al. Ectopic matrix metalloproteinase-9 expression in human brain tumor cells enhances oncolytic HSV vector infection. Gene therapy. 2010; 17:1200-5.
23. Zhang Y, Chao T, Li R, Liu W, Chen Y, Yan X, et al. MicroRNA-128 inhibits glioma cells proliferation by targeting transcription factor E2F3a. J Mol Med. 2009; 87:43-51.
24. Shi L, Cheng Z, Zhang J, Li R, Zhao P, Fu Z, et al. hsa-mir-181a and hsa-mir-181b function as tumor suppressors in human glioma cells. Brain Res. 2008; 1236:185-93.
25. Silber J, Lim DA, Petritsch C, Persson AI, Maunakea AK, Yu M, et al. miR-124 and miR-137 inhibit proliferation of glioblastoma multiforme cells and induce differentiation of brain tumor stem cells. BMC Med. 2008; 6:14.
26. Maiorano NA, Mallamaci A. The pro-differentiating role of miR-124: indicating the road to become a neuron. RNA Biol.7:528-33.
27. Lavon I, Zrihan D, Granit A, Einstein O, Fainstein N, Cohen MA, et al. Gliomas display a микроРНК expression profile reminiscent of neural precursor cells. Neuro Oncol.12:422-33.
28. Karpowicz P, Willaime-Morawek S, Balenci L, DeVeale B, Inoue T, van der Kooy D. E-Cadherin regulates neural stem cell self-renewal. J Neurosci. 2009; 29:3885-96.
29. Katoh Y, Katoh M. Hedgehog signaling, epithelial-to-mesenchymal transition and miRNA (review). Int J Mol Med. 2008; 22:271-5.
30. Ocana OH, Nieto MA. A new regulatory loop in cancer-cell invasion. EMBO Rep. 2008; 9:521-2.
31. Verhaak RG, Hoadley KA, Purdom E, Wang V, Qi Y, Wilkerson MD, et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer cell.17:98-110.
32. Nduom EK, Hadjipanayis CG, Van Meir EG. Glioblastoma cancer stem-like cells: implications for pathogenesis and treatment. Cancer journal. 2012; 18:100-6.
33. He J, Liu Y, Lubman DM. Targeting glioblastoma stem cells: cell surface markers. Current medicinal chemistry. 2012; 19:6050-5.
34. Kambara H, Okano H, Chiocca EA, Saeki Y. An oncolytic HSV-1 mutant expressing ICP34.5 under control of a nestin promoter increases survival of animals even when symptomatic from a brain tumor. Cancer research. 2005; 65:2832-9.
35. Xia H, Cheung WK, Ng SS, Jiang X, Jiang S, Sze J, et al. Loss of brain-enriched miR-124 микроРНК enhances stem-like traits and invasiveness of glioma cells. The Journal of biological chemistry. 2012; 287:9962-71.
36. Macdonald SJ, Mostafa HH, Morrison LA, Davido DJ. Genome sequence of herpes simplex virus 1 strain KOS. Journal of virology. 2012; 86:6371-2.
37. Kaji K, Norrby K, Paca A, Mileikovsky M, Mohseni P, Woltjen K. Virus-free induction of pluripotency and subsequent excision of reprogramming factors. Nature. 2009; 458:771-5.
38. Krisky DM, Marconi PC, Oligino T, Rouse RJ, Fink DJ, Glorioso JC. Rapid method for construction of recombinant HSV gene transfer vectors. Gene therapy. 1997; 4:1120-5.
Пример 2
В этом примере описан эксперимент, в котором используется нацеленный на опухоль вирус oHSV типа 1 с металлопротеиназой матрикса 9 для улучшения распределения этого вектора и повышения его киллерной активности.
Материалы и методы
Клеточные линии. Человеческую глиобластому SNB19, U251, U87 (любезно предоставленную Dr. H Okada, University of Pittsburgh), J/A, J/C, J/EGFR [9], клетки почек африканской зеленой мартышки Vero и клетки 7b [15] культивировали стандартными методами.
Клетки культивировали в модифицированной по способу Дульбекко среде Игла (Life technologies, Grand Island, NY), в которую была добавлена 10% фетальная бычья сыворотка (Sigma. St. Louis, MO). Клеточные линии первичной глиобластомы GBM169, OG2 (любезно предоставленные Dr. Balveen Kaur, Ohio State University) и GBM30 культивировали в виде сфероидов в нейробазальной среде, в которую были добавлены Glutamax, B27, человеческий β-FGF, EGF, гепарин и пенициллин-стрептомицин.
Плазмиды. KGw BAC получали из KGE-4:T124BAC путем инсерции кластера Gateway, амплифицированного из плазмиды pcDNA3.1GW с использованием праймеров 5'-TGCCCGTCGCGCGTGTTTGATGTTAATAAATAACACATAAATTTGGCTGGCCACTAGTCCAGTGTGGTGG-3' (SEQ ID NO:12) и 5'-CTGAAATGCCCCCCCCCCCTTGCGGGCGGTCCATTAAAGACAACAAACAAATCCCCAGCATGCCTGCTATTGT-3' (SEQ ID NO:13).
Плазмиду pEnCM получали путем клонирования промотора CAG из плазмиды pCAGH [10] в pEntr-MMP9. pEntr-MMP9 получали путем клонирования кДНК mmp9 из ранее описанной плазмиды pCMV6-XL4-MMP9 в плазмиду pEntr1A [13].
Конструирование генома HSV. Хромосома BAC KOS-37 [14], содержащая полноразмерный геном штамма HSV-1 KOS в бактериальном искусственном геноме (BAC), была любезно предоставлена David Leib (Dartmouth Medical School, NH). Двойная рекомбинация Red в E.coli [24] была использована для введения серии модификаций в BAC KOS-37, то есть, в полноразмерный геномный клон штамма HSV-1 KOS на бактериальной искусственной хромосоме (BAC) [14]. В продукте KGBAC (фиг. 5) была делетирована область внутренних повторов (стыка), содержащая одну копию каждого из диплоидных генов ICP0, ICP34.5, LAT и ICP4 вместе с промотором гена ICP47.
KGwG4:T124BAC (обозначенный KGw) получали из KGE-4:T124BAC (как обсуждается в примере 1) путем инсерции кластера Gateway (из pcDNA3.1GW) и последовательности полиаденилирования бычьего гормона роста в межгенную область UL3-UL4 с применением технологии рекомбинации Red/ET (Gene Bridges GmbH, Heidelberg). MMP9-экспрессирующий вектор KMMP9G4:T124BAC (обозначенный KMMP9) получали из KGwG4:T124BAC путем замены кластера GW кластером «промотор CAG-MMP9», происходящим от pEnCM, посредством клоназной реакции LR. Для продуцирования вирусов, клетки Vero 7b трансфецировали либо KGwG4:T124BAC, либо KMMP9G4:T124BAC. Все рекомбинантные векторы подтверждали посредством FIGE-картирования, ПЦР-анализа и секвенирования ДНК по всем релевантным модифицированным областям.
Размножение и очистка вируса. ДНКBAC была преобразована в инфекционный вирус путем трансфекции клеток Vero 7b с использованием реагента липофектамина (Lipofectamine™ LTX) (Invitrogen). Биологические титры вирусных штаммов (б.о.е./мл) стабилизировали на клетках Vero, а физические титры в геномных копиях (г.к.)/мл определяли с помощью количественной ПЦР (кол.ПЦР) в реальном времени для вирусного гена gD как описано ниже.
Кол.ПЦР для вирусных геномов. ДНК экстрагировали из вирусных штаммов с использованием набора DNeasy Blood & Tissue (Qiagen, Valencia, CA) в соответствии с инструкциями производителей. Затем строили стандартную кривую по данным кол.ПЦР для ДНК, происходящей от плазмиды pENTR1A (Invitrogen), содержащей полноразмерную последовательность, кодирующую gD HSV-1 (штамм KOS) (pE-gD18), в соответствии с протоколом, описанным в руководстве по проведению ПЦР-анализа в реальном времени Applied Biosystems StepOne™ и StepOnePlus™. Ниже представлены последовательности праймеров и зонда: прямой gD: 5'-CCCCGCTGGAACTACTATGACA-3' (SEQ ID NO:14); обратный gD: 5'-GCATCAGGAACCCCAGGTT-3' (SEQ ID NO:15); зонд: 5'-FAM-TTCAGCGCCGTCAGCGAGGA-TAMRA-3' (SEQ ID NO:16).
Вестерн-блот-анализ. Клетки лизировали в 1% буфере NP40, а затем лизаты подвергали электрофорезу на 10% ДСН-полиакриламидном геле, и белковые блоты подвергали реакции взаимодействия с поликлональным антителом к MMP-9 (разведение 1:1000) (Abcam, Cambridge, MA) или с антителом к gD (1:2000) (Santa Cruz, CA) и с ПХ-конъюгированным антикроличьим «вторым» антителом (Sigma, St. Louis, MO). Блоты проявляли с использованием хемилюминесцентного субстрата (Amersham Pharmacia, Piscataway, NJ). Нижнюю фракцию каждого блота подвергали реакции взаимодействия с поликлональным антителом против бета-тубулина (1:3000) (Sigma, St. Louis, MO) для выявления различий в загрузке. Затем блоты проявляли с использованием хемилюминесцентного субстрата SuperSignal West Dura (Thermo Scientific, Rockford, IL).
Зимография желатина. Образцы, перед их разделением на 10% ДСН-полиакриламидном геле, содержащем 0,2% желатин, не обрабатывали восстановителем и не нагревали. Гель промывали в промывочном буфере, используемом в зимографии (10 мМ трис, pH 7,5, 2,5% тритон X-100), инкубировали при 37°C в течение 16 часов в инкубирующем буфере (50 мМ трис, pH 7,5, 5 мМ CaCl2, 1 мкМ ZnCl2), окрашивали 1% кумасси бриллиантовым голубым R-250 и обесцвечивали обесцвечивающим буфером (4% метанол, 8% уксусная кислота) [13].
Анализ на связывание. Клетки J/A, J/C и J/EGFR инфицировали KMMP9, KGw или KG (экспрессирующими gD:wt) при плотности 10000, 1000 или 100 г.к./клетку в течение 6 часов и подвергали иммунологическому окрашиванию моноклональным мышиным антителом к ICP4 (1:300; Santa Cruz Biotechnology) и Cy3-конъюгированным овечьим антителом против мышиного IgG (1:400; Sigma) [9].
Анализ с использованием MTT. Клетки высевали в 48-луночные планшеты и инфицировали при 100 г.к./клетку (MOI 0,2) в течение 3 или 6 дней. Затем клетки обрабатывали 0,5 мг/мл раствора MTT (Sigma) при 37°C в течение 3 часов. После удаления раствора MTT добавляли 100% ДМСО, и OD570 регистрировали на микропланшет-ридере Wallac (Perkin Elmer, Waltham, MA). Процент жизнеспособных клеток вычисляли по формуле 100%×OD (инфицированных)/OD(неинфицированных).
Сфероидная культура и конфокальная визуализация. Сфероиды подвергали диссоцииации и подсчитывали. 3000 клеток культивировали по отдельности в суспензии в течение 2 дней до образования сфероидов. Каждый сфероид отдельно инфицировали 1000 б.о.е. или 4×107г.к./клетку KMMP9 или KGw в аналитических микропланшетах. Затем каждый день получали изображения eGFP на флуоресцентном микроскопе. Для осуществления конфокальной визуализации, сфероиды, после их инфицирования, переносили в чашки со стеклянным дном (Willco wells, Amsterdam, the Netherlands). Через 5 дней после инфицирования (dpi), сфероиды фиксировали в 4% параформальдегиде, обрабатывали заливочной средой с DAPI (Vector Laboratories, Burlingame, CA) и получали изображения Z-секций на конфокальной визуализирующей системе FV1000 (Olympus, Miami, FL).
Животные с моделью опухоли и их обработка. 3-4-недельных бестимусных nu/nu-мышей BALB/c закупали в лаборатории Charles River Laboratory (Wilmington, MA) и содержали в клетках для животных BSL2. Все эксперименты на животных осуществляли в соответствии с требованиями и рекомендациями, утвержденными специалистами по уходу и содержанию лабораторных животных (Institute for Laboratory Animal Research, 1985) и одобренными Комитетом по уходу и содержанию лабораторных животных при Университете Питсбурга (IACUC).
2×105 человеческих клеток GBM30 интракраниально имплантировали «голым» мышам как описано в [9]. Через 5 или 10 дней после инфицирования, 5,65×109 геномных копий KMMP9, KGw или PBS (n=3-4/group) инокулировали в тех же самых пространственных координатах, в которых вводили инъекции опухолевых клеток (на расстоянии 0,5 мм до брегмы, на 2 мм сбоку (справа) и на 3 мм вглубь брегмы), и эта процедура также описана в [9]. Затем оценивали состояние здоровья и поведение животных, и при появлении признаков болезненного состояния, животных подвергали эвтаназии.
МРТ-визуализация. Из каждой экспериментальной группы (KMMP9, KGw, PBS) было произвольно отобрано несколько мышей. Животных визуализировали за 1 день до обработки (через 9 дней после имплантации GBM30) и через 3, 6, 9 и 13 дней после обработки. Изображения получали с использованием магнита Bruker BioSpec 94/30 (Bruker BioSpin, Karlsruhe, Germany), снабженным только катушкой диаметром 2,0 см для мышиного головного мозга и градиентной объемной катушкой диаметром 70 мм. Анестезированным мышам внутрибрюшинно инъецировали 0,1 ммоль/кг Magnevist (Bayer Health Care Pharmaceuticals, Wayne, NJ) и получали T2-взвешенные изображения (время повторения=3,500 миллисекунд, эхо-задержка=12 миллисекунд, Rare-фактор=8, Navgs=4) фронтально по всей представляющей интерес области на приборе Bruker 400 МГц, снабженном горизонтальным рабочим магнитом с зазором Paravision 4.0 (Bruker Biospin, Billerica, MA).
Статистический анализ. Для данногоанализа применяли непарный t-критерий с поправкой Велча с использованием программы GraphPad Prism для Windows версии 6.01 (GraphPad Software, La Jolla, CA; www. graphpad.com). По данным выживаемости животных строили кривые Каплана-Мейера, и эти кривые сравнивали с применением log-рангового критерия Мантеля-Кокса с использованием той же самой программы.
Результаты
Конструирование и характеризация регуляторных miR-ретаргетированных векторов, экспрессирующих MMP9
Конструкция и дизайн вектора, используемого в этом исследовании, представлены на фиг. 5A в виде диаграммы, и этот вектор имеет множество модификаций, которые были введены во избежание изменения каких-либо литических функций вируса и, тем самым, для максимизации репликации и литической активности в опухолевых клетках, а также для предотвращения размножения вируса в нормальном головном мозге.
Кластер Gateway (Gw) и последовательность полиаденилирования бычьего гормона роста встраивали между локусами UL3 и UL4 KGE-4:T124 (описанного в примере 1) с получением вектора KGwG4:T124BAC (обозначенного здесь KGw, контрольного вектора), а онколитический вектор, экспрессирующий MMP9, получали путем замены кластера Gateway геном MMP9, индуцируемым промотором CAG (β-актина куриного CMV) (указанный вектор KMMP9G4:T124BAC обозначается здесь KMMP9).
Вестерн-блот-анализ клеток Vero, инфицированных KMMP9, подтвердил «правильную» экспрессию MMP9 (фиг. 5B). Зимография с использованием желатина показала желатиназную активность трех линий первичной МГБ, GBM30, GBM169 и OG2, инфицированных KMMP9, по сравнению с клетками, инфицированными контрольным вектором (фиг. 5C), и желатиназную активность в супернатанте KMMP9- инфицированных клеток Vero по сравнению с клетками Vero, инфицированными контролем (фиг. 5D).
Затем авторами настоящего изобретения было определено, может ли экспрессия MMP9 влиять на связывание вируса посредством распознавания рецептора эпидермального фактора роста (EGFR). Клеточные линии, тестируемые на связывание вируса, представляют собой EGFR-трансдуцированные клетки J1.1-2 (J/EGFR) (Nakano et al., Virol., 413: 12-18 (2011)), которые являются резистентными к HSV дикого типа, что обусловлено отсутствием рецепторов gD; клетки J/A, экспрессирующие человеческий HVEM (Uchida et al., J. Virol. 83: 2951-2961 (2009)), и клетки J/C, экспрессирующие человеческий нектин-1 (Frampton et al., J. Virol., 81: 10879-889 (2007)), где HVEM и нектин-1 представляют собой природные рецепторы для gD дикого типа. Связывание вируса детектировали путем иммунологического окрашивания на предранний белок ICP4 HSV через 6 часов после инфицирования. Как показано на фиг. 6A, внедрение EGFR-ретаргетированных вирусов KMMP9 и KGw в клетки J/EGFR было таким же эффективным, как и внедрение родительского вектора HSV-1, экспрессирующего gD:wt, в клетки J/A или J/C. Ни один из ретаргетированных вирусов не обнаруживал какого-либо детектируемого внедрения в клетки J/A или J/C даже при высокой вирусной загрузке (10000 г.к./клетку), что указывало на то, что экспрессия MMP9 не влияет на эффективность или специфичность инфицирования ретаргетированным вектором.
Авторами настоящего изобретения был также проведен анализ для того, чтобы определить, может ли экспрессия MMP9 влиять на репликацию вируса в клеточной культуре человеческих клеток МГБ. Эти результаты (фиг. 6B и 6C) показали, что KMMP9 KG4:T124 реплицировался с такой же кинетикой, как и KGw в сфероидах двух первичных глиобластомных линий, GBM169 и GBM30, и в любой момент времени, выходы этих 2 вирусов, определенные с помощью кол.ПЦР, значительно не отличались.
Для оценки онколитической активности KMMP9, HSV-пермиссивные человеческие глиомные линии, включая U87MG, SNB19 и GBM30, которые, как известно, экспрессируют EGFR, были инфицированы при MOI 0,005 (100 г.к./клетку), и жизнеспособность клеток определяли с помощью анализа, проводимого с использованием бромида 3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолия (MTT), через 3 дня (фиг. 7A) и через 7 дней после инфицирования (фиг. 7B). В этот последний промежуток времени, KMMP9 обнаруживал значительно более высокий уровень цитолиза, то есть, цитолиза 2 клеточных линий по сравнению с KGw, что позволяет предположить, что MMP9 может усиливать опосредуемый вектором онколизис.
MMP-9 повышает инфекционность HSV в сфероидной культуре.
Для оценки влияния повышенного уровня клеточной экспрессии MMP-9 на распределение HSV в сфероидах опухолевых клеток, клетки GBM30 и GBM169 культивировали в виде отдельных сфероидов и инфицировали вирусом KMMP9 или KGw (фиг. 8A). Через 5 дней после инфицирования, KMMP9 усиливал распределение экспрессируемого вектором eGFP по сравнению с KGw. Количественная оценка eGFP-позитивных клеток в каждом сфероиде указывала приблизительно на 1,5-кратное увеличение KMMP9-инфицированных сфероидов по сравнению с KGw-инфицированными сфероидами через 6 дней после инфицирования (фиг. 8B; P=0,006).
Для дополнительной оценки влияния MMP9 на HSV-инфекционность первичных опухолевых сфероидов, клетки GBM30 инфицировали KMMP9 или KGw, и eGFP, экспрессируемый вектором, визуализировали на конфокальном микроскопе, применяемом для оценки пенетрации и инфекционности вируса. 3D-преобразование 5 мкм Z-секционных моделей выявило повышенную относительную инфекционность KMMP9 внутри сфероидов по сравнению с KGw (фиг. 8C). Авторами настоящего изобретения были также оценены различия в инфиционности KMMP9 по сравнению с KGw в сфероидах (фиг. 8С). Авторами были также оценены различия в инфекционности 5 сегментов каждого сфероида по глубине графов на оси Z (снизу вверх: 0-20 мкм, 25-50 мкм, 55-80 мкм, 85-100 мкм, 105-120 мкм и 125-140 мкм). Хотя какого-либо различия в самых крайних сегментах не обнаруживалось (0-20 мкм) (фиг. 8D и 8E), однако, KMMP9 обнаруживал значительно более высокую инфиционность в более глубоких сфероидах, чем KGw (25-50 и 50-85 мкм, P<0,05), что позволяет предположить, что MMP9 усиливает распределение вектора по всем сфероидам. Значимое различие также наблюдалось при сравнении сегментов всех сфероидов (парный t-критерий, P=0,013).
Онколитический вектор MMP9 является в высокой степени эффективным для лечения МГБ у мышей.
Ранее авторами настоящего изобретения было показано, что GBM30 устойчиво стабилизирует летальную опухоль у «голых» мышей, что приводит к гибели животных через 20 дней после инокуляции опухолевых клеток [9]. Для создания агрессивных интракраниальных опухолей у «голых» мышей, авторами настоящего изобретения были использованы сферообразующие клетки GBM30, взятые у пациента [9]. Животных ежедневно наблюдали, а при появлении признаков болезненного состояния, умерщвляли. В соответствии с опубликованными авторами результатами, мыши, которым инъецировали PBS через 5 дней после инокуляции опухолевых клеток в тех же самых стереотактических координатах, погибали через несколько недель после имплантации опухолевых клеток (в среднем через 18 дней; фиг. 9). В противоположность этому, лечение опухоли с использованием MMP9-экспрессирующего вируса, KMMP9 или контрольного вируса KGw предупреждало развитие заболевания у половины животных по меньшей мере в течение 35 дней, а медианы продолжительности жизни для этих двух групп были сравнимыми (29 и 31,5 дней, соответственно; P=0,61, log-ранговый критерий). Эти результаты показали, что продолжительность жизни 50% MMP9-обработанных животных составляла до 35 дней, в отличие от необработанных животных, продолжительность жизни которых составляла 18 дней (фиг. 9).
В параллельном независимом эксперименте, противоопухолевую эффективность KMMP9 и KGw сравнивали с эффективностью контроля (PBS) у животных с ортотопической моделью ксенотрансплантата GBM30 путем инъекции векторов через 10 после инокуляции опухолей. Мышей визуализировали с помощью магнитной резонансной томографии (МРТ) на изменение объема опухоли за 1 день до обработки и через 3, 6, 9 и 13 дней после обработки. На фиг. 10A в качестве примеров представлены T2-взвешенные изображения для каждой группы. Сравнение отдельных животных каждой группы, которые имели сравнимые объемы опухоли в начале обработки, со всей очевидностью показало, что MMP9 дает более сильный онколитический эффект, чем вектор KGw (фиг.10B).
Библиография к примеру 2
1. Grossman, S.A., et al., Survival of patients with newly diagnosed glioblastoma treated with radiation and temozolomide in research studies in the United States. Clin Cancer Res, 2010. 16(8): p. 2443-9.
2. Assi, H., et al., Gene therapy for brain tumors: basic developments and clinical implementation. Neurosci Lett, 2012. 527(2): p. 71-7.
3. Friedman, G.K., et al., Herpes simplex virus oncolytic therapy for pediatric malignancies. Mol Ther, 2009. 17(7): p. 1125-35.
4. Mohyeldin, A. and E.A. Chiocca, Gene and viral therapy for glioblastoma: a review of clinical trials and future directions. Cancer J, 2012. 18(1): p. 82-8.
5. Campadelli-Fiume, G., et al., Rethinking herpes simplex virus: the way to oncolytic agents. Rev Med Virol, 2011. 21(4): p. 213-26.
6. Broberg, E.K. and V. Hukkanen, Immune response to herpes simplex virus and gammal34.5 deleted HSV vectors. Curr Gene Ther, 2005. 5(5): p. 523-30.
7. Aghi, M., et al., Oncolytic herpes virus with defective ICP6 specifically replicates in quiescent cells with homozygous genetic mutations in pl6. Oncogene, 2008. 27(30): p. 4249-54.
8. Navaratnarajah, C.K. et al., Targeted entry of еnveloped viruses: measles and herpes simplex virus I. Curr Opin Virol, 2012. 2(1): p. 43-9.
9. Uchida, H., et al., Effective treatment of an orthotopic xenograft model of human glioblastoma using an EGFR-retargeted oncolytic herpes simplex virus. Mol Ther, 2013. 21(3): p. 561-9.
10. Uchida, H., et al., A double mutation in glycoprotein gB compensates for ineffective gD-dependent initiation of herpes simplex virus type 1 infection. J Virol, 2010. 84(23): p. 12200-9.
11. Payne, L.S. and P.H. Huang, The pathobiology of collagens in glioma. Mol Cancer Res, 2013. 11(10): p. 1129-40.
12. Mok, W., Y. Boucher, and R.K. Jain, Matrix metalloproteinases-l and -8 improve the distribution and efficacy of an oncolytic virus. Cancer Res, 2007. 67(22): p. 10664-8.
13. Hong, C.S., et al., Ectopic matrix metalloproteinase-9 expression in human brain tumor cells enhances oncolytic HSV vector infection. Gene Ther, 2010. 17(10): p. 1200-5.
14. Gierasch, W.W., et al., Construction and characterization of bacterial artificial chromosomes containing HSV-1 strains 17 and KOS. J Virol Methods, 2006. 135(2): p. 197-206.
15. Krisky, D.M., et al., Deletion of multiple immediate-early genes from herpes simplex virus reduces cytotoxicity and permits long-term gene expression in neurons. Gene Ther, 1998. 5(12): p. 1593-603.
16. Szymczak, A.L. and D.A. Vignali, Development of 2 A peptide-based strategies in the design of multicistronic vectors. Expert Opin Biol Ther, 2005. 5(5): p. 627-38.
17. Miao, H., et al., EphA2 promotes infiltrative invasion of glioma stem cells in vivo through cross-talk with Akt and regulates stem cell properties. Oncogene, 2014.
18. Yin, A.A., et al., The treatment of glioblastomas: a systematic update on clinical Phase III trials. Crit Rev Oncol Hematol, 2013. 87(3): p. 265-82.
19. Wong, J., et al., Targeted oncolytic herpes simplex viruses for aggressive cancers. Curr Pharm Biotechnol, 2012. 13(9): p. 1786-94.
20. Wakimoto, H., et al., Effects of innate immunity on herpes simplex virus and its ability to kill tumor cells. Gene Ther, 2003. 10(11): p. 983-90.
21. McKee, T.D., et al., Degradation of fibrillar collagen in a human melanoma xenograft improves the efficacy of an oncolytic herpes simplex virus vector. Cancer Res, 2006. 66(5): p. 2509-13.
22. Yun, CO., Overcoming the extracellular matrix barrier to improve intratumoral spread and therapeutic potential of oncolytic virotherapy. Curr Opin Mol Ther, 2008. 10(4): p. 356-61.
23. Dmitrieva, N., et al., Chondroitinase ABC I-mediated enhancement of oncolytic virus spread and antitumor efficacy. Clin Cancer Res, 2011. 17(6): p. 1362-72.
24. Tischer, B.K., et al., Two-step red-mediated recombination for versatile high-efficiency markerless DNA manipulation in Escherichia coli. Biotechniques, 2006. 40(2): p. 191-7.
25. Ishida, D., et al., Enhanced cytotoxicity with a novel system combining the paclitaxel-2'-ethylcarbonate prodrug and an HSV amplicon with an attenuated replication-competent virus, Cancer Lett, 2010. 288(1): p. 17-27.
Все цитируемые здесь документы, включая публикации, патентные заявки и патенты, вводятся в настоящее описание посредством ссылки так, как если бы каждый из этих документов был отдельно и конкретно во всей своей полноте введен в настоящее описание посредством ссылки. Содержание заявки на патент США 13/641649 (Национальная фаза PCT/US2011/032923), которая была опубликована как US 2013/0096186 и WO 2011/130749, и в которой испрашивается приоритет предварительной заявки на патент США 61/325137, также во всей своей полноте вводится в настоящее описание посредством ссылки; причем особое внимание следует уделить параграфам [0039], [0040] и [0041] заявки US 2013/0096186. Работа Mazzacurati et al, Mol. Ther. 2014 Sep 9. doi: 10.1038/mt.2014.177 также во всей своей полноте вводится в настоящее описание посредством ссылки [публикациия Epub в настоящее время находится в печати].
Используемые в настоящем описании, артикли «a», «an» и «the» и аналогичные референты, соответствующие контексту описания изобретения (а в частности, контексту прилагаемой формулы изобретения) и употребляемые с существительными в единственном числе, могут относиться и к существительным во множественном числе, если из контекста описания не следует обратное, или если это явно не противоречит контексту описания изобретения. Используемые здесь термины «составляющий», «имеющий», «включающий» и «содержащий» имеют инклюзивное значение (то есть, «включают, но не ограничиваются ими»), если это не оговорено особо. Используемые здесь указания на интервалы величин служат лишь способом сокращенной записи каждой отдельной величины, входящей в этот интервал, если это не оговорено особо, и каждая такая отдельная величина вводится в описание настоящего изобретения так, как если бы она указывалась индивидуально. Все описанные здесь способы могут быть осуществлены в любом нужном порядке, если это не оговорено особо, или если это не противоречит контексту описания изобретения. Любые или все пояснительные слова или слово «например» (например, «такой как») приводятся здесь лишь для лучшего понимания настоящего изобретения и не ограничивают объема изобретения, если это не оговорено особо. Ни одно приводимое здесь словесное описание любого элемента не должно рассматриваться как указание на какой-либо незаявленный элемент, который является необходимым для осуществления настоящего изобретения.
В настоящей заявке описаны предпочтительные варианты осуществления изобретения, включая, по мнению авторов, наилучший вариант осуществления изобретения. Другие предпочтительные варианты осуществления изобретения будут очевидны для специалиста в данной области исходя из описания, приведенного выше. Авторы настоящего изобретения считают, что такие варианты, при необходимости, могут входить в объем настоящего изобретения, и могут быть использованы для осуществления изобретения, если это не оговорено особо. В соответствии с этим, настоящее изобретение включает все модификации и эквиваленты объектов изобретения, заявленных в прилагаемой формуле изобретения, если это входит в область применимости закона. Кроме того, в настоящем изобретении рассматривается любая комбинация описанных выше элементов во всех возможных их вариантах, если это не оговорено особо, или если это явно не противоречит контексту описания изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Zauderer, Maurice
<120> ПРИМЕНЕНИЕ МОЛЕКУЛ, СВЯЗЫВАЮЩИХ СЕМАФОРИН-4D, ДЛЯ ЛЕЧЕНИЯ АТЕРОСКЛЕРОЗА
<130> 1843.0750000/EJH/BNC
<140> To be assigned
<141> Herewith
<150> US 61/889,421
<151> 2013-10-10
<160> 48
<170> PatentIn version 3.3
<210> 1
<211> 862
<212> Белок
<213> Homo sapiens
<400> 1
Met Arg Met Cys Thr Pro Ile Arg Gly Leu Leu Met Ala Leu Ala Val
1 5 10 15
Met Phe Gly Thr Ala Met Ala Phe Ala Pro Ile Pro Arg Ile Thr Trp
20 25 30
Glu His Arg Glu Val His Leu Val Gln Phe His Glu Pro Asp Ile Tyr
35 40 45
Asn Tyr Ser Ala Leu Leu Leu Ser Glu Asp Lys Asp Thr Leu Tyr Ile
50 55 60
Gly Ala Arg Glu Ala Val Phe Ala Val Asn Ala Leu Asn Ile Ser Glu
65 70 75 80
Lys Gln His Glu Val Tyr Trp Lys Val Ser Glu Asp Lys Lys Ala Lys
85 90 95
Cys Ala Glu Lys Gly Lys Ser Lys Gln Thr Glu Cys Leu Asn Tyr Ile
100 105 110
Arg Val Leu Gln Pro Leu Ser Ala Thr Ser Leu Tyr Val Cys Gly Thr
115 120 125
Asn Ala Phe Gln Pro Ala Cys Asp His Leu Asn Leu Thr Ser Phe Lys
130 135 140
Phe Leu Gly Lys Asn Glu Asp Gly Lys Gly Arg Cys Pro Phe Asp Pro
145 150 155 160
Ala His Ser Tyr Thr Ser Val Met Val Asp Gly Glu Leu Tyr Ser Gly
165 170 175
Thr Ser Tyr Asn Phe Leu Gly Ser Glu Pro Ile Ile Ser Arg Asn Ser
180 185 190
Ser His Ser Pro Leu Arg Thr Glu Tyr Ala Ile Pro Trp Leu Asn Glu
195 200 205
Pro Ser Phe Val Phe Ala Asp Val Ile Arg Lys Ser Pro Asp Ser Pro
210 215 220
Asp Gly Glu Asp Asp Arg Val Tyr Phe Phe Phe Thr Glu Val Ser Val
225 230 235 240
Glu Tyr Glu Phe Val Phe Arg Val Leu Ile Pro Arg Ile Ala Arg Val
245 250 255
Cys Lys Gly Asp Gln Gly Gly Leu Arg Thr Leu Gln Lys Lys Trp Thr
260 265 270
Ser Phe Leu Lys Ala Arg Leu Ile Cys Ser Arg Pro Asp Ser Gly Leu
275 280 285
Val Phe Asn Val Leu Arg Asp Val Phe Val Leu Arg Ser Pro Gly Leu
290 295 300
Lys Val Pro Val Phe Tyr Ala Leu Phe Thr Pro Gln Leu Asn Asn Val
305 310 315 320
Gly Leu Ser Ala Val Cys Ala Tyr Asn Leu Ser Thr Ala Glu Glu Val
325 330 335
Phe Ser His Gly Lys Tyr Met Gln Ser Thr Thr Val Glu Gln Ser His
340 345 350
Thr Lys Trp Val Arg Tyr Asn Gly Pro Val Pro Lys Pro Arg Pro Gly
355 360 365
Ala Cys Ile Asp Ser Glu Ala Arg Ala Ala Asn Tyr Thr Ser Ser Leu
370 375 380
Asn Leu Pro Asp Lys Thr Leu Gln Phe Val Lys Asp His Pro Leu Met
385 390 395 400
Asp Asp Ser Val Thr Pro Ile Asp Asn Arg Pro Arg Leu Ile Lys Lys
405 410 415
Asp Val Asn Tyr Thr Gln Ile Val Val Asp Arg Thr Gln Ala Leu Asp
420 425 430
Gly Thr Val Tyr Asp Val Met Phe Val Ser Thr Asp Arg Gly Ala Leu
435 440 445
His Lys Ala Ile Ser Leu Glu His Ala Val His Ile Ile Glu Glu Thr
450 455 460
Gln Leu Phe Gln Asp Phe Glu Pro Val Gln Thr Leu Leu Leu Ser Ser
465 470 475 480
Lys Lys Gly Asn Arg Phe Val Tyr Ala Gly Ser Asn Ser Gly Val Val
485 490 495
Gln Ala Pro Leu Ala Phe Cys Gly Lys His Gly Thr Cys Glu Asp Cys
500 505 510
Val Leu Ala Arg Asp Pro Tyr Cys Ala Trp Ser Pro Pro Thr Ala Thr
515 520 525
Cys Val Ala Leu His Gln Thr Glu Ser Pro Ser Arg Gly Leu Ile Gln
530 535 540
Glu Met Ser Gly Asp Ala Ser Val Cys Pro Asp Lys Ser Lys Gly Ser
545 550 555 560
Tyr Arg Gln His Phe Phe Lys His Gly Gly Thr Ala Glu Leu Lys Cys
565 570 575
Ser Gln Lys Ser Asn Leu Ala Arg Val Phe Trp Lys Phe Gln Asn Gly
580 585 590
Val Leu Lys Ala Glu Ser Pro Lys Tyr Gly Leu Met Gly Arg Lys Asn
595 600 605
Leu Leu Ile Phe Asn Leu Ser Glu Gly Asp Ser Gly Val Tyr Gln Cys
610 615 620
Leu Ser Glu Glu Arg Val Lys Asn Lys Thr Val Phe Gln Val Val Ala
625 630 635 640
Lys His Val Leu Glu Val Lys Val Val Pro Lys Pro Val Val Ala Pro
645 650 655
Thr Leu Ser Val Val Gln Thr Glu Gly Ser Arg Ile Ala Thr Lys Val
660 665 670
Leu Val Ala Ser Thr Gln Gly Ser Ser Pro Pro Thr Pro Ala Val Gln
675 680 685
Ala Thr Ser Ser Gly Ala Ile Thr Leu Pro Pro Lys Pro Ala Pro Thr
690 695 700
Gly Thr Ser Cys Glu Pro Lys Ile Val Ile Asn Thr Val Pro Gln Leu
705 710 715 720
His Ser Glu Lys Thr Met Tyr Leu Lys Ser Ser Asp Asn Arg Leu Leu
725 730 735
Met Ser Leu Phe Leu Phe Phe Phe Val Leu Phe Leu Cys Leu Phe Phe
740 745 750
Tyr Asn Cys Tyr Lys Gly Tyr Leu Pro Arg Gln Cys Leu Lys Phe Arg
755 760 765
Ser Ala Leu Leu Ile Gly Lys Lys Lys Pro Lys Ser Asp Phe Cys Asp
770 775 780
Arg Glu Gln Ser Leu Lys Glu Thr Leu Val Glu Pro Gly Ser Phe Ser
785 790 795 800
Gln Gln Asn Gly Glu His Pro Lys Pro Ala Leu Asp Thr Gly Tyr Glu
805 810 815
Thr Glu Gln Asp Thr Ile Thr Ser Lys Val Pro Thr Asp Arg Glu Asp
820 825 830
Ser Gln Arg Ile Asp Asp Leu Ser Ala Arg Asp Lys Pro Phe Asp Val
835 840 845
Lys Cys Glu Leu Lys Phe Ala Asp Ser Asp Ala Asp Gly Asp
850 855 860
<210> 2
<211> 861
<212> Белок
<213> Murine sp.
<400> 2
Met Arg Met Cys Ala Pro Val Arg Gly Leu Phe Leu Ala Leu Val Val
1 5 10 15
Val Leu Arg Thr Ala Val Ala Phe Ala Pro Val Pro Arg Leu Thr Trp
20 25 30
Glu His Gly Glu Val Gly Leu Val Gln Phe His Lys Pro Gly Ile Phe
35 40 45
Asn Tyr Ser Ala Leu Leu Met Ser Glu Asp Lys Asp Thr Leu Tyr Val
50 55 60
Gly Ala Arg Glu Ala Val Phe Ala Val Asn Ala Leu Asn Ile Ser Glu
65 70 75 80
Lys Gln His Glu Val Tyr Trp Lys Val Ser Glu Asp Lys Lys Ser Lys
85 90 95
Cys Ala Glu Lys Gly Lys Ser Lys Gln Thr Glu Cys Leu Asn Tyr Ile
100 105 110
Arg Val Leu Gln Pro Leu Ser Ser Thr Ser Leu Tyr Val Cys Gly Thr
115 120 125
Asn Ala Phe Gln Pro Thr Cys Asp His Leu Asn Leu Thr Ser Phe Lys
130 135 140
Phe Leu Gly Lys Ser Glu Asp Gly Lys Gly Arg Cys Pro Phe Asp Pro
145 150 155 160
Ala His Ser Tyr Thr Ser Val Met Val Gly Gly Glu Leu Tyr Ser Gly
165 170 175
Thr Ser Tyr Asn Phe Leu Gly Ser Glu Pro Ile Ile Ser Arg Asn Ser
180 185 190
Ser His Ser Pro Leu Arg Thr Glu Tyr Ala Ile Pro Trp Leu Asn Glu
195 200 205
Pro Ser Phe Val Phe Ala Asp Val Ile Gln Lys Ser Pro Asp Gly Pro
210 215 220
Glu Gly Glu Asp Asp Lys Val Tyr Phe Phe Phe Thr Glu Val Ser Val
225 230 235 240
Glu Tyr Glu Phe Val Phe Lys Leu Met Ile Pro Arg Val Ala Arg Val
245 250 255
Cys Lys Gly Asp Gln Gly Gly Leu Arg Thr Leu Gln Lys Lys Trp Thr
260 265 270
Ser Phe Leu Lys Ala Arg Leu Ile Cys Ser Lys Pro Asp Ser Gly Leu
275 280 285
Val Phe Asn Ile Leu Gln Asp Val Phe Val Leu Arg Ala Pro Gly Leu
290 295 300
Lys Glu Pro Val Phe Tyr Ala Val Phe Thr Pro Gln Leu Asn Asn Val
305 310 315 320
Gly Leu Ser Ala Val Cys Ala Tyr Thr Leu Ala Thr Val Glu Ala Val
325 330 335
Phe Ser Arg Gly Lys Tyr Met Gln Ser Ala Thr Val Glu Gln Ser His
340 345 350
Thr Lys Trp Val Arg Tyr Asn Gly Pro Val Pro Thr Pro Arg Pro Gly
355 360 365
Ala Cys Ile Asp Ser Glu Ala Arg Ala Ala Asn Tyr Thr Ser Ser Leu
370 375 380
Asn Leu Pro Asp Lys Thr Leu Gln Phe Val Lys Asp His Pro Leu Met
385 390 395 400
Asp Asp Ser Val Thr Pro Ile Asp Asn Arg Pro Lys Leu Ile Lys Lys
405 410 415
Asp Val Asn Tyr Thr Gln Ile Val Val Asp Arg Thr Gln Ala Leu Asp
420 425 430
Gly Thr Phe Tyr Asp Val Met Phe Ile Ser Thr Asp Arg Gly Ala Leu
435 440 445
His Lys Ala Val Ile Leu Thr Lys Glu Val His Val Ile Glu Glu Thr
450 455 460
Gln Leu Phe Arg Asp Ser Glu Pro Val Leu Thr Leu Leu Leu Ser Ser
465 470 475 480
Lys Lys Gly Arg Lys Phe Val Tyr Ala Gly Ser Asn Ser Gly Val Val
485 490 495
Gln Ala Pro Leu Ala Phe Cys Glu Lys His Gly Ser Cys Glu Asp Cys
500 505 510
Val Leu Ala Arg Asp Pro Tyr Cys Ala Trp Ser Pro Ala Ile Lys Ala
515 520 525
Cys Val Thr Leu His Gln Glu Glu Ala Ser Ser Arg Gly Trp Ile Gln
530 535 540
Asp Met Ser Gly Asp Thr Ser Ser Cys Leu Asp Lys Ser Lys Glu Ser
545 550 555 560
Phe Asn Gln His Phe Phe Lys His Gly Gly Thr Ala Glu Leu Lys Cys
565 570 575
Phe Gln Lys Ser Asn Leu Ala Arg Val Val Trp Lys Phe Gln Asn Gly
580 585 590
Glu Leu Lys Ala Ala Ser Pro Lys Tyr Gly Phe Val Gly Arg Lys His
595 600 605
Leu Leu Ile Phe Asn Leu Ser Asp Gly Asp Ser Gly Val Tyr Gln Cys
610 615 620
Leu Ser Glu Glu Arg Val Arg Asn Lys Thr Val Ser Gln Leu Leu Ala
625 630 635 640
Lys His Val Leu Glu Val Lys Met Val Pro Arg Thr Pro Pro Ser Pro
645 650 655
Thr Ser Glu Asp Ala Gln Thr Glu Gly Ser Lys Ile Thr Ser Lys Met
660 665 670
Pro Val Ala Ser Thr Gln Gly Ser Ser Pro Pro Thr Pro Ala Leu Trp
675 680 685
Ala Thr Ser Pro Arg Ala Ala Thr Leu Pro Pro Lys Ser Ser Ser Gly
690 695 700
Thr Ser Cys Glu Pro Lys Met Val Ile Asn Thr Val Pro Gln Leu His
705 710 715 720
Ser Glu Lys Thr Val Tyr Leu Lys Ser Ser Asp Asn Arg Leu Leu Met
725 730 735
Ser Leu Leu Leu Phe Ile Phe Val Leu Phe Leu Cys Leu Phe Ser Tyr
740 745 750
Asn Cys Tyr Lys Gly Tyr Leu Pro Gly Gln Cys Leu Lys Phe Arg Ser
755 760 765
Ala Leu Leu Leu Gly Lys Lys Thr Pro Lys Ser Asp Phe Ser Asp Leu
770 775 780
Glu Gln Ser Val Lys Glu Thr Leu Val Glu Pro Gly Ser Phe Ser Gln
785 790 795 800
Gln Asn Gly Asp His Pro Lys Pro Ala Leu Asp Thr Gly Tyr Glu Thr
805 810 815
Glu Gln Asp Thr Ile Thr Ser Lys Val Pro Thr Asp Arg Glu Asp Ser
820 825 830
Gln Arg Ile Asp Glu Leu Ser Ala Arg Asp Lys Pro Phe Asp Val Lys
835 840 845
Cys Glu Leu Lys Phe Ala Asp Ser Asp Ala Asp Gly Asp
850 855 860
<210> 3
<211> 30
<212> ДНК
<213> Искусственная
<220>
<223> Полиниуклеотид CDR1 VH анти-CD100
<400> 3
ggctacagct tcagcgacta ctacatgcac 30
<210> 4
<211> 51
<212> ДНК
<213> Искусственная
<220>
<223> Полинуклеотид CDR2 VH анти-CD100
<400> 4
cagattaatc ctaccactgg cggcgctagc tacaaccaga agttcaaggg c 51
<210> 5
<211> 27
<212> ДНК
<213> Искусственная
<220>
<223> Полинуклеотид CDR3 VH анти-CD100
<400> 5
tattactacg gcagacactt cgatgtc 27
<210> 6
<211> 10
<212> Белок
<213> Искусственный
<220>
<223> Полипептид CDR1 VH анти-CD100
<400> 6
Gly Tyr Ser Phe Ser Asp Tyr Tyr Met His
1 5 10
<210> 7
<211> 17
<212> Белок
<213> Искусственный
<220>
<223> Полипептид CDR2 VH анти-CD100
<400> 7
Gln Ile Asn Pro Thr Thr Gly Gly Ala Ser Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 8
<211> 9
<212> Белок
<213> Искусственный
<220>
<223> Полипептид CDR3 VH анти-CD100
<400> 8
Tyr Tyr Tyr Gly Arg His Phe Asp Val
1 5
<210> 9
<211> 118
<212> Белок
<213> Искусственный
<220>
<223> Полипептид 2503 VH анти-CD100
<400> 9
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Ser Asp Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gln Ile Asn Pro Thr Thr Gly Gly Ala Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Ile Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Tyr Gly Arg His Phe Asp Val Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 10
<211> 118
<212> Белок
<213> Искусственный
<220>
<223> Полипептид 67 VH анти-CD100
<400> 10
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Ser Asp Tyr
20 25 30
Tyr Met His Trp Val Lys Gln Ser Pro Glu Asn Ser Leu Glu Trp Ile
35 40 45
Gly Gln Ile Asn Pro Thr Thr Gly Gly Ala Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Lys Ser Leu Thr Ser Glu Glu Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Tyr Tyr Tyr Gly Arg His Phe Asp Val Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 11
<211> 45
<212> ДНК
<213> Искусственная
<220>
<223> Полинуклеотид CDR1 VL анти-CD100
<400> 11
aaggccagcc aaagcgtgga ttatgatggc gatagctata tgaac 45
<210> 12
<211> 21
<212> ДНК
<213> Искусственная
<220>
<223> Полинуклеотид CDR2 VL анти-CD100
<400> 12
gctgcatcca atctggaaag c 21
<210> 13
<211> 27
<212> ДНК
<213> Искусственная
<220>
<223> Полинуклеотид CDR3 VL анти-CD100
<400> 13
cagcaaagca atgaggatcc ctacacc 27
<210> 14
<211> 15
<212> Белок
<213> Искусственнный
<220>
<223> Полипептид CDR1 VL анти-CD100
<400> 14
Lys Ala Ser Gln Ser Val Asp Tyr Asp Gly Asp Ser Tyr Met Asn
1 5 10 15
<210> 15
<211> 7
<212> Белок
<213> Искусственный
<220>
<223> Полипептид CDR2 анти-CD100
<400> 15
Ala Ala Ser Asn Leu Glu Ser
1 5
<210> 16
<211> 9
<212> Белок
<213> Искусственный
<220>
<223> Полипептид CDR3 VL анти-CD100
<400> 16
Gln Gln Ser Asn Glu Asp Pro Tyr Thr
1 5
<210> 17
<211> 111
<212> Белок
<213> Искусственный
<220>
<223> Полипептид VL 2503 анти-CD100
<400> 17
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 18
<211> 111
<212> Белок
<213> Искусственный
<220>
<223> Полипептид VL 67 анти-CD100
<400> 18
Asp Ile Val Met Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 19
<211> 354
<212> ДНК
<213> Искусственная
<220>
<223> Полиниуклеотид VH 2503 анти-CD100
<400> 19
caggtgcagc tggtgcagag cggcgctgag gtgaagaagc ctggcagcag cgtgaaggtc 60
tcctgcaagg ctagcggcta cagcttcagc gactactaca tgcactgggt gagacaggcc 120
cctggccaag gcctggagtg gatgggccag attaatccta ccactggcgg cgctagctac 180
aaccagaagt tcaagggcaa ggccaccatt accgtggaca aaagcaccag cacagcctac 240
atggagctga gcagcctgag aagcgaggac accgccgtgt attactgtgc cagatattac 300
tacggcagac acttcgatgt ctggggccaa ggcaccacgg tcaccgtctc ttca 354
<210> 20
<211> 354
<212> ДНК
<213> Искусственная
<220>
<223> Полинуклеотид VH 67 анти-CD100
<400> 20
caggtccagc tgcagcagtc tggacctgag ctggtgaagc ctggggcttc agtgaagata 60
tcctgcaagg cttctggtta ctcattcagt gactactaca tgcactgggt gaagcaaagt 120
cctgaaaata gtcttgagtg gattggacag attaatccta ccactggggg tgctagctac 180
aaccagaagt tcaagggcaa ggccacatta actgtagata aatcctccag cacagcctac 240
atgcagctca agagcctgac atctgaagag tctgcagtct attactgtac aagatattac 300
tacggtagac acttcgatgt ctggggccaa gggaccacgg tcaccgtttc ctca 354
<210> 21
<211> 333
<212> ДНК
<213> Искусственная
<220>
<223> полинуклеотид VL 2503 анти-CD100
<400> 21
gacatcgtga tgacccagag cccagacagc ctggctgtga gcctgggcga gagggccacc 60
atcaactgca aggccagcca aagcgtggat tatgatggcg atagctatat gaactggtac 120
cagcagaaac caggccagcc tcctaagctg ctgatttacg ctgcatccaa tctggaaagc 180
ggcgtgcctg acagattcag cggcagcggc agcggcacag atttcactct gaccatcagc 240
agcctgcagg ctgaagatgt ggcagtgtat tactgtcagc aaagcaatga ggatccctac 300
accttcggcc aagggaccaa gctcgagatc aaa 333
<210> 22
<211> 333
<212> ДНК
<213> Искусственная
<220>
<223> Полинуклеотид VL 67 анти-CD100
<400> 22
gacattgtga tgacccagtc tccagcttct ttggctgtgt ctctagggca gagggccacc 60
atctcctgca aggccagcca aagtgttgat tatgatggtg atagttatat gaactggtac 120
caacagaaac caggacagcc acccaaactc ctcatctatg ctgcatccaa tctagaatct 180
gggatcccag ccaggtttag tggcagtggg tctgggacag acttcaccct caacatccat 240
cctgtggagg aggaggatgc tgcaacctat tactgtcagc aaagtaatga ggatccgtac 300
acgttcggag gggggaccaa gctcgagatc aaa 333
<210> 23
<211> 2586
<212> ДНК
<213> Homo sapiens
<400> 23
atgaggatgt gcacccccat tagggggctg ctcatggccc ttgcagtgat gtttgggaca 60
gcgatggcat ttgcacccat accccggatc acctgggagc acagagaggt gcacctggtg 120
cagtttcatg agccagacat ctacaactac tcagccttgc tgctgagcga ggacaaggac 180
accttgtaca taggtgcccg ggaggcggtc ttcgctgtga acgcactcaa catctccgag 240
aagcagcatg aggtgtattg gaaggtctca gaagacaaaa aagcaaaatg tgcagaaaag 300
gggaaatcaa aacagacaga gtgcctcaac tacatccggg tgctgcagcc actcagcgcc 360
acttcccttt acgtgtgtgg gaccaacgca ttccagccgg cctgtgacca cctgaactta 420
acatccttta agtttctggg gaaaaatgaa gatggcaaag gaagatgtcc ctttgaccca 480
gcacacagct acacatccgt catggttgat ggagaacttt attcggggac gtcgtataat 540
tttttgggaa gtgaacccat catctcccga aattcttccc acagtcctct gaggacagaa 600
tatgcaatcc cttggctgaa cgagcctagt ttcgtgtttg ctgacgtgat ccgaaaaagc 660
ccagacagcc ccgacggcga ggatgacagg gtctacttct tcttcacgga ggtgtctgtg 720
gagtatgagt ttgtgttcag ggtgctgatc ccacggatag caagagtgtg caagggggac 780
cagggcggcc tgaggacctt gcagaagaaa tggacctcct tcctgaaagc ccgactcatc 840
tgctcccggc cagacagcgg cttggtcttc aatgtgctgc gggatgtctt cgtgctcagg 900
tccccgggcc tgaaggtgcc tgtgttctat gcactcttca ccccacagct gaacaacgtg 960
gggctgtcgg cagtgtgcgc ctacaacctg tccacagccg aggaggtctt ctcccacggg 1020
aagtacatgc agagcaccac agtggagcag tcccacacca agtgggtgcg ctataatggc 1080
ccggtaccca agccgcggcc tggagcgtgc atcgacagcg aggcacgggc cgccaactac 1140
accagctcct tgaatttgcc agacaagacg ctgcagttcg ttaaagacca ccctttgatg 1200
gatgactcgg taaccccaat agacaacagg cccaggttaa tcaagaaaga tgtgaactac 1260
acccagatcg tggtggaccg gacccaggcc ctggatggga ctgtctatga tgtcatgttt 1320
gtcagcacag accggggagc tctgcacaaa gccatcagcc tcgagcacgc tgttcacatc 1380
atcgaggaga cccagctctt ccaggacttt gagccagtcc agaccctgct gctgtcttca 1440
aagaagggca acaggtttgt ctatgctggc tctaactcgg gcgtggtcca ggccccgctg 1500
gccttctgtg ggaagcacgg cacctgcgag gactgtgtgc tggcgcggga cccctactgc 1560
gcctggagcc cgcccacagc gacctgcgtg gctctgcacc agaccgagag ccccagcagg 1620
ggtttgattc aggagatgag cggcgatgct tctgtgtgcc cggataaaag taaaggaagt 1680
taccggcagc attttttcaa gcacggtggc acagcggaac tgaaatgctc ccaaaaatcc 1740
aacctggccc gggtcttttg gaagttccag aatggcgtgt tgaaggccga gagccccaag 1800
tacggtctta tgggcagaaa aaacttgctc atcttcaact tgtcagaagg agacagtggg 1860
gtgtaccagt gcctgtcaga ggagagggtt aagaacaaaa cggtcttcca agtggtcgcc 1920
aagcacgtcc tggaagtgaa ggtggttcca aagcccgtag tggcccccac cttgtcagtt 1980
gttcagacag aaggtagtag gattgccacc aaagtgttgg tggcatccac ccaagggtct 2040
tctcccccaa ccccagccgt gcaggccacc tcctccgggg ccatcaccct tcctcccaag 2100
cctgcgccca ccggcacatc ctgcgaacca aagatcgtca tcaacacggt cccccagctc 2160
cactcggaga aaaccatgta tcttaagtcc agcgacaacc gcctcctcat gtccctcttc 2220
ctcttcttct ttgttctctt cctctgcctc tttttctaca actgctataa gggatacctg 2280
cccagacagt gcttgaaatt ccgctcggcc ctactaattg ggaagaagaa gcccaagtca 2340
gatttctgtg accgtgagca gagcctgaag gagacgttag tagagccagg gagcttctcc 2400
cagcagaatg gggagcaccc caagccagcc ctggacaccg gctatgagac cgagcaagac 2460
accatcacca gcaaagtccc cacggatagg gaggactcac agaggatcga cgacctttct 2520
gccagggaca agccctttga cgtcaagtgt gagctgaagt tcgctgactc agacgcagat 2580
ggagac 2586
<210> 24
<211> 13
<212> Белок
<213> Искусственный
<220>
<223> Пептидный эпитоп протеолипидного белка PLP(139-151)
<400> 24
His Ser Leu Gly Lys Trp Leu Gly His Pro Asp Lys Phe
1 5 10
<210> 25
<211> 118
<212> Белок
<213> Искусственный
<220>
<223> Полипептид VH 76 анти-CD100
<400> 25
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Trp Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Thr Gly Tyr Ser Asp Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Pro Tyr Gly Trp Thr Met Asp Ser Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 26
<211> 10
<212> Белок
<213> Искусственный
<220>
<223> Полипептид CDR1 VH 76 анти-CD100
<400> 26
Gly Tyr Thr Phe Thr Arg Tyr Trp Met His
1 5 10
<210> 27
<211> 17
<212> Белок
<213> Искусственный
<220>
<223> Полипептид CDR2 VH 76 анти-CD100
<400> 27
Tyr Ile Asn Pro Ser Thr Gly Tyr Ser Asp Tyr Asn Gln Lys Phe Lys
1 5 10 15
Asp
<210> 28
<211> 9
<212> Белок
<213> Искусственный
<220>
<223> Полипептид CDR3 VH 76 анти-CD100
<400> 28
Asp Pro Tyr Gly Trp Thr Met Asp Ser
1 5
<210> 29
<211> 107
<212> Белок
<213> Искусственный
<220>
<223> Полипептид VL 76 анти-CD100
<400> 29
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Thr Ile Thr Ile Thr Cys His Ala Ser Gln Asn Ile Asn Val Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Ile Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gly Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Gln Ser Tyr Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 30
<211> 11
<212> Белок
Искусственный
<220>
<223> Полипептид CDR1 VL 76 анти-CD100
<400> 30
His Ala Ser Gln Asn Ile Asn Val Trp Leu Ser
1 5 10
<210> 31
<211> 7
<212> Белок
<213> Искусственный
<220>
<223> Полипептид CDR2 VL 76 анти-CD100
<400> 31
Lys Ala Ser Asn Leu His Thr
1 5
<210> 32
<211> 9
<212> Белок
<213> Искусственный
<220>
<223> Полипептид CDR3 VL 76 анти-CD100
<400> 32
Gln Gln Gly Gln Ser Tyr Pro Tyr Thr
1 5
<210> 33
<211> 354
<212> ДНК
Искусственная
<220>
<223> Полинуклеотид VH 76 анти-CD100
<400> 33
caggtccagc tgcagcagtc tggggctgaa ctggcaaaac ctggggcctc agtgaagatg 60
tcctgcaagg cttctggcta cacctttact aggtactgga tgcactgggt aaaacagagg 120
cctggacagg gtctggaatg gattggatac attaatccta gcactggtta ttctgattac 180
aatcagaagt tcaaggacaa ggccacattg actgcagaca aatcctccag cacagcctac 240
atgcaactga gcagcctgac atctgaggac tctgcagtct attactgtgc aagagacccc 300
tacggctgga ctatggactc ctggggccaa gggactctgg tcaccgtctc ctca 354
<210> 34
<211> 30
<212> ДНК
<213> Искусственная
<220>
<223> Полинуклеттид CDR1 VH 76 анти-CD100
<400> 34
ggctacacct ttactaggta ctggatgcac 30
<210> 35
<211> 51
<212> ДНК
<213> Искусственная
<220>
<223> Полинуклеотид CDR2 VH 76 анти-CD100
<400> 35
tacattaatc ctagcactgg ttattctgat tacaatcaga agttcaagga c 51
<210> 36
<211> 27
<212> ДНК
<213> Искусственная
<220>
<223> Порлинуклеотид CDR3 VH 76 анти-CD100
<400> 36
gacccctacg gctggactat ggactcc 27
<210> 37
<211> 321
<212> ДНК
<213> Искусственная
<220>
<223> Полинуклеотид VL 76 анти-CD100
<400> 37
gacatccaga tgacccagtc tccatccagt ctgtctgcat cccttggaga cacaattacc 60
atcacttgcc atgccagtca gaacattaat gtttggttaa gctggtacca gcagaaacca 120
ggaaatattc ctaaactatt gatctataag gcttccaact tgcacacagg cgtcccatca 180
aggtttagtg gcagtggatc tggaacaggt ttcacattaa ccatcagcag cctgcagcct 240
gaagacattg ccacttacta ctgtcaacag ggtcaaagtt atccgtacac gttcggaggg 300
gggaccaagc tcgagatcaa a 321
<210> 38
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Полинуклеотид CDR1 VL 76 анти-CD100
<400> 38
catgccagtc agaacattaa tgtttggtta agc 33
<210> 39
<211> 21
<212> ДНК
<213> Искусственная
<220>
<223> Полинуклеотид CDR2 VL 76 анти-CD100
<400> 39
aaggcttcca acttgcacac a 21
<210> 40
<211> 27
<212> ДНК
<213> Искусственная
<220>
<223> Полинуклеотид CDR3 VL 76 анти-CD100
<400> 40
caacagggtc aaagttatcc gtacacg 27
<210> 41
<211> 51
<212> ДНК
<213> Искусственная
<220>
<223> Эпитоп 1
<400> 41
ctgaaggtgc ctgtgttcta tgcactcttc accccacagc tgaacaacgt g 51
<210> 42
<211> 17
<212> Белок
<213> Искусственный
<220>
<223> Эпитоп 1
<400> 42
Leu Lys Val Pro Val Phe Tyr Ala Leu Phe Thr Pro Gln Leu Asn Asn
1 5 10 15
Val
<210> 43
<211> 45
<212> ДНК
<213> Искусственная
<220>
<223> Эпитоп 2
<400> 43
aaatggacct ccttcctgaa agcccgactc atctgctccc ggcca 45
<210> 44
<211> 15
<212> Белок
<213> Искусственный
<220>
<223> Эпитоп 2
<400> 44
Lys Trp Thr Ser Phe Leu Lys Ala Arg Leu Ile Ala Ser Arg Pro
1 5 10 15
<210> 45
<211> 42
<212> ДНК
<213> Искусственная
<220>
<223> Эпитоп 3
<400> 45
gagtttgtgt tcagggtgct gatcccacgg atagcaagag tg 42
<210> 46
<211> 14
<212> Белок
<213> Искусственный
<220>
<223> Эпитоп 3
<400> 46
Glu Phe Val Phe Arg Val Leu Ile Pro Arg Ile Ala Arg Val
1 5 10
<210> 47
<211> 113
<212> Белок
<213> Искусственная последовательность
<220>
<223> Домен 2282 VL
<400> 47
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Ser Pro Gly
1 5 10 15
Glu Pro Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Phe Asn Ser
20 25 30
Gly Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Leu Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Gly Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Asn
85 90 95
Asp His Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile
100 105 110
<210> 48
<211> 122
<212> Белок
<213> Искусственная последовательность
<220>
<223> Домен 2282 VH
<400> 48
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Thr Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Val Asn Pro Tyr His Gly Tyr Ala Thr Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Glu Asn Ser Tyr Asp Gly Tyr Tyr Gly Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<---
Реферат
Изобретение относится к биотехнологии. Предложен рекомбинантный онколитический вирус простого герпеса (oHSV), содержащий не-HSV-лиганд, специфичный к молекуле (белковой, липидной или углеводной детерминанте), присутствующей на поверхности клетки (такой как раковая клетка), и одну или более копий одной или более последовательностей-мишеней микроРНК, встроенных в один или более локусов гена HSV, а предпочтительно, в один или более генов HSV, необходимых для репликации HSV в нормальных (то есть, не-раковых) клетках. Изобретение может быть использовано в медицине для уничтожения опухолевых клеток с использованием oHSV. 11 н. и 39 з.п. ф-лы, 10 ил., 1 табл., 2 пр.
Комментарии