Способ микробиологического получения аминокислот семейства аспартатов и/или глутаматов и используемые в способе средства - RU2231552C2
Код документа: RU2231552C2
Чертежи








Описание
Изобретение относится к способу микробиологического получения аминокислот семейства аспартатов и/или глутаматов по п.п.1-17 формулы изобретения, генам пируваткарбоксилазы по п.п.18-23 формулы изобретения, генным структурам по п.24 формулы изобретения, векторам по п.25 формулы изобретения, трансформированным клеткам по п.п.26-31 формулы изобретения, а также к их применению по п.п.32-37 формулы изобретения.
Аминокислоты представляют большой экономический интерес, причем использование аминокислот многостороннее. Так, например, L-лизин, а также L-треонин, L-метионин и L-триптофан необходимы в качестве кормовой добавки, L-глутамат необходим в качестве пряной добавки, L-изолейцин и L-тирозин требуются в фармацевтической промышленности, L-аргинин и L-изолейцин пригодны в качестве лекарственного средства или L-глутамат, L-аспартат и L-фенилаланин пригодны в качестве исходного вещества для тонкого химического синтеза.
Предпочтительным способом получения этих различных аминокислот является биотехнологическое получение с помощью микроорганизмов, так как таким путем получают непосредственно биологически эффективную и оптически активную форму соответствующей кислоты, и при этом можно использовать простое и недорогостоящее сырье. В качестве микроорганизмов используют, например, Corynebacterium glutamicum и родственные ей ssp. flavum и ssp. lactofermentum (Liebel и др. Int. J.System Bacteriol., 41, 255-260 [1991]), а также Escherichia coli и родственные бактерии.
Эти бактерии нормальным образом продуцируют аминокислоты, однако, только в необходимом для роста количестве, так что не образуются и не выделяются никакие избыточные аминокислоты. Это основано на том, что в клетке биосинтез аминокислот многократно контролируется. Вследствие этого уже известны самые различные способы повышения продуктивности путем "выключения" механизмов контроля. В случае этих процессов используют, например, аналоги аминокислот, чтобы исключить эффективную регуляцию биосинтеза. Так, например, описан способ, в котором используют штаммы Corynebacterium, которые устойчивы по отношению к аналогам L-тирозина и L-фенилаланина (патенты Японии 19037/1976 и 39517/1978). Также описаны способы, в случае которых используют бактерии, устойчивые по отношению к аналогам L-лизина или L-треонина, для преодоления механизмов контроля (европейский патент ЕР 0205849, патент Великобритании GB 2152509).
Далее, также известны сконструированные с использованием рекомбинантной ДНК микроорганизмы, в которых также ликвидирована регуляция биосинтеза путем клонирования и экспримирования генов, которые кодируют более не ингибируемые путем обратной связи ключевые ферменты. Так, например, известна рекомбинантная, продуцирующая L-лизин бактерия с кодируемой с помощью плазмиды, устойчивой к обратной связи аспартаткиназой (европейский патент ЕР 0381527). Также описана рекомбинантная, продуцирующая L-фенилаланин бактерия с устойчивой к обратной связи префенатдегидрогеназой (патент Японии JP 123475/1986, европейский патент ЕР 0488424).
Кроме того, путем сверхэкспрессии генов, которые не кодируют чувствительные к обратной связи ферменты аминокислотного синтеза, повышают выход аминокислот. Так, например, образование лизина улучшается путем повышенного синтеза дигидродипиколинатсинтазы (европейский патент ЕР 0197335). Точно также путем повышенного синтеза треониндегидратазы достигают улучшенного образования изолейцина (европейский патент ЕР 0436886).
Дальнейшие попытки повышения продуцирования аминокислот направлены на улучшенную подготовку первичных клеточных метаболитов центрального обмена веществ. Так, известно, что достигаемое за счет рекомбинантных методов сверхпродуцирование транскетолазы делает возможным улучшенное образование L-триптофана, L-тирозина или L-фенилаланина (европейский патент ЕР 0600463). Далее, снижение активности фосфоенолпируваткарбоксилазы в Corynebacterium) приводит к улучшенному образованию ароматических аминокислот (европейский патент ЕР 03331145), между тем как повышение активности фосфоенолпируваткарбоксилазы в Corynebacterium приводит к повышенному выделению аминокислот семейства аспартатов (европейский патент ЕР 0358940).
Во время роста и особенно в условиях продуцирования аминокислот цикл трикарбоновой кислоты должен непрерывно и эффективно пополняться соединениями с 4 атомами углерода, например, с помощью оксалацетата, чтобы заменить промежуточные продукты, расходующиеся при биосинтезе аминокислот. Вплоть до недавнего времени считали, что за эти так называемые анаплеротические функции в Corynebacterium ответственна фосфоенолпируваткарбоксилаза (Kinoshita, Biology of industrial micro-organisms, 115-142 [1985], Benjamin/Cummings Publishing Company, London; Liebl, The procaryotes II, 1157-1171 [1991], Springer Verlag N.Y.; Vallino и Stephanopoulos, Biotechnol. Bioeng., 41, 633-646 [1993]). Было показано, что отрицательные мутанты фосфоенолпируваткарбоксилазы по сравнению с соответствующими исходными штаммами одинаково растут во всех испытанных средах (Peters-Wendisch и др., FEMS Microbiology Letters, 112, 269-274 [1993]; Gubler и др., Appl. Microbiol. Biotechnol., 40, 857-863 [1994]). Этот результат показывает, что фосфоенолпируваткарбоксилаза не существенна для роста и либо не играет роли, либо играет только второстепенную роль в анаплеротических реакциях. Далее, вышеуказанный результат свидетельствует о том, что в Corynebacterium должен быть другой фермент, который ответственен за синтез оксалацетата, необходимого для роста. Недавно действительно была обнаружена пируваткарбоксилазная активность в проницаемых клетках Corynebacterium glutamicum (Peters-Wendisch и др. Microbiology, 143, 1095-1103 [1997]). Этот фермент эффективно ингибируется аденозин-5'-монофосфатом (АМР), тригидратом тринатриевой соли аденозин-5'-дифосфата (ADP) и ацетилкоферментом А и в присутствии лактата в качестве источника углерода образуется в повышенном количестве. Наличие пируваткарбоксилазы в Corynebacterium glutamicum доказано и подтверждено другими исследователями с помощью13 С-ЯМР-спектроскопии и газоадсорбционной хроматографии-масс-спектрометрии (Park и др., Appl. Microbiol. BiotechnoL, 47, 430-440 [1997]). Так как нужно было исходить из того, что этот фермент является ответственным при росте в первую очередь за пополнение цикла трикарбоновой кислотой, следовало ожидать, что повышение экспресссии гена или соответственно ферментной активности либо не приводит ни к какому, либо, в крайнем случае, ведет к незначительному повышению содержания аминокислот, относящихся к семейству аспартатов. Далее, ожидали, что повышение экспрессии гена или соответственно ферментной пируваткарбоксилазной активности также не окажет никакого влияния на продуцирование аминокислот других семейств.
В настоящее время неожиданно обнаружено, что после повышения пируваткарбоксилазной активности за счет генетического изменения фермента и/или после повышения экспрессии гена пируваткарбоксилазы повышается выработка микробами аминокислот семейства аспартатов и/или глутаматов. Оказалось, что в особенности штаммы с повышенным числом копий гена пируваткарбоксилазы выделяют в культуральную среду примерно на 50% больше лизина, на 40% больше треонина и на 150% больше гомосерина. Далее, показано, что неожиданно также значительно увеличивается продуцирование глутаматов (см. пример 6 и таблицу 4).
Генетическое изменение пируваткарбоксилазы с целью повышения ферментной активности предпочтительно осуществляют путем мутации эндогенного гена. Такого рода мутации можно осуществлять либо ненаправленно обычными методами, как, например, путем УФ-облучения или с помощью вызывающих мутацию химикалиев, либо направленно посредством гентехнологических методов, как делеция (делеции), инсерция (инсерции) и/или обмен нуклеотидов (обмены нуклеотидов).
Экспрессия гена пируваткарбоксилазы повышается за счет увеличения числа копий гена и/или путем усиления регуляторных факторов, которые положительно влияют на экспрессию гена. Так, усиления регуляторных элементов, предпочтительно на уровне транскрипции, можно достигать в особенности увеличением сигналов транскрипции. Это можно осуществлять, например, благодаря тому, что путем изменения промоторной последовательности, предварительно включенной в структурный ген, повышают активность промотора, или тем, что промотор полностью заменяют более активными промоторами. Усиления транскрипции также можно достигать путем соответствующего влияния на ген-регулятор, соединенный с геном пируваткарбоксилазы. Далее, в случае необходимости, за счет мутации регуляторной генной последовательности, можно оказывать влияние на эффективность связывания регуляторного протеина с ДНК регулируемого гена пируваткарбоксилазы таким образом, что благодаря этому усиливается транскрипция и тем самым повышается экспрессия гена. Далее, с геном пируваткарбоксилазы в качестве регуляторных последовательностей также могут быть связаны так называемые "энхансеры", которые за счет улучшенного взаимодействия между РНК-полимеразой и ДНК также вызывают повышенную экспрессию гена пируваткарбоксилазы. Наряду с этим также возможно усиление трансляции, например, за счет повышения стабильности мРНК.
Для увеличения числа копий гена ген пируваткарбоксилазы встраивают в генно-инженерную конструкцию или соответственно в вектор. Генно-инженерная конструкция содержит в особенности связанные с геном пируваткарбоксилазы регуляторные последовательности, предпочтительно усиливающие экспрессию гена. Для встраивания гена пируваткарбоксилазы в генно-инженерную конструкцию ген выделяют предпочтительно из штамма микроорганизмов рода Corynebacterium и трансформируют в продуцирующий аминокислоту штамм микроорганизмов, в особенности Corynebacterium или в Escherichia coli или Settaria marcescens. Для предлагаемого согласно изобретению способа пригодны в особенности гены С. glutamicum или С. glutamicum ssp. flavum или С. glutamicum ssp. lactofermentum. После выделения гена и рекомбинации с известными векторами in vitro (см., например, Simon и др., Bio/Technology, 1, 784-791 [1983]; Fikmanns и др., Gene, 102, 93-98 [1991]) осуществляют трансформацию в продуцирующие аминокислоты штаммы путем электропорации (Liebel и др., FEMS Microbiology Letters, 65, 299-304 [1991]) или конъюгации (Schafer и др., J. BacterioL, 172, 1663-1666 [1990]). В качестве штаммов-хозяев используют предпочтительно такие аминокислотные продуценты, которые дерегулированы в синтезе соответствующей аминокислоты и/или обладают повышенной активностью экскреционного носителя для соответствующей аминокислоты. Далее, предпочтительны такие штаммы, которые содержат повышенное количество таких метаболитов центрального обмена веществ, которые принимают участие в синтезе соответствующей аминокислоты, и/или штаммы, которые содержат уменьшенное количество не участвующих в синтезе соответствующей кислоты метаболитов центрального обмена веществ, в особенности метаболитов, которые компетентны в конкурирующих реакциях, то есть предпочтительны такие штаммы, в случае которых биосинтез, конкурирующий с соответствующим биосинтезом аминокислот, протекает с пониженной активностью. Так, в особенности пригоден коринеобразный штамм микроорганизмов, устойчивый по отношению к β-метиловому эфиру L-аспарагиновой кислоты (АМЕ), с пониженной цитратсинтазной активностью (европейский патент ЕР 0551614).
После выделения получают гены пируваткарбоксилазы с нуклеотидными последовательностями, которые кодируют аминокислотную последовательность, указанную как SEQ ID №2, или ее аллельные варианты или соответственно которые имеют нуклеотидную последовательность от нуклеотида 165 до нуклеотида 3587 согласно последовательности SEQ ID №1 или по существу функционально эквивалентную (равнодействующую) ДНК-последовательность. Далее, получают гены с предварительно введенным промотором с нуклеотидной последовательностью от нуклеотида 20 до нуклеотида 109 согласно последовательности SEQ ID №1 или с функционально эквивалентной ДНК-последовательностью. Аллельные варианты, соответственно равнодействующие ДНК-последовательности включают в особенности функциональные производные, которые получают путем делеции (делеций), инсерции (инсерций) и/или замещения нуклеотидами из соответствующих последовательностей, причем ферментная активность, или ферментная функция, сохраняется или даже повышается. Эти гены пируваткарбоксилазы предпочтительно используют в способе, предлагаемом согласно изобретению.
В ген пируваткарбоксилазы с предварительно введенным промотором или без него или с присоединенным геном-регулятором или без него могут быть предварительно и/или после включены одна или несколько ДНК-последовательностей таким образом, что ген находится в генной структуре.
В ген пируваткарбоксилазы предпочтительно предварительно вводят tac-промотор (lacIQ-ген), причем к нему предпочтительно присоединяют регуляторные последовательности.
Путем клонирования гена пируваткарбоксилазы получают плазмиды, которые содержат ген и пригодны для трансформации аминокислотного продуцента. Получаемые путем трансформации клетки, в случае которых речь идет предпочтительно о трансформированных клетках Corynebacterium, содержат ген в реплицируемой форме, то есть в дополнительных копиях на хромосоме, причем копии гена интегрируются путем рекомбинации на любых свитах генома, и/или на плазмиде или векторе.
Пример
1. Клонирование гена пируваткарбоксилазы из Corynebacterium glutamicum.
Исходя из консервативных областей всех до сих пор известных (рус)генов пируваткарбоксилазы Saccharomyces cerevisiae (J.Biol. Chem., 263, 11493-11497 [1988]; Mol. Gen. Genet., 229, 307-315 [1991]), человека (Biochim. Biophys. Acta, 1227, 46-52 [1994]), мыши (Proc. Natl. Acad. Sci., USA, 90, 1766-1770 [1993]), Aedes aegypti (банк генов EMBL, номер по каталогу L36530), а также Mycobacterium tuberculosis (банк генов EMBL, номер по каталогу U00024), синтезировали праймеры полимеразной цепной реакции (ПЦР-праймеры)(MWG Biotech). Праймеры соответствовали основаниям 810-831 и 1015-1037 рус-гена М.tuberculosis. С помощью этих праймеров посредством полимеразной цепной реакции (ПЦР) по стандартному методу Innis и др. (PCR protocols, A guide to methods and applications, 1990, Academic Press) для недегенерированных гомологичных праймеров амплифицировали фрагмент размером примерно 200 пар оснований, выделенный из хромосомной ДНК С. glutamicum АТСС 13032 (Американская коллекции типовых культур, номер по каталогу 13032), как описано у Eikmfnns и др. (Microbiology, 140, 1817-1828 [1994]). Размер 200 пар оснований соответствовал ожидаемому для рус-гена. Продукт ПЦР секвенировали, как описано у Sanger и др. (Proc. Natl. Acad. Sci, USA, 74, 5463-5467 [1977]). Секвенирование осуществляли с помощью флуоресцентномаркированных дидезоксинуклеозидтрифосфатов (ddNTPs) при использовании автоматического аппарата для секвенирования ДНК (Applied Biosystems).
Исходя из этого фрагмента ДНК С. glutamicum, получали следующие гомологичные олигонуклеотиды:
рус 1: 5'-CGTCTTCATCGAAATGAAC-3'
рус 2: 5'-ACGGTGGTGATCCGGCACT-3'
Олигонуклеотиды использовали в качестве ПЦР-праймеров для выделения зонда к (рус)гену пируваткарбоксилазы из С. glutamicum. Праймеры использовали в полимеразной цепной реакции с хромосомной ДНК С. glutamicum и нуклеотидами, маркированными дигоксигенином. Реакцию осуществляли по методике "PCR DIG Labeling Kits" фирмы Берингер Маннхайм. С помощью этой смеси амплифицировали маркированный дигоксигенином фрагмент ДНК, который соответствовал ожидаемому размеру примерно 200 пар оснований. Таким образом полученный рус-зонд затем использовали для идентификации путем саузернблоттинг-гибридизации фрагмента ДНК в хромосомной ДНК С.glutamicum, в котором локализован рус-ген. Для этой цели соответственно 2-5 мкг хромосомной ДНК С. glutamicum дикого типа расщепляли с помощью ферментов рестрикции HindIII, SphI, Sail, Dral, EcoRI и BamHI, полученные фрагменты ДНК разделяли соответственно их размеру путем гель-электрофореза в 0,8%-ном геле агарозы в течение 16 часов при напряжении 20 вольт. Находящиеся в геле агарозы фрагменты ДНК денатурировали по методу Саузерна (Southern, J.Mol. Biol., 98, 503-517 [1975]) и при поддерживании вакуума с помощью аппарата VacuGene Blot фирмы Pharmacia LKB (Uppsala, Швеция) переносили из матрицы геля на найлоновую мембрану (Nytran N13, фирма Schleicher и Schull, Dassel, Швейцария), иммобилизировали и обнаруживали дигоксигениновую маркировку посредством NBT/X-фосфатного превращения с помощью щелочной фосфатазы. Таким образом смогли обнаружить следующие, гибридизирукяциеся с рус-ДНК-зондом хромосомные фрагменты: фрагмент HindIII размером 17 т.п.н. (тысяч пар нуклеотидов), фрагмент Sail размером 6,5 т.п.н. и фрагмент EcoRI размером 1,35 т.п.н.
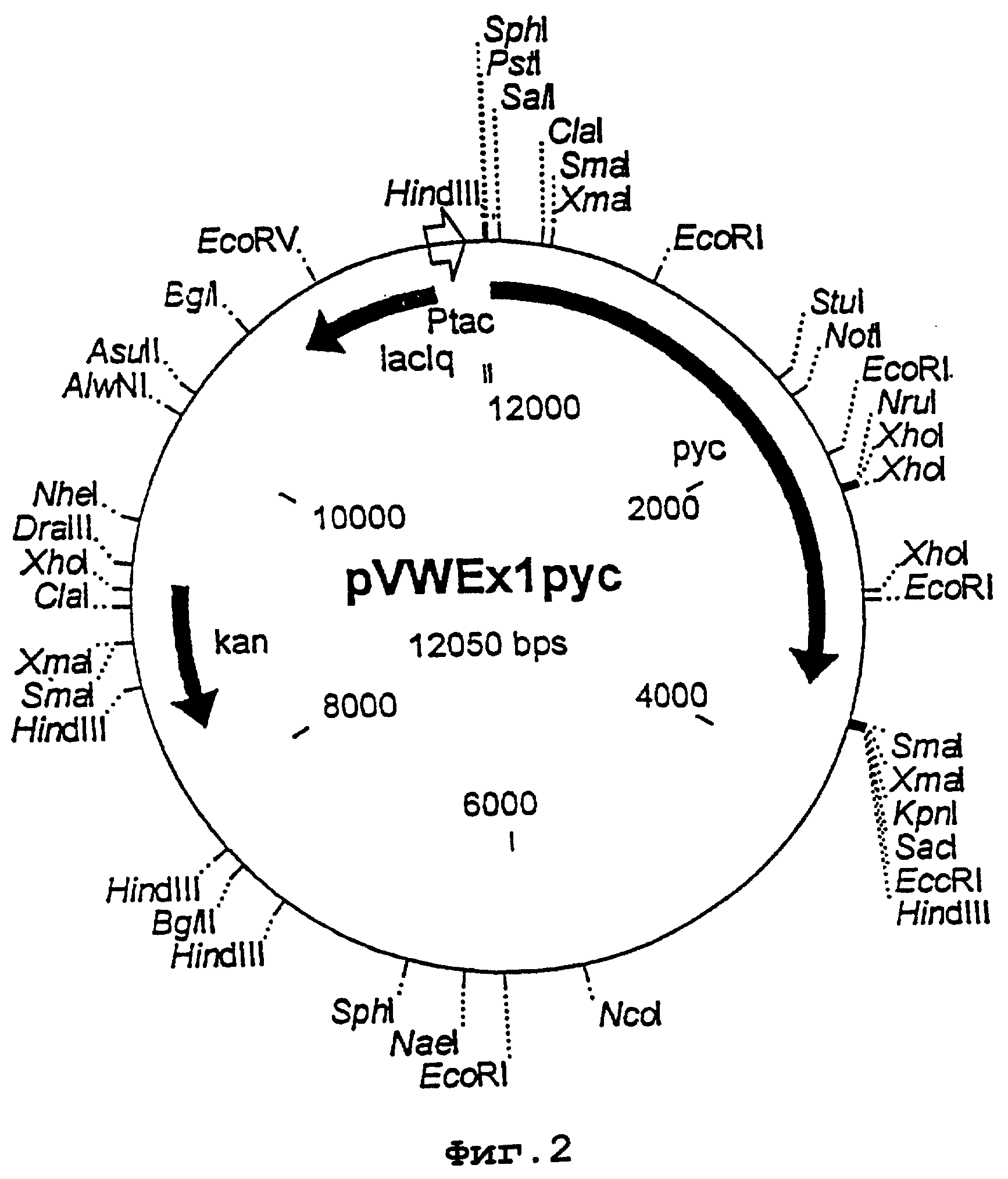
Фрагмент HindIII размером 17 т.п.н. выделяли и субклонировали. Для этого использовали банк космидных генов из хромосомной ДНК C.glutamicuia в космиде рНС79, который на 99% представлен геномом С.glutamicum (Mol. Microbiol., 6, 317-326 [1992]). Штамм E.coli DH5α трансформировали посредством СаСl2-метода Sambrook и др. (Molecular Cloning, A laboratory manual, 1989, Cold Spring Habour Laboratory Press) и по метолу агаровых слоев получали примерно вплоть до 300 колоний на агаровую пластинку при использовании бульона Луриа с 50 мкг/л канамицина (в целом 5000 колоний). Затем полученные трансформанты переносили на фильтр из Nitran N13 и для щелочного лизиса клеток и денатурации ДНК инкубировали их в течение 5 минут на пропитанной с помощью 0,5 М раствора NaOH и 1,5 М раствора NaCl ватмановской бумаге. Последующую нейтрализацию осуществляли с помощью 1 М буфера Tris/HCl, рН 7,5, и 1,5 М раствора NaCl. После инкубации фильтра в растворе 2×SSC высвободившуюся ДНК фиксировали на фильтре путем УФ-облучения при длине волны 366 нм. Затем остаточные "обломки" клеток удаляли путем встряхивания в растворе 3×SSC с 0,1% додецилсульфата натрия при температуре 50°С. Фильтры в этой форме использовали для гибридизации со специфическим рус-зондом, как описано у Саузерна (Southern, J.Mol. Biol., 98, 503-517 [1975]). Идентифицировали 3 трансформанта, которые были гибридизированы с рус-зондом. Из этих трансформантов выделяли космидную ДНК с помощью плазмидного препарата по методу щелочного лизиса согласно Birnboim (Meth. EnzyrooL, 100, 243-255 [1983]) и затем тестировали путем рестрикции и саузернблоттинг-анализа на наличие фрагмента HindIII. Космида рНС79-10, которая содержала вставку размером 40 т.п.н., включала полностью фрагмент HindIII размером 17 т.п.н. и ее анализировали далее. Оказалось, что также после рестрикции с помощью эндонуклеаз Sail и EcoRI получали такие же гибридизирующиеся фрагменты, как и в хромосомной ДНК, то есть фрагмент Sail размером 6,5 т.п.н. и фрагмент TcoRI размером 1,35 т.п.н. Фрагмент HindIII размером 17 т.п.н. выделяли из космиды путем рестрикции с помощью HindIII и лигировали с E. coli-вектором pUC18, который также расщепляли с помощью HindIII. В полученном в результате векторе pUCpyc проводили рестрикционный анализ фрагмента. Физическое картирование фрагмента представлено на фигуре 1.
2. Секвенирование гена пируваткарбоксилазы.
В следующих стадиях субклонирования путем рестрикции с помощью соответствующих рестриктаз из плазмиды pUCpyc выделяли фрагмент SalI-EcoRI размером 0,85 т.п.н., фрагмент EcoRI размером 1,35 т.п.н., фрагмент EcoRI-EcoRI-StuI размером 1,6 т.п.н., а также фрагмент Clal размером 1,6 т.п.н., который частично перекрывался с фрагментом SalI-EcoRI размером 0,85 т.п.н. Путем лигирования фрагменты клонировали в каждом случае в соответственно рестриктированном векторе pUC18 и затем секвенировали согласно Sanger и др. (Proc. Natl. Acad. Sci., 74, 5463-5467 [1977]), как описано выше. Полученные нуклеотидные последовательности анализировали с помощью пакета программ HUSAR (версия 3.0) Немецкого онкологического центра (Гейдельберг). Путем секвенирования фрагментов получили непрерывную открытую рамку считывания размером 3576 пар оснований, которая кодирует протеиновую последовательность из 1140 аминокислот. Сравнение выведенной протеиновой последовательности с банком данных по генам EMBL (Гейдельберг) выявило сходство со всеми известными пируваткарбоксилазами. Самая высокая идентичность (62%) найдена для предполагаемой пируваткарбоксилазы из Mycobacterium tuberculosis (банк генов EMBL: номер по каталогу U00024). Сходство составило, при учете сохраненного аминокислотного обмена, 76%. Сравнение с пируваткарбоксилазами других организмов показало 46-47% идентичных и 64-65% сходных аминокислот (Gene, 191, 47-50 [1997]; J.Bacteriol., 178, 5960-5970 [1996]; Proc. Natl. Acad. Sci., USA, 90, 1766-1770 [1993]; Biochem. J., 316, 631-637 [1996]; банк генов EMBL: номер по каталогу L36530; J. Biol. Chem., 263, 11493-11497 [1988]; Mol. en. Genet., 229, 307-315 [1991]). На основании этих результатов был сделан вывод, что клонированный фрагмент содержит ген пируваткарбоксилазы из C.glutamicum. Нуклеотидная последовательность гена представлена как последовательность SEQ ID №1 и соответствующая аминокислотная последовательность представлена как SEQ ID №2.
3. Сверхпродуцирование гена пируваткарбоксилазы.
Для сверхпродуцирования гена пируваткарбоксилазы из C.glutamicum ген из плазмиды pUCpyc в виде фрагмента SspI-Scal размером 6,2 т.п.н. клонировали в шаттл-векторе Е.соli - С.glutamicum pEKO (Gene, 102, 93-98 [1991]), который подвергали рестрикции с помощью рестриктаз EcoRI и Pstl. Путем обработки с помощью полимеразы Кленова выступающие концы дополняли (EcoRI), соответственно укорачивали (Pstl) до тупых концов, и линеаризированный вектор лигировали с фрагментом Sspl-Scal размером 6,2 т.п.н. Полученную конструкцию рЕКОрус сначала трансформировали в штамме E.coli DH5α, из полученных трансформантов выделяли плазмидную ДНК и контролировали правильность вставки путем рестрикции. ДНК затем вводили в штамм SP733 путем электропорации (FEMS Microbiol. Lett., 65, 299-304 [1989]). В случае этого штамма речь идет о мутанте рестрикционно-отрицательного штамма С. glutamicum R127 (Dechema Biotechnology Conference, 4, 323-327 [1990], Verlag Chemie), который был получен путем химического мутагенеза и отличается тем, что он не может расти в минимальной среде с пируватом и лактатом в качестве единственного источника углерода (Microbiology, 143, 1095-1103 [1997]). Этот фенотип получается за счет дефекта в пируваткарбоксилазе и он может быть дополнен за счет введения гена пируваткарбоксилазы из C.glutamicum, то есть штамм, который несет плазмиду рЕКОрус, в противоположность исходному штамму снова обладает способностью расти в минимальной среде с лактатом в качестве единственного источника углерода. Этим также было доказано, что ген кодирует функциональную пируваткарбоксилазу.
Сверх того, плазмиду рЕКОрус трансформировали в C.glutamicum дикого типа АТСС 13032 путем электропорации. Полученный в результате штамм дикого типа (рЕКОрус) исследовали по сравнению с диким типом АТСС 13032 в отношении его пируваткарбоксилазной активности. Штаммы культивировали в комплексной среде (Luria-Bertani, Molecular Cloning, A laboratory manual, 1989, Cold Spring Harbour Laboratory Press) с 0,5% лактата и в минимальной среде с 2% лактата, соответственно 4% глюкозы, и тестирование пируваткарбоксилазы осуществляли соответственно методу, описанному Peters-Wendisch и др. (Microbiology, 143, 1095-1103 [1997]). Результат анализа (таблица 1) показал, что пируваткарбоксилазная активность в несущем рЕКОрус штамме была в четыре раза выше, чем в исходном штамме.
4. Повышенная аккумуляция лизина за счет сверхпродуцирования гена пируваткарбоксилазы в штамме C.glutamicum DG52-5.
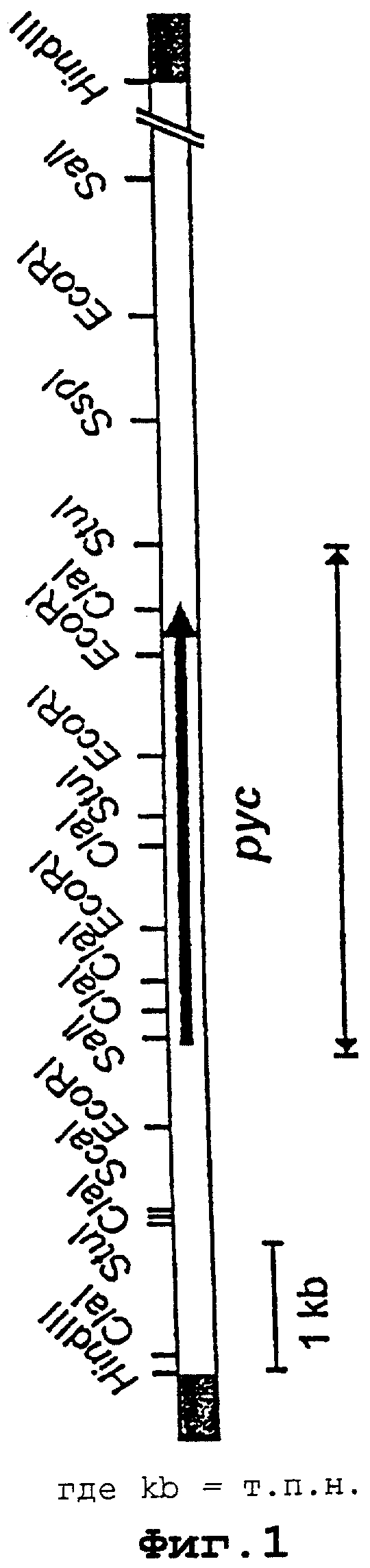
Для исследования воздействия сверхпродуцирования гена пируваткарбоксилазы в продуцирующем лизин штамме DG52-5 (J.Gen. MicrobioL, 134, 3221-3229 [1988]) использовали экспрессионный вектор pVWEXl, который позволяет осуществлять экспрессию, индуцируемую изопропилтиогалактозидом (IPTG). В этом векторе клонировали рус-ген в отсутствие промотора. Для этого сначала синтезировали праймеры полимеразной цепной реакции (праймер 1=положение 112-133; праймер 2=положение 373-355 в нуклеотидной последовательности №1) и посредством ПЦР амплифицировали 261 пару оснований начальной области гена пируваткарбоксилазы, не содержащей промотора. Праймеры выбирали так, чтобы праймер 1 опосредовал место расщепления PstI, а праймер 2 опосредовал место расщепления BamHI. После ПЦР выделяли полученный ПЦР-продукт размером 274 пары оснований, лигировали до конкатемера и затем расщепляли с помощью ферментов рестрикции PstI и BamHI. Смесь после рестрикции концентрировали путем осаждения этанолом и затем лигировали расщепленным Pst-BamHI вектором pVWEXl. Полученную конструкцию pVWEXl-ПЦР тестировали путем рестрикции. Концевую область рус-гена выделяли из вектора рЕКОрус путем обработки с помощью Real-фрагмент Кленова-SalI и лигировали обработанным с помощью BamHI-фрагмент Кленова-SalI вектором pVWEXl-ПЦР. Полученную конструкцию pVWEXIpyc анализировали путем рестрикционного картирования. Физическая карта плазми-ды представлена на фигуре 2.
Плазмиду вводили в штамм С.glutamicum DG52-5 путем электропорации. В качестве контроля штамм DG52-5 трансформировали с помощью вектора pVWEXl без вставки и сравнивали выделение L-лизина соответственно тремя различными трансформантами. Для этого DG52-5 (pVWEXl) 1, 2 и 3, а также DG52-5 (pVWEXlpyc) 3, 4 и 6 культивировали в комплексной среде (2×TY; Molecular Cloning, A laboratory manual, 1989, Cold Spring Harbour Laboratory Press; с 50 мкг/л канамицина) и сбраживаемую среду CGXlI (J. BacterioL, 175, 5595-5603 [1993]) в каждом случае отдельно инфицировали из предкультур. Среда содержала дополнительно канамицин, чтобы поддерживать стабильными плазмиды. Соответственно получали две параллельные смеси, причем в одну колбу добавляли 200 мкг изопропилтиогалактозида на 1 мл, в то время как вторая колба не содержала изопропилтиогалактозида. После культивирования в течение 48 часов при температуре 30°С на ротационном аппарате для встряхивания со 120 оборотами в минуту определяли аккумулированное в среде количество лизина. Определение концентрации аминокислоты осуществляли путем высокоэффективной жидкостной хроматографии (J. Chromat, 266, 471-482 [1983]). Результат ферментации представлен в таблице 2, причем указанные величины представляют собой средние значения из соответственно трех экспериментов с различными клонами. Оказалось, что сверхпродуцирование гена пируваткарбоксилазы приводит к повышенной на 50% аккумуляции лизина в среде. Таким образом, использование открытого и описанного гена анаплеротического фермента пируваткарбоксилазы представляет собой способ существенного улучшения образования L-лизина.
5. Повышенная аккумуляция треонина и гомосерина за счет сверхпродуцирования гена пируваткарбоксилазы в штамме С. glutamicum DM368-3.
Аналогично экспериментам в отношении образования L-лизина также исследовали аккумуляцию треонина в культуральном супернатанте за счет сверхпродуцирования гена пируваткарбоксилазы. Для этого, как описано в пункте 4, продуцирующий треонин штамм C.glutamicum DM368-3 (Degussa AG) трансформировали с помощью плазмиды pVWEXIpyc, а также для контроля с помощью плазмиды pVWEXl, и исследовали выделение треонина в каждом случае тремя различными трансформантами. Для этого DM368-3(pVWEXl) 1, 2 и 3, а также DM368-3(pVWEXlpyc) 1, 2 и 3 культивировали в комплексной среде (2×ТY с 50 мкг/л канамицина) и сбраживаемую среду CGX1I (J. Bacteriol., 175, 5595-5603 [1993]) в каждом случае отдельно инфицировали из предкультур. Среда содержала дополнительно канамицин, чтобы поддерживать стабильными плазмиды. Получали две параллельные смеси, причем в одну колбу добавляли 200 мкг изопропилтиогалактозида на 1 мл, в то время как вторая колба не содержала изопропилтиогалактозида. После культивирования в течение 48 часов при температуре 30°С на ротационной качалке со 120 оборотами в минуту определяли аккумулированное в среде количество треонина. Определение концентрации аминокислоты осуществляли также путем высокоэффективной жидкостной хроматографии (J.Chromat, 266, 471-482 [1983]). Результат ферментации представлен в таблице 3, причем указанные величины представляют собой средние значения из соответственно трех экспериментов с различными клонами. Оказалось, что сверхпродуцирование гена пируваткарбоксилазы приводит к повышению примерно на 40% аккумуляции треонина в среде. Таким образом, использование открытого и описанного гена анаплеротического фермента пируваткарбоксилазы представляет собой способ существенного улучшения образования L-треонина.
Далее, определение концентрации аминокислоты показало, что неожиданно штамм со сверхпродуцированным геном пируваткарбоксилазы, кроме того, выделяет примерно на 150% больше гомосерина в среду, чем штамм с геном, не подвергнутым сверхпродуцированию. Соответствующие результаты представлены также в таблице 3. Они отчетливо показывают, что благодаря предлагаемому согласно изобретению способу можно существенно улучшать как образование треонина, так и образование гомосерина.
6. Повышенная аккумуляция глутамата за счет сверхпродуцирования гена пируваткарбоксилазы в диком типе С.glutamicum.
Аналогично экспериментам в отношении образования L-лизина, L-треонина и L-гомосерина (см. выше в пп.4 и 5) также исследовали аккумуляцию глутамата в культуральном супернатанте за счет сверхпродуцирования гена пируваткарбоксилазы. Для этого, как описано в пункте 4, дикий тип С.glutamicum АТСС 13032 трансформировали с помощью плазмиды pVWEXlpyc, а также для контроля с помощью плазмиды pVWEXl, и исследовали выделение глутамата соответственно двумя различными трансформантами. Для этого C.glutamitum АТСС 13032 (pVWEXlpyc) D1 и D2, а также C.glutamitum АТСС 13032 (pVWEXlpyc) 1 и 2 культивировали в комплексной среде (2xTY с 50 мкг/л канамицина) и сбраживаемую среду CGXlI (J.Bacteriol., 175, 5595-5603 [1993]) в каждом случае отдельно инфицировали из предкультур. Среда содержала дополнительно канамицин, чтобы поддерживать стабильными плазмиды. Для индуцирования выделения глутамата в среду спустя примерно 6 часов после инокуляции добавляли 25 мг твина 60 на 1 мл. Получали две параллельные смеси, причем в одну колбу добавляли 200 мкг изопропилтиогалактозида на 1 мл, в то время как вторая колба не содержала изопропилтиогалактозида. После культивирования в течение 48 часов при температуре 30°С на ротационной качалке со 120 оборотами в минуту определяли аккумулированное в среде количество треонина. Определение концентрации аминокислоты осуществляли также путем высокоэффективной жидкостной хроматографии (J.Chromat, 266, 471-482 [1983]). Результат ферментации представлен в таблице 4, причем указанные величины представляют собой средние значения из соответственно трех экспериментов с различными клонами. Оказалось, что сверхпродуцирование гена пируваткарбоксилазы приводит вплоть до 500%-ного повышения концентрации глутамата в среде. Таким образом, использование открытого и описанного гена анаплеротического фермента пируваткарбоксилазы представляет собой способ существенного улучшения образования L-глутамата.




Во всех таблицах IPTG означает изопропилтиогалактозид.
Реферат
Настоящее изобретение относится к способу микробиологического получения аминокислот семейства аспартатов и/или глутаматов. Пируваткарбоксилазную активность повышают путем генетического изменения фермента и/или экспрессии гена пируваткарбоксилазы микроорганизма, продуцирующего соответствующую аминокислоту. Заявлен ген пируваткарбоксилазы с последовательностью, приведенной в описании, генная конструкция и плазмидный вектор, используемые в предлагаемом способе. Изобретение позволяет повысить выработку микробами аминокислот семейства аспартатов и/или глутаматов. 4 с. и 27 з.п.ф-лы, 2 ил., 4 табл.
Комментарии