Ген, придающий устойчивость к phytophthora infestans (фитофторозу) в семействе solanaceae - RU2361920C2
Код документа: RU2361920C2
Чертежи
































Описание
Фитофтороз, вызываемый относящимся к классу оомицетов патогеном Phytophthora infestans, является распространенной по всему свету наиболее деструктивной болезнью для культивирования картофеля. Эта болезнь является также угрозой культуре томата. Настоятельность получения устойчивых сортов (культиваров) интенсифицировалась по мере быстрого возникновения вирулентных, специализированных в отношении определенных культур и устойчивых к пестицидам штаммов этого патогена.
Способом предупреждения неурожаев или уменьшенных урожаев является применение фунгицидов, которые предотвращают или излечивают инфекцию P. infestans. Однако считается, что применение этих защищающих посев агентов является нагрузкой для окружающей среды. Так в нескольких западных странах законодательство становится более ограничивающим и частично запрещающим применение специфических фунгицидов, делая более трудной химическую борьбу с данной болезнью. Альтернативным подходом является применение сортов, которые несут частичную или полную устойчивость к фитофторозу. Два типа устойчивости к фитофторозу были описаны и использованы в селекции картофеля. Один тип придается несколькими основными доминантными генами, которые делают хозяина несовместимым со специфическими расами этого патогена (раса-специфическая устойчивость). Одиннадцать таких R-генов (R1-R11) были идентифицированы, и считается, что они происходят из дикого вида картофеля Solanum demissum, который является природным в Мексике, где обнаружена наибольшая генетическая изменчивость этого патогена. Некоторые из этих R-генов были картированы на генетической карте картофеля (обзор Gebhardt and Valkonen, 2001 Annu. Rev. Phytopathol. 39: 79-102). R1 и R2 локализованы на хромосомах 5 и 4 соответственно. R3, R6 и R7 локализованы на хромосоме 11. Неизвестные R-гены, придающие раса-специфическую устойчивость к фитофторозу, были описаны также в S. tuberosum ssp. andigena и S. berthaultii (Ewing et al., 2000 Mol. Breeding 6: 25-36). Вследствие высокого уровня устойчивости и легкости переноса многие сорта содержат происходящую из S. demissum устойчивость. К сожалению, происходящая из S. demissum раса-специфическая устойчивость хотя и является почти полной, не является продолжительной. Как только заново выведенные сорта выращивают в крупном масштабе в коммерческих полях, возникают новые вирулентности в P. infestans, которые делают этот патоген способным преодолевать приобретенную интрогрессией устойчивость. Второй тип устойчивости, называемый полевой устойчивостью и часто встречающийся в природе, считается не специфическим в отношении расы и более продолжительным. Полевая устойчивость к фитофторозу может быть обнаружена в некоторых видах Solanum в Мексике и Средней и Южной Америке (Rossi et al., 1986 PNAS 95:9750-9754).
Диплоидный S. bulbocastanum из Мексики и Гватемалы является одним из имеющих клубни видов, который известен из-за его высоких уровней полевой устойчивости к фитофторозу (Niederhauser and Mills, 1953 Phytopathology 43: 456-457). Несмотря на различия в балансовых числах в эндосперме интрогрессия признака устойчивости S. bulbocastanum была успешной. Манипуляции с плоидностью и ряд трудоемких bridge-скрещиваний привели к полученной из S. bulbocastanum, P. infestans-устойчивой зародышевой плазме (Hermsen and Ramanna, 1969 Euphytica 18:27-35; 1973 Euphytica 22:457-466; Ramanna and Hermsen, 1971 Euphytica 20:470-481; Hermsen and De Boer, 1971 Euphytica 20:171-180). Однако спустя почти 40 лет после получения первых гибридов и интенсивной и непрерывной селекционной работы селекционеров, занимающихся селекцией картофеля в Нидерландах с этой зародышевой плазмой, устойчивые к фитофторозу сорта все еще не были выведены на рынок. Успешное получение соматических гибридов S. bulbocastanum и S. tuberosum также сообщалось (Thieme et al., 1997 Euphytica 97(2):189-200; Helgeson et al., 1988 Theor Appl. Genet 96:738-742). Было обнаружено, что некоторые из этих гибридов и полученная обратным скрещиванием зародышевая плазма являются высокоустойчивыми к фитофторозу, даже при экстремальной нагрузке этой болезни. Несмотря на сообщения о супрессии рекомбинации устойчивость в полученном обратным скрещиванием материале находится, по-видимому, на хромосоме 8 в интервале приблизительно 6 сМ между маркерами RFLP СР53 и СТ64 (Naess et al., 2000 Theor. Appl Genet 101:697-704). Маркер CAPS, полученный из RFLP-зонда СТ88 томатов, корасщеплялся с устойчивостью. Супрессия рекомбинации между хромосомами S. bulbocastanum и S. tuberosum образует потенциальное препятствие для успешного восстановления повторно культивируемой зародышевой плазмы картофеля до уровня, который мог бы удовлетворять стандартам для новых селекционированных сортов картофеля. Выделение генов, которые кодируют устойчивость, обнаруженную в S. bulbocastanum, и последующая трансформация существующих сортов этими генами были бы гораздо более прямым и более быстрым подходом в сравнении использующей интрогрессию селекцией.
Клонирование и молекулярная характеристика многочисленных генов R растений, придающих устойчивость к болезням, вызываемым бактериями, грибами, вирусами, нематодами и насекомыми, идентифицировали несколько структурных признаков, характерных для генов R растений (обзор Dangl and Jones, 2001 Nature 411, 826-833). Большинство из них являются членами тесно связанных мультигенных семейств, и все гены R, охарактеризованные до сих пор, за исключением Pto, кодируют богатые лейцином повторы (LRR), структуры, которые, как было показано, участвуют в межбелковых взаимодействиях. LRR-содержащие гены R могут быть подразделены на два класса на основе присутствия предположительного состоящего из трех частей нуклеотидсвязывающего сайта (NBS). Гены R класса NBS-LRR содержат мотивы, которые являются общими с регуляторными белками апоптоза животных (van der Biezen et al., 1998 Curr. Biol. 8, 226-227; Aravind et al., 1999 Trends Biochem. Sci. 24, 47-53) и могут быть подразделены на две подгруппы на основе N-концевого домена, который либо проявляет сходство последовательности с белком Toll Drosophila и доменом рецептора интерлейкина-1 млекопитающего (TIR-NBS-LRR), либо содержит потенциальную лейциновую молнию или домен скрученной спирали (СС-NBS-LRR; Pan et al., 2000 Genetics, 155:309-22). Гены LRR-R без NBS кодируют трансмембранные белки, внеклеточный N-концевой район которых состоит из LRR (Jones et al., 1994 Adv. Bot. Res. 24, 89-167). Эти гены могут быть подразделены на две подгруппы на основе присутствия цитозольного домена серин/треонинкиназы (Song et al., 1995 Science, 270, 1804-1806). В настоящее время были клонированы четыре гена R из картофеля. Все четыре, в том числе полученный из S. demissum ген R1, придающий раса-специфическую устойчивость к фитофторозу, принадлежат к классу СС-NBS-LRR генов R растений (Bendahmane et al., 1999 Plant Cell 11, 781-791; Bendahmane et al., 2000 Plant J. 21, 73-81; van der Vossen et al., 2000 Plant Journal 23, 567-576; Ballvora et al., 2002 Plant Journal 30, 361-371).
Данное изобретение обеспечивает выделенную или рекомбинантную нуклеиновую кислоту, содержащую нуклеиновую кислоту, кодирующую аминокислотную последовательность фиг.8, или ее функциональный фрагмент или гомолог. Белок с указанной аминокислотой был обнаружен в качестве члена семейства белков, кодируемых кластером генов, идентифицируемых анализом филогенетического дерева, которое до сих пор состоит из белков, кодируемых Rpi-blb, RGC1-blb, RGC3-blb и RGC4-blb (далее называемыми кластером генов Rpi-blb) фиг.9.
Анализ филогенетического дерева проводят следующим образом. Сначала выполняют сопоставление множественных последовательностей - последовательностей нуклеиновых кислот и/или предпочтительно расшифрованных аминокислотных последовательностей генов, которые должны быть анализированы, с использованием программы CLUSTALW (http://www2.ebi.ac.uk/clustalw), которая рутинно используется в данной области. ClustalW дает.dnd file, который может быть прочтен с использованием программы TREEVIEW (http://taxonomy.zoology.gla.ac.uk/rod/rod.html). Филогенетическое дерево, изображенное на фиг.9А, является филограммой.
Филогенетические исследования расшифрованных аминокислотных последовательностей Rpi-blb, RGC1-blb, RGC3-blb и RGC4-blb и последовательностей большинства сходных генов из данной области (определенных с использованием BLASTX), полученных из отличающихся видов, с использованием способа Neighbour-Joining Saitou and Nei (1987 Molecular Biology and Evolution 4, 406-425), показывает, что соответствующие гены или их функциональные фрагменты кластера генов Rpi-blb могут быть помещены в отдельную ветвь (фиг.9А).
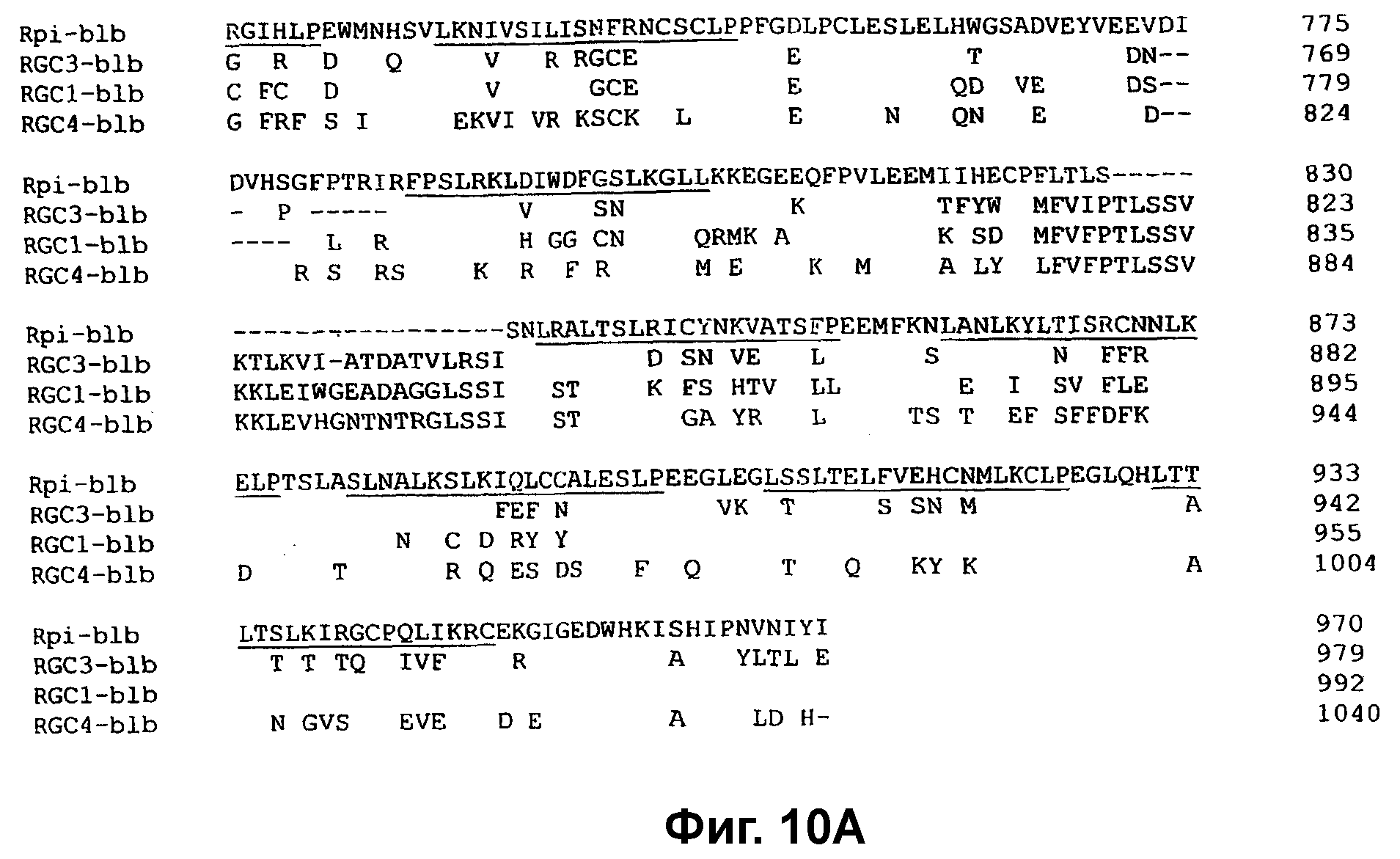
Сравнение последовательностей между четырьмя членами кластера генов Rpi-blb, идентифицированными на ВАС-клоне 8005-8 SPB-4, показывает, что гомология последовательностей в кластере генов Rpi-blb варьируется между 70% и 81% на уровне последовательностей аминокислот. Расшифрованная аминокислотная последовательность Rpi-blb имеет наивысшую общую гомологию с RGC3-blb (81% идентичность аминокислотной последовательности; таблица 4). При сравнении различных доменов видно, что эффекторные домены, присутствующие в N-концевых половинах этих белков (домены скрученных спиралей и NBS-ARC), имеют более высокую степень гомологии (91% идентичность последовательности), чем С-концевые половины этих белков, которые, как считается, содержат домены узнавания (LRR; 71% идентичность последовательности). Сравнение всех четырех аминокислотных последовательностей выявило в целом 104 Rpi-blb-специфических аминокислотных остатков (фиг.10). Большинство из них локализованы в LRR-районе (80/104). В этом последнем районе эти специфические остатки сконцентрированы в LRR-субдомене xxLxLxxxx. Относительная частота этих специфических аминокислотных остатков в этом LRR-субдомене является более, чем в 2 раза, более высокой (28,3%), чем частота, наблюдаемая в остальной части LRR-домена (12,3%). Считается, что остатки, расположенные около этих двух консервативных остатков лейцина в консенсусе xxLxLxxxx, экспонированы (обнажены) растворителем и, следовательно, участвуют, по-видимому, в создании/поддержании специфичности узнавания этого белка устойчивости.
Последовательности дополнительных членов кластера генов Rpi-blb могут быть получены скринингом библиотек геномной ДНК или инсертов, например, ВАС-библиотек, праймерами на основе сигнатурных последовательностей гена Rpi-blb. Скрининг различных ВАС-библиотек Solanum наборами праймеров А и/или В (таблица 2 и фиг.7) идентифицировал многочисленные гомологи Rpi-blb, полученные из различных видов Solanum. Сопоставление этих дополнительных последовательностей с последовательностями, представленными на фиг.10, поможет идентифицировать дополнительные члены кластера генов Rpi-blb и специфические аминокислотные остатки в их белках, ответственные за специфичность устойчивости к P. infestans. Кроме того, испытание дополнительных последовательностей в вышеописанных анализах филогенетического дерева, например, с использованием способа Neighbour-Joining Saitou and Nei (1987 Molecular Biology and Evolution 4, 406-425), обеспечивает дополнительную идентификацию генов, принадлежащих к кластеру генов Rpi-blb.
Данное изобретение обеспечивает развитие интраспецифического картирования популяции S. bulbocastanum, которая расщепляется в отношении раса-неспецифической устойчивости к фитофторозу. Эта устойчивость картирована на хромосоме 8, в районе, локализованном на расстоянии 0,3 сМ от СТ88. Благодаря раса-неспецифическому характеру этой устойчивости, всегда считалось, что устойчивость к фитофторозу S. bulbocastanum является независимой от гена R. Однако в данном изобретении авторы демонстрируют впервые, что раса-неспецифическая устойчивость S. bulbocastanum придается фактически геном, имеющим сходство с геном R NBS-LRR-типа.
Далее, данное изобретение обеспечивает молекулярный анализ этого геномного района и выделение с использованием клонирования на основе карты ДНК-фрагмента устойчивого родителя, который несет ген R, обозначенный Rpi-blb. Этот ДНК-фрагмент был субклонирован из клона бактериальной искусственной хромосомы (ВАС) длиной приблизительно 80 т.п.н., которая содержала четыре полных R-ген-подобных последовательности в кластер-подобном размещении. Трансформация чувствительного сорта картофеля Agrobacterium tumefaciens выявила, что одна из этих четырех R-ген-подобных последовательностей соответствует Rpi-blb, что обеспечивает раса-неспецифическую устойчивость к фитофторозу. Характеристика этого гена Rpi-blb показала, что он является членом класса NBS-LRR R-генов растений. Самыми близкими функционально охарактеризованными последовательностями предыдущего уровня являются члены семейства генов устойчивости I2 в томатах. Эти последовательности имеют общую идентичность аминокислотной последовательности приблизительно 32% с последовательностью Rpi-blb.
Таким образом, в первом варианте осуществления данное изобретение обеспечивает выделенную или рекомбинантную нуклеиновую кислоту, причем указанная нуклеиновая кислота кодирует генный продукт, имеющий последовательность Rpi-blb, или ее функциональный фрагмент, который способен обеспечивать член семейства Solanaceae раса-неспецифической устойчивостью против патогена из класса оомицетов.
Обеспеченное данным изобретением выделение гена, который кодирует желаемый признак устойчивости против фитофтороза, и последующая трансформация существующих сортов картофеля и томата этим геном обеспечивает теперь гораздо более прямой и более быстрый подход в сравнении с использующей интрогрессию селекцией. Обеспеченные здесь результаты предоставляют возможности дальнейшего исследования молекулярной основы взаимодействия растение-патоген, экологической роли R-генов в диком виде Мексиканского картофеля и являются применимыми для развития устойчивых сортов картофеля и томата с использованием генетической модификации.
В отличие от R-генов, клонированных и описанных до настоящего времени, ген, описанный здесь авторами данного изобретения, является первым выделенным R-геном из вида Solanum, который обеспечивает раса-неспецифическую устойчивость против патогена из класса оомицетов. Примечательно, что данное изобретение обеспечивает здесь нуклеиновую кислоту, которая обеспечивает желаемую устойчивость видам Solanum, среди которых находится S. tuberosum. В частности, этот ген является первым геном, который был выделен из филогенетически отличающегося родственного вида культивируемого картофеля, S. bulbocastanum, для которого было показано с использованием анализов на комплементацию, что он является функциональным в S. tuberosum. Эти данные предполагают, что ген Rpi-blb может быть легко использован в получении картофеля без необходимости отнимающей много времени и трудоемкой селекции с использованием интрогрессии.
Для терминов, используемых в описании и примерах, даются следующие определения.
- Нуклеиновая кислота: двухцепочечная или одноцепочечная молекула ДНК или РНК.
- Олигонуклеотид: короткая одноцепочечная молекула нуклеиновой кислоты.
- Праймер: термин праймер относится к олигонуклеотиду, который может праймировать (служить затравкой) синтез нуклеиновой кислоты.
- Гомология: гомология является термином, используемым в отношении сходства или идентичности биологической информации последовательности. Гомология может быть обнаружена на уровне нуклеотидной последовательности и/или на уровне кодируемой аминокислотной последовательности. Для расчета процентной идентичности может быть использован алгоритм BLAST (Altschul et al., 1997 Nucl. Acids Res. 25:3389-3402), использующий параметры по умолчанию, или, альтернативно, алгоритм GAP (Needleman and Wunsch, 1970 J. Mol. Biol. 48:443-453), использующий параметры по умолчанию, которые включены в пакет программ Wisconsin Genetics, Genetics Computer Group (GCG), 575 Science Dr., Madison, Wisconsin, USA. BLAST-поиски предполагают, что белки могут моделироваться в виде случайных последовательностей. Однако многие реальные белки содержат районы неслучайных последовательностей, которые могут быть гомополимерными участками, короткими периодическими повторами или районами, обогащенными одной или несколькими аминокислотами. Такие районы низкой сложности могут быть сопоставлены между неродственными белками даже в том случае, когда другие районы данного белка являются полностью несходными. Ряд программ-фильтров низкой сложности могут быть использованы для уменьшения таких сопоставлений низкой сложности. Например, фильтры низкой сложности SEG (Wooten and Federhen, 1993 Comput. Chem. 17:149-163) и XNU (Claverie and States, 1993 Comput. Chem. 17:191-201) могут использоваться по отдельности или в комбинации.
В применении здесь, термины «идентичность последовательности» или «идентичность» в контексте двух белковых последовательностей (или нуклеотидных последовательностей) включают в себя ссылку на остатки в двух последовательностях, которые являются одинаковыми при сопоставлении на максимальное соответствие на протяжении указанного окна сравнения. При использовании термина процент идентичности последовательности в отношении белков считается, что положения остатков, которые являются неидентичными, часто различаются консервативными аминокислотными заменами, где аминокислоты заменены другими аминокислотными остатками со сходными химическими свойствами (например, зарядом или гидрофобностью) и, следовательно, не изменяют функциональные свойства данной молекулы. Когда последовательности отличаются по консервативным заменам, процентная идентичность последовательности может корректироваться в сторону повышения для коррекции на консервативный характер этих замен. О последовательностях, которые отличаются такими консервативными заменами, говорят, что они имеют «сходство последовательности» или «сходство». Способы выполнения подобных коррекций хорошо известны лицам с квалификацией в данной области. Обычно они включают в себя оценку в баллах (очках) консервативной замены как частичного, а не полного ошибочного спаривания, что увеличивает процентную идентичность последовательности. Так, например, если идентичной аминокислоте дается оценка 1, а неконсервативной замене дается оценка 0, консервативной замене дается оценка между 0 и 1. Оценку в баллах консервативных замен рассчитывают, например, в соответствии с алгоритмом Meyers and Miller (Computer Applic. Biol. Sci. 4:11-17, 1988).
В применении здесь, «процентная идентичность последовательности» означает величину, определенную сравнением двух оптимально сопоставленных последовательностей на протяжении окна сравнения, где часть аминокислотной последовательности или нуклеотидной последовательности в окне сравнения может содержать добавления или делеции (т.е. гэпы) в сравнении со ссылочной последовательностью, для оптимального сопоставления этих двух последовательностей. Процент рассчитывают определением числа положений, в которых идентичные остатки аминокислот или нуклеотидов встречаются в обеих последовательностях, с получением числа соответствующих (правильно спаренных) положений, делением числа правильно спаренных положений на общее число положений в окне сравнения и умножением результата на 100 для получения процентной идентичности последовательности. Предпочтительно, аминокислотная последовательность белка данного изобретения имеет по меньшей мере 82% или более высокую гомологию с последовательностью, изображенной на фиг.8. Как показано в таблице 4, самая близкая функционально охарактеризованная последовательность предыдущего уровня (члены кластера генов I2 Fusarium в томате) имеет гораздо более низкий уровень идентичности аминокислотной последовательности, чем указанный уровень (32% относительно Rpi-blb). Гомология в кластере генов данного изобретения варьируется между 70% и 81% на уровне аминокислотной последовательности.
Гомологичные последовательности нуклеиновых кислот являются последовательностями нуклеиновых кислот, кодирующими определенный выше гомологичный белок. Одним примером такой нуклеиновой кислоты является последовательность, приведенная на фиг.6А. Однако имеются многочисленные последовательности, которые кодируют белок, являющийся на 100% идентичным белку, изображенному на фиг.8. Это обусловлено «качелями» в триплетах нуклеотидов, где одну и ту же аминокислоту может кодировать не один триплет, а несколько («гипотеза качелей», т.е. нестрогого соответствия в третьем положении триплета). Таким образом, даже без влияния на аминокислотную последовательность белка нуклеотидная последовательность, кодирующая этот белок, может существенно варьироваться. Общепризнанным является то, что нуклеотидные последовательности, кодирующие аминокислотные последовательности, которые не являются на 100% идентичными указанному белку, могут содержать даже еще больше вариаций. Таким образом, процентная идентичность на уровне последовательностей нуклеиновых кислот может варьироваться в более широких пределах, без отклонения от данного изобретения.
Промотор: термин «промотор» означает короткую ДНК-последовательность, с которой связываются РНК-полимераза и/или другие факторы инициации транскрипции перед транскрипцией ДНК, с которой функционально соединен этот промотор, позволяя иметь место транскрипции (т.е. запуская транскрипцию). Промотор обычно расположен слева (5') от кодирующей последовательности. В более широком смысле, термин «промотор» включает в себя сайт связывания РНК-полимеразы, а также регуляторные элементы последовательности, локализованные в нескольких сотнях нуклеотидов, иногда даже более далеко расположенные от стартового сайта транскрипции. Такими регуляторнными последовательностями являются, например, последовательности, которые участвуют в связывании белковых факторов, регулирующих эффективность инициации транскрипции в ответ на физиологические условия. Район промотора должен быть функциональным в клетке-хозяине и предпочтительно соответствует району природного промотора гена устойчивости Rpi-blb. Однако может быть использован любой гетерологичный промоторный район, пока он является функциональным в клетке-хозяина, в которой экспрессия является желательной. Гетерологичный промотор может быть либо конститутивным, либо регулируемым, тканеспецифическим, либо не специфическим в отношении конкретной ткани. Конститутивный промотор, такой как промотор CaMV 35S или промотор Т-ДНК, хорошо известные специалистам с квалификацией в данной области, является промотором, который по существу не подвергается регуляции, такой как индукция или репрессия, но делает возможной постоянную и по существу неизменную транскрипцию ДНК-последовательности, с которой он функционально связан во всех активных клетках организма, при условии, что другие требования для того, чтобы транскрипция имела место, являются удовлетворенными. Можно использовать тканеспецифический промотор, который запускает экспрессию в тех частях растения, которые имеют тенденцию к заражению патогеном. В случае Phytophthora может быть использован промотор, который запускает экспрессию в листьях, например промотор ферредоксина. Регулируемым промотором является промотор, функция которого регулируется одним или несколькими факторами. Эти факторы могут быть либо такими, которые своим присутствием гарантируют экспрессию релевантной ДНК-последовательности, либо могут, альтернативно, быть такими, которые подавляют экспрессию этой ДНК-последовательности, так что их отсутствие заставляет экспрессироваться данную ДНК-последовательность. Таким образом, промотор и необязательно связанная с ним регуляторная последовательность могут активироваться присутствием или отсутствием одного или нескольких факторов для влияния на транскрипциию ДНК-последовательностей генетической конструкции данного изобретения. Подходящие промоторные последовательности и способы получения увеличенной транскрипции и экспрессии известны специалистам с квалификацией в данной области.
Терминатор: терминатор транскрипции служит для терминации транскрипции ДНК в РНК и предпочтительно выбран из группы, состоящей из терминаторных последовательностей транскрипции растений, терминаторных последовательностей транскрипции бактерий и терминаторных последовательностей вирусов растений, известных специалистам с квалификацией в данной области.
Ген: термин “ген” используется для указания ДНК-последовательности, которая участвует в получении полипептидной цепи и которая включает в себя районы, предшествующие кодирующему району и следующие за кодирующим районом (расположенные 5'- (слева) и 3' (справа) от кодирующего района последовательности)), а также промежуточные последовательности, так называемые интроны, которые расположены между отдельными кодирующими сегментами (так называемыми экзонами) или в 5'- или 3'-районе. 5'-район (выше по ходу транскрипции) может содержать регуляторную последовательность, которая регулирует экспрессию гена, обычно промотор. 3'-район (ниже по ходу транскрипции) может содержать последовательности, которые участвуют в терминации транскрипции гена, и необязательно последовательности, ответственные за полиаденилирование транскрипта, и 3'-нетранслируемый район. Термин “ген устойчивости” обозначает выделенную нуклеиновую кислоту данного изобретения, причем указанная нуклеиновая кислота кодирует генный продукт, который способен обеспечивать устойчивость растения против патогена, причем более конкретно, указанное растение является членом семейства Solanaceae (семейства пасленовых), более конкретно, картофелем или томатом, причем указанный патоген является, более конкретно, патогеном из класса оомицетов, более конкретно Phytophthora, еще более конкретно Phytophthora infestans, и указанная нуклеиновая кислота предпочтительно содержит последовательность, изображенную на фиг.8, или ее часть, или гомологичную последовательность, имеющую по существу одинаковые функциональные и структурные характеристики. Функционально эквивалентный фрагмент такого гена или нуклеиновой кислоты устойчивости, обеспеченный данным изобретением, кодирует фрагмент полипептида, имеющего аминокислотную последовательность, изображенную на фиг.8, или ее часть, или гомологичный и/или функционально эквивалентный полипептид, причем указанный фрагмент проявляет свойство обеспечения по меньшей мере частичной устойчивости к инфекции оомицетов, такой как инфекция, вызываемая P. infestans, при включении и экспрессии в растении или клетке растения.
Продукт гена устойчивости: обозначает полипептид, имеющий аминокислотную последовательность, изображенную на фиг.8, или ее часть, или гомологичный и/или функционально эквивалентный полипептид, проявляющий свойство обеспечения по меньшей мере частичной устойчивости к инфекции оомицетов, такой как инфекция, вызываемая P. infestans, при включении и экспрессии в растении или клетке растения.
Функциональными эквивалентами белка данного изобретения являются белки, которые являются гомологичными белку, изображенному на фиг.8, или полученными из белка, изображенного на фиг.8, заменой, добавлением и/или делецией одной или более аминокислот, но все еще сохраняющие их активность в придании устойчивости к патогену. Такие эквиваленты могут быть легко получены конструированием белка in vivo, например, изменением открытой рамки считывания, способной кодировать этот белок, таким образом, что при этом происходит действие на эту аминокислотную последовательность. Пока эти изменения в аминокислотных последовательностях не уничтожают полностью эту активность данного белка, такие эквиваленты включены в данное изобретение. Далее, должно быть понятно, что эквиваленты должны быть получаемыми из белка, изображенного на фиг.8, при сохранении биологической активности, т.е. все или большая часть промежуточных продуктов между эквивалентным белком и белком, изображенным на фиг.8, должна иметь активность устойчивости к патогену. Большая часть означает 30% или более промежуточных продуктов, предпочтительно 40% или более, более предпочтительно 50% или более, еще более предпочтительно 60% или более, более предпочтительно 70% или более, более предпочтительно 80% или более, более предпочтительно 90% или более, более предпочтительно 95% или более, еще более предпочтительно 99% или более.
Предпочтительными эквивалентами являются эквиваленты, в которых богатый лейцинами повторяющийся район является высокогомологичным с LRR-районом, показанным на фиг.8. Другими предпочтительными эквивалентами являются эквиваленты, в которых N-концевой эффекторный домен является по существу одинаковым с эффекторным доменом Rpi-blb.
Белок данного изобретения содержит особый N-концевой эффекторный домен и богатый лейцином повторяющийся домен. Авторы считают, что консервативность этих районов является существенной для функции этого белка, хотя допустима некоторая изменчивость. Однако другие части этого белка являются менее важными для этой функции и могут быть более восприимчивыми к изменению.
Для обеспечения быстрого и простого испытания, способны ли модифицированные белки и/или генные конструкции к экспрессии указанных модифицированных белков, описанных здесь, или могут ли любые новые конструкции, которые являются очевидными для лиц с квалификацией в данной области после прочтения этой заявки, приводить к реакции устойчивости, специалист в данной области может выполнять быстрый тест транзиторной экспрессии, известный под названием АТТА (тест транзиторной экспрессии с использованием Agrobacterium tumefaciens). В этом анализе (подробное описание которого может быть найдено в Van den Ackerveken, G., et al., Cell 87, 1307-1316, 1996) нуклеотидную последовательность, кодирующую модифицированный белок, который должен быть испытан, помещают под контроль промотора CaMV 35S и вводят в штамм Agrobacterium, который используют также в протоколах для стабильной трансформации. После инкубирования этих бактерий с ацетосирингоном или любым другим фенольным соединением, о котором известно, что оно усиливает перенос Т-ДНК Agrobacterium, 1 мл культуры Agrobacterium инфильтрируют в лист растения in situ (например, из растения табака или картофеля) инъекцией, после чего эти растения помещают в оранжерею и инфицируют патогеном, предпочтительно P. infestans. Спустя 2-5 дней листья оценивают на появление симптомов устойчивости.
В данном изобретении авторы идентифицировали и выделили ген устойчивости Rpi-blb, который придает раса-неспецифическую устойчивость к P. infestans. Этот ген клонировали из генотипа Solanum bulbocastanum, который является устойчивым к P. infestans. Этот выделенный ген устойчивости данного изобретения может быть перенесен в чувствительное растение-хозяина с использованием опосредованной Agrobacterium трансформации или любым известным способом трансформации, и он может участвовать в придании этому растению-хозяину устойчивости к патогенам растений, в частности, к P. infestans. Растением-хозяином может быть картофель, томат или любое другое растение, в частности, член семейства Solanaceae, которое может быть инфицировано таким патогеном растения. Данное изобретение обеспечивает также последовательность нуклеиновой кислоты, кодирующую этот белок или его функциональный эквивалент, предпочтительно содержащий ген Rpi-blb, который изображен на фиг.6.
С использованием белка устойчивости Rpi-blb или его функционально эквивалентного фрагмента данного изобретения специалист имеет эффективное средство борьбы против патогенов растений, так как ген, кодирующий этот белок, может быть использован для трансформации чувствительных генотипов растений с получением таким образом генетически трансформированных растений, имеющих уменьшенную чувствительность или предпочтительно являющихся устойчивыми к патогену растения. В частности, растение, генетически трансформированное геном устойчивости Rpi-blb данного изобретения, имело уменьшенную чувствительность к P. infestans.
В предпочтительном варианте ген устойчивости Rpi-blb содержит кодирующую последовательность, приведенную на фиг.6А, или любую гомологичную последовательность или ее часть, перед которой находится промоторный район и/или за которой следует терминаторный район. Этот промоторный район должен быть функциональным в клетках растений и предпочтительно соответствовать природному промоторному району гена Rpi-blb. Однако может быть также использован промоторный район, который является функциональным в клетках-растениях, вместе с кодирующими последовательностями.
Кроме того, данное изобретение относится также к белку устойчивости Rpi-blb, который кодируется геном Rpi-blb данного изобретения и который имеет аминокислотную последовательность, приведенную на фиг.8, или ее функциональный вариант.
Сигнал, который запускает экспрессию гена устойчивости в S. bulbocastanum дикого типа или в трансгенных растениях данного изобретения, обусловлен, возможно, присутствием патогена, более конкретно, патогена P. infestans. Такие системы известны для других взаимодействий патоген-растение (Klement, Z., In: Phytopathogenic Prokaryotes, Vol. 2, eds.: Mount, M.S. and Lacy, G.H., New York, Academic Press, 1982, pp. 149-177), и эта система может быть использована для увеличения применимости белка устойчивости с получением устойчивости к большему числу патогенов (см. ЕР 474857). Эта система позволяет использовать соединение-«элиситор», полученное из патогена, соответствующий ген устойчивости, причем этот ген устойчивости при активации в присутствии элиситора мог бы приводить к локальной смерти клеток (реакция гиперчувствительности). В случае гена устойчивости данного изобретения соответствующий элиситорный компонент еще не был обнаружен, но авторы считают, что это может быть достигнуто специалистом в данной области. После выделения этого элиситорного компонента можно будет трансформировать ген, кодирующий указанный элиситор, вместе с геном, кодирующим белок устойчивости, в растение, посредством чего один из этих генов находится под контролем индуцируемого патогеном промотора. Эти промоторы хорошо известны в данной области (например, prp1, Fis1, Bet v 1, Vst1, gstA1 и промотор сесквитерпенциклазы, но может быть использован любой индуцируемый патогеном промотор, который включается после инфицирования патогеном). Если трансгенное растение содержит такую систему, то атака патогена, которая может запускать индуцируемый патогеном промотор, будет вызывать продуцирование компонента, который находится под контролем указанного промотора, и это, вместе с другим компонентом, экспрессируемым конститутивно, будет вызывать возникновение реакции устойчивости.
Можно также мутировать белок устойчивости, заставляя его находиться в активированном состоянии (см. ЕР 1060257). Поскольку это могло бы перманентно приводить к возникновению реакции устойчивости, которая в конечном счете ведет к локальной смерти клеток, следует заботиться о том, чтобы не произошла конститутивная экспрессия белка устойчивости. Это может выполняться помещением мутированного белка устойчивости под контроль индуцируемого патогеном промотора, который будет не только позволять экспрессию активного белка устойчивости только в моменты атаки патогена, но сделает возможным также более широкий спектр патогенов для индукции гиперчувствительной реакции. Мутация остатков треонина и серина в остатки аспарагиновой кислоты и глутаминовой кислоты часто приводит к активации, как показано в случае многих белков, активность которых модулируется фосфорилированием, например, в cлучае MAPK-активируемого белка (Engel et al., 1995, J. Biol. Chem. 270, 27213-27221) и в случае МАР-киназа-киназного белка (Huang et al., 1995 Mol. Biol. Cell 6, 237-245). С- и N-концевые, а также внутренние делеционные мутанты этих белков могут быть также испытаны на подходящие мутанты.
Более косвенный путь идентификации представляющих интерес мутантов, конститутивная активность которых является индуцированной, использует размножение кодирующей этот белок ДНК в так называемых «мутаторных» штаммах E. coli.
Быстрым способом тестирования всех полученных мутантов на их пригодность для индукции гиперчувствительной реакции является использование так называемого АТТА-анализа (Van den Ackerveken, G., et al., Cell 87, 1307-1316, 1996). Многочисленные мутанты могут быть подвергнуты скринингу без значительных усилий для идентификации мутантов, которые будут индуцировать HR после экспрессии.
Данное изобретение обеспечивает также вектор, содержащий нуклеиновую кислоту, описанную здесь, причем указанная нуклеиновая кислота кодирует генный продукт, который способен обеспечивать член семейства Solanaceae устойчивостью против патогена из класса оомицетов, или функционально эквивалентную выделенную или рекомбинантную нуклеиновую кислоту, в частности, когда указанный член включает в себя S. tuberosum или Lycopersicon esculentum.
Данное изобретение обеспечивает также клетку-хозяина, содержащую нуклеиновую кислоту или вектор данного изобретения. Пример указанной клетки-хозяина обеспечен здесь в подробном описании. В конкретном варианте указанная клетка-хозяин содержит клетку растения. В качестве клетки-растения предпочтительной является клетка, полученная из члена семейства Solanaceae, в частности, когда указанный член включает в себя S. tuberosum или Lycopersicon esculentum. Из такой клетки или протопласта, может возникать трансгенное растение, такое как трансгенное растение картофеля или растение томата, с устойчивостью против инфекции оомицетов. Таким образом, данное изобретение обеспечивает также растение или клубневидный корень, плод или семена или часть или потомство, полученные из них, содержащие клетку в соответствии с данным изобретением.
Кроме того, данное изобретение обеспечивает белковое вещество, проявляющее свойство обеспечения по меньшей мере частичной устойчивости к инфекции оомицетов, например, вызываемой P. infestans, при включении и экспрессии в растении или клетке-растения. В частности, обеспечено такое белковое вещество, которое кодируется нуклеиновой кислотой данного изобретения. В предпочтительном варианте данное изобретение обеспечивает белковое вещество, содержащее аминокислотную последовательность, изображенную на фиг.8, или ее функциональный компонент. Предпочтительно, такой функциональный компонент будет содержать одну или более из последовательностей, которые являются относительно уникальными относительно Rpi-blb в сравнении с RGC3-blb, RGC-blb и RGC4-blb. Такие последовательности могут быть обнаружены в сопоставлении (см. фиг.10А) и являются последовательностями RPLLGEM, AKMEKEKLIS, KHSYTHMM, FFYTLPPLEKFI, GDSTFNK, NLYGSGMRS, LQYCTKLC, GSQSLTCM, NNFGPHI, TSLKIYGFRGIH, IIHECPFLTLS, RICYNKVA и KYLTISRCN. Авторы считают, что одна или несколько из этих последовательностей обеспечивает функциональные свойства белка Rp1-blb.
Кроме того, данное изобретение обеспечивает связывающую молекулу, направленную на нуклеиновую кислоту данного изобретения. Например, ген Rpi-blb может быть использован для конструирования олигонуклеотидов, комплементарных одной цепи ДНК-последовательности, изображенной на фиг.7 и в таблице 2. Такие олигонуклеотиды, обеспеченные здесь, применимы в качестве зондов для скрининга библиотек, гибридизационных зондов для анализа по Саузерну/Нозерн-анализа, праймеров для ПЦР, для применения в диагностическом наборе для детектирования устойчивости к болезни и т.д. Такие олигонуклеотиды являются ценными фрагментами выделенной или рекомбинантной нуклеиновой кислоты, обеспеченной здесь, причем указанная нуклеиновая кислота кодирует генный продукт, который способен обеспечивать член семейства Solanaceae устойчивостью против гриба из класса оомицетов, или функционально эквивалентной выделенной или рекомбинантной нуклеиновой кислоты, в частности, когда указанный член включает в себя S. tuberosum или Lycopersicon esculentum. Они могут быть легко выбраны из последовательности, изображенной на фиг.6, или ее части. Конкретная точка узнавания содержит домен LRR, идентифицированный здесь. Такую связанную молекулу данного изобретения используют в качестве зонда или праймера, например, обеспеченного меткой, в частности, когда указанная метка содержит возбуждаемую часть, которая делает ее применимой в детектировании присутствия указанной связывающей молекулы.
Кроме того, данное изобретение обеспечивает способ для отбора растения или растительного материала или его потомства на его чувствительность или устойчивость к инфекции оомицетов, предусматривающий тестирование по меньшей мере части указанного растения или растительного материала или его потомства на присутствие или отсутствие нуклеиновой кислоты, причем указанная нуклеиновая кислота кодирует генный продукт, который способен обеспечивать член семейства Solanaceae устойчивостью против гриба из класса оомицетов, или на присутствие указанного генного продукта, причем указанный способ предпочтительно предусматривает контактирование по меньшей мере части указанного растения или растительного материала или его потомства со связывающей молекулой данного изобретения и определение связывания указанной молекулы с указанной частью. Указанный способ особенно применим, когда указанный оомицет включает в себя P. infestans, что позволяет отбирать растения или растительный материал на устойчивость против фитофтороза, например, когда указанное растение или материал включает в себя S. tuberosum. Авторы считают, что посредством анализа филогенетического дерева, обсуждаемого выше, белки, которые являются высокогомологичными относительно Rpi-blb и которые могли бы придавать устойчивость против патогенов растений, могут быть легко идентифицированы. Примером этого является детектирование трех высокогомологичных белков RGC1-blb, RGC3-blb и RGC4-blb, в отношении которых еще не было показано, что они придают устойчивость к P. infestans, но тем не менее считается, что они участвуют в устойчивости к патогенам в растениях.
Данное изобретение обеспечивает также применение нуклеиновой кислоты или вектора или клетки или вещества или связывающей молекулы данного изобретения в способе обеспечения растения или его потомства по меньшей мере частичной устойчивостью против инфекции оомицетов, в частности, когда указанный оомицет включает в себя P. infestans, особенно, когда указанное растение включает в себя S. tuberosum, причем указанный способ обеспечивает растение или его потомство по меньшей мере частичной устойчивостью против инфекции оомицетов, предусматривающий обеспечение указанного растения или его части геном, кодирующим белок устойчивости, или его функциональным фрагментом, содержащим нуклеиновую кислоту, причем указанный белок устойчивости способен обеспечивать член семейства Solanaceae устойчивостью против гриба из класса оомицетов, или обеспечение указанного растения или его части нуклеиновой кислотой или вектором или клеткой или веществом данного изобретения.
Кроме того, данное изобретение обеспечивает выделенный S. bulbocastanum или его часть, такую как клубень или семена, чувствительные к инфекции оомицетов, вызываемой P. infestans.
Далее данное изобретение представлено в виде подробного описания ниже.
ОПИСАНИЕ ФИГУР
Фиг.1. Географическая карта Мексики, указывающая происхождение поступлений S. bulbocastanum, используемых для выделения гена Rpi-blb. Буквы а, b и с указывают относительные географические происхождения используемых поступлений S. bulbocastanum.
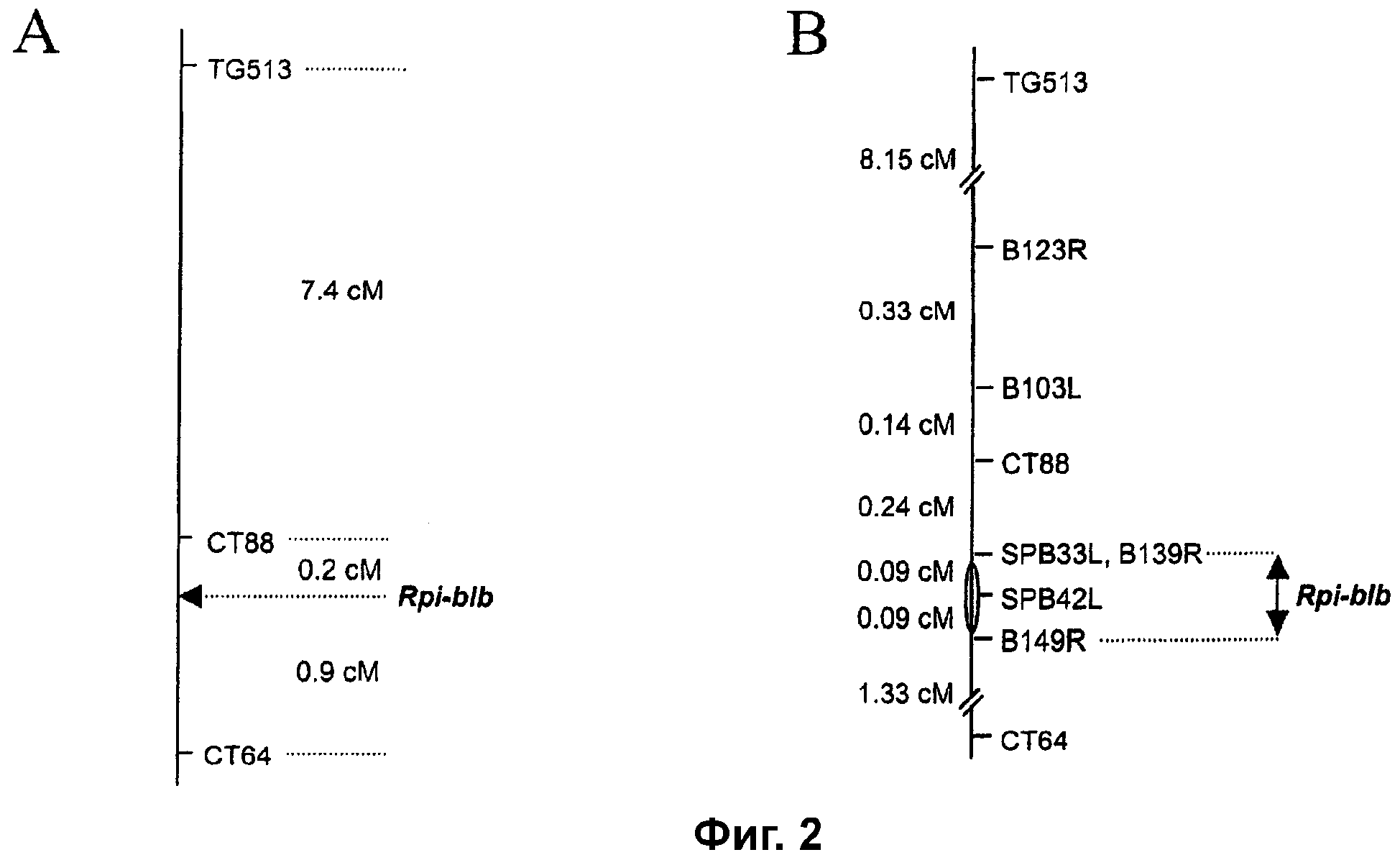
Фиг.2. Генетические карты сцепления локуса Rpi-blb на хромосоме 8 S. bulbocastanum. Горизонтальные линии указывают относительные положения маркеров, связанных с устойчивостью к фитофторозу. Расстояния между маркерами показаны в сантиморганах. А. Генетическое положение локуса Rpi-blb относительно маркеров TG513, СТ88 и СТ64 (n=508 генотипов). В. Генетическая карта сцепления высокой плотности локуса Rpi-blb (n=2109 генотипов).
Фиг.3. Физическая карта локуса Rpi-blb. А. Генетическая и физическая карта геномного района S. bulbocastanum, содержащего Rpi-blb. Вертикальные стрелки указывают относительные положения маркеров, сцепленных с устойчивостью. Числа выше горизонтальной линии указывают число рекомбинантов, идентифицированных между фланкирующими маркерами в 2109 растениях-потомках. Прямоугольники представляют клоны бактериальной искусственной хромосомы (ВАС). В. Относительные положения кандидатных генов для устойчивости к фитофторозу на ВАС SPB4. С. Схематическое представление структуры гена Rpi-blb. Горизонтальные линии указывают экзоны. Белые (открытые) блоки представляют кодирующую последовательность. Линии, идущие под углом вниз, указывают положение интронной последовательности длиной 678 нуклеотидов.
Фиг.4. Саузерн-блот-анализ ВАС-контига, охватывающего локус Rpi-blb. Названия над каждой дорожкой представляют названия ВАС-клонов. Показаны названия рестрикционных ферментов, используемых для расщепления ВАС-ДНК перед блоттингом по Саузерну.
Фиг.5. Анализы болезни на отделенных листьях. А. Устойчивый (левый), промежуточный (центральный) и чувствительный (правый) фенотипы, обнаруженные в картировании популяции В8 S. bulbocastanum спустя 6 дней после инокуляции (d.p.i.) капельками спорангиоспор P. infestans. В. Генетическая комплементация в отношении устойчивости к фитофторозу в картофеле. Характерные фенотипы болезни листьев, полученных из трансгенных растения картофеля, несущих RGC1-blb, RGC2-blb, RGC3-blb или RGC4-blb спустя 6 дней после инокуляции (d.p.i.) капельками спорангиоспор P. infestans. Генетические конструкции, несущие RGC, переносили в чувствительный сорт Impala опосредованной Agrobacterium трансформацией. С. Генетическая комплементация для устойчивости к фитофторозу в томате. Характерный фенотип болезни листьев, полученных из трансгенных растения томата, несущих Rpi-blb, спустя 6 дней после инокуляции (d.p.i.) капельками спорангиоспор P. Infestans (левая панель). Генетическую конструкцию, несущую Rpi-blb, переносили в чувствительный сорт томата Moneymaker опосредованной Agrobacterium трансформацией.
Фиг.6. Последовательности нуклеиновых кислот членов кластера генов Rpi-blb. А. Кодирующая последовательность нуклеиновой кислоты гена Rpi-blb. В. Кодирующая последовательность нуклеиновой кислоты гена Rpi-blb, включающая в себя интронную последовательность (положения 428-1106). С. Последовательность ВАС SPB4 геномного ДНК-фрагмента ScaI 5,2 т.п.н., генетической конструкции, используемой для генетической комплементации в отношении устойчивости к фитофторозу. Этот геномный фрагмент несет ген Rpi-blb, включающий в себя природные регуляторные элементы, необходимые для правильной экспрессии этого гена. Инициирующий кодон (положение ATG 1191-1193) и терминирующий кодон (положение ТАА 4781-4783) являются подчеркнутыми. D. Кодирующая последовательность нуклеиновой кислоты RGC1-blb, включающая в себя интронную последовательность (положение 428-708). Е. Кодирующая последовательность нуклеиновой кислоты RGC3-blb, включающая в себя интронную последовательность (положение 428-1458). F. Кодирующая последовательность нуклеиновой кислоты RGC4-blb, включающая в себя интронные последовательности (положения 434-510, 543-618 и 743-1365).
Фиг.7. Относительные положения праймеров. Горизонтальная черта представляет кодирующую последовательность гена Rpi-blb. Числа представляют положения нуклеотидов. Горизонтальные стрелки указывают относительные положения и ориентации праймеров. GSP1 и GSP2 представляют вмонтированные ген-специфические праймеры, используемые для 3'-RACE-экспериментов. GSP3 и GSP4 представляют вмонтированные ген-специфические праймеры, используемые для 5'-RACE-экспериментов. А(F), А(R), В(F) и В(R) являются праймерами, используемыми для амплификации гомологов Rpi-blb. Указано положение сайта рестрикции NsiI, используемого для получения обменов доменами между гомологами Rpi-blb.
Фиг.8. Расшифрованная последовательность белка Rpi-blb. Эта аминокислотная последовательность, расшифрованная из ДНК-последовательности Rpi-blb, разделена на три домена (А-С), как описано в примере 6. Гидрофобные остатки в домене А, которые образуют первый и четвертый остатки гептадных повторов потенциальных скрученных спиральных доменов. Консервативные мотивы в белках R написаны строчными буквами и курсивными буквами в домене В. Остатки, соответствующие консенсусу цитоплазматического LRR, показаны жирным шрифтом в домене С. Точки в этой последовательности были введены для сопоставления этой последовательности с консенсусной LRR-последовательностью цитоплазматических LRR.
Фиг.9. Анализ филогенетического дерева. А. Филогенетическое дерево последовательностей существующего уровня знаний, которые имеют некоторую степень гомологии с расшифрованной аминокислотной последовательностью Rpi-blb и членов кластера ее генов RGC1-blb, RGC3-blb и RGC4-blb. Это дерево было получено по способу Nieghbour-Joining Saitou and Nei (1987 Molecular Biology and Evolution 4, 406-425). Звездочка указывает, что этот ген был приписан функции. Кластер генов Rpi-blb показан в виде блока. В. Филогенетическое дерево последовательностей существующего уровня знаний, которые имеют некоторую степень гомологии с расшифрованной аминокислотной последовательностью Rpi-blb. В этот анализ включены Rpi-blb-гомологичные последовательности В149-blb, SH10-tub, SH20-tub и Т118-tar, последовательности, идентифицированные ПЦР-амплификацией с использованием праймеров, специфических для кластера генов Rpi-blb. С. Относительные положения ДНК-последовательностей существующего уровня знаний, которые обнаруживают существенную гомологию с частями последовательности гена Rpi-blb. Горизонтальные линии представляют относительные положения гомологичных последовательностей. Степень гомологии показана справа от каждой линии. Длина гомологичной последовательности указана над каждой линией.
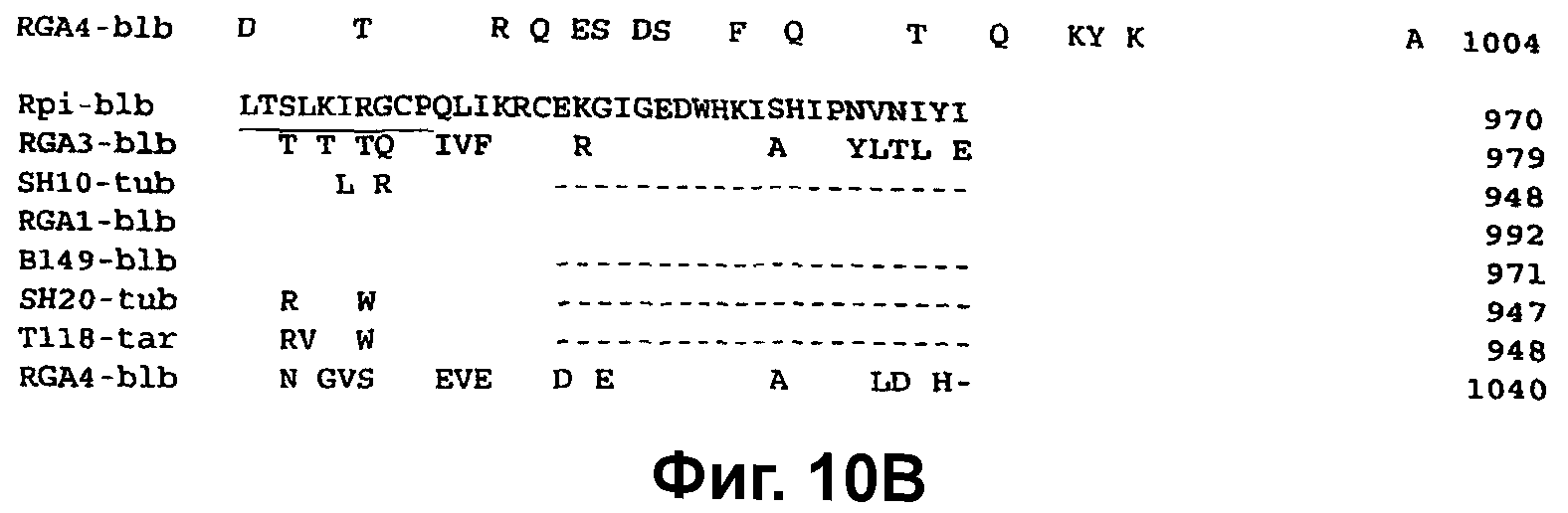
Фиг.10. Сопоставление предсказанного продукта гена Rpi-blb с предсказанными белковыми последовательностями гомологов Rpi-blb. А. Сопоставление расшифрованных белковых продуктов, кодируемых Rpi-blb, RGC1-blb, RGC3-blb и RGC4-blb. Показана полная аминокислотная последовательность Rpi-blb и аминокислотные остатки из RGC1-blb, RGC3-blb и RGC4-blb, которые отличаются от соответствующего аминокислотного остатка в Rpi-blb. Пунктиры указывают гэпы, вставленные для сохранения оптимального сопоставления. Аминокислотные остатки, которые являются специфическими для Rpi-blb при сравнении с остатками в соответствующих положениях в RGC1-blb, RGC3-blb и RGC4-blb, выделены жирным шрифтом. Районы LRR, которые соответствуют L..L..L.L..C/N/S..a..aP, подчеркнуты. Консервативные мотивы в NBS-домене показаны строчными буквами. В. Сопоставление расшифрованных белковых продуктов, кодируемых Rpi-blb, RGC1-blb, RGC3-blb, RGC4-blb, В149-blb, SH10-tub, SH20-tub и Т118-tar.
Фиг.11. Схематический обзор обменов доменами, выполненных между Rpi-blb и гомологами RGC1-blb и RGC3-blb. Вертикальная линия в виде точек указывает положение сайта NsiI, используемого для выполнения этих обменов. R и S указывают, являются ли трансгенные растения, несущие специфические химерные конструкции, устойчивыми или чувствительными к фитофторозному заражению соответственно.
Экспериментальная часть
Для картирования гена устойчивости была создана популяция для интраспецифического картирования S. bulbocastanum. Решающей стадией в этом процессе была идентификация чувствительных генотипов S. bulbocastanum. Для этой цели несколько поступлений S. bulbocastanum, происходящих из различных кластеров/зон в Мексике, анализировали на устойчивость или чувствительность к P. infestans в анализе на отделенных листьях (таблица 1 и фиг.1). Подвергнутые скринингу поступления BGRC 8008 и BGRC 7999 не содержали чувствительных генотипов. Однако в поступлениях BGRC 8005, BGRC 8006 и BGRC 7997 была обнаружена чувствительность в 9%, 7% и 14% анализируемых проростков соответственно. Затем был отобран чувствительный к P. infestans клон поступления BGRC 8006 и скрещен с устойчивым клоном поступления BGRC 8005. Полученную популяцию F1 использовали для картирования локуса Rpi-blb и в дальнейшем описании ее называют популяцией В8.
Первоначальный скрининг 42 генотипов В8 на устойчивость к P. infestans в анализе на отделенных листьях позволил предположить, что устойчивость к P. infestans в поступлении S. bulbocastanum 8005 могла быть обусловлена единственным доминантным геном R или тесно сцепленным кластером генов. Из 42 испытанных генотипов 22 оценивали как устойчивые и 16 как чувствительные в повторенном эксперименте. Фенотипы устойчивости остальных 4 проростков остаются неясными. Для определения хромосомного положения этой устойчивости S. bulbocastanum генотипы В8 с не вызывающим сомнений фенотипом использовали для маркерного анализа. Было обнаружено, что специфический маркер TG330 хромосомы 8 (таблица 2) является сцепленным в фазе отталкивания с устойчивым фенотипом, так как только один рекомбинант был получен между этим маркером и геном устойчивости в 12 генотипах В8. Кроме того, было обнаружено, что маркер СТ88 хромосомы 8 (таблица 2) был полностью сцепленным в фазе отталкивания с устойчивостью, что указывало на то, что локус, ответственный за устойчивость, обозначаемый как Rpi-blb, был локализован в этом районе хромосомы 8. По этой причине специфические для хромосомы 8 маркеры томата, которые картируются проксимально и дистально относительно СТ88 (TG513 и СТ64; Tanksley et al., 1992 Genetics 132: 1141-1160; таблица 2) делали маркерами CAPS и испытывали в 512 генотипах В8 с известными фенотипами устойчивости. В этом скрининге идентифицировали в целом пять СТ64-СТ88-рекомбинантных генотипов и 41 СТ88-TG513-рекомбинантный генотип (фиг.2А). Локус устойчивости Rpi-blb был картирован 1 событием рекомбинации, дистальным относительно маркера СТ88 (фиг.2А).
Картирование высокого качества локуса Rpi-blb проводили с маркерами CAPS, полученными из левой (L) и правой (R) пограничными последовательностями ВАС-клонов, выделенных из ВАС-библиотеки, полученной из устойчивого генотипа S. bulbocastanum BGRC 8005-8. ВАС-библиотеку сначала подвергали скринингу с использованием маркеров СТ88 и СТ64. ВАС-клоны, идентифицированные с использованием этих маркеров, использовали в качестве затравочных ВАС для последующей прогулки по хромосоме к локусу Rpi-blb. В целом 2109 генотипов В8 подвергали скринингу на рекомбинацию между маркерами TG513 и СТ64. Затем все рекомбинантные генотипы (219 из 2109) подвергали скринингу с использованием всех доступных маркеров в генетическом интервале СТ88-СТ64. Эти данные вместе с данными по устойчивости к болезни каждого рекомбинанта, полученными посредством анализов на отделенных листьях, позиционировали локус Rpi-blb между маркерами SPB33L и B149R, в генетическом интервале 0,1 сМ (4 из 2109 рекомбинантов), физически охватываемом перекрывающимися ВАС-клонами SPB4 и В49 (фиг.2b и 3). В пределах этого интервала устойчивость корасщеплялась с маркером SPB42L конца ВАС, последовательность которого была высокогомологичной частичным NBS-фрагментам из томата (например, Q194, Q137, Q152, Q153; Pan et al., 2000 Genetics 155: 309-322). Анализы по Саузерну ВАС-клонов, охватывающих интервал SP33L-B149R, с использованием32Р-меченых ПЦР-фрагментов маркера SPB42L в качестве зонда выявили присутствие по меньшей мере 4 копий этой подобной гену R последовательности в Rpi-blb-интервале (фиг.4). Кроме того, все из этих копий присутствовали на ВАС SPB4. Секвенирование и аннотирование полного инсерта этого ВАС-клона фактически идентифицировали четыре полных кандидата гена R (RGC1-blb, RGC2-blb, RGC3-blb и RGC4-blb) класса NBS-LRR генов R растений. ПЦР-маркер, который был локализован между RGC1-blb и RGC4-blb, выявил рекомбинацию между устойчивостью к P. infestans и RGC4-blb, исключающую возможность того, что RGC4-blb является Rpi-blb. Несмотря на это открытие, все четыре RGC были отобраны для анализа на комплементацию.
Геномные фрагменты приблизительно 10 т.п.н., несущие RGC1-blb, RGC2-blb, RGC3-blb или RGC4-blb, субклонировали из ВАС SPB4 в бинарный трансформирующий вектор pBINPLUS (van Engelen et al., 1995 Trans. Res. 4, 288-290) и переносили в чувствительный сорт картофеля при помощи стандартных способов трансформации. Первичные трансформанты испытывали на устойчивость к P. infestans, как описано в примере 1. Только генетическая конструкция, несущая RGC2-blb, была способна комплементировать чувствительный фенотип; 86% первичных трансформантов, несущих RGC2-blb, были устойчивыми (таблица 3), тогда как первичные трансформанты, содержащие RGC1-blb, RGC3-blb и RGC4-blb, были полностью чувствительными к P. infestans. Устойчивые трансформанты, содержащие RGC2-blb, обнаружили сходные фенотипы устойчивости с устойчивым родителем S. bulbocastanum (фиг.5). Таким образом, RGC2-blb был назван геном Rpi-blb, ДНК которого приведена на фиг.6.
ПРИМЕР 1: АНАЛИЗ БОЛЕЗНИ
Фенотип S. bulbocastanum и трансгенные генотипы S. tuberosum испытывали на устойчивость к P. infestans при помощи анализов на отделенных листьев. Листья из растений, выращиваемых в течение 6-12 недель в оранжерее, помещали в кусочках насыщенной водой применяемой флористами пены, приблизительно 35х4х4 см, и помещали в лоток (ширина 40 см, длина 60 см и высота 6 см) с перфорированным дном. Каждый лист инокулировали двумя капельками или более (каждая 25 мкл) раствора спорангиоспор на абаксиальной стороне. Затем этот лоток помещали в пластиковом мешке на верхнюю часть лотка, в который помещали насыщенную водой фильтровальную бумагу, и инкубировали в климатической камере при 17°С с фотопериодом день/ночь 16 ч/8 ч с флуоресцентным светом (Philips TLD50W/84HF). Спустя 6 дней эти листья оценивали на развитие симптомов болезни, вызываемой P. infestans. Растения с листьями, показавшими явно спорулирующие повреждения спустя 6 дней после инокуляции, считали растениями, имеющими чувствительный фенотип, тогда как растения с листьями, не обнаруживающими видимых симптомов или некроза на стороне инокуляции в отсутствие явной споруляции, считали устойчивыми. Этот анализ выполняли с комплексным изолятом 655-2А P. infestans (раса 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11), который получали из Plant Research International BV (Wageningen, The Netherlands).
ПРИМЕР 2: КАРТИРОВАНИЕ ЛОКУСА УСТОЙЧИВОСТИ Rpi-blb
Растительный материал
Для получения популяции интраcпецифического картирования, которая расщеплялась в отношении гена устойчивости к P. infestans, присутствующего в поступлении BGRC 8005 S. bulbocastanum (CGN 17692, PI 275193), требовался чувствительный генотип S. bulbocastanum. Несколько поступлений S. bulbocastanum, происходящих из различных кластеров/зон в Мексике, анализировали на устойчивость или чувствительность к P. infestans в анализе на отделенных листьях (таблица 1 и фиг.1). В поступлении BGRC 8008 и BGRC 7999 не обнаруживали чувствительности. В поступлении BGRC 8005, BGRC 8006 и BGRC 7997 чувствительность присутствовала только в 9%, 7% и 14% анализированных проростков соответственно. Таким образом, было получено только небольшое число чувствительных генотипов S. bulbocastanum.
Популяцию интраспецифического картирования S. bulbocastanum (В8) получали скрещиванием чувствительного клона поступления BGRC 8006 с устойчивым клоном поступления BGRC 8005. ДНК из 2109 растений-потомков экстрагировали из молодых листьев согласно Doyle and Doyle (1989 Focus 12, 13-15).
Анализ с маркерами CAPS
Для ПЦР-анализа готовили реакционные смеси по 15 мкл, содержащие 0,5 мкг ДНК, 15 нг каждого из праймеров, 0,2 мМ каждый из dNTP, 0,6 единиц Taq-полимеразы (15 Е/мкл, SphaeroQ, Leiden, The Netherlands), 10 мМ Трис-HCl рН 9, 1,5 мМ MgCl2, 50 мМ KCl, 0,1% Тритон Х-100 и 0,01% (мас./об.) желатин. ПЦР выполняли с использованием следующего профиля циклов: 25-секундная денатурация ДНК при 94°С, 30-секундный отжиг (см. таблицу 1) и 40-секундное удлинение при 72°С. В качестве первой стадии в ПЦР-амплификации ДНК денатурировали в течение 5 минут при 94°С и заканчивали дополнительной 5-минутной стадией элонгации при 72°С. Реакции амплификации проводили в термоциклере Biometra® T-Gradient или Biometra® Uno-II (Westburg, Leusden, The Netherlands). В зависимости от маркера, ПЦР-продукт расщепляли подходящей рестриктазой. Обзор маркеров, а также праймерных последовательностей, температур отжига и рестриктаз, приведен в таблице 2. Затем (расщепленные) ПЦР-продукты анализировали электрофорезом в агарозном или акриламидном геле. Для анализа в акриламидном геле использовали набор для анализа ДНК CleanGel DNA Analysis kit и набор для окрашивания ДНК серебром DNA Silver Staining kit (Amersham Pharmacia Biotech Benelux, Roosendaal, the Netherlands).
Генетическое картирование локуса Rpi-blb
Сначала небольшую группу из 42 растений-потомков популяции В8 подвергали скринингу на устойчивость к P. infestans в анализе на отделенных листьях. Растения с листьями, показавшими явно спорулирующие повреждения спустя 6 дней после инокуляции, считали растениями, имеющими чувствительный фенотип, тогда как растения с листьями, не обнаруживающими видимых симптомов или некроза на стороне инокуляции в отсутствие явной споруляции, считали устойчивыми. Из этих 42 проростков 22 были оценены как устойчивые и 16 как чувствительные. Фенотип остальных 4 проростков оставался неясным в этой начальной фазе. Эти данные показали, что устойчивость могла быть обусловлена единственным доминантным геном или тесно сцепленным кластером генов. Для определения хромосомного положения проростки с не вызывающим опасений фенотипом использовали для маркерного анализа. Было обнаружено, что маркер TG330 хромосомы 8 является сцепленным в фазе отталкивания с устойчивым фенотипом, так как только один рекомбинант был получен между этим маркером и геном устойчивости в 12 проростках В8. Кроме того, было обнаружено, что маркер СТ88 хромосомы 8 был полностью сцепленным в фазе отталкивания с устойчивостью, что указывало на то, что ген устойчивости был локализован на хромосоме 8.
Затем специфические маркеры хромосомы 8, которые были картированы проксимально и дистально относительно СТ88 (Tanksley et al., 1992 Genetics 132: 1141-1160), были разработаны в маркеры CAPS. Для более точного картирования этих маркеров другие 512 индивидуумов В8-популяции подвергали скринингу на устойчивость к фитофторозу при помощи анализа на отделенных листьях. Одновременно растения оценивали на маркеры СТ64, СТ88 и TG513. Для 5 проростков обнаруживали рекомбинацию между маркерами СТ64 и СТ88, тогда как 41 проросток были рекомбинантами между маркерами СТ88 и TG513 (фиг.2А). Ген устойчивости Rpi-blb был картирован между маркерами СТ64 и СТ88. На этой стадии позиционирование СТ88 проксимально относительно Rpi-blb было основано только на результате для одного рекомбинированного проростка.
Для более точного определения положения Rpi-blb относительно этих доступных маркеров выращивали другие 1555 проростков В8-популяции и анализировали на рекомбинацию между маркерами TG513 и СТ64. Таким образом, всего 2109 индивидуальных клонов потомства В8-популяции были подвергнуты скринингу. Рекомбинацию между маркерами TG513 и СТ64 обнаруживали в 219 из этих проростков (10,4 сМ). Все эти рекомбинанты подвергали скринингу с маркером СТ88 и фенотипировали на признак устойчивости с использованием анализа на отделенных листьях. В соответствии с более ранними результатами, ген Rpi-blb был картирован между маркерами СТ88 и СТ64 (фиг.2В).
ПРИМЕР 3: КОНСТРУИРОВАНИЕ ВАС-БИБЛИОТЕКИ S. bulbocastanum И КОНСТРУИРОВАНИЕ НЕПРЕРЫВНОГО ВАС-КОНТИГА, ОХВАТЫВАЮЩЕГО ЛОКУС Rpi-blb
Конструирование ВАС-библиотеки
Устойчивый клон поступления BGRC 8005 S. bulbocastanum (blb) (CGN 17692, PI 275193), гетерозиготный в отношении локуса Rpi-blb, использовали в качестве источника ДНК для конструирования геномной ВАС-библиотеки, далее называемой ВАС-библиотекой 8005-8. Получение высокомолекулярной ДНК и конструирование ВАС-библиотеки проводили, как описано в Rouppe van der Voort et al. (1999 MPMI 12:197-206). В конечном счете, получили 130000 клонов со средним размером инсерта 100 т.п.н., что соответствует 15 геномным эквивалентам. В целом приблизительно 83000 индивидуальных клонов хранили в 216 384-луночных микротитрационных планшетах (Invitrogen, The Netherlands), содержащих LB-буфер для замораживания (36 мМ К2НРО4, 13,2 мМ КH2PO4, 1,7 мМ цитрат, 0,4 мМ MgSO4, 6,8 мМ (NH4)2SO4, 4,4% (об./об.) глицерин, 12,5 мкг/мл хлорамфеникола в среде LB), при -80оС. Другие 50000 клонов хранили в виде бактериальных пулов, содержащих ~1000 белых колоний. Их получали соскребанием колоний из чашек с агаром в LB-среде, содержащей 18% глицерин и 12,5 мкг/мл хлорамфеникола, при помощи стерильного стеклянного спредера. Эти так называемые суперпулы также хранили при -80°С.
Скрининг ВАС-библиотеки и конструирование физической карты локуса Rpi-blb
ВАС-библиотеку 8005-8 сначала подвергали скринингу с использованием маркеров CAPS CТ88 и СТ64. Это выполняли следующим образом. Для первой части этой библиотеки из приблизительно 83000 клонов, хранящихся в 384-луночных микротитрационных планшетах, выделяли плазмидную ДНК с использованием стандартного протокола щелочного лизиса (Sambrook et al., 1989 in Molecular Cloning: A Laboratоry Manual, 2nd Edition, Cold Spring Harbor Press, Cold Spring Harbor, New York) из пулов бактерий каждого планшета с получением 216 планшетных пулов. Для идентификации индивидуальных ВАС-клонов, несущих маркеры CAPS, эти планшетные пулы подвергали скринингу при помощи ПЦР. После идентификации индивидуального планшетного пула как положительного в отношении конкретного маркера CAPS положительный ряд и положительный столбец в планшете идентифицировали с использованием двухмерного ПЦР-скрининга. Для этой цели «материнский» 384-луночный планшет реплицировали дважды на LB-среде, содержащей хлорамфеникол (12,5 мкг/мл). После выращивания колоний в течение 16 часов при 37оС один планшет использовали для выскребания 24 колоний из каждого ряда вместе, а другой планшет для выскребания 16 колоний из каждого столбца вместе. Бактерии каждого ряда или столбца ресуспендировали в 200 мкл ТЭ-буфера. Затем проводили анализ на маркеры CAPS на 5 мкл этих бактериальных суспензий, что приводило к идентификации отдельных положительных ВАС-клонов. Для второй части этой библиотеки, хранящейся в виде 50 пулов приблизительно 1000 клонов, плазмидную ДНК выделяли из каждого пула клонов с использованием стандартного протокола щелочного лизиса и проводили ПЦР для идентификации положительных пулов. Бактерии, соответствующие положительным пулам, разбавляли и высевали на чашки с LB-агаром, содержащим хлорамфеникол (12,5 мкг/мл). Затем индивидуальные белые колонии выскребали в 384-луночный планшет и затем отдельные положительные ВАС-клоны идентифицировали, как описано выше. Названия ВАС-клонов, выделенных из этих суперпулов, несут префикс SP (например, SPB33).
Размеры инсертов ВАС-клонов приближенно определяли следующим образом. Положительные ВАС-клоны анализировали выделением плазмидной ДНК из 2 мл ночных культур (LB-среда, дополненная 12,5 мг/мл хлорамфеникола) с использованием минипреп-протокола стандартного щелочного лизиса и ресуспендировали в 20 мкл ТЭ. Плазмидную ДНК (10 мкл) расщепляли 5 Е NotI в течение 3 часов при 37оС для отделения геномной ДНК от вектора pBeloBAC11. Расщепленную ДНК разделяли электрофорезом CHEF в 1% агарозном геле в 0,5х TBE при 4°С с использованием системы BIORAD CHEF DR II (Bio-Rad Laboratories, USA) при 150 В с временем постоянного импульса 14 с в течение 16 ч.
Скрининг ВАС-библиотеки 8005-8 маркером СТ88 идентифицировал два положительных ВАС-клона: В139 и В180, с инсертами ДНК картофеля 130 и 120 т.п.н. соответственно (фиг.3А). Расщепление ПЦР-продукта СТ88, полученного из этих ВАС-клонов и нескольких устойчивых и чувствительных растений-потомков популяции В8 для картирования, рестриктазой MboI, обнаружило, что ВАС139 нес аллель СТ88, который был сцеплен in cis с устойчивостью. Для идентификации относительного положения в геноме ВАС139, конструировали пары ПЦР-праймеров на основе последовательности правого (R) и левого (L) концов этого инсерта. Секвенирование ВАС-концов проводили, как описано в примере 4, с использованием 0,5 мкг ДНК ВАС в качестве матрицы. Были созданы полиморфные CAPS-маркеры расщеплением ПЦР-продуктов двух исходных генотипов популяции В8 и двух устойчивых и двух чувствительных генотипов-потомков с несколькими отрезающими 4 основания рестриктазами (таблица 2). Скрининг 37 СТ88-СТ164-рекомбинантных генотипов В8 картировал 5 из 7 СТ88-Rpi-blb-рекомбинантов между СТ88 и В139R, что свидетельствует о том, что маркер В139R был относительно более близким к локусу Rpi-blb, чем маркер СТ88. Скрининг 216 планшетных пулов с использованием В139R не приводил к идентификации положительные ВАС-клоны SPB33 и SPB42 с ДНК-инсертами 85 и 75 т.п.н. соответственно (фиг.3А). Скрининг полной ВАС-библиотеки с использованием SPB33L идентифицировал положительные ВАС-клоны В149 и SPB4. ВАС-клон SPB4 содержал аллель SPB33L, который был сцеплен in cis с устойчивостью, тогда как ВАС-клон В149 не содержал этот аллель. Однако скрининг панели СТ88-СТ64-рекомбинантов с использованием В149R обнаружил, что эта ВАС включала в себя локус Rpi-blb. В149R отделяли от локуса Rpi-blb двумя событиями рекомбинации (фиг.3А). Скрининг ВАС-библиотеки 8005-8 с использованием В149R идентифицировал ВАС-клон В49 как имеющий аллель В149R, который был сцеплен in cis с устойчивостью. Таким образом, этот ВАС-клон вместе с ВАС-клоном SPB4 образовывали ВАС-контиг, который включал в себя локус Rpi-blb (фиг.3).
ПРИМЕР 4: АНАЛИЗ ПОСЛЕДОВАТЕЛЬНОСТИ ВАС SPB4 И ИДЕНТИФИКАЦИЯ КАНДИДАТОВ ГЕНОВ УСТОЙЧИВОСТИ В ЛОКУСЕ Rpi-blb
В пределах интервала SPB33L-В149R устойчивость корасщеплялась с маркером SPB42L конца ВАС, последовательность которого была высокогомологична частичным NBS-фрагментам из томата (например, Q194, Q137, Q97, Q152, Q153; Pan et al., 2000 Genetics 155: 309-322). Анализы по Саузерну ВАС-клонов, охватывающих интервал SP33L-B149R, с использованием32Р-меченого ПЦР-фрагмента маркера SPB42L в качестве зонда выявили присутствие по меньшей мере 4 копий этой подобной гену R последовательности в Rpi-blb-интервале (фиг.4). Кроме того, все из этих копий присутствовали на ВАС SPB4. Таким образом, ДНК-последовательность ВАС-клона SPB4 определяли анализом последовательности по способу дробовика. Готовили ряд случайных клонов со средним размером инсерта 1,5 т.п.н. 10 мкг очищенной при помощи CsCl ДНК подвергали срезающему усилию в течение 6 секунд на льду при 6 микронах амплитуды в 200 мкл ТЭ с использованием ультразвукового дезинтегратора MSE soniprep 150. После осаждения этанолом и ресуспендирования в 20 мкл ТЭ концы этого ДНК-фрагмента репарировали посредством инкубирования с ДНК-полимеразой Т4 при 11°С в течение 25 минут в 50 мкл реакционной смеси, содержащей 1х буфер для ДНК-полимеразы Т4 (New England BioLabs, USA), 1 мМ ДТТ, 100 мкМ всех 4 dNTP и 25 Е ДНК-полимеразы Т4 (New England BioLabs, USA), с последующим инкубированием при 65°С в течение 15 минут. Затем обработанную срезающим усилием ДНК разделяли электрофорезом на 1% агарозном геле SeaPlaque LMP (FMC). Фракцию с размером 1,5-2,5 т.п.н. вырезали из геля и диализовали против 50 мл ТЭ в течение 2 часов при 4°С. Диализованные срезы агарозы переносили затем в пробирку Эппендорфа на 1,5 мл, расплавляли при 70оС в течение 5 минут, расщепляли 1 единицей GELASE (Epicentre Technologies, USA) на 100 мг агарозного геля в течение 1 часа при 45оС и затем осаждали ДНК. Фрагменты 1,5-2,5 т.п.н. лигировали при 16°С в расщепленный EcoRV и дефосфорилированный вектор pBlueScript SK+ (Stratagene Inc.). Затем эту смесь для лигирования использовали для трансформации ElectroMAX компетентных клеток E. coli DH10B (Life Technologies, UK) элекропорацией с использованием BioRad Gene Pulser. Установки на BioRad Gene Pulser были такими, как рекомендуемые для E. coli изготовителем. Клетки высевали распределением на чашках с агаром на бульоне Луриа-Бертани (LB), содержащем ампициллин (100 мкг/мл), 5-бром-4-хлор-3-индолил-β-D-галактозид (Xgal) (64 мкг/мл) и изопропил-1-тио-β-D-галактозид (IPTG) (32 мкг/мл). Чашки инкубировали при 37°С в течение 24 часов. Отдельные белые колонии выращивали в 96-луночных плоскодонных блоках (1,5 мл среды Terrific Broth, содержащей 100 мкг/мл ампициллина).
Плазмидную ДНК выделяли с использованием минипреп-системы QIAprep 96 Turbo вместе с BioRobot™ 9600 (QIAGEN) в соответствии с инструкциями изготовителя. Реакции секвенирования выполняли с использованием набора для секвенирования ABI PRISM BigDye™ Terminator cycle (Stratagene) в соответствии с инструкциями изготовителя. Все клоны секвенировали двунаправленно с использованием универсальных праймеров. Продукты секвенирования разделяли капиллярным электрофорезом на анализаторе ДНК Perkin Elmer ABI 3700.
Автоматическую сборку считываний (ДНК-фрагментов), полученных по способу “дробовика”, проводили с использованием программ Phred-Phrap (Ewing and Green, 1998 Genome Research 8, 186-194; Ewing et al., 1998 Genome Research 8, 175-185). Общее число 835 считываний обеспечивало полный охват ВАС-последовательности, равный 5х. Гэпы между контигами закрывали праймерной прогулкой или с использованием подхода комбинаторной ПЦР. Наконец, эту последовательность редактировали при качестве Phred 40 (1 ошибка на каждые 10000 нт) мануальным обследованием сборки с использованием редактора Gap4-contig editor и повторно секвенировали все районы низкого качества. Полная последовательность инсерта ВАС SPB4 состояла из 77283 нуклеотидов.
Анализ непрерывной последовательности ВАС SPB4 с использованием компьютерной программы GENSCAN (Burge and Karlin, 1997 J. Mol. Biol. 268, 78-94), GENEMARK (Lukashin and Borodovsky, 1998 NAR 26, 1107-1115) и BLASTX (Altschul et al., 1990 J. Mol. Biol. 215, 403-410) идентифицировал четыре полные кандидатные последовательности генов R (RGC1-blb, RGC2-blb, RGC3-blb и RGC4-blb), принадлежащие к классу NBS-LRR генов R растений. Маркер CAPS, сконструированный между RGC1-blb и RGC4-blb, маркер RGC4-blb, выявил рекомбинацию между устойчивостью к P. infestans и RGC1-4, что исключает возможность того, что RGC4-blb является Rpi-blb (фиг.3А и В). Несмотря на это открытие, все четыре RGC были отобраны для анализа на комплементацию.
ПРИМЕР 5: АНАЛИЗ НА КОМПЛЕМЕНТАЦИЮ
Субклонирование кандидатных генов и трансформациия в Agrobacterium tumefaciens
Геномные фрагменты приблизительно 10 т.п.н., несущие RGC1-blb, RGC2-blb, RGC3-blb или RGC4-blb, субклонировали из ВАС-клона SPB4 в бинарный трансформирующий вектор pBINPLUS (van Engelen et al., 1995 Trans. Res. 4, 288-290) и последующий отбор по размеру проводили следующим образом. Аликвоты ~ 1 мкг ДНК расщепляли 1 Е, 0,1 Е или 0,01 Е рестриктазы Sau3AI в течение 30 минут. Эту частично расщепленную ВАС-ДНК подвергали CHEF-электрофорезу при 4оС в 0,5 х ТВЕ с использованием линейно увеличивающегося времени импульса 1-10 секунд и силы поля 6 В/см в течение 16 часов. После электрофореза агарозный гель окрашивали бромидом этидия для локализации района геля, содержащего ДНК-фрагменты с размером приблизительно 10 т.п.н. Этот район вырезали из геля с использованием покровного стекла и диализовали против 50 мл ТЭ в течение 2 часов при 4оС. Затем диализованные срезы агарозы переносили в пробирку Эппендорфа на 1,5 мл, расплавляли при 70°С в течение 5 минут и расщепляли 1 единицей GELASE (Epicentre Technologies, USA) на 100 мг агарозного геля в течение 1 часа при 45оС. Лигирование отобранной по размеру ДНК с расщепленной BamHI и дефосфорилированной плазмидой pBINPLUS и последующую трансформацию ElectroMAX компетентных клеток E. coli DH10B (Life Technologies, UK) этой лигированной ДНК проводили, как описано в примере 5, с использованием для электропорации BioRad Gene Pulser. Клетки высевали распределением на чашках с агаром на бульоне Луриа-Бертани (LB), содержащем канамицин (50 мкг/мл), Xgal (64 мкг/мл) и IPTG (32 мкг/мл). Чашки инкубировали при 37°С в течение 24 часов. Отдельные белые колонии выращивали в 96-луночных планшетах (100 мкл LB-среды, содержащей 50 мкг/мл канамицина). В целом 480 клонов подвергали ПЦР-скринингу на присутствие RGC с использованием праймеров SPB42LF и SPB42LR или RGC4F и RGC4R (таблица 2). Положительные клоны отбирали для выделения плазмид и дополнительной характеристики. Идентификацию клонов, несущих RGC1-blb, RGC2-blb, RGC3-blb или RGC4-blb, проводили секвенированием ПЦР-фрагментов SPB42L, полученных из положительных клонов. Относительное положение этих RGC в субклоне определяли секвенированием концов этого клона и последующим сравнением этих последовательностей с полной последовательностью ВАС-инсерта. В конечном счете отобрали четыре бинарных плазмиды, рRGC1-blb, рRGC2-blb, рRGC3-blb и рRGC4-blb и их переносили в штаммы Agrobacterium tumefaciens AGL0 (Lazo et al., 1991 Bio/Technology 9, 963-967), LBA4404 (Hoekema et al., 1983 Nature 303: 179-180) или UIA143 (Farrand et al., 1989 J. of Bacteriology 171, 5314-5321) либо электропорацией с использованием BioRad Gene Pulser, либо конъюгацией. Установки на BioRad Gene Pulser были такими, как рекомендуемые для А. tumefaciens изготовителем. Конъюгацию проводили, как описано Simon et al. (1983 Bio/Tech. 1, 784-791). Клетки распределяли на чашках с агаром на бульоне Луриа-Бертани (LB), содержащем канамицин (100 мг/л) и рифампицин (50 мг/л). Чашки инкубировали при 28°С в течение 48 часов. Культуры небольшого масштаба из отобранных колоний выращивали в среде LB, содержащей канамицин (100 мг/л) и рифампицин (50 мг/л). Плазмидную ДНК выделяли, как описано выше, и целостность плазмид подтверждали рестрикционным анализом после повторного выделения из A. tumefaciens и последующей трансформации в E. coli. Культуры A. tumefaciens, несущие плазмиду с правильным распределением ДНК, использовали для трансформации чувствительного генотипа картофеля.
Трансформация чувствительного сорта картофеля
Штаммы A. tumefaciens выращивали в течение 2 дней при 28°С в 20 мл LB-среды, дополненной 50 мг/л рифампицина и 25 мг/л канамицина. Затем 0,2 мл культуры A. tumefaciens разбавляли в 10 мл LB-среды, содержащей те же самые антибиотики, и выращивали в течение ночи (28°С). Ночную культуру центрифугировали (30 минут, 2647 х g) и осадок ресуспендировали в 50 мл MS-среды (Murashige and Skoog, 1962 Physiol. Plant. 15, 473-497), дополненной 30 г/л сахарозы (MS30).
Сертифицированный семенной картофель сорта Impala очищали и поверхность стерилизовали в течение 30 минут в 1% растворе гипохлорита натрия, содержащем 0,1% Твина-20. Затем клубни промывали интенсивно большими объемами стерильной дистиллированной воды (4 раза, 10 минут). Диски с толщиной приблизительно 2 мм и диаметром 7 мм нарезали из цилиндров ткани клубней, полученных с использованием сверла для пробок. Диски клубней переносили в жидкую среду МS30, содержащую A. tumefaciens, и инкубировали в течение 15 минут. После удаления раствора A. tumefaciens диски клубней переносили на регенерационную среду, содержащую MS30, 0,9% мг/л IAA, 3,6 мг/л зеатинрибозида и 8 г/л агара (Hoekema et al., 1989 Bio/Technology 7, 273-278). Чашки инкубировали при 24°С, при длине дня 16 часов (Philips TLD50W/84HF). После 48 часов кокультивирования диски клубней промывали в течение 5 минут в жидкой среде MS30, содержащей антибиотики, 200 мг/л ванкомицина, 250 мг/л цефотаксима и 75 мг/л канамицина, и переносили на регенерационную среду, дополненную теми же самыми антибиотиками. Чашки инкубировали при 24°С, при длине дня 16 часов (Philips TLD50W/84HF). Каждые три недели диски клубней переносили на свежую среду. Укоренившиеся проростки размножали in vitro и тестировали на отсутствие клеток A. tumefaciens инкубированием среза стебля в 3 мл LB-среды (3 недели, 37°С, 400 об/мин). Одно растение каждого трансформированного регенеранта переносили в оранжерею.
Комплементация чувствительного фенотипа в картофеле
Первичные трансформанты испытывали на устойчивость к P. infestans, как описано в примере 1. Только генетическая конструкция, несущая RGC2-blb, была способна комплементировать чувствительный фенотип; 15 из 18 содержащих RGC2-blb первичных трансформантов были устойчивыми (таблица 3), тогда как все содержащие RGC1-blb, RGC3-blb и RGC4-blb первичные трансформанты были полностью чувствительными к P. infestans. Устойчивые трансформанты RGC2-blb показали сходные фенотипы устойчивости с устойчивым родителем S. bulbocastanum (фиг.5). Таким образом, RGC2-blb был назван геном Rpi-blb, ДНК-последовательность которого представлена на фиг.6.
Трансформация чувствительного томата
Семена чувствительной линии томата Moneymaker промывали в 70% этаноле до растворения оболочки семян и промывали стерильной водой. Затем эти семена поверхностно стерилизовали в 1,5% гипохлорите натрия в течение 15 минут, промывали три раза в стерильной воде и помещали в контейнеры, содержащие 140 мл MS-среды рН 6,0 (Murashige and Skoog, 1962 Physiol. Plant. 15, 473-497), дополненной 10 г/л сахарозы (MS10) и 160 мл вермикулита. Семена оставляли для прорастания на 8 дней при 25°С и освещении 0,5 В/м2.
Восьмидневные эксплантаты семядолей предварительно культивировали в течение 24 часов в чашках Петри, содержащих двухнедельные слои-фидеры клеток суспензии табака, посеянных на среду для кокультивации (MS30 рН 5,8, дополненную витаминами Nitsch (Duchefa Biochemie BV, Haarlem, The Netherlands), 0,5 г/л MES-буфера и 8 г/л агара Daichin).
Ночные культуры A. tumefaciens центрифугировали и осадок ресуспендировали в среде для суспензии клеток (MS30 рН 5,8, дополненной витаминами Nitsch, 0,5 г/л MES-буфера, рН 5,8), содержащей 200 мкМ ацетосирингон до конечной O.D.600 0,25. Затем эти эксплантаты инфицировали разбавленной ночной культурой штамма A. tumefaciens UIA143 (Farrand et al., 1989 J. of Bacteriology 171, 5314-5321), содержащего хелперную плазмиду pCH32 (Hamilton et al., 1996 PNAS 93, 9975-9979) и pRGC2-blb, в течение 25 минут, блоттировали (промакали) досуха на стерильной фильтровальной бумаге и кокультивировали в течение 48 часов на исходных чашках со слоями-фидерами. Условия культивирования были такими, как описанные выше.
После кокультивирования эксплантаты семядолей переносили на чашки Петри с селективной индуцирующей побеги средой (MS30 рН 5,8, дополненной 10 г/л глюкозы, включающей в себя витамины Nitsch, 0,5 г/л MES-буфера, 5 г/л агарового геля, 2 мг/л зеатинрибозида, 400 мг/л карбенициллина, 100 мг/л канамицина, 0,1 мг/л IAA) и культивировали при 25°С с освещением 3-5 В/м2. Эксплантаты субкультивировали каждые 3 недели на свежую среду. Появляющиеся побеги иссекали из лежащего ниже каллуса и переносили в контейнеры с селективной индуцирующей корни средой (MS10 рН 5,8, дополненной витаминами Nitsch, 0,5 г/л MES-буфера, 5 г/л агарового геля, 0,25 мг/л IBA, 200 мг/л карбенициллина и 100 мг/л канамицина).
Комплементация чувствительного фенотипа в томате
Для исследования, может ли Rpi-blb комплементировать чувствительный фенотип в томате, первичные трансформанты Moneymaker, несущие конструкцию гена Rpi-blb, сначала заражали полученными из картофеля изолятами P. infestans IPO655-2A и IPO428. Семь из девяти первичных трансформантов были устойчивыми (таблица 3). В связи с наблюдением, что испытанные изоляты P. infestans картофеля были менее вирулентными на томате, чем на картофеле, эти первичные трансформанты испытывали также с использованием изолята P. infestans, собранного из чувствительных растений томата приусадебного сада. Даже хотя этот изолят был значительно более вирулентным на Moneymaker, чем испытанные ранее изоляты, все 7 первичных трансформантов оставались устойчивыми. Эти результаты иллюстрируют потенциальную эффективность гена Rpi-blb не только против комплексных изолятов, полученных из картофеля, но также против изолятов, специализированных в отношении томата.
Молекулярный анализ первичных трансформантов
ОТ-ПЦР-анализ
Для получения кДНК смесь из 19 мкл, содержащая 1 мкг тотальной или полиА-РНК, 0,25 мМ каждого из dNTP, 50 мМ Трис-HCl рН 8,3, 75 мМ KCl,
3 мМ MgCl2, 10 мМ ДТТ и 530 нг олиго d(T)-праймера, GCTGTCAACGATACGCTACGTAACGGCATGACAGTG(T)18, денатурировали (1 мин 83°С). Затем эту смесь помещали при 42°С и добавляли 1 мкл обратной транскриптазы (M-MLV, Promega Benelux b.v., Leiden, The Netherlands). Спустя 60 минут смесь нагревали в течение 1 минуты при 99°С и переносили на лед. 2 мкл кДНК использовали для стандартной ПЦР.
Быстрая амплификация концов кДНК
5'- и 3'-концы кДНК Rpi-blb определяли быстрой амплификацией концов кДНК (RACE) с использованием набора GeneRacer™ (Invitrogen™, The Netherlands). 3'-RACE проводили с праймерами GSP1 (5'-GAGGAATCCATCTCCCAGAG) и GSP2 (5'-GTGCTTGAAGAGATGATAATTCACGAG) в комбинации с 3'-праймером GeneRacer™ и 3'-вмонтированным праймером GeneRacer™. 5'-RACE проводили на кДНК, синтезированной с праймером GSP3 (5'-GTCCATCTCACCAAGTAGTGG), с использованием праймеров GSP4 (5'-GAAATGCTCAGTAACTCTCTGG) и GSP5 (5'-GGAGGACTGAAAGGTGTTGG) в комбинации с 5'-праймером GeneRacer™ и 5'-вмонтированным праймером GeneRacer™ (фиг.7).
ПРИМЕР 6: СТРУКТУРА ГЕНА Rpi-blb И СООТВЕТСТВУЮЩЕГО БЕЛКА
Размер и структуру гена Rpi-blb определяли сравнением геномных последовательностей, полученных из инсерта pRGC2-blb, с кДНК-фрагментами, полученными 5'- и 3'-быстрой амплификацией концов кДНК. RACE идентифицировала 5'- и 3'-Rpi-blb-специфические кДНК-фрагменты единственного вида соответственно, что предполагает, что этот геномный клон кодирует единственный Rpi-blb-специфический транскрипт. Кодирующая последовательность транскрипта Rpi-blb равна 2913 нуклеотидов. Приближенно определено, что предположительный Rpi-blb-транскрипт имеет 3138 нуклеотидов (нт) и содержит 5'- и 3'-нетранслируемый район (UTR) длиной 44 и 181 нт соответственно. Ген Rpi-blb содержит единственный интрон 678 нт, начинающийся при 428 нт после стартового кодона трансляции ATG этого гена (фиг.3C).
Расшифрованная открытая рамка считывания гена Rpi-blb кодирует предсказанный полипептид из 970 аминокислот с приближенно оцениваемой молекулярной массой 110,3 кД (фиг.8). Несколько функциональных мотивов, присутствующих в генах R класса NBS-LRR генов R растений, являются очевидными в этом кодируемом белке, которые могут быть подразделены на 3 домена (А, В и С; фиг.8). N-концевая часть этого белка содержит потенциальные скрученные спиральные домены, гептадные повторы, в которых первый и четвертый остатки являются обычно гидрофобными (домен А). Домен В несет NBS и другие мотивы, которые составляют домен NB-ARC (ARC обозначает Apaf-1, R-белок и CED-4) белков R и регуляторов смерти клеток в животных (van der Biezen and Jones, 1998). Этот домен включает в себя мотивы Ар-АТФазы, присутствующие в белках эукариотического и прокариотического происхождения (Aravind et al., 1999 Trends Biochem. Sci. 24, 47-53). С-концевая половина Rpi-blb содержит группу из 19-20 неравномерных LRR (домен С). Эти LRR могут быть выстроены в соответствии с консенсусной последовательностью LxxLxxLxLxxC/N/SxxLxxLPxxa, где х обозначает любой остаток, а «а» обозначает положения алифатических аминокислот с последующим районом варьирующейся длины. Этот формат повторов приближается к консенсусу для цитоплазматических LRR (Jones and Jones, 1997 Adv. Bot. Res. 24, 89-167).
ПРИМЕР 7: ПРИРОДНЫЕ ГОМОЛОГИ И СИНТЕТИЧЕСКИЕ ВАРИАНТЫ ГЕНА Rpi-blb
Природные гомологи
Поиски гомологии с использованием алгоритма BLASTN с кодирующей ДНК-последовательностью гена Rpi-blb идентифицировали ряд последовательностей со значительной гомологией относительно коротких сегментов гена Rpi-blb (фиг.9С). Нуклеотиды 549-1245 кодирующей последовательности гена Rpi-blb имеют 81-90% идентичность последовательности с частичными NBS-фрагментами из томата (например, Q194, Q137, Q198 и Q199; (Pan et al., 2000 Genetics, 155:309-22). Эти гомологичные последовательности варьируются по их длине между 525 и 708 нуклеотидами и являются ПЦР-фрагментами, которые были идентифицированы системным сканированием генома томата с использованием (вырожденных) пар праймеров на основе повсеместных NBS-мотивов (Pan et al., 2000 Genetics, 155:309-22; Leister et al., 1996 Nat Genet. 14:421-429). Другой район гена Rpi-blb, который имеет значительную гомологию с последовательностью существующего уровня знаний, содержит нуклеотиды 76-805 этой кодирующей последовательности. Эта последовательность длиной 729 нт имеет 91% идентичность последовательности с EST из картофеля (номер по каталогу поступлений базы данных EMBL BG890602; фиг.9С). Последовательность гена Rpi-blb справа от нуклеотида 1245, содержащая район LRR, не имеет значимой гомологии ни с одной из искусственных последовательностей. Поиски гомологии BLASTX с кодирующей последовательностью гена Rpi-blb выявили, что гомология аминокислотной последовательности с различными искусственными генами не превышает 36% идентичности последовательности (таблица 4). Наилучшая оценка BLASTX была получена с геном NBS-LRR, полученным из Oryza sativa (36,5% идентичность аминокислотной последовательности). Гены NBS-LRR, имеющие общую гомологию последовательности (27-36% идентичность аминокислотной последовательности) с Rpi-blb, могут быть обнаружены, среди прочих, в Arabidopsis thaliana, Phaseolus vulgaris, Lycopersicon esculentum (кластер генов I2 Fusarium; Ori et al., 1997 Plant Cell, 9, 521-532; Simons et al., 1998 Plant Cell 10, 1055-1068), Zea mays, Hordeum vulgare и Lactuca sativa. Филогенетические исследования расшифрованных аминокислотных последовательностей Rpi-blb, RGC1-blb, RGC3-blb, RGC4-blb и последовательностей гомологичных генов существующего уровня знаний (определенных BLASTX), полученных из отличающихся видов, с использованием способа Neighbour-Joining Saitou and Nei (1987 Molecular Biology and Evolution 4, 406-425), показывают, что члены кластера генов Rpi-blb могут быть помещены в отдельную ветвь (фиг.9).
Сравнения последовательностей четырех RGC кластера генов Rpi-blb, идентифицированных на ВАС-клоне 8005-8 SPB-4, показывают, что гомология последовательностей в кластере генов Rpi-blb варьируется между 70% и 81% на уровне последовательностей аминокислот. Расшифрованная аминокислотная последовательность Rpi-blb имеет наивысшую общую гомологию с RGC3-blb (81% идентичность аминокислотной последовательности; таблица 4). При сравнении различных доменов ясно, что N-концевые половины этих белков (домены скрученных спиралей и NB-ARC) имеют более высокую степень гомологии (91% идентичность аминокислотной последовательности), чем С-концевые половины этих белков (LRR; 71% идентичность аминокислотной последовательности). N-конец белков NBS-LRR влияет на требование сигнальных компонентов, находящихся справа (ниже по ходу транскрипции), и, следовательно, считается, что он является предположительным эффекторным доменом (Feys and Parker, 2000 Trends Genet 16:449-55). С-концевой LRR-район участвует в соответствии с генетическими исследованиями в специфичности узнавания элиситора (Ellis et al., 2000 Trends Plant Sci. 5:373-379; Dodds et al., 2001 Plant Cell 13:163-78).
Сравнение всех четырех аминокислотных последовательностей выявило в целом 104 Rpi-blb-специфических аминокислотных остатков (фиг.10А). Большинство из них локализованы в LRR-районе (80/104). В этом последнем районе эти специфические остатки сконцентрированы в LRR-субдомене xxLxLxxxx. Относительная частота этих специфических аминокислотных остатков в этом LRR-субдомене является более чем в 2 раза более высокой (28,3%), чем частота, наблюдаемая в остальной части LRR-домена (12,3%). Считается, что остатки, расположенные около этих двух консервативных остатков лейцина в консенсусе xxLxLxxxx, экспонированы (обнажены) растворителем и, следовательно, участвуют, по-видимому, в создании/поддержании специфичности узнавания этого белка устойчивости.
Последовательности дополнительных гомологов гена Rpi-blb могут быть получены скринингом библиотек геномной ДНК или инсертов, например, ВАС-библиотек, праймерами на основе сигнатурных последовательностей гена Rpi-blb. Скрининг различных ВАС-библиотек Solanum наборами праймеров А и/или В (таблица 2 и фиг.7) идентифицировал другие гомологи Rpi-blb, полученные из различных видов S. bulbocastanum (В149-blb), S. tuberosum (SH10-tub и SH20-tub) и S. tаrijense (T118-tar). Сравнение всех 8 белковых последовательностей уменьшает число специфических аминокислотных остатков Rpi-blb до 51 (51/970; 5,25%) (фиг.10В). Большинство из них локализованы в LRR-районе (42/51; 82%). Относительная частота этих специфических аминокислотных остатков в субдомене LRR xxLxLxxxx в 3,3 раза выше, чем частота, наблюдаемая в остальном домене LRR (18,8% против 5,7% соответственно). Эти данные ясно предполагают, что эволюция специфичности устойчивости к P. infestans в кластере генов Rpi-blb происходит в основном посредством смещений в LRR-специфических остатках Rpi-blb.
Включение этих дополнительных гомологов Rpi-blb в вышеописанные анализы филогенетического дерева, с использованием способа Neighbour-Joining Saitou and Nei (1987 Molecular Biology and Evolution 4, 406-425), дополнительно оправдывает анализ филогенетического дерева в качестве способа для определения гомологичных последовательностей Rpi-blb (фиг.9В). Любой функциональный продукт R-гена, который имеет по меньшей мере 70% идентичность последовательности на уровне аминокислот, будет попадать в ту же самую ветвь, что и генные продукты кластера генов Rpi-blb, и, следовательно, может быть определен как гомолог Rpi-blb.
Синтетические варианты
Для определения роли различных последовательностей в устойчивости к P. infestans могут проводиться обмены доменами между различными гомологами. Например, рестриктаза NsiI, которая узнает ДНК-последовательность ATGCAT, присутствующую в консервативном мотиве MHD, может быть использована для обмена полного LRR-домена Rpi-blb с доменом RGC1-blb или RGC3-blb с использованием способов, известных специалистам с квалификацией в данной области. Получали химерные варианты гена Rpi-blb, которые кодируют N-концевую половину Rpi-blb и С-концевую половину RGC1-blb или RGC3-blb и наоборот, т.е. N-концевую половину RGC1-blb или RGC3-blb и С-концевую половину Rpi-blb (фиг.11). Эти варианты трансформировали в чувствительный генотип картофеля Impala и испытывали на устойчивость к P. infestans. Химерные гены RGC3-blb, содержащие LRR-домен Rpi-blb, были устойчивыми к P. infestans, что свидетельствует о том, что специфичность гена Rpi-blb кодируется этой частью данного гена.
Реферат
В растение или его часть вводят нуклеиновую кислоту, которая кодирует продукт, способный обеспечивать растение семейства Solanaceae и его потомство устойчивостью к поражению оомицетным грибком Phytophthora infestans. Это позволяет получать сорта картофеля и томата, устойчивые к фитофторозу. 11 н. и 17 з.п. ф-лы, 19 ил., 4 табл.
Комментарии