Новая мутантная карбамоилфосфатсинтетаза и способ продукции соединений - производных карбамоилфосфата - RU2264459C2
Код документа: RU2264459C2
Чертежи

Описание
Предпосылки к созданию изобретения
Область техники
Настоящее изобретение относится к микробиологической промышленности, в частности к способу продукции соединений - производных карбамоилфосфата. Более точно, настоящее изобретение касается использования нового фермента, устойчивого к ингибированию по типу обратной связи, вовлеченного в биосинтез аргинина и пиримидинов штаммами Е. coli - продуцентами соединений - производных карбамоилфосфата, таких как аргинин, цитруллин и пиримидины, включающие оротовую кислоту, уридин, уридин 5'-монофосфат (UMP), цитидин и цитидин 5'-монофосфат (СМР).
Описание предшествующего уровня техники
Карбамоилфосфатсинтетаза (CPSase) из E.coli катализирует комплексный синтез карбамоилфосфата (СР) из бикарбоната, глутамина и двух молекул Mg-ATP с высвобождением глутамата, фосфата и двух молекул Mg-ADP (Meister A., Advan. Enzymol. Mol. Biol., v.62, p.315-374, 1989). Синтез СР является промежуточным для двух путей биосинтеза, а именно для синтеза пиримидиновых нуклеотидов и синтеза аргинина. В первом пути СР связывается с аспартаткарбамоилтрансферазой (ATCase) с образованием в ходе двухстадийного процесса оротата. Оротат является важным промежуточным метаболитом для биосинтеза пиримидиновых производных, влючающих пиримидины, такие как урацил; пиримидиновые нуклеозиды, такие как оротидин, уридин и цитидин; и пиримидиновые нуклеотиды, такие как оротидин 5'-монофосфат (ОМР), UMP и СМР. Ранее было показано, что присутствие в питательной среде оротата в ходе ферментации широкого круга бактерий в значительной степени способствует продукции и накоплению пиримидиновых производных, в частности урацила (патент США 3214344). Во втором пути СР связывается с орнитином посредством орнитинкарбамоилтрансферазы (OTCase), включаясь в шестиступенчатый путь (начиная с глутамата) биосинтеза аргинина. CPSase активируется орнитином и инозинмонофосфатом (IMP) - предшественником пуриновых-нуклеотидов и ингибируется UMP. Карбамоилфосфатсинтетаза состоит из двух субъединиц.
Известно, что в коринеформных бактериях (ЕР 1026247 А1) и бактериях, принадлежащих к родам Escherichia и Bacillus, эти субъединицы кодируются генами carA и carB. Транскрипция оперона carAB кумулятивно репрессируется конечными продуктами обоих путей биосинтеза (Charlier D., et al., J.Mol.Biol., vol. 226, p. 367-386, 1992; Wang H., et al., J.Mol.Biol., vol. 277, p. 805-824, 1998; GlansdorffN., et al., Paths to Pyrimidines, vol. 6, p. 53-62, 1998). Природная CPSase является гетеродимером, состоящим из малой субъединицы весом 41,270 Да и большой субъединицы 117,70 Да, кодируемых генами carA и carB соответственно. Малая субъединица катализирует гидролиз глутамина и отвечает за перенос NH3 к большой субъединице, где, собственно, и происходит синтез СР. Большая субъединица содержит места связывания бикарбоната, аммиака, два различных места связывания Mg-ATP и 18 кДа С-концевую область, составляющую регуляторный домен (Rubio V., et al., Biochemistry, vol. 30, р. 1068-1075, 1991; Cervera J., et al., Biochemistry, vol. 35, p. 7247-7255, 1996). Недавно была описана кристаллическая структура аллостерически активированной формы CPSase (Thoden J., et al.. Biochemistry, vol. 36, p. 6305-6316, 1997; Thoden J., et al., Acta Crystallogr.Sec.D., vol. 55, p. 8-24, 1999). Первые три отчетливых домена большой субъединицы, обозначеные как А, В, С, являются очень похожими по структуре, но четвертый домен совершенно отличается от остальных. Домен D (остатки 937-1073) отвечает за связывание и аллостерическую регуляцию эффекторами: IMP, UMP и орнитин. Также было показано, что два остатка, серин 948 и треонин 1042, представляются ключевыми в аллостерической регуляции CPSase (Delannay S., et al., J.Mol.Biol., vol. 286, p. 1217-1228, 1999). Когда серин 948 замещается на фенилаланин, фермент становится нечувствительным к UMP и IMP, но все еще подвергается регуляции орнитином. Фермент с мутацией Т 10421 обладает пониженной чувствительностью к регуляции орнитином.
Как правило, ферменты с фенотипом устойчивости к ингибированию по типу обратной связи (fbr) возникают в результате замены аминокислотного остатка в одном или нескольких места аминокислотной последовательности и подобные замены приводят к понижению активности фермента. Например, замена Met-256 в сериновой ацетилтрансферазе (SAT) из E.coli (ген cysE) остальными 19 аминокислотными остатками в большинстве случаев приводит в возникновению fbr фенотипа, но мутантные белки SAT не обладают уровнем активности природной SAT (Nakamuri S. et al. AEM, vol.64, p. 1607-1611, 1998). Таким образом, недостатком мутантных ферментов, полученных подобным образом, является пониженная активность этих мутантных ферментов в сравнении с активностью природных ферментов.
Краткое описание изобретения
В настоящем изобретении описывается конструирование фермента, устойчивого к ингибированию по типу обратной связи и обладающего высокой активностью, играющего ключевую роль в биосинтезе пиримидинов и аргинина или цитруллина в Е. coli.
В настоящем изобретении предлагается новый процесс получения большого количества мутантных генов carB путем полной рандомизации фрагмента гена carB. Случайные замены некоторых аминокислотных остатков фрагмента аминокислотной последовательности, в котором могут находиться fbr мутации, могут привести к образованию мутантных белков с уровнем активности, близким к природному, вследствие более точного соответствия трехмерной структуры фермента. Таким образом было осуществлено настоящее изобретение, описанное ниже.
Что предоставляет настоящее изобретение:
(1) крбамоилфосфатсинтетаза, в которой последовательность аминокислот, соответствующая положениям с 947 по 951 в последовательности SEQ ID NO:20, заменена любой из последовательностей аминокислот SEQ ID NO:1 - SEQ ID NO:9, и в которой ингибирование UMP по типу обратной связи ослаблено;
(2) карбамоилфосфатсинтетаза по п.1, которая является карбамоилфосфатсинтетазой из Escherichia coli;
(3) карбамоилфосфатсинтетаза по п.1, в которой последовательность аминокислот в положении с 947 по 951 в последовательности SEQ ID NO:20 заменена любой из последовательностей аминокислот SEQ ID NO:1 - SEQ ID NO:9, и в которой ингибирование UMP по типу обратной связи ослаблено;
(4) карбамоилфосфатсинтетаза по п.1, которая включает делеции, замены, вставки или добавления одной или нескольких аминокислот в одном или нескольких положениях, кроме положений с 947 по 951, и в которой ингибирование UMP по типу обратной связи ослаблено;
(5) ДНК, кодирующая карбамоилфосфатсинтетазу по любому из пп.1-4, в которой ингибирование UMP по типу обратной связи ослаблено;
(6) бактерия, принадлежащая к роду Escherichia, которая трансформирована с помощью ДНК по п.5;
(7) бактерия по п.6, которая обладает способностью к продукции соединения, выбранного из группы, состоящей из L-аргинина, цитруллина и производных пиримидина;
(8) бактерия по 7, где производными пиримидина являются оротовая кислота, уридин, UMP, цитидин и СМР;
(9) способ продукции соединения, выбранного из группы, состоящей из L-аргинина, цитруллина и производных пиримидина, который включает стадии выращивания бактерии по любому из пп.6-8 в питательной среде с целью продукции и накопления указанного соединения в питательной среде, и выделения указанного соединения из культуральной жидкости; и
(10) способ по п.9, где производными пиримидина являются оротовая кислота, уридин, UMP, цитидин и СМР.
CPSase, содержащая любую из fbr мутаций, описанных выше, может упоминаться как «мутантная CPSase», ДНК, кодирующая мутантную CPSase, может упоминаться как «мутантные гены carA и carB», CPSase, не содержащая мутации, может упоминаться как «природная CPSase».
Далее настоящее изобретение будет разъяснено более детально.
<1> Мутантная CPSase и мутантные гены carA и car В.
Последовательная селекция и отбор рекомбинантных клонов, несущих мутантные гены carB, клонированные в экспрессирующем векторе как оперон carAB, позволяют выбрать fbr варианты мутантной CPSase с различным уровнем их биологической активности.
В соответствии с данными, полученными Delannay S. et al. (Delannay S., et al., J.Mol.Biol., v.286, 1217-1228, 1999), мутант (S948F) карбамоилфосфатсинтетазы из E.coli нечувствителен к UMP. Основываясь на этих данных, в качестве области для модификации была выбрана область CPSase, включающая позицию 948.
Мутантная CPSase и мутантный ген carB были получены в процессе мутагенеза путем рандомизации определенного фрагмента. Чтобы получить множественные мутации в гене carB, была проведена рандомизация фрагмента гена carB из 15 нуклеотидов, кодирующего область от Leu947 до Glu951 в последовательности аминокислот SEQ ID NO:20 (смотри ниже). Фрагмент из 15 нуклеотидов со случайной последовательностью может дать 412 или около 1,5·107 различных последовательностей ДНК, которые кодируют 4·105 различных остатков аминокислот в 5-мерном пептаде. Вероятность того, что в рамке считывания в этой последовательности не образуется стоп-кодон, составляет 0.95 или 95%. Таким образом, рандомизация фрагмента гена carB, кодирующего пептид от 947 по 951 аминокислотный остаток, должна дать приблизительно 4·105 различных последовательностей аминокислот со всем возможным многообразием этого пептидного фрагмента в структуре CPSase. Последовательная селекция и отбор рекомбинантных клонов, содержащих мутантные гены carB, клонированные в экспрессирующий вектор, позволяют отобрать fbr варианты мутантной CPSase с различным уровнем их биологической активности.
Последовательности аминокислот мутантной CPSase с подходящим fbr фенотипом были выявлены в настоящем изобретении. Вследствие этого, мутантная CPSase может быть получена на основе указанных последовательностей путем введения мутаций в природный ген car В с использованием традиционных методов. В качестве природного гена car В может быть упомянут ген car В из E.coli (нуклеотиды с 10158 по 133 79 в последовательности Genbank с инвентарным номером АЕ000113 U00096: SEQ ID NO: 19).
Последовательностью аминокислот с 947 по 951 в мутантной CPSase согласно настоящему изобретению является любая из последовательностей с SEQ ID NO:1 no 9. Соответствующая последовательность аминокислот известной fbr CPSase, в которой серин в положении 948 заменен на фенилаланин, а также природная CPSase из E.coli представлены в Таблице 1. Примеры последовательностей нуклеотидов, кодирующих указанные последовательности аминокислот, также приведены в Таблице 1.
Мутантная CPSase может содержать делеции, замены, вставки и добавления одной или нескольких аминокислот в одном или нескольких положениях, кроме положений с 947 по 951, при условии, что активность CPSase не нарушается. Термин «активность CPSase» означает активность по катализу реакции комплексного синтеза карбамоилфосфата из бикарбоната, глутамина и двух молекул Mg-ATP. Число «нескольких» аминокислот различно в зависимости от положения или типа остатка аминокислоты в трехмерной структуре белка. Это объясняется следующими причинами. Например, некоторые аминокислоты являются в достаточной степени взаимозаменяемыми и отличия в этих аминокислотах не влияют в значительной степени на трехмерную структуру белка. Следовательно, мутантной CPSase согласно настоящему изобретению может быть мутантная CPSase, у которой степень гомологии не ниже чем 30-50%, предпочтительно 50-70%, по отношению ко всем остаткам аминокислот, составляющим CPSase согласно настоящему изобретению, и которая обладает fbr активностью CPSase.
В настоящем изобретении «последовательность аминокислот, соответствующая положениям с 947 по 951», означает последовательность аминокислот, соответствующую последовательности аминокислот в положении с 947 по 951 в последовательности аминокислот SEQ ID NO:20. Положение остатка аминокислоты может быть изменено. Например, если какой-либо остаток аминокислоты добавлен в N-концевой участок, то остаток аминокислоты, находившийся ранее в положении 947, оказывается в положении 948. В таком случае остаток аминокислоты, соответствующий первоначальному положению 947, рассматривается как остаток аминокислоты в положении 947 согласно настоящему изобретению.
Фраза «ингибирование UMP по типу обратной связи ослаблено» означает, что степень ингибирования по типу обратной связи снижена. Снижение степени ингибирования по типу обратной связи может быть определено путем измерения снижения активности CPSase в присутствии UMP и последующего его сравнения со снижением активности белка, обладающего последовательностью аминокислот SEQ ID NO:20. Более того, «ингибирование UMP по типу обратной связи ослаблено» означает, что существенное ослабление ингибирования является достаточным и полная утрата чувствительности не является необходимой.
ДНК, кодирующая практически такой же белок, как мутантная CPSase, описанная выше, может быть получена, например, путем модификации последовательности нуклеотидов методом сайт-специфического мутагенеза таким образом, что белок, кодируемый подобной ДНК, будет в определенном положении содержать делеции, замены, вставки или добавления одного или нескольких остатков аминокислот. - ДНК, модифицированная описанным выше способом, может быть получена традиционным способом мутагенеза. К мутагенезу относится метод обработки ДНК, содержащей ген carB, in vitro, например, с помощью гидроксиламина, и метод обработки микроорганизма, например, бактерии, принадлежащей к роду Escherichia и содержащей ген carB, УФ-светом или мутирующим агентом, таким как N-метил-N'-нитро-N-нитрозогуанидин (NTG) и азотистая кислота, обычно используемым для подобного рода обработок.
К делениям, заменам, вставкам или добавлениям нуклеотидов, описанным выше, относятся мутации, которые встречаются в природных условиях (мутанты или варианты), например, в случае индивидуальных или родовых и видовых различий бактерий, содержащих CPSase.
ДНК, кодирующая практически такой же белок, как мутантная CPSase, может быть получена путем выделения ДНК, которая гибридизуется с ДНК, содержащей последовательность известного гена carB или его часть, используемой в качестве зонда в жестких условиях, и которая кодирует белок, обладающий активностью CPSase, из клетки, содержащей мутантную CPSase, являющейся объектом мутагенеза.
Термин «жесткие условия» обозначает условия, при которых образуется так называемый специфический гибрид (дуплекс), а неспецифический - не образуется. Четко описать эти условия с помощью численных значений довольно трудно. Однако, например, жесткими условиями являются такие условия, при которых молекулы ДНК, обладающие высокой гомологией, например, ДНК, обладающие гомологией друг с другом не менее 50%, гибридизуются, а ДНК с меньшей гомологией - не гибридизуются. В качестве альтернативы, примером жестких условий являются условия, при которых ДНК гибридизуются друг с другом при концентрации солей, соответствующей обычной концентрации при отмывке в ходе гибридизации по Саузерну, т.е. 60°С, 1×SSC, 0.1% SDS, предпочтительно 0.1×SSC, 0.1% SDS.
В число генов, которые гибридизуются в описанных выше условиях, включаются гены, содержащие стоп-кодон внутри кодирующего участка гена, а также те, которые кодируют неактивный белок вследствие мутаций в активном центре. Однако подобные затруднения могут быть легко разрешены путем лигирования гена в коммерчески доступный вектор для экспрессии и изучения в экспрессированном белке активности CPSase.
<2> Бактерия, принадлежащая к роду Escherichia, согласно настоящему изобретению.
Бактерией, принадлежащей к роду Escherichia, согласно настоящему изобретению является бактерия, принадлежащая к роду Escherichia, в которую введен мутантный ген carB, описанный выше. Примером бактерии, принадлежащей к роду Escherichia, является Е. coli. Мутантный ген carB может быть введен, например, путем трансформации бактерии, принадлежащей к роду Escherichia, рекомбинантной плазмидой, содержащей вектор, функционирующий в бактерии, принадлежащей к роду Escherichia, и мутантный ген carB. Мутантный ген carB также может быть введен заменой гена carB на мутантный ген car В в хромосоме.
Примерами векторов, которые можно использовать для введения мутантного гена car В, являются плазмидные векторы, такие как pBR322, pMW118, pUC19 или подобные им, фаговые векторы, такие как 11059,1BF101, M13mp9 или подобные им, и транспозоны, такие как Mu, Tn10, Тn5 или подобные им.
Введение ДНК в бактерию, принадлежащую к роду Escherichia, может быть осуществлено, например, по методу D.A. Morrison (Methods in Enzymology, 68, 326 (1979)) или методом, в котором бактериальная клетка - реципиент обрабатывается хлоридом кальция для увеличения проницаемости для ДНК (Mandel, M. and Higa, A., J.Mol.Biol. 53, 159 (1970)) или подобным им методом.
Количество продуцируемого соединения - производного карбамоилфосфата, такого как L-аргинин, цитруллин и производные пиримидина, может быть увеличено путем введения мутантного ген carB в бактерию - продуцент, принадлежащую к роду Escherichia. Кроме того, способность к продукции соединения - производного карбамоилфосфата, такого как L-аргинин, цитруллин и производные пиримидина, может быть придана бактерии, в которую мутантный ген carB уже введен. Производными пиримидина являются оротовая кислота, уридин, UMP, цитидин и СМР.
Примерами бактерий, принадлежащих к роду Escherichia, обладающих способностью к продукции L-аргинина, являются штаммы АJ1 1531 и AF11538 (JP56-106598А2), АJ11593 (FERM P-5616) и АJ1 1594 (FERM P-5617) (выложенная заявка Японии №57-5693), ВКПМ В-7925 (Российская патентная заявка№2000117677). Штамм ВКПМ В-7925 был депонирован во Всероссийскую коллекцию промышленных микроорганизмов (ВКПМ) 10 апреля 2000.
В настоящее время не известны примеры бактерий, принадлежащих к роду Escherichia, и обладающих способностью к продукции L-цитруллина и уридин-5'-монофосфата (UMP).
Примерами бактерий, принадлежащих к роду Bacillus, обладающих способностью к продукции L-цитруллина, являются штаммы B.siibtilis К-Х-1 А-1 (АТСС No 15561) и К-К-1 А-9 (АТСС No 15562) (патент США 3,282,794), штамм Bacillus sp. cit-70 (выложенная заявка Японии №08-089269). Примерами бактерий, принадлежащих к роду Brevibacterium, обладающих способностью к продукции L-цитруллина, являются штаммы Brevibacterium flavum AJ3408 (FERM Р-1645) (патент США 5164307) и АJ1 1677 (выложенная заявка Японии №57-163488). Примером коринеформной бактерии, обладающей способностью к продукции L-цитруллина, является штамм Corynebacterium glutamicum АJ1 1588 (FERM P-5643) (патент США 5164307).
Примерами коринеформных бактерий, обладающих способностью к продукции UMP, являются штаммы Corynebacterium ammoniagenes LK40-2 (ВКПМ В-7811), LK75-15 (ВКПМ В-7812), и LK75-66 (ВКПМ В-7813) (Российская патентная заявка №99122774).
<3> Способ продукции L-аргинина, цитруллина и производных пиримидина.
Такие соединения, как L-аргинин, цитруллин и производные пиримидина, могут быть получены с высокой эффективностью при выращивании бактерии, в которую введен мутантный ген carB и которая обладает способностью к продукции указанных соединений, в питательной среде, продукции и накоплении указанных соединений в питательной среде и выделения указанных соединений из культуральной жидкости. Производными пиримидина являются оротовая кислота, уридин, UMP, цитидин и СМР.
В способе согласно настоящему изобретению выращивание бактерии, принадлежащей к роду Escherichia, накопление и выделение указанных соединений из культуральной жидкости может быть осуществлено способом, подобным способу, традиционно используемому для продукции L-аргинина методом ферментации с использованием бактерий. Питательная среда для выращивания может быть как синтетической, так и натуральной, при условии, что она содержит источники углерода и азота, минеральные соединения и, если необходимо, питательные добавки в количестве, необходимом для роста бактерии. Источники углерода включают в себя различные углеводы, такие как глюкоза и сахароза, и различные органические кислоты, в зависимости от способности к их усвоению используемыми бактериями. Могут быть использованы спирты, такие как этанол и глицерин. В качестве источников азота используются аммиак, различные соли аммония, такие как сульфат аммония, другие соединения азота, такие как амины, природные источники азота, такие как пептон, гидролизат соевых бобов и микробный ферментализат. В качестве минеральных соединений используются однозамещенный фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, карбонат кальция.
Выращивание проводят предпочтительно в аэробных условиях, таких как взбалтывание и аэрация с перемешиванием. Температура культуры обычно поддерживается от 20°С до 40°С, предпочтительно от 30°С до 38°С. Значение рН находится в пределах от 5 до 9, предпочтительно в пределах от 6.5 до 7.2. рН среды может быть скорректировано аммиаком, карбонатом кальция, различными кислотами, основаниями и буферами. Обычно выращивание в течение от 1 до 3 дней приводит к накоплению соединения в культуральной жидкости.
Выделение соединения может быть осуществлено путем удаления из питательной среды после выращивания нерастворимых компонент, таких как клетки, методом центрифугирования или фильтрации на мембране, с последующим выделением и очисткой соединения методами ионного обмена, концентрации и фракционной кристаллизации или подобными им.
Краткое описание чертежей.
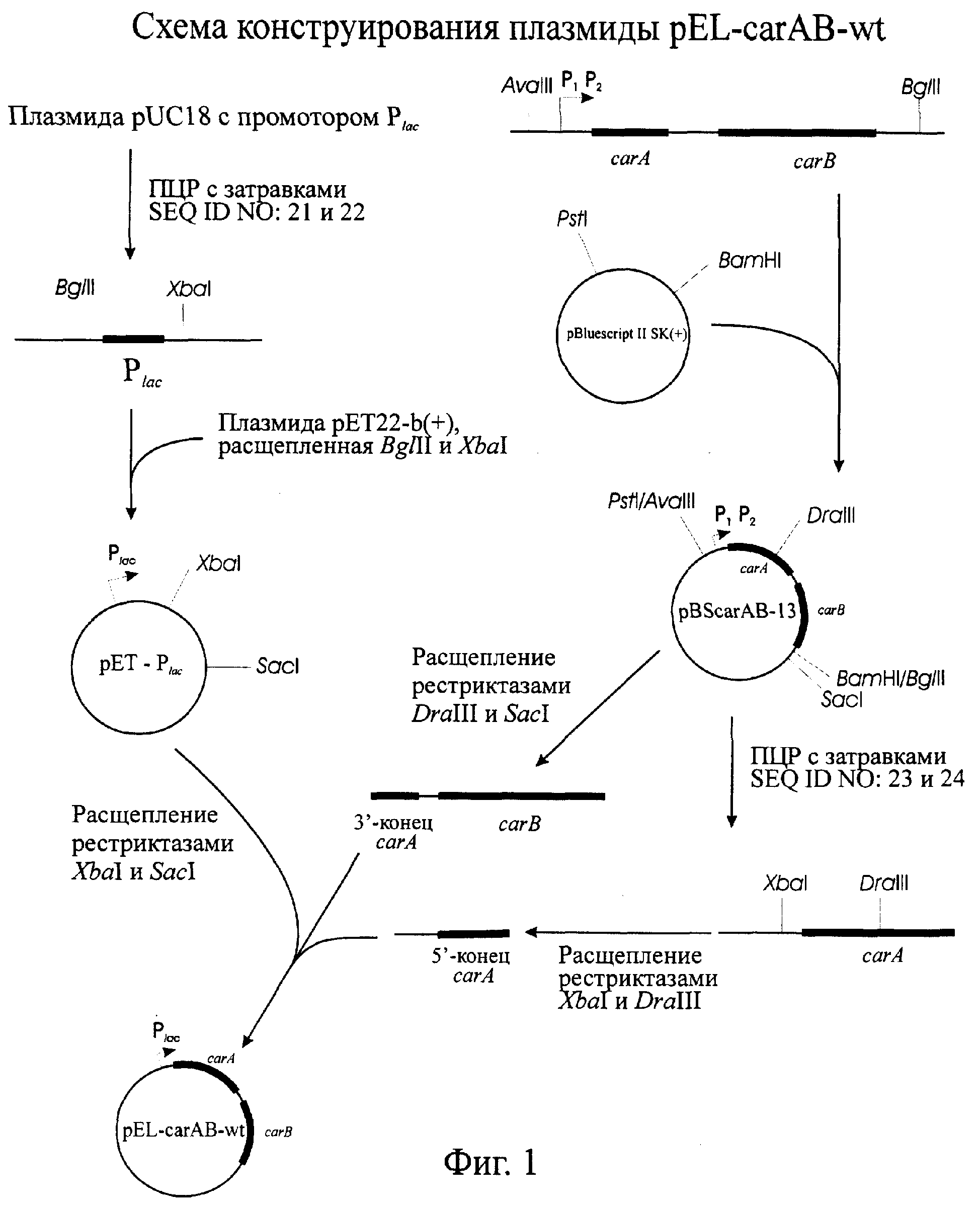
На Фиг.1 показана схема конструирования плазмиды pEL-carAB-wt.
На Фиг.2 показана схема конструирования набора мутантных генов carB.
Наилучший способ осуществления изобретения.
Настоящее изобретение будет детальнее разъяснено со ссылкой на следующие примеры и последовательности, которые приведены в конце описания.
Пример 1.
Плазмида pBScarAB-13, содержащая природные гены carAB из E.coli, была сконструирована путем клонирования AvaIII-BglII фрагмента ДНК (4911 п.o.) из хромосомы Е. coli в вектор pBluescript II SK(+) (Fermentas, Литва), предварительно обработанный рестриктазами BamHI и PstI.
Плазмида рЕТ22-b(+) (Novagen, США) была модифицирована путем замены промотора Т7 на lac промотор. lac промотор был получен методом амплификации в ходе ПЦР с использованием плазмиды pUC18 в качестве матрицы и олигонуклеотидов 5'-accagatctgcgggcagtgagcgcaacgc-3' (SEQ ID NO: 21) и 5'-gtttctagatcctgtgtgaaattgttatccgc-3'(SEQ ID NO: 22) в качестве затравок. Полученный фрагмент (0.14 kb), содержащий lac промотор, был расщеплен рестриктазами BglII и XbaI и слонирован в вектор рЕТ22-b(+), предварительно обработанный теми же рестриктазами. Полученная плазмида pET-Plac использовалась для клонирования генов сагАВ без промотора из плазмиды pBScarAB-13.
5'-концевой фрагмент гена car A (1.18 kb) был получен путем амплификации в ходе ПЦР с использованием плазмиды pBScarAB-13 в качестве матрицы и олигонуклеотидов 5'-cctctagaaataaagtgagtgaatattc-3'(SEQ ID NO: 23) и 5'-cttagcggttttacggtactgc-3'(SEQ ID NO:24) в качестве затравок. Полученный фрагмент был расщеплен рестриктазами XbaI и DraIII u XbaI - DraIII фрагмент (0.61 kb), содержащий 5'-концевую последовательность гена carA, был очищен методом электрофогеза в агарозном геле. Смесь двух фрагментов, XbaI-DraIII. и DraII-SacI фрагмента плазмиды pBScarAB-13 (содержащей 3'-концевую последовательность гена carA и ген carB) и вектора pET-Plac, расщепленного рестриктазами XbaI и SacI, была лигирована и использована при трансформации клеток E.coli TG1. Полученная рекомбинантная плазмида рЕТ-carAB-wt содержала последовательность природного оперона carAB под контролем lac промотора.
Для амплификации в ходе ПЦР использовали ДНК полимеразу TaKaRa La, полученную от Takara Shuzo Co. (Япония), в условиях, рекомендованных производителем.
<1> Мутагенез путем рандомизации определенного фрагмента.
В качестве матрицы для ПЦР была использована плазмида pBScarAB-13, в качестве 5'-затравки был использован пример Р1: 5'-ggtcgtgcgctgNN(T/C)N(T/C)CNN(T/C)NNN(G/A)NNggcgataaagaacgcgtggtg -3' (SEQ ID NO:25) (48 нуклеотидов), сконструированный на основании последовательности нуклеотидов гена car В, в качестве 3'-затравки - стандартная затравка М13. Фиксированный 5'-участок из 12-нуклеотидов и фиксированный 3'-участок из 21 нуклеотида затравки Р1 являются гомологичными последовательности гена car В перед кодоном Leu947 и после кодона Glu951 соответственно.
Фрагмент ДНК длиной 0,75 т.п.о. (3'-конец гена carB) был синтезирован в процессе первого раунда ПЦР (15 циклов) с использованием затравки Р1 с областью из 15 нуклеотидов со случайной последовательностью.
Первый раунд ПЦР был осуществлен следующим образом. 100 нг плазмиды pBScarAB-13 были добавлены в качестве матрицы в раствор для ПЦР (50 мкл), содержащий обе затравки в концентрации 10 пмоль. Было проведено 15 циклов ПЦР (94°С в течение 15 сек, 52°С в течение 20 сек, 72°С в течение 1 мин) на термоциклере ДНК модели 2400 (Perkin-Elmer Co., Foster City, USA).
Во втором раунде амплификации были проведены следующие 15 циклов (94°С в течение 1 мин, 35°С в течение 1 мин, 72°С в течение 2 мин), в ходе которых (-)-цепь полученного фрагмента функционировала в качестве «затравки» для того, чтобы нарастить этот фрагмент до полной последовательности гена.
В третьем раунде ПЦР 10 мкл полученной реакционной смеси были добавлены к 90 мкл свежего раствора, содержащего 100 пмоль 5'-затравки - М13, и затравки Р2: 5'-ccacttcctcgatgacgcgg-3' (SEQ ID NO: 26) в качестве 3'-затравки, и затем были проведены дополнительные 15 циклов (94°С в течение 0.5 мин, 55°С в течение 20 сек, 72°С в течение 2 мин).
Фрагменты ДНК длиной 1,32 т.п.о., кодирующие набор мутантных вариантов 3"-концевого фрагмента гена carB, были очищены методом электрофореза в агарозном геле, затем расщеплены рестриктазами AflII и SacI, а затем лигированы в вектор pEL-carAB-wt, предварительно расщепленный теми же рестриктазами.
Около 150 нг лигированной ДНК были использованы для трансформации клеток -реципиентов E.coli TG1 (supE hsdΔ5 thi Δ(lac-proAB) F'[traD36 proAB+ lacIq lacZΔM15]) (J.Sambrook et al., Molecular Cloning, 1989) в ряде последовательных экспериментов, что дало около 2000 рекомбинантных клонов в каждом случае. Набор рекомбинантных плазмид (pEL-carAB-NN) был очищен и ими был трансформированы клетки Е. coli ВКПМ В-6969 (саr5::Тn10), использовавшиеся для отбора рекомбинантных плазмид pEL-carAB-NN, кодирующих активные ферменты.
<2> Сайт-специфический мутагенез
Для введения точечной мутации Ser948Phe была осуществлена ПЦР с использованием плазмиды pBScarAB-13 в качестве матрицы, 5'-концевая затравка 5'-cgtgcgctgcttttcgtgcgcgaaggcgataaag - 3' (34 bases) (SEQ ID NO: 27), сконструированная на основе последовательности нуклеотидов гена car В, и затравки М13 в качестве 3'-концевой затравки. Амплификация в ходе ПЦР и клонирование фрагментов были осуществлены, как это описано выше.
Фрагмент ДНК длиной 1,32 т.п.о., кодирующий 3'-концевой фрагмент гена carB с единичной мутацией, был очищен методом электрофореза в агарозном геле, расщеплен рестриктазами AflII и SacI, а затем лигирован в вектор pET-carAB-wt, предварительно расщепленный теми же рестриктазами.
Около 100 нг полученной ДНК было использовано для трансформации клеток E.coli ВКПМ В-6969 и была отобрана плазмида pEL-carAB-6, кодирующая активные ферменты CarAB с единичной заменой Ser948Phe.
Пример 2. Выделение новых мутантов carB и влияние замен аминокислотных остатков в CPSase на каталитические свойства.
Сначала активность фермента CarAB и его устойчивость к UMP по типу обратной связи были оценены в реакции биосинтеза цитруллина из орнитина, катализируемой ферментами CarAB и ArgI (орнитинкарбамоилтрансфераза), в сорока рекомбинантных клонах B-6969(pEL-carAB-NN).
Схема реакции следующая:

В этой реакции карбамоилфосфатсинтетаза использует в качестве субстрата свободный NH4+.
Белковые экстракты из сорока штаммов B-6969(pEL-carAB-NN) и клеток TGl(pUC-argi) были приготовлены из неочищенного клеточного экстракта клеток, разрушенных ультразвуком, путем осаждения (NH4)2SO4 (75% насыщенности). Осадки белков были растворены в буфере следующего состава:Tris-HCl (50 мМ), рН 7.5, 2-меркаптоэтанол (2 мМ).
Тест-система включала в себя белковые экстракты из штаммов B-6969(pEL-carAB-NN) и клеток TGl(pUC-argI), а также следующие реагенты: АТР (8 мМ), MgSO4 (8 мМ), (NH4)2SO4 (200 мМ), Na2СО3 (8 мМ) и орнитин (1 мМ), (рН 7.5). Содержание орнитина и цитруллина в реакционной смеси анализировалось в помощью ТСХ с использованием подвижной фазы следующего состава: изопропанол/этилацетат/гидроксид аммония/Н2О=40/20/13/27 (v/v).
9 клонов, экспрессирующих активные и устойчивые к ингибированию UMP по типу обратной связи мутантные CPSase, и один клон, экспрессирующий мутантную CPSase с точечной заменой Ser948Phe, были использованы для измерения активности мутантных ферментов.
Плазмиды из указанных 10 клонов были очищены и была определена последовательность фрагментов генов carB со случайной последовательностью методом Сенгера с использованием дидезоксинуклеотидов (Таблица 1).
Затем белковые экстракты из указанных 9 клонов B-6969(pEL-carAB-NN) и клона В-6969(pEL-carAB-6) были использованы для оценки активности и fbr мутантов CPSase в реакции синтеза карбамоилфосфата (СР) из глутамина или аммиака.
Неочищенные клеточные экстракты клеток (20 мг влажных осадков клеток), разрушенных ультразвуком, были растворены в 0.5 мл буфера А (200 мМ К2HPO4/КН2PO4, рН 8.0, 1 мМ EDTA, 1 мМ PMSF, 1 мМ DTT) и обработаны твердым сульфатом аммония до достижения 65% насыщения. После инкубирования в течение 10 мин при 4°С суспензия была отцентрифугирована в течение 10 мин при 13000 об/мин и полученные осадки растворены в 1 мл буфера В (20 мМ К2HPO4/КН2PO4, рН 8.0, 50 мМ KCl, 1 мМ PMSF, 1 мМ DTT). Аликвоты полученных белковых экстрактов были использованы для оценки активности CPSase. Схемы реакций следующие:
I. Gln+СО2+2MgATP+H2O→NH2СООРО32-+2MgADP+Glu+Pi
II. NH3 +CO2+2MgATP+H2O→NH2СООРО32-+2MgADP+Pi.
50 мкл каждой реакционной смеси содержали:
реакция I - 20 мМ Tris-HCl, рН 8.0, 100 мМ KCl, 5 мМ Na2CO3, 10 мМ АТР, 10 мМ MgCl2, 5 mM глутамина, 10 мкл белкового экстракта;
реакция II - 20 мМ tris-HCl, рН 8.0, 100 мМ KCl, 5 мМ Na2СО3, 10 мМ АТР, 10 мМ MgCl2, 200 мМ (NH4)2SO4, 10 мкл белкового экстракта.
Также, для оценки уровня ингибирования CPSase по типу обратной связи часть реакций I проводилась в присутствии 10 мМ UTP.
После инкубирования в течение 10 мин при 37°С реакции были остановлены путем добавления равного объема этилового спирта, охлажденного при -20°С в течение 10 мин, и полученная смесь центрифуговалась в течение 1 мин при 13000 об/мин при комнатной температуре. Супернатанты были охлаждены при -20°С.
Содержание СР в реакционной смеси было проанализировано методом капиллярного электрофореза. Разделение осуществлялось на системе капиллярного электрофореза Quanta 4000Е ("Waters", США) в режиме непрямого УФ-детектирования при 254 нм. Инъекция осуществлялась гидростатически в течение 25 сек. Разделение осуществлялось на не модифицированном кварцевом капилляре (75u I.D.* 60см, эффективная длина 53 см) при напряжении -25kV. Температура поддерживалась при 20°С. Буфер для разделения состоял из 50 мМ Tris-base, 25 мМ бензойной кислоты (для непрямого детектирования), рН 8.5, 0.25 мМ ТТАВ (бромид тетрадецилтриметиламмония) (для обращения электроосмотического потока).
В Таблице 2 приведены экспериментальные данные измерения активности и fbr мутантных CPSase в реакции синтеза СР.
Таким образом, мутантные CPSase в значительной степени не чувствительны к UMP, но точечная мутация значительно снижает активность фермента. Полученные результаты указывают на то, что фрагмент белка с 947 по 951 остаток аминокислоты отвечает за ингибирование CPSase UMP по типу обратной связи, а также за уровень каталитической активности мутантных CPSase.
Гены, кодирующие природную CarAB и мутантную СагАВ-34, были клонированы в плазмиду pMWl 19. Для этой цели плазмиды pEL-carAB-wt и pEL-carAB-34 были расщеплены рестриктазами SacI и XbaI (проводилось неполное расщепление, так как указанные плазмиды содержали два места узнавания XbaI) и фрагменты, кодирующие гены carAB, были клонированы в вектор pMW119, предварительно расщепленный теми же рестриктазами. В результате были сконструированы низкокопийные плазмиды pMW119carAB-wt и pMW119carAB-34, содержащие гены сагАВ под контролем lac промотора.
Пример 3. Продукция оротовой кислоты с использованием штаммов, содержащих мутантные гены carAB.
Штамм 311 был получен из штамма E.coli К 12, содержащего вставку Тn10 в ген argA (ВКПМ В-3853), как мутантный штамм, устойчивый к 6-азаурацилу (1 мг/мл). Штамм 311 был депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) в соответствии с Будапештским договором под инвентарным номером ВКПМ В-8085 5 марта 2001 г.
Штамм 311 был трансформирован плазмидами pMW119carAB-wt и pMWl 19carAB-34 и была проверена продукция оротовой кислоты полученными рекомбинантными штаммами в присутствии различных концентраций уридина.
Условия выращивания в ходе ферментации в пробирках были следующими: 1/20 разбавленной ночной культуры, 60 г/л глюкозы, 25 г/л сульфата аммония, 2 г/л КН2PO4, 1 г/л MgSO4, 0.1 мг/л тиамина, 5 г/л дрожжевой экстракт Difco, 25 г/л мела в 1 л воды.
Глюкоза и мел стерилизовались раздельно. 2 мл питательной среды были помещены в пробирки, выращивание проводилось при 32°С в течение 3 дней на качалочной мешалке. Уровень продукции оротовой кислоты был определен методом ВЭЖХ (Таблица 3).
Как видно из Таблицы 3, штамм 311 (pMWcarAB-34), содержащий мутантный ген carAB, продуцировал больше оротовой кислоты, чем родительский штамм 311(pMW119) и штамм 311 (pMWcarAB-wt), содержащий природный ген carAB.
Пример 4. Продукция аргинина и/или цитруллина с использованием штаммов, содержащих мутантные гены carAB.
Штаммы 333 и 374 - продуценты аргинина были получены путем селекции из штамма - производного от штамма E.coli 57 (ВКПМ В-7386), содержащего вставку транспозона Тn5 в ген ilvA, как мутанты, устойчивые к 6-азаурацилу (1 мг/мл). Штаммы 333 и 374 были депонированы во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) 5 марта 2001 г. в соответствии с Будапештским договором под инвентарными номерами ВКПМ В-8084 и ВКПМ В-8086 соответственно.
Штаммы 333 и 374 были трансформированы плазмидами pMW119carAB-wt и pMW119carAB-34, и была проверена продукция аргинина и цитруллина полученными рекомбинантными штаммами.
Ферментация в пробирках была осуществлена таким же способом, как в Примере 3. Уровень продукции аргинина и/или цитруллина штаммами 333(pMWcarAB-wt) и 333(pMWcarAB-34) в синтетической питательной среде, содержащей 100 мг/л уридина, показан в Таблице 4.
Уровень продукции цитруллина штаммами 374(pMWcarAB-wt) и 374(pMWcarAB-34) в синтетической питательной среде, содержащей 100 мг/л уридина, показан в Таблице 5.
Как видно из Таблицы 4, штамм 333(pMWcarAB-34), содержащий мутантный ген carAB, продуцировал больше аргинина и цитруллина, чем штамм, содержащий природный ген carAB. Как видно из Таблицы 5, штамм 374(pMWcarAB-34), содержащий мутантный ген carAB, продуцировал больше цитруллина, чем штамм, содержащий природный ген carAB.








Реферат
Изобретение относится к биотехнологии. Предложена мутантная карбамоилфосфатсинтетаза из Escherichia coli, в которой последовательность аминокислот, соответствующая положениям 947-951 в природной карбамоилфосфатсинтетазе, заменена любой из последовательностей аминокислот SEQ ID NO:1 - SEQ ID NO:9. Описана также аналогичная мутантная карбамоилфосфатсинтетаза из Escherichia coli, содержащая любые делеции, замены, вставки или добавления одной или нескольких аминокислот в одном или множестве положений, кроме 947-951. Предложен фрагмент ДНК, кодирующий указанные мутантные карбамоилфосфатсинтетазы, а также способ продукции производного карбамоилфосфата с использованием штамма Escherichia coli, трансформированного указанным фрагментом ДНК. В изобретении описываются также различные штаммы Escherichia coli, каждый из которых трансформирован предложенным фрагментом ДНК: штамм Escherichia coli 311 (ВКПМ В-8085)-продуцент оротовой кислоты, штамм Escherichia coli 333 (ВКПМ В-8084) - продуцент L-аргинина и цитруллина, штамм Escherichia coli 374 (ВКПМ В-8086) - продуцент цитруллина. Использование изобретения позволяет получать повышенные количества производных карбамоилфосфата, таких как L-аргинин, цитруллин, производных пиримидина, по сравнению с природными штаммами Escherichia coli. 7 н. и 3 з.п. ф-лы, 2 ил., 5 табл.
Комментарии