Композиции для индукции гаплоидии и способы их применения - RU2771141C2
Код документа: RU2771141C2
Чертежи


















































































































































































Описание
ИСПРАШИВАНИЕ ПРИОРИТЕТА
[0001] Настоящая заявка испрашивает приоритет в соответствии с §120 раздела 35 U.S.C. по предварительной заявке на патент США №62/256902, поданной 18 ноября 2015 г., и предварительной заявке на патент США №62/300507, поданной 26 февраля 2016 г., содержание которых включено в данный документ посредством ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] К настоящей заявке прилагается перечень последовательностей под названием 80906-PCT_ST25.txt, созданный 16 ноября 2016 г., размер которого составляет примерно 392 килобайта. Этот перечень последовательностей включен в данный документ посредством ссылки во всей своей полноте. Этот перечень последовательностей подается вместе с настоящей заявкой посредством EFS-Web и соответствует §1.824(а)(2)-(6) и (b) раздела 37 C.F.R.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[0003] Раскрытый в данном документе объект изобретения относится к диагностическому выявлению индукции гаплоидии ("HI") или ее отсутствия и/или присутствия у растений, которые являются индукторами гаплоидии или не являются таковыми. Более конкретно, раскрытый в данном документе объект изобретения относится к нуклеиновым кислотам, которые можно использовать для индуцирования HI у растений, и/или видам биологической активности, которые можно модифицировать для получения или предупреждения HI у растения, которое в противном случае будет демонстрировать HI, либо у растения, которое в противном случае не будет демонстрировать HI. Еще более конкретно, раскрытый в данном документе объект изобретения относится к молекуле нуклеиновой кислоты, которая кодирует биологически активную молекулу, а также к способам применения ее для регулирования HI у растений.
[0004] В данном документе предусматриваются серия независимых индуцированных человеком мутаций, обнаруживаемых по меньшей мере в одном гене пататин-подобной фосфолипазы А2α ("PLA") маиса; растения маиса, имеющие эти мутации по меньшей мере в одном из их генов PLA; и способ создания и идентификации сходных и/или дополнительных мутаций в гене PLA посредством скрининга объединенных и/или отдельных растений маиса. Растения маиса по настоящему изобретению индуцируют гаплоидию в результате нетрансгенных мутаций по меньшей мере в одном из их генов PLA. Также предусматриваются способы индуцирования de novo индукции гаплоидии посредством нанесения химических веществ на репродуктивные ткани растений. Также предусматриваются способы увеличения уровня завязывания семян и уровня индукции гаплоидии ("HIR", определенного в данном документе как количество выживших гаплоидных зерен по сравнению с общим количеством зерен после опыления початка пыльцой, индуцирующей гаплоидию) посредством нанесения химических веществ на репродуктивные ткани растения во время опыления.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[0005] Опыление является сложным процессом. Пыльцевые зерна покрытосеменных состоят из крупной вегетативной клетки и двух мужских гамет (спермиев). После попадания на дыхальце из зерна прорастает пыльцевая трубка, которая демонстрирует быстрый концевой рост по мере перемещения вниз по женским передающим путям под воздействием хемоаттрактантов, секретируемых двумя синергидными клетками на микропилярном конце зародышевого мешка. Во время прохождения вниз по трубке спермин соединены друг с другом и с вегетативным ядром посредством тяжистой цитоплазмы, называемой мужской зародышевой единицей. Вскоре после контакта с одной из двух синергид пыльцевая трубка разрывается, и два спермия продвигаются через цитоплазму умирающей синергидной клетки, чтобы независимо слиться с яйцеклеткой и центральными клетками зародышевого мешка, завершая двойное оплодотворение. Даже после первоначального контакта неудачные события оплодотворения могут быть исправлены второй пыльцевой трубкой, которая оплодотворяет зародышевый мешок посредством взаимодействия с персистентной синергидной клеткой.
[0006] Селекционеры скрещивают инбредные родительские линии, при этом одна выступает в качестве мужской особи, а одна - в качестве женской особи, для образования гибридных семян. Процесс разработки инбредных родительских линий, которые являются практически гомозиготными, обычно требует выбора гибридного продукта скрещивания и его самоопыления (самооплодотворения) в течение множества поколений для того, чтобы он стал почти гомозиготным. Этот процесс является времязатратным и дорогостоящим. Для сокращения времени для разработки гомозиготных инбредных растений маиса, риса, пшеницы, ячменя и других сельскохозяйственных культур селекционеры могут предпочесть использовать линию, индуцирующую гаплоидию, для индукции образования гаплоидных семян у гибридной родительской особи. Число хромосом гаплоидных растений затем удваивают, например, посредством средства, удваивающего число хромосом, такого как колхицин, с образованием двойных гаплоидных гомозиготных инбредных линий.
[0007] Индукция гаплоидии ("HI") представляет собой класс явлений у растений, характеризующийся утратой хромосом индуктора в ходе развития зародыша. В WO 2012/030893, включенном в данный документ посредством ссылки, описана область хромосомы 1 маиса, которая может отвечать за индукцию гаплоидии. Идентифицированные маркеры в этой области, увеличивающие индукцию гаплоидии, описаны как находящиеся в положениях 48249509-51199249, которые соответствуют имеющемуся в открытом доступе маркеру umc1169, имеющему физическое положение (60213661). Эта область, по-видимому, не выравнивается с областью индукции гаплоидии у Stock 6. В Dong et al., (2013) Theor. Appl. Genet. 126: 1713-1720 описан QTL, расположенный в секции 1.04, который объясняет до 66% генотипической дисперсии включительно для HIR.
[0008] Индукцию гаплоидии наблюдали у множества видов растений, таких как сорго, ячмень, пшеница и другие злаковые растения. У маиса HI, по-видимому, является результатом реаранжировки, мутаций и/или рекомбинаций, вставки или делеций в пределах области хромосомы 1 (с существенным исключением для индукции гаплоидии ig-типа, которая является результатом мутации в гене INDETERMINATE GAMETOPHYTE1 в хромосоме 3). Предусматриваемые линии HI были изучены и приближенно идентифицированы. Однако не были представлены экспериментальные данные, демонстрирующие генетический причинный фактор HI у маиса. Также ранее не были идентифицированы перечисленные в данном документе маркеры, которые ассоциированы с этим признаком.
[0009] У маиса гаплоидные семена или зародыши, в частности, образуются посредством осуществления скрещиваний между мужской особью, индуцирующей гаплоидию (т.е. "пыльцой, индуцирующей гаплоидию"), и практически любым выбранным початком, при этом початок может быть получен из любой инбредной, гибридной или другой зародышевой плазмы. Гаплоиды образуются, если ДНК пыльцы, индуцирующей гаплоидию, не полностью передается и/или сохраняется на протяжении первых клеточных делений зародышей. Полученный в результате фенотип не является полностью пенетрантным, при этом некоторые семязачатки содержат гаплоидные зародыши, а другие содержат диплоидные зародыши, анеуплоидные зародыши, химерные зародыши или недоразвитые зародыши. Гаплоидные зерна имеют зародыши, которые содержат только материнскую ДНК вместе с нормальным триплоидным эндоспермом. После индукции гаплоидии гаплоидные зародыши или семена, как правило, отделяют от диплоидных и анеуплоидных сибсов с помощью скрининга по фенотипическим или генетическим маркерам, и из них выращивают или культивируют гаплоидные растения. Эти растения затем превращают естественным путем либо посредством химической манипуляции (т.е. с помощью колхицина) в двойные гаплоидные (DH) растения, которые затем образуют инбредные семена.
[0010] Линии HI содержат локус количественного признака ("QTL") в хромосоме 1, отвечающий по меньшей мере за 66% изменчивости индукции гаплоидии. QTL вызывает индукцию гаплоидии на различных уровнях, если он интрогрессирован в различные окружения. Все линии, индуцирующие гаплоидию, применяемые в промышленном семеноводстве, являются производными линии-основателя HI, известной как Stock6, и все они имеют мутацию в QTL хромосомы 1, индуцирующую гаплоидию. В данном документе авторы настоящего изобретения раскрывают ключевую мутацию в QTL, при комплементации которой восстанавливается нормальное размножение. Несмотря на то, что причины этой мутации являются неясными, она возникает во всех индуцирующих линиях, в том числе Stock6.
[0011] Селекция растений облегчается посредством применения двойных гаплоидных (DH) растений. Получение DH-растений позволяет селекционерам-растениеводам получать инбредные линии без инбридинга в нескольких поколениях, сокращая таким образом время, необходимое для получения гомозиготных растений. DH-растения обеспечивают бесценный инструмент для селекционеров-растениеводов, в частности для получения инбредных линий, картирования QTL, цитоплазматических превращений, интрогрессии признаков и скрининга F2 в отношении высокопроизводительного улучшения признака. Экономится большое количество времени, поскольку гомозиготные линии образуются по существу за одно поколение с исключением необходимости в традиционном инбридинге в нескольких поколениях. В частности, поскольку DH-растения являются полностью гомозиготными, они хорошо поддаются количественным генетическим исследованиям. Образование гаплоидных семян является критически важным для процесса селекции двойных гаплоидов. Гаплоидные семена образуются в материнской зародышевой плазме при оплодотворении пыльцой от гиногенетического индуктора, такого как Stock 6.
[0012] Растения маиса, индуцирующие гаплоидию, образуют пыльцу, которая при скрещивании с неиндуцирующей зародышевой плазмой обуславливает гиногенетическое развитие гаплоидных семян. К сожалению, в результате этого процесса часто получают низкую частоту гаплоидных зерен. Неэффективная частота индукции гаплоидии является ограничивающим фактором в программах селекции двойных гаплоидов маиса.
КРАТКОЕ ОПИСАНИЕ
[0013] Высокий HIR обеспечивает большую частоту образования гаплоидных семян родительским растением, представляющим интерес. Родительские растения можно подвергнуть предварительному скринингу с использованием генетических маркеров, ассоциированных с необходимыми признаками, или фенотипически наблюдаемых признаков для обогащения генетического потенциала родительских растений. Если эти желаемые родительские растения опыляют индуктором гаплоидии, который характеризуется более высоким HIR, то получают более высокий потенциал получения желаемых двойных гаплоидов с необходимым генотипом и фенотипом.
[0014] Несмотря на то, что процесс селекции двойных гаплоидов приводил в результате к более быстрому получению гомозиготных инбредных растений, объем получаемых двойных гаплоидных инбредных растений может быть ограничен. Известные индуцирующие линии включают без ограничения Stock 6, MHI (молдавский индуктор гаплоидии), линию с мутацией неопределенного гаметофита ("ig"), KEMS, RWK, ZEM, ZMS и KMS. Все из них характеризуются относительно низким HIR. Stock 6, например, индуцирует образование только 1-3% гаплоидных семян. В силу этого индукция образования гаплоидов представляет собой ограничивающую уровень стадию в процессе получения двойных гаплоидных линий.
[0015] Авторы настоящего изобретения изобрели способ индукции получения гаплоидов и/или увеличения уровня индукции гаплоидии у растений посредством обработки растений липидным соединением, ингибитором фосфолипаз и ингибитором десатураз жирных кислот. Одна такая группа способов включает нанесение специфических химических веществ на репродуктивные ткани при скрещивании с пыльцой дикого типа (не индуцирующей гаплоидию). Авторы настоящего изобретения впервые химическим путем вызвали индукцию гаплоидии de novo. Это осуществляется посредством введения ингибитора фосфолипаз метил-альфа-линоленоилфторфосфоната (MALFP) в определенной концентрации в цветок во время опыления, что приводит к высокому уровню индукции гаплоидии, составляющему до 9% HIR включительно. Кроме того, авторы настоящего изобретения вызвали индукцию гаплоидии de novo посредством введения арахидонилфторфосфоната (MAFP) в определенной концентрации в цветок во время опыления. Кроме того, авторы настоящего изобретения вызвали индукцию гаплоидии de novo посредством введения 1,2-дистеароил-sn-глицеро-3-фосфатидилхолина (также известного как дистеарилфосфатидилхолин; "DSPC") в определенной концентрации в цветок во время опыления. Авторы настоящего изобретения также вызвали индукцию гаплоидии de novo посредством введения альфа-линоленовой кислоты в определенной концентрации в цветок во время опыления. Авторы настоящего изобретения также вызвали индукцию гаплоидии de novo посредством введения легкодоступных соединений, в том числе кукурузного масла и льняного масла, а также химически синтезированных линолевой кислоты, этилового сложного эфира олеиновой кислоты (ОАЕЕ), метилового сложного эфира арахидоновой кислоты (ААМЕ) и ингибитора фосфолипаз маноалида. Авторы настоящего изобретения также увеличили уровень индукции гаплоидии при скрещивании растений с линией, индуцирующей гаплоидию (например, RWK, Stock 6 или ZMS), посредством введения ингибитора фосфолипаз метил-альфа-линоленоилфторфосфоната (MALFP) в определенной концентрации. При концентрациях 2% MALFP+смесь поверхностно-активных веществ 91, эмульгированных буферным раствором DML, авторы настоящего изобретения удвоили уровень индукции гаплоидии в опылениях с применением RWK в качестве опылителя. Типичный уровень индукции для RWK составляет приблизительно 10-18%. При нанесении MALFP уровень индукции увеличивается до 20-35%. Авторы настоящего изобретения также повышали уровень индукции гаплоидии посредством нанесения линолевой кислоты (LLE), этилового сложного эфира линолевой кислоты (LLAEE) и ингибитора фосфолипаз, называемого метиларахидонилфторфосфонатом (MAFP), в определенных концентрациях на цветок во время опыления. Кроме того, при выполнении этого таким образом также снижается уровень недоразвитости зародышей и увеличивается уровень образования зерен для MALFP, LLAEE, MAFP и LLE. В совокупности эти эффекты приводят к увеличению общего количества гаплоидных зародышей, воспроизводящихся на початке после опыления.
[0016] Авторы настоящего изобретения также клонировали ген, отвечающий за индукцию гаплоидии у маиса, и определили его характеристики. Ген имеет название PLA2 и характеризуется экспрессией, специфической для пыльцы. Белок PLA2, по-видимому, локализуется в цитоплазме спермия, возможно, в эндоплазматическом ретикулуме или тельцах Гольджи. Идентификация гена привела к изобретениям нескольких новых методик улучшения процесса индукции гаплоидии, определяемых в качестве действий по получению гаплоидных зародышей, зерен, семян или растений посредством скрещивания любого початка с пыльцой, индуцирующей гаплоидию. Идентификация гена также привела к изобретениям новых способов индукции образования гаплоидов. Другая группа включает способы создания новых линий, индуцирующих гаплоидию, путем изменения последовательности причинного гена посредством направленного мутагенеза, TILLING или CRISPR/Cas9. Экспрессия белка PLA2 может быть подавлена с помощью RNAi или посредством применения направленного мутагенеза в промоторе, 3'-UTR, 5'-UTR или сайтах сплайсинга.
[0017] С учетом идентификации этой мутации авторы настоящего изобретения изобрели способы модификации и улучшения процесса индукции гаплоидии, и авторы настоящего изобретения впервые раскрывают способы получения гаплоидов de novo посредством химической обработки во время опыления. Авторы настоящего изобретения демонстрируют способы увеличения уровня индукции гаплоидии ("HIR," т.е. процентного количества гаплоидных зародышей, обнаруживаемых на указанном початке, подвергнутом индукции гаплоидии), а также способы увеличения уровня выживаемости зерен в ходе индукции гаплоидии. Авторы настоящего изобретения также обсуждают способы создания новых линий, индуцирующих гаплоидию, с применением стратегий генетической модификации ("GM") или направленного мутагенеза.
[0018] Процесс индукции гаплоидии можно улучшить посредством множества способов. Во-первых, можно стремиться улучшить средний HIR. HIR имеет ограниченный уровень при крупномасштабном получении двойных гаплоидных ("DH") растений, поскольку HIR является относительно низким. См. фигуру 1. За редкими исключениями, HIR обычно составляет менее 25% и наиболее часто находится в диапазоне 10-20%, что означает, что 75-90% зерен являются диплоидными. С некоторой низкой частотой в ходе индукции гаплоидии также образуются анеуплоидные зародыши. Примеры соединений, которые, как было показано, увеличивают количество образующихся гаплоидов и/или уровень образования гаплоидов в ходе скрещиваний с индуктором гаплоидии без увеличения уровня недоразвитости зародышей, включают метил-альфа-линоленоилфторфосфонат (MALFP), метиларахидонилфторфосфонат (MAFP), линолевую кислоту (LLA) и этиловый сложный эфир линолевой кислоты (LLAEE). Во-вторых, можно уменьшить уровень недоразвитости зерен и частоту неудачного оплодотворения в ходе индукции гаплоидии, что приводит к образованию большего количества зерен на один початок. Недоразвитость зерна возникает после того, как происходит успешное оплодотворение, но при этом не могут развиться функциональные зародыш или эндосперм, и в результате вырастает небольшое, бесцветное зерно, не имеющее внутри зрелого зародыша. Уровень недоразвитости зерен является высоким в процессах индукции гаплоидии - 10-50% оплодотворенных семязачатков в початке. Это ограничивает количество гаплоидных зерен, которое может воспроизводиться на один початок. Неудачное оплодотворение происходит, если семязачаток неспособен быть оплодотворен пыльцевым зерном, и характеризуется отсутствием какого-либо развития зерен после опыления. Этот уровень в большинстве случаев оказывается высоким в ходе индукции гаплоидии - 10-70% зерен в некоторых початках, подвергнутых индукции гаплоидии, неспособны быть оплодотворены, в зависимости от синхронности созревания мужского и женского цветков, типа осуществляемого скрещивания и генетических характеристик мужской и женской особей. Примеры соединений, которые, как было показано, увеличивают уровень образования жизнеспособных зерен посредством снижения частоты неудачного оплодотворения и уровня недоразвитости зародышей, включают те же четыре соединения, упомянутые выше: MALFP, MAFP, LLA и LLAEE. Эти четыре молекулы при нанесении на пыльцу, метелки или другие части цветка в ходе скрещиваний для индукции гаплоидии, увеличивают количество образующихся гаплоидов посредством увеличения как уровня индукции гаплоидии, так и числа зерен.
[0019] И наконец, можно улучшить процесс индукции гаплоидии с исключением необходимости в мужских особях, индуцирующих гаплоидию, посредством обеспечения возникновения индукции гаплоидии de novo на початке или в семязачатке при его самоскрещивании или скрещивании с пыльцой от линии, не индуцирующей гаплоидию. Эти способы индукции гаплоидии de novo, в том числе путем нанесения фосфолипидов, таких как DSPC, жирных кислот, таких как линоленовая кислота (LNA), широко известных смесей триглицеридов, таких как кукурузное масло и льняное масло, или ингибиторов фосфолипаз, таких как MALFP, можно применять при любом ауткроссе или самоопылении для индукции образования гаплоидов у кукурузы. В данном документе авторы настоящего изобретения описывают примеры, которые относятся к одной или нескольким из этих категорий, представляющие улучшения процесса индукции гаплоидии или новые изобретения в нем.
[0020] Настоящее изобретение относится к способу индуцирования образования гаплоидных зародышей при скрещивании между двумя родительскими растениями. Это осуществляют путем изменения экспрессии фосфолипазы в одном из родительских растений. Это изменение можно осуществлять несколькими путями: либо вызывая экспрессию мутантной фосфолипазы одним из родительских растений; либо посредством введения малой интерферирующей РНК в одно или оба родительских растения, что вызывает подавление экспрессии фосфолипазы; либо посредством трансформации одного из родительских растений мутантной фосфолипазой; либо посредством редактирования фосфолипазы одного из родительских растений, например, посредством сайт-направленного мутагенеза, такого как технологии на основе CRISPR или TALEN. Если экспрессия фосфолипазы в одном из родительских растения изменена посредством одной из этих методик, то в случае, если это родительское растение применяется в скрещивании, образуется по меньшей мере один гаплоидный зародыш.
[0021] В одном варианте осуществления способа фосфолипаза представляет собой пататин-подобную фосфолипазу. В другом варианте осуществления пататин-подобная фосфолипаза является ортологом pPLAIIα, которая кодируется нуклеотидной последовательностью, содержащей SEQ ID NO: 1, или последовательностью, по меньшей мере на 70% идентичной SEQ ID NO: 1. Нуклеотидная последовательность, кодирующая пататин-подобную фосфолипазу, может быть подвергнута мутации, и в одном варианте осуществления нуклеотидная последовательность имеет мутацию сдвига рамки считывания, при которой создается искусственный стоп-кодон. Последовательность с мутацией сдвига рамки считывания содержит SEQ ID NO: 3 или последовательность, по меньшей мере на 70% идентичную SEQ ID NO: 3.
[0022] Другие мутации являются возможными и находятся в пределах объема настоящего изобретения. Сайт-направленный мутагенез можно применять для создания большего количества мутаций фосфолипазы. Применение CRISPR/Cas9, TALEN, "цинковых пальцев" и мегануклеаз является способами осуществления сайт-направленного мутагенеза в соответствии с вариантами осуществления настоящего изобретения.
[0023] Настоящее изобретение является применимым при многих типах скрещиваний между растениями. В одном варианте осуществления родительские растения, применяемые в скрещивании, являются однодольными растениями, такими как маис, рис, ячмень и пшеница. Родительские растения могут относиться к одному и тому же виду однодольных, или же они могут относиться к разным видам. В другом варианте осуществления родительские растения, применяемые в скрещивании, представляют собой двудольные растения, такие как соя, подсолнечник, томат, перец, сахарная свекла или разновидности брюссельской капусты. В предпочтительном варианте осуществления родительские растения представляют собой растения маиса или риса. В пределах объема настоящего изобретения находятся гаплоидный зародыш, полученный с помощью данного способа, гаплоидное семя, содержащее гаплоидный зародыш, и гаплоидное растение, выращенное из гаплоидного семени. Также в пределах объема настоящего изобретения находится двойной гаплоид, полученный посредством воздействия на гаплоидный зародыш средства, удваивающего число хромосом, такого как колхицин или трифлуралин.
[0024] Настоящее изобретение относится к кДНК, содержащей SEQ ID NO: 3, или последовательность, ортологичную SEQ ID NO: 3, или последовательность, на 70% идентичную SEQ ID NO: 3. В предпочтительных вариантах осуществления последовательность, ортологичная SEQ ID NO: 3, охватывает пататин-подобные фосфолипазы из маиса, риса, пшеницы, сои и подсолнечника. В частности, последовательность, ортологичная SEQ ID NO: 3, содержит ген Os03g27610 риса. В более предпочтительном варианте осуществления последовательность, ортологичная SEQ ID NO: 3, охватывает SEQ ID NO: 23 и 73-81.
[0025] Настоящее изобретение относится к растению, содержащему индуцированную человеком нетрансгенную мутацию в его гене пататин-подобной фосфолипазы. В предпочтительном варианте осуществления ген пататин-подобной фосфолипазы представляет собой pPLAIIα. В другом варианте осуществления мутация вызывает кодирование преждевременного стоп-кодона в гене. В более предпочтительном варианте осуществления растение представляет собой любое однодольное или любое двудольное растение, но особенно предпочтительными являются маис или рис.
[0026] Настоящее изобретение также относится к способу индуцирования образования гаплоидных зародышей и семян посредством обработки репродуктивных тканей растения соединением, включающим в себя липид или ингибитор фосфолипаз. В одном варианте осуществления обработку осуществляют до, во время или непосредственно после опыления. Обрабатываемые растения могут представлять собой любые однодольные или двудольные растения, но в предпочтительных вариантах осуществления растения представляют собой маис, рис, пшеницу, сою, подсолнечник и сахарную свеклу. В другом варианте осуществления липид или ингибитор фосфолипаз выбраны из группы, указанной в таблице 7. В предпочтительных вариантах осуществления соединение для обработки включает в себя метил-альфа-линоленоилфторфосфонат ("MALFP"), этиловый сложный эфир линолевой кислоты ("LLAEE"), линолевую кислоту ("LLA"), кукурузное масло, дистеарилфосфатидилхолин ("DSPC") или метиларахидонилфторфосфонат ("MAFP").
[0027] В другом варианте осуществления липид или ингибитор фосфолипаз соответствуют следующей формуле (I):
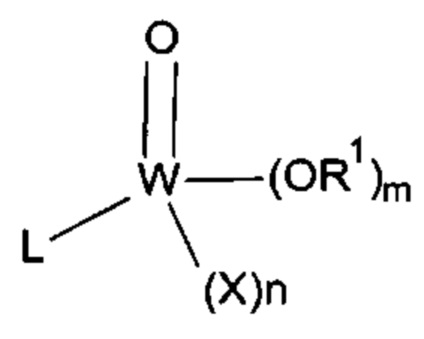
[0028] В предпочтительном варианте осуществления X представляет собой F, Cl, CF3, CCl3, CF2H, CCl2H, CF2CF3, CCl2CCl3, CF2Cl, CF2CH3, С(O)СН3 или CN. В более предпочтительном варианте осуществления галоген представляет собой F или Cl.
[0029] Важно отметить, что в R1 C1-С6алкил включает в себя линейные, разветвленные и циклические алкильные группы. В предпочтительном варианте осуществления R1 представляет собой C1-С6алкил, замещенный одной-шестью гидроксильными группами.
[0030] В одном варианте осуществления каждый L в формуле (I) независимо представляет собой углеродную цепь С2-C30, в том числе разветвленные цепи, которые могут быть насыщенными, ненасыщенными или полиненасыщенными. В предпочтительном варианте осуществления углеродная цепь L содержит от одной до четырех групп, независимо выбранных из алкенила, алкинила, фенила и гетероарила. Ненасыщенность представлена в виде двойных или тройных связей. Алкенил или алкинил могут находиться в пределах углеродной цепи или на конце углеродной цепи. Фенильные и/или гетероарильные кольца могут быть присоединены к углеродной цепи в орто-, мета- или пара-положениях или могут находиться на конце углеродной цепи. Арильные кольца могут быть необязательно замещены. В предпочтительном варианте осуществления углеродная цепь прерывается одним-шестью атомами кислорода. Как используется в данном документе, "прерывается" означает то, что углеродная цепь содержит по меньшей мере два атома углерода в последовательности, за которыми расположен атом кислорода. Например, -СН2-СН2-О-СН2-СН2-СН3 представляет собой углеродную цепь, прерываемую атомом кислорода. В предпочтительном варианте осуществления углеродная цепь прерывается одним-двумя атомами кислорода.
[0031] Примеры подходящих углеродных цепей, соответствующих требованиям L, включают (СН2)8-(СН)2-СН2-(СН)2-СН2-(СН)2-СН2-СН3; (СН2)3-(СН)2-СН2-(СН)2-СН2-(СН)2-СН2-(СН)2-(СН2)4-СН3; (СН2)7-(СН)2-(СН2)7-СН3; (СН2)8-(СН)2-СН2-фенил-СН2-(СН)2-СН2-СН3; (СН2)8-(СН)2-(СН2)2-O-СН2-(СН)2-СН2-СН3 и (СН2)8-(СН)2-СН2-фенил-O-(СН2)3-СН3.
[0032] Обработка этими соединениями осуществляется посредством нанесения соединений с помощью любой из следующих методик: погружения, инъекции, местного применения с использованием пульверизатора, распыления, местного применения с использованием пипетки и местного применения с использованием кисти, а также любого другого вида местного применения. В предпочтительных вариантах осуществления применяют аэрозоль или распылитель.
[0033] Настоящее изобретение дополнительно относится к способу увеличения завязывания семян и снижения недоразвитости зародышей у растений в ходе индукции гаплоидии, включающему обработку репродуктивных тканей растения, таких как рыльца, метелки, пыльца, початки, зерна или другие цветущие ткани, соединением в подходящей концентрации до, во время или после опыления. В одном варианте осуществления соединение выбрано из группы, состоящей из членов из таблицы 7. В другом варианте осуществления соединение представляет собой метил-альфа-линоленоилфторфосфонат (MALFP). В другом варианте осуществления соединение представляет собой линолевую кислоту (LLA), этиловый сложный эфир линолевой кислоты (LLAEE), линоленовую кислоту (LNA), дистеароилфосфатидилхолин (DSPC) или метиларахидонилфторфосфонат (MAFP).
[0034] Настоящее изобретение дополнительно относится к способу увеличения уровня индукции гаплоидии у растения, включающему нанесение липидной композиции на ткани растения непосредственно до, во время или непосредственно после опыления. В одном варианте осуществления растение является однодольным или двудольным растением; или растение представляет собой растение маиса или растение риса. В другом варианте осуществления липид выступает в качестве ингибитора фосфолипаз и/или ингибитора десатураз жирных кислот. В другом варианте осуществления липид представляет собой жирную кислоту (например, LLA) или сложный эфир жирной кислоты (например, LLAEE) с определенной длиной цепи и степенью насыщенности (восемнадцать атомов углерода и две двойные связи), относящиеся к классу длин цепей жирных кислот, не представленному в пыльце, индуцирующей гаплоидию. В целях иллюстрации, а не ограничения, липид представляет собой, например, ингибитор фосфолипаз метил-альфа-линоленоилфторфосфонат (MALFP), растворенный в буферном растворе DMSO при концентрациях MALFP от 0,0001 мг/мл до 1 г/мл или растворенный в составе на основе поверхностно-активных веществ и затем эмульгированный в буферном растворе диметилацетамида (DML) при концентрациях MALF от 0,0001 мг/мл до 1 г/мл. В целях дополнительной иллюстрации, а не ограничения, липидную композицию наносят посредством погружения, инъекции, опрыскивания, разбрызгивания, распыления, полива, нанесения кистью или любого другого способа нанесения на репродуктивные ткани растения. В одном варианте осуществления липидную композицию объединяют с пыльцой в смесь, и эту смесь затем наносят на ткани растения. В другом варианте осуществления смесь наносят на репродуктивные ткани растения, например, на пыльцу или рыльца растения маиса.
[0035] Настоящее изобретение относится к способу индуцирования de novo индукции гаплоидии у растения, включающему введение липидного соединения по меньшей мере в репродуктивную ткань растения во время опыления, перед опылением или после опыления. В одном варианте осуществления растение выбрано из группы, состоящей из однодольных и двудольных растений. В другом варианте осуществления растение выбрано из группы, состоящей из риса, маиса, пшеницы, сорго, томата, сахарной свеклы, проса, ячменя, сои, подсолнечника, хлопчатника, разновидностей овса, табака, овощных культур, плодовых культур и любых других культурных растений.
[0036] В соответствии с одним иллюстративным вариантом осуществления настоящее изобретение включает растение маиса или риса, способное индуцировать гаплоидию в результате индуцированной человеком мутации в гене пататин-подобной фосфолипазы AIIα ("PLA"), а также семена, пыльцу, части растения и потомство этого растения.
[0037] В соответствии с еще одним другим иллюстративным вариантом осуществления настоящее изобретение включает растение маиса или риса, способное индуцировать образование гаплоидов, созданное посредством стадий получения растительного материала из родительского растения маиса или риса, индуцирования по меньшей мере одной мутации по меньшей мере в одной копии гена PLA растительного материала посредством обработки растительного материала мутагеном с созданием растительного материала, подвергнутого мутагенезу, культивирования растительного материала, подвергнутого мутагенезу, с получением растений-потомков риса или маиса, анализа растений-потомков риса или маиса для выявления по меньшей мере одной мутации по меньшей мере в одной копии гена PLA, отбора растений-потомков риса или маиса, которые обладают способностью индуцировать образование гаплоидов, по сравнению с родительским растением риса или маиса; и повторения цикла культивирования растений-потомков риса или маиса с получением дополнительных растений-потомков, обладающих способностью индуцировать образование гаплоидов.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ В ПЕРЕЧНЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0038] SEQ ID NO: 1 представляет собой последовательность кДНК немутантной фосфолипазы, обнаруживаемую в GRMZM2G471240-NIL. Немутантный аллель гена фосфолипазы в данном документе переименован в MATRILINEAL.
[0039] SEQ ID NO: 2 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 1.
[0040] SEQ ID NO: 3 представляет собой нуклеотидную последовательность кДНК мутантной фосфолипазы, обнаруживаемую в GRMZM2G471240-mtl, содержащую вставку из 4 пар оснований. Мутантный аллель гена фосфолипазы в данном документе переименован в matrilineal.
[0041] SEQ ID NO: 4 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 3.
[0042] SEQ ID NO: 5 представляет собой праймер GRMZM2G471240_nil.F1.
[0043] SEQ ID NO: 6 представляет собой праймер GRMZM2G471240_nil.R1.
[0044] SEQ ID NO: 7 представляет собой праймер GRMZM2G471240_rwk.F1.
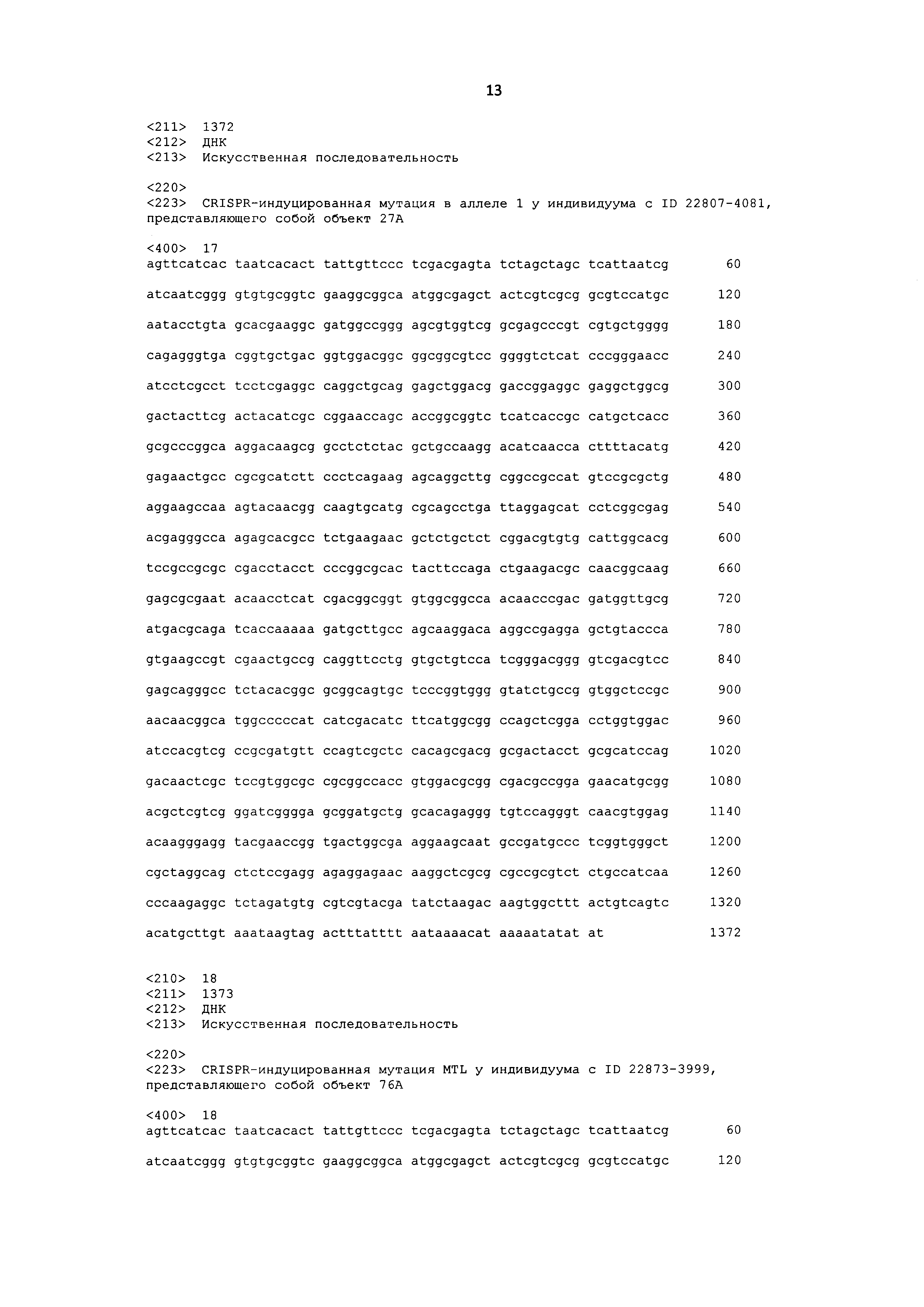
[0045] SEQ ID NO: 8 представляет собой праймер GRMZM2G471240_rwk.R1.
[0046] SEQ ID NO: 9 представляет собой нуклеотидную последовательность с TALEN-индуцированной мутацией MTL в аллеле 1 у индивидуума Т1 с ID 22808-3954, представляющего собой объект 39А.
[0047] SEQ ID NO: 10 представляет собой нуклеотидную последовательность с TALEN-индуцированной мутацией MTL в аллеле 1 у индивидуума Т1 с ID 22808-3924, представляющего собой объект 23А.
[0048] SEQ ID NO: 11 представляет собой нуклеотидную последовательность с TALEN-индуцированной мутацией MTL у индивидуума Т1 с ID 22808-3932, представляющего собой объект 81А, индивидуума с ID 22808-3317, представляющего собой объект 81А, и индивидуума с ID 22808-3303, представляющего собой объект 81А.
[0049] SEQ ID NO: 12 представляет собой нуклеотидную последовательность с TALEN-индуцированной мутацией MTL в аллеле 2 у объекта 39А с ID 22808-3954.
[0050] SEQ ID NO: 13 представляет собой нуклеотидную последовательность с TALEN-индуцированной мутацией MTL в аллеле 2 у объекта 23А с ID 22808-3924.
[0051] SEQ ID NO: 14 представляет собой нуклеотидную последовательность с TALEN-индуцированной мутацией MTL в аллеле 1 у индивидуума Т1 с ID 22808-4108, представляющего собой объект 38А.
[0052]SEQ ID NO: 15 представляет собой нуклеотидную последовательность с CRISPR-индуцированной мутацией MTL у индивидуума Т1 с ID 22807-4016, представляющего собой объект 18А.
[0053] SEQ ID NO: 16 представляет собой нуклеотидную последовательность с CRISPR-индуцированной мутацией MTL в аллеле 1 у индивидуума Т1 с ID 22807-4073, представляющего собой объект 27А.
[0054] SEQ ID NO: 17 представляет собой нуклеотидную последовательность с CRISPR-индуцированной мутацией MTL в аллеле 1 у индивидуума Т1 с ID 22807-4081, представляющего собой объект 27А.
[0055] SEQ ID NO: 18 представляет собой нуклеотидную последовательность с CRISPR-индуцированной мутацией MTL у индивидуума Т1 с ID 22873-3999, представляющего собой объект 76А.
[0056] SEQ ID NO: 19 представляет собой нуклеотидную последовательность с CRISPR-индуцированной мутацией MTL у индивидуума Т1 с ID 22873-3991, представляющего собой объект 32А.
[0057] SEQ ID NO: 20 представляет собой нуклеотидную последовательность направляющей CRISPR-РНК.
[0058] SEQ ID NO: 21 представляет собой геномную нуклеотидную последовательность Os03g27610, ортолога PLA2 риса.
[0059]SEQ ID NO: 22 представляет собой последовательность кДНК для SEQ ID NO: 21.
[0060] SEQ ID NO: 23 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 22.
[0061] SEQ ID NO: 24 представляет собой нуклеотидную последовательность немутантного GRMZM2G471240-B73.
[0062] SEQ ID NO: 25 представляет собой нуклеотидную последовательность немутантного GRMZM2G471240-RWK.
[0063] SEQ ID NO: 26 представляет собой нуклеотидную последовательность немутантного GRMZM2G471240-ST6.
[0064] SEQ ID NO: 27 представляет собой аминокислотную последовательность, ткодируемую SEQ ID NO: 24.
[0065] SEQ ID NO: 28 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 25.
[0066] SEQ ID NO: 29 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 26.
[0067] SEQ ID NO: 30 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 22466, содержащую MATRILINEAL дикого типа.
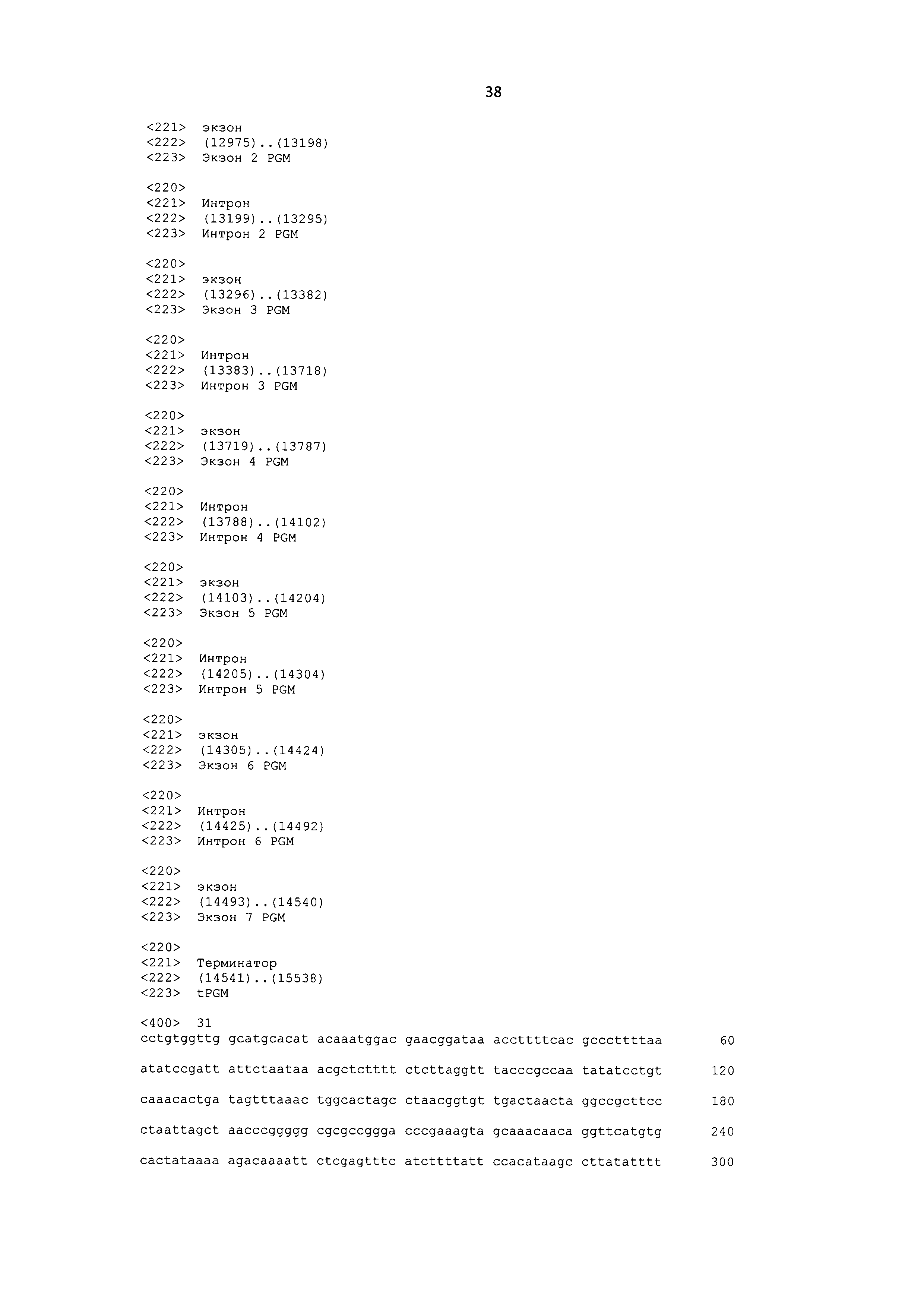
[0068] SEQ ID NO: 31 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 22467, содержащую ген фосфоглицератмутазы дикого типа.
[0069] SEQ ID NO: 32 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 22503, содержащей последовательность, кодирующую структуру типа "стебель-петля", нацеленную на экзон 2 MATRILINEAL.
[0070] SEQ ID NO: 33 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 22513, содержащую последовательность, кодирующую структуру типа стебель-петля, нацеленную на экзон 4 MATRILINEAL.
[0071] SEQ ID NO: 34 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 22807, содержащую последовательности, кодирующие аппарат редактирования CRISPR/Cas9, нацеленный на MATRILINEAL в NP2222.
[0072] SEQ ID NO: 35 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 22808, содержащую последовательности, кодирующие аппарат редактирования CRISPR/Cas9, нацеленный на MATRILINEAL в NP2222.
[0073]SEQ ID NO: 36 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 22873, содержащую последовательности, кодирующие аппарат редактирования CRISPR/Cas9, нацеленный на MATRILINEAL в NP2222.
[0074] SEQ ID NO: 37 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 23123, содержащую последовательности, кодирующие аппарат редактирования TALEN, нацеленный на MATRILINEAL в NP2222.
[0075] SEQ ID NO: 38 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 23501 с двумя направляющими последовательностями gRNA, нацеленными на экзон 4 риса.
[0076] SEQ ID NO: 39 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 23501 с одной направляющей последовательностью gRNA, нацеленной на экзон 4 риса.
[0077] SEQ ID NO: 40 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 23501 с двумя направляющими последовательностями gRNA, нацеленными на экзон 1 риса.
[0078] SEQ ID NO: 41 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 23501 с одной направляющей последовательностью gRNA, нацеленной на экзон 1 риса.
[0079] SEQ ID NO: 42 представляет собой нуклеотидную последовательность с TALEN-индуцированной мутацией MTL в аллеле 2 у объекта 38А с ID 22808-4108.
[0080] SEQ ID NO: 43 представляет собой нуклеотидную последовательность с CRISPR-индуцированной мутацией MTL в аллеле 2 у объекта 27А с ID 22807-4073.
[0081] SEQ ID NO: 44 представляет собой нуклеотидную последовательность с CRISPR-индуцированной мутацией MTL в аллеле 2 у объекта 27А с ID 22807-4081.
[0082] SEQ ID NO: 45 представляет собой нуклеотидную последовательность линии 1139, полученной с помощью TILLING.
[0083] SEQ ID NO: 46 представляет собой нуклеотидную последовательность линии 3594, полученной с помощью TILLING.
[0084] SEQ ID NO: 47 представляет собой нуклеотидную последовательность линии 0505, полученной с помощью TILLING.
[0085] SEQ ID NO: 48 представляет собой нуклеотидную последовательность линии 2658, полученной с помощью TILLING.
[0086] SEQ ID NO: 49 представляет собой нуклеотидную последовательность линии 1983, полученной с помощью TILLING.
[0087] SEQ ID NO: 50 представляет собой нуклеотидную последовательность линии 2732, полученной с помощью TILLING.
[0088] SEQ ID NO: 51 представляет собой нуклеотидную последовательность линии 2414, полученной с помощью TILLING.
[0089] SEQ ID NO: 52 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 45.
[0090] SEQ ID NO: 53 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 46.
[0091] SEQ ID NO: 54 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 47.
[0092] SEQ ID NO: 55 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 48.
[0093] SEQ ID NO: 56 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 49.
[0094] SEQ ID NO: 57 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 50.
[0095] SEQ ID NO: 58 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 51.
[0096] SEQ ID NO: 59 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 9.
[0097] SEQ ID NO: 60 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 10.
[0098] SEQ ID NO: 61 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 11.
[0099] SEQ ID NO: 62 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 12.
[00100] SEQ ID NO: 63 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 13.
[00101] SEQ ID NO: 64 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 14.
[00102] SEQ ID NO: 65 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 15.
[00103] SEQ ID NO: 66 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 16.
[00104] SEQ ID NO: 67 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 17.
[00105] SEQ ID NO: 68 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 18.
[00106] SEQ ID NO: 69 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 19.
[00107] SEQ ID NO: 70 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 42.
[00108] SEQ ID NO: 71 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 43.
[00109] SEQ ID NO: 72 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 44.
[00110] SEQ ID NO: 73 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Sorghum bicolor.
[00111] SEQ ID NO: 74 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Setaria italica.
[00112] SEQ ID NO: 75 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Hordeum vulgare.
[00113] SEQ ID NO: 76 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Brachypodium distachyon.
[00114] SEQ ID NO: 77 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Oryza sativa v. indica.
[00115] SEQ ID NO: 78 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Triticum aestivum.
[00116] SEQ ID NO: 79 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Musa acuminata.
[00117] SEQ ID NO: 80 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Elaeis guineensis.
[00118] SEQ ID NO: 81 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Arabidopsis thaliana.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[00119] На фигуре 1 представлена схема картирования, применяемая для картирования признака индукции гаплоидии в RWK.
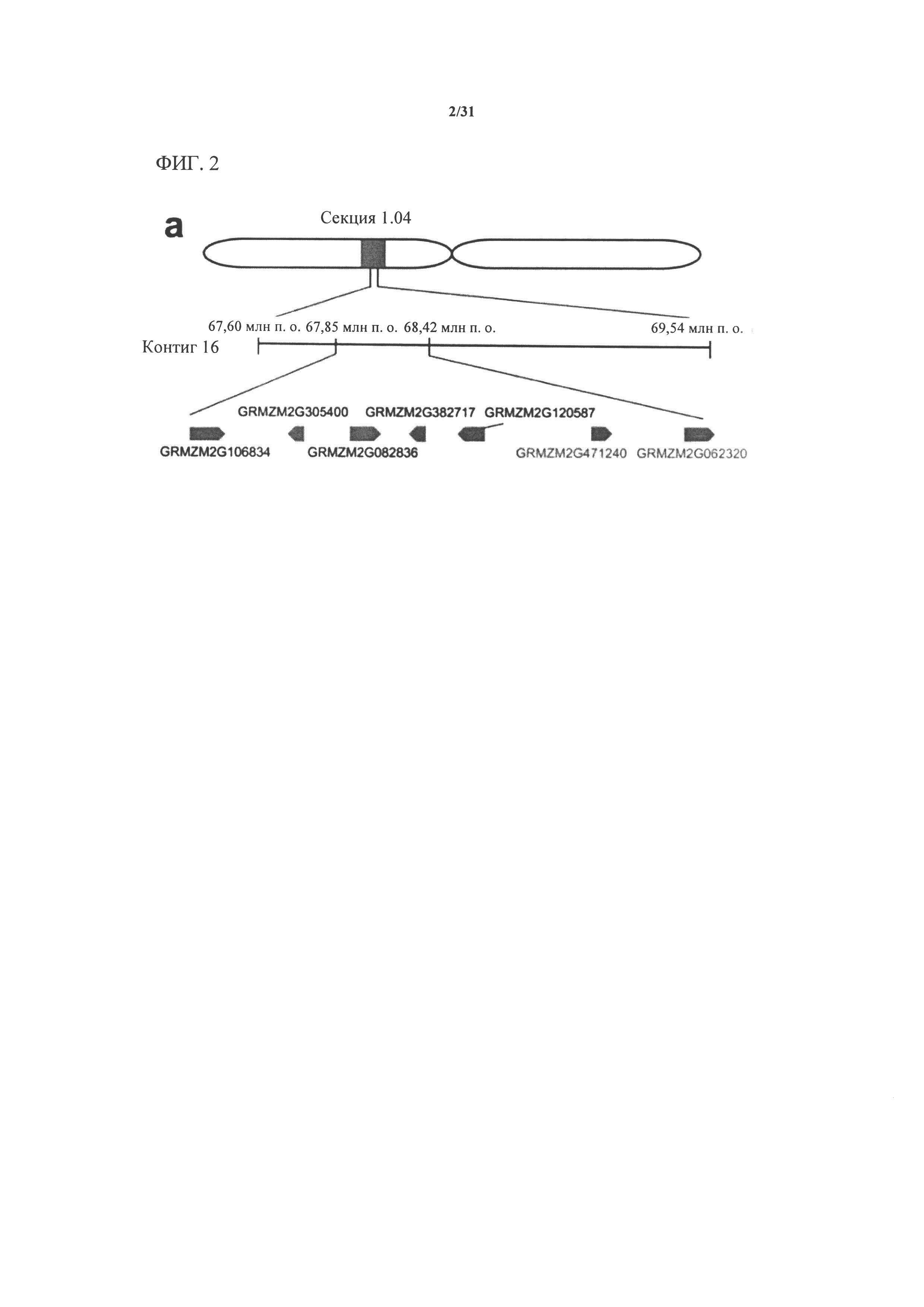
[00120] На фигуре 2 показано тонкое картирование, ограничивающее главный QTL очень небольшим интервалом в секции 1.04 от 67,85 млн п.о. до 68,42 млн п.о. Эта область имеет семь аннотированных генов. Авторы настоящего изобретения секвенировали и собирали гены в этом интервале в нескольких линиях. Два гена с наиболее значительными мутациями в линиях, индуцирующих гаплоидию, показаны в правой нижней части (GRMZM2G471240 и GRMZM2G062320).
[00121] На фигуре 3 показаны различия в экспрессии GRMZM2G471240 в пыльце, индуцирующей и не индуцирующей гаплоидию, и пыльцевых мешках после цветения (спорофитной ткани с удаленными пыльцевыми зернами). Этот ген, в частности, экспрессируется в мужском гаметофите.
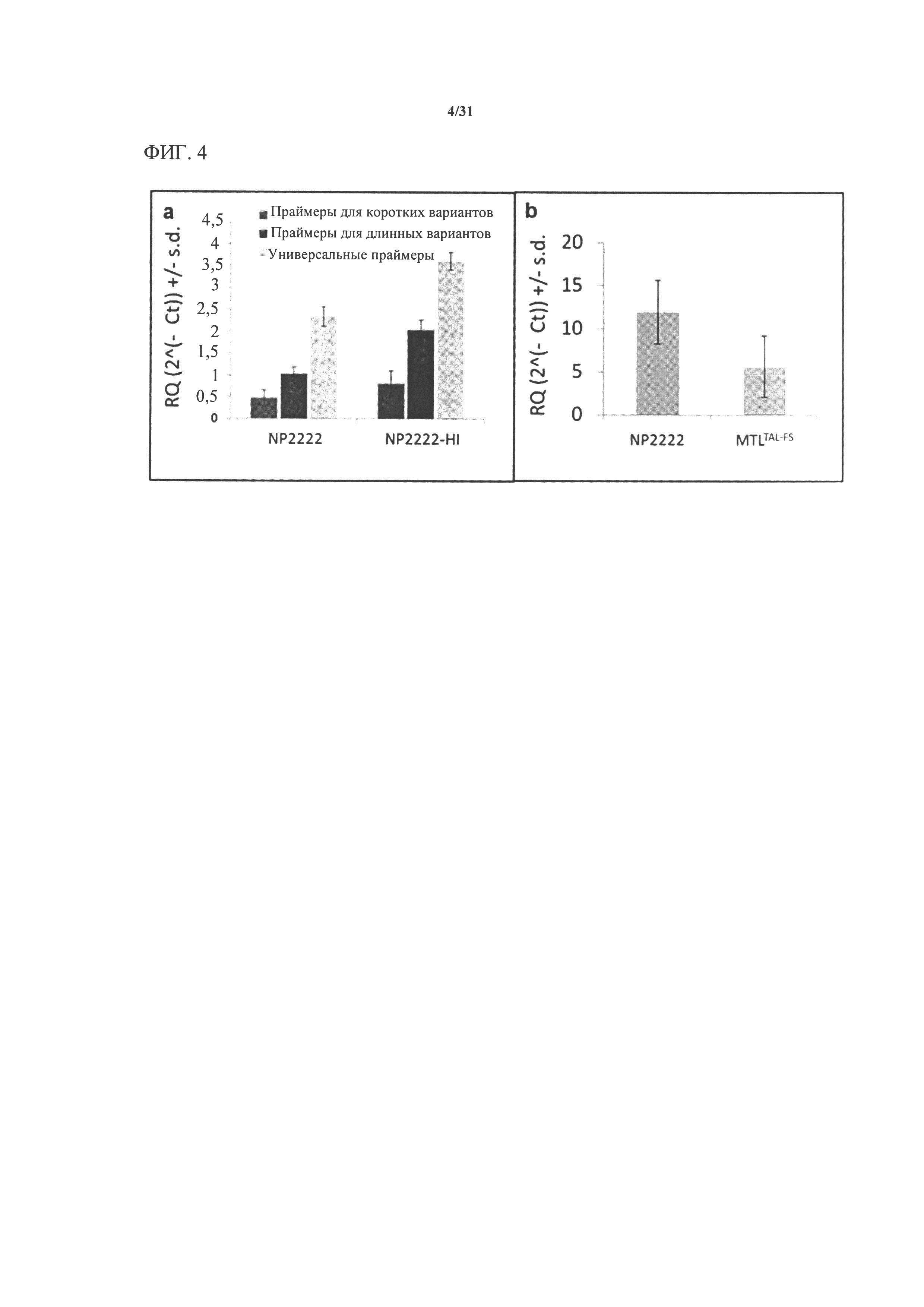
[00122] На фигуре 4а показаны результаты сплайс-специфической qRT-PCR для GRMZM2G471240. Три биологические реплики пыльников на стадии R1 тестировали в трех технических повторностях, и для каждой реакции рассчитывали среднее значение Ct и среднеквадратическое отклонение. Относительное количество каждого типа транскрипта сравнивали с эндогенным контролем с применением Iog2-регрессии дельта Ct. Два набора праймеров применяли для оценки относительной представленности каждого из двух аннотированных сплайс-вариантов по сравнению с набором праймеров, инвариантным относительно сплайс-вариантов. Более короткий вариант транскрипта по сравнению с длинным транскриптом характеризуется относительно низкой представленностью в генотипах как NP2222 (дикий тип), так и NP2222-HI (индуктор гаплоидии). Экспрессия мутантных копий гена в NP2222-HI была значительно более высокой для всех трех тестируемых пар праймеров.
[00123] На фигуре 4b показано пять биологических реплик свежей пыльцы от растений NP2222 и MTLTAL-FS (растений Т1, которые являются гомозиготными по отредактированным mtl-подобным аллелям), которые тестировали в трех технических повторах с универсальным праймером, и для каждой реакции рассчитывали среднее значение Ct и среднеквадратическое отклонение. Относительное количество каждого типа транскрипта сравнивали с эндогенным контролем с применением log2-регрессии дельта Ct. Пыльца MTLTAL-FS характеризуется более низкой представленностью транскриптов, чем пыльца NP2222 (дикого типа).
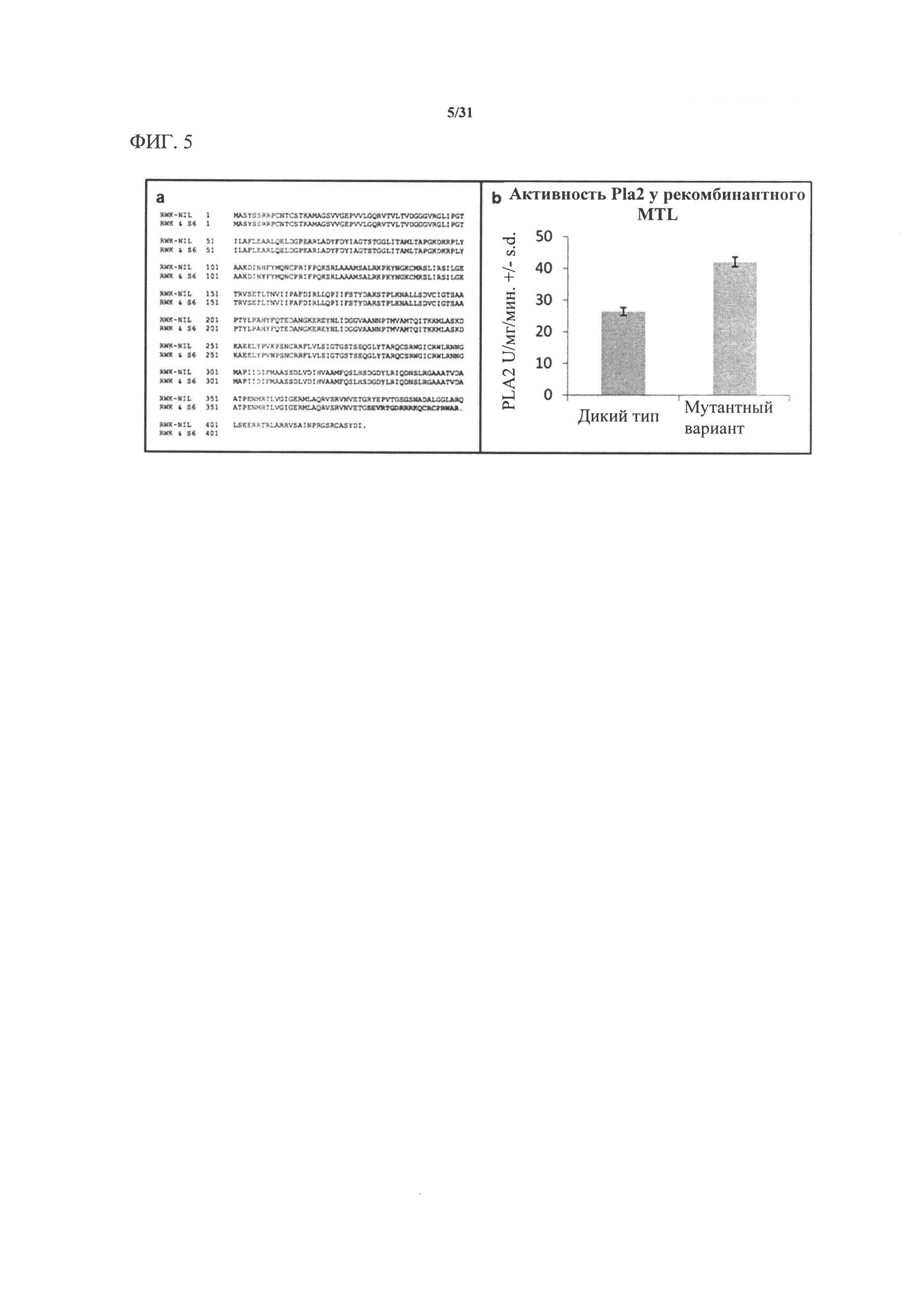
[00124] На фигуре 5а показано выравнивание предсказанной аминокислотной последовательности белка В73, кодируемой длинным сплайс-вариантом гена GRMZM2G471240 в В73 и RWK-NIL, с предсказанной последовательностью, кодируемой аллелем mtl, обнаруживаемым в RWK и Stock 6 (S6). Аминокислоты, которые отличаются, указаны красным цветом; аминокислоты, которые совпадают, указаны обычным серым текстом, а стоп-кодоны указаны точкой. Две точечные мутации приводят в результате к аминокислотным заменам: гистидина (Н) на тирозин (Y), а лизина (K) на аргинин (N). Эти изменения не являются консервативными; возможно, что одно или оба из них модифицируют фенотип индукции гаплоидии - что позволяет предположить, что аллельная серия может быть обнаружена при дополнительном исследовании вариантов.
[00125] На фигуре 5b показано, что MTL дикого типа и мутантный (усеченный) MTL, кодируемый аллелем mtl, характеризуются фосфолипазной активностью in vitro. Фосфолипазную активность PLA2 измеряли посредством флуоресцентного липосомного анализа рекомбинантного очищенного белка, полученного с применением кДНК MTL и mtl. Планки погрешностей указывают на среднеквадратическую погрешность, вычисленную по среднему значению для четырех повторностей.
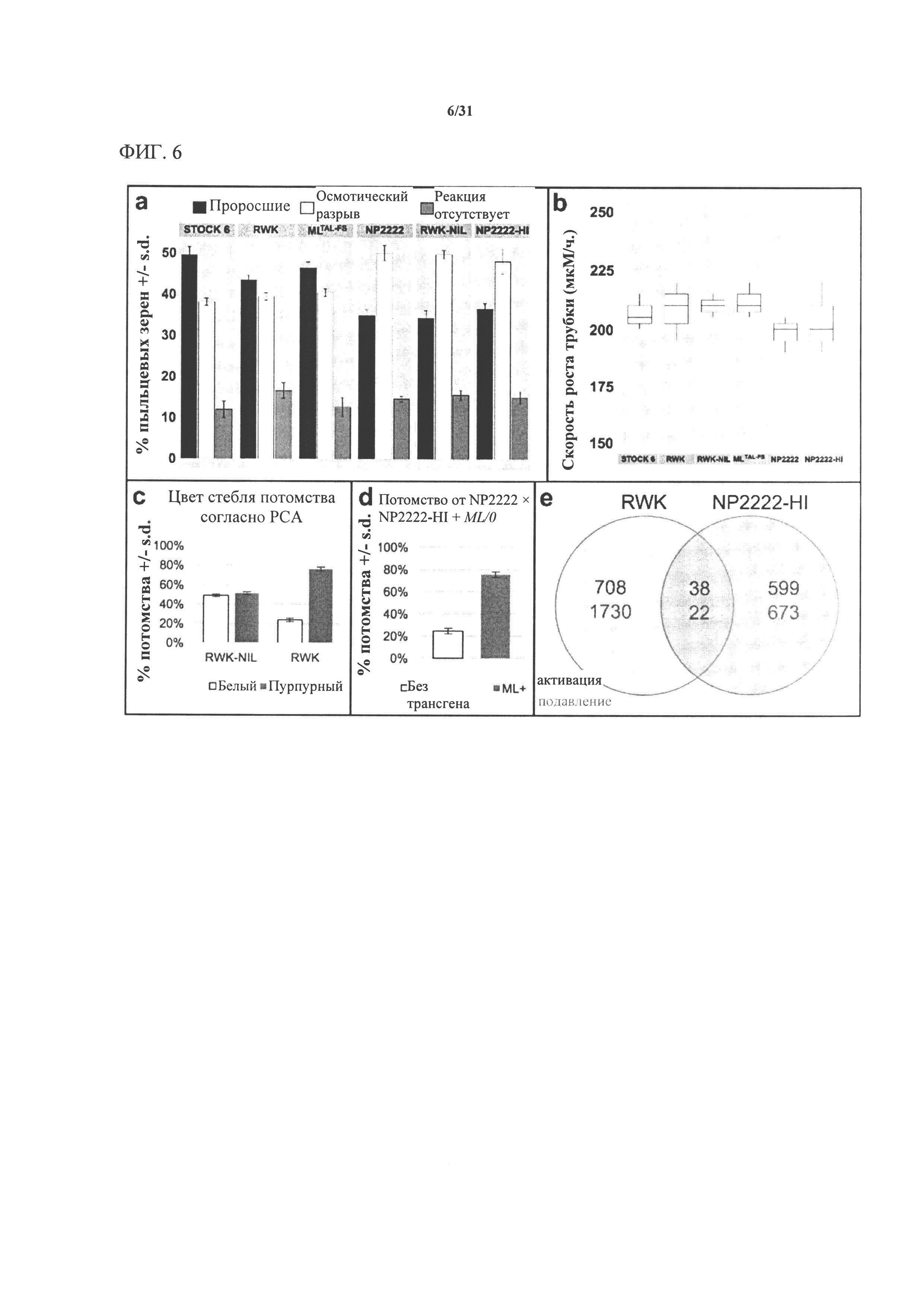
[00126] На фигуре 6 показано, что mtl отвечает за плейотропные фенотипы, ассоциированные с индукцией гаплоидии. 6А: скорость прорастания пыльцевой трубки была сходной у индукторов и неиндукторов (n=200). 6В: первоначальное удлинение пыльцевой трубки также было сходным (n=25). 6С: RWK, но не RWK-NIL, подвергается нарушению сегрегации (SD), как следует из низкой (25%) степени передачи признаков прорастающему потомству (n=300). 6D: Комплементационные линии MTL/0 также демонстрируют SD в отношении mtl у прорастающего потомства (n=400). 6Е: диаграмма Венна, на которой показаны результаты профилирования двух пар почти изогенных линий, индуцирующих гаплоидию, методом секвенирования РНК (левая часть - RWK по сравнению с RWK-NIL; правая часть - NP2222-HI по сравнению с NP2222; красный текст -активация; зеленый текст - подавление). Было обнаружено, что только 60 генов значительно изменились в одном и том же направлении.
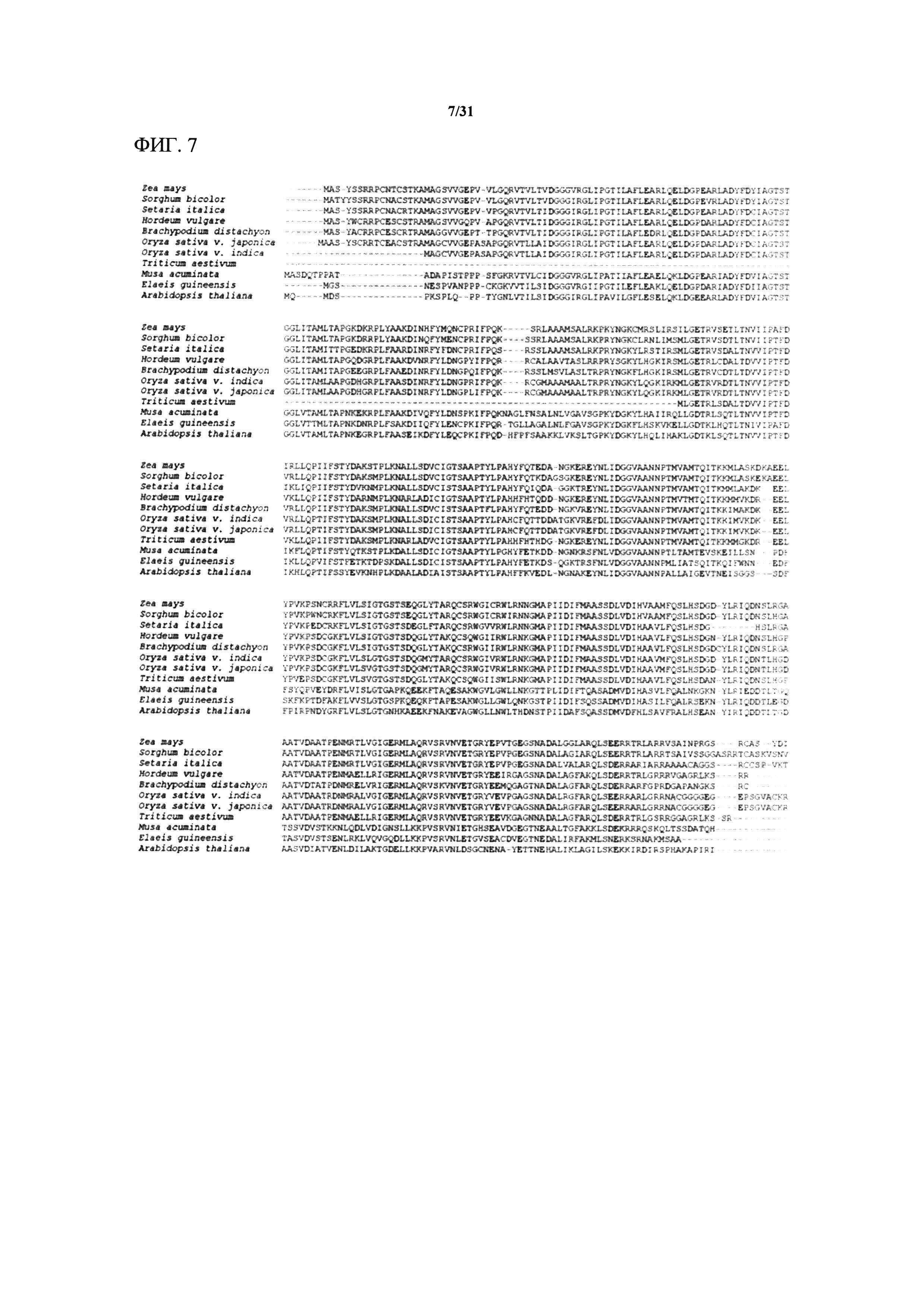
[00127] На фигуре 7 показано выравнивание аминокислотных последовательностей, кодируемых геном MTL маиса и имеющимися в открытом доступе ортологами MTL восьми видов злаковых растений, двух однодольных растений, отличных от злаковых, и Arabidopsis (резуховидки Таля). Это выравнивание охватывает маис (Zea mays), сорго (Sorghum bicolor, 92% идентичность последовательностей с MTL), итальянское просо (Setaria italica, 85% идентичность), ячмень (Hordeum vulgare, 78% идентичность), Brachypodium distachyon (78% идентичность), индийский и японский сорта риса (Oryza sativa v. indica и japonica, Os3g27610, 78% и 79% идентичность соответственно), пшеницу обыкновенную (Triticum aestivum, 55% идентичность), банан (Musa acuminata, 57% идентичность), масличную пальму (Elaeis guineensis, 56% идентичность) и Arabidopsis thaliana (52% идентичность).
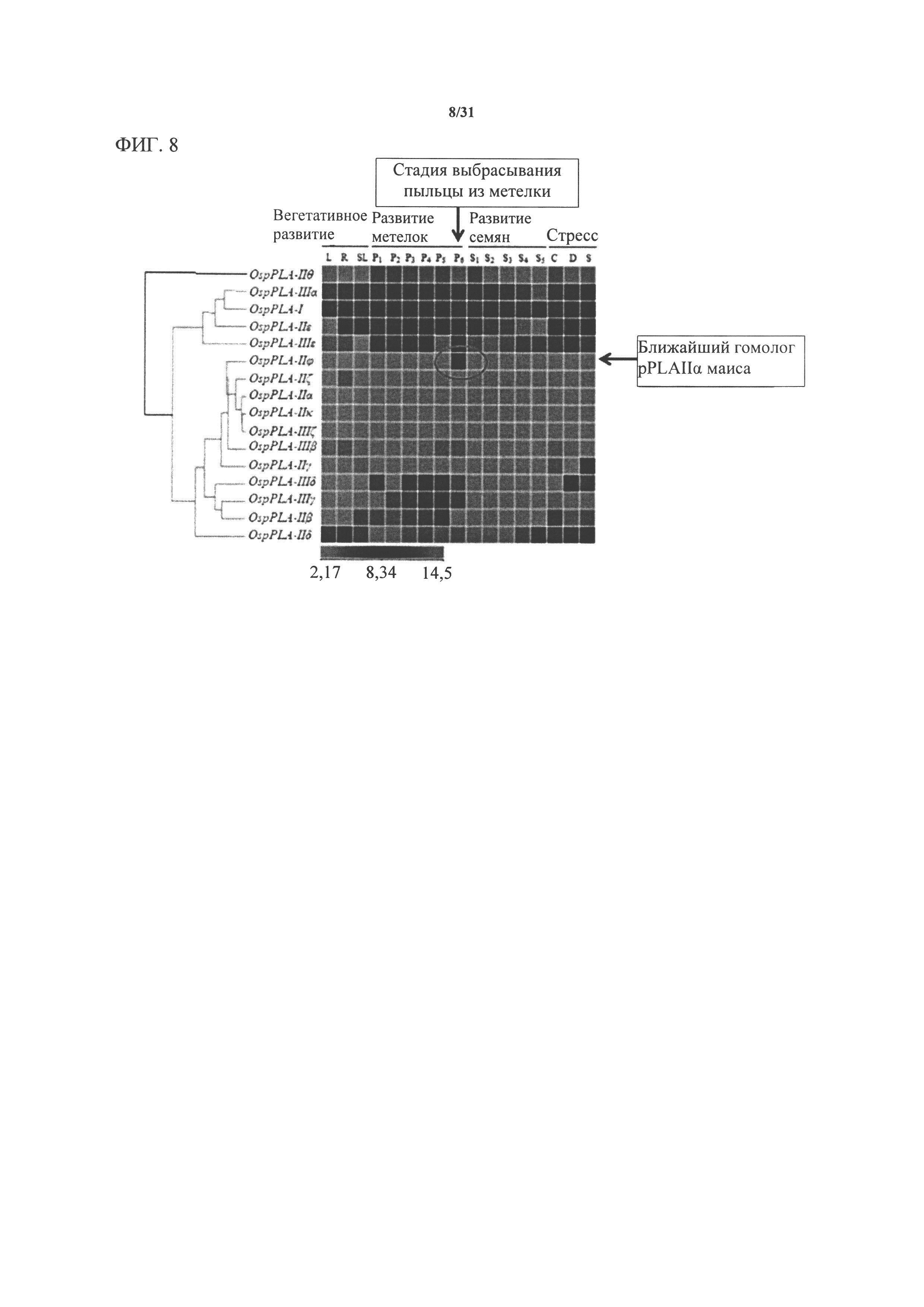
[00128] Фигура 8. Профиль экспрессии фосфолипаз риса (по материалам Singh, A., et al., Rice phospholipase A superfamily: organization, phylogenetic and expression analysis during abiotic stresses and development, PLoS One 7: e30947 (2012)). Ближайшим гомологом MTL является ген риса OspPLAIIϕ Os3g27610).
[00129] Фигура 9. Схема, на которой показан путь редактирования Os3g27610 для получения линий, индуцирующих гаплоидию. Можно целенаправленно воздействовать на любую часть гена (в данном документе показано целенаправленное воздействие на первый и четвертый экзоны) и ожидать создания мутаций сдвига рамки считывания, которые приведут к нокауту и утрате функции гена, что приведет к индукции гаплоидии.
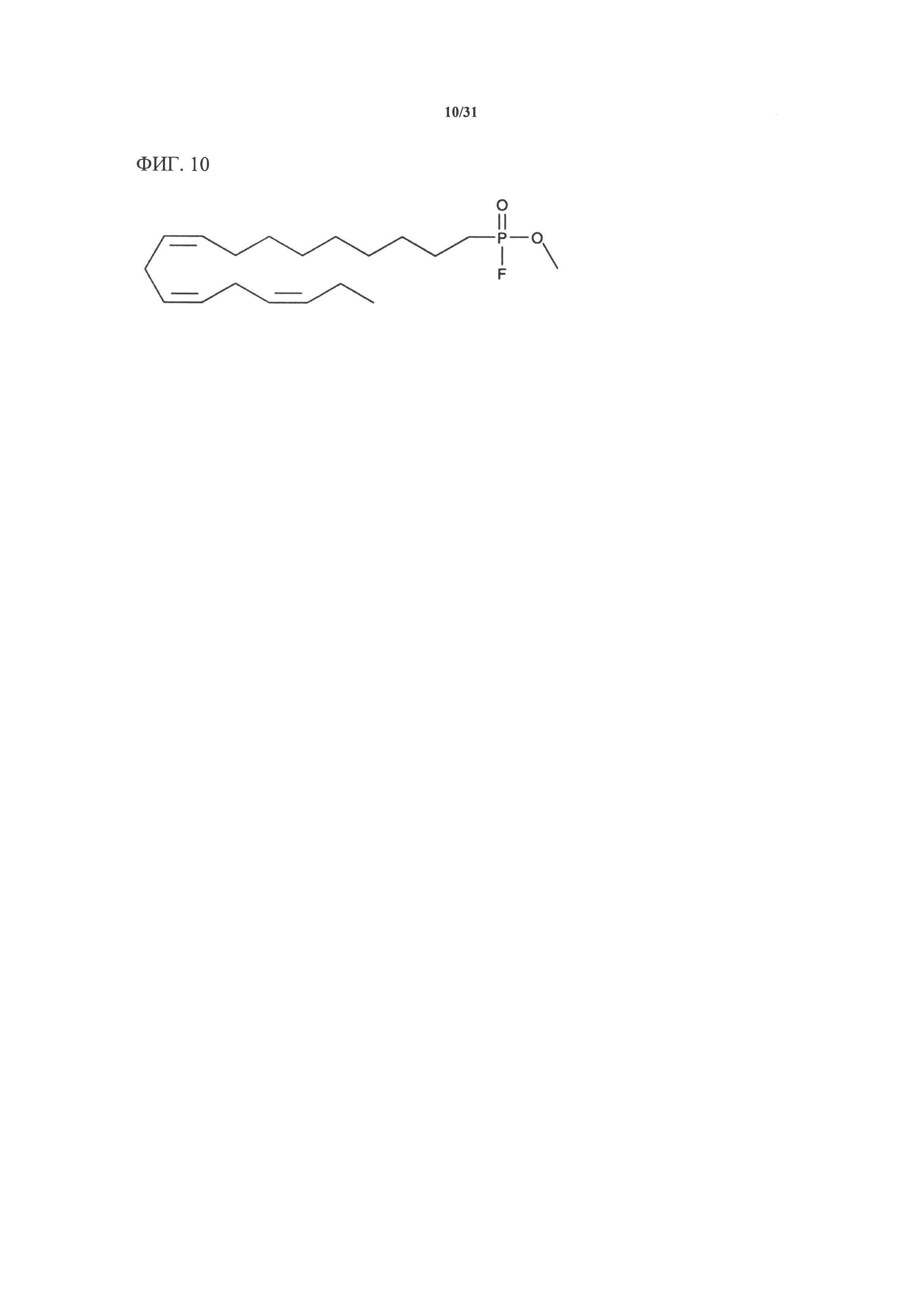
[00130] На фигуре 10 показана атомная структура метил-альфа-линоленоилфторфосфоната (MALFP).
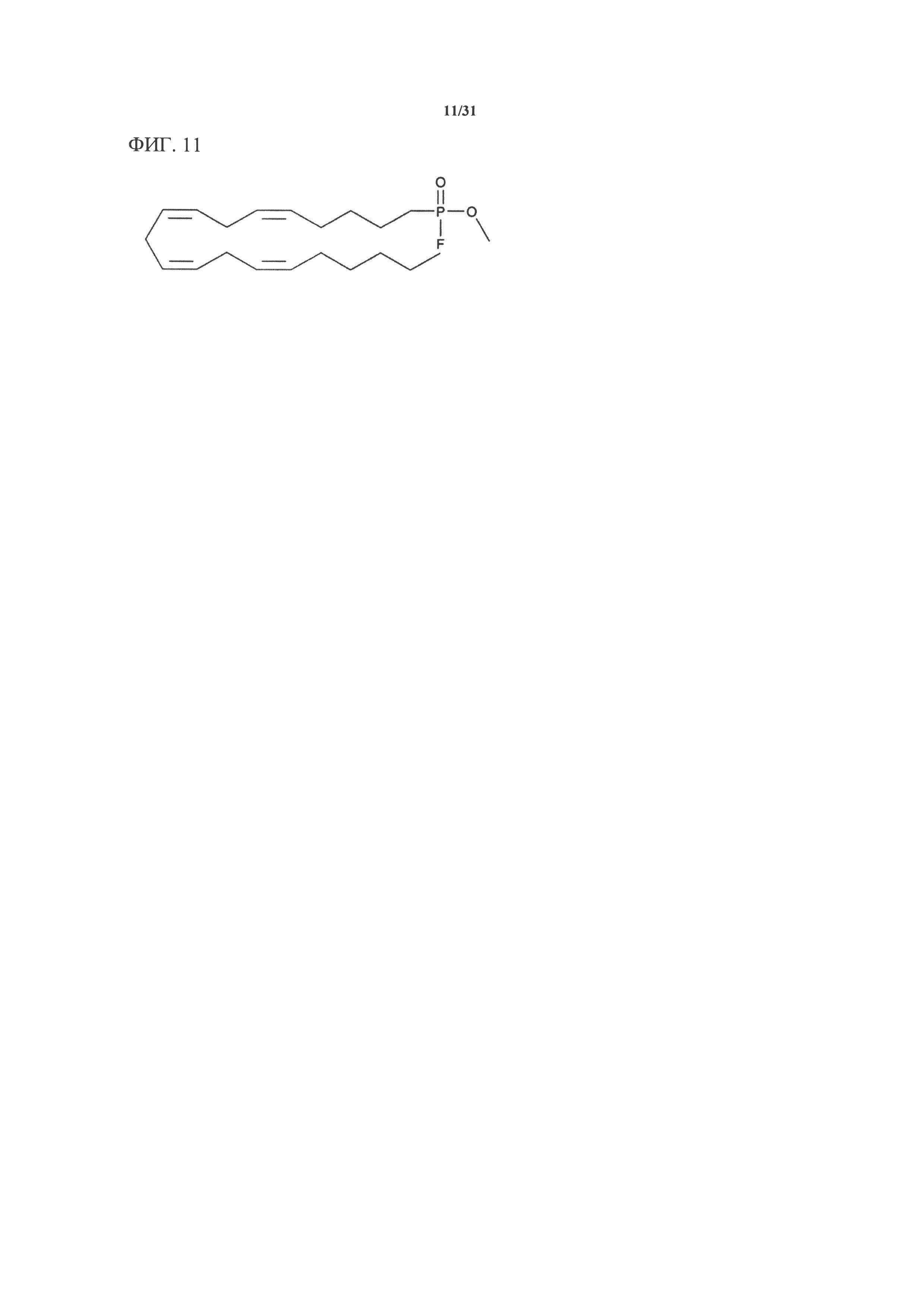
[00131] На фигуре 11 показана атомная структура метиларахидонилфторфосфоната (MAFP).
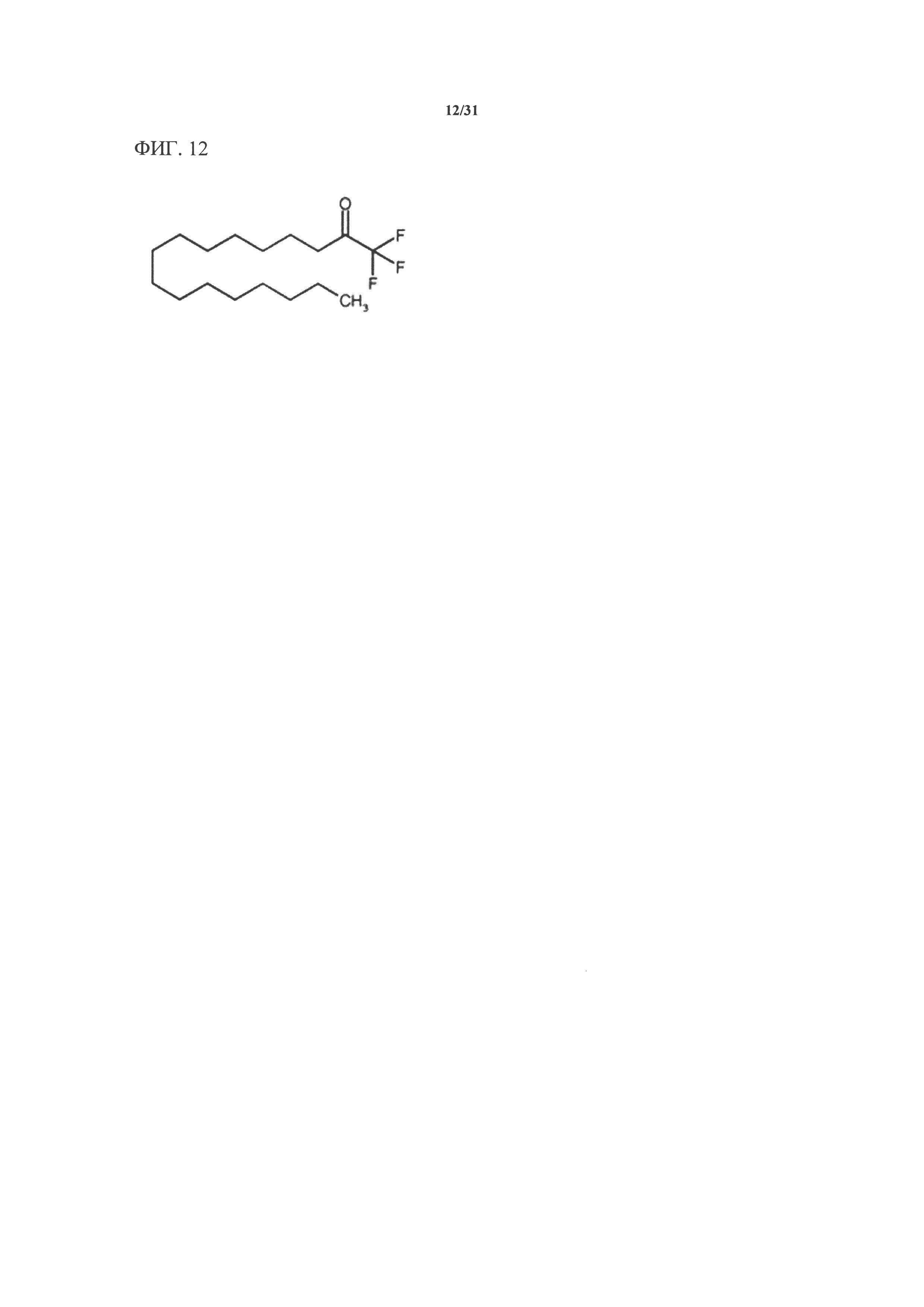
[00132] На фигуре 12 показана атомная структура пальмитилтрифторметилкетона (PACOCF3).
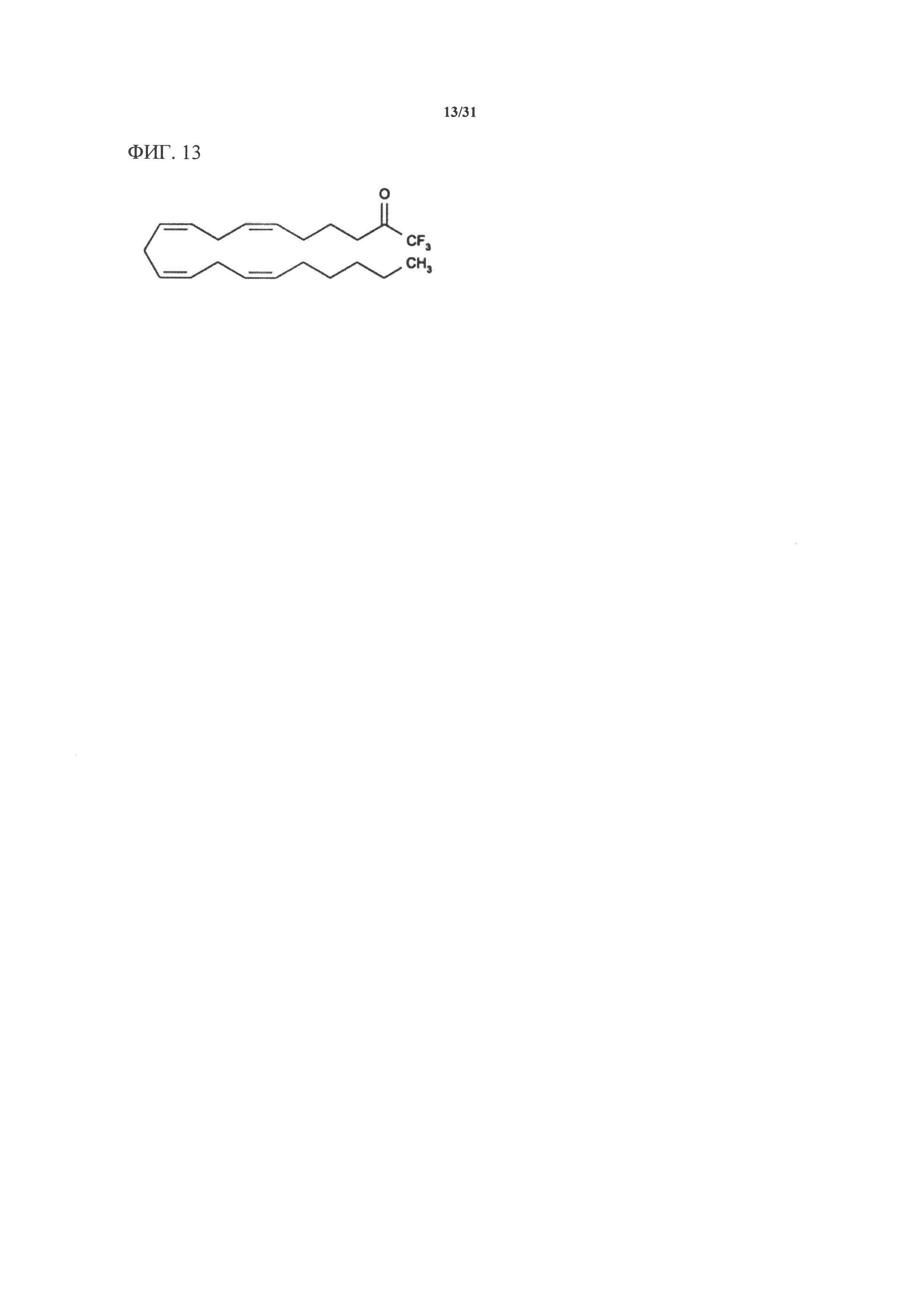
[00133] На фигуре 13 показана атомная структура арахидонилтрифторметилкетона (AACOCF3).
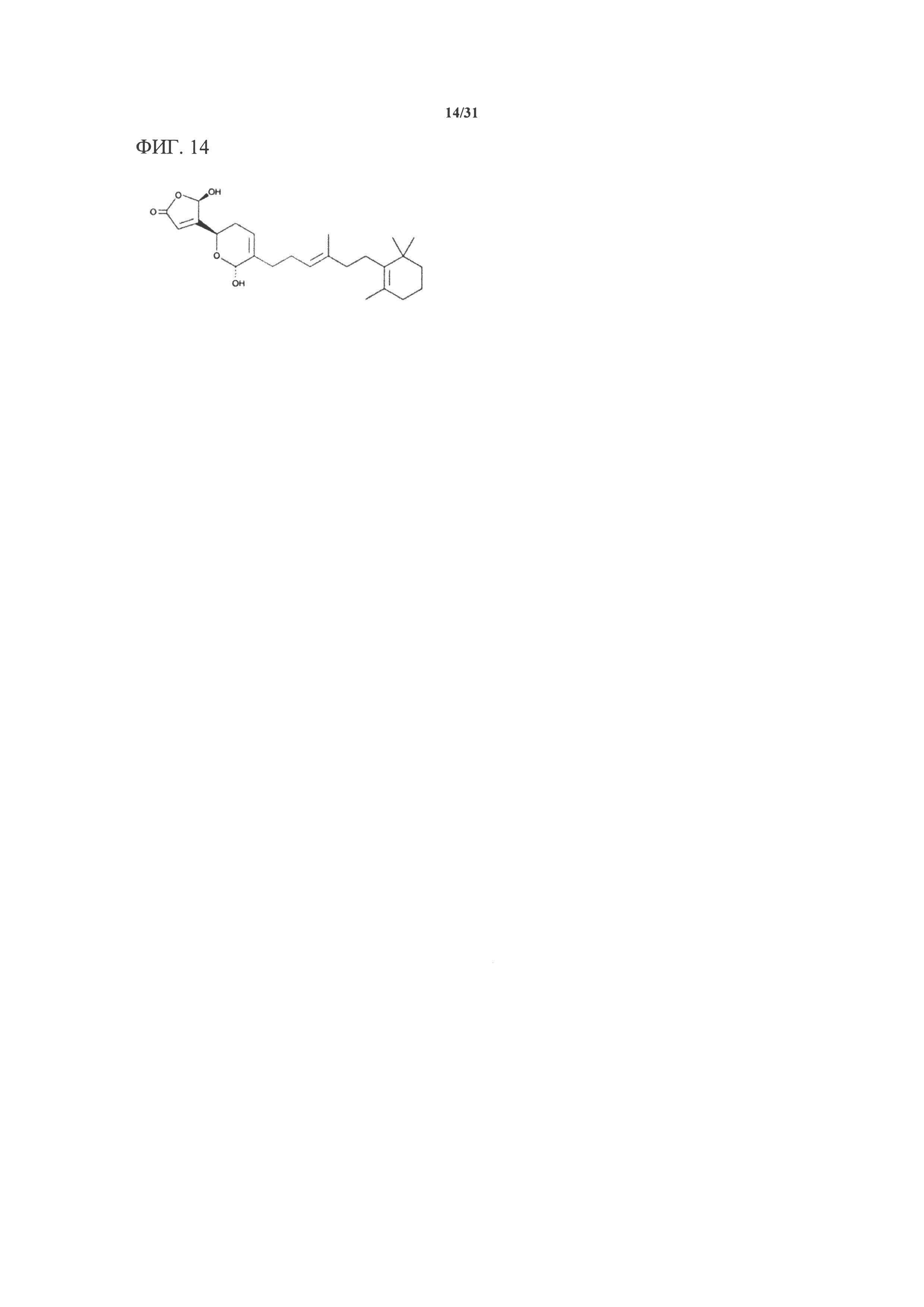
[00134] На фигуре 14 показана атомная структура маноалида.
[00135] На фигуре 15 показана атомная структура этилового сложного эфира линолевой кислоты (LLAEE).
[00136] На фигуре 16 показана атомная структура этилового сложного эфира линоленовой кислоты (LNAEE).
[00137] На фигуре 17 показана атомная структура метилового сложного эфира арахидоновой кислоты (ААМЕ).
[00138] На фигуре 18 показана атомная структура метилового сложного эфира олеиновой кислоты (ОАМЕ).
[00139] На фигуре 19 показана атомная структура этилового сложного эфира олеиновой кислоты (ОАЕЕ).
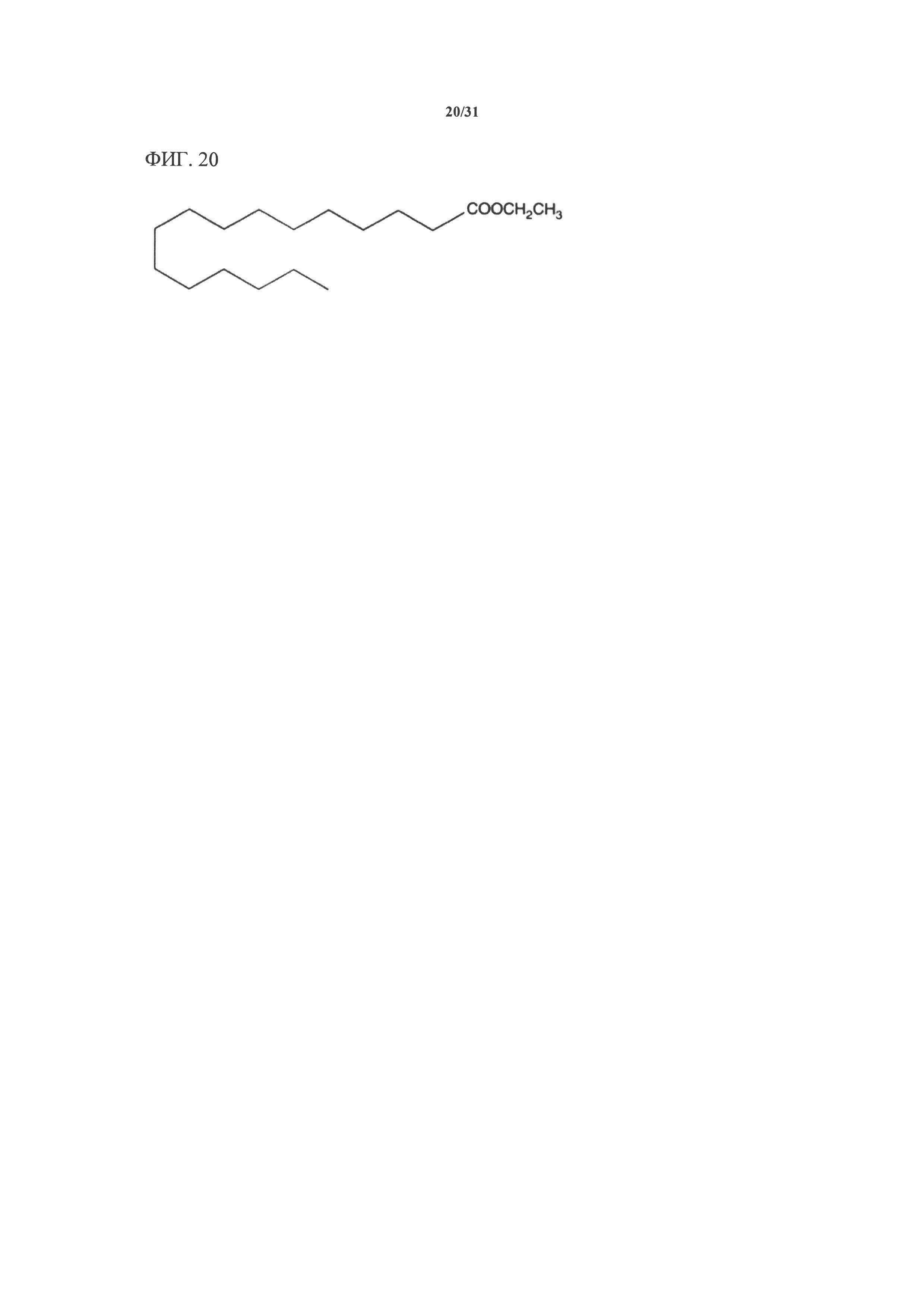
[00140] На фигуре 20 показана атомная структура этилового сложного эфира пальмитиновой кислоты (РАЕЕ).
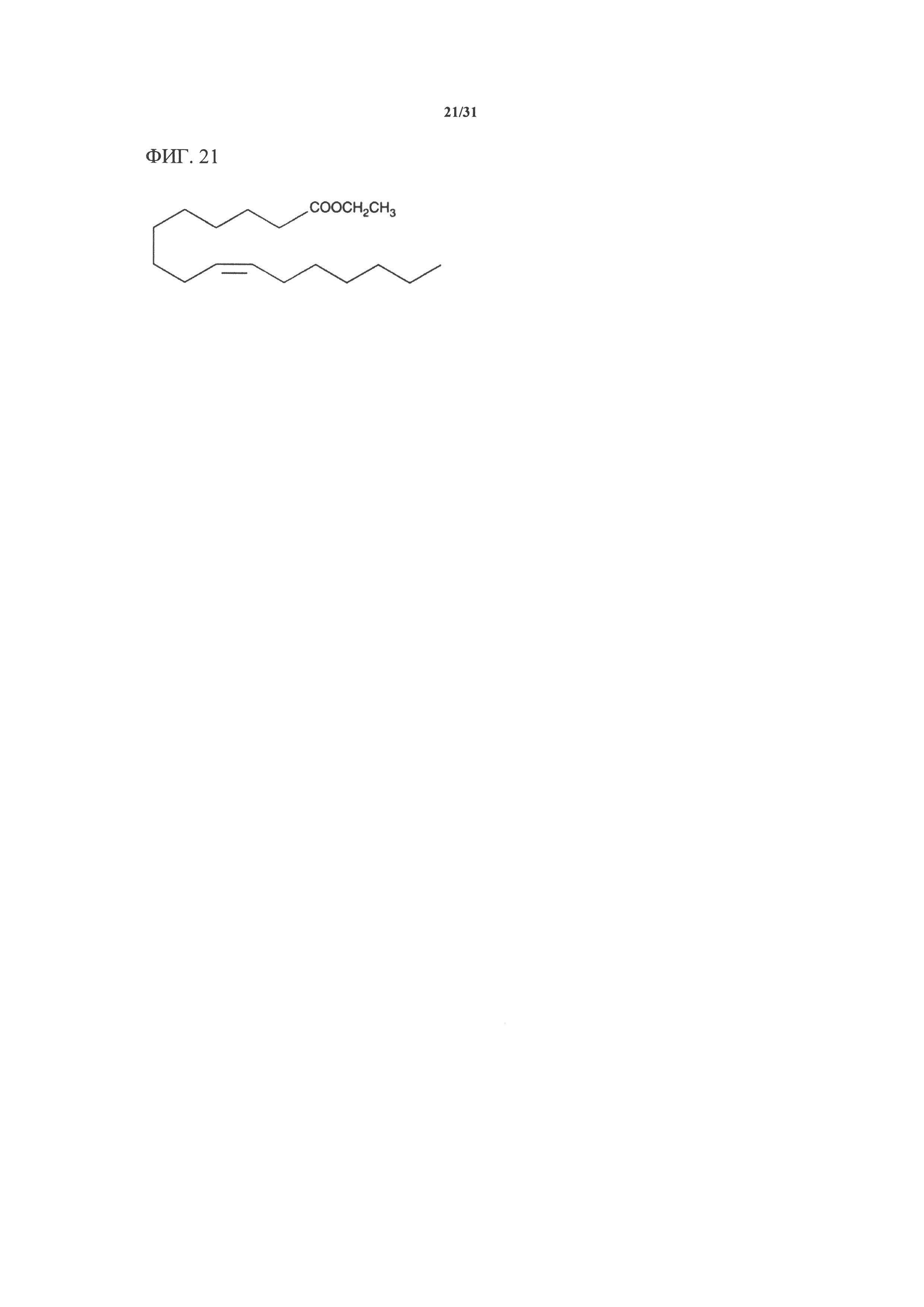
[00141] На фигуре 21 показана атомная структура этилового сложного эфира пальмитолеиновой кислоты (PLAEE).
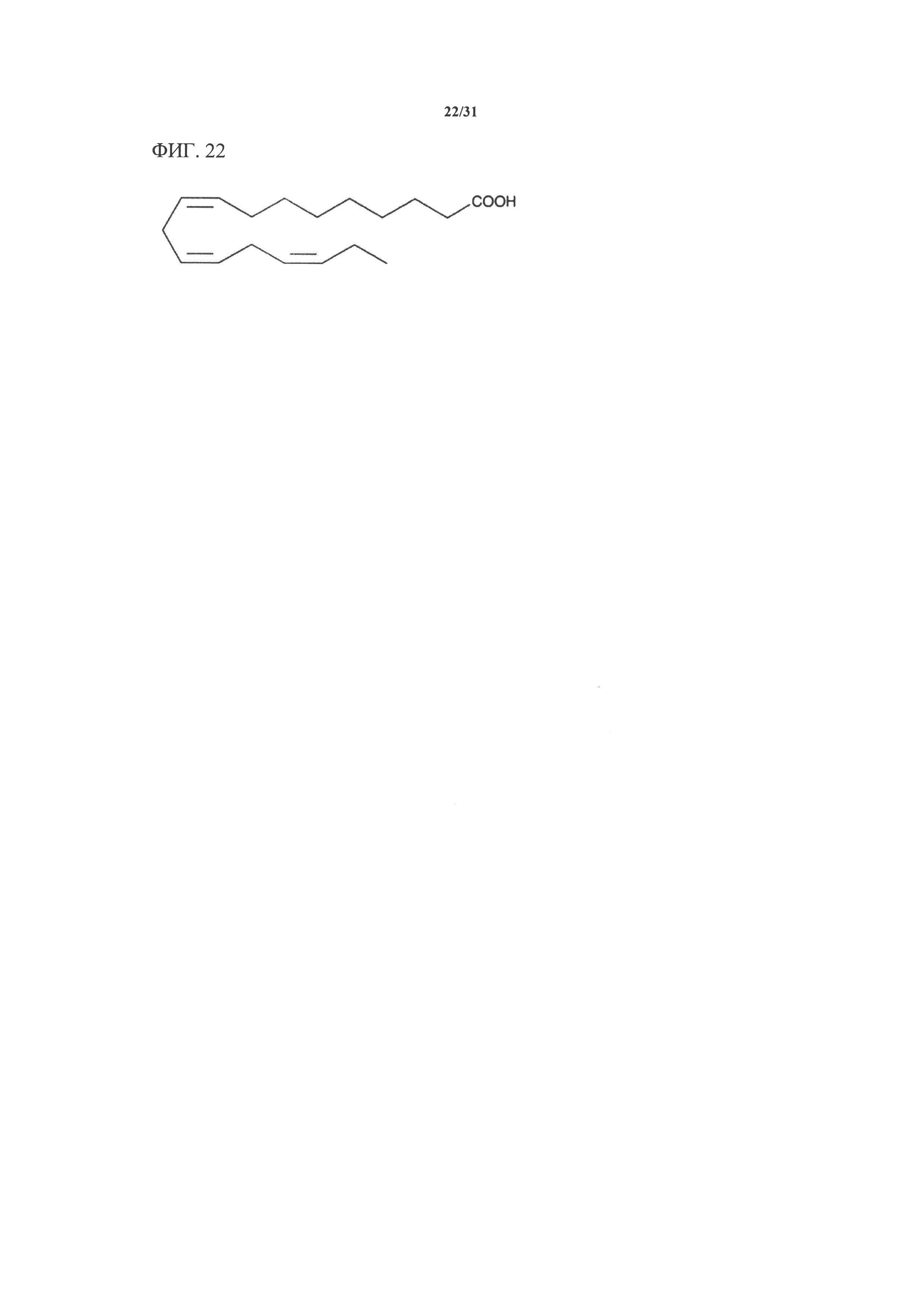
[00142] На фигуре 22 показана атомная структура альфа-линоленовой кислоты (aLNA).
[00143] На фигуре 23 показана атомная структура гамма-линоленовой кислоты (gLNA).
[00144] На фигуре 24 показана атомная структура олеиновой кислоты.
[00145] На фигуре 25 показана атомная структура линолевой кислоты.
00146] На фигуре 26 показана атомная структура арахидоновой кислоты.
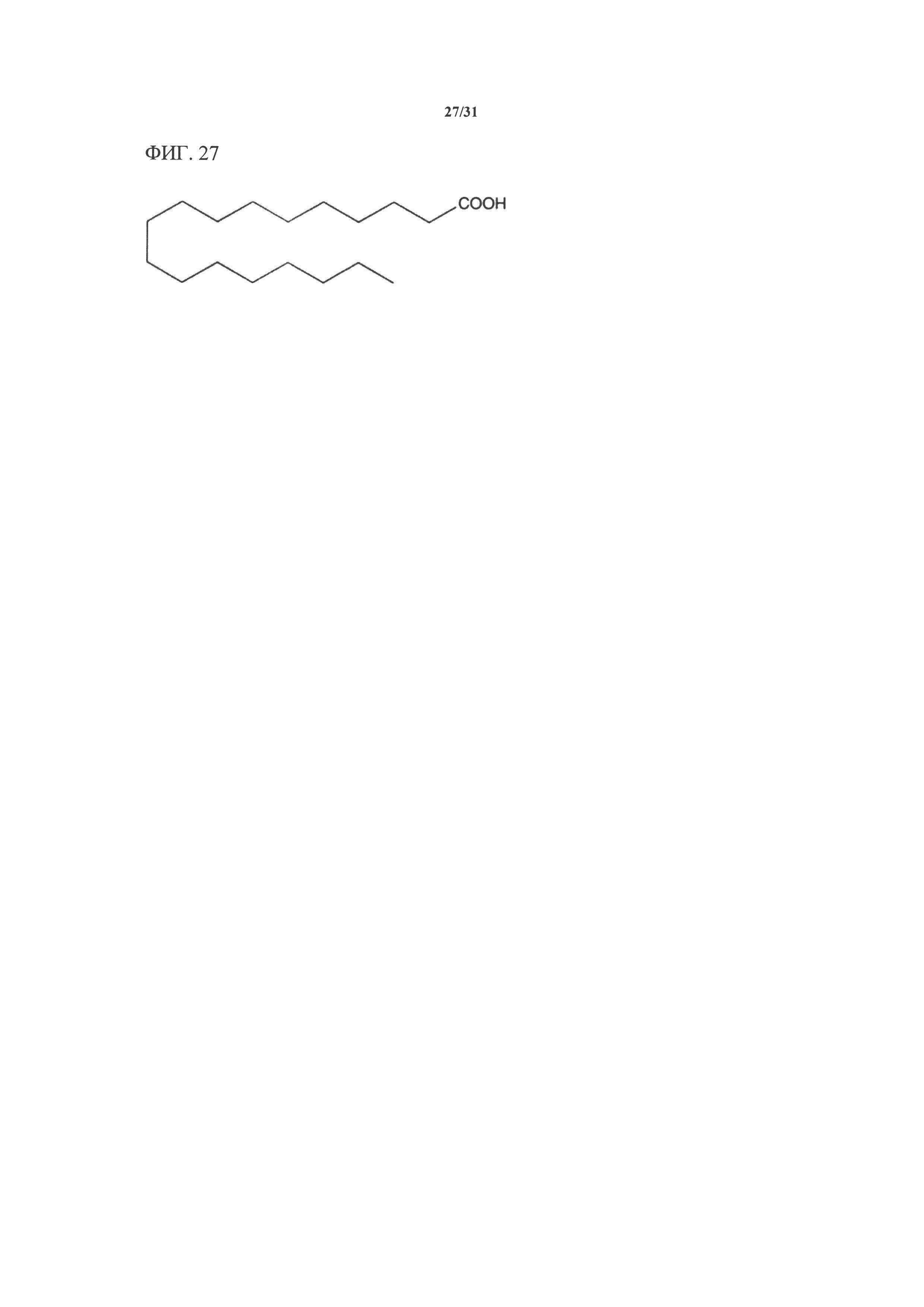
[00147] На фигуре 27 показана атомная структура стеариновой кислоты.
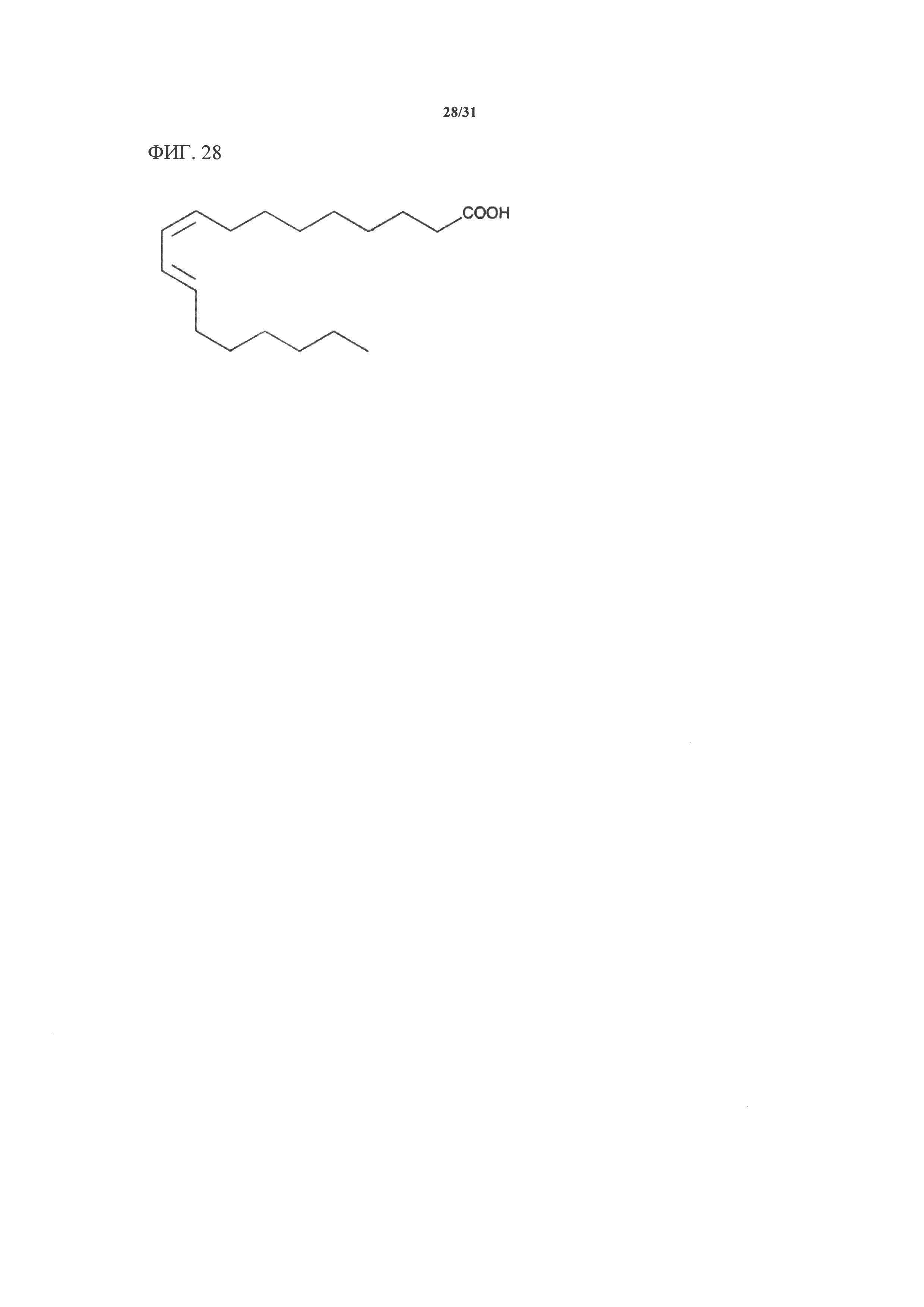
[00148] На фигуре 28 показана атомная структура 9(Z)-11(Е)-сопряженной линолевой кислоты.
[00149] На фигуре 29 показана атомная структура дистеароилфосфатидилхолина (DSPC).
[00150] На фигуре 30 показана атомная структура 2-олеоил-1-пальмитоил-sn-глицеро-3-фосфоэтаноламина.
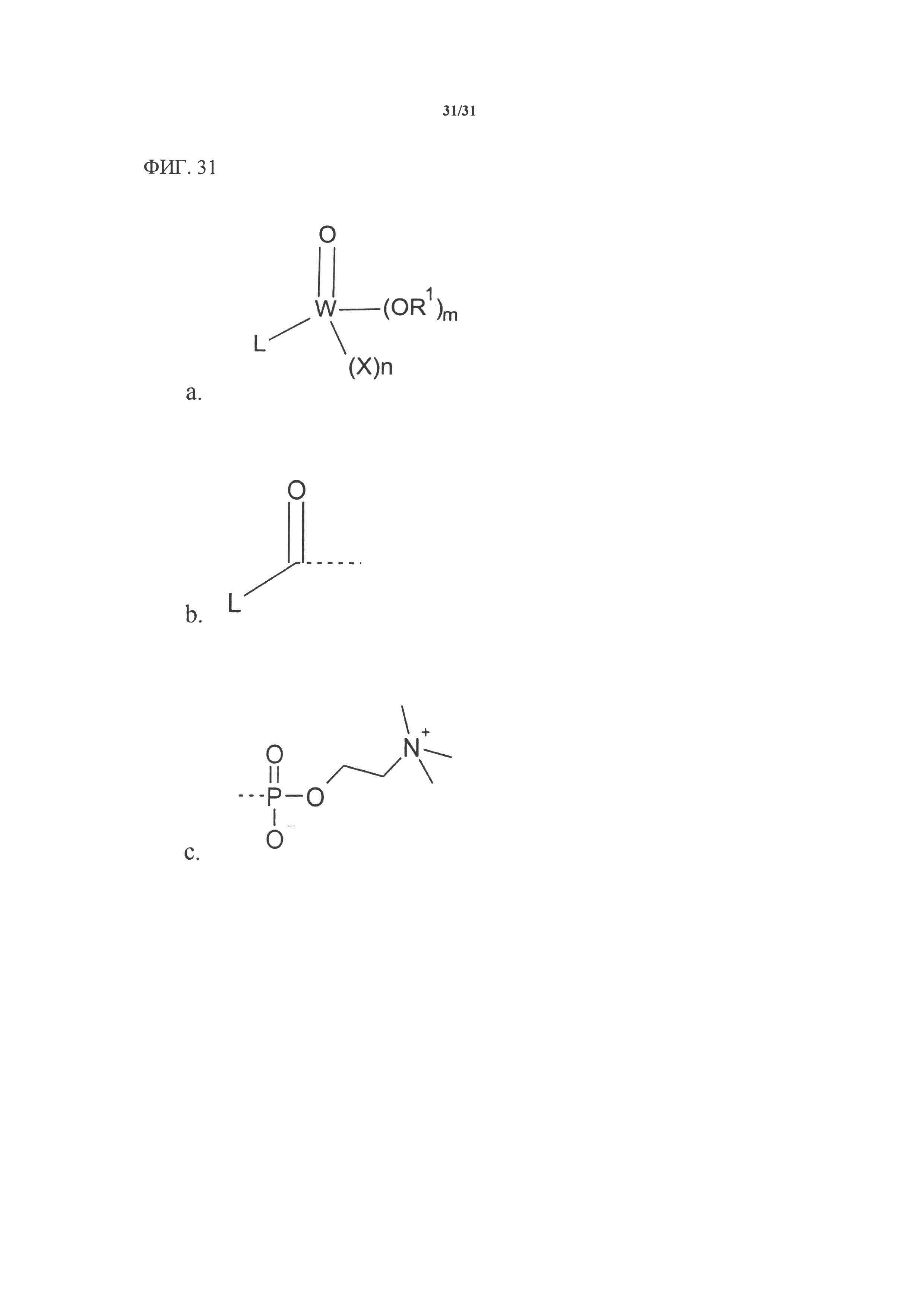
[00151] На фигуре 31 показана общая атомная структура молекул, пригодных к использованию в заявленном изобретении.
ОПРЕДЕЛЕНИЯ
[00152] Хотя следующие термины, как предполагается, хорошо понятны среднему специалисту в данной области, следующие определения приведены для облегчения пояснения раскрытого в данном документе объекта изобретения.
[00153] Все используемые в данном документе технические и научные термины, если ниже не указано иное, предполагаются как имеющие такое же значение, которое обычно понятно среднему специалисту в данной области. Ссылки на используемые в данном документе методики предполагаются как относящиеся к методикам, общепринятым в данной области техники, в том числе к изменениям этих методик и/или заменам на эквивалентные методики, которые будут очевидны специалистам в данной области. Хотя следующие термины, как предполагается, хорошо понятны среднему специалисту в данной области, следующие определения приведены для облегчения пояснения раскрытого в данном документе объекта изобретения.
[00154] Согласно устоявшейся традиции в патентном праве формы существительного единственного числа относятся к "одному или нескольким" при использовании в настоящей заявке, в том числе в формуле изобретения. Например, фраза "клетка" относится к одной или нескольким клеткам и в некоторых вариантах осуществления может относиться к ткани и/или органу. Аналогичным образом, фраза "по меньшей мере один" при использовании в данном документе для обозначения объекта относится, например, к 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40, 45, 50, 75, 100 или большему количеству этих объектов, включая без ограничения все целочисленные значения от 1 до 100, а также целые числа больше 100.
[00155] Если не указано иное, все числа, выражающие количества ингредиентов, условия реакции и т.д., используемые в настоящем описании и формуле изобретения, следует понимать как модифицированные во всех случаях термином "приблизительно". Термин "приблизительно", используемый в данном документе в отношении измеряемого значения, такого как величина массы, веса, времени, объема, концентрации или процентного значения, предназначено для охвата изменений определенного количества, составляющих в некоторых вариантах осуществления ±20%, в некоторых вариантах осуществления ±10%, в некоторых вариантах осуществления ±5%, в некоторых вариантах осуществления ±1%, в некоторых вариантах осуществления ±0,5% и в некоторых вариантах осуществления ±0,1%, поскольку такие изменения подходят для осуществления раскрытых способов и/или использования раскрытых композиций, нуклеиновых кислот, полипептидов и т.д. Соответственно, если не указано обратное, числовые параметры, изложенные в настоящем описании и прилагаемой формуле изобретения, являются приблизительными величинами, которые могут варьироваться в зависимости от желаемых свойств, которых стремятся достичь посредством раскрытого в данном документе объекта изобретения.
[00156] Используемый в данном документе термин "аллель" относится к варианту или альтернативной форме последовательности в генетическом локусе. У диплоидов отдельный аллель наследуется индивидуумом-потомком от каждой родительской особи отдельно в каждом локусе. Два аллеля данного локуса, присутствующие в диплоидном организме, занимают соответствующие места в паре гомологичных хромосом, хотя специалист в данной области понимает, что аллели в любом конкретном индивидууме необязательно представляют все аллели, которые присутствуют у данного вида.
[00157] Используемый в данном документе термин "и/или" при использовании применительно к перечню объектов относится к объектам, присутствующим в отдельности или в комбинации. Таким образом, например, фраза "А, В, С и/или D" включает А, В, С и D отдельно, но также включает все возможные комбинации и подкомбинации А, В, С и D (например, АВ, AC, AD, ВС, BD, CD, ABC, ABD и BCD). В некоторых вариантах осуществления один или несколько элементов, к которым относится "и/или", может также отдельно присутствовать в однократных или многократных случаях в комбинации(комбинациях) и/или подкомбинации(подкомбинациях).
[00158] Используемая в данном документе фраза "ассоциированный с" относится к распознаваемой и/или анализируемой взаимосвязи между двумя объектами. Например, фраза "ассоциированный с HI" относится к признаку, локусу, гену, аллелю, маркеру, фенотипу и т.д. или их экспрессии, присутствие или отсутствие которых может влиять на меру и/или степень, в которой растение или его потомство демонстрируют HI. Соответственно, маркер является "ассоциированным с" признаком, если он сцеплен с ним и если наличие маркера является индикатором того, будут ли и/или в какой степени будут желаемый признак или форма признака встречаться у растения/в зародышевой плазме, содержащих маркер. Аналогичным образом, маркер является "ассоциированным с" аллелем, если он сцеплен с ним и если наличие маркера является индикатором того, присутствует ли аллель у растения/в зародышевой плазме, содержащих маркер. Например, "маркер, ассоциированный с HI", относится к маркеру, наличие или отсутствие которого можно применять для предсказания того, будет ли и/или в какой степени будет растение проявлять индукцию гаплоидии.
[00159] Термин "содержащий", который является синонимом "включающего", "вмещающего" и "характеризующегося", является включающим или открытым и не исключает дополнительных, не упомянутых элементов и/или стадий способа. "Содержащий" является термином из уровня техники, который означает, что названные элементы и/или стадии присутствуют, но могут быть добавлены другие элементы и/или стадии, которые при этом по-прежнему находятся в пределах объема соответствующего объекта изобретения.
[00160] Используемая в данном документе фраза "состоящий из" исключает любой элемент, стадию или ингредиент, не упомянутые конкретно. Если фраза "состоит из" появляется в основной части пункта формулы изобретения, а не сразу после ограничительной части, то она ограничивает только элемент, изложенный в этой части; другие элементы не исключаются из пункта формулы изобретения в целом.
[00161] Используемая в данном документе фраза "состоящий фактически из" ограничивает объем связанного раскрытия или пункта формулы изобретения указанными материалами и/или стадиями, а также материалами и/или стадиями, которые существенно не влияют на основную(основные) и новую(новые) характеристику(характеристики) раскрытого и/или заявленного объекта изобретения.
[00162] Что касается терминов "содержащий", "состоящий фактически из" и "состоящий из", если в данном документе используется один из этих трех терминов, то раскрытый в данном документе и заявленный объект изобретения может включать в некоторых вариантах осуществления применение любого из двух других терминов. Например, это справедливо, если объект изобретения относится в некоторых вариантах осуществления к нуклеиновым кислотам, кодирующим полипептиды, содержащие аминокислотные последовательности, которые по меньшей мере на 95% идентичны SEQ ID NO: 2 или 3. Следует понимать, что раскрытый объект изобретения, таким образом, также охватывает нуклеиновые кислоты, кодирующие полипептиды, которые в некоторых вариантах осуществления состоят фактически из аминокислотных последовательностей, по меньшей мере на 95% идентичных этим SEQ ID NO: 2 или 3, а также нуклеиновые кислоты, кодирующие полипептиды, которые в некоторых вариантах осуществления состоят из аминокислотных последовательностей, по меньшей мере на 95% идентичных этим SEQ ID NO: 2 или 3. Аналогичным образом, также следует понимать, что в некоторых вариантах осуществления способы согласно раскрытому объекту изобретения включают стадии, раскрытые в данном документе, в некоторых вариантах осуществления способы согласно раскрытому в данном документе объекту изобретения состоят фактически из раскрытых стадий, и в некоторых вариантах осуществления способы согласно раскрытому в данном документе объекту изобретения состоят из стадий, раскрытых в данном документе.
[00163] Используемый в данном документе термин "индукция гаплоидии de novo" относится к вызыванию индукции гаплоидии посредством введения средства, индуцирующего спонтанную гаплоидию. Такое введение можно осуществлять посредством местного опрыскивания, опыления ручным способом, мутагенеза или трансгенных способов. Термины "индукция гаплоидии de novo", "HI de novo" и "de novo индукция гаплоидии" используются взаимозаменяемо во всем настоящем описании.
[00164] Используемый в данном документе термин "ген" относится к единице наследственности, содержащей последовательность ДНК, которая занимает определенное местоположение в хромосоме и которая содержит генетическую инструкцию для определенной характеристики или признака, свойственных организму.
[00165] "Генетическая карта" представляет собой описание взаимосвязей при генетическом сцеплении между локусами в одной или нескольких хромосомах у данного вида, обычно изображаемое в форме схемы или таблицы.
[00166] Как используется в данном документе, растение, называемое "гаплоидным", имеет одинарный набор (геном) хромосом, и уменьшенное количество хромосом (n) в гаплоидном растении равно количеству хромосом в гамете. Как используется в данном документе, растение, называемое "двойным гаплоидным", развивается посредством удвоения гаплоидного набора хромосом. Растение или семя, которые получают из двойного гаплоидного растения, самооплодотворяемого в течение любого количества поколений, можно по-прежнему идентифицировать как двойное гаплоидное растение. Двойное гаплоидное растение считается гомозиготным растением. Растение считается двойным гаплоидным, если оно является фертильным, даже если вся вегетативная часть растения состоит из клеток не с двойным набором хромосом; то есть растение будет считаться двойным гаплоидным, если оно содержит жизнеспособные гаметы, даже если оно является химерным.
[00167] Используемый в данном документе термин "индуцированная человеком мутация" относится к любой мутации, появляющейся в результате непосредственного либо опосредованного действия человека. Этот термин включает без ограничения, мутации, получаемые посредством любого способа направленного мутагенеза.
[00168] Используемые в данном документе термины "маркерный зонд" и "зонд" относятся к нуклеотидной последовательности или молекуле нуклеиновой кислоты, которые можно применять для выявления наличия или отсутствия последовательности в пределах большей последовательности посредством гибридизации нуклеиновых кислот, например, к зонду на основе нуклеиновой кислоты, комплементарному всему маркеру или маркерному локусу или его части. Для гибридизации нуклеиновых кислот можно применять маркерные зонды, содержащие приблизительно 8, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90, 100 или более смежных нуклеотидов.
[00169] Используемый в данном документе термин "молекулярный маркер" можно использовать для обозначения генетического маркера, определенного выше, или кодируемого им продукта (например, белка), применяемого в качестве исходной точки при идентификации наличия/отсутствия HI-ассоциированного локуса. Молекулярный маркер может быть получен из геномных нуклеотидных последовательностей или из экспрессируемых нуклеотидных последовательностей (например, из РНК, кДНК и т.д.). Данный термин также относится к нуклеотидным последовательностям, комплементарным маркерным последовательностям или фланкирующим их, таким как нуклеотидные последовательности, применяемые в качестве зондов и/или праймеров, способных амплифицировать маркерную последовательность. Нуклеотидные последовательности являются "комплементарными", если они специфически гибридизируются в растворе (например, согласно правилам спаривания оснований Уотсона-Крика). Данный термин также относится к генетическим маркерам, которые указывают на признак посредством отсутствия нуклеотидных последовательностей, комплементарных маркерным последовательностям или фланкирующих их, таких как нуклеотидные последовательности, применяемые в качестве зондов и/или праймеров, способных амплифицировать маркерную последовательность.
[00170] Используемые в данном документе термины "нуклеотидная последовательность", "полинуклеотид", "последовательность нуклеиновой кислоты", "молекула нуклеиновой кислоты" и "фрагмент нуклеиновой кислоты" относятся к полимеру РНК или ДНК, который является одно- или двухнитевым, необязательно содержащему синтетические, неприродные и/или измененные нуклеотидные основания. "Нуклеотид" представляет собой мономерную единицу, из которой сконструированы полимеры ДНК или РНК и которая состоит из пуринового или пиримидинового основания, пентозы и группы фосфорной кислоты. Нуклеотиды (обычно обнаруживаемые в форме их 5'-монофосфата) называют их однобуквенным обозначением следующим образом: "А" в случае аденилата или дезоксиаденилата (соответственно в случае РНК или ДНК), "С" в случае цитидилата или дезоксицитидилата, "G" в случае гуанилата или дезоксигуанилата, "U" в случае уридилата, "Т" в случае дезокситимидилата, "R" в случае пуринов (А или G), "Y" в случае пиримидинов (С или Т), "K" в случае G или Т, "Н" в случае А, или С, или Т, "I" в случае инозина и "N" в случае любого возможного нуклеотида.
[00171] Используемый в данном документе термин "идентичность нуклеотидных последовательностей" относится к наличию идентичных нуклеотидов в соответствующих положениях двух полинуклеотидов. Полинуклеотиды имеют "идентичные" последовательности в случае, если последовательность нуклеотидов в двух полинуклеотидах является одной и той же при выравнивании с максимальным соответствием (например, в окне сравнения). Сравнение последовательностей двух или более нуклеотидов обычно осуществляют путем сравнения частей двух последовательностей в окне сравнения для выявления и сравнения локальных областей сходства последовательностей. Как правило, окно сравнения составляет от приблизительно 20 до 200 смежных нуклеотидов. "Процентное значение идентичности последовательностей" для полинуклеотидов, как, например, приблизительно 50-, 55-, 60-, 65-, 70-, 75-, 80-, 85-, 90-, 95-, 98-, 99-или 100-процентную идентичность последовательностей, можно определять путем сравнения двух оптимально выровненных последовательностей в окне сравнения, где часть полинуклеотидной последовательности в окне сравнения может содержать добавления или делеции (т.е. гэпы) по сравнению с эталонной последовательностью для оптимального выравнивания двух последовательностей. В некоторых вариантах осуществления процентное значение рассчитывают путем (а) определения числа положений, в которых идентичное основание нуклеиновой кислоты встречается в обеих последовательностях; (b) деления числа совпадающих положений на общее число положений в окне сравнения и (с) умножения результата на 100. Оптимальное выравнивание последовательностей для сравнения можно также проводить с помощью компьютерных реализаций известных алгоритмов или путем визуального осмотра. Общедоступными алгоритмами сравнения последовательностей и множественного выравнивания последовательностей являются соответственно средство поиска основного локального выравнивания (BLAST) и программы ClustalW/ClustalW2/Clustal Omega, доступные в Интернете (например, на веб-сайте EMBL-EBI). Другие подходящие программы включают без ограничений GAP, BestFit, PlotSimilarity и FASTA, которые являются частью пакета программ Accelrys GCG, доступного от Accelrys, Inc. Сан-Диего, Калифорния, Соединенные Штаты Америки. См. также Smith & Waterman, 1981; Needleman & Wunsch, 1970; Pearson & Lipman, 1988; Ausubel et al, 1988 и Sambrook & Russell, 2001.
[00172] Одним примером алгоритма, который подходит для определения процентного значения идентичности последовательностей и сходства последовательностей, является алгоритм BLAST, который описан в Altschul et al., 1990. В некоторых вариантах осуществления процентное значение идентичности последовательностей относится к идентичности последовательностей по всей длине одной из сравниваемых последовательностей gDNA, кДНК или предсказываемого белка в наибольшей ORF SEQ ID NO: 1. В некоторых вариантах осуществления при расчете с целью определения процентного значения идентичности последовательностей нуклеиновых кислот не включаются в расчет какие-либо положения нуклеотидов, в которых любая из сравниваемых нуклеиновых кислот содержит "N" (т.е. где в данном положении может находиться любой нуклеотид).
[00173] Термин "открытая рамка считывания" (ORF) относится к последовательности нуклеиновой кислоты, которая кодирует полипептид. В некоторых вариантах осуществления ORF содержит кодон инициации трансляции, кодон терминации трансляции (т.е. стоп-кодон) и последовательность нуклеиновой кислоты между ними, которая кодирует аминокислоты, присутствующие в полипептиде. Термины "инициирующий кодон" и "терминирующий кодон" относятся к единице из трех прилегающих нуклеотидов (т.е. кодону) в кодирующей последовательности, которая определяет соответственно инициацию и терминацию цепи в синтезе белка (трансляции мРНК).
[00174] Пататин-подобная фосфолипаза А2α может также быть известна как PLA, pPLA, pPLAIIA, pPLAIIα, PLA2-альфа или PLA2 или другие сходные варианты. Пататин-подобную фосфолипазу AIIα также называют MATRILINEAL. Данные термины везде используются взаимозаменяемо. Ген MATRILINEAL, содержащий мутацию сдвига рамки считывания, охватывающую четыре пары оснований, в данном документе называется matrilineal.
[00175] Используемые в данном документе термины "фенотип", "фенотипический признак" или "признак" относятся к одному или нескольким признакам растения или растительной клетки. Фенотип можно наблюдать невооруженным глазом или с помощью любых других средств оценки, известных в данной области техники, например, микроскопии, биохимического анализа или электромеханического анализа. В некоторых случаях фенотип непосредственно контролируется одним геном или генетическим локусом (т.е. соответствует "признаку, определенному одним геном"). В случае индукции гаплоидии применение маркеров цвета, таких как R-Navajo, и других маркеров, в том числе трансгенов, визуализируемых по наличию или отсутствию цвета в семени, дает свидетельство того, является ли семя семенем с индуцированной гаплоидией. Применение R-Navajo в качестве маркера цвета и применение трансгенов хорошо известно из уровня техники в качестве способов выявления индукции образования гаплоидных семян на женском растении. В других случаях фенотип является результатом взаимодействий между несколькими генами, и в некоторых вариантах осуществления он также являются результатом взаимодействия растения и/или растительной клетки с их окружающей средой.
[00176] Используемый в данном документе термин "растение" может относится к целому растению, любой его части или культуре клеток или тканей, полученным из растения. Таким образом, термин "растение" может относится к любому из целых растений, компонентов или органов растения (например, листьев, стеблей, корней и т.д.), растительных тканей, семян и/или растительных клеток.
[00177] Растительная клетка представляет собой клетку растения, взятую из растения или полученную посредством культивирования из клетки, взятой из растения. Таким образом, термин "растительная клетка" включает без ограничения клетки в семенах, суспензионных культурах, зародышах, областях меристемы, каллюсной ткани, листьях, побегах, гаметофитах, спорофитах, пыльце и микроспорах. Фраза "часть растения" относится к части растения, в том числе к отдельным клеткам и клеточным тканям, таким как растительные клетки, являющиеся интактными в растениях, скопления клеток и культуры тканей, из которых можно регенерировать растения. Примеры частей растения включают без ограничения отдельные клетки и ткани из пыльцы, семязачатков, листьев, зародышей, корней, кончиков корней, пыльников, цветков, плодов, стеблей, побегов и семян; а также привоев, подвоев, протопластов, каллюсов и т.п.
[00178] Используемый в данном документе термин "праймер" относится к олигонуклеотиду, который способен отжигаться с нуклеиновой кислотой-мишенью (в некоторых вариантах осуществления специфически отжигаться с нуклеиновой кислотой-мишенью), обеспечивая присоединение к ней ДНК-полимеразы и/или обратной транскриптазы и выступая таким образом в качестве точки инициации синтеза ДНК, будучи помещенным в условия, в которых индуцируется синтез продукта удлинения праймера (например, в присутствии нуклеотидов и средства для полимеризации, такого как ДНК-полимераза, и при подходящих значениях температуры и рН). В некоторых вариантах осуществления для амплификации нуклеиновых кислот растения используют одну или несколько совокупностей праймеров (например, с применением полимеразной цепной реакции; ПЦР).
[00179] Используемый в данном документе термин "зонд" относится к нуклеиновой кислоте (например, однонитевой нуклеиновой кислоте или нити двухнитевой нуклеиновой кислоты или нуклеиновой кислоты более высокого порядка или их подпоследовательности), которая может образовывать стабилизированный водородными связями дуплекс с комплементарной последовательностью в последовательности нуклеиновой кислоты-мишени. Как правило, зонд имеет достаточную длину для образования стабильной и специфичной для последовательности дуплексной молекулы с комплементарной ему последовательностью, и, следовательно, может использоваться в некоторых вариантах осуществления для выявления последовательности, представляющей интерес, присутствующей во множестве нуклеиновых кислот.
[00180] Используемые в данном документе термины "потомство" и "растение-потомок" относятся к растению, полученному в результате вегетативного или полового размножения от одного или нескольких родительских растений. При индукции гаплоидии семена на женской родительской особи являются гаплоидными и, следовательно, не являются потомством линии, индуцирующей гаплоидию. Потомство гаплоидных семян не является единственным желаемым потомством. Существуют также семена HI и последующие растения- и семена-потомки растения, индуцирующего гаплоидию. Как гаплоидные семена, так и семена HI могут являться потомством. Растение-потомка можно получать путем клонирования или самооплодотворения одного родительского растения или путем скрещивания двух или более родительских растений. Например, растение-потомка можно получать путем клонирования или самооплодотворения одного родительского растения или путем скрещивания двух родительских растений и включает продукты самооплодотворения, а также F1 или F2 или более далекие поколения. F1 представляет собой потомство первого поколения, полученное от родительских особей, по меньшей мере одну из которых впервые используют в качестве донора признака, тогда как потомство второго поколения (F2) или последующих поколений (F3, F4 и т.п.) представляет собой образцы, полученные в результате самооплодотворений, перекрестных опылений, возвратных скрещиваний и/или других скрещиваний F1, F2 и т.п. F1, таким образом, может представлять собой (и в некоторых вариантах осуществления представляет собой) гибрид, полученный в результате скрещивания двух родительских особей из чистых линий (т.е. каждая из родительских особей из чистых линий является гомозиготной по признаку, представляющему интерес, или его аллелю), тогда как F2 может представлять собой (и в некоторых вариантах осуществления представляет собой) потомство, полученное в результате самоопыления гибридов F1.
[00181] Используемая в данном документе фраза "рекомбинация" относится к обмену фрагментами ДНК между двумя молекулами ДНК или хроматидами парных хромосом ("кроссинговеру") в области сходства или идентичности нуклеотидных последовательностей. "Событие рекомбинации" в данном документе понимают как относящееся в некоторых вариантах осуществления к мейотическому кроссинговеру.
[00182] Используемый в данном документе термин "эталонная последовательность" относится к определенной нуклеотидной последовательности, применяемой в качестве основы для сравнения нуклеотидных последовательностей. В некоторых вариантах осуществления любая из SEQ ID NO: 1-4, 22-23 или 73-81 может выступать в качестве эталонной последовательности для сравнения с другими последовательностями, полученными из растений.
[00183] Используемый в данном документе термин "регенерировать" и его грамматические варианты относятся к получению растения из культуры тканей.
[00184] Используемая в данном документе фраза "жесткие условия гибридизации" относится к условиям, при которых полинуклеотид гибридизируется со своей подпоследовательностью-мишенью, как правило, в сложной смеси нуклеиновых кислот, но фактически не делает этого с другими последовательностями. Жесткие условия зависят от последовательности и могут различаться при разных обстоятельствах.
[00185] Более длинные последовательности, как правило, специфически гибридизируются при более высоких температурах. Исчерпывающее руководство по гибридизации нуклеиновых кислот можно найти в Sambrook & Russell, 2001. Как правило, жесткие условия выбирают так, чтобы температура была на приблизительно 5-10°С ниже температурной точки плавления (Tm) для конкретной последовательности при определенных ионной силе и рН. Tm представляет собой температуру (при определенных ионной силе, рН и концентрации нуклеиновой кислоты), при которой 50% зондов, комплементарных мишени, гибридизируются с последовательностью-мишенью в равновесном состоянии (поскольку последовательности-мишени присутствуют в избытке, то при Tm в равновесном состоянии занято 50% зондов). Иллюстративными жесткими условиями являются условия, при которых концентрация солей составляет менее чем приблизительно 1,0 М ионов натрия, как правило, концентрация ионов натрия (или других солей) составляет приблизительно 0,01-1,0 М при рН 7,0-8,3, а температура составляет по меньшей мере приблизительно 30°С для коротких зондов (например, 10-50 нуклеотидов) и по меньшей мере приблизительно 60°С для длинных зондов (например, более 50 нуклеотидов).
[00186] Жестких условий также можно достичь путем добавления дестабилизирующих средств, таких как формамид. Дополнительные иллюстративные жесткие условия гибридизации включают инкубирование в 50% формамиде, 5х SSC и 1% SDS при 42°С; или инкубирование в SSC, 1% SDS при 65°С; с одной или несколькими стадиями отмывки в 0,2х SSC и 0,1% SDS при 65°С. Для ПЦР температура, составляющая приблизительно 36°С, является типичной для амплификации в условиях низкой жесткости, хотя значения температуры отжига могут варьироваться от приблизительно 32°С до 48°С (или выше) в зависимости от длины праймера. Дополнительные руководства по определению параметров гибридизации представлены во множестве литературных источников (см. например, Ausubel et al., 1999).
[00187] Используемый в данном документе термин "признак" относится к фенотипу, представляющему интерес, гену, вносящему вклад в фенотип, представляющий интерес, а также к последовательности нуклеиновой кислоты, ассоциированной с геном, вносящим вклад в фенотип, представляющий интерес. Например, "признак HI" относится к фенотипу индукции гаплоидии, а также к гену, который вносит вклад в индукцию гаплоидии, и последовательности нуклеиновой кислоты (например, продукту гена, ассоциированному с HI), ассоциированной с наличием или отсутствием фенотипа индукции гаплоидии.
[00188] Используемый в данном документе термин "трансген" относится к молекуле нуклеиновой кислоты, введенной в организм или одного или нескольких его предков посредством некоторой формы методики искусственного переноса. Посредством методики искусственного переноса, таким образом, создают "трансгенный организм" или "трансгенную клетку". Следует понимать, что методику искусственного переноса можно осуществлять в организме-предке (или его клетке и/или клетке, из которой может развиться организм-предок), и тем не менее любой индивидуум-потомок, который имеет искусственно перенесенную молекулу нуклеиновой кислоты или ее фрагмент, по-прежнему считается трансгенным объектом, если в результате одного или нескольких естественных и/или вспомогательных скрещиваний искусственно перенесенная молекула нуклеиновой кислоты присутствует в индивидууме-потомке.
[00189] Используемый в данном документе термин "направленный мутагенез" или "стратегия мутагенеза" относится к любому способу мутагенеза, который приводит в результате к преднамеренному мутагенезу выбранного гена. Направленный мутагенез включает способы с использованием CRISPR, TILLING, TALEN и другие способы, которые еще не обнаружены, но которые можно применять для достижения такого же результата.
[00190] Используемый в данном документе термин "уровень индукции гаплоидии" ("HIR") означает количество выживших гаплоидных зерен по сравнению с общим количеством зерен после опыления початка пыльцой, индуцирующей гаплоидию.
[00191] Конкретными проблемами, преследующими эту индукцию гаплоидии, являются увеличенные уровни недоразвитости зародышей и увеличенные частоты неудачного оплодотворения (пониженные уровни завязывания семян). В силу описанных причин существует потребность в успешном определении причины HI и применении этого знания для определения способов создания гаплоидных растений в стабильном или увеличивающемся объеме при одновременном снижении частоты неудачного оплодотворения и недоразвитости зародышей.
[00192] В частности, предполагается, что промотор можно подвергнуть мутагенезу, чтобы потенциально улучшить пригодность элементов для экспрессии трансгенов у растений. Мутагенез этих элементов можно осуществлять случайным образом, и подвергнутые мутагенезу промоторные последовательности можно подвергать скринингу в отношении активности по методу проб и ошибок. В качестве альтернативы, можно идентифицировать определенные последовательности, которые придают промотору желаемые характеристики экспрессии или придают промотору активность усиления экспрессии, и эти или сходные последовательности можно вводить в промотор посредством мутации. Дополнительно предполагается, что эти последовательности можно подвергнуть мутагенезу для усиления экспрессии трансгенов с их помощью у определенного вида. Способы осуществления мутагенеза сегмента ДНК, кодирующего промоторную последовательность по настоящему изобретению, хорошо известны специалистам в данной области. Как указано, модификации промотора или другого регуляторного элемента можно осуществлять посредством процедур случайного или сайт-специфического мутагенеза. Промотор и другой регуляторный элемент можно модифицировать путем изменения их структуры посредством добавления или делеции одного или нескольких нуклеотидов из последовательности, которая кодирует соответствующие немодифицированные последовательности.
[00193] Мутагенез можно осуществлять в соответствии с любыми методиками, известными из уровня техники, такими как без ограничения синтез олигонуклеотида, имеющего одну или несколько мутаций в пределах последовательности в конкретной регуляторной последовательности. В частности, сайт-специфический мутагенез представляет собой методику, применимую в получении мутантных промоторов посредством специфического мутагенеза лежащей в их основе ДНК. Также можно применять РНК-управляемые эндонуклеазы ("RGEN", например, CRISPR/Cas9). Методика дополнительно обеспечивает легкодоступную возможность получения и тестирования вариантов последовательностей, например, с учетом одного или нескольких из вышеуказанных соображений, посредством введения одного или нескольких изменений нуклеотидной последовательности в ДНК. Сайт-специфический мутагенез обеспечивает получение мутантных форм посредством применения специфических олигонуклеотидных последовательностей, которые кодируют последовательность ДНК с желаемой мутацией, а также достаточного количества прилегающих нуклеотидов, для получения последовательности праймера, имеющей достаточные размер и сложность последовательности для образования стабильного дуплекса по обе стороны от пересекаемой границы делеции. Как правило, предпочтительным является праймер, составляющий в длину от приблизительно 17 до приблизительно 75 нуклеотидов или более, имеющий от приблизительно 10 до приблизительно 25 или более остатков по обе стороны от границы в последовательности, которую изменяют.
[00194] В случае, если клон, содержащий промотор, был выделен в соответствии с настоящим изобретением, можно пожелать определить границы существенных областей промотора в пределах клона. Один эффективный направленный способ для получения подвергнутых мутагенезу промоторов основан на идентификации предполагаемых регуляторных элементов в промоторной последовательности. Ее можно начать со сравнения с промоторными последовательностями, которые, как известно, характеризуются сходными тканеспецифическими или уникальными для стадии развития профилями экспрессии. Последовательности, которые являются общими среди промоторов со сходными профилями экспрессии, являются вероятными кандидатами для связывания с факторами транскрипции и, таким образом, являются вероятными элементами, обеспечивающими профили экспрессии. Подтверждение этих предполагаемых регуляторных элементов можно осуществлять посредством делеционного анализа каждой предполагаемой регуляторной последовательности с последующим функциональным анализом каждой делеционной конструкции посредством анализа репортерного гена, функционально присоединенного к каждой конструкции. Таким образом, после получения исходной промоторной последовательности можно легко получить любую из ряда различных делеционных мутантных форм исходного промотора.
[00195] В настоящем изобретении, раскрытом в данном документе, предусматриваются молекулы полинуклеотидов, содержащие фрагменты регуляторных элементов, которые можно применять при конструировании новых химерных регуляторных элементов. Новые комбинации, содержащие фрагменты этих молекул полинуклеотидов и по меньшей мере один другой регуляторный элемент или фрагмент, можно конструировать и тестировать у растений, и они считаются находящимися в пределах объема настоящего изобретения. Таким образом, разработка, конструирование и применение химерных регуляторных элементов составляют один вариант осуществления настоящего изобретения. Промоторы по настоящему изобретению включают в себя гомологи цис-действующих элементов, которые, как известно, влияют на регуляцию экспрессии генов, демонстрирующие гомологию с промоторными последовательностями по настоящему изобретению.
[00196] Функциональные эквивалентные фрагменты одной из нуклеиновых кислот, регулирующих транскрипцию, описанных в данном документе, содержат по меньшей мере 50, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950 или 1000 пар оснований нуклеиновой кислоты, регулирующей транскрипцию. Из эквивалентных фрагментов нуклеиновых кислот, регулирующих транскрипцию, которые получают посредством делеции области, кодирующей 5'-нетранслируемую область мРНК, можно затем получить только (нетранскрибируемую) промоторную область. 5'-Нетранслируемую область можно легко определить с помощью способов, известных из уровня техники (таких как анализ 5'-RACE). Соответственно, некоторые из нуклеиновых кислот, регулирующих транскрипцию, описанных в данном документе, являются эквивалентными фрагментами других последовательностей.
[00197] Как указано выше, также можно случайным образом получать и затем анализировать делеционные мутантные формы промотора по настоящему изобретению. Следуя этой стратегии, получают серию конструкций, каждая из которых содержит отличную от других часть промотора (субклон), и затем эти конструкции подвергают скринингу в отношении активности. Подходящими способами для осуществления скрининга в отношении активности являются присоединение промоторной или интронной конструкции с делецией, которая содержит сегмент, подвергнутый делеции, к селектируемому или подвергаемому скринингу маркеру, и выделение только тех клеток, которые экспрессируют маркерный ген. Таким путем идентифицируют ряд различных промоторных конструкций с делециями, которые по-прежнему сохраняют желаемую или даже повышенную активность. Благодаря этому идентифицируют наименьший сегмент, требуемый для обеспечения активности, посредством сравнения выбранных конструкций. Этот сегмент можно затем применять для конструирования векторов для экспрессии экзогенных генов.
[00198] Кассета экспрессии, описанная в данном документе, может содержать дополнительные регуляторные элементы. Этот термин в данном контексте следует понимать в широком значении, включающем все последовательности, которые могут влиять на конструкцию или функцию кассеты экспрессии. Регуляторные элементы могут, например, модифицировать транскрипцию и/или трансляцию у прокариотических или эукариотических организмов. Кассета экспрессии, описанная в данном документе, может находиться ниже (в 3'-направлении) последовательности нуклеиновой кислоты, подлежащей экспрессии, и необязательно может содержать дополнительные регуляторные элементы, такие как транскрипционные или трансляционные энхансеры. Каждый дополнительный регуляторный элемент может быть функционально связан с последовательностью нуклеиновой кислоты, подлежащей экспрессии (или нуклеотидной последовательностью, регулирующей транскрипцию). Дополнительные регуляторные элементы могут включать в себя дополнительные промоторы, минимальные промоторы, промоторные элементы или транспозонные элементы, которые могут модифицировать или усиливать свойства регуляции экспрессии. Кассета экспрессии может также содержать один или несколько интронов, один или несколько экзонов и один или несколько терминаторов.
[00199] Кроме того, предполагается, что могут быть применимыми промоторы, в которых объединены элементы из более чем одного промотора. Например, в патенте США №5491288 раскрывается объединение промотора вируса мозаики цветной капусты с гистоновым промотором. Таким образом, элементы из промоторов, раскрытых в данном документе, можно объединять с элементами из других промоторов. Промоторы, применимые для экспрессии трансгенов у растений, включают индуцируемые, вирусные, синтетические, конститутивные (Odell Nature 313: 810-812 (1985)), регулируемые во времени, регулируемые в пространстве, тканеспецифические и регулируемые в пространстве и времени промоторы. Применяя регуляторные элементы, описанные в данном документе, можно экспрессировать множество агрономически значимых генов в трансформированных растениях. Более конкретно, растения можно создавать методами генной инженерии для экспрессии различных фенотипов, представляющих агрономический интерес.
[00200] Соединения по настоящему изобретению могут существовать в виде различных геометрических или оптических изомеров (диастереоизомеров и энантиомеров) или таутомерных форм. Настоящее изобретение охватывает все такие изомеры и таутомеры и их смеси во всех количественных соотношениях, а также изотопные формы, такие как дейтерированные соединения. Настоящее изобретение также охватывает все соли, N-оксиды и металлоидные комплексы соединений по настоящему изобретению.
[00201] Каждый алкильный фрагмент в отдельности либо как часть большей группы (такой как алкокси, алкоксикарбонил, алкилкарбонил, алкиламинокарбонил, диалкиламинокарбонил) имеет линейную или разветвленную цепь и представляет собой, например, метил, этил, н-пропил, н-бутил, н-пентил, н-гексил, изопропил, н-бутил, втор-бутил, изобутил, трет-бутил или неопентил. Алкильные группы включают в себя С1-С6алкил, С1-С4алкил и С1-С3алкил.
[00202] Термин "алкенил", используемый в данном документе, представляет собой алкильный фрагмент, имеющий по меньшей мере одну углерод-углеродную двойную связь, например, С2-С6алкенил. Конкретные примеры включают винил и аллил. Алкенильный фрагмент может представлять собой часть большей группы (такой как алкенокси, алкеноксикарбонил, алкенилкарбонил, алкениламинокарбонил, диалкениламинокарбонил).
[00203] Термин "ацетокси" относится к -ОС(=O)СН3.
[00204] Термин "алкинил", используемый в данном документе, представляет собой алкильный фрагмент, имеющий по меньшей мере одну углерод-углеродную тройную связь, например, С2-С6алкинил. Конкретные примеры включают этинил и пропаргил. Алкинильный фрагмент может представлять собой часть большей группы (такой как алкинокси, алкиноксикарбонил, алкинилкарбонил, алкиниламинокарбонил, диалкиниламинокарбонил).
[00205] Галоген представляет собой фтор (F), хлор (Cl), бром (Br) или йод (I).
[00206] Галогеналкильные группы (в отдельности либо как часть большей группы, такой как галогеналкокси или галогеналкилтио) являются алкильными группами, замещенными одним или несколькими одинаковыми или разными атомами галогенов, и представляют собой, например, -CF3, -CF2Cl, -CH2CF3 или -CH2CHF2.
[00207] Гидроксиалкильные группы являются алкильными группами, замещенными одной или несколькими гидроксильными группами, и представляют собой, например, -CH2OH, -CH2CH2OH или -СН(ОН)СН3.
[00208] Алкоксиалкильные группы представляют собой алкоксигруппу, связанную с алкилом (R-O-R'), например, -(CH2)rO(CH2)sCH3, где r равняется 1-6, и s равняется 1-5.
[00209] В контексте настоящего описания термин "арил" относится к кольцевой системе, которая может быть моно-, би- или трициклической. Примеры таких колец включают фенил, нафталинил, антраценил, инденил или фенантренил.
[00210] Если не указано иное, алкенил и алкинил сами по себе или как часть другого заместителя могут иметь линейную или разветвленную цепь и могут содержать от 2 до 6 атомов углерода и в соответствующих случаях могут находиться либо в (Е)-, либо (Z)-конфигурации. Примеры включают винил, аллил, этинил и пропаргил.
[00211] Если не указано иное, циклоалкил может быть моно- или бициклическим, может быть необязательно замещен одной или несколькими С1-С6алкильными группами и содержит от 3 до 7 атомов углерода. Примеры циклоалкила включают циклопропил, 1-метилциклопропил, 2-метилциклопропил, циклобутил, циклопентил и циклогексил.
[00212] Термин "гетероциклил" относится к кольцевой системе, содержащей от одного до четырех гетероатомов, выбранных из N, О и S, где атомы азота и серы необязательно окислены, и атом(атомы) азота необязательно кватернизирован(кватернизированы). Гетероциклил включает в себя гетероарил, его насыщенные аналоги и, кроме того, их ненасыщенные или частично ненасыщенные аналоги, такие как 4,5,6,7-тетрагидробензотиофенил, 9Н-флуоренил, 3,4-дигидро-2Н-бензо-1,4-диоксепинил, 2,3-дигидробензофуранил, пиперидинил, 1,3-диоксоланил, 1,3-диоксанил, 4,5-дигидроизоксазолил, тетрагидрофуранил и морфолинил. Кроме того, термин "гетероциклил" включает гетероциклоалкил, неароматическое моноциклическое или полициклическое кольцо, содержащее атомы углерода и водорода и по меньшей мере один гетероатом, выбранный из атомов азота, кислорода и серы, например, азоксетанил или тиетанил. Моноциклический гетероциклоалкил может содержать 3-7 членов.
[00213] Термин "гетероарил" относится к ароматической кольцевой системе, содержащей от одного до четырех гетероатомов, выбранных из N, О и S, где атомы азота и серы необязательно окислены, например, имеющей 5, 6, 9 или 10 членов и состоящей из одного кольца либо из двух или более конденсированных колец. Отдельные кольца могут содержать не более трех гетероатомов, а бициклические системы - не более четырех гетероатомов, которые предпочтительно будут выбраны из атомов азота, кислорода и серы. Примеры таких групп включают пиридил, пиридазинил, пиримидинил, пиразинил, фуранил, тиенил, оксазолил, изоксазолил, оксадиазолил, тиазолил, изотиазолил, тиадиазолил, пирролил, пиразолил, имидазолил, триазолил и тетразолил.
ПРИМЕРЫ
I. Идентификация мутации сдвига рамки считывания в PLA
[00214] В настоящем изобретении определены серия независимых индуцированных человеком мутаций, обнаруживаемых по меньшей мере в одном гене пататин-подобной фосфолипазы AIIα (pPLAIIα) маиса; растения маиса, имеющие эти мутации по меньшей мере в одном из их генов PLA; и способ создания и идентификации сходных и/или дополнительных мутаций в гене PLA посредством скрининга объединенных и/или отдельных растений риса и маиса. Растения риса и маиса по настоящему изобретению индуцируют гаплоидию в результате нетрансгенных мутаций по меньшей мере в одном из их генов PLA.
[00215] Более конкретно, с помощью настоящего изобретения получают новые линии маиса, индуцирующие гаплоидию. Существует ряд известных линий маиса, индуцирующих гаплоидию, в том числе без ограничения Stock 6, MHI (молдавский индуктор гаплоидии), линия с мутацией неопределенного гаметофита ("ig"), KEMS, ZEM, ZMS, KMS, RWS и RWK. Настоящее изобретение относится к способу идентификации и/или отбора зародышевой плазмы, которая способна или не способна индуцировать образование гаплоидов. Настоящее изобретение также относится к увеличению количества и дополнительной разработке отобранной зародышевой плазмы, индуцирующей гаплоидию. Настоящее изобретение дополнительно относится к способу улучшения зародышевой плазмы, индуцирующей гаплоидию, для увеличения индукции образования гаплоидов у родительской особи, образующей семена.
[00216] Первоначальная стадия в получении гаплоидных семян от гибридного или сегрегантного материнского родительского растения начинается с опыления початка растения, образующего семена, пыльцой индуктора гаплоидии. Результатом этого процесса гибридизации является получение диплоидных и материнских гаплоидных (1n) зерен. Между зернами с индуцированной гаплоидией (1n) и диплоидными семенами часто проводят различие посредством применения маркеров цвета, которые указывают на плоидность зародыша. Диплоидные семена обычно отбрасывают, тогда как гаплоидные зерна или зародыши часто подвергают процессу удвоения числа хромосом с получением двойных гаплоидных растений. Более конкретно, гаплоидный генетический материал обрабатывают одним или несколькими средствами, блокирующими митоз, для обеспечения образования гомологичных пар в гаплоидном (1n) наборе хромосом в одной или нескольких клетках. После процедуры химической обработки химическое(химические) вещество(вещества), удваивающее(удваивающие) число хромосом, удаляют. Маису, являющемуся теперь двойным гаплоидом, позволяют созревать, и полученные в результате двойные гаплоидные семена при посадке будут давать гомозиготные растения (также называемые инбредными растениями или линиями). Эти инбредные линии являются материалами, которые селекционеры используют для проведения своих программ выведения гибридов.
[00217] Локус признака индукции гаплоидии был подвергнут тонкому картированию. Хотя главный QTL в хромосоме 1, отвечающий за индукцию гаплоидию, был картирован и опубликован в Dong et al. Theor. Appl. Genet (2013) 126: 1713-1720, точный ген/генетический элемент, отвечающий за процесс индукции, до сегодняшнего дня не был идентифицирован. Чтобы прояснить генетические механизмы развития, лежащие в основе индукции гаплоидии, производную Stock 6 RWK (~13% HIR) получали из Гогенгеймского университета в 2006 г., скрещивали с инбредными линиями NP2460 и NP2391 и впоследствии подвергали возвратному скрещиванию с RWK для образования картирующих популяций. См. фигуру 1.
[00218] Повышенный HIR в обоих популяциях, косегрегировал с маркером SM020SDQ в секции 1.04, что согласуется с недавними сообщениями о QTL, называемом qhir1. См. Prigge, et al., New Insights into the Genetics of in Vivo Induction of Maternal Haploids, the Backbone of Doubled Haploid Technology in Maize, Genetics (2012) 190:781-793 (в которой обсуждается главный QTL для HI в секции 1.04 [qhir1] и минорные QTL для HI в секциях 3.02 [qhir2], 3.06 [qhir3], 4.03 [qhir4], 5.01 [qhir5], 5.04 [qhir6], 7.01 [qhir7] и 9.01 [qhir8]); Liu, et al., Fine mapping of qhir8 affecting in vivo haploid induction in maize, Theor. Appl. Genet. (2015) 128:2507-2515 (в которой описывается тонкое картирование тридцати пяти генов с отнесением их к qhir8); Hu, et al., Genetic Basis of Haploid Induction in Maize Identified with a Novel Genome-Wide Association Method, genetics (2016) 202:1267-1276 (в которой утверждается, что qhir1 представляет собой два QTL: qhir11 и qhir12, и описывается тонкое картирование qhir6 с отнесением его к области размером 1,1 млн п.о.). Авторы настоящего изобретения провели несколько циклов тонкого картирования и ограничили QTL областью размером примерно 0,57 млн п.о. между 67,85 млн п.о. и 68,42 млн п.о., которая лежит в пределах qhir11. Эта область имеет семь аннотированных генов (фигура 2).
[00219] С помощью Illumina HiSeq2000 авторы настоящего изобретения секвенировали RWK, Stock 6 и неиндуцирующую линию BC3F5 "RWK-NIL", которая является почти изогенной RWK, но имеет гаплотипы NP2391 в интервале qhir11. Путем сравнения индуцирующей и неиндуцирующей зародышевой плазмы было определено, что вставка из четырех нуклеотидов, присутствующая у индукторов гаплоидии, которая сдвигает рамку считывания для кодирования аминокислот в GRMZM2G471240, не присутствует в неиндуцирующей зародышевой плазме. Таким образом, в настоящем изобретении был идентифицирован ген с мутацией сдвига рамки считывания в индуцирующей зародышевой плазме в качестве отвечающего за индукцию гаплоидии у маиса. Ген-кандидат, соответствующий генной модели GRMZM2G471240, кодирует пататин-подобную фосфолипазу AIIα (pPLAIIα), которую авторы настоящего изобретения переименовали в MATRILINEAL (MTL) для представления аллеля дикого типа, а аллель со сдвигом рамки считывания называется matrilineal (mtl).
[00220] Последовательность ДНК получали для каждого гена-кандидата из двух индуцирующих линий (Stock6 и RWK) и одной неиндуцирующей линии (RWK-NIL). Кроме того, имеющиеся в открытом доступе данные о геноме В73 использовали в качестве второй неиндуцирующей линии. Информацию из генной модели сравнивали с данными о EST/кДНК для подтверждения структуры каждого гена. Аннотированные данные о последовательностях сравнивали с различиями по каталогу между четырьмя аллелями каждого гена.
[00221] Сравнения последовательностей выявили, что аллели В73 и RWK-NIL являлись сходными друг с другом, и аллели RWK и Stock 6 являлись сходными друг с другом. Большинство различий между последовательностями представляли собой однонуклеотидные полиморфизмы, которые не изменяют последовательность, кодирующую белок. Наблюдались некоторые вставки и некоторые делеции, большинство из которых находились в последовательности, не кодирующей белок.
[00222] В таблице 1 ниже перечислены наименования генов в показанном интервале. Эта информация относится к хромосоме 1, см., например, Patrick S. Schnable et al., The B73 Maize Genome: Complexity, Diversity, and Dynamics, 326 Science 1112-15 (2009), включенный в данный документ посредством ссылки во всей своей полноте, и включает краткое описание других кодируемых белков из генов в пределах локуса, индуцирующего гаплоидию.
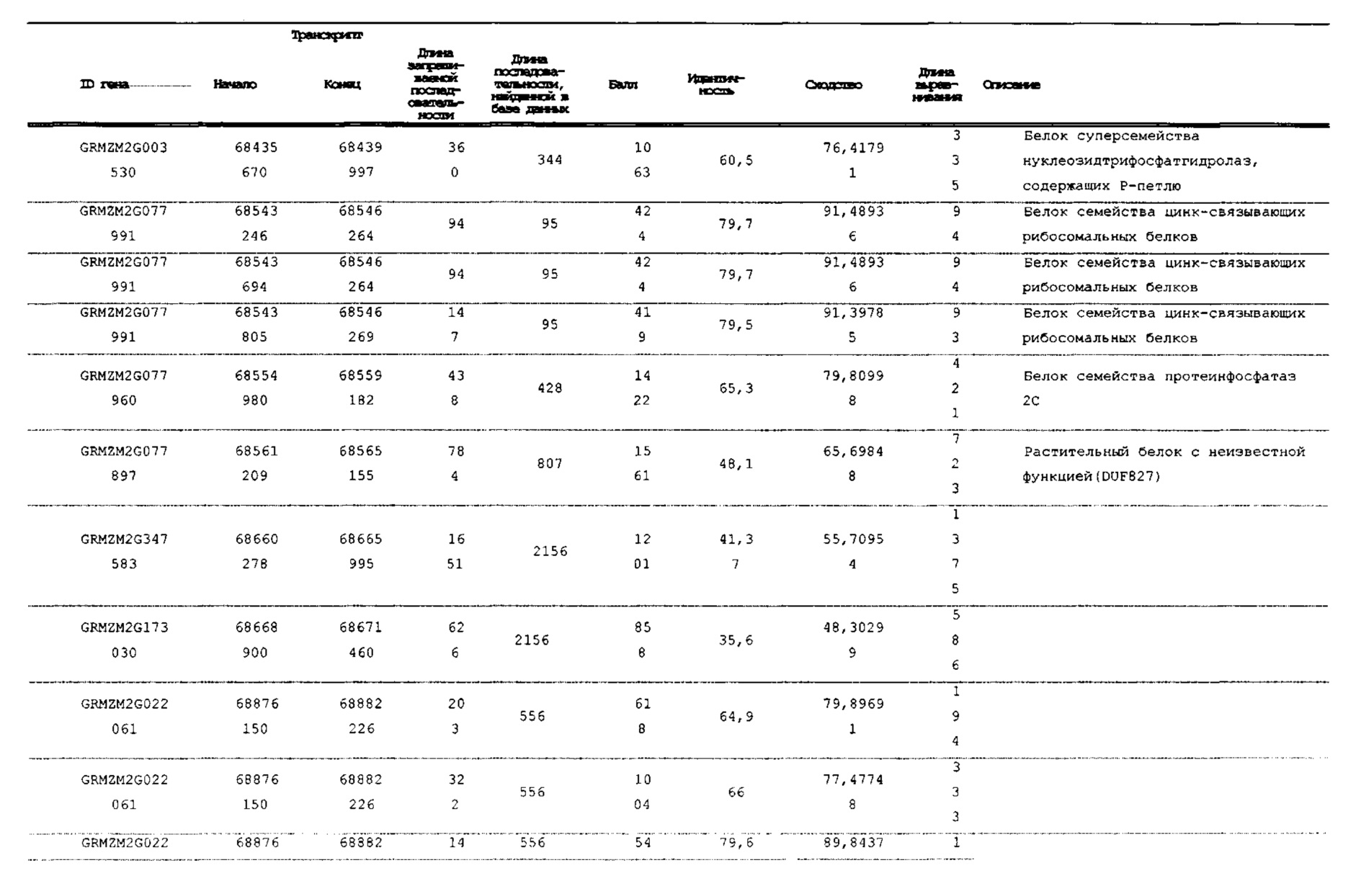
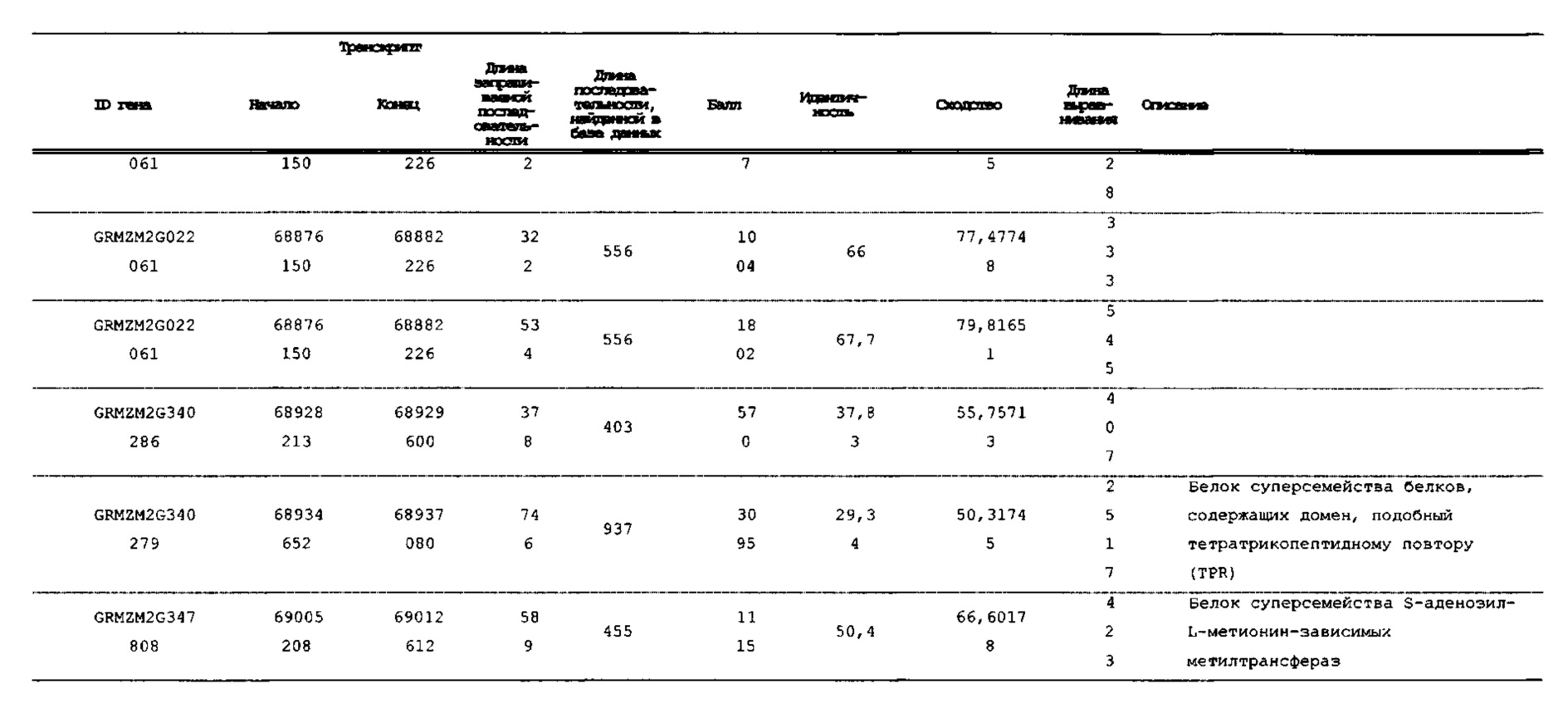
[00223] После завершения тонкого картирования признака индукции гаплоидии с отнесением его к интервалу, содержащему только семь генов, авторы настоящего изобретения сосредоточились на них при сборке и анализе последовательностей. Последовательности семи генов были почти идентичными в сравнении между В73 и RWK-NIL, но в RWK и Stock 6 отсутствовал GRMZM2G062320, ген фосфоглицератмутазы (PGM), и имелась вставка из 4 пар оснований ("п. о.") в четвертом экзоне GRMZM2G471240, гена пататин-подобной фосфолипазы AIIα (pPLAIIα) (фигура 2). Авторы настоящего изобретения обнаружили, что как RWK, так и Stock6 имели одну и ту же вставку из 4 п. о. в четвертом экзоне pPLAIIα, и что этот ген специфически экспрессируется в пыльце (см. maizegdb.org/gene_center/gene?id=GRMZM2G471240, включенный в данный документ посредством ссылки). Немутантный GRMZM2G471240 представлен под SEQ ID NO: 1. GRMZM2G471240, содержащий вставку из 4 п. о. в четвертом экзоне, представлен под SEQ ID NO: 3.
[00224] Большинство гаплоидов, которые были идентифицированы, были обнаружены с помощью маркерного теста Taqman. В данном маркерном тесте используются преимущества различия в гене pPLAIIα между скрещиваемыми RWK и NP2222. В скрещиваниях, где авторы настоящего изобретения применяли RWK в качестве женской особи, а NP2222 в качестве мужской особи, родительская особь RWK является гомозиготной по аллелю mtl, тогда как NP2222 является гомозиготной по аллелю MTL. Диплоидное потомство представляет собой MTL/mtl, а гиногенетическое гаплоидное потомство представляет собой mtl/Q. Таким образом, при выполнении этого теста результаты Taqman показывают 1 копию аллеля mtl и одну копию аллеля MTL у диплоидного потомства, а также 1 копию mtl и отсутствие копий MTL у гаплоидного потомства. При осуществлении этого типа скрещивания початки собирают в период 12-21 дня после опыления, зародыши извлекают, и небольшой образец зародышей отбирают для маркерного анализа Taqman. В качестве альтернативы, зародыши высевают на твердые среды и проращивают в темноте с тем, чтобы можно было отобрать более крупный образец удлиненного побега или корня 2-10 дней спустя для маркерного анализа. В то же время некоторую часть ткани сохраняют для анализа плоидности. В этом последнем случае после применения молекулярного теста более крупные образцы гаплоидов можно анализировать на анализаторе плоидности CyFlow Space и подтверждать в качестве гаплоидов. В большинстве случае это приводит в результате к положительной идентификации гаплоидов. В нескольких более редких случаях это приводит в результате к опровержению ложноположительных результатов маркерного анализа и коррекции сигнала как диплоидного.
[00225] Другим способом, которым авторы настоящего изобретения проводили тестирование на наличие гаплоидов, является анализ доминантных маркеров. В данном случае применяли мужских особей линии Х26. Эта линия является гомозиготной в отношении маркера, который действует доминантным образом. В таком скрещивании в качестве женской особи можно применять растение любой линии, при условии, что оно не имеет маркера или каких-либо генов или аллелей, функционирующих как подавляющие маркерный фенотип. Линия Х26 является неиндуцирующей и гомозиготной по MTL. При применении такой линии потомство срезали в период 12-21 дня после опыления и оценивали в отношении наличия маркера, или его исследовали непосредственно на початке, или высушенные зерна собирали и оценивали в отношении наличия маркера. Диплоидное потомство демонстрирует маркерный фенотип, поскольку оно имеет одну копию маркерного гена от мужской родительской особи Х26, тогда как гиногенетическое гаплоидное потомство не демонстрирует маркерный фенотип. Пенетрантность маркера и уровень спонтанной индукции гаплоидии у Х26 тестировали во множестве контрольных скрещиваний. С применением этой системы авторы настоящего изобретения осуществляли скрининг в отношении гаплоидов и затем тестировали их на анализаторе плоидности для подтверждения того, что они действительно являются гаплоидами.
[00226] Авторы настоящего изобретения разработали ПЦР-тесты для специфического выявления аллелей "дикого типа" и "мутантных" аллелей для скрининга девятнадцати индукторов, происходящих от Stock 6, в том числе индуктора гаплоидии NP2222 (NP2222-HI), линию ВС3, полученную в результате интрогрессии RWK в стандартную трансформируемую инбредную линию Syngenta NP2222. Авторы настоящего изобретения также подвергли скринингу девять неиндуцирующих контрольных линий.
[00227] Для разработки ПЦР-теста, с помощью которого можно было бы провести различие между гаплотипами RWK/Stock6 и RWK-NIL, разрабатывали две пары праймеров: одна пара должна амплифицировать аллель со сдвигом рамки считывания RWK/Stock6, тогда как другая должна амплифицировать аллель B73/RWK-NIL. Эти пары функционировали согласно ожидаемому для ДНК RWK-NIL, RWK и Stock6: gDNA RWK-NIL амплифицировалась только парой праймеров RWK-NIL. gDNA RWK и Stock6 амплифицировалась только парой праймеров RWK/Stock6, которая специфически выявляет аллель со сдвигом рамки считывания. Продукты ПЦР секвенировали, и последовательности были идентичными последовательностям, определенным в результате полногеномного секвенирования. SNP, которые идентифицировали в рамках полногеномного секвенирования, подтверждали в продуктах ПЦР. Ниже, на фигуре 2, ДНК, применяемая в каждой реакции, указана прописными буквами. Праймерами являются "nil.F1/R1" и "rwk.F1/R1."
GRMZM2G471240_nil.F1: GTACGCCGTGCGCTAACA (SEQ ID NO: 5).
GRMZM2G471240_nil.R1: TCGTACCTCCCTGTCTCCAC (SEQ ID NO: 6).
GRMZM2G471240_rwk.F1: TACGCCGTGCGCTAACATA (SEQ ID NO: 7).
GRMZM2G471240_rwk.R1: GTACCTCGCTCCCTGTCTCC (SEQ ID NO: 8).
[00228] Пары праймеров "rwk.F1/R1" и "nil.F1/R1" применяли для генотипирования панели сильных, слабых индукторов и неиндукторов. Авторы настоящего изобретения обнаружили, что все 19 линий, индуцирующих гаплоидию, в том числе, среди прочих, Stock6 (3% уровень индукции гаплоидии ["HIR"]), RWK (линия, полученная из запасов Гогенгеймского университета, 10-15% HIR), RWS и Z22, имели вставку из 4 п. о. В отличие от этого, во всех девяти линиях, не индуцирующих гаплоидию (средний HIR 0,1%), был обнаружен аллель дикого типа. Данные указывают на то, что гомозиготность по аллелю со сдвигом рамки считывания коррелирует с индукционной способностью: 12/12 сильных и 7/7 слабых индукторов амплифицировались в анализе сдвига рамки считывания, но не в анализе дикого типа, тогда как 9/9 неиндукторов амплифицировались в анализе дикого типа, но не сдвига рамки считывания. Это указывает на то, что индукционная способность коррелирует с мутацией GRMZM2G471240, и что pPLAIIα лежит в основе qhir11 и является основной мутацией, отвечающей за индукцию гаплоидии в этих линиях.

[00229] Авторы настоящего изобретения также идентифицировали ряд однонуклеотидных полиморфизмов ("SNP") между аллелем со сдвигом рамки считывания и аллелем RWK-NIL. Для многих из этих SNP последовательности STOCK6 и RWK согласовывались с другими инбредными растениями, которые авторы настоящего изобретения секвенировали, и, следовательно, по всей вероятности представляют собой естественную изменчивость. Действительно, большинство из этих SNP не изменяло аминокислотную последовательность и, следовательно, по всей вероятности не вносит вклад в фенотип индукции гаплоидии. Два SNP приводили в результате к аминокислотным изменениям (H107Y; K232N), и они не являются высококонсервативными изменениями, поэтому они могут вносить небольшой вклад в фенотип, но в большинстве случаев они не влияют на фенотип, поскольку сдвиг рамки считывания вызывает потерю функции.
[00230] Авторы настоящего изобретения переименовали pPLAIIα в "MATRILINEAL" (MTL; т. е. SEQ ID NO: 1) и нативный аллель со вставкой из 4 п. о. в "matrilineal" (mtl; т. е. SEQ ID NO: 3). В соответствии с предсказанной последовательностью белка вставка из 4 п. о. вызывает сдвиг открытой рамки считывания белка в положении аминокислоты ("АА") 352 из 401. Сдвиг рамки считывания приводит к появлению преждевременного стоп-кодона.
[00231] После обнаружения нокаут-мутации сдвига рамки считывания авторы настоящего изобретения непосредственно тестировали эффект, который она оказывала на индукцию гаплоидии, посредством комплементации линии, индуцирующей гаплоидию, трансгеном pPLAIIα дикого типа. Гетерологичная комплементация NP2222-HI (10,2% HIR) копией MATRILINEAL дикого типа практически устраняла индукцию гаплоидии и недоразвитость зерна. По сравнению с контролями HIR уменьшался в 50 раз, от 10% до 0,23%. Она также уменьшала уровень недоразвитости зародышей до 0,65%. Также получали линии с полноразмерными функциональными репортерами с помощью трансгенных слияний гена MTL дикого типа с GFP, а также мутантного аллеля mtl с GFP для того, чтобы как визуализировать субклеточную локализацию MTL дикого типа, так также и увидеть, правильно ли локализуется мутантная версия белка или вырабатывается ли она вообще. Эти линии также служили в качестве дополнительного материала для тестирования комплементации. Материал, индуцирующий гаплоидию (NP2222-HI), который являлся гомозиготным по трансгену MTL-GFP, также не демонстрировал фенотип индукции гаплоидии. Уровень индукции NP2222-HI падает до 0,60%, если он является гомозиготным по MTL-GFP. Кроме того, трансген MTL-GFP также снижал недоразвитость зародышей до 4,86%. И наконец, авторы настоящего изобретения тестировали, обеспечивает ли мутантный аллель mtl, слитый с GFP, комплементацию фенотипа индукции гаплоидии, и было обнаружено, что этого не происходит. Уровни индукции гаплоидии и недоразвитости зародышей были весьма сходными у NP2222-HI по сравнению с NP2222-HI, который являлся гомозиготным по мутантному слитому трансгену mtl-GFP. См. таблицу 3. Это представляет убедительное доказательство того, что сдвиг рамки считывания в MATRILINEAL отвечает за индукцию гаплоидии и необходим для этого. Для применения этих знаний авторы настоящего изобретения демонстрируют, что осуществление мутации или модулирование экспрессии pPLAIIα в линии дикого типа приводит к созданию новых линий с индуцированной гаплоидией.
Таблица 3. Репродуктивные характеристики линий, индуцирующих гаплоидию, комплементированных и отредактированных линий. В этой таблице показаны уровни индукции гаплоидии и недоразвитости зерна у индуцирующих линий в окружении NP2222. Вначале приведено количество анализирующих скрещиваний ("початки"), а затем приведены статистические данные о зернах и зародышах. Недоразвитость зародышей и индукция гаплоидии обычно наблюдаются одновременно на одном и том же початке. По этой причине уровень недоразвитости зародышей является таким высоким в початке, скрещенном с мужской особью NP2222-HI, которая характеризуется уровнем индукции гаплоидии (HIR) 10,17%. Оба трансгена WT, в том числе один без GFP и один, слитый с GFP, комплементировали фенотип индукции гаплоидии. В то же время мутантный mtl, слитый с GFP, не комплементировал его.

[00232] Несколько mtl-подобных аллелей создавали в инбредном растении NP2222 посредством введения небольших делеций в MTL вблизи сайта вставки из 4 п. о. в mtl, с применением эффекторных нуклеаз, подобных активаторам транскрипции (TALEN) (Boch, J. et al., Breaking the code of DNA binding specificity of TAL-type III effectors, Science 326:1509-1512 (2009), включенный в данный документ посредством ссылки). Несколько мутантных объектов подвергали самоопылению, и растения Т1, лишенные вставки Т-ДНК, введенной с помощью TALEN, но гомозиготные по небольшим делециям в MTL, подвергали ауткроссингу с NP2222. Отредактированные линии, гомозиготные по делециям со сдвигом рамки считывания в MTL (далее в данном документе называемые MTLTAL-FS), демонстрировали HIR, составляющий 4,0-12,5% (в среднем 6,65%) (таблица 4). Статус плоидности 118/127 предполагаемых гаплоидов подтверждали посредством проточной цитометрии, и фенотипические оценки указывали на то, что эти растения являлись гаплоидами. Эти результаты доказывают, что сдвиг рамки считывания в MTL является достаточным для индуцирования высоких уровней индукции гаплоидии. Картировали другие факторы, вносящие вклад в фенотип, в том числе соседний qhir12 (см. Liu, 2016), который может являться причиной различия в HIR между MTLTAL-FS и NP2222-HI. Разумно сделать вывод о том, что уровни завязывания семян, HIR и недоразвитости зерна устанавливаются благодаря mtl посредством специфических взаимодействий отцовского и материнского генотипа.
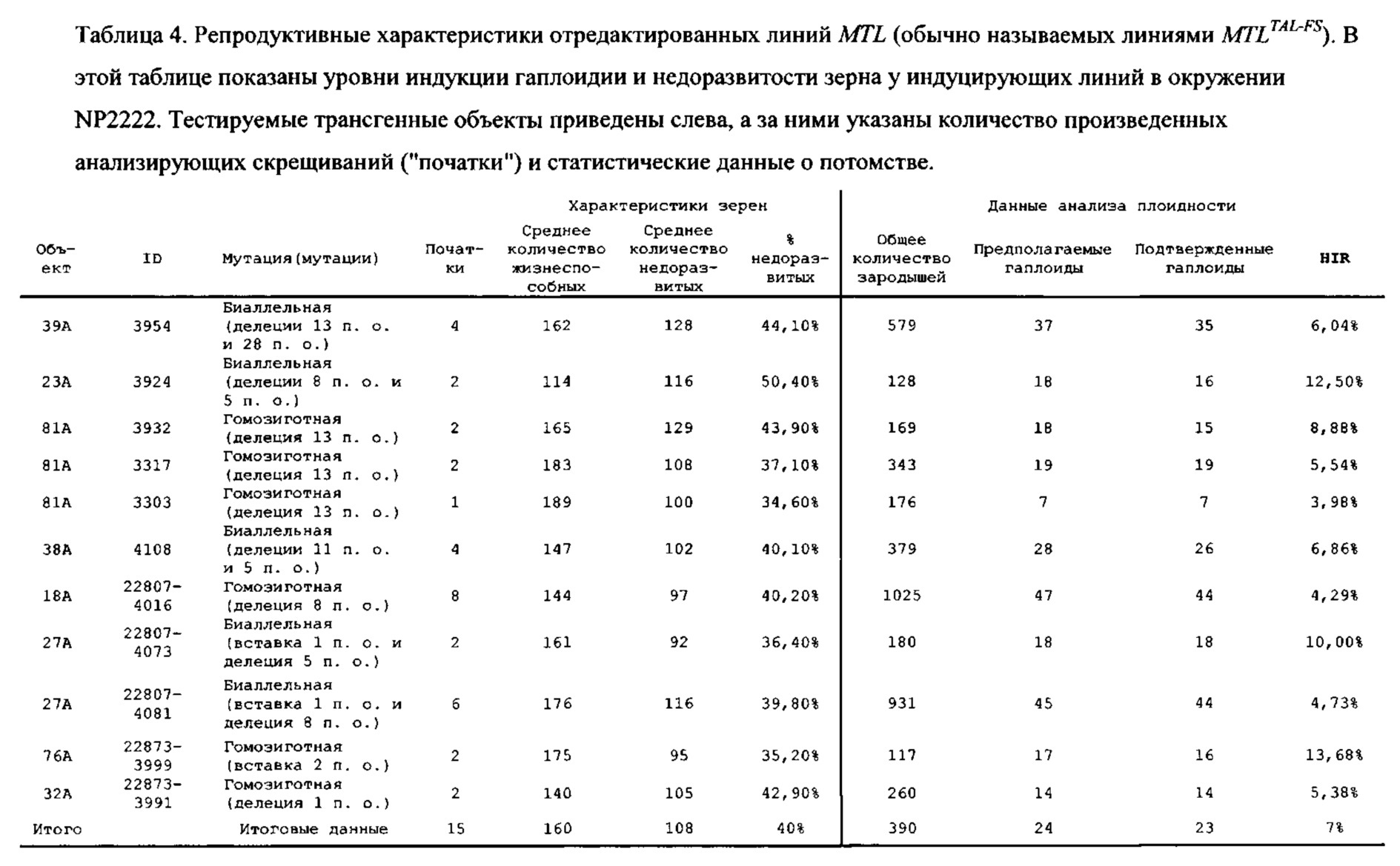
[00233] Образование гаплоидных семян у маиса имеет постзиготический характер и вызывается дефектом в мужском гаметофите. Этот факт отражен в данных об экспрессии MTL. Имеющиеся в открытом доступе профили секвенирования РНК указывают на то, что транскрипт MTL дикого типа является специфическим для пыльников на стадии цветения (см. Sekhon, R. S., et al. Genome-wide atlas of transcription during maize development, Plant Journal, 66, 553-563 (2011), включенный в данный документ посредством ссылки), что согласуется с профилем развития, обнаруживаемым исключительно в пыльниках до вскрывания (см. Zhai, J., et al. Spatiotemporally dynamic, cell-type-dependent premeiotic and meiotic phasiRNAs in maize anthers, PNAS 112, 3146-3151 (2015), включенный в данный документ посредством ссылки). Авторы настоящего изобретения обнаружили, что пыльца дикого типа имела в 18 раз больше транскриптов MTL, чем пыльцевые мешки после цветения, что указывало на то, что ген является специфическим для мужского гаметофита (фигура 3). Попытки осуществления нокдауна MTL посредством RNAi привели к повышению уровней образования гаплоидов только для MTLRNAi (таблица 5). Существует два аннотированных сплайс-варианта MTL, отображающие альтернативный 81 нуклеотид сайта сплайсинга перед 3'-концом экзона 2. По сравнению с NP2222 уровень Mtl был повышен в пыльце NP2222-HI, но не MTLTAL-FS (фигура 4), тогда как представленность двух аннотированных сплайс-вариантов была единообразной.
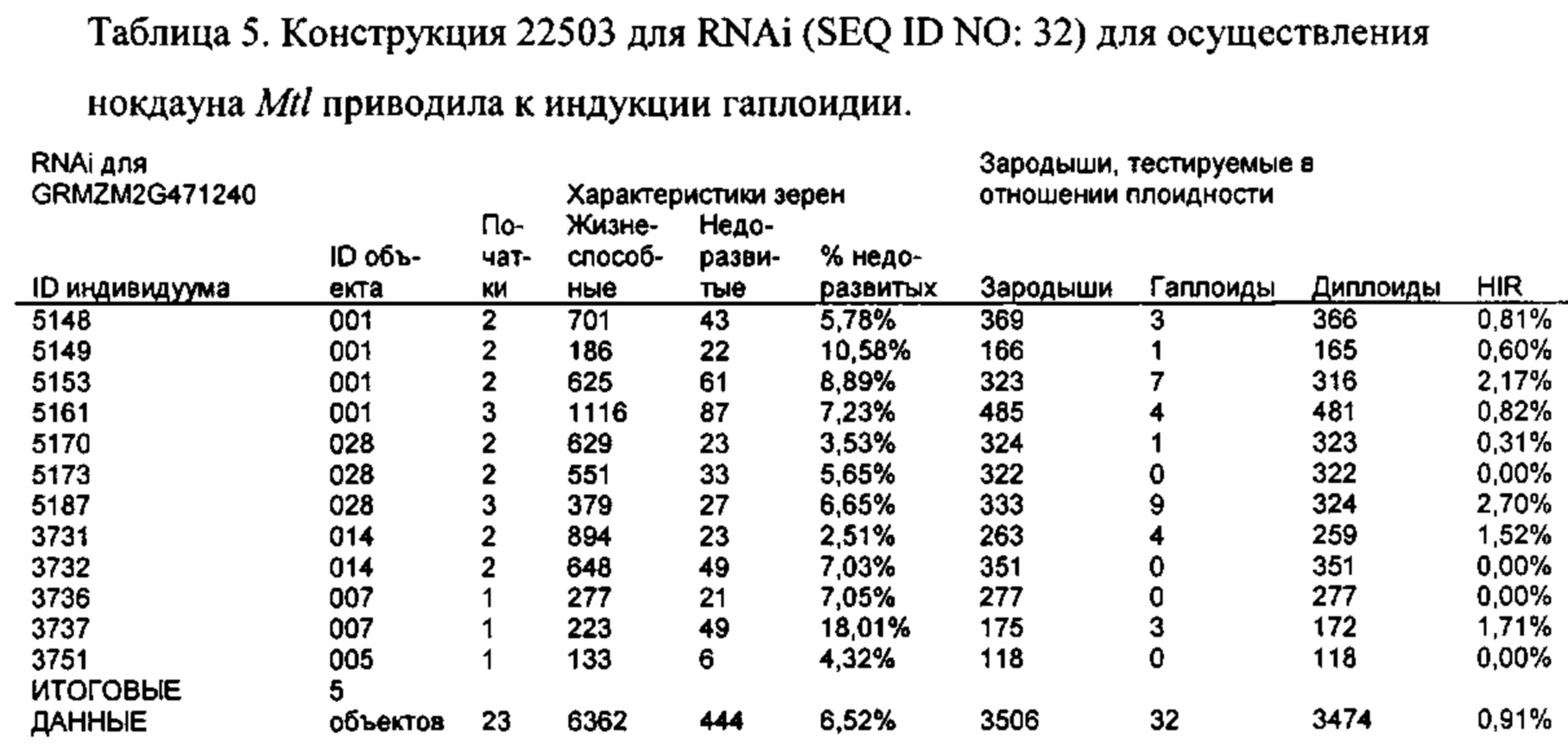
[00234] Сдвиг рамки считывания в mtl появляется в положении аминокислоты 380, что приводит к изменению 20 аминокислот с появлением расположенного после них преждевременного стоп-кодона, который усекает белок на 29 аминокислот (фигура 5а). Белок MTL дикого типа был обнаружен в профилях LS-MS пыльцы RWK-NIL и NP2222, но его уровень находился ниже предела выявления в трех из трех образцов RWK и 3 из трех 3 образцов NP2222-HI (таблица 6). Это демонстрирует, что, несмотря на наличие мутантного транскрипта mtl, образующегося в пыльце, белок не выявляется, что подтверждает наличие мутации потери функции. Как мутантные формы, так и формы дикого типа рекомбинантного белка MTL проявляют фосфолипазную активность in vitro во флуоресцентных анализах расщепления липосом pPLAIIα-подобным белком (фигура 5b). Это демонстрирует, что функциональная аннотация гена MTL (т.е. то, что он кодирует белок фосфолипазу) является правильной.
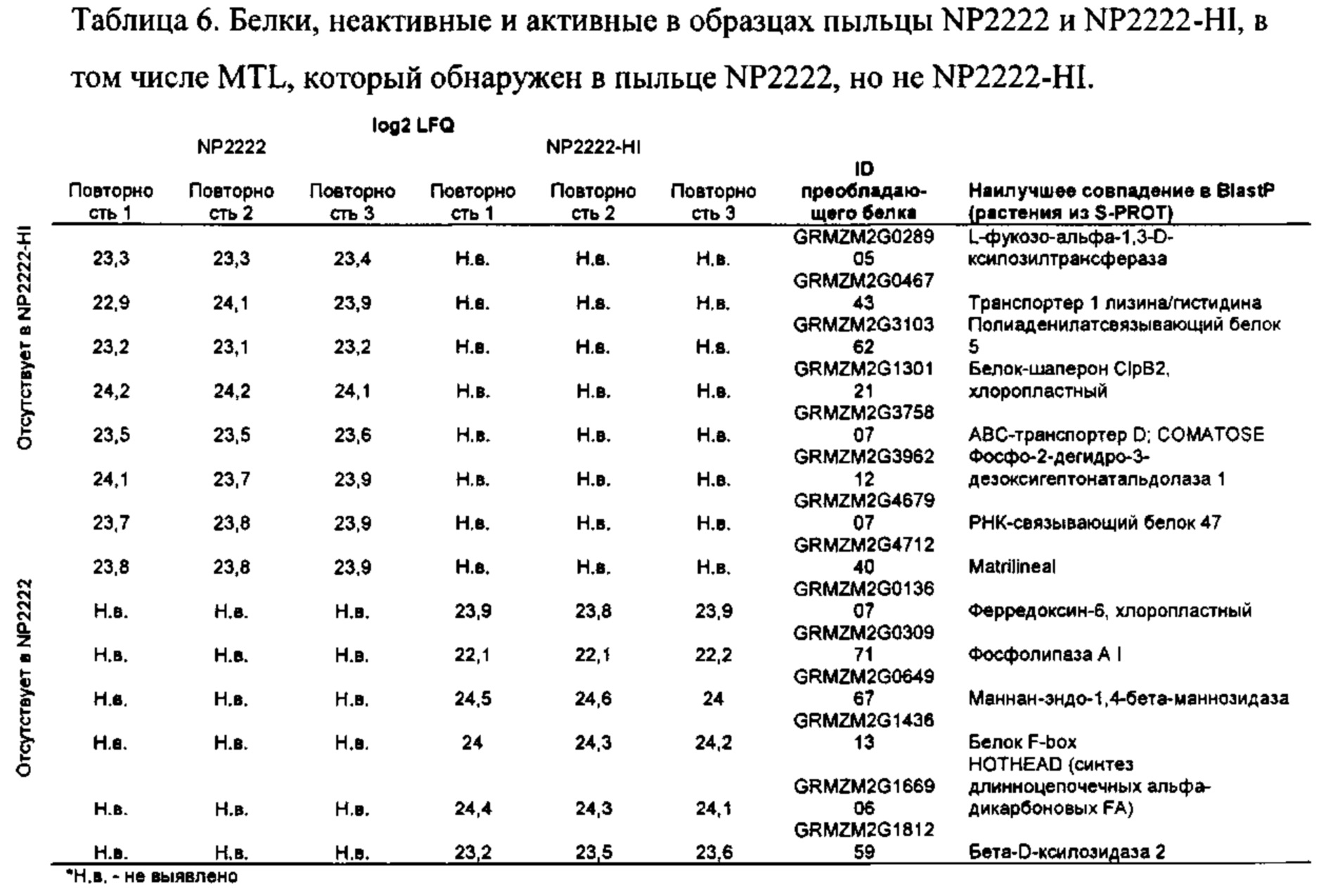
II. Эксперименты по проращиванию пыльцы и определению локализации
[00235] Линии с полноразмерными функциональными репортерами использовали для определения характеристик локализации MTL. Сигнал не был обнаружен в пыльце NP2222 или NP2222 + mtl-GFP/mtl-GFP. В отличие от этого, пыльца NP2222 + MTL-GFP/MTL-GFP демонстрировала сильный сигнал в цитоплазме двух спермиев. Этот сигнал был обнаружен в тяжистой цитоплазме гамет в пределах проросших пыльцевых трубок. Зародышевые мешки NP2222, фиксированные через 18 часов после опыления пыльцой MTL-GFP, давали сигнал в области дегенерирующей синергиды, согласующийся с сигналом SC, доставляемых в ходе оплодотворения. Это указывает на то, что MIL является частью мужской зародышевой единицы, которая откладывается в зародышевом мешке после разрыва пыльцевой трубки. MTL-GFP, но не mtl-GFP устранял индукцию гаплоидии в NP2222-HI (таблица 3). Совместно эти данные указывают на то, что MTL является фосфолипазой, специфической для цитоплазмы SC, и что сдвиг рамки считывания в mtl нарушает локализацию или стабильность MTL в пыльце, индуцирующей гаплоидию..
[00236] Идентификация MTL в качестве причинного гена при индукции гаплоидии у маиса позволила провести анализ плейотропных фенотипов, традиционно ассоциированных с признаком. Мутации фосфолипазы ассоциированы с замедленным прорастанием пыльцы и ростом трубки (см. Kim, Н.J., et al. Endoplasmic reticulum- and golgi-localizedphospholipase A2 plays critical roles in Arabidopsis pollen development and germination, Plant Cell 23,94-110 (2011)), но они являлись нормальными в линиях RWK, Stock 6 и MTLTAL-FS (фигура 6А, В). Початки, опыленные пыльцой NP2222-HI и MTLTAL-FS, демонстрируют ~10-25% частоту неудачного оплодотворения, и конкурентный анализ пыльцы показал, что RWK подвергается нарушению сегрегации (SD) (фигура 6С), что согласуется с предыдущими сообщениями (см. Xu, X., et al. Gametophytic and zygotic selection leads to segregation distortion through in vivo induction of a maternal haploid in maize, J. exp. ВОТ. 64, 1083-1096 (2013)). Скрещивания с гемизиготной пыльцой NP2222-HI + MTL/0 давали пропорциональное смещение в сторону потомства MTL + (фигура 6D), что указывало на то, что индукция SD может быть отнесена на счет mtl. Недоразвитость зародышей, неизменный побочный результат индукции гаплоидии, связанный с неудачной пролиферацией эндосперма, имела место на сходных уровнях у нативного индуктора и индуктора MTLTAL-FS (таблица 4). Совместно эти данные подразумевают участие mtl в каждом репродуктивном дефекте, ассоциированном с индукцией гаплоидии. Двумя механизмами, которые, как правило, предлагают для объяснения образования гаплоидов, являются одинарное оплодотворение и постзиготическая элиминация генома (см. Sarkar, K.R. & Сое, Е.Н, A genetic analysis of the origin of maternal haploids in maize, genetics 54, 453-464 (1966); Zhang, Z., et al., Chromosome elimination and in vivo haploidproduction induced by Stock 6-derived inducer line in maize (Zea mays L.), Plant CELL REP. 27, 1851-1860 (2008); и Barret, P., Brinkmann, M., & Beckert, M., A major locus expressed in the male gametophyte with incomplete penetrance is responsible for in situ gynogenesis in maize, theor. APPL. genet. 117, 581-594 (2008)). В первом случае гаплоиды образуются в результате оплодотворения центральной клетки, но не яйцеклетки, которая впоследствии развивается посредством партеногенеза. В последнем случае двойное оплодотворение предшествует элиминации мужских хромосом. Выяснение точного механизма потребует тщательных эмбриологических исследований после опылений MTLTAL-FS наряду с получением количественных данных для отслеживания редкой стойкости мужской ДНК у гаплоидов маиса.
[00237] Индукцию гаплоидии недавно осуществляли в Arabidopsis посредством манипуляции с геном центромерного варианта гистона Н3, который вызывает однородительскую элиминацию генома посредством постзиготического дисбаланса центромер между гибридизированными геномами. Попытка повторить это у маиса была успешной (см. Ravi, М. & Chan, S. W. L. Haploid plants produced by centromere-mediated genome elimination, Nature 464, 615-618 (2010)), но эта публикация является первым случаем описания системы индукции гаплоидии, запускаемой цитоплазматическим белком, который не связывается с хроматином. Таким образом, данная работа подчеркивает важность неядерных компонентов спермия в репродуктивном успехе и точном переносе генома. Консервативность MTL у злаковых растений (см. фиг. 7), в частности у риса, где ближайший гомолог является специфическим для пыльцы и также обнаружен в спермин, позволяет предположить, что эти полученные результаты приведут к разработке новых внутривидовых линий важных культурных растений, индуцирующих гаплоидию.
III. Химические аэрозоли для индукции HI de novo
[00238] После обнаружения того, что фосфолипаза MTL (и в частности, мутация потери функции со сдвигом рамки считывания в mtl) вызывает индукцию гаплоидию, авторы настоящего изобретения тестировали, могут ли ингибиторы фосфолипаз, жирные кислоты или другие липидные соединения выступать в качестве химических индукторов гаплоидии.
[00239] Авторы настоящего изобретения успешно достигли индукции гаплоидии de novo посредством нанесения различных химических веществ на цветки, рыльца, початки, метелки и пыльцу. Все эти химические вещества принадлежат к классу липидов или ингибиторов фосфолипаз. В таблице 7 в общих чертах изложены эти химические вещества и уровни индукции гаплоидии, которые были получены в результате их нанесения. Также в таблице 7 представлена информация о составе и информация о способе нанесения. Из этой таблицы понятно, что множество липидных соединений индуцирует образование гаплоидов de novo при нанесении до, во время или после опыления на рыльца, которые представляют собой рыльцевую ткань, специфичную для растения маиса, пыльцу, метелки (которые содержат мужские цветки, содержащие пыльцу до выбрасывания пыльцы) и початки. Такие соединения включают смешанные масла, жирные кислоты, сложные эфиры жирных кислот, фосфолипиды и ингибиторы фосфолипаз.
[00240] Авторы настоящего изобретения тестировали много ингибиторов фосфолипаз, в том числе маноалид, ингибитор фосфолипаз, не имеющий каких-либо цепей жирных кислот. Это химическое вещество было способно индуцировать образование гаплоидов de novo. Имея этот результат, авторы настоящего изобретения начали тестировать больше соединений. Авторы настоящего изобретения обнаружили, что наиболее активным соединение для индукции de novo является метил-альфа-линоленоилфторфосфонат (MALFP), сильнодействующий ингибитор фосфолипаз и ингибитор других липид-модифицирующих ферментов. Это соединение содержит три цепи жирной кислоты линолената (восемнадцать атомов углерода с тремя тройными связями), а также метальную группу и атом фтора в положении группы головки. Другим высокоактивным соединением, индуцирующим гаплоидию de novo, является метиларахидонилфторфосфонат (MAFP), другой ингибитор фосфолипаз с несколько отличающейся структурой цепи жирной кислоты (20 атомов углерода и четыре двойные связи). Не желая ограничиваться какой-либо теорией, полагают, что MALFP и MAFP ингибируют фосфолипазы, располагаясь в связывающем кармане для цепей жирных кислот и в некоторых случаях катализируя необратимое фосфорилирование аминокислоты серина в активном центре (см. Lio Y.C., Reynolds L.J., Balsinde J., и Dennis E.A. Irreversible inhibition of Ca(2+)-independent phospholipase A2 by methyl arachidonyl fluorophosphonate. BIOCHIM BIOPHYS ACTA 1302:55-60 (1996)). Некоторые жирные кислоты, такие как линолевая кислота и линоленовая кислота, могут также неконкурентно ингибировать фосфолипазы, располагаясь в связывающем кармане для цепей жирных кислот без ковалентной модификации активного центра, однако эффект этого ингибирования является более слабым. Ballou, L.R., и Cheung, W.Y. Inhibition of human platelet phospholipase A2 activity by unsaturated fatty acids PROC Natl Acad Sci 82 (2): 371-375 (1985)). Эти литературные источники могут помочь объяснить основания для полученных авторами настоящего изобретения результатов, заключающихся в том, что индукция гаплоидии de novo вызывается не только сильными ингибиторами фосфолипаз, такими как MALFP и MAFP, но также и сложными эфирами жирных кислот, в том числе метиловым сложным эфиром арахидоновой кислоты (ААМЕ), который имеет 20 атомов углерода и четыре двойные связи (сложный эфир полиненасыщенной жирной кислоты из 20 атомов углерода), а также линолевой кислотой (LLE), которая имеет 18 атомов углерода и две двойные связи (18:2), и линоленовой кислотой (LNA), которая имеет 18 атомов углерода и три двойные связи (18:3), и этиловым сложным эфиром олеиновой кислоты (ОАЕЕ), и арахидоновой кислотой (которая имеет 20 атомов углерода и четыре двойные связи). Опять же, полагают, что эти цепи ненасыщенных жирных кислот, будь то в их форме свободной жирной кислоты, в их форме сложного эфира (которые синтезируются в лаборатории и могут быть способны проникать в ткани) или в некоторой другой форме, могут выступать в качестве ингибиторов фосфолипаз. Несмотря на то, что, как правило, не признается, что эти соединения в форме сложного эфира могут ингибировать фосфолипазы (см. Ballou, выше), авторы настоящего изобретения обнаружили, что такие метиловые и этиловые сложные эфиры жирных кислот могут в действительности приводить к индукции гаплоидии de novo. См. таблицу 7. Следует отметить, что тестировали насыщенные жирные кислоты и их производные, в том числе этиловый сложный эфир стеариновой кислоты и пальмитиновой кислоты. В предшествующем уровне техники указано, что молекулы, которые содержат боковые цепи насыщенных жирных кислот, неспособны или не будут способны конкурентно ингибировать фосфолипазу, находясь в ее активном центре. См. Ballou, выше. Считалось, что такие соединения не обладают способностью ингибировать фосфолипазы. Однако, авторы настоящего изобретения неожиданно смогли применять молекулы, содержащие боковые цепи насыщенных жирных кислот, для индукции образования гаплоидов de novo. Например, этиловый сложный эфир пальмитиновой кислоты (РАЕЕ) индуцирует образование гаплоидов. См. таблицу 7.
[00241] Авторы настоящего изобретения также решили протестировать, могут ли определенные масла и фосфолипиды, в том числе широко известные растительные масла, вызывать индукцию гаплоидии de novo. Авторы настоящего изобретения обнаружили, что одна высокоактивная липидная смесь, индуцирующая гаплоидию de novo, представляет собой кукурузное масло, натуральный растительный экстракт, который можно приобрести в продуктовом магазине. Другая высокоактивная липидная смесь представляет собой льняное масло, которое можно приобрести в магазине хозяйственных товаров. Также будет действовать масло канолы, растительное масло, арахисовое масло, кунжутное масло или любое другое масло при нанесении правильным образом. Другое высокоактивное соединение представляет собой 1,2-дистеарил-sn-глицерофосфатидилхолин (DSPC) (также известный как PC 18:0/18:0), фосфолипид с фосфатидилхолиновой группой головки и двумя цепями насыщенной жирной кислоты из 18 атомов углерода. Этот широко известный фосфолипид присутствует во многих биологических материалах, поскольку он является одним из основных и наиболее распространенных компонентов фосфолипидных бислоев, которые составляют клеточные мембраны всех живых существ на планете. Другое соединение, индуцирующее гаплоидию, представляет собой фосфатидилэтаноламин с одной стеарильной цепью и одной олеоильной цепью (также известный как 1-стеарил-2-олеоил-sn-фосфатидилэтаноламин), другой широко известный фосфолипид. Другие фосфолипиды, а также лизофосфолипиды и другие триацилглицериды, диацилглицерины, лизофосфолипиды, сложные эфиры тритерпеноидов или глицеролипиды, будут выступать в качестве индукторов гаплоидии de novo. Эти масла и фосфолипиды не являются общеизвестными ингибиторами фосфолипаз, но они могут являться исходным материалом для получения побочных продуктов фосфолипазной активности, в том числе тех самых жирных кислот, которые ингибируют фосфолипазы. Тот факт, что эти соединения выступают в качестве химических веществ, индуцирующих гаплоидию de novo, таким образом, указывает на то, что эти масла и фосфолипиды могут ингибировать фосфолипазы опосредованно, или, возможно, они вызывают дисбаланс или изменение состава липидов или мембраны пыльцы, что приводит к индукции гаплоидии. В любом случае из данной работы ясно, что огромный ряд липидосодержащих соединений, в дополнение к нелипидным ингибиторам фосфолипаз, при нанесении на цветки может выступать в качестве химических индукторов образования гаплоидных семян.
[00242] Авторы настоящего изобретения также тестировали, могут ли буферы, которые авторы настоящего изобретения использовали для нанесения этих соединений на цветки, индуцировать образование гаплоидов сами по себе. Эти буферы вариативно содержали определенные смеси поверхностно-активных веществ, и/или соль, и/или DMSO (диметилсульфоксид), и/или другие близкие добавки. Эти буферы не могли индуцировать образование гаплоидов, что можно увидеть в строках 121-123 таблицы 7. Кроме того, индукция гаплоидии de novo без добавления каких-либо соединений является чрезвычайно редкой у маиса, что можно увидеть в строке 1. Среди 3073 тестированных потомков было обнаружено два гаплоида (Kelliher, et al. Maternal haploids are preferentially induced by CENH3-tailswap transgenic complementation in maize, Frontiers in Plant Science 7:414 (2016)). Таким образом, было обнаружено, что контрольный уровень индукции составляет примерно 0,065%, что попадает в диапазон гиногенетической индукции гаплоидии, о котором сообщается в предшествующих литературных источниках (0,05-0,1%) (Chang, М.-Т., и Сое, Е. "Doubled haploids", в Molecular Genetic Approaches to Maize Improvement, eds A.L. Kriz и В.A. Larkins (Heidelberg: Springer), 127-142).
[00243] Способ нанесения этих различных липидных соединений и соединений-ингибиторов липаз может являться достаточно вариативным и при этом по-прежнему обеспечивать образование гаплоидов. Один такой способ нанесения заключается в растворении соединений в солевом растворителе с добавлением 1% DMSO. Другой такой растворитель представляет собой смесь поверхностно-активных веществ (таблица 8). Ткань для нанесения может также изменяться, что видно из таблицы 7, где нанесение как на пыльцу, так и на рыльца во время опыления, а также на метелки за несколько часов или дней до опыления может привести в результате к образованию гаплоидов. Кроме того, соединения могут индуцировать образование гаплоидов в большом разнообразии концентраций от 20 мкМ до 100 мМ или от 0,2 мг/мл до 50 мг/мл включительно. На самом деле образование гаплоидов способен индуцировать намного более широкий диапазон, чем этот, что можно увидеть из таблицы 7.
[00244] В двух случаях авторы настоящего изобретения также разработали новые смеси поверхностно-активных веществ, чтобы обеспечить способность конкретных классов липидов к эмульгированию и образованию микроэмульсии (водно-органической смеси с липидными каплями, имеющими размер, меньший определенного размера - по меньшей мере меньше 10 микрон в диаметре и во многих случаях меньше 1 микрона в диаметре). См. таблицу 8 для состава двух смесей поверхностно-активных веществ. Смесь "91" в идеальном варианте применяют с жирными кислотами в концентрации 6,5 части смеси поверхностно-активных веществ на 1 часть жирной кислоты. Однако также могут действовать несколько близких концентраций и близких смесей поверхностно-активных веществ. Смесь поверхностно-активных веществ "92" в идеальном варианте применяют со сложными эфирами жирных кислот, а также маслами в чистой форме либо в виде смеси триглицеридов. В идеальном варианте их смешивают в соотношении 10 частей смеси поверхностно-активных веществ на 1 часть сложного эфира или масла. Любая из этих смесей или близких смесей соединений может являться подходящей для надлежащего растворения или образования микроэмульсии с близкими молекулярными классами липидов помимо жирных кислот, сложных эфиров и масел. Они могут включать в себя фосфолипиды, диацилглицерины, лизофосфолипиды, сложные эфиры тритерпеноидов или глицеролипиды.
[00245] Эти смеси поверхностно-активных веществ, как правило, смешивают с активным ингредиентом и затем смешивают в определенном процентном соотношении с водным буфером с получением эмульсии и в идеальном варианте микроэмульсии. При получении эмульсии процентное содержание смеси поверхностно-активных веществ + активного ингредиента в водном буфере может иметь любое значение в пределах 0,01-50%. Водный буфер может состоять из любого количества элементов. Авторы настоящего изобретения применяли 1x PBS + 1% DML и PBS + 50% DML, но другие составы также действуют. "PBS" означает фосфатно-солевой буферный раствор; "DML" означает диметиллактамид.
[00246] Применение этих буферов в отдельности не было способно привести к индукции гаплоидии de novo, что можно увидеть в строках 121-123 таблицы 7.
[00247] Способ нанесения соединений может иметь любое количество форм, включая погружение цветочной почки, инъекцию в цветочную почку, микроинъекцию, замачивание пыльцы, разбрызгивание на пыльцу, опрыскивание пыльцы, разбрызгивание на цветочную почку, опрыскивание цветочной почки, окунание рыльца, опрыскивание рыльца и т. д. Авторы настоящего изобретения чаще всего применяли опрыскивания метелок, пыльцы и рыльца до, во время или после опыления. Авторы настоящего изобретения достигали образования гаплоидов de novo посредством DSPC и линоленовой кислоты в случае опрыскивания метелок за два дня до опыления. Авторы настоящего изобретения также достигали образования гаплоидов de novo посредством DSPC в случае опрыскивания метелок всего за несколько минут до сбора пыльцы и последующего опыления. Аналогичным образом, авторы настоящего изобретения достигали образования гаплоидов de novo посредством DSPC в случае опрыскивания пыльцы непосредственно до или во время опыления, а также в случае опрыскивания рылец до, во время или после опыления, в том числе в период до двух дней включительно до опыления и до шести часов включительно после опыления.
[00248] Авторы настоящего изобретения также продемонстрировали образование гаплоидов de novo при применении различных аппликаторов, в том числе дешевого пластикового пульверизатора, который можно приобрести в магазине товаров повседневного спроса, и распылителя, который представляет собой медицинское устройство, применяемое для пероральной доставки лекарственных препаратов в форме очень маленьких водных капель. При том, что пульверизатор, как правило, образует капли размером 50-150 микрон в диаметре, распылитель способен образовывать капли размером менее 10 микрон. Это является преимущественным для нанесения на пыльцу, поскольку размеры пыльцы у большинства растений находятся в диапазоне 20-200 микрон в диаметре (примерно 70 микрон у маиса; примерно 50 микрон у риса), и если одна из капель из обычного пульверизатора попадет на зерно, то капля будет больше, чем зерно. Пыльца чрезвычайно чувствительна к влажности и осмотическому шоку. Если пыльцевое зерно вступает в контакт с каплей слишком большого размера, то из этого зерна неспособна прорасти успешная растущая пыльцевая трубка. То же самое может быть и в случае, если зерно попадает на дыхальце или рыльце, которое является слишком влажным - например, если это рыльце или дыхальце получило слишком много наносимого липида (будь то микроэмульсия или просто липидные капли или мицеллы, образовавшиеся в водном растворе). Если бы авторы настоящего изобретения наносили более 3 мл аэрозольного липида на пыльцу или рыльца с помощью пульверизатора или распылителя, авторы настоящего изобретения часто получали бы очень низкий уровень завязывания семян.
[00249] Таким образом, способ нанесения имеет решающее значение, и, в частности, наиболее важно попытаться нанести как можно меньший объем аэрозоля на пыльцу, или дыхальце, или рыльца, или на цветок в целом, и наилучшим способом распределения активного ингредиента, в частности, липида, в как можно меньшем объеме является получение микроэмульсии этого липида в водном буфере, где размеры липидных капель находятся на субмикронном уровне, а затем дозирование этой микроэмульсии с помощью распылителя или аналогичного устройства, способного образовывать капли, имеющие размер в диапазоне 1 -2 микрона. Это приводит в результате к доставке миллионов капель активного ингредиента в "парообразной" форме в соответствующие ткани, при этом размер капли по объему в > 100000 раз меньше размера самого пыльцевого зерна (если капля имеет размер 1 микрон, а пыльцевое зерно имеет размер 50 микрон, то диаметр капли в 50 раз меньше диаметра зерна). А 503 = 125000, таким образом, объем капли в > 105000 раз меньше объема пыльцевого зерна. При использовании микроэмульсий размер липидных капель может находиться в диапазоне примерно 20-1000 нм в диаметре, таким образом, в зависимости от концентрации смеси поверхностно-активных веществ вместе с активным ингредиентом в водном растворе в каждой капле, доставляемой к каждому пыльцевому зерну или рыльцу, может находиться одна или несколько связок липидов. Кроме того, принимая во внимание то, что некоторые молекулы липидного типа или ингибиторы фосфолипазы можно растворить в водном растворе вместо доставки в виде микроэмульсии, авторы настоящего изобретения обнаружили, что соединения могут индуцировать образование гаплоидов в большом разнообразии концентраций от 20 мкг/мл до 50 мг/мл включительно. Возможно, в более высоких концентрациях соединения будут индуцировать образование гаплоидов de novo.
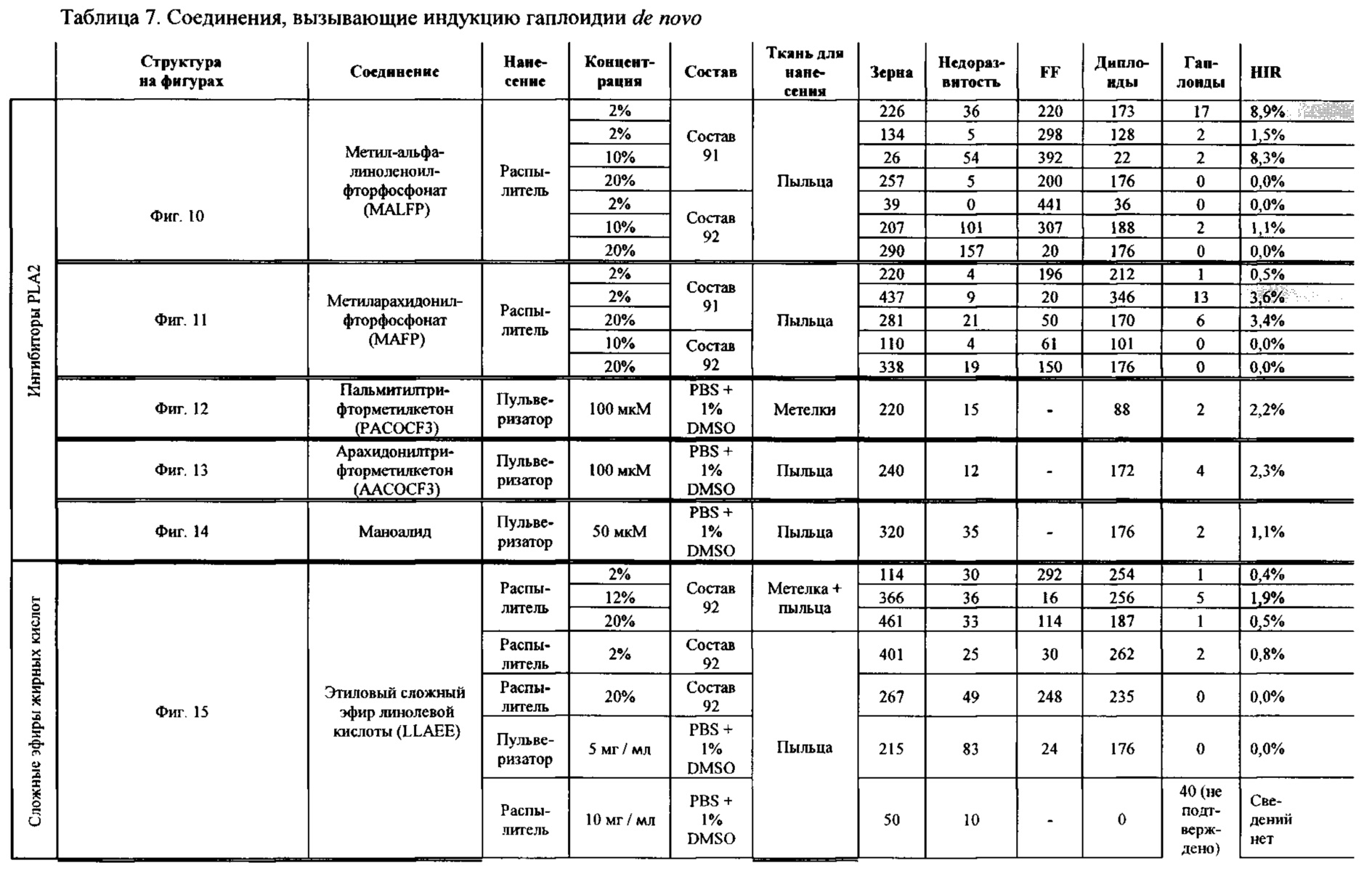
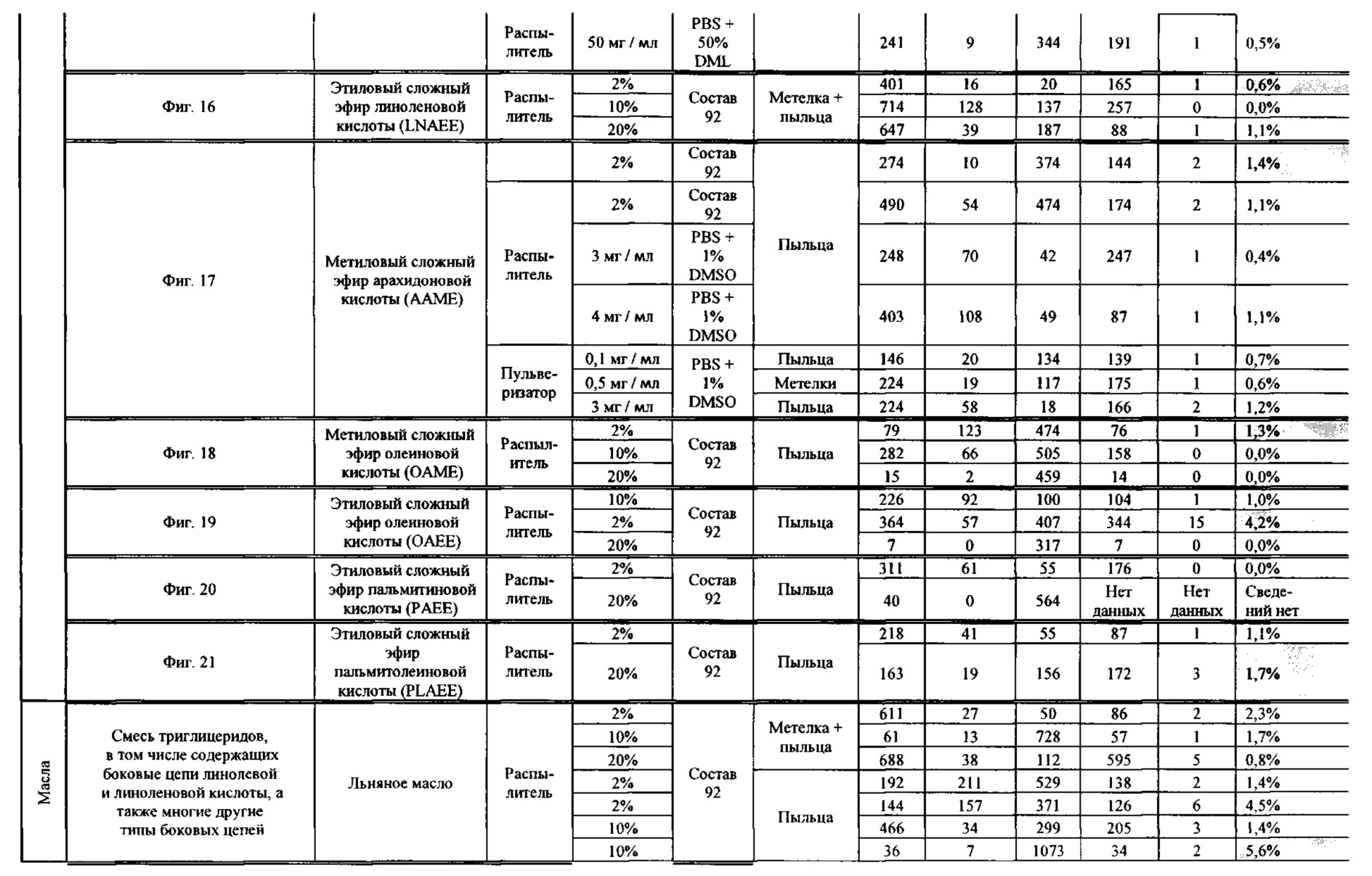
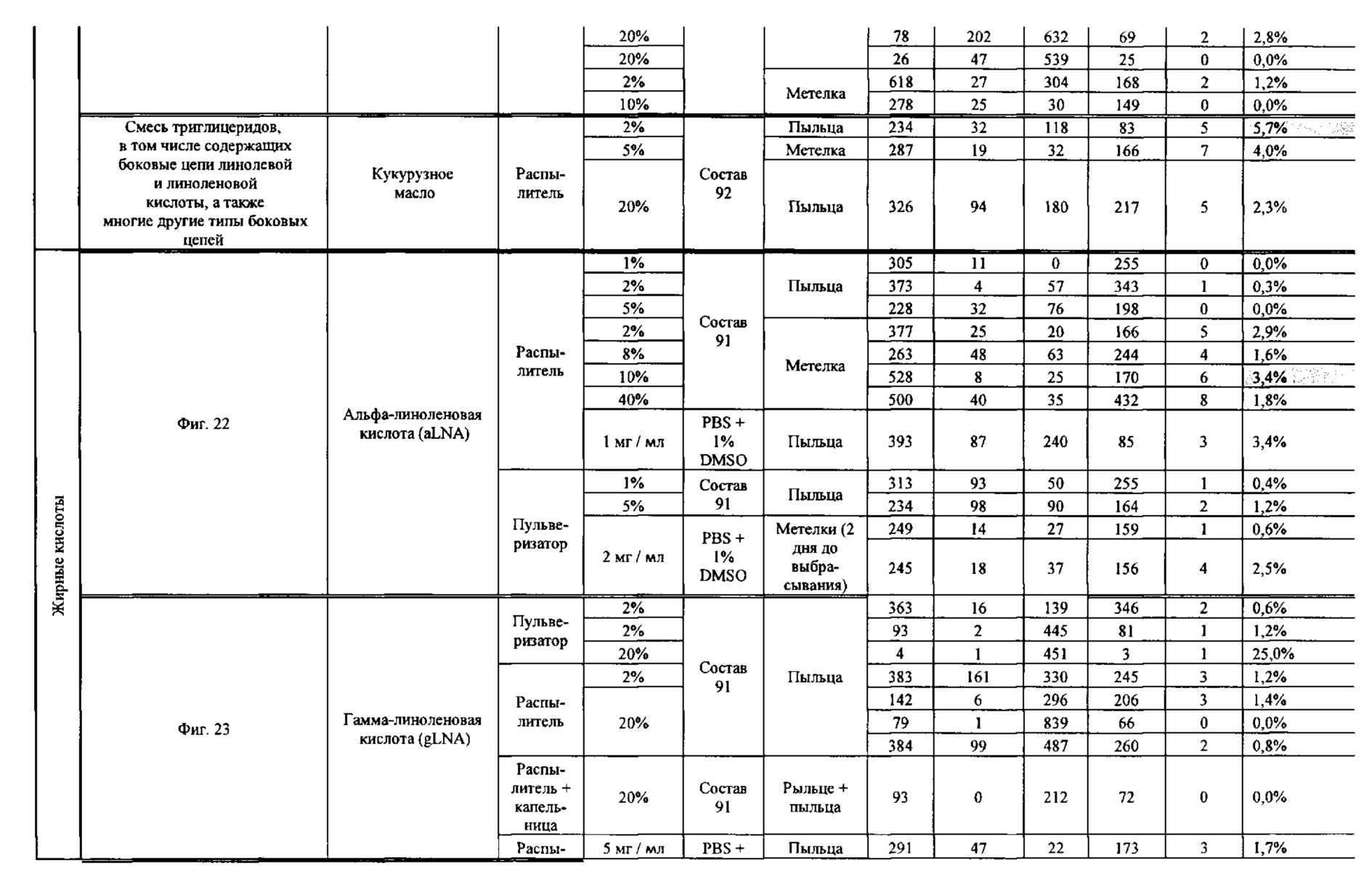
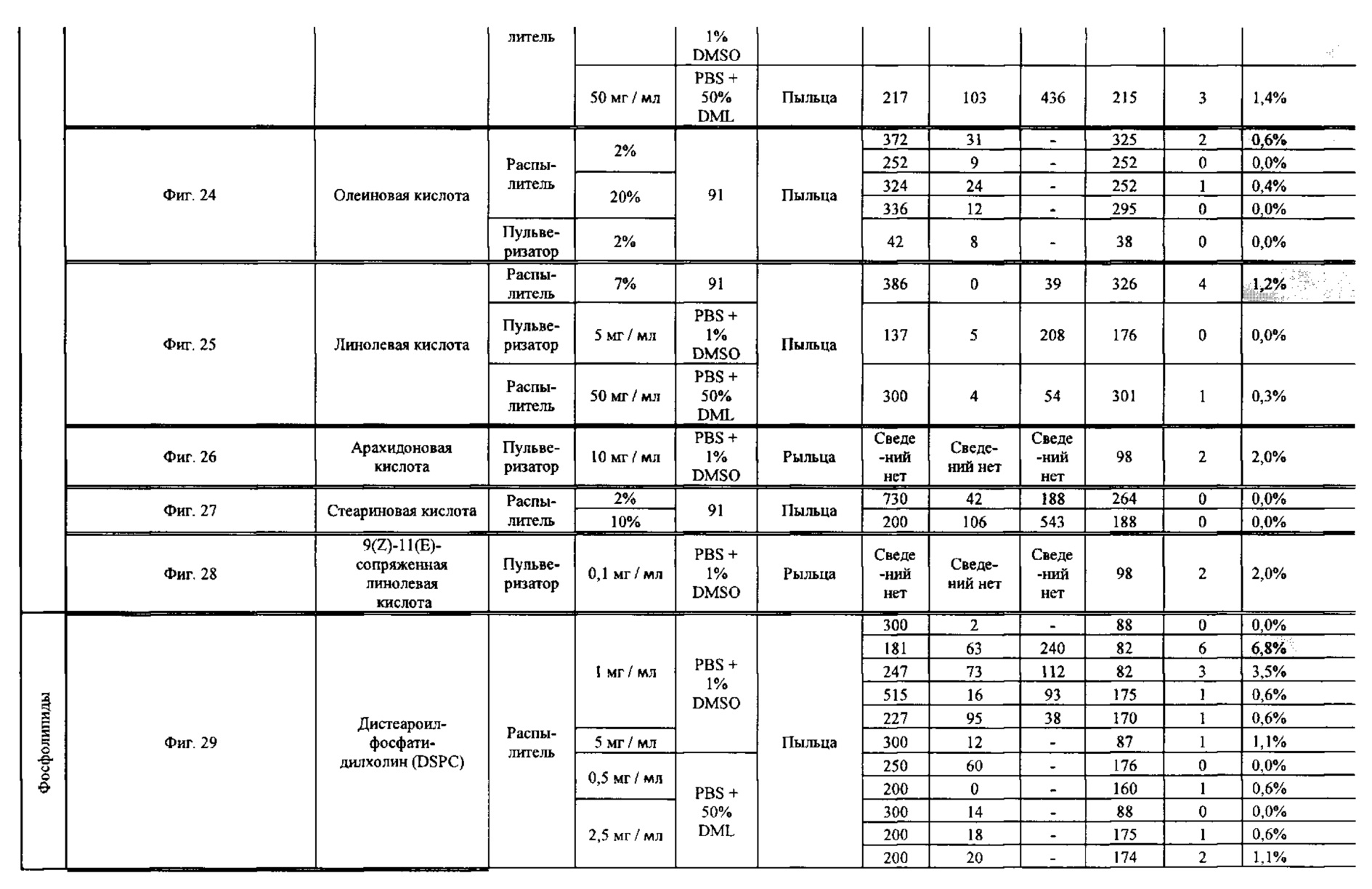
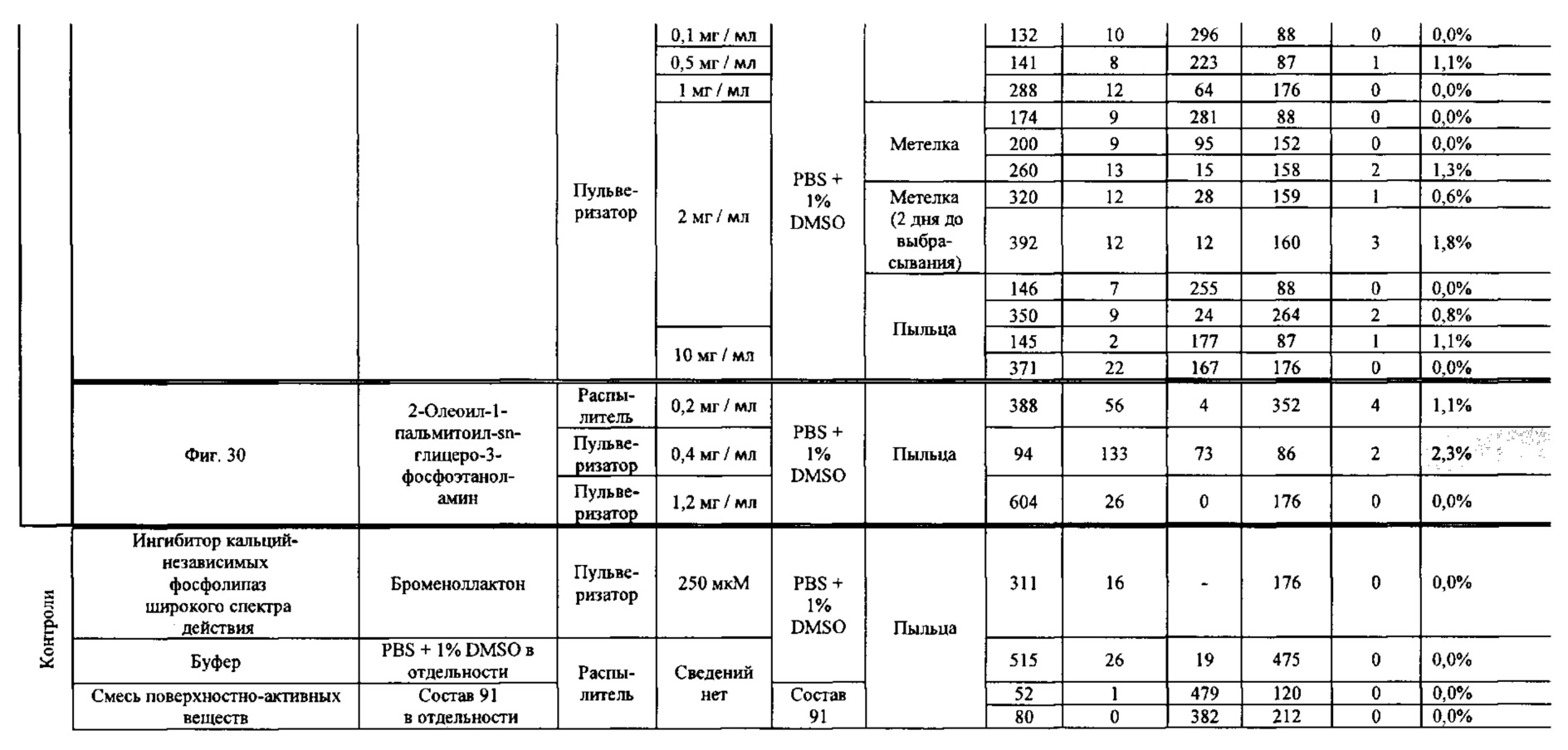
Таблица 8. Рецепты состава 91 ("F91") и состава 92 ("F92"), которые способствуют эмульгированию активных ингредиентов, таких как жирные кислоты, масла, сложные эфиры и фосфолипиды, в виде очень маленьких липидных капель. Состав 91 представляет собой смесь поверхностно-активных веществ, подлежащую смешиванию с жирной кислотой, в оптимальном варианте при соотношении 1 часть жирной кислоты на шесть с половиной частей состава 91. Смесь предназначена для образования эмульсий с водным буфером при концентрациях 0,001-50%. Состав 92 также представляет собой смесь поверхностно-активных веществ, которую можно смешивать со сложными эфирами или маслами, в оптимальном варианте при соотношении 1 часть активного ингредиента на 10 частей состава 92, и затем эмульгировать в водном буфере при концентрациях 0,001-50%.
IV. Увеличение HIR и восстановление завязывания семян посредством нанесения липидного соединения
[00250] Молекулярное профилирование, в том числе метаболомическое и липомическое профилирование, пыльцы, индуцирующей и не индуцирующей гаплоидию (сравнительно), показывает, что пыльца, индуцирующая гаплоидию, является, в частности, дефицитной по определенным типам классов липидов и избыточной по другим. Этот дефицит особенно выражен в классе молекул липидов с цепью из 18 атомов углерода, хотя он также наблюдается в классах с цепью из 20 атомов углерода. Дефицит, в частности, выражен для липидов с цепью из 18 атомов углерода с одной и двумя двойными связями (так называемых олеатов и линолеатов), а избыток был обнаружен, в частности, для липидов с цепью из 18 атомов углерода с тремя двойными связями (так называемых линоленатов). Что касается типов липидов, измененных в пыльце, индуцирующей гаплоидию, то их спектр является очень широким и включает триглицериды, диацилглицериды, свободные жирные кислоты, лизофосфолипиды и фосфолипиды. Изменения содержания липидов в пыльце, индуцирующей гаплоидию, также варьируются среди жирных кислот с различными уровнями насыщения. После рассмотрения этих данных авторы настоящего изобретения решили наносить липидные соединения на цветки во время опыления пыльцой, индуцирующей гаплоидию, чтобы увидеть, может ли простое добавление определенных типов липидов повлиять на уровень образования гаплоидов или число зерен. Авторы настоящего изобретения были особенно заинтересованы в нанесении соединений, которые имели в своей молекулярной структуре некоторые насыщенные жирные кислоты с цепями из 18 или 20 атомов углерода, находившиеся среди тех, представленность которых, как было замечено, является более низкой в пыльце, индуцирующей гаплоидию (например, олеат и линолеаты).
[00251] Нанесение липидов может также привести к увеличению уровня образования гаплоидов, завязывания семян и снижению частоты недоразвития зерен, что указано в таблицах 10-13. В таблице 10 показано девять отдельных контролируемых экспериментов с применением соединения метил-альфа-линоленоилфторфосфоната (MALFP). Каждый эксперимент предусматривал скрещивание двух початков с одним и тем же мужским родительским растением, индуцирующим гаплоидию, в один и тот же день почти точно в одно и то же время (в пределах 5 минут друг от друга) из точно одной и той же популяции пыльцы, собранной с одной и той же родительской мужской особи. Эти два початка получали наносимый контрольный буфер либо буфер вместе с активным ингредиентом. Как мужские, так и женские тестируемые растения выращивали рядом друг с другом в одних и тех же условиях в одной и той же теплице. В среднем, и по сравнению с контрольными процедурами нанесения буфера без MALFP, в случае нанесения MALFP уровень индукции гаплоидии увеличивался на 3,6%, количество зерен увеличивалось почти на 42 на один початок, и общее количество воспроизводящихся гаплоидов на один початок увеличивалось на 11,8 гаплоидов на один початок, а частота недоразвития зародышей уменьшалась на 1,1% (таблица 9). Все это представляет улучшения схемы получения индуцированных гаплоидов. Улучшения были особенно очевидными, если состав 91 применяли в качестве смеси поверхностно-активных веществ для растворения MALFP. Механизм, посредством которого эти ингибиторы оказывают аддитивный эффект вместе с mtl, приводящий к более высоким уровням индукции, может заключаться в том, что они разрушают другие фосфолипазы или липид-модифицирующие белки.
[00252] Тот же тип данных собирали для этилового сложного эфира жирной кислоты LLAEE, который дает увеличение уровня индукции гаплоидии на 2,4%, увеличение образования зерен более чем на 43 на один початок, увеличение образования гаплоидов более чем на 8 на один початок и уменьшение частоты недоразвития зародышей на 3,7%. Хотя многие из этих видов обработки предусматривали различные количества активного ингредиента, в большинстве из этих экспериментов LLAEE давал повышение этих статистических показателей образования гаплоидов и завязывания семян. См. таблицу 10.
[00253] Тот же тип данных собирали для жирной кислоты линолевой кислоты (LLA) (таблица 11), которая давала увеличение уровня индукции гаплоидии на 1,5%, увеличение образования жизнеспособных зерен на 41 на один початок и гаплоидов на 7,6 на один початок и уменьшение уровня недоразвитости зародышей на 2,9%. Это является результатом 22 отдельных экспериментов, при этом опять же каждый эксперимент состоит из опыления одного початка контрольным буфером и опыления одного початка буфером вместе с активным ингредиентом (LLE). Интересно, что представленность 18:2 снижена в пыльце, индуцирующей гаплоидию (что показывают липомические данные). Не желая ограничиваться какой-либо теорией, разумно выдвинуть гипотезу, что это может вызывать проблемы оплодотворения и недоразвитость зародышей или вносить вклад в них. Если авторы настоящего изобретения добавляли их обратно посредством нанесения LLA и LLAEE путем опрыскивания, эти молекулы могли ингибировать фосфолипазы, что приводило к более высокому HIR, и восполнять запасы 18:2, что приводило к большему количеству зерен.
[00254] И наконец, аналогичные данные собирали для MAFP, и были отмечены аналогичные результаты, в том числе повышение уровня индукции гаплоидии на 1,9%, увеличение количества воспроизводящихся гаплоидов на один початок на 6,6 и общего количества зерен на 28, а также уменьшение уровня недоразвитости зародышей на 2,2% (таблица 12). Эти соединения, а также другие липиды, можно применять для повышения частоты образования гаплоидных зародышей в рассадниках для индукции гаплоидии и также можно применять для увеличения уровня завязывания семян в случае с самоопыляемыми линиями, индуцирующими гаплоидию, для увеличения запаса семян этих линий.

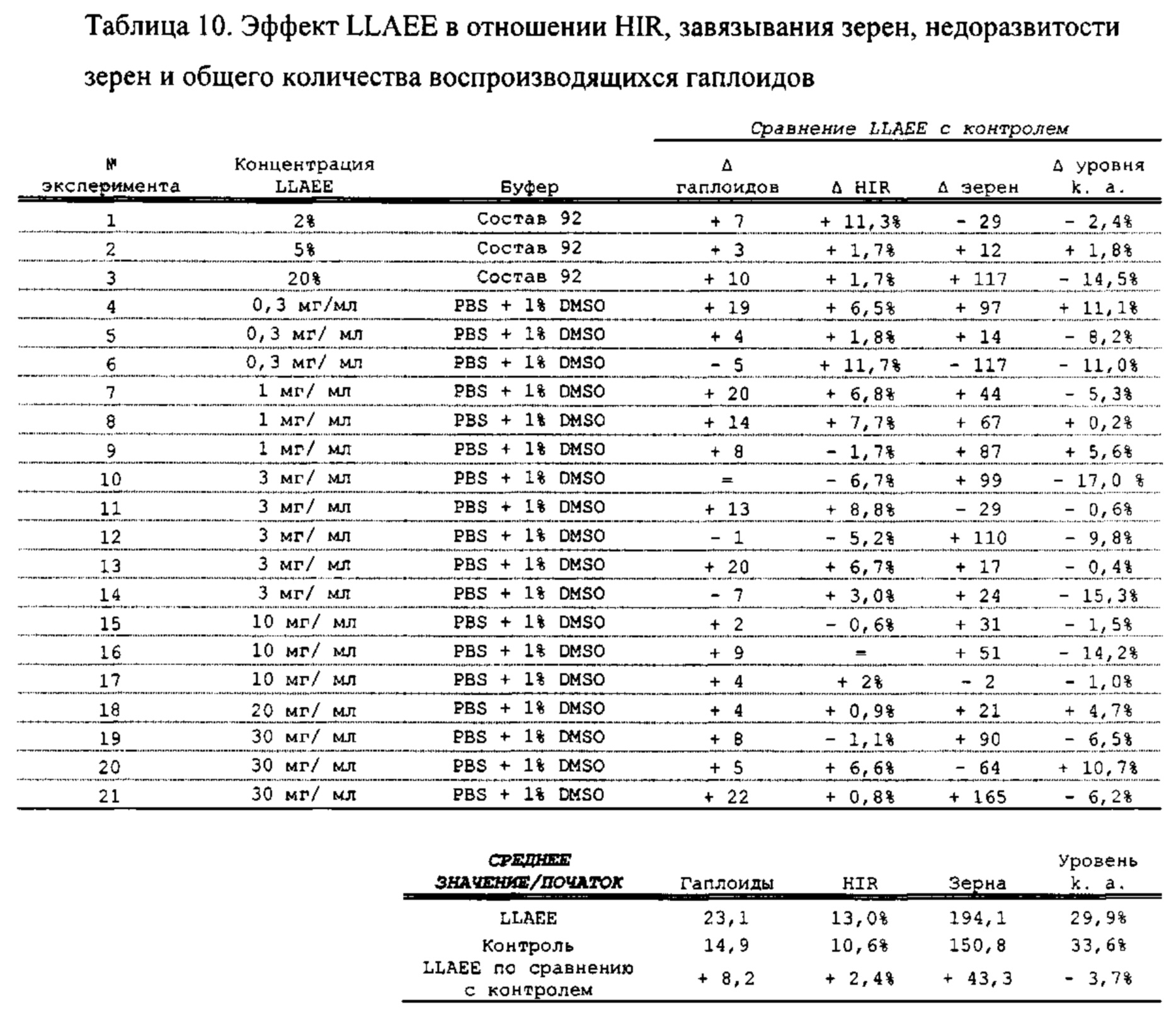
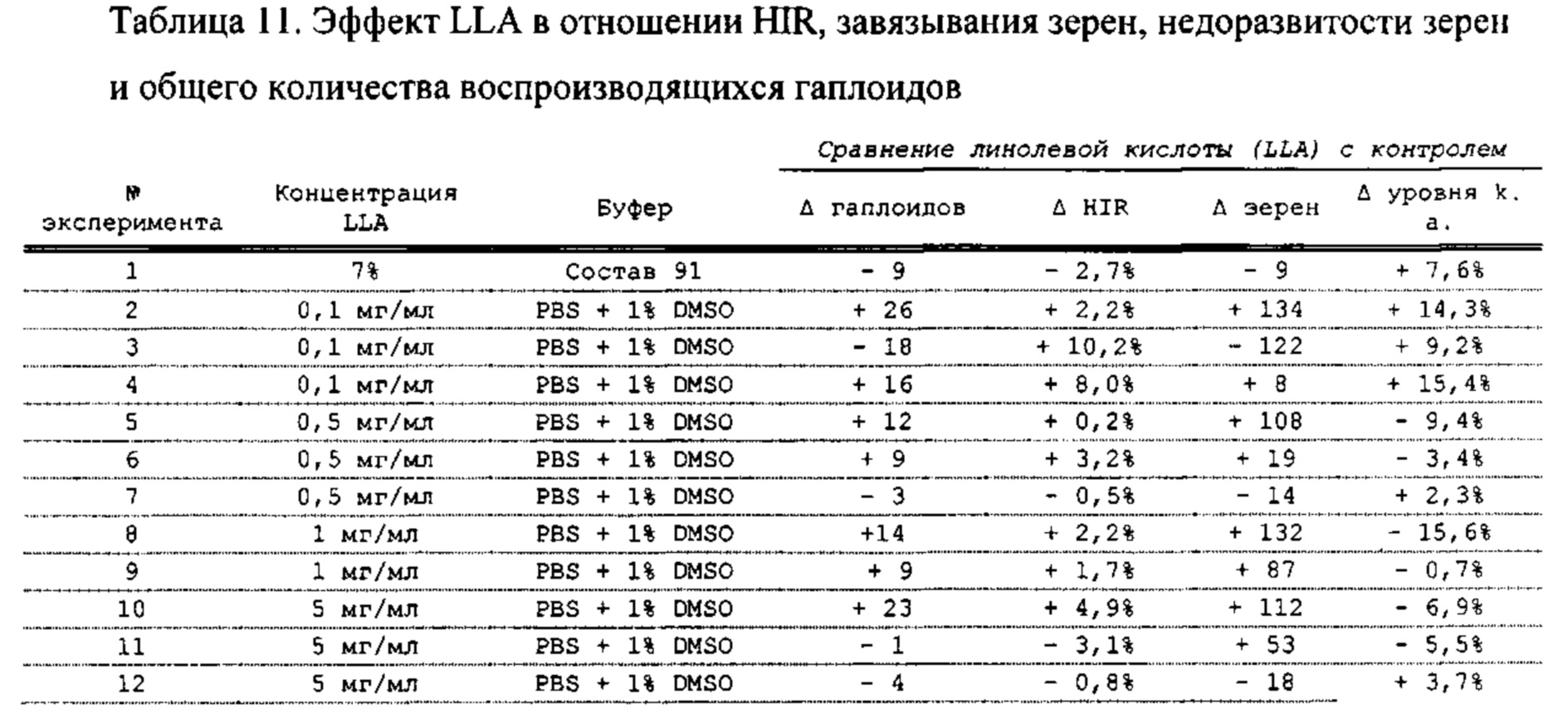
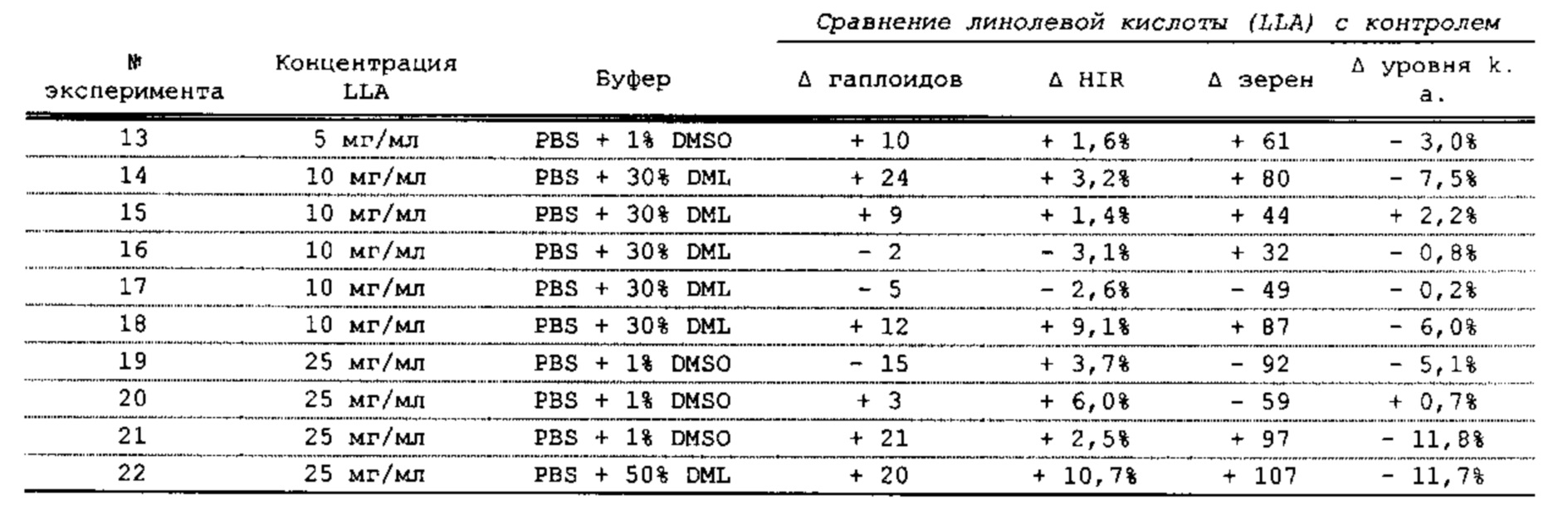
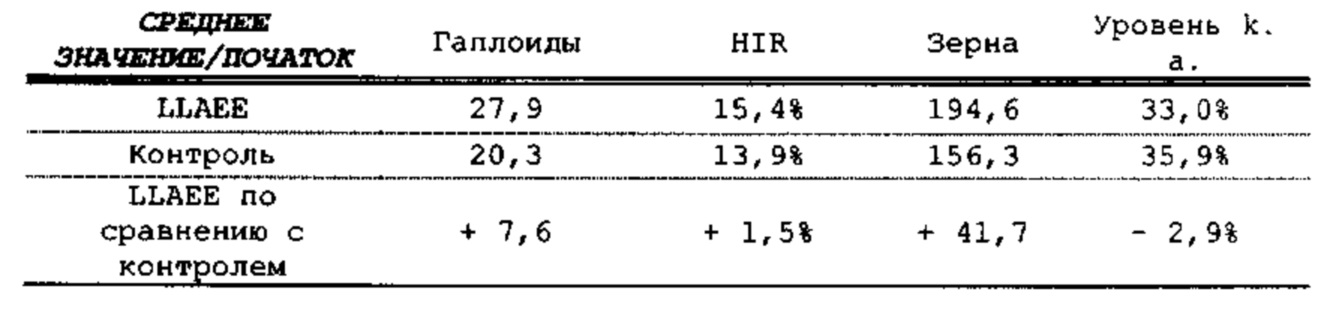
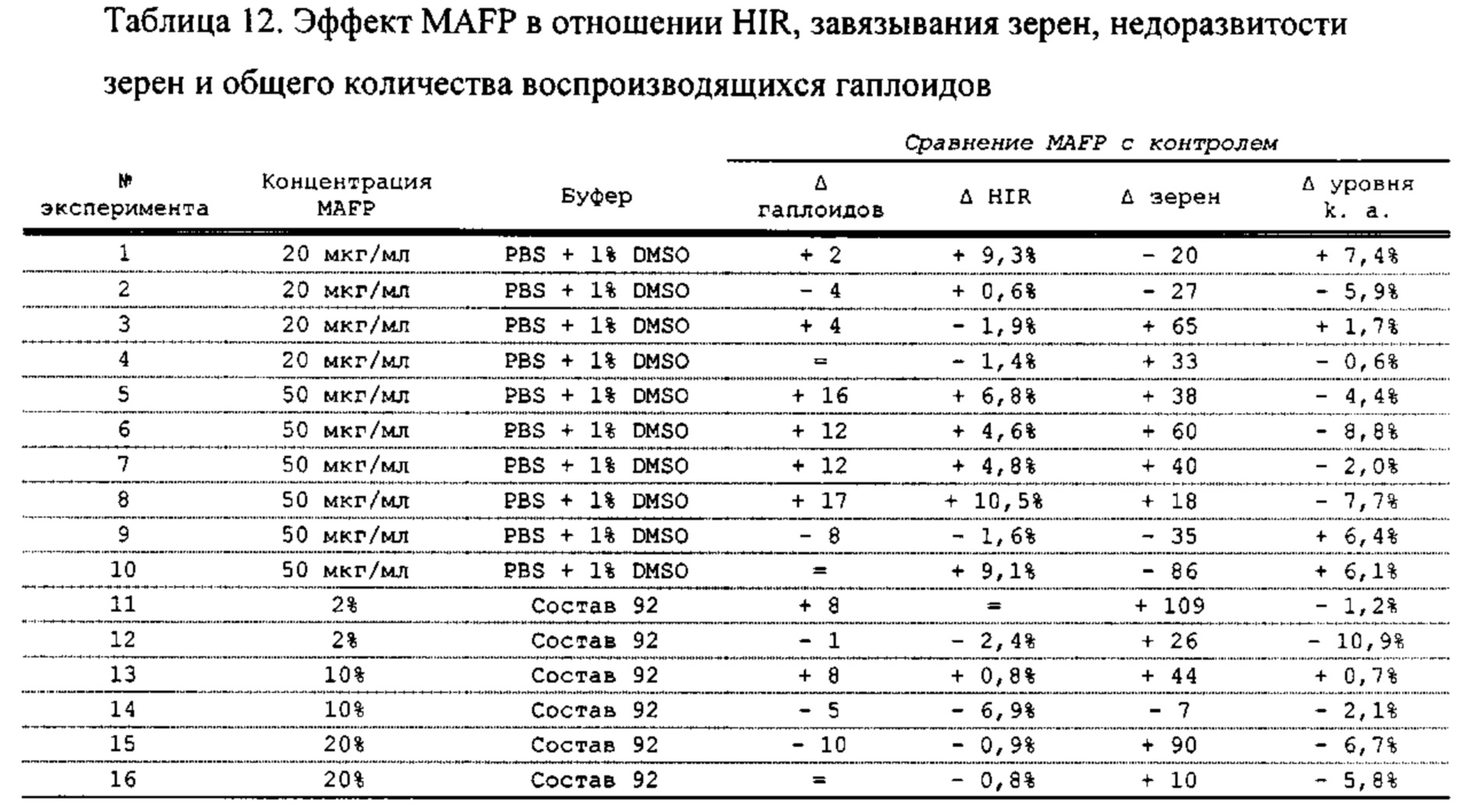
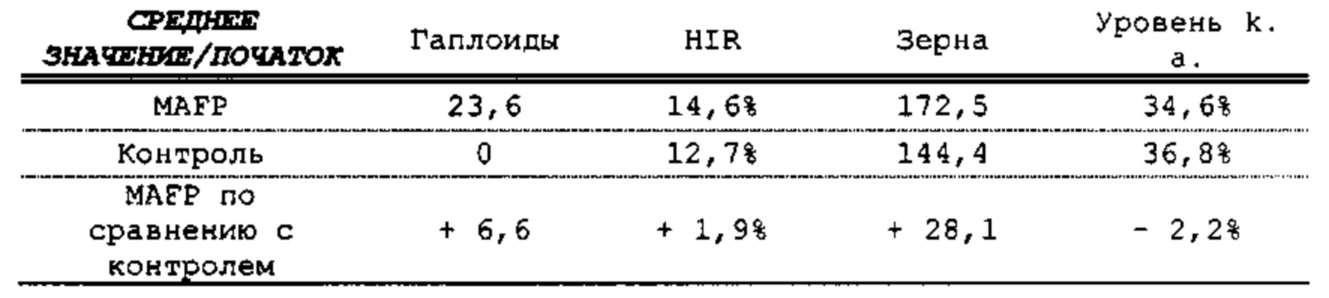
V. Мутагенез/нокаутные варианты PLA
[00255] В попытке изменить уровень индукции гаплоидии или уменьшить уровень недоразвитости зародышей в ходе скрещиваний для индукции гаплоидии авторы настоящего изобретения создавали или получали несколько мутантных линий посредством нескольких способов, в том числе GM-линии, полученные с помощью RNAi, линии, полученные с помощью TILLING, линии, полученные с помощью CRISPR, и линии, полученные с помощью TALEN. Вначале авторы настоящего изобретения искали доказательства того, что направленный мутагенез pPLAIIa является осуществимой стратегией для создания новых линий, индуцирующих гаплоидию. Таким образом, авторы настоящего изобретения тестировали стратегию направленных мутаций маиса с использованием как CRISPR/CAS9, так и TALEN, направленную на одну и ту же последовательность, которая содержит сдвиг рамки считывания в мутантном аллеле, индуцирующем гаплоидию. Это приводило к образованию линий с новыми мутациями, которые авторы настоящего изобретения тестировали в отношении индукции гаплоидии.
[00256] Существует три ключевых компонента для CRISPR-опосредованного процесса. См. патент США №8697359 В1, включенный в данный документ посредством ссылки во всей своей полноте. Первым ключевым компонентом является последовательность-мишень. Вторым является Cas9, которая представляет собой эндонуклеазу. Третьим ключевым компонентом является направляющая РНК ("gRNA"), которая является комплементарной последовательности-мишени и отвечает за привлечение Cas9 в необходимое местоположение. Последовательность-мишень составляет 18-20 п. о. в длину и необязательно должна располагаться сразу после мотива, прилегающего к протоспейсеру ("РАМ"), в 5'-направлении относительно него в геноме растения. Для Cas9 из Streptococcus pyogenes последовательность РАМ должна представлять собой 5'-NGG-3'. Транскрипция gRNA может управляться промотором Pol III U3 (если РНК начинается с А) или U6 (если РНК начинается с G). gRNA должна нести последовательность-мишень на 5'-конце непосредственно после А (U3) или G (U6). Cas9 будет образовывать двухнитевой разрыв ("DSB") в последовательности-мишени на расстоянии в три пары оснований в 5'-направлении относительно последовательности РАМ. Аминокислотная последовательность Cas9 является такой же, как у Cas9 из Streptococcus pyogenes штамма SF370, с двумя аминокислотными изменениями L1164V и I1179V в PI-домене (1099-1368) в NUC-доле. Активность Cas9, как было показано в экспериментах по трансформации, обуславливает примерно 90% частоту мутаций тестируемой последовательности-мишени у кукурузы. Как правило, рекомендуется идентифицировать несколько кандидатов среди РАМ и последовательностей-мишеней в области-мишени, а затем отыскать наилучшую, рассматривая то, какая из последовательностей является уникальной в геноме мишени. Растение-мишень представляет собой растение маиса, риса или любое однодольное растение.
[00257] Этой стратегии следовали для идентификации последовательностей-мишеней для CRISPR, которые перекрываются с существующей мутацией сдвига рамки считывания. Точный сайт разрезания находится всего в двух парах оснований от точки вставки в место сдвига рамки считывания. Конструкции, содержащие как Cas9, так и gRNA, вводили в растения маиса путем трансформации. Как правило, из множества образованных объектов воспроизводимыми являются биаллельные или гомозиготные мутантные растения, но гетерозиготные мутантные растения также являются применимыми. Гетерозиготные растения подвергали самооплодотворению, затем семена Т1 выращивали, подвергали скринингу в отношении гомозиготности мутации и подвергали ауткроссингу. Гомозиготных или биаллельных мутантных трансформантов Т0 просто подвергали самооплодотворению и ауткроссингу с трансформированной NP2222. Всех полученных в результате ауткроссинга зародышей выделяли для анализа плоидности для обнаружения гаплоидов.
[00258] Три различные созданные конструкции для направленного мутагенеза: CRISPR/CAS9 I, CRISPR/CAS9 II и TALEN. Различие между CRISPR/CAS9 I и II является незначительным. Локус сайта-мишени для всех трех конструкций представлял собой ту же область, где был обнаружен сдвиг рамки считывания в линиях, индуцирующих гаплоидию. Для конструкций CRISPR последовательность направляющей РНК начинается с нуклеотида +1560: -GTCAACGTGGAGACΤAGGG-(т. е. SEQ ID NO: 20). Сайт -AGG- РАМ SEQ ID NO: 20 подчеркнут и выделен курсивом. Вставка четырех пар оснований в линиях, индуцирующих гаплоидию, находится точно в том же сайте в нуклеотиде +1576. После трансформации несколько различных объектов, полученных с помощью CRISPR I (содержащих экспрессионную конструкцию, находящуюся под SEQ ID NO: 34), объектов, полученных с помощью CRISPR II (содержащих экспрессионную конструкцию, находящуюся под SEQ ID NO: 36), и объектов, полученных с помощью TALEN (содержащих экспрессионную конструкцию, находящуюся под SEQ ID NO: 35), были отобраны, выращены до зрелости и завязывали жизнеспособные семена. В поколении Т0 авторы настоящего изобретения осуществляли ПЦР в сайте-мишени и секвенировали продукты ПЦР после субклонирования. Авторы настоящего изобретения идентифицировали много уникальных мутаций среди этих объектов (и многие из объектов являлись химерами или имели несколько аллелей).
[00259] Многие растения являлись химерными, о чем свидетельствует появление нескольких различных последовательностей в поколении Т1. После самоопыления Т0 растения Т1 сегрегировали 1:2:1 по конструкции для направленного мутагенеза, и многие из них имели новые мутации в локусе-мишени в биаллельном либо в гомозиготном состоянии. Авторы настоящего изобретения осуществляли скрининг проростков на уровне ДНК с применением маркеров TAQMAN, идентифицировали биаллельные объекты, у которых отсутствовали трансгены, введенные с помощью Cas9 или TALEN, и осуществляли ПЦР-секвенирование с получением продукта ПЦР, прочитываемого от +1494 до +1691 пары оснований в последовательности гена GRMZM2G471240. Авторы настоящего изобретения затем тестировали гомозиготные мутантные формы в отношении способности индуцировать гаплоидию. См. SEQ ID NO: 9-19 и 42-44 в отношении последовательностей новых растений Т1 в гене mtl.
[00260] HIR измеряли для новых предполагаемых линий. См. таблицу 4, выше. Эти данные HIR получены из скрещиваний, где мужская особь относилась к предполагаемой линии, индуцирующей гаплоидию, а женская особь относилась к собственной стандартной инбредной трансформируемой линии NP2222. Предполагаемые линии, индуцирующие гаплоидию, создавали посредством направленной мутации локуса pPLAIIα, опосредованной TALEN либо CRISPR/CAS9. Среди показанных в данном документе существует одиннадцать различных предполагаемых индуцирующих растений, содержащих восемь различных объектов из трех отдельных трансформационных конструкций. Объект 39А представлял собой объект, полученный с помощью TALEN. Объекты 18А и 27А представляли собой объекты, полученные с помощью CRISPR. Последний представлял собой химеру, используемую в качестве растения Т0, и после его самоопыления в популяции Т1, в том числе в "биаллельных" растениях (под биаллельными авторы настоящего изобретения подразумевают то, что при секвенировании области pPLAIIα, которая была подвергнута мутации, авторы настоящего изобретения обнаружили два различных новых аллеля - так что очевидно, что обе копии гена дикого типа были подвергнуты мутации, но они были подвергнуты мутации различным образом, поэтому они представляют собой два новых аллеля), было обнаружено несколько мутаций. Каждое из этих одиннадцати отдельных растений, таким образом, имело различные комбинации мутаций в pPLAIIα. Общим у них всех было то, что ни одно из одиннадцати растений не имело копии pPLAII дикого типаα. Таким образом, все они являются "гомозиготными мутантными" по гену pPLAIIα. Все мутации представляли собой сдвиги рамки считывания в экзоне 4, имитирующие исходную мутацию в нативных линиях, индуцирующих гаплоидию. Используя эти пять растений в качестве мужских особей, авторы настоящего изобретения скрещивали их с одним либо несколькими женскими початками с образованием нескольких тысяч зародышей. Авторы настоящего изобретения срезали это потомство, осуществляли в его отношении анализ плоидности и обнаружили, что каждый из наборов потомков имел по меньшей мере 3,98% гаплоидов при максимальном содержании 12,5% гаплоидов. Это демонстрирует то, что образование мутаций в pPLAIIα приводит к индукции гаплоидии. Авторы настоящего изобретения думают, что другие типы мутаций, помимо сдвигов рамки считывания, также приводят к индукции гаплоидии. Эти мутации могут находиться в любом месте в гене, и они могут представлять собой точечные мутации, или вставки, или делеции, или другие типы мутаций.
[00261] Для образования линий, индуцирующих гаплоидию, также применяли RNAi. Для RNAi получали две шпилечные конструкции; одну, соответствующую границе между экзоном 1 и 2, и другую, соответствующую экзону 4 (фигуры 8 и 9). Конструкции вводили в растения дикого типа путем трансформации, и растения Т0 подвергали самооплодотворению. Семена Т1 из трех объектов на одну конструкцию выращивали, подвергали скринингу в отношении гомозиготности по трансгену и подвергали ауткроссингу с несколькими початками в качестве тестеров индукции гаплоидии. После исследования более 1500 зерен, полученных в результате этих ауткроссингов, авторы настоящего изобретения обнаружили, что оба объекта индуцировали гаплоидию на уровне, составляющем примерно 1%-2%. Наиболее высокий уровень индукции гаплоидии, полученный на одном початке, составлял 4,3%. Этот початок содержал приблизительно 300 зерен, так что авторы настоящего изобретения могут сделать вывод, что уровень недоразвитости зародышей также был ниже, чем у обычной линии, являющейся сильным индуктором. Эта работа демонстрирует то, что стратегию RNAi + GM можно применять для создания новых линий, индуцирующих гаплоидию, из линий, в иных отношениях обычно являющихся линиями дикого типа, путем изменения экспрессии pPLAIIα.
[00262] Для создания и идентификации мутаций фосфолипазы и маиса по настоящему изобретению также применяли способ мутагенеза TILLING. Публикации, в которых описан TILLING, доступны для культурных растений, таких как рис: Till et al., ВМС Plant Biology 7:19 (2007), томат: Rigola et al. PLOS ONE March 13, 2009, и маис: Till et al. BMC Plant Biol. 2004 Jul 28;4:12 (2004), все из которых включены в данный документ посредством ссылки. В базовой методике TILLING растительный материал, такой как семена, подвергают химическому мутагенезу, в результате которого создается серия мутаций в геномах клеток семян. Из семян, подвергнутых мутагенезу, выращивают взрослые растения M1, и их подвергают самоопылению. Образцы ДНК из полученных в результате этого растений М2 объединяют и затем подвергают скринингу в отношении мутаций в гене, представляющем интерес. После идентификации мутации в гене, представляющем интерес, из семян растений М2, несущих эту мутацию, выращивают взрослые растения М3, и их подвергают скринингу в отношении фенотипических характеристик, ассоциированных с геном, представляющим интерес.
[00263] Любой культивар маиса, имеющий по меньшей мере один ген фосфолипазы со значительной гомологией с SEQ ID NO: 1, можно применять в соответствии с настоящим изобретением. Используемое в данном документе выражение "значительная гомология" означает, что последовательность ДНК гена является достаточно сходной с SEQ ID NO: 1 на нуклеотидном уровне, чтобы кодировать белок, эквивалентный кодируемому SEQ ID NO: 1, при возможном наличии аллельных различий между культиварами. В соответствии с одним аспектом иллюстративного варианта осуществления настоящего изобретения "значительная гомология" может иметь место уже тогда, когда гомология между геном фосфолипазы и SEQ ID NO: 1 составляет приблизительно 85%, при условии, что гомология в консервативных областях гена является более высокой (например, по меньшей мере приблизительно 90%). Предпочтительно, процентная идентичность в кодирующей области составляет 85-90%, более предпочтительно 90-95%, и в оптимальном варианте она составляет более 95%. Специалист в данной области может предпочитать культивар маиса, имеющий коммерческую популярность или имеющий конкретные желаемые характеристики, из которого можно создать маис с мутантной фосфолипазой. В качестве альтернативы, специалист в данной области может предпочитать культивар маиса, имеющий незначительное количество полиморфизмов, такой как инбредный культивар, для облегчения скрининга в отношении мутаций в пределах локусов генов фосфолипаз.
[00264] В соответствии с одним аспектом иллюстративного варианта осуществления настоящего изобретения семена риса и маиса подвергали мутагенезу, и затем из них выращивали растения M1. Затем обеспечивали самоопыление растений M1, и из семян растений M1 выращивали растения М2, которые затем подвергали скринингу в отношении мутаций в их локусе гена фосфолипазы. Хотя скринингу в отношении мутаций можно подвергать и растения M1, преимущество осуществления скрининга растений М2 заключается в том, что все соматические мутации соответствуют герминативным мутациям. Специалисту в данной области будет понятно, что множество растительных материалов маиса, в том числе без ограничения семена, пыльцу, растительную ткань или растительные клетки, можно подвергнуть мутагенезу для создания маиса с мутантной фосфолипазой по настоящему изобретению. Однако тип растительного материала, подвергаемого мутагенезу, может оказывать влияние при осуществлении скрининга растительной ДНК в отношении мутаций. Например, если пыльцу подвергают мутагенезу до опыления не подвергнутого мутагенезу растения, то из семян, полученных в результате этого опыления, выращивают растения M1. Каждая клетка растений M1 будет содержать мутаций, созданные в пыльце, поэтому эти растения M1 можно затем подвергать скринингу в отношении мутации фосфолипазы вместо того, чтобы ждать получения поколения М2.
[00265] Для создания мутаций по настоящему изобретению можно применять мутагены, которые создают главным образом точечные мутации и короткие делеции, вставки, трансверсии и или транзиции (от приблизительно 1 до приблизительно 5 нуклеотидов), такие как химические мутагены или радиоактивное излучение. Мутагены, соответствующие способу по настоящему изобретению, включают в себя без ограничения этилметансульфонат (EMS), метилметансульфонат (MMS), N-этил-N-нитрозомочевину (ENU), триэтилмеламин (ТЕМ), N-метил-N-нитрозомочевину (MNU), прокарбазин, хлорамбуцил, циклофосфамид, диэтилсульфат, мономерный акриламид, мелфалан, азотистый иприт, винкристин, диметилнитрозамин, N-метил-N'-нитронитрозогуанидин (MNNG), нитрозогуанидин, 2-аминопурин, 7,12-диметилбенз(а)антрацен (DMBA), этиленоксид, гексаметилфосфорамид, бисульфан, диэпоксиалканы (диэпоксиоктан (DEO), диэпоксибутан (ВЕВ) и т.п.), дигидрохлорид 2-метокси-6-хлор-9[3-(этил-2-хлорэтил)аминопропиламино]акридина (ICR-170) и формальдегид. Спонтанные мутации в области ядрышкового организатора ("NOR"), которые могут не быть непосредственно вызваны мутагеном, можно также идентифицировать в соответствии с различными вариантами осуществления настоящего изобретения.
[00266] Любой подходящий способ получения растительной ДНК, который известен на данный момент или будет разработан в будущем, можно применять для получения ДНК растения маиса для скрининга в отношении мутации фосфолипазы. Например, см. Chen и Ronald, Plant Molecular Biology Reporter 17:53-57, 1999; Stewart и Via, Bio Techniques 14:748-749, 1993. Кроме того, доступно несколько коммерческих наборов, в том числе наборы от Qiagen (Валенсия, Калифорния) и Qbiogene (Карлсбад, Калифорния).
[00267] В соответствии с одним аспектом иллюстративного варианта осуществления настоящего изобретения образцы ДНК из отдельных растений маиса получали и затем объединяли для ускорения скрининга в отношении мутаций в фосфолипазе во всей популяции растений, происходящих из растительной ткани, подвергнутой мутагенезу. Размер объединенной группы может зависеть от чувствительности применяемого способа скрининга. В соответствии с одним аспектом иллюстративного варианта осуществления настоящего изобретения отдельные растения маиса объединяли в группы из четырех или более растений.
[00268] В соответствии с другим аспектом иллюстративного варианта осуществления после объединения образцов ДНК пулы подвергали методикам амплификации, специфичным в отношении последовательности фосфолипазы, таким как полимеразная цепная реакция (ПЦР). В отношении общего обзора ПЦР см. PCR Protocols: A Guide to Methods and Applications (Innis, Gelfand, Sninsky, J., и White, eds.), Academic Press, San Diego, 1990, который включен в данный документ посредством ссылки. Любой праймер, специфичный в отношении локуса гена фосфолипазы или последовательностей, непосредственно прилегающих к локусу гена фосфолипазы, можно использовать для амплификации последовательностей фосфолипазы в объединенном образце ДНК. Праймер предпочтительно разрабатывают для амплификации областей локуса гена фосфолипазы, где с наибольшей вероятностью возникнут полезные мутации. Праймер наиболее предпочтительно разрабатывают для выявления мутаций в кодирующей области гена фосфолипазы. Кроме того, предпочтительно, чтобы праймер избегал известных полиморфных сайтов для облегчения скрининга в отношении точечных мутаций. Для облегчения выявления продуктов ПЦР в геле праймер для ПЦР можно метить с помощью традиционного или разработанного в будущем способа мечения.
[00269] В соответствии с одним аспектом иллюстративного варианта осуществления настоящего изобретения продукты ПЦР-амплификации можно подвергнуть скринингу в отношении мутаций фосфолипазы с помощью любого способа, обеспечивающего идентификацию нуклеотидных различий между последовательностью дикого типа и мутантной последовательностью. Он может включать без ограничения секвенирование, денатурирующую жидкостную хроматографию высокого давления (dHPLC), капиллярный электрофорез с постоянной концентрацией денатурирующего средства (CDCE), капиллярный электрофорез с температурным градиентом (TGCE) (см. Li et al., Electrophoresis 23(10): 1499-1511, 2002) или фрагментацию с помощью ферментативного расщепления, как, например, применяемого в высокопроизводительном способе, описанном в Colbert et al., Plant Physiology 126:480-484, 2001. Продукты ПЦР-амплификации предпочтительно инкубируют с эндонуклеазой, которая преимущественно расщепляет участки ошибочного спаривания оснований в гетеродуплексах между последовательностью дикого типа и мутантной последовательностью. В соответствии с другим аспектом иллюстративного варианта осуществления продукты расщепления подвергают электрофорезу с помощью автоматизированного устройства для электрофореза в секвенирующем геле, и изображения геля анализируют с помощью стандартной коммерческой программы для обработки изображений.
[00270] Авторы настоящего изобретения определили, что для достижения индукции гаплоидии у маиса желаемыми являются мутации, которые изменяют функцию фосфолипазы. Предпочтительные мутации включают миссенс-мутации, нонсенс-мутации и изменения границы сплайсинга, в том числе мутации, проявляющиеся в преждевременном завершении трансляции белка фосфолипазы с информационной РНК, как, например, такие мутации, которые создают стоп-кодон в пределах кодирующих областей гена фосфолипазы. Такие мутации включают вставки, повторения последовательностей, модификации открытых рамок считывания (ORF) и, наиболее предпочтительно, точечные мутации.
[00271] В соответствии с еще одним другим аспектом иллюстративного варианта осуществления настоящего изобретения после идентификации растения М2, имеющего мутантную последовательность фосфолипазы, мутации анализируют для определения их эффектов в отношении экспрессии, трансляции и/или активности белка. В соответствии с одним иллюстративным вариантом осуществления фрагмент фосфолипазы, содержащий мутацию, секвенируют с помощью стандартных методик секвенирования для определения точного местоположения мутации относительно всей последовательности фосфолипазы. Каждую мутацию оценивают для того, чтобы предсказать ее влияние на функцию белка (т.е. от полной переносимости до потери функции), с помощью биоинформатических инструментов, таких как SIFT (рассортировка непереносимых и переносимых мутаций; Ng et al., Nucleic Acids Research 31:3812-3814, 2003), PSSM (позиционно-специфическая матрица замен; Henikoff и Henikoff, Computer Applications in the Biosciences 12:135-143, 1996) и PARSESNP (Taylor и Greene, Nucleic Acids Research 31:3808-3811, 2003). Например, балл SIFT, который составляет менее 0,05, и значительное изменение балла PSSM (например, примерно на 10 или более) указывает на мутацию, которая по всей вероятности оказывает пагубный эффект в отношении функции белка.
[00272] В соответствии с дополнительным аспектом иллюстративного варианта осуществления в случае, если первоначальная оценка мутации у растения М2 указывает на то, что она имеет полезную природу и находится в полезном положении в гене фосфолипазы, то проводят дополнительный фенотипический анализ растения маиса, содержащего эту мутацию. Вначале растение М2 дважды подвергают возвратному скрещиванию или ауткроссингу для создания растения ВС1 с целью устранения фоновых мутаций. Затем полученное в результате возвратного скрещивания или ауткроссинга растение ВС1 подвергают самоопылению для создания растения BC1F2, которое является гомозиготным по мутации фосфолипазы.
[00273] Оценивают несколько физических характеристик этих гомозиготных по фосфолипазе мутантных растений для определения того, приводит ли мутация в результате к полезному фенотипическому изменению у маиса. Мутантный по фосфолипазе маис оценивают в отношении индукции гаплоидии в сравнении с нормальным родительским маисом (например, дикого типа) или с контрольным сибсом маиса дикого типа. В таблице 13 показаны новые мутации, полученные с помощью TILLING.
Таблица 13. Новые мутации pPLAIIα, полученные с помощью TILLING, и их HIR. В столбце с нуклеотидными изменениями представлены положения от начала последовательности кДНК (последовательность №1), и измененный нуклеотид указан заглавной буквой в рамках его кодона. Затем указано аминокислотное изменение, а за ним - влияние этого изменения (переносимое или непереносимое). Из двух аллелей, для которых наблюдалась непереносимость, один индуцировал образование гаплоидов на уровне 1,04% (3/288).

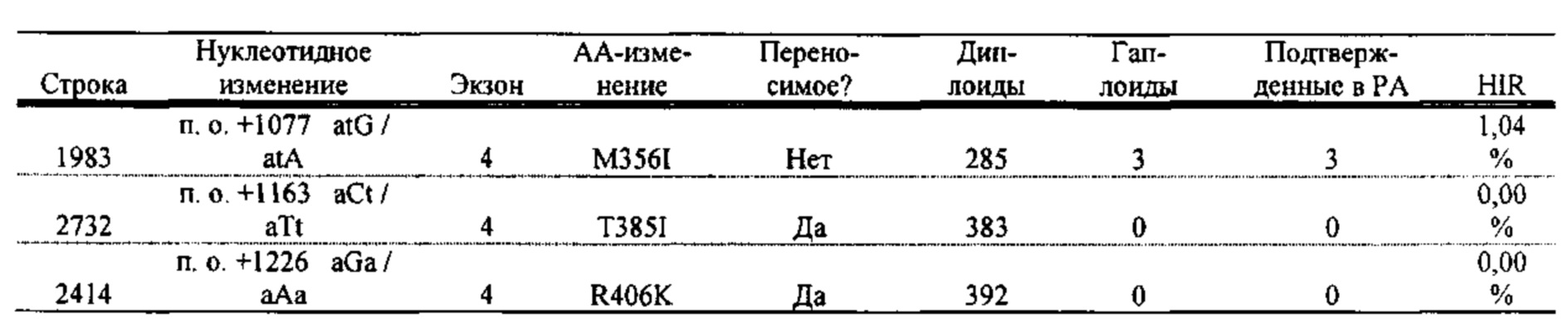
[00274] С помощью номенклатуры, применяемой в таблице 13, указываются нуклеотид или аминокислота дикого типа, за которыми указано их положение в соответствии с эталонной SEQ ID NO: 1, за которыми указаны измененные нуклеотид или аминокислота в этом положении, с применением стандартной терминологии генетического кода.
[00275] В случае с маисом в результате TILLING гена pPLAIIα маиса образуются новые аллели, которые характеризуются низкими уровнями индукции гаплоидии. Это позволяет создавать аллельные серии, в том числе нокаутные варианты, GRMZM2G471240. Последовательность двух сегментов этого гена (максимум 1,5 т. п. о., что равняется 20 ампликонам на один ген) подвергают скринингу в отношении мутаций. Эти последовательности включали в себя геномную последовательность, содержащую интроны, а также предсказанную последовательность кДНК и кодирующие последовательности для двух сплайс-вариантов. На основании этих последовательностей разрабатывают соответствующие и уникальные последовательности ампликонов, и скрининг в отношении мутаций осуществляют в существующей общей популяции кукурузы М2. Идентифицированные мутантные формы характеризуют в категориях последовательности ДНК и последствий в отношении транслируемой последовательности белка. Семена М3 выращивают и подвергают самооплодотворению с получением линий М4 с сегрегирующими предполагаемыми мутантными гомозиготными индивидуумами. Этих индивидуумов идентифицируют посредством ПЦР-секвенирования и подвергают ауткроссингу и самооплодотворению для тестирования способности этих мутантных линий индуцировать образование гаплоидов.
[00276] Для осуществления анализирующих скрещиваний новые линии выращивают рядом с маркерной линией, которая является гомозиготной рецессивной по нелетальному гену маркера цвета. Реципрокные скрещивания применяют для тестирования специфичности индукции в отношении передачи по мужской и женской линии посредством оценки полученных в результате растений в отношении наличия гаплоидов, которые демонстрируют фенотип цвета. Положительные хиты подтверждают посредством анализа плоидности, описанного выше.
[00277] Индивидуумов, которые являются гомозиготными в отношении мутаций SNP, скрещивали в качестве мужской особи с женской особью маркерной линии, что в некоторых случаях приводило к образованию гаплоидов на низком уровне (см. таблицу 18). Положительные хиты подтверждают посредством анализа плоидности, описанного выше. В частности, линия, которая давала образование гаплоидов, имела мутацию замены G на А в паре оснований 1077 последовательности кДНК. Эта мутация вызывает аминокислотную замену метионина (М) на изолейцин (I) в положении аминокислоты 356. Это является неконсервативным аминокислотным изменение, которое может нарушать активность белка, что приводит к низкому уровню образования гаплоидов. Среди 288 тестированных потомков авторы настоящего изобретения обнаружили трех гаплоидов при уровне индукции 1% (3/288).
VI. Создание линий риса, индуцирующих гаплоидию
[00278] У риса ближайшим гомологом pPLAIIα маиса является Os03g27610, пататин-подобная фосфолипаза риса (OspPLAIIϕ) со сходной аннотацией, структурой гена и профилем экспрессии, т. е. она экспрессируется в пыльце и отсутствует в других частях растений (фигура 10). SEQ ID NO: 21 содержит последовательность геномной ДНК Os03g27610, SEQ ID NO: 22 содержит последовательность кДНК, и SEQ ID NO: 23 содержит аминокислотную последовательность. Близкое совпадение этих признаков, наряду с коротким эволюционным расстоянием между этими двумя злаковыми растениями, позволяет предположить, что мутация в гене риса может также способствовать образованию линии, индуцирующей гаплоидию. В недавней публикации белок риса был выявлен в ядрах спермиев пыльцевых зерен (Abiko et al., 2013), что позволяет предположить участие этого белка в оплодотворении и/или развитии зиготы.
[00279] Для улучшения уровня индукции гаплоидии у маиса и создания первых линий риса, индуцирующих гаплоидию, применяли подход "обратной генетики" TILLING для получения новых индивидуумов, мутантных по гену GRMZM2G471240 маиса и гену Os03g27610 риса. См. McCallum CM et al. (2000) Targeting induced local lesions IN genomes (TILLING) for plant functional genomics, Plant Physiol. 123:439-42, включенный в данный документ посредством ссылки. TILLING представляет собой беспристрастный подход для образования новых мутантных форм, поскольку исследователь не может контролировать, где мутаген этилметансульфонат (EMS) будет создавать новые мутации. Получали новые аллели в определенном разнообразии и представленности, и их тестировали в отношении уровня индукции гаплоидии.
[00280] С помощью TILLING получали тринадцать различных линий М3. См. таблицу 14. В столбце "Положение в геноме" указано нуклеотидное положение мутации и изменение (например, G803A указывает на то, что пара оснований G в положении 803 была изменена на А). Эффектом является изменение аминокислоты или другое изменение белка в результате мутации (например, А209Т указывает на то, что аланин в положении аминокислоты 209 был изменен на треонин). Балл BLOSUM является предсказательной оценкой силы эффекта, который изменение аминокислоты будет оказывать на конформацию или сворачивание белка (чем она более отрицательная, тем сильнее эффект). "Тип" указывает на тип изменения аминокислоты ("NSM" означает немолчащую мутацию; "PSM" означает частично молчащую мутацию; "молчащая" означает молчащую мутацию; "сплайс" означает мутацию сайта сплайсинга, приводящую в результате к аберрантному сплайсингу; "интрон" означает, что мутация происходит в интроне). Наконец, № в GSOR представляет собой ID линии в коллекции генетического фонда Oryza в USDA.
[00281] Эти тринадцать линий подвергали самооплодотворению с получением М4, и семена М4 выращивали и тестировали в отношении гомозиготности. Гомозиготных мутантных индивидуумов подвергали самооплодотворению и ауткроссингу для тестирования способности индуцировать гаплоидию. Полученное в результате потомство исследовали в отношении содержания ДНК на одну клетку (плоидности) с помощью анализатора плоидности.
[00282] Неконсервативные изменения, такие как изменения в сайте сплайсинга и изменения с наиболее отрицательными баллами BLOSUM, имеют наибольший потенциал индукции гаплоидии. Они должны оказывать более выраженные дестабилизирующие эффекты на белковый продукт и поэтому в сравнении с другими индуцирующими гаплоидию аллелями, полученными с помощью TILLING, превосходят их, способствуя образованию большего количества гаплоидов на одно скрещивание для индукции гаплоидии и по всей вероятности обуславливая частично нарушенное завязывание семян. В действительности авторы настоящего изобретения уже начали наблюдать это в некоторых самоопылениях Т4. Линия с наиболее низким уровнем завязывания семян содержала мутацию G153A в сайте сплайсинга, при этом воспроизводилось только 29 семян на 12 скрещенных гомозиготных мутантных растений М4. Другие линии имели более 100 воспроизводящихся семян.
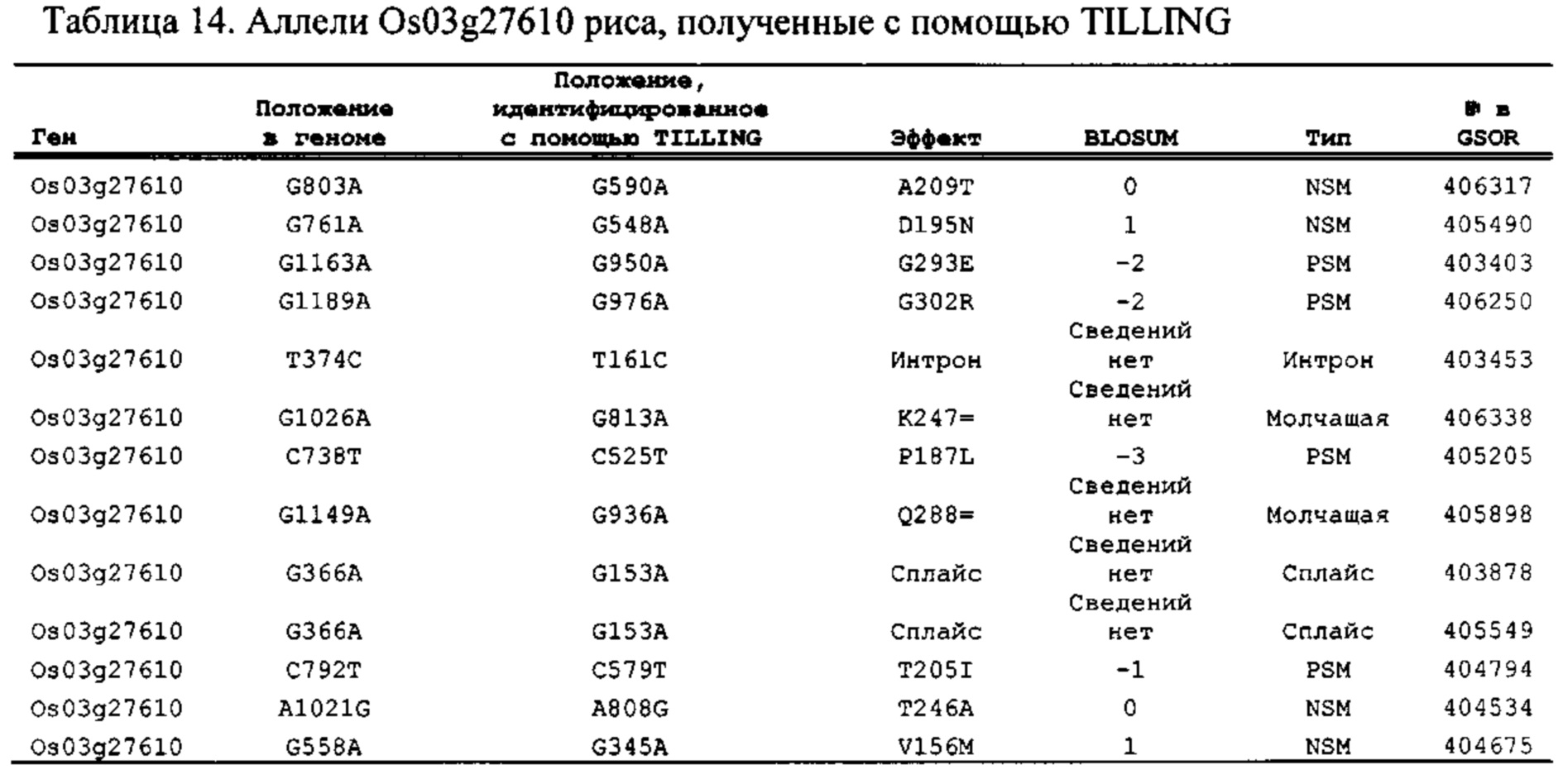
[00283] В качестве альтернативы, ген фосфолипазы риса, обнаруженный в Os03g27610, может быть отредактирован с помощью способов CRISPR/Cas9. Как указано выше, существует три ключевых компонента для CRISPR-опосредованного процесса. Первым ключевым компонентом является последовательность-мишень. Вторым является Cas9, которая представляет собой эндонуклеазу. Третьим ключевым компонентом является направляющая РНК ("gRNA"), которая является комплементарной последовательности-мишени и отвечает за привлечение Cas9 в необходимое местоположение. Направляющие РНК могут находиться в форме одинарной направляющей РНК (sgRNA) или двойной направляющей РНК (dgRNA). Для риса авторы настоящего изобретения создали четыре конструкции, целенаправленно воздействующие на ген фосфолипазы риса. SEQ ID NO: 38 содержит кассету экспрессии, которая обеспечивает нацеливание dgRNA на экзон 4 Os03g27610 очень близко к тому месту, где в гомологе маиса расположена природная мутация, затрагивающая четыре пары оснований. В гене риса сайтом-мишенью для направляющей РНК является GAGACCGGCAGGTACGTCGAGG. SEQ ID NO: 39 содержит кассету экспрессии, которая обеспечивает нацеливание sgRNA на тот же сайт-мишень для gRNA в экзоне 4 Os03g27610, на который нацеливается SEQ ID NO: 38. Ожидается, что в случае как с SEQ ID NO: 38, так и с SEQ ID NO: 39 мутации сдвига рамки считывания появятся там, где помещена вертикальная черта между G и Т в последовательности CAGGTACG | TCGAGG (в паре оснований +1150 последовательности gDNA в SEQ ID NO: 22). Таким образом, как ожидается, обе из этих конструкций будут обеспечивать образование мутаций, индуцирующих гаплоидию, расположенных всего на семь пар оснований ниже места, где у маиса расположена вставка, индуцирующая гаплоидию. Эти мутации в большинстве случаев будут представлять собой мутации сдвига рамки считывания, которые индуцируют небольшие вставки или делеции, например, делецию G или Т в сайте разрезания, или любую другую аналогичную мутацию. SEQ ID NO: 40 содержит кассету экспрессии, которая обеспечивает нацеливание dgRNA на Os03g27610. SEQ ID NO: 41 содержит кассету экспрессии, которая обеспечивает нацеливание sgRNA на Os03g27610. Обе они содержат направляющие РНК, которые нацеливаются на последовательность CCTCGCCGATTACTTCGACTGCA в экзоне 1. В результате этого должен образовываться нокаут большей части кодирующей последовательности гена. Образующаяся мутация должна появиться в сайте разрезания, где помещена вертикальная черта между С и С в последовательности CCTCGC | CGATTAC (в паре оснований +215 последовательности кДНК в SEQ ID NO: 22). Таким образом, конструкции как 40, так и 41, как ожидается, будут обеспечивать высокую частоту образования растений, содержащих нокаут-мутации гена, что должно также привести к высоким уровням индукции гаплоидии у риса.
[00284] Растения риса трансформируют с помощью трансформационной конструкции, содержащей последовательность, выбранную из группы, состоящей из SEQ ID NO: 38-41. Посредством аппарата CRISPR/Cas9, закодированного в трансформационной конструкции, у трансформантов, т.е. растений риса Т0, образуются новые аллели гена фосфолипазы. Растения риса Т0 выращивают и скрещивают (т.е. подвергают самоопылению) для создания растений Т1. Растения риса Т1 тестируют в отношении гомозиготности по новому аллелю гена фосфолипазы. Гомозиготные растения риса Т1 скрещивают с линией риса, и полученное в результате потомство тестируют в отношении гаплоидии с помощью анализатора плоидности. Гаплоидные зародыши, не содержащие выявляемой ДНК Т1, идентифицируют и подсчитывают, и измеряют HIR. В результате скрещивания образуется по меньшей мере один гаплоидный зародыш, и HIR является повышенным. HIR предпочтительно составляет по меньшей мере 5%. По меньшей мере одного гаплоидного зародыша обрабатывают средством, удваивающим число хромосом, например, колхицином, и из него выращивают двойное гаплоидное растение.
Реферат
Изобретение относится к области биохимии, в частности к способу индукции образования гаплоидных зародышей. Изобретение позволяет эффективно получать растения, индуцирующие гаплоидию. 6 з.п. ф-лы, 31 ил., 14 табл., 6 пр.
Комментарии