Химерные рецепторы антигенов - RU2745705C2
Код документа: RU2745705C2
Чертежи









































Описание
Область техники
Настоящее изобретение относится к химерным рецепторам антигенов (CAR), включающим спейсеры на основе рецептора фактора роста нервов с низким сродством (LNGFR).
Уровень техники
Иммунотерапия на основе адоптивного переноса иммунных клеток (например, Т-клеток) в организм пациента может играть важную роль в лечении заболеваний, в частности рака. Среди множества различных типов иммунотерапевтических агентов, один из самых перспективных терапевтических методов предполагает использование химерных рецепторов антигенов (CAR). CAR представляют собой сконструированные с помощью генной инженерии рецепторы, которые предназначены для нацеливания на специфический антиген, такой как опухолевый антиген (Sadelain et al., Cancer Discovery 2013. 3(4):388-98). Например, Т-клетки трансдуцируют с использованием CAR, так что Т-клетки, экспрессирующие CAR, уничтожают опухоли через антиген-мишень.
CAR включают экстраклеточный лигандсвязывающий домен, обычно одноцепочечный вариабельный фрагмент моноклонального антитела (scFv), связанный с внутриклеточными, передающими сигналы компонентами, обычно CD3ζ отдельно или в сочетании с одним или более костимулирующих доменов. Между экстраклеточным антигенсвязывающим доменом и трансмембранным фрагментом часто добавляют спейсер для оптимизации взаимодействия с мишенью.
Обычно Fc-домен, в состав которого входят шарнирная область-СН2-СН3 константной области иммуноглобулина IgG, используется в качестве спейсерного домена. Этот спейсер используется для отбора и отслеживания клеток, экспрессирующих CAR. Однако спейсер на основе IgG1 может также связываться с поверхностными Fc-гамма-рецепторами для IgG, представленными на врожденных иммунных клетках, подобных макрофагам и клеткам природным киллерам (Hombach et al. Gene Ther 2000, Jun;7(12):1067-75). Это связывание активирует как сконструированные Т-клетки, так и врожденные иммунные клетки независимо от специфичности связывающего домена CAR, приводя к нежелательной, нецелевой иммунной реакции.
Существует потребность в CAR, которые не вызывают нецелевые иммунные реакции, и не устраняются преждевременно иммунной системой хозяина. Существует также потребность в CAR, включающих спейсерные единицы, которые облегчают отбор клеток, сконструированных с помощью генной инженерии для экспрессии CAR. Настоящее изобретение направлено на удовлетворение этих потребностей.
Сущность изобретения
В соответствии с первым аспектом настоящего изобретения обеспечивается химерный рецептор антигена (CAR), включающий экстраклеточный спейсер, который включает по крайней часть экстраклеточного домена рецептора фактора роста нервов с низким сродством (LNGFR) человека или ее производное.
CAR может включать по крайней мере фрагмент экстраклеточного домена рецептора фактора роста нервов с низким сродством (LNGFR) человека или его производное.
Предпочтительно, когда по крайней мере часть LNGFR подходит для облегчения иммуноселекции клеток, трансдуцированных указанным CAR.
Предпочтительно, когда в спейсере отсутствует внутриклеточный домен LNGFR.
Предпочтительно, когда экстраклеточный спейсер включает первые три TNFR-Cys домена LNGFR или их фрагменты или производные.
В одном варианте осуществления спейсер включает все четыре TNFR-Cys домена LNGFR или их фрагменты или производные.
В другом варианте осуществления спейсер включает четвертый домен TNFR-Cys (TNFR-Cys 4), но при этом следующие аминокислоты удалены из указанного домена: NHVDPCLPCTVCEDTERQLRECTRW. Предпочтительно последовательность NHVDPCLPCTVCEDTERQLRECTRW заменена следующей аминокислотной последовательностью ARA.
В другом варианте осуществления спейсер включает богатую серином/треонином ножку LNGFR.
В другом варианте осуществления в спейсере отсутствует богатая серином/треонином ножка LNGFR.
Спейсер может включать последовательность, выбираемую из группы, состоящей из SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7, или последовательность, которая по меньшей мере на 80, 85, 90, 95, 96, 97, 98 или 99% идентична ей.
В другом варианте осуществления спейсер может состоять из последовательности, выбираемой из группы, состоящей из SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7, или последовательности, которая по меньшей мере на 80, 85, 90, 95, 96, 97, 98 или 99% идентична ей.
SEQ ID NO:1, SEQ ID NO:3 и SEQ ID NO:5 являются предпочтительными спейсерными элементами.
В соответствии с одним аспектом настоящего изобретения обеспечивается химерный рецептор антигена (CAR), включающий
(i) антигенспецифический таргетирующий (нацеливающий) домен;
(ii) экстраклеточный спейсерный домен, определенный здесь;
(iii) трансмембранный домен;
(iv) необязательно по крайней мере один костимулирующий домен; и
(v) внутриклеточный, передающий сигналы домен.
Предпочтительно, когда антигенспецифический таргетирующий домен включает антитело или его фрагмент, более предпочтительно одноцепочечный вариабельный фрагмент.
Предпочтительно, когда мишенью антигенспецифического таргетирующего домена является опухолевый антиген. Примеры таких антигенов включают CD44, CD19, CD20, CD22, CD23, CD123, CS-1, ROR1, мезотелин, c-Met, PSMA, Her2, GD-2, CEA, MAGE A3 TCR.
Предпочтительно, когда опухолевым антигеном является изоформа 6 CD44 (CD44v6).
Примеры трансмембранных доменов включают трансмембранный домен дзета-цепи Т-клеточного рецепторного комплекса, CD28 и CD8a.
Примеры костимулирующих доменов включают костимулирующий домен из CD28, CD137 (4-1BB), CD134 (OX40), DaplO, CD27, CD2, CD5, ICAM-1, LFA-1, Lck, TNFR-I, TNFR-II, Fas, CD30 и CD40.
Примеры внутриклеточных, передающих сигналы доменов включают дзета-цепь CD3 человека, FcγRIII, FcsRI, цитоплазматический хвост Fc-рецептора и несущие иммунорецепторный тирозиновый активирующий мотив (ITAM) цитоплазматические рецепторы.
В предпочтительном варианте осуществления мишенью антигенспецифического таргетирующего домена CAR является CD44v6, трансмембранный домен CAR включает трансмембранный домен CD28, внутриклеточный, передающий сигналы домен CAR включает внутриклеточный, передающий сигналы домен дзета-цепи CD3 человека, а костимулирующий домен CAR включает эндо-костимулирующий домен CD28.
В другом аспекте настоящего изобретения обеспечивается полинуклеотид, кодирующий CAR настоящего изобретению и определенный здесь.
Предпочтительно, когда полинуклеотид кодирует спейсерный домен, который включает последовательность SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:8, или последовательность, которая по меньшей мере на 80, 85, 90, 95, 96, 97, 98 или 99% идентична ей.
В одном варианте осуществления полинуклеотид кодирует спейсерный домен, который состоит из последовательности SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:8, или последовательности, которая по меньшей мере на 80, 85, 90, 95, 96, 97, 98 или 99% идентична ей.
В другом аспекте настоящего изобретения обеспечивается вектор, включающий полинуклеотид настоящего изобретения.
В одном варианте осуществления вектор представляет собой вирусный вектор.
В другом аспекте настоящего изобретения обеспечивается клетка, включающая CAR, полинуклеотид или вектор настоящего изобретения. Предпочтительно, когда клетка представляет собой Т-клетку.
В другом аспекте настоящего изобретения обеспечивается фармацевтическая композиция, включающая клетку настоящего изобретения.
В другом аспекте настоящего изобретения обеспечивается CAR, полинуклеотид, вектор или клетка настоящего изобретения для применения в терапии, предпочтительно терапии рака.
В другом аспекте настоящего изобретения обеспечивается CAR настоящего изобретения, мишенью антигенспецифического таргетирующего домена которого является CD44v6 для применения при лечении опухолей, которые экспрессируют CD44.
В другом аспекте настоящего изобретения обеспечивается способ лечения, включающий введение CAR, полинуклеотида, вектора или клетки настоящего изобретения субъекту, нуждающемуся в таком введении.
Примеры CAR представлены на фиг. 10-17.
Описание чертежей
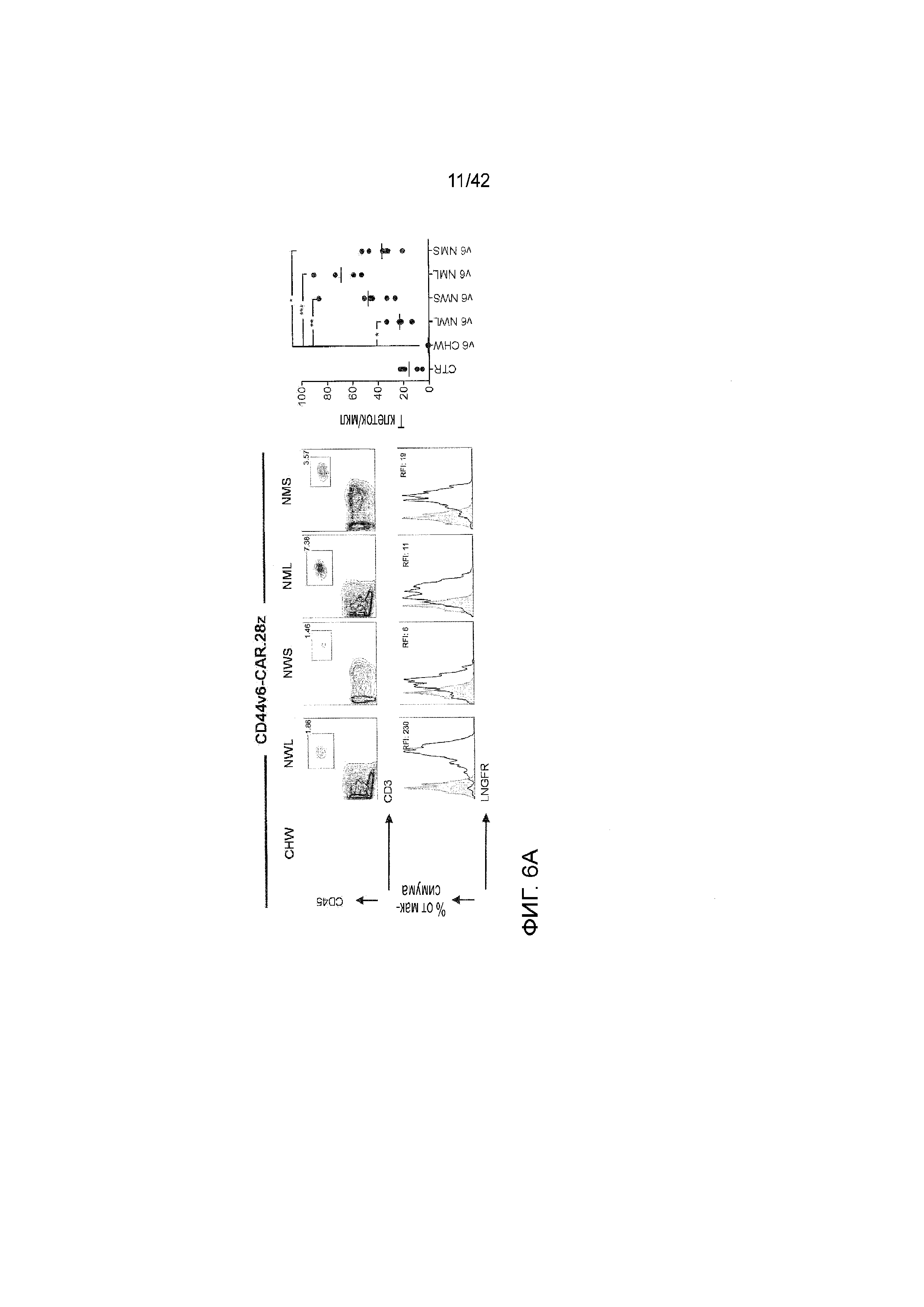
Фиг. 1. Обоснование создания конструкций CD44v6-CAR.28z с различными спейсерами на основе LNGFR. А. Схема, объясняющая ограничения Т-клеток, экспрессирующих CAR, содержащий спейсер CH2CH3 IgG1. B. Структура экстраклеточной части рецептора фактора роста нервов с низким сродством (LNGFR) и 4-х новых конструкций CAR, которые были созданы. Также включены CD44v6-CAR.28z, содержащие спейсер CH2CH3 IgG1 дикого типа или мутированный (mCH2CH3). CHW: CD44v6-CAR.28z, содержащий спейсер CH2CH3 дикого типа. CHM: CD44v6-CAR.28z, содержащий мутированный спейсер CH2CH3. NWL: CD44v6-CAR.28z, содержащий длинный спейсер на основе LNGFR дикого типа (включающий 4 TNFR-Cys домена и ножку). NWS: CD44v6-CAR.28z, содержащий короткий спейсер на основе LNGFR дикого типа (включающий только 4 TNFR-Cys домена). NML: CD44v6-CAR.28z, содержащий мутированный длинный спейсер на основе LNGFR (включающий 4 TNFR-Cys домена с делецией в четвертом домене и ножку). NMS: CD44v6-CAR.28z, содержащий мутированный короткий спейсер на основе LNGFR (включающий 4 TNFR-Cys домена с делецией в четвертом домене и ножку). Фигурные скобки указывают длину спейсера, представленную в аминокислотах. Серый цвет: scFv. Белый цвет: костимулирующий домен CD28; Черный цвет: CD3ζ.
Фиг. 2. Экспрессирующие CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клетки могут быть отсортированы с использованием мАт против LNGFR, эффективно размножаться in vitro и сохранять рано дифференцированный фенотип. Т-клетки активировали с использованием CD3/CD28-сфер, трансдуцировали ретровирусными векторами (RV), кодирующими CD44v6.CAR28z с различными спейсерами на основе LNGFR, и культивировали с IL-7/IL-15. А. Идентификация CAR на поверхности Т-клеток с использованием специфического для LNGFR мАт C40-1457 (верхние графики). Идентификация CAR на поверхности Т-клеток с использованием LNGFR-специфического мАт ME20.4 (нижние графики) B. Слева: Т-клетки, экспрессирующие CD44v6-CAR.28z с различными спейсерами на основе LNGFR, после сортировки с использованием мАт C40-1457 и анти-PE сфер. Справа: кинетика увеличения количества отсортированных экспрессирующих CD44v6-CAR.28z со спейсером CH2CH3 и спейсерами на основе LNGFR Т-клеток, представленная как кратное увеличение. С. Фенотип функциональной дифференцировки Т-клеток, экспрессирующих CD44v6-CAR.28z с различными спейсерами на основе LNGFR, через 15 дней после активации. CD45RA+/CD62L+ стволовые Т-клетки памяти, CD45RA-/CD62L+ центральные Т-клетки памяти, CD45RA-/CD62L- эффекторные Т-клетки памяти, CD45RA+/CD62L- эффекторные Т-клетки памяти RA. Графики и кривые соответствуют n=4 независимым экспериментам.
Фиг. 3. Экспрессирующие CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клетки специфически распознают CD44v6+ опухолевые клетки in vitro. А. После сортировки экспрессирующие CD44v6-CAR.28z с различными спейсерами на основе LNGFR Т-клетки (NWL, NWS, NML, NMS), экспрессирующие CD44v6-CAR со спейсером CH2CH3 Т-клетки (CHW, CHM) и Т-клетки, несущие нерелевантный CAR, культивировали с CD44v6+ клетками миеломы MM1.S, CD44v6+ лейкозными клетками ТНР-1 или CD44v6- лимфобластоидными клетками BV-173 при различных соотношениях Е:Т. Спустя 4 дня, остаточные опухолевые клетки подсчитывали и анализировали с помощью FACS. Представлен индекс элиминации (смотрите пример «Методы») с помощью экспрессирующих CD44v6-CAR.28z Т-клеток при различных соотношениях E:T. В. В экспрессирующие CD44v6-CAR.28z Т-клетки вводили краситель CFSE, и их стимулировали с использованием линий облученных опухолевых клеток в соотношении Е:Т=1:5. Спустя 6 дней, пролиферацию Т-клеток анализировали с помощью FACS, выражая в виде разбавляющих CFSE клеток. Графики и кривые соответствуют n=4 независимым экспериментам.
Фиг. 4. У экспрессирующих CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клеток отсутствует FcRg-опосредованное распознание. А. После сортировки экспрессирующие CD44v6-CAR.28z с различными спейсерами на основе LNGFR Т-клетки (NWL, NWS, NML, NMS), экспрессирующие CD44v6-CAR со спейсером CH2CH3 Т-клетки (CHW, CHM) и Т-клетки, несущие нерелевантный CAR, от n=4 здоровым донорам культивировали с CD44v6+/FcRg+ лейкозными клетками ТНР-1 или CD44v6-/FcRg+ лейкозными клетками HL-60 при различных соотношениях Е:Т. Спустя 4 дня, остаточные опухолевые клетки подсчитывали и анализировали с помощью FACS. Представлен индекс элиминации (смотрите пример «Методы») с помощью экспрессирующих CD44v6-CAR.28z Т-клеток при различных соотношениях E:T. В. В экспрессирующие CD44v6-CAR.28z Т-клетки вводили краситель CFSE, и их стимулировали с использованием линий облученных клеток THP1, HL60 или CD44v6-/FcRg- лимфобластоидных клеток BV-173. Через 6 дней пролиферацию Т-клеток анализировали с помощью FACS и представляли в виде разбавляющих CFSE клеток. Графики и кривые соответствуют n=4 независимым экспериментам.
Фиг. 5. CD44v6-CAR.28z со спейсерами на основе LNGFR не стимулируются с помощью растворимого NGF. А. После подвергания в течение 24 ч воздействию человеческого рекомбинантного NGF в различных концентрациях, LNGFR+ нервные клетки PC-12 анализировали в отношении образования дендритов с помощью оптической микроскопии. B. После сортировки в экспрессирующие CD44v6-CAR.28z с различными спейсерами на основе LNGFR Т-клетки (NWL, NWS, NML, NMS) и экспрессирующие CD44v6-CAR.28z со спейсером CH2CH3 Т-клетки (CHW, CHM) вводили краситель CFSE, и их подвергали воздействию различных концентраций NGF. Через 4 дня пролиферацию Т-клеток анализировали с помощью FACS и представляли в виде разбавляющих CFSE клеток. Разведение CFSE после сокультивирования с CD44v6+ клетками миеломы MM1.S или CD44v6- лимфобластоидными клетками BV-173 представлено для сравнения. Фотографии и кривые соответствуют n=2 независимым экспериментам. Графики показывают среднее значение ± стандартное отклонение от двух экспериментов.
Фиг. 6. 3. Экспрессирующие CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клетки лучше размножаются, сохраняются и опосредуют превосходные противолейкозные эффекты в модели минимальной остаточной болезни. Мышам NSG вводили путем инфузии CD44v6+ лейкозные клетки ТНР-1, и спустя 3 дня их подвергали лечению экспрессирующими CD44v6-CAR.28z с различными спейсерами на основе LNGFR Т-клетками (NWL, NWS, NML, NMS), экспрессирующими CD44v6-CAR со спейсером CH2CH3 Т-клетками (CHW) или Т-клетками, экспрессирующими нерелевантный CAR (CTR), всех их сортировали до составляющей >95% степени чистоты. A. Репрезентативные графики (слева) и включающий все график (справа), на котором представлены циркулирующие, экспрессирующие CD44v6-CAR.28z Т-клетки у каждой мыши через три дня после инфузии. CD44v6-CAR.28z с различными спейсерами отслеживались с помощью FACS после окрашивания поликлональным антителом против IgG (CTR и CHW) или LNGFR-специфическим мАт, C40-1457 мАт. В. Кинетика увеличения количества и персистенции со временем экспрессирующих CD44v6-CAR28z Т-клеток. C. Вес печени с инфильтрующими клетками Thp1 у подвергнутых лечению мышей при умерщвлении (7 недель). Пунктирная зона изображает диапазон нормального веса печени от подобранных по возрасту/полу нормальных мышей NSG. Результаты однофакторного дисперсионного анализа представлены в случае статистической значимости (*Р<0,05, **Р<0,01, ***Р<0,001).
Фиг. 7. Экспрессирующие CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клетки лучше размножаются, сохраняются и опосредуют превосходные противомиеломные эффекты в модели устойчивого заболевания. Мышам NSG вводили путем инфузии CD44v6+ клетки MM1.S, и спустя 5 недель их подвергали лечению экспрессирующими CD44v6-CAR.28z с различными спейсерами на основе LNGFR Т-клетками (NWL, NWS, NMS), экспрессирующими CD44v6-CAR.28z со спейсером CH2CH3 Т-клетками (CHW) или Т-клетками, экспрессирующими нерелевантный CAR (CTR), всех их сортировали до составляющей >95% степени чистоты. A. Включающий все график (справа), на котором представлены циркулирующие, экспрессирующие CD44v6-CAR.28z Т-клетки у каждой мыши через три дня после инфузии. CD44v6-CAR.28z с различными спейсерами отслеживались с помощью FACS после окрашивания поликлональным антителом против IgG (CTR и CHW) или LNGFR-специфическим мАт, C40-1457 мАт. В. Кривые выживаемости Каплана-Мейера подвергнутых лечению мышей. Представлены результаты проверки по критерию Кокса, сравнивающей различные условия (ns: незначимое, *Р<0,05, ***Р<0,001).
Фиг. 7 BIS. 3. Экспрессирующие CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клетки опосредуют превосходные противомиеломные эффекты в модели устойчивого заболевания. Мышам NSG вводили путем инфузии CD44v6+ клетки MM1.S, экспрессирующие люциферазу, и через 26 дней их подвергали лечению экспрессирующими CD44v6-CAR.28z со спейсером на основе LNGFR Т-клетками (NMS), экспрессирующими CD44v6-CAR со спейсером CH2CH3 Т-клетками (CHW) или Т-клетками, экспрессирующими нерелевантный CAR (CTR), всех их сортировали до составляющей >95% степени чистоты. А. Циркулирующее количество опухолевых клеток оценивали в виде относительных световых единиц (RLU) в указанные моменты времени. B. Кривые выживаемости Каплана-Мейера подвергнутых лечению мышей. Представлены результаты проверки по критерию Кокса, сравнивающей различные условия (**P<0,01).
Фиг. 8. Последовательность LNGFR человека.
Фиг. 9. Последовательность CD44v6CAR.28z. Представлены последовательности scFv, CH2CH3, CD28 и дзета-цепи.
Фиг. 10. Приводимая в качестве примера последовательность CD44v6CAR.28z со спейсером LNGFR дикого типа длинным (NWL) (SEQ ID NO:21)
Фиг. 11. Приводимая в качестве примера последовательность CD44v6-CAR.28z со спейсером LNGFR дикого типа коротким (NWS) (SEQ ID NO:22)
Фиг. 12. Приводимая в качестве примера последовательность CD44v6-CAR.28z со спейсером LNGFR мутированным длинным (NML) (SEQ ID NO:23)
Фиг. 13. Приводимая в качестве примера последовательность CD44v6-CAR.28z со спейсером LNGFR мутированным коротким (NMS) (SEQ ID NO:24)
Фиг. 14. Приводимая в качестве примера последовательность CD44v6CAR.28z со спейсером LNGFR дикого типа длинным (NWL) (SEQ ID NO:25)
Фиг. 15. Приводимая в качестве примера последовательность CD44v6-CAR.28z со спейсером LNGFR дикого типа коротким (NWS) (SEQ ID NO:26)
Фиг. 16. Приводимая в качестве примера последовательность CD44v6CAR.28z со спейсером LNGFR мутированным длинным (NML) (SEQ ID NO:27)
Фиг. 17. Приводимая в качестве примера последовательность CD44v6-CAR.28z со спейсером LNGFR мутированным коротким (NMS) (SEQ ID NO:28)
Фиг. 18. Приводимая в качестве примера последовательность CD44v6-4GS2-CAR28z со спейсером LNGFR дикого типа длинным (NWL) (SEQ ID NO:32)
Фиг. 19. Приводимая в качестве примера последовательность CD44v6-4GS2-CAR28z со спейсером LNGFR дикого типа коротким (NWS) (SEQ ID NO:33)
Фиг. 20. Приводимая в качестве примера последовательность CD44v6-4GS2-CAR28z со спейсером LNGFR мутированным длинным (NML) (SEQ ID NO:34)
Фиг. 21. Приводимая в качестве примера последовательность CD44v6-4GS2-CAR28z со спейсером LNGFR мутированным коротким (NMS) (SEQ ID NO:35)
Фиг. 22. Создание CAR с различными спейсерами на основе LNGFR. Структура экстраклеточной части рецептора фактора роста нервов с низким сродством (LNGFR) и различных конструкций CAR, мишенью которых являются CD19 и СЕА, которые были созданы. Также включены CD19/CEA-CAR.28z, содержащие спейсер CH2CH3 IgG1 дикого типа (CH2CH3). NWL: CD19/CEA-CAR.28z, содержащий длинный спейсер на основе LNGFR дикого типа (включающий 4 TNFR-Cys домена и ножку). NMS: CD19/CEA-CAR.28z, содержащий мутированный короткий спейсер на основе LNGFR (включающий 4 TNFR-Cys домена с делецией в четвертом домене). Фигурные скобки указывают длину спейсера, представленную в аминокислотах. Серый цвет: scFv.
Фиг. 23. Экспрессирующие CD19/CEA-CAR.28z со спейсерами на основе LNGFR Т-клетки могут быть окрашены мАт против LNGFR. Т-клетки активировали с использованием CD3/CD28-сфер, трансдуцировали ретровирусными векторами (RV), кодирующими CD19/CEA.CAR28z с различными спейсерами на основе LNGFR, культивировали с IL-7/IL-15 и отбирали с использованием мАт C40-1457 и анти-PE сфер. В качестве положительного контроля, экспрессирующие CD44v6-4GS2.CAR28z Т-клетки были получены в тех же самых условиях. Представлена идентификация CAR на поверхности Т-клеток с использованием LNGFR-специфического мАт C40-1457.
Фиг. 24. Экспрессирующие CD19/CEA-CAR.28z со спейсерами на основе LNGFR Т-клетки специфически распознают антигенэкспрессирующие опухолевые клетки in vitro. А. После сортировки экспрессирующие CD19/CEA/CD44v6-4GS2-CAR.28z с различными спейсерами на основе LNGFR Т-клетки (NWL, NWS) и экспрессирующие CD19/CEA-CAR со спейсером CH2CH3 Т-клетки (CHW) Т-клетки культивировали с лейкозными клетками ALL-CM и HL60, лимфобластоидными клетками BV-173 и клетками карциномы BXPC3 при соотношении Е:Т=1:10. Спустя 4 дня, остаточные опухолевые клетки подсчитывали и анализировали с помощью FACS. Представлен индекс элиминации (смотрите пример «Методы») с помощью различных CAR.28z+ Т-клеток. В. В. Супернатанты от сокультур, описанных в A, собирали через 24 часа и анализировали на продукцию цитокинов с помощью анализа СВА (Biolegend). Представлен выброс IFNα, IL-2 и TNFα при распознании клеток-мишеней.
Фиг. 25. Экспрессирующие CD19-CAR.28z со спейсерами на основе LNGFR Т-клетки опосредуют противолейкозные эффекты. Мышей NSG подвергали инфузии CD19+ лейкозных клеток ALL-CM и через три дня подвергали лечению экспрессирующими CD19-CAR.28z с различными спейсерами на основе LNGFR Т-клетками (19 NWL и 19 NMS). Т-клетки, экспрессирующие неродственную конструкцию CD44v6-4GS2-CAR.28z (v6 NWL и v6 NMS), использовали для инфузии в качестве контроля. Все CAR+ Т-клетки сортировали до составляющей >95% степени чистоты перед инфузией. График показывает наличие опухолевых клеток ALL-см в костном мозге (ВМ) каждой мыши в момент умерщвления. Опухолевые клетки отслеживали с помощью FACS после окрашивания с использованием мАт против hCD45 и hCD19. Результаты проверки по критерию Стьюдента представлены в случае статистической значимости (*Р<0,05, **Р<0,01, ***Р<0,001).
Фиг. 26. Полинуклеотидная последовательность CD44v6-4GS2-CAR28z, со спейсером LNGFR дикого типа длинным (NWL) (SEQ ID NO: 37)
Фиг. 27. Полинуклеотидная последовательности CD44v6-4GS2-CAR28z, со спейсером LNGFR дикого типа коротким (NWS) (SEQ ID NO:38)
Фиг. 28. Полинуклеотидная последовательность CD44v6-4GS2-CAR28z со спейсером LNGFR мутированным длинным (NML) (SEQ ID NO:39)
Фиг. 29. Полинуклеотидная последовательность CD44v6-4GS2-CAR28z со спейсером LNGFR мутированным коротким (NMS) (SEQ ID NO:40)
Подробное описание изобретения
Различные предпочтительные признаки и варианты осуществления настоящего изобретения будут теперь описаны в качестве неограничивающих примеров.
При осуществлении на практике настоящего изобретения будут использоваться, если не указано иное, обычные методы химии, биохимии, молекулярной биологии, микробиологии и иммунологии, которые находятся в пределах возможностей специалиста со средним уровнем компетентности в данной области техники. Такие методы объяснены в литературе. Смотрите, например, Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989) Molecular Cloning: A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory Press; Ausubel, F.M. et al. (1995 и приложения к периодическому изданию) Current Protocols in Molecular Biology, Ch. 9, 13 and 16, John Wiley & Sons; Roe, B., Crabtree, J., and Kahn, A. (1996) DNA Isolation and Sequencing: Essential Techniques, John Wiley & Sons; Polak, J.M., and McGee, J.O'D. (1990) In Situ Hybridization: Principles and Practice, Oxford University Press; Gait, M.J. (1984) Oligonucleotide Synthesis: A Practical Approach, IRL Press; и Lilley, D.M., and Dahlberg, J.E. (1992) Methods in Enzymology: DNA Structures Part A: Synthesis and Physical Analysis of DNA, Academic Press. Каждый из этих общих текстов включен сюда посредством ссылки.
Химерные рецепторы антигенов
«Химерный рецептор антигена» или «CAR», как здесь используется, относится к сконструированным рецепторам, которые могут обеспечивать специфичность в отношении антигена клеткам (например, Т-клеткам, такими как необученные Т-клетки, центральные Т-клетки памяти, эффекторные Т-клеток памяти или их комбинации). CAR также известные как искусственные Т-клеточные рецепторы, химерные Т-клеточные рецепторы или химерные иммунорецепторы. Предпочтительно, когда CAR настоящего изобретения включают антигенспецифический таргетирующий район, экстраклеточный домен, трансмембранный домен, необязательно один или более костимулирующих доменов и внутриклеточный, передающий сигналы домен.
Антигенспецифический таргетирующий домен
Антигенспецифический таргетирующий домен обеспечивает CAR способностью к связыванию с представляющим интерес антигеном-мишенью. Предпочтительно мишенью антигенспецифическего таргетирующего домена является представляющий клинический интерес антиген, против которого был бы желательным вызов эффекторного иммунного ответа, который приводит к уничтожению опухоли.
Антигенспецифическим таргетирующим доменом может быть любой белок или пептид, который обладает способностью к специфическому распознанию и связыванию с биологической молекулы(ой) (например, рецептора клеточной поверхности или опухолевого белка, или его компонента). Антигенспецифический таргетирующий домен включает любой встречающийся в природе, синтетический, полусинтетический или рекомбинантно полученный партнер по связыванию для биологической молекулы, представляющей интерес.
Иллюстративные антигенспецифические таргетирующие домены включают антитела или фрагменты или производных антител, экстраклеточные домены рецепторов, лиганды для молекул/рецепторов клеточной поверхности или их связывающиеся с рецепторами домены, и связывающиеся с опухолью белки.
В предпочтительном варианте осуществления аАнтигенспецифический таргетирующий домен представляет собой, или является происходящим из, антитело(а). Происходящий из антитела таргетирующий домен может быть фрагментом антитела или полученным методами генной инженерии продуктом одного или более фрагментов антитела, при этом фрагмент участвует в связывании с антигеном. Примеры включают вариабельную область (Fv), определяющий комплементарность участок (CDR), Fab, одноцепочечное антитело (scFv), вариабельную область тяжелой цепи (VH), вариабельную область легкой цепи (VL) и антитело семейства верблюдовые (VHH). В предпочтительном варианте осуществления связывающий домен представляет собой одноцепочечное антитело (scFv). ScFv может быть мышиным, человеческим или гуманизированным scFv.
«Определяющий комплементарность район» или «CDR», в отношении антитела или его антигенсвязывающего фрагмента, относится к высокой степени вариабельному петлевому участку в вариабельной области тяжелой цепи или легкой цепи антитела. CDR может взаимодействовать с конформацией антитела и в значительной степени определять связывание с антигеном (хотя некоторые каркасные области, как известно, участвуют в связывании). Каждая из вариабельной области тяжелой цепи и вариабельной области легкой цепи содержит 3 CDR.
«Вариабельная область тяжелой цепи» или «VH» относится к фрагменту тяжелой цепи антитела, который содержит три CDR, расположенные между фланкирующими участками, известными как каркасные области, которые являются в более высокой степени консервативными, чем CDR, и образуют каркас для поддержки CDR.
«Вариабельная область легкой цепи» или «VL» относится к фрагменту легкой цепи антитела, который содержит три CDR, расположенные между каркасными областями.
«Fv» относится к наименьшему фрагменту антитела, который несет полный антигенсвязывающий сайт. Fv-фрагмент состоит из вариабельной области одной легкой цепи, связанной с вариабельной областью одной тяжелой цепи.
«Одноцепочечное Fv-антитело» или «scFv» относится к сконструированному антителу, состоящему из вариабельной области легкой цепи и вариабельной области тяжелой цепи, которые соединены друг с другом непосредственно или через последовательность пептидного линкера.
Антитела, которые специфически связываются с молекулой на поверхности опухолевых клеток, могут быть получены, используя методы, хорошо известные в данной области техники. Такие методы включают фаговый дисплей, методы получения человеческих или гуманизированных антител, или методы с использованием трансгенного животного или растения, созданного для продуцирования антител человека. Библиотеки фагового дисплея частично или полностью синтетических антител доступны и могут быть подвергнуты скринингу на антитело или его фрагмент, которое(ый) может связываться с молекулой-мишенью. Библиотеки фагового дисплея антител человека также доступны. После идентификации аминокислотная последовательность или полинуклеотидная последовательность, кодирующая антитело, может быть изолирована и/или определена.
Примеры антигенов, на которые могут быть нацелены CAR настоящего изобретения, включают, но без ограничения ими, антигены, представленные на раковых клетках, и антигены, представленные на клетках, связанных с различными гематологическими заболеваниями, аутоиммунными заболеваниями, воспалительными заболеваниями и инфекционными заболеваниями.
Что касается таргетирующих доменов, мишенью которых являются раковые антигены, выбор таргетирующего домена будет зависеть от типа рака, подлежащего лечению, и он может быть нацелен на опухолевые антигены. Образец опухоли от субъекта может быть охарактеризован в отношении наличия определенных биомаркеров или маркеров клеточной поверхности. Например, клетки рака молочной железы у субъекта могут быть позитивными или негативными по каждому из Her2Neu, эстрогенового рецептора и/или рецептора прогестерона. Выбирают молекулу опухолевого антигена или клеточной поверхности, которая встречается на опухолевых клетках отдельного субъекта. Предпочтительно, когда мишенью антигенспецифического таргетирующего домена является молекула клеточной поверхности, которая встречается на опухолевых клетках и по существу не встречается в нормальных тканях, или ее экспрессия ограничивается не являющимися необходимыми для жизни нормальными тканями.
Дополнительные специфические для рака антигены, на которые может быть нацелен CAR настоящего изобретения, включают, но без ограничения, любой один или более из карциноэмбрионального антигена (СЕА), специфического для простаты антигена, PSMA, Her2/neu, эстрогенового рецептора, рецептора прогестерона, эфрина B2, ROR1, мезотелина, c-Met, GD-2, и MAGE А3 TCR, 4-1ВВ, 5T4, антигена аденокарциномы, альфа-фетопротеина, BAFF, клетки В-лимфомы, антигена С242, СА-125, карбоангидразы 9 (СА-IX), CCR4, CD152, CD200, CD22, CD19, CD22, CD123, CD221, CD23 (рецептора IgE), CD28, CD30 (TNFRSF8), CD33, CD4, CD40, CD44, CD44 v6, CD51, CD52, CD56, CD74, CD80, CS-1, CEA, CNT0888, CTLA-4, DR5, EGFR, EpCAM, CD3, FAP, дополнительного домена-B фибронектина, рецептора 1 фолиевой кислоты, GD2, ганглиозида GD3, гликопротеина 75, GPNMB, HGF, рецепторной киназы фактора роста гепатоцитов человека, рецептора IGF-1, IGF-1, IgG1, L1-САМ, IL-13, IL-6, рецептора инсулиноподобного фактора 1 роста, интегрина α5β1, интегрина ανβ3, MORAb-009, MS4A1, MUC1, муцинового CanAg, N-гликолилнейраминовой кислоты, NPC-1C, PDGF-Rα, PDL192, фосфатидилсерина, клеток карциномы простаты, RANKL, RON, SCH 900105, SDC1, SLAMF7, TAG-72, тенасцина C, TGF бета 2, TGF-β, TRAIL-R1, TRAIL-R2, опухолевого антигена CTAA16.88, VEGF-A, VEGFR1, VEGFR2 или виментина.
Специфические для воспалительных заболеваний антигены, на которые может быть нацелен CAR настоящего изобретения, включают, но без ограничения, любой один или более из AOC3 (VAP-1), CAM-3001, CCL11 (эотаксина-1), CD125, CD147 (басигина), CD154 (CD40L), CD2, CD20, CD23 (рецептора IgE), CD25 (α-цепи рецептора IL-2), CD3, CD4, CD5, IFN-α, IFN-γ, IgE, IgE, Fc-фрагмента IgE, IL-1, IL-12, IL-23, IL-13, IL-17, IL-17A, IL-22, IL-4, IL-5, IL-5, IL-6, рецептора IL-6, интегрина α4, интегрина α4β7, шерсти ламы, LFA-1 (CD11а), MEDI-528, миостатина, ОХ-40, rhuMAb β7, склеросцина, SOST, TGFβ1, TNF-а или VEGF-A.
Специфические для нейрональных нарушений антигены, на которые может быть нацелен CAR настоящего изобретения, включают, но без ограничения, любой один или более из бета-амилоида или MABT5102A.
Специфические для диабета антигены, на которые может быть нацелен CAR настоящего изобретения, включают, но без ограничения, любой один или более из L-1β или CD3. Другие антигены, специфические для диабета или других нарушений обмена веществ, будут очевидны квалифицированным специалистам в данной области.
Специфические для сердечнососудистых заболеваний антигены, на которые может быть нацелен CAR настоящего изобретения, включают, но без ограничения, любой один или более из C5, сердечного миозина, CD41 (интегрина альфа-IIb), фибрина II, бета-цепи, ITGB2 (CD18) и сфингазин-1-фосфата.
Предпочтительно антигенспецифический связывающий домен специфически связывается с опухолевым антигеном. В конкретном варианте осуществления полинуклеотид кодирует одноцепоченый Fv, который специфически связывается с CD44v6.
Иллюстративным антигенспецифическим таргетирующим доменом является CD44v6-специфический одноцепочечный фрагмент (scFv), например, описанный в Casucci M et al. Blood, 2013, Nov 14; 122(20):3461-72). Такая последовательность представлена ниже:
CD44v6-специфический одноцепочечный фрагмент (scFv)
MEAPAQLLFLLLLWLPDTTGEIVLTQSPATLSLSPGERATLSCSASSSINYIYWLQQKPGQAPRILIYLTSNLASGVPARFSGSGSGTDFTLTISSLEPEDFAVYYCLQWSSNPLTFGGGTKVEIKRGGGGSGGGGSGGGGSGGGGSEVQLVESGGGLVKPGGSLRLSCAASGFTFSSYDMSWVRQAPGKGLEWVSTISSGGSYTYYLDSIKGRFTISRDNAKNSLYLQMNSLRAEDTAVYYCARQGLDYWGRGTLVTVSS(SEQ ID NO:17)
В одном варианте осуществления CD44v6-специфический одноцепочечный фрагмент по меньшей мере на 85, 90, 95, 97, 98 или 99% идентичен последовательности SEQ ID NO:17.
В еще одном предпочтительном варианте осуществления вариабельная область легкой цепи и вариабельная область тяжелой цепи CD44v6-специфического одноцепочечного фрагмента соединены друг с другом через пептидный линкер, имеющий следующую последовательность GGGGSGGGGS (4GS2). Такой CD44v6-специфический одноцепочечный фрагмент (CD44v6-4GS2) имеет следующую последовательность:
MEAPAQLLFLLLLWLPDTTGEIVLTQSPATLSLSPGERATLSCSASSSINYIYWLQQKPGQAPRILIYLTSNLASGVPARFSGSGSGTDFTLTISSLEPEDFAVYYCLQWSSNPLTFGGGTKVEIKRGGGGSGGGGSEVQLVESGGGLVKPGGSLRLSCAASGFTFSSYDMSWVRQAPGKGLEWVSTISSGGSYTYYLDSIKGRFTISRDNAKNSLYLQMNSLRAEDTAVYYCARQGLDYWGRGTLVTVSS (SEQ ID NO:31)
Костимулирующий домен
CAR настоящего изобретения может также включать один или более костимулирующих доменов. Этот домен может усиливать пролиферацию клеток, выживаемость клеток и развитие клеток памяти.
Каждый костимулирующий домен включает костимулирующий домен любого одного или более из, например, членов суперсемейства TNFR, CD28, CD137 (4-1ВВ), CD134 (OX40), DaplO, CD27, CD2, CD5, ICAM-1, LFA-1, Lck, TNFR-1, TNFR-II, Fas, CD30, CD40 или их комбинаций. Костимулирующие домены из других белков могут быть также использованы с CAR настоящего изобретения. Дополнительные костимулирующие домены будут очевидны квалифицированным специалистам в данной области техники.
В одном варианте осуществления как трансмембранный, так и костимулирующий домены получены из CD28. В одном варианте осуществления трансмембранный и внутриклеточный костимулирующий домены включают последовательность, приведенную ниже:
Трансмембранная и внутриклеточная часть CD28 человека (UNIPROT: P10747, CD28_HUMAN, положения 153-220)
FWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRS (SEQ ID NO:18)
В одном варианте осуществления трансмембранный и внутриклеточный, передающий сигналы домен по меньшей мере на 85, 90, 95, 97, 98 или 99% идентичен последовательности SEQ ID NO:18.
В одном варианте осуществления трансмембранный домен CD28 включает последовательность FWVLVVVGGVLACYSLLVTVAFIIFWV (SEQ ID NO:29).
В одном варианте осуществления внутриклеточный костимулирующий домен CD28 включает последовательность RSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRS (SEQ ID NO:30).
Внутриклеточный, передающий сигналы домен
CAR настоящего изобретения может также включать внутриклеточный, передающий сигналы домен. Этот домен может быть цитоплазматическим и может передавать эффекторный функциональный сигнал и направлять клетки на выполнение своей специальной функции. Примеры внутриклеточных, передающих сигналы доменов включают, но без ограничения ими, ζ-цепь Т-клеточного рецептора или любой из ее гомологов (например, η-цепь, FcεR1γ и β-цепи, MB1 (lgα) цепь, B29 (lgβ) цепь и т.д.), полипептиды CD3 (Δ, δ и ε), семейство Syk тирозинкиназ (Syk, ZAP 70 и т.д.), семейство scr тирозинкиназ (Lck, Fyn, Lyn, и т.д.) и другие молекулы, участвующие в преобразовании сигналов в Т-клетках, такие как CD2, CD5 и CD28. Внутриклеточный, передающий сигналы домен может быть дзета-цепью CD3 человека, FcγRIII, FcsRI, цитоплазматическими хвостами Fc-рецепторов, несущими иммунорецепторный тирозиновый активирующий мотив (ITAM) цитоплазматическими рецепторами или их комбинациями.
Предпочтительно, когда внутриклеточный, передающий сигналы домен включает внутриклеточный, передающий сигналы домен дзета-цепи CD3 человека.
В одном варианте осуществления внутриклеточный, передающий сигналы домен дзета-цепи CD3 человека включает следующую последовательность:
UNIPROT: P20963, CD3Z_HUMAN, положения 31-143
RVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPQRRKNPQEG LYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO:20)
В одном варианте осуществления внутриклеточный, передающий сигналы домен по меньшей мере на 85, 90, 95, 97, 98 или 99% идентичен последовательности SEQ ID NO:20.
Дополнительные внутриклеточные, передающие сигналы домены будут очевидны квалифицированным специалистам в данной области техники и могут быть использованы в связи с альтернативными вариантами осуществления настоящего изобретения.
Трансмембранный домен
CAR настоящего изобретения может также включать трансмембранный домен. Трансмембранный домен может включать трансмембранную последовательность из любого белка, который имеет трансмембранный домен, в том числе любого из трансмембранных белков типа I, типа II или типа III. Трансмембранный домен CAR настоящего изобретения может также включать искусственную гидрофобную последовательность. Трансмембранные домены CAR настоящего изобретения могут быть выбраны таким образом, чтобы не подвергаться димеризации. Дополнительные трансмембранные домены будут очевидны квалифицированным специалистам в данной области техники. Примеры трансмембранных (ТМ) районов, используемых в конструкциях CAR, являются: 1) ТМ-район CD28 (Pule et al. Mol Ther, 2005, Nov;12(5):933-41; Brentjens et al. CCR, 2007, Sep 15;13(18 Pt 1):5426-35; Casucci et al. Blood, 2013, Nov 14;122(20):3461-72.); 2) ТМ-район OX40 (Pule et al. Mol Ther, 2005, Nov;12(5):933-41); 3) ТМ-район 41BB (Brentjens et al. CCR, 2007, Sep 15;13(18 Pt 1):5426-35); 4) ТМ-район CD3 дзета (Pule et al. Mol Ther, 2005, Nov; 12(5):933-41; Savoldo B, Blood, 2009, Jun 18;113(25):6392-402); 5) ТМ-район CD8a (Maher et al. Nat Biotechnol, 2002, Jan; 20(1):70-5.; Imai C, Leukemia, 2004, Apr;18(4):676-84; Brentjens et al. CCR, 2007, Sep 15;13(18 Pt 1):5426-35; Milone et al. Mol Ther, 2009, Aug;17(8):1453-64).
В одном варианте осуществления как трансмембранный, так и внутриклеточный, передающий сигналы домены получены из CD28. В одном варианте осуществления трансмембранный и внутриклеточный, передающий сигналы домен включает последовательность, приведенную ниже:
Трансмембранная и внутриклеточная часть CD28 человека (UNIPROT: P10747, CD28_HUMAN, положения 153-220)
FWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPR DFAAYRS (SEQ ID NO:18)
В одном варианте осуществления трансмембранный и внутриклеточный, передающий сигналы домен по меньшей мере на 85, 90, 95, 97, 98 или 99% идентичен последовательности SEQ ID NO:18.
Спейсерный домен - рецептор фактора роста нервов с низким сродством (LNGFR)
CAR настоящего изобретения включает экстраклеточный спейсерный домен. Экстраклеточный спейсерный домен присоединен к антигенспецифическому таргетирующему району и трансмембранному домену.
CAR настоящего изобретения включает экстраклеточный спейсер, который включает по крайней мере часть экстраклеточного домена рецептора фактора роста нервов человека с низким сродством (LNGFR) или ее производное.
LNGFR не экспрессируется в большинстве гемопоэтических клеток человека, так что делается возможным количественный анализ экспрессии трансдукцированных генов с помощью иммунофлуоресценции, с разрешающей способностью=одной клетке. Таким образом, анализ экспрессии LNGFR с использованием сортера клеток с активацией флуоресценции может быть выполнен на трансдуцированных клетках для изучения экспрессии гена. Более подробную информацию об анализе с использованием LNGFR можно найти в Mavilio 1994, Blood 83, 1988-1997.
Последовательность LNGFR человека представлена на фиг. 8 (SEQ ID NO:14).
В настоящем изобретении в одном варианте осуществления используется усеченный LNGFR (также известный как ΔLNGFR). Предпочтительно, когда LNGFR, используемый в настоящем изобретении, является усеченным в его внутрицитоплазматическом домене. Такое усечение описано в Mavilio 1994.
Таким образом, предпочтительно спейсер LNGFR настоящего изобретения включает по крайней мере часть экстраклеточного домена, или ее производное, но в нем отсутствует внутриклеточный домен LNGFR. Экстраклеточный домен может включать аминокислоты 29-250 из LNGFR или его производного.
Экстраклеточный домен LNGFR человека (UNIPROT #P08138, TNR16_HUMAN, положения 29-250)
KEACPTGLYTHSGECCKACNLGEGVAQPCGANQTVCEPCLDSVTFSDVVSATEPCKPCTECVGLQSMSAPCVEADDAVCRCAYGYYQDETTGRCEACRVCEAGSGLVFSCQDKQNTVCEECPDGTYSDEANHVDPCLPCTVCEDTERQLRECTRWADAECEEIPGRWITRSTPPEGSDSTAPSTQEPEAPPEQDLIASTVAGVVTTVMGSSQPVVTRGTTDN (SEQ ID NO:19)
Предпочтительно, когда в LNGFR отсутствует сигнальный пептид.
В одном варианте осуществления спейсер включает по крайней мере часть белка, идентичного по меньшей мере на 85, 90, 95, 96, 97, 98 или 99% экстраклеточному домену LNGFR (например, SEQ ID NO:19). В одном варианте осуществления спейсер включает по крайней мере часть белка, идентичного по меньшей мере на 85, 90, 95, 96, 97, 98 или 99% аминокислотам 29-250 белка LNGFR.
LNGFR включает 4 TNFR-Cys домена (TNFR-Cys 1, TNFR-Cys 2, TNFR-Cys 3 и TNFR-Cys 4). Последовательности доменов проиллюстрированы ниже:
TNFR-Cys 1, SEQ ID NO:9
ACPTGLYTHSGECCKACNLGEGVAQPCGANQTVC
TNFR-Cys 2, SEQ ID NO:10
PCLDSVTFSDVVSATEPCKPCTECVGLQSMSAPCVEADDAVC
TNFR-Cys 3, SEQ ID NO:11
RCAYGYYQDETTGRCEACRVCEAGSGLVFSCQDKQNTVC
TNFR-Cys 4. SEQ ID NO:12
ECPDGTYSDEANHVDPCLPCTVCEDTERQLRECTRWADAEC
В одном варианте осуществления спейсер включает TNFR-Cys 1, 2 и 3 домены или их фрагменты или производные. В другом варианте осуществления спейсер включает TNFR-Cys 1, 2, 3 и 4 домены или их фрагменты или производные.
В одном варианте осуществления спейсер включает последовательность, идентичную по меньшей мере на 80, 85, 90, 95, 96, 97, 98, 99% или 100% TNFR-Cys 1 (SEQ ID NO:9), последовательность, идентичную по меньшей мере на 80, 85, 90, 95, 96, 97, 98, 99% или 100% TNFR-Cys 2 (SEQ ID NO:10), или последовательность, идентичную по меньшей мере на 80, 85, 90, 95, 96, 97, 98, 99% или 100% TNFR-Cys 3 (SEQ ID NO:11). Спейсер может, кроме того, включать последовательность, идентичную по меньшей мере на 80, 85, 90, 95, 96, 97, 98, 99% или 100% TNFR-Cys 4 (SEQ ID NO:12).
Вместо того, чтобы включать в себя полный TNFR-Cys 4 домен, спейсер может включать TNFR-Cys 4 домен со следующими удаленными из указанного домена аминокислотами: NHVDPCLPCTVCEDTERQLRECTRW. В одном варианте осуществления аминокислоты NHVDPCLPCTVCEDTERQLRECTRW заменены следующими аминокислотами ARA.
В одном варианте осуществления в спейсере отсутствует богатая серином/треонином ножка LNGFR. В другом варианте осуществления спейсер включает богатую серином/треонином ножку LNGFR.
Спейсер может включать или состоять из последовательность(и) SEQ ID NO:1 или последовательность(и), идентичную по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98% или 99% SEQ ID NO:1.
Спейсер может включать или состоять из последовательность(и) SEQ ID NO:3 или последовательность(и), идентичную по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98% или 99% SEQ ID NO:3.
Спейсер может включать или состоять из последовательность(и) SEQ ID NO:5 или последовательность(и), идентичную по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98% или 99% SEQ ID NO:5.
Спейсер может включать или состоять из последовательность(и) SEQ ID NO:7 или последовательность(и), идентичную по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98% или 99% SEQ ID NO:7.
Спейсер может придавать CAR свойства из условия, чтобы делалась возможной иммуноселекция клеток, предпочтительно Т-клеток, экспрессирующих указанный CAR.
CAR настоящего изобретения (включающий спейсер, упоминаемый здесь) предпочтительно дает возможность Т-клеткам, экспрессирующим CAR, пролиферироваться в присутствии клеток, экспрессирующих антиген, для которого разработан CAR.
CAR настоящего изобретения (включающий спейсер, упоминаемый здесь) предпочтительно дает возможность Т-клеткам, экспрессирующим CAR, опосредовать терапевтически значимые противораковые эффекты против рака, который CAR предназначен делать мишенью.
CAR настоящего изобретения (включающий спейсер, упоминаемый здесь) предпочтительно пригоден для облегчения иммуноселекции клеток, трансдуцированных указанным CAR.
CAR настоящего изобретения, включающий спейсер на основе LNGFR, позволяет избежать активации нежелательных и потенциально токсичных нецелевых иммунных реакций и позволяет CAR-экспрессирующим Т-клеткам продолжать существовать in vivo, не будучи преждевременно устраненными иммунной системой хозяина.
Как упомянуто ниже, настоящее изобретение также охватывает применение вариантов, производных, гомологов и фрагментов спейсерных элементов, описанных здесь.
Производные и фрагменты
Помимо специфических белков, пептидов и нуклеотидов, упомянутых здесь, настоящее изобретение также охватывает применение их производных и фрагментов.
Термин «производное», используемый здесь, в отношении белков или полипептидов настоящего изобретения, включает любое замещение, изменение, модификацию, замену, делецию и/или добавление одного (или более) аминокислотных остатков из или в последовательности при условии, что полученный в результате белок или полипептид сохраняет желаемую функцию.
Как правило, аминокислотные замены могут быть составлены, например, из 1, 2 или от 3 до 10 или 20 замен при условии, что модифицированная последовательность сохраняет требуемую активность или способность. Замены аминокислот могут включать использование не встречающихся в природе аналогов.
Белки или пептиды, используемые в настоящем изобретении, могут также иметь делеции, вставки или замены аминокислотных остатков, которые вызывают молчащие изменения и дают в результате функционально эквивалентный белок. Преднамеренные аминокислотные замены могут быть осуществлены на основе сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатков, пока сохраняется эндогенная функция. Например, отрицательно заряженные аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту; положительно заряженные аминокислоты включают лизин и аргинин; и аминокислоты с незаряженными полярными концевыми группами, имеющие сходные величины гидрофильности, включают аспарагин, глутамин, серин, треонин и тирозин.
Консервативные замены могут быть осуществлены, например, в соответствии с приведенной ниже таблицей. Аминокислоты в одном и том же блоке во втором столбце и предпочтительно в той же строке в третьем столбце могут быть заменены друг на друга:
Производное может быть гомологом. Термин «гомолог», используемый здесь, означает структуру, имеющую определенную степень гомологии с аминокислотной последовательностью дикого типа и нуклеотидной последовательностью дикого типа. Термин «гомология» может быть приравнен к «идентичности».
Гомологичная последовательность может включать аминокислотную последовательность, которая может быть по меньшей мере на 50%, 55%, 65%, 75%, 85% или 90% идентична, предпочтительно по меньшей мере на 95% или 97% или 99% идентична рассматриваемой последовательности. Как правило, гомологи будут включать такие же активные центры и т.д., как и рассматриваемая аминокислотная последовательность. Хотя гомологию можно также рассматривать с точки зрения сходства (т.е. аминокислотные остатки, имеющие сходные химические свойства/функции), в контексте настоящего изобретения предпочтительно выражать гомологию в терминах идентичности последовательности.
Гомологичная последовательность может включать нуклеотидную последовательность, которая может быть по меньшей мере на 50%, 55%, 65%, 75%, 85% или 90% идентична, предпочтительно по меньшей мере на 95% или 97% или 99% идентична рассматриваемой последовательности. Хотя гомологию также можно рассматривать с точки зрения сходства, в контексте настоящего изобретения предпочтительно выражать гомологию в терминах идентичности последовательности.
Сравнения гомологии могут проводиться на глаз или, чаще всего, с помощью легкодоступных программ сравнения последовательностей. Эти коммерчески доступные компьютерные программы могут рассчитать процент гомологии или идентичности между двумя или более последовательностей.
Процент гомологии может быть рассчитан на протяжении прилегающих последовательностей, т.е. одну последовательность совмещают с другой последовательностью, и каждую аминокислоту в одной последовательности непосредственно сравнивают с соответствующей аминокислотой в другой последовательности, один остаток за один раз. Это называется совмещением, «не содержащим разрывы». Как правило, такие не содержащие разрывы совмещения выполняют только на протяжении относительно короткого числа остатков.
Несмотря на то, что это очень простой и согласующийся метод, он не в состоянии принять во внимание, что, например, в идентичной во всем остальном паре последовательностей одна вставка или делеция в нуклеотидной последовательности может привести к тому, что следующие кодоны будут выведены из совмещения, что таким образом может потенциально привести к значительному уменьшению процента гомологии при выполнении общего совмещения. Следовательно, большинство методов сравнения последовательностей разработано для получения оптимальных совмещений, которые принимают во внимание возможные вставки и делеции без неоправданного ухудшения общей оценки гомологии. Это достигается за счет вставки «разрывов» при совмещении последовательностей, чтобы попытаться максимизировать локальную гомологию.
Однако эти более сложные методы назначают «штрафы за разрывы» каждому разрыву, который возникает при совмещении, так что, для одного и того же числа идентичных аминокислот, совмещение последовательностей с наименьшим по возможности количеством разрывов, что отражает более высокую степень родства между двумя сравниваемыми последовательностями, получит более высокую оценку, чем совмещение со множеством разрывов. Как правило, используются «аффинные штрафы за разрывы», которые взимают относительно высокий штраф за существование разрыва и меньший штраф за каждый последующий остаток в разрыве. Это является наиболее часто используемой системой подсчета очков за разрывы. Высокие штрафы за разрывы будут, конечно, давать оптимизированные совмещения с меньшим количеством разрывов. Большинство программ для совмещений позволяет корректировать штрафы за разрывы. Однако предпочтительно использовать значения по умолчанию при использовании такого программного обеспечения для сравнения последовательностей. Например, при использовании пакета программ GCG Wisconsin Bestfit штраф за разрыв по умолчанию для аминокислотных последовательностей составляет -12 за разрыв и -4 за каждое расширение.
По этой причине для расчета максимального процента гомологии прежде всего требуется получения оптимального совмещения, принимая во внимание штрафы за разрывы. Подходящей компьютерной программой для проведения такого совмещения является пакет программ GCG Wisconsin Bestfit (University of Wisconsin, USA; Devereux et al. (1984) Nucleic Acids Res. 12: 387). Примеры другого программного обеспечения, которое может выполнять сравнение последовательностей, включают, но без ограничения ими, пакет программ BLAST (смотрите Ausubel et al. (1999) там же главу 18), FASTA (Atschul et al. (1990) J. Mol. Biol. 403-410) и набор инструментов для сравнения GENEWORKS. Как BLAST, так и FASTA доступны для поиска в режиме оффлайн (вне Интернета) и онлайн (смотрите Ausubel et al. (1999) там же, страницы с 7-58 до 7-60). Однако для некоторых применений предпочтительно использовать программу GCG Bestfit. Еще один инструмент, называемый BLAST 2 Sequences, также доступен для сравнения белковых и нуклеотидных последовательностей (смотрите FEMS Microbiol Lett (1999) 174: 247-50; FEMS Microbiol Lett (1999) 177: 187-8).
Хотя конечный процент гомологии можно измерить в терминах идентичности, сам процесс совмещения обычно не основывается на попарном сравнении по принципу «все или ничего». Вместо этого, обычно используется матрица дифференцированных оценок сходства, которая присваивает оценки каждому попарному сравнению на основе химического сходства или эволюционного расстояния. Примером такой обычно используемой матрицы является матрица BLOSUM62 - матрица по умолчанию для пакета программ BLAST. Программы GCG Wisconsin обычно используют либо общие значения по умолчанию или таблицу сравнения пользовательских символов, если предоставляется (смотрите инструкцию для получения более подробной информации). Для некоторых применений предпочтительно использовать общие значения по умолчанию для пакета программ GCG или, в случае другого программного обеспечения, матрицу по умолчанию, такую как BLOSUM62.
После того как программа произвела оптимальное совмещение, можно вычислить процент гомологии, предпочтительно процент идентичности последовательностей. Программное обеспечение, как правило, делает это как часть сравнения последовательностей и выдает численный результат.
Фрагменты обычно относятся к выбранной области полипептида или полинуклеотида, которая представляет интерес функционально. «Фрагмент», таким образом, относится к аминокислотной последовательности, которая представляет собой часть полноразмерного полипептида, или последовательности нуклеиновой кислоты, которая представляет собой часть полноразмерного полинуклеотида. Поскольку фрагменты представляют интерес функционально, например, сохраняют желаемую функциональность, они, следовательно, исключают например, одну аминокислоту или одну нуклеиновую кислоту.
Такие производные и фрагменты могут быть получены, используя стандартные методы рекомбинантной ДНК, такие как сайт-направленный мутагенез. Если вставки должны быть сделаны, синтетическая ДНК, кодирующая вставку вместе с 5' и 3' фланкирующими районами, соответствующими встречающейся в природе последовательности с той или другой стороны от места вставки, могут быть созданы. Фланкирующие районы будут содержать удобные сайты рестрикции, соответствующие сайтам во встречающейся в природе последовательности, так что последовательность может быть разрезана с помощью соответствующего фермента(ов), и синтетическая ДНК лигирована в место разреза. Затем ДНК экспрессируют в соответствии с настоящим изобретением для выработки кодируемого белка. Эти методы являются только иллюстрацией многочисленных стандартных методов, известных в данной области техники для манипулирования последовательностями ДНК, и другие известные методы могут быть также использованы.
Полинуклеотиды
Полинуклеотиды настоящего изобретения могут включать ДНК или РНК. Они могут быть одноцепочечными или двухцепочечными. Квалифицированному специалисту будет понятно, что многочисленные различные полинуклеотиды могут кодировать один и тот же полипептид в результате вырожденности генетического кода. Кроме того, следует понимать, что квалифицированный специалист может, используя обычные методы, произвести замены нуклеотидов, которые не влияют на последовательность полипептида, кодируемую полинуклеотидами настоящего изобретения, чтобы отразить частоту использования кодонов любого конкретного организма хозяина, в котором полипептиды настоящего изобретения должны быть экспрессированы.
Полинуклеотиды могут быть модифицированы любым способом, доступным в данной области техники. Такие модификации могут быть выполнены для того, чтобы увеличить in vivo активность или продолжительность существования полинуклеотидов настоящего изобретения.
Полинуклеотиды, такие как ДНК-полинуклеотиды, могут быть получены рекомбинантным способом, синтетически или любыми способами, доступными квалифицированным специалистам в данной области техники. Они также могут быть клонированы с помощью стандартных методов.
Более длинные полинуклеотиды, как правило, получают, используя рекомбинантные способы, например, используя методы клонирования с использованием полимеразной цепной реакции (ПЦР). Они будут включать в себя получение пары праймеров (например, приблизительно от 15 до 30 нуклеотидов), фланкирующих последовательность-мишень, которую желательно клонировать, приведение праймеров в контакт с мРНК или кДНК, полученной из клетки животного или человека, выполнение полимеразной цепной реакции в условиях, которые вызывают амплификацию желаемого района, выделение амплифицированного фрагмента (например, путем очистки реакционной смеси с использованием агарозного геля) и выделение амплифицированной ДНК. Могут быть разработаны праймеры, которые содержат подходящие сайты узнавания ферментами рестрикции, так что амплифицированная ДНК может быть клонирована в подходящий вектор.
Оптимизация частоты использования кодонов
Полинуклеотиды, используемые в настоящем изобретении, могут быть оптимизированы в отношении частоты использования кодонов. Оптимизация частоты использования кодонов ранее была описана в WO 1999/41397 и WO 2001/79518. Различные клетки отличаются использованием в них отдельных кодонов. Это предпочтение кодонов соответствует смещению относительной численности отдельных тРНК в этом типе клеток. Изменяя кодоны в последовательности таким образом, чтобы подогнать их к соответствию с относительным содержанием соответствующих тРНК, можно увеличить экспрессию. К тому же, можно уменьшить экспрессию путем намеренного выбора кодонов, для которых соответствующие тРНК, как известно, являются редкими в конкретном типе клеток. Таким образом, доступна дополнительная степень трансляционного контроля.
Векторы
Вектор представляет собой инструмент, который делает возможным или облегчает перенос структуры из одной среды в другую. В соответствии с настоящим изобретением, и в качестве примера, некоторые векторы, используемые в методах рекомбинантных нуклеиновых кислот, делают возможным перенос таких структур, как сегмент нуклеиновой кислоты (например, сегмент гетерологичной ДНК, такой как сегмент гетерологичной кДНК), в клетки-мишени. Векторы могут быть невирусными или вирусными. Примеры векторов, используемых в методах рекомбинантных нуклеиновых кислот, включают, но без ограничения ими, плазмиды, молекулы мРНК (например, in vitro транскрибированные мРНК), хромосомы, искусственные хромосомы и вирусы. Вектор может также быть, например, «голой» нуклеиновой кислотой (например, ДНК). В своей простейшей форме, сам вектор может быть представляющим интерес нуклеотидом.
Векторы, используемые в настоящем изобретении, могут представлять собой, например, плазмиды, мРНК или вирусные векторы и могут включать промотор для экспрессии полинуклеотида и необязательно регулятор промотора.
Векторы, включающие полинуклеотиды настоящего изобретения, могут быть введены в клетки с использованием различных методов, известных в данной области техники, таких как трансформация и трансдукция. В данной области техники известно несколько методов, например, инфицирование рекомбинантными вирусными векторами, такими как ретровирусные, лентивирусные, аденовирусные, аденоассоциированные вирусные, бакуловирусные векторы и векторы на основе вируса простого герпеса; прямая инъекция нуклеиновых кислот и баллистическая трансформация.
Невирусные системы доставки включают, но без ограничения ими, методы трансфекции ДНК. Здесь, трансфекция включает процесс с использованием невирусного вектора для доставки гена в клетку-мишень.
Типичные способы трансфекции включают электропорацию, баллистическую трансфекцию ДНК, трансфекцию с использованием липидов, трансфекцию с использованием упакованной ДНК, липосомы, иммунолипосомы, липофектин, трансфекцию с использованием катионных агентов, катионные поверхностные амфифилы (CFA) (Nat Biotechnol (1996) 14: 556) и их комбинации.
Ретровирусные векторы
В одном варианте осуществления вектор, используемый в настоящем изобретении, представляет собой вектор на основе ретровируса, которой был сконструирован с использованием методов генной инженерии таким образом, что он не может реплицироваться и давать потомство инфекционных вирусных частиц, как только вирус вошел в клетку-мишень. Существует множество ретровирусов, которые широко используются для доставки генов как в условиях культивирования тканей, так и в живые организмы. Примеры включают, и не ограничиваются ими, вирус лейкоза мышей (MLV), вирус иммунодефицита человека (ВИЧ-1), вирус инфекционной анемии лошадей (EIAV), вирус опухоли молочной железы мышей (MMTV), вируса саркомы Рауса (RSV), вирус саркомы Фудзинами (FuSV), вирус лейкоза Молони мышей (Mo-MLV), вирус остеосаркомы FBR мышей (FBR МСВ), вирус саркомы Молони мышей (Мо-МСВ), вирус лейкоза Абельсона мышей (А-MLV), вирус-29 миелоцитоматоза птиц (MC29) и вирус эритробластоза птиц (AEV) и все остальные вирусы семейства Retroviridiae, включая лентивирусы. Подробный список ретровирусов можно найти в Coffin et al., 1997, ʺretrovirusesʺ, Cold Spring Harbour Laboratory Press Eds: JM Coffin, SM Hughes, HE Varmus pp 758-763.
Основной структурой генома ретровирусов является 5' LTR и 3' LTR, между или внутри которых расположены сигнальная последовательность упаковки, которая делает возможной упаковку генома, сайт связывания праймера, сайты интеграция, которые делают возможной интеграцию в геном клетки-хозяина, и гены gag, pol и env, кодирующие компоненты упаковки - они представляют собой полипептиды, необходимые для сборки вирусных частиц. Более сложные ретровирусы имеют дополнительные признаки, такие как последовательности rev и RRE у ВИЧ, которые делают возможным эффективный экспорт РНК-транскриптов с интегрированного провируса из ядра в цитоплазму инфицированной клетки-мишени.
В провирусе, эти гены фланкированы на обоих концах участками, называемыми длинными концевыми повторами (LTR). LTR ответственны за интеграцию провируса и транскрипцию. LTR также служат в качестве энхансерных-промоторных последовательностей и могут контролировать экспрессию вирусных генов. Инкапсулирование ретровирусных РНК происходит благодаря последовательности psi, расположенной на 5'-конце вирусного генома.
Сами LTR представляют собой одинаковые последовательности, которые можно разделить на три элемента, которые называются U3, R и U5. U3 происходит от последовательности, уникальной для 3' конца РНК. R происходит от последовательности, повторяющейся на обоих концах РНК, и U5 происходит от последовательности, уникальной для 5' конца РНК. Размеры трех элементов могут значительно варьировать среди различных ретровирусов.
В геноме дефектного ретровирусного вектора gag, pol и env могут отсутствовать или не быть функциональными. R-районы на обоих концах РНК представляют собой повторяющиеся последовательности. U5 и U3 представляют собой уникальные последовательности на 5' и 3'-концах РНК-генома, соответственно.
Более предпочтительно, когда вирусный вектор является таргетирующим вектором, то есть он имеет тканевой тропизм, который изменен по сравнению со встречающимся в природе вирусом, так что вектор является нацеленным на конкретные клетки. Это может быть достигнуто путем изменения белка Env ретровируса. Предпочтительно белок оболочки представляет собой нетоксичную оболочку или оболочку, которая может продуцироваться в нетоксичных количествах в первичных клетках-мишенях, такую как, например, амфотропная оболочка MMLV или модифицированная амфотропная оболочка.
Предпочтительно, когда оболочкой является оболочка, которая делает возможной трансдукцию клеток человека. Примеры подходящих генов env включают, но без ограничения ими, VSV-G, амфотропный env MLV, такой как env 4070A, env вируса лейкоза кошек RD114 или гемагглютинин (HA) из вируса гриппа. Белок Env может быть таким, который способен связываться с рецептором на ограниченном количестве типов клеток человека, и может быть сконструированным белком оболочки, содержащим таргетирующие составляющие. Кодирующие последовательности env и gag-pol транскрибируются с промотора и необязательно энхансера, являющего активным в выбранной пакующей линии клеток, и единица транскрипции терминируется сигналом полиаденилирования. Например, если пакующая клетка представляет собой клетку человека, подходящей комбинацией промотор-энхансер является таковая из основного немедленно-раннего гена цитомегаловируса человека (hCMV-MIE), и может быть использован сигнал полиаденилирования из вируса SV40. Другие подходящие промоторы и сигналы полиаденилирования известны в данной области техники.
MLV
Предпочтительно, когда ретровирусный вектор, используемый в настоящем изобретении, представляет собой вектор в виде вируса лейкоза мышей (MLV). Ретровирусные векторы, полученные исходя из амфотропного вируса лейкоза Молони мышей (MLV-A), обычно используются в клинических протоколах по всему миру. Эти вирусы используют рецепторы-транспортеры фосфата на клеточной поверхности для вхождения, а затем надолго интегрируются в хромосомы пролиферирующихся клеток. Эти гены затем сохраняются в течение времени существования клетки. Активность генов в конструкциях на основе MLV легко контролируется и может быть эффективной в течение длительного времени. Клинические испытания, проведенные с этими системами на основе MLV, показали, что они хорошо переносятся без каких-либо неблагоприятных побочных эффектов.
Пример вектора MLV для использования в настоящем изобретении представляет собой вектор, происходящий от SFCMM-3, который несет как ген «самоубийства» HSV-tk, так и маркерный ген ΔLNGFR (Verzeletti 98, Human Gene Therapy, 9:2243). Первоначальным вектором, использованным для получения SFCMM-3, является LXSN (Miller et al. Improved retroviral vectors for gene transfer and expression. BioTechniques 7:980-990, 1989) (номер доступа в Genebank 28248). Вектор LXSN был модифицирован путем вставки гена HSV-tk в уникальный сайт HpaI (расщепление с образованием «тупых концов»), удаления гена neo путем расщепления HindIII и NaeI и вставки кДНК, кодирующей ΔLNGFR, в этот сайт.
Лентивирусный вектор
В одном варианте осуществления вектор настоящего изобретения может представлять собой лентивирусный вектор. Лентивирусные векторы являются частью большой группы ретровирусных векторов. Подробный список лентивирусов можно найти в Coffin et al. (ʺRetrovirusesʺ 1997 Cold Spring Harbour Laboratory Press Eds: JM Coffin, SM Hughes, HE Varmus pp 758-763). Вкратце, лентивирусы можно разделить на группы лентивирусов приматов и не приматов. Примеры лентивирусов приматов включают, но без ограничения ими: вирус иммунодефицита человека (ВИЧ), возбудитель синдрома приобретенного иммунодефицита человека (СПИД) и обезьяний вирус иммунодефицита (SIV). Группа лентивирусов не приматов включает вирус висна-маеди (VMV) прототипа «медленный вирус», а также родственный вирус артрита-энцефалита коз (CAEV), вирус инфекционной анемии лошадей (EIAV) и недавно описанный вирус иммунодефицита кошачьих (FIV), и вирус иммунодефицита крупного рогатого скота (BIV).
Различие между семейством лентивирусов и другими типами ретровирусов заключается в том, что лентивирусы обладают способностью инфицировать как делящиеся, так и не делящиеся клетки. В противоположность этому, другие ретровирусы - такие как MLV - не способны инфицировать не делящиеся или медленно делящиеся клетки, такие как те, которые составляют, например, мышечную ткань, ткань головного мозга, легких и печени. Поскольку лентивирусы способны к трансдукции окончательно дифференцированных/первичных клеток, использование стратегии скрининга лентивирусов позволяет выбрать библиотеку в первичной, целевой не делящейся или медленно делящейся клетке-хозяине.
Аденовирусные векторы
В другом варианте осуществления вектором настоящего изобретения может быть аденовирусный вектор. Аденовирус представляет собой содержащий двухцепочечную, линейную ДНК вирус, который не проходит через стадию РНК-посредника. Существует более 50 различных серотипов аденовируса человека, которые делятся на 6 подгрупп на основе гомологии генетических последовательностей. Естественной мишенью аденовируса является эпителий дыхательных путей и желудочно-кишечного тракта, как правило, порождая лишь легкие симптомы. Серотипы 2 и 5 (с 95% гомологией последовательности) чаще всего используются в аденовирусных векторных системах и, как правило, связаны с инфекциями верхних дыхательных путей у молодых.
Аденовирусы представляют собой правильные икосаэдры без оболочки. Типичным аденовирусом является вирус с заключенной в капсид ДНК диаметром 140 нм. Икосаэдральная симметрия вируса состоит из 152 капсомеров: 240 гексонов и 12 пентонов. Ядро частицы содержит линейную дуплексную ДНК размером 36 т.п.о., которая ковалентно связана на 5'-концах с терминальным белком (ТР), который действует в качестве праймера для репликации ДНК. ДНК содержит инвертированные концевые повторы (ITR), и их длина меняется в зависимости от серотипа.
Аденовирус представляет собой содержащий двухцепочечную ДНК вирус без оболочки, который способен к in vivo и in vitro трансдукции широкого ряда типов клеток человека и нечеловеческого происхождения. Эти клетки включают эпителиальные клетки дыхательных путей, гепатоциты, мышечные клетки, кардиомиоциты, синовиоциты, первичные эпителиальные клетки молочных желез и постмитотически окончательно дифференцированные клетки, такие как нейроны.
Аденовирусные векторы также способны к трансдукции не делящихся клеток. Это очень важно для таких заболеваний, как муковисцидоз, в случае которого пораженные клетки в эпителии легкого имеют низкую скорость кругооборота. На самом деле, несколько испытаний ведутся с использованием аденовирус-опосредованного переноса белка транспортера муковисцидоза (CFTR) в легкие взрослых пациентов, больных муковисцидозом.
Аденовирусы были использованы в качестве векторов для генной терапии и для экспрессии гетерологичных генов. Большой (36 т.п.о.) геном может вместить вплоть до 8 т.п.о. вставки чужеродной ДНК и способен эффективно реплицироваться в линии дополняющих клеток с продукцией очень высоких титров вплоть до 1012. Таким образом, аденовирус является одной из лучших систем для исследования экспрессии генов в первичных не размножающихся клетках.
Для экспрессии вирусных или чужеродных генов с генома аденовируса не требуется размножающаяся клетка. Аденовирусные векторы входят в клетки с помощью опосредованного рецептором эндоцитоза. Оказавшись внутри клетки, аденовирусный вектор редко интегрируется в хромосому хозяина. Вместо этого, он функционирует эписомально (независимо от генома хозяина) в виде линейного генома в ядре хозяина. Следовательно, применение рекомбинантного аденовируса устраняет проблемы, связанные со случайной интеграцией в геном хозяина.
Поксвирусные векторы
Поксвирусные векторы могут быть использованы в соответствии с настоящим изобретением, так как крупные фрагменты ДНК легко клонировать в их геном, и были описаны рекомбинантные аттенюированные варианты вируса осповакцины (Meyer et al. 1991; Smith and Moss, 1983).
Примеры поксвирусных векторов включают, но без ограничения ими, лепорипоксвирус: Upton et al. 1986, (вирус фиброма Шоупа); каприпоксвирус: Gershon et al., 1989, (вирус оспы овец-1 из Кении); ортопоксвирус: Weir et al., 1983, (вирус коровьей оспы); Esposito et al., 1984, (вирус оспы обезьян и натуральной оспы); Hruby et al., 1983, (вирус коровьей оспы); Kilpatrick et al., 1985, (вирус опухоли обезьян Яба); авипоксвирус: Binns et al., (1988) (вирус птичьей оспы); Boyle et al., 1987, (вирус оспы кур); Schnitzlein et al., 1988, (вирус оспы кур, вирус оспы перепелов); энтомопоксвирус (Lytvyn et al., 1992).
Поксвирусные векторы широко используются в качестве векторов для экспрессии интересующих генов в эукариотических клетках. Легкость их клонирования и размножения во множестве клеток-хозяев привела, в частности, к широкому использованию поксвирусных векторов для экспрессии чужеродного белка и в качестве средств доставки антигенов вакцины.
Векторы на основе вируса коровьей оспы
Вектор настоящего изобретения может представлять собой вектор на основе вируса коровьей оспы, такой как MVA или NYVAC. Наиболее предпочтительным является штамм модифицированного вируса коровьей оспы Анкара (MVA) или штамм, происходящий от него. Альтернативные варианты векторов на основе вируса коровьей оспы включают векторы на основе авипоксвирусов, такие как вирус птичьей оспы или оспы канареек, известный как ALVAC, и производные от него штаммы, которые могут инфицировать и экспрессировать рекомбинантные белки в клетках человека, но не способны реплицироваться.
Клетки
Настоящим изобретением также обеспечиваются сконструированные с помощью генной инженерии клетки, которые включают и стабильно экспрессируют CAR настоящего изобретения.
Антигенспецифические таргетирующие домены могут быть способны к специфическому связыванию, MHC-неограниченным образом, с антигеном, который обычно не связывается с Т-клеточным рецептором таким образом. В одном варианте осуществления антигенспецифические таргетирующие районы включают специфические в отношении мишени антитела или их функциональные эквиваленты или их фрагменты или производные. Антигенспецифическое антитело может представлять собой Fab-фрагмент антитела или одноцепочечный вариабельный фрагмент (scFv) антитела.
Сконструированные с помощью генной инженерии клетки, которые могут включать и экспрессировать CAR настоящего изобретения, включают, но без ограничения ими, Т-клетки, необученные Т-клетки, стволовые Т-клетки памяти, центральные Т-клетки памяти, эффекторные Т-клетки памяти, клетки-природные киллеры, гемопоэтические стволовые клетки и/или клетки, способные давать начало терапевтически соответствующему потомству. В одном варианте осуществления сконструированные с помощью генной инженерии клетки являются аутологичными клетками. В качестве примера, отдельные Т-клетки настоящего изобретения могут быть CD4+/CD8-, CD4-/CD8+, CD4-/CD8- или CD4+/CD8+. Т-клетки могут представлять собой смешанную популяцию CD4+/CD8- и CD4-/CD8+ клеток или популяцию одного клона.
Генетически модифицированные клетки могут быть получены путем стабильной трансфекции клеток с использованием ДНК, кодирующей CAR настоящего изобретения.
Различные методы дают стабильные трансфектанты, которые экспрессируют CAR настоящего изобретения. В одном варианте осуществления способ стабильной трансфекции и перенацеливания клеток осуществляют путем электропорации с использованием голой ДНК. При использовании голой ДНК, время, необходимое для создания перенацеленных клеток, может быть значительно уменьшено. Дополнительные методы генной инженерии клеток с использованием голой ДНК, кодирующей CAR настоящего изобретения, включают, но без ограничения, химические методы трансформации (например, с использованием фосфата кальция, дендримеров, липосом и/или катионных полимеров), нехимические методы трансформации (например, электропорацию, оптическую трансформацию, электроперенос генов и/или гидродинамическую доставку) и/или методы на основе частиц (например, импалефекция, используя генную пушку, и/или магнетофекцию). Трансфецированные клетки, демонстрирующие присутствие одного интегрированного не перестроенного вектора и экспрессию CAR, могут быть размножены ех vivo. В одном варианте осуществления клетки, выбранные для ех vivo размножения, являются CD8+ и демонстрируют способность специфически распознавать и лизировать антигенспецифические клетки-мишени.
Вирусные методы трансдукции также могут быть использованы для создания перенацеленных клеток, которые экспрессируют CAR настоящего изобретения.
Стимуляция Т-клеток с помощью антигена при соответствующих условиях приводит к пролиферации (размножению) клеток и/или продукции IL-2. Клетки, включающие CAR настоящего изобретения, будут увеличиваться в количестве в ответ на связывание одного или более антигенов с антигенспецифическими таргетирующими районами CAR. Настоящим изобретением также обеспечивается способ получения и увеличения количества клеток, экспрессирующих CAR. Способ может включать трансфекцию или трансдукцию клеток вектором, экспрессирующим CAR, после стимуляции клеток: 1) поликлональными стимулами, такими как бесклеточные каркасы, предпочтительно сферы с оптимальным размером, содержащие по крайней мере активирующий полипептид, предпочтительно антитело, специфическое для CD3, и активирующий полипептид, предпочтительно антитело, специфическое для CD28; 2) опухолевыми клетками, экспрессирующими антиген-мишень; 3) естественными искусственными антигенпрезентирующими клетками, и культивирование их вместе с цитокинами, включающими IL-2, IL-7, IL-15, IL-21 отдельно или в комбинации.
Терапевтические методы и фармацевтические композиции
Здесь обеспечиваются способы лечения заболевания, связанного с антигеном, являющегося мишенью CAR настоящего изобретения, у субъекта, нуждающегося в этом. Способ включает введение эффективного количества CAR, полинуклеотида или вектора, кодирующего CAR, или клетки, экспрессирующей указанный CAR, для осуществления лечения заболевания, связанного с этим антигеном, у субъекта.
Также обеспечивается фармацевтическая композиция, включающая CAR настоящего изобретения. CAR настоящего изобретения в композиции может быть любым одним или более из полинуклеотида, кодирующего CAR, вектора, кодирующего CAR, белка, включающего CAR, или генетически модифицированных клеток, включающих CAR.
Фармацевтическая композиция представляет собой композицию, которая включает или состоит из терапевтически эффективное(го) количество(а) фармацевтически активного агента. Она предпочтительно включает фармацевтически приемлемый носитель, разбавитель или наполнитель (в том числе их комбинации). Приемлемые носители или разбавители для терапевтического применения хорошо известны в фармацевтической области и описаны, например, в Remington's Pharmaceutical Sciences, Mack Publishing Co. (A. R. Gennaro edit. 1985). Выбор фармацевтического носителя, наполнителя или разбавителя может быть осуществлен с учетом предполагаемого пути введения и стандартной фармацевтической практики. Фармацевтические композиции могут включать в качестве - или в дополнение к - носителя(ю), наполнителя или разбавителя любое подходящее связующее вещество(а), смазочный материал(ы), суспендирующий агент(ы), обволакивающее средство(а) или солюбилизатор(ы).
Примеры фармацевтически приемлемых носителей включают, например, воду, солевые растворы, спирт, силикон, воски, вазелин, растительные масла, полиэтиленгликоли, пропиленгликоль, липосомы, сахара, желатин, лактозу, амилозу, стеарат магния, тальк, поверхностно-активные вещества, кремниевую кислоту, вязкий парафин, парфюмерное масло, моноглицериды и диглицериды жирных кислот, сложные эфиры жирных кислот, растворенные в петролейном эфире, гидроксиметилцеллюлозу, поливинилпирролидон и тому подобное.
Примеры
Пример 1 - Методы
Создание конструкций CD44v6-CAR.28z со спейсерами на основе LNGFR
Последовательности спейсеров на основе LNGFR происходили из экстраклеточной части рецептора фактора роста нервов с низким сродством (LNGFR), за исключением сигнального пептида (P08138, TNR16_HUMAN). Длинная конструкция дикого типа (NWL) содержит как четыре богатых цистеином домена TNFR, так и богатую серином/треонином ножку. Короткая конструкция дикого типа (NWS) содержит только четыре богатых цистеином домена TNFR. Мутированная длинная конструкция (NML) содержит четыре богатых цистеином домена TNFR, богатую серином/треонином ножку и включает специфическую модификацию в четвертом домене во избежание связывания с NGF (Yan et al. J Biol Chem, 1991, Jun 25;266(18):12099-104). Мутированная короткая конструкция (NMS) содержит только четыре богатых цистеином домена TNFR, включая специфическую модификацию в четвертом домене. Спейсеры были синтезированы GENEART, фланкированы специфическими сайтами рестрикции (BamHI и PflMI), чтобы сделать возможным клонирование в принадлежащую авторам настоящего изобретения, оригинальную CD44v6-специфическую конструкцию CAR второго поколения (фиг. 9, SEQ ID NO:15), вместо спейсера CH2CH3 IgG1. Все конструкции были оптимизированы в отношении частоты использования кодонов для экспрессии в организме человека. Все конструкты были экспрессированы в основах SFG-RV (широко используемом ретровирусном векторе на основе MoMLV, который опирается на сплайсинг для экспрессии генов (Riviere et al., PNAS, 1995, Jul 18;92(15):6733-7)).
Спейсер LNGFR дикого типа длинный (NWL):
Последовательность белка (SEQ ID NO:1)
KEACPTGLYTHSGECCKACNLGEGVAQPCGANQTVCEPCLDSVTFSDVVSATEPCKPCTECVGLQSMSAPCVEADDAVCRCAYGYYQDETTGRCEACRVCEAGSGLVFSCQDKQNTVCEECPDGTYSDEANHVDPCLPCTVCEDTERQLRECTRWADAECEEIPGRWITRSTPPEGSDSTAPSTQEPEAPPEQDLIASTVAGVVTTVMGSSQPVVTRGTTDN.
Нуклеотидная последовательность (SEQ ID NO:2):
AAAGAGGCCTGCCCCACCGGCCTGTACACCCACAGCGGAGAGTGCTGCAAGGCCTGCAACCTGGGAGAGGGCGTGGCCCAGCCTTGCGGCGCCAATCAGACCGTGTGCGAGCCCTGCCTGGACAGCGTGACCTTCAGCGACGTGGTGTCCGCCACCGAGCCCTGCAAGCCTTGCACCGAGTGTGTGGGCCTGCAGAGCATGAGCGCCCCCTGCGTGGAAGCCGACGACGCCGTGTGTAGATGCGCCTACGGCTACTACCAGGACGAGACAACCGGCAGATGCGAGGCCTGTAGAGTGTGCGAGGCCGGCAGCGGCCTGGTGTTCAGTTGTCAAGACAAGCAGAATACCGTGTGTGAAGAGTGCCCCGACGGCACCTACAGCGACGAGGCCAACCACGTGGACCCCTGCCTGCCCTGCACTGTGTGCGAGGACACCGAGCGGCAGCTGCGCGAGTGCACAAGATGGGCCGACGCCGAGTGCGAAGAGATCCCCGGCAGATGGATCACCAGAAGCACCCCCCCTGAGGGCAGCGACAGCACCGCCCCTAGCACCCAGGAACCTGAGGCCCCTCCCGAGCAGGACCTGATCGCCTCTACAGTGGCCGGCGTGGTGACAACCGTGATGGGCAGCTCTCAGCCCGTGGTGACACGGGGCACCACCGACAAT.
Спейсер LNGFR дикого типа короткий (NWS):
Последовательность белка (SEQ ID NO:3):
KEACPTGLYTHSGECCKACNLGEGVAQPCGANQTVCEPCLDSVTFSDVVSATEPCKPCTECVGLQSMSAPCVEADDAVCRCAYGYYQDETTGRCEACRVCEAGSGLVFSCQDKQNTVCEECPDGTYSDEANHVDPCLPCTVCEDTERQLRECTRWADAECEE.
Нуклеотидная последовательность (SEQ ID NO:4):
AAAGAGGCCTGCCCCACCGGCCTGTACACCCACAGCGGAGAGTGCTGCAAGGCCTGCAACCTGGGAGAGGGCGTGGCCCAGCCTTGCGGCGCCAATCAGACCGTGTGCGAGCCCTGCCTGGACAGCGTGACCTTCAGCGACGTGGTGTCCGCCACCGAGCCCTGCAAGCCTTGCACCGAGTGTGTGGGCCTGCAGAGCATGAGCGCCCCCTGCGTGGAAGCCGACGACGCCGTGTGTAGATGCGCCTACGGCTACTACCAGGACGAGACAACCGGCAGATGCGAGGCCTGTAGAGTGTGCGAGGCCGGCAGCGGCCTGGTGTTCAGTTGTCAGGACAAGCAGAACACCGTGTGTGAAGAGTGCCCCGACGGCACCTACAGCGACGAGGCCAACCACGTGGACCCCTGCCTGCCCTGCACTGTGTGCGAGGACACCGAGCGGCAGCTGCGCGAGTGCACAAGATGGGCCGACGCCGAGTGCGAGGAA.
Спейсер LNGFR мутированный длинный (NML):
Последовательность белка (SEQ ID NO:5):
KEACPTGLYTHSGECCKACNLGEGVAQPCGANQTVCEPCLDSVTFSDVVSATEPCKPCTECVGLQSMSAPCVEADDAVCRCAYGYYQDETTGRCEACRVCEAGSGLVFSCQDKQNTVCEECPDGTYSDEAARAADAECEEIPGRWITRSTPPEGSDSTAPSTQEPEAPPEQDLIASTVAGVVTTVMGSSQPVVTRGTTDN.
Нуклеотидная последовательность (SEQ ID NO:6):
AAAGAGGCCTGCCCCACCGGCCTGTACACCCACAGCGGAGAGTGCTGCAAGGCCTGCAACCTGGGAGAGGGCGTGGCCCAGCCTTGCGGCGCCAATCAGACCGTGTGCGAGCCCTGCCTGGACAGCGTGACCTTCAGCGACGTGGTGTCCGCCACCGAGCCCTGCAAGCCTTGCACCGAGTGTGTGGGCCTGCAGAGCATGAGCGCCCCCTGCGTGGAAGCCGACGACGCCGTGTGTAGATGCGCCTACGGCTACTACCAGGACGAGACAACCGGCAGATGCGAGGCCTGTAGAGTGTGCGAGGCCGGCAGCGGCCTGGTGTTCAGTTGTCAAGACAAGCAGAATACCGTGTGTGAAGAGTGCCCCGACGGCACCTACAGCGACGAAGCCGCCAGAGCCGCCGACGCCGAGTGCGAAGAGATCCCCGGCAGATGGATCACCAGAAGCACCCCCCCTGAGGGCAGCGACAGCACCGCCCCTAGCACCCAGGAACCTGAGGCCCCTCCCGAGCAGGACCTGATCGCCTCTACAGTGGCCGGCGTGGTGACAACCGTGATGGGCAGCTCTCAGCCCGTGGTGACACGGGGCACCACCGACAAT.
Спейсер LNGFR мутированный короткий (NMS):
Последовательность белка (SEQ ID NO:7):
KEACPTGLYTHSGECCKACNLGEGVAQPCGANQTVCEPCLDSVTFSDVVSATEPCKPCTECVGLQSMSAPCVEADDAVCRCAYGYYQDETTGRCEACRVCEAGSGLVFSCQDKQNTVCEECPDGTYSDEAARAADAECEE.
Нуклеотидная последовательность (SEQ ID NO:8):
AAAGAGGCCTGCCCCACCGGCCTGTACACCCACAGCGGAGAGTGCTGCAAGGCCTGCAACCTGGGAGAGGGCGTGGCCCAGCCTTGCGGCGCCAATCAGACCGTGTGCGAGCCCTGCCTGGACAGCGTGACCTTCAGCGACGTGGTGTCCGCCACCGAGCCCTGCAAGCCTTGCACCGAGTGTGTGGGCCTGCAGAGCATGAGCGCCCCCTGCGTGGAAGCCGACGACGCCGTGTGTAGATGCGCCTACGGCTACTACCAGGACGAGACAACCGGCAGATGCGAGGCCTGTAGAGTGTGCGAGGCCGGCAGCGGCCTGGTGTTCAGTTGTCAGGACAAGCAGAACACCGTGTGTGAAGAGTGCCCCGACGGCACCTACAGCGACGAGGCCGCCCGGGCCGCCGACGCCGAGTGCGAGGAA.
Условные обозначения:
С подчеркиванием: богатый цистеином домен c номером 1 TNFR.
Жирный шрифт: богатый цистеином домен c номером 2 TNFR.
Жирный шрифт с подчеркиванием: богатый цистеином домен c номером 3 TNFR.
Курсив: богатый цистеином домен c номером 4 TNFR.
Курсив с подчеркиванием: богатая серином/треонином ножка
Трансдукция и условия культивирования
Т-клетки активировали с использованием CD3/CD28-сфер клеточного размера (ClinExVivo, Invitrogen) плюс IL-7/IL-15 (5 нг/мл, Peprotech) и RV-трансдуцировали путем двух циклов центрифужной инокуляции в день 2 и 3 после стимуляции. В день 6, сферы удаляли, и Т-клетки культивировали в среде RPMI 1640 (Gibco-BRL), 10% FBS (BioWhittaker) в присутствии IL-7 и IL-15. Поверхностная экспрессия CD44v6-специфических конструкций CAR со спейсером CH2CH3 (CHW и CHM) была обнаружена с помощью мАт, специфических для спейсера CH2CH3 IgG1 (Jackson Laboratories), в то время как поверхностную экспрессию CD44v6-специфических конструкций CAR со спейсерами на основе LNGFR (NWL, NWS, NML и НМС) анализировали с использованием LNGFR-специфических мАт от BD Bioscience (клон: C40-14579) или от Miltenyi (клон: ME20.4). Между днем 9 и днем 15 с момента активации, экспрессирующие CD44v6-CAR.28z со спейсером CH2CH3 IgG1 Т-клетки были FACS-отсортированы, используя поликлональные, специфические для CH2CH3 IgG1 мАт, в то время как экспрессирующие CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клетки окрашивали РЕ-конъюгированным, LNGFR-специфическим мАт C40-14579 и сортировали с помощью колонок, используя анти-PE парамагнитные сферы (Miltenyi). После сортировки увеличение количества Т-клеток представляли как кратное увеличение: число Т-клеток в день х/число Т-клеток после сортировки.
In vitro анализы для анализа специфического распознавания
В анализах сокультивирования, CAR-отсортированные Т-клетки культивировали с клетками-мишенями при различных соотношениях Е:Т. Спустя 4 дня выжившие клетки подсчитывали и анализировали с помощью FACS. Т-клетки, трансдуцированные нерелевантным CAR (CD19), всегда использовались в качестве контроля. Индекс элиминации рассчитывали следующим образом: 1-(количество остаточных клеток-мишеней в присутствии CD44v6.CAR28z+ Т-клеток)/(число остаточных клеток-мишеней в присутствии CTR.CAR28z+ Т-клеток). В анализах разведения CFSE, в CAR-отсортированные Т-клетки вводили CFSE, и их стимулировали с использованием облученных (10000 рад) опухолевых клеток в соотношении Е:Т=1:5 или биологически активных концентраций NGF. Спустя 6 дней, пролиферацию Т-клеток измеряли с помощью FACS, анализируя процент клеток, которые разбавили краситель CFSE.
Модели противоопухолевой эффективности с использованием ксенотрансплантата
Экспериментальные протоколы были одобрены Комитетом по уходу за животными и их использованием (IACUC). Для модели минимальной остаточной болезни, мышам NSG (Jackson) внутривенно вводили путем инфузии 1,5×106 лейкозных клеток Thp1/мышь. Через три дня мышей подвергали лечению путем внутривенного введения 5×106 отсортированных, экспрессирующих CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клеток, экспрессирующих CD44v6-CAR.28z со спейсером CH2CH3 Т-клеток или Т-клеток, несущих нерелевантный CAR (CD19). Приживление Т-клеток контролировали еженедельно с помощью кровопускания и анализа FACS. Через 7 недель мышей умерщвляли, и их печень анализировали с помощью гистопатологии и FACS на наличие клеток ТНР-1. Для модели устойчивого заболевания, мышам NSG вводили внутривенно 2×106 клеток миеломы MM1.S/мышь. Через пять недель после этого мышей подвергали лечению путем внутривенного введения 5×106 отсортированных, экспрессирующих CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клеток, экспрессирующих CD44v6-CAR.28z со спейсером CH2CH3 Т-клеток или Т-клеток, несущих нерелевантный CAR (CD19). Приживление Т-клеток и прогрессирование миеломы контролировали еженедельно с помощью кровопускания и анализа FACS (клетки миеломы будут отличать от Т-клеток по отличному CD45/CD3 человека фенотипу). Когда циркулирующие клетки MM1.S превышали 30 клеток/мкл, и/или мыши проявляли явные признаки связанного с опухолью страдания (паралич или >10% потери веса), мышей подвергали эвтаназии.
Проточная цитометрия
Для анализа FACS были использованы конъюгированные с FITC, PE, PerCP, PE-Cy7, APC, APC-Cy7 и Pacific Blue антитела, направленные против человеческого CD44v6, CD4 (е-Bioscience), CD123, CD19, CD14, CD3, CD8, CD45RA, CD62L, CXCR4, CD127, CD33, CD38, CD45, LNGFR, мышиного CD45, 7AAD (BD Biosciences) и CH2CH3 IgG1 (Jackson Laboratories). Клетки (2×105) инкубировали с антителами в течение 15 минут при 4οС и промывали PBS с 1% FBS. Образцы пропускали через проточный цитометр FACS Canto II (BD Biosciences), и данные были проанализированы с помощью программного обеспечения Flow Jo (Tree star Inc). Относительная интенсивность флуоресценции (RFI) была рассчитана следующим образом: средняя интенсивность флуоресценции образца/среднюю интенсивность флуоресценции соответствующего изотипа в качестве контроля.
Пример 2 - Создание конструкций CD44v6-CAR.28z со спейсерами на основе LNGFR.
Недавно авторы настоящего изобретения сконструировали CD44v6-специфический CAR на основе цепи CD3ζ в сочетании с эндокостимулирующим доменом CD28 (Casucci e al, Blood 2013, Nov 14;122(20):3461-72). В экстраклеточный спейсерный район этого CAR, спейсер CH2CH3 IgG1 был вставлен для лучшего таргетинга антигена CD44v6 и для обеспечения возможности отбора и in vivo отслеживания трансдуцированных Т-клеток. Однако серьезным недостатком CAR со спейсером CH2CH3 является их взаимодействие с Fcγ-рецепторами (FcγR) (Hombach et al. Gene Ther 2000, Jun;7(12):1067-75), что потенциально ведет к неспецифическому таргетингу клеток, экспрессирующих эти рецепторы, (например, моноцитов/макрофагов) и/или in vivo устранению трансдуцированных Т-клеток (фиг. 1). Чтобы обойти эту проблему, авторы настоящего изобретения заменили первоначальный спейсер CH2CH3 различными экстраклеточными доменами из рецептора фактора роста нервов с низким сродством (LNGFR). Усеченный вариант LNGFR, в котором отсутствуют внутриклеточные, передающие сигналы компоненты, уже использовался в клинике для генной маркировки Т-клеток (Bonini et al., Nat Med, 2003, Apr;9(4):367-9;Ciceri et al., Lancet Oncol, 2009,May;10(5):489-500). Экстраклеточная часть LNGFR состоит из 4 богатых цистеином районов TNFR и богатой серином/треонином ножки (фиг. 1В). Прежде всего, авторы настоящего изобретения создали две конструкции CD44v6-CAR.28z: одну со спейсером в виде всей экстраклеточной части LNGFR (LNGFR дикого типа длинным или NWL), а другую только с 4 богатыми цистеином районами TNFR (LNGFR дикого типа короткий или NWS). Для исключения возможности антиген-независимой активации конструкции со спейсером на основе LNGFR с помощью природного лиганда NGF, авторы настоящего изобретения создали две дополнительные конструкции CD44v6-CAR.28z, несущие делецию четвертого богатого цистеином домена TNFR, который, как известно, аннулирует передачу сигнала от NGF (Yan et al., J Biol Chem, 1991, Jun 25; 266(18):12099-104), создавая мутированную по LNGFR длинную изоформу или NML и мутированную по LNGFR короткую изоформу или NMS, соответственно. В качестве контроля, авторы настоящего изобретения создали также конструкцию CD44v6-CAR.28z, включающую мутированный вариант первоначального спейсера CH2CH3 (CHM), который не способен распознавать FcγRI (Hombach et al. Gene Ther 2000, Jun;7(12):1067-75). Примечательно, что как FcγRII, так и FcγRIII могут использовать остатки помимо этого общего набора, что позволяет предположить, что эта мутация не полностью аннулирует связывание (Shields et al. J Biol Chem, 2001, Mar 2;276(9):6591-604. Armour et al. Mol Immunol, 2003, Dec; 40(9):585-93).
Пример 3 - Конструкции CD44v6-CAR.28z со спейсерами на основе LNGFR могут быть использованы для отбора и отслеживания трансдуцированных Т-клеток
Различные конструкции CD44v6-CAR.28z со спейсерами на основе LNGFR были клонированы в ретровирусные векторы (RV) для трансдукции первичных Т-клеток. Для трансдукции, Т-клетки активировали с использованием CD3/CD28-сфер плюс IL-7/IL-15, в соответствии с протоколом, который лучше сохраняет их функциональный фенотип (Kaneko et al. Blood, 2009, Jan 29;113(5):1006-15. Bondanza et al. Blood 2011, Jun 16;117(24):6469-78. Cieri et al. Blood, 2013, Jan 24;121(4):573-84). После трансдукции, все конструкции могли быть идентифицированы на поверхности Т-клеток, используя направленное против LNGFR мАт C40-1457 (фиг. 2А), что указывало на то, что они правильно процессировались, закреплялись на клеточной мембране и, самое главное, распознавались направленными против NGFR мАт. Как следствие, экспрессирующие CD44v6-CAR.28z с различными спейсерами на основе LNGFR Т-клетки могли быть отсортированы с использованием иммуномагнитных сфер (фиг. 2В). При ближайшем рассмотрении, авторы настоящего изобретения обнаружили, что только изоформа со спейсером NWL связывалась с другим направленным против LNGFR мАт, ME20.4, что позволяет предположить, что конформационные изменения, определяемые спейсерами на основе LNGFR различных длин, могут контролировать доступность эпитопа для ME20.4. Важно отметить, что кинетика увеличения количества экспрессирующих конструкции с различными спейсерами на основе LNGFR клеток была схожа с таковой экспресссирующих CD44v6-CAR.28z со срейсером СН2СН3 Т-клеток (фиг. 2В), что исключает потенциальное пролиферативное преимущество, вызванное экстраклеточными последовательностями LNGFR, установленными в CAR. В конце культивирования получаемая в результате популяция была обогащена в отношении рано дифференцированных Т-клеток (фиг. 2С), что указывает на отсутствие нарушения пути функциональной дифференцировки активированных с помощью сфер Т-клеток в присутствии IL-7/IL-15.
Пример 4 - Экспрессирующие CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клетки сохраняют CD44v6-специфическое распознавание, потеряв при этом неспецифическое распознавание, опосредованное взаимодействием с FcγR.
Для проверки сохранения CD44v6-специфического распознавания после замены первоначального спейсера CH2CH3 спейсерами на основе LNGFR, экспрессирующие CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клетки были исследованы в экспериментах сокультивирования с CD44v6-экспрессирующими опухолевыми клетками. Подобно экспрессирующим CD44v6-CAR.28z со спейсером CH2CH3 Т-клеткам, экспресссирующие CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клетки эффективно устраняли CD44v6+ (линии клеток MM1S и ТНР-1), но не CD44v6- (линии клеток BV173) опухолевые клетки (фиг. 3А). Кроме того, CD44v6-специфическое распознавание было связано с сильным увеличением количества Т-клеток (фиг. 3B), что означает полное сохранение терапевтического потенциала экспрессирующих CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клеток. Следовательно, в результате получается, что CAR со спейсерами на основе LNGFR в соответствии с настоящим изобретением являются эффективными против опухолевых моделей, экспрессирующих специфический антиген, на который они нацелены.
Чтобы продемонстрировать отсутствие неспецифического распознавания, опосредованного взаимодействием с FcRγ, экспрессирующие CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клетки сокультивировали с CD44v6+/FcγRs+ лейкозными клетками Thp1 или с CD44v6-/FcγRs+ лейкозными клетками HL-60. В этой системе, в то время как экспрессирующие CD44v6-CAR.28z со спейсером СН2СН3 Т-клетки устраняли как CD44v6+ клетки Thp1, так и CD44v6- клетки HL-60, экспрессирующие CD44v6-CAR.28z CAR со спейсерами на основе LNGFR Т-клетки специфически устраняли CD44v6+ клетки ТНР-1, но не CD44v6- клетки HL-60 (фиг.4А). Соответственно, экспрессирующие CD44v6-CAR.28z CAR со спейсерами на основе LNGFR Т-клетки пролиферировали в ответ на CD44v6+ клетки ТНР-1, но не на CD44v6- клетки HL-60 (фиг. 4В). В обеих системах, поведение экспрессирующих конструкций со спейсерами на основе LNGFR клеток перекрывалось с таковым экспрессирующих CD44v6-CAR.28z CAR с мутированным спейсером CH2CH3 Т-клеток, демонстрируя отмену FcγR-опосредованных эффектов.
Следовательно, из-за отсутствия Fc-домена, в состав которого входят шарнирная область-СН2-СН3 константной области иммуноглобулина IgG, в качестве спейсера, CAR, содержащие происходящий от LNGFR спейсер в соответствии с настоящим изобретением, не связываются с Fc-гамма-рецепторами для IgG, что, таким образом, позволяет избежать активации нежелательной и потенциально токсичной нецелевой иммунной реакции. Соответственно, CAR со спейсерами на основе LNGFR являются более безопасными, чем те, которые содержат шарнирную область-СН2-СН3 IgG.
И, наконец, чтобы исключить антиген-независимую стимуляцию с помощью растворимого NGF, экспресссирующие CD44v6-CAR28.z со спейсерами на основе LNGFR Т-клетки культивировали in vitro с NGF. Даже при сверхфизиологических концентрациях NGF, которые, как известно, подталкивает дифференциацию линии LNGFR-экспрессирующих нервных клеток РС12 (фиг. 5А), экспрессирующие CD44v6-CAR.28z CAR со спейсерами на основе LNGFR Т-клетки не были стимулированы к пролиферации (фиг. 5В), что указывает на отсутствие передачи сигнала с помощью растворимого NGF.
Пример 5 - Экспрессирующие CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клетки лучше сохраняются in vivo и опосредуют превосходные противоопухолевые эффекты
После демонстрации эффективного и специфического распознавания in vitro, экспрессирующие CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клетки испытывали на противоопухолевую активность in vivo, сначала в модели минимальной остаточной болезни, а затем в модели устойчивого заболевания (WED). В первой модели, мышей NSG подвергали инфузии лейкозных клеток ТНР-1 и через три дня подвергали лечению экспрессирующими CD44v6-CAR.28z со спейсером СН2СН3 или различными спейсерами на основе LNGFR Т-клетками. Экспрессирующие CD44v6-CAR.28z с различными спейсерами на основе LNGFR Т-клетки лучше размножались (фиг. 6А) и сохранялись (фиг. 6В), чем экспрессирующие CD44v6-CAR.28z со спейсером СН2СН3 Т-клетки. Соответственно, экспрессирующие CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клетки, по-видимому, опосредует более сильные противоопухолевые эффекты, на что указывала лучшая нормализация веса печени с инфильтратом клеток Thp1 по сравнению с мышами, которым осуществлена инфузия экспрессирующих CD44v6-CAR.28z со спейсером CH2CH3 Т-клеток (фиг. 6В). Во второй модели устойчивого заболевания, мышей NSG подвергали инфузии CD44v6-экспрессирующих миеломных клеток ММ1.S, а через 5 недель, когда опухоль уже колонизировала костный мозг, подвергали лечению экспрессирующими CD44v6-CAR.28z со спейсером СН2СН3 или различными спейсерами на основе LNGFR Т-клетками. Экспрессирующие CD44v6-CAR.28z Т-клетки, несущие изоформу NML, не были включены. В этой более точной модели, в то время как экспрессирующие CD44v6-CAR.28z со спейсером СН2СН3 Т-клетки едва приживлялись и не опосредовали какой-либо значительный противоопухолевый эффект, экспрессирующие CD44v6-CAR.28z с различными спейсерами на основе LNGFR Т-клетки размножались (Фиг. 7А), сохранялись и приводили к поразительный противоопухолевой активности (фиг. 7В).
Способность экспрессирующих CD44v6-CAR.28z со спейсерами на основе LNGFR Т-клеток опосредовать превосходную противоопухолевую активность была в дальнейшем подтверждена с использованием модели устойчивой миеломы с использованием CD44v6+ клеток ММ1.S, экспрессирующих секретируемую люциферазу. Наличие этой люциферазы позволяет контролировать изо дня в день количество циркулирующих опухолевых клеток ММ1.S у мышей, подвергнутых лечению экспрессирующими CD44v6-CAR.28z со спейсером CH2CH3 (v6 CHW) или со спейсером NMS LNGFR (v6 NMS) Т-клетками. В этой сложной модели, в то время как экспрессирующие CD44v6-CAR.28z со спейсером CH2CH3 Т-клетки продемонстрировали одинаковую противоопухолевую активность с экспрессирующими неродственный CAR Т-клетками (CTR), экспрессирующие CD44v6- CAR.28z со спейсером NMS Т-клетки способны держать под контролем количество циркулирующих опухолевых клеток вплоть до 21 дней (фиг. 7BIS A) и значительно увеличивать общую выживаемость (фиг. 7BIS B).
Пример 6 - Методы
Создание конструкций CD19-CAR.28z и CEA-CAR.28z со спейсерами на основе LNGFR
Стратегию, аналогичную той, которая описана в примере 1, использовали для создания CD19-специфических и СЕА-специфических конструкций CAR (фиг. 22). Были созданы следующие конструкции:
CD19-CAR.28z: содержащая CD19-специфический таргетирующий домен, цепь CD3ζ в сочетании с эндокостимулирующим доменом CD28 и спейсером CH2CH3 IgG1 дикого типа (CH2CH3)
NWL: CD19-CAR.28z, содержащая длинный спейсер на основе LNGFR дикого типа (включающий 4-TNFR-Cys домена и ножку)
NMS: CD19-CAR.28z, содержащая мутированный короткий спейсер на основе LNGFR (в том числе 4-TNFR-Cys домена с делецией в четвертом домене)
CEA-CAR.28z: содержащая СЕА-специфический, таргетирующий домен, цепь CD3ζ в сочетании с эндокостимулирующим доменом CD28 и спейсером CH2CH3 IgG1 дикого типа (CH2CH3)
NWL: CEA-CAR.28z, содержащая длинный спейсер на основе LNGFR дикого типа (включающий 4-TNFR-Cys домена и ножку)
NMS: СЕА-CAR.28z, содержащая мутированный короткий спейсер на основе LNGFR (включающий 4-TNFR-Cys домена с делецией в четвертом домене)
Трансдукция и условия культивирования
Т-клетки активировали с использованием CD3/CD28-сфер клеточного размера (ClinExVivo, Invitrogen) плюс IL-7/IL-15 (5 нг/мл, Peprotech) и RV-трансдуцировали путем двух циклов центрифужной инокуляции в день 2 и 3 после стимуляции. В день 6, сферы удаляли, и Т-клетки культивировали в среде RPMI 1640 (Gibco-BRL), 10% FBS (BioWhittaker) в присутствии IL-7 и IL-15. Поверхностная экспрессия CD19- и СЕА-специфических конструкций CAR со спейсером CH2CH3 (CHW) была обнаружена с помощью мАт, специфических для спейсера CH2CH3 IgG1 (Jackson Laboratories), в то время как поверхностную экспрессию конструкций CAR со спейсерами на основе LNGFR (NWL и NWS) анализировали с использованием LNGFR-специфических мАт от BD Bioscience (клон: C40-14579). Между днем 9 и днем 15 с момента активации, экспрессирующие CAR.28z со спейсером CH2CH3 Т-клетки были FACS-отсортированы, используя поликлональные, специфические для CH2CH3 IgG1 мАт, в то время как экспрессирующие CAR.28z со спейсерами на основе LNGFR Т-клетки окрашивали РЕ-конъюгированным, LNGFR-специфическим мАт C40-14579 и сортировали с помощью колонок, используя анти-PE парамагнитные сферы (Miltenyi).
In vitro анализы для анализа специфического распознавания
В анализах сокультивирования, CAR-отсортированные Т-клетки культивировали с клетками-мишенями в соотношении Е:Т=1:10. Спустя 4 дня выжившие клетки подсчитывали и анализировали с помощью FACS. Индекс элиминации рассчитывали следующим образом: 1-(количество остаточных клеток-мишеней в присутствии CD44v6- 4GS2.CAR28z+ Т-клеток, CD19.CAR28z+ Т-клеток и CEA.CAR28z+ Т-клеток)/(число оставшихся клеток-мишеней в присутствии CD19.CAR28z+ Т-клеток). Супернатант от сокультур собирали через 24 часа инкубации и анализировали на продукцию цитокинов (IFNγ, IL-2 и TNFα) с помощью анализа CBA (BD BioLegend).
Модели противоопухолевой эффективности с использованием ксенотрансплантата
Для модели минимальной остаточной болезни, мышам NSG (Jackson) внутривенно вводили путем инфузии 1,5×106 лейкозных клеток ALL-CM/мышь. Через три дня мышей подвергали лечению путем внутривенного введения 5×106 отсортированных, экспрессирующих CD19-CAR.28z или CD44v6-4GS2.CAR.28 со спейсерами на основе LNGFR (NWL, NMS). Приживление Т-клеток контролировали еженедельно с помощью кровопускания и анализа FACS. Через 7 недель, мышей умерщвляли, и их костный мозг (ВМ) анализировали с помощью FACS на наличие клеток ALL-CM с использованием направленных против hCD45 и hCD19 мАт.
Пример 7 - Конструкции CAR.28z со спейсерами на основе LNGFR могут быть использованы для отбора и отслеживания трансдуцированных Т-клеток
Конструкции CAR.28z с различными спейсерами на основе LNGFR были клонированы в ретровирусные векторы (RV) для трансдукции первичных Т-клеток. Для трансдукции, Т-клетки активировали с использованием CD3/CD28-сфер плюс IL-7/IL-15, в соответствии с протоколом, который лучше сохраняет их функциональный фенотип (Kaneko et al. Blood, 2009, Jan 29;113(5):1006-15. Bondanza et al. Blood 2011, Jun 16;117(24):6469-78. Cieri et al. Blood, 2013, Jan 24;121(4):573-84). После трансдукции, Т-клетки можно было отсортировать с помощью иммуномагнитных сфер (фиг. 23), что указывает на то, что, как это показано для CAR, мишенью которых является антиген CD44v6, происходящие от LNGFR спейсеры правильно процессировались и закреплялись на клеточной мембране также в контексте двух других CAR, специфических для антигенов CD19 и CEA.
Пример 8 - Экспрессирующие CD19-CAR.28z, СЕА-CAR.28z и CD44v6-4GS2.CAR.28z со спейсерами на основе LNGFR Т-клетки сохраняют антигенспецифическое распознавание, потеряв при этом неспецифическое распознавание, опосредованное взаимодействием с FcγR.
Для проверки сохранения CD19- и СЕА-специфического распознавания после замены первоначального спейсера CH2CH3 спейсерами на основе LNGFR, экспрессирующие CD19-CAR.28z и СЕА-CAR.28z со спейсерами на основе LNGFR Т-клетки были исследованы в экспериментах сокультивирования с антигенэкспрессирующими опухолевыми клетками. Подобно экспрессирующим конструкцию со спейсером CH2CH3 Т-клеткам, экспресссирующие CD19-CAR.28z, СЕА-CAR.28z со спейсерами на основе LNGFR Т-клетки эффективно устраняли CD19+, CEA+ и CD44v6+ опухолевые клетки, соответственно, сберегая негативные по антигену опухолевые клетки (фиг. 24А). В частности, экспрессирующие CD19-CAR.28z CAR со спейсером на основе LNGFR Т-клетки специфически устраняли CD19+ клетки ALL-CM и BV-173, но не CD19- клетки HL-60 и BXPC3 (фиг. 24А). Точно так же, экспресссирующие CEA-CAR.28z со спейсерами на основе LNGFR Т-клетки специфически устраняли СЕА+ клетки BXPC3, но не CEA- клетки HL-60, ALL-CM и BV-173 (фиг. 24A), а экспрессирующие CD44v6-4GS2.CAR.28z Т-клетки специфически устраняли CD44v6+ клетки BXPC3 клетки, но не CD44v6- клетки ALL-СМ, BV173 и HL-60 (фиг. 24А). Сравнимые результаты были получены при оценке антигенспецифического высвобождения цитокинов (IFNγ, IL-2 и TNFα) (фиг. 24В).
CAR, содержащие LNGFR в качестве спейсера в соответствии с настоящим изобретением, как получается в результате, сохраняют специфичность и противоопухолевый эффект с различными антигенспецифическими таргетирующими доменами.
Чтобы продемонстрировать отсутствие неспецифического распознавания, опосредованного взаимодействием с FcRγ, экспрессирующие CD19-CAR.28z, CEA-CAR.28z CD44v6-4GS2.CAR.28z со спейсерами на основе LNGFR Т-клетки сокультивировали с FcγRs+ CD19- CEA- клетками HL-60. В этой системе только экспрессирующие CD19-CAR.28z и СЕА-CAR.28z со спейсером СН2СН3 Т-клетки способны устранять клетки HL-60-мишени, тем самым подтверждая, что использование спейсера на основе LNGFR позволяет избежать активации нежелательного врожденного иммунного ответа.
Пример 12 - Экспрессирующие CD19-CAR.28z со спейсерами на основе LNGFR Т-клетки опосредуют противоопухолевые эффекты in vivo
После демонстрации эффективного и специфического распознавания in vitro, экспрессирующие CD19-CAR.28z со спейсерами на основе LNGFR Т-клетки испытывали на противоопухолевую активность in vivo, в модели минимальной остаточной болезни. Мышей NSG подвергали инфузии лейкозных клеток ALL-CM и через три дня подвергали лечению экспрессирующими CD19-CAR.28z с различными спейсерами на основе LNGFR (NWL и NMS) Т-клетками. В этом случае, экспрессирующие CD44v6-4GS2.CAR.28z со спейсерами на основе LNGFR (NWL и NMS) Т-клетки были использованы в качестве отрицательного контроля, поскольку лейкозные клетки ALL-CM не экспрессируют антиген CD44v6 (фиг. 25). Экспрессирующие CD19-CAR.28z с обоими спейсерами на основе LNGFR Т-клетки, по-видимому, опосредует противоопухолевые эффекты, на что указывала более низкая концентрация клеток ALL-CM, инфильтрующих костный мозг, по сравнению с мышами, которым осуществлена инфузия экспрессирующих CD44v6-CAR.28z Т-клеток (фиг. 25).
Все публикации, упомянутые в вышеприведенном описании, включены сюда посредством ссылки. Различные модификации и вариации описанных CAR, полинуклеотидов, векторов, клеток и композиций настоящего изобретения будут очевидны квалифицированным специалистам в данной области техники, не выходя за рамки объема и сущности настоящего изобретения. Хотя настоящее изобретение было описано в связи с конкретными предпочтительными вариантами осуществления, следует понимать, что заявленное изобретение не должно быть чрезмерно ограничено такими конкретными вариантами осуществления. В самом деле, различные модификации описанных способов осуществления настоящего изобретения, которые очевидны специалистам в области биохимии и биотехнологии или в смежных областях, как предполагается, находятся в пределах объема нижеследующей формулы изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> MOLMED SpA
<120> Химерные рецепторы антигенов
<130> P105739PCT
<150> EP 14184838.2
<151> 2014-09-15
<160> 40
<170> Патент в версии 3.5
<210> 1
<211> 222
<212> Белок
<213> Искусственная последовательность
<220>
<223> Спейсер LNGFR дикого типа длинный (NWL)
<400> 1
Lys Glu Ala Cys Pro Thr Gly Leu Tyr Thr His Ser Gly Glu Cys Cys
1 5 10 15
Lys Ala Cys Asn Leu Gly Glu Gly Val Ala Gln Pro Cys Gly Ala Asn
20 25 30
Gln Thr Val Cys Glu Pro Cys Leu Asp Ser Val Thr Phe Ser Asp Val
35 40 45
Val Ser Ala Thr Glu Pro Cys Lys Pro Cys Thr Glu Cys Val Gly Leu
50 55 60
Gln Ser Met Ser Ala Pro Cys Val Glu Ala Asp Asp Ala Val Cys Arg
65 70 75 80
Cys Ala Tyr Gly Tyr Tyr Gln Asp Glu Thr Thr Gly Arg Cys Glu Ala
85 90 95
Cys Arg Val Cys Glu Ala Gly Ser Gly Leu Val Phe Ser Cys Gln Asp
100 105 110
Lys Gln Asn Thr Val Cys Glu Glu Cys Pro Asp Gly Thr Tyr Ser Asp
115 120 125
Glu Ala Asn His Val Asp Pro Cys Leu Pro Cys Thr Val Cys Glu Asp
130 135 140
Thr Glu Arg Gln Leu Arg Glu Cys Thr Arg Trp Ala Asp Ala Glu Cys
145 150 155 160
Glu Glu Ile Pro Gly Arg Trp Ile Thr Arg Ser Thr Pro Pro Glu Gly
165 170 175
Ser Asp Ser Thr Ala Pro Ser Thr Gln Glu Pro Glu Ala Pro Pro Glu
180 185 190
Gln Asp Leu Ile Ala Ser Thr Val Ala Gly Val Val Thr Thr Val Met
195 200 205
Gly Ser Ser Gln Pro Val Val Thr Arg Gly Thr Thr Asp Asn
210 215 220
keacptglyt hsgecckacn lgegvaqpcg anqtvcepcl dsvtfsdvvs atepckpcte
cvglqsmsap cveaddavcr caygyyqdet tgrceacrvc eagsglvfsc qdkqntvcee
cpdgtysdea nhvdpclpct vcedterqlr ectrwadaec eeipgrwitr stppegsdst
apstqepeap peqdliastv agvvttvmgs sqpvvtrgtt dn
<210> 2
<211> 666
<212> Белок
<213> Искусственная последовательность
<220>
<223> Спейсер LNGFR дикого типа длинный (NWL)
<400> 2
aaagaggcct gccccaccgg cctgtacacc cacagcggag agtgctgcaa ggcctgcaac 60
ctgggagagg gcgtggccca gccttgcggc gccaatcaga ccgtgtgcga gccctgcctg 120
gacagcgtga ccttcagcga cgtggtgtcc gccaccgagc cctgcaagcc ttgcaccgag 180
tgtgtgggcc tgcagagcat gagcgccccc tgcgtggaag ccgacgacgc cgtgtgtaga 240
tgcgcctacg gctactacca ggacgagaca accggcagat gcgaggcctg tagagtgtgc 300
gaggccggca gcggcctggt gttcagttgt caagacaagc agaataccgt gtgtgaagag 360
tgccccgacg gcacctacag cgacgaggcc aaccacgtgg acccctgcct gccctgcact 420
gtgtgcgagg acaccgagcg gcagctgcgc gagtgcacaa gatgggccga cgccgagtgc 480
gaagagatcc ccggcagatg gatcaccaga agcacccccc ctgagggcag cgacagcacc 540
gcccctagca cccaggaacc tgaggcccct cccgagcagg acctgatcgc ctctacagtg 600
gccggcgtgg tgacaaccgt gatgggcagc tctcagcccg tggtgacacg gggcaccacc 660
gacaat 666
<210> 3
<211> 162
<212> Белок
<213> Искусственная последовательность
<220>
<223> Спейсер LNGFR дикого типа короткий (NWS)
<400> 3
Lys Glu Ala Cys Pro Thr Gly Leu Tyr Thr His Ser Gly Glu Cys Cys
1 5 10 15
Lys Ala Cys Asn Leu Gly Glu Gly Val Ala Gln Pro Cys Gly Ala Asn
20 25 30
Gln Thr Val Cys Glu Pro Cys Leu Asp Ser Val Thr Phe Ser Asp Val
35 40 45
Val Ser Ala Thr Glu Pro Cys Lys Pro Cys Thr Glu Cys Val Gly Leu
50 55 60
Gln Ser Met Ser Ala Pro Cys Val Glu Ala Asp Asp Ala Val Cys Arg
65 70 75 80
Cys Ala Tyr Gly Tyr Tyr Gln Asp Glu Thr Thr Gly Arg Cys Glu Ala
85 90 95
Cys Arg Val Cys Glu Ala Gly Ser Gly Leu Val Phe Ser Cys Gln Asp
100 105 110
Lys Gln Asn Thr Val Cys Glu Glu Cys Pro Asp Gly Thr Tyr Ser Asp
115 120 125
Glu Ala Asn His Val Asp Pro Cys Leu Pro Cys Thr Val Cys Glu Asp
130 135 140
Thr Glu Arg Gln Leu Arg Glu Cys Thr Arg Trp Ala Asp Ala Glu Cys
145 150 155 160
Glu Glu
keacptglyt hsgecckacn lgegvaqpcg anqtvcepcl dsvtfsdvvs atepckpcte
cvglqsmsap cveaddavcr caygyyqdet tgrceacrvc eagsglvfsc qdkqntvcee
cpdgtysdea nhvdpclpct vcedterqlr ectrwadaec ee
<210> 4
<211> 486
<212> Белок
<213> Искусственная последовательность
<220>
<223> Спейсер LNGFR дикого типа корткий (NWS)
<400> 4
aaagaggcct gccccaccgg cctgtacacc cacagcggag agtgctgcaa ggcctgcaac 60
ctgggagagg gcgtggccca gccttgcggc gccaatcaga ccgtgtgcga gccctgcctg 120
gacagcgtga ccttcagcga cgtggtgtcc gccaccgagc cctgcaagcc ttgcaccgag 180
tgtgtgggcc tgcagagcat gagcgccccc tgcgtggaag ccgacgacgc cgtgtgtaga 240
tgcgcctacg gctactacca ggacgagaca accggcagat gcgaggcctg tagagtgtgc 300
gaggccggca gcggcctggt gttcagttgt caggacaagc agaacaccgt gtgtgaagag 360
tgccccgacg gcacctacag cgacgaggcc aaccacgtgg acccctgcct gccctgcact 420
gtgtgcgagg acaccgagcg gcagctgcgc gagtgcacaa gatgggccga cgccgagtgc 480
gaggaa 486
<210> 5
<211> 200
<212> Белок
<213> Искусственная последовательность
<220>
<223> Спейсер LNGFR мутированный длинный (NML)
<400> 5
Lys Glu Ala Cys Pro Thr Gly Leu Tyr Thr His Ser Gly Glu Cys Cys
1 5 10 15
Lys Ala Cys Asn Leu Gly Glu Gly Val Ala Gln Pro Cys Gly Ala Asn
20 25 30
Gln Thr Val Cys Glu Pro Cys Leu Asp Ser Val Thr Phe Ser Asp Val
35 40 45
Val Ser Ala Thr Glu Pro Cys Lys Pro Cys Thr Glu Cys Val Gly Leu
50 55 60
Gln Ser Met Ser Ala Pro Cys Val Glu Ala Asp Asp Ala Val Cys Arg
65 70 75 80
Cys Ala Tyr Gly Tyr Tyr Gln Asp Glu Thr Thr Gly Arg Cys Glu Ala
85 90 95
Cys Arg Val Cys Glu Ala Gly Ser Gly Leu Val Phe Ser Cys Gln Asp
100 105 110
Lys Gln Asn Thr Val Cys Glu Glu Cys Pro Asp Gly Thr Tyr Ser Asp
115 120 125
Glu Ala Ala Arg Ala Ala Asp Ala Glu Cys Glu Glu Ile Pro Gly Arg
130 135 140
Trp Ile Thr Arg Ser Thr Pro Pro Glu Gly Ser Asp Ser Thr Ala Pro
145 150 155 160
Ser Thr Gln Glu Pro Glu Ala Pro Pro Glu Gln Asp Leu Ile Ala Ser
165 170 175
Thr Val Ala Gly Val Val Thr Thr Val Met Gly Ser Ser Gln Pro Val
180 185 190
Val Thr Arg Gly Thr Thr Asp Asn
195 200
keacptglyt hsgecckacn lgegvaqpcg anqtvcepcl dsvtfsdvvs atepckpcte
cvglqsmsap cveaddavcr caygyyqdet tgrceacrvc eagsglvfsc qdkqntvcee
cpdgtysdea araadaecee ipgrwitrst ppegsdstap stqepeappe qdliastvag
vvttvmgssq pvvtrgttdn
<210> 6
<211> 600
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Спейсер LNGFR мутированный длинный (NML)
<400> 6
aaagaggcct gccccaccgg cctgtacacc cacagcggag agtgctgcaa ggcctgcaac 60
ctgggagagg gcgtggccca gccttgcggc gccaatcaga ccgtgtgcga gccctgcctg 120
gacagcgtga ccttcagcga cgtggtgtcc gccaccgagc cctgcaagcc ttgcaccgag 180
tgtgtgggcc tgcagagcat gagcgccccc tgcgtggaag ccgacgacgc cgtgtgtaga 240
tgcgcctacg gctactacca ggacgagaca accggcagat gcgaggcctg tagagtgtgc 300
gaggccggca gcggcctggt gttcagttgt caagacaagc agaataccgt gtgtgaagag 360
tgccccgacg gcacctacag cgacgaagcc gccagagccg ccgacgccga gtgcgaagag 420
atccccggca gatggatcac cagaagcacc ccccctgagg gcagcgacag caccgcccct 480
agcacccagg aacctgaggc ccctcccgag caggacctga tcgcctctac agtggccggc 540
gtggtgacaa ccgtgatggg cagctctcag cccgtggtga cacggggcac caccgacaat 600
<210> 7
<211> 140
<212> Белок
<213> Искусственная последовательность
<220>
<223> Спейсер LNGFR мутированный короткий (NMS)
<400> 7
Lys Glu Ala Cys Pro Thr Gly Leu Tyr Thr His Ser Gly Glu Cys Cys
1 5 10 15
Lys Ala Cys Asn Leu Gly Glu Gly Val Ala Gln Pro Cys Gly Ala Asn
20 25 30
Gln Thr Val Cys Glu Pro Cys Leu Asp Ser Val Thr Phe Ser Asp Val
35 40 45
Val Ser Ala Thr Glu Pro Cys Lys Pro Cys Thr Glu Cys Val Gly Leu
50 55 60
Gln Ser Met Ser Ala Pro Cys Val Glu Ala Asp Asp Ala Val Cys Arg
65 70 75 80
Cys Ala Tyr Gly Tyr Tyr Gln Asp Glu Thr Thr Gly Arg Cys Glu Ala
85 90 95
Cys Arg Val Cys Glu Ala Gly Ser Gly Leu Val Phe Ser Cys Gln Asp
100 105 110
Lys Gln Asn Thr Val Cys Glu Glu Cys Pro Asp Gly Thr Tyr Ser Asp
115 120 125
Glu Ala Ala Arg Ala Ala Asp Ala Glu Cys Glu Glu
130 135 140
keacptglyt hsgecckacn lgegvaqpcg anqtvcepcl dsvtfsdvvs atepckpcte
cvglqsmsap cveaddavcr caygyyqdet tgrceacrvc eagsglvfsc qdkqntvcee
cpdgtysdea araadaecee
<210> 8
<211> 420
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Спейсер LNGFR мутированный короткий (NMS)
<400> 8
aaagaggcct gccccaccgg cctgtacacc cacagcggag agtgctgcaa ggcctgcaac 60
ctgggagagg gcgtggccca gccttgcggc gccaatcaga ccgtgtgcga gccctgcctg 120
gacagcgtga ccttcagcga cgtggtgtcc gccaccgagc cctgcaagcc ttgcaccgag 180
tgtgtgggcc tgcagagcat gagcgccccc tgcgtggaag ccgacgacgc cgtgtgtaga 240
tgcgcctacg gctactacca ggacgagaca accggcagat gcgaggcctg tagagtgtgc 300
gaggccggca gcggcctggt gttcagttgt caggacaagc agaacaccgt gtgtgaagag 360
tgccccgacg gcacctacag cgacgaggcc gcccgggccg ccgacgccga gtgcgaggaa 420
<210> 9
<211> 34
<212> Белок
<213> Homo sapiens
<400> 9
Ala Cys Pro Thr Gly Leu Tyr Thr His Ser Gly Glu Cys Cys Lys Ala
1 5 10 15
Cys Asn Leu Gly Glu Gly Val Ala Gln Pro Cys Gly Ala Asn Gln Thr
20 25 30
Val Cys
<210> 10
<211> 42
<212> Белок
<213> Homo sapiens
<400> 10
Pro Cys Leu Asp Ser Val Thr Phe Ser Asp Val Val Ser Ala Thr Glu
1 5 10 15
Pro Cys Lys Pro Cys Thr Glu Cys Val Gly Leu Gln Ser Met Ser Ala
20 25 30
Pro Cys Val Glu Ala Asp Asp Ala Val Cys
35 40
<210> 11
<211> 39
<212> Белок
<213> Homo sapiens
<400> 11
Arg Cys Ala Tyr Gly Tyr Tyr Gln Asp Glu Thr Thr Gly Arg Cys Glu
1 5 10 15
Ala Cys Arg Val Cys Glu Ala Gly Ser Gly Leu Val Phe Ser Cys Gln
20 25 30
Asp Lys Gln Asn Thr Val Cys
35
<210> 12
<211> 41
<212> Белок
<213> Homo sapiens
<400> 12
Glu Cys Pro Asp Gly Thr Tyr Ser Asp Glu Ala Asn His Val Asp Pro
1 5 10 15
Cys Leu Pro Cys Thr Val Cys Glu Asp Thr Glu Arg Gln Leu Arg Glu
20 25 30
Cys Thr Arg Trp Ala Asp Ala Glu Cys
35 40
<210> 13
<211> 25
<212> Белок
<213> Homo sapiens
<400> 13
Asn His Val Asp Pro Cys Leu Pro Cys Thr Val Cys Glu Asp Thr Glu
1 5 10 15
Arg Gln Leu Arg Glu Cys Thr Arg Trp
20 25
<210> 14
<211> 427
<212> Белок
<213> Homo sapiens
<400> 14
Met Gly Ala Gly Ala Thr Gly Arg Ala Met Asp Gly Pro Arg Leu Leu
1 5 10 15
Leu Leu Leu Leu Leu Gly Val Ser Leu Gly Gly Ala Lys Glu Ala Cys
20 25 30
Pro Thr Gly Leu Tyr Thr His Ser Gly Glu Cys Cys Lys Ala Cys Asn
35 40 45
Leu Gly Glu Gly Val Ala Gln Pro Cys Gly Ala Asn Gln Thr Val Cys
50 55 60
Glu Pro Cys Leu Asp Ser Val Thr Phe Ser Asp Val Val Ser Ala Thr
65 70 75 80
Glu Pro Cys Lys Pro Cys Thr Glu Cys Val Gly Leu Gln Ser Met Ser
85 90 95
Ala Pro Cys Val Glu Ala Asp Asp Ala Val Cys Arg Cys Ala Tyr Gly
100 105 110
Tyr Tyr Gln Asp Glu Thr Thr Gly Arg Cys Glu Ala Cys Arg Val Cys
115 120 125
Glu Ala Gly Ser Gly Leu Val Phe Ser Cys Gln Asp Lys Gln Asn Thr
130 135 140
Val Cys Glu Glu Cys Pro Asp Gly Thr Tyr Ser Asp Glu Ala Asn His
145 150 155 160
Val Asp Pro Cys Leu Pro Cys Thr Val Cys Glu Asp Thr Glu Arg Gln
165 170 175
Leu Arg Glu Cys Thr Arg Trp Ala Asp Ala Glu Cys Glu Glu Ile Pro
180 185 190
Gly Arg Trp Ile Thr Arg Ser Thr Pro Pro Glu Gly Ser Asp Ser Thr
195 200 205
Ala Pro Ser Thr Gln Glu Pro Glu Ala Pro Pro Glu Gln Asp Leu Ile
210 215 220
Ala Ser Thr Val Ala Gly Val Val Thr Thr Val Met Gly Ser Ser Gln
225 230 235 240
Pro Val Val Thr Arg Gly Thr Thr Asp Asn Leu Ile Pro Val Tyr Cys
245 250 255
Ser Ile Leu Ala Ala Val Val Val Gly Leu Val Ala Tyr Ile Ala Phe
260 265 270
Lys Arg Trp Asn Ser Cys Lys Gln Asn Lys Gln Gly Ala Asn Ser Arg
275 280 285
Pro Val Asn Gln Thr Pro Pro Pro Glu Gly Glu Lys Leu His Ser Asp
290 295 300
Ser Gly Ile Ser Val Asp Ser Gln Ser Leu His Asp Gln Gln Pro His
305 310 315 320
Thr Gln Thr Ala Ser Gly Gln Ala Leu Lys Gly Asp Gly Gly Leu Tyr
325 330 335
Ser Ser Leu Pro Pro Ala Lys Arg Glu Glu Val Glu Lys Leu Leu Asn
340 345 350
Gly Ser Ala Gly Asp Thr Trp Arg His Leu Ala Gly Glu Leu Gly Tyr
355 360 365
Gln Pro Glu His Ile Asp Ser Phe Thr His Glu Ala Cys Pro Val Arg
370 375 380
Ala Leu Leu Ala Ser Trp Ala Thr Gln Asp Ser Ala Thr Leu Asp Ala
385 390 395 400
Leu Leu Ala Ala Leu Arg Arg Ile Gln Arg Ala Asp Leu Val Glu Ser
405 410 415
Leu Cys Ser Glu Ser Thr Ala Thr Ser Pro Val
420 425
<210> 15
<211> 2046
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность конструкции CD44v6-специфического CAR второго поколения, CD44v6-CAR.28z
<400> 15
atggaagccc ctgcccagct gctgttcctg ctgctgctgt ggctgcccga caccaccggc 60
gagatcgtgc tgacacagag ccccgccacc ctgtctctga gccctggcga gagagccacc 120
ctgagctgta gcgccagcag cagcatcaac tacatctact ggctgcagca gaagcccggc 180
caggccccca gaatcctgat ctacctgacc agcaacctgg ccagcggcgt gcccgccaga 240
ttttctggca gcggcagcgg caccgacttc accctgacca tcagcagcct ggaacccgag 300
gacttcgccg tgtactactg cctgcagtgg tccagcaacc ccctgacctt cggcggaggc 360
accaaggtgg aaatcaagcg gggtggtggt ggttctggtg gtggtggttc tggcggcggc 420
ggctccggtg gtggtggatc tgaggtgcag ctggtggaaa gcggcggagg cctggtcaag 480
cctggcggca gcctgagact gagctgtgcc gccagcggct tcaccttcag cagctacgac 540
atgagctggg tccgacaggc tccaggcaag ggactggaat gggtgtccac catcagcagc 600
ggcggcagct acacctacta cctggacagc atcaagggcc ggttcaccat cagccgggac 660
aacgccaaga acagcctgta cctgcagatg aacagcctgc gggccgagga caccgccgtc 720
tactactgtg cccggcaggg cctcgactac tggggcagag gcaccctggt caccgtgtcc 780
agcggggatc ccgccgagcc caaatctcct gacaaaactc acacatgccc accgtgccca 840
gcacctgaac tcctgggggg accgtcagtc ttcctcttcc ccccaaaacc caaggacacc 900
ctcatgatct cccggacccc tgaggtcaca tgcgtggtgg tggacgtgag ccacgaagac 960
cctgaggtca agttcaactg gtacgtggac ggcgtggagg tgcataatgc caagacaaag 1020
ccgcgggagg agcagtacaa cagcacgtac cgtgtggtca gcgtcctcac cgtcctgcac 1080
caggactggc tgaatggcaa ggagtacaag tgcaaggtct ccaacaaagc cctcccagcc 1140
cccatcgaga aaaccatctc caaagccaaa gggcagcccc gagaaccaca ggtgtacacc 1200
ctgcccccat cccgggatga gctgaccaag aaccaggtca gcctgacctg cctggtcaaa 1260
ggcttctatc ccagcgacat cgccgtggag tgggagagca atgggcaacc ggagaacaac 1320
tacaagacca cgcctcccgt gctggactcc gacggctcct tcttcctcta cagcaagctc 1380
accgtggaca agagcaggtg gcagcagggg aacgtcttct catgctccgt gatgcatgag 1440
gctctgcaca accactacac gcagaagagc ctctccctgt ctccgggtaa aaaagatccc 1500
aaattttggg tgctggtggt ggttggtgga gtcctggctt gctatagctt gctagtaaca 1560
gtggccttta ttattttctg ggtgaggagt aagaggagca ggctcctgca cagtgactac 1620
atgaacatga ctccccgccg ccccgggccc acccgcaagc attaccagcc ctatgcccca 1680
ccacgcgact tcgcagccta tcgctccaga gtgaagttca gcaggagcgc agacgccccc 1740
gcgtaccagc agggccagaa ccagctctat aacgagctca atctaggacg aagagaggag 1800
tacgatgttt tggacaagag acgtggccgg gaccctgaga tggggggaaa gccgagaagg 1860
aagaaccctc aggaaggcct gtacaatgaa ctgcagaaag ataagatggc ggaggcctac 1920
agtgagattg ggatgaaagg cgagcgccgg aggggcaagg ggcacgatgg cctttaccag 1980
ggtctcagta cagccaccaa ggacacctac gacgcccttc acatgcaggc cctgccccct 2040
cgctaa 2046
<210> 16
<211> 681
<212> Белок
<213> Искусственная последовательность
<220>
<223> Белок CD44v6-CAR.28z
<400> 16
Met Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1 5 10 15
Asp Thr Thr Gly Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Ile Asn Tyr Ile Tyr Trp Leu Gln Gln Lys Pro Gly Gln Ala Pro Arg
50 55 60
Ile Leu Ile Tyr Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln Trp Ser Ser
100 105 110
Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys
145 150 155 160
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
165 170 175
Ser Ser Tyr Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
180 185 190
Glu Trp Val Ser Thr Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Leu
195 200 205
Asp Ser Ile Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
210 215 220
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
225 230 235 240
Tyr Tyr Cys Ala Arg Gln Gly Leu Asp Tyr Trp Gly Arg Gly Thr Leu
245 250 255
Val Thr Val Ser Ser Gly Asp Pro Ala Glu Pro Lys Ser Pro Asp Lys
260 265 270
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
275 280 285
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
290 295 300
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
305 310 315 320
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
325 330 335
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
340 345 350
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
355 360 365
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
370 375 380
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
385 390 395 400
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
405 410 415
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
420 425 430
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
435 440 445
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
450 455 460
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
465 470 475 480
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
485 490 495
Lys Lys Asp Pro Lys Phe Trp Val Leu Val Val Val Gly Gly Val Leu
500 505 510
Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val
515 520 525
Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr
530 535 540
Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro
545 550 555 560
Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser
565 570 575
Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu
580 585 590
Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg
595 600 605
Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln
610 615 620
Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr
625 630 635 640
Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp
645 650 655
Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala
660 665 670
Leu His Met Gln Ala Leu Pro Pro Arg
675 680
<210> 17
<211> 261
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD44v6-специфический одноцепочечный фрагмент
<400> 17
Met Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1 5 10 15
Asp Thr Thr Gly Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Ile Asn Tyr Ile Tyr Trp Leu Gln Gln Lys Pro Gly Gln Ala Pro Arg
50 55 60
Ile Leu Ile Tyr Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln Trp Ser Ser
100 105 110
Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys
145 150 155 160
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
165 170 175
Ser Ser Tyr Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
180 185 190
Glu Trp Val Ser Thr Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Leu
195 200 205
Asp Ser Ile Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
210 215 220
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
225 230 235 240
Tyr Tyr Cys Ala Arg Gln Gly Leu Asp Tyr Trp Gly Arg Gly Thr Leu
245 250 255
Val Thr Val Ser Ser
260
<210> 18
<211> 68
<212> Белок
<213> Homo sapiens
<400> 18
Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu
1 5 10 15
Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser
20 25 30
Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly
35 40 45
Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala
50 55 60
Ala Tyr Arg Ser
65
<210> 19
<211> 222
<212> Белок
<213> Homo sapiens
<400> 19
Lys Glu Ala Cys Pro Thr Gly Leu Tyr Thr His Ser Gly Glu Cys Cys
1 5 10 15
Lys Ala Cys Asn Leu Gly Glu Gly Val Ala Gln Pro Cys Gly Ala Asn
20 25 30
Gln Thr Val Cys Glu Pro Cys Leu Asp Ser Val Thr Phe Ser Asp Val
35 40 45
Val Ser Ala Thr Glu Pro Cys Lys Pro Cys Thr Glu Cys Val Gly Leu
50 55 60
Gln Ser Met Ser Ala Pro Cys Val Glu Ala Asp Asp Ala Val Cys Arg
65 70 75 80
Cys Ala Tyr Gly Tyr Tyr Gln Asp Glu Thr Thr Gly Arg Cys Glu Ala
85 90 95
Cys Arg Val Cys Glu Ala Gly Ser Gly Leu Val Phe Ser Cys Gln Asp
100 105 110
Lys Gln Asn Thr Val Cys Glu Glu Cys Pro Asp Gly Thr Tyr Ser Asp
115 120 125
Glu Ala Asn His Val Asp Pro Cys Leu Pro Cys Thr Val Cys Glu Asp
130 135 140
Thr Glu Arg Gln Leu Arg Glu Cys Thr Arg Trp Ala Asp Ala Glu Cys
145 150 155 160
Glu Glu Ile Pro Gly Arg Trp Ile Thr Arg Ser Thr Pro Pro Glu Gly
165 170 175
Ser Asp Ser Thr Ala Pro Ser Thr Gln Glu Pro Glu Ala Pro Pro Glu
180 185 190
Gln Asp Leu Ile Ala Ser Thr Val Ala Gly Val Val Thr Thr Val Met
195 200 205
Gly Ser Ser Gln Pro Val Val Thr Arg Gly Thr Thr Asp Asn
210 215 220
<210> 20
<211> 113
<212> Белок
<213> Homo sapiens
<400> 20
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Gln Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln
50 55 60
Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu
65 70 75 80
Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr
85 90 95
Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro
100 105 110
Arg
<210> 21
<211> 669
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD44v6-CAR28z со спейсером LNGFR дикого типа длинным (NWL)
<400> 21
Met Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1 5 10 15
Asp Thr Thr Gly Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Ile Asn Tyr Ile Tyr Trp Leu Gln Gln Lys Pro Gly Gln Ala Pro Arg
50 55 60
Ile Leu Ile Tyr Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln Trp Ser Ser
100 105 110
Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys
145 150 155 160
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
165 170 175
Ser Ser Tyr Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
180 185 190
Glu Trp Val Ser Thr Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Leu
195 200 205
Asp Ser Ile Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
210 215 220
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
225 230 235 240
Tyr Tyr Cys Ala Arg Gln Gly Leu Asp Tyr Trp Gly Arg Gly Thr Leu
245 250 255
Val Thr Val Ser Ser Gly Asp Pro Lys Glu Ala Cys Pro Thr Gly Leu
260 265 270
Tyr Thr His Ser Gly Glu Cys Cys Lys Ala Cys Asn Leu Gly Glu Gly
275 280 285
Val Ala Gln Pro Cys Gly Ala Asn Gln Thr Val Cys Glu Pro Cys Leu
290 295 300
Asp Ser Val Thr Phe Ser Asp Val Val Ser Ala Thr Glu Pro Cys Lys
305 310 315 320
Pro Cys Thr Glu Cys Val Gly Leu Gln Ser Met Ser Ala Pro Cys Val
325 330 335
Glu Ala Asp Asp Ala Val Cys Arg Cys Ala Tyr Gly Tyr Tyr Gln Asp
340 345 350
Glu Thr Thr Gly Arg Cys Glu Ala Cys Arg Val Cys Glu Ala Gly Ser
355 360 365
Gly Leu Val Phe Ser Cys Gln Asp Lys Gln Asn Thr Val Cys Glu Glu
370 375 380
Cys Pro Asp Gly Thr Tyr Ser Asp Glu Ala Asn His Val Asp Pro Cys
385 390 395 400
Leu Pro Cys Thr Val Cys Glu Asp Thr Glu Arg Gln Leu Arg Glu Cys
405 410 415
Thr Arg Trp Ala Asp Ala Glu Cys Glu Glu Ile Pro Gly Arg Trp Ile
420 425 430
Thr Arg Ser Thr Pro Pro Glu Gly Ser Asp Ser Thr Ala Pro Ser Thr
435 440 445
Gln Glu Pro Glu Ala Pro Pro Glu Gln Asp Leu Ile Ala Ser Thr Val
450 455 460
Ala Gly Val Val Thr Thr Val Met Gly Ser Ser Gln Pro Val Val Thr
465 470 475 480
Arg Gly Thr Thr Asp Asn Pro Lys Phe Trp Val Leu Val Val Val Gly
485 490 495
Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile
500 505 510
Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met
515 520 525
Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro
530 535 540
Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe
545 550 555 560
Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu
565 570 575
Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp
580 585 590
Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Gln Arg Arg
595 600 605
Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met
610 615 620
Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly
625 630 635 640
Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp
645 650 655
Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
660 665
<210> 22
<211> 609
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD44v6-CAR28z со спейсером LNGFR дикого типа коротким (NWS)
<400> 22
Met Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1 5 10 15
Asp Thr Thr Gly Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Ile Asn Tyr Ile Tyr Trp Leu Gln Gln Lys Pro Gly Gln Ala Pro Arg
50 55 60
Ile Leu Ile Tyr Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln Trp Ser Ser
100 105 110
Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys
145 150 155 160
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
165 170 175
Ser Ser Tyr Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
180 185 190
Glu Trp Val Ser Thr Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Leu
195 200 205
Asp Ser Ile Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
210 215 220
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
225 230 235 240
Tyr Tyr Cys Ala Arg Gln Gly Leu Asp Tyr Trp Gly Arg Gly Thr Leu
245 250 255
Val Thr Val Ser Ser Gly Asp Pro Lys Glu Ala Cys Pro Thr Gly Leu
260 265 270
Tyr Thr His Ser Gly Glu Cys Cys Lys Ala Cys Asn Leu Gly Glu Gly
275 280 285
Val Ala Gln Pro Cys Gly Ala Asn Gln Thr Val Cys Glu Pro Cys Leu
290 295 300
Asp Ser Val Thr Phe Ser Asp Val Val Ser Ala Thr Glu Pro Cys Lys
305 310 315 320
Pro Cys Thr Glu Cys Val Gly Leu Gln Ser Met Ser Ala Pro Cys Val
325 330 335
Glu Ala Asp Asp Ala Val Cys Arg Cys Ala Tyr Gly Tyr Tyr Gln Asp
340 345 350
Glu Thr Thr Gly Arg Cys Glu Ala Cys Arg Val Cys Glu Ala Gly Ser
355 360 365
Gly Leu Val Phe Ser Cys Gln Asp Lys Gln Asn Thr Val Cys Glu Glu
370 375 380
Cys Pro Asp Gly Thr Tyr Ser Asp Glu Ala Asn His Val Asp Pro Cys
385 390 395 400
Leu Pro Cys Thr Val Cys Glu Asp Thr Glu Arg Gln Leu Arg Glu Cys
405 410 415
Thr Arg Trp Ala Asp Ala Glu Cys Glu Glu Pro Lys Phe Trp Val Leu
420 425 430
Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val
435 440 445
Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His
450 455 460
Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys
465 470 475 480
His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser
485 490 495
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
500 505 510
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
515 520 525
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
530 535 540
Pro Gln Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln
545 550 555 560
Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu
565 570 575
Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr
580 585 590
Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro
595 600 605
Arg
<210> 23
<211> 647
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD44v6-CAR28z со спейсером LNGFR мутированным длинным (NML)
<400> 23
Met Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1 5 10 15
Asp Thr Thr Gly Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Ile Asn Tyr Ile Tyr Trp Leu Gln Gln Lys Pro Gly Gln Ala Pro Arg
50 55 60
Ile Leu Ile Tyr Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln Trp Ser Ser
100 105 110
Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys
145 150 155 160
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
165 170 175
Ser Ser Tyr Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
180 185 190
Glu Trp Val Ser Thr Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Leu
195 200 205
Asp Ser Ile Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
210 215 220
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
225 230 235 240
Tyr Tyr Cys Ala Arg Gln Gly Leu Asp Tyr Trp Gly Arg Gly Thr Leu
245 250 255
Val Thr Val Ser Ser Gly Asp Pro Lys Glu Ala Cys Pro Thr Gly Leu
260 265 270
Tyr Thr His Ser Gly Glu Cys Cys Lys Ala Cys Asn Leu Gly Glu Gly
275 280 285
Val Ala Gln Pro Cys Gly Ala Asn Gln Thr Val Cys Glu Pro Cys Leu
290 295 300
Asp Ser Val Thr Phe Ser Asp Val Val Ser Ala Thr Glu Pro Cys Lys
305 310 315 320
Pro Cys Thr Glu Cys Val Gly Leu Gln Ser Met Ser Ala Pro Cys Val
325 330 335
Glu Ala Asp Asp Ala Val Cys Arg Cys Ala Tyr Gly Tyr Tyr Gln Asp
340 345 350
Glu Thr Thr Gly Arg Cys Glu Ala Cys Arg Val Cys Glu Ala Gly Ser
355 360 365
Gly Leu Val Phe Ser Cys Gln Asp Lys Gln Asn Thr Val Cys Glu Glu
370 375 380
Cys Pro Asp Gly Thr Tyr Ser Asp Glu Ala Ala Arg Ala Ala Asp Ala
385 390 395 400
Glu Cys Glu Glu Ile Pro Gly Arg Trp Ile Thr Arg Ser Thr Pro Pro
405 410 415
Glu Gly Ser Asp Ser Thr Ala Pro Ser Thr Gln Glu Pro Glu Ala Pro
420 425 430
Pro Glu Gln Asp Leu Ile Ala Ser Thr Val Ala Gly Val Val Thr Thr
435 440 445
Val Met Gly Ser Ser Gln Pro Val Val Thr Arg Gly Thr Thr Asp Asn
450 455 460
Pro Lys Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr
465 470 475 480
Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys
485 490 495
Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg
500 505 510
Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp
515 520 525
Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp Ala
530 535 540
Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu
545 550 555 560
Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp
565 570 575
Pro Glu Met Gly Gly Lys Pro Gln Arg Arg Lys Asn Pro Gln Glu Gly
580 585 590
Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu
595 600 605
Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu
610 615 620
Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His
625 630 635 640
Met Gln Ala Leu Pro Pro Arg
645
<210> 24
<211> 587
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD44v6-CAR28z со спейсером LNGFR мутированным коротким (NMS)
<400> 24
Met Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1 5 10 15
Asp Thr Thr Gly Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Ile Asn Tyr Ile Tyr Trp Leu Gln Gln Lys Pro Gly Gln Ala Pro Arg
50 55 60
Ile Leu Ile Tyr Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln Trp Ser Ser
100 105 110
Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys
145 150 155 160
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
165 170 175
Ser Ser Tyr Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
180 185 190
Glu Trp Val Ser Thr Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Leu
195 200 205
Asp Ser Ile Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
210 215 220
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
225 230 235 240
Tyr Tyr Cys Ala Arg Gln Gly Leu Asp Tyr Trp Gly Arg Gly Thr Leu
245 250 255
Val Thr Val Ser Ser Gly Asp Pro Lys Glu Ala Cys Pro Thr Gly Leu
260 265 270
Tyr Thr His Ser Gly Glu Cys Cys Lys Ala Cys Asn Leu Gly Glu Gly
275 280 285
Val Ala Gln Pro Cys Gly Ala Asn Gln Thr Val Cys Glu Pro Cys Leu
290 295 300
Asp Ser Val Thr Phe Ser Asp Val Val Ser Ala Thr Glu Pro Cys Lys
305 310 315 320
Pro Cys Thr Glu Cys Val Gly Leu Gln Ser Met Ser Ala Pro Cys Val
325 330 335
Glu Ala Asp Asp Ala Val Cys Arg Cys Ala Tyr Gly Tyr Tyr Gln Asp
340 345 350
Glu Thr Thr Gly Arg Cys Glu Ala Cys Arg Val Cys Glu Ala Gly Ser
355 360 365
Gly Leu Val Phe Ser Cys Gln Asp Lys Gln Asn Thr Val Cys Glu Glu
370 375 380
Cys Pro Asp Gly Thr Tyr Ser Asp Glu Ala Ala Arg Ala Ala Asp Ala
385 390 395 400
Glu Cys Glu Glu Pro Lys Phe Trp Val Leu Val Val Val Gly Gly Val
405 410 415
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
420 425 430
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
435 440 445
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
450 455 460
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg
465 470 475 480
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
485 490 495
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
500 505 510
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Gln Arg Arg Lys Asn
515 520 525
Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu
530 535 540
Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly
545 550 555 560
His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr
565 570 575
Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
580 585
<210> 25
<211> 668
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD44v6-CAR28z со спейсером LNGFR дикого типа длинным (NWL)
<400> 25
Met Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1 5 10 15
Asp Thr Thr Gly Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Ile Asn Tyr Ile Tyr Trp Leu Gln Gln Lys Pro Gly Gln Ala Pro Arg
50 55 60
Ile Leu Ile Tyr Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln Trp Ser Ser
100 105 110
Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys
145 150 155 160
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
165 170 175
Ser Ser Tyr Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
180 185 190
Glu Trp Val Ser Thr Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Leu
195 200 205
Asp Ser Ile Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
210 215 220
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
225 230 235 240
Tyr Tyr Cys Ala Arg Gln Gly Leu Asp Tyr Trp Gly Arg Gly Thr Leu
245 250 255
Val Thr Val Ser Ser Gly Asp Pro Lys Glu Ala Cys Pro Thr Gly Leu
260 265 270
Tyr Thr His Ser Gly Glu Cys Cys Lys Ala Cys Asn Leu Gly Glu Gly
275 280 285
Val Ala Gln Pro Cys Gly Ala Asn Gln Thr Val Cys Glu Pro Cys Leu
290 295 300
Asp Ser Val Thr Phe Ser Asp Val Val Ser Ala Thr Glu Pro Cys Lys
305 310 315 320
Pro Cys Thr Glu Cys Val Gly Leu Gln Ser Met Ser Ala Pro Cys Val
325 330 335
Glu Ala Asp Asp Ala Val Cys Arg Cys Ala Tyr Gly Tyr Tyr Gln Asp
340 345 350
Glu Thr Thr Gly Arg Cys Glu Ala Cys Arg Val Cys Glu Ala Gly Ser
355 360 365
Gly Leu Val Phe Ser Cys Gln Asp Lys Gln Asn Thr Val Cys Glu Glu
370 375 380
Cys Pro Asp Gly Thr Tyr Ser Asp Glu Ala Asn His Val Asp Pro Cys
385 390 395 400
Leu Pro Cys Thr Val Cys Glu Asp Thr Glu Arg Gln Leu Arg Glu Cys
405 410 415
Thr Arg Trp Ala Asp Ala Glu Cys Glu Glu Ile Pro Gly Arg Trp Ile
420 425 430
Thr Arg Ser Thr Pro Pro Glu Gly Ser Asp Ser Thr Ala Pro Ser Thr
435 440 445
Gln Glu Pro Glu Ala Pro Pro Glu Gln Asp Leu Ile Ala Ser Thr Val
450 455 460
Ala Gly Val Val Thr Thr Val Met Gly Ser Ser Gln Pro Val Val Thr
465 470 475 480
Arg Gly Thr Thr Asp Asn Pro Lys Phe Trp Val Leu Val Val Val Gly
485 490 495
Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile
500 505 510
Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met
515 520 525
Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro
530 535 540
Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe
545 550 555 560
Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu
565 570 575
Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp
580 585 590
Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys
595 600 605
Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala
610 615 620
Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys
625 630 635 640
Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr
645 650 655
Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
660 665
<210> 26
<211> 608
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD44v6-CAR28z со спейсером LNGFR дикого типа коротким (NWS)
<400> 26
Met Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1 5 10 15
Asp Thr Thr Gly Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Ile Asn Tyr Ile Tyr Trp Leu Gln Gln Lys Pro Gly Gln Ala Pro Arg
50 55 60
Ile Leu Ile Tyr Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln Trp Ser Ser
100 105 110
Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys
145 150 155 160
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
165 170 175
Ser Ser Tyr Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
180 185 190
Glu Trp Val Ser Thr Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Leu
195 200 205
Asp Ser Ile Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
210 215 220
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
225 230 235 240
Tyr Tyr Cys Ala Arg Gln Gly Leu Asp Tyr Trp Gly Arg Gly Thr Leu
245 250 255
Val Thr Val Ser Ser Gly Asp Pro Lys Glu Ala Cys Pro Thr Gly Leu
260 265 270
Tyr Thr His Ser Gly Glu Cys Cys Lys Ala Cys Asn Leu Gly Glu Gly
275 280 285
Val Ala Gln Pro Cys Gly Ala Asn Gln Thr Val Cys Glu Pro Cys Leu
290 295 300
Asp Ser Val Thr Phe Ser Asp Val Val Ser Ala Thr Glu Pro Cys Lys
305 310 315 320
Pro Cys Thr Glu Cys Val Gly Leu Gln Ser Met Ser Ala Pro Cys Val
325 330 335
Glu Ala Asp Asp Ala Val Cys Arg Cys Ala Tyr Gly Tyr Tyr Gln Asp
340 345 350
Glu Thr Thr Gly Arg Cys Glu Ala Cys Arg Val Cys Glu Ala Gly Ser
355 360 365
Gly Leu Val Phe Ser Cys Gln Asp Lys Gln Asn Thr Val Cys Glu Glu
370 375 380
Cys Pro Asp Gly Thr Tyr Ser Asp Glu Ala Asn His Val Asp Pro Cys
385 390 395 400
Leu Pro Cys Thr Val Cys Glu Asp Thr Glu Arg Gln Leu Arg Glu Cys
405 410 415
Thr Arg Trp Ala Asp Ala Glu Cys Glu Glu Pro Lys Phe Trp Val Leu
420 425 430
Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val
435 440 445
Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His
450 455 460
Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys
465 470 475 480
His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser
485 490 495
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
500 505 510
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
515 520 525
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
530 535 540
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
545 550 555 560
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
565 570 575
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
580 585 590
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
595 600 605
<210> 27
<211> 646
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD44v6-CAR28z со спейсером LNGFR мутированным длинным (NWL)
<400> 27
Met Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1 5 10 15
Asp Thr Thr Gly Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Ile Asn Tyr Ile Tyr Trp Leu Gln Gln Lys Pro Gly Gln Ala Pro Arg
50 55 60
Ile Leu Ile Tyr Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln Trp Ser Ser
100 105 110
Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys
145 150 155 160
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
165 170 175
Ser Ser Tyr Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
180 185 190
Glu Trp Val Ser Thr Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Leu
195 200 205
Asp Ser Ile Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
210 215 220
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
225 230 235 240
Tyr Tyr Cys Ala Arg Gln Gly Leu Asp Tyr Trp Gly Arg Gly Thr Leu
245 250 255
Val Thr Val Ser Ser Gly Asp Pro Lys Glu Ala Cys Pro Thr Gly Leu
260 265 270
Tyr Thr His Ser Gly Glu Cys Cys Lys Ala Cys Asn Leu Gly Glu Gly
275 280 285
Val Ala Gln Pro Cys Gly Ala Asn Gln Thr Val Cys Glu Pro Cys Leu
290 295 300
Asp Ser Val Thr Phe Ser Asp Val Val Ser Ala Thr Glu Pro Cys Lys
305 310 315 320
Pro Cys Thr Glu Cys Val Gly Leu Gln Ser Met Ser Ala Pro Cys Val
325 330 335
Glu Ala Asp Asp Ala Val Cys Arg Cys Ala Tyr Gly Tyr Tyr Gln Asp
340 345 350
Glu Thr Thr Gly Arg Cys Glu Ala Cys Arg Val Cys Glu Ala Gly Ser
355 360 365
Gly Leu Val Phe Ser Cys Gln Asp Lys Gln Asn Thr Val Cys Glu Glu
370 375 380
Cys Pro Asp Gly Thr Tyr Ser Asp Glu Ala Ala Arg Ala Ala Asp Ala
385 390 395 400
Glu Cys Glu Glu Ile Pro Gly Arg Trp Ile Thr Arg Ser Thr Pro Pro
405 410 415
Glu Gly Ser Asp Ser Thr Ala Pro Ser Thr Gln Glu Pro Glu Ala Pro
420 425 430
Pro Glu Gln Asp Leu Ile Ala Ser Thr Val Ala Gly Val Val Thr Thr
435 440 445
Val Met Gly Ser Ser Gln Pro Val Val Thr Arg Gly Thr Thr Asp Asn
450 455 460
Pro Lys Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr
465 470 475 480
Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys
485 490 495
Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg
500 505 510
Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp
515 520 525
Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp Ala
530 535 540
Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu
545 550 555 560
Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp
565 570 575
Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu
580 585 590
Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile
595 600 605
Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr
610 615 620
Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met
625 630 635 640
Gln Ala Leu Pro Pro Arg
645
<210> 28
<211> 586
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD44v6-CAR28z со спейсером LNGFR мутированным коротким (NWL)
<400> 28
Met Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1 5 10 15
Asp Thr Thr Gly Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Ile Asn Tyr Ile Tyr Trp Leu Gln Gln Lys Pro Gly Gln Ala Pro Arg
50 55 60
Ile Leu Ile Tyr Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln Trp Ser Ser
100 105 110
Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys
145 150 155 160
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
165 170 175
Ser Ser Tyr Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
180 185 190
Glu Trp Val Ser Thr Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Leu
195 200 205
Asp Ser Ile Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
210 215 220
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
225 230 235 240
Tyr Tyr Cys Ala Arg Gln Gly Leu Asp Tyr Trp Gly Arg Gly Thr Leu
245 250 255
Val Thr Val Ser Ser Gly Asp Pro Lys Glu Ala Cys Pro Thr Gly Leu
260 265 270
Tyr Thr His Ser Gly Glu Cys Cys Lys Ala Cys Asn Leu Gly Glu Gly
275 280 285
Val Ala Gln Pro Cys Gly Ala Asn Gln Thr Val Cys Glu Pro Cys Leu
290 295 300
Asp Ser Val Thr Phe Ser Asp Val Val Ser Ala Thr Glu Pro Cys Lys
305 310 315 320
Pro Cys Thr Glu Cys Val Gly Leu Gln Ser Met Ser Ala Pro Cys Val
325 330 335
Glu Ala Asp Asp Ala Val Cys Arg Cys Ala Tyr Gly Tyr Tyr Gln Asp
340 345 350
Glu Thr Thr Gly Arg Cys Glu Ala Cys Arg Val Cys Glu Ala Gly Ser
355 360 365
Gly Leu Val Phe Ser Cys Gln Asp Lys Gln Asn Thr Val Cys Glu Glu
370 375 380
Cys Pro Asp Gly Thr Tyr Ser Asp Glu Ala Ala Arg Ala Ala Asp Ala
385 390 395 400
Glu Cys Glu Glu Pro Lys Phe Trp Val Leu Val Val Val Gly Gly Val
405 410 415
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
420 425 430
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
435 440 445
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
450 455 460
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg
465 470 475 480
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
485 490 495
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
500 505 510
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
515 520 525
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
530 535 540
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
545 550 555 560
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
565 570 575
Ala Leu His Met Gln Ala Leu Pro Pro Arg
580 585
<210> 29
<211> 27
<212> Белок
<213> Homo sapiens
<400> 29
Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu
1 5 10 15
Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val
20 25
<210> 30
<211> 41
<212> Белок
<213> Homo sapiens
<400> 30
Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr
1 5 10 15
Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro
20 25 30
Pro Arg Asp Phe Ala Ala Tyr Arg Ser
35 40
<210> 31
<211> 251
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD44v6-4GS2, CD44v6-специфический одноцепочечный фрагмент
<400> 31
Met Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1 5 10 15
Asp Thr Thr Gly Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Ile Asn Tyr Ile Tyr Trp Leu Gln Gln Lys Pro Gly Gln Ala Pro Arg
50 55 60
Ile Leu Ile Tyr Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln Trp Ser Ser
100 105 110
Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
130 135 140
Gly Gly Gly Leu Val Lys Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala
145 150 155 160
Ala Ser Gly Phe Thr Phe Ser Ser Tyr Asp Met Ser Trp Val Arg Gln
165 170 175
Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Thr Ile Ser Ser Gly Gly
180 185 190
Ser Tyr Thr Tyr Tyr Leu Asp Ser Ile Lys Gly Arg Phe Thr Ile Ser
195 200 205
Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg
210 215 220
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Gly Leu Asp Tyr
225 230 235 240
Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
245 250
<210> 32
<211> 658
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD44v6-4GS2-CAR28z со спейсером LNGFR дикого типа длинным (NWL)
<400> 32
Met Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1 5 10 15
Asp Thr Thr Gly Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Ile Asn Tyr Ile Tyr Trp Leu Gln Gln Lys Pro Gly Gln Ala Pro Arg
50 55 60
Ile Leu Ile Tyr Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln Trp Ser Ser
100 105 110
Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
130 135 140
Gly Gly Gly Leu Val Lys Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala
145 150 155 160
Ala Ser Gly Phe Thr Phe Ser Ser Tyr Asp Met Ser Trp Val Arg Gln
165 170 175
Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Thr Ile Ser Ser Gly Gly
180 185 190
Ser Tyr Thr Tyr Tyr Leu Asp Ser Ile Lys Gly Arg Phe Thr Ile Ser
195 200 205
Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg
210 215 220
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Gly Leu Asp Tyr
225 230 235 240
Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Gly Asp Pro Lys Glu
245 250 255
Ala Cys Pro Thr Gly Leu Tyr Thr His Ser Gly Glu Cys Cys Lys Ala
260 265 270
Cys Asn Leu Gly Glu Gly Val Ala Gln Pro Cys Gly Ala Asn Gln Thr
275 280 285
Val Cys Glu Pro Cys Leu Asp Ser Val Thr Phe Ser Asp Val Val Ser
290 295 300
Ala Thr Glu Pro Cys Lys Pro Cys Thr Glu Cys Val Gly Leu Gln Ser
305 310 315 320
Met Ser Ala Pro Cys Val Glu Ala Asp Asp Ala Val Cys Arg Cys Ala
325 330 335
Tyr Gly Tyr Tyr Gln Asp Glu Thr Thr Gly Arg Cys Glu Ala Cys Arg
340 345 350
Val Cys Glu Ala Gly Ser Gly Leu Val Phe Ser Cys Gln Asp Lys Gln
355 360 365
Asn Thr Val Cys Glu Glu Cys Pro Asp Gly Thr Tyr Ser Asp Glu Ala
370 375 380
Asn His Val Asp Pro Cys Leu Pro Cys Thr Val Cys Glu Asp Thr Glu
385 390 395 400
Arg Gln Leu Arg Glu Cys Thr Arg Trp Ala Asp Ala Glu Cys Glu Glu
405 410 415
Ile Pro Gly Arg Trp Ile Thr Arg Ser Thr Pro Pro Glu Gly Ser Asp
420 425 430
Ser Thr Ala Pro Ser Thr Gln Glu Pro Glu Ala Pro Pro Glu Gln Asp
435 440 445
Leu Ile Ala Ser Thr Val Ala Gly Val Val Thr Thr Val Met Gly Ser
450 455 460
Ser Gln Pro Val Val Thr Arg Gly Thr Thr Asp Asn Pro Lys Phe Trp
465 470 475 480
Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val
485 490 495
Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Leu
500 505 510
Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr
515 520 525
Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr
530 535 540
Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln
545 550 555 560
Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu
565 570 575
Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly
580 585 590
Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu
595 600 605
Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly
610 615 620
Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser
625 630 635 640
Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro
645 650 655
Pro Arg
<210> 33
<211> 598
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD44v6-4GS2-CAR28z со спейсером LNGFR дикого типа коротким (NWS)
<400> 33
Met Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1 5 10 15
Asp Thr Thr Gly Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Ile Asn Tyr Ile Tyr Trp Leu Gln Gln Lys Pro Gly Gln Ala Pro Arg
50 55 60
Ile Leu Ile Tyr Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln Trp Ser Ser
100 105 110
Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
130 135 140
Gly Gly Gly Leu Val Lys Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala
145 150 155 160
Ala Ser Gly Phe Thr Phe Ser Ser Tyr Asp Met Ser Trp Val Arg Gln
165 170 175
Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Thr Ile Ser Ser Gly Gly
180 185 190
Ser Tyr Thr Tyr Tyr Leu Asp Ser Ile Lys Gly Arg Phe Thr Ile Ser
195 200 205
Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg
210 215 220
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Gly Leu Asp Tyr
225 230 235 240
Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Gly Asp Pro Lys Glu
245 250 255
Ala Cys Pro Thr Gly Leu Tyr Thr His Ser Gly Glu Cys Cys Lys Ala
260 265 270
Cys Asn Leu Gly Glu Gly Val Ala Gln Pro Cys Gly Ala Asn Gln Thr
275 280 285
Val Cys Glu Pro Cys Leu Asp Ser Val Thr Phe Ser Asp Val Val Ser
290 295 300
Ala Thr Glu Pro Cys Lys Pro Cys Thr Glu Cys Val Gly Leu Gln Ser
305 310 315 320
Met Ser Ala Pro Cys Val Glu Ala Asp Asp Ala Val Cys Arg Cys Ala
325 330 335
Tyr Gly Tyr Tyr Gln Asp Glu Thr Thr Gly Arg Cys Glu Ala Cys Arg
340 345 350
Val Cys Glu Ala Gly Ser Gly Leu Val Phe Ser Cys Gln Asp Lys Gln
355 360 365
Asn Thr Val Cys Glu Glu Cys Pro Asp Gly Thr Tyr Ser Asp Glu Ala
370 375 380
Asn His Val Asp Pro Cys Leu Pro Cys Thr Val Cys Glu Asp Thr Glu
385 390 395 400
Arg Gln Leu Arg Glu Cys Thr Arg Trp Ala Asp Ala Glu Cys Glu Glu
405 410 415
Pro Lys Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr
420 425 430
Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys
435 440 445
Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg
450 455 460
Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp
465 470 475 480
Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp Ala
485 490 495
Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu
500 505 510
Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp
515 520 525
Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu
530 535 540
Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile
545 550 555 560
Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr
565 570 575
Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met
580 585 590
Gln Ala Leu Pro Pro Arg
595
<210> 34
<211> 636
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD44v6-4GS2-CAR28z со спейсером LNGFR мутированным длинным(NML)
<400> 34
Met Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1 5 10 15
Asp Thr Thr Gly Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Ile Asn Tyr Ile Tyr Trp Leu Gln Gln Lys Pro Gly Gln Ala Pro Arg
50 55 60
Ile Leu Ile Tyr Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln Trp Ser Ser
100 105 110
Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
130 135 140
Gly Gly Gly Leu Val Lys Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala
145 150 155 160
Ala Ser Gly Phe Thr Phe Ser Ser Tyr Asp Met Ser Trp Val Arg Gln
165 170 175
Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Thr Ile Ser Ser Gly Gly
180 185 190
Ser Tyr Thr Tyr Tyr Leu Asp Ser Ile Lys Gly Arg Phe Thr Ile Ser
195 200 205
Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg
210 215 220
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Gly Leu Asp Tyr
225 230 235 240
Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Gly Asp Pro Lys Glu
245 250 255
Ala Cys Pro Thr Gly Leu Tyr Thr His Ser Gly Glu Cys Cys Lys Ala
260 265 270
Cys Asn Leu Gly Glu Gly Val Ala Gln Pro Cys Gly Ala Asn Gln Thr
275 280 285
Val Cys Glu Pro Cys Leu Asp Ser Val Thr Phe Ser Asp Val Val Ser
290 295 300
Ala Thr Glu Pro Cys Lys Pro Cys Thr Glu Cys Val Gly Leu Gln Ser
305 310 315 320
Met Ser Ala Pro Cys Val Glu Ala Asp Asp Ala Val Cys Arg Cys Ala
325 330 335
Tyr Gly Tyr Tyr Gln Asp Glu Thr Thr Gly Arg Cys Glu Ala Cys Arg
340 345 350
Val Cys Glu Ala Gly Ser Gly Leu Val Phe Ser Cys Gln Asp Lys Gln
355 360 365
Asn Thr Val Cys Glu Glu Cys Pro Asp Gly Thr Tyr Ser Asp Glu Ala
370 375 380
Ala Arg Ala Ala Asp Ala Glu Cys Glu Glu Ile Pro Gly Arg Trp Ile
385 390 395 400
Thr Arg Ser Thr Pro Pro Glu Gly Ser Asp Ser Thr Ala Pro Ser Thr
405 410 415
Gln Glu Pro Glu Ala Pro Pro Glu Gln Asp Leu Ile Ala Ser Thr Val
420 425 430
Ala Gly Val Val Thr Thr Val Met Gly Ser Ser Gln Pro Val Val Thr
435 440 445
Arg Gly Thr Thr Asp Asn Pro Lys Phe Trp Val Leu Val Val Val Gly
450 455 460
Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile
465 470 475 480
Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met
485 490 495
Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro
500 505 510
Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe
515 520 525
Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu
530 535 540
Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp
545 550 555 560
Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys
565 570 575
Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala
580 585 590
Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys
595 600 605
Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr
610 615 620
Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
625 630 635
<210> 35
<211> 576
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD44v6-4GS2-CAR28z со спейсером LNGFR мутированным коротким (NMS)
<400> 35
Met Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1 5 10 15
Asp Thr Thr Gly Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Ile Asn Tyr Ile Tyr Trp Leu Gln Gln Lys Pro Gly Gln Ala Pro Arg
50 55 60
Ile Leu Ile Tyr Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln Trp Ser Ser
100 105 110
Asn Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
130 135 140
Gly Gly Gly Leu Val Lys Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala
145 150 155 160
Ala Ser Gly Phe Thr Phe Ser Ser Tyr Asp Met Ser Trp Val Arg Gln
165 170 175
Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Thr Ile Ser Ser Gly Gly
180 185 190
Ser Tyr Thr Tyr Tyr Leu Asp Ser Ile Lys Gly Arg Phe Thr Ile Ser
195 200 205
Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg
210 215 220
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Gly Leu Asp Tyr
225 230 235 240
Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Gly Asp Pro Lys Glu
245 250 255
Ala Cys Pro Thr Gly Leu Tyr Thr His Ser Gly Glu Cys Cys Lys Ala
260 265 270
Cys Asn Leu Gly Glu Gly Val Ala Gln Pro Cys Gly Ala Asn Gln Thr
275 280 285
Val Cys Glu Pro Cys Leu Asp Ser Val Thr Phe Ser Asp Val Val Ser
290 295 300
Ala Thr Glu Pro Cys Lys Pro Cys Thr Glu Cys Val Gly Leu Gln Ser
305 310 315 320
Met Ser Ala Pro Cys Val Glu Ala Asp Asp Ala Val Cys Arg Cys Ala
325 330 335
Tyr Gly Tyr Tyr Gln Asp Glu Thr Thr Gly Arg Cys Glu Ala Cys Arg
340 345 350
Val Cys Glu Ala Gly Ser Gly Leu Val Phe Ser Cys Gln Asp Lys Gln
355 360 365
Asn Thr Val Cys Glu Glu Cys Pro Asp Gly Thr Tyr Ser Asp Glu Ala
370 375 380
Ala Arg Ala Ala Asp Ala Glu Cys Glu Glu Pro Lys Phe Trp Val Leu
385 390 395 400
Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val
405 410 415
Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His
420 425 430
Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys
435 440 445
His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser
450 455 460
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
465 470 475 480
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
485 490 495
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
500 505 510
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
515 520 525
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
530 535 540
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
545 550 555 560
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
565 570 575
<210> 36
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пептидный линкер
<400> 36
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 37
<211> 8331
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CD44v6-4GS2-CAR.28Z С NGFR ДИКОГО ТИПА ДЛИННЫМ (V6 NWL)
<400> 37
aagctttgct cttaggagtt tcctaataca tcccaaactc aaatatataa agcatttgac 60
ttgttctatg ccctaggggg cggggggaag ctaagccagc tttttttaac atttaaaatg 120
ttaattccat tttaaatgca cagatgtttt tatttcataa gggtttcaat gtgcatgaat 180
gctgcaatat tcctgttacc aaagctagta taaataaaaa tagataaacg tggaaattac 240
ttagagtttc tgtcattaac gtttccttcc tcagttgaca acataaatgc gctgctgagc 300
aagccagttt gcatctgtca ggatcaattt cccattatgc cagtcatatt aattactagt 360
caattagttg atttttattt ttgacatata catgtgaatg aaagacccca cctgtaggtt 420
tggcaagcta gcttaagtaa cgccattttg caaggcatgg aaaaatacat aactgagaat 480
agaaaagttc agatcaaggt caggaacaga tggaacagct gaatatgggc caaacaggat 540
atctgtggta agcagttcct gccccggctc agggccaaga acagatggaa cagctgaata 600
tgggccaaac aggatatctg tggtaagcag ttcctgcccc ggctcagggc caagaacaga 660
tggtccccag atgcggtcca gccctcagca gtttctagag aaccatcaga tgtttccagg 720
gtgccccaag gacctgaaat gaccctgtgc cttatttgaa ctaaccaatc agttcgcttc 780
tcgcttctgt tcgcgcgctt atgctccccg agctcaataa aagagcccac aacccctcac 840
tcggggcgcc agtcctccga ttgactgagt cgcccgggta cccgtgtatc caataaaccc 900
tcttgcagtt gcatccgact tgtggtctcg ctgttccttg ggagggtctc ctctgagtga 960
ttgactaccc gtcagcgggg gtctttcatt tgggggctcg tccgggatcg ggagacccct 1020
gcccagggac caccgaccca ccaccgggag gtaagctggc cagcaactta tctgtgtctg 1080
tccgattgtc tagtgtctat gactgatttt atgcgcctgc gtcggtacta gttagctaac 1140
tagctctgta tctggcggac ccgtggtgga actgacgagt tcggaacacc cggccgcaac 1200
cctgggagac gtcccaggga cttcgggggc cgtttttgtg gcccgacctg agtcctaaaa 1260
tcccgatcgt ttaggactct ttggtgcacc ccccttagag gagggatatg tggttctggt 1320
aggagacgag aacctaaaac agttcccgcc tccgtctgaa tttttgcttt cggtttggga 1380
ccgaagccgc gccgcgcgtc ttgtctgctg cagcatcgtt ctgtgttgtc tctgtctgac 1440
tgtgtttctg tatttgtctg aaaatatggg cccgggctag cctgttacca ctcccttaag 1500
tttgacctta ggtcactgga aagatgtcga gcggatcgct cacaaccagt cggtagatgt 1560
caagaagaga cgttgggtta ccttctgctc tgcagaatgg ccaaccttta acgtcggatg 1620
gccgcgagac ggcaccttta accgagacct catcacccag gttaagatca aggtcttttc 1680
acctggcccg catggacacc cagaccaggt ggggtacatc gtgacctggg aagccttggc 1740
ttttgacccc cctccctggg tcaagccctt tgtacaccct aagcctccgc ctcctcttcc 1800
tccatccgcc ccgtctctcc cccttgaacc tcctcgttcg accccgcctc gatcctccct 1860
ttatccagcc ctcactcctt ctctaggcgc ccccatatgg ccatatgaga tcttatatgg 1920
ggcacccccg ccccttgtaa acttccctga ccctgacatg acaagagtta ctaacagccc 1980
ctctctccaa gctcacttac aggctctcta cttagtccag cacgaagtct ggagacctct 2040
ggcggcagcc taccaagaac aactggaccg accggtggta cctcaccctt accgagtcgg 2100
cgacacagtg tgggtccgcc gacaccagac taagaaccta gaacctcgct ggaaaggacc 2160
ttacacagtc ctgctgacca cccccaccgc cctcaaagta gacggcatcg cagcttggat 2220
acacgccgcc cacgtgaagg ctgccgaccc cgggggtgga ccatcctcta gactgccatg 2280
gaagcccctg cccagctgct gttcctgctg ctgctgtggc tgcccgacac caccggcgag 2340
atcgtgctga cacagagccc cgccaccctg tctctgagcc ctggcgagag agccaccctg 2400
agctgtagcg ccagcagcag catcaactac atctactggc tgcagcagaa gcccggccag 2460
gcccccagaa tcctgatcta cctgaccagc aacctggcca gcggcgtgcc cgccagattt 2520
tctggcagcg gcagcggcac cgacttcacc ctgaccatca gcagcctgga acccgaggac 2580
ttcgccgtgt actactgcct gcagtggtcc agcaaccccc tgaccttcgg cggaggcacc 2640
aaggtggaaa tcaagcgggg tggtggtggt tctggtggtg gtggttctga ggtgcagctg 2700
gtggaaagcg gcggaggcct ggtcaagcct ggcggcagcc tgagactgag ctgtgccgcc 2760
agcggcttca ccttcagcag ctacgacatg agctgggtcc gacaggctcc aggcaaggga 2820
ctggaatggg tgtccaccat cagcagcggc ggcagctaca cctactacct ggacagcatc 2880
aagggccggt tcaccatcag ccgggacaac gccaagaaca gcctgtacct gcagatgaac 2940
agcctgcggg ccgaggacac cgccgtctac tactgtgccc ggcagggcct cgactactgg 3000
ggcagaggca ccctggtcac cgtgtccagc ggggatccca aagaggcctg ccccaccggc 3060
ctgtacaccc acagcggaga gtgctgcaag gcctgcaacc tgggagaggg cgtggcccag 3120
ccttgcggcg ccaatcagac cgtgtgcgag ccctgcctgg acagcgtgac cttcagcgac 3180
gtggtgtccg ccaccgagcc ctgcaagcct tgcaccgagt gtgtgggcct gcagagcatg 3240
agcgccccct gcgtggaagc cgacgacgcc gtgtgtagat gcgcctacgg ctactaccag 3300
gacgagacaa ccggcagatg cgaggcctgt agagtgtgcg aggccggcag cggcctggtg 3360
ttcagttgtc aagacaagca gaataccgtg tgtgaagagt gccccgacgg cacctacagc 3420
gacgaggcca accacgtgga cccctgcctg ccctgcactg tgtgcgagga caccgagcgg 3480
cagctgcgcg agtgcacaag atgggccgac gccgagtgcg aagagatccc cggcagatgg 3540
atcaccagaa gcaccccccc tgagggcagc gacagcaccg cccctagcac ccaggaacct 3600
gaggcccctc ccgagcagga cctgatcgcc tctacagtgg ccggcgtggt gacaaccgtg 3660
atgggcagct ctcagcccgt ggtgacacgg ggcaccaccg acaatcccaa attttgggtg 3720
ctggtggtgg ttggtggagt cctggcttgc tatagcttgc tagtaacagt ggcctttatt 3780
attttctggg tgaggagtaa gaggagcagg ctcctgcaca gtgactacat gaacatgact 3840
ccccgccgcc ccgggcccac ccgcaagcat taccagccct atgccccacc acgcgacttc 3900
gcagcctatc gctccagagt gaagttcagc aggagcgcag acgcccccgc gtaccagcag 3960
ggccagaacc agctctataa cgagctcaat ctaggacgaa gagaggagta cgatgttttg 4020
gacaagagac gtggccggga ccctgagatg gggggaaagc cgagaaggaa gaaccctcag 4080
gaaggcctgt acaatgaact gcagaaagat aagatggcgg aggcctacag tgagattggg 4140
atgaaaggcg agcgccggag gggcaagggg cacgatggcc tttaccaggg tctcagtaca 4200
gccaccaagg acacctacga cgcccttcac atgcaggccc tgcctcctcg ctaagcatgc 4260
aacctcgatc cggattagtc caatttgtta aagacaggat atcagtggtc caggctctag 4320
ttttgactca acaatatcac cagctgaagc ctatagagta cgagccatag ataaaataaa 4380
agattttatt tagtctccag aaaaaggggg gaatgaaaga ccccacctgt aggtttggca 4440
agctagctta agtaacgcca ttttgcaagg catggaaaaa tacataactg agaatagaga 4500
agttcagatc aaggtcagga acagatggaa cagctgaata tgggccaaac aggatatctg 4560
tggtaagcag ttcctgcccc ggctcagggc caagaacaga tggaacagct gaatatgggc 4620
caaacaggat atctgtggta agcagttcct gccccggctc agggccaaga acagatggtc 4680
cccagatgcg gtccagccct cagcagtttc tagagaacca tcagatgttt ccagggtgcc 4740
ccaaggacct gaaatgaccc tgtgccttat ttgaactaac caatcagttc gcttctcgct 4800
tctgttcgcg cgcttctgct ccccgagctc aataaaagag cccacaaccc ctcactcggg 4860
gcgccagtcc tccgattgac tgagtcgccc gggtacccgt gtatccaata aaccctcttg 4920
cagttgcatc cgacttgtgg tctcgctgtt ccttgggagg gtctcctctg agtgattgac 4980
tacccgtcag cgggggtctt tcacacatgc agcatgtatc aaaattaatt tggttttttt 5040
tcttaagtat ttacattaaa tggccatagt acttaaagtt acattggctt ccttgaaata 5100
aacatggagt attcagaatg tgtcataaat atttctaatt ttaagatagt atctccattg 5160
gctttctact ttttctttta tttttttttg tcctctgtct tccatttgtt gttgttgttg 5220
tttgtttgtt tgtttgttgg ttggttggtt aatttttttt taaagatcct acactatagt 5280
tcaagctaga ctattagcta ctctgtaacc cagggtgacc ttgaagtcat gggtagcctg 5340
ctgttttagc cttcccacat ctaagattac aggtatgagc tatcattttt ggtatattga 5400
ttgattgatt gattgatgtg tgtgtgtgtg attgtgtttg tgtgtgtgac tgtgaaaatg 5460
tgtgtatggg tgtgtgtgaa tgtgtgtatg tatgtgtgtg tgtgagtgtg tgtgtgtgtg 5520
tgtgcatgtg tgtgtgtgtg actgtgtcta tgtgtatgac tgtgtgtgtg tgtgtgtgtg 5580
tgtgtgtgtg tgtgtgtgtg tgtgtgttgt gaaaaaatat tctatggtag tgagagccaa 5640
cgctccggct caggtgtcag gttggttttt gagacagagt ctttcactta gcttggaatt 5700
cactggccgt cgttttacaa cgtcgtgact gggaaaaccc tggcgttacc caacttaatc 5760
gccttgcagc acatccccct ttcgccagct ggcgtaatag cgaagaggcc cgcaccgatc 5820
gcccttccca acagttgcgc agcctgaatg gcgaatggcg cctgatgcgg tattttctcc 5880
ttacgcatct gtgcggtatt tcacaccgca tatggtgcac tctcagtaca atctgctctg 5940
atgccgcata gttaagccag ccccgacacc cgccaacacc cgctgacgcg ccctgacggg 6000
cttgtctgct cccggcatcc gcttacagac aagctgtgac cgtctccggg agctgcatgt 6060
gtcagaggtt ttcaccgtca tcaccgaaac gcgcgatgac gaaagggcct cgtgatacgc 6120
ctatttttat aggttaatgt catgataata atggtttctt agacgtcagg tggcactttt 6180
cggggaaatg tgcgcggaac ccctatttgt ttatttttct aaatacattc aaatatgtat 6240
ccgctcatga gacaataacc ctgataaatg cttcaataat attgaaaaag gaagagtatg 6300
agtattcaac atttccgtgt cgcccttatt cccttttttg cggcattttg ccttcctgtt 6360
tttgctcacc cagaaacgct ggtgaaagta aaagatgctg aagatcagtt gggtgcacga 6420
gtgggttaca tcgaactgga tctcaacagc ggtaagatcc ttgagagttt tcgccccgaa 6480
gaacgttttc caatgatgag cacttttaaa gttctgctat gtggcgcggt attatcccgt 6540
attgacgccg ggcaagagca actcggtcgc cgcatacact attctcagaa tgacttggtt 6600
gagtactcac cagtcacaga aaagcatctt acggatggca tgacagtaag agaattatgc 6660
agtgctgcca taaccatgag tgataacact gcggccaact tacttctgac aacgatcgga 6720
ggaccgaagg agctaaccgc ttttttgcac aacatggggg atcatgtaac tcgccttgat 6780
cgttgggaac cggagctgaa tgaagccata ccaaacgacg agcgtgacac cacgatgcct 6840
gtagcaatgg caacaacgtt gcgcaaacta ttaactggcg aactacttac tctagcttcc 6900
cggcaacaat taatagactg gatggaggcg gataaagttg caggaccact tctgcgctcg 6960
gcccttccgg ctggctggtt tattgctgat aaatctggag ccggtgagcg tgggtctcgc 7020
ggtatcattg cagcactggg gccagatggt aagccctccc gtatcgtagt tatctacacg 7080
acggggagtc aggcaactat ggatgaacga aatagacaga tcgctgagat aggtgcctca 7140
ctgattaagc attggtaact gtcagaccaa gtttactcat atatacttta gattgattta 7200
aaacttcatt tttaatttaa aaggatctag gtgaagatcc tttttgataa tctcatgacc 7260
aaaatccctt aacgtgagtt ttcgttccac tgagcgtcag accccgtaga aaagatcaaa 7320
ggatcttctt gagatccttt ttttctgcgc gtaatctgct gcttgcaaac aaaaaaacca 7380
ccgctaccag cggtggtttg tttgccggat caagagctac caactctttt tccgaaggta 7440
actggcttca gcagagcgca gataccaaat actgtccttc tagtgtagcc gtagttaggc 7500
caccacttca agaactctgt agcaccgcct acatacctcg ctctgctaat cctgttacca 7560
gtggctgctg ccagtggcga taagtcgtgt cttaccgggt tggactcaag acgatagtta 7620
ccggataagg cgcagcggtc gggctgaacg gggggttcgt gcacacagcc cagcttggag 7680
cgaacgacct acaccgaact gagataccta cagcgtgagc attgagaaag cgccacgctt 7740
cccgaaggga gaaaggcgga caggtatccg gtaagcggca gggtcggaac aggagagcgc 7800
acgagggagc ttccaggggg aaacgcctgg tatctttata gtcctgtcgg gtttcgccac 7860
ctctgacttg agcgtcgatt tttgtgatgc tcgtcagggg ggcggagcct atggaaaaac 7920
gccagcaacg cggccttttt acggttcctg gccttttgct ggccttttgc tcacatgttc 7980
tttcctgcgt tatcccctga ttctgtggat aaccgtatta ccgcctttga gtgagctgat 8040
accgctcgcc gcagccgaac gaccgagcgc agcgagtcag tgagcgagga agcggaagag 8100
cgcccaatac gcaaaccgcc tctccccgcg cgttggccga ttcattaatg cagctggcac 8160
gacaggtttc ccgactggaa agcgggcagt gagcgcaacg caattaatgt gagttagctc 8220
actcattagg caccccaggc tttacacttt atgcttccgg ctcgtatgtt gtgtggaatt 8280
gtgagcggat aacaatttca cacaggaaac agctatgacc atgattacgc c 8331
<210> 38
<211> 8151
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CD44V6-4GS2-CAR.28Z С NGFR ДИКОГО ТИПА КОРОТКИМ (V6 NWS)
<400> 38
aagctttgct cttaggagtt tcctaataca tcccaaactc aaatatataa agcatttgac 60
ttgttctatg ccctaggggg cggggggaag ctaagccagc tttttttaac atttaaaatg 120
ttaattccat tttaaatgca cagatgtttt tatttcataa gggtttcaat gtgcatgaat 180
gctgcaatat tcctgttacc aaagctagta taaataaaaa tagataaacg tggaaattac 240
ttagagtttc tgtcattaac gtttccttcc tcagttgaca acataaatgc gctgctgagc 300
aagccagttt gcatctgtca ggatcaattt cccattatgc cagtcatatt aattactagt 360
caattagttg atttttattt ttgacatata catgtgaatg aaagacccca cctgtaggtt 420
tggcaagcta gcttaagtaa cgccattttg caaggcatgg aaaaatacat aactgagaat 480
agaaaagttc agatcaaggt caggaacaga tggaacagct gaatatgggc caaacaggat 540
atctgtggta agcagttcct gccccggctc agggccaaga acagatggaa cagctgaata 600
tgggccaaac aggatatctg tggtaagcag ttcctgcccc ggctcagggc caagaacaga 660
tggtccccag atgcggtcca gccctcagca gtttctagag aaccatcaga tgtttccagg 720
gtgccccaag gacctgaaat gaccctgtgc cttatttgaa ctaaccaatc agttcgcttc 780
tcgcttctgt tcgcgcgctt atgctccccg agctcaataa aagagcccac aacccctcac 840
tcggggcgcc agtcctccga ttgactgagt cgcccgggta cccgtgtatc caataaaccc 900
tcttgcagtt gcatccgact tgtggtctcg ctgttccttg ggagggtctc ctctgagtga 960
ttgactaccc gtcagcgggg gtctttcatt tgggggctcg tccgggatcg ggagacccct 1020
gcccagggac caccgaccca ccaccgggag gtaagctggc cagcaactta tctgtgtctg 1080
tccgattgtc tagtgtctat gactgatttt atgcgcctgc gtcggtacta gttagctaac 1140
tagctctgta tctggcggac ccgtggtgga actgacgagt tcggaacacc cggccgcaac 1200
cctgggagac gtcccaggga cttcgggggc cgtttttgtg gcccgacctg agtcctaaaa 1260
tcccgatcgt ttaggactct ttggtgcacc ccccttagag gagggatatg tggttctggt 1320
aggagacgag aacctaaaac agttcccgcc tccgtctgaa tttttgcttt cggtttggga 1380
ccgaagccgc gccgcgcgtc ttgtctgctg cagcatcgtt ctgtgttgtc tctgtctgac 1440
tgtgtttctg tatttgtctg aaaatatggg cccgggctag cctgttacca ctcccttaag 1500
tttgacctta ggtcactgga aagatgtcga gcggatcgct cacaaccagt cggtagatgt 1560
caagaagaga cgttgggtta ccttctgctc tgcagaatgg ccaaccttta acgtcggatg 1620
gccgcgagac ggcaccttta accgagacct catcacccag gttaagatca aggtcttttc 1680
acctggcccg catggacacc cagaccaggt ggggtacatc gtgacctggg aagccttggc 1740
ttttgacccc cctccctggg tcaagccctt tgtacaccct aagcctccgc ctcctcttcc 1800
tccatccgcc ccgtctctcc cccttgaacc tcctcgttcg accccgcctc gatcctccct 1860
ttatccagcc ctcactcctt ctctaggcgc ccccatatgg ccatatgaga tcttatatgg 1920
ggcacccccg ccccttgtaa acttccctga ccctgacatg acaagagtta ctaacagccc 1980
ctctctccaa gctcacttac aggctctcta cttagtccag cacgaagtct ggagacctct 2040
ggcggcagcc taccaagaac aactggaccg accggtggta cctcaccctt accgagtcgg 2100
cgacacagtg tgggtccgcc gacaccagac taagaaccta gaacctcgct ggaaaggacc 2160
ttacacagtc ctgctgacca cccccaccgc cctcaaagta gacggcatcg cagcttggat 2220
acacgccgcc cacgtgaagg ctgccgaccc cgggggtgga ccatcctcta gactgccatg 2280
gaagcccctg cccagctgct gttcctgctg ctgctgtggc tgcccgacac caccggcgag 2340
atcgtgctga cacagagccc cgccaccctg tctctgagcc ctggcgagag agccaccctg 2400
agctgtagcg ccagcagcag catcaactac atctactggc tgcagcagaa gcccggccag 2460
gcccccagaa tcctgatcta cctgaccagc aacctggcca gcggcgtgcc cgccagattt 2520
tctggcagcg gcagcggcac cgacttcacc ctgaccatca gcagcctgga acccgaggac 2580
ttcgccgtgt actactgcct gcagtggtcc agcaaccccc tgaccttcgg cggaggcacc 2640
aaggtggaaa tcaagcgggg tggtggtggt tctggtggtg gtggttctga ggtgcagctg 2700
gtggaaagcg gcggaggcct ggtcaagcct ggcggcagcc tgagactgag ctgtgccgcc 2760
agcggcttca ccttcagcag ctacgacatg agctgggtcc gacaggctcc aggcaaggga 2820
ctggaatggg tgtccaccat cagcagcggc ggcagctaca cctactacct ggacagcatc 2880
aagggccggt tcaccatcag ccgggacaac gccaagaaca gcctgtacct gcagatgaac 2940
agcctgcggg ccgaggacac cgccgtctac tactgtgccc ggcagggcct cgactactgg 3000
ggcagaggca ccctggtcac cgtgtccagc ggggatccca aagaggcctg ccccaccggc 3060
ctgtacaccc acagcggaga gtgctgcaag gcctgcaacc tgggagaggg cgtggcccag 3120
ccttgcggcg ccaatcagac cgtgtgcgag ccctgcctgg acagcgtgac cttcagcgac 3180
gtggtgtccg ccaccgagcc ctgcaagcct tgcaccgagt gtgtgggcct gcagagcatg 3240
agcgccccct gcgtggaagc cgacgacgcc gtgtgtagat gcgcctacgg ctactaccag 3300
gacgagacaa ccggcagatg cgaggcctgt agagtgtgcg aggccggcag cggcctggtg 3360
ttcagttgtc aggacaagca gaacaccgtg tgtgaagagt gccccgacgg cacctacagc 3420
gacgaggcca accacgtgga cccctgcctg ccctgcactg tgtgcgagga caccgagcgg 3480
cagctgcgcg agtgcacaag atgggccgac gccgagtgcg aggaacccaa attttgggtg 3540
ctggtggtgg ttggtggagt cctggcttgc tatagcttgc tagtaacagt ggcctttatt 3600
attttctggg tgaggagtaa gaggagcagg ctcctgcaca gtgactacat gaacatgact 3660
ccccgccgcc ccgggcccac ccgcaagcat taccagccct atgccccacc acgcgacttc 3720
gcagcctatc gctccagagt gaagttcagc aggagcgcag acgcccccgc gtaccagcag 3780
ggccagaacc agctctataa cgagctcaat ctaggacgaa gagaggagta cgatgttttg 3840
gacaagagac gtggccggga ccctgagatg gggggaaagc cgagaaggaa gaaccctcag 3900
gaaggcctgt acaatgaact gcagaaagat aagatggcgg aggcctacag tgagattggg 3960
atgaaaggcg agcgccggag gggcaagggg cacgatggcc tttaccaggg tctcagtaca 4020
gccaccaagg acacctacga cgcccttcac atgcaggccc tgcctcctcg ctaagcatgc 4080
aacctcgatc cggattagtc caatttgtta aagacaggat atcagtggtc caggctctag 4140
ttttgactca acaatatcac cagctgaagc ctatagagta cgagccatag ataaaataaa 4200
agattttatt tagtctccag aaaaaggggg gaatgaaaga ccccacctgt aggtttggca 4260
agctagctta agtaacgcca ttttgcaagg catggaaaaa tacataactg agaatagaga 4320
agttcagatc aaggtcagga acagatggaa cagctgaata tgggccaaac aggatatctg 4380
tggtaagcag ttcctgcccc ggctcagggc caagaacaga tggaacagct gaatatgggc 4440
caaacaggat atctgtggta agcagttcct gccccggctc agggccaaga acagatggtc 4500
cccagatgcg gtccagccct cagcagtttc tagagaacca tcagatgttt ccagggtgcc 4560
ccaaggacct gaaatgaccc tgtgccttat ttgaactaac caatcagttc gcttctcgct 4620
tctgttcgcg cgcttctgct ccccgagctc aataaaagag cccacaaccc ctcactcggg 4680
gcgccagtcc tccgattgac tgagtcgccc gggtacccgt gtatccaata aaccctcttg 4740
cagttgcatc cgacttgtgg tctcgctgtt ccttgggagg gtctcctctg agtgattgac 4800
tacccgtcag cgggggtctt tcacacatgc agcatgtatc aaaattaatt tggttttttt 4860
tcttaagtat ttacattaaa tggccatagt acttaaagtt acattggctt ccttgaaata 4920
aacatggagt attcagaatg tgtcataaat atttctaatt ttaagatagt atctccattg 4980
gctttctact ttttctttta tttttttttg tcctctgtct tccatttgtt gttgttgttg 5040
tttgtttgtt tgtttgttgg ttggttggtt aatttttttt taaagatcct acactatagt 5100
tcaagctaga ctattagcta ctctgtaacc cagggtgacc ttgaagtcat gggtagcctg 5160
ctgttttagc cttcccacat ctaagattac aggtatgagc tatcattttt ggtatattga 5220
ttgattgatt gattgatgtg tgtgtgtgtg attgtgtttg tgtgtgtgac tgtgaaaatg 5280
tgtgtatggg tgtgtgtgaa tgtgtgtatg tatgtgtgtg tgtgagtgtg tgtgtgtgtg 5340
tgtgcatgtg tgtgtgtgtg actgtgtcta tgtgtatgac tgtgtgtgtg tgtgtgtgtg 5400
tgtgtgtgtg tgtgtgtgtg tgtgtgttgt gaaaaaatat tctatggtag tgagagccaa 5460
cgctccggct caggtgtcag gttggttttt gagacagagt ctttcactta gcttggaatt 5520
cactggccgt cgttttacaa cgtcgtgact gggaaaaccc tggcgttacc caacttaatc 5580
gccttgcagc acatccccct ttcgccagct ggcgtaatag cgaagaggcc cgcaccgatc 5640
gcccttccca acagttgcgc agcctgaatg gcgaatggcg cctgatgcgg tattttctcc 5700
ttacgcatct gtgcggtatt tcacaccgca tatggtgcac tctcagtaca atctgctctg 5760
atgccgcata gttaagccag ccccgacacc cgccaacacc cgctgacgcg ccctgacggg 5820
cttgtctgct cccggcatcc gcttacagac aagctgtgac cgtctccggg agctgcatgt 5880
gtcagaggtt ttcaccgtca tcaccgaaac gcgcgatgac gaaagggcct cgtgatacgc 5940
ctatttttat aggttaatgt catgataata atggtttctt agacgtcagg tggcactttt 6000
cggggaaatg tgcgcggaac ccctatttgt ttatttttct aaatacattc aaatatgtat 6060
ccgctcatga gacaataacc ctgataaatg cttcaataat attgaaaaag gaagagtatg 6120
agtattcaac atttccgtgt cgcccttatt cccttttttg cggcattttg ccttcctgtt 6180
tttgctcacc cagaaacgct ggtgaaagta aaagatgctg aagatcagtt gggtgcacga 6240
gtgggttaca tcgaactgga tctcaacagc ggtaagatcc ttgagagttt tcgccccgaa 6300
gaacgttttc caatgatgag cacttttaaa gttctgctat gtggcgcggt attatcccgt 6360
attgacgccg ggcaagagca actcggtcgc cgcatacact attctcagaa tgacttggtt 6420
gagtactcac cagtcacaga aaagcatctt acggatggca tgacagtaag agaattatgc 6480
agtgctgcca taaccatgag tgataacact gcggccaact tacttctgac aacgatcgga 6540
ggaccgaagg agctaaccgc ttttttgcac aacatggggg atcatgtaac tcgccttgat 6600
cgttgggaac cggagctgaa tgaagccata ccaaacgacg agcgtgacac cacgatgcct 6660
gtagcaatgg caacaacgtt gcgcaaacta ttaactggcg aactacttac tctagcttcc 6720
cggcaacaat taatagactg gatggaggcg gataaagttg caggaccact tctgcgctcg 6780
gcccttccgg ctggctggtt tattgctgat aaatctggag ccggtgagcg tgggtctcgc 6840
ggtatcattg cagcactggg gccagatggt aagccctccc gtatcgtagt tatctacacg 6900
acggggagtc aggcaactat ggatgaacga aatagacaga tcgctgagat aggtgcctca 6960
ctgattaagc attggtaact gtcagaccaa gtttactcat atatacttta gattgattta 7020
aaacttcatt tttaatttaa aaggatctag gtgaagatcc tttttgataa tctcatgacc 7080
aaaatccctt aacgtgagtt ttcgttccac tgagcgtcag accccgtaga aaagatcaaa 7140
ggatcttctt gagatccttt ttttctgcgc gtaatctgct gcttgcaaac aaaaaaacca 7200
ccgctaccag cggtggtttg tttgccggat caagagctac caactctttt tccgaaggta 7260
actggcttca gcagagcgca gataccaaat actgtccttc tagtgtagcc gtagttaggc 7320
caccacttca agaactctgt agcaccgcct acatacctcg ctctgctaat cctgttacca 7380
gtggctgctg ccagtggcga taagtcgtgt cttaccgggt tggactcaag acgatagtta 7440
ccggataagg cgcagcggtc gggctgaacg gggggttcgt gcacacagcc cagcttggag 7500
cgaacgacct acaccgaact gagataccta cagcgtgagc attgagaaag cgccacgctt 7560
cccgaaggga gaaaggcgga caggtatccg gtaagcggca gggtcggaac aggagagcgc 7620
acgagggagc ttccaggggg aaacgcctgg tatctttata gtcctgtcgg gtttcgccac 7680
ctctgacttg agcgtcgatt tttgtgatgc tcgtcagggg ggcggagcct atggaaaaac 7740
gccagcaacg cggccttttt acggttcctg gccttttgct ggccttttgc tcacatgttc 7800
tttcctgcgt tatcccctga ttctgtggat aaccgtatta ccgcctttga gtgagctgat 7860
accgctcgcc gcagccgaac gaccgagcgc agcgagtcag tgagcgagga agcggaagag 7920
cgcccaatac gcaaaccgcc tctccccgcg cgttggccga ttcattaatg cagctggcac 7980
gacaggtttc ccgactggaa agcgggcagt gagcgcaacg caattaatgt gagttagctc 8040
actcattagg caccccaggc tttacacttt atgcttccgg ctcgtatgtt gtgtggaatt 8100
gtgagcggat aacaatttca cacaggaaac agctatgacc atgattacgc c 8151
<210> 39
<211> 8265
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CD44v6-4GS2-CAR.28Z с NGFR МУТИРОВАННЫМ ДЛИННЫМ (V6 NML)
<400> 39
aagctttgct cttaggagtt tcctaataca tcccaaactc aaatatataa agcatttgac 60
ttgttctatg ccctaggggg cggggggaag ctaagccagc tttttttaac atttaaaatg 120
ttaattccat tttaaatgca cagatgtttt tatttcataa gggtttcaat gtgcatgaat 180
gctgcaatat tcctgttacc aaagctagta taaataaaaa tagataaacg tggaaattac 240
ttagagtttc tgtcattaac gtttccttcc tcagttgaca acataaatgc gctgctgagc 300
aagccagttt gcatctgtca ggatcaattt cccattatgc cagtcatatt aattactagt 360
caattagttg atttttattt ttgacatata catgtgaatg aaagacccca cctgtaggtt 420
tggcaagcta gcttaagtaa cgccattttg caaggcatgg aaaaatacat aactgagaat 480
agaaaagttc agatcaaggt caggaacaga tggaacagct gaatatgggc caaacaggat 540
atctgtggta agcagttcct gccccggctc agggccaaga acagatggaa cagctgaata 600
tgggccaaac aggatatctg tggtaagcag ttcctgcccc ggctcagggc caagaacaga 660
tggtccccag atgcggtcca gccctcagca gtttctagag aaccatcaga tgtttccagg 720
gtgccccaag gacctgaaat gaccctgtgc cttatttgaa ctaaccaatc agttcgcttc 780
tcgcttctgt tcgcgcgctt atgctccccg agctcaataa aagagcccac aacccctcac 840
tcggggcgcc agtcctccga ttgactgagt cgcccgggta cccgtgtatc caataaaccc 900
tcttgcagtt gcatccgact tgtggtctcg ctgttccttg ggagggtctc ctctgagtga 960
ttgactaccc gtcagcgggg gtctttcatt tgggggctcg tccgggatcg ggagacccct 1020
gcccagggac caccgaccca ccaccgggag gtaagctggc cagcaactta tctgtgtctg 1080
tccgattgtc tagtgtctat gactgatttt atgcgcctgc gtcggtacta gttagctaac 1140
tagctctgta tctggcggac ccgtggtgga actgacgagt tcggaacacc cggccgcaac 1200
cctgggagac gtcccaggga cttcgggggc cgtttttgtg gcccgacctg agtcctaaaa 1260
tcccgatcgt ttaggactct ttggtgcacc ccccttagag gagggatatg tggttctggt 1320
aggagacgag aacctaaaac agttcccgcc tccgtctgaa tttttgcttt cggtttggga 1380
ccgaagccgc gccgcgcgtc ttgtctgctg cagcatcgtt ctgtgttgtc tctgtctgac 1440
tgtgtttctg tatttgtctg aaaatatggg cccgggctag cctgttacca ctcccttaag 1500
tttgacctta ggtcactgga aagatgtcga gcggatcgct cacaaccagt cggtagatgt 1560
caagaagaga cgttgggtta ccttctgctc tgcagaatgg ccaaccttta acgtcggatg 1620
gccgcgagac ggcaccttta accgagacct catcacccag gttaagatca aggtcttttc 1680
acctggcccg catggacacc cagaccaggt ggggtacatc gtgacctggg aagccttggc 1740
ttttgacccc cctccctggg tcaagccctt tgtacaccct aagcctccgc ctcctcttcc 1800
tccatccgcc ccgtctctcc cccttgaacc tcctcgttcg accccgcctc gatcctccct 1860
ttatccagcc ctcactcctt ctctaggcgc ccccatatgg ccatatgaga tcttatatgg 1920
ggcacccccg ccccttgtaa acttccctga ccctgacatg acaagagtta ctaacagccc 1980
ctctctccaa gctcacttac aggctctcta cttagtccag cacgaagtct ggagacctct 2040
ggcggcagcc taccaagaac aactggaccg accggtggta cctcaccctt accgagtcgg 2100
cgacacagtg tgggtccgcc gacaccagac taagaaccta gaacctcgct ggaaaggacc 2160
ttacacagtc ctgctgacca cccccaccgc cctcaaagta gacggcatcg cagcttggat 2220
acacgccgcc cacgtgaagg ctgccgaccc cgggggtgga ccatcctcta gactgccatg 2280
gaagcccctg cccagctgct gttcctgctg ctgctgtggc tgcccgacac caccggcgag 2340
atcgtgctga cacagagccc cgccaccctg tctctgagcc ctggcgagag agccaccctg 2400
agctgtagcg ccagcagcag catcaactac atctactggc tgcagcagaa gcccggccag 2460
gcccccagaa tcctgatcta cctgaccagc aacctggcca gcggcgtgcc cgccagattt 2520
tctggcagcg gcagcggcac cgacttcacc ctgaccatca gcagcctgga acccgaggac 2580
ttcgccgtgt actactgcct gcagtggtcc agcaaccccc tgaccttcgg cggaggcacc 2640
aaggtggaaa tcaagcgggg tggtggtggt tctggtggtg gtggttctga ggtgcagctg 2700
gtggaaagcg gcggaggcct ggtcaagcct ggcggcagcc tgagactgag ctgtgccgcc 2760
agcggcttca ccttcagcag ctacgacatg agctgggtcc gacaggctcc aggcaaggga 2820
ctggaatggg tgtccaccat cagcagcggc ggcagctaca cctactacct ggacagcatc 2880
aagggccggt tcaccatcag ccgggacaac gccaagaaca gcctgtacct gcagatgaac 2940
agcctgcggg ccgaggacac cgccgtctac tactgtgccc ggcagggcct cgactactgg 3000
ggcagaggca ccctggtcac cgtgtccagc ggggatccca aagaggcctg ccccaccggc 3060
ctgtacaccc acagcggaga gtgctgcaag gcctgcaacc tgggagaggg cgtggcccag 3120
ccttgcggcg ccaatcagac cgtgtgcgag ccctgcctgg acagcgtgac cttcagcgac 3180
gtggtgtccg ccaccgagcc ctgcaagcct tgcaccgagt gtgtgggcct gcagagcatg 3240
agcgccccct gcgtggaagc cgacgacgcc gtgtgtagat gcgcctacgg ctactaccag 3300
gacgagacaa ccggcagatg cgaggcctgt agagtgtgcg aggccggcag cggcctggtg 3360
ttcagttgtc aagacaagca gaataccgtg tgtgaagagt gccccgacgg cacctacagc 3420
gacgaagccg ccagagccgc cgacgccgag tgcgaagaga tccccggcag atggatcacc 3480
agaagcaccc cccctgaggg cagcgacagc accgccccta gcacccagga acctgaggcc 3540
cctcccgagc aggacctgat cgcctctaca gtggccggcg tggtgacaac cgtgatgggc 3600
agctctcagc ccgtggtgac acggggcacc accgacaatc ccaaattttg ggtgctggtg 3660
gtggttggtg gagtcctggc ttgctatagc ttgctagtaa cagtggcctt tattattttc 3720
tgggtgagga gtaagaggag caggctcctg cacagtgact acatgaacat gactccccgc 3780
cgccccgggc ccacccgcaa gcattaccag ccctatgccc caccacgcga cttcgcagcc 3840
tatcgctcca gagtgaagtt cagcaggagc gcagacgccc ccgcgtacca gcagggccag 3900
aaccagctct ataacgagct caatctagga cgaagagagg agtacgatgt tttggacaag 3960
agacgtggcc gggaccctga gatgggggga aagccgagaa ggaagaaccc tcaggaaggc 4020
ctgtacaatg aactgcagaa agataagatg gcggaggcct acagtgagat tgggatgaaa 4080
ggcgagcgcc ggaggggcaa ggggcacgat ggcctttacc agggtctcag tacagccacc 4140
aaggacacct acgacgccct tcacatgcag gccctgcctc ctcgctaagc atgcaacctc 4200
gatccggatt agtccaattt gttaaagaca ggatatcagt ggtccaggct ctagttttga 4260
ctcaacaata tcaccagctg aagcctatag agtacgagcc atagataaaa taaaagattt 4320
tatttagtct ccagaaaaag gggggaatga aagaccccac ctgtaggttt ggcaagctag 4380
cttaagtaac gccattttgc aaggcatgga aaaatacata actgagaata gagaagttca 4440
gatcaaggtc aggaacagat ggaacagctg aatatgggcc aaacaggata tctgtggtaa 4500
gcagttcctg ccccggctca gggccaagaa cagatggaac agctgaatat gggccaaaca 4560
ggatatctgt ggtaagcagt tcctgccccg gctcagggcc aagaacagat ggtccccaga 4620
tgcggtccag ccctcagcag tttctagaga accatcagat gtttccaggg tgccccaagg 4680
acctgaaatg accctgtgcc ttatttgaac taaccaatca gttcgcttct cgcttctgtt 4740
cgcgcgcttc tgctccccga gctcaataaa agagcccaca acccctcact cggggcgcca 4800
gtcctccgat tgactgagtc gcccgggtac ccgtgtatcc aataaaccct cttgcagttg 4860
catccgactt gtggtctcgc tgttccttgg gagggtctcc tctgagtgat tgactacccg 4920
tcagcggggg tctttcacac atgcagcatg tatcaaaatt aatttggttt tttttcttaa 4980
gtatttacat taaatggcca tagtacttaa agttacattg gcttccttga aataaacatg 5040
gagtattcag aatgtgtcat aaatatttct aattttaaga tagtatctcc attggctttc 5100
tactttttct tttatttttt tttgtcctct gtcttccatt tgttgttgtt gttgtttgtt 5160
tgtttgtttg ttggttggtt ggttaatttt tttttaaaga tcctacacta tagttcaagc 5220
tagactatta gctactctgt aacccagggt gaccttgaag tcatgggtag cctgctgttt 5280
tagccttccc acatctaaga ttacaggtat gagctatcat ttttggtata ttgattgatt 5340
gattgattga tgtgtgtgtg tgtgattgtg tttgtgtgtg tgactgtgaa aatgtgtgta 5400
tgggtgtgtg tgaatgtgtg tatgtatgtg tgtgtgtgag tgtgtgtgtg tgtgtgtgca 5460
tgtgtgtgtg tgtgactgtg tctatgtgta tgactgtgtg tgtgtgtgtg tgtgtgtgtg 5520
tgtgtgtgtg tgtgtgtgtg ttgtgaaaaa atattctatg gtagtgagag ccaacgctcc 5580
ggctcaggtg tcaggttggt ttttgagaca gagtctttca cttagcttgg aattcactgg 5640
ccgtcgtttt acaacgtcgt gactgggaaa accctggcgt tacccaactt aatcgccttg 5700
cagcacatcc ccctttcgcc agctggcgta atagcgaaga ggcccgcacc gatcgccctt 5760
cccaacagtt gcgcagcctg aatggcgaat ggcgcctgat gcggtatttt ctccttacgc 5820
atctgtgcgg tatttcacac cgcatatggt gcactctcag tacaatctgc tctgatgccg 5880
catagttaag ccagccccga cacccgccaa cacccgctga cgcgccctga cgggcttgtc 5940
tgctcccggc atccgcttac agacaagctg tgaccgtctc cgggagctgc atgtgtcaga 6000
ggttttcacc gtcatcaccg aaacgcgcga tgacgaaagg gcctcgtgat acgcctattt 6060
ttataggtta atgtcatgat aataatggtt tcttagacgt caggtggcac ttttcgggga 6120
aatgtgcgcg gaacccctat ttgtttattt ttctaaatac attcaaatat gtatccgctc 6180
atgagacaat aaccctgata aatgcttcaa taatattgaa aaaggaagag tatgagtatt 6240
caacatttcc gtgtcgccct tattcccttt tttgcggcat tttgccttcc tgtttttgct 6300
cacccagaaa cgctggtgaa agtaaaagat gctgaagatc agttgggtgc acgagtgggt 6360
tacatcgaac tggatctcaa cagcggtaag atccttgaga gttttcgccc cgaagaacgt 6420
tttccaatga tgagcacttt taaagttctg ctatgtggcg cggtattatc ccgtattgac 6480
gccgggcaag agcaactcgg tcgccgcata cactattctc agaatgactt ggttgagtac 6540
tcaccagtca cagaaaagca tcttacggat ggcatgacag taagagaatt atgcagtgct 6600
gccataacca tgagtgataa cactgcggcc aacttacttc tgacaacgat cggaggaccg 6660
aaggagctaa ccgctttttt gcacaacatg ggggatcatg taactcgcct tgatcgttgg 6720
gaaccggagc tgaatgaagc cataccaaac gacgagcgtg acaccacgat gcctgtagca 6780
atggcaacaa cgttgcgcaa actattaact ggcgaactac ttactctagc ttcccggcaa 6840
caattaatag actggatgga ggcggataaa gttgcaggac cacttctgcg ctcggccctt 6900
ccggctggct ggtttattgc tgataaatct ggagccggtg agcgtgggtc tcgcggtatc 6960
attgcagcac tggggccaga tggtaagccc tcccgtatcg tagttatcta cacgacgggg 7020
agtcaggcaa ctatggatga acgaaataga cagatcgctg agataggtgc ctcactgatt 7080
aagcattggt aactgtcaga ccaagtttac tcatatatac tttagattga tttaaaactt 7140
catttttaat ttaaaaggat ctaggtgaag atcctttttg ataatctcat gaccaaaatc 7200
ccttaacgtg agttttcgtt ccactgagcg tcagaccccg tagaaaagat caaaggatct 7260
tcttgagatc ctttttttct gcgcgtaatc tgctgcttgc aaacaaaaaa accaccgcta 7320
ccagcggtgg tttgtttgcc ggatcaagag ctaccaactc tttttccgaa ggtaactggc 7380
ttcagcagag cgcagatacc aaatactgtc cttctagtgt agccgtagtt aggccaccac 7440
ttcaagaact ctgtagcacc gcctacatac ctcgctctgc taatcctgtt accagtggct 7500
gctgccagtg gcgataagtc gtgtcttacc gggttggact caagacgata gttaccggat 7560
aaggcgcagc ggtcgggctg aacggggggt tcgtgcacac agcccagctt ggagcgaacg 7620
acctacaccg aactgagata cctacagcgt gagcattgag aaagcgccac gcttcccgaa 7680
gggagaaagg cggacaggta tccggtaagc ggcagggtcg gaacaggaga gcgcacgagg 7740
gagcttccag ggggaaacgc ctggtatctt tatagtcctg tcgggtttcg ccacctctga 7800
cttgagcgtc gatttttgtg atgctcgtca ggggggcgga gcctatggaa aaacgccagc 7860
aacgcggcct ttttacggtt cctggccttt tgctggcctt ttgctcacat gttctttcct 7920
gcgttatccc ctgattctgt ggataaccgt attaccgcct ttgagtgagc tgataccgct 7980
cgccgcagcc gaacgaccga gcgcagcgag tcagtgagcg aggaagcgga agagcgccca 8040
atacgcaaac cgcctctccc cgcgcgttgg ccgattcatt aatgcagctg gcacgacagg 8100
tttcccgact ggaaagcggg cagtgagcgc aacgcaatta atgtgagtta gctcactcat 8160
taggcacccc aggctttaca ctttatgctt ccggctcgta tgttgtgtgg aattgtgagc 8220
ggataacaat ttcacacagg aaacagctat gaccatgatt acgcc 8265
<210> 40
<211> 8085
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CD44v6-4GS2-CAR.28Z с NGFR МУТИРОВАННЫМ КОРОТКИМ (V6 NMS)
<400> 40
aagctttgct cttaggagtt tcctaataca tcccaaactc aaatatataa agcatttgac 60
ttgttctatg ccctaggggg cggggggaag ctaagccagc tttttttaac atttaaaatg 120
ttaattccat tttaaatgca cagatgtttt tatttcataa gggtttcaat gtgcatgaat 180
gctgcaatat tcctgttacc aaagctagta taaataaaaa tagataaacg tggaaattac 240
ttagagtttc tgtcattaac gtttccttcc tcagttgaca acataaatgc gctgctgagc 300
aagccagttt gcatctgtca ggatcaattt cccattatgc cagtcatatt aattactagt 360
caattagttg atttttattt ttgacatata catgtgaatg aaagacccca cctgtaggtt 420
tggcaagcta gcttaagtaa cgccattttg caaggcatgg aaaaatacat aactgagaat 480
agaaaagttc agatcaaggt caggaacaga tggaacagct gaatatgggc caaacaggat 540
atctgtggta agcagttcct gccccggctc agggccaaga acagatggaa cagctgaata 600
tgggccaaac aggatatctg tggtaagcag ttcctgcccc ggctcagggc caagaacaga 660
tggtccccag atgcggtcca gccctcagca gtttctagag aaccatcaga tgtttccagg 720
gtgccccaag gacctgaaat gaccctgtgc cttatttgaa ctaaccaatc agttcgcttc 780
tcgcttctgt tcgcgcgctt atgctccccg agctcaataa aagagcccac aacccctcac 840
tcggggcgcc agtcctccga ttgactgagt cgcccgggta cccgtgtatc caataaaccc 900
tcttgcagtt gcatccgact tgtggtctcg ctgttccttg ggagggtctc ctctgagtga 960
ttgactaccc gtcagcgggg gtctttcatt tgggggctcg tccgggatcg ggagacccct 1020
gcccagggac caccgaccca ccaccgggag gtaagctggc cagcaactta tctgtgtctg 1080
tccgattgtc tagtgtctat gactgatttt atgcgcctgc gtcggtacta gttagctaac 1140
tagctctgta tctggcggac ccgtggtgga actgacgagt tcggaacacc cggccgcaac 1200
cctgggagac gtcccaggga cttcgggggc cgtttttgtg gcccgacctg agtcctaaaa 1260
tcccgatcgt ttaggactct ttggtgcacc ccccttagag gagggatatg tggttctggt 1320
aggagacgag aacctaaaac agttcccgcc tccgtctgaa tttttgcttt cggtttggga 1380
ccgaagccgc gccgcgcgtc ttgtctgctg cagcatcgtt ctgtgttgtc tctgtctgac 1440
tgtgtttctg tatttgtctg aaaatatggg cccgggctag cctgttacca ctcccttaag 1500
tttgacctta ggtcactgga aagatgtcga gcggatcgct cacaaccagt cggtagatgt 1560
caagaagaga cgttgggtta ccttctgctc tgcagaatgg ccaaccttta acgtcggatg 1620
gccgcgagac ggcaccttta accgagacct catcacccag gttaagatca aggtcttttc 1680
acctggcccg catggacacc cagaccaggt ggggtacatc gtgacctggg aagccttggc 1740
ttttgacccc cctccctggg tcaagccctt tgtacaccct aagcctccgc ctcctcttcc 1800
tccatccgcc ccgtctctcc cccttgaacc tcctcgttcg accccgcctc gatcctccct 1860
ttatccagcc ctcactcctt ctctaggcgc ccccatatgg ccatatgaga tcttatatgg 1920
ggcacccccg ccccttgtaa acttccctga ccctgacatg acaagagtta ctaacagccc 1980
ctctctccaa gctcacttac aggctctcta cttagtccag cacgaagtct ggagacctct 2040
ggcggcagcc taccaagaac aactggaccg accggtggta cctcaccctt accgagtcgg 2100
cgacacagtg tgggtccgcc gacaccagac taagaaccta gaacctcgct ggaaaggacc 2160
ttacacagtc ctgctgacca cccccaccgc cctcaaagta gacggcatcg cagcttggat 2220
acacgccgcc cacgtgaagg ctgccgaccc cgggggtgga ccatcctcta gactgccatg 2280
gaagcccctg cccagctgct gttcctgctg ctgctgtggc tgcccgacac caccggcgag 2340
atcgtgctga cacagagccc cgccaccctg tctctgagcc ctggcgagag agccaccctg 2400
agctgtagcg ccagcagcag catcaactac atctactggc tgcagcagaa gcccggccag 2460
gcccccagaa tcctgatcta cctgaccagc aacctggcca gcggcgtgcc cgccagattt 2520
tctggcagcg gcagcggcac cgacttcacc ctgaccatca gcagcctgga acccgaggac 2580
ttcgccgtgt actactgcct gcagtggtcc agcaaccccc tgaccttcgg cggaggcacc 2640
aaggtggaaa tcaagcgggg tggtggtggt tctggtggtg gtggttctga ggtgcagctg 2700
gtggaaagcg gcggaggcct ggtcaagcct ggcggcagcc tgagactgag ctgtgccgcc 2760
agcggcttca ccttcagcag ctacgacatg agctgggtcc gacaggctcc aggcaaggga 2820
ctggaatggg tgtccaccat cagcagcggc ggcagctaca cctactacct ggacagcatc 2880
aagggccggt tcaccatcag ccgggacaac gccaagaaca gcctgtacct gcagatgaac 2940
agcctgcggg ccgaggacac cgccgtctac tactgtgccc ggcagggcct cgactactgg 3000
ggcagaggca ccctggtcac cgtgtccagc ggggatccca aagaggcctg ccccaccggc 3060
ctgtacaccc acagcggaga gtgctgcaag gcctgcaacc tgggagaggg cgtggcccag 3120
ccttgcggcg ccaatcagac cgtgtgcgag ccctgcctgg acagcgtgac cttcagcgac 3180
gtggtgtccg ccaccgagcc ctgcaagcct tgcaccgagt gtgtgggcct gcagagcatg 3240
agcgccccct gcgtggaagc cgacgacgcc gtgtgtagat gcgcctacgg ctactaccag 3300
gacgagacaa ccggcagatg cgaggcctgt agagtgtgcg aggccggcag cggcctggtg 3360
ttcagttgtc aggacaagca gaacaccgtg tgtgaagagt gccccgacgg cacctacagc 3420
gacgaggccg cccgggccgc cgacgccgag tgcgaggaac ccaaattttg ggtgctggtg 3480
gtggttggtg gagtcctggc ttgctatagc ttgctagtaa cagtggcctt tattattttc 3540
tgggtgagga gtaagaggag caggctcctg cacagtgact acatgaacat gactccccgc 3600
cgccccgggc ccacccgcaa gcattaccag ccctatgccc caccacgcga cttcgcagcc 3660
tatcgctcca gagtgaagtt cagcaggagc gcagacgccc ccgcgtacca gcagggccag 3720
aaccagctct ataacgagct caatctagga cgaagagagg agtacgatgt tttggacaag 3780
agacgtggcc gggaccctga gatgggggga aagccgagaa ggaagaaccc tcaggaaggc 3840
ctgtacaatg aactgcagaa agataagatg gcggaggcct acagtgagat tgggatgaaa 3900
ggcgagcgcc ggaggggcaa ggggcacgat ggcctttacc agggtctcag tacagccacc 3960
aaggacacct acgacgccct tcacatgcag gccctgcctc ctcgctaagc atgcaacctc 4020
gatccggatt agtccaattt gttaaagaca ggatatcagt ggtccaggct ctagttttga 4080
ctcaacaata tcaccagctg aagcctatag agtacgagcc atagataaaa taaaagattt 4140
tatttagtct ccagaaaaag gggggaatga aagaccccac ctgtaggttt ggcaagctag 4200
cttaagtaac gccattttgc aaggcatgga aaaatacata actgagaata gagaagttca 4260
gatcaaggtc aggaacagat ggaacagctg aatatgggcc aaacaggata tctgtggtaa 4320
gcagttcctg ccccggctca gggccaagaa cagatggaac agctgaatat gggccaaaca 4380
ggatatctgt ggtaagcagt tcctgccccg gctcagggcc aagaacagat ggtccccaga 4440
tgcggtccag ccctcagcag tttctagaga accatcagat gtttccaggg tgccccaagg 4500
acctgaaatg accctgtgcc ttatttgaac taaccaatca gttcgcttct cgcttctgtt 4560
cgcgcgcttc tgctccccga gctcaataaa agagcccaca acccctcact cggggcgcca 4620
gtcctccgat tgactgagtc gcccgggtac ccgtgtatcc aataaaccct cttgcagttg 4680
catccgactt gtggtctcgc tgttccttgg gagggtctcc tctgagtgat tgactacccg 4740
tcagcggggg tctttcacac atgcagcatg tatcaaaatt aatttggttt tttttcttaa 4800
gtatttacat taaatggcca tagtacttaa agttacattg gcttccttga aataaacatg 4860
gagtattcag aatgtgtcat aaatatttct aattttaaga tagtatctcc attggctttc 4920
tactttttct tttatttttt tttgtcctct gtcttccatt tgttgttgtt gttgtttgtt 4980
tgtttgtttg ttggttggtt ggttaatttt tttttaaaga tcctacacta tagttcaagc 5040
tagactatta gctactctgt aacccagggt gaccttgaag tcatgggtag cctgctgttt 5100
tagccttccc acatctaaga ttacaggtat gagctatcat ttttggtata ttgattgatt 5160
gattgattga tgtgtgtgtg tgtgattgtg tttgtgtgtg tgactgtgaa aatgtgtgta 5220
tgggtgtgtg tgaatgtgtg tatgtatgtg tgtgtgtgag tgtgtgtgtg tgtgtgtgca 5280
tgtgtgtgtg tgtgactgtg tctatgtgta tgactgtgtg tgtgtgtgtg tgtgtgtgtg 5340
tgtgtgtgtg tgtgtgtgtg ttgtgaaaaa atattctatg gtagtgagag ccaacgctcc 5400
ggctcaggtg tcaggttggt ttttgagaca gagtctttca cttagcttgg aattcactgg 5460
ccgtcgtttt acaacgtcgt gactgggaaa accctggcgt tacccaactt aatcgccttg 5520
cagcacatcc ccctttcgcc agctggcgta atagcgaaga ggcccgcacc gatcgccctt 5580
cccaacagtt gcgcagcctg aatggcgaat ggcgcctgat gcggtatttt ctccttacgc 5640
atctgtgcgg tatttcacac cgcatatggt gcactctcag tacaatctgc tctgatgccg 5700
catagttaag ccagccccga cacccgccaa cacccgctga cgcgccctga cgggcttgtc 5760
tgctcccggc atccgcttac agacaagctg tgaccgtctc cgggagctgc atgtgtcaga 5820
ggttttcacc gtcatcaccg aaacgcgcga tgacgaaagg gcctcgtgat acgcctattt 5880
ttataggtta atgtcatgat aataatggtt tcttagacgt caggtggcac ttttcgggga 5940
aatgtgcgcg gaacccctat ttgtttattt ttctaaatac attcaaatat gtatccgctc 6000
atgagacaat aaccctgata aatgcttcaa taatattgaa aaaggaagag tatgagtatt 6060
caacatttcc gtgtcgccct tattcccttt tttgcggcat tttgccttcc tgtttttgct 6120
cacccagaaa cgctggtgaa agtaaaagat gctgaagatc agttgggtgc acgagtgggt 6180
tacatcgaac tggatctcaa cagcggtaag atccttgaga gttttcgccc cgaagaacgt 6240
tttccaatga tgagcacttt taaagttctg ctatgtggcg cggtattatc ccgtattgac 6300
gccgggcaag agcaactcgg tcgccgcata cactattctc agaatgactt ggttgagtac 6360
tcaccagtca cagaaaagca tcttacggat ggcatgacag taagagaatt atgcagtgct 6420
gccataacca tgagtgataa cactgcggcc aacttacttc tgacaacgat cggaggaccg 6480
aaggagctaa ccgctttttt gcacaacatg ggggatcatg taactcgcct tgatcgttgg 6540
gaaccggagc tgaatgaagc cataccaaac gacgagcgtg acaccacgat gcctgtagca 6600
atggcaacaa cgttgcgcaa actattaact ggcgaactac ttactctagc ttcccggcaa 6660
caattaatag actggatgga ggcggataaa gttgcaggac cacttctgcg ctcggccctt 6720
ccggctggct ggtttattgc tgataaatct ggagccggtg agcgtgggtc tcgcggtatc 6780
attgcagcac tggggccaga tggtaagccc tcccgtatcg tagttatcta cacgacgggg 6840
agtcaggcaa ctatggatga acgaaataga cagatcgctg agataggtgc ctcactgatt 6900
aagcattggt aactgtcaga ccaagtttac tcatatatac tttagattga tttaaaactt 6960
catttttaat ttaaaaggat ctaggtgaag atcctttttg ataatctcat gaccaaaatc 7020
ccttaacgtg agttttcgtt ccactgagcg tcagaccccg tagaaaagat caaaggatct 7080
tcttgagatc ctttttttct gcgcgtaatc tgctgcttgc aaacaaaaaa accaccgcta 7140
ccagcggtgg tttgtttgcc ggatcaagag ctaccaactc tttttccgaa ggtaactggc 7200
ttcagcagag cgcagatacc aaatactgtc cttctagtgt agccgtagtt aggccaccac 7260
ttcaagaact ctgtagcacc gcctacatac ctcgctctgc taatcctgtt accagtggct 7320
gctgccagtg gcgataagtc gtgtcttacc gggttggact caagacgata gttaccggat 7380
aaggcgcagc ggtcgggctg aacggggggt tcgtgcacac agcccagctt ggagcgaacg 7440
acctacaccg aactgagata cctacagcgt gagcattgag aaagcgccac gcttcccgaa 7500
gggagaaagg cggacaggta tccggtaagc ggcagggtcg gaacaggaga gcgcacgagg 7560
gagcttccag ggggaaacgc ctggtatctt tatagtcctg tcgggtttcg ccacctctga 7620
cttgagcgtc gatttttgtg atgctcgtca ggggggcgga gcctatggaa aaacgccagc 7680
aacgcggcct ttttacggtt cctggccttt tgctggcctt ttgctcacat gttctttcct 7740
gcgttatccc ctgattctgt ggataaccgt attaccgcct ttgagtgagc tgataccgct 7800
cgccgcagcc gaacgaccga gcgcagcgag tcagtgagcg aggaagcgga agagcgccca 7860
atacgcaaac cgcctctccc cgcgcgttgg ccgattcatt aatgcagctg gcacgacagg 7920
tttcccgact ggaaagcggg cagtgagcgc aacgcaatta atgtgagtta gctcactcat 7980
taggcacccc aggctttaca ctttatgctt ccggctcgta tgttgtgtgg aattgtgagc 8040
ggataacaat ttcacacagg aaacagctat gaccatgatt acgcc 8085
<---
Реферат
Изобретение относится к области биотехнологии, конкретно к химерным антигенным рецепторам (CAR), и может быть использовано в медицине для противоопухолевой терапии. Сконструирован CAR, который включает экстраклеточный спейсер, который включает по меньшей мере первые три TNFR-Cys экстраклеточного домена рецептора фактора роста нервов с низким сродством (LNGFR) человека. Изобретение обеспечивает облегчение иммуноселекции и идентификации клеток, трансдуцированных указанным CAR. 7 н. и 18 з.п. ф-лы, 29 ил., 1 табл., 12 пр.
Формула
Документы, цитированные в отчёте о поиске
Т-клетки с трансдуцированным в них антигеном, применяемые в качестве системы доставки антигенов
Комментарии