Синтез днк - RU2714253C2
Код документа: RU2714253C2
Чертежи








Описание
Область техники
Настоящее изобретение имеет отношение к усовершенствованному способу синтеза ДНК, РНК, белков и подобных молекул, в частности, бесклеточному ферментативному синтезу ДНК предпочтительно в большом масштабе.
Уровень техники
Амплификация дезоксирибонуклеиновой кислоты (ДНК) может осуществляться с использованием процессов, основанных на клетках, например, путем культивирования бактерий в биореакторах, и таким образом обеспечивая амплификацию ДНК. Также описаны бесклеточные ферментативные процессы для амплификации ДНК из исходной матрицы, включая полимеразную цепную реакцию и реакции с замещением цепей.
В прошлом амплификация ДНК в экспериментальном масштабе осуществлялась с использованием прибора на основе микротитрационных планшетов и пипеток, контролируемых с помощью робота и предназначенных для добавления компонентов реакции при необходимости. Такие устройства и процессы пригодны для производства небольших количеств молекул ДНК для экспериментальных целей, однако, они не обеспечивают достаточных количеств ДНК для других целей. Крупномасштабная амплификация и производство определенных нуклеиновых кислот и белков в основном осуществляется с помощью процессов на клеточной основе. Такие методы, как правило, являются эффективными для производства сверхбольших объемов продукта, однако их организация является дорогостоящей.
Существует также множество доступных устройств, специально приспособленных для амплификации образцов ДНК с использованием термоциклического метода для проведения полимеразной цепной реакции (ПЦР). Эти устройства идеально подходят для проведения такой реакции, однако они являются негибкими и не могут быть адаптированы для проведения других реакций. Пример такого устройства раскрывается в США 8,163,489.
Известны способы крупномасштабного синтеза ДНК с использованием химического синтеза, например, фосфорамидитные методы, однако у них есть свои недостатки. Такая реакция в большинстве случаев должна осуществляться в органических растворителях, многие из которых являются токсичными или опасными по другим причинам. Другой недостаток химического синтеза заключается в том, что он является малопродуктивным, так как после каждого добавления нуклеотида, по меньшей мере, 2 процента наращиваемых олигонуклеотидных цепей кэппируется, приводя к потере в выходе продукта. Потеря общего выхода синтезируемой нуклеотидной цепи, таким образом, увеличивается с каждым нуклеотидом, добавленным к последовательности. Эта характерная для химического синтеза олигонуклеотидов неэффективность в конечном итоге ограничивает длину олигонуклеотида, который может быть успешно получен, до олигонуклеотидов, имеющих 50 остатков нуклеиновых кислот или меньше.
В настоящее время биологический катализатор (фермент), такой как полимераза, обычным путем не может использоваться для производства ДНК-продуктов в промышленном масштабе, и реакции в основном ограничиваются объемами в масштабе микролитров. Увеличение масштаба процессов с использованием ферментативного синтеза ДНК признано проблематичным, что не в последнюю очередь связано с неоправдывающим ожидания выходом ДНК-продукта.
Следовательно, существует необходимость в процессах, которые могут использоваться для синтеза ДНК и подобных молекул в значительном масштабе.
Раскрытие изобретения
Настоящее изобретение имеет отношение к процессу выработки ДНК в бесклеточной системе. Этот процесс дает возможность повышенной выработки ДНК по сравнению с существующими методиками с использованием замкнутых периодических процессов. Этот процесс значительно повышает производительность и в то же время уменьшает стоимость синтеза ДНК, в частности, в больших масштабах. В общем, настоящее изобретение имеет отношение к изотермическим методам амплификации ДНК, при использовании которых не требуется циклическое изменение температуры путем нагревания и охлаждения.
Соответственно, предоставляется способ синтеза ДНК в бесклеточной системе, включающий контактирование матрицы ДНК, по меньшей мере, с одной полимеразой в присутствии одного или более праймеров и нуклеотидов для образования реакционной смеси при условиях, способствующих амплификации указанной матрицы путем замещения реплицированных цепей посредством репликации другой цепи с замещением цепей, при этом дополнительные нуклеотиды контролируемым образом доставляются в реакционную смесь.
Таким образом, бесклеточный способ предполагает амплификацию матрицы посредством репликации с замещением цепи. Предоставляется бесклеточный процесс синтеза ДНК, включающий контактирование ДНК-матрицы, по меньшей мере, с одной полимеразой в присутствии одного или более праймеров и нуклеотидов для образования реакционной смеси при условиях, способствующих амплификации указанной матрицы путем репликации с замещением цепей, при этом дополнительные нуклеотиды контролируемым образом доставляются в реакционную смесь.
Соответственно, также предоставляется бесклеточный способ синтеза ДНК, включающий контактирование ДНК-матрицы, по меньшей мере, с одной полимеразой в присутствии нуклеотидов для образования реакционной смеси при условиях, способствующих амплификации указанной матрицы путем замещения реплицированных цепей посредством репликации с замещением другой цепи, при этом дополнительные нуклеотиды контролируемым образом доставляются в реакционную смесь.
Таким образом, бесклеточный способ предполагает амплификацию матрицы посредством репликации с замещением цепей. Предоставляется бесклеточный способ синтеза ДНК, включающий контактирование ДНК матрицы, по меньшей мере, с одной полимеразой в присутствии нуклеотидов для образования реакционной смеси при условиях, способствующих амплификации указанной матрицы посредством репликации с замещением цепи, при этом дополнительные нуклеотиды контролируемым образом доставляются в реакционную смесь.
Таким образом, обеспечение контролируемой подачи дополнительных нуклеотидов в реакционную смесь полезно, поскольку такая рациональная стратегия контроля может обеспечить улучшение скорости синтеза ДНК и/или улучшенный выход ДНК. Такое усовершенствование сравнивается с аналогичной реакционной смесью, когда все нуклеотиды добавляются в реакционную смесь в начале. Контролируемая подача дополнительных нуклеотидов может обеспечить возможность повышенной продуктивности и/или процессивности фермента полимеразы.
Альтернативно, предоставляется бесклеточный способ синтеза ДНК, включающий контактирование ДНК матрицы, по меньшей мере, с одной полимеразой в присутствии одного или более праймеров, нуклеотидов и катионов металлов для образования реакционной смеси при условиях, способствующих амплификации указанной матрицы путем замещения реплицированных цепей посредством репликации другой цепи с замещением цепей, при этом дополнительные катионы металлов контролируемым образом подаются в реакционную смесь.
Таким образом, бесклеточный процесс предполагает амплификацию матрицы посредством репликации с замещением цепей. Предоставляется бесклеточный способ синтеза ДНК, включающий контактирование ДНК матрицы, по меньшей мере, с одной полимеразой в присутствии одного или более праймеров, нуклеотидов и катионов металлов для образования реакционной смеси при условиях, способствующих амплификации указанной матрицы путем репликации с замещением цепей, при этом дополнительные ионы металлов контролируемым образом подаются в реакционную смесь.
Альтернативно, предоставляется бесклеточный способ синтеза ДНК, включающий контактирование ДНК матрицы, по меньшей мере, с одной полимеразой в присутствии нуклеотидов и катионов металлов для образования реакционной смеси при условиях, способствующих амплификации указанной матрицы путем замещения реплицированных цепей посредством репликации другой цепи с замещением, при этом дополнительные катионы металлов контролируемым образом подаются в реакционную смесь.
Бесклеточный способ, таким образом, подразумевает амплификацию матрицы посредством репликации с замещением цепей. Предоставляется бесклеточный способ синтеза ДНК, включающий контактирование ДНК матрицы, по меньшей мере, с одной полимеразой в присутствии нуклеотидов и катионов металлов для образования реакционной смеси при условиях, способствующих амплификации указанной матрицы посредством репликации с замещением цепей, при этом дополнительные катионы металлов контролируемым образом подаются в реакционную смесь.
Таким образом, обеспечение контролируемой подачи дополнительных катионов металлов в реакционную смесь полезно, поскольку такая рациональная стратегия регулирования может обеспечить улучшение скорости синтеза ДНК. Такое усовершенствование сравнивается с аналогичной реакционной смесью, когда все катионы металлов подаются в реакционную смесь в начале. Контролируемая подача дополнительных катионов металлов может обеспечить возможность повышенной продуктивности и/или процессивности фермента полимеразы.
Также предоставляется бесклеточный способ синтеза ДНК, включающий контактирование матрицы ДНК, по меньшей мере, с одной полимеразой в присутствии одного или более праймеров, нуклеотидов и катионов металлов для образования реакционной смеси при условиях, способствующих амплификации указанной матрицы путем замещения реплицированных цепей посредством репликации другой цепи с замещением цепей, при этом дополнительные нуклеотиды и катионы металлов контролируемым образом подаются в реакционную смесь.
Бесклеточный способ, таким образом, подразумевает амплификацию матрицы путем репликации с замещением цепей. Предоставляется бесклеточный способ синтеза ДНК, включающий контактирование ДНК матрицы, по меньшей мере, с одной полимеразой в присутствии одного или более праймеров, нуклеотидов и ионов металлов для образования реакционной смеси при условиях, способствующих амплификации указанной матрицы путем репликации с замещением цепей, при этом дополнительные нуклеотиды и дополнительные катионы металлов контролируемым образом подаются в реакционную смесь.
Также предоставляется бесклеточный способ синтеза ДНК, включающий контактирование матрицы ДНК, по меньшей мере, с одной полимеразой в присутствии нуклеотидов и катионов металлов для образования реакционной смеси при условиях, способствующих амплификации указанной матрицы путем замещения реплицированных цепей посредством репликации другой цепи с замещением цепей, при этом дополнительные нуклеотиды и катионы металлов контролируемым образом подаются в реакционную смесь.
Бесклеточный способ, таким образом, подразумевает амплификацию матрицы путем репликации с замещением цепей. Предоставляется бесклеточный способ синтеза ДНК, включающий контактирование ДНК матрицы, по меньшей мере, с одной полимеразой в присутствии нуклеотидов и ионов металлов для образования реакционной смеси при условиях, способствующих амплификации указанной матрицы путем репликации с замещением цепей, при этом дополнительные нуклеотиды и дополнительные катионы металлов контролируемым образом подаются в реакционную смесь.
Дополнительные нуклеотиды и дополнительные катионы металлов могут подаваться вместе или по отдельности. Предпочтительно нуклеотиды и ионы металлов не смешиваются до того, как добавляются в реакционную смесь. Дополнительные нуклеотиды и дополнительные катионы металлов могут добавляться в реакционную смесь независимо друг от друга по мере того, как каждый из них требуется.
Таким образом, обеспечение контролируемой подачи дополнительных нуклеотидов и дополнительных катионов металлов в реакционную смесь полезно, поскольку такая рациональная стратегия контроля может обеспечить улучшение скорости синтеза ДНК и/или улучшенный выход ДНК. Такое усовершенствование сравнивается с аналогичной реакционной смесью, когда все нуклеотиды и катионы металлов подаются в реакционную смесь в начале. Контролируемая подача дополнительных нуклеотидов может обеспечить повышенную продуктивность и/или процессивность фермента полимеразы.
Согласно каждому из нескольких дополнительных аспектов изобретения предоставляется устройство, созданное для осуществления способа в соответствии с первым, вторым или третьим аспектом настоящего изобретения. Данное устройство может использоваться для осуществления способа согласно любому аспекту данного изобретения.
Согласно дополнительному аспекту изобретения матрица ДНК может содержать один или более сайтов-мишеней процессирующего фермента. Реакционная смесь в любой момент времени может входить в контакт с одним или более процессирующими ферментами. Указанные ферменты могут доставляться в указанную реакционную смесь контролируемым образом. Предпочтительно процессирующим ферментом является фермент протеломераза.
Независимо от того, необязательным признаком любого аспекта изобретения может быть: ДНК матрица может быть кольцевой. Амплификация с замещением цепей указанной ДНК матрицы может осуществляться путем амплификации по типу катящегося кольца (RCA). ДНК полимераза может представлять собой Phi29 или ее вариант. Амплификация ДНК может быть изотермической амплификацией. Один или более праймеров могут быть случайными праймерами. Синтезированная ДНК может содержать конкатамеры, включающие тандемные повторы последовательности ДНК, амплифицированной из ДНК матрицы. ДНК матрица может быть замкнутой линейной ДНК; предпочтительно ДНК матрица инкубируется в денатурирующих условиях для образования замкнутой кольцевой одноцепочечной ДНК. ДНК матрица может содержать, по меньшей мере, одну последовательность-мишень для процессирующего фермента, предпочтительно последовательность-мишень рекомбиназы или последовательность-мишень протеломеразы. Матрица ДНК может содержать экспрессионную кассету, содержащую эукариотический промотор, функционально связанный с представляющей интерес кодирующей последовательностью, и необязательно эукариотическую последовательность терминации транскрипции. Экспрессионная кассета может быть фланкирована с обеих сторон последовательностью-мишенью процессирующего фермента, предпочтительно последовательностью-мишенью рекомбиназы или последовательностью-мишенью протеломеразы. Амплифицированная ДНК может входить в контакт с ферментом рекомбиназой или протеломеразой, по необходимости.
Количество ДНК, которое может быть синтезировано, равно или выше, чем 1 грамм на литр реакционной смеси.
Дополнительные нуклеотиды и/или дополнительные ионы металлов могут доставляться в реакционную смесь в некотором количестве аликвот. Дополнительные нуклеотиды и/или дополнительные ионы металлов могут доставляться с равными промежутками на всем протяжении процесса, необязательно по меньшей мере каждые 30 минут. По меньшей мере, 3, 4, 5, 6, 7, 8, 9 или 10 аликвот может быть добавлено в реакционную смесь. Дополнительные нуклеотиды и/или дополнительные ионы металлов могут непрерывно добавляться в реакционную смесь. Дополнительные ионы металлов и/или дополнительные нуклеотиды могут подаваться в реакционную смесь, необязательно из внешнего источника при помощи насоса или подаваться в реакционную смесь с помощью осмотического насоса.
Нуклеотиды или дополнительные нуклеотиды могут включать биологически неактивные нуклеотиды; необязательно эти биологически неактивные нуклеотиды доставляются в реакционную смесь посредством активации, предпочтительно с помощью химических или физических способов. Биологически неактивные нуклеотиды могут быть «запертыми» нуклеотидами, которые активируются фотолизом.
Дополнительные нуклеотиды и/или дополнительные ионы металлов и/или процессирующий фермент может доставляться по сигналу, связанному с концентрацией ДНК в реакционной смеси, мониторинг которой необязательно может проводиться путем измерения разницы давления реакционной смеси, или ее части, которая вероятно может генерировать сигнал. Сигнал, показывающий концентрацию ДНК, может генерироваться путем измерения разницы давления, создаваемой насосом поршневого типа, который удаляет и возвращает часть реакционной смеси, необязательно с помощью датчика давления.
Катионы металлов или дополнительные катионы металлов могут включать один или более металлов, выбранных из списка, состоящего из Mg2+, Be2+, Ca2+, Sr2+, Li+, Na+, K+, Mn2+ или Zn2+, предпочтительно Mg2+. Дополнительные катионы металлов могут доставляться вместе с дополнительными нуклеотидами. Соотношение между катионами металлов и нуклеотидами может поддерживаться около 3:1 в реакционной смеси. Концентрация ионов металлов, обсуждаемая в описании, в частности концентрация свободных ионов металлов, имеет отношение к конкретному представляющему интерес катиону металла (т.е. Mg2+), а не к концентрации всех видов ионов металлов в реакционной смеси.
Реакционная смесь может содержать, по меньшей мере, один нуклеотид. Затем может быть добавлен один или более дополнительных нуклеотидов. Нуклеотиды или дополнительные нуклеотиды являются дезоксирибонуклеозид трифосфатами (дНТФ) или их производными или модифицированными вариантами. Нуклеотиды или дополнительные нуклеотиды являются одним или более из числа дезоксиаденозин трифосфата (дАТФ), дезоксигуанозин трифосфата (дГТФ), дезоксицитидин трифосфата (дЦТФ), дезокситимидин трифосфата (дТТФ) и их производных. Нуклеотиды или дополнительные нуклеотиды предоставляются в виде одной или более свободных кислот, их солей или их хелатов. Соли и хелаты могут включать один или более из числа следующих ионов металлов: Mg2+, Be2+, Ca2+, Sr2+, Li+, Na+, K+, Mn2+ или Zn2+. Концентрация нуклеотидов в реакционной смеси может поддерживаться в пределах от 0.001мM до 6мM, необязательно около 3мM.
Способ может представлять собой периодический процесс или непрерывный производственный процесс. Дополнительные необязательные признаки описаны в зависимых пунктах формулы изобретения.
Дополнительные преимущества описаны далее.
Краткое описание чертежей
Настоящее изобретение будет описываться далее со ссылкой на характерные варианты осуществления и прилагаемые чертежи.
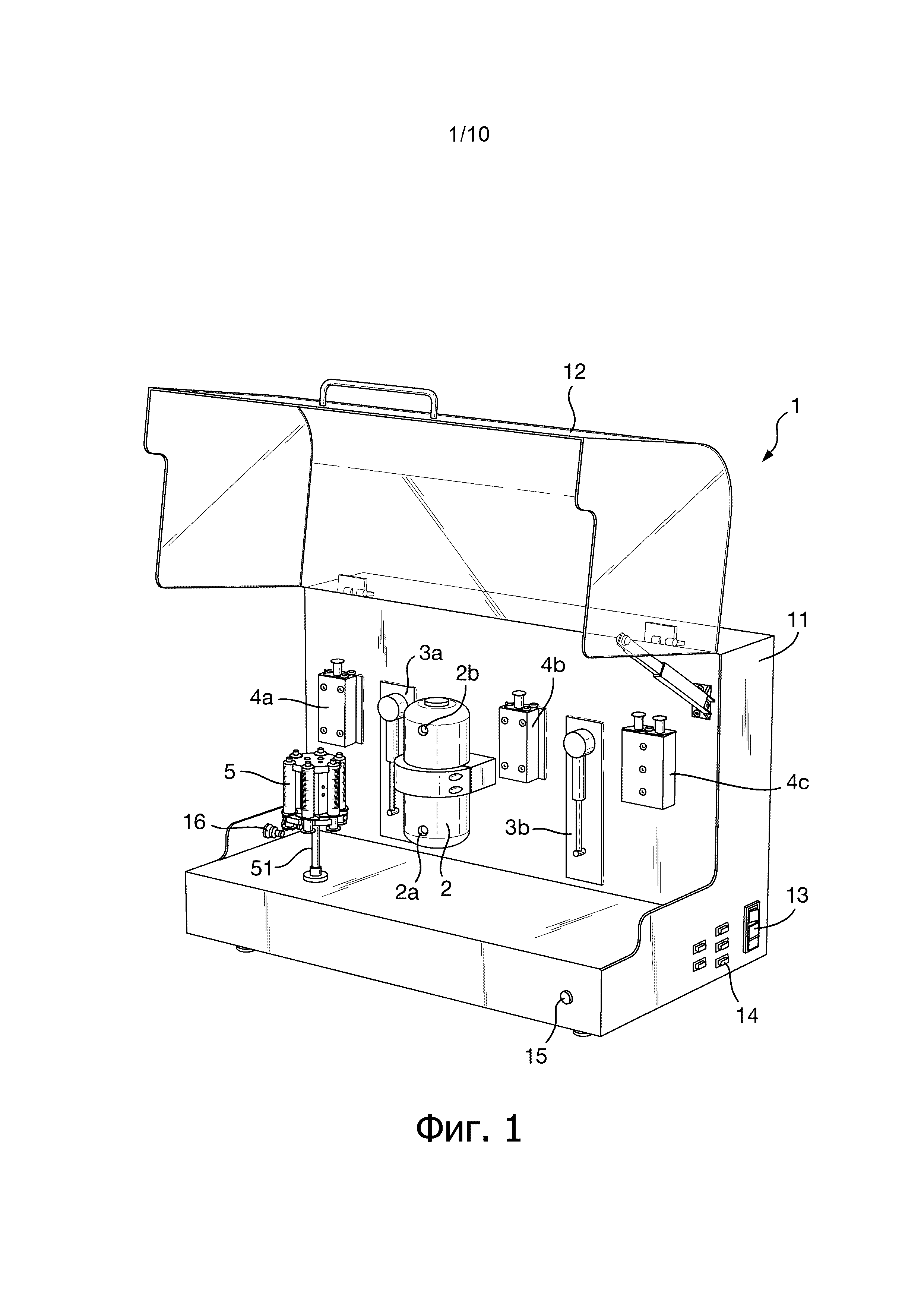
Фигура 1 - изображение в перспективе устройства для синтеза, используемого в соответствии с одним вариантом осуществления изобретения. Для облегчения понимания и чтобы сделать главные компоненты более очевидными, некоторые составные части и соединительные трубки (каналы) опущены на этой фигуре;
фигура 2 - схематический чертеж устройства, показанного на фигуре 1;
фигура 3 - вид в поперечном разрезе реакционного сосуда, используемого в соответствии с одним вариантом осуществления изобретения;
фигура 4 - график, показывающий результаты сравнения данных, полученных в контрольной реакции с водой (квадраты), с данными, полученными в результате стандартной реакции амплификации ДНК с использованием Phi29 ДНК полимеразы (ромбы) в зависимости от времени (как описано в таблице 1). Единицы разности давления являются произвольными, а время показано в минутах;
фигура 5 - график разности давления в зависимости от концентрации ДНК, измеренной с помощью флуориметрического анализа Qubit® BR dsДНК. Единицы разности давления являются произвольными, а концентрация ДНК показана в мкг/мл;
фигура 6 - фотография гель-электрофореза ковалентно замкнутых линейных ДНК продуктов (2.6 т.о. и 2 т.о.) в реакции ДНК амплификации с исходными концентрациями дНТФ 2мM, 4мM и 8мM;
фигура 7 представляет график измерений разности давления в зависимости от времени (минут) для реакции с исходной концентрацией дНТФ 2мМ (без добавления дополнительного количества дНТФ) (незакрашенные кружки), 2мМ дНТФ с подачей дополнительных 2мМ дНТФ (треугольники), 4мМ дНТФ с начала реакции (без добавления дополнительных дНТФ) (ромбы) и 2мМ дНТФ с подачей дополнительных дНТФ в концентрации 2мМ три раза (крестики). Стрелки показывают время добавления дНТФ для каждой реакции;
фигура 8 представляет график начальной скорости реакции (разность давлений единиц/минуту) в зависимости от отношения ионов магния к дНТФ для реакции с исходной концентрацией 2мМ дНТФ и дополнительной подачей 2мМ дНТФ трижды;
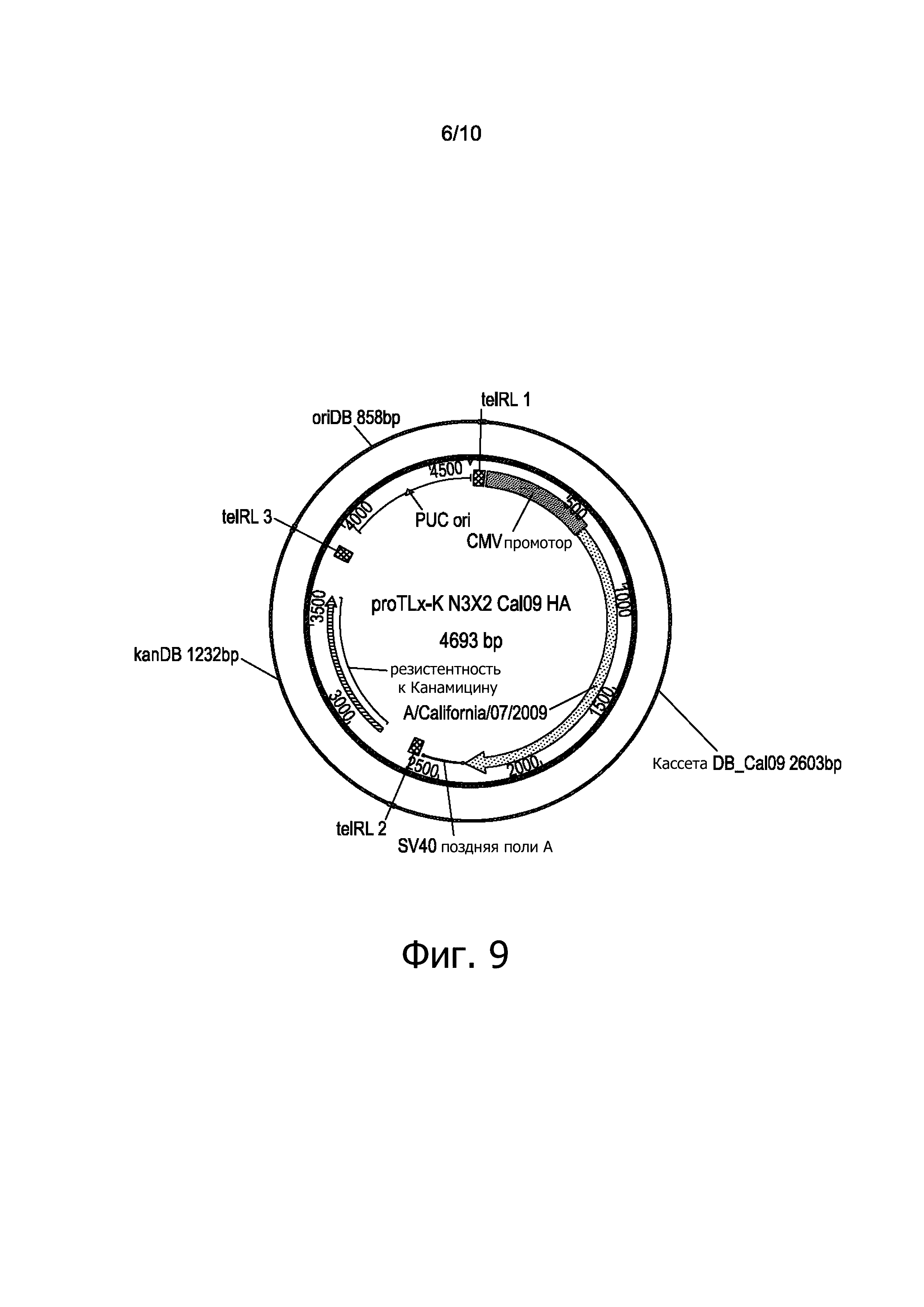
фигура 9 представляет матрицу ДНК proTLx-N3X2 Cal 09 HA;
фигура 10 представляет матрицу ДНК proTLx B5X4 Cal 09 HA;
фигура 11 представляет матрицу ДНК proTLx-K N3X2 ova;
фигура 12 представляет матрицу ДНК proTLx-K N3X2 lux; и
фигура 13 представляет график измерений разности давления в зависимости от времени (минут) для реакции, первоначально содержащей 2мМ дНТФ и 7.5мМ Mg2+, в которую было сделано контролируемое добавление (“Feed”) нуклеотидов и Mg2+ в указанных точках (300мин, 600мин, 900мин, 1080мин, 1230мин и 1440 мин – белые треугольники), с 2мМ дНТФ и достаточным количеством Mg2+ для поддержания отношения 3:1 Mg2+: дНТФ после первого добавления), к которому контролируемое добавление протеломеразы сделано в момент времени 1050 мин (2мкМ TelN), как указано (квадратик). Стрелки показывают время каждого добавления дНТФ сразу после добавления Mg2+ и добавления TelN.
Подробное описание
Настоящее изобретение имеет отношение к бесклеточным способам синтеза ДНК. Способы изобретения могут обеспечить синтез ДНК с высокой пропускной способностью.
Дезоксирибонуклеиновая кислота (ДНК), синтезированная в соответствии с настоящим изобретением может быть любой молекулой ДНК. ДНК может быть одноцепочечной или двухцепочечной. ДНК может быть линейной. ДНК может подвергаться процессингу с образованием колец, в частности миниколец, одноцепочечных замкнутых колец, двухцепочечных замкнутых колец, двухцепочечных открытых колец или замкнутых линейных двухцепочечных ДНК. ДНК может образовать или подвергнуться процессингу с образованием определенной вторичной структуры, такой как, но без ограничения, петли «шпильки» («петли на стебле»), неполные петли шпильки, псевдоузлы или любой из различных типов двойной спирали (A-ДНК, B-ДНК, или Z-ДНК). Кроме того, ДНК может образовывать «шпильки».
Синтезированная ДНК может иметь любую подходящую длину. При использовании способов изобретения возможна длина вплоть до или превышающая 77 тысяч оснований. Конкретнее, длина ДНК, которая может быть синтезирована согласно данному способу, может быть порядка вплоть до 60 тысяч оснований, или до 50 тысяч оснований, или до 40 тысяч оснований, или до 30 тысяч оснований. Предпочтительно синтезированная ДНК может иметь от 100 нуклеотидов до 60 тысяч оснований, от 200 нуклеотидов до 20 тысяч оснований, более предпочтительно от 200 нуклеотидов до 15 тысяч оснований, наиболее предпочтительно от 2 тысяч оснований до 15 тысяч оснований.
Количество ДНК, синтезированной в соответствии со способами настоящего изобретения, может превышать 10г/л реакционной смеси. Предпочтительно, что количество синтезированной ДНК, достигает 10, 9, 8, 7, 6 или 5 г/л. Предпочтительное количество синтезированной ДНК составляет от 0.2 до 5г/л, предпочтительно от 1 до 5г/л. Количество произведенной ДНК может быть охарактеризовано как промышленное или коммерческое количество, производство в крупном масштабе или массовое производство. ДНК, произведенная при использовании данного способа, может быть однородной по качеству, а именно по длине и последовательности ДНК. Таким образом, процессы могут быть пригодны для крупномасштабного синтеза ДНК.
При использовании существующих методов синтеза ДНК производится в небольшом или лабораторном масштабе, реакции происходят в объемах микролитров. Попытки увеличения масштаба ПЦР по-прежнему приводят к низкому выходу ДНК, например, приблизительно 20-40мг на литр реакционной смеси (Vandalia), поскольку реакция проходит только с исходным количеством нуклеотидов, при этом дополнительные нуклеотиды не подаются, и образование продукта ингибирует реакцию.
Матрица ДНК может быть любой подходящей матрицей. ДНК-матрица может быть одноцепочечной (ss) или двухцепочечной (ds). ДНК-матрица может быть линейной или кольцевой. Двухцепочечная матрица ДНК может быть открытой кольцевой ds ДНК, замкнутой кольцевой ds ДНК, открытой линейной ds ДНК или замкнутой линейной ds ДНК. Одноцепочечная ДНК матрица может быть линейной ss ДНК или замкнутой кольцевой ss ДНК. Последняя ДНК матрица может быть получена путем денатурации замкнутой линейной ds ДНК. Замкнутая линейная ДНК, т.е. линейные ds ковалентно замкнутые молекулы ДНК, как правило, содержат ковалентно замкнутые концы, т.е. шпилькообразные концы, где спаривание оснований между комплементарными цепями ДНК отсутствует. Петля «шпилька» соединяет концы комплементарных цепей ДНК. Петли сами могут содержать комплементарные последовательности, в частности, если петли содержат часть последовательности-мишени протеломеразы. Замкнутая линейная ds ДНК может быть плазмидой.
Матрица ДНК может содержать любую последовательность, или природную или искусственную. ДНК-матрица может содержать, по меньшей мере, одну последовательность-мишень процессирующего фермента, например, один, два, три, четыре или более сайтов-мишеней для процессирующего фермента. Такая последовательность-мишень создает возможность процессинга ДНК в ряде случаев также после синтеза. Процессирующий фермент представляет собой фермент, который распознает свой сайт-мишень и осуществляет процессинг ДНК. Последовательность-мишень процессирующего фермента может быть последовательностью-мишенью для рестрикционного фермента. Рестрикционный фермент, т.е. рестрикционная эндонуклеаза, связывается с последовательностью-мишенью и расщепляет ее в конкретном месте. Последовательность-мишень для процессирующего фермента может быть мишенью для рекомбиназы. Рекомбиназа направленно катализирует реакции замещения (цепей) ДНК между короткими (30–40 нуклеотидов) последовательностями сайтов-мишеней, которые специфичны для каждой рекомбиназы. Примеры рекомбиназ включают Cre рекомбиназу (с loxP в качестве последовательности-мишени) и FLP рекомбиназу (с короткими распознаваемыми флиппазой (FRT) сайтами-мишенями). Последовательность-мишень для процессирующего фермента может быть мишенью для сайт-специфичной интегразы, такой как phiC31 интеграза. Последовательность-мишень процессирующего фермента может быть мишенью фермента протеломеразы. Последовательность-мишень для протеломеразы представляет собой любую последовательность ДНК, присутствие которой в матрице ДНК обеспечивает ее превращение в замкнутую линейную ДНК при помощи ферментативной активности протеломеразы. Другими словами, последовательность-мишень для протеломеразы требуется для расщепления и повторного лигирования двухцепочечной ДНК протеломеразой с образованием ковалентно замкнутой линейной ДНК. Как правило, последовательность-мишень для протеломеразы содержит какую-либо палиндромную последовательность, т.е. какую-либо двухцепочечную последовательность ДНК, имеющую двустороннюю симметрию, также описанную в данном документе как инвертированный повтор. Длина инвертированного повтора различается в зависимости от конкретного организма. Палиндромный или инвертированный повтор может быть полным или неполным. Последовательность-мишень для протеломеразы предпочтительно содержит двухцепочечную палиндромную (инвертированный повтор) последовательность, по меньшей мере, 14 нуклеотидов в длину. Подходящие сайты-мишени для протеломеразы известны в данной области техники и обсуждаются в EP2,391,731, включенном в описание путем отсылки. Подходящим ферментом протеломеразой может быть TelN из фага N15 Escherichia coli. Предпочтительно сайт-мишень для процессирующего фермента представляет собой сайт-мишень протеломеразы.
Последовательность-мишень процессирующего фермента может быть мишенью-последовательностью для РНК полимеразы, в результате чего ДНК становится матрицей для синтеза полипептидов. В этом случае сайт-мишень процессирующего фермента является промотором, предпочтительно эукариотическим промотором.
Данный процесс может включать дополнительную стадию контактирования реакционной смеси с процессирующим ферментом. Реакционная смесь может вступать в контакт с процессирующим ферментом в любой момент времени. Предпочтительно реакционная смесь вступает в контакт с процессирующим ферментом, как только синтез ДНК завершается. Процессирующий фермент может быть РНК полимеразой, рекомбиназой, рестрикционной эндонуклеазой или протеломеразой. Предпочтительно процессирующий фермент является протеломеразой.
Протеломераза, используемая в изобретении, представляет собой какой-либо полипептид, способный расщеплять и снова соединяться с матрицей, содержащей сайт-мишень протеломеразы для того, чтобы продуцировать ковалентно замкнутую линейную молекулу ДНК. Таким образом, протеломераза обладает функцией расщепления и лигирования ДНК. Ферменты, обладающие протеломеразным типом активности, также описаны как резольвазы теломеров (например, в Borrelia burgdorferi). Типичным субстратом для протеломеразы является кольцевая двухцепочечная ДНК. Если эта ДНК содержит сайт-мишень для протеломеразы, фермент может разрезать ДНК в этом сайте и лигировать концы с образованием линейной двухцепочечной ковалентно замкнутой ДНК молекулы. Требования к сайтам-мишеням протеломеразы обсуждаются выше. Как уже было указано ранее, способность данного полипетида катализировать производство замкнутой линейной ДНК из матрицы, содержащей сайт-мишень протеломеразы, может быть определена с помощью любого подходящего метода анализа, описанного в данной области техники.
Для процессинга ДНК может требоваться использование, по меньшей мере, одной протеломеразы. Процесс изобретения может включать использование более чем одной протеломеразы, например, двух, трех, четырех, пяти или более разных протеломераз. Примеры подходящих протеломераз включают протеломеразы бактериофагов, такие как phiHAP-1 из Halomonas quamarina, PY54 из Yersinia enterolytica, phiK02 из Klebsiella oxytoca и VP882 из Vibrio sp., и N15 из Escherichia coli (SEQ ID NO: 15), или варианты любой из них. Особенно предпочтительным является использование протеломеразы бактериофага N15 или ее варианта. Этот фермент называется также TelN. Эти ферменты также описаны в WO2012/017210, включенном в данное описание путем отсылки.
Таким образом, процесс может осуществляться на матрице ДНК, включающей процессирующий фермент, а процессирующий фермент может подаваться в реакционную смесь контролируемым образом. Процессирующий фермент может подаваться в любое подходящее время в реакционную смесь; это может происходить в ответ на сигнал. Может быть сделано одно или более добавлений процессирующего фермента. Предпочтительно, чтобы процессирующий фермент добавлялся в аликвотах, которые по отдельности добавляются в реакционную смесь. 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более аликвот процессирующего фермента может быть добавлено в реакционную смесь. Может быть целесообразным добавление процессирующего фермента непосредственно перед внесением аликвоты необходимых дополнительных нуклеотидов и/или дополнительных ионов металлов. Альтернативно, процессирующий фермент может быть добавлен вместе с или после добавления аликвоты дополнительных нуклеотидов в реакционную смесь. Процессирующий фермент может быть добавлен в ответ на сигнал о том, что ДНК в реакционной смеси достигла желаемой концентрации. Эта концентрация ДНК может быть установлена, как описано в данном документе.
Матрица ДНК может включать экспрессионную кассету, содержащую, состоящую или в основном состоящую из эукариотического промотора, функционально связанного с последовательностью, включающей в себя представляющий интерес белок, и необязательно эукариотическую последовательность терминации транскрипции. Необязательно экспрессионная кассета может быть минимальной кассетой, у которой отсутствует одна или более бактериальных или векторных последовательностей, как правило, выбранных из группы, состоящей из: (i) бактериальной точки начала репликации; (ii) бактериальных селективных маркеров (таких как гены антибиотикорезистентности) и (iii) неметилированных CpG мотивов. "Промотор" представляет собой последовательность нуклеотидов, которая инициирует и регулирует транскрипцию полинуклеотида. Промоторы могут включать индуцибельные промоторы (когда экспрессия последовательности полинуклеотида, функционально связанного с промотором, индуцируется веществом-аналитом, кофактором, регуляторным белком и т.д.), репрессируемые промоторы (когда экспрессия последовательности полинуклеотида, функционально связанного с промотором, подавляется веществом-аналитом, кофактором, регуляторным белком и т.д.) и конститутивные промоторы. Подразумевается, что термин "промотор" или "контролирующий элемент" включает полноразмерные промоторные участки и функциональные (например, контролирующие транскрипцию или трансляцию) сегменты этих участков. "Функционально связанный" относится к расположению элементов, при котором описанные таким образом компоненты располагаются так, чтобы выполнять свою обычную функцию. Таким образом, конкретный промотор, функционально связанный с нуклеиновокислотной последовательностью, является способным к осуществлению экспрессии такой последовательности в присутствии надлежащего фермента. Промотор может и не быть смежным с данной последовательностью, поскольку его функция заключается в управлении ее экспрессией. Таким образом, например, промежуточные нетранслируемые и все-таки транскрибируемые последовательности могут присутствовать между промоторной последовательностью и нуклеиновокислотной последовательностью, при этом промоторная последовательность может, тем не менее, считаться "функционально связанной" с кодирующей последовательностью. Таким образом, термин "функционально связанный" включает любое расположение в пространстве или ориентацию промоторного элемента и интересующей последовательности ДНК, которая обеспечивает инициацию транскрипции интересующей последовательности ДНК после узнавания промотора транскрипционным комплексом.
ДНК матрица может быть любой подходящей длины. В частности, матрица ДНК может достигать 60 тысяч оснований, или 50 тысяч оснований, или 40 тысяч оснований, или 30 тысяч оснований. Предпочтительно матрица ДНК может иметь от 100 нуклеотидов до 60 тысяч оснований, от 200 тысяч оснований до 20 тысяч оснований, более предпочтительно от 200 нуклеотидов до 15 тысяч оснований, наиболее предпочтительно от 2 тысяч оснований до 15 тысяч оснований.
ДНК-матрица может обеспечиваться в количестве, достаточном для использования в процессе, с помощью любого, известного в данной области техники метода. Например, матрица может продуцироваться с помощью ПЦР.
Полная ДНК-матрица или выбранный участок ДНК матрицы может амплифицироваться в данном процессе.
ДНК-матрица может содержать последовательность ДНК для экспрессии. ДНК может предназначаться для экспрессии в клетке (т.е. трансфицированной клетке in vitro или in vivo) или может предназначаться для экспрессии в бесклеточной системе (т.е. синтез белка). ДНК последовательность, предназначенная для экспрессии, может требоваться для терапевтических целей, т.е. генотерапии с помощью ДНК-вакцины. Последовательность, предназначенная для экспрессии, может быть геном, и указанный ген может кодировать ДНК-вакцину, терапевтический белок и тому подобное. Последовательность ДНК может содержать последовательность, которая транскрибируется в активную форму РНК, т.е. небольшую интерферирующую РНК молекулу (siRNA).
Матрицу ДНК вводят в контакт, по меньшей мере, с одной полимеразой. Может использоваться одна, две, три, четыре или пять различных полимераз. Полимераза может быть любой подходящей полимеразой, синтезирующей полимеры ДНК. Полимераза может быть ДНК-полимеразой. Может использоваться любая ДНК полимераза, включая любую коммерчески доступную ДНК-полимеразу. Можно использовать две, три, четыре, пять или более разных ДНК полимераз, например, одна из которых обладает корректирущей функцией, и одну или более других, которые не обеспечивают исправление ошибок. Могут использоваться ДНК полимеразы, имеющие различные механизмы, например, тип полимераз, функционирующих по принципу замещения цепи, и ДНК полимеразы, реплицирующие ДНК с помощью других способов. Подходящим примером ДНК полимеразы, которая не обладает активностью замещения цепи, является T4 ДНК полимераза.
Полимераза может быть высокоустойчивой, так что ее активность существенно не уменьшается при длительной инкубации в условиях процесса. Следовательно, фермент предпочтительно имеет длительный период полужизни в некотором диапазоне условий процесса, включая, но не ограничиваясь этим, температуру и значение pH. Также предпочтительно, что полимераза имеет одну или более характеристик, подходящих для производственного процесса. Полимераза предпочтительно обладает высокой точностью, например, благодаря тому, что обладает корректирующей активностью. Более того, предпочтительно, что полимераза демонстрирует высокую процессивность, высокую активность по замещению цепи и низкую Km для дНТФ и ДНК. Полимераза может быть способна использовать кольцевую и/или линейную ДНК в качестве матрицы. Полимераза может быть способна использовать dsДНК или ssДНК в качестве матрицы. Предпочтительно, что полимераза не проявляет активность ДНК экзонуклеазы, которая не связана с ее корректирующей активностью.
Специалист может определить, демонстрирует или нет данная полимераза описанные выше характерные свойства, путем сравнения со свойствами, которые проявляют коммерчески доступные полимеразы, например, Phi29 (New England Biolabs, Inc., Ipswich, MA, US), Deep Vent® (New England Biolabs, Inc.), Bacillus stearothermophilus (Bst) ДНК полимераза I (New England Biolabs, Inc.), фрагмент Кленова ДНК полимеразы I (New England Biolabs, Inc.), обратная транскриптаза M-MuLV (New England Biolabs, Inc.), VentR®(exo-minus) ДНК полимераза (New England Biolabs, Inc.), VentR® ДНК полимераза (New England Biolabs, Inc.), Deep Vent® (экзо-)ДНК полимераза (New England Biolabs, Inc.) и большой фрагмент Bst ДНК полимеразы (New England Biolabs, Inc.). Когда речь идет о высокой процессивности, она, как правило, показывает среднее количество нуклеотидов, добавленных с помощью фермента полимеразы за одну ассоциацию/диссоциацию с матрицей, т.е. величину удлинения праймера, полученную в результате одного случая ассоциации.
Полимеразы, функционирующие по принципу замещения цепи, являются предпочтительными. Предпочтительными полимеразами такого типа являются Phi 29, Deep Vent и Bst ДНК полимераза I или любые их варианты. "Замещение цепи" описывает способность полимеразы замещать комплементарные цепи при обнаружении участка двухцепочечной ДНК в ходе синтеза. Матрица, таким образом, амплифицируется путем замещения комплементарных цепей и синтеза новой комплементарной цепи. Таким образом, во время репликации с замещением цепи только что реплицированная цепь будет замещаться, чтобы дать возможность полимеразе реплицировать также комплементарную цепь. Реакция амплификации инициируется при отжиге праймера или 3’ свободного конца одноцепочечной матрицы с комплементарной последовательностью на матрице (и то и другое являются инициирующими событиями). Когда синтез ДНК переходит к следующему этапу, и если происходит обнаружение еще одного праймера или другой цепи, присоединенных к матрице, полимераза вытесняет их и продолжает элонгацию своей цепи. Замещение цепи приводит к образованию вновь синтезированной одноцепочечной ДНК, которая снова может служить в качестве матрицы для инициирующих событий. Праймирование вновь синтезированной ДНК приводит к повышенной разветвленности и высокому выходу продуктов. Следует понимать, что методы амплификации с замещением цепей отличаются от методов на основе ПЦР тем, что циклы денатурации не являются необходимыми для эффективной ДНК амплификации, так как двухцепочечная ДНК не является препятствием для продолжения синтеза новых цепей ДНК. Для амплификации с замещением цепей может требоваться только один первоначальный цикл нагревания, чтобы денатурировать исходную матрицу, если она является двухцепочечной, для того, чтобы произошло присоединение (отжиг) праймера к сайту связывания праймера, в случае его использования. После этого амплификация может описываться как изотермическая, поскольку дальнейшее нагревание или охлаждение не требуется. В противоположность этому, ПЦР методы требуют циклов денатурации (т.е. увеличения температуры до 94°С или выше) во время процесса амплификации, чтобы «расплавить» двухцепочечную ДНК и обеспечить новые одноцепочечные матрицы. В ходе «замещения цепей» полимераза будет замещать цепи уже синтезированной ДНК. Кроме того, она будет использовать вновь синтезированную ДНК в качестве матрицы, обеспечивая быструю амплификацию ДНК.
Замещающая цепи полимераза, используемая в процессе изобретения, предпочтительно имеет процессивность, по меньшей мере, 20 тысяч оснований, более предпочтительно по меньшей мере 30 тысяч оснований, по меньшей мере, 50 тысяч оснований, или по меньшей мере 70 тысяч оснований или больше. В одном варианте осуществления замещающая цепи ДНК полимераза имеет процессивность, сравнимую с или превышающую процессивность phi29 ДНК полимеразы.
Репликация с замещением цепей является, следовательно, предпочтительной. В ходе репликации с замещением цепей матрица амплифицируется путем замещения уже реплицированных цепей, которые были синтезированы под действием полимеразы, в свою очередь замещающих другую цепь, которая может быть исходной комплементарной цепью двухцепочечеой матрицы, или вновь синтезированной комплементарной цепью, последняя синтезируется под действием полимеразы на прежнем праймере, отожженном к матрице. Таким образом, амплификация матрицы может происходить путем замещения реплицированных цепей посредством репликации другой цепи с замещением цепей. Этот процесс может быть описан как амплификация с замещением цепей или репликация с замещением цепей.
Предпочтительным процессом репликации с замещением цепей является опосредованная образованием петель изотермическая амплификация (петлевая изотермическая амплификация или LAMP). LAMP, как правило, использует 4-6 праймеров, узнающих 6-8 разных участков матрицы ДНК. Коротко, замещающая цепи ДНК полимераза инициирует синтез и пара праймеров формируют петлеобразные структуры, чтобы облегчить последующие циклы амплификации. Внутренний праймер, содержащий последовательности смысловой (кодирующей) и антисмысловой нитей целевой ДНК, инициирует LAMP. Последующий синтез ДНК с замещением цепи, инициированный внешним праймером, высвобождает одноцепочечную ДНК. Она служит в качестве матрицы для синтеза ДНК, инициированного вторым внутренним и внешним праймерами, которые гибридизируются с другим концом мишени, что приводит к образованию структуры ДНК «петля-на-стебле». В последующем LAMP цикле один внутренний праймер гибридизируется с петлей на продукте и инициирует замещающий синтез ДНК с образованием исходной ДНК типа «стебель-петля» и новой ДНК типа «стебель-петля» со стеблем вдвое длиннее.
Предпочтительным процессом репликации с замещением цепей является амплификация по типу катящегося кольца (RCA). Термин RCA описывает способность полимераз RCA-типа непрерывно продвигаться вокруг кольцевой цепи ДНК матрицы при удлинении гибридизованного праймера. Это приводит к образованию линейных одноцепочечных продуктов с множественными повторами амплифицированной ДНК. Последовательность кольцевой матрицы (отдельная структурная единица) многократно повторяется в пределах линейного продукта. Что касается кольцевой матрицы, исходным продуктом амплификации с замещением цепей является одноцепочечный конкатамер, который является смысловым или антисмысловым в зависимости от полярности матрицы. Эти линейные одноцепочечные продукты служат основой для многократной гибридизации, удлинения праймера и замещения цепи, приводя к образованию конкатамерных двухцепочечных ДНК продуктов, снова включающих множественные повторы амплифицированной ДНК. Таким образом, существует множество копий каждой амплифицированной "отдельной структурной единицы" ДНК в конкатамерных двухцепочечных ДНК продуктах. В частности, RCA полимеразы являются предпочтительными для использования в процессах настоящего изобретения. Продукты процессов репликации с замещением цепи RCA-типа могут требовать процессинга для высвобождения отдельных структурных единиц ДНКs. Это является желательным, если требуются отдельные структурные единицы ДНК.
Для обеспечения амплификации ДНК матрица также входит в контакт с одним или более праймерами. Праймеры могут быть неспецифическими (т.е. со случайной последовательностью) или могут быть специфическими для одной или более последовательностей, содержащихся в пределах ДНК матрицы. Если праймеры являются праймерами со случайной последовательностью, они обеспечивают неспецифическую инициацию в каком-либо сайте на ДНК матрице. Это обеспечивает высокую эффективность амплификации посредством множества реакций инициации на каждой цепи матрицы. Примерами случайных праймеров являются гексамеры, гептамеры, октамеры, нонамеры, декамеры или более длинные последовательности, например, состоящие из 12, 15, 18, 20 или 30 нуклеотидов в длину. Случайный праймер может иметь от 6 до 30, от 8 до 30 или от 12 до 30 нуклеотидов в длину. Случайные праймеры обычно предоставляются в виде смеси олигонуклеотидов, которые соответствуют всем возможным комбинациям, например, гексамеров, гептамеров, октамеров или нонамеров в матрице ДНК.
В одном варианте осуществления праймеры или один или более из праймеров являются специфическими. Это означает, что они имеют последователность, комплементарную последовательности в матрице ДНК, откуда желательна инициация амплификации. В этом варианте осуществления пара праймеров может использоваться для того, чтобы специфически амплифицировать участок ДНК матрицы, который находится между сайтами связывания двух праймеров. Альтернативно, может использоваться один специфический праймер.
В одном варианте осуществления ДНК матрица включает одну или более последовательностей-мишеней для протеломеразы. Такие последовательности-мишени являются палиндромными по природе. В этом варианте осуществления возможно использование специфического праймера, который связывается с палиндромной последовательностью в пределах сайта связывания протеломеразы, и может, таким образом, инициировать амплификацию на обеих цепях ДНК матрицы. Такой процесс подробно описывается в WO2012/017210. Таким образом, одного праймера достаточно для амплификации.
Праймеры могут быть немеченными или могут содержать одну или более меток, например, радионуклиды или флуоресцентные красители. Праймеры также могут содержать химически модифицированные нуклеотиды. Например, праймер может быть кэппирован для того, чтобы препятствовать инициации синтеза ДНК до тех пор, пока «кэп» не будет удален, т.е. с помощью химических или физических способов. Длина праймеров/последовательностей, как правило, может быть выбрана с учетом температурных условий, т.е. так, чтобы праймер был способен связываться с матрицей при используемой температуре на стадии амплификации.
Контактирование ДНК матрицы с полимеразой и одним или более праймерами может происходить при условиях, способствующих отжигу праймеров на матрице ДНК. Условия включают присутствие одноцепочечной ДНК, создающей возможность гибридизации праймеров. Условия также включают температуру и буфер, обеспечивающий отжиг праймера на матрице. Подходящие условия отжига/условия гибридизации могут быть выбраны в зависимости от природы праймера. Пример предпочтительных условий отжига, используемых в настоящем изобретении, включает буфер 30мМ Tris-HCl pH 7.5, 20мМ KCl, 8мМ MgCl2. Отжиг может проводиться после денатурации c использованием нагревания с постепенным охлаждением до желательной температуры реакции.
Однако, амплификация с использованием репликации с замещением цепи также может происходить без праймера, и, таким образом, не требовать гибридизации и удлинения праймера. Вместо этого, одноцепочечная ДНК матрица самоинициируется путем образования «шпилек», которые имеют свободный 3’-конец, доступный для удлинения. Остальные стадии амплификации остаются одинаковыми.
Матрицу ДНК и полимеразу также приводят в контакт с нуклеотидами. Объединение матрицы ДНК, полимеразы и нуклеотидов образует реакционную смесь. Реакционная смесь также может содержать один или более праймеров. Реакционная смесь также может независимо включать один или более катионов металлов.
Нуклеотид является мономером, или структурной единицей нуклеиновых кислот, а нуклеотиды состоят из азотистого основания, пятиуглеродного сахара (рибозы и дезоксирибозы), и по меньшей мере, одной фосфатной группы. Может использоваться любой подходящий нуклеотид.
Нуклеотиды могут присутствовать в виде свободных кислот, их солей или хелатов, или смеси свободных кислот и/или солей или хелатов.
Нуклеотиды могут присутствовать в виде солей, образованных с ионами одновалентных металлов или ионами двухвалентных металлов. Соли могут включать соли с ионами двухвалентных металлов, таких как, но без ограничения, магний (Mg2+), марганец (Mn2+), кальций (Ca2+), бериллий (Be2+), цинк (Zn2+) и стронций (Sr2+), или соли с ионами одновалентных металлов, включая, но не ограничиваясь этим, литий (Li+), натрий (Na+) или калий (K+).
Азотистое основание может быть аденином (A), гуанином (G), тимином (T), цитозином (C) и урацилом (U). Азотистое основание также может быть модифицированным основанием, таким как 5-метилцитозин (m5C), псевдоуридин (Ψ), дигидроуридин (D), инозин (I) и 7-метилгуанозин (m7G).
Предпочтительно, чтобы пятиуглеродный сахар являлся дезоксирибозой, в результате чего нуклеотид будет дезоксинуклеотидом.
Нуклеотид может быть в виде дезоксинуклеозид трифосфата, обозначаемого дНТФ. Это является предпочтительным вариантом осуществления настоящего изобретения. Подходящие дНТФ могут включать дАТФ (дезоксиаденозин трифосфат), дГТФ (дезоксигуанозин трифосфат), дТТФ (дезокситимидин трифосфат), dUTP (дезоксиуридин трифосфат), дЦТФ (дезоксицитидин трифосфат), dITP (дезоксиинозин трифосфат), dXTP (дезоксиксантозин трифосфат), и их производные и модифицированные варианты. Предпочтительно, чтобы дНТФ содержали один или более из дАТФ, дГТФ, дТТФ или дЦТФ, или их модифицированных вариантов или производных. Предпочтительным является использование смеси дАТФ, дГТФ, дТТФ и дЦТФ или их модифицированных вариантов.
Нуклеотиды могут находиться в растворе или предоставляться в лиофилизированной форме. Предпочтительным является раствор нуклеотидов.
Нуклеотиды могут включать модифицированные нуклеотиды, причем эти модифицированные нуклеотиды могут быть в биологически неактивной форме. Модифицированный нуклеотид, таким образом, может быть биологически неактивным нуклеотидом. Биологически инактивированный нуклеотид может иметь легкоудаляемый фрагмент, защищающий группу нуклеотида, такую как атом кислорода у 3′ атома углерода или кислород концевого фосфата. Биологически неактивный нуклеотид может быть «запертым» нуклеотидом или блокированным нуклеотидом. Подходящие фрагменты включают, но не ограничиваются этим фотоактивируемую («запирающую») группу, включая α-карбокси-2-нитробензил (CNB), 1-(2-нитрофенил)этил (NPE), 4,5-диметокси -2-нитробензил (DMNB), 1-(4,5-диметокси -2-нитрофенил)этил (DMNPE) и 5-карбоксиметокси-2-нитробензил (CMNB) (Molecular Probes). Эти группы, как правило, присоединяются посредством атома кислорода концевого фосфата. Функциональная группа может быть удалена любым подходящим способом. Например, если функциональная группа является фотолабильной, импульсный фотолиз функциональной группы под действием ультрафиолетового света приводит к быстрому и весьма локализованному высвобождению биологически активного нуклеотида в месте освещения. Функциональная группа может являться термолабильной, и, таким образом, биологически активный нуклеотид может высвобождаться под действием нагревания. Если для биологической активации нуклеотида используется нагревание, необходимо принимать во внимание требования полимеразы к температуре, и это находится в пределах компетенции квалифицированного специалиста. Например, терминальный фосфат может быть этерифицирован «запирающей» или блокирующей функциональной группой.
Нуклеотиды могут предоставляться в смеси, содержащей одно или более подходящих оснований, предпочтительно одно или более из числа аденина (A), гуанина (G), тимина (T), цитозина (C). Два, три или предпочтительно все четыре нуклеотида (A, G, T, и C) используются в процессе синтеза ДНК.
Все нуклеотиды могут быть природными нуклеотидами (т.е. немодифицированными), они могут быть модифицированными нуклеотидами, которые функционируют как природные нуклеотиды и являются биологически активными (т.е. LNA нуклеотидами, LNA – запертая нуклеиновая кислота), они могут быть модифицированными и биологически неактивными или они могут быть смесью немодифицированных и модифицированных нуклеотидов, и/или смесью биологически активных и биологически неактивных нуклеотидов. Каждый тип (т.е. основание) нуклеотида может предоставляться в одной или более формах, т.е. немодифицированной и модифицированной, или биологически активной и биологически неактивной.
В одном аспекте изобретения нуклеотиды включаются в реакционную смесь, а доставка дополнительных нуклеотидов в реакционную смесь контролируется. Согласно этому аспекту нуклеотиды могут присутствовать в реакционной смеси в исходной концентрации менее чем 8мМ, менее чем 7мМ, менее чем 6мМ, менее чем 5мМ, менее чем 4мМ, менее чем 3мМ, менее чем 2мМ или менее чем 1мМ. Таким образом, нуклеотиды добавляются, чтобы образовать такую реакционную смесь, чтобы исходная концентрация нуклеотидов в реакционной смеси составляла 8мМ или менее, предпочтительно 5мМ или менее, более предпочтительно 4мМ или менее. Исходная концентрация относится к общей концентрации нуклеотидов (всех оснований) в реакционной смеси в начале реакции.
Нуклеотиды могут присутствовать в реакционной смеси в исходной концентрации, достаточной для того, чтобы синтезировать, по меньшей мере, 0.3г/л ДНК. Предпочтительно исходная концентрация является достаточной для того, чтобы синтезировать 0.5г/л ДНК, более предпочтительно 0.6г/л или 0.65г/л ДНК. Данный объем представляет собой объем реакционной смеси. Исходной концентрации нуклеотидов будет недостаточно для того, чтобы синтезировать желательное количество ДНК из реакционной смеси, т.е. ее может быть недостаточно для достижения конечного выхода. Исходная концентрация нуклеотидов может быть определена, исходя из наличия одной или более полимераз в реакционной смеси.
Альтернативно или в дополнение к этому, исходная концентрация нуклеотидов может быть определена как отношение концентрации полимеразы к концентрации нуклеотидов. Отношение числа молекул нуклеотидов к каждому ферменту полимеразы в начале может достигать 500,000:1, 300,000:1 или 200,000:1 или в пределах от 5,000:1 до 100,000:1, предпочтительно от 20,000:1 до 90,000:1, более предпочтительно от 30,000:1 до 80,000:1, более предпочтительно от 40,000:1 до 60,000:1. Отношение числа молекул нуклеотидов к каждой полимеразе предпочтительно выше, чем 5,000:1. В предпочтительном аспекте отношение нуклеотиды: полимераза вначале составляет 50,000:1.
Реакционную смесь инкубируют при условиях, способствующих амплификации указанной матрицы. Амплификация с замещением цепей является предпочтительной. Предпочтительно условия способствуют амплификации указанной матрицы путем замещения реплицированных цепей посредством репликации другой цепи с замещением цепей. Условия включают использование любой температуры, дающей возможность амплификации ДНК, обычно в пределах от 20 до 90°С. Предпочтительный диапазон температуры составляет примерно от 20 до 40°С или примерно от 25 до 35°С. Предпочтительная температура для LAMP амплификации составляет примерно от 50 до 70°С.
Как правило, соответствующую температуру выбирают, исходя из температуры, при которой специфическая полимераза обладает оптимальной активностью. Эта информация обычно доступна и составляет часть общих знаний квалифицированного специалиста. Например, когда используется phi29 ДНК полимераза, подходящий температурный диапазон будет составлять примерно от 25 до 35°С, предпочтительно около 30°С. Специалист в рабочем порядке способен определить подходящую температуру для эффективной амплификации согласно процессу изобретения. Например, процесс может проводиться в некотором диапазоне температур, причем можно контролировать выход амплифицированной ДНК с целью установления оптимального диапазона температур для данной полимеразы. Амплификация может осуществляться при постоянной температуре, причем изотермический процесс является предпочтительным. Поскольку амплификация с замещением цепей является предпочтительной, отсутствует необходимость изменения температуры для разделения цепей ДНК. Таким образом, процесс может быть изотермическим процессом.
Другие условия, способствующие амплификации ДНК матрицы, включают присутствие подходящих буферных веществ /pH и других факторов, которые требуются для активности и устойчивости фермента. Подходящие условия включают любые известные в данной области техники условия, используемые для обеспечения активности полимеразы.
Например, pH реакционной смеси может находиться в пределах от 3 до 10, предпочтительно от 5 до 8 или около 7, например, около 7.5. Значение pH может поддерживаться в этом диапазоне при помощи одного или более буферных веществ. Такие буферы включают, но без ограничения, MES, Bis-Tris, ADA, ACES, PIPES, MOBS, MOPS, MOPSO, Bis-Tris пропан, BES, TES, HEPES, DIPSO, TAPSO, Trizma, HEPPSO, POPSO, TEA, EPPS, трицин, Gly-Gly, бицин, HEPBS, TAPS, AMPD, TABS, AMPSO, CHES, CAPSO, AMP, CAPS, CABS, фосфат, лимонная кислота - динатрийфосфат, лимонная кислота-цитрат натрия, натрий ацетат-уксусная кислота, имидазол и натрий карбонат-натрий бикарбонат.
В то время как применение нагревания (экспозиция при 95°C в течение нескольких минут) используется для денатурации двухцепочечной ДНК, можно использовать другие подходы, более подходящие для синтеза ДНК. Двухцепочечная ДНК может быть легко денатурирована при воздействии высокого или низкого значения pH окружающей среды или в случаях, когда катионы отсутствуют или присутствуют в очень низких концентрациях, например, как в деионизированной воде. Полимераза нуждается в связывании короткой последовательности олигонуклеотидного праймера с одноцепочечным участком ДНК матрицы, чтобы инициировать репликацию. Устойчивость этого взаимодействия и, следовательно, эффективность ДНК амплификации может, в частности, находиться под влиянием концентрации катионов металлов и, в частности, двухвалентных катионов, таких как ионы Mg2+, которые могут быть рассмотрены как неотъемлемая часть процесса.
Реакционная смесь также может содержать ионы металлов. Реакционная смесь также может содержать соли металлов, такие как, но без ограничения, соли двухвалентных ионов металлов: магния (Mg2+), марганца (Mn2+), кальция (Ca2+), бериллия (Be2+), цинка (Zn2+) и стронция (Sr2+), или соли одновалентных ионов металлов, включая, но не ограничиваясь этим, литий (Li+), натрий (Na+) или калий (K+). Соли могут включать хлориды, ацетаты и сульфаты. Другие соли, которые могут быть включены, являются солями аммония, в частности, сульфатом аммония. Специалист должен иметь в виду, что подача нуклеотидов в форме солей с ионами металлов будет влиять на концентрацию ионов металлов в реакционной смеси, и может учитывать этот дополнительный источник ионов металлов, при необходимости. Общая концентрация ионов металлов в реакционной смеси предпочтительно включает все источники ионов металлов.
В реакционные смеси также могут быть включены детергенты. Примеры подходящих детергентов включают Тритон X-100, Tween 20 и производные любого из них. В реакционную смесь также могут быть включены стабилизирующие агенты. Может использоваться любой подходящий стабилизирующий агент, в частности, бычий сывороточный альбумин (BSA) и другие стабилизирующие белки. Условия реакции также могут быть улучшены добавлением агентов, которые распрямляют ДНК и облегчают денатурацию матрицы. Такие агенты включают, например, диметилсульфоксид (DMSO), формамид, глицерин и бетаин. Вещества, вызывающие конденсацию ДНК, также могут быть включены в реакционную смесь. Такие агенты включают, например, полиэтиленгликоль или катионный липид или катионные полимеры.
Должно быть понятно, что специалисты способны модифицировать и оптимизировать условия амплификации и инкубации для процесса изобретения с использованием дополнительных компонентов и условий на основе их общих знаний. Аналогичным образом, конкретные концентрации определенных веществ могут быть выбраны на основе предшествующих примеров в данной области техники и дополнительно оптимизированы на основе общих знаний. В качестве примера, количество полимеразы, присутствующей в реакционной смеси может быть оптимизировано. Под этим может подразумеваться дополнительное добавление полимеразы в реакционную смесь в ходе синтеза ДНК. В качестве еще одного примера, количество ДНК матрицы может быть оптимизировано. Под этим может подразумеваться дополнительное добавление ДНК матрицы в реакционную смесь в ходе синтеза ДНК. Дополнительная подача фермента полимеразы и/или ДНК матрицы может быть непрерывной или прерывистой, предпочтительно прерывистой (периодической).
В качестве примера, подходящим реакционным буфером, используемым в методах на основе RCA в данной области техники, является 50мМ Tris HCl, pH 7.5, 10мМ MgCl2, 20мМ (NH4)2SO4, 5% глицерин, 0.2мМ BSA, 1мМ дНТФ. Предпочтительным реакционным буфером, используемым в RCA амплификации изобретения, является 30мМ Tris-HCl pH 7.4, 30мМ KCl, 7.5мМ MgC12, 10мМ (NH4)2SO4, 4мМ DTT, 2мМ дНТФ. Этот буфер является особенно пригодным для использования вместе с Phi29 RCA полимеразой.
Реакционная смесь также может включать один или более дополнительных белков. ДНК матрица может быть амплифицирована в присутствии, по меньшей мере, одной пирофосфатазы, такой как дрожжевая неорганическая пирофосфатаза. Может быть использовано две, три, четыре, пять или более различных пирофосфатаз. Эти ферменты способны разрушать пирофосфат, образованный полимеразой из дНТФ во время репликации цепи. Образование пирофосфата в реакции может вызывать ингибирование ДНК полимераз и уменьшать скорость и эффективность ДНК амплификации. Пирофосфатазы могут расщеплять пирофосфат с образованием неингибирующего фосфата. Примером подходящей пирофосфатазы для использования в процессе настоящего изобретения является Saccharomyces cerevisiae пирофосфатаза, коммерчески доступная от компании New England Biolabs, Inc.
Любой связывающий одноцепочечную ДНК белок (SSBP) может использоваться в процессе изобретения для того, чтобы стабилизировать одноцепочечную ДНК. SSBP являются необходимыми компонентами живых клеток и участвуют во всех процессах, которые затрагивают ssДНК, таких как ДНК репликация, репарация и рекомбинация. В этих процессах SSBP связываются с временно образованной ssДНК и помогают стабилизировать ssДНК структуру. Примером подходящего SSBP для использования в процессе настоящего изобретения является белок, кодируемый геном 32 бактериофага T4, коммерчески доступный от New England Biolabs, Inc.
Реакционная смесь может быть проинкубирована при условиях, способствующих амплификации ДНК матрицы, как описано выше. Амплификация предпочтительно является репликацией с замещением цепи. Реакционная смесь содержит нуклеотиды, предпочтительно в исходной концентрации или в исходном отношении (к полимеразе), которое обсуждалось ранее. Реакционная смесь может дополнительно содержать ионы металлов.
Для того, чтобы синтезировать ДНК, требуется подача дополнительных нуклеотидов в соответствии с одним аспектом изобретения. Доставка нуклеотидов в реакционную смесь контролируется в соответствии с первым аспектом изобретения. Таким образом, все нуклеотиды, необходимые для синтеза ДНК, не добавляются в реакционную смесь в начале процесса. Дополнительные нуклеотиды могут подаваться или предоставляться в любое подходящее время в реакционную смесь. Временной режим подачи дополнительных нуклеотидов в реакционную смесь и/или количество нуклеотидов, доставленных в реакционную смесь, контролируется или управляется. Таким образом, сама реакция может контролироваться точной подачей нуклеотидов. Этот контроль может основываться на теоретическом расчете ферментативной кинетики, или контроль может основываться на измеряемых параметрах реакционной смеси, так что дополнительные нуклеотиды добавляют, поскольку они нужны для работы полимеразы.
Дополнительные нуклеотиды могут быть добавлены сразу, как только реакционная смесь образована и проинкубирована при условиях, способствующих амплификации ДНК матрицы. Дополнительные нуклеотиды могут быть добавлены после того, как амплификация начинается или инициируется. В одном варианте осуществления дополнительная подача нуклеотидов начинается, по меньшей мере, через 20 минут или, по меньшей мере, 30 минут или, по меньшей мере, 40 минут после начала добавления нуклеотидов. Однако, подача может быть начата в начале реакции, в частности, если подача нуклеотидов является непрерывной.
В одном варианте осуществления концентрация или количество нуклеотидов в реакционной смеси не превышает верхнего порога во время процесса. Подача нуклеотидов контролируется так, что концентрация или количество нуклеотидов не превышает верхнего порога. Верхний порог может составлять 8мМ, или 7мМ или 6мМ или 5мМ или 4мМ или 3мМ или 2мМ нуклеотидов в реакционной смеси. Предпочтительно верхний порог составляет 4мМ нуклеотидов в реакционной смеси. Аналогичным образом, в одном варианте осуществления концентрация или количество нуклеотидов в реакционной смеси не может опускаться ниже нижнего порога в ходе процесса. Подача нуклеотидов контролируется таким образом, что концентрация или количество нуклеотидов не опускается ниже нижнего порога. Этот нижний порог может составлять 0.1мкМ, 0.5мкМ, 1мкМ, 5мкМ или 10мкМ нуклеотидов в реакционной смеси. Предпочтительно, чтобы концентрация нуклеотидов в реакционной смеси поддерживалась между верхним и нижним порогами в ходе процесса. Таким образом, концентрация нуклеотидов в реакционной смеси может поддерживаться в пределах между 0.0001мМ и 8мМ, предпочтительно между 0.001мМ и 6мМ, наиболее предпочтительно между 0.01мМ и 5мМ. Концентрация нуклеотидов может поддерживаться путем контролирования подачи дополнительных нуклеотидов в реакционную смесь. В одном варианте осуществления концентрация нуклеотидов в реакционной смеси может быть оценена с помощью ионной хроматографии. Альтернативно, приблизительная концентрация нуклеотидов может быть вычислена посредством оценки количества синтезированной ДНК, как обсуждается в данном документе.
Альтернативно, верхний и нижний пороги могут быть описаны в связи с отношением количества нуклеотидов к полимеразе, присутствующей в реакционной смеси. Таким образом, верхний порог может составлять 500,000:1 (нуклеотиды: полимераза), 200,000:1, 150,000:1, 100,000:1 или 80,000:1. Нижний порог может составлять 1,000:1, 1,500:1 или 2,000:1. Таким образом, отношение нуклеотидов к полимеразе может поддерживаться в пределах от 1,000:1 до 500,000:1, предпочтительно от 2,000:1 до 450,000:1, более предпочтительно от 5,000:1 до 400,000:1, наиболее предпочтительно в пределах от 10,000:1 до 100,000:1. Наиболее предпочтительно отношение поддерживается в пределах от 40,000:1 до 70,000:1, примерно 50,000:1.
В одном варианте осуществления дополнительные нуклеотиды доставляются в реакционную смесь в виде одной или более отдельных добавок, т.е. в виде аликвот. Любое число добавок или аликвот находится в рамках этого варианта осуществления. Таким образом, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17 , 18, 19, 20 или более, т.е. 20-25, 25-30, 35-40 добавок или аликвот может быть добавлено в реакционную смесь. Таким образом, 1-50, 1-40, 1-30 или 1-20 аликвот может использоваться для доставки нуклеотидов. Аликвота может быть частью общего количества нуклеотидов, необходимых для процесса или добавленных к процессу. Объем и/или концентрация нуклеотидов в аликвоте может варьировать в зависимости от потребностей процесса. Аликвота может содержать небольшое количество нуклеотидов, т.е. количество, достаточное для того, чтобы гарантировать, что определенная концентрация или количество нуклеотидов присутствует в реакционной смеси. Нуклеотиды могут быть добавлены в реакционную смесь в виде аликвот через равные промежутки времени (т.е. каждые 30 минут, каждые 60 минут, каждые 90 минут, каждые 120 минут, каждые 180 минут или каждые 240 минут или любой другой подходящий период времени). Нуклеотиды могут быть добавлены в реакционную смесь в виде аликвот с нерегулярными интервалами. Например, период времени между каждым добавлением нуклеотидов может быть первоначально более коротким, а затем может увеличиваться, по мере того, как протекает процесс или наоборот. Аликвоты могут предоставляться для поддержания постоянной концентрации нуклеотидов в реакционной смеси, или могут предоставляться, чтобы пополнить концентрацию нуклеотидов до исходной концентрации. В предпочтительном варианте осуществления нуклеотиды добавляют так, что концентрация в реакционной смеси не превышает 8мM или 7мM, или 6мM, или 5мM, или 4мM. В предпочтительном варианте осуществления нуклеотиды добавляют так, чтобы гарантировать, что концентрация или количество нуклеотидов в реакционной смеси попадает в пределы верхнего и нижнего порогов.
Предпочтительно, чтобы аликвота не доставлялась с единственной целью возместить объем реакционной смеси, взятый для проведения анализа или тестирования, но она доставляется, поскольку для реакции требуются дополнительные нуклеотиды. Предпочтительно, чтобы тестирование реакционной смеси являлось неинвазивным, и что материал не извлекается из реакционной смеси до завершения реакции.
Дополнительные нуклеотиды могут доставляться в случае необходимости наличия большего количества нуклеотидов в реакционной смеси. Это может определяться несколькими параметрами, включая, но не ограничиваясь этим, скорость синтеза ДНК, концентрацию синтезированной ДНК, вязкость или объем реакционной смеси. Скорость синтеза ДНК может быть вычислена, исходя из концентрации синтезированной ДНК в течение некоторого времени. По мере того как скорость синтеза ДНК начинает снижаться, могут добавляться дополнительные нуклеотиды. Альтернативно, концентрацию синтезированной ДНК можно контролировать с помощью методов, которые обсуждаются далее. Когда концентрация ДНК возрастает до порогового уровня, могут быть добавлены дополнительные нуклеотиды. Может быть измерена вязкость реакционной смеси, и увеличение вязкости может указывать на необходимость в дополнительных нуклеотидах. Вязкость реакции может измеряться с помощью вискозиметра.
В одном варианте осуществления дополнительные нуклеотиды доставляются в реакционную смесь постоянно, т.е. они добавляются в реакционную смесь продолжительно. Альтернативно, нуклеотиды могут подаваться по каплям или постоянно. Доставка дополнительных нуклеотидов согласно этому варианту осуществления может происходить в достаточно небольших количествах таким образом, что доставка нуклеотидов соответствует «потребности» полимеразы в дополнительных нуклеотидах. В этом варианте осуществления постоянная доставка нуклеотидов поддерживает приблизительно постоянную концентрацию нуклеотидов в реакционной смеси. Предпочтительно, чтобы постоянная доставка дополнительных нуклеотидов в реакционную смесь гарантировала, что концентрация нуклеотидов в реакционной смеси поддерживается в пределах верхнего и нижнего порогов.
Дополнительные нуклеотиды могут физически доставляться в реакционную смесь, т.е. они находятся вне реакционной смеси. Таким образом, доставка может происходить из внешнего источника, например, резервуара. Этот резервуар может быть соединен с реакционной смесью. Нуклеотиды могут накачиваться в реакционную смесь. Таким образом, нуклеотиды и реакционная смесь являются физически раздельными, а доставка дополнительных нуклеотидов является физической доставкой нуклеотидов. Альтернативно, нуклеотиды могут доставляться с помощью осмотического насоса. Осмотический насос обеспечивает способ контролирования и непрерывной доставки нуклеотидов.
Нуклеотиды могут альтернативно доставляться в реакционную смесь путем активации биологически инактивированных нуклеотидов, обсужденных ранее. Для того, чтобы доставить нуклеотиды в реакционную смесь, биологически неактивные нуклеотиды активируют подходящими способами. Таким образом, дополнительные нуклеотиды могут включать любые подходящие биологически неактивные нуклеотиды. Биологически неактивные нуклеотиды могут быть включены в реакционную смесь в начале, а доставляться в реакционную смесь посредством контролируемой активации. Альтернативно, биологически неактивные нуклеотиды могут быть физически добавлены в реакционную смесь в виде аликвот или непрерывно, а впоследствии активированы, чтобы доставить нуклеотиды в реакционную смесь. Активация может быть физической (т.е. нагревание или освещение) или химической, и это и является контролем активации, которая контролирует доставку нуклеотида.
Доставка нуклеотидов в реакционную смесь может контролироваться до тех пор, пока синтез ДНК не завершен, например, когда ДНК матрица исчерпывается и дополнительное включение невозможно. Альтернативно, реакция может быть завершена, как только был синтезирован ожидаемый или желательный выход ДНК. Такой выход может основываться на количестве добавленных нуклеотидов. Синтез ДНК может быть завершен, когда синтезировано необходимое количество ДНК, например, 1г на литр реакционной смеси 2г/л, 3г/л, 4г/л, 5г/л или вплоть до или превышающее 10г/л. Синтез ДНК может быть завершен, когда скорость синтеза ДНК понижается, а дополнительная доставка нуклеотида не повышает эту скорость, реакционная смесь истощает запас другого компонента (такого как, но не ограничиваясь этим, праймер), фермент полимераза инактивируется, неорганическая пирофосфатаза инактивируется, отсутствует дополнительная доступная одноцепочечная матрица, необходимые кофакторы, такие как катионы металлов, исчерпываются, вязкость реакционной смеси достигает определенного порога, объем реакционной смеси достигает определенного порога, или полимераза ингибируется.
Выход реакции имеет отношение к количеству синтезированной ДНК. Ожидаемый выход из процесса согласно настоящему изобретению может превышать 10 грамм/литр реакционной смеси или 10, 9, 8, 7, 6, 5 4, 3, 2 или 1г/л. Настоящее изобретение увеличивает выход, возможный в результате ферментативного синтеза ДНК. Цель настоящего изобретения - улучшить выход бесклеточного ферментативного процесса синтеза ДНК, так чтобы можно было синтезировать ДНК в крупном масштабе экономически эффективно. Настоящее изобретение дает возможность экономичного производства/синтеза ДНК в промышленном масштабе с использованием ферментативного процесса, катализируемого полимеразой. Настоящий процесс обеспечивает возможность эффективного включения нуклеотидов в ДНК-продукт. Есть основания полагать, что процесс изобретения позволит увеличить масштаб реакционных смесей до нескольких литров, включая десятки литров. Улучшенный выход, скорость синтеза ДНК, продуктивность или процессивность можно сравнить с аналогичной реакционной смесью, когда все нуклеотиды добавляются в начале процесса. Есть основания полагать, что выход может быть увеличен посредством контролирования доставки дополнительных нуклеотидов в реакционную смесь, как продемонстрировано в Примерах.
Скорость синтеза ДНК имеет отношение к количеству или концентрации ДНК, синтезированной за минуту процесса. В Примерах будет видно, что скорость синтеза ДНК постепенно замедляется по мере того, как процесс продвигается вперед.
Продуктивность процесса или скорость выхода ДНК на единицу доставленного нуклеотида, может быть увеличена путем использования процесса или процессов изобретения.
В одном варианте осуществления настоящее изобретение имеет отношение к процессу увеличения синтеза ДНК. Это увеличение можно сравнить с идентичной реакционной смесью, с тем исключением, что все необходимые компоненты, включая нуклеотиды, добавляются в начале процесса. Дополнительные доставки в эту контрольную реакционную смесь не делаются.
В одном аспекте, предоставляется бесклеточный способ синтеза ДНК, включающий контактирование ДНК матрицы, по меньшей мере, с одной полимеразой в присутствии одного или более праймеров, нуклеотидов и катионов металлов с образованием реакционной смеси, при условиях, способствующих амплификации указанной матрицы путем замещения реплицированных цепей посредством репликации другой цепи с замещением цепей, при этом дополнительные катионы металлов доставляются контролируемым образом в реакционную смесь.
Альтернативно, амплификация происходит путем репликации матрицы с замещением цепей. Таким образом, изобретение предоставляет бесклеточный способ синтеза ДНК, включающий контактирование ДНК матрицы, по меньшей мере, с одной полимеразой в присутствии одного или более праймеров, нуклеотидов и катионов металлов с образованием реакционной смеси, при условиях способствующих репликации с замещением цепи указанной матрицы, при этом дополнительные нуклеотиды и дополнительные катионы металлов контролируемым образом добавляются в реакционную смесь.
Предпочтительно, чтобы скорость реакции бесклеточного способа синтеза ДНК управляется контролируемой доставкой дополнительных катионов в реакционную смесь.
Доставка катионов металлов в реакционную смесь может контролироваться в соответствии с первым аспектом изобретения. Считается, что такой контроль улучшает скорость синтеза ДНК, в частности, в крупномасштабном процессе. Один или более катионов металлов может присутствовать в реакционной смеси в начале. Таким образом, реакционная смесь также может содержать один или более катионов металлов, предпочтительно доставленных в виде солей. Доставка дополнительных катионов металлов в реакционную смесь может контролироваться. Может доставляться один или более катионов металлов. Подходящие катионы металлов включают двухвалентные ионы металлов: магния (Mg2+), марганца (Mn2+), кальция (Ca2+), бериллия (Be2+), цинка (Zn2+) и стронция (Sr2+), или одновалентные ионы металлов, включая, но без ограничения, ионы лития (Li+), натрия (Na+) или калия (K+). Катионы металлов предпочтительно доставляются в виде солей. Соли могут включать хлориды, ацетаты и сульфаты, и предпочтительно, чтобы катионы металлов доставлялись в виде хлористых солей. Другие соли, которые могут быть включены, являются солями аммония, в частности сульфатом аммония. Предпочтительно, катионы металлов являются катионами магния или марганца, наиболее предпочтительно магния. В наиболее предпочтительном варианте осуществления катионы металлов предоставляются в виде хлорида магния.
Для того, чтобы синтезировать ДНК, требуется доставка дополнительных катионов металлов в соответствии с одним аспектом изобретения. Доставка катионов металлов в реакционную смесь контролируется в соответствии со вторым аспектом изобретения. Таким образом, все катионы металлов, необходимые для синтеза ДНК, добавляются в реакционную смесь в начале процесса. Дополнительные катионы металлов могут доставляться или предоставляться в реакционную смесь в любое подходящее время. Момент подачи дополнительных катионов металлов в реакционную смесь и/или количество ионов металлов, доставленных в реакционную смесь, контролируется или направляется. Таким образом, реакция сама может контролироваться точной подачей ионов металлов. Этот контроль может быть основан на теоретических расчетах ферментативной кинетики, или контроль может основываться на измеряемых параметрах реакционной смеси, в результате этого дополнительные ионы металлов доставляются как раз в то время, когда они требуются. Доставка дополнительных катионов металлов может контролироваться параллельно или в сочетании с контролируемой доставкой дополнительных нуклеотидов. Доставка может контролироваться независимо или контролироваться совместно.
Дополнительные катионы металлов могут быть добавлены, как только реакционная смесь образована и проинкубирована при условиях, способствующих амплификации ДНК матрицы. Дополнительные катионы металлов могут быть добавлены после того, как амплификация началась или была инициирована. В одном варианте осуществления дополнительная доставка катионов металлов начинается, по меньшей мере, через 20 минут или, по меньшей мере, 30 минут или, по меньшей мере, 40 минут после обеспечения исходной концентрации катионов металлов. Однако, доставка может быть начата в начале реакции, в частности, если доставка катионов металлов является непрерывной.
В одном варианте осуществления концентрация или количество свободных или несвязанных катионов металлов в реакционной смеси не превышает верхний порог во время процесса. Доставка катионов металлов контролируется так, что концентрация или количество свободных или несвязанных катионов металлов не превышает самый высокий порог. Самый высокий порог может составлять 4мM, 3мM или 2мM в реакционной смеси. Предпочтительно самый высокий порог составляет около 3мM в реакционной смеси. Подобным образом, в одном варианте осуществления концентрация или количество свободных катионов металлов в реакционной смеси не может опускаться ниже самого низкого порога во время процесса. Доставка катионов металлов контролируется так, что концентрация или количество свободных катионов металлов не опускается ниже самого низкого порога. Этот самый низкий порог может составлять 0.03мкM, 0.15мкM, 0.3мкM, 1.5мкM или 3мкM в реакционной смеси. Предпочтительно, чтобы концентрация свободных катионов металлов в реакционной смеси поддерживается между самым высоким и самым низким порогами во время процесса. Таким образом, концентрация свободных катионов металлов в реакционной смеси может поддерживаться в пределах от 0.03мкM до 4мM, или от 0.5мкM до 3мM, или от 1мкM до 2мM, или от 10мкM до 1мM. Предпочтительно концентрация свободных катионов металлов в реакционной смеси поддерживается в пределах от 1мM до 1.5мM.
Альтернативно, самый высокий порог может составлять 10мM, 8мM, 6мM, 4мM, 3мM или 2мM в реакционной смеси. Предпочтительно самый высокий порог составляет около 3мM в реакционной смеси. Подобным образом в одном варианте осуществления концентрация или количество свободных катионов металлов в реакционной смеси не может опускаться ниже самого низкого порога во время процесса. Доставка катионов металлов контролируется так, что концентрация или количество свободных катионов металлов не опускается ниже самого низкого порога. Этот самый низкий порог может составлять 0.03мкM, 0.15мкM, 0.3мкM, 1.5мкM, 3мкM и 5мкM в реакционной смеси. Предпочтительным является, что концентрация свободных катионов металлов в реакционной смеси поддерживается в пределах между самым высоким и самым низким порогами во время процесса. Таким образом, концентрация свободных катионов металлов в реакционной смеси может поддерживаться в пределах от 0.03µM до 10мМ, или от 0.15мкM до 8мM, или от 0.3мкM до 6мM, или от 1.5мкM до 3мM. Предпочтительно концентрация свободных катионов металлов в реакционной смеси поддерживается в пределах от 0.15мкM до 5мкM.
Концентрация свободных катионов металлов может поддерживаться путем контролирования подачи дополнительных ионов металлов в реакционную смесь. В одном варианте осуществления концентрация свободных ионов металлов в реакционной смеси может быть оценена с помощью атомно-абсорбционной спектроскопии, ионообменной хроматографии и других методов, известных в данной области техники. Альтернативно, приблизительная концентрация свободных катионов металлов может быть вычислена посредством оценки количества синтезированной ДНК, как обсуждалось в данном документе. Во избежание недоразумений уточняем, катионы металлов, такие как, ионы магния, могут быть связаны с разными субстанциями в реакционной смеси, включая, но не ограничиваясь этим, пирофосфат, фосфат и ДНК. Эти ионы являются связанными, и не являются свободными.
В одном варианте осуществления дополнительные катионы металлов доставляются в реакционную смесь в виде одного или более отдельных добавлений, т.е. в виде аликвот. Любое число добавлений или аликвот входит в рамки этого варианта осуществления. Таким образом, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более, т.е. 20-25, 25-30, 35-40 добавлений может быть сделано или добавлено аликвот в реакционную смесь. Таким образом, 1-50, 1-40, 1-30 или 1-20 аликвот может использоваться для доставки катионов металлов. Аликвота может быть частью общего количества катионов металлов, необходимого для процесса или добавленного к процессу. Объем и/или концентрация катионов металлов в аликвоте может варьировать в зависимости от требований процесса. Аликвота может содержать небольшое количество катионов металлов, т.е. количество, достаточное для того, чтобы гарантировать присутствие определенной концентрации или количества катионов металлов в реакционной смеси. Катионы металлов могут доставляться в реакционную смесь в виде аликвот через равные промежутки времени (т.е. каждые 30 минут, каждые 60 минут, каждые 90 минут, каждые 120 минут, каждые 180 минут или каждые 240 минут или любой другой промежуток времени). Катионы металлов могут доставляться в реакционную смесь в виде аликвот через неравные промежутки времени. Например, период времени между каждой подачей катионов металлов сначала может быть короче, а затем может увеличиваться, в зависимости от того, как протекает процесс, или наоборот. Аликвоты могут доставляться, чтобы поддержать постоянную концентрацию катионов металлов в реакционной смеси, или могут доставляться, чтобы пополнить концентрацию катионов металлов до исходной концентрации, предпочтительно оптимальной концентрации свободных ионов металлов.
В предпочтительном варианте осуществления катионы металлов подаются таким образом, чтобы гарантировать, что концентрация или количество свободных катионов металлов в реакционной смеси попадает в пределы самого высокого и самого низкого порога.
Дополнительные катионы металлов могут доставляться в реакционную смесь в ответ на такую необходимость. Это может определяться некоторыми параметрами, как обсуждалось ранее.
В одном варианте осуществления дополнительные катионы металлов доставляются в реакционную смесь непрерывно.
Дополнительные катионы металлов могут физически доставляться в реакционную смесь, т.е. они находятся вне реакционной смеси.
Предпочтительно, чтобы доставка дополнительных нуклеотидов также контролировалась.
В следующем аспекте предоставляется бесклеточный способ синтеза ДНК, включающий контактирование матрицы ДНК, по меньшей мере, с одной полимеразой в присутствии одного или более праймеров, нуклеотидов и катионов металлов с образованием реакционной смеси при условиях, способствующих амплификации указанной матрицы путем замещения реплицированных цепей посредством репликации другой цепи с замещением цепей, при этом дополнительные нуклеотиды и катионы металлов контролируемым образом доставляются в реакционную смесь.
Также предоставляется бесклеточный способ синтеза ДНК, включающий контактирование матрицы ДНК, по меньшей мере, с одной полимеразой в присутствии нуклеотидов и катионов металлов для образования реакционной смеси при условиях, способствующих амплификации с замещением цепи указанной матрицы, при этом дополнительные нуклеотиды и дополнительные катионы металлов контролируемым образом доставляются в реакционную смесь.
Дополнительные нуклеотиды и дополнительные ионы металлов могут доставляться вместе или по отдельности. Контролируемая доставка дополнительных нуклеотидов или дополнительных катионов металлов может быть достигнута так, как обсуждалось ранее в отношении этих частиц.
Доставка катионов металлов или дополнительных катионов металлов может происходить параллельно с доставкой нуклеотидов, или может происходить в разное с доставкой нуклеотидов время.
Предпочтительным является то, что дополнительные катионы металлов могут подаваться в реакционную смесь вместе с подачей нуклеотидов, в частности, когда катионы металлов являются одновалентными. Таким образом, катионы металлов и нуклеотиды могут доставляться в виде смеси. Таким образом, доставка катионов металлов и нуклеотидов может быть объединена, хотя предпочтительно они хранятся отдельно.
Доставка дополнительных катионов металлов и/или дополнительных нуклеотидов может быть, как описано, индивидуальной для каждой из этих субстанций.
В предпочтительном варианте осуществления изобретения катионы металлов и нуклеотиды доставляются так, что отношение (количество) катионов металлов к нуклеотидам находится в пределах от 9:1 до 1:9 или от 6:1 до 1:6, или 3:1, или 1:3, или 1:1. Предпочтительно отношение между их количествами в реакционной смеси находится в пределах от 3:1 до 1:3, предпочтительно около 3:1. Эти отношения являются особенно предпочтительными, когда катион металла является катионом двухвалентного металла, такого как, но не ограничиваясь этим, магний (Mg2+), марганец (Mn2+), кальций (Ca2+), бериллий (Be2+), цинк (Zn2+) и стронций (Sr2+).
Есть основания полагать, что поддержание отношения катионы металлов: нуклеотиды, предпочтительно катионов магния, может быть важным для контролирования скорости синтеза ДНК. Отношение нуклеотид: полимераза также может поддерживаться, как обсуждалось ранее. Таким образом, контролирование этих двух отношений посредством контроля доставки нуклеотидов и катионов металлов может контролировать синтез ДНК.
В отношении любого аспекта изобретения, если нуклеотиды доставляются в виде солей катионов металлов, тогда катионы металлов, доставленные в реакционную смесь в этих солях, будут включаться в концентрацию катионов металлов в реакционной смеси. Специалисту известно, что доставка нуклеотидов в форме солей с ионами металлов будет влиять на концентрацию катионов металлов в реакционной смеси, и он может учитывать это дополнительное количество.
Нуклеозид трифосфаты (ATP, GTP, CTP и TTP) вследствие их химических свойств обладают потенциальной возможностью вмешиваться в ДНК амплификацию разными путями, если для увеличения выхода ДНК-продукта используются высокие концентрации. Высокими концентрациями могут считаться концентрации дНТФ выше 4мM в реакционной смеси. Фосфатные группы дНТФ способны взаимодействовать с одно- и двухвалентными катионами металлов и обладают сильнейшей аффинностью к Mg2+. Например, аффинность ATP к Mg2+ в 700 раз выше, чем к Na+, и примерно в 400 раз выше, чем к Li+.
В ходе амплификации полимеразы высвобождают пирофосфат из нуклеотидов, которые встраиваются в растущую цепь ДНК. Пирофосфат обладает аффинностью связывания с ионами Mg2+сходной сATP, и поэтому свободные ионы Mg2+ не высвобождаются при этом процессе. Результатом использования высоких исходных концентраций нуклеотидов в ходе амплификации будет уменьшение уровней свободных ионов Mg2+. Поскольку Mg2+ ионы могут требоваться для каталитической активности полимеразы, субоптимальные (недостаточные) уровни, вызванные взаимодействием с фосфатами или фосфатными группами, вероятно, оказывают негативное воздействие на эффективную амплификацию. Однако, наоборот, показано, что высокие концентрации ионов Mg2+ влияют на точность работы некоторых ДНК полимераз. Концентрация ионов Mg2+ в реакции может по этой причине считаться критической для выхода и качества ДНК.
Расщепление пирофосфата может катализироваться ферментом и, как предполагается, может высвобождать связанные ионы Mg2+ из пирофосфата. Однако, Mg2+ ионы связываются с ионами PO43- с образованием практически нерастворимого Mg3(PO4)2 (растворимость 0.01мM в воде при 20°C).
Mg2+ ионы также будут связываться с синтезированной ДНК.
Уровень свободных ионов Mg2+ в любой катализируемой ДНК полимеразой реакции в любой конкретный момент времени будет зависеть от исходной концентрации, общей концентрации молекул, способных связывать катион, и pH реакции. Поскольку процессы находятся в равновесии, всегда наблюдается наличие некоторого количества свободного Mg2+. В одном варианте осуществления уровень свободных Mg2+ управляется путем контроля доставки свободных Mg2+ ионов, предпочтительно в виде солей магния, в реакционную смесь.
Полимеразы, способные амплифицировать кольцевые ДНК матрицы по механизму замещения цепи по типу катящегося кольца, также способны продуцировать длинные линейные конкатамерные повторы матрицы. Такие полимеразы, осуществляющие замещение цепи по типу катящегося кольца, являются предпочтительными. Например, сообщается, что Phi29 ДНК полимераза может амплифицировать круговые ДНК матрицы с получением линейных повторов до 77 килобаз в длину. Эти очень длинные молекулы ведут себя подобно полимерам и могут значительно изменять реологические свойства реакции путем увеличения вязкости. Это увеличение вязкости можно измерить по изменению давления, которое может напрямую коррелировать с концентрацией синтезированной ДНК. Такие измерения могут проводиться постоянно или в назначенное время, чтобы получить сигнал, предпочтительно сигнал в режиме онлайн, который говорит о состоянии реакции. Доставка дополнительных нуклеотидов может управляться по сигналу датчика давления. Такая доставка нуклеотидов в ответ на сигнал, указывающий на параметр реакционной смеси, может быть сделана с целью увеличения выхода и/или продуктивности реакционной смеси.
Измерения разницы давлений для оценки ДНК концентраций имеют важное преимущество по сравнению с анализами, основанными на связывании флуоресцентных химических реагентов, таких как красители Syto. В отличие от этих реагентов, которые связываются только с двухцепочечной ДНК, измерения разницы давлений не дают ошибочных оценок ДНК в образцах, содержащих одноцепочечные формы.
В одном варианте осуществления подача нуклеотидов управляется по сигналу. Сигналом может быть любой подходящий сигнал и может быть сигналом в режиме онлайн. Предпочтительно, чтобы сигнал имел отношение к концентрации ДНК, в частности, основывается на оценке общей концентрации ДНК в реакционной смеси. Сигнал может альтернативно иметь отношение к объему реакционной смеси. Доставка дополнительных ионов металлов может аналогичным образом управляться по сигналу. Сигнал может быть тем же самым сигналом или другим сигналом.
Предпочтительным сигналом является сигнал, который относится к вязкости реакционной смеси. Вязкость реакционной смеси может использоваться для оценки концентрации ДНК в реакционной смеси. Вязкость может быть измерена с помощью любого подходящего способа, включая вискозиметр. Предпочтительно, чтобы вязкость измерялась по изменению давлений. Разница давлений может быть измерена с помощью датчика давления, датчика избыточного давления, индикаторов давления, пьезометров и манометров. В одном варианте осуществления разница давлений может создаваться с помощью насоса поршневого типа, который извлекает и возвращает часть реакционной смеси. Предпочтительно, чтобы реакция контролировалась в режиме реального времени неинвазивным образом, как указано в описании.
Процесс изобретения может осуществляться как периодическая реакция. В этом варианте осуществления реакционная смесь находится в реакторе. Дополнительные нуклеотиды и/или дополнительные катионы металлов могут подаваться в реактор. Таким образом, процесс может осуществляться в реакторе с периодической загрузкой. Альтернативно, процесс может проводиться в реакторе с непрерывным потоком. Такие реакторы находятся в пределах знаний квалифицированного специалиста. Особенно предпочтительный формат представлен ниже.
В одном варианте осуществления один или более процессирующих ферментов добавляют в реакционную смесь в виде одного или более отдельных добавлений, т.е. в виде аликвот. Любое число добавок или аликвот находится в рамках этого варианта осуществления. Таким образом, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более, т.е. 20-25, 25-30, 35-40 добавлений или аликвот может быть добавлено в реакционную смесь. Таким образом, 1-50, 1-40, 1-30 или 1-20 аликвот может использоваться для доставки процессирующих ферментов. Аликвота может быть частью всего количества процессирующего фермента, необходимого для процесса или добавленного к процессу. Объем и/или концентрация процессирующего фермента в аликвоте может варьировать в зависимости от потребностей процесса. Аликвота может содержать небольшое количество процессирующего фермента, т.е. количество, достаточное, чтобы гарантировать, что определенная концентрация или количество процессирующего фермента присутствует в реакционной смеси. Процессирующий фермент может добавляться в реакционную смесь в виде аликвот через равные промежутки времени (т.е. каждые 30 минут, каждые 60 минут, каждые 90 минут, каждые 120 минут, каждые 180 минут или каждые 240 минут или любой другой подходящий период времени). Процессирующий фермент может добавляться в реакционную смесь в виде аликвот с нерегулярными интервалами. Например, период времени между каждым добавлением процессирующего фермента может быть первоначально более длинным, а затем может уменьшаться, по мере того, как протекает процесс или наоборот.
Процессирующие ферменты могут доставляться в реакционную смесь в ответ на необходимость. Эта необходимость может быть определена по нескольким параметрам, как обсуждалось ранее, включая концентрацию ДНК в реакционной смеси и/или скорость синтеза ДНК.
Процессирующие ферменты могут физически доставляться в реакционную смесь, т.е. они находятся вне реакционной смеси. Эти ферменты могут находиться в отдельном резервуаре и высвобождаться в реакционную смесь контролируемым образом.
Предпочтительно, чтобы процессирующий фермент имел одно обновление, как, например, в случае с протеломераза, поскольку тогда контролирование количества процессирующего фермента напрямую контролирует количество ДНК, процессированной ферментом. Процессирующий фермент может добавляться вместе с дополнительными нуклеотидами, или после дополнительных нуклеотидов. В случае, если также требуются дополнительные ионы металлов, может быть предпочтительно, чтобы они добавлялись после добавления процессирующего фермента, так что процессинг ДНК происходит до добавления дополнительных ионов металлов.
Процесс может проводиться в любом подходящем приборе. А теперь будет описано отдельное устройство, пригодное для осуществления такого процесса. Устройство для синтеза 1 выполнено в автономном корпусе 11 и снабжено открываемой передней крышкой 12, чтобы дать возможность доступа к основным компонентам. Передняя крышка 12 желательно сделана из прозрачного материала, но может быть тонирована, чтобы защитить компоненты реакции от избыточного уровня освещенности. Крышка также служит для ограничения проникновения загрязнений. Устройство снабжено внешне доступным выключателем питания и входом питания 13, например, сбоку устройства. Кроме того, имеются порты ввода/вывода 14, которые дают возможность устройству поддерживать связь с и/или быть под контролем компьютера общего назначения, снабженного пакетом программ, обеспечивающим автоматизацию исследований, такого как, например, Labview™, поставляемый корпорацией National Instruments Corporation, Техас, США. Световой индикатор работы 15 располагается спереди устройства в таком положении, чтобы быть видимым, и показывает, находится ли устройство в действии.
Центральным компонентом устройства для синтеза является заключенный в кожух реакционный сосуд 2, который может иметь объем более, чем 30 мл, например, 50 мл, 75 или 120 мл. Объем реакционного сосуда может быть менее 300 мл. Объем реакционного сосуда может быть выбран в соответствии с предполагаемым использованием устройства и количеством продукта, которое должно быть получено за один раз, или в соответствии со скоростью производства в непрерывном процессе. Совершенно ясно, что размер реакционного сосуда определяет верхний предел количества продукта, которое может быть получено за одну загрузку.
Отношение высоты и ширины реакционного сосуда не может быть больше, чем 8:1, или не больше, чем 7:1, 6:1 или 5:1. Если реакционный сосуд слишком высокий, тогда становится труднее осуществлять тщательное перемешивание содержимого путем извлечения и возвращения реакционной смеси. Если реакционный сосуд слишком широкий, тогда становится труднее обеспечивать равномерную температуру во всем объеме, в частности, при использовании небольших объемов реакции. Перемешивание смеси может способствовать получению желаемой температуры во всей смеси, в частности, если реакционная смесь нагревается до 95°C. Реакционные сосуды с отношением высоты и ширины в указанных выше пределах можно адаптировать к различным количествам компонентов реакции.
Реакционный сосуд 2, который может использоваться в варианте осуществления изобретения, показан в поперечном сечении на фигуре 3. Реакционный сосуд 2 включает внутренний сосуд 23, окруженный кожухом 25. Жидкость, обеспечивающая поддержание температуры, циркулирует по пространству 26 между внутренним сосудом 23 и кожухом 25 для того, чтобы контролировать температуру содержимого внутреннего сосуда 23. Нижний порт 27 и верхний порт 28 обеспечивают соединение внутреннего сосуда с избирательными клапанами (переключателями) 32b, 32a, соответственно.
Внутренний сосуд 23 может иметь основную часть, которая имеет форму цилиндра диаметром D1, например, в пределах от 20 до 50 мм, предпочтительно от 25 до 35 мм, и конусообразную часть 24, соединяющую цилиндрическую часть с нижним портом 27. Общая высота H1 внутреннего сосуда может находиться в пределах от 150 мм до 300 мм, желательно от 200 мм до 250 мм. Высота конусообразной части 24 может составлять в пределах от 5 до 20 мм, предпочтительно от 10 до 15 мм. Угол боковых стенок конусообразной части может быть в пределах от 30 до 60° относительно оси внутреннего сосуда, предпочтительно около 45°. Диаметр D2 нижнего порта 27 может быть в пределах от 5 до 10 мм.
Реакционный сосуд может иметь непостоянное поперечное сечение, например, за счет конусности на всем протяжении высоты или центрального днища.
Температура реакционного сосуда может контролироваться потоком жидкости, регулирующей температуру, через внешний кожух к реакционному сосуду. Жидкость, регулирующая температуру, может доставляться через входное отверстие 2a и удаляться из выходного отверстия 2b с помощью рециркулирующего регулятора температуры 22. Рециркулирующий регулятор температуры контролирует температуру жидкости, регулирующей температуру, путем нагревания и/или охлаждения жидкости. Реакционный сосуд может поддерживаться при постоянной температуре, например, несмотря на возникновение эндотермических или экзотермических реакций, или может регулироваться так, чтобы следовать желательному температурному профилю. Жидкость, регулирующая температуру, может быть водой или маслом. Жидкость, регулирующая температуру, может доставляться через порты 16 на корпусе устройства. Подходящим рециркулирующим регулятором температуры является система обеспечения температурного режима Presto A30, произведенная компанией Julabo USA, Пенсильвания, США. В одном варианте осуществления изобретения рециркулирующий регулятор температуры 22 содержит омические нагревательные элементы и/или устройства Пельтье. В одном варианте осуществления температура реакционного сосуда поддается регулированию в пределах от 4°C до 95°C с точностью до 0,01°C.
Для компонентов реакции обеспечиваются терморегулируемые резервуары 4a-4c, которые используются для проведения реакции синтеза. Компоненты реакции содержатся в шприцах 42a-42c, например, одноразовых полипропиленовых шприцах, которые вмонтированы в регуляторы температуры 41a-41c. Регуляторы температуры в этом варианте осуществления содержат блоки из электропроводного материала, например, алюминия, имеющие отверстия, предназначенные для того, чтобы плотно удерживать шприцы 42a-42c. Можно использовать теплопроводящую мастику, если желательно увеличить теплопроводность между блоком и шприцами. Нагревательные/охлаждающие устройства, например, омические нагревательные элементы или устройства Пельтье, прикрепляются к теплопроводящим блокам, для того, чтобы контролировать их температуру и, следовательно, компонентов реакции, содержащихся в резервуарах 42a-42c.
Регуляторы температуры 41a-41c могут использоваться или для нагревания, или для охлаждения содержащихся компонентов реакции. Некоторые компоненты реакции желательно содержать при низкой температуре, например, около 4°C, для того, чтобы предотвратить их деградацию. Праймеры могут храниться при повышенной температуре для того, чтобы предотвратить и/или уменьшить димеризацию праймеров. Резервуары также могут быть нагреты для того, чтобы денатурировать или подвергнуть отжигу матрицы и праймеры до добавления в реакционный сосуд. Как и в случае с основной регулировкой температуры реакционного сосуда, терморегулируемые резервуары могут поддерживаться при постоянной температуре или регулироваться так, чтобы следовать желательному температурному профилю, например, чтобы нагревать компонент реакции перед добавлением в реакционный сосуд. В одном варианте осуществления температуры терморегулируемых резервуаров независимо поддаются регулированию в пределах от 4°C до 95°C с точностью до 0,01°C.
Один или более проточных нагревателей (не показано) может предоставляться для повышения температуры, например, до 95°C, компонента реакции, например, праймера, до добавления в реакционный сосуд. Нагреватель располагается на канале между резервуаром 4a-c, 5a-g и переключателем 32c и/или между переключателем 32a и реакционным сосудом 2.
В этом воплощении, один из терморегулируемых резервуаров 4c содержит два резервуара-шприца 42c и 42c’ в термическом контакте с одним блоком температурного контроля 41c. Это бывает полезно, когда два компонента реакции должны поддерживаться при одинаковой температуре или когда больший объем отдельного компонента реакции требуется для желательной реакции.
Устройство также содержит несколько дополнительных резервуаров 5a-5g, которые снабжаются устройствами, регулирующими температуру. В этом варианте осуществления имеется семь таких резервуаров, в других вариантах осуществлениях предоставляется больше или меньше таких резервуаров. Дополнительные резервуары 5a-5g также могут содержать шприцы, например, стеклянные или полипропиленовые шприцы, которые могут быть удалены и утилизированы в опорожненном виде. Использование шприцов является предпочтительным, поскольку объем резервуара автоматически уменьшается по мере того, как компоненты реакции удаляются. Подобный эффект может быть достигнут при использовании складных (сжимаемых) пакетов или трубок. В случае использования жесткого контейнера, желательным является наличие фильтра в любом месте выхода в атмосферу, для того, чтобы предотвратить проникновение загрязнений.
В целях удобства подставка резервуаров 5 снабжается поворотной стойкой 51. Если требуются большие количества компонента реакции, устройство может быть снабжено портами для соединения с внешним резервуаром. Желательно, устройство спланировано таким образом, чтобы минимизировать длину канала, необходимого для соединения резервуаров с переключателями. Каналы могут быть отсоединены от резервуаров и затем могут быть присоединены к коллектору, который присоединяется для доставки очистительной жидкости или воды для промывки. Такими образом, все устройство может быть очищено и промыто in situ, т.е. не будучи демонтировано.
Резервуары 4a-c и 5a-g могут иметь вместимость от 3 до 10 мл, например, около 5 мл. Может быть предусмотрено наличие одного или более больших резервуаров, например, для буферных растворов, с вместимостью от 10 до 500 мл, например, около 250 мл.
Подача компонента реакции в реакционный сосуд может осуществляться возвратно-поступательным шприцевым насосом 3a. Этот насос называется подающим насосом. Он содержит шприц 31a, например, стеклянный или полипропиленовый шприц, приводимый в движение электромагнитом 33a и соединенный с управляемым переключателем 32a, чтобы выборочно соединять электромагнит с каждым из резервуаров и реакционным сосудом. Чтобы добавить контролируемое количество компонента реакции в реакционный сосуд, сначала переключатель 32a используется для соединения шприцевого насоса с соответствующим резервуаром, затем электромагнит срабатывает так, чтобы втянуть в шприц соответствующий объем компонента реакции. Затем, переключатель 32a используется для соединения шприца 31a с реакционным сосудом 2 и электромагнит меняет направление на обратное, чтобы вытолкнуть содержимое шприца в реакционный сосуд. При такой конфигурации можно быстро и последовательно добавить необходимые количества компонентов реакции в реакционный сосуд, без использования множества шприцевых насосов и множества входов в реакционный сосуд. Однако, если желательно добавить некоторое количество компонентов реакции одновременно в реакционный сосуд, можно обеспечить несколько подающих насосов 3a. Это также может быть желательно, если необходимо держать отдельные компоненты реакции абсолютно отдельно до их добавления в реакционный сосуд.
Кроме того, второй возвратно-поступательный шприцевой насос 3b может использоваться для перемешивания содержимого реакционного сосуда, а также для удаления реакционной смеси или продукта или в конце реакции или с целью отбора проб в ходе процесса реакции. Этот насос называется откачивающим насосом и включает шприц 31b, приводимый в движение электромагнитом 33b. Второй шприцевой насос 3b соединен со стоком на дне реакционного сосуда через второй переключатель 32b. Чтобы перемешать содержимое реакционного сосуда, используется второй переключатель 32b для соединения шприца 31b с реакционным сосудом, при этом некоторое количество реакционной смеси удаляется из реакционного сосуда, а затем возвращается туда. Количество реакционной смеси, которое удаляется, и скорость с которой она возвращается, может регулироваться, для того, чтобы контролировать качество осуществленного перемешивания. Такое перемешивание может осуществляться непрерывно в ходе реакции, периодически или в определенные моменты времени, имеющие отношение к добавлению компонентов реакции.
Альтернативно или дополнительно, второй возвратно-поступательный шприцевой насос 3b также может соединяться с резервуаром 4a и использоваться для доставки компонента реакции, содержащегося в этом резервуаре, в реакционный сосуд 2.
Датчик давления 82 может располагаться на коротком выступе, соединенном с каналом между реакционным сосудом 2 и переключателем 32b или между переключателем 32b и откачивающим насосом 3b. Во время удаления и возвращения материала из или в реакционный сосуд 2, давление, измеряемое датчиком 82, показывает вязкость переносимой смеси: более высокое давление, более высокая вязкость. В частности, амплитуда изменения давления в ходе циклического удаления и возвращения материала показывает вязкость реакционной смеси. Точное соотношение между давлением и вязкостью зависит от различных факторов, включая скорость удаления или возвращения материала, диаметр используемых каналов и форму реакционного сосуда. Это соотношение может определяться теоретически или путем калибровки. В одном варианте осуществления точное соотношение не требуется, а сигналы от датчика давления 82 могут использоваться просто для обнаружения изменений в реакционной смеси, без знания ее точной вязкости. В такой реакции, когда вязкость увеличивается в результате образования продукта, сигнал от датчика давления может использоваться для управления реакцией. На основе сигнала о давлении и путем активизирования доставки с помощью насоса одного или более соответствующих реагентов из одного или более резервуаров с надлежащей скоростью или в соответствующем количестве, можно увеличить выход продукта. В других случаях сигнал о давлении может указывать на завершение реакции или стадии в реакции и потому может использоваться для инициации сбора продукта или добавления компонентов для следующей стадии.
Сигнал от датчика давления 82 может использоваться для управления скоростью извлечения и/или возвращения смеси в реакционный сосуд, чтобы удостовериться, что ДНК материал в смеси не поврежден сдвиговыми силами. Скорость извлечения и/или возвращения смеси может контролироваться, чтобы удостовериться, что предельное давление не превышено. Предельное давление может зависеть от вязкости.
Чтобы удалить материал, например, продукт реакции, из реакционного сосуда откачивающий насос 3b сначала соединяется с реакционным сосудом с помощью второго переключателя 32b и электромагнита, работающего так, чтобы оттягивать поршень шприца с целью извлечения материала из реакционного сосуда 2. Затем откачивающий насос соединяется с выходным портом 7 с помощью второго переключателя и электромагнита 33b, приведенного в действие так, чтобы выпустить собранный материал через выходной порт 7.
Выходной порт 7 может соединяться, например, с контейнером для продукта реакции, контейнером для проб, измерительным устройством или любым другим прибором. Также может быть, что образец, который был извлечен с целью измерения, возвращается в реакционный сосуд после измерения.
Откачивающий насос 3b может иметь бóльшую вместимость, чем подающий насос 3a, для того, чтобы, например, весь реакционный сосуд мог опорожняться за одну операцию отсасывания. Такая большая вместимость откачивающего насоса 3b также желательна, чтобы принимать участие в очистке устройства посредством прокачки больших объемов очищающего раствора и/или деионизированной воды через всю систему. В этих целях устройство может быть оснащено дополнительными большими резервуарами для хранения достаточных количеств очищающего раствора и деионизированной воды.
Для доставки компонентов реакции, для перемешивания реакционной смеси и извлечения реакционной смеси и/или продукта может использоваться один насос.
Подающий насос 3a и/или откачивающий насос 3b могут представлять собой другую форму насоса, например, пульсирующий насос.
Устройство также может содержать сливной сосуд 6, который соединяется с атмосферой через отверстие 62, которое может снабжаться одноходовым клапаном. Сливной сосуд 6 соединяется с верхней частью реакционного сосуда 2, для того, чтобы обеспечить отверстие, и вследствие этого предотвратить повышение давления в реакционном сосуде 2. Также предоставляются каналы для соединения сливного сосуда 6 с переключателями 32a и 32b для того, чтобы обеспечить выгрузку продукта из реакционного сосуда и любых неиспользованных и нежелательных компонентов реакции. Сливной сосуд также может использоваться для принятия очищающих и промывающих жидкостей в ходе in-situ очистки и промывки, а также небольших количеств компонентов реакции, являющихся результатом предпусковой подготовки каналов с целью создания условий для точной доставки необходимых количеств в реакционный сосуд.
Сливной сосуд 6 может отсутствовать. В этом случае желательно обеспечить предохранительный клапан в реакционном сосуде 2 и обеспечить соединения от переключателей 32a, 32b до внешней канализации.
Работа прибора управляется регулирующим устройством (контроллером) 8, который соединен электрически с регуляторами температуры 41a-c, с набором шприцов 32a-b, электромагнитами 33a-b, с регулятором температуры 22 и датчиком давления 82 для того, чтобы управлять этими компонентами прибора. Контроллер 8 также содержит интерфейс 81, обеспечивающий связь с внешним компьютером 9 для полного контроля процесса, который должен быть осуществлен.
Контроллер 8 может быть предварительно запрограммирован на выполнение конкретных широкоиспользуемых режимов работы, таких как предварительное заполнение, отбор проб, сбор продукта, промывка и очистка. В одном варианте осуществления контроллер 8 просто передает команды от внешнего компьютера 9 к разным компонентам устройства. Упомянутые общепринятые режимы работы могут осуществляться под контролем внешнего компьютера или вручную.
Предпусковая подготовка устройства может включать извлечение компонентов реакции из каждого из резервуаров для того, чтобы каналы между резервуарами и переключателями заполнились соответствующими компонентами реакции. Это необходимо для того, чтобы увеличить точность количеств компонентов реакции, доставляемых в реакционный сосуд, и уменьшить возможность загрязнения. В одном варианте осуществления отбор проб из реакционной смеси включает извлечение заранее определенного количества компонента реакции во внешний сосуд или датчик во время реакции. В некоторых случаях возможно возвращение образца в реакционный сосуд. Сбор продукта может включать извлечение некоторых или всех продуктов реакции в конце периодического процесса или в соответствующее время в непрерывном процессе. Промывка может включать промывку использованных каналов и резервуаров чистой водой. Очистка устройства может включать промывку каналов и резервуаров очищающим раствором.
В одном варианте осуществления ДНК матрица, по меньшей мере, одна полимераза, один или более праймеров и нуклеотидов добавляется в реакционный сосуд для образования реакционной смеси. Часть или порция реакционной смеси извлекается из реакционного сосуда и возвращается туда, для того, чтобы перемешать реакционную смесь и обеспечить ее смешивание. В конце реакции ДНК извлекается из реакционного сосуда. В одном варианте осуществления реакционная смесь извлекается из порта, находящегося в нижней части реакционного сосуда, предпочтительно в самой нижней точке реакционного сосуда. В одном варианте осуществления часть реакционной смеси, которая извлекается, составляет 50% или менее от всей реакционной смеси, желательно 40% или менее, предпочтительно 30% или менее. В одном варианте осуществления часть реакционной смеси, которая извлекается, составляет до 5% или более от всей реакционной смеси, желательно 10% или более, предпочтительно 20% или более.
В одном варианте осуществления исходный объем реакционной смеси составляет от 10 мл до 10 литров, предпочтительно от 10 до 100 мл, от 100 мл до 1000 мл или от 1 до 10 литров.
Теперь изобретение будет описано со ссылкой на некоторые неограничивающие примеры.
Примеры
Материалы и методы
Амплификация ДНК в следующих примерах была проведена с использованием Phi29 ДНК полимеразы, действующей на ds кольцевую матрицу, содержащую сайты-мишени для протеломеразы TelN. Phi29 полимераза продуцирует длинные линейные повторы кольцевой матрицы (конкатамеры), которые могут процессироваться протеломеразой TelN (Touchlight Genetics) в желаемый замкнутый линейный ДНК-продукт. Создание конкатамерной ДНК с помощью Phi29 ДНК полимеразы приводит к увеличению вязкости реакции вследствие их длины (по имеющимся сведениям до 77 килобаз), тогда как процессинг двухцепочечной конкатамерной ДНК с использованием протеломеразы TelN приводит к уменьшению вязкости вследствие их более короткой длины (в пределах от 2 до 3 килобаз в данных примерах).
В Примерах 1 и 3 ДНК образуется в устройстве, описанном в данном документе (фигуры 1-3).
Пример 1. Количественное определение в режиме онлайн полученной в ходе реакции ДНК с помощью измерений разницы давлений
Первый прибор очистили и дезинфицировали. Трубки отсоединили от 10 резервуаров для реагентов (42a, 42b, 42c’, 42c, 5a - 5g) и повторно соединили через фитинговые соединения Люэра с коллектором в положении 10, который снабжается от 120мл резервуара, содержащего 10% раствор гипохлорита натрия. Аналогично, 60мл резервуар с 10% раствором гипохлорита натрия был присоединен к выходному порту 7 от переключателя 32b. Путем регулирования положений переключателей 32a и 32b и действий электромагнитов 33a и 33b на 5мл стеклянные шприцы 31a и 31b, соответственно, вся система, включая сам реакционный сосуд 2, была полностью заполнена раствором гипохлорита натрия без «мертвых пространств». Минимум 5мл раствора было вытянуто через каждую трубку. Реакционный сосуд (вместимость 120мл) нагревали до 50°C под регулирующим воздействием термоциркулятора 22, и система поддерживалась в этом состоянии в течение 30 минут. Результатом применения этой процедуры было полное разрушение всех загрязняющих ДНК во всех подающих трубках и в реакционном сосуде.
Аналогично, при помощи соответствующей программы, управляющей переключателями и электромагнитами, описанными выше, всю систему опорожнили от раствора гипохлорита натрия, сбросив его в сливной сосуд 6. Повторение описанного выше процесса 5 раз после замены раствора гипохлорита натрия в резервуарах на деионизированную воду гарантировало полное удаление любых остатков гипохлорита натрия из всей системы, делая систему готовой к эксплуатации.
Затем трубки были отсоединены от манифольда, вновь подсоединены к резервуарам для реагентов 10, содержащим стерильные одноразовые полипропиленовые шприцы, с помощью наконечников Люэра (вместимость от 5 мл до 20 мл). В случае, когда указано ниже, эти шприцы содержали отдельные компоненты реакции в минимальном объеме 2 мл. В других случаях они оставались пустыми, при этом поршни шприцов были полностью опущены. Резервуар 5d состоял из двух 60мл шприцов одновременно, чтобы давать полный объем деионизированной воды 120 мл. После заполнения шприцов из каждого цилиндра шприцов, содержащих реагенты, выгнали воздух, вручную опуская поршень до тех пор, пока жидкость не начинала вытекать из выходного отверстия шприца. В ходе этой процедуры выходное отверстие шприца держали в вертикальном положении выше поршня шприца, чтобы обеспечить удаление всего воздуха из цилиндра шприца.
Таблица 1
Ферменты, Phi29 ДНК полимераза и пирофосфатаза, в резервуарах 42c и 42c’ соответственно, содержались в стабильных условиях при 4°C при помощи регулятора температуры 41c. Все другие реагенты хранились при комнатной температуре.
Для осуществления точно дозированной подачи реагентов в реакционный сосуд 2, трубки между резервуарами с реагентами и переключателями 32a и 32b предварительно заполнили, последовательно выпустив очень небольшие количества реагентов в сливной сосуд 6.
Для того, чтобы денатурировать ДНК матрицу и связать олигонуклеотидный праймер, реагенты из резервуаров 5a, 5b и 5c были распределены через переключатель 32a (под воздействием электромагнита 33a на шприце 31a) в закрытый пустой шприц 5мл (резервуар 42b). Температура этого резервуара находилась под контролем регулятора температуры 41b. Регулирование с помощью 41b использовали для повышения его температуры до 95°C в течение 3 минут и затем охлаждения до 30°C. Затем объединенные в резервуаре 42b компоненты реакции были отправлены в реакционный сосуд 2 через переключатель 32a. Оставшиеся 5 компонентов реакции затем были распределены через переключатель 32a в порядке, показанном в таблице 1, так что конечный объем в реакционном сосуде 2 составлял 100мл. Температура реакционного сосуда 2 была установлена 30°C при помощи регулирования на термоциркуляторе 22. Компоненты реакции не подвергались действию поверхностных температур, превышавших больше чем на 2°C заданное значение.
Смешивание компонентов реакции осуществлялось путем автоматического регулирования работы электромагнита 33b и движений смешивающего шприца 31b. Работа шприца управлялась так, чтобы он извлекал из сосуда и возвращал в реакционный сосуд 2,5мл компонентов реакции. Смешивание было периодическим – шприц работал в течение 1 минуты с интервалами 29 минут со скоростью извлечения/распределения 55мл/мин.
Изменение давления в канале между реакционным сосудом 2 и шприцом 31b было измерено с помощью датчика вакуума и давления (Model PX409-2.5CGUSBH, Omega Engineering Limited, Manchester, M44 5BD, UK). Этот датчик измерял разницу давлений от -2.5 до + 2.5 psi и был присоединен через T-образный разветвитель с помощью короткого отрезка тефлоновой трубки диаметром ⅛ дюйма. Выходной сигнал от сенсора контролировался в режиме реального времени и регистрировался на компьютере, контролирующем реактор с помощью программного обеспечения, поставляемого производителем. Возвратно-поступательное движение смешивающего шприца генерирует выходной сигнал датчика давления, который колеблется между положительными и отрицательными значениями (двухтактное движение «пулл-пуш» смешивающего шприца). В течение минуты данные о давлении регистрируется в каждый момент смешивания, показатели минимального и максимального давления вычислялись по 3 наборам по 20 секунд. Минимальное значение вычитается из максимального значения для каждого набора и вычисляется среднее. Притом, что фиксировались все другие переменные, такие как скорость шприца, материал трубок, внутренний диаметр и длина и размер избирательного крана (переключателя), любые изменения в высоте пика могли быть только следствием изменения вязкости компонентов реакции. Очень небольшие изменения давления, которые могли быть отнесены за счет окружающей температуры и колебаний давления, не компенсировались подсчетами разницы давления. Первую экспериментальную точку, используемую для анализа, снимали приблизительно через 45 минут, чтобы гарантировать, что система полностью уравновешена.
Фигура 4 явно показывает увеличение разницы давления в зависимости от времени в течение реакции амплификации ДНК, катализируемой Phi29 ДНК полимеразой. Образцы, взятые из этой реакции, также были проанализированы напрямую в отношении ДНК с использованием набора для флуориметрического анализа QubitTMBR dsДНК (Life Technologies, Пейсли, Великобритания.). В этом методе измеряется только двухцепочечная ДНК. Конкатамерная ДНК, полученная в данной реакции, была подвергнута процессированию протеломеразой TelN с получением трех ковалентно замкнутых линейных молекул ДНК. Процессированную ДНК осадили PEG 8000 и NaCl в конечных концентрациях 5% и 2M, соответственно. Осажденную ДНК центрифугировали при 14,000 g в течение 20 минут и ресуспендировали в эквивалентном объеме не содержащей нуклеазу деионизированной воды.
Фигура 5 явно показывает, что увеличение сигнала разницы давлений в ходе реакции прямо коррелирует с увеличением концентрации ДНК. Прямая линейная зависимость между давлением и измеренной ДНК также указывает, что в течение этого периода реакции ДНК существует преимущественно в двухцепочечной форме.
Пример 2. Эффект исходных концентраций дНТФ на эффективность их включения в ДНК
В этом эксперименте реакции ДНК амплификации (объем 5мл) проводили в 50мл пропиленовых центрифужных пробирках. Компоненты последовательно добавляли в пробирки в порядке, показанном в таблице 2, с получением указанных конечных концентраций. Эффекты исходных концентраций дНТФ на ДНК амплификацию были протестированы при концентрациях 2мМ, 4мМ и 8мМ.
Таблица 2
Все пробирки инкубировали при 30°C в орбитальном инкубаторе при минимальном перемешивании в течение 61.5 часов. Этот период времени соответствовал приблизительно 10-кратному периоду времени, который требуется для завершения реакции ДНК амплификации при обеспечении 2мМ дНТФ. В конце реакции конкатамерная ДНК, полученная при каждом условии, подвергалась процессированию протеломеразой TelN с получением двух разных ковалентно связанных линейных молекул ДНК. Процессированную ДНК осаждали с помощью PEG 8000 и NaCl в окончательных концентрациях 5% и 2M, соответственно. Осажденную ДНК центрифугировали при 14,000 g в течение 20 минут и ресуспендировали в 5мл деионизированной воды, не содержащей нуклеазу. Образец этого ДНК продукта разводили 1 к 50 в деионизированной воде и нагревали до 75°C в течение 1 минуты. Образец 10мкл этой разбавленной ДНК смешивали с 2мкл загрузочного красителя (Novel Juice, Newmarket Scientific Ltd., UK.) и 10мкл образца наносили на 0.8% агарозный гель и проводили электрофорез в стандартных условиях до полного разделения компонентов ДНК (фигура 6).
Гель-электрофорез на фигуре 6 явно демонстрирует значительное снижение выхода продукта в виде двух ковалентно связанных линейных ДНК, когда исходная дНТФ концентрация составляла 8мМ. Поскольку реакция проводилась в течение длительного периода времени, это указывает на очень сильный ингибирующий эффект на амплификацию ДНК. Образцы, взятые в конце реакции, были также исследованы в отношении ДНК путем измерений оптической плотности при 260нм, результаты представлены в таблице 3.
Таблица 3. Концентрации общей процессированной ДНК в конце реакции были вычислены на основании измерений оптической плотности при 260нм и выражены в виде процентов от максимального теоретического выхода ДНК за счет разных уровней дНТФ
Таблица 3 подтверждает результаты электрофореза, показанные на фигуре 6, что исходные дНТФ концентрации свыше 4мМ могут значительно уменьшать выход ДНК в реакциях, катализируемых Phi 29 ДНК полимеразой. Также было очевидно, что при использовании исходной дНТФ концентрации 8мМ также наблюдалось значительное снижение чистоты продукта, подтвержденное размыванием вокруг двух основных полос продукта (фигура 6).
Наблюдаемое уменьшение выхода ДНК при увеличении исходных концентраций дНТФ может быть отнесено за счет целого ряда воздействий на реакцию. Сюда включается, но без ограничения: субстратное ингибирование фермента; эффекты Li+ или нуклеотидов на устойчивость взаимодействий цепи ДНК преимущественно взаимодействий праймера и матрицы и/или уровень свободных ионов Mg2+, доступных для активности фермента вследствие связывания Mg2+ с дНТФ. Ингибирование ДНК амплификации может быть вызвано одним фактором или множеством факторов, действующих прямым или косвенным образом. Очевидно, что увеличение выхода ДНК в реакции до уровней, считающихся приемлемыми для промышленного производства, не может быть достигнуто просто путем увеличения исходных концентраций дНТФ.
Пример 3. Эффект подачи дНТФ в реакцию ДНК-амплификации, катализируемую Phi29 ДНК полимеразой
Были проведены эксперименты для сравнения эффекта добавления дополнительных количеств дНТФ в реакции ДНК-амплификации, которые катализировал фермент Phi29. Реакции осуществлялись, как описано в Примере 1, но концентрации ДНК были вычислены по результатам измерения оптической плотности при 260нм. Были сделаны сравнения между реакцией с 2мМ дНТФ и без добавления дополнительных дНТФ (Реакция 1), реакцией с 2мМ дНТФ и добавлением дополнительных 2мМ дНТФ на момент времени 530 минут (Реакция 2), реакцией с 4мМ дНТФ с самого начала (без добавления дополнительных дНТФ) и реакцией с 2мМ дНТФ с троекратным добавлением дНТФ в концентрации 2мМ (в моменты времени 500, 1430 и 4340 минут). Инициирующим событием для подачи любых дополнительных дНТФ было выравнивание сигнала разности давлений. Это активировало добавление соответствующего объема раствора дНТФ из резервуара, так что концентрация дНТФ в реакторе увеличивалась на 2мМ.
На фигуре 7 показан график разницы давлений в зависимости от времени для каждого из этих условий реакции.
Из представленных на фигуре 7 данных видно, что начальные скорости были вычислены до добавления, между добавлениями и после добавления дНТФ, в зависимости от конкретного случая. Результаты показаны в таблице 4. Кроме того, в конце каждой реакции был оценен окончательный выход ДНК и вычислен как процент от максимального теоретического выхода, исходя из добавленной концентрации дНТФ. Результаты показаны в таблице 5.
Таблица 4: Исходные уровни амплификации ДНК для реакций 1 – 4, оцененные по измерению разности давлений
Таблица 5. Конечный выход ДНК для реакций 1 - 4 был вычислен, исходя из измерений оптической плотности при 260нм
Таблица 4 ясно показывает, что увеличение концентрации дНТФ значительно снижает скорость, с которой ДНК амплифицируется Phi29 ДНК полимеразой. Эффект на скорость реакции является кумулятивным и не зависит от того, когда были добавлены дополнительные дНТФ.
В Примере 2 было показано, что реакция с исходной дНТФ концентрацией 8 мМ давала только 15% от максимального теоретического выхода, указывая на чрезвычайно медленную реакцию. В таблице 5, реакция, к которой 8 мМ дНТФ добавляются в виде 2мМ аликвот (Реакция 4), достигает выхода ДНК 54% от максимального теоретического выхода, что является высоко значимым улучшением. Наряду с тем, что постепенная подача дНТФ, по-видимому, ведет к постепенному снижению скорости реакции, общая скорость быстрее и выход выше, чем при введении высокой концентрации дНТФ в начале реакции.
Таблица 4 также показывает, что в процессе подачи дНТФ молярное отношение ионов Mg2+ к фосфатным группам дНТФ (отношение Mg2+/PO43-) значительно снижается и соответствует наблюдаемому снижению скорости реакции. Это проиллюстрировано на фигуре 8.
Таким образом, подача дНТФ может увеличить выработку ДНК до приемлемых с точки зрения промышленного производства уровней в реакциях, катализируемых ферментами. Однако, для достижения более высоких скоростей реакции, судя по всему, молярное отношение ионов Mg2+ к PO43- группам дНТФ нуждается в поддержании на оптимальном уровне путем доставки ионов Mg2+ вместе с дНТФ.
Пример 4. Действие концентрация ионов магния на эффективность включения дНТФ в ДНК
В этом эксперименте реакции амплификации ДНК (объемы 5мл) проводили в 50мл полипропиленовых центрифужных пробирках. Компоненты были предварительно нагреты до 30°C и потом добавлены в пробирки в порядке, показанном в таблице 6, с получением указанных конечных концентраций. Исходные концентрации Mg2+ 2.24мМ, 4.52мМ, 9мМ и 11.24 мМ в сочетании с исходной концентрацией дНТФ 3мМ давали отношение Mg2+/PO43- 0.25, 0.5, 1 и 1.25, соответственно. Пробирки инкубировали при 30°C в течение 19 часов до добавления дополнительных аликвот дНТФ и MgCl2 для увеличения концентрации в реакции дНТФ до 6мМ и концентраций Mg2+до 4.48мМ, 9.04мМ, 18мМ и 22.48мМ, чтобы сохранить Mg2+/PO43- отношения 0.25, 0.5, 1 и 1.25. Реакции позволили продолжаться в течение дополнительных 24 часов, ДНК продукт, произведенный за это время, собрали и оценили его концентрацию. ДНК была обработана достаточным количеством протеломеразы TelN, ДНК осадили с помощью 6% PEG 8000 в буфере, содержащем 500мМ NaCl, 100мМ MgCl2. ДНК ресуспендировали в объеме 4 мл и количественно оценили ДНК путем измерения оптической плотности при 260нм.
Таблица 6. Компоненты реакционной смеси и порядок добавления
Таблица 7. Выход ДНК в реакциях с добавлением дНТФ с сохранением фиксированных отношений Mg2+/PO43-
Результаты в таблице 7 показывают воздействие разных отношений Mg2+/PO43- на выход ДНК через 43 часа в реакциях с добавлением дНТФ. Низкий выход ДНК, полученной за это время, при отношении Mg2+/PO43- 0.25 указывает на медленную скорость реакции или ингибирование реакции из-за недостаточной концентрации ионов Mg2+. В любом случае результаты показывают, что существует оптимальное отношение Mg2+/PO43-, которое нужно поддерживать для эффективной реакции амплификации ДНК для промышленного использования. Полученные данные показывают, что оптимальное отношение может быть в пределах от 0.5 до 1.25. Данные также показывают, что если дНТФ поступают в реакцию, добавление дополнительных ионов Mg2+ для сохранения оптимального отношения Mg2+/PO43- (в идеале 1) обеспечивает эффективное преобразование в ДНК.
Пример 5. эффект добавления протеломеразы TelN на реакцию амплификации ДНК, катализируемую полимеразой Phi29, в сочетании с добавлением нескольких аликвот дНТФ
Эксперименты проводили с целью сравнения эффекта добавления Phi29 катализируемых реакций амплификации ДНК на плазмиде ProTLx-K N3X2 Lux (фигура 13) с добавлением дополнительных количеств дНТФ и протеломеразы TelN. Реакции проводили в основном как описано в Примере 1, но с использованием резервуара 42b, содержащего 25мкМ TelN раствор в 12мМ Tris-HCl (pH7.4), 90мМ NaCl, 0.1мМ EDTA. 1мМ DTT, 50% глицерин, поддерживаемый при 4°C, и резервуар 5г, содержащий 100мМ MgCl2 раствора в деионизированной воде. Исходная концентрация матрицы ProTLx-K_N3X2 Lux была 5мкг/мл.
В соответствии с порционной фазой реакции аликвоты дНТФ добавляли в реакционный сосуд 2 через шприц 31a для достижения увеличения концентраций на 2мМ с выбранными интервалами времени 300мин, 600мин, 900мин, 1080мин, 1230мин и 1440 мин. С теми же самыми интервалами и сразу после добавления дНТФ были добавлены аликвоты MgCl2 для достижения отношения 3:1 [Mg2+] : [дНТФ]. В момент времени 1050 мин в реакционный сосуд 2 была подана протеломераза TelN с получением конечной концентрации 2мкМ.
В результате первоначальной порционной фазы реакции (до 300 мин) наблюдается постоянное увеличение измерения перепада давления, соответствующее синтезу конкатамерной ДНК. После добавления протеломеразы TelN в реакцию на момент времени 1050 минут наблюдается 3.6-кратное увеличение скорости изменения давления, соответствующее значительному увеличению синтеза ДНК. Есть основания полагать, что одно добавление TelN во время фазы подачи дНТФ/Mg2+ приводит к выработке замкнутой линейной ДНК, которая может действовать как матрица для дополнительной ДНК амплификации. Дополнительная доставка Mg2+/дНТФ после этой точки имеет очень большое значение для обеспечения наблюдаемой повышенной скорости синтеза ДНК.
Реферат
Изобретение относится к области биотехнологии и молекулярной биологии. Предложен способ бесклеточного синтеза ДНК, включающий контактирование матрицы ДНК по меньшей мере с одной полимеразой, функционирующей по принципу замещения цепи, в присутствии нуклеотидов и катионов одного или нескольких металлов с образованием реакционной смеси, где изотермическая амплификация матрицы ДНК происходит посредством репликации с замещением цепи, при этом дополнительные нуклеотиды подаются в реакционную смесь непрерывно или с интервалами во время синтеза ДНК и дополнительно подаются катионы металлов, таким образом, что соотношение катионы металлов : нуклеотиды в реакционной смеси сохраняется между 9:1 и 1:9. Способ позволяет улучшить выход ДНК и скорость ее синтеза, что может быть использовано в медицине и биотехнологиях. 2 н. и 24 з.п. ф-лы, 13 ил., 7 табл., 5 пр.
Комментарии