Микрофлюидизированная композиция для индукции специфичного цитотоксического т-лимфоцитного иммунного ответа и ее использование для лечения заболеваний - RU2201253C2
Код документа: RU2201253C2
Чертежи


















Описание
Находящиеся на рассмотрении заявка США 08/919787, поданная 24 июля, 1992, и заявка США 07/735069, поданная 25 июля 1991, озаглавленные "Индукция цитотоксических Т-лимфоцитных иммунных ответов", Syamal Raychaudhuri и William H. Rastetter (в настоящее время аннулированные), включаются в качестве ссылки во всей их полноте в данное описание. Это изобретение относится к способам и композициям, полезным для индуцирования опосредованных цитотоксическими Т-клетками иммунных ответов у людей и домашних или сельскохозяйственных животных.
Считают, что цитотоксические Т-лимфоциты (ЦТЛы) являются основным механизмом защитных сил организма в ответ на разнообразные вирусные инфекции и опухолевый или злокачественный рост. Эти клетки уничтожают инфицированные или трансформированные клетки распознаванием фрагментов антигенов в ассоциации с разнообразными молекулами (названы молекулами МНС класса I) на инфицированных или трансформированных клетках. ЦТЛ можно индуцировать экспериментально цитоплазматической загрузкой определенных растворимых антигенов в специфические клетки. Иммунизация только растворимым антигеном обычно недостаточна для индукции специфичных цитотоксических Т-лимфоцитов.
Один способ, которым можно индуцировать иммунный ЦТЛ-ответ, включает использование способов рекомбинантной инженерии для введения критических компонентов рассматриваемого антигена в геном мягкого инфекционного агента. Целью такой стратегии является генерирование антиген-специфичных цитотоксических Т-лимфоцитных иммунных ответов на желаемый эпитоп путем введения хозяину слабой, самоограничивающейся инфекции. Были описаны химерные векторы с использованием коровьей оспы, вируса полиомиелита, адено- и ретровирусов, а также бактерий, например Listeria и БСЖ. Например, Takahashi et al., 85 Proc. Natl. Acad. Sci. , USA 3105, 1988, описывают использование рекомбинантного вируса коровьей оспы, экспрессирующего ген белка в оболочке gr160 ВИЧ, в качестве возможного средства для индукции цитотоксических Т-лимфоцитов.
Второй способ, которым можно индуцировать опосредованный клетками иммунный ответ, включает использование адъювантов. Хотя в данной области знания продолжаются дискуссии об использовании адъювантов, до сих пор неясно, был ли индуцирован иммунитет, опосредованный (медиированный) клетками, и включал ли такой медиированный клетками иммунитет цитотоксический Т-лимфоцитный иммунный ответ. Тем не менее, далее представляются разные публикации в этой области.
Stover et al. , 361 Nature 456, 1991 (не допускается, чтобы эта работа была предшествующим уровнем данной заявки) описывает иммунный ЦТЛ-ответ на β-галактозидазу с использованием рекомбинантной бактерии БСЖ, содержащей ген β-галактозидазы. Такой ответ не был обнаружен при использовании неполного адъюванта Фрейнда и β-галактозидазы.
Mitchell et al., 8 J. Clinical Oncology 856, 1990 (не допускается, чтобы эта работа рассматривалась как предшествующий уровень данной заявки) описывает лечение пациентов с метастатической меланомой адъювантом, названным "DETOKC", и лизатами аллогенной меланомы, введенными пять раз в течение периода шести недель. У небольшой части пациентов наблюдали повышение цитотоксических Т-клеток. Авторы описывают потребность в повышении уровня продуцирования цитотоксических Т-лимфоцитов и предлагают комбинированную терапию адъювантом с интерлейкином-2, а также в качестве предварительного лечения циклофосфамидом для уменьшения уровня специфичных для опухоли Т-супрессорных клеток, которые могут иметь место. Детокс включает детоксицированный эндотоксин (монофосфориллипид А) из Salmonella minnesota, каркасы клеточных стенок Mycobacterium phlei, скваленовое масло и эмульгатор.
Allison и Gregoriadis, 11, Immynology Today 427, 1990 (не допускается, чтобы эта работа была предшествующим уровнем данного изобретения) указывают, что единственным адъювантом, "разрешенным для использования" в вакцинах человека, являются алюминиевые соли (квасцы), которые не вызывают стойкого медиированного клетками иммунитета. Allison и Gregoriadis заявляют: "здесь имеется, следовательно, потребность в разработке адъювантов с эффективностью полного адъюванта Фрейнда, но без его различных побочных действий, например гранулем". Они продолжают заявлять, что существует три возможные стратегии, например использование липосом; использование адъювантов, названных иммуностимулирующими комплексами (ИСКОМы, которые включают сапонин или Квил A (Quil А) (тритерпеноид с двумя углеводными цепями), холестерин и фосфатидилхолин), которые авторизованы для использования в вакцине против гриппа для лошадей (Morein et al., Immunological Adjuvants and Vaccines, Plenum Press, 153); и использование эмульсии (SAF) сквалена или сквалана (с плуроновым средством или без него) и мурамилдипептида (МДП). Говорят, что SAF вызывает медиированный клетками иммунитет у мышей, хотя "считали, что субъединичные антигены не могут вызывать цитотоксические Т-клеточные (ЦТЛ) иммунные реакции".
Takahashi et al., 344 Nature 873, 1990, описывают рестриктированный помощник (класса II) и цитотоксическую Т-лимфоцитную индукцию путем использования ИСКОМов в однократной подкожной иммунизации у мышей. Они заявляют, что адъювант Фрейнда, неполный адъювант Фрейнда и забуференный фосфатом соляной раствор не индуцируют цитотоксическую Т-лимфоцитную активность против мишеней, в которых они были заинтересованы. Они заявляют, что, в противоположность результатам с другими формами экзогенного растворимого белкового антигена, ими показано, что возможно примировать антигенспецифические МНС (класса 1)-рестриктированные CD8+ СD4--ЦТЛ иммунизацией экзогенным интактным белком, используя ИСКОМЫ. Они заявляют также, что описанные эксперименты допускают, что возможно вызывать индукцию ЦТЛ человека использованием ИСКОМов, содержащих белки ВИЧ, и что с использованием вакцин на основе ИСКОМов можно достичь долго достигаемой цели, т.е. индукции как ЦТЛ, так и антител очищенным белком.
Byars и Allison, 5 Vaccines 223, 1987, описывают использование SAF-1, который включает твин 223, плуроник L121 и сквален или сквалан с мурамилдипептидом или без него, и предполагают, что их данные указывают на то, что композиция с мурамилдипептидом будет полезна для приготовления человеческих и ветеринарных вакцин. Бустер-инъекции адъюванта вводили без мурамилдипептида. Говорят, что мурамилдипептид значительно повышает продуцирование антител по сравнению с использованием адъюванта без мурамилдипептида. Медиированный клетками иммунитет измеряли как гиперчувствительность замедленного типа кожными тестами для определения индукции Т-клеток-хелперов. Такая гиперчувствительность была сильнее и более отсроченная, когда мурамилдипептид присутствовал в адъюванте. Аналогичные адъюванты описываются Allison et al., патент США 4770874 (где заявляется, что комбинация мурамилдипептида и плуронового полиола существенна для вызывания сильной медиированной клетками и гуморальной иммунной реакции против яичного альбумина); Allison et al., патент США 4772466; Murphy-Corb et al., 246, Science 1293, 1989 (где указано, что использование комбинированных адъювантов с мурамилдипептидом может повысить индукцию как гуморального, так и клеточного плеч иммунного ответа); Allison и Byars, 87, Vaccines 56, 1987 (где указано, что медиированный клетками иммунитет вызывается SAF (с мурамилдипептидом), как показано гиперчувствительностью замедленного типа, пролиферативными ответами Т-клеток на антиген, продуцированием интерлейкина-2 и специфическим генетически рестриктированным лизисом клеток-мишеней, несущих иммунизирующий антиген); Allison и Byars, Immynopharmacology of Infections Diseases: Vaccine Adjuvants and Modulators of Non-Specific Resistance 191-201, 1987; Morgan et al., 29 J. Medical Virology 74, 1989; Kenney et al., 121 J. Immynological Methods 157, 1989; Allison и Byars, 95 J. Immynological Methods, 157, 1986 (где было показано, что эмульсии алюминиевых солей и минерального масла повышают образование антител, но не медиированный клетками иммунитет, и было показано, что композиции мурамилдипептида вызывают медиированный клетками иммунитет); Byars et al. , 8 Vaccine 49, 1990 (не допускается, чтобы эта работа была предшествующим уровнем заявки), где указано, что их композиция адъюванта заметно повышает гуморальный иммунный ответ и в меньшей степени усиливает медиированные клетками иммунные ответы на гемагглютининовый антиген гриппа); Allison and Byars, 28 Molecular Immynology 279, 1991 (не допускается, чтобы эта работа была предшествующим уровнем заявки; которые заявляют, что функцией мурамилдипептида является индуцирование экспрессии цитокинов и повышение экспрессии генов главного комплекса гистосовместимости (ГКГ), и что получали лучшие клеточные ответы и ответы антител, чем с другими адъювантами; и что они надеются выяснить, эффективна ли аналогичная стратегия для людей); Allison and Byars, Technology Advances in Vaccine Development 401, 1988 (которые описывают медиированный клетками иммунитет с использованием SAF); Epstein et al., 4 Advance Drug Delivery Reviews 223, 1990 (которые дают обзор разных адъювантов, используемых для получения вакцин); Allison and Byars, 95 J. Immynological Methods 157, 1986 (которые заявляют, что добавление мурамилдипептида к адъюванту заметно повышает медиированные клетками иммунные ответы на различные антигены, включая моноклональные иммуноглобулины и вирусные антигены); и Morgan et al., 29 J. Medical Virology 74, 1989 (которые описывают использование SAF-1 для получения вакцины для вируса Эпштейн-Барра.
Kwak et al., Idiotype Networks in Biology and Medicine. Elsevier Science Publisher, p. 163, 1990 (не допускается, чтобы эта работа была предшествующим уровнем заявки) описывают использование SAF без мурамилдипептида в качестве адъюванта для идиотипа В-клеточной лимфомы у человека. В частности, эмульсию плуроника L121, сквалана и 0,4% твина 80 в забуференном фосфатом солевом растворе вводили с идиотипом. Они заявляют, что "добавление адъюванта должно далее повышать... гуморальные иммунные ответы и, кроме того, может облегчить индукцию клеточных иммунных ответов".
Другие иммунологические препараты включают липосомы (Allison et al., патенты США 4053585 и 4117113); циклические пептиды (Dreesman et al., патент США 4778784); полный адъювант Фрейнда (Asherson et al., 22 Immynology 465, 1972; Berman et al., 2 International J. Cancer 539, 1967; Allison, 18 Immynopotentiation 73, 1973; and Allison, Non specific Factors Influencing Host Resistance 247, 1973); ИСКОМы (Letvin et al., 87 Vaccines 209, 1987); адъюванты, содержащие неионные блок-сополимерные агенты, образованные с минеральным маслом, поверхностно-активное вещество и твин 80 (Hunter and Bennett, 133 J. Immunology 3167, 1984; and Hunter et al., 127 J. Immunology 1244, 1981); адъюванты, составленные из минерального масла и эмульгирующего средства с убитыми микобактериями или без них (Sanchez-Pescador et al., 141 J. Immunology 1720, 1988) и другие адъюванты, такие как липофильное производное мурамилтрипептида и мурамилдипептида, ковалентно конъюгированный с рекомбинантым белком (id.).
Краткое изложение существа изобретения
Заявитель обнаружил безопасный и выгодный способ и композиции, которыми у людей и домашних или важных для сельского
хозяйства животных можно индуцировать иммунный ЦТЛ-ответ. Способ включает
использование антигенной композиции, которая слаботоксична или нетоксична для животных и не содержит иммуностимулирующего
пептида (например мурамилдипептида), присутствие которого будет понижать
желаемый клеточный иммунный ответ. Кроме того, эта методология проста в использовании и не требует экстенсивной работы in vivo
для изменения существующих клеток способами рекомбинантной ДНК, чтобы
сделать их более антигенными. Это открытие неожиданное, поскольку не ожидали, что такие иммунные ЦТЛ-ответы могут быть
индуцированы путем использования такой антигенной композиции, не содержат
иммуностимулирующих пептидов или их эквивалентов. Открытие заявителя позволяет использовать такие антигенные композиции при
болезненных состояний широкого спектра или в качестве профилактического
средства. Введение такой антигенной композиции можно использовать, например, для лечения вирусных болезней, в которых важен
иммунный ЦТЛ-ответ, например при лечении инфекции ВИЧ или гриппа;
использование ее можно также расширить до использования при лечении бактериальных инфекций, рака, паразитарных инфекций и тому
подобных. В качестве профилактического средства антигенная композиция,
комбинированная с подходящим антигеном, полезна для предупреждения инфекции вирусами, ответственными за вышеуказанные вирусные
болезни, в частности профилактики ВИЧ-инфекции, а также для профилактики
пациентов с риском рака, например после резекции первичной опухоли.
Таким образом, первым аспектом изобретения является представление способа индукции иммунного ЦТЛ-ответа у человека или домашнего (например кошки или собаки) или важного для сельского хозяйства животного (например козы, коровы или свиньи) к антигену, отличному от антигена В-клеточной лимфомы или яичного альбумина. Способ включает стадии получения антигена, для которого желателен иммунный ЦТЛ-ответ, и получения нетоксичной антигенной композиции, которая включает стабилизирующий детергент, мицеллообразующий агент и биоразрушаемое и биосовместимое масло или состоит или по существу состоит из этих компонентов. Эта антигенная композиция предпочтительно не содержит иммуностимулирующий пептидный компонент или имеет достаточно низкий уровень такого компонента, чтобы не уменьшился требуемый клеточный иммунный ответ. Эту композицию предпочтительно предоставляют в виде стабильной эмульсии типа масло-в-воде. То есть каждый из различных компонентов выбирают так, чтобы эта эмульсия оставалась в состоянии эмульсии в течение периода по меньшей мере одного месяца, предпочтительно в течение более чем одного года, без разделения фаз. В этом способе антиген и антигенную композицию смешивают для образования смеси (предпочтительно микрофлюидизацией) и эту смесь вводят животному в количестве, достаточном для индуцирования иммунного ЦТЛ-ответа у этого животного. Такое введение требуется проводить только один раз.
Термин "стабилизирующий детергент" означает детергент, который позволяет компонентам эмульсии сохраняться в виде стабильной эмульсии. Такие детергенты включают полисорбат, твин 80 (сорбитан-моно-9-октадеканоат-поли(окси-1,2-этандиил; производимый ICI Americas, Wilmington, DE), твин 40, твин 20, твин 60, цвиттергент 3-12, типол НВ7 и спан 85. Эти детергенты обычно применяют в количестве приблизительно 0,05-0,5%, предпочтительно около 0,2%.
Термин "мицеллообразующий агент" обозначает агент, который способен стабилизировать эмульсию, образованную с другими компонентами, так чтобы образовывалась мицеллоподобная структура. Такие агенты предпочтительно вызывают некоторое раздражение в месте инъекции, чтобы привлечь макрофаги для повышения клеточного иммунного ответа. Примеры таких агентов включают полимерные поверхностно-активные вещества, описанные в публикациях BASF Wyadotte, например Schmolka, 54 J. Am. Oil. Chem. Soc., 110, 1977, и Hunter et al. , 129 J. Immunol. 1244, 1981, причем обе эти публикации включены в качестве ссылок, плуроник L62LF, L101 и L64, ПЭГ1000 и тетроник 1501, 150R1, 701, 901 1301 и 130R1. Химические структуры таких агентов хорошо известны в данной области. Предпочтительно выбирают агент, который имеет гидрофильно-липофильный баланс (ГЛБ) между 0 и 2, как определено Hunter and Bennett, 133 Journal of Immunology 3167, 1984. Этот агент предпочтительно применяют в количестве от 0,5 до 10%, предпочтительно в количестве от 1,25 до 5%.
Выбирают масло, которое промотирует удерживание антигена в эмульсии масло-в-воде, то есть обеспечивает носитель для требуемого антигена, и предпочтительно имеет температуру плавления менее чем 65oС, так что эмульсия образуется либо при комнатной температуре (около от 20 до 25oС), либо один раз температуру эмульсии снижают до комнатной температуры. Примеры таких масел включают сквален, сквалан, эйкозан, тетратетраконтан, глицерин и арахисовое масло или другие растительные масла. Масло предпочтительно применяют в количестве от 1 до 10%, наиболее предпочтительно от 2,5 до 5%. Важно, чтобы масло было биоразлагаемым и биосовместимым, так чтобы организм мог разрушать масло в течение некоторого времени и чтобы при использовании такого масла не были очевидными отрицательные действия, например гранулемы.
В указанной выше композиции важно, чтобы в ней не содержался пептидный компонент, в особенности мурамилдипептид (МДП). Такой пептид будет мешать индукции иммунного ЦТЛ-ответа, если он присутствует в количестве, большем, чем около 20 микрограмм, на одно обычное введение композиции человеку. Предпочтительно, чтобы такие пептиды полностью отсутствовали в антигенной композиции, несмотря на их очевидную стимуляцию гуморального компартмента иммунной системы. То есть заявитель нашел, что, хотя такие пептиды могут усиливать гуморальную иммунную реакцию, они невыгодны, когда требуется цитотоксический Т-лимфоцитный иммунный ответ.
Другие связанные аспекты изобретения заключаются в том, что антигенную композицию образуют только из двух из указанных выше трех компонентов и используют с любым требуемым антигеном (этот термин включает белки, полипептиды и их фрагменты, которые иммуногенные), за исключением яичного альбумина (или других альбуминов, например человеческого сывороточного альбумина (HSA), бычьего сывороточного альбумина (BSA) и овальбумина (ОВА), для индукции иммунного ЦТЛ-ответа у указанных выше животных или людей.
Заявитель считает, что указанные выше композиции значительно выгоднее, чем предшествующие композиции (включая ИС-КОМы, детокс и SAF) для использования для людей. В отличие от таких композиций данная композиция как включает мицеллообразующий агент, так и не имеет пептидов, каркасов клеточных стенок или компонентов бактериальных клеток. Данная композиция также индуцирует иммунный ЦТЛ-ответ, который либо не имеет место при использовании предшествующих композиций, либо значительно повышается по сравнению с этими композициями.
Термин "нетоксичный" означает, что наблюдается слабое побочное действие антигенной композиции или не наблюдается такого действия у обработанного такой композицией животного или человека. Обычные специалисты в области медицины или ветеринарии должны признать, что этот термин имеет широкое значение. Например, по существу, у здорового животного или человека может допускаться только слабая токсичность, тогда как у человека, страдающего от иммунной болезни, может допускаться значительно более высокая токсичность.
В предпочтительных воплощениях антигенная композиция состоит, по существу, из двух или трех компонентов: детергента, агента и масла, и способ состоит, по существу, в однократном введении смеси (антиген плюс антигенная композиция) человеку или животному; человека или животного инфицируют вирусом и он страдает от одного или нескольких симптомов (обычно определяемых врачами в соответствующей области) инфекции вирусом; и антигенная композиция нетоксична для человека или животного.
В других предпочтительных воплощениях антиген выбирают из антигенных частей ВИЧ-антигенов: gр160, gаg, pol, Nef, Tat и Rev; антигенов малярии: белка спорозоита (CS) и поверхностного белка 2 спорозоита; поверхностных антигенов гепатита В: Pre-S1, Pre-S2, HBc Аg и НВе Аg; антигенов гриппа: НА, NP и NA; поверхностных антигенов гепатита А; антигенов вируса герпеса: gр340 EBV, gр85 EBV, gВ HSV, gD HSV, gH HSV, раннего белкового продукта HSV, антигенов вируса папилломы человека (например HPV-антигенов, таких как антигены L1, Е4, Е6, Е7, в частности антигенов Е6 и Е7 из HPV16 и 18, двух наиболее обычных типов HPV, связанных с цервикальной карциномой, Е4 и L1, полученных из HPV6 и HPV11, двух наиболее обычных типов HPV, связанных с остроконечной кондиломой; антигена предстательной железы, gВ цитомегаловируса, gН цитомегаловируса и белка gР72 1Е; антигенов респираторно-синцитиального вируса: белка F, белка G и белка N; и опухолевых антигенов карциномы СЕА, связанной с муцином карциномы, Р21 карциномы, Р53 карциномы, MPG меланомы, р97 меланомы и онкогенного продукта Neu карциномы, продукта гена р53 карциномы, антигена меланомы, названного MAGE и мутированного белка р21 ras, присутствующего в различных злокачественных опухолях.
В другом аспекте изобретение характеризуется композицией, содержащей, состоящей или по существу состоящей из антигена, смешанного с описанной выше композицией, и антиген выбирают из перечисленных антигенных частей.
В другом близком аспекте изобретение характеризуется способами лечения пациента, инфицированного ВИЧ-вирусом, страдающего малярией, страдающего гриппом, страдающего гепатитом, страдающего раком, инфицированного вирусом герпеса, страдающего цервикальным раком, страдающего остроконечной кондиломой (остроконечные бородавки) или инфицированного респираторно-синцитиальным вирусом, введением композиции, включающей подходящий антиген (например, выбранный из перечисленных выше антигенов), смешанный с одной из выше указанных композиций антигена. Эти антигены и способы лечения с их использованием являются только примерными антигенами, которые можно использовать в антигенных композициях-объектах данного изобретения.
Другие признаки и преимущества данного изобретения будут очевидными из следующего описания предпочтительных воплощений его и из формулы изобретения.
Описание предпочтительных воплощений
Сначала будут кратко описаны
чертежи.
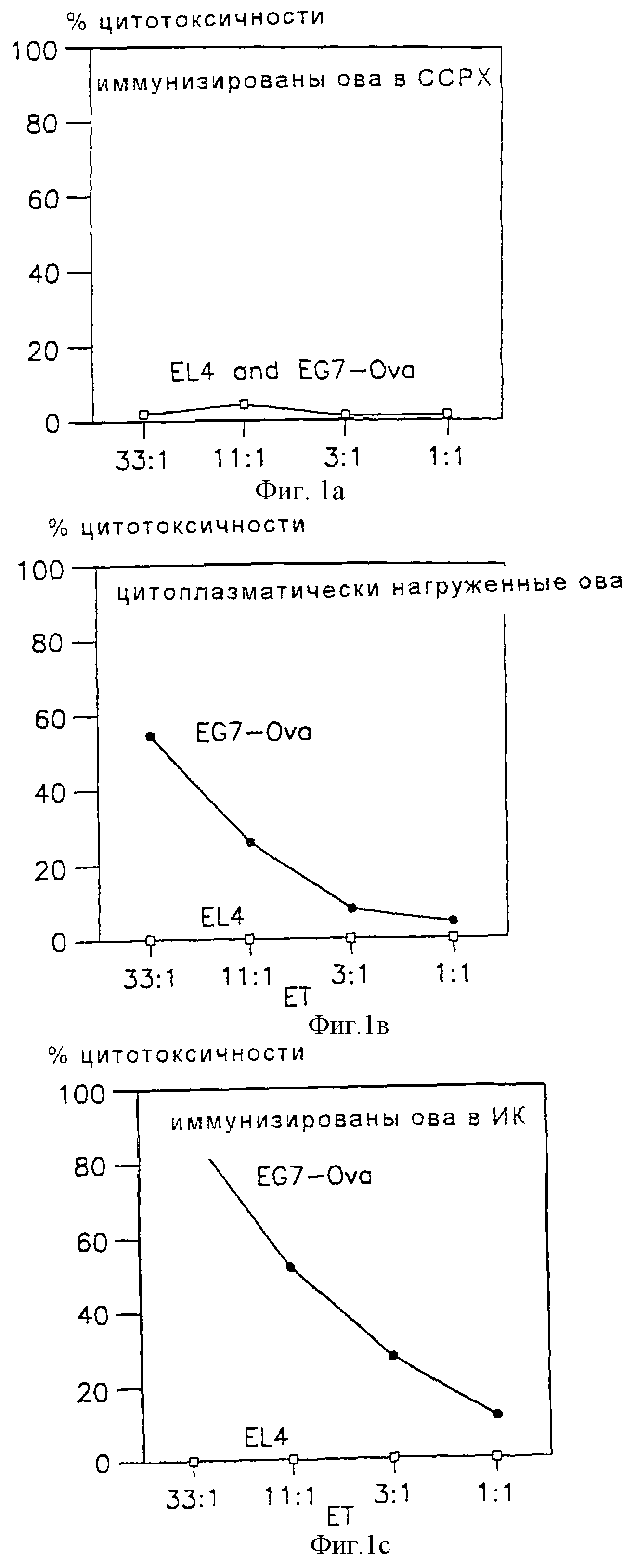
Фиг. 1a-1c и 4a-4c являются графическими представлениями данных, сравнивающих индукцию ЦТЛ различными композициями овальбумина; Е:Т представляет отношение эффектора к мишени на всех чертежах.
Фиг. 2a и 2b являются графическими представлениями данных, сравнивающих индукцию ЦТЛ разными композициями β-галактозидазы.
Фиг. 3 является графическим представлением данных, сравнивающих индукцию ЦТЛ овальбумином в липосоме и в антигенной композиции.
Фиг. 5 и 6 являются графическими представлениями данных, показывающих влияние истощения клеток CD4 и CD8 на индукцию ЦТЛ.
Фиг. 7 является графическим представлением данных, показывающих индукцию ЦТЛ белком gр120.
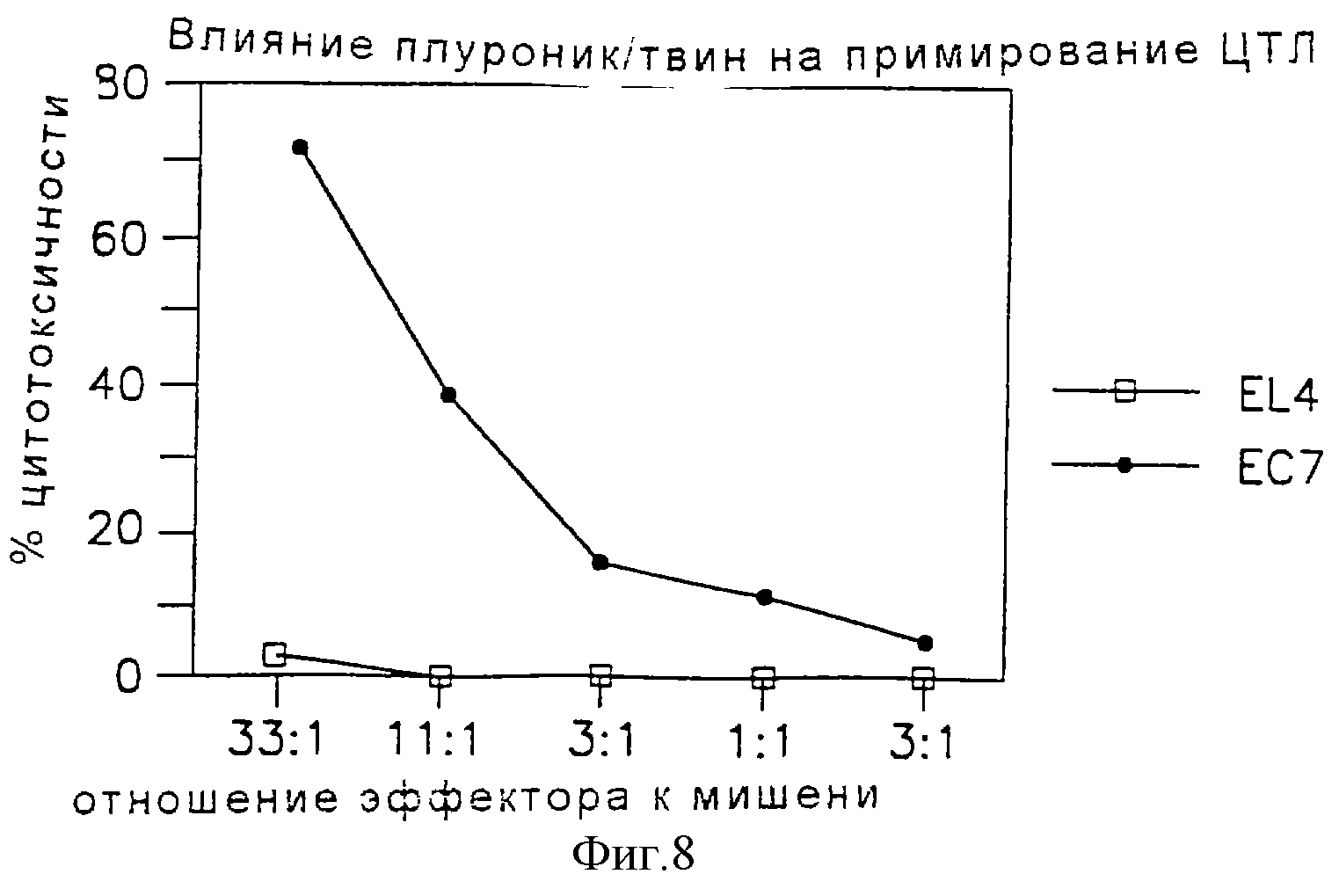
Фиг. 8 является графическим представлением данных, показывающих индукцию ЦТЛ смесью плуроника и твина и антигена.
Фиг. 9 является графическим представлением данных, показывающих индукцию ЦТЛ смесью сквалана и плуроника и антигена.
Фиг.10 является графическим представлением данных, показывающих индукцию ЦТЛ смесью сквалана и плуроника и антигена.
Фиг. 11 является графическим представлением влияния овальбумина (ова) с разными антигенными композициями на иммунный ЦТЛ-ответ.
Фиг. 12 является графическим представлением индукции антител против gp120IIIb у обезьян различными антигенными композициями.
Фиг.13 изображает противоопухолевую активность клеток НОРЕ2 через десять дней после одной иммунизации растворимым белком Е7 в адъюванте.
Фиг. 14 изображает противоопухолевую активность клеток НОРЕ2 на 10, 19 день после двух иммунизаций растворимым белком Е7 в адъюванте.
Антигенная
композиция
Антигенные композиции, полезные в этом изобретении, в общих чертах описываются
выше. Специалисты в данной области должны признать, что эквивалентные композиции легко получаются и,
как можно ожидать, имеют эквивалентные свойства в индукции иммунного ЦТЛ-ответа. Такие композиции
легко испытывать на их свойства с использованием методик, эквивалентных методикам, описанным в
приведенных ниже примерах.
Здесь даются примеры изобретения с использованием антигенной композиции (АК), состоящей из около 2,5% сквалана, 5% плуроновой кислоты и твина 80 в забуференном фосфатом солевом растворе. В частности, эмульсия АК включала: 15 мг сквалана, 37,5 мг полоксамера 401 (плуроник L121), 6 мг полисорбата 80 (твин 80), 0,184 мг хлорида калия, 0,552 мг первичного кислого фосфата калия, 7,36 мг хлорида натрия, 3,3 мг вторичного кислого фосфата натрия (безводного) на 1 мл воды, рН 7,4, Эту эмульсию микрофлюидизировали, используя стандартные способы (модель M110F Microfluidics) с модулем обратного давления при 0,773-0,984 атм с постепенным возвращением к атмосферному давлению, охлаждение и упаковку в мокрый лед.
В других примерах антиген смешивали с микрофлюидизированной смесью сквалана (С), плуроника (П) и твина 80 (Т) для достижения конечной концентрации 0,2% твина 80, 1,25% плуроника и 5% сквалана соответственно. Для определения субкомпонентов, необходимых для индукции антигенспецифичного иммунного ответа, получали смеси сквалан-твин 80, плуроник-твин 80 или сквалан-плуроник с такой же концентрацией, как и для трехкомпонентной смеси. Плуроник, сквалан или твин 80 готовили также индивидуально для определения влияния индивидуального компонента на индукцию ЦТЛ. Проводили также замену твина 20, твина 40 или цвиттергента на твин 80 для определения влияния разных производных твина на индукцию ЦТЛ в ова-системе. Сквалан в трехкомпонентной композиции заменяли на эйкозан или триаконтан и сополимерный плуроник в такой же трехкомпонентной системе заменяли на ПЭГ 1000, плуроник L62LF и тетроники 1501 и 150R1. В качестве двухкомпонентных композиций разные аналоги в разных комбинациях смешивали и испытывали на ова-специфическую индукцию ЦТЛ. Они являются смесью холестерин-твин 80, сквалан-твин 20, пристан-твин 80 или оливковое масло-твин 80. Для изучения стабилизации микрофлюидизированную смесь сквалан-твин 80 смешивали с декстрозой до конечной концентрации 5%. Во всех случаях комбинации наполнителей смешивали в микрофлюидизаторе для получения стабильной эмульсии. В некоторых экспериментах двухкомпонентные композиции смешивали с разными концентрациями МДП для индукции иммунного ЦТЛ-ответа и гуморального иммунного ответа. В таблице 1 приводится исчерпывающий список различных композиций, используемых в этом исследовании.
Композицию адъюванта синтекс (микрофлюидизированная; SAFm) использовали в качестве адъювантного контроля, она состоит из двух частей. Часть I состоит из забуференного фосфатом солевого раствора, содержащего как конечную концентрацию 5% сквалана, 1,25% плуроника и 0,2% твина 80 (наполнитель или I-SAF). Часть II состоит из N-ацетилмурамил-L-треонил-D-изо-глутамина (Тhr-МДП), производного компонента стенок клеток микобактерий. Для целей иммунизации антиген смешивают с микрофлюидизированным наполнителем (часть I) для получения гомогенной эмульсии. В приготовленную микрофлюидизированную композицию адъюванта синтекс (SAFm) добавляют МДП и смесь недолго интенсивно перемешивают. Концентрацию МДП в смеси изменяют для определения, была ли достигнута оптимальная его концентрация для ЦТЛ-индукции. В качестве адъювантного контроля мышей также иммунизировали растворимыми антигенами, смешанными с квасцами в соответствии с инструкцией производителя (Pierce Chemical, Rockford, IL) или с полным адъювантом Фрейнда (ПАФ).
Эту композицию антигена используют для индукции цитотоксического Т-лимфоцитного иммунного ответа у мышей. Обычные специалисты в данной области должны признать, что такая мышиная модель является показателем того, что эквивалентные эксперименты или способы лечения будут подобным образом индуцировать цитотоксический Т-лимфоцитный иммунный ответ у людей, домашних или сельскохозяйственных животных. Количество композиции антигена и антигена, полезное для продуцирования желательной клеточного иммунного ответа можно определить эмпирически стандартными способами, хорошо известными специалистам в данной области, без дополнительного экспериментирования. Таким образом, если желательно снизить до минимума побочное действие при лечении такой смесью, специалисты в данной области могут определить минимальный уровень такой смеси для введения человеку, домашнему или сельскохозяйственному животному для вызывания иммунного ЦТЛ-ответа и, таким образом, индуцирования иммунитета к желаемому антигену. При обычном использовании такую смесь нужно инъецировать любым одним из ряда стандартных методов, но, в частности, предпочтительна локальная внутримышечная инъекция, которая позволит эмульсии оставаться в стабильной форме в течение периода нескольких дней или нескольких недель.
Способы
В представленных ниже примерах использовали следующие
материалы и способы, если нет других указаний:
Мыши
Самок мышей С57В/6
(Н-2d) и BALB/c (Н-2d) покупали у Harlen Sprague (San Diego, California).
Антигены
Овальбумин (ова, сорт VII, Sigma Chemical Co., St. Louis, МО)
использовали в нативной форме, β-галактозидазу (β-гал, сорт VIII; BRL) использовали в нативной форме и
после кипячения в 1 М NaOH в течение 2 мин для проведения щелочного гидролиза.
Рекомбинантный gp120 покупали у American Biotechnology.
Опухолевые клетки и трансфектанты
Использованные опухолевые клетки были Iа--линиями EL4 (C57BL/6, тимома
Н-2b) и Р815 (DBA/2, мастоцитома Н-2d). Получение производного ова-продуцирующего трансфектанта
EL4, ЕG7-ова, описано ранее Moore et al., 54 Cell 777, 1988.
β
-Гал-продуцирующий трансфектант, Р13.1, получали электропорацией 107 Р815-клеток в 1 мл забуференного фосфатом
соляного раствора с 10 мг линеаризованного при помощи PstI pCH110 (Pharmacia
LKB Biotechnology Inc., Piscatway, NJ) и 1 мг линеаризованного при помощи Pvul pSV2 neo (Southern et al. , 1 J. Mol. Appl.
Genet. 327, 1982) с последующей селекцией в 400 мкг/мл антибиотика G418.
С3-4-Трансфектант получали из гибридомы IgM 662 BALB/c трансфекцией плазмидой, кодирующей ген β-гал, слитый с третьим и
четвертым экзоном тяжелой цепи IgM (Rammensee et al., 30 Immunogenetics
296. 1989). Фибробласт 3Т3, экспрессирующий gp160IIIb, 15-12, был получен Dr. Germain NIH (Bethesda, MD). Kb
-трансфектированная L-клеточная линия была предоставлена Dr. Carbone, Monash
University, Australia. Dd и Ld-трансфектированные L-клеточные линии были предоставлены Dr. Ted
Hensen, Washington University, St. Louis.
Иммунизация
Мышей иммунизировали внутривенно 200 мкл суспензии 25•106 спленоцитов после цитоплазматической
загрузки, как описано Moore et al., смотри выше, и Carbone et al. , J. Exp. Med.
169:603, 1989). Для иммунизации композицией ова-антиген или композицией β-гал-антиген 30 мкг каждого белкового
антигена инъецировали каждой мыши подкожно в подушечку лапы и основу хвоста.
Каждая инъекция состояла из 67 мкл микрофлюидизированной антигенной композиции (приготовлена в соответствии со
стандартными способами) и 30 мкг белкового антигена в конечном объеме 200 мкл. Конечный
объем устанавливали при помощи ССРХ (сбалансированный солевой раствор Хенка), смотри руководство Whittaker
(Welkersville, MD). МДП вводили в концентрациях от 0 до 300 мкг. Там, где сообщается, мышей
иммунизировали растворимыми антигенами в ПАФ или квасцах в общем объеме 200 мкл.
Стимуляция
in vitro популяций клеток-эффекторов
Клетки селезенки (30•106)
нормальных или иммунизированных мышей, которые были примированы по меньшей мере за 14 дней ранее,
инкубировали с 1,5•106 ЕG7-ова (облученные 20000 рад) для ова-иммунных ответов или с
1,5•106 С3-4-клетками (облученные 20000 рад) для иммунного β-гал-ответа
в планшетах на 24 лунки при 37oС в среде 7% СO2/воздух. Все культивирования
тканей выполняли в полной среде, состоящей из среды Дульбенко, модифицированной по способу Исков
(IMDM), смотри Whittaker Manual (Welkersville, MD), дополненной 10% фетальной телячьей сывороткой (FSC),
2 мМ глутамина, гентамицином и 2•10-5 М 2-меркаптоэтанола. Для экспериментов
с истощением in vitro праймированные in vivo или стимулированные in vitro клетки селезенки обрабатывали
моноклональными антителами (mAbs) RL.172 (анти-СD4) или mABs 3,168 (анти-СD8) для удаления
CD4+ или СD+-Т-клеток (Sarmiento et al., 125 J. Jmmunol. 2665, 1980, and Ceredig et al.,
314 Nature 98, 1985). mAb RL.172 и mAb 3,168 получали от Dr. Jonathan Sprent et Scripps
Clinic and Research Foundation, La Jolla, CA.
Клетки селезенки (30•106)
нормальных или иммунизированных мышей, которые были праймированы по меньшей мере на 21 дней
ранее, инкубировали с 1,5•106 клетками 15-12 (обработанные 200 мкг митомицина С в течение
45 минут на 108 клеток) или с
500 мкг пептида 18IIIb, содержащего
доминантный эпитоп ЦТЛ в мышах Balb/c, в полной IMDM-среде (Irvine Scientific, Santa Ana, CA), содержащей 10%
предварительно подвергнутой скринингу FCS (ICN Flow; ICN Biochemicals, Inc., Costa Mesa,
CA), 2 мМ глутамина, гентамицина и 2•105 М 2-меркаптоэтанола. Для стимуляции in vitro
пептидами клетки селезенки культивировали в полной IMDM-среде, содержащей 5% супернатанта
конканавалина А (СоnА).
Для экспериментов с истощением праймированные in vitro или стимулированные in vitro клетки селезенки обрабатывали mAbs RL.172 (анти-СD4) или mAbs 3.168 (анти-CD8) в присутствии малотоксичного кроличьего комплемента (Cederlane Laboratories, Ltd., Hornby Ontario, Canada) для удаления CD4+ или СD8+-Т-клеток (22, 23). (mAbs) RL.172 и mABs 3.168 были подарены Dr. Jonathan Sprent et Scripps Clinic and Research Foundation, La Jolla, CA).
Испытания на цитотоксичность
Клетки-мишени (1•106)
метили 100 мкКи [51Сr]-хромата натрия в течение 60 мин. Для "обученных" пептидом мишеней
(комитированных лимфоцитов) во время мечения мишеней51Сr добавляли 50 мкл пептидного
раствора 1 мг/мл в ССРХ. После промывания 104 меченых мишеней и продукты серийного
разведения клеток-эффекторов инкубировали в 200 мкл RP10 в течение 4 ч при 37oС. Собирали 100
мкл супернатанта и специфический лизис определяли следующим образом: Процент специфического
лизиса = 100 х {(выделение посредством ЦТЛ - самопроизвольное выделение)/(максимальное выделение
- самопроизвольное выделение)}. Самопроизвольное выделение в отсутствие цитотоксических Т-лимфоцитов
(ЦТЛ) было <25% максимального выделения детергентом во всех экспериментах.
Определение гуморального иммунного ответа у мышей и обезьян
Каждую лунку планшетов на 90
лунок с U-образным дном (Costar, Cambridge, МА) покрывали 150 нг ova или gr120 в 50 мкл ССРХ и
инкубировали в течение ночи при 4oС. Для определения гуморальных иммунных ответов против gp120
и против ova у мышей планшеты блокировали 1% бычьим сывороточным альбумином BSA в течение 1
часа. Серийно разведенные сыворотки добавляли в объеме 25 мкл на лунку и инкубировали в течение 2 часов.
Планшеты промывали и добавляли на лунку 50 мкл разведенного 1:1000 антимышиного козлиного IgG,
конъюгированного с HRPO (SBT, Alabama), в 1% BSA. После 1 часа инкубирования планшеты промывали и на
лунку добавляли 100 мкл субстрата. OD405 проводили через 10-15 минут. Для определения
гуморального иммунного ответа как образование обезьяньих антител против gр120 все указанные стадии, за
исключением как блокирования планшетов, так и разбавления сыворотки, проводили в 5% нормальной
козьей сыворотке в сбалансированном солевом растворе Хэнкса.
Синтез пептидов
Синтетические пептиды, соответствующие аминокислотным последовательностям 253-276
(Последовательность 1: EQLESIINFEKLTEWTSSNVMEER; где стандартный однобуквенный код используют для обозначения каждой
аминокислоты) овальбумина (ova 253-276), аминокислотным последовательностям 84-102
миелинового базового белка (МВР 84-102) (Последовательность 2: DENPVVHFFKNIVTPRTPP) и синтетические пептиды,
соответствующие аминокислотным последовательностям 308-322 (18IIIb-последовательность)
gp120IIIb, были "собраны" твердофазным пептидным синтезом с использованием синтезатора Applied Biosystems 430A.
Аминокислоты связывали через предварительно образованные симметричные ангидриды, за
исключением аспарагина, глутамина и аргинина, которые связывали в виде сложных эфиров гидроксибензотриазола.
Эффективность связывания контролировали нингидриновой реакцией по методу Kaiser et al., 34
Anal. Biochim. 595, 1970. Пептиды выделяли из подложки при помощи HF после процедуры "низкий-высокий",
описанной Tam, et al., 21 J. Am. Chem. Soc. 6442, 1983, и экстрагировали из смолы 10% уксусной
кислотой. После лиофилизации пептиды обессоливали на колонке с сефадексом G-25 и пробы пептидов затем
очищали ЖХВР хроматографией с обращенной фазой на препаративной колонке С-18 Vydac. Очищенные
пептиды (98%) растворяли в ССРХ при конечной концентрации 10 мг/мл и разбавляли до желаемой концентрации в
полной среде.
Гидролиз CNBr
Пробы белка (например, β
-галактозидазы) обрабатывали 100-кратным молярным избытком бромида циана в растворе 100 мМ трифторуксусной кислоты.
Реакцию проходила в течение 18 часов при комнатной (около 20oС)
температуре при перемешивании. После прохождения предписанного времени реакции пептидные фрагменты отделяли от реагента,
используя аппаратуру SEP-PAK С-18 (Waters), элюировали 95% ацетонитрилом и
лиофилизовали.
Гидролиз щелочью
Образцы белков (например, β-галактозидазы) обрабатывали 1 н
NaOH и кипятили в течение 2 минут и получаемые пептидные фрагменты отделяли
от реагентов, используя аппаратуру С-18 SEP-PAK (Waters), и элюировали 95% ацетонитрилом и лиофилизовали.
Пример 1: Примирование рестриктированных (по классу) I ЦТЛ
Moore et
al., 113 UCLA Symp. Mol. Cell. Biol. 1989 and Carbone and Bevan, 171 J. Exp. Medicine 377, 1990, показывают, что мыши,
иммунизированные клетками селезенки, заполненными цитоплазматически растворимым
овальбуцином, были праймированы для ова-специфического иммунного ответа в виде рестриктированных по классу I ЦТЛ.
Ова-экспрессирующий EL4-трансфектант EG7-ova использовали для стимуляции in vitro
примированных in vivo лимфоцитов селезенки и использовали также в качестве мишени для ова-специфического
ЦТЛ-медиированного киллинга. Это исследование показывает также, что СD8+-эффекторы,
индуцированные ЕG7-ова-трансфектантом или клетками селезенки, цитоплазматически загруженными ова,
распознают детерминанту, картированную пептидом ова 258-276 в контексте H-2Kb, лизируют
EG7-ova и нейтрализуют также клетки EL4, покрытые ова 258-276. Таким образом, чтобы оценить, может ли
эндогенный путь по рестриктированным (по классу I) CD88-T-клeткaм быть индуцирован
растворимым антигеном, указанную выше систему использовали для определения, можно ли обычные антигенные
композиции использовать для введения растворимого антигена в рестриктированный путь класса
I.
а) ова (ova)
Мышей C57BL иммунизировали один раз различными количествами ova (30
мкг - 1 мг на мышь) с использованием антигенной композиции или без нее. Мышей инъецировали
подкожно и в основание хвоста. Клетки селезенки отбирали у иммунизированных мышей по меньшей мере через две
недели после иммунизации и in vitro стимулировали трансфектантами EG7-ova. Концентрация до
30 мкг была так же эффективна, как доза 1 мг. Поэтому исследования ЦТЛ, как обычно, проводили с клетками
селезенки мышей, примированных от 30 мкг ova. После культивирования с EG7-ova in vitro в
течение пяти дней примирование оценивали по присутствию ova-специфических эффекторов, способных лизировать
EG7-ova.
Мыши, инъецированные растворимым ova в ССРХ вплоть до 1 мг, не продемонстрировали доказательства примирования ЦТЛ (фиг.1a). Однако мыши, иммунизированные 30 мкг ova в антигенной композиции, описанной выше (на фигурах показана как АК), показали значительный трансфектант-специфический иммунный ЦТЛ-ответ (фиг.1c). Кроме того, степень киллинга EG7-ova клетками селезенки, иммунизированными ova-AK, была сравнима со степенью киллинга ova-наполненными клетками селезенки иммунизированных мышей (фиг.1b).
То что специфичность ЦТЛ-примирования in vivo была антиген-специфичной, было показано неспособностью клеток селезенки иммунизированных β-галактозидазой мышей проявлять вторичный иммунный ЦТЛ-ответ in vitro при стимуляции EG7-ova. Не наблюдали индукцию ова-специфических ЦТЛ.
b) β-галактозидаза
Похожие результаты получали с использованием другого растворимого белкового антигена, β-гал. Для
анализа β-гал-специфичного иммунного ЦТЛ-ответа используемой мишенью был полученный из
мышей BALB/c β-гал-экспрессирующий С3-4-трансфек-тант. Иммунизация мышей BALB/c растворимым β
-гал давал фоновый иммунный ЦТЛ-ответ. Поэтому для определения специфического иммунного
ЦТД-ответа сбор был отсрочен по меньшей мере на восемь недель до того, как лимфоциты селезенки были собраны и
культивированы в течение пяти дней в присутствии облученных С3-4-трансфектантов.
Фиг. 2b показывает, что 30 мкг β-галактозидазы в АК индуцировали сильный специфический иммунный ЦТЛ-ответ против трансфектанта. При отношении эффектора к мишени (Э:М) 3:1 иммунизированные β-гал-АК мыши показали около 80% специфического киллинга С3-4. Тем не менее, только 20% киллинга той же мишени достигали эффекторами, выделенными из мышей, иммунизированных β-гал в ССРХ, при таком же отношении Э:М (фиг.2a). Поскольку ни EL4, ни Р815 не экспрессируют продукты гена МНС класса II и лизис показывает сингенную рестрикцию, эти ova- и β -гал-специфические эффекторы являются рестриктированными ГКГ класса I.
Для демонстрации полезности антигенной композиции мышей иммунизировали растворимым ова, капсулированным в два типа липосом, один из которых был рН-чувствительной липосомой. Через одну неделю клетки селезенки стимулировали in vitro, как описано выше, и испытывали по сравнению с51Сr-меченым EG7-ova или EL4. Фиг.3 показывает характерный результат, демонстрирующий, что ova в липосоме не может примировать мышей для заметной индукции иммунного ЦТЛ-ответа. Похожие результаты наблюдали, когда для иммунизации использовали ova в квасцах.
Пример 2: Распознавание эпитопа ЦТЛ
Carbone and Bevan, смотри выше,
показали, что ЦТЛ, индуцированные в мышах C57BL/6 трансфектантом
ЕG7-ова и цитоплазматически ова-нагруженными спленоцитами, распознают клетки EL4, покрытые пептидом ова 258-276. Для определения того,
индуцирует ли растворимый овальбумин в АК похожие иммунные
ЦТЛ-ответы, клетки селезенки получали от иммунизированных мышей и стимулировали in vitro ЕG7-ова. Эффекторы испытывали против клеток EL4,
покрытых пептидом 253-276 ова или контрольным пептидом,
полученным из базального белка миелина (ББМ 84-102). Результаты показывают, что ова-АК праймировали ЦТЛ со специфичностью, подобной
специфичности ЦТЛ, праймированных трансфектантами, или
цитоплазмически наполненных ова (фиг.1a, 1b и 1c). Клетки-эффекторы, праймированные ова-АК, эффективно лизировали ЕG7-ова и нетрансфектированные
EL4-клетки, покрытые 50 мкг/108 клеток
ова-пептида, но не лизировали EL4-клетки, покрытые 50 мкг/108 клеток ББМ-пептида.
Carbone and Bevan, смотри выше, указывали, что в β-галактозидазной системе β -гал-экспрессирующий трансфектант и спленоциты, цитоплазматически нагруженные растворимой β-галактозидазой, индуцировали ЦТЛ, которые лизировали β-гал-экспрессирующий трансфектант и нетрансфектантные клетки Р815, покрытые гидролизованной щелочью β-галактозидазой. Растворимая β-галактозидаза индуцирует ЦТЛ, имеющие специфичность, схожую со специфичностью ЦТЛ при иммунизации в АК (фиг.2).
Пример 3: Эффекторы ЦТЛ являются CD88-T-клeткaми
То что растворимые белковые антигены в АК
индуцируют СD8+-эффекторы-Т-клетки,
было показано следующим образом. Спленоциты от иммунизированных мышей культивировали в течение пяти дней с облученными трансфектантами in vitro. После
этого времени клетки собирали и истощали на
СD4+- или CD8+-T-клeтки, используя моноклональные антитела против CD4 или CD8 плюс комплемент. Истощенные популяции испытывали по
сравнению с51Сr-ЕG7-ова в
ова-системе или51Cr-P13.1 в β-гал-системе. Данные, показанные на фиг.4, указывают, что в ова-системе истощение Т-клеток на CD8
отменяет цитотоксическую активность,
присвоенную всей популяцией клеток-эффекторов. Тем не менее, истощение популяции Т-клеток на CD4+ не оказывает действия на лизис ЕG7-ова.
Аналогично в β-гал-системе обеднение Т-клеток на CD8+ отменяет цитолитическую активность клеток селезенки, иммунизированных β-гал-антигенной композицией.
Пример 4: Растворимый ова в AF
примирует СD8+-Т-клетки
Для демонстрации того, что система ова-АК примирует популяции СD8+-Т-клеток in vivo и что она критическая для
вторичной иммунной реакции in
vitro, селезенки иммунизированных ова-АК мышей и нативных мышей истощали на популяции CD44 и CD88. Эти обработанные популяции затем стимулировали in
vitro только ЕG7-ова или
ЕG7-ова в комбинации с CD4+ и CD8+-T-клетками мышей, иммунизированных ова-АК, или с различными комбинациями CD4+ или CD8+-T-клеток
мышей, иммунизированных
ова-АК, с CD4+ и CD8+-клeткaми нативных мышей. Фиг. 5 показывает, что праймированные CD8+-клeтки важны для проявления вторичного иммунного
ЦТЛ-ответа in vitro. Эти
данные показывают также, что для эффективного вторичного иммунного ЦТД-ответа in vitro требуются CD4+-T-клeтки. CD4+-T-клетки не нужны для примирования.
Аналогично, CD8+-T-клeтки были нужны для демонстрации β-гал-специфического вторичного иммунного ЦТЛ-ответа in vitro.
Приведенные выше примеры показывают влияние антигенной композиции на индукцию иммунного ответа в виде рестриктированных по классу I ЦТЛ против растворимых белковых антигенов. Медиированный антигенной композицией растворимый антиген индуцировал примирование ЦТЛ и оно сходно по активности с примированием, индуцированным трансфектантами и спленоцитами, цитоплазматически нагруженными растворимым ова или β-гал. В овальбуминовой системе, ЕG7-ова, цитоплазматически нагруженные ова спленоциты и ова-АК индуцировали: (а) рестриктированные СD8+-ЦТЛ класса I; (b) ЦТЛ, которые распознают мишень, сенсибилизированную синтетическим пептидом ова 253-276, и (с) долгоживущие ЦТЛ только после одной иммунизации. В β-галактозидазной системе β-гал-АК индуцировал ЦТЛ, которые распознают β-гал-экспрессирующий трансфектант С3-4 и также нетрансфектированные клетки Р815, сенсибилизированные β-гал, гидролизованный щелочью. То есть аналогично тому, что наблюдали с ЦТЛ, индуцированными иммунизацией клетками селезенки, цитоплазматически загруженными β-галактозидазой. Индукция ова-специфических ЦТЛ антигенной композицией необычна, поскольку ни ова, капсулированный в рН-чувствительную липосому, ни квасцы не могут индуцировать примирование ЦТЛ in vivo.
Эти примеры показывают, что используемая выше антигенная композиция и ее эквиваленты, полезные в терапии человека и в разработке вакцины для индукции ЦТЛ в разных видах рака и вирусных болезнях.
Пример 5
Это специфический пример для показа использования вышеуказанной АК для продуцирования
рестриктированных ЦТЛ
класса I, праймируемых растворимым gp120 из ВИЧ.
Экспрессирующая gр120 IIIB-клеточная линия (15-12) была продуцирована в полученной из фибробластов мышей Balb/c клеточной линии 3Т3. Ее получали от Drs. Ron Germain and Jay Berzofsky, National Institute of Health, Bethesda, M. D. Экспрессирующую gр160 клеточную линию использовали для стимуляции in vitro примированных in vivo лимфоцитов селезенки и использовали также в качестве мишени для индукции gр160-специфичных ЦТЛ. Мышей Balb/c иммунизировали один раз 10 мкг gр160 на мышь с АК или без нее. Мышей инъецировали в подушечки лапок и основание хвоста подкожно. Клетки селезенки брали у иммунизированных мышей через две недели после иммунизации и стимулировали in vitro облученными gр160-трансфектантами. После пяти дней культивирования in vitro примировали оценивали по присутствию специфических эффекторов, способных лизировать gр160-трансфектанты, но не нетрансфектированные клеточные линии. Результаты приводятся на фиг.7, где иммунные ЦТЛ-ответы потенциировали АК и gr120.
Следующий пример демонстрирует использование антигенных композиций этого изобретения с использованием только одного или двух компонентов. Эти примеры показывают, что иммунные ЦТЛ-ответы можно индуцировать только двумя из указанных выше трех компонентов.
Пример 6:
Определение критических
компонентов, необходимых для индукции ЦТЛ
Для определения того, все ли указанные выше компоненты необходимы для индукции антигенспецифичных ЦТЛ, мышей иммунизировали
овальбумином в
микрофлюидизированной композиции различных комбинаций двух из трех компонентов, присутствующих в указанных выше АК. Используемые комбинации двух компонентов были следующими:
сквалан/твин в PBS,
сквалан/плуроник в PBS или плуроник/твин в фосфатно-буферном растворе. Включали другую серию групп, где мышей иммунизировали ова в однокомпонентной композиции, т.е. композиции
только сквалана в
фосфатно-буферном растворе, плуроника в PBS или твина в фосфатно-буферном растворе. Указанную выше трехкомпонентную антигенную композицию модифицировали для исключения одного
компонента, одновременно
заменяя его на фосфатно-буферный раствор.
Указанные выше антигенные композиции состоят из:
0,300 г твина 80 (Aldrich, WI), 1,875 г плуроника L121
(BASF, NJ) и 7,5 г сквалана
(Aldrich, WI), объем композиции доводили до 50 мл добавлением фосфатно-буферного раствора.
Двухкомпонентные композиции были:
Сквалан/твин: 0,300 г
твина 80 и 7,5 г сквалана,
объем композиции доводили до 50 мл добавлением фосфатно-буферного раствора.
Плуроник/твин: 1,875 г плуроника L121 и 0,300 г твина 80, объем композиции доводили до 50 мл добавлением фосфатно-буферного раствора.
Плуроник/сквалан: 1,875 г плуроника L121 и 7,5 г сквалана, объем композицию доводили до 50 мл добавлением фосфатно-буферного раствора.
Образцы затем пропускали через микрофлюидизатор, модель 110Т, Microfluidics соrр., разливали по сосудам и хранили при 4oС до использования.
Овальбумин (Sigma, МО) взвешивали и получали раствор его концентрации 0,3 мг/мл в ССРХ (Whittaker, смотри выше). Исходный раствор 0,3 мг/мл комбинировали с двухкомпонентной композицией в следующих количествах: 5 частей раствора овальбумина концентрации 0,3 мг/мл, 3,3 части двухкомпонентной композиции и 1,7 части ССРХ.
Композицию интенсивно перемешивали и выдерживали на льду до инъецирования. Все растворы смешивали непосредственно перед инъекцией.
Каждая мышь получала 200 мкл одной композиции, содержащей 30 мкл ова, инъекцией в подушечки обеих задних лап и любое оставшееся количество раствора инъецировали подкожно в основание хвоста. Мышей оставляли для отдыха в течение от двух до четырех недель до отбора клеток селезенки.
Через две недели после иммунизации получали клетки селезенки и стимулировали in vitro облученным ЕG7-ова. Через пять дней после культивирования присутствие ова-специфических ЦТЛ измеряли испытанием по сравнению с51Сr-ЕG7-ова или51Cr-EL4 в 4-часовом анализе на выделение51Сr. Результаты, показанные на фиг.8-10, демонстрируют, что овальбумин в композиции микрофлюидизированной двухкомпонентной системы может примировать ова-специфические ЦТЛ in vivo.
Мы далее оценивали относительный вклад индивидуальных компонентов в их способность индуцировать ЦТЛ при комбинировании с белковыми антигенами. Для целей иммунизации растворимый антиген смешивали с микрофлюидизированными наполнителями для получения стабильной гомогенной эмульсии с пределами размера частиц 250-300 нм. Для дальнейшего определения компонентов композиции сквалан-твин 80-плуроник (СТП), ответственных за индукцию ЦТЛ, иммунизировали мышей ова в смеси сквалан-твин 80 (СТ), смеси плуроник-твин 80 (ПТ) или смеси сквалан-плуроник (СП) и в качестве контроля в сквалане (С), твине 80 (Т) или плуронике (П). Мышей также иммунизировали ова-SAFm (содержал 70 мг МДП) или ова-квасцы в качестве адъювантного контроля. Для положительного контроля мышей иммунизировали клетками селезенки, цитоплазматически наполненными растворимым ова. Использовали также другие комбинации и замещения, результаты приводятся в таблице 1.
Для определения примирования ЦТЛ мышей иммунизировали один раз. Через две недели после иммунизации клетки смешивали с облученными EG7-oвa (ова-экспрессирующими EL4-клетками) в течение пяти дней и тестировали по сравнению с51Сr-ЕG7-ова или51Cr-EL4-клетками. Результаты (фиг.11) показывают, что 30 мкг ова в комбинации с СТП или СТ примируют иммунный ответ рестриктированных по классу I ЦТЛ в мышах. Примирование ова-специфических ЦТЛ ова в СТП или ова в СТ, по-видимому, лучше, чем примирование, индуцированное клетками селезенки, цитоплазматически загруженными растворимым ова. Ова в ПТ или в СП может индуцировать ова-специфичный иммунный ЦТЛ-ответ у мышей, но ответ нестойкий и недостаточный. Подобно SAFm, добавление МДП в композицию СТ не компрометирует ова-специфическую индукцию ЦТЛ у мышей (таблица 2). Никакой ова-специфической индукции ЦТЛ не было, когда мышей индуцировали ова в смеси с индивидуальными компонентами С, П или Т, ни когда мышей иммунизировали ова-SAFm или ова-квасцы. Мыши, иммунизированные вплоть до 1 мг ова в (а) ССРХ, в (b) SAFm или (с) адсорбированного на квасцах, не примировали ова-специфичные ЦТЛ.
Пример 7: Компоненты, необходимые для продуцирования ова-специфических антител
Мыши были иммунизированы три раза с интервалами две недели
30 мкг ова в ССРХ, СТП, СТ, ПТ или СП. В качестве положительного контроля мыши были иммунизированы ова-SAFm, так как известно, что SAFm индуцирует сильный гуморальный иммунный ответ. Через семь дней
после второй и третьей иммунизации мышам делали кровопускание и сыворотку испытывали на ова-специфичный гуморальный иммунный ответ. Результаты приводятся в таблице 3. Они показывают, что мыши,
иммунизированные ова в СТП, СР или в SAFm, показывают похожие гуморальные иммунные ответы против ова после двух иммунизаций.
Пример 8: Индукция ЦТЛ, специфических для gр120 ВИЧ
Gp120 IIIB ВИЧ использовали в качестве второй антигенной системы для определения индукции ЦТЛ в СТП, СТ или в МП-Т. Мыши были иммунизированы 1 мкг gp120 IIIb в ССРХ, СТП, РТ или в СТ. В
качестве
контрольных опытов мыши были иммунизированы 1 мкг gp120 IIIb в SAFm или ПАФ (полный адъювант Фрейнда) или в RIBI-адъювантной системе, содержащей МФЛ (монофосфориллипид А) и ДМТ (димиколит
трегалозы).
Через три недели после иммунизации приготовляли клетки селезенки и стимулировали in vitro обработанными митомицином клетками-трансфектантами 15-12 или пептидом 18IIIb. После пятидневного
культивирования получаемые клетки-эффекторы испытывали по сравнению с коровьей оспой: gр160 IIIB или родоначальными инфицированными коровьей оспой клетками Р815 в качестве мишеней. Результаты
показывают, что композиция gр120-сквалан-твин 80 и композиция не gр120-сквалан-твин 80-плуроник или gр120-ССРХ индуцировали gр120-специфический иммунный ЦТЛ-ответ у мышей (таблица 4).
Пример 9: Индукция gr120-специфического гуморального иммунного ответа у мышей
Для индукции gр12-специфических гуморальных иммунных ответов мыши были иммунизированы 1 мкг gр120IIIb три раза с
интервалами две недели. Животным пускали кровь и испытывали на присутствие антител IgG, детектирующих gp120IIIb, анализом ELISA. Результаты показывают, что gp120-СТ является лучшим иммуногеном, чем
gp120-ССРХ, gp120-SAFm (таблица 5) или gр120-СТП.
Пример 10: gp120-специфические гуморальные иммунные ответы у обезьян
Обезьяны (две на группу) были иммунизированы gp120-SAFm,
gр120-СПТ, gp120-СТ или gр120-ССРХ. В качестве контроля группу обезьян иммунизировали рекомбинантной коровьей оспой, содержащей gp160 IIIb. Обезьян иммунизировали с интервалами две недели и кровь
отбирали через две недели и три недели после второй иммунизации. Пре- и иммунную сыворотки каждой обезьяны серийно разбавляли и анализировали на активность против gр120 по ELISA, как описано в
материалах и способах. Данные (фиг.12) показывают, что обезьяны, иммунизированные gр120-СТП или gp120-SAFm, индуцировали схожие иммунные ответы. Одна обезьяна, иммунизированная gр120-СТ, индуцировала
гуморальный иммунный ответ против gр120, похожий на ответ группы, иммунизированной gp120-SAFm или gр120-СПТ. Одна обезьяна, иммунизированная gр120-СТ, индуцировала сильный гуморальный иммунный ответ
против gр120 после двух иммунизаций.
Пример 11: Активность АК in vivo в комбинации с белком Е7 16 ВИЧ
1. Генерация рекомбинантного белка Е7 16 ВИЧ для иммунизации
a)
PCR и клонирование гена Е7
Ген Е7 16 ВИЧ клонировали из плазмиды, полученной от Dr. Raren Vousden (Ludwig Institute), кодирующей ген Е7, полученной из клеточной линии карциномы CaSki.
Кодирующие области были амплифицированы PCR с использованием праймеров, которые кодируют 5'- и 3'-концы генов, фланкированных клонирующими сайтами Bam HI и Sal I. Продукт Е7 PCR лигировали в
экспрессирующий вектор pGEX - 4Т-1 (Pharmacia Biotech), получая экспрессирующую плазмиду pGEX. E7. Штамм Е. coli XLI - голубой (стратаген) трансфектировали экспрессирующей плазмидой pGEX. E7.
Последовательность E7 получали из плазмид образующихся колоний, она была идентична последовательности E7, полученной из клеток CaSki.
b) Получение и очистка бактериально
экспрессированного Е7
Бактериальная экспрессирующая плазмида pGEX. E7 кодирует слитый белок глутатион-S-трансферазы (GST), состоящий из GST у аминоконца, сайта расщепления тромбинпротеазы и
белка E7 у карбоксиконца. Белок E7 получали и очищали, как описано в информационной литературе к продукту от производителя вектора pGEX-4T-1 (Pharmacia Biotech). Говоря кратко, бактерии, содержащие
вектор экспрессии pGEX. E7, индуцировали для экспрессии слитого белка добавлением в культуральную среду изопропил-β-D-тиогалактозидазы. Клетки собирали и лизировали мягкой ультразвуковой
обработкой. Лизат вводили в глутатионсефарозу 4В (Pharmacia Biotech). После связывания слитого белка с матрицей смолу промывали для удаления неспецифически связанных белков. Связанный слитый белок
гидролизовали тромбином для выделения белка E7 из слитого белка глутатион-S-трансферазы (GST).
Препарат белка E7 анализировали SDS-PAGE и концентрацию белка E7 определяли анализом Bradforda (BioRad). На литр бактериальной культуры получали 9 мг растворимого белка E7.
2. Генерация трансфектанта E7 Х21
Кодирующие последовательности для белка HPV16 E7
(смотри выше) были встроены в патентованную IDEC эукариотическую экспрессирующую плазмиду INPEP4. В пределах этого вектора экспрессия E7 контролировалась промотор/энхансерными транскрипциональными
элементами цитомегаловируса. Кроме того, первые три нуклеотида кодирующей последовательности E7 были удалены и заменены лидерной последовательностью легкой цепи иммуноглобулина, помещенной
непосредственно против входа (слева) и в рамки считывания с кодирующим районом Е7. После трансфекции в мышиную клеточную линию Х21 отдельные G418-резистентные клоны испытывали при помощи
нозерн-блоттингов на образование матрицы Е7. Каждый клон обнаруживал детектируемую матрицу Е7. Затем выполняли вестерн-блоттинг клеточных лизатов двух из этих клонов, 4Е7 и 1С7 (НОРЕ1 и НОРЕ2
соответственно) и показали образование белка Е7.
3. Активность in vivo иммунизации растворимым антигеном Е7/АК
В этих исследованиях использовали самок мышей фона С3Н (H2к/к, Harlan Sprague Dawley). Животных содержали в соответствии с "Руководством для наблюдения за лабораторными животными и их использованию" (DHHS Publication No. NIH 86-23, Bethesda, MD:NIH,
1985), они получали корм и воду по потребности. В этих исследованиях использовали трансфекцированную Е7 клеточную линию (НОРЕ2 H2к/к). Опухолевую клеточную линия сохраняли серийным
пассированием in vitro.
Было показано, что эта клеточная линия сохраняет Е7-цитоплазматическую антигенную экспрессию, определяемую анализом вестерн-блоттингом, после повторения пассивировании in vitro. Опухоли инициировали у сингенных мышей С3Н подкожной инъекцией 150000 пассированных in vitro клеток.
Опухоли измеряли в двух перпендикулярных направлениях с
интервалами в две недели. Объем опухоли (V) рассчитывали в соответствии со следующей формулой:
V(мм3) = (L x W2), деленная на 2,
где L - длина самой длинной
оси,
измеренной в мм; W - длина перпендикулярной оси (мм).
Данные в таблице 6 представляют как опухолевые мыши (число имеющих опухоли животных по сравнению с общим числом инъецированных животных). Данные на фиг.13 и 14 представляют как средний размер опухоли (мм3) каждой обработанной или контрольной группы. Каждую обработанную группу сравнивали с контрольной группой, которая не получала лечения. Терапию начинали через 10 дней после инокуляции клеток НОРЕ2, когда основная часть опухолей была пальпируемой (приблизительно 50-75 мм2 ). Терапию инициировали иммунизацией мышей растворимым белком Е7 в АК или квасцах-адъювантах (подкожно при общем объеме 0,2 мл). Непосредственно перед иммунизацией АК смешивали в течение 60 секунд с белком Е7 в сбалансированном солевом растворе Хэнкса (ССРХ), так чтобы каждая мышь получала либо 30 мкг, либо 90 мкг белка Е7 в 0,2 мл. Квасцы (Pierce Chemical Со.) смешивали с белком Е7 в соответствии с инструкциями по приготовлению, так чтобы каждое животное получало 90 мкг белка Е7 в 0,2 мл. Животные в группе второй обработки получали вторую иммунизацию через 9 дней (19 дней после инокуляции опухолевых клеток). Бустер-иммунизацию проводили непосредственно перед инокуляцией, как описано выше.
В этом примере (таблица 6: Хр #233) через 41 день после инокуляции опухолевых клеток только 4/8 и 5/8 мышей, получивших одну инъекцию растворимого Е7 в АК (30 мкг или 90 мкг соответственно), имели измеримые опухоли. Напротив, все из мышей, иммунизированных белком Е7 в квасцах (8/8) имели активно растущие опухоли. Дополнительно, как показано на фиг.13, значительное ингибирование роста опухоли наблюдали только в группах обработки, иммунизированных белком Е7 в АК, по сравнению с контрольными (необработанными) или обработанными квасцами группами. Ингибирование роста опухоли (фиг. 13) или повышенные скорости регресса опухоли (таблица 6) не наблюдали у мышей, которые получали одну инъекцию Е7 в квасцах.
Похожие результаты наблюдали также, используя группы обработки, которые получали две иммунизации, на 10 и 19 день после контрольного заражения опухолью (таблица 6 и фиг. 14), хотя некоторое замедление роста опухоли наблюдали у мышей, получивших две инъекции Е7 в квасцах.
Результаты показывают, что значительную противоопухолевую активность, измеряемую пониженным числом имеющих опухоли мышей, и ингибирование роста опухоли наблюдали после иммунизации растворимым Е7 в АК. Наоборот, все животные, иммунизированные одной или двумя инъекциями растворимого белка Е7 в квасцах, имели растущие опухоли. Говоря кратко, иммунизация растворимым белком Е7 в АК привела к значительному ингибированию роста опухолевых клеток, что не наблюдали при использовании иммунизации растворимым Е7 в квасцах.
Реферат
Изобретение относится к области иммунологии, в частности иммунотерапии. Для решения данной задачи используют антиген папилломавируса, выбранного из группы антигенов HPV16 E6, HPV16 E7, HPV18 E6, HPV18 E7, HPV6 E4, HPV6 L1, HPV11 E4 и HPV11 L1. Помимо антигена композиция может включать стабилизирующий детергент, мицеллообразующий агент, биоразлагаемое и биосовместимое масло. Композиция используется для лечения цервикального рака, остроконечной кондиломы. Использование композиции позволяет повысить иммунный ответ на антиген папилломавируса. 3 с. и 5 з.п.ф-лы, 6 табл., 19 ил.
Комментарии