Усиление иммунной реакции, медиатором которой является слитый протеин антитело-цитокин, при помощи комбинированного лечения агентами, увеличивающими поглощение иммуноцитокина - RU2272644C2
Код документа: RU2272644C2
Чертежи











Описание
Связанные заявки
Данная заявка предоставляет приоритет и использует информацию, раскрытую в заявке 60/215038, зарегистрированной 29 июня 2000 г., сведения из которой здесь введены посредством ссылок.
Область изобретения
Настоящее изобретение относится к слитым протеинам антитело-цитокин, полезным для целевой иммунотерапии. В общем случае изобретение относится к применению агентов, увеличивающих поглощение иммуноцитокина, в комбинированной терапии, направленной на усиление иммунной реакции, медиатором которой является слитый протеин антитело-цитокин, против предварительно выбранной мишени, например клеток в опухоли. В частности, изобретение относится к назначению слитых протеинов антитело-цитокин в комбинации с химиотерапевтическими препаратами, такими, как таксаны и/или алкилирующие агенты, для обработки клеток опухоли и других раковых или больных клеток.
Обоснование изобретения
Эффективное лечение таких заболеваний, как рак, требует усиления иммунных реакций одного или более типов эффекторных клеток, таких, как природные убийцы (NK), макрофаги и Т-лимфоциты. У животных и людей с опухолями иммунная система не способна эффективно реагировать на рост опухоли, главным образом, из-за того, что в опухоли задействованы специфические механизмы для подавления иммунной реакции. Во многих случаях потенциально способные разрушать опухоль моноцитные клетки, т.е. макрофаги, мигрируют к месту роста опухоли, но клетки опухоли секретируют такие факторы, как простагландины, TGF-β и IL-10, которые снижают их цитотоксическую активность (см., например, Sharma et al., 1999, J. IMMUNOL. 163:5020-5028). Аналогичным образом лимфоцитные клетки, такие, как NK и Т-клетки, мигрирующие внутрь опухолей, могут подавляться факторами, которые секретирует опухоль, так же, как и взаимодействиями с рецепторами, которые экспрессируются на поверхности опухолевых клеток и активируют апоптоз иммунных клеток (см., например, Villunger et al., 1997, BLOOD 90: 12-20). У этих лимфоцитов, которые подвергаются иммуноподавляющему воздействию моноцитных клеток в окрестности опухоли, может в дальнейшем уменьшаться способность к эффективной антиопухолевой реакции.
Попытки преодолеть иммуноподавляющее воздействие микроокружения опухоли включают направленную иммуностимуляцию, такую, как лечение слитым протеином антитело-цитокин, обладающим специфическим сродством к опухоли. Эффективное лечение на основе этого подхода продемонстрировано на нескольких моделях метастазов опухолей у мышей, однако это лечение становится значительно менее эффективным при увеличении размера опухоли. Вероятно, это обусловлено повышением, при большей массе опухоли, уровня ее секреции подавляющих факторов, а также и другими причинами, такими, как увеличение давления внутритканевой жидкости в опухоли (Griffon-Etienne et al. 1999, CANCER RES. 59: 3776-3782), которое препятствует проникновению в твердую опухоль терапевтических агентов.
В то время как для лечения большинства раковых пациентов по-прежнему назначают один или более курсов химиотерапии, хорошо известно, что цитотоксическая терапия рака создает угрозу иммунной системе. В человеческом организме иммунные клетки делятся быстрее, чем подавляющее большинство других типов клеток, и любое лечение, которое убивает размножающиеся клетки, будет убивать также и иммунные клетки. Следовательно, все варианты лечения, включающего облучение, повреждающие ДНК химические вещества, ингибиторы синтеза ДНК и ингибиторы микротубулярной функции, разрушают иммунную систему. Пересадка костного мозга как дополнение терапии рака требуется именно потому, что необходимо восстанавливать обусловленные противораковыми препаратами нарушения иммунной системы. Метотрексат и другие противораковые лекарства часто используют в качестве иммунодепрессантов. Очевидно также, что противораковые назначения могут специфически ингибировать функцию клеток Т. Например, пациенты, которых лечили от болезни Ходжкина путем облучения всего тела, страдают от постоянной потери простых клеток Т (Watanabe et al., 1997, Blood 90: 3662).
На основе современного уровня знаний становится очевидным, что обычные способы лечения (химиотерапия и облучение) и местная иммуностимуляция не могут составить полезный комбинированный подход к эффективному лечению рака. Следовательно, имеется необходимость в разработке способов усиления иммунной реакции, медиатором которой является слитый протеин антитело-цитокины, направленной против предварительно выбранных типов клеток, например клеток опухоли, и композиций, которые можно применять с этой целью.
Краткое описание изобретения
Обнаружено, что при назначении слитого протеина антитело-цитокин (иммуноцитокин) млекопитающим с опухолью или опухолевыми метастазами можно вызывать более сильную антиопухолевую реакцию в случаях, когда его назначают до, одновременно или после лечения агентом, увеличивающим поглощение иммуноцитокина, который повышает или усиливает терапевтическое действие слитого протеина антитело-цитокин за счет усиления или увеличения его поглощения опухолью. Было обнаружено, что в число полезных агентов, увеличивающих поглощение иммуноцитокина, входят алкилирующие химиотерапевтические агенты и таксаны, такие, как паклитаксел. В частности, было обнаружено, что такие комбинации полезны как посредники иммунного разрушения предварительно выбранных типов клеток, таких, как клетки опухоли вирусно-инфицированные клетки.
В одном из аспектов воплощения изобретение предоставляет способ стимулирования у млекопитающих убивающей клетки иммунной реакции против предварительно выбранных типов клеток. Способ включает назначение млекопитающим (i) иммуноцитокина, содержащего связывающий антитело сайт, который может связывать клетки предварительно выбранного типа, и цитокин, который может индуцировать такую иммунную реакцию против клеток предварительно выбранного типа, и (ii) агент, увеличивающий поглощение иммуноцитокина, в количестве, достаточном для усиления иммунной реакции, аналогичной той иммунной реакции, которую стимулирует сам иммуноцитокин, применяемый самостоятельно.
В предпочтительном воплощении изобретения клетками предварительно выбранного типа могут быть раковые клетки, присутствующие, например, в твердой опухоли, более предпочтительно в твердой опухоли большего размера (т.е. больше 100 мм3). Альтернативно клетками предварительно выбранного типа могут быть раковые клетки, присутствующие в виде мелких метастазов.
В другом предпочтительном воплощении изобретения агент, увеличивающий поглощение иммуноцитокина, можно назначать одновременно с иммуноцитокином. Альтернативно агент, увеличивающий поглощение иммуноцитокина, можно назначать до назначения иммуноцитокина. Более того, предполагается, что иммуноцитокин можно назначать вместе с множеством различных агентов, увеличивающих поглощение иммуноцитокина. Альтернативно предполагается, что агент, увеличивающий поглощение иммуноцитокина, можно назначать вместе с множеством различных иммуноцитокинов.
В другом аспекте изобретение предлагает композицию для стимулирования у млекопитающих убивающей клетки иммунной реакции. Эта композиция включает комбинацию: (i) иммуноцитокин, содержащий связывающий антитело сайт, который может связывать клетки предварительно выбранного типа, и цитокин, который может индуцировать такую иммунную реакцию против клеток предварительно выбранного типа у млекопитающих, и (ii) агент, увеличивающий поглощение иммуноцитокина, в количестве, достаточном для усиления убивающей клетки реакции, которую стимулирует иммуноцитокин из комбинации, аналогичной убивающей клетки реакции, которую иммуноцитокин стимулирует при самостоятельном назначении.
В предпочтительном воплощении изобретения связывающий антитело сайт иммуноцитокина предпочтительно включает высокомолекулярную цепочку иммуноглобулина или ее фрагмент, связывающий антиген. Высокомолекулярная цепочка иммуноглобулина предпочтительно включает, на конце с аминогруппой в направлении конца с карбоксигруппой, домен вариабельной области (VH) иммуноглобулина, способный связывать предварительно выбранный антиген, высокомолекулярный домен постоянной области иммуноглобулина 1 (СН1), высокомолекулярный домен постоянной области иммуноглобулина 2 (СН2) и может, кроме того, включать высокомолекулярный домен постоянной области иммуноглобулина 3 (СН3). В более предпочтительном воплощении изобретения иммуноцитокин представляет собой слитый протеин, включающий высокомолекулярную цепочку иммуноглобулина или ее фрагмент, связывающий антиген, слитый с цитокином через полипептидную связь. Следовательно, предпочтительный слитый протеин антитело-цитокин включает, на амино-конце в направлении карбокси-конца, (i) связывающий антитело сайт, который содержит вариабельную область иммуноглобулина, которая может связывать антиген поверхности клетки предварительно выбранного типа клетки, домен СН1 иммуноглобулина, домен СН2 иммуноглобулина (возможно, домен СН3), и (ii) цитокин. Способы получения и применения таких слитых протеинов подробно описаны в работе Gillies et al. (1992) Proc. Natl. Acad. Sci. USA 89: 1428-1432; Gillies et al. (1998) J. Immunol. 160: 6195-6203, и US Patent No. 5650150.
Домены постоянной области иммуноглобулина (т.е. домены СН1, СН2 и/или СН3) могут быть доменами постоянной области, которые обычно связаны с доменом вариабельной области встречающихся в природе антител. Альтернативно один или более доменов постоянной области иммуноглобулина можно извлечь из антител, отличных от антител, которые используют как источник домена вариабельной области. Иными словами, домены вариабельной и постоянной областей иммуноглобулина можно извлекать из различных антител, например антител, выделенных из различных биологических видов. См., например, US Patent No. 4816567. Более того, вариабельные области иммуноглобулина могут включать последовательности области рамки (FR), извлеченные из других видов, например человека, и последовательности области, определяющей дополнительность (CDR), помещенные между последовательностями FR, извлеченные из второго, отличного вида, например мыши. Способы получения и применения таких химерических вариабельных областей иммуноглобулина раскрыты, например, в патентах US №5225539 и 5585089.
Основанные на антителе иммуноцитокины предпочтительно включают, кроме того, низкомолекулярную цепочку иммуноглобулина, которая предпочтительно ковалентно связана с высокомолекулярной цепочкой иммуноглобулина посредством, например, дисульфидной связи. Вариабельные области связанных высокомолекулярной и низкомолекулярной цепочек иммуноглобулина вместе определяют единственный и полный связывающий сайт для связывания предварительно выбранного антигена. В других воплощениях изобретения иммуноцитокины включают две химерических цепочки, каждая из которых содержит по крайней мере часть высокомолекулярной цепочки иммуноглобулина, слитого с цитокином. Две химерические цепочки предпочтительно связаны друг с другом ковалентно, например, одной или более внутрицепочечными дисульфидными связями.
Изобретение, таким образом, предлагает слитые протеины, в которых специфичность связывания антигена и активность антитела объединены с высокой биологической активностью цитокина. Слитый протеин по настоящему изобренению можно применять для избирательной доставки цитокина к клетке-мишени in vivo, таким образом, чтобы цитокин мог оказывать локализованное биологическое действие в окрестности клетки-мишени. В предпочтительном воплощении изобретения компонент антитела слитого протеина специфически связывает антиген, который находится на поверхности или внутри раковой клетки, и в результате слитый протеин проявляет локализованную противораковую активность. В альтернативном предпочтительном воплощении изобретения компонент антитела слитого протеина специфически связывает вирусно-инфицированные клетки, такие, как ВИЧ-инфицированные клетки, и в результате слитый протеин проявляет локализованную антивирусную активность.
В число цитокинов, которые могут быть включены в иммуноцитокины по данному изобретению, входят, например, факторы некроза опухоли, интерлейкины, колониестимулирующие факторы и лимфокины, а также другие известные в настоящее время цитокины. Предпочтительная группа факторов некроза опухоли включает, например, фактор некроза тканей α (TNFα). Предпочтительная группа интерлейкинов включает, например, интерлейкин-2 (IL-2), интерлейкин-4 (IL-4), интерлейкин-5 (IL-5), интерлейкин-7 (IL-7), интерлейкин-12 (IL-12), интерлейкин-15 (IL-15) и интерлейкин-18 (IL-18). Предпочтительные колониестимулирующие факторы включают, например, гранулоцит-макрофаг-колониестимулирующий фактор (GM-CSF) и макрофаг-колониестимулирующий фактор (M-CSF). В группу предпочтительных лимфокинов входит, например, лимфотоксин (LT). В число других полезных цитокинов входят интерфероны, включая IFN-α, IFN-β и IFN-γ, каждый из которых проявляет иммунологическое действие, так же, как и антиангиогенное действие, которые не зависят от их антивирусных активностей.
Было обнаружено, что некоторые типы химиотерапевтических агентов являются эффективными агентами для увеличения поглощения иммуноцитокина. В частности, в число полезных агентов, увеличивающих поглощение иммуноцитокина, входят таксаны и алкилирующие химиотерапевтические агенты. Некоторые таксаны известны в этой области (см. Bissery и Lavelle, 1997, в Cancer Therapeutics: Experimental and Clinical Agents, Chapter 8, В. Teicher, ed.). В предпочтительном воплощении изобретения таксаном является Taxol, известный также под названием паклитаксел. Другие воплощения изобретения включают полусинтетический таксан, доцетаксел, который в некоторых моделях опухолей и при некоторых клинических показаниях оказался более эффективным, чем паклитаксел. Дальнейшие воплощения изобретения включают дополнительные производные таксана, такие, как выделенный из природного исходного материала 10-деацетил Baccatin III, экстрагированный из хвои Европейского Тиса. Одним из таких примеров является соединение для орального приема IDN5109, которое представляет собой также бедный субстрат для Р-гликопротеина и в общем случае более активно действует против опухолей, проявляющих высокую устойчивость к множеству лекарственных препаратов. В дополнение к его пригодности для орального приема, это соединение толерантно при более высоких дозах и обладает меньшими нейротоксическими побочными эффектами (Polizzi et al., 1999, Cancer Res. 59: 1036-1040).
В изобретении предлагаются также предпочтительные дозировки и курсы назначений для назначения иммуноцитокинов в комбинации с агентами, увеличивающими поглощение иммуноцитокина.
Описание чертежей:
фиг.1 - схематическое представление цитокина;
фиг.2 - влияние паклитаксела и иммуноцитокина на изменение объема опухоли LLC/KSA с течением времени;
фиг.3 - влияние многократных доз паклитаксела и иммуноцитокина на изменение среднего объема опухоли с течением времени;
фиг.4 - влияние паклитаксела и иммуноцитокина на вес опухоли по результатам анализа метастазов в легких;
фиг.5 - влияние паклитаксела и иммуноцитокина на изменение объема опухоли CT26/KSA с течением времени;
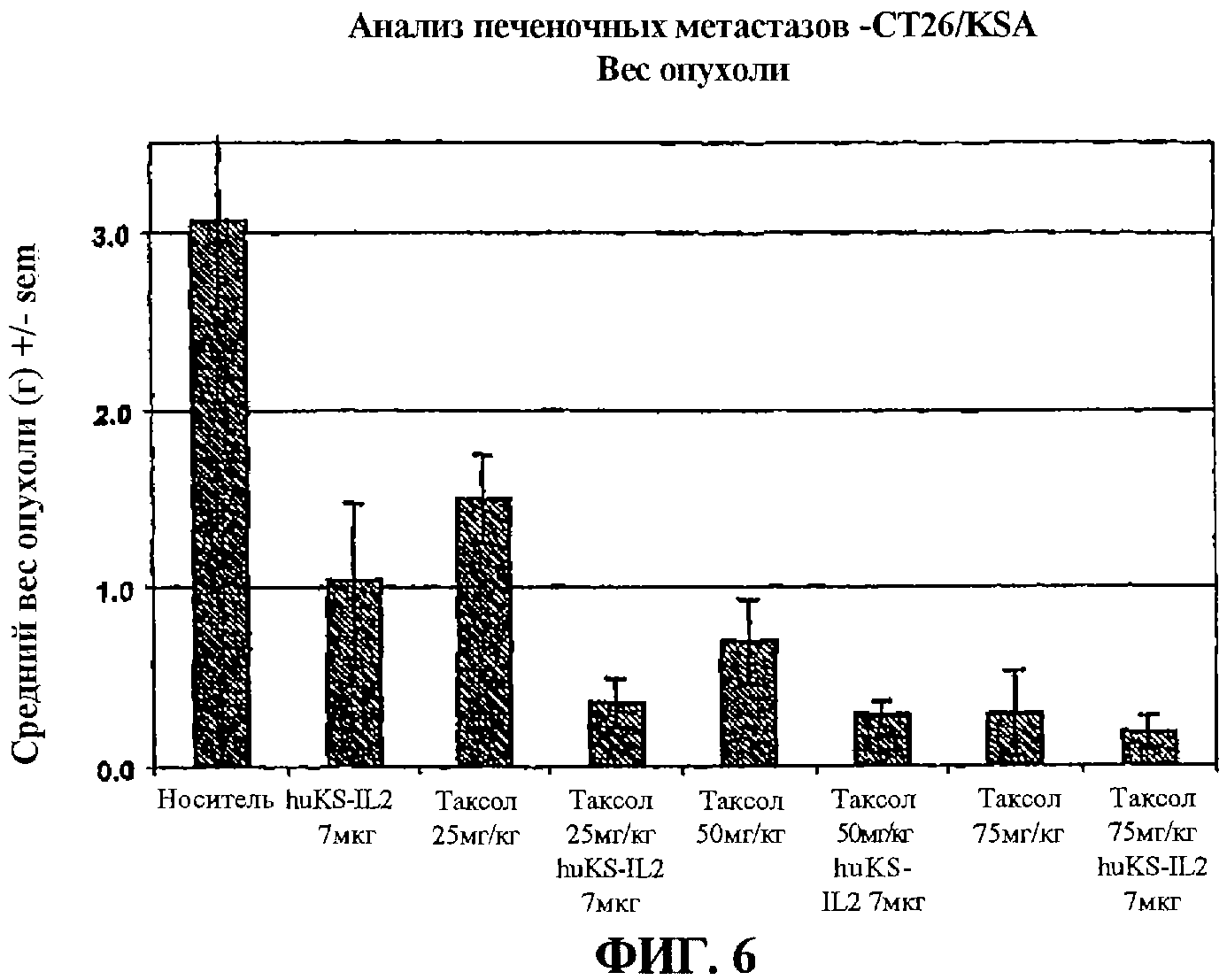
фиг.6 - влияние паклитаксела и иммуноцитокина на вес опухоли по результатам анализа метастазов в печени;
фиг.7А и 7Б - влияние паклитаксела на поглощение иммуноцитокина в опухоли;
фиг.8 - влияние циклофосфамида на поглощение иммуноцитокина в опухоли;
фиг.9А - влияние циклофосфамида и иммуноцитокина на вес опухоли по результатам анализа метастазов в легких;
фиг.9Б - влияние циклофосфамида и иммуноцитокина на объем опухоли по результатам анализа роста опухоли;
фиг.9В: влияние циклофосфамида и иммуноцитокина на объем опухоли по результатам анализа роста опухоли;
фиг.10: влияние карбоплатина и иммуноцитокина на объем опухоли по результатам анализа роста опухоли.
Подробное описание изобретения
Исследования свидетельствуют, что твердые опухоли большого размера значительно более устойчивы по отношению к терапевтическому вмешательству посредством антител и к иммунотерапии в целом, чем рассредоточенные метастатические образования (Sulitzeanu et al. (1993) Adv. Cancer Res. 60: 247-267). Считается, слабая реакция на терапию, основанную на антителах, частично обусловлена тем, что опухоли генерируют иммуноподавляющие факторы.
Хотя механизм уничтожения опухоли еще не известен полностью, предполагается, что реакция цитотоксических Т-лимфоцитов (CTL) может приводить к разрушению раковых клеток и обеспечивать иммунную память. Более того, предполагается, что при определенных обстоятельствах клетки природные убийцы (NK) ответственны за уничтожение опухоли в отсутствие CTL. Различные иммунные реакции могут быть следствием факта, что определенные опухоли генерируют различные типы и количества веществ, способных подавлять Т-клетки. Это более верно для твердых опухолей, чем для микрометастатических образований, т.е. для таких, которые достигли критической массы и способны генерировать и секретировать иммуноподавляющие факторы на уровнях, достаточных для снижения иммунной реакции против опухоли.
Теперь обнаружено, что иммунные реакции, приводящие к уничтожению клеток опухоли, инициированные иммуноцитокином против предварительно выбранных типов клеток, могут быть значительно усилены путем назначения иммуноцитокина вместе с агентом, усиливающим поглощение иммуноцитокина. Комбинированная терапия особенно эффективна в плане содействия иммунному разрушению больных тканей, таких, как упрочившаяся опухоль. Без попыток создать связь с теорией предполагается, что агент, увеличивающий поглощение иммуноцитокина, увеличивает проникновение иммуноцитокина в микроокружение опухоли, что позволяет ему преодолевать иммуноподавляющее действие и быть более эффективным в плане активирования клеточной иммунной реакции против опухоли. Аналогичным образом предполагается, что такой способ может быть полезньм для лечения определенных вирусных заболеваний, при которых аналогичный иммуноподавляющий механизм предотвращает эффективное проявление клеточного иммунитета, например, при инфекции ВИЧ. Предполагается, что агент, увеличивающий поглощение иммуноцитокина, вместе с иммуноцитокином оказывает синергическое действие как медиатор иммунного разрушения больных тканей, таких, как упрочившаяся опухоль, или вирусно инфицированных клеток. В настоящем изобретении описаны также способы изготовления и применения полезных иммуноцитокинов, а также анализ, пригодный для тестирования в доклиническом моделировании на животных in vivo их фармакокинетических активностей при комбинировании с подходящими агентами, увеличивающими поглощение иммуноцитокина.
Используемый здесь термин «агент, увеличивающий поглощение иммуноцитокина» следует понимать как любой агент, который усиливает иммунную реакцию, приводящую к уничтожению клеток опухоли, инициируемую иммуноцитокином против предварительно выбранного типа клеток. Более конкретно, предпочтительный агент, увеличивающий поглощение иммуноцитокина, это агент, который увеличивает поглощение опухолью иммуноцитокина и, таким образом, увеличивает его проникновение внутрь опухоли. Примеры агентов, увеличивающих поглощение иммуноцитокина, включают, но не ограничиваются ими, химиотерапевтические агенты, такие, как таксаны, агенты, разрушающие ДНК, в том числе алкилирующие химиотерапевтические агенты, агенты лучевой терапии и агенты, снижающие кровяное давление. Предпочтительными таксанами являются таксол, доцетаксел, 10-деацетил Baccatin III и их производные. Предпочтительными алкилирующими агентами являются циклофосфамид, карбоплатин, цисплатин и их производные. Предпочтительным видом облучения является гамма-излучение. Предпочтительным агентом, снижающим кровяное давление, является агонист ангиотенсина II, такой, как сам ангиотенсин II, предпочтительно назначаемый периодически в соответствии с общими принципами, описанными в работах Netti et al. (Cancer Research [1995] 55: 5451-8) и Netti et al. (Proc. Nat. Acad. Sci. [1999] 96: 3137-3142). Иммунную реакцию можно определять по известным методикам и/или по описанному здесь способу.
Используемый здесь термин "иммунная реакция, приводящая к уничтожению клеток опухоли" следует понимать как любую иммунную реакцию в млекопитающих либо гуморальной, либо клеточной природы, которая стимулируется иммуноцитокином и которая либо убивает, либо иным способом уменьшает жизнеспособность предварительно выбранного типа клеток в млекопитающих. Иммунная реакция может включать один или более типов клеток, в том числе Т-клетки, клетки NK и макрофаги.
Используемый здесь термин "иммуноцитокин" означает слияние (i) связывающего антитело сайта, обладающего специфичностью к связыванию предварительно выбранного антигена и способностью его связывать, например, специфического антигена клеточного типа, и (ii) цитокина, способного инициировать или стимулировать иммунную реакцию, приводящую к уничтожению клеток опухоли, обычно - против раковых или вирусно инфицированных клеток. В число примеров предварительно выбранных входят антигены поверхности клеток, таких, которые присутствуют на раковых клетках или вирусно инфицированных клетках, и нерастворимые внутриклеточные антигены, например, некротических клеток, которые могут оставаться присоединенньми к клеточной мембране. Предпочтительными антигенами являются целевые антигены, характерные для клеток опухоли, такие, как специфические к опухоли антигены. Следовательно, иммуноцитокин может селективно доставлять цитокин к мишени (которой обычно является клетка) in vivo таким образом, что цитокин может служить медиатором локализованной иммунной реакции против целевой клетки. Например, если компонент антитела иммуноцитокина селективно связывает антиген на раковой клетке, такой, как раковая клетка в твердой опухоли, в частности в большей твердой опухоли, размер которой превышает 100 мм, иммуноцитокин усиливает локализованную антираковую активность, если компонент антитела иммуноцитокина селективно связывает антиген на вирусно инфицированной клетке, такой, как клетка, инфицированная HIV, иммуноцитокин усиливает локализованную антивирусную активность.
Используемый здесь термин "сайт, связывающий антитело" следует понимать как по крайней мере часть высокомолекулярной цепочки иммуноглобулина, например вариабельную область иммуноглобулина, способную связывать предварительно выбранный антиген типа клеточного антигена. Сайт, связывающий антитело, также предпочтительно включает по крайней мере часть постоянной области иммуноглобулина, в том числе, например, домен CH1, домен СН2, и, возможно, домен СН3, или по крайней мере домен СН2, или одну или более их частей. Более того, высокомолекулярная цепочка иммуноглобулина может быть связана либо ковалентно, либо нековалентно с низкомолекулярной цепочкой иммуноглобулина, включая, например, вариабельную область низкомолекулярной цепочки иммуноглобулина и, возможно, постоянную область низкомолекулярной цепочки. Соответственно, предполагается, что сайт, связывающий антитело, может содержать целое антитело или его фрагмент либо отдельную цепочку антитела, способную связывать предварительно выбранный антиген.
Относительно иммуноцитокина предполагается, что фрагмент антитела может быть связан с цитокином множеством широко известных способов. Например, сайт, связывающий антитело, предпочтительно связан через полипептидную связь или линкер с цитокином в структуре слитого протеина. Альтернативно сайт, связывающий антитело, может быть химически спарен с цитокином через химически активные группы, например сульфгидрильные группы, внутри боковых цепочек аминокислот, присутствующих в сайте, связывающем антитело и цитокин.
Используемый здесь термин "цитокин" следует понимать как любой протеин или пептид, их аналог или функциональный фрагмент, который способен стимулировать или инициировать иммунную реакцию, приводящую к уничтожению клеток против предварительно выбранного типа клеток, например раковой клетки или вирусно инфицированной клетки, в млекопитающих. Следовательно, предполагается, что в иммуноцитокины по данному изобретению может быть включено множество цитокинов. В группу полезных цитокинов входят, например, факторы некроза опухоли (TNF), интерлейкины (IL), лимфокины (L), колониестимулирующие факторы (CSF), интерфероны (IFN), включая варианты видов и их усеченные аналоги, которые способны стимулировать или инициировать иммунную реакцию, приводящую к уничтожению клеток. В число подходящих факторов некроза опухоли входят, например, TNFα. В число подходящих лимфокинов входят, например, LT. В число подходящих колониестимулирующих факторов, входят, например, GM-CSF и M-CSF. В число подходящих интерлейкинов входят, например, IL-2, IL-4, IL-5, IL-7, IL-12, IL-15 и IL-18. В число подходящих интерферонов входят, например, IFN-α, IFN-β и IFN-γ. Ген, кодирующий цитокин, который представляет особый интерес, может быть клонирован de novo, получен из доступных источников или синтезирован в стандартном синтезе ДНК из известных нуклеотидных последовательностей. Например, последовательность ДНК LT известна (см., например, Nedwin et al. (1985) Nucleic acids res. 13: 6361), известны также и последовательности для IL-2 (см., например, Taniguchi et al. (1983) NATURE 302: 305-318), GM-CSF (см., например, Gasson et al. (1984) Science 266: 1339-1342) и TNF α (см., например, Nedwin et al. (1985) NUCLEIC ACIDS RES. 13: 6361).
В предпочтительном воплощении изобретения иммуноцитокины представляют собой рекомбинантные слитые протеины, полученные в рамках традиционных методологий рекомбинантных ДНК, т.е. путем формирования структуры нуклеиновой кислоты, кодирующей химерический иммуноцитокин. Конструирование рекомбинантного слитого протеина антитело-цитокин было описано в прототипе. См., например, Gillies et al. (1992) Proc. Natl. Acad. Sci. USA 89: 1428-1432; Gillies et al. (1998) J. Immunol. 160: 6195-6203, и патент US №5650150. Предпочтительно генная конструкция, кодирующая иммуноцитокин по данному изобретению, включает, в направлении от 5' к 3', сегмент ДНК, кодирующий домен вариабельной области высокомолекулярной цепочки иммуноглобулина, сегмент ДНК, кодирующий домен постоянной области высокомолекулярной цепочки иммуноглобулина и ДНК, кодирующую цитокин. Слитый ген монтируют или вставляют в вектор экспрессии для трансфекции в подходящую клетку-реципиент, в которой слитый ген экспрессируют. Гибридную полипептидную цепочку предпочтительно комбинируют с низкомолекулярной цепочкой иммуноглобулина таким образом, чтобы вариабельная область высокомолекулярной цепочки (VH) иммуноглобулина и вариабельная область низкомолекулярной цепочки (VL) иммуноглобулина вместе образовывали единый и полный сайт для связывания предварительно выбранного антигена. В предпочтительном воплощении изобретения высокомолекулярная и низкомолекулярная цепочки иммуноглобулина ковалентно связаны, например, через внутрицепочечную дисульфидную связь. Более того, высокомолекулярные цепочки двух иммуноглобулинов, один или оба из которых слиты с цитокином, могут быть ковалентно связаны, например, посредством одной или более внутрицепочечных дисульфидных связей.
Следовательно, способы по данному изобретению пригодны для повышения антиопухолевой активности иммуноцитокина, который применяется в терапевтическом методе лечения опухоли, включая иммуноцитокиновые композиции и способы, раскрытые в WO 99/29732, WO 99/43713, WO 99/52562, WO 99/53958 и WO 01/10912, и основаны на антителе слитых протеинов с измененной аминокислотной последовательностью в области связывания. В одном из воплощений способы по данному изобретению полезно применять в комбинации со слитыми протеинами Fc, такими, как Fc-интерферон-α.
На фиг.1 показано схематическое представление типичного иммуноцитокина 1. В этом воплощении молекулы цитокина 2 и 4 пептидно связаны с карбокси-концами 6 и 8 областей СН3 10 и 12 высокомолекулярных цепочек антитела 14 и 16. Области 26 и 28 VL показаны спаренными с областями 18 и 20 VH в обычной конфигурации IgG, что обеспечивает наличие двух связывающих антиген сайтов 30 и 32 на амино-концах иммуноцитокина 1 и двух связывающих рецептор цитокина сайтов 40 и 42 на карбокси-концах иммуноцитокина 1. Разумеется, в более широких аспектах нет необходимости спаривать иммуноцитокины, как это показано, или требуется сливать с молекулой цитокина только одну из двух высокомолекулярных цепочек иммуноглобулина.
Иммуноцитокины по данному изобретению можно рассматривать как химерические в силу двух аспектов их структуры. Во-первых, иммуноцитокин является химерическим в том, что включает высокомолекулярную цепочку иммуноглобулина, содержащую связывающую антиген специфичность, привязанную к данному цитокину. Во-вторых, иммуноцитокин по данному изобретению может быть химерическим в том смысле, что он включает вариабельную область (V) иммуноглобулина и постоянную область (С) иммуноглобулина, каждая из которых извлечена из различных антител, так что получающийся протеин является химерой V/C. Например, вариабельная и постоянная области могут быть извлечены из молекул встречающихся в природе антител, которые можно выделить из различных биологических видов. См., например, патент US 4816567. Кроме того, приемлемыми являются конструкции, в которых любая из двух или обе вариабельные области иммуноглобулина включают последовательности области рамки (FR) и последовательности области, определяющей дополнительность (CDR), извлеченные из различных биологических видов. Такие конструкции раскрыты, например, в работах Jones et al. (1986) Nature 321: 522-525, Verhoyen et al. (1988) SCIENCE 239: 1534-1535, и в патентах US №№5225539 и 5585089. Кроме того, предполагается, что последовательности вариабельной области могут быть получены скринированием библиотек, например библиотек фагов, для последовательностей вариабельной области, которые связывают предварительно выбранный антиген с желательной степенью сродства. Способы создания и библиотек фагов раскрыты, например, в работах Huse et al. (1989) Science 246: 1275-1281, и Kang et al. (1991) Proc. Natl. Acad. Sci. USA 88: 11120-11123.
Домены постоянной области высокомолекулярной цепочки иммуноглобулина иммуноцитокина могут быть выбраны из любого из пяти классов иммуноглобулина, на которые ссылаются как на IgA (Igα), IgD (Igδ), IgE (Igε), IgG (Igγ) и IgM (Igμ). Тем не менее постоянные области из высокомолекулярной цепочки иммуноглобулина класса IgG являются предпочтительными. Кроме того, предполагается, что высокомолекулярные цепочки иммуноглобулина можно извлекать из любого из подклассов антител IgG, которые известны как IgG1, IgG2, IgG3 и IgG4. Как известно, каждая постоянная область высокомолекулярной цепочки иммуноглобулина включает четыре или пять доменов. Домены называются в следующей последовательности: СН1-шарнир-СН2-СН3-(-СН4). СН4 присутствует в IgM, который не содержит областей шарнира. Последовательности ДНК доменов высокомолекулярной цепочки имеют пересекающуюся гомологию среди классов иммуноглобулина, например, домен СН2 из IgG является гомологическим домену СН2 из IgA и IgD и домену СН3 из IgM и IgE. Низкомолекулярные цепочки иммуноглобулина могут иметь постоянную цепочку либо каппа (κ), либо лямбда (λ). Последовательности и расположение последовательностей этих областей иммуноглобулина хорошо известны (см., например, Kabat et al., "Sequences of Proteins Immunological Interest, "U.S. Department of Health и Human Services, третье издание 1983, четвертое издание 1987, и Huck et al. (1986) NUC. ACIDS RES. 14: 1779-1789).
В предпочтительных воплощениях данного изобретения вариабельные области извлекают из антител, специфических по отношению к антигену поверхности предварительно выбранной клетки (антигену, связанному с больными клетками, такими, как раковая клетка или вирусно инфицированная клетка), а постоянные области включают домены СН1 и СН2 (и возможно, СН3) из антитела, которое является таким же или отличным от антитела, которое является источником вариабельной области. В практическом осуществлении данного изобретения относящаяся к антителу часть иммуноцитокина предпочтительно является неиммуногенной или слабо иммуногенной в намеченном реципиенте. Соответственно, относящуюся к антителу часть, максимально насколько это возможно, предпочтительно извлекают из тех же самых биологических видов, что и намеченный реципиент. Например, если иммуноцитокин предполагается назначать людям, домены постоянной области предпочтительно получают из человеческого источника. См., например, патент US №4816567. Более того, если вариабельную область иммуноглобулина извлекают из видов, иных, чем намеченный реципиент, например, когда источником последовательностей вариабельной области являются мыши, а намеченный реципиент - человек, то вариабельная область предпочтительно включает последовательности FR человека и мышиные последовательности CDR, помещенные между последовательностями FR, так, чтобы получалась химерическая вариабельная область, которая обладает специфичностью к связыванию предварительно выбранного антигена, но по-прежнему минимизирует иммунную активность намеченного хозяина. Конструирование и синтез таких химерических вариабельных областей описаны в работах Jones et al. (1986) Nature 321: 522-525, Verhoyen et al. (1988) SCIENCE 239: 1534-1535, и патенты US №№5225539 и 5585089. Клонирование и экспрессия приспособленного для человека слитого протеина антитело-цитокин, KS-1/4 анти-ЕрСАМ антитело -EL-12 слитый протеин, а также его способность разрушать упрочившуюся колонию метастазов карциномы была описана в работе Gillies et al. (1998) J. Immunol. 160: 6195-6203.
Ген, кодирующий цитокин, присоединен либо прямо, либо через линкер, например, при помощи линкера (Gly4-Ser)3, кодирующего ДНК в рамке, к концу 3' гена, кодирующего постоянную область иммуноглобулина (т.е. экзоны СН2 или СН3). В определенных воплощениях данного изобретения линкер может включать нуклеотидную последовательность, кодирующую сайт протеолитического расщепления. Этот сайт, если он помещен между постоянной областью иммуноглобулина и цитокином, может быть спроектирован таким образом, чтобы обеспечивать протеолитическое высвобождение цитокина на сайте мишени. Например, хорошо известно, что плазмин и трипсин отщепляются после радикалов лизина и аргинина на сайгах, которые являются подходящими для протеаз. Многие другие специфические к сайту эндопротеазы и аминокислотные последовательности, которые они отщепляют, хорошо известны. Предпочтительные сайты протеолитического расщепления и протеолитические ферменты, проявляющие активность с такими сайтами расщепления, раскрыты в патентах US №№5541087 и 5726044.
Структура нуклеиновой кислоты необязательно может включать эндогенный промотор и энхансер для гена, кодирующего вариабельную область, чтобы регулировать экспрессию химерической цепочки иммуноглобулина. Например, гены, кодирующие вариабельную область, могут быть получены в виде фрагментов ДНК, включающих лидирующий пептид, ген VJ (функционально перегруппированные вариабельные области (V) с присоединением сегмента (J)) для низкомолекулярной цепочки, или ген VDJ для высокомолекулярной цепочки, и эндогенный промотор и энхансер для этих генов. Альтернативно ген, кодирующий вариабельную область, может быть получен без регулирующих эндогенных элементов, и его можно применять в векторе экспрессии, который обеспечивает эти элементы.
Гены вариабельной области можно получать по обычной методике клонирования ДНК из клеток, которые генерируют желательные антитела. Скринирование библиотеки генов для специфической функционально перегруппированной вариабельной области можно произвести при помощи подходящих образцов ДНК, таких, как сегменты ДНК, содержащие область J последовательности ДНК и дальше отстоящие последовательности. Идентификация и подтверждение правильных клонов достигаются секвенированием клонированных генов и сравнением последовательности с соответствующей последовательностью полной длины, по сути, сплайсингованной mРНК.
Антиген-мишень может быть антигеном поверхности клетки опухоли, поверхности раковой клетки, вирусно инфицированной клетки или другой больной клетки. Антиген-мишень может также быть нерастворимым внутриклеточным антигеном некротической клетки (см., например, патент US №5019368). Гены, кодирующие подходящие вариабельные области, можно в общем случае получать из генерирующих иммуноглобулин колоний лимфоидных клеток, происходящих от одного общего предка. Например, колонии клеток гибридом, генерирующих иммуноглобулин, специфический для ассоциированных с опухолью антигенов, или вирусных антигенов, могут быть получены по хорошо известной стандартной методике гибридизации соматических клеток (см., например, патент US №4196265). Эти генерирующие иммуноглобулин колонии обеспечивают источник генов вариабельной области в функционально перегруппированной форме. Гены вариабельной области обычно бывают мышиного происхождения, поскольку эта мышиная система приспосабливается к генерированию широкого многообразия иммуноглобулинов желательной специфичности. Кроме того, последовательности вариабельной области можно получать путем скринирования библиотек, например библиотек фагов, для последовательностей вариабельной области, которые связывают предварительно выбранный антиген с регулируемым сродством. Способы создания и скринирования образцов библиотек фагов описаны, например, в работах Huse et al. (1989) Science 246: 1275-1281, и Kang et al. (1991) Proc. Natl. Acad. Sci. USA 88: 11120-11123.
Фрагмент ДНК, кодирующий функционально активный ген вариабельной области, связан с фрагментом ДНК, содержащим ген, кодирующий желательную постоянную область (или ее часть). Постоянные области иммуноглобулина (высокомолекулярная и низкомолекулярная цепочки) можно получать из генерирующих антитела клеток по обычным методикам клонирования генов. Были клонированы гены для двух классов низкомолекулярных цепочек человека (κ и λ) и пяти классов высокомолекулярных цепочек человека (α, δ, ε, γ и μ), и, таким образом, постоянные области человеческого происхождения легко можно получать из этих клонов.
Слитый ген, кодирующий гибридную высокомолекулярную цепочку иммуноглобулина, is монтируют или вставляют в вектор экспрессии для введения в клетку-реципиент. Введение генной структуры в плазмидные векторы можно осуществлять по обычным методикам сплайсинга генов. Химерическую высокомолекулярную цепочку иммуноглобулина можно ко-экспрессировать в той же клетке, с соответствующей низкомолекулярной цепочкой иммуноглобулина, так что полный иммуноглобулин можно экспрессировать и монтировать одновременно. С этой целью структуры высокомолекулярной и низкомолекулярной цепочек в один и тот же или в различные векторы.
Колонии клеток-реципиентов представляют собой в общем случае лимфоидные клетки. Предпочтительной клеткой-реципиентом является миелома (или гибридома). Миеломы могут синтезировать, монтировать и секретировать иммуноглобулины, кодированные трансфекцированными генами, и они способны гликозилировать протеины. В особо предпочтительную группу реципиентов или клеток-хозяев входят миелома Sp2/0, которая в обычном состоянии не генерирует эндогенный иммуноглобулин, и мышиная миелома NS/0.
После трансфекции клетка производит только иммуноглобулин, кодированный трансфекцированной генной структурой. Трансфекцированные миеломы можно выращивать в культуре или в брюшине мыши, где секретированный иммуноцитокин можно извлекать из асцитной жидкости. Другие лимфоидные клетки, такие, как лимфоциты В, можно применять в качестве клеток-реципиентов.
Существует несколько способов трансфекции лимфоидных клеток векторами, содержащими структуры нуклеиновых кислот, кодирующих химерическую цепочку иммуноглобулина. Например, векторы можно вводить в лимфоидные клетки путем слияния сферобластов (см., например. Gillies et al. (1989) BIOTECHNOL. 7: 798-804). В число других подходящих методов входят электропорация или осаждение фосфатом кальция (см., например, Sambrook et al. eds (1989) "Molecular Cloning: A Laboratory Manual" Cold Spring Harbor Press).
Другие подходящие методы получения иммуноцитокинов включают приготовление последовательности РНК, кодирующей структуру, и ее трансляцию в подходящую систему экспрессии in vivo или in vitro. Предполагается, что методологии рекомбинантной ДНК для синтеза генов, кодирующих слитые протеины антитело-цитокин, для внедрения генов в клетки-хозяева, для экспрессии генов в хозяине и для сбора получающихся слитых протеинов, хорошо известны и подробно описаны в специальной литературе. Специфические протоколы описаны, например, в работе Sambrook et al. eds (1989) "Molecular Cloning: A. Laboratory Manual" Cold Spring Harbor Press.
Понятно, что химически спаренные иммуноцитокины можно получать с применением широкого многообразия хорошо известных методик. Например, антитело или фрагмент антитела можно химически связать с цитокином при помощи химически активных боковых цепочек аминокислот в антителе или фрагменте антитела и цитокине. Боковые цепочки аминокислот могут быть ковалентно связанными, например, через дисульфидные связи или при посредстве гомо- или гетеробифункциональных структурообразующих реагентов, включая, например, N-сукцинимидил 3(-2-пиридиилдитио)пропионат, m-малеимидобензоил-N-гидроксисукцинатовый эфир, m-малеимидобензоил-N-гидроксисульфосукцинимидовый эфир и 1,4-ди-[3'-(2'-пиридилтио)пропионамидо]бутан, каждый из которых можно приобрести у компании Pierce, Rockford, IL.
По способу по данному изобретению комбинирование иммуноцитокинов с агентами, увеличивающими поглощение иммуноцитокина, пригодно для усиленного стимулирования иммунной системы, и, таким образом, приводит к цитотоксической реакции клеток целевого типа, например, клеток опухоли или других больных клеток. Следует ожидать, что комбинация иммуноцитокина и агента, увеличивающего поглощение иммуноцитокина, не будет приводить к совместному или синергическому антиопухолевому эффекту in vitro, поскольку в отдельности иммуноцитокин не является цитотоксичным.
Безотносительно к какой-либо конкретной теории предполагается, что эффекты комбинированной терапии in vivo могут включать повышенное поглощение одного из агентов в результате действия других, что приводит к проявлению одного из двух либо сразу обоих эффектов: (1) повышению химиотерапевтической цитотоксичности (если иммуноцитокин повышает поглощение химиотерапевтического агента, увеличивающего поглощение иммуноцитокина в клетках опухоли) и/или (2) усилению иммуностимуляции (если агент, увеличивающий поглощение иммуноцитокина, некоторым образом повышает поглощение иммуноцитокина опухолью). В отношении механизма (1) проведенные ранее исследования показали, что можно повысить поглощение антител с радиоактивной меткой (и предположительно, маленьких молекул лекарственных препаратов) опухолью при помощи предварительной обработки высокими дозами иммуноконъюгата антитело-IL2, которое инициирует локальную проницаемость сосудов (см., например, Homick et al., 1999, CLIN CANCER RES. 5: 51-60). Если этот конкретный механизм задействован в комбинированной терапии иммуноцитокинами и агентами, увеличивающими поглощение иммуноцитокина, то должна возникать необходимость сначала обрабатывать животное-носитель опухоли иммуноцитокином. Тем не менее, если единственная доза агента, увеличивающего поглощение иммуноцитокина, введенная перед лечением иммуноцитокином, приводит к синергическому эффекту в отношении противоопухолевой активности, то такой механизм не может быть задействованным. Скорее, более правдоподобным окажется объяснение, что лечение агентом, увеличивающим поглощение иммуноцитокина, повышает поглощение иммуноцитокина по механизму (2). Эта гипотеза может быть далее подкреплена демонстрацией факта, что совместное назначение с агентом, увеличивающим поглощение иммуноцитокина, повышает поглощение опухолью иммуноцитокина с радиоактивной меткой.
По способу по данному изобретению преимущество комбинированной терапии состоит в том, что назначение иммуноцитокина усиливает цитотоксическое действие химиотерапевтического агента, который действует как агент, увеличивающий поглощение иммуноцитокина. Следовательно, пациенту можно назначать меньшие дозы химиотерапевтического агента. Соответственно, можно уменьшить подавление некоторых аспектов иммунной системы пациента, связанное с лечением с применением химиотерапевтического агента. В одном из воплощений данного изобретения единственную дозу химиотерапевтического агента, увеличивающего поглощение иммуноцитокина, назначают пациенту до назначения иммуноцитокина. Химиотерапевтический агент, увеличивающий поглощение иммуноцитокина, назначают предпочтительно за срок от примерно 4 дней до примерно 4 часов, наиболее предпочтительно за примерно 24-48 часов до назначения иммуноцитокина. В другом воплощении данного изобретения перед назначением иммуноцитокина пациенту назначают несколько доз химиотерапевтического агента, увеличивающего поглощение иммуноцитокина. В других воплощениях изобретения химиотерапевтический агент, увеличивающий поглощение иммуноцитокина, можно назначать до, одновременно и/или после иммуноцитокина.
Паклитаксел является примером химиотерапевтического агента, увеличивающего поглощение иммуноцитокина, который может подавлять или подвергать опасности аспекты иммунной системы пациента. В то время как большая часть иммуностимулирующего действия паклитаксела осуществляется через медиаторы - клетки макрофаг/моноцит, многочисленные исследования лимфоцитной функции свидетельствуют о вредном воздействии паклитаксела на эту подсистему. Например, было обнаружено, что лечение паклитакселом подвергает серьезной опасности способность лимфоцитов к пролиферации как у нормальной мыши, так и у мыши-носителя опухоли (Mullins et al., 1998, IMMUNOPHARMACOL IMMUNOTOXICOL 20: 473-492) и ослабляет как цитотоксичность клеток NK, так и генерирование активированной лимфокином цитотоксичности в культурах клеток, содержащих IL-2 (Chuang et al., 1993, GYNECOL ONCOL 49: 291-298). Фактически имеющиеся данные свидетельствуют, что лимфоцитная подсистема клеток является важнейшим движущим фактором популяции в антиопухолевой активности иммуноцитокинов (Lode et al, 1998, PHARMACOL THER 80: 277-292). Экспериментальные данные, которые содержатся в настоящем изобретении, приводят к некоторым новым выводам, которые не могли быть предсказаны в прототипе, особенно относительно порядка назначения лекарственных препаратов.
Таксаны можно назначать одновременно с иммуноцитокином или назначать отдельно, с различными способами приема. Композиции по настоящему изобретению можно назначать при любом способе приема, совместимом с конкретными молекулами. Таким образом, если это уместно, назначение может быть оральным или парентеральным, включая внутривенные и внутрибрюшинные способы назначения.
Композиции по настоящему изобретению можно вводить животным любым подходящим способом, прямо (т.е. локально, как при инъекции, имплантации или местном назначении в очаге ткани) или системно (т.е. парентерально или орально). Если композиции вводят парэнтерально, например, при внутривенном, подкожном, офтальмологическом, внутрибрюшинном, внутримышечном, ротовом, ректальном, внутривлагалищном, внутриглазном, внутримозговом, внутричерепном, внутриспинномозговом, внутрижелудочковом, внутриоболочковом, внутриполостном, интракапсулярном, внутриносовом или аэрозольном назначении, то композиции предпочтительно включают водную составную часть или физиологически совместимую жидкую суспензию или раствор. Следовательно, носитель или переносчик является физиологически приемлемым, так что дополнительно к доставке пациенту желательной композиции он, в других отношениях, не оказывает неблагоприятного на электролитический и/или объемный баланс пациента. Таким образом, жидкая среда для агента может включать обычный физиологический раствор (т.е. 9,85% водный раствор NaCl, 0,15 М, рН 7-7,4). Для многих таксанов рецептуры значительно более сложны, за счет их в общем случае неблагоприятных характеристик в отношении растворимости. Например, обычно для паклитаксела используется следующая рецептура: 10% кремофора, 10% этанола и 80% физиологического раствора (0,9% NaCl), а рецептура для доцетаксела включает смесь этанол : эфир полиоксиэтиленовой жирной кислоты 80 в соотношении 1:1, которую затем перед назначением разбавляют до 1:10 раствором 5% глюкозы (Bissery и Lavelle, 1999). Тем не менее могут быть известны и другие рецепты, включающие таксаны и их внось синтезированные аналоги, либо специалистам нетрудно их разработать.
Предпочтительные дозы при назначении иммуноцитокина находятся в пределах 0,1 - 100 мг/м2, более предпочтительно в интервале 1 - 20 мг/м2 и наиболее предпочтительно в интервале 2 - 6 мг/м2. Предпочтительные дозы агента, увеличивающего поглощение иммуноцитокина, зависят в общем случае от типа применяемого агента, увеличивающего поглощение иммуноцитокина, тем не менее оптимальные дозы можно определить при помощи стандартных экспериментов. Назначение иммуноцитокина и/или агента, увеличивающего поглощение иммуноцитокина, можно осуществлять путем периодического введения порций лекарства или путем непрерывного внутривенного или внутрибрюшинного назначения из внешнего резервуара (например, из пакета для внутривенного вливания) или внутреннего (например, из биоразмываемого импланта). Кроме того, предполагается, что иммуноцитокин по данному изобретению можно также назначать намеченному пациенту вместо с множеством различных агентов, увеличивающих поглощение иммуноцитокина. Предполагается тем не менее, что оптимальная комбинация иммуноцитокинов и агентов, увеличивающих поглощение иммуноцитокина, способы назначения и дозы можно определить по широко известным стандартным экспериментальным методикам.
Чтобы оценить эффективность воздействия на иммунные реакции комбинированной терапии с применением слитых протеинов антитело-цитокин и агентов, увеличивающих поглощение иммуноцитокина, можно использовать множество различных методик. Например, описанные в приведенных ниже примерах модели на животных или другие подходящие модели на животных можно использовать для тестирования, которое позволит определить, какие из агентов, увеличивающих поглощение иммуноцитокина, или комбинации агентов, увеличивающих поглощение иммуноцитокина, наиболее эффективны в отношении синергического действия с иммуноцитокином (например, слитый протеин антитело-IL2) на усиление иммунного разрушения упрочившейся опухоли. Агент, увеличивающий поглощение иммуноцитокина, или комбинацию агентов, увеличивающих поглощение иммуноцитокина, можно назначать до или одновременно с курсом терапии иммуноцитокином и влияние на опухоль можно легко отслеживать посредством измерений объема. Далее, после идентификации новых агентов, увеличивающих поглощение иммуноцитокина, специалисты смогут применять описанные здесь методы для оценки потенциала этих новых соединений в плане усиления или другого модифицирования антираковой активности слитых протеинов антитело-цитокин.
Альтернативно после терапии опухоли можно вырезать, секционировать и окрашивать по стандартным гистологическим методикам или при помощи специфических иммуногистологических реагентов, чтобы оценить воздействие комбинированной терапии на иммунную реакцию. Например, простое окрашивание гематоксолином и эозином может выявить различия в инфильтрации лимфоцитов в твердые опухоли, что является показателем клеточной иммунной реакции. Более того, иммуноокрашивание срезов антителами для специфических классов иммунных клеток может выявить природу вызываемой реакции. Например, антитела, которые связаны с CD45 (общий маркер лейкоцитов), CD4 и CD8 (для идентификации подкласса Т-клеток) и NK1.1 (маркер клеток NK) можно использовать для оценки типа иммунной реакции, медиаторами которой являются иммуноцитокины по данному изобретению.
Альтернативно тип иммунной реакции, медиаторами которой являются иммуноцитокины, можно оценить по обычному способу удаления подклассов клеток, описанному, например, в работе Lode et al. (1998) Blood 91: 1706-1715. Примеры исчерпывающихся антител включают те антитела, которые вступают в реакцию с маркерами Т-клеток CD4 и CD8, а также и те, которые образовывают связи с маркерами NK - NK1.1 и асиало GM. Коротко говоря, эти антитела вводят животному до начала лечения протеином антитело-цитокин, в довольно высоких дозах (например, при дозах около 0,5 мг/мышь), и впоследствии дают с недельными интервалами до завершения эксперимента. Эта методика позволяет идентифицировать типы клеток, необходимые для определения наблюдаемой иммунной реакции в млекопитающих.
При другом подходе цитотоксическую активность спленоцитов, выделенных из животных, которые были подвергнуты комбинированной терапии, можно сопоставить с цитотоксическую активностью спленоцитов, выделенных из других групп животных, которые подвергались другой обработке. Культуры спленоцитов приготавливают путем механического измельчения выздоровевшей стерильной селезенки по стандартной методике, описанной во многих руководствах для иммунологических лабораторий. См., например, Coligan et al. (eds) (1988) "Current Protocols in Immunology" John Wiley & Sons, Inc. Полученные клетки затем культивируют в подходящей для культуры клеток среде (например, DMEM от фирмы GIBCO), содержащей сыворотку, антибиотики и низкие концентрации IL-2 (˜10 U/мл). Например, чтобы сопоставить активность NK, оптимальный срок культивирования обычно составляет 3 дня, однако, чтобы сопоставить цитотоксическую активность Т-клеток, оптимально проводить культивирование в течение 5 дней. Цитотоксическую активность можно измерить при помощи радиоактивной маркировки опухолевых клеток-мишеней (например, клеток LLC) изотопом51Cr в течение 30 мин. После удаления избытка радиоактивного маркера меченые клетки смешивают в различных концентрациях с культивированными клетками селезенки на 4 часа. В конце инкубирования выделившийся из клеток51Cr измеряют при помощи счетчика гамма частиц и эти данные затем используют для количественной оценки степени лизиса клеток, вызванного иммунными клетками. По этой методике традиционно измеряют активность цитотоксичных Т-лимфоцитов (или CTL).
Дальше изобретение проиллюстрировано на приведенных ниже не ограничивающих его примерах.
Пример 1. Моделирование на животных.
Для изучения эффекта от комбинирования иммуноцитокинов и таксанов на их действие как медиаторов эффективной цитотоксической реакции против опухоли были разработаны модели рака мыши. Иммуноцитокины, использованные в приведенных ниже примерах, связывали ЕрСАМ, антиген опухоли человека, который обнаруживается на большинстве извлеченных эпителиальных опухолей (см. Perez и Walker (1989) J. Immunol. 142: 3662-3667). Для тестирования эффективности в иммуночувствительной мышиной модели необходимо было экспрессировать антиген человека на поверхности клетки опухоли мыши, который был бы сингенным с мышью-хозяином. Клетки карциномы легких Lewis (LLC), широко известной культуры клеток рака легких мыши, были первой культурой клеток, выбранной для этой цели. Об этой линии клеток известно, что она генерирует высокие уровни ингибиторов иммунной системы и инициирует производство IL-10 из иммунных клеток в микроокружении опухоли, что приводит к локализованному подавлению иммунной реакции (Sharma et al., 1999, J. IMMUNOL 163: 5020-5028). Антиген человеческой опухоли, ЕрСАМ (который называют также KSA), был экспрессирован на поверхности клеток LLC таким образом, что на него можно было in vivo нацеливать иммуноцитокины, выделенные из мышиного антитела анти-ЕрСАМ, KS-1/4. Это осуществляли путем преобразования последовательности сДНК ЕрСАМ с рекомбинантным ретровирусным вектором по известной методике (Gillies, заявка на патент US 09/293042), что приводило к получению целевой линии клеток LLC/KSA. Эти клетки выдерживали в DMEM, дополненном инактивированной теплом 10% эмбриональной бычьей сывороткой, L-глутамином, смесью пенициллин/стрептомицин и генетицин (GIBCO) при 37°С и 7,0% СО2.
Дополнительные линии клеток, представляющие карциному из различных тканевых источников, конструировали аналогичным образом. Неиммуногенную линию клеток карциномы молочной железы мыши 4Т1 предоставил Dr. Paul Sondel (Univ. of Wisconsin). Эта линия выращивается медленно и постепенно после подкожной имплантации, и самопроизвольно метастазирует во многие органы даже до хирургического удаления первичной опухоли. Можно также индуцировать экспериментальные метастазы в легкие посредством внутривенной инъекции. Колонию линии клеток мышиной карциномы СТ26, выделенную при помощи интраректальной инъекции N-нитрозо-N-метилуретана в мышах BALB/C, предоставил Dr. I.J.Fidler (MD Anderson Cancer Center, Houston, TX). Клетки 4Т1 и СТ26 были трансфекцированы Ер-САМ по известной методике (Gillies et al., 1998, J. IMMUNOL 160: 6195-6203). Клетки 4T1/KSA выдержали в RPMI, добавили инактивированную теплом 10% бычью сыворотку, L-глутамин, смесь пенициллин/стрептомицин и генетицин (GIBCO) при 37°С и 7,0% CO2. Клетки CT26/KSA выдержали в DMEM, добавили инактивированную теплом 10% бычью сыворотку, L-глутамин, витамины, натриевую соль пировиноградной кислоты, второстепенные аминокислоты, пенициллин/стрептомицин и генетицин (GIBCO, Gaithersberg, MD) при 37°С и 7,0% СО2. Генетицин добавили к клеткам, чтобы поддержать экспрессию KSA. Все трансфекцированные линии клеток росли постепенно, в виде кожных опухолей (после подкожной инъекции) или в виде метастазов (после внутривенной инъекции), и убивали мышей, хотя в них на поверхности клеток происходила экспрессия молекулы ЕрСАМ человека (потенциальный чужеродный антиген).
Для исследования роста опухоли опухоли LLC/KSA или CT26/KSA имплантировали подкожно на спинах мышей. В исследованиях LLC/KSA опухоли пересаживали из нескольких классов опухолей, которые вводили в одной клеточной суспензии из 1×106 клеток в 100 мкл PBS. Примерно через две недели опухоли асептически собирали и пропускали через сито с ячейкой 150 мкм. Затем клетки пропускали через шприц и иглу 23 размера два или три раза, дважды промывали и повторно суспендировали в PBS. Единственную клеточную суспензию из 1×106 клеток LLC/KSA в 100 мкл PBS вводили подкожно иглой размера 301/2 на спинах мышей. В исследованиях CT26/KSA клетки, которые росли в культуре экспоненциально, вводили как одну клеточную суспензию из 1×10 клеток в 100 мкл PBS. Когда опухоль упрочивалась, примерно через две недели после имплантации, в День 0 начинали давать лекарство. Опухоли измеряли кронциркулем в трех измерениях дважды в неделю. Объемы опухоли рассчитывали по следующему уравнению:
Объем=½×4/3π(L/2×W/2×Н), где L = длина, W = ширина и Н = высота опухоли.
Животных взвешивали и контролировали общее состояние здоровья на протяжении всего исследования. Когда опухоль становилась некротической или когда животные начинали погибать, их подвергали эвтаназии при помощи асфиксии СО2.
Данные представлены в графическом виде. Диаграммы отражают индивидуальный или средний объем опухолей (+/- SEM) на протяжении и после введения лекарства. Кроме того, данные выражены как процент наблюдаемых средних объемов опухоли у мышей, которые получали лекарство относительно данных для мышей, которым вводили носитель. Для определения значительных различий объемов индивидуальных опухолей производили студенческий тест t.
Для экспериментального исследования печеночных метастазов мышь анестезировали при помощи 80 мг/кг кетамина HCl (Fort Dodge Animal Health, Fort Dodge, IA) и 5 мг/кг ксилазина (Вауег, Shawnee Mission, KS). Единую клеточную суспензию из 1×105 клеток CT26/KSA в 100 мкл DMEM, содержащего 25 мМ HEPES (GIBCO), вводили иглой размера 27 ½ под селезеночной мембраной через 60 секунд в День 0. Еще через 2 минуты сосуды селезенки каутеризировали термокаутером (Roboz, Rockville, MD) и селезенку удаляли, животным накладывали швы при помощи автозажимов. Через три недели после инокуляции животных умертвляли; их печень извлекали и взвешивали. Затем эти органы фиксировали и окрашивали в растворе Bouin (Sigma, St. Louis МО).
Данные представлены в графическом виде. Диаграмма показывает средний вес опухоли (+/- SEM) на момент умерщвления. Вес опухоли определяли путем вычитания веса обычной печени из веса экспериментальных образцов. Кроме того, данные выражены как процент наблюдаемого среднего веса опухоли мышей, получавших лекарство, от среднего веса опухоли мышей, получавших носитель. Для определения значительных различий веса индивидуальных опухолей производили студенческий тест t.
Для экспериментального исследования метастазов в легких одну клеточную суспензию из 2,5×105 клеток 4T1/KSA в 100 мкл PBS медленно ввели иглой размера 27 ½ в боковое ответвление хвостовой вены в День 0. Через три недели после инокуляции животных умертвили; их легкие извлекли и взвесили. Затем легкие зафиксировали и окрасили в растворе Bouin (Sigma). Данные представлены в графическом виде. На диаграмме показан средний вес опухолей (+/- SEM) на момент умерщвления. Вес опухоли определяли вычитанием веса нормального легкого из веса экспериментальных образцов. Кроме того, данные выражены как процент наблюдаемого среднего веса опухоли мышей, получавших лекарство от среднего веса опухоли мышей, получавших носитель. Для определения значительных различий веса индивидуальных опухолей производили студенческий тест t.
Пример 2. Приготовление слитых с антителом протеинов (иммуноцитокины).
Некоторые слитые протеины антитело-цитокин обсуждаются в приведенных ниже примерах.
huKS-huγ1-huIL2 (аббревиатура KS-IL2)
Ген, кодирующий слитый протеин huKS-huγ1-huIL2, приготовили и экспрессировали практически в полном соответствии с методикой, описанной в работах Gillies и др. (1998) J. Immunol. 160: 6195-6203, и патенте US №5650150. Поступали следующим образом: гуманизированные вариабельные области мышиного антитела KS1/4 (Varki и др., (1984) Cancer Res. 44: 681-687) моделировали по методике, описанной в работе Jones и др. (1986) Nature 321: 522-525, которая включает вложение CDR каждой вариабельной области KS 1/4 в структуру обобщающих типичных последовательностей человеческих вариабельных областей с высокой степенью гомологичности. Молекулярное моделирование при помощи Silicon Graphics Indigo с программой BioSym подтвердило, что формы CDR были сохранены. Затем протеиновые последовательности подвергли ретрансляции и сконструировали гены путем сшивки перекрывающихся олигонуклеотидов.
Полученные вариабельные области вставили в вектор экспрессии, содержащий постоянные области человеческой низкомолекулярной цепочки κ и человеческой высокомолекулярной цепочки Сγ1 придерживаясь, в основном, методики, описанной в работе Gillies et al. (1992) Proc. Natl. Acad. Sci. USA 89: 1428-1432, за исключением того, что промоторы металлотионеина и энхансеры высокомолекулярной цепочки иммуноглобулина были заменены промоторами/энхансерами CMV для экспрессии обеих цепочек. Слияние зрелых последовательностей IL-2 с карбокси-концами человеческих высокомолекулярных цепочек производили придерживаясь, в основном, методики, описанной в работе Gillies et al. (1992) Proc. Natl. Acad. Sci. USA 89: 1428-1432, за исключением того, что нетранслированные области 3' гена IL-2 были извлечены из поли(А) области SV40.
Слитый протеин IL-2 был экспрессирован трансфекцией полученной плазмиды в линию клеток миеломы NS/0 с выбором среды, содержащей 0,1 мкМ метотрексата (МТХ). Чтобы получить стабильно трансфекцированные клоны, ДНК плазмиды вводили в клетки NS/0 мышиной миеломы электропорацией. Клетки NS/0 выращивали в среде Dulbecco, модифицированной средой Eagle, дополненной 10% эмбриональной бычьей сывороткой. Примерно 5×106 клеток один раз промыли PBS и повторно суспендировали в 0,5 мл PBS. Затем 10 мкг линеаризованной ДНК плазмиды инкубировали с клетками в Gene Pulser Cuvette (межэлектродное расстояние 0,4 см, BioRad) на льду в течение 10 мин. Электропорацию производили при помощи аппарата Gene Pulser (BioRad, Hercules, CA), установленного на 0,25 В и 500 мкФ. Клеткам предоставили возможность восстанавливаться в течение 10 минут на льду, после этого их повторно суспендировали в среде выращивания и поместили на две пластинки с 96 колодцами каждая. Стабильно трансфекцированные клоны были отобраны путем выращивания в присутствии 100 нМ метотрексата, который ввели через два дня после трансфекции. Клетки подпитывали через каждые 3 дня в течение трижды большего времени, и МТХ-устойчивые появлялись в течение от 2 до 3 недель.
Экспрессирующие клоны были идентифицированы при помощи ELISA с Fc или цитокином, с использованием соответствующих антител (см., например, Gillies et al. (1989) Biotechnol. 7: 798-804). Полученный слитый протеин очистили связыванием с протеиновой A Sepharose (Pharmacia) и элюированием в соответствии с инструкциями изготовителя.
huKS-huγ4-huIL2
Ген, кодирующий слитый протеин huKS-huγ4-huIL2, сконструировали и экспрессировали, придерживаясь, в основном, методики, описанной в заявке U.S.S.N. 09/256156, зарегистрированной 24 февраля 1999 года, которая притязает на приоритет заявки U.S.S.N. 60/075887, зарегистрированной 25 февраля 1998 г.
Версию Igγ4 описанного выше слитого протеина huKS-huγ 1-huIL2 приготовили путем удаления фрагмента гена Сγ1 постоянной области иммуноглобулина из вектора экспрессии huKS-huγ1-huIL2 и его замены соответствующей последовательностью из человеческого гена Сγ4. Последовательности и порядок последовательностей постоянных областей Сγ1, Сγ2, Сγ3 и Сγ4 высокомолекулярной цепочки человека описаны в работе Huck et al. (1986) Nuc. ACIDS RES. 14: 1779-1789.
Обмен фрагментов Cγ1 и Сγ4 произвели путем переваривания исходной ДНК плазмиды, содержащей Cγ1, при помощи Hind III и Xho I и очистки большого фрагмента 7,8 kb агарозным гель-электрофорезом. Вторую ДНК плазмиды, содержащую ген Сγ4, дигетировали при помощи Hind III и Nsi I и очистили фрагмент размером 1, 75 kb. Третью плазмиду, содержащую человеческую сДНК IL-2 и полиА сайт SV40, слитую с карбоксильными концами человеческого гена Сγ1, дигетировали при помощи Xho I и Nsi I, и очистили маленький фрагмент 470 bp. Все три фрагмента связали вместе в примерно одинаковых молярных количествах. Продукт связывания использовали для трансформации соответствующей Е.coli и выбрали колонии путем выращивания на пластинках, содержащих ампициллин. Корректно смонтированные рекомбинантные плазмиды идентифицировали при помощи рестрикционного анализа препаратов ДНК плазмиды из выделенных трансформантов, а для дифференциации генных вставок Сγ1 (без Fsp I) и Сγ4 (один сайт) использовали дигетирование при помощи Fsp I.
Окончательный вектор, содержащий замену высокомолекулярной цепочки Сγ4-IL2, ввели в клетки миеломы мыши NS/0 при помощи электропорации (0,25 В и 500 мкФ) и трансфектанты выбрали путем выращивания в среде, содержащей метотрексат (0,1 мкМ). Идентифицировали клоны клеток, экспрессирующих высокие уровни слитых протеинов huKS-huγ4-huIL2, увеличили их и очистили слитый протеин из надосадочной жидкости культуры хроматографией на протеине A Sepharose. Чистоту и целостность слитого протеина Сγ4 определяли SDS-полиакриламидным гель-электрофорезом. Активность IL-2 измеряли при помощи анализа пролиферации Т-клеток (Gillis et al. (1978) J. Immunol. 120: 2027-2032) и обнаружили, что она идентична аналогичной характеристике структкры γ1.
huKS-muγ2a-muIL2
Ген, кодирующий слитый протеин huKS-muγ2a-muIL2, сконструировали путем замены по описанной выше методике постоянных областей человеческого антитела и человеческого IL-2 слитого протеина huKS-huγ1-huIL2 соответствующими последовательностями мыши. Конкретно, человеческую ДНК Cγ1-IL2 заменили фрагментом мышиной сДНК Сγ2а, слитым с мышиным IL-2, кодирующим ДНК. Область VH huKS соединили в рамке генетического кода с мышиной сДНК γ2а путем проведения перекрывающихся полимеразных цепных реакций (PCR) с использованием перекрывания олигонуклеотидных праймеров:
(сенс) 5' СС GTC ТСС ТСА GCC AAA АСА АСА GCC ССА TCG GTC (посл. ID №3);
(антисенс) 5' GG GGC TGT TGT TIT GGCTGA GGA GAC GGT GAC TGA CG (посл. ID №4);
(сенс) 5' С ТТА AGC CAG ATC CAG TTG GTG CAG (посл. ID №5) и
(антисенс) 5' СС CGG GGT CCG GGA GAA GCT CTT AGT С (посл. ID №6).
Олигонуклеотиды последовательностей ID №№3 и 4 были предназначены для гибридизации с соединением домена VH huKS и постоянной области мышиной сДНК γ2а (курсив). В первом цикле PCR провели две отдельные реакции. В одной реакции VH из ДНК huKS использовали в качестве шаблона, с олигонуклеотидами последовательностей ID №№4 и 5. Праймер последовательности ID №5 ввел рестрикционный сайт AfIII (CTTAAG) в начало последовательности, кодирующей зрелые амино-концы VНhuKS (жирный шрифт). В другой реакции мышиную сДНК γ2а использовали в качестве шаблона, с олигонуклеотидами последовательностей ID №№3 и 6. Праймер последовательности ID №6 гибридизировали с сДНК, кодирующей область вокруг С-концов γ2а, и ввели рестрикционный сайт XmaI (CCCGGG) для последующего сшивания с сДНК muIL2. Продукты PCR из этих двух реакций смешали и подвергли второму циклу PCR, с использованием олигонуклеотидов последовательностей ID №№5 и 6. Получившиеся в PCR продукты клонировали и, при верификации последовательности, фрагмент AfIII-XmaI, кодирующий VH huKS и постоянную область γ2а мыши использовали для сшивки с ДНК, кодирующей сигнальный пептид на сайте AfIII и сДНК muIL2 на сайте XmaI.
Мышиную сДНК IL2 клонировали из тРНК мононуклеарных клеток периферической крови мыши с использованием олигонуклеотидов, расположенных четвертыми в последовательностях ID №№7 и 8, а именно:
(сенс) 5' GGC CCG GGT AAA GCA CCC ACT TCA AGC TCC (посл. ID №7) и
(антисенс) 5' CCCTCGAGTTATTGAGGGCTTGTTG (посл. ID №8).
Праймер последовательности ID №7 приспосабливает muIL2 (последовательность в крови) к присоединению к muγ2a на рестрикционном сайте XmaI (CCCGGG). Праймер последовательности ID №8 ввели на рестрикционный сайт XhoI (CTCGAG) немедленно после прекращения трансляции кодона (антисенс в крови).
Аналогичным образом вариабельный низкомолекулярный (VL) домен huKS присоединили к последовательности сДНК mu κ при помощи PCR перекрывания. Использовали перекрывающиеся олигонуклеотиды, включая
(сенс) 5' G GAA ATA AAA CGG GCT GAT GCT GCA CCA ACT G (посл. ID №9);
(антисенс) 5' GC AGC ATC AGC CCGTT ТТА ТТТ CCA GCT TGG TCC (посл. ID №10);
(сенс) 5' С ТТА AGC GAG ATC GTG CTG ACC CAG (посл. ID №11) и
(антисенс) 5' СТС GAG СТА АСА СТС АТТ ССТ GTT GAA GC (посл. ID №12).
Олигонуклеотиды были предназначены для гибридизации к соединению VL huKS и постоянной области мышиной сДНК κ (курсив). В первом цикле PCR провели две раздельные реакции. В одной реакции VL ДНК huKS использовали в качестве шаблона, с олигонуклеотидами, которые располагаются четвертыми в последовательности ID №№10 и 11, который ввел рестрикционный сайт AfIII (CTTAAG) в начало последовательности, кодирующей зрелые амино-концы VL huKS (в крови). В другой реакции мышиную сДНК κ использовали в качестве шаблона, с олигонуклеотидами, которые располагаются четвертыми в последовательности ID №№9 и 12, который ввел рестрикционный сайт XhoI после прекращения трансляции кодона (антисенс в крови).
Продукты PCR этих двух реакций смешали и подвергли второму циклу PCR с использованием олигонуклеотидных праймеров, которые располагаются четвертыми в последовательности ID №№11 и 12. Полученный в результате PCR продукт клонировали и, при верификации последовательности, фрагмент AfIII-XhoI, кодирующий VL huKS и постоянную область мышиной κ, сшили с ДНК, кодирующей сигнальный пептид сайта AfIII.
Обе последовательности мышиных высокомолекулярной и низкомолекулярной цепочек были использованы для замены человеческих последовательностей в pdHL7. Полученный в результате вектор экспрессии антитела, содержащий маркерный ген, отличаемый dhfr, электропорировали (6,25 В, 500 мкФ) в клетки мышиной миеломы NS/0 и клоны выбрали путем культурирования в среде, содержащей 0,1 мкМ метотрексата. Затем, по стандартным методикам ELISA, произвели тестирование трансфекцированных клонов, устойчивых к метотрексату, на секрецию детерминант антитела. Слитые протеины очистили хроматографией на протеиновой A Sepharose в соответствии с инструкциями изготовителей.
huKS-muγ2a-muIL12
Ген, кодирующий слитый протеин huKS-muγ2a-muIL12, сконструировали и экспрессировали, придерживаясь, в основном, методики, описанной в заявке U.S.S.N. 08/986997, зарегистрированной 8 декабря 1997 года, и работе Gillies et al. (1998) J. Immunol. 160: 6195-6203. С этой целью произвели слияние субъединицы р35 IL-12 мышиной сДНК с предварительно приготовленной областью, кодирующей высокомолекулярную цепочку huKS-muγ2a. Затем полученный вектор трансфекцировали в линию клеток миеломы NS/0, предварительно трансфекцированную субъединицей р40 IL-12 и способную ее экспрессировать. Иными словами, линию клеток трансфекцировали одной р40 и отобрали стабильные клетки с высоким уровнем экспрессии, которые затем использовали в качестве реципиентов для трансфекции слитым протеином, содержащим р35 (т.е. последовательная трансфекция).
Мышиные субъединицы р35 и р40 IL-12 выделили при помощи PCR из тРНК, приготовленной из клеток селезенки, активированных Concanavalin A (5 мкг/мл в среде культуры в течение 3 дней). Праймеры PCR, использованные для выделения последовательности нуклеиновых кислот, кодирующей р35, которая, кроме того, адаптирована к сДНК р35 как к рестрикционному фрагменту XmaI-XhoI, включали
5' CCCCGGGTAGGGTCATTCCAGTCTCTGG (посл. ID №13) и
5' CTCGAGTCAGGCGGAGCTCAGATAGC (посл. ID №14).
Праймер PCR, использованный для выделения последовательности нуклеиновых кислот, кодирующей р40, включал
5' TCTAGACCATGTGTCCTCAGAAGCTAAC (посл. ID №15) и
5' CTCGAGCTAGGATCGGACCCTGCAG (посл. ID №16).
По известной методике (Gillies et al. J. Immunol. Methods 125: 191) сконструировали вектор плазмиды (pdHL7-huKS-muγ2a-p35), который содержал распознаваемый dhfr маркерный ген, транскрипционную единицу, кодирующую низкомолекулярную цепочку гуманизированного антитела KS, и транскрипционную единицу, кодирующую высокомолекулярную цепочку мыши, слитую с субъединицей р35 мышиной IL-12. Слияние осуществили при помощи сшивки XmaI с фрагментом XhoI сДНК, адаптированной к субъединице р35; к единому сайгу Xmal на конце экзона СН3 приготовленного заранее мышиного гена γ2а. Обе транскрипционные единицы цепочек Н и L включали промотор цитомегаловируса (CMV) (вместо промотора металлотионеина в оригинальной ссылке) на конце 5' и сайт полиаденилирования на конце 3'.
Аналогичный вектор (pNC-p40) был сконструирован для экспрессии свободных субъединиц р40, он включал распознаваемый маркерный ген (ген, устойчивый к неомицину), но по-прежнему использовал промотор CMV для транскрипции. В этом случае кодирующая область включала последовательность природного лидера субъединицы р40 для корректной транспортировки к эндоплазматической сети и монтажа со слитым протеином. Плазмиду pNC-p40 электропорировали в клетки, клетки поместили на пластинки и произвели отбор в среде, содержащей G418. В этом случае надосадочную жидкость культуры из устойчивых к лекарственным препаратам клонов подвергли тестированию ELISA для получения субъединиц р40.
По методике, описанной в работе Gillies et al. (1998) J. Immunol. 160: 6195-6203, вектор экспрессии pdHL7-huKS-muγ2a-p35 электропорировали в линию клеток NS/0, которая уже экспрессирует мышиную р40. Трансфекцированные клоны, устойчивые к мето-трексату, тестировали на секрецию детерминант антител и мышиной IL-12 по стандартной методике ELISA. Полученный в результате протеин очистили связыванием с протеиновой A Sepharose и элюированием в колонке в соответствии с инструкциями изготовителя.
Пример 3. Цитотоксическая активность комбинированной терапии in vitro.
Тестирование линий клеток, сконструированных для применения в животных моделях (пример 1), на их чувствительность к вызванной таксаном цитотоксичности в культуре клеток провели в присутствии и при отсутствии иммуноцитокина, основанного на L-2, состоящего из гуманизированной формы антитела KS-1/4, слитой на карбоксильным конце цепочки Н с человеческим IL-2 (huKS-huγ1-huIL2, ниже используется аббревиатура KS-IL2). Клетки равномерно распределили по 1000 клеток/колодец на пластинках с 96 плоскодонными колодцами и инкубировали в течение 24 часов при 37°С, 7% СО2. Паклитаксел при 2-кратных разбавлениях от 200 нг/мл до 3,125 нг/мл, KS-IL2 при 200 нг/мл и IL-2 при 33,3 нг/мл (эквивалентное количество IL-2 в KS-IL2) добавили в двух экземплярах к культуре клеток на пластинках и инкубировали в течение 6 дней при 37°С, 7% СО2. Колориметрический анализ MTS (Promega), позволяющий определять жизнеспособность клеток, основанный на клеточном превращении соли тетразолия, производили непосредственно на 96-колодезных пластинках. После считывания и регистрации пластинок жизнеспособные клейкие клетки окрасили при помощи Crystal violet (Sigma, St. Louis, МО). Окрашенные Crystal violet пластинки были использованы для верификации результатов анализа MTS. Полученные результаты представлены в табличном виде. IC50 обозначает концентрацию лекарственного препарата, которая приводит к снижению цитотоксичности до 50% уровня контрольных образцов.
Анализ цитотоксичности произвели при самостоятельном применении паклитаксела (от 3 до 200 нг/мл), а также при его комбинированном применении с KS-IL2 (200 нг/мл) или IL-2 (33,3 нг/мл, эквивалентное количество IL-2 в KS-IL2) против клеток CT26/KSA, LLC/KSA и 4T1/KSA. Наблюдалась низкая вплоть до отсутствия цитотоксичность применяемых индивидуально KS-IL2 или IL-2 к трем использованным в тестировании линиям клеток (от 81 до 101% от контрольных образцов, см. таблицу). Добавление как KS-IL2, так и IL-2 не влияло на цитотоксичность паклитаксела. Таким образом, поскольку ни KS-IL2, ни IL-2 не влияет на цитотоксичность паклитаксела, любое увеличение противоопухолевой активности в мышах в результате комбинированного лечения должно быть связано с другими механизмами, которые действуют только в животных-носителях опухоли.
Пример 4. Комбинированная терапия KS-IL2 и таксаном опухоли кожи LLC.
Регрессионный анализ роста опухоли производили с использованием быстро растущей опухоли LLC/KSA. Через неделю после введения единственной дозы паклитаксела (80 мг/кг) назначали в течение 5 дней внутривенные инъекции KS-IL2 (20 мкг) в хвостовую вену (фиг.2). Никакого эффекта при самостоятельном назначении как паклитаксела, так и KS-IL2 (в Дни 0-4) не наблюдалось. Тем не менее, когда KS-IL2 назначали через неделю после паклитаксела, наблюдалось значительное уменьшение среднего объема опухоли (41% от объема в контрольных экспериментах) и приостановка роста опухоли (TGD) в течение примерно 8 дней, что сильно отличается от варианта самостоятельного применения паклитаксела alone (p=0,023).
Никакой связанной с лекарством явной интоксикации не наблюдалось, за исключением потери веса <5% в группе, обработанной паклитакселом.
Далее, эффект от применения многократных доз паклитаксела, что обычно рассматривается как более эффективный курс химиотерапии, сопоставили с эффектом, полученным при введении единственной дозы паклитаксела в комбинации с KS-IL2, чтобы определить, как курс лечения влияет на улучшение. Обнаружилось, что самостоятельное применение KS-IL2 (20 мкг, Дни 0-4) опять не оказывает никакого эффекта на рост опухоли LLC/KSA, но самостоятельное применение паклитаксела, при назначении многократными дозами (50 мг/кг, через день), уменьшает средний объем опухоли до 63% наблюдаемого в контрольных экспериментах и приводит к 4-дневной приостановке роста опухоли (TGD) (фиг.3). Когда иммуноцитокин KS-IL2 назначали через одну неделю после лечения паклитакселом, наблюдалось уменьшение объема опухоли до 27% от наблюдавшегося в контрольной группе, а TGD составляла 10 дней, что значительно отличается от результатов, полученных при самостоятельном применении паклитаксела (р=0,016). Никакой связанной с лекарством явной интоксикации не наблюдалось, за исключением потери веса <5% в группе, обработанной паклитакселом. В группе, к которой применялась комбинированная терапия, потеря веса была даже меньшей. Эти положительные результаты комбинированной терапии тем более удивительны, что между химиотерапевтическим (и потенциально опасным для иммунной системы) лечением и началом лечения, основанного на возможности стимулировать пролиферацию и цитотоксичность лимфоцитов, проходило сравнительно короткое время.
Единственное объяснение эффекта комбинированного действия состоит в том, что вызванный таксаном апоптоз части растущей массы опухоли уменьшает внутритканевое давление, что, в свою очередь, увеличивает эффективное поглощение KS-IL2 в опухоли. Недавние исследования (Griffon-Etienne et al. 1999, CANCER RES. 59: 3776-3782) свидетельствуют, что действие единственной дозы паклитаксела состоит в эффективном снижении давления внутритканевой жидкости, причем максимальный эффект наблюдается от 24 до 48 часов (Griffon-Etienne et al. 1999, CANCER RES. 59: 3776-3782). Хотя этот эффект может приводить к созданию наилучших условий для поглощения иммуноцитокина в опухоли, это слишком короткий временной интервал после химиотерапии. Тем не менее мы обрабатывали KS-IL2 мышей-носителей опухоли LLC/KSA пять последующих дней, начиная точно через 24 часа после введения им единственной дозы паклитаксела. Результаты свидетельствуют, что возникает даже лучшая комбинированная реакция, если лечение иммуноцитокином начинать раньше, чем через неделю после введения единственной дозы паклитаксела как с этой линией опухоли, так и в случае колонии карциномы СТ26 (см. ниже).
Пример 5. Комбинированная терапия KS-IL2 и таксаном метастазов 4Т1.
Поскольку мы обнаружили, что лечебные интервалы между назначениями таксана и иммуноцитокина могут быть короче, чем ожидалось, было проведено тестирование комбинированных режимов, при которых таксан и иммуноцитокин вводили в один и тот же день, при этом сопоставляли эффект от введения единственной дозы (75 мг/кг) паклитаксела с действием фракционированной дозы (25 мг/кг × 3 дня), которые назначали одновременно с лечением KS-IL2 (15 мкг/доза × 3 дня, вводили через 4 часа после паклитаксела). В этих экспериментах использовали экспериментальную модель легочных метастазов, индуцированных клетками карциномы молочной железы 4T1/KSA. Дозы лекарственных препаратов выбирали таким образом, чтобы они были практически оптимальными сами по себе, так, что можно было наблюдать любое потенциальное улучшение или синергическую активность.
Каждый агент, введенный самостоятельно, значительно (р<0,02) уменьшал средний вес легких примерно в одинаковой мере: уменьшение на 43% наблюдалось при введении единственной дозы паклитаксела и уменьшение на 49% в случае многократных доз паклитаксела, который вводили самостоятельно, и уменьшение на 39% наблюдали при самостоятельном введении KS-IL2 (фиг.4). Комбинирование паклитаксела и KS-IL2 приводило к еще небольшому уменьшению легочных метастазов, но эффект был менее аддитивного: уменьшение на 58% наблюдалось в случае единственной дозы паклитаксела в комбинации с KS-IL2 и уменьшение на 68% имело место при многократном введении паклитаксела в комбинации с KS-IL2. Несмотря на то, что синергизма не наблюдалось, единственная доза паклитаксела в комбинации с KS-IL2 приводила к значительному отличию от варианта самостоятельного назначения паклитаксела (р=0,047).
Во всех группах наблюдалась потеря веса менее 10%, тем не менее самая большая потеря веса имела место в случае, когда 25 мг/кг паклитаксела вводили 3 раза, через день. На основании этих данных можно сделать вывод, что наилучшей программой в данном анализе легочных метастазов 4Т1 в плане наибольшего эффекта от комбинированной терапии оказался вариант введения единственной дозы паклитаксела с последующим назначением KS-IL2, как было и в случае регрессионной модели роста опухоли LLC/KSA. Поскольку дозировочный интервал в данном случае составлял только 4 часа, результаты могли не быть оптимальными для эффективного поглощения опухолью.
Пример 6. Комбинированная терапия KS-IL2 и таксаном кожной опухоли СТ26.
Результаты, описанные в примере 5, приводят к предположению, что временной интервал 4 часа между дозами двух агентов может быть слишком коротким. Возможно, что уровни паклитаксела, все еще остающиеся в животном на момент введения KS-IL2, могли служить прямым препятствием активации лимфоцитов, таким образом, снижая их потенциальную антиопухолевую активность при комбинированном введении препаратов. Кроме того, на момент через 4 часа после введения паклитаксела его максимальный эффект в аспекте внутритканевого давления еще не мог быть достигнут. Поэтому мы спланировали другой эксперимент, на этот раз с использованием упрочившейся кожной опухоли колонии карциномы CT26/KSA, в котором мы комбинировали единственную дозу паклитаксела (75 мг/кг) с 5-дневным курсом KS-IL2, который начинали через 24 часа после назначения таксана. При самостоятельном введении паклитаксел не оказывал влияния на рост опухоли (фиг.5). Лечение субоптимальными дозами KS-IL2 (10 мкг, Дни 1-5) приводило к объемам опухоли, которые составляли 71% от объемов опухолей в контрольной группе. Резкое и синергическое уменьшение объема опухоли - до 8% объема контрольных образцов - наблюдалось при комбинировании паклитаксела и KS-IL2; этот результат сильно отличается от варианта самостоятельного применения паклитаксела (р<0,001). Минимальная потеря веса ˜5% наблюдалась в обеих обработанных паклитакселом группах.
Второй эксперимент провели с использованием модели CT26/KSA, на этот раз произвели тестирование действия комбинированной терапии на упрочившиеся метастазы в печени и опять использовали перерыв 24 часа между введением паклитаксела и назначением KS-IL2. Мы также произвели сравнение эффектов в зависимости от дозы паклитаксела при комбинированной терапии. Мышам делали инъекции 25, 50 или 75 мг/кг паклитаксела в День 5 после внедрения метастазов, самостоятельно или с назначаемым днем позже лечением KS-IL2 (7 мкг) в течение 5 дней. Реакцию на дозу паклитаксела проверяли сначала при его самостоятельном применении, в этом эксперименте введение мышам 25, 50, 75 мг/кг препарата приводило к снижению веса опухоли соответственно до 49%, 23%, 10% веса опухоли контрольных образцов (фиг.6). Комбинирование паклитаксела с KS-IL2 обеспечивало дальнейшее уменьшение легочных метастазов - до 12%, 9% и 6% веса опухоли контрольных образцов соответственно при тех же дозах паклитаксела. Самая низкая доза паклитаксела (25 мг/кг) в комбинации с KS-IL2 приводила к самому большому и наиболее значительному (р<0,001) уменьшению веса опухоли по сравнению с более высокими дозами паклитаксела в комбинации с KS-IL2. Таким образом, комбинация KS-IL2 с предшествующим введением паклитаксела приведет к большему противоопухолевому эффекту, чем самостоятельное применение любого из этих агентов. Далее, самая низкая доза паклитаксела в комбинации с KS-IL2 давала такой же противоопухолевый эффект, что и самая высокая доза паклитаксела при его самостоятельном введении. Следовательно, применение меньших доз паклитаксела в комбинации с KS-IL2 должно обеспечивать снижение токсичности при сохранении хорошей эффективности.
Пример 7. Измерение поглощения KS-IL2 в опухолях.
Эффект от обработки единственной дозой цитотоксичного лекарственного препарата, введенного перед проведением терапии иммуноцитокином, состоит в снижении внутритканевого давления в опухоли и увеличении проницаемости опухолей, что должно поддаваться измерениям при помощи маркированного радиоактивным изотопом иммуноцитокина, т.е. KS-IL2. Маркировку очищенного KS-IL2 изотопом произвели по стандартной методике (ссылка) путем контакта с имеющимся в продаже125I (New England Nuclear, Billerica, MA). Кожные опухоли CT26/KSA имплантировали подкожно по методике, описанной в примере 1, и дали им разрастись до объемов порядка 100-200 мм3. Двум группам из 4 мышей каждая сделали инъекции либо паклитаксела (50 мг/кг) в носителе, либо одного носителя, после чего через 1 час (эксперимент 1) или через 24 часа (эксперимент 2) ввели 10 мкг125I-KS-IL2 (95 мкCi). Через шесть часов после инъекции маркированного радиоактивным изотопом иммуноцитокина мышей умертвили и их опухоли удалили хирургическим путем. В качестве контроля печени животных тоже собрали и все ткани взвесили, затем произвели измерения при помощи счетчика гамма частиц. Результаты были выражены как количество импульсов в минуту (СРМ) на грамм ткани путем деления суммарного СРМ в ткани на ее вес.
Когда маркированный KS-IL2 вводили через 1 час после обработки паклитакселом (фиг.7А), в опухолях, вырезанных у получивших лекарство животных, наблюдалось только небольшое увеличение радиоактивности. И наоборот, когда маркированный KS-IL2 вводили через 24 часа после обработки паклитакселом, наблюдалось резкое увеличение поглощения (>200%) по сравнению с контрольной группой, получившей носитель (фиг.7Б). Это большое различие между поглощением опухолью во временных точках 1 час и 24 часа находится в соответствии с данными по обусловленным таксаном изменениям внутритканевого давления (Griffon-Etienne et al. 1999, CANCER RES. 59: 3776-3782) и согласуется с данными, полученными в наших моделях опухоли, которые свидетельствуют, что начало лечения через 24 часа после паклитаксела более эффективно, чем если начать его раньше (4 часа).
Мы также провели тестирование других классов лекарственных препаратов, имея целью выяснить, могут ли они увеличивать поглощение маркированного иммуноцитокина твердой опухолью. В этом случае мышам делали инъекцию единственной дозы циклофосфамида (40 мг/кг) либо за 24 часа, либо за 3 дня до начала эксперимента. Инъекцию KS-IL2, маркированного125I, делали всем мышам, включая контрольную группу, предварительно PBS, и через 16 часов измеряли радиоактивность вырезанных опухолей. Результаты (фиг.8) свидетельствуют, что предварительная обработка циклофосфамидом увеличила поглощение KS-IL2 на 48% у мышей, предварительная обработка которых была произведена за 24 часа до начала эксперимента, и на 70% у мышей, предварительная обработка которых была произведена за 3 дня.
Пример 8. Комбинированная терапия huKS-huγ4-IL2 и таксаном
Недавно были описаны новые формы иммуноцитокинов, для которых характерны увеличенные периоды полувыведения и повышенная эффективность, что связано с уменьшенным сродством к рецепторам Fc (см. Gillies et al. 1999, CANCER RES. 59: 2159-2166). Было проведено тестирование одного представителя из этой группы улучшенных иммуноцитокинов IL-2, huKS-huγ4-IL2, в комбинированной терапии с единственной дозой паклитаксела. В этом случае тоже наблюдалось повышение эффективности, когда два лекарственных препарата последовательно вводили мышам-носителям кожной опухоли CT26/KSA.
Пример 9. Комбинированная терапия huKS-muγ2a-muIL12 и таксаном.
Чтобы проверить, ограничивается ли синергический терапевтический эффект только иммуноцитокинами, основанными на IL-2, мы сначала обработали большие упрочившиеся опухоли CT26/KSA паклитакселом (единственная доза 75 мг/кг), а через 24 после этой инъекции провели 5-дневный курс лечения препаратом huKS-muγ2a-muIL12 (5 мкг в день). Этот иммуноцитокин представляет собой продукт слияния мышиной формы антитела HuKS (т.е. мышиным С каппа и С гамма 2а были возвращены постоянные области) и мышиной IL-12. Необходимо было использовать мышиные последовательности IL-12, поскольку, в отличие от IL-2, этот цитокин в высокой мере специфичен к биологическому виду, и его человеческая форма проявляет слабую активность в мыши. Результаты свидетельствуют, что лечение одним паклитакселом оказывает весьма небольшое воздействие на рост опухоли. Лечение субоптимальными дозами huKS-muγ2a-muIL12 давало противоопухолевый эффект и он увеличивался у мышей, которых предварительно обрабатывали единственной дозой паклитаксела.
Пример 10. Комбинированная терапия huKS-IL2 и алкилирующим агентом.
i. Был продемонстрирован также повышенный терапевтический эффект комбинации huKS-IL2 с циклофосфамидом, химиотерапевтическим препаратом из группы алкилирующих агентов. За 3 дня до начала лечения иммунодостаточным мышам сделали инъекции клеток карциномы молочной железы 4Т1, чтобы спровоцировать легочные метастазы. Мышам ввели одну дозу циклофосфамида (15, 40 или 80 мг/кг), а через три дня после этого провели 5-дневный курс лечения huKS-IL2 (15 мкг/день). Несмотря на то, что две самые маленькие дозы циклофосфамида при самостоятельном применении приводили только к умеренному уменьшению веса легочных метастазов опухоли, комбинация с huKS-IL2 обеспечила значительно большее уменьшение веса опухоли по сравнению с самостоятельным применением циклофосфамида (р<0,05, фиг.9). Тем не менее при самой высокой дозе (80 мг/кг) синергии не наблюдалось.
ii. Повышенный терапевтический эффект комбинации huKS-IL2 с циклофосфамидом был также подтвержден анализом роста опухоли в иммунодостаточных мышах-носителях упрочившихся подкожных опухолей карциномы молочной железы. Мышей обрабатывали единственной дозой 80 мг/кг циклофосфамида либо самостоятельно, либо в комбинации с 5 девными дозами huKS-IL2 (30 мкг), который начинали вводить через 3 дня после введения циклофосфамида. Средние объемы опухолей в случаях раздельного лечения huKS-IL2 и одной дозой 80 мг/кг циклофосфамида уменьшались на 31% и 69% соответственно (фиг.9Б). Комбинированное лечение обеспечивало уменьшение среднего размера опухоли на 100% на День 25, что значительно отличалось от результатов самостоятельного применения как huKS-IL2, так и циклофосфамида (р<0,05), и полностью ликвидировало опухоли у шести мышей из восьми через двенадцать недель после начала лечения. Животные переносили это лечение хорошо, во всех группах наблюдалась потеря веса менее 10%.
iii. Повышенный терапевтический эффект комбинации huKS-IL2 с циклофосфамидом был подтвержден также анализом роста опухоли, проведенным на иммунодостаточных мышах-носителях упрочившихся подкожных опухолей карциномы легких. Мышей обрабатывали единственной дозой 80 мг/кг циклофосфамида либо самостоятельно, либо в комбинации с 5 дневными дозами huKS-IL2 (20 мкг), которые начинали вводить через 3 дня после лечения циклофосфамидом. При самостоятельном применении huKS-IL2 и 80 мг/кг циклофосфамида средние объемы опухоли уменьшались на 2% и 27% соответственно (фиг.9В). Комбинированное лечение уменьшает средние размеры опухолей 48% на День 20, что значительно отличается от результатов самостоятельного применения как huKS-IL2, так и циклофосфамида (р<0,05). Животные переносили это лечение хорошо, во всех группах наблюдалась потеря веса менее 10%.
Пример 11. Комбинированная терапия huKS-IL2 и алкилирующим агентом.
Продемонстрирован повышенный терапевтический эффект комбинации huKS-IL2 с карбоплатином, другим химиотерапевтическим препаратом из группы алкилирующих агентов. Мышей-носителей упрочившихся подкожных опухолей карциномы легких с немаленькими клетками (LLC/KSA) обработали карбоплатином (75 мг/кг) в День 0, а через три дня после этого провели 5-дневный курс лечения KS-IL2 (20 мкг в день). Раздельное лечение карбоплатином и KS-IL2 приводило к умеренному торможению роста опухоли, тем не менее только комбинированное лечение приводило к значительному уменьшению среднего объема опухолей на День 20 (р<0,05, фиг.10). Кроме того, рост опухолей, при котором мышей обрабатывали комбинацией названных препаратов, сильно отличался от случая самостоятельного применения карбоплатина (р<0,05).
Изобретение может быть воплощено в других конкретных формах без отклонений от его сущности или неотъемлемой специфики. Приведенные выше воплощения следует, таким образом, рассматривать во всех отношениях скорее как иллюстративные, чем как ограничивающие описанное изобретение. Широта данного изобретения более определена приведенной ниже формулой изобретения, чем изложенным выше его описанием, и все изменения значения и диапазона эквивалентности в формуле предназначены для охвата этой цели.
Каждый из патентных документов и научные публикации введены в текст описания изобретения посредством ссылок.
Реферат
Изобретение относится к области иммунологии. Сущность его заключается в разработке способа и композиции, способствующей повышению поглощения иммуноцитокинов в опухолях. Композиции основаны на комбинировании цитокина с агентом, увеличивающим поглощение иммуноцитокина. Технический результат - повышение эффективности терапии опухолей и метастазов у млекопитающих. 2 н. и 25 з.п. ф-лы, 13 ил., 1 табл.

Комментарии