Генерирование дендритных клеток из моноцитарных предшественников дендритных клеток с помощью gm-csf в отсутствие дополнительных цитокинов - RU2364625C2
Код документа: RU2364625C2
Чертежи













Описание
ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка претендует на приоритет предварительной Заявки на Патент Соединенных Штатов с номером 60/451015, поданной 27 февраля 2003 года, описание которой включено здесь полностью.
УРОВЕНЬ ТЕХНИКИ
Антигенпрезентирующие клетки (АПК) являются важными в индукции эффективного иммунного ответа. АПК не только презентируют антигены Т-клеткам с антигенспецифическими рецепторами, но также обеспечивают сигналы, необходимые для активации Т-клеток. Такие сигналы остаются не полностью определенными, но известно, что они включают различные молекулы клеточной поверхности, а также цитокины или факторы роста. Факторы, необходимые для активации «наивных» или непраймированных Т-клеток, могут отличаться от факторов, требуемых для реактивации ранее праймированных Т-клеток памяти. Хотя было показано, что моноциты и В-клетки являются компетентными АПК, их антигенпрезентирующие способности, являются, по-видимому, ограниченными повторной активацией ранее сенсибилизированных Т-клеток. Таким образом, они неспособны непосредственно активировать функционально наивные или непраймированные Т-клеточные популяции. С другой стороны, дендритные клетки способны активировать как наивные, так и ранее праймированные Т-клетки.
Дендритные клетки являются гетерогенной клеточной популяцией с характерной морфологией и широко распространенным распределением в тканях, в том числе в крови (см., например, Steinman, Ann. Rev. Immunol. 9:271-96 (1996)). Клеточная поверхность дендритных клеток является необычной, с характерными вуалеподобными выступами. Зрелые дендритные клетки обычно идентифицируют как CD3-, CD11c+, CD19-, CD83+, CD86+ и HLA-DR+.
Дендритные клетки процессируют и презентируют антигены и стимулируют реакции из "наивных" или непраймированных Т-клеток и Т-клеток памяти. В частности, дендритные клетки имеют высокую способность сенсибилизации МНС-рестриктированных Т-клеток и являются очень эффективными в презентации антигенов Т-клеткам, как аутоантигенов во время развития Т-клеток и Т-клеточной толерантности, так и чужеродных антигенов во время иммунной реакции. Кроме их роли в презентации антигенов, дендритные клетки также непосредственно сообщаются с нелимфатической тканью и осуществляют надзор за нелимфатической тканью в отношении сигнала повреждения (например, ишемии, инфекции или воспаления) или опухолевого роста. После передачи сигнала дендритные клетки инициируют иммунный ответ высвобождением цитокинов, которые стимулируют активность лимфоцитов и моноцитов.
Вследствие их эффективности в презентации антигенов существует растущий интерес к использованию дендритных клеток в качестве иммуностимуляторного агента как in vivo, так и in vitro. Однако применение выделенных дендритных клеток в качестве иммуностимуляторных агентов было ограниченным вследствие низкой частоты встречаемости дендритных клеток в периферической крови и низкой чистоты дендритных клеток, выделяемых прежними способами. В частности, частота встречаемости дендритных клеток в периферической крови человека была оценена при приблизительно 0,1% лейкоцитов. Подобным образом, существует ограниченная доступность дендритных клеток из других тканей, таких как лимфоидные органы. Эта низкая частота встречаемости дендритных клеток увеличивала интерес к выделению клеточной популяции, обогащенной предшественниками дендритных клеток, и культивированию этих предшественников ех vivo или in vitro для получения обогащенных популяций незрелых или зрелых дендритных клеток. Поскольку характеристики предшественников дендритных клеток остаются не полностью определенными, способы, обычно используемые для выделения предшественников дендритных клеток, не приводят к очищенным фракциям желаемых предшественников, а вместо этого обычно продуцируют смешанные популяции лейкоцитов, обогащенные предшественниками дендритных клеток. Были идентифицированы несколько типов клеток как имеющих потенциал функционирования в качестве предшественников дендритных клеток. Полученные из крови CD14+ моноциты, в частности, CD14+ моноциты, которые экспрессируют на своей поверхности рецептор для фактора роста, а именно колониестимулирующего фактора гранулоцитов-моноцитов (GM-CSF), являются известными предшественниками дендритных клеток. Другие происходящие из крови предшественники дендритных клеток могут быть выделены сначала удалением моноцитов и других "недендритных клеток-предшественников" (см., например, Патенты США с номерами 5994126 и 5851756). Другие известные предшественники дендритных клеток включают происходящие из костного мозга клетки, которые экспрессируют маркер клеточной поверхности CD34.
Клеточные популяции, обогащенные предшественниками дендритных клеток, получали различными способами, такими как, например, разделение в градиенте плотности, клеточный сортинг с активацией флуоресценции, иммунологические способы разделения клеток, например пэннинг, лизис комплемента, метод розеткообразования, магнитные способы разделения клеток, разделение с использованием найлоновой ваты и комбинации таких способов (см., например, O'Doherty et al., J. Exp. Med. 178:1067-76 (1993); Young and Steinman, J. Exp. Med. 171:1315-32 (1990); Freudenthal and Steinman, Proc. Natl. Acad. Sci. USA 87:7698-702 (1990); Macatonia et ai., Immunol. 67:285-89 (1989); Markowicz and Engleman, J. Clin. Invest. 85:955-61 (1990), все из которых полностью включены здесь в качестве ссылки). Способы для иммуноселекции дендритных клеток включают в себя, например, использование антител к маркерам клеточной поверхности, ассоциированным с предшественниками дендритных клеток, таких как анти-CD34-и/или анти-CD14-антитела, связанные с субстратом (см., например, Bernhard et al., Cancer Res. 55:1099-104 (1995); Caux et ai., Nature 360:258-61 (1992)).
В одном типичном способе-примере лейкоциты выделяют процедурой лейкафереза. Дополнительные способы обычно используют для дополнительной очистки для обогащения в отношении фракций клеток, которые, как предполагается, содержат дендритные клетки и/или предшественники дендритных клеток. Подобным образом, такие способы, как дифференциальное центрифугирование (например, выделение лейкоцитарной пленки), пэннинг с моноклональными антителами, специфическими в отношении определенных белков клеточной поверхности (например, положительный и отрицательный отбор), и фильтрование также дают неочищенную смесь лейкоцитов, содержащую предшественников дендритных клеток.
Другим сообщенным способом для выделения пролиферирующих предшественников дендритных клеток является использование коммерчески обработанного пластикового субстрата (например, гранул или магнитных гранул) для селективного удаления прикрепленных моноцитов и других «недендритных клеток-предшественников» (см., например, Патенты США с номерами 5994126 и 5851756). Прикрепленные моноциты и недендритные клетки-предшественники выбрасывают, тогда как неприкрепленные клетки сохраняют для культивирования и созревания ex vivo. В другом способе клетки афереза культивировали в пластиковых культуральных мешках, к которым были добавлены пластиковые, т.е. полистироловые или стироловые, гранулы-микроносители для увеличения площади поверхности этого мешка. Эти клетки культивировали в течение достаточного периода времени для прикрепления к этим гранулам и неприкрепленные клетки вымывали из этого мешка (Maffei, et al., Transfusion 40:1419-1420 (2000); WO 02/44338, включенные здесь в качестве ссылки).
После по существу всех сообщенных способов получения популяции клеток, обогащенной предшественниками дендритных клеток, эти популяции клеток обычно культивируют ех vivo или in vitro для дифференцировки предшественников дендритных клеток или поддержания и/или размножения этих дендритных клеток. Вкратце, дифференцировка ex vivo моноцитарных предшественников дендритных клеток включала культивирование популяции смешанных клеток, обогащенных предшественниками дендритных клеток, в присутствии комбинаций клеточных факторов роста, таких как цитокины. Например, моноцитарные предшественники дендритных клеток требуют колониестимулирующего фактора гранулоцитов/моноцитов (GM-CSF) в комбинации по меньшей мере с одним другим цитокином, например, выбранным из интерлейкина 4 (IL-4), интерлейкина 15 (IL-15), интерлейкина 13 (IL-13), интерферона α (IFN-α) и т.п., для дифференцировки этих клеток в оптимальное состояние для поглощения, процессинга и/или презентации антигена. Количества дендритных клеток из немоноцитарных предшественников дендритных клеток, таких как клетки, полученные удалением моноцитов и других предшественников недендритных клеток (адсобцией на пластиковой поверхности) или отбором на CD34+ клетки, также размножали культивированием в присутствии определенных цитокинов. Либо только GM-CSF, либо комбинацию GM-CSF и IL-4 использовали в способах получения популяций дендритных клеток из таких пролиферирующих предшественников дендритных клеток для терапевтического применения.
Однако эффективность таких дифференцировки, поддержания и/или размножения ех vivo была ограничена качеством исходной популяции, обогащенной дендритными клетками и предшественниками дендритных клеток. При некоторых условиях культивирования популяции дендритных клеток и предшественников дендритных клеток, которые в сильной степени загрязнены нейтрофилами, макрофагами и лимфоцитами или их комбинациями, могут быть уменьшены этими последними клетками, что приводит к низкому выходу дендритных клеток. Культуры дендритных клеток, содержащие большие количества нейтрофилов, макрофагов и лимфоцитов или их комбинации, являются менее пригодными для применения в качестве иммуностимуляторных препаратов.
Незрелые или зрелые дендритные клетки после получения обычно подвергали действию антигена-мишени (антигенов-мишеней) и агентов созревания для обеспечения активированных зрелых дендритных клеток. Обычно этот антиген добавляли в популяцию клеток, обогащенную незрелыми или зрелыми дендритными клетками, при подходящих условиях культивирования. В случае незрелых дендритных клеток этим клеткам затем позволяли в течение достаточного времени поглощать и процессировать антиген и экспрессировать антигенные пептиды на клеточной поверхности в ассоциации с маркерами либо МНС класса I, либо МНС класса II. Антиген может презентироваться незрелым дендритным клеткам на поверхности клеток в очищенной форме, в полуочищенной форме, такой как выделенный белок или слитый белок (например, слитый белок GM-CSF-антиген), в виде лизата мембран, в виде комплекса липосома-белок и другими способами. Кроме того, поскольку зрелые дендритные клетки не способны поглощать и процессировать антиген, к зрелым дендритным клеткам для презентации могут быть добавлены антигенные пептиды, которые связываются с молекулами МНС класса I или МНС класса II.
После получения активированных дендритных клеток их вводили пациенту для стимуляции иммунного ответа. Активированные дендритные клетки могут вводиться пациенту болюсной инъекцией, непрерывной инфузией, пролонгированным высвобождением из имплантатов или другими подходящими способами, известными в данной области. Активированные дендритные клетки могут также вводиться совместно с физиологически приемлемыми носителями, наполнителями, буферами и/или разбавителями. Кроме того, активированные дендритные клетки могут быть использованы для активации Т-клеток, например цитотоксических Т-клеток, ex vivo с использованием способов, хорошо известных квалифицированному специалисту. Затем антигенспецифические цитотоксические Т-клетки могут быть введены пациенту, например, для лечения растущей опухоли или бактериальной или вирусной инфекции. Эти композиции могут быть использованы сами по себе или в качестве адъюванта для других терапий, таких как, например, хирургическая резекция, химиотерапия, лучевая терапия и их комбинации, а также другие терапевтические способы воздействия, подходящие для подлежащего лечению состояния.
В данном изобретении обнаружено, что в противоположность предыдущим способам моноцитарные предшественники дендритных клеток могут быть дифференцированы в незрелые дендритные клетки и поддержаны в подходящем состоянии, которое является полностью компетентным для процессинга и презентации антигена, в присутствии только GM-CSF без дополнительных цитокинов. Эти способы предусматривают обеспечение популяции клеток, содержащих предшественники дендритных клеток, которые не были активированы, и культивирование этих клеток in vitro или ex vivo в среде для культивирования дендритных клеток, дополненной GM-CSF, без каких-либо дополнительных цитокинов. Способы, обычно используемые для обогащения клеточных популяций предшественниками дендритных клеток, могут активировать эти клетки-предшественники, инициируя конечную дифференцировку этих клеток, например, в макрофаг. Добавление других цитокинов, например IL-4, IL-13, IL-15 или TNF-α, противодействовало эффектам выделения, связанным с активацией этих клеток. Использование на практике способов данного изобретения обеспечивает простой и более экономичный способ получения и поддержания незрелых дендритных клеток с состоянии, оптимизированном для поглощения, процессинга и презентации выбранного антигена.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение обеспечивает способ дифференцировки и поддержания незрелых дендритных клеток ех vivo или in vitro в состоянии, оптимизированном для поглощения, процессинга и презентации выбранного антигена. Этот способ предусматривает обеспечение клеточной популяции, содержащей неактивированные моноцитарные предшественники дендритных клеток, т.е. моноциты, которые экспрессируют рецептор GM-CSF на своей поверхности, и другие такие предшественники дендритных клеток, и контактирование этих неактивированных предшественников дендритных клеток с культуральной средой для дендритных клеток, дополненной колониестимулирующим фактором гранулоцитов/макрофагов, в отсутствие дополнительных цитокинов. В противоположность предыдущим способам, эти дополнительные цитокины не требуются для генерирования дендритных клеток из выделенных неактивированных моноцитарных предшественников дендритных клеток.
Активация моноцитарных предшественников дендритных клеток может быть предотвращена, например, ингибированием или блокированием адгезии клеток-предшественников к твердой поверхности, с которой эти клетки контактируют во время обычного выделения и/или процесса обогащения или во время культивирования клеток. Твердой поверхностью может быть культуральный сосуд, такой как колба, бутыль или мешок для культуры клеток, используемый для получения или поддержания клеток ex vivo или in vitro. Этой твердой поверхностью может быть также любая поверхность сосуда или устройства, используемого в получении клеточной популяции, обогащенной предшественниками дендритных клеток, например, поверхность фильтра; гранула, используемая в очистке, такая как магнитная, стеклянная или пластиковая гранула; трубка, культуральный сосуд и т.п. Ингибирование адгезии неактивированных моноцитарных предшественников дендритных клеток может осуществляться добавлением высокой концентрации белка животного или человека к клеточной культуре или к среде для выделения. Высокая концентрация белка животного или человека, в данном контексте, включает приблизительно 1-10% мас./об. белка. Белок животного может включать альбумин, сыворотку, плазму, желатин, полиаминокислоту и т.п., пока они сами не активируют эти клетки. Активация моноцитарных предшественников дендритных клеток может быть также блокирована или ингибирована добавлением хелатора металла к клеточной культуре и/или среде для выделения. Хелатор металла может включать ЭДТА, ЭГТА и т.п. Считается, что добавление этих агентов дендритных клеток минимизирует активацию клеток-предшественников уменьшением концентрации двухвалентных катионных металлов в культуральной среде.
Активация моноцитарных предшественников дендритных клеток может быть также предотвращена или ингибирована выделением или обогащением и культивированием предшественников дендритных клеток в сосуде для культуры с низкой клеточной авидностью. Сосуды для культуры с низкой клеточной авидностью обычно содержат такие материалы, как полипропилен, Тефлон®, PFTE и т.п. Как и при добавлении белка животного или человека, уменьшение или блокирование адгезии предшественников дендритных клеток к твердой поверхности предотвращает активацию этих клеток и делает возможными дифференцировку и поддержание этих клеток в незрелые дендритные клетки в присутствии среды для культивирования дендритных клеток, дополненной GM-CSF, без каких-либо дополнительных цитокинов. Выполнение выделения клеток-предшественников при температурах ниже приблизительно 37°С, таких как комнатная температура, дополнительно уменьшает долю моноцитарных предшественников дендритных клеток, которая подвергается активации в клеточной популяции. Способы данного изобретения могут предусматривать комбинацию любых или всех этих агентов, материалов и/или условий. В одном конкретном варианте осуществления культуральной средой для дендритных клеток является бессывороточная среда, и белок животного, например сывороточный альбумин, добавляют для уменьшения авидности предшественников дендритных клеток в отношении поверхности культурального сосуда для предотвращения и/или уменьшения активации моноцитарных предшественников дендритных клеток.
Обычно клеточные популяции, которые содержат моноцитарные предшественники дендритных клеток, получают из периферической крови, продукта лейкафереза, продукта афереза, пуповинной крови, селезенки, лимфатического узла, тимуса или костного мозга. Эти клеточные популяции могут быть криоконсервированы до и после применения на практике данного изобретения. Далее эта клеточная популяция может быть дополнительно обогащена моноцитарными клетками-предшественниками фильтрацией с тангенциальным потоком, пэннигом антител, дифференциальным центрифугированием и т.п. При дополнительном обогащении этой клеточной популяции фильтрацией с тангенциальным потоком фильтр обычно содержит поры размером 5,5 мкм, скорость рециркуляции равна приблизительно 1400 мл/мин, скорость фильтрации равна приблизительно 15-21 мл/мин, обычно 17 мл/мин, и время фильтрации составляет приблизительно 60-90 мин (см., например, заявку WO 2004/000444, опубликованную 31 декабря 2003 года, включенную здесь в качестве ссылки).
Незрелые дендритные клетки, которые получали способами данного изобретения, могут быть проконтактированы с выбранным представляющим интерес антигеном в течение периода времени, достаточного для поглощения и процессинга этого антигена. После процессинга антиген презентируется на поверхности дендритных клеток. Далее незрелые дендритные клетки могут быть проконтактированы с агентом созревания дендритных клеток до контактирования с представляющим интерес антгеном, одновременно с контактированием или после контактирования с представляющим интерес антигеном. Агент созревания дендритных клеток может включать Bacillus Calmette-Guerin (BCG), липополисахарид (LPS), фактор некроза опухоли α (TNF-α), интерферон гамма (IFNγ) или их комбинации. В конкретных вариантах данного изобретения агент созревания дендритных клеток является комбинацией инактивированного BCG и IFNγ. Выбранные антигены, применимые в способах данного изобретения, включают, но не ограничиваются ими, опухолеспецифический антиген, опухоль-ассоциированный антиген, вирусный антиген, бактериальный антиген, опухолевые клетки, нуклеиновую кислоту, полученную из опухолевых клеток, бактериальные клетки, вирусные частицы, рекомбинантные клетки, экспрессирующие антиген, клеточный лизат, препарат мембран, рекомбинантно полученный антиген, пептид, произведенный из представляющего интерес антигена, или выделенный представляющей интерес антиген. На любой стадии, в том числе после контакта с выбранным антигеном, поглощения, процессинга и созревания дендритных клеток, эти клетки могут быть криоконсервированы для более позднего использования.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1 изображает гистограммы экспрессии CD14 и CD1a на поверхности дендритных клеток для оценки дифференцировки in vitro моноцитарных предшественников дендритных клеток в CD1а+-дендритные клетки в присутствии только GM-CSF без дополнительных цитокинов и в присутствии блокирующего агента (3% сывороточного альбумина человека; САЧ), который предотвращает прочное связывание с поверхностью контейнера клеточной культуры.
Фигура 2 изображает измерение экспрессии CD1a и CD14 при дифференцировке моноцитов в дендритные клетки (ДК) после культивирования in vitro в отсутствие IL-4. Дифференцировка показана реципрокной экспрессией маркеров CD1a и CD14 на "живых" CD11с+ клетках. Фигура 2А демонстрирует повышение экспрессии CD1a на ДК в день 5 относительно его экспрессии на поверхности моноцитов-предшественников. Фигура 2В демонстрирует понижение экспрессии CD14 на ДК относительно его уровня на моноцитах-предшественниках. Данные показывают соответствующие культуры после электронной установки дискриминационного окна на клетках моноцитарной линии дифференцировки (CD11c+) для (i) субпопуляций и (ii) относительной экспрессии, измеряемой по средней интенсивности флуоресценции (mfi). Фоновое окрашивание, наблюдаемое с контрольными антителами релевантного типа, вычитали. Эти данные представляют средние величины из моноцитов, выделенных и культивируемых из двух независимых доноров.
Фигура 3 изображает количественное определение секреции IL-12 р70 из моноритарных предшественников дендритных клеток, которым давали плотно прикрепляться или рыхло прикрепляться к субстрату перед культивированием либо в GM-CSF и IL-4, либо только GM-CSF.
Фигура 4 изображает экспрессию типичных маркеров дендритных клеток для моноцитарных предшественников дендритных клеток, культивируемых только в GM-CSF.
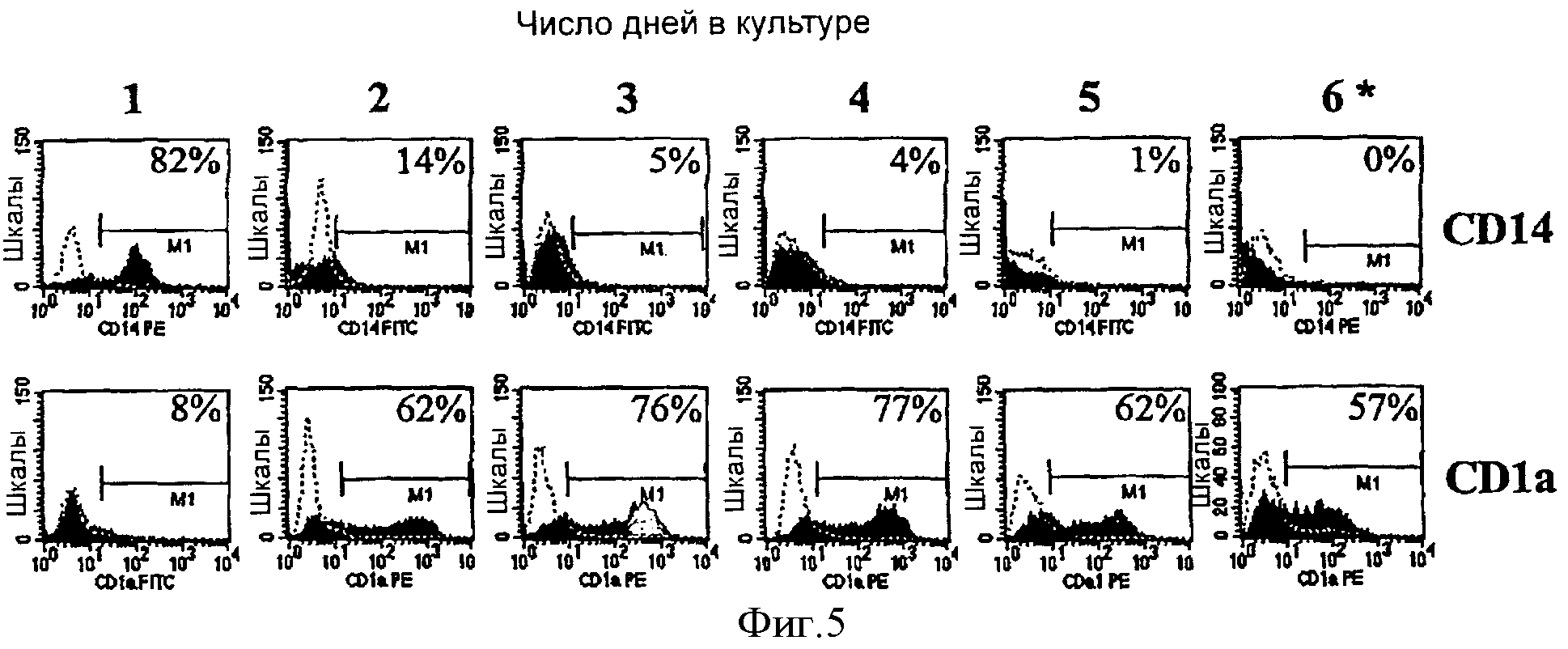
Фигура 5 изображает кинетику in vitro дифференцировки в дендритные клетки неактивированных моноцитов в клеточной культуральной среде, дополненной только GM-CSF, определяемую экспрессией CD1a и CD14.
Фигуры 6А-6Е изображают сравнение фенотипа неактивированных моноцитов относительно дендритных клеток, культивируемых в мешках из Тефлона® или пластиковых колбах для культуры ткани в клеточной культуральной среде, дополненной только GM-CSF или GM-CSF плюс IL-4, в присутствии или в отсутствие фактора созревания дендритных клеток. Фигура 6А изображает процент клеток, которые были CD1a-положительными. Фигура 6В изображает процент клеток, которые были CD83-положительными. Фигура 6С изображает относительный уровень экспрессии (mfi) CD80. Фигура 6D изображает относительный уровень экспрессии (mfi) CD86. Фигура 6Е изображает относительный уровень экспрессии (mfi) HLA-DR.
Фигуры 7А и 7В изображают генерируемую прикреплением к покрытым стеклом гранулам-микроносителям антигенспецифическую Т-клеточную реакцию дендритных клеток, культивируемых в присутствии либо только GM-CSF, либо GM-CSF и IL-4, и затем контактированных либо с контрольным антигеном гемоцианина фиссуреллы, либо с 40-мерным пептидом М1-А4 вируса гриппа А и агентом созревания дендритных клеток. Фигура 7А изображает анализ антигенспецифических цитотоксических Т-клеток для клеток, выделенных из донора Р016, и фигура 7В-подобный анализ для донора Р052.
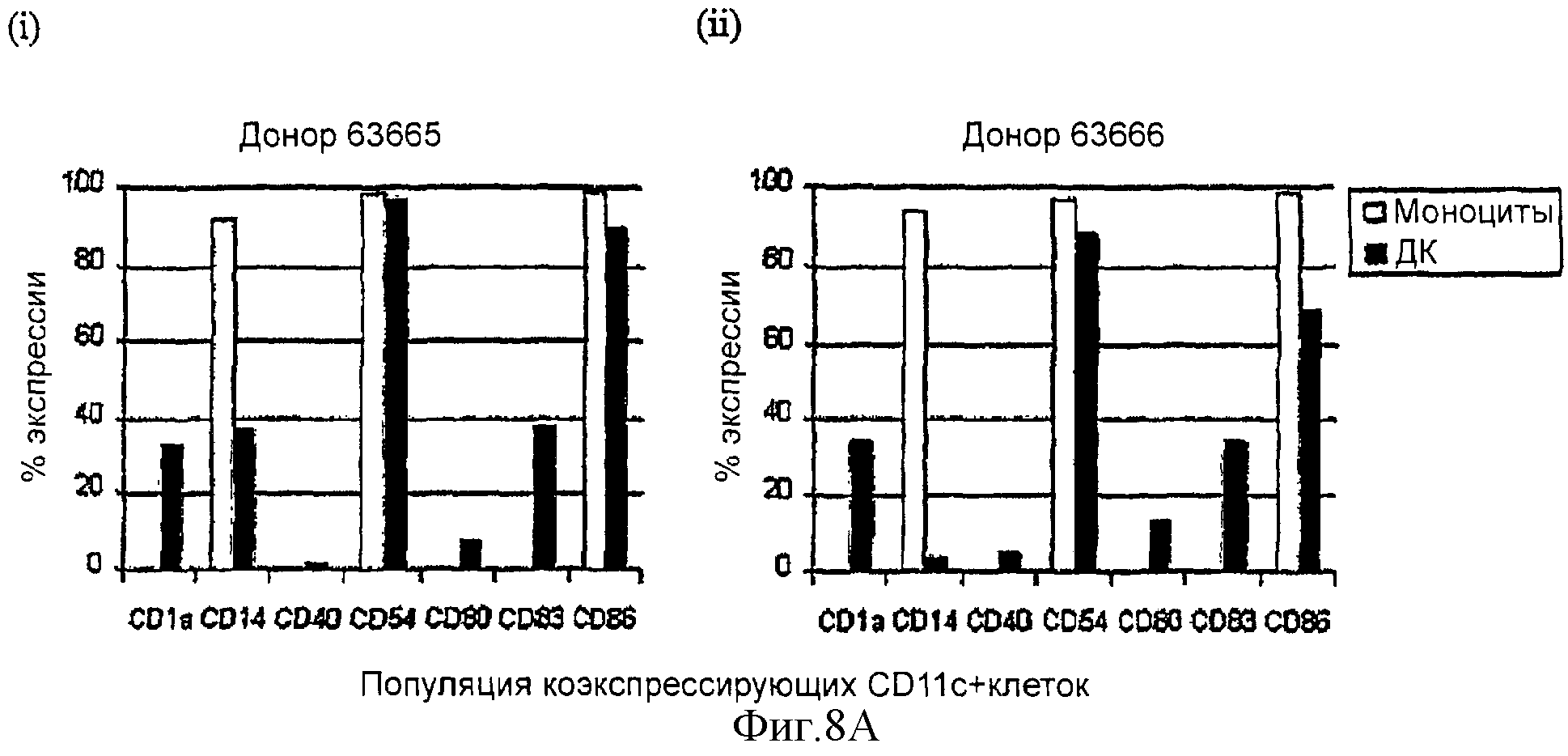
Фигуры 8А и 8В изображают фенотипические профили на клетках моноцитарной линии дифференцировки, которые дифференцировались в дендритные клетки (ДК) после культивирования in vitro в отсутствие IL-4. Маркеры на всех субпопуляциях и их соответствующие уровни экспрессии (mfi) показаны на «живых» CD11c+ клетках. Фигура 8А изображает процент CD11c+ клеток, которые коэкспрессируют специфические маркеры на моноцитах и ДК дня 5. Фигура 8В изображает относительную экспрессию фенотипических маркеров. Показаны результаты для независимых культур из двух различных доноров, обозначенных 63665 и 63666, после электронной установки дискриминационного окна на клетках моноцитарной линии дифференцировки (CD11с+) для (i) субпопуляций и (ii) относительной экспрессии, измеряемой по средней интенсивности флуоресценции (mfi). Фоновое окрашивание, наблюдаемое с контрольными антителами релевантного типа, вычитали.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Данное изобретение обеспечивает способы дифференцировки моноцитарных предшественников дендритных клеток в незрелые дендритные клетки (ДК). Моноцитарные предшественники дендритных клеток, которые не были активированы, могут быть проконтактированы с культуральной средой для дендритных клеток, дополненной GM-CSF, в качестве единственного цитокина для индукции дифференцировки и поддержания этих клеток в виде незрелых дендритных клеток. Способы, которые требуют только добавления единственного цитокина, являются менее дорогими и являются более эффективными, чем способы, используемые ранее, которые требуют добавления других цитокинов для предотвращения дифференцировки моноцитарных предшественников дендритных клеток в другие типы клеток, в том числе, например, макрофаг и т.п.
Незрелые дендритные клетки, получаемые способами данного изобретения, являются фенотипически и функционально сходными с клетками, получаемыми предыдущими способами, которые используют более сложные условия культивирования, и могут быть затем проконтактированы с фактором созревания дендритных клеток, таким как BCG и IFNγ, и необязательно с заранее определенным антигеном в подходящих условиях созревания. Антиген может быть проконтактирован с незрелыми дендритными клетками данного изобретения либо во время, либо до созревания.
Моноцитарные предшественники дендритных клеток и незрелые дендритные клетки
Моноцитарные предшественники дендритных клеток, в данном контексте, включают моноциты, которые имеют GM-CSF-рецептор на их поверхности, и другие миелоидные клетки-предшественники, которые отвечают на GM-CSF. Эти клетки могут быть получены из любой ткани, в которой они находятся, в частности из лимфоидных тканей, таких как селезенка, костный мозг, лимфатические узлы и тимус. Моноцитарные предшественники дендритных клеток могут быть также выделены из системы кровотока. Периферическая кровь является легкодоступным источником моноцитарных предшественников дендритных клеток. Кровь пупочного канатика является другим источником моноцитарных предшественников дендритных клеток. Моноцитарные предшественники дендритных клеток могут быть выделены из различных организмов, в которых может быть вызван иммунный ответ. Такие организмы включают, например, людей и животных, таких как приматы, млекопитающие (в том числе собаки, кошки, мыши и крысы), птицы (в том числе куры), а также их трансгенные виды.
В некоторых вариантах осуществления моноцитарные предшественники дендритных клеток и/или незрелые дендритные клетки могут быть выделены из здорового субъекта или из субъекта, нуждающегося в иммуностимуляции, такого как пациент с раком или другой субъект, для которого клеточная иммуностимуляция может быть полезной или желательной (т.е. субъект, имеющий бактериальную, вирусную или паразитарную инфекцию и т.п.). Моноцитарные предшественники дендритных клеток и/или незрелые дендритные клетки могут быть также получены из HLA-соответствующего здорового индивидуума для превращения в незрелые дендритные клетки, созревания, активации и введения HLA-соответствующему субъекту, нуждающемуся в иммуностимуляции.
Способы выделения неактивированных моноцитарных предшественников дендритных клеток и незрелых дендритных клеток из различных источников, описанных выше, в том числе из крови и костного мозга, могут выполняться рядом способов. Обычно клеточную популяцию собирают из индивидуума и обогащают неактивированными моноцитарными предшественниками дендритных клеток. Например, смешанная популяция клеток, содержащая неактивированные моноцитарные предшественники дендритных клеток, может быть получена из периферической крови лейкаферезом, аферезом, центрифугированием в градиенте плотности, дифференциальным лизисом, фильтрацией, пэннингом антител или получением лейкоцитарной пленки. Выбранный способ не должен активировать моноцитарные предшественники дендритных клеток. Например, если для обогащения клеточной популяции в отношении предшественников выбран пэннинг антител, выбранные антитела не должны активировать эти клетки, например, индукцией вхождения ионов кальция, которая может возникать как следствие сшивания этих молекул на поверхности, с которой связываются антитела. Обычно при пэннинге антител используют антитела, которые элиминируют макрофаги, В-клетки, природные клетки-киллеры, Т-клетки и т.п. Антитела могут быть также использованы для положительного отбора на моноцитподобные клетки, экспрессирующие CD14.
В одном варианте осуществления данного изобретения неактивированные моноцитарные предшественники дендритных клеток получают предотвращением прочного прикрепления популяции клеток, содержащей моноцитарные предшественники дендритных клеток, к сосуду для культуры клеток. Прочное прикрепление может предотвращаться, например, добавлением блокирующего агента к культуральной среде, используемой для поддержания предшественников дендритных клеток in vitro или ex vivo. Такие блокирующие агенты могут включать высокие концентрации белка, в том числе, например, и не в качестве ограничения, белка животного или белка человека, такого как альбумины, сыворотка, плазма, желатин, полиаминокислоты и т.п. В частности, используют альбумины из бычьего или человеческого источника. Обычно используют концентрацию блокирующего агента приблизительно 1-10% мас./об. В частности, альбумин сыворотки человека (САЧ) может быть использован при концентрации приблизительно 1%, 2% или до приблизительно 5% или более. Следует отметить, что должны выбираться блокирующие агенты, которые сами не активируют эти клетки. Культуральными средами могут быть любые среды, обычно используемые для культуры моноцитарных предшественников дендритных клеток, в том числе среды, которые не требуют сыворотки.
В другом варианте осуществления данного изобретения к культуральной среде может быть добавлен хелатор металла для дополнительного предотвращения или уменьшения активации моноцитарных предшественников дендритных клеток посредством хелатирования двухвалентных катионов, в том числе, но без ограничения, ионов кальция. Применение культуральных сосудов с низкой прикрепляемостью или с низким связыванием может также уменьшать авидность прикрепления или связывания предшественников дендритных клеток для предупреждения активации этих клеток. Особенно предпочтительные материалы низкого связывания включают, например, но не только, полипропилен, Тефлон®, PFTE и т.п. Хелатор металлов может быть использован в комбинации с вышеописанными блокирующими агентами.
Моноцитарные предшественники дендритных клеток и незрелые дендритные клетки могут быть также получены в замкнутой, асептической системе. В данном контексте термины «замкнутая, асептическая система» или «замкнутая система» относятся к системе, в которой подвергание действию нестерильного, окружающего или циркулирующего воздуха или других нестерильных условий является минимизированным или элиминированным. Замкнутые системы для изоляции предшественников дендритных клеток и незрелых дендритных клеток обычно исключают центрифугирование в градиенте плотности в открытых сверху пробирках, открытый перенос клеток, культивирование клеток в чашках для культуры ткани или негерметизированных колбах и т.п. В типичном варианте осуществления замкнутая система делает возможным асептический перенос предшественников дендритных клеток и незрелых дендритных клеток из исходного сосуда для сбора в герметизированный сосуд для культуры ткани без экспонирования нестерильного воздуха.
В некоторых вариантах неактивированные моноцитарные предшественники дендритных клеток выделяют частичным прикреплением к моноцитсвязывающему субстрату, как описано в заявке WO 03/010292, описание которой включено здесь в качестве ссылки. Например, популяция лейкоцитов (например, выделенная лейкаферезом) может быть проконтактирована с субстратом, приклеивающим моноцитарный предшественник дендритной клетки, например, покрытой стеклом микроносителем-гранулой, в присутствии блокирующего агента, который предотвращает неспецифическое связывание, а также уменьшает авидность связывания моноцитарных предшественников дендритных клеток. При контактировании этой популяции лейкоцитов с субстратом эти моноцитарные предшественники дендритных клеток в данной популяции лейкоцитов предпочтительно неплотно прикрепляются к субстрату. Другие лейкоциты (в том числе другие потенциальные предшественники дендритных клеток), например пролиферирующие предшественники дендритных клеток и т.п., обнаруживают уменьшенную аффинность связывания с субстратом, позволяя таким образом субпопуляции этих моноцитарных предшественников дендритных клеток преимущественно обогащаться на поверхности этого субстрата. Неплотная адгезия не активирует моноцитарные предшественники дендритных клеток. После связывания клеток и элюции неприкрепленных клеток субпопуляцию моноцитарных предшественников дендритных клеток элюируют из субстрата забуференным солевым раствором, который может быть дополнен нетоксичным хелатообразующим агентом. «Нетоксичный хелатообразующий агент» обозначает хелатообразующие агенты, которые по существу не уменьшают жизнеспособность моноцитарных предшественников дендритных клеток, например, но без ограничения, ЭДТА, ЭГТА и т.п.
Подходящие субстраты включают, например, субстраты, имеющие большую площадь поверхности относительно объемной доли, такие как стеклянные гранулы или покрытый стеклом микроноситель. Такой субстрат может быть, например, состоящим из частиц или волокнистым субстратом. Подходящие состоящие из частиц субстраты включают, например, стеклянные частицы, покрытые стеклом пластиковые частицы, покрытые стеклом полистироловые частицы и другие гранулы, подходящие для адсорбции белка. Подходящие волокнистые субстраты включают стеклянные или покрытые стеклом микрокапиллярные трубки и микроворсинчатую мембрану. Состоящий из частиц или волокнистый субстрат обычно позволяет элюировать прикрепленные моноцитарные предшественники дендритных клеток по существу без уменьшения жизнеспособности прикрепленных клеток. Состоящий из частиц или волокнистый субстрат может быть по существу непористым для облегчения элюции моноцитарных предшественников дендритных клеток или дендритных клеток из этого субстрата. «По существу непористый» субстрат обозначает субстрат, в котором по меньшей мере большинство пор, присутствующих в данном субстрате, являются меньшими, чем клетки, для мимнимизации улавливания клеток в этом субстрате.
Прикрепление моноцитарных предшественников дендритных клеток к субстрату без активации может быть необязательно модулировано добавлением связывающих сред. Подходящие связывающие среды включают культуральные среды для моноцитарных предшественников дендритных клеток (например, AIM-V®, RPMI 1640, DMEM, X-VIVO 15® и т.п.), дополненные, индивидуально или в любой комбинации, например, цитокинами (например, колониестимулирующим фактором гранулоцитов/макрофагов (GM-CSF), плазмой крови, сывороткой (например, сывороткой человека, такой как аутологичные или аллогенные сыворотки), очищенными белками, такими как сывороточный альбумин, двухвалентными катионами (например, ионами кальция и/или магния) и другими молекулами, которые способствуют специфическому прикреплению моноцитарных предшественников дендритных клеток к субстрату или которые предотвращают прикрепление немоноцитарных предшественников дендритных клеток к этому субстрату. В некоторых вариантах осуществления плазма или сыворотка крови может быть инактивирована нагреванием. Инактивированная нагреванием плазма может быть аутологичной или гетерологичной относительно лейкоцитов.
В другом способе обогащения клеточной популяции моноцитарными предшественниками дендритных клеток из пробы компонентов крови предусматривается фильтрация с тангенциальным потоком лейкоцитов от клеточного дебриса, эритроцитов и других клеток и частиц в пробе крови. Описание этого устройства и его использование описаны в заявке WO 2004/000444, полностью включенной здесь в качестве ссылки. Этот способ предусматривает (1) введение пробы крови в фильтрующий элемент с тангенциальным потоком (TFF), причем этот TFF содержит камеру поперечного потока, камеру фильтрата и фильтр, служащий для жидкостного сообщения камеры поперечного потока и камеры фильтрата, причем этот фильтр имеет размер пор приблизительно 1-10 мкм, обычно приблизительно 5,5 мкм; (2) рециркуляцию этой пробы через элемент TFF при заданной скорости введения, обычно приблизительно 1400 мл/мин, и заданной скорости фильтрации, обычно приблизительно 15-21 мл/мин, более предпочтительно приблизительно 17 мл/мин, причем заданная скорость введения по меньшей мере в 5 раз выше заданной скорости фильтрации; где заданная скорость фильтрации меньше, чем беспрепятственная скорость фильтрации для этого фильтра; и (3) выделение клеточной популяции, обогащенной в отношении лейкоцитов. Обычно время фильтрации равно приблизительно 60-90 мин. Этот способ может приводить к обогащенной клеточной популяции, которая является по существу свободной от нелейкоцитарных компонентов крови, в том числе плазмы, тромбоцитов и эритроцитов. Обогащенная клеточная популяция, полученная этим способом, может содержать по меньшей мере приблизительно 50% моноцитарных предшественников дендритных клеток и предпочтительно по меньшей мере приблизительно 70% моноцитарных предшественников дендритных клеток, которые не были активированы. Этот способ может дополнительно предусматривать сбор крови из субъекта и получение пробы из этой крови посредством лейкафереза, центрифугирования в градиенте плотности, дифференциального лизиса, фильтрации или получения лейкоцитарной пленки перед фильтрацией с тангенциальным потоком. Выполнение TFF-очистки моноцитарных предшественников ДК при комнатной температуре или при более низкой температуре (т.е. ниже 37°С) дополнительно способствует уменьшению активации этих клеток.
Клеточные популяции, обогащенные в отношении неактивированных моноцитарных предшественников дендритных клеток, культивируют ex vivo или in vitro для дифференцировки, созревания и/или размножения. (В данном контексте выделенные незрелые дендритные клетки, предшественники дендритных клеток, Т-клетки и другие клетки обозначают клетки, которые, благодаря рукам человека, существуют отдельно от их природного окружения и, следовательно, не являются продуктом природы. Выделенные клетки могут существовать в очищенной форме, в полуочищенной форме или в неприродном окружении.) Вкратце, дифференцировка ех vivo обычно включает культивирование неактивированных предшественников дендритных клеток или популяций клеток, содержащих неактивированные предшественники дендритных клеток, в присутствии одного или нескольких агентов дифференцировки. В частности, агент дифференцировки в данном изобретении является колониестимулирующим фактором гранулоцитов-макрофагов (GM-CSF), используемым отдельно, без других добавленных цитокинов, в частности без использования интерлейкина 4 (IL-4). В некоторых вариантах осуществления неактивированные моноцитарные предшественники дендритных клеток дифференцируются с образованием происходящих из моноцитов незрелых дендритных клеток, способных индуцировать активацию и пролиферацию значительного количества Т-клеток в популяции мононуклеарных клеток периферической крови.
Предшественники дендритных клеток могут быть дифференцированы и поддержаны в виде незрелых предшественников дендритных клеток в подходящих культуральных условиях. Подходящие среды для культуры ткани включают AIM-V®, RPMI 1640, DMEM, X-VIVO 15® и т.п., дополненные GM-CSF. Эти среды для культуры ткани могут быть дополнены сывороткой, аминокислотами, витаминами, двухвалентными катионами и т.п. для стимуляции дифференцировки этих клеток в дендритные клетки. В некоторых вариантах осуществления предшественники дендритных клеток могут культивироваться в бессывороточных средах. Такие условия культивирования могут необязательно включать любые произведенные из животных продукты. Обычно GM-CSF добавляют в культуральную среду при концентрации приблизительно 100-1000 единиц/мл или обычно 500 единиц/мл GM-CSF. Предшественники дендритных клеток при дифференцировке с образованием незрелых дендритных клеток демонстрируют типичный профиль экспрессии белков клеточной поверхности, наблюдаемых на незрелых моноцитарных дендритных клетках, например, эти клетки обычно являются CD14-и CD11c+, CD83-и экспрессируют низкие уровни CD86. Кроме того, незрелые дендритные клетки способны захватывать растворимые антигены посредством специализированных механизмов поглощения.
Незрелые дендритные клетки могут созревать с образованием зрелых дендритных клеток. Зрелые ДК утрачивают способность поглощать антиген, и эти клетки проявляют повышенную экспрессию костимуляторных молекул клеточной поверхности и секретируют различные цитокины. Конкретно, зрелые ДК зкспрессируют более высокие уровни антигенов МНС класса I и II и обычно идентифицируются как CD80+, CD83+ и CD86+. Более высокая экспрессия МНС приводит к увеличению плотности антигенов на поверхности ДК, тогда как повышенная экспрессия костимуляторных молекул CD80 и CD86 усиливает сигнал активации Т-клеток через дополняющие компоненты этих костимуляторных молекул, такие как CD28, на Т-клетках.
Зрелые дендритные клетки могут быть получены (т.е. сделаны зрелыми) контактированием незрелых дендритных клеток, которые культивировали в присутствии только GM-CSF, с эффективными количествами или концентрациями агента созревания дендритных клеток. Агенты созревания дендритных клеток могут включать, например, BCG, IFNγ, LPS, TNFα и т.п. Эффективные количества BCG обычно находятся в диапазоне приблизительно 105-107 КОЕ на миллилитр среды культуры ткани. Эффективные количества IFNγ обычно находятся в диапазоне приблизительно 100-1000 Е на миллилитр среды культуры ткани. Bacillus Calmette-Guerin (BCG) является авирулентным штаммом М. bovis. В данном контексте BCG относится к целому BCG, а также к компонентам клеточных стенок, полученным из BCG липоарабидоманнанам и другим компонентам BCG, которые связаны с индукцией иммунного ответа типа 2. BCG является необязательно инактивированным, например инактивированным нагреванием BCG, обработанным формалином BCG и т.п.
Незрелые ДК обычно контактируют с эффективными количествами BCG и IFNγ в течение приблизительно одного часа - приблизительно 48 часов. Эти незрелые дендритные клетки могут быть прокультивированы и сделаны зрелыми в подходящих условиях культуры для созревания. Подходящие среды для культуры ткани включают AIM-V®, RPMI 1640, DMEM, X-VIVO 15® и т.п. Эти среды для культуры ткани могут быть дополнены аминокислотами, витаминами, цитокинами, такими как GM-CSF, двухвалентными катионами и т.п. для стимуляции созревания этих клеток. Обычно используют приблизительно 500 единиц/мл GM-CSF.
Созревание дендритных клеток может быть подвергнуто мониторингу способами, известными в данной области для дендритных клеток. Маркеры клеточной поверхности могут быть детектированы в анализах, известных в данной области, таких как проточная цитометрия, иммуногистохимия и т.п. Эти клетки могут быть также подвергнуты мониторингу на продуцирование цитокинов (например, при помощи ELISA, другого иммуноанализа или с использованием массива олигонуклеотидов). Зрелые ДК данного изобретения также утрачивают способность поглощения антигена, что может анализироваться исследованиями на поглощение, известными специалисту с обычной квалификацией в данной области.
Антигены
Зрелые, праймированные дендритные клетки, полученные способами данного изобретения, могут презентировать антиген Т-клеткам. Зрелые, праймированные дендритные клетки могут быть образованы контактированием незрелых дендритных клеток с заданным антигеном перед созреванием или во время созревания.
Подходящие заданные антигены для применения в данном изобретении могут включать любой антиген, для которого желательна активация Т-клеток. Такие антигены могут включать, например, бактериальные клетки или другой препарат, содержащий бактериальные антигены, опухолеспецифические или опухоль-ассоциированные антигены (например, целые опухолевые или раковые клетки, лизат опухолевых клеток, препараты мембран опухолевых клеток, выделенные или частично выделенные антигены из опухолей, слитые белки, липосомы и т.п.), вирусные частицы или другие препараты, содержащие вирусные антигены, и любой другой антиген или фрагмент антигена, например пептидный или полипептидный антиген. В некоторых вариантах осуществления этот антиген может быть ассоциирован с раком предстательной железы, например, этот антиген может быть, но не только, специфическим мембранным антигеном предстательной железы (PSMA), кислой фосфатазой предстательной железы (РАР) или специфическим антигеном предстательной железы (PSA) (см., например, Pepsidero et al., Cancer Res. 40:2428-32 (1980); McCormack et al., Urology 45:729-44 (1995)). Этим антигеном может также быть бактериальная клетка, бактериальный лизат, мембранный фрагмент из клеточного лизата или любой другой источник, известный в данной области. Этот антиген может быть экспрессирован или продуцирован рекомбинантно или даже синтезирован химически. Рекомбинантный антиген может быть также экспрессирован на поверхности клетки-хозяина (например, бактерий, дрожжей, клеток насекомых, позвоночных или млекопитающих), может присутствовать в лизате или может быть очищен из этого лизата. Альтернативно, этот антиген может кодироваться нуклеиновыми кислотами, которые могут быть рибонуклеиновой кислотой (РНК) или дезоксирибонуклеиновой кислотой (ДНК), которые очищают или амплифицируют из опухолевой клетки.
Антиген может также присутствовать в пробе из субъекта. Например, проба ткани из гиперпролиферативного или другого состояния субъекта может быть использована в качестве источника антигена. Такая пробы может быть получена, например, биопсией или хирургической резекцией. Такой антиген может быть использован в виде лизата или в виде выделенного препарата. Альтернативно, мембранный препарат клеток из субъекта (например, ракового пациента) или установленная клеточная линия могут быть использованы в качестве антигена, или источника антигена, или нуклеиновой кислоты, кодирующей этот антиген.
В примерном варианте осуществления лизат опухолевых клеток, извлеченный из хирургических образцов, может быть источником антигена. Например, проба собственной опухоли ракового пациента, полученная при биопсии или хирургической резекции, может быть использована непосредственно для презентации антигена дендритным клеткам или для обеспечения клеточного лизата или нуклеиновых кислот для презентации антигена. Альтернативно, может быть использован мембранный препарат опухолевых клеток ракового пациента. Опухолевой клеткой может быть, например, клетка предстательной железы, легкого, яичника, ободочной кишки, головного мозга, меланомы или любой другой тип опухолевой клетки. Лизат и препарат мембран могут быть приготовлены из выделенных опухолевых клеток способами, известными в данной области.
В другом примерном варианте осуществления в качестве антигена может быть использован очищенный или полуочищенный специфический мембранный антиген предстательной железы (PSMA, также известный как антиген PSM), который специфически реагирует с моноклональным антителом 7Е11-С.5 (см. в общем Horoszewicz et al., Prog. Clin. Biol. Res. 37:115-32 (1983), Патент США №5162504; Патент США №5788963; Feng et al., Proc. Am. Assoc. Cancer Res. 32: (Abs. 1418) 238 (1991); описания которых включены здесь в качестве ссылки). Еще в одном примерном варианте осуществления в качестве антигена может быть использован антигенный пептид, имеющий аминокислотную последовательность Leu Leu His Glu Thr Asp Ser Ala Val (SEQ ID NO:1), (обозначенный PSM-P1), который соответствует аминокислотным остаткам 4-12 PSMA. Альтернативно, в качестве антигена может быть использован антигенный пептид, имеющий последовательность аминокислотных остатков Ala Leu Phe Asp lle Glu Ser Lys Val (SEQ ID NO:2), (обозначенный PSM-P2), который соответствует аминокислотным остаткам 711-719 PSMA.
В конкретном варианте осуществления в качестве антигена может быть использован антигенный пептид, имеющий последовательность аминокислотных остатков Хаа Leu (или Met) Xaa Хаа Хаа Хаа Хаа Хаа Val (или Leu), (обозначенный PSM-PX), где Хаа обозначает любой аминокислотный остаток. Этот пептид похож на мотив связывания HLA-A0201, т.е. мотив связывания из 9-10 аминокислотных остатков с «якорными остатками», лейцином и валином, обнаруживаемыми у HLA-А2-пациентов (см., например, Grey et al., Cancer Surveys 22:37-49 (1995)). Этот пептид может быть использован в качестве антигена для HLA-A2+ пациентов (см. Central Data Analysis Commitee "Allele Frequencies", Section 6.3, Tsuji, K. et al., (eds.), Tokyo University Press, pp.1066-1077). Подобным образом могут быть использованы пептиды, сходные с другими мотивами связывания HLA.
Обычно незрелые дендритные клетки, полученные способами данного изобретения, культивируют в присутствии агента созревания дендритных клеток, такого как BCG, IFNγ, LPS, TNFα, или их комбинаций и заданного антигена при подходящих условиях созревания, описанных выше. Необязательно, незрелые дендритные клетки могут быть смешаны с этим заданным антигеном в типичной культуральной среде для дендритных клеток с GM-CSF или без GM-CSF и/или агентом созревания. Спустя по меньшей мере приблизительно 10 минут - 2 дня культивирования с этим антигеном этот антиген может быть удален и могут быть добавлены культуральные среды, дополненные BCG и IFNγ. К средам для созревания может быть также добавлен GM-CSF без дополнительных цитокинов, таких как IL-4. Способы контактирования дендритных клеток с антигеном обычно известны в данной области (см. в общем Steel and Nutman, J. Immunol. 160:351-60 (1998); Tao et al., J. Immunol. 158:4237-44 (1997); Dozmorov and Miller, Cell Immunol. 178:187-96 (1997); Inaba et al., J Exp Med. 166:182-94 (1987); Macatonia et al., J. Exp Med. 169:1255-64 (1989); DeBruijn et al., Eur. J. Immunol. 22:3013-20 (1992); описания которых включены здесь в качестве ссылки).
Затем полученные зрелые, праймированные дендритные клетки коинкубируют с Т-клетками, такими как наивные Т-клетки. Т-клетки, или субпопуляция Т-клеток, могут быть получены из различных лимфоидных тканей для использования в качестве клеток-респондеров. Такие ткани включают, но не ограничиваются ими, селезенку, лимфатические узлы и/или периферическую кровь. Эти клетки могут быть культивированы совместно со зрелыми, праймированными дендритными клетками в виде популяции смешанных Т-клеток или в виде очищенной субпопуляции Т-клеток. Очистка Т-клеток может быть достигнута положительным или отрицательным отбором, в том числе, но не только, с использованием антител, направленных на CD2, CD3, CD4, CD8 и т.п.
Контактированием Т-клеток со зрелыми, праймированными дендритными клетками обеспечивают антигенреактивные или активированные, поляризованные Т-клетки или Т-лимфоциты. В данном контексте термин «поляризованные» относится к Т-клеткам, которые продуцируют высокие уровни IFNγ или в другом случае праймированы в отношении ответа типа 1 (Th-1). Такие способы обычно предусматривают контактирование дендритных клеток с BCG или IFNγ для получения зрелых, праймированных дендритных клеток. Незрелые дендритные клетки могут быть проконтактированы с заданным антигеном во время созревания или перед созреванием. Незрелые дендритные клетки могут культивироваться совместно с Т-клетками (например, наивными Т-клетками) во время созревания или культивироваться совместно с Т-клетками (например, наивными Т-клетками) во время созревания и праймирования дендритных клеток для индукции ответа типа 1. Далее незрелые дендритные клеток или зрелые дендритные клетки могут быть частично очищены, или обогащены, перед созреванием. Кроме того, Т-клетки могут быть обогащены из популяции лимфоцитов перед контактированием с дендритными клетками. В конкретном варианте осуществления обогащенные или очищенные популяции CD4+ Т-клеток контактируют со зрелыми, праймированными дендритными клетками. Совместное культивирование зрелых, праймированных дендритных клеток с Т-клетками приводит к стимуляции специфических Т-клеток, которые созревают в антигенреактивные CD4+ Т-клетки или антигенреактивные CD8+ Т-клетки.
В другом аспекте обеспечены способы для повторной стимуляции Т-клеток in vitro культивированием этих клеток в присутствии незрелых дендритных клеток или зрелых дендритных клеток, праймированных в отношении индукции Т-клеточного ответа типа 1 (Th-1). Такие Т-клетки необязательно могут культивироваться на клетках-фидерах. Незрелые дендритные клетки или зрелые, праймированные дендритные клетки необязательно могут быть облучены перед контактированием с Т-клетками. Подходящие условия культивирования могут включать в себя один или несколько цитокинов (например, очищенный IL-2, супернатант стимулированных Конканавалином А клеток селезенки, интерлейкин 15 (IL-15) и т.п., а также их комбинации). Такая in vitro повторная стимуляция Т-клеток может быть использована для стимуляции размножения Т-клеточной популяции.
Стабильная культура антигенспецифических, поляризованных Т-клеток или линии Т-клеток может поддерживаться in vitro в течение продолжительных периодов времени посредством периодической рестимуляции. Созданная таким образом Т-клеточная культура или Т-клеточная линия может храниться и при консервации (например, посредством криоконсерванта или замораживанием) применяться для повторной подачи активированных, поляризованных Т-клеток при желаемых интервалах для долгосрочного использования.
В некоторых вариантах осуществления активированные CD8+ или CD4+ Т-клетки могут быть генерированы в соответствии со способом данного изобретения. Обычно зрелые, праймированные дендритные клетки, используемые для генерирования антигенреактивных, поляризованных Т-клеток, являются сингенными относительно субъекта, которому они должны вводиться (например, получены из этого субъекта). Альтернативно, дендритные клетки, имеющие тот же самый HLA-гаплотип, что и предполагаемый реципиентный субъект, могут быть получены in vitro с использованием нераковых клеток (например, нормальных клеток) из HLA-соответствующего донора. В конкретном варианте осуществления антигенреактивные Т-клетки, в том числе CTL и Th-1 клетки, размножают in vitro в качестве источника клеток для лечения.
В соответствии с другим аспектом данного изобретения неактивированные моноцитарные предшественники дендритных клеток, незрелые дендритные клетки и зрелые праймированные дендритные клетки могут быть консервированы, например, криоконсервированием. Каждая популяция может быть извлечена перед продолжением описанных здесь процессов. Например, моноцитарные предшественники дендритных клеток могут быть получены из пациента в форме продукта лейкафереза или афереза перед культивированием в среде для культивирования дендритных клеток в присутствии блокирующего адгезию агента и GM-CSF для образования и поддержания незрелых дендритных клеток. После приготовления незрелых дендритных клеток эти клетки могут быть криоконсервированы либо перед подверганием действию антигена и созреванием, либо перед введением индивидууму, подлежащему лечению. Агенты криоконсервирования, которые могут быть использованы, включают, но не ограничиваются ими, диметилсульфоксид (ДМСО), глицерин, поливинилпирролидон, полиэтиленгликоль, альбумин, декстран, сахарозу, этиленгликоль, и-эритрит, D-рибит, D-маннит, D-сорбит, и-инозит, D-лактозу, холинхлорид, аминокислоты, метанол, ацетамид, моноацетат глицерина и неорганические соли. Различные криопротективные агенты и различные типы клеток обычно имеют различные оптимальные скорости охлаждения. Нагревание фазы слияния, в которой вода превращается в лед, обычно должно быть минимальным. Процедура охлаждения может проводиться, например, с использованием программируемого замораживающего устройства или процедуры с использованием метанольной бани. Программируемый аппарат замораживания позволяет определить оптимальные скорости охлаждения и облегчает стандартное воспроизводимое охлаждение. Программируемые морозильники с регулируемой скоростью, такие как Cryomed или Planar, позволяют настраивать режим замораживания относительно желаемой кривой скорости охлаждения.
После тщательного замораживания дендритные клетки могут быть быстро перенесены в резервуар долгосрочного криогенного хранения. В типичном варианте осуществления пробы могут храниться криогенно в жидком азоте (-196°С) или в его парах (-165°С). Рассмотрения и процедуры для манипуляции, криоконсервирования и долгосрочного хранения гемопоэтических стволовых клеток, в частности, из костного мозга или периферической крови в значительной степени приложимы к неактивированным дендритным клеткам данного изобретения. Обсуждение криоконсервирования для гемопоэтических стволовых клеток может быть найдено, например, в следующих ссылках, включенных здесь: Taylor et al., Cryobiology 27:269-78 (1990); Gorin, Clinics in Haematology 15:19-48 (1986); Bone-Marrow Conservation, Culture and Transplantation, Proceedings of a Panel, Moscow, Jul. 22-26, 1968, International Atomic Energy Agency, Vienna, pp.107-186.
Замороженные клетки обычно быстро оттаивают (например, на водяной бане, поддерживаемой при 37-41°С) и охлаждают немедленно после оттаивания. Может быть желательной обработка этих клеток для предотвращения образования комков при оттаивании. Для предотвращения образования комков могут быть использованы различные процедуры, в том числе, но не только, добавление до и/или после оттаивания ДНКазы (Spitzer et al., Cancer 45:3075-85 (1980)), низкомолекулярного декстрана и цитрата, гидроксиэтилкрахмала (Stiff et al., Cryobiology 20: 17-24 (1983)) и т.п. Криопротективные агенты, если они являются токсичными для людей, должны быть удалены перед терапевтическим использованием оттаянных дендритных клеток с низкой адгезией. Одним путем, которым может быть удален криопротективный агент, является разбавление до незначительной концентрации. После оттаивания и извлечения замороженных ДК они могут быть использованы для активации Т-клеток, как описано здесь в отношении незамороженных ДК.
Введение in vivo клеточной популяции
В другом аспекте данного изобретения обеспечены способы для введения зрелых, праймированных дендритных клеток или активированных поляризованных Т-клеток или популяции клеток, содержащей такие клетки, субъекту, нуждающемуся в иммуностимуляции. Такие клеточные популяции могут включать незрелые дендритные клетки, частично созревшие дендритные клетки, популяции зрелых праймированных дендритных клеток и/или популяции активированных, поляризованных Т-клеток. В некоторых вариантах осуществления эти способы выполняют посредством получения неактивированных дендритных клеток-предшественников или незрелых дендритных клеток, дифференцировкой этих клеток с использованием GM-CSF в отсутствие дополнительных цитокинов и созреванием этих клеток в присутствии агента созревания, такого как, например, BCG и/или IFNγ, и заданного антигена для образования популяции зрелых дендритных клеток, праймированных в отношении Th-1-ответа. Незрелые дендритные клетки могут быть проконтактированы с антигеном до или во время созревания. Такие зрелые, праймированные дендритные клетки могут вводиться непосредственно субъекту, нуждающемуся в иммуностимуляции.
В родственном варианте осуществления зрелые, праймированные дендритные клетки могут быть проконтактированы с лимфоцитами из субъекта для стимуляции Т-клеток в популяции лимфоцитов. Эти активированные, поляризованные лимфоциты, необязательно с последующим клональным размножением в клеточной культуре антигенреактивных CD4+ и CD8+ Т-клеток, могут быть введены субъекту, нуждающемуся в иммуностимуляции. В некоторых вариантах осуществления активированные, поляризованные Т-клетки являются аутологичными относительно субъекта.
В другом варианте осуществления дендритные клетки, Т-клетки и реципиентный субъект имеют один и тот же гаплотип МНС (HLA). Способы определения гаплотипа HLA субъекта известны в данной области. В родственном варианте осуществления дендритные клетки и/или Т-клетки являются аллогенными относительно реципиентного субъекта. Например, дендритные клетки могут быть аллогенными относительно Т-клеток и реципиента, которые имеют один и тот же гаплотип МНС (HLA). Эти аллогенные клетки обычно совпадают в отношении по меньшей мере одного аллеля МНС (например, имеют общий по меньшей мере один, но не все аллели МНС). В менее типичном варианте осуществления дендритные клетки, Т-клетки и реципиентный субъект являются, все, аллогенными относительно друг друга, но все имеют по меньшей мере один общий аллель МНС.
Согласно одному варианту осуществления Т-клетки получают из того же самого субъекта, из которого получают незрелые дендритные клетки. После созревания и поляризации in vitro аутологичные Т-клетки вводят субъекту для индукции и/или увеличения существующего иммунного ответа. Например, Т-клетки могут быть введены внутривенной инфузией, например, при дозах приблизительно 108-109 клеток/м2 площади поверхности тела (см., например, Ridell et al., Science 257:238-41 (1992), включенную здесь в качестве ссылки). Инфузия может повторяться при желаемых интервалах, например один раз в месяц. Реципиенты могут быть подвергнуты мониторингу во время и после инфузий Т-клеток на любое появление неблагоприятных эффектов.
В соответствии с другим вариантом осуществления дендритные клетки, полученные способом, описанным в данной заявке, выращивают только в присутствии GM-CSF, подвергают созреванию с BCG и IFNγ и в соответствии с данным изобретением могут быть введены инъекцией непосредственно в опухоль, участок, окружающий опухоль, или другую ткань, содержащую антиген-мишень. Такие зрелые клетки могут поглощать антиген и презентировать этот антиген Т-клеткам in vivo.
Следующие примеры приведены только в качестве иллюстрации различных аспектов данного изобретения и не должны рассматриваться как ограничивающие каким-либо образом данное изобретение.
Пример 1
В этом примере было продемонстрировано, что дифференцировка in vitro моноцитов в CD1a+ дендритные клетки в присутствии только GM-CSF требует, чтобы эти клетки не могли осуществлять начальное прилипание к культуральному сосуду.
Вкратце, CD4+CD1а-моноциты ресуспендировали либо в модифицированной Исковым среде Дульбекко (IMDM, BioWhittaker) плюс 2 мМ L-глутамин (Gibco BRL), либо в X-VIVO-15® (BioWhittaker) плюс 3% сывороточный альбумин человека (САЧ, Bayer). Клеточные суспензии переносили в культуральные колбы Т-25 (Greiner) и инкубировали в течение 30 мин в термостате с 6% СО2, 37°С. После инкубирования добавляли сывороточный альбумин человека (САЧ) и колониестимулирующий фактор гранулоцитов-макрофагов (GM-CSF, Immunex) для получения конечной концентрации 3% САЧ и 500 единиц/мл GM-CSF. Обе культуры инкубировали в течение 4 дней в термостате с 6% CO2, 37°C. Поверхностную экспрессию CD14 и CD1a анализировали с использованием меченых моноклональных антител, специфических в отношении этих молекул, и детектированием с использованием проточной флуорометрии. Пунктирные гистограммы представляют окрашивание контроля на изотип (фон) (фигура 1).
Клетки, моноциты, исходно инкубированные в среде без САЧ, показали прочное прилипание к пластику, как определено фазово-контрастной микроскопией, как видно по уплощению этих клеток на поверхности, тогда как клетки, инкубированные в среде с САЧ, показали уменьшенную степень адгезии. Это доказывалось сферической формой клеток, как определено с использованием фазово-контрастной микроскопии. После 4 дней в культуре первая культура сохраняла некоторую экспрессию CD14 и экспрессировала очень низкие уровни CD1a (фигура 1). В противоположность этому, большинство клеток, которым не давали прочно прикрепиться к поверхности культурального сосуда, были CD4-и CD1a+, что характерно для незрелых дендритных клеток.
В другом примере было показано, что моноциты могли дифференцироваться in vitro в CD1a+ дендритные клетки в присутствии только GM-CSF, когда эти клеткам не давали прилипать к культуральному мешку из Тефлона®.
Вкратце, выделенные моноциты из двух лейкаферезных доноров независимо ресуспендировали в среде X-VIVO-15® (BioWhittaker) плюс колониестимулирующий фактор гранулоцитов-макрофагов (GM-CSF, Immunex) и сывороточный альбумин человека (САЧ, Plasbumin™, Bayer) для получения конечной концентрации 500 единиц/мл GM-CSF и 2% САЧ, в мешках из Тефлона®. Клеточные суспензии в этих мешках переносили в термостат с 6% СО2, 37°C на 5 дней. В конце этого периода культивирования к культурам добавляли агенты созревания (разведение 1:400 инактивированного BCG (Organon-Teknika) и 500 Е/мл IFNγ (R&D Systems)). Событию созревания давали протекать в течение 4 часов. Поверхностную экспрессию CD14 и CD1a на «живых» клетках анализировали после пропускания сигналов рассеяния вперед (FS) и бокового рассеяния (SS) с мечеными моноклональными антителами, специфическими для этих молекул, с использованием проточного анализа клеток с возбуждением флуоресценции (фигуры 2А и 2В). Контрольные антитела изотипа использовали в качестве контролей на фоновую флуоресценцию, и они были IgG1 для антитела, специфического для CD1a, и IgG2b для антитела, специфического для CD14.
Клетки-предшественники, выделенные первоначально, экспрессировали высокие уровни CD14 и не экспрессировали CD1a или экспрессировали низкие уровни CD1a, типичные для моноцитов, (фигуры 2А и 2В). В отличие от этого, большинство клеток из конечной культуры со слабым прилипанием были CD14-и CD1a+, как это ожидалось для полученных из моноцитов дендритных клеток.
Пример 2
В этом примере незрелые дендритные клетки, полученные из моноцитов, которым не давали прикрепляться к поверхности культурального сосуда, были подвергнуты созреванию, и их испытывали на секрецию IL-12. Количество IL-12, секретированного из этих дендритных клеток, сравнивали со зрелыми дендритными клетками, выделенными обычными способами и культивируемыми в присутствии GM-CSF и IL-4.
Вкратце, криоконсервированные моноциты ресуспендировали при концентрации 1×106 клеток/мл в модифицированной Исковым среде Дульбекко (IMDM, BioWhittaker) плюс 2 мМ L-глутамин (Gibco BRL) в отсутствие или в присутствии 3% сывороточного альбумина человека (САЧ, Bayer). Клеточную суспензию переносили в две колбы для культуры ткани (Greiner) на одно условие (вариант) и инкубировали в течение 30 мин в увлажненном термостате с 6% CO2, 37°С. После этого периода инкубирования добавляли САЧ для получения конечной концентрации 3% САЧ во всех колбах. Колониестимулирующий фактор гранулоцитов-макрофагов (GM-CSF, Immunex) или GM-CSF и интерлейкин-4 (IL-4, R&D Systems) добавляли к каждой культуре для получения конечной концентрации 500 единиц/мл. Все культуры инкубировали в течение 30 мин в увлажненном термостате с 6% CO2, 37°С в течение 4 дней. В конце этого периода культивирования в колбу добавляли агенты созревания (разведение 1:400 инактивированного BCG (Organon-Teknika) и 500 Е/мл IFNγ (R&D Systems)). Событию созревания давали протекать в течение 18-24 часов. Культуральные супернатанты собирали и анализировали на секрецию IL-12 р70. Результаты из двух отдельных экспериментов сравнивали. В обоих экспериментах секреции IL-12 р70 детектировали в культурах, дополненных только GM-CSF или GM-CSF и IL-4. (фигура 3). Кроме того, моноциты, подвергнутые прочному первоначальному прилипанию с последующим культивированием только с GM-CSF, не секретировали IL-12 р70 в обоих экспериментах. В одном эксперименте секрецию IL-12 р70 детектировали в культуре, подвергнутой прочному начальному прилипанию, с последующим культивированием в течение 4 дней в присутствии GM-CSF м IL-4.
Пример 3
В этом примере экспрессию маркеров клеточной поверхности, типичных для дендритных клеток, анализировали в неактивированных моноцитах, культивируемых в присутствии только GM-CSF. Неактивированные моноциты, культивируемые только в GM-CSF, продемонстировали экспрессию маркеров клеточной поверхности, типичных для зрелых ДК.
Вкратце, криоконсервированные моноциты ресуспендировали при концентрации 1×106 клеток/мл в культуральной среде для ДК, содержащей X-VIVO-15® (BioWhittaker), 2% сывороточный альбумин человека (Bayer) и 500 Е/мл GM-CSF (Immunex). Эту клеточную суспензию переносили в колбу для культуры ткани (Greiner) и культивировали в течение 4 дней в увлажненном термостате с 6% СО2, 37°С. В конце этого периода культивирования в колбу добавляли агенты созревания (разведение 1:400 инактивированного BCG (Organon-Teknika) и 500 Е/мл IFNγ (R&D Systems)). Событию созревания давали протекать в течение 18-24 часов. Зрелые ДК собирали и характеризовали. Клетки реагировали с мечеными моноклональными антителами, специфическими для CD11c, CD1a, CD40, CD80, CD86, CD54 и CD83. Кроме того, эти клетки окрашивали иодидом пропидия. Метку определяли проточной цитометрией. Эти данные показали, что 80% извлеченных клеток были живыми ДК (CD11с+ и окрашивались иодидом пропидия). Кроме того, эти клетки экспрессировали типичные маркеры ДК, т.е. не экспрессировали CD4 и экспрессировали CD11с, CD1a, CD40, CD80, CD86, CD54 и CD83 (фигура 4).
Пример 4
В этом примере моноцитарные предшественники дендритных клеток, которые культивировали в присутствии блокирующего адгезию агента, испытывали в отношении кинетики дифференцировки in vitro в дендритные клетки в среде, дополненной только GM-CSF.
Вкратце, криоконсервированные моноциты ресуспендировали при концентрации 1×106 клеток/мл в культуральной среде для ДК, содержащей X-VIVO-15® (BioWhittaker), 2% сывороточный альбумин человека (Bayer) и 500 Е/мл GM-CSF (Immunex). Эту клеточную суспензию переносили в Тефлоновый® мешок (American Fluoroseal) и культивировали в течение 5 дней в увлажненном термостате с 6% СО2, 37°С. В день 5 в культуральный мешок добавляли агенты созревания (разведение 1:400 инактивированного BCG (Organon-Teknika) и 500 Е/мл IFNγ (R&D Systems)). Событию созревания давали протекать в течение 18 часов. Клетки собирали ежедневно из этого мешка для проточных цитометрических анализов экспрессии CD14 и CD1a. Результаты из этих анализов показали, что превращение из моноцитов
(CD14+ и CD1a-) в ДК (CD14-, CD1a+) начиналось между 1 и 2 днями после старта культуры (фигура 5). В день 3 фенотипические превращения завершались.
Пример 5
В этом примере сравнивали фенотип ДК, культивируемых либо в Тефлоновых мешках, либо в колбах при различных условиях культивирования. Клетки выращивали либо только в GM-CSF, либо с GM-CSF, дополненном IL-4. Сравнивали также фенотип клеток, которые были подвергнуты или не были подвергнуты действию агентов созревания.
Вкратце, моноциты ресуспендировали при 1×106 клеток/мл в X-VIVO-15® (BioWhittaker) и 2% САЧ (Bayer), дополненной 500 Е/мл только GM-CSF или GM-CSF в комбинации с 500 Е/мл IL-4. Суспензии клеток (мешки в двух повторностях для каждого из условий культивирования) культивировали в Тефлоновых мешках (American Fluoroseal) или колбах для культуры ткани (только комбинация GM-CSF/IL-4) в термостате с 6% СО2, 37°С. Спустя 5 дней в одну из двух повторных культур в Тефлоновых мешках добавляли агенты созревания (разведение 1:400 инактивированного BCG (Organon-Teknika) и 500 Е/мл IFNγ (R&D Systems)). В день 6 все культуры собирали. Их фенотипы анализировали с использованием окрашивания мечеными моноклональными антителами, специфическими для CD80, CD83, CD86 и HLA-DR, с детектированием проточной цитометрией.
Большинство ДК из всех пяти условий культивирования экспрессировали CD1a (89-97%) (фигура 6А). Как в культурах с GM-CSF, так и в культурах с GM-CSF/IL-4 наблюдали значимую экспрессию маркера созревания ДК, CD83 (фигура 6В) только в культурах, подвергнутых действию агентов созревания. Кроме того, наблюдали значимое увеличение поверхностной экспрессии костимуляторных молекул (CD80 и CD86, фигуры 6С и 6D), а также HLA-DR (фигура 6Е), так как эти ДК были зрелыми. Уровни экспрессии этих молекул были сходными во всех трех популяциях зрелых ДК.
Кроме того, функции по стимуляции Т-клеток ДК, генерированных из моноцитов, подвергнутых стадии начального прочного прилипания, сравнивали с ДК, генерируемыми в отсутствие прочного прилипания в присутствии блокирующего адгезию агента. В этом исследовании в качестве исходных популяций использовали два источника моноцитов. Для популяции прочно прилипших моноцитов мононуклеарные клетки периферической крови (РВМС) инкубировали в ОРТIМЕМ-1 (Gibco BRL) плюс 1% инактивированная нагреванием аутологичная плазма в течение 1 часа в колбе для культуры ткани (Greiner). После инкубирования неприкрепленные клетки удаляли, оставляя обогащенную прилипшую популяцию активированных моноцитов на поверхности колбы. Для получения популяции неактивированных моноцитов моноциты получали из колонки, содержащей стеклянные гранулы-микроносители, покрытые сывороточным альбумином человека (САЧ) (камеры для гранул).
Затем каждую из популяций моноцитов, активированных и неактивированных, инкубировали в X-VIVO-15® с 2% САЧ в присутствии только GM-CSF или в комбинации с IL-4 в течение 5 дней. Полученные незрелые ДК загружали 40-мерным пептидом вируса гриппа А М1-А4 или гемоцианином фиссуреллы (KLH) в течение одного часа до промывки и созревания с помощью BCG (разведение 1:400) и IFNγ (500 Е/мл). После сбора и промывки зрелых ДК устанавливали совместные культуры ДК с аутологичными РВМС при соотношении 1:10 ДК:РВМС в среде AIM-V® плюс 5% АВ-сыворотки человека (HuAB Sera), дополненной 20 нг/мл IL-2, со дня 2 по день 8. После восьми дней культивирования Т-клеточные линии собирали и анализировали на размножение М1-А4-специфических CD8 Т-клеток (Vβ17+ CD8+ Т-клеток) проточной цитометрией.
В сравнении с KLH-контролями ДК, генерированные прилипанием к пластику, требуют IL-4 для генерирования специфического ответа на грипп (М1-А4) (фигура 7А). Однако ДК, генерируемые из выделенных из камеры для гранул моноцитов (неактивированные), являются наиболее эффективными в инициации вторичного CD8 Т-клеточного ответа при использовании только GM-CSF во время их генерирования (фигура 7В).
Пример 6
В этом примере экспрессию поверхностных маркеров, типичных для дендритных клеток, анализировали в неактивированных моноцитах, которые были обогащены фильтрацией с тангенциальным потоком и культивировались в присутствии только GM-CSF. Дендритные клетки, культивируемые только в GM-CSF, демонстрировали экспрессию маркеров клеточной поверхности, типичных для созревающих ДК.
Вкратце, криоконсервированные моноциты предварительно выделяли процессом фильтрации с тангенциальным потоком из двух разных доноров крови. Этот процесс предусматривал TFF пробы моноцитов в устройстве, имеющем фильтр с размерами пор 5,5 мкм. Скорость рециркуляции (вхождения) была приблизительно 1400 мл/мин, скорость фильтрации была приблизительно 17 мл/мин, и время фильтрации было приблизительно 90 мин. Обогащенные моноцитарные предшественники дендритных клеток независимо культивировали при концентрации 1×106 клеток/мл в среде для культуры ДК, содержащей X-VIVO-15® (BioWhittaker), 2% сывороточный альбумин человека (Bayer) и 500 Е/мл GM-CSF (Immunex). Клеточные суспензии в Тефлоновых® мешках культивировали в течение 5 дней в увлажненном термостате с 6% CO2, 37°С. В конце периода культивирования в культуры добавляли агенты созревания (разведение 1:400 инактивированного BCG (Organon-Teknika) и 500 Е/мл IFNγ (R&D Systems). Событию созревания давали протекать в течение 4 часов. Созревающие ДК собирали и характеризовали. Клетки реагировали с мечеными моноклональными антителами, специфическими для CD11c, CD1a, CD40, CD80, CD86, CD54 и CD83. Поверхностную экспрессию на «живых» клетках анализировали после пропускания сигналов рассеяния вперед (FS) и бокового рассеяния (SS) с мечеными моноклональными антителами, специфическими для этих молекул, и детектирования с использованием проточной цитометрии. Кроме того, эти клетки окрашивали иодидом пропидия. Метку детектировали проточной цитометрией.
Более чем 80% извлеченных клеток были клетками моноцитарного направления дифференцировки, то есть экспрессирующими CD11c, и были "живыми" ДК (результаты с иодидом пропидия, не показаны). Важно, что клетки, дифференцированные в отсутствие IL-4, экспрессируют типичные маркеры ДК, т.е. имеют уменьшенную экспрессию CD14 и экспрессию CD1a, CD40, CD80, CD86, CD54 и CD83 (фигура 8А и 8В). Фоновую флуоресценцию измеряли с использованием контрольных антител на изотип, и эти антитела были IgG1, за исключением CD14, где контролем на изотип было антитело IgG2b.
Пример 7
В этом примере было определено, что моноциты, которые были подвергнуты действию пластиковых поверхностей (т.е. колбы для культуры ткани), становились активированными, если прочное взаимодействие не блокировалось добавлением блокирующего агента, такого как сывороточный альбумин человека (САЧ).
Моноциты (1×106/мл) высевали в колбы для культуры ткани в модифицированной Исковым среде Дульбекко (IMDM), с 3% (мас./об.) САЧ или без него, в течение 1 часа. После 1 часа инкубирования при 37°С 3% САЧ также добавляли к культуре, которую высевали исходно в отсутствие САЧ, и обе культуры инкубировали в течение ночи при 37°С. Затем супернатанты собирали и измеряли уровни различных цитокинов, которые обычно ассоциированы с активацией моноцитов. Концентрации этих цитокинов (нг/мл) показаны в таблице 1 ниже.
Предыдущие примеры приведены для иллюстрации, но не для ограничения объема заявленных изобретений. Другие варианты этих изобретений будут вполне очевидными специалистам с обычной квалификацией в данной области и включены в прилагаемую формулу изобретения. Все публикации, патенты, заявки на патенты и другие ссылки, процитированные здесь, включены тем самым в качестве ссылки.
Реферат
Изобретение относится к области биотехнологии, медицины и клеточной биологии. Предложен способ дифференцировки моноцитарных предшественников дендритных клеток в незрелые дендритные клетки, а также способ продуцирования зрелых дендритных клеток с использованием колониестимулирующего фактора гранулоцитов - макрофагов GM-CSF без других цитокинов при условиях, которые предотвращают адгезию клеток к сосуду. Изобретение может быть применено в иммунотерапии на основе дендритных клеток, например, для использования в лечении заболеваний, в том числе рака. 7 н. и 15 з.п. ф-лы, 15 ил., 1 табл.
Формула
a) использование популяции клеток, содержащей неактивированные моноцитарные предшественники дендритных клеток;
b) контактирование неактивированных моноцитарных предшественников дендритных клеток в культуральном сосуде со средой для культивирования дендритных клеток, дополненной колониестимулирующим фактором гранулоцитов-макрофагов GM-CSF в отсутствие дополнительных цитокинов при условиях, которые предотвращают адгезию неактивированных моноцитарных предшественников дендритных клеток к культуральному сосуду в течение периода времени, достаточного для дифференцировки моноцитарных предшественников дендритных клеток в незрелые дендритные клетки.
a) использование популяции клеток, содержащей неактивированные моноцитарные предшественники дендритных клеток;
b) контактирование неактивированных моноцитарных предшественников дендритных клеток в культуральном сосуде со средой для культивирования дендритных клеток, дополненной колониестимулирующим фактором гранулоцитов-макрофагов GM-CSF в отсутствие дополнительных цитокинов и хелаторов металла при условиях, которые предотвращают адгезию неактивированных моноцитарных предшественников дендритных клеток к культуральному сосуду в течение периода времени, достаточного для дифференцировки моноцитарных предшественников дендритных клеток в незрелые дендритные клетки.
a) использование популяции клеток, содержащей неактивированные моноцитарные предшественники дендритных клеток;
b) контактирование неактивированных моноцитарных предшественников дендритных клеток в культуральном сосуде со средой для культивирования дендритных клеток, дополненной колониестимулирующим фактором гранулоцитов-макрофагов GM-CSF, в отсутствие дополнительных цитокинов при условиях, которые предотвращают адгезию неактивированных моноцитарных предшественников дендритных клеток к культуральному сосуду;
где указанная популяция включает периферическую кровь, продукт лейкафереза, продукт афереза, пуповинную кровь, селезенку, лимфатический узел, тимус или костный мозг.
a) использование популяции клеток, содержащей неактивированные моноцитарные предшественники дендритных клеток;
b) обогащение неактивированных моноцитарных предшественников дендритных клеток посредством фильтрации с тангенциальным потоком;
c) контактирование неактивированных моноцитарных предшественников дендритных клеток в культуральном сосуде со средой для культивирования дендритных клеток, дополненной колониестимулирующим фактором гранулоцитов-макрофагов GM-CSF в отсутствие дополнительных цитокинов при условиях, которые предотвращают адгезию неактивированных моноцитарных предшественников дендритных клеток к культуральному сосуду в течение периода времени, достаточного для дифференцировки моноцитарных предшественников дендритных клеток в незрелые дендритные клетки.
a) использование популяции клеток, содержащей неактивированные моноцитарные предшественники дендритных клеток;
b) контактирование неактивированных моноцитарных предшественников дендритных клеток в культуральном сосуде со средой для культивирования дендритных клеток, дополненной колониестимулирующим фактором гранулоцитов-макрофагов GM-CSF в отсутствие дополнительных цитокинов при условиях, которые предотвращают адгезию неактивированных моноцитарных предшественников дендритных клеток к культуральному сосуду; и
c) контактирование дифференцированных предшественников дендритных клеток с представляющим интерес антигеном в течение периода времени, достаточного для поглощения антигена.
a) использование популяции клеток, содержащей неактивированные моноцитарные предшественники дендритных клеток;
b) контактирование неактивированных моноцитарных предшественников дендритных клеток в культуральном сосуде со средой для культивирования дендритных клеток, дополненной колониестимулирующим фактором гранулоцитов-макрофагов GM-CSF в отсутствие дополнительных цитокинов при условиях, которые предотвращают адгезию неактивированных моноцитарных предшественников дендритных клеток к культуральному сосуду;
c) контактирование дифференцированных предшественников незрелых дендритных клеток с представляющим интерес антигеном в течение периода времени, достаточного для поглощения антигена; и
d) контактирование дифференцированных предшественников незрелых дендритных клеток с агентом созревания дендритных клеток.
a) использование популяции клеток, содержащей неактивированные моноцитарные предшественники дендритных клеток; и
b) контактирование неактивированных моноцитарных предшественников дендритных клеток в культуральном сосуде со средой для культивирования дендритных клеток, дополненной колониестимулирующим фактором гранулоцитов-макрофагов GM-CSF в отсутствие дополнительных цитокинов и хелаторов металла при условиях, которые предотвращают адгезию неактивированных моноцитарных предшественников дендритных клеток к культуральному сосуду в течение периода времени, достаточного для дифференцировки моноцитарных предшественников дендритных клеток в незрелые дендритные клетки.
Комментарии