Мультизимы и их использование в получении полиненасыщенных жирных кислот - RU2517608C2
Код документа: RU2517608C2
Чертежи




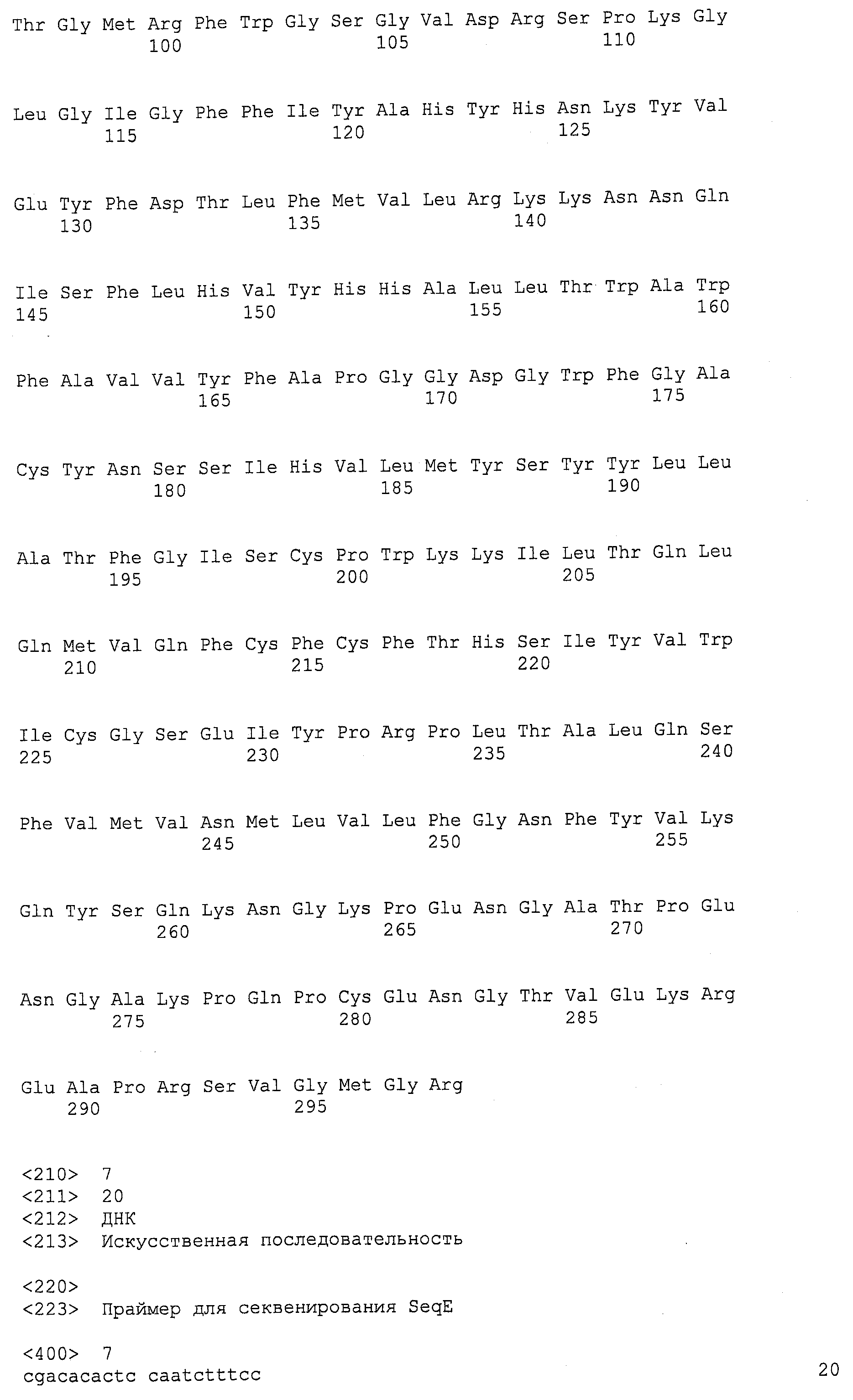





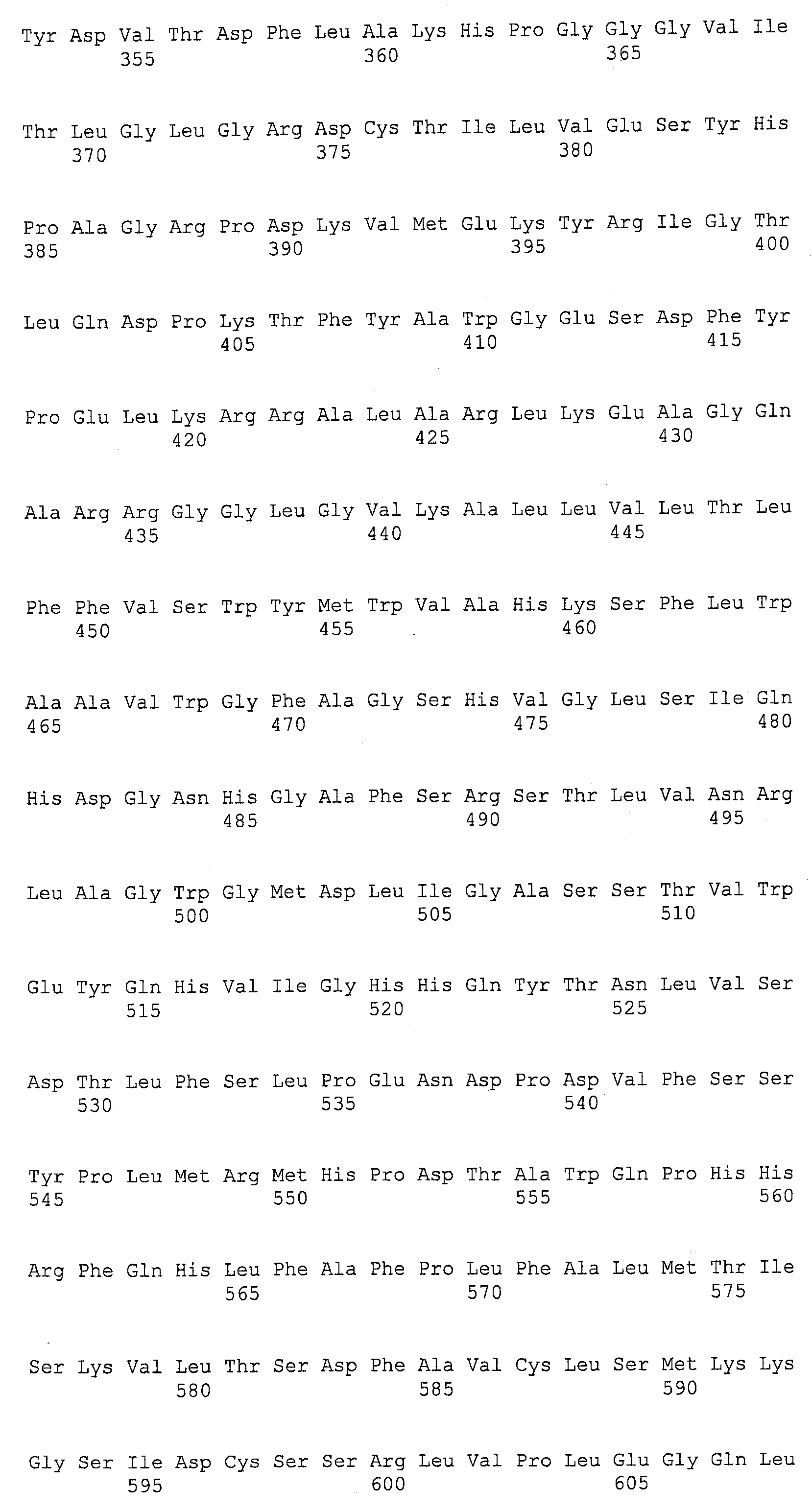







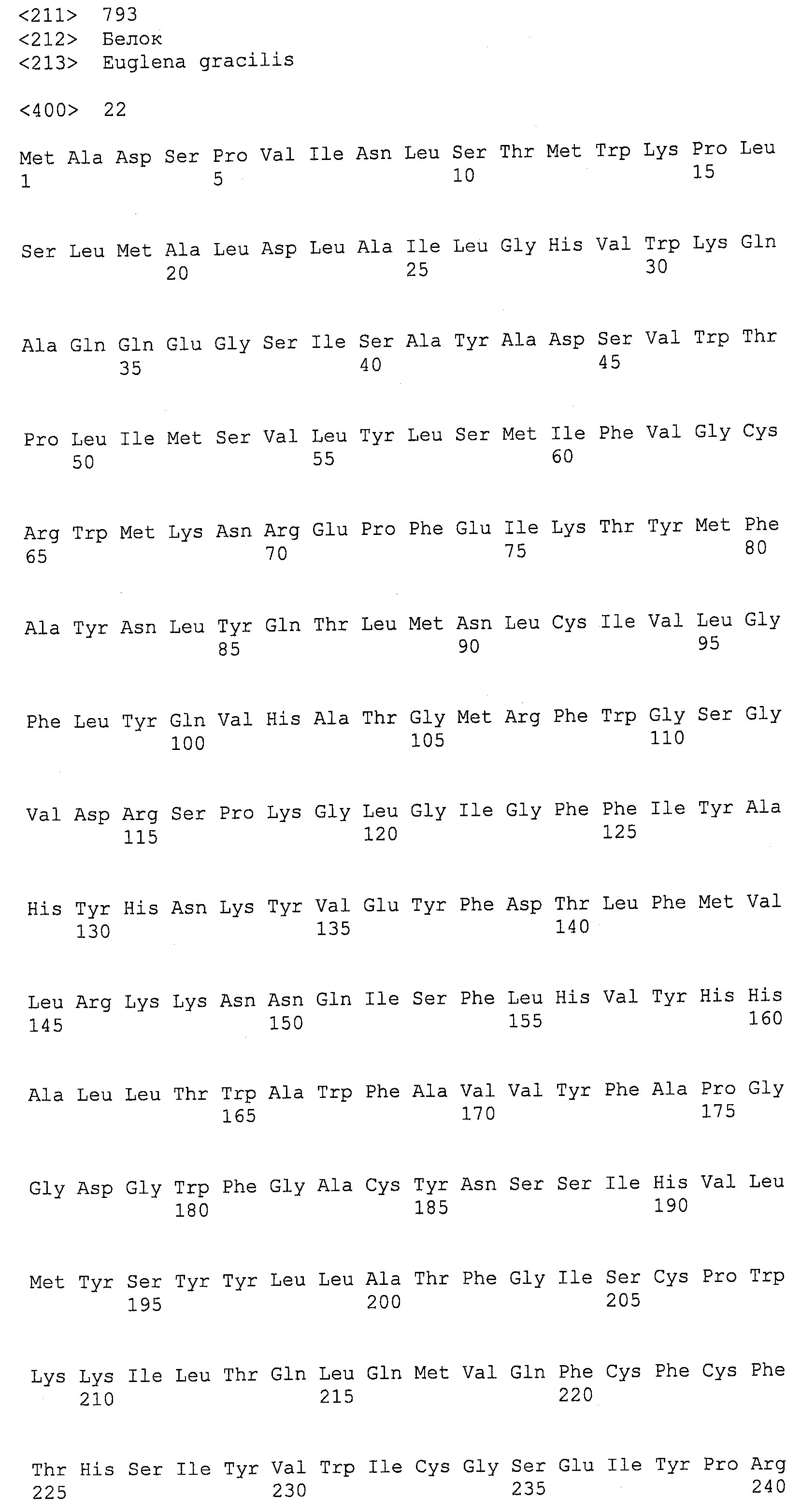

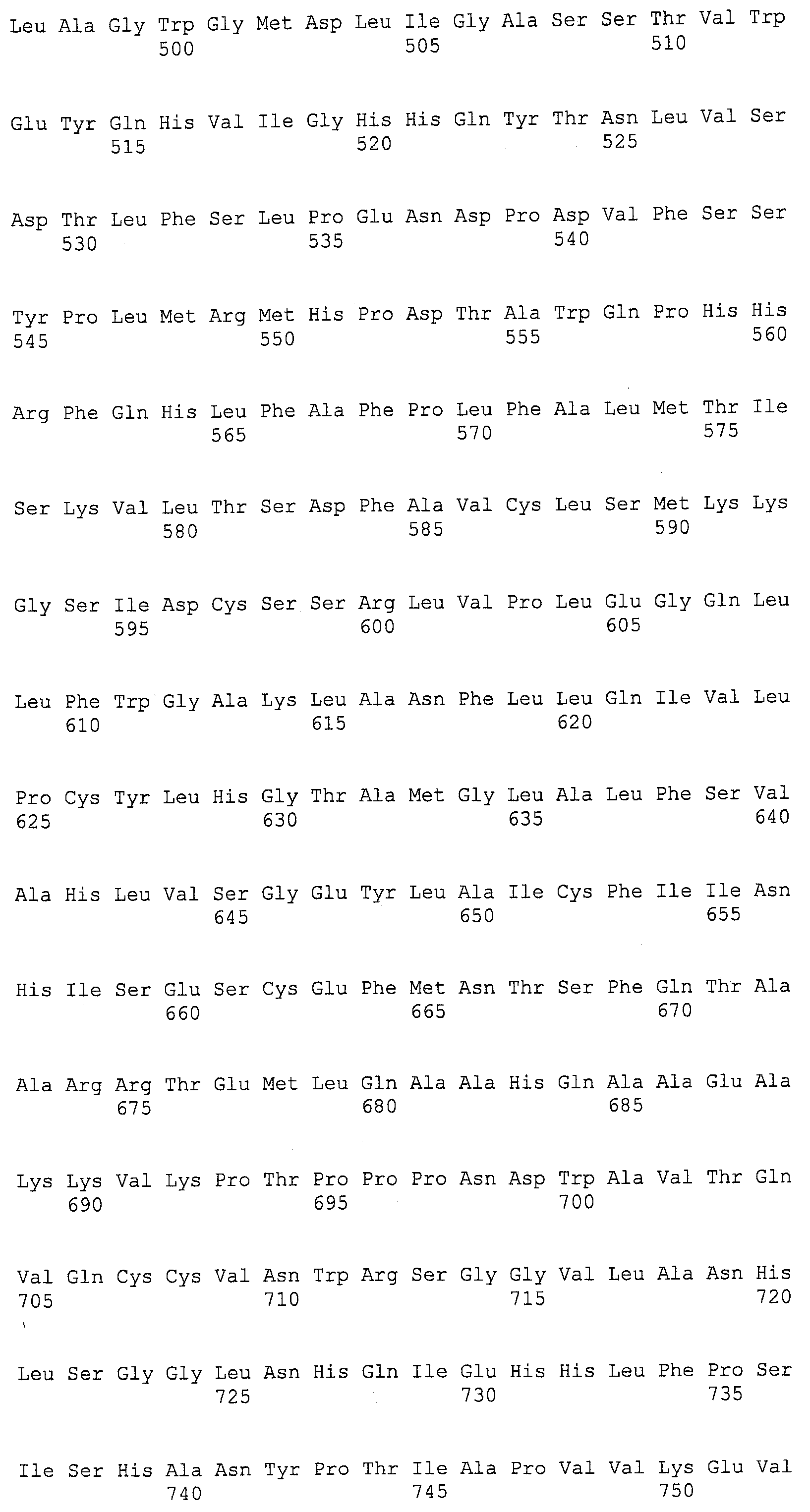



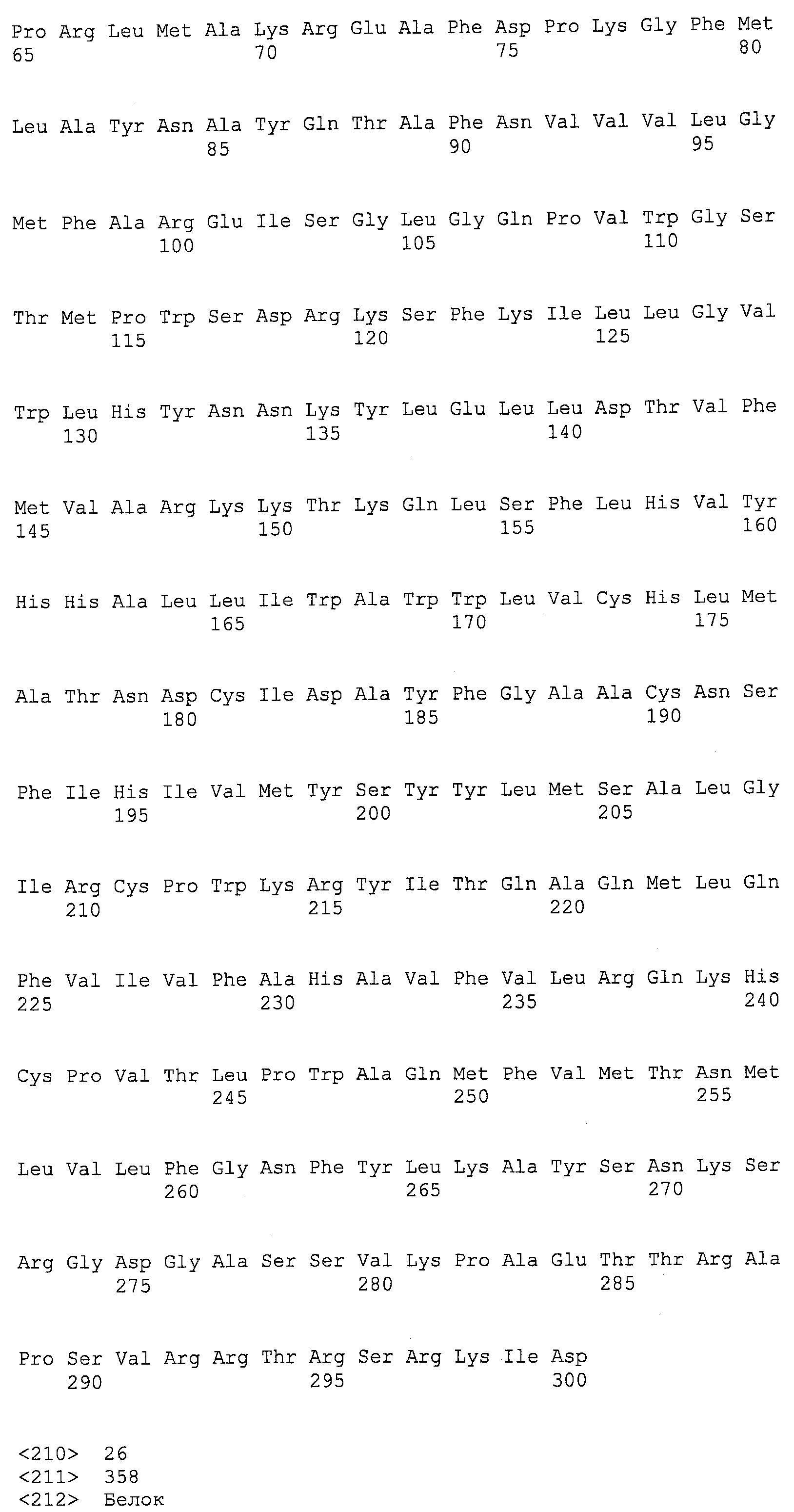
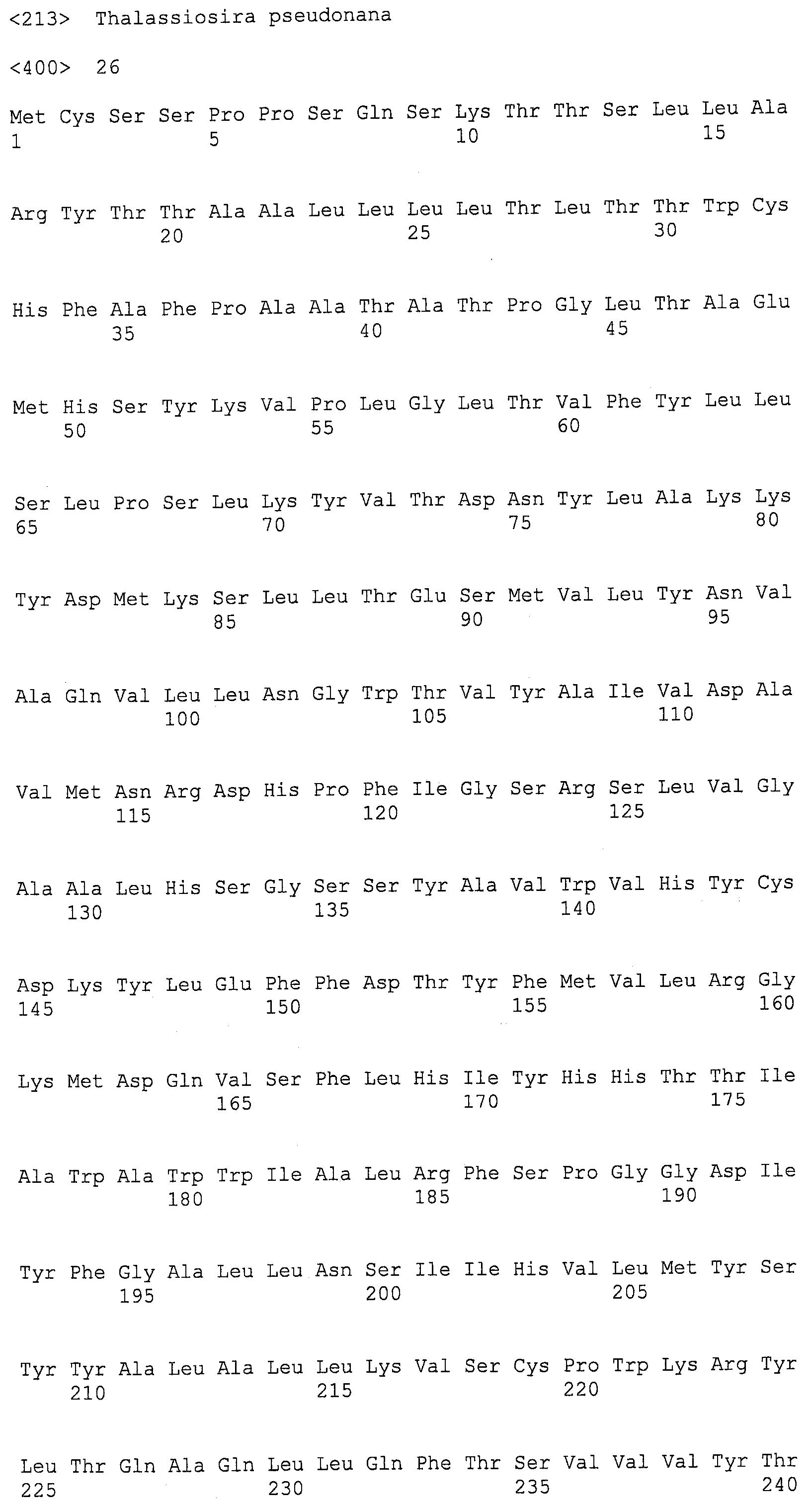



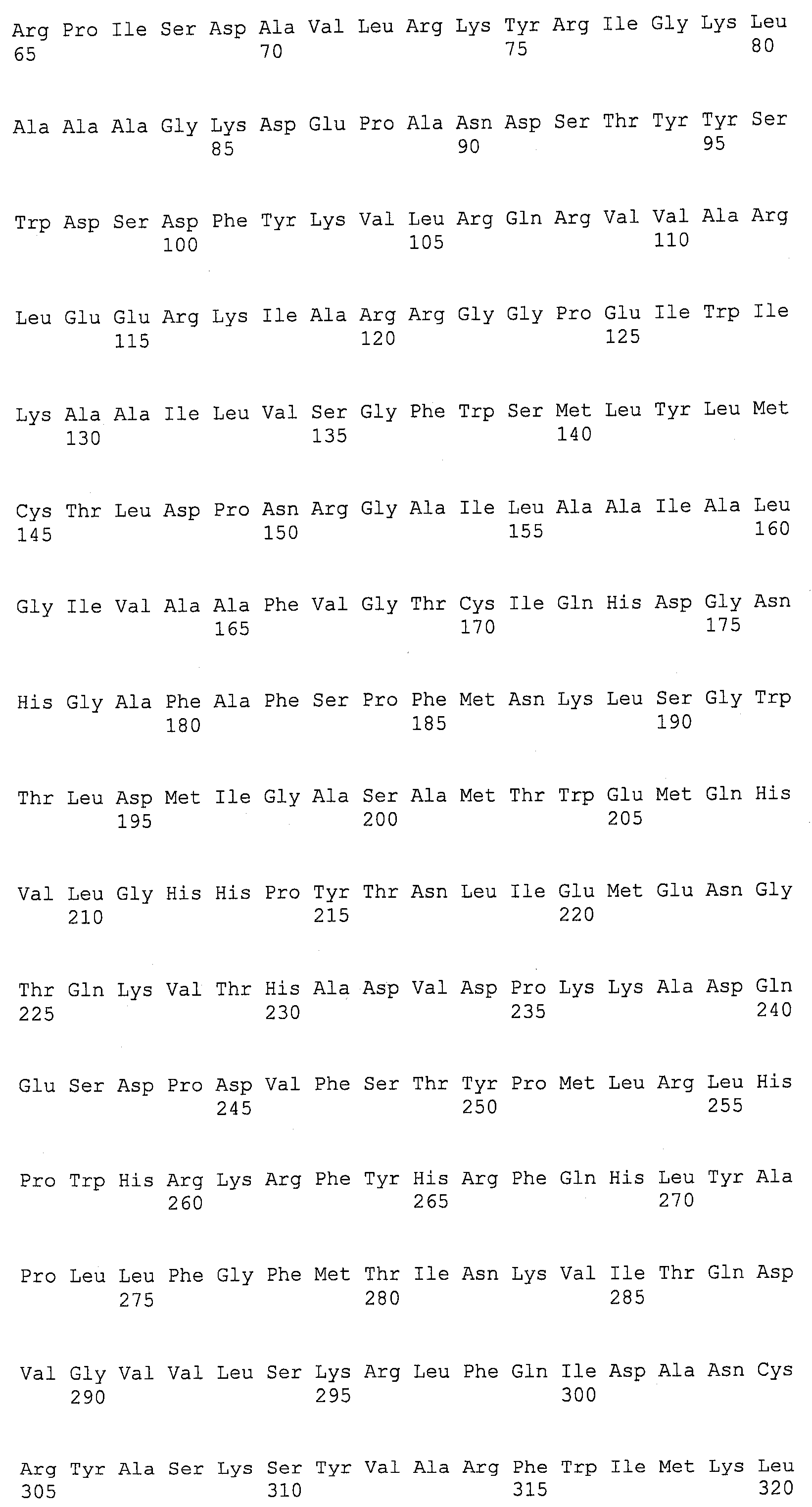
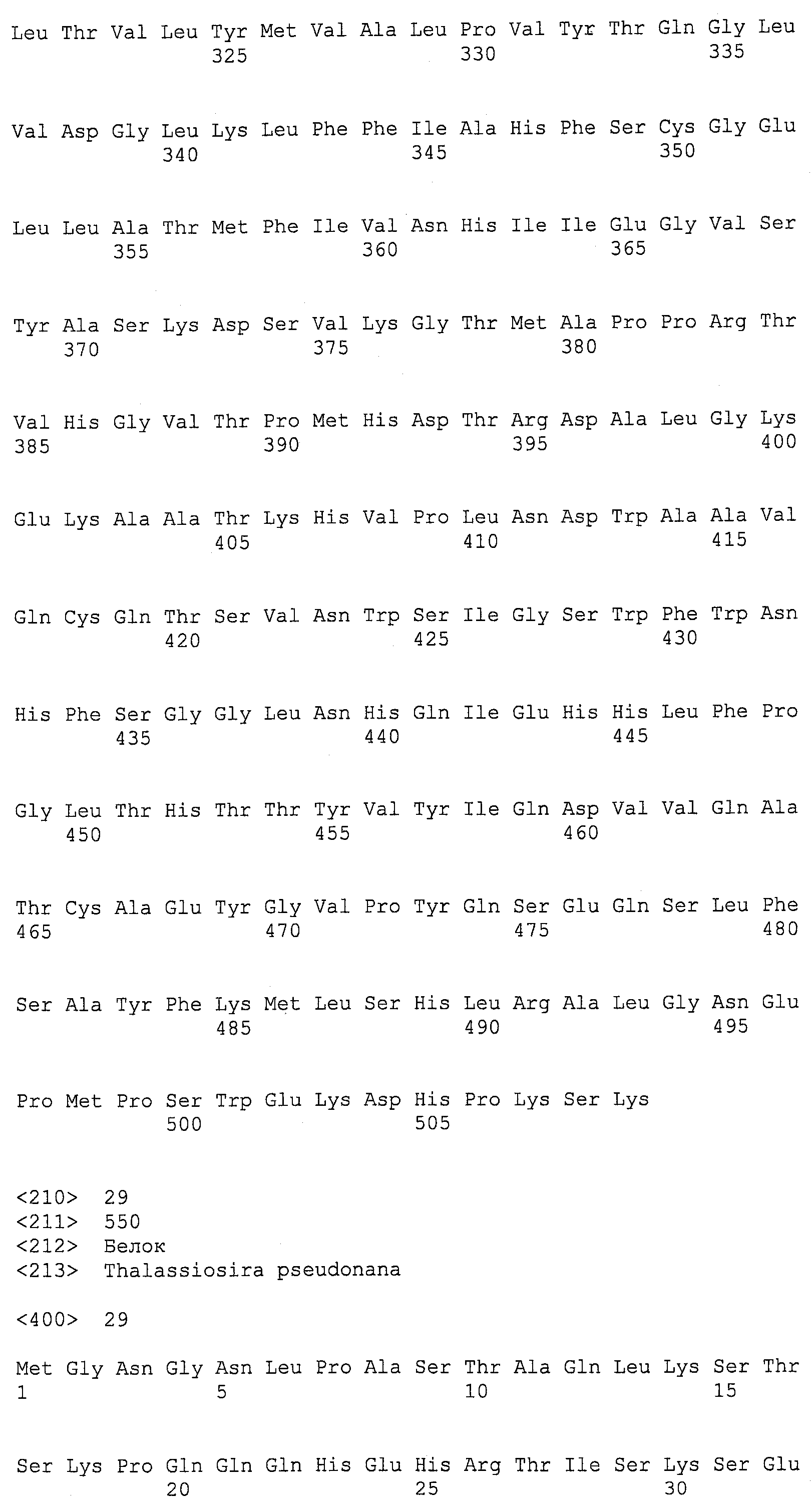











































































































































































































































































































































































































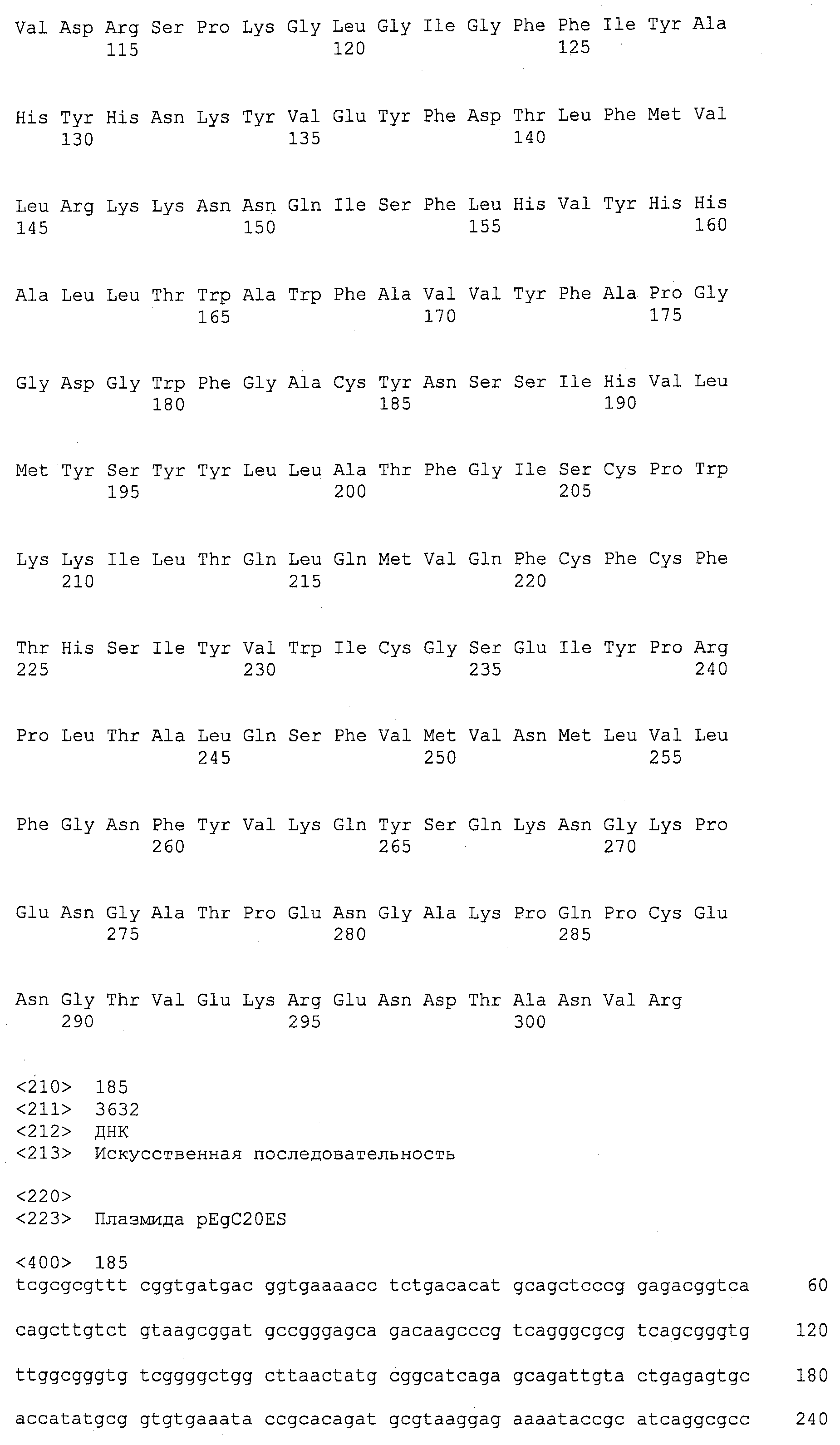







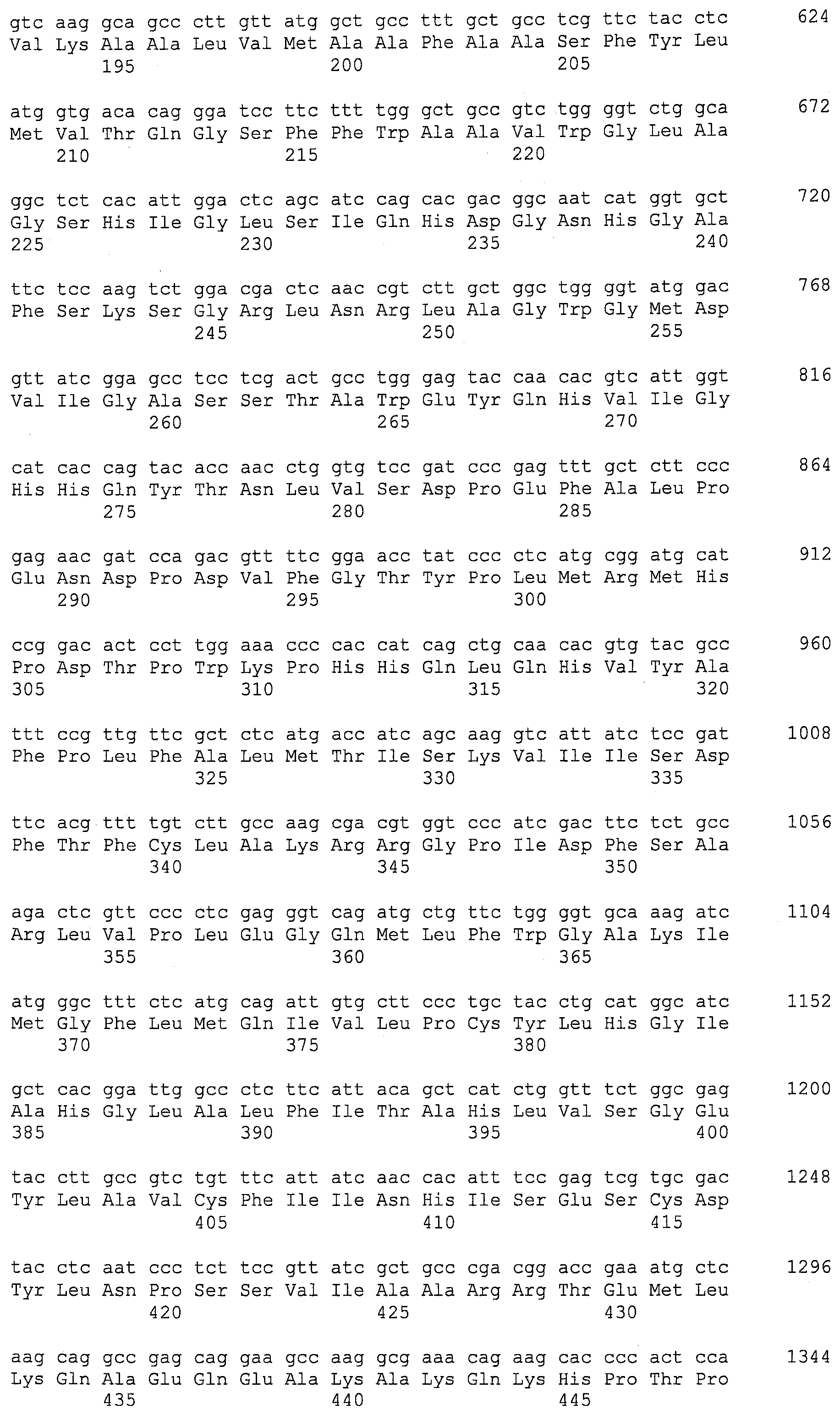



















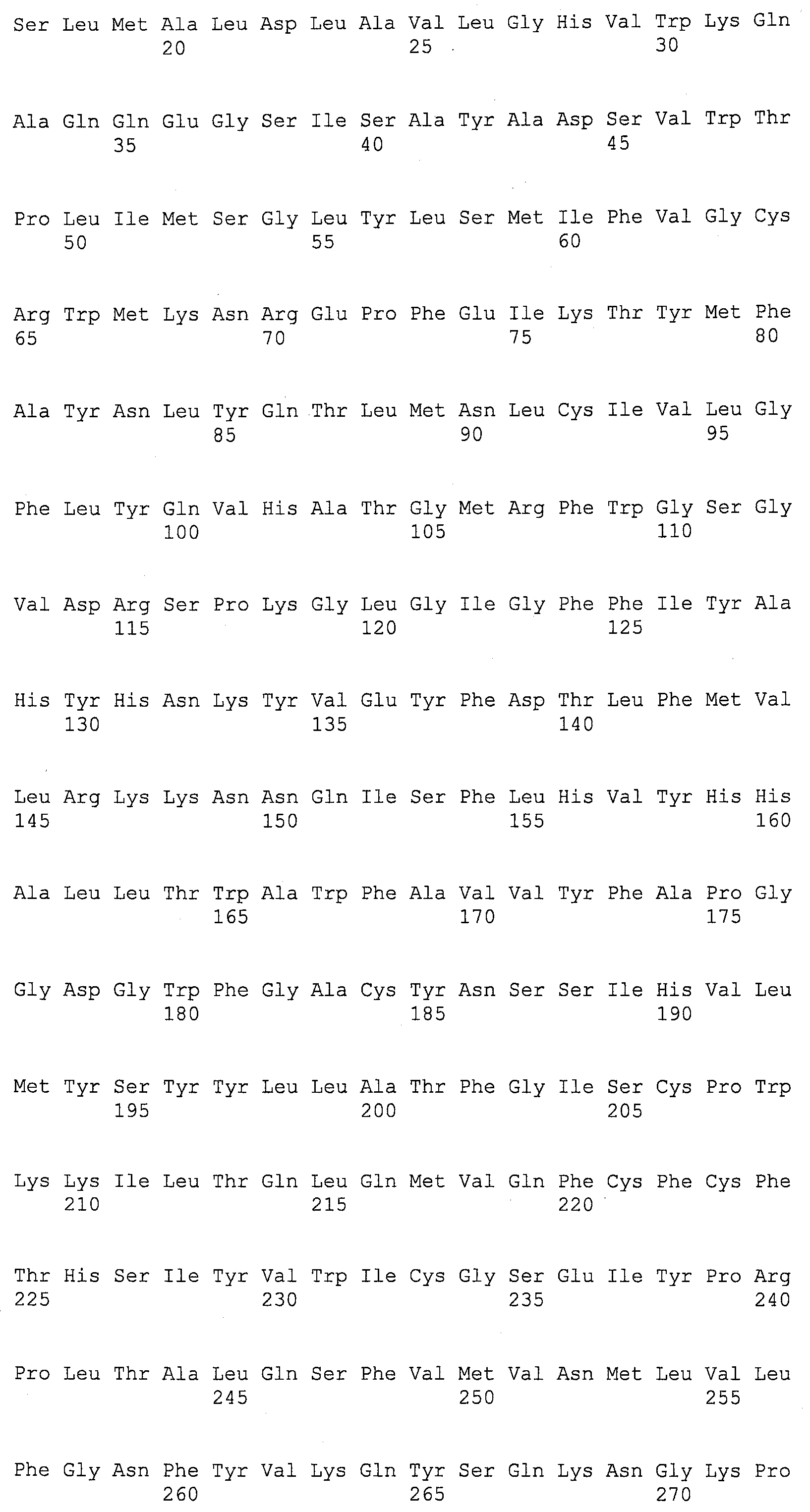













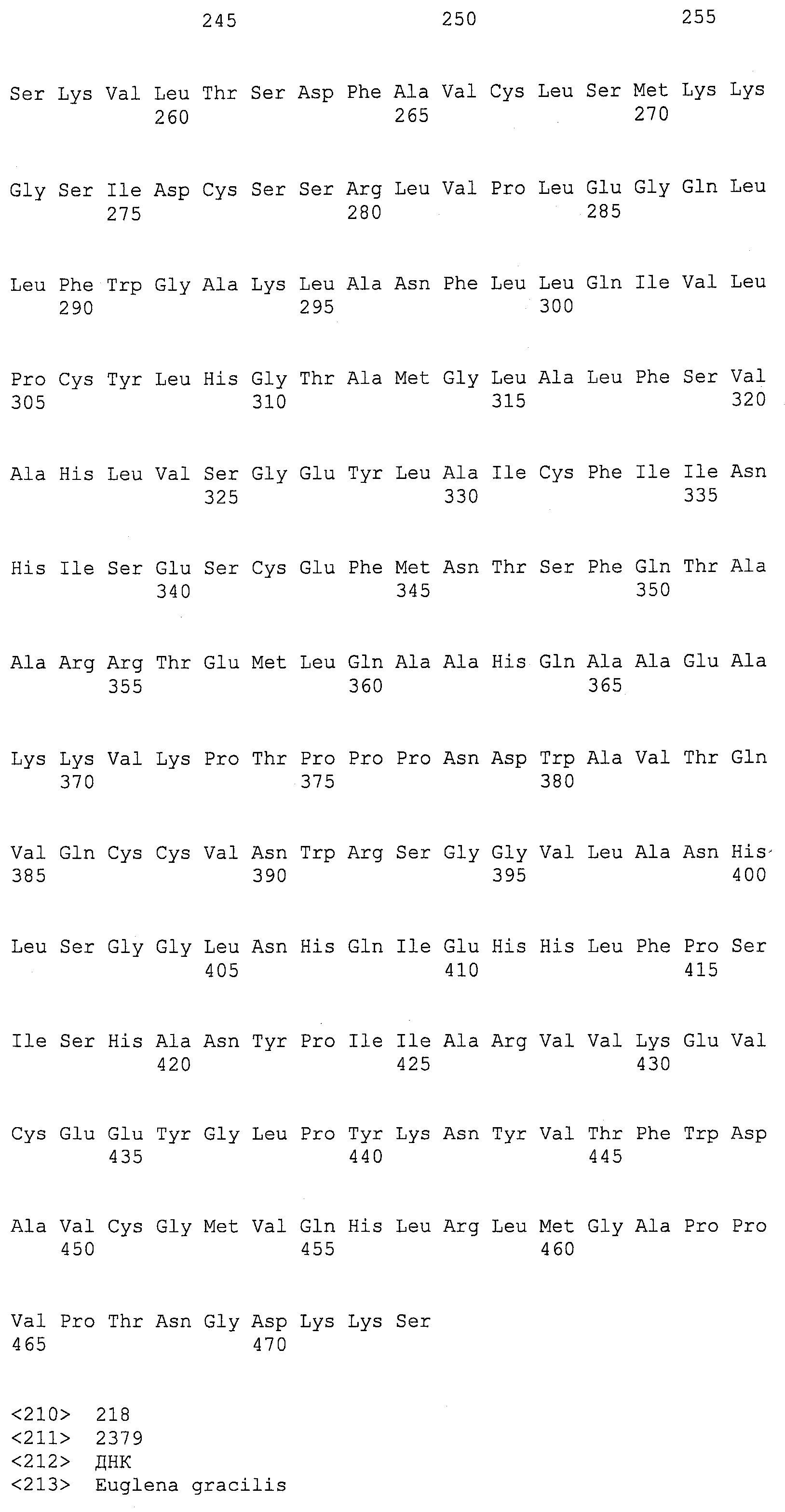














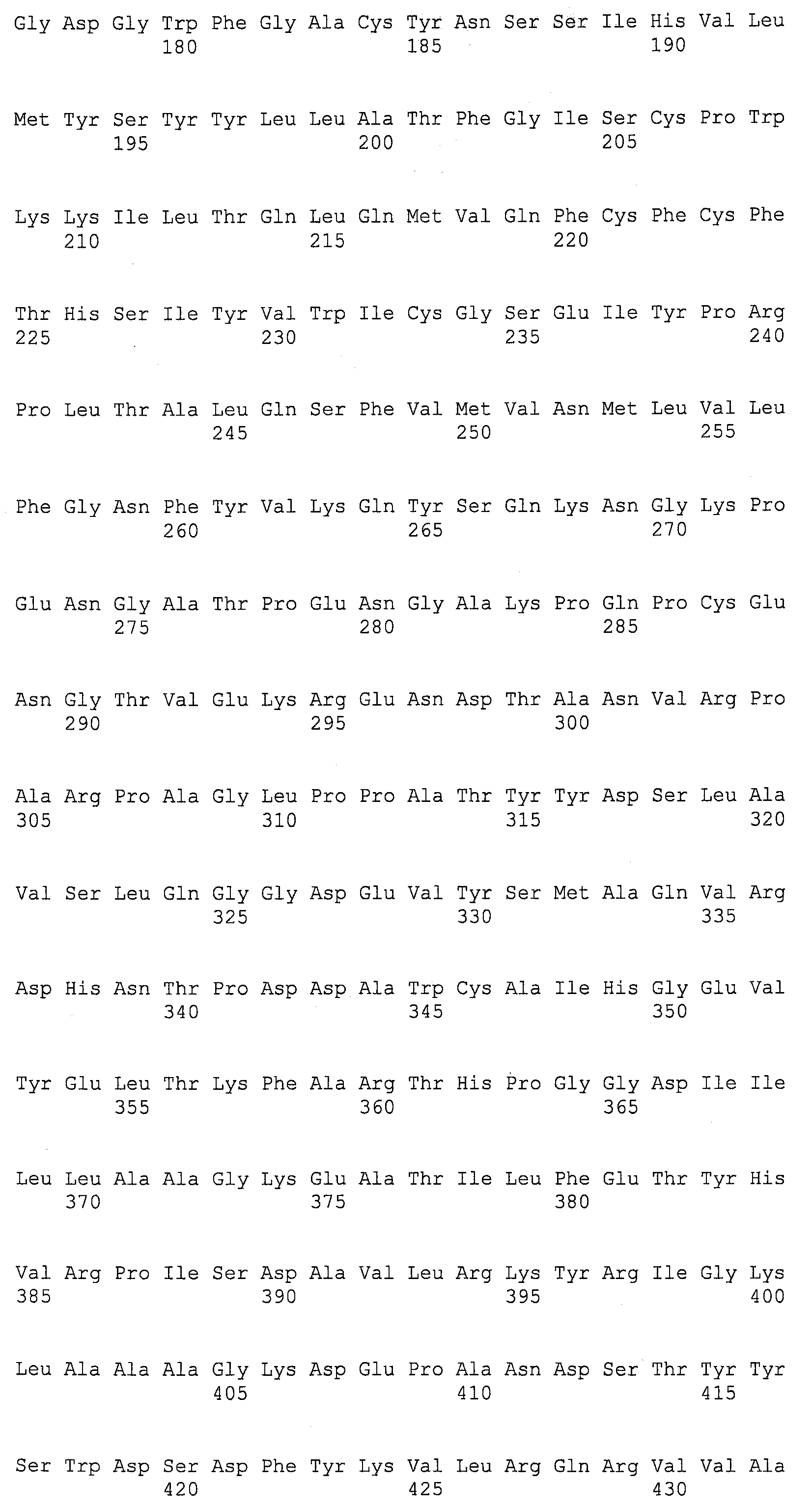
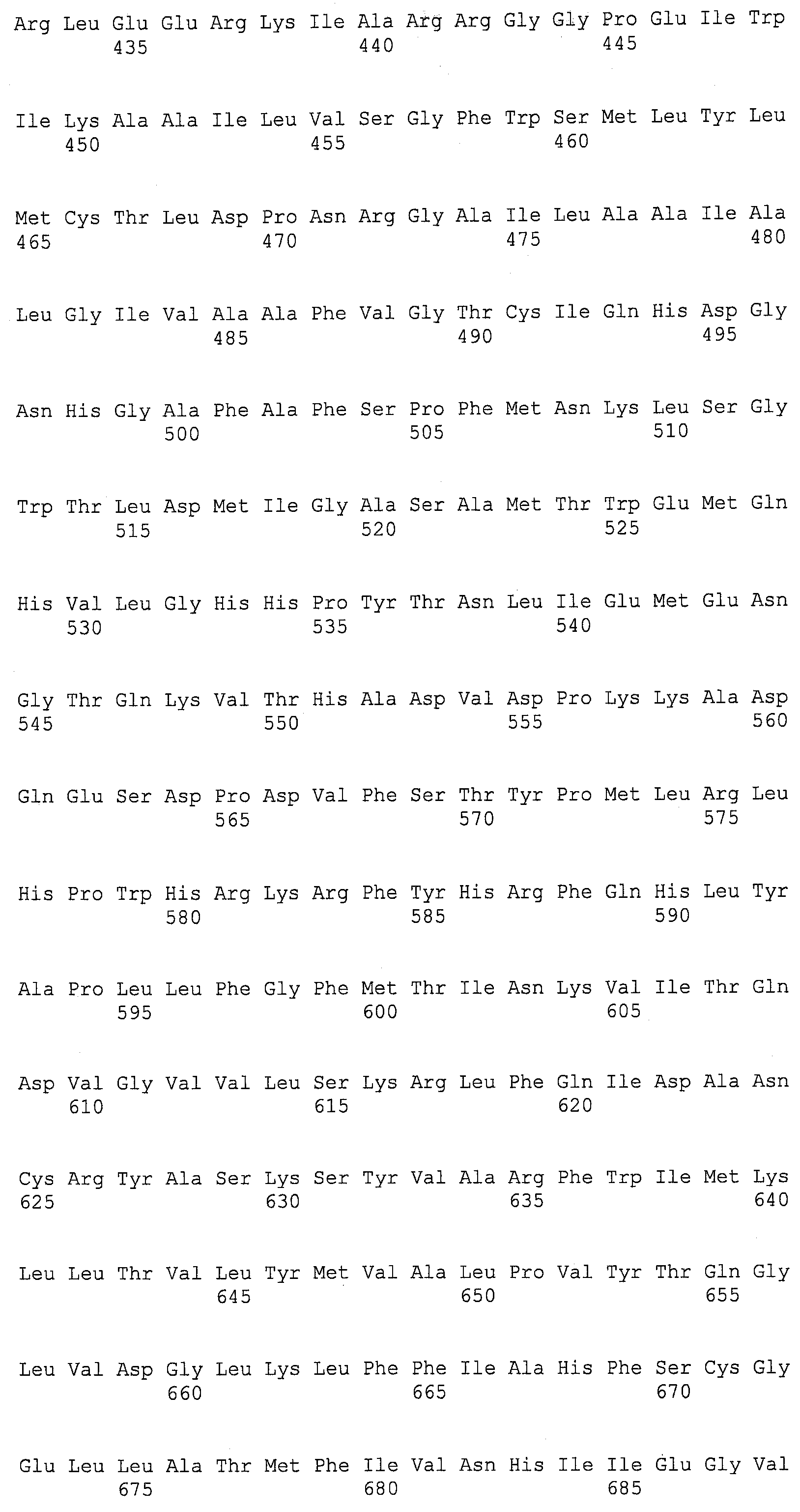






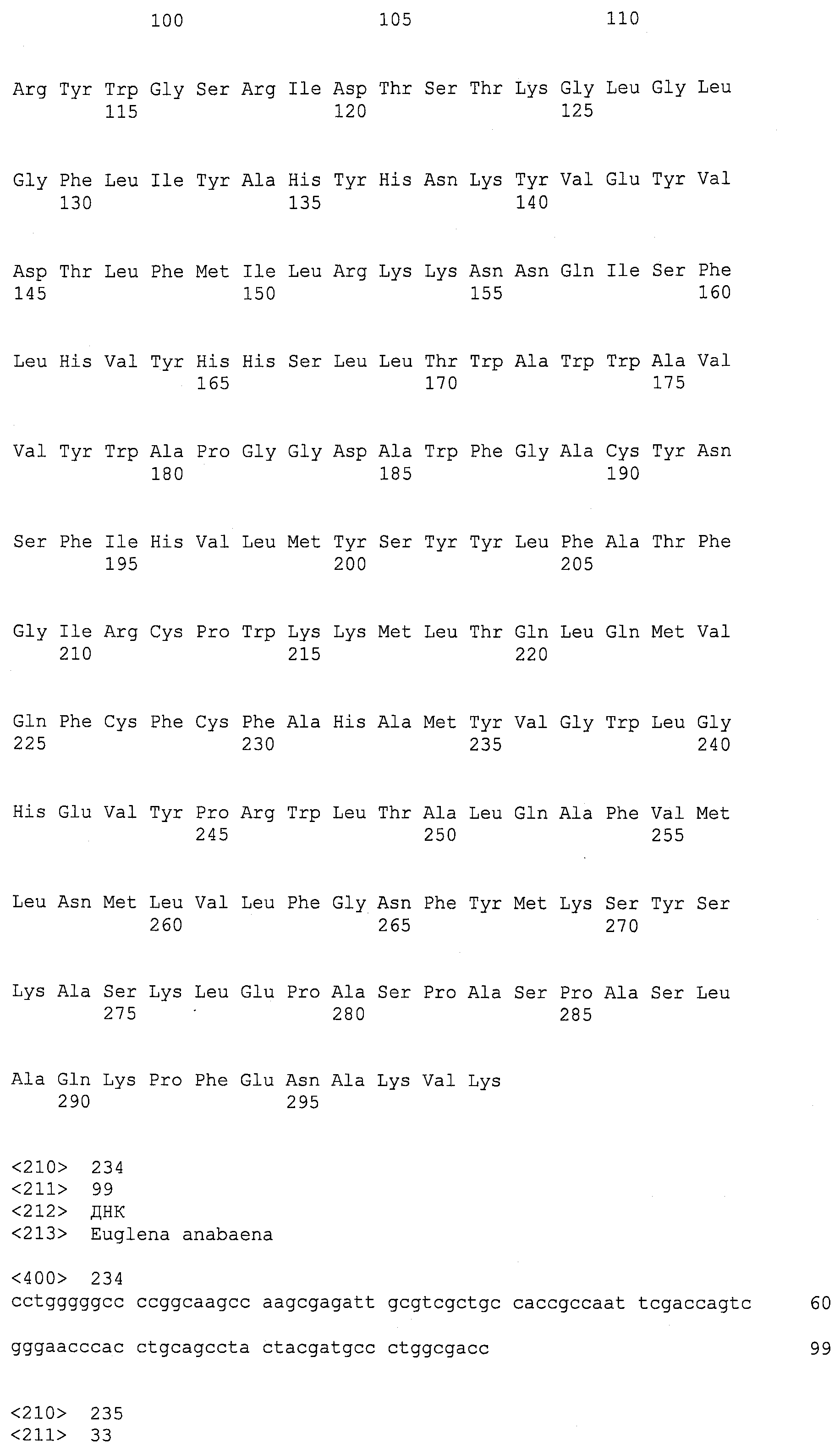



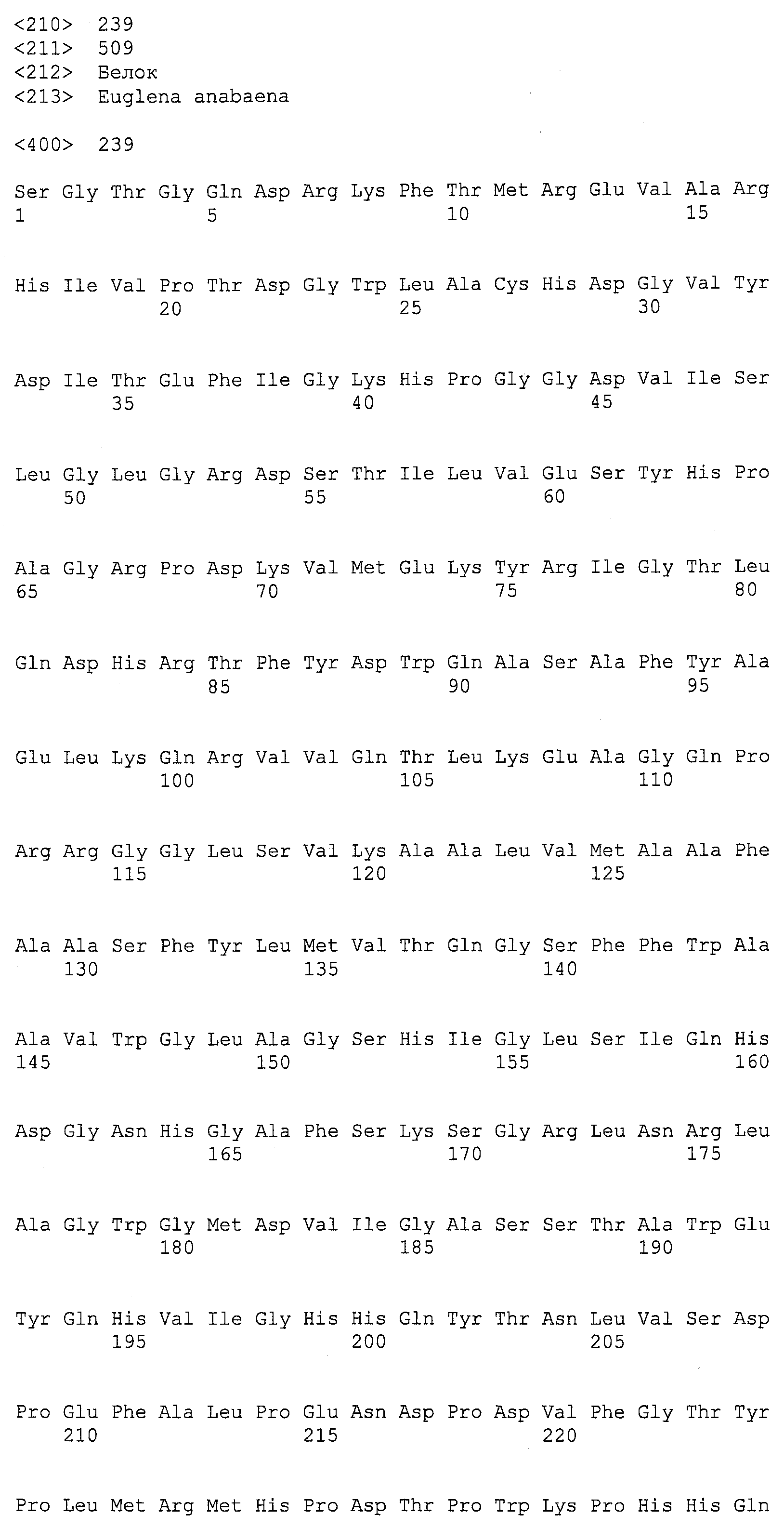













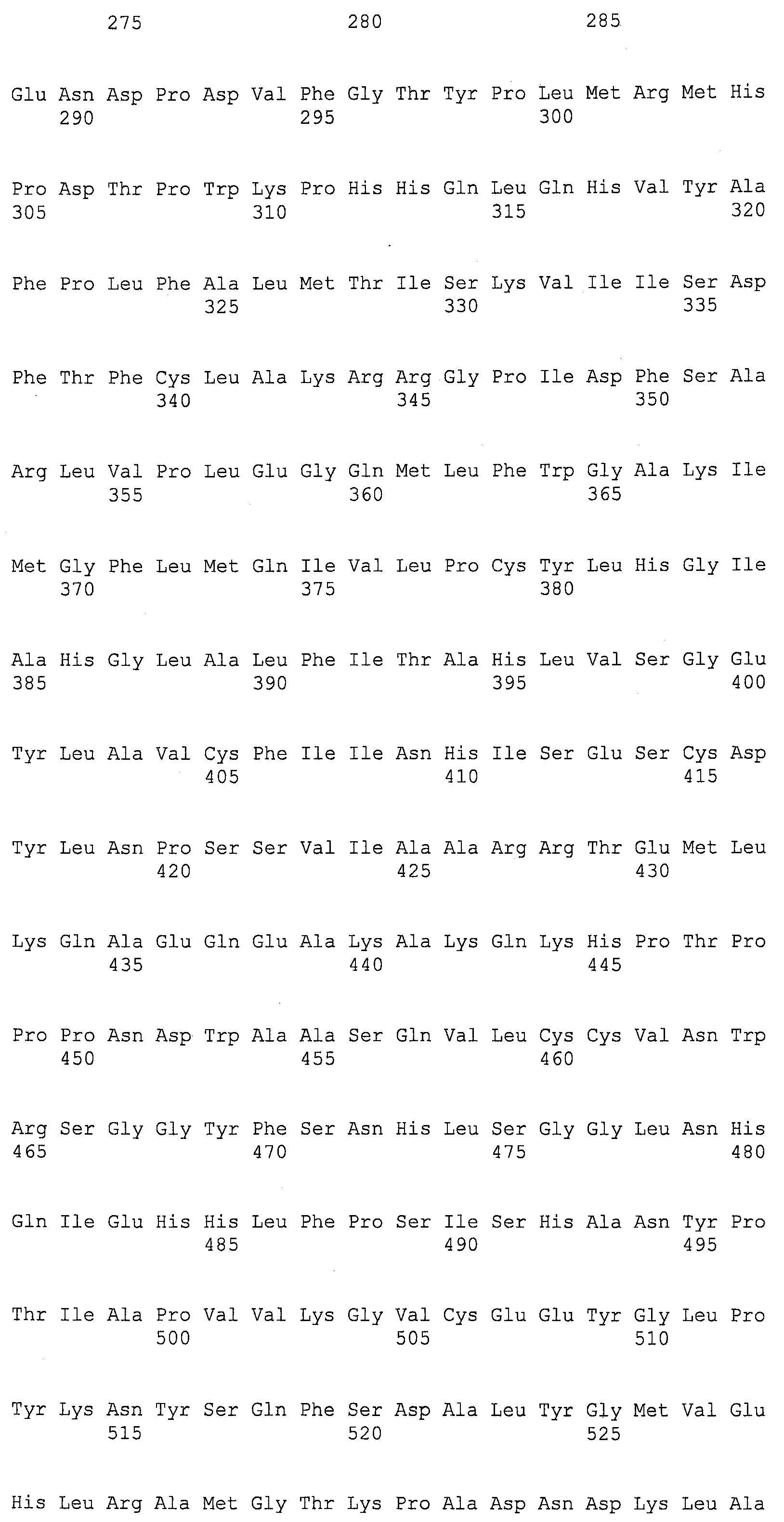

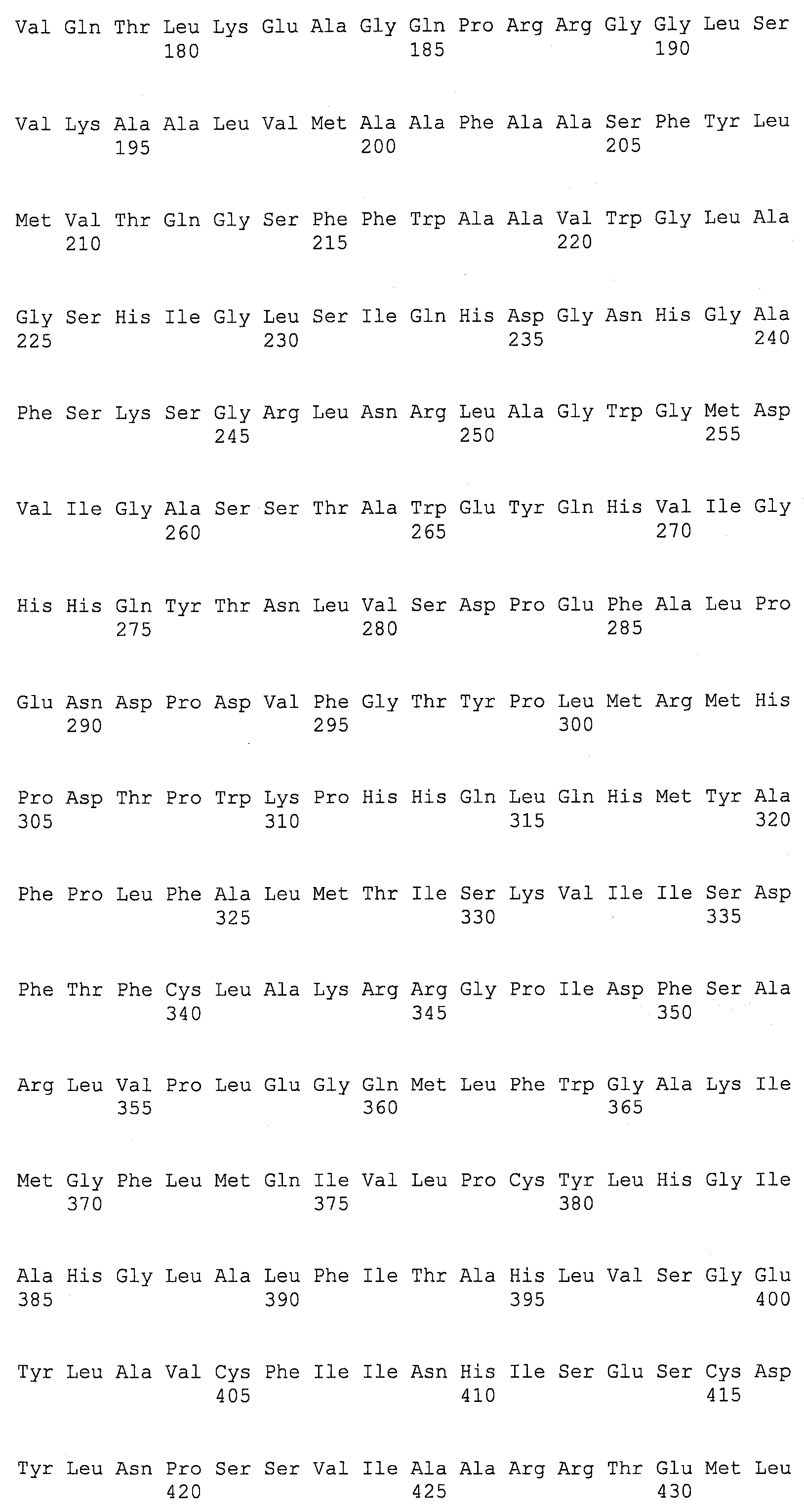

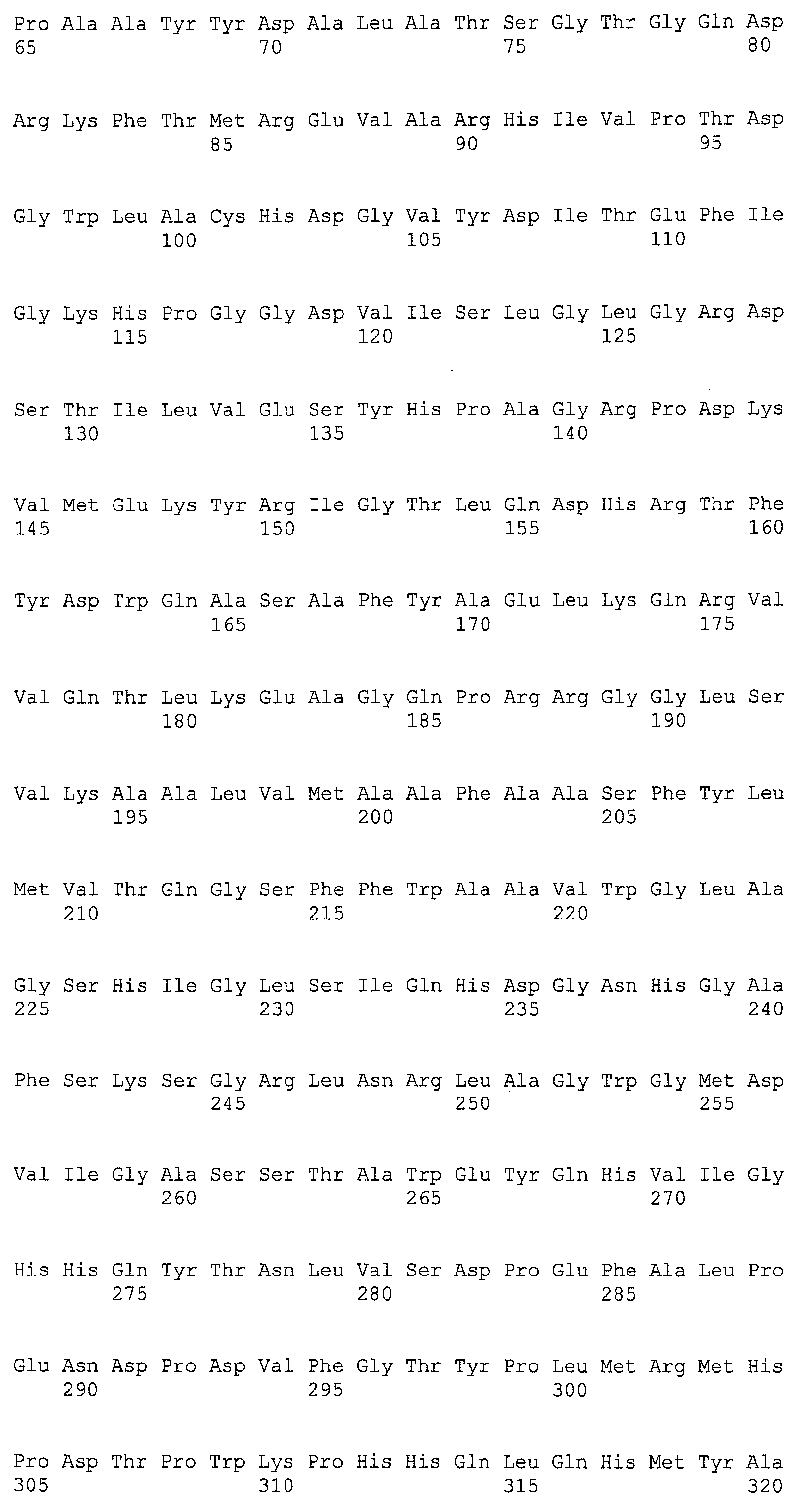
































































Описание
Данная заявка провозглашает приоритет предварительной заявки США № 60/909790 от 3 апреля 2007 года и предварительной заявки США № 61/027898 от 12 февраля 2008 года, на которые в данном тексте сделаны полные ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к области биотехнологии. Более конкретно, данное изобретение касается полинуклеотидных последовательностей, которые кодируют мультизимы, и их использования в синтезе длинноцепочечных полиненасыщенных жирных кислот (ПНЖК).
УРОВЕНЬ ТЕХНИКИ
Важность ПНЖК неоспорима. Например, некоторые ПНЖК являются важными биологическими компонентами здоровых клеток и признаны как: «незаменимые» жирные кислоты, которые не могут быть синтезированы de novo в млекопитающих, и вместо этого должны получаться или через пищу или путем дополнительного удлинения и десатурации линолевой кислоты (ЛК; 18:2 омега-6) или α-линоленовой кислоты (АЛК; 18:3 омега-3); составляющие плазматических клеточных мембран, где они могут быть найдены в таких формах как фосфолипиды или триацилглицерины; необходимые для соответствующего развития (особенно в развитии мозга младенца) и для формирования и восстановления ткани; и, предшественники нескольких биологически активных эйкозаноидов, важных для млекопитающих (например, простациклинов, эйкозаноидов, лейкотриенов, простагландинов). Кроме того, значительное потребление длинноцепочечных омега-3 ПНЖК продуцирует кардиоваскулярные защитные эффекты (Dyerberg et al., Amer. J. Clin. Nutr. 28:958-966 (1975); Dyerberg et al., Lancet. 2(8081):117-119 (1978); Shimokawa, H., World Rev. Nutr. Diet 88:100-108 (2001); von Schacky et al., World Rev. Nutr. Diet 88:90-99 (2001)). Многочисленые другие исследования свидетельствуют о значительной пользе для здоровья, которую приносит использование омега-3 и/или омега-6 ПНЖК против разнобразных симптомов и болезней (например, астмы, псориаза, экземы, диабета, рака).
В настоящее время исследуется множество различных хозяев, включая растения, водоросли, грибы и дрожжи, как средств для промышленного производства ПНЖК путем использования многочисленных разнообразных попыток. Хотя производительные способности организмов-хозяев в отношении природных ПНЖК иногда существенны для данной методологии, генная инженерия также показала, что природные способности некоторых хозяев (даже тех, которым присуща природная ограниченная способность к производству ЛК и АЛК жирных кислот) могут быть значительно изменены в плане повышения уровня производства различных длинноцепочечных омега-3/омега-6 ПНЖК. Независимо от того, является ли этот эффект результатом природных способностей или рекомбинантной технологии, арахидоновая кислота (АРК; 20:4 омега-6), эйкозапентаеновая кислота (ЭПК; 20:5 омега-3), и докозагексаеновая кислота (ДГК; 22:6 омега-3) требуют экспрессии либо дельта-9-элонгаза/дельта-8-десатуразного пути (который действует в некоторых организмах, таких как вид эвгленоидных, и который характеризуется выработкой эйкозадиеновой кислоты (ЭДК; 20:2 омега-6) и/или эйкозатриеновой кислоты (ЭТрК; 20:3 омега-3)) или дельта-6-десатураза/дельта-6-элонгазного пути (который находят, преимущественно, в водорослях, мхах, грибах, нематодах и человеческом организме, и который характеризуется выработкой гамма-линоленовой кислоты (ГЛК; 18:3 омега-6) и/или стеаридоновой кислоты (СТК; 18:4 омега-3)) (ФИГ.1). Дельта-6-элонгаза известна также как C18/20-элонгаза.
Идентифицированные до сих пор дельта-8-десатуразные ферменты обладают способностью к превращению как ЭДК в дигомо гамма-линоленовую кислоту (ДГЛК (известную также как ГГЛК); 20:3, n-6) так и ЭТрК в эйкозатетраеновую кислоту (ЭТК; 20:4, n-3). АРК и ЭПК потом синтезируются из ДГЛК и ЭТК, соответственно, по реакции с дельта-5-десатуразой. Однако синтез ДГК требует последующей экспрессии дополнительной C20/22-элонгазы и дельта-4-десатуразы. Большинство идентифицированных до сих пор C20/22-элонгаз обладают главной способностью к превращению ЭПК в ДПК, и вторичной активностью в отношении превращения арахидоновой кислоты (АРК; 20:4 омега-6) в докозатетраеновую кислоту (ДТК; 22:4 омега-6), тогда как большинство идентифицированных до сих пор дельта-4-десатуразных ферментов обладают главной способностью к превращению ДПК в ДГК, и вторичной активностью в отношении превращения докозатетраеновой кислоты (ДТК; 22:4 омега-6) в ω-6 докозапентаеновую кислоту (ДПКn-6; 22:5 омега-6).
Исходя из той роли, которую играют C20/22-элонгазный и дельта-4-десатуразный ферменты в синтезе ДГК, были предприняты значительные усилия для идентификации и анализа этих ферментов из различных источников. Как таковые, многочисленные C20/22-элонгазы были раскрыты как в открытой так и патентной литературе (например, Pavlova sp. CCMP459 (номер доступа GenBank AAV33630), Ostreococcus tauri (номер доступа GenBank AAV67798) и Thalassiosira pseudonana (номер доступа GenBank AAV67800)). Подобно этому, были раскрыты следующие дельта-4-десатуразы: Euglena gracilis (SEQ ID NO:13; номер доступа GenBank AAQ19605; Meyer et al., Biochemistry, 42(32):9779-9788 (2003)); Thalassiosira pseudonana (SEQ ID NO:29; номер доступа GenBank AAX14506; Tonon et al., FEBS J., 272(13):3401-3412 (2005)); Thraustochytrium aureum (SEQ ID NO:27; номер доступа GenBank AAN75707); Thraustochytrium sp. (номер доступа GenBank CAD42496; патент США № 7087432); Schizochytrium aggregatum (SEQ ID NO:28; PCT публикация № WO 2002/090493); Pavlova lutheri (номер доступа GenBank AAQ98793); and Isochrysis galbana (SEQ ID NO:30; номер доступа GenBank AAV33631; Pereira et al., Biochem. J., 384(2):357-366 (2004); PCT публикация № WO 2002/090493)].
Правопреемник заявителя имеет ряд патентных заявок, которые касаются производства ПНЖК в жировых дрожжах (т.е. Yarrowia lipolytica), включая, например: патенты США №№ 7238482 и 7125672; заявка США № 11/265761 (от 2 ноября 2005 года); заявка США № 11/264784 (от 1 ноября 2005 года); заявка США № 11/264737 (от 1 ноября 2005 года).
Подобным образом, PCT публикация № WO 2004/071467 (опубликована 26 августа 2004 года) касается производства ПНЖК в растениях, тогда как РСТ публикация № WO 2004/071178 (опубликована 26 августа 2004 года) касается аннексиновых промоторов и их использования в экспрессии трансгенов в растениях. Обе являются совместными заявками правопреемника заявителя, которые находятся на рассмотрении.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Данное изобретение касается мультизима, который включает отдельный полипептид, обладающий по меньшей мере двумя независимыми и раздельными ферментативными активностями.
Во втором варианте ферментативные активности данного мультизима могут выбираться из группы, которая состоит из элонгазы жирных кислот, десатуразы жирных кислот, ацилтрансфераз, ацил CoA синтаз и тиоэстераз. Более конкретно, ферментативные активности могут включать по меньшей мере одну элонгазу жирных кислот, связанную с по меньшей мере одной десатуразой жирных кислот.
В третьем варианте данный мультизим может включать первую ферментативную активность, связанную со второй ферментативной активностью, и указанная связь выбирается из группы, которая состоит из полипептидной связи, SEQ ID NO:198 (линкер EgDHAsyn1), SEQ ID NO:200 (линкер EgDHAsyn2), SEQ ID NO:235 (1 линкер EaDHAsyn), SEQ ID NO:438, SEQ ID NO:445, SEQ ID NO:472, и SEQ ID NO:504.
В четвертом варианте изобретение касается выделенного полинуклеотида, кодирующего ДГК синтазу, включая:
(a) нуклеотидную последовательность, кодирующую полипептид, который обладает ДГК синтазной активностью, где данный полипептид имеет по меньшей мере 80% аминокислотную идентичность, основываясь на методе группировки Clustal V, при сравнении с аминокислотной последовательностью, как это изложено в SEQ ID NO:12, SEQ ID NO:22, SEQ ID NO:95, SEQ ID NO:96, или SEQ ID NO:97;
(b) нуклеотидную последовательность, кодирующую полипептид, который обладает ДГК синтазной активностью, где данная нуклеотидная последовательность имеет по меньшей мере 80% идентичность последовательности, основываясь на сравнительном анализе первичной структуры согласно программе BLASTN, при сравнении с нуклеотидной последовательностью, как изложено в SEQ ID NO:11, SEQ ID NO:205, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93, или SEQ ID NO:410;
(c) нуклеотидную последовательность, кодирующую полипептид, который обладает ДГК синтазной активностью, где данная нуклеотидная последовательность гибридизуется при жестких условиях с нуклеотидной последовательностью, как изложено в SEQ ID NO:11, SEQ ID NO:205, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93, или SEQ ID NO:410; или
(d) комплемент нуклеотидной последовательности (a), (b) или (c), где данные комплемент и нуклеотидная последовательность состоят из одинакового количества нуклеотидов и являются на 100% комплементарными.
В пятом варианте данное изобретение касается полинуклеотида, кодирующего полипептид, который обладает ДГК синтазной активностью, где нуклеотидная последовательность включает SEQ ID NO:11, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93, или SEQ ID NO:410.
В шестом варианте данное изобретение касается полипетида данного изобретения, обладающего ДГК синтазной активностью, где аминокислотная последовательность данного полиппетида включает SEQ ID NO:12, SEQ ID NO:22, SEQ ID NO:95, SEQ ID NO:96, или SEQ ID NO:97.
В седьмом варианте данное изобретение касается выделенного полинуклеотида, кодирующего C20-элонгазу, включая:
(a) нуклеотидную последовательность, кодирующую полипептид, который обладает активностью C20-элонгазы, где данный полипептид имеет по меньшей мере 80% аминокислотную идентичность, основываясь на методе группировки Clustal V, при сравнении с аминокислотной последовательностью, как изложено в SEQ ID NO:12, SEQ ID NO:22, SEQ ID NO:95, SEQ ID NO:96, SEQ ID NO:97, SEQ ID NO:202 (EgDHAsyn1 домен C20-элонгазы), SEQ ID NO:204 (EgDHAsyn2 домен C20-элонгазы), SEQ ID NO:231 (EaDHAsyn1 домен C20-элонгазы), SEQ ID NO:232 (EaDHAsyn2 домен C20-элонгазы) или SEQ ID NO:233 (EaDHAsyn3 домен C20-элонгазы);
(b) нуклеотидную последовательность, кодирующую полипептид, который обладает активностью C20-элонгазы, где данная нуклеотидная последовательность имеет по меньшей мере 80% идентичность последовательности, основываясь на сравнительном анализе первичной структуры согласно программе BLASTN, при сравнении с нуклеотидной последовательностью, как изложено в SEQ ID NO:11, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93, SEQ ID NO:183, SEQ ID NO:188, SEQ ID NO:201 (EgDHAsyn1 домен C20-элонгазы), SEQ ID NO:206 (EgDHAsyn1* домен C20-элонгазы), SEQ ID NO:203 (EgDHAsyn2 домен C20-элонгазы), SEQ ID NO:227 (EaDHAsyn1 домен C20-элонгазы), SEQ ID NO:228 (EaDHAsyn2 домен C20-элонгазы), SEQ ID NO:229 (EaDHAsyn3 домен C20-элонгазы) или SEQ ID NO:230 (EaDHAsyn4 домен C20-элонгазы);
(c) нуклеотидную последовательность, кодирующую полипептид, который обладает активностью C20-элонгазы, где данный нуклеотид гибридизуется при жестких условиях с нуклеотидной последовательностью, как изложено в SEQ ID NO:11, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93, SEQ ID NO:183, SEQ ID NO:188, SEQ ID NO:201 (EgDHAsyn1 домен C20-элонгазы), SEQ ID NO:206 (EgDHAsyn1* домен C20-элонгазы), SEQ ID NO:203 (EgDHAsyn2 домен C20-элонгазы), SEQ ID NO:227 (EaDHAsyn1 домен C20-элонгазы), SEQ ID NO:228 (EaDHAsyn2 домен C20-элонгазы), SEQ ID NO:229 (EaDHAsyn3 домен C20-элонгазы) или SEQ ID NO:230 (EaDHAsyn4 домен C20-элонгазы); или
(d) комплемент нуклеотидной последовательности (a), (b) или (c), где данные комплемент и нуклеотидная последовательность состоят из одинакового количества нуклеотидов и являются на 100% комплементарными.
В восьмом варианте данное изобретение касается выделенного полинуклеотида, кодирующего дельта-4-десатуразу, включая:
(a) нуклеотидную последовательность, кодирующую полипептид, который обладает дельта-4-десатуразной активностью, где данный полипептид имеет по меньшей мере 80% аминокислотную идентичность, основываясь на методе группировки Clustal V, при сравнении с аминокислотной последовательностью, как изложено в SEQ ID NO:12, SEQ ID NO:22, SEQ ID NO:95, SEQ ID NO:96, SEQ ID NO:97, SEQ ID NO:193, SEQ ID NO:215, SEQ ID NO:217, SEQ ID NO:221, SEQ ID NO:239, SEQ ID NO:240, SEQ ID NO:241, SEQ ID NO:246, SEQ ID NO:247, SEQ ID NO:248, SEQ ID NO:249, SEQ ID NO:382, SEQ ID NO:384, SEQ ID NO:386, SEQ ID NO:388, SEQ ID NO:404, SEQ ID NO:406, или SEQ ID NO:408;
(b) нуклеотидную последовательность, кодирующую полипептид, который обладает активностью дельта-4-десатуразы, где данная нуклеотидная последовательность имеет по меньшей мере 80% идентичность последовательности, основываясь на сравнительном анализе первичной структуры согласно программе BLASTN, при сравнении с нуклеотидной последовательностью, как изложено в SEQ ID NO:11, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93, SEQ ID NO:192, SEQ ID NO:214, SEQ ID No:216, SEQ ID NO:220, SEQ ID NO:236, SEQ ID NO:237, SEQ ID NO:238, SEQ ID NO:242, SEQ ID NO:243, SEQ ID NO:244, SEQ ID NO:245, SEQ ID NO:381, SEQ ID NO:383, SEQ ID NO:385, SEQ ID NO:387, SEQ ID NO:403, SEQ ID NO:405, или SEQ ID NO:407;
(c) нуклеотидную последовательность, кодирующую полипептид, который обладает активностью дельта-4-десатуразы, где данная нуклеотидная последовательность гибридизуется при жестких условиях с нуклеотидной последовательностью, как изложено в SEQ ID NO:11, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93, SEQ ID NO:214, SEQ ID NO:216, SEQ ID NO:220, SEQ ID NO:236, SEQ ID NO:237, SEQ ID NO:238, SEQ ID NO:242, SEQ ID NO:243, SEQ ID NO:244, SEQ ID NO:245, SEQ ID NO: 381, SEQ ID NO:383, SEQ ID NO:385, SEQ ID NO:387, SEQ ID NO:403, SEQ ID NO:405, или SEQ ID NO:407; или
(d) комплемент нуклеотидной последовательности (a), (b) или (c), где данные комплемент и нуклеотидная последовательность состоят из одинакового количества нуклеотидов и являются на 100% комплементарными.
В девятом варианте данное изобретение касается выделенного полинуклеотида, который кодирует ДГК синтазу, указанный полинуклеотид включает последовательность, представленную в любой из SEQ ID NO:11, SEQ ID NO:205, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93, или SEQ ID NO:410.
В десятом варианте данное изобретение касается выделенного полинуклеотида, кодирующего C20-элонгазу, указанный фермент полинуклеотид кодирует C20-элонгазу, указанный полинуклеотид включает последовательность, представленную в любой из SEQ ID NO:183, SEQ ID NO:188, SEQ ID NO:201 (EgDHAsyn1 домен C20-элонгазы, SEQ ID NO:206 (EgDHAsyn1* домен C20-элонгазы), SEQ ID NO:203 (EgDHAsyn2 домен C20-элонгазы), SEQ ID NO:227 (EaDHAsyn1 домен C20-элонгазы), SEQ ID NO:228 (EaDHAsyn2 домен C20-элонгазы), SEQ ID NO:229 (EaDHAsyn3 домен C20-элонгазы) или SEQ ID NO:230 (EaDHAsyn4 домен C20-элонгазы).
В одиннадцатом варианте данное изобретение касается выделенного полинуклеотида, кодирующего дельта-4-десатуразу, указанный полинуклеотид включает последовательность, представленную в SEQ ID NO:192, SEQ ID NO:214, SEQ ID NO:220, SEQ ID NO:236, SEQ ID NO:237, SEQ ID NO:238, SEQ ID NO:242, SEQ ID NO:243, SEQ ID NO:244, SEQ ID NO:245, SEQ ID NO:381, SEQ ID NO:383, SEQ ID NO:385, SEQ ID No:387, SEQ ID NO:403, SEQ ID NO:405, или SEQ ID NO:407.
В двенадцатом варианте данное изобретение касается рекомбинантного конструкционного элемента, который включает любой из выделенных полинуклеотидов данного изобретения, связанный оперативно с по меньшей мере одной регуляторной последовательностью.
В тринадцатом варианте данное изобретение касается клетки-хозяина, которая содержит в своем геноме рекомбинантный конструкционный элемент данного изобретения. Более конкретно, клетка-хозяин представляет собой рекомбинантную микробную клетку-хозяина, включающую мультизим изобретения, где первой ферментативной активностью является дельта-9-элонгаза, и второй ферментативной активностью является дельта-8-десатураза. В другом аспекте первая ферментативная активность представляет собой С20-элонгазу, и вторая ферментативная активность представляет собой дельта-4-десатуразу.
В четырнадцатом варианте данное изобретение касается трансформированного вида Yarrowia, включающего рекомбинантный конструкционный элемент изобретения.
В пятнадцатом варианте данное изобретение касается способа трансформирования клетки, который включает трансформацию клетки рекомбинантным конструкционным элементом данного изобретения и селекцию этих клеток, трансформированных посредством указанного рекомбинантного конструкционного элемента.
В шестнадцатом варианте данное изобретение касается способа получения трансформированного растения, который включает трансформацию растительной клетки любым из полинуклеотидов данного изобретения и восстановление растения из трансформированной растительной клетки.
В семнадцатом варианте данное изобретение касается способа получения дрожжей, который включает трансформацию дрожжевой клетки любым из полинуклеотидов данного изобретения и выращивание дрожжей из трансформированной дрожжевой клетки.
В восемнадцатом варианте данное изобретение касается растения, которое содержит в своем геноме рекомбинантный конструкционный элемент изобретения. Также представляют интерес семена, полученные от таких растений, масло, полученное из таких семян, пища или корм, которые содержат такое масло, и напиток, который содержит масло данного изобретения.
В девятнадцатом варианте данное изобретение касается изолированной молекулы нуклеиновой кислоты, которая кодирует C20-элонгазу, как представлено в SEQ ID NO:183, где по меньшей мере 147 кодонов кодон-оптимизировано для экспрессии в Yarrowia.
В двадцатом варианте данное изобретение касается изолированной молекулы нуклеиновой кислоты, которая кодирует C20-элонгазу, как представлено в SEQ ID NO:188, где по меньшей мере 134 кодона кодон-оптимизированы для экспрессии в Yarrowia.
В двадцать первом варианте данное изобретение касается изолированной молекулы нуклеиновой кислоты, которая кодирует дельта-4-десатуразный фермент, как представлено в SEQ ID NO:192, где по меньшей мере 285 кодонов кодон-оптимизированы для экспрессии в Yarrowia.
В двадцать втором варианте данное изобретение касается способа получения мультизима, который включает:
(a) связывание первого полипетида с по меньшей мере вторым полипептидом, где каждый полиппетид обладает независимой и раздельной ферментативной активностью; и
(b) оценку продукта стадии (a) в отношении независимой и раздельной ферментативных активностей.
В двадцать третьем варианте данное изобретение касается способа изменения жирнокислотного профиля масличного растения, который включает:
a) трансформацию клетки масличного растения посредством рекомбинантного конструкционного элемента данного изобретения;
b) восстановление растения из трансформированной клетки масличного растения стадии (a), где данное растение имеет измененный жирнокислотный профиль.
В двадцать четвертом варианте данное изобретение касается выделенного полинуклеотида, кодирующего ДГЛК синтазу, включая:
(a) нуклеотидную последовательность, кодирующую полипептид, который обладает ДГЛК синтазной активностью, где данный полипептид представлен в SEQ ID NO:441, SEQ ID NO:454, SEQ ID NO:461, SEQ ID NO:464, SEQ ID NO:471, SEQ ID NO:515, SEQ ID NO:516, SEQ ID NO:517, SEQ ID NO:518, или SEQ ID NO:519;
(b) нуклеотидную последовательность, кодирующую полипептид, который обладает ДГЛК синтазной активностью, где данная нуклеотидная последовательность представлена в SEQ ID NO:440, SEQ ID NO:446, SEQ ID NO:453, SEQ ID NO:460, SEQ ID NO:463, SEQ ID NO:470, SEQ ID NO:492, SEQ ID NO:493, SEQ ID NO:494, SEQ ID NO:495, или SEQ ID NO:496;
(c) нуклеотидную последовательность, кодирующую полиппетид, котрый обладает ДГЛК синтазной активностью, где данная нуклеотидная последовательность гибридизуется при жестких условиях с нуклеотидной последовательностью, как представлено в SEQ ID NO:440, SEQ ID NO:446, SEQ ID NO:453, SEQ ID NO:460, SEQ ID NO:463, SEQ ID NO:470, SEQ ID NO:492, SEQ ID NO:493, SEQ ID NO:494, SEQ ID NO:495, или SEQ ID NO:496; или
комплемент нуклеотидной последовательности (a), (b) или (c), где данные комплемент и нуклеотидная последовательность состоят из одинакового количества нуклеотидов и являются на 100% комплементарными.
В двадцать пятом варианте данное изобретение касается способа превращения линолевой кислоты в дигомо гамма-линоленовую кислоту, которая включает:
a) предоставление рекомбинантной микробной клетки-хозяина, которая включает:
i) ДГЛК синтазу, включая:
1) по меньшей мере один полипептид, кодирующий дельта-9-элонгазу;
2) по меньшей мере один полипептид, кодирующий дельта-8-десатуразу; и
3) полипептидный линкер;
где данный линкер помещен между дельта-9-элонгазой и дельта-8-десатуразой; и
ii) источник линолевой кислоты; и
b) выращивание клетки-хозяина (a) при условиях, в которых образуется дигомо гамма-линоленовая кислота.
В двадцать шестом варианте данное изобретение касается способа превращения α-линоленовой кислоты в эйкозатриеновую кислоту, который включает:
a) предоставление рекомбинантной микробной клетки-хозяина, которая включает:
i) ДГЛК синтазу, включая:
1) по меньшей мере один полипептид, кодирующий дельта-9-элонгазу;
2) по меньшей мере один полипептид, кодирующий дельта-8-десатуразу; и
3) полипептидный линкер;
где данный линкер помещен между дельта-9-элонгазой и дельта-8-десатуразой; и
ii) источник α-линоленовой кислоты; и
b) выращивание клетки-хозяина (a) при условиях, в которых образуется эйкозатриеновая кислота.
В двадцать седьмом варианте данное изобретение касается способа превращения эйкозапентаеновой кислоты в докозагексаеновую кислоту, котрый включает:
a) предоставление рекомбинантной микробной клетки-хозяина, которая включает:
i) ДГК синтазу, включая:
1) по меньшей мере один полипептид, кодирующий C20-элонгазу;
2) по меньшей мере один полипептид, кодирующий дельта-4-десатуразу; и
3) полипептидный линкер;
где данный линкер помещен между C20-элонгазой и дельта-4-десатуразой; и
ii) источник ейкозапентаеновой кислоты; и
b) выращивание клетки-хозяина (a) при условиях, в которых образуется докозагексаеновая кислота.
В двадцать восьмом варианте данное изобретение касается способа превращения арахидоновой кислоты в докозапентаеновую кислоту, который включает:
a) предоставление рекомбинантной микробной клетки-хозяина, которая включает:
i) ДГК синтазу, включая:
1) по меньшей мере один полипептид, кодирующий C20-элонгазу;
2) по меньшей мере один полипептид, кодирующий дельта-4-десатуразу; и
3) полипептидный линкер;
где данный линкер помещен между C20-элонгазой и дельта-4-десатуразой; и
ii) источник арахидоновой кислоты; и
b) выращивание клетки-хозяина (a) при условиях, в которых образуется докозапентаеновая кислота.
В двадцать девятом варианте данное изобретение касается способа идентификации полипептида, обладающего улучшенной активностью дельта-4-десатуразы, который включает:
a) предоставление дельта-4-десатуразного полипептида дикого типа, выделенного из Euglena anabena, обладающего базисной активностью дельта-4-десатуразы;
b) усечение полипептида (a) дикого типа на приблизительно от 1 до приблизительно 200 аминокислот для создания усеченного мутантного полипептида, обладающего активностью дельта-4-десатуразы, которая повышена по сравнению с базисной активностью дельта-4-десатуразы.
В тридцатом варианте данное изобретение касается микробной клетки-хозяина, которая продуцирует полиненасыщенную жирную кислоту и экспрессирует полипептиды, кодирующие ферменты, на следующем последовательном пути:
1) дельта-9-десатураза,
2) дельта-12-десатураза,
3) дельта-9-элонгаза,
4) дельта-8-десатураза,
5) дельта-5-десатураза,
6) дельта-17-десатураза,
7) C20/22-элонгаза, и
8) дельта-4-десатураза;
где данные полипептиды включают по меньшей мере один мультизим, слияние, включая слияние между по меньшей мере одной смежной ферментной парой.
БИОЛОГИЧЕСКИЕ ДЕПОЗИТЫ
Следующие биологические материалы сохраняются в Американском собрании типовых культур (АСТК), 10801 University Boulevard, Manassas, VA 20110-2209, и имеют следующие обозначения, номера доступа и даты вложения (Таблица 1).
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ И ЛИСТИНГОВ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Данное изобретение может быть лучше понято из последующего подробного описания и сопроводительных рисунков, и листинга последовательностей, которые образуют часть данной заявки.
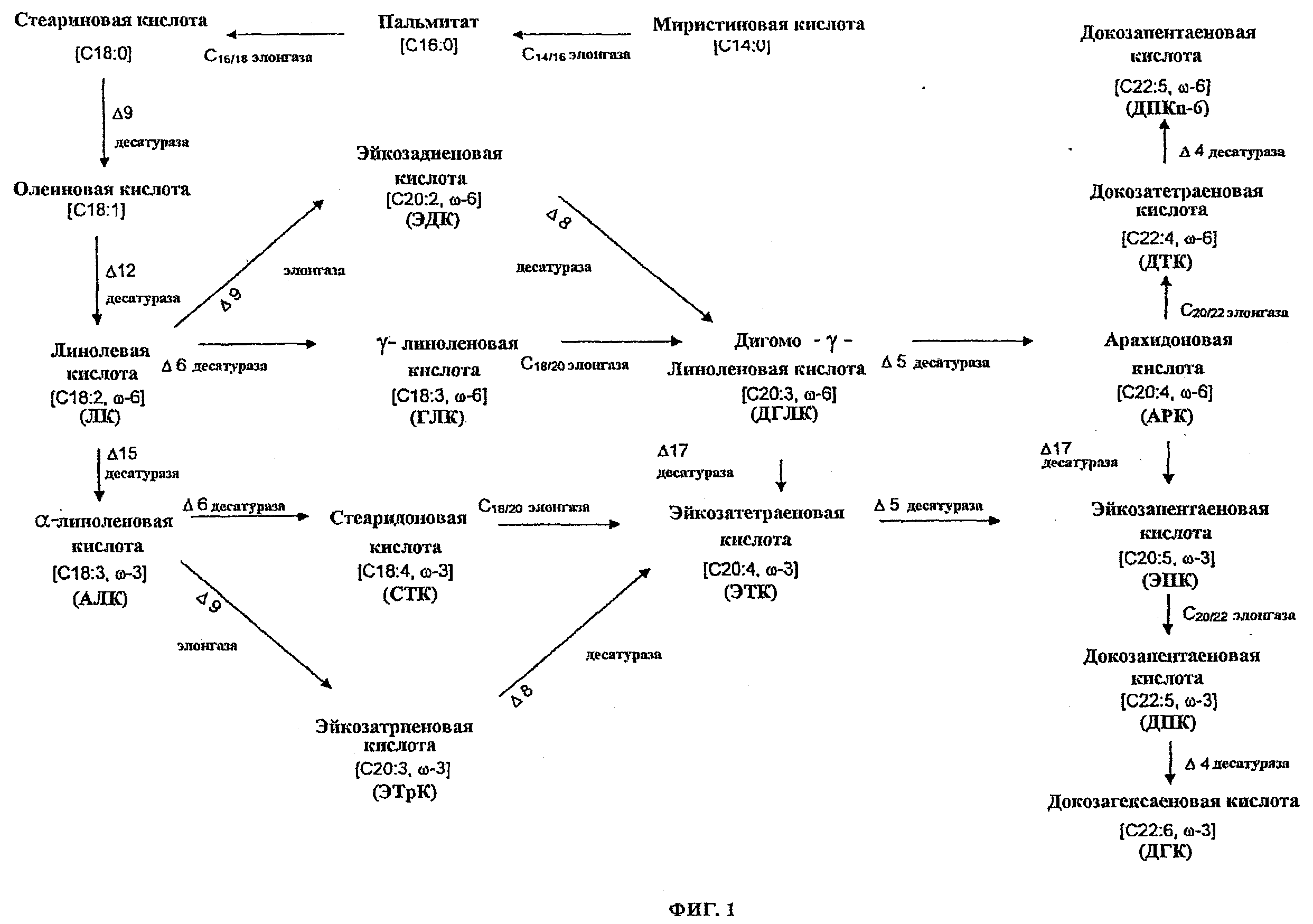
ФИГ.1 представляет собой репрезентативный омега-3 и омега-6 жирнокислотный путь, представленный для превращения миристиновой кислоты через разные интермедиаты в ДГК.
ФИГ.2 изображает группировку Clustal W между участком кодирующей последовательности EgDHAsyn2 (SEQ ID NO:21), кДНК последовательностью Euglena gracilis дельта-40-десатуразы (SEQ ID NO:23) (номер доступа НЦБИ AY278558 (GI 33466345), локус AY278558, Meyer et al., Biochemistry 42(32):9779-9788 (2003)), и кодирующей последовательностью Euglena gracilis дельта-4-десатуразы (SEQ ID NO:24) (Meyer et al., supra).
ФИГ.3A и 3B изображают группировку Clustal W между аминокислотной последовательностью EgDHAsyn1 (SEQ ID NO:12), EgDHAsyn2 (SEQ ID NO:22), и EgC20elo1 (SEQ ID NO:6).
ФИГ.4A и 4B изображают группировку Clustal W N-конца EgDHAsyn1 (SEQ ID NO:12) и N-конца EgDHAsyn2 (SEQ ID NO:22) ch EgC20elo1 (SEQ ID NO:6), Pavlova sp. CCMP459 C20-PUFA Elo (SEQ ID NO:2), Ostreococcus tauri ПНЖК элонгаза 2 (SEQ ID NO:25) (номер доступа НЦБИ AAV67798 (GI 55852396), локус AAV67798, CDS AY591336; Meyer et al., J. Lipid Res. 45(10):1899-1909 (2004)), и Thalassiosira pseudonana ПНЖК элонгаза 2 (SEQ ID NO:26) (номер доступа НЦБИ AAV67800 (GI 55852441), локус AAV67800, CDS AY591338; Meyer et al., J. Lipid Res. 45(10):1899-1909 (2004)).
ФИГ.5A , 5B, 5C и 5D изображают группировку Clustal W C-конца EgDHAsyn1 (EgDHAsyn1_CT; аминокислоты 253-793 SEQ ID NO:12; N-конец EgDHAsyn1 не показан и обозначен “…”) и C-конца EgDHAsyn2 (EgDHAsyn2_CT; аминокислоты 253-793 SEQ ID NO:22, N-конец EgDHAsyn2 не показан и обозначен “…”) с Euglena gracilis дельта-4-десатуразы жирных кислот (SEQ ID NO:13), Thraustochytrium aureum дельта-4-десатуразой (SEQ ID NO:27) (номер доступа НЦБИ AAN75707(GI 25956288), локус AAN75707, CDS AF391543), Schizochytrium aggregatum дельта-4-десатуразой (SEQ ID NO:28) (PCT публикация № WO 2002/090493), Thalassiosira pseudonana дельта-4-десатуразой (SEQ ID NO:29) (номер доступа НЦБИ AAX14506 (GI 60173017), локус AAX14506, CDS AY817156; Tonon et al., FEBS J. 272 (13):3401-3412 (2005)), и Isochrysis galbana дельта-4-десатуразой (SEQ ID NO:30) (номер доступа НЦБИ AAV33631 (GI 54307110), локус AAV33631, CDS AY630574; Pereira et al., Biochem. J., 384(2):357-366 (2004) и PCT публикация № WO 2002/090493).
ФИГ.6 изображает группировку внутренних фрагментов EgDHAsyn1 (EgDHAsyn1_ NCT; аминокислот 253-365 SEQ ID NO:12) и EgDHAsyn2 (EgDHAsyn2_NCT; аминокислот 253-365 SEQ ID NO:22), связывающую как участок C20-элонгазы, так и дельта-4-десатуразный домен (основываясь на гомологии), с C-концами C20-элонгаз (EgC20elo1_CT, аминокислот 246-298 SEQ ID NO:6; PavC20elo_CT, аминокислот 240-277 SEQ ID NO:2; OtPUFAelo2_CT, аминокислот 256-300 SEQ ID NO:25; TpPUFAelo2_CT, аминокислот 279-358 SEQ ID NO:26) и N-концы дельта-4-десатураз (EgD4_NT, аминокислот 1-116 SEQ ID NO:13; TaD4_NT, аминокислот 1-47 SEQ ID NO:27; SaD4_NT, аминокислот 1-47 SEQ ID NO:28; TpD4_NT, аминокислот 1-82 SEQ ID NO:29; IgD4_NT, аминокислот 1-43 SEQ ID NO:30).
ФИГ.7 представляет плазмидные карты для следующего: (A) pY115 (см. также SEQ ID NO:33); (B) Yarrowia lipolytica Gateway® дестинационный вектор pBY1 (см. также SEQ ID NO:34); (C) Yarrowia lipolytica Gateway® дестинационный вектор pY159 (см. также SEQ ID NO:38); и (D) pBY-EgC20elo1 (см. также SEQ ID NO:39).
ФИГ.8 представляет плазмидные карты для следующего: (A) pY132 (см. также SEQ ID NO:40); (B) pY161 (см. также SEQ ID NO:41); (C) pY164 (см.также SEQ ID NO:42); и (D) pY141 (см. также SEQ ID NO:49).
ФИГ.9 представляет плазмидные карты для следующего: (A) pY143 (см. также SEQ ID NO:52); (B) pY149 (см. также SEQ ID NO:55); (C) pY150 (см. также SEQ ID NO:62); и (D) pY156 (см. также SEQ ID NO:64).
ФИГ.10 представляет плазмидные карты для следующего: (A) pY152 (см. также SEQ ID NO:67); (B) pY157 (см. также SEQ ID NO:69); (C) pY153 (см. также SEQ ID NO:72); и (D) pY151 (см. также SEQ ID NO:76).
ФИГ.11 представляет собой карту pY160 (см. также SEQ ID NO:77).
ФИГ.12 представляет хроматограмму липидного профиля Euglena anabaena клеточного экстракта, как описано в Примерах.
ФИГ.13A, 13B и 13C изображают группировку Clustal W аминокислотных последовательностей для EaDHAsyn1 (SEQ ID NO:95), EaDHAsyn2 (SEQ ID NO:96), EaDHAsyn3 (SEQ ID NO:97), и EaDHAsyn4 (SEQ ID NO:98).
ФИГ.14 представляет плазмидные карты для следующего: (A) pY165 (см. также SEQ ID NO:99); (B) pY166 (см. также SEQ ID NO:100); (C) pY167 (см. также SEQ ID NO:101); и (D) pY168 (см. также SEQ ID NO:102).
ФИГ.15 представляет плазмидные карты для следующего: (A) pKR1061 (см. также SEQ ID NO:111); (B) pKR973 (см. также SEQ ID NO:128); (C) pKR1064 (см. также SEQ ID NO:132); и (D) pKR1133 (см. также SEQ ID NO:145).
ФИГ.16 представляет плазмидные карты для следующего: (A) pKR1105 (см. также SEQ ID NO:156); (B) pKR1134 (см. также SEQ ID NO:161); (C) pKR1095 (см. также SEQ ID NO:167); и (D) pKR1132 (см. также SEQ ID NO:170.
ФИГ.17 представляет собой карту KS373 (см. также SEQ ID NO:179).
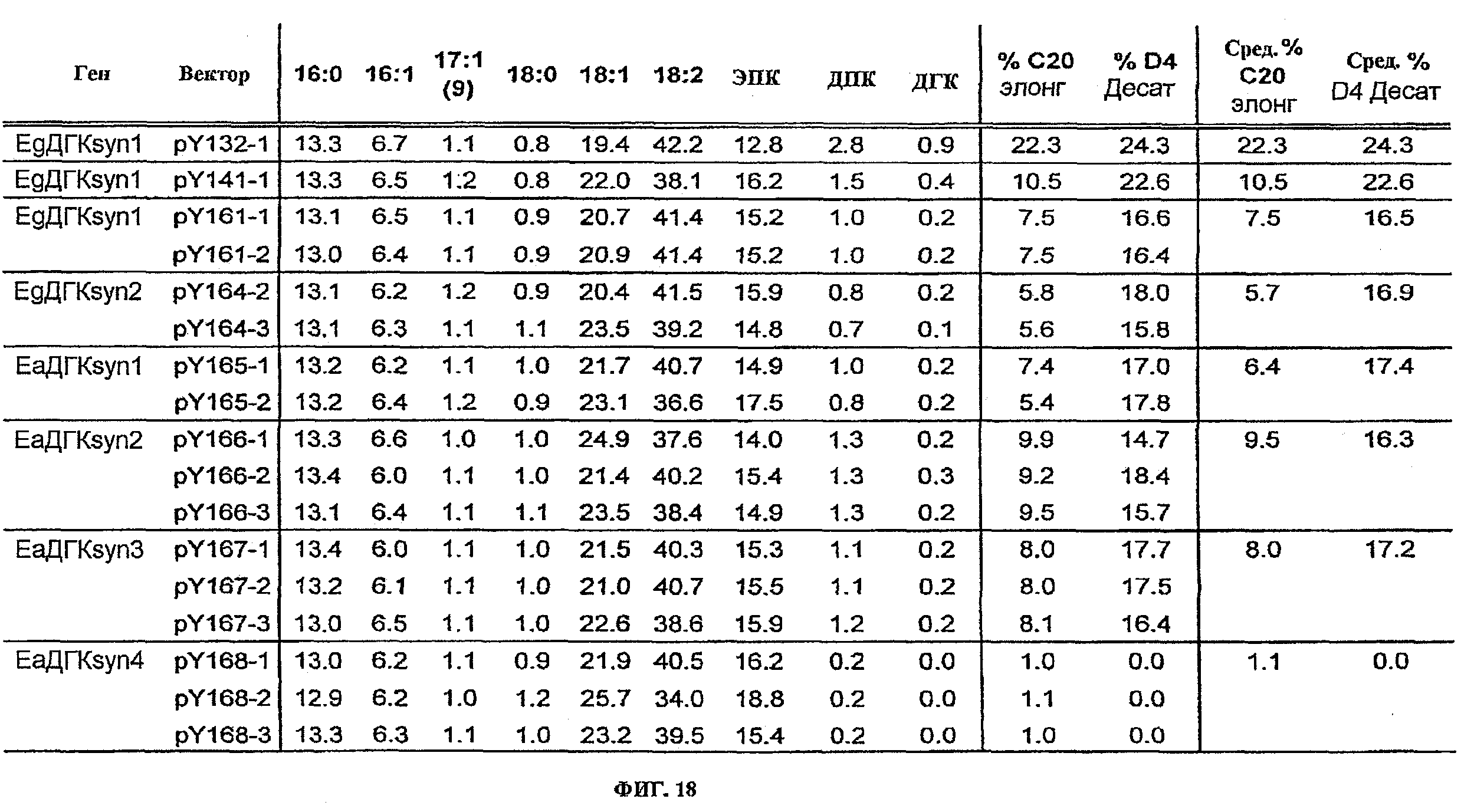
ФИГ.18 изображает жирнокислотные профили, вычисленные % элонгации, и вычисленные % десатурации для клонов (кроме pBY-EgC20elo1), показанных в Таблице 24ФИГ.19 изображает жирнокислотные профили, вычисленные % элонгации, и вычисленные % десатурации для подачи ЭПК только к контрольному вектору, pY141, pY143, pY149, pY156, pY157, и pY160.
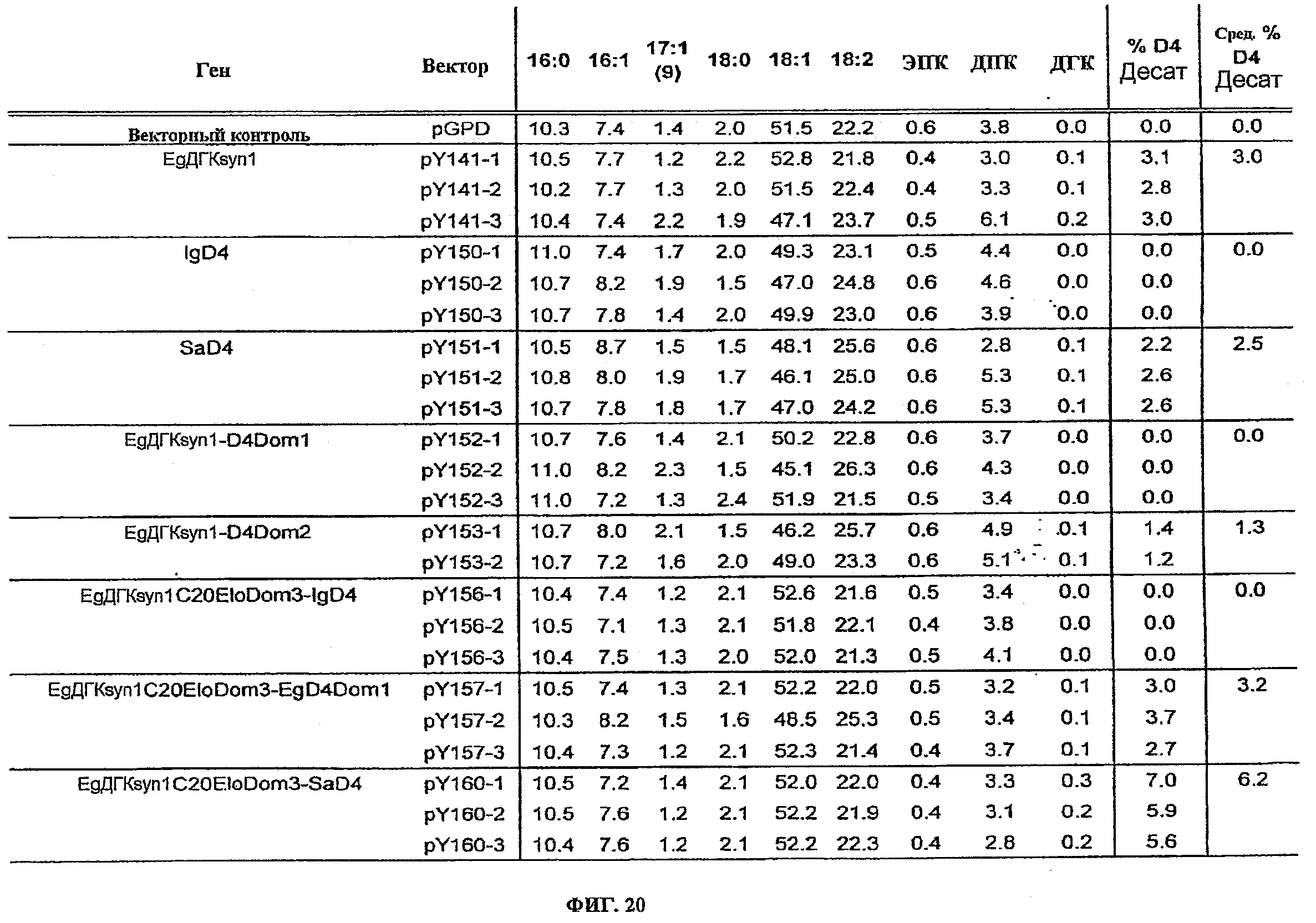
ФИГ.20 изображает жирнокислотные профили, вычисленные % элонгации и вычисленные % десатурации для подачи ДПК только к контрольному вектору, pY141, pY150, pY151, pY152, pY153, pY156, pY157, и pY160.
ФИГ.21 представляет схему относительной доменной структуры для каждого конструкционного элемента, описанного в Таблице 25.
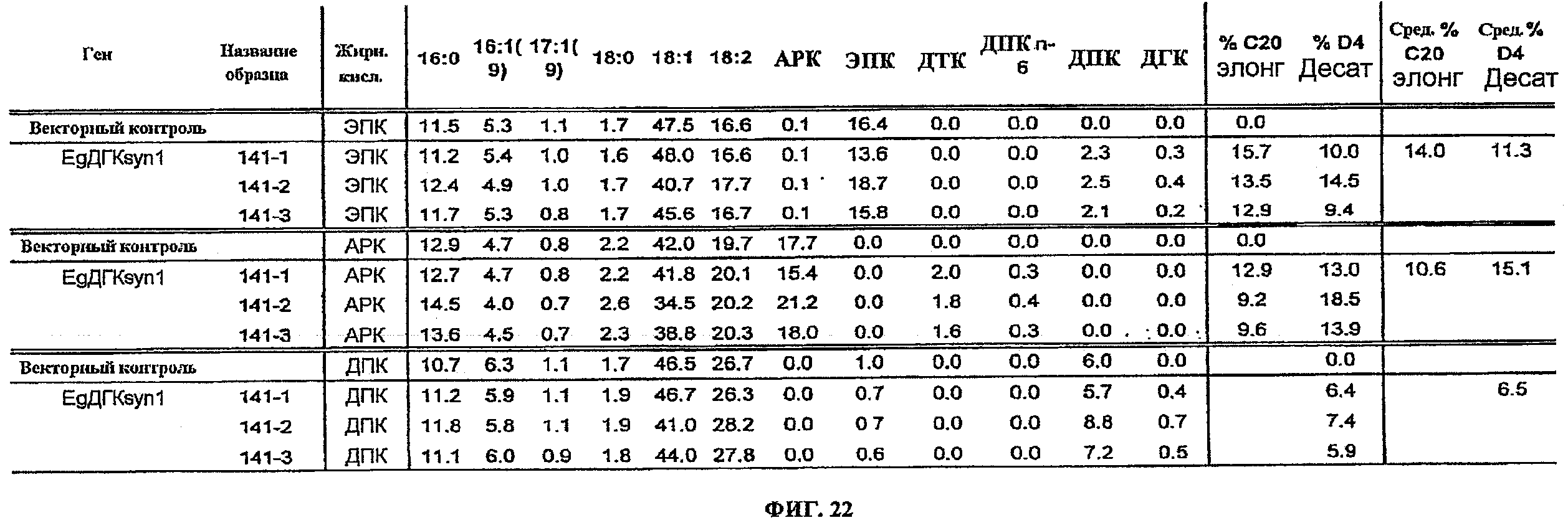
ФИГ.22 изображает жирнокислотные профили, вычисленные % элонгации и вычисленные % десатурации для подачи ЭПК, АРК и ДПК к клеткам Yarrowia, трансформированным pY141 (EgDHAsyn1; SEQ ID NO:49), и только к контрольному вектору.
ФИГ.23 изображает жирнокислотные профили для индивидуальных зародышей из репрезентативного события в соматических соевых зародышах, трансформированных соевыми экспрессирующими векторами pKR973 и pKR1064 (см. Таблицу 26).
ФИГ.24 изображает жирнокислотные профили из пяти наилучших элонгационных событий в соевых зародышах, трансформированных соевым экспрессирующим вектором KS373.
ФИГ.25 резюмирует данные BLASTP и значения процентов идентичности для EgC20elo1 (Пример 3), EgDHAsyn1 (Пример 4), и EgDHAsyn2 (Пример 5).
ФИГ.26 изображает жирнокислотные профили от питания соевых зародышей ЭПК. Данные соевые зародыши были выбраны из наилучших C20/дельта-5-элонгазной и дельта-4-десатуразной активностей в соевых зародышах, трансформированных соевым экспрессирующим вектором pKR1105.
ФИГ.27 изображает хроматограмму липидного профиля Euglena gracilis клеточного экстракта, как описано в данных Примерах.
ФИГ.28 представляет собой карту pKR1183 (см. также SEQ ID NO:266).
ФИГ.29 резюмирует Euglena anabaena ДГК синтазные доменные последовательности.
ФИГ.30 представляет собой карту pKR1253 (см. также SEQ ID NO:270).
ФИГ.31 представляет собой карту pKR1255 (см. также SEQ ID NO:275).
ФИГ.32 представляет собой карту pKR1189 (см. также SEQ ID NO:285).
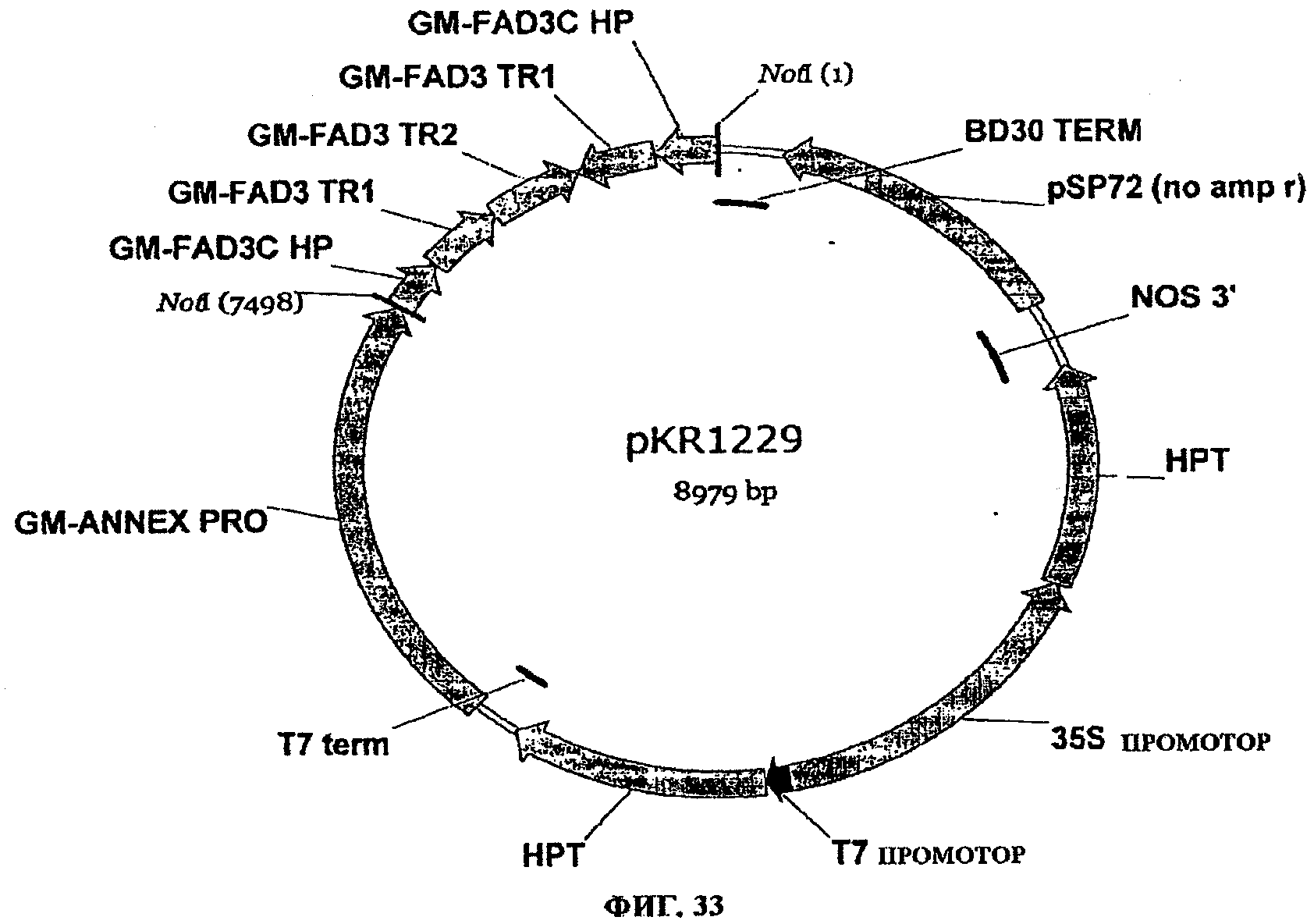
ФИГ.33 представляет собой карту pKR1229 (см. также SEQ ID NO:296).
ФИГ.34 представляет собой карту pKR1249 (см. также SEQ ID NO:297).
ФИГ.35 представляет собой карту pKR1322 (см. также SEQ ID NO:314).
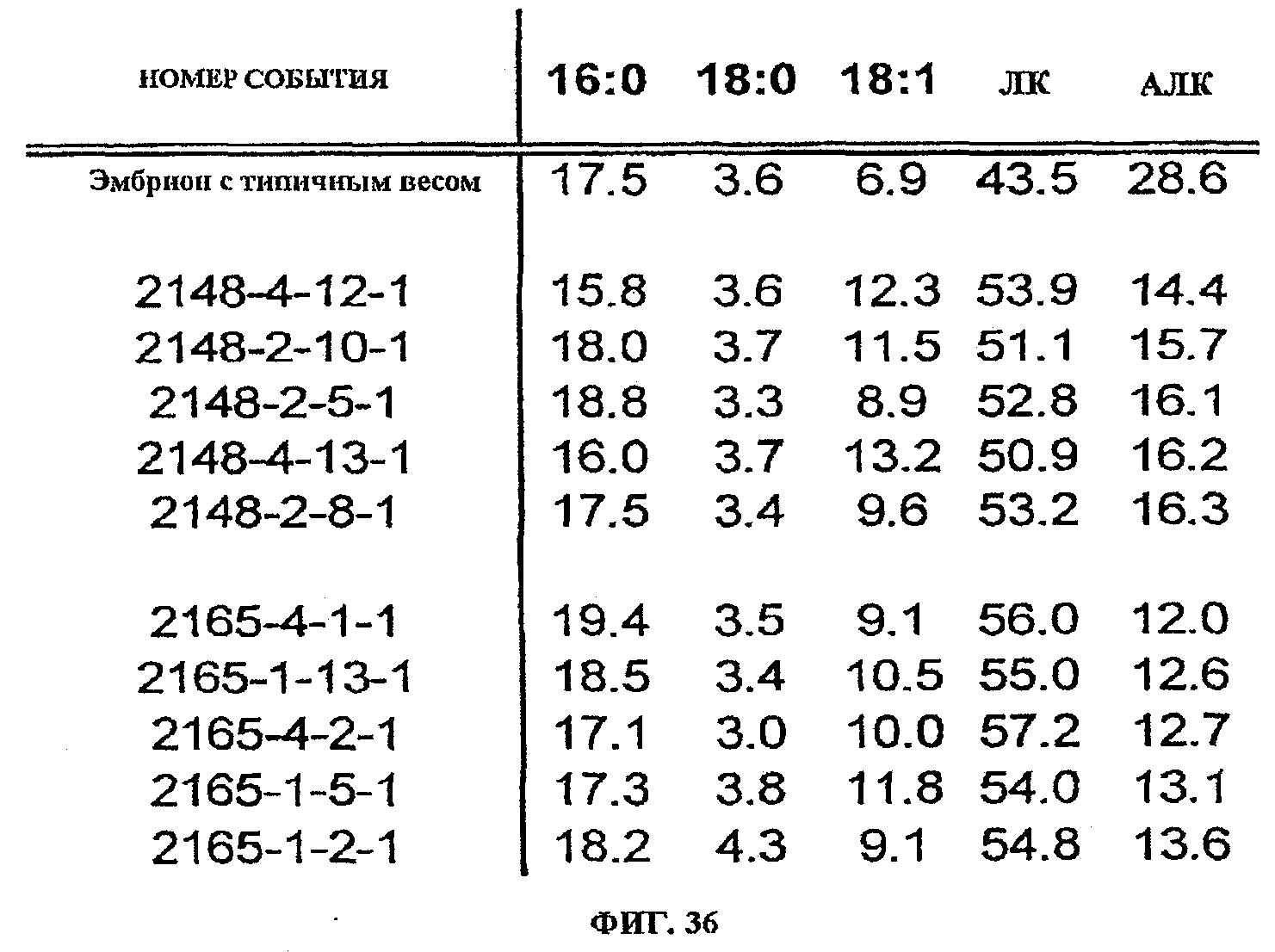
ФИГ.36 показывает жирнокислотные профили для пяти событий, трансформированных pKR1189, которые имеют самое низкое среднее содержание АЛК (среднее из 5 проанализированных соевых соматических зародышей), совместно с событием (2148-3-8-1), которое имеет жирнокислотный профиль, типичный для зародышей дикого типа для этого эксперимента. Жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК и АЛК. Составы жирных кислот выражены в виде весового процента (вес. %) от общего содержания жирных кислот.
ФИГ.37 показывает жирнокислотные профили для пяти событий, трансформированных pKR1183, которые имеют самое высокое среднее содержание ДГЛК (среднее из 5 проанализированных соевых соматических зародышей). Жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК, АЛК, ЭДК, ЭРК, ДГЛК и ЭТК. Составы жирных кислот выражены в виде весового процента (вес. %) от общего содержания жирных кислот.
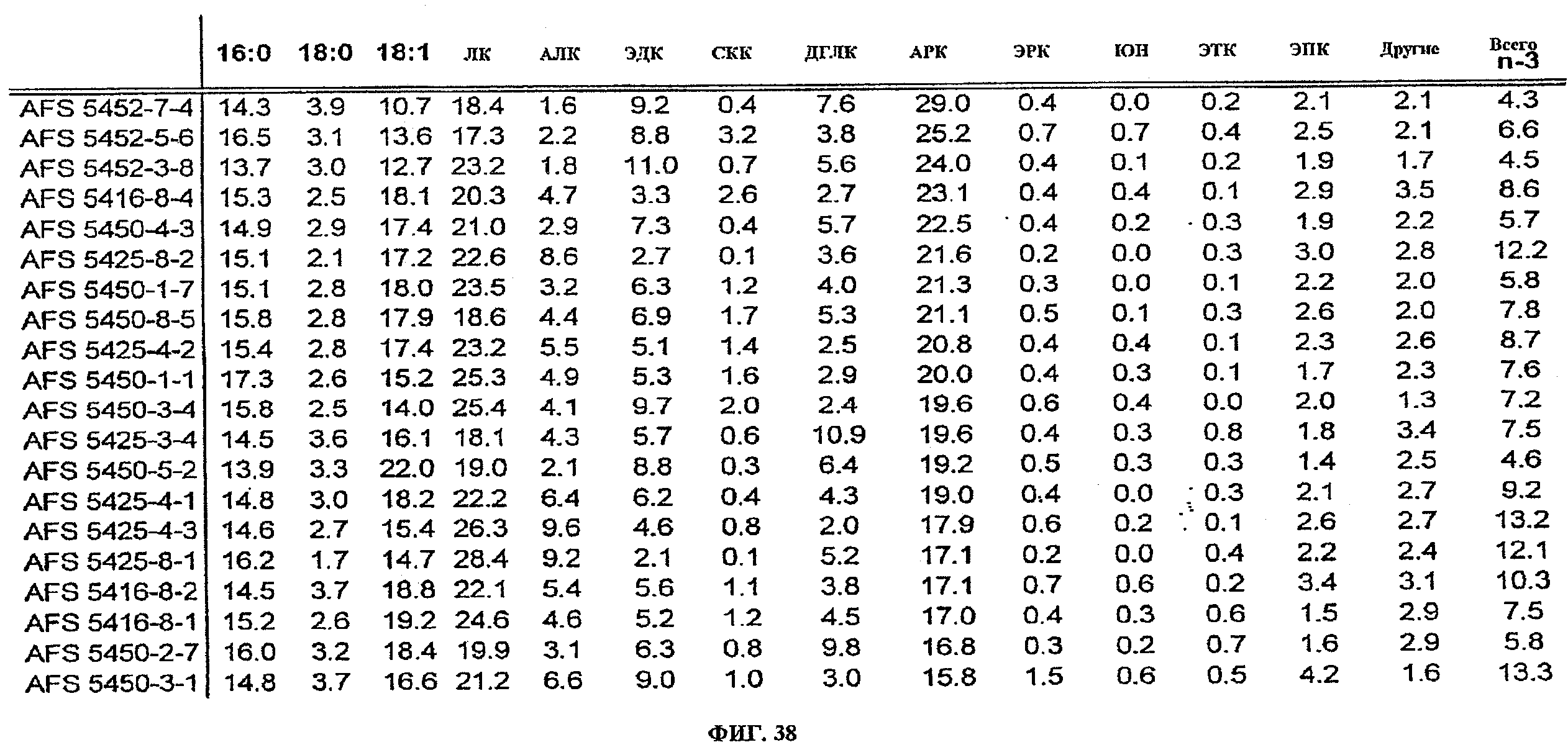
ФИГ.38 показывает усредненные жирнокислотные профили (среднее из 10 соевых соматических зародышей) для 20 событий, трансформированных pKR1249 и pKR1253, которые имеют наивысшее АРК. Жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК, АЛК, ЭДК, СКК, ДГЛК, АРК, ЭРК, ЮН, ЭТК и ЭПК. Составы жирных кислот выражены в виде весового процента (вес. %) от общего содержания жирных кислот. Жирные кислоты, перечисленные как "другие", включают: 18:2 (5,9), 18:3 (5,9,12), СТК, 20:0, 20:1(11), 20:2 (7,11) или 20:2 (8,11), и ДПК.
ФИГ.39 показывает фактические жирнокислотные профили для каждого соевого соматического зародыша из одного события (AFS 5416-8-1-1), со средним содержанием АРК 17,0% и средним содержанием ЭПК 1,5%. Жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК, АЛК, ЭДК, СКК, ДГЛК, АРК, ЭРК, ЮН, ЭТК и ЭПК. Составы жирных кислот выражены в виде весового процента (вес. %) от общего содержания жирных кислот. Жирные кислоты, перечисленные как "другие”, включают: 18:2 (5,9), 18:3 (5,9,12), СТК, 20:0, 20:1(11), 20:2 (7,11) или 20:2 (8,11), и ДПК.
ФИГ.40 показывает усредненные жирнокислотные профили (среднее из 9 или 10 соевых соматических зародышей) для 20 событий, тансформированных pKR1249 и pKR1255, которые имеют наивысшее АРК. Жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК, АЛК, ЭДК, СКК, ДГЛК, АРК, ЭРК, ЮН, ЭТК и ЭПК; составы жирных кислот выражены в виде весового процента (вес. %) от общего содержания жирных кислот. Жирные кислоты, перечисленные как "другие", включают: 18:2 (5,9), 18:3 (5,9,12), СТК, 20:0, 20:1(11), 20:2 (7,11) или 20:2 (8,11), и ДПК.
ФИГ.41 показывает жирнокислотные профили при питании зародышей ЭПК. Соевые зародыши выбирались из событий с наилучшими активностями C20-дельта-элонгазы и дельта-4-десатуразы в соевых зародышах, трансформированных соевым экспрессирующим вектором pKR1134. Жирные кислоты на ФИГ.41 идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК, АЛК, ЭПК, 22:0 (докозановая кислота), ДПК, 24:0 (тетракозановая кислота), ДГК и 24:1 (невоновая кислота). Составы жирных кислот, перечисленных на ФИГ.41, выражены в виде весового процента (вес. %) от общего содержания жирных кислот.
ФИГ.42 показывает жирнокислотные профили при питании соевых зародышей ЭПК. Соевые зародыши выбирались из событий с наилучшими активностями C20-дельта-5-элонгазы и дельта-4-десатуразы из 20 новых событий, проанализированных для сои, трансформированной pKR1105. Жирные кислоты на ФИГ.42 идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК, АЛК, ЭПК, 22:0 (докозановая кислота), ДПК, 24:0 (тетракозановая кислота), ДГК и 24:1 (невоновая кислота). Составы жирных кислот, перечисленных на ФИГ.42, выражены в виде весового процента (вес. %) от общего содержания жирных кислот.
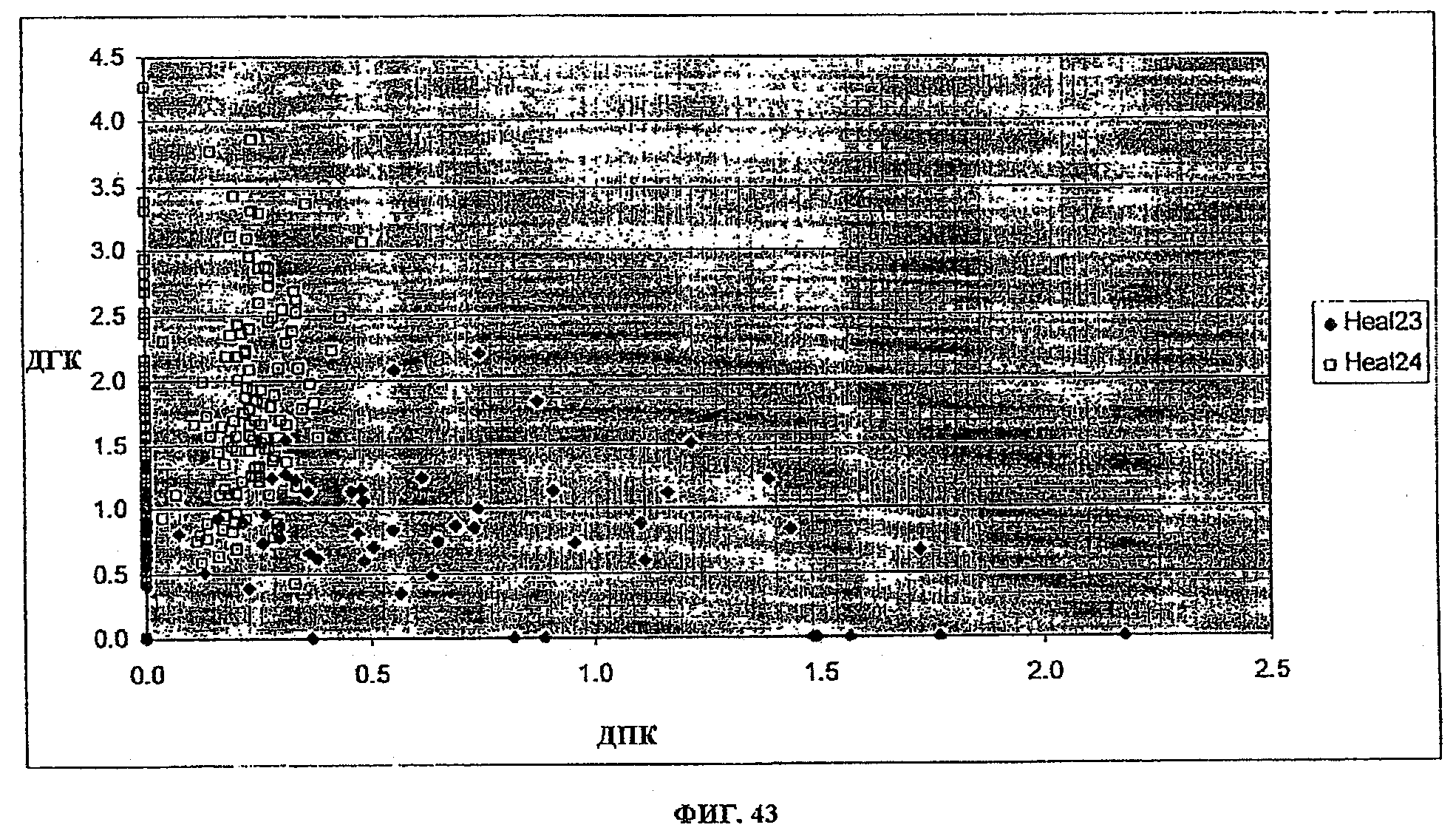
ФИГ.43 представляет собой график, изображающий относительные активности событий, трансформированных или pKR1105 (C20-элонгаза и дельта-4-десатуразы экспрессированы отдельно) или pKR1134 (C20-элонгаза и дельта-4-десатураза экспрессированы как гибрид), когда соевые зародыши питались ЭПК.
ФИГ.44 представляет схему создания штамма Y4305U3 Yarrowia lipolytica.
ФИГ.45 представляет плазмидные карты для следующего: (A) pZKLeuN-29E3 и (B) pY116.
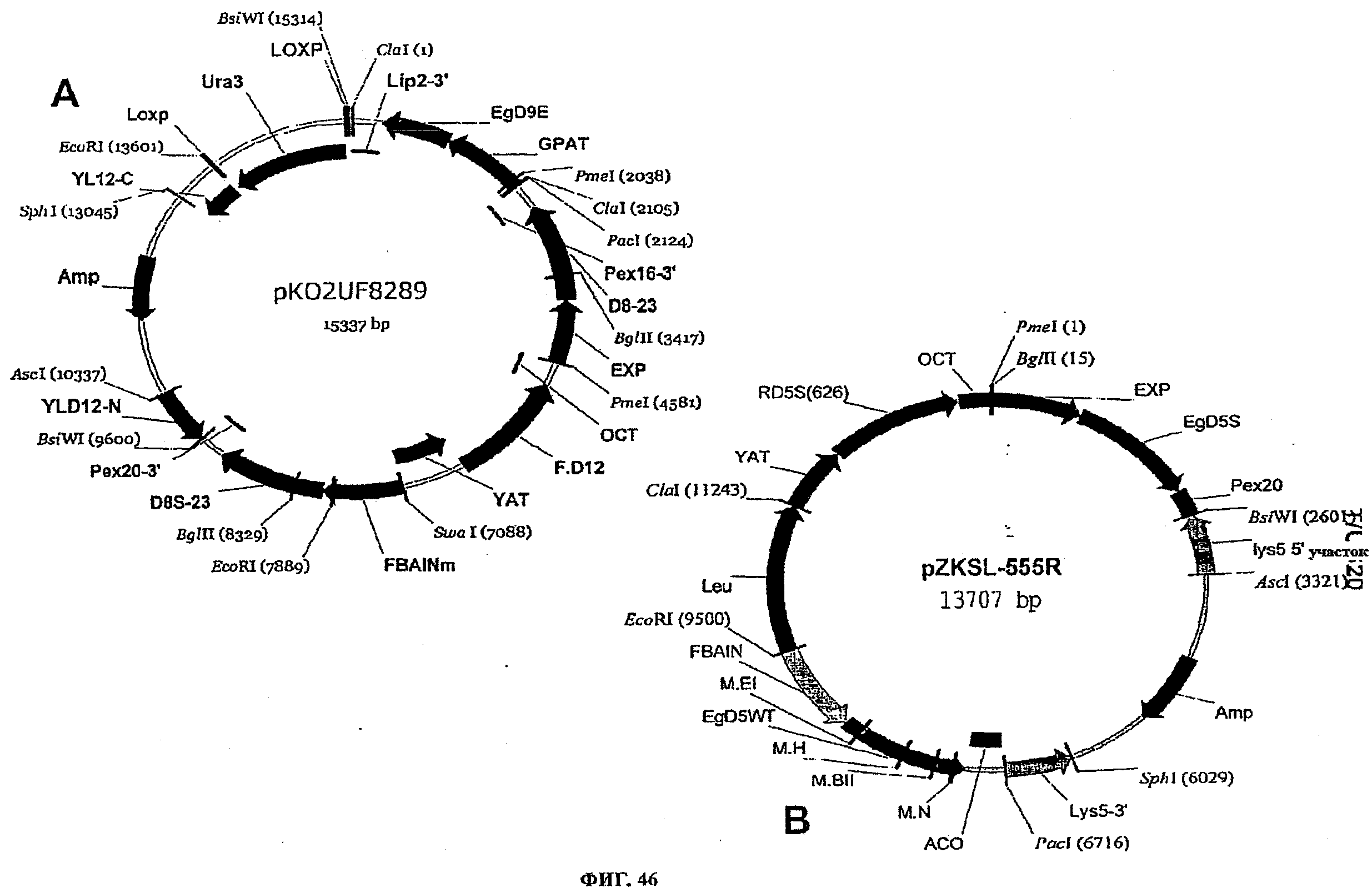
ФИГ.46 представляет плазмидные карты для следующего: (A) pKO2UF8289 и (B) pZKSL-555R.
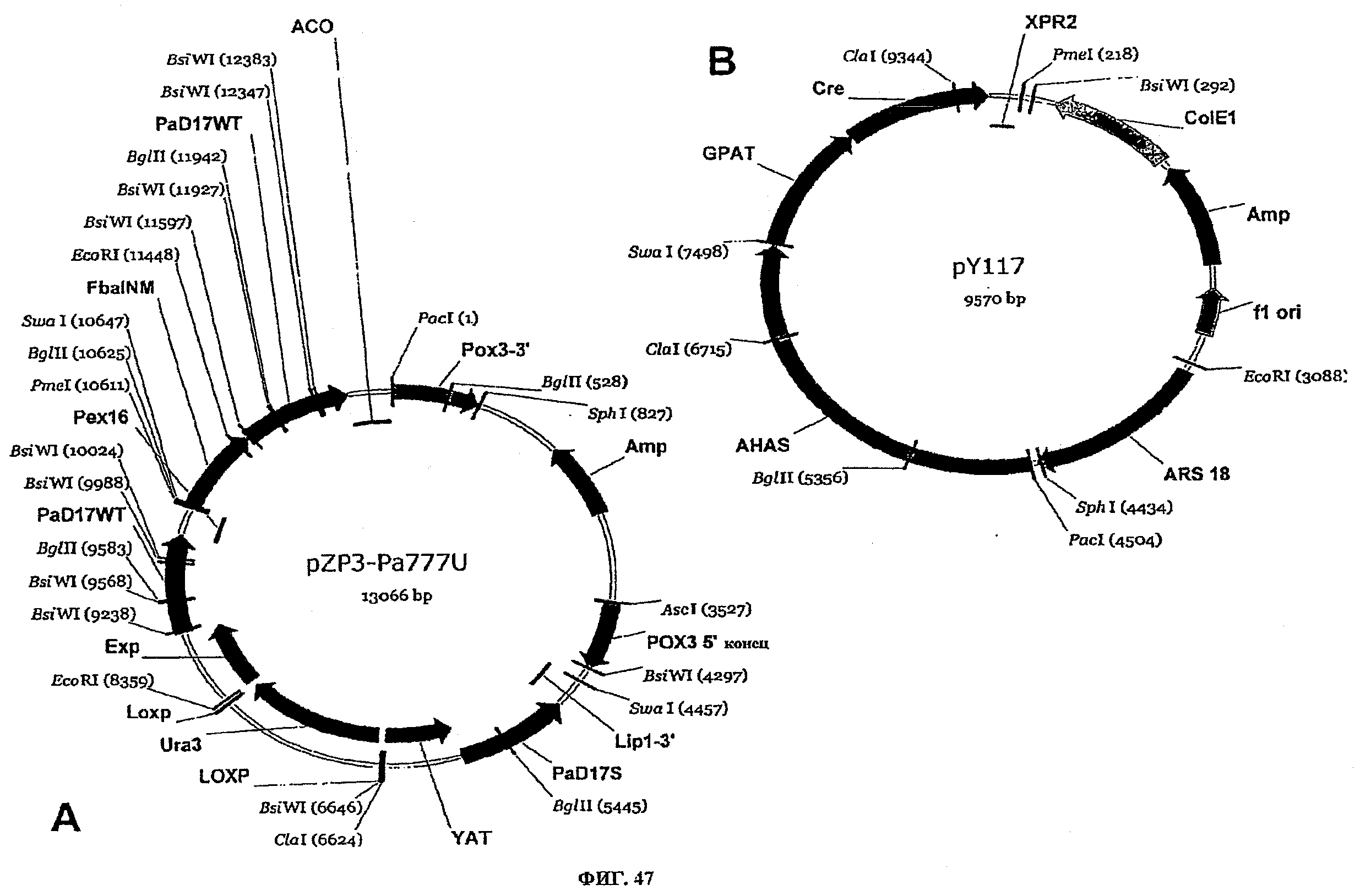
ФИГ.47 представляет плазмидные карты для следующего: (A) pZP3-Pa777U и (B) pY117.
ФИГ.48 представляет плазмидные карты для следующего: (A) pZP2-2988 и (B) pZKUE3S.
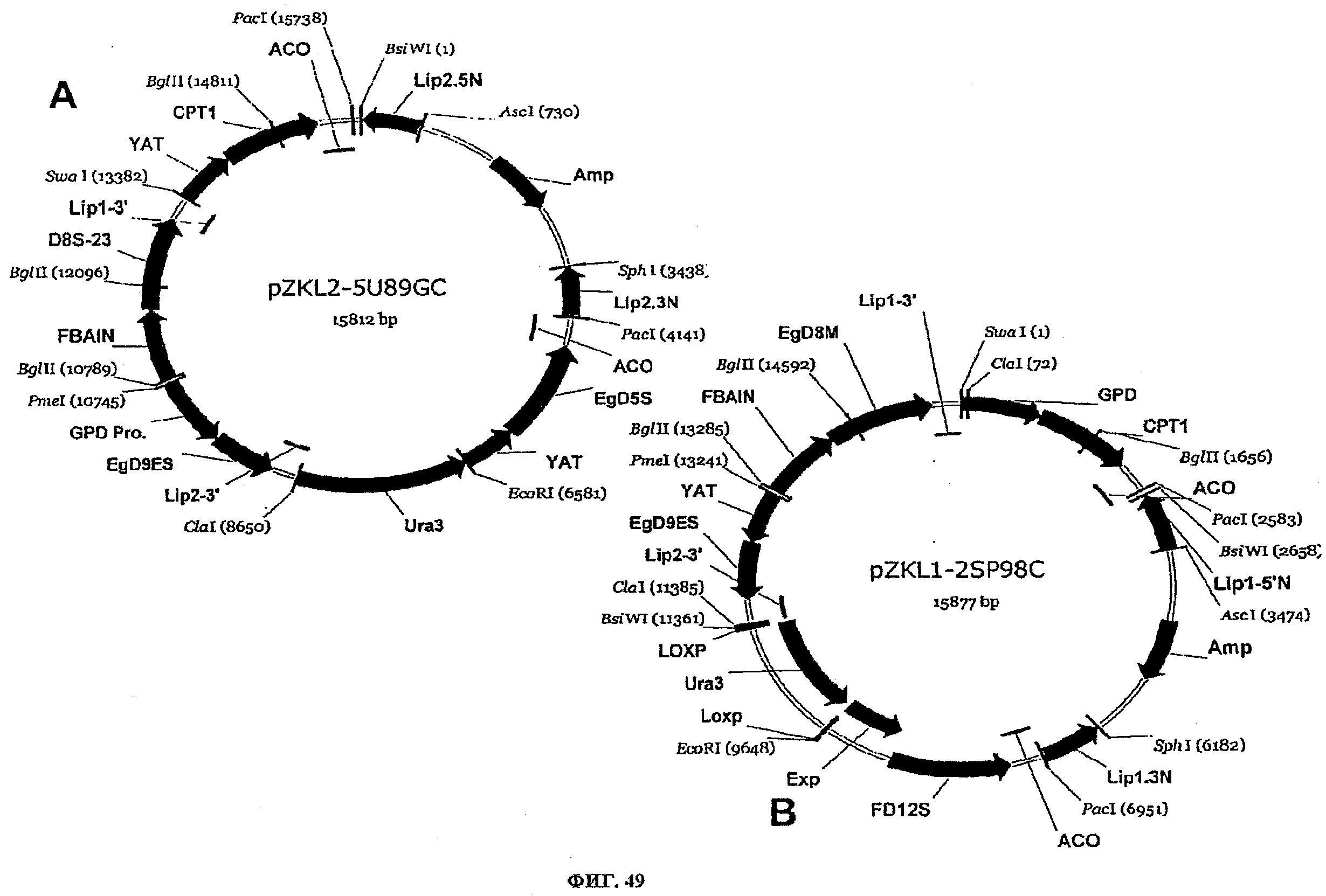
ФИГ.49 представляет плазмидные карты для следующего: (A) pZKL2-5U89GC и (B) pZKL1-2SP98C.
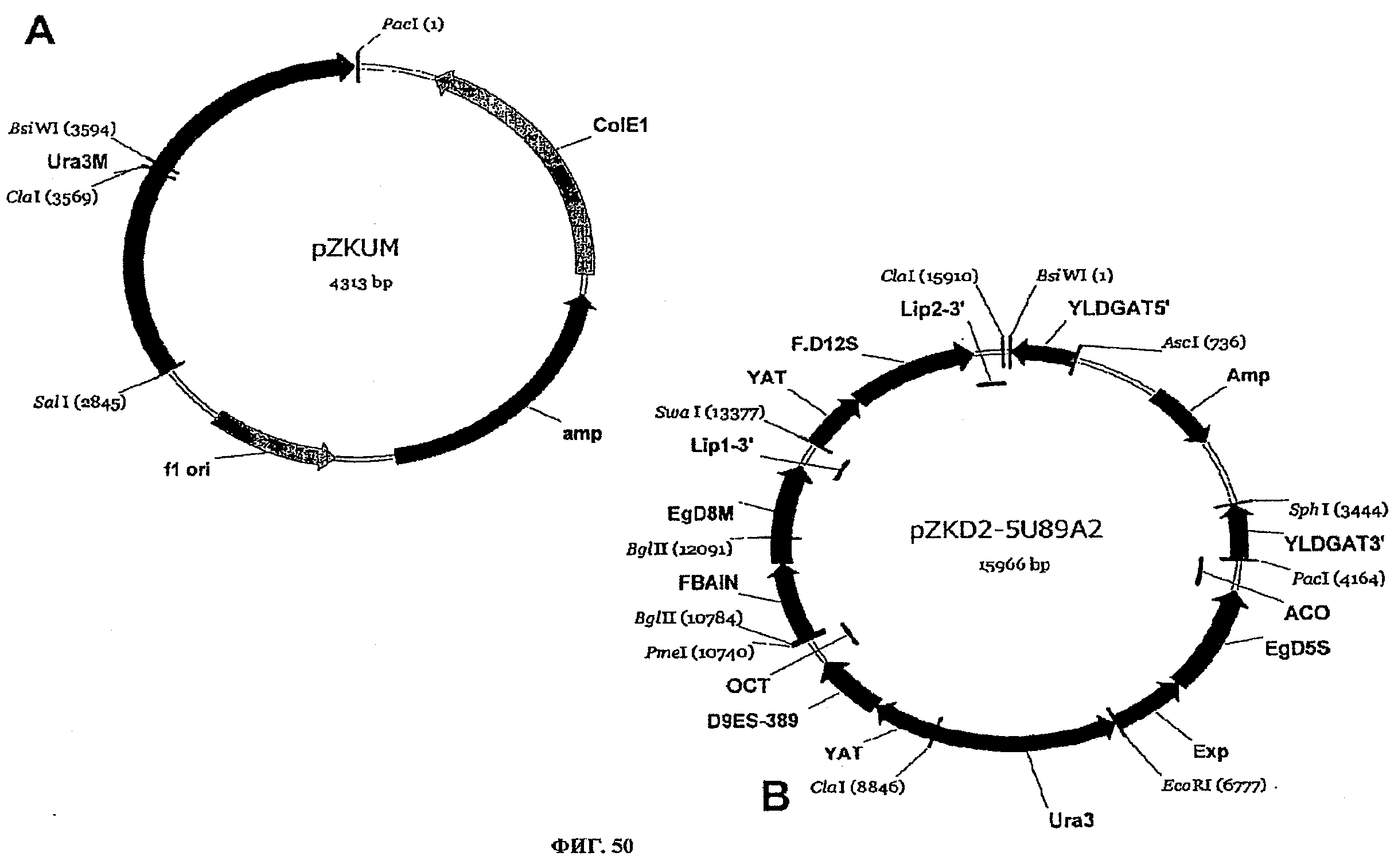
ФИГ.50 представляет плазмидные карты для следующего: (A) pZKUM и (B) pZKD2-5U89A2.
ФИГ.51A представляет схему создания штамма Y4184U Yarrowia lipolytica. ФИГ.51B представляет плазмидную карту для pEgC20ES.
ФИГ.52 представляет плазмидные карты для следующего: (A) pZUFmEgC20ES и (B) pZKL4-220EA4. ФИГ.52C представляет схему, показывающую перекрывание участка 3′ EaC20E домена (SEQ ID NO:231) участком 5′ EaD4 домена (SEQ ID NO:246) внутри EaDHAsyn1 (SEQ ID NO:95).
ФИГ.53A показывает группировку N-концов EaD4S (SEQ ID NO:193), EaD4S-1 (SEQ ID NO:382), EaD4S-2 (SEQ ID NO:384), и EaD4S-3 (SEQ ID NO:386). ФИГ.53B показывает группировку N-концов EgD4S (SEQ ID NO:388), EgD4S-1 (SEQ ID NO:404), EgD4S-2 (SEQ ID NO:406), и EgD4S-3 (SEQ ID NO:408).
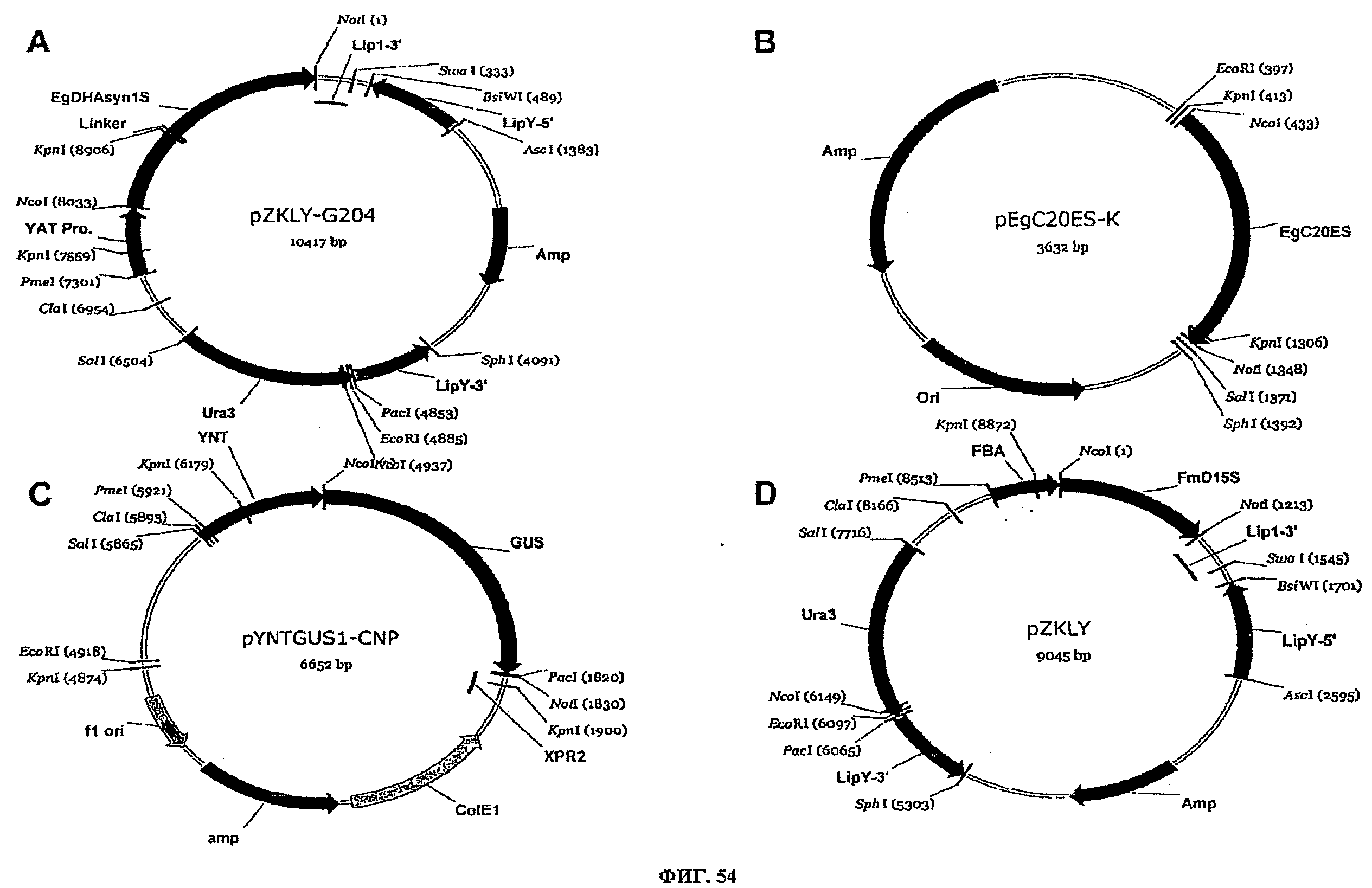
ФИГ.54 представляет плазмидные карты для следующего: (A) pZKLY-G204, (B) pEgC20ES-K, (C) pYNTGUS1-CNP, и (D) pZKLY.
ФИГ.55 представляет плазмидные карты для следующего: (A) pZUFmG9G8fu и (B) pZUFmG9A8.
ФИГ.56 представляет собой карту pKR1014.
ФИГ.57 представляет собой карту pKR1152.
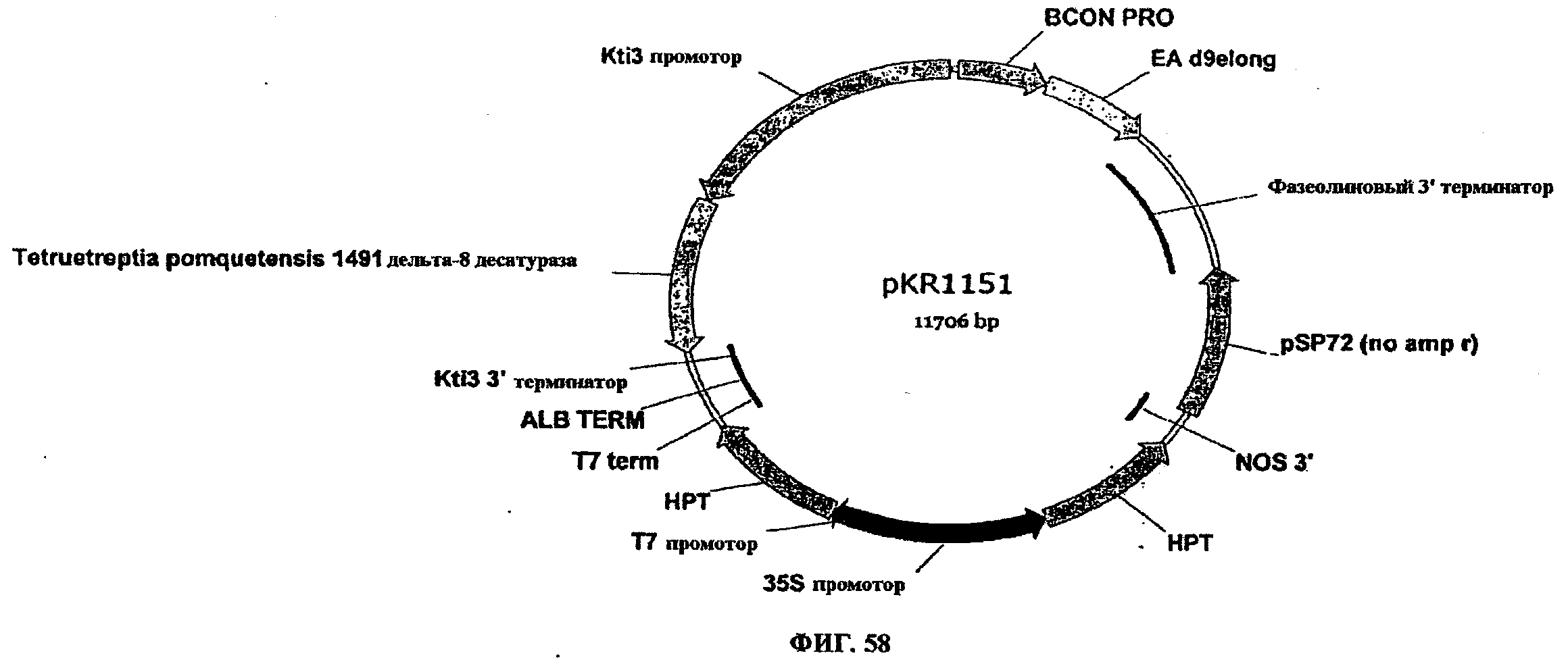
ФИГ.58 представляет собой карту pKR1151.
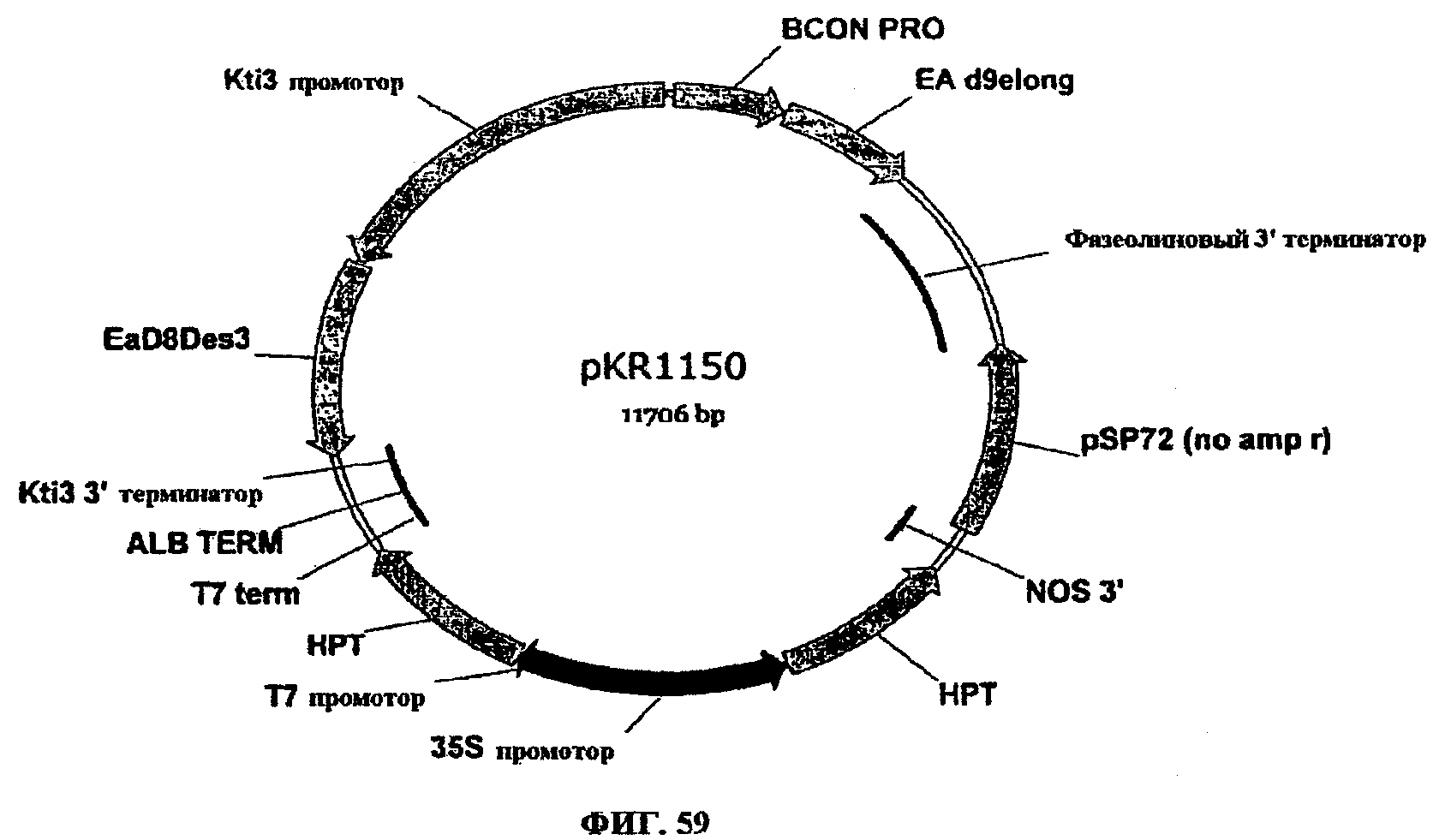
ФИГ.59 представляет собой карту pKR1150.
ФИГ.60 представляет собой карту pKR1199.
ФИГ.61 представляет собой карту pKR1200.
ФИГ.62 представляет собой карту pKR1184.
ФИГ.63 представляет собой карту pKR1321.
ФИГ.64 представляет собой карту pKR1326.
Для ФИГ.65-71, жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК, АЛК, ЭДК, ЭРК, ДГЛК и ЭТК, и составы жирных кислот выражены как весовой процент (вес. %) от общего количества жирных кислот. Кроме того, элонгационная активность выражена как % дельта-9-элонгации C18 жирных кислот (C18% дельта-9-элонг), вычисленный в соответствии со следующей формулой: ([продукт]/[субстрат+продукт])*100. Более конкретно, комбинированный процент элонгации для ЛК и АЛК определяется как: ([ДГЛК+ЭТК+ЭДК+ЭРК]/[ЛК+АЛК+ДГЛК+ЭТК+ЭДК+ЭРК])*100. Комбинированный процент десатурации для ЕДК и ЭРК, представленный как “C20 % дельта-8-десат”, определяется как: ([ДГЛК+ЭТК]/[ДГЛК+ЭТК+ЭДК+ЭРК])*100, и на него также ссылаются как на полный % десатурацииФИГ.65 представляет жирнокислотные профили для пяти событий, трансформированных pKR1014, которые имеют самое высокое среднее содержание ДГЛК (среднее из 5 проанализированных соевых соматических зародышей).
ФИГ.66 представляет жирнокислотные профили для пяти событий, трансформированных pKR1152, которые имеют самое высокое среднее содержание ДГЛК (среднее из 5 проанализированных соевых соматических зародышей).
ФИГ.67 представляет жирнокислотные профили для пяти событий, трансформированных pKR1151, которые имеют самое высокое среднее содержание ДГЛК (среднее из 5 проанализированных соевых соматических зародышей).
ФИГ.68 представляет жирнокислотные профили для пяти событий, трансформированных pKR1150, которые имеют самое высокое среднее содержание ДГЛК (среднее из 5 проанализированных соевых соматических зародышей).
ФИГ.69 представляет жирнокислотные профили для пяти событий, трансформированных pKR1199, которые имеют самое высокое среднее содержание ДГЛК (среднее из 5 проанализированных соевых соматических зародышей).
ФИГ.70 представляет жирнокислотные профили для пяти событий, трансформированных pKR1200, которые имеют самое высокое среднее содержание ДГЛК (среднее из 5 проанализированных соевых соматических зародышей).
ФИГ.71 представляет жирнокислотные профили для пяти событий, трансформированных pKR1184, которые имеют самое высокое среднее содержание ДГЛК (среднее из 5 проанализированных соевых соматических зародышей).
ФИГ.72 представляет сравнение индивидуально экспрессированных дельта-9-элонгаз и дельта-8-десатураз относительно эквивалентного дельта-9-элонгаза-дельта-8-десатуразного гибрида. Каждая точка данных представляет средний % ДГЛК или % ЭДК для 5-6 зародышей (как % от общего количества жирных кислот) для всех проанализированных событий, и построен график зависимости Средн. % ДГЛК от Средн. % ЭДК. На (A), EgTpom представляет EgD9e, коэкспрессированный с TpomD8 (pKR1014), и EgTpomfus представляет EgD9e/TpomD8 гибрид (pKR1199). На (B), EgEa представляет EgD9e, коэкспрессированный с EaD8 (pKR1152), и EgEafus представляет EgD9e/EaD8 гибрид (pKR1200). На (C), EaTpom представляет EaD9e, коэкспрессированный с TpomD8 (pKR1151), и EaTpomfus представляет EaD9e/TpomD8 гибрид (pKR1183). На ФИГ.(D), EaEa представляет EaD9e, коэкспрессированный с EaD8 (pKR1150), и EaEafus представляет EaD9e/EaD8 гибрид (pKR1200).
ФИГ.73 представляет жирнокислотные профили для пяти событий, трансформированных pKR1322 (Эксперимент MSE2274), которые имеют самое высокое среднее содержание АРК и ЭПК (среднее из 5 проанализированных зародышей). Жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), 18:2 (5,9), ЛК, АЛК, ЭДК, ЭРК, СКК, ДГЛК, ЮН (называемая также ЮП), ЭТК, АРК и ЭПК. Составы жирных кислот выражены как весовой процент (вес. %) от общего количества жирных кислот. Элонгационная активность выражена как % дельта-9-элонгации C18 жирных кислот (% Эло), вычисленный в соответствии со следующей формулой: ([продукт]/[субстрат+продукт])*100. Более конкретно, комбинированный процент элонгации для ЛК и АЛК определяется как: ([ДГЛК+ЭТК+ЭДК+ЭРК+ЭПА+АРК]/[ЛК+АЛК+ДГЛК+ЭТК+ЭДК+ЭРК+ЭПК+
АРК])*100. Комбинированный процент дельта-8-десатурации для ЭДК и ЭРК, представленный как “% D8”, определяется как: ([ДГЛК+ЭТК+ЭПК+АРК]/[ДГЛК+ЭТК+ЭДК+ЭРК+ЭПК+АРК])*100. На него ссылаются также как на полный % дельта-8-десатурации. Комбинированный процент дельта-5-десатурации для ДГЛК и ЭТК, представленный как “% D5”, определяется как: ([ЭПК+АРК]/[ДГЛК+ЭТК+ЭПК+АРК])*100. На него также ссылаются как на полный % дельта-5-десатурации.
ФИГ.74 представляет жирнокислотные профили для пяти событий, тансформированных pKR1326 (Эксперимент MSE2275), которые имеют самое высокое среднее содержание ДГЛК и ЭТК (среднее из 5 проанализированных зародышей). Жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК, АЛК, ЭДК, ЭРК, ДГЛК и ЭТК. Составы жирных кислот выражены как весовой процент (вес. %) от общего количества жирных кислот. Элонгационная активность выражена как % дельта-9-элонгации C18 жирных кислот (C18 % дельта-9 элонг), вычисленный в соответствии со следующей формулой: ([продукт]/[субстрат+продукт])*100. Более конкретно, комбинированный процент элонгации для ЛК и АЛК определяется как: ([ДГЛК+ЭТК+ЭДК+ЭРК]/[ЛК+АЛК+ДГЛК+ЭТК+ЭДК+ЭРК])*100. Комбинированный процент десатурации для ЭДК и ЭРК, представленный как “C20 % дельта-8-десат”, определяется как: ([ДГЛК+ЭТК]/[ДГЛК+ЭТК+ЭДК+ЭРК])*100. На него также ссылаются как на полный % десатурации.
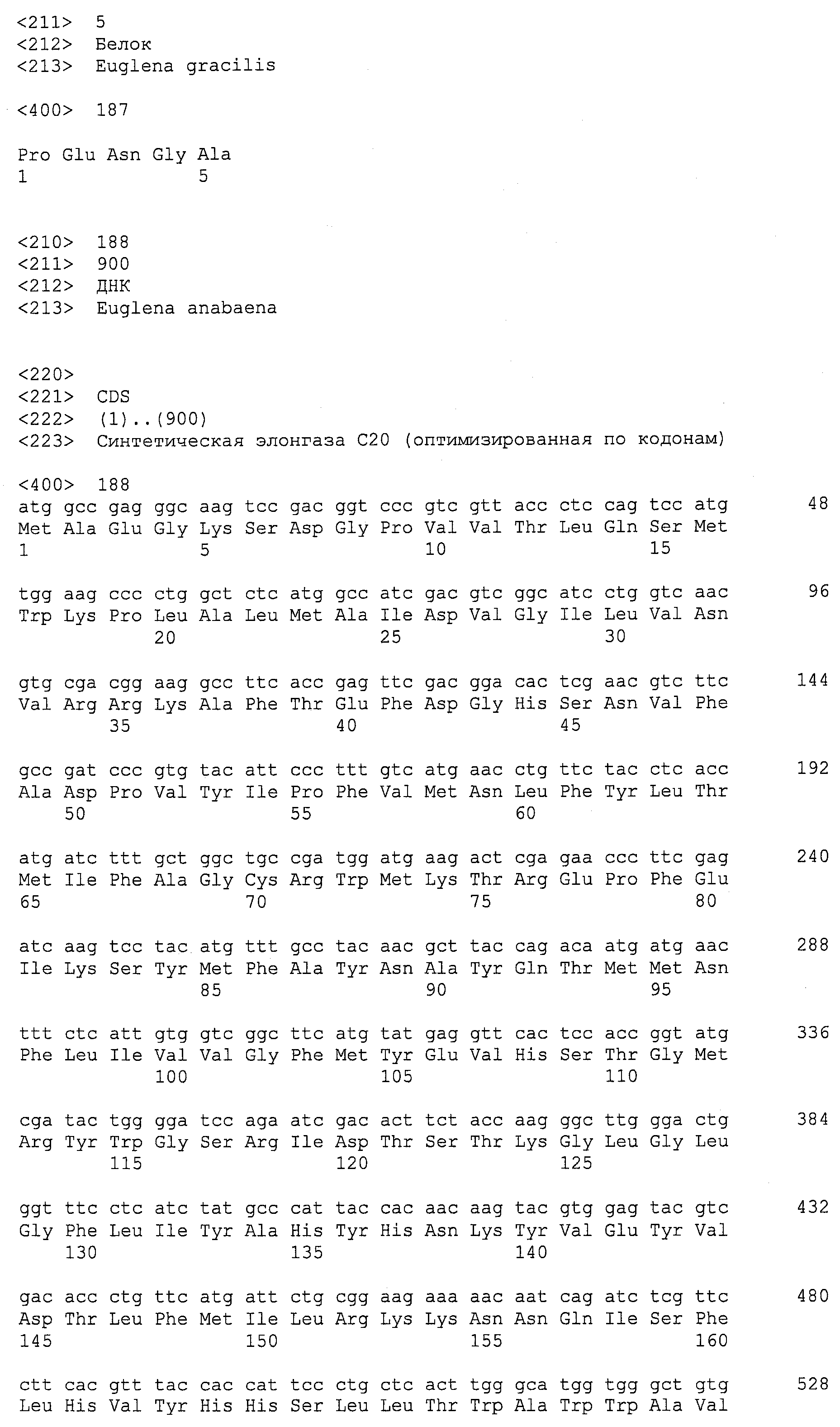
Описания последовательностей резюмируют Листинг последовательностей, который прилагается. Листинг последовательностей содержит однобуквенные коды для символов нуклеотидных последовательностей и одно- и трибуквенные коды для аминокислот, как определено в стандартах IUPAC-IUB, которые описаны в Nucleic Acids Research 13:3021-3030 (1985) и в Biochemical Journal 219(2):345-373 (1984).
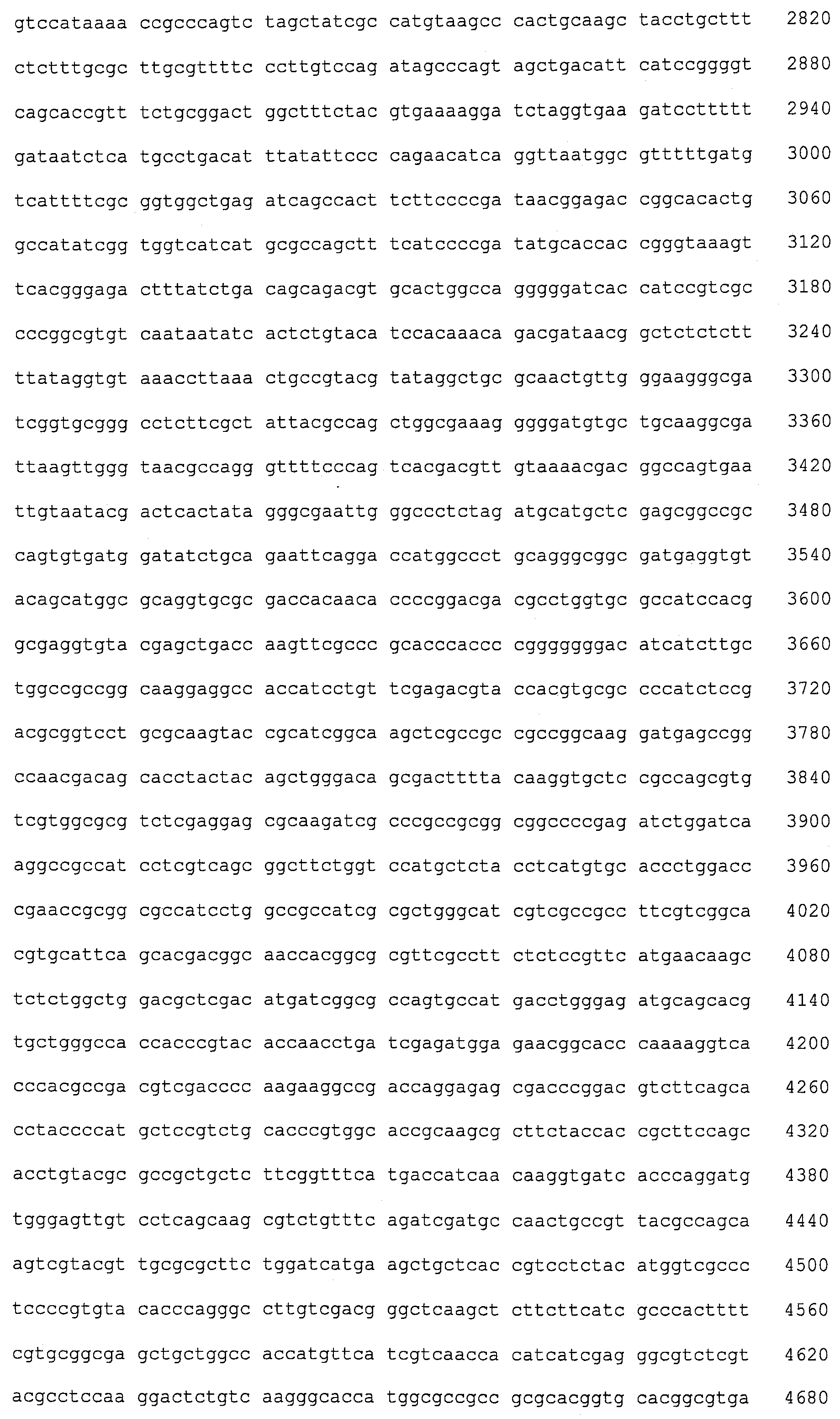
SEQ ID NOs:1-519 являются праймерами, ОРС кодируют гены, протеины (или их участки), или плазмиды, как показано в Таблице 2.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
На каждую из приведеннях в данном тексте работ и других материалов сделана полная ссылка.
Данное изобретение касается мультизимов, таких как ДГК синтаза. Они полезны, помимо всего прочего, для управления биохимическими путями в производстве целебных ПНЖК и, более конкретно, для производства докозагексаеновой кислоты (ДГК). Таким образом, представленное изобретение находит много применений. ПНЖК или их производные, полученные по раскрытой здесь методологии, могут быть использованы как пищевые заменители или добавки, в частности, как питательные смеси для младенцев, для пациентов, подвергающихся внутривенному кормлению, для предупреждения или лечения нарушений питания.
Как альтернатива, очищенные ПНЖК (или их производные) могут быть введены в кулинарные масла, жиры или маргарины, составленные таким образом, что при нормальном применении реципиент получает необходимое количество диетической добавки. ПНЖК могут также водиться в детские смеси, пищевые добавки или другие пищевые продукты и могут найти применение как противовоспалительные или снижающие холестерин агенты. При необходимости, данные композиции могут использоваться для фармацевтических целей (для человека или в ветеринарии). В этом случае ПНЖК применяются, в общем, перорально, но могут использоваться в соответствии с любой схемой, по которой они могут успешно абсорбироваться, например, парэнтерально (например, подкожно, внутримышечно или внутривенно), ректально, вагинально или местным образом (например, как мазь или лосьон для кожи).
Пополнение рационов человека или животного ПНЖК, произведенными рекомбинантными способами, может обеспечить повышенные уровни добавленных ПНЖК, так и их метаболитов. Например, лечение ЭПК может не только повысить уровни ЭПК, но также и уровни более низких продуктов ЭПК, таких как эйкозаноиды (то есть простагландины, лейкотриены, тромбоксаны). Сложные регуляторные механизмы могут сделать желательным комбинирование разных ПНЖК или использование различных конъюгатов ПНЖК с целью предупреждения, контроля или преодоления таких механизмов для достижения необходимых уровней специфических ПНЖК у данного субъекта.
Определения
Как используется в данном тексте и в прилагаемой формуле изобретения, сингулярные формы "a", "an", и "the" включают, если в контексте четко не указано иное, множественные ссылки. Так, например, ссылка на "a plant" включает множество таких растений, ссылка на "a cell" включает одну или несколько клеток и их эквивалентов, которые известны специалистам в данной области, и т.д.
Термин “изобретение” или “данное изобретение”, как используется в данном тексте, не предполагает ограничения каким-либо одним специфическим вариантом данного изобретения, а применяется, в общем, к любому и ко всем вариантам данного изобретения, как описано в пунктах формулы изобретения и спецификации.
В контексте данной заявки использован ряд терминов и сокращений. Введены следующие определения.
“Открытая рамка считывания” имеет сокращение ОРС.
“Полимеразная цепная реакция” имеет сокращение ПЦР.
“Американское собрание типовых культур” имеет сокращение АСТК.
“Полиненасыщенная жирная кислота(ы)” имеет сокращение ПНЖК.
“Триацилглицерины” имеют сокращение ТАГ.
Термины “снижать уровень регуляции или снижение уровня регуляции”, как тут используется, касаются снижения или ослабления уровня экспрессии гена или полинуклеотида.
Термин “мультизим” касается отдельного полипептида, который обладает по меньшей мере двумя независимыми и раздельными ферментативными активностями. Лучше, когда мультизим включает первую ферментативную активность, связанную со второй ферментативной активностью.
Термин “слитый протеин” употребляется взаимозаменяемо с термином “мультизим”. Так, “слитый протеин” касается отдельного полипептида, который обладает по меньшей двумя независимыми и раздельными ферментативними активностями.
Термин “слитый ген” касается полинуклеотида или гена, который кодирует мультизим. Слитый ген может быть сконструирован путем связывания по меньшей мере двух ДНК фрагментов, где каждый ДНК фрагмент кодирует независимую и раздельную ферментативную активность. Пример слитого гена описан здесь ниже в Примере 38, где Hybrid1-ГГЛК синтазный слитый ген был сформирован путем связывания Euglena anabaena дельта-9-элонгазы (EaD9Elo1; SEQ ID NO:252) и Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразы (TpomD8; SEQ ID NO:162) с использованием Euglena gracilis ДГК синтаза 1 обогащенного пролином линкера, (EgDHAsyn1Link; SEQ ID NO:197).
“Домен” или “функциональный домен” представляет собой обособленную, непрерывную часть или последовательность полипептида, которая может быть ассоциирована с функцией (например, ферментативной активностью). Как тут применяется, термин “домен” включает, но не ограничивается этим, жирнокислотные биосинтетические ферменты и участки жирнокислотных биосинтетических ферментов, котрые сохраняют ферментативную активность.
“ДГК синтаза” служит примером мультизима. Конкретно, ДГК синтаза включает C20-элонгазу, связанную с дельта-4-десатуразой с использованием любого из описанных здесь линкеров. Другим примером мультизима является отдельный полипептид, котрый содержит дельта-9-элонгазу, связанную с дельта-8-десатуразой как описано ниже.
Термин “связь” касается соединения или связывания по меньшей мере двух полипептидов, которые обладают независимыми и раздельными ферментативными активностями.
Термин “линкер” касается связи или сцепления между двумя или больше полипептидами, каждый из которых обладает независимыми и раздельными ферментативными активностями.
Связь, используемая для формирования мультизима, включает, как минимум, отдельную полипептидную связь. В другом аспекте, связь может включать один аминокислотный остаток, такой как пролин, или полипептид. Если данная связь является полипептидом, может быть желательно для данной связи иметь по меньшей мере один пролиновый аминокислотный остаток.
Пример линкера показан в SEQ ID NO:198 (EgDHAsyn1 обогащенный пролином линкер).
Термин “жирные кислоты” касается длинноцепочечных алифатических кислот (алкановых кислот) с варьирующими длинами цепи, от приблизительно C12 до C22 (хотя известны кислоты как с более длинными, так и более короткими длинами цепей). Преобладающие длины цепей лежат в области от C16 до C22. Дополнительные детали, которые касаются различий между “насыщенными жирными кислотами” и “ненасыщенными жирными кислотами”, “мононенасыщенными жирными кислотами” и “полиненасыщенными жирными кислотами ” (или “ПНЖК”), и “омега-6 жирными кислотами” (ω-6 или n-6) и “омега-3 жирными кислотами” (ω-3 або n-3), приведены в патенте США № 7238482.
Жирные кислоты описаны в данном тексте с помощью простой системы обозначений “X:Y”, где X представляет собой полное число углеродных атомов (C) в конкретной жирной кислоте, и Y - число двойных связей. Цифра после обозначения жирной кислоты указывает на положение двойной связи от карбоксильного конца жирной кислоты, с афиксом “c” для цис-конфигурации данной двойной связи (например, пальмитиновая кислота (16:0), стеариновая кислота (18:0), олеиновая кислота (18:1, 9c), петрозелиновая кислота (18:1, 6c), ЛК (18:2, 9c,12c), ГЛК (18:3, 6c,9c,12c) и АЛК (18:3, 9c,12c,15c)). Если не указано иное, 18:1, 18:2 и 18:3 касаются, соответственно, олеиновой, ЛК и АЛК жирных кислот. Если специально не указано иное, двойные связи имеют, как предполагается, цис-конфигурацию. Например, двойные связи в 18:2 (9,12), как предполагается, находятся в цис-конфигурации.
Номенклатура, которая используется для описания ПНЖК в данной заявке, приведена в Таблице 3 ниже. В колонке, обозначенной “Стенографическое обозначение”, используется система омега-ссылок для указания количества углеродных атомов, количества двойных связей и положения двойной связи, ближайшей к омега углеродному атому, считая от этого омега углеродного атома (который с этой целью обозначен номером 1). Остаток таблицы содержит общие названия омега-3 и омега-6 жирных кислот и их предшественников, сокращения, которые будут использованы далее в данной спецификации, и химическое название каждого соединения.
Метаболический или биосинтетический путь, в биохимическом смысле, может рассматриваться как ряд химических реакций, которые происходят внутри клетки, которые катализируются ферментами, или для образования метаболического продукта, который используется или сохраняется клеткой, или для инициации другого метаболического пути (который был потом назван стадией генерации потока). Многие из этих путей детально разработаны и включают пошаговую модификацию первичного вещества для его формовання в продукт, которых имеет необходитмую точную химическую структуру.
Термин “ПНЖК биосинтетический путь” касается метаболического процесса, котрый превращает олеиновую кислоту в ЛК, ЭДК, ГЛК, ДГЛК, АРК, ДТК, ДПКn-6, АЛК, СТК, ЭТрК, ЭТК, ЭПК, ДПК и ДГК. Этот процесс хорошо описан в литературе (например, смотри РСТ публикацию № WO 2006/052870). Упрощенно говоря, этот процесс включает элонгацию углеродной цепи путем добавления углеродных атомов и десатурации молекулы путем добавления двойных связей через ряд специальных десатурационных и элонгационных ензимов (т.е. “ферментов ПНЖК биосинтетического пути”), которые присутствуют в мембране эндоплазматичного ретикулума. Более конкретно, “ферменты ПНЖК биосинтетического пути” касаются любого из следующих ферментов (и генов, которые кодируют указанные ферменты), связанных с биосинтезом ПНЖК, включая: дельта-4-десатуразу, дельта-5-десатуразу, дельта-6-десатуразу, дельта-12-десатуразу, дельта-15-десатуразу, дельта-17-десатуразу, дельта-9-десатуразу, дельта-8-десатуразу, дельта-9-элонгазу, С14/16-элонгазу, С16/18-элонгазу, С18/20-элонгазу, С20/22-элонгазу, ДГК синтазу и/или мультизим настоящего изобретения.
Термин “омега-3/омега-6 жирнокислотный биосинтетический путь” касается набора генов, которые, при экспрессии в соответствующих условиях, кодируют ферменты, которые катализируют образование каждой или обеих омега-3 и омега-6 жирных кислот. Типично, гены, которые включены в биосинтетический омега-3/омега-6 жирнокислотный путь, кодируют ПНЖК ферменты биосинтетического пути. Репрезентативный путь иллюстрируется ФИГ.1, представляя конверсию миристиновой кислоты через различные интермедиаты в ДГК, что демонстрирует, каким образом обе омега-3 и омега-6 жирные кислоты могут образовываться из совместного источника. Данный путь природно разделяется на две части, где в одной части будут генерироваться омега-3 жирные кислоты а в другой части омега-6 жирные кислоты.
Термин “функциональный”, как здесь используется в контексте с омега-3/омега-6 жирнокислотным биосинтетическим путем, означает, что некоторые (или все) гены на данном пути экспрессируют активные ферменты, обеспечивая in vivo катализ или конверсию субстрата. Следует понимать, что “омега-3/омега-6 жирнокислотный биосинтетический путь” или “функциональный омега-3/омега-6 жирнокислотный биосинтетический путь” не предусматривает, что необходимы все ПНЖК ферментные гены биосинтетического пути, поскольку ряд жирнокислотных продуктов будет требовать только экспрессии подмножества генов этого пути.
Термин “дельта-6-десатураза/дельта-6-элонгазный путь” касается ПНЖК биосинтетического пути, котрый, как минимум, включает по меньшей мере одну дельта-6-десатуразу и по меньшей мере одну C18/20-элонгазу, обеспечивая тем самым биосинтез ДГЛК и/или ЭТК из ЛК и АЛК, соответственно, с ГЛК и/или СТК как промежуточными жирными кислотами. С экспрессией других десатураз и элонгаз могут быть также синтезированы АРК, ДТК, ДПКn-6, ЭПК, ДПК и ДГК.
Термин “дельта-9-элонгаза/дельта-8-десатуразный путь” касается ПНЖК биосинтетического пути, который, как минимум, включает по меньшей мере одну дельта-9-элонгазу и по меньшей мере одну дельта-8-десатуразу, обеспечивая таким образом биосинтез ДГЛК и/или ЭТК из ЛК и АЛК, соответственно, с ЭДК и/или ЭТрК как промежуточными жирными кислотами. С экспрессией других десатураз и элонгаз могут быть также синтезированы АРК, ДТК, ДПКn-6, ЭПК, ДПК и ДГК. Этот путь может иметь преимущества в некоторых вариантах, поскольку биосинтез ГЛК и/или СТК исключен.
Термин “промежуточная жирная кислота” касается любой жирной кислоты, которая получена на жирнокислотном метаболическом пути, которая может превращаться далее в необходимый жирнокислотный продукт этим путем под действием ферментов другого метаболического пути. Например, когда ЭПК производится с использованием дельта-9-элонгаза/дельта-8-десатуразного пути, могут образоваться ЭДК, ЭТрК, ДГЛК, ЭТК и АРК, и их считают “промежуточными жирными кислотами”, поскольку эти жирные кислоты могут быть конвертированы далее в ЭПК под действием ферментов другого метаболического пути.
Термин “побочная жирная кислота” касается любой жирной кислоты, которая получена на жирнокислотном метаболическом пути, которая не является ни продуктом назначения данного пути, ни “промежуточной жирной кислотой” данного пути. Например, когда ЭПК получается с использованием дельта-9-элонгаза/дельта-8 десатуразного пути, под действием дельта-5-десатуразы на ЭДК или ЭТрК, соответственно, могут также образоваться скиадоновая кислота (СКК) иа юнипероновая кислота (ЮП). Они считаются “побочными жирными кислотами”, поскольку не могут быть превращены далее в ЭПК под действием ферментов другого метаболического пути.
Термины “триацилглицерин”, “масло” и “ТАГ” касаются нейтральных липидов, которые состоят из трех жирных ацильных остатков, этерифицированных до молекулы глицерина (и такие термины будут употребляться взаимозаменяемо в данной заяке). Такие масла могут содержать длинноцепочечные ПНЖК, так же как и более короткие насыщенные и ненасыщенные жирные кислоты и насыщенные жирные кислоты с более длинными цепями. Таким образом, “масляный биосинтез” касается, в общем, синтеза ТАГ в клетке.
“Процент (%) ПНЖК в совокупных липидных и масляных фракциях” касается процента ПНЖК относительно общего количества жирных кислот в этих фракциях. Термины “совокупная липидная фракция” или “липидная фракция” оба касаются суммарного количества всех липидов (т.е. нейтральных и полярных) внутри маслянистого организма, включая, таким образом, те липиды, что локализованы в фосфатидилхолиновой (ФХ) фракции, фосфатидилэтаноламиновой (ФЕ) фракции и триацилглицериновой (ТАГ или масляной) фракции. Однако, термины “липид” и “масло” будут использоваться в настоящей спецификации взаимозаменяемо.
Термины “эффективность конверсии” и “процент конверсии субстрата” касаются эффективности, с которой конкретный фермент (например, десатураза) может превратить субстрат в продукт. Эффективность конверсии измеряется в соответствии со следующей формулой: ([продукт]/[субстрат+продукт])*100, где 'продукт′ включает промежуточный продукт и все продукты, полученные на этом пути.
“Десатураза” представляет собой полипептид, который может десатурировать, т.е. вводить двойную связь, в одну или несколько жирных кислот с образованием жирной кислоты или предшественника, которые представляют интерес. Несмотря на использование в данной спецификации системы омега-ссылок для ссылок на специфические жирные кислоты, удобнее определять активность десатуразы путем счета от карбоксильного конца субстрата с использованием дельта-системы. Особенный интерес представляют здесь дельта-8-десатуразы, которые десатурируют жирную кислоту между восьмым и девятым углеродным атомом, понумерованным от карбоксил-терминального конца молекулы, и которые могут, например, катализировать конверсию ЭДК в ДГЛК и/или ЭТрК в ЭТК. Другие полезные десатуразы жирных кислот включают, например, (1) дельта-5-десатуразы, которые катализируют конверсию ДГЛК в АРК и/или ЭТК в ЭПК; (2) дельта-6-десатуразы, которые катализируют конверсию ЛК в ГЛК и/или АЛК в СТК; (3) дельта-4-десатуразы, которые катализируют конверсию ДПК в ДГК и/или ДТК в ДПКn-6; (4) дельта-12-десатуразы, которые катализируют конверсию олеиновой кислоты в ЛК; (5) дельта-15-десатуразы, которые катализируют конверсию ЛК в АЛК и/или ГЛК в СТК; (6) дельта-17-десатуразы, которые катализируют конверсию АК в ЭПК и/или ДГЛК в ЭТК; и (7) дельта-9-десатуразы, которые катализируют конверсию пальмитата в пальмитолеиновую кислоту (16:1) и/или стеарата в олеиновую кислоту (18:1). В данной области на дельта-15 и дельта-17-десатуразы также иногда ссылаются как на “омега-3-десатуразы”, “w-3-десатуразы”, и/или “n-3-десатуразы”, исходя из их способности конвертировать омега-6 жирные кислоты в свои омега-3 аналоги (например, превращение ЛК в АЛК и АРК в ЭПК, соответственно). В некоторых вариантах наиболее предпочтительно эмпирически определять специфичность конкретной десатуразы жирных кислот путем превращения пригодного хозяина геном в десатуразе жирных кислот и определения ее влияния на жирнокислотный профиль данного хозяина.
Термин “дельта-4-десатураза” касается фермента, который будет десатурировать жирную кислоту между четвертым и пятым углеродным атомами, считая от карбоксил-терминального конца данной молекулы, и это может, например, катализировать конверсию ДПК в ДГК и/или ДТК в ДПКn-6. Для целей данной заявки термин “EgDHAsyn1” касается ДГК синтазного фермента (SEQ ID NO:12), выделенного из Euglena gracilis, кодируемого здесь SEQ ID NO:11. Термин “EgDHAsyn2” касается ДГК синтазного фермента (SEQ ID NO:22), выделенного из Euglena gracilis, кодируемого здесь SEQ ID NO:21. Термин “EaDHAsyn1” касается ДГК синтазного фермента (SEQ ID NO:95), выделенного из Euglena anabaena, кодируемого здесь SEQ ID NO:91. Термин “EaDHAsyn2” касается ДГК синтазного фермента (SEQ ID NO:96), выделенного из Euglena anabaena, кодируемого здесь SEQ ID NO:92. Термин “EaDHAsyn3” касается ДГК синтазного фермента (SEQ ID NO:97), выделенного из Euglena anabaena, кодируемого здесь SEQ ID NO:93. Термин “EaDHAsyn4” касается фермента (SEQ ID NO:98), выделенного из Euglena anabaena, кодируемого здесь SEQ ID NO:94.
Термин “элонгазная система” касается набора из четырех ферментов, которые ответственны за элонгацию жирнокислотной углеродной цепи с образованием жирной кислоты, которая на два углеродных атома длиннее, чем жирнокислотный субстрат, на который действует данная элонгазная система. Более конкретно, процесс элонгации происходит в ассоциации с жирнокислотной синтазой, соответственно с чем СоА служит ацильным носителем (Lassner et al., Plant Cell 8:281-292 (1996)). На первой стадии, которая, как было установлено, является субстрат-специфической и лимитированной скоростью, малонил-СоА конденсируется с длинноцепочечным ацил-СоА с образованием диоксида углерода (СО2) и β-кетоацил-СоА (где ацильная составляющая удлинена на два углеродных атома). Следующие реакции включают восстановление до β-гидроксиацил-СоА, дегидратацию до эноил-СоА и второе восстановление с образованием удлиненного ацил-СоА. Примерами реакций, которые катализируются элонгазными системами, являются конверсия ГЛК в ДГЛК, СТК в ЭТК, ЛК в ЭДК, АЛК в ЭТрК и ЭПК в ДПК.
Для целей данной заявки, на фермент, который катализирует первую реакцию конденсации (т.е. конверсию малонил-СоА и длинноцепочечную ацил-CvoA в β-кетоацил-СоА), будем ссылаться, в общем, как на ”элонгазу”. В общем, селективность элонгаз в отношении субстрата несколько широкая, но ограничивается как длиной цепи так и степенью ненасыщенности. Соответственно, элонгазы могут иметь разные специфичности. Например, С14/16-элонгаза будет утилизировать С14 субстрат (например, миристиновую кислоту), С16/18-элонгаза будет утилизировать С16 субстрат (например, пальмитат), С18/20-элонгаза будет утилизировать С18 субстрат (например, ГЛК, СТК) и С20/22-элонгаза будет утилизировать С20 субстрат (например, АРК, ЭПК). Подобным образом, “дельта-9-элонгаза” способна катализировать конверсию ЛК в ЭДК и/или АЛК в ЭТрК.
Важно отметить, что некоторые элонгазы имеют широкую специфичность, и, следовательно, отдельный ензим может быть способным катализировать несколько элонгазных реакций. Так, например, “дельта-9-элонгаза” может также действовать как С16/18-элонгаза, С18/20 элонгаза и/или С20/22 элонгаза, и может иметь измененные, однако не наилучшие, специфичности относительно дельта-5 и дельта-6 жирных кислот, таких как ЭПК и/или ГЛК, соответственно.
Термин “C20-элонгаза”, как здесь используется, касается фермента, который утилизирует C20 субстрат, такой как ЭПК или АРК, например. Термин “C20/дельта-5-элонгаза” касается фермента, который утилизирует C20 субстрат с дельта-5 двойной связью.
Подобно этому, термин “EgD9elo” или “EgD9e” касается дельта-9-элонгазы, выделенной из Euglena gracilis (см. SEQ ID NO:112; также см. патентную заявку США № 11/601563 (от 16 ноября 2006 г, которая опубликована как US-2007-0118929-A1 24 мая 2007 г.)).
Как здесь используется, термин “нуклеиновая кислота” означаєт полинуклеотид и включает одно- или двухцепный полимер деоксирибонуклеотидного или рибонуклеотидного оснований. Нуклеиновые кислоты могут также включать фрагменты и модифицированные нуклеотиды. Таким образом, термины “полинуклеотид”, “нуклеиновокислотная последовательность”, “нуклеотидная последовность” и “нуклеиновокислотный фрагмент” используются здесь взаимозаменяемо и представляют собой полимер РНК или ДНК, который является одно- или двухцепным, который содержит, при необходимости, синтетические, неприродные или измененные нуклеотидные основания. На нуклеотиды (которые обычно находятся в своей 5′-монофосфатной форме) ссылаются однобуквенным обозначением следующим образом: “A” касается аденилата или деоксиаденилата (для РНК или ДНК, соответственно), “C” касается цитидилата или деоксицитидилата, “G” касается гуанилата или деоксигуанилата, “U” касается уридата, “T” касается деокситимидилата, “R” касается пуринов (А або G), “Y” касается пиримидинов (С або Т), “K” касается G или Т, “H” касается А или С, или Т, “I” касается инозина, и “N” касается любого нуклеотида.
Термины “субфрагмент, который является функционально эквивалентным” и “функционально эквивалентный субфрагмент” используются здесь взаимозаменяемо. Эти термины касаются участка или субпоследовательности фрагмента изолированной нуклеиновой кислоты, в которой способность изменять экспрессию гена или создавать некий фенотип сохраняется, кодирует или не кодирует данный фрагмент или субфрагмент активный фермент. Например, данный фрагмент или субфрагмент может быть использован в конструировании химерных генов для создание желательного фенотипа в трансформированном растении. Химерные гены могут быть созданы для использования в супрессии путем связывания фрагмента нуклеиновой кислоты или ее субфрагмента, кодирует или не кодирует он активный фермент, в смысловой или антисмысловой ориентации относительно промоторной последовательности растения.
Термин “сохраненный домен” или “мотив” означает набор аминокислот, которые консервируются в специфических положениях вдоль выстроенной последовательности эволюционно родственных протеинов. Хотя аминокислоты в других положеннях могут варьировать между гомологичными протеинами, аминокислоты, которые сильно законсервированы в специфических положениях, отвечают аминокислотам, которые являются незаменимыми по структуре, стабильности или активности протеина. Термины “гомология”, “гомологический”, “подобный в значительной мере” и “такой, который отвечает в значительной мере” применяются здесь взаимозаменяемо. Они касаются фрагментов нуклеиновой кислоты, где изменения в одном или нескольких нуклеотидных основаниях не влияют на способность данного фрагмента нуклеиновой кислоты служить посредником в генной экспрессии или создавать определенный фенотип. Эти термины также касаются модификаций фрагментов нуклеиновой кислоты данного изобретения, таких как делеция или инсерция одного или нескольких нуклеотидов, которые значительно не изменяют функциональные свойства результирующего фрагмента нуклеиновой кислоты относительно первичного, немодифицированного фрагмента. Таким образом, специалистам в данной области понятно, что данное изобретение охватывает больше, чем специфические иллюстративные последовальности.
Кроме того, специалисту в данной области понятно, что в значительной мере схожие нуклеиновокислотные последовательности, которые охватываются данным изобретением, также определяются по их способности гибридизоваться (при умеренно жестких условиях, например, 0,5X SSC, 0,1% SDC, 60°C) с последовательностями, которые здесь приведены как пример, или с каким-либо участком раскрытых здесь нуклеотидных последовальностей, и которые функционально эквивалентны раскрытым здесь любым нуклеиновокислотным последовательностям. Условия жесткости могут быть отрегулированы для проведения скрининга умеренно сходных фрагментов, таких как гомологические последовательности от удаленно родственных организмов, сильно сходных фрагментов, таких как гены, которые дублируют функциональные ферменты из близко родственных организмов. Пост-гибридизационныеі промывки определяют условия жесткости.
Термин "селективно гибридизует" включает ссылку на гибридизацию, при жестких условиях гибридизации, нуклеиновокислотной последовательности с определенной нуклеиновокислотной последовательностью-мишенью в заметно большей мере (например, по меньшей мере с двукратным превышением относительно фона), чем ее гибридизация с нуклеиновокислотными последовальностями, которые не являются мишенями, и существенное исключение нуклеиновых кислот, которые не являются мишенями. Селективно гибридизованные последовательности, типично, имеют приблизительно по меньшей мере 80% идентичность последовальности, или 90% идентичность последовательности, включая до 100% идентичность последовательности (т.е. полную комплементарность) между собой.
Термин "жесткие условия" или “жесткие условия гибридизации” включает ссылки на условия, при которых зонд будет селективно гибридизован со своей последовательностью-мишенью. Условия жесткости зависят от последовательности и будут отличными при разных обстоятельствах. Путем контроля жесткости условий гибридизации и/или условий промывки могут быть идентифицированы последовательности-мишени, которые на 100% комплементарны с зондом (гомологичное зондирование). Как альтернатива, условия жесткости могут быть отрегулированы таким образом, что имеет место некоторое несоответствие в последовательностях, так что проявляются меньшие степени подобия (гетерологическое зондирование). В общем, зонд имеет длину меньше приблизительно 1000 нуклеотидов, при необходимости, менее 500 нуклеотидов.
Типично, жесткими условиями будут такие, где концентрация соли составляет менше приблизительно 1,5 M Na ионов, типично, приблизительно от 0,01 до 1,0 M Na ионов (или других солей) при pH от 7,0 до 8,3, и температура равняется по меньшей мере приблизительно 30°C для коротких зондов (например, 10-50 нуклеотидов) и по меньшей мере приблизительно 60°C для длинных зондов (например, больше 50 нуклеотидов). Жесткие условия могут также достигаться путем добавления дестабилизирующих агентов, таких как формамид. Иллюстративные условия низкой жесткости включают гибридизацию с буферным раствором от 30 до 35% формамида, 1 M NaCl, 1% НДС (натрий додецилсульфат) при 37°C, и промывку в 1X-2X SSC (20X SSC=3,0 M NaCl/0,3 M тринатрийцитрата) при 50-55°C. Иллюстративные условия умеренной жесткости включают гибридизацию в 40-45% формамида, 1 M NaCl, 1% НДС при 37°C, и промывку в 0,5X-1X SSC при 55-60°C. Иллюстративные условия высокой жесткости включают гибридизацию в 50% формамида, 1 M NaCl, 1% НДС при 37°C, и промывку в 0,1X SSC при 60-65°C.
Специфичность является, обычно, функцией пост-гибридизационных промывок, критическими факторами являются ионная сила и температура концевого промывочного раствора. Для ДНК-ДНК гибридов Tm можно аппроксимировать из уравнения Meinkoth et al., Anal. Biochem. 138:267-284 (1984): Tm=81,5°C+16,6 (log M)+0,41 (%GC)-0,61 (% форма)-500/L; где M отвечает молярности моновалентных катионов, %GC является процентом гуанозинового и цитозинового нуклеотидов в данной ДНК, % форма представляет собой процент формамида в гибридизующем растворе, и L отвечает длине гибрида в базисных парах. Tm является температурой (при определенных ионной силе и pH), при которой 50% комплементарной последовальности-мишени гибридизуется с полностью согласованным зондом. Tm снижается на приблизительно 1°C на кажлдый 1% несоответствия; таким образом, Tm, условия гибридизации и/или промывки могут быть отрегулированы таким образом, чтобы гибридизация осуществлялась до необходимой идентичности последовательностей. Например, если проводится поиск последовательностей с >90% идентичности, Tm может быть снижена на 10°C. В целом, условия жесткости выбираются таким образом, чтобы быть на приблизительно 5°C ниже, чем точка теплового плавления (Tm) для специфической последовательности и ее комплемента при определенных ионной силе и pH. Однако, при сильно жестких условиях могут использоваться гибридизация и/или промывка при температурах на 1, 2, 3, или 4°C ниже, чем тепловая точка плавления (Tm); при умеренно жестких условиях могут использоваться гибридизация и/или промывка при температурах на 6, 7, 8, 9, или 10°C ниже, чем тепловая точка плавления (Tm); при условиях низкой жесткости могут использоваться гибридизация и/или промывка при температурах на 11, 12, 13, 14, 15, или 20°C ниже, чем тепловая точка плавления (Tm). С использованием указаного уравнения, гибридизационных и промывочных композиций и желательной Tm рядовым специалистам в данной области будет понятно, каким способом достичь вариаций жесткости условий гибридизации и/или нужных условий промывки. Если необходимая мера несоответствия приводит к Tm меньше 45°C (водный раствор) или 32°C (формамидный раствор), предпочтение отдается тому, чтобы повысить концентрацию SSC, так что может быть использована более высокая температура. Подробным пособием по гибридизации нуклеиновых кислот служит работа Tijssen, Laboratory Techniques in Biochemistry and Molecular Biology--Hybridization with Nucleic Acid Probes, Part I, Chapter 2 "Overview of principles of hybridization and the strategy of nucleic acid probe assays", Elsevier, New York (1993); и работа Current Protocols in Molecular Biology, Chapter 2, Ausubel et al., Eds., Greene Publishing and Wiley-Interscience, New York (1995). Указанные условия гибридизации и/или промывки могут использоваться в течение по меньшей мере 10, 30, 60, 90, 120, или 240 минут.
“Идентичность последовательности” или “идентичность” в контексте нуклеиновокислотных или полипептидных последовательностей касается нуклеиновокислотных оснований или аминокислотных остатков в двух последовательностях, которые одинаковы при их группировке на максимальное соответствие по определенной области сравнения.
Таким образом, “процент идентичности последовательности” касается величины, которая определяется путем сравнения двух оптимально сгруппированных последовательностей по области сравнения, где участок полинуклеотидной или полипептидной последовательности в области сравнения может включать добавки или делеции (т.е., гэпы), в сравнении с эталонной последовательностью (которая не содержит дополнений или делеций), для оптимального группирования данных двух последовательностей. Процент вычисляется путем определения количества положений, в которых идентичное нуклеиновокислотное основание или аминокислотный остаток встречаются в обеих последовательностях, с получением количества положений, которые совпадают, деления количества положений, что совпадают, на общее число положений в области сравнения и умножением результатов на 100 с получением процента идентичности последовательности. Полезные примеры процента идентичности последовательности включают, однако не ограничиваясь этим, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% або 95%, или любой целый процент от 50% до 100%. Эти идентичности могут быть определены с использованием любых программ, которые здесь описаны.
Группирование последовательностей и процент идентичности или подобные расчеты могут быть проведены с использованием разнообразных методов сравнения, разработанных для выявления гомологических последовательностей, включая, однако не ограничиваясь этим, MegAlign™ программу вычислительного биоинформативного пакета LASERGENE (DNASTAR Inc., Madison, WI). В контексте данной заявки станет понятным, что когда для анализа используются программы секвенирования, результаты этого анализа будут базироваться на “значеннях по умолчанию” использованной программы, если не указано иное. Как употребляется в данном тексте, “значения по умолчанию” представляют собой любой набор значений или параметров, которые загружаются с программой при первой инициализации.
“Метод группировки Clustal V” отвечает методу группировки, маркированному Clustal V (который описан Higgins and Sharp, CABIOS. 5:151-153 (1989); Higgins, D.G. et al. (1992) Comput. Appl. Biosci. 8:189-191), который можно найти в программе MegAlign™ вычислительного биоинформативного пакета LASERGENE (DNASTAR Inc., Madison, WI). Для множественных группировок значения по умолчанию отвечают GAP PENALTY=10 и GAP LENGTH PENALTY=10. Параметрами по умолчанию для парных группировок и вычисления процента идентичности протеиновых последовательностей с использованием метода Clustal V являются KTUPLE 1, GAP PENALTY=3, WINDOW=5 и DIAGONALS SAVED=5. Для нуклеиновых кислот этими параметрами служат KTUPLE=2, GAP PENALTY=5, WINDOW=4 и DIAGONALS SAVED=4. После группирования последовательностей с использованием программы Clustal V можно получить “процент идентичности” путем визуализации таблицы “отклонений последовательностей” в той самой программе.
“Метод группировки Clustal W” отвечает методу группировки, маркированному Clustal W (который описан Higgins and Sharp, supra; Higgins, D.G. et al., supra), который можно найти в программе MegAlign™ v6.1 вычислительного биоинформативного пакета LASERGENE (DNASTAR Inc., Madison, WI). Для множественных группировок парметры по умолчанию отвечают GAP PENALTY=10 и GAP LENGTH PENALTY=0,2. Delay Divergen Seqs(%)=30, DNA Transition Weight=0,5, Protein Weight Matrix=Gonnet Series, DNA Weight Matrix=IUB. После группирования последовательностей с использованием программы Clustal W можно получить “процент идентичности” путем визуализации таблицы “отклонений последовательностей” в той самой программе.
Сравнительный анализ первичной структуры согласно программе BLASTN” представляет алгоритм, введенный Национальным центром биотехнологической информации (НЦБИ) для сравнения нуклеотидных последовательностей с использованием параметров по умолчанию.
Специалистам в данной области хорошо понятно, что полезными в идентификации полипептидов относительно других видов является множество уровней идентичности последовательности, где такие полипептиды имеют такую же или похожую функцию или активность. Полезные примеры процента идентичности включают, однако не ограничиваясь этим, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95%, или любой целый процент от 50% до 100%. Действительно, любая целочисленная аминокислотная идентичность от 50 до 100% может быть полезной в описании данного изобретения, такая как 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%. Кроме того, представляет интерес любой полномасштабный или частичный комплемент этого изолированного нуклеотидного фрагмента.
“Ген” касается фрагмента нуклеиновой кислоты, который экспрессирует специфический протеин, и может включать или один кодирующий участок или кодирующий участок, добавленный к регуляторным последовательностям, которые предшествуют (5′ некодирующие последовательности) и следуют за (3′ некодирующие последовательности) кодирующей последовательностью. “Нативный ген” касается гена, который существует в природе со своими собственными регуляторными последовательностями. “Химерный ген” касается любого гена, который не является нативным геном, который содержит регуляторную и кодирующую последовательности, которые вместе в природе не существуют. Соответственно, химерный ген может включать регуляторные последовательности и кодирующие последовательности, которые получаются из разных источников, или регуляторные последовательности и кодирующие последовательности, которые происходят из одного источника, но расположены таким образом, что отличаются от существующего в природе. “Эндогенный ген“ касается нативного гена в своем природном расположении в геноме организма. “Посторонний” ген касается гена, который обычно не существует в организме хозяина, но который вводится в организм хозяина путем переноса гена. Посторонние гены могут включать нативные гены, введенные в не-нативный организм, или химерные гены. “Трансген” представляет собой ген, который введен в геном путем процедуры трансформации.
Термин “геном”, как он применяется к растительным клеткам, охватывает не только хромосомную ДНК, найденную внутри ядра, но и органоидную ДНК, найденную внутри субклеточных компонентов (например, митохондриальных, пластидных) клеток.
“Кодон-оптимизированный ген” представляет собой ген, который имеет свою частоту использования кодона, созданный для имитации частоты предпочтительного использования кодона клетки хозяина.
“Аллель” представляет собой одну из нескольких альтернативных форм гена, занимающую данниый локус на хромосоме. Когда все аллели, присутствующие в данном локусе на хромосоме, одинаковы, это растение является гомозиготным в данном локусе. Если аллели, присутствующие в данном локусе на хромосоме, отличны, это растение является гетерозиготным в данном локусе.
“Кодирующая последовательность” касается ДНК последовальности, которая кодирует специфическую аминокислотную последовательность. “Регуляторные последовательности” касаются нуклеотидных последовательностей, расположенных вверху (5′ некодирующие последовательности), внутри или внизу (3′ некодирующие последовательности) от кодирующей последовательности, и которые влияют на транскрипцию, РНК процессинг или стабильность, или трансляцию ассоциированной кодирующей последовательности. Регуляторные последовательности могут включать, однако не ограничиваясь этим, промоторы, трансляционные сигнальные последовательности, интроны, полиаденилирующие распознающие последовательности, РНК процессинговые сайты, эффекторные связывающие сайты и структуры типа “стебель-петля”.
“Промотор” касается ДНК последовальности, способной контролировать экспрессию кодирующей последовательности или функциональной РНК. Промоторная последовательность состоит из ближайших и более удаленных “верхних” элементов, на последние элементы часто ссылаются как на энхансеры. Соответственно, “энхансер” представляет собой ДНК последовательность, которая может стимулировать промоторную активность, и может быть природным элементом промотора или гетерологическим элементом, который введен для усиления уровня или тканевой специфичности промотора. Промоторы могут быть получены в своей полноте из нативного гена, или могут состоять из разных элементов, полученных от разных промоторов, что существуют в природе, или даже включать сегменты синтетической ДНК. Специалистам в данной области понятно, что разные промоторы могут направлять экспрессию гена в разных тканях или типах клеток, или на разных стадиях развития, или действовать как реакция на разные условия окружающей среды. Кроме того, признано, что поскольку в большинстве случаев точные пределы регуляторных последовательностей определены не полностью, ДНК фрагменты некоторых вариаций могут иметь идентичную промоторную активность. На промоторы, которые вызывают экспрессию гена в большинстве типов клеток в большинства случаев, обычно ссылаются как на “конститутивные промоторы”. Новые промоторы разных типов, полезные в растительных клетках, постоянно открывают; множество примеров можно найти в компиляции Okamuro, J.K., та Goldberg, R.B. Biochemistry of Plants 15:1-82 (1989).
“Трансляционная лидерная последовательность” касается полинуклеотидной последовательности, которая локализована между промоторной последовательностью гена и кодирующей последовательностью. Трансляционная лидерная последовательность присутствует в подверженной полному процессингу мРНК вверх от трансляционной инициирующей (старт) последовательности. Трансляционная лидерная последовательность может влиять на процессинг первичного транскрипта до мРНК, мРНК стабильность или трансляционную эффективность. Примеры трансляционных лидерных последовательностей описаны в литературе (Turner, R. и Foster, G.D., Mol. Biotechnol. 3:225-236 (1995)).
“3′ некодирующие последовательности”, “транскрипционный терминатор” или “терминирующие последовательности” касаются ДНК последовательностей, расположенных вниз от кодирующей последовательности, и включают полиаденилирующие распознающие последовательности и другие последовательности, которые кодируют регуляторные сигналы, способные влиять на мРНК процессинг или экспрессию гена. Сигнал полиаденилирования обычно характеризуется влиянием на добавление трактов полиадениловой кислоты к 3′ концу мРНК предшественника. Использование разных 3′ некодирующих последовательностей иллюстрируется Ingelbrecht, I.L., et al. Plant Cell 1:671-680 (1989).
“РНК транскрипт” касается продукта, который является результатом катализированной РНК полимеразой транскрипции ДНК последовательности. Когда РНК транскрипт является совершенной комплементарной копией ДНК последовательности, на него ссылаются как на основной транскрипт. На РНК транскрипт ссылаются как на зрелую РНК, когда РНК последовательность получена в результате пост-транскрипционного процессинга основного транскрипта. “Матричная РНК” или “мРНК” касается РНК, которая не имеет интронов и которая может быть транслирована в протеин данной клеткой. “кДНК” касается ДНК, которая комплементарна к и синтезирована из мРНК матрицы с использованием ферментной ревертазы. кДНК может быть одноцепочечной или может быть конвертирована в двухцепочечную форму с использованием Кlenow фрагмента ДНК полимеразы І. “Смысловая” РНК касается РНК транскрипта, который включает мРНК и может быть транслирована в протеин внутри клетки или in vitro.
“Антисмысловая РНК” касается РНК транскрипта, который комплементарен ко всему или части таргетного первичного транскрипта или мРНК, и который блокирует экспрессию таргетного гена (U.S. Patent No. 5107065). Комплементарность антисмысловой РНК может быть с любой частью специфического генного транскрипта, т.е. 5′ некодирующей последовательностью, 3′ некодирующей последовательностью, интронами или кодирующей последовательностью. “Функциональная РНК” касается антисмысловой РНК, рибозимной РНК или другой РНК, которая не может транслироваться, но все же имеет влияние на клеточные процессы. Термины “комплемент” или “обратный комплемент” используются здесь взаимозаменяемо относительно мРНК транскриптов, и определяют, как представляется, антисмысловую РНК данного месседжа.
Термин “сцепленный действующим образом” касается ассоциации нуклеиновокислотных последовательностей на отдельном нуклеиновокислотном фрагменте, так что функция одной регулируется другой. Например, промотор сцеплен действующим образом с кодирующей последовательностью, когда он способен регулировать экспрессию этой кодирующей последовательности (т.е. данная кодирующая последовательность находится под транскрипционным контролем промотора). Кодирующие последовательности могут быть сцеплены действующим образом с регуляторными последовательностями в смысловой или антисмысловой ориентации. В другом примере участки комплементарной РНК данного изобретения могут быть сцеплены действующим образом прямо или опосредствованно, 5′ с таргетной мРНК, или 3′ с таргетной мРНК, или внутри таргетной мРНК, или первый комплементарный участок 5′ и его комплемент 3′ с таргетной мРНК.
Стандартная рекомбинантная ДНК и методы молекулярного клонирования, которые здесь использованы, хорошо изестны в данной области и более полно описаны в работе Sambrook, J., Fritsch, E.F. и Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY (1989). Способы превращения хорошо известны специалистам в данной области и описаны ниже.
“ПЦР” или “полимеразная цепная реакция” представляет собой метод синтеза больших количеств специфических ДНК сегментов и состоит из ряда повторных циклов (Perkin Elmer Cetus Instruments, Norwalk, CT). Типично, двухцепочечная ДНК подвергается тепловой денатурации, два праймера, комплементарные к 3′ границам данного таргетного сегмента, закаляются при низкой температуре и потом растягиваются при промежуточной температуре. Одна группа из таких трех последовательных шагов называется “циклом”.
Термин “рекомбинант” касается искусственной комбинации двух отделенных разным способом сегментов последовательности, например, путем химического синтеза или манипуляции изолированными сегментами нуклеиновых кислот методами генной инженерии.
Термины “плазмида” или “вектор” касаются экстрахромосомного элемента, который часто несет гены, которые не являются частью центрального метаболизма данной клетки, и, обычно, в форме круговых двухцепочечных ДНК фрагментов. Такие элементы могут быть автономными репликационными последовательностями, геномными интегрирующими последовательностями, фаговыми или нуклеотидными последовательностями, линейными или круговыми, одно- или двухцепочечных ДНК или РНК, полученных из любого источника, где ряд нуклеотидных последовательностей был объединен или рекомбинирован в уникальную конструкцию, которая способна вводить экспрессирующую кассету(ты) в клетку. “Экспрессирующая кассета” касается фрагмента ДНК, который содержит посторонний ген и имеет элементы в дополнение к постороннему гену, которые позволяют осуществлять усиленную экспрессию этого гена в постороннего хозяина. “Трансформирующая кассета” касается фрагмента ДНК, который содержит посторонний ген и имеет элементы в дополнение к постороннему гену, которые облегчают трансформацию конкретной клетки-хозяина.
Термины “рекомбинантная конструкция ”, “экспрессионный конструкция”, “химерный конструкция”, “конструкция” и “рекомбинантная ДНК конструкция” используются здесь взаимозаменяемо. Рекомбинантная конструкция включает искусственную комбинацию фрагментов нуклеиновой кислоты, например, регуляторную и кодирующую последовательности, которые вместе в природе не встречаются. Например, химерный конструкция может включать регуляторные последовательности и кодирующие последовательности, которые получены из разных источников, или регуляторные последовательности и кодирующие последовательности, полученные из одного источника, но расположеные способом, отличающимся от существующего в природе. Такая конструкция может использоваться сам по себе или в сочетании с вектором. Если используется вектор, выбор вектора зависит от способа, который будет использован для трансформации клеток-хозяев, как это хорошо известно специалистам в данной области. Например, может быть использован плазмидный вектор. Опытный специалист хорошо осведомлен относительно генетических элементов, которые должны присутствовать на векторе для успешной трансформации, селекции и размножения клеток хозяина, которые включают какой-либо из изолированных фрагментов нуклеиновой кислоты данного изобретения. Опытный специалист также признает, что разные независимые трансформационные события дадут разные уровни и виды экспрессии (Jones et al., EMBO J. 4:2411-2418 (1985); De Almeida et al., Mol. Gen. Genetics 218:78-86 (1989)), и потому эти множественные события должны быть подвергнуты скринингу для получения линий, которые проявляют желательный уровень экспрессии и вид. Такой скрининг может проводиться, среди прочего, методом саузерн анализа ДНК, норзерн анализа мРНК экспрессии, иммуноблоттингового анализа экспрессии протеина или фенотипного анализа.
Термин “экспрессия”, как здесь применяется, касается выработки функционального конечного продукта (например, мРНК или протеина [как предшественника, так и зрелого]).
Термин “введенный” означает введение нуклеиновой кислоты (например, экспрессионная конструкция) или протеина в клетку. «Введенный» включает ссылку на встраивание нуклеиновой кислоты в эукариотную или прокариотную клетку, где данная нуклеиновая кислота может быть встроена в геном клетки, и включает ссылку на временное введение нуклеиновой кислоты или протеина в данную клетку. «Введенный» включает ссылку на стабильные или переходные методы трансформации, так же как и на половое скрещивание. Таким образом, “введенный” в контексте введения фрагмента нуклеиновой кислоты (например, рекомбинантный ДНК конструкция/экспрессионная конструкция) в клетку означает “трансфекцию” или “трансформацию” или “трансдукцию”, и включает ссылку на встраивание фрагмента нуклеиновой кислоты в эукариотную или прокариотную клетку, где данный фрагмент нуклеиновой кислоты может быть встроен в геном данной клетки (например, хромосомную, плазмидную, пластидную или митохондриальную ДНК), конвертирован в автономный репликон, или временно экспрессирован (например, трансфектированная мРНК).
“Зрелый” протеин касается полипептида, который подвергался пост-трансляционному процессингу (т.е. такого, из которого были удалены любые пре- или пропептиды, присутствующие в первичном трансляционном продукте). “Предшествующий” протеин касается первичного продукта трансляции мРНК (т.е. с еще присутствующими пре- и пропептидами). Пре- и пропептиды могут быть, однако не ограничиваясь этим, сигналами внутриклеточной локализации.
“Стабильная трансформация” касается переноса фрагмента нуклеиновой кислоты в геном организма хозяина, включая ядерный и органелпярный геномы, что дает генетически устойчивую наследственность. В отличие от этого, “временная трансформация” касается переноса фрагмента нуклеиновой кислоты в ядро или органеллу, которая содержит ДНК, или организм хозяина, что дает экспрессию гена без интеграции или устойчивой наследственности. На организмы хозяина, которые содержат трансформированные фрагменты нуклеиновой кислоты, ссылаются как на “трансгенные” организмы.
Как здесь используется, "трансгенный" касается растения или клетки, которая содержит в своем геноме гетерологический полинуклеотид. Лучше, когда гетерологический полинуклеотид стабильно интегрирован в данном геноме, так что данный полинуклеотид передается последующим генерациям. Данный гетерологический полинуклеотид может быть интегрирован в геном отдельно или как часть экспрессирующей конструкции. «Трансгенный», как используется в данном тексте, включает любую клетку, клеточную линию, калюс, ткань, часть растения или растение, генотип которых изменен присутствием гетерологической нуклеиновой кислоты, включая как те трансгеники, которые изменены указанным способом, так и те, что были созданы путем половых скрещиваний или вегетативного размноження из первичного трансгеника. Термин "трансгенный”, как здесь используется, не охватывает смену генома (хромосомную или экстра-хромосомную) с помощью обычных методов селекции растений или путем природных событий, таких как случайное перекрестное оплодотворение, нерекомбинантная вирусная инфекция, нерекомбинантная бактериальная трансформация, нерекомбинантная транспозиция или спонтанная мутация.
“Антисмысловое ингибирование” касается создания антисмысловых РНК транскриптов, способных подвергать супрессии экспрессию таргетного протеина. “Косупрессия” касается продуцирования смысловых РНК трапнскриптов, способных подавлять экспрессию идентичных или в значительной мере схожих посторонних или эндогенных генов (патент США № 5231020). Косупрессионные конструкцияы в растениях были предварительно разработаны путем сосредоточения внимания на гиперэкспрессии нуклеиновокислотной последовательности, которая имеет гомологию с эндогенной мРНК, в смысловой ориентации, результатом чего явилась редукция всех РНК, которые гомологичны с гиперэкспрессированной последовательностью (Vaucheret et al., Plant J. 16:651-659 (1998); Gura, Nature 404:804-808 (2000)). Полная эффективность этого феномена низкая, и степень РНК редукции варьирует в широких пределах. В недавней работе описано использование “шпильковых” структур, которые включают всю или часть мРНК кодирующей последовательности в комплементарной ориентации, что дает потенциальную структуру “стебель-петля” для экспрессированной РНК (РСТ публикация № WO 99/53050; РСТ публикация № WO 02/00904). Это повышает частоту косупрессии в восстановленных трансгенных растениях. Другая вариация описывает использование растительных вирусных последовательностей для направления супрессии или “заглушивания” ближайших мРНК кодирующих последовательностей (РСТ публикация № WO 98/36083). Оба эти косупрессивные явления не были выяснены механистически, хотя генетические факты начали раскрывать эту сложную ситуацию (Elmayan et al., Plant Cell 10:1747-1757 (1998)).
Термин “маслянистый” касается тех организмов, которые имеют тенденцию сохранять свой источник энергии в форме липида (Weete, In: Fungal Lipid Biochemistry, 2nd Ed., Plenum, 1980). На класс растений, которые идентифицируются как маслянистые, обычно ссылаются как на “маслянистые” растения. Примеры маслянистих растений включают, однако не ограничиваясь этим, сою (виды Glycine и Soja), лен (вид Linum), рапс (вид Brassica), маис, хлопок, сафлор (вид Carthamus) и подсолнечник (вид Helianthus).
В общем, содержание клеточного масла или ТАГ в этих микроорганизмах следует сигмоидной кривой, где концентрация липида возрастает до тех пор, пока не достигает максимума в поздней логарифмической или ранней стационарной фазе роста и потом постепенно снижается на протяжении поздней стационарной фазы и фазы отмирания (Yongmanitchai and Ward, Appl. Environ. Microbiol. 57:419-25 (1991)).
Термин “жировые дрожжи” касается тех микроорганизмов, которые классифицируются как дрожжи, которые вырабатывают масло. Для маслянистых микроорганизмов вполне обычным является накопление более приблизительно 25% сухого клеточного веса в виде масла. Примеры маслянистых дрожжей включают, однако не ограничиваясь этим, следующие роды: Yarrowia, Candida, Rhodotorula, Rhodosporidium, Cryptococcus, Trichosporon и Lipomyces.
Применяемый здесь термин “биомасса” касается конкретно использованного дрожжевого клеточного материала, образовавшегося в результате ферментации рекомбинантного хозяина-производителя, который вырабатывает ПНЖК в коммерчески значимых количествах, где предпочтительным хозяином-производителем является рекомбинантный штамм жировых дрожжей, Yarrowia lipolytica. Биомасса может быть в виде целых клеток, целоклеточных лизатов, гомогенизированных клеток, частично гидролизованного клеточного материала, и/или частично очищенного клеточного материала (например, выработанного микробами масла).
Термин “эвгленофиковые” (“Euglenophyceae”) касается группы одноклеточных бесцветных или фотосинтетических флагеллатов (“эвгленоидов”), которые найдены существующими в пресноводных, морских, почвенных и паразитных средах. Данный класс характеризуется отдельными одноклеточными организмами, где большинство является водоплавающим и имеет два жгутика (один из которых может быть невсплывающим), которые возникают из передней инвагинации, известной как резервуар. Фотосинтетические эвгленоиды содержат от одного до множества травянистозеленых хлоропластов, которые варьируют от крохотных дисков до расширенных пластинок или ленточек. Ассимиляция нутриента бесцветными эвгленоидами зависит от осмотрофии или фаготрофии. Приблизительно 1000 видов было описано и классифицировано на приблизительно 40 родов и 6 рядов. Примеры эвгленофиковых включают, однако не ограничиваясь этим, следующие роды: Eutreptiella и Tetruetreptia.
Термин “растение” касается целых растений, растительных органов, растительных тканей, семян, растительных клеток, семечек и их потомства. Растительные клетки включают, без ограничений, клетки из семечек, суспензионных культур, зародышей, меристематических участков, калюсной ткани, листвы, корней, побегов, гаметофитов, спорофитов, пыльцы и микроспор.
“Потомство” включает любую последующую генерацию растения.
Обзор: Микробный биосинтез жирных кислот и триацилглицеринов
В общем, аккумуляция липидов в маслянистых микроорганизмах запускается как реакция на имеющееся в ростовой среде общее соотношение углерода и азота. Этот процесс, ведущий к de novo синтезу свободного пальмитата (16:0) в маслянистых микроорганизмах, подробно описан в патенте США № 7238482. Пальмитат является предшественником производных более длинноцепочечных насыщенных и ненасыщенных жирных кислот, которые образуются под действием элонгаз и десатураз (ФИГ.1).
ТАГи (основные единицы хранения для жирных кислот) образуются в результате ряда реакций, которые включают: (1) этерификацию одной молекулы ацил-CoA в глицерин-3-фосфат через ацилтрансферазу с образованием лизофосфатидной кислоты; (2) этерификацию второй молекулы ацил-CoA через ацилтрансферазу с образованием 1,2-диацилглицеринфосфата (идентифицируемого обычно как фосфатидная кислота); (3) удаление фосфата фосфатидкислотной фосфатазой с образованием 1,2-диацилглицерина (ДАГ); и (4) добавление третьей жирной кислоты действием ацилтрансферазы с образованием ТАГ. Широкий спектр жирных кислот может быть введен в ТАГи, включая насыщенные и ненасыщенные жирные кислоты, и короткоцепные и длинноцепочечные жирные кислоты.
Биосинтез омега жирных кислот
Метаболический процесс, где олеиновая кислота конвертируется в длинноцепочечные омега-3/омега-6 жирные кислоты, включает элонгацию углеродной цепи путем добавления углеродных атомов и десатурацию данной молекулы путем добавления двойных связей. Это требует ряда специальных десатурационных и элонгационных ферментов, присутствующих в эндоплазматичной сеточной мембране. Однако, как видно из ФИГ.1 и как описано ниже, для этого часто существуют множественные альтернативные пути производства специфических длинноцепочечных омега-3/омега-6 жирных кислот.
Конкретно, все пути требуют первичной конверсии олеиновой кислоты в ЛК, первую из омега-6 жирных кислот, с помощью дельта-12-десатуразы. Потом, с использованием “дельта-9-элонгаза/дельта-8-десатуразного” пути и ЛК как субстрата длинноцепочечные омега-6 жирные кислоты получаются следующим образом: (1) ЛК конвертируется в ЭДК под действием дельта-9-элонгазы; (2) ЭДК конвертируется в ДГЛК под действием дельта-8-десатуразы; (3) ДГЛК конвертируется в АРК под действием дельта-5-десатуразы; (4) АРК конвертируется в ДТК под действием С20/22-элонгазы; и (5) ДТК конвертируется в ДПКn-6 под действием дельта-4-десатуразы. Как альтернатива, “дельта-9-элонгаза/дельта-8-десатуразный путь” может использовать АЛК как субстрат с образованием длинноцепочечных омега-3 жирных кислот следующим образом: (1) ЛК конвертируется в АЛК, первую из омега-3 жирных кислот, под действием дельта-15-десатуразы; (2) АЛК конвертируется в ЭТрК под действием дельта-9-элонгазы; (3) ЭТрК конвертируется в ЭТК под действием дельта-8-десатуразы; (4) ЭТК конвертируется в ЭПК под действием дельта-5-десатуразы; (5) ЭПК конвертируется в ДПК под действием С20/22-элонгазы; и (6) ДПК конвертируется в ДГК под действием дельта-4-десатуразы. При необходимости, омега-6 жирные кислоты могут быть конвертированы в омега-3 жирные кислоты; например, ЭТК и ЭПК образуются из ДГЛК и АРК, соответственно, под действием активности дельта-17-десатуразы.
Альтернативные пути для биосинтеза омега-3/омега-6 жирных кислот используют дельта-6-десатуразу и С18/20-элонгазу (известную также как дельта-6-элонгаза, эти термины могут использоваться взаимозаменяемо) (т.е. “дельта-6-десатураза/дельта-6-элонгазный путь”). Более конкретно, ЛК и АЛК могут конвертироваться в ГЛК и СТК, соответственно, с помощью дельта-6-десатуразы; потом С18/20-элонгаза превращает ГЛК в ДГЛК и/или СТК в ЭТК.
Предполагается, что конкретные функциональные группы, которые необходимо ввести в специфический организм-хозяин для выработки омега-3/омега-6 жирных кислот, будут зависеть от клетки-хозяина (и ее нативного ПНЖК профиля и/или десатураза/элонгазного профиля), наличия субстрата и желательного конечного продукта(ов). Например, экспрессия дельта-9-элонгаза/дельта-8-десатуразного пути может быть предпочтительной в некоторых вариантах, в противоположность экспрессии дельта-6-десатураза/дельта-6-элонгазного пути, поскольку ПНЖК, полученные с помощью первого пути, свободны от ГЛК.
Специалист в данной области сможет идентифицировать разные гены-кандидаты, которые кодируют каждый из ферментов, необходимых для омега-3/омега-6 жирнокислотного биосинтеза. Полезные десатуразные и элонгазные последовательности могут быть получены из любого источника, например, выделены из природного источника (из бактерий, водорослей, грибов, растений, животных и т.д.), получены по полусинтетической схеме или синтезированы de novo. Хотя конкретный источник десатуразных и элонгазных генов, введенных в хозяина, не является критичным, соображения по поводу выбора специфического полипептида, который обладает десатуразной или элонгазной активностью, включают: (1) специфичность субстрата данного полипептида; (2) является ли данный полипептид или его компонент ферментом, который лимитирует скорость; (3) является ли данная десатураза или элонгаза незаменимой для синтеза нужной ПНЖК; (4) кофакторы, необходимые для данного полипептида; и/или (5) является ли данный полипептид модифицированным после его получения (например, с помощью киназы или пренилтрнсферазы). Экспрессированный полипептид имеет, лучше, параметры, совместимые с биохимическим окружением его локации в клетке хозяина (в отношении дополнительных деталей см. патентСША № 7238482).
В дополнительных вариантах будет также полезным рассмотреть эффективность конверсии каждой конкретной десатуразы и/или элонгазы. Более конкретно, поскольку каждый фермент в редких случаях функционирует со 100% эффективностью в отношении конверсии субстрата в продукт, конечный липидный профиль неочищенных масел, образованных в клетке хозяина, будет, типично, представлять собой смесь разных ПНЖК, которая состоит из желательной омега-3/омега-6 жирной кислоты, так же как и разных верхних промежуточных ПНЖК. Таким образом, эффективность конверсии, которую обеспечивает каждый фермент, также представляет собой переменную при оптимизации биосинтеза необходимой жирной кислоты, которую следует принимать во внимание.
С учетом всех вышерассмотренных соображений могут быть идентифицированы гены-кандидаты, которые обладают соответствующими активностями десатуразы и элонгазы (например, дельта-6-десатуразы, C18/20-элонгазы, дельта-5-десатуразы, дельта-17-десатуразы, дельта-15-десатуразы, дельта-9-десатуразы, дельта-12-десатуразы, C14/16-элонгазы, C16/18-элонгазы, дельта-9-элонгазы, дельта-8-десатуразы, дельта-4-десатуразы, C20/22-элонгазы и ДГК синтазы), в соответствии с имеющейся доступной для общественности литературой (например, GenBank), патентной литературой, и экспериментальным анализом организмов, которые способны производить ПНЖК. Эти гены будут пригодны для введения в организм конкретного хозяина для обеспечения или усиления синтеза ПНЖК этим организмом.
Мультизимы и линкеры
В одном варианте данное изобретение касается мультизима, который содержит отдельный полипептид, обладающий по меньшей мере двумя независимыми и раздельными ферментативными активностями.
Примеры пригодных ферментативных активностей включают элонгазы, десатуразы жирных кислот, трансферазы, ацил CoA синтазы и тиоэстеразы. Например, подходящие десатуразы жирных кислот включают, однако не ограничиваясь этим: дельта-4-десатуразу, дельта-5-десатуразу, дельта-6-десатуразу, дельта-8-десатуразу, дельта-9-десатуразу, дельта-12-десатуразу, дельта-15-десатуразу, и/или дельта-17-десатуразу. Примеры подходящих элонгаз включают, однако, не ограничиваясь этим: дельта-9-элонгазу, C14/16-элонгазу, C16/18-элонгазу, C18/20-элонгазу и/или C20/22-элонгазу.
Примеры подходящих трансфераз включают, однако не ограничиваясь этим, ацил трансферазы, такие как глицерин-3-фосфат O-ацилтрансфераза (называемая также глицерин-фосфат ацил трансфераза или глицерин -3-фосфат ацил трансфераза; ГФАТ), 2-ацилглицерин O-ацилтрансфераза, 1-ацилглицерин-3-фосфат O-ацилтрансфераза (называемая также 1-ацилглицерин-фосфат ацилтрансфераза или лизо-фосфатидкислотная ацилтрансфераза; АГФАТ или ЛФКАТ идли ЛФАТ), 2-ацилглицерин-3-фосфат O-ацилтрансфераза, 1-ацилглицерофосфохолин O-ацилтрансфераза (называемая также лизо-лецитин ацилтрансфераза или лизо-фосфатидилхолин ацилтрансфераза; АГФХАТ или ЛЛАТ или ЛФХАТ), 2-ацилглицерофосфохолин O-ацилтрансфераза, диацилглицерин O-ацилтрансфераза (называемая также диглицерид ацил трансфераза; ДАГАТ или ДГАТ) и фосфолипид:диацилглицерин ацилтрансфераза (ФДАТ).
Пример подходящей ацил CoA синтазы включает, однако не ограничиваясь этим, длинноцепочечную-жирнокислотную-CoA лигазу (называемую также ацил-активирующий фермент или ацил-CoA синтаза).
Пример подходящей тиоэстеразы включает, однако не ограничиваясь этим, олеоил-[ацил-носитель-протеин] гидролаза (называемая такжек ацил-[ацил-носитель-протеин] гидролаза, ацил-АНП-гидролаза или ацил-АНП-тиоэстераза.
Лучше, когда мультизим данного изобретения обладает ферментативными активностями, которые включают по меньшей мере одну элонгазу жирных кислот, сцепленную с по меньшей мере одной десатуразой жирных кислот.
Связь, используемая для формирования мультизима, включает, как минимум, отдельную полипептидную связь. В другом аспекте, связь может включать один аминокислотный остаток, такой как пролин, или полипептид. Если данная связь является полипептидом, может быть желательным для данной связи иметь по меньшей мере один пролиновый аминокислотный остаток.
Лучше, когда мультизим настоящего изобретения обладает первой ферментативной активностью, связанной со второй ферментативной активностью, и данная связь выбирается из группы, которая состоит из полипептидной связи, SEQ ID NO:198 (линкер EgDHAsyn1), SEQ ID NO:200 (линкер EgDHAsyn2), SEQ ID NO:235 (линкер EaDHAsyn1), SEQ ID NO:472, SEQ ID NO:504, и модифицированных Yarrowia lipolytica линкеров (SEQ ID NOs:438 и 445).
Кроме того, в объем данного изобретения входит способ получения мультизима, который включает:
(a) связывание первого полипептида с по меньшей мере вторым полипептидом, где каждый полипептид обладает независимой и раздельной ферментативной активностью; и
(b) оценку продукта стадии (a) в отношении независимой и раздельной ферментативных активностей.
Как описывалось выше, ферментативные активности выбираются из группы, которая состоит из элонгазы жирных кислот, десатуразы жирных кислот, ацил трансфераз, ацил CoA синтаз и тиоэстераз. Лучше, когда ферментативные активности включают по меньшей мере одну элонгазу жирных кислот, связанную с по меньшей мере одной десатуразой жирных кислот.
Примеры подходящих десатураз, элонгаз и линкеров описаны выше.
Хотя выше описаны многочисленные примеры мультизимов, ДГК синтазы (включающие как активность C20-элонгазы, так и активность дельта-4-десатуразы) и ДГЛК синтазы (включающие как активности дельта-9-элонгазы, так и дельта-8-десатуразы) представляют особый интерес. Описанные выше данные подтверждают, что связывание двух доменов внутри каждой синтазы дает повышенную эффективность или поток, в сравнении с эффективностью или потоком, наблюдаемыми, когда ферментативные домены существуют как независимые сущности, т.е. не связанные вместе в мультизиме.
Например, когда мультизим, включающий Euglena gracilis C20домен элонгазы и Schizochytrium aggregatum дельта-4-десатуразу, был экспрессирован в Yarrowia lipolytica, активность дельта-4-десатуразы была приблизительно в 2-3 раза выше в слитой конструкции, в противоположность ее активности при отдельной экспрессии (Пример 28). Подобно этому, когда Euglena gracilis C20 домен элонгазы-Schizochytrium aggregatum дельта-4-десатуразный гибрид был экспрессирован как мультизим в сое, наблюдался повышенный поток ЭПК в ДГК, в отличие от ситуации, когда два указанных фермента были экспрессированы независимо (Пример 49).
Повышенная эффективность (или поток ЛК в ДГЛК) была также продемонстрирована в различных ДГЛК синтазах, которые были созданы. Ряд из шести дельта-9-элонгаза/дельта-8-десатуразных слитых конструкций был создан с использованием различных комбинаций дельта-9-элонгаз, полученных из E. gracillis, E. anabaena UTEX 373 и Eutreptiella sp. CCMP389, и дельта-8-десатураз, полученных из E. gracillis и E. anabaena UTEX 373; они были индивидуально экспрессированы в Yarrowia lipolytica (Примеры 55 и 56, соответственно). Во всех случаях слитый ген обладал более высокой активностью, чем один индивидуальный ген, экспрессированный в Yarrowia. Эти данные вновь предполагают, что продукт дельта-9-элонгазы может прямо каналироваться как субстрат дельта-8-десатуразы в слитый протеин. Специалист в данной области сможет употребить приведенные здесь рассуждения для создания разных других мультизимов, которые обладают повышенной эффективностью или потоком. Соответственно, данное изобретение касается любого мультизима, который создан с использованием линкера, полученного из последовательностей настоящего изобретения. Мультизимами, которым отдается предпочтение, являются те, которые объединяют различные гены ПНЖК биосинтетического пути.
Идентификация последовательностей новых ДГК синтаз
В настоящем изобретении нуклеотидные последовательности, кодирующие ДГК синтазы, были выделены из Euglena gracilis и Euglena anabaena, как резюмировано ниже в Таблице 4.
В некоторых вариантах текущие EgDHAsyn1, EgDHAsyn2, EaDHAsyn1, EaDHAsyn2 и EaDHAsyn3 DHA синтазные последовательности могут быть кодон-оптимизированы для экспрессии в конкретном организме хозяина. Как хорошо известно в данной области, это может послужить полезным средством для дополнительной оптимизации экспрессии данного фермента в измененном хозяине, поскольку использование предпочитаемых хозяином кодонов может значительно усилить экспрессию постороннего гена, кодирующего данный полипептид. EgDHAsyn1, например, был кодон-оптимизирован для экспрессии в Yarrowia lipolytica (Пример 54), с образованием EgDHAsyn1S (как указано в патенте США № 7238482 и патенте США № 7125672).
Специалист в данной области сможет использовать наработки данного изобретения для создания различных других кодон-оптимизированных ДГК синтазных протеинов, подходящих для оптимальной экспрессии в альтернативных хозяевах, основываясь на последовательностях дикого типа EgDHAsyn1, EgDHAsyn2, EaDHAsyn1, EaDHAsyn2 и/или EaDHAsyn3, описанных выше в Таблице 4. Соответственно, настоящее изобретение касается любого кодон-оптимизированного ДГК синтазного протеина, который получен из последовательности дикого типа данного изобретения. В некоторых вариантах, которым отдается предпочтение, может быть желательным модифицировать часть кодонов, кодирующих EgDHAsyn1, EgDHAsyn2, EaDHAsyn1, EaDHAsyn2 и/или, EaDHAsyn3, для усиления экспрессии данного гена в организме хозяина, включая, однако не ограничиваясь этим, растение или часть растения.
В другом варианте данное изобретение касается выделенного полинуклеотида, кодирующего ДГК синтазу, включая:
(a) нуклеотидную последовательность, кодирующую полипептид, обладающий ДГК синтазной активностью, где данный полипептид имеет по меньшей мере 80% аминокислотную идентичность, основываясь на методе группировки Clustal V, при сравнении с аминокислотной последовательностью, как представлено в SEQ ID NO:12, SEQ ID NO:22, SEQ ID NO:95, SEQ ID NO:96, или SEQ ID NO:97;
(b) нуклеотидную последовательность, кодирующую полипептид, обладающий ДГК синтазной активностью, где данная нуклеотидная последовательность имеет по меньшей мере 80% идентичность последовательности, основываясь на сравнительном анализе первичной структуры согласно программе BLASTN, при сравнении с нуклеотидной последовательностью, как представлено в SEQ ID NO:11, SEQ ID NO:205, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93, или SEQ ID NO:410;
(c) нуклеотидную последовательность, кодирующую полипептид, обладающий ДГК синтазной активностью, где данная нуклеотидная последовательность гибридизуется при жестких условиях с нуклеотидной последовательностью, как представлено в SEQ ID NO:11, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93, SEQ ID NO:205, или SEQ ID NO:410; или
(d) комплемент нуклеотидной последовательности (a), (b) или (c), где данные комплемент и нуклеотидная последовательность состоят из одинакового числа нуклеотидов и являются на 100% комплементарными.
Еще в другом аспекте, данное изобретение касается выделенного полинуклеотида, включая:
(a) нуклеотидную последовательность, кодирующую полипептид, который обладает ДГК синтазной активностью, где данный полипептид имеет по меньшей мере 80% аминокислотную идентичность, основываясь на методе группировки Clustal V, при сравнении с аминокислотной последовательностью, как представлено в SEQ ID NO:12, SEQ ID NO:22, SEQ ID NO:95, SEQ ID NO:96, SEQ ID NO:97, или SEQ ID NO:411;
(b) нуклеотидную последовательность, кодирующую полипептид, который обладает ДГК синтазной активностью, где данный полипептид имеет по меньшей мере 80% аминокислотную идентичность, основываясь на сравнительном анализе первичной структуры согласно программе BLASTN, при сравнении с аминокислотной последовательностью, как представлено в SEQ ID NO:11, SEQ ID NO:205, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93 или SEQ ID NO:410;
(c) нуклеотидную последовательность, кодирующую полипептид, обладающий ДГК синтазной активностью, где данная нуклеотидная последовательность гибридизуется при жестких условиях с нуклеотидной последовательностью, как представлено в SEQ ID NO:11, SEQ ID NO:205, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93 или SEQ ID NO:410; или
(d) комплемент нуклеотидной последовательности (a), (b) или (c), где данные комплемент и нуклеотидная последовательность состоят из одинакового числа нуклеотидов и являются на 100% комплементарными.
Лучше, когда выделенный полинуклеотид, кодирующий ДГК синтазу, содержит последовательность, представленную в какой либо из SEQ ID NO:11, SEQ ID NO:205, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93, или SEQ ID NO:410.
Идентификация и выделение гомологов
Любая из текущих ДГК синтазных последовательностей (т.е. EgDHAsyn1, EgDHAsyn2, EaDHAsyn1, EaDHAsyn2 и EaDHAsyn3) или их частей может быть использована для поиска гомологов ДГК синтазы в тех же самых или других видах бактерий, водорослей, грибов, эвгленоидов или растений с использованием программы анализа последовательностей. В общем, такая компьютерная программа подбирает схожие последовательности, приписывая разные степени гомологии различным субституциям, делециям и другим модификациям.
Как альтернатива, любая из текущих ДГК синтазных последовательностей или ее частей могут также быть использованы как реагенты гибридизации для идентификации ДГК синтазных гомологов. Базовые компоненты нуклеиновокислотного гибридизационного теста включают зонд, образец, который, как предполагается, содержит нужный ген или генный фрагмент и специфический метод гибридизации. Зонды настоящего изобретения, типично, представляют собой одноцепочечные нуклеиновокислотные последовательности, комплементарные к нуклеиновокислотным последовательностям, которые предстоит обнаружить. Зонды “гибридизуемы” с нуклеиновокислотными последовательностями, которые предстоит выявить. Хотя длина зонда может варьировать от 5 оснований до десятков тысяч оснований, обычно подходит длина зонда от приблизительно 15 оснований до приблизительно 30 оснований. Только часть молекулы зонда должна быть комплементарной к нуклеиновокислотной последовательности, которую предстоит обнаружить. Кроме того, комплементарность между зондом и таргетной последовательностью не обязательно должна быть совершенной. Гибридизация все же имеет место между несовершенно комплементарными молекулами с тем результатом, что некоторая доля оснований на участке гибридизации не спарена с соответствующим комплементарным основанием.
Методы гибридизации хорошо известны. Обычно зонд и образец должны быть смешаны при условиях, которые дают возможность осуществлять нуклеиновокислотную гибридизацию. Это включает контактирование зонда и образца в присутствии неорганической или органической соли при подходящих концентрации и температуре. Нуклеиновые кислоты зонда и образца должны находиться в контакте на протяжении достаточно длительного времени, чтобы между ними могла произойти любая возможная гибридизация. Концентрация зонда или мишени в смеси будет определять время, необходимое для протекания гибридизации. Чем выше концентрация зонда или мишени, тем меньше будет время инкубации гибридизации. При необходимости, может добавляться разобщающий агент (например, гуанидинхлорид, гуанидинтиоцианат, цианат натрия, тетрахлороацетат лития, перхлорат натрия, тетрахлороацетат рубидия, йодид калия, трифтороацетат цезия). При желании, к гибридизационной смеси может быть добавлен формамид, обычно 30-50% (объем/объем).
Могут применяться различные гибридизационные растворы. Обычно, они содержат от приблизительно 20 до 60% по объему, лучше, 30%, полярного органического растворителя. Обычный гибридизационный раствор содержит приблизительно 30-50% объем/объем формамида, приблизительно от 0,15 до 1 M хлорида натрия, приблизительно от 0,05 до 0,1 M буферов (например, цитрат натрия, Tris-HCl, PIPES или HEPES (область pH приблизительно 6-9)), приблизительно от 0,05 до 0,2% детергента (например, додецилсульфат натрия), или 0,5-20 мМ EDTA, FICOLL (объединение Фармация) (приблизительно 300-500 кДальтонов), поливинилпирролидон (приблизительно 250-500 кДальтонов), и сывороточный альбумин. Кроме того, в типичный раствор для гибридизации входят немеченые нуклеиновые кислоты-носители от приблизительно 0,1 до 5 мг/мл, фрагментированная нуклеиновая ДНК (например, ДНК из зобной железы теленка или ДНК из молок лососевых, или дрожжевая РНК), и при необходимости приблизительно от 0,5 до 2% вес/объем глицина. Могут включаться и другие добавки, такие как агенты, уменьшающие объем, которые включают ряд полярных водорастворимых или разбухающих агентов (например, полиэтиленгликоль), анионные полимеры (например, полиакрилат или полиметакрилат) и анионные сахаридные полимеры (например, декстрансульфат).
Нуклеиновокислотная гибридизация адаптируема к различным форматам образцов для анализа. Одним из наиболее подходящих является сэндвичевый формат. Сэндвичевая проба особенно приспособлена к гибридизации при неденатурирующих условиях. Основным компонентом пробы сэндвичевого типа является твердый носитель, к которому путем адсорбции или за счет ковалентных связей присоединен иммобилизированный нуклеиновокислотный зонд, котрый непомечен и комплементарен к одному участку данной последовательности.
В дополнительных вариантах любой из ДГК синтазных нуклеиновокислотных фрагментов, описанных здесь (или каких-либо их гомологов, идентифицированных здесь), может быть использован для выделения генов, которые кодируют гомологичные протеины, из тех же самых или других бактериальных, водорослевых, грибковых, эвгленоидных или растительных видов. Изоляция гомологичных генов с использованием зависимых от последовательности протоколов хорошо известна в данной оторасли. Примеры зависимых от последовательности протоколов включают, однако не ограничиваясь этим: (1) методы нуклеиновокислотной гибридизации; (2) методы ДНК и РНК амплификации, как иллюстрируется различными применениями нуклеиновокислотных технологий амплификации [например, полимеразная цепная реакция (ПЦР), Mullis et al., патент США № 4683202; лигазная цепная реакция (ЛЦР), Tabor et al., Proc. Acad. Sci. USA 82:1074 (1985); или амплификация со смещением цепочки (АСЦ), Walker et al., Proc. Natl. Acad. Sci. U.S.A., 89:392 (1992)]; и (3) методы построения библиотеки и скрининга путем комплементации.
Например, гены, кодирующие схожие протеины или полипептиды в мультизим или его индивидуальный домен (такие как ДГК синтазы), описанные в настоящей заявке, могли бы быть выделены непосредственно путем использования всех или части текущих нуклеиновокислотных фрагментов как ДНК гибридизационных зондов для скрининга библиотек из, например, любых необходимых дрожжей или грибков с использованием методологии, хорошо известной специалистам в данной области (где предпочтение отдается организмам, производящим ДТК, ДПКn-6, ДПК и/или ДГК). Специфические олигонуклеотидные зонды на основе текущих нуклеиновокислотных последовательностей могут быть сконструированы и синтезированы с использованием известных в данной области методов (Maniatis, supra). Кроме того, для синтеза ДНК зондов могут быть непосредственно использованы полные последовательности с применением методов, известных специалистам в данной области (например, ДНК мечение с использованием рассеянной затравки, “ник” трансляция или метод концевого мечения), или РНК зондов с использованием имеющихся in vitro транскрипционных систем. В дополнение, специфические праймеры могут быть созданы и использовагы для амплификации части (или всей длины) данных последовательностей. Результирующие продукты амплификации могут метиться непосредственно при реациях амплификации или метиться после реакций амплификации и использоваться как зонды для выделения фрагментов ДНК полной длины в условиях подходящей жесткости.
Обычно, в технике амплификации типа ПЦР праймеры имеют отличные последовательности и некомплементарны один к другому. В зависимости от желательных условий теста последовательности праймеров должны быть сконструированы таким образом, чтобы обеспечить как эффективную, так и правильную репликацию нуклеиновой кислоты-мишени. Методы создания ПЦР праймеров носят общий характер и хорошо известны в данной области (Thein и Wallace, “The use of oligonucleotide as specific hybridization probes in the Diagnosis of Genetic Disorders”, in Human Genetic Diseases: A Practical Approach, K. E. Davis Ed., (1986) pp 33-50, IRL: Herndon, VA; and Rychlik, W., In Methods in Molecular Biology, White, B. A. Ed., (1993) Vol. 15, pp 31-39, PCR Protocols: Current Methods and Applications. Humania: Totowa, NJ).
В общем, два коротких сегмента текущих последовательностей могут быть использованы в ПЦР протоколах для амплификации более длинных нуклеиновокислотных фрагментов, кодирующих гомологичные гены из ДНК или РНК. ПЦР может также осуществляться на библиотеке клонированных нуклеиновокислотных фрагментов, где последовательность одного праймера получается из текущих нуклеиновокислотных фрагментов, и последовательность другого праймера использует преимущество присутствия трактов полиадениловой кислоты в 3' конце мРНК предшественника, кодирующего эукариотные гены.
Как альтернатива, последовательность второго праймера может основываться на последовательностях, полученных из клонирующего вектора. Например, опытный специалист может следовать протоколу RACE (Frohman et al., Proc. Natl Acad. Sci. U.S.A., 85:8998 (1988)) для генерации кДНК путем использования ПЦР для амплификации копий участка между отдельной точкой в транскрипте и 3′ или 5′ концом. Праймеры, ориентированные в направлениях 3′ и 5′ концов, могут быть созданы из текущих последовательностей. С использованием имеющихся в продаже 3′ RACE или 5′ RACE систем (Gibco/BRL, Gaithersburg, MD), могут быть выделены специфические 3′ или 5′ кДНК фрагменты (Ohara et al., Proc. Natl Acad. Sci. U.S.A., 86:5673 (1989); Loh et al., Science 243:217 (1989)).
В других вариантах любые из ферментов (например, мультизимы, ДГК синтазы или отдельные домены, описанные здеь) могут быть модифицированы. Как хорошо известно специалистам в данной области, для получения мутаций природных генов могут быть использованы in vitro мутагенез и селекция, химический мутагенез, методы “перетасовки генов” или другие меры. Как альтернатива, мультизимы могут быть синтезированы путем перестановки доменов, где функциональный домен из любого фермента может обмениваться на или добавляться к функциональному домену в альтернативном ферменте, что дает таким образом новый протеин.
Идентификация последовательностей новых C20-элонгаз
В настоящем изобретении нуклеотидные последовательности, кодирующие C20-элонгазы, были выделены из Euglena gracilis и Euglena anabaena, как показано ниже в Таблице 5.
Настоящее изобретение касается выделенного полинуклеотида, кодирующегл C20-элонгазу, включая:
(a) нуклеотидную последовательность, кодирующую полипептид, который обладает активностью C20-элонгазы, где данный полипептид имеет по меньшей мере 80% аминокислотную идентичность, основываясь на методе группировки Clustal V, при сравнении с аминокислотной последовательностью, как представлено в SEQ ID NO:12, SEQ ID NO:22, SEQ ID NO:95, SEQ ID NO:96, SEQ ID NO:97; SEQ ID NO:202, SEQ ID NO:204, SEQ ID NO:231, SEQ ID NO:232, или SEQ ID NO:233;
(b) нуклеотидную последовательность, кодирующую полипептид, который обладает активностью C20-элонгазы, где данная нуклеотидная последовательность имеет по меньшей мере 80% идентичность последовательности, основываясь на сравнительном анализе первичной структуры согласно программе BLASTN, при сравнении с аминокислотной последовательностью, как представлено в SEQ ID NO:11, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93, SEQ ID NO:183, SEQ ID NO:188, SEQ ID NO:201, SEQ ID NO:206, SEQ ID NO:203, SEQ ID NO:227, SEQ ID NO:228, SEQ ID NO:229, или SEQ ID NO:230; (c) нуклеотидную последовательность, кодирующую полипептид, который обладает активностью C20-элонгазы, где данная нуклеотидная последовательность гибридизуется при жестких условиях с нуклеотидной последовательностью, как представлено в SEQ ID NO:11, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93, SEQ ID NO:183, SEQ ID NO:188, SEQ ID NO:201, SEQ ID NO:206, SEQ ID NO:203, SEQ ID NO:227, SEQ ID NO:228, SEQ ID NO:229, SEQ ID NO:230; или
(d) комплемент нуклеотидной последовательности (a), (b) или (c), где данные комплемент и нуклеотидная последовательность состоят из одинакового числа нуклеотидов и являются на 100% комплементарными.
Лучше, когда фермент полинуклеотид, кодирующий C20-элонгазу, включает последовательность, представленную в любой из SEQ ID NO:183, SEQ ID NO:188, SEQ ID NO:201, SEQ ID NO:206, SEQ ID NO:203, SEQ ID NO:227, SEQ ID NO:228, SEQ ID NO:229, или SEQ ID NO:230.
Идентификация последовательностей новых дельта-4-десатураз
В настоящем изобретении нуклеотидные последовательности, кодирующие дельта-4-десатуразы, были выделены из Euglena gracilis и Euglena anabaena, как показано в Таблице 6.
В альтернативных вариантах текущие дельта-4-десатуразные доменные последовательности могут быть кодон-оптимизировавны для экспрессии в конкретном организме хозяина. Нпример, Euglena anabaena дельта-4-десатуразный домен EaDHAsyn2 был кодон-оптимизирован для экспрессии в Yarrowia lipolytica. Например, Euglena gracilis дельта-4-десатуразный домен EgDHAsyn1 был также кодон-оптимизирован для экспрессии в Yarrowia lipolytica. Специалист в данной области сможет применить приведенный здесь поучительный материал для создания различных других кодон-оптимизированных дельта-4-десатуразных протеинов, подходящих для оптимальной экспрессии в альтернативных хозяевах, исходя из дельта-4-десатуразных доменных последовательностей дикого типа EgDHAsyn1, EgDHAsyn2, EaDHAsyn1, EaDHAsyn2 и/или EaDHAsyn3 как описано выше в Таблице 6. Соответственно, настоящее изобретение касается любого из кодон-оптимизированного дельта-4-десатуразного протеина, который получен из последовательности дикого типа данного изобретения. В некоторых вариантах, которым отдается предпочтение, может быть желательным модифицировать часть кодонов, кодирующих дельта-4-десатуразные доменные последовательности EgDHAsyn1, EgDHAsyn2, EaDHAsyn1, EaDHAsyn2 и/или EaDHAsyn3 для усиления экспрессии данного гена в организме хозяина, включая, однако не ограничиваясь этим, растение или часть растения.
Более того, основываясь на наблюдении того, что C-терминальный участок C20-элонгазного домена ДГК синтазы, как представляется, перекрывается с N-терминальным участком дельта-4-десатуразного домена, были проведены функциональные анализы для определения оптимального функционального дельта-4-десатуразного доимена. Как описано в Примерах 51 и 53 ниже, с использованием кодон-оптимизированных протеиновых последовательностей, EaD4S (SEQ ID NO:193) и EgD4S (SEQ ID NO:388), были проведены исследования по делеции мутагенеза. Были получены следующие варианты: EaD4S-3 (SEQ ID NO:386), EaD4S-2 (SEQ ID NO:384), EaD4S-1 (SEQ ID NO:382), EgD4S-3 (SEQ ID NO:408), EgD4S-2 (SEQ ID NO:406) и EgD4S-1 (SEQ ID NO:404).
Специалисту в данной области должно быть понятно, что поскольку точные границы этих конкретных дельта-4-десатуразных последовательностей из Euglena gracilis и Euglena anabaena полностью не определены, фрагменты протеинов или полипептиды увеличеной или уменьшенной длины могут обладать сравнимой активностью дельта-4-десатуразы. Подобно этому, легко могут быть сделаны сравнимые усечения на основе дельта-4-десатуразных доменных последовательностей дикого типа EgDHAsyn1, EgDHAsyn2, EaDHAsyn1, EaDHAsyn2 и/или EaDHAsyn3, как описано выше в Таблице 6, с образованием дельта-4-десатуразы, обладающей достаточной степенью активности дельта-4-десатуразы, где предпочтение отдается эквивалентной или повышенной активности дельта-4-десатуразы.
Таким образом, настоящее изобретение касается также выделенного полинуклеотида, кодирующего дельта-4-десатуразу, включая:
(a) нуклеотидную последовательность, кодирующую полипептид, который обладает активностью дельта-4-десатуразы, где данный полипептид имеет по меньшей мере 80% аминокислотную идентичность, основываясь на методе группировки Clustal V, при сравнении с аминокислотной последовательностью, как представлено в SEQ ID NO:12, SEQ ID NO:22, SEQ ID NO:95, SEQ ID NO:96, SEQ ID NO:97, SEQ ID NO:215, SEQ ID NO:217, SEQ ID NO:221, SEQ ID NO:239, SEQ ID NO:240, SEQ ID NO:241, SEQ ID NO:246, SEQ ID NO:247, SEQ ID NO:248, SEQ ID NO:249, SEQ ID NO:193, SEQ ID NO:382, SEQ ID NO:384, SEQ ID NO:386, SEQ ID NO:388, SEQ ID NO:404, SEQ ID NO:406, или SEQ ID NO:408;
(b) нуклеотидную последовательность, кодирующую полипептид, который обладает активностью дельта-4-десатуразы, где данная нуклеотидная последовательность имеет по меньшей мере 80% идентичность последовательности, основываясь на сравнительном анализе первичной структуры согласно программе BLASTN, при сравнении с аминокислотной последовательностью, как представлено в SEQ ID NO:11, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93, SEQ ID NO:214, SEQ ID NO:216, SEQ ID NO:220, SEQ ID NO:236, SEQ ID NO:237, SEQ ID NO:238, SEQ ID NO:242, SEQ ID NO:243, SEQ ID NO:244, SEQ ID NO:245, SEQ ID NO:192, SEQ ID NO:381, SEQ ID NO:383, SEQ ID NO:385, SEQ ID NO:387, SEQ ID NO:403, SEQ ID NO:405 или SEQ ID NO:407;
(c) нуклеотидную последовательность, кодирующую полипептид, который обладает активностью дельта-4-десатуразы, где данная нуклеотидная последовательностьа гибридизуется при жестких условиях с нуклеотидной последовательностью, как представлено в SEQ ID NO:11, SEQ ID NO:21, SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93, SEQ ID NO:214, SEQ ID NO:216, SEQ ID NO:220, SEQ ID NO:236, SEQ ID NO:237, SEQ ID NO:238, SEQ ID NO:242, SEQ ID NO:243, SEQ ID NO:244, SEQ ID NO:245, SEQ ID NO:192, SEQ ID NO:381, SEQ ID NO:383, SEQ ID NO:385, SEQ ID NO:387, SEQ ID NO:403, SEQ ID NO:405 или SEQ ID NO:407; или
(d) комплемент нуклеотидной последовательности (a), (b) или (c), где данные комплемент и нуклеотидная последовательность состоят из одинакового числа нуклеотидов и являются на 100% комплементарными.
Лучше, когда фермент полинуклеотид, кодирующий дельта-4-десатуразу, включает последовательность, представленную в любой из SEQ ID NO:214, SEQ ID NO:220, SEQ ID NO:236, SEQ ID NO:237, SEQ ID NO:238, SEQ ID NO:242, SEQ ID NO:243, SEQ ID NO:244, SEQ ID NO:245, SEQ ID NO:192, SEQ ID NO:381, SEQ ID NO:383, SEQ ID NO:385, SEQ ID NO:387, SEQ ID NO:403, SEQ ID NO:405, или SEQ ID NO:407.
Эффект усечения Euglena anabaena дельта-4-десатуразы состоит в том, что ферментативная активность повышена, по сравнению с ферментативной активностью последовательности дикого типа. Этот результат неожиданный и непредвиденный, поскольку обычный специалист в данной области мог бы предполагать, что активность усеченной последовательности будет не лучше и возможно хуже, чем у последовательности дикого типа. Соответственно, данное изобретение также представляет новый способ получения дельта-4-десатуразы, обладающей более высокой активностью, чем последовательность дикого типа, данный способ включает: a) предоставление дельта-4-десатуразного полипептида дикого типа, выделенного из Euglena anabena, обладающего базисной активностью дельта-4-десатуразы; и б) усечение полипептида дикого типа (a) на приблизительно от 1 до приблизительно 200 аминокислот с образованием усеченного мутантного полипептида, обладающего активностью дельта-4-десатуразы, которая повышена по сравнению с базовой активностью дельта-4-десатуразы. “Базовая” активность, как используется в этом контексте, определяется как активность фермента дикого типа, измеренная или in vivo или in vitro, в соответствии со стандартными ферментативными протоколами, как описано в данной заявке.
В других вариантах любой из ферментов (например, мультизимы, ДГК синтазы, C20-элонгазы, дельта-4-десатуразы, и/или любой из гомологов), который здесь идентифицирован, может быть модифицирован для генерации новых и/или улучшенных ферментов ПНЖК биосинтетического пути. Как хорошо известно специалистам в данной области, для получения мутаций природных генов могут применяться методы in vitro мутагенеза и селекции, химического мутагенеза, “перетасовки” генов или другие меры. Как альтернатива, мультизимы могут быть синтезированы методом перестановки доменов, где функциональный домен из какого-либо фермента может обмениваться с или добавляться к функциональному домену в альтернативном ферменте с образованием, таким образом, нового протеина.
Способы получения различных омега-3 и/или омега-6 жирных кислот
Ожидается, что введение химерных генов, кодирующих ДГК синтазы, описанных в данном тексте (т.е. EgDHAsyn1, EgDHAsyn2, EaDHAsyn1, EaDHAsyn2 и EaDHAsyn3 или других мутантных ферментов, кодон-оптимизированных ферментов или их гомологов), под контролем подходящих промоторов обеспечит повышенное производство ДТК, ДПКn-6, ДПК и/или ДГК в трансформированном организме хозяина, соответственно. Как таковое, данное изобретение охватывает способ прямого производства ПНЖК, включающий экспонирование жирнокислотного субстрата (т.е. ЭПК или ДПК) относительно ДГК синтазных ферментов, описанных здесь (например, EgDHAsyn1, EgDHAsyn2, EaDHAsyn1, EaDHAsyn2 и EaDHAsyn3), такое, что данный субстрат конвертируется в необходимый жирнокислотный продукт (т.е. ДГК).
Более конкретно, настоящее изобретение касается способа трансформации клетки-хозяина, такого, что данная клетка-хозяин содержит в своем геноме рекомбинантная конструкция данного изобретения.
Примеры пригодных клеток-хозяев включают, однако не ограничиваясь этим, растения и дрожжи. Лучше, когда растительные клетки получаются из маслянистого растения, такого как соя и подобного, и дрожжевые клетки получаются из жировых дрожжей, таких как вид Yarrowia.
Кроме того, в объем данного изобретения входит способ получения трансформированного растения или дрожжей, включающий трансформацию растительной клетки или дрожжевой клетки с помощью любого из полинуклеотидов данного изобретения и восстановление растения из трансформированной растительной клетки или выращивание трансформированных дрожжевых клеток.
Более конкретно, предметом настоящего изобретения является представление способа выработки ДПКn-6 или ДГК в клетке-хозяине (например, растений, жировых дрожжей), где данная клетка-хозяин включает:
изолированную нуклеотидную молекулу, кодирующую полипептид, который обладает ДГК синтазной активностью, где данный полипептид имеет по меньшей мере 80% аминокислотную идентичность, основываясь на методе группировки Clustal V, при сравнении с аминокислотной последовательностью, представленной в SEQ ID NO:12, SEQ ID NO:22, SEQ ID NO:95, SEQ ID NO:96, или SEQ ID NO:97; и,
(ii) источник АРК или ЭПК;
где клетка-хозяин выращивается при таких условиях, что полипептид, обладающий ДГК синтазной активностью, экспрессирован, и АРК конвертирована в ДПКn-6 и/или ЭПК конвертирована в ДГК, и где ДПКn-6 или ДГК, при необходимости, восстановлена.
В альтернативных вариантах настоящее изобретение касается способа выработки ДТК или ДПК в клетке-хозяине (например, растений, жировых дрожжей), где данная клетка-хозяин включает:
изолированную нуклеотидную молекулу, кодирующую полипептид, который обладает активностью С20-элонгазы, где данный полипептид имеет по меньшей мере 80% аминокислотную идентичность, основываясь на методе группировки Clustal V, при сравнении с аминокислотной последовательностью, представленной в SEQ ID NO:12, SEQ ID NO:22, SEQ ID NO:95, SEQ ID NO:96, SEQ ID NO:97, SEQ ID NO:202, SEQ ID NO:204, SEQ ID NO:231, SEQ ID NO:232, or SEQ ID NO:233; и,
(ii) источник АРК или ЭПК;
где клетка-хозяин выращивается при таких условиях, что полипептид, обладающий активностью С20-элонгазы, экспрессирован, и АРК конвертирована в ДТК и/или ЭПК конвертирована в ДПК, и где ДТК или ДПК, при необходимости, восстановлена.
Дополнительно, данное изобретение представляет способ выработки ДПКn-6 или ДГК, где клетка-хозяин включает:
изолированную нуклеотидную молекулу, кодирующую полипептид, который обладает активностью дельта-4-десатуразы, где данный полипептид имеет по меньшей мере 80% аминокислотную идентичность, основываясь на методе группировки Clustal V, при сравнении с аминокислотной последовательностью, представленной в SEQ ID NO:12, SEQ ID NO:22, SEQ ID NO:95, SEQ ID NO:96, SEQ ID NO:97, SEQ ID NO:215, SEQ ID NO:221, SEQ ID NO:239, SEQ ID NO:240, SEQ ID NO:241, SEQ ID NO:246, SEQ ID NO:247, SEQ ID NO:248, SEQ ID NO:249, SEQ ID NO:382, SEQ ID NO:384, SEQ ID NO:386, SEQ ID NO:388, SEQ ID NO:404, SEQ ID NO:406, или SEQ ID NO:408; и,
(ii) источник ДТК или ДПК;
где клетка-хозяин выращивается при таких условиях, что полипептид, обладающий активностью дельта-4-десатуразы, экспрессирован, и ДТК конвертирована в ДПКn-6 и/или ДПК конвертирована в ДГК, и где ДПКn-6 или ДГК, при необходимости, восстановлена.
Источник субстрата(ов) АРК, ДТК, ЭПК или ДПК, использованных в каком-либо из методов, рассмотренных выше, может быть получен хозяином или природным путем или трансгенно, или может быть введен экзогенно.
Связывыание индивидуальных доменов с образованием мультизима может привести к снижению количества промежуточных жирных кислот. Например, связывание C20-элонгазы с дельта-4-десатуразой в мультизим, такой как ДГК синтаза, может привести к уменьшению количества промежуточной жирной кислоты ДПК при производстве ДГК. Подобно этому связывание дельта-9-элонгазы с дельта-8-десатуразой с использованием EgDHAsyn1 линкера с образованием мультизима, как здесь описано, может привести к выработке ДГЛК и ЭТК со снижением образования ЭДК и ЭРК интермедиатов.
Как альтернатива, каждый мультизимный ген, включающий ДГК синтазу и соответствующие ферментные продукты, описанные здесь, может быть использован косвенно для производства разных омега-6 и омега-3 ПНЖК, включая, например, ДТК, ДПКn-6, ДГЛК, ЭТК, АРК, ЭПК, ДПК и/или ДГК (ФИГ.1; см. патент США № 7238482). Косвенное образование омега-3/омега-6 ПНЖК имеет место, где жирнокислотный субстрат конвертируется косвенным образом в необходимый жирнокислотный продукт через промежуточную стадию(и) или интермедиат(ы) данного пути. Таким образом, предполагается, что ДГК синтазы, описанные здесь (т.е. EgDHAsyn1, EgDHAsyn2, EaDHAsyn1, EaDHAsyn2 и EaDHAsyn3, или другие мутантные ферменты, кодон-оптимизированные ферменты или их гомологи), могут быть экспрессированы в сочетании с дополнительными генами, кодирующими ферменты ПНЖК биосинтетического пути (например, дельта-6-десатуразы, C18/20-элонгазы, дельта-17-десатуразы, дельта-8-десатуразы, дельта-15-десатуразы, дельта-9-десатуразы, дельта-12-десатуразы, C14/16-элонгазы, C16/18-элонгазы, дельта-9-элонгазы, дельта-5-десатуразы, дельта-4-десатуразы, C20/22-элонгазы, ДГК синтазы), с достижением более высоких уровней выработки более длинноцепочечных омега-3/омега-6 жирных кислот (например, АРК, ДТК, ДПКn-6, ЭПК, ДПК и/или ДГК).
Специфические гены, включенные в конкретную экспрессирующую кассету, будут зависеть от клетки-хозяина (и ее ПНЖК профиля, и/или десатураза/элонгазного профиля), наличия субстрата и желательного конечного продукта(ов).
В некоторых случаях может быть желательным минимизировать побочные жирные кислоты. Относительное содержание побочных жирных кислот может быть снижено путем связывания индивидуальных путевых ферментов вместе с линкером с образованием мультизима. Например, присутствие скиадоновой кислоты (СКК) и/или юнипероновой кислоты (ЮП) [которые обычно находят в семенных липидах голосеменных растений (Wolff et al., Lipids 35(1):1-22 (2000)), таких как растения из семейства Pinaceae (сосна], могут рассматриваться как побочные жирные кислоты дельта-6-десатураза/дельта-6-элонгазного пути или дельта-9-елонгаза/дельта-8-десатуразного пути. Хотя эти жирные кислоты, как считается, сами по себе обладают различными оздоровительными свойствами (Nakane et al., Biol. Pharm. Bull. 23: 758-761 (2000)), их присутствие как побочных жирных кислот на разработанном методами генной инженерии ПНЖК пути, таком как в маслянистой культуре, может быть нежелательным, в зависимости от применения. Связывание дельта-9-элонгазы вместе с дельта-8-десатуразой с использованием линкера с образованием мультизима (ДГЛК и/или ЭТК синтазы), например, может дать увеличенный поток через эти стадии, что приведет к уменьшенному количеству ЭДК/ЭРК промежуточных жирных кислот на пути к дельта-5-десатуразе и, таким образом, к сниженным концентрациям СКК и ЮП.
Случайно, дельта-6-элонгаза может удлинить жирные кислоты, другие, чем предусмотренную жирную кислоту. Например, дельта-6-элонгаза преобразует, в общем, ГЛК в ДГЛК, однако некоторые дельта-6-элонгазы могут также превращать непредусмотренные субстраты, такие как ЛК или АЛК в ЭДК или ЭТрК, соответственно. На дельта-6-десатураза/дельта-6-элонгазном пути ЭДК и ЭТрК могут рассматриваться как “побочные жирные кислоты”. Добавление дельта-8-десатуразы к дельта-6-десатураза/дельта-6-элонгазному пути может предоставить средства для превращения “побочных жирных кислот” ЭДК и ЭТрК назад в “промежуточные жирные кислоты”, соответственно, ДГЛК и ЭТК.
В альтернативных вариантах может быть полезным разорвать нативную ДГК синтазу, С20-элонгазу или дельта-4-десатуразу организма хозяина, основываясь на полных последовательностях, описанных здесь, комплементе этих полных последовательностей, существенных участках этих последовательностей, кодон-оптимизированных десатуразах, полученных из них, и тех последовательностях, которые в значительной мере гомологичны к ним.
Растительные системы экспрессии, кассеты и векторы, и трансформация
В одном варианте данное изобретение касается рекомбинантного конструкционного элемента, который включает какой-либо из выделенных полинуклеотидов изобретения, связанный действующим образом с по меньшей мере одной регуляторной последовательностью, пригодной для экспрессии в клетке-хозяине, таком как растение. Промотор представляет собой ДНК последовательность, которая направляет клеточный механизм растения на выработку РНК из соседней кодирующей последовательности вниз (3′) от промотора. Промоторный участок влияет на скорость, стадию развития и тип клетки, в которой РНК транскрипт данного гена выработан. РНК транскрипт подвергается процессингу с образованием мРНК, которая служит матрицей для трансляции РНК последовательности в аминокислотную последовательность кодированного полипептида. 5′ нетранслированная лидерная последовательность представляет собой участок мРНК вверх от участка, который кодирует протеин, которая может играть роль в инициации и трансляции мРНК. 3′ транскрипционно-терминационная/полиаденилирующая сигнальная последовательность представляет собой нетранслированный участок вниз от кодирующего протеин участка, который функционирует в растительной клетке, обуславливая терминацию РНК транскрипта и добавление полиаденилатных нуклеотидов к 3′ концу РНК.
Происхождение промотора, выбранного для экспрессии кодирующей последовательности, не является важным, поскольку он обладает достаточной транскрипционной активностью для реализации данного изобретения путем экспрессии способной к трансляции мРНК для необходимых фрагментов нуклеиновой кислоты в необходимой ткани хозяина в корректный момент времени. Для практической реализации данного изобретения могут быть использованы как гетерологические, так и негетерологические (т.е. эндогенные) промоторы. Например, подходящие промоторы включают, однако не ограничиваясь этим, альфа первичную субъединицу бета конглицининового промотора, кунитц трипсин ингибиторный 3 промотор, аннексиновый промотор, глицининовый Gy1 промотор, бета субъединицу бета конглицининового промотора, P34/Gly Bd m 30K промотор, альбуминовый промотор, Leg A1 промотор и Leg А2 промотор.
Аннексиновый, или Р34 промотор описан в РСТ публикации под номером WO 2004/071178 (опубликована 26 августа 2004 года). Уровень активности аннексинового промотора сравним с уровнем многих известных сильных промоторов, таких как: (1) CaMV 35S промотор (Atanassova et al., Plant Mol. Biol. 37:275-285 (1998); Battraw и Hall, Plant Mol. Biol. 15:527-538 (1990); Holtorf et al., Plant Mol. Biol. 29:637-646 (1995); Jefferson et al., EMBO J. 6:3901-3907 (1987); Wilmink et al., Plant Mol. Biol. 28:949-955 (1995)); (2) Arabidopsis олеозиновые промоторы (Plant et al., Plant Mol. Biol. 25:193-205 (1994); Li, Texas Кандидатская дисскертация университета A&M, стр. 107-128 (1997)); (3) Arabidopsis убиквитиновые экстензионные протеиновые промоторы (Callis et al., J. Biol. Chem. 265(21):12486-93 (1990)); (4) томатный убиквитиновый генный промотор (Rollfinke et al., Gene. 211(2):267-76 (1998)); (5) соевый теплошоковый протеиновый промотор (Schoffl et al., Mol. Gen. Genet. 217(2-3):246-53 (1989)); и (6) маисовый Н3 гистонный генный помотор (Atanassova et al., Plant Mol. Biol. 37(2):275-85 (1989)).
Другой полезной особенностью аннексинового промотора является его профиль экспрессии в семенах, которые развиваются. Аннексиновый промотор наиболее активен в семенах, которые развиваются на ранних стадиях (до 10 суток после опыления), и в значительной мере спокойный на поздних стадиях. Профиль экспрессии аннексинового промотора отличается от профиля многих специфических в отношении семян промоторов, например, протеиновых промоторов хранения семян, которые часто представляют самую высокую активность на поздних стадиях развития (Chen et al., Dev. Genet. 10:112-122 (1989); Ellerstrom et al., Plant Mol. Biol. 32:1019-1027 (1996); Keddie et al., Plant Mol. Biol. 24:327-340 (1994); Plant et al., (supra); Li, (supra)). Аннексиновый промотор имеет более обычный профиль экспрессии, но остается отличным от других известных специфических в отношении семян промоторов. Таким образом, аннексиновый промотор будет очень привлекательным кандидатом, когда на ранней стадии развития необходима гиперэкспрессия или супрессия гена в зародыше. Например, может возникнуть проблема в гиперэкспрессии гена, который регулирует раннее развитие зародыша, или гена, задействованного в метаболизме до вызревания семян.
После идентификации соответствующего промотора, пригодного для экспрессии кодирующей последовательности, данный промотор потом связывают действующим образом в смысловой ориентации с использованием обычных средств, которые хорошо известны специалистам в данной области.
Стандартные методы рекомбинантной ДНК и молекулярного клонирования, которых здесь применены, хорошо известны в этой области и более полно описаны в работе Sambrook, J. Et al., In Molecular Cloning: A Laboratory Manual; 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, New York, 1989 (в дальнейшем, “Sambrook et al., 1989”) или Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A. и Struhl, K., Eds.; In Current Protocols in Molecular Biology; John Wiley and Sons: New York, 1990 (в дальнейшем, “Ausubel et al., 1990”). Например, гибридный ген может буть сконструирован путем сцепления по меньшей мере двух ДНК фрагментов в каркасе таким образом, чтобы не вводить терминирующий кодон (слияние в каркасе). Результирующий гибридный ген будет таким, что каждый ДНК фрагмент кодирует по меньшей мере одну независимую и раздельную ферментативную активность.
Когда рекомбинантный конструкционный элемент создан, он может быть потом введен в нужную растительную клетку с использованием методов, которые хорошо известны обычным специалистам в данной области (например, трансфекция, трансформация и электропорация). Растительными клетками, которым отдается предпочтение, являются маслянистые растительные клетки. Затем трансформированная растительная клетка культивируется и регенерируется при приемлемых условиях, которые дают возможность осуществлять экспрессию длинноцепочечной ПНЖК, которая потом, при необходимости, восстанавливается и очищается.
Рекомбинантные конструкции данного изобретения могут быть введены в одну растительную клетку; или, как альтернатива, каждый конструкция может быть введен в отдельные растительные клетки.
Экспрессия в растительной клетке может проводиться по временной или стабильной схеме как описано выше.
Необходимая длинноцепочечная ПНЖК может быть экспрессирована в семенах. Кроме того, в объем данного изобретения входят семена или части растения, полученные из таких трансформированных растений.
Части растений включают дифференциированные и недифференциированные ткани, включая, однако не ограничиваясь этим, следующие: корни, стебли, ростки, листья, пыльцу, семена, опухлевую ткань и разные формы клеток и культур (например, отдельные клетки, протопласты, зародыши и калюсную ткань). Данная растительная ткань может быть в растении или в растительном органе, ткани или клеточной культуре.
Термин “растительный орган” касается растительной ткани или группы тканей, которые составляют морфологично и функционально отличную часть растения. Термин “геном” касается следующего: (1) всего комплемента генетического материала (гены и некодирующие последовательности), присутствующего в каждой клетке организма или вируса, или органеллы; (2) полного набора хромосом, унаследованного как (гаплоидная) единица от одного предка.
Таким образом, данное изобретение также касается способа трансформации клеток, который включает трансформирование клетки рекомбинантным конструкционным элементом данного изобретения и селекцию этих клеток, трансформированных рекомбинантными конструкционными элементами, описанными в формуле изобретения.
Также представляет интерес способ продуцирования трансформированного растения, который включает трансформирование растительной клетки полинуклеотидами данного изобретения и регенерацию растения из трансформироованной растительной клетки.
Способы трансформирования двудольних растений (главным образом, путем использования Agrobacterium tumefaciens) и получения трансгенных растений опубликованы, среди прочих, для: хлопка (патент США № 5004863; патент США № 5159135); сои (патент США № 5569834; патент США № 5416011); Brassica (патент США № 5463174); арахиса (Cheng et al., Plant Cell Rep. 15:653-657 (1996); McKently et al., Plant Cell Rep. 14:699-703 (1995)); папайи (Ling, K. Et al., Bio/technology 9:752-758 (1991)); и гороха (Grant et al., Plant Cell Rep. 15;254-258 (1995)). В отношении обзора других общих методов превращения растений см. работу Newell, C.A. (Mol. Biotechnol. 16:53-65 (2000)). В одном из этих методов трансформации используется Agrobacterium rhizogenes (Tepfler, M. и Casse-Delbart, F. Microbiol. Sci. 4:24-28 (1987)). Трансформация сои с использованием прямой доставки ДНК и PEG слияния опубликована в РСТ публикации под номером WO 92/17598, електропорации (Chowrira, G.M. et al., Mol. Biotechnol 3:17-23 (1995); Christou, P. et al., Proc. Natl. Acad. Sci. U.S.A. 84:3962-3966 (1987)), микроинъекции и бомбардировки частичками (McCabe, D.E. et al., Bio/Technology 6:923 (1988); Christou et al., Plant Physiol. 87:671-674 (1988)).
Существует множество методов регенерации растений из растительной ткани. Конкретный метод регенерации будет зависеть от исходной растительной ткани и конкретного вида растения, которое должно регенерироваться. Регенерация, развитие и культивирование растений из отдельных растительных протопластных трансформантов или из разных трансформированных эксплантатов хорошо известны в данной области (Weissbach and Weissbach, In: Methods for Plant Molecular Biology, (Eds.), Academic: San Diego, CA (1988)). Эта регенерация и процесс роста обычно включают стадии селекции трансформированных клеток и культивирование данных индивидуальных клеток через обычные стадии эмбрионального развития через корневую проростковую стадию. Трансгенные зародыши и семена регенерируются подобным способом. Результирующие трансгенные корневые побеги затем выращиваются в соответствующей среде растительного роста, такой как почва. Лучше, когда регенерированные растения самоопыляются, давая гомозиготные трансгенные растения. По-другому, пыльца, полученная от регенерированных растений, скрещивается с растениями, которые выращены из семян агрономически важных линий. Напротив, пыльца от растений этих важних линий используется для опыления регенерированных растений. Трансгенное растение данного изобретения, которое содержит необходимый полипептид, культивируется с использованием методов, которые хорошо известны специалистам в данной области.
В дополнение к описанным выше процедурам, специалистам в данной области знакомы стандартные ресурсные материалы, которые описывают специфические условия и процедуры для конструирования, манипуляции и выделения макромолекул (например, ДНК молекул, плазмид и т.д.), генерации фрагментов рекомбинантной ДНК и рекомбинантных экспрессирующих конструкций, и скрининга и выделения клонов. См., например, Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor: NY (1989); Maliga et al., Methods in Plant Molecular Biology, Cold Spring Harbor: NY (1995); Birren et al., Genome Analysis: Detecting Genes, Vol.1, Cold Spring Harbor: NY (1998); Birren et al., Genome Analysis: Analyzing DNA, Vol.2, Cold Spring Harbor: NY (1998); Plant Molecular Biology: A Laboratory Manual, eds. Clark, Springer: NY (1997).
Примеры маслянистих растений включают, однако не ограничиваясь этим, сою, виды Brassica, подсолнечник, маис, хлопок, лен и сафлор красильный.
Примеры ПНЖК, которые имеют по меньшей мере двадцать углеродных атомов и четыре или более двойных связей углерод-углерод, включают, однако не ограничиваясь этим, омега-3 жирные кислоты, такие как ЭПК, ДПК и ДГК. Семена, полученные от таких растений, также подпадают под объем данного изобретения, так же как и масло, полученное из таких семян.
Таким образом, настоящее изобретение касается также способа изменения жирнокислотного профиля маслянистого растения, который включает:
a) трансформацию клетки маслянистого растения рекомбинантным конструкционныи элементом формулы данного изобретения; и
b) регенерацию растения из трансформированной клетки маслянистого растения (a), где данное растение имеет измененный жирнокислотный профиль.
Микробные системы экспрессии, кассеты и векторы
ДГК синтазные гены и генные продукты, описанные здесь (т.е. EgDHAsyn1, EgDHAsyn2, EaDHAsyn1, EaDHAsyn2 и EaDHAsyn3, или другие мутантные ферменты, кодон-оптимизированные ферменты или их гомологи), могут также образовываться в гетерологических микробных клетках-хозяевах, в частности, в клетках жировых дрожжей (например, Yarrowia lipolytica).
Микробные системы экспрессии и экспрессирующие векторы, содержащие регуляторные последовательности, которые направляют высокоуровневую экспрессию посторонних протеинов, хорошо известны специалистам в данной области. Любые из них могут быть использованы для конструирования химерных генов для выработки любого из генных продуктов текущих последовательностей. Эти химерные гены могут затем вводиться в соответствующие микроорганизмы путем трансформации, обеспечивая высокоуровневую экспрессию кодированных ферментов.
Векторы, полезные для трансформации подходящих микробных клеток-хозяев, хорошо известны в данной области. Специфический выбор последовательностей, присутствующих в данном конструкцияе, зависит от желательных продуктов экспрессии (supra), природы клетки-хозяина и предлагаемых средств отделения трансформированных клеток от нетрансформированных клеток. Однако, обычно вектор содержит по меньшей мере одну экспрессирующую кассету, селектируемый маркер и последовательности, позволяющие осуществлять автономную репликацию или хромосомную интеграцию. Подходящие экспрессирующие кассеты включают участок 5′ данного гена, который контролирует инициацию транскрипции (например, промотор), кодирующую ген последовательность, и участок 3′ ДНК фрагмента, который контролирует терминацию транскрипции (т.e. терминатор). Наиболее предпочтительно, когда оба контрольных участка получаются из генов от данной трансформированной микробной клетки-хозяина, хотя следует понимать, что нет потребности в том, чтобы такие контрольные участки получались из генов, нативных относительно специфических видов, выбранных в качестве хозяина-производителя.
Инициирующие контрольные участки промоторов, которые полезны для драйва экспрессии текущих мультизимов, таких как ДГК синтаза или отдельный домен ОРС, в желательную микробную клетку-хозяина, многочисленны и знакомы специалистам в данной области. В сущности, любой промотор, способный направлять экспрессию этих генов в выбранную клетку-хозяина, пригоден для настоящего изобретения. Экспрессия в микробную клетку-хозяина можут осуществляться по временной или постоянной схеме. Временная экспрессия может осуществляться путем индуцирования активности регулирующего промотора, связанного действующим образом с нужным геном. Постоянная экспрессия может достигаться путем использования конститутивного промотора, связанного действующим образом с нужным геном. Как пример, когда клетка-хозяин является дрожжевой, вводятся транскрипционные и трансляционные участки, функциональные в дрожжевых клетках, в частности, из хозяйских видов (например, см. патент США № 7238482 и PCT публикацию № WO 2006/052870 относительно предпочтительных транскрипционных инициирующих регуляторных участков для использования в Yarrowia lipolytica). Любая из ряда регуляторных последовательностей может быть использована, в зависимости от того, какая требуется транскрипция, конститутивная или индуцированная, эффективности данного промотора в экспрессии нужной ОРС, легкости конструирования и подобного.
Нуклеотидные последовательности, окружающие трансляционный инициирующий кодон ′ATG′, как было найдено, влияют на экспрессию в дрожжевых клетках. Если необходимый полипептид плохо экспрессирован в дрожжах, нуклеотидные последовательности экзогенных генов могут быть модифицированы для включения эффективной дрожжевой трансляционной инициирующей последовательности с достижением оптимальной экспрессии генов. Для экспрессии в дрожжах это может быть сделано путем сайт-направленного мутагенеза неэффективно экспрессированного гена путем слияния его в каркасе с эндогенным дрожжевым геном, лучше, сильноэкспрессированным геном. В качестве альтернативы, можно определить консенсусную трансляционную инициирующую последовательность в хозяине и методами генной инженерии внедрить эту последовательность в гетерологичные гены для их оптитмальной экспрессии в нужном хозяине.
Терминирующий участок может быть получен из 3' участка гена, из которого был получен инициирующий участок, или из другого гена. Известен целый ряд терминирующих участков, которые удовлетворительно функционируют в разных хозяевах (когда оба используются или в тех же самых родах или разных родах и видах, из которых они были получены). Терминируюший участок обычно выбирается, скорее, из соображений удобства, а не из-за какого-либо конкретного свойства. Терминирующие контрольные участки могут также получаться из различных генов, нативных для хозяев, которым отдается предпочтение. В альтернативных вариантах 3′-участок может также быть синтетическим, так как специалист в данной области может воспользоваться имеющейся информацией для конструирования и синтеза последовательности 3′-участка, что функционирует как транскрипционный терминатор. Когда это необходимо, можно обойтись без терминирующего сайта, однако, наибольшее предпочтение отдается его включению.
Как известно специалисту в данной области, простая инсерция гена в клонирующий вектор не гарантирует, что он будет успешно экспрессирован на необходимом уровне. Как реакция на потребность в высокой степени экспрессии, было создано множество специализированных экспрессирующих векторов путем манипулирования рядом разных генетических элементов, которые контролируют аспекты транскрипции, трансляции, стабильности протеина, кислородного ограничения и секреции из микробной клетки-хозяина. Более конкретно, некоторые молекулярные свойства, которыми манипулируют с целью контроля генной экспрессии, включают: природу релевантной транскрипционной промоторной и терминирующей последователностей; число копий клонированного гена; ограничен ли данный ген плазмидой или он интегрирован в геном клетки-хозяина; конечную клеточною локацию данного синтезированного постороннего протеина; эффективность трансляции и корректной укладки данного протеина в организме хозяина; внутреннюю устойчивость мРНК и протеина клонированного гена в клетке-хозяине; и использование кодона в клонированном гене, так что его частота приближается к частоте предпочтительного использования кодона клетки-хозяина. Каждый тип модификации входит в объем данного изобретения как средство для дальнейшей оптимизации экспрессии ДГК синтаз, описанных в данной заявке.
Трансформация микробных клеток-хозяев
Когда кассета, которая подходит для экспрессии в соответствующей клетке-хозяине, получена (например, химерный ген, который включает промотор, ОРС и терминатор), она помещается в плазмидный вектор, способный к автономной репликации в клетке-хозяине, или прямо интегрируется в геном клетки-хозяина. Интеграция экспрессирующих кассет может происходить случайным образом в геноме хозяина или может направляться путем использования конструкций, которые содержат участки гомологии с геномом хозяина, достаточные для правильного нацеливания рекомбинации внутри хозяйского локуса. Там, где конструкцияы нацелены на эндогенный локус, все или некоторые из транскрипционных и трансляционных регуляторных участков могут быть снабжены эндогенным локусом.
Когда два или более гена экспрессированы из разных репликационных векторов, желательно, чтобы каждый вектор имел различные средства селекции, и он не должен обладать гомологией с другим конструкцияом(ами) для сохранения стабильной экспрессии и предотвращения пересортировки элементов среди конструкций. Разумный выбор регуляторных участков, средств селекции и способа пропагации введенной конструкции(ий) могут быть экспериментально определены, так что все введенные гены экспрессированы на необходимых уровнях, обеспечивая синтез желательных продуктов.
Конструкционные элементы, содержащие необходимый ген(ы), могут быть введены в микробную клетку-хозяина с помощью любых стандартных способов. Эти способы включают трансформацию (например, литий ацетатную трансформацию [Methods in Enzymology, 194:186-187 (1991)]), протопластное слияние, баллистический удар, электропорацию, микроинъекцию или любой другой способ, с помощью которого нужный ген(ы) вводится в клетку-хозяина.
Для удобства, на клетку-хозяина, с которой проводились манипуляции с применением любого метода для поглощения ДНК последовательности (например, экспрессирующая кассета), будем в данной заявке ссылаться как на "трансформированную" или "рекомбинантную". Трансформированный хозяин будет иметь по меньшей мере одну копию экспрессирующей конструкции и может иметь две или больше, в зависимости от того, интегрирована ли экспрессирующая кассета в геном или присутствует на экстрахромосомном элементе, имеющем множественное число копий.
Трансформированная клетка-хозяин может быть идентифицирована посредством различных методов селекции, как описано в патентах США №№ 7238482 и 7259255, и PCT публикации № WO 2006/052870.
После трансформации субстраты, подходящие для данных ДНК синтаз (и, при необходимости, других ПНЖК ферментов, которые коэкспрессированы в клетке-хозяине), могут вырабатываться хозяином или природным путем или трансгенно, или могут вводиться экзогенным образом.
Предпочтительные микробные хозяева для рекомбинантной экспрессии
Клетки микробных хозяев для экспрессии текущих генов и фрагментов нуклеиновых кислот могут включать хозяев, которые растут на разном исходном сырье, включая простые или сложные углеводы, жирные кислоты, органические кислоты, масла и спирты, и/или углеводороды, в широкой области температур и значений pH. Исходя из потребностей правопреемника заявителя, гены, описанные в данном изобретении, будут экспрессироваться в жировых дрожжах (и, в частности, Yarrowia lipolytica); однако предполагается, что поскольку транскрипция, трансляция и аппарат для биосинтеза протеинов сильно законсервированы, любые бактерии, дрожжи, водоросли, эвгленоиды и/или грибки будут подходящими микробными хозяевами для экспрессии рассматриваеиых нуклеиновокислотных фрагментов.
Однако предпочтительными микробными хозяевами являются маслянистые организмы, такие как жировые дрожжи. Эти организмы способны природным путем синтезировать и накапливать масла, где данное масло может составлять более приблизительно 25% сухого веса клетки, лучше, больше приблизительно 30% сухого веса клетки, и еще лучше, больше приблизительно 40% сухого веса клетки. Роды, которые обычно идентифицируются как жировые дрожжи, включают, однако не ограничиваясь этим: Yarrowia, Candida, Rhodotorula, Rhodosporidium, Cryptococcus, Trichosporon и Lipomyces. Более конкретно, иллюстративные синтезирующие масло дрожжи включают: Rhodosporidium toruloides, Lipomyces starkeyii, L. lipoferus, Candida revkaufi, C. pulcherrima, C. tropicalis, C. utilis, Trichosporon pullans, T. cutaneum, Rhodotorula glutinus, R. graminis, и Yarrowia lipolytica (ранее классифицированная как Candida lipolytica).
Наиболее предпочтительными являются жировые дрожжи Yarrowia lipolytica; и, в дальнейшем варианте наиболее предпочтительными являются штаммы Y. Lipolytica, обозначенные как ATCC #20362, ATCC #8862, ATCC #18944, ATCC #76982 и/или LGAM S(7)1 (Papanikolaou S., и Aggelis G., Bioresour. Technol. 82(1):43-9 (2002)).
В историческом плане, различные штаммы Y. lipolytica использовались для получения и производства: изоцитратлиазы; липаз; полигидроксиалканоатов; лимонной кислоты; эритритола; 2-оксоглутаровой кислоты; гамма-декалактона; гамма-додекалатона; и пировиноградной кислоты. Конкретные пособия, применимые для трансформации жировых дрожжей (т.е. Yarrowia lipolytica), включают патент США № 4880741 и патент США № 5071764 и Chen, D. C. et al. (Appl. Microbiol. Biotechnol., 48(2):232-235 (1997)). Конкретными пособиями, подходящими для получения методами генной инженерии АРК, ЭПК и ДГК в Y. lipolytica,являются патентная заявка США № 11/264784 (PCT публикация No. WO 2006/055322), патентная заяска США № 11/265761 (PCT публикация No. WO 2006/052870) и патентная заявка США № 11/264737 (PCT публикация No. WO 2006/052871), соответственно.
Средства для синтеза и трансформации экспрессирующих векторов, содержащих C20-элонгазы и дельта-4-десатуразы, в жировых дрожжах (т.е. Yarrowia lipolytica), подробно рассмотрены в PCT публикации № WO 2006/052871. Предпочтительным методом экспрессии генов в Yarrowia lipolytica является интеграция линейной ДНК в геном хозяина. Интеграция в множественные локации внутри генома может быть особенно полезной, когда желательны высокие уровни экспрессии генов [например, в Ura3 локус (номер доступа GenBank AJ306421), Leu2 генный локус (номер доступа GenBank AF260230), Lys5 генный локус (номер доступа GenBank M34929), Aco2 генный локус (номер доступа GenBank AJ001300), Pox3 генный локус (Pox3: номер доступа GenBank XP_503244; или, Aco3: номер доступа GenBank AJ001301), дельта-12-десатуразный генный локус (патент США № 7214491), Lip1 генный локус (номер доступа GenBank Z50020), Lip2 генный локус (номер доступа GenBank AJ012632), и/или Pex10 генный локус (номер доступа GenBank CAG81606)].
Терминирующие участки, полезные в настоящей заявке для Yarrowia экспрессирующих векторов, включают, например: ~100 bp 3′ участка Yarrowia lipolytica экстрацеллюларной протеазы (XPR; номер доступа GenBank M17741); ацил-CoA оксидазные (Aco3: номер доступа GenBank AJ001301 и No. CAA04661; Pox3: номер доступа GenBank XP_503244) терминаторы; Pex20 (номер доступа GenBank AF054613) терминатор; Pex16 (номер доступа GenBank U75433) терминатор; Lip1 (номер доступа GenBank Z50020) терминатор; Lip2 (номер доступа GenBank AJ012632) терминатор; и 3-оксацил-CoA тиолазный (OCT; номер доступа GenBank X69988) терминатор.
Предпочтительными методами селекции для использования в Yarrowia lipolytica являются резистентность к канамицину, гигромицину и аминогликозиду G418, так же как и способность расти на средах, лишенных урацила, лейцина, лизина, триптофана или гистидина. В альтернативных вариантах для селекции дрожжевых Ura- мутантов используется 5-фторооротовая кислота (5-фтороурацил-6-карбоновая кислота, моногидрат; “5-FOA”). Данное соединение является токсичным к дрожжевым клеткам, которые обладают функционирующим URA3 геном, кодирующим оротидин 5′-монофосфат декарбоксилазу (ОМД декарбоксилазу); таким образом, на основе этой токсичности 5-FOA особенно полезна для селекции и идентификации Ura- мутантных дрожжевых штаммов (Bartel, P.L. и Fields, S., Двухгибридная система дрожжей, Оксфордский университет: Нью-Йорк, т. 7, стр. 109-147, 1997; см. также PCT публикацию № WO 2006/052870 для использования 5-FOA в Yarrowia). Более конкретно, сначала необходимо выбить нативный Ura3 ген для получения штамма, имеющего Ura- фенотип, где происходит селекция, основанная на 5-FOA резистентности. Затем кластер из множества химерных генов и новый Ura3 ген могут быть интегрированы в другой локус генома Yarrowia с образованием нового штамма, имеющего Ura+ фенотип. Последующая интеграция продуцирует новый Ura3- штамм (вновь идентифицируемый с использованием 5-FOA селекции), когда введенный Ura3 ген выбит. Таким образом, Ura3 ген (в комбинации с селекцией 5-FOA) может быть использован в качестве селекционного маркера в множественных раундах трансформации, позволяя легко интегрировать генетические модификации в Yarrowia геном.
Другие предпочтительные микробные хозяева включают маслянистые бактерии, водоросли, эвгленоиды и другие грибки; и, внутри этой широкой группы микробных хозяев особый интерес представляют микроорганизмы, которые синтезируют омега-3/омега-6 жирные кислоты (или те, что могут с помощью генетических методов послужить этой цели [например, другие дрожжи, такие как Saccharomyces cerevisiae]). Так, например, трансформация Mortierella alpina (которая используется в промышленности для производства АРК) с помощью любой из текущих ДГК синтазных генов под контролем индуцируемых или регулируемых промоторов может дать трансформантный организм, способный синтезировать повышенные количества ПНЖК. Способ трансформации M. alpina описан Mackenzie et al. (Appl. Environ. Microbiol., 66:4655 (2000)). Подобно этому, методы трансформации микроорганизмов Thraustochytriales раскрыты в патенте США № 7001772.
Может потребоваться подпитка субстрата.
Независимо от хозяина, выбранного для экспрессии мультизимов (например, ДГК синтаз), множественные трансформанты должны быть подвергнуты скринингу для получения штамма, проявляющего необходимый уровень экспрессии и структуру. Такой скрининг может быть проведен методом саузерн анализа ДНК блотов (Southern, J. Mol. Biol., 98:503 (1975)), нозерн анализа мРНК экспрессии (Kroczek, J. Chromatogr. Biomed. Appl., 618(1-2):133-145 (1993)), вестерн и/или Elisa анализов протеиновой экспрессии, фенотипического анализа или ГХ анализа ПНЖК продуктов.
Конечно, поскольку вырабатываемые природным путем ПНЖК в жировых дрожжах ограничиваются 18:2 жирными кислотами (т.е. ЛК), и не так часто 18:3 жирными кислотами (т.е. АЛК), в более предпочтительных вариантах настоящего изобретения жировые дрожжи будут подвергаться генетической обработке для экспрессии множества ферментов, необходимых для биосинтеза длинноцепочечных ПНЖК (обеспечивая тем самым производство, например, АРК, ЭПК, ДПК и ДГК, в дополнение к описанным здесь мультизимам.
В особо предпочтительных вариантах по меньшей мере один дополнительный рекомбинантный ДНК конструкция кодирует ДГЛК синтазу, так что данный мультизим обладает активностью дельта-9-элонгазы и активностью дельта-8-десатуразы. В некоторых вариантах дельта-9-элонгаза может быть изолирована или получена из Isochrysis galbana (номер доступа GenBank AF390174; IgD9e или IgD9eS), или же дельта-9-элонгаза может быть изолирована или получена из Euglena gracilis или Euglena anabaena. Например, см. ДГЛК синтазы, представленные как SEQ ID NO:441, SEQ ID NO:447, SEQ ID NO:454, SEQ ID NO:461, SEQ ID NO:464 и SEQ ID NO:471.
Метаболический инжениринг омега-3 и/или омега-6 жирнокислотного биосинтеза в микробах
Способы манипуляции биохимическими путями хорошо известны специалистам в данной области; и ожидается, что будет возможно проводить множественныеі манипуляции для максимизации биосинтеза омега-3 и/или омега-6 жирных кислот в жировых дрожжах, и в частности, в Yarrowia lipolytica. Это может потребовать метаболического инжениринга непосредственно внутри ПНЖК биосинтетического пути или дополнительных манипуляций путями, которые вносят углерод в ПНЖК биосинтетический путь. Методы, полезные для повышения уровня регуляции желательных биохимических путей и понижения уровня регуляции нежелательных биохимических путей, хорошо известны специалистам в данной области.
Например, биохимические пути, конкурирующие с омега-3 и/или омега-6 жирнокислотными биосинтетическими путями в отношении энергии углерода или нативных ферментов ПНЖК биосинтетического пути, которые мешают выработке конкретного ПНЖК конечного продукта, могут быть элиминированы путем разрыва гена или понижения уровня регуляции с помощью других средств (например, антисмысловой мРНК).
Подробное обсуждение манипуляций внутри ПНЖК биосинтетического пути как средства для увеличения выхода АРК, ЭПК или ДГК (и связанных с этим методов) представлено в PCT публикации № WO 2006/055322 [патентная публикация США № 2006-0094092-A1], PCT публикации № WO 2006/052870 [патентная публикация США № 2006-0115881-A1] и PCT публикации № WO 2006/052871 [патентная публикация США № 2006-0110806-A1], соответственно, как и необходимых манипуляций на ТАГ биосинтетическом пути и ТАГ деградационном пути (и связанных с этим методов).
В контексте настоящего изобретения может быть полезным модулировать экспрессию жирнокислотного биосинтетического пути с привлечением любой из стратегий, описанных выше. Например, настоящее изобретение предоставляет методы, посредством которых гены, кодирующие ключевые ферменты ПНЖК биосинтетического пути, вводятся в жировые дрожжи для производства омега-3 и/или омега-6 жирных кислот. Особенно полезным будет экспрессировать текущие ДГК синтазные гены в жировые дрожжи, которые не обладают природными омега-3 и/или омега-6 жирнокислотными биосинтетическими путями, и скоординировать экспрессию этих генов для максимизации производства предпочтительных ПНЖК продуктов с использованием различных средств для метаболического инжениринга хозяйского организма.
Микробные ферментационные процессы для производстап ПНЖК
Трансформированная клетка хозяина выращивается при условиях, которые оптимизируют экспрессию химерных генов и дают наибольший и самый экономичный выход ПНЖК, которым отдается предпочтение. В общем, условия среды, которые могут быть оптимизированы, включают тип и количество углеродного источника, тип и количество азотного источника, отношение углерод-азот, количество различных минеральных ионов, уровень кислорода, температуру выращивания, рН, продолжительность фазы производства биомассы, продолжительность фазы накопления масла и время и метод сбора клеток. Yarrowia lipolytica обычно выращиваются в комплексных средах (например, дрожжевой экстракт-пептон-декстрозный бульон (ДПД)) или определенных минимальных средах, в которых отсутствует компонент, необходимый для роста, вынуждая поэтому проводить селекцию необходимых экспрессирующих кассет (например, Yeast Nitrogen Base (Difco Laboratories, Detroit, Ml)).
Ферментационные среды в данном изобретении должны содержать подходящий источник углерода. Пригодные углеродные источники обсуждены в патенте США № 7238482. Хотя предполагается, что источник углерода, использованный в данном изобретении, может охватывать широкое разнообразие источников, которые содержат углерод, источниками углерода, которым отдается предпочтение, являются сахара, глицерин и/или жирные кислоты. Наибольшее предпочтение отдается глюкозе и/или жирным кислотам, которые содержат 10-22 углеродных атомов.
Азот может поставляться из неорганического (например, (NH4)2SO4) или органического (например, мочевина или глутамин) источника. В дополнение к углеродному и азотному источникам ферментационные среды должны также содержать подходящие минералы, соли, кофакторы, буферы, витамины и другие компоненты, известные специалистам в данной области, пригодные для выращивания маслянистого хозяина и промотирования ферментационных путей, необходимых для производства ПНЖК. Особенное внимание уделяется нескольким металлическим ионам (например, Fe+2, Cu+2, Mn+2, Co+2, Zn+2, Mg+2), которые промотируют синтез липидов и ПНЖК (Nakahara, T. et al., Ind. Appl. Single Cell Oils, авторы D.J. Kyle и R. Colin, стр. 61-97 (1992)).
Ростовыми средами, которым в данном изобретении отдается предпочтение, являются обычные, приготовляемые в промышленности среды, такие как Yeast Nitrogen Base (Difco Laboratoties, Detroit, Ml). Могут также использоваться и другие определенные или синтетические ростовые среды, и соответствующая среда для выращивания трансформантных клеток-хозяев должна быть известна специалистам в области микробиологии или ферментации. Подходящая область рН для ферментации составляет, обычно, 4,0-8,0, где области рН 5,5-7,5 отдается предпочтение как области условий для первичного роста. Ферментация может проводиться в аэробных или анаэробных условиях, где предпочтение отдается микроаэробным условиям.
Типично, накопление высоких уровней ПНЖК в клетках жировых дрожжей требует двухстадийного процесса, поскольку метаболическое состояние должно “балансировать” между ростом и синтезом/сохранением жиров. Таким образом, лучше всего, когда для производства ПНЖК в жировых дрожжах (например, Yarrowia lipolytica) используется двухстадийный ферментационный процесс. Этот подход описан в патенте США № 7238482, так же, как и разные приемлемые схемы ферментационного процесса (например, порционная, с подпиткой и непрерывная) и соображения относительно роста.
Очистка и обработка ПНЖК масел
ПНЖК могут быть найдены в микроорганизмах-хозяевах и растениях-хозяевах в виде свободных жирных кислот или в этерифицированных формах, таких как ацилглицерины, фосфолипиды, сульфолипиды или гликолипиды, и могут быть экстрагированы из клеток-хозяев с использованием множества средств, хорошо известных в данной области. Одним из обзоров способов экстракции, анализа качества и стандартов пригодности для дрожжевых липидов является обзор Z. Jacobs (Critical Reviews in Biotechnology, 12(5/6):463-491 (1992)). Краткий обзор нижнего процессинга имеется также в работе A. Singh и O. Ward (Adv. Appl. Microbiol., 45:271-312 (1997)).
В целом, средства для очистки ПНЖК могут включать экстракцию (например, патент США № 6797303 и патент США №5648564) органическими растворителями, обработку ультразвуком, экстракцию флюидом в сверхкритическом состоянии (например, с использованием диоксида углерода), омыление и физические средства, такие как прессование или их комбинации. Дополнительные подробности можно найти в патенте США № 7238482. Способы выделения масел из семян хорошо известны в данной области: (Young et al., Обработка масел и жиров, Руководство по липидам авторы, Gunstone et al., глава, Chapter 5 стр. 253-257; Chapman и Hall: Лондон (1994)). Например, соевое масло вырабатывается с использованием ряда шагов, включая экстракцию и очистку пищевого масляного продукта из маслянистых семян. Соевые масла и соевые побочные продукты производятся с использованием обобщенных шагов, представленных в Таблице 7.
Более конкретно, семена сои подвергаются очистке, темперированию, лущению и вальцеванию, повышая таким способом эффективность экстракции масла. Экстракция масла обычно осуществляется растворителем (например, гексаном), но может также достигаться с помощью комбинации физического давления и/или экстракции растворителем. Результирующее масло называется сырым маслом. Сырое масло может быть обессмолено путем гидрирования фосфолипидов и других полярных и нейтральных липидных комплексов, что облегчает их отделение от триглицеридной фракции, которая не гидрируется (соевое масло). Результирующие лецитиновые смолы могут дополнительно обрабатываться для приготовления коммерционно важных лецитиновых продуктов, которые используются в различных пищевых и промышленных продуктах, таких как эмульсификаторы и вещества, которые предупреждают слипание. Обессмоленное масло потом может подвергаться рафинированию для удаления примесей (главным образом, жирных кислот, пигментов и остаточных смол). Рафинирование проводится путем добавления каустического агента, который реагирует со свободной жирной кислотой с образованием мыла и гидратов фосфатидов и протеинов в сыром масле. Вода используется для вымывания следов мыла, которые образовались в процессе рафинирования. Побочный мыльный продукт может использоваться непосредственно в животных кормах или может подвергаться подкислению для восстановления свободных жирных кислот. От окраски избавляются путем адсорбции отбеливающей землей, которая удаляет большую часть хлорофилла и каротиновых соединений. Рафинированное масло может быть гидрировано, с образованием, в результате, жиров с разными свойствами плавления и текстурами. Фракционирование охлаждением может быть использовано для удаления стеарина из гидрированного масла путем кристаллизации при тщательно контролируемых условиях охлаждения. Дезодорирование (главным образом, путем паровой дистилляции в вакууме) является последним шагом и предназначено для удаления соединений, которые придают запах или аромат маслу. Другие ценные побочные продукты, такие как токоферолы и стерины, могут быть удалены во время процесса дезодорирования. Дезодорированный дистиллят, который содержит эти побочные продукты, может быть продан для производства натурального витамина Е и других высокоценных фармацевтических продуктов. Рафинированные, отбеленные (гидрированные, фракционированные) и дезодорированные масла и жиры могут упаковываться и прямо продаваться или дополнительно перерабатываться в более специализированные продукты. Более подробная ссылка на обработку соевых семян, производство соевого масла и использование побочных продуктов может быть найдена в работе Erickson, Practical Handbook of Soybean Processing and Utilization, The American Oil Chemists′ Society and United Soybean Board (1995). Соевое масло является жидким при комнатной температуре, поскольку оно имеет относительно низкое содержание насыщенных жирных кислот в сравнении с маслами, такими как кокосовое, пальмовое и какао.
Растительные и микробные масла, которые содержат ПНЖК, которые были рафинированы и/или очищены, могут быть подвергнуты гидрированию, давая в результате жиры с разными свойствами плавления и структурами. Многие переработанные жиры (включая спреды, кондитерские жиры, твердые масла, маргарины, хлебопекарные жиры и т.д.) требуют разных степеней твердости при комнатной температуре и могут вырабатываться только путем изменения физических свойств исходного масла. В наиболее общем случае это достигается путем каталитического гидрирования.
Гидрирование представляет собой химическую реакцию, в которой водород доставляется к двойным связям ненасыщенной жирной кислоты с помощью катализатора, такого как никель. Например, высокоолеиновое соевое масло содержит ненасыщенную олеиновую, ЛК и линоленовую жирные кислоты, и каждая из них может гидрироваться. Гидрирование имеет два главных эффекта. Во-первых, окислительная стойкость масла повышается в результате восстановления ненасыщенных жирных кислот. Во-вторых, физические свойства масла изменяются, поскольку жирнокислотные модификации повышают точку плавления, что дает полутвердый или твердый жир при комнатной температуре.
Существует много переменных, которые влияют на реакцию гидрирования, что в свою очередь изменяет состав конечного продукта. Рабочие условия, которые включают давление, температуру, тип катализатора и концентрацию, перемешивание и конструкцию реактора, находятся среди наиболее важных параметров, которые могут контролироваться. Условия селективного гидрирования могут быть использованы для гидрирования более насыщенных жирных кислот в сравнении с менее насыщенными кислотами. Очень легкое или щеточное гидрирование часто используется для повышения стабильности жидких масел. Дальнейшее гидрирование превращает жидкое масло в физически твердый жир. Степень гидрирования зависит от необходимой производительности и характеристик плавления, которые предполагаются для конечного продукта. Жидкие шортенинги (которые используются в производстве хлебопекарских продуктов, твердых жиров и шортенингов, применяемых для операций жарки и обжаривания в торговле) и основное сырье для производства маргарина находятся среди огромного множества возможных масляных и жировых продуктов, которые получаются путем гидрирования. Более детальное описание гидрирования и продуктов гидрирования можно найти в работе Patterson, H.B.W., Hydrogenation of Fats and Oils: Theory and Practice. The American Oil Chemists′ Society (1994).
Гидрированные масла стали в некоторой степени сомнительными в плане употребления из-за присутствия транс-жирнокислотных изомеров, которые возникают в результате процесса гидрирования. Употребление больших количеств транс-изомеров связывается с вредными для здоровья эффектами, включая повышенные отношения липопротеинов низкой плотности к липопротеинам высокой плотности в плазме крови и повышенный риск коронарной болезни сердца).
Масла, содержащие ПНЖК, для использования в пищевых продуктах, продуктах здорового питания и животных кормах
В настоящее время рынок поддерживает значительное разнообразие продуктов питания, которые включает омега-3 и/или омега-6 жирные кислоты (в частности, АЛК, ГЛК, АРК, ЭПК, ДПК и ДГК). Предполагается, что растительные/семенные масла, измененные семена и микробная биомасса и/или масла данного изобретения, которые содержат ПНЖК, будут действовать в пище и пищевых продуктах с пользой для здоровья. По сравнению с другими растительными маслами, масла данного изобретения, как представляется, действуют подобно другим маслам в пищевых приложениях с физической точки зрения (например, частично гидрированные масла, такие как соевое масло, широко применяются как ингредиенты для мягких спреев, маргарина и шортенингов для выпекания и жарки).
Растительные/семенные масла и измененные семенные масла, которые содержат омега-3 и/или омега-6 жирные кислоты, как тут описано, будут пригодны для употребления в различной пище и пищевых продуктах, включая, однако не ограничиваясь этим, пищевые аналоги, мясные продукты, крупяные продукты, мучные продукты, закуски и молочные продукты.
Кроме того, растительные/семенные масла, измененные семенные масла и микробная биомасса, и/или масла могут быть использованы в препаратах, которые действуют с пользой для здоровья в лечебном питании, включая медицинское питание, пищевые добавки, детские смеси, так же как и фармацевтические продукты. Специалисту в области обработки пищевых продуктов и составления пищевых рационов понятно, каким образом и в каком количестве и качестве к еде или пищевому продукту могут быть добавлены растительные и микробные масла данного изобретения. На такое количество будем здесь ссылаться как на “эффективное” количество, и оно будет зависеть от конкретной пищи или пищевого продукта, диеты, которую должен дополнять данный продукт, или медицинского состояния, которое предролагается скорректировать или вылечить с помощью медицинского питания.
Пищевые аналоги могут быть изготовлены с использованием способов, которые хорошо известны специалистам в данной области. Здесь можно отметить мясные аналоги, сырные аналоги, молочные аналоги и такое подобное. Мясные аналоги, изготовленные из сои, содержат соевый белок или тофу и другие инградиенты, смешанные для симуляции разных видов мяса. Эти мясные альтернативы продаются как замороженные, консервированные или высушенные пищевые продукты. Обычно, они могут быть использованы тем самым способом, что и пища, которую они заменяют. Мясные альтернативы, изготовленные из сои, служат прекрасными источниками протеина, железа и витаминов В. Примеры мясных аналогов включают, однако не ограничиваясь этим, аналоги ветчины, колбасные аналоги, аналоги бекона и такое подобное.
Пищевые аналоги могут быть классифицированы как имитация или заменители, в зависимости от их функциональных и составных характеристик. Например, имитационный сыр должен лишь походить на сыр, который он должен заменять. Однако, продукт может, в общем, называться заменителем сыра, только если он является пищевым эквивалентом сыра, который он заменяет и удовлетворяет минимумум требований по составу для этого сыра. Таким образом, заменитель сыра часто имеет более высокие уровни протеина, чем имитационные сыры, и должен быть подкреплен витаминами и минералами.
Молочные аналоги или немолочные пищевые продукты включают, однако не ограничиваясь этим, имитационное молоко и немолочные замороженные десерты (например, изготовленные из сои и/или соевого белка).
Мясные продукты охватывают широкое разнообразие продуктов. В США “мясо” включает “красное мясо”, полученное из крупного рогатого скота, свиней и овец. В дополнение к красному мясу имеются изделия из птицы, которые включают цыплят, индюков, гусей, рябчиков, уток и рыбу и моллюсков. Имеется широкий ассортимент высушенных и переработанных мясных продуктов: свежих, вяленых и обжареных, и вяленых и выпеченных. Колбаса и хотдоги служат примерами переработанных мясных продуктов. Таким образом, термин “мясные продукты”, как здесь используется, включает, однако не ограничиваясь этим, переработанные мясные продукты.
Крупяной пищевой продукт представляет собой пищевой продукт, полученный в результате переработки крупяного зерна. Крупяное зерно касается любого растения из семейства трав, которое дает съедобное зерно (семечки). Наиболее популярными являются ячмень, кукуруза, просо, овес, киноа, рис, рожь, сорго, тритикале, пшеница и канадский рис. Примеры крупяных продуктов включают, однако не ограничиваясь этим: целое зерно, измельченное зерно, крупу грубого помола, муку, высевки, семечки, хлопья для завтрака, экструдированные пищевые продукты, пасты и таму подобное.
Выпеченные продукты включают любой из крупяных продуктов, упомянутый выше, который был выпечен или переработан с помощью способа, подобного выпечке (например, подверженный действию тепла для высушивания и твердения). Примеры выпеченных продуктов включают, однако не ограничиваясь этим: хлеб, пироги, пончики, палочки, пасты, хлебное крошево, вареные закуски, мини-бисквиты, мини-крекеры, мини-булочки и мини-крендели. Как отмечалось выше, масла настоящего изобретения могут быть использованы в качестве ингредиента.
Закусочные пищевые продукты содержат любой из описанных выше или ниже пищевых продуктов.
Жареный пищевой продукт включает любой из описанных выше или ниже пищевых продуктов, который был поджарен.
Продукт здорового питания представляет собой любой пищевой продукт, который благотворно влияет на здоровье. Многие из пищевых продуктов, которые получены из маслянистых семян, могут считаться продуктами здорового питания.
Напиток может быть в жидкой или сухой порошковой форме.
Например, здесь можно упомянуть негазированные напитки, такие как фруктовые соки, свежие, замороженные, консервированные или концентраты; ароматизированные или простые молочные напитки и т.д. Питательные смеси для детей и взрослых хорошо известны в данной области и имеются в продаже (например, Similac®, Ensure®, Jevity® и Alimentum® от фирмы Ross Products Division, Abbott Laboratories).
Детские смеси являются жидкостями или восстановленными порошками, которыми кормят младенцев или маленьких детей. “Детская смесь” определена здесь как энтеральный пищевой продукт, которым можно заменить грудное молоко человека при кормлении младенцев и которая обычно состоит из необходимого процента жира, смешанного с необходимым процентом углеводов и белков в водном растворе (например, см. патент США № 4670285). Основываясь на мировых исследованиях состава и на уровнях, определенных экспертными группами, усредненное грудное молоко человека, типично, содержит приблизительно 0,20-0,40% совокупных жирных кислот (в допущении приблизительно 50% калорий из жира); и, в целом, отношение ДГК к АРК варьирует от приблизительно 1:1 до 1:2 (см., например, рецепты Enfamil LIPIL™ (Mead Johnson & Company) и Similac Advance™ (Ross Products Division, Abbott Laboratories)). Детские смеси должны играть специальную роль в детских диетах, поскольку они часто являются единственным источником питания для детей; и, хотя кормление грудью остается наилучшим способом кормления детей, детская смесь является важным компонентом не только для выживания, но и развития детей.
Молочный продукт является продуктом, полученным из молока. Аналог молока или немолочный продукт получается из источника, иного, чем молоко, например, соевого молока, как обсуждалось выше. Такие продукты включают, однако не ограничиваясь этим, несобранное молоко, собранное молоко, продукты из сброженного молока, такие как йогурт или кислое молоко, сливки, масло, сгущенное молоко, обезвоженное молоко, порошковый молочный заменитель для кофе, порошковый сливочный заменитель для кофе, мороженное, сыр и т.д.
Дополнительными пищевыми продуктами, в которые могут включаться масла данного изобретения, которые содержат ПНЖК, являются, например, жевательные резинки, конфеты и сахарная глазурь, желатины и пудинги, твердые и мягкие леденцы, джемы и желе, белый сахар-песок, заменители сахара, сладкие пюре, десерты и сиропы, и смешанные в сухом состоянии порошковые смеси.
Продуктом здорового питания является любой пищевой продукт, который оказывает благотворное влияние на здоровье человека и включает функциональную пищу, медицинскую пищу, лечебное питание и диетические добавки. Кроме того, растительные/семенные масла, измененные семена и микробные масла данного изобретения могут быть использованы в стандартных фармацевтических композициях (например, длинноцепочечные ПНЖК, которые содержат масла, могли бы быть легко введены в любой из упомянутых выше пищевых продуктов, образуя таким способом функциональную или медицинскую пищу). Более концентрированные препараты, которые содержат ПНЖК, включают капсулы, порошки, таблетки, мягкие гели, гелевые колпачки, жидкие концентраты и эмульсии, которые могут быть использованы как диетические добавки к питанию людей или животных.
Животный корм определяется здесь, в целом, как продукты, предназначенные для использования в виде корма или для смешивания с кормом для животных. Растительные/семенные масла, измененые семена и микробные масла данного изобретения могут использоваться как ингредиент в различных животных кормах.
Более конкретно, однако без каких-либо ограничений, ожидается, что масла настоящего изобретения могут использоваться в кормах для домашних животных, жвачных животных, птицы и аквакультур. Кормами для домашних животных служат те продукты, которые предназначены для кормления домашних любимцев (например, собак, котов, птиц, рептилий, грызунов). Эти корма могут включать крупяные продукты и продукты здорового питания, упомянутые выше, так же как и мясо и мясные побочные продукты, продукты из соевого белка, травяные продукты и продукты из сена (например, люцерна, тимофеевка луговая, овес или костер (Bromus), овощи). Корма для жвачных животных и птицы являются теми, которые предназначены для кормления этих животных (например, индюков, цыплят, крупного рогатого скота, свиней). Как и в случае кормов для домашних любимцев, эти корма могут включать крупяные продукты и продукты здорового питания, продукты из соевого белка, мясо и мясные побочные продукты, и травяные продукты, и продукты из сена, как перечислено выше. Корма для аквакультур (или “аквакорма”) являются кормами, которые предназначены для использования на аквафермах для разведення, культивирования или выращивания водных организмов и/или животных в пресной или морской водах.
ПРИМЕРЫ
Данное изобретение дополнительно определяется в следующих Примерах, где части и проценты являются весовыми, и температура приведена в градусах Цельсия, если не указано иное. Следует понимать, что эти Примеры, хотя они относятся к вариантам данного изобретения, которым отдается предпочтение, приведены лишь как иллюстрации. Исходя из приведенного выше обсуждения и этих Примеров, специалист в данной области сможет уяснить существенные особенности данного изобретения, и не отклоняясь от сути и объема изобретения, сможет сделать разные изменения и модификации изобретения с целью его адаптации к разным применениям и условиям. Так, разные модификации данного изобретения, в добавление к показанным и описанным здесь, будут очевидными для специалистов в данной области из предыдущего описания. Такие модификации также, как представляется, подпадают под объем пунктов формулы изобретения, которая прилагается.
Значения сокращений следующие: “sec” отвечает секундам (с), “min” отвечает минуте(ам), “h” отвечает часу(ам), “d” отвечает суткам, “µl” отвечает микролитру(ам), “mL” отвечает миллилитру(ам), “L” отвечает литру(ам), “µМ” отвечает микромолярной (концентрации), “mM” отвечает миллимолярной, “М” отвечает молярной, “mmol” отвечает миллимольному (количеству), “µmol” отвечает микромольной, “g” отвечает грамму(ам), “µg” отвечает микрограмму(ам), “ng” отвечает нанограмму(ам), “U” отвечает единице(ам), “bp” отвечает основной паре(ам) и “kB” отвечает тысяче пар (нуклеотидов).
ОБЩИЕ МЕТОДЫ:
Номенклатура для экспрессирующих кассет:
Структура экспрессирующей кассеты будет представлена простой системой обозначений “X::Y::Z”, где X описывает промоторный фрагмент, Y описывает фрагмент участка, кодирующего ген, и Z описывает терминирующий фрагмент, которые связаны между собой действующим образом.
Трансформация и культивирование Yarrowia lipolytica:
Штаммы Yarrowia lipolytica с номерами доступа ATCC 20362, 76982 и 90812 приобретали на фирме American Type Culture Collection (Rockville, MD) (АСТК - Американское собрание типовых культур) Штаммы Yarrowia lipolytica выращивали, типично, при 28-30оС в нескольких средах, в соответствии с рецептами, приведенными ниже. Агаровые пластинки готовили путем добавления 20 г/л агара в каждую жидкую среду, в соответствии со стандартной методологией.
YPD агаровая среда (на литр): 10 г дрожжевого экстракта [Difco], 20 г of Bacto пептона[Difco]; и 20 г глюкозы.
Базальная минимальная среда (МС) (на литр): 20 г глюкозы; 1,7 г дрожжевой азотной основы без аминокислот; 1,0 г пролина; и pH 6,1 (не скорректирован).
Минимальная среда+урацил (МС+урацил или МСУ (на литр): Приготовить МС среду как выше и добавить 0,1 г урацила и 0,1 г уридина.
Минимальная среда+урацил+сульфонилмочевина (МСУ+СМ) (на литр): Приготовить МСУ среду как выше и добавить 280 мг сульфонилмочевины.
Минимальная среда+лейцин (МС+лейцин или MCLeu) (на литр): Приготовить МС среду как выше и добавить 0,1 г лейцина.
Минимальная среда+лейцин+урацил (MCLeuUra) (на литр): Приготовить МС среду как выше и добавить 0,1 г лейцина, 0,1 г урацила и 0,1 г уридина.
Минимальная среда+лейцин+лизин (MCLeuLys) (на литр): Приготовить МС как выше и добавить 0,1 г лизина, 0,1 г лейцина.
Минимальная среда+5-фторооротовая кислота (МС+5-FOA) (на литр): 20 г глюкозы, 6,7 г Yeast Nitrogen основания, 75 мг урацила, 75 мг уридина и соответствующее количество FOA (Zymo Research Corp., Orange, CA), исходя из тестирования FOA активности в области концентраций от 100 мг/л до 1000 мг/л (поскольку для каждой порции, полученной от поставщика, отмечается расхождение).
Среда с высоким содержанием глюкозы (HGM) (на литр): 80 г глюкозы, 2,58 г KH2PO4 и 5,36 г K2HPO4, pH 7,5 (корректировка не требуется).
Трансформация Yarrowia lipolytica проводилась в соответствии с методом Chen, D.C. et al. (Appl. Microbiol. Biotechnol. 48(2):232-235 (1997)), если не указано иное. В кратком изложении, Yarrowia наносили штрихами на YPD пластинку и выращивали при 30°С на протяжении приблизительно 18 часов. Несколько больших петель клеток соскребали с пластинки и повторно суспендировали в 1 мл трансформационного буфера, котрый содержал 2,25 мл 50% PEG, средний МВ 3350; 0,125 мл 2М ацетата лития, рН 6,0; 0,125 мл 2М DTT; и 50 мкг гидродинамически фрагментированной ДНК молок лосося. Потом приблизительно 500 нг линеаризованной плазмидной ДНК инкубировали в 100 мкл ресуспендированных клеток и выдерживали при 39°С на протяжении 1 часа с вихревым перемешиванием при 15-минутных интервалах. Клетки помещали на пластинки с селективными средами и выдерживали при 30°С на протяжении 2-3 суток.
Анализ Yarrowia lipolytica на жирные кислоты:
Для анализа жирных кислот клетки собирали путем центрифугирования, и липиды экстрагировали как описано в работе Bligh, E.G. & Dyer, W.J. (Can. J. Biochem. Physiol. 37:911-917 (1959)). Метиловые эфиры жирных кислот готовили путем трансэтерификации липидного экстракта метоксидом натрия (Roughan, G. и Nishida, I., Arch Biochem Biophys. 276(1):38-46 (1990)) и потом анализировали с помощью Hewlett-Packard 6890 GC (газовый хроматограф), снабженного 30 м×0,25 мм (внутр. диам.) HP-INNOWAX (Hewlett-Packard) колонкой. Температура печи составляла от 170°С (25 минут выдержки) до 185°С при 3,5°С/минуту.
Для прямой трансэтерификации основания культуру Yarrowia (3 мл) собирали, промывали один раз в дистиллированной воде и высушивали под вакуумом в устройстве Speed-Vac на протяжении 5-10 минут. К образцу прибавляли метоксид натрия (100 мкл 1%), потом его подвергали вихревому перемешиванию и встряхиванию на протяжении 20 минут. После добавления 3 капель 1М NaCl и 400 мкл гексана образец подвергали вихревому перемешиванию и центрифугированию. Верхний слой удаляли и анализировали методом газовой хроматографии как описано выше.
Конструирование штамма Y4305U3 Yarrowia lipolytica:
Штамм Y4305U3 Y. lipolytica был использован как хозяин в Примерах 52, 53 и 54, infra. Последующее описание является сводкой конструирования штамма Y4305U3, полученного из Yarrowia lipolytica ATCC № 20362. Штамм Y4305U3 способен произвести приблизительно 53,2% ЭПК относительно всех липидов через экспрессию дельта-9-элонгаза/дельтаа-8-десатуразного пути (ФИГ.44).
Разработка штамма Y4305U3 потребовала конструирования штамма Y2224 (FOA резистентный мутант как результат автономной мутации Ura3 гена дикого типа Yarrowia штамма ATCC № 20362), штамма Y4001 (производящего 17% ЭДК с Leu- фенотипом), штамма Y4001U1 (производящего 17% ЭДК с Leu- и Ura- фенотипом), штамма Y4036 (производящего 18% ДГЛК с Leu- фенотипом), штамма Y4036U (производящего 18% ДГЛК с Leu- и Ura- фенотипом), штамма Y4070 (производящего 12% АРК с Ura- фенотипом), штамма Y4086 (производящего 14% ЭПК), штамма Y4086U1 (Ura3-), штамма Y4128 (производящего 37% ЭПК), штамма Y4128U3 (Ura-), штамма Y4217 (производящего 42% ЭПК), штамма Y4217U2 (Ura-), штамма Y4259 (производящего 46,5% ЭПК) и штамма Y4259U2 (Ura-).
Генерация штамма Y2224: Штамм Y2224 изолировали следующим способом: клетки Yarrowia lipolytica ATCC #20362 из YPD агаровой пластинки наносили штрихами на МС пластинку (по 75 мг/л урацила и уридина, 6,7 г/л YNB с сульфатом аммония, без аминокислот, и 20 г/л гдюкозы) с содержанием 250 мг/л 5-FOA (Zymo Research). Пластинки инкубировали при 28°C, и четыре из результирующих колоний намазывали раздельно на МС пластинки, содержащие 200 мг/мл 5-FOA и МС пластинки без урацила и уридина. Это проделывали для подтверждения ауксотрофии урацил Ura3.
Генерация штамма Y4001, производящего около 17% ЭДК из всех липидов: Штамм Y4001 создавался путем интеграции конструкции pZKLeuN-29E3 (ФИГ.45A). Эта конструкция, содержащая четыре химерных гена (т.е. дельта-12-десатураза, C16/18-элонгаза и две дельта-9-элонгазы), был интегрирован в Leu2 локусы штамма Y2224, давая таким образом возможность получить ЭДК.
Конструкция pZKLeuN-29E3, содержала компоненты, показанные ниже в Таблице 8.
Плазмиду pZKLeuN-29E3 расщепляли AscI/SphI и затем использовали для трансформации штамма Y2224Y. lipolytica (т.е. ATCC № 20362 Ura3-) в соответствии с разделом Общие методы. Трансформированные клетки помещали на пластинки со средой MCLeu, и пластинки выдерживали при 30°C в течение от 2 до 3 суток. Колонии собирали и наносили штрихами на МС и MCLeu селекционные пластинки. Колонии, которые могли расти на MCLeu пластинках, но не на МС пластинках, были выбраны в качестве Leu- штаммов. Отдельные колонии Leu- штаммов были использованы для инокуляции жидкой MCLeu, и жидкие культуры встряхивали при 250 оборотов/минуту в течение 2 суток при 30°C. Клетки собирали путем центрифугирования, и липиды экстрагировали. Жирнокислотные метиловые эфиры готовили путем трансэтерификации и потом анализировали с помощью газового хроматографа Hewlett-Packard 6890 GC.
Газо-хроматографические анализы показали наличие ЭДК в трансформантах, содержащих 4 химерных гена pZKLeuN-29E3, но не в контрольном штамме Yarrowia Y2224. Большиство из выбранных 36 Leu- штаммов продуцировали приблизительно от 12 до 16,9% ЭДК от всех липидов. Три штамма, обозначенных как Y4001, Y4002, и Y4003, давали приблизительно 17,4%, 17%, и 17,5% ЭДК от всех липидов, соответственно.
Отдельные колонии штаммов Y4001, Y4002, и Y4003 использовали для инокуляции жидкой MCLeu, и жидкие культуры встряхивали при 250 оборотов/минуту в течение 2 суток при 30°C. Клетки собирали путем центрифугирования, ресуспендировали в HGM, и потом встряхивали при 250 оборотов/минуту в течение 5 суток. Клетки собирали путем центрифугирования, и липиды экстрагировали. Жирнокислотные метиловые эфиры готовили путем трансэтерификации и потом анализировали на газовом хроматографе Hewlett-Packard 6890 GC. Анализы GC показали, что штаммы Y4001, Y4002, и Y4003 вырабатывали приблизительно 24% ЭДК от всех липидов.
Генерация штамма Y4001U (Leu-, Ura-): Штамм Y4001U создавали путем временной экспрессии Cre рекомбиназного фермента в плазмиде pY116 (ФИГ.45B) в штамме Y4001 с получением Leu- и Ura- фенотипа. Конструкция pY116 содержала следующие компоненты:
Плазмиду pY116 использовали для трансформации свежевыращенных Y4001 клеток в соответствии с разделом Общие методы. Трансформированные клетки помещали на MCLeuUra пластинки, содержащие 280 мкг/мл сульфонилмочевины (хлоримурон этил, E. I. duPont de Nemours & Co., Inc., Wilmington, DE), и пластинки выдерживали при 30°C в течение от 3 до 4 суток. Четыре колонии собирали и потом использовали для инокуляции в 3 мл жидкого YPD. Жидкие культуры встряхивали при 250 оборотов/минуту в течение 1 суток при 30°C. Культуры разбавляли до1:50000 жидкой MCLeuUra средой, и 100 мкл помещали на новые YPD пластинки. Данные пластинки выдерживали при 30°C в течение 2 суток. Колонии собирали и наносили штрихами на селекционные пластинки MCLeu и MCLeuUra. Колонии, которые могли расти на пластинках MCLeuUra, но не на пластинках MCLeu, выбирали и проанализировали методом газовой хроматографии, и подтвердили наличие C20:2 (ЭДК). Несколько штаммов, каждый из которых имел Leu- и Ura- фенотип, вырабатывали приблизительно 17% ЭДК от всех липидов и были коллективно обозначены как Y4001U. Один из этих штаммов был обозначен как Y4001U1.
Генерация Y4036 штамма, производящего около 18% ДГЛК от всех липидов: Конструкцию pKO2UF8289 (ФИГ.46A; SEQ ID NO:324) генерировали для интеграции четырех химерных генов (включающих дельта-12-десатуразу, одну дельта-9-элонгазу, и две мутантных дельта-8-десатуразы) в дельта-12-локусы штамма Y4001U1, обеспечивая тем самым получение ДГЛК. Конструкция pKO2UF8289 содержала следующие компоненты:
Плазмиду pKO2UF8289 расщепляли AscI/SphI и затем использовали для трансформации штамма Y4001U1 в соответствии с разделом Общие методы. Трансформированные клетки помещались на пластинки со средой MCLeu, и пластинки выдерживали при 30°C в течение от 2 до 3 суток. Колонии собирали и наносили штрихами на MCLeu селекционные пластинки при 30°С на протяжении 2 суток. Эти клетки потом использовали для инокуляции жидкой MCLeu среды, и жидкие культуры встряхивали при 250 оборотов/минуту в течение 2 суток при 30°C. Клетки собирали путем центрифугирования, и липиды экстрагировали. Жирнокислотные метиловые эфиры готовили путем трансэтерификации и потом анализировали с помощью газового хроматографа Hewlett-Packard 6890 GC.
Газо-хроматографические анализы показали наличие ДГЛК в трансформантах, содержащих 4 химерных гена pKO2UF8289, но не в родительском штамме Y4001U1. Большиство из выбранных 96 штаммов продуцировали приблизительно от 7 до 13% ДГЛК от всех липидов. Шесть штаммов, обозначенных как Y4034, Y4035, Y4036, Y4037, Y4038 и Y4039 давали приблизительно 15%, 13,8%, 18,2%, 13,1%, 15,6% и 13,9% ДГЛК от всех липидов, соответственно.
Генерация штамма Y4036U (Leu-, Ura3-): Конструкцию pY116 (ФИГ.45B; SEQ ID NO:323) использовали для временной экспрессии Cre рекомбиназного фермента в штамме Y4036. Это высвобождало LoxP сэндвичевый Ura3 ген из генома.
Плазмиду pY116 использовали для трансформации штамма Y4036 в соответствии с разделом Общие методы. Следуя трансформации, клетки помещали на MCLeuUra пластинки, и пластинки выдерживали при 30°C в течение от 2 до 3 суток. Отдельные колонии, выращенные на MCLeuUra пластинках, наносили штрихами в YPD жидкую среду. Жидкие колонии встряхивали при 250 оборотов/минуту в течение 1 суток при 30°C со структурированием pY116 плазмиды. Клетки из выращенных культур наносили штрихами на MCLeuUra пластинки. После двух суток при 30°C отдельные колонии повторно наносили на MCLeuUra, MCU и MCLeu пластинки. Те колонии, которые могли расти на MCLeuUra, но не на MCU или MCLeu пластинках, выбирали. Один штамм с Leu- и Ura- фенотипами был обозначен как Y4036U (Ura-, Leu-).
Генерация Y4069 и Y4070 штаммов, производящих около 12% АРК от всех липидов: Конструкцию pZKSL-555R (ФИГ.46B; SEQ ID NO:331) генерировали для интеграции трех дельта-5-десатуразных генов в Lys локусы штамма Y4036U, обеспечивая тем самым выработку АРК. pZKSL-555R плазмида содержала следующие компоненты:
Плазмиду pZKSL-555R расщепляли AscI/SphI и потом использовали для трансформации штамма Y4036U в соответствии с разделом Общие методы. Трансформированные клетки помещали на MCLeuLys пластинки, и эти пластинки выдерживали при 30°C в течение от 2 до 3 суток. Отдельные колонии затем наносили повторно штрихами на MCLeuLys пластинки, и результирующие колонии использовали для инокуляции жидкой MCLeuLys. Жидкие культуры потом встряхивали при 250 оборотв/минуту в течение 2 суток при 30°C. Данные клетки собирали путем центрифугирования, и липиды экстрагировали. Жирнокислотные метиловые эфиры готовили путем трансэтерификации и впоследствии анализировали на газовом хроматографе Hewlett-Packard 6890 GC.
Газо-хроматографический анализ показал наличие АРК в трансформантах, содержащих 3 химерных гена pZKSL-555R, но не в родительском Y4036U штамме. Большинство из выбранных 96 штаммов производили ~10% АРК от всех липидов. Четыре штамма, обозначненные как Y4068, Y4069, Y4070, и Y4071, производили около 11,7%, 11,8%, 11,9% и 11,7% АРК от всех липидов, соответственно. Дальнейший анализ показал, что данные три химерных гена pZKSL-555R не интегрировались в Lys5 сайт в Y4068, Y4069, Y4070 и Y4071 штаммов. Все штаммы обладали Lys+ фенотипом.
Конечный генотип штамма Y4070, по отношению к Yarrowia lipolytica ATCC #20362 дикого типа, был Ura-, неизвестен 1-, неизвестен 3-, Leu+, Lys+, GPD::FmD12::Pex20, YAT1::FmD12::OCT, YAT1::ME3S::Pex16, GPAT::EgD9e::Lip2, EXP1::EgD9eS::Lip1, FBAINm::EgD9eS::Lip2, FBAINm::EgD8M::Pex20, EXP1::EgD8M::Pex16, FBAIN::EgD5::Aco, EXP1::EgD5S::Pex20, YAT1::RD5S::OCT.
Генерация штамма Y4086, производящего около 14% ЭПК от всех липидов: Конструкцию pZP3-Pa777U (ФИГ.47A; SEQ ID NO:338) генерировали для трех дельта-17-десатуразных генов в Pox3 локусы (номер доступа GenBank AJ001301) штамма Y4070, обеспечивая тем самым выработку ЭПК. Плазмида pZP3-Pa777U содержала следующие компоненты:
Плазмиду pZP3-Pa777U расщепляли AscI/SphI и затем использовали для трансформации штамма Y4070 в соответствии с разделом Общие методы. Трансформированные клетки помещали на МС пластинки, и данные пластинки выдерживали при 30°C в течение от 2 до 3 суток. Затем отдельные колонии повторно наносили штрихами на МС пластинки, и результирующие колонии использовали для инокуляции жидкой MCLeuLys. Жидкие культуры встряхивали при 250 оборотов/минуту в течение 2 суток при 30°C. Данные клетки собирали путем центрифугирования, и липиды экстрагировали. Жирнокислотные метиловые эфиры готовили путем трансэтерификации и впоследствии анализировали с помощью газового хроматографа Hewlett-Packard 6890 GC.
ГХ (газо-хроматографический) анализы выявили присутствие ЭПК в трансформантах, содержащие 3 химерных гена pZP3-Pa777U, но не в родительском Y4070 штамме. Большинство из выбранных 96 штаммов производили 10-13% ЭПК от всех липидов. Два штамма, обозначенных как Y4085 и Y4086, вырабатывали около 14,2% и 13,8% ЭПК от всех липидов, соответственно.
Конечный генотип штамма Y4086, в отношении Yarrowia lipolytica ATCC #20362 дикого типа, был Ura3+, Leu+, Lys+, неизвестен 1-, неизвестен 2-, YALI0F24167g-, GPD::FmD12::Pex20, YAT1::FmD12::OCT, YAT1::ME3S::Pex16, GPAT::EgD9e::Lip2, EXP1::EgD9eS::Lip1, FBAINm::EgD9eS::Lip2, FBAINm::EgD8M::Pex20, EXP1::EgD8M::Pex16, FBAIN::EgD5::Aco, EXP1::EgD5S::Pex20, YAT1::RD5S::OCT, YAT1::PaD17S::Lip1, EXP1::PaD17::Pex16, FBAINm::PaD17::Aco.
Генерация штамма Y4086U1 (Ura3-): Штамм Y4086U1 создавали посредством временной экспрессии Cre рекомбинантного фермента в конструкции pY117 (ФИГ.47B; SEQ ID NO:343) в штамме Y4086 с образованием Ura- фенотипа. Это высвободило LoxP сэндвичевый Ura3 ген из генома. Мутантный Yarrowia AHAS фермент в плазмиде pY117 дал SUR, который использовали как позитивный скрининговый маркер.
Плазмиду pY117 получали из плазмиды pY116 (описанной supra, и в патентной заявке США № 11/635258) путем инсерции мутантного AHAS гена, фланкированного PacI-SwaI сайтами, в PacI-SwaI, расщепленный pY116, замещая таким образом LEU селективный маркер сульфонилмочевинным маркером. Конструкция pY117 содержала, таким образом, следующие компоненты:
Плазмиду pY117 использовали для трансформации штамма Y4086 в соответствии с разделом Общие методы. Следуя трансформации, клетки помещали на MCU+SU (280 мкг/мл сульфонилмочевины; известная также как хлоримурон этил, E. I. duPont de Nemours & Co., Inc., Wilmington, DE) пластинки, и данные пластинки выдерживали при 30°C в течение от 2 до 3 суток. Отдельные SUR колонии, выращенные на MCU+SU пластинках, собирали и вносили в YPD жидкую среду, и жидкие культуры встряхивали при 250 оборотов/минуту на протяжении 1 суток при 30°C для структурирования pY117 плазмиды. Клетки из выращенных культур наносили штрихами на MCU пластинки. После двух суток при 30°C отдельные колонии повторно наносили на МС и MCU пластинки. Те колонии, которые могли расти на MCU, но не на МС пластинках, отбирали. Два из этих штаммов с Ura- фенотипами были обозначены как Y4086U1 и Y4086U2 (Ura-).
Генерация штамма Y4128, производящего около 37% ЭПК от всех липидов: Конструкцию pZP2-2988 (ФИГ.48A; SEQ ID NO:345) генерировали для интеграции одного дельта-12-десатуразного гена, двух дельта-8-десатуразных генов, и одного дельта-9-элонгазного гена в Pox2 локусы (номер доступа GenBank AJ001300) штамма Y4086U1, обеспечивая тем самым высокий уровень выработки ЭПК. Плазмида pZP2-2988 содержала следующие компоненты:
Плазмиду pZP2-2988 расщепляли AscI/SphI и затем использовали для трансформации штамма Y4086U1 в соответствии с разделом Общие методы. Трансформированные клетки помещали на МС пластинки, и данные пластинки выдерживали при 30°C в течение от 2 до 3 суток. Отдельные колонии повторно наносили на МС пластинки, и результирующие колонии испоьзовали для инокуляции жидкой MCLeuLys. Жидкие культуры встряхивали при 250 оборотов/минуту в течение 2 суток при 30°C. Клетки собирали путем центрифугирования, ресуспендировали в HGM, и затем встряхивали при 250 оборотов/минуту в течение 5 суток. Данные клетки собирали путем цетрифугирования, и липиды экстрагировали. Жирнокислотные метиловые эфиры готовили путем трансэтерификации и впоследствии анализировали с помощью газового хроматографа Hewlett-Packard 6890 GC.
ГХ анализы выявили, что большинство из выбранных 96 штаммов производили 12-15,6% ЭРК от всех липидов. Два штамма, обозначенных как Y4128 и Y4129, вырабатывали приблизительно 37,6% и 16,3% ЭПК от всех липидов, соответственно.
Конечный генотип штамма Y4128, относительно Yarrowia lipolytica ATCC #20362 дикого типа, был: YALI0F24167g-, Pex10-, неизвестен 1-, неизвестен 2-, GPD::FmD12::Pex20, YAT1::FmD12::OCT, GPM/FBAIN::FmD12S::OCT, YAT1::ME3S::Pex16, GPAT::EgD9e::Lip2, EXP1::EgD9eS::Lip1, FBAINm::EgD9eS::Lip2, FBA::EgD9eS::Pex20, FBAINm::EgD8M::Pex20, EXP1::EgD8M::Pex16, GPDIN::EgD8M::Lip1, YAT1::EgD8M::Aco, FBAIN::EgD5::Aco, EXP1::EgD5S::Pex20, YAT1::RD5S::OCT, YAT1::PaD17S::Lip1, EXP1::PaD17::Pex16, FBAINm::PaD17::Aco. Штамм Yarrowia lipolytica Y4128 был депонирован в Американском собрании типовых культур 23 августа 2007 года и носит обозначение ATCC PTA-8614.
Генерация штаммов Y4128U: Для разрушения Ura3 гена в штамме Y4128 создали конструкцию pZKUE3S (ФИГ.48B; SEQ ID NO:351) для интеграции EXP1::ME3S::Pex20 химерного гена в Ura3 ген штамма Y4128. Плазмида ZKUE3S содержала следующие компоненты:
Плазмиду ZKUE3S расщепляли SphI/PacI и затем использовали для трансформации штамма Y4128 в соответствии с разделом Общие методы. Следуя трансформации, клетки помещали на МС+5-FOA селекционные пластинки, и данные пластинки выдерживли при 30°C в течение от 2 до 3 суток.
Всего собрали 24 трансформанта, которых выращивали на МС+5-FOA селекционных пластинках, их собирали и повторно наносили на свежеприготовленные МС+5-FOA пластинки. Данные клетки снимали с пластинок, и липиды экстрагировали. Жирнокислотные метиловые эфиры готовили путем трансэтерификации и впоследствии анализировали с помощью газового хроматографа Hewlett-Packard 6890 GC.
ГХ анализы выявили присутствие 10-15% ЭПК во всех трансформантах с pZKUE3S с пластинок. Штаммы, обозначеные как Y4128U1, Y4128U2, Y4128U3, Y4128U4, Y4128U5, и Y4128U6, вырабатывали 12,9%, 14,4%, 15,2%, 15,4%, 14%, и 10,9% ЭПК, соответственно (коллективно, Y4128U).
Расхождение в % ЭПК в Y4128 (37,6%) относительно Y4128U (среднее 13,8%) основано на разных условиях выращивания. Конкретно, предыдущая культура была проанализирована после 2 суток выращивания в жидкой культуре, тогда как последняя культура анализировалась после выращивания на агаровой пластинке. Заявители наблюдали 2-3 кратное возрастание % ЭПК, при сравнении результатов на агаровой пластинки с результатами в жидкой культуре. Таким образом, хотя данные результаты не являются прямо сопоставимыми, оба штамма, Y4128 и Y4128U, демонстрируют значительную выработку ЭПК.
Генерация штамма Y4217, производящего около 42% ЭПК от всех липидов: Конструкцию pZKL2-5U89GC (ФИГ.49A; SEQ ID NO:348) генерировали для интеграции одного дельта-9-элонгазного гена, одного дельта-8-десатуразного гена, одного дельта-5-десатуразного гена, и одного Yarrowia lipolytica диацилглицерин холинфосфотрансферазного гена (CPT1) в Lip2 локусы (номер доступа GenBank AJ012632) штамма Y4128U3, обеспечивая таким образом более высокие уровни производства ЭПК. Плазмида pZKL2-5U89GC одержала следующие компоненты:
Плазмиду pZKL2-5U89GC расщепляли AscI/SphI и затем использовали для трансформации штамма Y4128U3 в соответствии с разделом Общие методы. Трансформированные клетки помещали на МС пластинки, и эти пластинки выдерживали при 30°C в течение от 3 до 4 суток. Отдельные колонии повторно наносили на МС пластинки, и результирующие колонии потом использовались для инокуляции жидкой МС. Жидкие культуры встряхивали при 250 оборотов/минуту в течение 2 суток при 30°C. Клетки собирали путем центрифугирования, ресуспендировали в HGM, и потом встряхивали при 250 оборотов/минуту на протяжении 5 суток. Данные клетки собирали путем центрифугирования, и липиды экстрагировали. Жирнокислотные эфиры готовили путем трансэтерификации и затем анализировали с помощью газового хроматографа Hewlett-Packard 6890 GC.
ГХ анализы выявили, что большинство из выбранных 96 штаммов вырабатывали 32-39,9% ЭПК от всех липидов. Шесть штаммов, обозначенных как Y4215, Y4216, Y4217, Y4218, Y4219 и Y4220, вырабатывали около 41,1%, 41,8%, 41,7%, 41,1%, 41% и 41,1% ЭПК от всех липидов, соответственно. Конечный генотип каждого штамма, относительно Yarrowia lipolytica ATCC #20362 дикого типа, был: YALI0C18711g-, Pex10-, YALI0F24167g-, неизвестен 1-, неизвестен 3-, GPD::FmD12::Pex20, YAT1::FmD12::OCT, GPM/FBAIN::FmD12S::OCT, YAT1::ME3S::Pex16, EXP1::ME3S::Pex20, GPAT::EgD9e::Lip2, EXP1::EgD9eS::Lip1, FBAINm::EgD9eS::Lip2, FBA::EgD9eS::Pex20, GPD::EgD9eS::Lip2, FBAINm::EgD8M::Pex20, FBAIN::EgD8M::Lip1, EXP1::EgD8M::Pex16, GPDIN::EgD8M::Lip1, YAT1::EgD8M::Aco, FBAIN::EgD5::Aco, EXP1::EgD5S::Pex20, YAT1::EgD5S::Aco, YAT1::RD5S::OCT, YAT1::PaD17S::Lip1, EXP1::PaD17::Pex16, FBAINm::PaD17::Aco, YAT1::YlCPT1::ACO.
Генерация штамма Y4217U2 (Ura3-): Для разрушения Ura3 гена в штамме Y4217, конструкцию pZKUE3S (ФИГ.48B; SEQ ID NO:351) использовали для интеграции химерного EXP1::ME3S::Pex20 гена в Ura3 ген штамма Y4217. Следуя трансформации, клетки помещали на МС+5-FOA селекционные пластинки, и данные пластинки выдерживали при 30°C в течение от 3 до 4 суток.
Все из 6 трансформантов, выращенных на МС+5-FOA пластинках, собирали и повторно наносили штрихами на МС пластинки и МС+5-FOA пластинки. Все 6 штаммов имели Ura- фенотип (т.е. клетки могли расти на МС+5-FOA пластинках, но не на МС пластинках). Клетки соскребали с МС+5-FOA пластинок, и липиды экстрагировали. Жирнокислотные метиловые эфиры готовили путем трансэтерификации и впоследствии анализировали с помощью газового хроматографа Hewlett-Packard 6890 GC.
ГХ анализы выявили присутствие от 18,7% до 28,6% ЭПК во всех трансформантах с pZKUE3S, выращенных на МС+5-FOA пластинках. Два штамма, обозначенных как Y4217U1 и Y4217U2, вырабатывали 22,5% и 28,6% ЭПК, соответственно.
Генерация штамма Y4259, производящего около 46,5% ЭПК от всех липидов: Конструкцию pZKL1-2SP98C (ФИГ.49B; SEQ ID NO:352) генерировали для интеграции одного дельта-9-элонгазного гена, одного дельта-8-десатуразного гена, одного дельта-12-десатуразного гена, и одного Yarrowia lipolytica диацилглицерин холинфосфотрансферазного гена (CPT1) в Lip1 локусы (номер доступа GenBank Z50020) штамма Y4217U2, обеспечивая таким образом более высокие уровни выработки ЭПК. Плазмида pZKL1-2SP98C содержала следующие компоненты:
Плазмиду pZKL1-2SP98C расщепляли AscI/SphI и потом использовали для трансформации штамма Y4217U2 в соответствии с разделом Общие методы. Трансформированные клетки помещали на МС пластинки, и данные пластинки выдерживали при 30°C в течение от 3 до 4 суток. Отдельные колонии повторно наносили на МС пластинки, и результирующие колонии потом использовали для инокуляции жидкой МС. Жидкие культуры потом встряхивали при 250 оборотов/минуту на протяжении 2 суток при 30°C. Клетки собирали путем центрифугирования, ресуспендировали в HGM и потом встряхивали при 250 оборотов/минуту в течение 5 суток. Клетки собирали путем центрифугирования, и липиды экстрагировали. Жирнокислотные метиловые эфиры готовили путем транс-этерификации и впоследствии анализировали с помощью газового хроматографа Hewlett-Packard 6890 GC.
ГХ анализы показали, что большинство из выбранных 72 штаммов вырабатывали 40-44% ЭПК от всех липидов. Шесть штаммов, обозначенных как Y4259, Y4260, Y4261, Y4262, Y4263, и Y4264, производили около 46,5%, 44,5%, 44,5%, 44,8%, 44,5%, и 44,3% ЭПК от всех липидов, соответственно.
Конечный генотип штамма Y4259 относительно Yarrowia lipolytica ATCC #20362 дикого типа был: YALI0C18711g-, Pex10-, YALI0F24167g-, неизвестен 1-, неизвестен 3-, неизвестен 8-, GPD::FmD12::Pex20, YAT1::FmD12::OCT, GPM/FBAIN::FmD12S::OCT, EXP1::FmD12S::Aco, YAT1::ME3S::Pex16, EXP1::ME3S::Pex20 (2 копии), GPAT::EgD9e::Lip2, EXP1::EgD9eS::Lip1, FBAINm::EgD9eS::Lip2, FBA::EgD9eS::Pex20, GPD::EgD9eS::Lip2, YAT1::EgD9eS::Lip2, FBAINm::EgD8M::Pex20, FBAIN::EgD8M::Lip1 (2 копии), EXP1::EgD8M::Pex16, GPDIN::EgD8M::Lip1, YAT1::EgD8M::Aco, FBAIN::EgD5::Aco, EXP1::EgD5S::Pex20, YAT1::EgD5S::Aco, YAT1::RD5S::OCT, YAT1::PaD17S::Lip1, EXP1::PaD17::Pex16, FBAINm::PaD17::Aco, YAT1::YlCPT1::ACO, GPD::YlCPT1::ACO.
Генерация штамма Y4259U2 (Ura3-): Для разрушения Ura3 гена в штамме Y4259, конструкцию pZKUM (ФИГ.50A; SEQ ID NO:353) использовали для интеграции Ura3 мутантного гена в Ura3 ген штамма Y4259. Плазмида pZKUM содержала следующие компоненты:
Всего 3 трансформанта, выращенных на МС+5-FOA пластинках, собирали и повторно наносили на МС пластинки и МС+5-FOA пластинки. Все 3 штамма имели Ura- фенотип (т.е. клетки могли расти на МС+5-FOA пластинках, но не на МС пластинках). Клетки соскребали с МС+5-FOA пластинок, и липиды экстрагировали. Жирнокислотные метиловые эфиры готовили путем трансэтерификации и впоследствии анализировали с помощью газового хроматографа Hewlett-Packard 6890 GC.
ГХ анализы выявили присутствие 31,4%, 31% и 31,3% ЭПК в № 1, № 2 и № 3 трансформантах с pZKUM, выращенных на МС+5-FOA пластинках. Эти три штамма были обозначены как Y4259U1, Y4259U2 и Y4259U3, соответственно (коллективно, Y4259U).
Генерация штамма Y4305, производящего около 53% ЭПК от всех липидов: Конструкцию pZKD2-5U89A2 (ФИГ.50B; SEQ ID NO:355) генерировали для интеграции одного дельта-9-элонгазного гена, одного дельта-5-десатуразного гена, одного дельта-8-десатуразного гена, и одного дельта-12-десатуразного гена в диацилглицерил ацилтрансферазные (DGAT2) локусы штамма Y4259U2, обеспечивая таким образом более высокие уровни выработки ЭПК. Плазмида pZKD2-5U89A2 содержала следующие компоненты:
Плазмиду pZKD2-5U89A2 расщепляли AscI/SphI и потом использовали для трансформации штамма Y4259U2 в соответствии с разделом Общие методы. Трансформированные клетки поимещались на МС пластинки, и данные пластинки выдерживали при 30°C в течение от 3 до 4 суток. Отдельные колонии повторно наносили на МС пластинки, и результирующие колонии использовали для инокуляции жидкой МС. Жидкие культуры встряхивали при 250 оборотов/минуту в течение 2 суток при 30°C. Клетки собирали путем центрифугирования, ресуспендировали в HGM, и потом встряхивали при 250 оборотов/минуту в течении 5 суток. Данные клетки собирали путем центрифугирования, и липиды экстрагировали. Жирнокислотные метиловые эфиры готовили путем трансэтерификации и впоследствии анализировали с помощью газового хроматографа Hewlett-Packard 6890 GC.
ГХ анализы показали, что большинство из выбранных 96 штаммов вырабатывали 40-46% ЭПК от всех липидов. Четыре штамма, обозначенные как Y4305, Y4306, Y4307 и Y4308, вырабатывали около 53,2%, 46,4%, 46,8%, и 47,8% ЭПК от всех липидов, соответственно. Полный липидный профиль Y4305 следующий: 16:0 (2,8%), 16:1 (0,7%), 18:0 (1,3%), 18:1 (4,9%), 18:2 (17,6%), АЛК (2,3%), ЭДК (3,4%), ДГЛК (2,0%), АРК (0,6%), ЭТК (1,7%), и ЭПК (53,2%). Полный липидный % от сухого веса клетки (dcw) составлял 27,5.
Конечный генотип штамма Y4305 относительно Yarrowia lipolytica ATCC #20362 дикого типа был SCP2- (YALI0E01298g), YALI0C18711g-, Pex10-, YALI0F24167g-, неизвестен 1-, неизвестен 3-, неизвестен 8-, GPD::FmD12::Pex20, YAT1::FmD12::OCT, GPM/FBAIN::FmD12S::OCT, EXP1::FmD12S::Aco, YAT1::FmD12S::Lip2, YAT1::ME3S::Pex16, EXP1::ME3S::Pex20 (3 копии), GPAT::EgD9e::Lip2, EXP1::EgD9eS::Lip1, FBAINm::EgD9eS::Lip2, FBA::EgD9eS::Pex20, GPD::EgD9eS::Lip2, YAT1::EgD9eS::Lip2, YAT1::E389D9eS::OCT, FBAINm::EgD8M::Pex20, FBAIN::EgD8M::Lip1 (2 копии), EXP1::EgD8M::Pex16, GPDIN::EgD8M::Lip1, YAT1::EgD8M::Aco, FBAIN::EgD5::Aco, EXP1::EgD5S::Pex20, YAT1::EgD5S::Aco, EXP1::EgD5S::ACO, YAT1::RD5S::OCT, YAT1::PaD17S::Lip1, EXP1::PaD17::Pex16, FBAINm::PaD17::Aco, YAT1::YlCPT1::ACO, GPD::YlCPT1::ACO.
Генерация штамма Y4305U3 (Ura3-): Для разрушения Ura3 гена в штамме Y4305, конструкцию pZKUM (ФИГ.50A; SEQ ID NO:353) использовали для интеграции Ura3 мутантного гена в Ura3 ген штамма Y4305. Всего 8 трансформантов, выращенных на МС+5-FOA пластинках, повторно наносили на МС пластинки и МС+5-FOA пластинки, раздельно. Все 8 штаммов имели Ura- фенотип (т.е. клетки могли расти на МС+5-FOA пластинках, но не на МС пластинках). Клетки соскребали с МС+5-FOA пластинок, и липиды экстрагировали. Жирнокислотные метиловые эфиры готовили путем трансэтерификации и впоследствии анализировали с помощью газового хроматографа Hewlett-Packard 6890 GC.
ГХ анализы выявили присутствие 37,6%, 37,3% и 36,5% ЭПК в pZKUM трансформантах №1, №6 и №7, выращенных на МС+5-FOA пластинках. Эти три штамма были обозначены как штаммы Y4305U1, Y4305U2 и Y4305U3, соответственоо (коллективно, Y4305U).
Конструирование Yarrowia lipolytica штамма Y4184U
Y. lipolytica штамм Y4184U был использован как хозяин в Примерах 32, 33, 34 и 51, infra. Штамм Y4184U был получен из Y. lipolytica ATCC #20362, и он способен вырабатывать около 31% ЭПК относительгно полного количества липидов посредством экспрессии дельта-9-элонгаза/дельта-8-десатуразного пути.
Разработка штамма Y4184U потребовала конструирования штамма Y2224, штамма Y4001, штамма Y4001U, штамма Y4036, штамма Y4036U и штамма Y4069 (supra). Дальнейшая разработка штамма Y4184U (см. схему на ФИГ.51A) потребовала конструирования штамма Y4084 (вырабатывающего 14% ЭПК), штамма Y4084U1 (Ura-), штамма Y4127 (вырабатывающего 18% ЭПК), штамма Y4127U2 (Ura-), штамма Y4158 (вырабатывающего 25% ЭПК), штамма Y4158U1 (Ura-), и штамма 4184 (вырабатывающего 30,7% ЭПК). Хотя детали, касающиеся трансформации и селекции ЭПК-производящих штаммов разрабатывались позднее, и штамм Y4069 здесь не конструировался, методология, использованная для изоляции штамма Y4084, штамма Y4084U1, штамма Y4127, штамма Y4127U2, штамма Y4158, штамма Y4158U1, штамма Y4184, и штамма Y4184U, отвечала описанной при конструировании штамма Y4305, supra.
Вкратце, конструкцию pZP3-Pa777U (ФИГ.47A; SEQ ID NO:338) использовали для интеграции трех дельта-17-десатуразных генов в Pox3 локусы (номер доступа GenBank AJ001301) штамма Y4069, в результате чего произошла изоляция штамма Y4084 (вырабатывающего 14% ЭПК). Штамм Y4084U1 был создан посредством временной экспрессии Cre рекомбиназного фермента в конструкцию pY117 (ФИГ.47B; SEQ ID NO:343) внутри штамма Y4084 с образованием Ura- фенотипа. Конструкцию pZP2-2988 (ФИГ.48A; SEQ ID NO:345) затем использовали для интеграции одного дельта-12-десатуразного гена, двух дельта-8-десатуразных генов, и одного дельта-9-элонгазного гена в Pox2 локусы (номер доступа GenBank AJ001300) штамма Y4084U1, что привело к изоляции штамма Y4127 (вырабатывающего 18% ЭПК). Yarrowia lipolytica штамм Y4127 был депонирован в Американское собрание типовых культур 29 ноября 2007 года и носит обозначение ATCC PTA-8802.
Штамм Y4127U2 был создан путем разрыва Ura3 гена в штамме Y4127 посредством конструкции pZKUE3S (ФИГ.48B; SEQ ID NO:351), включающей химерный EXP1::ME3S::Pex20 ген, нацеленный на Ura3 ген. Конструкцию pZKL1-2SP98C (ФИГ.49B; SEQ ID NO:352) использовали для интеграции одного дельта-9-элонгазного гена, одного дельта-8-десатуразного гена, одного дельта-12-десатуразного гена, и одного Yarrowia lipolytica диацилглицерин холинфосфотрансферазный гена (CPT1) в Lip1 локусы (номер доступа GenBank Z50020) штамма Y4127U2, что привело к изоляции штамма Y4158 (вырабатывающего 25% ЭПК). Потом была создана Ura- производная (т.е. штамм Y4158U1), посредством трансформации конструкцией pZKUE3S (ФИГ.48B; SEQ ID NO:351), включающей химерный EXP1::ME3S::Pex20 ген, нацеленный на Ura3 ген. Наконец, конструкция pZKL2-5U89GC (ФИГ.49A; SEQ ID NO:348) была использована для интеграции одного дельта-9-элонгазного гена, одного дельта-8-десатуразного гена, одного дельта-5-десатуразного гена, и одного Yarrowia lipolytica CPT1 в Lip2 локусы (номер доступа GenBank AJ012632) Y4158U1, что привело к изоляции штамма Y4184.
Полный липидный профиль штамма Y4184 следующий: 16:0 (3,1%), 16:1 (1,5%), 18:0 (1,8%), 18:1 (8,7%), 18:2 (31,5%), АЛК (4,9%), ЭДК (5,6%), ДГЛК (2,9%), АРК (0,6%), ЭТК (2,4%), и ЭПК (28,9%). Полный липидный % от веса сухой клетки (dcw) составлял 23,9.
Конечный генотип штамма Y4184 относительно Yarrowia lipolytica ATCC #20362 дикого типа был неизвестен 1-, неизвестен 2-, неизвестен 4-, неизвестен 5-, неизвестен 6-, неизвестен 7-, YAT1::ME3S::Pex16, EXP1::ME3S::Pex20 (2 копии), GPAT::EgD9e::Lip2, FBAINm::EgD9eS::Lip2, EXP1::EgD9eS::Lip1, FBA::EgD9eS::Pex20, YAT1::EgD9eS::Lip2, GPD::EgD9eS::Lip2, GPDIN::EgD8M::Lip1, YAT1::EgD8M::Aco, EXP1::EgD8M::Pex16, FBAINm::EgD8M::Pex20, FBAIN::EgD8M::Lip1 (2 копии), GPM/FBAIN::FmD12S::Oct, EXP1::FmD12S::Aco, YAT1::FmD12::Oct, GPD::FmD12::Pex20, EXP1::EgD5S::Pex20, YAT1::EgD5S::Aco, YAT1::Rd5S::Oct, FBAIN::EgD5::Aco, FBAINm::PaD17::Aco, EXP1::PaD17::Pex16, YAT1::PaD17S::Lip1, YAT1::YlCPT1::Aco, GPD::YlCPT1::Aco.
Для разрыва Ura3 гена в штамме Y4184 конструкцию pZKUM (ФИГ.50A; SEQ ID NO:353) использовали с целью интеграции Ura3 мутантного гена в Ura3 ген штамма Y4184.
Всего 11 трансформантов, выращенных на МС+5-FOA пластинках, собирали и повторно наносили на МС пластинки и МС+5-FOA пластинки, раздельно. Все 11 штаммов имели Ura- фенотип (т.е. клетки могли расти на МС+5-FOA пластинках, но не на МС пластинках). Данные клетки соскребали с МС+5-FOA пластинок; липиды экстрагировали; и жирнокислотные метиловые эфиры готовили путем трансэтерификации и впоследствии анализировали с помощью газового хроматографа Hewlett-Packard 6890 GC.
ГХ анализы выявили присутствие 11,2%, 10,6%, и 15,5% ЭПК в № 7, № 8 и № 10 трансформантах с pZKUM, выращенных на МС+5-FOA пластинках. Эти три штамма были обозначены как штаммы Y4184U1, Y4184U2 и Y4184U4, соответственно (коллективно, Y4184U).
ПРИМЕР 1
Условия выращивания Euglena gracilis, липидный профиль и выделение мРНК
Euglena gracilis получали из лаборатории Dr.Richard Triemer at Michigan State University (East Lansing, MI). Из 10 мл активно растущей культуры 1 мл аликвотного количества переносили в 250 мл Euglena gracilis (Eg) среды в 500 мл стеклянную бутылку. Eg среду готовили путем комбинирования 1 г ацетата натрия, 1 г мясного бульона (U126-01, Difco Laboratories, Detroit, MI), 2 г Bacto® триптона (0123-17-3, Difco Laboratories) и 2 г Bacto® дрожжевого экстракта (0127-17-9, Difco Laboratories) в 970 мл воды. После фильтрационной стерилизации 30 мл почвенно-водного супернатанта (15-3790, Carolina Biological Supply Company, Burlington, NC) добавляли асептическим образом с получением конечной Eg среды. Культуры Euglena gracilis выращивали при 23°С c 16 часовым световым, 8 часовым темновым циклом на протяжении 2 недель без перемешивания.
Через 2 недели 10 мл культуры извлекали для липидного анализа и центрифугировали при 1800xg в течение 5 минут. Таблетку промывали один раз водой и повторно центрифугировали. Результирующую таблетку высушивали в течение 5 минут под вакуумом, ресуспендировали в 100 мкл триметилсульфоний гидроксида (ТМСГ) и инкубировали при комнатной температуре на протяжении 15 минут при встряхивании. После этого добавляли 0,5 мл гексана, и бутылочки инкубировали в течение 15 минут при комнатной температуре при встряхивании. Жирнокислотные метиловые эфиры (5 мкл, выделенных из гексанового слоя) отделяли и анализировали количественно с использованием газового хроматографа марки Hewlett-Packard 6890 Gas Chromatograph, снабженного капиллярной колонкой из плавленного кварца Omegawax 320 (корпорация Supelco Inc., Cat. No. 24152). Температура печи была запрограммирована на выдержку при 220°С в течение 2,7 минут, повышение до 240°С при 20°С/минуту и последующую выдержку еще на протяжении 2,3 минут. Газ-носитель подавался с помощью водородного генератора Whatman. Время удерживания было сравнимо с таковым для метиловых эфиров стандартов, которые имеются в продаже (Nu-Chek Prep, Inc. Cat. No. U-99-A), и результирующая хроматограмма показана на ФИГ.27.
Оставшуюся двухнедельную культуру (240 мл) таблетировали путем центрифугирования при 1800xg на протяжении 10 минут, промывали один раз водой и подвергали повторному центрифугированию. Полную РНК экстрагировали из результирующей таблетки с использованием RNA STAT-60™ реагента (TEL-TEST, Inc., Friendswood, TX) и в соответствии с представленным протоколом производителя (использовать 5 мл реагента, растворенную РНК в 0,5 мл воды). Таким способом из таблетки получали 1 мг полной РНК (2 мг/л). мРНК выделяли из 1 мг полной РНК с использованием набора mRNA Purification Kit (Amersham Biosciences, Piscataway, NJ) в соответствии с протоколом производителя. Таким способом получили 85 мкг мРНК.
ПРИМЕР 2
Синтез Euglena gracilis кДНК, конструирование библиотеки и секвенирование
кДНК библиотеку генерировали с использованием набора Cloneminer™ cDNA Library Construction Kit (Cat. No. 18249-029, Invitrogen Corporation, Carlsbad, CA) и согласно представленного протокола производителя (Version B, 25-0508). С использованием метода не-радиомечения кДНК была синтезирована из 3,2 мкг мРНК (описано выше) с использованием Biotin -attB2-Oligo(dT) праймера. После синтеза первой и второй цепи добавляли и лигировали адаптер attB1, и кДНК фракционировали по размерам с использованием колонковой хроматографии. ДНК из фракций 7 и 8 (размеры варьировали от приблизительно 800 до 1500 bp) концентрировали, рекомбинировали в pDONR™222 и трансформировали в клетки E. coli ElectroMAX™ DH10B™ T1 Phage-Resistant (Invitrogen Corporation). Библиотека Euglena gracilis получила название eeg1c.
Для секвенирования клоны сначала восстанавливали из архивных глицериновых культур, выращенных/замороженных в 384-ячеечных планшетах с морозильной средой. С использованием автоматического QPix® сборщика колоний (Genetix) клетки собирали и потом использовали для инокуляции 96-ячеечных планшетов с глубокими ячейками, которые содержали LB+50 мкг/мл канамицина. После выращивания в течение 20 часов при 37°С клетки таблетировали путем цетрифугирования и сохраняли при температуре -20°С. Потом плазмиды выделяли на автомате Eppendorf 5Prime с использованием модифицированного “минипреп” (miniprep) метода щелочного лизиса в 96-ячеечном формате (Eppendorf PerfectPrep®). В кратком изложении, для облегчения удаления клеточных остатков после осаждения ацетатом применяли фильтр и вакуумный коллектор. Плазмидную ДНК потом связывали на втором фильтровальном планшете прямо из фильтрата, промывали, высушивали и элюировали.
Плазмиды подвергали конечному секвенированию в 384-ячеечных планшетах с использованием затравленного вектором M13F Universal праймера (SEQ ID NO: 1) и набора для секвенирования ABI BigDye version 3 Prism. Для реакции секвенирования использовали 100-200 нг матрицы и 6,4 пмоль праймера, и следующие условия реакции повторялись 25 раз: 96°С на протяжении 10 с, 50°С на протяжении 5 с и 60°С на протяжении 4 минут. После этанольной очистки продукты цикличной реакции секвенирования растворяли и анализировали на автоматических секвенаторах Perkin-Elmer ABI 3700.
ПРИМЕР 3
Идентификация C20-ПНЖК элонгирующих ферментных гомологов из Euglena gracilis кДНК библиотеки eeg1c
кДНК клоны, кодирующие С20-ПНЖК элонгирующие ферментные гомологи (т.е. “С20-ПНЖК Elo”), идентифицировали путем проведения поисков с помощью BLAST (Basic Local Alignment Search Tool; Altschul et al., J. Mol. Biol. 215:403-410 (1993)) на подобие относительно последовательностей, которые содержатся в базе данных BLAST “nr” (которая включает все не-избыточные GenBank CDS трансляции, последовательности, полученные из 3-мерной структуры Brookhaven Protein Data Bank, последний основной выпуск базы данных протеиновых последовательностей SWISS-PROT, базы данных EMBL и DDBJ). кДНК последовательности, полученные в Примере 2, проанализировали на подобие относительно всех доступных ДНК последовательностей, которые содержатся в “nr” базе данных, с использованием алгоритма BLASTN, представленного Национальным центром биотехнологической информации (НЦБИ). ДНК последовательности транслировали во всех рамках считывания и сравнивали на совпадение со всеми доступными протеиновыми последовательностями, которые содержатся в “nr” базе данных, с использованием алгоритма BLASTX (Gish и States, Nat. Genet. 3:266-272 (1993)), представленного НЦБИ. Для удобства, Р-значения (вероятность) наблюдения просто случайного совпадения кДНК последовательности с последовательностью, которая содержится в поисковых базах данных, определяется с помощью BLAST, здесь приводятся как “pLog” значения, которые представляют собой отрицательные значения логарифма приведенного Р-значения. Соответственно, чем больше значение pLog, тем больше вероятность того, что кДНК последовательность и “попадание” BLAST представляют гомологичные протеины.
BLASTX поиск с использованием нуклеотидной последовательности из клона eeg1c.pk005.p14.f выявил подобие протеина, кодированного кДНК, с C20-PUFA Elo из Pavlova sp. CCMP459 (SEQ ID NO:2) (номер доступа НЦБИ AAV33630 (GI 54307108), локус AAV33630, CDS AY630573; Pereira et al., Biochem. J. 384:357-366 (2004)). Последовательность участка кДНК инсерта из клона eeg1c.pk005.p14.f показана в SEQ ID NO:3 (5′ конец кДНК инсерта). Потом была получена полная последовательность инсерта (т.е. eeg1c.pk005.p14.f), и она показана в SEQ ID NO:4. Последовательность для кодирующей последовательности (CDS) приведена в SEQ ID NO:5. Последовательность для соответствующей выведенной аминокислотной последовательности показана в SEQ ID NO:6.
Секвенирование всего инсерта (FIS) проводили с использованием модифицированного транспозиционного протокола. Клоны, идентифицированные для FIS, восстанавливали из архивных глицериновых запасов как отдельные колонии, и плазмидная ДНК была изолирована путем щелочного лизиса. Плазмидные матрицы подвергали транспозиции через Template Generation System (TGS II) транспозиционный набор (Finnzymes Oy, Espoo, Finland) согласно протоколу изготовителя. Подвергнутые транспозиции ДНК трансформировали в ЕН10В электро-компетентные клетки (Edge BioSystems, Gaithersburg, MD) путем электропорации. Множественные трансформаниты произвольно выбирали из каждой транспозиционной реакции, была создана плазмидная ДНК, и матрицы подвергали секвенированию как и выше (ABI BigDye v3.1) наружу от сайта транспозиционного события, с использованием уникальных праймеров SeqE (SEQ ID NO:7) и SeqW (SEQ ID NO:8).
Данные по последовательностям собирали (ABI Prism Collections software (программа)) и составляли с использованием программы составления последовательностей Phrap (P. Green, University of Washington, Seattle). Результаты рассматриваются Consed sequence editor (D. Gordon, University of Washington, Seattle) для окончательного издания.
Аминокислотная последовательность, определенная в SEQ ID NO:6, оценивалась BLASTP и дала значение pLog 61,22 (значение Е составляет 6е-62) относительно Pavlova sp. CCMP459 C20-PUFA Elo (SEQ ID NO:2). Аминокислотная последовательность, определенная в SEQ ID NO:6, на 45,1% идентична с Pavlova sp. CCMP459 C20-PUFA Elo последовательностью (SEQ ID NO:2), с использованием метода Jotun Hein. Вычисления процента идентичности последовательностей, проведенные с помощью метода Jotun Hein (Hein, J. J., Meth. Enz. 183:626-645 (1990)), были проведены с использованием программы MegAlign™ v6.1 вычислительного биоинформационного пакета LASERGENE (DNASTAR Inc., Madison, WI) с параметрами по умолчанию для парной группировки (KTUPLE=2). Аминокислотная последовательность, представленная в SEQ ID NO:6, на 40,4% идентична с Pavlova sp. CCMP459 C20-PUFA Elo последовательностью (SEQ ID NO:2), с использованием метода Clustal V. Вычисления процента идентичности последовательности, которые проводились по методу Clustal V (Higgins, D.G. та Sharp, P.M., Comput. Appl. Biosci. 5:151-153 (1989); Higgins et al., Comput. Appl. Biosci. 8:189-191 (1992)), осуществлялись с использованием программы MegAlign™ v6.1 вычислительного биоинформационного пакета LASARGENE (supra) с параметрами по умолчанию для парной группировки (KTUPLE=1, GAP PENALTY=3, WINDOW=5 и DIAGONALS SAVED=5 и GAP LENGTH PENALTY=10). BLAST оценки и вероятности указывают на то, что фрагмент данной нуклеиновой кислоты (SEQ ID NO:5) кодирует весь Euglena gracilis C20-PUFA Elo ген, обозначенный здесь как EgC20elo1.
ФИГ.25 резюмирует BLAST данные и данные по процентам идентичности для EgC20elo1 (Пример 3), EgDHAsyn1 (Пример 4, infra), и EgDHAsyn2 (Пример 5, infra).
ПРИМЕР 4
Идентификация ДГК синтазы 1 (EgDHAsyn1) из Euglena gracilis кДНК библиотеки eeg1c
КДНК клоны, кодирующие дополнительные C20-ПНЖК Elo гомологи, идентифицировали путем проведения BLAST поисков в отношении подобия последовательностям, которые содержатся в базе данных BLAST “nr” как описано в Примере 3.
BLASTX поиск с использованием нуклеотидных последовательностей из клона eeg1c.pk016.e6.f (также называемого pKR1049) выявил подобие протеина, кодированного кДНК, с C20-PUFA Elo из Pavlova sp. CCMP459 (SEQ ID NO:2) (номер доступа НЦБИ AAV33630 (GI 54307108), локус AAV33630, CDS AY630573; Pereira et al., Biochem. J. 384:357-366 (2004)). Последовательность части кДНК инсерта из клона eeg1c.pk016.e6.f показана в SEQ ID NO:9 (5′ конец кДНК инсерта). Затем была получена последовательность всего инсерта (eeg1c.pk016.e6.f:fis) как описано в Примере 3 и показано в SEQ ID NO:10. Кодирующая последовательность приведена в SEQ ID NO:11; соответствующая выведенная аминокислотная последовательность представлена в SEQ ID NO:12.
Аминокислотная последовательность, представленная в SEQ ID NO:12, оценивалась BLASTP, как описано в Примере 3. Интересно отметить, что SEQ ID NO:12, как было найдено, была схожа как с C20-PUFA Elo, так и с дельта-4-десатуразой жирных кислот. N-окончание SEQ ID NO:12 (приблизительно от аминокислот 16-268) дает значение pLog 60,30 (E значение 5e-61; 124/258 идентичные аминокислоты; 48% идентичность), относительно Pavlova sp. CCMP459 C20-PUFA Elo (SEQ ID NO:2). C-окончание SEQ ID NO:12 (приблизительно от аминокислот 253-793) дает E значение 0,0 (535/541 идентичные аминокислоты; 98% идентичность), относительно дельта-4-десатуразы жирных кислот из Euglena gracilis (SEQ ID NO:13) (номер доступа НЦБИ AAQ19605 (GI 33466346), локус AAQ19605, CDS AY278558; Meyer et al., Biochemistry 42(32):9779-9788 (2003)). Данные и вероятности BLAST указывают на то, что текущий фрагмент нуклеиновой кислоты (SEQ ID NO:11) кодирует весь Euglena gracilis C20-PUFA Elo/delta-4-десатуразы жирных кислот слитый ген, названный здесь Euglena gracilis ДГК синтаза 1 (EgDHAsyn1).
Аминокислотная последовательность EgDHAsyn1 (SEQ ID NO:12) на 47,8% идентична C20-PUFA Elo из Pavlova sp. CCMP459 (SEQ ID NO:2) и на 98,9% идентична дельта-4-десатуразы жирных кислот из Euglena gracilis (SEQ ID NO:13), при использовании метода Jotun Hein, как описано в Примере 3. Аминоксилотная последовательность EgDHAsyn1 (SEQ ID NO:12) на 41,2% идентична C20-PUFA Elo из Pavlova sp. CCMP459 (SEQ ID NO:2) и на 98,9% идентична дельта-4-десатуразы жирных кислот из Euglena gracilis (SEQ ID NO:13), с использованием метода Clustal V, как описано в Примере 3.
ФИГ.27 резюмирует данные BLASTP и значения процентов идентичности для EgDHAsyn1 (Пример 4), EgC20elo1 (Пример 3, supra) и EgDHAsyn2 (Пример 5, infra).
ПРИМЕР 5
Идентификация ДГК синтазы 2 (EgDHAsyn2) из Euglena gracilis кДНК библиотеки eeg1c
Приблизительно 17000 клонов Euglena gracilis кДНК библиотеки eeg1c высевали на три больших квадратных (24 см×24 см) чашки Петри (Corning, Corning, NY), каждая из которых содержала LB+50 мкг/мл канамицин агаровой среды. Клетки выращивали в течение ночи при 37°C, и потом чашки охлаждали до комнатной температуры.
Лифтинг колоний:
Биодин B 0,45 мкм мембрану (Cat. No. 60207, Pall Corporation, Pensacola, FL) обрезали до приблизительных размеров 22 см×22 см и потом ее осторожно помещали на верхнюю часть агара для удаления пузырьков воздуха. После инкубирования в течение 2 минут при комнатной температуре мембрану пометили для ориентации, подняли с помощью пинцета и поместили верхней стороной колоний на фильтровальную бумагу, пропитанную 0,5 M гидроксида натрия и 1,5 M хлорида натрия. После денатурации в течение 4 минут гидроксид натрия нейтрализовали путем помещения мембраны на фильтровальную бумагу, пропитанную 0,5 M Tris-HCL (pH 7.5) и 1,5 M хлорида натрия, на 4 минуты. Эту стадию повторяли, и мембрану в течение непродолжительного времени ополаскивали в 2X SSC буфере (20X SSC представляет собой 3M хлорид натрия, 0,3 M цитрат натрия; pH 7,0) и высушивали на воздухе на фильтровальной бумаге.
Гибридизация:
Мембраны подвергали предварительной гибридизации при 65°C в 200 мл гибридизационного раствора в течение 2 часов. Гибридизационный раствор содержал 6X SSPE (20X SSPE представляет собой 3 M хлорид натрия, 0,2 M фосфат натрия, 20 мМ ЭДТА; pH 7,4), 5X реактив Денгардта (Denhardt) (100X реактив Денгардта представляет собой 2% (вес/объем) фикола, 2% (вес/объем) поливинилпирролидона, 2% (вес/объем) ацетилированного альбумина бычьей сыворотки), 0,5% натрия додецила сульфата (НДС), 100 мкг/мл ДНК срезанных молок лосося и 5% декстрана сульфата.
ДНК зонд сделали с использованием NcoI/NotI ДНК фрагмента, очищенного агарозным гелем, который содержал EgDHAsyn1*, из pY141 (см. Пример 10), меченый P32 dCTP с использованием системы для радиоактивного мечения RadPrime DNA Labeling System (Cat. No. 18428-011, Invitrogen, Carlsbad, CA), следуя инструкциям производителя. Невнедренный P32 dCTP отделяли с использованием колонки NICK (Cat. No. 17-0855-02, Amersham Biosciences, Piscataway, NJ) в соответствии с инструкциями производителя. Данный зонд денатурировали в течение 5 минут при 100°C, помещали на лед на 3 минуты, и его половину добавляли к гибридизационному раствору.
Указанную мембрану подвергали гибридизации с зондом в течение ночи при 65°C при легком встряхивании и на следующий день промывали два раза 2X SSC, который содержал 0,5% SDS (каждый раз по 5 минут), и два раза 0,2X SSC, который содержал 0,1% SDS (каждый раз по15 минут). После промывания гиперпленку (Cat. No. RPN30K, Amersham Biosciences, Piscataway, NJ) экспонировали на мембране на протяжении ночи при -80°C.
Исходя из расположения пластинок с экспонированной гиперпленкой, позитивные колонии собирали с использованием затупленного конца пипетки Пастера в 1 мл воды и встряхивали. Было сделано несколько разбавлений, и их высевали на небольшие круглые чашки Петри (82 мм), содержавшие LB среду плюс 50 мкг/мл канамицина, с получением около 100 хорошо изолированных колоний на отдельной чашке. Лифтинги были сделаны как описано выше, за исключением использования мембранных кружочков NytranN (Cat, No. 10416116, Schleicher & Schuell, Keene, NH), и гибридизация проводилась в 100 мл с использованием меченого радиоактивным изотопом зонда, который остался. Таким способом был идентифицирован один позитивный клон (обозначенный как eeg 1c-1). На плазмиду из eeg 1c-1 можно также ссылаться как на pLF116.
Индивидуальные позитивные клоны выращивали при 37°C в LB+50 мкг/мл канамициновой жидкостной среде, и плазмиду подвергали очистке с использованием набора QIAprep® Spin Miniprep Kit (Qiagen Inc.) согласно протоколу производителя. Плазмидный инсерт секвенировали как описано в Примере 2, с помощью набора для секвенирования ABI BigDye version 3 Prism с использованием затравленного вектором M13F Universal праймера (SEQ ID NO:1), затравленного вектором M13rev праймера (SEQ ID NO:14), и затравленного поли(A) хвостом праймера WobbleT олигонуклеотидов. Вкратце, WobbleT праймер является эквимолярной смесью 21mer поли(T)A, поли(T)C, и поли(T)G, использованной для секвенирования 3′ конца кДНК клонов. На основе первичных данных по секвенированию подобным образом была получена дополнительная последовательность внутреннего фрагмента с использованием олигонуклеотидов oEUGel4-1 (SEQ ID NO:15), EgEloD4Mut-5 (SEQ ID NO:16), oEUGel4-2 (SEQ ID NO:17), EgDHAsyn5' (SEQ ID NO:18), и EgDHAsyn3′ (SEQ ID NO:19). Таким способом была получена последовательность полного инсерта eeg1c-1, и она показана в SEQ ID NO:20. Кодирующая последовательность представлена в SEQ ID NO:21, и соответствующая выведенная аминокислотная последовательность представлена в SEQ ID NO:22.
Аминокислотную последовательность, представленную в SEQ ID NO:22, оценивали с помощью BLASTP, как описано в Примере 3. Как и в случае для EgDHAsyn1, SEQ ID NO:22, как было установлено, схожа как с C20-PUFA Elo, так и с дельта-4-десатуразы жирных кислот. N-окончание SEQ ID NO:22 (приблизительно от аминокислот 41-268) дает значение pLog 61,0 (E значение 1e-61; 118/231 идентичные аминокислоты; 51% идентичности) относительно Pavlova sp. CCMP459 C20-PUFA Elo (SEQ ID NO:2). C-окончание SEQ ID NO:22 (приблизительно от аминокислот 253-793) дает E значение 0,0 (541/541 идентичные аминокислоты; 100% идентичность), относительно аминокислотной последовательности дельта-4-десатуразы жирных кислот из Euglena gracilis (SEQ ID NO:13). BLAST данные и вероятности указывают, что текущий нуклеиновокислотный фрагмент (SEQ ID NO:21) кодирует весь Euglena gracilis C20-ПНЖК Elo/дельта-4-десатуразы жирных кислот слитый ген, названный здесь Euglena gracilis ДГК синтаза 2 (EgDHAsyn2).
Аминокислотная последовательность EgDHAsyn2 (SEQ ID NO:22) на 48,2% идентична C20-PUFA Elo из Pavlova sp. CCMP459 (SEQ ID NO:2) и на 100% идентична дельта-4-десатуразы жирных кислот из Euglena gracilis (SEQ ID NO:13), при использовании метода Jotun Hein, как описано в Примере 3. Аминокислотная последовательность EgDHAsyn2 (SEQ ID NO:22) на 41,2% идентична C20-PUFA Elo из Pavlova sp. CCMP459 (SEQ ID NO:2) и на 100% идентична дельта-4-десатуразы жирных кислот из Euglena gracilis (SEQ ID NO:13), при использовании метода Clustal V, как описано в Примере 3.
ФИГ.25 резюмирует данные BLASTP и значения процентов идентичности для EgDHAsyn2 (Пример 5), EgC20elo1 (Пример 3, supra) и EgDHAsyn1 (Пример 4, supra).
ПРИМЕР 6
Основной структурный анализ EgC20elo1, EgDHAsyn1 и EgDHAsyn2
При 100% аминокислотной идентичности между С-окончанием EgDHAsyn2 (SEQ ID NO:22) и Euglena gracilis дельта-4-десатуразой (SEQ ID NO:13) выполняли группирование нуклеотидной последовательности между кодирующей последовательностью EgDHAsyn2 (SEQ ID NO:21), кДНК последовательностью Euglena gracilis дельта-4-десатуразы (SEQ ID NO:23) (номер доступа НЦБИ AY278558 (GI 33466345), локус AY278558, Meyer et al., Biochemistry 42(32):9779-9788 (2003)), и кодирующей последовательностью Euglena gracilis дельта-4-десатуразы (SEQ ID NO:24) (Meyer et al., supra). Группировку последовательностей проводили с использованием метода Clustal W (с помощью программы MegAlign™ v6.1 вычислительного биоинформационного пакета LASERGENE (DNASTAR Inc.) с параметрами по умолчанию для множественной группировки (GAP PENALTY=10, GAP LENGTH PENALTY=0.2, Delay Divergen Seqs(%)=30, DNA Transition Weight=0.5, Protein Weight Matrix=Gonnet Series, DNA Weight Matrix=IUB). Группировка показана на ФИГ.2. Кодирующая последовательность Euglena gracilis дельта-4-десатуразы названа EgD4_CDS (SEQ ID NO:24); Euglena gracilis дельта-4-десатуразная кДНК последовательность названа EgD4_cDNA (SEQ ID NO:23); и Euglena gracilis ДГК синтаза 2 кодирующая последовательность названа EgDHAsyn2_CDS (SEQ ID NO:21).
5′ конец (где последовательности расходятся) и 3′ конец (где последовательности идентичны) данной группировки усечены с целью размещения группировки на одной странице. ФИГ.2 иллюстрирует, что последовательности сильно расходятся от начала Euglena gracilis дельта-4-десатуразной кДНК до 83 bp вверх от инициирующего сайта кодирующей последовательности (CDS). Из группировки очевидно, что нуклеотидные последовательности для EgD4_cDNA и EgDHAsyn2_CDS идентичны от 83 bp вверх от CDS инициирующего сайта Euglena gracilis дельта-4-десатуразной кДНК последовательности (SEQ ID NO:23), который эквивалентен нуклеотиду 674 EgDHAsyn2_CDS (SEQ ID NO:21), вплоть до конца данных последовательностей. В точной точке дивергенции NotI сайт может быть найден в Euglena gracilis кДНК последовательности (нуклеотиды 656-663 SEQ ID NO:23), и поскольку NotI линкеры были использованы в первичном клонировании Euglena gracilis дельта-4-десатуразной кДНК (см. Meyer et al., supra), вероятно, что то, что было клонировано, являлось незавершенным, с неполной длиной транскрипта для EgDHAsyn2.
Аминокислотную последовательность EgDHAsyn1 (SEQ ID NO:12) сравнивали с EgDHAsyn2 (SEQ ID NO:22) и EgC20elo1 (SEQ ID NO:6) с использованием метода Clustal W, как описано выше, и группировка показана на ФИГ.3A и 3B. В сравнении с EgDHAsyn1 и EgDHAsyn2, EgC20elo1 имеет делецию 7 аминокислот (т.е. A L D L A [V/I] L) и 2 других аминокислотных субституции (т.е. W47R, T48I; основываясь на нумерации для EgDHAsyn1) в N-окончании. После аминокислоты 289 EgC20elo1, данные последовательности сильно различаются при сравнении с ДГК синтазами. EgDHAsyn1 и EgDHAsyn2 имеют дополнительные 498 аминокислоты в своих C-терминальных концах с гомологией к дельта-4-десатуразы жирных кислот, тогда как EgC20elo1 заканчивается после лишь 9 дополнительных аминокислот. Аминокислотные последовательности EgDHAsyn1 (SEQ ID NO:12) и EgDHAsyn2 (SEQ ID NO:22) имеют 8 аминокислотных различий между данными 2 последовательностями (т.е. V25I, G54V, A305T, L310P, V380I, S491N, I744T, R747P; основываясь на нумерации для EgDHAsyn1). Последние четыре различия имеют место в дельта-4-десатуразнром домене.
ФИГ.4A и 4B изображают группировку Clustal W N-окончания EgDHAsyn1 (SEQ ID NO:12) и N-окончания EgDHAsyn2 (SEQ ID NO:22) с EgC20elo1 (SEQ ID NO:6), Pavlova sp. CCMP459 C20-PUFA Elo (SEQ ID NO:2), Ostreococcus tauri ПНЖК элонгазой 2 (SEQ ID NO:25) (номер доступа НЦБИ AAV67798 (GI 55852396), локус AAV67798, CDS AY591336; Meyer et al., J. Lipid Res. 45(10):1899-1909 (2004)), и Thalassiosira pseudonana ПНЖК элонгазой 2 (SEQ ID NO:26) (номер доступа НЦБИ AAV67800 (GI 55852441), локус AAV67800, CDS AY591338; Meyer et al., J. Lipid Res., supra). На ФИГ.4A и 4B, Pavlova, Ostreococcus, и Thalassiosira протеины помечены как PavC20elo, OtPUFAelo2, и TpPUFAelo2, соответственно.
ФИГ.5A, 5B, 5C, и 5D изображают группировку Clustal W C-окончания EgDHAsyn1 (EgDHAsyn1_CT; аминокислот 253-793 SEQ ID NO:12; N-окончание EgDHAsyn1 не показано и обозначено как “…”) и C-окончания EgDHAsyn2 (EgDHAsyn2_CT; аминокислот 253-793 of SEQ ID NO:22, N-окончание EgDHAsyn2 не показано и обозначено как “…”) с Euglena gracilis дельта-4-десатуразы жирных кислот (SEQ ID NO:13), Thraustochytrium aureum дельта-4-десатуразой (SEQ ID NO:27) (номер доступа НЦБИ AAN75707(GI 25956288), локус AAN75707, CDS AF391543), Schizochytrium aggregatum дельта-4-десатуразой (SEQ ID NO:28) (PCT публикация № WO 2002/090493), Thalassiosira pseudonana дельта-4-десатуразой (SEQ ID NO:29) (номер доступа НЦБИ AAX14506 (GI 60173017), локус AAX14506, CDS AY817156; Tonon et al., FEBS J. 272 (13):3401-3412 (2005)), и Isochrysis galbana дельта-4-десатуразой (SEQ ID NO:30) (номер доступа НЦБИ AAV33631 (GI 54307110), локус AAV33631, CDS AY630574; Pereira et al., Biochem. J. 384(2),:357-366 (2004) и PCT публикация № WO 2002/090493). На ФИГ.5A, 5B, 5C, и 5D, Euglena, Thraustochytrium, Thalassiosira, и Isochrysis протеины помечены как EgD4, TaD4, TpD4, и IgD4, соответственно.
ФИГ.6 изображает группировку внутренних фрагментов EgDHAsyn1 (помеченных как “EgDHAsyn1_ NCT.pro”; аминокислот 253-365 SEQ ID NO:12) и EgDHAsyn2 (помеченных как “EgDHAsyn2_NCT.pro”; аминокислот 253-365 SEQ ID NO:22), разделяющих как C20-элонгазный участок так и дельта-4-десатуразный домен (основываясь на гомологии), с C-окончаниями C20-элонгаз (EgC20elo1_CT.pro, аминокислот 246-298 SEQ ID NO:6; PavC20elo_CT.pro, аминокислот 240-277 SEQ ID NO:2; OtPUFAelo2_CT.pro, аминокислот 256-300 SEQ ID NO:25; TpPUFAelo2_CT.pro, аминокислот 279-358 SEQ ID NO:26) и N-окончанием дельта-4-десатураз (EgD4_NT.pro, аминокислот 1-116 SEQ ID NO:13; TaD4_NT.pro, аминокислот 1-47 SEQ ID NO:27; SaD4_NT.pro, аминокислот 1-47 of SEQ ID NO:28; TpD4_NT.pro, аминокислот 1-82 of SEQ ID NO:29; IgD4_NT.pro, аминокислот 1-43 of SEQ ID NO:30). Законсервированный мотив в C-окончании всех C20-элонгазных доменов (т.е. VLFXXFYXXXY (SEQ ID NO:180)) также присутствует в N-окончании EgD4 и дополнительно подтверждает то, что EgD4 является неполной ДГК синтазой.
В C-окончании C20-элонгазного домена для каждого из EgDHAsyn1, EgDHAsyn2, и EgC20elo1 имеется повторяющаяся последовательность, содержащая NG мотив (т.е. KNGK (SEQ ID NO:186), PENGA (SEQ ID NO:187), PENGA (SEQ ID NO:187), и PCENGTV (SEQ ID NO:191); названные NG повторами и обозначенные на ФИГ.6 линиями под последовательностью). Хотя данная структура встречается с высокой вероятностью появления, просмотр NG повторяющегося участка с использованием Prosite показывает последний NG мотив (т.е. NGTV) в этой области как потенциальный N-гликосилирующий сайт. После NG повторяющегося участка как EgDHAsyn1 так и EgDHAsyn2 содержат обогащенный пролином участок (помеченный как “Обогащенный пролином линкер” на ФИГ.6), который может действовать в качестве линкера между C20-элонгазным и дельта-4-десатуразным доменами. Данный линкер может играть роль в поддержании C20-элонгазного и дельта-4-десатуразного доменов в соответствующей структурной ориентации, позволяющей осуществлять эффективную конверсию ЭПК в ДГК. Хотя обогащенный пролином линкер показан на ФИГ.6 как тянущийся от P304 до V21 (основываясь на нумерации для EgDHAsyn1), NG повторяющийся участок также в некоторой степени обогащен пролином и может также играть роль в этой линкерной функции.
Нуклеотидная и соответствующая аминокислотная последовательности для обогащенного пролином линкера EgDHAsyn1, как определено на ФИГ.6, представлены в SEQ ID NO:197 и SEQ ID NO:198, соответственно. Нуклеотидная и соответствующая аминокислотная последовательности для обогащенного пролином линкера EgDHAsyn2, как определено на ФИГ.6, представлены в SEQ ID NO:199 и SEQ ID NO:200, соответственно.
Нуклеотидная и соответствующая аминокислотная последовательности для EgDHAsyn1 C20-элонгазного домена из EgDHAsyn1 представлены в SEQ ID NO:201 и SEQ ID NO:202, соответственно. Нуклеотидная и соответствующая аминокислотная последовательности для EgDHAsyn2 C20-элонгазного домена представлены в SEQ ID NO:203 и SEQ ID NO:204, соответственно.
ПРИМЕР 7
Конструирование pDMW263
Плазмида pY5-30 (которая была ранее описана в патенте США № 7259255 (на содержание которого здесь сделана ссылка)), представляет собой челночную плазмиду, которая может воспроизводиться как в E. coli так и в Yarrowia lipolytica. Плазмида pY5-30 содержит следующее: Yarrowia автономную репликационную последовательность (ARS18); ColE1 плазмидный источник репликации; ампициллин-резистентный ген (AmpR), для селекции в E. coli; Yarrowia LEU2 ген, для селекции в Yarrowia; и химерный TEF::GUS::XPR ген. Плазмиду pDMW263 (SEQ ID NO:31) была создана из pY5-30 путем замены TEF промотора Yarrowia lipolytica FBAINm промотором (патент США № 7202356), с использованием методов, хорошо известных специалистам в данной области. Вкратце, FBAIN промотор локализован в 5′ верхнем нетранслированном участке перед 'ATG′ инициирующим трансляцию кодоном фруктоза-бисфосфат альдолазного фермента (E.C. 4.1.2.13), кодированного fba1 геном. Этот промотор необходим для экспрессии и включает часть 5′ кодирующего участка, который содержит интрон. Модифицированный промотор, FBAINm, имеет 52 bp делецию между ATG инициирующим трансляцию кодоном и интроном FBAIN промотора (включая таким образом только 22 аминокислоты N-окончания) и новый трансляционный консенсусный мотив после данного интрона. Таблица 20 резюмирует компоненты pDMW263 (SEQ ID NO:31; описано также в PCT публикации № WO 2007/061845).
ПРИМЕР 8
Конструирование Yarrowia lipolytica экспрессирующего вектора pY115 и Gateway® дестинационных векторов pBY1 и pY159
NcoI/SalI ДНК фрагмент из pDMW263 (SEQ ID NO:31) (см. конструкцию в Примере 7), содержащий Yarrowia lipolytica FBAINm промотор, клонировали в NcoI/SalI ДНК фрагмент pDMW237 (SEQ ID NO:32), описанный ранее в PCT публикации № WO 2006/012325 (на содержание которой здесь сделана ссылка). pDMW237 содержит синтетический дельта-9-элонгазный ген, полученный из Isochrysis galbana и кодон-оптимизированный для экспрессии в Yarrowia lipolytica (IgD9e). Таким способом была создана плазмида pY115 (SEQ ID NO:33; ФИГ.7A). На ФИГ.7A модифицированный FBAINm промотор называется FBA1+Intron. На модифицированный промотор FBAINm на других фигурах ссылаются или как на FBA1+Intron или YAR FBA1 PRO+Intron; эти термины используются взаимозаменяемо с FBAINm.
Плазмиду pY115 (SEQ ID NO:33) расщепляли NcoI/NotI, и результирующие ДНК концы наполняли с использованием Klenow. После наполнения с образованием затупленных концов ДНК фрагменты обрабатывали телячьей кишечной щелочной фосфатазой и разделяли с использованием агарозного гель-электрофореза. 6989 bp фрагмент, содержащий Yarrowia lipolytica FBAINm промотор, подвергали эксцизии из агарозного геля и очищали с использованием набора QIAquick® Gel Extraction Kit (Qiagen Inc., Valencia, CA), согласно протоколу производителя. Очищенный 6989 bp фрагмент лигировали с кассетой rfA с использованием системы Gateway Vector Conversion System (Cat. No. 11823-029, Invitrogen Corporation), следуя протоколу производителя, с образованием Yarrowia lipolytica Gateway® дестинационного вектора pBY1 (SEQ ID NO:34; ФИГ.7B).
При конструировании pBY1 наполненный NcoI сайт представляет ATG исходное положение для инициирования трансляции. Таким образом, гены, перенесенные в этот экспрессирующий вектор, экспрессируются как слитые протеины и должны находиться в корректном каркасе после Gateway® клонирования. Также, 5′ нетранслированная последовательность приводит к добавлению к N-концу результирующего протеина дополнительных аминокислот. По этой причине был создан второй Gateway® дестинационный вектор с удаленным полученным из вектора ATG инициирующим кодоном, позволяя тем самым инициировать трансляцию от введенного гена.
FBAINm промотор амплифицировали из плазмиды pY115 (SEQ ID NO:33) с использованием ПЦР с олигонуклеотидными праймерами oYFBA1 (SEQ ID NO:35) и oYFBA1-6 (SEQ ID NO:36). Праймер oYFBA1 (SEQ ID NO:35) был создан для введения BglII сайта в 5′ конце промотора, и праймер oYFBA1-6 (SEQ ID NO:36) был создан для введения NotI сайта в 3′ конце данного промотора с удалением NcoI сайта и, следовательно, ATG инициирующего кодона. Результирующий ПЦР фрагмент был расщеплен BglII и NotI и клонирован в BglII/NotI фрагмент pY115, содержащий векторный остов, с образованием pY158 (SEQ ID NO:37).
Плазмиду pY158 (SEQ ID NO:37) расщепляли NotI, и результирующие ДНК концы наполняли. После наполнения с образованием тупых концов ДНК фрагменты обрабатывали телячьей кишечной щелочной фосфатазой и разделяли с использованием агарозгого гель-электрофореза. 6992 bp фрагмент, содержащий Yarrowia lipolytica FBAINm промотор, подвергали эксцизии из агарозного геля и очищали с использованием набора QIAquick® Gel Extraction Kit (Qiagen Inc., Valencia, CA), следуя протоколу производителя. Очищенный 6992 bp фрагмент лигировали с кассетой rfA с использованием системы Gateway Vector Conversion System (Cat. No. 11823-029, Invitrogen Corporation), следуя протоколу производителя, с образованием Yarrowia lipolytica Gateway® дестинационного вектора pY159 (SEQ ID NO:38; ФИГ.7C).
ПРИМЕР 9
Конструирование Yarrowia lipolytica экспрессирующих векторов pBY-EgC20elo1 (EgC20elo1), pY132 (EgDHAsyn1), pY161 (EgDHAsyn1) и pY164 (EgDHAsyn2)
Плазмиду очищали от клонов eeg1c.pk005.p14.f (Пример 3), eeg1c.pk016.e6.f (Пример 4), и eeg1c-1 (Пример 5) с использованием набора QIAprep® Spin Miniprep Kit (Qiagen Inc., Valencia, CA), следуя протоколу производителя. С использованием Gateway® LR Clonase™ II ферментной смеси (Cat. No. 11791-020, Invitrogen Corporation) и следуя протоколу производителя, кДНК инсерты из eeg1c.pk001.p14.f (включая EgC20elo1) и eeg1c.pk016.e6.f (включая EgDHAsyn1) были перенесены в pBY1 (SEQ ID NO:34; ФИГ.7B) с образованием pBY-EgC20elo1 (SEQ ID NO:39, ФИГ.7D) и pY132 (SEQ ID NO:40; ФИГ.8A), соответственно. КДНК инсерт из eeg1c-1 (включая EgDHAsyn2) не переносился в pBY1, поскольку это привело бы к экспрессии неправильного трансляционного каркаса.
C использованием Gateway® LR Clonase™ II ферментной смеси (Cat. No. 11791-020, Invitrogen Corporation) и следуя протоколу производителя, кДНК инсерты из eeg1c.pk016.e6.f и eeg1c-1 были перенесены в pY159 (SEQ ID NO:38; Пример 8) с образованием pY161 (SEQ ID NO:41, ФИГ.8B) и pY164 (SEQ ID NO:42; ФИГ.8C), соответственно.
ПРИМЕР 10
Конструирование Yarrowia lipolytica экспрессирующих векторов pY141 (EgDHAsyn1*), pY143 (EgDHAsyn1*C20EloDom1) и pY149 (EgDHAsyn1*C20EloDom2Linker)
EgDHAsyn1 амплифицировали из клона eeg1c.pk001.e6.f олигонуклеотидными праймерами EgEPAEloDom-5 (SEQ ID NO:43) и oEUG el4-3 (SEQ ID NO:44), с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S, Finnzymes Oy, Finland), следуя протоколу. Результирующий ДНК фрагмент клонировали в pCR-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), в соответствии с протоколом производителя, с образованием pKR1062 (SEQ ID NO:45).
Внутренний NcoI сайт при нуклеотидах 619-624 удаляли из EgDHAsyn1 в pKR1062 с использованием набора Quickchange® Site Directed Mutagenesis kit (Cat. No. 200518, Stratagene, La Jolla, CA), с олигонуклеотидами EgEloD4Mut-5 (SEQ ID NO:46) и EgEloD4Mut-3 (SEQ ID NO:47), следуя протоколу производителя. После экстенсивного секвенирования клон с удаленным NcoI сайтом (т.е. мутация ccatgg в ccttgg) и без каких-либо дополнительных изменений был выбран для дальнейших исследований. Этот клон был обозначен как pLF115-7 (SEQ ID NO:48). Нуклеотидная последовательность для EgDHAsyn1 с удаленным NcoI сайтом (EgDHAsyn1*) представлена в SEQ ID NO:205. Соответствующая аминокислотная последовательность идентична SEQ ID NO:12.
Конструирование плазмиды pY141, экспрессирующей EgDHAsyn1*: NcoI/NotI ДНК фрагмент из pLF115-7 (SEQ ID NO:48), содержащий EgDHAsyn1 (SEQ ID NO:205; без внутреннего NcoI сайта; при нуклеотиде 621 EgDHAsyn1 CDS; ccatgg в ccttgg), клонировали в NcoI/NotI DNA фрагмент из pY115, содержащий Yarrowia lipolytica FBAINm промотор, с образованием pY141 (SEQ ID NO:49; ФИГ.8D). Таким образом, плазмида pY141 содержит ген полной длины EgDHAsyn1* (помеченный как “EgDHAsyn1(-NcoI)” на фигуре), под контролем Yarrowia lipolytica FBAINm промотора (PCT публикация № WO 2005/049805; патент США № 7202356; помеченный как “Fba1+Intron” на фигуре), и Pex20 терминирующую последовательность из Yarrowia Pex20 гена (номер доступа GenBank AF054613).
Конструирование плазмиды, экспрессирующей EgDHAsyn1-C20EloDom1: Нуклеотидная последовательность для EgDHAsyn1* C20-элонгазного домена (EgDHAsyn1C20EloDom1) в pY141 представлена в SEQ ID NO:206 (идентична SEQ ID NO:201 но с удаленным сайтом NcoI). Соответствующая аминокислотная последовательность идентична SEQ ID NO:202.
EgDHAsyn1C20EloDom1 (SEQ ID NO:206) амплифицировали из pLF115-7 олигонуклеотидными праймерами EgEPAEloDom-5 (SEQ ID NO:43) и EgDPAEloDom-3 (SEQ ID NO:50) с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S, Finnzymes Oy, Finland) согласно протокола производителя. Результирующий ДНК фрагмент клонировали в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), следуя протоколу производителя, с образованием pHD16 (SEQ ID NO:51).
NcoI/NotI DNA фрагмент из pHD16 (SEQ ID NO:51), содержащий EgDHAsyn1C20EloDom1 (без внутреннего NcoI сайта), клонировали в NcoI/NotI ДНК фрагмент из pY115, содержащий Yarrowia lipolytica FBAINm промотор, с образованием pY143 (SEQ ID NO:52; ФИГ.9A). Плазмида pY143 содержит N-концевой домен EgDHAsyn1* (EgDHAsyn1C20EloDom1) и не включает обогащенный пролином линкер или дельта-4-десатуразный домен.
Конструирование плазмиды pY149, экспресирующей EgDHAsyn1-C20EloDom2Linker: EgDHAsyn1* домен C20-элонгазы (SEQ ID NO:206) и обогащенный пролином линкер (SEQ ID NO:197), амплифицировали из pLF115-7 (SEQ ID NO:48) олигонуклеотидными праймерами EgEPAEloDom-5 (SEQ ID NO:43) и oEUGsyn6-2 (SEQ ID NO:53) с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S, Finnzymes Oy, Finland) согласно протоколу производителя. Результирующий ДНК фрагмент клонировали в pCR-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), в соответствии с протоколом производителя, с образованием pKR1071 (SEQ ID NO:54).
NcoI/Ecl132II ДНК фрагмент из pKR1071 (SEQ ID NO:54) клонировали в NcoI/NotI ДНК фрагмент из pY115 (где NotI сайт был наполнен), содержащий Yarrowia lipolytica FBAINm промотор, с образованием pY149 (SEQ ID NO:55; ФИГ.9B). Плазмида pY149 содержит EgDHAsyn1C20EloDom1/обогащенный пролином линкер слитый ген (т.е. EgDHAsyn1C20EloDom2Linker; SEQ ID NO:207), но не содержит дельта-4-десатуразного домена. Аминокислотная последовательность EgDHAsyn1C20EloDom2Linker представлена в SEQ ID NO:208. В дополнение к аминокислотам из EgDHAsyn1* C20-элонгазного домена и обогащенного пролином линкера вводились 4 дополнительных аминокислоты (т.е. SCRT) после линкерного участка как результат применения данного способа синтеза и клонирования этого фрагмента.
ПРИМЕР 11
Конструирование Yarrowia lipolytica экспрессирующих векторов для генерации новых C20-элонгаза/дельта-4-десатуразных слитых протеинов
С целью синтеза новых C20-элонгаза/дельта-4-десатуразных слитых протеинов уникальный SbfI сайт добавляли к 3′ концу C20-элонгазного домена EgDHAsyn1* после участка обогащенного пролином линкера (EgDHAsyn1C20EloDom3Linker). EgDHAsyn1C20EloDom3 амплифицировали из pLF115-7 (SEQ ID NO:48) олигонуклеотидными праймерами EgEPAEloDom-5 (SEQ ID NO:43) и oEUGsyn6-3 (SEQ ID NO:56) с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S, Finnzymes Oy, Finland) в соответствии с протоколом производителя. Результирующий ДНК фрагмент клонировали в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), следуя протоколу производителя, с образованием pKR1091 (SEQ ID NO:57).
NcoI/Ecl136II ДНК фрагмент из pKR1091 (SEQ ID NO:57), содержащий EgDHAsyn1C20EloDom3Linker, клонировали в NcoI/NotI ДНК фрагмент из pY115 (где NotI был наполнен с образованием затупленного конца), содержащий Yarrowia lipolytica FBAINm промотор, с образованием pY155 (SEQ ID NO:58).
С целью синтеза новых C20-элонгаза/дельта-4-десатуразных слитых протеинов, уникальный SbfI сайт добавляли к 5′ концу разных дельта-4-десатураз. В каждом случае SbfI сайт локализовался после ATG инициирующего сайта каждой кодирующей последовательности и обусловливал добавление и/или замену нескольких аминокислот в N-окончание дельта-4-десатуразы, кодированной этими генами.
Конструирование плазмиды pY156, экспрессирующей EgDHAsyn1-C20EloDom3-IgD4*: Isochrysis galbana дельта-4-десатураза (SEQ ID NO:209; IgD4) амплифицировали из pRIG6 (ранее описана в PCT публикации № WO 2002/090493) олигонуклеотидами oRIG6-1 (SEQ ID NO:59) и oRIG6-2 (SEQ ID NO:60) с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S) согласно протокола производителя. Результирующий ДНК фрагмент, который содержит IgD4 CDS и который идентичен SEQ ID NO:209, за исключением того, что SbfI сайт добавлялся в 5′ конце после инициирующего кодона (IgD4*; SEQ ID NO:210), клонировали в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), следуя протоколу производителя, с образованием pKR1067 (SEQ ID NO:61). Аминокислотная последовательность для IgD4* из pKR1067 представлена в SEQ ID NO:211 и является идентичной IgD4 (SEQ ID NO:30), за исключением того, что первые 4 аминокислоты (т.е. MCNA) были изменены на MALQ вследствие добавления SbfI сайта в нуклеотидную последовательность.
NcoI/NotI ДНК фрагмент из pKR1067 (SEQ ID NO:61), содержащий IgD4*, клонировали в NcoI/NotI ДНК фрагмент из pY115, содержащий Yarrowia lipolytica FBAINm промотор, с образованием pY150 (SEQ ID NO:62; ФИГ.9C). На ФИГ.9C, IgD4* помечен как “Ig d4 DS”. Таким способом IgD4* мог быть один экспрессирован в Yarrowia.
XbaI/SbfI ДНК фрагмент из pKR1091 (SEQ ID NO:57; supra), содержащий EgDHAsyn1C20EloDom3Linker, клонировали в XbaI/SbfI ДНК фрагмент из pKR1067 (SEQ ID NO:61), содержащий IgD4*, с образованием pKR1097 (SEQ ID NO:63). Таким образом было осуществлено слияние в каркасе между EgDHAsyn1C20EloDom3Linker и IgD4*, разделенных обогащенным пролином линкерным участком (названным EgDHAsyn1C20EloDom3-IgD4; SEQ ID NO:212). Аминокислотная последовательность для EgDHAsyn1C20EloDom3-IgD4 представлена в SEQ ID NO:213.
NcoI/NotI ДНК фрагмент из pKR1097 (SEQ ID NO:63), содержащий EgDHAsyn1C20EloDom3-IgD4, клонировали в NcoI/NotI ДНК фрагмент из pY115, содержащий Yarrowia lipolytica FBAINm промотор, с образованием pY156 (SEQ ID NO:64; ФИГ.9D). На ФИГ.9D EgDHAsyn1C20EloDom3-IgD4 помечен как “EGel-IGd4”.
Конструирование плазмиды pY152, экспрессирующей EgDHAsyn1-D4Dom1*: Участок C-окончания EgDHAsyn1* (SEQ ID NO:205), содержащий дельта-4-десатуразный домен (EgDHAsyn1D4Dom1; SEQ ID NO:214; соответствующая аминокислотная последовательность для EgDHAsyn1D4Dom1 представлена в SEQ ID NO:215), начинающийся как раз за концом участка линкера, обогащенного пролином, амплифицировали из pLF115-7 (как описано в Примере 10) олигонуклеотидами oEGslne6-1 (SEQ ID NO:65) и oEUGel4-3 (SEQ ID NO:44) с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S) в соответствии с протоколом производителя. Олигонуклеотид oEGslne6-1 (SEQ ID NO:65) вводил ATG инициирующий кодон в 5′ конце ПЦР продукта с последующим SbfI сайтом. Результирующий ДНК фрагмент клонировали в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), следуя протоколу производителя, с образованием pKR1069 (SEQ ID NO:66). Новые CDS и аминокислотные последовательности, содержащие EgDHAsyn1D4Dom1 из pKR1069 (т.е. EgDHAsyn1D4Dom1*), представлены в SEQ ID NO:216 и SEQ ID NO:217, соответственно. Аминокислотная последовательность для EgDHAsyn1D4Dom1* (SEQ ID NO:217) идентична таковой для EgDHAsyn1D4Dom1 (SEQ ID NO:215), за исключением того, что первые 2 аминокислоты (т.е. SG) были изменены на MAL вследствие добавления SbfI сайта в нуклеотидную последовательность.
NcoI/NotI ДНК фрагмент из pKR1069 (SEQ ID NO:66), содержащий EgDHAsyn1D4Dom1*, клонировали в NcoI/NotI ДНК фрагмент из pY115, содержащий Yarrowia lipolytica FBAINm промотор, с образованием pY152 (SEQ ID NO:67; ФИГ.10A). На ФИГ.10A EgDHAsyn1D4Dom1* помечен как “EUG d4 (fus test)”. Таким способом, EgDHAsyn1D4Dom1* мог быть один экспрессирован в Yarrowia.
Конструирование плазмиды pY157, экспрессирующей EgDHAsyn1-C20EloDom3-EgD4Dom1: XbaI/SbfI ДНК фрагмент из pKR1091 (SEQ ID NO:57), содержащий EgDHAsyn1C20EloDom3-Linker, клонировали в XbaI/SbfI ДНК фрагмент из pKR1069, содержащий EgDHAsyn1D4Dom1*, с образованием pKR1099 (SEQ ID NO:68). Таким способом было осуществлено каркасное слияние между EgDHAsyn1C20EloDom и EgDHAsyn1D4Dom1*, разделенных обогащенным пролином линкерным участком (названным EgDHAsyn1C20EloDom3-EgD4Dom1; SEQ ID NO:218). Аминокислотная последовательность EgDHAsyn1C20EloDom3-EgD4Dom1 (SEQ ID NO:219) почти идентична EgDHAsyn1, за исключением изменения одной аминокислоты (т.е. G323L, основываясь на нумерации для EgDHAsyn1), из-за SbfI клонирующего сайта и соединения путем слияния.
NcoI/NotI DNA фрагмент из pKR1099 (SEQ ID NO:68), содержащий EgDHAsyn1C20EloDom3-EgD4Dom1, клонировали в NcoI/NotI ДНК фрагмент из pY115, содержащий Yarrowia lipolytica FBAINm промотор, с образованием pY157 (SEQ ID NO:69; ФИГ.10B). На ФИГ.10B EgDHAsyn1C20EloDom3-EgD4Dom1 помечен как “EGel-EGd4 fus”.
Конструирование плазмиды pY153, экспрессирующей EgDHAsyn1*D4Dom2: Участок C-окончания EgDHAsyn1, содержащий дельта-4-десатуразный домен и часть C20-элонгазного домена (EgDHAsyn1D4Dom2; SEQ ID NO:220; соответствующая аминокислотная последовательность для EgDHAsyn1D4Dom2 представлена в SEQ ID NO:221), которая соответствует аминокислотной последовательности, идентифицированной как EgD4 (SEQ ID NO:13; Meyer et al., Biochemistry 42(32):9779-9788 (2003)), амплифицировали из pLF115-7 (как описано в Примере 10) олигонуклеотидами oEUGel4-4 (SEQ ID NO:70) и oEUGel4-3 (SEQ ID NO:44) с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S) в соответствии с протоколом производителя. Результирующий ДНК фрагмент клонировали в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), следуя протоколу производителя, с образованием pKR1073 (SEQ ID NO:71).
PciI/NotI ДНК фрагмент из pKR1073 (SEQ ID NO:71), содержащий EgDHAsyn1D4Dom2, клонировали в NcoI/NotI ДНК фрагмент из pY115, содержащий Yarrowia lipolytica FBAINm промотор, с образованием pY153 (SEQ ID NO:72; ФИГ.10C). На ФИГ.10C EgDHAsyn1D4Dom2 помечен как EUG d4 (HZ). Таким способом EgDHAsynD4Dom2 мог быть один экспрессирован в Yarrowia.
Конструирование плазмиды pY160, экспрессирующей EgDHAsyn1-C20EloDom3-SaD4*: Schizochytrium aggregatum дельта-4-десатуразу (SEQ ID NO:222; SaD4) амплифицировали из pRSA-1 (ранее описана в PCT публикации № WO 2002/090493) олигонуклеотидами oRSA1-1 (SEQ ID NO:73) и oRSA1-2 (SEQ ID NO:74) с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S) в соответствии с протоколом производителя. Результирующий ДНК фрагмент, который содержит SaD4 CDS и идентичен SEQ ID NO:222, за исключением того, что SbfI сайт добавляли в 5′ конце после инициирующего кодона (SaD4*; SEQ ID NO:223), клонировали в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), следуя протоколу производителя, с образованием pKR1068 (SEQ ID NO:75). Аминокислотная последовательность для SaD4* из pKR1068 представлена в SEQ ID NO:224 и идентична таковой для SaD4 (SEQ ID NO:28), за исключением того, что первые 3 аминокислоты (т.е. MTV) были изменены на MALQ из-за добавления SbfI сайта в нуклеотидную последовательность.
NcoI/NotI ДНК фрагмент из pKR1068 (SEQ ID NO:75) (частично расщепленный для исключения внутреннего NcoI сайта), содержащий SaD4*, клонировали в NcoI/NotI ДНК фрагмент из pY115, содержащий Yarrowia lipolytica FBAINm промотор, с образованием pY151 (SEQ ID NO:76; ФИГ.10D). На ФИГ.10D, SaD4* помечен как “RSA d4 DS”. Таким способом, SaD4* мог быть один экспрессирован в Yarrowia.
SbfI/NotI DNA фрагмент из pKR1068 (SEQ ID NO:75), содержащий SaD4*, клонировали в SbfI/NotI ДНК фрагмент из pY157 (SEQ ID NO:69), содержащий EgDHAsyn1C20EloDom3Linker, с образованием pY160 (SEQ ID NO:77; ФИГ.11). Таким способом, было осуществлено каркасное слияние между EgDHAsyn1C20EloDom3 и SaD4*, разделенных обогащенным пролином линкерным участком (т.е. EgDHAsyn1C20EloDom3-SaD4; SEQ ID NO:225). Аминокислотная последовательность для EgDHAsyn1C20EloDom3-SaD4 представлена в SEQ ID NO:226.
ПРИМЕР 12
Условия выращивания Euglena anabaena, липидный профиль и выделение мРНК
Euglena anabaena получали из лаборатории Dr.Richard Triemer at Michigan State University (East Lansing, MI). Приблизительно 2 мл культуры извлекали для липидного анализа и подвергали центрифугированию при 1800xg на протяжении 5 минут. Таблетку промывали один раз водой и повторно центрифугировали. Результирующую таблетку высушивали в течение 5 минут под вакуумом, ресуспендировали в 100 мкл триметилсульфоний гидроксида (ТМСГ) и инкубировали при комнатной температуре в течение 15 минут при встряхивании. После этого добавляли 0,5 мл гексана, и ампулы инкубировали в течение 15 минут при комнатной температуре при встряхивании. Жирнокислотные метиловые эфиры (5 мкл, вытянутых из гексанового слоя) отделяли и анализировали количественно с использованием газового хроматографа марки Hewlett-Packard 6890 Gas Chromatograph, снабженного капиллярной колонкой из плавленого кварца Omegawax 320 (корпорация Supelco Inc., Cat. No. 24152). Температура печи была запрограммирована на выдержку 170°С в течение 1,0 минуты, повышение до 240°С при 5°С/минуту и дополнительную выдержку в течение 1,0 минуты. Газ-носитель поставлялся водородным генератором Whatman. Время удерживания было сравнимо с таковым для стандартов метиловых эфиров, которые имеются в продаже (Nu-Chek Prep, Inc. Cat. No. U-99-A), и результирующая хроматограмма показана на ФИГ.12. Присутствие ЭПК и ДГК в жирнокислотном профиле дает возможность предположить, что Euglena anabaena может служить хорошим источником для длинноцепочечных ПНЖК биосинтетических генов, таких как, однако не оганичиваясь этим, С20-элонгазы, дельта-4-десатуразы и/или ДГК синтазы.
Оставшиеся 5 мл активно растущей культуры переносили в 25 мл среды AF-6 (Watanabe & Hiroki, NIES-Collection List of Strains, 5th ed., National Institute for Environmental Studies, Tsukuba, 127 pp (2004)) в 125 мл стеклянной колбе. Культуры Euglena anabaena выращивали при 22°С с 16 часовым световым, 8 часовым темновым циклом в течение 2 недель при очень осторожном перемешивании.
Через 2 недели культуру (25 мл) переносили в 100 мл среды AF-6 в 500 мл стеклянной бутылке, и данная культура выращивалась в течение 1 месяца как описано выше. После этого две 50 мл аликвоты переносили в две отдельные 500 мл стеклянные бутылки, которые содержали 250 мл среды AF-6, и данные культуры выращивались на протяжении двух месяцев как описано выше (давая, в общем, ~600 мл культуры). После этого данные культуры таблетировали путем центрифугирования при 1800xg в течение 10 минут, промывали один раз водой и подвергали повторному центрифугированию. Полную РНК экстрагировали из одной из результирующих таблеток с использованием RNA STAT-60™ реагента (TEL-TEST, Inc., Friendswood, TX) и следуя протоколу производителя (использовать 5 мл реагента, растворенную РНК в 0,5 мл воды). Таким способом из таблетки получали 340 мкг полной РНК (680 мкг/мл). Таблетку, которая осталась, заморозили в жидком азоте и хранили при -80°С. мРНК выделяли из 340 мкг полной РНК с использованием набора mRNA Purification Kit (Amersham Biosciences, Piscataway, NJ) согласно протоколу производителя. Таким образом получили 9,0 мкг мРНК.
ПРИМЕР 13
Синтез Euglena anabaena кДНК, конструирование библиотеки и идентификация ДГК синтаз из кДНК библиотеки eug1c
кДНК библиотеку генерировали с использованием набора Cloneminer™ cDNA Library Construction Kit (Cat. No. 18249-029, Invitrogen Corporation, Carlsbad, CA) и в соответствии с протоколом производителя (Version B, 25-0608). С использованием метода не-радиомечения кДНК синтезировали из 5,12 мкг мРНК (Пример 12) с использованием Biotin -attB2-Oligo(dT) праймера. После синтеза первой и второй цепочки добавляли адаптер attB1; проводили лигирование; и кДНК фракционировали по размерам с использованием колонковой хроматографии. ДНК из фракций концентрировали, рекомбинировали в pDONR™222 и трансформировали в клетки E. coli ElectroMAX™ DH10B™ T1 Phage-Resistant (Invitrogen Corporation). Библиотеку Euglena anabaena назвали eug1c.
Приблизительно 17000 клонов кДНК библиотеки eug1c высевали на три больших квадратных (24 см×24 см) чашки Петри (Corning, Corning, NY), каждая из которых содержала LB+50 мкг/мл канамицин агаровой среды. Клетки выращивали, переносили на Biodyne B мембрану и гибридизовывали с меченым Ncol/Notl ДНК фрагментом, содержащим EgDHAsyn1*, из pY141, в точности как описано в Примере 5. Таким способом идентифицировали 11 позитивных клонов (обозначенных как eug1c-1-eug1c-11).
Позитивные клоны выращивали, и ДНК очищали и секвенировали как описано в Примере 2 с использованием затравленного вектором M13F Universal праймера (SEQ ID NO:1), затравленного вектором M13-28rev праймера (SEQ ID NO:14), и затравленного поли(А) хвостом праймера WobbleT олигонуклеотидов. Исходя из первичных данных секвенирования, подобным способом была получена дополнительная последовательность внутреннего фрагмента с использованием олигонуклеотидов EaDHAsyn5′ (SEQ ID NO:78), EaDHAsyn5′2 (SEQ ID NO:79), EaDHAsyn5′3 (SEQ ID NO:80), EaDHAsyn5′4 (SEQ ID NO:81), EaDHAsyn3′ (SEQ ID NO:82), EaDHAsyn3′2 (SEQ ID NO:83), EaDHAsyn3′3 (SEQ ID NO:84), EaDHAsyn3′4 (SEQ ID NO:85), и EaDHAsyn3′5 (SEQ ID NO:86). Таким способом были получены полноинсертные последовательности eug1c клонов.
Последовательности группировали и сравнивали с использованием прибора Sequencher™ (Version 4.2, Gene Codes Corporation, Ann Arbor, MI), и таким способом клоны могли быть категоризированы на одну из четырех четких групп, основанных на инсертной последовательности (идентифицированы как EaDHAsyn1-EaDHAsyn4). Репрезентативные клоны, содержащие кДНК для каждого класса последовательности, выбирали для дальнейшего исследования, и последовательности для каждой представительной плазмиды (т.е. pLF117-1, pLF117-2, pLF117-3 и pLF117-4) показаны как SEQ ID NO: 87, SEQ ID NO:88, SEQ ID NO:89, и SEQ ID NO:90, соответственно. Последовательность pLF117-1, изображенная цепочкой NNNN, представляет участок полиА хвоста, который не подвергался секвенированию. Кодирующие последовательности для EaDHAsyn1, EaDHAsyn2, EaDHAsyn3, и EaDHAsyn4 показаны как SEQ ID NO:91, SEQ ID NO:92, SEQ ID NO:93, и SEQ ID NO:94, соответственно. Соответствующие аминокислотные последовательности для EaDHAsyn1, EaDHAsyn2, EaDHAsyn3, и EaDHAsyn4 представлены как SEQ ID NO:95, SEQ ID NO:96, SEQ ID NO:97, и SEQ ID NO:98, соответственно.
Аминокислотные последовательности для EaDHAsyn1 (SEQ ID NO:95), EaDHAsyn2 (SEQ ID NO:96), EaDHAsyn3 (SEQ ID NO:97), и EaDHAsyn4 (SEQ ID NO:98) оценивали с помощью BLASTP как описано в Примере 3 и, как и в случае для EgDHAsyn1 (SEQ ID NO:12) и EgDHAsyn2 (SEQ ID NO:22), все четыре EaDHAsyn последовательности, как было найдено, также были схожи, как с C20-PUFA Elo, так и с дельта-4-десатуразы жирных кислот. N-окончания EaDHAsyn1 (SEQ ID NO:95), EaDHAsyn2 (SEQ ID NO:96), EaDHAsyn3 (SEQ ID NO:97), и EaDHAsyn4 (SEQ ID NO:98) все дали значение pLog 58,5 (E значение 3e-59; 114/247 идентичные аминокислоты; 46% идентичности) относительно Pavlova sp. CCMP459 C20-ПНЖК Elo (SEQ ID NO:2). C-окончания EaDHAsyn1 (SEQ ID NO:95), EaDHAsyn2 (SEQ ID NO:96), EaDHAsyn3 (SEQ ID NO:97), и EaDHAsyn4 (SEQ ID NO:98) дали E значения 0,0 (378/538 идентичные аминокислоты; 70% идентичности), 0,0 (378/538 идентичные аминокислоты; 70% идентичности), 0,0 (379/538 идентичные аминокислоты; 70% идентичности), и 0,0 (368/522 идентичные аминокислоты; 70% идентичности), соответственно, относительно аминокислотной последовательности дельта-4-десатуразы жирных кислот из Euglena gracilis (SEQ ID NO:13). Данные BLAST и вероятности указывают, что текущие нуклеиновокислотные фрагменты кодируют все Euglena anabaena C20-ПНЖК Elo/дельта-4-десатуразы жирных кислот.
Аминокислотные последовательности для EaDHAsyn1 (SEQ ID NO:95), EaDHAsyn2 (SEQ ID NO:96), EaDHAsyn3 (SEQ ID NO:97), и EaDHAsyn4 (SEQ ID NO:98) сравнивали с использованием метода Clustal W как описано в Примере 6, и группировки показаны на ФИГ.13A, 13B, и 13C, соответственно. Интересно отметить, что вследствие делеции одной bp в нуклеотидной последовательности C-окончание результирующей аминокислотной последовательности для EaDHAsyn4 (приблизительно последние 35 аминоксилот) сильно расходится и меньше, чем другие три EaDHAsyn протеины.
При сравнении с аминокислотнолй последовательностью EgDHAsyn1 (SEQ ID NO:12) с использованием BLASTP, аминокислотные последовательности EaDHAsyn1 (SEQ ID NO:95), EaDHAsyn2 (SEQ ID NO:96), EaDHAsyn3 (SEQ ID NO:97), и EaDHAsyn4 (SEQ ID NO:98) были на 70% (558/791), 70% (558/791), 70% (559/791) и 70% (548/775) идентичны, соответственно.
Как было в случае для EgDHAsyn1 (SEQ ID NO:12) и EgDHAsyn2 (SEQ ID NO:22), все четыре EaDHAsyn последовательности имеют обогащенный пролином линкерный участок (приблизительно от P300 до T332, основываясь на нумерации для EaDHAsyn1). Данный линкер, как представляется, несколько длиннее, чем таковой для EgDHAsyn1 (SEQ ID NO:12) или EgDHAsyn2 (SEQ ID NO:22). Для всех четырех EaDHAsyn последовательностей характерно отсутствие NG повторяющегося мотива, найденного вверху от обогащенного пролином мотива EgDHAsyn1 и EgDHAsyn2; однако, этот участок, как было в случае для EgDHAsyn1 и EgDHAsyn2, также несколько обогащен пролином во всех четырех EaDHAsyn последовательностях и может играть роль в линкерной функции.
Нуклеотидные последовательности для C20-элонгазных доменов EaDHAsyn1, EaDHAsyn2, EaDHAsyn3, и EaDHAsyn4 представлены в SEQ ID NO:227, SEQ ID NO:228, SEQ ID NO:229, и SEQ ID NO:230, соответственно. Аминоксилотные последовательности для C20-элонгазных доменов EaDHAsyn1, EaDHAsyn2, и EaDHAsyn3 представлены в SEQ ID NO:231, SEQ ID NO:232, и SEQ ID NO:233, соответственно. Аминокислотная последовательность C20-элонгазного домена EaDHAsyn4 идентична таковой для EaDHAsyn1.
Нуклеотидные и аминокислотные последовательности для обогащенного пролином линкера EaDHAsyn1 представлены в SEQ ID NO:234 и SEQ ID NO:235, соответственно. Нуклеотидные и аминокислотные последовательности для обогащенных пролином линкеров EaDHAsyn2, EaDHAsyn3, и EaDHAsyn4 идентичны такой для EaDHAsyn1.
Нуклеотидные последовательности для дельта-4-десатуразного домена 1 каждого из EaDHAsyn1, EaDHAsyn2, и EaDHAsyn4 представлены в SEQ ID NO:236, SEQ ID NO:237, и SEQ ID NO:238, соответственно. Аминокислотные последовательности для дельта-4-десатуразных доменов EaDHAsyn1, EaDHAsyn2, и EaDHAsyn4 представлены в SEQ ID NO:239, SEQ ID NO:240, и SEQ ID NO:241, соответственно. Нуклеотидная и аминокислотная последовательностьТ дельта-4-десатуразного домена 1 EaDHAsyn3 идентична таковой для EaDHAsyn1.
Нуклеотидные последовательности для дельта-4-десатуразного домена 2 EaDHAsyn1, EaDHAsyn2, EaDHAsyn3, и EaDHAsyn4, включая обогащенный пролином линкер и часть 3′ конца C20-элонгазного домена, представлены в SEQ ID NO:242, SEQ ID NO:243, SEQ ID NO:244, и SEQ ID NO:245, соответственно. Аминокислотные последовательности для дельта-4-десатуразных доменов EaDHAsyn1, EaDHAsyn2, EaDHAsyn3, и EaDHAsyn4 представлены в SEQ ID NO:246, SEQ ID NO:247, SEQ ID NO:248, и SEQ ID NO:249, соответственно.
ФИГ.29 резюмирует Euglena anabaena ДГК синтазные доменные последовательности.
ПРИМЕР 14
Конструирование Yarrowia lipolytica экспрессирующих векторов pY165, pY166, pY167 и pY168
С использованием Gateway® LR Clonase™ II ферментной смеси (Cat. No. 11791-020, Invitrogen Corporation) и следуя протоколу производителя, кДНК инсерты из pLF117-1 (SEQ ID NO:87), pLF117-2 (SEQ ID NO:88), pLF117-3 (SEQ ID NO:89), и pLF117-4 (SEQ ID NO:90) переносили в pY159 (SEQ ID NO:38; Пример 8) с образованием pY165 (SEQ ID NO:99, ФИГ.14A), pY166 (SEQ ID NO:100; ФИГ.14B), pY167 (SEQ ID NO:101; ФИГ.14C), и pY168 (SEQ ID NO:102; ФИГ.14D), соответственно. Таким образом, каждая плазмида содержит ген полной длины EaDHAsyn, под контролем Yarrowia lipolytica FBAINm промотора (PCT публикация № WO 2005/049805; патент США № 7202356; помеченный как “Yar Fba1 Pro+Intron” на фигуре) и Pex20 терминирующей последовательности из Yarrowia Pex20 гена (номер доступа GenBank AF054613).
ПРИМЕР 15
Конструирование экспрессирующего вектора сои pKR1061 для коэкспрессии Euglena gracilis ДГК синтазы 1 (EgDHAsyn1) с Saprolegnia diclina дельта-17-десатуразой (SdD17)
Настоящий пример описывает конструирование соевого вектора для коэкспрессии EgDHAsyn1 (SEQ ID NO:12) с SdD17 и гигромицин фосфотрансферазным селективным маркером (гфт).
EgDHAsyn1 амплифицировали из pKR1049 (клон eeg1c.pk016.e6.f) олигонуклеотидными праймерами oEGel2-1 (SEQ ID NO:103) и oEUG el4-3 (SEQ ID NO:44), с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S, Finnzymes Oy, Finland) в соответствии с протоколом производителя. Результирующий ДНК фрагмент встраивали в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), следуя протоколу производителя, с образованием pKR1055 (SEQ ID NO:104).
Исходная плазмида pKR72 (номер доступа АСТК PTA-6019; SEQ ID NO:105, 7085 bp последовательность), производная pKS123, которая была ранее описана в PCT публикации № WO 2002/008269 (на содержание которой в данном тексте сделана ссылка), содержит гигромицин B фосфотрансферазный ген (ГФТ) (Gritz, L. и Davies, J., Gene 25:179-188 (1983)), фланкированный T7 промотором и транскрипционным терминатором (T7prom/HPT/T7term кассета), и бактериальный источник репликации (ori) для селекции и репликации в бактериях (например, E. coli). Кроме того, pKR72 также содержит ГФТ, фланкированный 35S промотором (Odell et al., Nature 313:810-812 (1985)) и NOS 3′ транскрипционным терминатором (Depicker et al., J. Mol. Appl. Genet. 1:561-570 (1982)) (35S/HPT/NOS3′ кассета), для селекции в растениях, таких как соя. pKR72 также содержит NotI рестрикционный сайт, фланкированный данным промотором для α′ субъединицы β-конглицинина (Beachy et al., EMBO J. 4:3047-3053 (1985)), и 3′ транскрипционный терминирующий участок фазеолинового гена (Doyle et al., J. Biol. Chem. 261:9228-9238 (1986)), что позволяет осуществлять сильную, специфическую к тканям экспрессию в семенах сои генов, клонированных в NotI сайт.
βcon/NotI/Phas3′ кассету в плазмиде pKR72 (SEQ ID NO:105, с номером доступа АСТК PTA-6019) амплифицировали с использованием олигонуклеотидных праймеров oCon-1 (SEQ ID NO:106) и oCon-2 (SEQ ID NO:107), с использованием VentR® DNA Polymerase (Catalog No. M0254S, New England Biolabs Inc., Beverly, MA) в соответствии с протоколом производителя. Результирующий ДНК фрагмент расщепляли XbaI и клонировали в XbaI сайт pUC19, с образованием pKR179 (SEQ ID NO:108).
EgDHAsyn1 избавляли от pKR1055 (SEQ ID NO:104) путем расщепления NotI и встраивали в NotI сайт плазмиды pKR179 (SEQ ID NO:108) с образованием pKR1057 (SEQ ID NO:109).
SbfI фрагмент pKR1057 (SEQ ID NO:109), содержащий βcon/EgDHAsyn1/Phas3′ кассету, клонировали в SbfI сайт pKR328 (SEQ ID NO:110; которая описана в PCT публикации № WO 2004/071467 и на содержание которой в данной заявке сделана ссылка), содержащий SdD17, с образованием вектора pKR1061 (SEQ ID NO:111). Схематическое изображение pKR1061 представлено на ФИГ.15A.
ПРИМЕР 16
Конструирование экспрессирующего вектора сои pKR973 для коэкспрессии Pavlova lutheri дельта-8-десатуразы (PavD8) с Euglena gracilis дельта-9-элонгазой (EgD9elo) и Mortierella alpina дельта-5-десатуразой (MaD5)
Euglena gracilis дельта-9-элонгаза (EgD9elo):
Клон из Euglena кДНК библиотеки (eeg1c), названный eeg1c.pk001.n5f, содержащий Euglena gracilis дельта-9-элонгазу (EgD9elo; SEQ ID NO:112; которая описана в патентной заявке США № 11/601563 (от 16 ноября 2006 года, которая опубликована 24 мая 2007 года как US-2007-0118929-A1; реестр уполномоченного No. BB-1562) на содержание которой в данной заявке сделана ссылка), использовали как матрицу для амплификации EgD9elo олигонуклеотидными праймерами oEugEL1-1 (SEQ ID NO:113) и oEugEL1-2 (SEQ ID NO:114) с использованием VentR® DNA Polymerase (Cat. No. M0254S, New England Biolabs Inc., Beverly, MA) в соответствии с протоколом производителя. Результирующий ДНК фрагмент встраивали в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), следуя протоколу производителя, с образованием pKR906 (SEQ ID NO:115).
Плазмиду KR906 расщепляли NotI, и фрагмент, содержащий Euglena gracilis дельта-9-элонгазу, клонировали в плазмиду pKR132 (SEQ ID NO:116; которая описана в PCT публикации № WO 2004/071467), с образованием pKR953 (SEQ ID NO:117).
Mortierella alpina дельта-5-десатураза (MaD5):
Вектор pKR287 (SEQ ID NO:118; которая описана в PCT публикации № WO 2004/071467, опубликованной 26 августа 2004 года; на содержание которой в данном тексте сделана ссылка), содержит Mortierella alpina дельта-5-десатуразу (MaD5; SEQ ID NO:119, которая описана в патенте США № 6075183 и PCT публикациях №№ WO 2004/071467 и WO 2005/047479, на содержание которых в данной заявке сделана ссылка), фланкировали соевым глицининовым Gy1 промотором и терминирующим участком легумина горохаА2 3′ (Gy1/MaD5/legA2 кассета). Вектор pKR287 расщепляли SbfI/BsiWI, и фрагмент, содержащий Gy1/MaD5/legA2 кассету, клонировали в SbfI/BsiWI фрагмент pKR277 (SEQ ID NO:120; которая описана в PCT публикации № WO 2004/071467, на содержание которой здесь сделана ссылка) с образованием pK952 (SEQ ID NO:121).
Вектор pKR457 (SEQ ID NO:122), описанный ранее в PCT публикации № WO 2005/047479 (на содержание которой в данной заявке сделана ссылка), содержит NotI сайт, фланкированный соевым трипсиновым ингибиторным промотором Kunitz (КТи) (Jofuku et al., Plant Cell 1:1079-1093 (1989)) и КТи 3′ терминирующим участком, выделение которого описано в патенте США № 6372965, за которым идет соевый альбуминовый транскрипционный терминатор, описанный ранее в PCT публикации № WO 2004/071467 (Kti/NotI/Kti3′Salb3′ кассета). Через ряд стадий субклонирования к 5′ и 3′ концам кассеты Kti/NotI/Kti3′Salb3′ добавляли последовательности, содержащие Asp718 рестрикционные сайты, с образованием SEQ ID NO:123.
Pavlova lutheri дельта-8-десатураза (PavD8):
Pavlova lutheri (CCMP459) получали из культуры морского фитопланктона (Culture of Marine Phytoplankton) (CCMP, West Boothbay Harbor, ME) и выращивали в 250 мл колбах, содержащих 50 мл F/2-Si среды (приготовленной с использованием набора F/2 Family Medium Kit-KIT20F2 и Filtered Seqwater-SEA2 из CCMP) при 26°C и встряхивании при 150 оборотов/минуту. Культуры переносили в новую среду еженедельно с использованием 1:4 (старая культура:новая среда) разведения.
Культуры из 28 колб (1400 мл) объединяли, и клетки таблетировали путем центрифугирования при 1800xg в течение 10 минут, промывали один раз водой и центрифугировали повторно. Полную РНК экстрагировали из результирующей таблетки с использованием реагента RNA STAT-60™ (TEL-TEST, Inc., Friendswood, TX), следуя протоколу производителя. Таким способом из таблетки получали 2,6 мг полной РНК (2,6 мг/мл). мРНК выделяли из 1,25 мг полной РНК с использованием набора mRNA Purification Kit (Amersham Biosciences, Piscataway, NJ), следуя протоколу производителя. Таким способом получали 112 мкг мРНК.
КДНК синтезировали из 224 нг мРНК с использованием системы синтеза SuperScript™ First-Strand Synthesis System для набора RT-PCR Kit (Invitrogen™ Life Technologies, Carlsbad, CA) с предоставленным олиго(dT) праймером, согласно протокола производителя. После РНКаза H оброботки согласно протокола, Pavlova lutheri дельта-8-десатураза (PavD8; SEQ ID NO:124; которая описана в патентной заявке США № 11/737772 (от 20 апреля 2007 года; реестр уполномоченного № BB-1566) на содержание которой в данном тексте сделана ссылка) амплифицировали из результирующей кДНК олигонуклеотидными праймерами PvDES5′Not-1 (SEQ ID NO:125) и PvDES3′Not-1 (SEQ ID NO:126) с использованием условий, описанных ниже.
кДНК (2 мкл) из описанной выше реакции, объединяли с 50 пмоль PvDES5′Not-1 (SEQ ID NO:125), 50 пмоль PvDES3′Not-1 (SEQ ID NO:126), 1 мкл ПЦР нуклеотидной смеси (10 мМ, Promega, Madison, WI), 5 мкл 10X ПЦР буфера (Invitrogen Corporation), 1,5 мкл MgCl2 (50 мМ, Invitrogen Corporation), 0,5 мкл Taq полимеразы (Invitrogen Corporation), и воды до 50 мкл. Условиями реакции были 94°C в течение 3 минут с последующими 35 циклами при 94°C на протяжении 45 c, при 55°C в течение 45 c, и при 72°C в течение 1 минуты. ПЦР завершали при 72°C в течении 7 минут, и затем выдерживали при 4°C. ПЦР реакцию анализировали методом агарозного гель-электрофореза на 5 мкл, и при этом наблюдалась ДНК полоса с молекулярным весом около 1,3 kb. Оставшийся продукт отделяли методом агарозного гель-электрофореза, и ДНК очищали с использованием набора Zymoclean™ Gel DNA Recovery Kit (Zymo Research, Orange, CA), согласно протоколу производителя.
PavD8, фланкированную NotI сайтами, клонировали в NotI сайт модифицированной Kti/NotI/Kti3′Salb3′ кассеты (SEQ ID NO:123), и затем ДНК фрагмент расщепляли Asp718 и встраивали в SbfI сайт pKR952 (SEQ ID NO:121) с образованием pKR970 (SEQ ID NO:127).
Плазмиду pKR953 (SEQ ID NO:117) расщепляли PstI, и фрагмент, содержащий Euglena gracilis дельта-9-элонгазу, клонировали в SbfI сайт pKR970 (SEQ ID NO:127) с образованием pKR973 (SEQ ID NO:128, ФИГ.15B
Таким образом, Pavlova lutheri дельта-8-десатураза могла быть коэкспрессирована с Mortierella alpina дельта-5-десатуразой и Euglena gracilis дельта-9-элонгазой за сильными, специфичными к семенам промоторами.
ПРИМЕР 17
Конструирование экспрессирующего вектора сои pKR1064 для коэкспрессии Euglena gracilis ДГК синтазы 1 (EgDHAsyn1) с Saprolegnia diclina дельта-17-десатуразой (SdD17)
Данный пример описывает конструирование соевого вектора для коэкспрессии EgDHAsyn1 с SdD17 и ацетолактатным синтазным (АЛС) селективным маркером.
PstI фрагмент, содержащий Ann/Sdd17/BD30 кассету из pKR271 (SEQ ID NO:129; которая описана в PCT публикации № WO 2004/071467 и на содержание которой в данном тексте сделана ссылка), встраивали в SbfI сайт pKR226 (SEQ ID NO:130; которая также описана в PCT публикации № WO 2004/071467) с образованием вектора pKR886r (SEQ ID NO:131). Таким способом, Saprolegnia diclina дельта-17-десатуразу (SdD17) клонировали за аннексиновым промотором, который является сильным и специфическим к семенам.
SbfI фрагмент pKR1057 (SEQ ID NO:109), содержащий βcon/EgDHAsyn1/Phas3′ кассету, клонировали в SbfI сайт pKR886r (SEQ ID NO:131), содержащий SdD17, с образованием вектора pKR1064 (SEQ ID NO:132). Схематическое изображение pKR1064 представлено на ФИГ.15C.
ПРИМЕР 18
Конструирование экспрессирующего вектора сои pKR1133 для коэкспрессии Euglena gracilis ДГК синтазы 1 (EgDHAsyn1) с Euglena gracilis дельта-9-элонгазой (EgD9elo) и Mortierella alpina дельта-5-десатуразой (MaD5)
Глицининовый Gy1 промотор ПЦР амплифицировали из pZBL119 (SEQ ID NO:133; которая описана в PCT публикации № WO 2004/071467 и на содержание которой в данной заявке сделана ссылка) с использованием праймеров oSGly-2 (SEQ ID NO:134) и oSGly-3 (SEQ ID NO:135). Результирующий ПЦР фрагмент субклонировали в промежуточный клонирующий вектор ПЦР-Script AMP SK(+) (Stratagene), в соответствии с протоколом производителя, с образованием плазмиды pPSgly32 (SEQ ID NO:136).
PstI/NotI фрагмент плазмиды pSGly32 (SEQ ID NO:136), содержащий Gy1 промотор, клонировали в PstI/NotI фрагмент из плазмиды pKR142 (SEQ ID NO:137; которая описана в PCT публикации № WO 2004/071467 и на содержание которой в данной заявке сделана ссылка), содержащий легуминА2 3′ транскрипционный терминирующий участок, ампициллин резистентный ген, и бактериальный источник репликации (ori), с образованием pKR264 (SEQ ID NO:138). Таким образом, вектор pKR264 содержит NotI сайт, фланкированный промотором для глицининового Gy1 гена и легуминА2 3′ транскрипционным терминирующим участком (Gy1/NotI/legA2 кассета).
EgDHAsyn1 высвобождали из pKR1055 (SEQ ID NO:104; Пример 15) путем расщепления NotI и клонировали в NotI сайт плазмиды pKR264 (SEQ ID NO:138), с образованием pKR1128 (SEQ ID NO:139).
NotI фрагмент pKS129 (SEQ ID NO:140; которая описана в PCT публикации № WO 2004/071467 и на содержание которой в настоящей заявке сделана ссылка), содержащий MaD5, клонировали в NotI сайт pKR457 (SEQ ID NO:122; Пример 16), с образованием pKR606 (SEQ ID NO:141).
Вектор pKR606 (SEQ ID NO:141) расщепляли BsiWI и после наполнения для затупления концов фрагмент, содержащий Gy1/MaD5/legA2 кассету, встраивали в наполненный NgoMI сайт pKR277 (SEQ ID NO:120) с образованием pKR804 (SEQ ID NO:142).
BsiWI фрагмент из pKR1128 (SEQ ID NO:139), содержащий Gy1/EgDHAsyn1/legA2 кассету, клонировали в BsiWI сайт pKR804 (SEQ ID NO:142) с образованием pKR1130 (SEQ ID NO:143).
Плазмиду pKR953 (SEQ ID NO:117) расщепляли BsiWI; концы затупляли путем наполнения; и pKR953 потом расщепляли BamHI. Наполненный BsiWI/BamHI фрагмент pKR953, содержащий Salb/EgD9Elo/Phas3′ кассету, клонировали в PmeI/BamHI сайты pNEB193 (New England Biolabs, Ipswich, MA) с образованием pKR1131 (SEQ ID NO:144).
Плазмиду pKR1131 (SEQ ID NO:144) расщепляли PstI, и фрагмент, содержащий Euglena gracilis дельта-9-элонгазу, встраивали в SbfI сайт pKR1130 (SEQ ID NO:143) с образованием pKR1133 (SEQ ID NO:145, ФИГ.15D).
Таким образом, Euglena gracilis ДГК синтаза 1 могла быть коэкспрессирована с Mortierella alpina дельта-5-десатуразой и Euglena gracilis дельта-9-элонгазой за сильными, специфическими по отношению к семенам промоторами.
ПРИМЕР 19
Конструирование экспрессирующего вектора сои pKR1105 для коэкспрессии Euglena gracilis ДГК синтаза 1 C20-элонгазного домена (EgDHAsyn1C20EloDom1) с Schizochytrium aggregatum дельта-4-десатуразой (SaD4)
βcon/NotI/Phas кассету ПЦР амплифицировали из pKS123 (SEQ ID NO:146; которая описана в PCT публикации № WO 2004/071467 и на содержание которой в настоящей заявке сделана ссылка) с использованием праймеров oKti5 (SEQ ID NO:147) и oKti6 (SEQ ID NO:148). Результирующий ПЦР фрагмент расщепляли BsiWI и клонировали в BsiWI сайт pKR124 (SEQ ID NO:149; которая описана в PCT публикации № WO 2004/071467 и на содержание которой в настоящем изобретении сделана ссылка), содержащий бактериальный источник репликации и селекции, с образованием плазмиды pKR193 (SEQ ID NO:150).
EgDHAsyn1C20Elodom1 выделяли из pHD16 (SEQ ID NO:51; Пример 10) путем расщепления NotI и был встраивали в NotI сайт плазмиды pKR193 (SEQ ID NO:150) с образованием pKR1103 (SEQ ID NO:151).
BsiWI фрагмент, содержащий EgDHAsyn1C20Elodom1, выделяли из pKR1103 (SEQ ID NO:151) и клонировали в BsiWI сайт pKR226 (SEQ ID NO:130; Пример 17) с образованием вектора pKR1104 (SEQ ID NO:152).
Вектор pKR300 (SEQ ID NO:153; которая описана в PCT публикации № WO 2004/071467, опубликованной 26 августа 2004 года; на содержание которой в настоящем изобретении сделана ссылка), содержит Schizochytrium aggregatum дельта-4-десатуразу (SaD4), которая описана в патенте США № 7045683 и PCT публикацию № WO 02/090493, на содержание которых в настоящем изобретении сделаны ссылки), фланкированную NotI рестрикционными сайтами. AscI сайт, присутствующий в SaD4, удаляли без воздействия на соответствующую аминокислотную последовательность с образованием новой последовательности (SEQ ID NO:154), которая остается фланкированной NotI сайтами. NotI фрагмент (SEQ ID NO:154) клонировали в NotI сайт плазмиды pKR457 (SEQ ID NO:122; Пример 16) с образованием pKR1102 (SEQ ID NO:155).
Плазмиду pKR1102 (SEQ ID NO:155) расщепляли PstI, и фрагмент, содержащий SaD4, клонировали в SbfI сайт pKR1104 (SEQ ID NO:152) с образованием pKR1105 (SEQ ID NO:156; ФИГ.16A). Таким способом, Euglena gracilis ДГК синтаза 1 домен C20-элонгазы мог быть коэкспрессирован с Schizochytrium aggregatum дельта-4-десатуразой за сильными, специфическими по отношению к семенам промоторами.
ПРИМЕР 20
Конструирование экспрессирующего вектора сои pKR1134 для экспрессии Euglena gracilis ДГК синтаза 1 C20-элонгазного домена/Schizochytrium aggregatum дельта-4-десатуразного гибрида (EgDHAsyn1C20EloDom3-SaD4)
EgDHAsyn1C20EloDom3 амплифицировали из pKR1091 олигонуклеотидными праймерами EgEPAEloDom-5 (SEQ ID NO:43) и oEUGsyn6-4 (SEQ ID NO:157) с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S, Finnzymes Oy, Finland) в соответствии с протоколом производителя. Результирующий ДНК фрагмент встраивали в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), следуя протоколу производителя, с образованием pKR1107 (SEQ ID NO:158).
Плазмиду pKR1107 (SEQ ID NO:158) расщепляли NotI, и фрагмент, содержащий EgDHAsyn1C20EloDom3, религировали с образованием pKR1112 (SEQ ID NO:159).
XbaI/PstI ДНК фрагмент из pKR1112 (SEQ ID NO:159), содержащий EgDHAsyn1C20EloDom3, клонировали в XbaI/SbfI ДНК фрагмент из pKR1068 (SEQ ID NO:75; Пример 11), содержащий SaD4, с образованием pKR1115 (SEQ ID NO:160). Таким образом, EgDHAsyn1C20Elodom3-SaD4 повторно создали без внутреннего SbfI сайта, но кодирует идентичную аминокислотную последовательность, как это описано в Примере 11.
EgDHAsyn1C20Elodom3-SaD4 выделяли из pKR1115 (SEQ ID NO:160) путем расщепления NotI и клонировали в NotI сайт плазмиды pKR1104 (SEQ ID NO:152), содержащей ALS селектируемый маркер, с образованием pKR1134 (SEQ ID NO:161; ФИГ.16B).
ПРИМЕР 21
Конструирование экспрессирующего вектора сои pKR1095 для коэкспрессии Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразы (TpomD8) с Saprolegnia diclina дельта-17-десатуразой (SdD17)
Настоящий пример описывает конструирование соевого вектора для коэкспрессии TpomD8 с SdD17 и гигромициновым фосфотрансферазным селектируемым (гфт) маркером.
Tetruetreptia pomquetensis CCMP1491 клетки (из 1 литра культуры) приобретали на фирме Provasoli-Guillard National Center for Culture of Marine Phytoplakton (CCMP) (Bigelow Laboratory for Ocean Sciences, West Boothbay Harbor, Maine). Полную РНК выделяли с использованием тризолового реактива (Invitrogen, Carlsbad, CA), согласно протоколу производителя. Клеточную таблетку ресуспендировали в 0,75 мл тризолового реактива, смешанного с 0,5 мл 0,5 мм стеклянных шариков, и гомогенизировали в приборе Biospec mini beadbeater (Bartlesville, OK) при установке самых высоких параметров в течение 3 минут. Данную смесь центрифугировали на центрифуге Eppendorf в течение 30 с при 14000 оборотов/минуту для удаления остатков и стеклянных шариков. Супернатант экстрагировали 150 мкл смеси 24:1 хлороформ:изоамиловый спирт. Верхнюю водную фазу использовали для РНК излоляции.
Для выделения РНК водную фазу смешивали с 0,375 мл изопропилового спирта, и ее инкубировали при комнатной температуре в течение 5 минут. Осажденную РНК собирали путем центрифугирования при 8000 оборотов/минуту и выдерживали при 4°C в течение 5 минут. Таблетку промывали один раз 0,7 мл 80% этанола и высушивали на воздухе. Таким образом получали 95 мкг полной РНК из Tetruetreptia pomquetensis CCMP1491.
Полную РНК (0,95 мкг полной РНК в 1 мкл) использовали как матрицу для синтеза двухцепочечной кДНК. Использовали набор CreatorTM SMARTTM cDNA Library Construction Kit от фирмы BD Bioscience Clontech (Palo Alto, CA). Полную РНК (1 мкл) смешивали с 1 мкл SMART IV олигонуклеотида (SEQ ID NO:181), 1 мкл адаптерного праймера из набора Invitrogen 3′-RACE kit (SEQ ID NO:182), и 2 мкл воды. Данную смесь нагревали до 75°C на 5 минут и потом охлаждали на льду в течение 5 минут. К данной смеси добавляли: 2 мкл 5х буфера первой цепочки, 1 мкл 20 мМ DTT, 1 мкл смеси dNTP (10 мМ каждого из dATP, dCTP, dGTP и dTTP), и 1 мкл PowerScript обратной транскриптазы. Образец инкубировали при 42°C в течение 1 часа. Результирующие первоцепочечные кДНК затем использовали как матрицы для амплификации.
Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразу (TpomD8; SEQ ID NO:162; которая описана в патентной заявке США № 11/876115 (от 22 октября 2007 года; реестр уполномоченного No. BB-1574), на содержание которой в настоящей заявке сделана ссылка) амплифицировали из кДНК олигонуклеотидными праймерами TpomNot-5 (SEQ ID NO:163) и TpomNot-3 (SEQ ID NO:164) с использованием Taq полимеразы (Invitrogen Corporation) согласно протокола производителя.
Tetruetreptia pomquetensis CCMP1491 cDNA (1 мкл) комбинировали с 50 пмоль TpomNot-5 (SEQ ID NO:163), 50 пмоль TpomNot-3 (SEQ ID NO:164), 1 мкл ПЦР нуклеотидной смесью (10 мМ, Promega, Madison, WI), 5 мкл 10х ПЦР буфера (Invitrogen Corporation), 1,5 мкл MgCl2 (50 мМ, Invitrogen Corporation), 0,5 мкл Taq полимеразы (Invitrogen Corporation) и воды до 50 мкл. Условиями реакции были 94°C в течение 3 минут с последующими 35 циклами при 94°C на протяжении 45 c, при 55°C в течение 45 c, и при 72°C в течение 1 минуты. ПЦР завершали при 72°C в течении 7 минут, и затем выдерживали при 4°C. 5 мкл ПЦР реакционной смеси анализировали методом агарозного гель-электрофореза, и при этом наблюдалась ДНК полоса с молекулярным весом около 1,3 kb.
Оставшийся продукт отделяли методом агарозного гель-электрофореза, и ДНК очищали с использованием набора Zymoclean™ Gel DNA Recovery Kit (Zymo Research, Orange, CA), следуя протоколу производителя. Результирующую ДНК клонировали в pGEM®-T Easy Vector (Promega), следуя протоколу производителя, с образованием pLF114-10 (SEQ ID NO:165).
TpomD8 выделяли из pLF114-10 (SEQ ID NO:165) путем расщепления NotI и клонировали в NotI сайт плазмиды pKR179 (SEQ ID NO:108; Пример 15) с образованием pKR1002 (SEQ ID NO:166).
PstI фрагмент pKR1002 (SEQ ID NO:166), содержащий βcon/TpomD8/Phas3′ кассету, клонировали в SbfI сайт pKR328 (SEQ ID NO:110; Пример 15), содержащий SdD17, с образованием вектора pKR1095 (SEQ ID NO:167). Схематическое изображение pKR1095 представлено на ФИГ.16C.
ПРИМЕР 22
Конструирование экспрессирующего вектора сои pKR1132 для коэкспрессии Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразы (TpomD8) с Euglena gracilis дельта-9-элонгазой (EgD9elo) и Mortierella alpina дельта-5-десатуразой (MaD5)
TPomD8 выделяли из pLF114-10 (SEQ ID NO:165; Пример 21) путем расщепления NotI и клонировали в сайт плазмиды pKR264 (SEQ ID NO:138; Пример 18) с образованием pKR1127 (SEQ ID NO:168).
BsiWI фрагмент из pKR1127 (SEQ ID NO:168), содержащий Gy1/TPomD8/legA2 кассету, клонировали в BsiWI сайт pKR804 (SEQ ID NO:142; Пример 18) с образованием pKR1129 (SEQ ID NO:169).
Плазмиду pKR1131 (SEQ ID NO:144; Пример 18) расщепляли PstI, и фрагмент, содержащий Euglena gracilis дельта-9-элонгазу, клонировали в SbfI сайт pKR1129 (SEQ ID NO:169) с образованием pKR1132 (SEQ ID NO:170, ФИГ.16D). Таким способом Tetruetreptia pomquetensis дельта-8-десатураза могла быть коэкспрессирована с Mortierella alpina дельта-5-десатуразой и Euglena gracilis дельта-9-элонгазой за сильными, специфическими в отношении семян промоторами.
ПРИМЕР 23
Конструирование экспрессирующего вектора сои KS373 для экспрессии Euglena gracilis дельта-9-элонгаза/Euglena gracilis ДГК синтаза 1 линкер/Pavlova lutheri дельта-8-десатуразного гибрида (EgD9elo-EgDHAsyn1Link-PavD8)
Внутрикаркасный гибрид Euglena gracilis дельта-9-элонгазы (EgD9elo; Пример 16; SEQ ID NO:112), Euglena gracilis ДГК синтаза 1 обогащенного пролином линкера (EgDHAsyn1Link; SEQ ID NO:171; описана в Примере 6 и показана на ФИГ.6), и Pavlova lutheri дельта-8-десатуразы (PavD8; Пример 16; SEQ ID NO:124) создавали при описанных ниже условиях.
Был создан первичный внутрикаркасный гибрид между EgD9elo и EgDHAsyn1Link (EgD9elo-EgDHAsyn1Link), фланкированный NcoI сайтом в 5′конце и NotI сайтом в 3′ конце, путем ПЦР амплификации. EgD9elo (SEQ ID NO:112) амплифицировали олигонуклеотидами MWG507 (SEQ ID NO:172) и MWG509 (SEQ ID NO:173), с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S, Finnzymes Oy, Finland), следуя протоколу производителя. EgDHAsyn1Link (SEQ ID NO:171) амплифицировали схожим способом олигонуклеотидами MWG510 (SEQ ID NO:174) и MWG511 (SEQ ID NO:175). Два результирующих ПЦР продукта объединяли и повторно амплифицировали с использованием MWG507 (SEQ ID NO:172) и MWG511 (SEQ ID NO:175) с образованием EgD9elo-EgDHAsyn1Link. Последовательность EgD9elo-EgDHAsyn1Link представлена в SEQ ID NO:176. EgD9elo-EgDHAsyn1Link не содержит внутрикаркасного терминирующего кодона вверху NotI сайта в 3′ конце, и поэтому ДНК фрагмент, клонированный в NotI site, может дать начало внутрикаркасному слиянию с EgD9elo-EgDHAsyn1Link.
Плазмида KS366 (SEQ ID NO:177) содержит уникальные NcoI и NotI рестрикционные сайты, фланкированные промотором для α′ субъединицы β-конглицинина (Beachy et al., EMBO J. 4:3047-3053 (1985)) и 3′ транскрипционным терминирующим участком фазеолинового гена (Doyle et al., J. Biol. Chem. 261:9228-9238 (1986)). За исключением замены уникального NotI сайта в pKR72 (SEQ ID NO:105) уникальным NcoI/NotI составным клонирующим сайтом, Bcon/NcoINotI/Phas3′ кассета в KS366 идентична найденной в pKR72 (SEQ ID NO:105), кроме того, что фланкирующие HindIII сайты были замещены BamHI сайтами. Bcon/NcoINotI/Phas3′ кассету KS366 встраивали в BamHI сайт pBluescript II SK(+) вектор (Stratagene).
NcoI/NotI ДНК фрагмент, содержащий EgD9elo-EgDHAsyn1Link (SEQ ID NO:176), клонировали в NcoI/NotI ДНК фрагмент из KS366 (SEQ ID NO:177), содержащий промотор для α′ субъединицы β-конглицинина, с образованием KS366-EgD9elo-EgDHAsyn1Link (SEQ ID NO:178).
NotI фрагмент, содержащий PavD8 (генерированный как описано в Примере 16), клонировали в NotI фрагмент KS366-EgD9elo-EgDHAsyn1Link (SEQ ID NO:178) с образованием KS373 (SEQ ID NO:179; ФИГ.17).
ПРИМЕР 24
Конструирование дополнительных экспрессирующих векторов сои для экспрессии ДГК синтаз, C20-элонгазных доменов, дельта-4-десатуразных доменов, синтетических C20-элонгаза/дельта-4-десатуразных слитых протеинов и других синтетических элонгаза/десатуразных слитых протеинов
В дополнение к генам, промоторам, терминаторам и генным кассетам, описанным здесь, специалисту в данной области понятно, что другие промотор/ген/терминаторные кассетные комбинации могут быть синтезированы способом, схожим, однако не ограничиваясь этим, с описанным здесь для экспрессии EgDHAsyn1. Подобно этому, может возникнуть необходимость в экспрессии других ПНЖК генов (таких как те, что описаны в Таблице 23) для коэкспрессии с любой из ДГК синтаз настоящего изобретения или ДГК синтазными доменами (т.е. домен C20-элонгазы или дельта-4-десатуразный домен экспрессированы индивидуально). Кроме того, могут быть осуществлены и экспрессированы синтетические слияния между элонгазным доменом и десатуразным доменом, разделенными подходящим линкерным участком. Например, может быть осуществлено и экспрессировано синтетическое слияние между C20-элонгазой (или C20-элонгазным доменом из ДГК синтазы) и подходящей дельта-4-десатуразой (или дельта- десатуразным доменом из ДГК синтазы). Как альтернатива, могут быть использованы другие элонгазы или десатуразы, такие как, однако не ограничиваясь этим, для описанного здесь синтетического слияния между дельта-9-элонгазой и дельта-8-десатуразой, которые разделены линкером из ДГК синтазы (т.е. Пример 23).
Например, PCT публикации №№ WO 2004/071467 и WO 2004/071178 описывают выделение ряда промоторных и транскрипционных последовательностей для использования в эмбрион-специфической экспрессии в сое. Кроме того, PCT публикации №№ WO 2004/071467, WO 2005/047479 и WO 2006/012325 описывают синтез множества промотор/ген/терминаторных кассетных комбинаций путем лигирования индивидуальных промоторов, генов и транскрипционных терминаторов в уникальные комбинации. В общем, NotI сайт, фланкированный подходящим промотором (таким как те, что перечислены, но не ограничиваясь этим, в Таблице 21) и транскрипционным терминатором (таким, как те, что перечислены, но не ограничиваясь этим, в Таблице 22), используется для клонирования желательного гена. NotI сайты могут добавляться к требуемому гену, как это перечислено, однако не ограничиваясь этим, в Таблице 23, с использованием ПЦР амплификации олигонуклеотидами. созданными для введения NotI сайтов в 5′ и 3′ концы данного гена. Затем результирующий ПЦР продукт расщепляется NotI и встраивается в подходящую промотор/NotI/терминирующую кассету.
В дополнение, PCT публикации №№ WO 2004/071467, WO 2005/047479 и WO 2006/012325 описывают дальнейшее связывание индивидуальных генных кассет в уникальные комбинации, совместно с подходящими селективными маркерными кассетами, для получения необходимой фенотипической экспрессии. Хотя это осуществляется, главным образом, с использованием разных рестрикционных ферментных сайтов, специалисту в данной области понятно, что для получения необходимой промотор/ген/транскрипционной терминаторной комбинации может быть использован ряд методов. При осуществлении этого может быть достигнута любая комбинация эмбрион-специфических промотор/ген/транскрипционных терминирующи кассет. Специалисту в данной области также понятно, что эти кассеты могут быть локализованы на отдельных ДНК фрагментах или на множественных фрагментах, где выходом котрансформации множественных ДНК фрагментов является коэкспрессия генов.
ПРИМЕР 25
Продуктивная и модельная системная трансформация соматических соевых эмбриональных культур соевыми экспресирующими векторами
Условия культивирования:
Соевые эмбриогенные суспензионные культуры (cv. Jack) выдерживали в 35 мл жидкой среды SB196 (ниже) на роторном шейкере, 150 оборотов/минуту, 26°С, при условиях освещения холодным белым люминесцентным излучением при 16:8 часовом фотопериоде день/ночь и интенсивности света 60-85 мкЕ/м2/с. Культуры пересеивали каждые 7 суток до двух недель путем инокуляции приблизительно 35 мг ткани в 35 мл свежей жидкости SB196 (интервал пересеивания, которому отдается предпочтение, составляет 7 суток).
Соевые эмбриогенные суспензионные культуры трансформировали соевыми экспрессирующими плазмидами по методу пушечной бомбардировки частицами (Klein et al., Nature, 327:70 (1987)) с использованием инструмента (гелиевая модификация) DuPont Biolistic PDS1000/HE для всех трансформаций.
Инициация соевых эмбриогенных суспензионных культур:
Соевые культуры подвергали инициации два раза в месяц с интервалами 5-7 суток между каждой инициацией. Стручки с недозрелыми семенами от наличных растений сои собирали через 45-55 суток после высаживания. Освобождали от шелухи и помещали в стерилизированную фуксиновую камеру. Семена сои стерилизовали, встряхивая их в течение 15 минут в 5% растворе Clorox с одной каплей дентинового мыла (т.е. 95 мл кипяченой дистиллированной воды плюс 5 мл Clorox и одна капля мыла, хорошо перемешанных). Семена ополаскивали, с использованием двух 1-литровых бутылок стерильной дистиллированной воды, и семена, меньшие 4 мм, помещали на отдельные предметные стекла. Малый конец семечка отрезали, и котиледоны выдавливали из оболочки семени. Когда культуры готовили для продуктивной трансформации, котиледоны переносили на плашки, содержащие SB1 среду (25-30 котиледонов на плашку). Плашки оборачивали лентой из фибры и сохраняли при 26°С в условиях освещения холодным белым люминесцентным излучением при 16:8 часовом фотопериоде день/ночь и интенсивности света 60-80 мкЕ/м2/с в течение восьми недель, со сменой сред через 4 недели. Во время подготовки культур к исследованию модельной системы, котелидоны переносили на плашки, содержищие среду SB199 (25-30 котелидонов на плашку) на 2 недели и затем переносили в SB1 на 2-4 недели. Условия температуры и освещения оставались как и описано выше. После инкубации на SB1 среде вторичные эмбрионы вырезали и помещали в SB196 жидкую среду на 7 суток.
Приготовление ДНК к бомбардировке:
Для бомбардировки использовали или нетронутую плазмиду или ДНК плазмидный фрагмент, содержащий нужные гены и селективный маркерный ген. Фрагменты от соевых экспрессирующих плазмид, конструкция которых здесь описана, получали путем гель-изоляции расщепленных плазмид. В каждом случае 100 мкг плазмидной ДНК использовали в 0,5 мл специальной ферментной смеси, которая описана ниже. Плазмиды подвергали расщеплению Ascl (100 единиц) в NEBuffer 4 (20 мМ Tris-ацетата, 10 мМ ацетата магния, 50 мМ ацетата калия, 1 мМ дитиотреитола, рН 7,9), 100 мкг/мл BSA и 5 мМ бета-меркаптоэтанола при 37оС в течение 1,5 часов. Результирующие ДНК фрагменты разделяли по методу гель-электрофореза на 1% SeaPlaque GTG агарозе (BioWhitaker Molecular Application), и ДНК фрагменты, которые содержали генные кассеты, вырезали из агарозного геля. ДНК очищали от агарозы с использованием GELase расщепляющего фермента в соответствии с протоколом производителя.
50 мкл аликвотного количества стерильной дистиллированной воды, которая содержала 3 мг частиц золота (3 мг золота), добавляли к 30 мкл 10 нг/мкл ДНК раствора (или нетронутая плазмида или ДНК фрагмент, полученный как описано выше), 25 мкл 5М CaCl2 и 20 мкл 0,1М спермидина. Данную смесь встряхивали в течение 3 минут на уровне 3 вихревого шейкера и центрифугировали в течение 10 с в лабораторной микроцентрифуге. Супернатант удаляли с последующей промывкой 400 мкл 100% этанола и путем еще одного короткого центрифугирования. 400 мкл этанола удаляли, и таблетку ресуспендировали в 40 мкл 100% этанола. По пять мкл ДНК суспензии дозировали в каждый подвижный диск дискового инструмента Biolistic PDS 1000/HE. Каждые 5 мкл аликвотного количества содержали приблизительно 0,375 мг золота на одну бомбардировку (т.е. на диск).
Для модельных системных трансформаций протокол идентичен, за исключением некоторых незначительных изменеий (например, 1 мг частиц золота прибавляли к 5 мкл раствора ДНК 1 мкг/мкл: 50 мкл 2,5 М CaCl2 и гранулы полностью ресуспендировали в 85 мкл 100% етанола получая при этом 0,058 мг золота за каждую).
Приготовление ткани и бомбардировка ДНК:
Приблизительно 150-200 мг эмбриогеных суспензионных культур возрастом 7 суток помещали в пустую, стерильную 60×15 мм чашку Петри, и последнюю накрывали пластиковой сеткой. Камеру вакуумировали до давления 27-28 дюйм рт.ст., и ткань бомбардировали 1 или 2 ударами на пластинку при давлении разрыва мембраны, установленном на 1100 фунт/кв.дюйм. Ткань помещали приблизительно за 3,5 дюйма от удерживающего/останавливающего экрана. Условия для мдельных системных трансформаций идентичны, за исключением использования эмбриогенной ткани в количестве 100-150 мг; устанавливали давление разрыва 650 фунт/кв.дюйм; и ткань помещали приблизительно на расстоянии 2,5 от удерживающего экрана.
Селекция трансформированных зародышей:
Трансформированные зародыши подвергали селекции с использованием гигромицина (когда селективный маркер используется как гигромицин В фосфотрансферазный (ГФТ) ген) или хлоросульфурона (когда селективный маркер используется как ацетолактатсинтазный (АЛС) ген).
После бомбардировки ткань помещали в свежую SB196 среду и культивировали как описано выше. Через шесть-восемь суток после бомбардировки SB196 обменивали на свежую SB196, которая содержала либо 30 нг/л, либо 100 нг/л хлоросульфурона, в зависимости от использованного селективного маркера. Селекционные среды обновляли каждую неделю. Через четыре-шесть недель после селекции можно было наблюдать зеленую, трансформированную ткань, которая росла из нетрансформированных, некротических эмбриогенных кластеров.
Вызревание зародышей:
Для продуктивных трансформаций изолированная, зеленую ткань удаляли и инокулировали в многоячеечные планшеты для генерации новых, клонально размножаемых, трансформированных эмбриогенных суспензионных культур. Трансформированные эмбриогенные кластеры культивировали в течение четырех-шести недель в многоячеечных планшетах при 26°С в SB196 под белыми люминесцентными лампами холодного излучения (Phillips cool white Econowatt F40/CW/RS/EW) и Agro (Phillips F40 Agro) (40 Вт) при 16:8 часовом фотопериоде и интенсивности света 90-120 мкЕ/м2/с. После завершения этого срока зародышевые кластеры помещали в твердую агарозную среду, SB166, на 1-2 недели и потом пересеивали в среду SB103 на 3-4 недели для созревания зародышей. После вызревания на планшетах в SB103 отдельные эмбрионы удаляли из кластеров, высушивали и подвергали скринингу относительно изменений в их составах жирных кислот как описано supra.
Для модельных системных трансформаций эмбрионы созревали в соевой жидкой среде для гистодифференциации и вызревания (жидкие среды SHaM; Schmidt et al., Cell Biology and Morphogenesis 24:393 (2005)) с использованием модифицированной процедуры. Вкратце, после 4 недель селекции в SB196, как описано выше, эмбрионные кластеры переносили в 35 мл SB228 (жидкие среды SHaM) в 250 мл колбе Эрленмейера. Ткань выдерживали в жидкой среде SHaM на роторном шейкере при 130 оборотов/минуту и 26°С под белыми люминесцентными лампами холодного излучения при 16:8 часовом фотопериоде день/ночь и интенсивности света 60-85 мкЕ/м2/с в течение 2 недель созревания зародышей. Эмбрионы, выращенные на протяжении 2 недель в жидкой среде SHaM, были эквивалентны по размерам и содержанию жирных кислот эмбрионам, культивированным на SB166/SB103 в течение 5-8 недель.
После вызревания в жидкой среде SHaM отдельные эмбрионы удаляли из кластеров, высушивали и подвергали скринингу относительно изменений в их составах жирных кислот как описано supra.
Рецепты сред:
SB 196 - FN Lite жидкая пролиферационная среда (на литр)
FN Lite маточные растворы
SB1 твердая среда (на литр)
1 упаковка MS солей (Gibco/BRL-Cat. No. 11117-066)
1 мл B5 витаминов 1000х маточный раствор
31,5 г глюкозы
2 мл 2,4-D (20 мг/л конечная концентрация)
pH 5,7
8 г TC агара
SB199 твердая среда (на литр)
1 упаковка MS солей (Gibco/BRL-Cat. No. 11117-066)
1 мл B5 витаминов 1000×маточный раствор
30 г сахарозы
4 мл 2,4-D (40 мг/л конечная концентрация)
pH 7,0
2 г гельрита
SB 166 твердая среда (на литр)
1 упаковка MS солей (Gibco/BRL-Cat. No. 11117-066)
1 мл B5 витаминов 1000х маточный раствор
60 г мальтозы
750 мг MgCl2 гексагидрата
5 г активированного угля
pH 5,7
2 г гельрита
SB 103 твердая среда (на литр)
1 упаковка MS солей (Gibco/BRL-Cat. No. 11117-066)
1 мл B5 витаминов 1000х маточный раствор
60 г мальтозы
750 мг MgCl2 гексагидрата
pH 5,7
2 г гельрита
SB 71-4 твердая среда (на литр)
1 бутылка Gamborg′s B5 солей w/сахароза (Gibco/BRL-Cat. No. 21153-036)
pH 5,7
5 г TC агара
2,4-D маточный раствор
Получен предварительно приготовленным от Phytotech Cat. No. D 295 - концентрациія 1 мг/мл
B5 витаминный маточный раствор (на 100 мл)
Сохранять аликвотные количества при -20°C
10 г мио-инозитола
100 мг никотиновой кислоты
100 мг пиридоксина HCl
1 г тиаминау
Если раствор не растворяется достаточно быстро, подвести низкий уровень тепла через нагревательную пластину.
SB 228- гистодифференциация и вызревание сои (SHaM) (на литр)
FN-lite макро для SHAM 10X- маточный раствор № 1 (на литр)
MS микро 1000х-маточный раствор №2 (на литр)
FeEDTA 100х-маточный раствор №3 (на литр)
Ca 100х-маточный раствор №4 (на литр)
CaCl2*2H20 (кальция хлорид дигидрат) 44 г
Довести до объема
Автоклав
B5 витамин 1000х-маточный раствор №5 (на литр)
4% глутамин- маточный раствор № 6 (на литр)
ПРИМЕР 26
Хлоросульфуроновая селекция (ALS) и регенерация растений
Хлоросульфуроновая (ALS) селекция:
После бомбардировки ткань разделяли между двумя колбами со свежей SB196 средой и культивировали как описано в Примере 25. Через шесть-семь суток после бомбардировки SB196 обменивали на свежую SB196, содержащую селекционный агент из 100 нг/мл хлоросульфурона (хлоросульфуроновый маточный раствор представляет собой 1 мг/мл в 0,01 N гидроксиде аммония). Селекционную среду обновляли еженедельно. Через четыре-шесть недель после селекции можно было наблюдать зеленую, трансформированную ткань, растущую из нетрансформированных, некротических эмбриогенных кластеров. Выделенную зеленую ткань удаляли и инокулировали в многоячеечные планшеты, содержащие SB196, и эмбрионы созревали как описано в Примере 25.
Регенерация соевых соматических эмбрионов в растения:
Чтобы получить целые растения из эмбриогенных суспензионных культур, данная ткань должна быть регенерирована. Эмбрионы созревали как описано в Примере 25. После пересева на среде SB103 в течение 3 недель отдельные зародыши могли быть удалены из кластеров и подвергали скринингу в отношении их жирнокислотного состава, как здесь описано. Следует отметить, что на этой стадии мог быть идентифицирован любой обнаружимый фенотип, проистекающий из экспрессии необходимых генов. Это включает, однако не ограничиваясь этим, изменения в жирнокислотном профиле, протеиновом профиле и содержимом, содержании углеводов, скорости роста, жизнеспособности или способности к нормальному развитию в соевое растение.
Созревшие отдельные эмбрионы обезвожиали путем помещения их в пустую, маленькую чашку Петри (35×10 мм) приблизительно на 4-7 суток. Чашки герметизировали фибровой лентой (образуя камеру с небольшой влажностью). Осушенные эмбрионы высаживали в SB71-4 среду, где их оставляли до прорастания при тех же самых условиях культивирования, которые описаны выше. Проросшие ростки удаляли из среды для проращивания и ополаскивали тщательно водой, и затем высаживали в 24-ячеечный укутанный лоток Redi-Earth, накрытый прозрачным пластиковым куполом. Через 2 недели купол снимали, и растения закаляли еще в течение недели. Если ростки выглядели крепкими, их пересаживали в 10” горшочки Readi-Earth приблизительно по 3 ростка на горшочек. Через 10-16 недель зрелые семена собирали, расщепляли и анализировали на жирные кислоты.
ПРИМЕР 27
Функциональный анализ Euglena gracilis и Euglena anabaena ДГК синтаз в Yarrowia lipolytica
Каждый из описанных ниже в Таблице 24 экспрессирующих векторов трансформировали в Yarrowia lipolytica штамм Y2224 (урацил ura3 ауксотрофный штамм Yarrowia lipolytica) как описано в разделе Общие методы, выше.
Отдельные колонии трансформанта Yarrowia lipolytica, содержащего соответствующий Yarrowia экспрессирующий вектор (см. Таблицу 24), выращивали в 3 мл МС без урацила с добавкой 0,2% тергитола при 30°C в течение 1 суток. После этого 0,05 мл переносили в 3 мл той же самой среды, дополненной любой жирной кислотой или ЭПК до 0,175 мМ. Эти растворы инкубировали в течение 16 часов при 30°C, 250 оборотов/минуту, и потом центрифугированием получали таблетки. Клетки промывали один раз водой, таблетировали путем центрифугирования и высушивали на воздухе. Таблетки подвергали трансэтерификации (Roughan, G. and Nishida, I., Arch. Biochem. Biophys. 276(1):38-46 (1990)) 500 мкл 1% натрий метоксида в течение 30 минут при 50°C, после чего добавляли 500 мкл 1 M хлорида натрия и 100 мкл гептана. После тщательного перемешивания и центрифугирования жирнокислотные метиловые эфиры (ЖКМЭ) анализировали методом ГХ, как описано supra (см. раздел Общие методы).
Жирнокислотный профиль для Yarrowia, экспрессирующей pBY-EgC20elo1, не выявил элонгации ЭПК до ДПК. Жирнокислотные профили, вычисленные % элонгации и вычисленные % десатурации для оставшихся клонов приведены на ФИГ 18. Процент C20-элонгации (% C20-Elong) вычисляля путем деления суммы весовых процентов (вес. %) для ДПК и ДГК на сумму вес.% для ЭПК, ДПК и ДГК и умножения на 100 для выражения в виде %. Подобным образом, процент дельта-4-десатурации (% D4 Desat) вычисляли путем деления вес. % для ДГК на сумму вес. % для ДПК и ДГК и умножения на 100 для выражения в виде %. Средние значения обозначены как Ave с последующим соответствующим заголовком.
Как резюме ФИГ.18, все из ДГК синтаз, за исключением EaDHAsyn4, функционировали как обе С20-элонгазы (элонгирующие ЭПК в ДПК) и как дельта-4-десатуразы (десатурирующие ДПК в ДГК) в Yarrowia. EaDHAsyn4, которая содержала существенно отличную аминокислотную последовательность в С-окончании вследствие сдвига каркаса в нуклеотидной последовательности, имела значительно сниженную функцию элонгации, и десатуразной активности не обнаруживалось. Экспрессия EgDHAsyn1 в pY132 последовательно приводила к более высокой активности в Yarrowia, при сравнении с другими EgDHAsyn1 конструкциями, вероятно, вследствие того факта, что EgDHAsyn1 экспрессировали как внутрикаркасное слияние между некоторой векторной последовательностью, 5′ UTR EgDHAsyn1, и EgDHAsyn1 кодирующей последовательностью. Созданный результирующий гибрид может привести к возникновению усиленной активности из-за усиленной экспрессии в Yarrowia или из-за прирожденного увеличения активности к самому ферменту. Когда экспрессирована лишь кодирующая последовательность EgDHAsyn1* (т.е. без 5′UTR; см. pY141), активность выше, чем когда 5′UTR присутствует, но не транслирована как слияние (т.е. см. pY161). Это наблюдение, вероятно, обусловлено снижением экспрессии EgDHAsyn1 из-за присутствия 5′UTR.
ПРИМЕР 28
Функциональный анализ EgDHAsyn1 независимого C20-элонгазного и дельта-4-десатуразного доменов и сравнение с гетерологическими слияниями
Каждый из экспрессирующих векторов, описанных ниже в Таблице 25 (и вектор только контроля) был трансформирован в Yarrowia lipolytica штамм Y2224, как описано в Примере 27. ЭПК и/или ДПК подавали в качестве подпитки к трансформированным Yarrowia клеткам. Схема, изображающая относительную доменную структуру для каждой конструкции в Таблице 25, приведена на ФИГ.21.
Жирнокислотные профили трансформантных клеток впоследствии анализировали как описано в Примере 27.
Результаты для подачи ЭПК к вектору только для контроля, pY141, pY143, pY149, pY156, pY157 и pY160 показаны на ФИГ.19. Жирнокислотные профили для Yarrowia, экспрессирующей pY150, pY151, pY152 и pY153, не выявили элонгации ЭПК в ДПК и на ФИГ.19 не показаны.
Результаты для подачи ДПК к вектору только контроля, pY141, pY150, pY151, pY152, pY153, pY156, pY157 и pY160 показаны на ФИГ.20. Жирнокислотные профили для Yarrowia, экспрессирующей pY143 и pY149, не выявили десатурации ДПК в ДГК и на ФИГ,20 не показаны.
Процент C20-элонгации (% C20 Elong), процент дельта-4-десатурации (% D4 Desat) и средние значения вычислены как описано в Примере 27.
Как резюме ФИГ.19 и ФИГ.20, когда EgDHAsyn1C20Elo домен экспрессирован один (с или без линкера; т.е. pY143 и pY149), средний процент C20-элонгации возрастает приблизительно на 40%, по сравнению с нативным EgDHAsyn1* (pY141; SEQ ID NO:49). Противоположное имеет место с EgDHAsyn1 дельта-4-десатуразным доменом, где активность с EgDHAsyn1D4Dom1 (pY152; SEQ ID NO:67; см. ФИГ.20) отсутствует, и приблизительно на 50% меньше с EgDHAsyn1D4Dom2 (pY153; SEQ ID NO:72; см. ФИГ.20), когда он экспрессирован один, в сравнении с EgDHAsyn1* (pY141; SEQ ID NO:49; см. ФИГ.20), питаемым ДПК. IgD4 не обладает активностью дельта-4-десатуразы, когда он экспрессирован один (pY150; SEQ ID NO:62; см. ФИГ.20) или как слияние (pY156; SEQ ID NO:64; см. ФИГ.19 и 20) и даже обусловливает приблизительно 50% снижение элонгационной активности при слиянии с EgDHAsyn1C20-элонгазным доменом (pY156; SEQ ID NO:64; см. ФИГ.19), возможно, из-за некорректного складывания. В противоположность этому, SaD4, экспрессированный один (pY151; SEQ ID NO:76; см. ФИГ.20), обладает приблизительно такой же самой активностью дельта-4-десатуразы, что и нативный EgDHAsyn1* (pY141; SEQ ID NO:49; см. ФИГ.20). Интересно, что активность дельта-4-десатуразы SaD4 возрастает приблизительно двукратно при слиянии с EgDHAsyn1 C20-элонгазным доменом (pY160; SEQ ID NO:77; см. ФИГ.20). При слиянии с EgDHAsyn1 C20-элонгазным доменом и подпитке ЭПК (pY160; SEQ ID NO:77; см. ФИГ.19), активность дельта-4-десатуразы приблизительно в три раза выше, чем при подпитке ДПК (pY160; SEQ ID NO:77; см. ФИГ.20), что предполагает, что связывание двух данных доменов дает повышенную эффективность или поток, возможно из-за каналирования субстрата.
ПРИМЕР 29
Специфичность субстрата EgDHAsyn1*
ГЛК, СТК, ЭДК, ЭРК, ДГЛК, ЭТК, АРК, ЭПК и ДПК подавали к Yarrowia клеткам, трансформированным pY141 (EgDHAsyn1*; SEQ ID NO:49), и проводили анализ векторов только для контроля и жирнокислотных профилей как описано в Примере 27.
Результаты для подпитки ЭПК, АРК и ДПК показаны на ФИГ.22. Жирнокислотные профили для Yarrowia, подпитываемой ГЛК, СТК, ЭДК, ЭРК, ДГЛК и ЭТК, не выявили элонгации и на ФИГ.22 не показаны. Процент C20-элонгации (% C20 Elong) и процент дельта-4-десатурации (% D4 Desat), и средние значения вычисляли, как описано в Примере 27, когда подпитку осуществляли ЭПК или ДПК. При подпитке АРК процент C20-элонгации (% C20-Elong) вычисляли путем деления суммы вес.% для докозатетраеновой кислоты [ДТК; 22:4 (7,10,13,16)] и омега-6 докозапентаеновой кислоты [ДПКn-6; 22:5(4,7,10,13,16)] на сумму вес. % для АРК, ДТК и ДПКn-6 и умножения на 100 для выражения в виде %. Подобно этому, процент дельта-4-десатуразы (% D4-Desat) при подпитке АРК высисляли путем деления вес. % для ДПКn-6 на сумму вес.% для ДТК и ДПКn-6 и умножения на 100 для выражения в виде %.
Как резюме ФИГ.22, EgDHAsyn1* элонгирует как АРК так и ЭПК, хотя небольшое преимущество (примерно на 40% больше активности) отмечается для ЭПК. Продукт элонгации АРК (т.е. ДТК) также десатурируется в дельта-4 положении EgDHAsyn1 с образованием ДПКn-6, и его активность приблизительно на 40% выше для ДТК, чем для ДПК.
ПРИМЕР 30
Коэкспрессия Euglena gracilis ДГК синтазы 1 с Pavlova lutheri дельта-8-десатуразой, Mortierella alpina дельта-5-десатуразой, Saprolegnia diclina дельта-17-десатуразой и Euglena gracilis дельта-9-элонгазой в соевых эмбрионах, трансформированных экспрессирующими векторами сои pKR973 и pKR1064
Зрелые соматические зародыши сои служат хорошей моделью для зиготных зародышей. Пребывая в глобулярном эмбриональном состоянии в жидкой культуре, соматические зародыши сои содержат очень низкие количества триацилглицерина или накопленных протеинов, что типично для созревающих зиготных зародышей сои. На этой стадии развития отношение суммарных триацилглицеридов к суммарным полярным липидам (фосфолипидам и гликолипидам) составляет приблизительно 1:4, что типично для зиготных зародышей сои на стадии развития, с которой была инициирована соматическая эмбриональная культура. В той же мере, на глобулярной стадии, мРНК для заметных семечковых протеинов, α′-субъединицы β-конглицинина, трипсинового ингибитора 3 kunitz и семечкового лецитина в значительной мере отсутствуют. После перенесения в свободную от гормонов среду с предоставлением возможности дифференциации до созревательного соматического эмбрионального состояния триацилглицерин становится наиболее изобильным липидным классом. В той же мере, мРНК для α′-субъединицы β-конглицинина, трипсинового ингибитора 3 kunitz и семечкового лецитина становятся весьма изобильными мессиджами в общей мРНК популяции. На этой основе, соматическая соевая эмбриональная система ведет себя очень схожим образом с созревающими зиготными соевыми эмбрионами in vivo, и поэтому является хорошей и быстрой модельной системой для анализа фенотипических эффектов модифицирования экспрессии генов на жирнокислотном биосинтетическом пути (см. РСТ публікацію № WO 2002/00904, Пример 3). Наиболее важно, что данная модельная система является также предсказательной в отношении составов жирных кислот семян от растений, полученных из трансгенных зародышей.
Анализ на жирные кислоты трансгенных соматических соевых зародышей, экспрессирующих pKR973 и pKR1064:
Соевые эмбриогенные суспензионные культуры (cv. Jack) трансформировали AscI фрагментами pKR973 и pKR1064 (фрагментами, содержащими экспрессирующие кассеты), как описано для производства в Примере 25 и как обобщено в Таблице 26.
Подмножество соевых эмбрионов, генерированных для каждого события (десять эмбрионов на событие), собирали и вносили в стеклянные ГХ ампулы, и жирнокислотные метиловые эфиры готовили путем трансэтерификации. Для трансэтерификации 50 мкл триметилсульфоний гидроксида (ТМСГ) и 0,5 мл гексана добавляли к зародышам в стеклянных ампулах и инкубировали в течение 30 минут при комнатной температуре при встряхивании. Жирнокислотные метиловые эфиры (5 мкл извлекали из гексанового слоя) разделяли и анализировали количественно с использованием газового хроматографа Hewlett-Packard 6890 Gas Chromatograph, снабженного капиллярной колонкой из плавленого кварца Omegawax 320 (Cat. No. 24152, Supelco Inc.). Температура печи была запрограммирована на выдержку при 220°C в течение 2,6 минут, повышение до 240°C при 20°C/минуту и затем дополнительную выдержку в течение 2,4 минут. Газ-носитель поставлялся водородным генератором Whatman. Величины времени сравнивали с величинами времени удерживания для стандартов метиловых эфиров, которые имеются в продаже (Nu-Chek Prep, Inc.). События с хорошим фенотипом были повторно анализированы методом ГХ с использованием идентичных условий, исключая то, что температура печи выдерживали при 150°C в течение 1 минуты и потом повышали до 240°C при 5°C/минуту.
Жирнокислотные профили для индивидуальных эмбрионов от репрезентативного события показаны на ФИГ.23. Жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), 18:2(ЛК), ГЛК, 18:3 (АЛК), ЭДК, ДГЛК, АРК, ЭРК, ЮН, ЭПК, 22:3(10,13,16) (докозатриеновая кислота), ДТК, ДПК и ДГК; и, составы жирных кислот, перечисленных на ФИГ.23, выражены в виде весового процента (вес. %) от всех жирных кислот.
Активность EgDHAsyn1 выражена как процент C20 элонгации (% C20-Elong) и/или процент дельта-4-десатурации (% D4-Desat), вычисленный в соответствии со следующей формулой: ([продукт]/[субстрат+прдукт])*100.
Более конкретно, процент элонгации для ЭПК представлен как % C20-Elong, определяемый как: ([ДПК+ДГК]/[ЭПК+ДПК+ДГК])*100. Процент дельта-4-десатурации для ДПК показан как % D4-Desat, определенный как ([ДГК]/[ДПК+ДГК])*100. Другие жирные кислоты, которые могут удлиняться или десатурироваться, не были включены в эти вычисления.
Кроме элонгационных и десатурационных продуктов для ЭПК и АРК, представляется, что в сое ДГЛК также элонгируется EgDHAsyn1, поскольку было получено значительное количество жирной кислоты 22:3(10,13,16). Данную жирную кислоту идентифицировали как 22:3(10,13,16), поскольку, как было найдено, она имела массу для 22:3, определенную методом ГХ-МС, и имела МС профиль, который согласуется с таким для 22:3(10,13,16).
ПРИМЕР 31
Экспрессия Euglena gracilis дельта-9-элонгаза/Pavlova lutheri дельта-8-десатуразного гибрида (EgD9elo-EgDHAsyn1Link-PavD8) в соевых эмбрионах, трансформированных экспрессирующими векторами сои KS373
Соевую эмбриогенную суспензионную культуру (cv. Jack) трансформировали KS373 (SEQ ID NO:179; ФИГ.17) и KS120 (описано в PCT публикации № WO 2004/071467, на содержание которой в настоящей заявке сделана ссылка) как описано для модельной системы в Примере 25. KS120 содержит гигромициновую селекцию. KS373, полученный в Примере 23, позволял осуществлять экспрессию слитого протеина, включающего Euglena gracilis дельта-9-элонгазу и Pavlova lutheri дельта-8-десатуразу, где данные два домена были сцеплены Euglena gracilis ДГК синтаза 1 линкером (т.е. EgDHAsyn1Link).
Жирнокислотные профили для пяти индивидуальных эмбрионов от 31 события получали как описано в Примере 30. Результаты пяти наилучших элонгационных событий приведены на ФИГ.24. Жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), 18:2 (ЛК), ГЛК, 18:3 (АЛК), ЭДК, ДГЛК, ЭРК и ЭТК; и, составы жирных кислот, перечисленные на ФИГ.24, выражены в виде весового процента (вес.%) от всех жирных кислот.
Активность EgD9elo-EgDHAsyn1Link-PavD8 выражена как процент дельта-9-элонгации (% D9-Elong) и/или процент дельта-8-десатурации (% D8-Desat), вычисленный согласно следующей формулы: ([продукт]/[субстрат+продукт])*100.
Более конкретно, процент дельта-9-элонгации представлен как % D9-Elong, определяемый как: ([ЭДК+ЭРК+ДГЛК+ЭТК]/[ЛК+АЛК+ЭДК+ЭРК+ДГЛК+ЭТК])*100. Процент дельта-8-десатурации представлен как % D8 Desat, определяемый как ([ДГЛК+ЭТК]/[ЭДК+ЭРК+ДГЛК+ЭТК])*100.
Наилучшее % D9 Elong событие имело среднюю элонгацию 22,1% при среднем % D8 Desat 92,7%. Элонгация несколько меньше, чем наблюдаемая, когда дельта-9-элонгаза экспрессирована одна в соевых эмбрионах, хотя это может быть обусловлено малым числом анализируемых событий. В противоположность этому, десатурация существенно выше, когда PavD8 слита с EgD9elo и EgDHAsyn1Link, чем когда PavD8 экспрессирована одна в соевых эмбрионах, достигая почти 100% конверсии в некоторых событиях. Эта усиленная конверсия дельта-8-десатуразой может быть обусловлена повышенной эффективностью или потоком, возможно, из-за каналирования субстрата.
ПРИМЕР 32
Синтез и функциональный анализ кодон-оптимизированного C20-элонгазного гена (EgC20ES), из Euglena gracilis в Yarrowia lipolytica
Использование кодона C20-элонгазного домена EgDHAsyn1 (EgDHAsyn1C20EloDom1) Euglena gracilis (т.е. соответствующего аминокислотам 1-303 SEQ ID NO:12) оптимизировали для экспрессии в Yarrowia lipolytica, способом, подобным описанному в PCT публикации № WO 2004/101753 и патенте США № 7125672. Конкретно, был создан кодон-оптимизированный C20-элонгазный ген (обозначенный как “EgC20ES” и имеющий нуклеотидную последовательность, как представлено в SEQ ID NO:183, и аминокислотную последовательность, как представлено в SEQ ID NO:184), основанный на кодирующей последовательности C20-элонгазного домена EgDHAsyn1 (SEQ ID NO:201), в соответствии со схемой использования Yarrowia кодона (PCT публикация № WO 2004/101753), консенсусной последовательностью вокруг ′ATG′ трансляционного инициирующего кодона и общими правилами РНК стабильности (Guhaniyogi, G. и J. Brewer, Gene, 265(1-2):11-23 (2001)). В добавок к модификации трансляционного инициирующего сайта, были модифицированы 163 bp 909 bp кодирующего участка (17,9%), и 147 кодонов было оптимизировано (48,5%). Ни одна из модификаций кодон-оптимизированного гена не изменила аминоксислотную последовательность кодированного протеина (т.е. SEQ ID NO:184 на 100% идентична по последовательности аминокислотам 1-303 SEQ ID NO:12). Созданный EgC20ES ген (SEQ ID NO:183) был синтезирован корпорацией GenScript Corporation (Piscataway, NJ) и клонирован в pUC57 (номер доступа GenBank Y14837) с генерацией pEgC20ES (ФИГ.51B; SEQ ID NO:185).
Для анализа функции кодон-оптимизированного EgC20ES гена сконструировали плазмиду pZuFmEgC20ES (ФИГ.52A; SEQ ID NO:360), включающую химерный FBAINm::EgC20ES::Pex20 ген. Плазмида pZuFmEgC20ES содержала следующие компоненты:
Плазмиду pZuFmEgC20ES трансформировали в Yarrowia lipolytica штамм Y4184U4, как описано в разделе Общие методы. Трансформанты отбирали на МС пластинках. После роста в течение 2 суток при 30°C, 10 трансформантов, выращенных на МС пластинках, извлекали и вновь наносили штрихами на свежие МС пластинки. После выращивания 10 штаммов инокулировали отдельно в 3 мл жидкую МС при 30°C и встряхивали при 250 оборотов/минуту в течение 2 суток. Клетки собирали путем центрифугирования; липиды экстрагировали; и жирнокислотные метиловые эфиры готовили путем трансэтерификации, и впоследствии анализировали с помощью газового хроматографа Hewlett-Packard 6890 GC.
ГХ анализы показали наличие около 6,2% ЭПК и 2,8% ДПК от всех липидов, выработанных во всех десяти трансформантах, где эффективность конверсии ЭПК в ДПК в этих 10 штаммах, как было найдено, составляла около 31% (вычислено как описано в Примере 27). Таким образом, эти экспериментальные данные продемонстрировали, что синтетическая Euglena gracilis C20-элонгаза, кодон-оптимизированная для экспрессии в Yarrowia lipolytica (т.е. EgC20ES, как представлено в SEQ ID NO:183), активно конвертирует ЭПК в ДПК.
ПРИМЕР 33
Синтез и функциональный анализ кодон-опимизированного C20-элонгазного гена (EaC20ES) из Euglena anabaena в Yarrowia lipolytica
Использование кодона C20-элонгазного домена EaDHAsyn2 (SEQ ID NO:228) Euglena anabaena (т.е. соответствующего аминокислотам 1-299 SEQ ID NO:96), оптимизировали для экспрессии в Yarrowia lipolytica, способом, подобным описанному в PCT публикации № WO 2004/101753, патенте США № 7125672 и выше в Примере 32. Конкретно, был создан кодон-оптимизированный C20-элонгазный ген (обозначенный как “EgC20ES” и имеющий нуклеотидную последовательность, как представлено в SEQ ID NO:188, и аминокислотную последовательность, как представлено в SEQ ID NO:189), основанный на кодирующей последовательности C20-элонгазного домена EaDHAsyn2 (SEQ ID NO:92), в соответствии со схемой использования Yarrowia кодона (PCT публикация № WO 2004/101753), консенсусной последовательностью вокруг ′ATG′ трансляционного инициирующего кодона и общими правилами РНК стабильности (Guhaniyogi, G. и J. Brewer, Gene, 265(1-2):11-23 (2001)). В добавок к модификации трансляционного инициирующего сайта, модифицировали 143 bp 897 bp кодирующего участка (15,9%), и 134 кодона оптимизировали (44,8%). Ни одна из модификаций кодон-оптимизированного гена не изменила аминоксислотную последовательность кодированного протеина (т.е. SEQ ID NO:189 на 100% идентична по последовательности аминокислотам 1-299 SEQ ID NO:96). Созданный EaC20ES ген (SEQ ID NO:188) был синтезирован корпорацией GenScript Corporation (Piscataway, NJ) и клонирован в pUC57 (номер доступа GenBank Y14837) с генерацией pEaC20ES (SEQ ID NO:190).
Для анализа функции кодон-оптимизированного EaC20ES гена сконструировали плазмиду pZuFmEaC20ES (SEQ ID NO:361), включающую химерный FBAINm::EaC20ES::Pex20 ген. Плазмида pZuFmEaC20ES (SEQ ID NO:361) была идентична по конструкции таковой для плазмиды pZuFmEgC20ES (SEQ ID NO:360; ФИГ.52A), исключая то, что EaC20ES (SEQ ID NO:188) использовался вместо EgC20ES (SEQ ID NO:183).
Плазмиду pZuFmEaC20ES (SEQ ID NO:361) трансформировали в Yarrowia lipolytica штамм Y4184U4, как описано в разделе Общие методы. Данные трансформанты отбирали на МС пластинках. После роста в течение 2 суток при 30°C 20 трансформантов, выращенных на МС пластинках, извлекали и повторно наносили на свежие МС пластинки. После выращивания 20 штаммов инокулировали отдельно в 3 мл жидкую МС при 30°C и встряхивали при 250 оборотов/минуту в течение 2 суток. Клетки собирали путем центрифугирования; липиды экстрагировали; и жирнокислотные метиловые эфиры готовили путем трансэтерификации, и впоследствии анализировали с помощью газового хроматографа Hewlett-Packard 6890 GC.
ГХ анализы показали наличие около 7,4% ЭПК и 1% ДПК от всех липидов, выработанных во всех 20 трансформантах, где эффективность конверсии ЭПК в ДПК в этих 20 штаммах, как было найдено, составляла около 11% (вычислено как описано в Примере 27). Таким образом, эти экспериментальные данные продемонстрировали, что синтетическая Euglena anabaena C20-элонгаза, кодон-оптимизированная для экспрессии в Yarrowia lipolytica (т.е. EaC20ES, как представлено в SEQ ID NO:188), активно конвертирует ЭПК в ДПК.
ПРИМЕР 34
Синтез и функциональный анализ кодон-оптимизированного дельта-4-десатуразного гена (EaD4S) из Euglena anabaena в Yarrowia lipolytica
Использование кодона дельта-4-десатуразного домена EaDHAsyn2 (SEQ ID NO:243) Euglena anabaena (т.е. соответствующего аминокислотам 259-841 SEQ ID NO:96), оптимизировали для экспрессии в Yarrowia lipolytica, способом, подобным описанному в PCT публикации № WO 2004/101753, патенте США № 7125672 и выше в Примерах 32 и 33. Конкретно, был создан кодон-оптимизированный дельта-4-десатуразный ген (обозначенный как “EaD4S” и имеющий нуклеотидную последовательность, как представлено в SEQ ID NO:192, и аминокислотную последовательность, как представлено в SEQ ID NO:193), основанный на кодирующей последовательности дельта-4-десатуразного домена EaDHAsyn2 (SEQ ID NO:92), который также представлен как SEQ ID NO:194 (нуклеотид) и SEQ ID NO:195 (аминокислота). В дополнение к модификации трансляционного инициирующего сайта, 307 bp из 1752 (включая TAA терминирующий кодон) bp кодирующего участка были модифицированы (17,5%), и 285 кодонов были оптимизированы (48.8%). Кроме того, NcoI сайт вводили вокруг трансляционного инициирующего кодона путем изменения второй аминокислоты дикого типа дельта-4-десатуразного домена (т.e. аминокислотного остатка 260 из SEQ ID NO:96 или аминокислотного остатка 2 из SEQ ID NO:195) от лейцинового к валиновому остатку в синтетическом EaD4S гене; таким образом, аминокислотная последовательность EaD4S представлена в SEQ ID NO:193. Сконструированный EaD4S ген (SEQ ID NO:192) был синтезирован корпорацией GenScript Corporation (Piscataway, NJ) и клонирован в pUC57 (номер доступа GenBank Y14837) с генерацией pEaD4S (SEQ ID NO:196).
Для анализа функции кодон-оптимизированного EaD4S гена сконструировали плазмиду pZKL4-220EA4 (ФИГ.52B; SEQ ID NO:362) с интеграцией двух химерных C20-элонгазных генов и химерного EaD4S гена в липазе 4 подобный локус (номер доступа GenBank XM_503825) Yarrowia lipolytica штамм Y4184U4. Плазмида pZKL4-220EA4 содержала следующие компоненты:
Плазмиду pZKL4-220EA4 расщепляли AscI/SphI, и потом трансформировали в Yarrowia lipolytica штамм Y4184U4, как описано в разделе Общие методы. Трансформанты отбирали на МС пластинках. После роста в течение 5 суток при 30°C 8 трансформантов, выращенных на МС пластинках, извлекали и повторно наносили штрихами на свежие МС пластинки. После выращивания эти штаммы инокулировали отдельно в 3 мл жидкую МС при 30°C и встряхивали при 250 оборотов/минуту в течение 2 суток. Данные клетки собирали путем центрифугирования, ресуспендировали в HGM и затем встряхивали при 250 оборотов/минуту в течение 5 суток. Клетки собирали путем центрифугирования, липиды экстрагировали, и жирнокислотные метиловые эфиры готовили путем трансэтерификации, и затем анализировали с помощью газового хроматографа Hewlett-Packard 6890 GC.
ГХ анализы показали наличие, в среднем, 0,4% ДГК и 10,2% ДПК от всех липидов, выработанных во всех 8 трансформантах, где эффективность конверсии ДПК в в этих восьми штаммах, как было найдено, составляла около 4,2% (вычислено как описано в Примере 27). Таким образом, эти экспериментальные данные продемонстрировали, что Euglena anabaena дельта-4-десатураза, кодон-оптимизированная для экспрессии в Yarrowia lipolytica (т.е. EaD4S, как представлено в SEQ ID NO:192), активна, но функционирует с относительно низкой эффективностью.
ПРИМЕР 35
Коэкспрессия Euglena gracilis ДГК синтаза 1 C20-элонгазного домена (EgDHAsyn1C20EloDom1) с Schizochytrium aggregatum дельта-4-десатуразой (SaD4) в соевых зародышах, трансформированных экспрессирующим вектором сои pKR1105
Следующий пример описывает генерацию трансгенных соевых событий, экспрессирующих EgDHAsyn1C20EloDom1 и SaD4, которые, при генерации в растениях, могут быть скрещены с ЭПК-продуцирующими соевыми событиями с генерацией ДГК-продуцирующих растений.
Соевую эмбриогенную суспензионную культуру (cv. Jack) трансформировали AscI фрагментом pKR1105 (SEQ ID NO:156; ФИГ.16A; фрагмент, содержащий экспрессирующую кассету) и зародыши созревали, как это описано для производства в Примере 25, но со следующим изменением. После созревания на SB103 в течение 10-12 суток отдельный кластер эмбрионов для каждого события переносился в 4 мл SB148 жидкой среды (рецепт приведен ниже), содержащей 0,02% тергитола и 0,33 мМ ЭПК, в шестиячеечный титрационный микропланшет.
SB 103 твердая среда (на литр)
1 упаковка MS солей (Gibco/BRL - Cat. No. 11117-066)
1 мл B5 витаминов 1000х маточный раствор
60 g maltose
pH 5,7
Кластеры осторожно разбивали для освобождения отдельных зародышей, и титрационный микропланшеты встряхивали на роторном шейкере при 150 оборотов/минуту и 26°C под белыми люминесцентными лампами холодного излучения при 16:8 часовом фотопериоде день/ночь и интенсивности света 60-85 мкЕ/м2/с в течение 48 часов. Через 48 часов зародыши ополаскивали водой, высушивали, и пять эмбрионов на событие собирали в стеклянные ГХ ампулы. Жирнокислотные метиловые эфиры готовили путем трансэтерификации TMSH и количественно анализировали на газовом хроматографе Hewlett-Packard 6890 Gas Chromatograph, снабженном капиллярной колонкой из плавленого кварца Omegawax 320 (Cat. No. 24152, Supelco Inc.), как это описано в Примере 30. Температуру печи программировали на выдержку при 150°C в течение 1 минуты и потом повышали до 240°C при 5°C/минуту. Величины времени сравнивали с величинами времени удерживания для стандартов метиловых эфиров, которые имеются в продаже (Nu-Chek Prep, Inc.).
Таким способом было проанализировано 122 события, трансформированных pKR1105. Из 122 проанализированных событий 49 были идентифицированы как такие, что элонгировали ЭПК (активность C20-дельта-5-элонгазы), и из них 41 событие было идентифицировано как такое, что десатурировало ДПК (активность дельта-4-десатуразы) с образованием ДГК. События с наилучшими активностями C20-дельта-5-элонгазы и дельта-4-десатуразы были успешными, и жирнокислотные профили от зародышей, подпитывавшихся ЭПК, показаны на ФИГ.26.
Жирные кислоты на ФИГ.26 идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК, АЛК, ЭПК, 22:0 (докозановая кислота), ДПК, 24:0 (тетракозановая кислота), ДГК и 24:1 (невоновая кислота); и, составы жирных кислот, перечисленные на ФИГ.26, выражены в виде весового процента (вес. %) от всех жирных кислот. Активность EgDHAsyn1C20EloDom1 выражали как процент C20-дельта-5-элонгации (% C20-delta-5 elong), вычисленный в соответствии со следующей формулой: ([продукт]/[субстрат+продукт])*100. Более конкретно, объединенный процент элонгации для ЭПК представлен как “% C20-delta-5 elong”, определяемый как: ([ДПК+ДГК]/[ЭПК+ДПК+ДГК])*100.
Активность SaD4 выражали в виде процента дельта-4-десатурации (% delta-4 desat), вычисленного в соответствии со следующей формулой: ([продукт]/[субстрат+продукт])*100. Более конкретно, объединенный процент десатурации для ДПК представляли как “% delta-4 desat”, определяемый как: (ДГК/[ДПК+ДГК])*100.
ПРИМЕР 36
Идентификация дельта-9-элонгазы из Euglena anabaena UTEX 373
Настоящий пример описывает идентификацию дельта-9-элонгаз из Euglena anabaena UTEX 373 кДНК библиотеки. Эта работа также описана в предварительной заявке США № No. 60/911925 (от 16 апреля 2007 года; реестр уполномоченного № BB-1613; на содержание которой в настоящей заявке сделана ссылка).
Выращивание Euglena anabaena UTEX 373 и приготовление РНК
Амплифицированную кДНК библиотеку eug1c высевали, и колонии подвергали лифтингу как описано в Примере 13. ДНК зонд был сделан с использованием очищенного агарозным гелем NcoI/NotI ДНК фрагмента, содержащего Euglena gracilis дельта-9-элонгазный ген, из pKR906 (SEQ ID NO:115; Пример 16 и WO 2007/061845, которая опубликована 31 мая 2007 года; реестр уполномоченного № BB-1562; на содержание которой в настоящей заявке сделана ссылка), меченый P32 dCTP с использованием системы RadPrime DNA Labeling System (Cat. No. 18428-011, Invitrogen, Carlsbad, CA) согласно инсрукциям производителя.
Лифтинги колоний зондировали, и позитивы идентифицировали и подтверждали как описано в Примере 13. Плазмиду ДНК выделяли и секвенировали в точности с тем как описано в Примере 2, и последовательности группировали и сравнивали с использованием Sequencher™ (Version 4.2, Gene Codes Corporation, Ann Arbor, MI). Таким способом клоны категоризовали в одну из двух четких групп на основе инсерционных последовательностей (обозначенных как EaD9Elo1 и EaD9Elo2). Репрезентативные клоны, содержащие кДНК для каждого класса последовательности, выбирали для дальнейшего исследования, и данные последовательности для каждой репрезентативной плазмиды (pLF121-1 и pLF121-2) показанв в SEQ ID NO:250 и SEQ ID NO:251, соответственно. Последовательность, изображенная цепочкой NNNN, представляет участок полиА хвоста, который не секвенировался. Кодирующие последовательности для EaD9Elo1 и EaD9Elo2 представлены в SEQ ID NO:252 и SEQ ID NO:253, соответственно. Соответствующие аминокислотные последовательности для EaD9Elo1 и EaD9Elo2 представлены в SEQ ID NO:254 и SEQ ID NO:255, соответственно.
ПРИМЕР 37
Идентификация дельта-5-десатуразы из Euglena anabaena UTEX 373
Настоящий пример описывает идентификацию дельта-5-десатуразы из Euglena anabaena UTEX 373. Эта работа также описана в предварительной заявке США № 60/915733 (от 3 мая 2007 года; реестр уполномоченного № BB-1614; на содержание которой в настоящей заявке сделана ссылка).
Амплифицированную кДНК библиотеку eug1c высевали, и колонии подвергали лифтингу как описано в Примере 13. ДНК зонд был сделан с использованием очищенного агарозным гелем NcoI/NotI ДНК фрагмента, содержащего Euglena gracilis дельта-5-десатуразный ген (EgD5; SEQ ID NO:267) из pDMW367, описанный ранее в РСТ публикации № WO 2007/136877 (которая опубликована 29 ноября 2007 года; реестр уполномоченного № BB-1629; на содержание которой в настоящей заявке сделана ссылка), меченый P32.
Лифтинги колоний зондировали, и позитивы идентифицировали и подтверждали как описано в Примере 13. Плазмиду ДНК выделяли и секвенировали в точности с тем как описано в Примере 2, и последовательности группировали и сравнивали с использованием Sequencher™ (Version 4.2, Gene Codes Corporation, Ann Arbor, MI).
Репрезентативный клон, содержащий кДНК (pLF119), представлен в SEQ ID NO:256, и ген, содержащийся внутри кДНК, был назван EaD5Des1. Кодирующая последовательность для EaD5Des1 представлена в SEQ ID NO:257. Соответствующая аминокислотная последовательность для EaD5Des1 представлена в SEQ ID NO:258.
ПРИМЕР 38
Конструирование экспрессирующего вектора сои pKR1183 для экспрессии Euglena anabaena дельта-9-элонгаза-Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразного слитого гена (Hybrid1-HGLA Synthase)
Внутрикаркасный гибрид Euglena anabaena дельта-9-элонгазы (EаD9Еlo 1; SEQ ID NO:252, Пример 36), Euglena gracilis ДГК синтаза 1 обогащенного пролином линкера (EgDHAsyn1Link; SEQ ID NO:197; Пример 6) и Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразы (TpomD8; SEQ ID NO:162; Пример 21; см. также совместную заявку правопреемника заявителя, которая находится на рассмотрении, с патентной заявкой США № 11/876115 (от 22 октября 2007 года; реестр уполномоченного № ВВ-1574)) был создан при описанных ниже условиях.
Был создан первичный внутрикаркасный гибрид между EаD9Еlo 1 и EgDHAsyn1Link (EаD9elo-EgDHAsyn1Link), фланкированный NcoI сайтом в 5′ конце и NotI сайтом в 3′ конце, путем ПЦР амплификации. EаD9Еlo 1 (SEQ ID NO:252) амплифицировали из pLF121-1 (SEQ ID NO:250) олигонуклеотидами EaD9-5Bbs (SEQ ID NO:259) и EaD9-3fusion (SEQ ID NO:260), с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S, Finnzymes Oy, Finland), следуя протоколу производителя. EgDHAsyn1Link (SEQ ID NO:197) амплифицировали схожим способом из pKR1049 (Пример 4) олигонуклеотидами EgDHAsyn1Link-5fusion (SEQ ID NO:261) и MWG511 (SEQ ID NO:175). Два результирующих ПЦР продукта объединяли и повторно амплифицировали с использованием EaD9-5Bbs (SEQ ID NO:259) и MWG511 (SEQ ID NO:175) с образованием EaD9Elo1-EgDHAsyn1Link. Последовательность EaD9Elo1-EgDHAsyn1Link представлена в SEQ ID NO:262. EaD9Elo1-EgDHAsyn1Link не содержит внутрикаркасного терминирующего кодона вверху NotI сайта в 3′ конце, и поэтому ДНК фрагмент, клонированный в NotI site, может дать начало внутрикаркасному слиянию с EgD9elo1-EgDHAsyn1Link, если выбран корректный каркас. EaD9Elo1-EgDHAsyn1Link клонировался в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), следуя протоколу производителя, с образованием pLF124 (SEQ ID NO:263).
BbsI/NotI ДНК фрагмент pLF124 (SEQ ID NO:263), содержащий EaD9Elo1-EgDHAsyn1Link, встраивали в NcoI/NotI ДНК фрагмент из KS366 (SEQ ID NO:177; Пример 23), содержащий промотор для α′ субъединицы β-конглицинина, с образованием pKR1177 (SEQ ID NO:264).
BamHI ДНК фрагмент pKR1177 (SEQ ID NO:264), содержащий EaD9Elo1-EgDHAsyn1Link, встраивали в BamHI ДНК фрагмент pKR325, описанный ранее в PCT публикации №. WO 2006/012325 (на содержание которого в настоящей заявке сделана ссылка) с образованием pKR1179 (SEQ ID NO:265).
NotI фрагмент из pLF114-10 (Пример 21; SEQ ID NO:165), содержащий TpomD8, клонировали в NotI фрагмент pKR1179 (SEQ ID NO:265) с образованием pKR1183 (SEQ ID NO:266; ФИГ.28). На ФИГ.28 слитый ген (Hybrid1-HGLA synthase) называется EAd9ELONG-TPOMd8DS.
ПРИМЕР 39
Конструирование экспрессирующего вектора сои pKR1253 для экспрессии Euglena anabaena дельта-9-элонгаза-Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразного слитого гена (Hybrid1-HGLA Synthase) с Euglena gracilis дельта-5-деспатуразой
Через ряд стадий субклонирования NotI сайт добавляли к 5′ концу Euglena gracilis дельта-5-десатуразы (EgD5; SEQ ID NO:267) из pDMW367, и этот NotI фрагмент, содержащий EgD5, встраивали в NotI сайт pKR457 (SEQ ID NO:122; Пример 16) с образованием pKR1237 (SEQ ID NO:268).
AscI фрагмент pKR1183 (SEQ ID NO:266; Пример 38), содержащий Hybrid1-HGLA синтазу, клонировали в AscI фрагмент pKR277 (SEQ ID NO:120, которая была ранее описана в PCT публикации № WO 2004/071467 и опубликована 8/26/2004; реестр уполномоченного No. BB-1538 (на содержание которой в настоящей заявке сделана ссылка) с образованием pKR1252 (SEQ ID NO:269).
BsiWI фрагмент pKR1237 (SEQ ID NO:268), содержащий EgD5 ген, клонировали в BsiWI сайт pKR1252 (SEQ ID NO:269) с образованием pKR1253 (SEQ ID NO:270; ФИГ.30).
ПРИМЕР 40
Конструирование соевого вектора pKR1139 для экспрессии Euglena anabaena дельта-5-десатуразы
Настоящий пример описывает клонирование дельта-5-десатуразы из Euglena anabaena UTEX 373 в экспрессирующий вектор сои. Эта работа также описана в предварительной заявке США №. 60/915733 (от 3 мая 2007 года; реестр уполномоченного No. BB-1614 (на содержание которой в настоящей заявке сделана ссылка)).
EaD5Des1 (SEQ ID NO:257) амплифицировали из pLF119 (SEQ ID NO:256, Пример 37) oEAd5-1-1 (SEQ ID NO:271) и oEAd5-1-2 (SEQ ID NO:272), с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S, Finnzymes Oy, Finland) в соответствии с протоколом производителя. Результирующий ПЦР продукт клонировали в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), в соответствии с протоколом производителя, с использованием pKR1136 (SEQ ID NO:273).
NotI фрагмент для pKR1136 (SEQ ID NO:273), содержащий EaD5Des1, клонировали в NotI фрагмент pKR974, описанный ранее в PCT публикации № WO 2007/136877, опубликованной 29 ноября 2007 года (реестр уполномоченного No. BB-1629 (на содержание которой в настоящей заявке сделана ссылка)), с образованием pKR1139 (SEQ ID NO:274).
ПРИМЕР 41
Конструирование экспрессирующего вектора сои pKR1255 для экспрессии Euglena anabaena дельта-9-элонгаза-Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразного слитого гена (Hybrid1-HGLA Synthase) с Euglena gracilis дельта-5-десатуразой и Euglena anabaena дельта-5-десатуразой
Плазмиду pKR1139 (SEQ ID NO:274; Пример 40) расщепляли SbfI, и фрагмент, содержащий EaD5Des1, клонировали в SbfI сайт pKR1253 (SEQ ID NO:270; Пример 39) с образованием pKR1255 (SEQ ID NO:275; ФИГ.31).
ПРИМЕР 42
Конструирование экспрессирующего вектора сои pKR1189 для снижения уровня регуляции экспрессии сои Fad3
Настоящий пример описывает соевый экспрессирующий вектор, созданный для снижения fad3 экспрессии в сое.
Стартовый вектор pKR561 (SEQ ID NO:276) собирали путем внедрения BsiWI фрагмента pKR268 (описанного ранее в PCT публикации № WO 04/071467), содержащего аннексиновый промотор, в BsiWI сайт pKR145, который описан в PCT публикации № WO 04/071467.
Плазмида XF1, описанная в PCT публикации № WO 93/11245 (которая была опубликована 10 июня 1993 года; также патент США № 5952544; на содержание которых в настоящей заявке сделана ссылка), содержит соевый дельта-15-десатуразный (fad3) ген (SEQ ID NO:277; номер доступа GenBank L22964; называемый также GmFAD3B).
Часть 5′ конца fad3 гена амплифицировали из XF1 Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S, Finnzymes Oy, Finland), следуя протоколу производителя, с использованием HPfad3-1 (SEQ ID NO:278) и HPfad3-2 (SEQ ID NO:279) с образованием ДНК фрагмента, названного HPfad3AB (SEQ ID NO:280).
Часть 3′ конца fad3 гена амплифицировали из XF1 Phusion™ High-Fidelity DNA Polymerase, с использованием HPfad3-3 (SEQ ID NO:281) и HPfad3-1 (SEQ ID NO:278) с образованием ДНК фрагмента, названного HPfad3A′-2 (SEQ ID NO:282).
HPfad3AB и HPfad3A′-2 скомбинировали и амплифицировали с использованием Phusion™ High-Fidelity DNA Polymerase HPfad3-1 (SEQ ID NO:278) с образованием HPfad3ABA′-2 (SEQ ID NO:283).HPfad3ABA′-2 (SEQ ID NO:283) имеет NotI сайт как в 5′ так и 3′ конце ДНК фрагмента. Результирующий ПЦР продукт клонировали в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), в ссответствии с протоколом производителя, с образованием pLF129 (SEQ ID NO:284).
NotI фрагмент для pLF129 (SEQ ID NO:284), содержащего fad3 шпильку, клонировали в NotI фрагмент pKR561 (SEQ ID NO:276) с образованием pKR1189 (SEQ ID NO:285; ФИГ.32). На ФИГ.32, A и A′ домены для fad3 обозначены как TR1, тогда как B домен обозначен TR2.
ПРИМЕР 43
Конструирование экспрессирующего вектора сои pKR1249 для снижения уровня регуляции сои Fad3 и сои Fad3c
NotI/HindIII фрагмент pLF129 (SEQ ID NO:284), содержащий TR1 и TR2 домены fad3, как указано на ФИГ.32, клонировали в NotI/HindIII каркасный фрагмент pLF129 (SEQ ID NO:284) с образованием pKR1209 (SEQ ID NO:286).
Кодирующая последовательность GmFad3C (номер доступа GenBank AY204712) (Bilyeu et al., Crop Sci. 43:1833-1838 (2003); Anai et al., Plant Sci. 168:1615-1623 (2005)) представлена в SEQ ID NO:287, и соответствующая аминокислотная последовательность представлена в SEQ ID NO:288. Часть fad3c гена амплифицировали из соевой кДНК библиотеки, описанной в PCT публикации № WO 93/11245 (которая опубликована 10 июня 1993 года; также патент США № 5952544) (на содержание которых в настоящей заявке сделана ссылка), Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S, Finnzymes Oy, Finland) в соответствии с протоколом производителя, с использованием fad3c-5 (SEQ ID NO:289) и fad3c-3 (SEQ ID NO:290). Результирующий ДНК фрагмент клонировали в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), согласно протоколу прооизводителя, с образованием pKR1213 (SEQ ID NO:291).
EcoRV/XhoI фрагмент pKR1213 (SEQ ID NO:291), содержащий фрагмент fad3c, клонировали в NotI(filled)/XhoI сайт pKR1209 (SEQ ID NO:286) с образованием pKR1218 (SEQ ID NO:292).
NotI/HindIII фрагмент pLF129 (SEQ ID NO:284), содержащий TR1 домен только из fad3, как указано на ФИГ.32, клонировали в NotI/HindIII каркасный фрагмент pLF129 (SEQ ID NO:284) с образованием pKR1210 (SEQ ID NO:293).
EcoRV/XhoI фрагмент pKR1213 (SEQ ID NO:291), содержащий фрагмент fad3c, клонировали в NotI(filled)/XhoI сайт pKR1210 (SEQ ID NO:293) с образованием pKR1219 (SEQ ID NO:294).
XhoI(filled)/HindIII фрагмент pKR1218 (SEQ ID NO:292), содержащий фрагмент fad3c, так же, как и fad3 TR1 и TR2 домены, клонировали в MluI(filled)/HindIII сайт pKR1219 (SEQ ID NO:294), содержащий фрагмент fad3c, так же само как и fad3 TR1 единственный домен, с образованием pKR1225 (SEQ ID NO:295). Таким способом сформировали новую шпильку, включающую fad3 и fad3c и фланкированная NotI сайтами.
NotI фрагмент для pKR1225 (SEQ ID NO:295), содержащий новую шпильку, включая fad3 и fad3c, клонировали в NotI фрагмент pKR561 (SEQ ID NO:276; Пример 42) с образованием pKR1229 (SEQ ID NO:296; FIG. 33). Таким образом, fad3/fad3c шпилька может быть экспрессирована из сильного, специфического в отношении семян промотора с гигромициновой селекцией в растениях.
BsiWI фрагмент для pKR1225 (SEQ ID NO:295), содержащий новую шпильку, включая fad3 и fad3c, клонировали в BsiWI фрагмент pKR226 (SEQ ID NO:130; Пример 17) с образованием pKR1249 (SEQ ID NO:297; ФИГ.34). На ФИГ.34, pKR1249 помечен как pKR1249_PHP33240. Таким образом, fad3/fad3c шпилька может быть экспрессирована из сильного, специфического в отношении семян промотора с хлоросульфуроновой (ALS) селекцией в растениях.
ПРИМЕР 44
Конструирование экспрессирующего вектора сои pKR1322 для экспрессии Euglena anabaena дельта-9-элонгаза-Tetruetreptia pomquetensis CCMP1491 дельта-8-десатураза-Euglena anabaena дельта-5-десатуразного гена (EaD9Elo1-TpomD8-EaD5Des1 fusion)
Настоящий пример описывает конструирование внутрикаркасного слитого гена между Euglena anabaena дельта-9-элонгазой (EaD9Elo1; SEQ ID NO:252, Пример 36), Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразой (TpomD8; SEQ ID NO:162; Пример 21) и Euglena anabaena дельта-5-десатуразой (EaD5Des1; SEQ ID NO:257; Пример 37). Каждый домен, отделенный EgDHAsyn1 линкером (EgDHAsyn1Link; SEQ ID NO:197; Пример 6).
EaD9Elo1-EgDHAsyn1Link (SEQ ID NO:262; Пример 38), амплифицировался из pLF124 (SEQ ID NO:263) олигонуклеотидами oEAd9el1-1 (SEQ ID NO:298) и oLINK-1(SEQ ID NO:299), с использованиием Phusion™ полимеразы ДНК высокого качества (Cat. No. F553S, Finnzymes Oy, Finland) в соответствии с протоколом производителя. EaD9Elo1-EgDHAsyn1Link фланкировали NotI в 5′ конце и EagI в 5′ и 3′ концах и не содержит внутрикаркасного терминирующего кодона вверху EagI сайта в 3′ конце. Поэтому ДНК фрагмент, клонированный в EagI сайт, может усилить внутрикаркасное слияние с EgD9elo-EgDHAsyn1Link, если выбран корректный каркас. Результирующий ДНК фрагмент, содержащий EaD9Elo1-EgDHAsyn1Link, клонировали в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), следуя протоколу производителя, с образованием pKR1298 (SEQ ID NO:300).
Путем ПЦР амплификации проводили внутрикаркасное слияние между TpomD8 и EgDHAsyn1Link (TpomD8-EgDHAsyn1Link), которое содержало NotI сайт в 5′ конце и EagI сайты в 5′ и 3′ концах. TpomD8 (SEQ ID NO:162) амплифицировали из pLF114-10 (SEQ ID NO:165) олигонуклеотидами oTPd8-1 (SEQ ID NO:301) и oTPd8fus-1 (SEQ ID NO:302), с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S, Finnzymes Oy, Finland) в соответствии с протоколом производителя. EgDHAsyn1Link (SEQ ID NO:197) амплифицировали подобным способом из pKR1049 (Пример 4) олигонуклеотидами oLINK-2 (SEQ ID NO:303) и oLINK-1 (SEQ ID NO:304). Два результирующих ПЦР продукта комбинировали и повторно амплифицировали с использованием oTPd8-1 (SEQ ID NO:301) и oLINK-1 (SEQ ID NO:304) с образованием TpomD8-EgDHAsyn1Link (SEQ ID NO:305). TpomD8-EgDHAsyn1Link встраивали в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), согласно протоколу производителя, с образованием pKR1291 (SEQ ID NO:306).
The EagI фрагмент pKR1291, содержащий TpomD8-EgDHAsyn1Link, клонировали в NotI сайт pBluescript II SK(+) вектор (Stratagene) с образованием либо pKR1301 (SEQ ID NO:307) в одной ориентации либо pKR1301R (SEQ ID NO:308) в противоположной ориентации.
Плазмиду pKR1301 (SEQ ID NO:307) расщепляли MfeI/BamHI, ДНК фрагмент, содержащий TpomD8-EgDHAsyn1Link, был полностью наполнен, и результирующий ДНК фрагмент религировали с образованием pKR1311 (SEQ ID NO:309).
Плазмиду pKR1301R (SEQ ID NO:308) расщепляли EcoRI, и фрагмент, содержащий 5′ конец TpomD8-EgDHAsyn1Link (названный TPOMD8TR2), и векторный каркас религировали с образованием pKR1304 (SEQ ID NO:310).
EagI сайт pKR1298 (SEQ ID NO:300), содержащий EaD9Elo1-EgDHAsyn1Link, клонировали в EagI сайт pKR1304 (SEQ ID NO:310) с образованием pKR1309 (SEQ ID NO:311).
NotI сайт pKR1298 (SEQ ID NO:300), содержащий EaD9Elo1-EgDHAsyn1Link, клонировали в EagI сайт pKR1304 (SEQ ID NO:310) с образованием pKR1309 (SEQ ID NO:311).
NotI фрагмент для pKR1136 (SEQ ID NO:273; Пример 40), содержащий EaD5Des1, клонировали в EagI сайт pKR1311 (SEQ ID NO:309) с образованием pKR1313 (SEQ ID NO:312).
MfeI/Ecl136II фрагмент pKR1313 (SEQ ID NO:312) клонировали в EcoRV/MfeI сайты pKR1309 (SEQ ID NO:311) с образованием pKR1315 (SEQ ID NO:313).
NotI фрагмент pKR1315 (SEQ ID NO:313) клонировали в NotI сайт pKR72 (SEQ ID NO:105; Пример 15) с образованием pKR1322 (SEQ ID NO:314; ФИГ.35).
На ФИГ.35 EaD9Elo1-TpomD8-EaD5Des1 слияние помечено как EAd9el+TPd8ds+EAd5ds слияние.
ПРИМЕР 45
Снижение уровня регуляции соевых fad3 и fad3c генов в соевых соматических зародышах путем трансформации pKR1189 или pKR1229
Настоящий пример описывает трансформацию и экспрессию в соевых соматических зародышах pKR1189 (SEQ ID NO:285, Пример 42), содержащих fad3 шпильковый конструкция или pKR1229 (SEQ ID NO:296; Пример 43), содержащий fad3 и fad3c шпильковую конструкцию. Обе конструкции также имеют гигромицин фосфотрансферазный ген для селекции на гигромицин.
Соевую эмбриогенную суспензионную культуру (cv. Jack) трансформировали pKR1189 (SEQ ID NO:285) или pKR1229 (SEQ ID NO:296), и зародыши созревали в соевой жидкой среде для гистодифференциации и созревания (жидкие среды SHaM; Schmidt et al., Cell Biology and Morphogenesis, 24:393 (2005)), как описано в Примере 25 и ранее описывалось в PCT публикации № WO 2007/136877, опубликованной 29 ноября 2007 года (на содержание которой в настоящей заявке сделана ссылка).
После созревания в жидкой среде SHaM, отдельные эмбрионы удаляли из кластеров, высушивали и подвергали скринингу на изменения в их жирнокислотных составах, как описано в Примере 1. В каждом случае подмножество соевых эмбрионов (т.е. пять зародышей на событие), тансформированных либо pKR1189 (SEQ ID NO:285) либо pKR1229 (SEQ ID NO:296), собирали и анализировали.
Таким способом проанализировали 41 событие, трансформированное pKR1189 (SEQ ID NO:285; эксперимент 2148) или pKR1229 (SEQ ID NO:296; эксперимент 2165). Жирнокислотные профили для данных пяти событий, имеющих самое низкое среднее содержание АЛК (среднее из 5 проанализированных эмбрионов), совместно с событием (2148-3-8-1), имеющим жирнокислотный профиль, типичный для зародышей дикого типа для этого эксперимента, показаны на ФИГ.36. На ФИГ.36, жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК и АЛК. Составы жирных кислот выражены в виде весового процента (вес. %) от всех жирных кислот.
Содержание АЛК в соматических зародышах, экспрессирующих либо fad3 шпильковую конструкцию (номер события 2148, ФИГ.36) или fad3c шпильковую конструкцию (номер события 2165, ФИГ.36), показало, по меньшей мере, 50% снижение, по сравнению с типичными зародышами дикого типа (ФИГ.36). Это в значительной мере указывает на то, что любая из шпильковых конструкций функциональна в плане снижения содержания АЛК в соевых зародышах.
ПРИМЕР 46
Соевые соматические зародыши, трансформированные pKR1183, для экспрессии Euglena anabaena дельта-9-элонгаза-Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразного слитого гена (Hybrid1-HGLA Synthase)
Настоящий пример описывает трансформацию и экспрессию в соевых соматических зародышах pKR1183 (SEQ ID NO:266; Пример 38), содержащих Euglena anabaena дельта-9-элонгаза-Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразный слитый ген (Hybrid1-HGLA Synthase) и гигромицин фосфотрансферазный ген для селекции на гигромицин.
Соевую эмбриогенную суспензионную культуру (cv. Jack) трансформировали pKR1183 (SEQ ID NO:266), и зародыши созревали в соевой жидкой среде для гистодифференциации и созревания (жидкие среды SHaM; Schmidt et al., Cell Biology and Morphogenesis, 24:393 (2005)), как описано в Примере 25 и ранее описывалось в PCT публикации № WO 2007/136877, опубликованной 29 ноября 2007 года (на содержание которой в настоящей заявке сделана ссылка).
После созревания в жидкой среде SHaM, подмножество соевых эмбрионов (т.е. четыре эмбриона на событие), тансформированных pKR1183 (SEQ ID NO:266), собирали и анализировали, как описано выше.
Таким способом проанализировали 20 событий, трансформированных pKR1183 (SEQ ID NO:266; эксперимент 2145). Жирнокислотные профили для данных пяти событий, имеющих самое высокое среднее содержание ДГЛК (среднее из 5 проанализированных эмбрионов), показаны на ФИГ.37. На ФИГ.37, жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК, АЛК, ЭДК, ЭРК, ДГЛК и ЭТК. Составы жирных кислот выражены в виде весового процента (вес. %) от всех жирных кислот.
ПРИМЕР 47
Соевые зародыши, трансформированные экспрессирующими векторами сои pKR1253 для экспрессии Euglena anabaena дельта-9-элонгаза-Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразного слитого гена (Hybrid1-HGLA Synthase) с Euglena gracilis дельта-5-десатуразой и pKR1249 для снижения уровня регулирования сои Fad3 и сои Fad3c
Соевую эмбриогенную суспензионную культуру (cv. Jack) трансформировали AscI фрагментами pKR1249 (SEQ ID NO:297; Пример 43) и pKR1253 (SEQ ID NO:270; Пример 39) как описано в Примере 25. Подмножество соевых эмбрионов, генерированных из каждого события (десять эмбрионов на событие), собирали, переносили в стеклянные ГХ ампулы, и жирнокислотные метиловые эфиры (ЖКМЭ) готовили путем трансэтерификации и анализировали методом газовой хроматографии как описано в Примере 1. Величины времени сравнивали с величинами времени удерживания для стандартов метиловых эфиров, имеющихся в продаже (Nu-Chek Prep, Inc.).
Таким способом проанализировали 142 события, трансформированных pKR1249 (SEQ ID NO:297) и pKR1253 (SEQ ID NO:270) (эксперимент под названием Heal 25). Из 142 проанализированных событий 90 идентифицировали как те, что вырабатывали АРК по меньшей мере в одном зародыше из десяти проанализированных при относительном содержании больше 1,0% от всех жирных кислот. Из них 64 идентифицировали как те, что вырабатывали АРК в по меньшей мере одном зародыше из десяти проанализированных при относительном содержании более 10,0% от всех жирных кислот. Из них 44 события идентифицировали как те, что вырабатывали АРК в по меньшей мере одном зародыше из десяти проанализированных при относительном содержании более 20,0% от всех жирных кислот.
Усредненные жирнокислотные профили (среднее из 10 эмбрионов) для 20 событий с наибольшим содержанием АРК показаны на ФИГ.38. Жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК, АЛК, ЭДК, СКК, ДГЛК, АРК, ЭРК, ЮН, ЭТК и ЭПК; и составы жирных кислот, перечисленных на ФИГ.38, выражены в виде весового процента (вес. %) от всех жирных кислот. Для ФИГ.38, жирные кислоты, перечисленные как “другие”, включают: 18:2 (5,9), 18:3 (5,9,12), СТК, 20:0, 20:1(11), 20:2 (7,11) или 20:2 (8,11) и ДПК. Каждая из этих жирных кислот присутствует при относительном содержании менее 2,0% от всех жирных кислот. Среднее содержание всех омега-3 жирных кислот (Всего n-3) является суммой средних содержаний всех омега-3 жирных кислот).
Фактические жирнокислотные профили для каждого зародыша из одного события (AFS 5416-8-1-1), имеющего среднее содержание АРК 17,0% и среднее содержание ЭПК 1,5%, показаны на ФИГ.39. Жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК, АЛК, ЭДК, СКК, ДГЛК, АРК, ЭРК, ЮН, ЭТК и ЭПК; и составы жирных кислот, перечисленных на ФИГ.39, выражены в виде весового процента (вес. %) от всех жирных кислот. Для ФИГ.39, жирные кислоты, перечисленные как “другие", включают: 18:2 (5,9), 18:3 (5,9,12), СТК, 20:0, 20:1(11), 20:2 (7,11) или 20:2 (8,11) и ДПК. Каждая из этих жирных кислот присутствует при относительном содержании менее 2,0% от всех жирных кислот. Среднее содержание всех омега-3 жирных кислот (Всего n-3) является суммой средних содержаний всех омега-3 жирных кислот).
Поскольку содержание АЛК обычно в 1,5-3 раза выше в соевых соматических эмбрионах, чем в семенах (т.е. 15%-30% в эмбрионах (см., например, типичный эмбрион дикого типа на ФИГ.36), в зависимости от условий созревания и времени, в сравнении с 7-10% в семенах (Bilyeu et al., 2005, Crop Sci. 45:1830-1836), можно предполагать, что содержание омега-3 в целом и содержание ЭПК в частности, будет значительно ниже в семенах, чем в соматических зародышах.
ПРИМЕР 48
Соевые зародыши, трансформированные экспрессирующими векторами сои pKR1255, для экспрессии Euglena anabaena дельта-9-элонгаза-Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразного слитого гена (Hybrid1-HGLA Synthase) с Euglena gracilis дельта-5-десатуразой и Euglena anabaena дельта-5-десатуразой и pKR1249 для снижения уровня регулирования сои Fad3 и сои Fad3c
Соевую эмбриогенную суспензионную культуру (cv. Jack) трансформировали AscI фрагментами pKR1249 (SEQ ID NO:297; Пример 43) и pKR1255 (SEQ ID NO:275; Пример 41) как описано в Примере 25. Подмножество соевых эмбрионов, генерированных из каждого события (десять эмбрионова на событие), собирали, переносили в стеклянные ГХ ампулы, и жирнокислотные метиловые эфиры (ЖКМЭ) готовили путем трансэтерификации и анализировали методом ГХ, как описано в Примере 1. Величины времени сравнивали с величинами времени удерживания для стандартов метиловых эфиров, которые имеются в продаже (Nu-Chek Prep, Inc.).
Таким способом проанализировали 197 событий, трансформированных pKR1249 (SEQ ID NO:297) и pKR1255 (SEQ ID NO:275) (эксперимент под названием Heal 26). Из 197 проанализированных событий 128 идентифицировали как те, что вырабатывали АРК в по меньшей мере одном зародыше из десяти проанализированных при относительном содержании, большем 1,0% от всех жирных кислот. Из них 105 идентифицировали как те, что вырабатывали АРК в по меньшей мере одном зародыше из десяти проанализированны при относительном содержании, большем 10,0% от всех жирных кислот. И из них 83 события идентифицировали как те, что вырабатывали АРК в по меньшей мере одном зародыше из десяти проанализированных при относительном содержании, большем 20,0% от всех жирных кислот.
Усредненные жирнокислотные профили (среднее из 9 или10 эмбрионов) для 20 событий с наибольшим содержанием АРК показаны на ФИГ.40. Жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК, АЛК, ЭДК, СКК, ДГЛК, АРК, ЭРК, ЮН, ЭТК и ЭПК; и составы жирных кислот, перечисленных на ФИГ.40, выражены в виде весового процента (вес. %) от всех жирных кислот. Для ФИГ.40, жирные кислоты, перечисленные как “другие”, включают: 18:2 (5,9), 18:3 (5,9,12), СТК, 20:0, 20:1(11), 20:2 (7,11) или 20:2 (8,11) и ДПК. Каждая из этих жирных кислот присутствует при относительном содержании менее 2,0% от всех жирных кислот. Среднее содержание всех омега-3 жирных кислот (Всего n-3) является суммой средних содержаний всех омега-3 жирных кислот).
ПРИМЕР 49
Экспрессия Euglena gracilis ДГК синтаза 1 C20 домен элонгазы/Schizochytrium aggregatum дельта-4-десатуразного гибрида (EgDHAsyn1C20EloDom3-SaD4), трансформированного экспрессирующим вектором сои pKR1134
Соевую эмбриогенную суспензионную культуру (cv. Jack) трансформировали AscI фрагментом pKR1134 (SEQ ID NO:161; Пример 20; фрагмент, содержащий экспрессирующую кассету), и зародыши созревали как описано для производства в Примере 25. Подпитку субстрата ЭПК и анализ проводили как описано в Примере 35.
Таким способом проанализировали 198 событий, трансформированных pKR1134 (эксперимент под названием Heal24). Из 198 проанализированных событий 193 идентифицировали как те, что элонгировали ЭПК (активность C20-дельта-5-элонгазы), и все они десатурировали ДПК (активность дельта-4-десатуразы) с образованием некоторого количества ДГК. События с наилучшими активностями C20-дельта-5-элонгазы и дельта-4-десатуразы были успешными, и жирнокислотные профили от эмбрионов с подпиткой ЭПК показаны на ФИГ.41.
Жирные кислоты на ФИГ.41 были идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК, АЛК, ЭПК, 22:0 (докозановая кислота), ДПК, 24:0 (тетракозановая кислота), ДГК и 4:1 (невоновая кислота); и составы жирных кислот, перечисленные на ФИГ.41, выражены в виде весового процента (вес. %) от всех жирных кислот. Активность EgDHAsyn1C20EloDom1 выражена как процент C20-дельта-5-элонгации (% C20-delta-5 elong), вычисленный в соответствии со следующей формулой: ([продукт]/[субстрат+продукт])*100. Более конкретно, комбинированный процент элонгации для ЭПК представлени как “% C20-delta-5 elong”, определяемый как: ([ДПК+ДГК]/[ЭПК+ДПК+ДГК])*100.
Активность SaD4 выражена в виде процента дельта-4-десатурации (% delta-4 desat), вычисленного в соответствии со следующей формулой: ([продукт]/[субстрат+продукт])*100. Более конкретно, комбинированный процент десатурации для ДПК представлен как “% delta-4 desat”, определяемый как: (ДГК/[ДПК+ДГК])*100.
В дополнение к 122 событиям, проанализированным для сои, трансформированной pKR1105 (эксперимент под названием Heal23), как описано в Примере 35, проанализировали еще 20 событий, поскольку Пример 35 написали с доведением общего числа проанализированных событий до 142. Из 20 новых проанализированных событий 18 идентифицировали как те, что элонгировали ЭПК (активность C20-дельта-5-элонгазы) и 17 из них также десатурировали ДПК (активность дельта-4-десатуразы) с образованием некоторого количества ДГК. События с наилучшими активностями C20-дельта-5-элонгазы и дельта-4-десатуразы от 20 новых событий, проанализированных для сои, трансформированной pKR1105, были успешными, и жирнокислотные профили от зародышей с подпиткой ЭПК показаны на ФИГ.42.
Относительные активности событий, трансформированных либо pKR1105 (C20-элонгаза и дельта-4-десатураза экспрессированы отдельно) либо pKR1134 (C20-элонгаза и дельта-4-десатураза экспрессированы как слияние), сравнивали путем построения графика зависимости % ДГК (вес. %) от % ДПК (вес. %) для всех событий, где эмбрионы подпитывали ЭПК. Результаты представлены на ФИГ.43. На ФИГ.43, Heal23 является названием эксперимента, где трансформировали pKR1105, и Heal24 представляет эксперимент, где трансформировали pKR1134. Из ФИГ.43 очевидно, что, в целом, концентрации ДГК возросли, тогда, как концентрации ДПК снизили (до необнаружимого уровня), когда C20-элонгаза была слита с дельта-4-десатуразой. Таким образом, слияние 2 независимых ферментов вместе с образованием одного слитого протеина, разделенного линкерным участком, усиливало поток от ЭПК к ДГК.
ПРИМЕР 50
Экспрессия Euglena anabaena дельта-9-элонгаза-Tetruetreptia pomquetensis CCMP1491 дельта-8-десатураза-Euglena anabaena дельта-5-десатуразного слитого гена (EaD9Elo1-TpomD8-EaD5Des1 fusion)
Соевую эмбриогенную суспензионную культуру (cv. Jack) трансформировали pKR1322 (SEQ ID NO:314), и эмбрионы созревали и их анализировали на жирнокислотные профили как здесь описано.
Соевые соматические эмбрионы, трансформированные pKR1322 (SEQ ID NO:314), элонгируют ЛК до ЭДК, ЭДК десатурируется до ДГЛК, и ДГЛК десатурируется далее до АРК. Поскольку соя дикого типа также содержит АЛК, некоторая часть АЛК элонгируется до ЭРК, ЭРК будет десатурироваться до ЭТК, и ЭТК будет десатурироваться далее до ЭПК.
Соевые растения, экспрессирующие слитый ген из pKR1322, могут быть регенерированы из зародышей, как описано в Примере 26, и могут быть получены семена.
В средах, которые имеют высокий уровень АЛК, или где дельта-15-десатураза и или дельта-17-десатураза коэкспрессированы, ЭПК будет доминировать. Наоборот, АРК может быть обогащена путем использования среды с низким содержанием АЛК (например, путем скрещивания с растением с низким содержанием АЛК или низким содержанием лин) или путем “нокаутирования” эндогенного fad3 гена(ов), как здесь описано. Промежуточные жирные кислоты (т.е. ЭДК и ДГЛК или ЭРК и ЭТК) будут в меньшем количестве, когда применяется слияние, чем когда отдельные активности трансформируются независимо (т.е. без слияния).
Другие генные комбинации (включая комбинации линкеров) могут быть слиты вместе схожим образом, как описано в Примере 24. Подобно этому, могут быть осуществлены другие промотор/слитый ген/терминаторные комбинации.
ПРИМЕР 51
Определение функционального домена в синтетической дельта-4-десатуразе, полученной из Euglena anabaena и кодон-оптимизированной для экспрессии в Yarrowia lipolytica (EaD4S)
Как схематически показано на ФИГ.52C, С-концевая часть C20-элонгазного домена EaDHAsyn1 (помеченного на фигуре как “EaC20E”) представляется перекрывающейся с N-концевой частью дельта-4-десатуразного домена EaDHAsyn1 (помеченного на фигуре как “EaD4”). Это предполагается на основе сравнения последовательностей.
Для определения функционального дельта-4-десатуразного домена в EaD4S (SEQ ID NO:192; Пример 34) генерировали три EaD4S* мутанта с разными N-концевыми усечениями. Конкретно, сконструировали pZuFmEaD4S (SEQ ID NO:364) путем замены NcoI/NotI фрагмента pZuFmIgD9ES (SEQ ID NO:365) фрагментом NcoI/NotI EaD4S pEaD4S (SEQ ID NO:196; Пример 34). NcoI сайт введили в pZuFmEaD4S (SEQ ID NO:364) путем сайт-направленного мутагенеза с использованием праймерных пар YL921 и YL922 (SEQ ID NOs:366 и 367, соответственно), YL923 и YL924 (SEQ ID NOs:368 и 369, соответственно) и YL925 и YL926 (SEQ ID NOs:370 и 371, соответственно) с генерацией pZuFmEaD4S-M1 (SEQ ID NO:372), pZuFmEaD4S-M2 (SEQ ID NO:373) и pZuFmEaD4S-M3 (SEQ ID NO:374), соответственно. pZuFmEaD4S-M1, pZuFmEaD4S-M2 и pZuFmEaD4S-M3 плазмиды расщепляли NcoI, и потом самолигировались с генерацией pZuFmEaD4S-1 (SEQ ID NO:375), pZuFmEaD4S-2 (SEQ ID NO:376) и pZuFmEaD4S-3 (SEQ ID NO:377) конструкций. NcoI/NotI фрагменты, содержащие разные усечения EaD4S из pZuFmEaD4S-1, pZuFmEaD4S-2, pZuFmEaD4S-3, использовали для продуцирования pZKL4-220EA4-1 (SEQ ID NO:378), pZKL4-220EA4-2 (SEQ ID NO:379) и pZKL4-220EA4-3 (SEQ ID NO:380) конструкций. Эти три конструкции были точно такими же как pZKL4-220EA4 (SEQ ID NO:362 и ФИГ.52B, как описано в Таблице 28 Примера 34), за исключением того, что кодирующий участок EaD4S был усечен в N-концевой области. Конкретно, вместо кодирующей последовательности длиной 583 аминокислоты EaD4S (SEQ ID NO:193), усеченный EaD4S* полипептид был длиной 547 аминокислот в pZKL4-220EA4-1 (т.е. SEQ ID NO:382), длиной 527 аминокислот в pZKL4-220EA4-2 (т.е. SEQ ID NO:384) и длиной 512 аминокислот в pZKL4-220EA4-3 (т.е. SEQ ID NO:386). N-концевые участки этих полипептидов (соответствующие аминокислотам 1-90 SEQ ID NO:193) сгруппированы на ФИГ.53A.
Плазмиды pZKL4-220EA4 (SEQ ID NO:362), pZKL4-220EA4-1 (SEQ ID NO:378), pZKL4-220EA4-2 (SEQ ID NO:379) и pZKL4-220EA4-3 (SEQ ID NO:380) расщепляли AscI/SphI, и затем трансформировали в Yarrowia lipolytica штамм Y4184U4, как описано в разделе Общие методы. Трансформанты отбирали на МС пластинках. После 5 суток роста при 30°C, 5 трансформантов, выращенных на МС пластинках из каждой конструкции, отбирали и повторно наносили штрихами на свежие МС пластинки. После выращивания эти штаммы инокулировали индивидуально в 3 мл жидкой МС при 30°C и встряхивали при 250 оборотов/минуту в течение 2 суток. Клетки собирали путем центрифугирования, ресуспендировали в HGM и потом встряхивали при 250 оборотв/минуту в течение 5 суток. Данные клетки собирали путем центрифугирования, липиды экстрагировали, и жирнокислотные метиловые эфиры готовили путем трансэтерификации, и впоследствии анализировали на газовом хроматографе Hewlett-Packard 6890 GC.
ГХ результаты показаны ниже в Таблице 29. Составы ДПК и ДГК представлены в виде % от всех жирных кислот. Эффективность конверсии (“Conv. Effic.”) измеряли в соответствии со следующей формулой: ([ДГК]/[ДПК+ДГК])*100. Активность дельта-4-десатуразы каждого усеченного EaD4S* (т.е. полипетидов из 547 аминоксилот, 527 аминокислот или 512 аминокислот в Yarrowia трансформантах pZKL4-220EA4-1, pZKL4-220EA4-2 и pZKL4-220EA4-3, соответственно) сравнивали с таковой для EaD4S (т.е. Yarrowia трансформантов pZKL4-220EA4) в колонке, обозначенной как “% Delta-4 Activity” (“% дельта-4 активности”.
Эти данные продемонстрировали, что N-концевые 37 аминокислот EaD4S (т.е. аминоксилоты 1-37 SEQ ID NO:193) оказывают отрицательное влияние на активность дельта-4-десатуразы. Повышенная активность дельта-4-десатуразы отмечается относительно EaD4S в каждом из данных EaD4S* усеченных протеинов, хотя EaD4S* полипептид из 547 аминокислот (SEQ ID NO:382) превосходит по активности EaD4S* полипептиды без дополнительных аминоксислот из их N-окончания (т.e. SEQ ID NOs:384 и 386).
Пример 52
Синтез и функциональный анализ кодон-оптимизированного дельта-4-десатуразного гена (EgD4S) из Euglena gracilis в Yarrowia lipolytica
Использование кодона дельта-4-десатуразного домена EgDHAsyn1 (SEQ ID NO: 221) Euglena gracilis (соответствующего аминокислотам 253-793 SEQ ID NO:12) оптимизировали для экспрессии в Yarrowia lipolytica, способом, подобным описанному в PCT публикации № WO 2004/101753, патенте США № 7125672 и Примерах 32, 33 и 34 данной заявки. Конкретно, создали кодон-оптимизированный дельта-4-десатуразный ген (обозначенный как “EgD4S”; SEQ ID NO:387), основанный на кодирующей последовательности дельта-4-десатуразного домена EgDHAsyn1, в соответствии со схемой использования Yarrowia кодона (PCT публикация № WO 2004/101753), консенсусной последовательностью вокруг ′ATG′ трансляционного инициирующего кодона и общими правилами РНК стабильности (Guhaniyogi, G. и J. Brewer, Gene, 265(1-2):11-23 (2001)). В дополнение к модификации трансляционного инициирующего сайта, модифицировали 282 bp 1623 bp кодирующего участка (17,4%), и 270 кодонов оптимизировали (49,9%). Таким способом кодон-оптимизированный кодирующий участок EgS4S длиной 1623 bp кодирует полипептид из 540 аминокислот (SEQ ID NO: 388). Таким образом, EgD4S на одну аминокислоту короче, чем дельта-4-десатуразный домен дикого типа EgDHAsyn1 (SEQ ID NO:221; конкретно, лейциновый остаток, соответствующий аминокислотному положению 2 SEQ ID NO:13 , был удален в EgD4S (SEQ ID NO:388). Созданный EgD4S ген (SEQ ID NO:387) был синтезирован корпорацией GenScript Corporation (Piscataway, NJ) и клонирован в pUC57 (номер доступа GenBank Y14837) с генерацией pEgD4S (SEQ ID NO:389).
Для анализа функции кодон-оптимизированного EgD4S гена сконструировали плазмиду pZKL4-220Eg4 (SEQ ID NO:390) для интеграции двух химерных C20-элонгазных генов и химерного EgD4S гена в липаза 4 подобный локус (номер доступа GenBank XM_503825) Yarrowia lipolytica штамм Y4305U3. Плазмида pZKL4-220Eg4 (SEQ ID NO:390) идентична по структуре с плазмидой pZKL4-220Ea4 (SEQ ID NO:362; ФИГ.52B; Таблица 28 Примера 34), с тем исключением, что EgD4S (SEQ ID NO:387) использовали вместо EaD4S (SEQ ID NO:192).
Плазмиду pZKL4-220Eg4 расщепляли AscI/SphI, и затем трансформировали в Yarrowia lipolytica штамм Y4305U3, как описано в разделе Общие методы. Трансформанты отбирали на МС пластинках. После 5 суток роста при 30°C, 14 трансформантов, выращенных на МС пластинках, отбирали и повторно наносили штрихами на свежие МС пластинки. После выращивания эти штаммы инокулировали индивидуально в 3 мл жидкой МС при 30°C и встряхивали при 250 оборотов/минуту в течение 2 суток. Клетки собирали путем центрифугирования, ресуспендировали в HGM и потом встряхивали при 250 оборотв/минуту в течение 5 суток. Данные клетки собирали путем центрифугирования, липиды экстрагировали, и жирнокислотные метиловые эфиры готовили путем трансэтерификации, и впоследствии анализировали на газовом хроматографе Hewlett-Packard 6890 GC.
ГХ анализы показали, что в среднем во всех 14 трансформантах было выработано 4,9% ДГК и 17,8% ДПК от всех липидов, где эффективность конверсии ДПК в ДГК составляла, как было найдено, около 21,5% (вычислено как описано в Примере 27). Таким образом, эти экспериментальные данные продемонстрировали, что синтетическая Euglena gracilis дельта-4-десатураза, кодон-оптимизированная для экспрессии в Yarrowia lipolytica (т.е. EgD4S, как представлено в SEQ ID NO:387), была активной в плане превращения ДПК в ДГК.
ПРИМЕР 53
Определение функционального домена в синтетической дельта-4-десатуразе, полученной из Euglena gracilis и кодон-оптимизированной для экспрессии в Yarrowia lipolytica (EgD4S)
Как и в случае, наблюдаемом в EaDHAsyn1 (ФИГ.52C), С-концевая часть C20-элонгазного домена EgDHAsyn1 представляется перекрывающейся с N-концевой частью дельта-4-десатуразного домена EgDHAsyn, как можно судить из сравнения последовательностей.
С целью определения функционального дельта-4-десатуразного домена в EgD4S (SEQ ID NO:387) генерировали три EgD4S мутанта с разными N-концевыми усечениями. NcoI сайт вводили в pEgD4S (SEQ ID NO:389; Пример 52) путем сайт-направленного мутагенеза с использованием праймерных пар YL935 и YL936 (SEQ ID NOs:391 и 392, соответственно), YL937 и YL938 (SEQ ID NOs:393 и 394, соответственно) и YL939 и YL940 (SEQ ID NOs:395 and 396, соответственно) с генерацией pEgD4S-M1 (SEQ ID NO:397), pEgD4S-M2 (SEQ ID NO:398) и pEgD4S-M3 (SEQ ID NO:399) конструкций, соответственно. NcoI/NotI фрагменты, содержащие разные усечения EgD4S из pEgD4S-M1, pEgD4S-M2 и pEgD4S-M3, использовали для генерации pZKL4-220Eg4-1 (SEQ ID NO:400), pZKL4-220Eg4-2 (SEQ ID NO:401) и pZKL4-EgD4-3 (SEQ ID NO:402) конструкций. Данные три конструкции были в точности такими же, как pZKL4-220Eg4 (SEQ ID NO:390; Пример 52), исключая то, что кодирующий участок EgD4S был усечен в N-концевом участке. Конкретно, вместо кодирующей последовательности длиной в 540 аминокислот EgD4S (SEQ ID NO:388), усеченный EgD4S* полипептид имел длину в 513 аминоксилот в pZKL4-220Eg4-1 (т.е. SEQ ID NO:404), 490 аминокислот в in pZKL4-220Eg4-2 (т.е SEQ ID NO:406) и 474 аминоксилоты в pZKL4-220Eg4-3 (т.е. SEQ ID NO:408). N-концевые учвстки этих полипептидов (соответствующих аминокислотам 1-80 SEQ ID NO:388) сгруппированы на ФИГ.53B.
Плазмиды pZKL4-220Eg4 (SEQ ID NO:390), pZKL4-220Eg4-1 (SEQ ID NO:400), pZKL4-220Eg4-2 (SEQ ID NO:401) и pZKL4-EgD4-3 (SEQ ID NO:402) расщепляли AscI/SphI, и затем трансформировали в Yarrowia lipolytica штамм Y4305U3 индивидуально, как описанно в разделе Общие методы. Трансформанты отбирали на МС пластинках. После 5 суток роста при 30°C, 4 трансформанта, выращенных на МС пластинках из каждой конструкции, извлекали и повторно наносили на свежие МС пластинки. После выращивания эти штаммы инокулировали индивидуально 3 мл жидкой МС при 30°C и встряхивали при 250 оборотов/минуту в течение 2 суток. Клетки собирали путем центрифугирования, ресуспендировали в HGM и потом встряхивали при 250 оборотов/минуту в течение 5 суток. Эти клетки собирали путем центрифугирования, липиды экстрагировали, и жирнокислотные метиловые эфиры готовили путем трансэтерификации и потом анализировали на газовом хроматографе Hewlett-Packard 6890 GC.
ГХ результаты приведены ниже в Таблице 30. Составы ДПК и ДГК представлены в виде % от всех жирных кислот. Эффективность конверсии (“Conv. Effic.”) определяли в соответствии со следующей формулой: ([ДГК]/[ДПК+ДГК])*100. Активность дельта-4-десатуразы каждого усеченного EgD4S* (т.е. полипептидов из 513 аминокислот, 490 аминокислот или 474 аминокислоты в Yarrowia трансформантах pZKL4-220Eg4-1, pZKL4-220Eg4-2 и pZKL4-220Eg4-3, соответственно) сравнивали с таковой для EgD4S (т.е. Yarrowia трансформантами pZKL4-220Eg4) в колонке, обозначенной как “% Delta-4 Activity” (“% дельта-4 активности”).
Эти данные продемонстрировали, что N-концевые 28 аминоксислот EgD4S* не играют роли (т.е. см. pZKL4-220Eg4-1, где первые 28 аминокислот EgD4S были усечены, и усеченный протеин, представленный как SEQ ID NO:404, сохранял полную активность дельта-4-десатуразы). Уменьшенная активность дельта-4-десатуразы была найдена в трансформантах с pZKL4-220EgD4-2 и pZKL4-220EgD4-3, когда дополнительные аминокислоты были усечены от 5′ части EgD4S протеина, давая таким образом SEQ ID NO:406 и SEQ ID NO:408.
ПРИМЕР 54
Синтез и функциональный анализ кодон-оптимизированного EgDHAsyn1 гена (EgDHAsyn1S) из Euglena gracilis в Yarrowia lipolytica
Плазмиду pZKLY-G204 (ФИГ.54A; SEQ ID NO:409) сконструировали для интеграции химерного гена, содержащего кодон-оптимизированный EgDHAsyn1 кодирующий участок (т.е. EgDHAsyn1S, представленнный как SEQ ID NOs:410 и 411), в липазе 7 локус (номер доступа GenBank AJ549519) Yarrowia lipolytica штамм Y4305U3. В дополнение к модификации трансляционного инициирующего сайта, 417 bp 2382 bp кодирующего участка модифицировали (17,5%), и 391 кодон из всех 794 кодонов оптимизировали (49,2%). Аминокислотная последовательность кодон-оптимизированного EgDHAsyn1S (SEQ ID NO:411) на 100% идентична по последовательности таковой для EgDHAsyn1 (SEQ ID NO:12).
Для генерации pZKLY-G204, KpnI сайт вводили в pEgC20ES (SEQ ID NO:185; см. ФИГ.51B, Пример 32) с генерацией pEgC20ES-K (SEQ ID NO:412; ФИГ.54B) путем сайт-направленного мутагенеза с использованием олигонуклеотидов YL973 и YL974 (SEQ ID NOs:413 и 414, соответственно) как праймеров и pEgC20ES как матрицы. 732 bp PmeI/NcoI фрагмент, содержащий YAT1 промотор (патентная публикация США № 2006/0094102-A1) pYNTGUS1-CNP (ФИГ.54C; SEQ ID NO:415), 873 bp NcoI/KpnI фрагмент pEgC20ES-K, содержащий кодон-оптимизированный N-концевой участок EgDHAsyn1S и 1512 bp KpnI/NcoI фрагмент pEgD4S (SEQ ID NO:389; Пример 52), содержащий кодон-оптимизированный C-концевой участок EgDHAsyn1S, изолировали и затем использовали для замены PmeI/NotI фрагмента pZKLY (ФИГ.54D; SEQ ID NO:416) с генерацией pZKLY-G204 (ФИГ.54A). Таким образом, pZKLY-G204 содержала следующие компоненты:
Плазмиду pZKLY-G204 расщепляли AscI/SphI, и затем трансформировали в Yarrowia lipolytica штамм Y4305U3, как описано в разделе Общие методы. Трансформанты отбирали на МС пластинках. После роста в течение 5 суток при 30°C 8 трансформантов, выращенных на МС пластинках, извлекали и повторно наносили на свежие МС пластинки. После выращивания эти штаммы индивидуально инокулировали в 3 мл жидкую МС при 30°C и встряхивали при 250 оборотов/минуту в течение 2 суток. Клетки собирали путем центрифугирования, ресуспендировали в HGM и потом встряхивали при 250 оборотов/минуту в течение 5 суток. Данные клетки собирали путем центрифугирования, липиды экстрагировали, и жирнокислотные метиловые эфиры готовили путем трансэтерификации, и впоследствии анализировали на газовом хроматографе Hewlett-Packard 6890 GC.
ГХ анализы показали, что во всех 8 трансформантах присутствовало около 5,0% ДГК, 13,5% ДПК и 26,5% ЭПК от всех продуцированных липидов. Эффективность конверсии ЭПК в ДПК и ДГК составляла, как было найдено, около 41%; и, эффективность конверсии ДПК в ДГК составляла, как было определено, приблизительно 27% в этих восьми штаммах (вычислено как описано в Примере 27). Таким образом, эти экспериментальные данные продемонстрировали, что синтетическая Euglena gracilis ДГК синтаза, кодон-оптимизированная для экспрессии в Yarrowia lipolytica (т.е. EgDHAsyn1S, как представлено в SEQ ID NO:410), обладала активностью как C20-элонгазы так и активностью дельта-4-десатуразы. EgDHAsyn1S может использовать ЭПК как субстрат для выработки ДГК.
ПРИМЕР 55
Создание дельта-9-элонгаза/дельта-8-десатуразных генных слияний для экспрессии в Yarrowia lipolytica
Для улучшения ферментативной активности дельта-9-элонгазы и дельта-8-десатуразы в Yarrowia lipolytica в настоящем примере создали серию из шести дельта-9-элонгаза/дельта-8-десатуразных генных слияний (мультизимов) с использованием двухвариантных линкерных последовательностей (т.е. SEQ ID NO:438 [GPARPAGLPPATYYDSLAV] и SEQ ID NO:445 [GAGPARPAGLPPATYYDSLAVMGS]) полученных из EgDHAsyn1 обогащенного пролином линкера (SEQ ID NO:198; PARPAGLPPATYYDSLAV).
Данная работа потребовала: идентификации соответствующих дельта-9-элонгаз и дельта-8-десатураз для экспрессии в Yarrowia lipolytica; и, конструирования плазмиды pZUFmG9G8fu (включающей EgD9ES/EgD8M генное слияние), плазмиды pZuFmG9G8fu-B (включающей EgD9ES/EgD8M генное слияние), плазмиды pZUFmG9A8 (включающей EgD9ES/EaD8S генное слияние), плазмиды pZUFmA9G8 (включающей EaD9ES/EgD8M генное слияние), плазмиды pZUFmA9A8 (включающей EaD9ES/EaD8S генное слияние) и плазмиды pZUFmR9G8 (включающей E389D9eS/EgD8M генное слияние). Все плазмиды использовали совместно общий векторный каркас и поэтому отличались только генным слиянием, которое каждая из них включала.
Функциональный анализ активности каждого генного слияния тестировали infra, в Примере 56.
Описание синтетических дельта-9-элонгазного и дельта-8 десатуразного генов, кодон-оптимизированных для экспрессии в Yarrowia lipolytica
Заявители проделали множество анализов разных дельта-9-элонгаз и дельта-8-десатураз для определения ферментов, которые обладают оптимальной специфичностью субстрата и/или селективностью субстрата при экспрессии в Yarrowia lipolytica. Исходя из этих анализов, гены, описанные ниже в Таблице 32 и полученные из них кодон-оптимизированные гены (с использованием методологии патента США № 7125672), были определены как предпочтительные для экспрессии в Y. lipolytica. Те гены, которые выделены жирным шрифтом, впоследствии использовали для создания дельта-9-элонгаза/дельта-8 десатуразных генных слияний.
Об эффективности конверсии ЛК в ЭДК EgD9eS, E389D9eS и EaD9eS сообщается в каждой из заявок, цитированных выше. Вкратце, когда каждую дельта-9-элонгазу была экспрессировали как химерный ген в Yarrowia lipolytica штамме Y2224 (ФИГ.44), под контролем Yarrowia FBAINm промотора (PCT публикация № WO 2005/049805; патент США № 7202356) и Pex20 терминирующей последовательности из Yarrowia Pex20 гена (номер доступа GenBank AF054613), независимо получали следующие значения конверсии субстрата: EgD9eS (20,1%); EaD9eS (13%); и, E389D9eS (12%). Все синтетические кодон-оптимизированные гены функционировали с большей эффективностью конверсии субстрата, чем соответствующий ген дикого типа.
Патент США № 7256033 раскрывает E. gracilis дельта-8-десатуразу (“EgD8”), способную десатурировать ЭДК и ЭтрК в ДГЛК и ЭТК, соответственно, так же как и синтетическую дельта-8-десатуразу, полученную из EgD8 и кодон-оптимизированную для экспрессии в Yarrowia lipolytica (“EgD8S”). Несмотря на полезность EgD8 и EgD8S, синтетически полученная мутантная дельта-8-десатураза, идентифицированная здесь как EgD8M (SEQ ID NOs:327 и 328), используется в более предпочтительных вариантах для экспрессии в Yarrowia lipolytica. Как описано в патентной заявке США № 11/635258, “EgD8M” (идентифицированная здесь как “EgD8S-23”) создали путем множественных раундов направленных мутаций внутри EgD8S. Влияние каждой мутации на активность дельта-8-десатуразы результирующего мутанта было подвержено скринингу, чтобы убедиться в функциональной эквивалентности с активностью EgD8S дельта-8-десатуразы (SEQ ID NO:426). Как результат этой работы, мутантная EgD8M (SEQ ID NO:328) включает следующие 24 аминокислотные мутации в отношении к снтетической кодон-оптимизированной EgD8S последовательности, представленной как SEQ ID NO:426: 4S к A, 5K к S, 12T к V, 16T к K, 17T к V, 66P к Q, 67S к A, 108S к L, 117G к A, 118Y к F, 120L к M, 121M к L, 125Q к H, 126M к L, 132V к L, 133 L к V, 162L к V, 163V к L, 293L к M, 407A к S, 408V к Q, 418A к G, 419G к A и 422L к Q. Парное группирование EgD8M и EgD8S протеиновых последовательностей с использованием параметров по умолчанию Vector NTI®′s AlignX программы (Invitrogen Corporation, Carlsbad, CA) выявило 94,3% идентичность последовательности и 97,9% соответствия между данными двумя протеинами на длине 422 аминокислот. Среднее значение субстратной конверсии ЭДК в ДГЛК этим мутантом дельта-8-десатуразы составляло, как было найдено, 37%, когда EgD8M экспрессировали в Yarrowia lipolytica штамм Y4001 (ФИГ.44), под контролем Yarrowia FBAINm промотора (PCT публикация № WO 2005/049805; патент США № 7202356) и Pex20 терминирующей последовательности из Yarrowia Pex20 гена (номер доступа GenBank AF054613).
Когда EaD8S экспрессировали в Yarrowia lipolytica штамм Y4001U (ФИГ.44), под контролем Yarrowia FBAINm промотора (патент США № 7202356) и Pex20 терминирующей последовательности из Yarrowia Pex20 гена (номер доступа GenBank AF054613), имела место 41% эффективность конверсии в ДГЛК с эндогенной ЭДК в качестве субстрата.
Генерация конструкция pZUFmEgD9ES-Na, включающего EgD9ES
Плазмида pZuFmEgD9ES (SEQ ID NO:431), которая была ранее описана в патентной публикации США № 2007-0117190 A1, включает химерный FBAINm::EgD9ES::Pex20 ген, ColE1 плазмидный источник репликации, ампициллин-резистентный ген (AmpR) для селекции в E. coli,
Yarrowia автономную репликационную последовательность (ARS18; номер доступа GenBank A17608), и Yarrowia Ura 3 ген (номер доступа GenBank AJ306421).
Nar I сайт вводили в pZuFmEgD9ES с генерацией pZuFmEgD9ES-Na (SEQ ID NO:432) с использованием олигонуклеотидоа YL989 и YL990 (SEQ ID NOs:433 и 434, соответственно) как праймеров и pZuFmEgD9ES и как матрицы. Введенный Nar I сайт (т.е. GGCGCC) локализовали как раз перед трансляционным терминирующим кодоном EgD9ES; поэтому кодирующий участок EgD9ES расширили двумя дополнительными аминокислотами (т.е. глицином и аланином).
Генерация конструкции pZUFmG9G8fu, включающей EgD9ES/EgD8M генное слияние
N-концевую часть EgD8M (SEQ ID NO:327) амплифицировали ПЦР с использованием олигонуклеотидов YL991 и YL992 (SEQ ID NOs:435 и 436, соответственно) как праймеров pKO2UFm8A (SEQ ID NO:437) и как матрицы. Олигонуклеотид YL991, который содержал Nar I сайт в своем 5′ конце и ДНК последовательность, кодирующую модифицированный вариант EgDHAsyn1 обогащенного пролином линкера (т.е. GPARPAGLPPATYYDSLAV, как представлено в SEQ ID NO:438 в зависимости от PARPAGLPPATYYDSLAV, как представлено в SEQ ID NO:198). Этот линкер обладал дополнительным глицином в 5′ конце, относительно EgDHAsyn1 обогащенного пролином линкера (SEQ ID NO:198).
Nar I/Bgl II, расщепляющий ПЦР продукт, включающий 5′ участок EgD8M, и Bgl II/Not I, расщепляющий фрагмент pKO2UFm8A, включающий 3′ участок EgD8M, использовали для замены NarI/NotI фрагмента pZUFmEgD9ES-Na с генерацией pZUFmG9G8fu (ФИГ.55A), которая содержала, таким образом, следующие компоненты:
Генерация конструкции pZuFmG9G8fu-B, включающей EgD9ES/EgD8M генное слияние
BamH I сайт (т.е. GGATCC) введили в pZUFmG9G8fu с генерацией pZUFmG9G8fu-B (SEQ ID NO:442) с использованием олигонуклеотидов YL1043 и YL1044 (SEQ ID NOs:443 и 444, соответственно) как праймеров рZuFmG9G8fu и как матрицы. BamH I сайт локализовали как раз после трансляционного инициирующего кодона ATG и находился в рамке считывания EgD8M, что привело к инсерции двух аминокислот (т.е. глицина и аланина) между метиониновым аминокислотным остатком и оставшейся частью EgD8M полипептида. Эта модификация привела к тому, что линкерный участок между EgD9ES и EgD8M стал пептидом с последовательностью, представленной как SEQ ID NO:445 (т.е. GAGPARPAGLPPATYYDSLAVMGS); нуклеотидная и транслированная аминокислотная последовательности EgD9ES/EgD8M генного слияния полной длины представлены как SEQ ID NOs:446 и 447, соответственно.
Генерация конструкции pZUFmG9A8, включающей EgD9ES/EaD8S генное слияние
Плазмиду pEaD8S (SEQ ID NO:448) создали, когда EaD8S ген (SEQ ID NO:429) клонировали в pUC57 (номер доступа GenBank Y14837). Затем BamH I сайт вводили в pEaD8S с использованием олигонуклеотидов YL1059 и YL1060 (SEQ ID NOs:449 и 450, соответственно) как праймеров pEaD8S и как матрицы, с генерацией pEaD8S-B (SEQ ID NO:451). Введенный BamH I сайт (т.е. GGATCC) локализовали как раз перед трансляционным инициирующим кодоном EaD8S в pEaD8S-B, и находится в той же рамке считывания, что и EaD8S.
BamHI/NotI фрагмент pEaD8S-B, включающий EaD8S, использовали для замены BamH I/Not I фрагмента pZUFmG9G8fu-B (SEQ ID NO:442) с генерацией pZUFmG9A8 (SEQ ID NO:452; ФИГ.55B) (вводя тем самым EaD8S вместо EgD8M). Линкерный участок между EgD9ES и EaD8S представлял собой пептид с последовательностью, представленной в SEQ ID NO:445 (т.е. GAGPARPAGLPPATYYDSLAVMGS). Таким образом, плазмида pZUFmG9A8 содержала EgD9ES/EaD8S генное слияние, фланкированное Yarrowia lipolytica FBAINm промотором и Pex20 терминатором (номер доступа GenBank AF054613). Нуклеотидная и транслированная аминокислотная последовательности генного слияния EgD9ES/EaD8S полной длины представлены как SEQ ID NOs:453 и 454, соответственно.
Генерация конструкции pZUFmA9G8, включающей EaD9ES/EgD8M генное слияние
Плазмида pZUFmEaD9ES (SEQ ID NO:455) содержала EaD9ES ген, фланкированный Yarrowia lipolytica FBAINm промотором и Pex20 терминатором (номер доступа GenBank AF054613). Nar I сайт вводили в данную плазмиду с генерацией pZUFmEaD9ES-Na (SEQ ID NO:456) с использованием олигонуклеотидов YL1049 и YL1050 (SEQ ID NOs:457 и 458, соответственно) как праймеров и pZUFmEaD9ES как матрицы. Введенный Nar I сайт (т.е. GGCGCC) был локализован как раз перед трансляционным терминирующим кодоном EaD9ES, и находится в той же самой рамке считывания EaD9ES.
Nco I/Nar I фрагмент pZUFmEaD9ES-Na (SEQ ID NO:456), включающий EaD9ES, использовали для замены Nco I/Nar I фрагмента pZUFmG9G8fu-B (SEQ ID NO:442) с генерацией pZUFmA9G8 (SEQ ID NO:459) (вводя тем самым EaD9ES вместо EgD9ES). Линкерный участок между EaD9ES и EgD8M представлял собой пептид с последовательностью, представленной в SEQ ID NO:445 (т.е. GAGPARPAGLPPATYYDSLAVMGS). Таким образом, плазмида pZUFmA9G8 содержала EaD9ES/EgD8M генное слияние, фланкированное Yarrowia lipolytica FBAINm промоторм и Pex20 терминатором (номер доступа GenBank AF054613). Нуклеотидная и транслированная аминокислотная последовательности генного слияния EaD9ES/EgD8M полной длины представлены как SEQ ID NOs:460 и 461, соответственно.
Генерация конструкции pZUFmA9A8, включающей EaD9ES/EaD8S генное слияние
BamH I I/Not I фрагмент pEaD8S-B (SEQ ID NO:451), включающий EaD8S, использовали для замены BamH I/Not I фрагмента pZUFmA9G8 (SEQ ID NO:459) с генерацией pZUFmA9A8 (SEQ ID NO:462) (вводя тем самым EaD8S вместо EgD8M). Линкерный участок между EaD9ES и EaD8S представлял собой пептид с последовательностью, представленной в SEQ ID NO:445 (т.е. GAGPARPAGLPPATYYDSLAVMGS). Таким образом, плазмида pZUFmA9A8 содержала EaD9ES/EaD8S генное слияние, фланкированное Yarrowia lipolytica FBAINm промотором и Pex20 терминатором (номер доступа GenBank AF054613). Нуклеотидная и транслированная аминокислотная последовательности генного слияния EaD9ES/EaD8S полной длины представлены в SEQ ID NOs:463 и 464, соответственно.
Генерация конструкции pZUFmR9G8, включающей E389D9eS/EgD8M генное слияние
Плазмиду pE389S (SEQ ID NO:465) создали, когда E389D9eS ген (SEQ ID NO:358) клонировали в pUC57 (номер доступа GenBank Y14837). Затем NarI сайт введили в pE389S с генерацией pE389S-Na (SEQ ID NO:466) с использованием олигонуклеотидов YL1051 и YL1052 (SEQ ID NOs:467 и 468, соответственно) как праймеров и pE389S как матрицы. Введенный Nar I (т.е. GGCGCC) сайт локализовали как раз перед трансляционным терминирующим кодоном и находился в той же самой рамке считывания E389D9eS.
Nco I/Nar I фрагмент pE389S-Na, включающий E389D9eS, использовали для замены Nco I/Nar I фрагмента pZUFmG9G8fu-B, включая EgD9ES, с генерацией pZUFmR9G8 (SEQ ID NO:469) (вводя тем самым E389D9eS вместо EgD9ES). Линкерный участок между E389D9eS и EgD8M представлял собой пептид с последовательностью, представленной в SEQ ID NO:445 (т.е. GAGPARPAGLPPATYYDSLAVMGS). Таким образом, плазмида pZUFmR9G8 содержала E389D9eS/EgD8M генное слияние, фланкированное Yarrowia lipolytica FBAINm промоторм и Pex20 терминатором (номер доступа GenBank AF054613). Нуклеотидная и транслированная аминокислотная последовательности генного слияния E389D9eS/EgD8M полной длины представлены в SEQ ID NOs:470 и 471, соответственно.
Пример 56
Функциональный анализ дельта-9-элонгаза/дельта-8-десатуразных генных слияний в Yarrowia lipolytica штамме Y2224
Плазмиды из Примера 55 [т.е. pZUFmEgD9ES (SEQ ID NO:431), pZUFMEgD9ES-Na (SEQ ID NO:432), pZUFMG9G8fu (SEQ ID NO:439), pZUFmG9G8fu-B (SEQ ID NO:442), pZUFmG9A8 (SEQ ID NO:452), pZUFmA9G8 (SEQ ID NO:459), pZuFmA9A8 (SEQ ID NO:462) и pZUFmR9G8 (SEQ ID NO:469)] индивидуально трансформировали в Yarrowia lipolytica штамм Y2224, как описано в разделе Общие методы. Трансформанты отбирали на МС пластинках. После роста в течение 2 суток при 30°C восемь трансформантов из каждой реакции трансформации наносили штрихами на новые МС пластинки и инкубировали еще в течение 2 суток при 30°C. После выращивания эти штаммы инокулировали индивидуально в 3 мл жидкой МС при 30°C и встряхивали при 250 оборотов/минуту в течение 2 суток. Клетки собирали путем центрифугирования, супернатант удаляли, и прибавляли 3 мл HGM. Данные штаммы выращивали при 30°C в инкубаторе при встряхивании при 250 оборотов/минуту еще на протяжении 5 суток. Клетки собирали путем центрифугирования, липиды экстрагировали, и жирнокислотные метиловые эфиры готовили путем трансэтерификации, и впоследствии анализировали с помощью газового хроматографа Hewlett-Packard 6890 GC.
ГХ анализы показали, что активности дельта-9-элонгазы и дельта-8-десатуразы проявлялись во всех штаммах, имеющих дельта-9-элонгаза/дельта-8-десатуразное слияние. Сводка полученных результатов приведена ниже в Таблице 34. Активность дельта-9-элонгазы вычисляли путем деления суммы весовых процентов (вес. %) для ЭДК и ДГЛК на сумму весовых процентов для ЛК, ЭДК и ДГЛК и умножением на 100 для выражения в виде процента; схожим образом, активность дельта-8-десатуразы вычисляли путем деления вес. % для ДГЛК на сумму вес. % для ЭДК и ДГЛК и умножением на 100 для выражения в виде процента.

Активности дельта-9-элонгазы и дельта-8-десатуразы в Yarrowia, трансформированной различными генными слияниями
Как резюме Таблицы 34, данные показывают, что все шесть слитых генов обладают активностями дельта-9-элонгазы и дельта-8-десатуразы; таким образом, слитые протеины из слитых генов эффективно позволяют осуществлять экспрессию двух независимых и раздельных ферментативных активностей. Более важно, слияние двух независимых ферментов в один слитый протеин, разделенных линкерным участком, повышало поток от ЛК к ДГЛК. Во всех случаях слитый ген имел более высокую активность, чем по меньшей мере один из индивидуальных генов при отдельной экспрессии в Yarrowia. Эти данные предполагают, что продукт дельта-9-элонгазы мог быть прямо каналирован как субстрат дельта-8-десатуразы в слитом протеине.
Более конкретно, в случае EgD9ES-EgD8M генного слияния (т.е. pZuFmG9G8fu-B) и EgD9ES-EaD8S генного слияния (т.е. pZuFmG9A8), EgD9ES дельта-9-элонгаза (21% конверсия) действовала сравнимым образом с тем случаем, когда EgD9ES была экспрессирована одна (20% конверсия [pZuFmEgD9ES данные и Пример 55]). Однако в противоположность этому, EgD8M активность дельта-8-десатуразы в Yarrowia, экспрессирующей pZuFmG9G8-B, была на приблизительно 97% более эффективной, чем когда EgD8M была экспрессирована одна (73% относительно 37% конверсии [Пример 55]). Подобно этому, EaD8S активность дельта-8-десатуразы в Yarrowia, экспрессирующей pZuFmG9A8, была на приблизительно 63% больше, чем когда EaD8S была экспрессирована одна (67% относительно 41% конверсии [Пример 55]).
В случае EaD9ES/EgD8M генного слияния (т.е. pZuFmA9G8) и EaD9ES/EaD8S генного слияния (т.е. pZuFmA9A8), EaD9ES активность дельта-9-элонгазы была на приблизительно 15% и 38% более эффективной, соответственно, чем когда EaD9ES экспрессировали одну (15% и 18% конверсия, соответственно, относительно 13% конверсии [Пример 55]). Подобно этому, EgD8M активность дельта-8-десатуразы в Yarrowia, экспрессирующей pZuFmA9G8, была на приблизительно 46% более эффективной, чем когда EgD8M была экспрессирована одна (54% относительно 37% конверсии [Пример 55]). Схожим образом, EaD8S активность дельта-8-десатуразы в Yarrowia, экспрессирующей pZuFmA9A8, была на приблизительно 32% более эффективной, чем когда EaD8S была экспрессирована одна (58% относительно 41% конверсии [Пример 55]).
Наконец, в случае E389D9eS-EgD8M генного слияния (т.е. pZuFmR9G8), E389D9eS активность дельта-9-элонгазы была на приблизительно 50% более эффективной, чем когда E389D9eS была экспрессирована одна (18% относительно 12% конверсии [Пример 55]). Подобно этому, EgD8M активность дельта-8-десатуразы в Yarrowia, экспрессирующей pZuFmR9G8, была на приблизительно 89% более эффективной, чем когда EgD8M экспрессировалась одна (70% относительно 37% конверсии [Пример 55]).
Таблица 34 также демонстрирует, что модифицированый линкер GAGPARPAGLPPATYYDSLAVMGS (SEQ ID NO:445) был предпочтительным, в противоположность линкеру, представленному как SEQ ID NO:438 в Yarrowia lipolytica, при слиянии дельта-9-элонгазного и дельта-8-десатуразных генов.
Для специалистов в данной области будет очевидным, что другие ПНЖК десатуразный и элонгазный гены, которые предпочтительны для экспрессии в Yarrowia lipolytica (включая, например, любой из генов, описанных в Таблицах 8-19), могут быть слиты вместе с помощью способа, описанного выше, и экспрессированы в Yarrowia lipolytica. Предпочтительные промоторы и терминаторы, подходящие для конструирования экспрессирующей кассеты (где экспрессированная ОРС кодирует мультизим), могут быть выбраны из описанных в Таблицах 8-19. Гипотетически, повышенная эффективность или поток будут наблюдаться в слитом гене, в противоположность той ситуации, когда каждый (или оба) индивидуальный ген экспрессирован отдельно.
ПРИМЕР 57
Создание дельта-9-элонгаза/дельта-8-десатуразных генных слияний для экспрессии в сое
С целью описания мультизимных слияний между дельта-9-элонгазами и дельта-8-десатуразами в сое создали ряд дельта-9-элонгаза/дельта-8 мультизимов. Дельта-9-элонгазный и дельта-8-десатуразный домены разделяли EgDHAsyn1 обогащенным пролином линкером (SEQ ID NO:198). Для сравнения, также создали конструкции, которые коэкспрессировали индивидуальные дельта-9-элонгазные и дельта-8-десатуразные гены. Использованные дельта-9-элонгазы включают EgD9elo (Пример 16; SEQ ID NO:112; на которую здесь ссылаются также как на EgD9e и EgD9E , но они идентичны) и EaD9elo1 (SEQ ID NO:252; Пример 36; на которую здесь ссылаются также как на EaD9E и EaD9e, но все они идентичны). Использованные дельта-8-десатуразы включают TpomD8 (SEQ ID NO:162; Пример 21) и Euglena anabaena дельта-8-десатуразу (EaD8Des3; SEQ ID NO:427; на которую здесь также ссылаются как на EaD8, но они идентичны; описанную в предварительной заявке США № 60/910831(от 10 апреля 2007 года; реестр уполномоченного No. BB-1615).
В настоящем Примере и для Примера 23, который описывает синтез EgD9elo-EgDHAsyn1Link-PavD8 слияния, к 3' концу EgDHAsyn1 обогащенной пролином линкерной последовательности добавляли дополнительные нуклеотиды для обеспечения клонирования при создании слияний для всех конструкций. Таким образом, между концом EgDHAsyn1 обогащенного пролином линкера (SEQ ID NO:198; PARPAGLPPATYYDSLAV) и началом использованной дельта-8-десатуразы (т.е. SEQ ID NO:472; PARPAGLPPATYYDSLAVSGRT) включали дополнительные 4 аминокислоты.
Плазмида pKR1183 (SEQ ID NO:266), включающая Euglena anabaena дельта-9-элонгаза-Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразное слияние (Hybrid1-HGLA Synthase; называвемая также EaD9e/TpomD8), была описана в Примере 38.
Другие плазмиды, описанные в настоящем примере, включают: pKR1014 (описана в патентной заявке США № 11/876115 (от 22 октября 2007 года; реестр уполномоченного No. BB-1574; включающая EgD9e и TpomD8, экспрессированные индивидуально), pKR1152 (включающая EgD9e и EaD8, экспрессированные индивидуально), pKR1151 (включающая EaD9e и TpomD8, экспрессированные индивидуально), pKR1150 (включающая EaD9e и EaD8, экспрессированные индивидуально), pKR1184 (включающая EaD9e/EaD8 генное слияние), pKR1199 (включающая EgD9e/TpomD8 генное слияние), и pKR1200 (включающая EgD9e/EaD8 генное слияние). Сводка созданных конструкций и соответствующих тестированных генов совместно с SEQ ID NOs для нуклеотидной и аминокислотной последовательностей приведена в Таблице 35.
Функциональный анализ активности каждого генного слияния проводили infra, в Примере 60.
Конструирование pKR1014
Вектор pKR123r, который был ранее описан в PCT публикации № WO 2004/071467 (опубликованной 26 августа 2004 года), содержит NotI сайт, фланкированный соевым трипсиновым ингибиторным промотором Kunitz (KTi3) (Jofuku et al., Plant Cell 1:1079-1093 (1989)) и KTi 3′ терминаторным участком, выделение которого описано в патенте США № 6372965 (KTi3/NotI/KTi3′ кассета). TpomD8 (SEQ ID NO:162; Пример 21) выделяли из pLF114-10 (SEQ ID NO:165; Пример 21) путем расщепления NotI и клонировали в NotI сайт pKR123r с образованием pKR1007 (SEQ ID NO:473).
Вектор pKR912, который ранее был описан в US-2007-0118929-A1, опубликованной 24 мая 2007 года, содержит гигромицин B фосфотрансферазный ген, фланкированный 35S промотором (Odell et al., Nature 313:810-812 (1985)) и NOS 3′ транскрипционным терминатором (Depicker et al., J. Mol. Appl. Genet. 1:561-570 (1982)) (35S/hpt/NOS3′ кассета) для селекции в растениях, таких как соя. Вектор pKR912 также содержит EgD9e (SEQ ID NO:112), фланкированный промотором для α′ субъединицы β-конглицинина (Beachy et al., EMBO J. 4:3047-3053 (1985)) и 3′ транскрипционным терминирующим участком фазеолинового гена (Doyle et al., J. Biol. Chem. 261:9228-9238 (1986)), обеспечивая таким образом сильную специфическую к тканям экспрессию EgD9e в семенах сои.
Плазмиду pKR1007 (SEQ ID NO:473) расщепляли PstI, и фрагмент, содержащий Tetruetreptia pomquetensis дельта-8-десатуразу, клонировали в SbfI сайт pKR912, образуя pKR1014 (SEQ ID NO:474). Таким способом Tetruetreptia pomquetensis дельта-8-десатураза коэкспрессировали с Euglena gracilis дельта-9-элонгазой за сильными, специфическими к семенам промоторами. Схематическое изображение pKR1014 приведено на ФИГ.56. На ФИГ.56, TpomD8 называется Tetruetreptia pomquetensis 1491 дельта-8-десатуразой, и EgD9e называется eug el1.
Конструирование pKR1151
Для введения NotI и NcoI рестрикционных сайтов в 5′ конец кодирующих последовательностей и NotI сайта в 3′ конец кодирующих последовательностей, EaD8 (SEQ ID NO:427) амплифицировали олигонуклеотидными праймерами EaD8-5 (SEQ ID NO:475) и EaD8-3 (SEQ ID NO:476) с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S, Finnzymes Oy, Finland) в соответствии с протоколом производителя. Результирующий ДНК фрагмент клонировали в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), в соответствии с протоколом производителя, с образованием pLF120-3 (SEQ ID NO:477).
Вектор pLF120-3 (SEQ ID NO:477) расщепляли NotI, и фрагмент, содержащий EaD8, клонировали в NotI сайт pKR457 (SEQ ID NO:122; Пример 16), с образованием pKR1138 (SEQ ID NO:478).
Вектор pKR1138 (SEQ ID NO:478) расщепляли BsiWI, и фрагмент, содержащий EaD8, клонировали в BsiWI сайт pKR912 с образованием pKR1152 (SEQ ID NO:479). Схематическое изображение pKR1152 приведено на ФИГ.57. На ФИГ.57, EaD8 называется EaD8Des3, и EgD9e называется eug el1.
Конструирование pKR1152
Для введения NotI и NcoI рестрикционных сайтов в 5′ конец кодирующих последовательностей и NotI сайта в 3′ конец кодирующих последовательностей EaD9e был ПЦР амплифицирован из pLF121-1 (SEQ ID NO:250; Пример 36) олигонуклеотидными праймерами oEAd9el1-1 (SEQ ID NO:298; Пример 44) и oEAd9el1-2 (SEQ ID NO:480) с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S, Finnzymes Oy, Finland) в соответствии с протоколом производителя. Результирующие ДНК фрагменты клонировали в ПЦР-Blunt клонирующий вектор с использованием набора для клонирования Zero Blunt PCR Cloning Kit (Invitrogen Corporation), согласно протоколу производителя, с образованием pKR1137 (SEQ ID NO:481).
EaD9e выделяли из pKR1137 (SEQ ID NO:481) путем расщепления NotI и клонировали в NotI сайт pKR72 (SEQ ID NO:105; Пример 15) с образованием pKR1140 (SEQ ID NO:482).
TpomD8 выделяли из pLF114-10 (SEQ ID NO:165; Пример 21) путем расщепления NotI и клонировали в NotI сайт плазмиды pKR457 (SEQ ID NO:122; Пример 16) с образованием pKR1145 (SEQ ID NO:483).
Вектор pKR1145 (SEQ ID NO:483) расщепляли BsiWI, и фрагмент, содержащий TpomD8, клонировали в BsiWI сайт pKR1140 (SEQ ID NO:482) с образованием pKR1151 (SEQ ID NO:484). Схематическое изображение pKR1151 показано на ФИГ.58. На ФИГ.58, TpomD8 называется Tetruetreptia pomquetensis 1491 дельта-8-десатураза, и EaD9e называется EAd9elong.
Конструирование pKR1150
Вектор pKR1138 (SEQ ID NO:478) расщепляли BsiWI, и фрагмент, содержащий EaD8, клонировали в BsiWI сайт pKR1140 (SEQ ID NO:482) с образованием pKR1150 (SEQ ID NO:485). Схематическое изображение pKR1150 показано на ФИГ.59. На ФИГ.59, EaD8 называется EaD8Des3, и EaD9e называется EAd9elong.
Конструирование pKR1199
NcoI/NotI ДНК фрагмент KS373 (SEQ ID NO:179; Пример 23), содержащий EgD9elo-EgDHAsyn1Link, клонировали в NcoI/NotI ДНК фрагмент из pKR1177 (SEQ ID NO:264; Пример 38), содержащий промотор для α′ субъединицы β-конглицинина, с образованием pKR1190 (SEQ ID NO:486).
NotI фрагмент из pLF114-10 (SEQ ID NO:165; Пример 21), содержащий TpomD8, клонировали в NotI фрагмент pKR1190 (SEQ ID NO:486) с образованием pKR1195 (SEQ ID NO:487).
BamHI ДНК фрагмент pKR1195 (SEQ ID NO:487), содержащий EgD9e/TpomD8 слитый ген, клонировали в BamHI ДНК фрагмент pKR325, описанный ранее в РСТ публикации № WO 2006/012325 с образованием pKR1199 (SEQ ID NO:488). Схематическое изображение pKR1199 показано на ФИГ.60. На ФИГ.60, EgD9e/TpomD8 называется EGd9elong-TPOMd8DS.
Конструирование pKR1200
NotI фрагмент из pLF120-3 (SEQ ID NO:477), содержащий EaD8, клонировали в NotI фрагмент pKR1190 (SEQ ID NO:486) с образованием pKR1196 (SEQ ID NO:489).
BamHI ДНК фрагмент pKR1196 (SEQ ID NO:489), содержащий EgD9e/EaD8 слитый ген, клонировали в BamHI ДНК фрагмент pKR325, описанный ранее в PCT публикации № WO 2006/012325, с образованием pKR1200 (SEQ ID NO:490). Схематическое изображение pKR1200 приведено на ФИГ.61. На ФИГ.61, EgD9e/EaD8 называется EGd9ELONG-EAd8DS.
Конструирование pKR1184
NotI фрагмент из pLF120-3 (SEQ ID NO:477), содержащий EaD8, клонировали в NotI фрагмент pKR1179 (SEQ ID NO:265) с образованием pKR1184 (SEQ ID NO:491). Схематическое изображение pKR1184 показано на ФИГ.62. На ФИГ.62, EaD9e/EaD8 называется EAd9ELONG-EAd8DS.
ПРИМЕР 58
Конструирование экспрессирующего вектора сои pKR1321 для экспрессии Tetruetreptia pomquetensis CCMP1491 дельта-8-десатураза-Euglena anabaena дельта-9-элонгазного слитого гена (TpomD8-EaD9Elo1 fusion)
Настоящий пример описывает конструирование внутрикаркасного слитого гена между Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразой (TpomD8; SEQ ID NO:162; Пример 21) и Euglena anabaena дельта-9-элонгазой (EaD9e; SEQ ID NO:252, Пример 36). Каждый домен отделен EgDHAsyn1 линкером с дополнительными 4 аминокислотами, включенными между концом EgDHAsyn1 обогащенного пролином линкера и началом EaD9e, как описано в Примере 57 (т.е. SEQ ID NO:472; PARPAGLPPATYYDSLAVSGRT).
Плазмиду pKR1301 (SEQ ID NO:307; Пример 44) расщепляли EcoRI, и ДНК фрагмент, содержащий 3′ конец TpomD8-EgDHAsyn1Link (называемый TpomD8+L1TR1), религировали с образованием pKR1303 (SEQ ID NO:497).
NotI фрагмент pKR1137 (SEQ ID NO:481; Пример 57), содержащий EaD9e, клонировали в EagI сайт pKR1303 (SEQ ID NO:497) с образованием pKR1308 (SEQ ID NO:498). Таким способом EaD9e была слита с 3′ концом TpomD8.
Gy1/Pavelo/legA2 кассету выделили из плазмиды pKR336 (описанной в PCT публикации № WO 04/071467; на содержание которой в настоящей заявке сделана ссылка) путем расщепления PstI/BamHI и клонировали в PstI/BamHi сайт pKR268 (описанный в PCT публикации № WO 04/071467) с образованием pKR393 (SEQ ID NO:499). Pavelo ген был выделен из pKR393 (SEQ ID NO:499) путем расщепления NotI, и данный вектор религировали с образованием pKR407 (SEQ ID NO:500).
TpomD8 выделяли из pLF114-10 (SEQ ID NO:165; Пример 21) путем расщепления NotI и клонировали в NotI сайт плазмиды pKR407 (SEQ ID NO:500) с образованием pKR1018 (SEQ ID NO:501).
Плазмиду pKR1018 (SEQ ID NO:501) расщепляли HindIII/EcoRI, и фрагмент, содержащий 5′ конец Tpomd8, клонировали в HindIII/EcoRI сайт pKR1308 (SEQ ID NO:498) с образованием pKR1312 (SEQ ID NO:502). Таким способом TpomD8 последовательность восстановили, и сформировали TpomD8/EaD9e слияние.
NotI фрагмент pKR1312 (SEQ ID NO:502), содержащий TpomD8/EaD9e слияние, клонировали в NotI сайт pKR72 (SEQ ID NO:105; Пример 23) с образованием pKR1321 (SEQ ID NO:503). Схематическое изображение pKR1321 показано на ФИГ.63. На ФИГ.63, TpomD8/EaD9e называется TPd8ds-EAd9el слиянием.
ПРИМЕР 59
Конструирование экспрессирующего вектора сои pKR1326 для экспрессии Euglena anabaena дельта-9-элонгаза-Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразного слитого гена с использованием Euglena anabaena DHAsyn1 обогащенного пролином линкера
Настоящий пример описывает конструирование внутрикаркасного слитого гена между Euglena anabaena дельта-9-элонгазой (EaD9e; SEQ ID NO:252, Пример 36) и Tetruetreptia pomquetensis CCMP1491 дельта-8-десатуразой (TpomD8; SEQ ID NO:162; Пример 21). Каждый домен отделен EaDHAsyn1 обогащенным пролином линкером (SEQ ID NO:235), но с дополнительными 3 аминокислотами, включенными между концом EaDHAsyn1 обогащенного пролином линкера (EaDHAsyn1Link) и началом EaD9e (т.е. SEQ ID NO:504; PGGPGKPSEIASLPPPIRPVGNPPAAYYDALATGRT). Клонирование осуществляли подобно описанному в Примере 57.
Первичное внутрикаркасное слияние между EaD9e и EaDHAsyn1Link (EaD9elo-EgDHAsyn1Link) проводили путем ПЦР амплификации и фланкировали NotI и NcoI сайтом в 5′ конце и NotI сайтом в 3′ конце. EaD9e (SEQ ID NO:252) амплифицировали из pLF121-1 (SEQ ID NO:250) олигонуклеотидами oEAd9el1-1 (SEQ ID NO:298) и EaLink1 (SEQ ID NO:505), с использованием Phusion™ High-Fidelity DNA Polymerase (Cat. No. F553S, Finnzymes Oy, Finland) в соответствии с протоколом производителя. EaDHAsyn1Link (SEQ ID NO:234) амплифицировали схожим способом из pLF117-1 (SEQ ID NO:87; Пример 13) олигонуклеотидами EaLink2 (SEQ ID NO:506) и EaLink3 (SEQ ID NO:507). Два результирующих ПЦР продукта скомбинировали и повторно амплифицировали с использованием oEAd9el1-1 (SEQ ID NO:298) и EaLink3 (SEQ ID NO:507) с образованием EaD9e-EaDHAsyn1Link. Последовательность EaD9e-EaDHAsyn1Link представлена в SEQ ID NO:508. EaD9e-EaDHAsyn1Link клонировали в ПЦР-Blunt® клонирующий вектор с использованием набора для клонирования Zero Blunt® PCR Cloning Kit (Invitrogen Corporation), следуя протоколу производителя с образованием pKR1305 (SEQ ID NO:509).
EagI ДНК фрагмент pKR1305 (SEQ ID NO:509), содержащий EaD9e-EaDHAsyn1Link, клонировали в NotI сайт pKR1304 (SEQ ID NO:310; Пример 44) с образованием pKR1317 (SEQ ID NO:510). Таким способом 5′ конец TpomD8 был слит с EaD9e-EaDHAsyn1Link.
The EcoRI/Asp718 фрагмент pKR1127 (SEQ ID NO:168; Пример 22), содержащий 3′ конец TpomD8, клонировали в EcoRI/Asp718 фрагмент pKR1317 (SEQ ID NO:510), содержащий EaD9e-EaDHAsyn1Link, с образованием pKR1320 (SEQ ID NO:511).
NotI фрагмент из pKR1320 (SEQ ID NO:511), содержащий данное слияние, клонировали в NotI фрагмент pKR72 (SEQ ID NO:105; Пример 15) с образованием pKR1326 (SEQ ID NO:512). Схематическое изображение pKR1326 показано на ФИГ.64. На ФИГ.64, EaD9e/TpomD8 с EaDHAsyn1 обогащенным пролином линкером называется EAd9el-TPOMd8ds L2fusion.
ПРИМЕР 60
Функциональные анализы дельта-9-элонгаза/дельта-8-десатуразных генных слияний в сое
Настоящий пример описывает трансформацию и экспрессию в соевых соматических зародышах pKR1014 (SEQ ID NO:474), pKR1152 (SEQ ID NO:479), pKR1151 (SEQ ID NO:484), pKR1150 (SEQ ID NO:485), pKR1199 (SEQ ID NO:488), pKR1200 (SEQ ID NO:490), и pKR1184 (SEQ ID NO:491), синтез которых был ранее описан в Примере 57. Функциональные анализы pKR1183 (SEQ ID NO:266) и KS373 (SEQ ID NO:179) были ранее описаны в Примерах 46 и 31, соответственно.
Соевую эмбриогенную суспензионную культуру (cv. Jack) трансформировали каждым из вышеприведенных векторов, и эмбрионы созревали в соевой жидкой среде для гистодиффеернциации и созревания (жидкие среды SHaM; Schmidt et al., Cell Biology and Morphogenesis, 24:393 (2005)), как описано в Примере 25 и ранее описано в PCT публикации № WO 2007/136877, опубликованной 29 ноября 2007 года (на содержание которой в настоящей заявке сделана ссылка).
После созревания в SHaM жидкой среде подмножество трансформированных соевых эмбрионов (т.е. 5-6 эмбрионов на событие) собирали и анализировали как здесь описано.
Таким способом проанализировали приблизительно 30 событий, трансформированных pKR1014 (SEQ ID NO:474), pKR1152 (SEQ ID NO:479), pKR1151 (SEQ ID NO:484), pKR1150 (SEQ ID NO:485), pKR1199 (SEQ ID NO:488), pKR1200 (SEQ ID NO:490), или pKR1184 (SEQ ID NO:491). Пять событий с наивысшим средним содержанием ДГЛК (среднее из 5 проанализированных эмбрионов) представлены на ФИГ.65, 66, 67, 68, 69, 70, или 71, соответственно. На ФИГ.65-71 жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК, АЛК, ЭДК, ЭРК, ДГЛК и ЭТК. Составы жирных кислот выражены в виде весового процента (вес. %) от всех жирных кислот. В Таблице 36 представлена сводка данных по векторам, использованным генам, номеру эксперимента (MSE#) и соответствующей ФИГ.
На ФИГ.65-71 элонгационная активность выражена в виде % дельта-9-элонгации C18 жирных кислот (C18 % delta-9 elong), вычисленного в соответствии со следующей формулой: ([продукт]/[субстрат+продукт])*100. Более конкретно, комбинированный процент элонгации для ЛК и АЛК определяется как: ([ДГЛК+ЭТК+ЭДК+ЭРК]/[ЛК+АЛК+ДГЛК+ЭТК+ЭДК+ЭРК])*100.
На ФИГ.65-71 комбинированный процент десатурации для ЭДК и ЭРК показан как “C20 % delta-8 desat”, определяемый как: ([ДГЛК+ЭТК]/[ДГЛК+ЭТК+ЭДК+ЭРК])*100. На него также ссылаются как на полный % десатурации.
Сравнение индивидуально экспрессированных дельта-9-элонгаз с дельта-8-десатуразами относительно эквивалентных дельта-9-элонгаза-дельта-8-десатуразных слияний приведено на ФИГ.72. На ФИГ.72 каждая точка данных представляет средний % ДГЛК или % ЭДК для 5-6 эмбрионов (как % от всех жирных кислот) для всех проанализированных событий, и средн. % ДГЛК представлен на графике в зависимости от средн. % ЭДК. На ФИГ.72A EgTpom представляет EgD9e, коэкспрессированный с TpomD8 (pKR1014), и EgTpomfus представляет EgD9e/TpomD8 слияние (pKR1199). На ФИГ.72B EgEa представляет EgD9e, коэкспрессированный с EaD8 (pKR1152), и EgEafus представляет EgD9e/EaD8 слияние (pKR1200). На ФИГ.72C EaTpom представляет EaD9e, коэкспрессированный с TpomD8 (pKR1151), и EaTpomfus представляет EaD9e/TpomD8 слияние (pKR1183). На ФИГ.72D EaEa представляет EaD9e, коэкспрессированный с EaD8 (pKR1150), и EaEafus представляет EaD9e/EaD8 слияние (pKR1200).
ПРИМЕР 61
Функциональные анализы дельта-9-элонгаза/дельта-8-десатураза/дельта-5-десатуразного генного слияния
Настоящий пример описывает трансформацию и экспрессию в соевых соматических эмбрионах pKR1322 (SEQ ID NO:314; Пример 50), включая EaD9Elo1-TpomD8-EaD5Des1 тройное слияние (также называемое EaD9e/TpomD8/EaD5). Каждый домен отделен EgDHAsyn1 линкером с дополнительными 4 аминокислотами (т.е. SEQ ID NO:472; PARPAGLPPATYYDSLAVSGRT).
Соевую эмбриогенную суспензионную культуру (cv. Jack) трансформировали pKR1322, и зародыши созревали в соевой жидкой среде для гистодифференциации и созревания (жидкие среды SHaM; Schmidt et al., Cell Biology and Morphogenesis, 24:393 (2005)) как описано в Примере 25 и ранее описано в PCT публикации № WO 2007/136877, опубликованной 29 ноября 2007 года (на содержание которой в настоящей заявке сделана ссылка).
После созревания в жидкой среде SHaM подмножество трансформированных соевых эмбрионов (т.е. 5 эмбрионов на событие) собирали и анализировали как здесь описано.
Таким способом проанализировали приблизительно 30 событий, трансформированных pKR1322 (эксперимент MSE2274), и пять событий с наивысшим средним содержанием АРК и ЭПК (среднее из 5 проанализированных зародышей) представлены на ФИГ.73. На ФИГ.73 жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), 18:2 (5,9), ЛК, АЛК, ЭДК, ЭРК, СКК, ДГЛК, ЮН (называемая также ЮП), ЭТК, АРК и ЭПК. Составы жирных кислот выражены в виде весового процента (вес. %) от всех жирных кислот.
На ФИГ.73 элонгационная активность выражена как % дельта-9-элонгации C18 жирных кислот (%Elo), вычисленный в соответствии со следующей формулой: ([продукт]/[субстрат+продукт])*100. Более конкретно, комбинированный процент элонгации для ЛК и АЛК определяется как: ([ДГЛК+ЭТК+ЭДК+ЭРК+ЭПК+АРК]/[ЛК+АЛК+ДГЛК+ЭТК+ЭДК+ЭРК+ЭПК+АРК])*100.
На ФИГ.73 комбинированный процент дельта-8-десатурации для ЭДК и ЭРК представленный как “% D8”, определялся как: ([ДГЛК+ЭТК+ЭПК+АРК]/[ДГЛК+ЭТК+ЭДК+ЭРК+ЭПК+АРК])*100. На него также ссылаются как на полный % дельта-8-десатурации.
На ФИГ.73 комбинированный процент дельта-5-десатурации для ДГЛК и ЭТК представлен как “% D5”, и определяется как: ([ЭПК+АРК]/[ДГЛК+ЭТК+ЭПК+АРК])*100. На него также ссылаются как на полный % дельта-5-десатурации.
Как резюме ФИГ.73, все три домена функциональны. На это слияние можно также ссылаться как на ЭПК синтазу или АРК синтазу.
ПРИМЕР 62
Функциональные анализы дельта-9-элонгаза/дельта-8-десатуразных и дельта-8-десатураза/дельта-9-элонгазных генных слияний
Настоящий пример описывает трансформацию и экспрессию в соевых соматических зародышах либо pKR1326 (SEQ ID NO:512), включая EaD9Elo1-TpomD8 слияние (называемое также EaD9e/TpomD8), отделенное EaDHAsyn1 линкером с дополнительными 3 аминокислотами (т.е. SEQ ID NO:504; PGGPGKPSEIASLPPPIRPVGNPPAAYYDALATGRT), либо pKR1321 (SEQ ID NO:503; Пример 58), включая TpomD8/EaD9e слияние, отделенное EgDHAsyn1 линкером.
Соевую эмбриогенную суспензионную культуру (cv. Jack) трансформировали pKR1326 или pKR1321, и зародыши созревали в соевой жидкой среде для гистодифференциации и созревания (жидкие среды SHaM; Schmidt et al., Cell Biology and Morphogenesis, 24:393 (2005)), как описано в Примере 25 и ранее описано в PCT публикации № WO 2007/136877, опубликованной 29 ноября 2007 года (на содержание которой в настоящей заявке сделана ссылка).
После созревания в жидкой среде SHaM подмножество трансформированных соевых эмбрионов (т.е.5 эмбрионов на событие) собирали и анализировали как здесь описано.
Таким способом проанализировали приблизительно 30 событий, трансформированных pKR1326 (эксперимент MSE2275), и пять событий с наивысшим средним содержанием ДГЛК и ЭТК (среднее из 5 проанализированных зародышей) представлены на ФИГ.74. На ФИГ.74 жирные кислоты идентифицированы как 16:0 (пальмитат), 18:0 (стеариновая кислота), 18:1 (олеиновая кислота), ЛК, АЛК, ЭДК, ЭРК, ДГЛК и ЭТК. Составы жирных кислот выражены в виде весового процента (вес. %) от всех жирных кислот.
На ФИГ.74 элонгационная активность выражена в виде % дельта-9-элонгации C18 жирных кислот (C18 % delta-9-elong), вычисленный в соответствии со следующей формулой: ([продукт]/[субстрат+продукт])*100. Более конкретно, комбинированный процент элонгации для ЛК и АЛК определяется как: ([ДГЛК+ЭТК+ЭДК+ЭРК]/[ЛК+АЛК+ДГЛК+ЭТК+ЭДК+ЭРК])*100.
На ФИГ.74 комбинированный процент десатурации для ЭДК и ЭРК, представленный как “C20 % delta-8-desat”, определялся как: ([ДГЛК+ЭТК]/[ДГЛК+ЭТК+ЭДК+ЭРК])*100. На него также ссылаются как на полный % десатурации.
Как резюме ФИГ.74, EaDHAsyn1 линкерные функции схожи с таковыми для EgDHAsyn1 линкера. Никакой активности для любого из событий, трансформированных pKR1321, где TpomD8 был слит с EaD9e EgDHAsyn1 линкером, не обнаруживалось.
Реферат
Изобретение относится к области биотехнологии и может быть использовано в сельском хозяйстве и пищевой промышленности. Сконструированы рекомбинантные нуклеотидные последовательности (НП), кодирующие полипептиды (ДГЛК-синтазы), которые состоят из дельта-9-элонгазы и дельта-8-десатуразы, независимо и раздельно проявляющих присущие им ферментативные активности. Предложены содержащие эти НП генетические конструкции и трансформированные указанными конструкциями клетки-хозяева, предпочтительно клетки масличных растений и дрожжей, которые экспрессируют активный фермент ДГЛК-синтазу и могут быть использованы для получения в указанных клетках длинноцепочечных полиненасыщенных жирных кислот, в частности промышленно значимых дигомо-гамма-линоленовой и эйкозатетраеновой. 10 н. и 8 з.п. ф-лы, 119 ил., 36 табл., 62 пр.
Формула
а) получение рекомбинантной микробной клетки-хозяина по п.2;
б) обеспечение линолевой кислоты в клетке-хозяине; и
в) выращивание клетки-хозяина в условиях, обеспечивающих превращение линолевой кислоты в дигомо-гамма-линоленовую кислоту.
а) получение рекомбинантной микробной клетки-хозяина по п.2;
б) обеспечение альфа-линоленовой кислоты в клетке-хозяине; и
в) выращивание клетки-хозяина в условиях, обеспечивающих превращение линоленовой кислоты в эйкозатетраеновую кислоту.
а) нуклеотидной последовательностью, кодирующей полипептид, который обладает ДГЛК-синтазной активностью, где данный полипептид представлен SEQ ID NO:441, SEQ ID NO:447, SEQ ID NO:454, SEQ ID NO:461, SEQ ID NO:464, SEQ ID NO:471, SEQ ID NO:515, SEQ ID NO:516, SEQ ID NO:517, SEQ ID NO:518 и SEQ ID NO:519; или
б) нуклеотидной последовательностью, кодирующей полипептид, который обладает ДГЛК-синтазной активностью, где данная нуклеотидная последовательность представлена SEQ ID NO:440, SEQ ID NO:446, SEQ ID NO:453, SEQ ID NO:460, SEQ ID NO:463, SEQ ID NO:470, SEQ ID NO:492, SEQ ID NO:493, SEQ ID NO:494, SEQ ID NO:495 и SEQ ID NO:496.