Способ получения двухнулевых линий-восстановителей фертильности brassica napus, имеющих хорошее агрономическое качество - RU2373698C2
Код документа: RU2373698C2
Чертежи
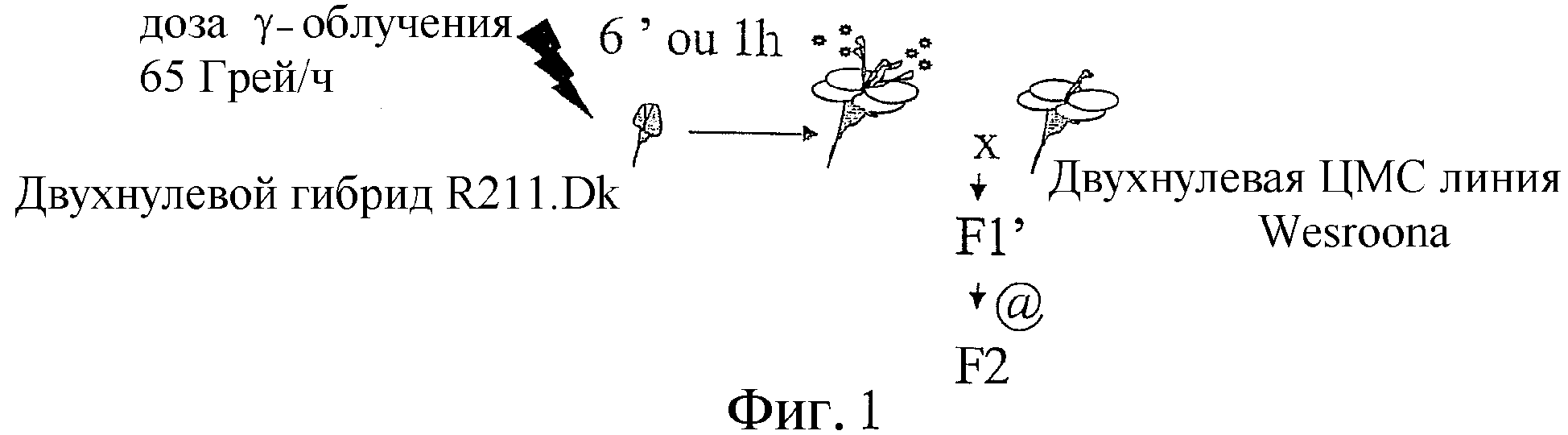























Описание
Изобретение относится к способу получения двухнулевых линий-восстановителей фертильности рапса Brassica napus с цитоплазматической мужской стерильностью (ЦМС) Ogura, представляющего собой интрогрессию редьки, несущую ген-восстановитель фертильности Rfo, вырезанный из аллели Pgi-2 редьки и рекомбинированный с геном Pgi-2 Brassica oleracea, имеющих хорошее агрономическое качество, отличающееся женской фертильностью, хорошим уровнем переноса Rfo и высокой вегетационной мощностью. Изобретение также относится к способу получения гибридных семян Brassica napus и их потомства, а также использования маркеров для селекции.
Линии-восстановители фертильности для размножения системы мужской цитоплазматической стерильности (ЦМС) Ogu-INRA рапса (Brassica napus L.) были основой целью исследований в течение последних нескольких лет. Широкое обратное скрещивание и селекция были необходимы для улучшения их женской фертильности и получения двухнулевых линий-восстановителей фертильности. Так называемые «double low-двухнулевые» разновидности - это те, которые содержат низкий уровень эруковой кислоты, а также низкий уровень глюкозинолатов в шроте после экстракции масла. Однако в размножении этих линий все еще могут встречаться трудности (перестройки интрогрессии, возможная связь с негативными особенностями) вследствие большой величины интрогрессии редьки.
Поэтому авторы изобретения поставили себе цель получить новую улучшенную двухнулевую линию-восстановитель фертильности с хорошим агрономическим качеством.
Эта цель была достигнута новым способом получения рекомбинантной двухнулевой линии-восстановителя фертильности для ЦМС Ogu-INRA рапса. Первая задача данного изобретения относится к способу получения двухнулевых линий-восстановителей фертильности Brassica napus для мужской цитоплазматической стерильности (ЦМС) Ogura, представляющего собой интрогрессию редьки, несущую ген-восстановитель фертильности Rfo, лишенный аллели Pgi-2 редьки и рекомбинированный с геном Pgi-2 из Brassica oleracea; которые имеют хорошее агрономическое качество, отличающееся женской фертильностью, хорошим уровнем переноса Rfo и высокой вегетационной мощностью, упомянутый способ включает в себя следующие стадии:
а) скрещивание двухнулевых линий ЦМС ярового рапса Brassica napus с вырезанной вставкой генома редьки с двухнулевой линией ярового рапса сорта Drakkar для формирования гетерозиготного растения Brassica napus с восстановленной фертильностью,
b) облучение перед мейозом гетерозиготных растений с восстановленной фертильностью, полученных на стадии а), гамма-лучами,
с) перекрестное опыление цветков, полученных на стадии b), с двухнулевой яровой линией ЦМС Wesroona,
d) тестирование потомства на мощность, женскую фертильность и уровень переноса гена ЦМС,
e) селекция линий потомства.
В данном изобретении термин «линия(и)» означает растение, которое в основном гомозиготно и воспроизводится самоопылением.
Способ по п.1, в котором доза облучения на стадии b) составляет 65 Грэй в течение 6 минут.
В соответствии с одной из предпочтительных форм реализации способа по данному изобретению двухнулевая линия ЦМС ярового Brassica napus на стадии а) является линией R211.
R211 представляет собой яровую линию-восстановитель фертильности INRA. Drakkar - это зарегистрированная яровая французская разновидность. Wesroona - это зарегистрированная яровая австралийская разновидность.
В соответствии с одной из предпочтительных форм реализации способа по данному изобретению тестирование выполняется с комбинацией из пяти маркеров, избранных из PGIol, PGIUNT, PGIint, BolJon и СР418.
Другая задача данного изобретения относится к двухнулевым линиям-восстановителям фертильности Brassica napus для ЦМС Ogura, представляющим собой делецию вставки Rfo в аллели Pgi-2 редьки, рекомбинированной с геном Pgi-2 Brassica oleracea и имеющим хорошие агрономические качества, отличающееся женской фертильностыо, хорошим уровнем переноса Rfo и высокой вегетационной мощностью.
В соответствии с одной из предпочтительных форм реализации двухнулевые линии-восстановители фертильности представляют собой уникальную комбинацию пяти маркеров, избранных из PGIol, PGIUNT, PGIint, BolJon и СР418.
Другая задача данного изобретения относится к способу формирования гибридных растений Brassica napus и их потомства, полученных следующими стадиями:
а) получение линий-восстановителей фертильности, продуцированных по пункту 1, и скрещивание их для получения гомозигот,
b) использование упомянутой линии-восстановителя фертильности на делянке для получения гибрида как опылителя,
с) использование ЦМС стерильных растений для получения гибридов на делянке в качестве растений, продуцирующих гибридные семена и
d) сбор гибридных семян с мужских стерильных растений.
Другая задача данного изобретения относится к семенам растений Brassica, полученных способом по данному изобретению.
Еще одна задача изобретения относится к семенам Brassica napus, депонированным в NCIMB Limited (Национальные Коллекции индустриальных пищевых и морских бактерий), 23 St Machar Drive, Aberdeen, Scotland, AB24 3RY, UK, 4 июля , 2003 под номером NCIMB41183.
Другая задача данного изобретения относится к использованию по меньшей мере четырех маркеров PGIol, PGIint, BolJon и CP418 или какой-либо их части, образующей по меньшей мере один полиморфный сайт, для характеризации рекомбинированных линий-восстановителей фертильности Brassica napus для ЦМС Ogura, имеющих делецию вставки Rfo в аллели Pgi-2 редьки и рекомбинированных с геном Pgi-2 Brassica oleracea и обладающих хорошим агрономическим качеством, отличающимся женской фертильностью, хорошим уровнем переноса Rfo и высокой вегетационной мощностью.
В предпочтительном варианте реализации комбинация образована пятью маркерами PGIol, PGIUNT, PGIint, BolJon и CP418.
В данном изобретении выражение "какая-либо часть их, образующая по меньшей мере один полиморфный сайт" означает какую-либо часть последовательности, демонстрирующую по меньшей мере отличие между последовательностью типа B.oleracea и последовательностью типа B.rapa.
Такие маркеры представлены в нижеследующих чертежах и списках последовательностей линии R2000.
В соответствии с одной из предпочтительных форм реализации данное изобретение относится к:
- маркеру PGIol, который амплифицирован с использованием праймеров PGIol U и PGIol L
(PGIol U: 5TCATTTGATTGTTGCGCCTG3',
PGIol L: 5TGTACATCAGACCCGGTAGAAAA3');
- маркеру PGIint, который амплифицирован с использованием праймеров PGIint U и PGIint L
(PGIint U: 5'CAGCACTAATCTTGCGGTATG3',
PGIint L: 5'CAATAACCCTAAAAGCACCTG3');
- маркеру PGIUNT, который амплифицирован с использованием праймеров PGIol U и PGIint L
(PGIol U: 5'TCATTTGATTGTTGCGCCTG3',
PGIint L: 5'CAATAACCCTAAAAGCACCTG3');
- маркеру BolJon, который амплифицирован с использованием праймеров BolJon U и BolJon L
(BolJon U: 5'GATCCGATTCTTCTCCTGTTG3',
BolJon L: 5'GCCTACTCCTCAAATCACTCT3');
- маркеру CP418, который амплифицирован с использованием праймеров SG129 U и pCP418 L
(SG129 U: cf Giancola et al., 2003. Theor Appl. Genet (in press);
pCP418 L: 5'AATTTCTCCATCACAAGGACC3').
Другая задача данного изобретения относится к маркерам PGIol, PGIUNT, PGIint, BolJon и CP418 со следующими последовательностями:
маркер PGIol R2000:

Маркер PGIUNT R2000:

Маркер PGIint R2000:


Маркер BolJon R2000:

Маркер CP418L R2000:

В прилагающихся нижеследующих чертежах использованы следующие сокращения:
Фиг.1 иллюстрирует облучение гамма-лучами и продукцию F2.
Фиг.2 иллюстрирует результаты посадки семян "R211" и "R2000".
Фиг.3 иллюстрирует число семян на стручок в различных линиях.
Фиг.4 иллюстрирует локализацию праймера PGIol в сегменте последовательности PGI из базы данных. На этой фигуре:
PGIol: - праймер PGIolU (именуемый в SGAP (Society for Growing Australian Plants - Общество австралийских растений): BnPGIch 1 U);
- праймер PGIol L (именуемый в SGAP: Bn PGIch 1 L);
PGIint: - праймер PGIint U;
- праймер PGIint L (за пределами приведенной последовательности).
Фиг.5 иллюстрирует гель-электрофорез гена PGI-2 (PGIol), маркера ПЦР (полимеразная цепная реакция) и SG34, маркера ПЦР, близкого к Rfo.
Фиг.6 иллюстрирует сегмент ДНК Pgi-2, амплифицированный с помощью ПЦР с праймерами PGIol.
Фиг.7 иллюстрирует переваривание продукта ПЦР PGIol с помощью Msel. На этой фигуре:
Sam и Darm представлены полосой 75 bp (пар оснований).
Drak, R21 l.Dk и R2000 представлены полосой 70 bр (Акриламид 15%).
8 - сходно с Samourai (75 bp); смесь с Drakkar (70 bр) позволяет визуализацию двух полос.
Фиг.8 иллюстрирует электрофорез маркера PGIUNT в агарозном геле.
На этой фигуре:
полоса PGIUNT (около 980 bp) представлена в B.oleracea, B.rapa cv Asko, поддерживающей и восстанавливающей фертильность в линиях за исключением "R211".
Амплификация в редьке и Arabidopsis отсутствует.
В различных генотипах Brassica амплифицировалась только одна полоса. Размер полосы сходен, но последовательности различны.
Фиг.9 иллюстрирует гель-электрофорез маркера ПЦР PGIint.
На этой фигуре PGIint линии 7 редьки имеет около 950 bp. Это такая же полоса, как в восстановленных RRH1 и R113. Она не обнаружена в R211. Ее также нет в R2000. Однако полоса PGIint имеет сходный размер около 870 bp в различных видах рода Brassica, но последовательности отличаются.
Фиг.10 иллюстрирует электрофорез маркера ПЦР BolJon в агарозном геле.
Фиг.11 иллюстрирует электрофорез маркера CP418 в агарозном геле.
На этой фигуре полоса CP418 (около 670 bp) специфична для генома B.oleracea. Она представлена у B.ol*, B.napus (Samourai, Drakkar, Pactol и гетерозиготного R2111*Dk). Она отсутствует у рапса с восстановленной фертильностью (RRH, R113 и R211). Она присутствует у гомозиготной R2000.
Фиг.12 иллюстрирует суммарную таблицу маркеров.
Фиг.13 (13(a), 13(b)) иллюстрируют сравнительный анализ первичной структуры маркера PGIol с Arabidopsis, редькой, B.rapa, B.oleracea и R2000.
Фиг.14 (14(a), 14(b), 14(c), 14(d)) иллюстрируют сравнительный анализ первичной структуры маркера PGlint-UNT с Arabidopsis, редькой, B.rapa, B.oleracea и R2000.
Фиг.15 (15(a), 15(b), 15(c)) иллюстрируют сравнительный анализ первичной структуры маркера CP418L с Arabidopsis, редькой, B. rapa, B.oleracea и R2000.
Фиг.16 (16 и 16bis) иллюстрирует маркеры BolJon у Arabidopsis, редьки и B.rapa. Их первичная структура сравнивается с последовательностями DB Arabidopsis (AC007190 концевой участок - AC011000 начальный участок), концевым участком EMBH959102 и начальным участком EMBH448336 B.oleracea и репрезентативной консенсусной последовательностью полос 1 и 2 маркеров SG129 в B.napus (в линиях Drakkar и Samourai соответственно).
Начиная с 836-й bp, последовательности ACG7190 - AC11000 и GCPATpBOJ не проявляют близкой гомологии с последовательностями Brassica.
Последовательности редьки и B.rapa (GCPconsen RsRf BOJ и BR) все еще близко гомологичны таковым B.napus с 858-й bp до 900-й bp и 981-й bp соответственно.
Кроме того, в редьке обнаружена только частичная гомология с последовательностью Brassica.
У вида B.rapa cv Asko остаток последовательности BolJon может быть после делеции 78 bp снова сравнен по структуре с таковыми B.oleracea и B.rapa в B.napus, начиная с 1057 bp до праймера BolJon L.
Фиг.17 (17 и 17bis) иллюстрируют локализацию праймеров Pgi-2 в последовательности th MJB21.12 Arabidopsis.
Фиг.18 иллюстрирует локализацию праймеров BolJon в гене mipsAtl62850 и перекрывающиеся участки клонов th AC007190 и AC011000 Arabidopsis. Представлено сравнение его первичной структуры с продуктом ПЦР BolJon в Arabidopsis (740 bp). Следует понимать, однако, что примеры приводятся как способ иллюстрации задачи изобретения, но никоим образом не образуют ограничений изобретения.
Пример I: Способ получения двухнулевой линии-восстановителя фертильности Brassica napus с цитоплазматической мужской стерильностью (ЦМС) Ogura, представляющего собой интрогрессию редьки, несущую ген-восстановитель фертильности Rfo, вырезанный из аллели Pgi-2 редьки, рекомбинированный с геном Pgi-2 Brassica oleracea, имеющих хорошее агрономическое качество, отличающееся женской фертильностью, хорошим уровнем переноса Rfo и высокой вегетационной мощностью.
Материалы и методы:
Генотипы: линию «R211» с вырезанной вставкой редьки скрещивали с яровым низкоглюкозинолятным (GLS) рапсом «Drakkar» для получения потомства Fl ('R211*Dk'). Яровая низкоглюкозинолятная ЦМС линия «Wesroona» (австралийского происхождения) использовалась для последующих скрещиваний. В качестве контроля в молекулярном анализе использовались: озимая линия с восстановленной фертильностью, происходящая от линии «Samourai» и несущая полную («RRHl») или неполную («R113») интрогрессию, а также европейская линия 7 редьки, азиатская редька с восстановленной фертильностью D81, гибрид Brassica napus*дикая редька, Brassica oleracea и B.rapa cv Asko, Arabidopsis thaliana.
Гамма-облучение: цветущие растения целиком были обработаны гамма-лучами от источника60Cо на контролируемой делянке. Сублетальная доза в 65 Грэй давалась перед мейозами.
Скрещивания и получение [поколения] F2: облученные растения переносили в защищенную от насекомых оранжерею после удаления цветочных почек, размер которых превышает 2 мм. Облученное потомство Fl использовали для опыления линии ЦМС «Wesroona» вручную. Восстановленным растениям F1 позволяли дать урожай семей растений F2, который собирали индивидуально и тщательно высевали на полевые испытания параллельно с необлученным контролем (Фиг.1).
Фенотипическая селекция: при полевых испытаниях оценивали три визуальных критерия (по шкале от 1 до 5) в течение 2 лет на 1200 растениях потомства F2 плюс 44 контрольных (82 330 оцененных растений):
1 - вегетационная мощность,
2 - нормальность соотношения фертильных/стерильных растений в расщеплении F2 и
3 - женская фертильность (развитие стручка и высев семян).
Последующие самоопыленные поколения избранных семей получали как на делянке, так и в оранжерее и продуцировали гомозиготные линии (F4) для дальнейшего анализа. Изозимный анализ выполняли, как описано в Delourme R. and Eber F. 1992. Theor Appl Genet 85: 222-228, развитие маркера (Fourmann M et al. 2002. Theor Appl Genet 105:1196-1206.): ПЦР-продукты оценены по секвенированию. Проверка первичной структуры выполнена с использованием программы Blast NCBI (Национальный Центр Биотехнологической информации США) и Uk Crop Net Brassica DB, а также программного обеспечения Multialin INRA Toulouse.
Способ:
Была выбрана одна низкоглюкозинолятная яровая гомозиготная линия с восстановленной фертильностью «R211», для которой уже показаны делеции в интрогрессии (Delourme R. and Eber F. 1992. Tlieor Appl Genet 85: 222-228. Delourme R et al. 1998. Theor Appl Genet 97: 129-134. Delourme R. et al. 1999.10 Int. Rapeseed Congress, Canberra.). Несколько молекулярных маркеров утрачены на обеих сторонах Rfo, такие как spATCHIA (Fourmann M et al. 2002. Theor Appl. Genet 105:1196-1206), spSG91 (Giancola S et al. 2003. Theor Appl Genet (in press)). "R211" утратила изозимную экспрессию аллели Pgi-2 гена редьки, но также и одну из аллелей Pgi-2 генома B.oleracea (1,2). Далее, гомозиготная "R211" демонстрирует сцепленные негативные черты, такие как низкая вегетационная мощность и очень малое количество семян. Было предположено, что эти растения утратили хромосомный сегмент рапса. Соотношение фертильности в поколении F2, полученном из этого материала, ниже ожидаемого (64% вместо 75%). Была начата программа, в которой попытались добиться рекомбинации между Rfo, несущим интрогрессию из этой вырезанной линии, и гомологичной хромосомой из двухнулевой линии B.napus.
Известно, что ионизирующая радиация индуцирует хромосомные перестройки в результате двухнитевых разрывов и последующих аберрантных соединений концов. Облучение гамма-лучами использовалось на гетерозиготных растениях F1, полученных от линии "R211", для индуцирования разрывов хромосом непосредственно перед мейозом, с целью добиться рекомбинации вырезанной интрогрессии редьки в геноме рапса.
Результаты:
Очень малое количество семей из 1200 тестированных семей F2 имели лучшие оценки по трем критериям.
Только одна, «R2000», оказалась способной продуцировать нормальное соотношение фертильных растений у самоопыленного потомства со стабильным восстановлением хороших агрономических характеристик, таких как хорошая женская фертильность при нормальном числе семян по сравнению с линией «R211» (Фиг.2 и 3). Эта семья была получена в результате 6-минутной обработки облучением при дозе 65 Грэй в час.
Анализ глюкозинолятов подтвердил их низкое содержание.
На Фиг.2 (растения «R211» и «R2000») R2000 демонстрирует нормальное цветение с нормальной внешней архитектурой.
На Фиг.3 (число семян на стручок) мы видим:
- показатели лучших семей F4 «R2000» при самоопылении и при тестовых скрещиваниях;
- показатели ЦМС-линии "Pactol" рапса и контрольных "R211".
Пример II: селекция маркеров в гене Pgi-2
Изозимный анализ PGI: потомство от «R2000» экспрессирует аллель Pgi-2 рапса из генома B.oleracea, исходно утраченного в «R211».
Были определены три ПЦР-маркера для того, чтобы охарактеризовать семейство R2000 по сравнению с известными восстановителями фертильности рапса RRH1 и R113.
1) Маркер PGIol получен из последовательностей BrassicaDB, специфичных для генома Brassica. Амплификация отсутствует как в редьке, так и в Arabidopsis th., но присутствует в Brassica в виде одной полосы 248 bp.
2) Маркер PGIint амплифицировал более длинную часть гена Pgi-2, позволяя четкое различение между различными испытанными видами Brassica, Raphanus и Arabidopsis. Виды B.rapa и B.oleracea различались не по размеру полосы в агарозном геле, а по последовательности их полосы PGINT.
3) Маркер PGIUnt представляет собой комбинацию праймеров PGI ol U и PGI int L. Этот маркер обладал специфичностью маркера PGIol, но амплифицировал более длинный участок, чем маркер PGIint.
II.1 Маркер PGIol
С праймерами PGIol родительская линия «R211» демонстрирует отсутствие амплификации, тогда как в протестированных озимых линиях присутствует полоса 248 bp. Ее последовательность ДНК гомологична последовательности PGI-2 из Crop Net UK DB для вида Brassica и последовательности из предыдущей работы авторов (наименованной последовательностями SGAP) (Локализация праймеров SGPGI chou, Фиг.4).
Это ортолог клона MJB21-12 на хромосоме V (34543 bp) у Arabidopsis (NCBI DB).
PGIol плюс SG34 для проведения теста на гомозиготность:
комбинированное использование двух наборов праймеров в смешанной ПЦР: PGIol, маркирующего ген Pgi-2, отсутствующего в гомозиготном растении с восстановленной фертильностью, и SG34 (из S. Giancola et al., Giancola S et al. 2003. Theor Appl. Genet. (in press)) - маркера, очень близкого к гену Rfo, было направлено на различение гомозиготных и гетерозиготных растений среди фертильных растений в расщеплении потомства F2, полученного из "R211". Вместо использования SG34 возможно использование какого-либо другого маркера, близкого к гену Rfo или входящего в его состав.
Только одно семейство "R2000" продемонстрировало отсутствие различий между гомозиготным и гетерозиготным потомством:
ген Pgi-2 присутствует в гомозиготе "R2000" при том, что он отсутствует в родительской гомозиготной R211.
На Фиг.5 (маркеры ПЦР PGIol и SG34):
гомозиготное семейство "R2000" восстанавливает полосу PGIol.
Последовательность ДНК полосы подтверждает гомологию с известными последовательностями Pgi-2 в Arabidopsis и Brassica. Контрольные генотипы (Drakkar, Pactol, и Samourai, Darmor) имеют такой же паттерн на геле. Последовательность этой общей полосы позволяет подтвердить высокую степень их гомологии, поскольку они практически сходны, за исключением замены одного основания.
У гомозиготного семейство «R2000» восстанавливается полоса PGIol типа Brassica oleracea. Это отличает ее от известного восстановителя фертильности группы Samourai.
Эта амплифицированная часть Pgi-2 очень консервативна, и трудно обнаружить какие-либо отличия между различными генотипами. Более длинная часть гена Pgi-2 была исследована.
II.2 Маркеры PGIUNT и PGIint
Паттерн электрофореза продуктов ПЦР:
маркер PGIUNT: второй обратный праймер PGIint L был сконструирован по большей части последовательности Pgi-2, чтобы амплифицировать и консервативные, и вариабельные участки гена.
При использовании праймера PGIol U амплифицируется полоса 980 bp только в геноме Brassica.
В «R211» не наблюдается каких-либо полос, а в гомозиготной «R2000» наблюдается полоса PGIUNT, как и в родительской линии Drakkar.
На Фиг.8 (маркер PGIUNT):
маркер PGIint амплифицировал сегмент PGIUNT. Верхний праймер PGIint позволяет амплификацию у всех тестируемых видов, что дает возможность провести ясное различие между Arabidopsis, редькой и Brassica. B.rapa и B.oleracea не различались по величине полосы в агарозном геле, но различались по своим последовательностям PGIint. Все протестированные генотипы с восстановленной фертильностью, за исключением линии «R211», демонстрируют полосу европейской редьки и одну полосу Brassica, гомологичную таковой B.rapa.
Гомозиготная «R2000» не демонстрирует полосу PGIint редьки, как в вырезанной родительской линии «R211», но демонстрирует полосу Brassica, гомологичную таковой B.oleracea.
Электрофорез маркера PGIint представлен на Фиг.9.
Анализ последовательности:
сравнение последовательностей PGI из баз данных.
Известен сегмент PGI величиной около 490 bp.
Последовательности сегмента величиной около 490 bp из различных генотипов (B.oleracea, B.rapa, B.napus) были изучены в лаборатории авторов, и некоторые последовательности переданы в Brassica Crop Net DB: EMAF 25875 до 25788 (M.Fouramnn) (4). Эти последовательности очень консервативны.
Сравнение последовательностей PGI B.rapa и B.oleracea (Фиг.13 и 14):
сравнение между последовательностями PGI мы получили из протестированных генотипов видов B.oleracea и B.rapa, оно показало, что эти последовательности различаются заменами 21 основания. Эти замены позволили различить последовательности PGIint из каждого из других протестированных генотипов рапса, гомологичных либо B.rapa cv Asko (RRH1 и R113) или B.oleracea (Drakkar, R211*DK, но также R2000).
Пример III: отбор маркера в области, близкой к Rfo
Окружающие ген Rfo во вставке редьки маркеры определены, чтобы облегчить клонирование гена (Desloires S. et al. 2003. EMBO reports 4, 6:588-594). Один из них, маркер ПЦР SGI 29, располагается очень близко к Rfo (Giancola S. et al. 2003. Theor Appl. Genet. (in press)): он коамплифицировал различающиеся полосы геномов B.oleracea и B.rapa у B.napus, однако полосу редьки очень трудно разглядеть на агарозном геле.
Мишенью последовательности SG129 был ортолог клона (AC011000, в локусе F16P17) у Arabidopsis thaliana. Этот клон перекрывается с соседним примыкающим (контиг) клоном Arabidopsis (AC07190).
В базе данных Brassica Crop Net DB авторы обнаружили один клон B.oleracea (EMBH448336, 764 bp), близкий по первичной структуре к начальному участку A011000, а второй клон B.oleracea (EMBH53971), который на карте генома Arabidopsis удален на приблизительно 300 bp, близкий к концевому участку AC07190.
Мы сконструировали новый ПЦР-маркер BolJon, занимающий место между двумя клонами B.oleracea. Мы удостоверились в том, что этот маркер позволяет амплифицировать специфические полосы ПЦР в различных генотипах, сравниваемых в данной заявке.
На Фиг.16 (гель-электрофорез ПЦР-продуктов BolJon):
- У Arabidopsis амплифицировалась полоса BolJon 815 bp, гомологичная перекрывающемуся сегменту примыкающих участков.
- В диплоидных видах Brassiceae маркер BolJon демонстрирует различающиеся полосы: одна - 950 bp у B.oleracea и одна - 870 bp у B.rapa. Это показывает, что два клона B.oleracea (EMBH53971 и EMBH448336) представляют собой непрерывность в геноме Brassica, как это имеет место в ортологической последовательности у Arabidopsis.
- У B.napus эти две полосы коамплифицируются в поддерживающих линиях Samourai или Drakkar.
- В линии 7 редьки амплифицировалась одна полоса BolJon длиной около 630 bp. Полоса у редьки с восстановленной фертильностью cmsRd81 была немного меньше. У всех линий с восстановленной фертильностью одна из полос BolJon была такой же величины, как у линии 7 редьки. BolJon - это маркер интрогрессии редьки.
- Гомозиготные линии рапса с восстановленной фертильностью «RRH1», «R113», а также «R211» демонстрируют только полосу B.rapa и полосу редьки 630 bp, что предполагает, что ортолог B.oleracea гена-мишени отсутствует или он был модифицирован, когда сегмент хромосомы редьки был вставлен в исходный геном рапса B.oleracea.
Гомозиготные растения «R2000» демонстрируют присутствие маркера ПЦР редьки BolJon плюс две полосы BolJon Brassica, которые снова восстанавливают полосу B.oleracea, утраченную у «R211» и в других линиях-восстановителях фертильности.
Мы сконструировали праймер pCP418L, специфический для генома B.oleracea у тестируемых видов. С праймером SG129U он амплифицирует только одну полосу ПЦР (670 bp) у вида B.oleracea species (Фиг.17).
Амплификации ни у B.rapa, ни у редьки, ни у Arabidopsis не было, но присутствовала ясная полоса CP418 у поддерживающих линий B.napus. Ее последовательность строго гомологична последовательности EMBH448336. Этот маркер представляет собой очень консервативную последовательность ДНК, не допускающую полиморфизма между генотипами, за исключением присутствия или отсутствия.
У RRH1, R113 и у R211 отсутствовала полоса CP418, что показывает, как в предыдущем случае, что ортолог гена-мишени B.oleracea отсутствует или был модифицирован вследствие вставки сегмента генома редьки.
Гомозиготные растения «R2000» демонстрируют полосу CP418, восстанавливающую таковую, специфичную для B.oleracea.
В данном изобретении новая рекомбинантная низкоглюкозинолятная линия-восстановитель фертильности селекционирована с хорошей женской фертильностью. Низкое качество линии «R211» позволяет селекцию в поле редких рекомбинантных событий и характеризации семьи «R2000».
Гомозиготная линия «R2000» представляет собой уникальную комбинацию маркеров PGIol, PGIUNT, PGIint и BolJon по сравнению с до сих пор анализированными восстановителями фертильности рапса:
маркер PGIinT демонстрирует, что у гомозиготных линий рапса с восстановленной фертильностью RRH1 и R113 присутствует полоса европейской редьки плюс одна полоса Brassica, гомологичная геному B.rapa. «R2000» демонстрирует отсутствие полосы редьки, утраченной, как и в родительской вырезанной линии R211, но демонстрирует одну полосу Brassica, гомологичную B.oleracea. Ортологическая последовательность PGIint в собственном геноме B.rapa не амплифицируется с этим маркером в генетическом окружении линий R211 и Drakkar.
Последовательности маркера PGIol и маркера PGIUNT в линиях с восстановленной фертильностью RRH1 и R113 были гомологичны таковой B.rapa cv Asko. У «R2000» последовательность PGIUNT гомологична B.oleracea. Ортологическая последовательность в ее геноме B.rapa не амплифицируется с этим маркером в генетическом окружении линий R211 и Drakkar.
Маркер BolJon показал, что гомозиготные линии рапса с восстановленной фертильностью, включая "R211", демонстрируют присутствие полосы европейской редьки плюс только полосу B.rapa. 'R2000' демонстрирует две полосы «R211» плюс восстановленную полосу BolJon B.oleracea.
Маркер CP418 показал, что у «R2000» восстановился консервативный сегмент B.oleracea.
Гипотеза авторов состоит в том, что событие рекомбинации имело место в опыленной материнской клетке, которая дала растения «R2000». Вырезанная интрогрессия редьки затем интегрировалась в нормальный гомологичный сегмент хромосомы, несущий ген Pgi-2 типа B.oleracea, а последовательность - мишень BolJon, охарактеризованная этими маркерами, вероятно, происходит из двухнулевого генома Drakkar «00», присутствующего в облученном гетерозиготном гибриде «R211*DK».
Паттерн, наблюдавшийся для BolJon, предполагает, что событие рекомбинации привело в результате к формированию в семействе «R2000» особого сдвоенного участка, одна часть которого происходит от редьки, а другая - от B.oleracea.



Реферат
Получают двухнулевую линию-восстановитель фертильности Brassica napus для мужской цитоплазматической стерильности (ЦМС) Ogura, представляющую собой интрогрессию редьки, несущую ген-восстановитель фертильности Rfo, вырезанную из аллели Pgi-2 редьки и рекомбинированную с геном Pgi-2 из Brassica oleracea, характеризующейся женской фертильностью, хорошим уровнем переноса Rfo и высокой вегетационной мощностью. Для характеризации полученной линии-восстановителя фертильности используют комбинацию маркеров PGIol, PGIUNT, PGIint, BolJon и СР418. Линия отличается хорошим агрономическим качеством. 2 н. и 5 з.п. ф-лы, 24 ил.
Формула
маркер PGIol амплифицируется с использованием праймеров PGIol U и PGIol L, имеющих следующие последовательности:
PGIol U: 5'TCATTTGATTGTTGCGCCTG3';
PGIol L: 5'TGTACATCAGACCCGGTAGAAAA3';
маркер PGIint амплифицируется с использованием праймеров PGIint U и PGIint L, имеющих следующие последовательности:
PGImtU: 5'CAGCACTAATCTTGCGGTATG3';
PGIint L: 5'CAATAACCCTAAAAGCACCTG3';
маркер PGIUNT амплифицируется с использованием праймеров PGIol U и PGIint L, имеющих следующие последовательности:
PGIol U: 5'TCATTTGATTGTTGCGCCTG3';
PGIint L: 5'CAATAACCCTAAAAGCACCTG3';
маркер BolJon амплифицируется с использованием праймеров BolJon U и BolJon L, имеющих следующие последовательности:
BolJon U: 5'ATCCGATTCTTCTCCTGTTG5';
BolJon L: 5'GCCTACTCCTCAAATCACTCT3';
маркер СР418 амплифицируется с использованием праймера рСР418 L, имеющего следующую последовательность:
рСР418 L: 5'AATTTCTCCATCACAAGGACC3'.
Комментарии