Способ направления (индукции) иммунного ответа, днк, вектор, способ лечения - RU2161044C2
Код документа: RU2161044C2
Чертежи








































Описание
Настоящее изобретение относится к функциональным T клеточным рецепторам, Fc рецепторам или B клеточным рецепторам химер, которые способны менять направление деятельности иммунной системы. Более конкретно, оно относится к регулированию лимфоцитов, макрофагов, природных киллерных клеток или гранулоцитов за счет экспрессии в указанные клетки химер, которые заставляют клетки реагировать на мишени, распознаваемые химерами. Настоящее изобретение относится также к функциональным T клеточным рецепторам, Fc рецепторам или B клеточным рецепторным химерам, которые способны управлять терапевтическими клетками для специфического распознавания и разрушения либо клеток, инфицированных специфическим инфицирующим агентом, самого инфицирующего агента, опухолевой клетки, или автоиммунно-генерированной клетки. Более конкретно, настоящее изобретение относится к продуцированию T клеточных рецепторов или Fc рецепторов химер, способных заставить цитотоксичные T лимфоциты специфически распознать и лизировать клетки, экспрессирующие протеины HIV оболочки. Поэтому настоящее изобретение предоставляет лечение от таких заболеваний, как AIDS /синдром приобретенного иммунодефицита, СПИД/, которое вызывается вирусом HIV.
Предпосылки изобретения
Распознавание T клетками антигена за счет рецептора
T клетки является основой иммунологии. T клетки управляют тем, что называется клеточным иммунитетом. Он включает разрушение клетками иммунной системы чужеродных тканей или инфицированных клеток.
Существует целый ряд T клеток, включая "хелперные" и "супрессорные" клетки, которые формируют иммунную реакцию, цитотоксичные /или "киллеры"/ клетки, которые непосредственно убивают ненормальные
клетки.
T клетки, которые распознают и связывают уникальные антигены, расположенные на поверхности других клеток, становятся активированными; тогда они могут умножаться, и, если это цитотоксичные клетки, они могут убить связанную клетку.
Автоиммунное заболевание характеризуется продуцированием либо антител, которые реагируют с тканями хозяина, или иммунных эффекторных T клеток, которые являются автореактивными. В некоторых случаях автоантитела могут возникнуть за счет нормальной T- и B-клеточной реакции, активированной чужими веществами или организмами, которые содержат антигены, которые перекрестно реагируют с аналогичными соединениями в тканях организма. Примерами клинических автоантител являются антитела против ацетилхолиновых рецепторов в Myasthenia gravis; и анти-ДНК, анти-эритроцитные и анти-тромбоцитные антитела при системном lupus erythematorus.
HIV и иммунопатогенез
В 1984 г. было обнаружено, что HIV
является этиологическим агентом AIDS. С того времени определение СПИД было пересмотрено много раз с точки зрения того, какие критерии должны быть включены в диагноз. Однако, несмотря на флуктуации
параметров диагностики, простым общим определителем СПИДа является инфицирование HIV и последующее развитие стойких конституциональных симптомов, и определяемые СПИД заболевания, такие, как вторичные
инфекции, неоплазмы и неврологические заболевания. Harrizon's Principles of Internal Medicine 12е изд., Mc. Graw Hill (1991).
HIV является ретровирусом человека группы лентивирусов. Четыре распознаваемых ретровируса человека принадлежат к двум различным группам: T лимфотропические /или лейкемия / ретровирусы человека HTLV-1 и HTLV-2, и вирусы иммунодефицита человека, HIV-1 и HIV-2. Первые представляют собой трансформирующиеся вирусы, тогда как последние являются цитопатическими вирусами.
HIV-1 был идентифицирован как наиболее часто вызывающий СПИД во всем мире. Соответствие гомологии между HIV-2 и HIV-1 составляет около 40%, причем HIV-2 более близок к некоторым членам группы вирусов иммунодефицита /SJV/. См. Cuman J. et al. Science 329:1357-1359 /1985/; Wеiss R. et al., Nature, 324:572-575 /1986/.
HIV содержит обычные ретровирусные гены /env, gag и pol/, а также шесть дополнительных генов, вовлеченных в репликацию и другие биологические активности вирусов. Как было указано ранее, общее определение СПИД состоит в глубокой иммунодепрессии, главным образом, клеточного иммунитета. Подавление иммунитета ведет к различным возможным заболеваниям, особенно к некоторым инфекциям и неоплазмам.
Основной причиной иммунодефицита при СПИДЕ, как было обнаружено, является количественный и качественный дефицит субкомплекта лимфоцитов, продуцируемых в тимусе /T/, T4 популяции. Этот субкомплект клеток фенотипически определяется по наличию CD4 поверхностной молекулы, которая как было продемонстрировано, является клеточным рецептором для HIV. Dalgleish et al., Nature, 312: 763 /1984/. Хотя именно T клетки являются основным типом клеток, инфицируемых HIV, важно, что любые клетки человека, которые экспрессируют CD4 молекулу на своей поверхности, способны связываться с HIV и быть инфицированы им.
Традиционно, CD4+ T клеткам была приписана роль хелпера/индуктора, что указывало на функцию в обеспечении активирующего сигнала на B клетки, или индуцируя T лимфоциты, которые несут реципрокный CD8 маркер и становятся цитотоксичными/супрессорными клетками. Peilihess and Schlossman Cell, 19: 821-827 /1980/ Goldstein et al., Immunol, Rev. 68:5-42 /1982/.
HIV связывается специфически и с высоким сродством, за счет участка секвенирования аминокислот в оболочке вируса /gp 120/, с частью VI участка CD4 молекулы, расположенной вблизи ее N-конца. После связывания, вирус сливается с мембраной мишеневой клетки и интернализируется. После интернализации он использует энзим обратной транскриптазы для транскрипции своей геномной РНК в ДНК, которая внедряется в клеточную ДНК, где и существует в течение жизни клетки в качестве "провируса".
Провирус может оставаться латентным или может быть активирован до транскрипции мРНК и геномной РНК, что ведет к синтезу протеинов, сборке, образованию нового вируса и бадингу вируса с поверхности клетки. Хотя точный механизм, за счет которого вирус вызывает гибель клетки, не установлен, по-видимому, основным механизмом является массивный вирусный бадинг с поверхности клетки, который приводит к разрушению плазмы мембраны и в результате к нарушению осмотического равновесия.
Во время инфицирования организм хозяина вырабатывает антитела против протеинов вирусов, включая основные гликопротеины оболочки gp120 и gp 41. Несмотря на этот гуморальный иммунитет, заболевание прогрессирует, что приводит к летальной иммуносуппрессии, которая характеризуется множественными различными инфекциями, паразитемией, слабоумием и смертью. Неспособность анти-вирусных антител хозяина остановить развитие заболевания и представляет один из наиболее тревожных и настораживающих аспектов заболевания и является плохим предзнаменованием для попыток вакцинации на основании обычных приближений.
Два фактора могут играть роль в эффективности гуморальной реакции на вирус иммунодефицита. Во-первых, также как и остальные РНК вирусы /и подобно ретровирусам, в частности/, вирусы иммунодефицита демонстрируют высокую степень мутации в ответ на иммунный контроль хозяина. Во-вторых, сами гликопротеины оболочки являются плохо гликозилируемыми молекулами, представляющими несколько эпитопов, подходящих для связывания с высоко афинными антителами. Плохая антигенная мишень, которую представляют оболочки, предоставляет хозяину мало возможностей для ограничения вирусной инфекции за счет продуцирования специфических антител.
Клетки, инфицированные вирусами HIV, экспрессируют gp 120 гликопротеин на своей поверхности. Gp 120 способствует актам слияния между CD4+ клетками за счет реакции, аналогичной той, с помощью которой вирус проникает в неинфицированные клетки, что приводит к образованию коротко-живущих многоядерных гигантских клеток. Образование синцития от непосредственного взаимодействия gp 120 гликопротеина оболочки с CD4 протеином. Dalgleish et al., Supra Klatrman D. et all Nature 312:763 (1984); Mc Dougal J.S. et al., Science 231: 382 (1986); Sodroski J. et al., Nature, 322:470 (1986); Zifson J.D. et al., Nature, 323:725 (1986); Sodroski, J. et al., Nature, 321:412 (1986).
Доказательством того, что CD4-gp 120 связывание ответственно за вирусную инфекцию клеток, несущих CD4 антиген, включает обнаружение того, что между gp120 и CD4 образуется специфический комплекс. Mc Dougal et al., Supra. Другие исследователи показали, что клеточные линии, которые были неинфективны для HIV, были превращены в инфицируемые клеточные линии для HIV, с последующей трансфекцией и экспрессией CD4 кДНК гена человека. Madolon et al. Cell, 46:333-348 /1968/.
Терапевтические программы на основании растворимого CD4 как пассивного агента, который помешал бы вирусной адсорбции и осуществляемой через синцитий клеточной трансмиссии, были предложены и с успехом продемонстрированы in vitro рядом исследовательских групп /Deen at al., 3321:82-84 /1988/; Fisher et al. , Nature, 331: 76-78 /1988/; Hussey et al., Nature 331:78-81 /1988/; Smith et al., Science, 238:1704-1707 /1987/; Franecker et al., Nature, 331: 84-86 /1988 /, а CD4 протеины слияния иммуноглобулина с увеличенным сроком полужизни и умеренной биологической активностью были последовательно разработаны /Nature, 337:525-531/1989/; Fraunecker et al., Nature, 339: 68-70 /1989/; Byrn et al., Nature, 344:667-670 /1990/; Zcttlemeissl at al. , DNA Cell Biol. 9:347-353 /1990/. Хотя конъюгаты CD4 иммунотоксина или протеины слияния демонстрируют потенциальную цитотоксичность для инфицированных in vitro клеток /Chaudhary et al. , Nature, 335:369-372 /1988/; Fill et. al., Science, 242:1166-1168 /1988//, латентность синдрома иммунодефицита делает маловероятным эффективность какой-либо разовой терапии по исключению основы вирусов, а антигенность чужих протеинов слияния, по-видимому, ограничит их применимость при лечении требуемыми повторными дозами. Эксперименты на обезьянах, зараженных SIV, показали, что растворимый CD4, если его вводить животным без видимой CD4 цитопении, может понизить титр SIV и повысить in vitro степень миелоидного потенциала /Watanabe et al., Nature, 337:267-270 /1989 /. Однако, после окончания лечения наблюдалось немедленное повторное появление вируса, что предполагает необходимость пожизненного введения препарата для предотвращения прогрессирующего вырождения иммунной системы.
T клетки и Fc рецепторы
Поверхностная клеточная экспрессия большинства многочисленных форм T клеточного антигенного рецептора /TCR/ требует совместной экспрессии, по крайней мере, 6
различных полипептидных цепей /Weiss et al., J. Exp. Med. 160:1284-1299 /1984/; Orloffhashi et al., Nature 316:606-609 /1985/; Berghout et al., J. Biol. Chem., 263:8528 - 8536 /1988/; Sussman et al.,
Cell, 54: 85-95 /1988/ /; α / β антиген связывающих цепей, трех полипептидов CD3 комплекса и z. Если отсутствует любая из этих цепей, не происходит стабильной экспрессии остальных членов
комплекса. z является лимитирующим полипептидом для поверхностной экспрессии полного комплекса /Suss man et al., Cell, 52: 85-95 /1988/ и, как известно, осуществляет, по крайней мере, часть клеточных
активационных программ, которые включаются рецептором распознавания лиганда /Weiss man et al., EMBO J. 8:3651 - 3656 /1989/; Frank et al., Science, 249: 174 - 177 /1990//. 32 кДа типа I интегральный
мембранный гомодимер, z /зета/ имеет 9 остаточных внеклеточных доменов, причем нет участков для присоединения I-связанных гликанов, и 112 остатков /мышиных/ или 113 остатков
/человеческих/ внутриклеточных доменов /Weiss man et al., Science, 238:1018 - 1020 /1988a/; Weiss man et al., Proc. Natl. Acad. Sci. USA, 85:9709 - 9713 /1988b//. Изоформа z, называемая η
/эта/ /Baniyash et al., J. Biol. Chem., 263: 9874 - 9878 /1988/; Orloff et al., J. Biol. Chem. 264:14812 - 14817 /1989//, которые образуются в результате другого направления иРНК сплайсинга /Jin et
al., Proc. Natl. Acad. Sci. USA, 87:3319 - 3323 /1990/, присутствуют в пониженных количествах в клетках, экспрессирущих рецептор антигена. z - η гетеродимеры, как обнаружено, осуществляют
образование инозитолфосфатов, а также инициируемую рецепторами запрограммированную гибель клеток, называемую апоптозисом /Mercep et al., Science, 242:571 - 574 /1988/, Mercep et al., Science, 246:1162
- 1165 /1989//.
Подобно z и η, связанная с Fc рецептором γ цепь экспрессируется в комплексы клеточной поверхности дополнительными полипептидами, некоторые из которых осуществляют распознавание лиганда, а другие имеют неопределенные функции.
γ /гамма/ имеет гомодимерную структуру и вся ее организация весьма схожа с организацией z, и она является компонентой как маст клетки /базофила с высоким сродством с IgE рецептором Fc ε RI, который состоит из, по крайней мере, трех различных полипептидных цепей /Blank et al., Nature, 337:187 - 189 /1989/; Ra et al., Nature, 241:752 - 754 /1989//, так и одного из рецепторов с низким сродством к IgG, представленным у мышей Fc γ RII α /Ra et al., J. Biol. Chem J. Biol. Chem. 264:1523 - 1527 /1989//, и у человека CD16 субтипом экспрессии макрофагов и природных киллерных клеток, CD6TM /CD16 трансмембрана/ /Zanier et al., Nature, 342:803 - 805 /1989/; Andersonet al, Proc. Natl. Acad. Sci. USA. 87:2274 - 2278 /1990//, и с полипептидом неидентифицированной функции /Anderson et al., Proc. Natl. Acad. Sci. USA. 87: 2274-2278 /1990/. Недавно сообщалось, что γ экспрессируют T клеточные линии мышей, CTL, в которых он образует гомодимеры, также, как γ - z и γ - η гетеродимеры /Orloff et al., Nature, 347:189 - 191 /1990//.
Fc рецепторы определяют патоцитолиз иммунных комплексов, трансцитолиз и зависимую от антител клеточную цитотоксичность /АДСС/ /Pavetch and Kinet Annu. Rev. Immunol. 9:457 - 492 /1991/; Unkeless et al., Annu, Rev. Immunol. 6: 251 - 281 /1988/; and Mellman, Curr. Opin. Immunol. 1:16-25 /1988/. Недавно было показано, что одна из изоформ мышиных с низким сродством Fc рецепторов /Fc R γ IIIBI/ определяет интернализацию Ig-покрытых мишеней в покрытые клатрином литы, и что другой рецептор с низким сродством /Fc γ IIA/ осуществляет ADCC за счет ассоциации с одной или более из мембран небольшого семейства "триггерных молекул" /Miettineu et al., Cell 58:317-327. /1989/; и Hunziker and Mellman J. Cell, Biol. 109:3291 - 3302 /1989/. Эти триггерные молекулы, T клеточный рецептор /TCR/ z цепи, TCR η цепи и Fc рецептор γ цепи, взаимодействуют с доменами распознавания лиганда различных рецепторов иммунных систем и могут автономно инициировать клеточные эффекторные программы, включая цитолиз, с последующей аггрегацией /Samelson et al., Cell 43: 223-231 (1985); Weissman et al., Science 239:1018-1020 (1988): Jin et al., Proc. Natl. Acad. Sci. USA 87:3319-3323 (1990); Blank et al., Nature 337: 187-189 (1989); Lanier et al., Nature 342:803-805 (1989); Kurosaki and Ravetch, Nature 342:805-807 (1989); Hibbs et al., Science 246:1608-1611 (1989); Anderson et al., Proc. Natl. Acad. Sci USA 87:2274-2278 (1990); and Irving and Weiss, Cell 64:891-901 (1991).
Проводя параллели между семействами Fc рецепторов с низким сродством мышей и людей, однако, стало очевидным, что Fc γ IIA и C изоформы человека не содержат мышиных дубликатов. Отчасти из-за этого их функции до сих пор не определены.
Благодаря тому, что гуморальные агенты на основе CD4 могут иметь ограниченное применение in vivo, авторы изобретения начали использовать возможность придания клеточного иммунитета HIV. В результате они сообщают о получении протеиновых химер, в которых внеклеточный домен CD4 слит с трансмембранным и/или внутриклеточным доменами T клеточного рецептора, J gG Fc рецептора, или B клеточного рецепторного трансдукторного сигнального элемента. Цитолитические T клетки, экспрессирующие химеры, которые включают внеклеточный CD4 домен, демонстрируют потенциальное MHC-независящее разрушение клеточных мишеней, экспрессирующих протеины оболочки HIV. Крайне важный и новый компонент этого приближения состоит в идентификации одиночного T клеточного рецептора, или Fc рецептора, и B клеточного рецептора цепей, аггрегации которых достаточно для инициирования клеточной реакции.
Одним из особенно ценных применений этого приближения было изобретение химер между CD4 и Z, η или γ, которые заставляют цитолитические T лимфоциты распознавать и убивать клетки, экспрессирующие HIV gp120.
Краткое содержание изобретения
Хотя нативные T клетки, В клетки и Fc
рецепторы являются, или могут быть, весьма сложными многомерными структурами, не позволяющими проводить с ними обычные манипуляции, настоящее изобретение демонстрирует легкость создания химер между
внутриклеточным доменом любой из многочисленных молекул, которые способны выполнять задачу распознавания мишени. В частности, создание химер, состоящих из внутриклеточной части T клетки/ Fc
рецепторных зета, эта или гамма цепей, присоединенных к внеклеточной части подходящим образом сконструированных молекул антител, позволяет специфически изменить направление мишеневого потенциала
распознавания клеток иммунной системы на распознавание антигена за счет внеклеточного участка антитела. Таким образом, имея участки антител, способные распознавать некоторые детерминанты на
поверхности патогенов, клетки иммунной системы, снабженные химерами, будут реагировать на патоген с эффекторной программой соответствующей им линии дифференцировки, например, хелперные T лимфоциты
будут реагировать цитотоксичной активностью против мишени, а В лимфоциты будут активизированы на синтез антител. Макрофаги и гранулоциты будут осуществлять свод эффекторные программы, включая
выделение цитокина, фагоцитов и вырабатывая реакционноспособный кислород. Аналогично, имея участок антител, способный распознавать опухолевые клетки, реакция иммунной системы на опухоль будет с
выгодой усилена. Имея антитела, способные распознавать иммунные клетки, обладающие вредной реакционной способностью с самодетерминантами, автореактивные клетки могут селективно направляться на
разрушение. Хотя эти примеры, описывающие использование химер антител в качестве удобного описательного инструмента, настоящее изобретение не ограничивается объемом химер антител, и, естественно,
использование специфических неантительных внеклеточных доменов может дать важные преимущества. Так, например, внеклеточная часть, то есть рецептор для вируса, бактерии или паразита, клетки, снабженные
химерами, будут специфически поражать клетки, экспрессирующие вирусные, бактериальные или паразитические детерминанты. Выгода такого приближения по сравнению с антителами состоит в том, что нативный
рецептор для патогена может иметь уникально высокую селективность или сродство с патогеном, обеспечивая большую степень точности в получении иммунной реакции. Аналогично, для исключения клеток
иммунной системы, с неправильными реакциями с самим антигеном, может оказаться достаточным соединить антиген /или его интактный протеин в случае терапии делеции B клеток/ с внутриклеточными зета, эта
или гамма цепями, и за счет этого обеспечить специфическое поражение клеток, неправильно реагирующих на свои детерминанты.
Другим применением химер является контроль за популяциями клеток in vivo, следующими за другими формами генетического конструирования. Так, например, использование опухолевых инфильтрующих лимфоцитов или природных киллерных клеток для переноса цитотоксинов в определенные участки опухоли, было предложено. Настоящее изобретение обеспечивает удобные средства для регулирования количества и активности таких лимфоцитов и клеток, не удаляя их из организма пациента для амплификации in vivo. Так, благодаря тому, что внутриклеточные домены химерических рецепторов осуществляют пролиферативные реакции клеток, координирование внеклеточных доменов за счет различных стимулов аггрегации, специфичных для внеклеточных доменов /например, антител, специфичных для внеклеточных доменов/ приведет к пролиферации клеток, несущих химеры.
Хотя конкретные варианты настоящего изобретения включают химеры между зета, эта и гамма цепями или их активными фрагментами /например, описываемыми далее/, любая рецепторная цепь с аналогичными функциями к этим молекулам, например, гранулоцитам или B лимфоцитам, может быть использована для раскрытых здесь целей. Отличительные характеристики целевых иммунных триггерных молекул включают способность быть автономно экспрессированными /например, как отдельная цепь/, способность сливаться с внеклеточным доменом так, чтобы полученная химера присутствовала на поверхности терапевтической клетки, и способность инактивировать клеточные эффекторные программы после аггрегации после столкновения с мишеневым лигандом.
В настоящее время наиболее удобным способом доставки химер в иммунную клеточную систему является способ, включающий некоторые формы генетической терапии. Однако, реконструирование клеток иммунной системы химерическими рецепторами за счет смешения клеток с подходящим солюбилизированным очищенным химерическим протеином также привело бы в результате к образованию популяций сконструированных клеток, способных реагировать на мишени, распознаваемые внеклеточными доменами химер. Аналогичное приближение можно использовать, например, для введения интактного HIV рецептора, CD4, в эритроциты с терапевтическими целями. В этом случае сконструированные клетки не будут способны к самообновлению.
Настоящее изобретение относится к функционально упрощенным T клеточным рецепторным, B клеточным рецепторным и Fc клеточным рецепторным химерам, которые способны изменять направление действия иммунной системы. Более конкретно, оно относится к регулированию лимфоцитов, макрофагов, природных киллерных клеток или гранулоцитов за счет экспрессии в указанные клетки химер, которые вызывают клеточную реакцию на мишени, распознаваемые химерами. Настоящее изобретение относится также к способу направления клеточной реакции на инфицирующий агент, опухолевые или канцерогенные клетки, или на клетки, вырабатываемые автоиммунно. Способ управления клеточной реакцией у млекопитающих включает введение эффективного количества терапевтических клеток указанному млекопитающему, причем указанные клетки способны распознавать и разрушать указанный инфицирующий агент, опухолевые, раковые клетки или автоиммунно генерированные клетки.
В другом варианте способ направления клеточной реакцией на инфицирующие клетки включает введение терапевтических клеток, которые способны распознавать и разрушать указанный агент, причем агентом является специфический вирус, бактерия, простейшее или грибок. Еще более конкретно, способ направлен против таких агентов, как HIV и Pneumocystis carinii.
Специфически настоящее изобретение предлагает способ направления клеточной реакции на клетки, инфицированные HIV.
Способ включает введение пациенту эффективного количества цитотоксичных T лимфоцитов, причем указанные лимфоциты способны специфически распознавать и подвергать лизису клетки, инфицированные HIV.
Таким образом, в одном из вариантов, настоящее изобретение представляет способ направления клеточной реакции на клетки, инфицированные HIV, включающий введение пациенту эффективного количества цитотоксичных T лимфоцитов, которые способны специфически распознавать и подвергать лизису клетки, инфицированные HIV.
В еще одном варианте изобретения предложены химерические рецепторные протеины, которые направляют цитотоксичные T лимфоциты на распознавание и лизис клеток, инфицированных HIV. Еще один вариант изобретения включает клетки хозяина, трансформированные вектором, содержащим химерические рецепторы.
В еще одном варианте настоящее изобретение предлагает антитела против химерических рецепторов настоящего изобретения.
Для получения цитотоксичных T лимфоцитов, которые специфически связывают и разрушают клетки, инфицированные HIV, авторы изобретения испробовали рецепторные химеры. Эти химерические рецепторы функционально активны и обладают экстраординарной способностью специфически связывать и разрушать клетки, экспрессирующие gp120.
Еще одной целью настоящего изобретения стало создание способа лечения индивидуумов, инфицированных HIV. В настоящем изобретении предложен ряд важных достижений в лечении СПИДа.
Эти и другие нелимитирующие варианты настоящего изобретения будут очевидны специалистам из последующего подробного описания изобретения.
В нижеследующем подробном описании будут сделаны ссылки на различные методологии, известные специалистам в области молекулярной биологии и иммунологии. Публикации и другие материалы, представленные далее для таких методик, на которые сделаны ссылки, включены сюда по ссылкам на их содержание.
Стандартные работы, в которых суммированы
общие принципы технологии рекомбинантных ДНК, включают:
Watson, J. D, et al., Molecular Biology of the Gene, Volumes I and II, the Bеnjamin/Cummings Publishing Company, Inc., publisher, Menlo
Park, CA (1987); Darnell, J. E. et al., Molecular Cell Biology. Scientific American Books, Inc. , Publisher, New York, N.Y. (1986); Lewin, B.M., Genes II, John Wiley & Sons, publishers, New
York, N.Y, (1985); Old, R.W., et al,. Principles of Gene Manipulation: An Introduction to Genetic Engineering, 2d edition, University of California Press, publisher, Berkeley, CA (1981); Maniatis, T. ,
et al., Molecular Cloning: A Laboratory Manual, 2nd Ed. Cold Spring Harbor Laboratory, publisher, Cold Spring Harbor, NY (1989); and Current Protocols in Molecular Biology, Ausubel et al., Wiley
Press, New York, NY (1989).
Определения
Под "клонированием" подразумевают in vitro рекомбинантную методику включения конкретного гена или другой ДНК последовательности в
векторную молекулу. Для успешного клонирования целевого гена, необходимо использовать способы создания ДНК фрагментов для соединения этих фрагментов с векторной молекулой, для введения композитной ДНК
молекулы в клетки хозяина, в которых они могут реплицироваться, и для отбора клона, содержащего мишеневый ген из реципиентных клеток хозяина.
Под "кДНК" подразумевают комплемент или копию ДНК, полученную из РНК темплата под действием РНК зависимой ДНК полимеразы /обратной транскриптазы/. Так, "кДНК клон" означает дуплексную ДНК последовательность, комплементарную РНК молекуле вставки, содержащей клонирующий вектор.
Под "кДНК библиотекой" подразумевают коллекцию рекомбинантных ДНК молекул, содержащих кДНК вставки, которые содержат ДНК копии иРНК, которые были экспрессированы клеткой во время создания кДНК библиотеки. Такую кДНК библиотеку можно получить способами, известными специалистам и описанными, например, в Maniatis et al., Molecular Cloning: A Laboratory Manual supra. Обычно РНК вначале выделяют из клеток организма, из генома которого предстоит клонировать конкретный ген. Предпочтительными для целей настоящего изобретения являются клеточные линии лимфоцитов млекопитающих и особенно человека. Предпочтительным вектором для этой цели является штамм вакцины вируса WR.
Под "вектором" подразумевают молекулу ДНК, полученную, например, из плазмид, бактериофагов или вирусов человека или насекомых, в которую можно включить или клонировать фрагменты ДНК. Вектор должен содержать один или более из рестрикционных сайтов и может быть способен автономно реплицироваться в определенном организме хозяина или носителя, таким образом, чтобы клонированная последовательность была репродуцируема. Так, под термином "вектор экспрессии ДНК" подразумевают автономный элемент, способный направлять синтез рекомбинантных пептидов. Такие ДНК векторы экспрессии ДНК включают бактериальные плазмиды и плазмиды и вирусы млекопитающих и насекомых.
Под "практически чистым" понимают соединение, например, протеин, полипептид или антитела, которые практически не содержат компонентов, которые в природе их сопровождают. Обычно соединение называют практически чистым, если, по крайней мере, 60%, более предпочтительно, по крайней мере, 75%, и наиболее предпочтительно, по крайней мере, 90% всего материала составляет представляющее интерес соединение. Чистоту определяют соответствующими способами, например, с помощью хроматографии на колоннах, электрофореза в полиакриламидном геле или с помощью высокоэффективной жидкостной хроматографии /HPLC = ВЖХ/. Если это касается нуклеиновых кислот, термин "практически чистый" относится к последовательности нуклеиновой кислоты, сегменту ее или фрагменту, который не содержит генов, который фланкирует ее в ее природно-встречающемся состоянии /например, не содержит последовательностей, которые фланкируют нуклеиновую кислоту в ее нативном геномном положении/. Под "функциональным производным" подразумевают "фрагменты", "варианты", "аналоги" или "химические производные" молекулы. "Фрагмент" молекулы, например, кДНК последовательности настоящего изобретения, означают, что этот термин относится к любому нуклеотидному субсету молекулы. "Вариант" такой молекулы относится к природно встречающейся молекуле, практически аналогичной либо целой молекуле, либо ее фрагменту, "Аналог" молекулы относится к не-активной молекуле, практически аналогичной либо всей молекуле, либо ее фрагменту. Говорят, что молекула "практически аналогична" другой молекуле, если последовательность аминокислот в обеих молекулах практически одинакова. Практически аналогичные молекулы аминокислот будут обладать аналогичными биологическими активностями. Так, при условии, что две молекулы обладают аналогичными активностями, их считают вариантами, так как этот термин используют даже, если одна молекула содержит дополнительно /или меньше/ аминокислотные остатки, которые не найдены в другой, или если последовательность аминокислотных остатков не идентична. В том смысле, как здесь использовано, говорят, что молекула является "химическим производным" другой молекулы, если она содержит дополнительные химические фрагменты, которые обычно не составляют части этой молекулы. Такие фрагменты могут повысить растворимость молекул, адсорбцию, биологическую полужизнь и т.д. Эти фрагменты в другом варианте могут снизить токсичность молекулы, исключить или ослабить любые нежелательные побочные эффекты молекулы и т.д. Фрагменты, способные осуществить такие эффекты, раскрыты, например, в Reminston's Pharmaceutical Sciences, 16th ed. Mack Publishing Co., Easton, Peuu (1980).
Аналогично, "функциональное производное" рецептора химеры настоящего изобретения подразумевает "фрагменты", "варианты" или "аналоги" гена, который может быть "практически аналогичен" нуклеотидной последовательности и который кодирует молекулу, обладающую аналогичной активностью с, например, T клеточными, B клеточными или Fc рецепторными химерами.
Так, в том смысле, как здесь использовано, T клеточные, B клеточные или Fc рецепторные химерические протеины также включают любые функциональные производные, варианты, аналоги или химические производные, которые могут быть существенно аналогичны "дикого типа" химерам и которые обладают аналогичной активностью, то есть, наиболее предпочтительно 90%, более предпочтительно 70%, предпочтительно 40% или, по крайней мере, 10% активности дикого типа рецепторных химер. Активность функциональных химерических рецепторных производных включает специфическое связывание /с их внеклеточной частью/ с мишеневым агентом и в результате разрушение /направляемое его внутриклеточной или трансмембранной частью/ агента или клетки; причем такую активность можно определить, используя любые аналитические способы, описанные далее.
ДНК последовательность, кодирующую T клеточные, B клеточные или Fc рецепторные химеры настоящего изобретения, или их функциональные производные, можно рекомбинировать вектором ДНК в соответствия с обычными способами, включая тупые концы или болтающиеся концы для лигирования, расщепления рестрикционным энзимом до получения соответствующих концов, заполняя их липкими концами соответствующим образом, обрабатывая щелочной фосфатазой во избежание нежелательных присоединений, и лигирования соответствующей лигазой. Методики таких манипуляций раскрыты Maniatis T., et al: supra, и хорошо известны специалистам.
Говорят, что молекулы нуклеиновых кислот, такие как ДНК, "способны к экспрессии" полипептида, если они содержат нуклеотидную последовательность, которая содержит транскрипционную и трансляционную регуляторную информацию, и такие последовательности являются "операбельно связанными" с нуклеотидными последовательностями, которые кодируют полипептид. Операбельной связью является такая связь, в которой регуляторные ДНК последовательности и ДНК последовательности, которые нужно экспрессировать, связаны таким образом, чтобы обеспечить экспрессию гена. Точная природа регуляторных участков, необходимых для экспрессии гена, может меняться от организма к организму, но должна, вообще, включать промоторный участок, который, у прокариотов, содержит оба промотора /которые руководят инициированием РНК транскрипции/, а также ДНК последовательности, которые будучи транскрибированы в РНК, дадут сигнал начала синтеза протеина, такие участки обычно включают те 5'-некодирующие последовательности, которые связаны с инициированием транскрипции и трансляции, такие как TATA, CAAT последовательности и т.п.
При желании, некодирующий 3'-участок генной последовательности, кодирующий протеин, можно получить вышеописанными способами. Этот участок можно сохранить для его трансляционной терминационной регуляторной последовательности, например, для терминации и полиаденилирования. Так, сохраняя 3'-участок, смежный с ДНК последовательностью, кодирующей протеин, можно получить сигналы окончания транскрипции. Если сигналы окончания транскрипции функционируют недостаточно при экспрессии клеток хозяев, тогда 3'-участок, функциональный в клетках хозяина, можно заменить.
Считают, что две ДНК последовательности /такие как последовательность промотерного участка и T клеточный рецептор, B клеточный рецептор или Fc рецепторная химера, кодирующая последовательность/ операбельно связаны, если природа связи между двумя ДНК последовательностями /1/ не приводит к введению мутации со сдвигом рамки, /2/ не мешает способности промотерного участка последовательности управлять транскрипцией рецепторной химерической генной последовательности, или /3/ не мешает способности рецепторной химерической генной последовательности быть транскрибированной промотерным участком последовательности. Промотерный участок должен быть операбельно связан с ДНК последовательностью, если промотер был способен осуществлять транскрипцию ДНК последовательности. Таким образом, для того чтобы осуществить экспрессию протеина, необходимы транскрипционный и трансляционные сигналы, распознаваемые соответствующим хозяином.
Настоящее изобретение охватывает экспрессию T клеточного рецептора, B клеточного рецептора или Fc рецепторного химерического протеина /или их функциональных производных/ либо в прокариотические, либо в эукариотические клетки, хотя предпочтительна экспрессия в эукариотические /и особенно, лимфоцитов человека/ клетки.
Антитела по способу настоящего изобретения можно получить любым из множества различных способов. Так, например, клетки, экспрессирующие рецепторный химерический протеин, или его функциональные производные, можно ввести животным для того, чтобы вызвать продуцирование сыворотки, содержащей поликлональные антитела, которые способны связывать химеры.
В предпочтительном способе изобретения, антитела являются моноклональными антителами. Такие моноклональные антитела можно получить, используя технологию гибридом /Kohler et al., Nature 256:495 (1975); Kohler et al., Eur J. Immunol, 6:511 (1976); Kohler et al., Eur. J. Immunol. 6:292 (1976); Hammerling et al., In Monoclonal Antibodies and T-Cell Hybridomas, Elsevier, N.J. p. 563-684 (1981)/. Вообще, такие процедуры включают иммунизацию животного T клеточным рецепторным, B клеточным рецепторным или Fc рецепторным химерическим антигеном. Спленоциты таких животных экстрагируют и сливают с подходящей миеломной клеточной линией. В способе настоящего изобретения можно использовать любые подходящие клеточные линии миелом. После слияния полученные гибридомные клетки селективно сохраняют в HAT среде, а затем клонируют ограниченным разбавлением, как описано у Wands J.R. et al. /Gastroehterology 80: 225 - 232 /1981//. Гибридомные клетки, полученные после такой селекции, затем анализируют для идентификации клонов, которые выделяют антитела, способные связывать химеры.
Антитела по способу настоящего изобретения также могут быть поликлональными или, предпочтительно, региоспецифическими поликлональными антителами.
Антитела против T клеточных рецепторов, B клеточных рецепторов или Fc рецепторов химер по способу настоящего изобретения можно использовать для слежения за количеством химерических рецепторов /или химерические рецепторы содержащих клеток/ у пациента. Такие антитела прекрасно приспособлены для использования в стандартном иммунодиагностическом анализе, известны специалистам, включают такие иммунометрические или "сэндвичевые" анализы, как опережающий сэндвичевый, обратный сэндвичевый и одновременный сэндвичевый анализы. Антитела можно использовать в любой из их комбинаций, которые могут определить специалисты, без ненужных экспериментов для осуществления иммуноанализа приемлемой специфичности, чувствительности и точности.
Стандартные работы, в которых изложены общие принципы иммунологии, включают: 13, Elsevier, Publisher, Amsterdam (1984); Klein, J., Immunology: The Science of Self-Nonself Discrimination, John Wiley & Sons, Publisher, New York (1982); and Kennett, R., et al., eds., Monoclonal Antibodies, Hybridoma: A New Dimension In Biological Analyses, Plenum Press, Publisher, New York (1980). Roitt, I., Essential Immunology, Sixth Ed., Blackwell Scientific Publications, Publisher, Oxford (1988); Kimball, J. W., Introduction to Immunology, Second Ed. , Macmillan Publishing Co. , Publisher, New York (1986); Roitt. I., et al., Immunology, Gower Medical Publishing Ltd,, Publisher, London, (1985); Campbell, A., "Monoclonal Antibody Technology", in, Burdon, R., et al., eds., Laboratory Techniques in Biochemistry and Molecular, Biology, Volume.
Под "детектированием" подразумевают определение наличия или отсутствия вещества или количественное определение его содержания. Этот термин относится к использованию материалов, композиций и способов настоящего изобретения для качественных и количественных определений.
Выделение других гибридом, секретирующих моноклональные антитела той же специфичности, что и у описанных ранее, можно осуществить методами анти-идиотинического скринирования /Potocmjak. et al., Science 215:1637 /1982//. Коротко, анти-идиотипическое антитело это такое антитело, которое распознает уникальные детерминанты, присутствующие на антителах, продуцируемых представляющим интерес клоном. Анти-идиотипические антитела получают за счет иммунизации животного тем же штаммом, который был использован в качестве источника моноклональных антител, моноклональными антителами, представляющими интерес. Иммунизованное животное будет распознавать и реагировать на идиотипические детерминанты иммунизирующих антител выработкой антител против этих идиотипических детерминант /анти-идиотипические антитела/.
Для репликации гибридные клетки можно культивировать как in vitro, так и in vivo. Высокая продуктивность in vivo делает этот способ предпочтительным способом культивирования. Короче, клетки из отдельных гибридных штаммов вводят внутрибрюшинно мыши pristane - primed BALB/с для получения асцитной жидкости, содержащей высокие концентрации целевых моноклональных антител. Моноклональные антитела изотипов IgM или IgG можно очистить из надосадочной жидкости культуры, используя хроматографические способы, хорошо известные специалистам.
Антитела по способу настоящего изобретения особенно пригодны для использования в иммуноанализах, где их можно использовать в жидкой фазе, или связанными с твердой фазой носителя. Кроме того, антитела в этих иммуноанализах можно детектируемо метить различными способами.
Существует множество меток и способов мечения, известных специалистам. Примеры такого типа меток, которые можно использовать в способе настоящего изобретения, включают, хотя и не ограничиваются ими, ферменты, радиоизотопы, флюоресцирующие соединения, хемилюминесцирующие соединения, биолюминесцирующие соединения и халаты металлов. Специалистам известны и другие возможные метки для связывания с антителами, или можно определить то же самое с использованием рутинных экспериментов. Более того, связывание таких меток с антителами можно осуществить, используя стандартные методики, обычно известные специалистам.
Одним из таких способов настоящего изобретения, с помощью которого можно детектируемо пометить антитело, является способ
связывания его с энзимом. Этот энзим, в свою очередь, будучи позднее экспонирован субстрату, будет реагировать с субстратом таким образом, что приведет к образованию химического фрагмента, который
можно детектировать, например, спектроскопическими или флуорометрическими способами. Примеры энзимов, которые можно использовать для детектируемой метки антител, включают малатдегидрогеназу,
стафилококкалнуклеазу, дельта- γ -стероидизомеразу, дрожжевую спиртовую дегидрогеназу, альфа-глицерофосфатдегидрогеназу, триозфосфатизомеразу, биотинавидинпероксидазу, пероксидазу хрена,
щелочную фосфатазу, аспарагиназу, глюкозооксидазу, бета-галактозидазу, рибонуклеазу, уреазу, каталазу,
глюкозо-VI-фосфатдегидрогеназу, глюкоамилазу и ацетилхолинэстеразу.
Наличие детектируемо меченых антител можно также определить, пометив антитела радиоактивными изотопами, которые затем определяют с помощью гамма-счетчиков или сцинтилляционных счетчиков. Изотопы, наиболее пригодные для целей настоящего изобретения:3H,125I,32P,35S,14C,51Cr,36Cl,57Co,58Co,59Fe и75Se.
Возможно также определять связывание детектируемо меченых антител путем введения метки флуоресцентного соединения в антитело. Если флуоресцентно меченое антитело экспонируют свету соответствующей длины волны, его присутствие можно определить за счет флуоресценции красителя. Среди наиболее часто используемых флуоресцентно меченых соединений находятся флуоресцин, изотиоциан, родамин, фикоэретрин, фикоцианин, аллофикоцианин орто-фтальдегид и флуоресцамин.
Антитела настоящего изобретения можно также детектируемо пометить, используя такие флуоресцирующие металлы, как152Eu, или другие лантаноиды. Эти металлы можно присоединить к антителам, используя такие хелатирующие группы, как диэтилэнтериаминпентауксусная кислота /DTPA/ или этилендиаминтетрауксусная кислота /EDTA/.
Антитела можно также детектируемо пометить, соединяя их с хемилюминесцентными соединениями. Наличие хемилюминесцентных антител определяют затем, детектируя наличие люминесценции, возникающей в процессе химической реакции. Примеры наиболее подходящих хемилюминесцентных метящих соединений включают люминал, изолюминол, сложный эфир тероматикакридиния, имидазол, соли акридиния, сложный оксалатный эфир и диоксэтан.
Аналогично, для того чтобы пометить антитела по способу настоящего изобретения, можно использовать биолюминесцентные соединения. Биолюминесценция представляет собой тип хемилюминесценции, возникающей в биологических системах, в которых каталитический протеин повышает эффективность реакции хемилюминесценции. Наличие биолюминесцентных антител определяют, детектируя наличие люминесценции. Важные биолюминесцентные соединения для целей введения меток включают люциферин, люциферин, люциферазаекворин.
Антитела и практически очищенные антигены настоящего изобретения идеально подходят для приготовления набора. Такой набор может содержать контейнер, разделенный на отделения, для размещения ампул, трубок и т.п., причем каждый контейнер содержит отдельные элементы, которые используют при анализах.
Типы анализов, для которых можно использовать этот набор, включают, например, конкурентный и неконкурентный анализы. Типичные примеры анализов, в которых можно использовать антитела настоящего изобретения, включают радиоиммуноанализ /RIA/, энзимативный иммуноанализ /EIA /, иммуносорбентный со связанным энзимом анализ /ELISA/ и иммунометрический, или сэндвичевый иммуноанализ.
Под термином "иммунометрический анализ" или "сэндвичевый иммуноанализ" подразумевают, что он одновременно включает сэндвичевый, опережающий сэндвичевый и обратный сэндвичевый иммуноанализы. Эти термины хорошо знакомы специалистам. Специалистам понятно также, что антитела по способу настоящего изобретения будут полезны в различных вариантах и формах анализов, которые уже известны или которые можно разработать в будущем. Они также включены в объем изобретения.
В предпочтительном варианте осуществления анализа важно, чтобы в инкубируемой среде присутствовали определенные "блокеры" /которые обычно добавляют с меченым растворимым антителом/. Такие "блокеры" добавляют для того, чтобы неспецифические протеины, протеазы или антитела человека к мышиным иммуноглобулинам, присутствующим в экспериментальном образце, не сшивали или не разрушали антитела на твердой фазе носителя или радиомеченные индикаторные антитела, и не давали бы неверные положительные или неверные отрицательные результаты. Поэтому выбор "блокеров" существенно добавляет специфичности к анализам, рассматриваемым в настоящем изобретении.
Было обнаружено, что ряд нерелевантных /то есть, неспецифичных/ антител того же класса или подкласса /изотипа/, что и те, которые используют в анализе /например, IgG1, IgG2, IgM и т. д./, можно использовать в качестве "блокеров". Концентрация "блокеров" /обычно 1 - 100 мкг/мл/ важна для поддержания соответствующей чувствительности, и все еще ингибирует любые нежелательные вмешательства за счет обычно встречающихся перекрестно-реакционноспособных протеинов в сыворотке человека. Кроме того, буферная система, содержащая "блокеры", нуждается в оптимизации. Предпочтительными буферам являются буфера на основе органических кислот, такие как имидазол, HEPPS, MOPS, TES, ADA, ACES, HEPES, PIPES, TRIS и тому подобные в интервале физиологических значений pH. Несколько менее предпочтительными буферами являются такие неорганические буферы, как фосфаты, бораты или карбонаты, и наконец, известные ингибиторы протеазы необходимо добавить /обычно 0,01 - 10 мкг/мл/ к буферу, который содержит "блокеры".
Существует множество иммуноадсорбентов, которые можно использовать и которые можно применить в способе настоящего изобретения. Хорошо известные иммуноадсорбенты включают стекло, полистирол, полипропилен, декстран, найлон или другие материалы в форме трубочек, шариков и микропластиков, изготовленных из или покрытых такими материалами и т.п. Иммобилизованные антитела можно либо ковалентно либо физически связать с твердой фазой иммуноадсорбента методиками, известными, например, ковалентным связыванием через амидную или сложноэфирную связь, или адсорбцией. Специалистам известно множество других подходящих твердофазных адсорбентов и способов иммобилизации на них антител, или они могут выполнить таковые, используя всего лишь рутинные эксперименты.
Для диагнозов in vivo, in vitro или in situ метки, такие как радионуклиды, можно связать с антителами по способу настоящего изобретения либо непосредственно, либо используя промежуточные функциональные группы. Промежуточной группой, которую часто используют для связывания радиоизотопов, которые существуют в виде катионов металлов, с антителами, является диэтилентриаминпентауксусная кислота /DTPA/. Типичные примеры катионов металлов, которые связывают таким образом, включают99мTc,123I,111In,131I, 9797Ru,67Cu,67Ga и68Ga. Антитела настоящего изобретения можно также метить нерадиоактивными изотопами для диагностических целей. Элементы, которые наиболее подходят в таком случае:157Gd,55Mn,162Dy,53Cr и 5656Fe.
Антиген настоящего изобретения можно выделить в практически чистом виде, используя антитела по способу настоящего изобретения. Так, в одном из вариантов настоящего изобретения, предложенном для практически чистых T клеточного рецептора, B клеточного рецептора или Fc рецептора химеры, указанный антиген, отличающийся тем, что его распознают и связывают антитела настоящего изобретения. В другом варианте настоящее изобретение обеспечивает способ выделения или очистки рецепторного химерического антигена за счет образования комплекса указанного антигена с одним или более из антител, направленных против рецепторной химеры.
Практически чистые антигены T клеточного рецептора, B клеточного рецептора или Fc рецептора химеры настоящего изобретения можно, в свою очередь, использовать для определения или измерения антител к химерам в образце, например, в сыворотке или в моче. Таким образом, один из вариантов настоящего изобретения включает способ определения наличия или количества антител к рецепторному химерическому антигену в образце, который включает контактирование образца, содержащего антитела, к химерическому антигену, с детектируемо меченной рецепторной химерой, и детектирование указанной метки. Следует учитывать, что иммунореактивные фракции и иммунореактивные аналоги химер также можно использовать. Под термином "иммунореактивные фракции" подразумевают любую часть химерического антигена, которая демонстрирует эквивалентную иммунную реакцию на антитела, направленные против рецепторных химер. Под термином "иммунореактивные аналоги" подразумевают протеины, которые отличаются от протеинов рецепторных химер одной или более аминокислотами, но которые демонстрируют эквивалентную иммунную реакцию на антитела настоящего изобретения.
Под выражением "специфически распознают и связывают" подразумевают антитела, которые распознают и связывают химерический рецепторный полипептид, но которые практически не распознают и не связывают другие молекулы в образце, например, в биологическом образце, который включает рецепторный полипептид.
Под выражением "автоиммунно-генерированная клетка" подразумевают клетки, продуцирующие антитела, которые реагируют с тканями хозяина или иммуноэффекторные клетки, которые являются автореактивными; такие клетки включают антитела против ацетилхолиновых рецепторов /ведущих, например, к myasthehia gravis/ или анти-ДНК, анти-эритроцит и анти-тромбоцитные антитела /ведущие, например, к lupus erythematosus/.
Под термином "терапевтические клетки" подразумевают клетки, которые были трансформированы химерами настоящего изобретения таким образом, что они способны распознавать и разрушать специфический инфицирующий агент, клетки, инфицированные специфическим агентом, опухолевые или канцерогенные клетки, или автоиммунно-генерированные клетки; предпочтительно такими терапевтическими клетками являются клетки системы гомепоэза /кроветворные/.
Под термином "внеклеточный" подразумевают содержащий, по крайней мере, часть молекулы, экспонированной на поверхности клетки. Под термином "внутриклеточный" подразумевают содержащий, по крайней мере, часть молекулы, экспонированной терапевтическим клеткам цитоплазмы. Под термином "трансмембранный" подразумевают содержащий, по крайней мере, часть молекулы, пронизывающей плазму мембраны. "Внеклеточная часть" и "внутриклеточная часть" и также "трансмембранная часть" в том смысле как здесь использованы, могут включать фланкирующие аминокислотные последовательности, которые простираются в прилегающие отделы клетки.
Под термином "олигомеризованный" подразумевают комплекс с другими протеинами с образованием димеров, тримеров, тетрамеров или других высших олигомеров. Такие олигомеры могут быть гомо-олигомерами или гетероолигомерами. Олигомеризующий участок представляет собой тот участок молекулы, который управляет образованием комплекса /то есть, олигомера/.
Под термином "цитолитический" подразумевают способный разрушить клетку /например, клетку, инфицированную патогеном, опухолевую или канцерогенную клетку, или автоиммуно-генерированную/ или способный разрушить инфицирующий агент /например, вирус/.
Под "вирусом иммунодефицита" подразумевают ретровирусы, которые в форме дикого типа способны инфицировать T клетки хозяина-примата, и обладают вирусным морфогенезом и морфологическими характеристиками подсемейства лентивирусов. Этот термин включает, без ограничений, все варианты HIV и SIV, включая HIV-1, HIV-2, SIVmac, SIVagm, SIVmnd, SIVSmm, SIVman, SIVmand и SIVcpz.
Под термином "МНС-зависимый" подразумевают, что клеточная цитолитическая реакция не требует присутствия какого-либо МНС класса II антигена на поверхности мишеневой клетки.
Под термином "функциональное цитолитическое сигнально-трансдукторное производное" подразумевают функциональное производное /как было определено ранее/, которое способно управлять, по крайней мере, 10%, предпочтительно, 40%, более предпочтительно, 70%, и наиболее предпочтительно, по крайней мере, 90% биологической активности дикого типа молекулы. В том смысле, как здесь использован, "функциональное цитолитическое сигнал-трансдукторное производное" может посылать непосредственно сигнал терапевтической клетке разрушить связанный с рецептором агент или клетку /например, в случае внутриклеточной химерической рецепторной части/ или может действовать косвенно, промотируя олигомеризацию цитолитического сигнального трансдукторного протеина терапевтической клетки /например, в случае трансмембранного домена/. Такие производные можно тестировать на эффективность, например, используя in vitro анализ, описанный здесь.
Под "функциональным производным связывающим оболочку HIV" подразумевают функциональное производное /как определено ранее/, которое способно связывать любой протеин оболочки HIV. Функциональные производные можно идентифицировать, используя например, in vitro анализ, описанный здесь.
Терапевтическое введение
Трансформированные клетки настоящего изобретения можно использовать для лечения ряда заболеваний. Текущие
способы введения таких трансформированных клеток включают адаптационную иммунотерапию или терапию переноса клеток. Эти способы позволяют вернуть иммунотрансформированные системы клеток в поток крови.
Roseuberg, S.A. Scientific Amerian, 62 (May 1990); Roseuberg, et al., The New Englang Journal of Medicine, 323 (9): 570 (1990).
Фармацевтические композиции настоящего изобретения можно вводить любому животному, которое может извлечь благоприятное воздействие от соединений настоящего изобретения. Прежде всего, среди животных следует назвать человека, хотя настоящее изобретение этим не ограничивается.
Клинические данные Cell Genesys по первоначальным испытаниям фазы II, в ходе которых исследовали пациентов с определяемым, несмотря на проводимую противоретровирусную лекарственную терапию содержанием ВИЧ в крови, свидетельствуют о безопасности проводимого лечения и сохранности генетически модифицированных T-клеток в циркуляторном русле, по крайней мере, в течение 100 суток после введения клеток. Были отмечены предварительные признаки противовирусной активности с тенденцией к снижению содержания ВИЧ в лимфоидной ткани желудочно-кишечного тракта, основного резервуара ВИЧ-инфицированных клеток, у четырех из пяти исследуемых пациентов".
Впоследствии та же рабочая группа сообщила о дальнейших проведенных клинических испытаниях Фазы II. В указанных последующих испытаниях участвовали 40 пациентов. 20 из них получали лечение в виде однократного введения экспрессирующих химерный рецептор T-клеток, а 20 контрольным пациентам вводили немодифицированные T-клетки. Результаты оценивали спустя 6 месяцев. В результате данных исследований выяснилось, что рецидив заболевания наблюдался лишь у 5 из 20 пациентов, которым вводили экспрессирующие химерный рецептор T-клетки, по сравнению с 10 из 20 в контрольной группе. Далее, у пациентов, получавших лечение в виде однократного введения экспрессирующих химерный рецептор T-клеток, по прошествии 6 месяцев наблюдалось как среднее 0,4 log отрицательное отклонение количества ВИЧ, высылаемого из циркулирующих клеток крови, от контрольных значений (p=0,02), так и среднее log отрицательное отклонение содержания ДНК ВИЧ, определяемой в биоптатах прямой кишки (p= 0,007), от контрольных значений. У пациентов, получавших контрольные немодифицированные T-клетки, существенные отклонения от контрольных значений в обоих исследованиях не наблюдались.
Подробное описание изобретения
Вначале описывается содержание чертежей.
Краткое описание чертежей
Фиг. 1 Характеристика CD4 химер. На фиг. 1A представлена аминокислотная последовательность вокруг сайта слияния между CD4 /остатки 1-369/ и различными рецепторными
цепями. Подчеркнутая последовательность показывает положение аминокислот, закодированных внутри BamHI сайта, использованного для конструкции слияния. Начало трансмембранного домена отмечено
вертикальной чертой. η последовательность идентична Z последовательности у аминоконца, но расходится у карбоксильного конца /Jin et al., Proc. Natl. Acad. Sci. USA, 87:3319 - 3323 /1990//. На
фиг. 1B представлены данные цитометрического анализа в потоке поверхностной экспрессии CD4, CD4:Z, CD4: γ и CD4 η в CVI клетках. Клетки были инфицированы вирусом, экспрессирующим CD4
химеры или CD16PI, инкубированы в течение 9 часов при 37oC и окрашены фикоэритрин-конъюгированными анти-CD4 Mab Leu 3A. На фиг. 1C представлено иммуноосаждение меченых CD4:Z,
CD4: η , или нативных CD4, экспрессированных в CVI клетки. Опыты проводили со снижением /R/ или без снижения /R/ агента. Молекулярные массы стандартно выражены в кД и указаны слева.
Фиг. 2 Поверхностная экспрессия CD16TM с последующей коинфекцией CD16TM отдельно /пунктир/ или совместной инфекцией с вирусом, экспрессирующим CD4: γ /штрихи/ или CD4 Z /сплошная линия/. Раздельные точки-клетки, инфицированные CD4: Z /отдельно, окрашенные 3G8/ Fleit et al., Proc. Natl. Acad. Sci. USA, 79:3275 - 3279 /1982 / /анти-CD16 Mab/.
Фиг. 3 Мутант CD4:Z химерического рецептора без ZASp-15 не поддерживает совместную экспрессию CD16TM. Фиг. 3A представляет собой авторадиограмму иммуноосажденных химер, обработанных электрофорезом либо с восстановлением /R/, либо без восстановления /NR/. Фиг. 3B детализирует поверхностную экспрессию CD16ТМ с последующим совместным инфицированием вирусом, экспрессирующим CD16TM и последующими Z химерами: CD4:Z (жирная линия) СД4:Z C11G /сплошная линия/, CD4: Z /пунктирная линия/, CD4:Z C11G /D15G /жирные точки/, без коинфицирования /CD16TM отдельно, редкие точки/. Клетки инкубируют с анти-CD16 Mab 3G8 и полиэритрин-конъюгированными Fab'2 козлиными антителами к мышиному IgG. Уровень экспрессии Z химер практически идентичен для различных анализированных мутантов, и коинфицирование клеток вирусами, экспрессирующими CD16TM, и химерами заметно не изменяет поверхностную экспрессию химер /не показано/.
Фиг. 4 Повышению внеклеточных свободных ионов кальция соответствует сшивка мутантных Z химер в T клеточной линии. Jurkat E6 клетки /Weiss et al. , J. Immunol 133:123-128 /1984/ /были идентифицированы с рекомбинантными вирусами вакцины и проанализированы с помощью цитометрии в потоке. Полученные результаты представлены для открытой CD4+ популяции, так что только клетки, экспрессирующие соответствующий химерический протеин, были анализированы. Среднее отношение фиолетовой к синей Индо-1 флуоресценции отражает внеклеточные концентрации свободного кальция в популяции как целом, а процент соответствующих клеток отражает долю клеток, которые превышают заранее определенное пороговое отношение /установленное таким образом, что положительными оказываются 10% непрореагировавших клеток/. Фиг. 4A и 4B изображают Jurkat клетки, экспрессирующие CD4: Z /сплошная линия/ или CD16:Z /пунктирная линия/, которые были экспонированы анти-CD4 Mab Leu3a /фикоэритриновый конъюгат/ с последующей сшивкой с козлиными антителами к мышиному IgG. Пунктирная линия показывает реакцию неинициированных клеток на анти-CD3 Mab OKT3. Фиг. 4C и 4D показывают Jurkat клетки, экспрессирующие CD4:Z D15G /сплошная линия; CD4: Z C11G /D15G /пунктир/; или CD4:Z C11G /точки/, которые были обработаны и проанализированы как на фиг. 4A и 4B.
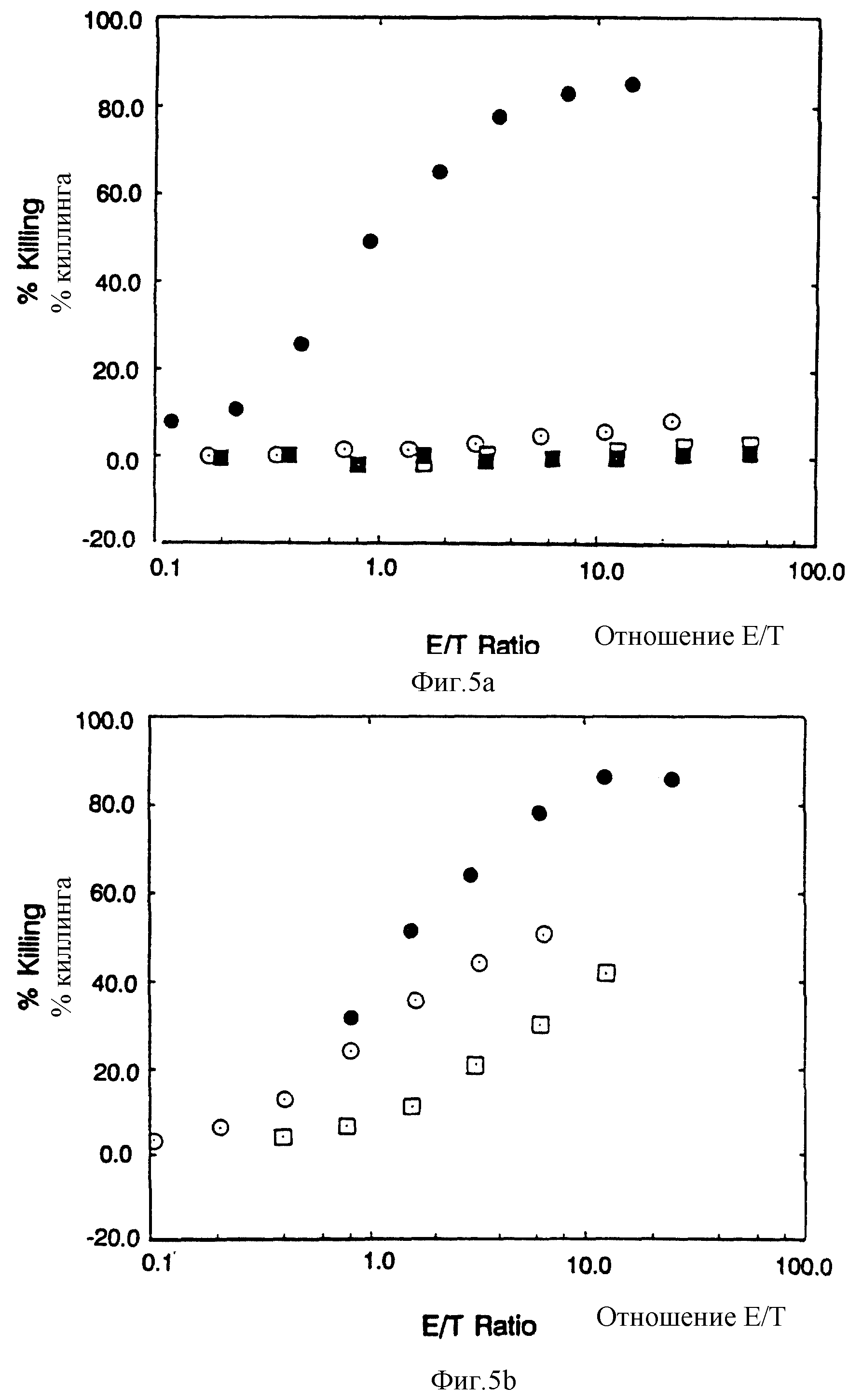
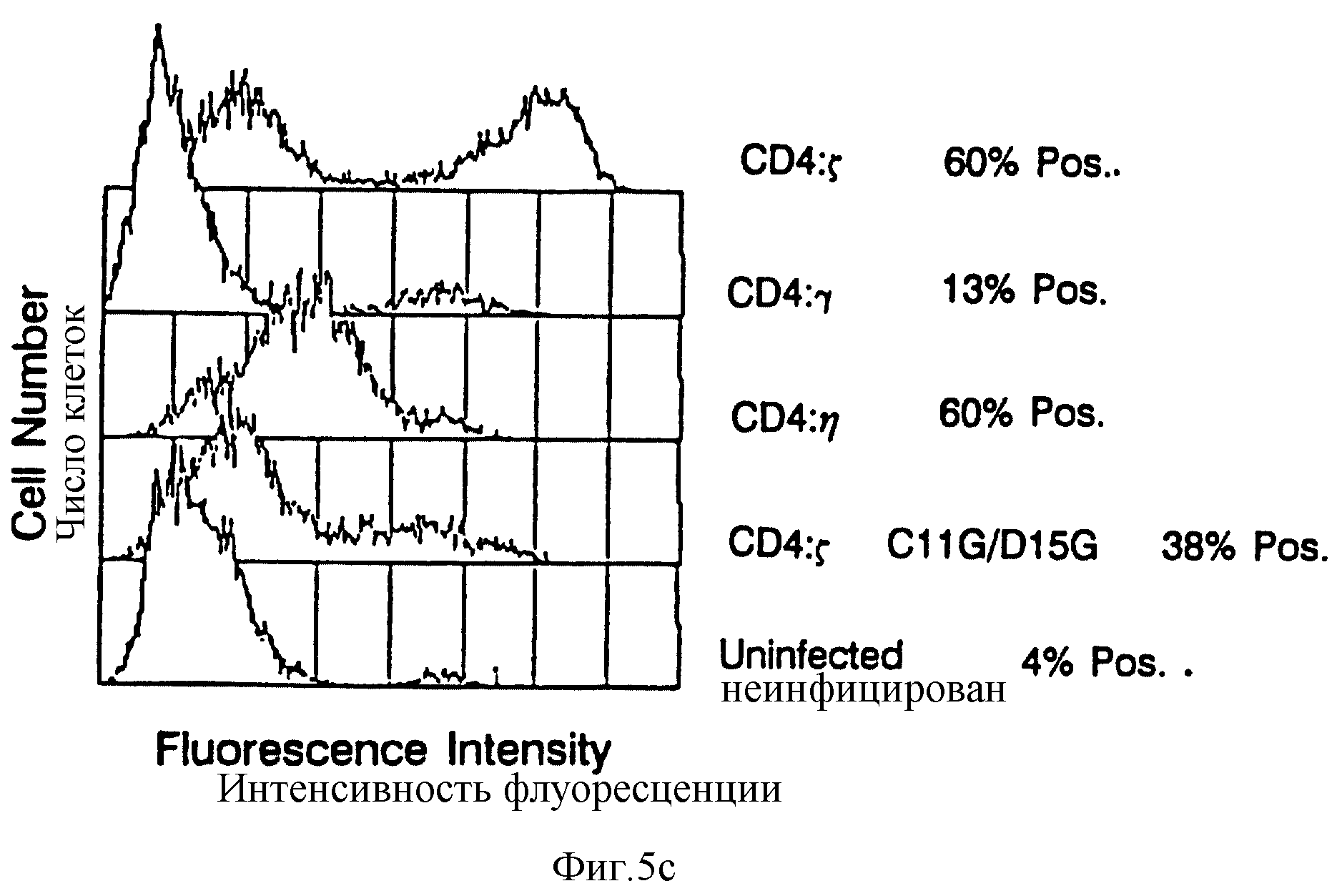
Фиг. 5 CD4: Z, CD4: η и CD4: γ рецепторы позволяют цитолитическим T лимфоцитам /CTL/ убивать мишени, экспрессирующие HIV-1 gp120/41. Фиг. 5a: сплошные кружочки, CTL, экспрессирующие CD4:Z, инкубированные с клетками HeLa, экспрессирующими gp120/41; пустые кружочки, CTZ экспрессирующие CD4:Z, инкубированные с неинфицированными клетками HeLa, сплошные квадраты, неинфицированные CTL, инкубированные с клетками HеLa, экспрессирующими gp120/41; пустые квадраты, неинфицированные CTL, инкубированные с неинфицированными клетками HeLa. Фиг. 5B: сплошные кружочки, CTL экспрессирующие CD4: η , инкубированные с клетками HeLa, экспрессирующими gp120/41; пустые кружочки, CTL экспрессирующие CD4: γ, инкубированные с клетками HeLa, экспрессирующими gp120/41; пустые квадраты, CTL экспрессирующие C 11G/D15G двойной мутант CD4: Z химера, инкубированные с клетками HeLa, экспрессирующими gp120/41, Фиг. 5C: Цитометрический анализ в потоке CD4 экспрессии за счет CTL, использованных на фиг. 5B. Для исправления мишени до эффекторных отношений процент клеток, экспрессирующих CD4 химеры, был определен вычитанием отрицательных /неинфицированных/ популяций наложением гистограмм; для сравнения на фиг. 5С неинфицированные клетки были приписаны предполагаемому пороговому значению, что дало приблизительно ту же часть, положительную для других клеточных популяций, что и на вычитании гистограмм.
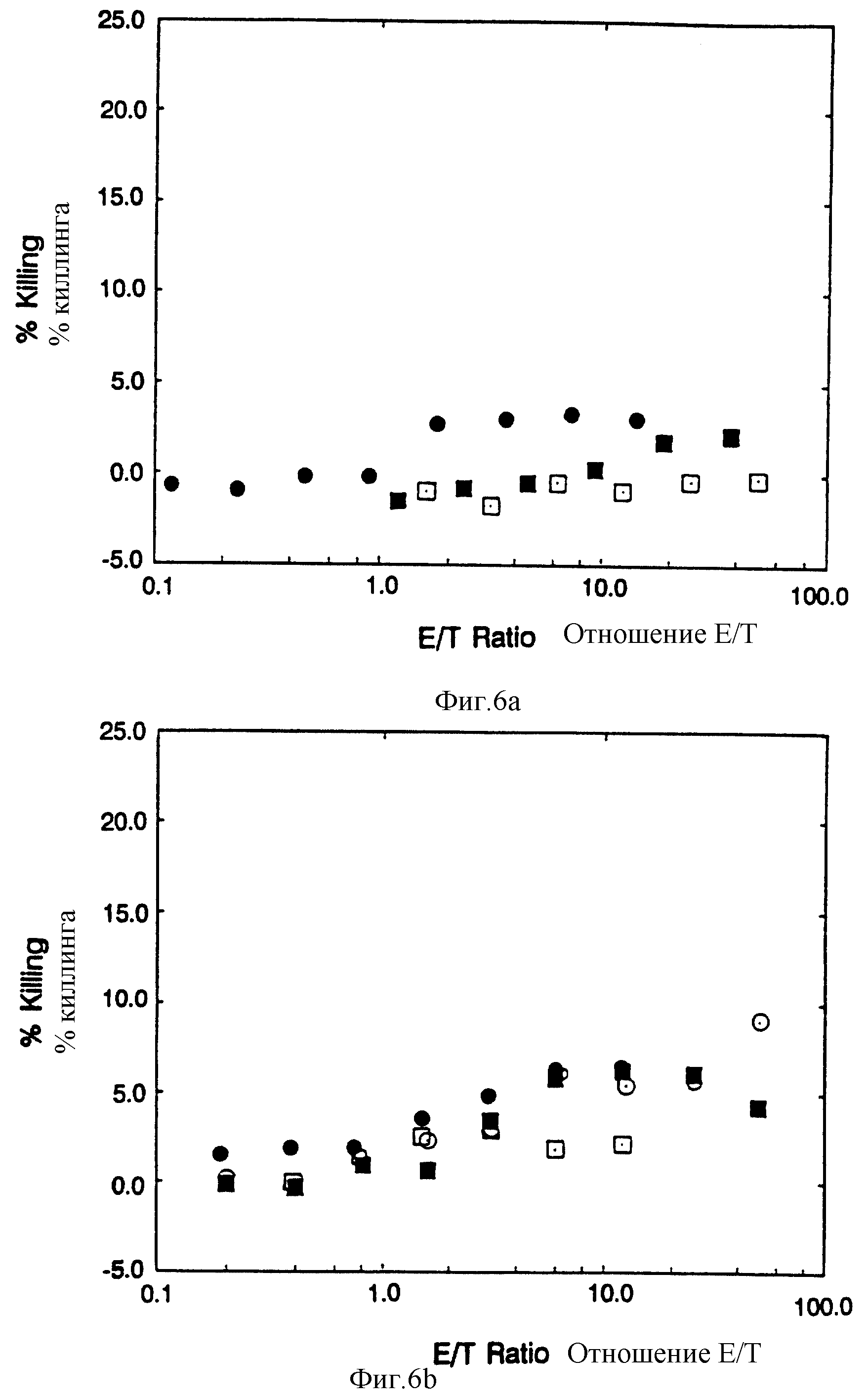
Фиг. 6 Специфичность цитолиза под управлением CD4:Z. Фиг. 6A: сплошные кружочки CTL экспрессирующие CD4:Z, инкубированные с клетками HeLa, экспрессирующими CD16PI ; пустые кружочки, CTL экспрессирующие CD4, инкубированные с клетками HeLa, экспрессирующими gp120; сплошные квадраты, CTL, экспрессирующие CD: Z, инкубированные с клетками HeLa, экспрессирующими gp120/41; пустые квадраты, CTL экспрессирующие CD16PI инкубированные с HeLa клетками, экспрессирующими gp120/41. Фиг. 6B: сплошные кружки, CTL экспрессирующие CD4: Z, инкубированные с Raji /МНС класс II+/ клетками; пустые кружки, неинфицированные CTL клетки, инкубированные с RJ 2.2.5 /МНС класс II- Raji мутант/ клетками; сплошные квадраты, неинфицированные CTL, инкубированные с Raji /МНС класс II+/ клетками; пустые квадраты, CTL экспрессирующие CD4:Z, инкубированные с RJ 2,2,5 /МНС класс II-/ клетками. Шкала ординат расширена.
Фиг. 7 Характеристика CD16:Z химерического рецептора. Фиг. 7A является схематической диаграммой CD16: Z слитого протеина. Внеклеточная часть фосфатидилинозитол-связанной формы мономерной CD16 была соединена с димерным Z как раз снаружи трансмембранного домена. Протеиновая последовательность в месте слияния представлена внизу. Фиг. 7B представляет цитометрический анализ в потоке мобилизации кальция с последующей сшивкой CD16:Z химеры с либо TCR или TCR отрицательной клеточной линией. Представлено среднее отношение фиолетовой к синей флуоресценции /мера относительной концентрации ионов кальция /среди клеточной популяции, обработанной антителами в момент времени 0. Сплошные квадраты, реакция Jurkat клеток на анти-CD3 Mab OKT3 оплошные треугольники, реакция CD16:Z на анти-CD16 Mab 3G8, сшивающие в REX33A TCR- мутант; пустые квадраты, реакция на CD16:Z сшивающие в Jurkat TCR- мутант линии PT3. T3.5; пустые треугольники, реакция на CD16:Z сшивающие Jurkat клетки; крестики, реакция на нехимерические CD16 в Jurkat клетках и точки, реакция на нехимерические CD16 в REX33A TCR- линии клеток.
Фиг. 8. Делеционный анализ цитолитического потенциала. Фиг. 8A показывает расположение конечных точек Z делеции. Здесь и всюду мутанты в Z представлены исходным остатком-расположением-мутантным остатком, так что D66*, например, обозначает замену ASp-66 терминационным кодоном. На фиг. 8B представлен цитолитический анализ в результате не подвергавшейся делеции CD16:Z и выступающей Z делеции. Гибридомные клетки, экспрессирующие поверхностные антитела к CD16, были загружены51Cr и инкубированы с повышающимся числом цитолитичеcких лимфоцитов человека /CTL/, инфицированных вакциной рекомбинантных экспрессирующих CD16:Z химеры. Процент51Cr, который высвобождается, откладывают как функцию клеточного отношения эффектора /CTL/ к мишени /гибридоме/е/т/. Сплошные кружки, цитолиз посредством клеток, экспрессирующих CD16:Z /mfi 18,7/; сплошные квадраты, цитолиз посредством клеток, экспрессирующих CD16:Z ASp66* /mfi/ 940,2/; пустые квадраты, цитолиз посредством клеток, экспрессирующих CD16: Z Glu60* /mfi 16,0/; пустые кружки, цитолиз посредством клеток, экспрессирующих CD16: Z Tyr51* /mfi 17,4 /; сплошные треугольники, цитолиз посредством клеток, экспрессирующих CD16:Z Phe34* /mfi 17,8/; и пустые треугольники, цитолиз, осуществляемый за счет клеток, экспрессирующих нехимерические CD16 /mfi 591/. Хотя в этих экспериментах экспрессия CD16: Z ASp66* не была такой же, как для других слитых протеинов, цитолиз клетками, экспрессирующими CD16:Z при одинаковых уровнях в тех же самых экспериментах дал результаты, практически идентичные тем, которые демонстрируют клетки, экспрессирующие CD16:Z ASp66* /не показано/.
Фиг. 9. Исключение потенциала для трансмембранного взаимодействия выявляет короткий сегмент, способный быть медиатором цитолиза. Фиг. 9A представляет схему мономерных химер, состоящих из двух или трех частей. Наверху представлен CD16:Z, усеченный по остатку 65 и не содержащий трансмембранных Cys и Asp остатков. Ниже представлены CD16:CD5:Z и CD16:CD7:Z конструкции и родственные контроли. Пептидные последовательности внутриклеточных доменов представлены ниже. На фиг. 9B представлена цитолитическая активность мономерных химерических мутантов делеции. Цитолитическая активность клеток, экспрессирующих CD16: Z /сплошные кружки; mfi 495/ сравнивается с активностью клеток, экспрессирующих CD16:Z Asp66* /сплошные квадраты; mfi 527/ или мутантов CD16: Z Cys11Gly /Asp66*, /пустые квадраты, mfi 338/ и CD16:Z Cys11Gly /Asp15-Gly/Glu60* /сплошные треугольники; mfi 259/. На фиг. 9C представлена цитолитическая активность через посредство слитых протеинов, состоящих из трех частей. Сплошные треугольники, CD16:Z Asp66*, пустые квадраты, CD16:5:Z /48 - 65/, сплошные квадраты CD16:7:Z /48 - 65/, пустые треугольники, CD16: 7: Z /48-59/, пустые кружки CD16:5, сплошные кружки, CD16:7. На фиг. 9 представлена мобилизация кальция мутантом и тройными химерами в TCR негативной Jurkat JRT3. T3.5. мутантной клеточной линии. Пустые кружки соответствуют клеточной экспрессии димерной CD16:Z Asp66*, сплошные квадраты соответствуют клеткам, экспрессирующим CD16:Z Cys11Gly/Asp15Gly/Asp66*, пустые квадраты, реакция клеток, экспрессирующих CD16:Z Cys11Gly/Asp15Gly/Glu60*, сплошные треугольники, реакция клеток, экспрессирующих CD16:7:Z /48-65/, и пустые треугольники, реакция клеток, экспрессирующих CD16:Z /48-59/.
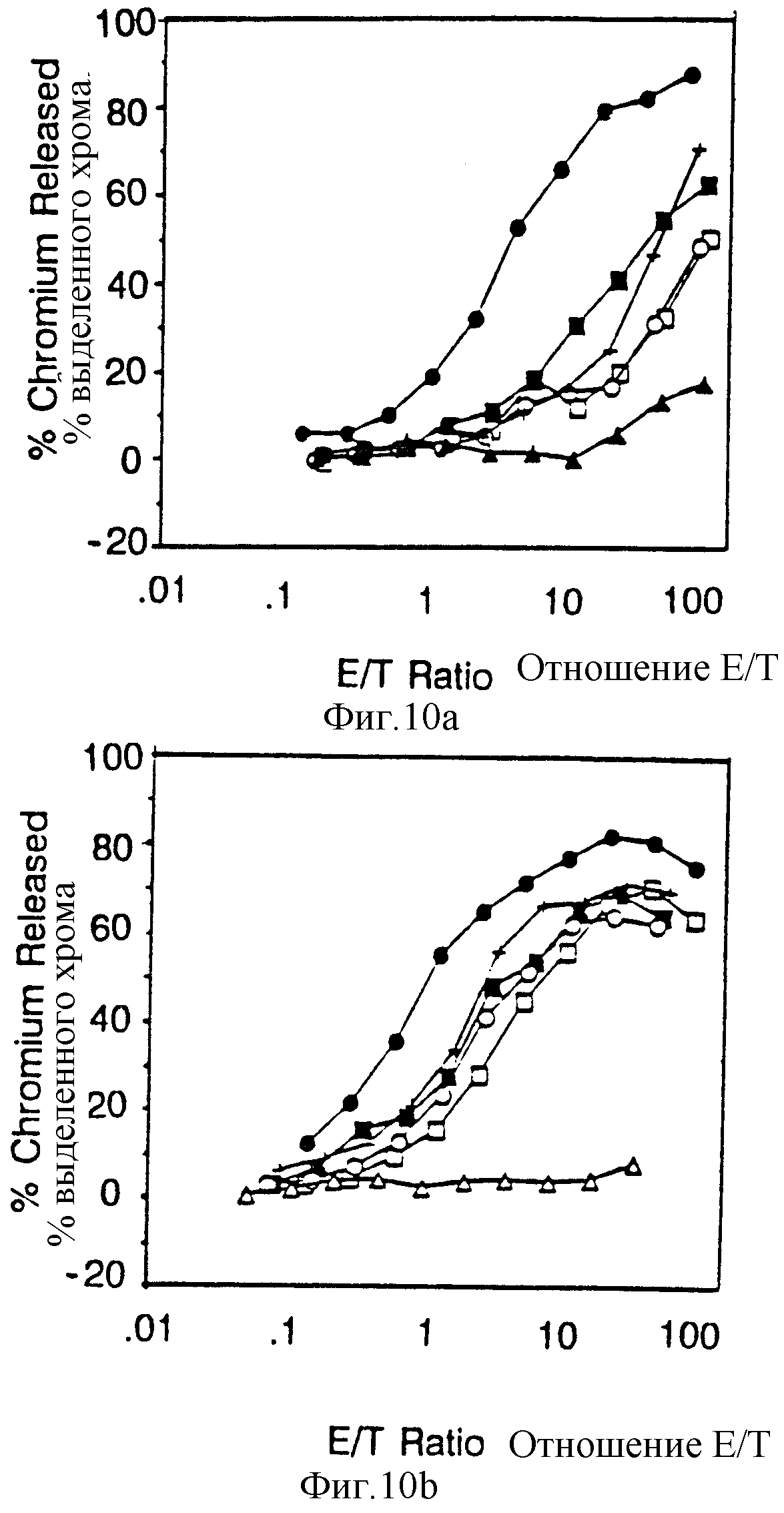
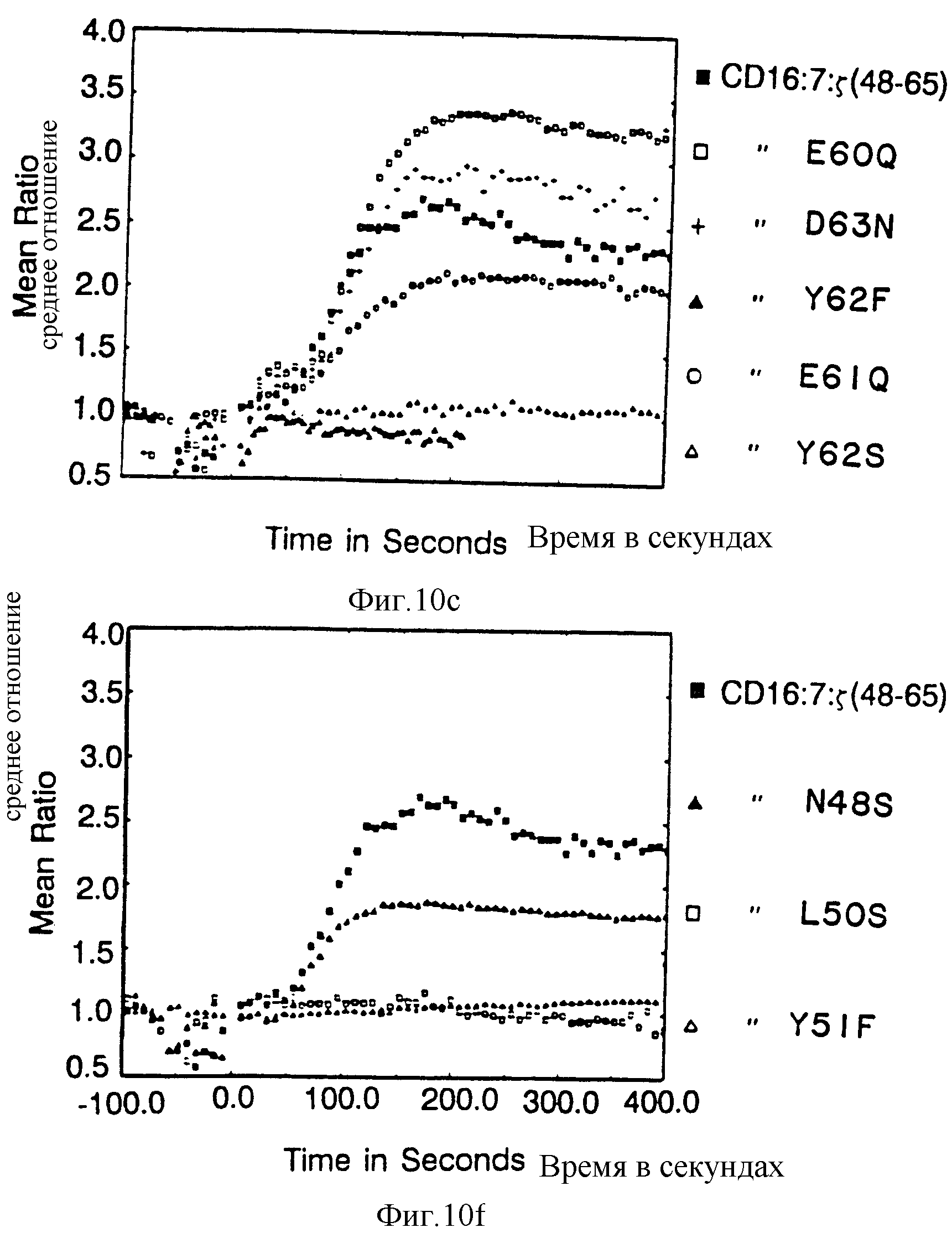
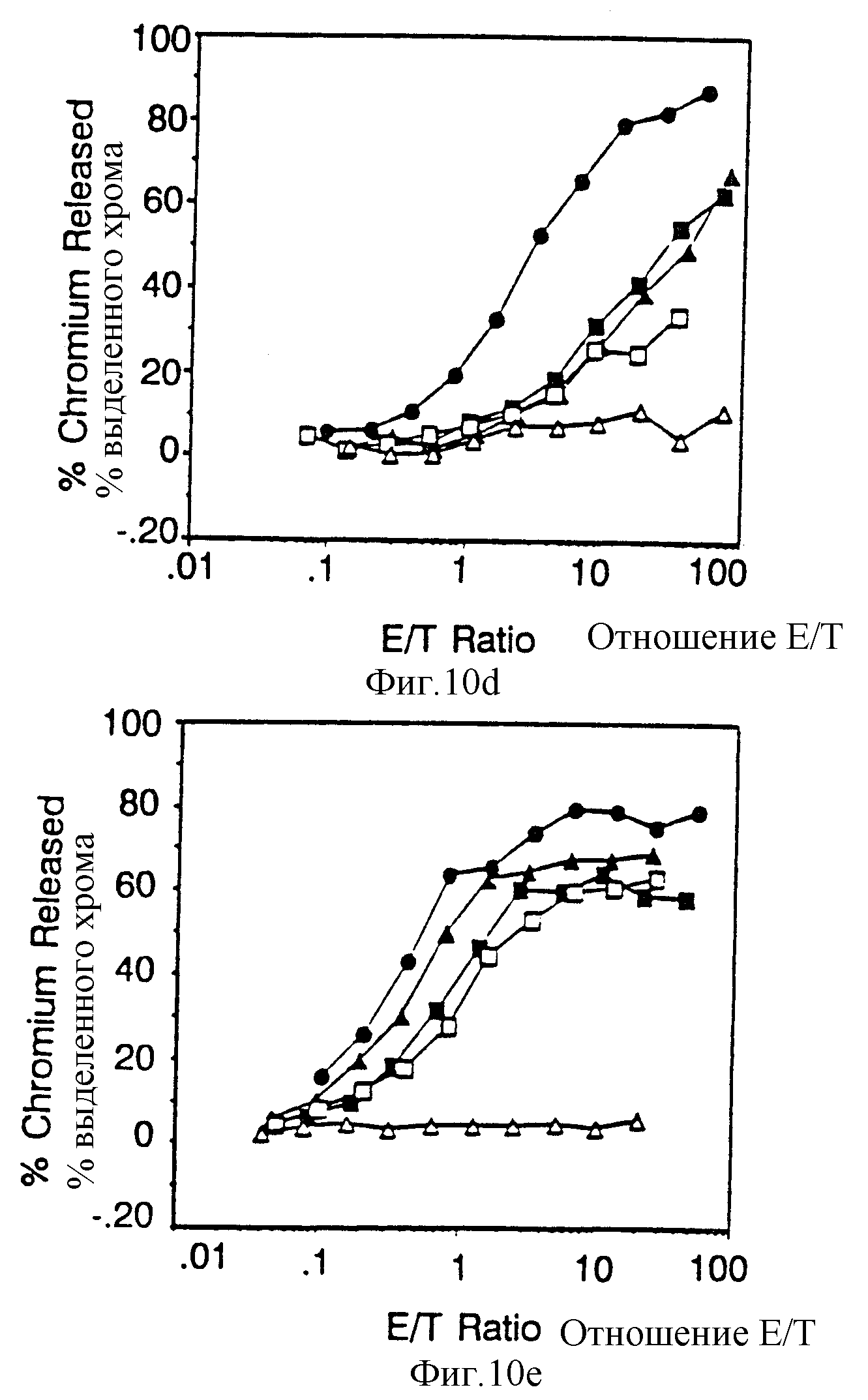
Фиг. 10 Вклад отдельных аминокислот в активность цитолитического передающего сигнал фрагмента из 18 остатков. Фиг. 10A и 10B показывают цитолитическую активность, а фиг. 10c показывает мобилизацию ионов кальция посредством химер, несущих точечные мутации вблизи карбоксильного терминального тирозина /V62/. Фиг. 10A и 10B представляют данные, полученные на клетках, экспрессирующих низкие и высокие количества, соответственно, CD16:Z протеинов слияния. Идентичные символы использованы для анализов по мобилизации кальция и цитолиза и представлены в однобуквенном коде справа. Сплошные кружки, клетки, экспрессирующие CD16:Z /mfi в A,21, B,376/; сплошные квадраты, клетки, экспрессирующие CD16:7:Z /48-65/ /mif A,31, B,82/, пустые квадраты CD16:7:Z /48-65/ GL60Gн /mfi A,33; B,92/, крестики, CD16:7:Z /48-65/ Asp63A н /mfi A, 30; B,74/, сплошные треугольники CD16:7:Z /48-65 /Tyr62Phe /mfi A,24, B,88/, пустые кружки, CD16: 7:Z /48-65/Gl61GIн /mfi A, 20, B, 62/, и пустые треугольники. CD16: 7:Z /48-65/ Tyr62 Serr /mfi B, 64/. Фиг. 10D и 10E демонстрируют цитолитическую активность, а на фиг. 10F показана мобилизация ионов кальция химерами, несущими точечные мутации вблизи аминотерминального тирозина /V51/. Идентичные символы использованы для анализов по мобилизации кальция и цитолизу и представлены справа. Сплошные кружки, клетки, экспрессирующие CD16: Z /mfi в D,21,2. в E, 672/, сплошные квадраты, клетки, экспрессирующие CD16: 7: Z /48-65/ /mfi D,31,3, E, 179/, оплошные треугольники, CD16: 7:Z /48-65/ /Asn48Ser /mfi D, 22,4, E, 209/, пустые квадраты, CD16:7:Z /48-65/ Leu50Ser/ D, 26,0, E, 142/ и пустые треугольники, CD16:7:Z /48-65/ Tyr51Phe /mfi D, 32,3, Е, 294/.
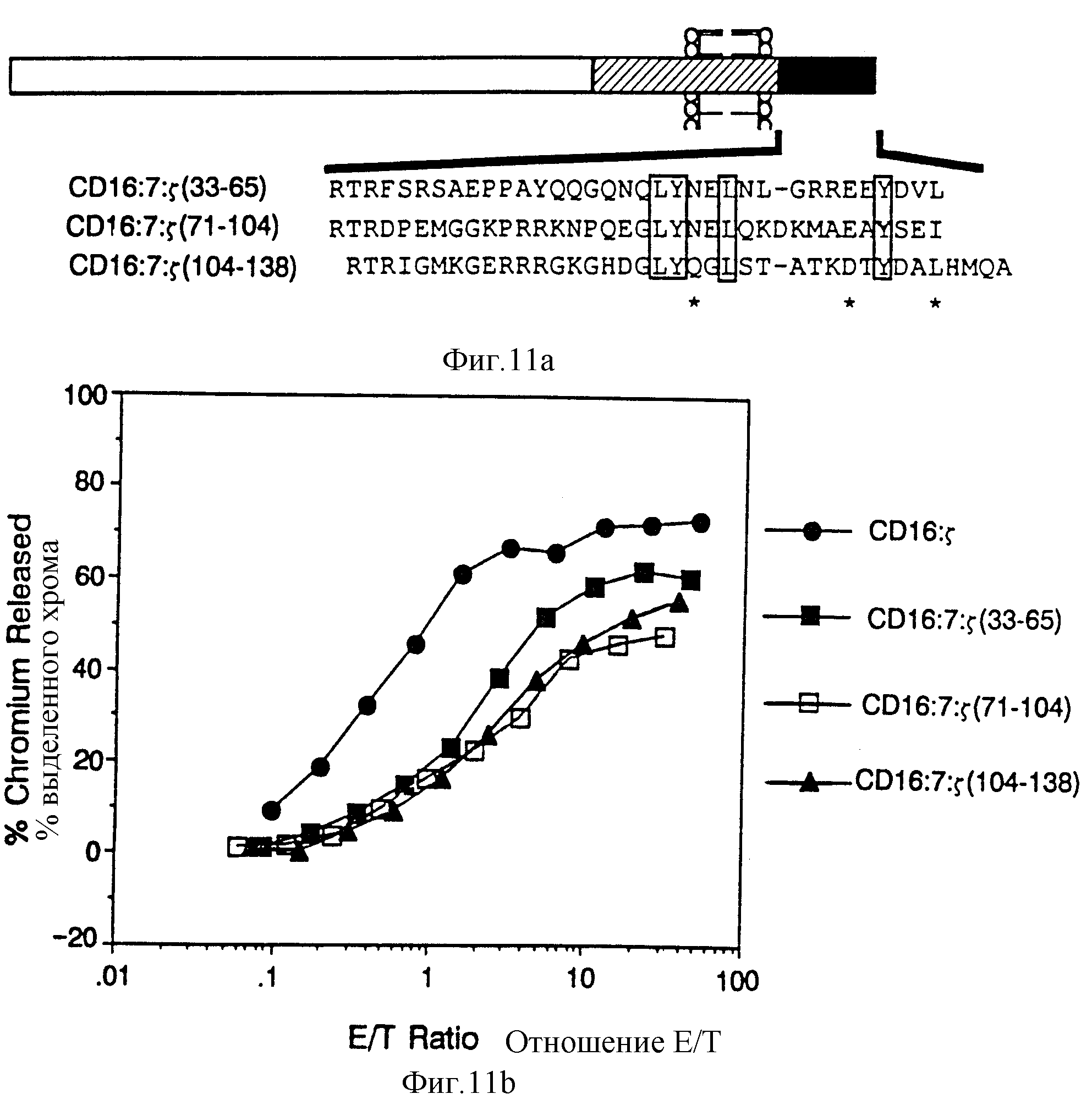
Фиг. 11 Соответствие внутренних повторов Z, и сравнение их способности поддерживать цитолиз. Фиг. 11A представляет схематическую диаграмму химер, полученных делением Z внутриклеточного домена на трети и присоединения их к трансмембранному домену CD16: 7 химер. Последовательности внутриклеточных доменов показаны ниже, причем нужные остатки заключены в прямоугольники, а родственные остатки помечены звездочками. Фиг. 11B показывает цитолитическую способность трех Z субдоменов. Сплошные кружки, клетки, экспрессирующие CD16: Z /mfi 476/, сплошные квадраты, CD16:7:Z /33-65/ /mfi 68/, пустые квадраты, CD16: 7: Z /71-104/ /mfi 114/ и сплошные треугольники, CD16:7:Z /104-138/ /mfi 104/.
Фиг. 12 представляет схематическую диаграмму химер CD16:FcR γ 11.
Фиг. 13. Мобилизация кальция с последующей сшивкой CD4:FcR γ 11 и CD16: FcR γ 11 химер. Фиг. 13A представляет отношение фиолетовой к синей флуоресценции, испускаемой клетками, загруженными чувствительным к кальцию флуорофор Indo-1, представленное как функция времени с последующей сшивкой CD16 внеклеточного домена с антителами. Фиг. 13B показывает аналогичный анализ повышения отношения фиолетовой к синей флуоресценции клеток, несущих CD4:FcR γ 11 химеры, с последующей сшивкой с антителами.
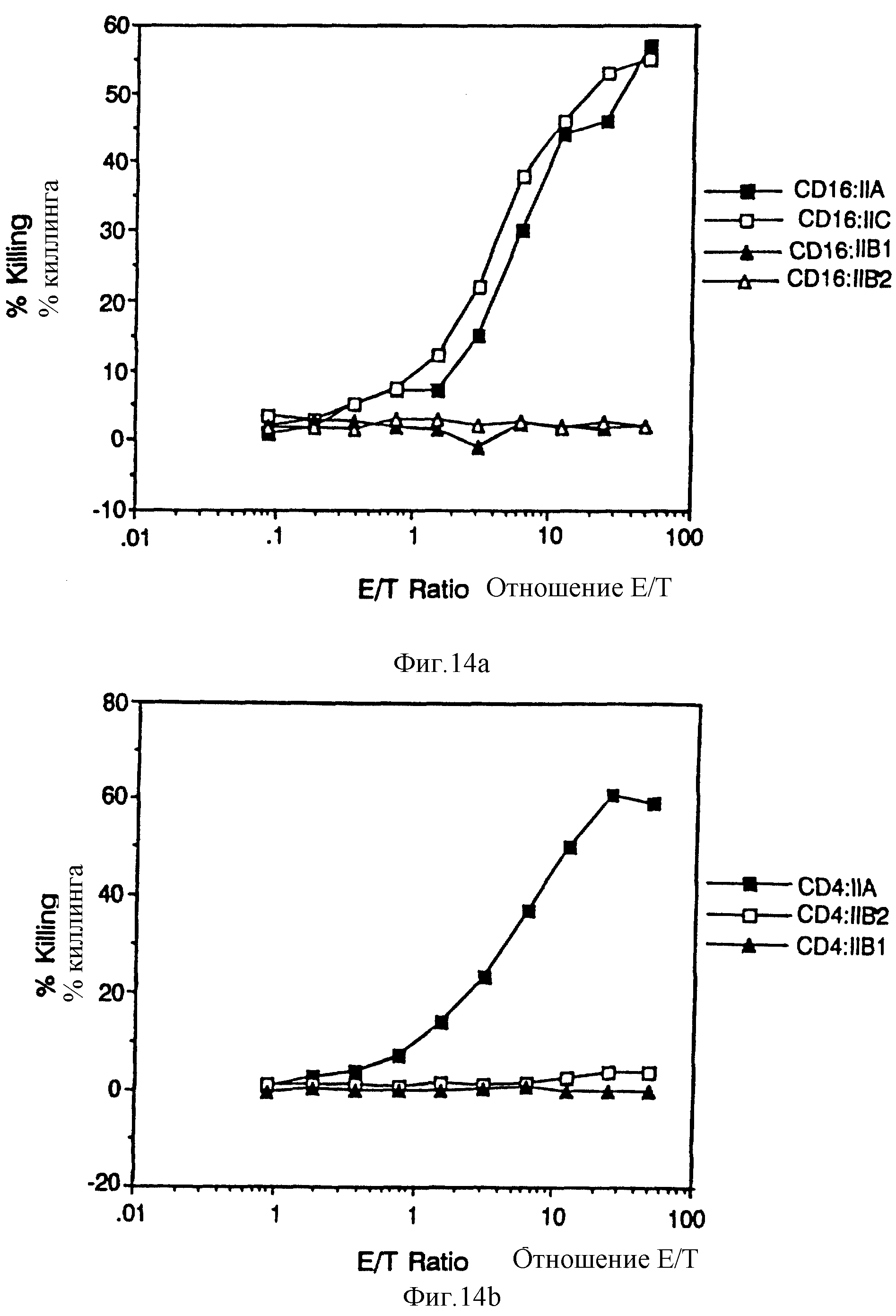
Фиг. 14. Цитолитический анализ CD4:FcR γ 11 и CD16:FcR γ 11 химер. Фиг. 14A показывает процент51Cr высвобождаемого из анти-CD16 гибридомных /мишеневых/ клеток, когда эти клетки экспонируют возрастающему количеству цитолитических T лимфоцитов, экспрессирующих CD16:FcR γ 11 химеры /эффекторные клетки/. Фиг. 14B показывает аналогичный анализ цитотоксичности через посредство CD4:FcR γ 11 химер против мишеневых клеток, экспрессирующих гликопротеины оболочки HIV.
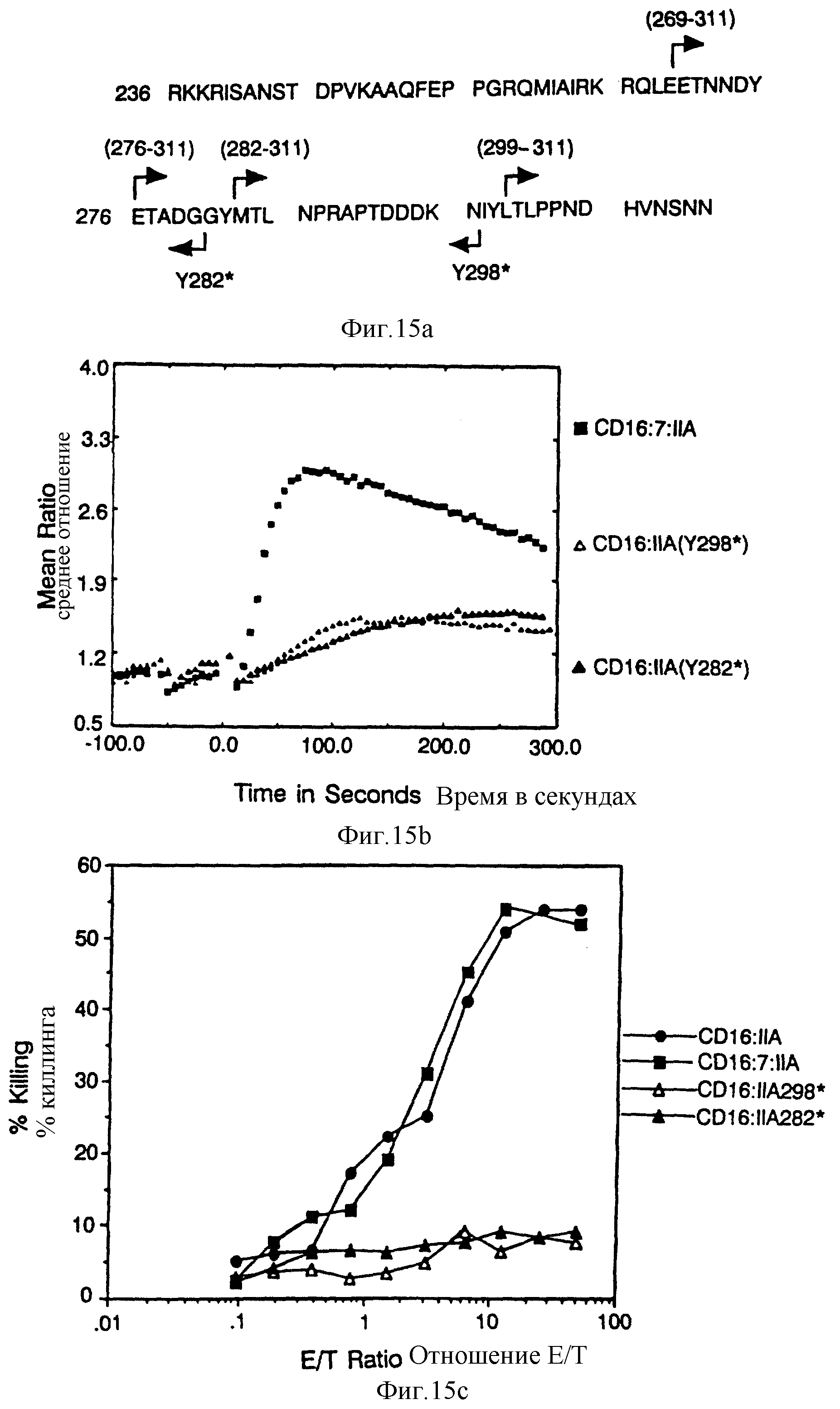
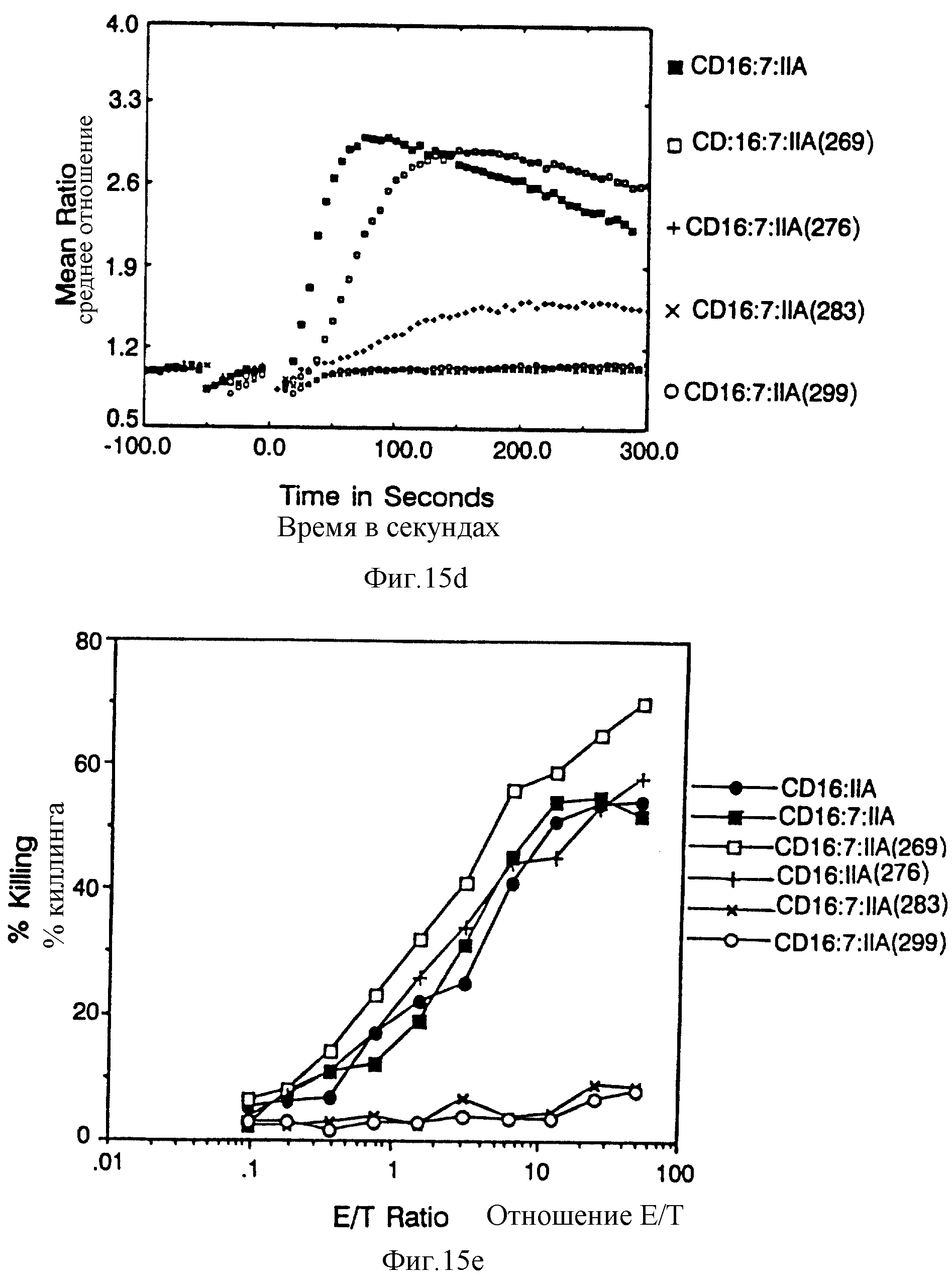
Фиг. 15. Идентификация остатков в FcR γ 11A хвосте, который важен для цитолиза, фиг. 15A представляет собой схематическую диаграмму делеций конструкций. Фиг. 15B и 15C демонстрируют мобилизацию кальция и цитолиз за счет карбоксил-терминальных делеций вариантов CD16:FcR γ 11A. Фиг. 15D и 15E показывают мобилизацию кальция и цитолиз химерами, состоящими из трех частей, содержащими прогрессивно меньше аминоконцов внутриклеточного хвоста CD16:FcR γ 11A.
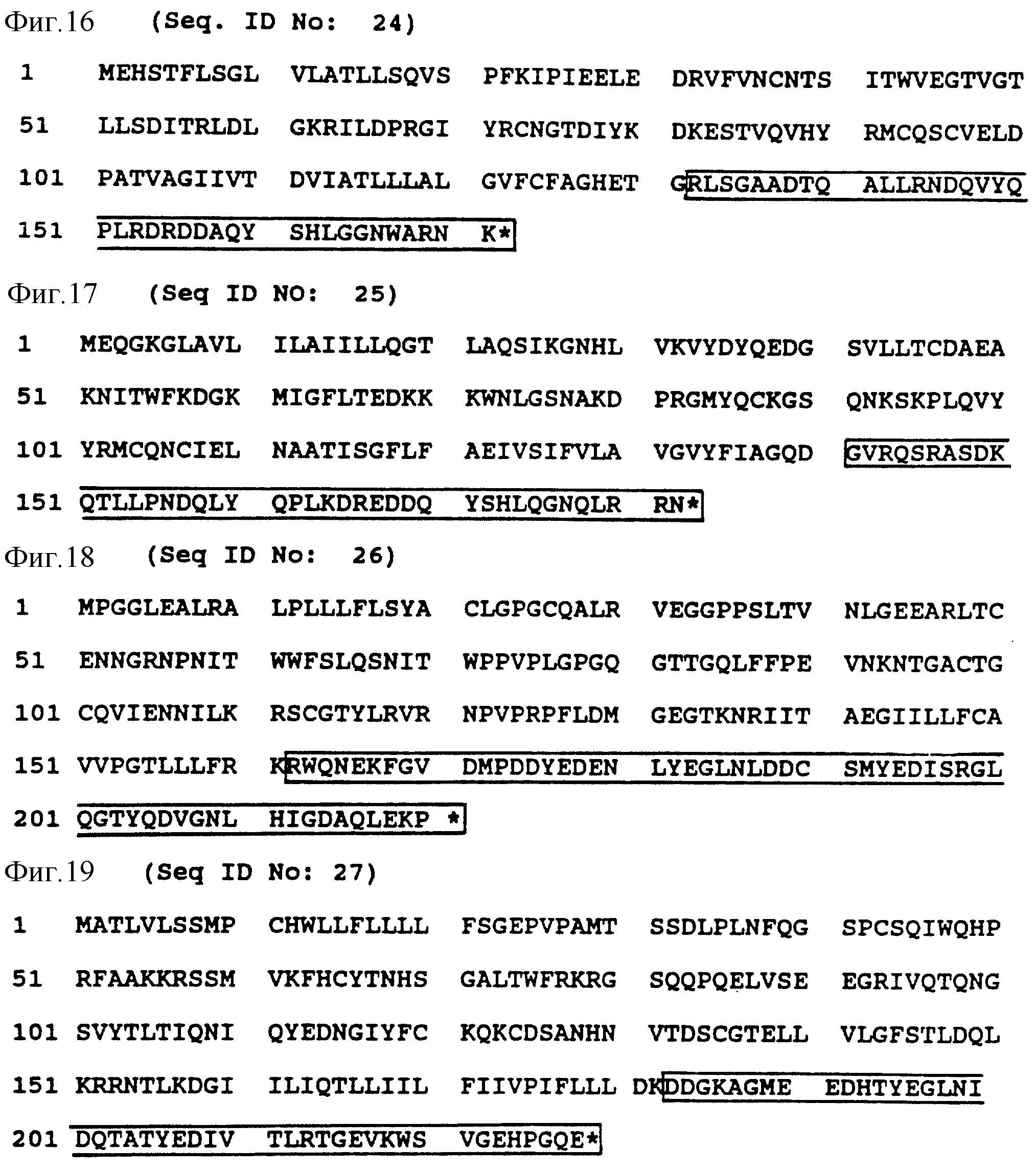
Фиг. 16. /SEQ ID N0:24/ изображает аминокислотную последовательность CD3 дельта рецепторного протеина; в рамке последовательность, которая представляет предпочтительную часть цитолитического сигнала трансдукции.
Фиг. 17 /SEQ ID N0:24/ представляет аминокислотную последовательность T3 гамма рецепторного протеина; заключенная в рамку последовательность представляет предпочтительную часть цитолитического сигнала трансдукции.
Фиг. 18 /SEQ ID N0:26/ представляет аминокислотную последовательность mbl рецепторного протеина; заключенная в рамку последовательность представляет предпочтительную часть цитолитического сигнала трансдукции.
Фиг. 19 /SEQ ID N0:27/ представляет аминокислотную последовательность B29 рецепторного протеина; заключенная в рамку последовательность представляет предпочтительную часть цитолитического сигнала трансдукции.
Пример 1
Конструирование IgG1 человека: рецепторные химеры
Получают тяжелые цепные последовательности человеческого IgG1, соединяя
последовательности в CH3 домене с кДНК фрагментом, полученным из 3' конца трансмембранной формы иРНК антител. 3' концевой фрагмент получают в цепной полимеразной реакции, используя
библиотеку кДНК из миндалин в качестве субстрата, и олигонуклеотиды, с последовательностями:
CGC GGG GTG ACC GTG CCC TCC AGC AGC TTC GGC /SEQ ID N0:7/ и
CGC GGG GAT CCG TCG TCC AGA
GCC CGT CCA GCC CGT CCT GGG CCT CA /SEQ ID N0: 8/, соответствующие 5' и 3' концам целевых фрагментов ДНК, соответственно. 5' олигокомплементарен сайту в CHI домене человеческого IgC1, а 3'
комплементарен сайту 5' последовательности, кодирующей мембранный спанинговый домен. PCR продукт переваривают с BStXI и BamHI и лигируют между CStXI и BamHI сайтами полусинтетического гена антитела
IgG1, несущего переменные и постоянные участки. После включения фрагмента BStXI до BamHI, амплифицирующую часть конструкции заменяют вплоть до Smal сайта в CH3 рестрикционным фрагментом
взаимообмена, так, что только участок между Smal сайтом и 3' олиго получают из PCR реакции.
Для создания человеческого IgC1: η химерического рецептора тяжелый цепной ген, оканчивающийся в ВамHI сайте, соединяют с ВамHI сайтом описанной далее химеры η , так что последовательность антител образует внеклеточную часть. Цитометрия в потоке COS клеток, трансфектированных плазмидой, кодирующей химеру, показывает высокий уровень экспрессии детерминант антител, если плазмида экспрессии, кодирующая легкие цепи кДНК, котрансфектируется, и умеренную экспрессию детерминант антител, если плазмида экспрессии легкой цепи отсутствует.
Аналогичные химеры, включая человеческий IgG1 слитый с η или γ /см. далее/, или любой сигнальный-трансдукторный участок T клеточного рецептора или Fc рецепторного протеина, можно конструировать обычно, как описано ранее, используя стандартные методики молекулярной биологии.
Для создания отдельной транскрипционной единицы, которая позволила бы экспрессировать как тяжелую, так и легкую цепи, из единого промотера, создают плазмиду, кодирующую bicistronic иРНК из
последовательностей, кодирующих тяжелые и легкие цепи, и 5' нетрансляционной части иРНК, кодирующей 78 кД глюкозо регуляторный протеин, известный иначе как grp78, или BiP. grp78 последовательность
получают за счет PCR человеческой геномной ДНК, используя праймеры, имеющие последовательности:
CGC GGG CGG CCG CGA CGC CGG CCA AGA CAG CAC /SEQ ID N0:9/ и
CGC GTT GAC GAG CAG CCA
GTT GGG CAC CAG /SEQ ID N0:10/
по 5' и 3' концам, соответственно. Полимеразные цепные реакции с этими олигомерами проводят в присутствии 10% диметилсульфоксида. Фрагмент, полученный за счет
PCR, переваривают с Wotl и Hincll и включают между Notl и Hpal сайтами после последовательности, кодирующей IgG1 человека. Последовательности, кодирующие кДНК легкой цепи IgG1 каппа человека, включают
затем после grp78 лидера, используя Hincll сайт и другие сайты в векторе. Плазмида экспрессии, полученная в результате этих манипуляций, состоит из синтетического гена тяжелой цепи с последующей grp78
лидерной последовательностью, с последующей последовательностью кДНК легкой цепи каппа, с последующими сигналами полиаденилирования, полученными из SV40 ДНК фрагмента. Трансфекция COS клеток
экспрессионной плазмидой дает заметно повышенную экспрессию детерминант тяжелой цепи, по сравнению с трансфекцией плазмид, кодирующих только детерминанты тяжелой цепи.
Для создания bicistronic гена, содержащего тяжелую цепь/рецепторную химеру, и легкую цепь, расположенные ранее, последовательности тяжелой цепи можно заменить любым химерическим тяжелая цепь/рецепторным геном, здесь описанным.
Пример 2
Конструирование CD4 рецепторных химер
Человеческие Z (Weissman et al., Proc. Natl. Acad. Sci. USA, 85: 9709-9713 (1998b)) и η
(Kuster et al., J. Biol. Chem. 265: 6448-6452 (1990).
кДНК выделяют в реакции полимеразной цепи из библиотеки, полученной из клеточной линии HPB-ALL опухоли /Aruffo et al. Proc. Natl. Acad. Sci. USA 84: 8573-8577 /1987// и из природных киллерных клеток человека, тогда как η ДНК /Jin et al. Proc. Natl. Acad. Sci. USA, 87:3319 - 3323 /1990// выделяют из библиотеки тимоцитов мышей, Z, η и γ кДНК соединяют с внеклеточным доменом сконструированной формы CD4, содержащей BamHI сайт сразу перед мембранным спанинговым доменом /Aruffo et al. Proc. Natl. Acad. Sci. USA 84:8573 - 8577 /1987/; Zeltlmeissl et al. DNA Cell Biol. 9347 - 353 /1990//, который присоединяют к BamHI сайту, природно встречающемуся в Z и η кДНК в аналогичном положении на несколько остатков ранее мембранного спанингового домена /SEQ ID NOS: 1, 3, 4 и 6/. Для получения слияния протеина с γ BamHI сайт включают в последовательность в том же самом /приблизительно/ месте /фиг. 1/ последовательности SEQ ID N0: 2 и 5/. Слитые гены вводят в вирус вакцины, экспрессирующий плазмиду, несущую E.coli gpt ген в качестве селектируемого маркера /M. Amiot and B.S., неопубликовано/ и включают в геном штамма WP вакцины за счет гомологической рекомбинации и отбора по росту в микофенолевой кислоте /Falkner et al., J. virol 62:1849 - 1854 /1988/, Boyle et al., Gene, 65:123 - 128 /1988/ /. Цитометрический анализ в потоке показывает, что рекомбинанты вакцины управляют обильным продуцированием CD4:Z и CD4 γ протеинами слияния на поверхности клеток, тогда как экспрессия CD4: η существенно ниже /фиг. 1/. Последнее открытие согласуется с недавним сообщением о том, что трансфекция η кДНК экспрессирующей плазмиды в гибридомную клеточную линию мышей дает существенно более низкую экспрессию, нежели трансфекция сравнимой Z экспрессионной плазмиды /Clayton et al. J. Exp. Med. 172:1243 - 1253 /1990//. Иммуноосаждение клеток, инфицированных рекомбинантами вакцины, показывает, что слитые протеины образуют ковалентные димеры, в отличие от природно встречающихся CD4 антигенов /фиг. 1/. Молекулярные массы мономерных CD4: Z и CD4: γ протеинов слияния и нативных CD4 оказались 63, 55 и 53 кД, соответственно. Более крупные массы протеинов слияния приблизительно соответствуют большей длине внеклеточной части, которая превышает нативные CD4 на 75 /CD4:Z/ или на 5 /CD4 γ / остатков.
Пример III
CD4 химеры можно ассоциировать с другими рецепторными цепями
Клеточная поверхностная экспрессия макрофаг
/природная киллерная клеточная форма Fc γ RIII /CD16TM/ человека на трансфектанты облегчается за счет котрансфекции с мышиным /Kurosaki et al., Nature, 342:805 - 807 /1989// или
человеческим /Hibbs et al., Science, 246: 1608 - 1611 /1989/, а также с человеческим /Lauier et al., Nature, 342:803 - 805 /1989//.
В согласии с этими сообщениями экспрессия химерами также обеспечивает поверхностную экспрессию /CD16TM/, если подается на мишеневые клетки либо за счет котрансфекции или за счет коинфекции рекомбинантными вирусами вакцины /фиг. 2/. Промотирование /CD16TM/ поверхностной экспрессии за счет Z более выражено, нежели промотирование за счет γ /фиг. 2/ в исследованных клеточных линиях, тогда как нативные CD4 /данные не приведены/ не превышают CD16TM поверхностной экспрессии.
Пример IV
Asp Z мутанты не ассоциируются совместно с Fc рецептором
Для создания химер, которые
не ассоциировались бы с существующими антигенами или Fc рецепторами, мутантный Z слитый протеин, который не содержит ни внутримембранного Asp, ни внутримембранного Cys остатка, был получен. Цитометрия
в потоке показывает, что интенсивность экспрессии клеточной поверхности за счет различных мутантных химер заметно не отличается от не мутантов-предшественников /данные не приведены/, а эксперименты по
иммуноосаждению показали, что полная экспрессия за счет химер была такой же /фиг. 3/. Как и ожидалось, мутантные химеры, не содержащие трансмембранного остатка цистеина, как было обнаружено, не
образуют димеров с дисульфидной связью /фиг. 3/. Две мутантные химеры, не содержащие Asp, были неспособны поддержать поверхностную экспрессию CD16, тогда как мономерные химеры, не содержащие Cys, но
несущие Asp, обеспечили совместную экспрессию CD16TM, но с более низкой эффективностью, нежели родительский димер /фиг. 3/.
Пример V
Мутантные рецепторы сохраняют
способность инициировать реакцию кальция
Для того чтобы определить, позволит ли сшивка слитых протеинов обеспечить аккумуляцию внутриклеточного кальция таким же образом, который известен для
T клеточных антигенных рецепторов, клетки линии T клеточной лейкемии человека, Jurkat E6 3ATCC Access or Number TIB 152, American Tyрe Culture Collection, Rockviell, MD/, инфицировали рекомбинантами
вакцины, и измеряли относительную концентрацию кальция в цитоплазме после сшивки внеклеточного домена с антителами. Осуществляли цитометрические измерения в потоке на клетках, с введенным
чувствительным к кальцию красителем lnoo-1 /Grynkievich et al., J. Biol. Chem. 260:3340-3450 /1985/; Rabinovitch et al., J. Immunol, 137: 952 - 961 /1986/ /. На фиг. 4 представлены результаты по
кальцию в экспериментах в потоке с клетками, инфицированными CD4:Z и Asp- и Gys- мутантами Z.
Сшивка химер воспроизводимо повышает внутриклеточный кальций. CD4: η и CD4: γ аналогичным образом позволяют аккумулировать внутриклеточный кальций в инфицированных клетках /данные не приведены/, Jurkat клетки экспрессируют низкие уровни CD4 на поверхности клеток, однако, сшивка нативных CD4 в присутствии или без CD16: Z /C. R. и B.S., не опубликовано/ /фиг. 4 и не приведенные данные/ не влияет на уровни содержания внутриклеточного кальция.
Пример VI
CD4: Z, η и γ химеры - посредники цитолиза мишеней, экспрессирующих HIV gp120/41
Для определения того факта, будут ли химерические
рецепторы включать цитолитическую эффекторную программу, была создана модель мишень:эффекторная система на основе CD4 распознавания комплекса HIV оболочка gp120/gp41. HeLa клетки были инфицированы
рекомбинантными вирусами вакцины, экспрессирующими gp120/gp41 /Chakrobarti et al., Nature 320:535 - 537/1986/; Earl et al., J. Virol, 64:2449 - 2451 /1990/ и помечены51Cr. Меченые клетки
инкубировали с клетками человеческой аллоспецифической /CD8+, CD4-/ клеточной линии цитотоксических T лимфоцитов, которые были инфицированы рекомбинантами вакцины,
экспрессирующими CD4: Z, CD4: η , или CD4: γ химеры, или CD4:Z Gys11Gly: Asp15Gly двойными мутантными химерами. На фиг. 5 показано, что HeLa клетки, экспрессирующие gp120/41, были
специфически разрушены цитотоксичными T лимфоцитами /CTL/, экспрессирующими CD4 химеры. Неинфицированные HeLa клетки не были поражены CTL, снабженными CD4:Z химерами, а HeLa клетки, экспрессирующие
gp120/41, не были распознаны неинфицированными CTL. Для сравнения эффективности различных химер внесли поправку в отношения эффектор/мишень на долю CTL, экспрессирующих CD4 химеры, и на долю HeLa
клеток, экспрессирующих gp120/41 по данным цитометрии в потоке. На фиг. 5C показаны данные цитометрического анализа CD4 экспрессии CTL, использованных в цитолитических экспериментах, представлены на
фиг. 4A и 4B. Хотя средняя плотность поверхностных CD4: Z значительно превышает среднюю плотность CD4: η , цитолитическая эффективность клеток, экспрессирующих любую из форм, была одинакова.
Поправки на долю мишеней, экспрессирующих gp120, эффективность цитолиза посредством CD4: Z и CD4: η протеинов по сравнению с наивысшей эффективностью, известной для специфических T клеточных
рецепторных пар мишень : эффектор /среднее отношение эффектор: мишень для 50% высвобождения T клетками, экспрессирующими CD4: Z, составило 1,9 ± 0,99, н=10/. CD4: γ слияния были менее
активны по сравнению с CD4:Z слияниями, в которых отсутствовали трансмембранные Asp и Cys остатки. Однако, в обоих случаях наблюдался значительный цитолиз /фиг. 5/.
Для того чтобы проконтролировать возможность того, что заражение вакциной может промотировать артефактуальное распознавание за счет CTL, были проведены аналогичные эксперименты по цитолизу с мишеневыми клетками, инфицированными рекомбинантами вакцины, экспрессирующими фосфатидилинозитольную связанную форму CD16 /CD16PI/ и меченые51Cr, и с CTL инфицированными контрольными рекомбинантами, экспрессирующими либо CD16PI, либо CD16:Z. Фиг. 16 показывает, что T клетки, экспрессирующие не CD4 химеры, не распознают нативные HeLa клетки или HeLa клетки, экспрессирующие gp120/41, и аналогично, что T клетки, экспрессирующие CD4 химеры, не распознают HeLa клетки, экспрессирующие другие закодированные вакциной поверхностные протеины. Кроме того, CTL, экспрессирующие не химерические CD4, заметно не разрушают HeLa клетки, экспрессирующие gp120/41 /фиг. 6A/.
Пример VII
Клетки, содержащие МНС, класс II, не поражаются химерами
Известно, что CD4
взаимодействует с неполиморфной последовательностью, экспрессируемой антигеном МНС класса II /Gay et al., Nature, 328:626 - 629 /1987/, Sfeckman et al., Nature, 328:351 - 353 /1987//. Хотя
специфическое взаимодействие между CD4 и антигеном класса II никогда не было подтверждено на очищенных протеинах, в некоторых условиях адгезия между клетками, экспрессирующими CD4, и клетками,
экспрессирующими молекулы класса II, может быть продемонстрирована /Doyle et al., Nature 330:256 - 259 /1987/; Claybou et al., J. Exp. Med. 172-1243 - 1253 /1990/; Lamarre et al. Science 245:743 - 746
/1989//. Следующими исследованиями были исследования вопроса, можно ли детектировать убийство клеток, несущих класс II. На фиг. 6B показано, что нет специфического цитолиза направляемого CD4: η
против Raji B клеточной линии, которая экспрессирует обильно антиген класса II. Хотя наблюдается умеренный /около 5%/ цитолиз, негативный мутант класса II Raji, RJ 2.2.5 /Accolla, P. S. J. Exp. Med.
157:1053 - 1058 /1983/ доказывает аналогичную подверженность, что и Raji клетки, инкубированные с неинфицированными T клетками.
Пример VIII
Требования последовательности для
индукции цитолиза T клеточным антигеном/Fc рецептором зета цепи
Хотя химеры между CD4 и Z можно снабдить цитотоксичными T лимфоцитами /CTL/ для убийства мишеневых клеток, экспрессирующих HIV
gp120, ищут альтернативу CD4 для однозначного сравнения свойств зета химер, введенных в T клеточные линии человека. Такие линии могут экспрессировать CD4, затрудняя специфическое определение связи
между
типом или степенью мобилизации кальция и цитотоксическим потенциалом различных химер. Для осуществления этого, создают химеры между Z и CD16, в которых внеклеточный домен CD16
присоединен к трансмембранной и внутриклеточной последовательности Z /фиг. 7A/. Слитые гены вводят в вирус вакцины, экспрессирующий плазмиду, несущую ген E.coli gpt в качестве селектируемого маркера,
и включают в геном вакцины штамма WR путем гомологической рекомбинации и селекции по росту в микофенолевой кислоте /Falkner and Moss, J. Virol, 62:1849 /1988/; Boyle and Coupar, Cene 65:123
/1988//.
T клеточную линию инфицируют рекомбинантами вакцины, и определяют относительную концентрацию цитоплазмически свободных ионов кальция с последующей сшивкой внеклеточных доменов с антителами. Были проведены как спектрофотометрические /объем популяции/, так и цитометрические в потоке /одиночное клетки/ измерения с клетками, меченными красителем Indo-1 (Grynkiewicz et al. , J. Biol. Chem. 260:3440 /1985/; Rabinovitch et al., J. Immunol 37:952 /1986//. На фиг. 7B представлены результаты анализа данных, полученных для клеток линии Jurkat Т клеток лейкемии человека, инфицированными рекомбинантами вакцины, экспрессирующими CD16: η протеины слияния. Сшивка химер воспроизводимо повышает содержание внутриклеточного кальция, хотя аналогичная обработка клеток, экспрессирующих нехимерические CD16, не оказывает влияния, или оно мало. Если химера была экспрессирована в мутантную клеточную линию без рецептора антигена, либо PEX33A /Breitmeyer et al., J. Immunol 138:726 /1987/; Sauco et al., J. Biol. Chem. 264:20760 /1989/ либо Jurkat мутанта JRT3, T3.5 /Wess et al., J. Immunol. 135:123 /1984/; наблюдается сильная реакция на сшивку CD16 антител. Аналогичные данные были получены на REX20A /Breitmeyer et al., supra., 1987; Blumbery et al., J. Biol. Chem. 265:14036 /1990// мутантной клеточной линии и на CD3/Ti негативном мутанте Jurkat клеточной линии, установленной в этой лаборатории /данные не представлены/. Заражение рекомбинантами, экспрессирующими CD16:Z, не сохраняет реакцию на анти-CD3 антитела, что показывает, что слитый протеин не действует, освобождая внутриклеточные CD3 комплексные цепи /данные не приведены/.
Для оценки способности химер изменять направление иммунитета через клетки-медиаторы, CTL были инфицированы рекомбинантами вакцины, экспрессирующими CD16 химеры, и использованы для специфически разрушенных гибридомных клеток, экспрессирущих связанные с мембраной анти-CD16 антитела /см. далее/. Этот анализ является продолжением анализа цитотоксичности гибридомы, вначале разработанный для анализа эффекторного механизма клеток, несущих Fc рецепторы /Craziano and Fanger, J. Immunol, 138:945, /1987/; Graziano and Fanger, J. Immunol, 139: 35 - 36 /1987/ Shem et al., Mol. Immunol 26:959 /1989/; Fauger et al. , Immunol Today 10:92 /1989/. На фиг. 8B показано, что экспрессия CD16:Z в цитотоксические T лимфоциты позволяет снабдить CTL для убийства 3G8 /анти-CD16, Fleit et al. , Proc. Natl. Acad. Sci. USA, 79:3275 /1982/ гибридомных клеток, тогда как CTL, экспрессирующие связанную фоcфатидилинозитолом форму CD16, являются неактивными. CTL, снабженная CD16:Z, также не убивает гибридомные клетки, экспрессирующие не интересующие антитела /данные не приведены/.
Для идентификации минимальной Z последовательности, необходимой для цитолиза, приготовили серии делеционных мутантов, в которых последовательно была удалена большая часть Z внутриклеточного домена с карбоксильного конца /фиг. 8A/. Большую часть внутриклеточного домена затем можно удалить с незначительными последствиями для цитолитического потенциала, химера полной длины CD16: Z была практически равна по эффективности химере с делецией до остатка 65, CD16:Z Asp66* /фиг. 8B/. Существенное снижение цитотоксичности наблюдали при делеции до Z остатка 59 /химера CD16:Z Glu60* /, а дальнейшая делеция до остатка 50 приводит к несколько меньшей активности. Однако, полной потери активности не наблюдалось, даже когда внутриклеточный домен был сокращен до трех остатков трансмембранного якоря /фиг. 8B/.
Так как Z является дисульфидно связанным димером, единственным объяснением сохранения цитолитической активности оказался тот факт, что эндогенный Z формировал гетеродимер с химерической Z делецией, за счет чего восстановил активность. Для проверки этой идеи, Z остатки 11 и 15 заменили в Asp и Gys соответственно на Gly/Gys11Gly/Asp15Gly/ и иммуноосаждение провели следующим образом. Приблизительно 2 • 106 CVI клеток инфицировали в течение одного часа в DME среде, не содержащей сыворотки, рекомбинантной вакциной при коэффициенте инфекции /moi/, по крайней мере 10. Спустя 6-8 часов после инфицирования, клетки смыли с пластин PBS/1 мМ EDTA и поверхностно пометили 0,2 мк Кюри125I на 2 • 106 клеток, используя лактопероксидазу и H2O2 по способу Clark and Einfeld /Zeukocyte Typing 11 pp 155-167, Springer-Verlag, NY, 1986/. Меченые клетки собрали центрифугированием и подвергли лизису в 1% NP-40, 0,1% SDS, 0,15 М NaCl, 0,05 М Трис, pH 8,0, 5 мМ MgCl2, 5 мM KCl, 0,2 М иодацетамида и 1 мМ PMSF. Ядра удалили центрифугированием, а CD16 протеины иммуноосадили антителами 3G8 /Fleit, Sypra 1982, Medarex/ и анти-мышиной IgG агарозой /Cappel, Durham, NC/. Образцы обрабатывали электрофорезом через 8% полиакриламид/SDS гель в невоccтановительных условиях, или через 10% гель в восстановительных условиях. Эти иммуноосаждения подтвердили, что CD16:Z Cys11-Gly/Asp15Gly химера не образует димерные структуры с дисульфидной связью.
Была проверена также цитолитическая активность мутантных рецепторов. Провели делецию мутантной химеры до остатка 65 /CD15:Z Gys11Gly/Asp66*/, и она оказалась в зависимости от условий анализа в 2 - 8 раз менее активной в цитолитическом анализе, нежели сравнимая не подвергшаяся мутации химера /CD16: Z Asp66*/, которая имеет обычно коэффициент 2 или неразличима в активности от CD16:Z /фиг. 9B/. Снижение активности у мутантных химер сравнимо со снижением, которое наблюдают для CD4 химер аналогичной структуры /см. выше /, и наиболее вероятно связано с более низкой эффективностью Z мономеров по сравнению c димерами. Напротив Asp-, Cys- подвергшиеся мутации химеры, усеченные до остатка 59, не обладают цитолитической активностью /фиг. 9B/, что подтверждает гипотезу о том, что ассоциация с другими цепями, осуществляемая трансмембранным Cys и/или Asp остатками, ответственна за слабое проявление цитолитической активности в делеции скорее аминотерминала, нежели остатка 65.
Цитометрические исследования в потоке показывают, что делеционные мутанты, не содержащие трансмембранных Asp и Gys остатков, все еще могут промотировать повышение содержания ионов свободного внутриклеточного кальция в ответ на антитела, сшивающие в TCR- мутант Jurkat клеточную линию /фиг. 9D/. Аналогичные результаты были получены для химер, экспрессированных в родительскую Jurkat клеточную линию /не показано/. В случае CD16:Z Gys11Gly/Asp15Gly/Glu60* эти данные демонстрируют то, что способность передавать отвечаемость кальция можно мутационно отделить от способности поддерживать цитолиз.
Для определенного исключения возможного вклада Z трансмембранных остатков трансмембрану и первые 17 цитоплазмических, остатков Z заменили последовательностью, кодирующей мембранный спанинг, и первыми 14 или первыми 17 цитоплазмическими остатками CD5 или CD7 антигена, соответственно /фиг. 9A/. Полученные слитые из трех частей протеины CD16:5:Z /48 - 65/ и CD16:7:Z /48 - 65/ не образуют димеров с дисульфидной связью как более простые CD16:Z химеры, так как у них нет цистеинового остатка в Z трансмембранном домене. Обе состоящие из трех частей химеры были способны мобилизовать кальций в Jurkat и TCR негативных клеточных линиях /фиг. 9D/ и увеличить цитолитическую реакцию в CTL /фиг. 9C и не представленные данные/. Однако, усечение Z части до остатка 59 в химере CD16:7:Z /48 - 59/ аннулирует способность тройного слияния управлять отвечаемостью кальция в TCR позитивных или негативных Jurkat клетках или цитолизом в зрелых CTL /фиг. 9C и 9D и не представленные данные/.
Для определения вклада отдельных остатков в 18-остаточный фрагмент, приготовили ряд мутантных вариантов за счет направленного в сайты мутагенеза, и оценили их способность посредничать в направляемом рецепторами киллинге в условиях низкой /фиг. 10A и 10D /или высокой /фиг. 10B и 10E/ экспрессии химерических рецепторов. Фиг. 10 показывает, что хотя ряд относительно консервативных замещений /то есть замена кислотных остатков их родственными амидами, или тирозина фенилаланином/, которые соединяют остатки 59 - 63, приводит к умеренному компромиссу цитолитической эффективности, обычно варианты сохраняют способность к мобилизации кальция. Однако, коллективно, эти остатки содержат важный субфрагмент, постольку поскольку их делеция исключает цитолитическую активность. Превращение Tyr62 либо в Phe, либо в Ser исключает как цитотоксичность, так и кальциевую реакцию. На аминокислотном конце сегмента из 18 остатков замена Tyr51 на Phe упраздняет как мобилизацию кальция, так и цитолитическую активность, тогда как замена Leu на Ser в положении 50 исключает реакцию кальция, но лишь частично влияет на цитолиз. Не связывая себя с определенной гипотезой, можно предположить, что неспособность Leu50Ser мутанта мобилизовывать кальций в кратковременном цитометрическом анализе в потоке, полностью не отражает его способность вызывать существенное повышение количества ионов свободного внутриклеточного кальция в более длительном цитолитическом анализе. Однако, как сообщалось, не чувствительная к кальцию цитолитическая активность для некоторых цитолитических клеточных линий и возможность того, что аналогичное явление подчеркивает описанные здесь результаты, не следует отбрасывать. Замена Asn48 на Ser частично придает цитотоксичность в некоторых экспериментах, хотя и оказывает мало влияния в других.
Для определения возможной роли излишней последовательности элементов, внутриклеточный домен Z разделяли на три сегмента, соединяя остатки 33 - 65, 71 до 104 и 104 до 138. Каждый из этих сегментов был присоединен к CD16:CD7 химере с помощью Mlu1 сайта, введенного как раз на периферию основной мембранной якорной последовательности внутриклеточного домена CD7 /см. далее, фиг. 11A/. Сравнение цитолитической эффективности трех элементов показало, что они практически обладают одинаковой эффективностью /фиг. 11B/. Сравнение последовательностей /фиг. 11A/ показало, что второй фрагмент несет 11 остатков между тирозинами, тогда как первый и третий содержат только десять.
Хотя точный учет процесса T клеточной активации и не был проделан, очевидно, что аггрегация антигенного рецептора и рецептора химер, которые содержат Z внутриклеточную последовательность, включает мобилизацию кальция, высвобождение цитокина и гранул и появление маркеров активации клеточной поверхности. Активный сайт Z, короткая линейная пептидная последовательность, вероятно, слишком мала, чтобы обладать собственной энзимативной активностью, по-видимому взаимодействует с одним или более из нескольких протеинов для создания клеточной активации. Ясно также, что одной только мобилизации свободного кальция недостаточно для клеточной активации, так как способность вызывать цитолиз можно мутационным способом отделить от способности вызывать накопление кальция.
Как было показано, добавление 18 остатков из внутриклеточного домена Z к трансмембранному и внутриклеточному доменам двух неродственных протеинов позволяет полученным химерам изменять направление цитолитической активности против мишеневых клеток, которые связаны с внеклеточной частью слитых протеинов. Хотя химеры, содержащие фрагмент из 18 остатков, приблизительно в восемь раз менее активны, нежели химеры, основанные на полной длине Z, пониженную активность можно приписать потере трансмембранного взаимодействия, которое обычно позволяет дикому типу Z образовывать димеры с дисульфидной связью. То есть, Z делеционные конструкции, которые имеют те же самые карбоксильные концы, что и фрагмент, и не имеют трансмембранных Gys и Asp остатков, обычно демонстрируют несколько меньшую активность, нежели химеры, содержащие только фрагмент из 18 остатков.
Элемент цитолитической компетентности, на котором мы сосредоточили внимание, имеет два тирозина и не содержит серина или треонина, что ограничивает возможный вклад в активность фосфорилирования. Мутация любого тирозина нарушает активность, однако, хотя предварительные эксперименты и не указали на существенное тирозиновое фосфорилирование с последующей сшивкой химерических поверхностных антигенов, содержащих фрагмент из 18 остатков; возможное участие такого фосфорилирования при низких уровнях нельзя исключить. Кроме указанных эффектов, отмеченных для двух тирозиновых остатков, ряд замен аминокислот у аминокислотного и карбоксильного концов фрагмента ослабляет активность в условиях низкой плотности рецепторов.
Последовательности, аналогичные Z активному фрагменту, можно найти в цитоплазмических доменах некоторых других трансмембранных протеинов, включая CD3 δ и γ молекулы, поверхностные IgM связанные протеины mbl и B29, а также β и γ цепи высокой афинности IgE рецепторы, Fc ε R1 /Ret, Nature, 338:383, 1989/. Хотя функции этих последовательностей не определены, если они эффективно экспрессированы, каждая может быть способна к автономной T клеточной активации, и также активности можно объяснить остаточной TCR отвечаемостью, которую можно наблюдать в зета-негативной мутантной клеточной линии /Sucsman et al., Cell, 52:85, 1988/.
Сам Z содержит три последовательности, приблизительно равной величины, и примерно три секции внутриклеточного домена показывают, что каждый способен инициировать цитолитическую реакцию Z, сплайс изоформа Z /Jin et al., supra, 1990; Clayton et al., Proc. Natl. Acad. Sci. USA 88:5202, 1991/ не содержит карбоксильной половины третьего фрагмента. Так как удаление карбоксильной половины первого фрагмента уничтожает активность, можно предположить, что основная биологическая эффективность Z может быть приписана первым двум фрагментам. Хотя при различных измерениях Z столь же активно как Z в промотировании осуществляемого антигеном высвобождения цитокина /Bauer et al., Proc. Natl, Acad. Sсi. USA, 88:384, 1991/ или в изменении направления цитолиза /см, выше/, Z не фосфорилируется в ответ на рецепторную стимуляцию /Bauer et al. , Supra, 1991/. Таким образом, либо наличие всех трех фрагментов необходимо для фосфорилирования, либо третий фрагмент представляет благоприятный субстрат для неидентифицированной тирозинкиназы.
Пример IX
Трансдукция цитолитического сигнала Fc рецептором человека
Для оценки действий различных человеческих Fc
рецепторных субтипов, были созданы химерические молекулы, в которых внеклеточный домен антигенов человека CD4, CD5 или CD16 был соединен с трансмембранными и внутриклеточными доменами FcR11 γ A,
B1, B2 и C субтипов /номенклатура Ravetchand kinet, Ann, Rev, Immunol. 9: 457, 1991/. Специфически, кДНК последовательности, соответствующие трансмембранным и цитоплазмическим доменам ранее описанных
FcR11A, B1 и B2 изоформ, были амплифицированы из предшественника клона PC23 или из библиотеки кДНК миндалин человека /сконструированной по стандартным методикам/, используя олигонуклеотидные
праймеры.
CCC GGA TCC CAG CAT GGG CAG CTC TT (SEQ ID N0: 18; FcRIIA forward)
CGC GGG GCG GCC GCT TTA GTT ATT ACT GTT GAC ATG GTC GTT (SEQ ID N0: 19; FcR11A reverse);
GCG GGG GGA TCC CAC TGT CCA AGC TCC CAG CTC TTC ACC G (SEQ ID N0: 20; FcR11B1 and FcR11B2 forward); and
GCG GGG GCG GCC GCC TAA ATA CGG TTC TGG TC (SEQ ID N0: 21; FcR11B1 and FcR11B2
reverse).
TCA GAA AGA GAC AAC CTG AAG AAA CCA ACA A (SEQ ID N0: 22) and
TTG TTG GTT TCT TCA GGT TGT GTC TTT CTG A (SEQ ID N0: 23).
Эти праймеры содержат
сайты расщепления для энзимов BamHI и Notl, соответственно, отступив на 6 остатков от 5' конца. Сайт Notl следует сразу за противосмысловым стоп кодоном либо CTA, либо TTA. Все праймеры содержат 18
или более остатков, комплементарных 5' концу и 3' концу целевого фрагмента. кДНК фрагмент, соответствующий FcR11 γ C цитоплазмическому домену, который отличается от 11A изоформы только одним
аминокислотным остатком /Z для P у остатка 268/, создается за счет управляемого сайтом мутагенеза за счет перекрывания PGR, используя праймерные последовательности:
TCA GAA AGA GAC AAC CTG
AAG AAA CCA ACA A /SEQ ID N0: 22/ и
TTG TTG GTT TCT TCA GGT TGT GTC TTT CTG A /SEQ ID N0: 23/.
PCR фрагменты были включены в векторы экспрессии вируса вакцины, которые содержали CD16 или CD4 внеклеточные домены, соответственно, а затем были включены в дикого типа вакцину за счет рекомбинации по тимидинкиназному локусу, с использованием селекции по коинтеграции E.coli gpt для облегчения идентификации целевых рекомбинант. Идентификация всех изоформ /представлены на фиг. 12/ была подтверждена дидеоксисеквенированием.
Получение химерических рецепторных протеинов было далее подтверждено исследованиями иммуноосаждения. Приблизительно 107 JRT3.T3.5 клеток было инфицировано в течение одного часа в среде IMDM не содержащей сыворотки, рекомбинантной вакциной при умножаемости инфекции, по крайней мере, 10. Спустя 12 часов после заражения, клетки собрали, и поверхности пометили 0,5 мКюри125I на 107 клеток, используя способ лектопероксидаза /глюкозооксидаза /Clark and Einfeld, supra/. Меченые клетки собрали центрифугированием и лизировали 1% NP-40, 0,1 мM MgCl2, 5 мМ KCl, 0,2 М иодоацетамида и 1 мМ PMSF. Ядра удалили центрифугированием, а CD16 слитые протеины иммуноосадили антителами 4G8 и анти-мышиной IgG агарозой. Образцы обработали электрофоретически при мягких условиях. Все иммуноосажденные химерические рецепторные молекулы были ожидаемой молекулярной массы.
Для проверки способности химерических рецепторов осуществлять повышение содержания цитоплазмических свободных ионов кальция, использовали рекомбинантные вирусы для заражения TCR- мутантной Jurkat клеточной линии JRT3.T3-5 /как показано далее/, и содержание цитоплазмического свободного иона кальция определяли в клетках /как здесь описано/ с последующей сшивкой рецепторных внеклеточных доменов с моноклональными антителами 3G8 или Leu-3A /как здесь описано/.
Эти эксперименты показали, что внутриклеточные домены FcR γ 11A и C были способны осуществлять повышение содержания цитоплазмических свободных ионов кальция после сшивки внеклеточных доменов, тогда как внутриклеточные домены FcR γ 11B1 и B2 были неактивны в аналогичных условиях /фиг. 13A и 13B/. CD4, CD5 и CD16 гибриды FcR γ 11A демонстрируют практически равную способность промотировать реакцию кальция /фиг. 13 и не представление данные/. Другие клеточные линии как моноцитного так и лимфоцитного происхождения, оказались способны реагировать на сигналы, инициируемые сшивкой внеклеточных доменов /данные не представлены/.
Для изучения вовлечения различных FcR γ 11 внутриклеточных доменов в цитолиз, цитотоксические T лимфоциты человека /CTL/ были инфицированы рекомбинантами вакцины, экспрессирующими CD16:FcR γ 11A, B1, B2 и C химеры. Инфицированные клетки затем совместно культивировали с гибридомными клетками с 51Cr /то есть 3G8 10 - 2 клетки/, которые экспрессировали клеточную поверхность антител к CD16. В этом анализе CTL содержащие CD16 химеры, убитые гибридомными мишеневыми клетками /что позволило высвободиться51Cr/, если CD16 внеклеточный домен химеры был соединен с внутриклеточным сегментом, способным активировать лимфоцитную эффекторную программу; этот цитолиз-анализ подробно описан далее. Фиг. 14A показывает, что CTL снабженная CD16:FcR γ 11A и C, но не FcR γ 11 B1 или B2, способны к лизису мишеневых клеток, экспрессирующих клеточную поверхность анти-CD16-антитела.
Для исключения возможности того, чтобы специфический цитолиз каким-либо образом был связан с взаимодействием с CD16 фрагментом, проводили эксперименты по цитолизу, в которых FcR11 внутриклеточные домены были присоединены к CD4 внеклеточным доменам. В этом случае мишеневыми клетками были HeLa клетки, экспрессирующие протеины gp120/41 оболочки HIV /специфично, клетки HeLa, инфицированные вектором VPE16 вакцины /доступны от National Institute of Aplergy and Infection Disease AIDS Depository, Bethesda, MD/. Как и в CD16 системе, мишеневые клетки, экспрессирующие оболочку HIV, подвергали лизису T клетками, экспрессирующими CD4:FcR γ 11A химеры, но не FcR γ 11B1 или B2 /фиг. 14B/.
Внутриклеточные домены FcR γ 11A и C не образовывали никаких подходящих последовательностей, гомологичных с любым другим протеином, включая члены расширенного семейства FcR γ /TCRZ. Для определения последовательности элементов, ответственных за возбуждение цитолиза, 5' и 3' делеции последовательностей, кодирующих внутриклеточный домен /описанные далее и представленные на фиг. 15A/, получили и оценили на эффективность мобилизации кальция и в цитолитическом анализе /как здесь описано/. В тех экспериментах, в которых аминотерминальная часть внутриклеточного домена была удалена, трансмембранный домен FcR γ 11 был заменен трансмембранным доменом неродственного CD7 антигена для исключения возможного вклада взаимодействий за счет связанного с мембраной домена.
На фиг. 15B и 15C показано удаление 14 карбокси-терминальных остатков, включая тирозин 298, что привело к полной потере цитолитической способности и существенному снижению потенциала мобилизации кальция. Дальнейшая делеция непосредственно тирозином 282 дает идентичный фенотип /фиг. 15B и 15C/. Делеция с N-конца внутриклеточного домена до остатка 268 не оказывает существенного действия ни на кальциевый профиль, ни на цитолитическую способность, тогда как делеция до остатка 275 заметно увеличивает высвобождение кальция, но оказывает слабое воздействие на цитолиз /фиг. 15D и 15E/. Дальнейшая делеция до остатка 282 дает FcR γ 11 хвосты, которые не обладают способностью ни к мобилизации кальция, ни к включению цитолиза /фиг. 15D и 15E/. "Активный элемент", определенный этими грубыми измерениями, относительно велик /36 аминокислот/ и содержит два тирозина, разделенные 16 остатками.
Пример X
Другие внутриклеточные и
трансмембранные сигнальные домены трансдукции по способу настоящего изобретения можно получить из протеинов T клеточных рецепторов, CD3 дельта и T3 гамма, и протеинов B клеточных рецепторов, mb1 и
B29.
Аминокислотные последовательности этих протеинов представлены на фиг. 16 /CD3 дельта, SEQ ID N0:24/, фиг. 17 /T3 гамма, SEQ ID N0:25/, фиг. 18 /mb1, SEQ ID N0: 26/ и фиг. 19 /B29, SEQ ID N0: 27/. Участки последовательностей, достаточные для трансдукции цитолитического сигнала /и поэтому, предпочтительно, включенные в химерический рецептор настоящего изобретения/, представлены в рамках. Химерические рецепторы, которые включают эти протеиновые домены, сконструированы и использованы в способе лечения настоящего изобретения, как описано ранее.
Пример XI
Экспериментальные методы
Инфицирование вакциной и радиоиммуноосаждение
Приблизительно 5 • 106 CVI клеток инфицировали в течение часа в DME среде не содержащей
сыворотки, с рекомбинантной вакциной с умножением инфекции /moi/, по крайней мере, 10 /титр, измеренный на CVI клетках/. Клетки помещают в свежую среду после того, как они инфицированы и метаболически
мечены 200 мк Кюри/мл35S-метионин плюс цистеином /Тган35S-метка, ICN, Costo Mesa CA/ в метионине, и не содержащем цистеина DMEM /Gibco, Grand Island, NY/ в течение 6 часов.
Меченые клетки отделили PBS, содержащим 1 мМ EDTA, собрали центрифугированием и провели лизис в 1% NP-40, 0,1% SDS, 0,15 М NaCl, 0,05 М Tris pH 8.0, 5 мМ EDTA и 1 мМ PMSF. Ядра удалили
центрифугированием, а CD4 протеины иммуноосадили OKT4 антителами и анти-мышь IgG агарозой /Cappel, Durham, NC/. Образцы обработали электрофоретически на 8% полиакриламид /SDS гелях в
невосстанавливающих /NR/ и в восстанавливающих /R/ условиях. Гели, содержащие35S-меченые образцы, были пропитаны En3 Hance /New England Nuclear, Boston, MA/ перед
авторадиографией. Облегченная экспрессия трансмембранной формы CD16, CD16TM, была измерена по сравнению с ее экспрессией в CVI клетки, просто инфицированные CD16TM, с экспрессией
в клетки коинфицированных вирусами, кодирующими CD16TM и Z или γ химеры. После инфицирования и инкубирования в течение 6 часов или более, клетки удалили с пластин PBS, 1 мМ EDTA и
определили экспрессию CD16TM или зимер по данным косвенной иммунофлуоресценции и цитометрии в потоке.
FI х анализ кальция
Клетки сублинии Jurkat E6 /Weiss et al.,
J. Immunol, 133:123-128 /1984// инфицировали рекомбинантными вирусами вакцины в течение одного часа в не содержащей сыворотки среде IMDM при moi = 10, и инкубировали в течение 3-9 часов в IMDM, 10%
FBS. Клетки собирали центрифугированием и повторно суспендировали при 3 • 106 клеток/мл в полной среде, содержащей 1 мМ Indo-1 ацетометоксиэфира /Grynkiewier et al., J. Biol. Chem.
260:3440-3450 /1985/ /Molecular Probes/, и инкубировали при 37oC в течение 45 минут. Клетки с введенным Indo-I таблетировали и повторно суспендировали при 1 • 106 мл в не
содержащей сыворотки IMDM, и хранили при комнатной температуре в темноте. Клетки анализировали на содержание свободных ионов кальция при одновременном измерении фиолетовой с синей флюоресценции с
помощью цитометрии в потоке /Rabinovitch et al. , J. Immunol, 137:952 - 961 /1986//. Для инициирования истечения кальция либо фикоэритрин /PE/-конъюгированный Leu-3A /анти-CD4/ /Beston Dickinson,
Lincoln Park NY/ при 1 мкг/мл добавляли к клеточной суспензии, с последующим добавлением 10 мкг/мл неконъюгированного козлиного анти-мышиного IgG во время 0, или неконъюгированные 3G8
/анти-CD16/ моноклональные антитела добавляли к клеточной суспензии при 1 мкг/мл с последующим добавлением 10 мкг/мл PE-конъюгированного Fab2 козлиного анти-мышиного IgG в момент времени 0.
Гистограммы отношения фиолетовой/синей эмиссии снимали для популяции РЕ позитивных /инфицированных/ клеток, которые обычно составляли 40 - 80% от всех клеток. Реакцию рецептора T клеточного антигена в
неинфицированных клетках включали антителами OKTЗ, без сшивки. Для экспериментов, включавших CD16 химерические рецепторы, образцы, у которых был дрейф базовой линии относительно низшего
внутриклеточного кальция /без антител/, исключали из анализа. Затем данные гистограммы анализировали, превращая бинарные данные в ASCII, используя программы Writl Hand Man /Cooper City, FL/, с
последующим анализом в коллекции программ FORTRAN. Отношение эмиссий фиолетовой/синей перед добавлением реагентов второго антитела использовали для установления нормального начального отношения,
приравнивали его к единице, а остальное пороговое отношение устанавливали таким образом, чтобы 10% оставшейся популяции превышало бы порог.
Анализ цитолиза
T Клеточную линию
WH3 человека, CD8+, CD4- HLA B44 рестриктированную цитолитическую линию выдерживали в IMDM, 10% сыворотке человека со 100U/мл IL-2, и периодически стимулировали либо
неспецифически, облученными /1000 рад/ HLA несовместимыми лимфоцитами периферической крови и 1 мкг/мл фитогемагглютинина, либо специфически, облученными B44-содержащими моноядерными клетками. Спустя
один день неспецифической стимуляции, PHA разбавляют до 0,5 мкг/мл, добавляя свежую среду, и спустя три дня среду меняют. Клетки выращивают в течение, по крайней мере, 10 дней с последующей
стимуляцией перед использованием в анализе на цитотоксичность. Клетки инфицируют рекомбинантной вакциной при умножении инфекции, по крайней мере, 10, в течение одного часа в не содержащей сыворотки
среде, с последующим инкубированием в полной среде в течение трех часов. Клетки собирают центрифугированием и повторно суспендируют при плотности 1 • 107 клеток/мл. 100 мкл добавляют
в каждую ячейку микротитровальной пластины, содержащую 100 мкл/ячейку полной среды. Клетки разбавляют поэтапно кратно 2. По две ячейки для каждого образца не содержат лимфоцитов, чтобы обеспечить
спонтанное высвобождение хрома и определить полный расход хрома. Меченые клетки из сублинии HeLa S3 инфицируют в 6,0 или 10,0 см пластинах при приблизительно moi 10 в течение одного часа в среде, не
содержащей сыворотки, с последующим инкубированием в полной среде в течение трех часов. Их удаляют из чашек с помощью PBS, 1 мМ EDTA, и считают. Аликвоты в 10' мишеневых клеток /HeLa, Raji или RJ2.2.5
клетки для экспериментов с CD4 химерическими рецепторами и 3G8 10-2 клетки, Shen et al., Mol. Immunol, 26:959 /1989/ для CD16 химерических рецепторных экспериментов / центрифугируют и повторно
суспендируют в 50 мкл стерильного51Cr-натрийхромата /1 м Кюри/мл, Дюпон, Уилмингтон, DE/ в течение одного часа при 37oC с перемежающимся перемешиванием, затем промывают трижды
PBS. 100 мкл меченых клеток снова суспендируют в среде при 105 клеток/мл, которые добавляют в каждую ячейку. Мишеневые клетки Raji и RJ2.2.5 мечены также как и HeLa клетки.
Микротитровальные пластины вращают при 750g в течение 1 минуты и инкубируют в течение 4 часов при 37oC. К концу периода инкубирования клетки в каждой из ячеек снова ресуспендируют,
осторожно взбалтывая пипеткой, образцы отбирают для определения полного количества включений, а микротитровальные пластины снова вращают при 750g в течение 1 минуты. 100 мкл аликвоты надосадочной
жидкости отбирают и считают в сцинтилляционном счетчике гамма лучей. В процент киллинга вносят поправку на долю инфицированных мишеневых клеток /обычно 50 - 90%/ по данным цитометрии в потоке. Для не
инфицированных эффекторных клеток отношение эффектор:мишень было исправлено на процент клеток, инфицированных /обычно 20 - 50% для экспериментов с CD4 химерическими рецепторами, и более 70% для
экспериментов с CD16 химерическими рецепторами/.
In vitro мутагенез Z последовательности
Для создания точечных мутаций в аминокислотных остатках 11 и/или 15Z
последовательности, были получены синтетические олигонуклеотидные праймеры, простирающиеся от BamHI сайта в обратном направлении от Z трансмембранного домена, и превращающие нативный Z остаток 11 от
Cys в Gly /C11G/ или остаток 15 от Asp в Gly /D15G/ или оба /C11G /D15G/, и используют в PCR реакциях для создания мутированных фрагментов, которые снова вставлены в дикого типа CD4: Z конструкции.
Для создания Z делеций, Z ДНК последовательности амплифицируют PCR, используя синтетические олигонуклеотидные праймеры, сконструированные для создания стоп кодона /UAG/ после остатка 50, 59 или 65. Праймеры, содержащие сайты расщепления для энзима Notl, вырезают 5 или 6 остатков с 5' конца, обычно в последовательности CGC GGG CGG CCG CTA /SEQ ID N0: 11/, где последние три остатка соответствуют стоп антикодону. За последовательностями Notl и стоп антикодона следуют 18 или более остатков, комплементарных целевому 3' концу фрагмента. Полученные химеры обозначены CD16:Z V51*, CD16:Z E60* и CD16:Z D66*, соответственно. BamHI сайт в обратном направлении от трансмембранного домена и Notl сайт используют для создания фрагментов, которые заново вводят в дикого типа CD16:Z конструкцию. Мономерные Z химеры создают, высвобождая Z трансмембранную и мембранную проксимальную внутриклеточные последовательности перевариванием BamHI и Sacl Asp- и Cys- CD4:Z конструкцию, описанную ранее, и включая фрагмент в CD16:Z E60* и CD16:Z D66* конструкции соответственно.
CD16: 7:Z /48-65/ и CD16:7:Z /48-59/ химерические конструкции, состоящие из трех частей.
Для получения конструкции CD16:Z D66*, Z кДНК последовательность, соответствующую трансмембранному домену, и последующих остатков цитоплазмического домена заменяют соответствующими трансмембранным и цитоплазмическим доменом, полученными из CD5 и CD7 кДНК. Фрагменты CD5 и CD7 создают в PCR реакции, используя форвардные олигонуклеотиды, включающие BamHI рестрикционный сайт расщепления и соответствующие участку непосредственно перед трансмембранным доменом CD5 и CD7, соответственно, с последующим обратным олигонуклеотидным перекрыванием CD5 и CD7 последовательностей, соответственно, и Z последовательности, которая содержит Sacl рестрикционный сайт расщепления.
CD5: Z: CGC GGG
CTC GTT ATA GAG CTG GTT CTG GCG CTG CTT CTT CTG /SEQ ID N0: 12/
CD7: Z: CGC GGG GAG CTC GTT ATA GAG CTC GTT TGG CGC CGA ATT CTT ATC CCG /SEQ ID N0: 13/.
CD5 и CD7 PCR продукты были переварены BamHI и Sacl и лигированы с BamHI и Sacl переваренными CD16:Z E60*, и заменяют Z последовательность из BamHI до Sacl фрагментом CD7. Для создания конструкций CD16:CD5 и CD16:CD17, фрагменты CD5 и CD7 были получены с помощью PCR, используя олигонуклеотиды, содержащие Notl рестрикционный сайт расщепления и кодирующий стоп кодон /UAA/ после остатка Gln416 и Ala193 CD5 и CD7, соответственно, CD5 и CD7 PCR фрагменты были переварены BamHI и Notl и включены в CD16:Z Asp66* конструкцию.
In vitro мутагенез N-терминальных остатков с
Z цитолитическим сигнальным фрагментом трансдукции
Синтетические олигонуклеотидные праймеры, простирающиеся от Sacl сайта внутрь Z фрагмента и превращающие нативный остаток 48 из Asn в Ser
/48S/, остаток 50 из Leu в Ser /LS 50S/, и остаток 51 из Туг в Phe /V52F/ были синтезированы и использованы в PCR реакции для создания фрагментов, которые снова включили в дикого типа CD16:7:Z
/48-65/ конструкцию.
In vitro мутагенез C-терминальных остатков с Z цитолитическим сигнальным фрагментом трансдукции
Синтетические олигонуклеотидные праймеры, простирающиеся
от Notl сайта 3' до стоп кодона и превращающие нативный остаток 60 из Glu в Gln /E60Q/, остаток 61 от Glu в Gln /E61Q/, остаток 62 из Tyr в Phe /V62F или V62S/, и остаток 63 из Asp в An /D63N/ были
синтезированы и использованы для создания фрагментов, которые были субклонированы в дикого типа CD16:Z D66* конструкцию от BamHI сайта до Notl сайта.
CD16: 7: Z /33-65/,
CD16:7:Z /71-104/, CD16:7:Z /104-137/ химерические конструкции
CD7 трансмембранный фрагмент, содержащий Mlul и Notl сайты по соединению между трансмембранным и внутриклеточным доменами, был
получен в результате PCR с использованием олигонуклеотида со следующей последовательностью: CGC GGG GCG GCC ACG CGT CCT CGC CAG CAC ACA /SEQ ID N0: 14/. Полученный PCR фрагмент был переварен BamHI и
Notl и снова включен в CD16:7:Z /48-65/ конструкцию. Z фрагменты, кодирующие остатки 33-65, 71-104 и 104-137, получают в результате реакции PCR, с использованием пар драйверов, содержащих Mlul сайты у
5' конца форвардных праймеров и стоп кодоны с последующими Notl сайтами у 5' конца обратных праймеров. В каждом случае рестрикционные сайты были включены на шесть остатков от 5' конца праймера для
обеспечения расщепления рестрикционным энзимом.
S33: CGC GGG ACG CGT TTC AGC CGT CCT CGC CAG CAC ACA (SEQ ID N0: 15),
S71: CGC GGG ACG CGT GAC CCT GAG ATG GGG GGA AAG (SEQ IS
N0: 16);
S104: CGC GGG ACG CGT ATT GGG ATG AAA GGC GAG CGC (SEQ ID N0: 17).
Конструирование FcR 11A делеционных мутантов
Карбоксильные терминальные FcR 11A
делеционные мутанты были сконструированы в результате PCR таким же способом, как и для полной длины конструкций, превращая последовательности, кодирующие тирозин, в положении 282 и 298 в стоп кодоны
/TAA/. N-терминальные делеции были созданы за счет амплификации фрагментов, кодирующих последовательно без внутриклеточного домена за счет PCR, с использованием олигонуклеотидов, которые позволяют
включать полученные фрагменты между Mlul и Notl рестрикционными сайтами в ранее сконструированные плазмиды экспрессии, кодирующие CD16 внеклеточный домен, слитый с CD7 трансмембранным доменом, причем
последний оканчивается в Mlul сайте и соединяет трансмембранный и внутриклеточный домены.
Другие варианты
Описанные ранее примеры демонстрируют тот факт, что аггрегации Z,
η или γ химер могут инициировать цитолитическую эффекторную клеточную реакцию в T клетках. Известный интервал экспрессии Z, η и γ , который включает T лимфоциты, природные
киллерные клетки, базофильные гранулоциты, макрофаги и мест-клетки, предполагает, что сохраненные фрагменты последовательности могут взаимодействовать с сенсорным аппаратом, общим для клеток
кроветворного происхождения, и что важный компонент защиты хозяина в иммунной системе может быть передан посредством рецепторной аггрегации.
Потенция цитолитической реакции и отсутствие реакции на мишеневые клетки, содержащие МНС класса II рецепторы, демонстрирует тот факт, что химеры на основе Z, η и γ создают основу для генетической интервенции для AIDS за счет адаптационной иммунотерапии. Широкое распределение эндогенных Z и η и доказательство того, что Fc рецепторы, связанные с γ передаваемой токсичностью в различных типах клеток /Fanger et al., Immunol, Today, 10: 92-99 /1989//, позволяет рассматривать для этой цели различные клетки. Так, например, нейтрофильные гранулоциты, которые имеют очень короткий срок полужизни /примерно 4 часа/ в циркуляции, и являются интенсивно цитолитическими, представляют привлекательные мишеневые клетки для экспрессии химер. Инфекция нейтрофилов HIV, по-видимому, не приводит к высвобождению вируса, и обилие этих клеток /наиболее превалирующие из лейкоцитов/ должно облегчить защиту хозяина. Другой привлекательной возможностью для клеток хозяина являются зрелые T клетки, популяция в настоящее время доступная для конструирования ретровирусов /Rosenberg, S.A. Sci. Am. 262:62-69 /1990//. С целью рекомбинации IL-2. Популяции T клеток можно размножить в культуре с относительной легкостью, и размноженная популяция обычно имеет ограниченный период полужизни /Rosenberg et al., N. England J. Med. 323:570-578 /1990//.
В соответствующих условиях HIV узнавание клетками, экспрессирующими CD4 химеры, должно также обеспечивать митогенный стимул, который предоставляет возможность популяции клеток с плечами динамично реагировать на вирусное поражение. Хотя мы сфокусировали внимание на поведении протеинов слияния в цитолитических T лимфоцитах, экспрессия химер в хелперные лимфоциты может обеспечить HIV-мобилизованный источник цитокинов, которые могут противодействовать коллапсу субсета хелперных клеток в AIDS. Недавнее описание нескольких схем конструирования устойчивости к инфекции на стадиях, отличных от проникновения вируса /Friedman et al., Nature 335; 452-454 (1988); Green et al., Cell, 58:215-223 (1989); Malim et al., Cell 58:205-214 (1989); Trono et al. , Cell 59:113-120 (1989); Buonocore et al., Nature, 345:625-628 (1990), предполагает, что клетки, содержащие CD4 химеры, можно сконструировать, и что они могут помешать продуцированию вирусов за счет экспрессии соответствующих агентов, содержащих внутриклеточный сайт действия.
Способность передавать сигналы T лимфоцитам через автономные химеры также обеспечивает способность для регуляции ретровирусно созданных лимфоцитов in vivo. Сшивая стимулы посредством, например, специфических IgM антител, сконструированных для удаления комплемент-связывающих доменов, можно дать возможность таким лимфоцитам увеличить свое количество in situ, тогда как обработкой аналогичными специфическими IgG антителами /например, распознавая аминокислотные варианты, созданные в химерической цепи/, можно селективно исчерпать сконструированную популяцию.
Кроме того, анти-CD4 IgM антитела не требуют дополнительной сшивки для мобилизации кальция в Jurkat клетках, экспрессирующих CD4:Z химеры /данные не представлены/. Способность регулировать клеточную популяцию без обращения в повторной экстракорпореальной амплификации может существенно расширить круг и эффективность текущих применений, предполагаемых для генетического конструирования T клеток.
Для создания других химер, содержащих Z, η или γ внутриклеточные последовательности, кДНК или геномные последовательности, кодирующие внеклеточный домен рецептора, можно обеспечить рестрикционным сайтом, вводимым в локус, как раз предшествующий выбранному трансмембранному домену. Фрагмент внеклеточного домена, заканчивающий его в рестрикционном сайте, можно затем соединить с Z, η или γ последовательностями. Типичные внеклеточные домены можно получить из рецепторов, которые распознают комплемент, углеводороды, вирусные протеины, бактерии, простейшие или metazoan паразиты, или продуцируемые ими протеины. Аналогично, лиганды или рецепторы, экспрессируемые патогенами или опухолевыми клетками, можно присоединить к Z, η или γ последовательностям для направления иммунной реакции против клеток, содержащих рецепторы, распознающие эти лиганды.
Хотя изобретение было описано в связи с его специфическими воплощениями, следует учитывать, что его можно в дальнейшем модифицировать, и такие применения должны охватывать варианты, использования или адаптации настоящего изобретения и включать такие отклонения от настоящего раскрытия, которые могут придти в голову специалистам, которым это изобретение предназначено, и как могут быть использованы его отличительные черты, изложенные ранее, как следует из объема прилагаемой формулы изобретения.
ПЕРЕЧИСЛЕНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
(2) Информация для последовательности N 1:
(i) Характеристики
последовательности:
(A) длина: 1728 пар оснований
(B) тип: нуклеиновая кислота
(C) тип цепей: двойная
(D) топология: линейная
(ii) тип молекулы: ДНК
(геномная)
(iii) Описание последовательности: SEQ ID N0: 1:
Реферат
Изобретение относится к биотехнологии, иммунологии и медицине и может быть использовано для направления клеточного иммунного ответа на инфекционный агент. Млекопитающему вводят трансформированные иммунные клетки, которые экспрессируют химерный рецептор, содержащий внеклеточную часть, способную распознавать инфекционный агент, внутриклеточную часть, происходящую из полипептида CD3, ζ или η Т - клеточного рецептора, или полипептида В-клеточного рецептора, или полипептида Fc-рецептора, которая обладает способностью передавать клетке сигнал на разрушение. Включение в химерный рецептор внеклеточной части CD4, распознающей и связывающей ВИЧ, и внутриклеточной части, происходящей из полипептида CD3, ζ или η Т - клеточного рецептора, позволяет передавать клетке сигнал на разрушение ВИЧ или ВИЧ-инфицированной клетки. 12 с. и 20 з.п. ф-лы, 45 ил.

Комментарии