Способ получения эндотелиальных клеток из эмбриональных стволовых клеток человека (варианты) - RU2359030C1
Код документа: RU2359030C1
Чертежи










Описание
Изобретение относится к биотехнологии, а именно к получению эндотелиальных клеток из эмбриональных стволовых клеток (ЭСК) человека и их применению. Человеческие эмбриональные стволовые клетки (обозначение, принятое в мире, hESCs) нужно рассматривать как ценный источник для регенеративной медицины из-за их способности дифференцироваться во все типы клеток. Полученные в результате дифференцировки эндотелиальные клетки могут найти широкое применение в медицинской практике и в изучении механизмов дифференцировки. Эндотелиальные клетки не только выстилают стенки кровеносного сосуда, но и также способны мигрировать в потоке крови к различным местам. Таким образом, они могли использоваться не только для регенерации сосудов с помощью их интеграции или секреции ростовых факторов, но и служить транспортным средством для терапевтического гена или доставки препарата. Однако потенциальное клиническое применение дифференцированных in vitro клеток требует подтверждения, что желательный клеточный фенотип будет поддержан без возвращения к плюрипотентному состоянию.
На стадии развития бластоцисты происходит формирование внутренней клеточной массы бластоцисты (ВКМ), из которой и получают линии ЭСК. Способность hESCs дифференцироваться в некоторые специфические типы клеток в настоящее время интенсивно изучается. Общая цель таких исследований обычно состоит в том, чтобы получать специализированные функциональные клетки. Однако дальнейшая возможность применения этих клеток для лечения человека может быть реализована только при условии обеспечения устойчивой клеточной специализации. Таким образом, нужно рассматривать не только морфологические, иммунологические, или функциональные свойства клетки, но также и генетические и эпигенетические.
Дифференцировка ЭСК по определенному пути сопровождается изменением паттерна экспрессии генов от генов, характерных для плюрипотентного состояния, к тканеспецифическим генам. Изменение паттерна экспрессии осуществляется за счет затухания экспрессии ключевых транскрипционных факторов, участвующих в поддержании плюрипотентности, и экспрессией тканеспецифических транскрипционных факторов. Параллельно работают эпигенетические механизмы регуляции экспрессии генов, которые репрессируют гены, необходимые для плюрипотентного состояния, и активируют тканеспецифические. Так как ЭСК - это in vitro аналог клеток эпибласта, искусственно поддерживаемых в недифференцированном состоянии, при изменении условий культивирования ЭСК спонтанно дифференцируются в различные типы клеток. In vitro спонтанная дифференцировка ЭСК человека наблюдается как при длительном культивировании прикрепленных колоний, так и в процессе роста эмбриоидных телец (ЭТ) в суспензии. В отличие от ЭСК мыши не все линии ЭСК человека легко образуют эмбриоидные тельца и ни одна линия ЭСК не образует эмбриоидные тельца из одиночных клеток.
Для ЭСК человека была показана способность к дифференцировке во многие типы клеток: нейроны и глия, эндотелий, кератиноциты, трофобласты, кардиомиоциты, остеобласты, клетки крови, гепатоциты, инсулин-продуцирующие клетки и некоторые другие.
Производные энтодермы получить оказалось труднее всего. В недавно опубликованных сообщениях о дифференцировке ЭСК человека в инсулин-секретирующие клетки [1] и гепатоциты [2] было показано, что по белковым и генетическим маркерам они совпадают с зрелыми инсулин-секретирующими клетками или гепатоцитами. Однако факторы, определяющие дифференцировку в энтодерму, плохо охарактеризованы, и почти отсутствуют маркеры ранней энтодермальной дифференцировки.
В отличие от ЭСК мыши, которые легко дифференцируются в кардиомиоциты в отсутствие leukemia inhibitory factor LIF, спонтанная дифференцировка ЭСК человека наблюдается у разных линий ЭСК в разном проценте случаев - от спонтанно сокращающихся участков в 70% ЭТ до полного отсутствия спонтанной дифференцировки [3]. Показано, что кардиомиоциты, дифференцированные из ЭСК человека, экспрессируют транскрипционные факторы Nkx 2.5, GATA4, а также специфические маркеры кардиомиоцитов - тропонин 1 и тяжелую цепь альфа-миозина. Недавно опубликована работа по дифференцировке кардиомиоцитов в бессывороточной среде, минуя стадию эмбриоидных телец [4]. При длительном культивировании ЭСК могут подвергаться искусственной случайной селекции. Кроме того, как оказалось, ЭСК, идентичные по основным морфологическим характеристикам и экспрессии основных генов, могут отличаться эпигенетическим статусом в регуляторных областях некоторых генов. Эти факторы могут приводить к тому, что клеточные линии ЭСК будут иметь преимущественные пути дифференцировки. То есть, например, ЭСК будут легко дифференцироваться в нейроны и плохо в кардиомиоциты. Возможно, именно разница в эпигенетическом статусе ЭСК мыши и человека может объяснить столь различный потенциал дифференцировки.
При нормальном развитии эмбриона сосудистая ткань формируется из мезодермы. Вскоре после гаструляции мезодерма формируется как отдельный слой между экто- и эндодермой. Предполагается, что эндотелиальные и гематопоэтические клетки происходят от общего мезодермального предшественника - гемангиобласта [5]. Клетки-предшественники, потенциально способные образовывать гематопоэтические и эндотелиальные клетки, были идентифицированы на модели мышиных ЭСК [6, 7, 8] и лишь позже in vivo при изучении гаструляции мышиного эмбриона [9]. Существует три основных технических подхода для изучения дифференцировки ЭСК в клетки эндотелия и гематопоэтические клетки. Первый - дифференцировка ЭСК в эндотелиальные или гематопоэтические клетки через стадию ЭТ [10-15]. Второй - совместное культивирование ЭСК со стромальными клетками [16, 17]. Третий - дифференцировка ЭСК в двумерных условиях [18, 8].
Дифференцировка ЭСК через образование ЭТ - это всегда спонтанная ненаправленная дифференцировка. Как уже упоминалось, ЭТ содержит клетки, происходящие из разных зародышевых листков, и поэтому доля клеток требуемого типа мала. Процент клеток эндотелия в эмбриоидных тельцах составляет около 2%. Кроме того, этот подход обладает еще несколькими ограничениями. Например, трудно наблюдать клетки в микроскоп, проводить анализ дифференцировки единичных клеток, трудно разделять клетки из агрегатов клеток в ЭТ. Однако на сегодняшний день в большом количестве публикаций используется именно этот подход. Это обусловлено, в частности, тем, что направленная дифференцировка клеток требует специального подбора условий, а ЭТ - грубый модельный аналог развивающегося эмбриона, в котором присутствуют различные клетки, в том числе и необходимые исследователю.
Нами было проверено, есть ли эндотелиальные клетки в ЭТ, образованных из линий ЭСК человека, имеющихся в распоряжении. Для анализа были взяты линии hESM01 и hESM03. Эти линии, выделенные в лаборатории ранее, проявляли характерную для данного типа клеток морфологию, экспрессировали соответствующие маркеры и имели нормальный кариотип [19]. При переводе в суспензионную культуру ЭСК формировали ЭТ с характерной морфологией для данного типа структур. При иммуногистохимическом анализе эмбриоидных телец после 10-12 дней культивирования были обнаружены в небольшом количестве (не более 1-5%) эндотелиоцитоподобные клетки, экспрессирующие CD31 (фиг.1).
Дифференцировка ЭСК человека в клетки эндотелия в двумерных условиях (минуя стадию образования ЭТ) использовалась только одной группой исследователей [20]. Известный способ выбран в качестве ближайшего аналога. Способ предусматривает дифференцировку ЭСК человека в присутствии человеческого фактора роста эндотелия сосудов (VEGF) на подложке из коллагена IV. Селекция эндотелиальных клеток осуществлялась по размеру клеток с помощью фильтрации через фильтр с диаметром пор 40 мкм. Из работы не ясно, какую долю составляли клетки эндотелия после селекции. Однако, полагая, что эндотелиальные клетки связаны с гематопоэтическими клетками и происходят от общего предшественника гемангиобласта, механический выбор не мог обеспечить желательную чистоту клеток. Скорее всего, данный метод селекции не стоит использовать, если необходимо получить чистую популяцию клеток эндотелия.
Была поставлена задача разработать эффективный и воспроизводимый метод дифференцировки hESCs в чистую популяцию функциональных эндотелиальных клеток, в которой изменения паттерна генной регуляции соответствуют зрелым эндотелиальным клеткам. В отличие от дифференцировки через стадию ЭТ прямая дифференцировка ЭСК проста для наблюдения за клетками и манипуляций с ними, однако требует предварительных исследований по определению условий дифференцировки. Кроме того, контролируемая дифференцировка, во-первых. должна приводить преимущественно к желаемому клеточному фенотипу. Во-вторых, должна существовать возможность селекции нужных популяций клеток, причем сам процесс селекции не должен существенным образом влиять на жизнеспособность клеток и их дифференцировку. Следующей поставленной перед нами задачей явилась разработка метода сепарации предшественников эндотелия и эндотелиальных клеток из дифференцированных ЭСК человека.
Отсутствие в литературе данных по дифференцировке ЭСК человека в двумерной системе без формирования ЭТ в эндотелиальные клетки потребовало определение условий и сред для дифференцировки. Было испробовано более 15 способов культивирования с различными сочетаниями сред, сывороток и ростовых факторов. Для этого колонии ЭСК перемещали в чашки Петри с ростовыми средами различного состава (табл.1).
В процессе дифференцировки были обнаружены отдельные клетки, экспрессирующие ранний маркер эндотелия CD31 и маркер позднего дифференцированного эндотелия - фактор Вон Виллибрандта (vWF).
После применения различных композиций сред в различных комбинациях с факторами роста были выявлены ростовые факторы, обеспечивающие эффективную дифференцировку ЭСК человека в эндотелиальные клетки. К таким ростовым факторам белковой природы относятся stem cell factor SCF (производитель Chemicon), человеческий фактор роста эндотелия сосудов VEGF (производитель Chemicon), bone morphogenic protein BMP4, basic fibroblast growth factor bFGF (производитель Chemicon), tumor growth factor TGFbeta.
Количественные соотношения данных ростовых факторов зависят от состава среды, наличия или отсутствия в среде компонентов неопределенного происхождения, а также могут варьировать в зависимости от используемых линий ЭСК. Количество ростовых факторов может быть подобрано экспериментальным путем.
В течение дифференцировки в среде, содержащей компоненты, необходимые для направленной дифференцировки ЭСК человека в эндотелиальные клетки, мы могли наблюдать изменения и в морфологии клеток и специфической экспрессии маркеров эндотелия. Сначала на 4 день после индукции дифференцирования были обнаружены CD31 эндотелиальные маркеры. Появление сосудистоподобных структур совпало с экспрессией vWF. На день 6-7 почти 60% клеток экспрессировали CD31 молекулу и к концу процедуры дифференцирования (12-й день), приблизительно 50% клеток экспрессировали CD 105 антиген (табл.2). При этом стоит заметить, что экспрессия CD 133 молекулы была высока в недифференцированных hESCs и понижалась в течение дифференцировки в эндотелиальных предшественниках. Довольно высокий процент дифференцировки в эндотелиальные клетки позволял нам проводить тест на формирования сосудов без селекции клеток.
CD31 положительные клетки представлены не только в составе сосудистоподобных структур, но и в виде монослоя (фиг.2). По данным исследований баланс между сосудистоподобными клетками и монослоем зависит от концентрации VEGF.
Для сепарации дифференцированных из ЭСК человека эндотелиальных клеток нами был выбран метод иммунологической селекции с использованием поверхностных или внутренних маркеров, специфических для клеток эндотелия. Например, в данном случае использовался метод иммуномагнитной сепарации MACS (magnetic activated cell sorting) - эффективная технология магнитного сортинга. основанная на разделении клеток по поверхностным антигенам (см. www.miltenyibiotec.com). Подобный метод позволяет быстро и эффективно (с чистотой 98% и более) провести селекцию нужной популяции клеток по поверхностным маркерам. Однако может быть использован и другой метод, например флуоресцентно-активированный клеточный сортинг (FACS) или любые другие, позволяющие разделить клетки в зависимости от поверхностных или внутренних маркеров.
Для селекции эндотелия, полученного из ЭСК в двумерной системе, мы воспользовались маркером CD31, но могут быть использованы и другие маркеры эндотелия, как CD34, фактор vWF или другие.
Селекцию на CD31 проводили на 5-7 день дифференцировки. Замечено, что жизнеспособность клеток, отобранных на 5 день, была выше, чем отобранных в день 7 после начала дифференцировки (фиг.3). Этот факт можно объяснить тем. что на 7 день дифференцировки клетки эндотелия уже формируют структуры, которые достаточно трудно разрушить ферментативной обработкой, а увеличение времени обработки и/или разрушение уже сформировавшихся межклеточных контактов ведет к существенному снижению жизнеспособности. Следовательно, проведение селекции на 5 день дифференцировки в наших условиях можно считать оптимальным. При использовании для селекции других маркеров селекцию можно проводить на 3-9 дни дифференцировки.
Выделенные клетки высевали на чашки, покрытые коллагеном, в среду для эндотелиальной дифференцировки. Клетки высевали в высокой плотности (не менее 50000 клеток на 1 см2 и более), так как посев в более низкой плотности негативно влиял на жизнеспособность клеток.
Нами предложено 2 варианта способа получения эндотелиальных клеток из ЭСК человека.
Сущность первого варианта способа получения эндотелиальных клеток из ЭСК человека заключается в следующем:
1. Проведение дифференцировки ЭСК человека на подложке и в синтетической среде, которые обеспечивают направленную прямую дифференцировку ЭСК человека в эндотелиальные клетки, с VEGF, SCF, ВМР4, bFGF, TGFbeta. Использование в качестве основы синтетической среды смеси жидкой DMEM/F12 с жидким КО заменителем сыворотки в количестве (об.%): жидкая DMEM/F12 - 75-95%, жидкая КО заменитель сыворотки - 5-25%. Причем TGFbeta вносят в синтетическую среду на 3-6-й день дифференцировки.
2. Проведение иммунологической селекции с использованием маркеров, специфических для клеток эндотелия, на 3-9 дни дифференцировки.
3. Культивирование эндотелиальных клеток при плотности посева не менее 50000 клеток на 1 см2
Сущность второго варианта способа получения эндотелиальных клеток из ЭСК человека заключается в следующем:
1. Проведение дифференцировки ЭСК человека на подложке и в среде, которые обеспечивают направленную прямую дифференцировку ЭСК человека в эндотелиальные клетки, с VEGF, SCF, bFGF. Использование в качестве основы среды смеси жидкой DMEM/F12 и фетальной бычьей сыворотки в количестве (об.%): жидкая DMEM/F12 - 75-95%, жидкая фетальная бычья сыворотка - 5-25%.
2. Проведение иммунологической селекции с использованием маркеров, специфических для клеток эндотелия, на 3-9 дни диференцировки.
3. Культивирование эндотелиальных клеток при плотности посева не менее 50000 клеток на 1 см2
В наших способах были опробованы линии клеток hESM01, hESM03, hESM02, 04, HI, HUES7, 8, 9, при этом полученные результаты были близкими, что подтверждает возможность его применения для дифференцировки любых ЭСК человека.
Соотношение компонентов среды может оптимизироваться для каждой клеточной линии в отдельности для достижения максимальной эффективности. В качестве среды для культивирования клеток могут быть использованы и другие синтетические среды полностью известного и охарактеризованного состава.
Заявленные способы позволяют получить высокий выход клеток в конце процедуры дифференцировки, имеющих иммунологические маркеры эндотелия, большинство из которых способно участвовать в формировании трубчатой сети. Даже больше, дифференцировка была воспроизведена с использованием заменителя сыворотки вместо характеризованной фетальной бычьей сыворотки (FBS), что значительно лучше существующих методов дифференцировки. Использование синтетической среды, не содержащей компонентов животного происхождения, делает метод воспроизводимым и независимым от состава сыворотки. Однако даже в этом случае приблизительно половина клеток была представлена неизвестным типом клеток или незначительной популяцией недифференцированных клеток. Важно отметить, что даже маленький процент недифференцированных клеток или клеток, которые дифференцировались в различные типы, может загрязнить клеточную культуру. Эти сторонние популяции могут иметь большое воздействие на безопасную и функциональную эффективность культуры для терапевтического применения или дальнейшего изучения. Таким образом, селекция специфической популяции клеток важна.
CD31 признан лучшим иммунологическим маркером эндотелиальных клеток, и в то же самое время его экспрессия была обнаружена очень рано - на день 6 hESCs дифференцировки, что позволило его использовать в методе иммуномагнитного сортинга. Однако использование других маркеров (например, CD34, фактор vWF) и других методов селекции (например, FACS) тоже давало хорошие результаты.
Возможность осуществления способа подтверждается следующими примерами его реализации, которые показывают лишь частные случаи его выполнения.
Пример 1. Культивирование ЭСК человека.
Линии ЭСК человека культивировали в среде, содержащей 80% KnockOut DMEM, 20% КО заменителя сыворотки, глутамин 1 мМ, 1% заменимые аминокислоты, 50 пенициллина ЕД/мл, 50 мкг/мл стрептомицина (производитель Invitrogen), 0,1 мМ β-меркаптоэтанола (Sigma), и 4 мкг/мл bFGF (Chemicon) на ингибированных митомицином С (10 мкг/мл, Sigma) мышиных эмбриональных фибробластах (МИФ) при плотности 104клеток/см2 в культуральных чашках, покрытых 0,1%-ным желатином (Merck). Фибробласты приготавливали из 12-дневных мышиных эмбрионов Fl (C57BL/6J×CBA/Ca). hESC колонии пассировали каждые 5-6 дней культивирования обработкой коллагеназы типом IV (200U/ml, Invitrogen) и механической диссекцией. Чашки с недифференцированными колониями hESC, выращенными в течение 8 дней, обрабатывали коллагеназой IV в течение 10 минут, смывали дважды ростовой средой и собрали колонии с фидера механически наконечником микропипетки вручную. Плюропотентные линии клеток, изолированные и выращенные на фидере, показывали типичную ЭСК морфологию клетки, экспрессировали иммунологические маркеры, характерные для недифференцированных клеток, и поддерживали нормальный кариотип. Мы не наблюдали даже маленький процент CD31 положительных клеток в недифференцированных колониях.
Пример 2. Дифференцировка ЭСК человека в эндотелиальные клетки.
Среда для дифференцировки.
Для дифференцировки использовали синтетическую среду следующего состава: основа 85% жидкой DMEM/F12 (Hyclone или аналог другой фирмы), 15% жидкого КО заменитель сыворотки (Invitrogen или аналог другой фирмы) с добавлением 1% заменимых аминокислот, 2 мМ L-глутамина, 50 ед/мл пенициллина, 50 мг/мл стрептомицина, 50 нг/мл SCF, 10 нг/мл ВМР4, 50 нг/мл VEGF, активина 10 нг/мл, 20 нг/мл bFGF, 2 нг/мл TGFbeta. TGF beta вносят на 4 день дифференцировки. Концентрации и сочетания ростовых факторов могут отличаться от указанных на 50-100% в зависимости от клеточной линии и индивидуальных условий культивирования.
Способ приготовления среды
Жидкая среда DMEM/F12 смешивалась с жидким заменителем сыворотки, антибиотиками и глутамином и для дальнейшего использования хранилась при +4°С.
Все остальные компоненты (ростовые факторы и пр.) добавлялись в среду непосредственно перед добавлением к клеткам. Полная среда не хранилась.
Методика проведения дифференцировки.
Чашки с недифференцированными колониями hESC, выращенными в течение 8 дней (фиг.4А) обрабатывали коллагеназой IV в течение 10 минут, смывали дважды ростовой средой и колонии собрали вручную наконечником микропипетки, разрезая на фрагменты размером 300-500 клеток. Колонии ЭСК для проведения дифференцировки были перемещены в 35 мм чашки Петри, покрытые коллагеном IV (70 нг/мл) в среде, указанного выше состава (фиг.4Б). На 4 день дифференцировки в среду добавляли 2 нг/мл TGFbeta 1. На следующий после добавления день клетки приобретали морфологию дифференцированных производных (фиг.4В). На 6-10 день культивирования колонии содержали кластеры сосудистоподобных структур (фиг.4Г).
Пример 3. Дифференцировка ЭСК человека в эндотелиальные клетки. Отличается от примера 1 использованием среды на основе смеси жидких DMEM/F12 (Hyclone) и фетальной бычьей сыворотки (Hyclone characterized).
Среда для дифференцировки.
Для дифференцировки использовали среду следующего состава: основа 85% жидкой DMEM/F12 (Hyclone или аналог другой фирмы) и 15% фетальной бычьей сыворотки (Hyclone, characterized или аналог другой фирмы), 2 мМ L-глутамина, 1% заменимые аминокислоты, 50 единиц/мл пенициллина, 50 мг/мл стрептомицина, (все от Invitrogen или аналог другой фирмы), 20 нг/мл SCF (Chemicon или аналог другой фирмы), от 10 до 50 нг/мл VEGF (Chemicon или аналог другой фирмы), 5 нг/мл bFGF (Chemicon или аналог другой фирмы) Ростовые факторы могут варьировать, как указано в примере 1. Среду готовят, как описано в примере 1.
Клетки также на 3-6 день приобретали морфологию дифференцированных производных. На 6-10 день культивирования колонии содержали кластеры сосудистоподобных структур.
Пример 4. Сепарация эндотелиальных клеток.
Для CD31+ клеточной селекции клетки на 5-7 день дифференцировки были отделены обработкой коллагеназы IV (Invitrogen или аналог другой фирмы) при 37°С в течение 10 минут и смыты дважды забуференным фосфатом физиологическим раствором (phosphate bufferd saline PBS), содержащим 0,5% альбумина бычьей сыворотки (BSA). Полученные клетки инкубировали в течение 30 минут с антителами мыши против CD31 человека в порции 10 мкг на 106 клеток и затем с IgG козы против мыши, пришитыми к магнитным шарикам (Miltenyi Biotec или аналог другой фирмы) в течение 15 мин при 8°С. CD31 положительные клетки были изолированы, проходя через фильтр и колонку MS, прикрепленную к магнитному блоку Mini-MACS separation unit (Miltenyi Biotech или аналог другой фирмы), чтобы получить сохраненную магнитом фракцию очищенных CD31+ клеток. После элюции CD31 положительные клетки были помещены на покрытые коллагеном IV плашки с плотностью посева 50000 клеток на 1 см2 и клетки были выращены в эндотелий-поддерживающей среде (фиг.5А).
Пример 5. Анализ полученных эндотелиальных клеток.
Чистоту выделенной популяции определяли по иммуноцитохимическому анализу. Чтобы узнать, являются ли CD31+ клетки, элюированные с колонки, клетками эндотелия или их предшественниками, часть клеток подвергали функциональному тесту на принадлежность клеток к эндотелию - тесту на матригеле. Анализ экспрессии генов, характерных для эндотелия, проводили методом полуколичественного ОТ-ПЦР (Обратной транскрипции ПЦР).
Иммуноцитохимический анализ.
Клетки фиксировали 100%-ным ледяным метанолом в течение 5 минут или 4%-ным парафармальдегидом в течение 10 минут при комнатной температуре. Слайды были промыты в трех PBS, 0,1% Tween 20 и инкубировались со вторичными антителами: Alexa Fluor 546-conjugated IgG козы против мыши, Alexa Fluor 488-conjugated IgG козы против мыши в разведении 1:800. Первичные используемые антитела были античеловеческим CD31, античеловеческим vWF, античеловеческим CD 105. античеловеческим CD34, античеловеческим CD133, античеловеческим Flk-1. античеловеческим CD 14, античеловеческим CD45. Клетки были также окрашены 4,6-диамидино-2-фенилондолом (DAPI). После этого иммуномеченые клетки или фрагменты были исследованы с помощью флюоресценцентного микроскопа. Чтобы избегать артефактов в обнаружении, мы использовали антитела с различной спецификой от используемых для селекции клеток. Чистота популяции клеток была больше 90% (фиг.5Б).
Испытание на способность образовывать сосудистоподобные структуры.
Испытание было проведено, как описано в [21]. Матригель добавлялся в 24 лунки планшета. (0,5-2)×104 клеток были помещены в лунки в 0,5 мл ростовой среды и инкубировались при 37С и 5% СО2. Морфология клеток анализировалась в фазовом контрасте на инвертированном световом микроскопе. Как видно из фиг.5В, клетки формируют характерную для эндотелия ячеистую сосудоподобную сеть.
Обратной транскрипции ПЦР (ОТ-ПЦР).
Тотальная РНК была изолирована от недифференцированных hESCs. CD31 селектированых эндотелиальных клеток и HUVECs (эндотелиальные клетки вены человеческой пуповины) использованием RNeasy Mini Kit (Qiagen). Обратная реакция транскрипции была выполнена, используя РНК (1-2 мкг), случайные шестинуклеотидные праймеры (Promega), RevertAid M-MuLV обратную транскриптазу и реактивы (Promega) согласно рекомендациям изготовителя. Кодирующая ДНК была растворена в конечном объеме 25 мл. Определенное количество кДНК (1-5 мл) было подвергнуто ПЦР амплификации, используя Taq ДНК полимеразу и реактивы (Fermentas). Все образцы РНК были способны привести к равной амплификации глицеральдегид-3-фосфатдегидрогеназы (GAPDH) как внутренний стандарт. Последовательности праймеров для ОТ-ПЦР указанны в Таблице 3. Продукты амплификации были сепарированы в 1,5% агарозном геле.
Факторы транскрипции играют важную роль в течение эмбрионального развития в направлении дифференцировки и поддержании фенотипа. Особенно эндотелий-специфичные факторы транскрипции GATA-2 и 3, которые почти полностью выключены в недифференцированных hESCs, хотя их экспрессия в полученной из hESCs чистой популяции эндотелиальных клеток была сопоставима с их экспрессией в HUVECs. Ген eNOSa не экспрессировался недифференцированными hESCs, но после дифференцировки в эндотелий его экспрессия была обнаружена в чистой популяции эндотелиальных клеток на том же самом уровне, как в HUVCs.
Таким образом. Анализ экспрессии генов, характерных для эндотелия, методом полуколичественного ОТ-ПЦР показал, что уровень экспрессии этих генов в клетках эндотелия из ЭСК сравним с экспрессией в HUVEC (фиг.6)
Геномное бисульфитное секвенирование.
Были выделены геномные ДНК от недифференцировынных hESCs. CD31 отобранных hESCs-полученных эндотелиальных клеток и HUVEC. Геномная ДНК была подвергнута обработке натрием бисульфитом используя CpGenome™ Fast DNA Modification Kit, Chemicon. Аликвоты ДНК, модифицированной бисульфитом натрия (25-50 нг), были использованы для ПЦР при 45 циклах амплификации. Праймеры для ПЦР в интересущих участках были подобраны для натрия бисульфит модифицированной матрицы (Таблица 4). Конечные ПЦР продукты были клонированы с помощью набора pGEM-T Easy (A1360, Promega) и клоны плазмиды (10 для каждого участка) были секвенированы использованием автоматизированной ABI Prism 377 ДНК Sequencer (Applied Biosystems).
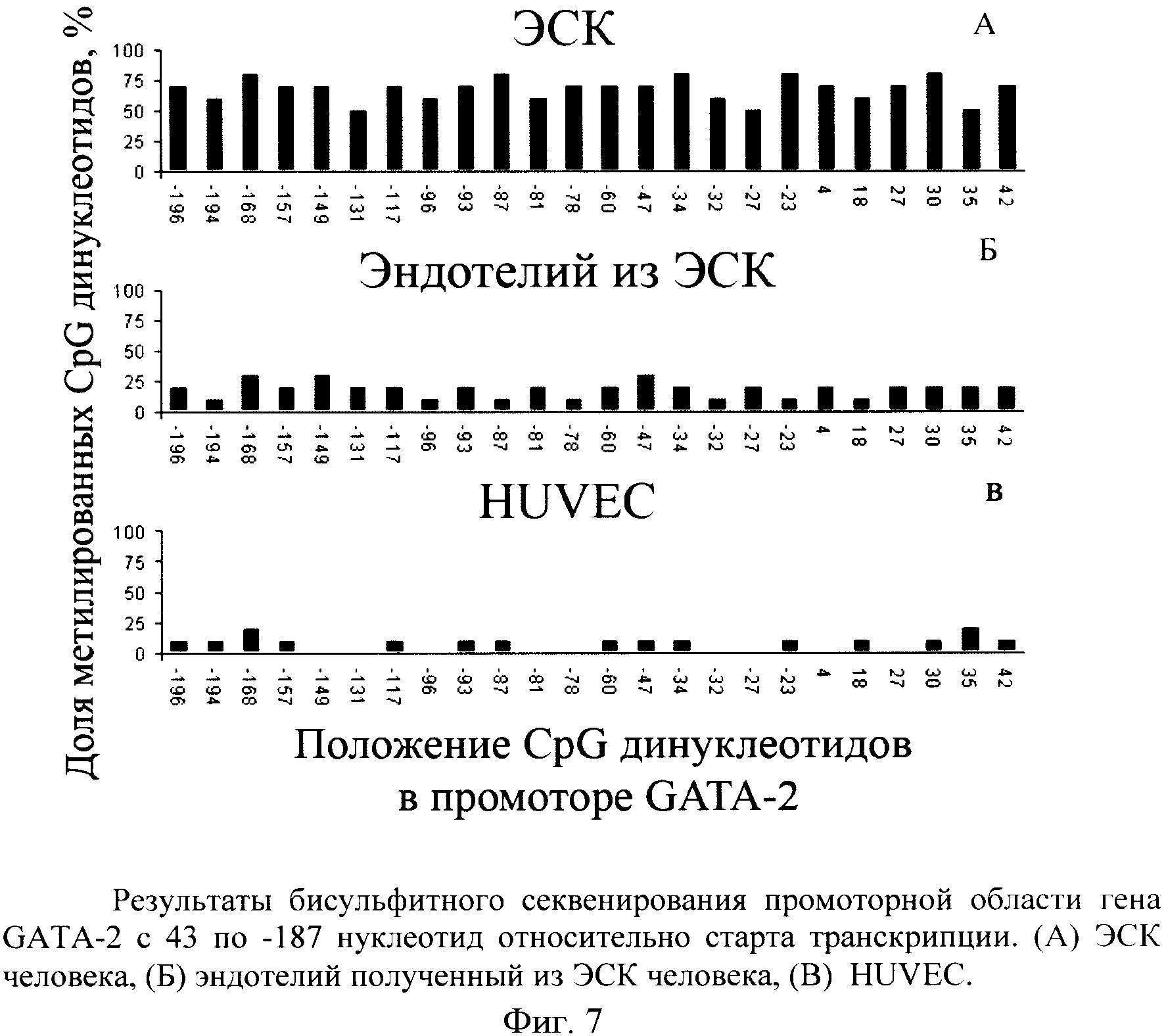
Чтобы подтвердить, что эндотелиальные клетки, дифференцированные из hESCs, имеют правильное проявление эпигенетического статуса, мы сравнили профили метилирования регулирующих участков GATA-2,-3 и генов eNOCa в hESCs эндотелиальных клеток, дифференцированных из hESCs и HUVECs. Мы исследовали уровень метилирования каждого CpG в области, соответствующей селективному промотору гена GATA-2 (с 43 по 187 нуклеотид относительно старта транскрипции, определяемого как +1) с помощью бисульфитного секвенирования. В недифференцированных hESCs эта область была высоко метилирована почти до 80% метилирования (фиг.7). Уровни метилирования селективного промотора в HUVEC и эндотелиальных клетках, дифференцированных из hESCs, были низкими и варьировали от 20% в эндотелиальных клетках, дифференцированных из hESCs, до 10%в HUVECs.
CpG, расположенные в области от 228-115 нуклеотидов, были высоко метилированы в недифференцированных клетках (до 90%), но в полученных из hESC клетках эндотелия и в HUVECs метилирование обозначенной области понижалось до 15-20% (фиг.8). Таким образом, несмотря на гиперметилирование проксимального промотора ген GATA-3 был слабо экспрессирован в недифференцированных hESCs, хотя гипометилированный промотор значительно увеличил его экспрессию.
Согласно данным Chan [22] в эндотелиальных клетках самые существенные эпигенитические модификации происходят в области промотора eNOSa самой близкой к сайту начала транскрипции. Эта область гена eNOSa между 13 и 237 нуклеотидами была выбрана для анализа бисульфитного секвенирования, который показал, что область была гиперметилирована в недифференцированных hESCs приблизительно на том же самом уровне, как регулирующие области GATA-2 и GATA-3 генов от 70 до 90% по каждому CpG положению. После hESCs дифференцировки в эндетелиальные происхождения метилирование этой области уменьшилось до 20% в функциональных эндотелиальных клетках, отобранных из hESCs, и не превышал 10% в HUVECs (фиг.9).
Бисульфитное секвенирование позволило выявить, что область селективного промотора GATA-2 гиперметилирована в hESCs и гипометилирована в hESCs-полученных эндотелиальных клетках. Таким образом, мы показали, что статус метелирования GATA-2 селективного промотора оказывает влияние на экспрессию этого фактора транскрипции.
Приняв во внимание корреляции экспрессии GATA-2 и его промотора метилирования в hESCs-полученных эндотелиальных клетках, мы могли признать важную роль селективного промотора GATA-2 в эндотелий-дифференцировке. Несмотря на гиперметилирование GATA-3 проксимального промотора GATA-3 ген был слабо экспрессирован в недифференцированных hESCs. Однако мы не могли исключить, что в недифференцированных hESCs GATA-3 ген запущен альтернативным промотором.
Проанализировав все полученные данные, мы можем заключить, что in vitro внешние сигналы вызывают сложную комбинацию молекулярных механизмов. заканчивающихся индукцией определенных факторов транскрипции экспрессии. Даже больше, полагая, что ДНК деметилирование является заключительным шагом эпигенетических событий, ведущих к активации гена, эта индукция постоянна и необратима. Несомненно, стволовые клетки имеют большой потенциал, однако этот потенциал должен управляться in vitro, иначе нежелательные последствия могут иметь место после неконтролируемой в естественных условиях дифференцировки.
Согласно нашим данным GATA-2 и eNOS промоторы подвергаются гипометилированию в ходе hESCs дифференцировки в эндотелий. Следовательно, мы можем заключить, что эндотелий-специфичная экспрессия eNOSa регулируется через двойной негативный эпигенетический контроль. В недифференцированных hESCs не только промотор eNOSa, но также и промотор его регулирующего фактора транскрипции GATA-2 был блокирован метилированием.
Наши исследования демонстрируют существенные различия в метилировании областей промоторов трех генов от hESCs к из hESCs-полученным эндотелиальным клеткам. Однако мы наблюдали некоторые различия между областями промоторов метилирования трех генов, изученных в из hESCs-полученных эндотелиальных клетках и HUVEC. В полученных из hESCs эндотелиальных клетках области промоторов изученных генов были немного больше метилированы, чем соответствующие области в HUVEC, хотя это не кончалось подавлением транскрипции. Возможно, это отражает, что происходящие из hESCs эндотелиальные клетки не были эндотелиальными клетками вены пуповины, а скорее представляли смесь специализированных подтипов эндотелиальных клеток (например, капиллярные, аортальные эндотелиальные клетки, легочные эндотелиальные клетки), где уровень метилирования является переменным.
Полученные эндотелиальные клетки могли быть подвергнуты заморозке и хранению. При этом клетки сохраняли жизнеспособность и свои свойства.
Криоконсервацию и оттаивание эндотелиальных клеток осуществляли следующим образом.
Среда для заморозки имела следующий состав: 90% фетальная бычья сыворотка (инактивированная при 56°С в течение 45 минут) (Invitrogen или аналог другой фирмы), 10% DMSO (ICN Biomedicals, Inc. или аналог другой фирмы).
С культивируемых клеток удаляли среду, промывали дважды PBS, добавляли трипсин/ЭДТА 2 мл на 100 мм чашку или 75 см2 флакон. Инкубировали при 37°С 5 минут, нейтрализовали трипсин/ЭДТА 5 мл культуральной среды и пипетировали, чтобы получить одноклеточную суспензию. Полученную суспензию переносили в 25 мл пробирку и центрифугировали 5 минут при 1200 rpm. Осадок суспендировали в 1 мл среды для заморозки, затем переносили в криовиалу и сразу помещали на 24 часа на -70°С. На следующий день криовиалы переносили в жидкий азот для продолжительного хранения. В одной криовиале замораживали 1,5-3 млн клеток.
Криовиалы оттаивали на водяной бане при 37°С, стерилизовали 95% этанолом. Содержимое виалы переносили в 15 мл пробирку и постепенно добавляли 9 мл культуральной среды. Центрифугировали 5 минут при 1000 rpm, надосадочную жидкость удаляли, осадок суспендировали в 1 мл среды и переносили в культуральную чашку или флакон со средой. Клетки равномерно распределяли по поверхности и помещали в инкубатор при 37°С и 5% CO2
Методы, способы, применение полученных клеток эндотелия (эндотелеоциты).
1. Полученные описанные выше способом клетки эндотелия могут быть применены для формирования сосудистой сети (коллатералей) de novo за счет неоангиогенеза путем введения 104, 105, 106 или большего количества клеток в вену или артерию млекопитающих, включая человека. На фиг.10 приведено характерное изменение плотности кровеносных сосудов через 30 дней после введения клеток эндотелия (фиг.10б)
2. Полученные клетки могут быть применены для васкуляризации имплантата путем заключения их в биосовместимый материал, такой как желатин, хитозан. коллаген, матригель и другие либо их комбинации, обеспечивающие формирование трехмерной структуры, в которую кроме клеток эндотелия могут входить такие клетки, как фибробласты, кардиомиоциты, гепатоциты, кератиноциты, нейроны, мультипотентные клетки костного мозга, глиальные клетки, хондроциты. остеоциты и другие клеточные типы или сочетания типов клеток и имплантирования (введения) в организм млекопитающих, включая человека. Васкуляризации имплантата может осуществляться либо за счет привлечения эндотелиальных клеток организма, либо за счет васкуляризации имплантированными клетками, содержащимися в трехмерной структуре, для обеспечения импланта (введенной тканевой или клеточной композиции) кровоснабжением. На фиг.11а приведен матригель, удаленный из экспериментальных животных через 8 дней и не содержащий клеток эндотелия. На фиг.11б представлен матригель, удаленный из экспериментальных животных через 8 дней и содержащий клетки эндотелия.
3. Полученные клетки эндотелия могут быть использованы для изготовления трубчатой структуры, покрытой эндотелиоцитами, путем нанесения их на двумерную подложку из коллагена, мтаригеля, хитозана, фибронектинов и/или других биосовместимых материалов в сочетании с другими типами клеток или без них, которая может быть использована в качестве трубки при операциях на сосудах для восстановления нарушенного или недостаточного кровообращения, путем замены части, фрагментиа или полного сосуда или его стентирования.
4. Полученные стандартизованным методом из стандартной возобновляемой клеточной культуры клетки эндотелия могут быть использованы для тестирования токсичности химических или биологических субстанций или могут быть использованы для определения эффективности веществ стимулировать рост (пролиферацию) клеток эндотелия путем определения пролиферации или гибели клеток эндотелия с помощью известных методов.
5. Клетки эндотелия могут быть использованы для диагностических или терапевтических целей для лечения генетических заболеваний или для доставки генов токсинов либо самих токсинов в места активного ангиогенеза (роста сосудов) для блокирования последнего (например, в случае роста сосудов в опухоли) с целью прекращения кровоснабжения органа или ткани и ее последующей гибелью путем их генетической модификации конструкциями, содержащими гены-репортеры, например зеленый флуоресцентный белок или аналогичные, или генетическими конструкциями, содержащими гены токсинов, или генетическими конструкциями. содержащими гены для восстановления работы нарушенных генов и введения модифицированных клеток эндотелия в организм млекопитающих, в частности человека. Эндотелиальные клетки, модифицированные генетическими конструкциями, кодирующими ростовые факторы, могут быть векторами для стимуляции роста определенной ткани, клеток, группы клеток.
6. Сочетания перечисленных выше методов использования.
Таким образом, наша двумерная система дифференцировки позволила нам производить эндотелиальные клетки с высокой эффективностью, выбирать чистую популяцию функциональных эндотелиальных клеток, которые приобретали тот же самый эпигенетический статус ключевых факторов транскрипции и гомеостаза поддержания генов, как их аналоги, дифференцирующиеся в эмбрионе in vivo. Полученные эндотелиальные клетки могут найти широкое применение в медицинской практике.
Отобранные hESC-полученные эндотелиальные клетки экспрессировали эндотелий-специфические гены, формировали сосудоподобные структуры и были морфологически неотличимы от первичных эндотелиальных клеток. CD31 отобранные эндотелиальные клетки, дифференцированные из hESCs, были способны культивироваться in vitro, по крайней мере, до 4 пассажей.
Используя наш способ дифференцировки, мы наблюдали CD31 и VE-cadherin экспрессию на 6 день. Несомненно, это могло произойти из-за различий между линиями клеток, хотя условия культивирования (то есть среда культивирования, факторы роста, обработка) имеют большое воздействие на свойства дифференцировки. Интересно то, что мы обнаружили VE cadherin экспрессию на 6 день hESCs дифференцировки, в то время как в естественных условиях его экспрессия наблюдалась только в 8-ю неделю беременности. Таким образом, маркер дифференцированных эндотелиальных клеток VE-cadherin был обнаружен очень рано in vitro.
Список литературы.
1. Baharvand, Н., Jafary, H., Massumi, M., Ashtiani, S.K. Generation of insulin-secreting cells from human embryonic stem cells. Dev Growth Differ. 2006, 48: 323-332.
2. Hay, D.C., Zhao, D., Ross, A., Mandalam, R., Lebkowski, J., Cui, W. Direct Differentiation of Human Embryonic Stem Cells to Hepatocyte-like Cells Exhibiting Functional Activities. Cloning Stem Cells. 2007, 9: 51-62.
3. Heng, B.C., Haider, H.Kh., Sim, E.K., Cao, Т., Ng, S.C. Strategies for directing the differentiation of stem cells into the cardiomyogenic lineage in vitro. Cardiovasc Res. 2004, 62: 34-42.
4. Xu, С., Не, J.Q., Kamp, T.J., Police, S., Hao, X., O'Sullivan, C., Carpenter, M.K.Lebkowski, J., Gold JD. Human embryonic stem cell-derived cardiomyocytes can be maintained in defined medium without serum. Stem Cells Dev. 2006, 15: 931-941.
5. Lacaud, G., Keller, G., Kouskoff, V. Tracking mesoderm formation and specification to the hemangioblast in vitro. Trends Cardiovasc Med. 2004, 14, 314-317.
6. Choi, К., Kennedy, M., Kazarov. A., Papadimitriou, J.C., Keller, G. A common precursor for hematopoietic and endothelial cells. Development. 1998, 125, 725-732.
7. Chung, Y.S., Zhang, W.J., Arentson, E., Kingsley, P.D., Palis, J., Choi, K. Lineage analysis of the hemangioblast as defined by FLK1 and SCL expression. Development. 2002, 129, 5511-5520.
8. Nishikawa, S.I., Nishikawa, S., Hirashima, M., Matsuyoshi, N., Kodama, H. Progressive lineage analysis by cell sorting and culture identifies FLK1+VE-cadherin + cells at a diverging point of endothelial and hemopoietic lineages. Development. 1998, 125, 1747-1757.
9. Huber, T.L., Kouskoff, V., Fehling, H.J., Palis, J., Keller, G. Haemangioblast commitment is initiated in the primitive streak of the mouse embryo. Nature. 2004, 432, 625-630.
10. Cerdan, С., Rouleau, A., Bhatia, M., VEGF-A165 augments erythropoietic development from human embryonic stem cells. Blood. 2004, 103, 2504-12.
11. Chadwick, К., Wang, L., Li, L., Menendez, P., Murdoch, В., Rouleau, A., Bhatia, M. Cytokines and BMP-4 promote hematopoietic differentiation of human embryonic stem cells. Blood. 2003, 102, 906-915
12. Levenberg, S., Golub, J.S., Amit, M., Itskovitz-Eldor, J., Langer, R. Endothelial cells derived from human embryonic stem cells. Proc Nati Acad Sci USA. 2002, 99: 4391-4396.
13. Ng, E.S., Davis, R.P., Azzola, L., Stanley, E.G., Elefanty, A.G. Forced aggregation of defined numbers of human embryonic stem cells into embryoid bodies fosters robust. reproducible hematopoietic differentiation. Blood. 2005, 106, 1601-1603.
14. Zambidis, E.T., Peault, В., Park, T.S., Bunz, F., Civin, C.I., Hematopoietic differentiation of human embryonic stem cells progresses through sequential hematoendothelial, primitive, and definitive stages resembling human yolk sac development. Blood. 2005, 106, 860-870.
15. Zhan, X., Dravid, G., Ye, Z., Hammond, H., Shamblott, M., Gearhart, J., Cheng, L. Functional antigen-presenting leucocytes derived from human embryonic stem cells in vitro. Lancet. 2004, 364, 163-171
16. Kaufman, D.S., Hanson, E.T., Lewis, R.L., Auerbach, R., Thomson, J.A. Hematopoietic colony-forming cells derived from human embryonic stem cells. Proc Nati Acad Sci USA. 2001, 98, 10716-10721.
17. Vodyanik, M.A., Bork, J.A., Thomson, J.A., Slukvin, I.I. Human embryonic stem cell-derived CD34+ cells: efficient production in the coculture with OP9 stromal cells and analysis of lymphohematopoietic potential. Blood. 2005, 105, 617-626.
18. Hirashima, M., Kataoka, H., Nishikawa, S., Matsuyoshi, N., Nishikawa, S. Maturation of embryonic stem cells into endothelial cells in an in vitro model of vasculogenesis. Blood. 1999, 93, 1253-1263.
19. Lagarkova, M.A., Volchkov, P.Y., Lyakisheva, A.V., Philonenko. E.S., Kiselev, S.L. Diverse epigenetic profile of novel human embryonic stem cell lines. Cell Cycle. 2006, 5: 416-420.
20. Gerecht-Nir, S., Ziskind, A., Cohen, S., Itskovitz-Eldor, J. Human embryonic stem cells as an in vitro model for human vascular development and the induction of vascular differentiation. Lab Invest. 2003, 83: 1811-1820.
21. Bompais, H. et al. Human endothelial cells derived from circulating progenitors display specific functional properties compared with mature vessel wall endothelial cells. Blood 103, 2577-2584 (2004)
22. Chan, Y., Fish, J.E., D'Abreo, C. The cell-specific expression of endothelial nitric-oxide synthase: a role for DNA methylation. J Biol Chem. 2004, 279: 35087-35100.
Реферат
Изобретение относится к биотехнологии. Предложен способ получения эндотелиальных клеток из эмбриональных стволовых клеток человека, который предусматривает дифференцировку ЭСК человека на подложке и в синтетической среде. Способ позволяет производить эндотелиальные клетки с высокой эффективностью, выбирать чистую популяцию функциональных эндотелиальных клеток, которые приобретали тот же самый эпигенетический статус ключевых факторов транскрипции и гомеостаза поддержания генов, как их аналоги, дифференцирующиеся в эмбрионе in vivo. Полученные эндотелиальные клетки могут найти широкое применение в медицинской практике. 2 н.п. ф-лы, 11 ил, 4 табл.
Комментарии