Способ получения l-аминокислот - RU2614258C2
Код документа: RU2614258C2
Чертежи








Описание
ОБЛАСТЬ ТЕХНИКИ
По настоящей заявке испрашивается приоритет по патентной заявке Кореи № 10-2013-0121090, зарегистрированной 11 октября 2013 года, и патентной заявке Кореи № 10-2014-0091307, зарегистрированной 18 июля 2014 года, в Korean Intellectual Property Office, описание которых включено в настоящее описание в качестве ссылок в полном объеме.
Один или несколько вариантов осуществления настоящего изобретения относятся к способу получения L-аминокислот с использованием способа ингибирования транскрипции генов.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
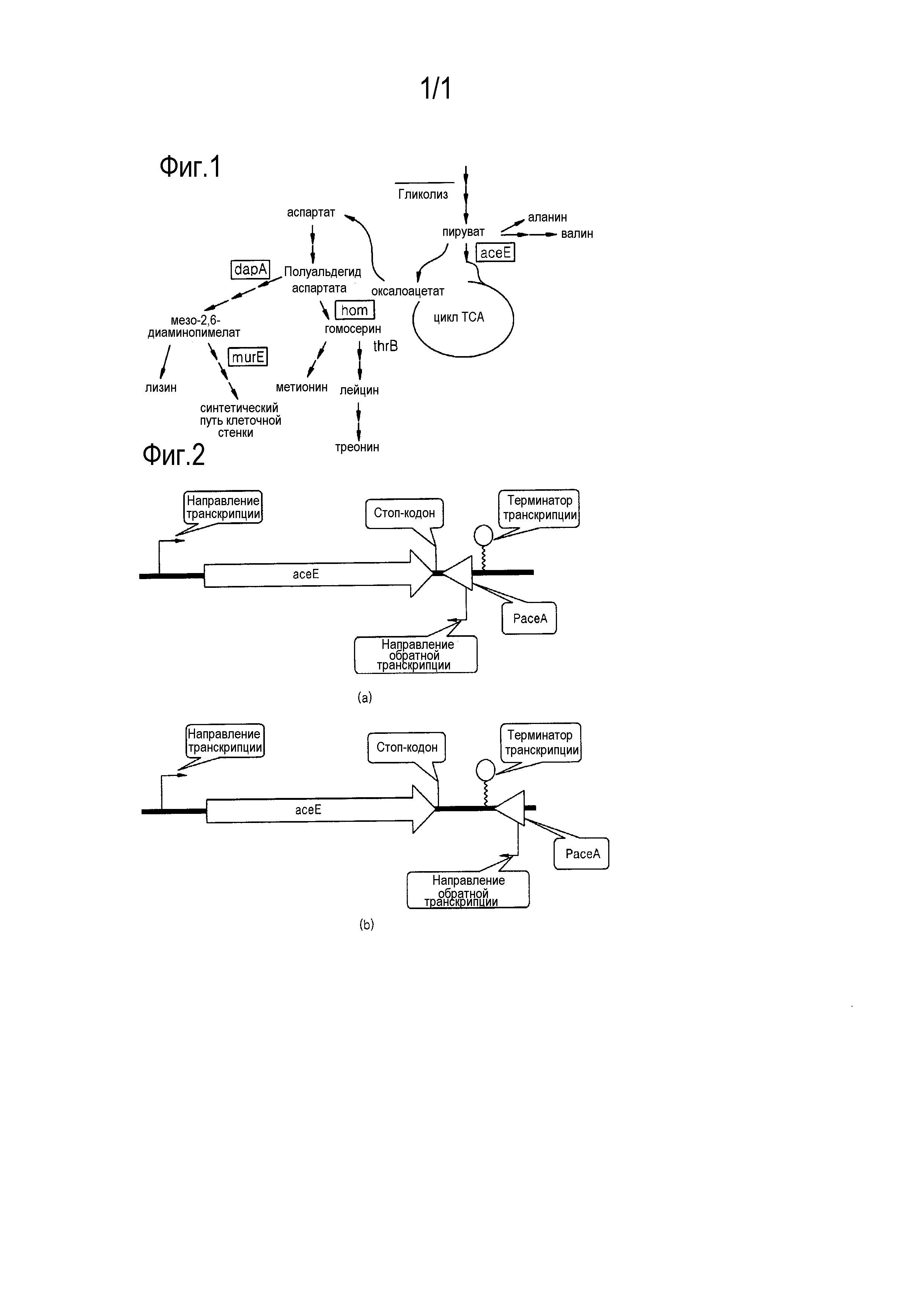
Пируват, получаемый посредством гликолиза различных источников углерода в коринеформных микроорганизмах, превращается в аспартат через оксалоацетат. Аспартат превращается в аминокислоты, такие как треонин, метионин, изолейцин и лизин, с помощью различных путей биосинтеза (фиг. 1). Таким образом, экспрессию генов, находящихся в каждой узловой точке процессов биосинтеза аминокислот, можно ингибировать для снижения образования побочных продуктов и повышения продукции целевых аминокислот.
Как описано выше, для получения штамма микроорганизма, способного к высокоактивной продукции целевых материалов, с использованием генетической инженерии и метаболической инженерии необходимо избирательно контролировать экспрессию генов, связанных с различными метаболическими процессами микроорганизма. Недавно сообщали о технологии ослабления экспрессии генов, названной "искусственная конвергентная транскрипция" (Krylov et al., J Mol Microbiol Biotechnol, 18:1-13, 2010). Искусственная конвергентная транскрипция является технологией ослабления экспрессии гена-мишени посредством инсерции промотора в нижележащую область терминатора транскрипции гена-мишени таким образом, что противоположное направление промотора вызывает столкновение комплексов РНК-полимеразы, происходящих из каждого промотора в течение транскрипции.
Авторы настоящего изобретения разрабатывали технологию избирательного ингибирования экспрессии гена-мишени в присутствии ацетата посредством инсерции ацетат-индуцибельного промотора в направлении, противоположном транскрипции гена-мишени, и эффективно применяли технологию для ингибирования экспрессии генов, находящихся в узловых точках в коринеформном микроорганизме. Затем авторы настоящего изобретения подтверждали получение коринеформного микроорганизма, продуцирующего L-аминокислоту с высоким выходом, с использованием технологии и осуществляли настоящее изобретение.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
ТЕХНИЧЕСКАЯ ПРОБЛЕМА
Целью настоящего изобретения является предоставление способа получения L-аминокислот с использованием ацетат-индуцибельного промотора для ингибирования транскрипции гена-мишени.
Один из вариантов осуществления настоящего изобретения относится к способу получения L-аминокислот, включающему
1) культивирование рекомбинантного коринеформного микроорганизма, способного продуцировать L-аминокислоты, где рекомбинантный коринеформный микроорганизм трансформируют посредством инсерции ацетат-индуцибельного промотора ниже стоп-кодона гена-мишени в хромосоме; и
2) добавление ацетата при культивировании для ослабления экспрессии гена-мишени и усиления способности рекомбинантного коринеформного микроорганизма продуцировать L-аминокислоты.
РЕШЕНИЕ ПРОБЛЕМЫ
Далее будут подробно описаны варианты осуществления, примеры которых проиллюстрированы в сопутствующих чертежах, где на всем протяжении подобные номера позиций относятся к подобным элементам. В связи с этим настоящие варианты осуществления могут иметь различные формы, и их не следует истолковывать как ограничение описаний, изложенных в настоящей заявке. Таким образом, варианты осуществления лишь описаны ниже со ссылкой на фигуры для объяснения аспектов настоящего описания. Выражения, такие как "по меньшей мере один", предшествующие списку элементов, модифицируют весь список элементов и не модифицируют отдельные элементы списка.
Далее в настоящем описании подробно описывают настоящее изобретение.
Один из вариантов осуществления настоящего изобретения относится к способу получения L-аминокислот, включающему
1) культивирование рекомбинантного коринеформного микроорганизма, способного продуцировать L-аминокислоты, где рекомбинантный коринеформный микроорганизм трансформируют посредством инсерции ацетат-индуцибельного промотора ниже стоп-кодона гена-мишени в хромосоме; и
2) добавление ацетата при культивировании для ослабления экспрессии гена-мишени и усиления способности рекомбинантного коринеформного микроорганизма продуцировать L-аминокислоты.
Термин "ацетат-индуцибельный промотор", используемый в настоящем описании, относится к промотору, имеющему активность, индуцирующую экспрессию генов в присутствие ацетата.
В коринеформном микроорганизме ацетат превращается ацетаткиназой (ackA, NCgl2656) и фосфотрансацетилазой (pta, NCgl2657) или сукцинил-КоА:ацетат-КоА-трансферазой (actA, NCgl2480) в ацетил-КоА, а затем метаболизируется изоцитратлиазой (aceA, NCgl2248) в глиоксилатном цикле. У Escherichia coli ацетат превращается в ацетил-КоА ацетил-КоА-синтазой (acs, b4069) (Gerstmeir et al., J Biotechnol, 104:99-122, 2003). Экспрессия указанных генов, участвующих в метаболизме ацетата, индуцируется в присутствие ацетата. Таким образом, при использовании промоторов генов экспрессию гена можно специально индуцировать в присутствие ацетата.
Ацетат-индуцибельные промоторы включают промотор гена, кодирующего изоцитратлиазу (aceA, NCgl2248), или промотор оперона гена, кодирующего ацетаткиназу (ackA, NCgl2656), и гена, кодирующего фосфотрансацетилазу (pta, NCgl2657), являющегося вышележащим промотором гена pta. Более конкретно, среди описываемых выше ацетат-индуцибельных промоторов, промотор гена aceA представлен нуклеотидной последовательностью SEQ ID NO: 1 и включает 486 пар оснований выше гена aceA и 36 пар оснований от N-конца открытой рамки считывания (ORF).
Вышележащий промотор гена pta, являющийся другим ацетат-индуцибельным промотором, представлен нуклеотидной последовательностью SEQ ID NO: 2 и включает 340 пар оснований выше гена pta.
Кроме того, очевидно, что в объем настоящего изобретения можно включать любой промотор, способный индуцировать экспрессию гена-мишени посредством ацетата. Например, ацетат-индуцибельные промоторы могут включать нуклеотидную последовательность, включающую нуклеотидную последовательность SEQ ID NO: 1 или 2 или включающую консервативную последовательность нуклеотидной последовательности SEQ ID NO: 1 или 2, и один или множество нуклеотидов (конкретно - от 2 до 20, более конкретно - от 2 до 10, еще более конкретно - от 2 до 5 нуклеотидов в зависимости от стерической конформации аминокислотных остатков белка), подвергнутых замене, делеции, инсерции, добавлению или инверсии в одном или нескольких положениях. При условии, что функцию индуцибельного промотора сохраняют или усиливают, он может включать нуклеотидную последовательность, имеющую более 80% гомологии по отношению к нуклеотидной последовательности SEQ ID NO: 1 или 2, конкретно - более 90%, более конкретно - более 95%, еще более конкретно - более 97%. При условии, что функцию индуцибельного промотора сохраняют, подвергнутая замене, делеции, инсерции, добавлению или инверсии нуклеотидная последовательность может включать спонтанно-мутантную последовательность или даже искусственную мутантную последовательность.
Термин "гомология", используемый в настоящем описании, относится к идентичности между двумя различными нуклеотидными последовательностями. Гомологию можно определять известным в этой области способом с использованием программного обеспечения BLAST 2.0, с помощью которого вычисляют такие параметры, как балл, идентичность и сходство. Однако способ определения гомологии не ограничен указанным выше.
Если в настоящем описании не указано иначе, термин "вышележащий" относится к 5'-направлению и термин "нижележащий" относится к 3'-направлению. Как правило, направлением транскрипции является 5'-3' таким образом, что положение промотора, как правило, находится выше (5') гена-мишени.
В настоящем описании ген-мишень на хромосоме может являться геном, участвующим в процессе биосинтеза аминокислот, таких как треонин, метионин, изолейцин и лизин, из различных источников углерода, в частности, геном, находящимся в узловой точке пути биосинтеза.
Например, что касается лизина, в узловых точках пути биосинтеза находятся ген субъединицы E1 пируватдегидрогеназы (aceE, NCgl2167), участвующей в превращении пирувата в ацетил-КоА, ген гомосериндегидрогеназы (hom, NCgl1136), продуцирующей гомосерин из аспартата, и ген УДФ-N-ацетилмурамоилаланил-D-глутамат-2,6-диаминопимелатлигазы (murE, NCgl2083), использующей мезо-2,6-диаминопимелат, являющийся предшественником лизина в соматическом синтезе.
Кроме того, что касается треонина, в узловой точке находится ген дигидродипиколинатсинтазы (dapA, NCgl1896), участвующей в продукции лизина из аспартата, и, что касается метионина, в узловой точке находится ген гомосеринкиназы (thrB, NCgl1137), участвующей в продукции треонина из гомосерина. Что касается аланина и валина, являющихся аминокислотами, образующимися из пирувата, в узловой точке находится ген субъединицы E1 пируватдегидрогеназы (aceE, NCgl2167), участвующей в превращении пирувата в ацетил-КоА.
Таким образом, ген-мишень, в качестве неограничивающих примеров, можно выбирать из группы, состоящей из гена, кодирующего субъединицу E1 пируватдегидрогеназы (aceE, NCgl2167), гена, кодирующего гомосериндегидрогеназу (hom, NCgl1136), гена, кодирующего УДФ-N-ацетилмурамоилаланил-D-глутамат-2,6-диаминопимелатлигазу (murE, NCgl2083), и гена, кодирующего дигидродипиколинатсинтазу (dapA, NCgl1896).
Конкретно, субъединица E1 пируватдегидрогеназы (aceE, NCgl2167) является одной из белковых субъединиц комплекса пируватдегидрогеназы (PDHC), участвующей в поступлении пирувата, являющегося конечным метаболитом гликолиза цикла трикарбоновых кислот (цикла TCA). Таким образом, ослабление экспрессии гена aceE может снижать поступление источников углерода в цикл TCA и повышать поступление источников углерода в путь биосинтеза лизина для повышения продукции лизина.
Гомосериндегидрогеназа (hom, NCgl1136) является ферментом, синтезирующим гомосерин из полуальдегида аспартата. Т.к. полуальдегид аспартата является одним из промежуточных предшественников в пути биосинтеза лизина, ослабление активности гена hom может снижать поступление источников углерода в путь биосинтеза гомосерина и повышать поступление источников углерода в путь биосинтеза лизина для повышения продукции лизина.
УДФ-N-ацетилмурамоилаланил-D-глутамат-2,6-диаминопимелатлигаза (murE, NCgl2083) использует мезо-2,6-диаминопимелат для соматического синтеза. Т.к. мезо-2,6-диаминопимелат также используется в качестве предшественника для биосинтеза лизина, ослабление активности гена murE может снижать поступление источников углерода в соматический синтез и повышать поступление источников углерода в путь биосинтеза лизина для повышения продукции лизина.
Дигидродипиколинатсинтаза (dapA, NCgl1896) является ферментом, участвующим в продукции лизина с использованием полуальдегида аспартата. Т.к. полуальдегид аспартата является одним из промежуточных предшественников в пути биосинтеза лизина, ослабление активности гена dapA может снижать поступление источников углерода в путь биосинтеза лизина и повышать поступление источников углерода в путь биосинтеза треонина для повышения продукции треонина.
Термин "стоп-кодон", используемый в настоящем описании, относится к кодонам, не кодирующим аминокислоту в мРНК, а действующим в качестве сигнала для терминации синтеза белка. Общепринято в качестве стоп-кодонов используют три кодона, включающих UAA, UAG и UGA.
Термин "терминатор транскрипции", используемый в настоящем описании, относится к GC-богатой последовательности инвертированного повтора. Терминатор транскрипции образует петлю-шпильку для терминации транскрипции генов.
В настоящем изобретении для ослабления экспрессии гена-мишени, как описано выше, можно встраивать ацетат-индуцибельный промотор ниже стоп-кодона гена-мишени, конкретно, между стоп-кодоном и выше терминатора транскрипции. Можно использовать ацетат-индуцибельный промотор для вызывания инверсии транскрипции гена-мишени таким образом, что комплексы РНК-полимеразы могут сталкиваться друг с другом с ослаблением экспрессии гена-мишени.
Экспрессию гена-мишени можно ослаблять при каждом культивировании. Более конкретно, экспрессию гена-мишени можно ослаблять до культивирования или в течение него.
Термин "трансформация", используемый в настоящем описании, относится к встраиванию вектора, включающего полинуклеотид, кодирующий ген-мишень, в клетку-хозяина таким образом, что в клетке-хозяине может экспрессироваться белок, кодируемый полинуклеотидом. При условии, что встраиваемый полинуклеотид может экспрессироваться в клетке-хозяине, полинуклеотид можно встраивать в хромосому клетки-хозяина, или он может находиться вне хромосомы. Кроме того, полинуклеотид включает ДНК или РНК, кодирующую белок-мишень. При условии, что полинуклеотид можно встраивать и экспрессировать в клетке-хозяине, полинуклеотид можно встраивать в любой форме.
В варианте осуществления изобретения коринеформный микроорганизм может включать микроорганизмы рода Corynebacterium, рода Brevibacterium, рода Arthrobacter sp. и рода Microbacterium sp. Примеры коринеформного микроорганизма включают Corynebacterium glutamicum, Corynebacterium thermoaminogenes, Brevibacterium flavum, Brevibacterium lactofermentum и полученные из них варианты, продуцирующие L-аминокислоты. Конкретно, коринеформный микроорганизм может являться, в качестве неограничивающих примеров, Corynebacterium glutamicum.
Более конкретно, коринеформные микроорганизмы в настоящем изобретении могут включать Corynebacterium glutamicum KCCM11016P (прежний регистрационный №: KFCC10881, ссылка на патент Кореи №: 10-0159812), Corynebacterium glutamicum KCCM10770P (ссылка на патент Кореи №: 10-0924065), и Corynebacterium glutamicum KCCM11347P (прежний регистрационный №: KFCC10750, ссылка на патент Кореи №: 10-0073610).
В настоящем изобретении коринеформные микроорганизмы также могут включать Corynebacterium glutamicum CJ3P. Согласно статье Binder et al. (Binder et al., Genome Biology, 13:R40, 2012), можно получать CJ3P, имеющий способность продуцировать лизин, посредством встраивания мутации в три гена, вносящих свой вклад в эффективность продукции лизина (pyc(P458S), hom(V59A) и lysC(T311I)), в родительском штамме Corynebacterium glutamicum дикого типа (ATCC13032).
Кроме того, в настоящем изобретении другим коринеформным микроорганизмом может являться Corynebacterium glutamicum KCCM11222P, являющийся L-треонин-продуцирующим штаммом (ссылка на патент Кореи №: 10-1335853).
По одному из вариантов осуществления настоящего изобретения во всех коринеформных микроорганизмах, в которые встраивали промотор, представленный нуклеотидной последовательностью SEQ ID NO: 1 или SEQ ID NO: 2, повышали продукцию L-лизина или L-треонина по сравнению с таковой у родительского штамма.
Что касается способа, представленного в настоящем изобретении, культивирование коринеформного микроорганизма можно осуществлять с использованием любых условий культивирования и способа культивирования, известных в этой области.
Среда для культивирования, которую можно использовать в культивировании штамма коринеформного микроорганизма, может являться, например, средами для культивирования, описываемыми в Manual of Methods for General Bacteriology, American Society for Bacteriology (Washington D.C., USA, 1981).
Источники углерода, которые можно использовать в среде для культивирования, могут включать углевод, такой как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал и целлюлоза, масло или липид, такой как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло, жирную кислоту, такую как пальмитиновая кислота, стеариновая кислота и линолевая кислота, спирт, такой как глицерин и этанол, и органическую кислоту, такую как уксусная кислота. Эти вещества могут находиться отдельно или в виде смеси.
Источники азота, которые можно использовать в среде для культивирования, могут включать пептон, дрожжевой экстракт, говяжий экстракт, солодовый экстракт, кукурузный экстракт, сою и мочевину и неорганический источник азота, такой как сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Эти источники азота также могут находиться отдельно или в виде смеси.
Источники фосфора, которые можно использовать в среде для культивирования, могут включать дигидрофосфат калия, гидрофосфат дикалия и их натрийсодержащую соль. Кроме того, среда для культивирования может включать соль металла, такую как сульфат магния и сульфат железа, необходимые для роста. Помимо описываемых выше веществ, можно использовать вещества, необходимые для роста, такие как аминокислоты и витамины. Кроме того, в среде для культивирования можно использовать подходящие предшественники. В культуральный раствор можно добавлять сырьевые материалы периодическим способом или непрерывным способом в течение культивирования.
В течение культивирования микроорганизма можно корректировать pH среды для культивирования посредством добавления основного соединения, такого как гидроксид аммония, гидроксид калия и аммиак, или кислого соединения, такого как фосфорная кислота и серная кислота, в среду для культивирования подходящим способом. Кроме того, можно подавлять образование пузырьков с использованием пеногасителя, такого как сложный эфир жирной кислоты и полигликоля. Для поддержания аэробных условий в среду для культивирования можно вводить кислород или кислородсодержащий газ (например, воздух). Температура среды для культивирования, как правило, может составлять от приблизительно 20°C до приблизительно 45°C, конкретно - от приблизительно 25°C до приблизительно 40°C. Можно продолжать культивирование до получения желаемого количества L-аминокислоты, но подходящее время культивирования может составлять от приблизительно 10 до 160 часов.
Что касается способа, представленного в настоящем изобретении, культивирование можно осуществлять непрерывным способом или периодическим способом, таким как периодический способ, периодический способ с подпиткой и периодический способ с повторной подпиткой. Эти способы культивирования известны в этой области, и можно использовать любой из способов культивирования.
Термин "культивирование", используемый в настоящем описании, может включать получение среды для культивирования и время в течение роста микроорганизмов.
Что касается способа, представленного в настоящем изобретении, он может включать дополнительный этап очистки или выделения. Целевую L-аминокислоту можно очищать или выделять из культурального раствора с использованием подходящего способа, известного в этой области, в соответствии со способом, таким как периодическое культивирование, непрерывное культивирование и периодическое культивирование с подпиткой.
L-аминокислота, продуцируемая при культивировании по настоящему изобретению, может являться аминокислотой, выбранной из группы, состоящей из треонина, метионина, изолейцина, лизина, валина и аланина, конкретно, лизина или треонина.
Краткое описание чертежей
Эти и/или другие аспекты станут очевидными и более понятными из следующего описания вариантов осуществления, представленных в комбинации с сопутствующими чертежами, в которых:
На фиг. 1 показаны узловые точки процесса биосинтеза аминокислот в коринеформном микроорганизме; и
Фиг. 2 является схематическим изображением, на котором показано ингибирование экспрессии гена aceE посредством инсерции промотора гена aceA (a) между стоп-кодоном и терминатором транскрипции выше гена aceE или (b) ниже терминатора транскрипции в направлении, противоположном направлению транскрипции гена aceE.
Способ по изобретению
Далее в настоящем описании настоящее изобретение будет более подробно описано со ссылкой на примеры. Эти примеры представлены исключительно в иллюстративных целях и не ограничивают объем настоящего изобретения.
Примеры
Пример 1: Выбор ацетат-индуцибельного промотора
Изоцитратлиаза (aceA, NCgl2248) является ключевым ферментом глиоксилатного цикла, и ген, кодирующий изоцитратлиазу, экспрессируется в присутствие ацетата. Кроме того, ацетаткиназа (ackA, NCgl2656) и фосфотрансацетилаза (pta, NCgl2657), являющиеся ферментами, участвующими в процессе метаболизма ацетата, образуют оперон, и их экспрессия усиливается в присутствие ацетата. Промоторные области гена aceA и оперон pta-ackA уже известны (Gerstmeir et al., J Biotechnol, 104, 99-122, 2003).
В примере 1 для ингибирования транскрипции гена-мишени в присутствие ацетата выбирали промотор гена aceA и промотор оперона pta-ack, являющийся вышележащей промоторной областью гена pta. На основе гена aceA, зарегистрированного в US NIH GenBank (регистрационный № NCBI: NCgl2248), получали нуклеотидную последовательность (SEQ ID NO: 1), включающую 486 пар оснований выше гена aceA и 36 пар оснований от N-конца открытой рамки считывания (ORF). Кроме того, на основе гена pta, зарегистрированного в US NIH GenBank (регистрационный № NCBI: NCgl2657), получали нуклеотидную последовательность (SEQ ID NO: 2), включающую 340 пар оснований выше гена pta.
Пример 2: Получение вектора для ингибирования экспрессии гена aceE
Субъединица E1 пируватдегидрогеназы (aceE, NCgl2167) является одной из белковых субъединиц комплекса пируватдегидрогеназы (PDHC), участвующей в поступлении пирувата, являющегося конечным метаболитом гликолиза в цикле TCA. Таким образом, ослабление экспрессии гена aceE может снижать поступление источников углерода в цикл TCA и повышать поступление источников углерода в путь биосинтеза лизина для повышения продукция лизина (Blombach et al., Appl Microbiol Biotechnol, 76(3):615-23, 2007).
Для избирательного ингибирования экспрессии гена aceE в присутствие ацетата промотор гена aceA встраивали ниже гена aceE таким образом, что транскрипция с этого промотора может происходить в направлении, противоположном исходному направлению транскрипции гена aceE (фиг. 2).
Сначала для прогнозирования терминатора транскрипции гена aceE использовали программное обеспечение CLC main workbench (CLC Bio, Denmark). Терминатор транскрипции является GC-богатой последовательностью инвертированного повтора и образует петлю-шпильку для терминации транскрипции генов. Результат прогнозирования терминатора транскрипции гена aceE свидетельствует о том, что 36 пар оснований, с 21-ой пары оснований до 56-ой пары оснований ниже стоп-кодона гена aceE, образуют петлю-шпильку в качестве терминатора транскрипции. С учетом этого результата получали два вектора для встраивания промотора aceE выше или ниже, соответственно, терминатора транскрипции гена aceE таким образом, что транскрипция с этого промотора может происходить в направлении, противоположном исходному.
<2-1> Получение вектора pDZ-aceE1-PaceA для ингибирования экспрессии гена aceE
Вектор получали посредством инсерции промотора гена aceA ниже стоп-кодона гена aceE, находящегося между стоп-кодоном и вышележащим терминатором транскрипции.
Для получения фрагмента гена aceE Corynebacterium glutamicum использовали хромосомную ДНК Corynebacterium glutamicum KCCM11016P в качестве матрицы для синтеза праймеров (SEQ ID NO: 3 и 4), сконструированных содержащими участок распознавания фермента рестрикции XbaI на 5'-конце фрагмента и участок распознавания фермента рестрикции SpeI на 3'-конце фрагмента. Осуществляли ПЦР с использованием синтезированных праймеров для получения фрагмента ДНК, включающего 296 пар оснований от 2474-го нуклеотида от инициаторного кодона гена aceE до 2769-го нуклеотида, стоп-кодона гена aceE. Кроме того, синтезировали праймеры (SEQ ID NO: 5 и 6), сконструированные содержащими участок распознавания SpeI на 5'-конце фрагмента и участок распознавания XbaI на 3'-конце фрагмента, и осуществляли ПЦР с использованием праймеров для получения фрагмента ДНК, включающего 300 пар оснований ниже стоп-кодона гена aceE. В качестве полимеразы использовали ДНК-полимеразу PfuUltra™ High-Fidelity (Stratagene) и осуществляли ПЦР с 30 циклами денатурации при 95°C в течение 30 секунд; отжига при 55°C в течение 30 секунд и полимеризации при 72°C в течение 30 секунд, а затем полимеризацией при 72°C в течение 7 минут.
Два продукта ПЦР-амплификации и вектор pDZ (ссылка на патент Кореи №: 10-0924065) для встраивания в хромосому, уже полученный посредством расщепления ферментом рестрикции XbaI, клонировали с использованием набора In-fusion Cloning Kit (TAKARA, JP) для получения вектора pDZ-aceE1.
SEQ ID NO: 3:
aceE-P1F 5'-ccggggatcctctagacctccggcccatacgttgc-3'
SEQ ID NO: 4: aceE-P1R 5'-ttgagactagttattcctcaggagcgtttg-3'
SEQ ID NO: 5: aceE-P2F 5'-gaataactagtctcaagggacagataaatc-3'
SEQ ID NO: 6:
aceE-P2R 5'-gcaggtcgactctagagaccgaaaagatcgtggcag-3'
Для получения фрагмента промотора гена aceA Corynebacterium glutamicum синтезировали праймеры, сконструированные содержащими участок распознавания фермента рестрикции SpeI на 5'-конце и на 3'-конце фрагмента (SEQ ID NO: 7 и 8). Осуществляли ПЦР с использованием хромосомной ДНК Corynebacterium glutamicum KCCM11016P в качестве матрицы и синтезированных праймеров для амплификации промоторной области из приблизительно 500 пар оснований, представленных нуклеотидной последовательностью SEQ ID NO: 1. Продукт ПЦР-амплификации и фрагмент ДНК, полученный посредством обработки pDZ-aceE1 ферментом рестрикции SpeI, клонировали с использованием набора In-fusion Cloning Kit (TAKARA, JP) для получения вектора pDZ-aceE1-PaceA.
SEQ ID NO: 7:
PaceA-P3F 5'-gtcccttgagactagtagcactctgactacctctg-3'
SEQ ID NO: 8:
PaceA-P3R 5'-ctgaggaataactagtttcctgtgcggtacgtggc-3'
<2-2> Получение вектора pDZ-aceE2-PaceA для ингибирования экспрессии генов aceE
Вектор получали посредством инсерции промотора гена aceA ниже терминатора транскрипции гена aceE.
Для получения фрагмента гена aceE Corynebacterium glutamicum в качестве матрицы использовали хромосомную ДНК Corynebacterium glutamicum KCCM11016P и использовали праймеры, сконструированные содержащими участок распознавания фермента рестрикции XbaI на 5'-конце фрагмента и участок распознавания фермента рестрикции SpeI на 3'-конце фрагмента (SEQ ID NO: 9 и 10). Осуществляли ПЦР с использованием синтезированных праймеров для получения фрагмента ДНК, включающего 294 пар оснований от 2538-го нуклеотида от инициаторного кодона гена aceE до 62-го нуклеотида ниже стоп-кодона. Кроме того, праймеры, сконструированные содержащими участок распознавания фермента рестрикции SpeI на 5'-конце фрагмента и участок распознавания фермента рестрикции XbaI на 3'-конце фрагмента (SEQ ID NO: 11 и 12), использовали для осуществления ПЦР для получения фрагмента ДНК, включающего 294 пар оснований от 69-го нуклеотида ниже стоп-кодона гена aceE до 362-го нуклеотида. В качестве полимеразы использовали ДНК-полимеразу PfuUltra™ High-Fidelity (Stratagene) и осуществляли ПЦР с 30 циклами денатурации при 95°C в течение 30 секунд; отжига при 55°C в течение 30 секунд и полимеризации при 72°C в течение 30 секунд, а затем полимеризацией при 72°C в течение 7 минут.
Два продукта ПЦР-амплификации и вектор pDZ для встраивания в хромосому, уже полученный посредством расщепления ферментом рестрикции XbaI, клонировали с использованием набора In-fusion Cloning Kit (TAKARA, JP) для получения вектора pDZ-aceE2.
SEQ ID NO: 9:
aceE-P4F 5'-ccggggatcctctagaggtcccaggcgactacacc-3'
SEQ ID NO: 10:
aceE-P4R 5'-gagctactagtacgacgaatcccgccgccagacta-3'
SEQ ID NO: 11:
aceE-P5F 5'-gtcgtactagtagctctttttagccgaggaacgcc-3'
SEQ ID NO: 12:
aceE-P5R 5'-gcaggtcgactctagacatgctgttggatgagcac-3'
Для получения фрагмента промотора гена aceA Corynebacterium glutamicum синтезировали праймеры, сконструированные содержащими участок распознавания фермента рестрикции SpeI на 5'-конце и на 3'-конце фрагмента (SEQ ID NO: 13 и 14). ПЦР осуществляли с использованием хромосомной ДНК Corynebacterium glutamicum KCCM11016P в качестве матрицы и синтезированных праймеров для амплификации промоторной области из приблизительно 500 пар оснований, представленных нуклеотидной последовательностью SEQ ID NO: 1. Продукт ПЦР-амплификации и фрагмент ДНК, полученный посредством обработки вектора pDZ-aceE2 ферментом рестрикции SpeI, клонировали с использованием набора In-fusion Cloning Kit (TAKARA, JP) для получения вектора pDZ-aceE2-PaceA.
SEQ ID NO: 13:
PaceA-P6F 5'-aaaaagagctactagtagcactctgactacctctg-3'
SEQ ID NO: 14:
PaceA-P6R 5'-gattcgtcgtactagtttcctgtgcggtacgtggc-3'
Пример 3: Получение штаммов, в которых промотор гена aceA встраивали ниже гена aceE
Векторы pDZ-aceE1-PaceA и pDZ-aceE2-PaceA, полученные в примере 2, встраивали соответствующим образом посредством электропорации в Corynebacterium glutamicum KCCM11016P, являющийся L-лизин-продуцирующим штаммом (способ трансформации описан в Van der Rest et al., Appl Microbiol Biotechnol, 52:541-545, 1999). Выбирали соответствующие штаммы, в которых промотор гена aceA встраивали ниже стоп-кодона гена aceE на хромосоме таким образом, что транскрипция с этого промотора может происходить в направлении, противоположном исходному направлению, осуществляя ПЦР для получения L-лизин-продуцирующего штамма. Выбранные штаммы называли Corynebacterium glutamicum KCCM11016P::aceE1-PaceA и Corynebacterium glutamicum KCCM11016P::aceE2-PaceA, соответственно. Corynebacterium glutamicum KCCM11016P::aceE1-PaceA находился на международном депонировании под названием Corynebacterium glutamicum CA01-2271 в Korean Culture Center of Microorganism (KCCM) на 12 июня 2013 года с регистрационным № KCCM11432P. Проверяли полученные штаммы, анализируя нуклеотидную последовательность полученной области-мишени посредством ПЦР с использованием SEQ ID NO: 3 и SEQ ID NO: 6 в качестве праймеров для KCCM11016P::aceE1-PaceA и SEQ ID NO: 9 и SEQ ID NO: 12 в качестве праймеров для KCCM11016P::aceE2-PaceA.
Пример 4: Сравнение продукции лизина штаммами, в которых промотор гена aceA встраивали ниже гена aceE
Штамм Corynebacterium glutamicum KCCM11016P, используемый в качестве родительского штамма, и Corynebacterium glutamicum KCCM11016P::aceE1-PaceA и Corynebacterium glutamicum KCCM11016P::aceE2-PaceA, являющиеся L-лизин-продуцирующими штаммами, полученными в примере 3, культивировали описываемым ниже способом.
Corynebacterium glutamicum KCCM11016P, KCCM11016P::aceE1-PaceA и KCCM11016P::aceE2-PaceA инокулировали соответствующим образом в 25 мл описываемой ниже среды для посева в колбах с дефлекторами емкостью 250 мл с последующим культивированием со встряхиванием при 200 об./мин при 30°C в течение 20 часов. 1 мл раствора посевной культуры добавляли в колбу с дефлекторами емкостью 250 мл, содержащую 24 мл описываемой ниже продукционной среды, с последующим культивированием со встряхиванием при 200 об./мин при 30°C в течение 72 часов. Соответствующие композиции среды для посева и продукционной среды описывают ниже.
<Среда для посева (pH 7,0)>
Глюкоза 20 г, пептон 10 г, дрожжевой экстракт 5 г, мочевина 1,5 г, KH2PO4 4 г, K2HPO4 8 г, MgSO4⋅7(H2O) 0,5 г, биотин 100 мкг, тиамин HCl 1000 мкг, пантотенат кальция 2000 мкг, никотинамид 2000 мкг (на 1 л дистиллированной воды).
<Продукционная среда (pH 7,0)>
Глюкоза 100 г, (NH4)2SO4 40 г, соевый белок 2,5 г, твердый кукурузный экстракт 5 г, мочевина 3 г, KH2PO4 1 г, MgSO4⋅7(H2O) 0,5 г, биотин 100 мкг, тиамин HCl 1000 мкг, пантотенат кальция 2000 мкг, никотинамид 3000 мкг, CaCO3 30 г (на 1 л дистиллированной воды).
После культивирования с использованием ВЭЖХ измеряли концентрацию L-лизина. Для случаев, когда не добавляли ацетат, концентрация L-лизина в культуральных растворах Corynebacterium glutamicum KCCM11016P, KCCM11016P::aceE1-PaceA и KCCM11016P::aceE2-PaceA представлена в таблице 1.
Кроме того, штаммы культивировали тем же способом, за исключением, что для сравнения продукции L-лизина в продукционную среду добавляли 5 г/л ацетата. Концентрация L-лизина в культуральных растворах представлена в таблице 2.
Как показано в таблице 1, в отсутствие ацетата продукция L-лизина штаммами KCCM11016P::aceE1-PaceA и KCCM11016P::aceE2-PaceA не отличалась от родительского штамма KCCM11016P.
Однако, как показано в таблице 2, в присутствие ацетата продукция L-лизина штаммом KCCM11016P::aceE1-PaceA была на 3,6% выше, чем у родительского штамма KCCM11016P, а штаммом KCCM11016P::aceE2-PaceA - на 2,5% выше, чем у родительского штамма KCCM11016P.
Кроме того, сравнение штамма KCCM11016P::aceE1-PaceA и штамма KCCM11016P::aceE2-PaceA показало, что продукция L-лизина штаммом KCCM11016P::aceE1-PaceA, в котором промотор aceA встраивали выше терминатора транскрипции гена aceE, находящегося между стоп-кодоном и вышележащей областью терминатора транскрипции, являлась более эффективной. Это свидетельствует о том, что область между стоп-кодоном и вышележащей областью терминатора транскрипции можно использовать для более эффективного ингибирования экспрессия генов.
Пример 5: Получение вектора pDZ-aceE-Ppta для ингибирования экспрессии гена aceE
Ацетаткиназа (ackA, NCgl2656) и фосфотрансацетилаза (pta, NCgl2657), являющиеся ферментами, участвующими в процессе метаболизма ацетата, образуют оперон, и их экспрессия усиливается в присутствие ацетата. Таким образом, при использовании промоторов генов экспрессию гена можно специально индуцировать в присутствие ацетата.
В примере 5 для ингибирования экспрессии гена aceE в присутствие ацетата получали вектор для использования промотора оперона pta-ack, являвшегося вышележащей промоторной областью гена pta.
Для ингибирования экспрессии гена aceE конструировали вектор, который может содержать промотор гена pta ниже стоп-кодона гена aceE, находящегося между стоп-кодоном и вышележащей областью терминатора транскрипции, таким образом, что транскрипция с промотора гена pta может происходить в направлении, противоположном исходному направлению транскрипции гена aceE.
Для получения фрагмента гена aceE Corynebacterium glutamicum в качестве матрицы использовали хромосомную ДНК Corynebacterium glutamicum KCCM11016P для получения вектора pDZ-aceE1 тем же способом, что и в примере 2.
Для получения фрагмента промотора гена pta Corynebacterium glutamicum синтезировали праймеры, сконструированные содержащими участок распознавания фермента рестрикции SpeI на 5'-конце и на 3'-конце фрагмента (SEQ ID NO: 15 и SEQ ID NO: 16). Для осуществления ПЦР для амплификации промоторной области из приблизительно 340 пар оснований, представленной нуклеотидной последовательностью SEQ ID NO: 2, использовали хромосомную ДНК Corynebacterium glutamicum KCCM11016P в качестве матрицы и синтезированные праймеры. Продукт ПЦР-амплификации и фрагмент ДНК, полученный посредством обработки вектора pDZ-aceE1 ферментом рестрикции SpeI, клонировали с использованием набора In-fusion Cloning Kit (TAKARA, JP) для получения вектора pDZ-aceE1-Ppta.
SEQ ID NO: 15:
Ppta-P7F 5'-gtcccttgagactagtctttgctggggtcagatttg-3'
SEQ ID NO: 16:
Ppta-P7R 5'-ctgaggaataactagtacatcgcctttctaatttc-3'
Пример 6: Получение штаммов, в которых промотор гена pta встраивали ниже гена aceE, и сравнение продукции ими лизина
Corynebacterium glutamicum KCCM11016P трансформировали с использованием вектора pDZ-aceE1-Ppta, полученного в примере 5, тем же способом, что и в примере 3. Выбирали штамм, в котором промотор гена pta встраивали ниже стоп-кодона гена aceE на хромосоме таким образом, что транскрипция может происходить в направлении, противоположном исходному направлению транскрипции гена aceE, осуществляя ПЦР для получения L-лизин-продуцирующего штамма, названного KCCM11016P::aceE1-Ppta. Проверяли полученный штамм KCCM11016P::aceE1-Ppta, анализируя полученную нуклеотидную последовательность области-мишени посредством ПЦР с использованием SEQ ID NO: 3 и SEQ ID NO: 6 в качестве праймеров.
Полученный штамм культивировали тем же способом, что и в примере 4, и измеряли концентрацию L-лизина, выделенного из культурального раствора. Для случаев, когда не добавляли ацетат, концентрация L-лизина в культуральном растворе представлена в таблице 3.
Кроме того, культивировали штамм тем же способом, за исключением того, что для сравнения продукции L-лизина в продукционную среду добавляли 5 г/л ацетата. Концентрация L-лизина в культуральном растворе представлена в таблице 4.
Как показано в таблице 3, в отсутствие ацетата продукция L-лизина штаммом KCCM11016P::aceE1-Ppta не отличалась от продукции родительским штаммом KCCM11016P.
Однако, как показано в таблице 4, в присутствие ацетата продукция L-лизина штаммом KCCM11016P::aceE1-Ppta была на 2,4% выше, чем у родительского штамма KCCM11016P.
Кроме того, т.к. в присутствие ацетата продукция L-лизина штаммом KCCM11016P::aceE1-PaceA являлась более высокой, чем у штамма KCCM11016P::aceE1-Ppta, экспрессию гена-мишени можно более эффективно ингибировать с использованием промотора гена aceA, чем с использованием промотора гена pta.
Пример 7: Получение штаммов, в которых промотор гена aceA встраивали ниже гена aceE
Три L-лизин-продуцирующих штамма, являвшихся Corynebacterium glutamicum KFCC10750, KCCM10770P и CJ3P, соответствующим образом трансформировали с использованием вектора pDZ-aceE1-PaceA, полученного в примере 2, тем же способом, что и в примере 3. Выбирали штаммы, в которых промотор гена aceA встраивали ниже стоп-кодона гена aceE на хромосоме таким образом, что транскрипция может происходить в направлении, противоположном исходному направлению транскрипции гена aceE, осуществляя ПЦР. Полученными тремя L-лизин-продуцирующими штаммами являлись KFCC10750::aceE1-PaceA, KCCM10770P::aceE1-PaceA и CJ3P::aceE1-PaceA. Проверяли полученные штаммы, анализируя полученную нуклеотидную последовательность области-мишени посредством ПЦР с использованием SEQ ID NO: 3 и SEQ ID NO: 6 в качестве праймеров.
Полученные штаммы культивировали тем же способом, что и в примере 4, и измеряли концентрацию L-лизина, выделенного из культуральных растворов. Для случаев, когда не добавляли ацетат, концентрация L-лизина в культуральных растворах представлена в таблице 5.
Кроме того, штаммы культивировали тем же способом, за исключением того, что для сравнения продукции L-лизина в продукционную среду добавляли 5 г/л ацетата. Концентрация L-лизина в культуральных растворах представлена в таблице 6.
Как показано в таблице 5, в отсутствие ацетата продукция L-лизина тремя штаммами KFCC10750::aceE1-PaceA, KCCM10770P::aceE1-PaceA, CJ3P::aceE1-PaceA не отличалась от продукции родительским штаммом.
Однако, как показано в таблице 6, в присутствие ацетата продукция L-лизина штаммом KFCC10750::aceE1-PaceA была на 5% выше, чем у родительского штамма, продукция штаммом KCCM10770P::aceE1-PaceA была на 2,8% выше, чем у родительского штамма, и продукция штаммом CJ3P::aceE1-PaceA была на 15% выше, чем у родительского штамма.
Пример 8: Получение вектора для ингибирования экспрессии гена hom
Для повышения продукции L-лизина можно ослаблять путь биосинтеза L-треонина, использующий тот же субстрат, что и путь биосинтеза L-лизина. Примером способов ослабления пути биосинтеза L-треонина является снижение ферментативной активности гомосериндегидрогеназы (hom, NCgl1136), синтезирующей гомосерин из аспартата.
У Corynebacterium glutamicum ген hom образует оперон hom-thrB с геном thrB, и терминатор транскрипции гена hom находится ниже гена thrB. Сообщают, что промотор находится выше оперона hom-thrB, находящегося выше гена hom. Кроме того, сообщают о том, что второй промотор находится выше оперона гена thrB (Mateos et al., J Bacteriol, 176:7362-7371, 1994). Таким образом, для избирательного ингибирования экспрессии гена hom в присутствие ацетата промотор гена aceA встраивали ниже стоп-кодона гена hom таким образом, что транскрипция с этого промотора может происходить в направлении, противоположном исходному направлению транскрипции гена hom. Для сохранения экспрессии гена thrB добавляли последовательность второго промотора выше ORF гена thrB.
В примере 8 рекомбинантный вектор получали посредством инсерции промотора гена aceA между нижележащим стоп-кодоном гена hom и вышележащим геном thrB.
Для получения фрагмента гена hom Corynebacterium glutamicum в качестве матрицы использовали хромосомную ДНК Corynebacterium glutamicum KCCM11016P и синтезировали праймеры, сконструированные содержащими участок распознавания фермента рестрикции XbaI на 5'-конце фрагмента и участок распознавания фермента рестрикции SpeI на 3'-конце фрагмента (SEQ ID NO: 17 и 18). ПЦР осуществляли с использованием синтезированных праймеров для получения фрагмента ДНК, включающего 300 пар оснований между 1039-ым нуклеотидом от инициаторного кодона гена hom и 1338-ым нуклеотидом, являющимся стоп-кодоном. Даже при встраивании aceA между оперонами hom-thrB необходимо сохранять экспрессию гена thrB. Таким образом, когда получали фрагмент ДНК, включающий 300 пар оснований ниже стоп-кодона гена hom, синтезировали праймеры, сконструированные для дополнительного включения последовательности промотора thrB из 32 пар оснований с 5'-стороны (SEQ ID NO: 19 и 20). ПЦР осуществляли с использованием этих праймеров (SEQ ID NO: 19 и 20) для получения фрагмента ДНК из 334 пар оснований, содержащего участок распознавания фермента рестрикции SpeI на 5'-конце фрагмента и участок распознавания фермента рестрикции XbaI на 3'-конце фрагмента. ПЦР осуществляли в тех же условиях, что и в примере 2.
Два продукта ПЦР-амплификации и вектор pDZ для встраивания в хромосому, уже полученный посредством расщепления ферментом рестрикции XbaI, клонировали с использованием набора In-fusion Cloning Kit (TAKARA, JP) для получения вектора pDZ-hom.
SEQ ID NO: 17:
hom-h1F 5'-ccggggatcctctagaccaggtgagtccacctacg-3'
SEQ ID NO: 18:
hom-h1R 5'-gaggcggatcactagtttagtccctttcgaggcgg-3'
SEQ ID NO: 19: hom-h2F 5'-actagtgatccgcctcgaaagggac-3'
SEQ ID NO: 20:
hom-h2R 5'-gcaggtcgactctagagactgcggaatgttgttgtg-3'
Для получения фрагмента промотора ген aceA Corynebacterium glutamicum синтезировали праймеры, сконструированные содержащими участок распознавания фермента рестрикции SpeI на 5'-конце и на 3'-конце фрагмента (SEQ ID NO: 21 и 22). ПЦР осуществляли с использованием хромосомной ДНК Corynebacterium glutamicum KCCM11016P в качестве матрицы и синтезированных праймеров для амплификации промоторной области из приблизительно 500 пар оснований, представленных нуклеотидной последовательностью SEQ ID NO: 1. Продукт ПЦР-амплификации и фрагмент ДНК, полученный посредством обработки вектора pDZ-hom ферментом рестрикции SpeI, клонировали с использованием набора In-fusion Cloning Kit (TAKARA, JP) для получения вектора pDZ-hom-PaceA.
SEQ ID NO: 21:
PaceA-h3F 5'-gaggcggatcactagtagcactctgactacctctg-3'
SEQ ID NO: 22:
PaceA-h3R 5'-aagggactaaactagtttcctgtgcggtacgtggc-3'
Пример 9: Получение штаммов, в которых промотор гена aceA встраивали ниже гена hom, и сравнение продукции ими лизина
Corynebacterium glutamicum KCCM11016P, являющийся L-лизин-продуцирующим штаммом, трансформировали с использованием вектора pDZ-hom-PaceA, полученного в примере 8, тем же способом, что и в примере 3. Выбирали штамм, в котором промотор гена aceA встраивали ниже стоп-кодона гена hom на хромосоме таким образом, что транскрипция может происходить в направлении, противоположном исходному направлению транскрипции гена hom, осуществляя ПЦР для получения L-лизин-продуцирующего штамма, названного KCCM11016P::hom-PaceA. Проверяли полученный штамм KCCM11016P::hom-PaceA, анализируя нуклеотидную последовательность полученной области-мишени посредством ПЦР с использованием SEQ ID NO: 17 и SEQ ID NO: 20 в качестве праймеров.
Штамм Corynebacterium glutamicum KCCM11016P, используемый в качестве родительского штамма, и полученный штамм KCCM11016P::hom-PaceA культивировали тем же способом, что и в примере 4, и измеряли концентрацию L-лизина, выделенного из культуральных растворов. Для случаев, когда не добавляли ацетат, концентрация L-лизина в культуральных растворах штамма Corynebacterium glutamicum KCCM11016P и KCCM11016P::hom-PaceA представлена в таблице 7.
Кроме того, штаммы культивировали тем же способом, за исключением того, что для сравнения продукции L-лизина в продукционную среду добавляли 5 г/л ацетата. Концентрация L-лизина в культуральных растворах представлена в таблице 8.
Как показано в таблице 7, в отсутствие ацетата продукция L-лизина штаммом KCCM11016P::hom-PaceA не отличалась от продукции родительским штаммом KCCM11016P.
Однако, как показано в таблице 8, в присутствие ацетата продукция L-лизина штаммом KCCM11016P::hom-PaceA была на 2,6% выше, чем у родительского штамма KCCM11016P.
Пример 10: Получение вектора для ингибирования экспрессии гена murE
УДФ-N-ацетилмурамоилаланил-D-глутамат-2,6-диаминопимелатлигаза (murE, NCgl2083) использует мезо-2,6-диаминопимелат, являющийся предшественником для биосинтеза лизина, в соматическом синтезе. Ослабление активности гена murE может снижать поступление источников углерода в путь соматического синтеза и повышать поступление источников углерода в путь биосинтеза лизина для повышения продукции лизина.
У Corynebacterium glutamicum ген murE (NCgl2083) образует оперон с семью генами от NCgl2076 до NCgl2082. Транскрипция оперона начинается с NCgl2083 murE в направлении гена NCgl2076. Таким образом, терминатор транскрипции находится ниже гена NCgl2076. Таким образом, для избирательного ингибирования экспрессии гена murE в присутствие ацетата промотор гена aceA встраивали ниже стоп-кодона гена murE таким образом, что транскрипция может происходить в направлении, противоположном исходному направлению транскрипции гена murE. Для поддержания экспрессии других семи генов за исключением гена murE, локализованного в первой области оперона, дополнительно включали промотор оперона murE выше ORF гена NCgl2082.
В примере 10 рекомбинантный вектор получали посредством инсерции промотора гена aceA ниже стоп-кодона гена murE.
Для получения фрагмента гена murE Corynebacterium glutamicum в качестве матрицы использовали хромосомную ДНК Corynebacterium glutamicum KCCM11016P и синтезировали праймеры, сконструированные содержащими участок распознавания фермента рестрикции XbaI на 5'-конце фрагмента и участок распознавания фермента рестрикции XhoI на 3'-конце фрагмента (SEQ ID NO: 23 и 24). ПЦР осуществляли с использованием синтезированных праймеров для получения фрагмента ДНК, включающего 300 пар оснований между 1267-ым нуклеотидом от инициаторного кодона гена murE и 1566-ым нуклеотидом, являвшимся стоп-кодоном. Кроме того, синтезировали праймеры, сконструированные содержащими участок распознавания фермента рестрикции XhoI на 5'-конце фрагмента и участок распознавания фермента рестрикции XbaI на 3'-конце фрагмента (SEQ ID NO: 25 и 26). ПЦР осуществляли с использованием синтезированных праймеров для получения фрагмента ДНК, включающего 292 пар оснований от 10-го нуклеотида ниже стоп-кодона гена murE. ПЦР осуществляли в тех же условиях, что и в примере 2. Два продукта ПЦР-амплификации и вектор pDZ для встраивания в хромосому, уже полученный посредством расщепления ферментом рестрикции XbaI, клонировали с использованием набора In-fusion Cloning Kit (TAKARA, JP) для получения вектора pDZ-murE.
SEQ ID NO: 23:
mur-m1F 5'-ccggggatcctctagaaaccctcgttcagaggtgc-3'
SEQ ID NO: 24:
mur-m1R 5'-ttgtgatcatctcgagctatccttcttccgtagtaag-3'
SEQ ID NO: 25: mur-m2F 5'-agctcgagatgatcacaatgacccttgg -3'
SEQ ID NO: 26:
mur-m2R 5'-gcaggtcgactctagacatgagcataaatgtcagc-3'
Для получения фрагмента гена aceA Corynebacterium glutamicum синтезировали праймеры, сконструированные содержащими участок распознавания фермента рестрикции XhoI на 5'-конце фрагмента (SEQ ID NO: 27 и 28). Для осуществления ПЦР для амплификации промоторной области из приблизительно 500 пар оснований, представленной нуклеотидной последовательностью SEQ ID NO: 1, использовали хромосомную ДНК Corynebacterium glutamicum KCCM11016P в качестве матрицы и синтезированные праймеры. Кроме того, для получения промоторной области оперона murE Corynebacterium glutamicum в качестве матрицы использовали хромосомную ДНК Corynebacterium glutamicum KCCM11016P и синтезировали праймеры, сконструированные содержащими участок распознавания фермента рестрикции XhoI на 3'-конце фрагмента (SEQ ID NO: 29 и 30). ПЦР осуществляли с использованием синтезированных праймеров для получения фрагмента ДНК, включающего 300 пар оснований выше ORF гена murE.
Два продукта ПЦР-амплификации и фрагмент ДНК, полученный посредством обработки вектора pDZ-murE ферментом рестрикции XhoI, клонировали с использованием набора In-fusion Cloning Kit (TAKARA, JP) для получения вектора pDZ-murE-PaceA-PmurE.
SEQ ID NO: 27: mur-m3F 5'-tcatcagcagcactctgactacctctg-3'
SEQ ID NO: 28:
mur-m3R 5'-agaaggatagctcgagttcctgtgcggtacgtggc-3'
SEQ ID NO: 29: mur-m4F 5'-agagtgctgctgatgatcctcgatttg-3'
SEQ ID NO: 30:
mur-m4R 5'-ttgtgatcatctcgagggttttctctcctccacagg-3'
Пример 11: Получение штаммов, в которых промотор гена aceA встраивали ниже гена murE, и сравнение продукции ими лизина
Corynebacterium glutamicum KCCM11016P трансформировали с использованием вектора pDZ-murE-PaceA-PmurE, полученного в примере 10, тем же способом, что и в примере 3.
Выбирали штамм, в котором промотор гена aceA встраивали ниже стоп-кодона гена murE на хромосоме таким образом, что транскрипция может происходить в направлении, противоположном исходному направлению транскрипция гена murE, осуществляя ПЦР для получения L-лизин-продуцирующего штамма, названного KCCM11016P::murE-PaceA-PmurE. Проверяли полученный штамм KCCM11016P::murE-PaceA-PmurE, анализируя нуклеотидные последовательности полученной области-мишени посредством ПЦР с использованием SEQ ID NO: 23 и SEQ ID NO: 26 в качестве праймеров.
Штамм Corynebacterium glutamicum KCCM11016P, используемый в качестве родительского штамма, и полученный штамм KCCM11016P::murE-PaceA-PmurE культивировали тем же способом, что и в примере 4, и измеряли концентрацию L-лизина, выделенного из культуральных растворов. Для случаев, когда не добавляли ацетат, концентрация L-лизина в культуральных растворах штамма Corynebacterium glutamicum KCCM11016P и штамма KCCM11016P::murE-PaceA-PmurE представлена в таблице 9.
Кроме того, штаммы культивировали тем же способом, за исключением того, что для сравнения продукции L-лизина в продукционную среду добавляли 5 г/л ацетата. Концентрация L-лизина в культуральных растворах представлена в таблице 10.
Как показано в таблице 9, в отсутствие ацетата продукция L-лизина штаммом KCCM11016P::murE-PaceA-PmurE не отличалась от продукции родительским штаммом KCCM11016P.
Однако, как показано в таблице 10, в присутствие ацетата продукция L-лизина штаммом KCCM11016P::murE-PaceA-PmurE была на 2,8% выше, чем у родительского штамма KCCM11016P.
Пример 12: Получение вектора для ингибирования экспрессии гена dapA
Для повышения продукции L-треонина можно ослаблять путь биосинтеза L-лизина, использующий тот же субстрат, что и путь биосинтеза L-треонина. Примером способов ослабления пути биосинтеза L-лизина является снижение ферментативной активности дигидродипиколинатсинтазы (dapA, NCgl1896), участвующей в продукции лизина из аспартата.
У Corynebacterium glutamicum ген dapA образует оперон dapA-ORF4 с геном ORF4 (NCgl1895), и, таким образом, терминатор транскрипции гена dapA находится ниже гена ORF4. Кроме того, сообщают, что промотор находится выше оперона dapA-ORF4, находящегося выше гена dapA, и второй промотор находится выше гена ORF4 (Patek et al., Biotechnology letters, 19:1113-1117, 1997). Таким образом, для избирательного ингибирования экспрессии гена dapA в присутствие ацетата промотор гена aceA встраивали ниже стоп-кодона гена dapA таким образом, что транскрипция может происходить в направлении, противоположном исходному направлению транскрипции гена dapA. Для сохранения экспрессии гена ORF4 последовательность промоторной области из приблизительно 100 пар оснований в вышележащем гене ORF4 добавляли в ORF вышележащего гена ORF4.
В примере 12 рекомбинантный вектор получали посредством инсерции промотора гена aceA ниже стоп-кодона гена dapA.
Для получения фрагмента гена dapA Corynebacterium glutamicum в качестве матрицы использовали хромосомную ДНК Corynebacterium glutamicum KCCM11016P и синтезировали праймеры, сконструированные содержащими участок распознавания фермента рестрикции XbaI на 5'-конце фрагмента и участок распознавания фермента рестрикции SpeI на 3'-конце фрагмента (SEQ ID NO: 31 и 32). ПЦР осуществляли с использованием синтезированных праймеров для получения фрагмента ДНК, включающего 301 пар оснований между 606-ым нуклеотидом от инициаторного кодона гена dapA и 906-ым нуклеотидом, являвшимся стоп-кодоном. Кроме того, синтезировали праймеры, сконструированные содержащими участок распознавания фермента рестрикции SpeI на 5'-конце фрагмента и участок распознавания фермента рестрикции XbaI на 3'-конце фрагмента (SEQ ID NO: 33 и 34). С помощью ПЦР получали фрагмент ДНК, дополнительно включающий последовательность промоторной области из приблизительно 100 пар оснований между 809-ым нуклеотидом от инициаторного кодона гена dapA и 2-ым нуклеотидом в нижележащем стоп-кодоне для сохранения экспрессии гена ORF4 и 213 пар оснований ниже стоп-кодона гена dapA. ПЦР осуществляли в тех же условиях, что и в примере 2. Два продукта ПЦР-амплификации и вектор pDZ для встраивания в хромосому, уже полученный посредством расщепления ферментом рестрикции XbaI, клонировали с использованием набора In-fusion Cloning Kit (TAKARA, JP) для получения вектора pDZ-dapA.
SEQ ID NO: 31:
dapA-d1F 5'-ccggggatcctctagatgtttggcttgctttgggc-3'
SEQ ID NO: 32:
dapA-d1R 5'-gttgatgcactagtttatagaactccagcttt-3'
SEQ ID NO: 33:
dapA-d2F 5'-ttctataaactagtgcatcaacgtaggagatcc-3'
SEQ ID NO: 34:
dapA-d2R 5'-gcaggtcgactctagacgttctgggaaccctgag-3'
Для получения фрагмента промотора гена aceA Corynebacterium glutamicum синтезировали праймеры, сконструированные содержащими участок распознавания фермента рестрикции SpeI на 5'-конце и на 3'-конце фрагмента (SEQ ID NO: 35 и 36). ПЦР осуществляли с использованием хромосомной ДНК Corynebacterium glutamicum KCCM11016P в качестве матрицы и синтезированных праймеров для амплификации промоторной области из приблизительно 500 пар оснований, представленной нуклеотидной последовательностью SEQ ID NO: 1. Продукт ПЦР-амплификации и фрагмент ДНК, полученный посредством обработки вектора pDZ-dapA ферментом рестрикции SpeI, клонировали с использованием набора In-fusion Cloning Kit (TAKARA, JP) для получения вектора pDZ-dapA-PaceA.
SEQ ID NO: 35:
PaceA-d3F 5'-acgttgatgcactagtagcactctgactacctctg-3'
SEQ ID NO: 36:
PaceA-d3R 5'-agttctataaactagtttcctgtgcggtacgtggc-3'
Пример 13: Получение штаммов, в которых промотор гена aceA встраивали ниже гена dapA, и сравнение продукции ими треонина
Для проверки эффекта ингибирования экспрессии гена dapA в L-треонин-продуцирующем штамме штамм Corynebacterium glutamicum KCCM11222P (патент Кореи №: 2013-0061570), являющийся L-треонин-продуцирующим штаммом, трансформировали с использованием вектора pDZ-dapA-PaceA, полученного в примере 12, тем же способом, что и в примере 3. Осуществляя ПЦР, выбирали штамм, в котором промотор гена aceA встраивали ниже стоп-кодона гена dapA на хромосоме таким образом, что транскрипция может происходить в направлении, противоположном исходному направлению транскрипции гена dapA, названный KCCM11222P::dapA-PaceA. Проверяли полученный штамм KCCM11222P::dapA-PaceA, анализируя нуклеотидную последовательность полученной области-мишени посредством ПЦР с использованием SEQ ID NO: 31 и SEQ ID NO: 34 в качестве праймеров.
Штамм Corynebacterium glutamicum KCCM11222P, используемый в качестве родительского штамма, и полученный штамм KCCM11222P::dapA-PaceA культивировали описываемым ниже способом.
Каждый из штаммов соответствующим образом инокулировали в 25 мл среды для посева в колбы с дефлекторами емкостью 250 мл с последующим культивированием со встряхиванием при 200 об./мин при 30°C в течение 20 часов. Затем 1 мл раствора посевной культуры добавляли в колбу с дефлекторами емкостью 250 мл, содержащую 24 мл продукционной среды, с последующим культивированием со встряхиванием при 200 об./мин при 30°C в течение 48 часов. Соответствующие композиции среды для посева и продукционной среды описаны ниже.
<Среда для посева (pH 7,0)>
Глюкоза 20 г, пептон 10 г, дрожжевой экстракт 5 г, мочевина 1,5 г, KH2PO4 4 г, K2HPO4 8 г, MgSO4⋅7(H2O) 0,5 г, биотин 100 мкг, тиамин HCl 1000 мкг, пантотенат кальция 2000 мкг, никотинамид 2000 мкг (на 1 л дистиллированной воды).
<Продукционная среда (pH 7,0)>
Глюкоза 100 г, (NH4)2SO4 20 г, соевый белок 2,5 г, твердый кукурузный экстракт 5 г, мочевина 3 г, KH2PO4 1 г, MgSO4⋅7(H2O) 0,5 г, биотин 100 мкг, тиамин HCl 1000 мкг, пантотенат кальция 2000 мкг, никотинамид 3000 мкг, CaCO3 30 г (на 1 л дистиллированной воды).
После культивирования концентрацию L-треонина в культуральном растворе измеряли с помощью ВЭЖХ. Для случаев, когда не добавляли ацетат, концентрация L-треонина в культуральных растворах Corynebacterium glutamicum KCCM11222P и KCCM11222P::dapA-PaceA представлена в таблице 11.
Кроме того, штаммы культивировали тем же способом, за исключением того, что для сравнения продукции L-треонина в продукционную среду добавляли 5 г/л ацетата. Концентрация L-треонина в культуральных растворах представлена в таблице 12.
Как показано в таблице 11, в отсутствие ацетата продукция L-треонина штаммом KCCM11222P::dapA-PaceA не отличалась от продукции родительским штаммом KCCM11222P.
Однако, как показано в таблице 12, в присутствие ацетата продукция L-треонина штаммом KCCM11222P::dapA-PaceA была на 50% выше, чем у родительского штамма KCCM11222P.
[Регистрационный №]
Депозитарное учреждение: Korean Culture Center of Microorganisms (International Depositary Authority)
Регистрационный №: KCCM11432P
Дата регистрации: 12 июня 2013 года
Как описано выше, согласно одному или нескольким из указанных выше вариантов осуществления настоящего изобретения можно использовать ацетат-индуцибельный промотор для эффективного получения L-аминокислот, т.к. целевую L-аминокислоту можно получать с высоким выходом, ослабляя экспрессию гена-мишени посредством добавления ацетата в соответствующий момент времени.
Следует понимать, что примеры вариантов осуществления, представленные в настоящем описании, необходимо истолковывать исключительно в описательном смысле, а не в целях ограничения. Описание признаков или аспектов в каждом варианте осуществления, как правило, следует считать пригодным для других аналогичных признаков и аспектов в других вариантах осуществления.
Хотя один или несколько вариантов осуществления настоящего изобретения описаны со ссылкой на фигуры, специалистам в этой области будет понятно, что можно осуществлять различные изменения формы и подробностей без отклонения от сущности и объема настоящего изобретения, определенных в прилагаемой формуле изобретения.
Реферат
Изобретение относится к области биотехнологии. Представлен способ получения L-лизина с использованием рекомбинантного коринеформного микроорганизма, способного продуцировать L-лизин, где рекомбинантный коринеформный микроорганизм трансформируют посредством инсерции ацетат-индуцибельного промотора ниже стоп-кодона гена-мишени на хромосоме для ослабления экспрессии гена-мишени и усиления способности рекомбинантного коринеформного микроорганизма продуцировать L-лизина, где ген-мишень представляет собой ген, расположенный в узловой точке пути биосинтеза L-лизина. Изобретение позволяет увеличить продукцию L-лизина по сравнению с родительским штаммом микроорганизма. 3 з.п. ф-лы, 2 ил., 12 табл., 13 пр.
Комментарии