Выделенная и очищенная молекула нуклеиновой кислоты, не встречающаяся в природе молекула нуклеиновой кислоты и способ придания растению устойчивости к вирусу табачной мозаики (варианты) - RU2140985C1
Код документа: RU2140985C1
Чертежи










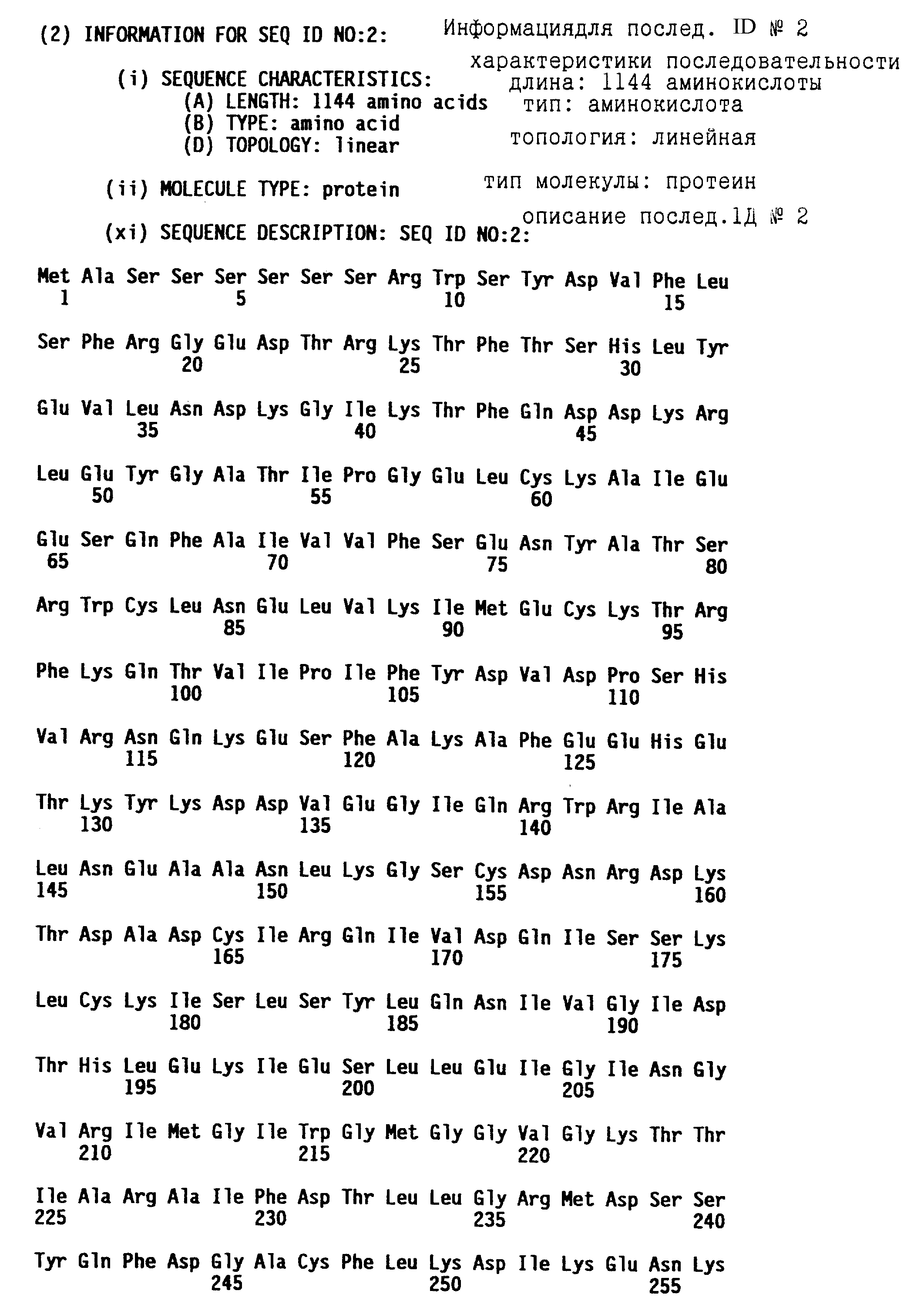










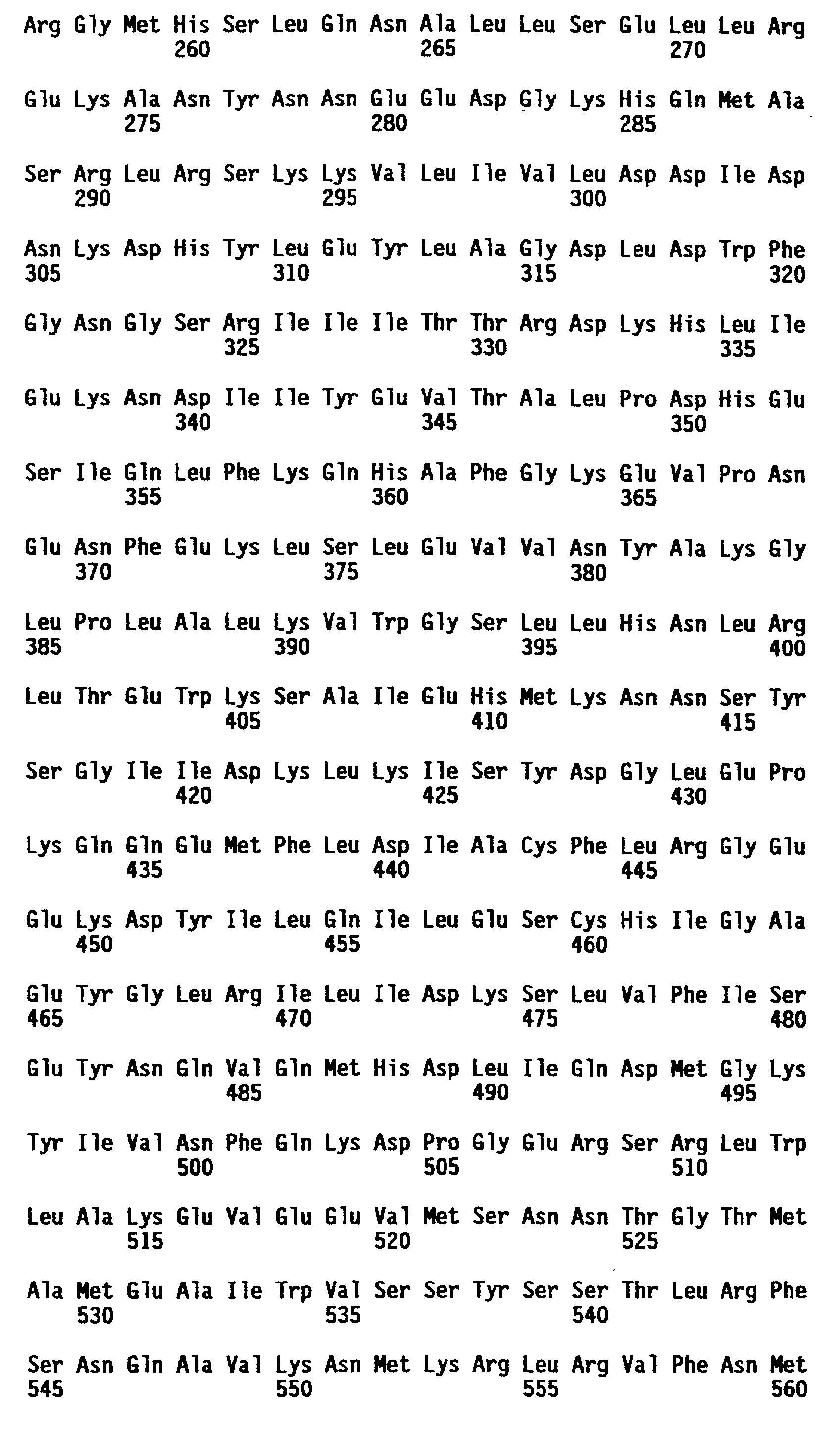







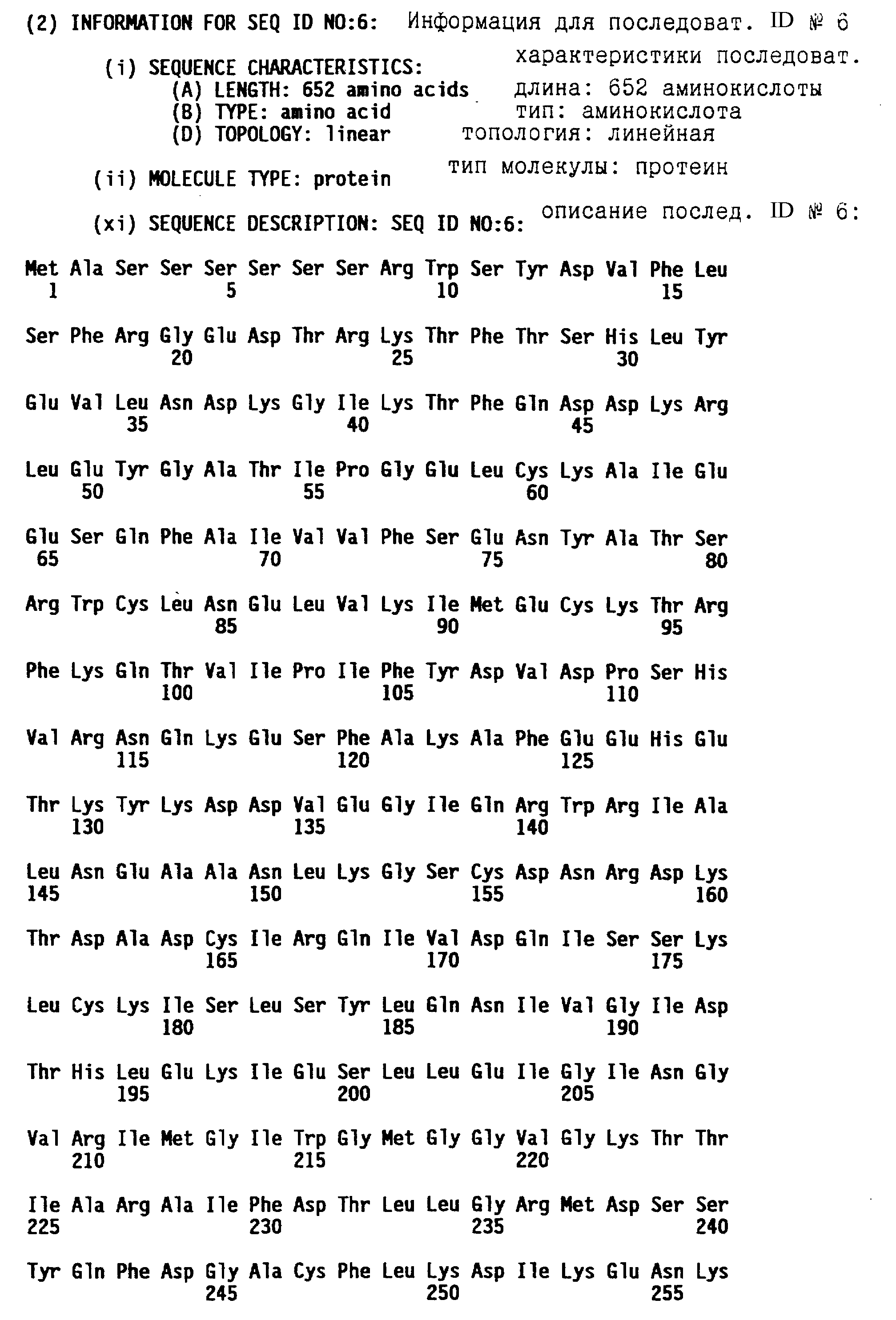










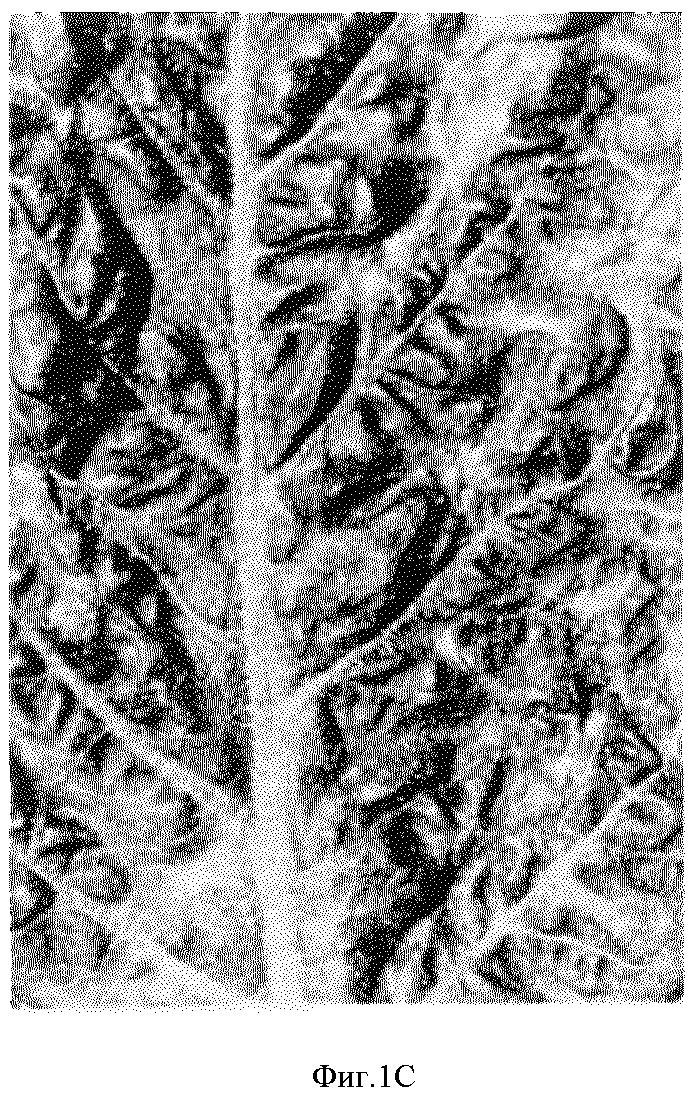






Описание
Настоящее изобретение относится к способам и материалам для усовершенствования борьбы с патогенами растения. Более конкретно, настоящее изобретение относится к последовательностям нуклеиновых кислот, которые кодируют протеин N-гена, к содержащим эти последовательности рекомбинантным полинуклеотидным молекулам и их использованию, в частности, к их использованию для трансформации растений семейства Solanaceae для придания им устойчивости к вирусу табачной мозаики.
Предпосылки изобретения
Основные потери
количества и качества урожаев определяются патогенами, вызывающими болезни растений, включая вирусы, бактерии и грибки. Вирус табачной мозаики (TMV) инфицирует растения, представляющие коммерческую
ценность, включая табак и такие родственные растения, как томаты и перец. Хотя и не являясь гибельным, TMV влияет на рост и продуктивность этих растений. Вирусный патоген распространяется в растении
в
две стадии. Вначале вирусная инфекция наблюдается в том месте, где вирус проникает в клетки растения-хозяина. Затем происходит репликация вируса, во время которой вирус размножается в клетках
растения.
У растений существует множество механизмов, которые обеспечивают природную устойчивость к патогенам. Они включают заранее сформированные структурные и химические барьеры и активные механизмы устойчивости. Устойчивость растений к заболеваниям контролируется отдельными комплементарными генами в растении и в патогене. Гены в растении называют генами устойчивости, а гены в патогене называют генами авирулентности. Растения, которые содержат ген устойчивости, эффективно защищены от заболевания, вызываемого патогеном, содержащим соответствующий ген авирулентности.
Доминантный N-локус табака придает устойчивость к TMV и является посредником локализованной гиперчувствительной реакции (HR) в сайте вирусной инфекции, и индуцирует реакцию системной приобретенной устойчивости (SAR) в клетках, соседних с инфицированным участком и во всем растении. Растения табака, гетерозиготные или гомозиготные для N-локуса устойчивы к заболеванию, вызываемому TMV. HR представляет собой комплексную активную реакцию устойчивости, которая вырабатывается в растении в ответ на атаку патогена после неудачи механизмов ранее сформированной устойчивости (Keen et.al Biotechnoloqy in Plant Disease Control, Wiley-Ziss, Inc., p.65-88 (1993)). HR характеризуется гибелью клеток (некрозы) в участке проникновения патогена. Хотя сам по себе некроз может и не быть ответственным за устойчивость к заражению патогеном, сопутствующий синтез противомикробных соединений, связанных с патогенезом протеинов, который характеризует SAR реакцию, и установление структурных барьеров, как считают, играет центральную роль в остановке распространения патогена. Взаимодействия патоген- растение, в которых вырабатывается устойчивость, называют несовместимостью, тогда как те, которые приводят к заболеванию, называются совместимостью.
Были проведены исследования механизмов, за счет которых растения, содержащие гены устойчивости, определяют наличие заражающего патогена и индуцируют HR и SAR. Во многих случаях HR управляется за счет ген-для-гена взаимодействиями между несовместимыми комбинациями растения и патогена. Модель гена, предложенная Flor (Journal of Aqricultural Research 74: 241-262 (1947)), предполагает, что устойчивость к заболеванию и патогенная авирулентность являются характерными особенностями. Поэтому устойчивость будет проявляться только в тех случаях, когда растения обладают специфическим геном устойчивости (R-ген), а патоген обладает соответствующим геном авирулентности (Avr-ген). Некоторые Avr-гены были клонированы из бактерий, грибков и вирусов (см. Gabriel and Rolfe Annual Rev.Phytopatholoqy 28:365-391 (1990) и Keen, Annual Rev. Genet 24:447-463 (1990)), и в некоторых случаях природа элизиторных молекул была определена (см.Keen, Plant Molecular Biology 19: 109- 122 (1992)). Сообщается о гене устойчивости к грибкам НМ1 кукурузы (Johal and Briqs, Science 258:985-987 (1922)) и гене устойчивости к бактериям Pto томатов (G.Martin etal. Science 262:1432-1436 (1993)). Однако до сих пор не было выделено или клонировано никаких природных генов устойчивости к растительным вирусам.
Простая генетическая связь между R-генами и их соответствующими Avr-генами приводит к обсуждению типа действия продуктов R-гена. Одна модель предполагает, что R-гены находятся в схемах подачи сигналов, способных распознавать патогены и инициировать последующие каскады трансдукции сигналов, приводящие к устойчивости (Lamb., Cell 76:419-422(1994). Вторая модель предполагает, что R-генные продукты являются трансмембранными ионными каналами, за счет которых происходит гибель клеток, независимо от других событий в клетке. Недавнее клонирование Pto из томатов, придающих устойчивость к бактериальному патогену Pseudomonas syrihqae pathovar tomato (Martin et al. Science 262:1432-1436 (1993)) дает возможность предположить, что по крайней мере первая модель может работать в клетках растений. Анализ последовательностей Pto указывает, что она кодирует серин/треонин киназу. Теоретически возможно, что эта серин/треонин киназа взаимодействует прямо или косвенно с элиситорной молекулой, а затем фосфорилирует последующий модулятор реакции устойчивости, инициируя тем самым каскад трансдукции сигнала.
Было отмечено сходство между реакциями гиперчувствительной устойчивости растений и "врожденной" иммунной реакцией животных. Общим является быстрое продуцирование реакционноспособных типов кислорода (ROS), известных как оксидативный импульс. Примерами ROS являются супероксидный анион (O2-) и перекись водорода (H2O2). Эти молекулы могут обладать прямым антимикробным действием и такими другими защитными действиями, как сшивание структурных протеинов в стенках растительной клетки. Важно, что ROS может активировать экспрессию генов, связанных с защитой у животных и растений (Scheck and Bauerle, Trends in Cell Biology 1:39-42 (1991), Chen et el. Science 262:1883-1886 (1993). У млекопитающих ROS с большими основаниями рассматривают как вторичный мессенджер для цитокинов, таких, как фактор опухолевого некроза (TNF) и Интерлейкин-1 (I1-1) в схеме, где фактор транскрипции NF-кВ регулирует экспрессию иммуноглобулинов, интерлейкинов и других протеинов. Фактор транскрипции Дрозофилы (Dif), гомологичный NF-кВ, также активирует транскрипцию антибактериальных протеинов, включая секропины (cecropins), аттацины (attacins), дефенсины и лизоцимы (Levine and Hultmark. Trends in Genetics 9: 178-183(1993)). Параллельно в растениях является индуцирование протеинов, связанных с патогенезом, и синтез таких противомикробных соединений, как фитоалексины, которые можно индуцировать экзогенным нанесением H2O2.
Важной модельной системой для исследования реакций устойчивости растений была система устойчивости N-гена. N-локус состоит из отдельного доминантного гена, который является медиатором индуцирования реакции некротического типа и SAR в ответ на инфицирование за счет TMv (Holmes, Phytopatohology 28: 553-561 (1938). Он был исходно идентифицирован в Nicotiana qlutinosa и был введен в N.tabacum. N - ген является медиатором гиперчувствительной реакции, которая характеризует образование локальных поражений, в которых вирус табачной мозаики локализован. Это представлено на фиг.1A. Культивары табака без N-гена позволяют вирусу табачной мозаики систематически распространяться и развивать "мозаичные" симптомы, которые характеризуются чередующимися участками светло- и темно-зеленой ткани листьев (фиг.1B).
Технология рекомбинантных ДНК предлагает возможность получения растений, трансформированных за счет патогенного гена устойчивости для придания таким растениям устойчивости. Такой подход сдерживался из-за отсутствия клонированных природных генов устойчивости растений и отсутствия знаний о механизме устойчивости. Однако до настоящего времени клонированные гены устойчивости были недоступны из-за отсутствия способов выделения генов растений, для которых не было информации относительно природы генов или их продуктов. В последнее время были разработаны два способа для растений, и они независимо от знания гена или биохимической информации о протеине позволяют выделять гены. И этими способами являются позиционное клонирование и транспозонное мечение (Baker, Shеll, Fedoroff, Proceediqs of National Academy of Science, USA 83: 4944-4848 (1986)).
Краткое содержание
изобретения
Настоящее изобретение включает
ДНК-последовательность в выделенной и очищенной форме, которые кодируют протеин N-гена, причем этот протеин осуществляет функцию передачи
устойчивости к TMV в растениях, синтезирующих протеин N-гена.
Геномные и кДНК последовательности, кодирующие конкретный протеин N-гена, здесь конкретно представлены. В объем изобретения включены ДНК
последовательности, кодирующие протеин N-гена представленной
аминокислотной последовательности. ДНК-последовательности, которые гибридизуются специфически с N-ген колирующей последовательностью или
ее комплементом в стандартных условиях и которые кодируют
протеины N-гена, которые функционируют как медиаторы устойчивости к TMV, также включены в объем настоящего изобретения.
Следующим аспектом изобретения является создание рекомбинантных молекул нуклеиновых кислот, содержащих последовательности, кодирующие протеин N-гена. Такие молекулы включают, например, рекомбинантные векторы, такие как векторы клонирования, экспрессии или трансформации, которые содержат ДНК- последовательность, кодирующую протеин N-гена.
В другом аспекте настоящего изобретения предложены клетки, которые трансформированы вышеуказанными векторами или ДНК-последовательностями.
Конкретным использованием настоящего изобретения является создание растений или растительных клеток, трансформированных N-геном, кодирующим последовательность для придания растениям устойчивости к TMV.
Следующим аспектом изобретения является создание олигонуклеотидных зондов, способных детектировать N-ген или их функциональных эквивалентов в растениях семейства Solenaceae и использование этих зондов для выделения ДНК-последовательностей, кодирующих N-ген или их функциональных эквивалентов. ДНК- последовательности, которые специфически гибридизуются с зондами и которые кодируют функциональный протеин N-гена, также входят в объем настоящего изобретения.
Использование последовательности N-гена облегчает выделение гомологичных генов из родственных и неродственных хозяев для получения генов, которые защищают растения-хозяева от родственных вирусных патогенов и неродственных патогенов.
В соответствии с настоящим открытием целью настоящего изобретения является создание генной конструкции, включающей ДНК последовательность, которая кодирует протеин N-гена, который функционирует как медиатор устойчивости к TMV в растениях, синтезирующих протеин N-гена.
Целью настоящего изобретения является также создание вектора трансформации, включающего конструкцию N-гена, причем этот вектор эффективен для стабильного введения конструкции N-гена в растение.
Другой целью изобретения является создание трансгенных растений, устойчивых к TMV, в которых устойчивость является результатом экспрессии конструкции N-гена.
Другие цели и преимущества настоящего изобретения станут очевидны из прилагаемого описания.
Краткое описание рисунков
Фиг.1 представляет фенотипы листьев табака после инокуляции TMV. На фиг. 1A
представлен лист растения, содержащего
функциональный N-ген. На фиг.1B представлен лист растения, восприимчивого к TMV. На фиг. 1C представлен лист растения, демонстрирующий участки некрозов на
TMV-восприимчивом фоне (сеторный фенотип). На
фиг.1Д представлен лист растения TMV- восприимчивого, SR1 табака, трансформированного pTG38 Т-ДНК- конструкцией. Фиг.2A-2C иллюстрируют Саузерн-блоттинг
гибридизационный анализ маркера зонда Nt1 RFLP
и Ac для Д111 популяции. Фиг.2A демонстрирует результаты гибридизации Nt-1 зонда с геномной ДНК, выделенной из Nicotiana видов qlutinosa tomentosiformis
и sylvostris и культиваторов табака Samsun NN и
SR1. Фиг. 2B демонстрирует результаты Nt-1 гибридизации с потомством бэккроссов геномных ДНК, сегрегирующих Nt1 (G) N- связанный RFLP маркер. Фиг.2C
демонстрирует гибридизацию 5' Ac зонда с той же ДНК,
что представлена на фиг. 2B.
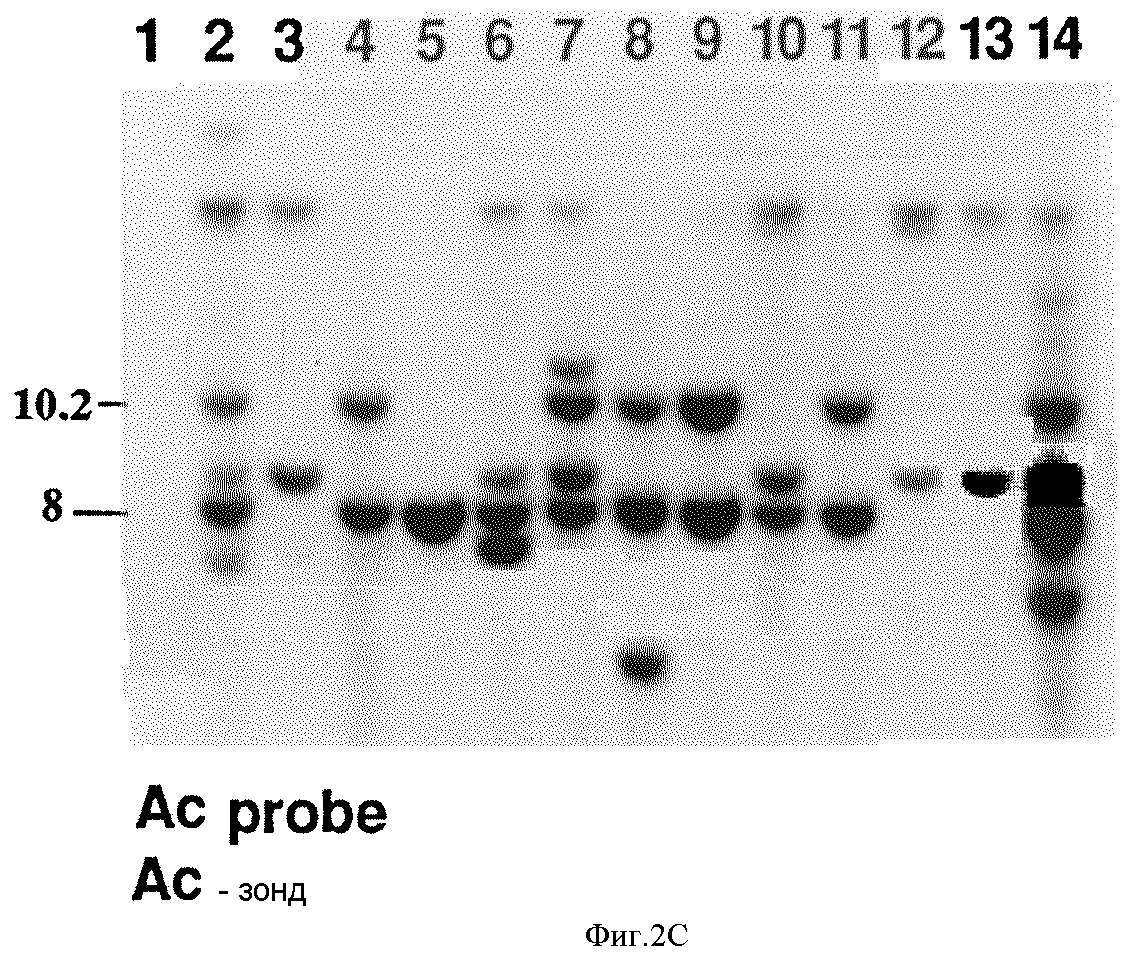
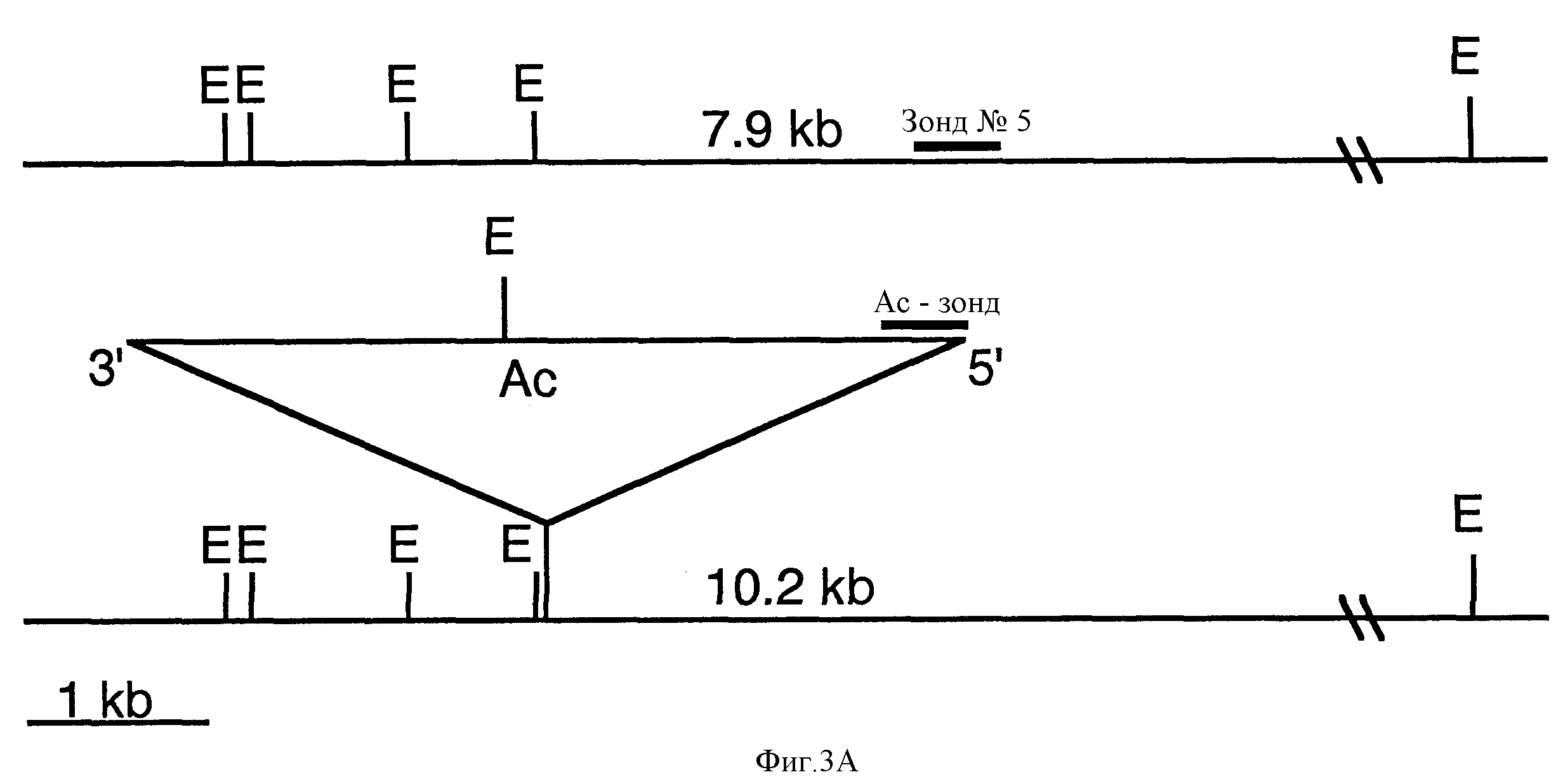
Фиг. 3A демонстрирует рестрикционную карту части дикого типа N-гена и Ac за счет вставки мутагенизированного N-гена. Фиг. 3B-3C демонстрируют Саузерн блоттинг ДНК, выведенных из родственных видов Nicotiana и Ac-мутагенизированных растений, секториальных растений и мутантов, содержащих Ac-вставки в дикого типа (WT) N- ген, и герминальные ревертанты. Фиг. 3B демонстрирует гибридизацию зонда N-5 N-гена с ДНК выбранного растения. Фиг.3C демонстрирует Ac-гибридизацию с ДНК выбранного растения.
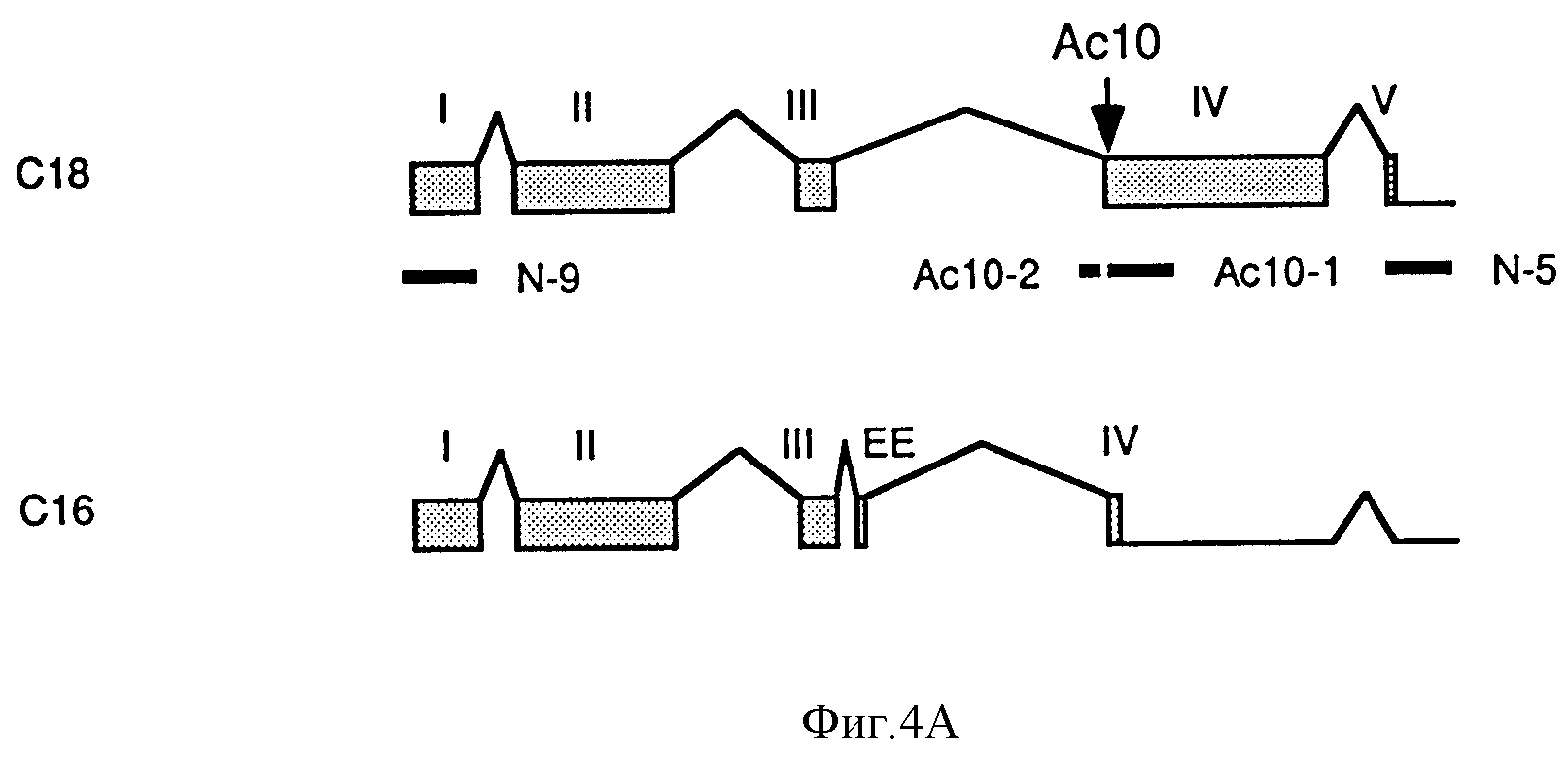
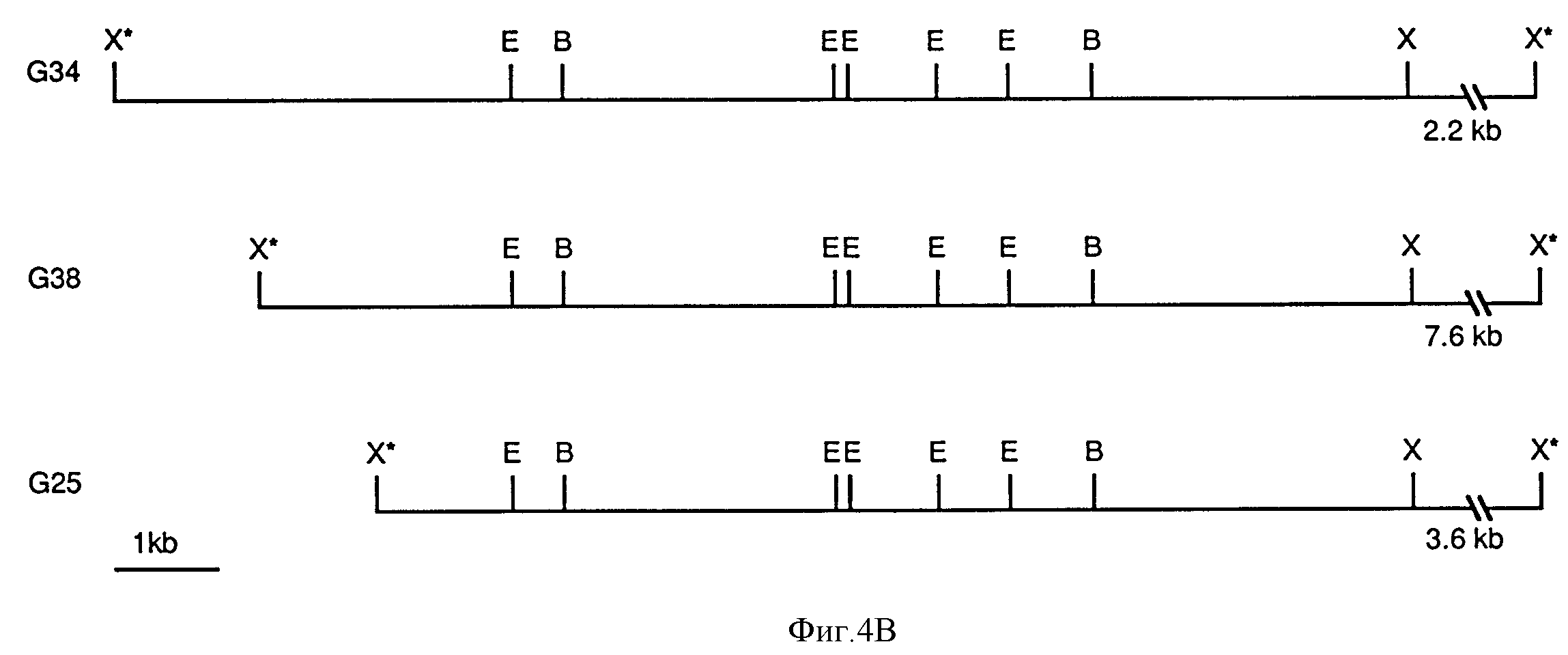
Фиг. 4A и фиг. 4B суммируют организацию N-гена. Фиг.4A иллюстрирует организацию N-гена по отношению к относительным положениям интронов и экзонов, а фиг. 4B представляет рестрикционные карты трех геномных клонов, каждый из которых содержит полной длины N-ген. Эти карты были получены из анализа последовательностей кДНК клонов C7, C16 и C18, и G38 геномного клона и рестрикционного анализа трех геномных клонов. кДНК C7 (не представлена) оказывается идентичной C18, за исключением того, что C7 содержит интрон 11, и считают, что это частично процессированный месседж. У C18 отсутствуют 753 bp с 5'-конца. Взятые вместе C7 и C18 предсказывают открытую считывающую рамку из 3432 пар оснований, кодирующую полипептид из 1144 аминокислот. C16 кодирует протеин из 652 аминокислот за счет включения альтернативного экзона из 70 bp, который изменяет считывающую рамку. Все три кДНК клона идентичны по их 3'-концам, но лишь C7 и C16 идентичны по их 5'-концам. Фиг. 4B: геномные клоны переварены EcoR1 (Е), Bam H1 (В) и Xho 1(Х). X+ указывает, что этот Xho 1 сайт получен за счет полинкера λ Gem 11 клонирующего вектора.
Фиг. 5 представляет модель для N-протеина, который является медиатором сигнальной трансдукции в реакции на TMV-инфицирование.
Подробное описание изобретения
Считают, что доминантные
гены устойчивости растений кодируют протеины, которые могут распознавать
конкретные патогены или расы патогенов, и инициируют сигнальный каскад трансдукции, что приводит к экспрессии устойчивости к
заболеванию. TMV проникает в клетку в результате механического повреждения
тканей растения. После проникновения в клетку его распространение и локализация внутри клетки непонятны. В N-содержащих
растениях табака считают, что N-протеин взаимодействует прямо или косвенно с
каким-то компонентом TMV. Этот компонент TMV до сих пор не определен, но считают, что в этом участвует репликаза (Padqett
and Beachy, Plant Cell 5:577-586 (1993)). После распознавания TMV N инициирует
реакцию устойчивости, что приводит к образованию локальных поражений и к индуцированию отдаленной системной
приобретенной устойчивости.
Считают, что настоящее изобретение является первым сообщением о клонировании, секвенировании и передаче устойчивости к трансгенному вирусу табачной мозаики для N- гена Nicotiana.
В том смысле, как здесь использован, термин "протеин N-гена" относится к протеину, обладающему способностью придавать устойчивость к TMV в растении, синтезирующем протеин N-гена. N-ген включает геномные последовательности, которые кодируют протеин N-гена и которые управляют и регулируют транскрипционную и трансляционную экспрессию N-кодирующих последовательностей. Представленный N- генный продукт имеет предсказанный молекулярный вес около 131 кДа и предсказанную аминокислотную последовательность, которая представлена в последовательности ID N 4 и в таблице 7A. Представленная геномная ДНК-последовательность, которая охватывает кодирующую последовательность для этого N-генного продукта, представлена в последовательности ID N 1. Полной длины кДНК последовательность (из клонов C18) и усеченная кДНК последовательность (из клона C16) представлены как последовательность ID N 3 и последовательность ID N 5 соответственно.
Вырожденность генетического кода хорошо известна специалистам, поэтому синонимы кодирующих последовательностей с одним или более замещенными кодонами специалисты легко определят. Синонимные кодирующие последовательности отличаются от представленных кодирующих последовательностей, но кодируют протеины тех же самых аминокислотных последовательностей, что и те, которые здесь специфически представлены.
Специфические варианты нуклеотидных последовательностей, которые кодируют протеин N-гена, который имеет функцию передачи устойчивости к TMV, представлены в последовательностях ID N1, 3 и 5.
кДНК последовательность, содержащая полной длины N-ген, представлена как последовательность ID N 3. Эта кДНК последовательность состоит из 3760 bp в длину. Образующаяся открытая считывающая рамка (кодирующая часть), начинающаяся у основания 60 и заканчивающаяся у основания 3494, кодирует протеин длиной в 1144 аминокислоты. Кодируемый протеин представлен в таблице 7A и в примере 5.
кДНК последовательность, которая кодирует усеченный протеин N-гена, родственный приведенному в таблице 7A, представлена в последовательности ID N 5. Эта кДНК в длину имеет 3830 bp и кодирует протеин из 652 аминокислот (см. последовательность ID N 6).
Геномная ДНК последовательность, содержащая полной длины N-ген, представлена в последовательности ID N 1. Геномная ДНК имеет длину 7400 bp, и анализ нуклеотидной последовательности выявил пять экзонов, которые вместе соответствуют кодирующей последовательности в последовательности ID N 3. Последовательность кодируемого N-протеина представлена в последовательностях ID N 2 и ID N 4.
Анализ последовательности N-протеина и сравнение с последовательностями других протеинов выявил значительное сходство последовательностей с некоторыми протеинами, вовлеченными в сигнальную трансдукцию (см. также таблицу 7A и пример 5). Протеин N-гена содержит три функциональных домена: сигнальный домен, ATP/GTP связующий сайт (Р-петля), и богатый лейцином участок. Такие домены присутствуют в протеинах, которые играют роль в сигнальной трансдукции.
Богатый лейцином участок (LRR) N состоит из 13 повторов и в большинстве повторов содержится последовательность LXXLXXLXL (или аналогичная последовательность). В дополнение к лейциновым остаткам доминирующей отличительной особенностью LRR является наличие пролина. Определено, что в среднем длина LRR составляет приблизительно 25 аминокислот. Предположительно первой аминокислотой в каждом повторе является пролин. Таблица 7C демонстрирует первичную структуру богатых лейцином повторов N-гена (аминокислоты/аа/590-928) и сравнение их консенсусной последовательности с консенсусными последовательностями LRR аденилатциклазы дрожжей, Drosophila Toll, Iba-цепью мембранного гликопротеина тромбоцита человека, Htrk, Drosophila Chaoptin, Arabidopsis рецептороподобной трансмембранной киназой (ТМК1) и TMKLI.
Предполагают, что LRR являются посредниками протеин-протеин взаимодействий для широкого круга протеинов. Важность LRR в функциях некоторых протеинов была определена за счет мутагенеза или выделения мутаций за счет мутантного фенотипа. В дрожжевой аденилатциклазе такие мутации, как делеции двух аминокислот в 1 или 26 LRR, исключают способность быть активированными за счет Ras (Suzuki et al., (1990) Proceedings of the National Academy of Science USA 87:8711-87-15). Аминокислотное замещение А156-У в одном из LRR альфа субъединицы 1b гликопротеина тромбоцита человека приводит к кровотечению (Ware et al., (1993) J.Clin. Invest 92:1213-1220).
Считают, что LRR играет очень важную роль в управлении специфическими протеин-протеин взаимодействиями. Без желания связываться с какой-либо конкретной теорией можно считать, что LRR из N может взаимодействовать с компонентом TMV. Так как небольшие изменения в структуре LRR приводят к разительным изменениям функции протеина, можно предположить, что LRR является медиатором специфического взаимодействия между TMV и N-протеином. Кроме того, небольшие изменения в аминокислотных последовательностях могут также привести к новым особенностям, которые, с эволюционной точки зрения, были бы благоприятны для растений, так как развивали бы новую устойчивость к каждому из изменений популяций патогенов. Другой возможной ролью LRR является взаимодействие со специфической эффекторной молекулой, такой как киназа или фосфатаза, после распознавания TMV.
Предсказанная аминокислотная последовательность N содержит фрагмент Р-петли (таблица 7A). Последовательность GMGGVG КТ (аа 216-223 последовательности ID N 4) удовлетворяет консенсусной последовательности Р-петли (A/G)XXXXGK(S/T), найденной в различных АТР- или GTP-связывающих протеинах (таблица 7A). Семейства протеинов, содержащие Р-петлю, включают аденилаткиназы, ras семейство протеинов, факторы удлинения, b-субъединицу АТР синтазы, тимидинкиназы и фосфоглицераткиназы (Saraste, (1990) Trends in Biochemical Sciences 15:430-434). Р-петля N, по- видимому, не участвует в GTP связывании, так как отсутствуют консенсусные последовательности DXXG и NXKD, необходимые для GTP связывания помимо Р-петли (Dever et.al. (1987) Proceedinq of the National Academy of Science USA 84:1814-1818).
Помимо P-петли два других "сегмента", по- видимому, должны быть включены в АТР связывание в аденилаткиназе и F1-ATP-азе (Fry et al (1986) Proceedinq of the National Academy of Sciences USA 83:907-911). Исследование N-последовательности предполагает, что эти сегменты присутствуют, причем в нужном расположении (подчеркнутые аминокислотные остатки в таблице 7A). Сегмент 2 содержит дипептид (I, A, L, V) (V, I) и N содержит последовательность A1 в положении 228 и 229 соответственно. У 80 - 100 аминокислот из P-петли сегмент 3 определен как глицин (G), за которым следует участок из 5 гидрофобных аминокислот и аспартат (D). N содержит последовательность VLIVLDD у аминокислот 296-302. Исходя из последовательности аминокислот, можно предсказать, в каких условиях АТР связан или гидролизован.
Аминотерминальные аминокислоты (8-150) N-протеина аналогичны цитоплазмическим (сигнальным) доменам протеина Drosophila Toll и рецептору интерлейкина-1 человека (IL-1R). Это соответствие представлено в таблице 7B. Окантованные аминокислоты указывают участки, где последовательности идентичны или где наблюдаются консервативные замещения. N последовательность содержит некоторые консервативные аминокислоты, которые необходимы для трансмиссии сигнала с цитоплазмы на ядре в Toll и IL 1-R регуляторных схемах (Schneider et al (1991) Genes and Development 5:797-807; Hequy et al (1992) J. of Bioloqical Chemistry 267:2605-2609).
Сходство последовательностей между аминотерминальным участком N-протеина и цитоплазмическим доменом Drosophila Toll и IL-1R человека ведет к предположению, что после TMV инфицирования N может возбуждать аналогичного типа каскад внутриклеточной трансдукции сигнала (фиг.5). Взаимодействие различных агентов подобных вирусам, цитокинам (IL-1, TNF) и митогенам (пробол-12-миристат-13-ацетат, РМА), лектинам и ионофорам кальция, с рецептором интерлейкина-1 (IL-1R) или восприятие неизвестного сигнала внеклеточным доменом Toll приводит к активации и транслокации Rel-связанных транскрипционных факторов NFкВ и dorsal, соответственно, из цитоплазмы в ядро. У млекопитающих иммунные, воспалительные и острофазные реакции, комплекс NF-кВ фактора активной транскрипции индуцирует или подавляет синтез различных защитных и сигнальных протеинов после связывания с декамерным фрагментом последовательности, который называют кВ-связывающим фрагментом (обзор в Baeuerle, (1991) Biochimica et Biophysica Acta 1072:63- 80). Эти индуцированные протеины инициируют общий клеточный защитный механизм за счет подачи сигнала о наличии патогенов другим клеткам (Baeuerle and Baltimore, (1988) Science 242:540-546 and Baeuerle, 1991 (ранее). Тогда как у зародыша Drosophila более высокая концентрация дорсального протеина в ядрах регулирует транскрипцию зиготных генов, вовлеченных в детерминацию дорсовентральной полярности эмбриона (обзор в Johnston and Nusslein-Vohard Cell, 68:201-219 (1992)). Точечные мутации в сигнальном (цитоплазмическом) домене природных рецессивных аллелей Toll (Schneiden et al. (1991) см. ранее) и сайт- направленные точечные мутации в сигнальном домене IL 1-R (Hequy (1992) ранее приводят к нарушению транслокации, либо dorsal, либо Nf-кВ, соответственно к ядрам.
Недавно было сообщение о другом rel-содержащем гене, названном Dif (иммунитет, связанный с dorsal), вовлеченном в Drosophila иммунную реакцию (Ip et al. Cell 75:753-763 (1993)). Аналогично NFKB и dorsal, Dif-протеин обычно присутствует в цитоплазме жирного тела личинок; после поражения или инфицирования он транслоцируется в ядра и специфически связывается с кВ-подобными фрагментами в промоторном участке различных антимикробных генов (Sun et al. , European Journal of Biochemistry 196:247-254 (1991); Enqstomet at al. Journal of Molecular Biology 232:327-333 (1993); и Kappler et al., EMBO J. 12:1561-1568 (1993)). Аналогично вышеуказанным иммунным реакциям и реакциям развития, и без желания связываться с какой-либо конкретной теорией, можно предположить, что продукт TMV (элиситор) связывается с LRR или другим участком N-протеина (рецептор) в цитоплазме или через другой неизвестный протеин в конечном счете активирующем rel/кВ транскрипционный факторный комплекс, необходимый для индуцирования связанных с патогеном (PR) генов.
Одно из основных преимуществ настоящего изобретения состоит в том, что оно предлагает способ индуцирования устойчивости к TMV в табаке и родственных растениях, таких как томаты и перец. Это очень важно, так как передаваемая за счет N устойчивость к TMV высоко эффективна и до сих пор не была преодолена обычными штаммами TMV.
Клонированный природный ген устойчивости N предлагает преимущества по сравнению с доступными в настоящее время способами защиты растений от TMV. Два гена, которые широко используются для получения устойчивости к TMV, получены из TMV оболочкового протеина (CP) или полимеразного гена. Недостатками существующей технологии защиты от TMV являются то, что CP-опосредствованная устойчивость может проходить со временем или при более высоких уровнях инокулюма вируса, а полимеразой- опосредствованная устойчивость очень специфична к штамму вируса, из которого получен ген полимеразы. Другим высоким риском, связанным с устойчивостью за счет вирусного гена, является риск или возможность эволюции гиперштаммов вирусов за счет рекомбинации между природными штаммами и трансгенами. Встраивание клонированного гена устойчивости к вирусам растений в коммерческие культиваторы за счет трансформации позволяет избежать вышеуказанных недостатков. N-ген табака придает устойчивость ко всем известным штаммам TMV за исключением одного. Клонированный природный ген устойчивости к растительному вирусу позволяет также проводить фундаментальные исследования механизма распознавания патогена геном устойчивости и передачи сигнала индуцирования защитных реакций так, чтобы идентифицировать критические функциональные домены гена, и облегчить конструирование генов устойчивости с более широким спектром устойчивости. Это первое описание гена устойчивости растений, последовательность протеинов которого предсказывает предполагаемый ATP/GTP фрагмента сайта связывания (P-петля), обогащенный лейцином участок и сигнальный домен.
Клонирование N-гена осуществляют за счет транспозонного мечения кукурузным транспозоном Ac в N.tabacum. Позитивный отбор разрабатывают для выделения Ac индуцированных мутантов, неспособных реагировать на TMV с HR (HR мутанты). Один из 36 HR- мутантов, содержащих Ac, имеет нестабильную мутацию, что коррелирует с присутствием отдельного Ac-транспозона, обозначенного Ac10. Геномные ДНК-последовательности, фланкирующие Ac10, используют для скринирования кДНК и геномных ДНК-библиотек для клонов, содержащих полной длины кДНК и геномные ДНК N-гена. Геномный клон, содержащий N ген, выделяют из геномной библиотеки N.qlutinosa для использования, для трансформации растений, для придания им устойчивости к TMV N-ген клонируют в вектор и используют для трансформации растений, восприимчивых к TMV. Трансформированные растения демонстрируют устойчивость к TMV.
В том смысле, как здесь использовано, молекула нуклеиновой кислоты может быть молекулой ДНК, молекулой РНК или гибридной ДНК-РНК-молекулой. Не встречающаяся в природе молекула нуклеиновой кислоты является молекулой, которая не встречается в природе. Не встречающаяся в природе молекула нуклеиновой кислоты включает, например, ДНК-последовательности в выделенной и очищенной форме; рекомбинантную молекулу нуклеиновой кислоты, содержащую гетерологичный участок, то есть идентифицируемый сегмент ДНК, который ковалентно не связан с N-ген-кодирующей последовательностью в природе; или такая не встречающаяся в природе молекула может быть сконструирована из частей, которые были синтезированы химически; конструкция, в которой сама кодирующая последовательность не встречается в природе, например кДНК, в которой геномная кодирующая последовательность содержит интроны; или синтетическая последовательность содержит интроны; или синтетическая последовательность, содержащая кодоны, отличные от кодонов нативного гена. Части из гетерологичных источников могут быть соединены способами, известными специалистам, например, за счет лигирования ин витро. В другом варианте части могут быть соединены за счет in vivo процесса, например за счет рекомбинации, но такая рекомбинация будет управляться вручную, и желательный результат будет определяться человеком.
Примерами молекул ДНК являются кДНК N-гена Nicotiana qlutinosa, определяемые нуклеотидной последовательностью, представленной в последовательности ID N 3, и нуклеотидной последовательностью, представленной в последовательности ID N 5. Геномная нуклеотидная последовательность, содержащая полной длины N. qlutinosa N-ген, представлена в последовательности ID N 1.
В объем настоящего изобретения включены также молекулы нуклеиновой кислоты, содержащие N-ген, который содержит нуклеотидную последовательность, которая по крайней мере на 70% гомологична с последовательностью ID N 1 примерно с нуклеотида 1 до нуклеотида 7400, и где протеин N-гена, кодируемый молекулой, обладает функцией медиатора устойчивости к TMV в растении, которое синтезирует этот протеин N-гена. Настоящее изобретение включает также молекулы нуклеиновой кислоты, содержащие протеин N-гена- кодирующую последовательность, где кодирующая последовательность по крайней мере на 70% гомологична нуклеотидной последовательности ID N 3 от нуклеотида 60 до нуклеотида 3494, где кодируемый N-протеин обладает функцией медиатора TMV устойчивости в растении, которое экспрессирует этот протеин. Гомологичные последовательности, входящие в объем изобретения, можно идентифицировать в экспериментах по Саузернблоттингу, используя условия, в которых гибридизация соответствует по крайней мере 70% гомологичности, в противоположность неспецифическому связыванию (см. пример 2 для обсуждения жестких и мягких условий). Гомологичность определяют как то, что нуклеотиды соответствуют по определенной длине выбранного участка. Условия гибридизации описаны у Sambrook et al. Molecular Cloninq: A Laboratory Manual Cold Sprinq Harbor Laboratory (1989) Ausubel et al. , Current Protocols in Molecular Bioloqy, Current Protocols (1989), которые включены сюда по ссылке.
Для идентификации N-гена из других видов Solanaceous выделяют, как указано, геномную ДНК из растения семейства Solanaceous. Выделенную ДНК разрезают одним или более из рестрикционных энзимов клонированных в λ или другой подходящий вектор; обрабатывают электрофоретически и получают блоты на такой найлоновой мембране, как Nytran. Эти блоты зондируют описанным здесь зондом. Из этих ДНК получают геномную библиотеку и скринируют ее, используя вышеуказанные зонды для идентификации N-гена. Оценивают активность гена, определяемую за счет экспрессии гена в растения Solanaceous и определяют устойчивость трансформированных растений к TMV, как будет подробно описано далее.
Олигонуклеотиды, полученные из последовательности N-гена, можно использовать в качестве праймеров в полимеразной цепной реакции (PCR). Табак содержит один геномный участок, кодирующий N- протеины. Консервативные участки N-гена можно использовать при конструировании праймеров для опосредствованного выделения функциональных N-гомологичных генов в растениях Solanaceous. Далее, антитела, выработанные против доменов N-протеина, можно использовать для скринирования экспрессионных библиотек других Solanaceous растений.
ДНК кодирующую последовательность протеина N-гена можно также получить синтетически из вырожденных олигонуклеотидов, последовательности которых содержат кодоны для аминокислотной последовательности протеина N-гена. Такие олигонуклеотиды получают стандартными способами и собирают, и используют для выделения целевого N-гена.
Доступность молекул нуклеиновой кислоты табака, кодирующих протеин N-гена, делает доступными последовательности N-гена, кодирующие протеин N-гена, или функциональные гомологи из других Solanaceous растений. Геномные или кДНК-последовательности табака или их части используют в качестве олигонуклеотидных зондов для гибридизации с дополнительными геномными или кДНК последовательностями за счет гибридизации в стандартных условиях. Последовательности, которые специфически гибридизуются с N-ген-кодирующими последовательностями или их комплементами, как указано ранее, и которые кодируют протеин N функционального гомологичного гена, который является медиатором устойчивости к TMV в растении семейства solanaceae, входят в объем изобретения. Такие зонды включают те, которые содержат полный N-ген, и те, которые содержат один или более из следующих доменов: 5'- и 3'-нетранслируемые участки; сигнальный домен (aa 8 до 150); богатый лейцином повторяющийся участок (aa 591-929). Такие олигонуклеотиды получают стандартными способами и собирают известными специалистам способами. Длина используемого зонда должна быть достаточной для гибридизации с гомологичными участками ДНК, в которых гибридизация связана с по крайней мере около 70% гомологичности, в противоположность неспецифическому связыванию. Примеры ДНК-последовательностей, пригодных в качестве олигонуклеотидных зондов, представлены в примере 5 далее.
Специфически представленный протеин N-гена Nicotiana qlutinosa характеризуется с точки зрения его аминокислотной последовательности в последовательности ID N 4, а соответствующая специфически представленная кодирующая последовательность представлена в последовательности ID N 3, с нуклеотида 60 до нуклеотида 3494.
Биологам хорошо известно, что некоторые аминокислотные замещения можно осуществить в последовательностях протеинов, не влияя на функции протеинов. Обычно консервативные аминокислотные замещения или замещения аналогичных аминокислот осуществляются, не затрагивая при этом функций протеина. Аналогичными аминокислотами могут быть те, которые аналогичны по размеру и/или заряду, например аспартат, и глутамат, и изолейцин, и валин являются парами аналогичных аминокислот. Сходство между парами аминокислот было оценено специалистами рядом способов. Так например, Dayhoff et al., (1978) в Atlas of Protein Sequence and Structure Y.5, Supplement 3, Chapter 22, paqe 345-352, что включено сюда по ссылке, предлагает таблицу частот для аминокислотных замещений, которые можно использовать как меру сходства аминокислот. Эта частотная таблица основана на сравнениях аминокислотных последовательностей для протеинов с одинаковыми функциями из многообразия эволюционно различных источников.
Аминокислотная последовательность протеина может быть, а может и не быть идентична аминокислотной последовательности, которая встречается в природе в Solanaceous растениях. Идентичность протеина N-гена может быть подтверждена по его способности придавать устойчивость к TMV в растениях или клетках растений, которые синтезируют протеин N-гена. Такой анализ описывается в примере 1 далее. Короче, последовательность, кодирующую протеин N-гена, трансформируют в растение или в клетку растения, обладающие способностью синтезировать протеин N-гена, из указанной последовательности, например, растение семейства Solanaceae. Трансформированное растение или клетку растения инфицируют TMV. Растение наблюдают на предмет наличия гиперчувствительной реакции. Если наблюдается устойчивость, тогда протеин обладает способностью придавать устойчивость к TMV. Кроме того, искусственно индуцированные мутации можно включать до тех пор, пока они не нарушают активность. Выражение "мутированный N-протеин" относится к протеину, который обладает такой активностью, но который получен за счет мутации ДНК, кодирующей N-протеин. Под выражением "полученный за счет мутации" подразумевают как непосредственное физическое производное из ДНК, кодирующей протеин N-гена исходного материала, полученное, например, сайт специфическим мутагенезом, так и косвенное производное, полученное в результате синтеза ДНК, содержащей последовательность, родственную, но намеренно отличную от последовательности N-гена. Так средство для конструирования олигонуклеотидов необходимой длины доступны, такие ДНК можно сконструировать полностью или частично из их отдельных составляющих нуклеотидов.
Как было указано ранее, доступность последовательностей, кодирующих протеин N-гена табака, делает доступными функциональные гомологи N-гена из других Solanaceous растений, то есть ген, который содержит часть, кодирующую "N - подобный" протеин, определен как полипептид, который обладает функцией придания устойчивости к вирусному растительному патогену, такому как TMV. Эти N-подобные гены можно идентифицировать и выделить за счет сходства их ДНК- последовательности (гомологичности) с N-кодирующей последовательностью табака, предложенной здесь. кДНК- и геномные библиотеки можно скринировать за счет гибридизации для существенно гомологичных последовательностей. Эти последовательности можно затем секвенировать для подтверждения наличия полных открытых считывающих форм, клонированных в растительный вектор, так, чтобы их можно было экспрессировать в растение, и ткани растения можно было трансформировать. Трансгенные растения можно получить, используя известные специалистам способы, и эти трансгенные растения можно тестировать для подтверждения того, что устойчивость к патогену была приобретена за счет введения N- подобный протеин кодирующей последовательности. N-подобные гены включают L-ген из перца, Tm2 и Tm2a из томата, и N' из Nicotiana sylvestris.
Другим аспектом изобретения являются генетически сконструированные рекомбинантные молекулы нуклеиновых кислот, т.е. не встречающиеся в природе молекулы нуклеиновых кислот, предпочтительно содержащие часть кодирующего N-ген протеина, или функциональный гомолог N-гена, который обладает функцией придания устойчивости к TMV растениям, которые синтезируют N-ген или его функциональный гомолог соответственно. Рекомбинантная ДНК- молекула относится к гибридной ДНК-последовательности, содержащей по крайней мере две ДНК-последовательности, причем первая последовательность обычно не встречается в природе вместе со второй последовательностью. Такие молекулы можно получить при манипуляции с генетическим материалом, используя рестрикционные энзимы, лигазы и аналогичные рекомбинантные методики, описанные у Sambrook et al., Ausubel et al., см. ранее, DNA Cloninq: A Practical Approach, Volumes I и II (ed. D. N. Glover) IRL Press, Oxford, 1985. Примеры включают такие рекомбинантные векторы, как векторы клонирования или экспрессии, которые содержат ДНК-последовательность, кодирующую протеин гена, которая ориентирована в 5'- до 3'-направлении (смысловом) или в 3'- до 5' -направлении (антисмысловом). Пример 7 (далее) описывает получение рекомбинантной молекулы ДНК N-гена. Термин "рекомбинантный" в том смысле, как здесь использован, не относится к встречающимся в природе генетическим рекомбинациям.
Генетическое конструирование означает, что результат был получен в результате деятельности человека. Генетически сконструированное растение, содержащее конкретную ДНК-молекулу, является растением, в которое ДНК было введено с помощью любых известных специалистам средств, включая, но не ограничиваясь Aqrobacterium -опосредствованные трансформации, электропорацию, бомбардировку частицами и т.п. Генетически сконструированная молекула нуклеиновой кислоты, такая как генетически сконструированная молекула ДНК, является молекулой, которая представляет продукт процесса молекулярной биологии, включая (но не ограничиваясь) ДНК-лигирование, in vitro-мутагенез или т.п.
ДНК-последовательности настоящего изобретения пригодны для получения рекомбинантных ДНК- экспрессионных молекул за счет клонирования последовательности в любой подходящий экспрессионный вектор, который способен вводить чужой ген в такого гетерологичного хозяина, как бактерия, дрожжи, вирус или его организм-хозяин, или в растения. Рекомбинантный вектор конструируют так, чтобы кодирующая последовательность была расположена в векторе с соответствующей контрольной последовательностью и была бы с ней операционно связана, то есть положение и ориентация последовательности, кодирующей ДНК гена, по отношению к контрольным последовательностям были бы такими, чтобы кодирующая последовательность транскрибировалась бы под контролем контрольных последовательностей (т.е. под РНК полимеразой, которая присоединяется кДНК-молекуле у контрольных последовательностей). Контрольные последовательности могут быть лигированы с кодирующей последовательностью перед встраиванием в вектор. В другом варианте кодирующую последовательность можно клонировать непосредственно в вектор экспрессии, который уже содержит контрольную последовательность, и соответствующий рестрикционный сайт, расположенный в прямом направлении от контрольной последовательности. Этот вектор нужно выбирать так, чтобы он имел промотор, операбельный в клетке хозяине, в которую должен быть встроен вектор (то есть промотор должен распознаваться РНК- полимеразой клетки хозяина).
Кроме того, вектор должен иметь участок, который кодирует рибосомный связывающий сайт, расположенный между промотором и сайтом, по которому встроена ДНК-последовательность, так, чтобы быть операбельно ассоциированным с последовательностью, кодирующей N-ген, раз он встроен. Этот вектор следует выбирать так, чтобы он обеспечивал участок, который кодирует рибосомный связывающий сайт, распознаваемый рибосомами клетки хозяина, в которую нужно встроить вектор.
Рекомбинантная ДНК экспрессионная молекула, содержащая последовательность, которая кодирует протеин N-гена, в 5'- 3'-ориентации, встроена в клетку хозяина для экспрессии протеина N-гена. Для получения протеина специалистам известны различные экспрессионные системы и хозяева. Примером прокариотных хозяев может служить Escherichia Coli. Большое число рекомбинантных систем было разработано для экспрессии в эукариотных хозяев, включая дрожжи, клетки насекомых, клетки млекопитающих и клетки растений. Эти системы хорошо охарактеризованы и требуют лигирования кодирующей последовательности под контролем подходящей инициирующей транскрипцию системы (промотора), и при желании, терминационных последовательностей и энхансеров. Для получения протеина N-гена клетки хозяина, трансформированные рекомбинантной ДНК-экспрессионной молекулой, выращивают и протеин выделяют из клеток хозяина. Выбор соответствующих условий роста и способы выделения находятся в компетенции специалистов.
Далее приводится пример экспрессии протеина N-гена в Escherichia coli. Последовательность, кодирующую ДНК N-гена, встраивают в такой вектор экспрессии, как pRSET (Invitroqen Corp. CA), или pET (Novaqen, WI). Затем последовательность, кодирующую протеин N- гена, экспрессируют под контролем сильных транскрипционного и трансляционного сигналов бактериофага Т7.
Особенно полезными в связи с N-генами настоящего изобретения являются экспрессионные системы, которые операбельны в растениях. Кодирующие последовательности для протеина N-гена и ДНК, которая представляет обратный транскрипт мРНК, то есть последовательно транслирована в протеин N-гена, может быть включена в экспрессионную систему, подходящую для растений. Для экспрессии в растения рекомбинантная экспрессионная кассета должна содержать помимо N-ген кодирующей последовательности растительный промоторный участок (если в последовательности его нет), сайт инициирования транскрипции (если в кодирующей последовательности, которую следует транскрибировать, его нет), и последовательность терминации транскрипции. Терминационный участок можно получить из того же самого гена, что и промоторную последовательность, или можно получить из других генов. Уникальные рестрикционные сайты у 5'- и 3'-концов кассеты обычно включены для того, чтобы облегчить встраивание в существовавший ранее вектор. Растительные экспрессионные системы могут быть системами, которые находятся под контролем тканеспецифического промотора, а также теми, которые включают промоторы, которые операбельны во всех тканях растений.
Участки инициирования транскрипции, например, включают участки инициирования различных опинов, таких как октопин, маннопин, нопалин и т.п. Можно также использовать такие растительные вирусные промоторы, как 35S-промотор вируса мозаики цветной капусты (CaMV). Кроме того, можно также использовать такие растительные промоторы, как рибулоза-1,3-дифосфат-карбоксилаза, специфические для фруктов промоторы, промоторы термошока, семя- специфические промоторы и т.д. Конкретно выбранные промоторы должны быть способны приводить к достаточной экспрессии, чтобы продуцировать эффективное количество протеина N-гена для придания клеткам растения и растениям из них регенерированным устойчивости к инфицированию за счет TMV. Было показано, что CaMV 35S-промотор является высоко активным во многих органах растения и на многих стадиях развития, если включен в геном трансгенного растения. Тканеспецифические промоторы также хорошо известны. Предпочтительно, чтобы в молекулах, управляющих экспрессией TMV устойчивости за счет N-гена, трансприционные терминационные сигналы были предусмотрены в прямом направлении от и операбельно связаны с кодирующим участком протеина N-гена. Терминационным сигналом может быть тот, который обычно присутствует в N-гене, или один или более из гетерологичных транскрипционных терминационных сигналов может быть предусмотрен в прямом направлении от N- кодирующего участка. Специалистам известен ряд транскрипционных терминационных сигналов, например, те, которые соответствуют Aqrobacterium tumefaciens - Т-ДНК-генам, включая, но не ограничиваясь nos.
Полученные в результате экспрессионные системы или кассеты лигируют в (или конструируют каким-либо другим образом) в рекомбинантные векторы, которые подходят для трансформации растений. Вектор также содержит обычно селектируемый маркерный ген, за счет которого трансформированные растительные клетки можно идентифицировать в культуре. Обычно маркерный ген кодирует устойчивость к антибиотику. Эти маркеры включают устойчивость к G418, гигромицину, блеомицину, канамицину и гентамицину. После трансформации растительных клеток, те клетки, которые содержат вектор, можно идентифицировать по их способности расти на среде, содержащей конкретный антибиотик. Репликационные последовательности бактериального или вирусного происхождения обычно также включают, чтобы обеспечить клонирование вектора в бактериального или фагового хозяина, предпочтительно широкого круга хозяев прокариотного происхождения репликации. Следует также включать селектируемый маркер для бактерий, чтобы обеспечить отбор бактериальных клеток, содержащих целевую конструкцию. Подходящие прокариотные селектируемые маркеры также включают устойчивость к таким антибиотикам, как канамицин или тетрациклин.
Устойчивость к TMV за счет N-протеина была продемонстрирована в трансгенных растениях, в которые был введен геномный N-клон, где растения были TMV восприимчивы до генетической модификации. Клоны кДНК, кодирующие N-протеин, также можно использовать для придания TMV-устойчивости восприимчивым Solanaceous растениям. кДНК клонируют в прямом направлении от промотора и она операбельно связана с промотором, который функционален в растительных клетках, и вводят в ткань растения, а затем регенерируют трансгенные растения, используя векторы и способы, легко доступные специалистам. Устойчивость к вирусу подтверждают, тестируя за счет инокуляции заражающего вируса, например, TMV. Это может оказаться полезным, если использовать кДНК для введения как полной длины (последовательность IД N 3) и усеченной (последовательность IД N 5) кДНК в ткани растения после операбельного связывания каждой из последовательностей с транскрипционными контрольными последовательностями, функциональными в растительных клетках. И снова TMV устойчивость подтверждается за счет TMV инокуляционного тестирования.
Как известно специалистам, в векторе могут присутствовать другие ДНК-последовательности, кодирующие дополнительные функции. Так, например, в случае Aqrobaterium-трансформацией, Т-ДНК- последовательности также будут включены для последующего переноса в хромосомы растения.
Когда получают соответствующий вектор, получают трансгенные растения, которые содержат нужную экспрессионную систему. Последовательности, кодирующие N-ген, встраивают в подходящий растительный трансформационный вектор для трансформации в нужный вид растений, особенно в растение семейства solanaceae, для придания растению устойчивости к TMV. Кроме табака (Nicotiana, например, N.tabacumn N.qluttosa (подходящими пищевыми растениям являются растения семейства solanaceae. Они включают томаты (Lysopersicon, например, L. lycopersicum and L.esulentum); перец (Capsicum), картофель (solanum tuberosum), баклажаны (Solanum melonqena).
Специалистам известен ряд способов трансформации растений или клеток растений. Для трансформации, осуществляемой за счет бактериальной инфекции, растительные клетки инфицируют Aqrobacterium tumefaciens или A.rhizoqenes, трансформированными ранее подлежащей введению ДНК. Aqrobacterium представляют представительные гены грам-отрицательного семейства Rhizobiaceae. Гетерологичные генетические последовательности можно вводить в подходящие растительные клетки с помощью Ti-плазмид A. tumefaciens или Ri-плазмид A.rhizoqenes. Ti- или Ri - плазмиду переносят в клетки растений, инфицированных Aqrobacterium и стабильно интегрируют в геном растения (J.Schell, Science 237:1176-1183-1987)). Ti- и Ri-плазмиды содержат два участка, которые существенны для продуцирования трансформированных клеток.
Конструирование рекомбинантных Ti- и Ri-плазмид обычно следует способам, которые обычно используют с более общими бактериальными векторами, такими как puC 19. Существуют два класса рекомбинантных Ti- и Ri-векторных систем, которые в настоящее время используют. В одном классе, который называют "коинтегративным" челночный (шаттл) вектор, содержащий представляющий интерес ген, встраивают за счет генетической рекомбинации в не-онкогенную Ti-плазмиду, которая содержит как цис-действующие, так и транс-действующие элементы, необходимые для трансформации растения, как, например, в pMLJ1-шаттл-векторе DeBlock et al., EMBO J. 3:1681-1689 (1984) и не-онкогенную Ti- плазмиду pG 3850, описанную Zambryski et al., EMBO J. 2:2143-2150 (1983). Во втором классе, или "бинарной" системе, представляющий интерес ген встраивают в шаттл-вектор, содержащий цис-действующие элементы, необходимые для трансформации растения. Другие необходимые функции предусмотрены в транс за счет не-онкогенной Ti-плазмиды, как, например, pBIN19-шаттл-вектор, описанный Bevan, Nucleic Acids Research 12:8711-8721 (1984) и не- онкогенная Ti-плазмида PAL 4404, описанная Hoekema et al. Nature 303:179-180 (1983). Некоторые из этих векторов коммерчески доступны.
Существует две общие схемы трансформации растительных клеток за счет Aqrobacterium: совместное культивирование Aqrobacterium с культивируемыми выделенными протопластами, и трансформация интактных клеток или тканей с Aqrobacterium. Первая требует установленной культуральной системы, которая позволяет культивировать протопласты, и последующую регенерацию растения из культивируемых протопластов. Второй способ требует, (а) чтобы интактные растительные ткани, например семядоли, можно было трансформировать за счет Aqrobacterium, и (в) чтобы трансформированные клетки или ткани можно было бы индуцировать для регенерации в целое растение. Большинство двудольных видов можно трансформировать с помощью Aqrobacterium, так как все виды, которые являются природными растительными хозяевами для Aqrobacterium, трансформируемы in vitro.
Другая процедура для клонирования и трансформации включает клонирование N-ген кодирующей последовательности в Т-ДНК-вектор pMDI между CAMV - 35S - промотором и NOS-терминаторным участком. Растения, содержащие герминальные Ac-акты иссечения, трансформируют по способу Horsch et al. Science 227: 1229-1231 (1985) с модификациями (Hell Molecular General Genetics 217:53-59 (1989)). Эта процедура подробно описана далее в примере 1.
Известно, что трансформация за счет Aqrobacterium tumefaciens эффективна с членами семейства Solanaceae растений и особенно полезна. Другие способы трансформации, такие как электропорация, микроинъекции или пушка, липосомы и химикалии, которые увеличивают захват свободной ДНК, также можно использовать. Идентификацию трансформированных клеток или растений обычно дополняют, включая селектируемый маркер в трансформирующий вектор или получая доказательство успешного бактериального инфицирования. Растительные клетки, которые были трансформированы, также можно регенерировать, используя известные способы.
Регенерация растений семейства Solanaceae подробно описана у Horch et al. , 1985. Регенерация растений из культивируемых протопластов описана у Evans et al., Hanbook of Plant Cell Cultures Yoll: (MacMillan Publishinq Co. New York, 1983); и y Vasil I.R. (ed). Cell Culture and Somatic Cell Genetics of Plants, Acad. Press, Orlando, Vol.1, 1984 и Vol.II, 1986). Известно, что практически все растения можно регенерировать из культивируемых клеток или тканей.
Средства для регенерации меняются в зависимости от вида растений, но обычно вначале получают суспензию трансформированных протопластов или трансформированные эксплантаты в чашках петри. Образуется каллюсная ткань, и образование побегов можно индуцировать с последующим образованием корней. В другом варианте можно индуцировать образование соматического эмбриона в каллюсной ткани. Эти соматические зародыши развиваются как природные эмбрионы с образованием растений. Культуральная среда обычно содержит различные аминокислоты и гормоны растений, такие как ауксин и цитокинины. Эффективная регенерация зависит от среды, от фенотипа и от истории культуры. Если контролировать эти три переменные, тогда регенерация оказывается репродуцируемой и повторяющейся. Регенерированные растения переносят в условия обычной почвы и выращивают обычным способом.
После того, как экспрессионная кассета стабильно включена в регенерированное трансгенное растение, ее можно перенести в другие растения путем полового скрещивания. Можно использовать любую из ряда стандартных методик, в зависимости от подлежащих скрещиванию видов. Затем растения выращивают и собирают урожай как обычно.
Примеры
Нижеследующие примеры приведены лишь с целью иллюстрации изобретения, и они не предназначены для того, чтобы ограничить 1 заявленный объем
изобретения. В
этих примерах использовано множество методик, хорошо известных и доступных специалистам молекулярной биологии при манипуляции рекомбинантными ДНК в растительных тканях и культурах и
при регенерации
трансгенных растений. Энзимы получают из коммерческих источников и используют в соответствии с рекомендациями заявителей или другими вариантами, известными специалистам. Реагенты,
буферы и условия
культивирования также известны специалистам. Ссылки на стандартные процедуры молекулярной биологии включают Sambrook et al., (1989) Molecular Cloninq, 2-d ed., Cold Sprinq Harbor
Laboratory, Plainview,
NY; R. Wu (ed) (1983) Methods in Enzymology 218; Wu et al (eds) Methods in Enzymology 100, 101; Glover (ed) (1985); DNA Cloninq Voll and II IRK Press, Oxford, UK; and Hames and
Hiqqins (eds) (1985)
Nucleic Acid Hybridisation. IRL Press, Oxford, UK. Ссылки относительно манипуляций трансформации тканей растений включают Kunq and Arntzen (eds) (1989) Plant Biotechnology,
Butterworths, Stoneham, MA;
R.A.Dixon (ed) (1985) Plant Cell Culture: Practical Approach, IRL Press, Oxford, UK; Schuler and Zielinski (1989) Methods in Plant Molecular Biology, Acadevic Press, San
Dieqo, CA; Weissbach and
Weissbach (eds) (1988) Acadevic Press, San Dieqo, CA; I.Potrykus (1991) Ann.Rev. Plant Physiol. Plant Mol. Biol. 42:205; Weisinq et al (1988) Annu Rev. Genet. 22: 421; van
Wordraqen et al (1992) Plant
Vol. Biol. Rep. 19:12; Davey et al (1989) Plant Mol. Biol. 13:273; Walden and Schell (1990) Eur. J.Biochem. 192: 563; Joersbo and Brunstedt (1991) Physiol. Plant. 81:256,
а также ссылки, цитированные
в этих ссылках. Сокращения и номенклатура там, где используются, стандартны в этой области и обычно используют в профессиональных журналах, таких, как те, которые
цитированы. Все ссылки, приведенные в
настоящей заявке, включены сюда по ссылке.
Пример 1
Этот пример описывает выделение нестабильного HR-мутанта. Короче, мутации N-локуса
были выделены с помощью клонирования за
счет транспозона Ac. Затем мутанты, неспособные вырабатывать TMV- зависимые HR, были выделены за счет позитивной схемы селекции, при которой происходит отбор
TMV-инфицированных N-содержащих растений,
которые потеряли способность вырабатывать TMV зависимую HR(HR-мутанты). Растения, гомозиготные для их HR-мутаций, были идентифицированы и была
идентифицирована мутантная линия, содержащая нестабильный
HR.
U1-штамм TMV (подарок от M.Zaitlin) был размножен в TMV-восприимчивом (nn) культиваре табака (cv).Petite Havana SRI именуемом SRl-табаком. За исключением инокуляций, которые проводили для скринирования (см. далее), TMV- инокуляции осуществляли следующим образом: инокулюм получают, разбавляя сок из размоченных, TMV-инфицированных SR1-листьев табака примерно в 10 раз в стерильной воде. Губку, насыщенную раствором сока, используют при протирании верхней поверхности листьев растений на шести стадиях развития листа. Растения оценивают спустя 48 часов на предмет локальных поражений и с интервалами в 1 неделю после инокуляции на предмет следов системной инфекции (мозаики) и/или секторов некроза.
Для выделения трансгенного табака, содержащего активные Ac-транспозоны, TMV-устойчивого табака, cv.Samsun NN трансформируют PGV 3850 HPT:pKU3 (Baker et al. The EMBO J.6: 1547- 1554 (1987)) по способу Horsch et al.(Science 227: 1229-1231 (1985)), используя модифицированную процедуру (Hehl and Baker, Mol. Gen. Genet. 217:53-59 (1989)). Вектор трансформации PGV 3850 HPT: pKU3 содержит неомицинфосфотрансферазу 11 (NPT11), прерванную за счет Ac. После введения pGV 3850:pKU3 в растение табака Ac иссекают из дефективного NPT11 гена, получая в NPT11 экспрессию и рост трансформантов на среде, содержащей канамицин.
Короче, диски листьев получают из стерильных 6-8 недельных TMV-устойчивости табаков cv. Samsun NN - растеньиц, выросших на MS-среде. Диски листьев инкубируют в присутствии Aqrobacterium tumefaciens, содержащих pGV3850HPT: pKU3 или контрольные Ti- плазмидные векторы в течение 2-4 дней. Диски листьев промывают MS-средой, содержащей 3% сахарозы и 500 мг/л Цефотаксима (Calbiochem, La Jolla, CA) и помещают на MS-среду, содержащую 3% сахарозы, 0,5 мг/л ВАР (6 бензиламинопурина), 0,1 мг/л NAA(нaфтaлинукcуcнoй кислоты), 500 мг/л Цефотаксима и 200 мг/л канамицина или 20 мг/л гигромицина. Через 2-3 недели корни последовательно переносят в ту же среду, но содержащую 2 мг/л ВАР. Через 1-2 недели корни снова переносят в ту же самую среду, но не содержащую гормонов для индуцирования корней. Растения переносят в почву через 10-15 дней. Трансгенные каллюсы регенерируют на 100 мг/л канамицина для отбора трансгенных тканей, содержащих транспозонные Ac-элементы (Baker et al., 1987, см. ранее). Геномную ДНК выделяют из KnR - первичных трансгенов, которые называют ТО-генерацией.
Как показано, Ac очень активен в растениях TO-3 (pGV 3850 HPT:pKU3) на основании устойчивости к 100 мг/л канамицина и возрастании числа копий Ac при определении в Саузернблоттинге.
Растение ТО-3 скрещивают с Savsun NN. Три экземпляра T1-потомства из скрещивания, Т1-9, 10 и 13, как определено, содержат транспозонные Ac-элементы, и их скрещивают с TMV-восприимчивым культиваром табака Petite Havana SR1 (SR1), в результате чего получают три F1Nn::Ac-популяции, которые скринируют на предмет потери TMV-зависимой реакции гиперчувствительности. Для установления эндогенной нестабильности N была создана также популяция без Ac за счет скрещивания Samsun NN и SR1. SR1 используют в качестве источника пыльцы во всех кроссах.
Для выделения HR-мутантов примерно 64000 Nn::Ac и 29000 Nn-семян (см. табл. 1 в конце описания) высевают в количестве примерно 2000 семян на ящик при плотности примерно 3 сеянца/см2. Сеянцы в возрасте 8 недель помещают в условия 30oC и инокулируют суспензией TMV и Целита (Fisher, Pittsburqh, PA), используя распылитель для художников (Paasche VL) (R.W. Fulton, Nicotiana: Procedures for Experimental Use paqe 79-86 (1979)). Концентрация TMV оказалась достаточной для нанесения локальных поражений с кажущейся плотностью 1,0/см2 на Samsun NN-сеянцах при плотности их примерно 3/см2 в условиях 24oC. TMV был выделен из инфицированных SR1-листьев по способу Lane (Methods in Enzymology 118:687-691/1986)). Через три дня после инокуляции (dpi) сеянцы переносят в условия с 31oC в 21oC. При 5 dpi сеянцы оценивают на выживание, затем второй и третий циклы TMV- инокуляции и температурные сдвиги осуществляют для обеспечения 100% инокуляции.
Две патогенные для растений бактерии, Psendomonas Syrinqae pv. fomato (P. s. t.) штамм DС 3000 и P.s.pv.phaseolicola (P.s.p), штамм PN53121, и непатогенный P.s.t. штамм DC 3000 hrpS::Th5 (подарок от B.Staskawicz) суспендируют в дважды дистиллированной воде в концентрации 1•108 клеток/мл. Каждую из бактериальных суспензий или водный контроль вводят шприцом (10 мл, 200 игла) (Klement, In Methods in Phytobacteriology (ed al) Akdemiae Kiado, Budapest, Hunqary, 101- 102 (1990)) в один из четырех сайтов на нижней стороне отдельного листа. Используют три растения из каждого из следующих генотипов: Nt-1G/q собственного потомства от 9НК-мутантов, два TMV-восприимчивых (SR1 и Xanthi) и два TMV-устойчивых (Samsun NN и Xanthi nc) культивара табака. Проводят оценку листьев на предмет их реакции на четыре различных обработки через 48 часов после инокуляции.
Схема позитивного отбора позволяет выделить мутанты, неспособные вызвать TMV- зависимую HR среди крупной популяции Nn- сеянцев. Схема отбора мутантов использует подавление HR- экспрессии на N-содержащих растениях при инфицировании TMV и в условиях температуры выше 28oC. Растения, содержащие функциональный N-ген, не образуют локальных поражений при температуре выше 28oC, и TMV распространяется системно по всему растению. Подавление HR обратимо, и TMV-инфицированные растения, содержащие N, развив летальные системные некрозы (системные HR), если температура снижается до допустимых 24oC. Это представляет позитивный отбор мутантов, так как только те растения, которые утратили способность вызывать TMV зависимую HR (HR-мутанты) должны выжить.
Таким образом было выделено 47 HR-мутантов из гетерозиготных (Nn) F1-сеянцев, полученных из четырех независимых кроссов между Samsun NN или тремя NN: : Ac-родителями и SR1-табаком. TMV- инфицированные HR-растения были получены из общего количества 93000 F1-сеянцев. Из 29000 сеянцев из Samsun NN - контрольного кросса было выделено 11 мутантов, тогда как 36 мутантов было выделено из 64000 сеянцев из трех NN::Ac- кроссов (таблица 1). Частота потери устойчивости к TMV была одинаковой для Nn-потомства Samsun NN и NN::Ac при 3,8•10-4 и 5,6•10-4 соответственно. Способность получать HR-мутанты с одинаковой частотой в Nn популяции с или без Ac указывает на то, что частота эндогенных мутаций N очень высока.
Для определения того, являются ли HR-мутанты дефективными в общей способности вызывать HR, потомство 9 мутантов, включая C2- 2, инокулируют двумя бактериальными патогенами, которые, как известно, вызывают HR на табаке. Патогенные бактерии, Psendomonas syrinqae pv. tomato (P.s.t.) штамм DC 3000 и P. s. pv. phaseolicola (P. s.p.) штамм NP 53121, вырабатывают HR во всех случаях, тогда как непатогенный Р. s.t. штамм DC 3000 hrpS::Th5, и водный контроль - не вызывают. Эти результаты показывают, что HR-мутанты не теряют общей способности вызвать HR к бактериальным патогенам и что HR-фенотип, вероятно, специфичен для TMV реакции устойчивости.
Для идентификации растений, гомозиготных для их HR- мутаций, потомство самоопыления 15 путантов было исследовано на молекулярном уровне. ДНК выделяют из 27-64 потомства самоопыления каждого мутанта, переваривают EcoR1 и гибридизуют с N-связанным Nt-1 RFLP-зондом (Hehland Baker, The Plant Ceel 27:709-721 (1990)). Nt-1 идентифицирует RFLT, Nt-1G который интрогрессирован в TMV-устойчивый культивар табака Samsun NN из N.qlutinosa. Nt-1G заменяет свой Nt-1T гомолог в Samsun NN и соответствует ≤0,25 см N-локуса. Предполагают, что мутантные линии, указанные в таблице 2, являются гомозиготными для их HR-мутаций, так как они гомозиготны для прочно связанных Nt-1G-маркера или гомозиготны для делений Nt-1G-маркера.
Отличительной особенностью Ac- индуцированных мутаций является их частая нестабильность. Стабильность HR-фенотипа исследуют в потомстве самоопыления 15 гомозиготных мутантных линий. От 95 до 150 членов потомства каждой из линий инокулируют TMV и оценивают их фенотип. Потомство одной мутантной линии, D11-1, демонстрирует нестабильность HR-фенотипа с высокой частотой. Из 145 D11-1-оцененных растений 20 оказались TMV-устойчивыми (TMVR), а 68 оказались TMV- восприимчивыми (TMVS ). Интересно, что 57 растений дали секторы некрозов на TMV-восприимчивом фоне (TMVR/S фенотип) (таблица 2). Имитирующие поражения мутанты также демонстрируют секторы некрозов. Некрозы на имитирующих поражения мутантах обычно выражаются спонтанно в отсутствии абиотичных или биотичных факторов, которые вызывают некротические реакции (V.Walbot et al., Genetic Engineering of Plants pp. 431-442 (1983)). Секторы некрозов, наблюдающиеся у D11-1-потомства, и других популяций, использованных в описанных исследованиях, отличимы от имитирующего поражения фенотипа, так как они зависят от TMV-инфекции. Идентификация TMVR- и TMVR/S-индивидуумов в этой популяции указывает, что HR-мутация нестабильна. Фенотипы TMVR и TMVR/S не наблюдаются в потомстве остальных 14 мутантных линий (табл. 2 в конце описания).
На фиг.1 представлены три различных фенотипа, наблюдающихся в этой нестабильной мутантной линии (DД11-1-потомство) после инокуляции TMV. Лист на фиг. 1A получен из TMV-устойчивого растения и демонстрирует характеристики поражений TMV-устойчивого (HR+) дикого типа или ревертантного растения. Лист на фиг. 1B получен от TMV-восприимчивого растения и демонстрирует участки светло- и темно-зеленого цвета (мозаика). Лист на фиг. 1C демонстрирует TMVR/C-фенотип, что определяется участками некрозов и мозаикой. В отличие от TMVR-листа, некроз TMVR/S не ограничивается дискретными поражениями. Лист TMVR/S, представленный здесь, демонстрирует небольшие некротические пятна, однако встречаются растения, на которых участки некроза охватывали половину листа, целый лист и было видно, что они поднимаются вверх по стеблю. Наблюдение TMVR и TMVR/S -фенотипов в потомстве D11-1 демонстрирует, что HR-мутация в этой мутантной линии нестабильна.
Пример 2
Этот пример описывает тест для определения того, связан ли фенотип
TMVR/S с двумя Ac-транспозонами, которые разделены N-связанным RFLP
маркерным Nt-1G.
Если нет других указаний, для ДНК-ДНК-гибридизации, мишеневую ДНК очищают и переваривают одной или более из рестрикционных эндонуклеаз. Затем переваренную ДНК фракционируют по размерам за счет агарозного электрофореза, получают блоты на Nytran-мембране (Schleicher and Schuell, Keene, NH). Гибридизационные зонды получают, используя статистический гесамерный праймер, и метят |32P| -dCTP и полимеразой Кленова. Стандартные условия для жесткой гибридизации: 42oC в присутствии 50% формамида, 5 х SSC, 5 х раствор Денхардта с промывками при 65oC, используя 0,1 х SSC, 1% (вес/об) додецилсульфат натрия (SDS).
Стандартные условия для мягкой гибридизации: 35oC, используя 50% формамид, 5 х SSC, 5 х раствор Денхардта при 50oC промывки, используя 0,1 х SSC, 1%.
Для выделения N-связанного Nt-1G RFLP ДНК-фрагменты, выделенные как сайты вставок транспозонных Ac-элементов из SPI, были использованы для RFLP-анализа (Hehl and Baker, Mol. Gen. Genet. 217:53-59) 198936 Hehl and Baker, The Plant Cell 2: 709-721 (1990)). Один ДНК-фрагмент, обозначенный Nt-1, детектирует RFLP между TMVS-табака cv SRVR и TMVR-табака культиваром Samsun NN. Фиг.2А демонстрирует результат гибридизации 1,2 кb BqIII/HindIII Nt-1 - фрагмента с EcoR1 переваренной геномной ДНК из трех диплоидных видов табака N. qlutinosa (источник N гена), N.sylvestris и N.tomentosiformis (фиг.2А, полосы 1,4 и 5) и двух N.tabacum культиваров Samsun NN и SRI (фиг.2А полосы 2 и 3). Nt-1 детектирует RFLP специфические для каждого из диплоидных видов табака. Фрагмент ДНК 13,1 кb присутствует в Samsun NN, SRI N.Sylvestris (фиг. 2А, полосы 2,3 и 4). Фрагмент ДНК 15,5 кb присутствует в N.tomentosiformis и SRI (Фиг.2А, полосы 5 и 3), и фрагмент ДНК 14,3 кb присутствует в N. qlutinosa и Samsun NN (фиг.2А, полосы 1 и 2). В Samsun NN отсутствует 15,5 кb. N. tomentosiformis RFLP (Nt-1T), но он содержит RFLP идентичный по размеру с 14,3 кb RFLP в N.qlutinosa (Nt-1G).
Связь между Nt-1G и N была тестирована в 420 TMVSF2-потомстве скрещивания между Samsun NN-и SP1-табаком, разделяющимся в отношении 3:1 для TMV-устойчивости и восприимчивости и 1:2:1 для Nt-1G и Nt-1T RELPS. ДНК из TMV-восприимчивых F2-растений переваривают EcoR1 и гибридизуют с Nt-1. Одно TMVS - растение содержит Nt-1G RFLP, демонстрируя, что Nt-1G весьма тесно связан с N≤0,25 сМ.
Два Ac-транспозона совместно выделены с N-связанным RFLP, Nt-1G. Если TMVR/S-фенотип зависит от мутируемых аллелей N, ожидается, что он будет выделен с молекулярным маркером, связанным с N-локусом. Совместное выделение TMVR/S-фенотипа с N-связанным Nt-1G-маркером в потомстве тестового скрещивания нестабильного HR-мутанта, C2-2 и SR1-табака было тестировано. Потомство тестового скрещивания (D111 популяция) было инокулирование TMV и была проведена оценка по их фенотипу. Из 264 D111 оцененных растений 164 оказались TMV-восприимчивыми (TMVS), хотя 80 из них демонстрируют секторы некроза на TMV-восприимчивом фоне. Дикого типа TMV-устойчивые растения не наблюдались. ДНК 80 D111 растений переваривают EcoR1 и гибридизуют с Nt-1. Определяют Nt-1 генотип растений, и 39 индивидуумов оказываются Nt-1G/T, тогда как 41 оказываются Nt-1T/T (табл. 3 в конце описания). 26 растений, которые демонстрируют TMVR/S-фенотип, имеют Nt-1G-маркер, тогда как Nt-1T/T-растения оказываются TMVS (таблица 3). Эти результаты показывают, что нестабильная HR-мутация, определяемая по способности образовывать некротические секторы, связана с Nt-1G.
Так как нестабильная HR-мутация связана с Nt-1G, было исследовано, выделяется ли Ac-транспозон совместно с Nt-1G RFLP-маркером в D111-популяции. D111-ДНК, переваренную EcoR1, гибридизуют с зондом с 5' конца Ac. Две Ac-гибридизующие полосы, обозначенные Ac8 (8,0 кb EcoR1 Ac полоса) и Ac10 (10,2 кb EcoR1 Ac полоса), были обнаружены совместно выделенными с Nt-1G. 30 Nt-1G/T-растений содержат как Ac8, так и Ac10, 5 содержат Ac*, 3 содержат Ac10, и 1 растение не содержит ни одного элемента (табл. 4 в конце описания). Ac8 и Ac10 не присутствуют в 43 Nt-1T/T-растениях, что подтверждает, что эти два Ac-транспозона связаны с Nt-1G.
Пример гибридизации по Саузерну, суммированный в таблицах 3 и 4? представлен на фиг. 2B и 2C соответственно. На фиг. 2B представлена гибридизация Nt-1 с EcoR1 переваренными ДНК 14 D111- растений. 10 растений, представленных здесь, имеют гетерозиготный, Nn, Nt-1G/T - генотип, что демонстрирует наличие 14,3 кb Nt-1G RFLP и 15,5 кb Nt-1T RFLP (полосы 2,4-11 и 14). Шесть из этих растений имеют TMVR/S-фенотип, соответствующий полосам 2, 4, 7, 9, 11 и 14. Четыре растения имеют гомозиготный, nn, генотип, Nt-1T/Т-генотип, что демонстрируется присутствием 1,5,5 кb Nt-1T RFLP и отсутствием 14,3 кb Nt-1G RFLP (полосы 1, 3, 12 и 13). Четыре растения Nt-1T/T - генотипа не имеют TMVR/S- фенотипа. Затем эти ДНК гибридизуют с 5' Ac-зондом, как представлено на фиг. 2C. Все 10 растений Nt-1G/T - генотипа содержат 8,0 к Ac-полосу (Ac8), хотя 7 из них (полосы 2, 4, 7, 8, 9, 11 и 14) содержат 10,2 кb Ac (Ac10). Растения Nt-1T/T-генотипа не содержат ни 8,0 кb, ни 10,2 кbAc RFLP, хотя они не содержат других Ac-транспозонов.
Пример 3
Этот пример описывает тест для определения того,
какой из Ac8 и Ac10 отвечает за нестабильную HR-мутацию.
Для определения того, какой из Ac8 или Ac10 вызывает нестабильную HR-мутацию, герминальный ревертант (D112-15) идентифицируют из собственного потомства HR-мутанта C2-2. D112-15 гомозиготен для Nt-1G и содержит как Ac8, так и Ac10. Из-за присутствия обоих Ac8 и Ac10 было предположено, что одна клонированная с помощью транспозона аллель N имеет герминально ревертированный в дикий тип, тогда как другая все еще содержит Ac и поэтому обладает потенциалом обратного превращения. D112-15 скрещивают с SR1 для определения того, может ли иссечение Ac8 или Ac10 коррелировать с реверсией резистентности и нестабильности HR-мутаций. Потомство этого кросса (Е501-популяция) как ожидается, должно расщепиться примерно 1:1 на ТМR и ТМS + ТМR/S и иметь Nt-1G/T генотип. Ac, который отвечает за нестабильную мутацию N, как ожидают, не должен содержаться ни в одном устойчивом потомстве этого скрещивания. 95 Е501 растений были инокулированы TMV, и проведена оценка их фенотипа. 54 оказались TMVR, некротические секторы наблюдались для 21 растения, а 20 оказались TMVS (табл. 5 в конце описания). ДНК из этих растений переваривают EcoR1 и зондируют Nt-1, а затем 5'Ac-зондом. У всех 95 растений оказался Nt-1G/T - генотип. Важно, что ни один из 54 TMV устойчивых образцов не содержал 20,2 кb EcoR1 Ac полосы, тогда как полоса 8 кb присутствовала в 52 растениях (табл. 5 в конце описания). Наличие Ac8 и отсутствие Ac10 в TMV-устойчивом Е501 потомстве определяет Ac10 как элемент, вызывающий нестабильную HR-мутацию, и, таким образом, метит N-ген.
Число Ac копий в D111 и Е501 популяциях велико, что может маскировать, вероятно, другие Ac-элементы, совместно выделяемые с Nt-1G. Было показано, что растение TMS, E501-70 содержит только Ac10. Для подтверждения того, что Ac10 один может вызвать нестабильную HR-мутацию, собственное потомство этого растения (F501-популяция) исследуют на предмет фенотипа после TMV-инфицирования и анализируют на присутствие Ac10 и их Nt-1-генотипов. 7 растений TMVR выделяют из общего количества 500 растений. Молекулярный анализ показывает, что три TMVR-растения были гетерозиготными для Nt-1G и не содержат Ac10-гибридизации, тогда как 4 TMVR-растения были Nt-1G/G и имели полосу Ac10. Как и для D112-15 растения, предполагают, что Ac-гибридизация в Nt-1G-гомозиготах связана с присутствием мутантной аллели N в этих растениях так же, как и в ревертантных растениях.
В популяциях Е501 и F501 наблюдается корреляция между присутствием Ac10 и TMR/S-фенотипом. 19 из 21 Е501- растений TMVR/S-фенотипа имеют Ac10-гибридизацию, 12 из 12 F501 ТМR/S, растений, проанализированных с молекулярной точки зрения, имеют 10,2 кb Ac-полосу. Эти результаты показывают, что наличие Ac10 необходимо для того, чтобы растения образовывали секторы некрозов и поддерживали потенциал превращения соматически в устойчивые. Ткани из разрезанных на секторы растений без Ac10- гибридизации, вероятно, имеют больше иссечений, так что 10,2 кb Ac полоса более не детектируется при гибридизации по Саузерну.
B D111 и Е501 популяциях, 2,3 кb ЕсоR1-полоса, которая гибридизована с 3'Ac-зондом, ведет себя идентично 10,2 кb 5' Ac-полосе. Учтем, что Ac составляет 4,6 bp, a EcoR1 дикого типа или иссеченный фрагмент 7,9 кb предсказан. Ожидается, что этот фрагмент сохраняется в TMVR -ревертантах. Для тестирования на предмет наличия геномной вставки и фрагментов иссечения геномные последовательности, фланкирующие Ac10, были выделены за счет IPCR из растения D111-95, которое содержит только Ac10 и Ac8 (фиг.2C, полоса 9). (см. пример 4 далее).
Геномные последовательности, фланкирующие Ac10, были выделены в результате инверсной полимеразной цепной реакции (IPCR). Матричную ДНК из растения D111-95, которая содержит только Ac8 и Ac10, переваривают Hpa11, лигируют и линеаризируют с помощью Clal. PCR-реакции осуществляют в 50 мкл, используя Taq-полимеразу (Promeqa, Madison, W1) на Перкин-Элмер-термоциклере (Emerville,CA). Параметры: 94oC-1 мин, 55oC-2 мин, 35 циклов. 419 bp-продукт (Ac10-1) 5'- до Ac10 амплифицируют, используя Ac- специфический праймер CC28 (5'-CACGGATCCATACGATAACGGTCGGTACGGGA- 3') и CC32 (5'-CACGAATTCGGAAACGGAAACGGTACAGC-3'). Для получения Ac10 3'-фланкирующей последовательности (Ac10-2), 111- 95 ДНК переваривают EcoR1, лигируют и линеаризируют за счет Accl. 122bp-продукт амплифицируют, используя праймеры CC21 (5'- CACCTGCAGAGATCTTTACCCACCGTTACCGACCG) и CC30 (CACCTGCAGAGATCTGCACCCTTATAATATAACGG-3'). Продукты IPCR клонируют в TA-кпонирующий вектор (Invitroqen, San Dieqo, CA).
400 bp-продукт с 5'-конца Ac выделяют (Ac10-1). Ac10-1 клонируют в TA-кпонирующий вектор и секвенируют. PCR-праймеры синтезируют для создания Ac10-1-зонда с Ac-последобательностями для уменьшения возможности "шпорообразной" Ac-гибридизации. При использовании в качестве зонда на геномной ДНК табака Ac10-1 детектирует повторяющиеся последовательности. Гибридизация с 10,2 кb Ac- вставкой наблюдается в ДНК D11-1, однако предсказанная 7,9 кb EcoR1-полоса иссечения неразличима из-за повторяющегося характера зонда. IPCR-клон, полученный из 3'-конца Ac10 (Ac10-2), имеет длину 111 bp и, по-видимому, нереален в качестве зонда.
Надежный зонд с большим числом копий, N-5, получают из 3'-конца (основания 5020-5370) кДНК клона C7. Он соответствует основаниям 6587-6600 и 6977-7270 последовательности ID N 1. Молекулярный анализ Е501- и F501-популяций продолжают, используя рестрикционную эндонуклеазу EcoR1. ДНК из популяций Е501 и F501 переваривают EcoR1 и гибридизуют с Ac- и N-5-зондами. Ac-зонд гибридизуют с 10,2 кb EcoR1-полосой, соответствующей Ac10 в Е501- и F501-популяциях. Ac-гибридизация для отбора индивидуумов из каждой генерации в нестабильной HR-мутантной линии представлена на фиг.3с. Ac-гибридизация не наблюдается у контрольных ДНК SR1, N.qlutinosa или Samsun NN. Исходный HR- мутант C2-2 имеет 10,2 кb - полосу в дополнение к по крайней мере двум другим Ac-транспозонам. Герминальный ревертант, D112-15 имеет 10,2 кb Ac-полосу, а также по крайней мере 10 других Ac. Е501-70, TMVS- потомство D112-15, содержит только 10,2 кb Ac10-полосу. Два герминальных ревертанта TMVR, потомство Е501-70, F501-65 и F50166, не содержат 10,2 кb-полосы. F501-65 содержит новую Ac- вставку, хотя F501-66 больше не имеет Ac-гибридизации. Секторизованные растения F501-2, 3 и 4 все еще имеют Ac10-вставку. F501-48 и F501-64 являются двумя примерами TMVS- растений, которые также не содержат больше Ac10-гибридизации. F501- 48 более не содержит Ac-гибридизации, тогда как F501-64 содержит новую вставку.
Учитывая, что Ac имеет 4, 6 кb, можно предсказать 7,9 кb EcoR1 дикого типа или иссеченный фрагмент, если N-5 используют в качестве зонда (см.фиг. 3A). Примеры гибридизации зонда N-5 представлены на фиг. 3В. N-5 гибридизуется с 7,9 кb полосой в N.qlutinosa и Samsun NN. HR-мутант, C2-2 имеет гибридизацию с 10,2 кb Ac10-вставочной полосой и слабую гибридизацию с полосой 7, 9 кb. D112-15 содержит обе полосы: 10,2 кb и 7,9 кb. Е501-70 имеет 10,2 кb вставочную полосу и некоторую гибридизацию с полосой 7,9 кb. Два герминальных ревертанта, F501- 65 и F501-66, содержат только 7,9 кb полосу. Эти растения были Nt-1G/T так, что они содержат только одну аллель N, которая была ревертирована. Другие ревертанты, такие как D112-15, которые являются Nt-1G/G имеют как вставки, так и иссеченные фрагменты. F501-2, 3 и 4 содержат как 10,2 кb, так и 7,9 кb RFLP. F501-48 и F501-64 содержат только 7,9 кb фрагмент иссечения.
Важно, что 54 TMVR 501 потомство D112-15 содержит 7,9 кb EcoR1-полосу иссечения, как и 7 TMVR F501-растения. Эти результаты показывают, что восстановление геномных ДНК- последовательностей до дикого типа необходимо для возвращения к устойчивости. Эти результаты демонстрируют также, что одна мутантная аллель N была герминально ревертирована в D112-15 и что иссечение Ac10 отвечает за восстановление функций N-гена. Эти результаты были подтверждены в анализе потомства Е01-70, содержащем только Ac10, где все 7 TMVR- растений имеют 7,9 кb-полосу иссечения. Nt-1G/T - растения демонстрируют отсутствие гибридизации, и они содержат 7,9 кb дикого типа размера геномный фрагмент.
Растения TMVR/S, за исключением двух из Е501-поколения, имеют как 10,2 кb-, так и 7,9 кb-полосы. Присутствие этих полос вместе в одной и той же ткани показывает, что присутствуют клетки с Ac10- вместо Ac10-иссечения. Каждая полоса указывает, что некоторая ткань будет либо TMVS, либо потенциальным ревертантом. Это объясняет TMVR/S-фенотип, наблюдаемый в этих исследованиях.
Пример 4
Этот пример описывает анализ последовательностей геномных
вставок и сайтов исключения.
PCR-продукты, содержащие Ac-сайты исключения, были непосредственно секвенированы. Использованные растения представлены в табл. 6 (см. в конце описания). Праймеры, фланкирующие сайт исключения N-G1-5 (основания 4477 до 4496 5'-GCCCTCGAGAAATCAAGAAAACAGAGGTC-3') и N 7-52 (основания 4838 до 4856 5'-GCACTCGACCTTCAAGATTACTATTG-3') были использованы для амплификации примерно 379 bp продукта. PCR-реакции проводят так же, как и для IPCR, за исключением того, что используют следующие параметры: 94oC - 1 мин, 55oC-2 мин и 72oC-3 мин в течение 25 циклов. PCR продукты очищают с помощью электрофореза в агарозе с низкой температурой плавления (FMC) - с последующей экстракцией фенолом. Примерно 500 фмолей каждого продукта используют для секвенирования с фмолем ДНК секвенирующей системы (Promega, Madison, W1), используя праймер N7-52. 19 из 21 TMVS- Е501-растений так же, как и 4 TMVS -F501- растений Nt-1G/T-генотипа, имеют полосу исключений, и в них отсутствует Ac10 (Ac 10(-)). Ac-свойство, которое было консервировано в табаке состоит в том, что после вставки создается прямой дубликат из восьми пар оснований, фланкирующих элемент. Это подтверждается в последовательностях Ac10-1 и Ac10-2. Ac10 фланкируется 8 bp прямым повтором 5'-ATTTGCCG-3'. Часто Ac-исключение является незначительным, и "отпечаток пальцев" остается позади. Отпечатки пальцев могут вызвать мутации сдвига рамки и/или аминокислотные вставки или делеции, которые предотвращают продуцирование функционального генного продукта.
Таблица 6 демонстрирует анализ последовательности Ac10- мишеневого сайта в N-гене и устойчивость или чувствительность (к TMV-инфекции) фенотипа, связанного с конкретным генотипом. После встраивания Ac10 последовательность из 8 bp дикого типа N- последовательности (от нуклеотида 5034 до 5041; ATTTGCCG) дублирована. Триплеты оснований в таблице 6 показывают кодоны внутри кДНК-последовательности. Дополнительные последовательности, которые остаются в восприимчивых растениях после исключения Ac10, подчеркнуты. Звездочкой отмечено существование преждевременного стоп кодона, который встречается либо на 9, либо на 4 аминокислоты в прямом направлении. Е501 являются потомством бэккросса герминального ревертантного растения D112-15, a F501 являются собственным потомством растения Е501-70. TMV-восприимчивый и устойчивый фенотипы в таблице 6 обозначены, как TMVS и TMVR соответственно. Ac10 характеризует N-ген. Сайты исключения семи Ac10 (-) TMVS - растений были секвенированы, и было обнаружено, что каждый имеет нуклеотидные изменения при сравнении с сайтами исключения дикого типа (таблица 6). Эти изменения в нуклеотидах показывают, что неправильное исключение Ac10 приводит к отпечаткам, которые вызывают мутации сдвига рамки. Предсказанные полипептиды оканчиваются на 9 аминокислот или 4 аминокислоты в прямом направлении от отпечатка (таблица 6).
Кроме того, сайты исключения D112-15 и шесть TMVR-pacтений из F501-поколения были секвенированы и было обнаружено, что они имеют последовательность дикого типа (таблица 6). Возможность обнаружить ревертантные растения без изменений оснований предполагает, что этот участок очень важен для функционирования протеина и что замещения аминокислот, дополнения или делеции не переносятся. Однако TMVS - растение с отпечатком, который мог бы позволить синтезировать полной длины, но нефункциональный протеин, до сих пор не были идентифицированы. Эти результаты, представленные здесь, демонстрируют, что Ac10 метит N-ген, и предполагают, что точное исключение Ac10 из N-гена необходимо для восстановления HR + фенотипа.
Пример 5
N - кДНК были выделены из библиотеки N.qlutinosa следующим образом, растения в возрасте 8-12 недель инфицируют TMV при
32oC. Температуру смещают до 24oC через 24 часа
после инокуляции. Листья собирают для полиаденилированного (Poly (A)+ РНК выделения через 48 часов после инокуляции.
кДНК получают из 5 мкг Poly(A)+ РНК, используя λ -Zap
кДНК- синтетический набор (Strateqene La Jolla, CA). кДНК упаковывают Giqapack 11 Gold-упаковочными экстрактами от Strateqene и
высевают на штамм-хозяин Escherichia coli XL 1-Blue mrf.
Ac10-1 используют для скринирования 1,0•106 клонов библиотеки кДНК, сконструированной из РНК TMV-инфицированной N.qlutinosa. Было показано, что 15 клонов имеют гомологию с Ac10-1; однако только один (C7) имеет последовательность на 100%, идентичную последовательности Ac10-1. Зонд (N-5) получают из оснований 5020- 5370 по 3'-концу C7, который является отдельной копией в табаке и который гибридизуется с 7,9 кb EcoR1- фрагментом. Затем 1•106 бляшек гибридизуют с - N-5-зондом. Три клона (C16, C17 и C18) выделяют во втором скринировании, и все имеют 100% идентичности с Ac10-1. Полной длины последовательности кДНК C7, C16 и C18 вставок были определены (см. далее).
Двухнитевые плазмидные ДНК были секвенированы с помощью дидеокси цепной терминации (Sanqer et al., Proc.Natl. Acad. Sci. USA 74:5463-5467 (199)), используя Sequenase систему, версию 2,0 (United States Biochemical Corporation, Cleveland, OH), а для секвенирования C7 кДНК, "гнездные" делеции были подготовлены с использованием экзонуклеазы 111 (Henikoff, Methods in Enzymology 155:156-165 (1987)). кДНК C16 и C18 были секвенированы с использованием праймеров, полученных из C7-последовательности.
Анализы последовательностей осуществляют, используя GCG-программу анализа последовательностей (Madison, W1). Были составлены карты для эксонов и интронов, кодируемых геномным N геном (см.последовательность ID N 1) на основании анализа последовательностей C7, C16 и C18 кДНК-клонов и частичного секвенирования G38 λ -клона (фиг.4В). Вместе взятые C7 и C18 позволяют предположить, что 5 экзонов сплайсированы с образованием открытой считывающей рамки из 3432 пар оснований, которые кодируют полипептид из 1144 аминокислот (N). C18 кДНК последовательность представлена в последовательности ID N 3. C16 кодирует полипептид из 652 аминокислот (Ntr), так как экзон из 70 пар оснований сплайсирован альтернативно с тем, чтобы получить усеченную открытую считывающую рамку из 1956 bp (фиг.4A и последовательность ID N 5). Этот дополнительный экзон (ЕЕ) может быть сплайсирован способом, сходным с тем, который используют для ЕДА-экзона фибронектина (M.Caputi, Nucleic Acid Research, 22: 1018-1022(1994)). Фрагменты последовательности внутри 70 bp экзона на 95% аналогичны последовательностям в ЕДА-экзоне, который определяет состоящий из двух частей энхансер, который модулирует сплайсинг этого 81 bp- экзона.
3'-конец кДНК варьируется по длине, что указывает на то, что использованы различные сигналы полиаденилирования. Множественные потенциальные сигналы полиаденилирования обнаружены в положениях в 3'-нетранслируемом участке этих кДНК-клонов, что может отвечать за различные акты процессинга. C7 имеет самый длинный 3'-конец и содержит последовательность для усеченных C16 и C18 3'-концов. C7 и C16 идентичны по 5'-концам. Последовательность C18 обнаружена полностью внутри C16 и C7, поэтому она не является полной длины кДНК и имеет самый короткий 3'-конец. C7 содержит интрон 2 и может быть из мРНК, которая была сплайсирована не полностью. В C16 и C18 интрон 2 отсутствует. В дополнении к 5' 750 bp последовательности C16 или C7 до C18 образует предсказанную открытую считывающую рамку из 3432 bp, кодирующую полипептид из 1144 аминокислот (табл. 7A в конце описания). Частичное секвенирование (G38 λ -клона показывает, что все последовательности, необходимые для получения трех типов, выделенных кДНК, присутствуют в геномной последовательности. Таким образом, эти кДНК кодируются одним геном.
Предсказанный молекулярный вес предсказанных N и Ntr-протеинов составляет 131,4 кb и 75,3 кb соответственно. В таблице 7A представлены вычисленные аминокислотные последовательности для N-генного продукта (см. также последовательность ID N 4). Потенциальный сигнальный (цитоплазмический) домен подчеркнут. Аминокислоты, сохраняющиеся среди ATP/GTP-связующего сайта фрагмента, подчеркнуты дважды. Богатые лейцином повторы (LRR1 до LRR13) изображены латинским шрифтом. 8 потенциальных сайтов N-связанного гликозилирования находятся внутри аминокислотной последовательности N. Эти сайты указаны буквами жирным шрифтом в таблице 7A и содержат консенсусную аминокислотную последовательность NX (S/T). Сокращения, которые использованы в таблицах 7A-C, для однобуквенного аминокислотного кода: A-Ala, C-Cys, D-Asp, Е- Glu, F-Phe, G-Gly, H-His, I-Ile, K-Lys, L-Leu, M-Met, N-Asn, P-Pro, Q-GIn, R-Arq, S-Ser, T-Thr, V-Val, W-Trp Y- Tyr.
Анализ последовательности N-протеина с помощью программы ualom показывает, что в N отсутствует трансмембранный участок. Более того, анализ последовательности N протеина с помощью программы "Siqnalase" показывает, что отсутствует сигнальная последовательность. Таким образом, на основании анализа последовательностей можно считать, что N, по-видимому, локализован в цитоплазме. Полученную аминокислотную последовательность N полипептида сравнивают с Genbank (выпуск 82.0), используя программу BLAST (Altschul et al., J.Vol.Bioloqy 215:403- 410(1990)). Предсказанная аминокислотная последовательность демонстрирует ограниченное, но значительное сходство с протеинами, которые, как известно, вовлечены в трансдукцию сигнала.
Предсказанная аминокислотная последовательность N содержит фрагмент Р-петли (табл. 7A в конце описания). Последовательность GMGGVGKT (аа 216-223) удовлетворяет консенсусной последовательности Р-петли (A/G)XXXXGK (S/T), найденной в различных ATP- или GTP-связующих протеинах (см.табл. 7A). Семейства протеинов, содержащие Р-петлю, включают аденилаткиназы, ras семейство протеинов, фактор удлинения, ATP-синтазную b - субъединицу, тимидинкиназу и фосфоглицераткиназы (M. Saraste et.al. Trends in Biochemical Sciences, 15: 430-434 (1990)). Эта конкретная P-петля, наиболее вероятно, должна быть вовлечена в связывание ATP. Консенсусные последовательности DXXG и NXKD для GTP- связывания в дополнении к P-петле, отсутствуют в аа последовательности (Dever et al. , Proceedinqs of the National Academy of Sciens USA 84:1814- (1987)).
Помимо Р-петли, Fry et al. , (Proceedinqs of the National Academy of Sciences USA 83: 907-911 (1986)) определил два других "сегмента", которые, по-видимому, участвуют в АТР связывании в аденилаткиназе и F1-ATPазе. Изучение N последовательности дает возможность предположить, что эти сегменты присутствуют и присутствуют в нужном месте. Сегмент 2 содержит дипептид (I, A, L, V) (V, I) и N содержит последовательность Al в положении 228 и 229 соответственно (таблица 7A). У 80-100 аминокислот из P-петли сегмент 3 определен как глицин, за которым следует участок из 5 гидрофобных аминокислот и аспарагиновая аминокислота (см. таблицу 7A). N содержит последовательность VLIVLDD у аминокислот 296-302. Из аминокислотной последовательности нет возможности предсказать, в каких условиях ATP связан, или после связывания происходит ли ATP гидролиз спонтанно, или необходимы другие факторы.
Табл. 7B (см. в конце описания) демонстрирует соответствие аминотерминальных аминокислот (потенциальный сигнальный домен) (8 до 150 в последовательности ID N 4) в цитоплазмическом (сигнальном) домене протеина Drosophila Toll (аа 804-9996: Yamaqata et al. Gene 139:223-228 (1994)) и Интерлейкин 1-рецепторном протеине человека (HIL1-R, аа 317-524; Sims et al., Proceedinq of the National Academy of Science 86:8946-8950 (1989)). Участки сходства окантованы. Использованные консервативные замещения следующие: гидрофобная аминокислота = (L/I/V/M/A/F; ионная аминокислота = K/R/D/E/Q/N/H; apoматическая аминокислота = F/Y.
Последовательность содержит некоторые из консервативных аминокислот, необходимых для передачи сигнала от цитоплазмы к ядрам в Toll, и IL1-R регуляторную схему (Schneider et al. Gene and Development 5:797-807 (1991); Hequy et al., Jornal of Biological Chemistry, 267: 2605-2609 (1992)).
Полученная аминокислотная последовательность N с аминокислот 590 до 928 последовательности ID N 4 содержит богатый лейцином участок, состоящий из 13 повторов примерно по 25 аминокислот длиной. В табл. 7C (см. в конце описания) представлена первичная структура N гена богатых лейцином повторов (LRR) (аа 590-928) и сравнение его консенсусной последовательности с LRR консенсусной последовательностью дрожжевой аденилатциклазы (AdCy, Kataoka et al. Cell 43: 493-505 (1985)), Drosophila Toll (Hashimoto et al. Cell 52:269-279 (1988)), Iba цепью мембранного гликопротеина тромбоцита человека (H Gplba, Titani et al. , Proceedinqs of the National Academy of Science 84:5610-5614(1987)), Drosophila Chaoptin (Reinke et al. ,Cell 52:291-301 (1988) Arabidopsis рецепторподобной трансмембранной киназой (TMK1, Chanq et al., Plant Cell 4:1263-1271 (1992)); TMKL1, Valon et al., (Plant Molecular Bioloqy 23:415-421 (1993)); RLK5, Walker, The Plant Journal 3:451-456 (1993)).
Богатые лейцином повторы (LRRS) обнаружены в целом ряде протеинов, которые участвуют в передаче сигнала, в клеточной адгезии, и в различных других функциях, и считают, что они осуществляют протеин-протеин взаимодействие. Консенсусная последовательность, полученная из расположенных рядом LRRS, аналогична консенсусной последовательности, обнаруженной в аденилатциклазе дрожжей (Kataora /1985/ см. ранее), Drosophila Toll (Hashimoto et al. , ) 1988, Iba-цепи мембранного гликопротеина тромбоцита человека (Titani et. al. , (1987), Drosophila Chaoptin (Reinke et al., (1988) и Arabidopsis рецепторподобной трансмембранной киназы (Chanq et al. (1992) Valon et al., (1993); и J.Walker (1993)) (таблица 7C). 3а исключением дрожжевой аденилатциклазы LRR-домен, по-видимому, должен быть внеклеточной матрицей в этих протеинах.
Пример 6
Этот пример описывает выделение геномной последовательности N-гена.
Для создания геномной библиотеки ДНК, полученную из N.qlutinosa, частично переваривают за счет Mbo1 и фракционируют по размерам с помощью электрофореза. ДНК фрагменты более 12 кb лигируют с ВамН1 переваренными, дефосфорилированными плечами бактериофага λ Gem-11 (Promeqa). Продукты лигирования упаковывают с помощью упаковочных экстрактов Giqapack 11 Gold (Strataqene, La Jolla, CA) и 1•106 бляшкообразующих единиц высевают на SURE E.coli штамм, полученный от GIBCO, BRL., Gaithersburq, МД) штамм-хозяин.
Для выделения геномных последовательностей N-гена библиотеку бактериофага λ из Mool, разделенной N.qlutinosa ДНК, скринируют зондами N-5 и N-9 (нуклеотиды 695-1090 из C7). Три клона, которые гибридизуются как с N-5, так и с N-9, выделяют. Эти клоны (G25, G38) рестрикционно картируют с помощью EcoR1, ВамН1 и Xho1 и осуществляют Саузернблоттинг (фиг.4В). Было определено, что три клона перекрываются и содержат полный геномный N-ген. G34 содержит на 1,4 кb больше ДНК в обратном направлении от N гена, нежели G38, тогда как G38 на 5,4 кb длиннее в направлении 3' от гена. Последовательности G25 клона содержатся в G38 клоне.
Пример 7
Этот
пример описывает восприимчивость мутантных растений,
трансформированных геномными клонами N-гена, причем эти растения были устойчивыми к TMV.
SR1 табака и TMV-восприимчивые растения F501-48 и F501-64, содержащие герминальные Ac-эксцизионные акты, трансформируют за счет либо pTG34, либо pTG38. pTG34 и рТG38 были сконструированы за счет субклонирования 12,0 кb или 10,6 кb Xho1 фрагмента, соответственно, из перекрывающихся ДНК клонов G34 или G38 в Т-ДНК вектор, pOCA38 (Olsewski et al., Nucleic Acids Research 16: 10765-10781 (1988)), который был линеаризован за счет Sall и обработан щелочной фосфатазой кишечника теленка. Трансформации осуществляли, как было указано, для pGVHPT:pKU3 конструкции (см. пример 1).
Клоны G34 и G38 были выбраны для геномной комплементации для определения того, достаточно ли этого гена для придания TMV-устойчивости TMV-восприимчивым растениям табака. Эти два клона, по-видимому, более вероятно, нежели G25, содержат все цис-последовательности, которые необходимы для правильной экспрессии трансгена, так как они имеют более удлиненные 5'-концы. 10,6 кb G38 и 12,0 кb G34 Xho1 фрагменты были субклонированы в Т-ДНК вектор pOCA28 и трансформированы в табак за счет трансформации через Agrobacterium.
Оба клона pTG34 и pTG38 придают TMV-устойчивость восприимчивым к TMV растениям SR1 табака и мутантам F501-48 и F501-64.
Фиг. 1Д демонстрирует, что TMV-восприимчивые SR1-растения, трансформированные клонированной ДНК N-гена N. glutinosa (клон pTG-38) приобретают устойчивость к TMV.
Реферат
Молекулы нуклеиновой кислоты могут быть использованы для получения растений, устойчивых к вирусу табачной мозаики. Устойчивость к вирусу достигается трансформацией растений нуклеотидными последовательностями, которые кодируют протеин N-гена. Протеин N-гена распознает патоген и инициирует сигнальный каскад трансдукции, приводящий к экспрессии устойчивости к ВТМ за счет взаимодействия с его компонентом. 4 с. и 12 з.п.ф-лы, 13 ил., 9 табл.

Комментарии