Аутопроцессирующиеся растения и части растений - RU2312144C2
Код документа: RU2312144C2
Чертежи











Описание
Связанные заявки
В настоящей заявке испрашивается приоритет заявки рег. №60/215281, поданной 27 августа 2001, которая вводится в настоящее описание посредством ссылки.
Область, к которой относится изобретение
В общих чертах, настоящее изобретение относится к молекулярной биологии растений, а более конкретно к созданию растений, которые экспрессируют процессирующий фермент, придающий нужные свойства растению или его продуктам.
Предшествующий уровень техники
Ферменты используются для обработки ряда сельскохозяйственных продуктов, таких как древесина, фрукты и овощи, крахмалы, соки т.п. Обычно процессирующие ферменты продуцируют и выделяют из различных источников в промышленном масштабе, например, путем микробной ферментации (под действием α-амилазы Bacillus) или выделения из растения (с помощью β-галактозидазы кофе или папаина из частей растения). Ферментные препараты используют в различных методах обработки путем смешивания фермента и субстрата при соответствующей влажности, температуре и в определенный период времени и механического перемешивания в целях осуществления ферментативной реакции в промышленных масштабах. Методы включают отдельные стадии продуцирования фермента, производство ферментного препарата, смешивания фермента и субстрата и помещение полученной смеси в подходящие условия, стимулирующие прохождение ферментативной реакции. При этом подходящим и предпочтительным является метод, который позволяет сэкономить время и энергию, сократить стадию смешивания, снизить финансовые затраты и/или стоимость продуцирования ферментов либо улучшить результаты или получить новые продукты. Одним из примеров процедуры, которая нуждается в усовершенствовании, является помол кукурузы.
В настоящее время кукурузу размалывают с получением кукурузного крахмала и других побочных продуктов, образующихся при размоле кукурузы, таких как кукурузный глютеновый продукт, кукурузная глютеновая мука или кукурузное масло. Крахмал, полученный таким способом, часто подвергают дальнейшей обработке с получением других продуктов, таких как дериватизированные крахмалы и сахара, или ферментации с получением различных продуктов, включая спирты или молочную кислоту. Обработка кукурузного крахмала часто предусматривает использование ферментов, а в частности ферментов, которые расщепляют крахмал и превращают его в ферментируемые сахара или фруктозу (под действием α- и глюкоамилазы, α-глюкозидазы, глюкозоизомеразы и т.п.). Современные промышленные способы требуют больших фмнансовых затрат, поскольку для осуществления обработки кукурузы в промышленном масштабе необходимо конструирование очень больших мельниц, что не отвечает приемлемому экономическому показателю "затраты - эффективность". Кроме того, способ требует отдельного производства гидролизующих или модифицирующих крахмал ферментов с последующим использованием оборудования для смешивания данного фермента и субстрата в целях продуцирования продуктов из гидролизованного крахмала.
Способ получения крахмала из кукурузного зерна хорошо известен и включает осуществление процесса мокрого помола. Мокрый помол кукурузы включает стадии замачивания кукурузного зерна, размола этого кукурузного зерна и отделения компонентов этого зерна. Зерно замачивают в чане для замачивания с обратным потоком воды примерно при 120°F, и зерно оставляют в чане для замачивания на 24-48 часов. Вода в чане для замачивания обычно содержит диоксид серы при концентрации примерно 0,2 мас.%. В этом способе диоксид серы используют для снижения микробного роста, а также для восстановления дисульфидных связей в белках эндосперма в целях более эффективного отделения крахмала от белка. Обычно на один бушель кукурузы используют примерно 0,59 галлонов воды для замачивания. Вода для замачивания идет в отходы и часто содержат нежелательные уровни остаточного диоксида серы.
Затем замоченное зерно обезвоживают и подвергают размолу в серии мельниц тонкого помола. В первой серии мельниц тонкого помола происходит дробление зерна, в результате чего из этого зерна высвобождается зародыш. Промышленные мельницы тонкого помола, подходящие для крупномасштабного мокрого помола, имеются в продаже под торговым знаком Bauer. Для отделения зародыша от остальной части зерна используют центрифугирование. Широко используемым промышленным центрифужным сепаратором является центрифужный сепаратор Merco. Мельницы тонкого помола и центрифужные сепараторы являются дорогостоящим оборудованием, которое потребляет много энергии.
В следующей стадии этого процесса оставшиеся компоненты зерна, включая крахмал, шелуху, волокно и клейковину, загружают на другую серию мельниц тонкого помола и пропускают через серию промывочных сит для отделения волокнистых компонентов от крахмала и клейковины (белка эндосперма). Крахмал и клейковина проходят через сита, а волокно не проходит через сита. Центрифугирование или третья стадия измельчения с последующим центрифугированием используется для отделения крахмала от белка эндосперма. В результате центрифугирования продуцируется крахмальная суспензия, которую обезвоживают, а затем промывают свежей водой и сушат до достижения примерно 12% влажности. В основном чистый крахмал обычно подвергают дополнительной обработке с использованием ферментов.
Отделение крахмала от других компонентов зерна осуществляют в целях удаления кожуры семян и белков зародыша и эндосперма, что способствует эффективному контактированию крахмала с процессирующими ферментами, а поэтому полученные продукты гидролиза, в основном, не содержат примесей других компонентов зерна. Такое разделение позволяет эффективно выделять другие компоненты зерна, которые могут затем поступать в продажу как вспомогательные продукты для повышения дохода, получаемого от использования мельницы.
После выделения крахмала в результате мокрого помола этот крахмал обычно подвергают таким стадиям обработки, как желатинизация, разжижение и декстринизация, в целях продуцирования мальтодекстрина, и последующим стадиям осахаривания, изомеризации и очистки в целях продуцирования глюкозы, мальтозы и фруктозы.
Желатинизация применяется при гидролизе крахмала, поскольку доступные в настоящее время ферменты не могут быстро гидролизовать кристаллический крахмал. Для того, чтобы сделать крахмал доступным для гидролитических ферментов, этот крахмал обычно приготавливают в виде водной суспензии (20-40% твердых веществ) и нагревают при подходящей температуре гелеобразования. Для кукурузного крахмала температура составляет 105-110°С. Желатинизированный крахмал обычно является очень вязким, а поэтому его разбавляют в следующей стадии, называемой разжижением. При разжижении некоторые связи между глюкозными молекулами крахмала разрываются, и это происходит либо под действием фермента, либо под действием кислоты. В этой стадии и в последующей стадии декстринизации используются термостабильные ферменты эндо-α-амилазы. Степень гидролиза регулируется в стадии декстринизации, в результате чего получают продукты гидролиза с нужным процентом декстрозы.
Последующий гидролиз декстриновых продуктов после стадии разжижения осуществляют с использованием различных экзоамилаз и деветвящих ферментов, в зависимости от нужного продукта. И наконец, если необходимо получить фруктозу, то обычно используют иммобилизованный фермент глюкозоизомеразу для превращения глюкозы во фруктозу.
Осуществление сухого помола для получения ферментируемых сахаров (например, с последующим получением этанола) из кукурузного крахмала способствует эффективному контактированию экзогенных ферментов с крахмалами. Этот способ требует меньших финансовых затрат, чем способ мокрого помола, однако было бы желательно получить еще большую экономическую выгоду, поскольку, в большинстве случаев, побочные продукты, образующиеся в указанных процессах, часто не являются такими ценными, как продукты, получаемые при мокром помоле. Так, например, при сухом помоле кукурузы зерно измельчается в порошок, что облегчает эффективный контакт крахмала с расщепляющими ферментами. После гидролиза кукурузной муки ферментами остаточные твердые вещества имеют определенную пищевую ценность, поскольку они содержат белки и некоторые другие компоненты. Недавно в работе Eckhoff, озаглавленной "Fermentation and costs of fuel ethanol from corn with quick-germ process" (Appl. Biochem. Biotechnol., 94:41 (2001)), были описаны возможности усовершенствования и существующие проблемы, связанные с сухим помолом. Метод "быстрого отделения зародыша" позволяет отделять богатый маслом зародыш от крахмала с использованием меньшего времени замачивания.
Одним из примеров растения, где регуляция и/или уровень эндогенных процессирующих ферментов может приводить к получению желаемого продукта, является сладкая кукуруза. Типичные сорта сладкой кукурузы отличаются от полевой кукурузы тем, что сладкая кукуруза не способна к биосинтезу нормальных уровней крахмала. Обычно, для ограничения биосинтеза крахмала, сорта сладкой кукурузы подвергают генетическим мутациям в генах, кодирующих ферменты, участвующие в биосинтезе крахмала. Такие мутации присутствуют в генах, кодирующих крахмалсинтазу и АДФ-глюкозопирофосфорилазу (такие как мутации в сахарных и суперсахарных сортах кукурузы). Фруктоза, глюкоза и сахароза, которые представляют собой простые сахара, необходимые для продуцирования приятного сладкого вкуса, являющегося предпочтительным для потребителя свежей кукурузы, аккумулируются в развивающемся эндосперме таких мутантов. Однако, если уровень накопления крахмала является слишком высоким, то есть если эту кукурузу оставляют на слишком длительный срок для созревания (поздний урожай), или если эта кукуруза, перед ее употреблением, хранится в течение очень длительного периода времени, то этот продукт теряет свою сладость и приобретает крахмалистый привкус и нежелательные органолептические свойства. Поэтому окно сбора урожая для сладкой кукурузы является слишком узким, и ее срок хранения ограничен.
Другая серьезная проблема, с которой сталкиваются фермеры, выращивающие сладкую кукурузу, связана с тем, что ценность этих сортов кукурузы ограничивается исключительно их использованием в пищевых продуктах. Если фермер захочет заранее, во время развития семян, собрать урожай сладкой кукурузы для ее использования в качестве пищевого продукта, то он понесет значительные убытки в урожае. Низкий выход и плохое качество зерна сладкой кукурузы можно объяснить двумя фундаментальными причинами. Первая причина заключается в том, что мутации в пути биосинтеза крахмала оказывают ингибирующее действие на механизмы биосинтеза крахмала, и зерно не наливается полностью, что приводит к снижению выхода и качества зерна. Во вторых, из-за высокого содержания сахаров в зерне и неспособности этих сахаров секвестироваться в качестве крахмалов влагостойкость семян снижается, что приводит к еще большему снижению срока сохранения питательных веществ в зерне. Эндоспермы семян сортов сладкой кукурузы сморщиваются и сплющиваются, но не подвергаются присущей им десикации и становятся восприимчивыми к болезням. Кроме того, низкое качество семян сладкой кукурузы связано с другими агрономическими проблемами, такими как низкая жизнеспособность семян, плохое прорастание семян, восприимчивость саженцев к болезням и плохая ранняя всхожесть, обусловленная комбинацией факторов, вызываемых неадекватным накоплением крахмала. Таким образом, плохое качество продуктов сладкой кукурузы экономически отражается на потребителях, фермерах/производителях, распространителях продукции и семеноводах.
Таким образом, что касается сухого помола, то необходимо разработать такой способ, который позволил бы повысить эффективность процесса и/или повысить ценность побочных продуктов. Что касается мокрого помола, то необходимо разработать такой способ обработки крахмала, который позволил бы избежать использования оборудования, требующегося для продолжительного замачивания, измельчения, помола и/или отделения компонентов зерна. Так, например, необходимо модифицировать или исключить стадию замачивания при мокром помоле, что позволит снизить количество сбросных вод, требующих удаления, и, тем самым, сэкономить энергию и время и увеличить производительность помола (время нахождения зерна в чанах для замачивания должно снижаться). Также необходимо исключить или усовершенствовать способ отделения крахмалсодержащего эндосперма от зародыша.
Краткое описание изобретения
Настоящее изобретение относится к самопроцессирующимся растениям и к частям таких растений, а также к способам их использования. Самопроцессирующиеся растения и части растения настоящего изобретения способны экспрессировать и активировать фермент(ы) (мезофильный, термофильный или гипертермофильный ферменты). После активации фермента(ов) (мезофильного, термофильного или гипертермофильного фермента) растение или часть растения обладает способностью к аутопроцессингу субстрата, что позволяет достичь нужного результата.
Настоящее изобретение относится к выделенному полинуклеотиду, (а) содержащему последовательность SEQ ID NO:2, 4, 6, 9, 19, 21, 25, 37, 39, 41, 43, 46, 48, 50, 52 или 59 или комплементарную последовательность, или к полинуклеотиду, который гибридизуется с последовательностью, комплементарной любой из последовательностей SEQ ID NO:2, 4, 6, 9, 19, 21, 25, 37, 39, 41, 43, 46, 48, 50, 52 или 59, в условиях низкой степени жесткости, и который кодирует полипептид, обладающий α-амилазной, пуллуланазной, α-глюкозидазной, глюкозоизомеразной или глюкоамилазной активностью, или (b) кодирующему полипептид, содержащий последовательность SEQ ID NO:10, 13, 14, 15, 16, 18, 20, 24, 26, 27, 28, 29, 30, 33, 34, 35, 36, 38, 40, 42, 44, 45, 47, 49 или 51, или его ферментативно активный фрагмент. Предпочтительно, выделенный полинуклеотид кодирует гибридный полипептид, содержащий первый полипептид и второй пептид, где указанный первый полипептид обладает α-амилазной, пуллуланазной, α-глюкозидазной, глюкозоизомеразной или глюкоамилазной активностью. Более предпочтительно, второй пептид содержит пептид сигнальной последовательности, который может нацеливать первый полипептид на вакуоль, эндоплазматический ретикулум, хлоропласт, крахмальные зерна, семена или клеточные стенки растения. Например, сигнальной последовательностью может быть N-концевая сигнальная последовательность, происходящая от восковидной кукурузы (waxy), N-концевая сигнальная последовательность, происходящая от γ-зеина, крахмалсвязывающий домен или С-концевой крахмалсвязывающий домен. Кроме того, настоящее изобретение охватывает полинуклеотид, который гибридизуется с последовательностью, комплементарной любой из последовательностей SEQ ID NO:2, 9 или 52, в условиях низкой степени жесткости, и кодирует полипептид, обладающий α-амилазной активностью; полинуклеотид, который гибридизуется с последовательностью, комплементарной любой из последовательностей SEQ ID NO:4 или 25, в условиях низкой степени жесткости, и кодирует полипептид, обладающий пуллуланазной активностью; полинуклеотид, который гибридизуется с последовательностью, комплементарной последовательности SEQ ID NO:6, и кодирует полипептид, обладающий α-глюкозидазной активностью; полинуклеотид, который гибридизуется с последовательностью, комплементарной любой из последовательностей SEQ ID NO:19, 21, 37, 39, 41 или 43, в условиях низкой степени жесткости, и кодирует полипептид, обладающий глюкозоизомеразной активностью; полинуклеотид, который гибридизуется с последовательностью, комплементарной любой из последовательностей SEQ ID NO:46, 48, 50 или 59, в условиях низкой степени жесткости, и кодирует полипептид, обладающий глюкоамилазной активностью.
Кроме того, настоящее изобретение относится к экспрессирующей кассете, содержащей полинуклеотид, (а) имеющий последовательность SEQ ID NO:2, 4, 6, 9, 19, 21, 25, 37, 39, 41, 43, 46, 48, 50, 52 или 59 или комплементарную последовательность, или полинуклеотид, который гибридизуется с последовательностью, комплементарной любой из последовательностей SEQ ID NO:2, 4, 6, 9, 19, 21, 25, 37, 39, 41, 43, 46, 48, 50, 52 или 59 или комплементарную последовательность в условиях низкой степени жесткости, и кодирует полипептид, обладающий α-амилазной, пуллуланазной, α-глюкозидазной, глюкозоизомеразной или глюкоамилазной активностью, или (b) кодирующий полипептид, содержащий последовательность SEQ ID NO:10, 13, 14, 15, 16, 18, 20, 24, 26, 27, 28, 29, 30, 33, 34, 35, 36, 38, 40, 42, 44, 45, 47, 49 или 51, или его ферментативно активный фрагмент. Предпочтительно, экспрессирующая кассета дополнительно содержит промотор, функционально присоединенный к полинуклеотиду, такой как индуцибельный промотор, тканеспецифический промотор или, предпочтительно, эндосперм-специфический промотор. Предпочтительным эндосперм-специфическим промотором является промотор γ-зеина кукурузы или промотор ADP-gpp кукурузы. В предпочтительном варианте осуществления изобретения промотор содержит последовательность SEQ ID NO:11 или SEQ ID NO:12. Кроме того, в другом предпочтительном варианте осуществления изобретения указанный полинуклеотид находится в смысловой ориентации по отношению к этому промотору. Экспрессирующая кассета согласно изобретению может дополнительно кодировать сигнальную последовательность, которая функционально присоединена к полипептиду, кодируемому указанным полинуклеотидом. Сигнальная последовательность предпочтительно нацеливает функционально присоединенный полипептид на вакуоль, эндоплазматический ретикулум, хлоропласт, крахмальные зерна, семена или на клеточные стенки растения. Предпочтительными сигнальными последовательностями являются N-концевая сигнальная последовательность, происходящая от восковидной кукурузы (waxy), N-концевая сигнальная последовательность, происходящая от γ-зеина, или крахмалсвязывающий домен.
Кроме того, настоящее изобретение относится к вектору или к клетке, содержащей экспрессирующие кассеты согласно изобретению. Клетка может быть выбрана из группы, состоящей из Agrobacterium, клетки однодольного растения, клетки двудольного растения, клетки растения семейства лилейных (Liliopsida), клетки растения семейства просовых (Panicoideae), клетки кукурузы и клетки злаков. Предпочтительной является клетка кукурузы.
Кроме того, настоящее изобретение охватывает растение, стабильно трансформированное векторами настоящего изобретения. Настоящее изобретение относится к растению, стабильно трансформированному вектором, содержащими α-амилазу, имеющую любую из аминокислотных последовательностей SEQ ID NO:1, 10, 13, 14, 15, 16, 33 или 35 или кодируемую полинуклеотидом, имеющим любую из последовательностей SEQ ID NO:2 или 9. Предпочтительной α-амилазой является гипертермофильная α-амилаза.
В другом варианте своего осуществления настоящее изобретение относится к растению, стабильно трансформированному вектором, содержащим пуллуланазу, имеющую любую из аминокислотных последовательностей SEQ ID NO:24 или 34 или кодируемую полинуклеотидом, содержащим последовательность SEQ ID NO:4 или 25. Кроме того, настоящее изобретение относится к растению стабильно трансформированному вектором, содержащими α-глюкозидазу, имеющую любую из аминокислотных последовательностей SEQ ID NO:26 или 27 или кодируемую полинуклеотидом, содержащим последовательность SEQ ID NO:6. Предпочтительной α-глюкозидазой является гипертермофильная α-глюкозидаза. Кроме того, настоящее изобретение относится к растению, стабильно трансформированному вектором, содержащим глюкозоизомеразу, имеющую любую из аминокислотных последовательностей SEQ ID NO:18, 20, 28, 29, 30, 38, 40, 42 или 44 или кодируемую полинуклеотидом, содержащим любую из последовательностей SEQ ID NO:19, 21, 37, 39, 41 или 43. Предпочтительной глюкозоизомеразой является гипертермофильная глюкозоизомераза. В другом варианте своего осуществления настоящее изобретение относится к растению, стабильно трансформированному вектором, содержащим глюкозоамилазу, имеющую любую из аминокислотных последовательностей SEQ ID NO:45, 47 или 49 или кодируемую полинуклеотидом, содержащим любую из последовательностей SEQ ID NO:46, 48, 50 или 59. Предпочтительно глюкозоамилаза является гипертермофильной.
Кроме того, настоящее изобретение относится к растительным продуктам, таким как семена, плоды или зерно, происходящее от стабильно трансформированных растений настоящего изобретения.
В другом варианте своего осуществления настоящее изобретение относится к трансформированному растению, геном которого пополнен за счет рекомбинантного полинуклеотида, кодирующего, по крайней мере, один процессирующий фермент, функционально присоединенный к промоторной последовательности, где последовательность указанного полинуклеотида оптимизирована для экспрессии в растении. Растением может быть однодольное растение, такое как кукуруза, или двудольное растение. Предпочтительным растением является злаковое растение или растение, культивируемое в коммерческих целях. Процессирующий фермент выбирают из группы, состоящей из α-амилазы, глюкоамилазы, глюкозоизомеразы, глюканазы, β-амилазы, α-глюкозидазы, изоамилазы, пуллуланазы, неопуллуланазы, изопуллуланазы, амилопуллуланазы, целлюлазы, экзо-1,4-β-целлобиогидролазы, экзо-1,3-β-D-глюканазы, β-глюкозидазы, эндоглюканазы, L-арабиназы, α-арабинозидазы, галактаназы, галактозидазы, маннаназы, маннозидазы, ксиланазы, ксилозидазы, протеазы, глюканазы, ксиланазы, эстеразы, фитазы и липазы. Предпочтительным процессирующим ферментом является крахмалпроцессирующий фермент, выбранный из группы, состоящей из α-амилазы, глюкоамилазы, глюкозоизомеразы, β-амилазы, α-глюкозидазы, изоамилазы, пуллуланазы, неопуллуланазы, изопуллуланазы и амилопуллуланазы. Более предпочтительно, указанный фермент выбирают из α-амилазы, глюкоамилазы, глюкозоизомеразы, α-глюкозидазы и пуллуланазы. Предпочтительно, чтобы процессирующий фермент был дополнительно гипертермофильным. В соответствии с этим аспектом настоящего изобретения ферментом может быть некрахмалразрушающий фермент, выбранный из группы, состоящей из протеазы, глюканазы, ксиланазы, эстеразы, фитазы и липазы. Кроме того, такие ферменты могут быть гипертермофильными. В предпочтительном варианте осуществления изобретения указанный фермент аккумулируется в вакуоли, эндоплазматическом ретикулуме, хлоропласте, крахмальных зернах, семенах или в клеточных стенках растения. Кроме того, в другом варианте осуществления изобретения геном растения может быть дополнительно пополнен вторым рекомбинантным полинуклеотидом, содержащим негипертермофильный фермент.
В другом своем аспекте настоящее изобретение относится к трансформированному растению, геном которого пополнен за счет рекомбинантного полинуклеотида, кодирующего, по крайней мере, один процессирующий фермент, выбранный из группы, состоящей из α-амилазы, глюкоамилазы, глюкозоизомеразы, α-глюкозидазы и пуллуланазы, и функционально присоединенному к промоторной последовательности, где последовательность указанного полинуклеотида оптимизирована для экспрессии в растении. Предпочтительно процессирующий фермент является гипертермофильным и происходит из кукурузы.
В другом своем варианте настоящее изобретение относится к трансформированному растению кукурузы, геном которого пополнен за счет рекомбинантного полинуклеотида, кодирующего, по крайней мере, один процессирующий фермент, выбранный из группы, состоящей из α-амилазы, глюкоамилазы, глюкозоизомеразы, α-глюкозидазы и пуллуланазы, и функционально присоединенному к промоторной последовательности, где последовательность указанного полинуклеотида оптимизирована для экспрессии в растении кукурузы. Предпочтительным процессирующим ферментом является гипертермофильный фермент.
Настоящее изобретение относится к трансформированному растению, геном которого пополнен за счет рекомбинантного полинуклеотида, имеющего последовательность SEQ ID NO:2, 9 или 52, и функционально присоединенному к промотору и к сигнальной последовательности. Кроме того, описано трансформированное растение, геном которого пополнен за счет рекомбинантного полинуклеотида, имеющего последовательность SEQ ID NO:4 или 25, и функционально присоединенному к промотору и к сигнальной последовательности. В другом своем варианте настоящее изобретение относится к трансформированному растению, геном которого пополнен за счет рекомбинантного полинуклеотида, имеющего последовательность SEQ ID NO:6, и функционально присоединенному к промотору и к сигнальной последовательности. Кроме того, описано трансформированное растение, геном которого пополнен за счет рекомбинантного полинуклеотида, имеющего последовательность SEQ ID NO:19, 21, 37, 39, 41 или 43. Описано трансформированное растение, геном которого пополнен за счет рекомбинантного полинуклеотида, имеющего последовательность SEQ ID NO:46, 48, 50 или 59.
В настоящей заявке также рассматриваются продукты трансформированных растений. Такими продуктами являются, например, семя, плод или зерно. Альтернативно, таким продуктом может быть процессирующий фермент, крахмал или сахар.
Описано также растение, полученное из стабильно трансформированных растений настоящего изобретения. В этом аспекте настоящего изобретения указанным растением может быть гибридное растение или инбредное растение.
В другом варианте осуществления изобретения крахмальная композиция включает, по крайней мере, один процессирующий фермент, которым является протеаза, глюканаза или эстераза. Предпочтительным ферментом является гипертермофильный фермент.
В другом варианте осуществления изобретения зерно содержит, по крайней мере, один процессирующий фермент, которым является α-амилаза, пуллуланаза, α-глюкозидаза, глюкоамилаза или глюкозоизомераза. Предпочтительным ферментом является гипертермофильный фермент.
В другом своем варианте настоящее изобретение относится к способу получения крахмальных зерен, включающему обработку зерна, которое содержит, по крайней мере, один не процессирующий крахмал фермент, в условиях, способствующих активации, по крайней мере, одного фермента; получение смеси, содержащей крахмальные зерна и некрахмальные продукты разложения, где указанное зерно получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один фермент; и выделение крахмальных зерен из смеси. Таким ферментом предпочтительно является протеаза, глюканаза, ксиланаза, фитаза или эстераза. Более того, указанный фермент, предпочтительно, является гипертермофильным. Зерно может быть раздроблено и/или обработано в условиях низкой или высокой влажности. Альтернативно, зерно может быть обработано диоксидом серы. Кроме того, настоящее изобретение может, предпочтительно, включать выделение некрахмальных продуктов из этой смеси. Далее также описаны крахмалопродукты и некрахмалопродукты, полученные этим способом.
В еще одном своем варианте настоящее изобретение относится к способу продуцирования сверхсладкой кукурузы, включающему в себя обработку трансформированного растения или его части, геном которого экспрессируется в эндосперме и пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один крахмалразрушающий или крахмализомеризирующий фермент, в условиях, способствующих активации по меньшей мере одного фермента и, тем самым, превращению полисахаридов в сахар в этой кукурузе, и получение сверхсладкой кукурузы. Экспрессирующая кассета предпочтительно дополнительно содержит промотор, функционально присоединенный к полинуклеотиду, кодирующему фермент. Промотором может быть, например, конститутивный промотор, семяспецифический промотор или эндоспермспецифический промотор. Предпочтительным ферментом является гипертермофильный фермент. Более предпочтительным ферментом является α-амилаза. Используемая здесь экспрессирующая кассета может дополнительно содержать полинуклеотид, который кодирует сигнальную последовательность, функционально присоединенную к полинуклеотиду, кодирующему по меньшей мере один фермент. Сигнальная последовательность может направлять гипертермофильный фермент, например, в апопласт или в эндоплазматический ретикулум. Фермент предпочтительно включает в себя любую из последовательностей SEQ ID NO:13, 14, 15, 16, 33 или 35.
В своем наиболее предпочтительном варианте настоящее изобретение относится к способу продуцирования сверхсладкой кукурузы, включающему в себя обработку трансформированного растения кукурузы или его части, геном которого пополнен за счет экспрессирующей кассеты, причем указанная кассета, кодирующая α-амилазу, экспрессируется в эндосперме в условиях, способствующих активации по меньшей мере одного фермента и, тем самым, превращению полисахаридов в сахар в этой кукурузе; и получение сверхсладкой кукурузы. Предпочтительным ферментом является гипертермофильный фермент, и эта гипертермофильная α-амилаза содержит аминокислотную последовательность SEQ ID NO:10, 13, 14, 15, 16, 33 или 35 или ее ферментативно активный фрагмент, обладающий α-амилазной активностью.
Описан способ получения раствора гидролизованного крахмалопродукта, включающий в себя обработку части растения, содержащей крахмальные зерна и по меньшей мере один процессирующий фермент в условиях, способствующих активации по меньшей мере одного фермента и, тем самым, процессингу крахмальных зерен, с образованием водного раствора, содержащего гидролизованный крахмалопродукт, где указанная часть растения получена из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один крахмалпроцессирующий фермент; и сбор водного раствора, содержащего гидрализованный крахмалопродукт. Гидрализованный крахмалопродукт может содержать декстрин, мальтоолигосахарид, глюкозу и/или их смеси. Предпочтительным ферментом является α-амилаза, α-глюкозидаза, глюкоамилаза, пуллуланаза, амилопуллуланаза, глюкозоизомераза или любая их комбинация. Более того, предпочтительным ферментом является гипертермофильный фермент. В другом аспекте настоящего изобретения геном части растения может быть дополнительно пополнен за счет экспрессирующей кассеты, кодирующей негипертермофильный крахмалпроцессирующий фермент. Негипертермофильный крахмалпроцессирующий фермент может быть выбран из группы, состоящей из амилазы, глюкоамилазы, α-глюкозидазы, пуллуланазы, глюкозоизомеразы или их комбинации. В еще одном аспекте настоящего изобретения процессирующий фермент, предпочтительно, экспрессируется в эндосперме. Предпочтительной частью растения является зерно, и этим зерном является зерно кукурузы, пшеницы, ячменя, ржи, овса, сахарного тростника или риса. При этом предпочтительно, чтобы по меньшей мере один процессирующий фермент был функционально присоединен к промотору и к сигнальной последовательности, которая нацеливает указанный фермент на крахмальные зерна или на эндоплазматический ретикулум, либо на клеточную стенку. Этот способ может дополнительно включать в себя выделение гидролизованного крахмалопродукта и/или ферментацию гидрализованного крахмалопродукта.
В другом аспекте настоящего изобретения описан способ получения гидролизованного крахмалопродукта, включающий обработку части растения, содержащей крахмальные зерна и, по крайней мере, один процессирующий фермент в условиях, способствующих активации, по крайней мере, одного фермента и, тем самым, процессингу крахмальных зерен с образованием водного раствора, содержащего гидролизованный крахмалопродукт, где указанная часть растения получена из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере одну α-амилазу; и сбор водного раствора, содержащего продукт гидролизованного крахмала. Предпочтительно гипертермофильная α-амилаза, а более предпочтительно гипертермофильная α-амилаза содержит любую из аминокислотных последовательностей SEQ ID NO:1, 10, 13, 14, 15, 16, 33 или 35 или ее активный фрагмент, обладающий α -амилазной активностью. При этом предпочтительно, чтобы экспрессирующая кассета содержала полинуклеотид, выбранный из последовательностей SEQ ID NO:2, 9, 46 или 52 или комплементарных последовательностей, или полинуклеотид, который гибридизуется с любой из последовательностей SEQ ID NO:2, 9, 46 или 52 в условиях низкой степени жесткости и кодирует полипептид, обладающий α -амилазной активностью. Кроме того, настоящее изобретение также относится к геному трансформированного растения, который дополнительно содержит полинуклеотид, кодирующий нетермофильный крахмалпроцессирующий фермент. Альтернативно, часть растения может быть обработана негипертермофильным крахмалпроцессирующим ферментом.
Настоящее изобретение также относится к части трансформированного растения, содержащей, по крайней мере, один крахмалпроцессирующий фермент, присутствующий в клетках этого растения, где часть растения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один крахмалпроцессирующий фермент. Предпочтительным ферментом является крахмалпроцессирующий фермент, выбранный из группы, состоящей из α-амилазы, глюкоамилазы, глюкозоизомеразы, β-амилазы, α-глюкозидазы, изоамилазы, пуллуланазы, неопуллуланазы, изопуллуланазы и амилопуллуланазы. Кроме того, указанный фермент является, предпочтительно, гипертермофильным. Растением может быть любое растение, предпочтительно кукуруза.
В другом своем варианте настоящее изобретение также относится к части трансформированного растения, содержащей по меньшей мере один крахмалпроцессирующий фермент, присутствующий в клеточных стенках или в клетках растения, где часть растения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один не процессирующий крахмал фермент или по меньшей мере один фермент, процессирующий полисахарид, не являющийся крахмалом. Этот фермент, предпочтительно, является гипертермофильным. Более того, не процессирующий крахмал фермент предпочтительно выбирают из группы, состоящей из протеазы, глюканазы, ксиланазы, эстеразы, фитазы и липазы. Частью растения может быть любая часть растения, а предпочтительно, колос, семя, плод, зерно, кукурузная солома, чешуя злаков или багасса.
Настоящее изобретение также относится к частям трансформированного растения. Так, например, описана часть трансформированного растения, содержащая α-амилазу, имеющую любую из аминокислотных последовательностей SEQ ID NO:1, 10, 13, 14, 15, 16, 33 или 35, или кодируемую полинуклеотидом, содержащим любую из последовательностей SEQ ID NO: 2, 9, 46 или 52; часть трансформированного растения, содержащая α-глюкозидазу, имеющую любую из аминокислотных последовательностей SEQ ID NO:5, 26 или 27 или кодируемую полинуклеотидом, содержащим последовательность SEQ ID NO:6; часть трансформированного растения, содержащая глюкозоизомеразу, имеющую любую из аминокислотных последовательностей SEQ ID NO:28, 29, 30, 38, 40, 42 или 44 или кодируемую полинуклеотидом, содержащим любую из последовательностей SEQ ID NO:19, 21, 37, 39, 41 или 43; часть трансформированного растения, содержащая глюкоамилазу, имеющую аминокислотную последовательность SEQ ID NO:45 или SEQ ID NO:47 или SEQ ID NO:49 или кодируемую полинуклеотидом, содержащим любую из последовательностей SEQ ID NO:46, 48, 50 или 59; часть трансформированного растения, содержащая пуллуланазу, кодируемую полинуклеотидом, содержащим любую из последовательностей SEQ ID NO:4 или 25.
В другом своем варианте настоящее изобретение относится к способу превращения крахмала в часть трансформированного растения, включающему активацию содержащегося в нем крахмалпроцессирующего фермента. Описаны также крахмал, декстрин, мальтоолигосахарид или сахар, продуцируемые в соответствии с этим способом.
Настоящее изобретение также относится к способу использования части трансформированного растения, содержащей, по крайней мере, один не процессирующий крахмал фермент в клеточной стенке или в клетках части растения, включающему обработку части трансформированного растения, содержащей, по крайней мере, один фермент, процессирующий полисахарид, не являющийся крахмалом в условиях, стимулирующих активацию по меньшей мере одного фермента и, тем самым, способствующих расщеплению полисахарида, не являющегося крахмалом, с образованием водного раствора, содержащего олигосахарид и/или сахара, где указанную часть растения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один фермент, процессирующий олигосахарид, не являющийся крахмалом; и сбор водного раствора, содержащего олигосахариды и/или сахара. Предпочтительный фермент, процессирующий полисахарид, не являющийся крахмалом, представляет собой гипертермофильный фермент.
Способ применения трансформированных семян, содержащих по меньшей мере один процессирующий фермент, включает в себя обработку трансформированных семян, содержащих по меньшей мере одну протеазу или липазу в условиях, стимулирующих активацию по меньшей мере одного фермента, с образованием водной смеси, содержащей аминокислоты и жирные кислоты, где семена получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один фермент; и сбор водной смеси. При этом предпочтительно выделяют аминокислоты, жирные кислоты или и те, и другие. Предпочтительно, чтобы по меньшей мере одна протеаза или липаза была гипертермофильной.
Настоящее изобретение относится к способу получения этанола, включающему в себя обработку части растения, содержащей по меньшей мере один полисахаридпроцессирующий фермент в условиях, стимулирующих активацию по меньшей мере одного фермента и, тем самым, способствующих расщеплению полисахарида, с образованием олигосахарида или ферментируемого сахара, где часть растения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один полисахаридпроцессирующий фермент; и инкубирование ферментируемого сахара в условиях, стимулирующих превращение ферментируемого сахара или олигосахарида в этанол. Предпочтительной частью растения является зерно, плод, семя, стебли, древесина, овощ или корень. Предпочтительно, часть растения получают из растения, выбранного из группы, состоящей из овса, ячменя, пшеницы, ягод, винограда, ржи, кукурузы, риса, картофеля, сахарной свеклы, сахарного тростника, ананаса, травянистых растений и деревьев. В другом предпочтительном варианте осуществления изобретения полисахаридпроцессирующий ферментом является α-амилаза, глюкоамилаза, α-глюкозидаза, глюкозоизомераза, пуллуланаза или их комбинации.
Настоящее изобретение относится к способу получения этанола, включающему обработку части растения, содержащей, по крайней мере, один фермент, выбранный из группы, состоящей из α-амилазы, глюкоамилазы, α-глюкозидазы, глюкозоизомеразы, пуллуланазы или их комбинаций, при нагревании в течение определенного периода времени и в условиях, стимулирующих активацию, по крайней мере, одного фермента и, тем самым, способствующих расщеплению полисахарида с образованием ферментируемого сахара, где часть растения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один фермент; и инкубирование указанного ферментируемого сахара в условиях, стимулирующих превращение ферментируемого сахара в этанол. Предпочтительно, чтобы, по крайней мере, один фермент был гипертермофильным или мезофильным.
В другом своем варианте настоящее изобретение относится к способу получения этанола, включающему обработку части растения, содержащей по крайней мере, один не процессирующий крахмал фермент в условиях, стимулирующих активацию, по крайней мере, одного фермента и, тем самым, способствующих расщеплению не являющегося крахмалом полисахарида в олигосахарид и ферментируемый сахар, где часть растения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один фермент; и инкубирование ферментируемого сахара в условиях, стимулирующих превращение ферментируемого сахара в этанол. Предпочтительным не процессирующим крахмал ферментом является глюканаза, ксиланаза или целлюлаза.
Настоящее изобретение относится к способу получения этанола, включающему обработку части растения, содержащей, по крайней мере, один фермент, выбранный из группы, состоящей из α-амилазы, глюкоамилазы, α-глюкозидазы, глюкозоизомеразы, пуллуланазы или их комбинаций, в условиях, стимулирующих активацию, по крайней мере, одного фермента и, тем самым, способствующих расщеплению полисахарида с образованием ферментируемого сахара, где часть растения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один фермент; и инкубирование указанного ферментируемого сахара в условиях, стимулирующих превращение ферментируемого сахара в этанол. Предпочтительно, чтобы указанный фермент был гипертермофильным.
Кроме того, описан способ получения подслащенного мучного пищевого продукта без добавления дополнительных подсластителей, включающий обработку части растения, содержащей, по крайней мере, один крахмалпроцессирующий фермент, в условиях, стимулирующих активацию, по крайней мере, одного фермента и, тем самым, способствующих процессированию крахмальных зерен в указанной части растения до сахаров с образованием подслащенного продукта, где часть растения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один фермент; процессинг подслащенного продукта, с получением мучного пищевого продукта. Мучной пищевой продукт может быть получен из подслащенного продукта и воды. Кроме того, указанный мучной пищевой продукт может содержать солод, отдушки, витамины, минералы, красители или любые их комбинации. Предпочтительно, чтобы, по крайней мере, один фермент был гипертермофильным. Фермент может быть выбран из α-амилазы, α-глюкозидазы, глюкоамилазы, пуллуланазы, глюкозоизомеразы или любой их комбинации. Растение может быть дополнительно выбрано из группы, состоящей из сои, ржи, овса, ячменя, пшеницы, кукурузы, риса и сахарного тростника. Предпочтительным мучным пищевым продуктом является зерновой пищевой продукт, пищевой продукт для завтрака, пищевой продукт, готовый к употреблению, и выпечка. Процессинг может включать выпекание, кипячение, нагревание, варку на пару, электрический разряд или любую их комбинацию.
Кроме того, настоящее изобретение относится к способу подслащивания крахмалсодержащего продукта без добавления дополнительный подсластителей, включающий обработку крахмала, содержащего, по крайней мере, один крахмалпроцессирующий фермент, в условиях, стимулирующих активацию, по крайней мере, одного фермента и, тем самым, способствующих расщеплению крахмала с образованием подслащенного крахмала, где указанный крахмал получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один фермент; и добавление подслащенного крахмала в продукт с получением пищевого продукта, содержащего подслащенный крахмал. Предпочтительно, трансформированное растение может быть выбрано из группы, состоящей из кукурузы, сои, ржи, овса, ячменя, пшеницы, риса и сахарного тростника.
Предпочтительно, чтобы, по крайней мере, один фермент был гипертермофильным. Более предпочтительно, чтобы, по крайней мере, один фермент представлял собой α-амилазу, α-глюкозидазу, глюкоамилазу, пуллуланазу, глюкозоизомеразу или любую их комбинацию.
Настоящее изобретение относится к мучному пищевому продукту и к продукту, содержащему подслащенный крахмал.
Настоящее изобретение также относится к способу подслащивания полисахаридсодержащих плодов или овощей, включающему обработку плодов или овощей, содержащих, по крайней мере, один полисахаридпроцессирующицй фермент в условиях, стимулирующих активацию, по крайней мере, одного фермента и, тем самым, способствующих процессингу полисахарида в плодах или овощах с образованием сахара и с получением подслащенных плодов или овощей, где плоды или овощи получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один полисахаридпроцессирующий фермент. Плоды или овощи выбирают из группы, состоящей из картофеля, томатов, бананов, тыквы, гороха и бобов. Предпочтительно, чтобы, по крайней мере, один фермент был гипертермофильным.
Настоящее изобретение, кроме того, относится к способу получения сахарсодержащего водного раствора, включающему в себя обработку крахмальных зерен, полученных из части растения в условиях, стимулирующих активацию, по меньшей мере, одного фермента и, тем самым, получение водного раствора, содержащего сахар.
В другом своем варианте настоящее изобретение относится к способу получения производных крахмалопродуктов из зерна, которое не подвергают мокрому или сухому помолу перед выделением крахмалопродуктов, включающему обработку части растения, содержащей крахмальные зерна, и, по крайней мере, один крахмалпроцессирующий фермент в условиях, стимулирующих активацию, по крайней мере, одного фермента и, тем самым, способствующих процессингу крахмальных зерен с образованием водного раствора, содержащего декстрины или сахара, где часть растения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один крахмалпроцессирующий фермент; и сбор водного раствора, содержащего производный крахмалопродукт. Предпочтительно, чтобы, по меньшей мере, один крахмалпроцессирующий фермент был гипертермофильным.
Настоящее изобретение также относится к способу выделения α-амилазы, глюкоамилазы, глюкозоизомеразы, α-глюкозидазы и пуллуланазы, включающему в себя культивирование трансформированного растения и выделения из него α-амилазы, глюкоамилазы, глюкозоизомеразы, α-глюкозидазы и пуллуланазы. Предпочтительно, чтобы указанный фермент был гипертермофильным.
Описан способ получения мальтодекстрина, включающий в себя смешивание трансгенного зерна с водой, нагревание указанной смеси, отделение твердого вещества от полученного декстринового сиропа и сбор мальтодекстрина. Предпочтительно, чтобы трансгенное зерно содержало, по меньшей мере, один крахмалпроцессирующий фермент. Предпочтительным крахмалпроцессирующим ферментом является α-амилаза, глюкоамилаза, α-глюкозидаза и глюкозоизомераза. Кроме того, настоящее изобретение относится к мальтодекстрину, полученному указанным способом, а также к композиции, полученной этим способом.
Настоящее изобретение относится к способу получения декстринов или сахаров из зерна, которое не подвергают механическому разрушению перед выделением крахмалсодержащего продукта, включающему обработку части растения, содержащей крахмальные зерна и, по крайней мере, один крахмалпроцессирующий фермент в условиях, стимулирующих активацию, по крайней мере, одного фермента и, тем самым, способствующих процессингу крахмальных зерен с образованием водного раствора, содержащего декстрины или сахара, где часть растения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один процессирующий фермент; и сбор водного раствора, содержащего сахар и/или декстрины.
Настоящее изобретение относится к способу получения ферментируемого сахара, включающему в себя обработку части растения, содержащей крахмальные зерна и, по меньшей мере, один крахмалпроцессирующий фермент в условиях, стимулирующих активацию по меньшей мере одного фермента и, тем самым, способствующих процессингу крахмальных зерен, с образованием водного раствора, содержащего декстрины или сахара, где часть растения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один процессирующий фермент; и сбор указанного водного раствора, содержащего ферментируемый сахар.
Кроме того, настоящее изобретение относится к растению кукурузы, стабильно трансформированному вектором, содержащим гипертермофильную α-амилазу. Так, например, настоящее изобретение охватывает, предпочтительно, растения кукурузы, стабильно трансформированные вектором, содержащим полинуклеотидную последовательность, которая кодирует α-амилазу и которая на 60% идентична последовательности SEQ ID NO:1 или SEQ ID NO:51.
Краткое описание графического материала
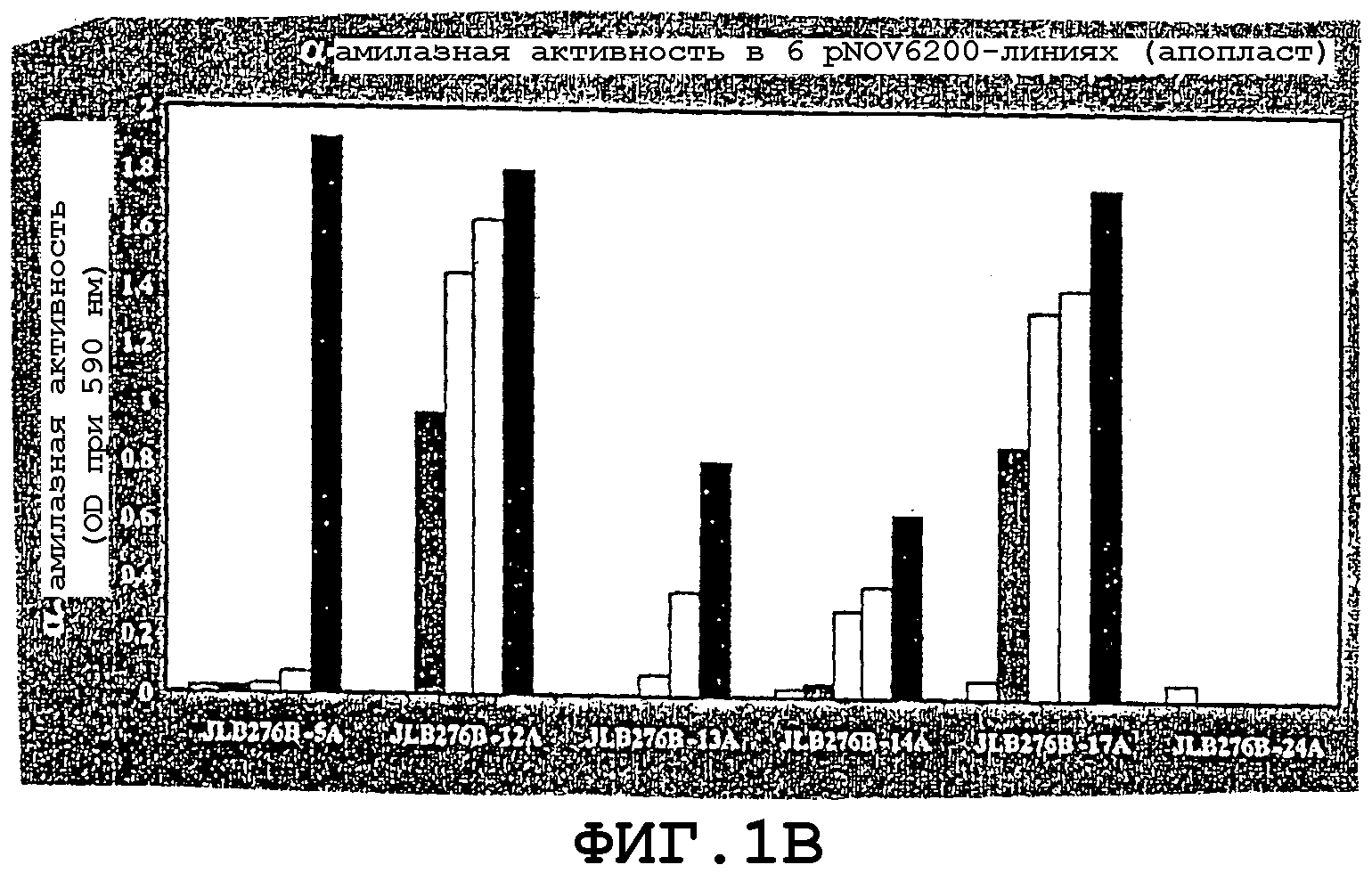
На фигурах 1А и 1В проиллюстрирована активность α-амилазы, экспрессируемой в кукурузном зерне и в эндосперме в результате сегрегации зерен Т1 из растений, трансформированных рNOV6201 и из шести рNOV6200-линий.
На фигуре 2 проиллюстрирована активность α-амилазы в сегрегирующихся зернах Т1 из линий рNOV6201.
На фигуре 3 показано количество этанола, продуцируемого при ферментации биомассы трансгенной кукурузы, содержащей термостабильную альфа-амилазу 797GL3 и подвергаемой разжижению в течение периода времени до 60 минут при 85°С и 95°С. На этой фигуре проиллюстрировано, что выход этанола после 72-часовой ферментации почти не изменялся во время разжижения, составляющего от 15 минут до 60 минут. Кроме того, было показано, что биомасса, полученная путем разжижения при 95°С, продуцировала большее количество этанола в любой момент времени, чем масса, полученная путем разжижения при 85°С.
На фигуре 4 показано количество остаточного крахмала (%) после ферментации биомассы трансгенной кукурузы, содержащей термостабильную альфа-амилазу и подвергаемую разжижению в течение периода времени до 60 минут при 85°С и 95°С. На этой фигуре проиллюстрировано, что выход этанола после 72-часовой ферментации почти не изменялся в течение периода времени разжижения, составляющего от 15 минут до 60 минут. Кроме того, было показано, что биомасса, полученная путем разжижения при 95°С, продуцировала большее количество этанола в любой момент времени, чем масса, полученная путем разжижения при 85°С.
На фигуре 5 проиллюстрирован выход этанола для биомассы трансгенной кукурузы, контрольной кукурузы и их различных смесей, полученных при 85°С и 95°С. На этой фигуре показано, что трансгенная кукуруза, содержащая α-амилазу, дает значительное увеличение продуцирования крахмала, доступного для ферментации, поскольку после ферментации наблюдается снижение остаточного крахмала.
На фигуре 6 показано количество остаточного крахмала, измеренного в осушенном перегонном кубе после ферментации биомассы трансгенного зерна, контрольного зерна и их различных смесей, полученных при 85°С и 95°С.
На фигуре 7 показаны выходы этанола в зависимости от времени ферментации образца, содержащего 3% трансгенной кукурузы, в течение 20-80 часов при различных рН в пределах от 5,2 до 6,4. На фигуре проиллюстрировано, что ферментация, осуществляемая при более низком рН, протекает быстрее, чем при рН 6,0 или выше.
На фигуре 8 показаны выходы этанола в процессе ферментации биомассы, содержащей различные массовые проценты трансгенной кукурузы от 0 до 12 мас.% при различных рН в пределах от 5,2 до 6,4. На этой фигуре показано, что выход этанола не зависит от количества трансгенного зерна, содержащегося в этом образце.
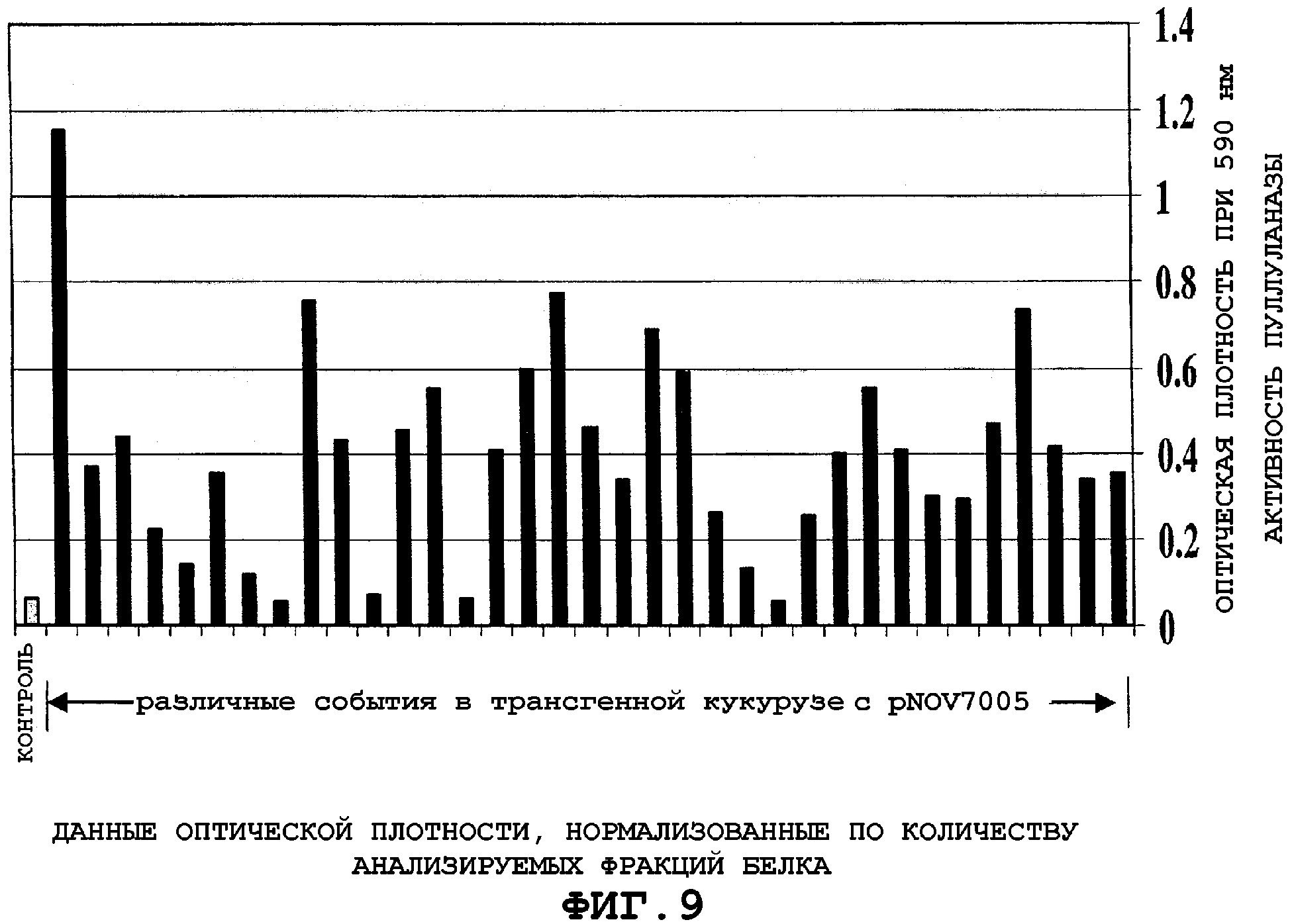
На фигуре 9 проиллюстрирован анализ семян Т2, полученных при различных экспериментах, трансформированных рNOV7005. В ряде случаев может быть детектирована высокая экспрессия пуллуланазной активности по сравнению с не-трансгенным контролем.
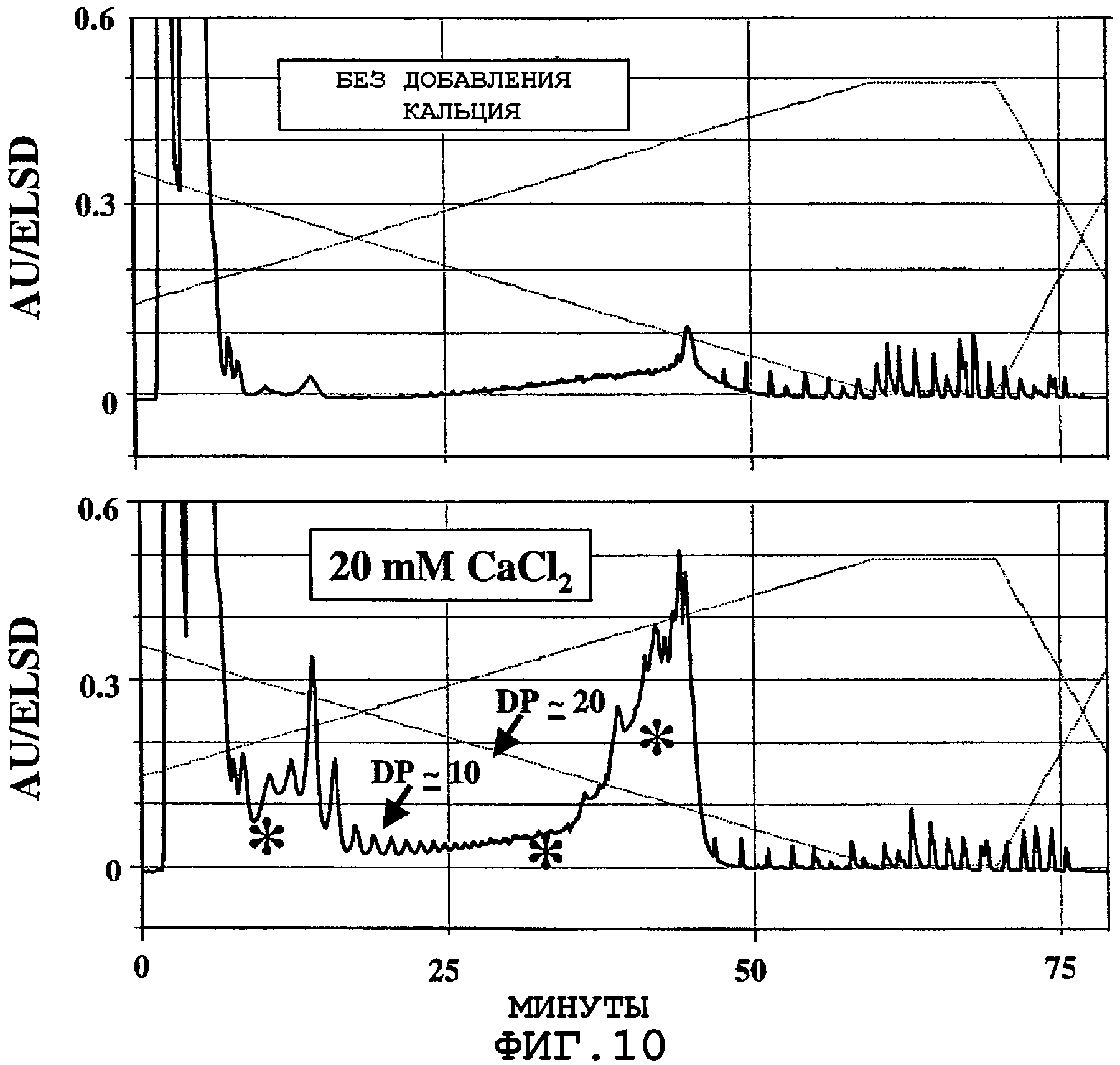
На фигурах 10А и 10В показаны результаты ВЭЖХ-анализа продуктов гидролиза, продуцируемых экспрессированной пуллуланазой из крахмала в муке трансгенной кукурузы. Инкубирование муки из кукурузы, экспрессирующей пуллуланазу, в реакционном буфере при 75°С в течение 30 минут приводило к продуцированию олигосахаридов со средней цепью (степень полимеризации (DP) ˜ 10-30) и с короткими амилозными цепями (DP ˜100-200) из кукурузного крахмала. На фигурах 10А и 10В также показано влияние добавления ионов кальция на активность пуллуланазы.
На фигурах 11А и 11В представлены данные ВЭЖХ-анализа продукта гидролиза крахмала из двух реакционных смесей. Первая реакционная смесь, указанная как "Амилаза", содержит смесь [1:1 (мас./мас.)] образцов кукурузной муки, полученных из трансгенной кукурузы, экспрессирующей α-амилазу, и не-трансгенной кукурузы А188; а вторая реакционная смесь "Амилаза + Пуллуланаза" содержит смесь [1:1 (мас./мас.)] образцов кукурузной муки, полученной из трансгенной кукурузы, экспрессирующей α-амилазу, и из трансгенной кукурузы, экспрессирующей пуллуланазу.
На фигуре 12 представлено количество сахарного продукта (мкг) в 25 мкл реакционной смеси для двух реакционных смесей. Первая реакционная смесь, указанная как "Амилаза", содержит смесь [1:1 (мас./мас.)] образцов кукурузной муки, полученный из трансгенной кукурузы, экспрессирующей α-амилазу, и не-трансгенной кукурузы А188; а вторая реакционная смесь "Амилаза + Пуллуланаза" содержит [1:1 (мас./мас.)] образцов кукурузной муки, полученный из трансгенной кукурузы, экспрессирующей α-амилазу, и из трансгенной кукурузы, экспрессирующей пуллуланазу.
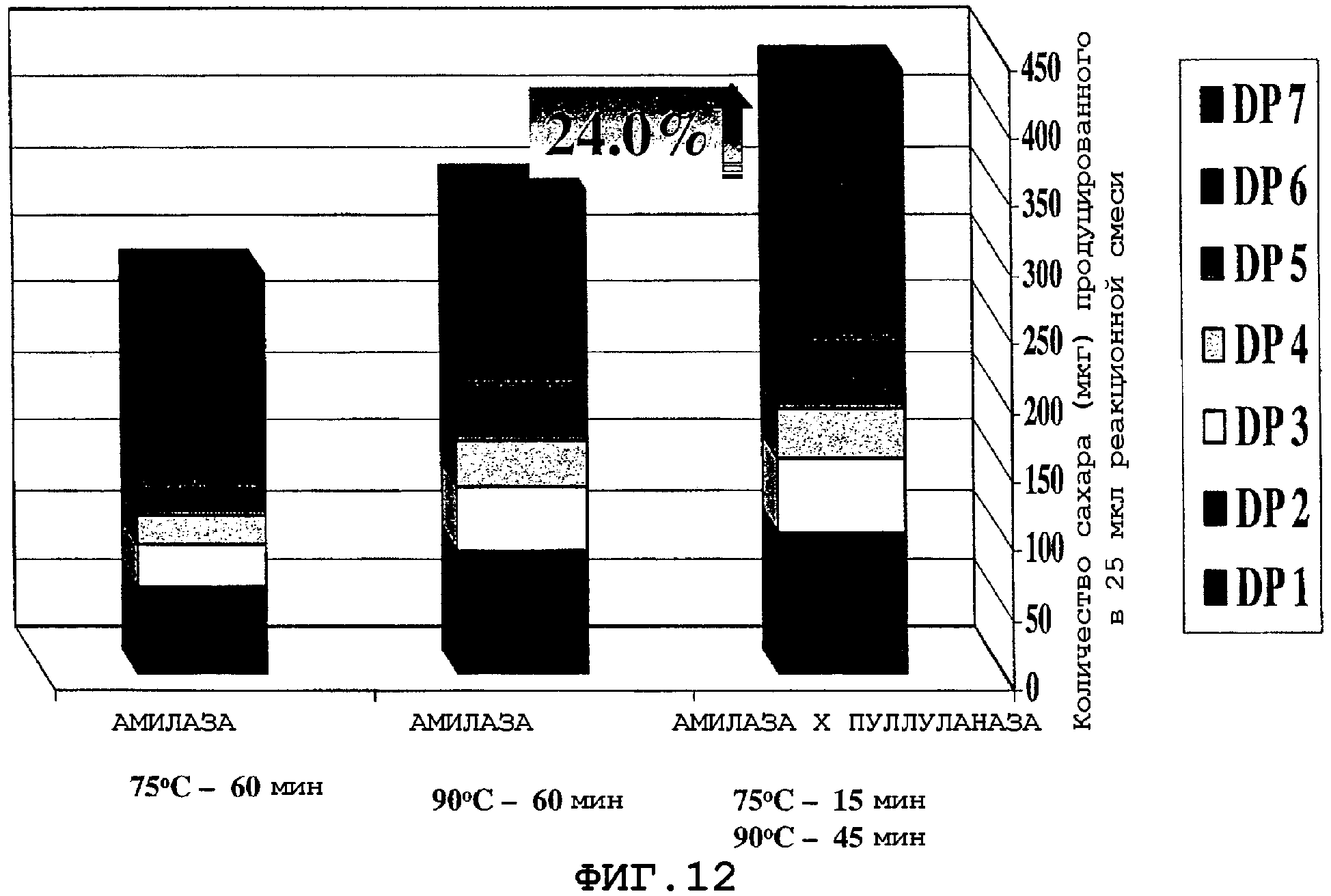
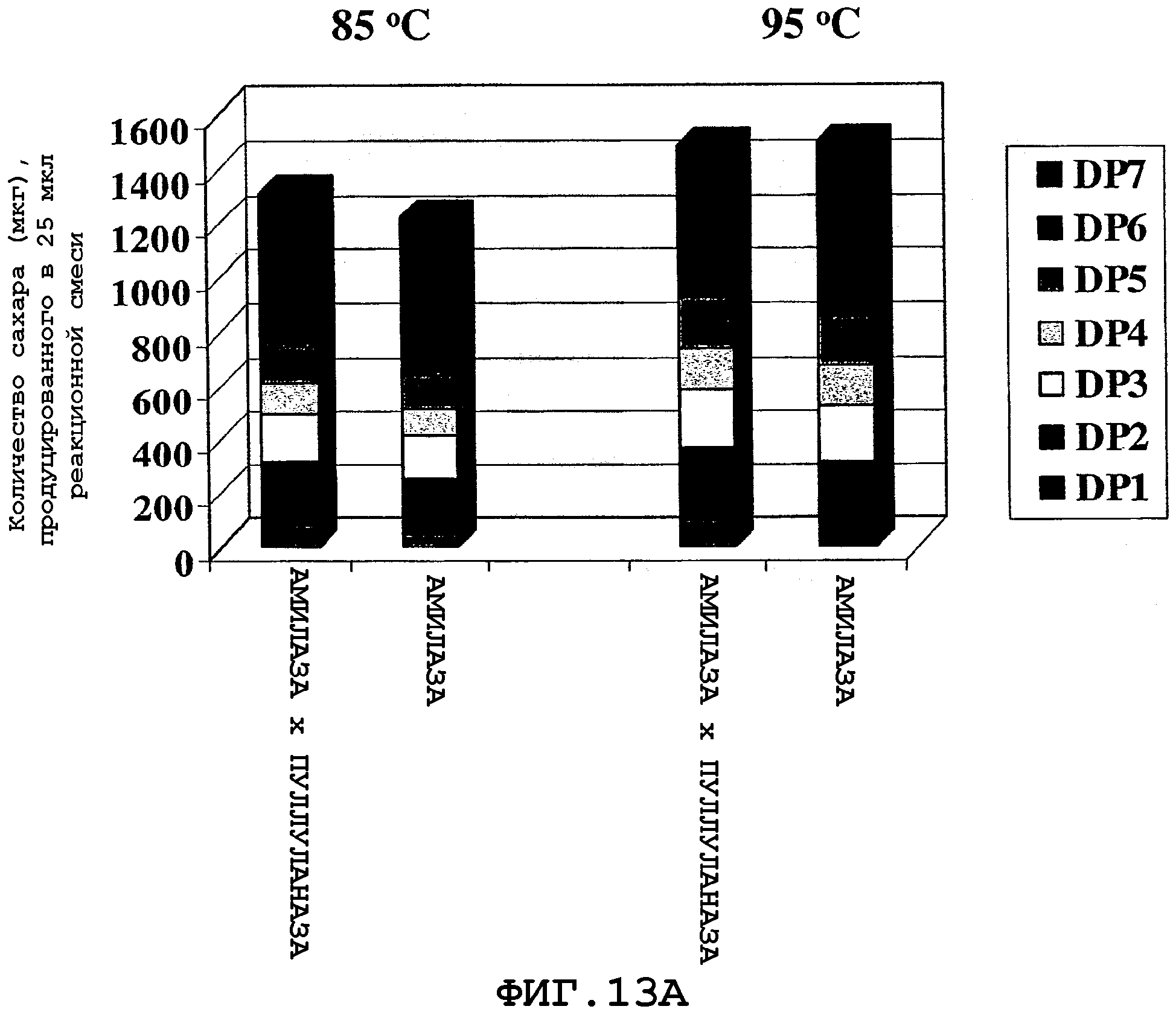
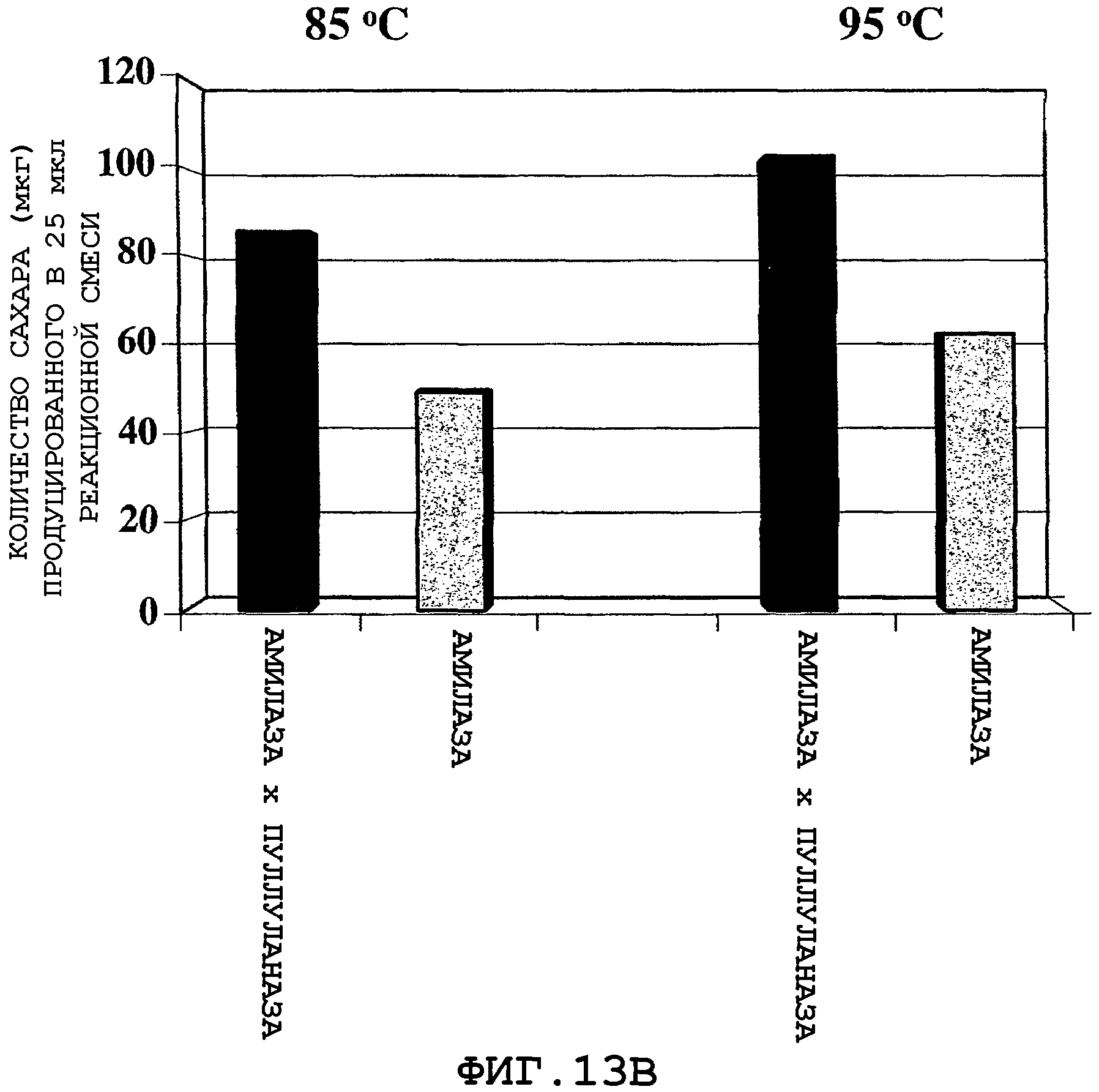
На фигурах 13А и 13В показан продукт гидролиза крахмала, полученного от двух серий реакционных смесей по окончании 30-минутного инкубирования при 85°С и 95°С. Для каждой серии были использованы две реакционные смеси, где первая реакционная смесь, указанная как "Амилаза × пуллуланаза", содержит муку из трансгенной кукурузы (полученной путем перекрестного опыления), экспрессирующей α-амилазу и пуллуланазу, а вторая реакционная смесь, указанная как "Амилаза", содержит смесь образцов кукурузной муки, полученных из трансгенной кукурузы, экспрессирующей α-амилазу, и из не-трансгенной кукурузы А188 в таком отношении, которое дает тот же самый уровень α-амилазной активности, как и уровень активности, наблюдаемый при перекрестном опылении ("Амилаза × Пуллуланаза").
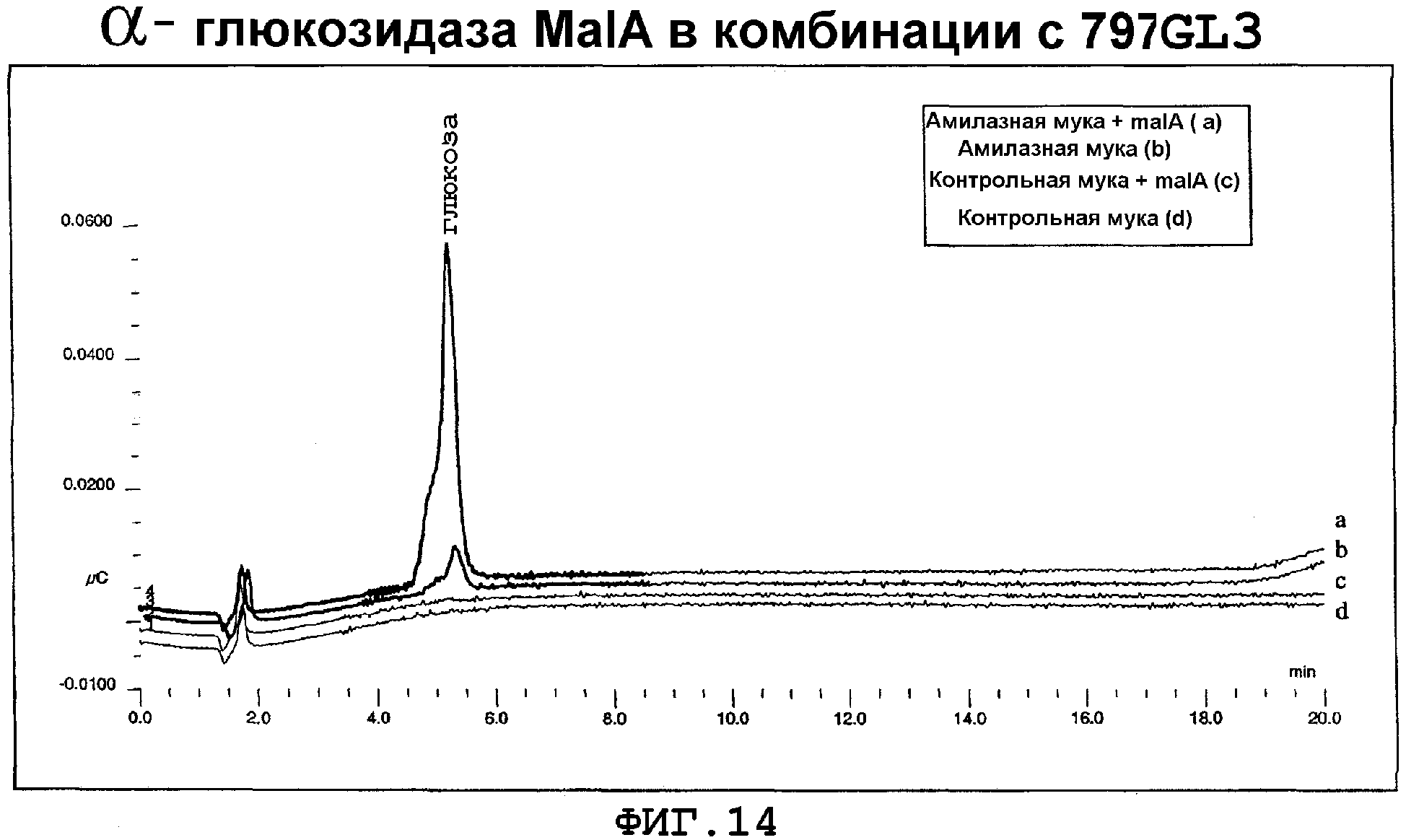
На фигуре 14 показано разложение крахмала до глюкозы с использованием семян не-трансгенной кукурузы (контроль), семян трансгенной кукурузы, содержащей α-амилазу 797GL3, и комбинацию семян трансгенной кукурузы с 797GL3 и семян с α-глюкозидазой MalA.
На фигуре 15 показано превращение исходного крахмала при комнатной температуре или при 30°С. На этой фигуре реакционные смеси 1 и 2 представляют собой комбинации воды и крахмала при комнатной температуре и при 30°С соответственно. Реакционные смеси 3 и 4 представляют собой комбинацию α -амилазы и крахмала ячменя при комнатной температуре и при 30°С соответственно. Реакционные смеси 5 и 6 представляют собой комбинации глюкоамилазы Thermoanaerobacterium и крахмала при комнатной температуре и при 30°С соответственно. Реакционные смеси 7 и 8 представляют собой комбинации α-амилазы ячменя (Sigma), глюкоамилазы Thermoanaerobacterium и крахмала при комнатной температуре и при 30°С соответственно. Реакционные смеси 9 и 10 представляют собой комбинации контрольной α-амилазы ячменя (Sigma) и крахмала при комнатной температуре и при 30°С соответственно. Показана степень полимеризации (DP) продуктов глюкоамилазы Thermoanaerobacterium.
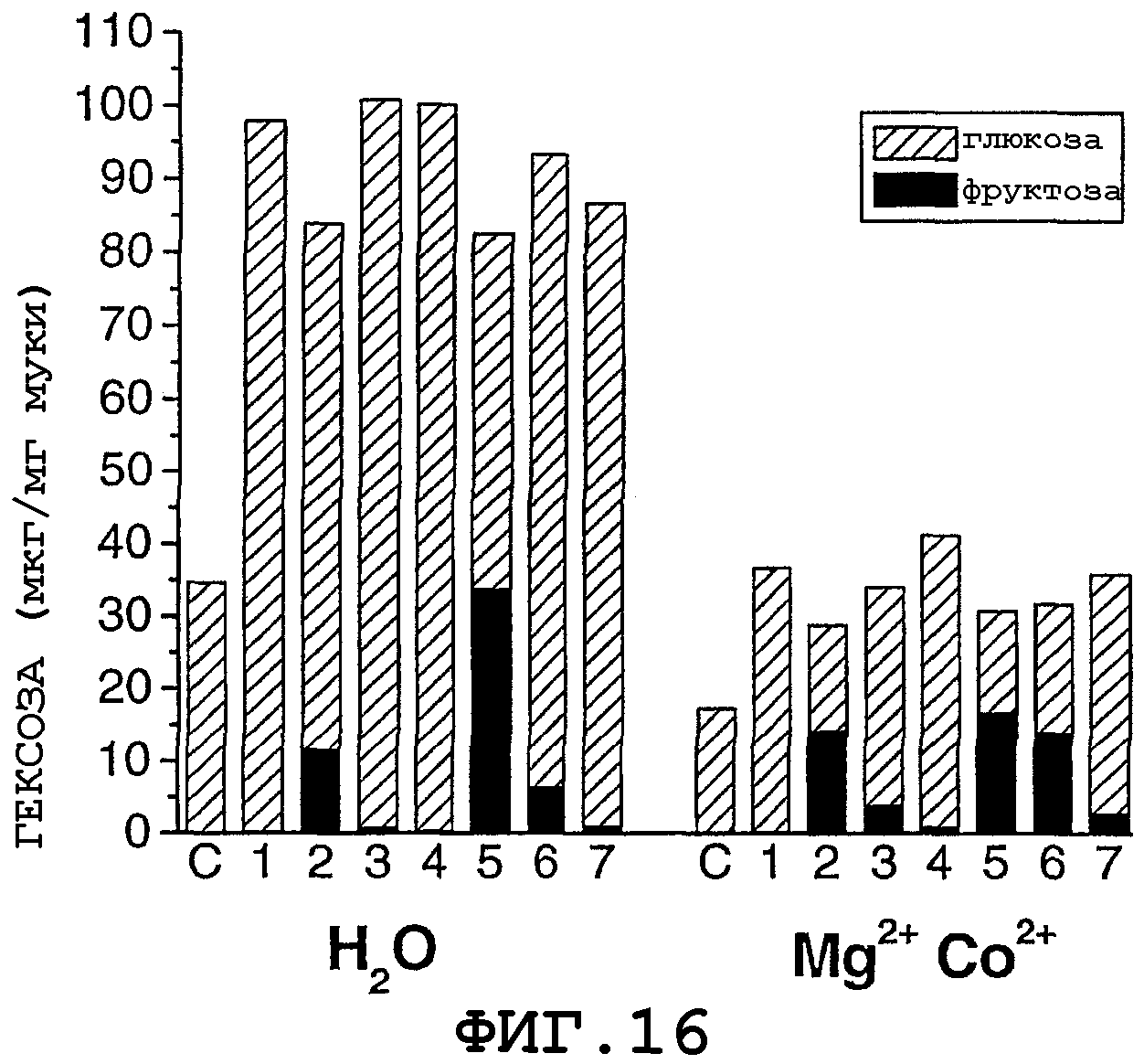
На фигуре 16 показано продуцирование фруктозы из муки амилазной трансгенной кукурузы с использованием комбинации альфа-амилазы, альфа-глюкозидазы и глюкозоизомеразы, как описано в примере 19. Кукурузную муку, содержащую амилазу, смешивали с ферментными растворами + вода или буфер. Все реакционные смеси содержали 60 мг муки с амилазой и всего 600 мкл жидкости и были инкубированы в течение 2 часов при 90°С.
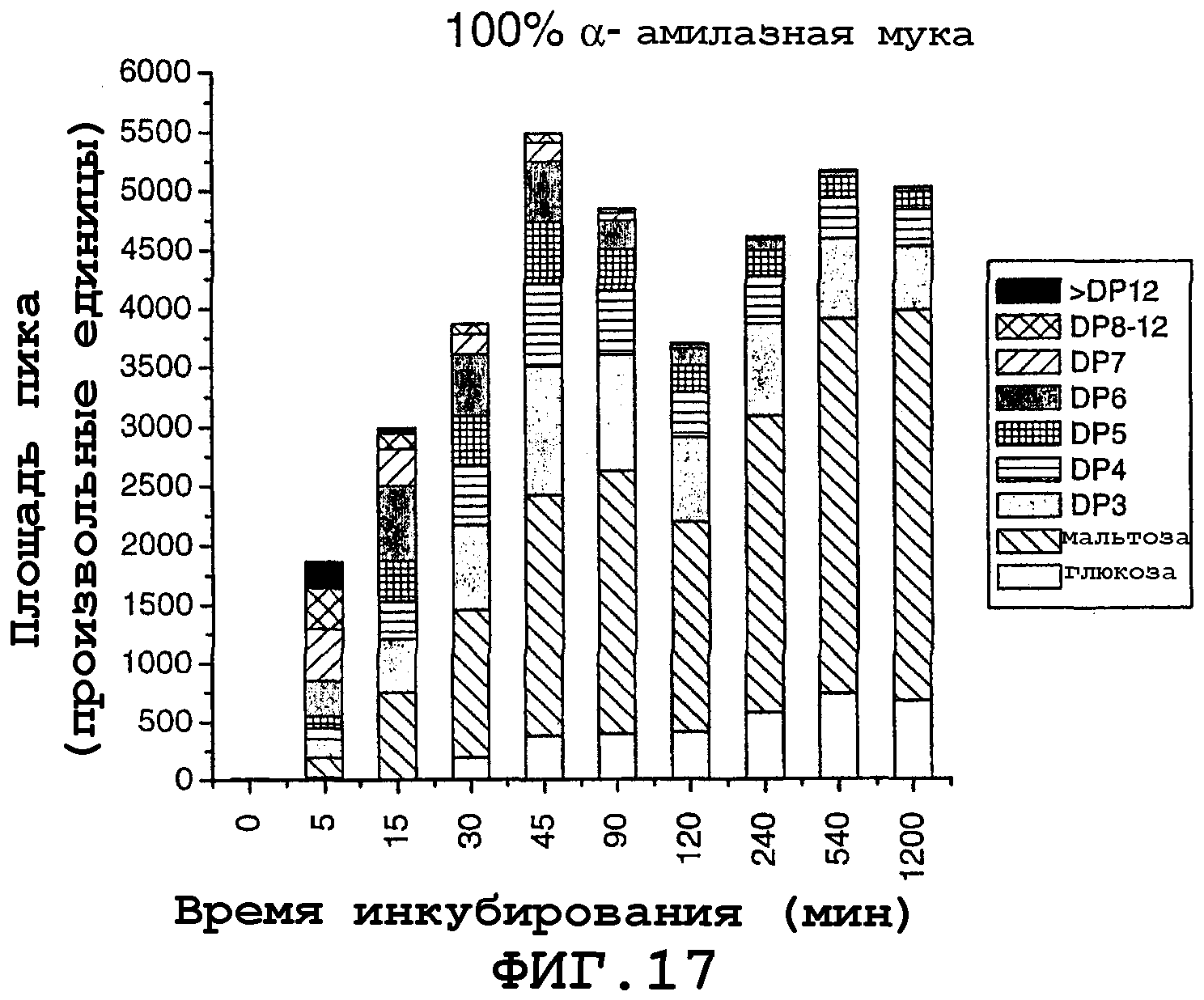
На фигуре 17 показаны площади пиков для продуктов реакции, продуцированных с использованием 100% амилазной муки, полученной из самопроцессирующегося зерна в зависимости от времени инкубирования в течение 0-1200 минут при 90°С.
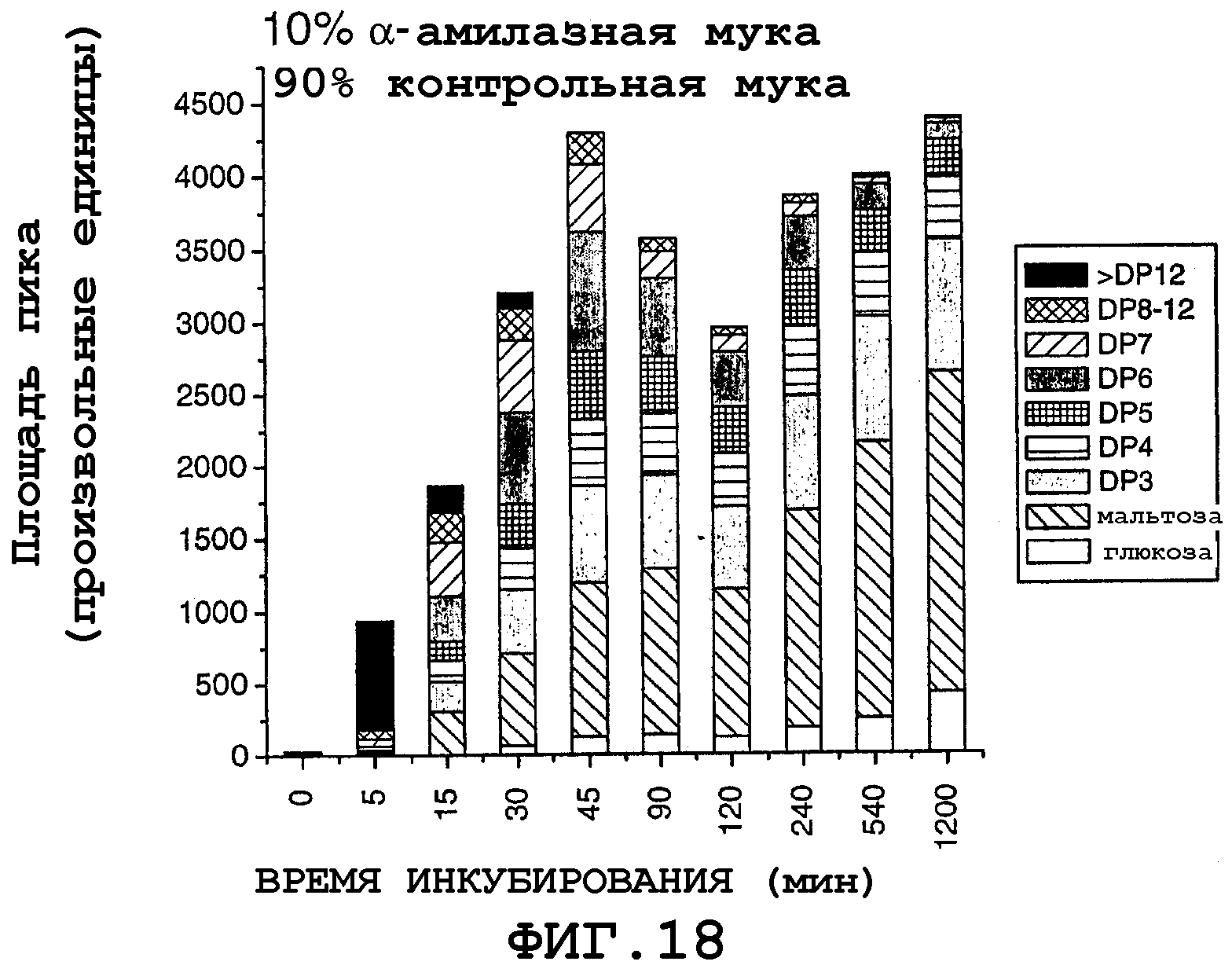
На фигуре 18 показаны площади пиков для продуктов реакции, продуцированных с использованием смеси, содержащей 10% трансгенной муки с амилазой, полученной из самопроцессирующегося зерна, и 90% контрольной кукурузной муки, в зависимости от времени инкубирования в течение 0-1200 минут при 90°С.
На фигуре 19 представлены результаты ВЭЖХ-анализа трансгенной муки с амилазой, инкубированной при 70°С, 80°С, 90°С или 100°С в течение периода времени вплоть до 90 минут, проводимого для оценки влияния температуры на гидролиз крахмала.
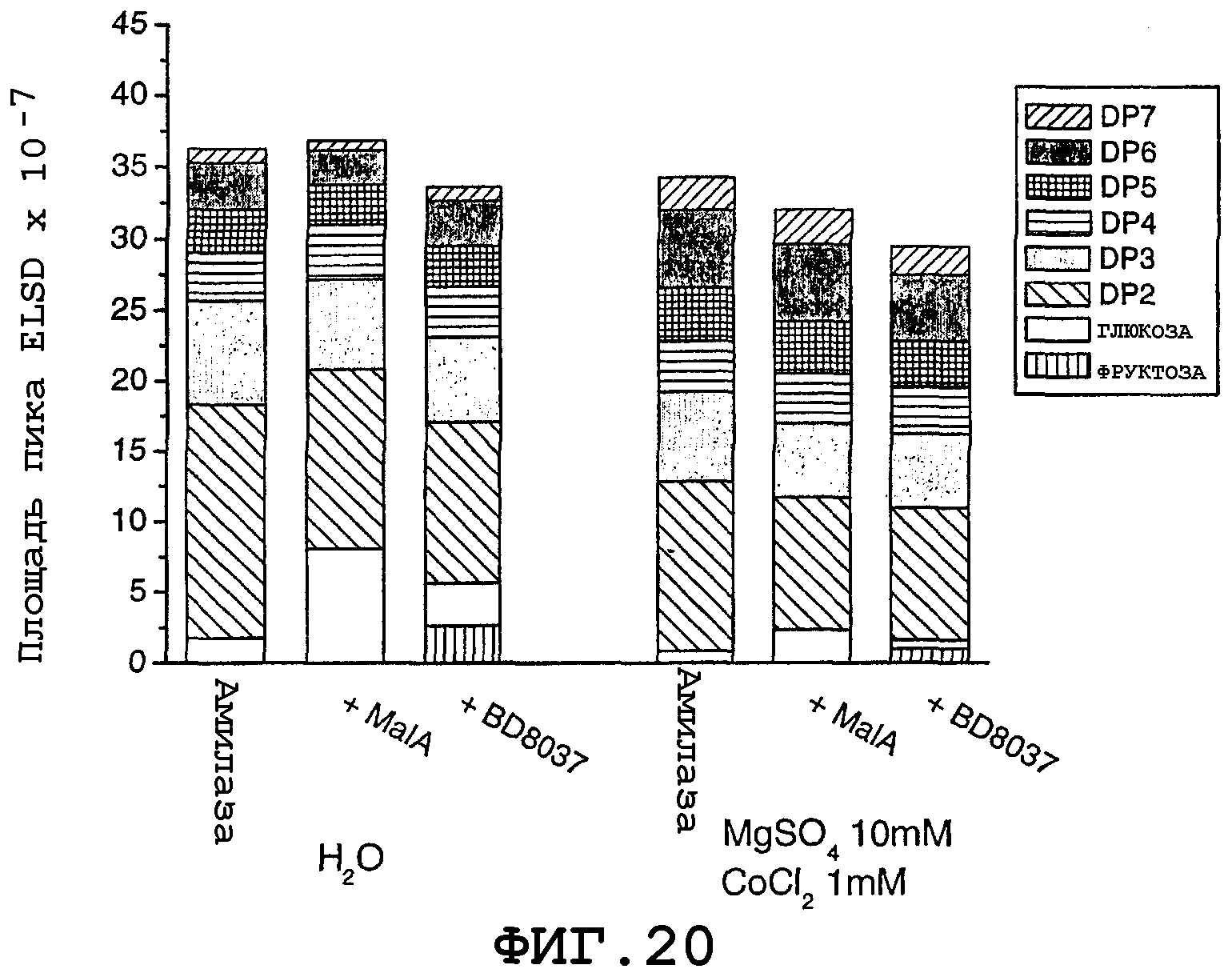
На фигуре 20 показана площадь пика для ELSD-образцов, содержащих 60 мг амилазной трансгенной муки, смешанной с ферментными растворами плюс вода или буфер в различных реакционных условиях. Одна серия реакционных смесей была забуферена 50 мМ MOPS, рН 7,0 при комнатной температуре, плюс 10 мМ MgSO4 и 1 мМ CoCl2; а во второй серии реакционных смесей металлсодержащий буферный раствор был заменен водой. Все реакционные смеси инкубировали в течение 2 часов при 90°С.
Подробное описание изобретения
В соответствии с настоящим изобретением "самопроцессирующееся растение" или часть растения содержит введенный в него (или в нее) выделенный полинуклеотид, кодирующий процессирующий фермент, способный к процессингу, например к модификации крахмалов, полисахаридов, липидов, белков и т.п., в растениях, где процессирующий фермент может быть мезофильным, термофильным или гипертермофильным, и он может быть активирован путем измельчения, добавления воды, нагревания или создания каких-либо других условий, благоприятных для функционирования данного фермента. Выделенный полинуклеотид, кодирующий процессирующий фермент, интегрируют в растение или в часть растения для экспрессии. После экспрессии и активации процессирующего фермента растение или часть растения настоящего изобретения способны к аутопроцессингу субстрата, после чего действует процессирующий фермент. Поэтому растение или части растения настоящего изобретения обладают способностью к аутопроцессингу субстрата фермента после активации содержащегося в них процессирующего фермента в отсутствие внешних источников или с пониженным количеством таких источников, которые обычно являются необходимыми для процессинга этих субстратов. Такие трансформированные растения, клетки трансформированного растения и части трансформированного растения обладают "присущими им" процессирующими способностями, которые позволяют процессировать нужные субстраты под действием введенных в них ферментов в соответствии с настоящим изобретением. При этом предпочтительно, чтобы полинуклеотид, кодирующий процессирующий фермент, был "генетически стабильным", то есть чтобы этот полинуклеотид стабильно сохранялся в трансформированном растении или в частях такого растения настоящего изобретения и стабильно передавался потомству последующих поколений.
В соответствии с настоящим изобретением способы, в которых используются эти растения и части растений, позволяют избежать необходимости помола или какого-либо другого физического нарушения целостности частей растения перед выделением крахмалопродуктов. Так, например, настоящее изобретение относится к усовершенствованным способам процессинга кукурузы и другого зерна для выделения крахмалопродуктов. Настоящее изобретение также относится к способу, позволяющему выделять крахмальные зерна, содержащие уровни крахмалразрушающих ферментов, которые находятся внутри этих зерен или на этих зернах, и которые являются адекватными для гидролиза специфических связей внутри крахмала и не требуют добавления экзогенно продуцируемых крахмалразрушающих ферментов. Настоящее изобретение также относится к усовершенствованным продуктам самопроцессирующегося растения или частей этого растения, полученных способами настоящего изобретения.
Кроме того, использование части "самопроцессирующегося" трансформированного растения, например зерна, и трансформированного растения позволяет избегать главные проблемы, связанные с существующей в настоящее время технологией, заключающиеся в том, что процессирующие ферменты, которые обычно продуцируются при ферментации микробов, требующей осуществления дорогостоящего выделения ферментов из супернатантов культуры, причем выделенные ферменты должны быть приготовлены для конкретных целей, и должны быть разработаны способы и оборудование для добавления, смешивания и осуществления реакции фермента с его субстратом. Трансформированное растение настоящего изобретения или его часть также служат источником самого процессирующего фермента, а также субстратов и продуктов этого фермента, таких как сахара, аминокислоты, жирные кислоты и полисахариды, являющиеся и не являющиеся крахмалом. Растение настоящего изобретения может быть также использовано для получения потомства растения, такого как гибридные и инбредные растения.
Процессирующие ферменты и кодирующие их полинуклеотиды
Полинуклеотид, кодирующий процессирующий фермент (мезофильный, термофильный или гипертермофильный), вводят в растение или в часть растения. Процессирующий фермент выбирают исходя из нужного субстрата, на который, как было обнаружено, он действует в растениях или в трансгенных растениях, и/или исходя из нужного конечного продукта. Так, например, указанным процессирующим ферментом может быть крахмалпроцессирующий фермент, такой как крахмалразрушающий фермент или крахмализомеризующий фермент, или не процессирующий крахмал фермент. Подходящими процессирующими ферментами являются, но не ограничиваются ими, крахмалразрушающие или крахмализомеризующие ферменты, включая, например, α-амилазу, эндо- или экзо-1,4- или 1,6-α-D-глюкоамилазу, глюкозоизомеразу, β-амилазы, α-глюкозидазы и другие экзо-амилазы, и крахмалдеветвящие ферменты, такие как изоамилаза, пуллуланаза, нео-пуллуланаза, изопуллуланаза, амилопуллуланаза и т.п., гликозилтрансферазы, такие как циклодекстрингликозилтрансферазы и т.п., целлюлазы, такие как экзо-1,4-β-целлобиогидролаза, экзо-1, 3-β-D-глюканаза, гемицеллюлаза, β-глюкозидаза и т.п., эндоглюканазы, такие как эндо-1,3-β-глюканаза и эндо-1,4-β-глюканаза и т.п., L-арабиназы, такие как эндо-1,5-α -L-арабиназа, α-арабинозидазы и т.п., галактаназы, такие как эндо-1,4-β-D-галактаназа, эндо-1,3-β-D-галактаназа, β-галактозидаза, α-галактозидаза и т.п., маннаназы, такие как эндо-1,4-β-D-маннаназа, β-маннозидаза, α-маннозидаза и т.п., ксиланазы, такие как эндо-1,4-β-ксиланаза, β-D-ксилозидаза, 1,3-β-D-ксиланаза и т.п., и пектиназы, и не процессирующие крахмал ферменты, включая протеазу, глюканазу, ксиланазу, тиоредоксин/тиоредоксин-редуктазу, эстеразу, фитазу и липазу.
В одном из вариантов осуществления изобретения процессирующим ферментом является крахмалразрушающий фермент, выбранный из группы, состоящей из α-амилазы, пуллуланазы, α-глюкозидазы, глюкоамилазы, амилопуллуланазы, глюкозоизомеразы или их комбинаций. В соответствии с этим вариантом настоящего изобретения крахмалразрушающий фермент позволяет самопроцессирующемуся растению или его части разлагать крахмал при активации фермента, содержащегося в этом растении или в части растения, как это будет описано ниже. Крахмалразрушающий(ие) фермент(ы) выбирают исходя из нужных конечных продуктов. Так, например, для превращения глюкозы (гексозы) во фруктозу может быть выбрана глюкозоизомераза. Альтернативно, фермент может быть выбран исходя из нужного конечного крахмалопродукта, имеющего цепи с различными длинами, в зависимости, например, от степени процессинга, или имеющего различные нужные типы ветвления. Так, например, α-амилаза, глюкоамилаза или амилопуллуназа могут быть использованы при небольшом периоде времени инкубирования для продуцирования декстриновых продуктов, и при более продолжительном времени инкубирования для продуцирования продуктов или сахаров с более короткой цепью. Пуллуланаза может быть использована, в частности, для гидролиза в точках ветвления крахмала с образованием высокоамилозного крахмала, а неопуллуланаза может быть использована для продуцирования крахмала с фрагментами, имеющими α-1,4-связи, перемежающиеся с α-1,6-связями. Глюкозидазы могут быть использованы для продуцирования предельных декстринов, а для получения других производных крахмала могут быть использованы комбинации ферментов.
В другом варианте осуществления изобретения, процессирующим ферментом является не процессирующий крахмал фермент, выбранный из протеазы, глюканазы, ксиланазы и эстеразы. Эти не процессирующие крахмал ферменты способны внедряться в целевую область самопроцессирующегося растения или в его часть, и при активации разрушать растение, оставляя, при этом, содержащиеся в нем крахмальные зерна интактными. Так, например, в предпочтительном варианте осуществления изобретения не разрушающие крахмал ферменты поступают в матрикс эндосперма клетки растения и, при активации, разрушают матрикс эндосперма, оставляя, при этом, содержащиеся в нем крахмальные зерна интактными и более легко выделяемыми из полученного материала.
Кроме того, в настоящем изобретении рассматриваются комбинации процессирующих ферментов. Так, например, крахмалпроцессирующие ферменты и не процессирующие крахмал ферменты могут быть использованы в комбинации. Комбинации процессирующих ферментов могут быть получены с использованием множества генных конструкций, кодирующих каждый из этих ферментов. Альтернативно, отдельные трансгенные растения, стабильно трансформированные ферментами, могут быть скрещены известными методами с получением растения, содержащего оба этих фермента. Другой метод включает использование экзогенного фермента(ов) и трансгенного растения.
Процессирующие ферменты могут быть выделены или получены из любого источника, и его соответствующие полинуклеотиды могут быть идентифицированы специалистом. Так, например, процессирующий фермент, предпочтительно α-амилазу, получают из Pyrococcus (например, Pyrococcus furiosus), Thermus, Thermococcus (например, Thermococcus hydrothermalis), Sulfolobus (например, Sulfolobus solfataricus), Thermotoga (например, Thermotoga maritima и Thermotoga neapolitana), Thermoanaerobacterium (например, Thermoanaerobacter tengcongensis), Aspergillus (например, Aspergillus shirousami and Aspergillus niger), Rhizopus (например, Rhizopus oryzae), Thermoproteales, Desulfurococcus (например, Desulfurococcus amylolyticus), Methanobacterium thermoautotrophicum, Methanococcus jannaschii, Methanopyrus kandleri, Thermosynechococcus elongatus, Thermoplasma acidophilum, Thermoplasma volcanium, Aeropyrum pernix и растений, таких как кукуруза, ячмень и рис.
Процессирующие ферменты настоящего изобретения могут быть активированы после их введения и экспрессии в геноме растения. Условия активации фермента определяют для каждого конкретного фермента и могут быть использованы различные условия, такие как соответствующая температура и рН, гидратация, присутствие металлов, активирующие соединения, инактивирующие соединения и т.п. Так, например, зависимыми от температуры ферментами могут быть мезофильные, термофильные и гипертермофильные ферменты. Мезофильные ферменты обычно имеют максимальную активность при температуре 20-65°С и являются инактивированными при температуре выше 70°С. Мезофильные ферменты обладают значительной активностью при 30-37оС, причем их активность при 30°С составляет, по крайней мере, 10% от максимальной активности, а более предпочтительно, по крайней мере, 20% от максимальной активности.
Термофильные ферменты обладают максимальной активностью при температуре от 50°С до 80°С и являются инактивированными при температуре выше 80°С. Термофильный фермент при 30°С предпочтительно обладает менее чем 20% от максимальной активности, а более предпочтительно менее чем 10% от максимальной активности.
"Гипертермофильный" фермент обладает активностью даже при более высоких температурах. Гипертермофильные ферменты обладают максимальной активностью при температуре выше 80°С и сохраняют активность при температуре, по крайней мере, 80°С, а более предпочтительно, если они сохраняют активность при температуре, по крайней мере, 90°С, а наиболее предпочтительно, если они сохраняют активность при температуре, по крайней мере, 95°С. Гипертермофильные ферменты также обладают пониженной активностью при низких температурах. Гипертермофильный фермент может обладать активностью при 30°С, т.е. менее чем 10% от максимальной активности, а предпочтительно менее чем 5% от максимальной активности.
Полинуклеотид, кодирующий процессирующий фермент, предпочтительно, модифицируют так, чтобы он включал кодоны, оптимизированные для экспрессии в выбранном микроорганизме, таком как растение (см., например, Wada et al., Nucl. Acids. Res., 18:2367 (1990), Murray et al., Nucl. Acids. Res., 17:477 (1989), патенты США №№5096825, 5625136, 5670356 и 5874304). Оптимизированные по кодонам последовательности являются синтетическими последовательностями, то есть они не встречаются в природе и, предпочтительно, кодируют идентичный полипептид (или ферментативно активный фрагмент полноразмерного полипептида, который обладает, в основном, такой же активностью, как и полноразмерный полипептид), кодируемый не оптимизированным по кодонам исходным полинуклеотидом, который кодирует процессирующий фермент. При этом предпочтительно, чтобы полипептид отличался по своим биохимическим свойствам или имел улучшенные биохимические свойства (что может быть достигнуто, например, путем рекурсивного мутагенеза ДНК, кодирующей конкретный процессирующий фермент), по сравнению с полипептидом родительского источника, так, чтобы он имел более высокую эффективность в этом способе применения. Предпочтительные полинуклеотиды оптимизированы для экспрессии в нужном растении-хозяине и кодируют процессирующий фермент. Способы получения этих ферментов включают мутагенез, например рекурсивный мутагенез и отбор. Методы мутагенеза и модификации нуклеотидной последовательности хорошо известны специалистам. См., например, Kunkel, Proc.Natl. Acad. Sci., USA, 82:488 (1985); Kunkel et al., Methods in Enzymol., 154:367 (1987); патент США №4873192; Walker and Gaastra, eds. (1983) Techniques in Molecular Biology (MacMillan Publishing Company, New York) и цитируемые там работы и Arnold et al., Chem. Eng. Sci. 51:5091 (1996)). Методы оптимизации экспрессии сегмента нуклеиновой кислоты в нужном растении или в микроорганизме хорошо известны специалистам. Для этого получают таблицу встречаемости кодонов, иллюстрирующую оптимальные кодоны, которые используются в организме-мишени, и выбирают оптимальные кодоны для их замены в нужном полинуклеотиде, а затем осуществляют химический синтез оптимизированной последовательности. Предпочтительные кодоны кукурузы описаны в патенте США №5625136.
Кроме того, рассматриваются комплементарные нуклеиновые кислоты полинуклеотидов настоящего изобретения. Примерами условий низкой степени жесткости для гибридизации комплементарных нуклеиновых кислот, которые имеют более чем 100 комплементарных остатков на фильтре в Саузерн- или Нозерн-блоте, являются использование 50% формамида, например проведение гибридизации в 50% формамиде, 1М NaCl, 1% ДСН при 37°С и промывка в 0,1 х SSC при 60°С-65°С. Примером условий гибридизации с низкой степенью жесткости является гибридизация с буферным раствором 30-35% формамида, 1М NaCl, 1% ДСН (додецилсульфат натрия) при 37°С и промывка в 1Х-2Х SSC (20X SSC=3,0М NaCl/0,3М тринатрийцитрат) при 50-55°С. Примерами условий гибридизации умеренной степени жесткости являются гибридизация в 40-45% формамиде, 1,0М NaCl, 1% ДСН при 37°С, и промывка в 0,5Х-1Х SSC при 55-60°С.
Кроме того, далее рассматриваются полинуклеотиды, кодирующие "ферментативно активный" фрагмент процессирующего фермента. Используемый здесь термин "ферментативно активный" фрагмент означает полипептидный фрагмент процессирующего фермента, который обладает, в основном, такой же биологической активностью, как и процессирующий фермент, и способен модифицировать субстрат, после чего процессирующий фермент обычно нормально функционирует в подходящих условиях.
В предпочтительном варианте, полинуклеотид настоящего изобретения представляет собой полинуклеотид, оптимизированный по кодонам кукурузы и кодирующий α-амилазу, такой как SEQ ID NO:2, 9, 46 и 52. В другом предпочтительном варианте осуществления изобретения полинуклеотидом является полинуклеотид, оптимизированный по кодонам кукурузы и кодирующий пуллуланазу, такой как SEQ ID NO:4 и 25. В еще одном предпочтительном варианте осуществления изобретения указанным полинуклеотидом является полинуклеотид, оптимизированный по кодонам кукурузы и кодирующий α -глюкозидазу, такой как SEQ ID NO:6. Другим предпочтительным полинуклеотидом является полинуклеотид, оптимизированный по кодонам кукурузы и кодирующий глюкозоизомеразу, такой как SEQ ID NO:19, 21, 37, 39, 41 или 43. В другом варианте осуществления изобретения предпочтительным является полинуклеотид, оптимизированный по кодонам кукурузы и кодирующий глюкоамилазу, такой как SEQ ID NO:46, 48 или 50. Кроме того, оптимизированным по кодонам кукурузы полинуклеотидом, кодирующим гибридный полипептид глюканазы/маннаназы, является полинуклеотид SEQ ID NO:57. Настоящее изобретение также относится к последовательностям, комплементарным таким полинуклеотидам, которые гибридизуются в условиях умеренной или, предпочтительно, низкой степени жесткости и которые кодируют полипептид, обладающий α -амилазной, пуллуланазной, α-глюкозидазной, глюкозоизомеразной, глюкоамилазной, глюканазной или маннаназной активностью, там, где это необходимо.
Термин "полинуклеотид" может быть использован наряду с терминами "нуклеиновая кислота" или "полинуклеиновая кислота" и означает дезоксирибонуклеотиды или рибонуклеотиды и их полимеры в одноцепочечной или в двухцепочечной форме, состоящие из мономеров (нуклеотидов), содержащих сахар, фосфат и основание, которым являются либо пурин, либо пиримидин. Если это не оговорено особо, термин охватывает нуклеиновые кислоты, содержащие известные аналоги природных нуклеотидов, которые обладают такими же свойствами связывания, как и исходная нуклеиновая кислота, и подвергаются метаболизму в соответствии с таким же механизмом, как и природные нуклеотиды. Если это не оговорено особо, то конкретная последовательность нуклеиновой кислоты также, безусловно, охватывает ее обратимо модифицированные варианты (например, замены вырожденных кодонов) и комплементарные последовательности, а также конкретно указанную последовательность. В частности, замены вырожденных кодонов могут быть осуществлены путем генерирования последовательностей, в которых один или более выбранных (или всех) кодонов в третьем положении заменен смешанными основаниями и/или дезоксиинозиновыми остатками.
Настоящее изобретение, кроме того, охватывает "варианты" или, в основном, аналогичные последовательности. Для нуклеотидных последовательностей вариантами являются последовательности, которые, из-за вырожденности их генетического кода, кодируют идентичную аминокислотную последовательность нативного белка. Природные аллельные варианты, такие как указанные здесь варианты, могут быть идентифицированы с использованием хорошо известных методов молекулярной биологии, например полимеразной цепной реакции (ПЦР), техники гибридизации и вторичной сборки путем лигирования. Вариантами нуклеотидных последовательностей также являются синтетические нуклеотидные последовательности, такие как последовательности, полученные, например, путем сайт-направленного мутагенеза и кодирующие природный белок, а также последовательности, кодирующие полипептид, имеющий аминокислотные замены. В общих чертах, варианты нуклеотидной последовательности настоящего изобретения, по крайней мере, на 40%, 50%, 60%, предпочтительно на 70%, более предпочтительно на 80%, еще более предпочтительно на 90%, а наиболее предпочтительно на 99% идентичны нативной нуклеотидной последовательности того же класса или отличаются от нее лишь одним нуклеотидом. Так, например, это сходство может составлять 71%, 72%, 73% и т.п., по крайней мере, до 90%. Такими вариантами могут быть также полноразмерный ген, соответствующий идентифицированному генному фрагменту.
Регуляторные последовательности: промоторы/сигнальные последовательности/селективные маркеры.
Полинуклеотидные последовательности, кодирующие процессирующий фермент настоящего изобретения, могут быть функционально присоединены к полинуклеотидным последовательностям, кодирующим сигнал локализации или сигнальную последовательность (у N- или С-конца полипептида), например, для нацеливания гипертермофильного фермента на конкретный компартмент в растении. Примерами таких мишеней являются, но не ограничиваются ими, вакуоль, эндоплазматический ретикулум, хлоропласт, амилопласт, крахмальные зерна или клеточные стенки или конкретная ткань, например семя. Экспрессия полинуклеотида, кодирующего процессирующий фермент, имеющий сигнальную последовательность в растении, а в частности, в сочетании с тканеспецифическим или индуцибельным промотором может давать высокие уровни локализованного процессирующего фермента в растении. Известно, что различные сигнальные последовательности влияют на экспрессию или на нацеливание полинуклеотида на конкретный компартмент или за пределы конкретного компартмента. Подходящие сигнальные последовательности и нацеливающие промоторы известны специалистам, и такими последовательностями являются, но не ограничиваются ими, описанные здесь последовательности.
Так, например, если желательно, чтобы экспрессия происходила в конкретных тканях или органах, то могут быть использованы тканеспецифические промоторы. В противоположность этому, если желательно, чтобы ген экспрессировался в ответ на стимулятор, то в качестве регуляторных элементов необходимо выбрать индуцибельные промоторы. Если желательна непрерывная экспрессия во всех клетках растения, то используют конститутивные промоторы. В экспрессионных конструкциях трансформирующих векторов могут присутствовать дополнительные регуляторные последовательности, расположенные выше и/или ниже от коровой промоторной последовательности, и эти последовательности будут обеспечивать различные уровни экспрессии гетерологичных нуклеотидных последовательностей в трансгенном растении.
Был описан ряд промоторов растения с различными экспрессионными свойствами. Примерами некоторых конститутивных промоторов, которые были описаны в литературе, являются актин 1 риса (Wang et al., Mol. Cell. Biol. 12:3399 (1992); патент США №5641876), CaMV 35S (Odell et al., Nature, 313:810 (1985)), CaMV 19S (Lawton et al., 1987), nos (Ebert et al., 1987), Adh (Walker et al., 1987), сахарозосинтаза (Yang & Russell, 1990) и промоторы убихитина.
Векторы, используемые для тканеспецифического нацеливания генов в трансгенных растениях, обычно включают тканеспецифические промоторы, и они могут также включать другие тканеспецифические регуляторные элементы, такие как энхансерные последовательности. Промоторы, которые регулируют специфическую или усиленную экспрессию в некоторых тканях растений, известны специалистам из описания настоящего изобретения. Такими промоторами являются, например, промотор rbcS, специфичный для зеленых тканей; промоторы ocs, nos и mas, которые обладают более высокой активностью в корнях или в ткани поврежденных листьев; усеченный промотор 35S (-90-(+8)), который обеспечивает усиленную экспрессию в корнях, ген α-тубулина, который обеспечивает экспрессию в корнях, и промоторы, происходящие от генов запасного белка зеина и обеспечивающие экспрессию в эндосперме.
Тканеспецифическая экспрессия может быть функционально осуществлена путем введения конститутивно экспрессируемого гена (во всех тканях) в комбинации с антисмысловым геном, который экспрессируется только в тех тканях, в которых продуцирование данного генного продукта нежелательно. Так, например, ген, кодирующий липазу, может быть введен вместе с промотором 35S вируса мозаики цветной капусты так, чтобы он экспрессировался во всех тканях. Экспрессия антисмыслового транскрипта гена липазы в зерне кукурузы с использованием, например, промотора зеина будет предотвращать накопление липазного белка в семенах. Поэтому белок, кодируемый указанным введенным геном, будет присутствовать во всех тканях, кроме зерен.
Кроме того, как уже сообщалось, в растениях имеется несколько тканеспецифических регуляторных генов и/или промоторов. Некоторыми из таких описанных тканеспецифических генов являются гены, кодирующие запасные белки семян (такие как напин, круциферин, бета-конглицинин и фазеолин), зеин или белки масляных телец (такие как олеозин) или гены, участвующие в биосинтезе жирных кислот (включая ацил-переносящий белок, стеароил-АСР-дезатуразу и дезатуразы жирных кислот (fad 2-1)), и другие гены, экспрессируемые в процессе развития зародыша (такие как Bce4, см. например, ЕР 255378 и Kridl et al., Seed Science Research, 1:209 (1991)). Примерами описанных тканеспецифических промоторов являются промоторы лектина (Vodkin, Prog. Clin. Biol. Res. 138:87 (1983); Lindstrom et al., Der. Genet., 11:160 (1990)), алкоголь-дегидрогеназы 1 кукурузы (Vogel et al., 1989, Dennis et al., Nucleic Acids. Res., 12:3983 (1984)), светособирающего комплекса кукурузы (Simpson, 1986; Bansal et al., Proc. Natl. Acad. Sci. USA, 89:3654 (1992)), белка теплового шока кукурузы (Odell et al., 1985; Rochester et al., 1986), карбоксилазы малой субъединицы RuBP гороха (Poulsen et al., 1986; Cashmore et al., 1983), маннопин-синтазы плазмиды Ti (Langridge et al., 1989), нопалин-синтазы плазмиды Ti (Langridge et al., 1989), халконизомеразы петунии (vanTunen et al., EMBO J. 7:1257 (1988)), богатого глицином белка 1 сои (Keller et al., Genes Dev. 3:1639 (1989)), усеченного CaMV 35s (Odell et al., Nature, 313:810 (1985)), пататина картофеля (Wenzler et al., Plant Mol. Biol. 13:347 (1989)), клеток корней (Yamamoto et al., Nucleic Acids., Res. 18:7449 (1990)), зеина кукурузы (Reina et al., Nucleic Acids. Res., 18:6425 (1990); Kriz et al., Mol.Gen.Genet., 207:90 (1987); Wandelt et al., Nucleic Acids Res., 17:2354 (1989), Langridge et al., Cell, 34:1015 (1983); Reina et al., Nucleic Acids Res., 17:7449 (1990)), глобулина-1 (Belanger et al., Genetics, 129:863 (1991)), α-тубулина, cab (Sullivan et al., Mol. Gen. Genet. 215:431 (1989)), РЕРСаза (Hudspeth & Grula, 1989), комплекс-ассоциированные промоторы гена R (Chandler et al., Plant Cell, 1:1175 (1989)) и промоторы халконсинтазы (Franken et al., EMBO J., 10:2605 (1991)). Особенно подходящим для семяспецифической экспрессии является промотор вицилина гороха (Czako et al., Mol. Gen. Genet., 235:33 (1992)). (См. также патент США №5625136, который вводится в настоящее описание посредством ссылки). Другими подходящими промоторами для экспрессии в распустившихся листьях являются промоторы, которые переключаются на начало процесса старения, такие как промотор SAG растения Arabidopsis (Gan et al., Science, 270:1986 (1995)).
Промоторы плод-специфического класса, экспрессируемые во время цветения или в течение периода времени от начала цветения до развития плода, по крайней мере, до начала его созревания, обсуждаются в патенте США №4943674, который вводится в настоящее описание посредством ссылки. Были выделены кДНК-клоны, которые экспрессируются преимущественно в волокне хлопчатника (John et al., Proc. Natl. Acad. Sci. USA, 89:5769 (1992)). Были также выделены и охарактеризованы кДНК-клоны томатов, в которых наблюдается дифференциальная экспрессия во время развития плода (Mansson et al., Gen. Genet., 200:356 (1985), Staler et al., Plant. Mol. Biol., 5:137 (1985)). Промотор гена полигалактуроназы является активным на стадии созревания плода. Ген полигалактуроназы описан в патенте США №4535060, в патенте США №4769061, в патенте США №4801590 и в патенте США №5107065, которые вводятся в настоящее описание посредством ссылки.
Другими примерами тканеспецифических промоторов являются промоторы, регулирующие экспрессию в клетках листьев после их повреждения (например, после поедания насекомыми), в клубнях (например, промотор гена пататина) и в клетках волокна (например, регулируемым на стадии развития клеток волокна белком является белок E6 (John et al., Proc. Natl. Acad. Sci. USA, 89:5769 (1992)). Ген Е6 обладает наибольшей активностью в волокне, хотя небольшие уровни его транскриптов обнаружены в листьях, семяпочках и в цветках.
Тканеспецифичность некоторых "тканеспецифических" промоторов не может быть абсолютной и может быть протестирована специалистом с использованием последовательности дифтерийного токсина. Тканеспецифическая экспрессия может быть также достигнута посредством "пиковой" экспрессии путем комбинирования различных тканеспецифических промоторов. (Beals et al., Plant Cell, 9:1527 (1997)). Другие тканеспецифические промоторы могут быть выделены специалистом в данной области (см. патент США №5589379).
В одном из вариантов осуществления изобретения генный продукт гидролиза полисахарида, такой как α-амилаза, может быть нацелен на конкретную органеллу, такую как апопласт, а не на цитоплазму. Это может быть достигнуто, например, с использованием N-концевой сигнальной последовательности γ-зеина кукурузы (SEQ ID NO:17), которая обеспечивает апопласт-специфическое нацеливание белков. Направление белка или фермента в конкретный компартмент позволяет ферменту локализоваться так, чтобы он не вступал в контакт с субстратом. Таким образом, ферментативное действие фермента не наступит, пока фермент не проконтактирует со своим субстратом. Контакт этого фермента с субстратом может быть осуществлен путем размалывания (физического разрушения целостности клеток) или нагревания клеток или тканей растения с нарушением физической целостности клеток или органов растения, содержащих фермент. Так, например, мезофильный крахмалразрушающий фермент может быть нацелен на апопласт или на эндоплазматический ретикулум, а поэтому он не будет вступать в контакт с крахмальными зернами в амилопласте. После помола зерна целостность этого зерна будет нарушаться, после чего крахмалгидролизующий фермент будет контактировать с крахмальными зернами. Таким образом можно избежать возможных негативных эффектов совместной локализации фермента и его субстрата.
В другом варианте осуществления изобретения тканеспецифическим промотором является эндосперм-специфические промоторы, такие как промотор γ-зеина кукурузы (представленной SEQ ID NO:12) или промотор ADP-gpp кукурузы (представленный SEQ ID NO:11 и включающий 5'-нетранслируемую последовательность и интронную последовательность). Таким образом, настоящее изобретение относится к выделенному полинуклеотиду, содержащему промотор, включающий последовательность SEQ ID NO:11 или 12, к полинуклеотиду, который гибридизуется с его комплементарной последовательностью в условиях низкой степени жесткости, или к его фрагменту, промоторная активность которого составляет, по крайней мере, 10%, а предпочтительно, по крайней мере, 50% от активности промотора, имеющего последовательность SEQ ID NO:11 или 12.
В другом варианте осуществления изобретения полинуклеотид кодирует гипертермофильный процессирующий фермент, который функционально присоединен к хлоропластному (амилопластному) транспортному пептиду (СТР) и крахмалсвязывающему домену, например кодируемому геном waxy. Характерный полинуклеотид в этом варианте настоящего изобретения кодирует последовательность SEQ ID NO:10 (α-амилазы, присоединенной к крахмалсвязывающему домену восковидной кукурузы). Другие характерные полинуклеотиды кодируют гипертермофильный процессирующий фермент, присоединенной к сигнальной последовательности, которая обеспечивает нацеливание указанного фермента на эндоплазматический ретикулум и секрецию в апопласт (например, полинуклеотид, кодирующий последовательность SEQ ID NO:13, 27 или 30, который содержит N-концевую последовательность γ-зеина кукурузы, функционально связанную с α-амилазой, α-глюкозидазой, глюкозоизомеразой соответственно); гипертермофильный процессирующий фермент, присоединенной к сигнальной последовательности, которая обеспечивает удерживание фермента в эндоплазматическом ретикулуме (например, полинуклеотид, кодирующий SEQ ID NO:14, 26, 28, 29, 33, 34, 35 или 36, которые содержат N-концевую последовательность γ-зеина кукурузы, функционально присоединенную к гипертермофильному ферменту, который функционально присоединен к SEKDEL, где ферментом являются α-амилаза, α-глюкозидаза malA, глюкозоизомераза T.maritima, глюкозоизомераза T.neapolitana); гипертермофильный процессирующий фермент, присоединенной к N-концевой последовательности, которая обеспечивает нацеливание фермента на амилопласт (например, полинуклеотид, кодирующий последовательность SEQ ID NO:15, которая содержит N-концевую последовательность, обеспечивающую нацеливание фермента на амилопласт восковидной кукурузы, и функционально присоединенную к последовательности α-амилазы); гипертермофильный гибридный полипептид, который обеспечивает нацеливание указанного фермента на крахмальные зерна (например, полинуклеотид, кодирующий последовательность SEQ ID NO:16, которая содержит N-концевую последовательность, обеспечивающую нацеливание фермента на амилопласт последовательность восковидной кукурузы, и функционально присоединенную к гибридному полипептиду α-амилазы/waxy, содержащему крахмалсвязывающий домен восковидной кукурузы); гипертермофильный процессирующий фермент, связанный с сигнальной последовательностью, обеспечивающей удерживание в ЭР (например, полинуклеотид, кодирующий SEQ ID NO:38 и 39). Кроме того, гипертермофильный процессирующий фермент может быть присоединен к сайту связывания с исходным крахмалом, имеющему аминокислотную последовательность (SEQ ID NO:53), где указанный полинуклеотид, кодирующий процессирующий фермент, присоединен к последовательности нуклеиновой кислоты, оптимизированной по кодонам кукурузы (SEQ ID NO:54) и кодирующей указанный сайт связывания.
Сообщалось также о нескольких индуцибельных промоторах. Многие из них описаны Gatz в работе Current Opinion in Biotechnology, 7:168 (1996) и Gatz, C., Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 48:89 (1997). Примерами таких промоторов являются система супрессии тетрациклина, система супрессии Lac, медь-индуцибельные системы, салицилат-индуцибельные системы (такие как система PR1a), глюкокортикоид-индуцибельная система (Aoyama T. et al., N-H Plant Journal, 11:605 (1997)) и экдизон-индуцибельная система. Другими индуцибельными промоторами являются АВА- и тургор-индуцибельные промоторы, промотор гена белка, связывающегося с ауксином (Schwob et al., Plant J. 4:423 (1993)), промотор гена UDP-глюкозофлавоноидгликозилтрансферазы (Ralston et al., Genetics, 119:185 (1988)), промотор ингибитора протеиназы MPI (Cordero et al., Plant. J., 6:141 (1994)), промотор гена глицеральдегид-3-фосфатдегидрогеназы (Kohler et al., Plant. Mol. Biol., 29:1293 (1995); Quigley et al., J. Mol. Evol. 29:412 (1989); Martinez et al., J. Mol. Biol. 208:551 (1989)). К этим промоторам также относятся бензолсульфонамид-индуцибельные системы (патент США №5364780) и спирт-индуцибельные системы (WO 97/06269 и WO 97/06268) и промоторы глутатион-S-трансферазы.
Проводились также другие исследования генов, индуцибельно регулируемых в ответ на стрессовые условия, создаваемые окружающей средой, или на стимуляторы, такие как повышение степени солености, засуха, наличие патогенов и повреждения (Graham et al., J. Biol. Chem. 260:6555 (1985); Graham et al., J. Biol. Chem. 260:6561 (1985), Smith et al. Planta, 168:94 (1986)). Накопление белка ингибитора металлокарбоксипептидазы наблюдалось в листьях поврежденных растений картофеля (Graham et al., Biochem. Biophys. Res. Comm. 101:1164 (1981)). Как уже сообщалось, другие гены растений индуцируются метилжасмонатом, стимуляторами, белками теплового шока, анаэробным стрессом или гербицидными препаратами.
Регулируемая экспрессия химерного белка, обладающего транс-активностью по отношению к репликации вируса, может дополнительно регулироваться другими методами генетической стратегии, например путем Cre-опосредованной активации генов (Odell et al., Mol. Gen. Genet., 113:369 (1990)). Таким образом, ДНК-фрагмент, содержащий 3'-регуляторную последовательность, связанную с lox-сайтами между промотором и последовательностью, кодирующей белок репликации и блокирующей экспрессию химерного гена репликации, начиная с промотора, может быть удален путем Cre-опосредованного вырезания, что приводит к экспрессии транс-активного гена репликации. В этом случае химерный ген Cre, химерный транс-активный ген репликации или оба этих гена могут находиться под контролем тканеспецифических и стадиеспецифических или индуцибельных промоторов. Альтернативная генная стратегия предусматривает использование гена-супрессора тРНК. Так, например, регулируемая экспрессия гена-супрессора тРНК может, в зависимости от условий, регулировать экспрессию последовательности, кодирующей транс-активный белок репликации и содержащей соответствующий кодон терминации (Ulmasov et al. Plant. Mol. Biol., 35:417 (1997)). И снова, химерный ген-супрессор тРНК, химерный транс-активный ген репликации или оба этих гена могут находиться под контролем тканеспецифических и стадиеспецифических или индуцибельных промоторов.
В случае многоклеточного организма промотор может также, что предпочтительно, быть специфическим к конкретной ткани, органу и стадии развития. Примерами таких промоторов являются, но не ограничиваются ими, промотор ADP-gpp Zea mays, промотор γ -зеина Zea mays и промотор глобулина Zea mays.
Экспрессия гена в трансгенном растении может оказаться желательной только в определенный период времени развития растения. Время развития часто коррелирует с тканеспецифической экспрессией гена. Так, например, экспрессия запасных белков зеинов инициируется в эндосперме примерно через 15 дней после опыления.
Кроме того, могут быть сконструированы векторы, которые могут быть использованы для внутриклеточного нацеливания специфического генного продукта на клетки трансгенного растения или для нацеливания белка на внеклеточное пространство. Это обычно достигается путем присоединения ДНК-последовательности, кодирующей транспортную или сигнальную пептидную последовательность, к кодирующей последовательности конкретного гена. Полученный транспортный или сигнальный пептид будет переносить данный белок в конкретный внутриклеточный или внеклеточный участок соответственно, а затем он будет посттрансляционно удален. Транспортные или сигнальные пептиды действуют посредством транспорта через внутриклеточные мембраны, например вакуоль, везикулу, пластидные и митохондриальные мембраны, тогда как сигнальные пептиды направляют белки через внеклеточную мембрану.
Сигнальная последовательность, такая как N-концевая сигнальная последовательность γ-зеина кукурузы, обеспечивающая нацеливание на эндоплазматический ретикулум и секрецию в апопласт, может быть функционально присоединена к полинуклеотиду, кодирующему гипертермофильный процессирующий фермент настоящего изобретения (Torrent et al., 1997). Так, например, последовательности SEQ ID NO:13, 27 и 30 представляют собой полинуклеотид, кодирующий гипертермофильный фермент, функционально присоединенный к N-концевой последовательности белка γ-зеина кукурузы. Другой сигнальной последовательностью является аминокислотная последовательность SEKDEL, обеспечивающая удерживание полипептидов в эндоплазматическом ретикулуме (Munro & Pelham, 1987). Так, например, полинуклеотид, кодирующий последовательности SEQ ID NO:14, 26, 28, 29, 33, 34, 35 или 36, которые содержат N-концевую последовательность γ-зеина кукурузы, функционально присоединенную к процессирующему ферменту, который функционально связан с SEKDEL. Полипептид может быть также нацелен на амилопласт путем присоединения к пептиду, обеспечивающему нацеливание на амилопласт восковидной кукурузы (Klosgen et al., 1986) или на крахмальные зерна. Так, например, полинуклеотид, кодирующий гипертермофильный процессирующий фермент, может быть функционально присоединен к хлоропластному (амилопластному) транспортному пептиду (СТР) и к крахмалсвязывающему домену, например, кодируемому геном waxy. Последовательность SEQ ID NO:10 представляет α-амилазу, присоединенную к крахмалсвязывающему домену восковидной кукурузы. Последовательность SEQ ID NO:15 представляет собой N-концевую последовательность, обеспечивающую нацеливание на амилопласт восковидной кукурузы и функционально присоединенную к α-амилазе. Кроме того, полинуклеотид, кодирующий процессирующий фермент, может быть присоединен для нацеливание на крахмальные зерна с использованием крахмалсвязывающего домена восковидной кукурузы. Так, например, последовательность SEQ ID NO:16 представляет собой гибридный полипептид, содержащий N-концевую амилопластную нацеливающую последовательность, обеспечивающую нацеливание на амилопласт восковидной кукурузы и функционально присоединенную к гибридному полипептиду α-амилазы/waxy, содержащему крахмалсвязывающий домен восковидной кукурузы.
Полинуклеотиды настоящего изобретения, помимо сигналов процессинга, могут, кроме того, включать другие регуляторные последовательности, известные специалистам. Термины "регуляторные последовательности" и "подходящие регуляторные последовательности" означают нуклеотидные последовательности, которые локализованы выше (5'-некодирующие последовательности), внутри или ниже (3'-некодирующие последовательности) кодирующей последовательности, которые влияют на транскрипцию, РНК-процессинг или стабильность, или на трансляцию ассоциированной кодирующей последовательности. Регуляторные последовательности включают энхансеры, промоторы, лидерные последовательности трансляции, интроны и сигнальные последовательности полиаденилирования. Такими последовательностями являются природные и синтетические последовательности, а также последовательности, которые могут представлять собой комбинацию синтетических и природных последовательностей.
В соответствии с настоящим изобретением могут быть также использованы селективные маркеры, которые позволяют проводить отбор трансформированных растений и тканей растения, как хорошо известно специалистам. В качестве экспрессируемого нужного гена или в дополнение к этому гену может оказаться желательным использование селективного или скринируемого маркерного гена. "Маркерные гены" представляют собой гены, которые придают клеткам, экспрессирующим этот маркерный ген, отличительный фенотипический признак и, тем самым, позволяют идентифицировать такие трансформированные клетки от клеток, которые не имеют такого маркера. Эти гены могут кодировать либо селективный, либо скринируемый маркер в зависимости от того, придает ли указанный маркер признак, по которому может быть проведен отбор химическими способами, то есть с использованием селективного агента (например, гербицида, антибиотика или т.п.), либо он просто является признаком, который можно идентифицировать посредством наблюдения или тестирования, то есть путем скрининга (например, признак R-локуса). Само собой разумеется, что известно много примеров подходящих маркерных генов, которые могут быть использованы для осуществления настоящего изобретения.
Термины "селективные" или "скринируемые" маркерные гены означают также гены, кодирующие "секретируемый маркер", секреция которого может быть детектирована методами идентификации или отбора на трансформированные клетки. Примерами могут служить маркеры, кодирующие секретируемый антиген, который может быть идентифицирован посредством взаимодействия с антителом, либо даже секретируемые ферменты, которые могут быть детектированы по их каталитической активности. Секретируемые белки принадлежат к ряду классов, включающих небольшие диффундируемые белки, детектируемые, например, с помощью ELISA; небольшие активные ферменты, детектируемые во внеклеточном растворе (например, α-амилаза, β -лактамаза, фосфинотрицин-ацетилтрансфераза); и белки, которые включаются в клеточные стенки или иммобилизуются на клеточных стенках (например, белки, которые включают лидерную последовательность, такую как последовательность, обнаруженную в экспрессионном элементе экстензина или PR-S табака).
Что касается селективных секретируемых маркеров, то особенно предпочтительно использовать ген, кодирующий белок, который становится секвестрированным в клеточной стенке и который включает уникальный эпитоп. Такой секретируемый антигенный маркер является идеальным из-за использования последовательности эпитопа, которая должна обеспечивать низкий фоновый уровень в ткани растения; промоторной-лидерной последовательности, которая должна обеспечивать эффективную экспрессию и прохождение через плазматическую мембрану и должна продуцировать белок, который связывается с клеточной стенкой и является к тому же доступным для антител. Обычно секретируемый белок клеточной стенки, модифицированный так, чтобы он включал уникальный эпитоп, должен удовлетворять всем указанным требованиям.
Одним из примеров белка, подходящего для такой модификации, является экстензин или богатый гидроксипролином гликопротеин (HPRG). Так, например, молекула HPRG кукурузы (Steifel et al., The Plant Cell, 2:785 (1990)) хорошо охарактеризована с точки зрения молекулярной биологии, экспрессии и структуры белка. Однако для создания скринируемого маркера любой белок из ряда экстензинов и/или богатых глицином белков клеточной стенки (Keller et al., EMBO Journal, 8:1309 (1989)) может быть модифицирован путем добавления области детерминанты.
а. Селективные маркеры
Возможными селективными маркерами, используемыми в настоящем изобретении, являются, но не ограничиваются ими, ген neo или nptII (Potrykus et al., Mol. Gen. Genet. 199:183 (1985)), которые придают резистентность к канамицину и могут быть выбраны для использования канамицина, G418 и т.п; ген bar, который придает резистентность к гербициду фосфинотрицину; ген, который кодирует модифицированный белок EPSP-синтазу (Hinchee et al., Biotech., 6:915 (1988)) и, тем самым, придает резистентность к глифосату; ген нитрилазы, такой как bxn от Klebsiella ozaenae, который придает резистентность к бромоксинилу (Stalker et al., Science, 242:419 (1988); мутантный ген ацетолактатсинтазы (ALS), который придает резистентность к имидазолинону, сульфонилмочевине или к другим ALS-ингибирующим химическим веществам (Европейская патентная заявка 154204, 1985); ген резистентности к метотрексату DHFR (Thillet et al., J. Biol. Chem. 263:12500 (1988)); ген далапондегалогеназы, который придает резистентность к гербициду далапону; ген фосфоманнозоизомеразы (PMI); мутированный ген антранилатсинтазы, который придает резистентность к 5-метилтриптофану; ген hph, который придает резистентность к антибиотику гигромицину; или ген маннозо-6-фосфатизомеразы (также называемый здесь геном фосфоманнозоизомеразы), который придает способность к метаболизму маннозы (патенты США №№5767378 и 5994629). Каждый специалист может выбрать подходящий селективный маркерный ген для использования в настоящем изобретении. При использовании мутированного гена EPSP-синтазы можно получить дополнительное преимущество от введения подходящего хлоропластного транспортного пептида СТР (Европейская патентная заявка 0218571, 1987).
Иллюстративным вариантом селективного маркерного гена, используемого в системах отбора трансформантов, являются гены, кодирующие фермент фосфинотрицин-ацетилтрансферазу, такие как ген bar от Streptomyces hygroscopicus или ген pat от Streptomyces viridochromogenes. Фермент фосфинотрицин-ацетилтрансфераза (РАТ) инактивирует активный ингредиент фосфинотрицин (РРТ) в гербициде биалафосе. РРТ ингибирует глутаминсинтетазу (Murakami et al., Mol. Gen. Genet. 205:42 (1986); Twell et al., Plant. Physiol., 91:1270 (1989)), что приводит к быстрому накоплению аммиака и к гибели клеток. Принимая во внимание тот факт, что главные трудности, как уже сообщалось, возникают при трансформации злаков, успешное использование этой селективной системы у однодольных растений было особенно неожиданным (Potrykus, Trends Biotech., 7:269 (1989)).
Если для осуществления настоящего изобретения необходимо использовать ген резистентности к биалафосу, то наиболее подходящими для этих целей являются гены bar или pat, которые могут быть получены из микроорганизмов вида Streptomyces (например, АТСС №21705). Клонирование гена bar описано в работах Murakami et al., Mol. Gen. Genet., 205:42 (1986) и Thompson et al., EMBO Journal, 6:2519 (1987), а также было описано использование гена bar для неоднодольных растений (De Block et al., EMBO Journal, 6:2513 (1987); De Block et al., Plant Physiol., 91:694 (1989)).
b. Скринируемые маркеры
Скринируемыми маркерами, которые могут быть использованы, являются, но не ограничиваются ими, ген β-глюкуронидазы или ген uidA (GUS), кодирующий фермент, для которого известны различные хромогенные субстраты; ген R-локуса, кодирующий продукт, который регулирует продуцирование антоцианиновых пигментов (красного цвета) в тканях растений (Dellaporta et al., Chromosome Structure and Function, pp.263-282 (1988)); ген β-лактамазы (Sutcliffe, PNAS USA, 75:3737 (1978)), кодирующий фермент, для которого известны различные хромогенные субстраты (например, PADAC, хромогенный цефалоспорин); ген xylE (Zukowsky et al., PNAS USA, 80:1101 (1983)), кодирующий катехол-диоксигеназу, который может превращать хромагенные катехолы; ген α-амилазы (Ikuta et al., Biotech, 8:241 (1990)); ген тирозиназы (Katz et al., J. Gen. Microbiol., 129:2703 (1983)), кодирующий фермент, способный окислять тирозин до DOPA и допахинона, который, в свою очередь, конденсируется с образованием легкодетектируемого соединения меланина; ген β-галактозидазы, кодирующий фермент, для которого имеются хромогенные субстраты; ген люциферазы (lux) (Ow et al., Science 234:856 (1986)), обеспечивающий биолюминесцентную детекцию; ген экворина (Prasher et al., Biochem. Biophys. Res. Comm., 126:1259 (1985)), который может быть использован для чувствительной к кальцию биолюминесцентной детекции, или ген белка, флуоресцирующего в зеленом диапазоне спектра (Niedz et al., Plant Cell Reports, 14:403 (1995)).
Считается, что гены R-генного комплекса кукурузы являются особенно ценными в качестве скринируемых маркеров. R-генный комплекс в кукурузе кодирует белок, который действует как регулятор продуцирования антоцианиновых пигментов в большинстве семян и тканей растения. Ген R-генного комплекса является подходящим для трансформации кукурузы, поскольку экспрессия этого гена в трансформированных клетках не оказывает негативного воздействия на эти клетки. Таким образом, R-ген, вводимый в такие клетки, будет приводить к экспрессии красного пигмента, и, при стабильном встраивании, он, в виде красного сектора, может обеспечивать визуальную оценку. Если линия растения кукурузы несет доминантные аллели генов, кодирующих ферментные промежуточные соединения в пути биосинтеза антоцианина (С2, А1, А2, Bz1 и Bz2), а также несет рецессивный аллель в R-локусе, то такая R-трансформация любой клетки этой линии будет приводить к образованию красного пигмента. Примерами таких линий являются линия Wisconsin 22, которая содержит аллель rg-Stedler и ТR112, и производное К55, которым является r-g, b, P1. Альтернативно, может быть использован любой генотип кукурузы, если С1- и R-аллели вводятся вместе. Другим скринируемым маркером, подходящим для использования в настоящем изобретении, является люцифераза огневки, кодируемая геном lux. Присутствие гена lux в трансформированных клетках может быть детектировано с использованием, например, рентгеновской пленки, сцинтилляционного счетчика, флуоресцентной спектрофотометрии, видеокамер, работающих при слабом освещении, счетчиков фотонов или мультиволновой люминометрии. Считается также, что может быть разработана система для скрининга популяций на биолюминесценцию, например, такая как планшеты для культивирования тканей или даже для скрининга целых растений.
Полинуклеотидами, используемыми для трансформации растений, могут быть, но не ограничиваются ими, ДНК, состоящей из растительных генов и нерастительных генов, таких как гены бактерий, дрожжей, животных или вирусов. Встроенная ДНК может включать модифицированные гены, части генов или химерные гены, включая гены от одного и того же или от различных генотипов кукурузы. Термины "химерный ген" или "химерная ДНК" означает ген или ДНК-последовательность или ее сегмент, содержащие, по крайней мере, две ДНК-последовательности или сегменты от видов, которые не объединяются в одну ДНК в природных условиях, либо ДНК-последовательности или сегменты, расположенные или присоединенные так, как это обычно не происходит в нативном геноме нетрансформированного растения.
Настоящее изобретение также относится к экспрессирующим кассетам, содержащим полинуклеотид, кодирующий гипертермофильный процессирующий фермент, предпочтительно оптимизированный по кодонам полинуклеотид. При этом предпочтительно, чтобы полинуклеотид в указанной экспрессирующей кассете (первый полинуклеотид) был функционально присоединен к регуляторным последовательностям, таким как промотор, энхансер, интрон, последовательность терминации или их любая комбинация, и, необязательно, ко второму полинуклеотиду, кодирующему сигнальную последовательность (N- или С-концевую), которая направляет фермент, кодируемый первым полинуклеотидом, в конкретный клеточный или субклеточный участок. Таким образом, промотор и одна или более сигнальных последовательностей могут обеспечивать высокие уровни экспрессии фермента в конкретных участках растения, тканях растения или клетках растения. Такими промоторами могут быть конститутивные промоторы, индуцибельные (условные) промоторы или тканеспецифические промоторы, например эндосперм-специфические промоторы, такие как промотор γ-зеина кукурузы (представленный SEQ ID NO:12) или промотор ADP-gpp кукурузы (представленный SEQ ID NO:11, которая включает 5′-нетранслируемую и интронную последовательность). Настоящее изобретение также относится к выделенному полинуклеотиду, содержащему промотор, включающий последовательность SEQ ID NO:11 или 12, к полинуклеотиду, который гибридизуется с его комплементом в условиях низкой степени жесткости, или к его фрагменту, промоторная активность которого составляет, по крайней мере, 10%, а предпочтительно, по крайней мере, 50% от активности промотора, имеющего последовательность SEQ ID NO:11 или 12. Настоящее изобретение также относится к векторам, которые содержат экспрессирующую кассету или полинуклеотид настоящего изобретения, и к трансформированным клеткам, содержащим полинуклеотид, экспрессирующую кассету или вектор согласно изобретению. Вектор согласно изобретению может содержать полинуклеотидную последовательность, кодирующую более чем один гипертермофильный процессирующий фермент настоящего изобретения, где указанная последовательность может находиться в смысловой или в антисмысловой ориентации, а трансформированная клетка может содержать один или более векторов настоящего изобретения. Предпочтительными векторами являются векторы, подходящие для встраивания нуклеиновых кислот в клетки растения.
Трансформация
Экспрессирующая кассета или векторная конструкция, содержащая экспрессирующую кассету, могут быть введены в клетку. Экспрессирующая кассета или векторная конструкция могут присутствовать в эписоме или они могут быть интегрированы в геном клетки. Затем трансформированная клетка может быть культивирована, с получением трансгенного растения. В соответствии с этим настоящее изобретение относится к продуктам трансгенного растения. Такими продуктами могут быть, но не ограничиваются ими, семена, плоды, потомство и продукты потомства трансгенного растения.
Для введения конструкции в клетку хозяина имеется ряд известных и доступных специалистам методов. Трансформация бактерий и многих эукариотических клеток может быть осуществлена с использованием полиэтиленгликоля, хлорида кальция, вирусной инфекции, фаговой инфекции, электропорации и других методов, известных специалистам. Методами трансформации растительных клеток или тканей являются трансформация с помощью ДНК, где в качестве трансформирующего средства используется A.tumefaciens или A.rhizogenes, электропорация, инжекция ДНК, бомбардировка микрочастицами, ускорение частиц и т.п. (см., например, ЕР 295959 и ЕР 138341).
В одном из вариантов осуществления изобретения бинарные векторы плазмид Ti и Ri Agrobacterium spp., то есть производные Ti-векторы, используются для трансформации широкого ряда высших растений, включая однодольные и двудольные растения, такие как соя, хлопчатник, рапс, табак и рис (Pacciotti et al. Bio/Technology, 3:241 (1985): Byrne et al. Plant Cell Tissue and Organ Culture, 8:3 (1987); Sukhapinda et al. Plant Mol. Biol., 8:209 (1987); Lorz et al. Mol, Gen. Genet., 199:178 (1985); Potrykus Mol. Gen. Genet., 199:183 (1985); Park et al., J. Plant Biol., 38:365 (1985): Hiei et al., Plant J., 6:271(1994)). Использование Т-ДНК для трансформации растительных клеток получило широкое распространение в научных исследованиях и подробно описано в литературе (EP 120516; Hoekema, In: The Binary Plant Vector_System. Offset-drukkerij Kanters B.V.; Alblasserdam (1985), Chapter V; Knauf, et al., Genetic Analysis of Host Range Expression By Agrobacterium In: Molecular Genetics of the Bacteria-Plant Interaction. Puhler, A. ed., Springer-Verlag, New York, 1983, p.245; and An. et al., EMBO J., 4:277 (1985)).
Специалистам известны и другие методы трансформации, такие как прямой захват чужеродной ДНК-конструкции (см. ЕР 295959), электропорация (Fromm et al., Nature (London), 319:791 (1986) или высокоскоростная баллистическая бомбардировка металлическими частицами, покрытыми конструкциями нуклеиновой кислоты (Kline et al., Nature (London), 327:70 (1987) и патент США №4945050). После трансформации клетки могут быть регенерированы методами, известными специалистам. Особенно подходящими являются недавно описанные способы трансформации чужеродных генов в коммерчески ценные культуры, такие как семена рапса (De Block et al., Plant Physiol. 91:694-701(1989)), подсолнечник (Everett et al., Bio/Technology. 5:1201(1987)), соя (McCabe et al., Bio/Technology. 6:923 (1988); Hinchee et al., Bio/Technology. 6:915 (1988); Chee et al., Plant Physiol., 91:1212 (1989); Christou et al., Proc. Natl. Acad. Sci USA, 86:7500 (1989) EP 301749), рис (Hiei et al., Plant J., 6:271 (1994)), и кукуруза (Gordon Kamm et al., Plant Cell, 2:603 (1990); Fromm et al., Biotechnology, 8:833, (1990)).
Экспрессирующие векторы, содержащие геномные или синтетические фрагменты, могут быть введены в протопласты, или в интактные ткани, или в выделенные клетки. Предпочтительно экспрессирующие векторы вводят в интактную ткань. Общие методы культивирования растительных тканей описаны, например, by Maki et al. "Procedures for Introducing Foreign DNA into Plants′" in Methods in Plant Molecular Biology & Biotechnology, Glich et al. (Eds.), pp.67-88 CRC Press (1993); and by Phillips et al. "Cell-Tissue Culture and In-Vitro Manipulation" in Corn & Com Improvement, 3rd Edition 10, Sprague et al. (Eds.) pp. 345-387, American Society of Agronomy Inc. (1988).
В одном из вариантов осуществления изобретения экспрессирующие векторы могут быть введены в кукурузу или в другие растительные ткани методами прямого переноса генов, такими как опосредуемое микрочастицами нацеливание, инжекция ДНК, электропорация и т.п. Экспрессирующие векторы вводят в растительные ткани путем бомбардировки микрочастицами с использованием биобаллистического устройства. См., например, Tomes et al. "Direct DNA transfer into intact plant cells via microprojectile bombardment" in Gamborg and Phillips (Eds.) Plant Cell, Tissue and Organ Culture: Fundamental Methods, Springer Verlag, Berlin (1995). Однако в настоящем изобретении рассматривается трансформация растений с использованием гипертермофильного процессирующего фермента в соответствии с известными методами трансформации. См. также Weissinger et al., Annual Rev. Genet., 22:421 (1988); Sanford et al., Particulate Science and Technology 5:27 (1987) (лук); Christou et al., Plant Physiol., 87:671 (1988) (соя); McCabe et al., Bio/Technology,. 6:923 (1988) (соя); Datta et al., Bio/Technology. 8:736 (1990) (рис); Klein et al., Proc. Natl. Acad. Sci. USA, 85:4305 (1988) (кукуруза); Klein et al., Bio/Technology, 6:559 (1988) (кукуруза); Klein et al., Plant Physiol., 91:440 (1988) (кукуруза); Fromm et al., Bio/Technology, 8:833 (1990) (кукуруза); and Gordon-Kamm et al., Plant Cell. 2, 603 (1990) (кукуруза); Svab et al., Proc. Natl. Acad. Sci. USA, 87:8526 (1990) (хлоропласт табака); Koziel et al., Biotechnology, 11:194 (1993) (кукуруза); Shimamoto et al., Nature. 338:274 (1989) (рис); Christou et al., Biotechnology, 9:957 (1991) (рис); Европейская патентная заявка EP 0332581 (ежа сборная и другие Мятликовые); Vasil et al., Biotechnology, 11:1553 (1993) (пшеница). Methods in Molecular Biology, 82. Arabidopsis Protocols Ed. Martinez-Zapater and Salinas 1998 Humana Press (Arabidopsis).
Трансформация может быть осуществлена одной ДНК-молекулой или множеством ДНК-молекул (то есть ко-трансформация), и оба этих метода являются подходящими для применения вместе с экспрессирующими кассетами и конструкциями согласно изобретению. Для трансформации растений имеется множество подходящих трансформирующих векторов, и экспрессирующие кассеты согласно изобретению могут быть использованы в сочетании с любым таким вектором. Выбор вектора зависит от предпочтительного метода трансформации и вида мишени для трансформации.
И наконец, наиболее предпочтительные ДНК-сегменты для введения в геном однодольного растения могут представлять собой гомологичные гены или семейство генов, которые кодируют нужный признак (например, гидролиз белков, липидов или полисахаридов), которые вводят под контролем новых промоторов или энхансеров и т.п., либо, возможно, даже под контролем гомологичных или тканеспецифичных (например, корнеспецифических, шейко/влагалищеспецифических, мутовкоспецифических, стеблеспецифических, початкоспецифических, зерноспецифических или листьеспецифичных) промоторов или регуляторных элементов. Действительно, предусматривается, что конкретное использование настоящего изобретения позволит осуществлять направленное нацеливание гена конститутивным или индуцибельным способом.
Примеры подходящих трансформирующих векторов
Многочисленные трансформирующие векторы, подходящие для трансформации растений, известны специалистам по трансформации растений, и гены настоящего изобретения могут быть использованы в сочетании с любыми векторами, известными специалистам. Выбор вектора будет зависеть от предпочтительного метода трансформации и вида мишени, используемой для трансформации.
а. Векторы, подходящие для трансформации Agrobacterium
Для трансформации с использованием Agrobacterium tumefaciens существует много векторов. Эти векторы обычно несут, по крайней мере, одну Т-ДНК-пограничную последовательность, и такими векторами являются рBIN19 (Bevan, Nucl. Acids. Res. (1984)). Ниже приводится конструирование двух типов векторов, подходящих для Agrobacterium-опосредованной трансформации.
рСIB200 и рСIB2001
Для конструирования рекомбинантных векторов с помощью Agrobacterium были использованы бинарные векторы рСIB200 и рСIB2001, которые были сконструированы следующим образом. рTJS75kan конструировали путем расщепления NarI рTJS75 (Schmidhauser & Helinski, J. Bacteriol., 164:446 (1985)), что позволяло вырезать ген резистентности к тетрациклину, с последующим встраиванием Асс1-фрагмента из плазмиды pUC4К, несущей NPTII (Messing & Vierra, Gene, 19:259 (1982): Bevan et al., Nature, 304:184 (1983): McBride et al., Plant Molecular Biology, 14:266 (1990)). XhoI-линкеры лигировали с EcoRV-фрагментом РСIB7, который содержит левые и правые пограничные Т-ДНК-последовательности, селективный химерный ген nos/nptII растения и полилинкер pUC (Rothstein et al., Gene, 53:153 (1987)), a XhoI-расщепленный фрагмент клонировали в SalI-расщепленный pTJS75kan с получением рСIВ200 (см. также ЕР 0332104, пример 19). рСIВ200 содержит нижеследующие уникальные рестрикционные сайты полилинкера: EcoRI, SstI, Kpnl, BglII, Xbal и Sail. pCIB2001 представляет собой производное рСIВ200, сконструированное путем встраивания в полилинкер дополнительных рестрикционных сайтов. Уникальными рестрикционными сайтами в полилинкере pCIB2001 являются EcoRI, SstI, Kpnl, BglII, Xbal, Sall, Mlul, Bc1l, AvrII, Apa1, Hpa1 и StuI. Полилинкер pCIB2001, помимо указанных уникальных рестрикционных сайтов, также содержит ген резистентности к канамицину для отбора в растениях и бактериях, левую и правую пограничные Т-ДНК-последовательности для Agrohacterium-опосредованной трансформации, функциональный ген trfA, происходящий от RK2, для активации в E.coli и других хозяевах, и функциональные гены OriT и OriV, также происходящие от RK2. Полилинкер pCIB2001 является подходящим для клонирования растительных экспрессирующих кассет, содержащих свои собственные регуляторные сигналы.
pCIB10 и его производные для отбора на гигромицин
Бинарный вектор pCIB10 содержит ген резистентности к канамицину для отбора в растениях и левую и правую пограничные Т-ДНК-последовательности и вводит последовательности, происходящие от плазмиды pRK252 широкого круга хозяев, что позволяет этому вектору реплицироваться как в E.coli, так и в Agrobacterium. Его конструирование описано Rothstein et al. (Gene., 53:153 (1987)). Были сконструированы различные производные рСIB10, которые вводят ген гигромицин В-фосфотрансферазы, описанный Gritz et al. (Gene, 25:179 (1983)). Эти производные позволяют проводить отбор трансгенных клеток растения только на гигромицин (рСIB743) или на гигромицин и канамицин (рСIB715, рСIB717).
b. Векторы, подходящие для трансформации, не опосредуемой Agrobacterium
Трансформация без использования Agrobacterium tumefaciens позволяет "обойти" требование присутствия Т-ДНК-последовательностей при выборе трансформирующего вектора, а поэтому помимо векторов, таких как описанные выше векторы, содержащие Т-ДНК-последовательности, могут быть использованы векторы, не имеющие этих последовательностей. Методы трансформации, которые не зависят от Agrobacterium, предусматривают осуществление трансформации путем бомбардировки микрочастицами, захват протопластами (например, ПЭГ и электропорация) и микроинжекцию. Выбор вектора, главным образом, зависит от предпочтительного отбора на трансформируемые виды растений. Далее приводятся неограничивающие примеры конструирования типичных векторов, подходящих для трансформации без использования Agrobacterium.
рСIB3064
рСIB3064 представляет собой вектор, происходящий от pUC и подходящий для осуществления методов прямого переноса генов в комбинации с методом отбора на гербицид basta (или фосфофинотрицина). Плазмида рСIB246 содержит промотор 35S CaMV, функционально присоединенный к гену GUS E.coli, и терминатор транскрипции 35S CaMV и описана в опубликованной РСТ-заявке WO 93/07278. Промотор 35S этого вектора содержит две последовательности ATG со стороны 5′-конца от сайта инициации. Эти сайты подвергали мутации с использованием стандартной ПЦР-техники в целях удаления ATG и генерирования рестрикционных сайтов Sspl и Pvul. Новые рестрикционные сайты находились на расстоянии 96 и 37 п.н. от уникального SalI-сайта и на расстоянии 101 и 42 п.н. от имеющегося сайта инициации. Полученное производное рСIВ246 обозначали рСIВ3205. Затем ген GUS вырезали из рСIВ3025 путем расщепления ферментами SalI и Sad, концы затупляли и снова лигировали с получением плазмиды рСIВ3060. Плазмида рJIТ82 может быть взята из Центра Джона Иннеса (Норвегия), а 400 п.н.-SmaI-фрагмент, содержащий ген bar от Streptomyces viridochromogenes, вырезали и встраивали в HpaI-сайт рСIВ3060 (Thompson et al., EMBO J., 6:2519 (1987)). Этот генерированный вектор рСIВ3064 содержит ген bar, находящийся под контролем промотора 35S CaMV и терминатора для отбора на гербицид, ген резистентности к ампициллину (для отбора в E.coli) и полилинкер с уникальными сайтами SphI, PstI, HindIII и BamHI. Этот вектор является подходящим для клонирования растительных экспрессирующих кассет, содержащих свои собственные регуляторные сигналы.
PSOG19 и pSOG35
Плазмида pSOG35 представляет собой трансформирующий вектор, который содержит ген дигидрофолатредуктазы E.coli (DHFR) в качестве селективного маркера, придающего резистентность к метотрексату. Для амплификации промотора 35S (-800 п.н.), интрона 6 гена Adh1 кукурузы (-550 п.н.) и 18 п.н. нетранслируемой лидерной последовательности GUS от рSOG10 использовали ПЦР. 250-п.н.-фрагмент, кодирующий ген гидрофолатредуктазы типа II E.coli, также амплифицировали с помощью ПЦР, и эти два ПЦР-фрагмента собирали с использованием SacI-PstI-фрагмента из вектора рВ1221 (Clontech), который содержит каркас вектора pUC19 и терминатор нопалин-синтазы. Сборка этих фрагментов приводит к генерированию вектора рSOG19, который содержит промотор 35S, присоединенный к последовательности интрона 6, лидерную последовательность GUS, ген DHFR и терминатор нопалин-синтазы. Замена лидерной последовательности GUS в рSOG19 на лидерную последовательность, происходящую от вируса хлорозной пятнистости кукурузы (MCMV), приводит к генерированию вектора рSOG35. Векторы рSOG19 и рSOG35 несут ген pUC резистентности к ампициллину и имеют HindIII-, SphI-, PstI- и EcoRI-сайты, доступные для клонирования чужеродного материала.
с. Вектор, подходящий для трансформации в хлоропластах
Для экспрессии нуклеотидной последовательности настоящего изобретения в пластидах растений использовали вектор для трансформации в пластидах рРН143 (WO 97/32011, пример 36). Нуклеотидную последовательность встраивали в рРН143 для замены кодирующей последовательности PROTOX. Затем этот вектор использовали для трансформации в пластидах и для отбора трансформантов на резистентность к спектиномицину. Альтернативно, нуклеотидную последовательность встраивали в рРН143 для замены гена aadH. В этом случае, трансформанты отбирали на резистентность к ингибиторам PROTOX.
Растения-хозяева, подвергаемые трансформации
Любая ткань растения, способная к клональному размножению, независимо от органогенеза или от эмбриогенеза, может быть трансформирована с помощью конструкции настоящего изобретения. Термин "органогенез" означает процесс, при котором побеги и корни последовательно развиваются из меристемных центров, а термин "эмбриогенез" означает процесс, при котором побеги и корни развиваются вместе и одновременно (не последовательно) либо из соматических клеток, либо из гамет. Выбор конкретной ткани может варьироваться в зависимости от систем клонального размножения, доступных и наиболее подходящих для конкретно трансформируемых видов. Примерами тканей-мишеней являются дифференцированные и недифференцированные ткани или растения, включая, но не ограничиваясь ими, диски листьев, корни, стебли, побеги, листья, пыльцу, семена, зародыши, семядоли, гипокотили, мегагаметофиты, каллусные ткани, присутствующую меристемную ткань (например, верхушечную меристему, пазушные почки и корневые меристемы) и индуцированную меристемную ткань (например, меристему семядоли и меристему гипокотиля), опухолевую ткань и различные формы клеток и культур, таких как одиночные клетки, протопласт, зародыши и каллусная ткань. Такая ткань растения может присутствовать в растении или в органе либо в тканевой или клеточной культуре.
Растения настоящего изобретения могут иметь различные формы. Такими растениями могут быть химеры трансформированных клеток и нетрансформированных клеток; растениями могут быть клональные трансформанты (например, все клетки, трансформированные так, чтобы они содержали экспрессирующую кассету); растения могут содержать трансплантаты трансформированных или нетрансформированных тканей (например, трансформированный корневой побег, привитый к нетрансформированному побегу цитрусового растения). Трансформированные растения могут быть размножены различными способами, такими как клональное размножение или классический способ естественного разведения. Так, например, первое поколение (или Т1) трансформированных растений может быть подвергнута самоопылению с получением второго поколения (или Т2) трансформированных растений, а растение Т2 может быть затем размножено классическими методами разведения растений. Для облегчения разведения доминантный селективный маркер (такой как npt II) может быть ассоциирован с экспрессирующей кассетой.
Настоящее изобретение может быть использовано для трансформации любых видов растений, а именно однодольных или двудольных растений, включая, но не ограничиваясь ими, кукурузу (Zea mays), Brassica sp.(например, B.napus, B.rapa, B.juncea), а в частности, виды растения Brassica, используемые в качестве источника для получения масличных семян, люцерна (Medicago sativa), рис (Oryza sativa), рожь (Secale cereale), сорго (Sorghum bicolor, Sorghum vulgare), просо (например, просо американское (Pennisetum glaucum), просо африканское (Panicum miliaceum), просо итальянское (Setaria italica), элевзине (Eleusine coracana)), подсолнечник (Helianthus annuus), сафлор (Carthamus tinctoris), пшеницу (Triticum aestivum), сою (Glycine max), табак (Nicotiana tabacum), картофель (Solanum tuberosum), арахис (Arachis hypogaea), хлопчатник (Gossypium barbadense, Gossypium hirsutum), батат (Ipomoea batatus), кассаву (Manihot esculenta), кофе (Cofea spp.), кокос (Cocos nucifera), ананас (Ananas comosus), цитрусовые (Citrus spp.), какао (Theobroma cacao), чай (Camellia sinensis), банан (Musa spp.), авокадо (Persea americana), инжир (Ficus casica), гуаву (Psidium guajava), манго (Mangifera indica), оливу (Olea europaea), папайю (Carica papaya), кешью (Anacardium occidentale), макадамию (Macadamia integrifolia), миндаль (Prunus amygdalus), сахарную свеклу (Beta vulgaris), сахарный тростник (Saccharum spp.), овес, ячмень, овощные растения, декоративные растения, древесные растения, такие как хвойные и листопадные деревья, тыква крупноплодная, тыква обыкновенная, конопля, цукини, яблоня, груша, айва, дыня, слива, вишня, персик, нектарин, абрикос, земляника, виноград, малина, ежевика, соя, сорго, сахарный тростник, рапс, клевер, морковь и резушка Таля (Arabidopsis thaliana).
Овощными растениями являются томаты (Lycopersicon esculentum), латук (например, Lactuca sativa), зеленые бобы (Phaseolus vulgaris), фасоль Лима (Phaseolus limensis), горох (Lathyrus spp.), цветная капуста, брокколи, турнепс, редька, шпинат, спаржа, лук, чеснок, перец, сельдерей и члены рода Cucumis, такие как огурец (C.sativus), канталупская дыня (C.cantalupensis) и мускусная дыня (С.melo). Декоративными растениями являются азалия (Rhododendron spp.), гортензия (Macrophylla hydrangea), гибискус (Hibiscus rosasanensis), розы (Rosa spp.), тюльпаны (Tulipa spp.), нарциссы (Narcissus spp.), петунии (Petunia hybrida), гвоздика (Dianthus caryophyllus), пуансетия (Euphorbia pulcherrima) и хризантема. Хвойными деревьями, которые могут быть использованы для осуществления настоящего изобретения, являются, например, сосны, такие как сосна ладанная (Pinus taeda), сосна Элиотта (Pinus elliotii), сосна желтая (Pinus ponderosa), сосна скрученная широкохвойная (Pinus contorta) и сосна лучистая (Pinus radiata), лжетсуга тиссолистная (Pseudotsuga menziesii); тсуга западная (Tsuga canadensis); ель ситхинская (Picea glauca); секвойя вечнозеленая (Sequoia sempervirens); пихты настоящие, такие как пихта белая (Abies amabilis) и пихта бальзамическая (Abies balsamea), и кедры, такие как туя (Thuja plicata) и кипарис желтый аляскинский (Chamaecyparis nootkatensis). Бобовыми растениями являются бобы и горох. Бобами являются хьюар, рожковое дерево, пажитник сеннной, соя, бобы посевные, вигна китайская, фасоль золотистая, фасоль Лима, кормовые бобы, чечевица, горох бараний и т.п. Бобовыми растениями являются, но не ограничиваются ими, Arachis, например, арахис, Vicia, например, вязель, вика мохнатая, фасоль лучистая, фасоль золотистая и горох бараний, Lupinus, например, люпин, клевер, Phaseolus, например, фасоль обыкновенная и фасоль Лима, Pisum, например, фасоль полевая, Melilotus, например, клевер, Medicago, например, люцерна, Lotus, например, трилистник, Lens, например, чечевица, и софора австралийская. Предпочтительными кормовыми и дернообразующими растениями, подходящими для использования в способах настоящего изобретения, являются люцерна, ежа сборная, овсянница высокая, плевел многолетний, полевица белая и полевица малая.
Предпочтительными растениями настоящего изобретения являются сельскохозяйственные растения, например кукуруза, люцерна, подсолнечник, капуста Brassica, соя, хлопчатник, сафлор, арахис, сорго, пшеница, просо, табак, ячмень, рис, томаты, картофель, тыква крупноплодная, дыни, бобовые культуры и т.п. Другими предпочтительными растениями являются Liliopsida и Panicoideae.
После трансформации нужной ДНК-последовательности в растения конкретного вида она может быть амплифицирована в растении того же вида либо введена в другой сорт растения того же самого вида, а в частности, в промышленные сорта, используемые в традиционных способах разведения.
Ниже описана трансформация двудольных и однодольных растений, а также репрезентативный способ трансформации в пластидах.
а. Трансформация двудольных растений
Методы трансформации двудольных растений хорошо известны специалистам, и такими методами являются методы с использованием Agrobacterium и методы без использования Agrobacterium. Методы без использования Agrobacterium предусматривают захват экзогенного генетического материала непосредственно протопластами или клетками. Это может быть осуществлено путем ПЭГ-опосредованного введения, введения путем электропорации, доставки путем бомбардировки микрочастицами или путем микроинжекции. Примеры таких методов описаны в литературе (Paszkowski et al., EMBO J. 3:2717 (1984), Potrykus et al. Mol. Gen. Genet., 199:169 (1985), Reich et al., Biotechnology, 4:1001 (1986), and Klein et al., Nature, 327:70 (1987)). В каждом случае трансформированные клетки регенерируют с получением целого растения стандартными методами, известными специалистам.
Agrobacterium-опосредованная трансформация является предпочтительным методом трансформации двудольных растений из-за его высокой эффективности трансформации и возможности использования для многих видов растений. Agrobacterium-трансформация обычно предусматривает перенос бинарного вектора, несущего представляющую интерес чужеродную ДНК (например, pCIB200 или pCIB2001) в соответствующий штамм Agrobacterium, и может зависеть от комплемента генов vir, присутствующих в штамме-хозяине Agrobacterium, либо присутствующих вместе на Ti-плазмиде, либо на хромосоме (например, штамм CIB542 для pCIB200 и pCIB2001 (Uknes et al., Plant Cell., 5:159 (1993)). Перенос рекомбинантного бинарного вектора в Agrobacterium осуществляют путем скрещивания трех родительских линий с использованием рекомбинантного бинарного вектора и хелперного штамма E.coli, который несет плазмиду, такую как рRK2013, и который способен к мобилизации этого рекомбинантного бинарного вектора в штамм-мишень Agrobacterium. Альтернативно, указанный рекомбинантный бинарный вектор может быть перенесен в Agrobacterium путем ее трансформации ДНК-последовательностью (Höfgen & Willmitzer, Nucl. Acids. Res., 16:9877 (1988)).
Трансформация растения нужного вида рекомбинантной агробактерией обычно предусматривает совместное культивирование Agrobacterium вместе с эксплантатом, взятым из растения, в соответствии с нижеследующими протоколами, хорошо известными специалистам. Трансформированную ткань регенерируют на селективной среде, несущей маркер резистентности к антибиотику или гербициду, присутствующему между Т-ДНК-фланкирующими последовательностями в бинарной плазмиде.
Векторы могут быть введены в растительные клетки известными методами. Клетками, предпочтительными для трансформации, являются Agrobacterium, клетки однодольного и клетки двудольного растения, включая клетки Liliopsida и Panicoideae. Предпочтительными клетками однодольного растения являются клетки зерновых растений, например маиса (кукурузы), ячменя и пшеницы, и крахмалзапасающих двудольных растений, например картофеля.
Другой способ трансформации клетки растения предусматривает "запуск" частиц, являющихся инертными или биологически активными в тканях и клетках растения. Этот способ описан в патентах США №№4945050, 5036006 и 5100792. В общих чертах, эта процедура предусматривает приведение в движение частиц, являющихся инертными или биологически активными в клетках, в условиях, эффективных для прохождения через внешнюю поверхность клетки и включение вовнутрь клетки. Если используется инертная частица, то вектор может быть введен в клетку путем покрытия частиц вектором, содержащим нужный ген. Альтернативно, клетка-мишень может быть окружена вектором так, чтобы этот вектор внедрялся в клетку с помощью данной частицы. Биологически активные частицы (например, осушенные дрожжевые клетки, осушенные бактерии или бактериофаг, которые содержат ДНК, предназначенную для введения) могут быть также внедрены в клетки и ткань растения.
b. Трансформация однодольных растений
В настоящее время трансформация большинства видов однодольных растений уже стала рутинной процедурой. Предпочтительными методами является прямой захват гена протопластами с использованием полиэтиленгликоля (ПЭГ) или техники электропорации, а также бомбардировки каллусной ткани частицами. Трансформация может быть осуществлена с использованием одного вида ДНК или множества видов ДНК (то есть ко-трансформация), и оба эти метода могут быть использованы в настоящем изобретении. Ко-трансформация может иметь то преимущество, что она позволяет избежать конструирования полного вектора и генерировать трансгенные растения с несцепленными локусами для нужного гена и селективного маркера и, тем самым, позволяет удалять этот селективный маркер в последующих поколениях, что должно рассматриваться как положительный фактор. Однако недостаток использования ко-трансформации заключается в том, что в этом случае отдельные виды ДНК интегрируются в геном с частотой менее чем 100% (Schocher et al., Biotechnology, 4:1093, 1986).
В патентных заявках ЕР 0292453, ЕР0392225 и WO 93/07278 описаны методы получения каллуса и протопластов из элитной имбредной линии кукурузы, трансформация протопластов с использованием ПЭГ или электропорации и регенерация растений кукурузы из трансформированных протопластов. В работе Gordon-Kamm et al. (Plant Cell, 2:603 (1990)) и Fromm et al. (Biotechnology, 8:833 (1990)) описаны методы трансформации линии растения кукурузы, происходящей от А188, посредством бомбардировки частицами. Кроме того, в WO 93/07278 и в работе Koziel et al. (Biotechnology, 11:194 (1993)) описаны методы трансформации элитных инбредных линий кукурузы путем бомбардировки частицами. В этом методе использовались незрелые зародыши кукурузы длиной 1,5-2,5 мм, вырезанные из початка кукурузы через 14-15 дней после опыления, и биобаллистическое устройство PDS-1000Не для бомбардировки.
Трансформация риса может быть также осуществлена методами прямого переноса гена с использованием протопластов или путем бомбардировки частицами. Опосредуемая протопластами трансформация была описана для видов Japonica и Indica (Zhang et al., Plant. Cell. Rep., 7:379 (1988); Shimamoto et al., Nature 338: 274 (1989); Datta et al., Biotechnology, 8:736 (1990)). Оба этих типа могут быть также рутинно трансформированы посредством бомбардировки частицами (Christou et al., Biotechnology, 9:957 (1991)). Кроме того, в WO 93/21335 описаны методы трансформации риса посредством электропорации. В патентной заявке ЕР 0332581 описаны методы генерирования, трансформации и регенерации протопластов растений семейства Мятликовых (Pooideae). Эти методы позволяют трансформировать Dactylis и пшеницу. Кроме того, трансформация пшеницы была описана в работе Vasil et al. (Biotechnology, 10:667 (1992)) посредством бомбардировки частицами клеток долгоживущего регенерируемого каллуса типа С, а также в работе Vasil et al. (Biotechnology 11:1553 (1993)) и Weeks et al. (Plant Physiol., 102:1077 (1993)), посредством бомбардировки частицами незрелых зародышей и каллуса, происходящего от незрелого зародыша. Однако предпочтительный метод трансформации пшеницы предусматривал трансформацию пшеницы посредством бомбардировки частицами незрелых зародышей и включает стадию культивирования с высоким содержанием сахарозы или высоким содержанием мальтозы перед доставкой гена. Перед бомбардировкой любое число зародышей (длиной 0,75-1 мм) высевали в среду MS с 3% сахарозой (Murashiga & Skoog, Physiologia Plantarum, 15:473 (1962)) и 3 мг/л 2,4-D для индуцирования соматических зародышей, которые оставляли в темноте. В день, выбранный для бомбардировки, зародыши удаляли из среды для индуцирования и помещали в условия осмоса (то есть в среду для индуцирования, содержащую сахарозу или мальтозу в нужной концентрации, обычно 15%). Зародыши оставляли на 2-3 часа для плазмолиза, а затем подвергали бомбардировке. Число в двадцать зародышей на данный планшет является типичным, но не критическим. Плазмиду, несущую соответствующий ген (такую как рСIB3064 или рSG35) осаждали на золотые частицы размером в несколько микрон с использованием стандартных процедур. Каждый планшет с зародышами простреливали с помощью гелиевого биобаллистического устройства Du Pont Biolistics® при давлении разрыва примерно 1000 фунт/кв.дюйм с использованием стандартного сита 80 меш. После бомбардировки зародыши помещали обратно в темноту примерно на 24 часа (пока еще в условия осмоса). Через 24 часа зародыши удаляли из среды в условиях осмоса и помещали обратно в среду для индуцирования, где они находились примерно один месяц перед регенерацией. Примерно через месяц эксплантаты зародышей с развивающимся эмбриогенным каллусом переносили в среду для регенерации (MS+1 мг/литр NAA, 5 мг/литр GA), которая дополнительно содержала подходящий селективный агент (10 мг/л basta в случае рСIB3064 и 2 мг/л метотрексата в случае рSOG35). Примерно через один месяц развившиеся побеги переносили в более крупные стерильные контейнеры, известные как "GA7s", которые содержали полуконцентрированную MS, 2% сахарозу и ту же самую концентрацию селективного агента.
Была также описана трансформация однодольных растений с использованием Agrobacterium. См. WO 94/00977 и патент США №5591616, которые вводятся в настоящее описание посредством ссылки.
с. Трансформация пластид
Семена Nicotiana tabacum, сорта "Xanthi nc", проращивали в количестве семь семян на планшет в 1-дюймовом кольцевом массиве на Т-агаровой среде и через 12-14 дней после высевания подвергали бомбардировке вольфрамовыми частицами размером в 1 мкм (М10, Biorad, Hercules, CA), покрытыми ДНК, происходящими от плазмид рРН143 и рРН145, в основном, как описано в литературе (Svab & Maliga, PNAS, 90:913 (1993)). Подвергнутые бомбардировке саженцы инкубировали на Т-среде в течение двух дней, после чего листья отрезали и помещали своей обратной стороной на яркий свет (350-500 мкмоль· фотон/м2/с) на планшеты со средой RMOP (Svab, Hajdukiewicz & Maliga, PNAS, 87:8526 (1990)), содержащей 500 мкг/мл дигидрохлорида спектиномицина (Sigma, St Louis, MO). Через три-восемь недель после бомбардировки резистентные побеги, находящиеся под обесцвеченными листьями, подвергали субклонированию на той же самой селективной среде, затем оставляли для образования каллуса и вторичные побеги отделяли и субклонировали. Полную сегрегацию копий трансформированного пластидного генома (гомоплазматичности) в независимых субклонах анализировали стандартными методами (Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor (1989)). Полную клеточную ДНК, расщепленную BamHI/EcoRI (Mettler I.J. Plant. Mol. Biol. Reporter, 5:346 (1987)), отделяли на агарозном геле с 1% трис-боратом (ТВЕ), переносили на найлоновые мембраны (Amersham) и зондировали32Р-меченными произвольно праймированными ДНК-последовательностями, соответствующими 0,7 т.п.н.-BamHI/HindIII-ДНК-фрагменту от плазмиды рС8, содержащей часть пластидной нацеливающей последовательности rps7/12. Гомоплазматические побеги асептически укореняли на среде MS/IBA, содержащей спектиномицин (McBride et al., PNAS, 91:7301 (1994)), и переносили в теплицу.
Продуцирование и характеризация стабильно трансформированных растений
Клетки трансформированного растения помещали в соответствующую селективную среду для отбора трансгенных клеток, которые затем выращивали до каллуса. Из каллуса вырастали побеги, и из этих побегов генерировали небольшие растения путем культивирования на среде для укоренения. Различные конструкции обычно присоединяли к маркеру для отбора в клетках растений. Как правило, маркер представляет собой ген резистентности к биоциду (а в частности, к антибиотику, такому как канамицин, G418, блеомицин, гигромицин, хлорамфеникол, гербицид или т.п.). Конкретно используемый маркер позволяет дифференцировать трансформированные клетки от клеток, не содержащих вводимой ДНК.
Компоненты ДНК-конструкций, включая транскрипционные/экспрессирующие кассеты согласно изобретению, могут быть получены из последовательностей, которые являются нативными (эндогенными) или чужеродными (экзогенными) для хозяина. Термин "чужеродный" означает, что последовательность не обнаруживается в хозяине дикого типа, в который вводят данную конструкцию. Гетерологичные конструкции будут содержать, по крайней мере, одну область, не являющуюся нативной для гена, от которого происходит область инициации транскрипции.
Для подтверждения присутствия трансгенов в трансгенных клетках и растениях может быть осуществлен Саузерн-блот-анализ методами, известными специалистам. Интеграция сегментов полинуклеиновой кислоты в геном может быть детектирована и количественно оценена с помощью Саузерн-блот-анализа, поскольку они могут быть легко дифференцированы от конструкций, содержащих указанные сегменты, с использованием подходящих рестриктирующих ферментов. Продукты экспрессии трансгенов могут быть детектированы любыми способами в зависимости от природы данного продукта, и такими способами являются Вестерн-блот-анализ и ферментный анализ. Одним из конкретно используемых способов количественной оценки экспрессии белка и детекции репликации в различных тканях растений является использование гена-репортера, такого как GUS. После получения трансгенных растений они могут быть культивированы для генерирования тканей или частей растения, имеющих нужный фенотип. Могут быть получены ткани растения или части растения и/или собраны семена. Семена могут служить источником для культивирования других растений с тканями или частями, имеющими нужные признаки.
Таким образом, настоящее изобретение относится к трансформированному растению или части растения, такой как колос, семена, плод, зерно, солома, чешуя злаков или багасса, содержащие по меньшей мере один полинуклеотид, экспрессирующую кассету или вектор согласно изобретению, к способу получения такого растения или к способам применения такого растения или его части. Указанное трансформированное растение или часть такого растения экспрессирует процессирующий фермент, необязательно присутствующий в конкретном клеточном или субклеточном компартменте некоторых тканей или в прорастающем зерне. Так, например, настоящее изобретение относится к трансформированному растению, содержащему по меньшей мере один крахмалпроцессирующий фермент, присутствующий в клетках указанного растения, где часть растения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один крахмалпроцессирующий фермент. Процессирующий фермент не действует на субстрат-мишень, если только он не активирован такими методами, как нагревание, измельчение, или другими методами, обеспечивающими контактирование указанного фермента с субстратом в условиях, где указанный фермент является активным.
Предпочтительные способы настоящего изобретения
Самопроцессирующиеся растения и части растений настоящего изобретения могут быть использованы в различных методах с применением процессирующих ферментов (мезофильных, термофильных или гипертермофильных), экспрессированных и активированных в этих растениях. В соответствии с настоящим изобретением часть трансгенного растения, полученная из трансгенного растения, геном которого пополнен, по крайней мере, на один процессирующий фермент, помещают в условия, при которых экспрессируется и активируется данный процессирующий фермент. После активации процессирующий фермент активируется и взаимодействует с субстратом, на который он обычно действует, с продуцированием нужного продукта. Так, например, крахмалпроцессирующие ферменты действуют на крахмал с последующим его разложением, гидролизом, изомеризацией или какой-либо другой модификации и с образованием нужного продукта при активации. Ферменты, не процессирующие крахмал, могут быть использованы для разрушения клеточной мембраны растения в целях стимуляции экстракции крахмала, липидов, аминокислот или других продуктов из растений. Кроме того, негипертермофильные и гипертермофильные ферменты могут быть использованы в комбинации с самопроцессирующимся растением или частями этого растения настоящего изобретения. Так, например, мезофильный фермент, не разрушающий крахмал, может быть активирован в целях разрушения клеточной мембраны растения для экстракции крахмала, а затем в этом самопроцессирующемся растении может быть активирован гипертермофильный крахмалразрушающий фермент для разложения крахмала.
Ферменты, экспрессированные в зерне, могут быть активированы путем помещения растения или части растения, содержащей эти ферменты, в условия, при которых стимулируется их активность. Так, например, может быть использован один или более из описанных ниже методов. Часть растения может контактировать с водой, которая является субстратом для гидролитического фермента, и, тем самым, активировать фермент. Часть растения может контактировать с водой, что способствует миграции фермента из компартмента, в котором он осаждается в процессе развития части растения, и, тем самым, ассоциируется с его субстратом. Миграция фермента возможна благодаря тому, что во время созревания, сушки зерна и регидратации происходит нарушение компартментализации. Целое или разрушенное зерно может контактировать с водой, позволяющей ферменту мигрировать из компартмента, в котором он осаждается в процессе развития части растения, и, тем самым, ассоциироваться с его субстратом. Ферменты могут быть также активированы путем добавления активирующего соединения. Так, например, кальцийзависимый фермент может быть активирован добавлением кальция. Другие активирующие соединения могут быть определены методами, известными специалистам. Ферменты могут быть активированы путем удаления инактиватора. Так, например, такими инактиваторами являются известные пептидные ингибиторы амилазных ферментов, где амилазы могут быть совместно экспрессированы с ингибитором амилазы, а затем активированы добавлением протеазы. Ферменты могут быть активированы путем изменения рН до значения, при котором указанный фермент становится наиболее активным. Ферменты могут быть также активированы путем повышения температуры. Активность фермента обычно увеличивается при достижении максимальной температуры для этого фермента. Активность мезофильного фермента будет увеличиваться, начиная с уровня его активности при комнатной температуре и до момента достижения температуры, при которой его активность будет снижаться, т.е., обычно, при температуре менее чем 70°С, или при температуре, равной 70°С. Аналогичным образом, термофильные и гипертермофильные ферменты могут быть также активированы путем повышения температуры. Термофильные ферменты могут быть также активированы путем повышения температуры до максимальной температуры их активности или стабильности. Для термофильного фермента максимальные температуры стабильности и активности обычно составляют в пределах от 70 до 85°С. Гипертермофильные ферменты будут иметь даже более высокую относительную активность, чем мезофильные или термофильные ферменты, благодаря их более сильному изменению потенциала при температуре от 25°С до 85°С-95°С или даже до 100°С. Повышенная температура может быть достигнута любым способом, таким как нагревание, например выпекание, кипячение, нагревание, обработка горячим паром, электрический разряд или любая их комбинация. Кроме того, в растениях, экспрессирующих мезофильный или термофильный фермент(ы), активация указанного фермента может быть осуществлена путем измельчения растения, что, тем самым, способствует контактированию указанного фермента с субстратом.
Оптимальные условия, например температура, гидратация, рН, и т.п., могут быть определены любым специалистом и могут зависеть от конкретно используемого фермента и от цели применения фермента.
Кроме того, настоящее изобретение относится к использованию экзогенных ферментов, которые могут стимулировать конкретный процесс. Так, например, самопроцессирующееся растение или часть растения настоящего изобретения могут быть использованы в комбинации с экзогенным ферментом для ускорения реакции. Например, трансгенная кукуруза, содержащая α-амилазу, может быть использована в комбинации с другими крахмалпроцессирующими ферментами, такими как пуллуланаза, α-глюкозидаза, глюкозоизомераза, маннаназа, гемицеллюлаза и т.п. для гидролиза крахмала или продуцирования этанола. Действительно, было обнаружено, что комбинации трансгенной кукурузы, содержащей α-амилазу, с такими ферментами приводили к неожиданно высоким уровням превращения крахмала по сравнению с использованием трансгенной кукурузы, содержащей только α-амилазу.
Ниже приводится пример подходящих способов, рассматриваемых в настоящей заявке.
а. Экстракция крахмала из растений
Изобретение относится к способу облегчения экстракции крахмала из растений. В частности, по крайней мере, один полинуклеотид, кодирующий процессирующий фермент, который разрушает физически ограничивающий матрикс эндосперма (клеточные стенки, не относящиеся к крахмалам полисахарид и белковый матрикс), вводят в растение так, чтобы данный фермент, предпочтительно, находился в непосредственной физической близости к крахмальным зернам в растении. В этом варианте осуществления изобретения трансформированные растения, предпочтительно, экспрессируют один или более ферментов, а именно протеазу, глюканазу, ксиланазу, тиоредоксин/тиоредоксинредуктазу, эстеразу и т.п., но не ферменты, которые обладают какой-либо крахмалразрушающей активностью, что способствует сохранению целостности крахмальных зерен. Таким образом, экспрессия этих ферментов в части растения, такой как зерно, приводит к улучшению функциональных свойства зерна. Процессирующий фермент может быть мезофильным, термофильным или гипертермофильным. В одном из примеров зерно трансформированного растения настоящего изобретения сушат нагреванием, что, вероятно, приводит к инактивации негипертермофильных процессирующих ферментов и к лучшему сохранению целостности семян. Зерно (или дробленое зерно) замачивают при низких температурах или при высоких температурах (где время играет важную роль) в условиях высокой или низкой влажности (см. руководство Primary Cereal Processing, Gordon & Willm, eds., pp.319-337 (1994), содержание которого вводится в настоящее описание посредством ссылки), в присутствии или в отсутствие диоксида серы. При повышения температуры, необязательно в условиях определенной влажности, целостность матрикса эндосперма нарушается посредством активации ферментов, например протеаз, ксиланаз, фитазы или глюканаз, которые разлагают белки и полисахариды, не являющиеся крахмалом и присутствующие в эндосперме, в результате чего крахмальные зерна остаются интактными и могут быть более легко выделены из полученного материала. Кроме того, выходящие белки и полисахариды, не являющиеся крахмалом, становятся, по крайней мере, частично расщепленными и высококонцентрированными, а поэтому они могут быть использованы для улучшенных кормов для животных, пищевых продуктов или в качестве компонентов сред, используемых для ферментации микроорганизмов. При этом предполагается, что выходящее вещество представляет собой жидкий кукурузный экстракт с улучшенным составом.
Таким образом, настоящее изобретение относится к способу получения крахмальных зерен. Способ включает в себя обработку зерна, например дробленого зерна, которое включает, по крайней мере, один не процессирующий крахмал фермент, в условиях, которые активируют по меньшей мере один фермент, с получением смеси, содержащей крахмальные зерна и не содержащие крахмал продукты разложения, например расщепленные продукты матрикса эндосперма. Не процессирующий крахмал фермент может быть мезофильным, термофильным или гипертермофильным. После активации фермента крахмальные гранулы выделяют из смеси. Зерно получают из трансформированного растения, геном которого содержит (пополнен) экспрессирующую кассету, кодирующую по меньшей мере один процессирующий фермент. Так, например, указанным процессирующим ферментом может быть протеаза, глюканаза, ксиланаза, фитаза, тиоредоксин/тиоредоксинредуктаза или эстераза. Указанный процессирующий фермент предпочтительно является гипертермофильным. Зерно может быть обработано в условиях низкой или высокой влажности, в присутствии или в отсутствие диоксида серы. В зависимости от активности и уровня экспрессии процессирующего фермента в зерне трансгенного растения трансгенное зерно может быть смешано с товарным зерном до или во время процессинга. Также рассматриваются продукты, полученные указанным способом, такие как крахмал, продукты, не содержащие крахмал; и вода для замачивания, содержащая, по крайней мере, один вспомогательный компонент.
b. Методы процессинга крахмала
Трансформированные растения или части растения настоящего изобретения могут содержать описанные здесь крахмалразрушающие ферменты, которые разрушают крахмальные зерна до декстринов, других модифицированных крахмалов или гексоз, (например, α-амилаза, пуллуланаза, α-глюкозидаза, глюкоамилаза, амилопуллуланаза) или превращают глюкозу во фруктозу (например, глюкозоизомераза). Предпочтительно, указанный крахмалразлагающий фермент выбирают из α-амилазы, α-глюкозидазы, глюкоамилазы, пуллуланазы, неопуллуланазы, амилопуллуланазы, глюкозоизомеразы и их комбинаций, и его используют для трансформации зерна. Кроме того, предпочтительно, чтобы указанный фермент был функционально присоединен к промотору или к сигнальной последовательности, которая нацеливает данный фермент на крахмальное зерно, амилопласт, апопласт или на эндоплазматический ретикулум. Более предпочтительно, если фермент экспрессируется в эндосперме, а в частности, в эндосперме кукурузы, и локализуется в одном или в более клеточных компартментах или внутри самого крахмального зерна. Предпочтительной частью растения является зерно. Предпочтительными частями растения являются части растений кукурузы, пшеницы, ячменя, ржи, овса, сахарного тростника или риса.
В соответствии с одним из способов разложения крахмала настоящего изобретения, трансформированное зерно, аккумулирующее крахмалразрушающий фермент в крахмальных зернах, замачивают, обычно, при 50°-60°С и подвергают мокрому помолу методом, известным специалистам. Предпочтительно, чтобы указанный крахмалразрушающий фермент был гипертермофильным. Благодаря субклеточному нацеливанию фермента на крахмальное зерно или благодаря ассоциации указанного фермента с крахмальным зерном посредством контактирования фермента и крахмального зерна в процессе мокрого помола при стандартных температурах процессирующий фермент подвергают совместной очистке с крахмальными зернами с получением смеси "крахмальные зерна/фермент". После получения смеси "крахмальные зерна/фермент", фермент активируют в условиях, благоприятствующих активации такого фермента. Так, например, обработка может быть осуществлена в различных условиях влажности и/или температуры для облегчения частичного (для получения дериватизированных крахмалов или декстринов) или полного гидролиза крахмала в гексозу. Таким же образом также получают сиропы с высоким содержанием эквивалентов декстрозы или фруктозы. Этот способ позволяет эффективно экономить время, энергию и стоимость фермента и повышать эффективность превращения крахмала в соответствующую гексозу и эффективность продуцирования продуктов, таких как вода для замачивания с высоким содержанием сахара и сиропы с повышенным содержанием эквивалентов декстрозы.
В другом варианте осуществления изобретения растение или продукт растения, такой как плоды, или зерно, или мука, изготовленная из зерна, которое экспрессирует указанный фермент, обрабатывают для активации указанного фермента и превращения экспрессируемых и содержащихся в растении полисахаридов в сахара. Этот фермент, предпочтительно, присоединяют к сигнальной последовательности, которая нацеливает данный фермент на описанные здесь крахмальные зерна, амилопласт, апопласт или эндоплазматический ретикулум. Продуцируемый сахар может быть затем выделен или экстрагирован из растения или продукта растения. В другом варианте осуществления изобретения процессирующий фермент, способный превращать полисахариды в сахара, помещают под контроль индуцибельного промотора методами, известными специалистам, и методами, описанными в настоящей заявке. Процессирующий фермент может быть мезофильным, термофильным или гипертермофильным. Растение культивируют до нужной стадии, и на этой стадии индуцируется промотор, обусловливающий экспрессию фермента и превращение полисахаридов в растении или в продукте растения в сахара. Фермент предпочтительно функционально присоединяют к сигнальной последовательности, которая нацеливает этот фермент на описанные здесь крахмальные зерна, амилопласт, апопласт или на эндоплазматический ретикулум. В другом варианте осуществления изобретения продуцируют трансформированное растение, которое экспрессирует процессирующий фермент, способный превращать крахмал в сахар. Фермент присоединяют к сигнальной последовательности, нацеливающей этот фермент на крахмальные зерна растения. После этого крахмал выделяют из трансформированного растения, которое содержит фермент, экспрессируемый трансформированным растением. Затем фермент, содержащийся в выделенном крахмале, может быть активирован для превращения крахмала в сахар. Этот фермент может быть мезофильным, термофильным или гипертермофильным. В настоящей заявке описаны примеры гипертермофильных ферментов, способных превращать крахмал в сахар. Описанные методы могут быть применены к любому растению, которое продуцирует полисахарид и которое может экспрессировать фермент, способный превращать полисахарид в сахара или гидролизованный крахмалопродукт, такой как декстрин, мальтоолигосахарид, глюкоза и/или их смеси.
Настоящее изобретение относится к способу продуцирования декстринов и модифицированных крахмалов из растения или продукта такого растения, которое было трансформировано процессирующим ферментом, гидролизующим некоторые ковалентные связи полисахарида с образованием полисахаридного производного. В одном из вариантов осуществления изобретения растение или продукт этого растения, такой как плод, или зерно, или мука, изготовленная из зерна, экспрессирующего указанный фермент, помещают в условия, благоприятные для активации фермента и превращения полисахаридов, содержащихся в этом растении, в полисахариды с более низкой молекулярной массой. Фермент, предпочтительно, присоединяют к сигнальной последовательности, которая нацеливает этот фермент на описанные здесь крахмальные зерна, амилопласт, апопласт или эндоплазматический ретикулум. Продуцированный декстрин или дериватизированный крахмал может быть затем выделен или экстрагирован из растения или из продукта растения. В другом варианте осуществления изобретения процессирующий фермент, способный превращать полисахариды в декстрины или модифицированные крахмалы, помещают под контроль индуцибельного промотора методами, известными специалистам и описанными в настоящей заявке. Растение культивируют до нужной стадии, после чего индуцируется промотор, инициирующий экспрессию фермента и превращение полисахаридов в растении или в продукте этого растения в декстрины или в модифицированные крахмалы. Предпочтительным ферментом является α-амилаза, пуллуланаза, изо- или нео-пуллуланаза, и этот фермент функционально присоединен к сигнальной последовательности, которая нацеливает данный фермент на описанные здесь крахмальные зерна, амилопласт, апопласт или эндоплазматический ретикулум. В одном из вариантов осуществления изобретения указанный фермент нацеливается на апопласт или на эндоплазматический ретикулум. В еще одном варианте осуществления изобретения получают трансформированное растение, экспрессирующее фермент, способный превращать крахмал в декстрины или в модифицированные крахмалы. Фермент присоединяют к сигнальной последовательности, которая нацеливает фермент на крахмальные зерна растения. Затем крахмал выделяют из трансформированного растения, которое содержит фермент, экспрессируемый трансформированным растением. Фермент, содержащийся в выделенном крахмале, может быть затем активирован в условиях, благоприятствующих превращению крахмала в декстрины или в модифицированные крахмалы. В настоящей заявке описаны примеры гипертермофильных ферментов, способных превращать крахмал в расщепленные крахмалопродукты. Описанные методы могут быть применены к любому растению, которое продуцирует полисахарид и которое может экспрессировать фермент, способный превращать полисахарид в сахар.
В другом варианте настоящего изобретения зерно, полученное от трансформированных растений настоящего изобретения, которые аккумулируют амилолитические ферменты, разрывающие связи в крахмальных зернах с образованием декстринов, модифицировнных крахмалов или гексозы (например, α-амилаза, пуллуланаза, α-глюкозидаза, глюкоамилаза, амилопуллуланаза), замачивают в условиях, благоприятствующих активации крахмалразрушающего фермента в течение определенного периода времени. Полученная смесь может содержать высокие уровни крахмалопродукта. Использование такого зерна (1) позволяет избежать необходимости помола зерна или какой-либо другой обработки зерна, обычно используемой для получения крахмальных зерен, (2) делает такой крахмал более доступным для ферментов благодаря присутствию ферментов непосредственно в ткани эндосперма зерна, и (3) позволяет избежать необходимости микробиологического продуцирования крахмалразрушающих ферментов. Таким образом, весь процесс мокрого помола до выделения гексозы может быть заменен простым нагреванием зерна, предпочтительно зерна кукурузы, в присутствии воды, которая стимулирует действие ферментов на крахмал.
Этот способ может быть также использован для получения этанола, сиропов с высоким содержанием фруктозы, ферментационной среды, содержащей гексозу (глюкозу), или для любого другого превращения крахмала, которое не требует очистки компонентов зерна.
Настоящее изобретение также относится к способу получения декстрина, мальтоолигосахаридов и/или сахара, включающему обработку части растения, содержащей крахмальные зерна или, по крайней мере, один крахмалпроцессирующий фермент в условиях, стимулирующих активацию, по крайней мере, одного фермента, с последующим ферментативным расщеплением крахмальных зерен и с получением водного раствора, содержащего сахара. Часть растения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один процессирующий фермент. Затем собирают водный раствор, содержащий декстрины, мальтоолигосахариды и/или сахар. В одном из вариантов осуществления изобретения процессирующим ферментом является α-амилаза, α-глюкозидаза, пуллуланаза, глюкоамилаза, амилопуллуланаза, глюкозоизомераза или любая их комбинация. Предпочтительно, чтобы указанный фермент был гипертермофильным. В другом варианте осуществления изобретения указанный способ дополнительно включает в себя выделение декстринов, мальтоолигосахаридов и/или сахара.
с. Улучшенные сорта кукурузы
Настоящее изобретение также относится к получению улучшенных сортов кукурузы (и сортов других культур), которые имеют нормальные уровни накопления крахмала и накапливают достаточные уровни амилолитического фермента(ов) в своем эндосперме, или в накапливающем крахмал органе, такие, чтобы после активации содержащегося в нем фермента, например при кипячении или нагревании указанного растения или его части, в случае гипертермофильного фермента, указанный(е) фермент(ы) активировался и стимулировал быстрое превращение крахмала в простые сахара. Эти простые сахара (главным образом, глюкоза) будут придавать обработанной кукурузе сладость. Полученное растение кукурузы будет представлять собой улучшенный сорт, предназначенный для двойного использования как в качестве зернопродуцирующего гибрида, так и в качестве сладкой кукурузы. Так, например, настоящее изобретение относится к способу продуцирования сверхсладкой кукурузы, включающему обработку трансформированной кукурузы или ее части, геном которой пополнен благодаря введению экспрессирующей кассеты и экспрессирует эту экспрессирующую кассету в эндосперме, где указанная кассета содержит промотор, функционально присоединенный к первому полинуклеотиду, кодирующему по меньшей мере один крахмалразрушающий фермент, в условиях, активирующих по меньшей мере один фермент, так, чтобы он превращал полисахариды кукурузы в сахар, с образованием сверхсладкой кукурузы. Промотором может быть конститутивный промотор, семяспецифический промотор или эндоспермспецифический промотор, который присоединен к полинуклеотидной последовательности, кодирующей процессирующий фермент, такой как α-амилаза, например, содержащий последовательность SEQ ID NO:13, 14 или 16. Предпочтительный фермент является гипертермофильным. В одном из вариантов осуществления изобретения экспрессирующая кассета дополнительно содержит второй полинуклеотид, который кодирует сигнальную последовательность, функционально присоединенную к ферменту, кодируемому первым полинуклеотидом. Сигнальные последовательности, характерные для этого варианта осуществления изобретения, нацеливают указанный фермент на апопласт, эндоплазматический ретикулум, крахмальные зерна или в амилопласт. Растение кукурузы культивируют до образования початков с зернами, после чего происходит индуцирование промотора, приводящее к экспрессии фермента и превращению полисахарида, содержащегося в этом растении, в сахар.
d. Самоферментирующиеся растения
В другом варианте осуществления изобретения растения, такие как кукуруза, рис, пшеница или сахарный тростник, конструируют так, чтобы в их клеточных стенках аккумулировались большие количества процессирующих ферментов, например ксиланазы, целлюлазы, гемицеллюлазы, глюканазы, пектиназы и т.п. (ферменты, разрушающие полисахариды, не являющиеся крахмалами). После сбора компонентов зерна (или сахара в случае сахарного тростника) в качестве источника фермента, который направляется для экспрессии и аккумуляции в клеточных стенках, и в качестве источника биомассы используют солому, чешую зерен или багассу. Солома (или другая ненужная ткань) используется в качестве исходного материала в способе выделения ферментативных сахаров. Способ получения таких ферментируемых сахаров заключается в активации фермента, разрушающего полисахариды, не являющиеся крахмалом. Так, например, активация может включать нагревание ткани растения в присутствии воды в течение периода времени, достаточного для расщепления не являющегося крахмалом полисахарида с образованием сахаров. Таким образом, самопроцессирующаяся солома продуцирует ферменты, которые необходимы для превращения полисахаридов в моносахариды, и которые, в основном, не требует дополнительных затрат, поскольку они являются компонентами исходного материала. Кроме того, температурозависимые ферменты не оказывают неблагоприятного действия на рост и развитие растения, и на нацеливание на клеточную стенку, и даже на нацеливание на полисахаридные микроволокна благодаря связывающим целлюлозу/ксилозу доменам, присоединенным к белку, что повышает доступность субстрата для данного фермента.
Таким образом, изобретение относится к способу использования части трансформированного растения, включающему нацеливание, по крайней мере, одного фермента, гидролизующего полисахарид, не являющийся крахмалом, на клеточную стенку клеток данной части растения. Способ включает обработку части трансформированного растения, содержащей, по крайней мере, один фермент, процессирующий полисахарид, не являющийся крахмалом, в условиях, способствующих активации, по крайней мере, одного фермента с последующим расщеплением крахмальных зерен и с образованием водного раствора, содержащего сахара, где часть растения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один фермент, процессирующий полисахарид, не являющийся крахмалом; и сбор водного раствора, содержащего сахара. Настоящее изобретение также относится к трансформированному растению или к части растения, содержащей по меньшей мере один фермент, который процессирует полисахарид, не являющийся крахмалом, и присутствует в клетке или в клеточной стенке клеток растения или части растения. Часть растения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один не процессирующий крахмал фермент, например ксиланазу, целлюлазу, глюканазу, пектиназу или любые их комбинации.
е. Высокое содержание водной фазы в белке и сахаре
В еще одном варианте осуществления изобретения протеазы и липазы конструируют так, чтобы они аккумулировались в семенах, например в семенах сои. После активации протеазы или липазы, например, путем нагревания эти ферменты в семенах гидролизуют липиды и запасные белки, присутствующие в сое во время процессинга. Таким образом, могут быть получены растворимые продукты, содержащие аминокислоты, которые могут быть использованы в качестве питательной, кормовой или ферментационной среды, и жирные кислоты. Полисахариды, обычно, присутствуют в нерастворимой фракции процессированного зерна. Однако, благодаря экспрессии полисахарид-разрушающего фермента и его аккумуляции в семенах, могут быть гидролизованы белки и полисахариды, присутствующие в водной фазе. Так, например, этим способом могут быть солюбилизированы зеины кукурузы и запасный белок и не являющиеся крахмалом полисахариды сои. Компоненты водной и гидрофобной фаз могут быть легко разделены путем экстракции органическим растворителем или надкритической двуокисью углерода. Таким образом, настоящее изобретение относится к способу получения водного экстракта зерна, которое содержит повышенные уровни белка, аминокислот, сахаров или сахаридов.
f. Ферментация самопроцессирующихся растений
Настоящее изобретение относится к способу продуцирования этанола, напитков или других продуктов, получаемых при брожении. Этот способ предусматривает получение растения, или продукта, или части растения, либо производного растения, такого как мука из зерна, в котором экспрессируется процессирующий фермент, превращающий полисахариды в сахар. Растение или его продукт обрабатывают так, чтобы при превращении полисахарида образовывался сахар, как описано выше. Затем сахара и другие компоненты растения подвергают ферментации с получением этанола, напитков или других продуктов, продуцируемых путем сбраживания методами, известными специалистам. См., например, патент США №4929452. Вкратце, сахар, продуцированный превращением полисахаридов, инкубируют с дрожжами в условиях, стимулирующих превращение сахара в этанол. Подходящими дрожжами являются дрожжевые штаммы, толерантные к высокому содержанию спирта и сахара, например, дрожжи S.cerevisiae АТСС №20867. Этот штамм был депонирован в Американской коллекции типовых культур, Rockville, MD, 17 сент., 1987 и зарегистрирован под номером АТСС №20867. Затем продукт или напиток, полученный при брожении, может быть подвергнут перегонке с выделением этанола или напитка, получаемого путем перегонки, либо этот продукт ферментации может быть выделен каким-либо иным способом. Растением, используемым в этом способе, может быть растение, содержащее полисахарид и способное экспрессировать фермент настоящего изобретения. В настоящей заявке описано много таких растений. Предпочтительным является растение, культивируемое в коммерческих целях. Более предпочтительным является растение, которое обычно используется для получения этанола либо продуктов или напитков, получаемых при брожении, например, такое растение как пшеница, ячмень, кукуруза, рожь, картофель, виноград или рис. Наиболее предпочтительным растением является кукуруза.
Способ включает обработку части растения, содержащей, по крайней мере, один полисахаридпроцессирующий фермент в условиях, стимулирующих активацию, по крайней мере, одного фермента и, тем самым, способствующих расщеплению полисахарида в указанной части растения с получением сбраживаемого сахара. Полисахаридпроцессирующий фермент может быть мезофильным, термофильным или гипертермофильным. Предпочтительным является гипертермофильный фермент. Часть растения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один полисахаридпроцессирующий фермент. В этом варианте осуществления изобретения частями растения являются, но не ограничиваются ими, зерно, плоды, семена, стебли, древесина, овощи или корни. Предпочтительными растениями являются, но не ограничиваются ими, овес, ячмень, пшеница, ягоды, виноград, рожь, кукуруза, рис, картофель, сахарная свекла, сахарный тростник, ананас, трава и деревья. Часть растения может быть объединена с товарным зерном или с другими коммерчески доступными продуктами, а источником субстрата для процессинга может быть источник, не являющийся самопроцессирующимся растением. Затем ферментируемый сахар инкубируют в условиях, стимулирующих превращение сбраживаемого сахара в этанол, например, дрожжами и/или другими микроорганизмами. В предпочтительном варианте осуществления изобретения часть растения происходит от кукурузы, трансформированной α-амилазой, что, как было обнаружено, позволяет сэкономить время ферментации и затраты на осуществление этой ферментации.
Было обнаружено, что количество остаточного крахмала снижается в том случае, когда трансгенная кукуруза, продуцируемая в соответствии с настоящим изобретением, экспрессирует термостабильную α-амилазу, например, при использовании в ферментации. Это указывает на то, что при ферментации солюбилизируется большее количество крахмала. При пониженном содержании остаточного крахмала в дистилляторе получают зерно с более высоким содержанием (по массе) белка и более высокой ценности. Более того, ферментация трансгенной кукурузы позволяет осуществлять разжижение при более низком рН, что дает возможность сократить затраты на химические вещества, используемые для коррекции рН при более высокой температуре, например при более чем 85°С, а предпочтительно более чем 90°С, более предпочтительно при 95°С или выше, что приводит к сокращению времени разжижения и к более полной солюбилизации крахмала и позволяет снижать время разжижения, в результате чего могут быть проведены эффективные реакции ферментации с более высокими выходами этанола.
Кроме того, было обнаружено, что контактирование соответствующих частей растения, даже с небольшой частью трансгенного растения, продуцированного в соответствии с настоящим изобретением, может приводить к снижению времени ферментации и затрат на ее осуществление. Таким образом, настоящее изобретение относится к способу снижения времени ферментации для растений, включающему использование части трансгенного растения, полученного от растения, содержащего полисахаридпроцессирующий фермент, который превращает полисахариды в сахар, вместо использования растения, которое не содержит полисахаридпроцессирующего фермента.
g. Ферменты, процессирующие исходный крахмал, и полинуклеотиды, их кодирующие
Полинуклеотид, кодирующий мезофильный процессирующий фермент(ы), вводят в растение или в часть растения. В предпочтительном варианте осуществления изобретения полинуклеотидом настоящего изобретения является полинуклеотид, оптимизированный по кодонам кукурузы, такой как полинуклеотид, имеющий последовательности SEQ ID NO:48, 50 и 59 и кодирующий глюкоамилазу, имеющую последовательность SEQ ID NO:47 и 49. В другом предпочтительном варианте осуществления изобретения полинуклеотидом настоящего изобретения является полинуклеотид, оптимизированный по кодонам кукурузы, такой как полинуклеотид, имеющий последовательность SEQ ID NO:52, кодирующий α-амилазу, имеющую последовательность SEQ ID NO:51. Кроме того, рассматриваются гибридные продукты процессирующих ферментов. В одном из предпочтительных вариантов осуществления изобретения полинуклеотидом настоящего изобретения является полинуклеотид, оптимизированный по кодонам кукурузы, такой как полинуклеотид, имеющий последовательность SEQ ID NO:46 и кодирующий гибрид альфа-амилазы и глюкоамилазы, имеющий последовательность SEQ ID NO:45. Кроме того, в настоящем изобретении рассматриваются комбинации процессирующих ферментов. Так, например, рассматривается комбинация крахмалпроцессирующих ферментов и не процессирующих крахмал ферментов. Такие комбинации процессирующих ферментов могут быть получены с использованием множества генных конструкций, кодирующих каждый из ферментов. Альтернативно, отдельные трансгенные растения, стабильно трансформированные ферментами, могут быть подвергнуты скрещиванию известными методами с получением растения, содержащего оба фермента. Другой метод предусматривает использование экзогенного фермента(ов) при получении трансгенного растения.
Источник крахмалпроцессирующего фермента и не процессирующего крахмал фермента может быть выделен или получен из любого материала, и его полинуклеотиды могут быть определены любым специалистом. Предпочтительно, α-амилазу получают из Aspergillus (например, Aspergillus shirousami и Aspergillus niger), Rhisopus (например, Rhizopus oryzae) и таких растений, как кукуруза, ячмень и рис. Предпочтительно, глюкоамилазу получают из Aspergillus (например, Aspergillus shirousami и Aspergillus niger), Rhisopus (например, Rhizopus oryzae) и Thermoanaerobacter (например, Thermoanaerobacter thermosaccharolyticum).
В другом варианте осуществления изобретения полинуклеотид кодирует мезофильный крахмалпроцессирующий фермент и функционально присоединен к оптимизированному по кодонам кукурузы полинуклеотиду, такому как SEQ ID NO:54, кодирующему связывающий исходный крахмал домен, такой как SEQ ID NO:53.
В другом варианте осуществления изобретения тканеспецифическими промоторами являются эндоспермспецифические промоторы, такие как промотор γ-зеина кукурузы (представленный SEQ ID NO:12) или промотор ADP-gpp кукурузы (представленный SEQ ID NO:11, который включает 5'-нетранслируемую и интронную последовательность). Таким образом, настоящее изобретение относится к выделенному полинуклеотиду, содержащему промотор, включающий SEQ ID NO:11 или 12; к полинуклеотиду, который гибридизуется с последовательностью, комплементарной последовательности в условиях низкой степени жесткости, или к его фрагменту, промоторная активность которого составляет, например, по крайней мере, 10%, а предпочтительно, по крайней мере, 50% от активности промотора, имеющего последовательность SEQ ID NO:11 или 12.
В одном из вариантов осуществления изобретения продукт гена гидролиза крахмала, такого как ген α-амилазы, глюкоамилазы или гибрида α-амилазы/глюкоамилазы, может быть нацелен на конкретную органеллу или на конкретный участок, такой как эндоплазматический ретикулум или апопласт, но не на цитоплазму. Это может быть, например, осуществлено с использованием N-концевой сигнальной последовательности γ-зеина кукурузы (SEQ ID NO:17), которая обеспечивает апопластспецифическое нацеливание белков, и использование N-концевой сигнальной последовательности γ-зеина (SEQ ID NO:17), функционально присоединенной к процессирующему ферменту, который, в свою очередь, функционально присоединен к последовательности SEKDEL для удерживания в эндоплазматическом ретикулуме. Нацеливание белка или фермента на конкретный компартмент обеспечивает такую локализацию указанного фермента, при которой он не может контактировать с субстратом. Таким образом, ферментативное действие указанного фермента не будет осуществляться до контакта этого фермента с субстратом. Фермент может контактировать со своим субстратом в процессе помола (физического разрушения целостности клетки) и гидратации. Так, например, мезофильный крахмалпроцессирующий фермент может быть нацелен на апопласт или на эндоплазматический ретикулум, а поэтому он не будет вступать в контакт с крахмальными зернами в амилопласте. Помол зерна будет разрушать целостность этого зерна, и крахмалразрушающий фермент будет контактировать с крахмальными зернами. Таким образом можно избежать потенциально негативных эффектов совместной локализации фермента и его субстрата.
h. Пищевые продукты без добавления подсластителя.
Настоящее изобретение также относится к способу получения подслащенного мучного пищевого продукта без добавления дополнительных подсластителей. Примерами мучных пищевых продуктов являются, но не ограничиваются ими, пищевой продукт для завтрака, пищевой продукт, готовый к употреблению, выпечка, паста и зерновые пищевые продукты, такие как зерновой завтрак. Способ включает обработку части растения, содержащей по крайней мере, один крахмалпроцессирующий фермент, в условиях, стимулирующих активацию крахмалпроцессирующего фермента и, тем самым, способствующих процессингу крахмальных зерен в указанной части растения до сахаров с образованием подслащенного продукта, например, относительно продукта, продуцируемого при процессинге крахмальных гранул из части растения, которая не содержит гипертермофильного фермента. Предпочтительным крахмалпроцессирующим ферментом является гипертермофильный фермент, который активируется при нагревании, например при выпекании, кипячении, нагревании, варке на пару, электрическом разряде или любой их комбинации. Часть растения получают из трансформированного растения, например из трансформированных растений сои, ржи, овса, ячменя, пшеницы, кукурузы, риса или сахарного тростника, геном которых пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один гипертермофильный крахмалпроцессирующий фермент, например α-амилазу, α-глюкозидазу, глюкоамилазу, пуллуланазу, глюкозоизомеразу или любую их комбинацию. Затем подслащенный продукт подвергают обработке с получением мучного пищевого продукта. Настоящее изобретение также относится к мучному пищевому продукту, например к зерновому пищевому продукту, пищевому продукту для завтрака, пищевому продукту, готовому к употреблению, или к выпечке, приготовленным указанным способом. Указанный мучной пищевой продукт может быть получен из подслащенного продукта и воды и может содержать солод, отдушки, витамины, минералы, красители или любые их комбинации.
Фермент может быть активирован так, чтобы он превращал полисахариды, содержащиеся в указанном растительном материале, в сахар перед включением растительного материала в зерновой продукт или при процессинге зернового продукта. В соответствии с этим полисахариды, содержащиеся в растительном материале, могут быть превращены в сахар путем активации материала, например путем нагревания в случае гипертермофильного фермента перед включением в мучной продукт. Затем растительный материал, содержащий сахар, продуцируемый путем превращения полисахаридов, добавляют в указанный продукт с получением подслащенного продукта. Альтернативно, полисахариды могут быть превращены в сахара под действием фермента во время процессинга мучного продукта. Примеры способов получения зерновых продуктов хорошо известны специалистам, и такими способами являются нагревание, выпекание, кипячение и т.п., как описано в патентах США №№6183788, 6159530, 6149965, 4988521 и 5368870.
Вкратце, тесто может быть приготовлено путем смешивания различных сухих ингредиентов вместе с водой и подвержения тепловой обработке для желатинизации крахмальных компонентов и развития вкуса перепастеризованных сливок. Этот термообработанный материал может быть затем подвергнут механической обработке с получением готового теста, такого как зерновое тесто. Сухие ингредиенты могут включать различные добавки, такие как сахара, крахмал, соль, витамины, минералы, красители, отдушки, соль и т.п. Помимо воды, могут быть добавлены различные жидкие ингредиенты, такие как кукурузный или солодовый сироп. Мучной продукт может включать зерно злаковых, дробленое зерно, зерно крупного помола или муку из пшеницы, риса, кукурузы, овса, ячменя, ржи или других злаков и их смеси, полученные из трансформированного растения согласно изобретению. Затем тесто может быть обработано, с получением нужной формы, таким способом, как экструзия или штамповка, с последующей тепловой обработкой с использованием таких средств, как варочный аппарат Джеймса, печи или устройства для подачи электрического разряда.
Кроме того, настоящее изобретение относится к способу подслащивания крахмалсодержащего продукта без добавления подсластителя. Способ включает обработку крахмала, содержащего, по крайней мере, один крахмалпроцессирующий фермент, в условиях, стимулирующих активацию, по крайней мере, одного фермента, и, тем самым, способствующих расщеплению крахмала до сахаров с образованием обработанного (подслащенного) крахмала, например, относительно продукта, продуцируемого при обработке крахмала, не содержащего гипертермофильного фермента. Крахмал настоящего изобретения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один процессирующий фермент. Предпочтительными ферментами являются α-амилаза, α-глюкозидаза, глюкоамилаза, пуллуланаза, глюкозоизомераза или любая их комбинация. Предпочтительно, чтобы указанный фермент был гипертермофильным и активировался нагреванием. Предпочтительными трансформированными растениями являются кукуруза, соя, рожь, овес, ячмень, пшеница, рис и сахарный тростник. Затем обработанный крахмал добавляют в продукт, с получением подслащенного крахмалсодержащего продукта, например мучного пищевого продукта. Настоящее изобретение также относится к подслащенному крахмалсодержащему продукту, полученному таким методом.
Кроме того, настоящее изобретение относится к способу подслащивания полисахаридсодержащих плодов или овощей, включающему обработку плодов или овощей, содержащих, по крайней мере, один полисахаридпроцессирующий фермент, в условиях, активирующих, по крайней мере, один фермент, с последующим превращением указанного полисахарида плодов или овощей в сахар и с получением более сладких плодов и овощей, например, по сравнению с плодами и овощами растения, не содержащего полисахаридпроцессирующий фермент. Плоды или овощи настоящего изобретения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один полисахаридпроцессирующий фермент. Предпочтительными плодами и овощами являются картофель, томаты, банан, тыква, горох и фасоль. Предпочтительными ферментами являются α-амилаза, α -глюкозидаза, глюкоамилаза, пуллуланаза, глюкозоизомераза или любая их комбинация. Фермент предпочтительно является гипертермофильным.
i. Подслащивание полисахаридсодержащего растения или растительного продукта.
Указанный способ предусматривает получение растения, которое экспрессирует полисахаридпроцессирующий фермент, превращающий полисахарид в сахар, как описано выше. В соответствии с этим фермент экспрессируется в растении и в продуктах растения, например в плодах или в овощах. В одном из вариантов осуществления изобретения фермент помещают под контроль индуцибельного промотора так, чтобы экспрессия этого фермента индуцировалась под влиянием внешних факторов. Такие индуцибельные промоторы и конструкции хорошо известны специалистам и описаны в настоящей заявке. Экспрессия фермента в растении или в продукте растения приводит к превращению полисахарида, содержащегося в растении или в его продукте, в сахар, и к подслащиванию этого растения или продукта. В другом варианте осуществления изобретения полисахаридпроцессирующий фермент экспрессируется конститутивно. Таким образом, растение или полученный из него продукт могут быть активированы в условиях, достаточных для активации фермента в целях превращения полисахаридов в сахар под действием фермента и с последующим подслащиванием растения или полученного из него продукта. В результате этого аутопроцессинг полисахарида в плодах или овощах с образованием сахара приводит к получению более сладких плодов или овощей по сравнению с плодами или овощами растения, не содержащего указанный полисахаридпроцессирующий фермент. Плоды или овощи настоящего изобретения получают из трансформированного растения, геном которого пополнен за счет экспрессирующей кассеты, кодирующей по меньшей мере один полисахаридпроцессирующий фермент. Предпочтительными плодами и овощами являются картофель, томаты, банан, тыква, горох и фасоль. Предпочтительными ферментами являются α-амилаза, α -глюкозидаза, глюкоамилаза, пуллуланаза, глюкозоизомераза или любая их комбинация. Предпочтительно, чтобы полисахаридпроцессирующий фермент являлся гипертермофильным.
j. Выделение крахмала из трансформированного зерна, которое содержит фермент, разрушающий матрикс эндосперма.
Настоящее изобретение относится к способу выделения крахмала из трансформированного зерна, где экспрессируется фермент, разрушающий матрикс эндосперма. Этот способ предусматривает получение растения, которое экспрессирует фермент, разрушающий матрикс эндосперма, путем модификации, например, клеточных стенок полисахаридов, не являющихся крахмалом, и/или белков. Примерами таких ферментов являются, но не ограничиваются ими, протеазы, глюканазы, тиоредоксин, тиоредоксинредуктаза и эстераза. Указанные ферменты не включают ферменты, обладающие крахмалразрушающий активностью, поскольку необходимо, чтобы сохранялась целостность крахмальных зерен. При этом предпочтительно, чтобы этот фермент был присоединен к сигнальной последовательности, нацеливающей этот фермент на крахмальные зерна. В одном из вариантов осуществления изобретения зерно подвергают тепловой сушке для активации фермента и для инактивации эндогенных ферментов, содержащихся в зерне. Тепловая обработка приводит к активации фермента, разрушающего матрикс эндосперма, который может быть затем легко выделен из крахмальных зерен. В другом варианте осуществления изобретения, зерно замачивают при низкой или высокой температуре, при высокой или низкой влажности и в присутствии или в отсутствие диоксида серы. Затем зерно подвергают тепловой обработке для разрушения матрикса эндосперма, что позволяет легко выделять из него крахмальные зерна. В другом варианте осуществления изобретения, создают условия с соответствующей температурой и влажностью, которые обеспечивают проникновение протеаз в крахмальные зерна и разложение белков, содержащихся в зернах. Такая обработка приводит к продуцированию крахмальных зерен с высоким выходом и с небольшим содержанием белков-примесей.
k. Сироп, имеющий высокое содержание сахарного эквивалента, и использование этого сиропа для продуцирования этанола или напитков, получаемых при брожении.
Указанный способ предусматривает получение растения, экспрессирующего полисахаридпроцессирующий фермент, который превращает полисахарид в сахар, как описано ниже. Растение или его продукт замачивают в водном потоке в условиях, при которых экспрессируемый фермент превращает полисахарид, содержащийся в растении или в его продукте, в декстрин, мальтоолигосахарид и/или в сахар. Затем водный поток, содержащий декстрин, мальтоолигосахарид и/или сахар, продуцированные посредством превращения полисахарида, выделяют для получения сиропа, имеющего высокое содержание сахарного эквивалента. Способ может включать и не включать дополнительную стадию мокрого помола растения или его продукта с получением крахмальных зерен. Примерами ферментов, которые могут быть использованы в способе, являются, но не ограничиваются ими, α-амилаза, глюкоамилаза, пуллуланаза и α-глюкозидаза. Предпочтительно, фермент является гипертермофильным. Сахарами, продуцированными таким способом, являются, но не ограничиваются ими, гексоза, глюкоза и фруктоза. Примерами растений, которые могут быть использованы в способе, являются, но не ограничиваются ими, кукуруза, пшеница или ячмень. Примерами продуктов растения, которые могут быть использованы, являются, но не ограничиваются ими, плоды, зерно и овощи. В одном из вариантов осуществления изобретения полисахаридпроцессирующий фермент помещают под контроль индуцибельного промотора. В соответствии с этим, перед замачиванием или в процессе замачивания, индуцируется промотор, обуславливающий экспрессию указанного фермента, который затем превращает полисахарид в сахар. Примеры индуцибельных промоторов и конструкций, содержащих эти промоторы, хорошо известны специалистам и описаны в настоящей заявке. Таким образом, если полисахаридпроцессирующим ферментом является гипертермофильный фермент, то замачивание осуществляют при комнатной температуре для активации гипертермофильного фермента и для инактивации эндогенных ферментов, содержащихся в растении или в его продукте. В другом варианте осуществления изобретения гипертермофильный фермент, способный превращать полисахарид в сахар, экспрессируется конститутивно. Фермент может быть (а может и не быть) нацелен на компартмент растения с использованием сигнальной последовательности. Растение или его продукт замачивают в условиях высокой температуры для инициации превращения полисахаридов, содержащихся в данном растении, в сахар.
Настоящее изобретение также относится к получению этанола или приготовлению напитка, получаемого при брожении, из сиропа, имеющего высокое содержание сахарного эквивалента. Способ предусматривает инкубирование сиропа с дрожжами в условиях, обеспечивающих превращение сахара, содержащегося в этом сиропе, в этанол, или в напиток, получаемый при брожении. Примерами таких напитков, получаемых при брожении, являются, но не ограничиваются ими, пиво и вино. Условия ферментации хорошо известны специалистам и описаны в патенте США №4929452 и в настоящей заявке. Предпочтительными дрожжами являются дрожжевой штамм, толерантный к высокому содержанию спирта и к высокому содержанию сахара, такой как S.cerevisiae, рег. АТСС №20867. Ферментированный продукт или напиток, получаемый при брожении, может быть подвергнут перегонке для выделения этанола или напитка, полученного перегонкой.
l. Аккумуляция гипертермофильного фермента в клеточных стенках растения.
Настоящее изобретение относится к способу аккумуляции гипертермофильного фермента в клеточной стенке растения. Способ предусматривает экспрессию в растении гипертермофильного фермента, который связан с сигналом, нацеливающим этот фермент на клеточную стенку, так, чтобы нацеливающий фермент аккумулировался в клеточной стенке. Предпочтительно, чтобы фермент был способен превращать полисахариды в моносахариды. Примерами нацеливающей последовательности являются, но не ограничиваются ими, домен, связывающий целлюлозу или ксилозу. Примерами гипертермофильных ферментов являются ферменты, представленные в SEQ ID NO:1, 3, 5,10, 13, 14, 15 или 16. В этом способе растительный материал, содержащий клеточные стенки, может быть добавлен как источник нужных ферментов для выделения сахаров из исходного пищевого материала или как источник ферментов для превращения полисахаридов, происходящих от других источников, в моносахариды. Кроме того, клеточные стенки могут служить источником, из которого могут быть выделены ферменты. Методы очистки ферментов хорошо известны специалистам, и такими методами являются, но не ограничиваются ими, гель-фильтрация, ионообменная хроматография, хроматофокусирование, изоэлектрическое фокусирование, аффинная хроматография, ЖЭХБ, ВЭЖХ, осаждение солью, диализ и т.п. В соответствии с этим настоящее изобретение также относится к очищенным ферментам, выделенным из клеточных стенок растений.
m. Способ получения и выделения процессирующих ферментов.
В соответствии с настоящим изобретением рекомбинантно продуцированные процессирующие ферменты настоящего изобретения могут быть получены путем трансформации растительной ткани или растительной клетки, содержащей процессирующий фермент настоящего изобретения, способный активироваться в растении, выбранном для получения трансформированной ткани или клетки; культивирования трансформированной ткани или клетки с получением трансформированного растения и выделения процессирующего фермента из указанного трансформированного растения или его части. Предпочтительным рекомбинантно продуцированным ферментом является α-амилаза, глюкоамилаза, глюкозоизомераза, α-глюкозидаза и пуллуланаза. Наиболее предпочтительно, чтобы указанный фермент кодировался полинуклеотидом, выбранным из последовательностей SEQ ID NO:2, 4, 6, 9, 19, 21, 25, 37, 39, 41, 43, 46, 48, 50, 52 или 59.
Настоящее изобретение, кроме того, описано в нижеследующих примерах, которые не должны рассматриваться как ограничение его объема.
Примеры
Пример 1
Конструирование оптимизированных по кодонам кукурузы генов для гипертермофильных крахмалпроцессирующих/изомеризующих ферментов
Ферменты, такие как α-амилаза, пуллуланаза, α-глюкозидаза и глюкозоизомераза, участвующие в разложении крахмала или в изомеризации глюкозы, были отобраны по их нужным профилям активности. Такими профилями являются, например, минимальная активность при комнатной температуре, активность/стабильность при высокой температуре и активность при низком рН. Затем конструировали соответствующие гены с использованием предпочтительных кодонов кукурузы, описанных в патенте США №5625136, и синтезировали в соответствии с общей технологией рекомбинантных ДНК (Integrated DNA Technologies, Inc. (Coralville, IA).
α-Амилазу 797GL3, имеющую аминокислотную последовательность SEQ ID NO:1, отбирали по ее гипертермофильной активности. Выводили последовательность нуклеиновой кислоты фермента и ее оптимизировали по кодонам кукурузы, как представлено в SEQ ID NO:2. Аналогичным образом отбирали пуллуланазу 6gp3, имеющую аминокислотную последовательность, представленную в SEQ ID NO:3. Последовательность нуклеиновой кислоты для пуллуланазы 6gp3 выводили и оптимизировали по кодонам кукурузы, как представлено в SEQ ID NO:4.
Аминокислотную последовательность α-глюкозидазы malA Sulfolobus solfataricus получали, как описано в литературе, J.Bact. 177:482-485 (1995); J.Bact. 180:1287-1295 (1998). На основе опубликованной аминокислотной последовательности белка (SEQ ID NO:5) конструировали синтетический ген, оптимизированный по кодонам кукурузы (SEQ ID NO:6) кодирующий α-глюкозидазу malA.
Было отобрано несколько ферментов глюкозоизомераз. Аминокислотная последовательность (SEQ ID NO:18) глюкозоизомеразы, происходящей от Thermotoga maritima, была предсказана исходя из опубликованной ДНК-последовательности, имеющей рег. №NC_000853, и был сконструирован синтетический ген, оптимизированный по кодонам кукурузы (SEQ ID NO:19). Аналогичным образом, аминокислотная последовательность (SEQ ID NO:20) глюкозоизомеразы, происходящей от Thermotoga neapolitana, была предсказана исходя из ДНК-последовательности, опубликованной в Appl. Envir. Microbiol. 61(5):1867-1875 (1995), рег.№L38994. Был сконструирован синтетический ген, оптимизированный по кодонам кукурузы и кодирующий глюкозоизомеразу Thermotoga neapolitana (SEQ ID NO:21).
Пример 2
Экспрессия гибрида α-амилазы 797GL3 и области инкапсуляции крахмала в E.coli
Конструкцию, кодирующую гипертермофильную α-амилазу 797GL3, присоединенную к области инкапсуляции крахмала (SER), происходящей от крахмал-синтазы, связанной с кукурузными зернами (waxy), встраивали и экспрессировали в E.coli. кДНК для связанной с кукурузными зернами крахмал-синтазы (SEQ ID NO:7), кодирующую аминокислотную последовательность (SEQ ID NO:8) (Klosgen R.B., et al.,1986), клонировали в качестве источника крахмалсвязывающего домена или области инкапсуляции крахмала (SER). Полноразмерную кДНК амплифицировали с помощью ОТ-ПЦР исходя из РНК, полученной из семян кукурузы, с использованием праймеров SV57: (5'-AGCGAATTCATGGCGGCTCTGGCCACGT-3') (SEQ ID NO:22) и SV58 (5'-AGCTAAGCTTCAGGGCGCGGCCACGTTCT-3') (SEQ ID NO:23), имеющихся в Genbank, рег. №Х03935. Полноразмерную кДНК клонировали в pBluescript в виде EcoRI/HindIII-фрагмента и плазмиду обозначали рNOV4022.
С-концевую часть (кодируемую п.н.919-1818) кДНК восковидной кукурузы, включая крахмалсвязывающий домен, амплифицировали из рNOV4022 и присоединяли в той же рамке считывания к 3'-концу полноразмерного гена 797GL3, оптимизированного по кодонам кукурузы (SEQ ID NO:2). Гибридный генный продукт, 797GL3/waxy, имеющий нуклеиновую кислоту SEQ ID NO:9 и кодирующий аминокислотную последовательность SEQ ID NO:10, клонировали в виде NcoI/XbaI-фрагмента в вектор рЕТ28b (NOVAGEN, Madison, WI), который разрезали NcoI/NheI. Ген 797GL3, взятый отдельно, также клонировали в вектор рЕТ28b в виде NcoI/XbaI-фрагмента.
Векторы рЕТ28/797GL3 и рЕТ28/797GL3/waxy трансформировали в клетки BL21/DЕ3 E.coli (NOVAGEN), культивировали и индуцировали в соответствии с инструкциями производителей. Анализ, проводимый путем электрофореза в ПААГ/окрашивания кумасси, выявил присутствие индуцированного белка в обоих экстрактах, соответствующих предсказанным размерам гибридной и негибридной амилазы соответственно.
Общие клеточные экстракты анализировали на активность гипертермофильной амилазы следующим образом: 5 мг крахмала суспендировали в 20 мкл воды, а затем разбавляли 25 мкл этанола. К указанной смеси добавляли стандартный амилазопозитивный контроль или образец, тестируемый на амилазную активность, после чего добавляли воду до конечного реакционного объема 500 мкл. Реакцию осуществляли при 80°С в течение 15-45 минут. Затем реакционную смесь охлаждали до комнатной температуры и добавляли 500 мкл о-дианизидина и смесь глюкозооксидазы/пероксидазы (Sigma). Полученную смесь инкубировали при 37оС в течение 30 минут. Для прекращения реакции добавляли 500 мкл 12 н. серной кислоты. Для определения количества глюкозы, высвобождаемой амилазой/образцом, измеряли оптическую плотность при 540 нм. Анализ экстрактов гибридной и негибридной амилазы показал аналогичные уровни активности гипертермофильной амилазы, тогда как контрольные экстракты были негативными по этой активности. Это свидетельствовало о том, что при присоединении к С-концевой части белка восковидной кукурузы амилаза 797GL3 еще оставалась активной (при высоких температурах).
Пример 3
Выделение фрагментов промотора для эндоспермспецифической экспрессии в кукурузе
Промотор и 5'-некодирующую область I (включая первый интрон), происходящую от крупной субъединицы ADP-gpp Zea mays (ADP-глюкозопирофосфорилаза), амплифицировали в виде фрагмента из 1515 пар нуклеотидов (SEQ ID NO:11), полученного из геномной ДНК кукурузы, с использованием праймеров, имеющихся в Genbank, рег. №М81603. Было показано, что промотор ADP-gpp является эндоспермспецифическим (Shaw & Hannah, 1992).
Промотор гена γ-зеина Zea mays амплифицировали в виде 673 п.н.-фрагмента (SEQ ID NO:12), происходящего от плазмиды pGZ27.3 (полученной от Dr. Brian Larkins). Было показано, что промотор γ-зеина был эндосперм-специфическим (Torrent et al., 1997).
Пример 4
Конструирование трансформирующих векторов для гипертермофильной α-амилазы 797GL3
Экспрессионние кассеты, предназначенные для экспрессии гипертермофильной амилазы 797GL3 в эндосперме кукурузы, конструировали с использованием различных сигналов для нацеливания следующим образом.
pNOV6200 (SEQ ID NO:13) содержит N-концевую сигнальную последовательность γ-зеина кукурузы (MRVLLVALALLALAASATS)(SEQ ID NO:17), присоединенную к синтетической амилазе 797GL3, полученной, как описано выше в примере 1, и предназначенную для нацеливания на эндоплазматический ретикулум и секреции на апопласт (Torrent et al., 1997). Гибрид клонировали в область, расположенную за промотором ADP-gpp кукурузы, для специфической экспрессии в эндосперме.
pNOV6201 (SEQ ID NO:14) содержит N-концевую сигнальную последовательность γ-зеина, присоединенную к синтетической амилазе 797GL3 с добавлением в С-конец последовательности SEKDEL для нацеливания и удерживания на эндоплазматическом ретикулуме (ЭР) (Munro & Pelham, 1987). Гибрид клонировали в область, расположенную за промотором ADP-gpp кукурузы, для специфической экспрессии в эндосперме.
pNOV7013 содержит N-концевую сигнальную последовательность γ-зеина, присоединенную к синтетической амилазе 797GL3 с добавлением в С-конец последовательности SEKDEL для нацеливания и удерживания на эндоплазматическом ретикулуме (ЭР). pNOV7013 был аналогичен pNOV6201, за исключением того, что для экспрессии этого гибрида в эндосперме, вместо промотора ADP-gpp кукурузы, использовали промотор γ-зеина кукурузы (SEQ ID NO:12).
pNOV4029 (SEQ ID NO:15) содержит амилопластный нацеливающий пептид восковидной кукурузы (Klosgen et al., 1986), присоединенный к синтетической амилазе 797GL3 для нацеливания на указанный амилопласт. Это гибрид клонировали в область, расположенную за промотором ADP-gpp кукурузы, для специфической экспрессии в эндосперме.
pNOV4031 (SEQ ID NO:16) содержит амилопластный нацеливающий пептид восковидной кукурузы, присоединенный к синтетическому гибридному белку 797GL3/waxy для нацеливания на крахмальные зерна. Гибрид клонировали в область, расположенную за промотором ADP-gpp кукурузы, для специфической экспрессии в эндосперме.
Были созданы дополнительные конструкции с использованием указанных гибридов, клонированных за промотором γ-зеина кукурузы, в целях получения более высоких уровней экспрессии фермента. Все экспрессирующей кассеты переносили в бинарный вектор для трансформации в кукурузу посредством Agrobacterium-инфекции. Бинарный вектор содержал ген фосфоманнозоизомеразы (PMI), который позволяет проводить отбор трансгенных клеток с использованием маннозы. Трансформированные растения кукурузы либо подвергали самоопылению, либо скрещивали, и полученные семена собирали для анализа.
Были созданы дополнительные конструкции с использованием описанных выше сигналов для нацеливания, присоединенных либо к пуллуланазе 6gр3, либо к α-глюкозидазе 340g12, точно так же, как было описано для α-амилазы. Эти гибриды клонировали в область, расположенную за промотором ADP-gpp кукурузы и/или за промотором γ-зеина кукурузы, и трансформировали в кукурузу, как описано выше. Трансформированные растения кукурузы либо подвергали самоопылению, либо скрещивали, и полученные семена собирали для анализа.
Комбинации ферментов могут быть продуцированы либо путем скрещивания растений, экспрессирующих отдельные ферменты, либо путем клонирования нескольких экспрессирующих кассет в тот же самый бинарный вектор для возможности ко-трансформации.
Пример 5
Конструирование векторов, предназначенных для трансформации растений в целях введения термофильной пуллуланазы 6GP3
Экспрессирующую кассету конструировали для экспрессии термофильной пуллуланазы 6GP3 в эндоплазматическом ретикулуме эндосперма кукурузы следующим образом.
pNOV7005 (SEQ ID NO:24 и 25) содержит N-концевую сигнальную последовательность γ-зеина кукурузы, присоединенную к синтетической пуллуланазе 6GP3, с добавлением в С-конец последовательности SEKDEL для нацеливания и удерживания на ЭР. Пептид SEKDEL присоединяли к С-концу указанных ферментов посредством ПЦР с использованием праймеров, сконструированных для амплификации синтетического гена, с одновременным добавлением 6 аминокислот у С-конца белка. Гибрид клонировали в область, расположенную за промотором γ-зеина кукурузы, для специфической экспрессии в эндосперме.
Пример 6
Конструирование векторов, предназначенных для трансформации растений, для гипертермофильной α -глюкозидазы malA
Экспрессирующие кассеты конструировали для экспрессии гипертермофильной α-глюкозидазы malA Sulfolobus solfataricus в эндосперме кукурузы с использованием различных сигналов для нацеливания следующим образом.
pNOV4831 (SEQ ID NO:26) содержит N-концевую сигнальную последовательность γ-зеина (MRVLLVALALLALAASATS) (SEQ ID NO:17), присоединенную к синтетической α-глюкозидазе malA с добавлением в С-конец последовательности SEKDEL для нацеливания и удерживания на эндоплазматическом ретикулуме (ЭР) (Munro & Pelham, 1987). Гибрид клонировали в область, расположенную за промотором γ-зеина кукурузы, для специфической экспрессии в эндосперме.
pNOV4839 (SEQ ID NO:27) содержит N-концевую сигнальную последовательность γ-зеина кукурузы (MRVLLVALALLALAASATS) (SEQ ID NO:17), присоединенную к синтетической α-глюкозидазе malA для нацеливания на эндоплазматический ретикулум и секреции в апопласт (Torrent et al., 1997). Гибрид клонировали в область, расположенную за промотором γ-зеина кукурузы, для специфической экспрессии в эндосперме.
pNOV4837 содержит N-концевую сигнальную последовательность γ-зеина кукурузы (MRVLLVALALLALAASATS) (SEQ ID N0:17), присоединенную к синтетической α-глюкозидазе malA, с добавлением в С-конец последовательности SEKDEL для нацеливания и удерживания на ЭР. Гибрид клонировали в область, расположенную за промотором ADP-gpp кукурузы, для специфической экспрессии в эндосперме. Аминокислотная последовательность клона была идентична последовательности pNOV4831 (SEQ ID NO:26).
Пример 7
Конструирование векторов, предназначенных для трансформации растений, в целях введения гипертермофильных глюкозоизомераз Thermotoga maritima и Thermotoga neapolitana
Ниже описано конструирование экспрессирующих кассет для экспрессии гипертермофильных глюкозоизромераз Thermotoga maritima и Thermotoga neapolitana в эндосперме кукурузы с использованием различных нацеливающих сигналов.
pNOV4832 (SEQ ID NO:28) содержит N-концевую сигнальную последовательность γ-зеина кукурузы (MRVLLVALALLALAASATS) (SEQ ID NO:17), присоединенную к синтетической глюкозоизомеразе Thermotoga maritima, с добавлением в С-конец последовательности SEKDEL для нацеливания и удерживания на ЭР. Гибрид клонировали в область, расположенную после промотора γ-зеина кукурузы, для специфической экспрессии в эндосперме.
рNOV4833 (SEQ ID NO:29) содержит N-концевую сигнальную последовательность γ-зеина кукурузы (MRVLLVALALLALAASATS)(SEQ ID NO:17), присоединенную к синтетической глюкозоизомеразе Thermotoga neapolitana с добавлением в С-конец последовательности SEKDEL для нацеливания и удерживания на ЭР. Гибрид клонировали в область, расположенную за промотором γ-зеина кукурузы, для специфической экспрессии в эндосперме.
рNOV4840 (SEQ ID NO:30) содержит N-концевую сигнальную последовательность γ-зеина кукурузы (MRVLLVALALLALAASATS)(SEQ ID NO:17), присоединенную к синтетической глюкозоизомеразе Thermotoga neapolitana для нацеливания на эндоплазматический ретикулум и секреции в апопласт (Torrent et al., 1997). Гибрид клонировали в область, расположенную за промотором γ-зеина кукурузы, для специфической экспрессии в эндосперме.
рNOV4838 содержит N-концевую сигнальную последовательность γ-зеина кукурузы (MRVLLVALALLALAASATS)(SEQ ID NO:17), присоединенную к синтетической глюкозоизомеразе Thermotoga neapolitana с добавлением в С-конец последовательности SEKDEL для нацеливания и удерживания на ЭР. Гибрид клонировали в область, расположенную за промотором ADP-gpp кукурузы, для специфической экспрессии в эндосперме. Аминокислотная последовательность клона была идентична последовательности рNOV4833 (SEQ ID NO:29).
Пример 8
Конструирование векторов, предназначенных для трансформации растений в целях экспрессии гипертермофильной глюканазы EglA
рNOV4800 (SEQ ID NO:58) содержит сигнальную последовательность альфа-амилазы AMY32b ячменя (MGKNGNLCCFSLLLLLLAGLASGHQ) (SEQ ID NO:31), присоединенную к последовательности зрелого белка EglA для локализации в апопласт. Гибрид клонировали в область, расположенную за промотором γ-зеина кукурузы, для специфической экспрессии в эндосперме.
Пример 9
Конструирование векторов, предназначенных для трансформации растений в целях экспрессии множества гипертермофильных ферментов
рNOV4841 содержит двойную генную конструкцию, состоящую из гибрида α-амилазы 797GL3 и гибрида пуллуланазы 6GP3. Гибрид 797GL3 (SEQ ID NO:33) и гибрид 6GP3 (SEQ ID NO:34) имели N-концевую сигнальную последовательность γ-зеина кукурузы и последовательность SEKDEL для нацеливания и удерживания на ЭР. Каждый гибрид клонировали в область, расположенную за отдельными промоторами γ-зеина кукурузы, для специфической экспрессии в эндосперме.
рNOV4842 содержит двойную генную конструкцию, состоящую из гибрида α-амилазы 797GL3 и гибрида α-глюкозидазы malA. Гибридный полипептид 797GL3 (SEQ ID NO:35) и гибридный полипептид α-глюкозидазы malA (SEQ ID NO:36) имели N-концевую сигнальную последовательность γ-зеина кукурузы и последовательность SEKDEL для нацеливания и удерживания на ЭР. Каждый гибрид клонировали в область, расположенную за отдельными промоторами γ-зеина кукурузы, для специфической экспрессии в эндосперме.
рNOV4843 содержит двойную генную конструкцию, состоящую из гибрида α-амилазы 797GL3 и гибрида α-глюкозидазы malA. Оба гибрида 797GL3 и α-глюкозидазы malA имели N-концевую сигнальную последовательность γ -зеина кукурузы и последовательность SEKDEL для нацеливания и удерживания на ЭР. Гибрид 797GL3 был клонирован в область, расположенную за промотором γ-зеина кукурузы, а гибрид malA был клонирован в область, расположенную за промотором ADP-gpp кукурузы, для специфической экспрессии в эндосперме. Аминокислотные последовательности гибрида 797GL3 и гибрида malA были идентичны последовательностям рNOV4842 (SEQ ID No:35 и 36 соответственно).
рNOV4844 содержит тройную генную конструкцию, состоящую из гибрида α-амилазы 797GL3, гибрида пуллуланазы 6GP3 и гибрида α-глюкозидазы malA. Все гибриды 797GL3, malA, и 6GP3 имели N-концевую сигнальную последовательность γ-зеина кукурузы и последовательность SEKDEL для нацеливания и удерживания на ЭР. Гибриды 797GL3 и malA были клонирован в область, расположенную за 2 отдельными промоторами γ-зеина кукурузы, а гибрид 6GP3 был клонирован в область, расположенную за промотором ADP-gpp кукурузы для специфической экспрессии в эндосперме. Аминокислотные последовательности гибридов 797GL3 и malA были идентичны последовательностям рNOV4842 (SEQ ID No:35 и 36 соответственно). Аминокислотная последовательность гибрида 6GP3 была идентична последовательности гибрида 6GP3 в рNOV4841 (SEQ ID NO:34).
Все экспрессирующие кассеты переносили в бинарный вектор pNOV2117 для трансформации в кукурузу посредством Agrobacterium-инфекции. pNOV2117 содержит ген фосфоманнозоизомеразы (PMI) для отбора трансгенных клеток с использованием маннозы. pNOV2117 представляет собой бинарный вектор с сайтами инициации репликации pVS1 и ColE1. Этот вектор содержит конститутивный ген VirG, происходящий из pAD1289 (Hansen G. et al., PNAS USA 91:7603-7607 (1994)), и ген резистентности к спектиномицину, происходящий из Tn7. Этот вектор, клонированный в полилинкер между его правой и левой границами, содержал промотор убихитина кукурузы, кодирующую область PMI и терминатор нопалинсинтазы pNOV117 (Negrotto D. et al., Plant Cell Report 19:798-803 (2000)). Трансформированные растения кукурузы либо подвергали самоопылению, либо скрещивали и полученные семена собирали для анализа. Комбинации различных ферментов могут быть получены либо путем скрещивания растений, экспрессирующих отдельные ферменты, либо путем трансформации растения кассетами, содержащими один ген, или мультигенными кассетами.
Пример 10
Конструирование бактериальных и Pichia экспрессирующих векторов
Ниже описано конструирование экспрессирующих кассет для экспрессии гипертермофильной α-глюкозидазы и глюкозоизромеразы либо в Pichia, либо в бактериях.
pNOV4829 (SEQ ID NO:37 и 38) содержит синтетический гибрид глюкозоизомеразы Thermotoga maritima с сигналом удерживания в ЭР в бактериальном экспрессирующем векторе рЕТ29а. Гибридный ген глюкозоизомеразы клонировали в NcoI- и SacI-сайты рЕТ29а, что приводило к присоединению у N-конца S-метки, используемой для очистки белка.
PNOV4830 (SEQ ID NO:39 и 40) содержит синтетический гибрид глюкозоизомеразы Thermotoga neapolitana с сигналом удерживания в ЭР в бактериальном экспрессирующем векторе рЕТ29а. Гибридный ген глюкозоизомеразы клонировали в NcoI- и SacI-сайты pET29a, что приводило к присоединению у N-конца S-метки, используемой для очистки белка.
рNOV4835 (SEQ ID NO:41 и 42) содержит синтетический ген глюкозоизомеразы Thermotoga maritima, клонированный в BamHI- и EcoRI-сайты бактериального экспрессирующего вектора рЕТ28С. В результате получали гибрид His-метки (для очистки белка) с N-концом глюкозоизомеразы.
рNOV4836 (SEQ ID NO:43 и 44) содержит синтетический ген глюкозоизомеразы Thermotoga neapolitana, клонированный в BamHI- и EcoRI-сайты бактериального экспрессирующего вектора рЕТ28С. В результате получали гибрид His-метки (для очистки белка) с N-концом глюкозоизомеразы.
Пример 11
Трансформацию незрелых зародышей кукурузы осуществляли, в основном, как описано в работе Negrotto D. et al., Plant Cell Reports 19:798-803. В этом примере все компоненты сред являются такими, как они были описаны у Negrotto D. et al., см. выше. Однако различные компоненты сред, описанные в литературе, могут быть заменены.
А. Трансформирующие плазмиды и селективный маркер
Гены, используемые для трансформации, клонировали в вектор, подходящий для трансформации кукурузы. Векторы, используемые в этом примере, содержали ген фосфоманнозоизомеразы (PMI) для отбора трансгенных линий (Negrotto et al. (2000), Plant Cell Reports 19:798-803).
В. Получение Agrobacterium tumefaciens
Штамм LВА4404 Agrobacterium (pSBI), содержащий плазмиду для трансформации растений, культивировали на твердой среде YEP (дрожжевой экстракт (5 г/л), пептон (10 г/л), NaCl (5 г/л), 15 г/л агара, рН 6,8) в течение 2-4 дней при 28°С. Приблизительно 0,8×109 агробактерий суспендировали в среде LS-inf, в которую было добавлено 100 мкМ As (Negrotto et al. (2000), Plant Cell Report 19:798-803). Бактерии предварительно индуцировали в среде в течение 30-60 минут.
С. Инокуляция
Незрелые зародыши от А188 или другого подходящего генотипа вырезали из 8-12-дневных початков и помещали в жидкую среду LS-inf+100 мкМ As. Зародыши один раз промывали свежей инфекционной средой. Затем добавляли раствор с агробактериями, зародыши встряхивали в течение 30 секунд и оставляли с бактериями на 5 минут для осаждения. Затем эти зародыши той стороной, где расположен щиток зародыша, переносили в среду LSA и культивировали в темноте в течение двух-трех дней. После этого примерно 20-25 зародышей на чашку Петри переносили на среду LSDs, в которую были добавлены цефотаксим (250 мг/л) и нитрат серебра (1,6 мг/л), и культивировали в темноте при 28°С в течение 10 дней.
D. Отбор трансформированных клеток и регенерация трансформированных растений
Незрелые зародыши, продуцирующие эмбриогенный каллус, переносили в среду LSD1М0.5S. Культуры отбирали на этой среде в течение 6 недель, при этом стадию субкультивирования проводили в течение 3 недель. Жизнеспособные каллусы переносили в среду Reg1, в которую была добавлена манноза. После культивирования на свету (в режиме день/ночь=16 часов/8 часов) зеленые ткани переносили на среду Reg2 в отсутствие регуляторов роста и инкубировали в течение 1-2 недель. Проростки переносили в вегетационные сосуды Magenta GA-7 (Magenta Corp, Chicago III), содержащие среду Reg3, и культивировали на свету. Через 2-3 недель растения тестировали на присутствие генов PMI и других нужных генов посредством ПЦР. Положительные растения, идентифицированные посредством ПЦР-анализа, переносили в теплицу.
Пример 12
Анализ семян Т1, полученных от растений кукурузы, экспрессирующих α-амилазу, нацеливаемую на апопласт или на ЭР
Получали семена Т1 от самоопыленных растений кукурузы, трансформированных либо рNOV6200, либо рNOV6201, как описано в примере 4. Было установлено, что аккумуляция крахмала в этих зернах была нормальной, на что указывала визуальная оценка и нормальное окрашивание иодным раствором в тесте на крахмал, проводимом до термической обработки. Незрелые зерна иссекали и очищенные эндоспермы отдельно помещали в микроцентрифужные пробирки и погружали в 200 мкл 50 мМ NaPO4-буфера. Пробирки помещали в водяную баню при 85°С на 20 минут, а затем охлаждали на льду. В каждую пробирку добавляли двадцать микролитров 1% иодного раствора и смешивали. Приблизительно 25% сегрегирующихся зерен давали положительную окраску на крахмал. Остальные 75% не давали окраску, что указывало на то, что этот крахмал разлагался на низкомолекулярные сахара, которые не окрашиваются иодом. Было обнаружено, что зерна Т1 с рNOV6200 и рNOV6201 представляли собой зерна с аутогидролизующимся кукурузным крахмалом. Какого-либо заметного снижения уровня крахмала после инкубирования при 37оС не наблюдалось.
Затем анализировали экспрессию амилазы путем выделения фракции гипертермофильного белка из эндосперма с последующим проведением электрофореза в ПААГ/окрашивания кумасси. Наблюдалась полоса сегрегирующегося белка с соответствующей молекулярной массой (50 кДа). Эти образцы подвергали анализу на α-амилазу с использованием коммерчески доступной окрашенной амилозы (AMYLAZYME, от Megazyme, Ireland). Высокие уровни активности гипертермофильной амилазы коррелировали с присутствием 50 кДа-белка.
Кроме того, было обнаружено, что крахмал в зернах, полученных от большинства растений трангсгенной кукурузы, которая экспрессировала гипертермофильную α-амилазу, транспортируемую в амилопласт, достаточно активно гидролизовался при комнатной температуре, если указанный фермент мог непосредственно контактировать с крахмальными зернами. Из восьмидесяти линий, содержащих гипертермофильную α-амилазу, нацеливаемую на амилопласт, были идентифицированы четыре линии, которые аккумулировали крахмал в зерне. Три из этих линий были проанализированы на термостабильную α-амилазную активность с использованием колориметрического анализа на амилазим (Megazyme). Анализ на амилазный фермент показал, что эти три линии имели низкие уровни термостабильной амилазной активности. При обработке очищенного крахмала, полученного от этих трех линий, в подходящих условиях влажности и нагревания, данный крахмал был гидролизован, что указывало на присутствие адекватных уровней α-амилазы, необходимых для обеспечения аутогидролиза крахмала, полученного из этих линий.
Были получены семена Т1 от множества независимых линий обоих трансформантов рNOV6200 и рNOV6201. Отдельные зерна от каждой линии рассекали и очищенные эндоспермы отдельно гомогенизировали в 300 мкл 50 мМ NaPO4-буфера. Аликвоты суспензий эндосперма анализировали на α-амилазную активность при 85°С. Приблизительно 80% линий сегрегировались по гипертермофильной активности (см. фигуры 1А, 1В и 2).
Зерна от растений дикого типа или от растений, трансформированных рNOV6201, нагревали при 100°С в течение 1, 2, 3 или 6 часов, а затем окрашивали на присутствие крахмала иодным раствором. Через 3 или 6 часов соответственно в зрелых зернах наблюдалось либо незначительное содержание крахмала, либо крахмал вообще отсутствовал. Таким образом, крахмал в зрелых зернах трансгенной кукурузы, которая экспрессировала гипертермофильную амилазу, нацеливаемую на эндоплазматический ретикулум, гидролизовался при инкубировании при высокой температуре.
В другом эксперименте частично очищенный крахмал, полученный от зрелых зерен Т1 растений с рNOV6201, которые замачивали при 50°С в течение 16 часов, гидролизовался после нагревания при 85°С в течение 5 минут. Это указывало на то, что α-амилаза, нацеливаемая на эндоплазматический ретикулум, связывается с крахмалом после измельчения зерна и способна гидролизовать крахмал после нагревания. Окрашивание иодом показало, что крахмал остается интактным в зрелых семенах после замачивания в течение 16 часов при 50°С.
В другом эксперименте сегрегирующиеся зрелые зерна, полученные от растений, трансформированных рNOV6201, нагревали при 95°С в течение 16 часов, а затем сушили. В семенах, экспрессирующих гипертермофильную α-амилазу, после сушки происходил гидролиз крахмала в сахар, на что указывал вид сморщенного зерна.
Пример 13
Анализ семян Т1, полученных из растений кукурузы, экспрессирующих α-амилазу, нацеливаемую на амилопласт
Получали семена Т1 от самоопыленных растений кукурузы, трансформированных либо рNOV4029, либо рNOV4031, как описано в примере 4. Аккумуляция крахмала в зернах от этих линий была, очевидно, аномальной. Все линии сегрегировались, с определенной степенью вариабельности, по фенотипу очень низкого содержания крахмала или отсутствия крахмала. Перед обработкой при высоких температурах эндосперм, очищенный из незрелых зерен, давал лишь слабую окраску иодом. После выдерживания в течение 20 минут при 85°С окраска исчезала. После высушивания початков зерно сморщивалось. Эта конкретная амилаза обладала, очевидно, достаточной активностью при температурах, создаваемых в теплице для гидролиза крахмала, если этот фермент непосредственно контактировал с крахмальными зернами.
Пример 14
Ферментация зерна растений кукурузы, экспрессирующих α-амилазу
100% трансгенное зерно, 85°С и 95°С, варьирующееся время разжижения
Трансгенная кукуруза (рNOV6201), которая содержала термостабильную α-амилазу, хорошо подвергалась ферментации без добавления экзогенной α -амилазы, требовала меньше времени для разжижения и давали более полную солюбилизацию крахмала. Ферментацию в лабораторных масштабах осуществляли в соответствии с протоколом, включающим следующие стадии (подробно описанные ниже): 1) измельчение, 2) анализ на влажность, 3) получение взвеси, содержащей измельченную кукурузу, воду, барду и α-амилазу, 4) разжижение и 5) одновременное осахаривание и ферментацию (SSF). В этом примере температура и время разжижения варьировались, как описано ниже. Кроме того, трансгенную кукурузу подвергали разжижению с использованием и без использования экзогенной α-амилазы и эффективность продуцирования этанола сравнивали с эффективностью контрольной кукурузы, обработанной коммерчески доступной α-амилазой.
Трансгенную кукурузу, используемую в данном примере, получали в соответствии с процедурами, описанными в примере 4, с использованием вектора, содержащего ген α-амилазы и селективный маркер PMI, а именно рNOV6201. Трансгенную кукурузу продуцировали путем опыления коммерчески доступного гибрида (N3030ВТ) пыльцой трансгенной линии, экспрессирующей высокий уровень термостабильной α -амилазы. Кукурузу сушили до состояния 11% влажности и хранили при комнатной температуре. Содержание α-амилазы в муке трансгенной кукурузы составляло 95 единиц/г, где 1 единица фермента генерирует 1 микромоль восстанавливающих концов за одну минуту в кукурузной муке при 85°С в буфере MES, рН 6,0. В качестве контроля была использована желтая зубовидная кукуруза, которая, как известно, хорошо продуцирует этанол.
1) Измельчение: трансгенную кукурузу (1180 г) измельчали в молотковой мельнице Perten 3100, снабженной 2,0 мм-ситом, и таким образом получали муку трансгенной кукурузы. Контрольную кукурузу измельчали в той же самой мельнице после тщательной очистки для предупреждения загрязнения трансгенной кукурузой.
2) Анализ на содержание влаги: образцы (20 г) трансгенной и контрольной кукурузы взвешивали в алюминиевых лодочках для взвешивания и нагревали при 100°С в течение 4 ч. Затем образцы снова взвешивали и содержание влаги вычисляли по потере массы. Содержание влаги в трансгенной муке составляло 9,26%, а в контрольной муке - 12,54%.
3) Получение взвесей: композицию взвесей приготавливали так, чтобы полученная биомасса содержала 36% твердых веществ в начале стадии SSF. Контрольные образцы приготавливали в пластиковых сосудах емкостью 100 мл, и эти образцы содержали 21,50 г муки контрольной кукурузы, 23 мл деионизованной воды, 6,0 мл барды (8% твердых веществ по массе) и 0,30 мл коммерчески доступной α-амилазы, разведенной 1/50 водой. Выбранная доза α-амилазы представляла собой дозу, обычно используемую в промышленном производстве. При анализе в условиях, описанных выше для анализа трансгенной α-амилазы, доза контрольной α-амилазы составляла 2 ед/г кукурузной муки. рН доводили до 6,0 добавлением гидроксида аммония. Трансгенные образцы приготавливали аналогичным образом, но из-за более низкого содержания влаги в трансгенной муке они содержали 20 г кукурузной муки. Взвеси трансгенной муки получали либо с α-амилазой в такой же дозе, как и контрольные образцы, либо без экзогенной α-амилазы.
4) Разжижение: сосуды, содержащие взвеси муки трансгенной кукурузы, погружали в водяную баню при 85°С или при 95°С на 5, 15, 30, 45 или 60 минут. Контрольные взвеси инкубировали в течение 60 минут при 85°С. Во время инкубирования при высокой температуре взвеси интенсивно размешивали вручную каждые 5 минут. После стадии обработки при высокой температуре взвеси охлаждали на льду.
5) Одновременное осахаривание и ферментация: биомассу, полученную путем разжижения, смешивали с глюкоамилазой (0,65 мл 1/50 разведения коммерчески доступной глюкоамилазы L-400), с протеазой (0,60 мл 1000-кратного разведения коммерчески доступной протеазы) и 0,2 мг лактоцида и мочевины (0,85 мл 10-кратного разведения 50% раствора мочевины). В крышке 100 мл-сосуда, содержащего указанную биомассу, делали отверстие для выпуска CO2 в целях вентиляции. Затем указанную биомассу инокулировали дрожжами (1,44 мл) и инкубировали в водяной бане при 90°F. После 24-часовой ферментации температуру понижали до 86°F, а через 48 часов ее доводили до 82°F.
Дрожжи для инокуляции размножали путем получения смеси, содержащей дрожжи (0,12 г), с 70 граммами мальтодекстрина, 230 мл воды и 100 мл барды, глюкомилазу (0,88 мл 10-кратного разведения коммерчески доступной глюкоамилазы), протеазу (1,76 мл 100-кратного разведения коммерчески доступного фермента), мочевину (1,07 граммов), пенициллин (0,67 мг) и сульфат цинка (0,13 г). Размножение культуры инициировали за один день до использования и инкубировали при перемешивании при 90°F.
Через 24, 48 и 72 часа образцы вынимали из каждого ферментера, фильтровали через 0,2 мкм-фильтры и анализировали посредством ВЭЖХ на этанол и сахара. Через 72 часа образцы анализировали на полное растворение твердых веществ и на остаточный крахмал.
ВЭЖХ-анализ осуществляли с использованием двойной градиентной системы, снабженной детектором показателя преломления, нагревательной колонной и колонной Bio-Rad Aminex НРХ-87Н. Систему уравновешивали 0,005 М Н2SO4 в воде при концентрации 1 мл/мин. Температура колонны составляла 50°С. Объем впрыска образца составлял 5 мкл; элюирование проводили в том же самом растворителе. Ответ RI калибровали путем инжекции известных стандартов. Уровень этанола и глюкозы измеряли для каждой инжекции.
Остаточный крахмал измеряли следующим образом. Образцы и стандарты сушили при 50°С в печи, а затем измельчали в порошок в мельнице для образцов. Порошок (0,2 г) взвешивали в градуированной центрифужной пробирке емкостью 15 мл. Порошок 3 раза промывали 10 мл водного этанола (80% об./об.) путем интенсивного перемешивания с последующим центрифугированием и удалением супернатанта. К осадку добавляли ДМСО (2,0 мл), а затем добавляли 3,0 мл термостабильной альфа-амилазы (300 единиц) в буфере MOPS. После интенсивного перемешивания, пробирки инкубировали в водяной бане при 85°С в течение 60 мин. В процессе инкубирования содержимое пробирок перемешивали четыре раза. Образцы охлаждали и добавляли 4,0 мл натрийацетатного буфера (200 мМ, рН 4,5), а затем 0,1 мл глюкоамилазы (20 ед.). Образцы инкубировали при 50°С в течение 2 часов, перемешивали, а затем центрифугировали в течение 5 минут при 3500 об./мин. Супернатант фильтровали через 0,2 мкм-фильтр и анализировали на глюкозу ВЭЖХ-методом, описанным выше. Для образцов с низким содержанием остаточного крахмала (<20% твердых веществ) объем впрыска составлял 50 мкл.
Результаты. Трансгенная кукуруза хорошо поддавалась ферментации без добавления α-амилазы. Выходы этанола через 72 часа были, в основном, идентичными, независимо от присутствия или отсутствия экзогенной α-амилазы, как показано в таблице 1. Эти данные также показали, что более высокий выход этанола достигается при более высокой температуре разжижения; фермент настоящего изобретения, экспрессируемый в трансгенной кукурузе, обладает активностью при более высоких температурах, чем другие коммерчески доступные ферменты, такие как α-амилаза Bacillus liquefaciens.
При изменении времени разжижения было обнаружено, что время разжижения, требуемое для эффективного продуцирования этанола, было гораздо меньше, чем один час, требуемый для осуществления стандартного процесса. На фигуре 3 показано, что выход этанола после 72-часовой ферментации почти не изменялся за период времени разжижения от 15 минут до 60 минут. Кроме того, разжижение при 95°С давало большее количество этанола в каждый период времени, чем разжижение при 85°С. Это наблюдение продемонстрировало повышение производительности процесса при использовании гипертермофильного фермента.
Контрольная кукуруза давала более высокий конечный выход этанола, чем трансгенная кукуруза, но была выбрана контрольная кукуруза, поскольку она обнаруживала высокоэффективную ферментацию. В противоположность этому, трансгенная кукуруза имела генетический фон, выбранный для стимуляции трансформации. Введение α-амилазного признака в зародышевую плазму элитной кукурузы путем хорошо известных способов скрещивания должно устранять это различие.
Оценка остаточных уровней крахмала в пиве, приготовленном через 72 часа (фигура 4), показала, что трансгенная α-амилаза давала значительное увеличение продуцирования крахмала, доступного для ферментации, то есть после ферментации оставалось гораздо меньшее количество крахмала.
С использованием уровней этанола и уровней остаточного крахмала оптимальное время разжижения составляло 15 минут при 95°С и 30 минут при 85°С. В настоящих экспериментах это время представляло собой общее время, в течение которого ферментеры находились в водяной бане, и, таким образом, включало период, за который температура образцов увеличивалась от комнатной температуры до 85°С или 95°С. При крупномасштабных промышленных процессах, при которых происходит быстрое нагревание биомассы с использованием такого оборудования, как варочный котел струйного типа, может быть оптимальным еще более короткое время разжижения. Стандартные промышленные процессы разжижения требуют таких чанов-сборников, которые позволяют инкубировать биомассу при высоких температурах в течение одного или более часов. Настоящее изобретение позволяет избежать необходимости использования таких чанов-сборников и повысить производительность оборудования для разжижения.
Одной из важных функций α-амилазы в процессах ферментации является снижение вязкости биомассы. При всех периодах времени образцы, содержащие муку трансгенной кукурузы, были заметно менее вязкими, чем контрольный образец. Кроме того, трансгенные образцы, очевидно, не проходили фазу желатинизации, которая наблюдалась для всех контрольных образцов и которая обычно происходит при варке кукурузных взвесей. Таким образом, присутствие α-амилазы, распределенной по всем фрагментам эндосперма, придает биомассе в процессе варки преимущественные физические свойства, предотвращающие образование большого количества геля, которое замедляет диффузию и повышает энергетические затраты на смешивание и подачу указанной биомассы.
Высокая доза α-амилазы в трансгенной кукурузе может также придавать указанной трансгенной биомассе предпочтительные свойства. При 85°С активность α -амилазы трансгенной кукурузы была во много раз больше, чем активность той же дозы экзогенной α-амилазы, используемой в контроле. Эта контрольная α-амилаза была выбрана как репрезентативная α-амилаза для промышленного применения.
Пример 15
Эффективная функция трансгенной кукурузы при смешивании с контрольной кукурузой
Муку трансгенной кукурузы смешивали с мукой контрольной кукурузы при различных уровнях муки трансгенной кукурузы от 5% до 100%. Эту смесь обрабатывали, как описано в примере 14. Биомассу, содержащую трансгенно экспрессированную α-амилазу, подвергали разжижению при 85°С в течение 30 минут и при 95°С в течение 15 минут, а контрольную биомассу получали, как описано в примере 14, и подвергали разжижению при 85°С в течение 30 или 60 минут (один раз каждую) или при 95°С в течение 15 или 60 минут (один раз каждую).
Данные, полученные для этанола, через 48 и 72 часа, и для остаточного крахмала приведены в таблице 2. Уровни этанола через 48 часов графически представлены на фигуре 5, а определение остаточного крахмала показано на фигуре 6. Эти данные показали, что трансгенно экспрессируемая термостабильная α-амилаза давала очень хороший уровень продуцирования этанола, даже когда трансгенное зерно составляло лишь небольшую часть (по крайней мере, 5%) от общего зерна в биомассе. Эти данные также показали, что уровень остаточного крахмала был заметно ниже, чем уровень в контрольной биомассе, даже когда трансгенное зерно составляло, по крайней мере, 40% от общего количества зерна.
Пример 16
Продуцирование этанола в зависимости от рН разжижения с использованием трансгенной кукурузы при уровнях 1,5-12% от общей кукурузы
Поскольку трансгенная кукуруза хорошо поддается ферментации на уровне 5-10% от общей кукурузы, была проведена дополнительная серия ферментаций, в которых трансгенная кукуруза составляла 1,5-12% от общей кукурузы. рН варьировался от 6,4 до 5,2, а фермент α-амилаза, экспрессируемый в трансгенной кукурузе, был оптимизирован по активности при более низком рН, чем это обычно используется в промышленности.
Эксперименты осуществляли, как описано в примере 15, со следующими исключениями:
1. Трансгенную муку смешивали с контрольной мукой, и процент трансгенной муки по общей сухой массе составлял в пределах 1,5%-12,0%.
2. Контрольной кукурузой была N3030ВТ, которая имела большее сходство с трансгенной кукурузой, чем контрольная кукуруза, используемая в примерах 14 и 15.
3. В образцы, содержащие трансгенную муку, не добавляли экзогенную α-амилазу.
4. Перед разжижением рН образцов доводили до 5,2, 5,6, 6,0 или 6,4. Для каждого рН получали, по крайней мере, 5 образцов, в которых содержание муки трансгенной кукурузы составляло 0%-12%.
5. Разжижение для всех образцов проводили при 85°С в течение 60 минут.
Изменение содержания этанола в зависимости от времени ферментации показано на фигуре 7. На этой фигуре представлены данные, полученные от образцов, содержащих 3% трансгенную кукурузу. При более низком рН ферментация протекает быстрее, чем при рН 6,0 и выше; при этом аналогичный характер зависимости наблюдался для образцов с другим содержанием трансгенного зерна. рН-профиль активности трансгенного фермента, наряду с высокими уровнями экспрессии, позволяет осуществлять разжижение при более низком рН, что будет приводить к ускорению ферментации и, тем самым, к увеличению производительности, чем это было возможно в стандартных процессах при рН 6,0.
Выходы этанола через 72 часа показаны на фигуре 8. Как можно видеть по выходам этанола, эти результаты свидетельствуют о незначительной зависимости от количества трансгенного зерна, содержащегося в образце. Таким образом, для повышения ферментативного продуцирования этанола зерно должно содержать избыточное количество амилазы. Это также указывает на то, что разжижение при более низком рН дает более высокий выход этанола.
Проводили мониторинг вязкости образцов после разжижения, и при этом наблюдалось, что при рН 6,0 для адекватного снижения вязкости было достаточно 6% трансгенного зерна. При рН 5,2 и 5,6 вязкость была эквивалентна вязкости контроля при 12% содержании трансгенного зерна, но не при более низком содержании трансгенного зерна.
Пример 17
Продуцирование фруктозы из кукурузной муки с использованием термофильных ферментов
Было показано, что кукуруза, которая экспрессирует гипертермофильную α-амилазу 797GL3, более эффективно продуцирует фруктозу при смешивании с α-глюкозидазой (MalA) и ксилозоизомеразой (XylA).
Семена от рNOV6201-трансгенных растений, экспрессирующих 797GL3, измельчали в муку в камере Kleco, и таким образом получали амилазную муку. Зерно не-трансгенной кукурузы измельчали аналогичным образом и получали контрольную муку.
α-Глюкозидазу, MalA (от S.solfataricus), экспрессировали в E.coli. Собранные бактерии суспендировали в 50 мМ фосфатно-калиевого буфера, рН 7,0, содержащего 1 мМ 4-(2-аминоэтил)бензолсульфонилфторид, а затем подвергали лизису в прессе Френча. Лизат центрифугировали при 23000×g в течение 15 минут при 4°С. Раствор супернатанта удаляли, нагревали до 70°С в течение 10 минут, охлаждали на льду в течение 10 минут, а затем центрифугировали при 34000×g в течение 30 минут при 4°С. Раствор супернатанта удаляли и MalA концентрировали до двухкратного снижения объема в центрифуге Centricon 10. Фильтрат, полученный в стадии центрифугирования на Centricon 10, оставляли для использования в качестве негативного контроля на MalA.
Ксилозо(глюкозо)изомеразу получали путем экспрессии гена ХуlА T.neapolitana в E.coli. Бактерии суспендировали в 100 мМ фосфата натрия, рН 7,0, и подвергали лизису путем пропускания через пресс Френча. После преципитации клеточного дебриса экстракт нагревали при 80°С в течение 10 минут, а затем центрифугировали. Раствор супернатанта обладал ХуlА-ферментативной активностью. Одновременно с экстрактом ХуlА получали экстракт "пустого" контрольного вектора.
Кукурузную муку (60 мг на образец) смешивали с буфером и с экстрактами от E.coli. Как показано в таблице 3, образцы содержали кукурузную муку с амилазой (амилазную) или контрольную кукурузную муку (контроль), 50 мкл либо экстракта (+) MalA, либо фильтрата (-), и 20 мкл либо экстракта ХуlА (+), либо контрольного "пустого" вектора (-). Все образцы также содержали 230 мкл 50 мМ MOPS, 10 мМ MgSO4 и 1 мМ CoCl2; рН буфера составлял 7,0 при комнатной температуре.
Образцы инкубировали при 85°С в течение 18 часов. По окончании времени инкубирования образцы разводили 0,9 мл воды при 85°С и центрифугировали для удаления нерастворенного материала. Затем фракцию супернатанта фильтровали путем пропускания через устройство для ультрафильтрации Centricon 3 и анализировали с помощью ВЭЖХ и ELSD-детекции.
Градиентная ВЭЖХ-система была снабжена колонкой Astec Polymer Amino Column, с размером частиц 5 микрон, 250×4,6 мм и детектором Alltech ELSD 2000. Эту систему предварительно уравновешивали смесью вода:ацетонитрил =15:85. Скорость потока составляла 1 мл/мин. Начальные условия поддерживались в течение 5 минут после инжекции, после чего в течение 20 минут проводили элюирование градиентом вода:ацетонитрил =50:50 и в течение 10 минут тем же самым растворителем. Систему промывали 20 минут смесью вода:ацетонитрил =80:20, а затем снова уравновешивали исходным растворителем. Фруктоза элюировалась за 5,8 минут, а глюкоза - за 8,7 минут.
Результаты ВЭЖХ также показали присутствие более высокого уровня мальтоолигосахаридов во всех образцах, содержащих α-амилазу. Эти результаты продемонстрировали, что три термофильных фермента, взятые вместе, могут продуцировать фруктозу из кукурузной муки при высокой температуре.
Пример 18
Амилазная мука с изомеразой
В другом примере амилазную муку смешивали с очищенной MalA, и с каждой из двух бактериальных ксилозоизомераз: ХуlА T.maritima и фермента, обозначенного BD8037 и полученного от Diversa. Амилазную муку получали, как описано в примере 18.
MalA S.solfataricus с 6His-меткой для очистки экспрессировали в E.coli. Клеточный лизат получали, как описано в примере 18, после чего этот лизат очищали до видимой гомогенности с использованием смолы, обладающий аффинностью по отношению к никелю (Probond, Invitrogen), в соответствии с инструкциями производителей для очистки нативного белка.
ХуlА T.maritima с добавленной S-меткой и с сигналом удерживания в ЭР экспрессировали в E.coli и получали таким же способом, как ХуlА T.neapolitana, описанным в примере 18.
Ксилозоизомеразу ВD8037 получали в виде лиофилизованного порошка и ресуспендировали в воде в объеме, составляющем 0,4 от исходного.
Амилазную кукурузную муку смешивали с ферментными растворами, содержащими воду или буфер. Все реакционные смеси содержали 60 мг амилазной муки и 600 мкл общего количества жидкости. Одну серию реакционных смесей забуферивали 50 мМ MOPS, рН 7,0, при комнатной температуре, с добавлением 10 мМ MgSO4 и 1 мМ CoCl2, а во второй серии реакционных смесей металлсодержащий буферный раствор заменяли водой. Количество изомеразного фермента варьировалось, как показано а таблице 4. Все реакционные смеси инкубировали в течение 2 часов при 90°С. Фракции реакционного супернатанта получали путем центрифугирования. Осадок еще раз промывали 600 мкл H2O и снова центрифугировали. Фракции супернатанта от каждой реакции объединяли, фильтровали через Centricon 10 и анализировали с помощью ВЭЖХ с ELSD-детекцией, как описано в примере 17. Количество полученной глюкозы и фруктозы графически представлено на фигуре 15.
В присутствии каждой изомеразы фруктоза продуцировалась из кукурузной муки в зависимости от дозы в том случае, когда в реакции присутствовали α-амилаза и α -глюкозидаза. Эти результаты продемонстрировали, что экспрессируемая зерном амилаза 797GL3 может функционировать с MalA и с рядом различных термофильных изомераз, в присутствие или в отсутствие ионов металла, с продуцированием фруктозы из кукурузной муки при высокой температуре. В присутствии добавленных двухвалентных ионов металла указанные изомеразы могут давать предсказанные уравновешенные уровни фруктозы:глюкозы при 90°С приблизительно с 55% содержанием глюкозы. Процесс может рассматриваться как усовершенствованный по сравнению с обычным процессом, в котором используются мезофильные изомеразы и который требует хроматографического разделения для увеличения концентрации фруктозы.
Пример 19
Экспрессия пуллуланазы в кукурузе
Трансгенные растения, которые являются гомозиготными либо по рNOV7013, либо по рNOV7005, скрещивали для получения семян трансгенной кукурузы, экспрессирующих как α-амилазу 797GL3, так и пуллуланау 6GP3.
Получали семена Т1 или Т2 от самоопыленных растений кукурузы, трансформированных либо рNOV7005, либо рNOV4093. рNOV4093 представляет собой гибрид синтетического гена 6GP3, оптимизированного по кодонам кукурузы (SEQ ID NO:3, 4), с амилопластной нацеливающей последовательностью (SEQ ID NO:7, 8), которая обеспечивает локализацию гибридного белка на амилопласте. Гибридный белок находится под контролем промотора ADP-gpp (SEQ ID NO:11) для специфической экспрессии в эндосперме. Конструкция рNOV7005 обеспечивает направленную экспрессию пуллуланазы в эндоплазматическом ретикулуме эндосперма. Локализация этого фермента в ЭР обеспечивает нормальную аккумуляцию крахмала в зернах. Кроме того, наблюдалось нормальное окрашивание крахмала иодным раствором перед какой-либо обработкой при высоких температурах.
Как было описано в случае α-амилазы, экспрессия пуллуланазы, нацеливаемой на амилопласт (рNOV4093), приводила к аномальному накоплению крахмала в зернах. При сушке кукурузных початков зерна сморщивались. Очевидно, что указанная термофильная пуллуланаза является достаточно активной при низких температурах и гидролизует крахмал в том случае, если происходит прямой контакт с крахмальными гранулами в эндосперме семян.
Получение фермента или экстракция фермента из кукурузной муки.
Фермент пуллуланазы экстрагировали из трансгенных семян путем их измельчения в мельнице Kleco с последующим инкубированием муки в 50 мМ NaOAc-буфере, рН 5,5, в течение 1 часа при комнатной температуре и при непрерывном встряхивании. Затем, после инкубирования, смесь центрифугировали в течение 15 минут при 14000 об/мин. Супернатант использовали как источник фермента.
Анализ на пуллуланазу: Аналитическую реакцию проводили в 96-луночном планшете. Фермент, экстрагированный из кукурузной муки (100 мкл), 10-кратно разводили 900 мкл 50 мМ NaOAc-буфера, рН 5,5, содержащего 40 мМ CaCl2. Смесь интенсивно перемешивали, а затем к каждой реакционной смеси добавляли 1 таблетку Limit-Dextrizyme (структурированный азурином пуллулан от Megazyme) и инкубировали при 75°С в течение 30 минут (или как было упомянуто выше). По окончании инкубирования реакционные смеси центрифугировали в течение 15 минут при 3500 об/мин. Супернатант 5-кратно разводили и переносили в плоскодонный 96-луночный планшет для измерения оптической плотности при 590 нм. Гидролиз структурированного азурином пуллуланового субстрата пуллуланазой приводил к продуцированию водорастворимых окрашенных фрагментов, и скорость их высвобождения (измеренная по оптической плотности при 590 нм) непосредственно коррелировала с ферментативной активностью.
На фигуре 9 проиллюстрирован анализ семян Т2 при различных случаях трансформации рNOV7005. В ряде случаев можно было детектировать высокую экспрессию пуллуланазной активности по сравнению с не-трансгенным контролем.
К измеренному количеству (˜100 мкг) сухой кукурузной муки, полученной от трансгенного растения (экспрессирующего пуллуланазу или амилазу или тот и другой фермент) и/или от контрольного (не-трансгенного) растения, добавляли 1000 мкл 50 мМ NaOAc-буфера, рН 5,5, содержащего 40 мМ CaCl2. Реакционные смеси интенсивно перемешивали и инкубировали на шейкере в течение 1 часа. Ферментативную реакцию инициировали путем переноса смесей для инкубирования в условия высокой температуры (75°С, оптимальной температуры реакции для пуллуланазы или как указано на фигурах) и инкубировали в течение периода времени, указанного на фигурах. Реакции прекращали путем охлаждения на льду. Затем реакционные смеси центрифугировали в течение 10 минут при 14000 об/мин. Аликвоту (100 мкл) супернатанта трехкратно разводили и фильтровали через 0,2-микронный фильтр для ВЭЖХ-анализа.
Образцы анализировали с помощью ВЭЖХ с использованием следующих условий:
Колонка: Alltech Prevail Carbohydrate ES, 5 микрон, 250×4,6 мм
Детектор: Alltech ELSD 2000
Насос: Gilson 322
Инжектор: инжектор/разбавитель Gilson 215
Растворители: ВЭЖХ, ацетонитрил высокого качества (Fisher Scientific) и вода (очищенная на системе Waters Millipore System)
Градиент, используемый для олигосахаридов низкой степени полимеризации (DP 1-15).
Градиент, используемый для сахаридов с высокой степенью полимеризации (DP 20-100 и выше).
Система, используемая для анализа данных: Gilson Unipoint Software System Version 3.2.
На фигурах 10А и 10В показан ВЭЖХ-анализ продуктов гидролиза крахмала в муке трансгенной кукурузы, генерированных экспрессированной пуллуланазой. Инкубирование муки из экспрессирующей пуллуланазу кукурузы в реакционном буфере при 75°С в течение 30 минут приводило к продуцированию олигосахаридов со средним размером цепи (DP ˜10-30) и амилозы с короткими цепями (DP ˜100-200) из кукурузного крахмала. На этой фигуре также продемонстрирована зависимость пуллуланазной активности от присутствия ионов кальция.
Трансгенная кукуруза, экспрессирующая пуллуланазу, может быть использована для продуцирования модифицированного крахмала/декстрина, который является неразветвленным (расщепленным по α -1-6-связям), а поэтому будет иметь высокий уровень амилозы/декстрина с прямой цепью. Кроме того, в зависимости от вида используемого крахмала (например, waxy, высокоамилозного и т.п.), распределение по длине цепи амилозы/декстрина, генерируемого пуллуланазой, будет варьироваться, а следовательно, будут варьироваться и свойства модифицированного крахмала/декстрина.
Был также продемонстрирован процессинг α-1-6-связей с использованием пуллулана в качестве субстрата. Пуллуланаза, выделенная из кукурузной муки, эффективно гидролизовала пуллулан. ВЭЖХ-анализ (описанный) продукта, генерированного по окончании инкубирования, указывал, как и ожидалось, на продуцирование мальтотриозы, что было обусловлено процессингом α-1-6-связей в молекулах пуллулана ферментом кукурузы.
Пример 20
Экспрессия пуллуланазы в кукурузе
Экспрессию пуллуланаз 6gp3 дополнительно анализировали путем экстракции из кукурузной муки с последующим электрофорезом в ПААГ и окрашиванием кумасси синим. Кукурузную муку получали путем измельчения семян в течение 30 сек на мельнице Kleco. Этот фермент экстрагировали примерно из 150 мг муки с использованием 1 мл 50 мМ NaOAc-буфера, рН 5,5. Смесь интенсивно перемешивали и инкубировали на шейкере при комнатной температуре в течение 1 часа, а затем инкубировали еще 15 минут при 70°С. Затем смесь центрифугировали (14000 об/мин в течение 15 минут при комнатной температуре) и супернатант использовали для анализа, проводимого путем электрофореза в ДСН-ПААГ. При этом наблюдалась полоса белка с соответствующей молекулярной массой (95 кДа). Эти образцы анализировали на пуллуланазу с использованием коммерчески доступных конъюгированных с красителем предельных декстринов (LIMIT-DEXTRIZYME, от Megazyme, Ireland). Высокие уровни термофильной пуллуланазной активности коррелировали с присутствием 95 кДа-белка.
Вестерн-блот-анализ и ELISA-анализ трансгенной кукурузы также продемонстрировал экспрессию ˜95 кДа-белка, который реагирует с антителом против пуллуланазы (экспрессированной в E.coli).
Пример 21
Увеличение скорости гидролиза крахмала и повышение выхода олигосахаридов с короткой цепью (ферментируемых) при добавлении кукурузы, экспрессирующей пуллуланазу
Данные, представленные на фигурах 11А и 11В, были получены исходя из вышеописанного ВЭЖХ-анализа на продукты расщепления крахмала от двух реакционных смесей. Первая реакционная смесь, указанная как "амилаза", содержала смесь [1:1 (мас./мас.)] образцов кукурузной муки для трансгенной кукурузы, экспрессирующей α-амилазу и приготовленной способом, например, описанным в примере 4, и для не-трансгенной кукурузы А188; а вторая реакционная смесь, указанная как "амилаза + пуллуланаза", содержала смесь [1:1 (мас./мас.)] образцов кукурузной муки для трансгенной кукурузы, экспрессирующей α-амилазу, и для трансгенной кукурузы, экспрессирующей пуллуланазу, и приготовленной способом, описанным в примере 19. Полученные результаты подтвердили преимущество использования пуллуланазы в комбинации с α -амилазой в процессе гидролиза крахмала. Это преимущество заключается в увеличении скорости гидролиза крахмала (фигура 11А) и увеличении выхода ферментируемых олигосахаридов с низкой DP (фигура 11В).
Было обнаружено, что для продуцирования мальтодекстрина (олигосахаридов с прямой или разветвленной цепью) может быть использована либо только α-амилаза, либо α -амилаза и пуллуланаза (либо любая другая комбинация крахмалразрушающих ферментов), экспрессируемые в кукурузе (фигуры 11А, 11В, 12 и 13А). В зависимости от условий реакций, типа гидролитических ферментов и их комбинаций и типа используемого крахмала, состав продуцируемых мальтодекстринов, а следовательно, и их свойства будут варьироваться.
На фигуре 12 представлены результаты эксперимента, проводимого способом, аналогичным способу, описанному на фигуре 11. На этой фигуре указаны различные температурные и временные режимы, используемые в процессе инкубирования реакционных смесей. Оптимальная температура реакции для пуллуланазы составляет 75°С, а для α-амилазы она составляет >95°С. Следовательно, указанные режимы соответствовали целям осуществления катализа пуллуланазой и/или α-амилазой при их соответствующих оптимальных температурах реакции. Из этих результатов можно сделать вывод, что комбинация α-амилазы и пуллуланазы давала большую эффективность в гидролизе кукурузного крахмала по окончании 60-минутного периода инкубирования.
ВЭЖХ-анализ, описанный выше (за исключением того, что в этих реакциях использовали ˜150 мг кукурузной муки) и проводимый для оценки продуктов гидролиза крахмала из двух серий реакционных смесей по окончании 30-минутного инкубирования, проиллюстрирован на фигуре 13А и 13В. Реакционные смеси в первой серии реакций инкубировали при 85°С, а реакционные смеси во второй серии инкубировали при 95°С. Для каждой серии были получены две реакционные смеси; первая реакционная смесь, указанная как "амилаза х пуллуланаза", содержит муку из трансгенной кукурузы (продуцированной путем перекрестного опыления), экспрессирующей как α -амилазу, так и пуллуланазу, и вторая реакционная смесь, указанная как "амилаза", представляет собой смесь образцов кукурузной муки из экспрессирующей α-амилазу трансгенной кукурузы и не-трансгенной кукурузы А188 в таком отношении, при котором α-амилазная активность присутствует в таком же количестве, как это наблюдается при перекрестной опылении (амилаза × пуллуланаза). Полный выход олигосахаридов с низкой DP был более высоким в случае кроссов "α-амилаза × пуллуланаза" по сравнению с кукурузой, экспрессирующей только α-амилазу, при инкубировании образцов кукурузной муки при 85°С. Инкубирование при температуре 95°С приводило к инактивации (по крайней мере, частично) фермента пуллуланазы, а поэтому между образцами "амилаза × пуллуланаза" и "амилаза" можно было наблюдать небольшие различия. Однако данные, полученные для обеих температур инкубирования, показали, что при использовании кукурузной муки, полученной от кроссов "амилаза × пуллуланаза", наблюдалось значительное увеличение количества продуцируемой глюкозы (фигура 13В) по окончании периода инкубирования, чем при использовании кукурузной муки, экспрессирующей только α-амилазу. Поэтому использование кукурузы, экспрессирующей как α-амилазу, так и пуллуланазу, может быть особенно предпочтительным в процессах, где желателен полный гидролиз крахмала в глюкозу.
Вышеуказанные примеры дают дополнительное подтверждение тому, что пуллуланаза, экспрессируемая в семенах кукурузы и используемая в комбинации с α-амилазой, совершенствует процесс гидролиза крахмала. Активность фермента пуллуланазы, являющейся специфичной к α-1-6-связям, гидролизует точки ветвления крахмала гораздо более эффективно, чем α-амилаза (фермент, специфичный к α-1-4-связям), что приводит к снижению количества разветвленных олигосахаридов (например, предельного декстрина, панозы, которые обычно не ферментируются) и к увеличению количества олигосахаридов с короткой прямой цепью (легко ферментируемых до этанола, и т.п.). Во-вторых, фрагментация молекул крахмала при катализируемом пуллуланазой гидролизе точек ветвления крахмала приводит к увеличению доступности субстрата для α-амилазы, а следовательно, и к увеличению эффективности реакций, катализируемых α -амилазой.
Пример 22
Для того, чтобы определить, может ли альфа-амилаза 797GL3 и альфа-глюкозидаза malA функционировать при тех же самых условиях рН и температуры с продуцированием повышенного количества глюкозы по сравнению с количеством глюкозы, продуцируемым любым из указанных ферментов, взятых отдельно, к раствору, содержащему 1% крахмал и крахмал, выделенный либо из семян не-трансгенной кукурузы (контроль), либо из семян трансгенной кукурузы 797GL3 (из семян кукурузы 797GL3, где альфа-амилаза выделяется вместе с крахмалом), добавляли приблизительно 0,35 мкг фермента альфа-глюкозидазы malA, продуцируемого в бактерии. Кроме того, крахмал, выделенный из не-трансгенных семян кукурузы и из семян трансгенной кукурузы 797GL3, добавляли к 1% кукурузному крахмалу в отсутствие какого-либо фермента malA. Смеси инкубировали при 90°С, рН 6,0, в течение 1 часа, центрифугировали для удаления любого нерастворившегося материала и растворимую фракцию анализировали посредством ВЭЖХ на уровни глюкозы. Как показано на фигуре 14, альфа-амилаза 797GL3 и альфа-глюкозидаза malA действуют при аналогичных рН и температуре, разлагая крахмал в глюкозу. В данном случае количество продуцируемой глюкозы было значительно выше, чем количество глюкозы, продуцируемой каким-либо одним ферментом.
Пример 23
Была определена эффективность глюкоамилазы Thermoanaerobacterium для гидролиза исходного крахмала. Как показано на фигуре 15, гидролитическое превращение исходного крахмала тестировали с использованием воды, α -амилазы ячменя (коммерчески доступный препарат Sigma), глюкоамилазы Thermoanaerobacterium и их комбинаций при комнатной температуре и при 30°С. Как было показано, комбинация α-амилазы ячменя с глюкоамилазой Thermoanaerobacterium способна гидролизовать исходный крахмал в глюкозу. Более того, количество глюкозы, продуцируемой амилазой ячменя и GA Thermoanaerobacter, значительно превышало количество глюкозы, продуцируемой каким-либо одним ферментом.
Пример 24
Оптимизированные по кодонам кукурузы гены и последовательности для гидролиза исходного крахмала и векторы для трансформации растений
Ферменты отбирали по их способности гидролизовать исходный крахмал при температурах приблизительно 20-50°С. Затем соответствующие гены или генные фрагменты конструировали с использованием кодонов кукурузы, предпочтительных для конструирования синтетических генов, как описано в примере 1.
Отбирали гибридный полипептид α-амилазы/глюкоамилазы Aspergillus shirousami (без сигнальной последовательности), который имеет аминокислотную последовательность, представленную в SEQ ID NO:45 и идентифицированную в Biosci. Biotech. Biochem. 56:884-889 (1992); Agric. Biol. Chem. 545:1905-14 (1990); Biosci. Biotechnol. Biochem. 56:174-79 (1992). Конструировали также нуклеиновую кислоту, оптимизированную по кодонам кукурузы, и эта нуклеиновая кислота представлена в SEQ ID NO:46.
Аналогичным образом отбирали глюкоамилазу Thermoanaerobacterium thermosaccharolyticum, имеющую аминокислотную последовательность SEQ ID NO:47, опубликованную в Biosci. Biotech. Biochem. 62:302-308 (1998). Конструировали нуклеиновую кислоту, оптимизированную по кодонам кукурузы (SEQ ID NO:48).
Отбирали глюкоамилазу Rhizopus oryzae, имеющую аминокислотную последовательность (без сигнальной последовательности) (SEQ ID NO:49), описанную в литературе (Agric. Biol. Chem. (1986) 50, pg. 957-964). Конструировали нуклеиновую кислоту, оптимизированную по кодонам кукурузы, и эта нуклеиновая кислота представлена в SEQ ID NO:50.
Кроме того, отбирали α-амилазу кукурузы, и ее аминокислотную последовательность (SEQ ID NO:51) и последовательность нуклеиновой кислоты (SEQ ID NO:52) получали из литературы. См., например, Plant Physiol. 105:759-760 (1994).
Конструировали экспрессирующие кассеты для экспрессии гибридного полипептида α-амилазы/глюкоамилазы Aspergillus shirousami из сконструированной и оптимизированной по кодонам кукурузы нуклеиновой кислоты, представленной в SEQ ID NO:46; глюкоамилазы Thermoanaerobacterium thermosaccharolyticum, из сконструированной и оптимизированной по кодонам кукурузы нуклеиновой кислоты, представленной в SEQ ID NO:48; глюкоамилазы Rhizopus oryzae, имеющей аминокислотную последовательность (без сигнальной последовательности) (SEQ ID NO:49), из сконструированной и оптимизированной по кодонам кукурузы нуклеиновой кислоты, представленной в SEQ ID NO:50, и α-амилазы кукурузы.
Плазмиду, содержащую N-концевую сигнальную последовательность γ-зеина кукурузы (MRVLLVALALLALAASATS) (SEQ ID NO:17), присоединяли к синтетическому гену, кодирующему фермент. Необязательно к С-концу синтетического гена присоединяли последовательность SEKDEL для нацеливания и удерживания на ЭР.
Гибрид клонировали в область, расположенную за промотором γ-зеина кукурузы, для специфической экспрессии в эндосперме в плазмиде для трансформации растений. Гибрид нацеливается на ткань кукурузы посредством Agrobacterium-трансфекции.
Пример 25
Для экспрессии ферментов конструировали экспрессирующие кассеты, содержащие выбранные ферменты. Плазмиду, содержащую последовательность сайта связывания с исходным крахмалом, присоединяли к синтетическому гену, кодирующему фермент. Сайт связывания с исходным крахмалом обеспечивал связывание фермента с нежелатинизированным крахмалом. Аминокислотную последовательность сайта связывания с исходным крахмалом (SEQ ID NO:53) определяли из литературных источников и последовательность нуклеиновой кислоты оптимизировали по кодонам кукурузы с получением SEQ ID NO:54. Последовательность нуклеиновой кислоты, оптимизированную по кодонам кукурузы, присоединяли к синтетическому гену, кодирующему фермент, в плазмиде для экспрессии в растении.
Пример 26
Конструирование генов, оптимизированных по кодонам кукурузы, и векторов для трансформации растений
Гены или генные фрагменты конструировали с использованием кодонов кукурузы, предпочтительных для создания синтетических генов, как описано в примере 1.
Отбирали EGLA, гипертермофильную эндоглюканазу Pyrococcus furiosus (без сигнальной последовательности), имеющую аминокислотную последовательность, представленную в SEQ ID NO:55 и идентифицированную в Journal of Bacteriology (1999) 181, pg.284-290). Конструировали нуклеиновую кислоту, оптимизированную по кодонам кукурузы и представленную в SEQ ID NO:56.
Отбирали ксилозоизомеразу Thermus flavus, имеющую аминокислотную последовательность, представленную в SEQ ID NO:57, и описанную в Applied Biochemistry and Biotechnology, 62:15-27 (1997).
Конструировали экспрессирующие кассеты для экспрессии EGLA (эндоглюканазы) Pyrococcus furiosus из оптимизированной по кодонам кукурузы нуклеиновой кислоты (SEQ ID NO:56) и ксилозоизомеразы Thermus flavus из оптимизированной по кодонам кукурузы нуклеиновой кислоты, кодирующей аминокислотную последовательность SEQ ID NO:57. Плазмиду, содержащую N-концевую сигнальную последовательность γ -зеина кукурузы (MRVLLVALALLALAASATS) (SEQ ID NO:17), присоединяли к синтетическому и оптимизированному по кодонам кукурузы гену, кодирующему фермент. Необязательно к С-концу синтетического гена присоединяли последовательность SEKDEL для нацеливания и удерживания на ЭР. Гибрид клонировали в область, расположенную за промотором γ-зеина кукурузы, для специфической экспрессии в эндосперме в плазмиде для трансформации растений. Гибрид нацеливается на ткань кукурузы посредством Agrobacterium-трансфекции.
Пример 27
Продуцирование глюкозы из кукурузной муки с использованием термофильных ферментов, экспрессируемых в кукурузе
Было показано, что экспрессия гипертермофильной α-амилазы, 797GL3, и α-глюкозидазы (MalA) приводит к продуцированию глюкозы при смешивании с водным раствором и инкубировании при 90°С.
Линию трансгенной кукурузы (линия 168А10В, pNOV4831), экспрессирующую фермент МаlА, идентифицировали путем измерения α-глюкозидаэной активности, на что указывал гидролиз п-нитрофенил-α-глюкозида.
Зерно кукурузы, полученное от трансгенных растений, экспрессирующих 797GL3, измельчали в муку в камере Kleco и получали муку, содержащую амилазу. Зерно кукурузы, полученное от трансгенных растений, экспрессирующих MalA, измельчали в муку в камере Kleco и получали муку, содержащую MalA. Зерно не-трансгенной кукурузы измельчали аналогичным способом и получали контрольную муку.
Буфер представлял собой 50 мМ MES-буфер, рН 6,0.
Реакции гидролиза кукурузной муки: Образцы получали, как описано ниже в таблице 5. Кукурузную муку (примерно 60 мг на образец) смешивали с 40 мл 50 мМ MES-буфера, рН 6,0. Образцы инкубировали в водяной бане при 90°С в течение 2,5 и 14 часов. В указанное время инкубирования брали образцы и анализировали на содержание глюкозы.
Образцы анализировали на глюкозу путем проведения анализа с использованием глюкозооксидазы/пероксидазы хрена. Реагент GOPOD содержал: 0,2 мг/мл о-дианизидина, 100 мМ трис, рН 7,5, 100 ед./мл глюкозооксидазы и 10 ед./мл пероксидазы хрена. 20 мкл образца или разведенного образца распределяли по лункам 96-луночного планшета вместе со стандартами глюкозы (которые варьировались от 0 до 0,22 мг/мл). В каждую лунку добавляли, при размешивании, 100 мкл реагента GOPOD и планшеты инкубировали при 37оС в течение 30 минут. Затем добавляли 100 мкл серной кислоты (9 М) и считывали оптическую плотность при 540 нм. Концентрацию глюкозы в образцах определяли по стандартной кривой, построенной с использованием эталона. Количество глюкозы, наблюдаемой в каждом образце, указано в таблице 5.
Эти данные продемонстрировали, что экспрессия гипертермофильной α-амилазы и α-глюкозидазы в кукурузе приводит к образованию кукурузного продукта, который будет продуцировать глюкозу при гидратации и нагревании в соответствующих условиях.
Пример 28
Продуцирование мальтодекстринов
Для получения мальтодекстринов использовали зерно, экспрессирующее термофильную α-амилазу. Проиллюстрированный способ не требует предварительного выделения крахмала и не требует добавления экзогенных ферментов.
Зерно кукурузы, полученное от трансгенных растений, экспрессирующих 797GL3, измельчали в муку в камере Kleco и получали "амилазную муку". Смесь 10% трансгенного/90% не-трансгенного зерна измельчали аналогичным способом и получали "10% амилазную муку".
Амилазную муку и 10% амилазную муку (приблизительно 60 мг/образец) смешивали с водой в соотношении 5 мкл воды на один мг муки. Полученные взвеси инкубировали при 90°С в течение периода времени до 20 часов, как указано в таблице 6. Реакции прекращали добавлением 0,9 мл 50 мМ EDTA при 85°С и смеси размешивали пипеткой. Брали 0,2 мл-образцы взвеси, центрифугировали для удаления нерастворившегося материала и разводили 3х в воде.
Образцы анализировали на сахара и мальтодекстрины с помощью ВЭЖХ с ELSD-детекцией. Градиентная ВЭЖХ-система была снабжена колонной Astec Polymer Amino Column, с размером частиц 5 микрон, 250×4,6 мм, и детектором Alltech ELSD 2000. Эту систему предварительно уравновешивали смесью вода:ацетонитрил =15:85. Скорость потока составляла 1 мл/мин. Начальные условия поддерживали в течение 5 минут после инжекции, после чего проводили элюирование в течение 20 минут градиентом вода:ацетонитрил =50:50, а затем в течение 10 минут тем же самым растворителем. Систему промывали 20 минут смесью вода:ацетонитрил =80:20, а затем снова уравновешивали исходным растворителем.
Полученные площади пиков нормализовали на объем и массу муки. Ответный фактор ELSD на один мкг углевода снижался по мере увеличения DP, и, таким образом, более высокая DP мальтодекстринов соответствует их более высокому процентному содержанию в целом, чем это было определено по площади пика.
Относительные площади пиков для продуктов реакции со 100% амилазной мукой показаны на фигуре 17. Относительные площади пиков для продуктов реакций с 10% амилазной мукой показаны на фигуре 18.
Эти данные продемонстрировали, что различные смеси мальтодекстринов могут быть продуцированы в различные периоды времени нагревания. Уровень α-амилазной активности может варьироваться при смешивании трансгенной кукурузы, экспрессирующей α -амилазу, с кукурузой дикого типа, что приводит к изменению профиля мальтодекстрина.
Продукты реакций гидролиза, описанных в этом примере, могут концентрированы и очищены в целях приготовления пищевого продукта и для других целей, с использованием ряда хорошо известных методов, включая: центрифугирование, фильтрацию, ионный обмен, гель-фильтрацию, ультрафильтрацию, нанофильтрацию, обратный осмос, обесцвечивание частицами углерода, сушку распылением и другие стандартные методы, известные специалистам.
Пример 29
Влияние времени и температуры на продуцирование мальтодекстрина
Состав мальтодекстриновых продуктов при аутогидролизе зерна, содержащего термофильную α-амилазу, может быть изменен в зависимости от времени и температуры реакции.
В другом эксперименте амилазную муку получали, как описано выше в примере 28, и смешивали с водой в соотношении 300 мкл воды на 60 мг муки. Образцы инкубировали при 70°С, 80°С, 90°С или 100°С в течение периода времени вплоть до 90 минут. Реакции прекращали добавлением 900 мкл 50 мМ EDTA при 90°С, центрифугировали для удаления нерастворившегося материала и фильтровали через найлоновые 0,45 мкм-фильтры. Фильтраты анализировали с помощью ВЭЖХ, как описано в примере 28.
Результат этого анализа представлен на фигуре 19. Число DP по номенклатуре означает степень полимеризации. DP2 относится к мальтозе, DP3 относится к мальтотриозе и т.п. Мальтодекстрины с более высокой DP элюировались с одним пиком почти в конце элюирования и были помечены ">DP12". Этот агрегат включал декстрины, которые проходили через 0,45 мкм-фильтры и через предохранительную колонку, и не включал каких-либо очень крупных фрагментов крахмала, удерживаемых на фильтре или на предохранительной колонке.
Этот эксперимент продемонстрировал, что состав мальтодекстринового продукта может изменяться в зависимости от температуры и времени инкубирования, в результате чего может быть получен нужный мальтоолигосахаридный или мальтодекстриновый продукт.
Пример 30
Продуцирование мальтодекстрина
Состав мальтодекстриновых продуктов, полученных от трансгенной кукурузы, содержащей термофильную α-амилазу, может быть изменен путем добавления других ферментов, таких как α-глюкозидаза и ксилозоизомераза, а также путем включения солей в водную смесь муки перед термообработкой.
В другом эксперименте амилазную муку, полученную, как описано выше, смешивали с очищенной MalA и/или бактериальной ксилозоизомеразой, обозначенной BD8037. MalA S. sulfotaricus с 6His-меткой для очистки экспрессировали в E.coli. Клеточный лизат получали, как описано примере 28, а затем лизат очищали до видимой гомогенности с использованием смолы, обладающий аффинностью по отношению к никелю (Probond, Invitrogen), в соответствии с инструкциями производителей для очистки нативного белка. Ксилозоизомеразу ВD8037 получали в виде лиофилизованного порошка от Diversa и ресуспендировали в воде в объеме, составляющем 0,4 от исходного.
Амилазную кукурузную муку смешивали с ферментными растворами, содержащими воду или буфер. Все реакционные смеси содержали 60 мг амилазной муки и 600 мкл общего количества жидкости. Одну серию реакционных смесей забуферивали 50 мМ MOPS, рН 7,0, при комнатной температуре, с добавлением 10 мМ MgSO4 и 1 мМ CoCl2, а во второй серии реакционных смесей металлсодержащий буферный раствор заменяли водой. Все реакционные смеси инкубировали в течение 2 часов при 90°С. Фракции супернатанта реакции получали путем центрифугирования. Осадок промывали еще 600 мкл H2O и снова центрифугировали. Фракции супернатанта от каждой реакции объединяли, фильтровали через Centricon 10 и анализировали с помощью ВЭЖХ с ELSD-детекцией, как описано выше.
Результаты графически представлены на фигуре 20. На этой фигуре продемонстрировано, что экспрессированная в зерне амилаза 797GL3 может функционировать вместе с другими термофильными ферментами в присутствии или в отсутствие ионов металла, в результате чего из кукурузной муки при высокой температуре продуцируется ряд мальтодекстриновых смесей. В частности, включение глюкоамилазы или α-глюкозидазы может приводить к образованию продукта с еще большим содержанием глюкозы и других продуктов с низкой DP. Включение фермента с глюкозоизомеразной активностью приводит к получению продукта, который содержит фруктозу, а поэтому должен быть более сладким, чем продукт, продуцируемый только амилазой или амилазой с α-глюкозидазной активностью. Кроме того, эти данные показали, что содержание мальтоолигосахаридов с DP5, DP6 и DP7 может быть увеличено путем включения двухвалентных катионных солей, таких как CoCl2 и MgSO4.
Другими способами изменения состава мальтодекстринов, продуцированных посредством описанной здесь реакции, являются: изменение рН реакции, изменение типа крахмала в трансгенном или не-трансгенном зерне, изменение соотношения твердых веществ или добавление органических растворителей.
Пример 31
Получение декстринов или сахаров из зерна, не подвергнутого механическому разрушению до выделения крахмалопродуктов
Сахара и мальтодекстрины получали посредством контактирования трансгенного зерна, экспрессирующего α -амилазу, 797GL3, с водой и нагревания до 90°С в течение ночи (>14 часов). Затем жидкость отделяли от зерна путем фильтрации. Жидкий продукт анализировали посредством ВЭЖХ методом, описанным в примере 15. В таблице 6 представлен профиль детектируемых продуктов.
Эти данные продемонстрировали, что сахара и мальтодекстрины могут быть получены путем контактирования интактного зерна, экспрессирующего α-амилазу, с водой и нагревания. Затем полученные продукты могут быть отделены от интактного зерна либо путем фильтрации или центрифугирования, либо путем гравитационного осаждения.
Пример 32
Ферментация исходного крахмала в кукурузе, экспрессирующей глюкоамилазу Rhizopus oryzae
Зерно трансгенной кукурузы собирали от трансгенных растений, полученных, как описано в примере 29. Зерно измельчали в муку. Зерно кукурузы экспрессирует белок, который содержит активный фрагмент глюкоамилазы Rhizopus oryzae (последовательность SEQ ID NO:49), нацеливаемый на эндоплазматический ретикулум.
Зерно кукурузы измельчали в муку, как описано в примере 15. Затем получали биомассу, содержащую 20 г кукурузной муки, 23 мл деионизованной воды и 6,0 мл барды (8% твердых веществ по массе). рН доводили до 6,0 добавлением гидроксида аммония. К указанной биомассе добавляли следующие компоненты: протеазу (0,60 мл 1000-кратного разведения коммерчески доступной протеазы), 0,2 мг лактоцида и мочевины (0,85 мл 10-кратного разведения 50% раствора мочевины). В крышке 100 мл-сосуда, содержащего указанную биомассу, делали отверстие для выпуска CO2 в целях вентиляции. Затем биомассу инокулировали дрожжами (1,44 мл) и инкубировали в водяной бане при 90°F. После 24-часовой ферментации температуру понижали до 86°F, а через 48 часов ее доводили до 82°F.
Дрожжи для инокуляции размножали, как описано в примере 14.
Образцы брали, как описано в примере 14, а затем анализировали методами, описанными в примере 14.
Пример 33
Пример ферментации исходного крахмала в кукурузе, экспрессирующей глюкоамилазу Rhizopus oryzae
Зерно трансгенной кукурузы собирали от трансгенных растений, полученных, как описано в примере 28. Зерно измельчали в муку. Зерно кукурузы экспрессирует белок, который содержит активный фрагмент глюкоамилазы Rhizopus oryzae (последовательность SEQ ID NO:49), нацеливаемый на эндоплазматический ретикулум.
Зерно кукурузы измельчали в муку, как описано в примере 15. Затем получали биомассу, содержащую 20 г кукурузной муки, 23 мл деионизованной воды и 6,0 мл барды (8% твердых веществ по массе). рН доводили до 6, 0 добавлением гидроксида аммония. К указанной биомассе добавляли следующие компоненты: протеазу (0,60 мл 1000-кратного разведения коммерчески доступной протеазы), 0,2 мг лактоцида и мочевины (0,85 мл 10-кратного разведения 50% раствора мочевины). В крышке 100 мл-сосуда, содержащего указанную биомассу, делали отверстие для выпуска CO2 в целях вентиляции. Затем указанную биомассу инокулировали дрожжами (1,44 мл) и инкубировали в водяной бане при 90°F. После 24-часовой ферментации температуру понижали до 86°F, а через 48 часов ее доводили до 82°F.
Дрожжи для инокуляции размножали, как описано в примере 14.
Образцы брали, как описано в примере 14, а затем анализировали методами, описанными в примере 14.
Пример 34
Ферментация исходного крахмала в цельном зерне кукурузы, экспрессирующей глюкоамилазу Rhizopus oryzae с добавлением экзогенной α-амилазы
Зерно трансгенной кукурузы собирали от трансгенных растений, полученных, как описано в примере 28. Зерно кукурузы экспрессирует белок, который содержит активный фрагмент глюкоамилазы Rhizopus oryzae (последовательность SEQ ID NO:49), нацеливаемый на эндоплазматический ретикулум.
Зерно кукурузы подвергали контакту с 20 г кукурузной муки, 23 мл деионизованной воды и 6,0 мл барды (8% твердых веществ по массе). рН доводили до 6,0 добавлением гидроксида аммония. Затем добавляли следующие компоненты: α-амилазу ячменя, закупленную у Sigma (2 мг), протеазу (0,60 мл 1000-кратного разведения коммерчески доступной протеазы), 0,2 мг лактоцида и мочевины (0,85 мл 10-кратного разведения 50% раствора мочевины). В крышке 100 мл-сосуда, содержащего указанную биомассу, делали отверстие для выпуска CO2 в целях вентиляции. Затем смесь инокулировали дрожжами (1,44 мл) и инкубировали в водяной бане при 90°F. После 24-часовой ферментации, температуру понижали до 86°F, а через 48 часов ее доводили до 82°F.
Дрожжи для инокуляции размножали, как описано в примере 14.
Образцы брали, как описано в примере 14, а затем анализировали методами, описанными в примере 14.
Пример 35
Ферментация исходного крахмала в кукурузе, экспрессирующей глюкоамилазу Rhizopus oryzae и амилазу Zea mays
Зерно трансгенной кукурузы собирали от трансгенных растений, полученных, как описано в примере 28. Зерно кукурузы экспрессирует белок, который содержит активный фрагмент глюкоамилазы Rhizopus oryzae (последовательность SEQ ID NO:49), нацеливаемый на эндоплазматический ретикулум. Зерно также экспрессирует амилазу кукурузы с доменом, связывающим исходный крахмал, как описано в примере 28.
Зерно кукурузы измельчали в муку, как описано в примере 14. Затем получали биомассу, содержащую 20 г кукурузной муки, 23 мл деионизованной воды и 6,0 мл барды (8% твердых веществ по массе). рН доводили до 6,0 добавлением гидроксида аммония. К указанной биомассе добавляли следующие компоненты: протеазу (0,60 мл 1000-кратного разведения коммерчески доступной протеазы), 0, 2 мг лактоцида и мочевины (0,85 мл 10-кратного разведения 50% раствора мочевины). В крышке 100 мл-сосуда, содержащего указанную биомассу, делали отверстие для выпуска CO2 в целях вентиляции. Затем биомассу инокулировали дрожжами (1,44 мл) и инкубировали в водяной бане при 90°F. После 24-часовой ферментации температуру понижали до 86°F, а через 48 часов ее доводили до 82°F.
Дрожжи для инокуляции размножали, как описано в примере 14.
Образцы брали, как описано в примере 14, а затем анализировали методами, описанными в примере 14.
Пример 36
Ферментация исходного крахмала в кукурузе, экспрессирующей глюкоамилазу Thermoanaerobacter thermosaccharolyticum
Зерно трансгенной кукурузы собирали от трансгенных растений, полученных, как описано в примере 28. Зерно кукурузы экспрессирует белок, который содержит активный фрагмент глюкоамилазы Thermoanaerobacter thermosaccharolyticum (последовательность SEQ ID NO:47), нацеливаемый на эндоплазматический ретикулум.
Зерно кукурузы измельчали в муку, как описано в примере 15. Затем получали биомассу, содержащую 20 г кукурузной муки, 23 мл деионизованной воды и 6,0 мл барды (8% твердых веществ по массе). рН доводили до 6,0 добавлением гидроксида аммония. К биомассе добавляли следующие компоненты: протеазу (0,60 мл 1000-кратного разведения коммерчески доступной протеазы), 0,2 мг лактоцида и мочевины (0,85 мл 10-кратного разведения 50% раствора мочевины). В крышке 100 мл-сосуда, содержащего указанную биомассу, делали отверстие для выпуска CO2 в целях вентиляции. Затем биомассу инокулировали дрожжами (1,44 мл) и инкубировали в водяной бане при 90°F. После 24-часовой ферментации температуру понижали до 86°F, а через 48 часов ее доводили до 82°F.
Дрожжи для инокуляции размножали, как описано в примере 14.
Образцы брали, как описано в примере 14, а затем анализировали методами, описанными в примере 14.
Пример 37
Ферментация исходного крахмала в кукурузе, экспрессирующей глюкоамилазу Aspergillus niger
Зерно трансгенной кукурузы собирали от трансгенных растений, полученных, как описано в примере 28. Зерно кукурузы экспрессирует белок, который содержит активный фрагмент глюкоамилазы Aspergillus niger (Fiil, N.P. "Glucoamylases G1 and G2 from Aspergillus niger are synthesized from two different but closely related mRNAs" EMBO J. 3(5), 1097-1102 (1984), рег. №Р04064). Нуклеиновая кислота, оптимизированная по кодонам кукурузы и кодирующая глюкоамилазу, имеет последовательность SEQ ID NO:59 и нацеливается на эндоплазматический ретикулум.
Зерно кукурузы измельчали в муку, как описано в примере 14. Затем получали биомассу, содержащую 20 г кукурузной муки, 23 мл деионизованной воды и 6,0 мл барды (8% твердых веществ по массе). рН доводили до 6,0 добавлением гидроксида аммония. К биомассе добавляли следующие компоненты: протеазу (0,60 мл 1000-кратного разведения коммерчески доступной протеазы), 0,2 мг лактоцида и мочевины (0, 85 мл 10-кратного разведения 50% раствора мочевины). В крышке 100 мл-сосуда, содержащего биомассу, делали отверстие для выпуска CO2 в целях вентиляции. Затем биомассу инокулировали дрожжами (1,44 мл) и инкубировали в водяной бане при 90°F. После 24-часовой ферментации температуру понижали до 86°F, а через 48 часов ее доводили до 82°F.
Дрожжи для инокуляции размножали, как описано в примере 14.
Образцы брали, как описано в примере 14, а затем анализировали методами, описанными в примере 14.
Пример 38
Пример ферментации исходного крахмала в кукурузе, экспрессирующей глюкоамилазу Aspergillus niger и амилазу Zea mays
Зерно трансгенной кукурузы собирали от трансгенных растений, полученных, как описано в примере 28. Зерно кукурузы экспрессирует белок, который содержит активный фрагмент глюкоамилазы Aspergillus niger (Fiil, N.P. "Glucoamylases G1 and G2 from Aspergillus niger are synthesized from two different but closely related mRNAs" EMBO J. 3(5), 1097-1102 (1984), рег. №Р04064) (SEQ ID NO:59, нуклеиновая кислота, оптимизированная по кодонам кукурузы) и нацеливается на эндоплазматический ретикулум. Зерно также экспрессирует амилазу кукурузы с доменом, связывающим исходный крахмал, как описано в примере 28.
Зерно кукурузы измельчали в муку, как описано в примере 14. Затем получали биомассу, содержащую 20 г кукурузной муки, 23 мл деионизованной воды и 6,0 мл барды (8% твердых веществ по массе). рН доводили до 6,0 добавлением гидроксида аммония. К биомассе добавляли следующие компоненты: протеазу (0,60 мл 1000-кратного разведения коммерчески доступной протеазы), 0,2 мг лактоцида и мочевины (0,85 мл 10-кратного разведения 50% раствора мочевины). В крышке 100 мл-сосуда, содержащего указанную биомассу, делали отверстие для выпуска CO2 в целях вентиляции. Затем биомассу инокулировали дрожжами (1,44 мл) и инкубировали в водяной бане при 90°F. После 24-часовой ферментации температуру понижали до 86°F, а через 48 часов ее доводили до 82°F.
Дрожжи для инокуляции размножали, как описано в примере 14.
Образцы брали, как описано в примере 14, а затем анализировали методами, описанными в примере 14.
Пример 39
Ферментация исходного крахмала в кукурузе, экспрессирующей глюкоамилазу Thermoanaerobacter thermosaccharolyticum и амилазу ячменя
Зерно трансгенной кукурузы собирали от трансгенных растений, полученных, как описано в примере 28. Зерно кукурузы экспрессирует белок, который содержит активный фрагмент глюкоамилазы Thermoanaerobacter thermosaccharolyticum (последовательность SEQ ID NO:47), нацеливаемый на эндоплазматический ретикулум. Зерно также экспрессирует ген amyl амилазы ячменя с низкой pI (Rogers, J.C. & Milliman, C. "Isolation and sequence analysis of a barley alpha-amylase cDNA clone" J. Biol. Chem. 258(13), 8169-8174 (1983)), модифицированный для направленной экспрессии указанного белка в эндоплазматический ретикулум.
Зерно кукурузы измельчали в муку, как описано в примере 14. Затем получали биомассу, содержащую 20 г кукурузной муки, 23 мл деионизованной воды и 6,0 мл барды (8% твердых веществ по массе). рН доводили до 6,0 добавлением гидроксида аммония. К биомассе добавляли следующие компоненты: протеазу (0,60 мл 1000-кратного разведения коммерчески доступной протеазы), 0,2 мг лактоцида и мочевины (0,85 мл 10-кратного разведения 50% раствора мочевины). В крышке 100 мл-сосуда, содержащего указанную биомассу, делали отверстие для выпуска CO2 в целях вентиляции. Затем биомассу инокулировали дрожжами (1,44 мл) и инкубировали в водяной бане при 90°F. После 24-часовой ферментации температуру понижали до 86°F, а через 48 часов ее доводили до 82°F.
Дрожжи для инокуляции размножали, как описано в примере 14.
Образцы брали, как описано в примере 14, а затем анализировали методами, описанными в примере 14.
Пример 40
Ферментация исходного крахмала в цельных зернах кукурузы, экспрессирующей глюкоамилазу Thermoanaerobacter thermosaccharolyticum и амилазу ячменя
Зерно трансгенной кукурузы собирали от трансгенных растений, полученных, как описано в примере 28. Зерно кукурузы экспрессирует белок, который содержит активный фрагмент глюкоамилазы Thermoanaerobacter thermosaccharolyticum (последовательность SEQ ID NO:47), нацеливаемый на эндоплазматический ретикулум. Зерно также экспрессирует ген amyl амилазы ячменя с низкой pI (Rogers, J.C. & Milliman, C. "Isolation and sequence analysis of a barley alpha-amylase cDNA clone" J. Biol. Chem. 258(13), 8169-8174 (1983)), модифицированный для направленной экспрессии белка в эндоплазматический ретикулум.
Зерно кукурузы подвергали контактированию с 20 г кукурузной муки, 23 мл деионизованной воды и 6,0 мл барды (8% твердых веществ по массе). рН доводили до 6,0 добавлением гидроксида аммония. К смеси добавляли следующие компоненты: протеазу (0,60 мл 1000-кратного разведения коммерчески доступной протеазы), и 0,2 мг лактоцида и мочевины (0,85 мл 10-кратного разведения 50% раствора мочевины). В крышке 100 мл-сосуда, содержащего биомассу, делали отверстие для выпуска CO2 в целях вентиляции. Затем смесь инокулировали дрожжами (1,44 мл) и инкубировали в водяной бане при 90°F. После 24-часовой ферментации температуру понижали до 86°F, а через 48 часов ее доводили до 82°F.
Дрожжи для инокуляции размножали, как описано в примере 14.
Образцы брали, как описано в примере 14, а затем анализировали методами, описанными в примере 14.
Пример 41
Ферментация исходного крахмала в кукурузе, экспрессирующей гибрид альфа-амилазы и глюкоамилазы
Зерно трансгенной кукурузы собирали от трансгенных растений, полученных, как описано в примере 28. Зерно кукурузы экспрессирует оптимизированный по кодонам кукурузы полинуклеотид, представленный в SEQ ID NO:46 и кодирующий гибрид альфа-амилазы и глюкоамилазы, представленный в SEQ ID NO:45, которые нацелены на эндоплазматический ретикулум.
Зерно кукурузы измельчали в муку, как описано в примере 14. Затем получали биомассу, содержащую 20 г кукурузной муки, 23 мл деионизованной воды и 6,0 мл барды (8% твердых веществ по массе). рН доводили до 6,0 добавлением гидроксида аммония. К биомассе добавляли следующие компоненты: протеазу (0,60 мл 1000-кратного разведения коммерчески доступной протеазы), 0,2 мг лактоцида и мочевины (0,85 мл 10-кратного разведения 50% раствора мочевины). В крышке 100 мл-сосуда, содержащего биомассу, делали отверстие для выпуска CO2 в целях вентиляции. Затем указанную биомассу инокулировали дрожжами (1,44 мл) и инкубировали в водяной бане при 90°F. После 24-часовой ферментации температуру понижали до 86°F, а через 48 часов ее доводили до 82°F.
Дрожжи для инокуляции размножали, как описано в примере 14.
Образцы брали, как описано в примере 14, а затем анализировали методами, описанными в примере 14.
Все публикации, патенты и патентные заявки вводятся в настоящее описание посредством ссылки. Хотя в вышеприведенном описании настоящего изобретения описаны некоторые предпочтительные варианты его осуществления и представлено много конкретных вариантов, имеющих иллюстративный характер, однако очевидно, что в настоящее изобретение могут быть внесены дополнительные варианты его осуществления, и что некоторые конкретные варианты могут быть модифицированы, если только это не выходит за рамки основной идеи настоящего изобретения.
Аминокислотная последовательность α-амилазы 797GL3 (SEQ ID NO:1)































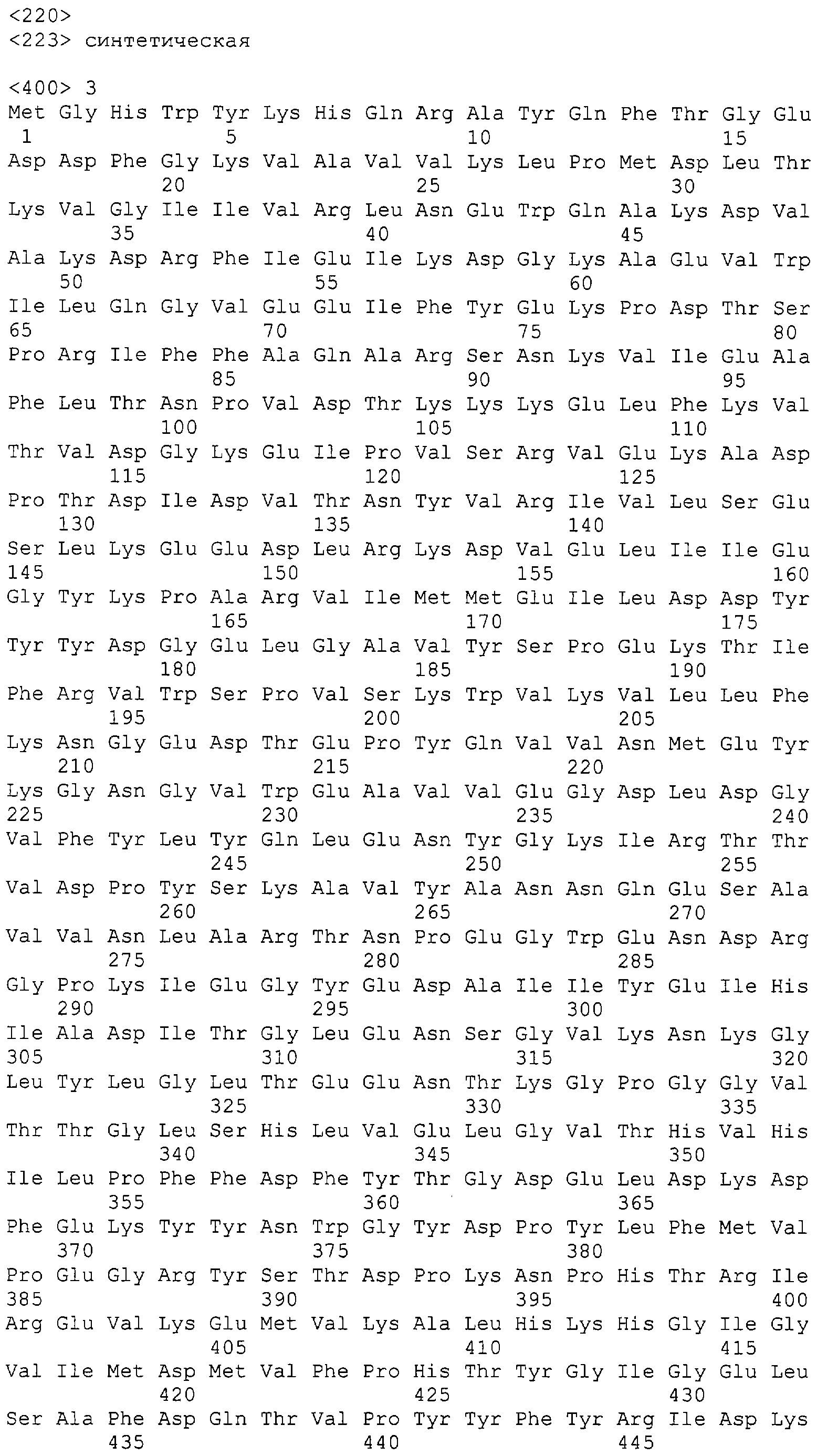


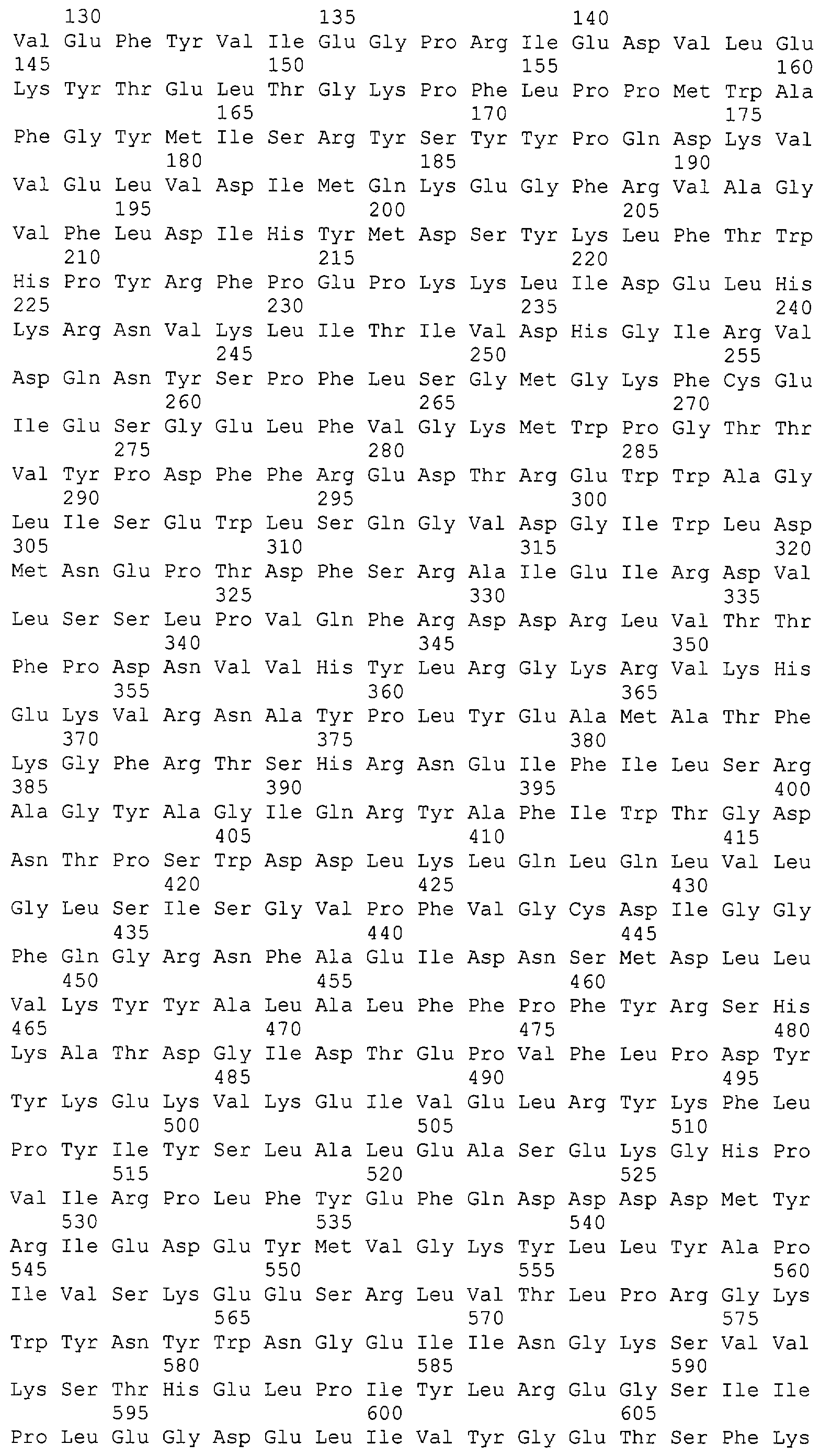
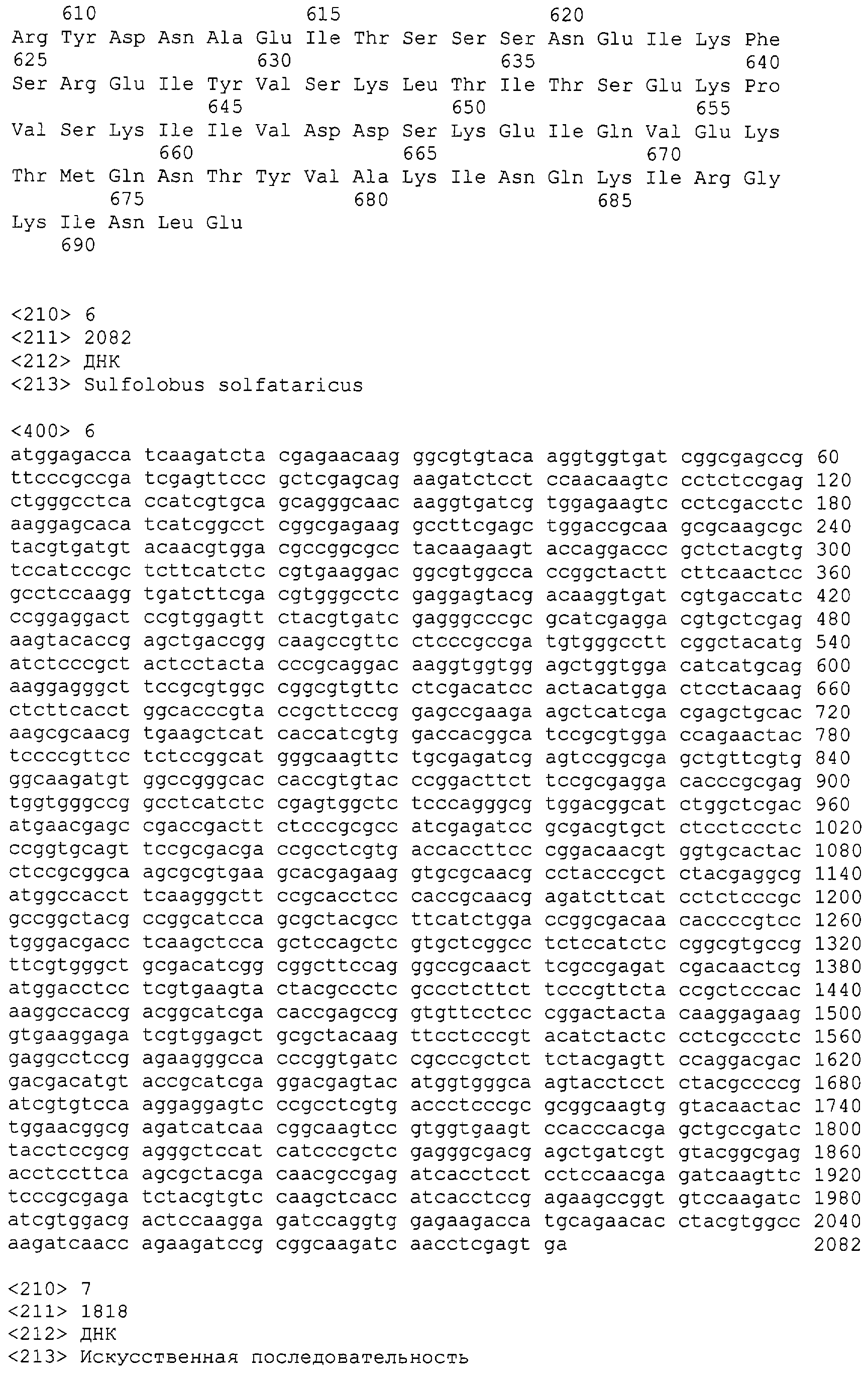





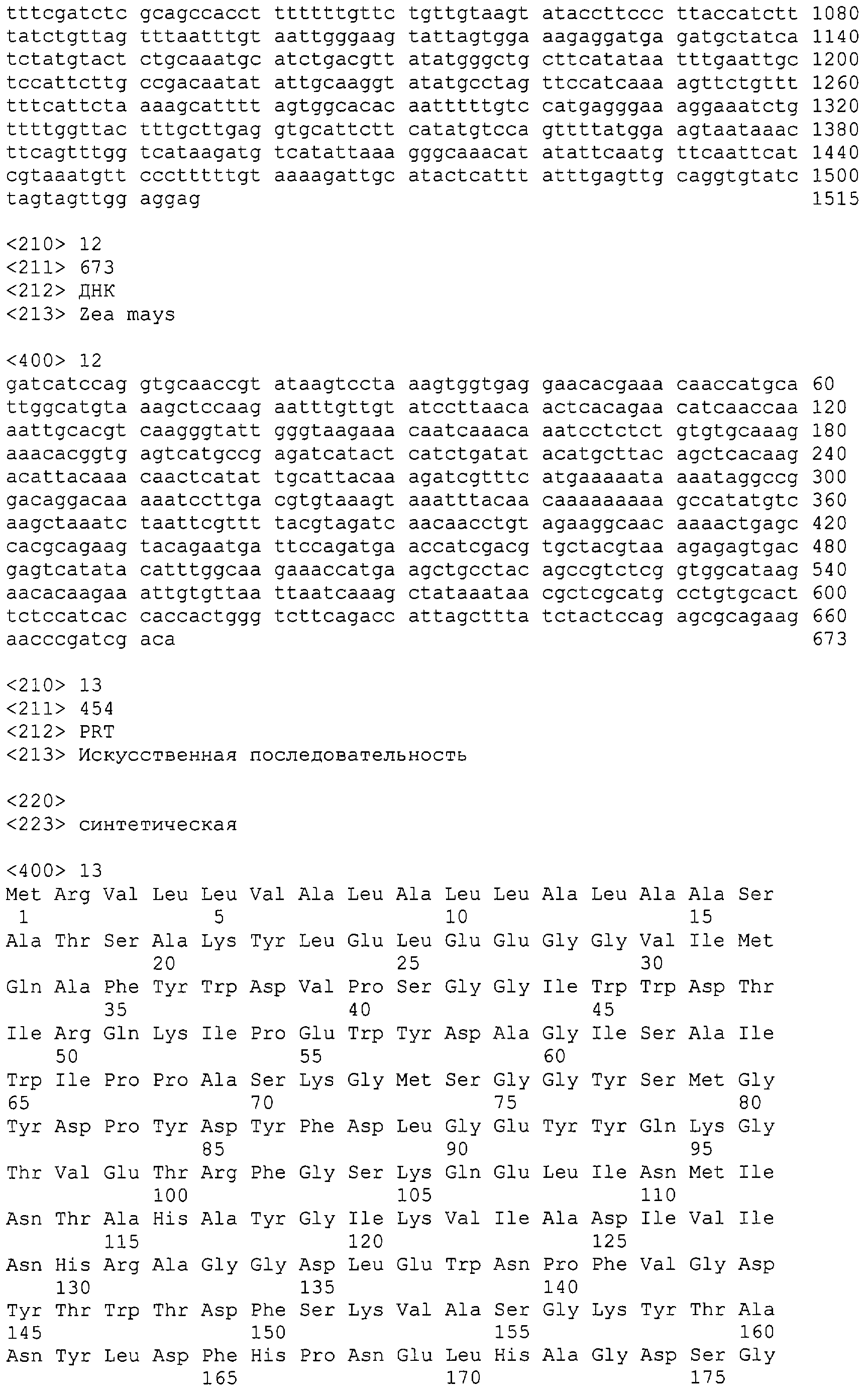

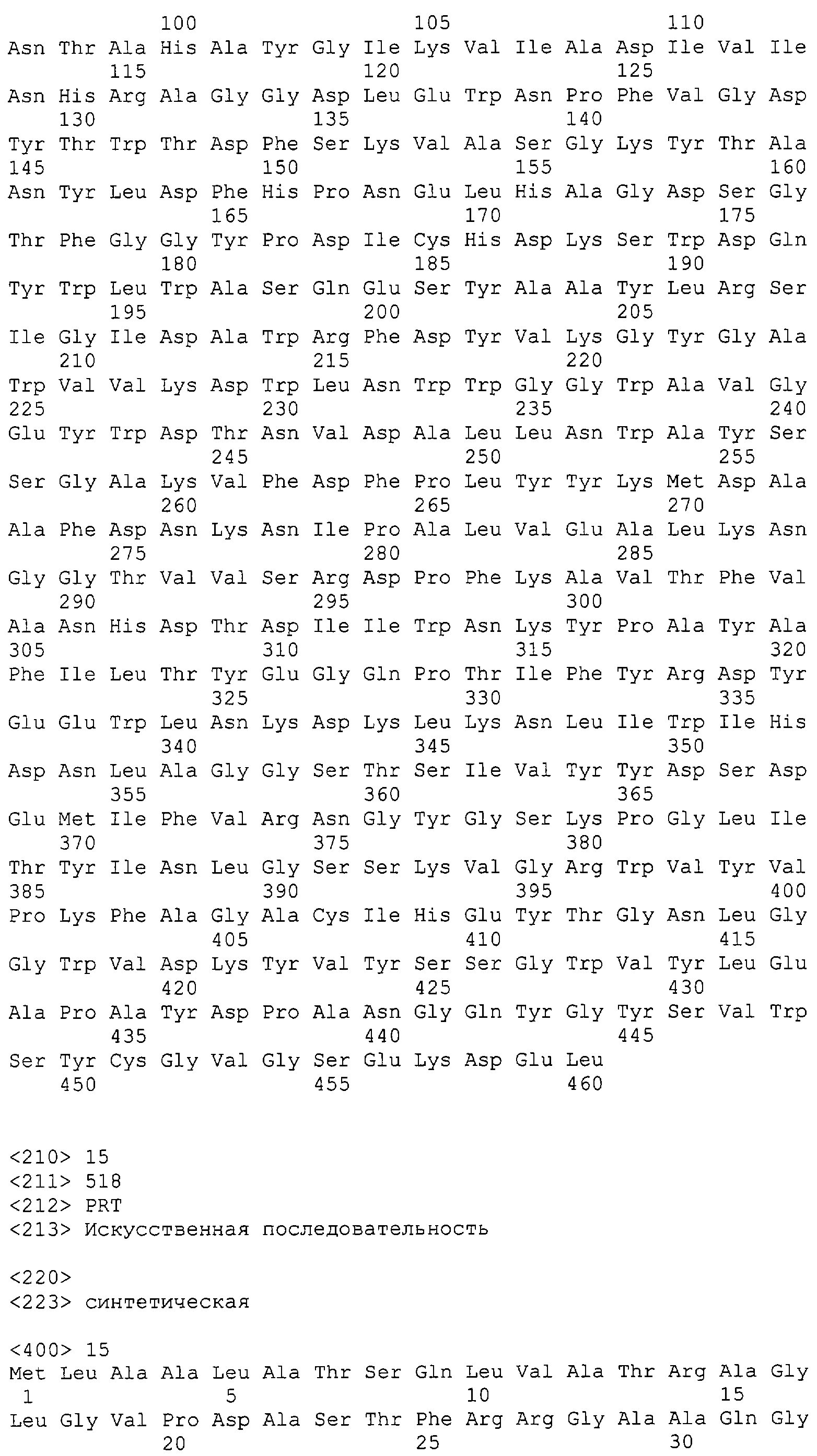









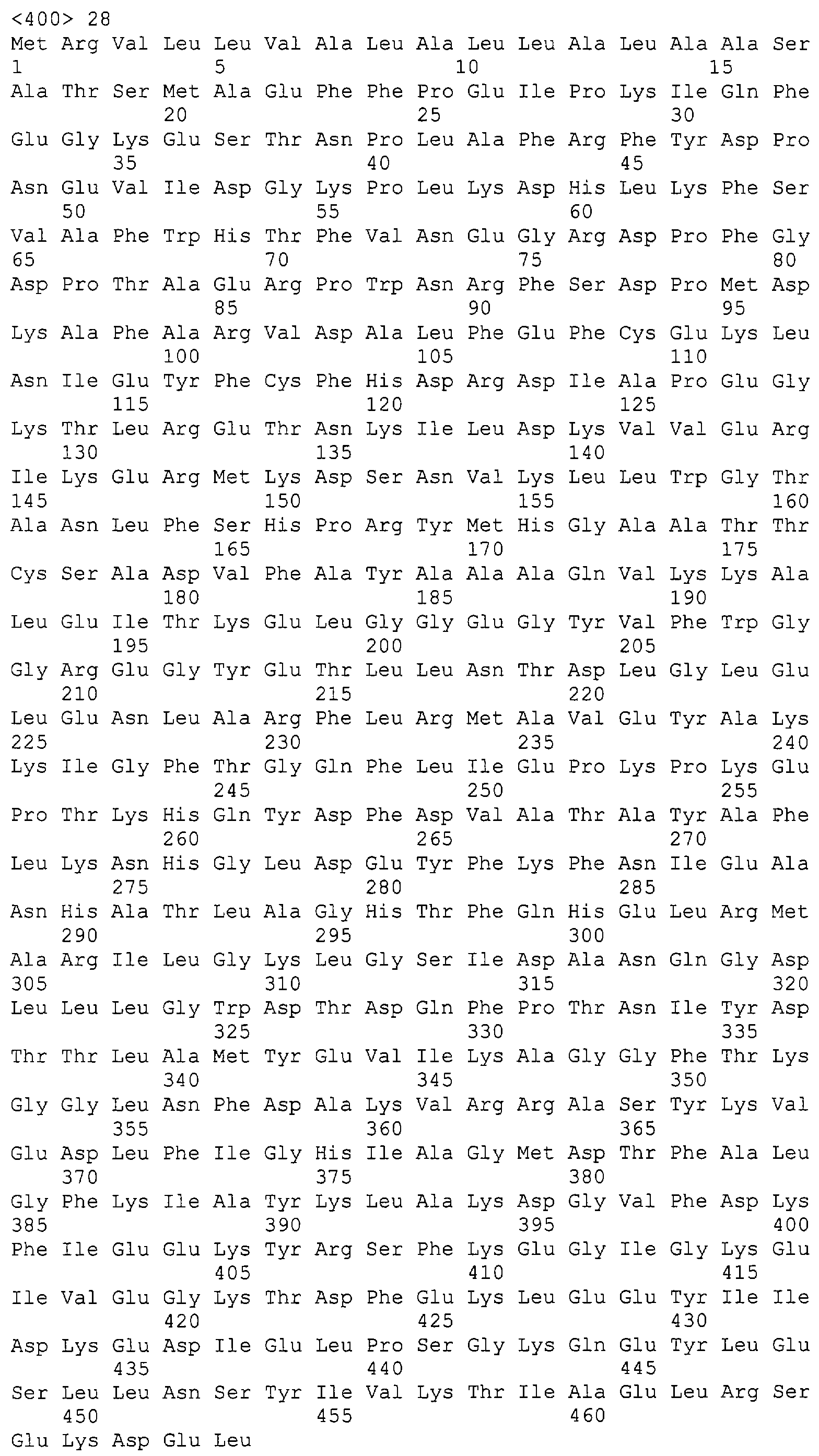
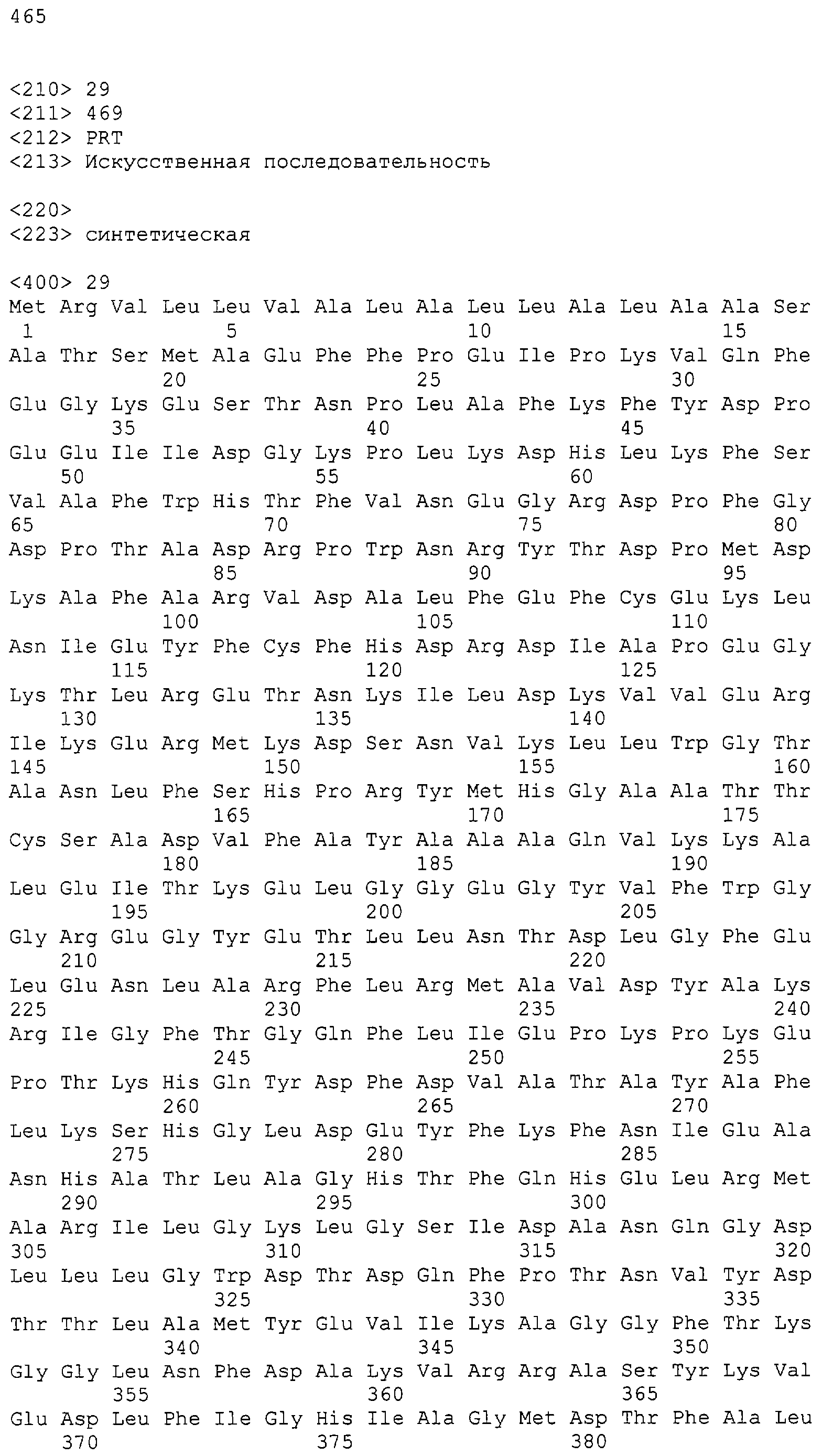





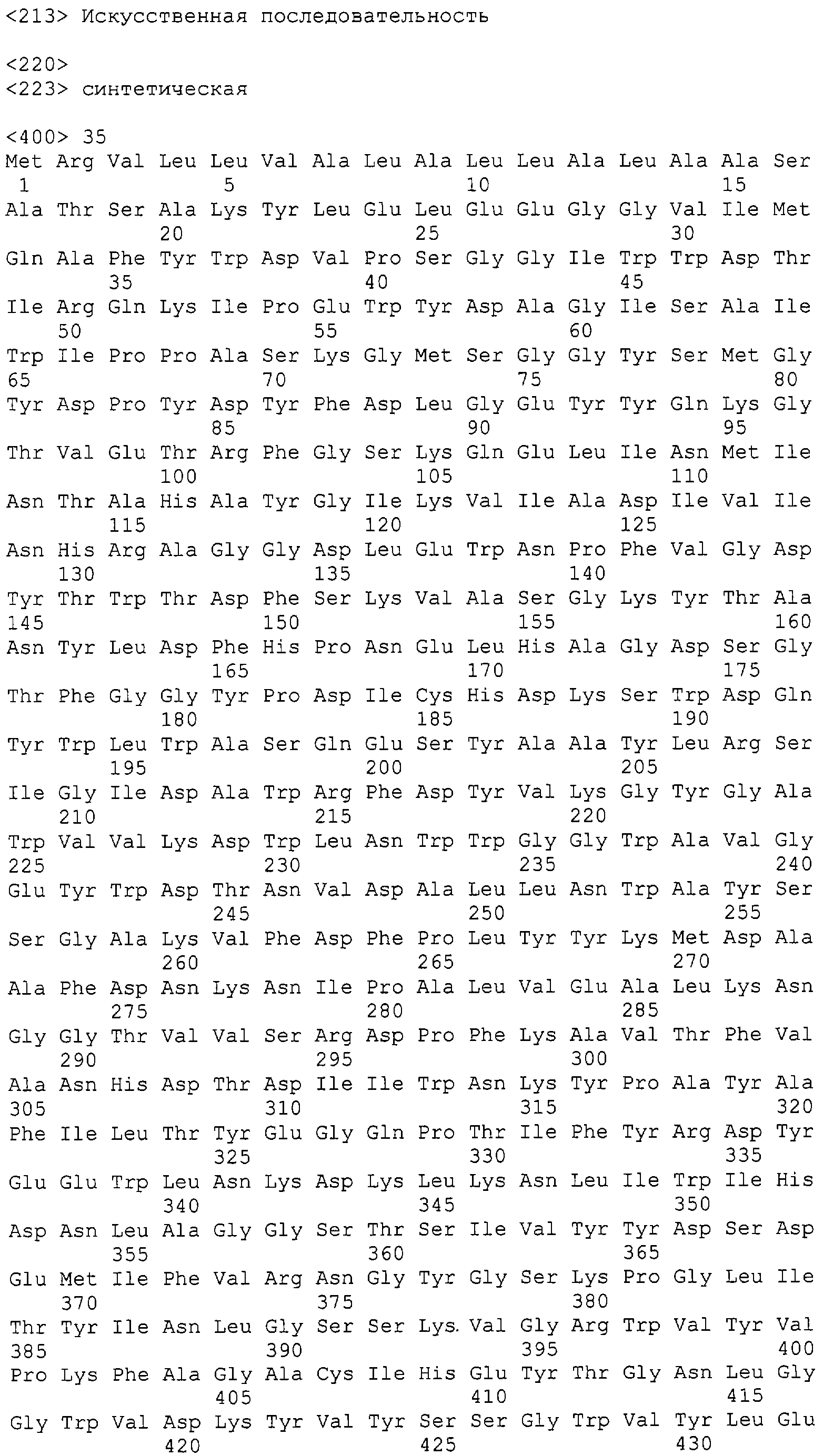
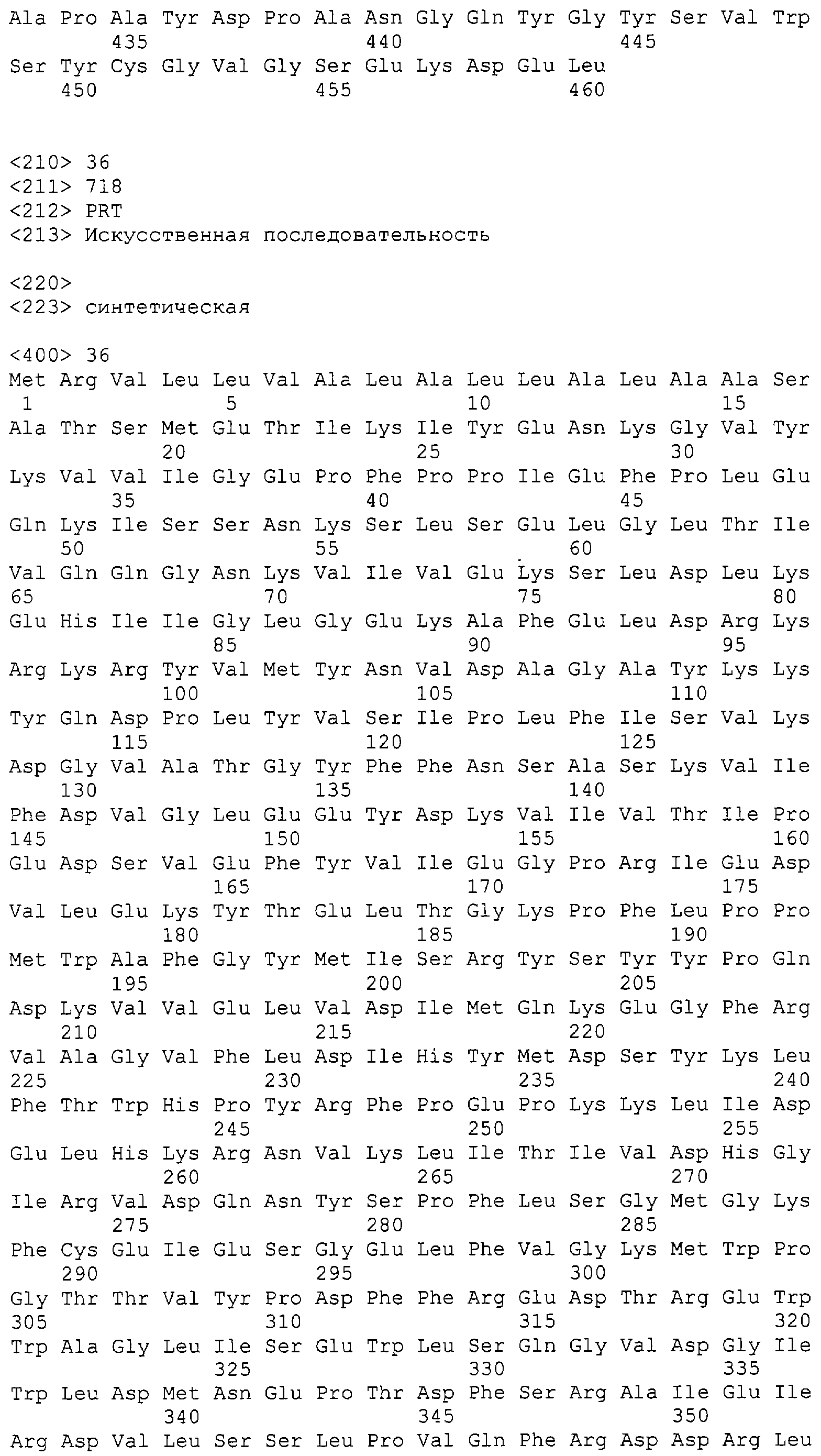




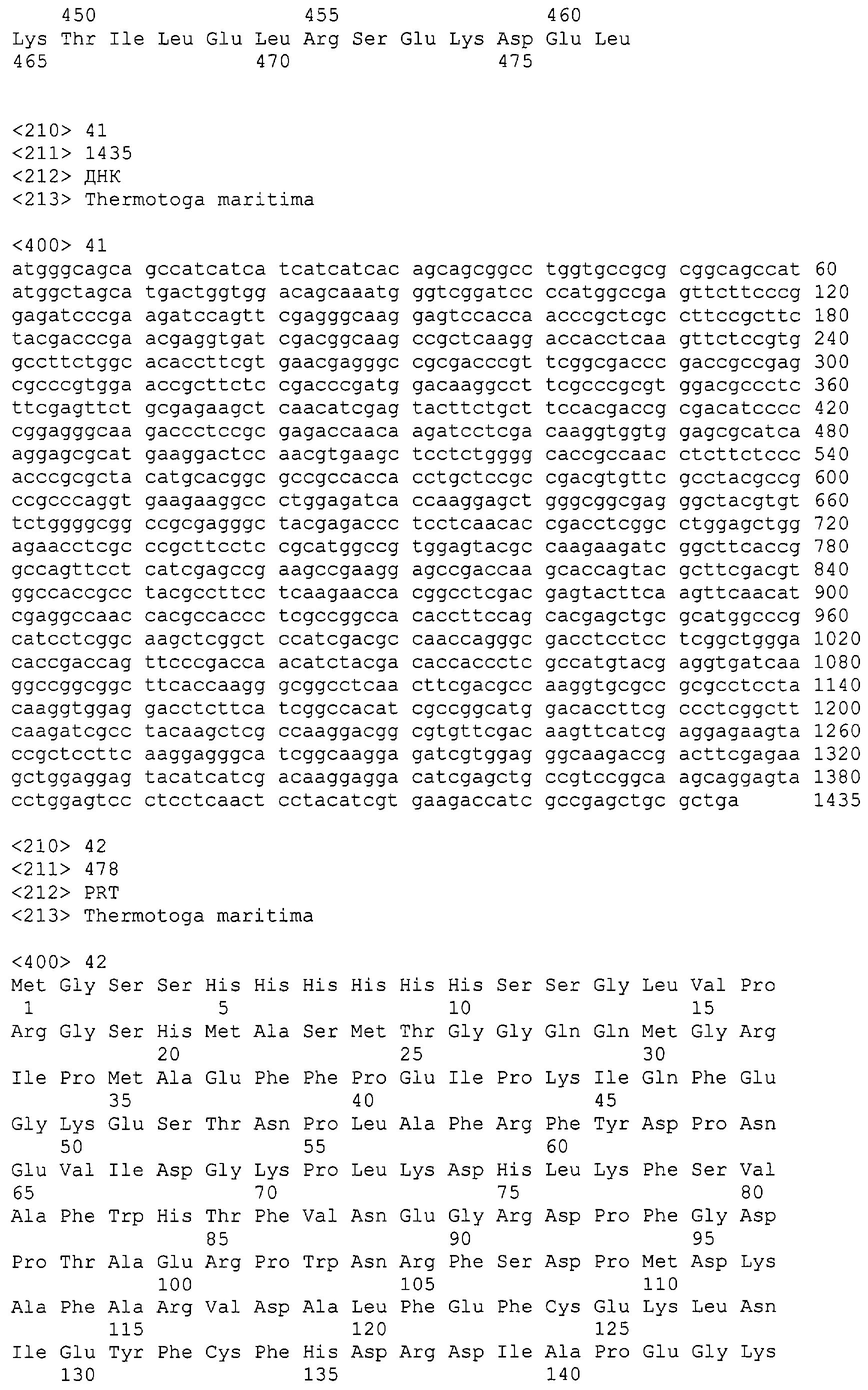


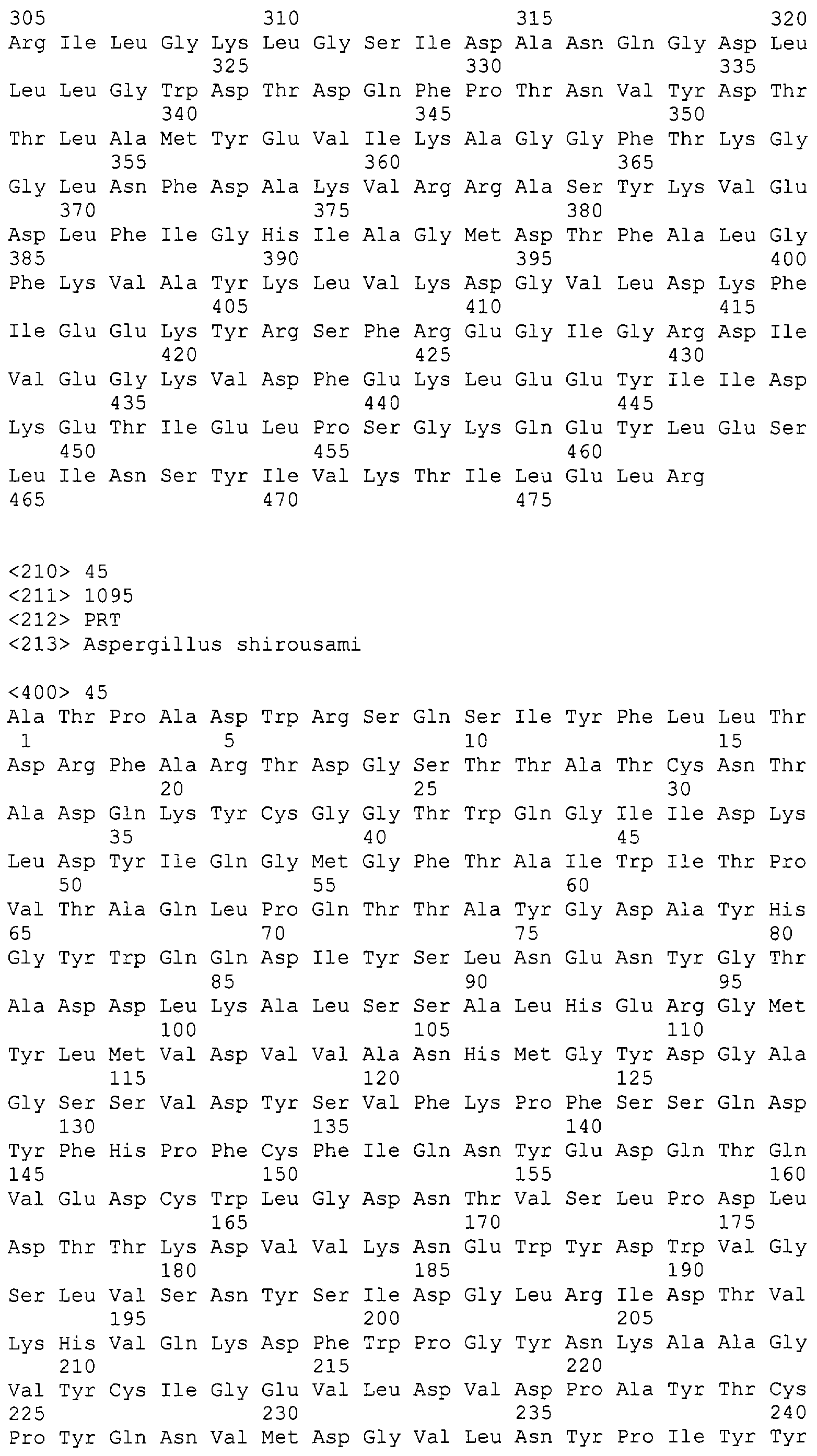
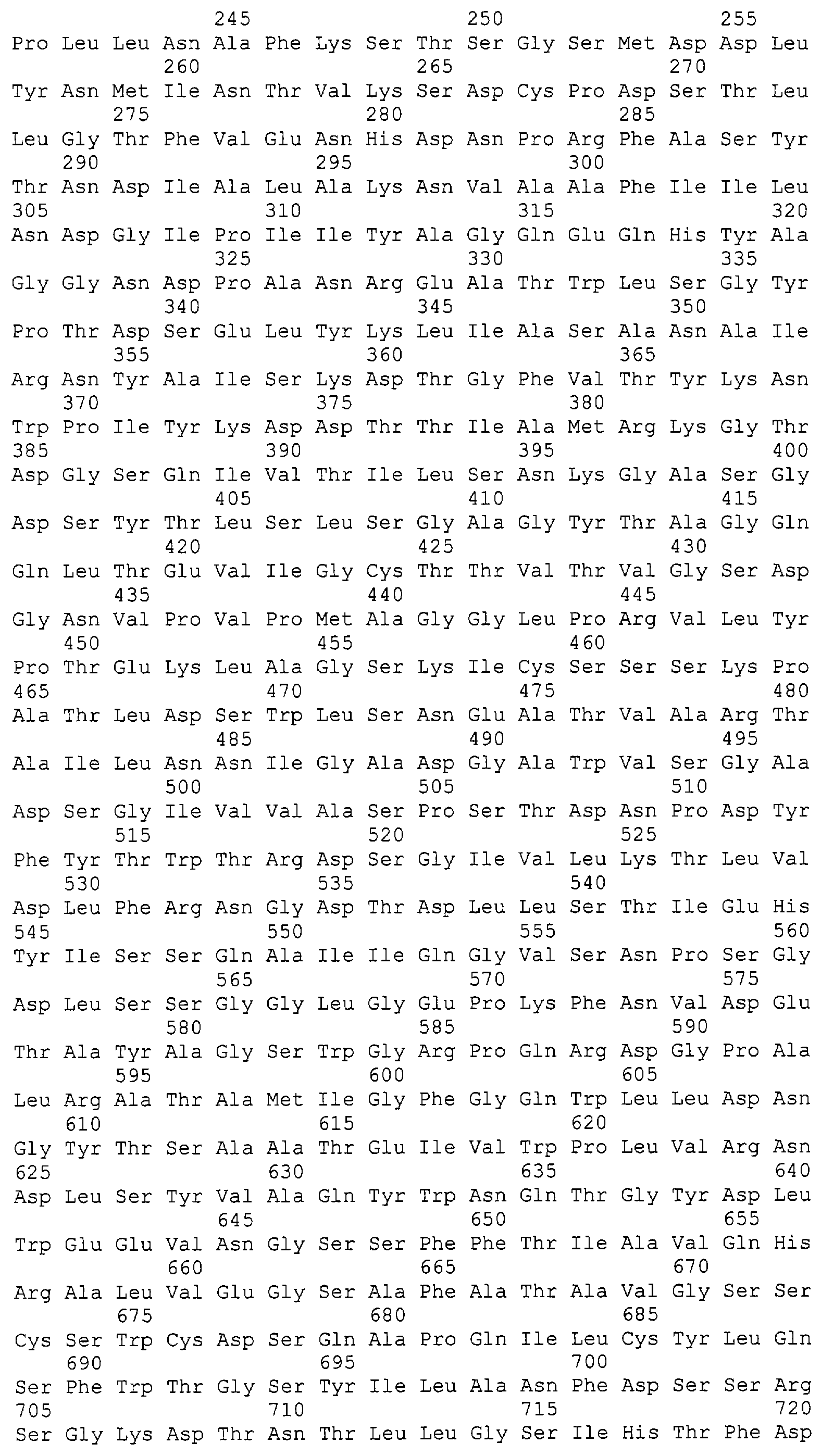


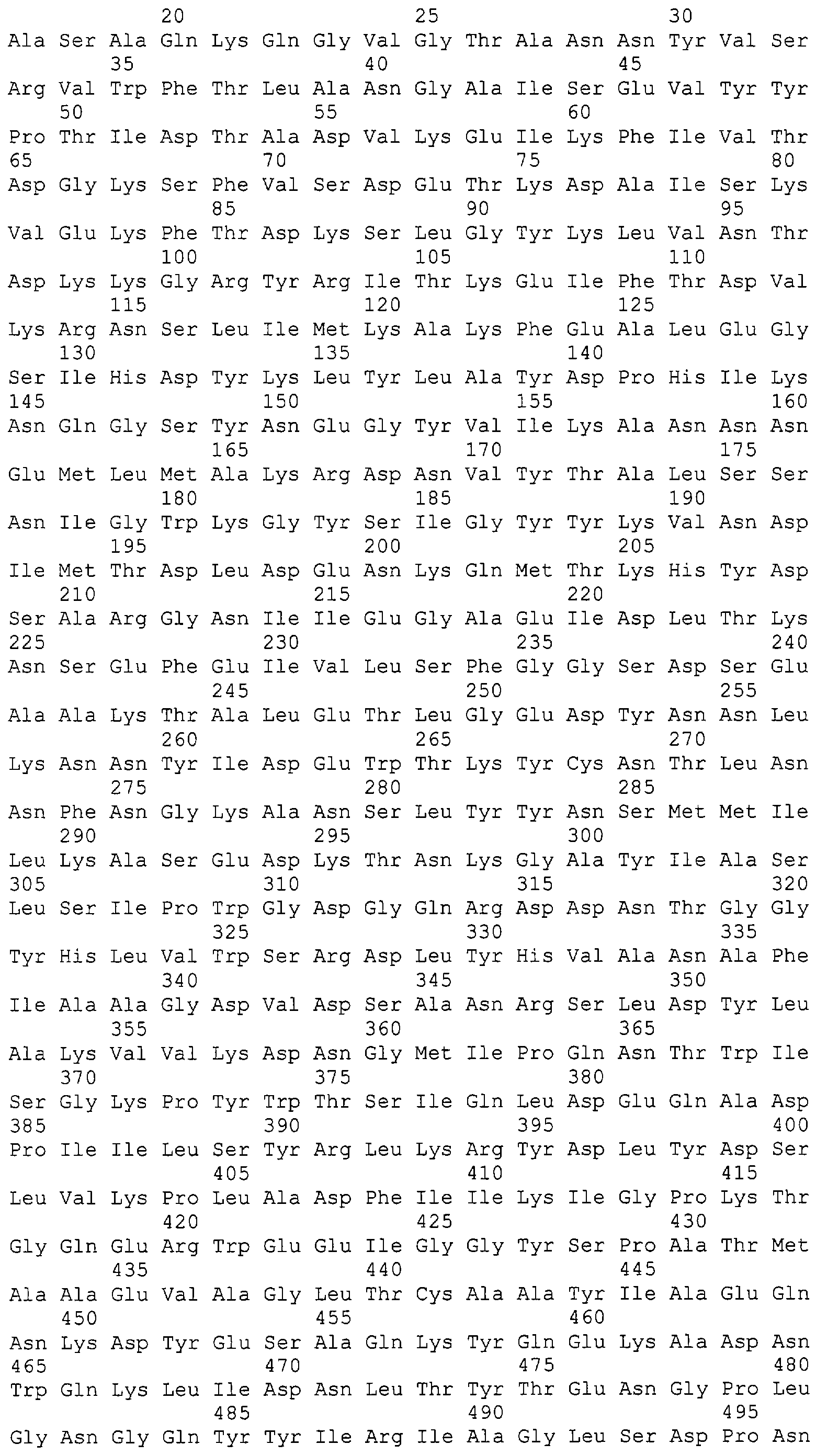







Реферат
Изобретение относится к полинуклеотидам, оптимизированным для экспрессии в растениях, кодирующим процессирующие ферменты. Полинуклеотиды кодируют мезофильные, термофильные или гипертермофильные процессирующие ферменты, которые активируются в условиях, подходящих для взаимодействия с нужным субстратом. Введение этих полинуклеотидов в растительный геном приводит к получению «самопроцессирующихся» трансгенных растений, части которых, например зерно, плод или семя, экспрессируют один или более указанных ферментов и имеют измененный состав. Аутопроцессирующиеся растения могут быть использованы, например, для получения пищевых продуктов, имеющих улучшенный вкус. 14 н. и 15 з.п. ф-лы, 23 ил., 6 табл.

Комментарии