Способ и композиции для клеточной иммунотерапии - RU2688185C2
Код документа: RU2688185C2
Чертежи










Описание
Настоящая заявка подана 23 марта 2012 года в виде международной заявки на выдачу патента PCT на имя Онкологического исследовательского центра имени Фреда Хатчинсона, национальной корпорации США, заявителя для случая указания всех стран, за исключением США, и Stanley R. Riddell, гражданина Канады, и Michael Hudecek, гражданина Германии, заявителей для случая указания только США, и испрашивает приоритет на основании заявки на выдачу патента США с регистрационным номером 61/466552, поданной 23 марта 2011 года, содержание которой включено в настоящее описание в виде ссылки в полном объеме.
Область техники, к которой относится изобретение
Настоящее изобретение относится к области биомедицины и, в частности, к способам, применимым для терапии злокачественных опухолей. В частности, варианты осуществления настоящего изобретения относятся к способам и композициям для проведения клеточной иммунотерапии.
Заявление относительно финансируемого из федерального бюджета исследования
Настоящее изобретение осуществлено при поддержке правительства в форме грантов R01CA18029 Департамента здравоохранения и социального обеспечения США, Национального института здравоохранения и гранта SCORE общества исследования лейкоза и лимфомы. Правительство Соединенных Штатов Америки имеет определенные права на изобретение.
Уровень техники
Исследования на грызунах показали, что адоптивная иммунотерапия с использованием специфичных для антигена T-клеток является эффективной в случае злокачественной опухоли и инфекций, и имеются данные о том, что такой способ обеспечивает терапевтическую активность у человека1-8. Для клинических применений необходимо выделить T-клетки с требуемой антигенной специфичностью или сконструировать T-клетки так, чтобы они экспрессировали рецепторы, которые нацелены на инфицированные или трансформированные клетки, и затем размножить такие клетки в культуре9-14. Перенос клонов T-клеток представляет интерес, поскольку он обеспечивает контроль специфичности и функции и облегчает оценку продолжительности жизни, токсичности и эффективности in vivo. Кроме того, при осуществлении аллогенной трансплантации стволовых клеток введение реципиентам T-клеточных клонов от донора, мишенью которых являются патогены или злокачественные клетки, может обеспечить возможность избегания болезни трансплантат-против-хозяина, которое возникает при инфузии не подвергнутых селекции T-клеток донора3,4,15. Однако из клинических исследований очевидно, что эффективность культивируемых T-клеток, особенно клонированных T-клеток CD8+, часто ограничена их неспособностью персистировать после адоптивного переноса16,17.
Пул лимфоцитов, из которого могут быть получены T-клетки для адаптивной иммунотерапии, может содержать наивные и долгоживущие контактировавшие с антигенами T-клетки памяти (TM). TM можно разделить на подгруппы центральных клеток памяти (TCM) и эффекторных клеток памяти (TEM), которые отличаются по фенотипу, свойствам хоминга и функции18. TCM CD8+ экспрессируют CD62L и CCR7 на клеточной поверхности, которые стимулируют миграцию в лимфатические узлы и быстро пролиферируют при повторной экспозиции с антигеном. TEM CD8+ не имеют CD62L на клеточной поверхности и предпочтительно мигрируют в периферические ткани и проявляют немедленную эффекторную функцию19. В ответ на антигенную стимуляцию TCM и TEM CD8+ дифференцируются в цитолитические эффекторные T-клетки (TE), которые экспрессируют высокий уровень гранзимов и перфорина, но являются короткоживущими20. Таким образом, короткое время жизни T-клеток в испытаниях по клинической иммунотерапии просто может быть результатом их дифференцировки во время культивирования in vitro в TE, гибель которых предопределена17,21,22. Существует необходимость в идентификации клеточных популяций и способах, которые обеспечивают повышенную жизнеспособность адоптивно переносимых T-клеток in vivo.
Сущность изобретения
В одном аспекте настоящее изобретение относится к способам и композициям для придания и/или усиления иммунных ответов, опосредованных клеточной иммунотерапией, такой как иммунотерапия посредством адоптивного переноса специфичных для опухоли, специфичных для подгруппы генетически модифицированных T-клеток CD4+, при этом T-клетки CD4+ придают и/или усиливают способность T-клеток CD8+ поддерживать противоопухолевую реактивность и увеличивать и/или максимизировать специфичную для опухоли пролиферацию.
В одном варианте настоящее изобретение относится к способу осуществления клеточной иммунотерапии у субъекта, имеющего заболевание или нарушение, посредством введения субъекту препарата генетически модифицированных цитотоксических T-лимфоцитов, который обеспечивает клеточный иммунный ответ, при этом препарат цитотоксических T-лимфоцитов содержит T-клетки CD8+, которые имеют химерный рецептор антигена с внеклеточным вариабельным доменом антитела, специфичным по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточным доменом передачи сигнала T-клеточных или других рецепторов, таким как костимулирующие домены; и препарата генетически модифицированных хелперных T-лимфоцитов, которые проявляют преобладающий Th1-фенотип, а также продуцируют другие цитокины, вызывают прямое распознавание опухоли и усиливают способность препаратов генетически модифицированных цитотоксических T-лимфоцитов опосредовать клеточный иммунный ответ, при этом препарат хелперных T-лимфоцитов содержит T-клетки CD4+, которые имеют химерный рецептор антигена, содержащий внеклеточный вариабельный домен антитела, специфичный по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного рецептора. Возможны различные модификации указанного выше способа. Например, химерные рецепторы антигена, модифицирующие T-клетку CD4+ и T-клетку CD8+, могут быть одинаковыми или разными. В альтернативных вариантах T-клетки могут быть модифицированы рекомбинантным рецептором T-клеток (TCR). TCR может быть специфичным по отношению к антигену, патогену или опухоли. Существуют TCR для многих опухолевых антигенов меланомы (например, MART1, gp100), лейкоза (например, WT1, минорные антигены гистосовместимости), рака молочной железы (например, her2, NY-BR1).
В другом варианте настоящее изобретение относится к композиции для адаптивной клеточной иммунотерапии, содержащей препарат генетически модифицированных цитотоксических T-лимфоцитов CD8+, который вызывает клеточный иммунный ответ, при этом препарат цитотоксических T-лимфоцитов содержит T-клетки CD8+, которые имеют химерный рецептор антигена с внеклеточным вариабельным доменом антитела, специфичным по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного или других рецепторов, такой как костимулирующий домен, и препарат генетически модифицированных хелперных T-лимфоцитов, которые проявляют преимущественный Th1-фенотип, а также продуцируют другие цитокины, вызывают прямое распознавание опухоли и усиливают способность препаратов генетически модифицированных цитотоксических T-лимфоцитов опосредовать клеточный иммунный ответ, при этом препарат хелперных T-лимфоцитов содержит T-клетки CD4+, которые имеют химерный рецептор антигена с внеклеточным вариабельным доменом антитела, специфичным по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного рецептора.
В еще одном варианте настоящее изобретение относится к композиции для адаптивной клеточной иммунотерапии, содержащей препарат модифицированных химерным рецептором антигена специфичных для опухоли цитотоксических T-лимфоцитов CD8+, который вызывает клеточный иммунный ответ, при этом препарат цитотоксических T-лимфоцитов содержит T-клетки CD8+, которые имеют химерный рецептор антигена, содержащий внеклеточное одноцепочечное антитело, специфичное по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного рецептора, и модифицированные антиген-реактивным химерным рецептором антигена наивные хелперные T-клетки CD4+, которые получены из CD45RO-негативных, CD62L-позитивных CD4-позитивных T-клеток, и фармацевтически приемлемый носитель.
В другом варианте настоящее изобретение относится к композиции для адаптивной клеточной иммунотерапии, содержащей препарат антиген-специфичных цитотоксических T-лимфоцитов CD8+, который вызывает клеточный иммунный ответ и содержит T-клетки CD8+, полученные от пациента, вместе с антиген-реактивными модифицированными химерным рецептором антигена хелперными T-клетками CD4+, которые вызывают цитокиновый ответ Th1-типа и усиливают иммунный ответ CD8+ на патогены, при этом препарат хелперных T-лимфоцитов содержит T-клетки CD4+, которые имеют химерный рецептор антигена с внеклеточным вариабельным доменом антитела, специфичным по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного рецептора.
В другом варианте настоящее изобретение относится к композиции для адаптивной клеточной иммунотерапии, содержащей антиген-реактивные модифицированные химерным рецептором антигена хелперные T-клетки CD4+, которые вызывают прямое распознавание опухоли и усиливают иммунный ответ CD8+ на патогены, при этом препарат хелперных T-лимфоцитов содержит T-клетки CD4+, которые имеют химерный рецептор антигена, содержащий внеклеточный вариабельный домен антитела, специфичный по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного рецептора.
В другом аспекте настоящее изобретение относится к способу производства композиции для адаптивной иммунотерапии посредством получения препарата модифицированных химерным рецептором антигена специфичных для опухоли цитотоксических T-лимфоцитов CD8+, который вызывает клеточный иммунный ответ, и антиген-реактивного химерного рецептора антигена, при этом препарат модифицированных цитотоксических T-лимфоцитов содержит T-клетки CD8+, которые имеют химерный рецептор антигена с внеклеточным вариабельным доменом антитела, специфичным по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный модуль передачи сигнала T-клеточного рецептора; и получения модифицированной наивной хелперной T-клетки CD4+, которая вызывает цитокиновый ответ Th1-типа, при этом препарат модифицированных хелперных T-лимфоцитов содержит клетки CD4+, которые имеют химерный рецептор антигена с внеклеточным вариабельным доменом антитела, специфичным по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного рецептора.
В другом варианте настоящее изобретение относится к способу производства композиции для адаптивной иммунотерапии посредством получения модифицированной наивной хелперной T-клетки CD4+, которая вызывает цитокиновый ответ Th1-типа, при этом препарат модифицированных хелперных T-лимфоцитов содержит T-клетки CD4+, которые имеют химерный рецептор антигена, содержащий внеклеточный вариабельный домен антитела, специфичный по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного рецептора, и объединения модифицированных наивных хелперных T-клеток CD4+ с препаратом антиген-специфичных цитотоксических центральных T-лимфоцитов памяти CD8+, которые имеют химерный рецептор антигена с внеклеточным вариабельным домен антитела, специфичный по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного или других рецепторов.
В одном варианте настоящее изобретение относится к способу осуществления клеточной иммунотерапии у субъекта, имеющего заболевание или нарушение, посредством введения субъекту препарата генетически модифицированных хелперных T-лимфоцитов, при этом препарат модифицированных хелперных T-лимфоцитов содержит T-клетки CD4+, которые имеют химерный рецептор антигена, содержащий внеклеточный вариабельный домен антитела, специфичный по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный модуль передачи сигнала T-клеточного рецептора.
Указанные и другие варианты осуществления изобретения описаны далее в сопровождающем описании, чертежах и формуле изобретения.
Краткое описание чертежей
Фиг. 1 показывает фенотип и анализ экспрессии химерного рецептора антигена (CAR) в CAR-трансдуцированной лентивирусом, кодирующим ROR1-CAR, и нетрансдуированной линии T-клеток CD8+ в качестве контроля. Кассета ROR1-CAR содержит укороченный EGFR, который служит в качестве маркера трансдукции и может быть выявлен по окрашиванию моноклональными анти-EGFR-антителами. Укороченный слитый белок Fc-ROR1 связывается непосредственно с антиген-связывающим доменом ROR1-CAR и избирательно окрашивает ROR1-CAR-трансдуцированную, но не окрашивает нетрансдуцированную контрольную T-клеточную линию. Экспрессию ROR1-CAR на клеточной поверхности T-клеток CD8+ измеряют непосредственно по связыванию со слитым белком ROR1-Fc и опосредованно по экспрессии укороченного EGFR, который закодирован в векторе ниже последовательности 2A.
Фиг. 2 показывает цитолитическую активность T-клеток CD8+, экспрессирующих ROR1-специфичный химерный рецептор антигена, против панели ROR1-позитивных линий опухолевых клеток человека (K562) и первичных опухолевых клеток (B-CLL) и аутологичных нормальных B-клеток в анализе высвобождения51Cr. В соответствии с равномерной экспрессией ROR1 на злокачественных, но не на зрелых нормальных B-клетках генетически модифицированные T-клетки CD8+ ROR1-CAR лизировали только опухолевые клетки ROR1+, но не лизировали зрелые нормальные B-клетки. T-клетки CD8+ ROR1-CAR проявляли специфичную литическую активность против ROR1-позитивных опухолевых клеток, включая первичные CLL, но не против нормальных B-клеток.
Фиг. 3 показывает фенотип и экспрессию CAR в ROR1-CAR-трансдуцированной и нетрансдуцированной T-клеточной линии CD4+ в качестве контроля. Экспрессию ROR1-CAR на клеточной поверхности T-клеток CD4+ измеряют на основании специфичного связывания со слитым белком ROR1-Fc. Укороченный слитый белок Fc-ROR1, но не белок Fc отдельно, непосредственно связывается с ROR1-CAR и избирательно окрашивает ROR1-CAR-трансдуцированную, но не окрашивает нетрансдуцированную контрольную T-клеточную линию CD4+, подтверждая экспрессию ROR1-CAR на клеточной поверхности и связывание с белком ROR1. Экспрессию ROR1-CAR на клеточной поверхности T-клеток CD4+ измеряют на основании специфичного связывания со слитым белком ROR1-Fc, но не с контрольным слитым белком Fc.
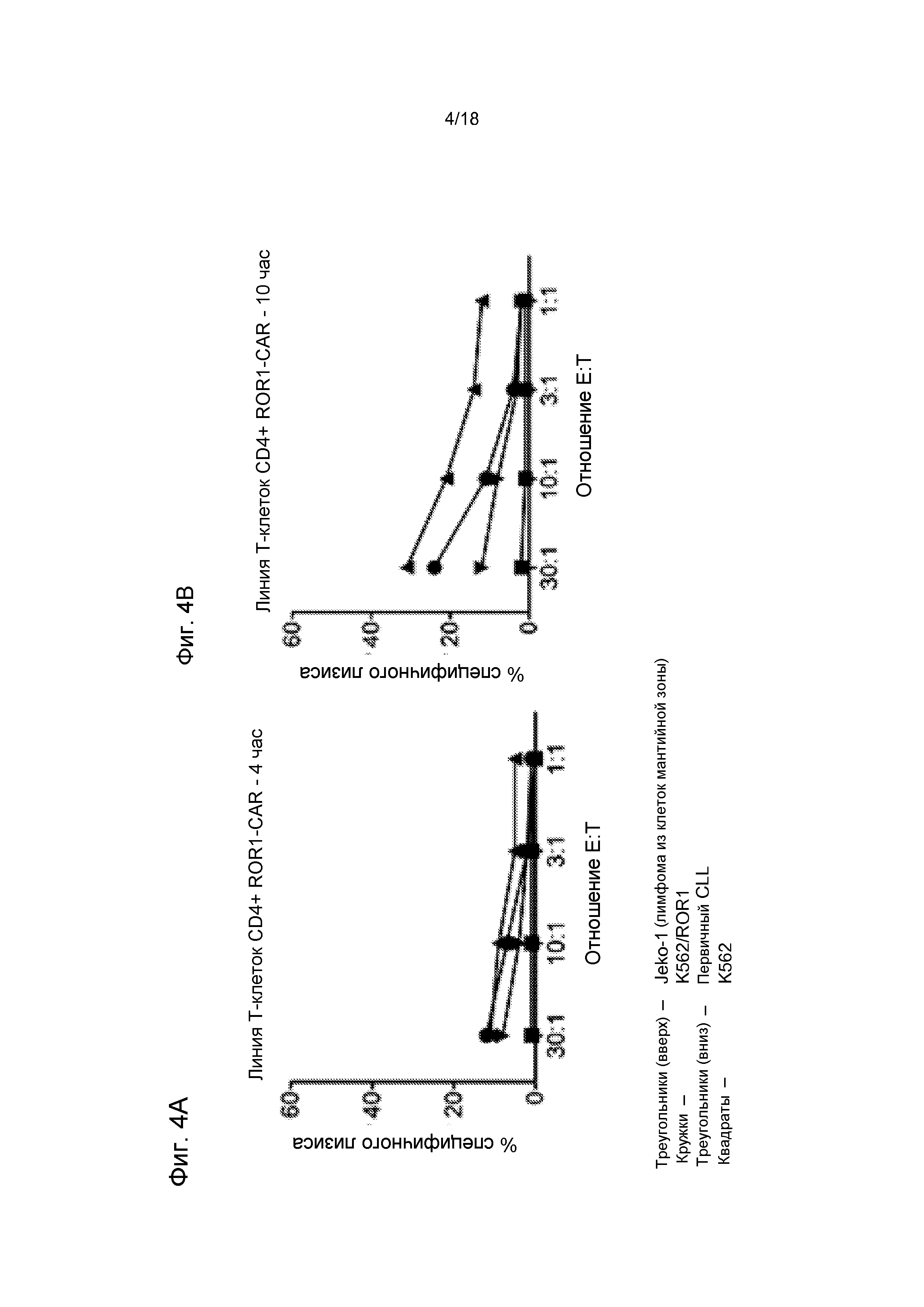
Фиг. 4 (т.е. Фиг. 4A-4B вместе) показывают слабую, но специфичную цитолитическую активность T-клеток CD4+ ROR1-CAR в анализе высвобождения51Cr против панели ROR1-позитивных опухолевых клеток, включая первичные CLL, линию лимфомы из клеток мантийной зоны Jeko-1, клетки K562, которые были стабильно трансфицированы ROR1 (K562/ROR1), но не наивные ROR1-негативные клетки K562. T-клетки CD4+ ROR1-CAR проявляют слабую, но специфичную литическую активность против ROR1-позитивных опухолевых клеток.
Фиг. 5 (т.е. Фиг. 5A-5B вместе) показывают результаты ELISA IFNγ (Фиг.5A) и мультиплексного анализа цитокинов (Фиг.5B). Секреция цитокинов в линиях T-клеток ROR1-CAR CD4+ и CD8+. T-клетки CD4+ ROR1-CAR и CD8 ROR1-CAR совместно инкубировали с опухолевыми клетками ROR1+ и измеряли уровни гамма-интерферона (IFNg) в ELISA (5A), и IFNg, TNFα, IL-2, IL-4, IL-10 и IL-17 измеряли в анализе Luminex (5B). ROR1-CAR-модифицированные T-клетки CD4+ специфично узнают ROR1-позитивные опухолевые клетки и линии опухолевых клеток и продуцируют большие количества Th1-цитокинов, включая IFN-γ, TNF-α и особенно IL-2, чем ROR1-CAR-модифицированные T-клетки CD8+. Полученные данные демонстрируют, что T-клетки CD4+ ROR1-CAR проявляют хелперные эффекторные функции после стимуляции с использованием ROR1-CAR и в дополнение к опосредованию прямой противоопухолевой реактивности также могут быть использованы для повышения способности ROR1-CAR-модифицированных T-клеток CD8+ опосредовать клеточный иммунный ответ.
На Фиг. 6 изображены результаты исследования пролиферации, показывающие, что T-клетки CD4+ ROR1-CAR индуцированы к пролиферации после стимуляции линиями ROR1-позитивных опухолевых клеток и первичными опухолевыми клетками (анализ CFSE) и что процент пролиферирующих клеток и количество клеточных делений, которым подвергалась подгруппа пролиферирующих клеток, были значимо выше по сравнению с ROR1-CAR-модифицированными T-клетками CD8+. T-клетки CD4+ ROR1-CAR сильнее пролиферируют после стимуляции ROR1-позитивными опухолевыми клетками (K562/ROR1, первичные CLL и Jeko MCL) по сравнению с CTL CD8+ ROR1-CAR.
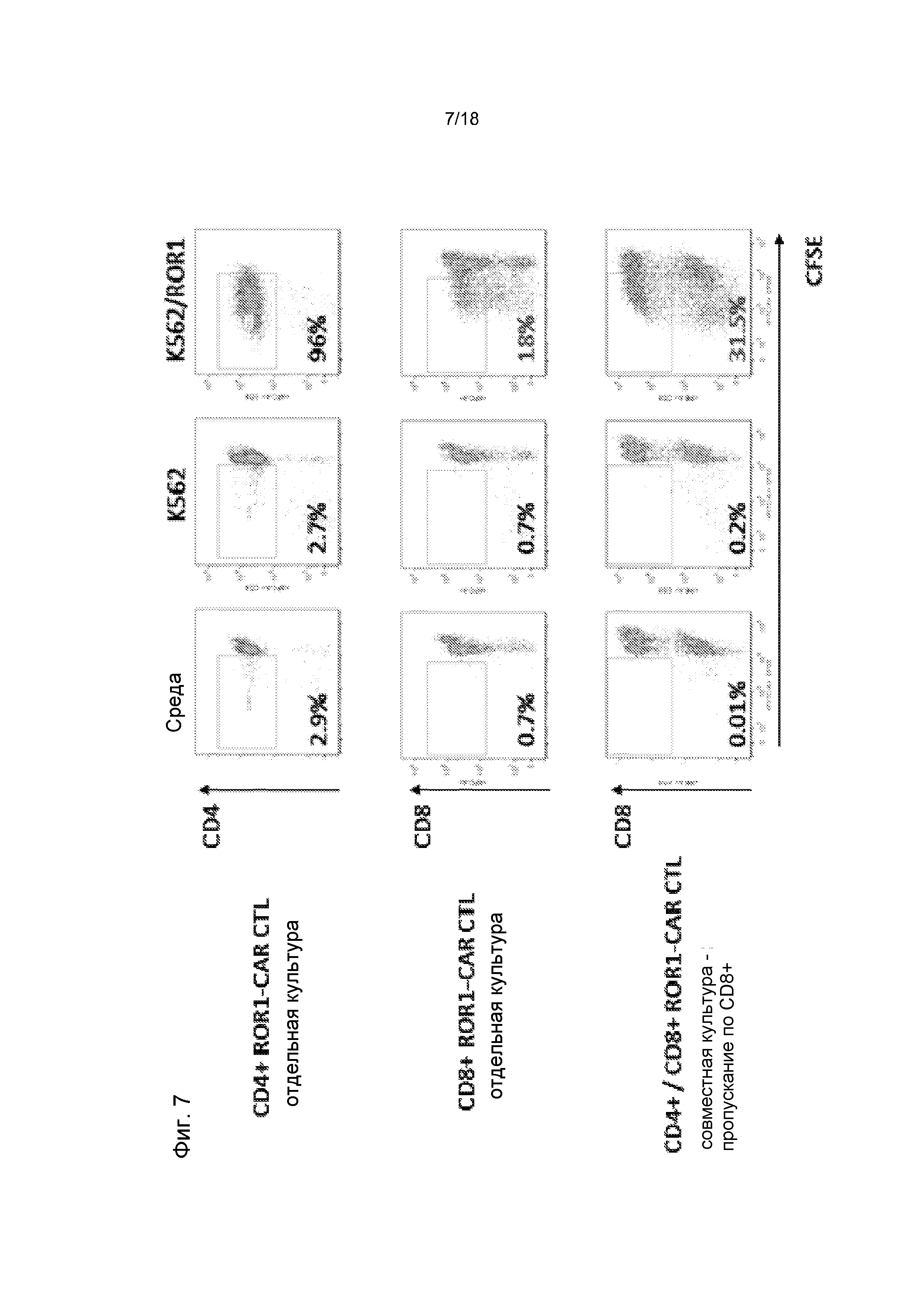
Фиг. 7: Поликлональные не подвергнутые селекции T-клетки CD4+ ROR1 CAR помогают CTL CD8+ ROR1-CAR, стимулируя их пролиферацию в ответ на опухоль. T-клетки CD4+ ROR1-CAR (полученные из общей массы T-клеток CD4+) значимо увеличивали пролиферацию поликлональных не подвергнутых селекции CTL CD8+ ROR1-CAR (18% в отдельной культуре → 31,5% после совместного культивирования с T-клетками CD4+ CAR).
Фиг. 8 (т.е. Фиг. 8A-8D вместе) показывает образование линий T-клеток CD4+ CAR из очищенных с использованием проточной сортировки подгрупп наивных, центральных и эффекторных клеток памяти CD4+ и анализ функции T-клеток. Профиль цитокинов и пролиферативная способность свидетельствуют, что T-клетки CD4+ ROR1-CAR, полученные из наивных T-клеток CD4+, могут быть наиболее подходящими в качестве помощников для CTL CD8+. Сходные данные получали в экспериментах, в которых сравнивали функцию T-клеточных линий CD4+ CAR, экспрессирующих CD19-специфичный CAR. На фиг. 8A показана очистка с использованием проточной сортировки наивных, центральных и эффекторных T-клеток памяти CD4+ на основе экспрессии CD45RA, CD45RO, CD62L. На фиг. 8B показан анализ пролиферации T-клеточных линий ROR1-CAR, которые были получены с использованием лентивирусной трансдукции очищенных сортировкой наивных, центральных и эффекторных T-клеток памяти CD4+ (анализ CFSE). На фиг. 8C показан анализ секреции цитокинов в T-клеточных линиях ROR1-CAR, полученных из очищенных сортировкой наивных, центральных и эффекторных T-клеток памяти CD4+ (анализ Luminex). На фиг. 8D показан анализ секреции цитокинов в T-клеточных линиях CD19-CAR, полученных из очищенных сортировкой наивных, центральных и эффекторных T-клеток памяти CD4+ (анализ Luminex). Профиль цитокинов, полученный в мультиплексном анализе цитокинов (Фиг. 8B), и пролиферативная способность на основании окрашивания CFSE (Фиг. 8C) показывают, что ROR1-CAR-модифицированные T-клетки CD4+, полученные из подгруппы наивных клеток, продуцировали более высокие уровни Th1-цитокинов и сильнее пролиферировали после стимуляции ROR1-позитивными опухолевыми клетками, что свидетельствует о том, что они могут быть наиболее подходящими для усиления CTL CD8+ ROR1-CAR. Анализ секреции цитокинов в T-клеточных линиях CD19-CAR, полученных из очищенных сортировкой наивных центральных и эффекторных T-клеток памяти CD4+ (анализ Luminex), демонстрирует, что активность подгрупп T-клеток CD4 может быть распространена на многие CAR.
Фиг. 9 показывает совместное культивирование ROR1-CAR-модифицированных T-клеток CD8+ с ROR1-CAR-модифицированными T-клетками CD4+ (но не с нетрансдуцированными контрольными T-клетками CD4+). Совместное культивирование CTL CD8+ ROR1-CAR и T-клеточных линий CD4+ ROR1-CAR, полученных из подгрупп наивных, центральных и эффекторных клеток памяти, для определения оптимального сочетания T-клеток CD8+ и CD4+, которое может обеспечить возможность максимальной пролиферации CTL CD8+ ROR1-CAR. Наивные T-клетки CD4 ROR1-CAR обеспечивают наибольшую пролиферацию центральных CTL-клеток памяти ROR1-CAR CD8. Совместное культивирование приводит к увеличению специфичной для опухоли пролиферации подгруппы CD8+, и такая максимальная пролиферация подгруппы CD8+ наблюдается после совместного культивирования с T-клетками CD4+ ROR1-CAR, полученными из наивных T-клеток CD4+, что свидетельствует о том, что наивные T-клетки CD4+ наилучшим образом подходят для усиления эффекторной функции CTL-клеток CD8+.
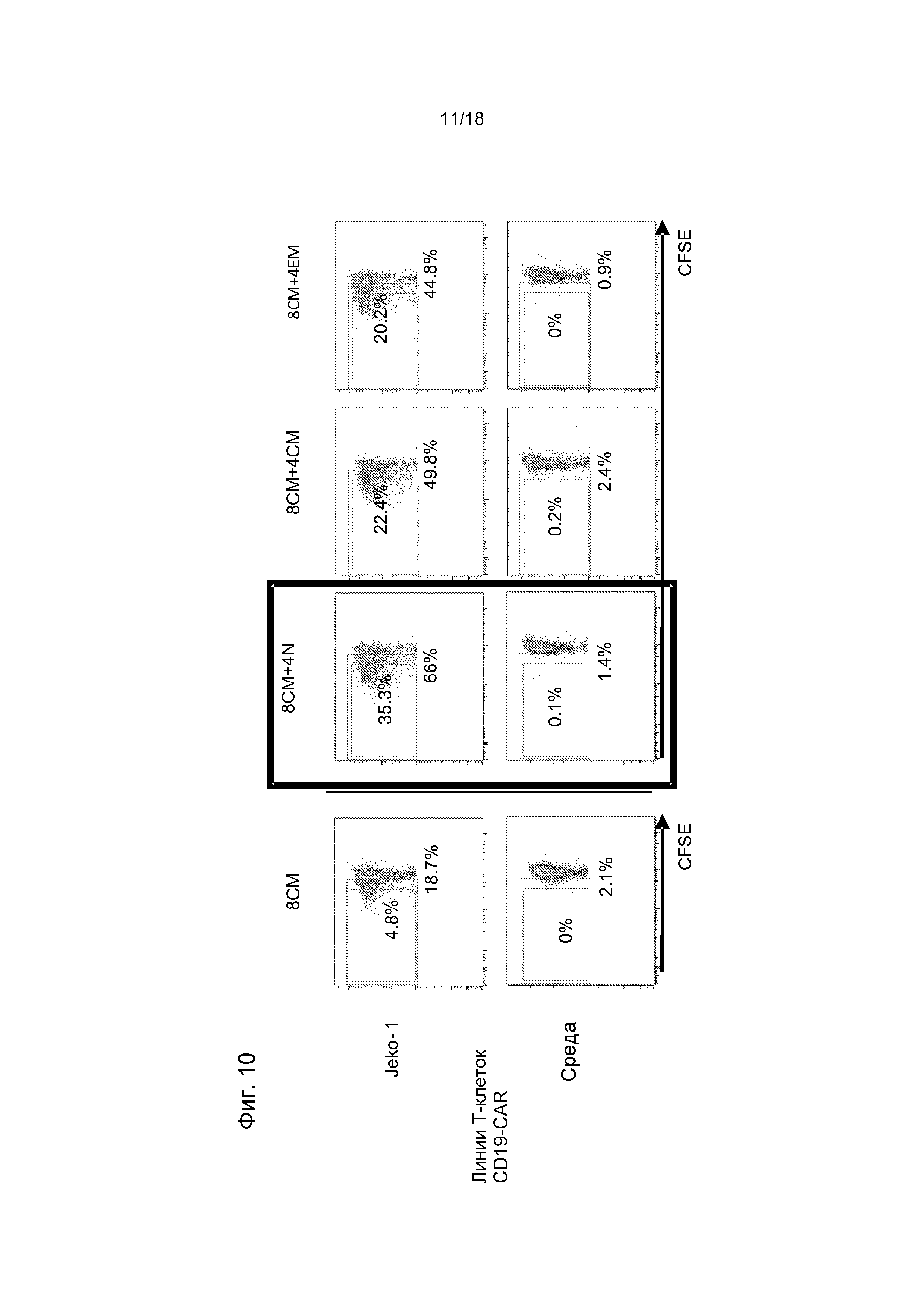
Фиг. 10 показывает превосходную способность T-клеточных линий CD4+ CAR, полученных из наивной подгруппы, усиливать специфичную для опухоли пролиферацию полученных из центральных CTL-клеток памяти CD8+ CAR в экспериментах по совместному культивированию с использованием CTL CD8+ CD19-CAR и T-клеточных линий CD4+ CD19-CAR, стимулированных линией лимфомы из клеток мантийной зоны CD19+ Jeko-1. Превосходная способность T-клеточных линий CD4+ CAR, полученных из наивной подгруппы, усиливать специфичную для опухоли пролиферацию полученных из центральных CTL-клеток памяти CD8+ CAR была подтверждена в экспериментах по совместному культивированию с использованием CTL CD8+ CD19-CAR и T-клеточных линий CD4+ CD19-CAR, стимулированных линий лимфомы из клеток мантийной зоны CD19+ Jeko-1.
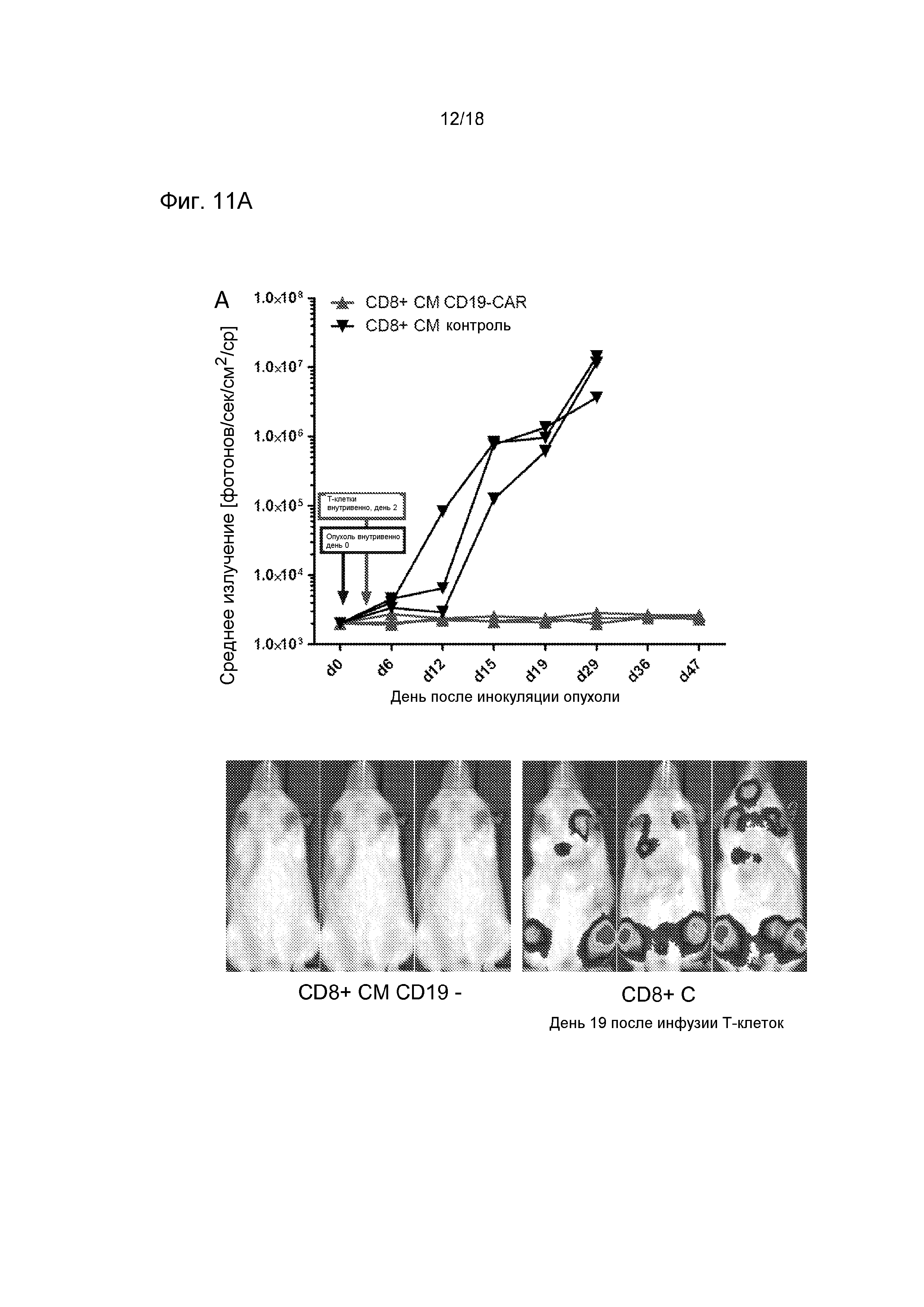
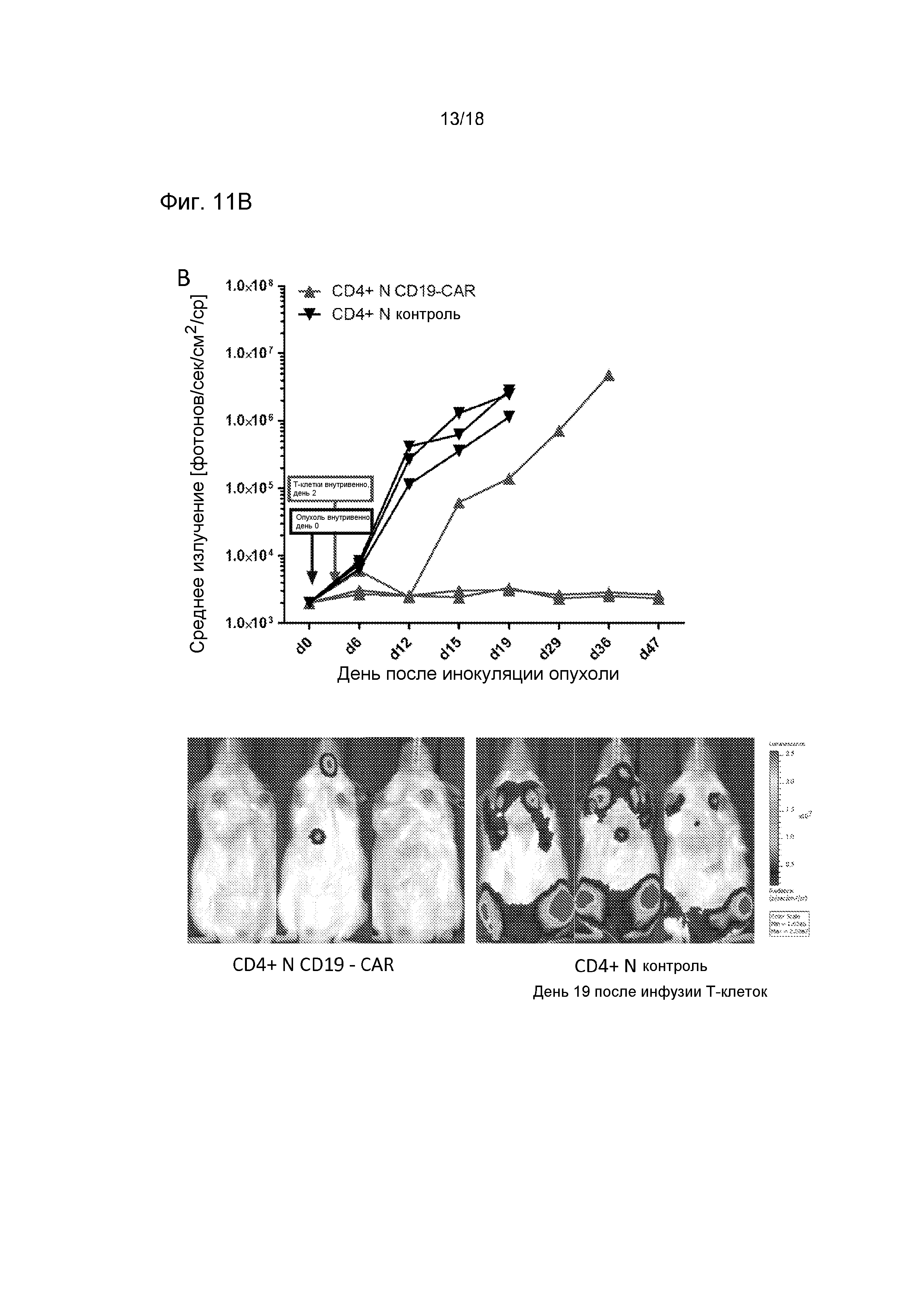
Фиг. 11 показывает, что T-клетки CD8+ CAR и T-клетки CD4+ CAR независимо обеспечивают прямую противоопухолевую эффективность в модели лимфомы у иммунодефицитных мышей (NOD/SCID-Raji). Группам мышей (n=3) инокулировали экспрессирующие люциферазу светляка опухолевые клетки Raji посредством инъекции в хвостовую вену и обрабатывали однократной дозой 10×106 T-клеток. Мыши получали либо CD19-CAR-трансдуцированные или контрольные ложно трансдуцированные T-клетки CD8+, полученные из центральных клеток памяти (A), либо CD19-CAR-трансдуцированные или контрольные ложно трансдуцированные T-клетки CD4+, полученные из наивных клеток (B). Опухолевую нагрузку и распределение анализировали, используя серийную визуализацию биолюминесценции.
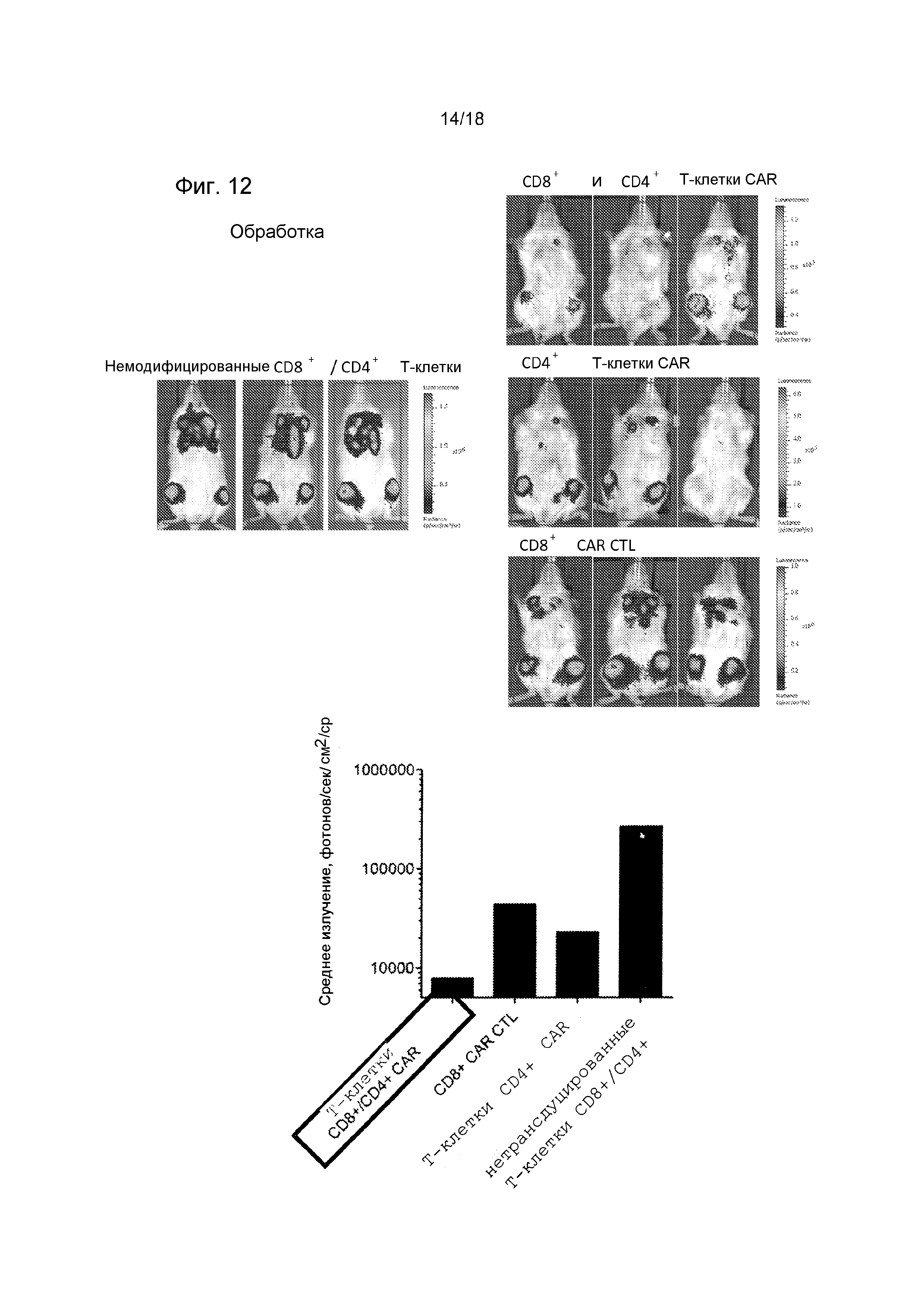
Фиг. 12 показывает стимуляцию и синергетическое действие ROR1-CAR-модифицированных T-клеток CD4+ на противоопухолевую эффективность CTL CD8+ROR1-CAR в мышиной опухолевой модели системной лимфомы из клеток мантийной зоны (NSG/Jeko-1-ffLuc). Противоопухолевая эффективность ROR1-CAR-модифицированных T-клеток CD8+ и CD4+ в мышиной опухолевой модели системной агрессивной лимфомы из клеток мантийной зоны (NSG/Jeko-1). Анализ опухолевой нагрузки с использованием визуализации биолюминесценции после адоптивного переноса CTL CD8+ ROR1-CAR, T-клеток CD4+ ROR1-CAR или сочетания T-клеток ROR1-CAR CD8+ и CD4+. Все мыши получали одинаковые общие дозы T-клеток CAR.
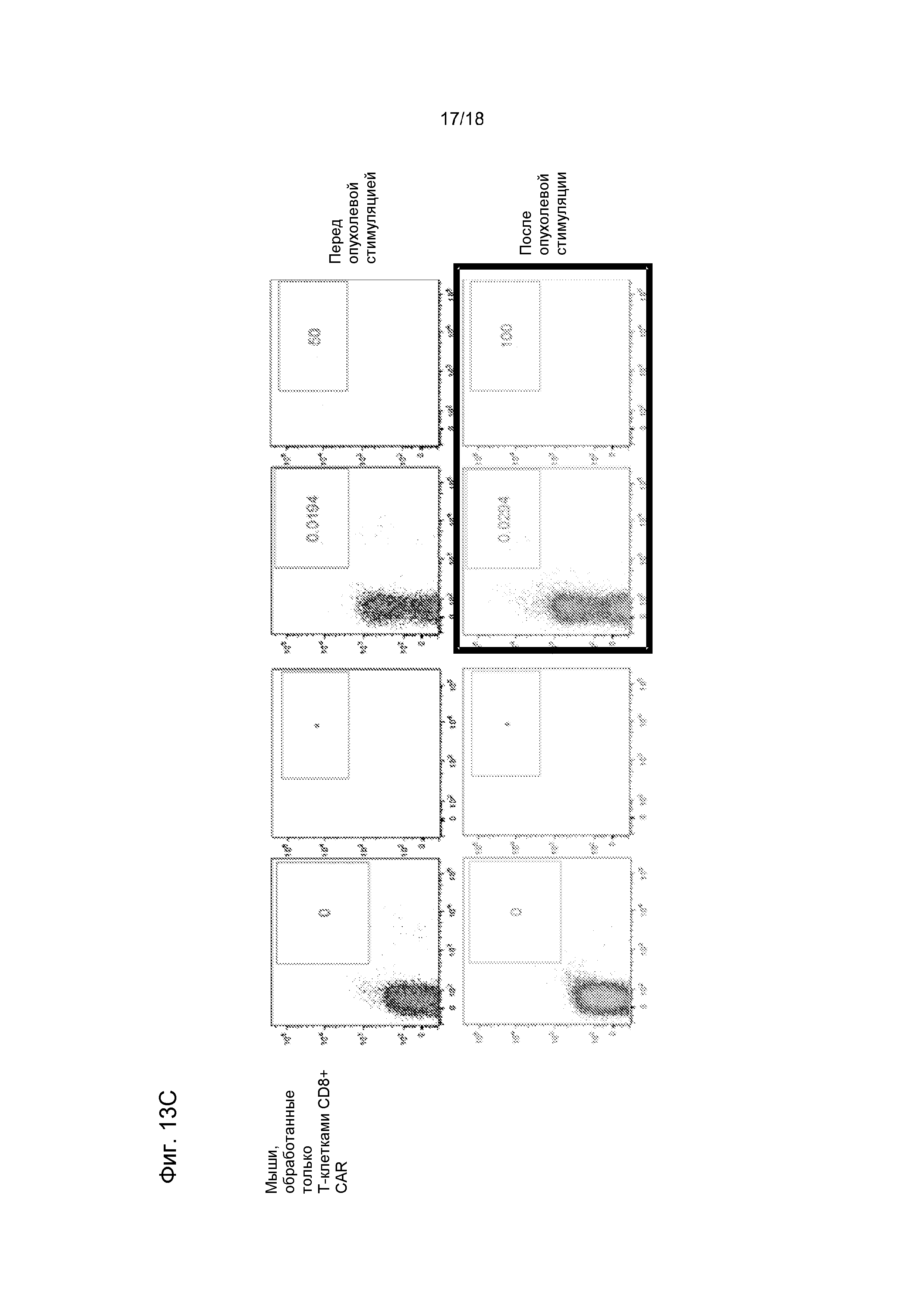
Фиг. 13 показывает синергию T-клеток CD19-CAR CD8+ и CD4+ в мышиной модели системной лимфомы (NSG/Raji). Мышам NSG инокулировали трансдуцированные люциферазой светляка опухолевые клетки Raji. Приживление опухоли Raji подтверждали, используя визуализацию биолюминесценции на 6-й день после инокуляции опухоли (перед обработкой) (схема обработки показана на Фиг. A, приживление опухоли на основании биолюминесценции показано на Фиг. B). Затем группы мышей (n=5) обрабатывали либо CD19-CAR-модифицированными T-клетками CD8+, либо объединенным T-клеточным продуктом, который содержал T-клетки CD19-CAR CD8+ и CD4+. Все мыши получали одну и ту же общую дозу T-клеток (10×106). Анализ опухолевой нагрузки с использованием визуализации биолюминесценции показал полное устранение опухолей Raji в когортах мышей, обработанных T-клетками CD8+ CD19-CAR, и у мышей, обработанных объединенным продуктом T-клеток CD19-CAR CD8+ и CD4+ (после обработки средние черный и серый столбики) B). Затем мышам вводили второй инокулят опухолевых клеток Raji и анализировали частоту встречаемости T-клеток CAR CD4+ и CD8+ в периферической крови и приживление опухоли. У мышей, обработанных объединенным продуктом T-клеток CAR CD8+ и CD4+ наблюдали значимо более высокие уровни T-клеток CAR CD8+ после инокуляции опухоли (нижние панели C) и полное отторжение инокулята Raji (после инокуляции опухоли правый серый столбик, B). Напротив, у мышей, которые получили только CTL CD8+ CD19-CAR, авторы не выявили увеличения количества T-клеток CAR после инокуляции опухоли (C), и опухолевые клетки Raji были способны приживаться (после инокуляции опухоли правый черный столбик, панель B).
Подробное описание предпочтительных вариантов осуществления
T-клетки, или T-лимфоциты, которые использованы в настоящем изобретении, могут быть получены от любого вида млекопитающего, предпочтительно приматов, включая мартышек, собак и человека. В некоторых вариантах T-клетки являются аллогенными (из того же самого вида, но другого донора) по отношению к субъекту-реципиенту; в некоторых вариантах T-клетки являются аутологичными (один и тот же субъект является и донором, и реципиентом); в некоторых вариантах T-клетки являются сингенными (донор и реципиенты являются разными, но идентичными близнецами).
Термин «цитотоксический T-лимфоцит» (CTL) в используемом в настоящем описании смысле относится к T-лимфоциту, который экспрессирует CD8 на своей поверхности (т.е. T-клетка CD8+). В некоторых вариантах такие клетки предпочтительно являются T-клетками «памяти» (клетки TM), которые подвергались контакту с антигеном.
Центральная T-клетка памяти (или TCM) в используемом в настоящем описании смысле относится к клетке CTL, которая подвергалась контакту с антигеном, которая экспрессирует CD62L и CD45RO на своей поверхности и не экспрессирует или имеет пониженную экспрессию CD45RA по сравнению с наивными клетками. В некоторых вариантах центральные клетки памяти являются позитивными в отношении экспрессии CD62L, CCR7, CD28, CD127, CD45RO и CD95 и имеют пониженную экспрессию CD54RA по сравнению с наивными клетками.
Эффекторная T-клетка памяти (или TEM) в используемом в настоящем описании смысле относится к клетке CTL, которая подвергалась контакту с антигеном, которая не экспрессирует или имеет пониженную экспрессию CD62L на своей поверхности по сравнению с центральными клетками памяти и не экспрессирует или имеет пониженную экспрессию CD45RA по сравнению с наивной клеткой. В некоторых вариантах эффекторные клетки памяти являются негативными в отношении экспрессии CD62L, CCR7, CD28, CD45RA и являются позитивными в отношении CD127 по сравнению с наивными клетками или центральными клетками памяти.
Термин «наивные» T-клетки в используемом в настоящем описании смысле относится к не подвергавшемуся контакту с антигеном T-лимфоциту, который экспрессирует CD62L и CD45RA и не экспрессирует или имеет пониженную экспрессию CD45RO- по сравнению с центральными клетками памяти. В некоторых вариантах наивные T-лимфоциты CD8+ характеризуются экспрессией фенотипических маркеров наивных T-клеток, включая CD62L, CCR7, CD28, CD3, CD127 и CD45RA.
Термин «эффекторные» T-клетки, «TE», в используемом в настоящем описании смысле относится к подвергавшимся контакту с антигеном цитотоксическим T-лимфоцитам, которые не экспрессируют или имеют пониженную экспрессию CD62L, CCR7, CD28 и являются позитивными в отношении гранзима B и перфорина по сравнению с центральными клетками памяти.
«Обогащенный» и «истощенный» в используемом в настоящем описании смысле при описании количеств типов клеток в смеси относится к случаю, когда смесь клеток подвергается процессу, или проходит стадию, которая приводит к увеличению количества «обогащенного» типа и уменьшению количества «подвергнутых истощению» клеток. Таким образом, в зависимости от источника исходной популяции клеток, подвергаемых процессу обогащения, смесь или композиция может содержать 60, 70, 80, 90, 95 или 99% или более (при оценке количества или импульсов) «обогащенных» клеток и 40, 30, 20, 10, 5 или 1% или менее (при оценке количества или импульсов) «истощенных» клеток.
Интерлейкин-15 известен и описан, например, в патенте США № 6344192.
«CAR» в используемом в настоящем описании смысле относится к химерному рецептору антигена, содержащему внеклеточный вариабельный домен антитела, специфичный по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного или других рецепторов, такой как костимулирующий домен.
Способы осуществления изобретения
T-лимфоциты CD4+ во время культивирования in vitro значимо увеличивали пролиферацию, продолжительность жизни и противоопухолевую реактивность специфичных для опухоли T-клеток CD8+ in vitro и in vivo. В некоторых вариантах наивные T-клетки CD4+ имеют присущую им программу, которая приводит к более высокой хелперной активности по сравнению с T-клетками CD4+, полученными из центральных и эффекторных T-клеток памяти или суммарных T-клеток CD4+.
В некоторых вариантах реактивные по отношению к опухоли T-клетки CD4+ модифицированы полученным из одноцепочечного антитела химерным рецептором антигена (CAR), специфичным по отношению к орфановому тирозинкиназному рецептору ROR1 или к молекуле CD19. ROR1 одинаково экспрессируется на клетках хронического лимфоцитарного лейкоза (CLL) и лимфомы из клеток мантийной зоны (MCL), и ROR1-специфичный CAR из моноклонального анти-ROR1-антитела (мАт) обеспечивает специфичное распознавание злокачественных, но не зрелых нормальных B-клеток, при экспрессии в цитотоксических T-клетках (CTL) CD8+. T-клетки ROR1-CAR из общей популяции и очищенных проточной сортировкой наивных центральных и эффекторных T-клеток памяти CD4+ получают из периферической крови здоровых доноров и пациентов с CLL. T-клетки CD4+ CAR обладают специфичной, но слабой цитолитической активностью, направленной против опухолей ROR1+, включая первичный CLL, MCL-линию Jeko-1 и клетки K562, трансфицированные ROR1. Мультиплексный анализ цитокинов выявляет высокий уровень продукции Th1-цитокинов со значимо более высокими уровнями IFNγ, TNFα и особенно IL-2 по сравнению с CTL CD8+ CAR. Окрашивание CFSE показывает существенно более высокую пролиферацию после стимуляции ROR1-позитивными опухолевыми клетками, при этом процент клеток, которые были индуцированы к пролиферации, и количество клеточных делений, которым подвергается пролиферирующая подгруппа, значимо выше, чем в случае CTL CD8+ CAR. T-клетки CD4+, полученные от здоровых доноров и пациентов с CLL, приобретают противоопухолевую реактивность после генетической модификации ROR1-специфичным CAR. Кроме того, способность пролиферировать в отсутствие экзогенных цитокинов и продуцировать высокие уровни Th1-цитокинов показывает, что T-клетки CD4+ CAR проявляют типичные хелперные функции после стимуляции посредством CAR, и свидетельствует, что в дополнение к приданию способности к прямым противоопухолевым эффектам они могут быть использованы для усиления специфичных для опухоли CTL CD8+.
Получают профиль цитокинов и определяют пролиферативную способность T-клеток ROR1-CAR, полученных из очищенных проточной сортировкой подгрупп наивных центральных и эффекторных клеток CD4+. T-клетки CD4+ CAR, полученные из подгруппы наивных клеток CD45RA+ CD45RO- CD62L+, продуцируют самые высокие уровни Th1-цитокинов, особенно IL-2 и пролиферируют в ответ на опухолевые клетки ROR1+. Действительно, в экспериментах по совместному культивированию добавление CAR-трансдуцированных, но не добавление нетрансдуцированных T-клеток CD4+ приводит к значимому увеличению специфичной для опухоли пролиферации CTL CD8+ CAR. В некоторых вариантах CAR-модифицированные T-клетки CD4+, полученные из наивной подгруппы, но не из подгрупп центральных и эффекторных клеток памяти, или общей массы T-клеток CD4+, приводят к усиленной пролиферации CTL CD8+ CAR.
Центральные T-клетки памяти CD8+ имеют присущую им программу, которая позволяет им выживать в течение продолжительных периодов времени после введения, что делает их предпочтительной подгруппой T-клеток CD8+ для иммунотерапии. В некоторых вариантах ROR1-CAR- или CD19-CAR-модифицированные CTL из очищенных сортировкой центральных T-клеток памяти CD8+ и наивных CAR-модифицированных T-клеток CD4+ обеспечивают усиленную пролиферацию подгруппы T-клеток CD8+. В некоторых вариантах специфичные для опухоли T-клетки CD4+ проявляют противоопухолевую реактивность и помогают специфичным для опухоли T-клеткам CD8+ in vitro и in vivo. В конкретном варианте используют специфичные для опухоли T-клетки CD4+ из наивной подгруппы.
В другом варианте T-клетки CD8+ и CD4+ могут быть модифицированы T-клеточным рецептором (TCR). TCR может быть специфичным по отношению к любому антигену, патогену или опухоли (существуют TCR ко многим опухолевым антигенам меланомы (например, MART1, gp100), лейкоза (например, WT1, минорные антигены гистосовместимости например), рака молочной железы (например, her2, NY-BR1).
Подробное описание
Композиции
Изобретение относится к композиции для адаптивной клеточной иммунотерапии, содержащей препарат генетически модифицированных хелперных T-лимфоцитов, которые усиливают способность препаратов генетически модифицированных цитотоксических T-лимфоцитов опосредовать клеточный иммунный ответ, при этом препарат хелперных T-лимфоцитов содержит T-клетки CD4+, которые имеют химерный рецептор антигена, содержащий внеклеточный вариабельный домен антитела, специфичный по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного рецептора или других рецепторов.
В некоторых вариантах композиция для адаптивной клеточной иммунотерапии дополнительно содержит препарат модифицированных химерным рецептором антигена специфичных для опухоли цитотоксических T-лимфоцитов CD8+, который вызывает клеточный иммунный ответ, при этом препарат цитотоксических T-лимфоцитов содержит T-клетки CD8+, которые имеют химерный рецептор антигена, содержащий внеклеточное одноцепочечное антитело, специфичное по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного рецептора.
В некоторых вариантах композиция для адаптивной клеточной иммунотерапии содержит препарат модифицированных химерным рецептором антигена специфичных для опухоли цитотоксических T-клеток CD8+, который вызывает клеточный иммунный ответ, при этом препарат цитотоксических T-лимфоцитов содержит T-клетки CD8+, которые имеют химерный рецептор антигена, содержащий внеклеточное одноцепочечное антитело, специфичное по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного рецептора, в сочетании с антиген-реактивными модифицированными химерным рецептором антигена наивными хелперными T-клетками CD4+, полученными из CD45RO-негативных, CD62L-позитивных, CD4-позитивных T-клеток, и фармацевтически приемлемый носитель.
В других вариантах композиция для адаптивной клеточной иммунотерапии содержит препарат антиген-специфичных цитотоксических T-лимфоцитов CD8+, которые вызывают клеточный иммунный ответ, полученные от пациента, в сочетании с антиген-реактивными модифицированными химерным рецептором антигена наивными хелперными T-клетками CD4+, которые усиливают иммунный ответ CD8+, при этом препарат хелперных T-лимфоцитов содержит T-клетки CD4+, которые имеют химерный рецептор антигена, содержащий внеклеточный вариабельный домен антитела, специфичный по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного рецептора.
В следующем варианте композиция для адаптивной клеточной иммунотерапии содержит антиген-реактивные модифицированные химерным рецептором антигена наивные хелперные T-клетки CD4+, которые усиливают иммунный ответ CD8+, при этом препарат хелперных T-лимфоцитов содержит T-клетки CD4+, которые имеют химерный рецептор антигена, содержащий внеклеточный вариабельный домен антитела, специфичный по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного рецептора.
В некоторых вариантах хелперный T-лимфоцит CD4+ выбран из группы, состоящей из наивных T-клеток CD4+, центральных T-клеток памяти CD4+, эффекторных T-клеток памяти CD4+ или общей популяции T-клеток CD4+. В некоторых вариантах клетка хелперного лимфоцита CD4+ представляет собой наивную T-клетку CD4+, при этом наивная T-клетка CD4+ включает T-клетку CD45RO-, CD45RA+, CD62L+, CD4+. В некоторых вариантах цитотоксический T-лимфоцит CD8+ выбран из группы, состоящей из наивных T-клеток CD8+, центральных T-клеток памяти CD8+, эффекторных T-клеток памяти CD8+ или общей популяции T-клеток CD8+. В некоторых вариантах клетка цитотоксического T-лимфоцита CD8+ представляет собой центральную T-клетку памяти, при этом центральная T-клетка памяти включает T-клетку CD45RO+, CD62L+, CD8+. В следующих вариантах цитотоксический T-лимфоцит CD8+ представляет собой центральную T-клетку памяти и хелперный T-лимфоцит CD4+ представляет собой наивную T-клетку CD4+.
В альтернативных вариантах T-клетки могут быть модифицированы рекомбинантным T-клеточным рецептором. TCR может быть специфичным по отношению к любому антигену, патогену или опухоли. Существуют TCR для многих опухолевых антигенов меланомы (например, MART1, gp100), лейкоза (например, WT1, минорные антигены гистосовместимости), рака молочной железы (например, her2, NY-BR1).
Селекция и сортировка популяций T-лимфоцитов
Композиции, описанные в настоящей публикации, обеспечивают антиген-реактивные T-лимфоциты CD4+ и CD8+.
T-лимфоциты могут быть собраны согласно известным методикам и обогащены или истощены известными способами, такими как аффинное связывание с антителами, такими как проточная цитометрия и/или иммуномагнитная селекция. После стадий обогащения и/или истощения размножение in vitro требуемых T-лимфоцитов может быть осуществлено в соответствии с известными методиками (включая без ограничения методики, описанные в патенте США № 6040177, Riddell et al.) или их вариантами, которые будут очевидны для специалистов в данной области.
Например, требуемая популяция или субпопуляция T-клеток может быть размножена добавлением исходной популяции T-лимфоцитов в культуральную среду in vitro и затем добавлением к культуральной среде питающих клеток, таких как неделящиеся мононуклеарные клетки периферической крови (PBMC), (например, так, чтобы полученная в результате популяция клеток содержала по меньшей мере приблизительно 5, 10, 20 или 40 или более питающих PBMC-клеток на каждый T-лимфоцит в исходной популяции, которую необходимо размножить); и инкубацией культуры (например, в течение периода времени, достаточного для увеличения количества T-клеток). Неделящиеся питающие клетки могут содержать облученные гамма-излучением питающие PBMC-клетки. В некоторых вариантах PBMC облучают гамма-лучами в диапазоне приблизительно от 3000 до 3600 рад. При желании порядок добавления T-клеток и питающих клеток в культуральные среды может быть обратным. Культуру, как правило, можно инкубировать в определенных условиях температуры и тому подобного, которые подходят для роста T-лимфоцитов. Например, для роста T-лимфоцитов человека температура обычно будет составлять по меньшей мере приблизительно 25ºС, предпочтительно по меньшей мере приблизительно 30°С, более предпочтительно приблизительно 37°С.
T-лимфоциты, подвергаемые размножению, включают цитотоксические T-лимфоциты (CTL) и хелперные T-лимфоциты, которые специфичны по отношению к антигену, присутствующему на опухоли человека, или патогену.
Необязательно, способ размножения может дополнительно включать стадию добавления неделящихся EBV-трансформированных лимфобластоидных клеток (LCL) в качестве питающих клеток. LCL могут быть облучены гамма-лучами в диапазоне от приблизительно 6000 до 10000 рад. Питающие LCL-клетки могут быть в любом подходящем количестве, так, чтобы соотношение между питающими LCL-клетками и исходными T-лимфоцитами составляло по меньшей мере приблизительно 10:1.
Необязательно, способ размножения может дополнительно включать стадию добавления моноклонального анти-CD3-антитела в культуральную среду (например, в концентрации по меньшей мере приблизительно 0,5 нг/мл). Необязательно, способ размножения может дополнительно включать стадию добавления IL-2 и/или IL-15 в культуральную среду (при этом концентрация IL-2 составляет, например, по меньшей мере приблизительно 10 единиц/мл).
После выделения T-лимфоцитов как цитотоксические, так и хелперные T-лимфоциты могут быть подвергнуты сортировке на субпопуляции наивных T-клеток, T-клеток памяти и эффекторных T-клеток либо до, либо после размножения.
Клетки CD8+ могут быть получены с применением стандартных способов. В некоторых вариантах клетки CD8+ дополнительно сортируют на наивные клетки, центральные клетки памяти и эффекторные клетки посредством идентификации антигенов клеточной поверхности, которые ассоциированы с каждым из указанных типов клеток CD8+. В некоторых вариантах T-клетки памяти представлены в обеих подгруппах CD62L+ и CD62L- лимфоцитов CD8+ периферической крови. PBMC сортируют на фракции CD62L-CD8+ и CD62L+CD8+ после окрашивания анти-CD8- и анти-CD62L-антителами. В некоторых вариантах экспрессия фенотипических маркеров центральных TCM памяти включает экспрессию CD45RO, CD62L, CCR7, CD28, CD3 и CD127, и они являются негативными в отношении гранзима B. В некоторых вариантах центральные T-клетки памяти являются T-клетками CD45RO+, CD62L+, CD8+. В некоторых вариантах эффекторные TE являются негативными в отношении CD62L, CCR7, CD28 и CD127 и позитивными в отношении гранзима B и перфорина. В некоторых вариантах наивные T-лимфоциты CD8+ характеризуются экспрессией фенотипических маркеров наивных T-клеток, включая CD62L, CCR7, CD28, CD3, CD127 и CD45RA.
Являются ли клетки или популяция клеток позитивными в отношении конкретного маркера клеточной поверхности, можно определить проточной цитометрией, используя окрашивание антителом, специфичным по отношению к маркеру поверхности, и совпадающее по изотипу контрольное антитело. Указание того, что популяция клеток является негативной в отношении маркера, означает отсутствие значимого окрашивания популяции клеток специфичным антителом больше, чем в случае контроля изотипа, является позитивной означает равномерное окрашивание популяции клеток больше, чем в случае контроля изотипа. В некоторых вариантах снижение экспрессии одного или нескольких маркеров относится к уменьшению на 1 log10 средней интенсивности флуоресценции и/или уменьшению процентного содержания клеток, на которых обнаруживают маркер по меньшей мере на 20% клеток, 25% клеток, 30% клеток, 35% клеток, 40% клеток, 45% клеток, 50% клеток, 55% клеток, 60% клеток, 65% клеток, 70% клеток, 75% клеток, 80% клеток, 85% клеток, 90% клеток, 95% клеток и 100% клеток и любой % от 20 до 100%, по сравнению с эталонной популяцией клеток. В некоторых вариантах популяция клеток, позитивных по одному или нескольким маркерам, относится к популяции, в которой процентное содержание клеток, на которых обнаруживают маркер, составляет по меньшей мере 50% клеток, 55% клеток, 60% клеток, 65% клеток, 70% клеток, 75% клеток, 80% клеток, 85% клеток, 90% клеток, 95% клеток и 100% клеток и любой % от 50 до 100% по сравнению с эталонной популяцией клеток.
Хелперные T-клетки CD4+ сортируют на наивные, центральные и эффекторные клетки памяти, идентифицируя популяции клеток, которые имеют антигены на клеточной поверхности. Лимфоциты CD4+ могут быть получены стандартными способами. В некоторых вариантах наивные T-лимфоциты CD4+ представляют собой T-клетки CD45RO-, CD45RA+, CD62L+ CD4+. В некоторых вариантах центральные клетки памяти CD4+ являются CD62L-позитивными и CD45RO-позитивными. В некоторых вариантах эффекторные клетки CD4+ являются CD62L- и CD45RO-негативными.
Популяции CD4+ и CD8+, которые являются антиген-специфичными, могут быть получены в результате стимуляции наивных или антиген-специфичных T-лимфоцитов антигеном. Например, могут быть созданы антиген-специфичные клоны T-клеток, специфичные по отношению к антигенам цитомегаловируса, посредством выделения T-клеток из организма инфицированных субъектов и стимуляции клеток in vitro таким же антигеном. Также можно использовать наивные T-клетки. Можно использовать любое количество антигенов из опухолевых клеток, злокачественных клеток или инфекционных агентов. Примеры таких антигенов включают антигены ВИЧ, антигены HCV, антигены HBV, антигены CMV, антигены паразитов и опухолевые антигены, такие как орфановый тирозинкиназный рецептор ROR1, tEGFR, Her2, L1-CAM, CD19, CD20, CD22, мезотелин и CEA. В некоторых вариантах композиции для адаптивной клеточной иммунотерапии применимы при лечении заболевания или нарушения, включая солидную опухоль, гематологическое злокачественное заболевание, меланому или вирусную инфекцию.
Модификация популяций T-лимфоцитов
В некоторых вариантах может быть требоваться введение функциональных генов в T-клетки, используемые для иммунотерапии согласно настоящему изобретению. Например, введенный ген или гены могут повышать эффективность терапии за счет повышения жизнеспособности и/или стимуляции функции переносимых T-клеток; или они могут обеспечивать генетический маркер для осуществления селекции и/или оценки жизнеспособности или миграции in vivo; или они могут включать функции, которые повышают безопасность иммунотерапии, например, делая клетки чувствительными к негативной селекции in vivo, как описано Lupton S.D. et al., Mol. and Cell Biol., 11: 6 (1991); и Riddell et al., Human Gene Therapy 3: 319-338 (1992); также см. публикации PCT/US91/08442 и PCT/US94/05601, Lupton et al., в которых описано применение бифункциональных селектируемых слитых генов, полученных в результате слияния доминантного позитивного селектируемого маркера с негативным селектируемым маркером. Такое введение можно осуществить согласно известным методикам (см., например, патент США № 6040177, Riddell et al., колонки 14-17) или их вариантам, которые будут очевидны для специалистов в данной области на основании настоящего описания.
В некоторых вариантах T-клетки модифицируют химерными рецепторами антигенов (CAR). В некоторых вариантах CAR содержат фрагмент одноцепочечного антитела (scFv), который получен из вариабельного домена тяжелой (VH) и вариабельного домена легкой (VL) цепи моноклонального антитела (мАт), связанный с цепью TCR CD3+, которая опосредует активацию и цитотоксичность T-клеток. Костимулирующие сигналы также могут быть обеспечены посредством CAR за счет слияния костимулирующего домена CD28 или 4-1BB с цепью CD3+. CAR специфичны по отношению к молекулам клеточной поверхности, независимым от HLA, таким образом преодолеваются ограничения TCR-распознавания, включая ограничения по HLA и низкие уровни экспрессии HLA на опухолевых клетках.
CAR могут быть сконструированы со специфичностью по отношению к любому маркеру клеточной поверхности с использованием антиген-связывающих фрагментов или вариабельных доменов антител, например, молекул антител. Антиген-связывающие молекулы могут быть связаны с одним или несколькими модулями передачи клеточных сигналов. В вариантах осуществления изобретения модули передачи клеточных сигналов включают трансмембранный домен CD3, внутриклеточные домены передачи сигналов CD3 и трансмембранные домены CD28. В вариантах осуществления изобретения внутриклеточный домен передачи сигнала содержит трансмембранный и передающий сигнал домен CD28, связанный с внутриклеточным доменом CD3. В некоторых вариантах CAR также может содержать маркер трансдукции, такой как tEGFR.
В вариантах осуществления изобретения внутриклеточный домен передачи сигнала цитотоксических T-клеток CD8+ является таким же, как и внутриклеточный домен передачи сигнала хелперных T-клеток CD4+. В других вариантах внутриклеточный домен передачи сигнала цитотоксических T-клеток CD8+ отличается от внутриклеточного домена передачи сигнала хелперных T-клеток CD4+.
В некоторых вариантах обе клетки, T-клетка CD8+ и T-клетка CD4+, генетически модифицированы доменом тяжелой цепи антитела, который специфично связывает антиген клеточной поверхности, специфичный для патогена. В вариантах осуществления изобретения CAR являются специфичными по отношению к экспрессируемым на клеточной поверхности антигенам, ассоциированным с патогенами, опухолями или злокачественными клетками. В некоторых вариантах CAR специфичен по отношению к антигенам ВИЧ, антигенам HCV, антигенам HBV, антигенам CMV, антигенам паразитов и опухолевым антигенам, таким как орфановый тирозинкиназный рецептор ROR1, tEGFR, Her2, L1-CAM, CD19, CD20, CD22, мезотелин и CEA. Способы получения CAR описаны в настоящей публикации, и их также можно найти в публикациях 6410319, Forman, и WO 2002/077029, 7446191, 2010/065818, 2010/025177, 2007/059298 и 7514537, Jensen et al., и они описаны Berger C. et al., J. Clinical Investigation, 118: 1 294-308 (2008), при этом все публикации включены в настоящее описание посредством ссылки.
В вариантах осуществления изобретения одинаковые или разные CAR могут быть введены в каждый из T-лимфоцитов CD4+ и CD8+. В вариантах осуществления изобретения CAR в каждой из таких популяций имеет антиген-связывающую молекулу, которая специфично связывается с одним и тем же антигеном. Молекулы клеточной передачи сигнала могут отличаться. В вариантах осуществления изобретения каждая из популяций T-лимфоцитов CD4 или CD8 перед трансдукцией может быть подвергнута сортировке на наивные, центральные клетки памяти, эффекторные клетки памяти или эффекторные клетки. В альтернативных вариантах каждая из популяций T-лимфоцитов CD4 или CD8 перед трансдукцией может быть подвергнута сортировке на наивные, центральные клетки памяти, эффекторные клетки памяти или эффекторные клетки.
В альтернативных вариантах T-клетки могут быть модифицированы рекомбинантным T-клеточным рецептором. TCR может быть специфичным по отношению к любому антигену, патогену или опухоли. Существуют TCR для многих опухолевых антигенов меланомы (например, MART1, gp100), лейкоза (например, WT1, минорные антигены гистосовместимости), рака молочной железы (например, her2, NY-BR1).
Были разработаны различные способы инфекции, в которых используют рекомбинантные инфекционные вирусные частицы для доставки генов. Такой способ представляет предпочтительный в настоящее время подход к трансдукции T-лимфоцитов согласно настоящему изобретению. Вирусные векторы, которые были использованы таким образом, включают вирусные векторы, полученные из вируса обезьян 40, аденовирусов, аденоассоциированного вируса (AAV), лентивирусных векторов и ретровирусов. Таким образом, способы переноса и экспрессии генов многочисленны, но основной их функцией является введение и экспрессия генетического материала в клетках млекопитающих. Несколько из указанных выше способов применяли для трансдукции гематопоэтических или лимфоидных клеток, включая трансфекцию с использованием фосфата кальция, слияние протопластов, электропорацию и инфекцию векторами на основе рекомбинантных аденовирусов, аденоассоциированных вирусов и ретровирусов. Первичные T-лимфоциты были успешно трансдуцированы электропорацией и инфекцией ретровирусами.
Ретровирусные векторы обеспечивают высокоэффективный способ переноса генов в эукариотические клетки. Кроме того, интеграция ретровирусов происходит контролируемым образом и приводит к стабильной интеграции одной или нескольких копий новой генетической информации в клетку.
Предполагается, что сверхэкспрессия стимулирующего фактора (например, лимфокина или цитокина) может быть токсичной для индивидуума, подвергаемого лечению. Поэтому в объем изобретения включены генные участки, которые вынуждают T-клетки согласно изобретению быть чувствительными к негативной селекции in vivo. Под негативной селекцией подразумевают, что инфузированная клетка может быть элиминирована в результате изменения состояния индивидуума in vivo. Негативный селектируемый фенотип может быть результатом инсерции гена, который придает чувствительность к вводимому агенту, например, соединению. Негативные селектируемые гены известны в данной области и включают наряду с прочими следующие гены: ген тимидинкиназы вируса простого герпеса типа I (TK HSV-I) (Wigler et al., Cell 11: 223, 1977), который придает чувствительность к ганцикловиру; клеточный ген гипоксантинфосфорибозилтрансферазы (HPRT), клеточный ген аденинфосфорибозилтрансферазы (APRT), ген бактериальной цитозиндезаминазы (Mullen et al., Proc. Natl. Acad. Sci. USA. 89: 33 (1992)).
В некоторых вариантах может быть полезным включение в T-клетки позитивного маркера, который делает возможным отбор клеток с негативным селектируемым фенотипом in vitro. Позитивный селектируемый маркер может представлять собой ген, который при введении в клетку-хозяина экспрессирует доминантный фенотип, обеспечивающий возможность позитивной селекции клеток, несущих ген. Гены такого типа известны в данной области и включают наряду с прочими ген гигромицин-B-фосфотрансферазы (hph), который придает резистентность к гигромицину B, ген аминогликозидфосфотрансферазы (neo или aph) из Tn5, который кодирует резистентность к антибиотику G418, ген дигидрофолатредуктазы (DHFR), ген аденозиндезаминазы (ADA) и ген множественной лекарственной резистентности (MDR).
Предпочтительно позитивный селектируемый маркер и негативный селектируемый элемент связаны так, чтобы утрата негативного селектируемого элемента также обязательно сопровождалась утратой позитивного селектируемого маркера. Еще более предпочтительно позитивные и негативные селектируемые маркеры сливают так, чтобы утрата одного обязательно приводила к утрате другого. Примером слитого полинуклеотида, который дает в качестве продукта экспрессии полипептид, который придает требуемые признаки как для позитивной, так и для негативной селекции, описанные выше, является слитый ген гигромицинфосфотрансферазы-тимидинкиназы (HyTK). Экспрессия такого гена дает полипептид, который придает резистентность к гигромицину B для позитивной селекции in vitro и чувствительность к ганцикловиру для негативной селекции in vivo. См. публикацию Lupton S.D. et al., Mol. and Cell. Biology 11: 3374-3378, 1991. Кроме того, в предпочтительных вариантах полинуклеотиды согласно изобретению, кодирующие химерные рецепторы, находятся в ретровирусных векторах, содержащих слитый ген, особенно таких, которые придают резистентность к гигромицину B для позитивной селекции in vitro и чувствительности к ганцикловиру для негативной селекции in vivo, например, ретровирусном векторе HyTK, описанном в публикации Lupton, S.D. et al. (1991), указанной выше. Также см. публикации PCT/US91/08442 и PCT/US94/05601, S.D. Lupton, описывающие применение бифункциональных селектируемых слитых генов, полученных в результате слияния доминантных позитивных селектируемых маркеров с негативными селектируемыми маркерами.
Предпочтительные позитивные селектируемые маркеры получают из генов, выбранных из группы, состоящей из hph, nco и gpt, и предпочтительные негативные селектируемые маркеры получают из генов, выбранных из группы, состоящей из цитозиндезаминазы, TK HSV-I, TK VZV, HPRT, APRT и gpt. Особенно предпочтительными маркерами являются бифункциональные селектируемые слитые гены, при этом позитивный селектируемый маркер получен из hph или neo, а негативный селектируемый маркер получен из цитозиндезаминазы или гена TK или селектируемого маркера.
Можно применять множество способов для трансдукции T-лимфоцитов, которые хорошо известны в данной области. Например, ретровирусные трансдукции могут быть осуществлены следующим образом: в 1-й день после стимуляции с использованием REM, как описано в настоящей публикации, клетки обеспечивают 20-30 единицами/мл IL-2; на 3-й день заменяют половину среды супернатантом ретровирусов, полученным согласно стандартным способам, и затем добавляют в культуры 5 мкг/мл полибрена и 20-30 единиц/мл IL-2; на 4-й день промывают клетки и помещают их в свежую культуральную среду с добавлением 20-30 единиц/мл IL-2; на 5-й день повторяют экспозицию с ретровирусом; на 6-й день помещают клетки в селективную среду (содержащую, например, антибиотик, соответствующий гену резистентности к антибиотикам, имеющемуся в ретровирусном векторе) с добавлением 30 единиц/мл IL-2; на 13-й день отделяют видимые живые клетки от мертвых клеток, используя разделение в градиенте плотности фиколл-гипак, и затем субклонируют живые клетки.
Клетки CD4+ и CD8+ могут быть модифицированы экспрессирующим вектором, кодирующим CAR. В вариантах осуществления изобретения такие клетки затем дополнительно сортируют на субпопуляции наивных, центральных клеток памяти и эффекторных клеток, как описано выше, за счет сортировки антигенов клеточной поверхности, уникальных для каждой из таких популяций клеток. Кроме того, популяции клеток CD4+ или CD8+ могут быть подвергнуты селекции на основе профиля их цитокинов или пролиферативных активностей. Например, могут быть отобраны T-лимфоциты CD4+, которые имеют повышенную продукцию таких цитокинов, как IL-2, IL-4, IL-10, TNFα и IFNγ по сравнению с ложно трансдуцированными клетками или трансдуцированными клетками CD8+. В других вариантах отбирают наивные T-клетки CD4+, которые имеют повышенную продукцию IL-2 и/или TNFα. Подобным образом отбирают клетки CD8+, которые имеют повышенную продукцию IFNγ по сравнению с ложно трансдуцированными клетками CD8+.
В вариантах осуществления изобретения отбирают клетки CD4+ и CD8+, которые пролиферируют в ответ на антиген. Например, отбирают клетки CD4+, которые энергично пролиферируют при стимуляции антигеном по сравнению с ложно трансдуцированными клетками или трансдуцированными клетками CD8+.
В некоторых вариантах отбирают клетки CD4+ и CD8+, которые являются цитотоксическими для клеток, несущих антиген. В вариантах осуществления изобретения предполагают, что CD4+ являются слабо цитотоксическими по сравнению с клетками CD8+.
В изобретении предполагается, что в композициях будут использованы сочетания T-клеток CD4+ и CD8+. В одном варианте сочетания CAR-трансдуцированные клетки CD4+ можно объединять с антиген-реактивными клетками CD8+ с такой же антигенной специфичностью, что и CAR. В других вариантах CAR-трансдуцированные клетки CD8+ клетки объединяют с антиген-реактивными клетками CD4+. В еще одном варианте объединяют CAR-модифицированные клетки CD4+ и CD8+.
Как описано в настоящей публикации, в изобретении предполагается, что клетки CD4+ и CD8+ могут быть дополнительно разделены на субпопуляции, такие как популяции наивных клеток, центральных клеток памяти и эффекторных клеток. Как описано в настоящей публикации, в некоторых вариантах наивные клетки CD4+ являются T-клетками CD45RO-, CD45RA+, CD62L+ CD4+. В некоторых вариантах центральные клетки памяти CD4+ являются CD62L-позитивными и CD45RO-позитивными. В некоторых вариантах эффекторные клетки CD4+ являются CD62L негативными и CD45RO-позитивными. Каждая из таких популяций независимо может быть модифицирована с использованием CAR.
Как описано в настоящей публикации, в вариантах осуществления изобретения T-клетки памяти присутствуют как в подгруппе CD62L+, так и в подгруппе CD62L- лимфоцитов CD8+ периферической крови. PBMC сортируют на фракции CD62L-CD8+ и CD62L+CD8+ после окрашивания анти-CD8- и анти-CD62L-антителами. В некоторых вариантах экспрессия фенотипических маркеров центральных клеток памяти TCM включает CD62L, CCR7, CD28, CD3 и CD127, и они являются негативными в отношении гранзима B. В некоторых вариантах центральные T-клетки памяти представляют собой T-клетки CD45RO+, CD62L+, CD8+. В некоторых вариантах эффекторные TE являются негативными в отношении CD62L, CCR7, CD28 и CD127 и позитивными в отношении гранзима B и перфорина. В некоторых вариантах наивные T-лимфоциты CD8+ характеризуются CD8+, CD62L+, CD45RO+, CCR7+, CD28+, CD127+ и CD45RO+. Каждая из таких популяций независимо может быть модифицирована CAR.
Каждая из субпопуляций клеток CD4+ и CD8+ может быть объединена с другой. В конкретном варианте модифицированные наивные клетки CD4+ объединяют с модифицированными центральными T-клетками памяти CD8+, чтобы получить синергетическое цитотоксическое действие на клетки, несущие антиген, такие как опухолевые клетки.
Способы
Изобретение относится к способам получения композиций для адаптивной иммунотерапии и применениям или способам применения таких композиций для осуществления клеточной иммунотерапии у субъекта, имеющего заболевание или нарушение.
В вариантах осуществления изобретения способ производства композиций включает получение модифицированной хелперной T-клетки CD4+, при этом препарат модифицированных хелперных T-лимфоцитов содержит T-клетки CD4+, которые имеют химерный рецептор антигена, содержащий внеклеточный вариабельный домен антитела, специфичный по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала.
В другом варианте способ дополнительно включает получение модифицированной цитотоксической T-клетки CD8+, при этом препарат модифицированных цитотоксических T-лимфоцитов содержит клетки CD8+, которые имеют химерный рецептор антигена, содержащий внеклеточный вариабельный домен антитела, специфичный по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного рецептора.
В другом варианте способ включает получение модифицированной цитотоксической T-клетки CD8+, при этом препарат модифицированных цитотоксических T-лимфоцитов содержит T-клетки CD8+, которые имеют химерный рецептор антигена, содержащий внеклеточный вариабельный домен антитела, специфичный по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного рецептора, и, кроме того, способ дополнительно включает объединение модифицированных цитотоксических T-клеток CD8+ с препаратом антиген-специфичных хелперных лимфоцитов CD4+.
Препарат клеток CD4+ и CD8+, которые модифицированы с использованием CAR, описаны выше, а также описаны в примерах. Антиген-специфичные T-лимфоциты могут быть получены от пациента, имеющего заболевание или нарушение, или могут быть получены стимуляцией T-лимфоцитов in vitro в присутствии антигена. Субпопуляции T-лимфоцитов CD4+ и CD8+ также могут быть выделены, как описано в настоящей публикации, и объединены при осуществлении способов производства.
Изобретение также относится к способам осуществления клеточной иммунотерапии у субъекта, имеющего заболевание или нарушение, включающим введение композиции по любому из пп. 1-19. В других вариантах способ включает введение субъекту препарата генетически модифицированных цитотоксических T-лимфоцитов, который обеспечивает клеточный иммунный ответ, при этом препарат цитотоксических T-лимфоцитов содержит T-клетки CD8+, которые имеют химерный рецептор антигена, содержащий внеклеточный вариабельный домен антитела, специфичный по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного или других рецепторов, и препарат генетически модифицированных хелперных T-лимфоцитов, который вызывает прямое распознавание опухоли и усиливает способность препаратов генетически модифицированных цитотоксических T-лимфоцитов опосредовать клеточный иммунный ответ, при этом препарат хелперных T-лимфоцитов содержит T-клетки CD4+, которые имеют химерный рецептор антигена, содержащий внеклеточный вариабельный домен антитела, специфичный по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный домен передачи сигнала T-клеточного рецептора.
В другом варианте способ осуществления клеточной иммунотерапии у субъекта, имеющего заболевание или нарушение, включает введение субъекту препарата генетически модифицированных хелперных T-лимфоцитов, при этом препарат модифицированных хелперных T-лимфоцитов содержит T-клетки CD4+, которые имеют химерный рецептор антигена, содержащий внеклеточный вариабельный домен антитела, специфичный по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный модуль передачи сигнала T-клеточного рецептора. В вариантах осуществления изобретения способ дополнительно включает введение субъекту препарата генетически модифицированных цитотоксических T-лимфоцитов, при этом препарат модифицированных цитотоксических T-лимфоцитов содержит CD8-позитивные клетки, которые имеют химерный рецептор антигена, содержащий внеклеточный вариабельный домен антитела, специфичный по отношению к антигену, ассоциированному с заболеванием или нарушением, и внутриклеточный модуль передачи сигнала T-клеточного рецептора.
Другой вариант описывает способ осуществления клеточной иммунотерапии у субъекта, имеющего заболевание или нарушение, включающий анализ биологического образца субъекта в отношении присутствия антигена, ассоциированного с заболеванием или нарушением, и введение композиций для адаптивной иммунотерапии, описанных в настоящей публикации, при этом химерный рецептор антигена специфично связывается с антигеном.
Получают CAR, который имеет компонент, который обеспечивает специфичное связывание с антигеном, ассоциированным с заболеванием или состоянием, таким как солидная опухоль, злокачественная опухоль, вирусная инфекция и инфекция паразитом. В вариантах осуществления изобретения внутриклеточный модуль передачи сигнала T-клеточного рецептора химерного рецептора антигена содержит трансмембранный домен, домен передачи сигнала CD28 и внутриклеточный домен передачи сигнала CD3 или другие домены костимулирующих молекул T-клеток. В некоторых вариантах молекула внутриклеточной передачи сигнала содержит внутриклеточный домен CD3, домен CD28, трансмембранный и передающий сигнал домен CD28, связанный с внутриклеточным доменом CD3, или другие домены костимулирующих молекул T-клеток.
В альтернативных вариантах T-клетки могут быть модифицированы рекомбинантным T-клеточным рецептором. TCR может быть специфичным по отношению к любому антигену, патогену или опухоли. Существуют TCR для многих опухолевых антигенов меланомы (например, MART1, gp100), лейкоза (например, WT1, минорные антигены гистосовместимости), рака молочной железы (например, her2, NY-BR1).
В некоторых вариантах хелперный T-лимфоцит CD4+ выбран из группы, состоящей из наивных T-клеток CD4+, центральных T-клеток памяти CD4+, эффекторных T-клеток памяти CD4+ или суммарной популяции T-клеток CD4+. В конкретном варианте хелперный лимфоцит CD4+ представляет собой наивную T-клетку CD4+, при этом наивная T-клетка CD4+ включает T-клетку CD45RO-, CD45RA+, CD62L+ CD4+. В следующих вариантах цитотоксический T-лимфоцит CD8+ выбран из группы, состоящей из наивных T-клеток CD8+, центральных T-клеток памяти CD8+, эффекторных T-клеток памяти CD8+ или суммарной популяции T-клеток CD8+. В конкретном варианте цитотоксический T-лимфоцит CD8+ представляет собой центральную T-клетку памяти, при этом центральная T-клетка памяти включает T-клетку CD45RO+, CD62L+, CD8+. В конкретном варианте цитотоксический T-лимфоцит CD8+ представляет собой центральную T-клетку памяти и хелперный T-лимфоцит CD4+ представляет собой наивную T-клетку CD4+.
В вариантах осуществления изобретения обе клетки, T-клетка CD8+ и T-клетка CD4+, генетически модифицированы CAR-содержащим доменом тяжелой цепи антитела, который специфично связывает патоген или специфичный для опухоли антиген клеточной поверхности. В других вариантах внутриклеточный домен передачи сигнала цитотоксических T-клеток CD8 является таким же, как внутриклеточный домен передачи сигнала хелперных T-клеток CD4. В следующих вариантах внутриклеточный домен передачи сигнала цитотоксических T-клеток CD8 отличается от внутриклеточного домена передачи сигнала хелперных T-клеток CD4.
Субъектами, которых можно лечить согласно настоящему изобретению, в общем, являются человек и, в ветеринарных целях, другие приматы, такие как мартышки и человекообразные обезьяны, Субъекты могут быть мужского или женского пола и любого подходящего возраста, включая новорожденных, детей, подростков, взрослых и пожилых субъектов.
Способы применимы для лечения, например, солидной опухоли, гематологического злокачественного заболевания, меланомы или инфекции, вызванной вирусом или другим патогеном. Инфекции, вызванные патогенами, включают инфекции ВИЧ, HCV, HBV, CMV и паразитарные заболевания. В некоторых вариантах антиген, ассоциированный с заболеванием или нарушением, выбран из группы, состоящей из орфанового тирозинкиназного рецептора ROR1, tEGFR, Her2, L1-CAM, CD19, CD20, CD22, мезотелина, CEA и поверхностного антигена гепатита B.
Субъекты, которых можно лечить, включают субъектов, пораженных злокачественной опухолью, включая, без ограничения, рак ободочной кишки, легкого, печени, молочной железы, простаты, яичника, кожи (включая меланому), кости и головного мозга и т.д. В некоторых вариантах ассоциированные с опухолью антигены известны, например, в случае меланомы, рака молочной железы, плоскоклеточной карциномы, рака ободочной кишки, лейкоза, миеломы, рака простаты и т.д. (в таких вариантах T-клетки памяти могут быть выделены или сконструированы путем введения генов T-клеточного рецептора). В других вариантах ассоциированные с опухолью белки могут быть мишенью генетически модифицированных T-клеток, экспрессирующих сконструированный иммунорецептор. Примеры включают, без ограничения, B-клеточную лимфому, рак молочной железы, рак простаты и лейкоз.
Субъекты, которых можно лечить, также включают субъектов, пораженных инфекционным заболеванием, или субъектов, для которых существует риск развития инфекционного заболевания, включая, без ограничения, вирусные, ретровирусные, бактериальные и протозойные инфекции и т.д. Субъекты, которых можно лечить, включают пациентов с иммунным дефицитом, пораженных вирусной инфекцией, включая, без ограничения, цитомегаловирус (CMV), вирус Эпштейна-Барр (EBV), аденовирус, полиомавирус BK у пациентов с трансплантатами и т.д.
Клетки, полученные, как описано выше, можно использовать в способах и композициях для адаптивной иммунотерапии согласно известным методикам или их вариантам, которые будут очевидны для специалистов в данной области на основании настоящего описания. См., например, публикацию заявки на выдачу патента США № 2003/0170238, Gruenberg et al.; также см. патент США № 4690915, Rosenberg.
В некоторых вариантах клетки готовят, сначала собирая их из их культуральной среды и затем промывая и концентрируя клетки в среде и системе для упаковки, подходящих для введения (фармацевтически приемлемый носитель) в эффективном для лечения количестве. Подходящей средой для инфузии может быть любой препарат изотонической среды, обычно можно использовать физиологический раствор соли, Normosol R (Abbott) или Plasma-Lyte A (Baxter), а также 5% декстрозу в воде или лактат Рингера. Среда для инфузии может быть дополнена человеческим сывороточным альбумином.
Эффективное для лечения количество клеток в композиции составляет по меньшей мере 2 клетки (например, 1 центральная T-клетка памяти CD8+ и 1 T-клетка подгруппы хелперов CD4+) или чаще более чем 102 клеток и до 106, вплоть до и включая 108 или 109 клеток, и может составлять более чем 1010 клеток. Количество клеток будет зависеть от конечного применения, для которого композиция предназначена, а также типа включенных в нее клеток. Например, если требуются клетки, которые специфичны для конкретного антигена, то популяция будет содержать более 70%, обычно более 80%, 85% и 90-95% таких клеток. Для применений, предлагаемых в настоящем изобретении, клетки обычно содержатся в объеме 1 л или менее, может быть 500 мл или менее, даже 250 мл или 100 мл или менее. Поэтому плотность требуемых клеток обычно более 106 клеток/мл и обычно более 107 клеток/мл, обычно 108 клеток/мл или более. Клинически подходящее количество иммунных клеток может быть распределено на несколько инфузий, и их количество в совокупности равно или превышает 109, 1010 или 1011 клеток.
В некоторых вариантах лимфоциты согласно изобретению могут быть использованы для придания иммунитета индивидуумам. Под «иммунитетом» подразумевают уменьшение одного или нескольких физических симптомов, ассоциированных с ответом на инфекцию, вызванную патогеном, или на опухоль, к которым направлен ответ лимфоцитов. Количество вводимых клеток обычно находится в диапазоне, имеющем место у нормальных индивидуумов, которые обладают иммунитетом к патогену. Таким образом, клетки обычно вводят посредством инфузии, при этом каждую инфузию осуществляют в диапазоне от 2 клеток до по меньшей мере 106-1010 клеток/м2, предпочтительно в диапазоне по меньшей мере от 107 до 109 клеток/м2. Клоны можно вводить в виде однократной инфузии или множества инфузий в течение определенного периода времени. Однако поскольку разные индивидуумы, как предполагают, отличаются по чувствительности, тип и количество инфузируемых клеток, а также количество инфузий и период времени, на протяжении которого проводят множественные инфузии, определяет лечащий врач, и определение может быть осуществлено при обычном экспериментировании. Создание достаточных уровней T-лимфоцитов (включая цитотоксические T-лимфоциты и/или хелперные T-лимфоциты) легко достижимо с использованием способа быстрого размножения согласно настоящему изобретению, который проиллюстрирован в настоящем описании. См., например, патент США № 6040177, Riddell et al., колонка 17.
Настоящее изобретение дополнительно проиллюстрировано в примерах, приведенных ниже.
ЭКСПЕРИМЕНТАЛЬНЫЙ РАЗДЕЛ
Пример 1: Трансдукция T-клеток и анализ экспрессии CAR
ROR1-специфичный CAR может быть экспрессирован в T-клетках CD8+ человека, и он обеспечивает специфичное распознавание B-клеточных опухолей ROR1+, но не зрелых нормальных B-клеток. Авторы сконструировали ROR1-специфичный химерный рецептор антигена, который при экспрессии в T-клетках от здоровых доноров или CLL-пациентов придавал способность к специфичному распознаванию первичного B-CLL и лимфомы из клеток мантийной зоны.
Материалы и способы
Линии клеток
Трансформированные вирусом Эпштейна-Барр B-клетки (EBV-LCL) получали, как описано (25). Линии опухолевых клеток Jeko-1 и BALL-1 предоставлены докторами Oliver Press и Jerald Radich (Онкологического исследовательского центра имени Фреда Хатчинсона). Все линии клеток поддерживали в RPMI, содержащей 10% фетальной сыворотки теленка, 0,8 мМ L-глутамин и 1% пенициллин-стерптомицин (среда LCL). Клетки K562 получали из Американской коллекции типов культур.
Трансфекция клеток K562 с использованием ROR1
Для основанной на полимеразной цепной реакции (ПЦР) амплификации ROR1-гена получали суммарную РНК из клеток B-CLL (набор RNeasyPlus; QIAGEN) и обратно транскрибировали в кДНК с использованием обратной транскриптазы M-MLV (Invitrogen). ПЦР осуществляли со специфичными праймерами
(ROR1-F: 5-XhoIAGAGGAGGAATGCACCGGCC-3 и
ROR1-R: 5-XhoI-CACAGAAGGTACTTGTTGCGATGT-3),
используя ДНК-полимеразу Herculase-II (Stratagene). ПЦР-продукт клонировали в ретровирусном векторе MIGR-1 (23) и последовательность подтверждали. Реагент для трансформации Effectene (QIAGEN) использовали для того, чтобы трансфицировать клетки Platinum-A (Cell Biolabs) MIGR-1/ROR1 и получить ROR1-кодирующий ретровирус. Клетки K562 трансдуцировали ретровирусом, используя центрифугирование при 2500 об/мин в течение 60 минут при 32°C, размножали и очищали ROR1-позитивную подгруппу, используя сортировку.
Количественная ПЦР в реальном времени
Первую нить кДНК B-CLL, нормальных неактивированных и активированных B-клеток и EBV-LCL получали, как описано в предыдущем абзаце. Первую нить кДНК из нормальных тканей (панели тканей человека I/II, фракции крови) получали из Clontech. Экспрессию мРНК ROR1 анализировали в двух повторах и нормализовали по отношению к GAPDH. Амплификации осуществляли на устройстве ABI Prism 7900 (Applied Biosystems) в 50 мкл реакционной смеси, состоящей из 25 мкл основной смеси для ПЦР Power SYBR Green (Applied Biosystems), 2,5 нг кДНК и 300 нМ специфичных для гена прямых и обратных праймеров:
ROR1-F 5-AGCGTGCGATTCAAAGGATT-3,
ROR1-R 5-GACTGGTGCCGACGATGACT-3,
GAPDH-F 5-GAAGGTGAAGGTCGGAGTC-3,
и GAPDH-R 5-GAAGATGGTGATGGGATTTC-3.
Пороговый цикл (Ct) определяли, используя компьютерную программу SDS v2.2.2 (Applied Biosystems), и уровень экспрессии генов вычисляли, используя способ сравнения Ct (2-(ΔΔCt)).
Конструирование векторов и создание лентивируса
Лентивирусные векторы, кодирующие CD20-CAR (CD20R-epHIV7) и зеленый флуоресцирующий белок (GFP), (GFP-epHIV7), описаны ранее (24). ROR1-CAR кодировали в таком же векторе. Мышиное мАт (клон 2A2), которое, как было показано, специфично связывается с ROR1 человека, экспрессируемым на первичных опухолевых линиях B-CLL и MCL, создавали, клонировании и характеризовали в предыдущем исследовании. Оптимизированную по кодонам нуклеотидную последовательность, кодирующую scFv, содержащий VL- и VH-цепи мАт 2A2, синтезировали (GENEART) и клонировали в CD20R-epHIV7, используя сайты рестрикции NheI и RsrII, чтобы заменить CD20-специфичный scFv. Лентивирус продуцировали в клетках 293T, котрансфицированных лентивирусным вектором и пакующими векторами pCHGP-2, pCMVRev2 и pCMV-G, используя Effectene (Qiagen). Среду меняли через 16 часов после трансфекции и лентивирусы собирали через 48 часов.
Лентивирусная трансдукция и выделение CAR-трансдуцированных T-клеточных клонов
PBMC от здоровых доноров и пациентов с B-CLL и очищенные сортировкой центральные T-клетки памяти (TCM) CD8+CD45RO+CD62L+ активировали, используя анти-CD3-мАт (30 нг/мл) (25), и трансдуцировали в супернатанте с лентивирусами с добавлением 1 мкг/мл полибрена (Sigma-Aldrich) и 50 ME(IU)/мл рекомбинантного интерлейкина-2 (IL-2) человека на 2-й и 3-й день после активации, центрифугируя при 2500 об./мин в течение 60 минут при 32°C. T-клетки размножали в RPMI, содержащей 10% сыворотки человека, 2 мМ L-глутамина и 1% пенициллина-стрептомицина (среда CTL) (25). После размножения аликвоту каждой линии трансдуцированных T-клеток красили конъюгированным с биотином анти-EGFR-мАт (против рецептора эпителиального фактора роста), стрептавидином-PE и анти-CD8-мАт. T-клетки EGFR+CD8+ очищали сортировкой и клонировали лимитирующим разведением (0,5 клеток/лунка) (25). ROR1-CAR-трансдуцированные T-клетки идентифицировали по окрашиванию биотинилированным рекомбинантным слитым белком Fc-внеклеточный домен ROR1 и стрептавидином-PE. Рекомбинантный ROR1-белок получали во временно трансфицированных клетках 293F (Invitrogen), очищали, как описано (26), и биотинилировали, используя набор BiotinTag (Sigma). GFP-трансдуцированные T-клетки CD8+ идентифицировали с использованием проточной цитометрии, очищали сортировкой и клонировали сходным образом.
Анализы высвобождения хрома и секреции цитокинов
Клетки-мишени метили51Cr (PerkinElmer) в течение ночи, промывали и инкубировали в трех повторах по 1-2×103 клеток/лунка с эффекторными T-клетками в разных соотношениях между эффекторными клетками и клетками-мишенями (E:T). Супернатанты собирали для подсчета гамма-излучения после 4-часовой инкубации и специфичный лизис вычисляли, используя стандартную формулу (25).
Результаты
Трансдуцированные T-клетки CD8+ очищали сортировкой, используя биотинилированное анти-EGFR-мАт и конъюгированные со стрептавидином красители. Экспрессию ROR1-CAR на поверхности очищенных сортировкой T-клеток оценивали окрашиванием клеток биотинилированным рекомбинантным слитым белком Fc-внеклеточный домен ROR1, который непосредственно связывается с scFv ROR1-CAR, и совместным окрашиванием конъюгатами стрептавидина. Белок Fc-ROR1 специфично окрашивал T-клетки CD8+, трансдуцированные лентивирусным вектором ROR1-CAR, но не окрашивал T-клетки CD8+, трансдуцированные контрольным лентивирусным вектором, кодирующим GFP (фиг. 1).
Авторы создали ROR1-CAR трансдуцированные (n=10) и контрольные GFP-трансдуцированные клоны T-клеток CD8+ (n=4), используя лимитирующее разведение, и подтвердили стабильную поверхностную экспрессию CAR после множества раундов размножения in vitro. Не было явного различия в росте ROR1-CAR-трансдуцированных по сравнению с нетрансдуцированными или GFP-трансдуцированными клонами T-клеток (данные не показаны).
ROR1-CAR трансдуцированные клоны T-клеток эффективно лизировали первичные B-CLL- и K562-клетки, которые были стабильно трансфицированы геном ROR1, но не лизировали наивные, ROR1-негативные клетки K562, что свидетельствует о специфичном распознавании ROR1 (фиг. 2).
Обсуждение
Адоптивная иммунотерапия, в которой используют CAR-модифицированные T-клетки, подвергается исследованию в клинических испытаниях в случае B-клеточных злокачественных заболеваний. Поверхностные молекулы, которые представляют собой мишени, являются специфичными для B-клеточной линии и включают CD19, который экспрессируется на нормальных клетках B-линии от стадии про-B-клеток до плазматических клеток, и CD20, который экспрессируется на нормальных B-клетках от стадии пре-B-клетки до B-клетки памяти. Таким образом, предполагаемым результатом эффективной терапии, нацеленной на такие молекулы, является истощение популяций нормальных B-клеток и B-клеточных предшественников. В исследованиях профиля экспрессии генов идентифицировали гены, которые преимущественно или исключительно экспрессировались злокачественными, но не экспрессировались нормальными B-клетками, и ROR1 выявлен в качестве характерного для CLL гена в 2 независимых анализах (27,28). Специфичные антитела к ROR1, вырабатываемые у пациентов с CLL после вакцинации аутологичными опухолевыми клетками, которые были модифицированы так, чтобы они экспрессировали CD154, и лечение леналидомидом без видимой токсичности для нормальных тканей, свидетельствуют о том, что такой опухолевый антиген может быть подходящей мишенью для иммунотерапии (29,30).
Исследования авторов показывают возможность целенаправленного воздействия на ROR1-позитивные злокачественные клетки сконструированными T-клетками, экспрессирующими ROR1-CAR. T-клетки CD8+ ROR1-CAR могут быть получены как от здоровых доноров, так и от пациентов с CLL после лентивирусной трансдукции либо общей популяции PBMC, либо очищенных сортировкой TCM, которые в животных моделях существуют в течение длительных периодов времени после адоптивного переноса (31). ROR1-CAR трансдуцированные T-клетки эффективно лизировали первичные B-CLL, но не лизировали нормальные неактивированные или активированные B-клетки. Такие T-клетки продуцировали эффекторные цитокины, включая TNF-α, IFNγ и IL-2, и были способны пролиферировать в ответ на ROR1-экспрессирующие опухолевые клетки.
Пример 2: Создание линий T-клеток CD4+ CAR и анализ эффекторной функции
T-клетки CD4+ ROR1-CAR могут быть получены из PBMC здоровых доноров/CLL-пациентов. ROR1-специфичный CAR может быть экспрессирован в T-клетках CD4+ человека и придает способность к специфичному распознаванию B-клеточных опухолей ROR1+, но не зрелых нормальных B-клеток.
Материалы и способы
Клеточные линии
Трансформированные вирусом Эпштейна-Барр B-клетки (EBV-LCL) создавали, как описано (25). Линии опухолевых клеток Jeko-1 и BALL-1 предоставлены докторами Oliver Press и Jerald Radich (Онкологического исследовательского центра имени Фреда Хатчинсона). Все линии клеток поддерживали в RPMI, содержащей 10% фетальной сыворотки теленка, 0,8 мМ L-глутамин и 1% пенициллин-стрептомицин (среда LCL). Клетки K562 и 293T получали из Американской коллекции типов культур и культивировали, как указано.
Трансфекция клеток K562 с использованием ROR1
Для основанной на полимеразной цепной реакции (ПЦР) амплификации ROR1-гена получали суммарную РНК из клеток B-CLL (набор RNeasyPlus; QIAGEN) и обратно транскрибировали в кДНК с использованием обратной транскриптазы M-MLV (Invitrogen). ПЦР осуществляли со специфичными праймерами
(ROR1-F: 5-XhoIAGAGGAGGAATGCACCGGCC-3 и
ROR1-R: 5-XhoI-CACAGAAGGTACTTGTTGCGATGT-3),
используя ДНК-полимеразу Herculase-II (Stratagene). ПЦР-продукт клонировали в ретровирусном векторе MIGR-1 (23) и последовательность подтверждали. Реагент для трансформации Effectene (QIAGEN) использовали для того, чтобы трансфицировать клетки Platinum-A (Cell Biolabs) MIGR-1/ROR1 и получить ROR1-кодирующий ретровирус. Клетки K562 трансдуцировали ретровирусом, используя центрифугирование при 2500 об/мин в течение 60 минут при 32°C, размножали и очищали ROR1-позитивную подгруппу, используя сортировку.
Конструирование вектора и создание лентивируса
Лентивирусные векторы, кодирующие CD20-CAR (CD20R-epHIV7) и зеленый флуоресцирующий белок (GFP), (GFP-epHIV7), описаны ранее (24). ROR1-CAR кодировали в таком же векторе. Мышиное мАт (клон 2A2), которое, как было показано, специфично связывается с ROR1 человека, экспрессируемым на первичных опухолевых линиях B-CLL и MCL, создавали, клонировали и характеризовали в предыдущем исследовании. Оптимизированную по кодонам нуклеотидную последовательность, кодирующую scFv, содержащий VL- и VH-цепи мАт 2A2, синтезировали (GENEART) и клонировали в CD20R-epHIV7, используя сайты рестрикции NheI и RsrII, чтобы заменить CD20-специфичный scFv. Лентивирус продуцировали в клетках 293T, котрансфицированных лентивирусным вектором и пакующими векторами pCHGP-2, pCMVRev2 и pCMV-G, используя Effectene (Qiagen). Среду меняли через 16 часов после трансфекции и лентивирусы собирали через 48 часов.
Лентивирусная трансдукция и выделение линий T-клеток CD4+ RoR1-CAR
T-клетки CD4+ выделяли из PBMC здоровых доноров и активировали моноклональными антителами против CD3 (30 нг/мл) (25) и трансдуцировали в супернатанте с лентивирусами с добавлением 1 мкг/мл полибрена (Sigma-Aldrich) и 50 IU/мл рекомбинантного интерлейкина-2 (IL-2) человека на 2-й и 3-й день после активации, центрифугируя при 2500 об./мин в течение 60 минут при 32°C. T-клетки размножали в RPMI, содержащей 10% сыворотки человека, 2 мМ L-глутамина и 1% пенициллина-стрептомицина (среда CTL) (25). После размножения аликвоту каждой линии трансдуцированных T-клеток красили конъюгированным с биотином анти-EGFR-мАт (против рецептора эпителиального фактора роста), стрептавидином-PE и анти-CD4-мАт. T-клетки EGFR+CD4+ очищали сортировкой и размножали. ROR1-CAR-трансдуцированные T-клетки идентифицировали по окрашиванию биотинилированным рекомбинантным слитым белком Fc-внеклеточный домен ROR1 и стерптавидином-PE. Рекомбинантный ROR1-белок получали во временно трансфицированных клетках 293 (Invitrogen), очищали, как описано (26), и биотинилировали, используя набор BiotinTag (Sigma). GFP-трансдуцированные T-клетки CD4+ идентифицировали с использованием проточной цитометрии, очищали сортировкой и клонировали сходным образом.
Анализы высвобождения хрома и секреции цитокинов
Клетки-мишени метили51Cr (PerkinElmer) в течение ночи, промывали и инкубировали в трех повторах по 1-2×103 клеток/лунка с эффекторными T-клетками в разных соотношениях между эффекторными клетками и клетками-мишенями (E:T). Супернатанты собирали для γ-подсчета после 4-часовой инкубации и специфичный лизис вычисляли, используя стандартную формулу (25). Для анализа секреции цитокинов клетки-мишени и эффекторные клетки трижды высевали в лунки в соотношении E/T 2:1, и интерферон INFγ, фактор некроза опухолей (TNF-α) и IL-2 измеряли в мультиплексном имуноанализе цитокинов (Luminex) в супернатанте, извлеченном после 24-часовой инкубации.
Анализ пролиферации с использованием CFSE
T-клетки метили с использованием 0,2 мкМ сложного карбоксифлуоресцеинсукцинимидилового эфира (CFSE; Invitrogen), промывали и высевали со стимулирующими клетками в соотношении 2:1 в среде CTL, содержащей 10 единиц/мл рекомбинантного IL-2 человека. После 72-часовой инкубации клетки метили, используя анти-CD4-мАт и йодид пропидия (PI), чтобы исключить мертвые клетки из анализа. Образцы анализировали проточной цитометрией и клеточное деление живых T-клеток CD4+ оценивали, используя разведение CFSE.
Анализ совместного культивирования
ROR1-CAR-трансдуцированные T-клетки CD4+ и ROR1-CAR-трансдуцированные цитотоксические T-лимфоциты CD8+ метили CFSE и культивировали совместно в соотношении 2:1, 1:1 и 1:2. Затем совместные культуры стимулировали клетками K562/ROR1 и контрольными клетками K562 и пролиферацию клеток измеряли в анализе с разведением красителя CFSE после 5 дней инкубации. Для проточного анализа образцы красили конъюгированным анти-CD8-мАт и анти-CD4-мАт, чтобы отличить подгруппы CD8+ и CD4+.
Результаты
Создание T-клеток CD4+ ROR1-CAR из PBMC здоровых доноров и CLL-пациентов
Авторы показали, что ROR1, онкофетальный тирозинкиназный рецептор, одинаково экспрессируется на CLL и MCL, и разработали ROR1-CAR из анти-ROR1-мАт, который придает способность к специфичному распознаванию злокачественных, но не зрелых нормальных B-клеток при экспрессии в T-клетках CD8+ (32). В настоящем изобретении авторы создали T-клетки CD4+ ROR1-CAR для анализа прямого распознавания опухоли и их способности усиливать CTL CD8+ ROR1-CAR. CAR-модифицированные T-клетки CD4+ могут быть легко созданы из общей популяции периферических T-клеток CD4+ от здоровых доноров (n=4) и CLL-пациентов (n=4) с использованием лентивирусного вектора, кодирующего ROR1-CAR. В таком векторе авторы кодировали укороченный домен EGFR (рецептора эпителиального фактора роста, tEGFR) ниже ROR1-CAR и самоотщепляемый элемент 2A, служащие как в качестве маркера трансдукции, так и для обогащения экспрессирующих трансген T-клеток с использованием анти-EGFR-мАт (фиг.3). Авторы определили частоту CAR-модифицированных T-клеток на 12-й день после однократной трансдукции лентивирусом, кодирующим ROR1-CAR (MOI=3), используя маркер tEGFR, и обнаружили стабильно более высокие эффективности трансдукции в линиях T-клеток CD4+ CAR по сравнению с линиями T-клеток CD8+ CAR, полученными от тех же индивидуумов. Чтобы подтвердить экспрессию ROR1-CAR на поверхности T-клеток CD4+, авторы использовали биотинилированный рекомбинантный слитый белок Fc-внеклеточный домен ROR1, который непосредственно связывается с scFv ROR1-CAR и специфично окрашивает T-клетки CD4+, трансдуцированные лентивирусом ROR1-CAR, но не нетрансдуцированные контрольные T-клетки CD4+ (фиг. 3). Авторы обогащали T-клетки CD4+, экспрессирующие трансген, используя маркер tEGFR, и размножали подгруппу CAR-позитивных T-клеток, стимулируя моноклональным анти-CD3-антителом. Может быть достигнуто более чем 3-log-увеличение численности T-клеток CD4+ CAR в конце 14-дневного цикла стимуляции, которое эквивалентно амплификации, наблюдаемой в CTL CD8+ CAR. После размножения клеток авторы подтверждали стабильную экспрессию ROR1-CAR на клеточной поверхности T-клеток CD4+ CAR (данные не показаны) и анализировали распознавание ROR1-позитивных опухолевых клеток.
T-клетки CD4+ ROR1-CAR специфично узнают ROR1-позитивные опухоли
Авторы анализировали эффекторную функцию T-клеток CD4+ ROR1-CAR против ROR1-позитивных первичных опухолевых клеток и линий опухолевых клеток. Авторы анализировали способность T-клеток CD4+ CAR обеспечивать прямую цитотоксичность, исследуемую в анализе высвобождения хрома (CRA), и выявили слабый, но специфичный лизис ROR1-позитивных клеток-мишеней в конце стандартной 4-часовой инкубации (фиг. 4). Авторы продлили CRA до 10 часов и наблюдали дополнительное увеличение специфичного лизиса, однако общая цитолитическая активность T-клеток CD4+ CAR все еще была ниже, чем у CTL CD8+ ROR1-CAR (фиг. 2, 4). T-клетки CD4+ ROR1-CAR как от здоровых доноров, так и от пациентов с CLL специфично узнавали первичные CLL-клетки, линии ROR1-позитивных опухолевых клеток Jeko-1 (MCL) и BALL-1 (B-ALL) и клетки K562, которые были стабильно трансфицированы ROR1-геном (K562/ROR1), но не нативные ROR1-негативные клетки K562, согласно ELISA IFN-γ, что свидетельствует о специфичном узнавании ROR1 на клеточной поверхности клеток-мишеней (фиг. 5A). Мультиплексный анализ цитокинов выявил продукцию других Th1-цитокинов, таких как TNF-α и IL-2 на значимо более высоких уровнях по сравнению с CTL CD8+ CAR, и продукцию IL-4, IL-10 и IL-17 (фиг. 5B).
Затем авторы оценивали пролиферацию T-клеток CD4+ CAR после стимуляции ROR1-позитивными опухолевыми клетками при окрашивании CFSE и использовали жесткие условия культивирования без добавления экзогенных цитокинов, чтобы исключить любой возможный неспецифичный стимул. В T-клетках CD4+ CAR наблюдали сильную и специфичную пролиферацию в ответ на ROR1-позитивные опухолевые клетки. Процентное содержание T-клеток, которые были индуцированы к пролиферации, и количество клеточных делений, которым подвергалась такая пролиферирующая подгруппа, были значимо выше в T-клетках CD4+ CAR по сравнению с T-клетками CD8+ CAR (фиг. 6). Полученные авторами данные в сочетании показывают, что T-клетки CD4+, полученные от здоровых доноров и пациентов с CLL, приобретают противоопухолевую реактивность после генетической модификации ROR1-специфичным CAR. Кроме того, способность пролиферировать в отсутствие экзогенных цитокинов и продуцировать высокие уровни Th1-цитокинов, свидетельствуют о том, что T-клетки CD4+ CAR проявляют типичные хелперные функции после стимуляции посредством CAR, и в дополнение к приданию способности к прямым противоопухолевым эффектам также могут быть использованы для усиления CTL CD8+ CAR.
CAR-модифицированные, но не нетрансдуцированные T-клетки CD4+ предоставляют помощь CTL CD8+ CAR
Чтобы проанализировать, способны ли T-клетки CD4+ CAR предоставлять помощь клеткам CD8+ CAR, авторы осуществляли эксперименты по совместному культивированию CAR-трансдуцированных и контрольных нетрансдуцированных поликлональных линий T-клеток CD4+ и CD8+, которые авторы получили из клеток здоровых доноров и пациентов с CLL. В качестве индикатора оказания помощи авторы определяли повышение специфичной для опухоли эффекторной функции CD8+ в присутствии T-клеток CD4+ по сравнению с T-клетками CD8+, культивируемыми отдельно. Авторы объединяли либо CAR-трансдуцированные, либо нетрансдуцированные контрольные T-клетки CD4+ с CTL CD8+ CAR в разных соотношениях CD4:CD8 (2:1, 1:1, 1:2), стимулировали их ROR1-позитивными опухолевыми клетками и измеряли пролиферацию, используя разведение красителя CFSE. Авторы обнаружили, что добавление CAR-трансдуцированных, но не добавление нетрансдуцированных T-клеток CD4+ к CTL CD8+ CAR значимо усиливало специфичную пролиферацию подгруппы CD8+ по сравнению с CTL CD8+ CAR отдельно (фиг.7). Усиление пролиферации было наиболее выраженным, когда в совместную культуру добавляли по меньшей мере эквивалентное количество T-клеток CD4+ CAR (соотношение CD4:CD8 2:1 или 1:1). Сочетание нетрансдуцированных T-клеток CD4+ с нетрансдуцированными T-клетками CD8+ служило в качестве дополнительного контроля и не индуцировало неспецифичной пролиферации в подгруппе CD8+ (данные не показаны).
Обсуждение
В исследованиях профиля экспрессии генов идентифицировали гены, которые предпочтительно или исключительно экспрессировались злокачественными, но не нормальными B-клетками, и ROR1 выступал в качестве гена, являющегося показателем CLL, в 2 независимых анализах (27,28). Исследования авторов показывают возможность целенаправленного воздействия на ROR1-позитивные злокачественные клетки сконструированными T-клетками, экспрессирующими ROR1-CAR. T-клетки CD8+ и CD4+ ROR1-CAR могут быть получены от здоровых доноров после лентивирусной трансдукции либо общей популяции PBMC, либо очищенных сортировкой T-клеток. ROR1-CAR-трансдуцированные T-клетки CD8+ эффективно лизировали первичные B-CLL, но не лизировали нормальные неактивированные или активированные B-клетки. ROR1-CAR-трансдуцированные T-клетки CD4+ слабо лизировали первичные B-CLL, но не лизировали нормальные неактивированные или активированные B-клетки. Такие T-клетки продуцировали эффекторные цитокины, включая TNF-α, IFNγ, IL-2, IL-4 и IL-10. CAR-трансдуцированные T-клетки CD4+ продуцировали значимо более высокие количества цитокинов, чем трансдуцированные клетки CD8+. Оба типа клеток были способны пролиферировать в ответ на ROR1-экспрессирующие опухолевые клетки. И в этом случае T-клетки CD4+ ROR1-CAR также пролиферировали в 2-3-раза сильнее, чем CTL CD8+ ROR1-CAR. Полученные результаты показывают, что трансдуцированные хелперные T-клетки CD4+ проявляют типичные хелперные функции, что свидетельствует о том, что их можно применять для усиления CTL CD8+ CAR.
Пример 3: Эффекторная функция T-клеток CD4+ ROR1-CAR, полученных из подгрупп наивных клеток, центральных и эффекторных клеток памяти
Сравнивали эффекторную функцию T-клеток CD4, полученных из подгрупп наивных клеток, центральных и эффекторных клеток памяти и затем модифицированных ROR1 CAR.
Материалы и способы
Очистка наивных клеток, центральных и эффекторных клеток памяти CD4 с использованием сортировки
T-клетки CD4+ выделяли из PBMC здорового донора, используя негативную селекцию на магнитных шариках (набор для выделения Miltenyi CD4), которая дает неактивные T-клетки CD4+. Фракцию CD4+ метили конъюгированными анти-CD45RA-, анти-CD45RO- и анти-CD62L-мАт и очищали посредством проточной сортировки, используя проточный сортировщик FACS Aria (BD Biosciences), и наивные T-клетки (CD45RA+ CD45RO- CD62L+), центральные T-клетки памяти (CD45RA- CD45RO+ CD62L+) и эффекторные T-клетки памяти (CD45RA- CD45RO+ CD62L-) CD4+ очищали на основании их экспрессии определенных маркеров.
Анализ пролиферации с использованием CFSE
T-клетки метили, используя 0,2 мкМ сложного карбоксифлуоресцеинсукцинимидилового эфира (CFSE; Invitrogen), промывали и высевали со стимулирующими клетками в соотношении 2:1 в среде CTL, содержащей 10 единиц/мл рекомбинантного IL-2 человека. После 72-часовой инкубации клетки метили, используя анти-CD8-мАт или анти-CD4-мАт и йодид пропидия (PI), чтобы исключить мертвые клетки из анализа. Образцы анализировали проточной цитометрией и клеточное деление живых T-клеток CD8+ и CD4+ оценивали, используя разведение CFSE.
Анализы цитокинов
Для анализов секреции цитокинов клетки-мишени и эффекторные клетки высевали в лунки в трех повторах в соотношении E/T 2:1, и интерферон INFγ, фактор некроза опухолей (TNF-α) и IL-2 измеряли в мультиплексном имуноанализе цитокинов (Luminex) в супернатанте, извлеченном после 24-часовой инкубации.
Результаты
Используя проточную сортировку авторы очищали T-клетки CD4+ N, центральные (CM) и эффекторные T-клетки памяти (EM) CD4+ из периферической крови 3 здоровых доноров на основании экспрессии CD45RA, CD45RO и CD62L (фиг. 8A) и сравнивали их эффекторную функцию после модификации ROR1-CAR. Авторы достигали сходных высоких эффективностей трансдукции в линиях T-клеток CAR, полученных из каждой из трех подгрупп. Многопараметровая проточная цитометрия после обогащения экспрессирующих трансген T-клеток показала наличие экспрессии CD45RO и утрату CD45RA в T-клеточной линии N CAR CD4+, что согласуется с активированным фенотипом после лентивирусной трансдукции. T-клеточные линии N, CM и EM CAR CD4+ сохраняли дифференциальную экспрессию CD62L, подтверждая, что начальная очистка проточной сортировкой была осуществлена с высокой чистотой.
Затем авторы анализировали распознавание опухоли, секрецию цитокинов и пролиферацию T-клеток CD4+ CAR, полученных из подгрупп N, CM и EM, и сравнивали их с линиями T-клеток CAR, созданными из общей популяции T-клеток CD4+. Авторы наблюдали специфичное распознавание ROR1-позитивных опухолевых клеток, используя ELISA IFN-γ, в каждой из клеточных линий. Мультиплексный анализ цитокинов показал, что T-клетки CD4+, полученные из N-подгруппы, явно продуцировали самые высокие уровни Th1-цитокинов, особенно IL-2 (фиг. 8C), и разведение красителя CFSE показало, что они пролиферировали наиболее энергично в ответ на стимуляцию ROR1-позитивными опухолевыми клетками (фиг. 8B).
Обсуждение
Исследования, проведенные авторами, иллюстрируют возможность целенаправленно воздействовать на ROR1-позитивные злокачественные клетки сконструированными T-клетками, экспрессирующими ROR1-CAR. T-клетки ROR1-CAR CD8+ и CD4+ могут быть получены от здоровых доноров после лентивирусной трансдукции либо общей популяции PBMC, либо очищенных сортировкой T-клеток из определенных подгрупп наивных T-клеток или T-клеток памяти. Наивные, центральные клетки памяти и эффекторные T-клетки CD4+ продуцировали эффекторные цитокины, включая TNFα, IFNγ, IL-2, IL-4 и IL-10. CAR-трансдуцированные клетки CD4+, полученные из подгруппы наивных клеток, продуцировали значимо более высокие количества TNFα и IL-2, чем полученные из центральных и эффекторных клеток памяти T-клетки CD4+ CAR, после передачи сигнала через CAR. Все типы CD4-клеток были способны пролиферировать в ответ на ROR1/K562, однако в CAR-трансдуцированных клетках CD4+, полученных из подгруппы наивных клеток, процентное содержание T-клеток, которые были индуцированы к пролиферации, и количество клеточных делений, которым подвергалась пролиферирующая подгруппа, были значимо выше. Оба показателя, профиль цитокинов и пролиферативная способность, свидетельствуют, что наивные T-клетки CD4+ ROR1-CAR лучше всего подходят для усиления CTL CD8+ ROR1-CAR.
Пример 4: Наивные T-клетки CD4+ являются более хорошими хелперами, чем T-клетки памяти CD4+
Трансдуцированные наивные, центральные памяти и эффекторные T-клетки CD4+ культивировали совместно с трансдуцированными цитотоксическими T-лимфоцитами CD8+ и измеряли пролиферативный ответ клеток в ответ на стимуляцию клетками K562/ROR1.
Материалы и способы
Совместное культивирование
Полученные из наивных клеток, центральных и эффекторных клеток памяти ROR1-CAR-трансдуцированные T-клетки CD4+ и ROR1-CAR-трансдуцированные цитотоксические T-лимфоциты CD8+, полученные из наивных и центральных T-клеток памяти CD8+, метили CFSE, и линии T-клеток CAR CD4+ и CD8+ культивировали совместно в соотношении 1:1. Затем совместные культуры стимулировали клетками K562/ROR1 и контрольными клетками K562 и измеряли пролиферацию клеток в анализе с разбавлением красителя CFSE после 5 дней инкубации. Для проточного анализа образцы красили конъюгированными анти-CD8-мАт и анти-CD4-мАт, чтобы отличить подгруппы CD8+ и CD4+.
Результаты
Наивные T-клетки CD4+ CAR обладают превосходной способностью усиливать эффекторную функцию CTL CD8+ CAR
Авторы сравнивали хелперную функцию линий T-клеток CD4+ CAR N, CM и EM, чтобы определить, можно ли благоприятный профиль цитокинов и пролиферативный потенциал T-клеток N CAR CD4+ также транслировать на наиболее сильный хелперный эффект по отношению к CTL CD8+ CAR. В предыдущей работе было показано, что имеют место существенные различия между T-клетками CD8+ N, CM и EM, которые влияют на их потенциальную применимость для адаптивной иммунотерапии. Авторы недавно показали, что полученные из CM, но не из EM, T-клетки CD8+ способны существовать в течение длительных периодов времени после адоптивного переноса, что делает их предпочтительной для иммунотерапии подгруппой T-клеток CD8+ (33, 34). Другие группы исследователей считают, что N-T-клетки CD8+ также обладают благоприятными признаками для применения в T-клеточной терапии (35, 36). Таким образом, авторы создали CTL CD8+ CAR из очищенных сортировкой N- и CM-T-клеток для определения оптимального сочетания подгрупп CAR-T-клеток CD8+ и CD4+. После лентивирусной трансдукции и обогащения CAR-трансдуцированных T-клеток CD8+ с использованием маркера tEGFR авторы подтвердили реактивность по отношению к опухоли N- и CM-CTL CD8+ CAR (данные не показаны) и осуществили эксперименты по совместному культивированию с T-клетками CD4+ CAR, как описано выше. Как и ожидалось, совместная культура N- и CM-CTL CD8+ CAR c N-T-клетками CD4+ CAR приводила к значимо более высокой специфичной для опухоли пролиферации подгруппы CD8+ по сравнению с совместной культурой с CM- и EM-T-клетками CD4+ CAR или с CTL CD8+ CAR отдельно (фиг. 9). Из всех сочетаний максимальную пролиферацию CTL CD8+ CAR в ответ на стимуляцию ROR1-позитивными опухолевыми клетками наблюдали после совместного культивирования N-T-клеток CD4+ CAR с CM-CTL CD8+ CAR (фиг. 9). Полученные авторами данные, вместе взятые, демонстрируют, что имеют место присущие клеткам различия между T-клетками CD4+ N, CM и EM в профиле их цитокинов и пролиферативном потенциале, при этом более высокую продукцию IL-2 и наиболее высокую пролиферацию наблюдали в N-T-клетках CD4+. Полученные авторами данные свидетельствуют, что очищенные сортировкой T-клетки CD4+ N, а не CM, EM или общая популяция T-клеток CD4+, лучше всего подходят для усиления эффекторной функции CTL CD8+, и дополняют предыдущую работу по T-клеткам CD8+ в том, что полученные из CM T-клетки CD8+, обладают благоприятными характеристиками для применения в адаптивной иммунотерапии.
Обсуждение
Полученные данные, взятые вместе, демонстрируют, что адоптивный перенос ROR1-CAR-модифицированных T-клеток CD4+ и CD8+ придает способность к мощным противоопухолевым ответам в модели in vivo агрессивной системной лимфомы, и представляют доказательство полезного и синергетического влияния T-клеток CD4+ CAR на противоопухолевую эффективность CTL CD8+ CAR. Полученные авторами данные иллюстрируют, каким образом анализ присущих клеткам качеств может давать информацию, необходимую для рациональной разработки клеточных продуктов, содержащих специфичные для опухоли T-клетки CD8+ и CD4+, чтобы улучшить результаты иммунотерапии злокачественной опухоли.
Пример 5: Мышиная опухолевая модель системной лимфомы из клеток мантийной зоны (NSG/Jeko-1-ffLuc)
Авторы исследовали влияние предоставления CD4-помощи в отношении противоопухолевой эффективности ROR1-CAR-модифицированных CTL CD8+ в модели in vivo агрессивной системной лимфомы из клеток мантийной зоны.
Материалы и способы
Сублетально облученным мышам NOD/SCID/гамма-/- (NSG) прививали посредством инъекции в хвостовую вену 5×105 клеток Jeko-1, которые были стабильно трансфицированы люциферазой светляка (Jeko-1/ffLuc), чтобы обеспечить возможность оценки опухолевой нагрузки и распределения с использованием визуализации биолюминесценции. Авторы подтвердили соответствующее приживление (частота приживления = 100%) и развитие быстро прогрессирующей диссеминированной лимфомы у мышей NSG в таких условиях. После прививки опухолей группы по 3 мыши в каждой получали либо CTL CD8+ CAR (группа 1), T-клетки CD4+ CAR (группа 2), сочетание ROR1-CAR-трансдуцированных T-клеток CD8+ и CD4+ (группа 3), нетрансдуцированные контрольные T-клетки (группа 4, 5, 6) посредством инъекции в хвостовую вену или не получали обработки (группа 7). Общее количество переносимых T-клеток составляло 10×106 во всех случаях. Авторы получали кровь из глаза мышей через 2 дня после адоптивного переноса и подтверждали присутствие ROR1-CAR-трансдуцированных или нетрансдуцированных T-клеток в периферической крови.
Результаты
На 6-й день после переноса T-клеток авторы осуществляли визуализацию биолюминесценции, чтобы оценить опухолевую нагрузку. Наибольший противоопухолевый эффект наблюдали у мышей, которые получали сочетание T-клеток ROR1-CAR CD8+ и CD4+, у которых происходило снижение сигнала биолюминесценции на >2 log по сравнению с контрольной группой (фиг. 10). Авторы также наблюдали мощный противоопухолевый эффект у мышей, которые получали либо CD8+, либо CD4+ ROR1-CAR-модифицированные T-клетки, при этом имело место снижение сигнала биолюминесценции на >1 log по сравнению с контролями (фиг.10). Важно, что уменьшение опухолевой нагрузки после введения сочетания CAR-T-клеток CD8+/CD4+ было больше, чем в случае объединяемых групп CTL CD8+ CAR и T-клеток CD4+ CAR, что свидетельствует о том, что T-клетки CD4+ CAR и CTL CD8+ CAR работают синергетически.
Обсуждение
Полученные данные, взятые вместе, демонстрируют, что адоптивный перенос ROR1-CAR-модифицированных T-клеток CD4+ и CD8+ придает способность к сильным противоопухолевым ответам в модели in vivo агрессивной системной лимфомы и представляют доказательство полезного и синергетического влияния T-клеток CD4+ CAR на противоопухолевую эффективность CTL CD8+ CAR. Полученные авторами данные иллюстрируют, каким образом анализ присущих клеткам качеств может давать информацию, необходимую для рациональной разработки клеточных продуктов, содержащих специфичные для опухоли T-клетки CD8+ и CD4+, чтобы улучшить результаты иммунотерапии злокачественной опухоли.
Пример 6: T-клетки CD19 CAR проявляют такую же синергию
Авторы исследовали влияние предоставления CD4-помощи в отношении противоопухолевой эффективности CD19-модифицированных CTL CD8+ в совместной культуре in vitro и в модели in vivo агрессивной системной лимфомы из клеток мантийной зоны.
Материалы и способы
T-клетки CD19 CAR можно получить, как описано в патенте US 2008/0131415, который включен в настоящее описание в виде ссылки.
Анализ совместной культуры
CD19-CAR-трансдуцированные T-клетки CD4+ и CD19-CAR-трансдуцированные цитотоксические T-лимфоциты CD8+ метили CFSE и совместно культивировали в соотношении 2:1, 1:1 и 1:2. Затем совместные культуры стимулировали клетками K562/ROR1 и контрольными клетками K562, и пролиферацию клеток измеряли в анализе с разведением красителя CFSE после 5 дней инкубации. Для проточного анализа образцы красили конъюгированным анти-CD8-мАт и анти-CD4-мАт, чтобы отличить подгруппы CD8+ и CD4+.
Модель in vivo
Сублетально облученным мышам NOD/SCID/гамма-/- (NSG) прививали посредством инъекции в хвостовую вену 5×105 клеток Jeko-1, которые были стабильно трансфицированы люциферазой светляка (Jeko-1/ffLuc), чтобы обеспечить возможность оценки опухолевой нагрузки и распределения с использованием визуализации биолюминесценции. Авторы подтвердили соответствующее приживление (частота приживления = 100%) и развитие быстро прогрессирующей диссеминированной лимфомы у мышей NSG в таких условиях. После прививки опухолей группы по 3 мыши в каждой получали либо CTL CD8+ CD19 CAR (группа 1), T-клетки CD4+ CD19 CAR (группа 2), сочетание CD19-CAR-трансдуцированных T-клеток CD8+ и CD4+ (группа 3), нетрансдуцированные контрольные T-клетки (группы 4, 5, 6) посредством инъекции в хвостовую вену или не получали обработки (группа 7). Общее количество переносимых T-клеток составляло 10×106 во всех случаях. Авторы получали кровь из глаза мышей через 2 дня после адоптивного переноса.
Результаты
На фиг. 10 показана превосходная способность линий T-клеток CD4+ CAR, полученных из подгруппы наивных клеток, усиливать специфичную для опухоли пролиферацию CTL CD8+ CAR, полученных из центральных клеток памяти, в экспериментах по совместному культивированию CTL CD8+ CD19-CAR и линий T-клеток CD4+ CD19-CAR, стимулированных линией CD19+ опухолевых клеток Jeko-1 лимфомы из клеток мантийной зоны. Хотя линии T-клеток CD4+ CAR, полученные из подгрупп центральных или эффекторных клеток памяти, усиливают специфичную для опухоли пролиферацию полученных из центральных клеток памяти CTL CD8+ CAR в намного меньшей степени.
Фиг. 11 показывает, что T-клетки CD8+ CAR и T-клетки CD4+ CAR независимо обеспечивают прямую противоопухолевую эффективность в модели лимфомы у иммунодефицитных мышей (NOD/SCID-Raji). Мыши получали либо CD19-CAR-трансдуцированные или контрольные ложно трансдуцированные T-клетки CD8+, полученные из центральных клеток памяти (A), либо CD19-CAR-трансдуцированные или контрольные ложно трансдуцированные T-клетки CD4+, полученные из наивных клеток (B).
Фиг. 12 показывает стимуляцию и синергетическое действие ROR1-CAR-модифицированных T-клеток CD4+ на противоопухолевую эффективность CTL CD8+ROR1-CAR в мышиной опухолевой модели системной лимфомы из клеток мантийной зоны (NSG/Jeko-1-ffLuc). Противоопухолевая эффективность ROR1-CAR-модифицированных T-клеток CD8+ и CD4+ в мышиной опухолевой модели системной агрессивной лимфомы из клеток мантийной зоны (NSG/Jeko-1) была усилена по сравнению либо с отдельной клеточной популяцией, либо по сравнению с нетрансдуцированными клетками.
Фиг. 13 показывает синергию T-клеток CD19-CAR CD8+ и CD4+ в мышиной модели системной лимфомы (NSG/Raji). Приживление опухоли Raji подтверждали, используя визуализацию биолюминесценции на 6-й день после инокуляции опухоли (перед обработкой) (схема обработки показана на A, приживление опухоли на основании биолюминесценции показано на B). Анализ опухолевой нагрузки с использованием визуализации биолюминесценции показал полное устранение опухолей Raji в когортах мышей, обработанных T-клетками CD8+ CD19-CAR, и у мышей, обработанных объединенным продуктом T-клеток CD19-CAR CD8+ и CD4+ (после обработки средние черный и серый столбики) B). Затем мышам вводили второй инокулят опухолевых клеток Raji и анализировали частоту встречаемости T-клеток CAR CD4+ и CD8+ в периферической крови и приживление опухоли. У мышей, обработанных объединенным продуктом T-клеток CAR CD8+ и CD4+ наблюдали значимо более высокие уровни T-клеток CAR CD8+ после инокуляции опухоли (нижние панели C) и полное отторжение инокулята Raji (после инокуляции опухоли правый серый столбик, B). Напротив, у мышей, которые получили только CTL CD8+ CD19-CAR, авторы не выявили увеличения количества T-клеток CAR после инокуляции опухоли (C), и опухолевые клетки Raji были способны приживаться (после инокуляции опухоли правый черный столбик, панель B).
Обсуждение
Полученные данные, вместе взятые, демонстрируют, что в случае трансдукции клеток другой конструкцией CAR, CD19, CD19-CAR-модифицированные T-клетки CD4+ и CD8+ обеспечивают сильные противоопухолевые ответы в модели in vivo агрессивной системной лимфомы и представляют доказательство полезного и синергетического влияния T-клеток CD4+ CAR на противоопухолевую эффективность CTL CD8+ CAR.
Вышесказанное является иллюстрацией настоящего изобретения, и его не следует рассматривать как ограничение изобретения. Изобретение определено следующей далее формулой изобретения, при этом включены эквиваленты формулы изобретения. Все ссылки и документы, упоминаемые в настоящем описании, включены в виде ссылки.
ССЫЛКИ



Реферат
Группа изобретений относится к медицине, а именно к онкологии, и может быть использована для адаптивной клеточной иммунотерапии. Композиция по изобретению содержит T-лимфоциты CD4и T-лимфоциты CD8, модифицированные химерным рецептором антигена. Применение композиции по изобретению предназначено для изготовления лекарственного средства для лечения рака. Способ изготовления композиции для адаптивной клеточной иммунотерапии включает стадию размножения in vitro, по отдельности, популяции T-лимфоцитов CD8, обогащенной центральными T-клетками памяти, и популяции T-лимфоцитов CD4и стадию модификации клеток. Использование изобретений позволяет усилить иммунный ответ за счет синергического действия модифицированных клеток. 3 н. и 16 з.п. ф-лы, 13 ил., 6 пр.
Комментарии