Ген липоксигеназы-1 ячменя, способ отбора ячменя, материалы для солодовых алкогольных напитков и способ получения солодовых алкогольных напитков - RU2348696C2
Код документа: RU2348696C2
Чертежи











Описание
Область техники, к которой относится изобретение
Данное изобретение относится к гену липоксигеназы-1 ячменя, способу отбора ячменя, материалам для солодовых алкогольных напитков и способу получения солодовых алкогольных напитков.
Уровень техники
Липоксигеназа-1 ячменя (далее "LOX-1") является ферментом, присутствующим в солоде, который окисляет образуемую солодом линолевую кислоту до 9-гидропероксиоктадекадиеновой кислоты во время затирания солода для получения солодовых алкогольных напитков (Kobayashi, N et al., J. Ferment. Bioeng., 76, 371-375, 1993). Далее, 9-гидропероксиоктадекадиеновая кислота превращается в тригидроксиоктадеценовую кислоту (THOD) пероксигеназа-подобной активностью (Kuroda, H., et al., J. Biosci. Bioeng., 93, 73-77, 2002). Известно, что THOD уменьшает пеностойкость пива, придает терпкий вкус и ухудшает однородность вкуса пива (Kobayashi, N., J. Am. Soc. Brew. Chem. 60: 37-41, 2002; и Kaneda, H. et al., J. Biosci. Bioeng., 92, 221-226, 2001), приводя к более низкому качеству солодовых алкогольных напитков. Кроме того, 9-гидропероксиоктадекадиеновая кислота превращается в транс-2-ноненаль, который является веществом, ответственным за неприятный привкус картона в выдержанных солодовых алкогольных напитках (Yasui, Journal of the Brewing Society of Japan, 96:94-99 (2001)).
В качестве стратегии ингибирования образования транс-2-ноненаля для улучшения стабильности вкуса солодовых алкогольных напитков был предложен способ получения солодовых алкогольных напитков с использованием солода с низкой активностью LOX-1 (Drost, J. Am. Soc. Brew. Chem. 48:124-131 (1990)).
Douma et al. индуцировали мутацию в ячмене мутагенной (химической) обработкой для создания индуцированной мутированной линии, обнаруживающей на 9% более низкую активность LOX-1 в сравнении с контролями, и пытались получать солодовые алкогольные напитки с использованием такого ячменя (WO 02/053721).
Однако даже при использовании такого ячменя уменьшенная концентрация транс-2-ноненаля у полученных солодовых алкогольных напитков является недостаточной, и стабильность вкуса не является достаточно улучшенной. Кроме того, абсолютно неопределенные результаты получали в отношении уменьшения THOD или улучшения пеностойкости.
Описание изобретения
Данное изобретение было выполнено в свете вышеуказанных проблем уровня техники, и его целью является обеспечение гена LOX-1 ячменя, который применим для получения солодовых алкогольных напитков, обнаруживающих улучшенную стабильность вкуса и стойкость пены, без манипуляций генами, способа отбора LOX-1-недостаточного ячменя, материалов для солодовых алкогольных напитков, получаемых из ячменя, полученного посредством этого способа отбора, и способа получения солодовых алкогольных напитков с использованием этих материалов для солодовых алкогольных напитков.
В результате очень тщательного исследования, проводимого для достижения вышеописанной цели, авторы данного изобретения завершили это изобретение обнаружением природной разновидности ячменя, которая полностью лишена активности LOX-1, и идентификацией нового мутантного гена LOX-1 из этой разновидности ячменя.
Конкретно, мутантный ген LOX-1 данного изобретения характеризуется тем, что гуанин в донорном сайте сплайсинга (5'-GT-3') 5-го интрона известного гена LOX-1 ячменя мутирован в другое основание. Предпочтительно этим другим основанием является аденин.
Способ отбора ячменя, лишенного LOX-1 ячменя, в соответствии с данным изобретением характеризуется распознаванием ячменя, лишенного LOX-1 ячменя, посредством определения, является или не является гуанин в донорном сайте сплайсинга 5-го интрона гена LOX-1 мутированным в другое основание. Предпочтительно этим другим основанием является аденин.
Способ отбора ячменя, лишенного LOX-1 ячменя, характеризуется также включением стадии экстракции геномной ДНК, в которой геномную ДНК экстрагируют из пробы ячменя; стадии амплификации ДНК-фрагмента, в которой ДНК-фрагмент, содержащий донорный сайт сплайсинга 5-го интрона гена LOX-1, амплифицируют из экстрагированной геномной ДНК; и стадии детектирования ДНК-фрагмента, в которой ДНК-фрагмент, содержащий донорный сайт сплайсинга 5-го интрона гена LOX-1, амплифицированный в стадии амплификации ДНК-фрагмента, расщепляют рестрикционным ферментом (рестриктазой), детектируют ДНК-фрагмент, имеющий предписанное количество оснований, и ячмень, лишенный LOX-1 ячменя, распознают посредством определения, является или не является гуанин в донорном сайте сплайсинга мутированным в другое основание.
Рестрикционным ферментом, используемым в стадии детектирования этого ДНК-фрагмента, является предпочтительно AfaI и/или RsaI, которые узнают нуклеотидную последовательность 5'-GTAC-3'.
Согласно данному изобретению сорт ячменя, имеющий признак недостаточности активности LOX-1, распознают на основании присутствия или отсутствия мутации гуанина в донорном сайте сплайсинга 5-го интрона гена LOX-1.
В результате, можно легко распознать разновидность ячменя, лишенную активности LOX-1, посредством анализа на генетическом уровне, без прямого измерения активности LOX-1. На активность фермента влияют индивидуальные стадии роста, среда и другие факторы, и, следовательно, ее трудно точно измерить, но данный способ позволяет распознать разновидность ячменя с отличающейся активностью LOX-1 способом, отличающимся от измерения активности фермента, и, следовательно, независимо от факторов окружающей среды и других факторов. Кроме того, в то время как активность фермента не может быть измерена, пока семена не созреют, скрининг ДНК может идентифицировать присутствие или отсутствие признака отсутствия этой активности на ранней стадии роста, так как его проводят перед цветением, и, следовательно, является эффективным для непрерывного обратного скрещивания.
Материал для солодовых алкогольных напитков данного изобретения характеризуется тем, что он является семенами, солодом, экстрактом солода, продуктом расщепления ячменя или обработанным ячменем, полученными из ячменя, имеющего мутантный ген LOX-1 в соответствии с данным изобретением.
Материал для солодовых алкогольных напитков данного изобретения характеризуется также тем, что он является семенами, солодом, экстрактом солода, продуктом расщепления ячменя или обработанным ячменем, полученными из ячменя, отобранного с использованием способа отбора лишенного LOX-1 ячменя в соответствии с данным изобретением.
Способ получения солодовых алкогольных напитков данного изобретения характеризуется использованием материала для солодовых алкогольных напитков в соответствии с данным изобретением.
Согласно данному изобретению можно получать солодовые алкогольные напитки с улучшенной стабильностью вкуса и стойкостью пены, так как LOX-1 не присутствует в этом материале, и, следовательно, не происходит легкого образования 9-гидропероксиоктадекадиеновой кислоты из линолевой кислоты, и, следовательно, не происходит также легкого образования THOD и транс-2-ноненаля в этом способе получения солодовых алкогольных напитков.
Кроме того, данное изобретение обеспечивает нуклеиновую кислоту, содержащую нуклеотидную последовательность от положения 1 до положения 1554, представленную в SEQ ID NO:10. Эта нуклеотидная последовательность представляет кодирующую область гена, кодирующего мутантный белок LOX-1, лишенный липоксигеназной активности белка LOX-1. Посредством детектирования присутствия или отсутствия этой нуклеиновой кислоты в пробе ячменя можно узнать, имеет или не имеет этот ячмень признак недостаточности активности LOX-1.
Далее, данное изобретение обеспечивает нуклеиновую кислоту, содержащую нуклеотидную последовательность, представленную в SEQ ID NO:11. Эта нуклеотидная последовательность представляет геномную последовательность гена, кодирующего мутантный белок LOX-1, лишенный липоксигеназной активности белка LOX-1. Посредством детектирования присутствия или отсутствия этой нуклеиновой кислоты в пробе ячменя можно узнать, имеет или не имеет этот ячмень признак недостаточности активности LOX-1.
Далее, данное изобретение обеспечивает нуклеиновую кислоту, содержащую нуклеотидную последовательность из 10-60 смежных оснований, в том числе основание 3178, в нуклеотидной последовательности, представленной в SEQ ID NO:11. Основание 3178 является полиморфизмом единственного нуклеотида, которое является G в случае аутентичного LOX-1 и А в случае мутантного LOX-1. Посредством детектирования присутствия или отсутствия нуклеиновой кислоты, включающей в себя этот полиморфный сайт, в пробе ячменя можно узнать, имеет или не имеет этот ячмень признак недостаточности активности LOX-1.
Кроме того, данное изобретение обеспечивает способ детектирования присутствия активности LOX-1 в ячмене, включающий стадию выделения геномной ДНК из пробы ячменя и стадию детектирования основания 3178 нуклеотидной последовательности, представленной в SEQ ID NO:11, причем присутствие этого основания является показателем присутствия активности LOX-1 в ячмене. Согласно этому способу можно узнать, имеет или не имеет испытуемый ячмень признак недостаточности активности LOX-1.
Семена, солод, экстракт солода, продукт расщепления ячменя или обработанный ячмень, полученные из ячменя, лишенного активности LOX-1, обнаруженного посредством этого способа, могут быть использованы в качестве сырьевого материала для получения солодовых алкогольных напитков с целью получения солодовых алкогольных напитков с улучшенной стабильностью вкуса и стойкостью пены.
Краткое описание фигур
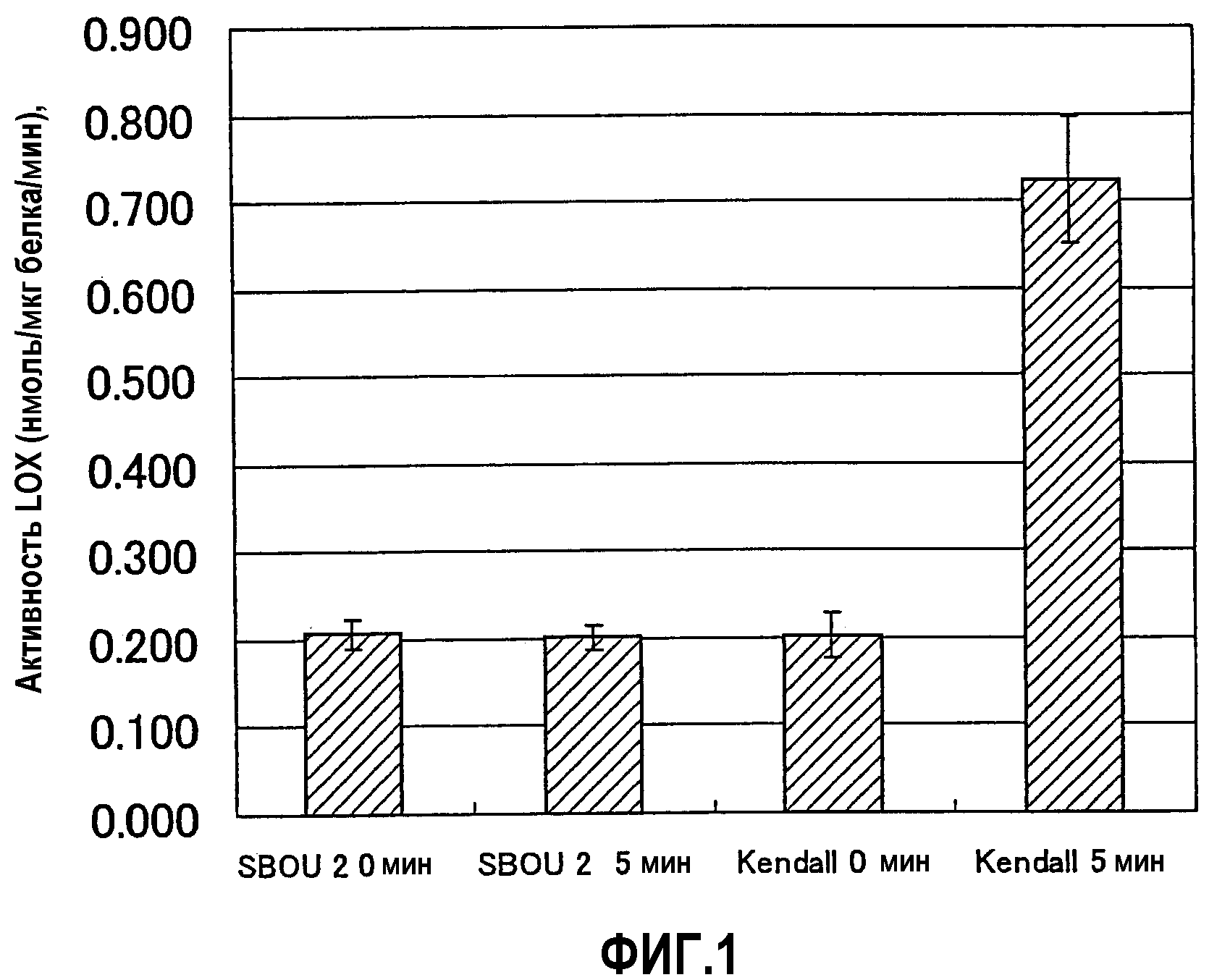
Фиг.1 является диаграммой, показывающей результаты активности LOX-1 в тесте поиска 1.
Фиг.2 является диаграммой, показывающей результаты ингибирующей LOX-1 активности в тесте верификации 1.
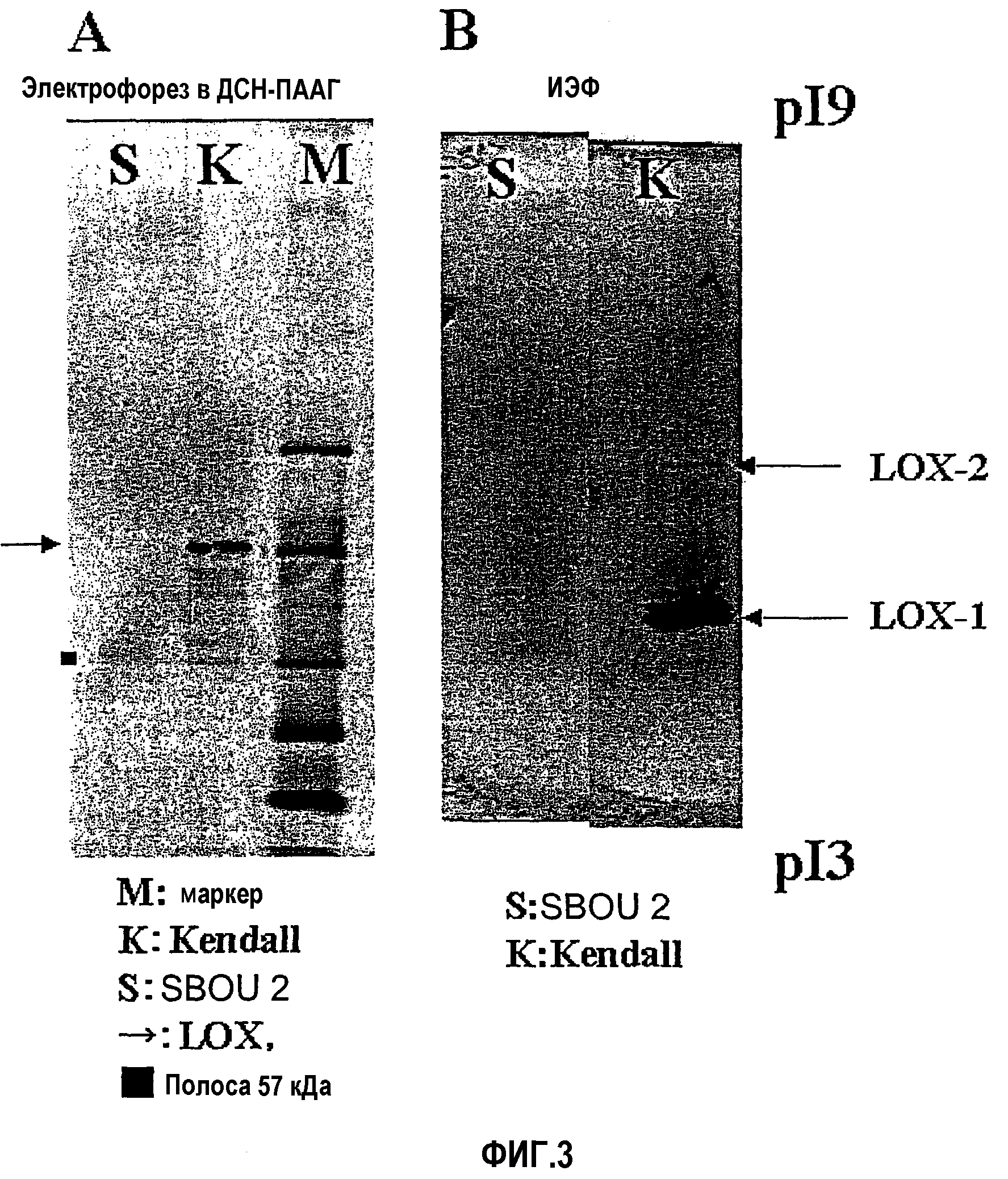
Фиг.3 является парой изображений электрофореза, показывающих результаты Вестерн-анализа белка LOX-1 семян ячменя в тесте верификации 2. Изображение А показывает результаты Вестерн-анализа после электрофореза в ПААГ, а В показывает результаты Вестерн-анализа после изоэлектрофокусирования (IEF).
Фиг.4 является изображением электрофореза, показывающим результаты анализа ОТ-ПЦР РНК семян ячменя в тесте верификации 3.
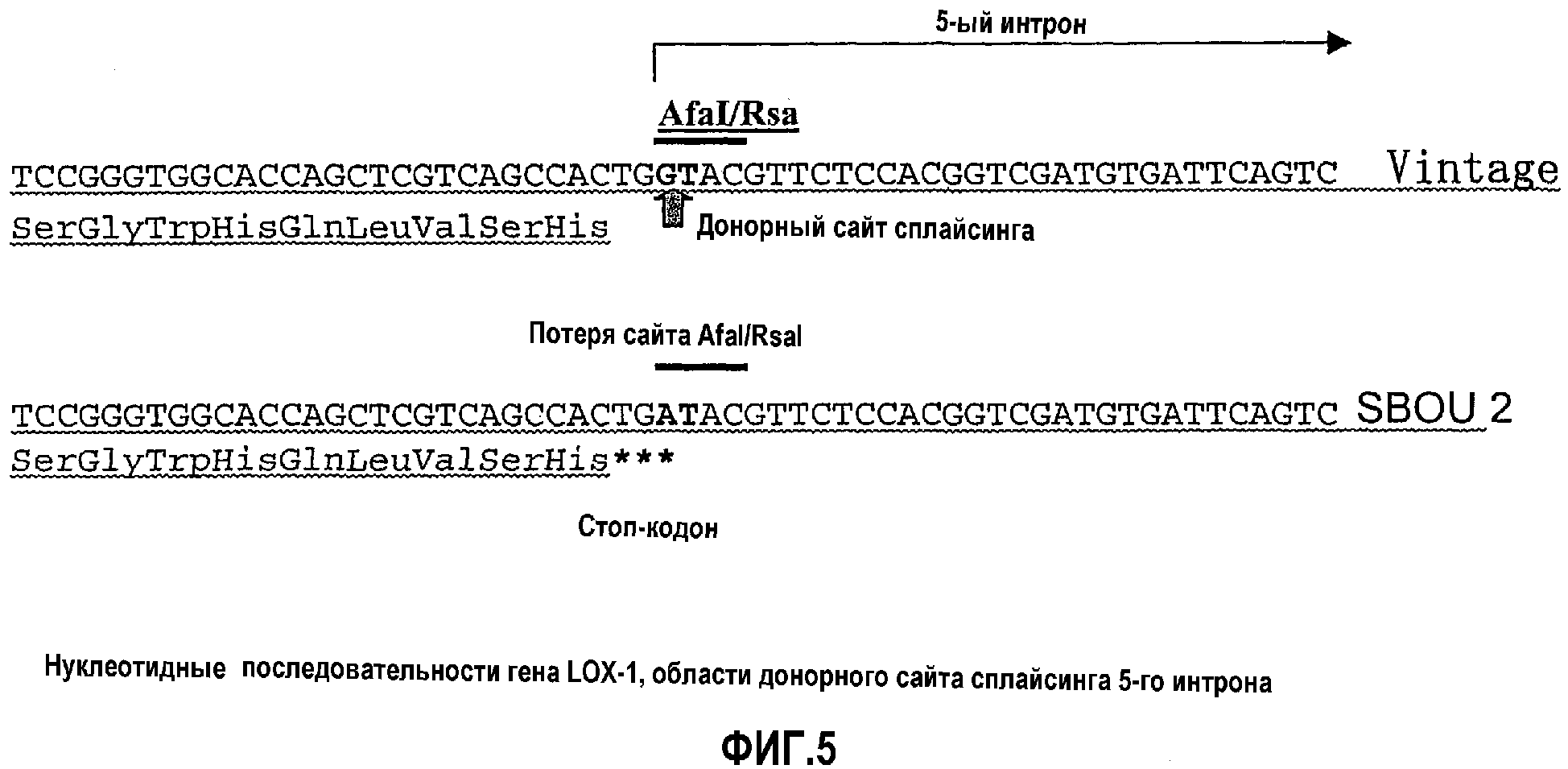
Фиг.5 является диаграммой, показывающей структуру донорного сайта сплайсинга 5-го интрона гена LOX-1 в тесте верификации 4.
Фиг.6 является парой изображений электрофореза, показывающих результаты анализа сплайсинга мутантного гена LOX-1 в тесте верификации 5. Изображение А является изображением электрофореза амплифицированных фрагментов, содержащих 3-й - 5-й интрон, а В является изображением электрофореза тех же самых фрагментов, что и в А, после расщепеления StuI.
Фиг.7 является изображением электрофореза, показывающим индуцированные экспрессией белки в E. coli в тесте верификации 7 и 8.
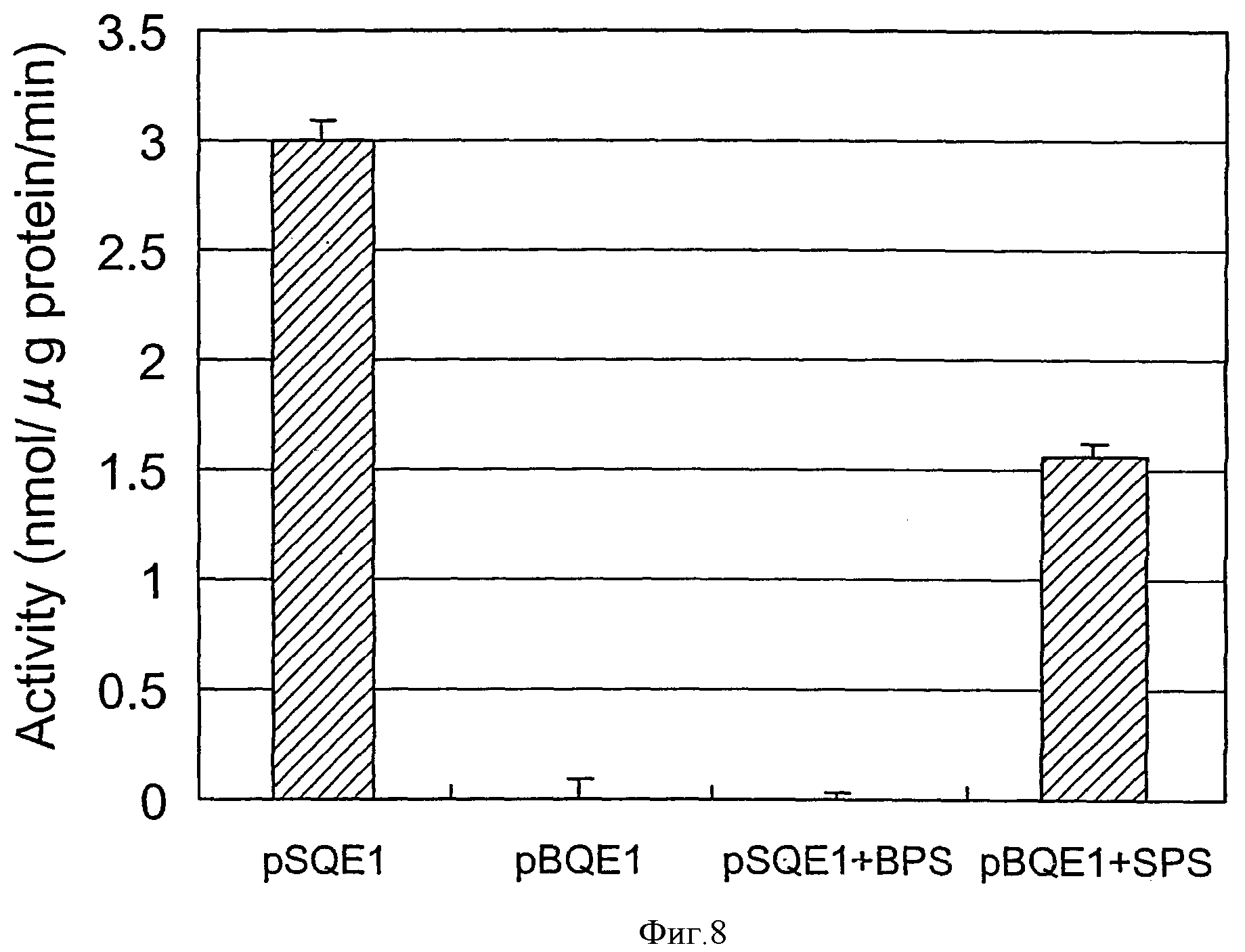
Фиг.8 является диаграммой, показывающей активность индуцированной экспрессией LOX-1 в E. coli в тесте верификации 1.
Фиг.9 является изображением электрофореза, показывающим полиморфизм ДНК во 2-ом дочернем поколении гибрида для Kendall × SBOU2 в тесте верификации 9.
Фиг.10 является таблицей, суммирующей полиморфизм ДНК в гибридном 2-м дочернем поколении и активность LOX-1 в 3-м дочернем поколении гибрида для Kendall x SBOU2 в тесте верификации 9.
Фиг.11 является изображением электрофореза, показывающим результаты анализа основного сорта/линии ячменя с использованием AfaI-способа в примере 1.
Фиг.12 является изображением электрофореза, показывающим результаты анализа LOX-1-недостаточного ячменя с использованием AfaI-способа в примере 2.
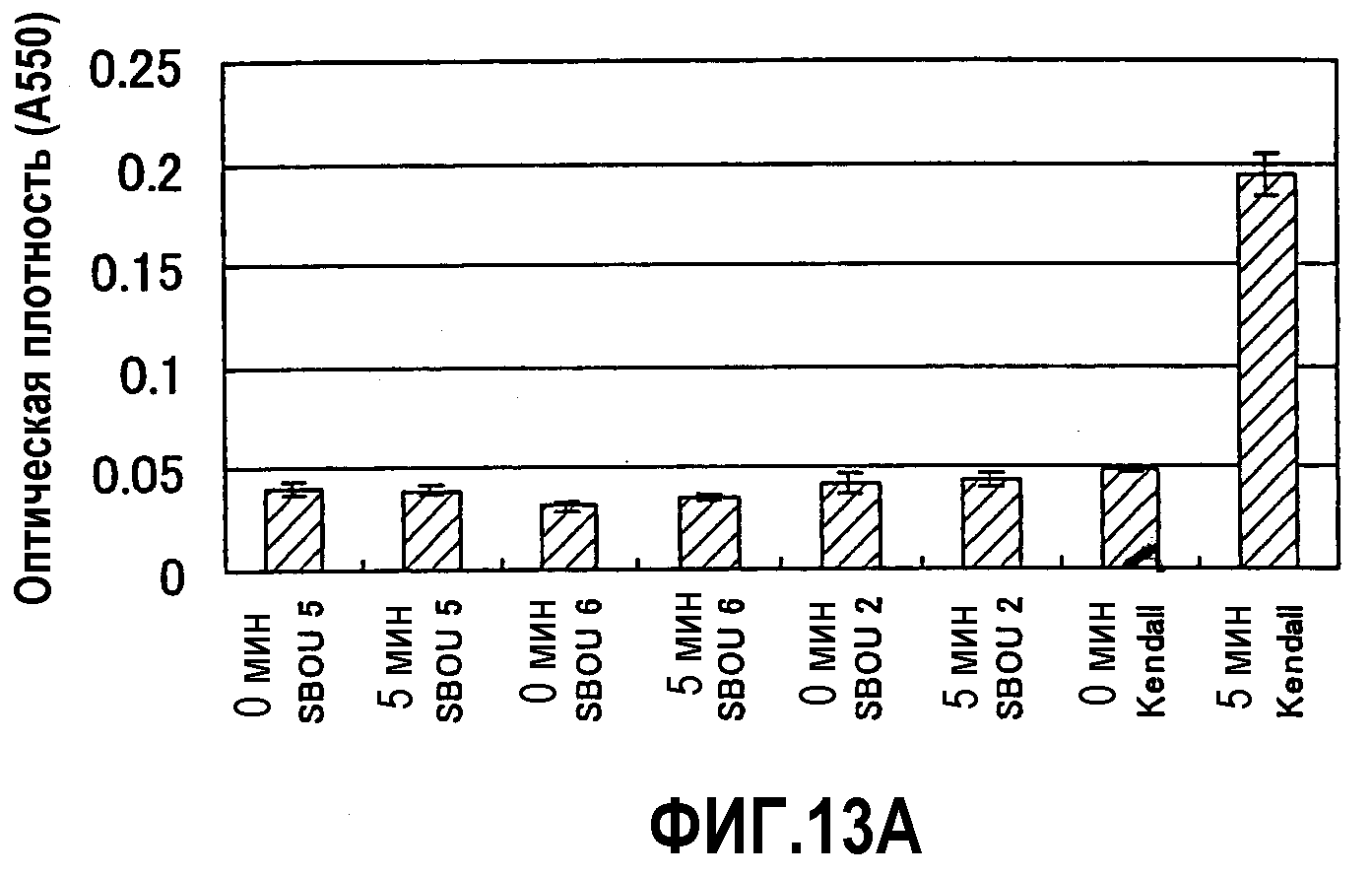
Фиг.13 является парой диаграмм, показывающих результаты активности LOX-1 LOX-1-недостаточных семян ячменя в примере 2. Диаграмма А показывает результаты при времени реакции фермента 5 минут, а диаграмма В показывает результаты при времени реакции 90 минут.
Фиг.14 является таблицей, показывающей результаты анализа солода семян из популяции LOX+F4 и LOX-F4 в примере 5.
Фиг.15 является точечной диаграммой, показывающей концентрации транс-2-ноненаля и потенциалы ноненаля для сусла в примере 5.
Наилучший способ проведения данного изобретения
Данное изобретение будет теперь объяснено более подробно.
Сначала будут даны объяснения по поводу мутантного гена LOX-1.
Мутантный ген LOX-1 данного изобретения является новым геном, обнаруженным авторами данного изобретения, и он характеризуется тем, что 60-е основание G известного гена LOX-1 ячменя (SEQ ID NO:1) заменено на А (SEQ ID NO:2). Поскольку основания 60-61 SEQ ID NO:1 составляют донорный сайт сплайсинга (5'-GT-3'), эта замена основания дает аберрацию в сплайсинге LOX-1, так что активный LOX-1 не может быть экспрессирован.
Нуклеотидная последовательность области 5-го интрона известного гена LOX-1 приведена в виде SEQ ID NO:1 в списке последовательностей, а нуклеотидная последовательность части мутантного гена LOX-1 данного изобретения, соответствующая области 5-го интрона гена LOX-1, приведена в списке последовательностей в виде SEQ ID NO:2.
Теперь будет объяснен способ отбора LOX-1-недостаточного ячменя в соответствии с данным изобретением.
Способ отбора LOX-1-недостаточного ячменя в соответствии с данным изобретением характеризуется распознаванием ячменя, лишенного LOX-1 ячменя, на основании того, является или не является гуанин в донорном сайте сплайсинга 5-го интрона гена LOX-1 мутированным в другое основание.
Способом отбора LOX-1-недостаточного ячменя, использующим вышеупомянутую мутацию основания, может быть, например, способ с использованием праймера, содержащего вышеупомянутый сайт мутации на 3'-конце последовательности праймера или внутри последовательности праймера, для амплификации ДНК, детектирования мутации основания на основании присутствия амплификации или эффективности амплификации и отбора LOX-1-недостаточного ячменя, или способ амплификации ДНК-фрагмента, содержащего вышеупомянутый сайт мутации, и отбора LOX-1-недостаточного ячменя.
Нет конкретных ограничений в отношении способа детектирования мутации нуклеотидной последовательности, пока данный способ позволяет детектировать ДНК-фрагменты, но подходящие способы, которые должны быть использованы, включают в себя электрофорез в агарозном геле и электрофорез в полиакриламидном геле. Когда подлежащая детектированию мутация ДНК основана на присутствии амплификации или эффективности амплификации, вместо электрофореза может быть использована количественная ПЦР, например способ TAQMAN.
Способ отбора LOX-1-недостаточного ячменя данного изобретения характеризуется включением предпочтительно стадии экстракции геномной ДНК, в которой геномную ДНК экстрагируют из пробы ячменя; стадии амплификации ДНК-фрагмента, в которой ДНК-фрагмент, содержащий донорный сайт сплайсинга 5-го интрона гена LOX-1, амплифицируют из экстрагированной геномной ДНК; и стадии детектирования ДНК-фрагмента, в которой ДНК-фрагмент, содержащий донорный сайт сплайсинга 5-го интрона гена LOX-1, амплифицированный в стадии амплификации ДНК-фрагмента, расщепляют рестрикционным ферментом (рестриктазой), детектируют ДНК-фрагмент, имеющий предписанное количество оснований, и лишенный LOX-1 ячменя ячмень распознают посредством определения, является или не является гуанин в донорном сайте сплайсинга мутированным в другое основание.
Сначала будет объяснена стадия экстракции геномной ДНК.
Нет особых ограничений в отношении способа экстракции геномной ДНК, и может быть использован любой широко известный способ. Конкретно, экстракцию можно проводить, например, способом CTAB (Murray et al., 1980, Nucleic Acids Res. 8:4321-4325) или способом с бромидом этидия (Varadarajan and Prakash 1991, Plant Mol. Biol. Rep. 9:6-12). Ткань, используемая для экстракции геномной ДНК не ограничивается семенами ячменя, но может быть также листьями, стеблями, корнями или т.п. Например, листья могут быть использованы для отбора из партии индивидуумов в генерациях обратного скрещивания.
Теперь будет объяснена стадия амплификации ДНК-фрагмента.
Нет особых ограничений в отношении способа амплификации ДНК-фрагмента, и, например, может быть использован способ ПЦР (полимеразной цепной реакции). Праймеры, используемые для способа ПЦР, не ограничены конкретно в их нуклеотидных последовательностях, пока они представляются областью, позволяющей амплифицировать ДНК-фрагмент, содержащий донорный сайт сплайсинга 5-го интрона гена LOX-1, и, конкретно, они являются предпочтительно 10-60 смежными основаниями и более предпочтительно 15-30 смежными основаниями гена LOX-1. Обычно нуклеотидная последовательность праймера будет предпочтительно иметь GC-содержание 40-60%. Предпочтительно также, чтобы различие в величинах Tm двух праймеров, используемых для способа ПЦР, было равно нулю или было очень малым. Эти праймеры предпочтительно не образуют вторичную структуру друг с другом.
Теперь будет объяснена стадия детектирования ДНК-фрагмента.
Мутантный ген LOX-1 в соответствии с данным изобретением имеет нуклеотидную последовательность, отличающуюся от известного LOX-1, как объяснено выше, и, следовательно, при использовании рестриктазы, которая узнает или расщепляет эту отличающуюся часть, для расщепления продукта амплификации, различие в размерах полученных ДНК-фрагментов будет очевидным. Нет особых ограничений в отношении рестриктазы, используемой для данного изобретения, пока она узнает или расщепляет отличающуюся часть, но предпочтительными являются рестриктазы AfaI и/или RsaI, которые, как уже было продемонстрировано, проявляют такую активность.
Другими словами, поскольку мутантный ген LOX-1 данного изобретения имеет гуанин в донорном сайте сплайсинга, мутированный в другое основание, он не содержит сайта расщепления для рестриктаз AfaI и RsaI (5'-GTAC-3': нуклеотидов 60-63 5-го интрона), который присутствует в известном гене LOX-1. В результате, его паттерн расщепления, при расщеплении продукта амплификации гена, содержащего этот сайт расщепления, рестриктазами AfaI и/или RsaI, будет отличаться от паттерна расщепления известного гена LOX-1, что позволяет идентифицировать мутантный ген LOX-1.
ДНК-фрагмент, имеющий предписанное количество оснований, не ограничивается в количестве оснований, пока он является ДНК-фрагментом, в котором присутствие отличающейся части приводит к различию в размере ДНК-фрагмента, получаемого расщеплением продукта амплификации рестриктазой.
Детектирование на этой стадии особо не ограничивается, пока оно является способом, делающим возможным детектирование ДНК-фрагмента, расщепляемого рестриктазой, и, конкретно, детектирование может выполняться, например, электрофорезом в агарозном геле или электрофорезом в полиакриламидном геле.
Теперь будет объяснен материал для солодовых алкогольных напитков данного изобретения.
Материал для солодовых алкогольных напитков данного изобретения характеризуется тем, что он является семенами, солодом, экстрактом солода, продуктом расщепления ячменя или обработанным ячменем, полученными из ячменя, имеющего мутантный ген LOX-1 в соответствии с данным изобретением, и тем, что он является семенами, солодом, экстрактом солода, продуктом расщепления ячменя или обработанным ячменем, полученными из ячменя, отобранного способом отбора LOX-1-недостаточного ячменя в соответствии с данным изобретением.
Солодовый экстракт является экстрактом из солода, и в качестве примеров может быть упомянут экстракт сахарных компонентов или белковых компонентов из солода. Продукт расщепления ячменя является продуктом расщепления ячменя ферментами или т.п., и он включает в себя ячменный затор и т.п. Обработанным ячменем называют дробленый ячмень, используемый в качестве добавки для солодовых алкогольных напитков.
Поскольку материал для солодовых алкогольных напитков по данному изобретению не содержит LOX-1, не происходит легкого образования 9-гидропероксиоктадекадиеновой кислоты из линолевой кислоты и, следовательно, не происходит легкого образования THOD и транс-2-ноненаля во время этого способа получения солодовых алкогольных напитков; таким образом, можно получать солодовые алкогольные напитки с улучшенной стабильностью вкуса и пеностойкостью.
Теперь будет объяснено получение солодовых алкогольных напитков.
Способ получения солодовых алкогольных напитков данного изобретения характеризуется использованием материала для солодовых алкогольных напитков в соответствии с данным изобретением.
Сначала будет объяснена стадия получения солода, характеризующаяся использованием LOX-1-недостаточного ячменя, и этот способ не является особо ограниченным и может быть любым широко известным способом. Более конкретно, например, замачивание до степени замачивания 40-45% сопровождается проращиванием при 10-20°С в течение 3-6 дней и печной сушкой для получения солода.
Теперь будет объяснена стадия затирания.
Стадия затирания в соответствии с данным изобретением является стадией получения сусла затиранием вышеупомянутого солода. Более конкретно, она состоит из четырех стадий.
Первой стадией является стадия затирания, в которой содержащий солод материал смешивают с водой, полученную смесь нагревают для затирания солода и получают сусло из осахаренного солода.
Солод, используемый для этой стадии, предпочтительно получают добавлением воды и воздуха к ячменю для проращивания с последующей сушкой для удаления зародышей. Этот солод является источником необходимых ферментов для получения сусла, а также основным источником крахмала в качестве материала для затирания. Высушенный печной сушкой пророщенный солод используют также для придания характерного вкуса и цвета солодовому алкогольному напитку. Кроме такого солода, могут быть добавлены добавки, такие как LOX-1-недостаточный ячмень по данному изобретению и/или обычный ячмень, кукурузный крахмал, кукурузная крупа, рис, сахариды или т.п.
В стадии получения сусла, описанной выше, продукт расщепления ячменя или обработанный ячмень, полученный из LOX-1-недостаточного ячменя данного изобретения и/или обычного ячменя, смешивают с заторной водой и добавляют, если необходимо, вышеупомянутые добавки для получения сусла.
Солод перемешивают после добавления заторной воды. При добавлении также добавок они могут быть также смешаны в этот момент. Сахариды могут добавляться перед кипячением, описанным ниже. Нет особых ограничений в отношении заторной воды, и может быть использована вода, которая пригодна для получаемых солодовых алкогольных напитков. Затирание проводят по существу при общепринятых условиях. После фильтрования солодового затора, полученного таким образом, добавляют материалы, которые придают аромат или горечь, такие как хмель или травы, и смесь кипятят и затем охлаждают с получением охлажденного сусла.
Второй стадией является стадия добавления дрожжей к охлажденному суслу для ферментации с целью получения промежуточных продуктов солодового алкогольного напитка.
Дрожжи, используемые в этой стадии, могут быть любыми пивными дрожжами для спиртового брожения, которые метаболизируют сахара в сусле, полученном затиранием солода, с образованием спирта и газообразного диоксида углерода, и, конкретно, могут быть упомянуты, например, Saccharomyces cerevisiae и Saccharomyces uvarum.
Сбраживание выполняют охлаждением сусла, полученного в стадии затирания, и добавлением к нему дрожжей. Условия сбраживания могут быть по существу такими же, что и условия для общепринятой ферментации, и, например, температура сбраживания будет обычно не выше 15оС и предпочтительно 8-10оС, тогда как период сбраживания составляет предпочтительно 8-10 дней.
Третьей стадией является стадия выдерживания, в которой промежуточные продукты солодового алкогольного напитка, полученного в стадии сбраживания, выдерживают.
В этой стадии сбраживаемый раствор, который завершил спиртовое брожение, переносят в герметизированный чан и выдерживают. Это вторичное сбраживание по существу является таким же, что и сбраживание при обычных условиях, и, например, температура выдерживания равна предпочтительно 0-2°С и время выдерживания равно приблизительно 20-90 дням. Выдерживание сброженного раствора делает возможными прохождение повторного брожения и созревания оставшегося экстракта.
Четвертой стадией является стадия фильтрования, в которой промежуточные продукты солодового алкогольного напитка, полученного на стадии выдерживания, фильтруют с получением солодового алкогольного напитка.
Условия фильтрования являются по существу такими же, что и общепринятые условия, и, например, используемым фильтрующим материалом могут быть диатомовая земля, поливинилпирролидон (PVPP), силикагель, порошок целлюлозы или т.п., и температура может быть 0±1°С.
Эта процедура дает солодовый алкогольный напиток. Затем отфильтрованный солодовый алкогольный напиток помещают в чан, разливают в бочки, разливают в бутылки или в консервные банки для транспортировки на рынок, либо сразу же, либо после стерильного фильтрования или тепловой обработки.
Доля солода, используемого для получения ячменного алкогольного напитка, особо не ограничивается, и алкогольный напиток может быть любым напитком, получаемым с использованием солода в качестве исходного материала. Конкретно могут быть упомянуты, например, пиво и искристый солодовый напиток. Безалкогольное пиво и безалкогольный искристый солодовый напиток также считаются солодовыми алкогольными напитками, так как используемый способ получения является сходным со способом получения таких солодовых алкогольных напитков, как пиво.
Поскольку материал по данному изобретению не содержит LOX-1, не происходит легкого образования 9-гидропероксиоктадекадиеновой кислоты из линолевой кислоты и, следовательно, ингибируется образование THOD и транс-2-ноненаля во время способа получения солодовых алкогольных напитков, и, следовательно, можно получать солодовые алкогольные напитки с улучшенной стабильностью вкуса и стойкостью пены.
Теперь будут объяснены нуклеиновая кислота и способ детектирования присутствия активности LOX-1 в ячмене в соответствии с данным изобретением.
Нуклеиновая кислота данного изобретения характеризуется тем, что она содержит нуклеотидную последовательность, представленную в SEQ ID NO:11. SEQ ID NO:11 представляет геномную последовательность для мутантного LOX-1 в сорте SDOU2 ячменя, лишенного активности LOX-1. То есть мутантный ген LOX-1 данного изобретения характеризуется тем, что он представлен последовательностью SEQ ID NO:11. Основание, соответствующее положению 3178, является G в аутентичном гене LOX-1, тогда как основание 3178 является мутированным в А в мутантном гене LOX-1. Это основание является также первым основанием 5-го интрона аутентичного гена LOX-1, и последовательность GT в виде последовательности оснований 3178-3179 соответствует донорному сайту сплайсинга (фиг.5). Однако в мутантном гене LOX-1 последовательность оснований 3178-3179, соответствующая донорному сайту сплайсинга, является АТ, и, следовательно, происходит аберрация сплайсинга, которая предотвращает сплайсинг. Кроме того, последовательность оснований 3176-3178 является TGA, которая является стоп-кодоном, и, следовательно, в этой точке трансляция заканчивается.
Поскольку мутантный белок LOX-1, экспрессируемый мутантным геном LOX-1 имеет только часть, соответствующую 5-му экзону, он лишен аминокислотных остатков на С-концевой стороне от 5-го экзона аутентичного белка LOX-1. Молекулярная масса аутентичного белка LOX-1 равна 95 кД, тогда как молекулярная масса мутантного белка LOX-1 равна 57 кД. Мутантный белок LOX-1 лишен липоксигеназной активности, и это коррелирует с известным фактом, что домен, соответствующий области экзона около 5-го интрона в аутентичном белке LOX-1, является активным центром LOX растений (Shibata and Axelrod (1995) J. Lipid Mediators and Cell Signaling 12:213-218).
Таким образом, если ячмень, имеющий мутантный ген LOX-1, используется в качестве сырьевого материала для получения солодовых алкогольных напитков, белок LOX-1 не будет присутствовать в этом сырьевом материале, и, следовательно, образование 9-гидропероксиоктадекадиеновой кислоты из линолевой кислоты во время этого способа получения солодовых алкогольных напитков будет уменьшено, так что в результате может быть достигнуто ингибирование образования THOD и транс-2-ноненаля с целью получения солодовых алкогольных напитков с улучшенными стабильностью вкуса и стойкостью пены. Таким образом, нуклеиновая кислота, содержащая нуклеотидную последовательность, представленную в SEQ ID NO:11, в соответствии с данным изобретением, является в высокой степени полезной для получения солодовых алкогольных напитков с улучшенными стабильностью вкуса и стойкостью пены.
Нуклеиновая кислота данного изобретения обеспечивает нуклеиновую кислоту, содержащую нуклеотидную последовательность из 10-60 смежных оснований, в том числе основание 3178, в нуклеотидной последовательности, представленной в SEQ ID NO:11. Эта нуклеиновая кислота может быть использована в качестве зонда для различения между аутентичной и мутантной формами гена LOX-1 ячменя. То есть, поскольку основание 3178 аутентичного гена LOX-1 является G, полученное ошибочное спаривание может быть использовано для различения аутентичной и мутантной форм на основании различия в гибридизации. Например, посредством образования гибридов между этими нуклеиновыми кислотами и нуклеиновой кислотой гена LOX-1, постепенного увеличения температуры и измерения температуры плавления гибридов можно легко различать между аутентичным геном LOX-1 и мутантным геном LOX-1, так как их температуры плавления будут различаться. Эта нуклеиновая кислота может быть также использована для различения между формами гена LOX-1 (аутентичной/мутантной формами) с использованием способов, известных специалистам с квалификацией в данной области. С точки зрения специфичности, эта нуклеиновая кислота предпочтительно содержит нуклеотидную последовательность из 20-50 смежных оснований, в том числе основание 3178, и предпочтительно она включает в себя основания 3178-3181. Эта нуклеиновая кислота может быть также мечена флуоресцентным веществом, радиоактивным изотопом или т.п.
Способ детектирования присутствия активности LOX-1 в ячмене в соответствии с данным изобретением предусматривает стадию выделения геномной ДНК из пробы ячменя и стадию детектирования основания 3178 нуклеотидной последовательности, представленной в SEQ ID NO:11, причем присутствие этого основания является показателем присутствия активности LOX-1 в ячмене. Согласно этому способу можно узнать, имеет или не имеет тестируемый ячмень признак недостаточности активности LOX-1.
Проба ячменя не ограничивается семенами ячменя, и она может быть листьями, стеблями корнями ячменя или т.п. Нуклеиновая кислота может быть выделена широко известными способами и, например, может быть использован способ CTAB или способ с использованием бромида этидия.
Детектирование основания 3178 нуклеиновой последовательности, представленной SEQ ID NO:11, может выполняться посредством способа, известного специалистам с квалификацией в данной области. Например, если необходимо, нуклеиновая кислота, содержащая основание 3178 нуклеотидной последовательности, представленной SEQ ID NO:11, может быть амплифицирована по способу амплификации нуклеиновых кислот, такому как ПЦР. Идентичность основания в положении 3178 выделенной нуклеиновой кислоты или амплифицированного фрагмента нуклеиновой кислоты можно различить, например, с помощью нуклеиновой кислоты, содержащей нуклеотидную последовательность, представленную SEQ ID NO:11, причем эта нуклеиновая кислота содержит последовательность из 10-60 смежных оснований, в том числе основание 3178, как описано выше.
Однако способ детектирования основания 3178 с использованием различия в основаниях 3178-3181 гена LOX-1 является более удобным и эффективным. Сайт оснований 3178-3181 является сайтом расщепления рестриктаз AfaI/RsaI в аутентичном гене LOX-1, но вследствие того, что основание 3178 является А в мутантном гене LOX-1, оно не образует сайта расщепления для рестриктаз AfaI/RsaI (фиг.5). Другими словами, при обработке выделенной нуклеиновой кислоты или амплифицированного фрагмента нуклеиновой кислоты рестриктазами AfaI/RsaI нуклеиновая кислота аутентичного LOX-1 будет расщепляться, тогда как нуклеиновая кислота мутантного LOX-1 не будет расщепляться. Электрофоретический анализ обработанной рестриктазами пробы нуклеиновой кислоты будет делать возможным различение формы гена LOX-1 (аутентичной или мутантной) на основании различия в картинах электрофореза, так что может быть различена идентичность основания в положении 3178. Кроме того, посредством использования нуклеиновой кислоты, содержащей нуклеотидную последовательность из 10-60 смежных оснований, в том числе оснований 3178-3181, в качестве зонда для гибридизации с обработанной рестриктазами нуклеиновой кислотой можно различить форму гена LOX-1 и, следовательно, сделать возможным узнавание идентичности основания 3178.
Если в результате распознавания основания 3178 таким образом обнаруживается, что это основание является G, то можно сделать вывод, что тестируемый ячмень имеет активность LOX-1 и не пригоден в качестве сырьевого материала для солодовых алкогольных напитков с улучшенными стабильностью вкуса и стойкостью пены. С другой стороны, если этим основанием является А, то можно сделать вывод, что тестируемый ячмень не имеет активности LOX-1 и, следовательно, пригоден в качестве сырьевого материала для солодовых алкогольных напитков с улучшенными стабильностью вкуса и стойкостью пены.
Нуклеиновая кислота данного изобретения характеризуется также тем, что она содержит нуклеотидную последовательность от положения 1 до положения 1554, представленную в SEQ ID NO:10. SEQ ID NO:10 представляет кДНК-последовательность мутантного LOX-1, экспрессируемого сортом ячменя SBOU2, лишенного активности LOX-1. Нуклеотидная последовательность от положения 1 до положения 1554 представляет кодирующую область. Как объяснено выше, мутантный белок LOX-1, кодируемый этой кДНК, не имеет аминокислотных остатков на С-концевой стороне от 5-го экзона аутентичного белка LOX-1, его молекулярная масса равна 57 кДа и он лишен липоксигеназной активности.
Таким образом, ячмень, экспрессирующий эту нуклеиновую кислоту, может быть использован в качестве сырьевого материала для получения солодовых алкогольных напитков с целью получения солодовых алкогольных напитков с улучшенными стабильностью вкуса и стойкостью пены, как упоминалось выше.
[Примеры]
Теперь данное изобретение будет объяснено более подробно посредством следующих примеров с пониманием того, что эти примеры никоим образом не являются ограничивающими в отношении данного изобретения.
Тест поиска 1 (поиск на LOX-1-недостаточный ячмень посредством измерения активности фермента LOX-1)
Активность фермента LOX-1 измеряли по способу, описанному ниже, а поиск на LOX-1-недостаточный ячмень проводили из ресурсов генов ячменя.
Сначала раствор неочищенного фермента экстрагировали из семян ячменя по следующему способу. Одно зрелое семя ячменя измельчали молотком и использовали 500 мкл буфера для экстракции (0,1 М натрий-ацетатный буфер (рН 5,5)) для экстракции со встряхиванием при 4°С в течение 30 минут. Полученный экстракт центрифугировали в течение 10 минут при 15000 об/мин и затем супернатант использовали в качестве раствора неочищенного фермента.
Затем 5 мкл раствора субстрата (40 мМ линолевая кислота, 1,0% (масса/объем) водный раствор Твина 20) и 85 мкл буфера для экстракции добавляли к 10 мкл раствора неочищенного фермента и смешивали вместе и затем реакцию проводили при 24°С в течение 5 минут. Реакцию останавливали добавлением и смешиванием 100 мкл стоп-раствора (80 мМ 2,6-ди-трет-бутил-п-крезол, раствор в метаноле). После стояния реакционной смеси при -20°С в течение 30 минут ее центрифугировали при 3000 об/мин в течение 20 минут и супернатант использовали в следующей реакции с развитием окраски. Часть 200 мкл образующего цвет раствора (4 мМ 2,6-ди-трет-бутил-п-крезол, 25 мМ серная кислота, 0,25 мМ гексагидрат сульфата аммония/железа(II), 100 мМ ксиленоловый оранжевый, 90% водный метанол) добавляли к 20 мкл полученного супернатанта и после стояния в течение 30 минут измеряли оптическую плотность при 550 нм. В качестве отрицательного контроля реакцию проводили таким же образом с использованием раствора неочищенного фермента, обработанного нагреванием при 100°С в течение 5 минут для инактивации LOX-1, тогда как в качестве положительного контроля использовали семена Kendall, сорта ячменя.
Как показано на фиг.1, поиск на ресурсы генов не обнаружил значимой активности LOX-1 в семенах сорта SBOU2. Поскольку SBOU2 является местным сортом, он является скорее спонтанным мутантом, чем искусственно мутагенезированной линией.
Тест верификации 1 (подтверждение отсутствия ингибирующей LOX-1 активности в растворах неочищенного фермента SBOU2)
Растворы неочищенного фермента SBOU2 испытывали на присутствие или отсутствие ингибирующей LOX-1 активности.
Растворы неочищенного фермента SBOU2 (10 мкл, 20 мкл, 50 мкл) добавляли к раствору неочищенного фермента, проявляющего активность LOX-1 (положительный контроль: PC), и исследовали изменения активности LOX-1. Активность LOX-1 не изменялась при добавлении раствора неочищенного фермента SBOU2, и, следовательно, ингибирующая LOX-1 активность не проявлялась растворами неочищенного фермента SBOU2 (фиг.2). Это позволяет предположить, что причина недостаточности активности LOX-1 в SBOU2 не была обусловлена ингибирующим активность LOX веществом.
Тест верификации 2 (подтверждение уровня экспрессии белка LOX-1 в семенах SBOU2)
Антитело анти-LOX-1 использовали для определения, экспрессируется или не экспрессируется белок LOX-1 в семенах SBOU2.
Сначала получали антитело анти-LOX-1. Белок LOX-1, используемый в качестве антигена, получали очисткой белка LOX-1, экспрессируемого E. coli (Kuroda et al. (2002) J. Bioscience and Bioengineering 93:73-77)). Этот очищенный белок использовали для иммунизации кроликов для получения антитела против LOX-1. Это антитело узнает как LOX-1, так и LOX-2.
Затем проводили Вестерн-блоттинг следующим образом для испытания экспрессии белка LOX-1 в семенах SBOU2. Часть 3 мкг общего растворимого белка, экстрагированного из SBOU2 с использованием 0,1 М натрий-ацетатного буфера (рН 5,5), фракционировали электрофорезом в ДСН-полиакриламидном геле (электрофорезом в ДСН-ПААГ) и затем блоттировали на мембрану PVDF (Millipore). Эту мембрану промывали TTBS (20 мМ Трис-HCl (рН 7,5), 0,15 М хлорид натрия, 0,05% (м/об.) Твин 20, 0,05% (м/об.) азид натрия) и затем использовали в реакции с раствором LOX-1-антитела (1000-кратное разведение/TTBS) в течение 30 минут. Мембрану промывали TTBS три раза в течение 5 минут каждый раз, после чего проводили реакцию в течение 30 минут с раствором меченного щелочной фосфатазой козьего антитела против кроличьего IgG (Santa Cruz Biotechnology, раствор в TTBS при 1000-кратном разведении). После промывания этой мембраны TTBS в течение 5 минут × 2 и затем АР9,5 (10 мМ Трис-HCl (рН 9,5), 0,1 М хлорид натрия, 5 мМ хлорид магния) в течение 5 минут × 1, проводили реакцию с раствором субстрата щелочной фосфатазы (1 мг/мл нитросинего тетразолия, 0,5 мг/мл BCIP, раствора в АР9,5) для развития окраски. В результате получали темную полосу с молекулярной массой около 95 кДа с контрольным сортом (Kendall), в то время как две очень слабые полосы детектировали в областях молекулярных масс около 95 кДа и около 57 кДа в случае семян сорта SBOU2 (фиг.3А).
Затем пробу этого экстракта подвергали изоэлектрическому фокусированию (IEF, pI 3-9) с использованием PhastSystem (Amersham Pharmacia) и подвергали Вестерн-анализу, как описано выше. В результате анализа с использованием SBOU2 детектировали полосу в положении pI LOX-2, но ясная полоса не присутствовала в положении pI LOX-1 (фиг.3В). Это предполагает, что полоса приблизительно 95 кД, обнаруживающаяся в случае экстрагированного из семян SBOU2 белка, была белком LOX-2.
Эти результаты подтвердили, что семена SBOU2 фактически не экспрессируют аутентичный белок LOX-1.
Тест верификации 3 (анализ РНК LOX-1 семян SBOU2)
ОТ-ПЦР проводили с использованием в качестве матрицы тотальной РНК, экстрагированной из приблизительно 4-недельных созревающих семян SBOU2 и 3-дневных проращиваемых семян. Реакцию проводили с использованием коммерчески доступных наборов (Roche Diagnostics, Perkin Elmer) в соответствии с руководствами наборов. Праймерами были 5'-GGAGAGGAGGCCAAGAACAAGATG-3' (SEQ ID NO:3) и 5'-GGTTGCCGATGGCTTAGAT-3' (SEQ ID NO:4), сконструированные на основе опубликованной последовательности (DNA Databank: Accession No. L35931). После инкубации при 94°С в течение 2 минут проводили ПЦР с реакцией при 94°С, 1 мин, 60°С, 2 мин и 72°С, 4 мин, повторяемой 31 раз, с последующей реакцией удлинения при 72°С, 7 мин.
При электрофорезе амплифицированной ДНК амплификацию полосы приблизительно 2,5 т.п.н. детектировали для РНК из созревающих и прорастающих семян, хотя слегка более низкое количество, чем в контроле (сорте Kendall) (фиг.4). Это показало, что ген LOX-1 был должным образом транскрибирован.
Поскольку вышеупомянутые результаты для семян SBOU2 показали, что (1) активность LOX-1 не была детектирована, (2) присутствовало только незначительное количество антигенного белка, реагирующего с антителом LOX-1, (3) детектировали присутствие белка с молекулярной массой приблизительно 57 кДа и (4) при детектировании мРНК LOX-1, был сделан вывод, что механизмом недостаточности активности LOX-1 в SBOU2 была аберрация после транскрипции.
Тест верификации 4 (структурный анализ интронных областей гена LOX-1 SBOU2)
Для анализа структуры областей интронов и экзонов гена LOX-1 выделяли геномную ДНК области, содержащей все эти экзоны. Тотальную ДНК из SBOU2 использовали в качестве матрицы. Праймеры (5'CACGTCGCCGTCCGATCCATC-3') (SEQ ID NO:5), 5'-GGTTCCGATGGCTTAGAT-3') (SEQ ID NO:4)) конструировали на основе известной последовательности (DNA Databank: Accession Nos. U83904, L35931). ПЦР проводили с реакцией при 94°С, 1 мин, 65°С, 2 мин и 72°С, 3 мин, повторяемой 31 раз, с последующей реакцией удлинения при 72°С, 7 мин. Полученный ДНК-фрагмент клонировали в pCR2.1 (pGLXABAL1) для применения в качестве матрицы для структурного анализа. Структурный анализ проводили с использованием ABI-секвенатора, а реакцию секвенирования проводили по способу терминации с красителем. Полная структура показана в SEQ ID NO:11 в списке последовательностей.
SEQ ID NO:1 списка последовательностей показывает структуру области сообщенной нуклеотидной последовательности гена LOX-1 (WO 02053721), содержащего 5-й интрон. Донорным сайтом сплайсинга является нуклеотидная последовательность 5'-GT-3' в положениях 60-61.
Нуклеотидная последовательность соответствующей области SBOU2, определенная из результатов анализа, находится в списке последовательностей в виде SEQ ID NO:2. В случае SBOU2 ясно, что гуанин в положении 60 в донорном сайте сплайсинга мутирован в аденин.
Замена 60-го основания SEQ ID NO:2 аденином образует новый стоп-кодон (нуклеотидная последовательность 5'-TGA-3' в положениях 58-60 SEQ ID NO:2), и, следовательно, предположительно трансляция белка LOX-1 заканчивается в этой точке, если этот сайт сплайсинга никогда не изменяется в направлении налево относительно 5'-конца (фиг.5).
Известно, что область экзона вблизи 5-го интрона является активным центром LOX растений (Shibata and Axelrod (1995), J. Lipid Mediators and Cell Signaling 12:213-228), и считается, что вышеупомянутая аберрация сплайсинга оказывает большое влияние на активность фермента LOX-1.
Тест верификации 5 (анализ сплайсинга в 5-м интроне)
ОТ-ПЦР-анализ проводили для подтверждения, что аберрация сплайсинга, действительно, имела место в 5-м интроне.
Тотальную РНК экстрагировали из прорастающих сортов SBOU2 и Kendall и для получения матричной ДНК использовали коммерчески доступный набор (Roche Diagnostics) для синтеза ДНК. ПЦР проводили с использованием двух различных праймеров (5'-CCATCACGCAGGGCATCCTG-3' (SEQ ID NO:6), 5'-GCGTTGATGAGCGTCTGCCG-3' (SEQ ID NO:7)), сконструированных для получения амплифицированного фрагмента, содержащего 3-й интрон (106 п.н.), 4-й интрон (132 п.н.) и 5-й интрон (79 п.н.) в последовательности геномной ДНК. ПЦР проводили с реакцией при 94°С, 1 мин, 65°С, 2 мин и 72°С, 3 мин, повторяемой 31 раз, с последующей реакцией удлинения при 72°С, 7 мин. Электрофорез в агарозном геле амплифицированных ДНК-фрагментов показал, что амплифицированный фрагмент SBOU2 был приблизительно на 80 п.н. больше, чем амплифицированный фрагмент Kendall (фиг.6А). Фрагмент приблизительно 1,2 т.п.н. амплифицировали из геномной ДНК SBOU2, что предполагает, что эти результаты ОТ-ПЦР были результатами для экспрессированной РНК.
Затем для исследования того, в каких интронах произошла аберрация сплайсинга (3-м интроне (106 п.н.), 4-м интроне (132 п.н.) или 5-м интроне (79 п.н.)), этот амплифицированный фрагмент расщепляли в сайте рестриктазы StuI, присутствующем в области экзона между 4-м интроном (132 п.н.) и 5-м интроном. В результате ДНК-фрагмент, содержащий 5-й интрон, имел подвижность, приблизительно эквивалентную подвижности одного из фрагментов амплификации из геномной ДНК, и, следовательно, явно произошла аберрация в сплайсинге 5-го интрона, которая либо предотвращала сплайсинг, либо смещала сайт сплайсинга к 3'-стороне (фиг.6В). То есть, очевидно, что стоп-кодон, новообразованный, как показано на фиг.5, останавливал трансляцию белка LOX-1 SBOU2 в этом кодоне, приводя к потере активности LOX-1.
Тест верификации 6 (структурный анализ кДНК LOX-1 SBOU2)
Для выяснения структуры LOX-1, полученной из SBOU2, кДНК выделяли тем же самым способом, который описан выше. Амплификацию проводили с использованием праймера, содержащего сайт BamHI и стартовый кодон (5'-GGATCCATGCTGCTGGGAGGGCTG-3' (SEQ ID NO:8)), и праймера, сконструированного для включения сайта HindIII и стоп-кодона (5'-AAGCTTTTAGATGGAGATGCTGTTG-3') (SEQ ID NO:9). Амплифицированный фрагмент клонировали в вектор рТ7 Blue T (Novagen) (pBDC1) и затем обеспечивали структурный анализ. Результаты структурного анализа давали нуклеотидную последовательность кДНК, показанную в виде SEQ ID NO:10. Этот кДНК-клон явно включает в себя всю область 5-го интрона (основания 1554-1632 SEQ ID NO:10).
Тест верификации 7 (трансформация E. coli и индуцированная экспрессия)
Для экспрессии LOX-1, происходящей из сорта Steptoe, сохраняющего LOX-1 дикого типа, полученную из Steptoe кДНК выделяли также вышеописанным способом и клонировали в E. coli (pSDC1), отдельно от SBOU2. BamHI-HindIII-фрагмент вырезали из каждого из клонов pSDC1 и pBDC1 и каждый из полученных фрагментов инсертировали в сайте BamHI-HindIII экспрессирующего вектора E. coli pQE80L (Qiagen) с получением экспрессирующих векторов E. coli (pSQE1 с инсерцией кДНК Steptoe) и pBQE1 (с инсерцией кДНК SBOU2)). Эти векторы использовали для трансформации E. coli JM109 и затем индуцировали экспрессию посредством IPTG в соответствии с инструкцией Qiagen. В результате индуцировалась экспрессия полосы приблизительно 95 кДа в pSQE1/JM109, тогда как экспрессия полосы приблизительно 57 кДа индуцировалась с pBQE1/JM109 (фиг.7). Клетки E. coli разрушали обработкой ультразвуком и анализировали активности LOX экстрактов неочищенных ферментов. В результате была обнаружена высокая активность LOX с pSQE1/JM109, тогда как активность LOX не была обнаружена с pBQE1/JM109 (фиг.8). Эти результаты вполне согласовались с результатами анализа активности LOX-1 и белка LOX-1 для растений SBOU2, свидетельствуя о том, что недостаточность LOX-1 SBOU2 может быть воспроизведена в E. coli.
Тест верификации 8 (тест обмена инсерций и экспрессии)
Затем PstI-StuI-фрагмент, содержащий мутацию в донорном сайте сплайсинга 5-го интрона pBQE1 (сайт StuI находится в основаниях 1502-1507 SEQ ID NO:10, а сайт PstI находится в основаниях 2048-2053 SEQ ID NO:10), подвергали взаимному обмену с PstI-StuI-фрагментом pSQE1 дикого типа (pSQE1+BPS, pBQE1+SPS) и экспрессию индуцировали в E. coli, как описано выше. В результате с pSQE1+BPS/JM109, имеющей происходящий из pBQE1 PstI-StuI-фрагмент, содержащий мутацию в донорном сайте сплайсинга 5-го интрона, встроенный в pSQE1, была индуцирована экспрессия белка приблизительно 57 кДа (фиг.7) и активность LOX-1 была утрачена (фиг.8), подобно pBQE1/JM109. Напротив, с pBQE1+SPS/JM109, имеющей происходящий из pSQE1 PstI-StuI-фрагмент, встроенный в pBQE1, была индуцирована экспрессия белка приблизительно 95 кДа (фиг.7) и была восстановлена активность LOX (фиг.8), что сходно с pSQE1/JM109. Нуклеотидная последовательность PstI-StuI-фрагмента pBQE1 была точно идентична гену LOX-1 дикого типа, за исключением донорного сайта сплайсинга 5-го интрона (G заменен на А в основании 1554 SEQ ID NO:10). Эти результаты ясно продемонстрировали, что присутствие или отсутствие мутации в донорном сайте сплайсинга 5-го интрона SBOU2 определяет, присутствует ли активность LOX-1.
Тест верификации 9 (картирование и отбор гибридного сорта ячменя с использованием мутации 5-го интрона)
В сообщенной нуклеотидной последовательности LOX-1, последовательность, содержащая донорный сайт сплайсинга 5-го интрона (нуклеотиды 60-63 SEQ ID NO:1: 5'GTAC-3'), может быть отщеплена рестриктазой AfaI (или RsaI), которая узнает последовательность GTAC. Дефектный ген LOX-1 был картирован с использованием того факта, что в последовательности этой области в линии SBOU2 присутствует мутация (нуклеотиды 60-63 SEQ ID NO:2: 5'-ATAC-3'), которая предотвращает расщепление рестриктазами AfaI и/или RsaI (фиг.5). Экстрагировали ДНК из листьев 144 индивидуумов 2-го дочернего поколения гибридов (F2) скрещивания между Kendall и линией SBOU2 и проводили ПЦР с использованием двух различных праймеров (5'-CCATCACGCAGGGCATCCTG-3' (SEQ ID NO:6), 5'-GCGTTGATGAGCGTCTGCCG-3' (SEQ ID NO:7)), сконструированных таким образом, что амплифицированный фрагмент должен содержать сайт AfaI. ПЦР проводили с реакцией при 94°С, 1 мин, 65°С, 2 мин и 72°С, 3 мин, повторяемой 31 раз, с последующей реакцией удлинения при 72°С, 7 мин. Каждый из амплифицированных фрагментов расщепляли AfaI и анализировали электрофорезом в 2,5% агарозном геле (далее эта процедура будет называться "способом AfaI"). В результате можно было легко выполнять различение между SBOU2, Kendall и гетеротипами (фиг.9). Кроме испытания полиморфизма по способу AfaI, полиморфизм в каждом сорте испытывали также с использованием маркера ДНК (JBC970) вблизи генного локуса LOXA хромосомы 4Н ячменя, несущей ген LOX-1 (фиг.10).
Зрелое семя каждого из индивидуумов F2 использовали для испытания активности LOX-1 семян. Для линий, не проявляющих активности LOX, активность измеряли с использованием нескольких (4-7) семян (фиг.10).
В результате испытания полиморфизма поколения F2 по способу AfaI и измерения активности LOX семян F3 расщепление признака дефектного LOX-1 SBOU2 точно совпадало с расщеплением полиморфизма по способу AfaI. Таким образом, эти серии генетических анализов ясно показали, что дефектный ген LOX-1 SBOU2 находится в генном локусе LOXA.
Эти результаты показали также, что применение способа AfaI в качестве примера способа отбора ячменя с использованием мутации ДНК позволяет проводить отбор LOX-1-недостаточных линий потомства на ранней стадии роста, что устраняет необходимость ожидания до времени созревания семян.
Пример 1
(Исследование полиморфизма AfaI других сортов ячменя)
Обычные сорта/линии ячменя использовали для исследования полиморфизма AfaI. В целом использовали 32 сорта/линии: Mikamo Golden, Golden Melon, Haruna Nijo, Myogi Nijo, Sakitama Nijo, Wasedori Nijo, Agurimochi, Harupin Nijo, Ryofu, Hokuiku 33, Hokuiku 35, Prior, Schooner, Sloop, Lofty Nijo, Franklin, Betzes, Harrington, Manley, B1251, CDC Kendall, CDC Stratus, CDC Copeland, Hanna, Merit, AC Metcalfe, TR145, Chariot, Stirling, Proctor, Koral и Heartland. В результате исследования полиморфизма по способу AfaI было определено, что испытанные сорта не были сортами типа SBOU2, но, вместо этого, расщеплялись в сайте рестриктазы AfaI, содержащем донорный сайт сплайсинга 5-го интрона (нуклеотиды 60-63 SEQ ID NO:1: 5'GTAC-3') (фиг.11). Это показало, что эти жизнеспособные линии не имеют мутации ДНК в сайте AfaI (нуклеотидах 60-63 SEQ ID NO:1: 5'GTAC-3'), и, следовательно, способ AfaI может быть эффективно использован для отбора дефектных генов LOX-1 среди линий потомства.
Пример 2
(Поиск в ресурсах генов)
Всемирно известные ресурсы генов ячменя (местного сорта), хранящиеся в Okayama University, исследовали с использованием способа AfaI. В результате были обнаружены пять новых линий (SBOU1, SBOU3, SBOU4, SBOU5 и SBOU6, хранящиеся в Okayama University), которые не расщеплялись в сайте рестрикатазы AfaI, содержащем донорный сайт сплайсинга 5-го интрона (нуклеотиды 60-63 SEQ ID NO:1: 5'GTAC-3') (фиг.12).
Активность LOX-1 семян этих линий измеряли по способу, описанному в тесте 1 поиска. Измерения активности для SBOU5 и SBOU6 проводили реакцией в течение 5 минут (фиг.13А), тогда как измерения активности для SBOU1, SBOU3 и SBOU4 проводили с продленным временем реакции 90 минут для точной идентификации активности (фиг.13В). В результате ни в одной из этих линий не была обнаружена значимая активность (фиг.13).
Это ясно продемонстрировало, что SBOU2, а также SBOU1, SBOU3, SBOU4, SBOU5 и SBOU6 (LOX-1-недостаточный ячмень типа SBOU2) были линиями ячменя, лишенными LOX-1. Поскольку все эти линии являются местными сортами (созданными улучшающим отбором), а не искусственно мутагенизированными линиями, они представляют спонтанные мутанты гена LOX-1.
Вышеописанные результаты показали, что способ AfaI, в качестве примера способа отбора ячменя с использованием мутации ДНК, является способом, делающим возможным не только отбор линий потомства LOX-1-недостаточного ячменя, но также эффективный отбор LOX-1-недостаточного ячменя из генных ресурсов ячменя.
Пример 3
(Выращивание ячменя для испытания варки напитков)
Сорт ячменя Taishomugi скрещивали с SBOU2 и полученное первое дочернее поколение (F1) самоопыляли для получения поколения F2. Признак недостаточности LOX-1 подтверждали посредством способа измерения активности фермента LOX-1, описанного в тесте поиска 1 и посредством способа AfaI, описанного в тесте верификации 9 выше, и группу LOX-1-недостаточных линий и группу LOX-1-сохраняющих линий обеспечивали в виде популяций для размножения семян, как описано ниже.
Размножение семян проводили для каждой линии (популяции) с использованием однообразной делянки или оранжереи, пока не получали семена F4. При определении активности фермента LOX-1 семян F4 было обнаружено, что соответствующие признаки активности LOX-1 индивидуумов F2 сохранялись, что свидетельствует о том, что признак недостаточности LOX-1 стабильно передается потомству.
Семена F4 использовали для следующего испытания получения солода и испытания получения солодовых алкогольных напитков.
Пример 4
(Получение солода для испытания варки напитков)
Получали популяцию F4 LOX-1-недостаточного ячменя (LOX-F4), содержащую семена ячменя, не имеющие активности LOX-1, и популяцию F4, сохраняющую активность LOX, (LOX+F4), из семян ячменя с LOX-1, причем обе популяции происходили из скрещивания Taishomugi × SBOU2, и использовали их для получения солода.
Получение солода проводили с использованием Automatic Micromalting System (Phoenix Systems) в условиях с температурой замачивания 16°С в течение в целом 82 часов (цикл 5 ч водяное замачивание/7 ч сухое состояние), проращивания при 15°С в течение 139 часов и печной сушки в течение 29 часов (55°С в течение 13,5 часов, 65°С в течение 8 часов, 75°С в течение 3,5 часов, 83°С в течение 4 часов).
Пример 5
(Анализ солода и конгрессного сусла)
Солод анализировали в соответствии со стандартным способом ЕВС (European Brewery Convention ed., Analytica EBC (4th Ed.), 1987). В результате, не обнаруживали значимого различия в результатах анализа солода между солодами, использующими LOX-F4 и LOX+F4, что свидетельствует о том, что они могут быть использованы для варки солодовых алкогольных напитков с целью сравнения присутствия или отсутствия активности LOX-1 (фиг.14).
Затем 50 г этого солода использовали для получения сусла в соответствии со стандартным конгрессным способом (European Brewery Convention ed., Analytica EBC (4th Ed.), 1987) и анализировали окисление липидов в конгрессном сусле.
Сначала измеряли количество транс-2-ноненаля в конгрессном сусле по следующему способу. Пробу сусла 8 мл помещали во флакон, добавляли 3 г NaCl и флакон закрывали крышкой. Затем в верхнее пространство этого флакона вставляли полидиметилсилоксановое волокно SPME (Supelco, Inc.) и флакон инкубировали при 40°С в течение 15 минут.
Затем это волокно вставляли в инжекционное отверстие прибора для газовой хроматографии/масс-спектрометрии, снабженного колонкой J&W DB-1 в качестве капиллярной колонки (30 м × 0,25 мм, толщина пленки: 1 мкм). В качестве газа-носителя использовали гелий (1 мл/мин) и условия печи были 60°С - 225°С (5°С в минуту), в режиме отбора ионов (m/z: 70). Количественное определение выполняли стандартным способом с использованием транс-2-ноненаля (Sigma) в качестве стандартной пробы.
В результате, концентрации транс-2-ноненаля конгрессных сусел, получаемых с использованием LOX-F4 и LOX+F4, были 0,36 млд.д. (миллиардных долей) и 3,85 млд.д. (миллиардных долей) соответственно. Таким образом, было показано, что образование сусла с использованием солода в соответствии с данным изобретением делает возможным такое ингибирование образования транс-2-ноненаля, как 1/10 или менее обычного сусла.
Потенциал ноненаля конгрессного сусла измеряли следующим способом. Сначала конгрессное сусло кипятили в течение 2 часов по способу Drost et al. (Drost, B.W., van den Berg, R., Freijee, F.J.M., van der Velde, E.G., and Hollemans, M., J. Am. Soc. Brew. Chem., 48, 124-131, 1990). Затем количество транс-2-ноненаля в пробе измеряли по способу измерения транс-2-ноненаля, описанному выше, и рассчитывали потенциал ноненаля конгрессного сусла.
В результате потенциалы ноненаля конгрессных сусел, использующих LOX-F4 и LOX+F4, были 2,74 млд.д. (миллиардных долей) и 11,9 млд.д. (миллиардных долей) соответственно. Поскольку потенциал ноненаля известен как показатель старения продукта (Drost, B.W., et al., J. Am. Soc. Brew. Chem., 48, 124-131, 1990; Ueda et al. (2001) EBC-proceedings 55:p3 28th Congress), солод, полученный из ячменя в соответствии с данным изобретением, может быть использован для варки солодовых алкогольных напитков для значительного улучшения стабильности вкуса солодовых алкогольных напитков.
Концентрации транс-2-ноненаля и потенциалы ноненаля в конгрессных суслах, полученных с использованием LOX-F4, показаны на фиг.15 в сравнении с концентрациями и потенциалами транс-2-ноненаля, полученными с использованием коммерчески доступного солода. Как видно из результатов, показанных на фиг.15, сусла данного изобретения обнаруживали величины, которые не были достигаемыми с обычным ячменем.
Эти результаты показали, что использование ячменя в соответствии с данным изобретением позволяет получать солод с улучшенным уровнем качества, не обнаруживаемым с обычными продуктами.
Концентрации THOD конгрессных сусел измеряли, используя метод высокоэффективной жидкостной хроматографии-масс-спектрометрии (ВЖХ-МС). Условия ВЖХ были следующими. Скорость тока подвижной фазы была 0,3 мл/мин, с использованием смешанного раствора 0,5% уксусной кислоты (Раствора А) и ацетонитрила (раствора В) в качестве подвижной фазы, с линейным градиентом раствор А:раствор В=35:65 (0 мин) - раствор А:раствор В=5:95 (30 мин). Использовали колонку Waters Asymmetry column (No. 106005; C18, 3,5 мкм: 2,1×150 мл), температура колонки была 50°С, и использовали систему Model 1100 HPLC (Hewlett-Packard) для отделения пробы 5 мкл сусла или солодовых алкогольных напитков. Анализ массы выполняли с использованием Waters ZQ, с мониторингом массы 329 в отрицательном режиме электрораспылительной (ES) ионизации. Исходным стандартным раствором THOD была проба экстракта пива (Kobayashi, N., et al., J. Biosci. Bioeng., 90, 69-73, 2000).
В результате, концентрации THOD конгрессных сусел, использующих LOX-F4 и LOX+F4, были 6,5 млд.д. (миллиардных долей) и 14,7 млд.д. (миллиардных долей) соответственно, что демонстрирует возможность ингибирования концентрации THOD сусла до

Как упоминалось выше, THOD образуется превращением из линолевой кислоты под действием LOX-1 солода и пероксигеназной активности солода на стадии затирания, но поскольку, как считают, активность пероксигеназы солода является ограничивающей скорость стадией для образования THOD (Kuroda, H., et al., J. Biosci. Bioeng., 93, 73-77, 2002), не было ясно, до какой степени уменьшенная активность LOX-1 ингибирует образование THOD. Однако результаты в примерах, обеспеченных в данном описании, показали, что использование солода, полученного из семян ячменя без активности LOX-1, уменьшает концентрацию THOD в сусле. Поскольку THOD присутствует до конечного продукта, не метаболизируясь дрожжами (Kobayashi, N., et al., J. Inst. Brew., 106, 107-110, 2000)), использование солода, полученного из ячменя в соответствии с данным изобретением, может явно приводить к получению солодовых алкогольных напитков с хорошими качеством вкуса и стойкостью пены.
Пример 6
(Испытание варки солодовых алкогольных напитков)
1. Получение и анализ сусла
Солод LOX-F4 и солод LOX+F4, полученные в примере 4 выше, затирали с использованием заторного аппарата 50-литрового масштаба в соответствии со стандартными способами затирания для Happoshu, алкогольного напитка с низким содержанием солода (содержание солода: 24%). Условия затирания были следующими.
Затирали 1,5 кг каждого солода по отдельности с 15 л заторной воды в соответствии со схемой 50°С в течение 20 мин, 65°С в течение 30 мин и 75°С в течение 3 мин. После затирания проводили фильтрование пивного сусла с использованием фильтровального чана. Получали 35 л отфильтрованных сусел.
Перед кипячением отфильтрованное сусло смешивали с 5 кг крахмального сиропа (75% сахаридов). В сусло добавляли 13 г гранул хмеля (87,0 BU (единиц горечи) (ЕВС)). После кипячения в течение 70 минут прокипяченное сусло охлаждали до 10°С. Содержание экстракта охлажденных сусел доводили добавлением воды до 11,6-11,8%.
Полученные сусла анализировали в соответствии со стандартными способами EBC (European Brewery Convention ed., Analytica EBC (4th Ed.), 1987). Результаты анализа показаны в таблице 1. Как видно из таблицы 1, не было обнаружено четкого различия между LOX-F4 и LOX+F4 в отношении этих показателей.
2. Получение солодового алкогольного напитка (Happoshu, алкогольного напитка с низким содержанием солода)
Сусло, полученное выше в 1, переносили в стерилизованный паром цилиндрически-конический чан на 30 л и затем добавляли дрожжи до начальной концентрации 30 миллионов клеток на мл для главного (собственно) брожения при 13°С. Когда содержание экстракта в сбраживаемой жидкости падало до 2,5%, ее переносили в новый подобный чан для стадии выдерживания. Стадию выдерживания проводили при 13°С в течение первых 6 дней и затем при 0°С в течение 2 недель после этого.
После завершения стадии выдерживания сбраживаемую жидкость подавали в аппарат фильтрования и розлива пива. Солодовый алкогольный напиток фильтровали и разливали в бутылки.
3. Анализ солодового алкогольного напитка
Солодовый алкогольный напиток, полученный выше в 2, анализировали следующим образом.
Анализ в соответствии со стандартным способом ЕВС показывает, что не было обнаружено значимого различия между LOX-F4 и LOX+F4 в отношении параметров общего анализа, других, чем параметры окисления липидов (таблица 2).
Стойкость пены солодового алкогольного напитка, полученного выше в 2, анализировали следующим способом.
Анализ стойкости пены проводили по способу NIBEM. При анализе стойкости пены с использованием тестера пеностойкости Haffmans (таблица 3) ячмень LOX-F4 имел явно более высокую стойкость пены, причем величина NIBEM была на 21 балл выше, чем в случае ячменя LOX+F4.
Так же, как и результат измерения концентрации THOD по способу, описанному в примере 5 выше, обнаружилось уменьшение содержания THOD LOX-F4 до менее чем половины содержания THOD LOX+F4 (таблица 3).
Эти результаты ясно показали, что способ получения солодовых алкогольных напитков данного изобретения позволяют получать солодовый алкогольный напиток с пониженным содержанием THOD и улучшенным сохранением пены.
Затем солодовый алкогольный напиток, полученный выше в 2, подвергали следующему сенсорному анализу, выполняемому 13 членами комиссии по оценке качества пищевых продуктов.
Сначала солодовые алкогольные напитки LOX-F4 и LOX+F4 хранили при 37°С в течение одной недели. Затем их наливали в чашки при обычной температуре принятия напитков и обеспечивали сенсорный анализ членами комиссии по оценке качества пищевых продуктов для оценки нестандартного вкуса и общей несвежести по шкале 0-4 (причем более высокий показатель представляет прогрессирующее старение) (таблицы 4А, В).
В результате 10 из 13 членов комиссии дали более низкие баллы LOX-F4 в отношении нестандартного вкуса, и, следовательно, LOX-F4 обнаруживал более низкий балл (среднее), чем LOX+F4. Было определено, что это различие в соответствии с парным t-критерием является значимым при 5% вероятности (таблица 4А).
В отношении общей несвежести, 11 из 13 членов комиссии дали более низкие баллы LOX-F4, и, следовательно, LOX-F4 обнаруживал более низкий балл (среднее), чем LOX+F4. Было определено, что это различие в соответствии с парным t-критерием является значимым при 5% вероятности (таблица 4В).
Описанные выше сенсорный анализ и статистический анализ показали, что LOX-F4 имел меньшие нестандартный вкус и общую несвежесть, чем LOX+F4.
В результате измерения содержания транс-2-ноненаля солодового алкогольного напитка, полученного выше в 2, LOX-F4 имел более низкое содержание транс-2-ноненаля до и после хранения при 37°С в течение одной недели. Содержание транс-2-ноненаля LOX-F4 уменьшалось до приблизительно 1/3 в сравнении с содержанием транс-2-ноненаля LOX+F4 после хранения (таблица 5).
Эти результаты сенсорного анализа и результаты анализа содержания транс-2-ноненаля в солодовых алкогольных напитках показали, что способ получения солодовых алкогольных напитков данного изобретения позволяет получать солодовые алкогольные напитки с улучшенной стабильностью вкуса.
Наконец, «тело» (экстрактивность) и однородность солодового алкогольного напитка, полученного выше в 2, анализировали сенсорным анализом и с использованием липидной мембраны в качестве датчика.
Сначала сенсорный анализ проводился 13 высококвалифицированными членами комиссии для сравнения качества вкуса. Солодовые алкогольные напитки LOX-F4 и LOX+F4 были обеспечены для сенсорного анализа, а "тело" (экстрактивность) и однородность оценивали по шкале 0-4 (причем более высокие баллы обозначают более наполненное "тело" и лучшую однородность) (таблица 6).
В отношении "тела" не было обнаружено значимого различия (уровень вероятности 5%) между LOX-F4 и LOX+F4 (таблица 6А).
Что касается однородности, 8 из 13 членов комиссии дали более высокие баллы LOX-F4 (таблица 6В). LOX-F4 имел более высокий балл (среднее), чем LOX+F4, и было определено, что различие в соответствии с парным t-критерием было значимым при 5% уровне вероятности.
Эти результаты показали, что варка солодовых алкогольных напитков с использованием LOX-F4 может улучшать однородность без влияния на "тело" (экстрактивность) напитка.
«Тело» и однородность оценивали также с использованием липидной мембраны в качестве датчика в соответствии со способом Kaneda et al. (Kaneda, H. et al., J. Biosci. Bioeng., 92, 221-226, 2001) (таблица 7).
«Тело» оценивали на основании адсорбции на липидную мембрану, и результаты не показали значимого статистического различия (при 5% уровне вероятности) между адсорбцией с LOX-F4 и LOX+F4 (таблица 7А).
Однородность оценивали на основании продолжительности адсорбции на липидную мембрану (с более высокой продолжительностью адсорбции, указывающей худшую однородность), и LOX-F4 обнаруживал остаток приблизительно

До сих пор не была обнаружена корреляция между активностью LOX-1 и уровнями образования THOD ячменя в стадиях затирания (Kobayashi, N. et al., (2000), J. Biosci. Bioeng. 90:69-73), и было неясно, до какой степени образование THOD уменьшается ингибированием LOX-1 ячменя. Кроме того, в предыдущем уровне техники было невозможно предсказать, может ли однородность усиливаться в результате такого ингибирования LOX-1 ячменя. Однако результаты сенсорного анализа и анализ «тела» и однородности продукта с использованием липидной мембраны в качестве датчика в примерах, приведенных в данном описании, показали впервые, что ячмень, содержащий заявленный здесь ген, может быть использован для увеличения однородности продукта без влияния на «тело» продукта.
Пример 7
(Испытание варки солодовых алкогольных напитков с использованием обработанного ячменя)
1. Получение и анализ сусла
С использованием обработанного ячменя LOX-F5 и LOX+F5, следующих поколений линий, полученных в примере 3 выше, дополнительное затирание проводили с использованием заторного аппарата в масштабе 50 л в соответствии со стандартными способами затирания для Happoshu, солодового алкогольного напитка с низким содержанием солода (содержание солода: 24%, содержание обработанного ячменя: 76%). Условия затирания были следующими.
1,2 кг коммерчески доступного солода для варки и 3,8 кг каждого обработанного ячменя затирали с 20 л заторной воды в соответствии со схемой: 50°С в течение 30 мин, 65°С в течение 60 мин и 75°С в течение 3 мин (добавляли ферменты, такие как α-амилаза и β-глюканаза, вследствие высокого содержания обработанного ячменя). После затирания проводили фильтрование сусла с использованием фильтровального чана. Получали 40 л отфильтрованных сусел.
В полученные отфильтрованные сусла добавляли 53 г гранул хмеля (единицы горечи: 25,6 BU (ЕВС)). После кипячения в течение 80 минут прокипяченное сусло охлаждали до 10°С. Содержание экстракта охлажденных сусел доводили добавлением воды до 7,5-7,6%.
Полученные сусла анализировали в соответствии со стандартным способом ЕВС. Величины анализа показаны в таблице 8. Как видно в таблице 8, не было обнаружено четкого различия между LOX-F5 и LOX+F5 в отношении этих показателей.
2. Получение солодового алкогольного напитка (Happoshu, алкогольного напитка с низким содержанием солода)
Сусло, полученное выше в 1, переносили в стерилизованный паром цилиндрически-конический чан на 30 л и затем добавляли дрожжи до начальной концентрации 30 миллионов клеток на мл для главного (собственно) брожения при 15°С. Когда содержание экстракта в сбраживаемой жидкости падало до 1,3%, ее переносили в новый подобный чан для стадии выдерживания. Стадию выдерживания проводили при 13°С в течение первых 5 дней и затем при 0°С в течение 2 недель после этого.
После завершения стадии выдерживания сбраживаемую жидкость подавали в аппарат фильтрования и розлива пива. Солодовый алкогольный напиток фильтровали и разливали в бутылки.
3. Анализ солодового алкогольного напитка
Солодовый алкогольный напиток, полученный выше в 2, анализировали следующим образом.
Анализ в соответствии со стандартным способом ЕВС показывает, что не было обнаружено значимого различия между LOX-F5 и LOX+F5 в отношении параметров общего анализа, других, чем параметры окисления липидов (таблица 9).
Стойкость пены солодового алкогольного напитка, полученного выше в 2, анализировали следующим способом.
Анализ стойкости пены проводили по способу NIBEM. При анализе стойкости пены с использованием тестера пеностойкости Haffmans (таблица 10) ячмень LOX-F5 имел явно более высокую стойкость пены, причем величина NIBEM была на 17 баллов выше, чем в случае ячменя LOX+F5.
Также, как и результат измерения концентрации THOD по способу, описанному в примере 5 выше, обнаруживалось уменьшение содержания THOD солодового алкогольного напитка LOX-F5 до менее чем половины содержания THOD LOX+F5.
Эти результаты ясно показали, что способ получения солодовых алкогольных напитков данного изобретения позволяет получать солодовый алкогольный напиток с пониженным содержанием THOD и улучшенным сохранением пены.
Затем солодовый алкогольный напиток, полученный выше в 2, подвергали следующему сенсорному анализу, выполняемому 13 членами комиссии по оценке качества пищевых продуктов. Конкретный способ этого сенсорного анализа является таким же, какой описан в примере 6.
В результате 11 из 13 членов комиссии дали более низкие баллы LOX-F5 в отношении нестандартного вкуса, и, следовательно, LOX-F5 проявлял более низкий балл (среднее), чем LOX+F5. Было определено, что это различие в соответствии с парным t-критерием является значимым при 5% вероятности (таблица 11А).
В отношении общей несвежести, 12 из 13 членов комиссии дали более низкие баллы LOX-F5, и, следовательно, LOX-F5 обнаруживал более низкий балл (среднее), чем LOX+F5. Было определено, что это различие в соответствии с парным t-критерием является значимым при 5% вероятности (таблица 11В).
Описанные выше сенсорный анализ и статистический анализ показали, что LOX-F5 имел меньшие нестандартный вкус и общую несвежесть, чем LOX+F5.
В результате измерения содержания транс-2-ноненаля солодового алкогольного напитка, полученного выше в 2, до и после хранения при 37°С в течение одной недели LOX-F5 имел сходное содержание транс-2-ноненаля в сравнении с LOX+F5 перед хранением. Содержание транс-2-ноненаля LOX-F5 уменьшалось до приблизительно1/2 в сравнении с содержанием транс-2-ноненаля LOX+F5 после хранения (таблица 12).
Пример 8.
(Испытание варки солодовых алкогольных напитков)
1. Получение и анализ сусла
Солод LOX-F4 и LOX+F4, полученные по способу, описанному в примере 4, затирали с использованием заторного аппарата 50-литрового масштаба в соответствии со стандартными способами затирания для пива (содержание солода: 71%). Условия затирания были следующими.
5,0 кг каждого испытуемого солода, указанного выше, и 2,0 кг добавки (кукурузного крахмала, кукурузной крупы и дробленого риса) затирали с 23 л заторной воды в соответствии со схемой: 50°С в течение 20 мин, 65°С в течение 40 мин и 75°С в течение 3 мин. После затирания проводили фильтрование пивного сусла с использованием фильтровального чана. Получали 40 л отфильтрованных сусел.
К полученным отфильтрованным суслам добавляли 40 г гранул хмеля (44,9 BU (единиц горечи) (ЕВС)). После кипячения в течение 90 минут прокипяченное сусло охлаждали до 10°С. Содержание экстракта охлажденных сусел доводили добавлением воды до 10,8-11,1%.
Полученные сусла анализировали в соответствии со стандартными способами EBC. Результаты анализа показаны в таблице 13. Как видно из таблицы 13, не было обнаружено четкого различия между LOX-F4 и LOX+F4 в отношении этих показателей.
2. Получение солодового алкогольного напитка (пива)
Сусло, полученное выше в 1, переносили в стерилизованный паром цилиндрически-конический чан на 30 л и затем добавляли дрожжи до начальной концентрации 15 миллионов клеток на мл для главного (собственно) брожения при 10,5°С. Когда содержание экстракта в сбраживаемой жидкости падало до 2,5%, ее переносили в новый подобный чан для стадии выдерживания. Стадию выдерживания проводили при 8°С в течение первых 8 дней и затем при 0°С в течение последующих 2 недель.
После завершения стадии выдерживания сбраживаемую жидкость подавали в аппарат фильтрования и розлива пива. Солодовый алкогольный напиток фильтровали и разливали в бутылки.
3. Анализ солодового алкогольного напитка
Стойкость пены солодового алкогольного напитка, полученного выше в 2, анализировали следующим способом. Анализ стойкости пены проводили по способу NIBEM. При анализе стойкости пены с использованием тестера пеностойкости Haffmans (таблица 14) ячмень LOX-F4 показал явно более высокую стойкость пены, причем величина NIBEM была на 30 баллов выше, чем в случае ячменя LOX+F4.
Так же, как и результат измерения концентрации THOD по способу, описанному в примере 5 выше, обнаруживалось уменьшение содержания THOD в солодовом алкогольном напитке LOX-F4 до менее, чем половины содержания THOD LOX+F4.
Эти результаты ясно показали, что способ получения солодовых алкогольных напитков данного изобретения позволяет получать солодовый алкогольный напиток с пониженным содержанием THOD и улучшенным сохранением пены.
Затем солодовый алкогольный напиток, полученный выше в 2, подвергали следующему сенсорному анализу, выполняемому 13 членами комиссии по оценке качества пищевых продуктов. Конкретный способ этого сенсорного анализа является таким же, какой описан в примере 6.
В результате 11 из 13 членов комиссии дали более низкие баллы LOX-F4 в отношении нестандартного вкуса, и, следовательно, LOX-F4 обнаруживал более низкий балл (среднее), чем LOX+F4. Было определено, что это различие в соответствии с парным t-критерием является значимым при 5% вероятности (таблица 15А).
В отношении общей несвежести, 12 из 13 членов комиссии дали более низкие баллы LOX-F4, и, следовательно, LOX-F4 обнаруживал более низкий балл (среднее), чем LOX+F4. Было определено, что это различие в соответствии с парным t-критерием является значимым при 5% вероятности (таблица 15В).
Описанные выше сенсорный анализ и статистический анализ показали, что LOX-F4 имел меньшие нестандартный вкус и общую несвежесть, чем LOX+F4.
Эти результаты сенсорного анализа и результаты анализа содержания транс-2-ноненаля в солодовых алкогольных напитках показали, что способ получения солодовых алкогольных напитков данного изобретения позволяет получать солодовые алкогольные напитки с улучшенной стабильностью вкуса.
Промышленная применимость
Может быть обеспечено следующее: мутантный ген, который применим для получения солодовых алкогольных напитков, обнаруживающих улучшенные стабильность вкуса и стойкость пены, без манипулирования генами, способ отбора LOX-1-недостаточного ячменя, материалы для солодовых алкогольных напитков, происходящие из ячменя, полученного этим способом отбора, и способ получения солодовых алкогольных напитков с использованием этих материалов для солодовых алкогольных напитков.








Реферат
Ген липоксигеназы-1 ячменя, в котором гуанин в донорном сайте сплайсинга 5-го интрона гена липоксигеназы-1 ячменя мутирован в аденин, кодирует белок, лишенный активности липоксигеназы-1. Ячмень, лишенный активности липоксигеназы-1 ячменя, распознают посредством определения, является или не является гуанин в донорном сайте сплайсинга 5-го интрона гена липоксигеназы ячменя мутированным в другое основание. Такой ячмень, его семена, солод, продукты экстракции, расщепления или обработки такого ячменя представляют собой материал для получения солодовых алкогольных напитков. 9 н. и 2 з.п. ф-лы, 16 ил., 20 табл.
Формула
Документы, цитированные в отчёте о поиске
Способ получения вкусовой основы, готовых блюд, вкусовая основа
Патенты аналоги
Способ получения вкусовой основы, готовых блюд, вкусовая основа
Комментарии