Способ репликации или амплификации кольцевой днк - RU2752611C2
Код документа: RU2752611C2
Чертежи

Описание
Область техники
[0001] Настоящее изобретение относится к способу репликации или амплификации кольцевой ДНК. Более конкретно, настоящее изобретение относится к способу, позволяющему осуществлять эффективную репликацию или амплификацию кольцевой ДНК в бесклеточной системе. Настоящее изобретение также относится к нуклеиновой кислоте, которая может быть использована в качестве функционального кластера для получения кольцевой ДНК.
Уровень техники
[0002] Технология клонирования ДНК, на которой основано развитие биотехнологии, представляет собой метод амплификации кольцевой ДНК, которая была получена путем разрезания и лигирования фрагментов ДНК в виде плазмиды в клетках E. coli и т.п. Применение технологии клонирования ДНК, в которой используются клетки для амплификации кольцевой ДНК, требует проведения трудоемких процедур, таких как культивирование клеток, экстракция/очистка амплифицированных продуктов и т.п. Кроме того, проведение экспериментов по такому клонированию ДНК ограничено условиями окружающей среды, поскольку для проведения клонирования ДНК с использованием клеток необходимо получить генетически модифицированные организмы.
[0003] Наиболее распространенным методом, применяемым для амплификации ДНК in vitro, является полимеразная цепная реакция (ПЦР). Однако, амплификация ДНК in vitro с помощью ПЦР не позволяет осуществлять амплификацию кольцевой ДНК. Методы амплификации кольцевой ДНК in vitro включают амплификацию по типу «катящегося кольца» (RCA) (NPL 1, PTL 1, PTL 2, PTL 3). Однако, при амплификации кольцевой ДНК по типу «катящегося кольца» каждый раз необходимо конструирование праймера, специфичного к ДНК-мишени. Кроме того, продуктом амплификации, который образуется непосредственно в результате амплификации по типу «катящегося кольца», является линейная ДНК, а поэтому необходимо проводить дополнительную стадию циклизации полученного продукта амплификации, такую как инкубирование с ферментом, осуществляющим рекомбинацию. Другим описанным в литературе методом является метод получения мономерного продукта репликации путем репликации минихромосомы E. coli (кольцевой ДНК oriC), а затем их разделения с получением мономерного кольцевого продукта репликации (NPL 2-5). Однако, что касается условий проведения реакций, описанных в этих публикациях, то было экспериментально продемонстрировано, что эффективность репликации молекул кольцевой ДНК при добавлении матричной ДНК составляет лишь приблизительно 15-40%, и таким образом, амплифицируемое количество не достигает даже двухкратного увеличения (NPL 3-6). Кроме того, размер кольцевой ДНК, используемой в качестве матрицы, как описано в этих публикациях, составляет лишь менее, чем 10 т.п.о.
[0004] Как показано выше, амплификация кольцевой ДНК посредством стандартной амплификации ДНК in vitro имеет те недостатки, что она требует конструирования праймеров, связанных с матричной ДНК, продуцирования линейной ДНК в качестве продукта амплификации и ограничения размера амплифицируемой ДНК несколькими т.п.о. Кроме того, существует проблема, заключающаяся в том, что при продуцировании кольцевого продукта амплификации с использованием системы репликации минихромосомы Escherichia coli, матричная кольцевая ДНК не может амплифицироваться даже с двухкратным увеличением.
Список цитируемых документов
Патентные документы
[0005] PTL 1: Публикация нерассмотренной заявки на патент Японии No. 2005-229950
PTL 2: Публикация нерассмотренной заявки на патент Японии No. 2008-161182
PTL 3: Публикация нерассмотренной заявки на патент Японии No. 2012-501173
Патентная литература
[0006] NPL 1: Fakruddin M et al., J Pharm Bioallied Sci. 2013, 5: 245-252
NPL 2: Peng H & Marians KJ. PNAS. 1993, 90: 8571-8575
NPL 3: Hiasa H & Marians KJ. J Biol Chem. 1994, 269: 32655-32659
NPL 4: Funnell B et al., J Biol Chem. 1986, 261: 5616-5624
NPL 5: Hiasa H et al., J Biol Chem. 1994, 269: 2093-2099
NPL 6: Hiasa H & Marians KJ. J Biol Chem. 1994, 269: 26959-26968
Сущность изобретения
Техническая проблема
[0007] Настоящее изобретение относится к способу эффективной репликации или амплификации кольцевой ДНК в бесклеточной системе. Настоящее изобретение также относится к нуклеиновой кислоте, которая может быть использована в качестве функционального кластера для получения кольцевой ДНК.
Решение проблемы
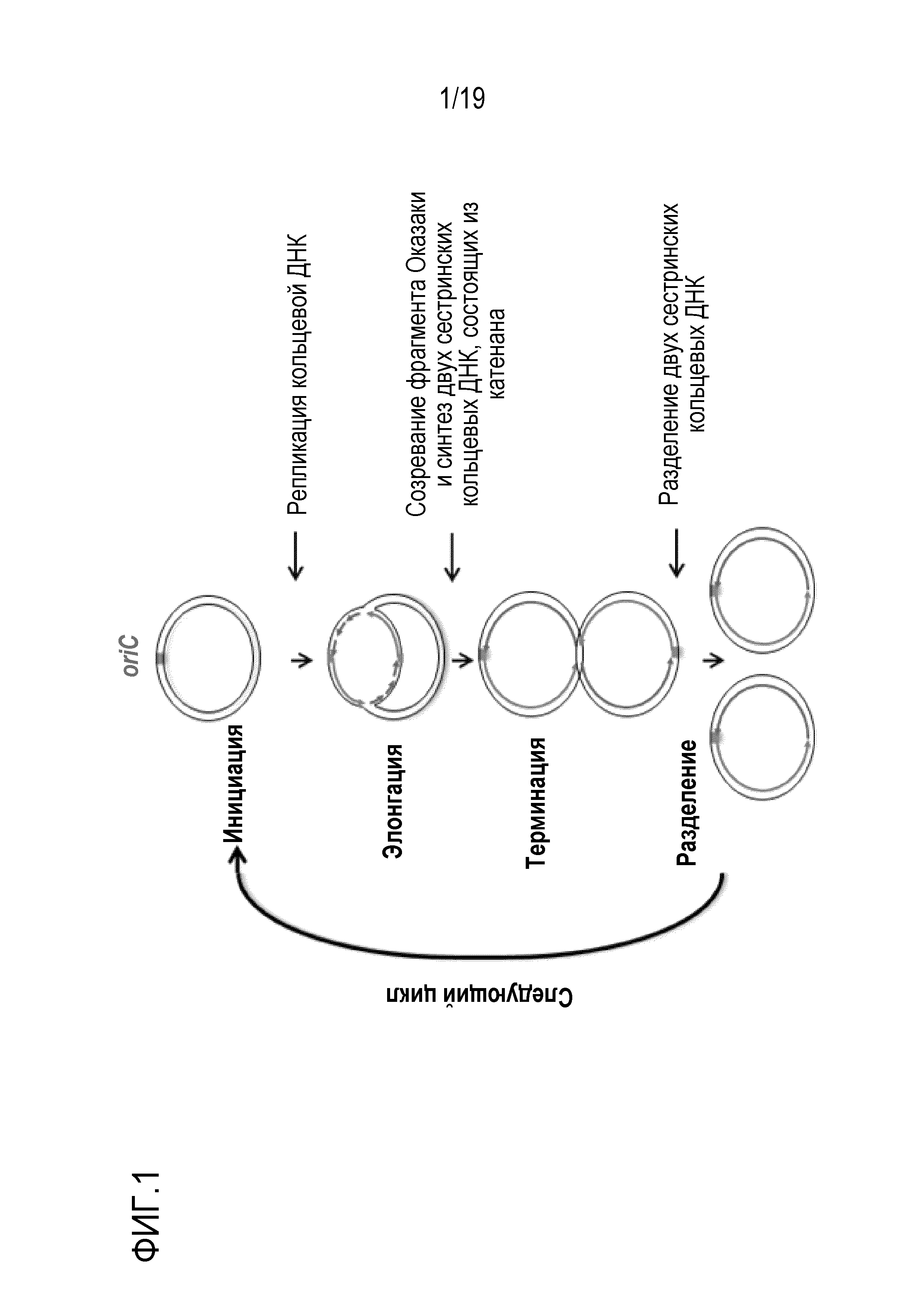
[0008] Авторами настоящего изобретения были проведены интенсивные исследования для решения вышеуказанной проблемы, и было обнаружено, что если кольцевая ДНК, имеющая последовательность ориджина репликации (ориджина хромосомы (oriC)), реплицируется или амплифицируется с использованием нижеследующих групп ферментов:
(1) первой группы ферментов, которые катализируют репликацию кольцевой ДНК;
(2) второй группы ферментов, которые катализируют созревание фрагмента Оказаки и синтезируют две сестринские кольцевые ДНК, состоящие из катенана; и
(3) третьей группы ферментов, которые катализируют разделение двух сестринских кольцевых ДНК, то образование ДНК-мультимера в качестве побочного продукта может подавляться по механизму терминации репликации с использованием ter-Tus, и/или по механизму разделения ДНК-мультимеров с использованием сайт-специфической системы рекомбинации, такой как dif-XerCD. Кроме того, авторы настоящего изобретения также обнаружили, что даже в случае, когда кольцевая ДНК, не содержащая oriC, присутствует в крайне низкой концентрации, то кольцевая ДНК может реплицироваться или амплифицироваться посредством введения oriC в кольцевую ДНК с использованием транспозона.
[0009] В настоящем изобретении, реакция репликации или амплификации кольцевой ДНК с использованием вышеописанных групп ферментов (1), (2) и (3), в некоторых случаях, будет обозначаться «RCR» (реакция цикла репликации)».
[0010] Кроме того, в настоящем описании, термин «ДНК-мультимер» означает мультимерную ДНК, полученную после репликации или амплификации кольцевой ДНК. В данном случае, мультимерная ДНК означает, что рассматриваемая ДНК является мультимеризованной, если кольцевая ДНК, используемая в качестве матрицы, определена как мономер. В настоящем описании, ДНК-мультимер иногда просто называется «мультимером».
[0011] Другими словами, настоящее изобретение охватывает нижеследующие аспекты, которые включают, но не ограничиваются ими:
[0012] [1] Способ репликации кольцевой ДНК в бесклеточной системе, включающий следующие стадии:
(1) образования реакционной смеси кольцевой ДНК в качестве матрицы с реакционным раствором, содержащим:
первую группу ферментов, которые катализируют репликацию кольцевой ДНК;
вторую группу ферментов, которые катализируют созревание фрагмента Оказаки и синтезируют две сестринские кольцевые ДНК, состоящие из катенана; и
третью группу ферментов, которые катализируют разделение двух сестринских кольцевых ДНК; и
(2) реакции взаимодействия реакционной смеси, полученной в стадии (1), где:
кольцевая ДНК включает последовательность ориджина репликации (ориджина хромосомы (oriC)), которая может связываться с ферментом, обладающим DnaA-активностью, а также включает пару последовательностей ter, каждая из которых была встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров, где:
если кольцевая ДНК имеет последовательности ter, то реакционный раствор в стадии (1) также включает белок, обладающий активностью ингибирования репликации посредством связывания с последовательностями ter, и если кольцевая ДНК имеет нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров, то реакционный раствор в стадии (1) также содержит фермент для разделения ДНК-мультимеров.
[0013] [2] Способ в соответствии с вышеуказанным [1], где ферментом для разделения ДНК-мультимеров является Cre или XerCD.
[0014] [3] Способ репликации кольцевой ДНК в бесклеточной системе, включающий следующие стадии:
(1) образования реакционной смеси кольцевой ДНК в качестве матрицы с реакционным раствором, содержащим:
первую группу ферментов, которые катализируют репликацию кольцевой ДНК;
вторую группу ферментов, которые катализируют созревание фрагмента Оказаки и синтезируют две сестринские кольцевые ДНК, состоящие из катенана; и
третью группу ферментов, которые катализируют разделение двух сестринских кольцевых ДНК; и
(2) реакции взаимодействия реакционной смеси, полученной в стадии (1), где:
кольцевая ДНК включает последовательность ориджина репликации (ориджина хромосомы (oriC)), которая может связываться с ферментом, обладающим DnaA-активностью, а также включает пару последовательностей ter, каждая из которых была встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую XerCD, где:
если кольцевая ДНК имеет последовательности ter, то реакционный раствор в стадии (1) также включает белок, обладающий активностью ингибирования репликации посредством связывания с последовательностями ter, и если кольцевая ДНК имеет нуклеотидную последовательность, распознаваемую XerCD, то реакционный раствор в стадии (1) также содержит белок XerCD.
[0015] [4] Способ в соответствии с любым из вышеуказанных [1]-[3], где пара последовательностей ter, каждая из которых встроена за пределами oriC, содержит: последовательность, включающую любую из последовательностей, представленных в SEQ ID NN: 1-14, которая встроена как одна последовательность ter с 5'-концевой стороны oriC; и последовательность, включающую последовательность, комплементарную любой из последовательностей, представленных в SEQ ID NN: 1-14, которая встроена как другая последовательность ter с 3'-концевой стороны oriC.
[0016] [5] Способ в соответствии с любым из вышеуказанных [1]-[4], где белок, обладающий активностью ингибирования репликации посредством связывания с последовательностями ter, представляет собой белок Tus или белок RTP.
[0017] [6] Способ в соответствии с вышеуказанными [2] или [3], где нуклеотидная последовательность, распознаваемая XerCD, представляет собой последовательность, содержащую любую из последовательностей, представленных в SEQ ID NN: 15-24, или комплементарную ей последовательность.
[0018] [7] Способ в соответствии с вышеуказанным [2], где нуклеотидная последовательность, распознаваемая Cre, представляет собой последовательность, содержащую любую из последовательностей, представленных в SEQ ID NN: 30-35 или комплементарную ей последовательность.
[0019] [8] Нуклеиновая кислота, которая представляет собой линейную ДНК, имеющую длину от 273 п.о. до 2,0 т.п.о., и содержащую oriC, и пару последовательносей ter, каждая из которых встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров.
[0020] [9] Нуклеиновая кислота, которая представляет собой линейную ДНК, имеющую длину от 273 п.о. до 2,0 т.п.о., и содержащую oriC, и пару последовательносей ter, каждая из которых встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую XerCD.
[0021] [10] Способ репликации кольцевой ДНК в бесклеточной системе, включающий следующие стадии:
(1) получения кольцевой ДНК, содержащей oriC, путем:
добавления транспозона и транспозазы oriC в буфер с образованием транспосомы oriC, где транспозон oriC представляет собой линейную ДНК, содержащую последовательность ориджина репликации (ориджина хромосомы (oriC)), которая может связываться с ферментом, обладающим DnaA-активностью, и внешние концевые последовательности (OE) у обоих концов; и
реакции взаимодействия транспосомы oriC с кольцевой ДНК, не содержащей oriC, в буфере для проведения реакции переноса,
(2) образования реакционной смеси кольцевой ДНК, содержащей oriC и полученной в стадии (1), с реакционным раствором, содержащим:
первую группу ферментов, которые катализируют репликацию кольцевой ДНК;
вторую группу ферментов, которые катализируют созревание фрагмента Оказаки и синтезируют две сестринские кольцевые ДНК, состоящие из катенана; и
третью группу ферментов, которые катализируют разделение двух сестринских кольцевых ДНК; и
(3) реакции взаимодействия реакционной смеси, полученной в стадии (2).
[0022] [11] Способ в соответствии с вышеуказанным [10], где последовательность OE содержит последовательность, представленную в SEQ ID NO: 25 (5'-CTGTCTCTTATACACATCT-3') и комплементарную ей последовательность, и где последовательность OE, содержащая последовательность, представленную в SEQ ID NO: 25, встроена у 5'-конца ДНК в стадии (1), и где последовательность OE, содержащая последовательность, комплементарную последовательности, представленной в SEQ ID NO: 25, встроена у 3'-конца линейной ДНК.
[0023] [12] Способ в соответствии с вышеуказанными [10] или [11], где кольцевая ДНК, содержащая oriC, также содержит пару последовательностей ter, каждая из которых встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров, где:
если кольцевая ДНК имеет последовательности ter, то реакционный раствор в стадии (2) также включает белок, обладающий активностью ингибирования репликации посредством связывания с последовательностями ter, и если кольцевая ДНК имеет нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров, то реакционный раствор в стадии (2) также содержит фермент для разделения ДНК-мультимеров.
[0024] [13] Способ в соответствии с вышеуказанными [10] или [11], где кольцевая ДНК, содержащая oriC, также содержит пару последовательностей ter, каждая из которых встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую XerCD, где:
если кольцевая ДНК имеет последовательности ter, то реакционный раствор в стадии (2) также включает белок, обладающий активностью ингибирования репликации посредством связывания с последовательностями ter, и если кольцевая ДНК имеет нуклеотидную последовательность, распознаваемую XerCD, то реакционный раствор в стадии (2) также содержит белок XerCD.
[0025] [14] Способ в соответствии с любым из вышеуказанных [10]-[13], где транспозон oriC в стадии (1) также содержит пару последовательностей ter, каждая из которых встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров, где:
если линейная ДНК имеет последовательности ter, то реакционный раствор в стадии (2) также включает белок, обладающий активностью ингибирования репликации посредством связывания с последовательностями ter, и если кольцевая ДНК имеет нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров, то реакционный раствор в стадии (2) также содержит фермент для разделения ДНК-мультимеров.
[0026] [15] Способ в соответствии с любым из вышеуказанных [10]-[13], где транспозон oriC в стадии (1) также содержит пару последовательностей ter, каждая из которых встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую XerCD, где:
если линейная ДНК имеет последовательности ter, то реакционный раствор в стадии (2) также включает белок, обладающий активностью ингибирования репликации посредством связывания с последовательностями ter, и если кольцевая ДНК имеет нуклеотидную последовательность, распознаваемую XerCD, то реакционный раствор в стадии (2) также содержит белок XerCD.
[0027] [16] Способ в соответствии с любым из вышеуказанных [10]-[15], также включающий:
(4) удаление транспозона oriC из кольцевой ДНК, реплицированной или амплифицированной в продукте реакции стадии (3).
[0028] [17] Нуклеиновая кислота, которая представляет собой линейную ДНК, имеющую длину от 311 п.о. до 2,0 т.п.о. и содержащую oriC и пару последовательносей ter, каждая из которых встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров, а также внешние концевые последовательности (OE) у обоих концов.
[0029] [18] Нуклеиновая кислота, которая представляет собой линейную ДНК, имеющую длину от 311 п.о. до 2,0 т.п.о. и содержащую oriC и пару последовательносей ter, каждая из которых встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую XerCD, а также внешние концевые последовательности (OE) у обоих концов.
[0030] [19] Набор для репликации кольцевой ДНК, включающий комбинацию:
первой группы ферментов, которые катализируют репликацию кольцевой ДНК;
второй группы ферментов, которые катализируют созревание фрагмента Оказаки и синтезируют две сестринские кольцевые ДНК, состоящие из катенана; и
третьей группы ферментов, которые катализируют разделение двух сестринских кольцевых ДНК;
транспозона oriC, который представляет собой линейную ДНК, содержащую последовательность ориджина репликации (ориджина хромосомы (oriC)), которая может связываться с ферментом, обладающим DnaA-активностью, и внешние концевые последовательности (OE) у обоих концов; и
транспозазы.
[0031] [20] Набор в соответствии с вышеуказанным [19], где транспозон oriC также содержит пару последовательностей ter, каждая из которых встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров.
[0032] [21] Набор в соответствии с вышеуказанным [20], также включающий:
белок, обладающий активностью ингибирования репликации посредством связывания с последовательностями ter; и/или
фермент для разделения ДНК-мультимеров.
[0033] [22] Набор в соответствии с вышеуказанным [19], где транспозон oriC также содержит пару последовательностей ter, каждая из которых встроена за пределами oriC и/или нуклеотидную последовательность, распознаваемую XerCD.
[0034] [23] Набор в соответствии с вышеуказанным [22], также включающий:
белок, обладающий активностью ингибирования репликации посредством связывания с последовательностями ter; и/или белок XerCD.
Предпочтительные эффекты изобретения
[0035] В соответствии со способом, описанным в настоящей заявке, если кольцевая ДНК, имеющая последовательность ориджина репликации (ориджина хромосомы (oriC)), реплицируется или амплифицируется с использованием нижеследующих групп ферментов:
(1) первой группы ферментов, которые катализируют репликацию кольцевой ДНК;
(2) второй группы ферментов, которые катализируют созревание фрагмента Оказаки и синтезируют две сестринские кольцевые ДНК, состоящие из катенана; и
(3) третьей группы ферментов, которые катализируют разделение двух сестринских кольцевых ДНК, то образование ДНК-мультимера в качестве побочного продукта может подавляться. Кроме того, кольцевая ДНК в крайне низкой концентрации может реплицироваться или амплифицироваться посредством введения oriC в кольцевую ДНК с использованием транспозона. Исходя из этих данных, можно сказать, что продукт репликации или продукт амплификации может быть эффективно получен способом, описанным в настоящей заявке.
Краткое описание чертежей
[0036] На фигуре 1 показана модель репликативного цикла кольцевой ДНК.
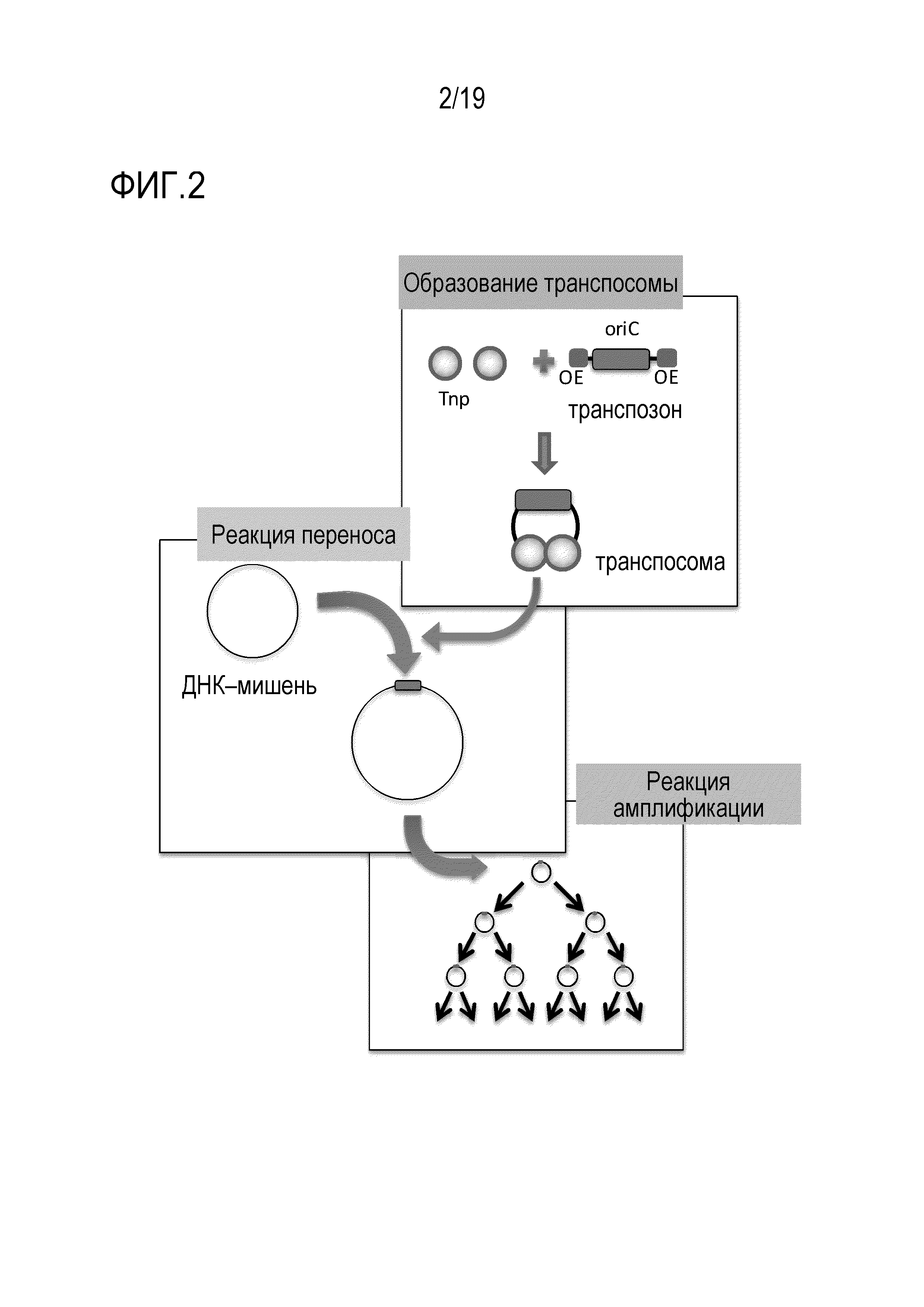
На фигуре 2 схематически представлено изображение, на котором показано введение кластера oriC с использованием транспозона и последующей реакции репликации или амплификации.
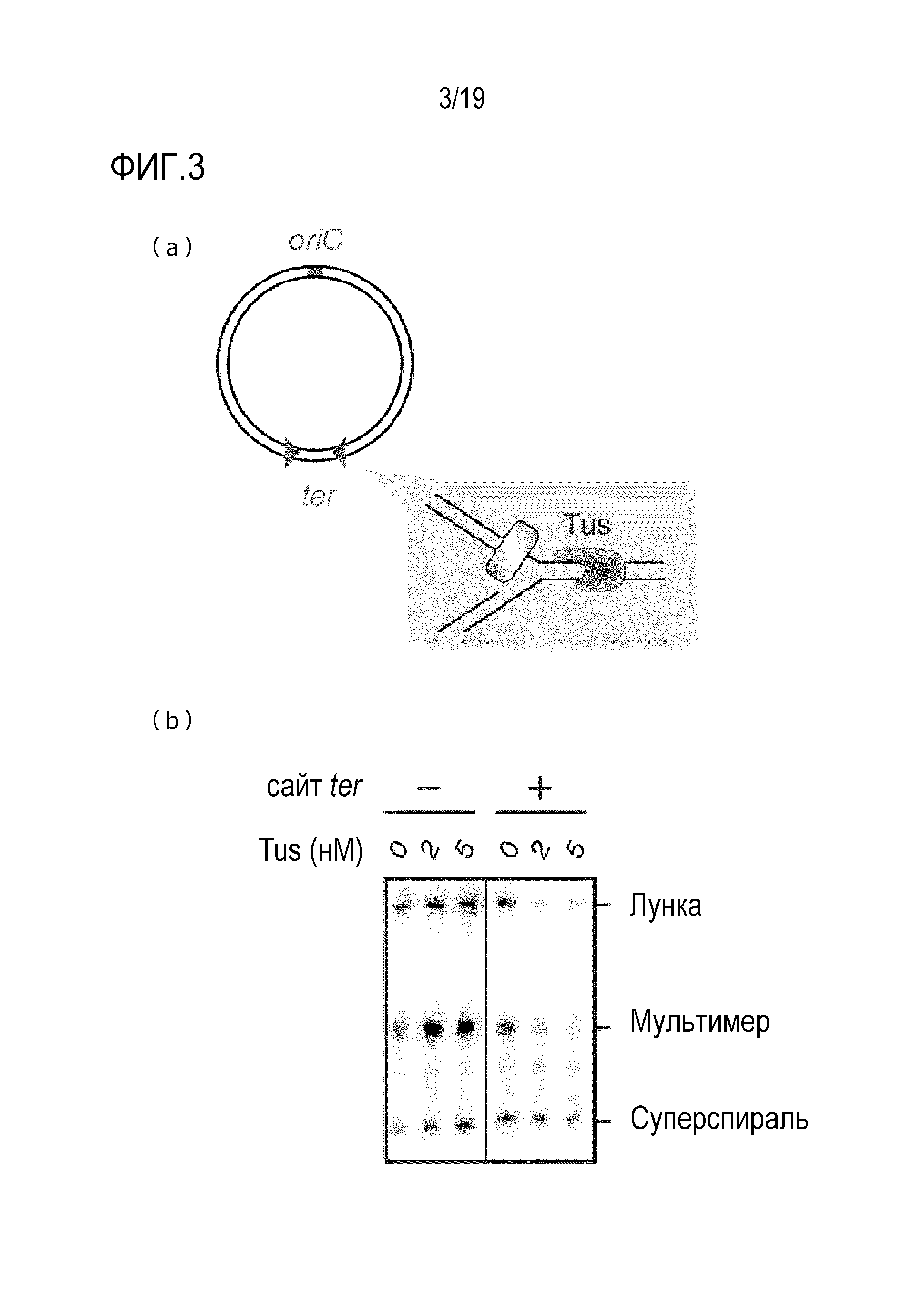
На фигуре 3 схематически представлено изображение (a), иллюстрирующее подавление образования ДНК-мультимера с использованием последовательности терминации ter и белка Tus, и фотография гель-электрофореза (b), иллюстрирующая полученные результаты.
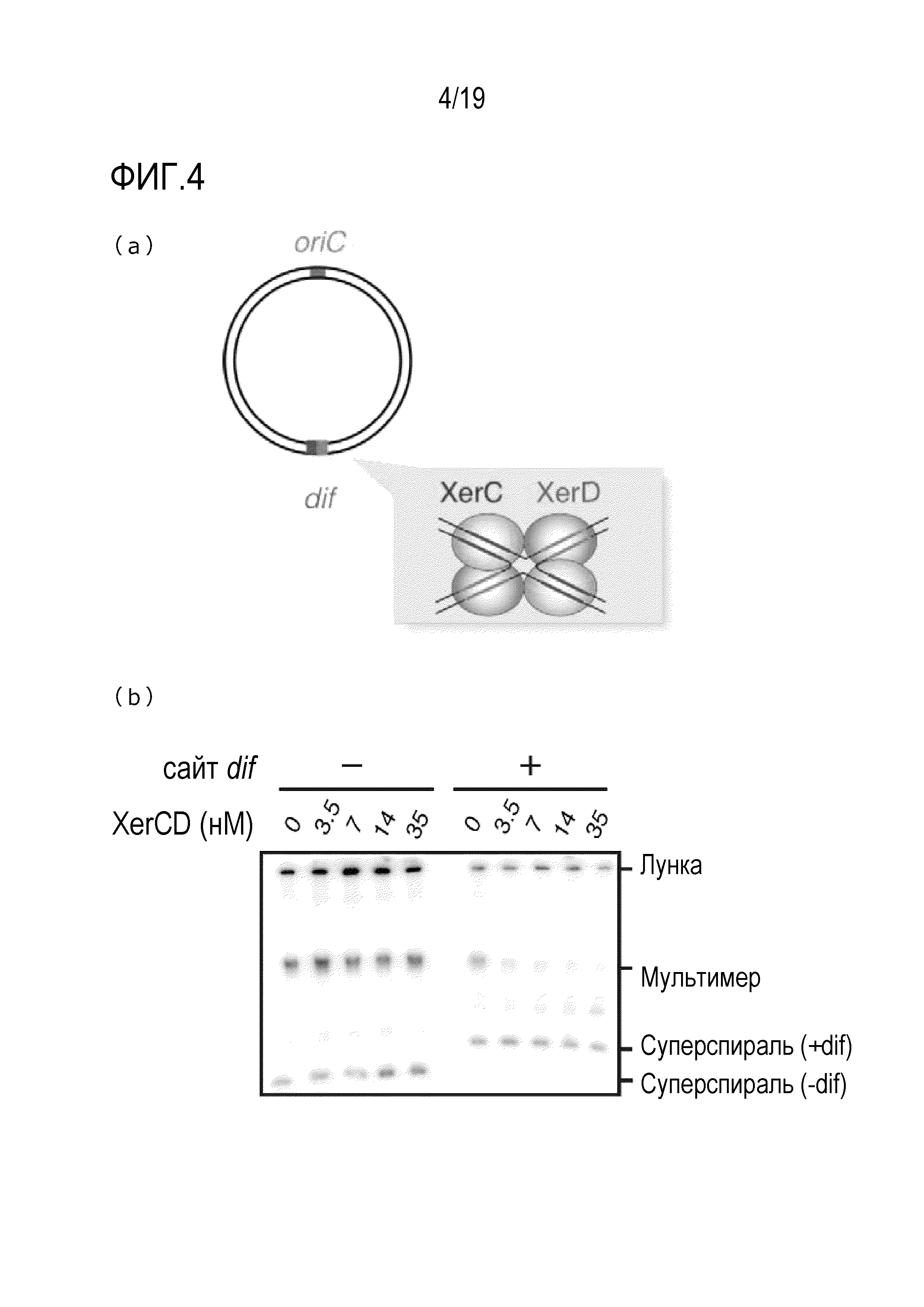
На фигуре 4 схематически представлено изображение (a), иллюстрирующее подавление образования ДНК-мультимера с использованием последовательностей сайт-специфической рекомбинации dif и XerCD, и фотография гель-электрофореза (b), иллюстрирующая полученные результаты.
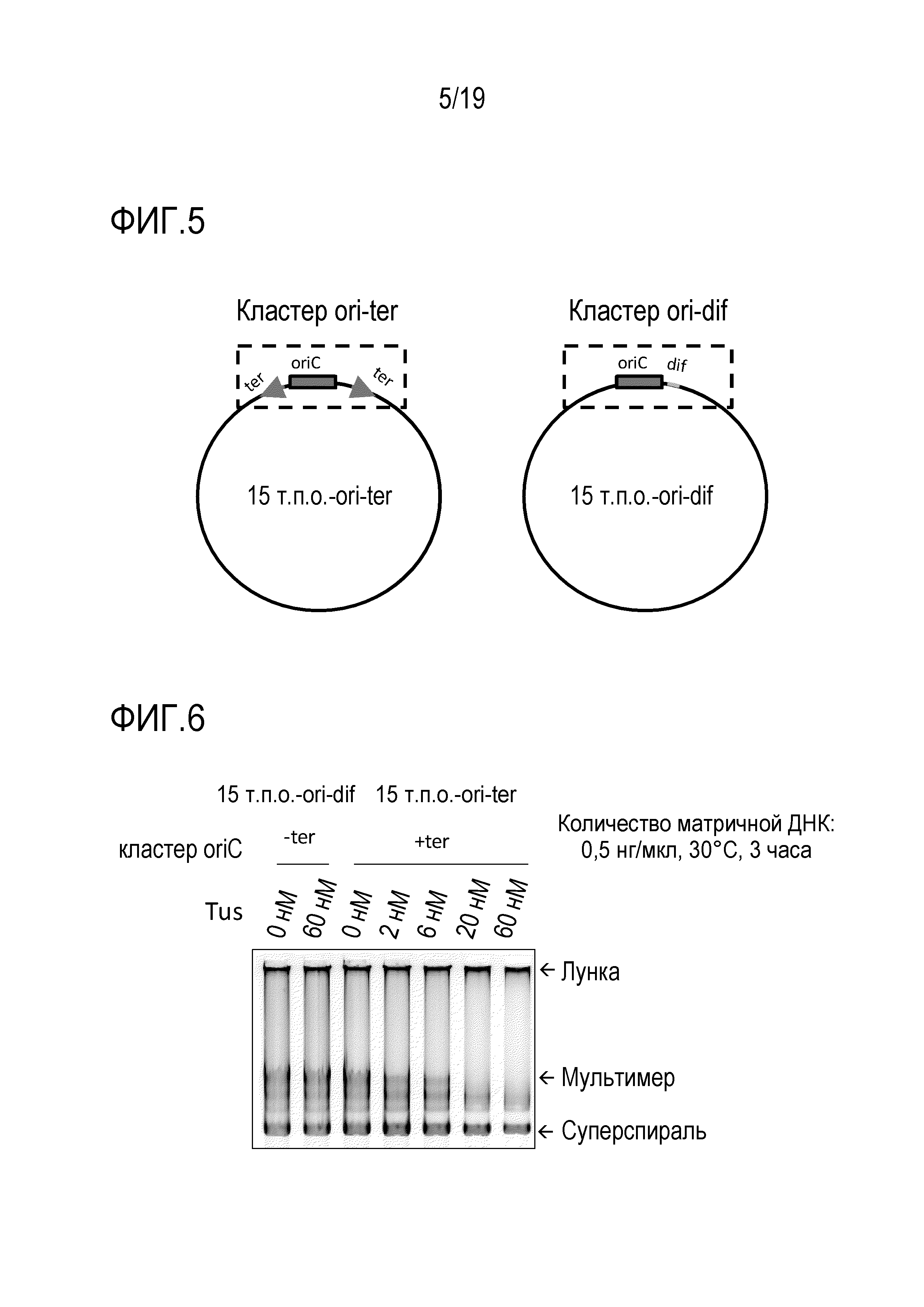
На фигуре 5 схематически представлено изображение, иллюстрирующее кольцевую 15 т.п.о. -ДНК ori-ter и кольцевую 15 т.п.о. -ДНК ori-dif.
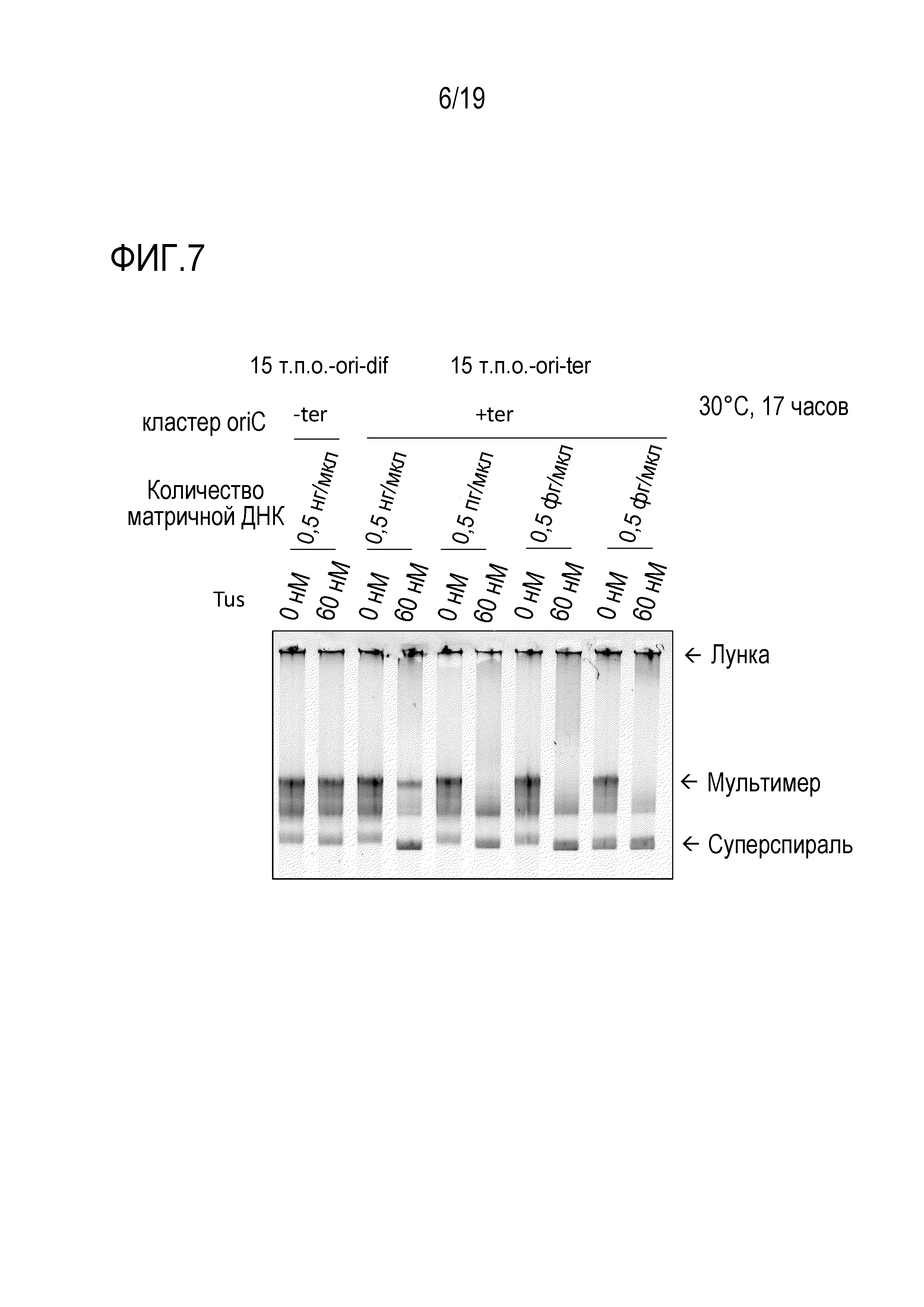
На фигуре 6 представлена фотография гель-электрофореза, иллюстрирующая результаты проведения титрования Tus на подавление образования ДНК-мультимера с использованием последовательности терминации ter и белка Tus.
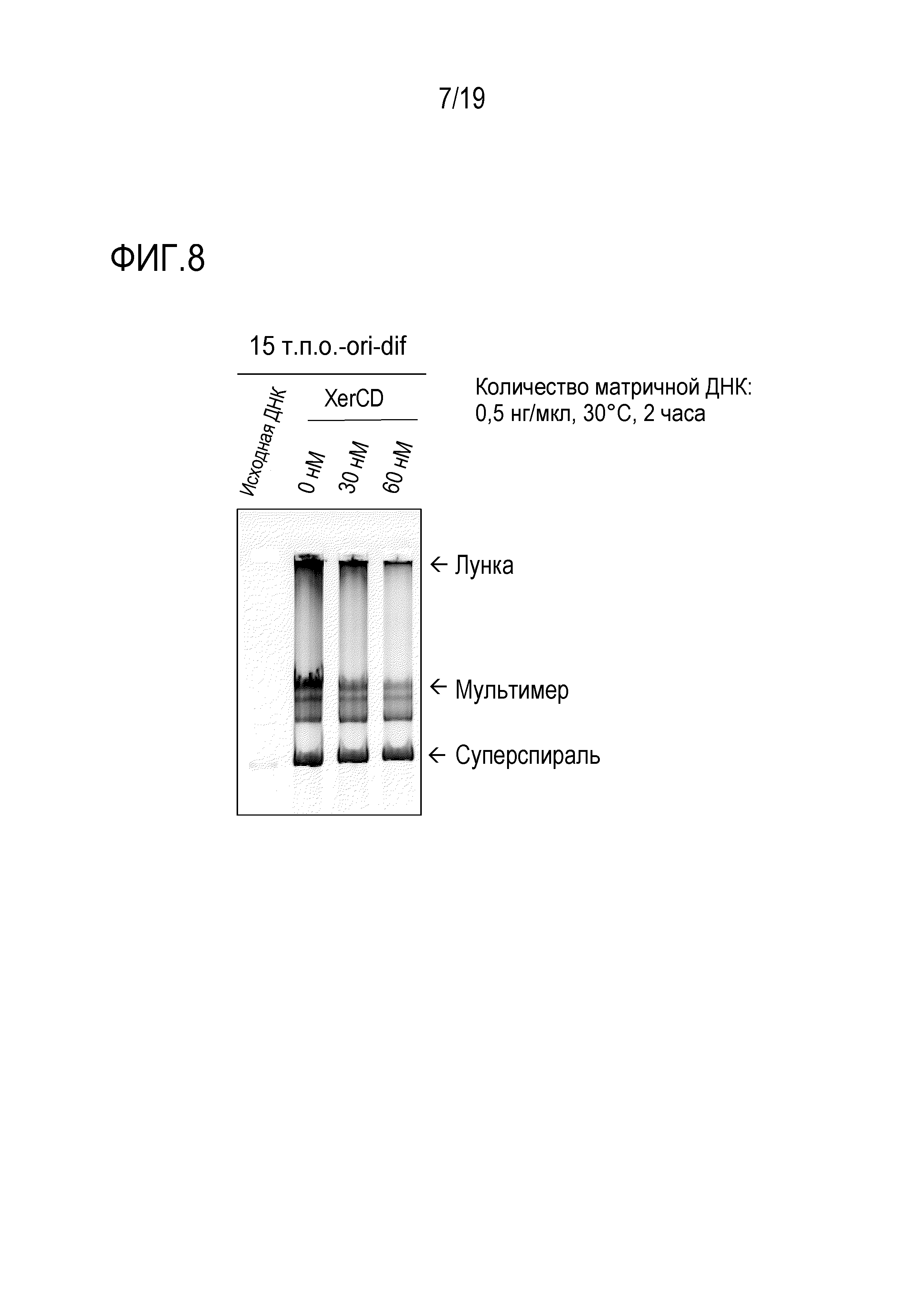
На фигуре 7 представлена фотография гель-электрофореза, иллюстрирующая результаты проведения титрования ДНК на подавление образования ДНК-мультимера с использованием последовательности терминации ter и белка Tus.
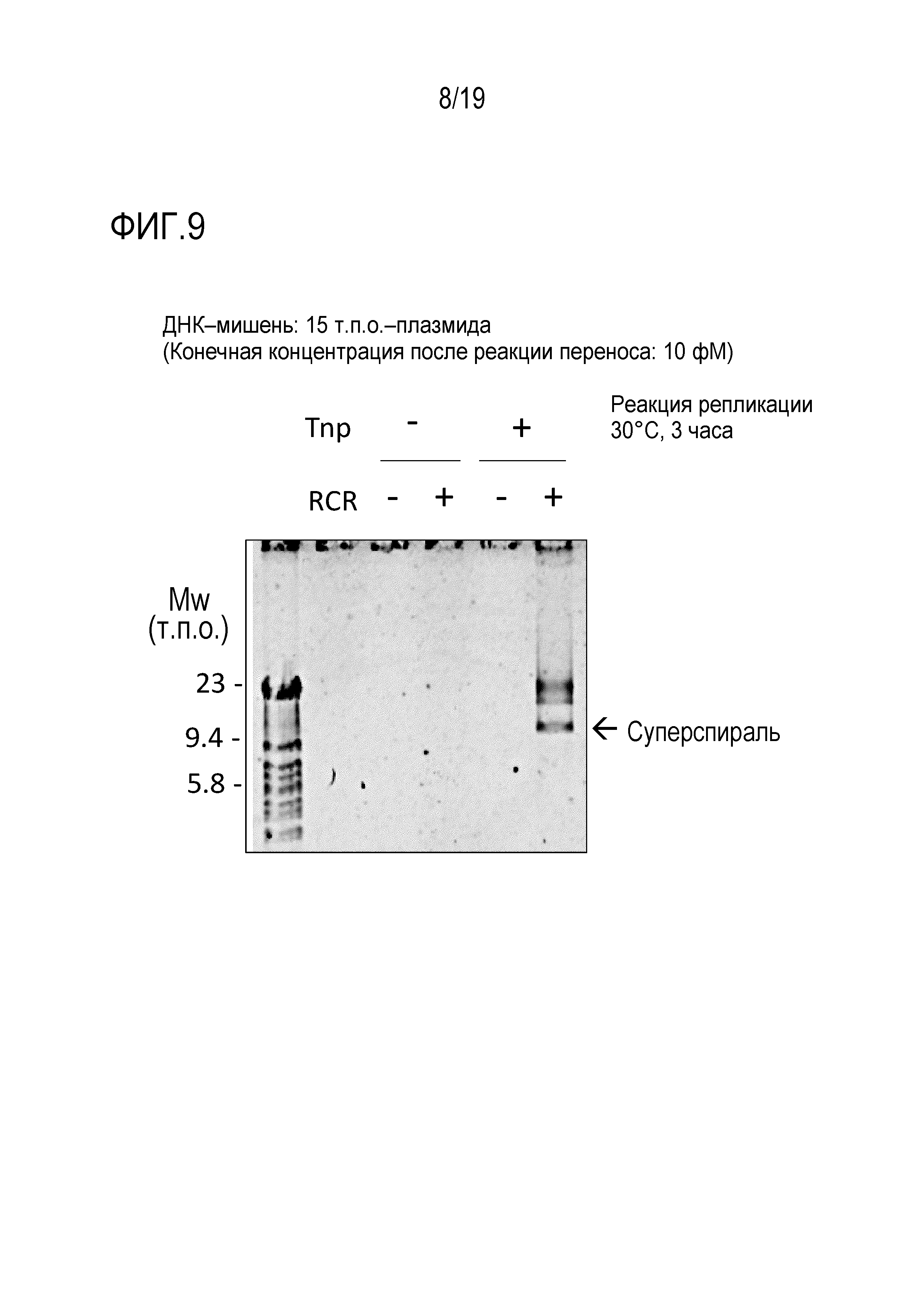
На фигуре 8 представлена фотография гель-электрофореза, иллюстрирующая результат подавления образования ДНК-мультимера посредством XerCD.
На фигуре 9 представлена фотография гель-электрофореза, иллюстрирующая результат амплификации 15 т.п.о. -плазмиды посредством переноса транспозона oriC.
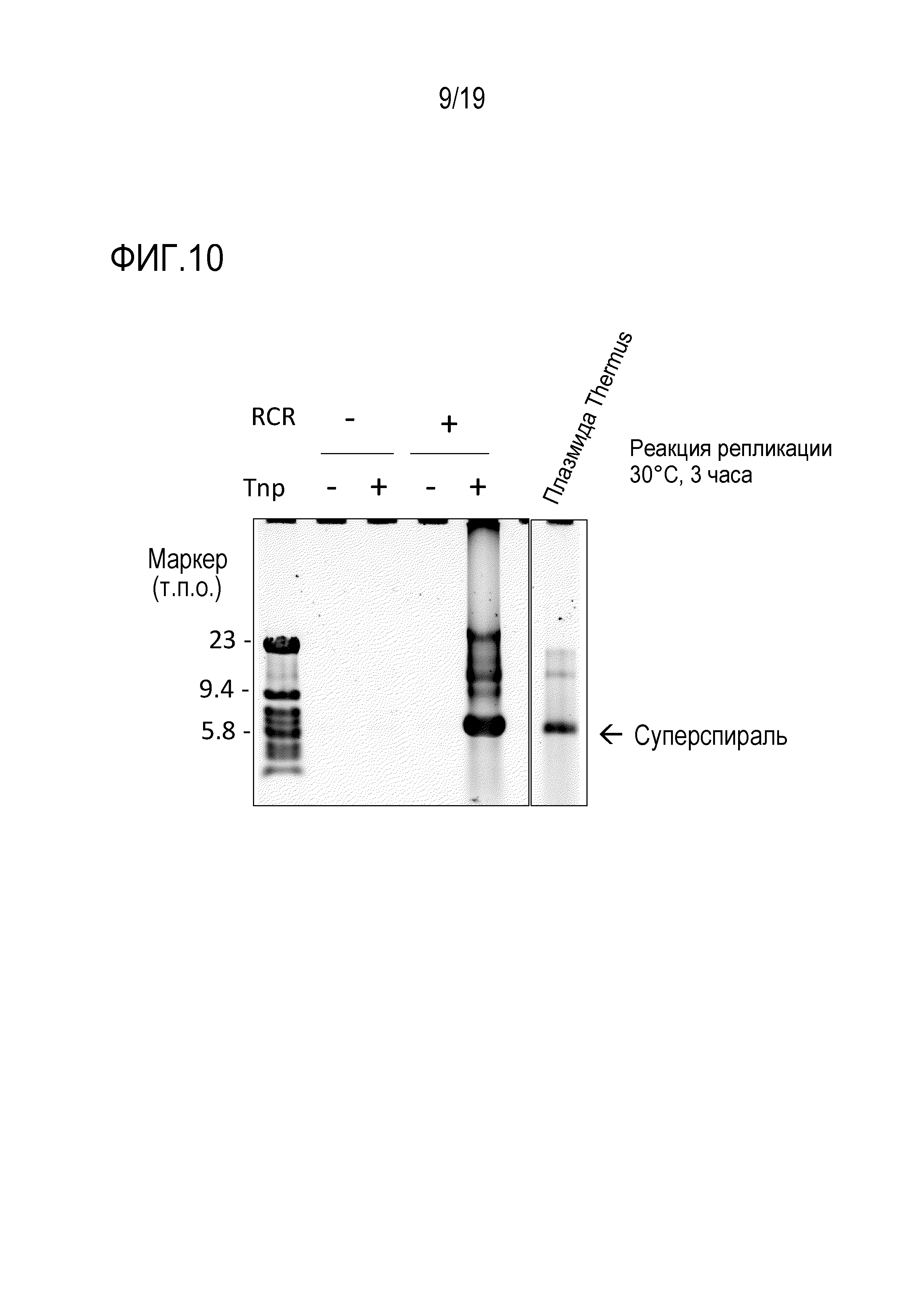
На фигуре 10 представлена фотография гель-электрофореза, иллюстрирующая результат амплификации происходящей от термофила 9,3 т.п.о. -плазмиды с высоким содержанием GC посредством переноса транспозона oriC.
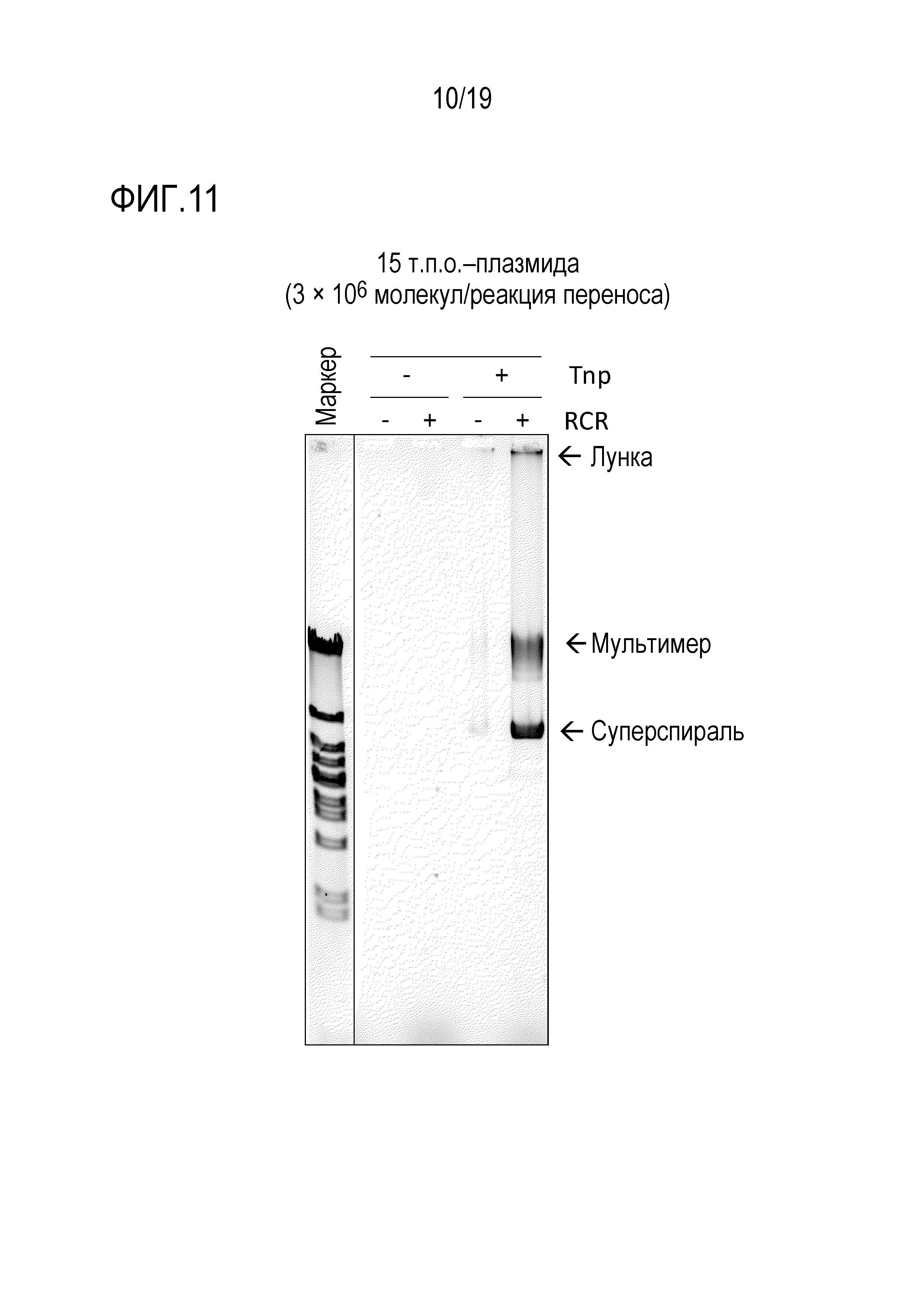
На фигуре 11 представлена фотография гель-электрофореза, иллюстрирующая результат амплификации 15 т.п.о. -плазмиды посредством переноса транспозона oriC.
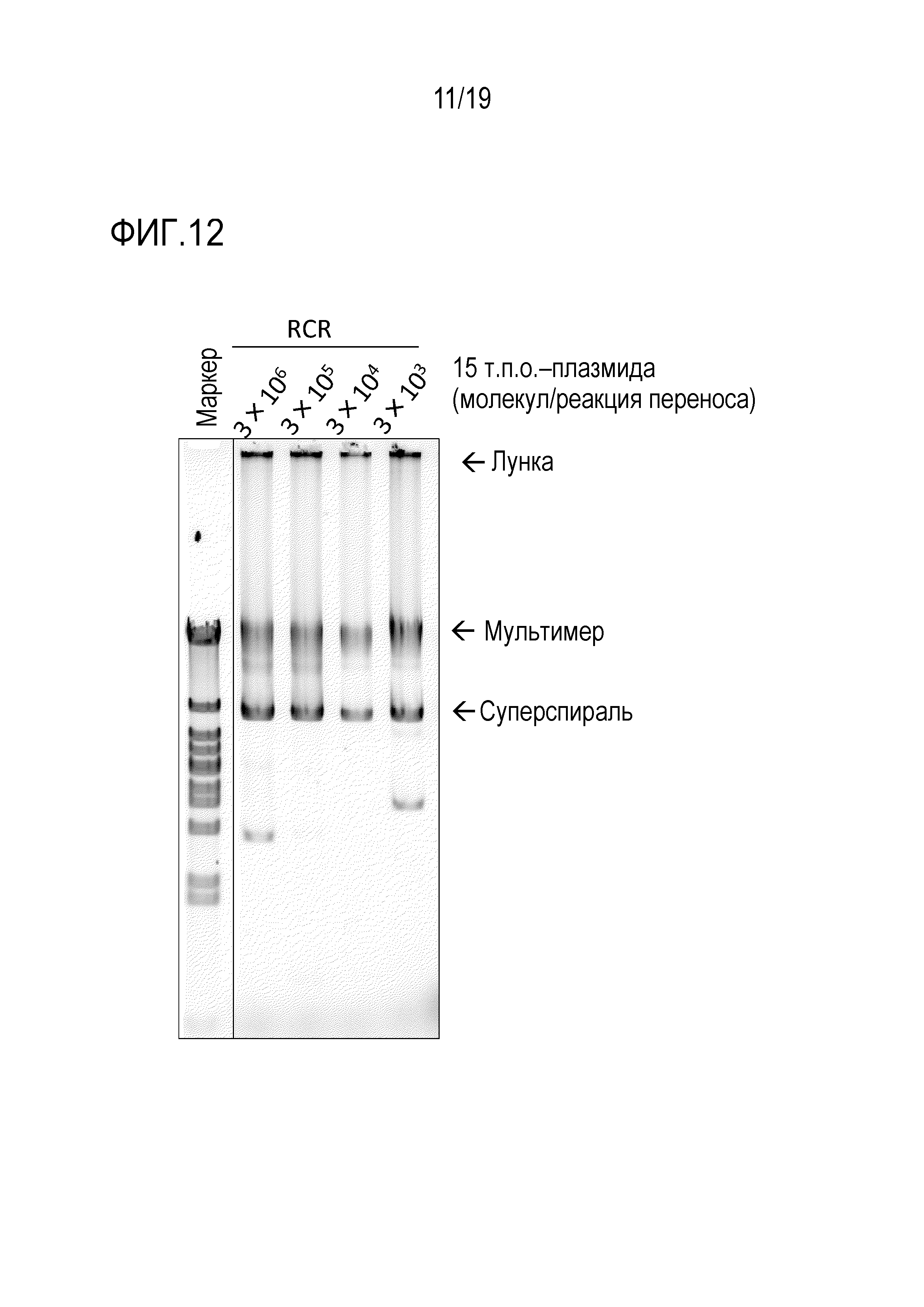
На фигуре 12 представлена фотография гель-электрофореза, иллюстрирующая результат амплификации 15 т.п.о. -плазмиды посредством переноса транспозона oriC в случае, если количество ДНК, используемой в реакции переноса транспозона oriC, было изменено.
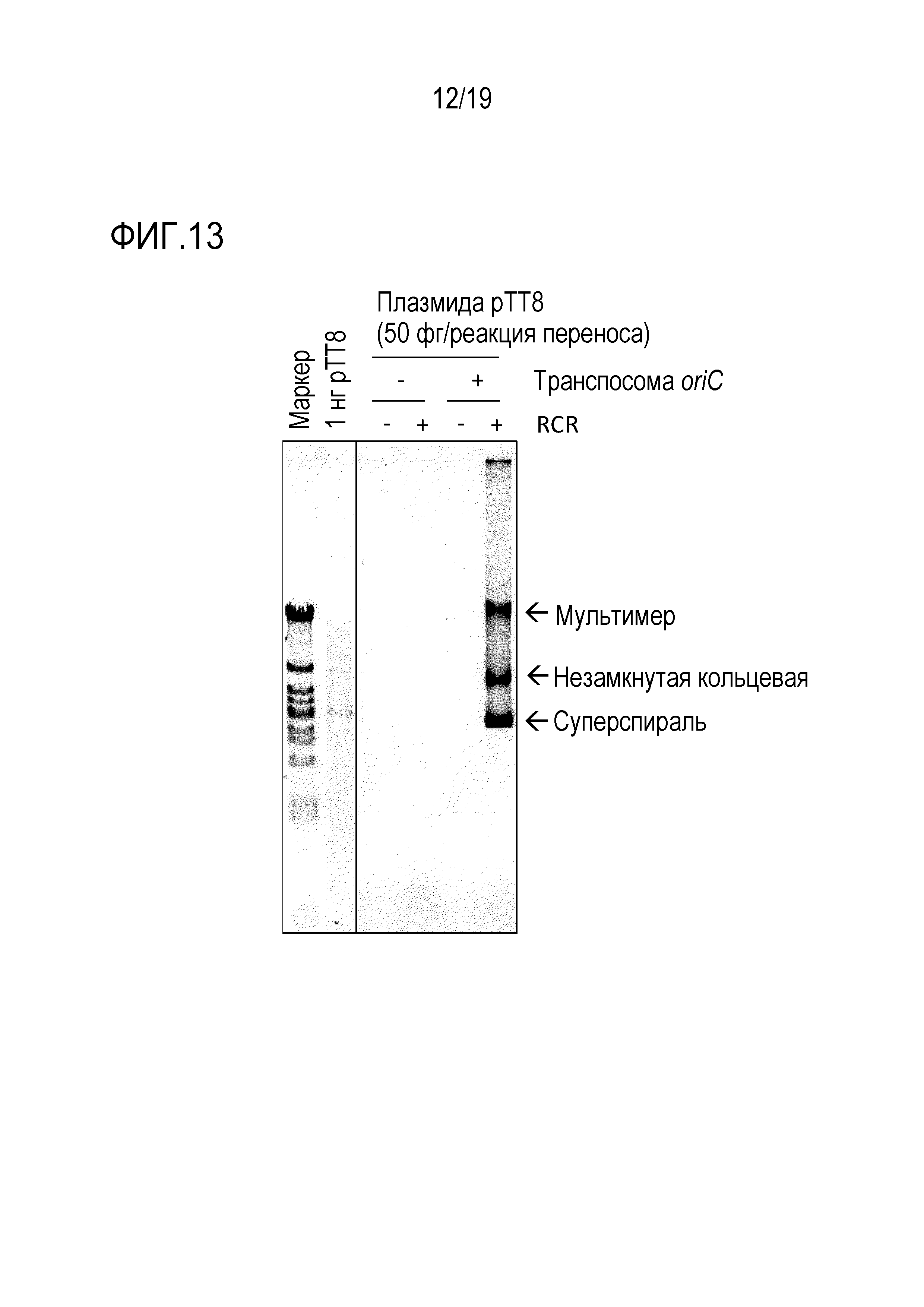
На фигуре 13 представлена фотография гель-электрофореза, иллюстрирующая результат амплификации происходящей от термофила 9,3 т.п.о. -плазмиды с высоким содержанием GC посредством переноса транспозона oriC.
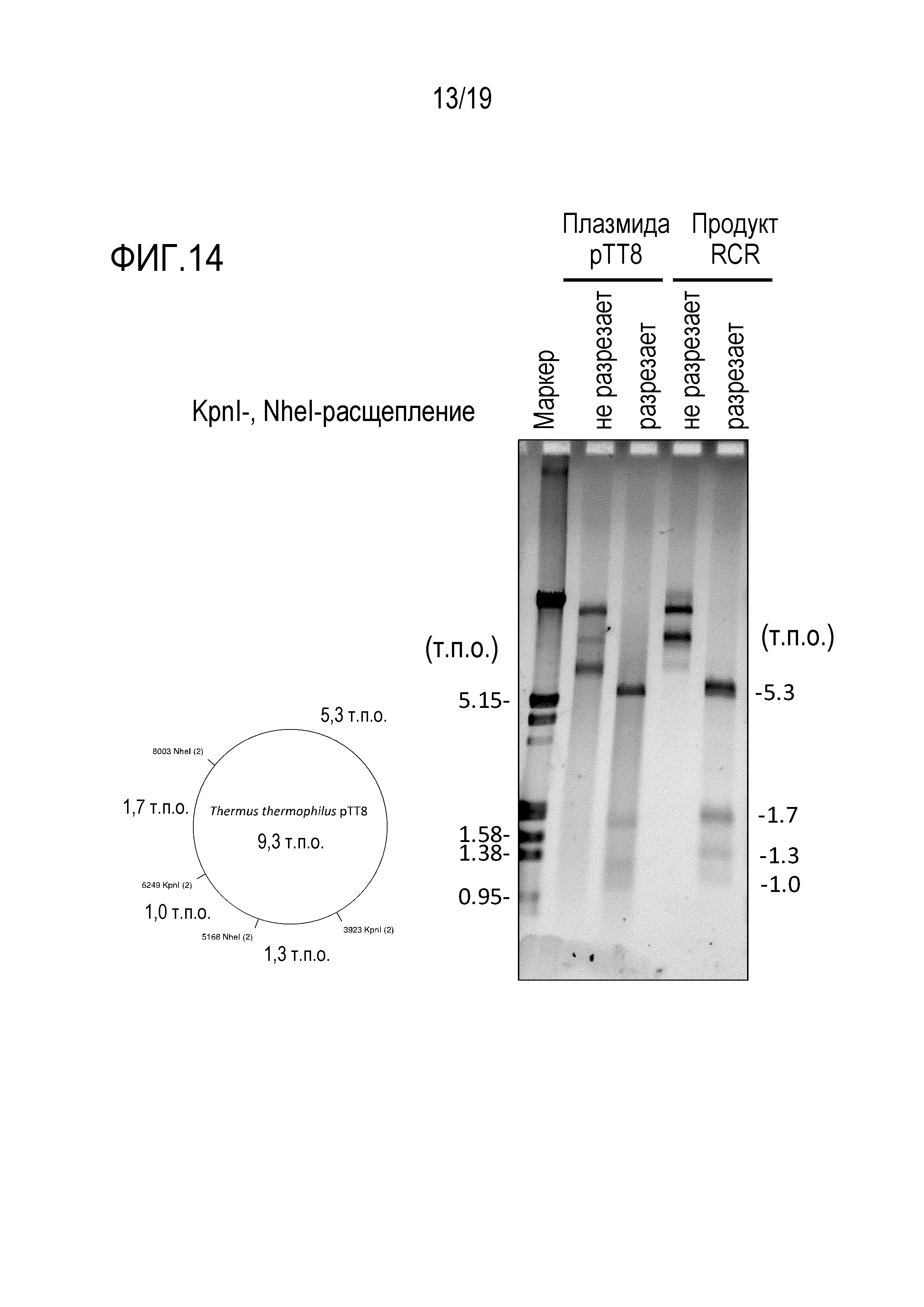
На фигуре 14 представлена фотография гель-электрофореза, иллюстрирующая результат проведения гидролиза рестриктирующими ферментами (KpnI и NheI) продукта амплификации происходящей от термофила 9,3 т.п.о. -плазмиды с высоким содержанием GC посредством переноса транспозона oriC.
На фигуре 15 представлена фотография гель-электрофореза, иллюстрирующая результат исследования циклизации λДНК.
На фигуре 16 представлена фотография гель-электрофореза, иллюстрирующая результат исследования циклизации λДНК и амплификации кольцевой ДНК посредством переноса транспозона oriC.
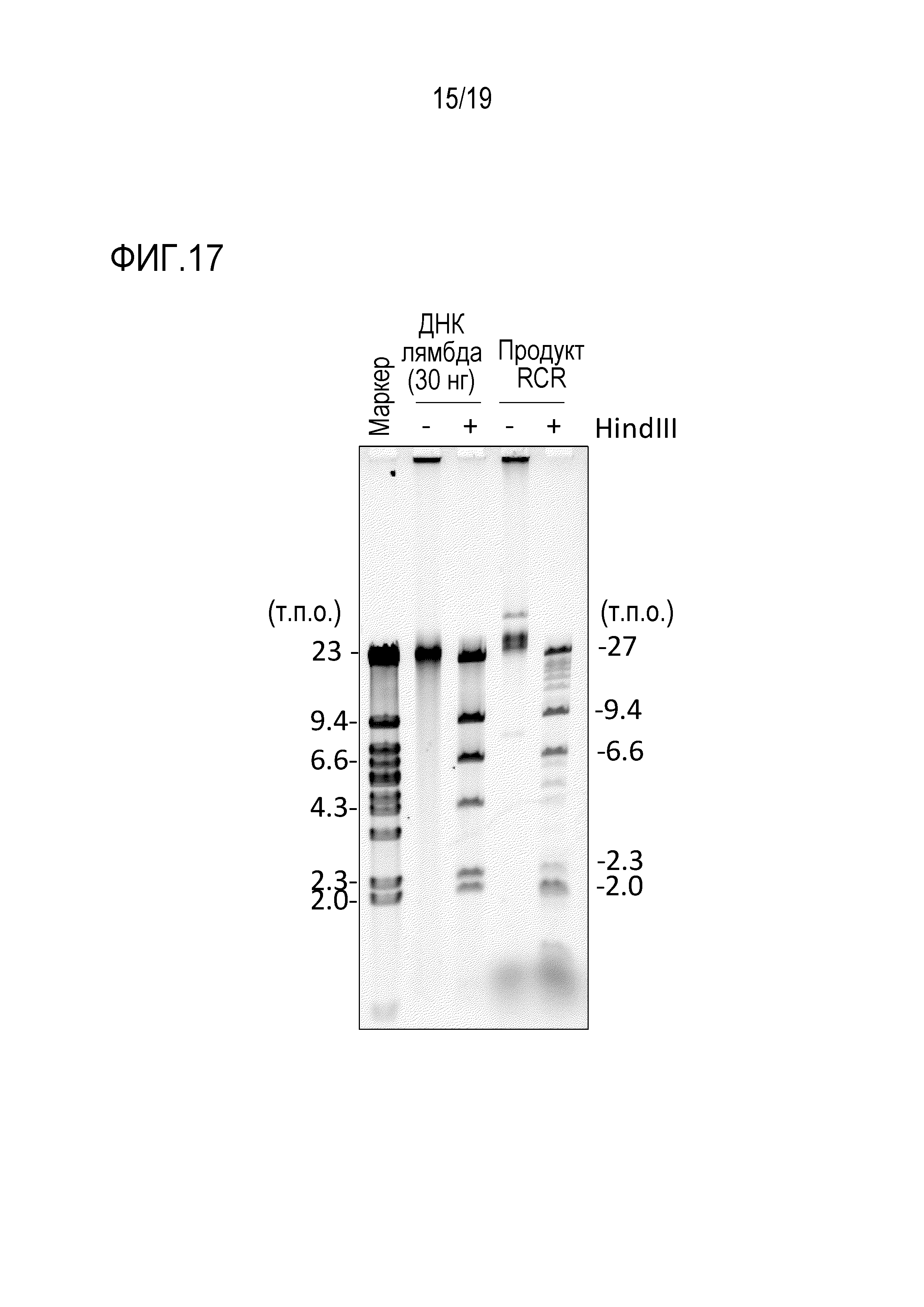
На фигуре 17 представлена фотография гель-электрофореза, иллюстрирующая результат циклизации λДНК и гидролиза рестриктирующим ферментом (HindIII) продукта амплификации кольцевой ДНК посредством переноса транспозона oriC.
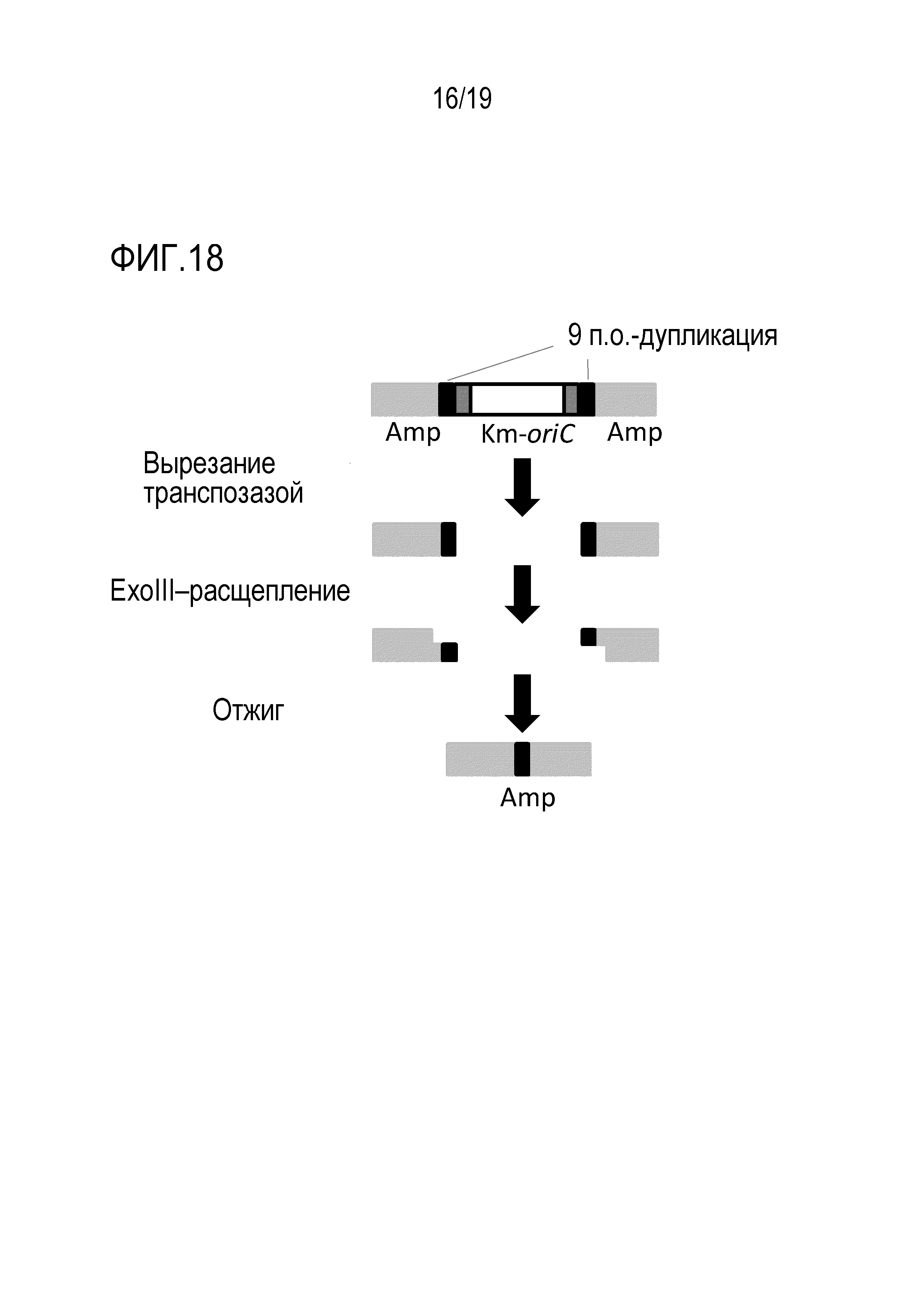
На фигуре 18 схематически представлено изображение, иллюстрирующее реакцию диссоциации транспозона oriC.
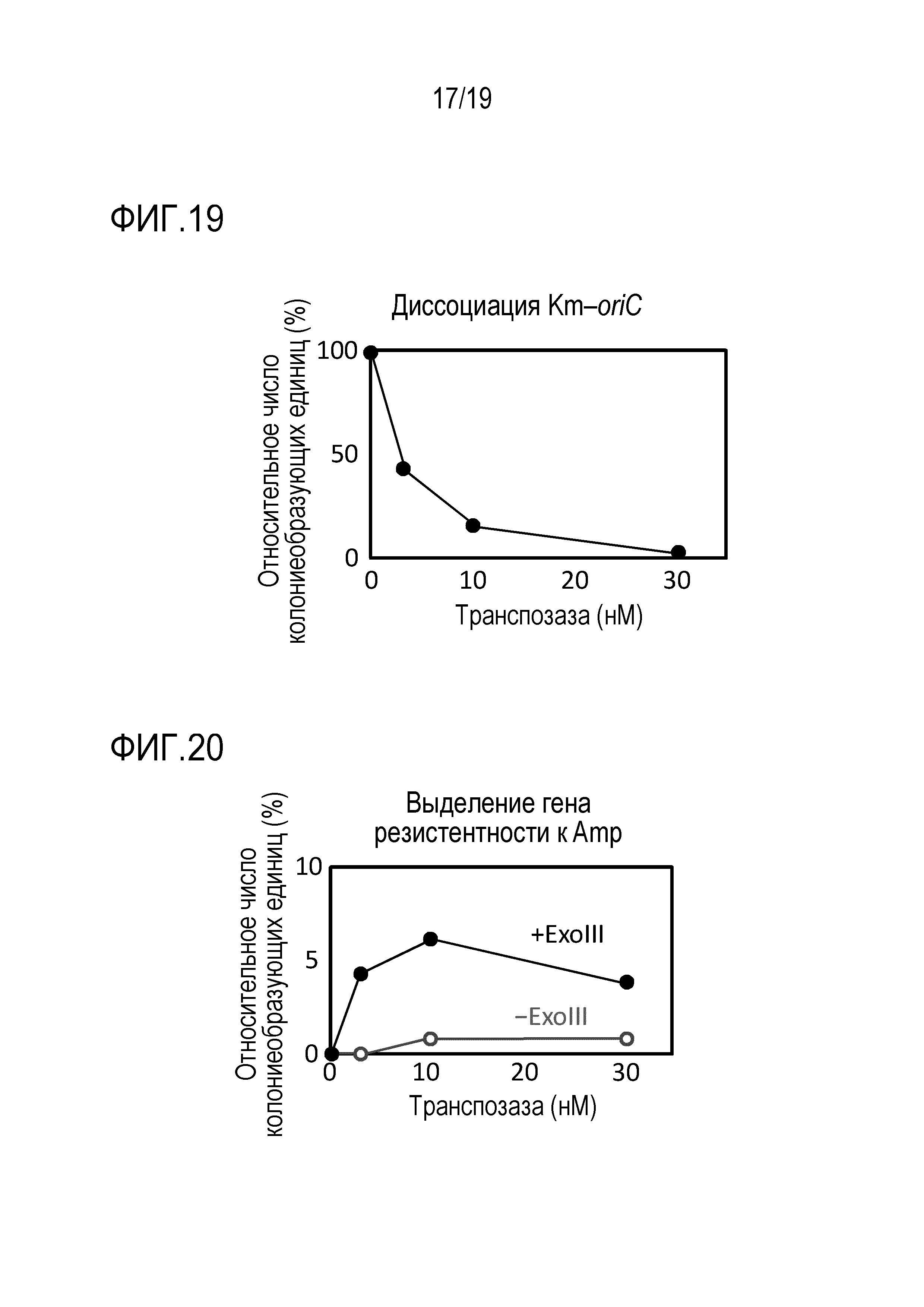
На фигуре 19 представлен график, иллюстрирующий оценку диссоциации транспозона oriC, содержащего Km-oriC.
На фигуре 20 представлен график, иллюстрирующий оценку гена резистентности к Amp, выделенного после диссоциации транспозона oriC.
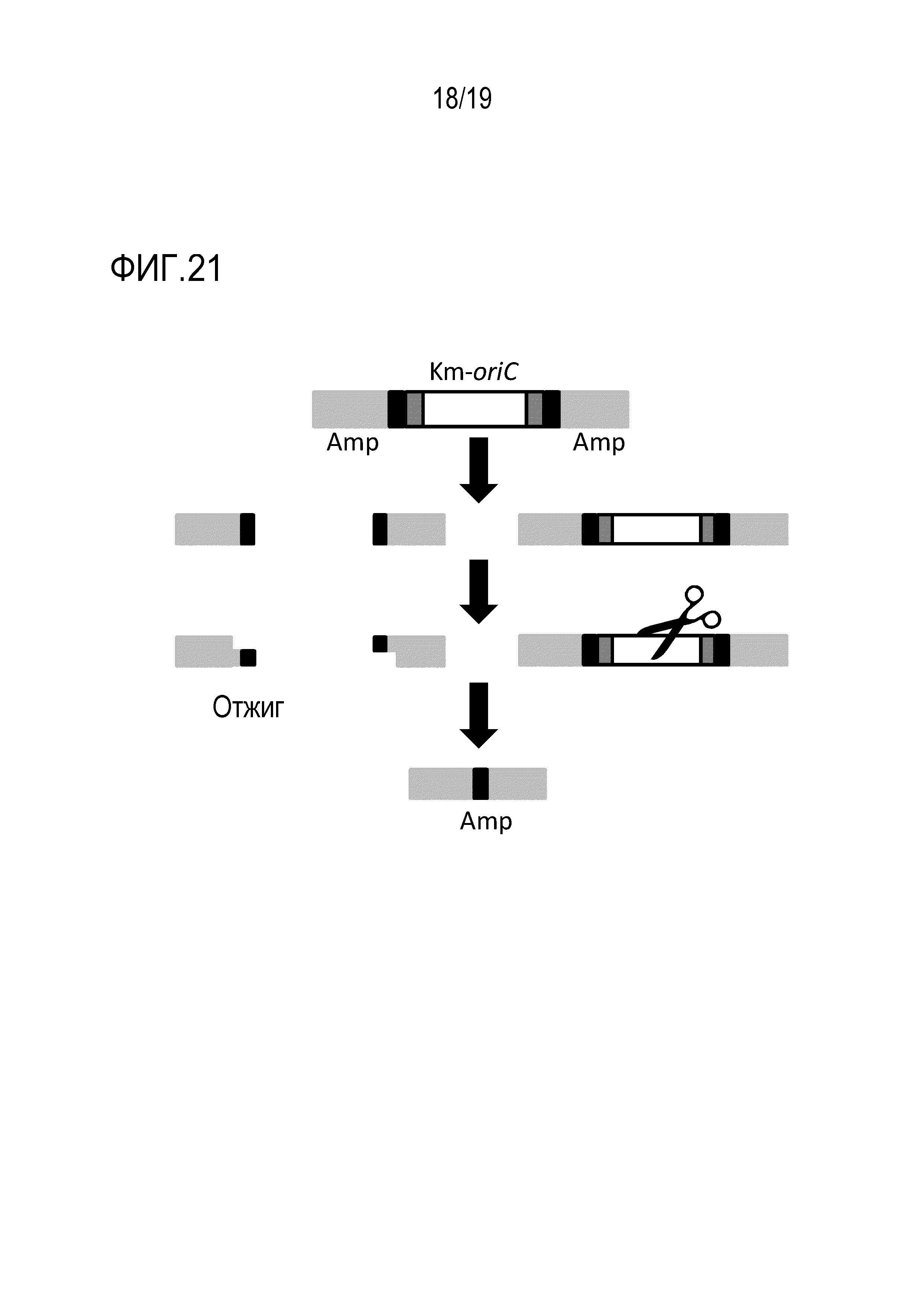
На фигуре 21 схематически представлено изображение, иллюстрирующее реакцию диссоциации транспозона oriC и реакцию удаления транспозона oriC посредством расщепления.
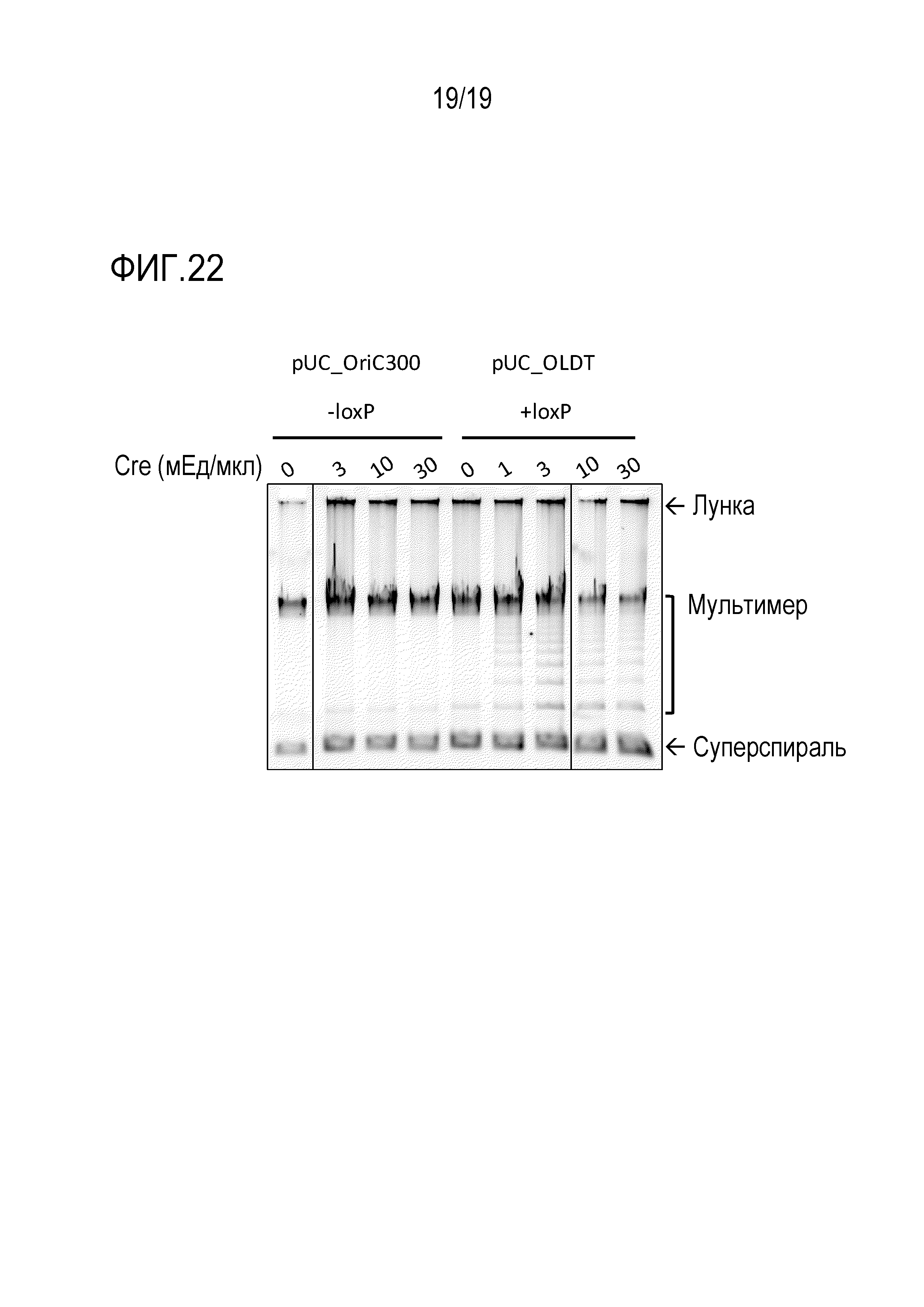
На фигуре 22 представлена фотография гель-электрофореза, иллюстрирующая результат подавления образования ДНК-мультимера посредством Cre.
Описание вариантов осуществления изобретения
[0037] Далее приводится подробное описание настоящего изобретения. Однако, настоящее изобретение не ограничивается таким описанием. Научные и технические термины, используемые в настоящем изобретении, имеют свои общепринятые значения, понятные специалистам в данной области, если это не оговорено особо.
[0038] <Кольцевая ДНК>
Кольцевая ДНК, используемая в качестве матрицы, предпочтительно, является двухцепочечной. Кольцевая ДНК, используемая в качестве матрицы, не имеет конкретных ограничений при условии, что она включает последовательность ориджина репликации (ориджина хромосомы (oriC)), которая может связываться с ферментом, обладающим DnaA-активностью, и примерами таких ДНК являются природная кольцевая ДНК, такая как кольцевая хромосома микроорганизмов; кольцевая ДНК, полученная путем лигирования природной кольцевой ДНК, которая была разрезана посредством ферментативного процессинга и т.п., с другим фрагментом ДНК с последующей циклизацией лигированного продукта; кольцевая ДНК, которая была получена путем обработки посредством циклизации ДНК, присутствующей в природе в линейной форме, и кольцевая ДНК, которая была получена путем синтеза. Что касается последовательностей ориджина репликации (oriC), которые могут связываться с ферментом, обладающим DnaA-активностью (эта последовательность может далее называться просто «последовательностью ориджина репликации» или «oriC»), то общеизвестные последовательности ориджина репликации, присутствующие в бактериях, таких как E. coli, Bacillus subtilis и т.п., могут быть взяты из общедоступной базы данных, такой как NCBI (http://www.ncbi.nlm.nih.gov/). Кроме того, последовательность ориджина репликации может быть получена путем клонирования фрагмента ДНК, который может связываться с ферментом, обладающим DnaA-активностью, с последующим анализом этой последовательности оснований.
[0039] Кольцевая ДНК, используемая в настоящем изобретении в качестве матрицы, может представлять собой кольцевую ДНК, которая изначально содержит последовательность ориджина репликации или кольцевую ДНК, которая по своей природе не содержит последовательности ориджина репликации, но в которую она может быть впоследствии введена.
[0040] В качестве метода получения кольцевой ДНК, используемой в качестве матрицы, путем введения последовательности ориджина репликации в кольцевую ДНК, которая по своей природе не содержит последовательности ориджина репликации, может быть применен любой метод, известный специалистам. В одном из вариантов осуществления изобретения, введение последовательности ориджина репликации в кольцевую ДНК, не содержащую такой последовательности ориджина репликации, может быть осуществлено путем присоединения ДНК транспозона, содержащей последовательность ориджина репликации, которая представляет собой фосфорилированную у 5'-конца линейную ДНК, содержащую последовательность ориджина репликации, а также содержащую внешние концевые (OE) последовательности у обоих концов, и добавления транспозазы в буфер с образованием транспосомы, содержащей последовательность ориджина репликации, с последующей реакцией взаимодействия транспосомы, содержащей последовательность ориджина репликации, с кольцевой ДНК, не содержащей такой последовательности ориджина репликации, в буфере для проведения реакции переноса.
[0041] Кольцевая ДНК, используемая в настоящем изобретении в качестве матрицы, может включать последовательности маркерных генов резистентности к лекарственным средствам, таким как канамицин, ампициллин, тетрациклин и т.п., в зависимости от цели их применения.
[0042] Кроме того, кольцевая ДНК, используемая в настоящем изобретении в качестве матрицы, может быть получена в очищенной форме, либо она может быть получена в форме суспензии после экстракции из бактерий, включающих кольцевую ДНК. В качестве матрицы может быть использована кольцевая ДНК одного типа, но в качестве матрицы может быть также использована смесь кольцевых ДНК нескольких типов, такая как библиотека ДНК в одной тест-пробирке.
[0043] Длина кольцевой ДНК, используемой в настоящем изобретении в качестве матрицы, не имеет конкретных ограничений и может составлять 1 т.п.о. (длиной 1000 оснований) или более, 5 т.п.о. (длиной 5000 оснований) или более, 8 т.п.о. (длиной 8000 оснований) или более, 10 т.п.о. (длиной 10000 оснований) или более, 50 т.п.о. (длиной 50000 оснований) или более, 100 т.п.о. (длиной 100000 оснований) или более, 200 т.п.о. (длиной 200000 оснований) или более, 500 т.п.о. (длиной 500000 оснований) или более, 1000 т.п.о. (длиной 1000000 оснований) или более или 2000 т.п.о. (длиной 2000000 оснований) или более.
[0044] <Первая, вторая и третья группы ферментов>
1. Первая группа ферментов
В настоящем описании, первая группа ферментов означает группу ферментов, которые катализируют репликацию кольцевой ДНК.
[0045] Примером первой группы ферментов, которые катализируют репликацию кольцевой ДНК, является группа ферментов, описанная Kaguni JM & Kornberg A. Cell. 1984, 38:183-90. В частности, примерами первой группы ферментов являются один или более ферментов или групп ферментов, выбранных из группы, состоящей из фермента, обладающего DnaA-активностью; нуклеоидного белка одного или более типов; фермента или группы ферментов, обладающих ДНК-гиразной активностью; белка, связывающегося с одноцепочечной ДНК (SSB); фермента, обладающего геликазной активностью DnaB-типа; фермента, обладающего ДНК-геликазной загрузочной активностью; фермента, обладающего ДНК-примазной активностью; фермента, обладающего ДНК-захватывающей активностью; и фермента или группы ферментов, обладающих активностью ДНК-полимеразы III*; и комбинаций всех вышеупомянутых ферментов или групп ферментов.
[0046] Фермент, обладающий DnaA-активностью, конкретно не ограничивается биологическим происхождением, при условии, что он обладает инициирующей активностью, аналогичной активности DnaA, который представляет собой инициирующий белок E. coli, при этом, предпочтительно, использовать DnaA, происходящий от E. coli. DnaA, происходящий от Escherichia coli, может содержаться в качестве мономера в реакционном растворе в количестве от 1 нМ до 10 мкМ, а предпочтительно, в количестве от 1 нМ до 5 мкМ, от 1 нМ до 3 мкМ, от 1 нМ до 1,5 мкМ, от 1 нМ до 1,0 мкМ, от 1 нМ до 500 нМ, от 50 нМ до 200 нМ или от 50 нМ до 150 нМ, но такое количество не ограничивается указанными значениями.
[0047] Нуклеоидным белком является белок в нуклеоиде. Нуклеоидный белок одного или более типов, используемый в настоящем изобретении, конкретно не ограничивается биологическим происхождением, при условии, что он обладает активностью, аналогичной активности нуклеоидного белка E. coli. Так, например, предпочтительно использовать IHF, происходящий от Escherichia coli, а именно, комплекс IhfA и/или IhfB (гетеродимер или гомодимер) или HU, происходящий от Escherichia coli, а именно, комплекс hupA и hupB. IHF, происходящий от Escherichia coli, может содержаться в виде гетеро/гомодимера в реакционном растворе в концентрации в пределах от 5 нМ до 400 нМ. Предпочтительно, IHF, происходящий от Escherichia coli, может содержаться в реакционном растворе в концентрации в пределах от 5 нМ до 200 нМ, от 5 нМ до 100 нМ, от 5 нМ до 50 нМ, от 10 нМ до 50 нМ, от 10 нМ до 40 нМ или от 10 нМ до 30 нМ, но такая концентрация не ограничивается указанными пределами. HU, происходящий от Escherichia coli, может содержаться в реакционном растворе в концентрации в пределах от 1 нМ до 50 нМ, а предпочтительно, он может содержаться в концентрации в пределах от 5 нМ до 50 нМ или от 5 нМ до 25 нМ, но такая концентрация не ограничивается указанными пределами.
[0048] Фермент или группа ферментов, обладающих ДНК-гиразной активностью, конкретно не ограничиваются биологическим происхождением, при условии, что они обладают активностью, аналогичной активности ДНК-гиразы E. coli. Так, например, предпочтительно, использовать комплекс GyrA и GyrB, происходящих от Escherichia coli. Такой комплекс GyrA и GyrB, происходящих от Escherichia coli, может содержаться в виде гетеротетрамера в реакционном растворе в концентрации в пределах от 20 нМ до 500 нМ, а предпочтительно, он может содержаться в концентрации в пределах от 20 нМ до 400 нМ, от 20 нМ до 300 нМ, от 20 нМ до 200 нМ, от 50 нМ до 200 нМ или от 100 нМ до 200 нМ, но такая концентрация не ограничивается указанными пределами.
[0049] Белок, связывающийся с одноцепочечной ДНК (SSB), конкретно не ограничивается биологическим происхождением, при условии, что он обладает активностью, аналогичной активности белка, связывающегося с одноцепочечной ДНК E. coli. Так, например, предпочтительно, использовать SSB, происходящий от Escherichia coli. Такой SSB, происходящий от Escherichia coli, может содержаться в виде гомотетрамера в реакционном растворе в концентрации в пределах от 20 нМ до 1000 нМ, а предпочтительно, он может содержаться в концентрации в пределах от 20 нМ до 500 нМ, от 20 нМ до 300 нМ, от 20 нМ до 200 нМ, от 50 нМ до 500 нМ, от 50 нМ до 400 нМ, от 50 нМ до 300 нМ, от 50 нМ до 200 нМ, от 50 нМ до 150 нМ, от 100 нМ до 500 нМ или от 100 нМ до 400 нМ, но такая концентрация не ограничивается указанными пределами.
[0050] Фермент, обладающий геликазной активностью DnaB-типа, конкретно не ограничивается биологическим происхождением, при условии, что он обладает активностью, аналогичной активности DnaB E. coli. Так, например, предпочтительно, использовать DnaB, происходящий от Escherichia coli. Такой DnaB, происходящий от Escherichia coli, может содержаться в виде гомогексамера в реакционном растворе в концентрации в пределах от5 нМ до 200 нМ, а предпочтительно, он может содержаться в концентрации в пределах от 5 нМ до 100 нМ, от 5 нМ до 50 нМ или от 5 нМ до 30 нМ, но такая концентрация не ограничивается указанными пределами.
[0051] Фермент, обладающий ДНК-геликазной загрузочной активностью, конкретно не ограничивается биологическим происхождением, при условии, что он обладает активностью, аналогичной активности DnaC E. coli. Так, например, предпочтительно, использовать DnaC, происходящий от Escherichia coli. Такой DnaC, происходящий от Escherichia coli, может содержаться в виде гомогексамера в реакционном растворе в концентрации в пределах от5 нМ до 200 нМ, а предпочтительно, он может содержаться в концентрации в пределах от 5 нМ до 100 нМ, от 5 нМ до 50 нМ или от 5 нМ до 30 нМ, но такая концентрация не ограничивается указанными пределами.
[0052] Фермент, обладающий ДНК-примазной активностью, конкретно не ограничивается биологическим происхождением, при условии, что он обладает активностью, аналогичной активности DnaG E. coli. Так, например, предпочтительно, использовать DnaG, происходящий от Escherichia coli. Такой DnaG, происходящий от Escherichia coli, может содержаться в виде мономера в реакционном растворе в концентрации в пределах от20 нМ до 1000 нМ, а предпочтительно, он может содержаться в концентрации в пределах от 20 нМ до 800 нМ, от 50 нМ до 800 нМ, от 100 нМ до 800 нМ, от 200 нМ до 800 нМ, от 250 нМ до 800 нМ, от 250 нМ до 500 нМ или от 300 нМ до 500 нМ, но такая концентрация не ограничивается указанными пределами.
[0053] Фермент, обладающий ДНК-захватывающей активностью, конкретно не ограничивается биологическим происхождением, при условии, что он обладает активностью, аналогичной активности DnaN E. coli. Так, например, предпочтительно, использовать DnaN, происходящий от Escherichia coli. Такой DnaN, происходящий от Escherichia coli, может содержаться в виде гомодимера в реакционном растворе в концентрации в пределах от10 нМ до 1000 нМ, а предпочтительно, он может содержаться в концентрации в пределах от 10 нМ до 800 нМ, от10 нМ до 500 нМ, от 20 нМ до 500 нМ, от 20 нМ до 200 нМ, от 30 нМ до 200 нМ или от 30 нМ до 100 нМ, но такая концентрация не ограничивается указанными пределами.
[0054] Фермент или группа ферментов, обладающих активностью ДНК-полимеразы III*, конкретно не ограничиваются биологическим происхождением, при условии, что они обладают активностью, аналогичной активности комплекса ДНК-полимеразы III* E. coli. Так, например, предпочтительно, использовать группу ферментов, содержащих любые из DnaX, HolA, HolB, HolC, HolD, DnaE, DnaQ и HolE, происходящих от Escherichia coli; предпочтительно, группу ферментов, содержащих комплекс DnaX, HolA, HolB и DnaE, происходящих от Escherichia coli; а более предпочтительно, фермент, содержащий комплекс DnaX, HolA, HolB, HolC, HolD, DnaE, DnaQ и HolE, происходящих от Escherichia coli. Такой комплекс ДНК-полимеразы III*, происходящий от Escherichia coli, может содержаться в виде гетеромультимера в реакционном растворе в концентрации в пределах от 2 нМ до 50 нМ, а предпочтительно, он может содержаться в концентрации в пределах от 2 нМ до 40 нМ, от 2 нМ до 30 нМ, от 2 нМ до 20 нМ, от 5 нМ до 40 нМ, от 5 нМ до 30 нМ или от 5 нМ до 20 нМ, но такая концентрация не ограничивается указанными пределами.
[0055] 2. Вторая группа ферментов
В настоящем описании, вторая группа ферментов означает группу ферментов, которые катализируют созревание фрагмента Оказаки и синтезируют две сестринские кольцевые ДНК, состоящие из катенана.
[0056] В настоящем изобретении, две сестринские кольцевые ДНК, состоящие из катенана, представляют собой две кольцевых ДНК, синтезированных посредством репликации ДНК с их последующим присоединением.
[0057] Примерами второй группы ферментов, которые катализируют созревание фрагмента Оказаки и синтезируют две сестринские кольцевые ДНК, состоящие из катенана, могут быть, например, один или более ферментов, выбранных из группы, состоящей из фермента, обладающего активностью ДНК-полимеразы I; фермента, обладающего ДНК-лигазной активностью; фермента, обладающего активностью РНКазы Н или комбинации этих ферментов.
[0058] Фермент, обладающий активностью ДНК-полимеразы I, конкретно не ограничивается биологическим происхождением, при условии, что он обладает активностью, аналогичной активности ДНК-полимеразы I E. coli. Так, например, предпочтительно, использовать ДНК-полимеразу I, происходящую от Escherichia coli. Такая ДНК-полимераза I, происходящая от Escherichia coli, может содержаться в виде мономера в реакционном растворе в концентрации в пределах от 10 нМ до 200 нМ, а предпочтительно, она может содержаться в концентрации в пределах от 20 нМ до 200 нМ, от 20 нМ до 150 нМ, от 20 нМ до 100 нМ, от 40 нМ до 150 нМ, от 40 нМ до 100 нМ или от 40 нМ до 80 нМ, но такая концентрация не ограничивается указанными пределами.
[0059] Фермент, обладающий ДНК-лигазной активностью, конкретно не ограничивается биологическим происхождением, при условии, что он обладает активностью, аналогичной активности ДНК-лигазы E. coli. Так, например, предпочтительно, использовать ДНК-лигазу, происходящую от Escherichia coli или ДНК-лигазу фага Т4. Такая ДНК-лигаза, происходящая от Escherichia coli, может содержаться в виде мономера в реакционном растворе в концентрации в пределах от 10 нМ до 200 нМ, а предпочтительно, она может содержаться в концентрации в пределах от 15 нМ до 200 нМ, от 20 нМ до 200 нМ, от 20 нМ до 150 нМ, от 20 нМ до 100 нМ или от 20 нМ до 80 нМ, но такая концентрация не ограничивается указанными пределами.
[0060] Фермент, обладающий активностью РНКазы Н, конкретно не ограничивается биологическим происхождением, при условии, что он обладает активностью в разложении цепи РНК гибрида РНК-ДНК. Так, например, предпочтительно, использовать РНКазу Н, происходящую от Escherichia coli. Такая РНКаза Н, происходящая от Escherichia coli, может содержаться в виде мономера в реакционном растворе в концентрации в пределах от 0,2 нМ до 200 нМ, а предпочтительно, она может содержаться в концентрации в пределах от 0,2 нМ до 200 нМ, от 0,2 нМ до 100 нМ, от 0,2 нМ до 50 нМ, от 1 нМ до 200 нМ, от 1 нМ до 100 нМ, от 1 нМ до 50 нМ или от 10 нМ до 50 нМ, но такая концентрация не ограничивается указанными пределами.
[0061] 3. Третья группа ферментов
В настоящем описании, третья группа ферментов означает группу ферментов, которые катализируют разделение двух сестринских кольцевых ДНК.
[0062] Примером третьей группы ферментов, которые катализируют разделение двух сестринских кольцевых ДНК, является группа ферментов, представленная, например, как группа ферментов, описанная Peng H & Marians KJ. PNAS. 1993, 90: 8571-8575. В частности, примерами третьей группы ферментов являются один или более ферментов, выбранных из группы, состоящей из фермента, обладающего активностью топоизомеразы IV; фермента, обладающего активностью топоизомеразы III; и фермента, обладающего геликазной активностью RecQ-типа или комбинации вышеупомянутых ферментов.
[0063] Фермент, обладающий активностью топоизомеразы III, конкретно не ограничивается биологическим происхождением, при условии, что он обладает активностью, аналогичной активности топоизомеразы III Escherichia coli. Так, например, предпочтительно, использовать топоизомеразу III, происходящую от Escherichia coli. Такая топоизомераза III, происходящая от Escherichia coli, может содержаться в виде мономера в реакционном растворе в концентрации в пределах от 20 нМ до 500 нМ, а предпочтительно, она может содержаться в концентрации в пределах от 20 нМ до 400 нМ, от 20 нМ до 300 нМ, от 20 нМ до 200 нМ, от 20 нМ до 100 нМ или от 30 нМ до 80 нМ, но такая концентрация не ограничивается указанными пределами.
[0064] Фермент, обладающий геликазной активностью RecQ-типа, конкретно не ограничивается биологическим происхождением, при условии, что он обладает активностью, аналогичной активности RecQ Escherichia coli. Так, например, предпочтительно, использовать RecQ, происходящий от Escherichia coli. Такой RecQ, происходящий от Escherichia coli, может содержаться в виде мономера в реакционном растворе в концентрации в пределах от 20 нМ до 500 нМ, а предпочтительно, он может содержаться в концентрации в пределах от 20 нМ до 400 нМ, от 20 нМ до 300 нМ, от 20 нМ до 200 нМ, от 20 нМ до 100 нМ или от 30 нМ до 80 нМ, но такая концентрация не ограничивается указанными пределами.
[0065] Фермент, обладающий активностью топоизомеразы IV, конкретно не ограничивается биологическим происхождением, при условии, что он обладает активностью, аналогичной активности топоизомеразы IV E. coli. Так, например, предпочтительно, использовать топоизомеразу IV, происходящую от Escherichia coli, то есть, комплекс ParC и ParE. Такая топоизомераза IV, происходящая от Escherichia coli, может содержаться в виде гетеротетрамера в реакционном растворе в концентрации в пределах от 0,1 нМ до 50 нМ, а предпочтительно, она может содержаться в концентрации в пределах от 0,1 нМ до 40 нМ, от 0,1 нМ до 30 нМ, от 0,1 нМ до 20 нМ, от 1 нМ до 40 нМ, от 1 нМ до 30 нМ, от 1 нМ до 20 нМ, от 1 нМ до 10 нМ или от 1 нМ до 5 нМ, но такая концентрация не ограничивается указанными пределами.
[0066] Первая, вторая и третья группы ферментов, описанных выше, могут представлять собой группу ферментов, которые являются коммерчески доступными или могут быть экстрагированы из микроорганизмов и очищены, если это необходимо. Экстракция и очистка ферментов из микроорганизмов могут быть осуществлены, если это необходимо, с применением методов, известных специалистам.
[0067] Если в качестве первой, второй и третей групп ферментов используются ферменты, не являющиеся вышеописанными ферментами, происходящими от Escherichia coli, то каждый из этих ферментов может использоваться как единица активности фермента в концентрации в пределах, соответствующих концентрации, определенной для вышеописанного фермента, происходящего от Escherichia coli.
[0068] Реакционный раствор, содержащий бесклеточные системы экспрессии белка, а именно, вышеупомянутых ферментов, может быть смешан с кольцевой ДНК, содержащей матрицу, с образованием реакционной смеси для репликации или амплификации кольцевой ДНК. Бесклеточная система экспрессии белка может представлять собой бесклеточную систему трансляции, которая включает общую РНК, содержащую РНК, состоящую из последовательности, которая комплементарна последовательности оснований генов, кодирующих вышеуказанные ферменты, мРНК или продукт трансляции in vitro в виде матричной РНК, либо она может представлять собой бесклеточную систему транскрипции/трансляции, которая содержит гены, кодирующие различные ферменты, или экспрессионные векторы, включающие гены, которые кодируют различные ферменты в виде матричной ДНК.
[0069] <Способ репликации кольцевой ДНК(А)>
В одном из своих аспектов, настоящее изобретение относится к способу репликации или амплификации кольцевой ДНК в бесклеточной системе, включающему следующие стадии:
(1) образования реакционной смеси кольцевой ДНК в качестве матрицы с реакционным раствором, содержащим:
первую группу ферментов, которые катализируют репликацию кольцевой ДНК;
вторую группу ферментов, которые катализируют созревание фрагмента Оказаки и синтезируют две сестринские кольцевые ДНК, состоящие из катенана; и
третью группу ферментов, которые катализируют разделение двух сестринских кольцевых ДНК; и
(2) реакции взаимодействия реакционной смеси, полученной в стадии (1), где:
кольцевая ДНК включает последовательность ориджина репликации (ориджина хромосомы (oriC)), которая может связываться с ферментом, обладающим DnaA-активностью, а также включает пару последовательностей ter, каждая из которых была встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую XerCD, где:
если кольцевая ДНК имеет последовательности ter, то реакционный раствор в стадии (1) также включает белок, обладающий активностью ингибирования репликации посредством связывания с последовательностями ter, и если кольцевая ДНК имеет нуклеотидную последовательность, распознаваемую XerCD, то реакционный раствор в стадии (1) также содержит белок XerCD (далее этот способ будет также называться «Способом (А)»).
[0070] Не ограничиваясь какой-либо конкретной теорией, авторы лишь отмечают, что в Способе (А), кольцевую ДНК подвергают репликации или амплификации путем проведения цикла репликации, показанного на фигуре 1, или повторения этого цикла репликации. В настоящем описании, репликация кольцевой ДНК означает, что была получена такая же молекула, как и кольцевая ДНК, используемая в качестве матрицы. Репликация кольцевой ДНК может быть подтверждена по такому феномену, как увеличение количества кольцевой ДНК в продукте реакции после завершения реакции по сравнению с количеством кольцевой ДНК, используемой в качестве матрицы, в начале реакции. Предпочтительно, репликация кольцевой ДНК означает, что количество кольцевой ДНК в продукте реакции было увеличено по меньшей мере в 2 раза, 3 раза, 5 раз, 7 раз или 9 раз по сравнению с количеством кольцевой ДНК в начале реакции. Амплификация кольцевой ДНК означает, что репликация кольцевой ДНК прогрессирует, а количество кольцевой ДНК в продукте реакции экспоненциально увеличивается по сравнению с количеством кольцевой ДНК, используемой в качестве матрицы в начале реакции. В соответствии с этим, амплификация кольцевой ДНК является одним из вариантов репликации кольцевой ДНК. В настоящем описанни, амплификация кольцевой ДНК означает, что количество кольцевой ДНК в продукте реакции увеличивалось по меньшей мере в 10 раз, 50 раз, 100 раз, 200 раз, 500 раз, 1000 раз, 2000 раз, 3000 раз, 4000 раз, 5000 раз или в 10000 раз по сравнению с количеством кольцевой ДНК, используемой в качестве матрицы в начале реакции.
[0071] В способе согласно изобретению, словосочетание «в бесклеточной системе» означает, что реакцию репликации не проводят в клетках. То есть, это означает, что способ согласно изобретению, проводимый в бесклеточной системе, осуществляют in vitro. То же самое относится и к «Способу (В)», описанному ниже.
[0072] Кольцевая ДНК, смешиваемая с реакционным раствором, описана выше в разделе «Кольцевая ДНК». Количество матричной ДНК, используемой в одной реакции, не имеет конкретных ограничений. Так, например, при инициации реакции, кольцевая ДНК может присутствовать в реакционном растворе в концентрации 10 нг/мкл или менее, 5 нг/мкл или менее, 1 нг/мкл или менее, 0,8 нг/мкл или менее, 0,5 нг/мкл, 0,1 нг/мкл или менее, 50 пг/мкл или менее, 5 пг/мкл или менее, 0,5 пг/мкл или менее, 50 фг/мкл или менее, 5 фг/мкл или менее или 0,5 фг/мкл или менее. Кроме того, при инициации реакции, одна молекула кольцевой ДНК в одной реакции может присутствовать в качестве матрицы, а поэтому она может быть использована для репликации или амплификации.
[0073] Кольцевая ДНК, используемая в качестве матрицы в способе (A), включает пару последовательностей ter, каждая из которых была встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую XerCD. Если эта кольцевая ДНК имеет последовательности ter, то реакционный раствор в стадии (1) также включает белок, обладающий активностью ингибирования репликации посредством связывания с последовательностями ter, и если кольцевая ДНК имеет нуклеотидную последовательность, распознаваемую XerCD, то реакционный раствор в стадии (1) также содержит белок XerCD.
[0074] В качестве белка, обладающего активностью ингибирования репликации посредством связывания с последовательностями ter и/или XerCD, может быть использован коммерчески доступный продукт, либо может быть также использован продукт, экстрагированный из микроорганизмов и т.п., а затем очищеннный, если это необходимо. Экстракция и очистка фермента из микроорганизмов могут быть осуществлены, если это необходимо, с применением методов, известных специалистам.
[0075] Комбинация последовательностей ter на ДНК и белке, обладающем активностью ингибирования репликации посредством связывания с последовательностями ter, представляет собой механизм терминации репликации. Этот механизм был обнаружен у бактерий множества типов, и, например, в Escherichia coli, этот механизм известен как система Tus-ter (Hiasa, H., and Marians, K. J., J. Biol. Chem., 1994, 269: 26959-26968; Neylon, C., et al., Microbiol. Mol. Biol. Rev., September 2005, p.501-526), а в бактериях Bacillus, этот механизм был известен как система RTP-ter (Vivian, et al., J. Mol. Biol., 2007, 370: 481-491). В способе согласно изобретению, благодаря этому механизму может подавляться образование ДНК-мультимера в качестве побочного продукта. Биологическое происхождение комбинации последовательностей ter на ДНК и белке, обладающем активностью ингибирования репликации посредством связывания с последовательностями ter, не имеет конкретных ограничений.
[0076] В предпочтительном варианте осуществления изобретения, в способе согласно изобретению используется комбинация последовательностей ter и белка Tus. Последовательность ter, используемая в комбинации с белком Tus, может представлять собой последовательность, содержащую 5'-GN[A/G][T/A]GTTGTAAC[T/G]A-3' (SEQ ID NO: 1), или более предпочтительно, 5'-G[T/G]A[T/A]GTTGTAAC[T/G]A-3' (SEQ ID NO: 2), 5'-GTATGTTGTAACTA-3' (SEQ ID NO: 3), 5'-AGTATGTTGTAACTAAAG-3' (SEQ ID NO: 4), 5'-GGATGTTGTAACTA-3' (SEQ ID NO: 5), 5'-GTATGTTGTAACGA-3' (SEQ ID NO: 6), 5'-GGATGTTGTAACTA-3' (SEQ ID NO: 7), 5'-GGAAGTTGTAACGA-3' (SEQ ID NO: 8), или 5'-GTAAGTTGTAACGA-3' (SEQ ID NO: 9). Происхождение белка Tus не имеет конкретных ограничений, но предпочтительно, чтобы белок Tus происходил от Escherichia coli. Белок Tus может содержаться в реакционном растворе в концентрации в пределах от 1 нМ до 200 нМ, и может предпочтительно, содержаться в концентрации в пределах от 2 нМ до 200 нМ, от 2 нМ до 100 нМ, от 5 нМ до 200 нМ, от 5 нМ до 100 нМ, от 10 нМ до 100 нМ, от 20 нМ до 100 нМ, или от 20 нМ до 80 нМ, но эти интервалы концентраций не ограничиваются указанными значениями.
[0077] В другом предпочтительном варианте осуществления изобретения, в способе согласно изобретению используется комбинация последовательностей ter и белка RTP. Последовательность ter, используемая в комбинации с белком RTP, представляет собой последовательность длиной в 23-30 нуклеотидов, содержащую 5'-AC[T/A][A/G]ANNNNN[C/T]NATGTACNAAAT-3' (SEQ ID NO: 10), или предпочтительно, 5'-ACTAATT[A/G]A[A/T]C[T/C]ATGTACTAAAT-3' (SEQ ID NO: 11), 5'-ACTAATT[A/G]A[A/T]C[T/C]ATGTACTAAATTTTCA-3' (SEQ ID NO: 12), 5'-GAACTAATTAAACTATGTACTAAATTTTCA-3' (SEQ ID NO: 13) или 5'-ATACTAATTGATCCATGTACTAAATTTTCA-3' (SEQ ID NO: 14). Если последовательность длиной в 23-30 нуклеотидов, содержащая любую из последовательностей, представленных в SEQ ID NN: 10-12, была отобрана как последовательность ter, то эта последовательность может иметь последовательность, которая по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, или по меньшей мере на 95% идентична SEQ ID NO: 13 или 14. Происхождение белка RTP не имеет конкретных ограничений, но предпочтительно, чтобы белок RTP происходил от бактерий Bacillus, а более предпочтительно, от Bacillus subtilis. Белок Tus может содержаться в реакционном растворе в концентрации в пределах от 1 нМ до 200 нМ, и может предпочтительно, содержаться в концентрации в пределах от 2 нМ до 200 нМ, от 2 нМ до 100 нМ, от 5 нМ до 200 нМ, от 5 нМ до 100 нМ, от 10 нМ до 100 нМ, от 20 нМ до 100 нМ, или от 20 нМ до 80 нМ, но эти интервалы концентраций не ограничиваются указанными значениями. Что касается последовательностей ter, то выражение «встроены за пределами oriC» означает, что последовательности ter встроены так, чтобы репликация, осуществляемая в направлении от oriC, достигалась под действием комбинации с белком, обладающим активностью ингибирования репликации посредством связывания с последовательностями ter, а репликация, осуществляемая в направлении к oriC, не достигалась и прекращалась. Стрелки в последовательностях ter, показанные на Фигуре 3(a) и на Фигуре 5, указывают на положение, в котором пара последовательностей ter встроена за пределами oriC. В соответствии с этим, что касается последовательности ter, то выражение «пара последовательностей ter, каждая из которых встроена за пределами oriC» означает, что последовательность, содержащая любую из последовательностей, представленных в SEQ ID NN: 1-14, встроена как одна последовательность ter с 5'-концевой стороны oriC, а последовательность, содержащая последовательность, комплементарную любой из последовательностей, представленных в SEQ ID NN: 1-14, встроена как другая последовательность ter с 3'-концевой стороны oriC.
[0078] Последовательности ter могут присутствовать в любых положениях, при условии, что каждая пара последовательностей ter встроена за пределами oriC. Так, например, пара последовательностей ter может присутствовать в области, противоположной oriC, либо она может присутствовать в области, расположенной рядом или смежной с обеими сторонами oriC. Если пара последовательностей ter присутствует в области, расположенной рядом или смежной с обеими сторонами oriC, то oriC и пара последовательностей ter могут быть получены в виде функционального кластера. В этом случае, преимущество заключается в облегчении введения oriC и пары последовательностей ter в ДНК и в снижении стоимости получения кольцевой ДНК, используемой в качестве матрицы.
[0079] Комбинация последовательности, распознаваемой XerCD на ДНК и белке XerCD, представляет собой механизм разделения ДНК-мультимеров (Ip, S. C. Y., et al., EMBO J., 2003, 22: 6399-6407). Белок XerCD представляет собой комплекс XerC и XerD. Такая последовательность, распознаваемая XerCD, известна как последовательность dif, последовательность cer и последовательность psi (Colloms, et al., EMBO J., 1996, 15(5): 1172-1181; Arciszewska, L. K., et al., J. Mol. Biol., 2000, 299: 391-403). В способе согласно изобретению, благодаря этому механизму может подавляться образование ДНК-мультимера в качестве побочного продукта. Биологическое происхождение комбинации последовательностей, распознаваемых XerCD на ДНК и белке XerCD, не имеет конкретных ограничений. Кроме того, стимулирующие факторы XerCD являются известными, и например, функция dif стимулируется белком FtsK (Ip, S. C. Y., et al., EMBO J., 2003, 22: 6399-6407). В одном из вариантов осуществления изобретения, такой белок FtsK может присутствовать в реакционном растворе, используемом в способе согласно изобретению.
[0080] Последовательность, распознаваемая XerCD, может представлять собой последовательность, содержащую 5'-GGTGCG[C/T][A/G][T/C]AANNNNNNTTATG[T/G]TAAA[T/C]-3' (SEQ ID NO: 15), 5'-GGTGCG[C/T]A[T/C]AANNNNNNTTATG[T/G]TAAAT-3' (SEQ ID NO: 16), 5'-GGTGCGC[A/G][T/C]AANNNNNNTTATGTTAAA[T/C]-3' (SEQ ID NO: 17), 5'-GGTGCG[C/T][A/G]CAANNNNNNTTATG[T/G]TAAA[T/C]-3' (SEQ ID NO: 18), 5'-GGTGCGCATAANNNNNNTTATGTTAAAT-3' (SEQ ID NO: 19), 5'-GGTGCGTACAANNNNNNTTATGGTAAAT-3' (SEQ ID NO: 20), 5'-GGTGCGCGCAANNNNNNTTATGTTAAAC-3' (SEQ ID NO: 21), 5'-GGTGCGCATAATGTATATTATGTTAAAT-3' (SEQ ID NO: 22/последовательность dif), 5'-GGTGCGTACAAGGGATGTTATGGTAAAT-3' (SEQ ID NO: 23/последовательность cer) или 5'-GGTGCGCGCAAGATCCATTATGTTAAAC-3' (SEQ ID NO: 24/последовательность psi), или последовательность, комплементарную любой из этих последовательностей. Нуклеотидная часть в положениях 1-11 в SEQ ID NN: 15-24 представляет собой XerC-связывающий сайт, а нуклеотидная часть в положениях 18-28 в SEQ ID NN: 15-24 представляет собой XerD-связывающий сайт. Поскольку нуклеотидная часть в положениях 12-17 в SEQ ID NN: 15-21 (то есть, часть из 6 нуклеотидов, состоящая из NNNNNN) не является областью, связывающейся с XerC или XerD, то их последовательность не имеет конкретных ограничений. Предпочтительно, последовательность нуклеотидов в положениях 12-17 в SEQ ID NN: 15-21 (то есть, часть из 6 нуклеотидов, состоящая из NNNNNN) может иметь последовательность, которая по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90% или по меньшей мере на 95% идентична последовательности нуклеотидов в положениях 12-17 в SEQ ID NN: 22-24.
[0081] Белок XerCD, предпочтительно, представляет собой белок XerCD, происходящий от Escherichia coli. Белок XerCD может содержаться в реакционном растворе в концентрации в пределах от 1 нМ до 200 нМ, а предпочтительно, в концентрации в пределах от 5 нМ до 200 нМ, от 5 нМ до 150 нМ, от 10 нМ до 200 нМ, от 10 нМ до 150 нМ, от 20 нМ до 200 нМ, от 20 нМ до 150 нМ или от 20 нМ до 100 нМ, но эти интервалы концентраций не имеют конкретных ограничений.
[0082] Последовательность, распознаваемая XerCD, может присутствовать в любом положении на кольцевой ДНК. Так, например, последовательность, распознаваемая XerCD, может присутствовать в области, противоположной oriC, либо она может присутствовать в области, расположенной рядом или смежной с oriC. Если последовательность, распознаваемая XerCD, присутствует в области, расположенной рядом или смежной с oriC, то oriC и последовательность, распознаваемая XerCD, могут быть получены в виде функционального кластера. Таким образом, преимущество заключается в облегчении введения oriC и последовательности, распознаваемой XerCD, в ДНК и в снижении стоимости получения кольцевой ДНК, используемой в качестве матрицы.
[0083] В описании настоящей заявки, идентичность (%) между двумя нуклеотидными последовательностями может быть определена путем визуального наблюдения и математического вычисления. Кроме того, идентичность (%) может быть также определена с использованием компьютерных программ. Примерами таких компьютерных программ для сравнения последовательностей могут быть программа BLASTN, которая имеется на web-сайте Национальной Медицинской Библиотеки США (http://www.ncbi.nlm.nih.gov/blast/bl2seq/bls.html) (Altschul et al. (1990) J. Mol. Biol. 215: 403-10); версия 2.2.7, или алгоритм WU-BLAST2.0. Что касается стандартных параметров по умолчанию для WU-BLAST2.0, то могут быть взяты параметры по умолчанию, описанные в нижеследующем интернет-сайте (http://blast.wustl.edu).
[0084] Первая, вторая и третья группы ферментов, содержащихся в реакционном растворе, описаны выше в разделе «Первая, вторая и третья группы ферментов».
[0085] В некоторых вариантах осуществления изобретения, первая группа ферментов, используемых в способе согласно изобретению, может включать комбинацию фермента, обладающего DnaA-активностью; одного или более нуклеоидных белков; фермента или группы ферментов, обладающих ДНК-гиразной активностью; белка, связывающегося с одноцепочечной ДНК (SSB); фермента, обладающего геликазной активностью DnaB-типа; фермента, обладающего ДНК-геликазной загрузочной активностью; фермента, обладающего ДНК-примазной активностью; фермента, обладающего ДНК-захватывающей активностью; и фермента или группы ферментов, обладающих активностью ДНК-полимеразы III*. В данном случае, один или более нуклеоидных белков могут представлять собой IHF или HU; фермент или группа ферментов, обладающих ДНК-гиразной активностью, могут представлять собой комплекс GyrA и GyrB; фермент, обладающий геликазной активностью DnaB-типа, может представлять собой геликазу DnaB; фермент, обладающий ДНК-геликазной загрузочной активностью, может представлять собой геликазный загрузочный фермент DnaC; фермент, обладающий ДНК-примазной активностью, может представлять собой примазу DnaG; фермент, обладающий ДНК-захватывающей активностью, может представлять собой ДНК-захватывающий фермент DnaN, а фермент или группа ферментов, обладающих активностью ДНК-полимеразы III*, могут представлять собой фермент или группу ферментов, содержащих любые из DnaX, HolA, HolB, HolC, HolD, DnaE, DnaQ и HolE.
[0086] В другом варианте осуществления изобретения, вторая группа ферментов, используемых в способе согласно изобретению, может включать комбинацию фермента, обладающего активностью ДНК-полимеразы I, и фермента, обладающего ДНК-лигазной активностью. Альтернативно, вторая группа ферментов может включать комбинацию фермента, обладающего активностью ДНК-полимеразы I, фермента, обладающего ДНК-лигазной активностью, и фермента, обладающего активностью РНКазы Н.
[0087] В другом варианте осуществления изобретения, третья группа ферментов, используемых в способе согласно изобретению, может включать фермент, обладающий активностью топоизомеразы III, и/или фермент, обладающий активностью топоизомеразы IV. Альтернативно, третья группа ферментов может включать комбинацию фермента, обладающего активностью топоизомеразы III, и фермента, обладающего геликазной активностью RecQ-типа. Альтернативно, третья группа ферментов может представлять собой комбинацию фермента, обладающего активностью топоизомеразы III, фермента, обладающего геликазной активностью RecQ-типа, и фермента, обладающего активностью топоизомеразы IV.
[0088] Реакционный раствор может содержать буфер, ATP, GTP, CTP, UTP, dNTP, источник иона магния и источник иона щелочного металла.
[0089] Буфер, содержащийся в реакционном растворе, не имеет конкретных ограничений, при условии, что он используется при pH в пределах от 7 до 9, а предпочтительно, при pH 8. Примерами буфера могут быть Трис-HCl, Hepes-KOH, фосфатный буфер, MOPS-NaOH и трицин-HCl. Предпочтительным буфером является Трис-HCl. Концентрация буфера может быть выбрана самим специалистом в данной области, и таким образом, не имеет конкретных ограничений. В случае Трис-HCl, выбранная концентрация может составлять, например, от 10 мМ до 100 мМ, от 10 мМ до 50 мМ или 20 мМ.
[0090] ATP означает аденозин-трифосфат. При инициации реакции, концентрация ATP, содержащегося в реакционном растворе, может составлять, например, в пределах от 0,1 мМ до 3 мМ, а предпочтительно, в пределах от 0,1 мМ до 2 мМ, от 0,1 мМ до 1,5 мМ или от 0,5 мМ до 1,5 мМ.
[0091] GTP, CTP и UTP означают гуанозин-трифосфат, цитидин-трифосфат и уридин-трифосфат, соответственно. При инициации реакции, концентрации GTP, CTP и UTP, содержащихся в реакционном растворе, могут независимо составлять, например в пределах от 0,1 мМ до 3,0 мМ, а предпочтительно, в пределах от 0,5 мМ до 3,0 мМ или от 0,5 мМ до 2,0 мМ.
[0092] dNTP представляет собой общий термин, означающий дезоксиаденозин-трифосфат (dATP), дезоксигуанозин-трифосфат (dGTP), дезоксицитидин-трифосфат (dCTP) и дезокситимидин-трифосфат (dTTP). При инициации реакции, концентрация dNTP, содержащегося в реакционном растворе, может составлять, например, в пределах от 0,01 до 1 мМ, а предпочтительно, в пределах от 0,05 мМ до 1 мМ или от 0,1 мМ до 1 мМ.
[0093] Источник иона магния представляет собой вещество, которое вводит ионы магния (Mg2+) в реакционный раствор. Примерами источника иона магния могут быть Mg(OAc)2, MgCl2 и MgSO4. Предпочтительным источником иона магния является Mg(OAc)2. При инициации реакции, концентрация источника иона магния, содержащегося в реакционном растворе, может представлять собой, например, концентрацию, необходимую для введения 5-50 мМ ионов магния в реакционный раствор.
[0094] Источник иона щелочного металла представляет собой вещество, которое вводит ионы щелочного металла в реакционный раствор. Примерами иона щелочного металла могут быть ионы натрия (Na+) и ионы калия (K+). Примерами источника иона щелочного металла могут быть глутамат калия, аспартат калия, хлорид калия, ацетат калия, глутамат натрия, аспартат натрия, хлорид натрия и ацетат натрия. Предпочтительным источником иона щелочного металла является глутамат калия. При инициации реакции, концентрация источника иона щелочного металла, содержащегося в реакционном растворе, может представлять собой концентрацию, необходимую для введения ионов щелочного металла в реакционный раствор в пределах от 100 мМ до 300 мМ, но эта концентрация не ограничивается указанными значениями. Как описано ранее, для достижения нужного баланса, концентрация 150 мМ может быть вычтена из концентрации вышеописанного источника иона щелочного металла.
[0095] Реакционный раствор может также включать неспецифический ингибитор адсорбции белка или неспецифический ингибитор адсорбции нуклеиновой кислоты. Предпочтительно, реакционный раствор может также включать неспецифический ингибитор адсорбции белка и неспецифический ингибитор адсорбции нуклеиновой кислоты. Благодаря присутствию в реакционном растворе такого неспецифического ингибитора адсорбции белка и/или неспецифического ингибитора адсорбции нуклеиновой кислоты, неспецифическая адсорбция между белками и/или между белком и кольцевой ДНК, или адгезия белка и кольцевой ДНК на поверхности сосуда могут подавляться, что позволяет повысить эффективность реакции.
[0096] Неспецифический ингибитор адсорбции белка представляет собой белок, который является не подходящим для реакции репликации или амплификации в способе согласно изобретению. Примерами такого белка могут быть альбумин бычьей сыворотки (BSA), лизозим, желатин, гепарин и казеин. Неспецифический ингибитор адсорбции белка может содержаться в реакционном растворе в концентрации в пределах от 0,02 до 2,0 мг/мл, а предпочтительно, от 0,1 до 2,0 мг/мл, от 0,2 до 2,0 мг/мл или от 0,5 до 2,0 мг/мл, но такая концентрация не ограничивается вышеуказанными пределами.
[0097] Неспецифический ингибитор адсорбции нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты или фактор, подобный нуклеиновой кислоте, которые являются не подходящими для реакции репликации или амплификации в способе согласно изобретению. Примерами такой молекулы нуклеиновой кислоты или фактора, подобного нуклеиновой кислоте, могут быть тРНК (транспортная РНК), рРНК (рибосомная РНК), мРНК (матричная РНК), гликоген, гепарин, олиго-ДНК, поли(I-C) (полиинозин-полицитидин), поли(dI-dC) (полидезоксиинозин-полидезоксицитидин), поли(A) (полиаденин) и поли(dA) (полидезоксиаденин). Неспецифический ингибитор адсорбции нуклеиновой кислоты может содержаться в реакционном растворе в концентрации в пределах от 1 до 500 нг/мкл, а предпочтительно, от 10 до 500 нг/мкл, от 10 до 200 нг/мкл или от 10 до 100 нг/мкл, но такая концентрация не ограничивается вышеуказанными пределами. Как было описано ранее, если тРНК была выбрана в качестве неспецифического ингибитора адсорбции нуклеиновой кислоты, то для достижения нужного баланса, концентрация 50 нг/мкл может быть вычтена из концентрации тРНК.
[0098] Реакционный раствор может также содержать экзонуклеазу, специфичную к линейной ДНК, или геликазу RecG-типа. Предпочтительно, реакционный раствор может также содержать экзонуклеазу, специфичную к линейной ДНК, или геликазу RecG-типа. Благодаря присутствию в реакционном растворе экзонуклеазы, специфичной к линейной ДНК, и/или геликазы RecG-типа, эти ферменты снижают количество линейной ДНК, образующейся в результате расщепления дуплекса или т.п. в процессе реакции репликации или амплификации, что приводит к увеличению выхода представляющего интерес суперспирализованного продукта.
[0099] Экзонуклеазой, специфичной к линейной ДНК, является фермент, который успешно гидролизует линейную ДНК у 5'-конца или 3'-конца. Экзонуклеаза, специфичная к линейной ДНК, конкретно не ограничивается типом или биологическим происхождением, при условии, что она обладает активностью, способствующей успешному гидролизу линейной ДНК у 5'-конца или 3'-конца. Так, например, могут быть использованы RecBCD, экзонуклеаза λ, экзонуклеаза III, экзонуклеаза VIII, экзонуклеаза T5, экзонуклеаза T7 и ATP-зависимая ДНКаза Plasmid-Safe™ (Epicentre). Предпочтительной экзонуклеазой, специфичной к линейной ДНК, является RecBCD. Экзонуклеаза, специфичная к линейной ДНК, может содержаться в реакционном растворе в концентрации в пределах от 0,01 до 1,0 ед./мкл, а предпочтительно, в пределах от 0,1 до 1,0 ед./мкл, но такая концентрация не ограничивается вышеуказанными пределами. Единицей ферментативной активности (ед.) экзонуклеазы, специфичной к линейной ДНК, является единица величины, достигаемой в том случае, когда количество фермента, необходимое для превращения 1 нмоль дезоксирибонеклеотида линейной ДНК в растворимую в кислоте ДНК в процессе реакции при 37°C в течение 30 минут было принято за 1 единицу.
[0100] Геликаза RecG-типа представляет собой фермент, который рассматривается как геликаза, разрушающая структуру ДНК, образующуюся как побочный продукт в результате взаимодействия между репликативными вилками по окончании реакции элонгации. Геликаза RecG-типа конкретно не ограничивается биологическим происхождением, при условии, что она обладает такой же активностью, как и RecG, происходящая от Escherichia coli. Так, например, предпочтительно использовать RecG, происходящую от Escherichia coli. RecG, происходящая от Escherichia coli, может содержаться в реакционном растворе в концентрации в пределах от 100 нМ до 800 нМ, а предпочтительно, в пределах от 100 нМ до 500 нМ, от 100 нМ до 400 нМ или от 100 нМ до 300 нМ, но такая концентрация не ограничивается вышеуказанными пределами. Геликаза RecG-типа, как единица ферментативной активности, может быть использована в концентрации в пределах, соответствующих пределам для вышеописанной RecG, происходящей от Escherichia coli.
[0101] Реакционный раствор может также содержать соль аммония. Примерами соли аммония могут быть сульфат аммония, хлорид аммония и ацетат аммония. Особенно предпочтительной солью аммония является сульфат аммония. Соль аммония может содержаться в реакционном растворе в концентрации в пределах от 0,1 мМ до 100 мМ, а предпочтительно, в пределах от 0,1 мМ до 50 мМ, от 1 мМ до 50 мМ или от 1 мМ до 20 мМ, но такая концентрация не ограничивается вышеуказанными пределами.
[0102] Если ДНК-лигаза, происходящая от Escherichia coli, которая представляет собой фермент, обладающий ДНК-лигазной активностью, используется в качестве фермента, принадлежащего ко второй группе ферментов, то в реакционном растворе содержится его кофактор NAD (никотинамид-аденин-динуклеотид). NAD может содержаться в реакционном растворе в концентрации в пределах от 0,01 мМ до 1,0 мМ, а предпочтительно, в пределах от 0,1 мМ до 1,0 мМ или от 0,1 мМ до 0,5 мМ, но такая концентрация не ограничивается вышеуказанными пределами.
[0103] Реакционный раствор, используемый в способе согласно изобретению, может также содержать восстановитель. Предпочтительными примерами восстановителя могут быть DTT, β-меркаптоэтанол и глутатион. Предпочтительным восстановителем является DTT.
[0104] Реакционный раствор, используемый в способе согласно изобретению, может также содержать фермент и субстрат, которые используются для регенерации ATP. Примерами комбинации фермента и субстрата в ATP-регенерирующей системе могут быть креатин-киназа и фосфат креатина, и пируват-киназа и фосфоенолпируват. Ферментом в ATP-регенерирующей системе является, например, миокиназа. Предпочтительной комбинацией фермента и субстрата в ATP-регенерирующей системе являются креатин-киназа и фосфат креатина.
[0105] Вышеописанная стадия (2) представляет собой стадию реакции взаимодействия реакционной смеси, полученной в стадии (1). Стадия (2) может представлять собой, например, стадию реакции взаимодействия реакционной смеси при температуре в пределах от 15°C до 80°C, от 15°C до 50°C или от 15°C до 40°C. Предпочтительно, стадия (2) может представлять собой стадию поддержания температуры в изотермических условиях. Такие изотермические условия не имеет конкретных ограничений, при условии, что они будут подходящими для прохождения реакции репликации ДНК. Так, например, изотермическими условиями могут быть постоянная температура в пределах от 20°C до 80°C, от 25°C до 50°C или от 25°C до 40°C, или приблизительно 30°C, которая является оптимальной температурой для ДНК-полимеразы. В настоящем описании, термины «поддержание в изотермических условиях» и «реакция в изотермических условиях» означают, что температура во время реакции поддерживается в пределах вышеописанных температур. Время поддержания температуры может быть определено, если это необходимо, в зависимости от количества продукта репликации или продукта амплификации представляющей интерес кольцевой ДНК. Время поддержания температуры может составлять, например, 1-24 часа.
[0106] Альтернативно, способ согласно изобретению может также включать, в качестве вышеописанной стадии (2), стадию инкубирования реакционной смеси, полученной в стадии (1), в термическом цикле повторения инкубирования при 30°C или выше и инкубирования при 27°C или ниже. Инкубирование при 30°C или выше не имеет конкретных ограничений, при условии, что температура будет составлять в пределах, подходящих для инициации репликации кольцевой ДНК, содержащей oriC. Так, например, температура может составлять от 30 до 80°C, от 30 до 50°C, от 30 до 40°C или 37°C. Инкубирование при 30°C или выше может быть проведено в течение периода времени от 10 секунд до 10 минут на цикл, хотя такое время не имеет конкретных ограничений. Инкубирование при 27°C или ниже не имеет конкретных ограничений, при условии, что такая температура будет подходящей для подавления инициации репликации и для прохождения реакции элонгации ДНК. Так, например, температура может составлять 10-27°C, 16-25°C или 24°C. Инкубирование при 27°C или ниже может быть, предпочтительно, проведено в зависимости от длины амплифицируемой кольцевой ДНК, но такая температура не имеет конкретных ограничений. Так, например, инкубирование может быть осуществлено в течение периода времени от 1 до 10 секунд на 1000 оснований за один цикл. Число термических циклов не имеет конкретных ограничений, но может составлять от 10 до 50 циклов, от 20 до 40 циклов, от 25 до 35 циклов или 30 циклов.
[0107] Способ согласно изобретению может также включать, после завершения стадии инкубирования реакционной смеси в изотермических условиях, стадию очистки продукта репликации или продукта амплификации кольцевой ДНК, необходимую для этой цели. Очистка кольцевой ДНК может быть осуществлена, если это необходимо, методами, известными специалистам.
[0108] Кольцевая ДНК, которая была реплицирована или амплифицирована с применением способа согласно изобретению, может быть использована для последующих целей, таких как трансформация, в форме реакционной смеси после реакции или в очищенной форме реакционной смеси.
[0109] <Способ репликации кольцевой ДНК (A')>
Известно, что, как и в случае комбинации XerCD и dif, и даже в случае использования комбинации Cre и его последовательности распознавания loxP, может быть осуществлено разделение ДНК-мультимеров (Ip, S. C. Y., et al., EMBO J., 2003, 22: 6399-6407). Авторами настоящего изобретения было обнаружено, что образование ДНК-мультимера в качестве побочного продукта может подавляться даже с использованием комбинации фермента для разделения ДНК-мультимеров и его распознающей последовательности вместо комбинации XerCD и dif в Способе (A).
[0110] В одном из своих вариантов, настоящее изобретение относится к способу репликации или амплификации кольцевой ДНК в бесклеточной системе, включающему следующие стадии:
(1) образования реакционной смеси кольцевой ДНК в качестве матрицы с реакционным раствором, содержащим:
первую группу ферментов, которые катализируют репликацию кольцевой ДНК;
вторую группу ферментов, которые катализируют созревание фрагмента Оказаки и синтезируют две сестринские кольцевые ДНК, состоящие из катенана; и
третью группу ферментов, которые катализируют разделение двух сестринских кольцевых ДНК; и
(2) реакции взаимодействия реакционной смеси, полученной в стадии (1), где:
кольцевая ДНК включает последовательность ориджина репликации (ориджина хромосомы (oriC)), которая может связываться с ферментом, обладающим DnaA-активностью, а также включает пару последовательностей ter, каждая из которых была встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров, где:
если кольцевая ДНК имеет последовательности ter, то реакционный раствор в стадии (1) также включает белок, обладающий активностью ингибирования репликации посредством связывания с последовательностями ter, и если кольцевая ДНК имеет нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров, то реакционный раствор в стадии (1) также содержит фермент для разделения ДНК-мультимеров (в настоящем описании, этот способ также называется «способом (А')).
[0111] Можно сказать, что Способ (A') представляет собой способ, в котором «XerCD» в Способе (A) относится к ферменту для разделения ДНК-мультимеров», а «нуклеотидная последовательность, распознаваемая XerCD» в Способе (A) относится к «нуклеотидной последовательности, распознаваемой ферментом для разделения ДНК-мультимеров». В соответствии с этим, объяснения, приведенные в разделе <Способ репликации кольцевой ДНК (A)> и относящиеся к отдельным аспектам Способа (A), также применимы к Способу (A').
[0112] Фермент для разделения ДНК-мультимеров представляет собой фермент, который инициирует генетическую рекомбинацию, в результате чего может достигаться разделение ДНК-мультимеров. Фермент сайт-специфической рекомбинации, который может распознавать специфическую нуклеотидную последовательность и может генерировать генетическую рекомбинацию в сайте нуклеотидной последовательности, может быть использован в качестве фермента для разделения ДНК-мультимеров. Специфическая нуклеотидная последовательность, распознаваемая ферментом для разделения ДНК-мультимеров, называется «нуклеотидной последовательностью, распознаваемой ферментом для разделения ДНК-мультимеров». При осуществлении генетической рекомбинации с использованием комбинации фермента для разделения ДНК-мультимеров и нуклеотидной последовательности, распознаваемой ферментом для разделения ДНК-мультимеров, может достигаться разделение ДНК-мультимеров. В Способе (A'), благодаря этому механизму может подавляться образование ДНК-мультимера в качестве побочного продукта. В качестве такого фермента для разделения ДНК-мультимеров может быть использован коммерчески доступный продукт, либо может быть использован фермент, экстрагированный из микроорганизмов и т.п., который затем очищают, если это необходимо. Экстракция и очистка фермента из микроорганизмов могут быть осуществлены, если это необходимо, методами, известными специалистам.
[0113] Примерами комбинации фермента для разделения ДНК-мультимеров и нуклеотидной последовательности, распознаваемой ферментом для разделения ДНК-мультимеров, могут быть, но не ограничиваются ими: последовательность XerCD и dif; последовательность Cre и loxP (Siegel, R. W., et al.., FEBS Lett., 2001, 499(1-2): 147-153; Araki, K., et al., Nucleic Acids Res.: 1997, 25(4): 868-872); последовательность рекомбинантного фермента FLP, происходящего от почкующихся дрожжей (Saccharomyces verevisiae), и последовательность FRT (Broach, J. R., et al., Cell, 1982, 29(1):227-234); последовательность рекомбинантного фермента DreO, происходящего от бактериофага D6, и последовательность rox (Anastassiadis, K., et al., Dis. Model. Mech., 2009, 2: 508-515); последовательность рекомбинантного ферментаR, происходящего от Zygosacchromyces rouxii, и последовательность RS (Araki, H., et al., J. Mol. Biol., 1985, 182(2): 191-203); и последовательности, принадлежащие к семейству серин-содержащих рекомбинантных ферментов (например, Gin, γδ, Tn3 и Hin) и разпознающие их последовательности (Smith, M. C., et al., Mol. Microbiol., 2002, 44: 299).
[0114] Последовательности XerCD и dif являются такими, как они были описаны выше в разделе <Способ репликации кольцевой ДНК (A)>.
[0115] Биологическое происхождение комбинации последовательности Cre и loxP не имеет конкретных ограничений. Cre, предпочтительно, представляет собой белок Cre, происходящий от бактериофага P1. Cre может содержаться в реакционном растворе в концентрации в пределах от 0,01 до 200 мЕд/мкл, и может, предпочтительно, содержаться в концентрации в пределах от 0,1 до 150 мЕд/мкл, от 0,1 до 100 мЕд/мкл, от 0,5 до 100 мЕд/мкл, от 0,5 до 80 мЕд/мкл, от 0,1 до 50 мЕд/мкл, от 1 до 50 мЕд/мкл, или от 1 до 30 мЕд/мкл, но эти концентрации не ограничиваются вышеуказанными интервалами.
[0116] Последовательность loxP, распознаваемая Cre, может представлять собой последовательность, содержащую 5'-ATAACTTCGTATAGCATACATTATACGAAGTTAT-3' (SEQ ID NO: 30), которая представляет собой консенсусную последовательность loxP, или 5'-ATAACTTCGTATAGtATACATTATACGAAGTTAT-3' (SEQ ID NO: 31/lox511), 5'-ATAACTTCGTATAGgATACtTTATACGAAGTTAT-3' (SEQ ID NO: 32/lox2272), 5'-ATAACTTCGTATAtacctttcTATACGAAGTTAT-3' (SEQ ID NO: 33/loxFAS), 5'-ATAACTTCGTATAGCATACATTATACGAAcggta-3' (SEQ ID NO: 34/lox RE), 5'-taccgTTCGTATAGCATACATTATACGAAGTTAT-3' (SEQ ID NO: 35/lox LE), которые представляют собой мутантные последовательности loxP (где строчные буквы означают мутантный нуклеотид, замененный на консенсусный нуклеотид), или последовательность, комплементарную любой из этих последовательностей.
[0117] Рекомбинантный фермент FLP, происходящий от почкующихся дрожжей (Saccharomyces verevisiae), может содержаться в реакционном растворе в концентрации в пределах от 1 нМ до 200 нМ, а предпочтительно, он может содержаться в концентрации в пределах от 5 нМ до 200 нМ, от 5 нМ до 150 нМ, от 10 нМ до 200 нМ, от 10 нМ до 150 нМ, от 20 нМ до 200 нМ, от 20 нМ до 150 нМ, или от 20 нМ до 100 нМ, но эти концентрации не ограничиваются вышеуказанными интервалами. Последовательность FRT, распознаваемая FLP, может представлять собой последовательность, содержащую 5'-GAAGTTCCTATTCTCTAGAAAGTATAGGAACTTC-3' (SEQ ID NO: 36), или комплементарную ей последовательность.
[0118] Рекомбинантный фермент DreO, происходящий от бактериофага D6, может содержаться в реакционном растворе в концентрации в пределах от 1 нМ до 200 нМ, а предпочтительно, он может содержаться в концентрации в пределах от 5 нМ до 200 нМ, от 5 нМ до 150 нМ, от 10 нМ до 200 нМ, от 10 нМ до 150 нМ, от 20 нМ до 200 нМ, от 20 нМ до 150 нМ, или от 20 нМ до 100 нМ, но эти концентрации не ограничиваются вышеуказанными интервалами. Последовательность rox, распознаваемая DreO, может представлять собой последовательность, содержащую 5'-TAACTTTAAATAATGCCAATTATTTAAAGTTA-3' (SEQ ID NO: 37), или комплементарную ей последовательность.
[0119] Рекомбинантный фермент R, происходящий от Zygosacchromyces rouxii, может содержаться в реакционном растворе в концентрации в пределах от 1 нМ до 200 нМ, а предпочтительно, он может содержаться в концентрации в пределах от 5 нМ до 200 нМ, от 5 нМ до 150 нМ, от 10 нМ до 200 нМ, от 10 нМ до 150 нМ, от 20 нМ до 200 нМ, от 20 нМ до 150 нМ, или от 20 нМ до 100 нМ, но эти концентрации не ограничиваются вышеуказанными интервалами. Последовательность RS, распознаваемая ферментом R, может представлять собой последовательность, содержащую последовательность, описанную Araki, H. et al. (J. Mol. Biol., 1985, 182(2): 191-203), или комплементарную ей последовательность.
[0120] Рекомбинантный фермент, принадлежащий к семейству серин-содержащих ферментов (γδ, Tn3, Gin и Hin), может содержаться в реакционном растворе в концентрации в пределах от 1 нМ до 200 нМ, а предпочтительно, он может содержаться в концентрации в пределах от 5 нМ до 200 нМ, от 5 нМ до 150 нМ, от 10 нМ до 200 нМ, от 10 нМ до 150 нМ, от 20 нМ до 200 нМ, от 20 нМ до 150 нМ, или от 20 нМ до 100 нМ, но эти концентрации не ограничиваются вышеуказанными интервалами. Рекомбинантный фермент, принадлежащий к семейству серин-содержащих ферментов γδ и Tn3, и каждая распознающая их последовательность res может представлять собой последовательность, содержащую последовательность, описанную Grindley N. D. F. et al. (Cell, 1982, 30: 19-27) или комплементарную ей последовательность. Рекомбинантный фермент, принадлежащий к семейству серин-содержащих ферментов Gin, и каждая распознающая их последовательность может представлять собой последовательность, содержащую последовательность, описанную Kahmann. R. et al. (Cell, 1985, 41: 771-780) или комплементарную ей последовательность. Рекомбинантный фермент, принадлежащий к семейству серин-содержащих ферментов Hin, и каждая распознающая их последовательность может представлять собой последовательность, содержащую последовательность, описанную Glasgow. A. C. et al. (J. Biol. Chem., 1989, 264: 10072-10082) или комплементарную ей последовательность.
[0121] Последовательность, распознаваемая ферментом для разделения ДНК-мультимеров, может присутствовать в любом положении на кольцевой ДНК. Так, например, последовательность, распознаваемая ферментом для разделения ДНК-мультимеров, может присутствовать в области, находящейся рядом или смежной с oriC, либо она может также присутствовать в области, противоположной oriC.
[0122] <Способ репликации кольцевой ДНК (B)>
В одном из своих вариантов, настоящее изобретение относится к способу репликации или амплификации кольцевой ДНК в бесклеточной системе, включающему следующие стадии:
(1) получения кольцевой ДНК, содержащей oriC, путем:
добавления транспозона и транспозазы oriC в буфер с образованием транспосомы oriC, где транспозон oriC представляет собой линейную ДНК, содержащую последовательность ориджина репликации (ориджина хромосомы (oriC)), которая может связываться с ферментом, обладающим DnaA-активностью, и внешние концевые последовательности (OE) у обоих концов; и
реакции взаимодействия транспосомы oriC с кольцевой ДНК, не содержащей oriC, в буфере для проведения реакции переноса,
(2) образования реакционной смеси кольцевой ДНК, содержащей oriC и полученной в стадии (1), с реакционным раствором, содержащим:
первую группу ферментов, которые катализируют репликацию кольцевой ДНК;
вторую группу ферментов, которые катализируют созревание фрагмента Оказаки и синтезируют две сестринские кольцевые ДНК, состоящие из катенана; и
третью группу ферментов, которые катализируют разделение двух сестринских кольцевых ДНК; и
(3) реакции взаимодействия реакционной смеси, полученной в стадии (2) (в настоящем описании, этот способ также называется «способом (В)).
[0123] Не ограничиваясь какой-либо конкретной теорией, авторы лишь отмечают, что в Способе (В), oriC вводят в кольцевую ДНК, не содержащую oriC, с использованием транспозона, с получением кольцевой ДНК, содержащей oriC, а затем кольцевую ДНК согласно изобретению, содержащую oriC, подвергают репликации или амплификации. Схематическое изображение представлено на Фигуре 2. Стадия, которая на Фигуре 2 указана под заголовком «Образование транспосомы» и «Реакция переноса», соответствует вышеописанной стадии (1). Что касается репликации или амплификации, то в вышеописанных стадиях (2) и (3), кольцевую ДНК подвергают репликации или амплификации за репликативный цикл, представленный на Фигуре 1, или путем повторения этого репликативного цикла. Определения репликации и амплификации кольцевой ДНК являются такими, как они были описаны выше.
[0124] Кольцевая ДНК, содержащая oriC и смешиваемая с реакционным раствором, является такой, как она описана выше в разделе <Кольцевая ДНК>. Количество кольцевой ДНК, содержащей oriC и используемой в одной реакции, является таким же, как оно указано выше и соответствует количеству матричной ДНК, используемой в Способе (A).
[0125] Кроме того, объяснение, относящееся к группам ферментов, содержащихся в реакционном растворе, и к другим компонентам, содержащимся, но необязательно, в реакционном растворе, является таким же, как оно приводится в Способе (A). Кроме того, вышеописанную стадию (3) осуществляют таким же способом, как и стадию (2) в Способе (A). Этот способ, также включающий стадию очистки продукта репликации или продукта амплификации кольцевой ДНК, и использование кольцевой ДНК, реплицированной или амплифицированной с применением способа согласно изобретению, является таким же, как и Способ (A).
[0126] Последовательности OE у обоих концов транспозона oriC могут представлять собой любые последовательности, при условии, что эти последовательности, как известно специалистам, могут распознаваться транспозазой и могут быть использованы в качестве последовательностей OE. В предпочтительном варианте осуществления изобретения, последовательность OE содержит последовательность, представленную в SEQ ID NO: 25 (5'-CTGTCTCTTATACACATCT-3') или комплементарную ей последовательность, и последовательность OE, содержащую последовательность, представленную в SEQ ID NO: 25, встраивают у 5'-конца линейной ДНК в стадии (1), а последовательность OE, содержащую последовательность, комплементарную последовательности, представленной в SEQ ID NO: 25, встраивают у 3'-конца линейной ДНК.
[0127] В вышеописанной стадии (1), концентрация транспозона oriC, используемого для получения транспосомы oriC, может составлять от 20 до 200 нМ, а предпочтительно, от 40 до 160 нМ.
[0128] Биологическое происхождение транспозазы не имеет конкретных ограничений, при условии, что она представляет собой фермент, который распознает последовательность OE, образует транспосому и переносит ДНК транспозона в кольцевую ДНК. Так, например, предпочтительно, может быть использована транспозаза, происходящая от Escherichia coli. Особенно предпочтительным является в высокой степени активный мутантный белок Tn5 (E54K, L372P) (Goryshin, I. Y., and Reznikoff, W. S., J. Biol. Chem., 1998, 273: 7367-7374). В качестве такой транспозазы может быть использован коммерчески доступный продукт, либо может быть также использован фермент, экстрагированый из микроорганизмов, а затем очищенный, если это необходимо. Экстракция и очистка ферментов из микроорганизмов могут быть осуществлены, если это необходимо, с применением методов, известных специалистам. Если в качестве такой транспозазы используется в высокой степени активный мутантный белок Tn5 (E54K, L372P), то концентрация белка, используемого для получения транспосомы oriC в вышеописанной стадии (1), может составлять от 50 до 200 нМ, а предпочтительно, от 80 до 150 нМ.
[0129] Буфер, используемый в стадии (1), не имеет конкретных ограничений, при условии, что он используется при pH 6-9, а предпочтительно, при pH 7,5. Примерами буфера могут быть Трис-уксусная кислота, Трис-HCl, Hepes-KOH, фосфатный буфер, MOPS-NaOH и трицин-HCl. Предпочтительным буфером является Трис-уксусная кислота или Трис-HCl. Концентрация буфера не имеет конкретных ограничений, и может быть выбрана самим специалистом в данной области. Если буфером является Трис-уксусная кислота или Трис-HCl, то его концентрация может быть выбрана, например, в пределах от 10 мМ до 100 мМ, от 10 мМ до 50 мМ или 20 мМ.
[0130] В стадии (1), стадию образования транспосомы oriC осуществляют путем инкубирования при температуре приблизительно 30°C в течение приблизительно 30 минут.
[0131] Реакцию переноса в стадии (1) осуществляют при оптимальной температуре транспозазы, которая составляет, например, 37°C. Время, необходимое для прохождения реакции переноса, может быть выбрано, если это необходимо, самим специалистом, и оно может составлять, например, приблизительно 15 минут. Кроме того, при проведении реакции переноса стадии (1) может быть добавлена тРНК. Что касается концентрации тРНК, добавленной при проведении реакции переноса в стадии (1), то такая концентрация может быть выбрана в пределах от 10 до 200 нг/мкл, от 30 до 100 нг/мкл или 50 нг/мкл.
[0132] В одном из вариантов осуществления изобретения, кольцевая ДНК, содержащая oriC в стадии (2), может также включать пару последовательностей ter, каждая из которых встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров, таким как XerCD или Cre. В этом случае, если кольцевая ДНК имеет последовательности ter, то реакционный раствор в стадии (2) также содержит белок, обладающий активностью ингибирования репликации посредством связывания с последовательностями ter, а если кольцевая ДНК имеет нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров, таким как XerCD или Cre, то реакционный раствор в стадии (2) также содержит фермент для разделения ДНК-мультимеров, такой как XerCD или Cre.
[0133] Кроме того, в другом варианте осуществления изобретения, пару последовательностей ter, каждая из которых встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров, таким как XerCD или Cre, получают так, чтобы они содержались в части транспозона oriC, и так, чтобы пара последовательностей ter и/или нуклеотидная последовательность, распознаваемая ферментом для разделения ДНК-мультимеров, таким как XerCD или Cre, могли быть также введены в кольцевую ДНК с использованием транспозона. В частности, в этом варианте осуществления изобретения, линейная ДНК в стадии (1) также содержит пару последовательностей ter, каждая из которых встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров, таким как XerCD или Cre; и если кольцевая ДНК имеет последовательности ter, то реакционный раствор в стадии (2) также содержит белок, обладающий активностью ингибирования репликации посредством связывания с последовательностями ter, а если кольцевая ДНК имеет нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров, таким как XerCD или Cre, то реакционный раствор в стадии (2) также содержит белок XerCD.
[0134] В настоящей заявке, определения и объяснения, относящиеся к паре последовательностей ter, каждая из которых встроена за пределами oriC, и/или к нуклеотидной последовательности, распознаваемой ферментом для разделения ДНК-мультимеров, таким как XerCD или Cre; и к белку, обладающему активностью ингибирования репликации посредством связывания с последовательностями ter, и/или к ферменту для разделения ДНК-мультимеров, такому как XerCD или Cre, являются такими, как они описаны выше в Способе (A) или в Способе (A').
[0135] В одном из вариантов осуществления изобретения, Способ (B) может также включать стадию (4) удаления транспозона oriC из кольцевой ДНК, реплицированной или амплифицированной в продукте реакции в стадии (3).
[0136] Стадия удаления транспозона oriC может включать обработку транспозазой в концентрации от 0,1 до 30 нМ, предпочтительно, от 1 до 20 нМ, а более предпочтительно, от 3 до 10 нМ, и обработку путем превращения конца ДНК в одноцепочечную ДНК с использованием одноцепочечной ДНК-экзонуклеазы, зависимой от линейной двухцепочечной ДНК, такой как ExoIII. Буфер, используемый для обработки транспозазой, может представлять собой буфер, используемый в стадии (1). Буфер, используемый для обработки одноцепочечной ДНК-экзонуклеазой, может представлять собой буфер любого состава, при условии, что одноцепочечная ДНК-экзонуклеаза будет действовать в этом буфере.
[0137] Кроме того, стадия удаления транспозона oriC может также включать обработку рестриктирующим ферментом, соответствующим сайту рестриктирующего фермента, содержащемуся в последовательности транспозона oriC. Такая обработка непосредственно направлена на специфическое расщепление транспозона oriC. Поэтому, в данном случае выбирают рестриктирующий фермент, соответствующий сайту рестриктирующего фермента, который содержится в транспозоне oriC, но не содержится в области, которая не является областью транспозона oriC в реплицированной и/или амплифицированной кольцевой ДНК. Для двухцепочечного расщепления, которое является специфичным к области, содержащейся в транспозоне oriC, вместо рестриктирующего фермента может быть использован CRISPR-Cas9. В этом случае, последовательность, специфичная к области, содержащейся в транспозоне oriC, называется руководящей РНК.
[0138] <Функциональный кластер (нуклеиновая кислота)>
В одном из своих аспектов, настоящее изобретение относится к нуклеиновой кислоте, содержащей oriC и пару последовательностей ter, каждая из которых встроена за пределами oriC, и/или к нуклеотидной последовательности, распознаваемой ферментом для разделения ДНК-мультимеров, таким как XerCD или Cre. Нуклеиновой кислотой, предпочтительно, является линейная ДНК, а более предпочтительно, двухцепочечная нуклеиновая кислота. Длина нуклеиновой кислоты не имеет конкретных ограничений, при условии, что такая нуклеиновая кислота может быть использован для получения кольцевой ДНК. В предпочтительном варианте осуществления изобретения, длина нуклеиновой кислоты составляет от 273 п.о. до 2,0 т.п.о., от 273 п.о. до 1,5 т.п.о. или от 273 п.о. до 1,0 т.п.о. Самая меньшая длина нуклеиновой кислоты составляет 273 п.о. Поскольку длина oriC составляет 245 п.о., а длина последовательности dif, которая представляет собой самую короткую последовательность из пары последовательностей ter и последовательности распознавания фермента для разделения ДНК-мультимеров, составляет 28 п.о., то длина в 273 п.о. представляет собой длину, полученную путем непосредственного связывания двух вышеуказанных последовательностей друг с другом.
[0139] Описанная выше нуклеиновая кислота может быть использована в качестве функционального кластера для получения кольцевой ДНК, используемой в качестве матрицы в Способе (A) согласно изобретению.
[0140] В другом своем варианте, настоящее изобретение относится к нуклеиновой кислоте, содержащей oriC и пару последовательностей ter, каждая из которых встроена за пределами oriC, и/или к нуклеотидной последовательности, распознаваемой ферментом для разделения ДНК-мультимеров, таким как XerCD или Cre, а также к внешним концевым последовательностям (ОЕ) у обоих концов. Нуклеиновой кислотой, предпочтительно, является линейная ДНК, а более предпочтительно, двухцепочечная нуклеиновая кислота. Длина нуклеиновой кислоты не имеет конкретных ограничений, при условии, что такая нуклеиновая кислота может быть использован для получения кольцевой ДНК. В предпочтительном варианте осуществления изобретения, длина нуклеиновой кислоты составляет от 311 п.о. до 2,0 т.п.о., от 311 п.о. до 1,5 т.п.о. или от 311 п.о. до 1,0 т.п.о. Самая меньшая длина нуклеиновой кислоты составляет 311 п.о. Длина oriC составляет 245 п.о., длина последовательности dif, которая представляет собой самую короткую последовательность из пары последовательностей ter и последовательности распознавания фермента для разделения ДНК-мультимеров, составляет 28 п.о., а длина двух последовательностей ОЕ составляет 38 п.о. Таким образом, длина 311 п.о. равна длине, полученной путем непосредственного связывания этих последовательностей друг с другом.
[0141] Описанная выше нуклеиновая кислота может быть использована в качестве функционального кластера, который действует как транспозон oriC в Способе (В) согласно изобретению.
[0142] В настоящей заявке, определения и объяснения, относящиеся к паре последовательностей ter, каждая из которых встроена за пределами oriC, и/или к нуклеотидной последовательности, распознаваемой ферментом для разделения ДНК-мультимеров, таким как XerCD или Cre; и к белку, обладающему активностью ингибирования репликации посредством связывания с последовательностями ter, и/или к ферменту для разделения ДНК-мультимеров, такому как XerCD или Cre, являются такими, как они описаны выше в Способе (A) или в Способе (A').
[0143] Преимущество вышеописанного функционального кластера заключается в возможном снижении стоимости получения кольцевой ДНК, содержащей oriC и используемой в качестве матрицы в Способах (A), (A') и (B).
[0144] <Набор>
В одном из своих аспектов, настоящее изобретение относится к набору для репликации или амплификации кольцевой ДНК, включающему комбинацию:
первой группы ферментов, которые катализируют репликацию кольцевой ДНК;
второй группы ферментов, которые катализируют созревание фрагмента Оказаки и синтезируют две сестринские кольцевые ДНК, состоящие из катенана; и
третьей группы ферментов, которые катализируют разделение двух сестринских кольцевых ДНК;
линейной ДНК, содержащей oriC и пару последовательностей ter, каждая из которых встроена за пределами oriC, и/или нуклеотидной последовательности, распознаваемой ферментом для разделения ДНК-мультимеров, таким как XerCD или Cre; и
если линейная ДНК имеет последовательности ter, то белок, обладает активностью ингибирования репликации посредством связывания с последовательностями ter, и/или если линейная ДНК имеет нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров, таким как XerCD или Cre, то фермент для разделения ДНК-мультимеров соответствует такой последовательности как XerCD или Cre (в настоящем описании, этот набор также называется «Набором (A))». Набор (A) представляет собой набор для осуществления Способа (A) или (A') согласно изобретению.
[0145] Конкретные ингредиенты и концентрации отдельных компонентов, включенных в Набор (A) согласно изобретению, являются такими же, как они описаны выше в разделах <Первая, вторая и третья группа ферментов>, <Способ амплификации кольцевой ДНК (A)> и <Способ амплификации кольцевой ДНК (A')>.
[0146] IB другом своем варианте, настоящее изобретение относится к набору для репликации или амплификации кольцевой ДНК, включающему комбинацию:
первой группы ферментов, которые катализируют репликацию кольцевой ДНК;
второй группы ферментов, которые катализируют созревание фрагмента Оказаки и синтезируют две сестринские кольцевые ДНК, состоящие из катенана; и
третьей группы ферментов, которые катализируют разделение двух сестринских кольцевых ДНК;
транспозона oriC, который представляет собой линейную ДНК, содержащую последовательность ориджина репликации (ориджина хромосомы (oriC)), которая может связываться с ферментом, обладающим DnaA-активностью, и внешние концевые последовательности (OE) у обоих концов; и
транспозазы (в настоящем описании, этот набор также называется «Набором (В))». Набор (В) представляет собой набор для осуществления Способа (В) согласно изобретению.
[0147] В некоторых вариантах осуществления изобретения, транспозон oriC, включенный в Набор (B), может также содержать пару последовательностей ter, каждая из которых встроена за пределами oriC, и/или нуклеотидную последовательность, распознаваемую ферментом для разделения ДНК-мультимеров, таким как XerCD или Cre. В этом случае, Набор (B) может также включать белок, обладающий активностью ингибирования репликации посредством связывания с последовательностями ter, и/или фермент для разделения ДНК-мультимеров, соответствующий последовательности, распознаваемой ферментом для разделения ДНК-мультимеров, встроенным в транспозон oriC.
[0148] Конкретные компоненты и концентрации каждого отдельного компонента, включенного в Набор (В) согласно изобретению, являются такими же, как они описаны выше в разделах <Первая, вторая и третья группа ферментов> и <Способ амплификации кольцевой ДНК (В)>.
[0149] Наборы (A) и (B) согласно изобретению могут представлять собой набор, содержащий все вышеописанные необходимые компоненты. Альтернативно, если набором согласно изобретению является набор, используемый в способе согласно изобретению, то он может и не содержать некоторых вышеописанных необходимых компонентов. Если набором согласно изобретению является набор, который не содержит некоторых вышеописанных необходимых компонентов, то специалист может добавить необходимые компоненты в набор во время амплификации в целях осуществления способов амплификации (А) и (В) согласно изобретению.
[0150] Наборы (А) и (В) согласно изобретению могут также включать дополнительные необходимые компоненты, включая один или более компонентов, выбранных из неспецифического ингибитора адсорбции белка; неспецифического ингибитора адсорбции нуклеиновой кислоты; экзонуклеазы, специфичной к линейной ДНК; геликазы RecG-типа, соли аммония; NAD; восстановителя; и комбинации фермента и субстрата в ATP-регенерирующей системе. Такие дополнительные необходимые компоненты могут быть включены в набор согласно изобретению в виде одного набора, либо они могут поставляться в виде другого набора, который впоследствии может быть использован вместе с набором согласно изобретению.
[0151] Наборы (А) и (В) согласно изобретению могут включать смесь вышеописанных необходимых компонентов, которые упакованы в виде одного продукта. В другом случае, набор согласно изобретению может включать вышеописанные необходимые компоненты, каждый из которых может быть упакован по отдельности, либо набор согласно изобретению может также включать необходимые компоненты нескольких типов, которые объединяют или смешивают, а затем упаковывают. Кроме того, наборы (А) и (В) согласно изобретению могут включать руководство, содержащее инструкции по осуществлению способов (А) и (В) амплификации кольцевой ДНК согласно изобретению.
Примеры
[0152] Конкретное объяснение настоящего изобретения приводится в Примерах. Следует отметить, что настоящее изобретение не ограничивается вариантами, представленными в нижеследующих Примерах.
[0153] Пример 1: Репликация кольцевой ДНК, ассоциированная с подавлением образования ДНК-мультимеров с использованием последовательности терминации ter и белка Tus
<Материалы и методы>
Кольцевую ДНК, используемую в качестве матрицы, получали следующим образом. Фрагмент oriC встраивали в плазмидный вектор M13mp18 с получением кольцевой ДНК размером 8,0 т.п.о. В область, противоположную oriC в этой кольцевой ДНК размером 8,0 т.п.о., встраивали ДНК-фрагмент, содержащий две последовательности ter (подчеркнуты), обращенные лицом друг к другу (5'-ACTTTAGTTACAACATACTTATT-N176-AATAAGTATGTTGTAACTAAAGT-3' (SEQ ID NO: 26)), с получением кольцевой ДНК размером 8,0 т.п.о. со встроенной ter (Фигура 3(a)). Эта кольцевая ДНК размером 8,0 т.п.о. со встроенной ter была использована в качестве матричной ДНК, а вышеупомянутая кольцевая ДНК размером 8,0 т.п.о. была использована в качестве контрольной ДНК, не содержащей последовательности ter.
[0154] Tus получали путем его продуцирования из Tus-экспрессирующего штамма Escherichia coli в соответствии со стадией, включающей аффинную колоночную хромотографию и колоночную гель-фильтрационную хромотографию.
[0155] Был получен реакционный раствор, имеющий состав, представленный в Таблице 1, и реакционный раствор, имеющий состав, представленный в Таблице 1, но с добавлением Tus до конечной концентрации 2 нМ или 5 нМ. К каждому из этих реакционных растворов добавляли матричную ДНК или контрольную ДНК до конечной концентрации 0,8 нг/мкл, а затем смешивали вместе на льду. После этого, полученную смесь инкубировали в инкубаторе при 30°C в течение 1 часа для реакции. Общий объем одной реакционной смеси составлял 10 микролитров. К реакционному раствору добавляли [α-32P]dATP, и после завершения реакции репликации ДНК, аликвоту реакционного раствора подвергали электрофорезу в агарозном геле (0,5% 1 × TAE, 150 В, 100 минут, 14°C). Затем, продукт с включенным32P детектировали на визуализирующем планшете BAS для подтверждения образования представляющей интерес суперспирализованной структуры.
[0156] [Таблица 1]
[0157] В этой таблице, SSB означает SSB, происходящий от E. coli; IHF означает комплекс IhfA и IhfB, происходящих от E. coli; DnaG означает DnaG, происходящий от E. coli; DnaN означает DnaN, происходящий от E. coli; PolIII* означает комплекс ДНК-полимеразы III*, состоящий из комплекса DnaX, HolA, HolB, HolC, HolD, DnaE, DnaQ и HolE; DnaB означает DnaB, происходящий от E. coli; DnaC означает DnaC, происходящий от E. coli; DnaA означает РНКазу H, происходящую от E. coli; лигаза представляет собой ДНК-лигазу, происходящую от E. coli; PolI означает ДНК-полимеразу I, происходящую от E. coli; GyrA означает GyrA происходящую от E. coli; GyrB означает GyrB, происходящую от E. coli; Topo IV означает комплекс ParC и ParE, происходящие от E. coli; Topo III означает топоизомеразу III, происходящую от E. coli; а RecQ означает RecQ, происходящую от E. coli.
[0158] SSB получали путем очистки штамма E. coli, экспрессирующего SSB, в стадиях, включающих преципитацию сульфатом аммония и ионообменную колоночную хроматографию.
[0159] IHF получали путем очистки штамма E. coli, коэкспрессирующего IhfA и IhfB, в стадиях, включающих преципитацию сульфатом аммония и аффинную колоночную хроматографию.
[0160] DnaG получали путем очистки штамма E. coli, экспрессирующего DnaG, в стадиях, которые включают преципитацию сульфатом аммония, анионообменную колоночную хроматографию и колоночную хроматографию методом гель-фильтрации.
[0161] DnaN получали путем очистки штамма E. coli, экспрессирующего DnaN, в стадиях, которые включают преципитацию сульфатом аммония и анионообменную колоночную хроматографию.
[0162] PolIII* получали путем очистки штамма E. coli, коэкспрессирующего DnaX, HolA, HolB, HolC, HolD, DnaE, DnaQ и HolE, в стадиях, которые включают преципитацию сульфатом аммония, аффинную колоночную хроматографию и колоночную хроматографию методом гель-фильтрации.
[0163] DnaB и DnaC получали путем очистки штамма E. coli, коэкспрессирующего DnaB и DnaC, в стадиях, которые включают преципитацию сульфатом аммония, аффинную колоночную хроматографию и колоночную хроматографию методом гель-фильтрации.
[0164] DnaA получали путем очистки штамма E. coli, экспрессирующего DnaA, в стадиях, которые включают преципитацию сульфатом аммония, преципитацию путем диализа и колоночную хроматографию методом гель-фильтрации.
[0165] GyrA и GyrB получали путем очистки смеси штамма E. coli, экспрессирующего GyrA, и штамма E. coli, экспрессирующего GyrB, в стадиях, которые включают преципитацию сульфатом аммония, аффинную колоночную хроматографию и колоночную хроматографию методом гель-фильтрации.
[0166] Topo IV получали путем очистки смеси штамма E. coli, экспрессирующего ParC, и штамма E. coli, экспрессирующего ParE, в стадиях, которые включают преципитацию сульфатом аммония, аффинную колоночную хроматографию и колоночную хроматографию методом гель-фильтрации.
[0167] Topo III получали путем очистки штамма E. coli, экспрессирующего Topo III, в стадиях, которые включают преципитацию сульфатом аммония и аффинную колоночную хроматографию.
[0168] RecQ получали путем очистки штамма E. coli, экспрессирующего RecQ, в стадиях, которые включают преципитацию сульфатом аммония, аффинную колоночную хроматографию и колоночную хроматографию методом гель-фильтрации.
[0169] Ферменты РНКаза H, лигаза и PolI были использованы в виде коммерчески доступных ферментов, происходящих от E. coli (Takara Bio Inc.).
[0170] <Результаты>
Результат детектирования продукта репликации представлен на Фигуре 3.
[0171] Было подтверждено, что если кольцевая ДНК размером 8,0 т.п.о. со встроенной ter была использована в качестве матрицы, а Tus содержалась в реакционном растворе, то кольцевая ДНК, имеющая представляющую интерес суперспирализованную структуру, была реплицирована или амплифицирована с подавлением образования мультимера как побочного продукта. С другой стороны, если кольцевая ДНК размером 8,0 т.п.о., не содержащая последовательности ter, была использована в качестве матрицы, и реакционный раствор не содержал Tus, то наблюдалось образование кольцевой ДНК, имеющей представляющую интерес суперспирализованную структуру, но в то же самое время также наблюдалось образование мультимера как побочного продукта.
[0172] Система Tus-ter представляет собой механизм терминации репликации кольцевой хромосомы. В соответствии с результатами эксперимента, представленными в данном примере, было подтверждено, что образование неспецифического ДНК-мультимера может подавляться путем включения этой системы в реакционную смесь для репликации или амплификации кольцевой ДНК.
[0173] Пример 2: Репликация кольцевой ДНК, ассоциированная с подавлением образования ДНК-мультимера с использованием последовательностей сайт-специфической рекомбинации dif и XerCD
<Материалы и методы>
Кольцевую ДНК размером 12 т.п.о. со встроенной dif, используемую в качестве матрицы, получали путем проведения реакции рекомбинации в клетках Escherichia coli так, чтобы последовательность dif (SEQ ID NO: 22) находилась в области, противоположной oriC кольцевой ДНК (Фигура 4(a)). В частности, с использованием клеток Escherichia coli, экспрессирующих группу белков рекомбинации фага λ, была проведена внутриклеточная реакция рекомбинации для получения кольцевой ДНК нужной длины, включающей кластер, содержащий oriC и ген резистентности к канамицину, и 4.2 т.п.о. -область, расположенную выше dif и 6.0 т.п.о. -область, расположенную ниже dif в хромосоме Escherichia coli.
[0174] В качестве контрольной ДНК, не содержащей последовательности dif, использовали кольцевую ДНК размером 8,0 т.п.о., описанную в Примере 1.
[0175] XerCD получали путем его очистки из штамма из штамма Escherichia coli, коэкспрессирующего XerC и XerD в соответствии со стадией, включающей преципитацию сульфатом аммония и аффинную колоночную хромотографию.
[0176] Был получен реакционный раствор, имеющий состав, представленный в Таблице 1 Примера 1, и реакционный раствор, имеющий состав, представленный в Таблице 1, но с добавлением XerCD до конечной концентрации 3,5 нМ, 7 нМ, 14 нМ или 35 нМ. К каждому из этих реакционных растворов добавляли матричную ДНК или контрольную ДНК до конечной концентрации 0,8 нг/мкл, а затем смешивали вместе на льду. После этого, полученную смесь инкубировали в инкубаторе при 30°C в течение 1 часа для реакции. Общий объем одной реакционной смеси составлял 10 микролитров. К реакционному раствору добавляли [α-32P]dATP, и после завершения реакции, побочный продукт детектировали способом, описанным в Пример 1. Затем подтверждали его структуру.
[0177] <Результаты>
Результат детектирования продукта репликации представлен на Фигуре 4(b).
[0178] Было подтверждено, что если кольцевая ДНК размером 12 т.п.о. со встроенной dif была использована в качестве матрицы, а XerCD содержался в реакционном растворе, то кольцевая ДНК, имеющая представляющую интерес суперспирализованную структуру, была реплицирована или амплифицирована с подавлением образования мультимера как побочного продукта. С другой стороны, если кольцевая ДНК размером 8,0 т.п.о., не содержащая последовательности dif, была использована в качестве матрицы, и реакционный раствор не содержал XerCD, то наблюдалось образование кольцевой ДНК, имеющей представляющую интерес суперспирализованную структуру, но в то же самое время также наблюдалось образование мультимера как побочного продукта.
[0179] Система XerCD-dif представляет собой механизм разделения кольцевой хромосомы на отдельные хромосомы. То есть, система XerCD-dif представляет собой механизм разделения ДНК-мультимеров. В соответствии с результатами эксперимента, представленными в данном примере, было подтверждено, что образование неспецифического ДНК-мультимера может подавляться путем включения этой системы в реакционную смесь для репликации или амплификации кольцевой ДНК.
[0180] Пример 3: Влияние положения последовательности ter или последовательности dif в кольцевой ДНК
В Примерах 1 и 2, матричную ДНК получали так, чтобы последовательность ter или последовательность dif находились в области, противоположной oriC в кольцевой ДНК. В Примере 3, реакцию репликации и/или амплификации осуществляли на кольцевой ДНК, в которой последовательность ter или последовательность dif находились в положении, близком или смежном с oriC.
[0181] <Материалы и методы>
Для конструирования кольцевой ДНК размером 15 т.п.о. с использованием генома Escherichia coli в качестве матрицы был амплифицирован и получен фрагмент ДНК размером 15 т.п.о., не содержащий oriC.
[0182] В качестве кольцевой ДНК, в которой последовательности ter были расположены рядом с oriC, кольцевую ДНК ori-ter размером в 15 т.п.о. получали следующим образом. Кластер ori-ter присоединяли к вышеописанному ДНК-фрагменту размером в 15 т.п.о., а затем подвергали циклизации с получением кольцевой ДНК ori-ter (Фигура 5). Последовательность кластера ori-ter (0,38 т.п.о.) указана ниже, а последовательности ter, расположенные по краям (подчеркнутые части), присутствовали с обоих концов кластера oriC (строчные буквы).
Кластер ori-ter: 5'-AGTATGTTGTAACTAAAGATAACTTCGTATAATGTATGCTATACGAAGTTATacagatcgtgcgatctactgtggataactctgtcaggaagcttggatcaaccggtagttatccaaagaacaactgttgttcagtttttgagttgtgtataacccctcattctgatcccagcttatacggtccaggatcaccgatcattcacagttaatgatcctttccaggttgttgatcttaaaagccggatccttgttatccacagggcagtgcgatcctaataagagatcacaatagaacagatctctaaataaatagatcttctttttaatacccaggatccATTTAACATAATATACATTATGCGCACCTTTAGTTACAACATACT-3' (SEQ ID NO: 27).
[0183] В качестве кольцевой ДНК, в которой последовательность dif была расположена рядом с oriC, кольцевую ДНК ori-dif размером в 15 т.п.о. получали следующим образом. Кластер ori-dif присоединяли к вышеописанному ДНК-фрагменту размером в 15 т.п.о., а затем подвергали циклизации с получением кольцевой ДНК ori-dif (Фигура 5). Последовательность кластера ori-dif (0,32 т.п.о.) указана ниже, а последовательность dif (подчеркнутая часть) присутствовала рядом слева от кластера oriC (строчные буквы).
Кластер ori-dif:
5'-ATTTAACATAATATACATTATGCGCACCAAGTATacagatcgtgcgatctactgtggataa
ctctgtCaggaagcttggatcaaccggtagttatccaaagaacaactgttgttcagtttttgagttgtgtataacccctcattctgat
cccAgcttatacggtccaggatcaccgatcattcacagttaatgatcctttccaggttgttgatcttaaaagccggatccttgttatc
cacagggcagtgcgatcctaataagagatcacaatagaacagatctctaaataaatagatcttctttttaatacccaggatcc-3' (SEQ ID NO: 28).
[0184] Для исследования подавления образования ДНК-мультимера, которое зависит от ter и Tus, к реакционному раствору, состав которого указан ниже в Таблице 2, добавляли Tus до конечной концентрации 0, 2, 6, 20 или 60 нМ, а затем к реакционной смеси добавляли кольцевую ДНК до конечной концентрации 0,5 нг/мкл, 5 пг/мкл, 50 фг/мкл или 0,5 фг/мкл. Полученную таким образом смесь подвергали реакции взаимодействия при 30°C в течение 3 часов или 17 часов.
[0185] Для исследования подавления образования ДНК-мультимера посредством XerCD, к реакционному раствору, состав которого указан ниже в Таблице 2, добавляли XerCD до конечной концентрации 0, 30 или 60 нМ, а затем к реакционной смеси добавляли кольцевую ДНК до конечной концентрации 0,5 нг/мкл. Полученную таким образом смесь подвергали реакции взаимодействия при 30°C в течение 2 часов.
[0186] Продукт реакции подвергали электрофорезу в агарозном геле (0,5% 1×TBE, 60 В, 60 минут), а затем окрашивали SybrGreen I (Takara Bio Inc.) для детектирования ДНК.
[0187] [Таблица 2]
[0188] Отдельные ферменты, указанные в Таблице, являются такими же, как они были описаны в Примере 1, и эти ферменты были получены или приобретены методами, описанными в Примере 1.
[0189] <Результат 1> Подавление образования ДНК-мультимера, которое зависит от ter и Tus
(1) Титрование Tus
Результат детектирования продукта репликации/амплификации представлен на Фигуре 6. Количество матричной ДНК составляло 0,5 нг/мкл, Tus использовали в количестве, указанном на Фигуре 6, а реакцию осуществляли при 30°C в течение 3 часов.
[0190] Если кольцевая ДНК ori-ter размером 15 т.п.о. была использована в качестве матрицы, а Tus содержался в реакционном растворе, то было подтверждено, что кольцевая ДНК, имеющая представляющую интерес суперспирализованную структуру, была реплицирована или амплифицирована с подавлением образования мультимера как побочного продукта. Кроме того, по мере увеличения концентрации Tus в реакционном растворе, эффект подавления образования мультимера усиливался. В частности, если Tus присутствовал в концентрации 20 нМ или 60 нМ, то образование мультимера было снижено до уровня, при котором образование мультимера уже почти не могло быть подтверждено.
[0191] С другой стороны, если кольцевая ДНК ori-dif размером 15 т.п.о. была использована в качестве матрицы, а Tus содержался в реакционном растворе, то эффект подавления образования мультимера не наблюдался. Этот результат показал, что система Tus-ter сообщает эффект подавления образования мультимера.
[0192] Кроме того, вышеупомянутый результат показал, что эффект подавления образования мультимера наблюдался даже в случае, когда последовательности ter присутствуют в положениях, находящихся рядом или смежных с oriC в кольцевой ДНК, используемой в качестве матрицы. То есть, это означает, что положения последовательностей ter, встроенных в кольцевую ДНК, не влияют на эффект подавления образования мультимера.
[0193] (2) Титрование ДНК
Результат детектирования продукта репликации/амплификации представлен на Фигуре 7. Матричная ДНК и Tus использовали в количествах, указанных на Фигуре 7, а реакцию осуществляли при 30°C в течение 17 часов.
[0194] Если кольцевая ДНК ori-ter размером 15 т.п.о. была использована в качестве матрицы, а Tus содержался в реакционном растворе, то было подтверждено, что кольцевая ДНК, имеющая представляющую интерес суперспирализованную структуру, была реплицирована или амплифицирована с подавлением образования мультимера как побочного продукта. В частности, было подтверждено, что этот эффект мог наблюдаться даже при снижении количества матричной ДНК до 0,5 фг/мкл.
[0195] <Результат 2> Подавление образования ДНК-мультимера посредством XerCD
Результат детектирования продукта репликации/амплификации представлен на Фигуре 8. Количество матричной ДНК составляло 0,5 нг/мкл, XerCD использовали в количестве, указанном на Фигуре 8, а реакцию осуществляли при 30°C в течение 2 часов.
[0196] Если кольцевая ДНК ori-dif размером 15 т.п.о. была использована в качестве матрицы, а XerCD содержался в реакционном растворе, то было подтверждено, что кольцевая ДНК, имеющая представляющую интерес суперспирализованную структуру, была реплицирована или амплифицирована с подавлением образования мультимера как побочного продукта. Кроме того, по мере увеличения концентрации XerCD в реакционном растворе, эффект подавления образования мультимера усиливался.
[0197] Кроме того, вышеупомянутый результат показал, что эффект подавления образования мультимера наблюдался даже в случае, когда последовательность dif присутствует в положении, находящемся рядом или смежном с oriC в кольцевой ДНК, используемой в качестве матрицы. То есть, это означает, что положение последовательности dif, встроенной в кольцевую ДНК, не влияет на эффект подавления образования мультимера.
[0198] Исходя из вышеупомянутых результатов 1 и 2, было обнаружено, что последовательности эффективно функционировали и подавляли образование ДНК-мультимера даже в том случае, когда последовательности ter и последовательность dif присутствовали в положении рядом или смежном с oriC. Последовательности ter и последовательность dif, которые могут находиться рядом с oriC, указывают на то, что эти последовательности, вместе с oriC, могут быть использованы в качестве функционального кластера для конструирования кольцевой ДНК.
[0199] Пример 4: Введение кластера oriC с использованием транспозона (1)
Для репликации или амплификации кольцевой ДНК способом согласно изобретению, oriC необходимо вводить в кольцевую ДНК, используемую в качестве матрицы. В Примере 4 описаны исследования по введению кластера oriC с использованием транспозона (Фигура 2).
[0200] <Материалы и методы>
В качестве транспозазы (Tnp) использовали в высокой степени активный мутантный белок Tn5 (E54K, L372P). Этот белок получали путем его очистки из штамма Escherichia coli, экспрессирующего этот белок, в соответствии со стадией, включающей преципитацию сульфатом аммония и аффинную колоночную хромотографию.
[0201] В качестве транспозона oriC использовали ДНК-фрагмент, состоящий из нижеследующей последовательности, в которой внешние концевые (OE) последовательности (подчеркнутые части) присутствовали у обоих концов последовательности, содержащей oriC, которая была затем 5'-фосфорилирована.
Транспозон oriC: 5'-CTGTCTCTTATACACATCTgaagatccggcagaagaatggctgggatcgtgggttaatttactcaaataagtatacagatcgtgcgatctactgtggataactctgtcaggaagcttggatcaaccggtagttatccaaagaacaactgttgttcagtttttgagttgtgtataacccctcattctgatcccagcttatacggtccaggatcaccgatcattcacagttaatgatcctttccaggttgttgatcttaaaagccggatccttgttatccacagggcagtgcgatcctaataagagatcacaatagaacagatctctaaataaatagatcttctttttaatacccaggatcccaggtctttctcaagccgacAGATGTGTATAAGAGACAG-3' (SEQ ID NO: 29).
[0202] Реакцию переноса oriC осуществляли путем инкубирования 116 нМ Tnp и 48 нМ транспозона oriC в буфере (10 мМ Трис-уксусной кислоты [pH 7,5], 15% глицерина, 50 мМ глутамата калия, 1 мМ DTT и 0,1 мМ EDTA) при 30°C в течение 30 минут с получением транспосомы oriC. Транспосому oriC (0,5 мкл) и ДНК-мишень (10 фМ) инкубировали в буфере (5 мкл; 10 мМ Трис-HCl [pH 7,5], 150 мМ глутамата калия и 10 мМ Mg(oAc)2) при 37°C в течение 15 минут для осуществления реакции переноса. В качестве такой ДНК-мишени использовали плазмиду для экспрессии гена Escherichia coli размером 15 т.п.о. (плазмиду pTT8), или плазмиду 9,3 т.п.о., экстрагированную из штамма HB8 Thermus thermophilus с высокой термофильностью. Затем проводили термоинактивацию при 70°C в течение 5 минут.
[0203] Аликвоту (0,5 мкл) реакционной смеси, полученной в результате проведения вышеописанной реакции переноса oriC, добавляли к реакционному раствору, состав которого указан в Таблице 2 Примера 3, и полученную таким образом смесь подвергали реакции взаимодействия при 30°C в течение 3 часов. Продукт реакции подвергали электрофорезу в агарозном геле (0,5% 1 × TBE, 60 В, 60 минут), а затем окрашивали SybrGreen I (Takara Bio Inc.) для детектирования ДНК.
[0204] <Результаты>
Результат, полученный с использованием плазмиды для экспрессии гена Escherichia coli размером 15 т.п.о. в качестве ДНК-мишени, представлен на Фигуре 9, а результат, полученный с использованием плазмиды размером 9,3 т.п.о. (плазмиды pTT8), экстрагированной из штамма HB8 Thermus thermophilus с высокой термофильностью и используемой в качестве ДНК-мишени, представлен на Фигуре 10.
[0205] Если аликвоту реакционной смеси, полученной в результате проведения реакции переноса oriC, в которой присутствовал Tnp, использовали в способе репликации или амплификации кольцевой ДНК, то была обнаружена суперспираль, которая представляет собой продукт репликации/продукт амплификации. С другой стороны, если Tnp не присутствует в реакции переноса oriC, то такой продукт репликации/продукт амплификации не обнаруживался даже после проведения способа репликации или амплификации кольцевой ДНК.
[0206] Кроме того, вышеупомянутый результат показал, что oriC мог быть эффективно введен в ДНК-мишень, в частности, в крайне низкой концентрации, которая составляла 10 фМ (0,1 пг/мкл), и что ДНК-мишень может быть амплифицирована способом репликации или амплификации кольцевой ДНК согласно изобретению. Этот результат показал, что введение oriC в ДНК-мишень может быть легко достигнуто с высокой эффективностью с использованием транспозона для кластера oriC, и что полученная таким образом кольцевая ДНК, содержащая oriC, может быть также эффективно амплифицирована способом репликации или амплификации кольцевой ДНК согласно изобретению.
[0207] Пример 5: Введение кластера oriC с использованием транспозона (2)
Были использованы та же самая транспозаза (Tnp) и тот же самый транспозон oriC как и в Примере 4.
[0208] Реакцию переноса oriC осуществляли путем инкубирования 116 нМ Tnp и 144 нМ транспозона oriC в буфере (10 мМ Трис-уксусной кислоты [pH 7,5], 15% глицерина, 50 мМ глутамата калия, 1 мМ DTT и 0,1 мМ EDTA) при 30°C в течение 30 минут с получением транспосомы oriC. Транспосому oriC (0,5 мкл), ДНК-мишень (1 пМ) (50 пг (3 × 106 молекул/5 мкл) и тРНК (50 нг/мкл) инкубировали в буфере (5 мкл; 10 мМ Трис-HCl [pH 7,5], 150 мМ глутамата калия и 10 мМ Mg(oAc)2) при 37°C в течение 15 минут для осуществления реакции переноса. В качестве такой ДНК-мишени использовали плазмиду для экспрессии гена Escherichia coli размером 15 т.п.о. Затем проводили термоинактивацию при 70°C в течение 5 минут.
[0209] Аликвоту (0,5 мкл) реакционной смеси, полученной в результате проведения вышеописанной реакции переноса oriC, добавляли к 5 мкл реакционного раствора, состав которого указан в Таблице 2 Примера 3, и полученную таким образом смесь подвергали реакции взаимодействия при 30°C в течение 4 часов. Продукт реакции подвергали электрофорезу в агарозном геле (0,5% 1 × TBE, 60 В, 55 минут), а затем окрашивали SybrGreen I (Takara Bio Inc.) для детектирования ДНК. Результат представлен на Фигуре 11. Как описано в Примере 4, если аликвоту реакционной смеси, полученной в результате проведения реакции переноса oriC, в которой присутствовал Tnp, использовали в реакции репликации или амплификации кольцевой ДНК, то была обнаружена суперспираль, которая представляет собой продукт репликации/продукт амплификации. С другой стороны, если Tnp не присутствует в реакции переноса oriC, то такой продукт репликации/продукт амплификации не обнаруживался даже после проведения реакции репликации или амплификации кольцевой ДНК.
[0210] Кроме того, дополнительное количество ДНК-мишени в реакции переноса oriC заменяли на 1 пМ (50 пг (3 × 106 молекул/5 мкл), 0,1 пМ (50 пг (3 × 105 молекул/5 мкл), 10 фМ (500 фг (3 × 104 молекул/5 мкл) и 1 фМ (50 фг (3 × 103 молекул/5 мкл), и проводили ту же самую реакцию, описанную выше. Результат представлен на Фигуре 12. Результат показал, что oriC мог быть эффективно введен в ДНК-мишень в крайне низкой концентрации, которая составляла 1 фМ (50 фг (3000 молекул/5 мкл), и что ДНК-мишень может быть амплифицирована способом репликации или амплификации кольцевой ДНК согласно изобретению.
[0211] Пример 6: Амплификация термофильной плазмиды посредством переноса транспозона oriC
Реакцию переноса oriC и реакцию репликации или амплификации кольцевой ДНК осуществляли как описано в Примере 5, за исключением того, что в реакции переноса oriC использовали плазмиду размером 9,3 т.п.о. (плазмиду pTT8), экстрагированную из штамма HB8 Thermus thermophilus с высокой термофильностью в количестве 50 фг в качестве ДНК-мишени. Результат представлен на Фигуре 13.
[0212] Если плазмиду pTT8 гидролизовали ферментами Kpn I и Nhe I, то получали фрагменты размером 5,3 т.п.о., 1,7 т.п.о., 1,3 т.п.о. и 1,0 т.п.о. как показано на плазмидной карте на Фигуре 14. Продукт репликации/продукт амплификации, полученный посредством вышеописанной реакции, гидролизовали рестриктирующими ферментами Kpn I и Nhe I. Результат представлен на Фигуре 14. Если продукт репликации/продукт амплификации, полученный посредством вышеописанной реакции, гидролизовали рестриктирующими ферментами Kpn I и Nhe I, то образование фрагментов размером 5,3 т.п.о., 1,7 т.п.о., 1,3 т.п.о. и 1,0 т.п.о. было подтверждено как и в случае плазмиды pTT8. Этот результат показал, что продукт репликации/продукт амплификации, полученный посредством вышеописанной реакции, представляет собой кольцевую ДНК, полученную путем переноса транспозона oriC на плазмиду pTT8 с последующей репликацией и/или амплификацией полученного продукта.
[0213] Эти результаты показали, что даже в случае использования плазмиды размером 9,3 т.п.о. с высоким процентом содержания GC (процент содержания GC составляет приблизительно 70%), которая происходила от гетерологичных клеток, oriC может быть эффективно введен в ДНК-мишень в крайне низкой концентрации как описано в Примере 5, и ДНК-мишень может быть амплифицирована способом репликации или амплификации кольцевой ДНК согласно изобретению.
[0214] Пример 7: Амплификация λДНК посредством переноса транспозона oriC
λДНК представляет собой линейную ДНК, происходящую от бактериофага. Эта ДНК была подвергнута циклизации, а затем в нее вводили oriC посредством переноса транспозона oriC с получением кольцевой ДНК. Затем, на кольцевой ДНК осуществляли способ репликации или амплификации кольцевой ДНК согласно изобретению.
[0215] (1) Реакция отжига и репарации гэпов
Реакцию отжига осуществляли следующим образом. 160 нг/мкл λДНК (48 т.п.о./Toyobo Co., Ltd.) добавляли в буфер (10 мМ Трис-HCl (pH 7,5), 50 мМ NaCl и 1 мМ EDTA) с получением 5 мкл раствора. Этот раствор инкубировали при 65°C в течение 5 минут, после чего охлаждали до 4°C при понижении температуры со скоростью -5°C/30 сек., и COS-сайты у обоих концов λДНК присоединяли друг к другу для циклизации.
[0216] Реакцию репарации гэпов осуществляли следующим образом. После завершения реакции отжига, 0,5 мкл полученного раствора добавляли к реакционному раствору, содержащему 50 нМ лигазы, 50 нМ Pol I, 20 мЕд/мкл Exo III, 5 нМ гиразы и 0,1 мг/мл BSA (где в качестве реакционного буфера использовали реакционный буфер, состав которого указан в Таблице 2 (то есть, реакционный буфер, не содержащий группы ферментов, указанной в Таблице 2)) и полученную таким образом смесь подвергали реакции взаимодействия при 30°C в течение 16 часов.
[0217] Продукт реакции, полученный в результате реакции репарации гэпов, подвергали электрофорезу в агарозном геле (0,5% 1 × TBE, 60 В, 55 минут), а затем окрашивали SybrGreen I (Takara Bio Inc.) для детектирования ДНК. Результат представлен на Фигуре 15. В продукте реакции, полученном в результате реакции репарации гэпов, наблюдалась полоса суперспирали, указывающая на присутствие кольцевой ДНК без гэпов.
[0218](2) Перенос транспозона oriC и амплификация λДНК
Продукт реакции (1 мкл), полученный в результате реакции репарации гэпов, использовали в качестве раствора, содержащего ДНК-мишень, а реакцию переноса oriC и реакцию репликации или амплификации кольцевой ДНК осуществляли как описано в Примере 5. В данном случае, для реакции репликации или амплификации кольцевой ДНК использовали реакционный раствор, состав которого показан в Таблице 2 Примера 3, и к которму также добавляли 60 нМ RecG и 0,5 Ед/мкл RecJf (NEB). RecG получали путем его продуцирования из RecG-экспрессирующего штамма Escherichia coli в соответствии со стадией, включающей преципитацию сульфатом аммония и аффинную колоночную хромотографию. Результат представлен на Фигуре 16.
[0219] Если λДНК гидролизовали рестриктирующим ферментом HindIII, то получали фрагменты размером 27 т.п.о., 9,4 т.п.о., 6,6 т.п.о., 2,3 т.п.о. и 2,0 т.п.о. Продукт репликации/продукт амплификации, полученный посредством вышеописанной реакции, гидролизовали рестриктирующим ферментом HindIII (37°C, 3 часа). Результат представлен на Фигуре 17. Если продукт репликации/продукт амплификации, полученный посредством вышеописанной реакции, гидролизовали ферментом HindIII, то фрагменты размером 7 т.п.о., 9,4 т.п.о., 6,6 т.п.о., 2,3 т.п.о. и 2,0 т.п.о. подтверждали как и в случае λДНК. Этот результат показал, что продукт репликации/продукт амплификации, полученный посредством вышеописанной реакции, представляет собой кольцевую ДНК, полученную путем переноса транспозона oriC на кольцевую λДНК с последующей репликацией и/или амплификацией полученной λДНК.
[0220] Эти результаты показали, что даже в случае линейной ДНК, реакция репликации или амплификации кольцевой ДНК согласно изобретению может быть осуществлена путем проведения реакции переноса oriC после циклизации линейной ДНК.
[0221] Пример 8: Удаление транспозона oriC (1)
Были проведены исследования для того, чтобы определить, может ли быть удален oriC, введенный в кольцевую ДНК путем реакции переноса oriC.
[0222] (1) Кольцевая ДНК
Транспозон oriC Примера 5 содержит ген резистентности к канамицину (Km) и oriC. С другой стороны, плазмида для экспрессии гена Escherichia coli размером 15 т.п.о. содержит ген резистентности к ампициллину (Amp). Затем, из плазмид, полученных путем реакции переноса oriC Примера 5, отбирали и выделяли кольцевую ДНК (обозначаемую «p15k::Km-oriC»), где транспозон oriC был перенесен в область, кодирующую ген резистентности к ампициллину плазмиды для экспрессии гена Escherichia coli размером 15 т.п.о.. В частности, плазмиды, полученные путем реакции переноса oriC Примера 5, переносили в Escherichia coli, и трансформанты, которые приобретали чувствительность к Amp и резистентность к Km, клонировали путем скрининга.
[0223] (2) Диссоциация транспозона oriC
Область, соответствующую транспозону oriC, перенесенному посредством реакции переноса oriC, диссоциировали с использованием транспозазы. Кроме того, области размером 9 п.о. образовывали дубликаты на обоих концах транспозона oriC, если транспозон oriC был перенесен путем реакции переноса oriC. Таким образом, после удаления транспозона oriC, необходимо обработать эти 9 п.о. -области после завершения присоединения так, чтобы эти области не могли образовывать дубликаты. Конец ДНК, содержащий 9 п.о. -область, полученную в результате экстракции транспозона oriC, превращали в одноцепочечный конец с использованием ExoIII, который представляет собой одноцепочечную ДНК-экзонуклеазу, зависимую от линейной двухцепочечной ДНК. Затем, с использованием дуплицированной части 9 п.о. -областей, одиночные цепи были присоединены друг к другу посредством отжига и подвергнуты циклизации. На Фигуре 18 представлено схематическое изображение такой процедуры.
[0224] В частности, была проведена нижеследующая реакция.
[0225] Реакцию диссоциации транспозона oriC осуществляли путем инкубирования 0-30 нМ (0 нМ, 3 нМ, 10 нМ и 30 нМ) транспозазы и 2 нг/мкл p15k::Km-oriC в буфере (5 мкл; 10 мМ Трис-HCl [pH 7,5], 150 мМ глутамата калия и 10 мМ Mg(oAc)2) при 37°C в течение 16 часов. Затем проводили термоинактивацию при 70°C в течение 5 минут.
[0226] Буфер Takara ExoIII (50 мМ Трис-HCl (pH 8,0), 5 мМ MgCl2 и 1 мМ DTT) и 20 мЕд/мкл ExoIII (Takara) добавляли к 1 мкл продукта реакции диссоциации транспозона oriC до конечного объема 5 мкл. Эту смесь подвергали реакции взаимодействия при 30°C в течение 10 минут, так, чтобы она была обработана ExoIII.
[0227] Продукт реакции, обработанный ExoIII, инкубировали при 65°C в течение 5 минут, а затем охлаждали до 4°C при снижении температуры со скоростью -0,5°C/30 секунд для отжига.
[0228] (3) Подтверждение диссоциации транспозона oriC посредством трансформации
Escherichia coli трансформировали с использованием 2 мкл образца, полученного в вышеописанной стадии (2), и химически компетентных клеток (Escherichia coli DH5α)50. Трансформированные клетки высевали на планшет, содержащий 100 мкг/мл ампициллина, 25 мкг/мл канамицина, а затем инкубировали при 37°C в течение ночи.
[0229] Образование колоний, если транспозазу не добавляли в вышеописанной стадии (2) (0 нМ), принимали за 100, а затем определяли относительное число колониеобразующих единиц (%) при изменении концентрации транспозазы.
[0230] (4) Результаты
Результаты представлены на Фигуре 19 и Фигуре 20.
[0231] Из результата, представленного на Фигуре 19, стало очевидным, что число трансформированных клеток, резистентных к канамицину, снижалось в зависимости от концентрации добавленной транспозазы. При добавлении 30 нМ транспозазы, трансформированные клетки, резистентные к канамицину, почти не присутствовали. Этот результат показал, что транспозон oriC может диссоциироваться при добавлении транспозазы.
[0232] Кроме того, из результата, представленного на Фигуре 20, стало очевидным, что ген ампициллина, после его деструкции, может быть восстановлен путем инсерции транспозона oriC в зависимости от ExoIII-обработки.
[0233] Пример 9: Удаление транспозона oriC (2)
Было проведено исследование удаления транспозона oriC путем проведения обработки с использованием рестриктирующего фермента, соответствующего сайту рестриктирующего фермента, содержащегося в последовательности транспозона oriC, одновременно с ExoIII-обработкой. Такая процедура схематически проиллюстрирована на Фигуре 21.
[0234] (1) Кольцевая ДНК
Была использована кольцевая ДНК, описанная в Примере 8.
[0235] (2) Диссоциация транспозона oriC
Реакцию диссоциации транспозона oriC осуществляли как описано в Примере 8, за исключением того, что использовали 10 нМ транспозазы.
[0236] Буфер Takara ExoIII (50 мМ Трис-HCl (pH 8,0), 5 мМ MgCl2 и 1 мМ DTT) и 20 мЕд/мкл одного ExoIII (Takara) или 20 мЕд/мкл ExoIII (Takara) и 0,6 Ед/мкл NheI (NEB) добавляли к 1 мкл продукта реакции диссоциации транспозона oriC до конечного объема 5 мкл. Эту смесь подвергали реакции взаимодействия при 30°C в течение 10 минут, так, чтобы она была обработана ExoIII.
[0237] Продукт реакции, обработанный ExoIII и NheI, инкубировали при 65°C в течение 5 минут, а затем охлаждали до 4°C при снижении температуры со скоростью -0,5°C/30 сек. для отжига.
[0238] (3) Подтверждение диссоциации транспозона oriC посредством трансформации
Трансформацию осуществляли способом, описанным в Примере 8. Затем определяли число колониеобразующих единиц.
[0239] (4) Результат
Результат представлен в Таблице 3.
[0240] [Таблица 3]
[0241] Этот результат показал, что проблема, связанная с тем, что плазмида, содержащая транспозоны, которые не были диссоциированы из кольцевой ДНК даже с использованием транспозазы, остается фоновой, может быть решена путем расщепления сайта ДНК транспозона рестриктирующим ферментом (NheI).
[0242] Пример 10: Подавление образования ДНК-мультимера посредством Cre
<Материалы и методы>
С использованием pUC19 (Takara Bio Inc.) в качестве матрицы была проведена ПЦР с использованием праймеров SUE1156: 5'-CTATGCGGCATCAGAGCAG-3' (SEQ ID NO: 38) и SUE1361: 5'-GTTAAGCCAGCCCCGACAC-3' (SEQ ID NO: 39), с получением фрагмента ДНК pUC размером 2,6 т.п.о.
[0243] В качестве кольцевой ДНК, в которой последовательность loxP находилась в положении рядом с oriC, получали кольцевую ДНК pUC19-OLDT следующим образом. Фрагмент ДНК pUC присоединяли к кластеру OLDT для циклизации с получением кольцевой ДНК pUC19-OLDT. Последовательность кластера OLDT (0,41 т.п.о.) представлена ниже, а кластер OLDT имеет последовательность loxP (подчеркнутая часть), смежную с кластером oriC и расположенную слева от него (эта часть показана строчными буквами).
Кластер OLDT:
5'-CGCGTCAGCGGGTGTTGGCGGGTGTCGGGGCTGGCTTAACAGTATGTT
GTAACTAAAGATAACTTCGTATAATGTATGCTATACGAAGTTATACAGA
TCGTGCgatctactgtggataactctgtcaggaagcttggatcaaccggtagttatccaaagaacaactgttgttcag
Tttttgagttgtgtataacccctcattctgatcccagcttatacggtccaggatcaccgatcattcacagttaatgatcctttcca
Ggttgttgatcttaaaagccggatccttgttatccacagggcagtgcgatcctaataagagatcacaatagaacagatctctaa
ataaatagatcttctttttaatacCCAGGATCCATTTAACATAATATACATTATGCGCACCTT
TAGTTACAACATACTATGCGGCATCAGAGCAGATTGTACTGAGAGTGCACCA
T-3' (SEQ ID NO: 40).
[0244] В качестве контрольной кольцевой ДНК, не содержащей такой последовательности loxP, была получена кольцевая ДНК pUC-OriC300 следующим образом. Фрагмент ДНК pUC присоединяли к кластеру oriC300 для циклизации с получением кольцевой ДНК pUC-oriC 300. Последовательность кластера oriC 300 (0,41 т.п.о.) представлена ниже, а кластер oriC 300 имеет кластер oriC (эта часть показана строчными буквами).
Кластер oriC 300:
5'-CGCGTCAGCGGGTGTTGGCGGGTGTCGGGGCTGGCTTAACAGTATGTTGTA
ACTAAAgatctactgtggataactctgtcaggaagcttggatcaaccggtagttatccaaagaacaactgttgttcagtttt
Gagttgtgtataacccctcattctgatcccagcttatacggtccaggatcaccgatcattcacagttaatgatcctttccaggttgtt
Gatcttaaaagccggatccttgttatccacagggcagtgcgatcctaataagagatcacaatagaacagatctctaaataaata
gatcttctttttaatacTTTAGTTACAACATACTATGCGGCATCAGAGCAGATTGTACTG
AGAGTGCACCAT-3' (SEQ ID NO: 41)
[0245] Был использован Cre, закупленный у NEB.
[0246] Был получен реакционный раствор, имеющий состав, представленный в Таблице 2 Примера 3, и реакционный раствор, имеющий состав, представленный в Таблице 3 Примера 3, в которые добавляли Cre до конечной концентрации 1 мЕд/мкл, 3 мЕд/мкл, 10 мЕд/мкл или 30 мЕд/мкл. В каждый из этих реакционных растворов добавляли кольцевую ДНК pUC19-OLDT или кольцевую ДНК pUC-OriC300 до конечной концентрации 0,01 нг/мкл, и эти растворы смешивали на льду. Затем полученную смесь инкубировали в инкубаторе при 33°C в течение 3 часов для реакции.
[0247] Продукт реакции подвергали электрофорезу в агарозном геле (0,5% 1×TBE, 60 В, 60 минут), а затем окрашивали SybrGreen I (Takara Bio Inc.) для детектирования ДНК.
[0248] <Результаты>
Результат детектирования продукта репликации/амплификации представлен на Фигуре 22.
[0249] Было подтверждено, что если кольцевая ДНК pUC-OLDT, в которой последовательность loxP расположена рядом с oriC, была использована в качестве матрицы, а Cre содержался в реакционном растворе, то кольцевая ДНК, имеющая представляющую интерес суперспирализованную структуру, была реплицирована или амплифицирована с подавлением образования мультимера как побочного продукта. В это время также наблюдалось появление промежуточного продукта в процессе разделения мультимера на мономеры. Кроме того, по мере увеличения концентрации Cre в реакционном растворе, эффект подавления образования мультимера усиливался.
[0250] При использовании кольцевой ДНК pUC19-OriC 300, не содержащей последовательности loxP, в качестве матрицы, эффект Cre не наблюдался.
Промышленное применение
[0251] В соответствии с настоящим изобретением может быть разработан способ простой и эффективной репликации или амплификации кольцевой ДНК, а в частности, длинноцепочечной кольцевой ДНК.
[0252] Список последовательностей в свободном формате
SEQ ID NO: 1: последовательность ter (консенсусная)
SEQ ID NO: 2: последовательность ter (консенсусная)
SEQ ID NO: 3: последовательность ter (terA, B, D, E или H)
SEQ ID NO: 4: последовательность ter (terA, B, D, E или H)
SEQ ID NO: 5: последовательность ter (terC)
SEQ ID NO: 6: последовательность ter (terF)
SEQ ID NO: 7: последовательность ter (terG)
SEQ ID NO: 8: последовательность ter (terI)
SEQ ID NO: 9: последовательность ter (tarJ)
SEQ ID NO: 10: последовательность ter (Bacillus, консенсусная)
SEQ ID NO: 11: последовательность ter (Bacillus subtilis, консенсусная)
SEQ ID NO: 12: последовательность ter (Bacillus subtilis, консенсусная)
SEQ ID NO: 13: последовательность ter (terVII)
SEQ ID NO: 14: последовательность ter (terIX)
SEQ ID NO: 15: последовательность, распознаваемая XerCD, (консенсусная)
SEQ ID NO: 16: последовательность, распознаваемая XerCD, (консенсусная, dif и cer)
SEQ ID NO: 17: последовательность, распознаваемая XerCD, (консенсусная, dif и psi)
SEQ ID NO: 18: последовательность, распознаваемая XerCD, (консенсусная, cer и psi)
SEQ ID NO: 19: последовательность dif
SEQ ID NO: 20: последовательность cer
SEQ ID NO: 21: последовательность psi
SEQ ID NO: 22: последовательность dif
SEQ ID NO: 23: последовательность cer
SEQ ID NO: 24: последовательность psi
SEQ ID NO: 25: Внешняя концевая (OE) последовательность
SEQ ID NO: 26: ДНК-фрагмент, содержащий пару последовательностей ter
SEQ ID NO: 27: кластер ori-ter
SEQ ID NO: 28: кластер ori-dif
SEQ ID NO: 29: транспозон oriC
SEQ ID NO: 30: консенсусная loxP
SEQ ID NO: 31: последовательность lox511
SEQ ID NO: 32: последовательность lox2272
SEQ ID NO: 33: последовательность loxFAS
SEQ ID NO: 34: последовательность lox RE
SEQ ID NO: 35: последовательность lox LE
SEQ ID NO: 36: последовательность FRT
SEQ ID NO: 37: последовательность rox
SEQ ID NO: 38: праймер SUE1156
SEQ ID NO: 39: праймер SUE1361
SEQ ID NO: 40: кластер OLDT
SEQ ID NO: 41: кластер OriC300.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Japan Science and Technology Agency
<120> Способ репликации или амплификации кольцевой ДНК
<130> FA4002-17267
<150> JP 2017-037489
<151> 2017-02-28
<160> 41
<170> PatentIn version 3.5
<210> 1
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ter (консенсусная)
<220>
<221> misc_feature
<222> (2)..(2)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (3)..(3)
<223> r представляет собой a или g
<220>
<221> misc_feature
<222> (4)..(4)
<223> w представляет собой a или t
<220>
<221> misc_feature
<222> (13)..(13)
<223> k представляет собой g или t
<400> 1
gnrwgttgta acka 14
<210> 2
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ter (консенсусная)
<220>
<221> misc_feature
<222> (2)..(2)
<223> k представляет собой g или t
<220>
<221> misc_feature
<222> (4)..(4)
<223> w представляет собой a или t
<220>
<221> misc_feature
<222> (13)..(13)
<223> k представляет собой g или t
<400> 2
gkawgttgta acka 14
<210> 3
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ter (terA, B, D, E или H)
<400> 3
gtatgttgta acta 14
<210> 4
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ter (terA, B, D, E или H)
<400> 4
agtatgttgt aactaaag 18
<210> 5
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ter (terC)
<400> 5
ggatgttgta acta 14
<210> 6
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ter (terF)
<400> 6
gtatgttgta acga 14
<210> 7
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ter (terG)
<400> 7
ggatgttgta acta 14
<210> 8
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ter (terI)
<400> 8
ggaagttgta acga 14
<210> 9
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ter (terJ)
<400> 9
gtaagttgta acga 14
<210> 10
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ter (Bacillus, консенсусная)
<220>
<221> misc_feature
<222> (3)..(3)
<223> w представляет собой a или t
<220>
<221> misc_feature
<222> (4)..(4)
<223> r представляет собой a или g
<220>
<221> misc_feature
<222> (6)..(10)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (11)..(11)
<223> y представляет собой t или c
<220>
<221> misc_feature
<222> (12)..(12)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (19)..(19)
<223> n представляет собой a, c, g или t
<400> 10
acwrannnnn ynatgtacna aat 23
<210> 11
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ter (Bacillus subtilis, консенсусная)
<220>
<221> misc_feature
<222> (8)..(8)
<223> r представляет собой a или g
<220>
<221> misc_feature
<222> (10)..(10)
<223> w представляет собой a или t
<220>
<221> misc_feature
<222> (12)..(12)
<223> y представляет собой c или t
<400> 11
actaattraw cyatgtacta aat 23
<210> 12
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ter (Bacillus subtilis, консенсусная)
<220>
<221> misc_feature
<222> (8)..(8)
<223> r представляет собой a или g
<220>
<221> misc_feature
<222> (10)..(10)
<223> w представляет собой a или t
<220>
<221> misc_feature
<222> (12)..(12)
<223> y представляет собой c или t
<400> 12
actaattraw cyatgtacta aattttca 28
<210> 13
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ter (terVII)
<400> 13
gaactaatta aactatgtac taaattttca 30
<210> 14
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ter (terIX)
<400> 14
atactaattg atccatgtac taaattttca 30
<210> 15
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, распознаваемая XerCD (консенсусная)
<220>
<221> misc_feature
<222> (7)..(7)
<223> y представляет собой c или t
<220>
<221> misc_feature
<222> (8)..(8)
<223> r представляет собой a или g
<220>
<221> misc_feature
<222> (9)..(9)
<223> y представляет собой c или t
<220>
<221> misc_feature
<222> (12)..(17)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (23)..(23)
<223> k представляет собой g или t
<220>
<221> misc_feature
<222> (28)..(28)
<223> y представляет собой c или t
<400> 15
ggtgcgyrya annnnnntta tgktaaay 28
<210> 16
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, распознаваемая XerCD (консенсусная, dif и cer)
<220>
<221> misc_feature
<222> (7)..(7)
<223> y представляет собой c или t
<220>
<221> misc_feature
<222> (9)..(9)
<223> y представляет собой c или t
<220>
<221> misc_feature
<222> (12)..(17)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (23)..(23)
<223> k представляет собой g или t
<400> 16
ggtgcgyaya annnnnntta tgktaaat 28
<210> 17
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, распознаваемая XerCD (консенсусная, dif и psi)
<220>
<221> misc_feature
<222> (9)..(9)
<223> y представляет собой c или t
<220>
<221> misc_feature
<222> (12)..(17)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (28)..(28)
<223> y представляет собой c или t
<400> 17
ggtgcgcrya annnnnntta tgttaaay 28
<210> 18
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, распознаваемая XerCD (консенсусная, cer и psi)
<220>
<221> misc_feature
<222> (7)..(7)
<223> y представляет собой c или t
<220>
<221> misc_feature
<222> (8)..(8)
<223> r представляет собой a или g
<220>
<221> misc_feature
<222> (12)..(17)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (23)..(23)
<223> k представляет собой g или t
<220>
<221> misc_feature
<222> (28)..(28)
<223> y представляет собой c или t
<400> 18
ggtgcgyrca annnnnntta tgktaaay 28
<210> 19
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность dif
<220>
<221> misc_feature
<222> (12)..(17)
<223> n представляет собой a, c, g, или t
<400> 19
ggtgcgcata annnnnntta tgttaaat 28
<210> 20
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность cer
<220>
<221> misc_feature
<222> (12)..(17)
<223> n представляет собой a, c, g, или t
<400> 20
ggtgcgtaca annnnnntta tggtaaat 28
<210> 21
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность psi
<220>
<221> misc_feature
<222> (12)..(17)
<223> n представляет собой a, c, g, или t
<400> 21
ggtgcgcgca annnnnntta tgttaaac 28
<210> 22
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность dif
<400> 22
ggtgcgcata atgtatatta tgttaaat 28
<210> 23
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность cer
<400> 23
ggtgcgtaca agggatgtta tggtaaat 28
<210> 24
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательнсоть psi
<400> 24
ggtgcgcgca agatccatta tgttaaac 28
<210> 25
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Внешняя концевая (OE) последовательность
<400> 25
ctgtctctta tacacatct 19
<210> 26
<211> 222
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК-фрагмент, содержащий пару последовательностей ter
<220>
<221> misc_feature
<222> (24)..(199)
<220>
<221> misc_feature
<222> (24)..(199)
<223> n представляет собой a, c, g, или t
<400> 26
actttagtta caacatactt attnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn 60
nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn 120
nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn 180
nnnnnnnnnn nnnnnnnnna ataagtatgt tgtaactaaa gt 222
<210> 27
<211> 383
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кластер ori-ter
<400> 27
agtatgttgt aactaaagat aacttcgtat aatgtatgct atacgaagtt atacagatcg 60
tgcgatctac tgtggataac tctgtcagga agcttggatc aaccggtagt tatccaaaga 120
acaactgttg ttcagttttt gagttgtgta taacccctca ttctgatccc agcttatacg 180
gtccaggatc accgatcatt cacagttaat gatcctttcc aggttgttga tcttaaaagc 240
cggatccttg ttatccacag ggcagtgcga tcctaataag agatcacaat agaacagatc 300
tctaaataaa tagatcttct ttttaatacc caggatccat ttaacataat atacattatg 360
cgcaccttta gttacaacat act 383
<210> 28
<211> 320
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кластер ori-dif
<400> 28
atttaacata atatacatta tgcgcaccaa gtatacagat cgtgcgatct actgtggata 60
actctgtcag gaagcttgga tcaaccggta gttatccaaa gaacaactgt tgttcagttt 120
ttgagttgtg tataacccct cattctgatc ccagcttata cggtccagga tcaccgatca 180
ttcacagtta atgatccttt ccaggttgtt gatcttaaaa gccggatcct tgttatccac 240
agggcagtgc gatcctaata agagatcaca atagaacaga tctctaaata aatagatctt 300
ctttttaata cccaggatcc 320
<210> 29
<211> 399
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК транспозона oriC
<400> 29
ctgtctctta tacacatctg aagatccggc agaagaatgg ctgggatcgt gggttaattt 60
actcaaataa gtatacagat cgtgcgatct actgtggata actctgtcag gaagcttgga 120
tcaaccggta gttatccaaa gaacaactgt tgttcagttt ttgagttgtg tataacccct 180
cattctgatc ccagcttata cggtccagga tcaccgatca ttcacagtta atgatccttt 240
ccaggttgtt gatcttaaaa gccggatcct tgttatccac agggcagtgc gatcctaata 300
agagatcaca atagaacaga tctctaaata aatagatctt ctttttaata cccaggatcc 360
caggtctttc tcaagccgac agatgtgtat aagagacag 399
<210> 30
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> консенсусная последовательность loxP
<400> 30
ataacttcgt atagcataca ttatacgaag ttat 34
<210> 31
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность lox 511
<400> 31
ataacttcgt atagtataca ttatacgaag ttat 34
<210> 32
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность lox 2272
<400> 32
ataacttcgt ataggatact ttatacgaag ttat 34
<210> 33
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность loxFAS
<400> 33
ataacttcgt atataccttt ctatacgaag ttat 34
<210> 34
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность lox RE
<400> 34
ataacttcgt atagcataca ttatacgaac ggta 34
<210> 35
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность lox LE
<400> 35
taccgttcgt atagcataca ttatacgaag ttat 34
<210> 36
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность FRT
<400> 36
gaagttccta ttctctagaa agtataggaa cttc 34
<210> 37
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность rox
<400> 37
taactttaaa taatgccaat tatttaaagt ta 32
<210> 38
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер SUE1156
<400> 38
ctatgcggca tcagagcag 19
<210> 39
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер SUE1361
<400> 39
gttaagccag ccccgacac 19
<210> 40
<211> 461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кластер OLDT
<400> 40
cgcgtcagcg ggtgttggcg ggtgtcgggg ctggcttaac agtatgttgt aactaaagat 60
aacttcgtat aatgtatgct atacgaagtt atacagatcg tgcgatctac tgtggataac 120
tctgtcagga agcttggatc aaccggtagt tatccaaaga acaactgttg ttcagttttt 180
gagttgtgta taacccctca ttctgatccc agcttatacg gtccaggatc accgatcatt 240
cacagttaat gatcctttcc aggttgttga tcttaaaagc cggatccttg ttatccacag 300
ggcagtgcga tcctaataag agatcacaat agaacagatc tctaaataaa tagatcttct 360
ttttaatacc caggatccat ttaacataat atacattatg cgcaccttta gttacaacat 420
actatgcggc atcagagcag attgtactga gagtgcacca t 461
<210> 41
<211> 378
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кластер oriC300
<400> 41
cgcgtcagcg ggtgttggcg ggtgtcgggg ctggcttaac agtatgttgt aactaaagat 60
ctactgtgga taactctgtc aggaagcttg gatcaaccgg tagttatcca aagaacaact 120
gttgttcagt ttttgagttg tgtataaccc ctcattctga tcccagctta tacggtccag 180
gatcaccgat cattcacagt taatgatcct ttccaggttg ttgatcttaa aagccggatc 240
cttgttatcc acagggcagt gcgatcctaa taagagatca caatagaaca gatctctaaa 300
taaatagatc ttctttttaa tactttagtt acaacatact atgcggcatc agagcagatt 360
gtactgagag tgcaccat 378
<---
Реферат
Настоящее изобретение относится к биотехнологии, в частности к способу репликации или амплификации кольцевой ДНК. Более конкретно, настоящее изобретение относится к способу подавления образования ДНК–мультимера в виде побочного продукта, если кольцевая ДНК, имеющая последовательность ориджина репликации (ориджина хромосомы (oriC)), была реплицирована или амплифицирована с использованием нижеследующих групп ферментов: (1) первой группы ферментов, которые катализируют репликацию кольцевой ДНК; (2) второй группы ферментов, которые катализируют созревание фрагмента Оказаки и синтезируют две сестринских кольцевых ДНК, состоящих из катенана; и (3) третьей группы ферментов, которые катализируют разделение двух сестринских кольцевых ДНК. Кроме того, настоящее изобретение также относится к способу, включающему введениеoriCв кольцевую ДНК с использованием транспозона. 4 н. и 10 з.п. ф-лы, 22 ил., 3 табл., 10 пр.
Комментарии