Двухцистронная бактериальная система экспрессии - RU2732148C2
Код документа: RU2732148C2
Чертежи


Описание
Область техники, к которой относится изобретение
Настоящее изобретение относится к двухцистронной независимой системе экспрессии в одном векторе, применяемой для получения представляющего интерес белка, включая рекомбинантные Fab-фрагменты антител или другой фрагмент антитела, пептиды и белки, экспрессируемые в виде нерастворимых телец включения, образующихся в бактериях E. сoli. Также настоящее изобретение предлагает способ экспрессии представляющего интерес белка с применением бицистронного вектора.
Предпосылки к созданию изобретения
Технология рекомбинантных ДНК (рДНК) полностью изменила способ получения терапевтических средств. В настоящее время требуемые белки получают внутри чужеродной клетки и осуществляют их очистку.
Белки с посттрансляционными модификациями (PTM) обычно экспрессируются в виде рекомбинантных молекул в дрожжевых системах или системах клеток млекопитающих. Дрожжевые системы экспрессии, такие как Pichia и Saccharomyces, являются наиболее близкими к системам млекопитающих в части PTM, но все же отличаются по типам гликозилирования, как, например, гликаны с высоким содержанием маннозы в случае Pichia, что делает их неподходящими для экспрессии рекомбинантных белков для использования у человека.
Моноклональные антитела (mAb), антитела, белки слияния, Fab-фрагменты mAb используют в качестве терапевтических средств. В технологии рДНК используют специальные векторы и системы экспрессии для получения терапевтических белков. Cистемы экспрессии включают, главным образом, бактериальные, дрожжевые системы экспрессии, системы экспрессии на основе клеток насекомых или млекопитающих. Изначально, большую часть рекомбинантных белков экспрессировали в бактериальной системе экспрессии с использованием E. coli в качестве хозяина. Существует ряд преимуществ от использования E. coli в качестве хозяина для экспрессии, таких как простота клонирования, простота экспрессии, сокращенные сроки, сокращенные периоды инкубации и высокие выходы. Таким образом, белки, не требующие каких-либо PTM, могут надежно экспрессироваться в E. coli.
Необходимость в экспрессии Fab, представляющих собой антигенсвязывающие фрагменты mAb, в системах млекопитающих отсутствует, поскольку они не содержат сайты гликозилирования, присутствующие в Fc-части антитела. Таким образом, экспрессию Fab обычно осуществляют в системе на основе E. coli. На протяжении 1980-90 несколько исследователей предпринимали попытки экспрессировать Fab в E. coli. Plückthun Aet. al.,1990 Behring Inst. Mitt. (87): 48-55 являются одними из самых первых исследователей, которые сообщили о секретировании Fab антитела из E. coli. Williamson R.A. et. al., 1991 Biochem J. 277 (Pt 2): 561-3 сообщили об использовании векторов на основе бактериофага-лямбда для экспрессии молекул Fab в E. coli. Система фагового дисплея используется для получения фрагментов Fab, бивалентного антитела или химерного антитела в E. coli.
Кроме того, Fab получали в E. coli в виде неправильно свернутых телец включения, а затем их подвергали рефолдингу с получением функциональной молекулы и, таким образом достигали 40% повышения выходов антител.
В большинстве упомянутых выше исследований использовали один промотор, т. e., phoA для управления экспрессией как тяжелых, так и легких цепей. Сайт связывания рибосом (rbs), присутствующий между последовательностями легких и тяжелых цепей, управляет транскрипцией и трансляцией второго гена.
В US5648237 также использовали аналогичную стратегию с одним промотором (phoA) для экспрессии генов Fab в E. coli для достижения секреции продукта. Основной недостаток упомянутой выше стратегии заключается в том, что уровни экспрессии второго гена обычно ниже, чем первого гена, что таким образом ограничивает выходы функционального Fab.
В патенте под № WO03018771 раскрывается способ получения антитела при помощи двух функциональных единиц трансляции, кодирующих, соответственно, легкую и тяжелую цепи определенного антитела или фрагмента, где обе цепи экспрессируются последовательно, со специфическим разделением тем самым получения легкой и тяжелой цепей и обеспечения возможности сборки легкой и тяжелой цепей.
В патенте № EP1356052B1 раскрывается способ получения полных антител в прокариотических клетках. В системе присутствует первый промотор и первый цистрон для получения легкой цепи иммуноглобулина, и второй промотор и второй цистрон для получения тяжелой цепи иммуноглобулина, при этом обе цепи подвергают фолдингу и сборке с образованием биологически активного иммуноглобулина.
Краткое описание изобретения
В одном варианте осуществления настоящее изобретение относится к двухцистронной независимой системе экспрессии с одним промотором, используемой для получения рекомбинантных белков и пептидов, экспрессируемых в бактериальных клетках в виде нерастворимых телец включения.
В другом варианте осуществления настоящее изобретение относится к способу получения двухцистронной независимой системы экспрессии в одном векторе, имеющем два разных промотора, для получения рекомбинантных белков и пептидов, экспрессируемых в виде нерастворимых телец включения в бактериальных клетках.
В другом варианте осуществления настоящее изобретение относится к двухцистронной независимой системе экспрессии в одном векторе, имеющем два разных промотора, используемой для получения фрагментов антител, экспрессируемых в виде нерастворимых телец включения в бактериальных клетках.
В другом варианте осуществления настоящее изобретение относится к двухцистронной независимой системе экспрессии в одном векторе, имеющем два разных промотора, для получения рекомбинантного Fab-фрагмента антител, экспрессируемых в виде нерастворимых телец включения в бактериальных клетках.
В другом варианте осуществления настоящее изобретение относится к двухцистронной независимой системе экспрессии в одном векторе, имеющем два разных промотора, используемой для получения рекомбинантных пептидов, экспрессируемых в виде нерастворимых телец включения в бактериальных клетках.
В другом варианте осуществления двухцистронная система экспрессии содержит
a) первый цистрон, содержащий промотор, функционально связанный с полинуклеотидной последовательностью, кодирующей представляющий интерес белок;
b) второй цистрон, содержащий промотор, функционально связанный с полинуклеотидной последовательностью, кодирующей представляющий интерес белок;
где первый и второй цистроны расположены в одном векторе и экспрессируют полинуклеотидную последовательность, кодирующую представляющий интерес белок в виде телец включения в бактериальной клетке.
В другом варианте осуществления настоящее изобретение относится к двухцистронному вектору, содержащему промотор, функционально связанный с сайтом множественного клонирования, содержащим представляющий интерес ген, сайт связывания рибосом и терминатор.
В еще одном варианте осуществления настоящее изобретение относится к способу получения представляющего интерес белка с использованием двухцистронной системы экспрессии.
Подробности одного или более вариантов осуществления настоящего изобретения, изложенные ниже, являются исключительно иллюстративными по своему характеру и не предназначены для ограничения объема настоящего изобретения. Другие признаки, цели и преимущества настоящего изобретения будут понятны из описания.
Краткое описание прилагаемых графических материалов
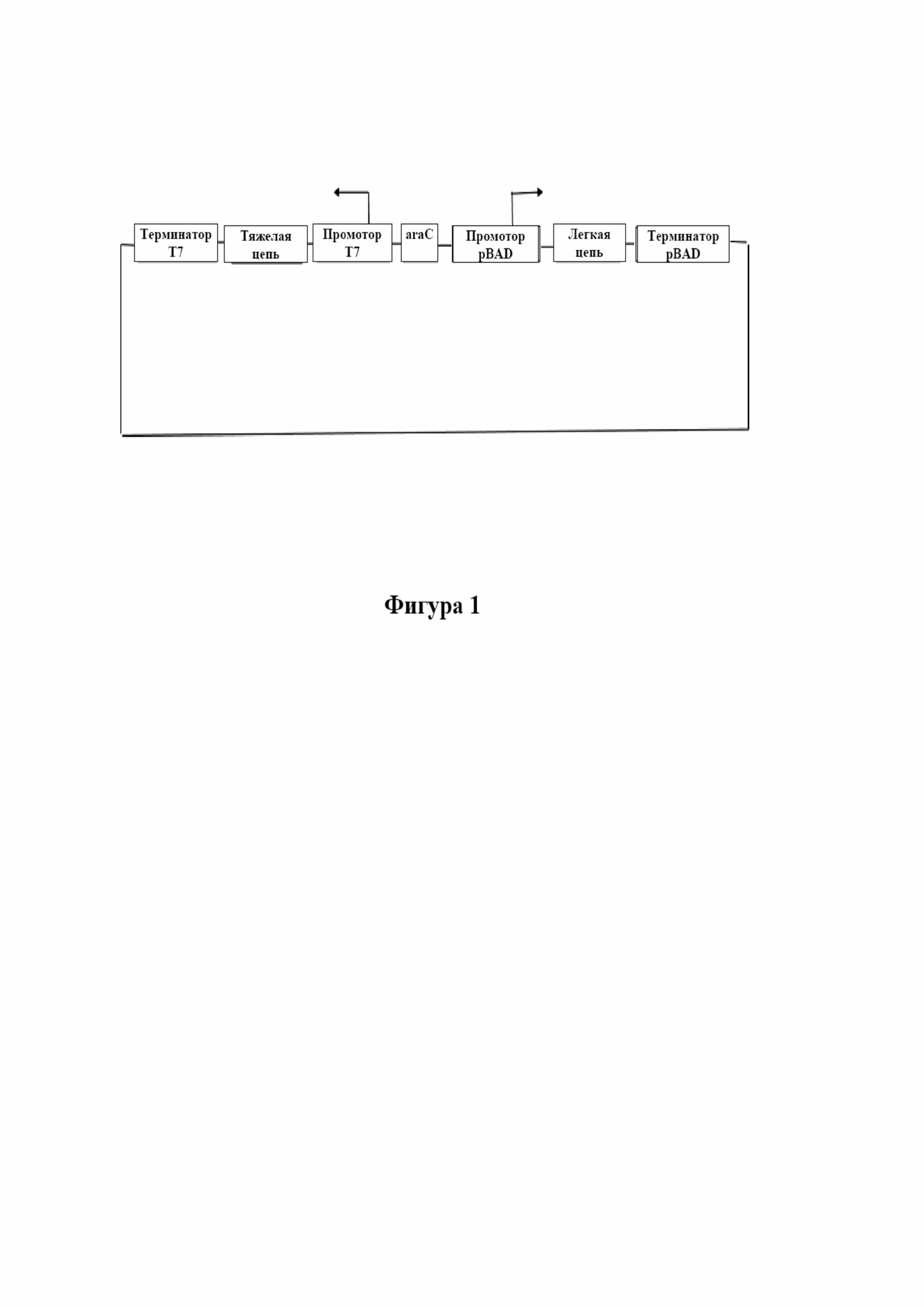
На фигуре 1 изображена схема бицистронного вектора.
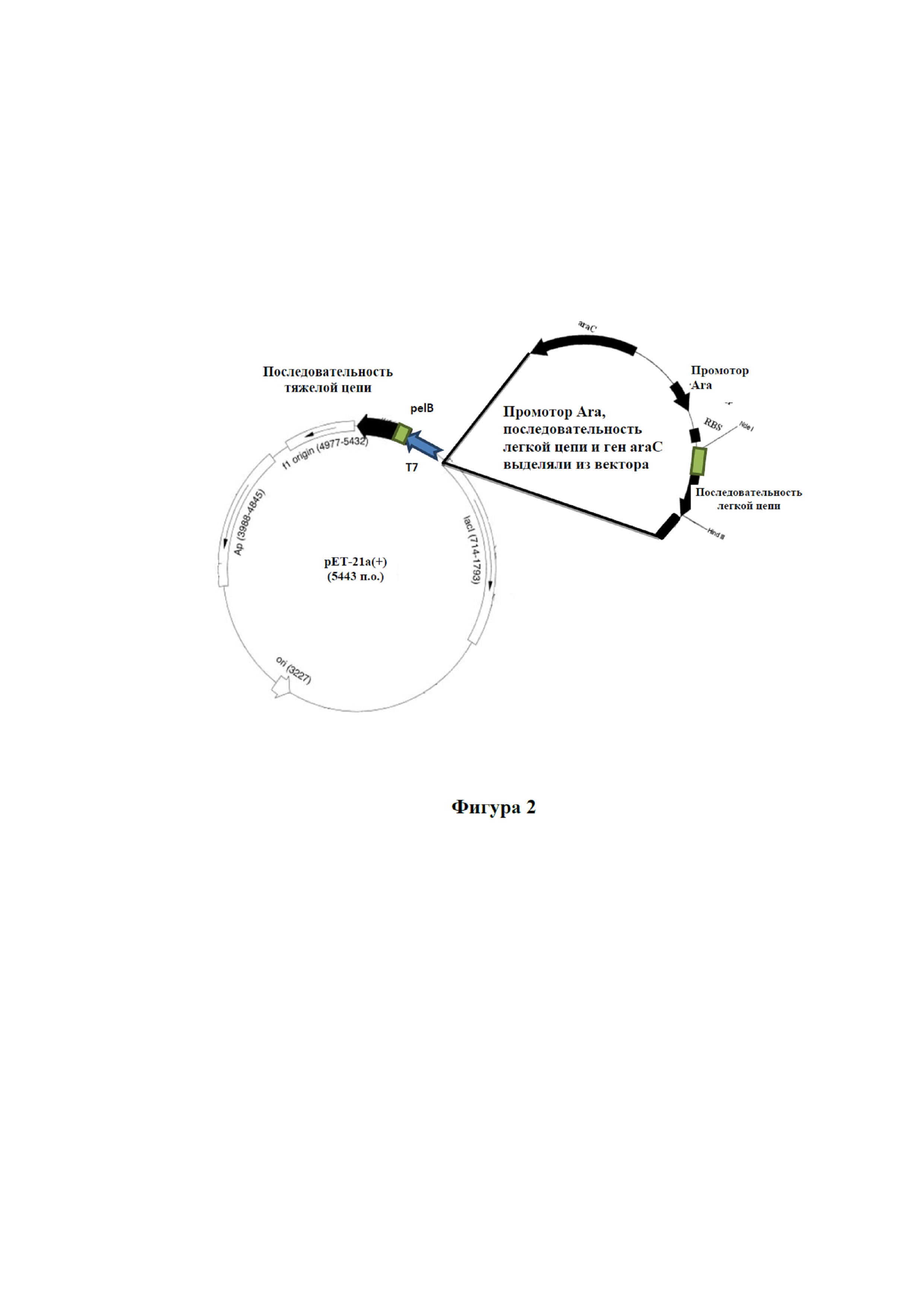
На фигуре 2 изображена карта вектора клона pET21a-HC-LC.
На фигуре 3 изображены результаты анализа в SDS PAGE нерастворимой осадочной фракции из клона E. coli BL21A1 вместе с контролями и эталонным продуктом.
На фигуре 4 изображены результаты анализа с помощью RP-HPLC для образцов растворимых IB с пиками LC и HC, отмечаемыми для клона, по сравнению с молекулой восстановленного Fab.
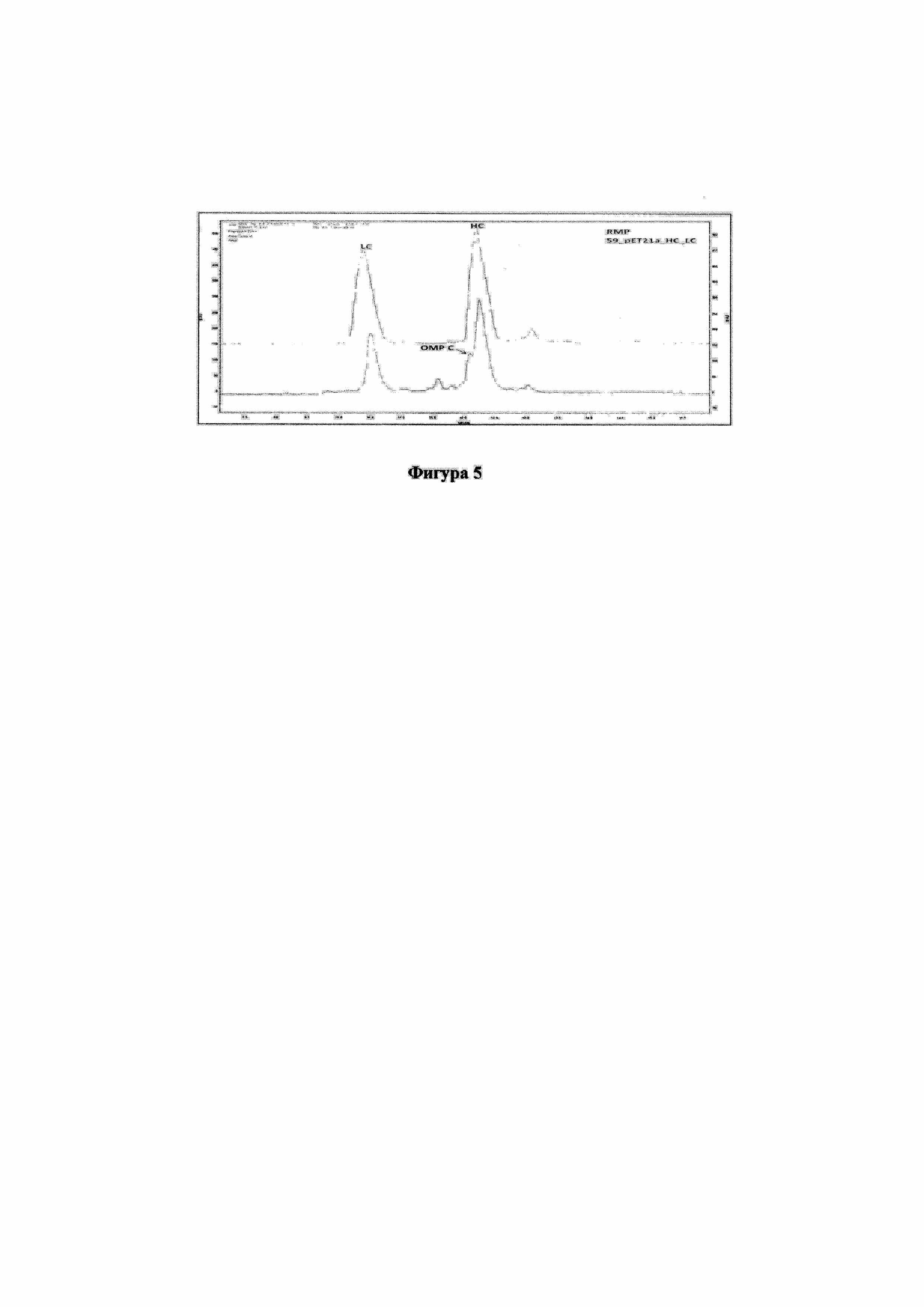
На фигуре 5 изображены циклы HPLC с отделенными пиками, соответствующими тяжелым цепям, от пиков, соответствующих другим белкам.
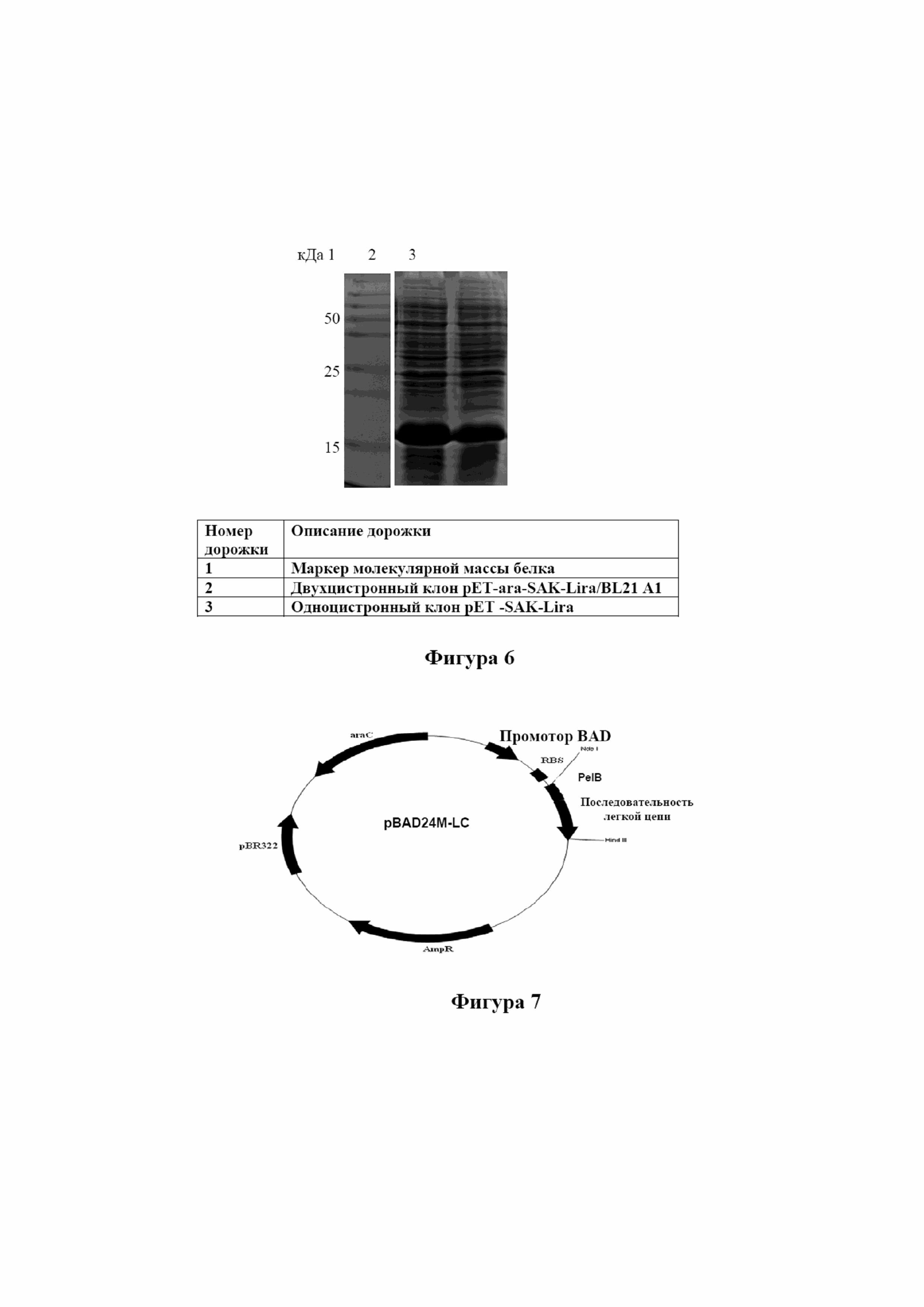
На фигуре 6 показано значимое повышение экспрессии клона SAK-Lira в двухцистронной конструкции по сравнению с одноцистронным клоном в линии клеток E. coliBL21 A1.
На фигуре 7 показана карта вектора pBAD24M-LC.
Подробное описание изобретения
Определения
Используемое в данном документе выражение «представляющий интерес белок» в данном документе относится к любому полипептиду, включая белок и пептиды, используемому в отрасли получения биотерапевтических средств или для диагностических, или для исследовательских целей.
Используемое в данном документе выражение «полинуклеотидная последовательность, кодирующая представляющий интерес белок», в данном документе включает ДНК, кодирующую ген, предпочтительно гетерологичный ген, экспрессирующий полипептид.
Используемые в данном документе термины «рекомбинантный белок и пептид» относятся к белку или пептиду, которые продуцируются в результате экспрессии рекомбинантной ДНК в живых клетках.
Используемые в данном документе термины «Fab» и «антитело» используют взаимозаменяемо, поскольку антитело содержит две части, т.e., Fab- и Fc-участок.
Используемый в данном документе термин «вектор» относится к молекуле ДНК, используемой в качестве носителя для искусственного переноса чужеродного генетического материала в бактериальную клетку, где он может реплицироваться и экспрессироваться.
Используемый в данном документе термин «цистрон» относится к части ДНК, которая содержит генетический код для одного полинуклеотида и функционирующей в виде функциональной единицы наследственности.
Используемое в данном документе выражение «двухцистронная независимая экспрессия» относится к двум отдельным цистронам, которые используют для независимой экспрессии двух одинаковых или двух разных белков.
Используемое в данном документе выражение «одинаковый» является взаимозаменяемым с идентичным или подобным.
Выражение «двухцистронная независимая система экспрессии», используемое в данном документе, включает полинуклеотидную последовательность, кодирующую подлежащий экспрессии полипептид, и последовательности, контролирующие ее экспрессию, такие как промотор и необязательно энхансерная последовательность. Промотор по настоящему изобретению либо функционально связан с подлежащим экспрессии геном, т. е. функциональной единицей транскрипции, либо отделен от нее промежуточным участком ДНК, таким как, например, 5'-нетранслируемый участок гетерологичного гена. Предпочтительно система экспрессии фланкирована одним или более подходящими сайтами рестрикции для обеспечения встраивания кассеты экспрессии в вектор и/или ее вырезания из вектора. Таким образом, система экспрессии в соответствии с настоящим изобретением может быть использована для конструирования вектора экспрессии, в частности, бактериального вектора экспрессии.
Используемый в данном документе термин «промоторы» относится к регуляторному участку ДНК, как правило располагающемуся против хода транскрипции гена, обеспечивающему контрольную точку регуляции транскрипции гена.
Используемое в данном документе выражение «функционально связанный» относятся к функциональной взаимосвязи между двумя или более сегментами ДНК в конкретных последовательностях подлежащих экспрессии генов и к последовательностям, контролирующим их экспрессию.
Используемый в данном документе термин «малые пептиды» или «пептиды» относится к пептидам с массой в диапазоне от 2 до 10 кДа, используемым в отрасли получения биотерапевтических средств и для диагностических или исследовательских целей, к таким как лираглутид, эксанетид, PTH и т. д.
Настоящее изобретение предлагает двухцистронную систему экспрессии для получения ряда представляющих интерес рекомбинантных белков. В определенном варианте осуществления двухцистронная система экспрессии содержит два цистрона, имеющие промотор, функционально связанный с полинуклеотидной последовательностью, кодирующей представляющий интерес белок, и с терминатором.
В определенном варианте осуществления двухцистронная система экспрессии содержит два цистрона, экспрессирующие полинуклеотидную последовательность, кодирующую представляющий интерес белок, которые расположены в одном векторе.
В одном варианте осуществления двухцистронная система экспрессии содержит:
a) первый цистрон, содержащий промотор, функционально связанный с полинуклеотидной последовательностью, кодирующей представляющий интерес белок;
b) второй цистрон, содержащий промотор, функционально связанный с полинуклеотидной последовательностью, кодирующей представляющий интерес белок;
где первый и второй цистроны расположены в одном векторе и экспрессируют полинуклеотидную последовательность, кодирующую представляющий интерес белок, в виде телец включения, образующихся в клетке-хозяине.
В одном варианте осуществления промотор может быть выбран из промотора T7, арабинозного промотора, промотора phoA, tac, lpp, lac-lpp, lac, trp, trc, а предпочтительно из промотора T7 и арабинозного промотора. В определенном варианте осуществления двухцистронная система экспрессии содержит два цистрона, экспрессирующие полинуклеотидную последовательность, кодирующую представляющий интерес белок, под контролем двух промоторов. В одном варианте осуществления оба промотора контролируют экспрессию полинуклеотидной последовательности, кодирующей один и тот же представляющий интерес белок. В другом варианте осуществления оба промотора контролируют экспрессию полинуклеотидной последовательности, кодирующей представляющий интерес белок, отличающийся по длине аминокислотной цепи или по физико-химическим свойствам.
В определенном варианте осуществления представляющий интерес белок может быть выбран из пептидов и белков.
В некоторых вариантах осуществления белки могут экспрессироваться с помощью бицистронного вектора. Белок представляет собой антитело или его фрагмент. Фрагмент антитела может экспрессироваться в бицистронной системе экспрессии. Фрагмент антитела может быть выбран из Fab-фрагмента тяжелой и легкой цепей антител или других фрагментов антител, таких как scFv, диатела, триатела, тетратела, бис-scFv, минитела Fab2 (биспецифические), Fab3 (триспецифические). В предпочтительном варианте осуществления в бицистронной системе экспрессии экспрессируется полинуклеотидная последовательность, кодирующая тяжелую и легкую цепь антитела, образующие Fab-фрагмент антитела. В таком варианте осуществления Fab-фрагмент антитела проявляет аффинность в отношении рецептора VEGF, при этом указанный Fab-фрагмент антитела представляет собой ранибизумаб.
В другом варианте осуществления белок может быть выбран без ограничения из G-CSF, IFN, эритропоэтина, инсулина и его вариантов, PTH (аминокислоты 1-84), FSH, LH, GH и протеиндисульфидизомеразы (PDI).
В некоторых вариантах осуществления пептид может экспрессироваться в бицистронном векторе. Пептиды содержат аминокислотную последовательность, выбранную из по меньшей мере менее 40 аминокислот, или предпочтительно менее 31 аминокислоты, или более предпочтительно менее 10 аминокислот. В определенном варианте осуществления молекулярная масса пептида выбрана из диапазона от приблизительно 2 до приблизительно 10 кДа. Пептид может быть выбран без ограничения из аналога пептида GLP-1, такого как лираглутид или эксендинор, тедуглутида, подобного пептиду GLP-2, и PTH (аминокислоты 1-34), и инсулина. В другом предпочтительном варианте осуществления в бицистронной системе экспрессии экспрессируется полинуклеотидная последовательность, кодирующая агонист рецепторов пептида GLP-1. В таком варианте осуществления пептид GLP-1 представляет собой лираглутид.
В другом варианте осуществления оба промотора независимо контролируют экспрессию разных представляющих интерес белков, таких как тяжелая цепь или легкая цепь антитела, которые отличаются по длине аминокислотной последовательности и физико-химическим свойствам.
В одном варианте осуществления двухцистронная система экспрессии содержит:
a) первый цистрон, содержащий промотор T7, функционально связанный с полинуклеотидной последовательностью, кодирующей тяжелую цепь антитела;
b) второй цистрон, содержащий арабинозный промотор, функционально связанный с полинуклеотидной последовательностью, кодирующей легкую цепь антитела;
где первый и второй цистроны расположены в одном векторе и экспрессируют тяжелую цепь и легкую цепь антитела в виде телец включения, образующихся в клетке-хозяине.
В таком варианте осуществления тяжелая цепь антитела и легкая цепь антитела предусматривает нуклеотидную последовательность с SEQ ID NO: 1 и SEQ ID NO: 2, или аминокислотную последовательность с SEQ ID NO: 3 и SEQ ID NO: 4. В некоторых вариантах осуществления положение первого и второго цистрона является взаимозаменяемым, при этом второй цистрон можно клонировать в вектор в положение первого цистрона, а первый цистрон можно поместить в положение второго цистрона. Тяжелая цепь и легкая цепь антитела независимо экспрессируются в виде телец включения и могут быть дополнительно обработаны с получением Fab-фрагмента антитела, который проявляет аффинность в отношении рецептора VEGF, при этом указанный Fab-фрагмент антитела представляет собой ранибизумаб.
В определенном варианте осуществления тяжелая цепь и легкая цепь антитела необязательно экспрессируются в комбинации с сигнальным пептидом, предпочтительно pelB. Сигнальный пептид управляет экспрессией белка в периплазматическом пространстве клетки-хозяина.
В одном варианте осуществления двухцистронная система экспрессии в одном векторе имеет два разных промотора, арабинозный промотор и промотор T7, регулирующие продуцирование рекомбинантных Fab-фрагментов тяжелой и легкой цепей, соответственно, при этом обе цепи имеют метку pelB, которые продуцируются в виде нерастворимых телец включения в периплазматическом пространстве E. coli.
В определенном варианте осуществления тяжелая цепь или легкая цепь антитела необязательно эксперссируется в комбинации с регулятором, предпочтительно геном AraC, для дополнительного повышения экспрессии белка.
Двухцистронная система экспрессии обеспечивает экспрессию представляющего интерес белка в эквимолярных количествах. Экспрессия в эквимолярном количестве является особенно желательной для получения представляющего интерес белка подходящего качества и в подходящем количестве. Она зависит от соотношения тяжелой и легкой цепи или соотношения функциональных единиц полипептида в случае клонирования нуклеотидной последовательности в вектор. В определенном варианте осуществления последовательности тяжелой и легкой цепи клонированы при подходящем соотношении, предусматривающем, что количество последовательностей тяжелой цепи равно или больше количества последовательностей легкой цепи, с достижением экспрессии тяжелой и легкой цепи в эквимолярном количестве. Последовательности тяжелой цепи и легкой цепи клонируют при соотношении, выбранном из от 1:5:0,7 до 1:1, которое включает 1:3:0,8, 1:2:0,9, 1:2:1 1:1.
В одном варианте осуществления двухцистронная система экспрессии содержит нуклеотидную последовательность, изложенную под SEQ ID NO: 19.
В другом варианте осуществления двухцистронная система экспрессии содержит:
a) первый цистрон, содержащий промотор T7, функционально связанный с полинуклеотидной последовательностью, кодирующей пептид;
b) второй цистрон, содержащий арабинозный промотор, функционально связанный с полинуклеотидной последовательностью, кодирующей пептид;
где первый и второй цистроны расположены в одном векторе и экспрессируют пептид в виде телец включения, образующихся в клетке-хозяине.
В таком варианте осуществления пептид представляет собой аналог GLP-1, содержащий нуклеотидную последовательность, изложенную под SEQ ID NO: 6, кодирующую агонист рецепторов пептида GLP-1, представляющий собой лираглутид, имеющий аминокислотную последовательность с SEQ ID NO: 7.
В определенном варианте осуществления пептид может быть необязательно экспрессирован с сигнальным пептидом или регулятором/энхансером, известным специалисту в данной области.
В определенном варианте осуществления пептид может быть необязательно экспрессирован с партнером для слияния или меткой слияния для предупреждения деградации пептида. Партнер для слияния представляет собой аминокислотную последовательность из от 30 аминокислот до 300 аминокислот. Партнер для слияния представляет собой аминокислотную последовательность, выбранную из приблизительно 50 аминокислот, 100 аминокислот, приблизительно 136 аминокислот, приблизительно 175 аминокислот, приблизительно 250 аминокислот, 300 аминокислот, предпочтительно приблизительно 136 аминокислот. Метка слияния может быть выбрана без ограничения из гистидиновой метки, глутатион-s-трансферазы (GST), мальтозосвязывающего белка, NusA, тиоредоксина (TRX), полигистидина (HIS), малого убиквитин-подобного модификатора (SUMO) и убиквитина (Ub), а также гена стафилокиназы (SAK). В предпочтительном варианте осуществления метка слияния представляет собой ген SAK. Подробности использования гена SAK в качестве метки слияния с представляющим интерес белком раскрываются в US8853380, который включен в данный документ посредством ссылки.
В некоторых вариантах осуществления бицистронная система экспрессии дополнительно включает в себя селективный маркер, который выбран из гена устойчивости к ампициллину, канамицину, предпочтительно из гена устойчивости к ампициллину.
В другом варианте осуществления настоящее изобретение предлагает способ получения представляющего интерес белка, предусматривающего стадии:
(i) трансформации клетки-хозяина одним вектором, включающим в себя, по сути, двухцистронную систему экспрессии;
(ii) культивирования трансформированной клетки в подходящей среде для экспрессии представляющего интерес белка, при этом первый и второй цистрон экспрессирует представляющий интерес белок в тельцах включения;
(iii) проведения солюбилизации телец включения;
(iv) проведения рефолдинга представляющего интерес белка.
В одном варианте осуществления бицистронной системой экспрессии трансфицируют подходящую бактериальную клетку-хозяина для того, чтобы экспрессировать представляющий интерес белок. Подходящая бактериальная клетка-хозяин представляет собой E. coli, в которой представляющий интерес белок экспрессируется в виде телец включения. Тельца включения представляют собой нерастворимые материалы, образовавшиеся в периплазме или цитоплазме E. coli. Тельца включения можно выделять, солюбилизировать, а представляющий интерес белок можно извлекать в активной форме посредством хорошо известных в данной области методик.
В одном варианте осуществления Fab-фрагмент тяжелой и легкой цепей антител или другие фрагменты антител, такие как scFv, диатела, триатела, тетратела, бис-scFv, минитела Fab2 (биспецифические), Fab3 (триспецифические) экспрессировали в виде нерастворимых телец включения в периплазматическом пространстве E. coli при помощи конструкций, созданных из двух независимых цистронов в одном векторе, имеющем два разных промотора, арабинозный промотор и промотор T7. Два разных промотора, т. e., промотор T7 и арабинозный промотор, обеспечивают экспрессию тяжелой и легкой цепей молекулы Fab-фрагмента, соответственно. Тяжелую и легкую цепи антитела получали в виде нефункциональных телец включения в бактериальной клетке, т. е, E. сoli,, которые в дальнейшем экстрагировали, подвергали рефолдингу и очищали.
В одном варианте осуществления по настоящему изобретению цистрон является таким, что каждый ген (тяжелой и легкой цепи) будет иметь свой собственный промотор и терминатор в одном векторе. Последовательность тяжелой цепи клонировали под контролем промотора T7, тогда как последовательность легкой цепи клонировали под контролем арабинозного промотора. Обе цепи начинались метками в виде сигнальных последовательностей pelB, используемых для получения продукта в периплазматическом пространстве бактериальной мембраны.
Преимущество бицистронной системы экспрессии заключается в том, что как арабинозный промотор, так и T7 являются сильными промоторами, причем высокий уровень экспрессии как легкой, так и тяжелой цепей, достигается в результате однократного цикла ферментации вместо отдельных циклов ферментации с клонами последовательностей легкой и тяжелой цепи. Двухцистронная система экспрессии облегчает характеристику и поддержание единого банка клеток вместо отдельных банков клеток для клонов легкой и тяжелой цепи. Кроме того, полученные таким образом тельца включения являются относительно чистыми, если их экстрагируют из периплазматического пространства бактериальных клеток. Высокий уровень экспрессии и более чистые формы легкой и тяжелой цепей, полученных в виде телец включения, обеспечивают возможность более легкого фолдинга в функциональные Fab ex vivo, значительно повышая тем самым выход продукта.
Другое преимущество данной системы заключается в том, что нуклеотидную последовательность представляющего интерес белка можно клонировать и экспрессировать под контролем арабинозного промотора и промотора T7, при этом может значительно повышаться уровень экспрессии белка.
Раскрытые далее примеры лишь иллюстрируют настоящее изобретение и не предназначены для ограничения.
Пример 1. Клонирование последовательности тяжелой цепи в вектор pET21a
Последовательность ДНК, используемая для клонирования последовательности тяжелой и легкой цепи Fab-фрагментов, приведена под SEQ ID NO: 1 и 2, соответственно. Вставку последовательности тяжелой цепи амплифицировали из синтетической ДНК с использованием специфических к генам праймеров. Праймеры конструировали в соответствии со способами, хорошо известными в данной области техники. ПЦР-продукт последовательности тяжелой цепи затем расщепляли с использованием ферментов NdeI-HindIII и лигировали в вектор pET21a, который расщепляли теми же самыми ферментами. Клоны подвергали скринингу посредством ПЦР для отбора колоний и их подтверждали с помощью рестрикционного анализа. Полученному в результате клону присвоили обозначение pET21a-HC. Рекомбинантный вектор вводили в линию клеток BL21A1 и проверяли клетки в отношении экспрессии тяжелой цепи.
Пример 2. Клонирование последовательности легкой цепи в вектор pBAD24M
Вставку последовательности легкой цепи амплифицировали из синтетической ДНК с использованием специфических к генам праймеров. Праймеры конструировали в соответствии со способами, хорошо известными в данной области техники. Амплифицированную последовательность легкой цепи расщепляли с использованием ферментов NdeI-HindIII и лигировали в расщепленный вектор pBAD24M (имеется в лаборатории) в те же самые сайты. Клоны подвергали скринингу посредством ПЦР для отбора колоний и их подтверждали с помощью рестрикционного анализа. Полученному в результате клону присвоили обозначение pBAD24M-LC. Рекомбинантный вектор вводили в линию клеток BL21A1 и проверяли клетки в отношении экспрессии легкой цепи.
Пример 3. Конструкция из двух независимых цистронов в одном и том же векторе
Праймеры конструировали для амплификации последовательности легкой цепи вместе с арабинозным промотором, терминатором и геном araC. Праймеры конструировали в соответствии со способами, хорошо известными в данной области техники. Данные праймеры присоединяли линкер с сайтом BglII к амплифицированному продукту. Вектор pET21a имел отдельный сайт BglII выше промотора T7. Кассету экспрессии с последовательностью легкой цепи амплифицировали из матрицы pBAD24M с использованием специфических к вектору праймеров и клонировали в клон pET21a-HC по сайту BglII. Клон подтверждали с помощью расщепления рестриктазами и подвергали секвенированию. Полученному в результате клону присваивали обозначение pET21a-HC-LC, а подходящие клоны вкратце перечислены с учетом экспрессии. Карта клона pET21a-HC-LC представлена на фигуре 2.
Полученный таким образом клон содержит все части, требующиеся для независимой регуляции и экспрессии как тяжелых, так и легких цепей.
Пример 4. Анализ экспрессии
Линию клеток E. coli BL21 A1 использовали в качестве хозяина для экспрессии. Помимо BL21 A1, использовали BL21 DE3 или какую-либо другую линию клеток, содержащую в своем геноме промотор T7. Клетки BL21 A1 трансформировали с использованием выбранного выше клона вместе с pET21a-HC и pBAD24M-LC в качестве контролей. Экспрессию тяжелой цепи индуцировали IPTG, тогда как экспрессию легкой цепи индуцировали арабинозой. Концентрация индуцирующего средства составляла 13 мM арабинозы и 1 мM IPTG, а индуцирование завершали тогда, когда OD600 культуры составляла ~1. Клетки собирали через 4 ч. после индуцирования. Исследование проводили во флаконах при встряхивании. Собранные клетки лизировали с помощью бус и центрифугировали для разделения растворимых и нерастворимых фракций. Образцы наносили на 12% гели для SDS PAGE с проведением электрофореза для проверки экспрессии. Анализ результатов SDS PAGE показан на фигуре 3. Восстановленный ранибизумаб вносили в лунку 5, что показано на фигуре 3, для подтверждения экспрессии восстановленных легкой и тяжелой цепей.
Анализ SDS PAGE показал экспрессию обеих цепей в нерастворимой осадочной фракции, и тот же самый результат подтвердили с помощью анализа RP-HPLC, где значения времени удерживания легкой и тяжелой цепей референтного продукта соответствовали значениям времени удерживания полученного в лаборатории продукта. Используемыми контролями были молекула восстановленного Fab (референтный продукт), и продукты клона pET21a-HC и клона pBAD24M. Таким образом, подтвердили экспрессию как тяжелой, так и легкой цепи в одном клоне. Анализ результатов RP-HPLC показан на фигурах 4 и 5. В ходе RP-HPLC солюбилизированные и восстановленные IB из двухцистронного клона сравнивали с восстановленным ранибизумабом (RMP) и клонами, экспрессирующими тяжелую и легкую цепи по отдельности, т. e. pET21-HC и pBAD24MLC.
Время удерживания (RT) основного пика солибилизированного IB из pBAD24MLC, экспрессирующего только легкую цепь, соответствовало RT легкой цепи восстановленного RMP. Пик примесей при RT в 13 минут соответствовал времени удерживания для тяжелой цепи, что было показателем аналогичной гидрофобности. Таким образом, примеси характеризовали посредством LC-MS/MS и в заключении установили, что это белок OMP C клетки-хозяина и легкой цепи с неотщепленной лидерной последовательностью при RT в 17 минут. Тяжелая цепь, экспрессируемая pET21a-HC, соответствовала референтной стандартной тяжелой цепи. На профиле также виден хвостовой пик на RT 19, который соответствовал тяжелой цепи с неотщепленной лидерной последовательностью. Клон pET21a_HC_LC с двумя цистронами, который экспрессирует как LC, так и HC, характеризуется 2 главными пиками, которые имеют значения время удерживания, эквивалентные значениям времени удерживания для LC и HC референтного стандарта. Но, поскольку OMP C элюировался одновременно с тяжелой цепью, способ анализа с обращенной фазой имел лучшую разрешающую способность, что показано на фигуре 4.
Существующий способ анализа на колонке Zorbax C8 RP модифицировали для Aeriswidepore C8 и проводили одновременное элюирование фрагментов с лучшей разрешающей способностью. Солюбилизированное IB из pET21a_HC_LC на Aeriswidepore C8 проявляло отчетливые пики, соответствующие LC, HC и OMP C, что давало возможность идентификации и точного количественного определения отдельных субъединиц в IB, что видно на фигуре 5.
Хотя некоторые варианты осуществления и примеры были подробно описаны выше, специалист в данной области техники сможет отчетливо понять, что в вариантах осуществления и примерах возможно много модификаций без отступления от их идей.
Пример 5. Клонирование нуклеотидной последовательности малого пептида (лираглутида) с меткой слияния в виде нуклеотидной последовательности стафилокиназы (SAK) в вектор pET24a
Гены SAK и лираглутида амплифицировали из синтетической ДНК с использованием специфических к генам праймеров. Праймеры сконструировали в соответствии со способами, хорошо известными в данной области техники, а ПЦР-продукты расщепляли ферментами NdeI-BamHI и BamHI-HindIII и лигировали в расщепленный вектор pET24a по сайтам NdeI-HindIII. Клоны подвергали скринингу посредством ПЦР для отбора колоний и их подтверждали с помощью рестрикционного анализа. Полученному в результате клону присвоили обозначение pET24a-SAK-Lira.
Пример 6. Клонирование нуклеотидной последовательности малого пептида (лираглутида) с меткой слияния в виде нуклеотидной последовательности стафилокиназы в вектор pBAD24M
Как описано в примере 5, нуклеотидную последовательность лираглутида с меткой SAK клонировали в вектор pBAD24M. Клону присвоили обозначение pBAD24M-SAK-Lira.
Пример 7. Конструкция из двух независимых цистронов в одном и том же векторе, причем оба цистрона экспрессируют пептидслияния SAK-Lira
Стратегию конструирования клона, используемую в примере 3, использовали для конструирования клона с двумя цистронами, экспрессирующего лираглутид, где ген слияния SAK-Lira вместе с кассетой экспрессии, контролируемой арабинозным промотором, амплифицировали из клона pBAD24M-SAK-Lira и клонировали в клон pET24a-SAK-Lira с получением конструкции, содержащей два цистрона. Клону присвоили обозначение pET-ara-SAK-Lira.
Пример 8. Анализ экспрессии двухцистронного клона с белком слияния SAK-Lira
Линию клеток E. coli BL21 A1 использовали в качестве хозяина для экспрессии. Помимо BL21 A1, использовали BL21 DE3 или какую-либо другую линию клеток, содержащую в своем геноме промотор T7. Клетки BL21 A1 трансформировали с использованием одно- или двухцистронной конструкций. Осуществляли индуцирование клонов с использованием IPTG и арабинозы. Концентрация индуцирующего средства составляла 13 мM арабинозы и 1 мM IPTG, а индуцирование завершали тогда, когда OD600 культуры составляла ~1. Клетки собирали через 4 ч. после индуцирования. Исследование проводили во флаконах при встряхивании. Собранные клетки лизировали с помощью бус и центрифугировали для разделения растворимых и нерастворимых фракций. Образцы наносили на 12% гели для SDS PAGE с проведением электрофореза для проверки экспрессии.
Анализ посредством SDS PAGE электрофореза четко показал повышение экспрессии белка-слияния SAK-Lira в двухцистронном клоне (фигура 6, дорожка 2) по сравнению с одноцистронным клоном pET24a-SAK-Lira (фигура 6, дорожка 3).
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЛЮПИН ЛИМИТЕД
<120> Двухцистронная бактериальная система экспрессии
<130> FPAA4613PCT
<150> 2245/MUM/2014
<151> 2014-07-09
<160> 19
<170> PatentIn версии 3.5
<210> 1
<211> 762
<212> ДНК
<213> искусственная
<220>
<223> Ранибизумаб, последовательность тяжелой цепи
<400> 1
atgaaatacc tgctgccgac agctgctgct ggtctgctgc tcctcgctgc ccagccggcg 60
atggccgaag tccaactggt cgaatcgggt ggtggtctgg tccaaccggg tggctctctg 120
cgtctgtcgt gtgctgcctc gggctatgat tttacccatt atggtatgaa ctgggtccgt 180
caggccccgg gtaaaggtct ggaatgggtg ggctggatta atacctacac gggtgaaccg 240
acctatgcgg ccgattttaa acgtcgcttt acgttctctc tggacacctc gaagagcacg 300
gcatatctgc agatgaacag tctgcgcgcg gaagataccg ccgtgtatta ctgcgcgaag 360
tacccgtatt actatggcac gtcccactgg tattttgacg tttggggcca aggtaccctg 420
gtcaccgtga gcagcgcgag caccaaaggc ccgagcgtgt tcccgctggc cccgagttcc 480
aagtctacca gtggcggtac ggcagctctg ggttgtctgg ttaaagatta ttttccggaa 540
ccggttaccg tctcctggaa cagcggcgca ctgacctctg gtgtgcatac gttcccggct 600
gttctgcagt catcgggcct gtacagcctg agcagcgtgg ttaccgttcc gagttcctca 660
ctgggtaccc aaacgtatat ctgcaacgtc aatcacaaac cgagcaatac caaagtggac 720
aaaaaagtgg aaccgaaatc gtgtgataaa acgcatctgt aa 762
<210> 2
<211> 711
<212> ДНК
<213> искусственная
<220>
<223> Ранибизумаб, легкая цепь: последовательность нуклеотидов 2295-3005
<400> 2
atgaaatacc tgctgccgac agctgctgct ggtctgctgc tcctcgctgc ccagccggcg 60
atggccgaca ttcaactgac gcaaagtccg agcagcctga gcgcatccgt gggcgaccgt 120
gtgacgatta cctgttccgc aagccaagac atctctaact atctgaattg gtaccagcaa 180
aaaccgggca aggcaccgaa agtcctgatt tattttacca gctctctgca ttccggcgtt 240
ccgtcacgtt ttagcggctc tggtagtggc accgatttca ccctgacgat cagttccctg 300
cagccggaag actttgctac gtattactgc cagcaataca gcaccgtgcc gtggacgttc 360
ggtcagggca ccaaggttga aattaaacgt acggttgcgg ccccgtctgt ctttatcttc 420
ccgccgagtg atgaacagct gaaatcgggt accgcaagcg tggtttgtct gctgaacaat 480
ttctatccgc gcgaagcaaa ggtccagtgg aaagtggaca acgctctgca gtccggcaat 540
tcacaagaat cggtgaccga acaagatagc aaggactcta cgtacagtct gtcatcgacc 600
ctgacgctgt ccaaagcgga ttatgaaaaa cacaaggttt acgcctgcga agtcacccat 660
caaggtctgt cgtctccggt taccaagagt ttcaatcgtg gcgaatgtta a 711
<210> 3
<211> 253
<212> БЕЛОК
<213> искусственная
<220>
<223> Ранибизумаб, аминокислотная последовательность тяжелой цепи
<400> 3
Met Lys Tyr Leu Leu Pro Thr Ala Ala Ala Gly Leu Leu Leu Leu Ala
1 5 10 15
Ala Gln Pro Ala Met Ala Glu Val Gln Leu Val Glu Ser Gly Gly Gly
20 25 30
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
35 40 45
Tyr Asp Phe Thr His Tyr Gly Met Asn Trp Val Arg Gln Ala Pro Gly
50 55 60
Lys Gly Leu Glu Trp Val Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro
65 70 75 80
Thr Tyr Ala Ala Asp Phe Lys Arg Arg Phe Thr Phe Ser Leu Asp Thr
85 90 95
Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
100 105 110
Thr Ala Val Tyr Tyr Cys Ala Lys Tyr Pro Tyr Tyr Tyr Gly Thr Ser
115 120 125
His Trp Tyr Phe Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser
130 135 140
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser
145 150 155 160
Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
165 170 175
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
180 185 190
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
195 200 205
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln
210 215 220
Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp
225 230 235 240
Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Leu
245 250
<210> 4
<211> 236
<212> БЕЛОК
<213> искусственная
<220>
<223> Ранибизумаб, аминокислотная последовательность легкой цепи
<400> 4
Met Lys Tyr Leu Leu Pro Thr Ala Ala Ala Gly Leu Leu Leu Leu Ala
1 5 10 15
Ala Gln Pro Ala Met Ala Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser
20 25 30
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Ser Ala Ser
35 40 45
Gln Asp Ile Ser Asn Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Val Leu Ile Tyr Phe Thr Ser Ser Leu His Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
100 105 110
Tyr Ser Thr Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
115 120 125
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
130 135 140
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
145 150 155 160
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
165 170 175
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
180 185 190
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
195 200 205
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
210 215 220
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 5
<211> 762
<212> ДНК
<213> искусственная
<220>
<223> Комплементарная последовательность тяжелой цепи 940-178
<400> 5
ttacagatgc gttttatcac acgatttcgg ttccactttt ttgtccactt tggtattgct 60
cggtttgtga ttgacgttgc agatatacgt ttgggtaccc agtgaggaac tcggaacggt 120
aaccacgctg ctcaggctgt acaggcccga tgactgcaga acagccggga acgtatgcac 180
accagaggtc agtgcgccgc tgttccagga gacggtaacc ggttccggaa aataatcttt 240
aaccagacaa cccagagctg ccgtaccgcc actggtagac ttggaactcg gggccagcgg 300
gaacacgctc gggcctttgg tgctcgcgct gctcacggtg accagggtac cttggcccca 360
aacgtcaaaa taccagtggg acgtgccata gtaatacggg tacttcgcgc agtaatacac 420
ggcggtatct tccgcgcgca gactgttcat ctgcagatat gccgtgctct tcgaggtgtc 480
cagagagaac gtaaagcgac gtttaaaatc ggccgcatag gtcggttcac ccgtgtaggt 540
attaatccag cccacccatt ccagaccttt acccggggcc tgacggaccc agttcatacc 600
ataatgggta aaatcatagc ccgaggcagc acacgacaga cgcagagagc cacccggttg 660
gaccagacca ccacccgatt cgaccagttg gacttcggcc atcgccggct gggcagcgag 720
gagcagcaga ccagcagcag ctgtcggcag caggtatttc at 762
<210> 6
<211> 96
<212> ДНК
<213> искусственная
<220>
<223> ДНК-последовательность лираглутида
<400> 6
catgcagaag gcacctttac gagtgatgtg agctcttatc tggaaggcca ggcggccaaa 60
gaatttattg cgtggctggt tcgtggccgt ggttaa 96
<210> 7
<211> 31
<212> БЕЛОК
<213> искусственная
<220>
<223> Аминокислотная последовательность лираглутида
<400> 7
His Ala Glu Gly Thr Phe Thr Ser Asp Val Ser Ser Tyr Leu Glu Gly
1 5 10 15
Gln Ala Ala Lys Glu Phe Ile Ala Trp Leu Val Arg Gly Arg Gly
20 25 30
<210> 8
<211> 456
<212> ДНК
<213> искусственная
<220>
<223> Нуклеотидная последовательность SAK (с сайтом EK)
<400> 8
catatgtcaa gttcattcga caaaggaaaa tataaaaaag gcgatgacgc gagttatttt 60
gaaccaacag gcccgtattt gatggtaaat gtgactggag ttgatggtaa aggaaatgaa 120
ttgctatccc ctcattatgt cgagtttcct attaaacctg ggactacact tacaaaagaa 180
aaaattgaat acctgcagga tgatgatgat aaatacgtag aatgggcatt agatgcgaca 240
gcatataaag agtttagagt agttgaatta gatccaagcg caaagatcga agtcacttat 300
tatgataaga ataagaaaaa agaagaaacg aagtctttcc ctataacaga aaaaggtttt 360
gttgtcccag atttatcaga gcatattaaa aaccctggat tcaacttaat tacaaaggtt 420
gttatagaaa agaaaggatc cgatgatgat gataaa 456
<210> 9
<211> 151
<212> БЕЛОК
<213> искусственная
<220>
<223> Аминокислотная последовательность SAK (с меткой EK)
<400> 9
Met Ser Ser Ser Phe Asp Lys Gly Lys Tyr Lys Lys Gly Asp Asp Ala
1 5 10 15
Ser Tyr Phe Glu Pro Thr Gly Pro Tyr Leu Met Val Asn Val Thr Gly
20 25 30
Val Asp Gly Lys Gly Asn Glu Leu Leu Ser Pro His Tyr Val Glu Phe
35 40 45
Pro Ile Lys Pro Gly Thr Thr Leu Thr Lys Glu Lys Ile Glu Tyr Leu
50 55 60
Gln Asp Asp Asp Asp Lys Tyr Val Glu Trp Ala Leu Asp Ala Thr Ala
65 70 75 80
Tyr Lys Glu Phe Arg Val Val Glu Leu Asp Pro Ser Ala Lys Ile Glu
85 90 95
Val Thr Tyr Tyr Asp Lys Asn Lys Lys Lys Glu Glu Thr Lys Ser Phe
100 105 110
Pro Ile Thr Glu Lys Gly Phe Val Val Pro Asp Leu Ser Glu His Ile
115 120 125
Lys Asn Pro Gly Phe Asn Leu Ile Thr Lys Val Val Ile Glu Lys Lys
130 135 140
Gly Ser Asp Asp Asp Asp Lys
145 150
<210> 10
<211> 17
<212> ДНК
<213> искусственная
<220>
<223> Промотор T7: 1011-1027
<400> 10
ctatagtgag tcgtatt 17
<210> 11
<211> 52
<212> ДНК
<213> искусственная
<220>
<223> Терминатор T7:26-72
<400> 11
ttcagcaaaa aacccctcaa gacccgttta gaggccccaa ggggttatgc ta 52
<210> 12
<211> 10
<212> ДНК
<213> искусственная
<220>
<223> RBS из pET21a: 947-957
<400> 12
tctccttctt 10
<210> 13
<211> 879
<212> ДНК
<213> искусственная
<220>
<223> Ген AraC: 1048-1927
<400> 13
ttatgacaac ttgacggcta catcattcac tttttcttca caaccggcac ggaactcgct 60
cgggctggcc ccggtgcatt ttttaaatac ccgcgagaaa tagagttgat cgtcaaaacc 120
aacattgcga ccgacggtgg cgataggcat ccgggtggtg ctcaaaagca gcttcgcctg 180
gctgatacgt tggtcctcgc gccagcttaa gacgctaatc cctaactgct ggcggaaaag 240
atgtgacaga cgcgacggcg acaagcaaac atgctgtgcg acgctggcga tatcaaaatt 300
gctgtctgcc aggtgatcgc tgatgtactg acaagcctcg cgtacccgat tatccatcgg 360
tggatggagc gactcgttaa tcgcttccat gcgccgcagt aacaattgct caagcagatt 420
tatcgccagc agctccgaat agcgcccttc cccttgcccg gcgttaatga tttgcccaaa 480
caggtcgctg aaatgcggct ggtgcgcttc atccgggcga aagaaccccg tattggcaaa 540
tattgacggc cagttaagcc attcatgcca gtaggcgcgc ggacgaaagt aaacccactg 600
gtgataccat tcgcgagcct ccggatgacg accgtagtga tgaatctctc ctggcgggaa 660
cagcaaaata tcacccggtc ggcaaacaaa ttctcgtccc tgatttttca ccaccccctg 720
accgcgaatg gtgagattga gaatataacc tttcattccc agcggtcggt cgataaaaaa 780
atcgagataa ccgttggcct caatcggcgt taaacccgcc accagatggg cattaaacga 840
gtatcccggc agcaggggat cattttgcgc ttcagccat 879
<210> 14
<211> 27
<212> ДНК
<213> искусственная
<220>
<223> Арабинозный промотор: 2203-2230
<400> 14
acgcttttta tcgcaactct ctactgt 27
<210> 15
<211> 425
<212> ДНК
<213> искусственная
<220>
<223> Арабинозный терминатор: 3011-3438
<400> 15
gctgttttgg cggatgagag aagattttca gcctgataca gattaaatca gaacgcagaa 60
gcggtctgat aaaacagaat ttgcctggcg gcagtagcgc ggtggtccca cctgacccca 120
tgccgaactc agaagtgaaa cgccgtagcg ccgatggtag tgtggggtct ccccatgcga 180
gagtagggaa ctgccaggca tcaaataaaa cgaaaggctc agtcgaaaga ctgggccttt 240
cgttttatct gttgtttgtc ggtgaacgct ctcctgagta ggacaaatcc gccgggagcg 300
gatttgaacg ttgcgaagca acggcccgga gggtggcggg caggacgccc gccataaact 360
gccaggcatc aaattaagca gaaggccatc ctgacggatg gcctttttgc gtttctacaa 420
actct 425
<210> 16
<211> 1084
<212> ДНК
<213> искусственная
<220>
<223> Последовательность LacI: 3810-4890
<400> 16
tgtgaaacca gtaacgttat acgatgtcgc agagtatgcc ggtgtctctt atcagaccgt 60
ttcccgcgtg gtgaaccagg ccagccacgt ttctgcgaaa acgcgggaaa aagtggaagc 120
ggcgatggcg gagctgaatt acattcccaa ccgcgtggca caacaactgg cgggcaaaca 180
gtcgttgctg attggcgttg ccacctccag tctggccctg cacgcgccgt cgcaaattgt 240
cgcggcgatt aaatctcgcg ccgatcaact gggtgccagc gtggtggtgt cgatggtaga 300
acgaagcggc gtcgaagcct gtaaagcggc ggtgcacaat cttctcgcgc aacgcgtcag 360
tgggctgatc attaactatc cgctggatga ccaggatgcc attgctgtgg aagctgcctg 420
cactaatgtt ccggcgttat ttcttgatgt ctctgaccag acacccatca acagtattat 480
tttctcccat gaagacggta cgcgactggg cgtggagcat ctggtcgcat tgggtcacca 540
gcaaatcgcg ctgttagcgg gcccattaag ttctgtctcg gcgcgtctgc gtctggctgg 600
ctggcataaa tatctcactc gcaatcaaat tcagccgata gcggaacggg aaggcgactg 660
gagtgccatg tccggttttc aacaaaccat gcaaatgctg aatgagggca tcgttcccac 720
tgcgatgctg gttgccaacg atcagatggc gctgggcgca atgcgcgcca ttaccgagtc 780
cgggctgcgc gttggtgcgg atatctcggt agtgggatac gacgataccg aagacagctc 840
atgttatatc ccgccgttaa ccaccatcaa acaggatttt cgcctgctgg ggcaaaccag 900
cgtggaccgc ttgctgcaac tctctcaggg ccaggcggtg aagggcaatc agctgttgcc 960
cgtctcactg gtgaaaagaa aaaccaccct ggcgcccaat acgcaaaccg cctctccccg 1020
cgcgttggcc gattcattaa tgcagctggc acgacaggtt tcccgactgg aaagcgggca 1080
gtga 1084
<210> 17
<211> 861
<212> ДНК
<213> искусственная
<220>
<223> bla (ген устойчивости к ампициллину): 7085-7942
<400> 17
ttaccaatgc ttaatcagtg aggcacctat ctcagcgatc tgtctatttc gttcatccat 60
agttgcctga ctccccgtcg tgtagataac tacgatacgg gagggcttac catctggccc 120
cagtgctgca atgataccgc gagacccacg ctcaccggct ccagatttat cagcaataaa 180
ccagccagcc ggaagggccg agcgcagaag tggtcctgca actttatccg cctccatcca 240
gtctattaat tgttgccggg aagctagagt aagtagttcg ccagttaata gtttgcgcaa 300
cgttgttgcc attgctgcag gcatcgtggt gtcacgctcg tcgtttggta tggcttcatt 360
cagctccggt tcccaacgat caaggcgagt tacatgatcc cccatgttgt gcaaaaaagc 420
ggttagctcc ttcggtcctc cgatcgttgt cagaagtaag ttggccgcag tgttatcact 480
catggttatg gcagcactgc ataattctct tactgtcatg ccatccgtaa gatgcttttc 540
tgtgactggt gagtactcaa ccaagtcatt ctgagaatag tgtatgcggc gaccgagttg 600
ctcttgcccg gcgtcaatac gggataatac cgcgccacat agcagaactt taaaagtgct 660
catcattgga aaacgttctt cggggcgaaa actctcaagg atcttaccgc tgttgagatc 720
cagttcgatg taacccactc gtgcacccaa ctgatcttca gcatctttta ctttcaccag 780
cgtttctggg tgagcaaaaa caggaaggca aaatgccgca aaaaagggaa taagggcgac 840
acggaaatgt tgaatactca t 861
<210> 18
<211> 1
<212> ДНК
<213> искусственная
<220>
<223> Последовательность инициации репликации 6323 в pBR322
<400> 18
t 1
<210> 19
<211> 8540
<212> ДНК
<213> искусственная
<220>
<223> Полная последовательность клона с двухцистронным вектором, включающим в себя последовательности тяжелой и легкой цепи fab
<400> 19
atccggatat agttcctcct ttcagcaaaa aacccctcaa gacccgttta gaggccccaa 60
ggggttatgc tagttattgc tcagcggtgg cagcagccaa ctcagcttcc tttcgggctt 120
tgttagcagc cggatctcag tggtggtggt ggtggtgctc gagtgcggcc gcaagctttt 180
acagatgcgt tttatcacac gatttcggtt ccactttttt gtccactttg gtattgctcg 240
gtttgtgatt gacgttgcag atatacgttt gggtacccag tgaggaactc ggaacggtaa 300
ccacgctgct caggctgtac aggcccgatg actgcagaac agccgggaac gtatgcacac 360
cagaggtcag tgcgccgctg ttccaggaga cggtaaccgg ttccggaaaa taatctttaa 420
ccagacaacc cagagctgcc gtaccgccac tggtagactt ggaactcggg gccagcggga 480
acacgctcgg gcctttggtg ctcgcgctgc tcacggtgac cagggtacct tggccccaaa 540
cgtcaaaata ccagtgggac gtgccatagt aatacgggta cttcgcgcag taatacacgg 600
cggtatcttc cgcgcgcaga ctgttcatct gcagatatgc cgtgctcttc gaggtgtcca 660
gagagaacgt aaagcgacgt ttaaaatcgg ccgcataggt cggttcaccc gtgtaggtat 720
taatccagcc cacccattcc agacctttac ccggggcctg acggacccag ttcataccat 780
aatgggtaaa atcatagccc gaggcagcac acgacagacg cagagagcca cccggttgga 840
ccagaccacc acccgattcg accagttgga cttcggccat cgccggctgg gcagcgagga 900
gcagcagacc agcagcagct gtcggcagca ggtatttcat atgtatatct ccttcttaaa 960
gttaaacaaa attatttcta gaggggaatt gttatccgct cacaattccc ctatagtgag 1020
tcgtattaat ttcgcgggat cgagatcttt atgacaactt gacggctaca tcattcactt 1080
tttcttcaca accggcacgg aactcgctcg ggctggcccc ggtgcatttt ttaaataccc 1140
gcgagaaata gagttgatcg tcaaaaccaa cattgcgacc gacggtggcg ataggcatcc 1200
gggtggtgct caaaagcagc ttcgcctggc tgatacgttg gtcctcgcgc cagcttaaga 1260
cgctaatccc taactgctgg cggaaaagat gtgacagacg cgacggcgac aagcaaacat 1320
gctgtgcgac gctggcgata tcaaaattgc tgtctgccag gtgatcgctg atgtactgac 1380
aagcctcgcg tacccgatta tccatcggtg gatggagcga ctcgttaatc gcttccatgc 1440
gccgcagtaa caattgctca agcagattta tcgccagcag ctccgaatag cgcccttccc 1500
cttgcccggc gttaatgatt tgcccaaaca ggtcgctgaa atgcggctgg tgcgcttcat 1560
ccgggcgaaa gaaccccgta ttggcaaata ttgacggcca gttaagccat tcatgccagt 1620
aggcgcgcgg acgaaagtaa acccactggt gataccattc gcgagcctcc ggatgacgac 1680
cgtagtgatg aatctctcct ggcgggaaca gcaaaatatc acccggtcgg caaacaaatt 1740
ctcgtccctg atttttcacc accccctgac cgcgaatggt gagattgaga atataacctt 1800
tcattcccag cggtcggtcg ataaaaaaat cgagataacc gttggcctca atcggcgtta 1860
aacccgccac cagatgggca ttaaacgagt atcccggcag caggggatca ttttgcgctt 1920
cagccatact tttcatactc ccgccattca gagaagaaac caattgtcca tattgcatca 1980
gacattgccg tcactgcgtc ttttactggc tcttctcgct aaccaaaccg gtaaccccgc 2040
ttattaaaag cattctgtaa caaagcggga ccaaagccat gacaaaaacg cgtaacaaaa 2100
gtgtctataa tcacggcaga aaagtccaca ttgattattt gcacggcgtc acactttgct 2160
atgccatagc atttttatcc ataagattag cggatcctac ctgacgcttt ttatcgcaac 2220
tctctactgt ttctccatac ccgttttttt gggctagaaa taattttgtt taactttaag 2280
aaggagatat acatatgaaa tacctgctgc cgacagctgc tgctggtctg ctgctcctcg 2340
ctgcccagcc ggcgatggcc gacattcaac tgacgcaaag tccgagcagc ctgagcgcat 2400
ccgtgggcga ccgtgtgacg attacctgtt ccgcaagcca agacatctct aactatctga 2460
attggtacca gcaaaaaccg ggcaaggcac cgaaagtcct gatttatttt accagctctc 2520
tgcattccgg cgttccgtca cgttttagcg gctctggtag tggcaccgat ttcaccctga 2580
cgatcagttc cctgcagccg gaagactttg ctacgtatta ctgccagcaa tacagcaccg 2640
tgccgtggac gttcggtcag ggcaccaagg ttgaaattaa acgtacggtt gcggccccgt 2700
ctgtctttat cttcccgccg agtgatgaac agctgaaatc gggtaccgca agcgtggttt 2760
gtctgctgaa caatttctat ccgcgcgaag caaaggtcca gtggaaagtg gacaacgctc 2820
tgcagtccgg caattcacaa gaatcggtga ccgaacaaga tagcaaggac tctacgtaca 2880
gtctgtcatc gaccctgacg ctgtccaaag cggattatga aaaacacaag gtttacgcct 2940
gcgaagtcac ccatcaaggt ctgtcgtctc cggttaccaa gagtttcaat cgtggcgaat 3000
gttaaaagct tggctgtttt ggcggatgag agaagatttt cagcctgata cagattaaat 3060
cagaacgcag aagcggtctg ataaaacaga atttgcctgg cggcagtagc gcggtggtcc 3120
cacctgaccc catgccgaac tcagaagtga aacgccgtag cgccgatggt agtgtggggt 3180
ctccccatgc gagagtaggg aactgccagg catcaaataa aacgaaaggc tcagtcgaaa 3240
gactgggcct ttcgttttat ctgttgtttg tcggtgaacg ctctcctgag taggacaaat 3300
ccgccgggag cggatttgaa cgttgcgaag caacggcccg gagggtggcg ggcaggacgc 3360
ccgccataaa ctgccaggca tcaaattaag cagaaggcca tcctgacgga tggccttttt 3420
gcgtttctac aaactcttag atctcgatcc tctacgccgg acgcatcgtg gccggcatca 3480
ccggcgccac aggtgcggtt gctggcgcct atatcgccga catcaccgat ggggaagatc 3540
gggctcgcca cttcgggctc atgagcgctt gtttcggcgt gggtatggtg gcaggccccg 3600
tggccggggg actgttgggc gccatctcct tgcatgcacc attccttgcg gcggcggtgc 3660
tcaacggcct caacctacta ctgggctgct tcctaatgca ggagtcgcat aagggagagc 3720
gtcgagatcc cggacaccat cgaatggcgc aaaacctttc gcggtatggc atgatagcgc 3780
ccggaagaga gtcaattcag ggtggtgaat gtgaaaccag taacgttata cgatgtcgca 3840
gagtatgccg gtgtctctta tcagaccgtt tcccgcgtgg tgaaccaggc cagccacgtt 3900
tctgcgaaaa cgcgggaaaa agtggaagcg gcgatggcgg agctgaatta cattcccaac 3960
cgcgtggcac aacaactggc gggcaaacag tcgttgctga ttggcgttgc cacctccagt 4020
ctggccctgc acgcgccgtc gcaaattgtc gcggcgatta aatctcgcgc cgatcaactg 4080
ggtgccagcg tggtggtgtc gatggtagaa cgaagcggcg tcgaagcctg taaagcggcg 4140
gtgcacaatc ttctcgcgca acgcgtcagt gggctgatca ttaactatcc gctggatgac 4200
caggatgcca ttgctgtgga agctgcctgc actaatgttc cggcgttatt tcttgatgtc 4260
tctgaccaga cacccatcaa cagtattatt ttctcccatg aagacggtac gcgactgggc 4320
gtggagcatc tggtcgcatt gggtcaccag caaatcgcgc tgttagcggg cccattaagt 4380
tctgtctcgg cgcgtctgcg tctggctggc tggcataaat atctcactcg caatcaaatt 4440
cagccgatag cggaacggga aggcgactgg agtgccatgt ccggttttca acaaaccatg 4500
caaatgctga atgagggcat cgttcccact gcgatgctgg ttgccaacga tcagatggcg 4560
ctgggcgcaa tgcgcgccat taccgagtcc gggctgcgcg ttggtgcgga tatctcggta 4620
gtgggatacg acgataccga agacagctca tgttatatcc cgccgttaac caccatcaaa 4680
caggattttc gcctgctggg gcaaaccagc gtggaccgct tgctgcaact ctctcagggc 4740
caggcggtga agggcaatca gctgttgccc gtctcactgg tgaaaagaaa aaccaccctg 4800
gcgcccaata cgcaaaccgc ctctccccgc gcgttggccg attcattaat gcagctggca 4860
cgacaggttt cccgactgga aagcgggcag tgagcgcaac gcaattaatg taagttagct 4920
cactcattag gcaccgggat ctcgaccgat gcccttgaga gccttcaacc cagtcagctc 4980
cttccggtgg gcgcggggca tgactatcgt cgccgcactt atgactgtct tctttatcat 5040
gcaactcgta ggacaggtgc cggcagcgct ctgggtcatt ttcggcgagg accgctttcg 5100
ctggagcgcg acgatgatcg gcctgtcgct tgcggtattc ggaatcttgc acgccctcgc 5160
tcaagccttc gtcactggtc ccgccaccaa acgtttcggc gagaagcagg ccattatcgc 5220
cggcatggcg gccccacggg tgcgcatgat cgtgctcctg tcgttgagga cccggctagg 5280
ctggcggggt tgccttactg gttagcagaa tgaatcaccg atacgcgagc gaacgtgaag 5340
cgactgctgc tgcaaaacgt ctgcgacctg agcaacaaca tgaatggtct tcggtttccg 5400
tgtttcgtaa agtctggaaa cgcggaagtc agcgccctgc accattatgt tccggatctg 5460
catcgcagga tgctgctggc taccctgtgg aacacctaca tctgtattaa cgaagcgctg 5520
gcattgaccc tgagtgattt ttctctggtc ccgccgcatc cataccgcca gttgtttacc 5580
ctcacaacgt tccagtaacc gggcatgttc atcatcagta acccgtatcg tgagcatcct 5640
ctctcgtttc atcggtatca ttacccccat gaacagaaat cccccttaca cggaggcatc 5700
agtgaccaaa caggaaaaaa ccgcccttaa catggcccgc tttatcagaa gccagacatt 5760
aacgcttctg gagaaactca acgagctgga cgcggatgaa caggcagaca tctgtgaatc 5820
gcttcacgac cacgctgatg agctttaccg cagctgcctc gcgcgtttcg gtgatgacgg 5880
tgaaaacctc tgacacatgc agctcccgga gacggtcaca gcttgtctgt aagcggatgc 5940
cgggagcaga caagcccgtc agggcgcgtc agcgggtgtt ggcgggtgtc ggggcgcagc 6000
catgacccag tcacgtagcg atagcggagt gtatactggc ttaactatgc ggcatcagag 6060
cagattgtac tgagagtgca ccatatatgc ggtgtgaaat accgcacaga tgcgtaagga 6120
gaaaataccg catcaggcgc tcttccgctt cctcgctcac tgactcgctg cgctcggtcg 6180
ttcggctgcg gcgagcggta tcagctcact caaaggcggt aatacggtta tccacagaat 6240
caggggataa cgcaggaaag aacatgtgag caaaaggcca gcaaaaggcc aggaaccgta 6300
aaaaggccgc gttgctggcg tttttccata ggctccgccc ccctgacgag catcacaaaa 6360
atcgacgctc aagtcagagg tggcgaaacc cgacaggact ataaagatac caggcgtttc 6420
cccctggaag ctccctcgtg cgctctcctg ttccgaccct gccgcttacc ggatacctgt 6480
ccgcctttct cccttcggga agcgtggcgc tttctcatag ctcacgctgt aggtatctca 6540
gttcggtgta ggtcgttcgc tccaagctgg gctgtgtgca cgaacccccc gttcagcccg 6600
accgctgcgc cttatccggt aactatcgtc ttgagtccaa cccggtaaga cacgacttat 6660
cgccactggc agcagccact ggtaacagga ttagcagagc gaggtatgta ggcggtgcta 6720
cagagttctt gaagtggtgg cctaactacg gctacactag aaggacagta tttggtatct 6780
gcgctctgct gaagccagtt accttcggaa aaagagttgg tagctcttga tccggcaaac 6840
aaaccaccgc tggtagcggt ggtttttttg tttgcaagca gcagattacg cgcagaaaaa 6900
aaggatctca agaagatcct ttgatctttt ctacggggtc tgacgctcag tggaacgaaa 6960
actcacgtta agggattttg gtcatgagat tatcaaaaag gatcttcacc tagatccttt 7020
taaattaaaa atgaagtttt aaatcaatct aaagtatata tgagtaaact tggtctgaca 7080
gttaccaatg cttaatcagt gaggcaccta tctcagcgat ctgtctattt cgttcatcca 7140
tagttgcctg actccccgtc gtgtagataa ctacgatacg ggagggctta ccatctggcc 7200
ccagtgctgc aatgataccg cgagacccac gctcaccggc tccagattta tcagcaataa 7260
accagccagc cggaagggcc gagcgcagaa gtggtcctgc aactttatcc gcctccatcc 7320
agtctattaa ttgttgccgg gaagctagag taagtagttc gccagttaat agtttgcgca 7380
acgttgttgc cattgctgca ggcatcgtgg tgtcacgctc gtcgtttggt atggcttcat 7440
tcagctccgg ttcccaacga tcaaggcgag ttacatgatc ccccatgttg tgcaaaaaag 7500
cggttagctc cttcggtcct ccgatcgttg tcagaagtaa gttggccgca gtgttatcac 7560
tcatggttat ggcagcactg cataattctc ttactgtcat gccatccgta agatgctttt 7620
ctgtgactgg tgagtactca accaagtcat tctgagaata gtgtatgcgg cgaccgagtt 7680
gctcttgccc ggcgtcaata cgggataata ccgcgccaca tagcagaact ttaaaagtgc 7740
tcatcattgg aaaacgttct tcggggcgaa aactctcaag gatcttaccg ctgttgagat 7800
ccagttcgat gtaacccact cgtgcaccca actgatcttc agcatctttt actttcacca 7860
gcgtttctgg gtgagcaaaa acaggaaggc aaaatgccgc aaaaaaggga ataagggcga 7920
cacggaaatg ttgaatactc atactcttcc tttttcaata ttattgaagc atttatcagg 7980
gttattgtct catgagcgga tacatatttg aatgtattta gaaaaataaa caaatagggg 8040
ttccgcgcac atttccccga aaagtgccac ctgaaattgt aaacgttaat attttgttaa 8100
aattcgcgtt aaatttttgt taaatcagct cattttttaa ccaataggcc gaaatcggca 8160
aaatccctta taaatcaaaa gaatagaccg agatagggtt gagtgttgtt ccagtttgga 8220
acaagagtcc actattaaag aacgtggact ccaacgtcaa agggcgaaaa accgtctatc 8280
agggcgatgg cccactacgt gaaccatcac cctaatcaag ttttttgggg tcgaggtgcc 8340
gtaaagcact aaatcggaac cctaaaggga gcccccgatt tagagcttga cggggaaagc 8400
cggcgaacgt ggcgagaaag gaagggaaga aagcgaaagg agcgggcgct agggcgctgg 8460
caagtgtagc ggtcacgctg cgcgtaacca ccacacccgc cgcgcttaat gcgccgctac 8520
agggcgcgtc ccattcgcca 8540
<---
Реферат
Настоящее изобретение относится к области иммунологии. Предложен способ получения антитела или его фрагмента. Настоящее изобретение может найти дальнейшее применение в производстве такого антитела как Ранибизумаб. 2 з.п. ф-лы, 7 ил., 8 пр.
Комментарии