Новый ген лизиндекарбоксилазы и способ получения l-лизина - RU2188235C2
Код документа: RU2188235C2
Чертежи









Описание
Область техники, к которой относится
изобретение
Изобретение относится к новому гену лизиндекарбоксилазы Escherichia coli, существенному для распада L-лизина, микроорганизму, относящемуся к роду Escherichia, с ограниченной
экспрессией указанного гена и/или другого гена лизиндекарбоксилазы, известного как cadA-ген, и способу получения L-лизина в результате использования указанного микроорганизма. В последнее время
потребность в L-лизине, как пищевой добавки, очень выросла.
Предшествующий уровень техники
Лизиндекарбоксилаза, которая катализирует реакцию образования кадаверина, в
результате декарбоксилирования L-лизина, известна как фермент Escherichia coli, расщепляющий L-лизин. Ранее уже сообщали о нуклеотидной последовательности ее гена, названного cadA, и аминокислотной
последовательности, кодируемой этим геном (Mend, S. and Bennet, G.N., J.Bacteriol., 174, 2659 (1992)). Имеются два сообщения по лизиндекарбоксилазе, кодируемой иным геном, чем cadA Escherichia coli, в
которых описывают ослабление активности, обнаруженное у мутантного штамма Escherichia coli (Goldemberg, S.H., J, Bacteriol. , 141, 1428 (1980); Wertheimer, S.J. and Leifer, Z., Biochem. Biophys. Res.
Соmmun. , 114, -882 (1983)). Однако относительно этой активности Goldemberg S.H. сообщал, что ферментативная активность снижалась приблизительно на 30% после тепловой обработки при 60o в
течение 4 минут, тогда как Wertheimer S.J. с соавт. сообщили о том, что такого явления не наблюдали. В соответствии с этим, наличие второй лизиндекарбоксилазы остается неопределенным.
С другой стороны, L-лизин получают известными способами, используя Escherichia coli, в том числе способ, включающий в себя культивирование мутантного штамма, резистентного к лизиновому аналогу, или рекомбинантного штамма, содержащего вектор с включенной дезоксирибонуклеиновой кислотой, которая переносит генетическую информацию, существенную для биосинтеза L-лизина (Японский открытый патент 56-18596). Однако во всех случаях не сообщается об образовании L-лизина с использованием микроорганизма, относящегося к общеизвестному роду Escherichia, с ограниченной экспрессией гена лизиндекарбоксилазы.
Раскрытие изобретения
Цель настоящего изобретения состоит в получении нового лизиндекарбоксилазного гена Escherichia coli, создания продуцирующего L-лизин
микроорганизма, относящегося к роду Escherichia, с ограниченной экспрессией указанного гена и/или гена cadA, и в разработке способа получения L-лизина путем культивирования микроорганизма,
относящегося к роду Escherichia.
Когда авторы настоящего изобретения создали штамм Escherichia coli, в котором разрушили ген cadA, известный как ген лизиндекарбоксилазы, было обнаружено, что кадаверин являющийся продуктом расщепления L-лизина лизиндекарбоксилазой, все еще продолжал образовываться в данном бактериальном штамме. Поэтому авторы настоящего изобретения сделали допущение, что у Escherichia coli должен был бы присутствовать новый ген лизиндекарбоксилазы и он, возможно, более всего влияет на ферментативное образование L-лизина, при использовании микроорганизма, относящегося к роду Escherichia. В результате попыток проклонировать указанный ген авторы настоящего изобретения добились получения нового гена лизиндекарбоксилазы, отличающегося от гена cadA. Обнаружили также, что его активность расщепления L-лизина очень снизилась или исчезла, а производительность L-лизина у микроорганизма Escherichia coli, продуцирующего L-лизин, существенно улучшилась в результате ограничения экспрессии данного гена и ограничения экспрессии известного гена cadA. Так осуществили настоящее изобретение.
То есть в настоящем изобретении описан новый ген, который кодирует лизиндекарбоксилазу, происходящий от Escherichia coli. Этот ген обозначили как "ldс"-ген.
Следующая цель настоящего изобретения - создать микроорганизм, относящийся к общеизвестному роду Escherichia, продуцирующий L-лизин при снижении или исчезновении в клетках активности лизиндекарбоксилазы.
Еще одна цель настоящего изобретения - разработать способ получения L-лизина, включающего в себя определенные стадии культивирования в жидкой среде указанного микроорганизма, относящегося к общеизвестному роду Escherichia coli, для получения и накопления L-лизина в культуральной среде и его сбора.
Этот микроорганизм, относящийся к роду Escherichia, включает в себя микроорганизм, в клетках которого активность лизиндекарбоксилазы снижена или исчезла в результате ограничения экспрессии указанного гена Idc и/или гена cadA.
Более подробно настоящее изобретение описывается ниже.
<1> Получение
фрагмента ДНК, содержащего новый ген лизиндекарбоксилазы
Фрагмент ДНК, содержащий новый ген (ldc) лизиндекарбоксилазы настоящего изобретения, можно получить следующим образом из
соответствующего штамма Escherichia coli, например штамма К-12, или штамма, произведенного из него.
Вначале ген cadA, который представляет собой известный ген лизиндекарбоксилазы, получали из хромосомной ДНК штамма W3110, происходящего из Escherichia coli K-12, методом полимеразной цепной реакции (везде далее употребляемой как "метод ПЦР"). Нуклеотидная последовательность гена cadA и аминокислотная последовательность, им кодируемая, показаны соответственно в SEQ ID NOS:5 и 6. Фрагменты ДНК, имеющие последовательности, сходные с геном cadA, клонировали из библиотеки хромосомной ДНК Escherichia coli W3110, согласно методу, использующему плазмидный вектор или фаговый вектор, чтобы подтвердить действительно или нет указанный новый ген лизиндекарбоксилазы содержится во фрагментах ДНК. Подтвердить факт, что ген действительно присутствует, можно методом Саузернгибридизации, используя зонд, полученный методом ПЦР.
Нуклеотидную последовательность указанного гена, содержащегося в полученном таким образом конкретном фрагменте ДНК, определяли следующим способом. Вначале этот фрагмент ДНК лигировали с плазмидным вектором, автономно реплицирующимся в клетках Escherlchia coli, чтобы получить рекомбинантную ДНК, которую интродуцировали в комплектные клетки Escherichia сoli. Полученный трансформант культивировали в жидкой среде, а рекомбинантную ДНК извлекали из пролиферированных клеток. Полную нуклеотидную последовательность фрагмента ДНК, содержащуюся в этой извлеченной рекомбинантной ДНК, определяли в соответствии с дидезокси-методом (Sanger, F. et al., Рrос. Natl. Acad. Sci. , 74, 5463 (1977)). Чтобы определить существующие положения промотора, оператора, SD-последовательности, кодона инициации, кодона терминации, открытой рамки считывания и так далее, определяли структуру ДНК.
Полученный новый ген лизиндекарбоксилазы настоящего изобретения имеет последовательность от 1005-1007-го ATG до 3141-3143-го GGA всей нуклеотидной последовательности указанного фрагмента ДНК, представленного в SEQ ID NO:3 в Списке последовательностей. Данный ген кодирует лизиндекарбоксилазу, обладающую аминокислотной последовательностью, представленной в SEQ ID NO:4 в Списке последовательностей. Установлено, что гомология между указанной новой лизиндекарбоксилазой и лизиндекарбоксилазой, кодируемой геном cadA, составляет 69,4%.
Полученный ген настоящего изобретения может быть геном, который кодирует лизиндекарбоксилазу, обладающей аминокислотной последовательностью, представленной в SEQ ID NO:4 в Списке последовательностей, нуклеотидной последовательностью, которая не ограничивается вышеописанной нуклеотидной последовательностью. Эта лизиндекарбоксилаза, кодируемая полученным геном настоящего изобретения, может иметь замену, делецию или инсерцию по одному или множеству аминокислотных остатков в вышеописанной аминокислотной последовательности, без существенного ухудшения ее лизиндекарбоксилазной активности. Гены, которые кодируют лизиндекарбоксилазу, имеющую такую делецию, инсерцию или замену можно получить из вариантов спонтанных мутантных штаммов или искусственных мутантных штаммов Escherichia coli, или из микроорганизмов, относящихся к общеизвестному роду Escherichia, но других, чем Escherichia coli. Такие мутантные гены, которые кодируют лнзиндекарбоксилазу, имеющую делецию, инсерцию или замену, можно получить в результате проведения мутационной обработки ин витро или сайт-направленного мутагенеза соответствующего гена, который кодирует лизиндекарбоксилазу, имеющую аминокислотную последовательность, представленную в SEQ ID NO:4. Эти мутационные обработки можно осуществить способами, хорошо известными специалистам в этой области, которые описаны ниже.
Однако соответствующий ген, который кодирует лизиндекарбоксилазу, имеющую замену, делецию или инсерцию по одному или множеству аминокислотных остатков, на что указывается здесь, включает в себя ген, который происходит из указанного "ldc-гена" и по существу может рассматриваться как тот же ldc-ген. Данное толкование не распространяется на гены, имеющие разное происхождение. Точно сослаться на определенный ряд "множественности" не представляется возможным. Однако специалистам в данной области понятно, что, например, указанный ген cadA, который кодирует белок, отличающийся от белка с аминокислотной последовательностью, представленный в SEQ ID NО:3, не менее чем по 200 аминокислотным остаткам, отличается от указанного гена настоящего изобретения, а гены, которые кодируют белки, обладающие эквивалентной активностью лизиндекарбоксилазы, включены в настоящее изобретение, даже если они отличаются от белка с аминокислотной последовательностью, представленной в SEQ ID NО:3, по двум или трем аминокислотным остаткам.
<2> Создание микроорганизма, относящегося к общеизвестному роду Echenchia, с
ограниченной экспрессией гена лизиндекарбоксцлазы
Соответствующий микроорганизм, относящийся к роду Escherichia настоящего изобретения представляет собой микроорганизм, относящийся к
общеизвестному роду Escherichia, в клетках которого активность лизиндекарбоксилазы снижена или исчезла. Этот микроорганизм, относящийся к общеизвестному роду Escherichia включает в себя Escherichia
сoli. Соответствующая активность лизиндекарбоксилазы в клетках снижается или исчезает, например, при ограничении экспрессии любого одного или обоих генов лизиндекарбоксилазы - нового гена (ldc) и
известного вышеуказанного гена cadA. Иначе в клетках эта лизиндекарбоксилазная активность может снижаться или исчезать в результате снижения или исчезновения удельных активностей ферментов
лизиндекарбоксилазы, кодируемых этими генами, вследствие модификации структуры этих ферментов.
Соответствующие способы по ограничению экспрессии указанного гена (ldc) и известного гена cadA включают в себя, например, способ по ограничению экспрессии этих генов на транскрипционном уровне путем замены, делеции, вставки, присоединения или инверсии одного или множества нуклеотидов в промоторных последовательностях этих генов, и снижения активностей промоторов (М. Rosenberd и D. Court, Ann. Rev. Genetics. 13 (1979) р. 319 и P. Youderian, и М. Susskind, Cell 30 (1982) р. 843-853). Экспрессию этих генов можно ограничить иначе, на трансляционном уровне, путем замены, делеции, вставки, присоединения или инверсии одного или множества нуклеотидов на участке между SD-последовательностью и инициирующим кодом (J.J. Dunn, E. Buzash-Pollert и F. W. Studier, Proc. nat, Acad. Sci. U.S.A., 75 (1978) р. 2743). Кроме того, для того чтобы снизить или убрать соответствующую удельную активность соответствующего фермента лизиндекарбоксилазы, существует способ, в котором соответствующую кодирующую область соответствующего гена лизиндекарбоксилазы модифицируют или разрушают путем замены, делеции, вставки, присоединения или инверсии одного или множества нуклеотидов в нуклеотидной последовательности кодирующей области.
Соответствующий ген, в котором происходит замена, делеция, вставка, присоединение или инверсия нуклеотида, помимо нового гена Idc и известного гена cadA, может относиться к генам Idc или генам cadA, обладающим заменой, делецией или вставкой одного или множества аминокислотных остатков, которые существенно не ухудшают активность кодируемой лизиндекарбоксилазы.
Соответствующий способ, вызывающий нуклеотидную замену, делецию, вставку или инверсию в соответствующем гене, включает в себя, в частности, способ сайт-направленного мутагенеза (Кramer, W. и Frits, H.J., Methods in Ezymology, 154, 350 (1987)), и способ обработки химическим агентом, таким как гипосульфит натрия и гидроксиламин (Shortle, D. и Nathans, D., Proc. Nati, Acad, Sci, U.S.A., 75, 270 (1978)).
Указанный способ сайт-направленного мутагенеза представляет собой способ с использованием синтетического олигонуклеотида, способ, позволяющий произвольно осуществлять замену, делецию, вставку, присоединение или инверсию относительно любой или конкретной нуклеотидной пары. Для того чтобы применить данный способ, во-первых, готовят одноцепочечную ДНК путем денатурации плазмиды, обладающей клонированным целевым геном с определенной нуклеотидной последовательностью. Затем синтезируют синтетический олигонуклеотид, комплементарный участку, в котором предполагается создать мутацию. Однако в данном методе соответствующий синтетический олигопуклеотид не представляет собой полностью комплементарную последовательность, но предназначен иметь произвольную нуклеотидную замену, делецию, вставку, добавку или инверсию. После этого соответствующую одноцепочечную ДНК отжигают с указанным синтетическим олигонуклеотидом, несущим произвольную нуклеотидную замену, делецию, вставку, добавку или инверсию. Полную двухцепочечную плазмиду синтезировали путем использования лигазы Т4 и фрагмента Кленова ДНК-полимеразы 1, которую интродуцировали в компетентные клетки Escherichia coli. Отдельные трансформанты, полученные таким образом, имели плазмиду, содержащую ген, в котором фиксировали произвольную нуклеотидную замену, делецию, вставку, добавку или инверсию. В качестве сходного метода интродуцирования мутации в ген, с целью его модификации или деструкции, можно упомянуть метод рекомбинатной ПЦР (Техника ПЦР, Stockton press (1989)).
Соответствующий способ с использованием химического агента представляет собой способ, в котором мутацию, представленную нуклеотидной заменой, делецией, вставкой, добавкой или инверсией, интродуцируют случайным образом во фрагмент ДНК путем обработки указанного фрагмента ДНК, содержащего целевой ген, непосредственно гипосульфитом натрия, гидроксиламином или им подобным.
Экспрессию указанного гена ldc и/или гена cadA в клетках можно ограничить в результате замещения нормального гена в хромосоме микроорганизма, относящегося к общеизвестному роду Escherichia, модифицированным или разрушенным геном, полученным в результате вышеописанной интродукции мутации. Указанный способ по замещению соответствующего гена включает в себя способы, использующие гомологичную рекомбинацию (Эксперименты в Молекулярной Генетике, Cold Spring Harbor Laboratory press (1972); Matsuyama, S. и Mizusbima, S. , J. Bacteriol., 162, 1196 (1985)). Такая гомологичная рекомбинация базируется на способности, обычно присущей микроорганизмам, относящимся к роду Escherichia. Когда плазмиду или ей подобную, обладающую гомологией с последовательностью в хромосоме, интродуцируют в клетки, рекомбинация происходит с определенной частотой на участке соответствующей последовательности, обладающей определенной гомологией, и вся интродуцированная плазмида инкорпорируется в данную хромосому. Если после этого рекомбинация происходит и далее на указанном участке последовательности, обладающей гомологией в хромосоме, указанная плазмида снова выпадает из хромосомы. Однако в течение этого процесса соответствующий ген с интродуцированной мутацией иногда фиксируется, предпочтительно в указанной хромосоме, в зависимости от позиции, на которой происходит рекомбинация, а первоначальный ген выпадает из хромосомы вместе с используемой плазмидой. Селекция таких бактериальных штаммов дает возможность получить бактериальный штамм, в котором нормальный ген в хромосоме замещен модифицированным или разрушенным геном, полученным в результате интродуцирования мутации, представленной нуклеотидной заменой, делецией, вставкой, добавкой или инверсией.
Соответствующий микроорганизм, относящийся к роду Escherichia, который подвергли замене гена, представляет собой микроорганизм, продуцирующий L-лизин. Указанный микроорганизм, относящийся к роду Escherichia, продуцирующий L-лизин, например бактериальный штамм Escherichia coli, можно получить в результате применения мутационной обработки штамма, не продуцирующего L-лизин, чтобы придать ему устойчивость к аналогу лизина, такому как S-(-2-аминоэтил)-L-цистеину (в дальнейшем именуемый как "АЕС"). Способы мутационной обработки включают в себя способы, в которых клетки Escherichia coli подвергают обработке химическим агентом, таким как N-метил-N'-нитро-N-нитрозогуанидин и азотистая кислота, или обработке облучением ультрафиолетовым светом, радиацией и т.п. Такой бактериальный штамм, в частности, включает в себя Escherichia coli AJ13069 (FERM Р-14690). Этот бактериальный штамм получили в результате придания АЕС-устойчивости штамму W3110, происходящему от Escherichia coli K-12. Escherichia coli AJ13069 депонировали в Национальном Институте Биологических и Прикладных наук Человека Агенства Промышленных наук и Технологий (почтовый код: 305, 1-3, Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Япония) под каталожным номером FERM Р-14690, 6 декабря 1994 г., переместили на международное храпение, на основании Будапештского Договора от 29 сентября 1995 г., и присвоили каталожный номер FERM BP-5252.
Бактериальный штамм Escheric. hia coli, продуцирующий L-лизин, можно также получить в результате интродуцирования и усиления ДНК, которая несет генетическую информацию, необходимую для биосинтеза L-лизина посредством генной рекомбинантной техники. Указанный вводимый ген представляет собой гены, которые кодируют ферменты биосинтетического пути L-лизина, такие как аспартаткиназа, дигидродипиколинатсинтетаза, дигидродипиколинатредуктаза, сукцинилдиаминопимелаттрансаминаза и сукцинилдиаминопимелатдезацилаза. Для случая гена фермента, который подвергается ингибированию по типу обратной связи L-лизином, такого как аспартаткиназа и дигидродипиколинатсинтетаза, желательно использовать мутантный тип гена, кодирующий фермент, у которого снижена чувствительность к такому ингибированию. Чтобы интродуцировать или усилить такой ген, пригоден способ, в котором этот ген лигирует с вектором, автономно реплицирующимся в клетках Escherichia coli, для получения рекомбинантной ДНК, с помощью которой трансформируют Escherichia coli. Альтернативно указанный ген можно также инкорпорировать в хромосому хозяина в соответствии со способом, использующим трансдукцию, транспозон (Berd, D.E. и Berd, C.M., Bio/Technol., 417 (1983), Mu-фага (открытый японский патент 2-109985) или гомологичную рекомбинацию (Эксперименты в Молекулярной Генетике, Cold Spring Harbor Lab. (1972)).
Другие способы получения микроорганизма, относящиеся к роду Escherichia, с уничтоженной функцией указанного гена, включает в себя способ, вызывающий генетическую мутацию в результате обработки клеток соответствующего микроорганизма, относящегося к роду Eschericha, имеющих указанный ген, химическим агентом, таким как N-метил-N'-нитро-N-нитрозогуанидин и азотистая кислота, или обработки облучением ультрафиолетовым светом, радиацией и т.п.
В нижеописанном Примере штамм Escherjchia coli, с уничтоженной функцией гена лизиндекарбоксилазы, создали путем делетирования части его кодирующей области и вставки вместо него гена лекарственной устойчивости, чтобы получить ген лизиндекарбоксилазы, который использовали для замены гена лизиндекарбоксилазы в хромосоме Escherichia. coli, в соответствии с вышеописанным способом применения гомологичной рекомбинации.
Представляется возможным в одном бактериальном штамме ограничить экспрессию любого одного гена - нового гена лизиндекарбоксилазы настоящего изобретения и cadA-гена, или ограничить экспрессию обоих. Экспрессию гена лизиндекарбоксилазы можно ограничить у микроорганизма, относящегося к роду Escherichia, продуцирующего L-лизин, или способность продуцировать L-лизин можно придать микроорганизму, относящемуся к роду Escherichia, с ограниченной экспрессией гена лизиндекарбоксилазы в соответствии с вышеописанным способом.
<3> Получение L-лизина при использовании микроорганизма, относящегося к роду Escherichia, с ограниченной экспрессией гена лизиндекарбоксилазы.
При культивировании микроорганизма, относящегося к роду Escherichia, с ограниченной экспрессией гена лизиндекарбоксилазы, полученного, как описано выше, в культуральной жидкости образуется и накапливается значительное количество L-лизина. Количество накопления L-лизина увеличивается только при ограничении экспрессии известного гена cadA. Однако более эффективное увеличение количества L-лизина связано с ограничением экспрессии нового гена лизиндекарбоксилазы настоящего изобретения. Наиболее предпочтительное образование лизина получают при использовании бактериального штамма, в котором ограничена экспрессия обоих генов - cadA-гена и нового гена лизиндекарбоксилазы настоящего изобретения.
Соответствующая среда, используемая для получения L-лизина, представляет собой обычную среду, содержащую источник углерода, источник азота, неорганические ионы и, необязательно, другие источники органических питательных веществ в следовых количествах. В качестве источника углерода можно использовать сахара, такие как глюкозу, лактозу, галактозу, фруктозу и гидролизованный крахмал; спирты, такие как глицерин и сорбитол; и органические кислоты, такие как фумаровая кислота и янтарная кислота. В качестве источника азота можно использовать аммониевые соли, такие как сульфат аммония, хлорид аммония и фосфат аммония; органические источники азота, такие как соевый гидролизат; газообразный аммиак и водоаммиачный раствор. В качестве источника неорганических ионов добавляли в небольших количествах фосфат калия, сульфат магния, ионы железа, ионы марганца и др. Помимо вышеуказанных веществ желательно присутствие витамина В1, дрожжевого экстракта или подобные им, в качестве органических источников в соответствующих следовых количествах, питательные вещества.
Культивирование предпочтительно осуществляли в аэробных условиях в течение 16-72 часов. Температуру культивирования поддерживали от 30oС до 45oС, а рН поддерживали во время культивирования на уровне 5-7. Для доведения рН использовали неорганические или органические вещества, кислоты и основания или газообразный аммиак и им подобные вещества.
После завершения культивирования выделение L-лизина из ферментной жидкости можно соответственно осуществить путем комбинирования методов ионного обмена на обычных смолах, способом осаждения и другими известными способами.
Краткое описание чертежей.
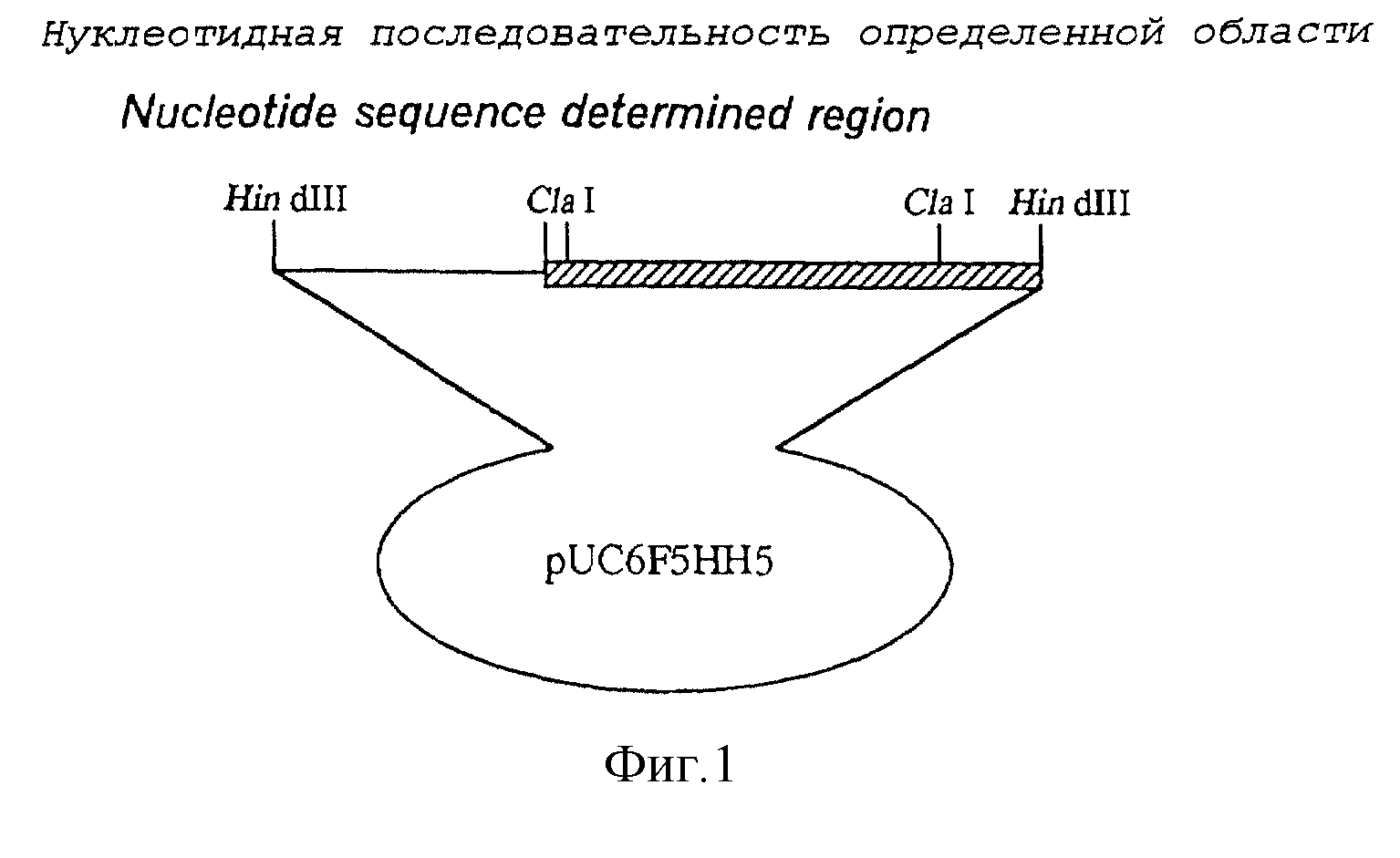
Фиг.1 показывает структуру плазмиды pUC6F5HH5, содержащей новый ген лизиндекарбоксилазы настоящего изобретения.
Фиг.2 показывает структуру температурочувствительной плазмиды pTS6F5HH5, содержащей новый ген лизиндекарбоксилазы настоящего изобретения и конструкцию плазмиды pTS6G5HH5, в которой часть указанного гена заменили фрагментом, содержащем ген, устойчивый к хлорамфениколу.
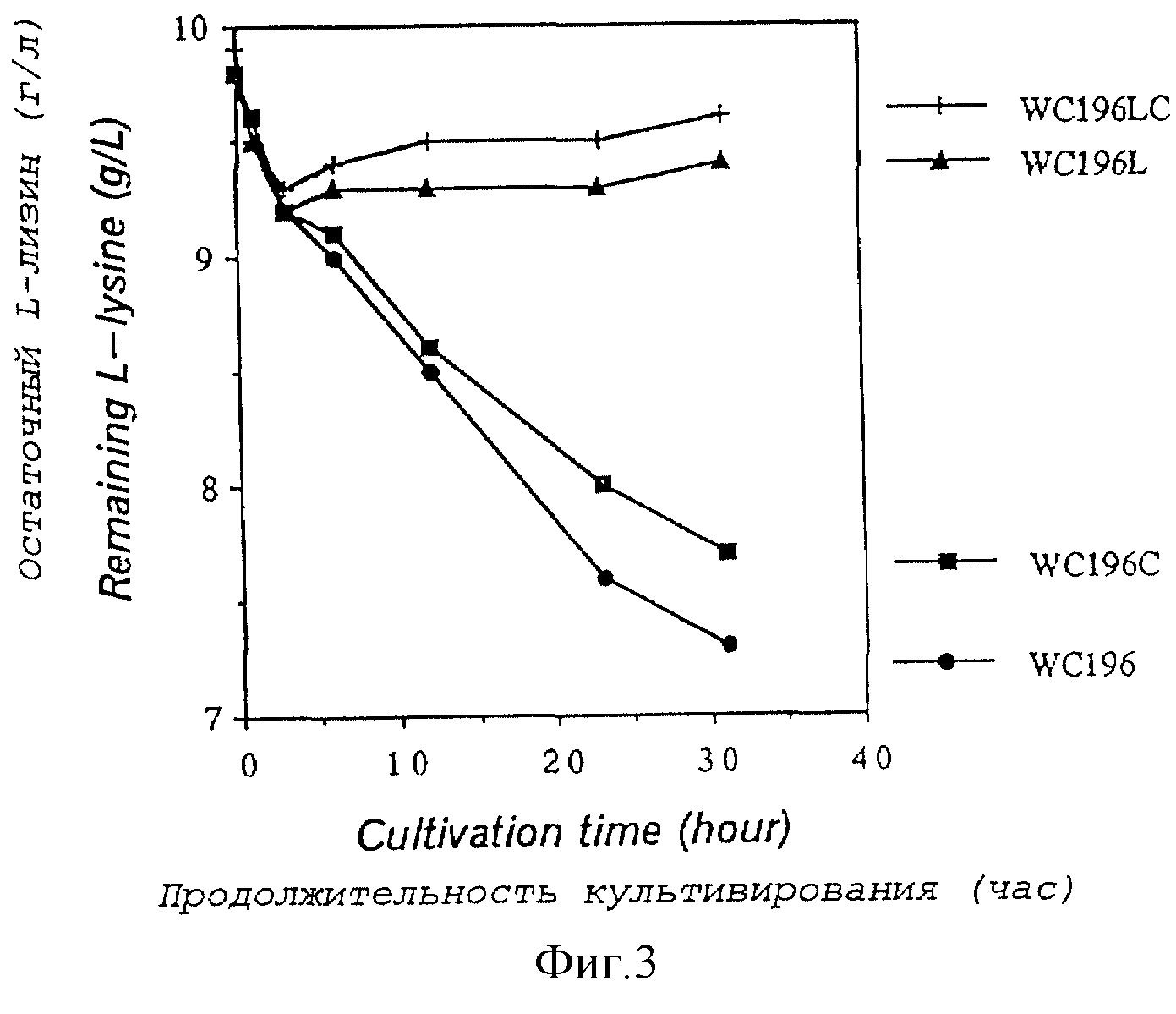
Фиг. 3 показывает сравнительные активности расщепления L-лизина в штамме WC196, несущего нормальный ген лизиндекарбоксилазы, и в штаммах WC196C, WC196L, WS196LC с разрушенными генами лизиндекарбоксилазы.
Предпочтительный способ осуществления настоящего изобретения
Более подробно настоящее изобретение объясняется ниже со ссылками на примеры.
Пример 1
(1)
Клонирование нового гена лизиндекарбоксилазы
Хромосомную ДНК экстрагировали стандартным способом из клеток Escherichia coli К-12 штамма W3110, полученного из Национального Института генетики
(Yata 1111, Misliima-shi, Shi-zuoka-ken, Япония). Кроме того, два синтетических праймера ДНК, которые показаны в SEQ ID NOS: 1 и 2, синтезировали стандартным способом на основании нуклеотидной
последовательности известного гена cadA (см. SEQ ID NO: 5), описанный у Meng, S. и Bennett, G.N., J, Bacteriol. , 174, 2659 (1992). Они имели последовательности, гомологичные, соответственно,
вышележащему 5'-концевому участку и 3'-концевому участку гена cadA. Эту хромосомную ДНК и эти ДНК-праймеры использовали для проведения ПЦР по методу Erlich с соавт. (Техника. ПЦР, Stockholm press
(1989)). Таким образом, был получен 2, 1 Kbp фрагмент ДНК, содержащий почти все части гена cadA. Данный фрагмент метили с помощью набора Kamdom Orimer Labeling Kit (производит Takara Shuzo) и [α
-32 PJdCTP (производит Amersham, Япония), чтобы получить гибридизационный зонд.
Далее гибридизацию осуществляли стандартным способом (Молекулярное клонирование (2-у издание), Cold Spring Harbor Laboratory press (1989)), используя приготовленный зонд и Escherichia coli мембрану для генного картирования (Geme mappimg Membrane, производит Takara Shuzo). Библиотеку Кохара и соавт. (библиотека фага лямбда хромосомной ДНК Escherichia coli: смотрите Kohara, Y. et al. Cell, 50, 495-508 (1987)) адсорбировали Escherichia coli/мембраной для генного картирования. Клоны фага лямбда, обладающие последовательностями, сходными с последовательностью гена cadA, скринировали после проведения гибридизации путем ослабления соответствующего условия отмывки данного зонда (2 х SSC, 55oС, 30 минут). В итоге, нам удалось обнаружить слабые сигналы из трех клонов Е2В8, 6F5H и 10F9, помимо сильных сигналов из клонов, содержащих соответствующую область гена cadA (21H11, 5G7). Инсерционные последовательности из трех клонов фага Е2В8, 6F5H и 10F9 продолжались в хромосоме Escherichia соli, хотя и перекрывали друг друга. В соответствии с этим, ДНК фага лямбда 6F5H, относящуюся к библиотеке Кохара и соавт. , (Kohara, Y. et al. Cell, 50, 495-508 (1987)), отделили стандартным способом, путем обработки различными ферментами рестрикации, чтобы осуществить Саузернблот-гибридизацию, используя вышеуказанный зонд, способом, близким описанному выше. В результате обнаружили, что последовательность, сходная с последовательностью гена cadA, присутствует в ДНК-фрагменте, с размером около 5 т.п.н., полученным путем обработки HindIII.
Итак, указанный фрагмент около 5 т.п.н., полученный в результате обработки ДНК фага лямбда HmdIII, лигировали с обработанной HindIII плазмидой pUC19 (производит Takara Shuzo), с использованием ДНК-лигазы фага Т4. Данную реакционную смесь использовали для трансформации Escherichia, coli JM109 (производит Takara Shuzo), чтобы получить устойчивые к ампициллину штаммы, растущие на полной среде для культивирования в бактериологических чашках (содержащей 10 г полипептона, 5 г дрожжевого экстракта и 5 г хлористого натрия в 1 л воды) с добавлением 50 мг/мл ампициллина. Посредством этого получили бактериальный штамм, который нес плазмиду со вставкой указанного фрагмента, размером около 5 т.п.н., полученного в результате обработки указанной ДНК фага лямбда HindIII. Из этих клеток экстрагировали плазмиду и получали плазмиду pUC6F5HH5. Фиг.1 показывает структуру полученной плазмиды pUC6F5HH5.
Escherichia coli JM109 pUC6F5HH5, несущую эту плазмиду, обозначили как AJ13068, и депонировали в Национальном Институте Биологических и Прикладных наук Человека Агенства Промышленных наук и Технологий под каталожным номером FERM Р-14689 6 декабря 1004 г., переместили на международное хранение на основании Будапештского Договора от 29 сентября 1995 г. и присвоили каталожный номер FERM BP-5251.
(2) Определение нуклеотидной последовательности нового гена лизиндекарбоксилазы
Нуклеотидную последовательность области между сайтами ферментов
рестрикации ClaI и HindIII полученной плазмиды pUC6F5HH5 определяли способом, описанным в Молекулярном Клониродании (2-е издание), Cold spring Harbor Laboratory press (1989). В результате обнаружено,
что кодирующей является указанная нуклеотидная последовательность, представленная в SEQ ID NO: 4 в Списке Последовательностей.
(3) Получение Escherichia coli, продуцирующей L-лизин,
Escherichia coli W3110 культивировали при 37oС в течение 4 часов в полной среде (содержащей 10 г полипептона, 5 г дрожжевого экстракта и 5 г хлористого натрия в 1 л воды), чтобы
получить бактериальные клетки, которые подвергали мутационной обработке при 37oС в течение 30 минут в растворе N-метил-N'-нитро-N-нитрозогуанидина при концентрации 200 мкг/мл, и затем
вносили в минимальную среду для культивирования в бактериологических чашках (содержащую 7 г двузамещенного натрия фосфорнокислого, 1 г хлористого аммония, 0,5 г хлористого натрия, 5 г глюкозы, 0,25 г
сернокислого магния семиводного и 15 г агара в 1 л воды) с добавлением 5 г/л АЕС. Штаммы, устойчивые к АЕС, получали отделением колоний, появляющихся после культивирования при 37oС в
течение 48 часов. Штамм WC196, как одна из них, продуцировал L-лизин. Штамм WC196 обозначали как AJ13069, депонировали в Национальном Институте Биологических и прикладных наук Человека Агенства
Промышленных наук и Технологий под каталожным номером FERM Р-14690 6 декабря 1994 г., переместили на международное хранение на основании Будапештского Договора от 29 сентября 1995
г. и присвоили
каталожный номер FERM ВР-5252.
(4) Создание штамма WC 196 с уничтоженной функцией нового гена лизиндекарбоксилазы
Вышеописанный фрагмент, размером около 5 т.п.н., получали
HindIII-обработкой ДНК фага лямбда из клона 6F5H, лигированного с НindIII-гидролизатом температурно-чувстви-тельной плазмиды рМАN031 (Yasueda, Н. et al., Appl Microbiol. Diotechnol., 36, 211 (1991)
ДНК-лигазой фага Т4. Данную реакционную смесь использовали для трансформации Escherichia coli JM 109 после культивирования при 37oС в течение 24 часов в полной среде, для культивирования в
бактериологических чашках, с добавлением 50 мг/ мл ампициллина для выращивания штаммов, устойчивых к ампициллину. Таким образом получали бактериальный штамм, который несет в себе плазмиду со вставкой
фрагмента, размером около 5 т.п.н., полученного в результате обработки рестриктазой HindIII ДНК фага лямбда из клона 6F5H. Из клеток данного штамма экстрагировали плазмиду и получали плазмиду
pTS6F5HH5. Полученную плазмиду pTS6F5HH5 обрабатывали EcoRV для удаления фрагмента, размером около 1 т.п.н. Затем использовали Т4-лигазу, чтобы вставить фрагмент, обладающий геном устойчивости к
хлорамфениколу, полученного в результате обработки pHSG399 (производит Takara Shuso) рестриктазой AccI. Так сконструировали плазмиду pTS6F5HH5Cm. В результате вышеописанной операции нам удалось
сконструировать плазмиду, несущую фрагмент ДНК с разрушенной функцией нового гена лизиндекарбоксилазы. Фиг.2 показывает структуру плазмиды pTS6F5HH5 и плазмиды pTS6F5HH4Cm.
Далее создали штамм, в котором новый ген лизиндекарбоксилазы в хромосоме штамма WC196 заменили фрагментом ДНК с разрушенной функцией нового гена лизиндекарбоксилазы, в соответствии с основной методикой рекомбинации (Matsuyama, S., J. Bacteriol., 162,- 1196 (1985)), используя свойство температурной чувствительности плазмиды pTS6F5HH5Cm. А именно, штамм WC196 трансформировали плазмидой pTS6F5HH5Cm, во-первых, чтобы получить штамм, который был бы устойчив к ампициллину и устойчив к хлорамфениколу при 30oС. Затем этот штамм использовали для получения штамма, который был бы устойчив к ампициллину и устойчив к хлорамфениколу при 42oС. Кроме того, этот штамм использовали для получения штамма, который был бы чувствителен к ампициллину и устойчив к хлорамфениколу при 30oС. Таким образом создали вышеописанный штамм, в котором новый ген лизиндекарбоксилазы в хромосоме штамма WC196 заменили фрагментом ДНК с разрушенной функцией нового гена лизиндекарбоксилазы. Данный штамм обозначили как WC196L-штамм.
(5) Создание штамма WC196 и штамма WC196L с дефицитом гена cadA
Сейчас известна Escherichia coli, включающая в себя, например, штамм
GNB10181, происходящий от Escherichia coli К-12 (см. Auger, E.A. et al., Mol. Microbiol., 3, 609 (1989), у которого cadA, известный как ген лизиндекарбоксилазы, разрушен; данный штамм доступен,
например, от Центра Генетических Штаммов Е. coii (Connecticut, USA)). Было обнаружено, что область указанного гена cadA отсутствует у данного бактериального штамма. Поэтому характерную нехватку гена
cadA, присущую штамму GNB10181, трансдуцировали в штамм WC196 в соответствии с основным способом, используя фаг Р1 (Краткий курс по генетике бактерий, Cold Spring Harbor Laboratory Press (1992)),
чтобы создать штамм WC196C. Нехватку указанного гена cadA в штамме WC196 подтвердили Саузерн-блот-гибридизацией. Кроме того, способом, подобным вышеописанному, из штамма WC196LC с нехваткой указанного
гена cadA.
Пример 2
(1) Подтверждение наличия у шаммов WC196, WC196L и WC196LC активностей расщепления L-лизина
Созданные вышеуказанные четыре штамма культивировали
при 37oС в течение 17 часов с использованием среды для образования L-лизина (содержащей 40 г глюкозы, 16 г сернокислого аммония, 1 г однозамещенного калия фосфорнокислого, 2 г дрожжевого
экстракта, 10 мг сернокислого марганца, четырех-пятиводного, и 10 мг сернокислого железа, двенадцативодного, в 1 л воды; рН доводили до 7,0 гидроксидом калия и затем отдельно добавляли 30 г
стерильного углекислого кальция). Выделенные бактериальные клетки дважды отмывали физиологическим раствором, суспендировали в среде для анализа распада L-лизина (содержащего 17 г натрия
фосфорнокислого двузамещенного, двенадцативодного, 3 г калия фосфорнокислого однозамещенного, 0,5 натрия хлористого и 10 г L-лизина гидрохлорида в 1 л воды), и культивировали при 37oС в
течение 31 часа.
Фиг.3 показывает изменения остаточного количества L-лизина в культуральных жидкостях в зависимости от времени. Уровень L-лизина определяли количественно, используя Biotech-анализатор AS-210 (производит Asahi Chemical Industry). Значительный распад L-лизина наблюдали для штамма WC196. Однако активность разложения L-лизина была несколько меньшей у штамма WC196C с нехваткой гена cadA, известного как ген лизиндекарбоксилазы. У штаммов WC196L и WC196LC, с разрушенной функцией нового гена лизиндекарбоксилазы не наблюдали распада L-лизина. Содержание остаточного L-лизина в культуральной жидкости снижалось на протяжении почти 3 часов культивирования для любого бактериального штамма. Однако это явление вызвано включением L-лизина в бактериальные клетки, а не его распадом.
(2) Образование L-лизина штаммами WC 196, WC196L и WC196LC.
Вышеуказанные четыре штамма культивировали при 37oС в течение 20 часов в вышеописанной среде для образования L-лизина. В культуральных жидкостях измеряли соответствующие количества L-лизина и кадаверина. Уровень L-лизина определяли количественно, используя Biotech-анализатор AS-210, как описано выше. Уровень кадаверина количественно определяли, используя жидкостную хроматографию высокого давления.
Результаты представлены в таблице 1. По сравнению со штаммом WC196, у штамма WC196C, с разрушенным геном cadA, и у штамма WC196L, с разрушенной функцией нового гена лизиндекарбоксилазы, по сравнению со штаммами WC196 и WC196C, уровень накопления L-лизина возрастал, а уровень накопления кадаверина, как продукта распада L-лизина, снижался. Кроме того, у штамма WC196LC, с разрушенной функцией обоих указанных генов лизиндекарбоксилазы, уровень накопления L-лизина возрастал, а накопление кадаверина, как продукта распада L-лизина, не обнаруживали.
Пример 3
Штамм WC196LC Escherichia coli, с отсутствием
активности разложения L-лизина, трансформировали плазмидой pUC6F5HH5, содержащей новый ген лизиндекарбоксилазы, чтобы получить штамм, устойчивый к ампициллину. Штамм WC196LC и штамм WC196LC/pUC6F5HH5
культивировали при 37oС в течение 16 часов в среде для образования L-лизина и измеряли получаемое количество кадаверина.
Результаты представлены в таблице 2. Штамм WC196LC не мог превращать L-лизин в кадаверин, тогда как штамм WC196LC/pUC6F5HH5 обладал способностью превращать L-лизин в кадаверин.
Промышленная применимость
Новый ген
лизиндекарбоксилазы настоящего изобретения участвует в разложении L-лизина у Escherichia coli. L-лизин можно получать недорого и эффективно путем культивирования бактерии, относящейся к общеизвестному
роду Escherichia, продуцирующей L-лизин при ограничении экспрессии вышеописанного гена настоящего изобретения и/или гена cadA.
Реферат
Изобретение относится к генной инженерии. Ген, который кодирует лизиндекарбоксилазу, имеет нуклеотидную последовательность, приведенную в описании. Аминокислотная последовательность лизиндекарбоксилазы приведена в описании. E. coli, обладающий ограниченной экспрессией вышеуказанного гена, культивируют в жидкой питательной среде. Целевой продукт собирают. Изобретение позволяет повысить скорость получения L-лизина. 2 с. и 1 з.п. ф-лы, 3 ил., 2 табл.
Комментарии