Химерные аденовирусы для применения для лечения злокачественного новообразования - RU2448157C2
Код документа: RU2448157C2
Чертежи





















































Описание
Настоящее изобретение имеет приоритет предварительной заявки на патент US 60/574,851, поданной 26 мая 2004 года, которая полностью включена в настоящее описание в качестве ссылки.
Область техники, к которой относится изобретение
Изобретение, описанное в настоящей заявке, в целом относится к молекулярной биологии, в частности к онколитическим аденовирусам, имеющим терапевтические применения.
Предпосылки создания изобретения
Злокачественные новообразования являются основной причиной смертности как в США, так и в других странах мира. В зависимости от типа злокачественного новообразования, для лечения обычно применяют хирургические методы, химиотерапию и/или лучевую терапию. Часто эти методы являются неэффективными и, следовательно, существует необходимость новых видов лечения, которые могут использоваться отдельно или в комбинации с классическими методами.
Одним из подходов является применение аденовирусов, отдельно или в качестве векторов, способных доставлять противораковые терапевтические белки к опухолевым клеткам. Аденовирусы представляют собой безоболочечные икосаэдрические вирусы с двухцепочечной молекулой ДНК, линейный геном которых содержит около 36 тыс. пар оснований. На каждом конце вирусного генома присутствует короткая последовательность, известная как инвертированный концевой повтор (или ITR), которая необходима для репликации вируса. Геномы всех аденовирусов человека, исследованных к настоящему времени, имеют одинаковую общую организацию; то есть гены, кодирующие специфические функции, расположены в одном и том же положении вирусного генома. Вирусный геном содержит пять ранних единиц транскрипции (Е1А, Е1В, Е2, Е3 и Е4), две отсрочено-ранние единицы (IX и Iva2) и одну позднюю единицу (главную позднюю), что приводит к образованию пяти семейств поздних мРНК (L1-L5). Белки, кодируемые ранними генами, вовлечены в репликацию, тогда как поздние гены кодируют вирусные структурные белки. Части вирусного генома легко могут быть замещены с помощью ДНК чужеродного происхождения и такие рекомбинантные аденовирусы являются структурно стабильными, а свойства, приобретаемые такими вирусами, потенциально делают их пригодными для генной терапии (см. Jolly, D. (1994) Cancer Gene Therapy 1:51-64).
В настоящее время попытки исследователей получить аденовирусы, пригодные для лечения, сосредоточены на серотипе аденовируса Ad5. Генетика этого аденовируса человека хорошо изучена, также описаны системы для его молекулярной манипуляции. Для поддержки клинического применения разработаны высокоэффективные способы получения и доступные некоторые опыты использования этого агента в клинике (см. Jolly, D. (1994) Cancer Gene Therapy, 1:51-64). Исследования, относящиеся к применению аденовирусов человека (Ad) для лечения злокачественного новообразования, сконцентрированы на разработке аденовирусов на основе Ad5, которые являются особенно эффективными в или предпочтительно нацелены на специфические типы опухолевых клеток, и существует потребность получения более эффективных онколитических вирусов для того, чтобы лечение с помощью аденовирусов нашло практическое применение в клинических условиях.
Ad5 является только одним из 51 известных в настоящее время аденовирусных серотипов, которые подразделяются на подгруппы A-F на основе различных характерных признаков, включая их гемагглютинирующие свойства (см. Shenk, "Adenoviridae: The Viruses and Their Replication," в Fields Virology, том 2, 4-е издание, Knipe, ea., Lippincott, Williams & Wilkins, cc.2265-2267 (2001)). Эти серотипы отличаются на различных уровнях, например, патологией, вызываемой у людей и грызунов, клеточными рецепторами, используемыми для прикрепления, но эти отличия в значительной степени игнорируются в качестве эффективных средств развития более сильнодействующих онколитических аденовирусов (за исключением изменений волокон, см. Stevenson и др. (1997) J.Virol. 71:4782-4790; Krasnykh и др. (1996) J. Virol. 70:6839-6846; Wickham и др. (1997) J.Virol. 71:8221-8229; Legrand и др. (2002) Curr. Gene Ther. 2:323-329; Barnett и др. (2002) Biochim. Biophys. Acta 1-3:1-14; заявка на патент US 2003/0017138).
Использование различий между серотипами аденовирусов может обеспечивать источник более эффективных лекарственных средств на основе аденовирусов при использовании новых аденовирусов с повышенной селективностью и активностью. Существует потребность в таком улучшенном лечении на основе аденовирусов.
Сущность изобретения
Настоящее изобретение обеспечивает новые химерные аденовирусы или их варианты или производные, пригодные для лечения с использованием вирусов. В частности, изобретение обеспечивает химерные аденовирусы или их варианты или производные, имеющие геном, содержащий участок Е2В,
где указанный участок Е2В содержит нуклеотидную последовательность, имеющую происхождение от первого аденовирусного серотипа, и нуклеотидную последовательность, имеющую происхождение от второго аденовирусного серотипа;
где указанные первый и второй аденовирусные серотипы каждый выбирают из подгрупп аденовирусов В, С, D, Е или F и отличаются друг от друга; и где указанный химерный аденовирус является онколитическим и проявляет повышенный терапевтический индекс в опухолевой клетке.
В одном варианте осуществления изобретения, химерный аденовирус дополнительно содержит участки, кодирующие фибриллярные, гексоновые и пентоновые белки, где нуклеиновые кислоты, кодирующие указанные белки, все получены из одного и того же аденовирусного серотипа. В другом варианте осуществления изобретения, химерный аденовирус по изобретению содержит модифицированный участок Е3 или Е4.
В другом варианте осуществления изобретения, химерный аденовирус проявляет повышенный терапевтический индекс в опухолевых клетках ободочной кишки, молочной железы, поджелудочной железы, легких, предстательной железы, яичника или опухолевых кроветворных клетках. В более предпочтительном варианте осуществления изобретения, химерный аденовирус проявляет повышенный терапевтический индекс в опухолевых клетках ободочной кишки.
В предпочтительном варианте осуществления изобретения, участок Е2В химерного аденовируса содержит SEQ ID NO: 3. В более предпочтительном варианте осуществления изобретения, химерный аденовирус содержит SEQ ID NO: 1.
Настоящее изобретение обеспечивает рекомбинантный химерный аденовирус или его вариант или производное, имеющий геном, содержащий участок Е2В,
где указанный участок Е2В содержит нуклеотидные последовательности, имеющие происхождение от первого аденовирусного серотипа, и вторую нуклеотидную последовательность, имеющую происхождение от второго аденовирусного серотипа;
где указанные первый и второй аденовирусные серотипы каждый выбирают из подгрупп аденовирусов В, С, D, Е или F и отличаются друг от друга;
а указанный химерный аденовирус является онколитическим и проявляет повышенный терапевтический индекс в опухолевой клетке; и
где указанному химерному аденовирусу придавали недостаточность репликации путем удаления одного или более аденовирусных участков, кодирующих белки, вовлеченные в репликацию аденовируса, выбранные из группы, включающей Е1, Е2, Е3 или Е4.
В одном варианте осуществления изобретения, химерный аденовирус по изобретению дополнительно содержит гетерологичный ген, который кодирует терапевтический белок, где указанный гетерологичный ген экспрессируется в клетке, инфицированной указанным аденовирусом. В предпочтительном варианте осуществления изобретения, терапевтический белок выбирают из группы, включающей цитокины и хемокины, антитела, ферменты, превращающие пролекарства, и иммунорегуляторные белки.
Настоящее изобретение обеспечивает способы применения химерных аденовирусов по изобретению для терапевтических целей. В одном варианте осуществления изобретения, химерные аденовирусы могут применяться для ингибирования роста злокачественных клеток. В предпочтительном варианте осуществления изобретения, химерный аденовирус, содержащий SEQ ID NO: 1, пригодный для ингибирования роста злокачественных клеток ободочной кишки.
В другом варианте осуществления изобретения, аденовирусы по изобретению пригодны в качестве векторов для доставки терапевтических белков в клетки.
Настоящее изобретение обеспечивает способ получения химерных аденовирусов по изобретению, который предусматривает
а) объединение аденовирусных серотипов, представляющих подгруппы аденовирусов B-F, таким образом получая смесь аденовирусов;
б) пассирование объединенной смеси аденовирусов со стадии (а) в активно растущей культуре опухолевых клеток при соотношении частичек на клетку, достаточно большом для стимуляции рекомбинации между серотипами, но не настолько большом, чтобы вызывать преждевременную гибель клеток;
в) сбор супернатанта со стадии (б);
г) инфицирование покоящейся культуры опухолевых клеток супернатантом, собранным на стадии (в);
д) сбор супернатанта клеточной культуры со стадии (г) до появления любых признаков СРЕ;
е) инфицирование покоящейся культуры опухолевых клеток супернатантом, собранным на стадии (д); и
з) выделение химерного аденовируса из супернатанта, собранного на стадии (е), путем очистки бляшки.
Краткое описание фигур
Фигура 1. Профили времени удерживания Ad на ВЭЖХ-колонке ТМАЕ. А) профили удерживания для отдельных серотипов Ad, используемых для получения оригинального исходного вирусного пула. Б) профили удерживания пулов 20 пассажа, имеющих происхождение от клеточных линий НТ-29, Panc-1, MDA-231 и РС-3, соответственно.
Фигура 2. Цитолитическая активность отдельных вирусных пулов. А) НТ-29, Б) MDA-231, В) Panc-1 и Г) PC-3 клетки инфицировали их соответствующими вирусными пулами при VP на соотношение клеток от 100 до 0,01. В различные дни после инфицирования проводили MTS-исследования (как указано), в зависимости от клеточной линии. Каждая точка данных на панели представляет собой исследование, осуществленное в четырех повторах, результаты выражали в виде средних значений +/- СО. На панели приведен один характерный пример и все вирусные пулы исследовали по меньшей мере три независимых раза на целевой клеточной опухолевой линии (обозначения на фигуре:


Фигура 3. Цитолитическая активность ColoAd1 и Ad5 на клеточных линиях опухолей человека. MTS-анализ осуществляли на А) широкой панели клеточных линий опухолей человека и Б) на панели клеточных линий рака ободочной кишки человека для определения возможной специфической активности. MTS-анализ осуществляли в различные дни в зависимости от клеточной линии. Каждая панель представляет собой типичный эксперимент, который повторяли по меньшей мере три раза. Каждая точка данных на панели представляет собой исследование, осуществленное в четырех повторах, результаты выражали в виде средних значений +/- СО (обозначения на фигуре:
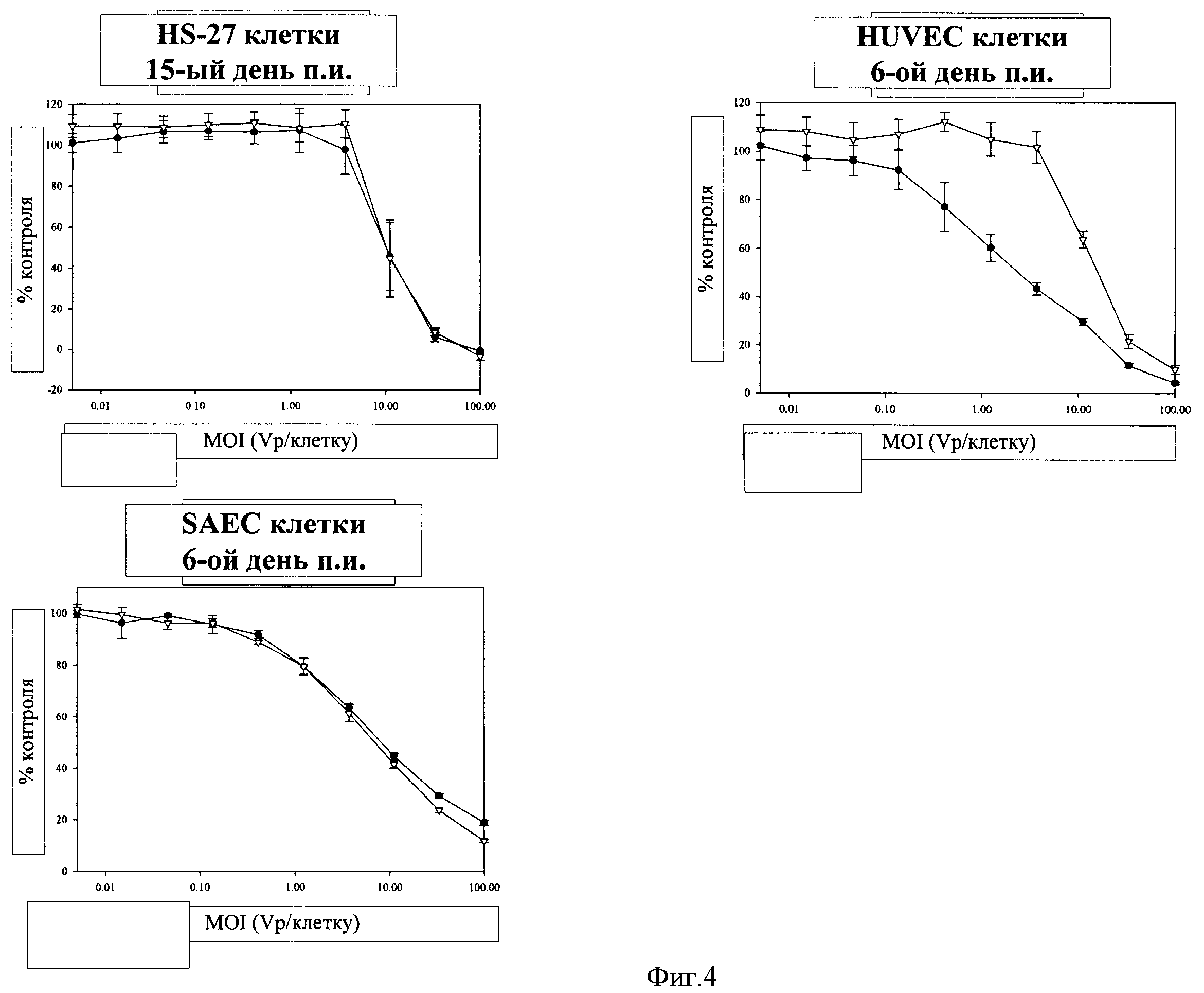
Фигура 4. Цитолитическая активность ColoAd1 и Ad5 на панели нормальных клеток. Клетки HS-27, HUVEC и SAEC (первичные фибробластные, эндотелиальные и эпителиальные клетки, соответственно) инфицировали ColoAd1 и Ad5 при VP на соотношение клеток от 100 до 0,01. MTS-анализ осуществляли в различные дни в зависимости от клетки и каждая панель представляет собой типичный эксперимент, который повторяли по меньшей мере три раза. Каждая точка данных на панели представляет собой исследование, осуществленное в четырех повторах, результаты выражали в виде средних значений +/- СО (обозначения на фигуре:
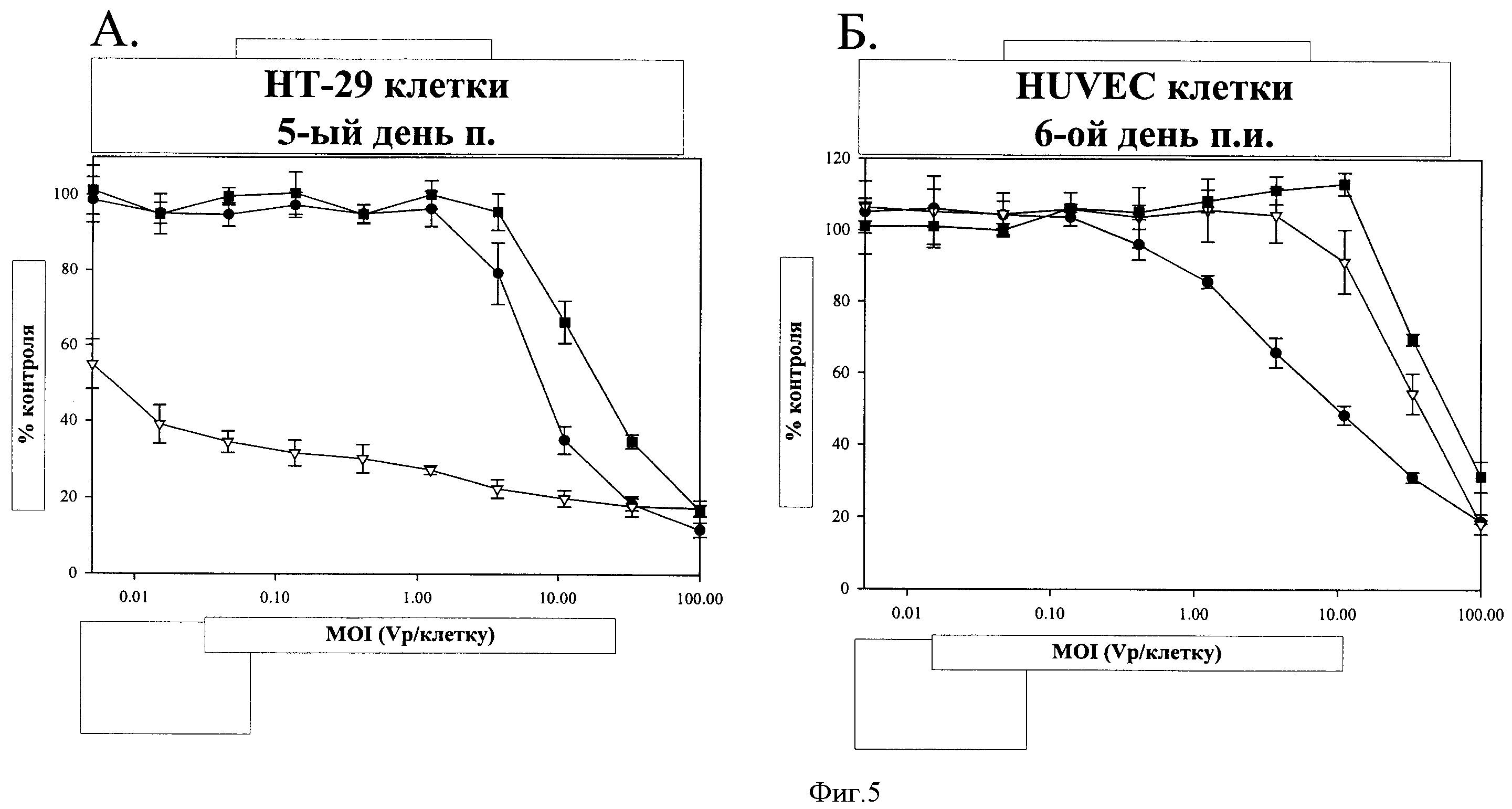
Фигура 5. Цитолитическая активность ColoAd1, Ad5 и ONYX-015 на первичных нормальных эндотелиальных клетках (HUVEC) и клеточной линии опухоли ободочной кишки (НТ-29). Каждая панель представляет собой типичный эксперимент, который повторяли по меньшей мере три раза. Каждая точка данных на панели представляет собой исследование, осуществленное в четырех повторах, результаты выражали в виде средних значений +/- СО (обозначения на фигуре:
Фигура 6. Цитолитическая активность ColoAd1, Ad11p и Ad5 на нормальной эпителиальной клеточной линии (SAEC) и клеточной линии рака ободочной кишки человека (НТ-29). Каждая панель представляет собой типичный эксперимент, который повторяли по меньшей мере три раза. Каждая точка данных на панели представляет собой исследование, осуществленное в четырех повторах, результаты выражали в виде средних значений +/- СО (обозначения на фигуре:
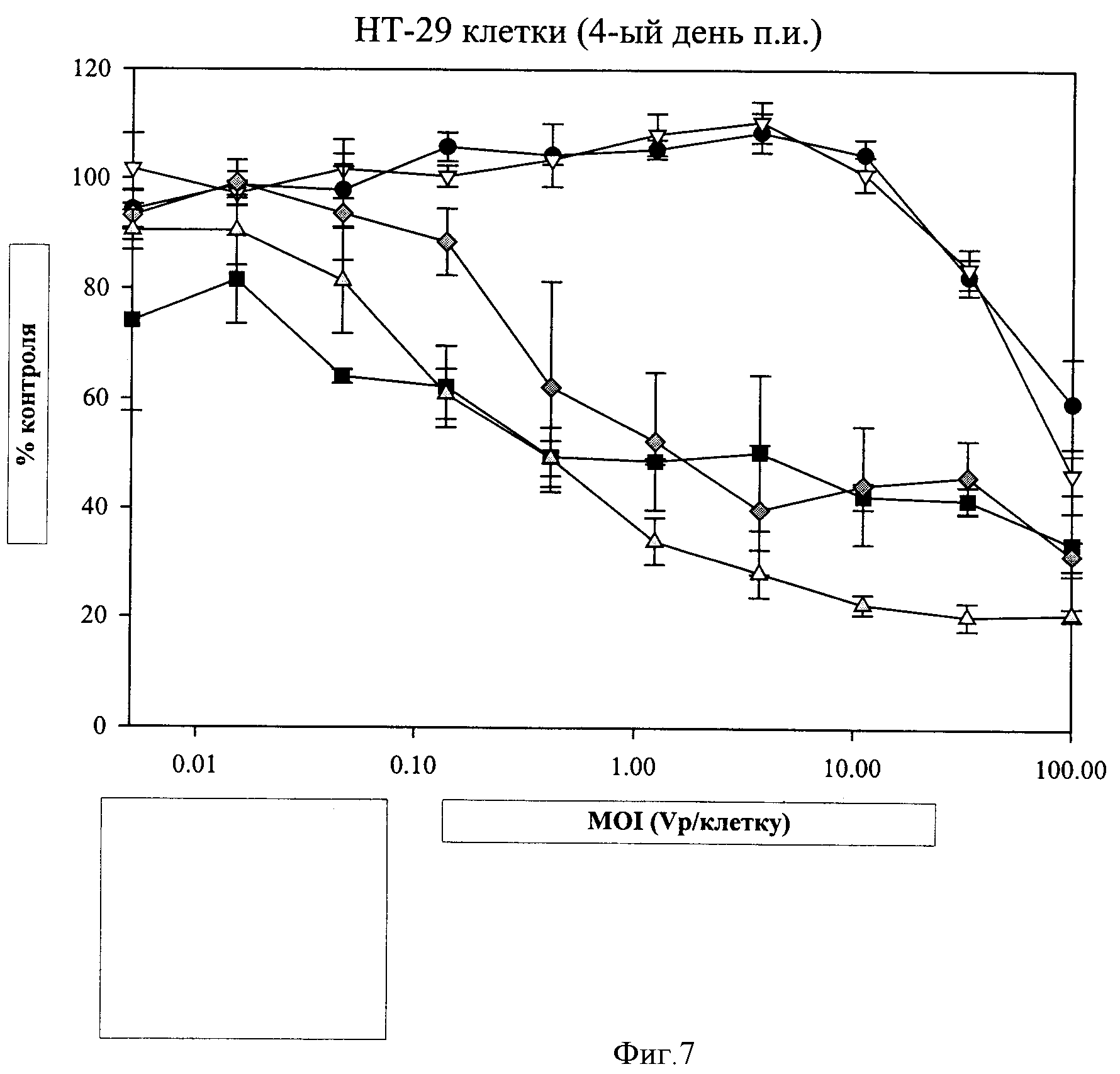
Фигура 7. Цитолитическая активность рекомбинантных вирусов. Конструировали рекомбинантные вирусы, представляющие четыре вирусные популяции (Adp11, ColoAd1, левый конец Ad11p/правый конец ColoAd1 (ColoAd1.1) и левый конец ColoAd1/правый конец Ad11p (ColoAd1.2)), как описано в примере 6. Определяли цитолитическую активность каждой популяции в НТ-29 клетках, как было описано ранее (обозначения на фигуре:

Подробное описание изобретения
Все публикации, включая патенты и патентные заявки, упомянутые в данном описании, включены в настоящее изобретение в качестве ссылки в таком объеме, если бы каждая отдельная публикация была специфически и отдельно полностью включена в качестве ссылки.
Определения
Если специально не указано иначе, то все технические и научные термины, используемые в настоящем изобретении, имеют такие же значения, что обычно подразумеваются средним специалистом в области техники, к которой относится данное изобретение. Как правило, номенклатура, используемая в настоящем изобретении, и лабораторные методики, описанные ниже, являются хорошо известными и обычно используемыми в данной области.
Как используется в настоящем изобретении, термин "аденовирус", "серотип" или "аденовирусный серотип" относится к любому из 51 аденовирусных серотипов человека, известных в настоящее время или выделенных в будущем (см., например, Strauss, "Adenovirus infections in humans," в The Adenoviruses, Ginsberg, ea., Plenum Press, Нью-Йорк, NY, cc.451-596 (1984)). Эти серотипы классифицируют на подгруппы A-F (см. Shenk, "Adenoviridae: The Viruses and Their Replication," в Fields Virology, том 2, 4-е издание, Knipe, ea., Lippincott Williams & Wilkins, cc.2265-2267 (2001)), как показано в таблице 1.
Как используется в настоящем изобретении, "химерный аденовирус" относится к аденовирусу, нуклеотидная последовательность которого содержит нуклеотидные последовательности по меньшей мере двух аденовирусных серотипов, описанных выше.
Как используется в настоящем изобретении, "родительский аденовирусный серотип" относится к аденовирусному серотипу, представляющему собой серотип, из которого имеет происхождение большинство геномов химерного аденовируса.
Как используется в настоящем изобретении, термин "гомологичная рекомбинация" относится к двум молекулам нуклеиновой кислоты, каждая имеющая гомологичные последовательности, где две молекулы нуклеиновой кислоты подвергаются кроссинговеру или рекомбинации в участке гомологии.
Как используется в настоящем изобретении, термин "эффективность" относится к литическому потенциалу вируса и характеризует его способность к репликации, лизису и распространению. Для целей настоящего изобретения, эффективность представляет собой величину, посредством которой сравнивается цитолитическая активность данного аденовируса по изобретению с таковой Ad5 в аналогичной клеточной линии, то есть эффективность = IC50 AdX/IC50 Ad5, где Х представляет собой определенный аденовирусный серотип, подвергаемый исследованию, и где для эффективности Ad5 заданным значением является 1.
Как используется в настоящем изобретении, термин "онколитический вирус" относится к вирусу, который предпочтительно убивает раковые клетки по сравнению с нормальными клетками.
Как используется в настоящем изобретении, термин "терапевтический индекс" или "терапевтическое окно" относится к числу, указывающему онколитический потенциал данного аденовируса, и определяется путем деления эффективности аденовируса в линии раковых клеток на эффективность того же самого аденовируса в обычной (то есть незлокачественной) клеточной линии.
Как используется в настоящем изобретении, термин "модифицированная" относится к молекуле нуклеотидной или аминокислотной последовательности, отличающейся от встречающейся в естественных условиях, например, дикого типа, нуклеотидной или аминокислотной последовательности. Модифицированная молекула может сохранять функцию или активность молекулы дикого типа, то есть модифицированный аденовирус может сохранять свою онколитическую активность. Модификации включают мутации нуклеиновых кислот, как описано ниже.
Как используется в настоящем изобретении, "мутация" по отношению к полинуклеотиду или полипептиду относится к встречающемуся в естественных условиях синтетическому, рекомбинантному или химическому изменению или отличию первичной, вторичной или третичной структуры полинуклеотида или полипептида по сравнению с полинуклеотидом или полипептидом соответственно, с которым проводят сравнение (например, по сравнению с полинуклеотидом или полипептидом дикого типа). Мутации включают такие изменения, как, например, делеции, вставки или замены. Полинуклеотиды и полипептиды, имеющие такие мутации, могут быть выделены или получены с помощью способов, хорошо известных в данной области.
Как используется в настоящем изобретении, "делеция" определяется как изменение в полинуклеотидных или в аминокислотных последовательностях, при котором один или несколько полинуклеотидов или аминокислотных остатков, соответственно, отсутствуют.
Как используется в настоящем изобретении, "инсерция" или "добавление" представляет собой такое изменение в полинуклеотидной или в аминокислотной последовательности, которое приводит к добавлению одного или нескольких полинуклеотидов или аминокислотных остатков, соответственно, по сравнению с встречающейся в естественных условиях полинуклеотидной или аминокислотной последовательностью.
Как используется в настоящем изобретении, "замена" является следствием замены одного(ой) или нескольких полинуклеотидов или аминокислот другими полинуклеотидами или аминокислотами, соответственно.
Как используется в настоящем изобретении, термин "производное аденовируса" относится к аденовирусу по изобретению, который модифицирован таким образом, что осуществлено добавление, делеция или замена к или в вирусном геноме, при этом полученное производное аденовируса проявляет эффективность и/или терапевтический индекс, превышающий таковой у родительского аденовируса, или в других отношениях является более терапевтически пригодным (то есть менее иммуногенно, с улучшенным профилем выведения). Например, производное аденовируса по изобретению может иметь делецию в одном из ранних генов вирусного генома, включая, но не ограничиваясь только ими, участок Е1А или Е2В вирусного генома.
Как используется в настоящем изобретении, "вариант" по отношению к полинуклеотиду или полипептиду относится к полинуклеотиду или полипептиду, которые могут отличаться по первичной, вторичной или третичной структуре, по сравнению с полинуклеотидом или полипептидом соответственно, с которым проводят сравнение (например, по сравнению с полинуклеотидом или полипептидом дикого типа). Например, аминокислотная или нуклеотидная последовательность может содержать мутацию или модификацию, которая отличается от аминокислотной или нуклеотидной последовательности, с которой проводят сравнение. В некоторых вариантах осуществления изобретения аденовирусный вариант может отличаться изоформой или полиморфизмом. Варианты могут представлять собой встречающиеся в естественных условиях, синтетические, рекомбинантные или химически модифицированные полинуклеотиды или полипептиды, выделенные или полученные с помощью способов, известных в данной области. Изменения в вариантах полинуклеотидной последовательности могут быть молчащими. Это происходит в том случае, если они не изменяют аминокислотную последовательность, кодируемую полинуклеотидом. Если вариации ограничены молчащими изменениями этого типа, то такой вариант будет кодировать полипептид, имеющий аминокислотную последовательность, идентичную последовательности, с которой проводят сравнение. Альтернативно, такие изменения в полинуклеотидной последовательности варианта могут изменять аминокислотную последовательность полипептида, кодируемого полинуклеотидом, с которым проводят сравнение, что приводит к консервативным или неконсервативным изменениям аминокислот, как описано ниже. Такие изменения в полинуклеотиде могут приводить к аминокислотным заменам, добавлениям, делециям, слияниям и усечениям аминокислот в полипептиде, кодируемым последовательностью, с которой проводят сравнение. Для оптимизации клонирования в плазмиде или вирусном векторе или экспрессирования в конкретной прокариотической или эукариотической системе могут осуществляться различные изменения кодонов, такие как молчащие изменения, образующие различные рестрикционные участки.
Как используется в настоящем изобретении, "аденовирусный вариант" относится к аденовирусу, полинуклеотидная последовательность которого отличается от полинуклеотида, с которым проводят сравнение, например аденовируса дикого типа, как описано выше. Отличия могут быть ограничены таким образом, что родительская полинуклеотидная последовательность и полинуклеотидная последовательность варианта являются в целом сходными и, в большинстве участков, идентичными. Как используется в настоящем изобретении, первая нуклеотидная или аминокислотная последовательность является "сходной" второй последовательности, если при сравнении двух последовательностей обнаружено, что они могут иметь несколько отличий в последовательности (то есть первая и вторая последовательности являются практически идентичными). Как используется в настоящем изобретении, различия полинуклеотидной последовательности у аденовирусного варианта и аденовируса, с которым проводят сравнение, не приводят к отличиям в эффективности и/или терапевтическом индексе.
Как используется в настоящем изобретении, термин "консервативный" относится к замене аминокислотного остатка на другой аминокислотный остаток, имеющий сходные химические свойства. Консервативные аминокислотные замены включают замену лейцина на изолейцин или валин, аспартата на глутамат или треонина на серин. Вставки или делеции обычно находятся в диапазоне приблизительно 1-5 аминокислот.
Как используется в настоящем изобретении, термин "неконсервативный" относится к замене аминокислотного остатка на другой аминокислотный остаток, который имеет другие химические свойства. Неконсервативные замены включают, но не ограничиваясь только ими, замену аспарагиновой кислоты (D) на глицин (G); аспарагина (N) на лизин (K) или аланина (А) на аргинин (R).
Однобуквенный код для аминокислотных остатков включает следующие обозначения: А = аланин, R = аргинин, N = аспарагин, D = аспарагиновая кислота, С = цистеин, Q = глутамин, Е = глутаминовая кислота, G = глицин, Н = гистидин, I = изолейцин, L = лейцин, K = лизин, М = метионин, F = фенилаланин, Р = пролин, S = серин, Т = треонин, W = триптофан, Y = тирозин, V = валин.
Следует принять во внимание, что полипептиды часто содержат аминокислоты, отличающиеся от тех 20-ти аминокислот, которые обычно обозначают как 20 аминокислот, встречающихся в естественных условиях, и что в данном полипептиде многие аминокислоты, включая концевые аминокислоты, могут быть модифицированы или в результате осуществления естественных процессов, таких как гликозилирование и другие посттрансляционные модификации, или с помощью методик химической модификации, которые хорошо известны в данной области техники. Даже общеизвестных модификаций, которые происходят в полипептидах, встречающихся в естественных условиях, очень много, и перечислить их здесь полностью невозможно, но они подробно описаны в основных пособиях и более подробных монографиях, а также в обширной исследовательской литературе, и они хорошо известны специалисту в данной области. Среди известных модификаций, которые могут присутствовать в полипептидах согласно настоящему изобретению, можно упомянуть, но только в качестве иллюстрации, ацетилирование, ацилирование, АДФ-рибозилирование, амидирование, ковалентное присоединение флавина, ковалентное присоединение молекулы гема, ковалентное присоединение полинуклеотида или производного полинуклеотида, ковалентное присоединение липида или производного липида, ковалентное присоединение фосфотидилинозитола, поперечное сшивание, циклизация, образование дисульфидной связи, деметилирование, образование ковалентных поперечных связей, образование цистина, образование пироглутамата, формилирование, гамма-карбоксилирование, глицирование, гликозилирование, образование GPI-якоря, гидроксилирование, йодирование, метилирование, миристоилирование, окисление, протеолитическая обработка, фосфорилирование, пренилирование, рацемизация, селеноирование, сульфатирование, опосредованное тРНК добавление аминокислот к белкам, такое как аргинилирование и убиквитинизация.
Такие модификации хорошо известны специалисту в данной области и очень подробно описаны в научной литературе. Некоторые предпочтительные распространенные модификации, гликозилирование, присоединение липидов, сульфатирование, гамма-карбоксилирование остатков глутаминовой кислоты, гидроксилирование и АДФ-рибозилирование, описаны, например, в большинстве основных пособий, таких как, например, I.E.Creighton, Proteins-Structure and Molecular Properties, 2-е изд., W.H.Freeman и Company, Нью-Йорк, 1993. По данному вопросу доступны различные подробные обзоры, такие как, например, подготовленные Wold, F., в Posttranslational Covalent Modification of Proteins, под ред. В.С.Johnson, Academic Press, New York, cc. 1-12, 1983; Seifter и др., Meth. Enzymol. 182: 626-646, 1990 и Rattan и др., Protein Synthesis: Posttranslational Modifications and Aging, Ann. N.Y. Acad. Sci. 663: 48-62, 1992.
Также следует принять во внимание, как хорошо известно и описано выше, что полипептиды не всегда являются полностью линейными. Например, полипептиды могут быть разветвленными вследствие убиквитинизации, и они могут быть кольцевыми с разветвлениями или без них, обычно вследствие пострансляционных воздействий, включая встречающиеся в естественных условиях воздействия и воздействия, осуществляемые людьми, которые не происходят в естественных условиях. Кольцевые, разветвленные и разветвленные кольцевые полипептиды могут быть синтезированы с помощью нетрансляционных естественных процессов, а также с помощью полностью синтетических способов.
Модификации могут осуществляться в любом участке полипептида, включая скелет пептида, боковые цепи аминокислот и амино- или карбоксильные концевые участки. В действительности наиболее частыми модификациями, которые обычно встречаются в естественных условиях или при синтезе полипептидов, являются блокирование аминогруппы или карбоксильной группы полипептида или обеих этих групп путем ковалентной модификации, и такие модификации могут присутствовать в полипептидах согласно настоящему изобретению. Например, аминоконцевой остаток полипептидов, полученных из E. coli, перед протеолитической обработкой практически всегда представляет собой N-формилметионин.
Модификации, которые происходят в полипептиде, очень часто будут зависеть от того, каким образом он был получен. Для полипептидов, получаемых путем экспрессирования клонируемого гена в организме-хозяине, например, природа и степень модификаций в основном будет определяться способностью клетки-хозяина к посттрансляционным модификациям и модификациями сигнальной последовательности, содержащейся в аминокислотной последовательности полипептида. Например, хорошо известно, что в бактериальных клетках-хозяевах, таких как Е. coli, гликозилирование очень часто не осуществляется. Следовательно, если желательно получить гликозилированный полипептид, то такой полипептид необходимо экспрессировать в хозяине, способном осуществлять гликозилирование, обычно в эукариотической клетке. Клетки насекомых очень часто обеспечивают такое же посттрансляционное гликозилирование, как и клетки млекопитающих, поэтому, в частности, для эффективной экспрессии белков млекопитающих, имеющих естественные схемы гликозилирования, развиваются экспрессионные системы на основе клеток насекомых. Аналогичные соображения применяются и для других модификаций.
Также следует принять во внимание, что аналогичный тип модификаций может иметь место в одинаковой или отличающейся степени в нескольких сайтах данного полипептида. Также данный полипептид может содержать модификации различных типов.
Как используется в настоящем изобретении, для описания соотношений последовательностей между двумя или более полинуклеотидными или аминокислотными последовательностями используют следующие термины: "последовательность, с которой проводят сравнение", "окно сравнения", "идентичность последовательностей", "процент идентичности последовательностей", "практически идентичная", "сходство" и "гомологичность". Под "последовательностью, с которой проводят сравнение" понимают последовательность, используемую как основу для сравниваемой последовательности; последовательность, с которой проводят сравнение, может представлять собой подмножество более длинной последовательности, например сегмент полноразмерной кДНК или последовательности гена, приведенных в перечне последовательностей, или она может включать полную кДНК или последовательность гена. Как правило, последовательность, с которой проводят сравнение, содержит по меньшей мере 18 нуклеотидов или 6 аминокислот, часто по меньшей мере 24 нуклеотида или 8 аминокислот, более предпочтительно 48 нуклеотидов или 16 аминокислот. Поскольку каждая из двух полинуклеотидных или аминокислотных последовательностей может содержать (1) последовательность (то есть часть полной полинуклеотидной или аминокислотной последовательности), которая является сходной для двух молекул, и (2) может содержать дополнительную последовательность, которая отличается у двух полинуклеотидных или аминокислотных последовательностей, то сравнивание последовательностей двух (или более) молекул обычно осуществляют путем сравнивания последовательностей двух молекул в "окне сравнения", идентифицируя и сравнивая локальные участки сходных последовательностей. Термин "окно сравнения", как он используется в настоящем описании, относится к абстрактному сегменту, включающему по меньшей мере 18 смежных нуклеотидов или 6 аминокислот, где полинуклеотидная последовательность или аминокислотная последовательность может быть сравнена с последовательностью, с которой проводят сравнение, состоящей по меньшей мере из 18 смежных нуклеотидов или 6 аминокислот, и где участок полинуклеотидной последовательности в окне сравнения может включать 20% или меньше вставок, делеций, замен и т.п. (то есть брешей), относительно последовательности, с которой проводят сравнение (которая не имеет вставок или делеций) для оптимального совмещения двух последовательностей. Оптимальное совмещение последовательностей для совмещения окна сравнения можно осуществлять, например, с помощью алгоритма локальной гомологии Smith и Waterman, Adv. Appl. Math. 2:482 (1981), с помощью алгоритма совпадения гомологии Needleman и Wunsch, J. Mol. Biol. 48:443 (1970), с помощью метода поиска сходства Pearson и Lipman, Proc. Natl. Acad. Sci. (USA) 85:2444 (1988), с помощью компьютерных реализаций этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA, входящих в пакет программ Wisconsin Genetics, версия 7,0 (Genetics Computer Group, 575 Science Dr., Мэдисон, Висконсин), VectorNTI от Informatix, Geneworks, или программного обеспечения MacVector) или путем проверки, и выбирают наилучшее совмещение (то есть дающее в результате наивысший процент гомологии в окне сравнения), полученное при осуществлении различных способов.
Как используется в настоящем изобретении, термин "идентичность последовательностей" обозначает, что две полинуклеотидные или аминокислотные последовательности являются идентичными (то есть по своим последовательно расположенным нуклеотидам или аминокислотным остаткам) в окне сравнения. "Процент идентичности последовательностей" рассчитывают путем сравнения двух оптимально совмещенных последовательностей в окне сравнения, определяя количество положений, в которых идентичные основания нуклеиновых кислот (например, А, Т, С, G, U или I) или аминокислотные остатки встречаются в обеих последовательностях, в результате чего получают количество совпадающих положений, деления количества совпавших положений на общее количество положений в окне сравнения (то есть на размер окна) и умножения результата на 100 для получения процента идентичности последовательностей. Термин "практически идентичная", как он используется в настоящем изобретении, относится к характеристике полинуклеотидной или аминокислотной последовательности и обозначает, что полинуклеотидная или аминокислотная последовательность имеет последовательность, которая по меньшей мере на 85%, предпочтительно по меньшей мере на 90-95%, более предпочтительно по меньшей мере на 99% идентична последовательности, с которой проводят сравнение, в окне сравнения, включающем по меньшей мере 18 положений нуклеотидов (6 аминокислот), часто в окне сравнения, включающем по меньшей мере 24-48 положений нуклеотидов (8-16 аминокислот), где процент идентичности последовательностей рассчитывают путем сравнивания последовательности, с которой проводят сравнения, с последовательностью, которая может включать делеции или вставки, составляющие в целом 20% или меньше последовательности, с которой проводят сравнение, в окне сравнения. Последовательность, с которой проводят сравнение, может представлять собой подмножество более длинной последовательности. Термин "сходство", когда он применяется для обозначения полипептида, определяется путем сравнивания аминокислотной последовательности и консервативных аминокислотных замен одного полипептида с последовательностью второго полипептида. Термин "гомологичный", когда он используется для описания полинуклеотида, указывает на то, что два полинуклеотида или их обозначенные последовательности при оптимальном совпадении и сравнении являются идентичными с соответствующими вставками или делециями нуклеотидов по меньшей мере для 70% нуклеотидов, обычно приблизительно для 75%-99% и наиболее предпочтительно по меньшей мере для 98-99% нуклеотидов.
Как используется в настоящем изобретении, "гомологичный", когда он используется для описания полинуклеотида, указывает на то, что два полинуклеотида или их обозначенные последовательности при оптимальном совпадении и сравнении являются идентичными с соответствующими вставками или делециями нуклеотидов по меньшей мере для 70% нуклеотидов, обычно приблизительно для 75%-99% и наиболее предпочтительно по меньшей мере для 98-99% нуклеотидов.
Как используется в настоящем изобретении, "полимеразная цепная реакция" или "ПЦР" относится к методике амплификации специфических участков ДНК, как описано в патенте US 4,683,195. Как правило, необходимо наличие информации о концевых последовательностях фрагмента полипептида, представляющего интерес, или о последовательностях, расположенных далее, с использованием таких сведений могут быть созданы олигонуклеотидные праймеры; затем эти праймеры направляют друг к другу и амплифицируют идентичные или подобные последовательности в противоположных цепях матрицы. 5'-Концевые нуклеотиды двух праймеров совмещают с концами амплифицируемого вещества. ПЦР может использоваться для амплификации специфических последовательностей ДНК из общей геномной ДНК, кДНК, транскрибируемой из общей клеточной РНК, плазмидных последовательностей и т.д. (более подробно см. Mullis и др., Cold Spring Harbor Symp. Quant. Biol., 51:263, 1987; под ред. Erlich, PCR Technology, Stockton Press, Нью-Йорк, 1989).
Как используется в настоящем изобретении, "строгость" обычно наблюдается в интервале приблизительно от Тпл (температура плавления) -5°С (5° ниже Тпл зонда) приблизительно до 20-25°С ниже Tm. Как очевидно для специалиста в данной области, строгая гибридизация может применяться для идентификации или определения идентичных полинуклеотидных последовательностей или для идентификации или определения сходных или родственных полинуклеотидных последовательностей. Как используется в настоящем изобретении, термин "строгие условия" обозначает, что гибридизация происходит только при наличии по меньшей мере 95% и предпочтительно по меньшей мере 97% идентичности между последовательностями.
Как используется в настоящем изобретении, термин "гибридизация", как он используется в настоящем изобретении, включает "любой процесс, с помощью которого полинуклеотидная цепь соединяется с комплементарной цепью путем спаривания оснований" (Coombs, J., Dictionary of Biotechnology, Stockton Press, Нью-Йорк, N.Y., 1994).
Как используется в настоящем изобретении, "терапевтически эффективная доза" или "эффективное количество" относится к количеству аденовируса, которое уменьшает интенсивность симптомов или облегчает болезненное состояние. Доза является терапевтически эффективной дозой для лечения рака или его метастаз, если рост опухоли или метастаз замедляется или останавливается, или если выявляется уменьшение размера опухоли или метастаз, что приводит к увеличению продолжительности жизни субъекта.
Аденовирусы по изобретению
Настоящее изобретение обеспечивает химерные аденовирусы или их варианты или производные, имеющие геном, в котором нуклеотидная последовательность участка Е2В химерного аденовируса содержит нуклеотидную последовательность, имеющую происхождение от по меньшей мере двух аденовирусных серотипов, каждый из которых выбирают из подгрупп аденовирусов В, С, D, Е и F и отличаются друг от друга. Химерный аденовирус по изобретению является онколитическим и проявляет повышенный терапевтический индекс в опухолевой клетке.
Выделение химерных аденовирусов
Химерные аденовирусы по изобретению или их варианты или производные могут быть получены с помощью модификации методики, на которую здесь ссылаются как "биоселекция", согласно которой аденовирус с требуемыми свойствами, такими как увеличенная онкогенность или специфичность к типу клеток, получают путем применения генетической селекции в контролируемых условиях (Yan и др. (2003) J. Virol. 77:2640-2650).
В настоящем изобретении, смесь аденовирусов различных серотипов объединяют и пассируют, предпочтительно по меньшей мере два раза, в субконфлюэнтной культуре опухолевых клеток при соотношении частичек на клетку, достаточно большом для стимуляции рекомбинации между серотипами, но не настолько большом, чтобы вызывать преждевременную гибель клеток. Предпочтительное соотношение частичек на клетку составляет приблизительно 500 частичек на клетку и легко может быть определено специалистом в данной области. Как используется в настоящем изобретении, "субконфлюэнтная культура" клеток относится к монослойной или суспензионной культуре, в которой клетки активно растут. Для роста клеток в виде монослоя примером может являться культура, в которой около 50-80% площади, доступной для роста клеток, покрыто клетками. Предпочтительной является культура, в которой около 75% ростовой площади покрыто клетками.
В предпочтительном варианте осуществления изобретения, смесью аденовирусов является смесь, которая включает аденовирусные серотипы, представляющие адновирусные подгруппы В, С, D, Е и F. Группа аденовирусов А не включена в смесь, так как они связаны с образованием опухолей у грызунов. Предпочтительными линиями опухолевых клеток, пригодными для биоселекции, являются, но не ограничиваясь только ими, те, которые имеют происхождение от молочной железы, ободочной кишки, поджелудочной железы, легкого и предстательной железы. Некоторыми примерами клеточных линий сóлидных опухолей, пригодных для "биоселективного" пассирования смеси аденовирусов, являются, но не ограничиваясь только ими, клетки MDA231, НТ-29, PAN-1 и РС-3. Гемопоэтические клеточные линии включают, но не ограничиваясь только ими, В-лимфоидные клетки Raji и Daudi, эритробластоидные клетки K562, миелоидные клетки U937 и Т-лимфоидные клетки HSB2.
Аденовирусы, получаемые при таких исходных пассажах, используют для инфицирования покоящихся опухолевых клеток при соотношении частичек на клетку ниже необходимого для возможности инфицирования клетки, но не больше чем один аденовирус. После осуществления вплоть до 20 пассажей в этих условиях супернатант из последнего пассажа собирают до видимого цитопатического действия (СРЕ, см. Fields Virology, том 2, четвертое изд., Knipe, ea., Lippincott Williams & Wilkins, cc.135-136) для повышения селекции сильнодействующих вирусов. Собранный супернатант может быть концентрирован с помощью методик, хорошо известных специалисту в данной области. Предпочтительным способом получения покоящихся клеток, то есть таких, у которых остановлен активный рост клеток, в монослойной культуре является предоставление возможности культуре расти в течение 3 дней с последующим слиянием, где слияние обозначает, что вся площадь, доступная для роста клеток, занята (покрыта клетками). Подобным образом суспензионные культуры могут расти до плотностей, характеризующих отсутствие активного роста клеток.
Профиль серотипа концентрированного супернатанта, который содержит биоселектируемый аденовирусный пул, можно исследовать путем измерения времени удерживания собранного вирусного пула на анионообменной колонке, где известно, что различные аденовирусные серотипы имеют характерные времена удерживания (Blanche и др. (2000) Gene Therapy 7:1055-1062); см. пример 3, фигуры 1А и В. Аденовирусы по изобретению могут быть выделены из концентрированного супернатанта путем разведения и очистки бляшек или других методик, хорошо известных в данной области, и роста для дальнейшей характеристики. Хорошо известные в данной области методики применяются для определения последовательностей выделенных химерных аденовирусов (см. пример 5).
Примером химерного аденовируса по изобретению является химерный аденовирус ColoAd1, который выделен с помощью клеток ободочной кишки НТ-29 в способе биоселекции. ColoAd1 имеет нуклеотидную последовательность SEQ ID NO: 1. Большая часть нуклеотидной последовательности ColoAd1 идентична нуклеотидной последовательности серотипа Ad11 (SEQ ID NO: 2) (Stone и др. (2003) Virology 309:152-165; Mei и др. (2003) J. Gen. Virology 84:2061-2071). В нуклеотидной последовательности ColoAd1 присутствуют две делеции по сравнению с Ad11, первая длиной 2444 пар оснований в пределах участка единицы транскрипции Е3 генома (пары оснований 27979-30423 SEQ ID NO: 2) и вторая, меньшая делеция, длиной 25 пар оснований (пары оснований 33164-33189 SEQ ID NO: 2), в пределах гена E4orf4. Участок единицы транскрипции Е2В (SEQ ID NO: 3) ColoAd1, который кодирует аденовирусные белки ДНК-полимеразу и терминальный белок, расположен между парами основ 5067 и 10354 SEQ ID NO: 1 и представляет собой область гомологичной рекомбинации между серотипами Ad11 и Ad3. В пределах этого участка ColoAd1 присутствует изменение 198 пар оснований по сравнению с последовательностью Ad11 (SEQ ID NO: 1). Изменения приводят к удлинению нуклеотидов в пределах участка Е2 В ColoAd1, которые являются гомологичными части участка Е2В Ad3 (SEQ ID NO: 8), с наибольшим удлинением гомологии между ColoAd1 и Ad3, имеющим длину 414 по. Участок Е2В ColoAd1 (SEQ ID NO: 3) предоставляет увеличенную эффективность аденовирусу ColoAd1 по сравнению с немодифицированным аденовирусом Ad11 (см. пример 6; фиг.7). В других вариантах осуществления изобретения, химерный аденовирус по изобретению может содержать нуклеотидные последовательности более чем из двух аденовирусных серотипов.
Химерный аденовирус по изобретению или его вариант или производное может быть исследован относительно его селективности в характерном типе опухоли путем оценки его литического потенциала на панели опухолевых клеток, имеющих происхождение от одной и той же ткани, где аденовирусный пул был подвержен исходному пассированию. Например, химерный аденовирус ColoAd1 (SEQ ID NO: 1), который исходно имел происхождение от аденовирусного пула, пассированного на клеточных линиях опухоли ободочной кишки НТ-29, повторно исследовали как на клетках НТ-29, так и на панели других клеточных линий, имеющих происхождение от опухоли ободочной кишки, включая DLD-1, LS174T, LS1034, SW403, НСТ116, SW48 и Colo320DM (см. фигуру 3Б). Любые доступные клеточные линии опухоли ободочной кишки равным образом могут быть пригодными для такой оценки. Аденовирусные клоны, выделенные из аденовирусных пулов, выбранных на клеточных линиях опухолей других типов, аналогично можно тестировать на подходящей панели опухолевых клеток, включая, но не ограничиваясь только ими, клеточные линии предстательной железы (например, клеточные линии DU145 и РС-3); клеточные линии поджелудочной железы (например, клеточная линия Panc-1); клеточные линии опухолей молочной железы (например, клеточная линия MDA231) и клеточные линии яичников (например, клеточная линия OVCAR-3). Другие доступные клеточные линии опухолей равным образом пригодны для выделения и идентификации аденовирусов по изобретению.
Химерные аденовирусы по изобретению имеют повышенный терапевтический индекс по сравнению с аденовирусными серотипами, производными которых они являются (см. фигуру 6, где сравнивается цитолитическая активность химерного аденовируса ColoAd1 с Ad11p).
Изобретение также охватывает химерные аденовирусы, которые сконструированы с помощью рекомбинантных методик, хорошо известных специалисту в данной области. Такие химерные аденовирусы содержат участок нуклеотидной последовательности, имеющий происхождение от одного аденовирусного серотипа, который встроен с помощью рекомбинантных методик в геном второго аденовирусного серотипа. Встроенная последовательность предоставляет родительскому аденовирусному серотипу свойство, например опухолевую специфичность или повышенную эффективность. Например, участок Е2В ColoAd1 (SEQ ID NO: 3) может быть встроен в геном Ad35 или Ad9.
Производные аденовирусов
Изобретение также охватывает химерный аденовирус по изобретению, который модифицирован для обеспечения других терапевтически пригодных химерных аденовирусов. Модификации включают, но не ограничиваясь только ими, те, которые описаны ниже.
Одной из модификаций является получение производных химерного аденовируса по изобретению с по существу отсутствующей способностью связывать р53, в результате мутации в аденовирусном гене, который кодирует белок Е1В-55K. В таких вирусах, как правило, удалены некоторые или все участки Е1В-55K (см. патент US 5,677,178). В патенте US 6,080,578 описаны, в частности, Ad5 мутанты, которые имеют делеции в участке белка Е1В-55K, отвечающего за связывание р53. Другими предпочтительными модификациями химерных аденовирусов согласно настоящему изобретению являются мутации в участке Е1А, как описано в патентах US 5,801,029 и 5,972,706. Эти типы модификаций обеспечивают производные химерных аденовирусов по изобретению с увеличенной селективностью к опухолевым клеткам.
Другим примером модификации, подпадающей под объем изобретения, является химерный аденовирус, который обладает повышенной степенью тканевой специфичности благодаря размещению репликации вируса под контролем тканеспецифичного промотора, как описано в патенте US 5,998,205. Репликация химерного аденовируса по изобретению также может находиться под контролем чувствительного элемента E2F, как описано в заявке на патент US 09/714,409. Эта модификация обеспечивает контролируемый механизм репликации вируса, основываясь на присутствии E2F, что приводит к повышенной специфичности к опухолевой ткани и отличается от контроля, осуществляемой тканеспецифичным промотором. В этих обоих вариантах осуществления изобретения тканеспецифичный промотор и чувствительный элемент E2F функционально связаны с аденовирусным геном, необходимым для репликации аденовируса.
Другой модификацией, подпадающей под объем изобретения, является применение химерного аденовируса по изобретению, например ColoAd1, в качестве основы получения новых аденовирусных векторов с недостаточностью репликации. Как описано в Lai и др. ((2002) DNA Cell Bio. 21:895-913), аденовирусные векторы с недостаточностью репликации могут использоваться для доставки и экспрессии терапевтических генов. Первое поколение (в котором удалены участки Е1 и Е3) и второе поколение (в котором дополнительно удален участок Е4) аденовирусных векторов, которые имеют происхождение от химерных аденовирусов по изобретению, оба обеспечиваются данным изобретением. Такие векторы легко получают с помощью методик, хорошо известных специалисту в данной области (см. Imperiale и Kochanek (2004) Curr. Top. Microbiol. Immunol. 273:335-357; Vogels и др. (2003) J. Virol. 77:8263-8271).
Дальнейшей модификацией, подпадающей под объем изобретения, является вставка гетерологичного гена, пригодного в качестве маркера или репортера для отслеживания эффективности инфицирования вирусом. Одним вариантом осуществления этого типа модификации является вставка гена тимидинкиназы (TK). Экспрессия TK в инфицированных клетках может использоваться для отслеживания уровня вирусов, оставшихся в клетках, после инфицирования вирусом с помощью радиоактивно меченных субстратов TK реакции (Sangro и др. (2002) Mol. Imaging Biol. 4:27-33).
Способы конструирования модифицированных химерных аденовирусов хорошо известны в данной области (см. Mittal, S.K. (1993) Virus Res. 28:67-90 и Hermiston, T. и др. (1999) Methods in Molecular Medicine: Adenovirus Methods and Protocols, под ред. W.S.M. Wold, Humana Press). В стандартных методиках используют методы рекомбинантной нуклеиновой кислоты, синтез полинуклеотидов и культивирование и трансформацию микроорганизмов (например, электропорацию, липофекцию). Обычно ферментативные реакции и этапы очистки осуществляют согласно рекомендациям производителя. Техники и методики осуществляют согласно методам, общепринятым в данной области, различным распространенным ссылкам (см. в основном Sambrook и др., Molecular Cloning: A Laboratory Manual, 2-е изд. (1989) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.), которые описаны в этом источнике. Номенклатура, используемая в настоящем описании, и лабораторные методики аналитической химии, химии органического синтеза и приготовления лекарственных средств являются хорошо известными и обычно используются в данной области.
Определение терапевтического потенциала
Химерные аденовирусы по изобретению или их варианты или производные могут быть оценены относительно их терапевтической полезности путем определения их литического потенциала на опухолевых клетках, имеющих происхождение от терапевтических мишеней, представляющих интерес. Линии опухолевых клеток, пригодные для тестирования таких аденовирусов, включают, но не ограничиваясь только ими, клеточные линии ободочной кишки, включая, но не ограничиваясь только ими, клеточные линии DLD-1, HCT116, НТ-29, LS1034 и SW48; клеточные линии предстательной железы, включая, но не ограничиваясь только ими, клеточные линии DU145 и РС-3; клеточные линии поджелудочной железы, включая, но не ограничиваясь только ими, клеточную линию Panc-1; клеточные линии опухолей молочной железы, включая, но не ограничиваясь только ими, клеточную линию MDA231, и клеточные линии яичника, включая, но не ограничиваясь только ими, клеточную линию OVCAR-3. Гемопоэтические клеточные линии включают, но не ограничиваясь только ими, В-лимфоидные клетки Raji и Daudi, эритробластоидные клетки K562, миелоидные клетки U937 и Т-лимфоидные клетки HSB2. Другие доступные клеточные линии опухолей равным образом пригодны для выделения и идентификации аденовирусов по изобретению для применения для лечения новообразования.
Цитолитическая активность аденовирусов по изобретению может быть определена в характерных опухолевых клеточных линиях и полученные при измерении данные превращены в эффективность, используя в качестве стандарта аденовирус, принадлежащий к подгруппе С, предпочтительно Ad5 (то есть данная эффективность равна 1). Предпочтительным способом определения цитолитической активности является MTS-анализ (см. пример 4, фигура 2).
Терапевтический индекс аденовируса по изобретению в конкретной опухолевой клеточной линии может быть определен путем сравнения эффективности данного аденовируса в опухолевой клеточной линии с эффективностью этого же аденовируса в незлокачественной клеточной линии. Предпочтительными незлокачественными клеточными линиями являются клетки SAEC (эпителиального происхождения) и клетки HUVEC (эндотелиального происхождения) (см. фигуру 4). Эти два типа клеток представляют нормальные клетки, имеющие происхождение от органов и сосудистой сети, соответственно, и характеризуют вероятные участки токсичности при осуществлении аденовирусной терапии, в зависимости от способа доставки аденовируса. Тем не менее, практическое осуществление по изобретению не ограничивается применением этих клеток, также можно использовать другие незлокачественные клеточные линии (например, В-клетки, Т-клетки, макрофаги, моноциты, фибробласты).
Химерные аденовирусы по изобретению дополнительно могут быть оценены относительно их способности нацеливать рост опухолевых клеток (то есть раковых) по их способности уменьшать опухолеобразование или нагрузку опухолевыми клетками у бесшерстых мышей, несущих трансплантированные опухолевые клетки, по сравнению с мышами, несущими опухолевые клетки в эквивалентной нагрузке, но не получающих лечения (см. пример 7).
Оценку аденовирусов по изобретению также можно осуществлять с помощью первичных эксплантатов опухолей человека (Lam и др. (2003) Cancer Gene Therapy; Grill и др. (2003) Mol. Therapy 6:609-614), которые обеспечивают тестируемые условия, присутствующие в опухоли, которые обычно не могут быть получены при исследовании ксенотрансплантированных опухолей.
Полезность для лечения
Настоящее изобретение обеспечивает применение химерных аденовирусов по изобретению для ингибирования роста опухолевых клеток, а также применение аденовирусных векторов, имеющих происхождение от этих химерных аденовирусов, для доставки терапевтически полезных белков, пригодных для лечения новообразований и других болезненных состояний.
Фармацевтические композиции и введение
Настоящее изобретение также относится к фармацевтическим композициям, которые содержат химерные аденовирусы по изобретению, включая их варианты и производные, приготовленные для терапевтического введения пациенту. Для терапевтического применения стерильную композицию, содержащую фармакологически эффективную дозу аденовируса, вводят человеку или в ветеринарии пациенту, кроме человека, для лечения, например, новообразования. Обычно композиция будет содержать около 1011 или больше частичек аденовируса в одной суспензии. Часто в таких стерильных условиях используют фармацевтически приемлемый носитель или наполнитель. Можно использовать различные водные растворы, например воду, буферизованную воду, 0,4% солевой раствор, 0,3% глицин и подобные. Эти растворы являются стерильными и практически свободны от других частичек, не представляющих собой требуемый аденовирусный вектор. Композиции могут содержать фармацевтически приемлемые вспомогательные вещества, необходимые для поддержания физиологических условий, такие как вещества для установления значения рН и буферные вещества, вещества, регулирующие токсичность, и подобные, например ацетат натрия, хлорид натрия, хлорид калия, хлорид кальция, лактат натрия и т.д. Можно включать наполнители, которые повышают инфицируемость клеток аденовирусом (см. патент US 6,392,069).
Аденовирусы по изобретению также могут доставляться к опухолевым клеткам с помощью липосом или иммунолипосом, такая доставка может быть селективно нацелена на опухолевые клетки на основе свойств клеточной поверхности, присутствующих в популяции опухолевых клеток (например, присутствие белка клеточной поверхности, который связывается с иммуноглобулином на липосоме). Обычно водную суспензию, содержащую вирионы, инкапсулируют в липосомы или иммунолипосомы. Например, суспензия аденовирусных вирионов может быть инкапсулирована в мицеллы с образованием липосом с помощью общепринятых способов (патент US 5,043,164, патент US 4,957,735, патент US 4,925,661; Connor и Huang (1985) J. Cell Biol. 101: 581; Lasic D.D. (1992) Nature 355: 279; Novel Drug Delivery (под ред. Prescott и Nimmo, Wiley, Нью-Йорк, 1989); Reddy и др. (1992) J. Immunol. 148:1585). Иммунолипосомы, содержащие антитело, которое специфически связывается с антигеном злокачественной клетки (например, CALLA, СЕА), находящимся на злокачественных клетках пациента, можно использовать для нацеливания вирионов на эти клетки (Fisher (2001) Gene Therapy 8:341-348).
Для дальнейшего повышения эффективности аденовирусов по изобретению их можно модифицировать для проявления повышенного тропизма к предпочтительным типам опухолевых клеток. Например, как показано в заявке PCT/US98/04964, протеин на наружной оболочке аденовируса можно модифицировать для выявления химического агента, предпочтительно полипептида, который связывается с рецептором, находящимся на опухолевых клетках, в большей степени, чем с нормальными клетками (см. также патенты US 5,770,442 и 5,712,136). Полипептид может представлять собой антитело, предпочтительным является одноцепочечное антитело.
Лечение с помощью аденовирусов
Аденовирусы по изобретению или их фармацевтические композиции могут вводиться для терапевтического лечения опухолевого заболевания или злокачественного новообразования. Для терапевтических показаний композиции вводят пациенту, у которого уже развилось опухолевое заболевание, в количестве, достаточном для лечения или по меньшей мере частично купирования состояния и его осложнений. Количество, достаточное для осуществления такого действия, определяется согласно настоящему изобретению как "терапевтически эффективная доза" или "действующая доза". Количество, эффективное для такого применения, будет зависеть от тяжести состояния, общего состояния пациента и пути введения.
В качестве примера, но не ограничиваясь только им, человека или млекопитающего, отличающегося от человека, имеющего сóлидное или гематологическое опухолевое заболевание (например, рак поджелудочной железы, ободочной кишки, яичников, легких или молочной железы, лейкоз или множественную миелому), можно подвергать лечению путем введения терапевтически эффективной дозы подходящего аденовируса по изобретению, то есть такого аденовируса, который проявляет улучшенный терапевтический индекс для этого типа ткани. Например, предпочтительным химерным аденовирусом для лечения рака ободочной кишки будет являться аденовирус ColoAd1 (SEQ ID NO: 1). Суспензии инфицирующих частиц аденовируса могут доставляться к опухолевой ткани различными путями, включая внутривенный, внутрибрюшинный, внутримышечный, подкожный и местный. Аденовирусная суспензия, содержащая около 103-1012 или больше частичек вириона на мл, может вводиться путем инфузии (например, в полость брюшины для лечения рака яичника, в воротную вену для лечения гепатокарциномы или метастаз в печени из других первичных опухолей непеченочного происхождения) или другим подходящим путем, включая прямую инъекцию в опухолевое образование (например, опухоль молочной железы), клизмы (например, рак ободочной кишки) или катетер (например, рак мочевого пузыря). Другие пути введения будут подходящими для карцином другого происхождения, то есть ингаляции в виде аэрозоля (например, для доставки в легкие при раке бронха, мелкоклеточном раке легкого, немелкоклеточном раке легкого, аденокарциноме легкого или раке гортани) или непосредственного введения в опухолевый участок (например, рак бронха, рак носоглотки, рак гортани, рак шейки матки).
Аденовирусную терапию с применением аденовирусов по настоящему изобретению можно комбинировать с другими противоопухолевыми протоколами, такими как общепринятая химиотерапия или лучевая терапия, для лечения конкретного злокачественного новообразования. Лечение может быть одновременным или последовательным. Предпочтительным химиотерапевтическим средством является цисплатин, и предпочтительная доза может быть выбрана лечащим врачом, исходя из природы злокачественного новообразования, подвергаемого лечению, и других факторов, которые обычно принимают во внимание при назначении цисплатина. Предпочтительно, цисплатин вводят внутривенно в дозе 50-120 мг/м2 в течение 3-6 часов. Более предпочтительно его вводят внутривенно в дозе 80 мг/м2 в течение 4 часов. Другим предпочтительным химиотерапевтическим средством является 5-фторурацил, который часто вводят в комбинации с цисплатином. Предпочтительная доза 5-фторурацила составляет 800-1200 мг/м2 в сутки в течение 5 последовательных дней.
Аденовирусную терапию с применением аденовирусов по настоящему изобретению в качестве аденовирусных векторов также можно комбинировать с другими генами, для которых известна полезность для лечения при генной терапии (см. патент US 5,648,478). В таких случаях, химерный аденовирус дополнительно содержит гетерологичный ген, который кодирует терапевтический белок, встроенный в вирусный геном, таким образом, что гетерологичный ген экспрессируется в инфицированной клетке. Терапевтический белок, как используется в настоящем изобретении, относится к белку, который, как предполагают, будет обеспечивать некоторые терапевтические преимущества при экспрессии в данной клетке.
В одном варианте осуществления изобретения, гетерологичный ген представляет собой пролекарство гена-активатора, такое как дезаминаза цитозина (CD) (см. патенты US 5,631,236; 5,358,866 и 5,677,178). В других вариантах осуществления изобретения, гетерологичный ген известен в качестве индуктора клеточной гибели, например апоптин или аденовирусный протеин гибели (ADP), или слитый белок, например сливающийся мембранный гликопротеин (Danen-Van Oorschot и др. (1997) Proc. Nat. Acad. Sci. 94:5843-5847; Tollefson и др. (1996) J. Virol. 70:2296-2306; Fu и др. (2003) Mol. Therapy 7: 48-754, 2003; Ahmed и др. (2003) Gene Therapy 10:1663-1671; Galanis и др. (2001) Human Gene Therapy 12(7): 811-821).
Дальнейшими примерами гетерологичных генов или их фрагментов являются те, которые кодируют иммуномодулирующие белки, такие как цитокины или хемокины. Примерами являются интерлейкин 2, патенты US 4,738,927 или 5,641,665; интерлейкин 7, патенты US 4,965,195 или 5,328,988; и интерлейкин 12, патент US 5,457,038; фактор некроза опухоли альфа, патент US 4,677,063 или 5,773,582; гамма-интерферон, патент US 4,727,138 или 4,762,791; или GM CSF, патент US 5,393,870 или 5,391,485, Mackensen и др. (1997) Cytokine Growth Factor Rev. 8:119-128. Дополнительные иммуномодуляторные белки, кроме того, включают макрофаговые воспалительные белки, включая MIP- 3. Также можно использовать моноцитный хемотоксический белок (МСР-3 альфа); предпочтительным вариантом осуществления гетерологичного гена является химерный ген, состоящий из гена, который кодирует белок, проходящий клеточные мембраны, например VP22 или ТАТ, слитого с геном, который кодирует белок, предпочтительно токсичный по отношению к раковым, но не нормальным клеткам.
Химерные аденовирусы по изобретению также можно использовать в качестве векторов для доставки генов, кодирующих терапевтически пригодные молекулы РНК, то есть миРНК (Dorsett и Tuschi (2004) Nature Rev Drug Disc 3:318-329).
В некоторых случаях, гены можно встраивать в химерный аденовирус по изобретению для дополнительного усиления способности онколитического вируса уничтожать опухоль, хотя он не обладает прямым воздействием на саму опухоль - такой вариант включает гены, кодирующие белки, которые нарушают презентацию МНС I класса (Hewitt и др. (2003,) Immunology 110: 163-169), блокируют комплемент, ингибируют интерфероны и механизмы, индуцированные интерфероном, хемокины и цитокины, гибель на основе NK клеток (Orange и др. (2002) Nature Immunol. 3: 1006-1012; Mireille и др. (2002) Immunogenetics 54: 527-542; Alcami (2003) Nature Rev. Immunol. 3: 36-50); понижают ответную иммунную реакцию (например, IL-10, TGF-бета, Khong и Restifo (2002) Nature Immunol. 3: 999-1005; 2002), и металлопротеазы, которые могут разрушать внеклеточный матрикс и усиливать распространение вирусов в опухоли (Bosman и Stamenkovic (2003) J. Pathol. 2000: 423-428; Visse и Nagase (2003) Circulation Res. 92: 827-839).
Наборы
Изобретение также относится к фармацевтическим упаковкам и наборам, которые состоят из одного или нескольких контейнеров, заполненных одним или несколькими компонентами указанных выше композиций, предлагаемых в изобретении. Такой(ие) контейнер(ы) должен(ны) быть снабжен(ы) вкладышем в форме, установленной правительственным органом, ответственным за производство, применение или продажу фармацевтических или биологических продуктов, в котором указано разрешение этого органа на производство, применение или продажу продукта для введения человеку.
В дальнейшем настоящее изобретение описано с помощью последующих примеров, которые иллюстрируют специфические варианты осуществления изобретения и их различные применения. Эти приведенные примеры, иллюстрирующие отдельные специфические варианты осуществления изобретения, не могут рассматриваться как такие, которые каким-либо образом ограничивают объем раскрытого изобретения.
Если специально не указано иначе, при практическом осуществлении изобретения используют общепринятые техники культуры клеток, молекулярной биологии, микробиологии, методики рекомбинантной ДНК, иммунологии, которые находятся в компетенции среднего специалиста в данной области. Такие методики подробно поясняются в литературе. См., например, Cell Biology: a Laboratory Handbook: J. Celis (ред.). Academic Press. N.Y. (1996); Graham, F.L. и Prevec, L. Adenovirus-based expression vectors and recombinant vaccines. B: Vaccines: New Approaches to Immunological Problems. R.W.Ellis (ред.) Butterworth. cc. 363-390; Grahan and Prevec Manipulation ofadenovirus vectors. B: Methods in Molecular Biology, том 7: Gene Transfer and Expression Techniques. E.J. Murray и J.M. Walker (ред.) Humana Press Inc., Clifton, N.J. cc. 109-128, 1991; Sambrook и др. (1989), Molecular Cloning, A Laboratory Manual, 2-е изд., Cold Spring Harbor Laboratory Press; Sambrook и др. (1989), и Ausubel и др. (1995), Short Protocols in Molecular Biology, John Wiley and Sons.
Примеры
Методы
Стандартные методики используются для методов рекомбинантной нуклеиновой кислоты, синтеза полинуклеотидов и культивирования и трансформации микроорганизмов (например, электропорация, липофекция). Обычно ферментативные реакции и этапы очистки осуществляют согласно рекомендациям производителя. Техники и методики осуществляют согласно методам, общепринятым в данной области, различным распространенным ссылкам (см., в основном, Sambrook и др., Molecular Cloning: A Laboratory Manual, 2-е издание (1989) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.), которые описаны в этом источнике. Номенклатура, используемая в настоящем описании, и лабораторные методики аналитической химии, химии органического синтеза и приготовления лекарственных средств и доставки и лечения пациентов. Способы конструирования мутантов аденовирусов хорошо известны в данной области. See, Mittal, S.K., Virus Res.,1993, том 28, cc.67-90; и Hermiston, Т. и др., Methods in Molecular Medicine: Adenovirus Methods and Protocols, под ред. W.S.M. Wold, Humana Press, 1999. Кроме того, геном аденовируса 5 зарегистрирован в банке генов 10 под №М73260, и вирус доступный из Американской коллекции типичных культур, Rockville, Maryland, USA, под регистрационным номером VR-5.
Вирусы и клеточные линии
Серотипы Ad Ad3 (штамм GB), Ad4 (штамм RI-67), Ad5 (штамм Adenoid 75), Ad9 (штамм Hicks), Ad11p (штамм Slobitski), Ad16 (штамм Ch. 79) и все клеточные линии, за исключением следующих, получали из АТСС: MDA231-mt1 (производное, выделенное Dr. Deb Zajchowski из быстро растущих подкожно ксенотрансплантированных клеток MDA231) и Panc 1-sct (полученных Dr. Sandra Biroc из быстро растущих подкожно ксенотрансплантированных клеток Panc 1), HUVEC (Vec Technologies, Rensselaer, NY), и SAEC (Clonetics, Walkersville, MD). Ad40 был любезно предоставлен Dr. William S.M. Wold из Сент-Луисского университета.
Пример 1 - Очистка и количественный анализ вирусов
Маточные вирусные растворы размножали на клетках 293 и очищали на градиентах CsCl (Hawkins и др., 2001). Способ, используемый для количественного анализа вирусов, основывался на способе Shabram и др. (1997) Human Gene Therapy 8:453-465, за исключением того, что анионообменный ТМАЕ Fractogel использовали вместо Resource Q. Вкратце, колонку 1,25 л загружали Fractogel EMD ТМАЕ-650 (S) (кат. №116887-7 ЕМ Science, Gibbstown, NJ 08027). Осуществляли ВЭЖХ-разделение на Agilent HP 1100 ВЭЖХ в следующих условиях: буфер А=50 мМ HEPES, рН 7,5; буфер В=1,0 М NaCl в буфере А; скорость потока 1 мл/мин. После уравновешивания колонки в течение не меньше 30 минут в буфере А приблизительно 109-1011 вирусных частиц образца загружали на колонку в 10-100 мкл объема, затем загружали 4 объема колонки буфером А. Использовали линейный градиент, простирающийся на 16 объемов колонки, и завершали 100% буфером В.
За вытекающими потоками из колонки наблюдали при А260 и А280 нм, площади пиков подсчитывали и определяли соотношение 260 к 280 нм. Вирусные пики идентифицировали как пики, имеющие соотношение А260/А280, близкое к 1,33. В каждую серию образцов включали вирусный стандарт. Количество вирусных частиц на мл стандарта определяли с помощью способа Lehmberg и др. (1999) J. Chrom. В, 732:411-423. В диапазоне используемых вирусных концентраций, площадь пика при А260 нм каждого образца была прямо пропорционально количеству вирусных частиц в образце. Количество вирусных частиц на мл в каждом тестируемом образце рассчитывали путем умножения известного количества вирусных частиц на мл в стандарте на соотношение площади вирусного пика образца при А260 нм к площади вирусного пика стандарта при А260 нм.
Колонку регенерировали после градиента каждого образца путем промывания двумя объемами колонки 0,5 н. NaOH, затем двумя объемами колонки 100% буфером А, 3 объемами колонки 100% буфером В и после этого 4 объемами колонки 100% буфером А.
Пример 2 - Биоселекция
Вирусные серотипы, представляющие подгруппы Ads B-F, объединяли и пассировали на субконфлюэнтных культурах целевых опухолевых клеточных линий при высоком соотношении частичек на клетку в течение двух циклов для способствования осуществления рекомбинации между серотипами. Затем супернатант (1,0; 0,1; 0,01; 0,001 мл) из второго цикла инфицирования при высоком соотношении вирусных частиц на клетку, субконфлюэнтные культуры, использовали для инфицирования серий сверхконфлюэнтных колб опухолевой культуры Т-75 целевых опухолевых клеточных линий РС-3, НТ-29, Panc-1 и MDA-231. Для достижения сверхконфлюэнтности каждую клеточную линию высевали при соотношениях расщепления, позволяющих достичь конфлюэнтности клеточными линиями в течение 24-40 часов после высевания, и клетки оставляли расти в целом в течение 72 часов после высевания перед инфицированием. Это осуществляли для максимизирования конфлюэнтности клеток для воспроизведения условий роста сóлидных опухолей у людей.
Супернатант клеточной культуры собирали из первой колбы в сериях с 10-кратным разведением, которые не проявляли никаких признаков СРЕ на 3-й или 4-й день после инфицирования (в случае НТ-29 и PC-3, их модифицировали для пассажей 10-20 для сбора из второй колбы, то есть собирали при 100-кратном разведении ниже разведения, при котором обнаруживается СРЕ на 3-й день после инфицирования). Каждый сбор служил в качестве исходного материала для последующего пассажа вируса. Этот процесс повторяли до тех пор, пока вирусный пул не достигал 20 биоселективных пассажей.
Характерные вирусы для каждого биоселектируемого пула выделяли путем осуществления очистки бляшек в два этапа на клетках А549 с помощью стандартных способов (Tollefson, A., Hermiston, T.W., и Wold, W.S.M.; "Preparation and Titration of CsCl-banded Adenovirus Stock" в Adenovirus Methods and Protocols, Humana Press, 1999, cc.1-10, под ред. W.S.M. Wold). Вкратце, для инфицирования клеток А549 использовали разведения супернатанта, собранного из 20-го пассажа, на каждой целевой опухолевой клеточной линии в стандартном анализе бляшкообразования. Хорошо видимые бляшки собирали и аналогичный метод бляшкообразования использовали для получения второго цикла характерных бляшек с этих сборов. Тщательно выделенные бляшки из второго цикла очистки бляшек считали чистыми, инфицированные культуры готовили с помощью этих очищенных бляшек и определяли онколитическую эффективность этих культуральных супернатантов с помощью MTS-анализа, как описано.
Пример 3 - Характеристика серотипа
Родительские аденовирусные серотипы, содержащие вирусные пулы или выделенный аденовирус ColoAd1, идентифицировали с помощью анионообменной хроматографии аналогично описанному в Shabram и др. (1997) Human Gene Therapy 8:453:465, за исключением того, что использовали анионообменную среду ТМАЕ Fractogel (ЕМ Industries, Gibbstown, NJ) вместо Resource Q, как описано в примере 1 (см. фигуру 1).
Аденовирус 5 типа элюировался приблизительно при 60% буфере В при градиенте. Другие серотипы (3, 4, 9, 11p, 16, 35 и 40) каждый элюировался при характерном времени удерживания, соответствующем временам удерживания на Q Sepharose XL, опубликованным Blanche и др. (2000) Gene Therapy 7:1055-1062.
Пример 4 - Цитолитический анализ
Литическую способность вирусов определяли с помощью модифицированного МТТ-анализа (Shen и др., 2001). Вкратце, MTS-анализ (Promega, CellTiter 96® водный нерадиоактивный анализ пролиферации клеток) использовали вместо МТТ-анализа, так как превращение клетками MTS в водном, растворимом формазане уменьшает время и позволяет избежать использования летучего органического растворителя, связанного с МТТ-анализом.
Для осуществления этого исследования клетки высевали при заданной плотности для каждой опухолевой клеточной линии таким образом, чтобы получить монослой в течение 24 часов. Этим плотно высеянным клеткам позволяли расти дополнительно в течение 2 дней перед обработкой тестируемым(ыми) вирусом(ами). Инфицирование как опухолевых, так и первичных нормальных клеток осуществляли в четырех повторах при серийных трехкратных разведениях вирусов, начиная с соотношения частичек на клетку 100 и заканчивая соотношением частичек на клетку 0,005. Инфицированные клетки инкубировали при 37°С и осуществляли MTS-анализ в моменты времени, указанные для конкретных линий первичных или опухолевых клеток. Ложно инфицированные клетки являлись отрицательным контролем и устанавливали точку 100% выживания в данном исследовании.
Пример 5 - Секвенирование ДНК
Секвенирование ДНК Ad11p (SEQ ID NO: 2) и ColoAd1 (SEQ ID NO: 1) геномных ДНК осуществляли следующим образом. Вкратце, очищенную кДНК аденовируса из ColoAd1 и Ad11p частично расщепляли с помощью рестрикционной эндонулкеазы Sau3Al и сделанный наугад фрагмент клонировали в плазмидном векторе pBluescript II (Stratagene, La Jolla, CA). Положительные клоны размножали и секвенировали с помощью праймеров M13R и KS (Stratagene, La Jolla, CA). Отдельные последовательности реакций обрезали, откорректировали и собирали с помощью Sequenchertm (Gene Codes Corp., Ann Arbor, Мичиган). Бреши в покрытии амлифицировали с помощью обычных олигонуклеотидних праймеров и секвенировали. Концы вирусных геномов отсеквенировали непосредственно с аденовирусной ДНК. В итоге, каждый геном секвенировали при 3Х+ покрытие и 431 оснований при 2Х покрытии.
Для определения начала Е2В участка ColoAd1 получали два набора праймеров, один к гену рТР Е2В (п.о. 9115, 5'GGGAGTTTCGCGCGGACACGG3' (SEQ ID NO: 4) и п.о. 9350, 5'GCGCCGCCGCCGCGGAGAGGT3' (SEQ ID NO: 5)) и один к гену ДНК-полимеразы (п.о. 7520 5'CGAGAGCCCATTCGTGCAGGTGAG3' (SEQ ID NO: 6) и п.о. 7982, 5'GCTGCGACTACTGCGGCCGTCTGT3' (SEQ ID NO: 7) и использовали ПЦР для выделения фрагментов ДНК из различных серотипов (Ad3, 4, 5, 9, 11p, 16 и 40), применяя реагенты из ПЦР-набора Advantage 2 PCR (Clonetics, Walkersville, MD; кат. № K1910-Y) и прогоняя в термоцикле РТС-200 от MJ Research (Watertown, МА). Затем эти фрагменты секвенировали вдоль последовательности ДНК Ad3, используя краситель терминатора секвенирования на генетическом анализаторе ABI 3100.
Участок Е2В Ad3 секвенировали с помощью выделенной ДНК Ad3 и перекрывающихся праймеров.
Информацию, полученную при секвенировании, анализировали с помощью программы Vector NTI (Informatix).
Пример 6 - Конструирование рекомбинантных вирусов
Геномные ДНК Ad11p (SEQ ID NO: 2) и ColoAd1 (SEQ ID NO: 1) очищали от вирусных частиц в ленточном градиенте CsCl. Геномные ДНК расщепляли с помощью PacI, которая разрезает каждую молекулу только один раз в пределах вирусного гена. Разрезание Рас1 происходит на основании 18141 в нуклеотидной последовательности ColoAd1 (SEQ ID NO: 1) и на основании 18140 в нуклеотидной последовательности Ad11 (SEQ ID NO: 2). Расщепленные ДНК смешивали в равных количествах и лигировали в присутствии ДНК-лигазы Т4 при 16°С в течение ночи. Эту лигирующую смесь трасфектировали в клетки А549 с помощью CaPO4 набора для трансфекции от Invitrogen, Carlsbad, СА (кат. № K2780-01). Выделенные бляшки снимали и скринировали путем расщепления рестрикционным ферментом и анализировали с помощью ПЦР для различения четырех вирусных популяций (Ad11p, ColoAd1, левый конец Ad11p/правый конец ColoAd1 (ColoAd1.1) и левый конец ColoAd1/правый конец Ad11p (ColoAd1.2)).
Литическую способность вирусов каждой популяции определяли в нескольких клеточных линиях, включая клеточные линии НТ-29 и HUVEC, как описано в примере 3. Полученные результаты свидетельствуют о наличии диапазона эффективности, от наименее сильнодействующих до наиболее сильнодействующих, как Ad11p, ColoAd1.2, ColoAd1.1, ColoAd1 (см. фигуру 7 для результатов в клетках НТ-29).
Также конструировали химерные аденовирусы pCJ144 и pCJ146, содержащие полноразмерный геном ColoAd1, в котором восстанавливали участки дикого типа Ad11p Е3 и Е4, соответственно. Эти модификации вносили путем гомологичной рекомбинации в BJ5183 Е. Coli (Chartier и др. (1996) J. Virol. 70:4805-4810). Оба эти химерных аденовируса демонстрировали уменьшенную литическую активность в клетках НТ-29 и HUVEC по сравнению с ColoAd1 или ColoAd1.2.
Пример 7 - Эффективность аденовирусов в условиях in vivo
В характерном исследовании на ксенотрансплантированной опухоли человека у бесшерстых мышей, животных инфицировали путем подкожного введения мышам 5×106 клеток в задний бок. При достижении опухолями размера 100-200 мкл им вводили наполнитель (ФСБР) или вирус в количестве 2×1010 частичек в течение пяти последовательных дней (всего 1×1011 частичек). Следует отметить уменьшение размера опухоли относительно ФСБР контроля и дополнительных контрольных вирусов (Ad5, ONYX-015).
Пример 8 - Селективность ColoAd1 на первичных эксплантатах опухоли человека
Образцы ткани, удаленные при хирургическом вмешательстве из колоректальных опухолей и соседних нормальных тканей, помещали в культуральную среду и инфицировали одинаковым количеством вирусов ColoAd1 или Ad5. Через 24 часа после инфицирования собирали культуральные супернатанты и определяли образованное количество вирусных частичек. ColoAd1 образовывали большее количество вирусных частичек относительно введенных частичек, чем Ad5, в ткани опухоли, но меньшее количество частичек относительно введенных частичек, чем Ad5, в нормальной ткани.
Реферат
Настоящее изобретение относится к онколитическим аденовирусам, имеющим терапевтические применения. Геном химерного аденовируса включает нуклеотидную последовательность SEQ ID NO:1, при условии, что участок транскрипции Е2В в ней содержит нуклеотидную последовательность SEQ ID NO:3. Аденовирус из настоящего изобретения проявляет повышенный терапевтический индекс. 3 н. и 8 з.п. ф-лы, 9 ил., 1 табл., 8 пр.
Комментарии