Способ детоксикации микотоксинов - RU2347810C2
Код документа: RU2347810C2
Чертежи





Описание
Настоящее изобретение относится к способу детоксикации микотоксинов, а также к способу продукции производных микотоксинов.
Целый комплекс близкородственных видов из рода Fusarium отвечает за деструктивные и экономически очень значимые болезни злаковых культур (фузариоз колосьев злаковых, FHB, поражающий пшеницу и ячмень) и кукурузы (гниль колосьев, вызываемая Fusarium). В те годы, когда климатические условия благоприятствуют развитию грибов, инфекции, вызываемые представителями рода Fusarium, могут достичь эпидемических показателей (1). Болезни, вызываемые растительными патогенами, не только существенно снижают урожай, но также приводят к контаминации зерна неприемлемо высокими количествами микотоксинов, создавая проблему глобального значения. Класс токсинов, которые вызывают особую озабоченность в контексте здоровья человека и животного, включает трихотеценам, эпоксиды сесквитерпеноидов, которые являются мощными ингибиторами синтеза белка у эукариотов. Более 180 соединений этого класса было выделено из природных источников, преимущественно из видов Fusarium (2). В зависимости от концентрации и от характера замещения трихотецена предпочтительно ингибируется инициация, элонгация или терминация транскрипции (3).
F. graminearum и F. culmorum представляют собой два наиболее важных агента, вызывающих FHB злаковых. В то время, как линии F. graminearum, распространенные в Европе и Северной Америке, продуцируют преимущественно дезоксиниваленол (DON) и его ацетилированные производные 3-ацетил-дезоксиниваленол (3-ADON) и 15-ацетил-дезоксиниваленол (15-DON), продуценты ниваленола (NIV), который содержит одну дополнительную гидроксильную группу (фиг.1А), доминируют в Азии (4). Несмотря на то что токсичность этих трихотеценов хорошо изучена на животных системах (5), очень мало известно о различиях в их фитотоксичности. Было показано, что DON (который также известен как вомитоксин) при своем воздействии на животных вызывает обратимые реакции в организме, при этом нервная и иммунная системы являются наиболее чувствительными мишенями (6). В отличие от продукции антител, для ингибирования которой необходимы высокие дозы DON, низкие дозы DON в сочетании с бактериальным липополисахаридом оказывают синергический эффект стимуляции провоспалительных процессов (6). Для защиты потребителей Администрация по пищевым и лекарственным продуктам США рекомендовала уровни для пищевых продуктов, а недавно в Европейском сообществе были разработаны рекомендации по предельным показателям действующих уровней DON (7).
Роль DON в развитии болезни растений все еще является предметом обсуждения (8), но большая часть данных указывает на его функцию в качестве фактора вирулентности. Мутанты Fusarium, содержащие нарушенный ген, участвующий в биосинтезе трихотеценов (триходиенсинтаза, Tri5), все еще сохраняют патогенность, хотя и демонстрируют сниженную вирулентность на пшенице (9) в связи с неспособностью распространяться от места инфекции (10). DON могут передвигаться вверх по грибам, инфицирующим растения, что указывает на их возможную роль с точки зрения кондиционирования ткани хозяйского организма, благоприятного для последующей колонизации (11). Результаты ряда исследований указывают на то, что резистентность in vitro разных сортов пшеницы в отношении DON коррелирует с резистентностью к FHB в полевых условиях (12), что также является основанием для сегрегации имеющихся экспериментальных популяций.
Повышенная резистентность к токсическим веществам может быть результатом действия ряда механизмов, охватывающих сниженное поглощение вплоть до включения обходных путей, суперэкспрессию или мутацию мишеней токсина. Кроме того, весьма важным процессом может быть метаболическая трансформация, которая часто является следствием компартментализации (13). Наблюдаемое снижение концентрации DON в пшенице, инфицированной Fusarium, в полевых условиях (14) позволяет полагать, что токсин может подвергаться метаболизации.
Было высказано предположение о том, что ацетилирование трихотеценов (например, DON) может быть использовано в рамках стратегии, направленной на достижение устойчивости (US 6346655 B1), однако получаемые ацетилированные трихотецены (например, ацетил-DON=ADON) эквивалентны по своей общей токсичности неацетилированной (то есть DON) форме, как было показано в работе Эудес с соавт. (Eudes et al.) в тесте по удлинению колеоптилей (53). В этой связи можно сказать, что процесс ацетилирования не приводит к снижению токсичности трихотеценов.
Два сорта пшеницы, различающиеся по резистентности в отношении Fusarium, демонстрируют различия по своей способности образовывать метаболит DON, который, как предполагается, является гликозидом (15). Сьювалд с соавт. (Sewald et al.) (16) в процессе инкубации суспензии культуры клеток кукурузы с радиоактивно меченым DON доказал, что он в первую очередь вступает в реакцию конъюгации с 3-R-D-глюкопиранозил-4-дезоксиниваленолом. Ранее не были описаны никакие растительные ферменты, способные модифицировать трихотецены или другие микотоксины за счет транспортировки сахарного фрагмента.
Исследования с использованием методики секвенирования генома показали наличие огромного числа генов в растениях, которые кодируют предположительные УДФ-глюкозилтрансферазы (UGT), которые, как предполагается, осуществляют конъюгацию малых молекул. Например, Arabidopsis thaliana содержит более 100 представителей данного мультигенного семейства (17), функции которого во многом неизвестны.
В этой связи объектом настоящего изобретения является разработка способов детоксикации микотоксинов.
Подходящие препараты микотоксинов или их производные, особенно производные микотоксинов в виде конъюгированных форм с сахарами, зачастую получить непросто. Поскольку стереоселективность является важнейшим фактором, определяющим качество такого рода продуктов, чисто химический синтез на основе объединенной смеси микотоксинов весьма затруднен, если не сказать, что невозможен, по меньшей мере, в промышленном масштабе. Другим объектом настоящего изобретения является, в этой связи, разработка способов синтеза производных микотоксинов в виде сахарных конъюгатов.
Исходя из этого, настоящее изобретение относится к способу детоксикации микотоксинов, в соответствии с которым микотоксин подвергают взаимодействию с глюкозилтрансферазой в присутствии активированной глюкозы.
Как указывалось выше, в литературе отсутствуют какие-либо сообщения относительно ферментов (растительных), способных модифицировать микотоксины за счет транспорта сахарной молекулы и, конечно, отсутствуют сообщения относительно ферментов, способных к детоксикации таких микотоксинов, то есть способных в значительной мере снижать токсичность микотоксинов, например, более чем на 80%, предпочтительно более чем на 90%, в особенности более чем на 95% при определении степени снижения в специфических тестах на микотоксины, таких как ингибирование синтеза белка in vitro.
В рамках настоящего изобретения было показано, что глюкозилтрансферазы представляют собой очень специфичный инструмент детоксикации микотоксинов. С одной стороны, специфичность определяется конкретной специфичностью данной глюкозилтрансферазы для ограниченного числа микотоксинов, а с другой стороны, специфичность также определяется способом детоксикации, а именно сайт-специфичного глюкозилирования микотоксинов. Поскольку указанная специфичность ограничивает область применения данной конкретной глюкозилтрансферазы, настоящее изобретение относится к инструментам, пригодным для простого определения специфичности любой глюкозилтрансферазы в отношении любого микотоксина, в особенности трихотеценовых микотоксинов. В этой связи в том случае, если конкретный фермент специфичен для данного набора микотоксинов, то на основе настоящего изобретения можно напрямую определить специфичности для других глюкозилтрансфераз и микотоксинов.
Поскольку уридиндифосфатглюкоза (УДФ-глюкоза, то есть глюкоза, которая образуется ферментами, такими как пирофосфорилаза) является предпочтительной активированной биологической формой глюкозы, предпочтительным вариантом настоящего изобретения является использование УДФ-глюкозилтрансферазы в качестве глюкозилтрансферазы и УДФ-глюкозы в качестве активированной глюкозы (= косубстрат), хотя любая другая форма глюкозы, которая может использоваться глюкозилтрансферазой (например, АДФ-глюкоза, ЦДФ-глюкоза или любая другая форма биологически и синтетически активированной глюкозы может быть приемлемым субстратом для осуществляемой глюкозилтрансферазами транспортировки к микотоксинам), может использоваться в соответствии с настоящим изобретением.
Глюкозилтрансфераза по настоящему изобретению определяется как естественный (природный) или синтетический (рекомбинантный), сконструированный или продуцируемый фермент, который способен (в качестве основного или в качестве побочного вида активности) к переносу глюкозного фрагмента к молекуле субстрата с использованием активированной глюкозы в качестве косубстрата, в результате чего образуется глюкозилированная молекула субстрата.
Предпочтительные микотоксины, подлежащие детоксикации по методу настоящего изобретения, включают, в связи с их экономической важностью, трихотецены, в особенности трихотецены, содержащие свободную гидроксигруппу в положении С3 и два атома водорода при С4. В особенности, если последние трихотецены включают специфическое глюкозилирование в положении 3 или по свободной гидроксильной группе в положении 7, при ее наличии, или в обоих указанных положениях. В этой связи предпочтительные микотоксины включают дезоксиниваленол, 15-ацетил-дезоксиниваленол, 15-ацетоксисцирпендиол или их смеси.
Предпочтительно способ осуществления настоящего изобретения осуществляется in vivo. Указанный способ может быть осуществлен на основе технологии рекомбинантной ДНК и/или с использованием соединений, повышающих экспрессию глюкозилтрансферазы. Предпочтительные примеры включают экспрессию глюкозилтрансферазы в клетке трансгенного растения (или в растительной ткани, или в целом растении), содержащей рекомбинантную глюкозилтрансферазу и/или рекомбинантный регуляторный участок для глюкозилтрансферазы, в особенности индуцируемый микотоксином промотор, в особенности глюкозилтрансферазы. Предпочтительные примеры соединений, повышающих экспрессию глюкозилтрансферазы, включают вещества, которые осуществляют позитивную регуляцию глюкозилтрансферазы, например токсины (если экспрессия трансферазы контролируется промоторами, индуцируемыми токсином). В принципе, любая клетка, которую можно подвергнуть генетической манипуляции, так чтобы она экспрессировала рекомбинантный ген (например, ген, который искусственно вводится в данную клетку, или его предшественник, который в природе не присутствует в данном сайте клеточного генома) растительной глюкозилтрансферазы может быть использована для данной цели. Предпочтительные варианты включают трансгенные бактерии, дрожжи, клетки насекомых, инфицированных бакуловирусом, и т.п., экспрессирующие ген глюкозилтрансферазы из растения.
При детоксикации микотоксинов в промышленном масштабе глюкозилтрансфераза может быть с успехом иммобилизована (в соответствии со стандартными методиками иммобилизации) на твердой поверхности, и раствор или суспензия, содержащие микотоксин, могут быть затем приведены в контакт с такой иммобилизованной глюкозилтрансферазой.
Предпочтительные примеры глюкозилтрансфераз, используемых в способе настоящего изобретения, включают УДФ-глюкозилтрансферазы, соответствующие подсемейству 73С из Arabidopsis thaliana, в особенности УДФ-глюкозилтрансферазы 73С4 и 73С5.
В соответствии с другим аспектом настоящее изобретение относится к способу, позволяющему создавать глюкозилированные микотоксины, который включает взаимодействие микотоксина с глюкозилтрансферазой в присутствии активированной глюкозы. При этом указанные выше особенности фермента могут быть использованы соответствующим образом для получения специфически глюкозилированных производных микотоксина. Если микотоксин глюкозилируется неферментативным способом («химическим»), то получают неочищенную смесь продуктов глюкозилирования (где каждая свободная гидроксигруппа в принципе является акцепторным сайтом для глюкозильных остатков). Способ ацетилирования в качестве средства защиты таких групп также имеет свои недостатки в плане специфичности (все гидроксигруппы могут подвергаться ацетилированию). Основываясь на использовании субстрат-специфичности и сайт-специфичности, авторы настоящего изобретения предлагают способ получения хорошо очищенных глюкозилированных производных. Если глюкозилированию подвергается более чем одна гидроксигруппа, или если более чем одно положение может быть глюкозилировано данной глюкозилтрансферазой, то такое ограниченное число продуктов (например, от 2 до 5) может быть легко разделено с использованием соответствующих способов разделения, таких как ВЭЖХ (например, DON, будучи глюкозилированным в положении 3, легко отделяется от DON, который глюкозилирован в положении 7).
Описанные выше предпочтительные стадии осуществления данного способа также применимы к этому аспекту настоящего изобретения.
В соответствии с другим аспектом настоящее изобретение относится к трансгенным растениям или трансгенным (растительным) клеткам, содержащим рекомбинантную глюкозилтрансферазу и/или рекомбинантный элемент, регулирующий экспрессию, в особенности промоторный участок глюкозилтрансферазы. Такие растения или (растительные) клетки могут использоваться в способе настоящего изобретения.
Настоящее изобретение также относится к генетически модифицированной клетке или организму, в особенности к растению или клетке растения, которые включают в своем геноме неприродную (трансгенную) глюкозилтрансферазу и обладают резистентностью к микотоксину, в особенности резистентностью к трихотецену, благодаря наличию такой глюкозилтрансферазы в виде трансгена. Альтернативно эндогенная глюкозилтрансфераза в клетке может вовлекаться в трансгенную экспрессию с использованием другого регуляторного участка или элемента, такого как промотор, ведущей к повышенной экспрессии эндогенной глюкозилтрансферазы (например, к повышенной активности экспрессии гомологичной глюкозилтрансферазы, определяемой наличием трансгенных промоторов). Кроме того, такие клетки демонстрируют повышенную резистентность к микотоксинам, как здесь будет далее описано. Такие клетки и растения могут быть получены при использовании стандартных способов и в соответствии с идеями настоящего изобретения (см., например, фиг.6В, а также примеры, например (54)).
Предпочтительные клетки согласно настоящему изобретению включают также клетки дрожжей. Поскольку клетки дрожжей могут обладать ограниченной способностью к поглощению, предпочтительные клетки дрожжей согласно настоящему изобретению могут быть разработаны или соответствующим образом отобраны, так чтобы достичь повышенной способности поглощать микотоксины в сравнении со штаммом дикого типа или с исходным штаммом. Конкретно предпочтительными согласно настоящему изобретению являются дрожжи, в которых один или более, предпочтительно три или более ABC-переносчиков обладают сниженной активностью или инактивированы по данной активности, особенно в отношении pdr5. Такие мутанты по АВС-переносчику характеризуются повышенным поглощением микотоксинов. Предпочтительно клетки согласно настоящему изобретению имеют делецию в pdr5, pdr10, pdr15, snq2, yor1, ayt1 или включают сочетания таких делеций.
Указанные клетки могут быть предпочтительно использованы для детоксикации растворов или суспензий, содержащих микотоксин, особенно для детоксикации трихотеценов в сельском хозяйстве и пивоварении, а также при получении глюкозилированных микотоксинов, в особенности глюкозилированных DON или 15-ADON.
Настоящее изобретение также может быть проиллюстрировано приведенными ниже примерами и чертежами, которые никоим образом не ограничивают настоящее описание.
Чертежи:
Фиг.1. Спектр устойчивости к трихотецену дрожжей, экспрессирующих УДФ-глюкозилтрансферазы в Arabidopsis thaliana из подсемейства UGT73C. А: Структура и нумерация кольцевой системы трихотеценов класса В, использованных для тестирования резистентности. В таблице показаны различные остатки в разных положениях R1-R4 (OAc: -OCOCH3, OBe: -OCOCH=CHCH3 (Z)). Релевантный R1 подчеркнут (OH в случае DON и уже блокированный ацетилированием в 3-ADON). Повышенная резистентность, придаваемая экспрессией DOGT1, указана знаком плюс, отсутствие защиты - знаком минус. B: Геномная организация генного кластера UGT в хромосоме II (подсемейство 73С), содержащего DOGT1 (UGT73C5, локус At2g36800). DON, детоксифицированные под действием DOGT1 и 73C4, представлены в виде затемненных серым цветом фигур. C: Дрожжевые штаммы наносят на планшеты со средой YPD, содержащей указанное количество токсинов (м.д.: мг/л). Используемые штаммы представляют собой штаммы дикого типа YZGA515 без плазмиды (wt), данный штамм подвергают трансформации пустым вектором (с), и показаны трансформанты, экспрессирующие myc-меченные 73С UGT (C5=DOGT1, подчеркнуто, наносимые в двойном повторе). Резистентность, придаваемая плазмидами, ведет к суперэкспрессии С5 и С4. D: Вестерн-блотт анализ дрожжевых штаммов, используемых для тестирования резистентности. Выявляют N-концевой с-Myc-меченый эпитоп, введенный в соответствующих представителей семейства 73С UGT (C5=DOGT1, подчеркнуто). Трансформант, содержащий пустой вектор экспрессии, используют в качестве контроля.
Фиг.2. Сопоставление аминокислотного состава растительных УДФ-глюкозилтрансфераз, имеющих высокую степень сходства по аминокислотному составу с DOGT1. Участки, вовлекаемые в субстрат-акцепторное связывание (ссылка 38) (указаны подчеркиванием в виде точечных линий), и последовательность консенсусного мотива в UGT (пунктирные линии) показаны внутри прямоугольников под последовательностями. Треугольник, приведенный над последовательностью в гипотетическом участке связывания акцептора, обозначает лизин 136 в DOGT1, который был заменен в ходе мутагенеза in vitro. В банке данных последовательностей (genbank) рассчитанные белки имеют следующие номера: ADGT-9, глюкозилтрансфераза-9 из Vigna angularis (AB070752); TOGT1, фенилпропаноид: глюкозилтрансфераза 1 из Nicotiana tabacum (AF346431); IS5a из Nicotiana tabacum (U32644); предположительная глюкозилтрансфераза из Oryza sativa (AP002523); Twi1 из Lycopersicon esculentum (x85138); Бетанидин-5-O-глюкозилтрансфераза из Dorothenathus bellidiformis (Y18871).
Фиг.3. Экспрессия DOGT1 регулируется и индуцируется в ходе развития под действием DON и соединений, имеющих отношение к стрессовой реакции. А: GUS-окрашивание проростков, гомозиготных по белку слияния промотора транскрипции DOGT1-GUS. Верхний ряд: через три дня после прорастания (3 DAG) экспрессия белка слияния ограничивается областью сосудистой структуры корня и гипокотиля и меристематическим участком корневой верхушки. Средний ряд: на более поздних фазах развития (10 DAG) степень окрашивания сосудистой структуры корня снижается, за исключением тех участков, где образуются боковые корешки. Нижний ряд: в надземных частях взрослых растений экспрессия DОGТ1 ограничивается лепестками цветов и зонами осыпания. В: Обработка DON (5 м.д. в течение 4 часов) проростков (14 DAG), экспрессирующих трансляционный GUS-содержащий белок слияния, индуцирует экспрессию GUS-репортера. Оба образца окрашивают в течение двух часов. С: полуколичественный анализ в рамках метода ПЦР с обратной транскриптазой явлений индукции экспрессии DOGT1, UGT73C4 и UGT73C6 с последующей обработкой DON (м.д.), СК (SA) (100 мкМ), ЖК (JA) (50 мкМ) и АЦК (ACC) (2 мкМ). UBQ5 используют в качестве внутреннего контроля.
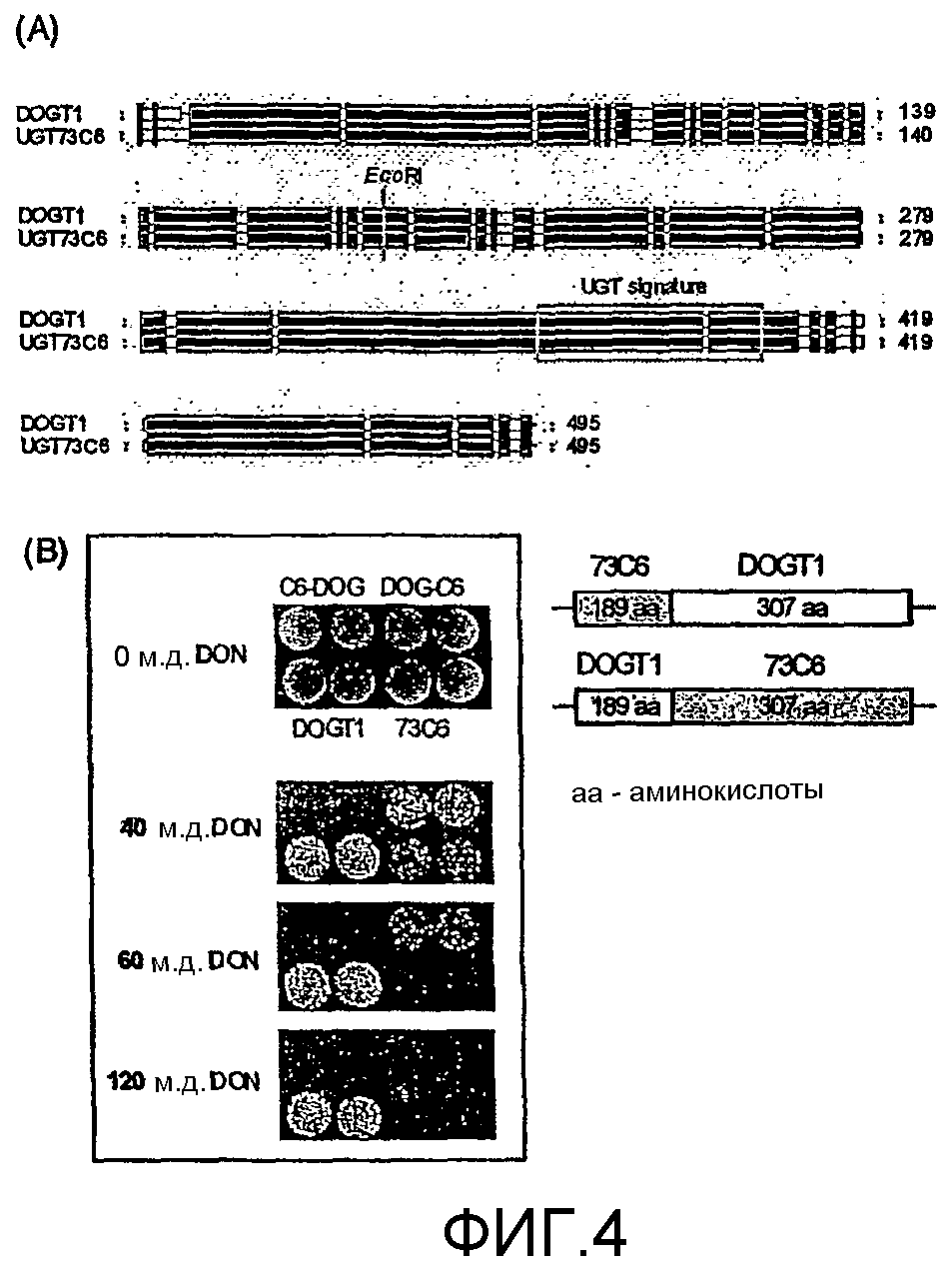
Фиг.4. N-терминальная часть DOGT1 необходима для проявления его способности к детоксикации DON. A: Сопоставление по аминокислотному составу DOGT1 и его ближайшего гомолога UGT73C6, который не имеет защиты против DON (черным цветом обозначена зона идентичности по аминокислотному составу). Консервативный сайт рестрикции EcoRI используют для создания химерных UGT, состоящих из N-конца одного и C-конца соответствующего другого гена. B: Дрожжевые трансформанты вносят в двойном повторе в среду YPD, содержащую указанные концентрации DON. Сверху, слева: трансформанты, экспрессирующие химерный фермент, состоящий из N-конца UGT73C6 и С-конца DOGT1 (короткий фрагмент: C6-DOG). Сверху, справа: штаммы, экспрессирующие химерный белок, состоящий из N-конца DOGT1 и С-конца UGT73C6 (DOG-С6). Нижний ряд: трансформанты, экспрессирующие DOGT1, придающий устойчивость, (слева) и неактивный 73С6 (справа). Гибрид, содержащий N-концевую часть из DOGT1, характеризуется резистентностью к более высоким концентрациям DON, чем штамм, экспрессирующий другой гибрид (С6 DOG) или UGT73C6.
Фиг.5. Картина фрагментации синтезированных DON-глюкозидов и DON-метаболита, образованного в дрожжевых клетках, экспрессирующих DOGT1, по результатам масс-спектрометрии. А: синтезированный DON-3-О-глюкозид дает фрагмент со значением 427,2 m/z, при тестировании в режиме линейного захвата ионов (MS/MS/MS). B: данный фрагмент не образуется при использовании образца синтезированного DON-15-О-глюкозида, поскольку отделению -CH2OH группы при C6 мешает глюкозный фрагмент, присутствующий при гидроксильной группе. C: DON-метаболит, образованный в дрожжах, экспрессирующих DOGT1, элюируется в ходе ВЭЖХ с тем же временем удерживания, что и DON-3-O-глюкозид, и демонстрируют картину фрагментации, соответствующую эталонному веществу DON-3-O-глюкозид.
Фиг.6. Глюкозилирование DON снижает токсичность in vitro и in vivo. A: Структура предполагаемого продукта реакции: DOGT1 катализирует перенос глюкозы от УДФ-глюкозы к 3-OH положению в DON. B: Сравнение степени ингибирования синтеза белка in vitro в рибосомах пшеницы под действием DON и DON-3-O-глюкозида, осуществляемое с использованием экстракта зародышей пшеницы в системе сопряженной транскрипции/трансляции. Измеряют интенсивность люминесценции и выражают полученный результат в виде процента от активности люциферазы в контрольных образцах без токсина. С: Вестерн-блотт анализ гомозиготных линий А. thaliana на наличие конструкции суперэкспрессии, кодирующей c-Myc-меченый DOGT1. Ниже схематически показан вектор трансформации растения (GENT: резистентность к гентамицину, используемый в качестве селектируемого маркера; дуплицированный промотор 2×35S для транскрипта 35S из цветной капусты (детали см. в разделе, описывающем процедуры проведения эксперимента)). Col-0 используют в качестве отрицательного контроля. Показаны 2 линии с высокими уровнями и 2 линии с низкими уровнями белка c-Myc-DOG1. D: Всхожесть семян в среде MS, содержащей DON. Сравнение трансгенных линий Arabidopsis, экспрессирующих высокие количества DOGT1 (например, линии 1319/2), с диким типом (Col-0), указывает на повышение резистентности.
ПРИМЕРЫ
Для иллюстрации настоящего изобретения ниже даны примеры с описанием клонирования UGT из Arabidopsis при функциональной экспрессии в дрожжах и ее характеристики. Данный фермент способен осуществлять детоксикацию микотоксина DON из Fusarium и его ацетилированного производного 15-ADON. Приведенные в данных примерах методики могут быть также использованы для характеристики других глюкозилтрансфераз и микотоксинов без проведения дополнительных экспериментальных процедур.
ЭКСПЕРИМЕНТАЛЬНЫЕ ПРОЦЕДУРЫ
Штаммы дрожжей. Штаммы дрожжей, используемые в настоящей работе получают из YPH499 (Mat a, ade2-101oc, his3-Δ200, leu2-Δ 1, lys2-801a, trp1-Δ1, ura3-52) (18). Релевантный генотип YZGA452 представляет собой pdr5Δ::TRP1, pdr10Δ::hisG, snq2Δ::hisG, yor1Δ::hisG. YZGA515 (pdr5Δ::TRP1, pdr10Δ::hisG, prd15Δ::loxP-KanMX-loxP, ayt1Δ::URA3) был сконструирован путем разрушения ацетилтрансферазного гена AYT1 в штамме YHW10515.
Растительный материал и условия выращивания. Эксперименты с A. thaliana проводят при использовании дикого экотипа Columbia-0 (Col-o). Для размножения семена стерилизуют, высевают на стандартную ростовую среду MS (19) с добавкой 1,0 % сахарозы и 1,0% фитагара (Life Technologies) и подвергают темновой обработке в течение 2 дней при температуре 4°С для синхронизации прорастания. Проростки растят в течение 2 недель в контролируемых условиях с циклом чередования светлой/темновой фаз 16 часов/8 часов (140 мкмоль.м-2·сек-1 при освещении белым светом) при температуре 22°С и затем переносят в почву и растят при температуре 20°С и уровне влажности 55% в условиях освещения белым светом.
Скрининг библиотеки кДНК из Arabidopsis thaliana в дрожжах. В штамме Saccharomyces cerevisiae YZGA452, который является дефицитным по кассете связывания с АТФ для переносчика (ABC) и характеризуется гиперчувствительностью к DON, трансформируют библиотекой кДНК из A. thaliana, способной конститутивно экспрессироваться под контролем промотора фосфоглюцераткиназы (PGK1) (20). С использованием минимальной среды, не содержащей урацил, отбирают в общей сложности 107 трансформантов и переносят их в среды, содержащие DON на уровне 180 м.д., достаточном для полного ингибирования роста дрожжей, трансформированных библиотекой пустых плазмид. Выделяют колонии, которые демонстрируют резистентность, и на отобранных вариантах, формирующих единичные колонии на средах, содержащих токсин, тестируют зависимость плазмиды от фенотипа с помощью препарата плазмидной ДНК и проводят повторную трансформацию YZGA452. Фрагмент NotI, содержащий вставку кДНК из отбираемого варианта (обозначенную как DON-глюкозилтрансфераза 1, DOGT1), подвергают субклонированию с использованием pBluescript SKII+ (Stratagene) с последующим секвенированием.
Конститутивная экспрессия и выявление иммунологическими методами DON-глюкозилтрансферазы (DOGT1) и ее близких гомологов в дрожжах. Не содержание интрон открытые рамки считывания (ORF) для DOGT1 (UGT73C5, локус At2g36800) и 5 ее ближайших гомологов (UGT73C1, At2g36750; UGT73C2, At2g36760; UGT73C3, At2g36780; UGT73C4, At2g36770; UGT73C6, At2g36790) амплифицируют при помощи ПЦР (система Triple Master PCR, Eppendorf) на основе геномной ДНК с использованием ген-специфичных праймеров, содержащих фланкирующие сайты рестрикции HindIII и NotI на 5'- и 3'-концах соответственно.
(DOGT1, fw: 5'-ACTAAGCTTGGAATCATGGTTTCCGAAACA-3',
rv: 5'-AAGCGGCCGCATACTCAATTATTGG-3';
73C1, fw: 5'-CTAAGCTTGGAATCATGGCATCGGAATTTCG-3', rv: 5'-TAGCGGCCGCATTCATTTCTTGGGTTGTTC-3';
73C2, fw: 5'-CTAAGCTTGGAATCATGGCTTTCGAGAAGACC-3', rv: 5'-TAGCGGCCGCATTCAACTCTTGGATTCTAC-3';
73C3, fw: 5'-CTAAGCTTGGAATCATGGCTACGGAAAAAACC-3', rv: 5'-TAGCGGCCGCATTCATTCTTGAATTGTGC-3'; 73C4, fw: 5'-CTAAGCTTGGAATCATGGCTTCCGAAAAATC-3', rv: 5'-TAGCGGCCGCATTCAGTTCTTGGATTTCA-3'; 73C6, 5'-CTAAGCTTGGAACATGTGTTCTCATGATCCT-3', rv: 5'-TAGCGGCCGCATTCAATTATTGGACTGTGC-3'). Продукты ПЦР клонируют в сайтах клонирования HindIII и NotI в векторе экспрессии дрожжей pYAK7 (PADH1-c-Myc-PDR5 LEU2 2 мк) с заменой PDR5 гена. Вектор pYAK7 конструируют путем первой вставки двуцепочечного линкера 5'-GGATGCCCGAACAAAAGTTAATTTCAGAAGAGGACTTATCAAAGCTTGAGGCCTCGCGA в сайт SmaI в векторе pAD4Δ (21), получая при этом N-концевой c-Myc эпитоп и сайт рестрикции HindIII, через который геномный фрагмент HindIII, содержащий дрожжевой PDR5, встраивают в рамку считывания.
Анализируют путем секвенирования меченые конструкции UGT и затем используют их для трансформации дрожжей штамм YZGA515. Пустой вектор (расщепленный HindIII + NotI и повторно лигированный с pYAK7) используют в качестве контроля. Трансформанты отбирают на минимальных средах без лейцина. Экспоненциально растущие культуры разбавляют до значения ОП600 0,05 и переносят на среды YPD, содержащие возрастающие концентрации различных трихотеценов. Используемые токсины включают: DON, 3-ADON, 15-ADON, NIV, трихотецин (TTC), T-2 токсин, HT-2 токсин, диацетоксисцирпенол (DAS) и веруккарин A (VA). За исключением DON, 3-ADON, NIV, которые были получены от компании Biopure Referenz-substanzen GmbH (Tulln, Austria), микотоксины приобретают от компании Sigma и хранят при температуре -20°С растворенными в 70% этаноле.
Для иммунологического анализа проводят экстракцию белков из дрожжевых клеток по методике, описанной Эгнером с соавт. (Egner et al.) (22). Вестерн-блотинг проводят с первичным антителом против с-Myc (1:5000, клон 9Е10, Invitrogen).
Перетасовка домена. ORF, включающую N-концевую метку c-Myc из DOGT1 и ее ближайшего гомолога UGT73C6, выделяют из векторов экспрессии в дрожжах путем расщепления с использованием SmaI + NotI, клонируют в векторе pBluescript SKII+ (который был разрезан с использованием XhoI, обработан ферментом Кленова и далее расщеплен с использованием NotI). Полученные плазмиды расщепляют с использованием HindIII, при этом в обоих генах обнаруживается консервативный сайт EcoRI, который расщепляет DOGT1 в нуклеотидном положении 565 (в случае 73С6 в 568). Гибриды конструируют путем лигирования N-концевой части одного гена с C-концевой частью другого. Полученные гены возвращают в вектор экспрессии в дрожжах pYAK7 с использованием для этого сайтов рестрикции HindIII и NotI. Дрожжевой штамм YZGA515 используют в качестве хозяйских клеток для тестирования конструкций, обладающих измененными способностями по детоксикации.
Сайт-направленный метагенез. Мутации создают по методу перекрывающейся ПЦР с удлинением (23) при использовании перекрывающихся мутантных праймеров DOG-K136E-fw (5'-TACAAGCGAAATCGCCAAGAAGTTCA-3') и DOG-K136E-rv (5'-CTTCTTGGCGATTTCGCTTGTATAAG-3') и фланкирующих праймеров DOGIpYAK7-fw-a (5'-ACTAAGCTTGGAATCATGGTTTCCGAAACA-3') и DOG-EcoRI-rv (5'-TCTTGTGAATTCAACTCTATC AGGA-3') для осуществления мутагенеза DOGT1, а также мутантных праймеров 73С6-E137K-fw (5'-TACAAGCAAAATCGCC AAGAAGTTCAA-3') и 73С6-Е137K-rv (5'-ACTTCTTGGCGATTTTGCTTGTATT-3') и фланкирующих праймеров 73С6pYAK7-fw (5'-TAAGCTTGGAATCATGTGTTCTCATGATCCT-3') и DOG-EcoRI-rv для мутагенеза 73С6. Полученные продукты ПЦР клонируют в виде фрагментов расщепления HindIII + EcoRI в соответствующих генах, присутствующих в векторе pBluescript SKII+. После секвенирования ORF перемещают в виде фрагментов расщепления HindIII + NotI назад в вектор экспрессии в дрожжах pYAK7 (с заменой гена PDR5) и полученные плазмиды трансформируют в штамм YZGA515 для анализа созданных при таких генетических манипулированиях UGT на их способность к детоксикации.
Синтез 3-β-D-глюкопиранозил-4-дезоксиниваленола и 15-β-D-глюкопиранозил-4-дезоксиниваленола. С целью получения эталонного материала для проведения ВЭЖХ и ТСХ в рамках 2-стадийных реакций синтезируют DON-3-глюкозид и DON-15-глюкозид. На первой стадии 15-ацетил-DON или 3-ацетил-DON модифицируют с использованием β-бром-1-дезокси-2,3,4,6-тетра-О-ацетил-α-D-глюкопиранозы (ацетобромглюкозы) в толуоле, содержащем CdCO3 в качестве катализатора, с образованием DON-глюкозидацетатов (24). На второй стадии проводят мягкий гидролиз ацетата до глюкозидов с использованием сильного основного анионообменника (DOWEX1x2-400, Aldrich, USA). После подтверждения прогрессирования реакции с помощью ТСХ (мобильная фаза: толуол/этилацетат, 1:1, объем/объем) DON-глюкозиды очищают методом флэш-хроматографии на силикагеле с использованием смеси 1-бутанол/1-пропанол/этанол/вода (2:3:3:1, объем/объем/объем/объем). Дальнейшую очистку веществ проводят по методу ВЭЖХ с использованием колонки RP-18 Aquasil (Keystone, Waltham, USA) и смеси ацетонитрил/вода (10:90, объем/объем) при температуре 22°С.
Синтезированные DON-производные анализируют с использованием интерфейсной системы электронапыления с отрицательными ионами (ESI). Анализ в рамках LC-MS/MS проводят в системе QTrap-LC-MS/MS (Applied Biosystems, Foster City, USA), снабженной ESI и установкой для проведения ВЭЖХ серии 1100 (Agilent, Waldbronn, Germany). Хроматографическое разделение проводят на колонке Aquasil RP-18 с размерами 150 мм × 4,6 мм (внутр.диам.), 3 мкм (Keystone, Waltham, USA) при температуре 22°С с использованием смеси метанол/вода (28:72, объем/объем). Скорость течения устанавливают на уровне 0,3 мл/мин. Интерфейс ESI используют в режиме регистрации отрицательных ионов при 400°С со значениями: CUR 20 psi, GS1 30 psi, GS2 75 psi, IS-4200 V, DP-46 V, EP- 9 V, CE-30 eV, CAD high, время заполнения LIT 50 миллисекунд, барьер входа Q3 8 V.
Выделение и анализ метаболитов DON in vivo. Для выяснения химической структуры метаболитов DON, образованных при ферментативной трансформации микотоксина под действием DOGT1, конструируют высоко толерантный штамм и экстрагируют DON-метаболит из клеток, обработанных токсином, и далее проводят анализ по методу ВЭЖХ.
Дрожжевой штамм YZGА515 трансформируют c-Myc-меченым DOGT1 под контролем конститутивного промотора ADH1 и с добавлением плазмиды pRM561. Данная плазмида содержит мутантный вариант рибосомального белка L3 (RPL3) из Saccharomyces cerevisiae, который существенно повышает резистентность к DON при экспрессии в дрожжах. Трансформанты отбирают на минимальных средах, не содержащих лейцин и аденин.
Полученный дрожжевой штамм растят до значения ОП600 0,7 в селективной среде (SC-LEU-ADE). Клетки переносят в среду YPD (10% глюкозы) с добавкой аденина, растят в течение 2 часов при температуре 30°С, собирают центрифугированием и разбавляют средой YPD до ОП600 3,0. Добавляют DON к 5 мл культуры, начиная с 200 м.д. DON. После 3 часов концентрацию повышают до 400 м.д., через 6 часов - до 600 м.д. и через 9 часов - до 1000 м.д. Клетки инкубируют еще в течение 15 часов при 30°С, после чего их собирают, промывают три раза охлажденной льдом водой, экстрагируют с помощью 2,5 мл смеси метанол/вода (4:1) и озвучивают. После центрифугирования супернатант фильтруют через стеклянный микроволокнистый фильтр (Whatman 1822 025). 500 мкл дрожжевых экстрактов концентрируют досуха в потоке азота, подаваемого с постоянной скоростью, и растворяют в 100 мкл воды категории ВЭЖХ.
Гетерологичная экспрессия DOGT1 в Escherichia coli. Белок DOGT1 экспрессируется в Escherichia coli XL1-blue в виде белка слияния с GST. Ген DOGT1 высвобождается из вектора экспрессии в дрожжах при расщеплении HindIII и заполнении ферментом Кленова с последующим расщеплением NotI. Полученный фрагмент клонируют в сайтах рестрикции SmaI + NotI гена GST в гибридном векторе pGEX-4T-3 (Amersham Pharmacia). Рекомбинантный белок слияния очищают с использованием сефарозы, связанной с глютатионом (Amersham Pharmacia), в соответствии с инструкциями производителя.
Для оценки влияния N-концевой GST-метки на активность ген, кодирующий белок слияния, амплифицируют по методу ПЦР с использованием ДНК-полимеразы с подтвержденной активностью по считыванию (Pfu-полимераза, MBI) и с использованием специфических праймеров для белка слияния GSTDOGpYAK7-fw (5'-TCACCCGGGAAACAGTAATCATGTCC-3') и GSTDOGpYAK7-rv (5'-CGAGGCAGATCGTCAGTCAGTC-3'). Продукт ПЦР клонируют с использованием HindIII+NotI в векторе экспрессии в дрожжах pYAK7. Способность к детоксикации DON определяют при экспрессии в YZGA515 на средах, содержащих токсин, как было описано выше.
Энзиматические тесты. Тест, используемый для определения активности глюкозилтрансферазы, включает смесь, содержащую 1 мкг рекомбинантного GST-содержащего белка слияния, 10 мМ 2-меркаптоэтанола, 50 мМ Трис/HCl, pH 7,0, 0,5 мМ радиоактивномеченой УДФ-[14C] глюкозы, (4,4*103 имп./мин, NEN Life Science Products, USA), 0,01% БСА и 1 мМ субстрата-акцептора (растворенного в ДМСО в 20 мМ основном растворе). Реакции проводят в объемах 20 мкл при температуре 30°С в течение 1 часа, останавливают добавлением 2 мкл трихлоруксусной кислоты (240 мг/мл), замораживают и хранят при температуре -20°С.
Анализ продуктов реакции проводят по методу ТСХ. Аликвоту каждого из образцов наносят в виде пятна на пластинку с силикагелем (Kieselgel 60; Merck) и проводят элюцию смесью 1-бутанол/1-пропанол/этанол/вода (2:3:3:1, объем/объем/объем/объем). Интенсивность каждого радиоактивномеченого пятна определяют с использованием системы фосфовизуализации (система STORM 860, Molecular Dynamics). Далее пластины дополнительно окрашивают п-анисовым альдегидом (0,5% в метаноле/H2SO4/уксусная кислота, 85:5:10, объем/объем/объем).
Ингибирование in vitro рибосом из пшеницы. Для определения, является ли 3-β-D-глюкопиранозил-4-дезоксиниваленол менее токсичным, чем его аглюкон, анализируют экстракт из зародышей пшеницы, введенный в систему транскрипции/трансляции in vitro (TNT Coupled Wheat Germ Extract, T3, Promega). После проведения реакций транскрипции/трансляции в течение 25 минут согласно инструкциям производителей в присутствии DON, очищенного DON-глюкозида (1 мкМ, 2,5 мкМ, 5 мкМ, 10 мкМ и 20 мкМ) или воды в качестве контроля определяют активность репортера люциферазы светлячков (Luciferase Assay System, Promega) с использованием люминометра (Victor 2, Wallac).
Обработка растений, для анализа характера экспрессии, различными соединениями, вовлекаемыми в стрессовую реакцию. Для анализа продуктов ПЦР с обратной транскрипцией (RT)-ПЦР с целью оценки экспрессии мРНК DOGT1 проростки обрабатывают DON, салициловой кислотой (СК), жасмоновой кислотой (ЖК) и 1-аминоциклопропилкарбоновой кислотой (АЦК) и растят в течение 2 недель в вертикальных чашках с MS (0,8% фитагара), после чего переносят в жидкие среды MS. Растения инкубируют в течение 48 часов на планетарной качалке (50 об/мин), после чего добавляют 5 м.д. DON, 200 мкМ СК, 2 мкМ АЦК или 50 мкМ ЖК. Соединения хранят в основных растворах при растворении их либо в 70% этаноле, либо в ДМСО. Обработка этанолом и ДМСО дает контрольные варианты. Растения собирают в разные временные точки, измельчают в жидком азоте и хранят при температуре -70°С до проведения экстракции РНК.
Анализ экспрессии мРНК DOGT1 и близких гомологов по методу ПЦР с обратной транскриптазой. Суммарную РНК выделяют из растительной ткани и измельчают в жидком азоте с использованием тризолового реагента в соответствии с инструкцией производителей (Gibco BRL Life Technologies). Количество РНК определяют фотометрически и визуально в геле с денатурированной РНК, используя для анализа 5 мкг суммарной РНК.
кДНК синтезируют на основе 1 мкг суммарной РНК (расщепленной ДНКазой I) с использованием 500 нг олиго(dt) из 18 остатков и обратной транскриптазы SuperScript (Gibco BRL Life Technologies). ПЦР проводят на основе примерно 2 мкл разбавленной 1:20 кДНК с использованием праймеров, получая после амплификации крупные фрагменты размером 200-400 н.п., локализованные в С-терминальной части генов, подлежащих анализу (DOGT1-fw: 5'-ATCCGGGGTTGAACAGCCT-3', DOGT-rv: 5'-TCAATTATTGGGTTCTGCC-3'; 73C4RT-fw: 5'-GGAGAAAATAGGAGTGTTA-3', 63C4RT-rv: 5'-TCAGTTCTTGGATTTCACT-3'; 73C6RT-fw: 5'-GAGAAACTGGTCGTACAA-3', 73C6RT-rv: TCAATTATTGGACTGTGCT-3'; UBQ5-U: 5'-GTCCTTCTTTCTGGTAAACGT-3', UBQ5-D: 5'-AACC CTTGAGGTTGAATCATC-3'). Для сравнения относительных количеств транскриптов в образцах фрагменты ДНК из UBQ5 вначале амплифицируют и объемы образцов, нормализованные по количеству продуктов, соответствующих транскриптам UBQ5, используют для проведения ПЦР.
Клонирование конструкций суперэкспрессии в растении и конструкций GUS-белок слияния. Для конститутивной суперэкспрессии с-Myc-меченного белка DOGT1 в Arabidopsis конструируют вектор pBP1319. Его получают на основе модифицированного варианта вектора экспрессии в растениях pPZP221 (25). Используют промоторную кассету, состоящую из двух копий 35S-промотора и сигнала полиаденилирования CaMV, штамм Cabb B-D из вектора р2RT, представляющего собой модифицированный вариант pRT100 (26).
C-Myc-меченный фрагмент DOGT1 высвобождают путем расщепления с использованием SmaI+NotI (при заполнении с использованием фермента Кленова) из вектора экспрессии в дрожжах и клонируют в pBluescript SKII+, который разрезают с использованием ClaI+SmaI и обрабатывают ферментом Кленова. На следующей стадии ген вырезают из полученной плазмиды в виде SalI+BamHI фрагмента и встраивают в XhoI+BamHI сайты в p2RT. Полученную кассету 2×35S c-Myc-DOGT1 выделяют путем расщепления PstI и клонируют в уникальном сайте PstI в pPZP221 после разрушения множественного сайта клонирования в данном векторе путем расщепления плазмиды с использованием EcoRI+SalI, заполняют сайты с помощью фермента Кленова и проводят повторное лигирование. В полученном векторе pBP1319 кассету 2×35S c-Myc-DOGT1 ориентируют в противоположном направлении относительно 2×35S маркера резистентности к гентамицину.
Для конструирования фактора транскрипции белка слияния DOGT1-GUS используют GUS-содержащий вектор pPZP-GUS.1, который получают из pPZP200 и который содержит GUS ген из pBI101.1 (вставленный с помощью HindIII+EcoRI в MCS) (27). Промоторный участок DOGT1 подвергают амплификации по методике ПЦР из геномной ДНК с использованием ДНК-полимеразы с подтвержденной активностью по считыванию (Pfu Polymerase, MBI) и специфических праймеров (DOGP-GUS-fw: 5'-GTTAAAAGCTTACATGTGCATTACGGTCTGTGTGAATA, DOGP-GUS-rv: 5'-TTTCGGATCCCATG ATTCAACCTTAGTAAGAAACTCTC). Полученный продукт клонируют в рамке считывания с сайтами HindIII+BamHI для GUS гена в векторе pPZP-GUS.1. Идентичность конструкции подтверждают секвенированием ДНК.
Создание и анализ трансгенной A. thaliana. Для проведения всех трансформаций растений используют recA-дефицитный штамм Agrobacterium tumefaciens, UIA143 (28), который содержит хелперную плазмиду pMP90 (29). Штамм A. thaliana трансформируют с использованием «флорал дип» методики (30). Проводят отбор потомства от 15 независимых трансформантов в течение трех поколений с получением гомозиготных линий.
Для иммунологического выявления c-Myc-меченного DOGT1 используют примерно 200 - 500 мг растительного материала, который гомогенизируют в жидком азоте. Добавляют 300 мкл экстрагирующего буфера (200 мМ Трис-HCl, pH 8,9; 200 мМ KCl; 35 мМ MgCl2; 12,5 мМ ЭГТА; 15 мМ ДТТ; 0,6 мМ сорбита) и 15 мкл ингибиторного коктейля для протеаз (Sigma, # 9599) к еще замороженным образцам и смесь инкубируют в течение 15 минут с энергичным встряхиванием при температуре 4°С. После центрифугирования (14000 об/мин в течение 15 минут при температуре 4°С) переносят по 200 мкл супернатантов в свежие пробирки и хранят при температуре -20°С. Берут эквивалентное количество белка (50 мкг) для проведения вестерн-блотт анализа, в котором используют очищенное первичное анти-c-Myc антитело из супернатанта от гибридом (клон 9Е10).
Для анализа DON-резистентности семена гомозиготных линий, демонстрирующих высокую экспрессию DOGT1, а также Col-0, в качестве контроля, проращивают на MS средах, содержащих разные концентрации DON (5-30 м.д.). Проростки растят в течение 5 недель и затем оценивают получаемый фенотип. Активность GUS анализируют при окрашивании проростков или органов взрослых растений в растворе X-Gluc в течение 2-4 часов при температуре 37°С (31).
РЕЗУЛЬТАТЫ
Выделение DON-глюкозилтрансферазы (DOGT1) в процессе гетерологичной экспрессии в дрожжах. Было проведено объективное функциональное тестирование на основе гетерологичной экспрессии кДНК в дрожжах с целью идентификации растительных генов, которые вносят вклад в генерирование резистентности против микотоксинов. Штамм Saccharomyces cerevisiae дикого типа характеризуется высокой резистентностью к дезоксиниваленолу (DON). Для снижения количества токсина, необходимого для оценки в рамках такого тестирования, создают штамм, дефицитный по четырем переносчикам ABC, которые в значительной степени ответственны за плейотропную резистентность к лекарственным препаратам у дрожжей (32). Штамм YZGA452 (snq2Δ::hisG pdr5Δ::TRP1 pdr10Δ::hisG yor1Δ::hisG) обладает гиперчувствительностью к широкому перечню различных ксенобиотиков и природных продуктов, включая DON. YZGA452 трансформируют библиотекой экспрессии кДНК из A. thaliana (20), где конститутивная экспрессия кДНК осуществляется под контролем промотора фосфоглюцераткиназы. Получают десять миллионов трансформантов и разбавленные пулы трансформантов вносят на чашки со средой, содержащей DON. После отбора DON-резистентных колоний дрожжей и подтверждения зависимости получаемого фенотипа от введенной плазмиды вставку подвергают субклонированию и секвенируют.
DOGT1 является представителем семейства УДФ-глюкозилтрансфераз из A. thaliana и демонстрирует высокое сходство с генами, индуцируемыми салициловой кислотой и повреждением, из других видов. кДНК, придающая резистентность, имеет размер 1,75 т.н.п. и содержит открытую рамку считывания длиной 1488 н.п., предположительно кодирующую уридиндифосфат (УДФ)-глюкозилтрансферазу. Идентифицированная DON-глюкозилтрансфераза (DOGT1) соответствует гену UGT73C5 и принадлежит к подсемейству 73С, как часть группы D из УДФ-глюкозилтрансфераз (UGT) из A. thaliana (33). UGT из Arabidopsis составляет очень крупное генное семейство, которое было разделено на 14 отдельных групп, которые, как считается, происходят от общих предков (17). DOGT1 расположен в кластере (фиг.1В) вместе с пятью другими представителями подсемейства 73С на хромосоме II (BAC клон F13K3, At2g36800). Все шесть тандемно повторяющихся генов не содержат интронов и характеризуются высоким сходством друг к другу (77-89% идентичности на аминокислотном уровне). Сходство является также очень высоким и на межгенных промоторных участках.
Поиски в базе данных по аминокислотной последовательности DOGT1 выявили высокое сходство с глюкозилтрансферазами из растений табака (TOGT1, ссылка 34; Is5a и Is10a, ссылка 35), из растений томатов (Twi-1, ссылка 36), экспрессия которых, как было показано, повышается после обработки салициловой кислотой (СК), элиситорами грибов или повреждением (34, 36, 37), а также с бетанидин-5-О-глюкозилтрансферазой из Dorotheanthus bellidiformis (38). Две предположительных неохарактеризованных глюкозилтрансферазы из Vigna angularis (ADGT-9) и из Oryza sativa, обладающие гомологией к DOGT1, были также включены в сопоставительный анализ на аминокислотном уровне, показанный на фиг.2. Участки с высокой степенью сходства наблюдались и на амино-, и на карбокси-терминальных доменах рассчитанных аминокислотных последовательностей. На фиг.2 показаны гипотетические участки связывания субстрата-акцептора (38) и консенсусная последовательность для UGT (33).
Экспрессия DOGT1 регулируется в процессе развития и индуцируется под действием DON и других соединений, связанных с реакцией на стресс. Для определения того, регулируется ли экспрессия DOGT1 по механизму, аналогичному описанному ранее для родственных генов у других видов растений, ORF репортерного гена β-глюкуронидазы помещают за промотором DOGT1 (PDOGT1-GUS). Тканеспецифичная экспрессия транскрипционного GUS-содержащего гибрида определяют гистохимически в трансгенной линии Arabidopsis, гомозиготной для данного гена слияния. Результаты, показанные на фиг.3, демонстрируют, что экспрессия DOGT1 регулируется в ходе развития и в целом является относительно низкой. В проростках активность GUS наблюдается специфично в корнях и гипокотиле, при этом наиболее сильная экспрессия отмечается в сосудистой системе, в меристематической ткани кончиков корешков (в первичном корне, а также в боковых корнях) и в сосудистой системе гипокотиля, сразу после прорастания. Окрашивание сосудистой сетки существенно снижается на более поздних фазах развития и при этом в эпидермальных клетках корней наблюдается очаговая картина окрашивания. У взрослых растений активность GUS выявляется на поздних стадиях развития цветов в лепестках и зонах осыпания (фиг.3А).
Показано, что экспозиция проростков с DON (5 м.д. в течение 4 часов) или с этиленовым предшественником аминоциклопропилкарбоновой кислоты (АЦК, 2мМ в течение 1 часа) индуцирует экспрессию PDOGT1-GUS (фиг.3). Не отмечается индукции экспрессии репортера при обработке СК (200 мкМ в течение 12 часов) или при обработке жасмоновой кислотой (ЖК, 50 мкМ в течение 1 часа). Был применен полуколичественный анализ по методу ПРЦ с обратной транскриптазой для подтверждения результатов, полученных при анализе GUS-репортера путем выявления изменений на уровне мРНК для DOGT1, 73С4 и 73С6 после обработки теми же концентрациями DON, СК, АЦК и ЖК, как указано выше.
Как показано на фиг.3С, результаты анализа по методу ПЦР с использованием обратной транскриптазы подтверждают наличие DON-индуцируемой экспрессии DOGT1, ранее выявленной с репортерной конструкцией, и в этой связи показывают, что две других UGT также могут индуцироваться под действием DON. Повышение количества транскриптов наблюдается сразу по прошествии 1 часа инкубации с токсином, достигая пика к 4 часам, и вновь снижается в период от 6 до 12 часов. Интересно отметить, что 73С6 демонстрирует более сильную индукцию экспрессии в ответ на воздействие DON, чем DOGT1 или 73С4, хотя приведенные ниже данные указывают на то, что он не может акцептировать токсин в качестве субстрата. Через 4 часа после обработки СК отмечается экспрессия DOGT1, 73С4 и 73С6 на низких уровнях, которая несколько возрастает к 12 часам. Было отмечено, что СК в концентрации 200 мкМ индуцирует экспрессию трех генов относительно слабо. Обработка жасмоновой кислотой и АЦК также ведет к слабой индукции экспрессии DOGT1 и UGT73С6, которая определяется уже через 1 час после обработки, но быстро снижается без накопления выявляемых транскриптов после 2 часов экспозиции с соединениями (фиг.3С).
Фенотипическое определение спектра трихотеценовой резистентности у дрожжей. Токсичность трихотеценов для животных зависит от характера гидроксилирования молекулы трихотецена, а также от локализации, количества и сложности явлений этерификации (5). Основная структура молекулы трихотецена с иллюстрацией системы нумерации атомов углерода в ней показаны на фиг.1А. Представители подкласса В (например, DON или NIV) содержат кетогруппу при углероде С-8, тогда как трихотецены типа А (например, высокотоксичный токсин Т-2, продуцируемый F. sporotrichoides) ее не имеют. Чрезвычайно токсичными являются макроциклические трихотецены, такие как веруккарин А, которые содержат макроциклическое кольцо со сложноэфирными мостиковыми связями между углеродом 4 и углеродом 15. Дрожжевые мутанты pdr5 обладают гиперчувствительностью ко всем исследованным трихотеценам, предоставляя возможность исследовать способность DOGT1 и других генов в кластере, придавать резистентность к разному числу трихотеценов.
Высокое сходство UGT73C1, C2, C3, C4, C5 (DOGT1) и С6 и их кластерная организация в структуры хромосомы позволяют полагать, что они вовлекались в процесс генной дупликации гена-предшественника и могут в этой связи иметь сходные ферментативные свойства. Для анализа возможной функциональной близости проводят реакции амплификации шести ORF со специфическими праймерами с последующей экспрессией в дрожжах в виде белков слияния с c-Myc-меченным по N-концу эпитопом. Сравнение трансформантов, экспрессирующих меченный или немеченный DOGT1, показывает, что эпитоп не мешает проявлению DON-протекторной активности. Дрожжевые трансформанты, содержащие полный генный набор кластера, вносят в среды, содержащие возрастающие концентрации различных трихотеценов. Трансформанты, содержащие пустой вектор экспрессии, используют в качестве контроля.
Результаты, приведенные на фиг.1, показывают, что UGT73C4, как и DOGT1, придает резистентность к DON. Хотя С4 характеризуется лишь вторым по степени выраженности сходством (79% идентичности к DOGT1), он обладает способностью к детоксикации DON и 15-ADON (фиг.1С), но не может придавать резистентность к 3-ADON, NIV, TTC, HT-2-токсину, Т2-токсину, DAS и VA. И наоборот, UGT73C6, который характеризуется идентичностью на аминокислотном уровне в 89% и демонстрирует в этой связи наивысший уровень сходства, не защищает ни от одного из исследованных трихотеценов. Аналогично 73С3 не способен придавать резистентность ко всем исследованным микотоксинам, хотя все четыре белка экспрессируются на близком уровне в дрожжах. Конструкции 73С1 и 73С2 не повышают резистентность к токсину, однако это также не определяется низким уровнем рекомбинантного белка, имеющегося в дрожжевых трансформантах (фиг.1D).
Специфичность по детоксикации DON локализована в N-терминальной части DOGT1. Различные характеристики близкородственных ферментов, несмотря на их высокое сходство по последовательности, дают возможность выяснить те особенности белков, которые являются решающими для распознавания DON. Сайт EcoRI, присутствующий как в DOGT1, так и в 73С6 (см. фиг.4А), был использован для конструирования гибридов, содержащих N-терминальную часть одного и C-терминальную часть соответствующего другого гена. Экспрессия в дрожжах и экспозиция с DON-содержащими средами показали, что N-конец DOGT1 (состоящий из первых 189 аминокислот) является обязательным для проявления способности к детоксикации DON (фиг.4В).
Лизиновый остаток в положении 136 в DOGT1 является консервативным для детоксифицирующего действия в случае 73С4, однако он замещается другими аминокислотами в двух других, не способных к детоксикации гомологах С3 и С6. Сайт-направленный мутагенез был использован для введения мутаций в DOGT1 и в UGT73C6, ведущих к замене лизинового остатка 136 в DOGT1 на глютаминовую кислоту, которая присутствует в соответствующем положении в С6 (аминокислота 137). Мутант DOGT1К136Е, имеющий тот же уровень защиты, что и DOGT1, иллюстрирует тот факт, что К136 не является обязательным для проявления его способности к детоксикации DON. Кроме того, обратная конструкция 73С6Е137К остается неактивной.
Анализ in vivo и in vitro подтверждает, что DOGT1 катализирует перенос глюкозы от УДФ-глюкозы специфично в 3-OH положение DON. Факты создания защиты против 15-ADON и невозможность достичь резистентности против 3-ADON позволяют полагать, что белок DOGT1 может катализировать образование DON-3-О-глюкозида. Для проверки этой гипотезы вначале был проведен химический синтез производных DON, в которых глюкозный фрагмент присоединен либо к С3-, либо к С15-гидроксильной группе DON. Оба продукта оценивают с помощью методов LC-MS/MS анализа. DON-3-О-глюкозид и DON-15-О-глюкозид элюируются на 12,43 минуте и 12,68 минуте соответственно. Масс-спектральный анализ глюкозидов демонстрирует наличие в них характерных различий по характеру фрагментации (фиг.5). Тогда как DON-3-О-глюкозид фрагментируется в данных условиях с образованием иона со значением 427,2 m/z, то этот ион не выявляется для DON-15-О-глюкозида. Потеря 30 единиц атомной массы может быть объяснена расщеплением -CH2OH группы при С6, что не происходит в том случае, когда гидроксильная группа конъюгирована с глюкозой в молекуле DON-15-О-глюкозида. Дальнейшее расщепление (MS) DON-глюкозида в системе линейного захвата ионов демонстрирует почти идентичный характер фрагментации, как и в случае [DON-H]- иона (295,3 m/z, не фрагментированный в Q2), что подтверждает наличие DON структур в реакционных продуктах.
Имея в руках указанные инструменты исследования, можно непосредственно определять, какой глюкозид образуется в дрожжевых клетках. Штамм дрожжей YZGA515, не способный к преобразованию DON в 3-DON из-за делетирования гена дрожжевой ацетилтрансферазы AYT1, трансформируют вектором экспрессии DOGT1 и плазмидой, содержащей ген, кодирующий нечувствительный к трихотецену мутантный рибосомальный белок L3, как мишень трихотеценов, для повышения DON-толерантности дрожжевых клеток. После инкубации полученного штамма с высокими количествами DON, достигающими в итоге таких концентраций в среде, как 1000 м.д., DON-метаболит экстрагируют из клеток и идентифицируют методами ВЭЖХ и масс-спектроскопии наличие прогнозируемого 3-О-глюкопиранозил-4-дезоксиниваленола (фиг.6А). На фиг.5 показаны характер фрагментации синтезированных эталонных веществ и пик продукта, характерные для дрожжей, экспрессирующих DOGT1. Как и ожидалось, данный метаболит не присутствует в контрольном штамме, который не демонстрирует DOGT1 активности.
Для дополнительного подтверждения субстратной специфичности конструируют гибрид GST-DOGT1. Слитый ген для GST-DOGT1 также экспрессируется в дрожжах, которым он придает резистентность к DON, как и в случае DOGT1 дикого типа. Генный продукт подвергают экспрессии в E. coli и очищают методами аффинной хроматографии. Реакционные продукты, получаемые in vitro в ходе инкубации либо белка слияния на основе DOGT1, либо GST с УДФ [14C]-глюкозой и DON, анализируют по методу ТСХ. Пятно с тем же самым значением Rf, что и синтезированный DON-глюкозид, выявляется в реакционной среде, содержащей белок слияния GST-DOGT1, но не в контроле.
Глюкозилирование 4-дезоксиниваленола существенно снижает его токсичность. Для анализа того, обладает ли 3-β-D-глюкопиранозил-4-дезоксиниваленол меньшей фитотоксичностью, чем DON, используют экстракт зародышей пшеницы, введенный в систему транскрипции/трансляции in vitro. Как показано на фиг.6В, наличие 1 мкМ DON в реакционной среде существенно ингибирует трансляцию белка, что снижает активность репортера для фермента до 36,8% в сравнении с контролем (принятым за 100%). Использование токсина в количестве 4 мкМ дает лишь 3,1% люминесценции, тогда как 20 мкМ синтезированного DON-3-О-глюкозида ингибирует люциферазную активность лишь на 8%. Данные результаты подтверждают, что глюкозилирование DON и есть процесс детоксикации.
Суперэкспрессия DOGT1 в Arabidopsis thaliana повышает резистентность к DON. Получают трансгенный Arabidopsis thaliana, конститутивно экспрессирующий DOGT1 под контролем тандемного 35S промотора, и определяют количество рекомбинантного белка в трансформантах по методу вестерн-блоттинга с использованием c-Myc-меченного по N-концу эпитопа. Семена гомозиготной линии 1319/2, которые, как было показано, характеризуются высоким уровнем экспрессии DOGT1 (фиг.6С), и штамм дикого типа Col-0, взятый в качестве контроля, проращивают на MS средах, содержащих 5-30 м.д. DON, и затем растят в течение 5 дней, после чего проводят фенотипический анализ.
Основным наблюдаемым фитотоксичным эффектом был низкий уровень прорастания растений дикого типа. Формирование корней в них не происходит вовсе, котиледоны развиваются, но начинают бледнеть еще до формирования истинных листьев. После 3 недель экспозиции с DON большая часть проростков дикого типа теряет практически все зеленое окрашивание и прекращает рост. Линии суперэкспрессии DOGT1 демонстрируют также явную задержку развития. В сравнении с вариантами дикого типа прорастание у них начинается раньше, корни формируются, котиледоны не обесцвечиваются и появляются истинные листья. Наблюдаемые различия по DON-чувствительности были самыми заметными в средах, содержащих 15 м.д. токсина (фиг.6D). Контрольные трансформанты с пустым вектором (содержащим только ген резистентности к гентамицину) и линии, экспрессирующие низкие количества DOGT1, были чувствительны в той же мере, что и линия дикого типа Col-0.
ОБСУЖДЕНИЕ
Биотехнологическая значимость. Болезни пшеницы и ячменя, вызываемые видами Fusarium, имеют огромное экономическое значение для стран во всем мире. Например, Соединенные Штаты были серьезно поражены эпидемией фузариоза за последние десятилетия. Непосредственные потери производителей пшеницы в США, связанные с FHB, исчисляются в среднем примерно 260 млн долларов в год, а в период 1998-2000 гг. общий экономический ущерб малых производителей зерновых исчислялся на уровне 2,7 млрд долларов (39). Дети относятся к группе населения, наиболее подверженной риску превышения допустимого уровня переносимого дневного поступления (TDI) DON. В проблемном в данном отношении 1998 г. у 80% детей до 1 года жизни в Нидерландах отмечалось превышение уровня TDI.
Высокая значимость болезней, вызываемых видами Fusarium, обосновывает необходимость работ, направленных на исследование способов инактивации микотоксинов, хотя резистентность против патогена является, скорее всего, полигенным фактором у культивируемых растений, тогда как резистентность к токсину представляет собой один из ее компонентов. Продукция трихотеценов является единственным фактором вирулентности грибного патогена, что было экспериментально подтверждено, за исключением плейотропных мутаций в MAP-киназе, которые воздействуют не только на вирулентность, но охватывают такие процессы, как образование конидий и перитеций, вегетативный рост и продукция микотоксинов (40, 41). Открытие авторами настоящего изобретения того факта, что суперэкспрессия DOGT1 ведет к повышенной резистентности к дезоксиниваленолу в трансгенных растениях Arabidopsis, открывает новые возможности для разработки биотехнологических подходов, направленных на антагонизацию фактора вирулентности грибов. Единственный опубликованный результат основывается на данных по ацетилтрансферазе Fusarium (42), которая превращает DON в 3-ADON и который примерно в 2 раза менее токсичен для лабораторных животных, так что при этом отсутствует потребность в детоксикации, понимаемой в контексте настоящего изобретения. Фенотип трансформированных дрожжей и Arabidopsis, а также данные, полученные с использованием систем трансляции in vitro, показывают, что DON-глюкозид демонстрирует сниженную токсичность. В основном, процесс глюкозилирования превращает реакционные и токсичные аглюконы в стабильные и нереактивные формы для хранения, ограничивая тем самым их взаимодействие с другими клеточными компонентами. Добавление сахара блокирует реакционную способность сайта для данного вещества и, следовательно, снижает его токсичность для растения. Такие модификации в этой связи рассматриваются, как возможность доступа к связанным с мембраной переносчикам, которые могут осуществить вывод из цитозоля, например, к клеточной стенке или к вакуоли (43).
Глюкозилирование трихотеценов отражает процесс детоксикации в растениях. Однако в пищеварительном тракте человека и животных микотоксинсодержащие глюкоконъюгаты могут быть легко гидролизованы с регенерацией токсина. Уровень транспортировки DON-глюкозида в вакуоль или апопласт в настоящее время не известен. Перед тем, как попытаться использовать трансгенные растения, осуществляющие суперэкспрессию DON-глюкозилтрансферазы, нужно понять, только лишь вакуолярный глюкозид или также токсин, связанный с материалом клеточной стенки в качестве «нерастворимого остатка», является источником «замаскированного микотоксина» (44), который не удается определить с помощью традиционных методик анализа и который может иметь значительно более серьезное токсикологическое воздействие, чем это в настоящее время предполагается. Аналитические инструменты и эталонные материалы, разработанные в настоящем изобретении, подходят для решения данных вопросов.
Инактивация токсинов путем глюкозилирования представляет собой, по всей видимости, важнейший природный механизм резистентности, позволяющий растениям взаимодействовать с огромным множеством токсичных микробных метаболитов, с которыми они могут сталкиваться в природе. DOGT1 представляет собой один из 118 UGT генов в A. thaliana, который, как предполагается, осуществляет конъюгацию малых молекул.
Экспрессия UGT генов в дрожжах представляет собой ценный подход в рамках комплементарной стратегии на основе широко используемой системы экспрессии в E. coli, в особенности в том случае, когда соответствующие химические вещества имеют мишени только в эукариотах. Одна из проблем заключается в том, что дрожжевые клетки дикого типа часто являются «непроницаемыми» для интересующих веществ. Инактивирование нескольких ABC-переносчиков в данном хозяйском штамме является исходной предпосылкой при выборе подхода, используемого для селекции кДНК, придающих резистентность. В случае трихотеценов это позволяет осуществлять фенотипическое выявление активности экспрессированных генов в простом чашечном тесте. В принципе, данный подход может быть адаптирован ко множеству других веществ.
Эволюция гена и субстратная специфичность. DOGT1 локализована в кластере (фиг.1В) вместе с 5 другими представителями семейства 73С на хромосоме II (At2g36800), указывая на генную дупликацию, осуществляемую за счет неоднородной рекомбинации (32). Можно предположить, что такая генная амплификация обеспечивает селективное преимущество в сравнении с токсиновым стрессом. DOGT1 и другие представители данного кластера имеют очень близкие белковые последовательности, что указывает на возможную избыточность ферментативной функции. Однако при тестировании гомологов с точки зрения детоксикации трихотеценов было показано, что штамм дрожжей, экспрессирующий ген с наивысшим уровнем сходства по последовательности с UGT73C6, был столь же чувствителен, как и ген дикого типа, ко всем исследованным токсинам (так же, как и UGT73C3), тогда как второй по уровню сходства гомолог UGT73C4 демонстрировал те же свойства, что и DOGT1. Ферментативная активность в отношении различных гидроксикумаринов была продемонстрирована для всех представителей кластера 73С (45), указывая на то, что UGT, не обладающие DON-протекторной активностью, не просто утеряли аллели для данной функции. Кластер в этой связи может рассматриваться в контексте данных, поддерживающих гипотезу, согласно которой дуплицированные UGT гены могут приобретать новые субстратные специфичности и функции в ходе эволюции.
Растительные УДФ-глюкозилтрансферазы структурно очень близки по своим карбокси-терминальным характерным мотивам с UGT млекопитающих, которые используют УДФ-глюкуроновую кислоту вместо УДФ-глюкозы в качестве субстрата-донора. Ферменты млекопитающих играют центральную роль в метаболизме и детоксикации таких химических веществ, как канцерогены или гидрофобные агенты. Было показано, что UGT высших растений осуществляют также параллельную группу активностей, способных модифицировать ксенобиотики, такие как гербициды и другие пестициды (46,47). [Один открытый вопрос заключается в том, являются ли эти реакции детоксикации побочной активностью UGT, осуществляющих конъюгацию эндогенных соединений растений. Хотя последние публикации поддерживают гипотезу, согласно которой UGT, осуществляющие преобразование эндогенных субстратов, могут также отвечать за способность растений детоксифицировать ксенобиотики (48), этот вопрос нуждается в дальнейших исследованиях для его окончательного разрешения].
Выявление того факта, что DOGT1 защищает против DON, но не против NIV, который содержит одну дополнительную гидроксильную группу в положении 4 остатка (фиг.1А), указывает на то, что вариация в характере гидроксилирования трихотеценового скелета представляет собой важный механизм, позволяющий грибам избежать такого рода реакций детоксикации. Неизвестно, могут ли растения также осуществлять реакцию конъюгации с NIV, который более токсичен, чем DON в животных системах (5). Отсутствие активности против NIV является серьезным аргументом против использования одного DOGT1-подобного гена в трансгенных растениях, поскольку NIV-продуцирующие хемотипы F. graminearum могут иметь селективное преимущество.
Возможность конструирования гибридных генов путем перетасовки N- и C-терминальных частей DOGT1 и UGTN73С6 и их экспрессия в дрожжах подтверждают тот факт, что наличие N-терминальных 189 аминокислот в DOGT1 обязательно для проявлений их способности к детоксикации DON, что согласуется с гипотезой, согласно которой связывание субстрата может происходить в консервативной гидрофобной области, локализованной между аминокислотами 130 и 150 (38). Сравнение двух генов, обладающих протекторным эффектом, и двух генов, не демонстрирующих такой активности, но со сходным уровнем экспрессии, дает основание полагать, что лизин, который присутствует в положении 136 аминокислоты в DOGT1 (и в соответствующем положении в UGT73С4), но который замещен в двух других UGT, не способных к детоксикации трихотеценов, может быть важен для детоксикации DON. Однако эта гипотеза была опровергнута при использовании сайт-направленного мутагенеза и экспрессии сконструированных белков в дрожжах.
Информация по специфическим доменам или аминокислотам, вовлекаемым при воздействии DOGT1 в связывание с акцептором, может быть потенциально полезной для поиска глюкозилтрансфераз, способных к детоксикации DON в зерновых растениях, таких как пшеница или кукуруза. Следует также отметить, что попытки объяснить функцию UGT исключительно на основе сходства на аминокислотном уровне ошибочны, как это было показано для случая 73С кластера. Тем не менее, функциональность предполагаемых генов может быть легко проверена при проведении их экспрессии в дрожжах. Одна возможность, которую нельзя исключить, связана с тем, что глюкозиды трихотеценов формируются в зерновых растениях при воздействии структурно весьма различающихся UGT. Например, UGT84В1, как было показано, демонстрирует наивысшую активность in vitro против ауксина, индол-3-уксусной кислоты (ИУК) из всех исследованных UGT A. thaliana, хотя этот фермент не демонстрирует наивысшего сходства по последовательности с функционально гомологичным геном iaglu в кукурузе (49).
Регуляция экспрессии гена. Было показано, что гены с наивысшим уровнем сходства по последовательности к DOGT1 из растений табака и томата, индуцируются салициловой кислотой или повреждающим воздействием (34, 36, 37). Анализ экспрессии DOGT1 и 73С6 после обработки СК и ЖК показывает, что они отвечают на СК, ЖК и этиленовый предшественник АЦК повышением через неделю уровня мРНК. Индуцибельность генной экспрессии под действием СК, ЖК или этилена рассматривается как индикатор возможной роли позитивно регулируемого генного продукта в растениях на стресс или в механизме защитной реакции (50).
При использовании анализа уровней мРНК и конструкции GUS-репортер оказалось возможным продемонстрировать, что транскрипция DOGT1 в A. thaliana дикого типа регулируется в ходе развития и быстро и в значительной мере индуцируется в ответ на воздействие DON. Индукция экспрессии микотоксином также наблюдалась для других представителей кластера 73С, независимо от наличия у них DON-протекторной активности. Было бы интересно выяснить, специфична ли такая индуцибельность для данного агента или она является общим феноменом, характерным для ингибиторов биосинтеза белка. «Риботоксичная реакция на стресс» в настоящее время активно изучается исследователями на клетках человека (51), и было показано, что микотоксин индуцирует экспрессию циклооксигеназы-2, ключевого фермента синтеза медиаторов воспалительной реакции через сигнальный механизм, опосредованный MAP-киназой. DON-индуцибельная экспрессия DOGT1 является привлекательной исходной точкой для изучения того, существуют ли подобные механизмы в растениях.
В настоящем изобретении было показано, что огромное генноесемейство глюкозилтрансфераз, в особенности, как показано в примерах, UGT играет важную роль во взаимодействии с растительным патогеном за счет участия их в детоксикации метаболитов, продуцируемых микробами, для повышения их вирулентности в отношении хозяйских организмов. Аналогично огромному семейству генов резистентности, относящихся к типу лицин-обогащенных повторов (52), вовлекаемых в распознавание патогенов, генная амплификация и соответствующий отбор UGT может привести к достижению широкого спектра защиты против грибных токсинов. Селективное давление, направленное на спасение от такого рода реакций глюкозилирования, может быть движущей силой в эволюции микробных биосинтетических реакций, ведущих к образованию широкого спектра токсиновых структур, как это было показано для трихотеценов.
СПИСОК ЦИТИРОВАННОЙ ЛИТЕРАТУРЫ
1. McMullen, M.P., Jones, R. and Gallenberg, D. (1997) Plant Dis. 81, 1340-1348
2. Grove J. F. (1996) Progress in the Chemistry of Organic Natural Products, pp. 1-70, Springer, Wien
3. Cundliffe, E. and Davies, J.E. (1977) Agents Chemother. 11, 491-499
4. O'Donnel, K., Kistler, H.C., Tacke, B.K. and Casper, H.H. (2000) PNAS 97, 7905-7910
5. Betina, V. (1989) Mycotoxins - chemical, biological and environmental aspects, pp. 192-241, Elsevier, Amsterdam
6. Canady, R.A., Coker, R.D., Egan, K., Krska, R., Kuiper-Good-man, T., Olsen M., Pestka, J., Resnik, S., & Schlatter, J. (2001) WHO Food Additives Series 47, 419-528
7. Codex Committee on Food Additives and Contaminants (2002) Joint FAO/WHO Food Standards Programme, CX/FAC 03/35, www.codex-alimentarius.net
8. McCormick, S.P. (2003) Fusarium head blight of wheat, pp.165-183, American Phytopathological Society, St. Paul, Minnesota, USA
9. Desjardins, A.E., Procter, R.H., Bai, G., McCormick, S.P., Shaner, G., Buechley, G. and Hohn, T. (1996) Mol. Plant-Microbe Interact. 9, 775-781
10. Bai, G.H, Desjardins, A.E. and Plattner R.D. (2002) My-copathologia 153, 91-98
11. Kang, Z. and Buchenauer, H. (1999) Physiol. Molec. Plant Pathol. 55, 275-288
12. Mesterhazy, A. (2003) Fusarium head blight of wheat, pp. 363-380, American Phytopathological Society, St. Paul, Minnesota, USA
13. Coleman, J.O.D., Blake-Kalff, M.M.A. and Davies, T.G.E. (1997) Trends Plant Sci. 2, 144-151
14. Scott, P.M., Nelson, K., Kanhere, S.R., Karpinski, K.F., Hayward, S., Neish, G.A. and Teich, A. (1984) Appl. Environ. Microbiol. 48, 884-886
15. Miller, J.D. and Arnison, P.G. (1986) Can J Plant Path 8, 147-150
16. Sewald, N., von Gleissenthall, J. L., Schuster, M., Müller,G. and Aplin, R.T. (1992) Tetrahedron Asymmetry 3, 953-960
17. Ross, J., Yi, L., Lim, E.-K. and Bowles, D. (2001) Genome Biol. 2, 3004.1-3004.6.
18. Sikorski, R.S. and Hieter, P. (1989) Genetics 122, 19-27
19. Murashige, T. and Skoog, F. (1962) Plant Physiol. 15, 473-497
20. Minet, M., Dufour, M.-E. and Lacroute, F. (1992) Plant J. 2, 417-422
21. Ballester, R., Michaeli, T., Ferguson, K., Xu, H.-P., Mc-Cormick F, and Wigler, M. (1989), Cell 59, 681-686
22. Egner, R., Mahe, Y., Pandjaitan, R. and Kuchler, K. (1995) Mol Cell Biol 15, 5879-5887
23. Pogulis, R.J., Vallejo, A.N. and Pease, L.R. (1996) Methods Mol Biol 57, 167-176
24. Savard, M. (1991) J. Agr. Food Chem., 39, 570-574
25. Hajdukiewicz, P., Svab, Z. and Maliga, P. (1994) Plant Mol Biol 25, 989-994
26. Töpfer, R., Matzeit, V., Groneborn, B., Schell, J. and Steinbiss H.-H. (1987) Nucleic Acid Research 14, 5890
27. Diener, A.C., Li, H., Zhou, W., Whoriskey, W.J., Nes, W.D. and Fink, G.R. (2000) Plant Cell 12, 853-870
28. Farrand, S.K., O'Morchoe, S.P. and McCutchan, J. (1989) J Bacteriol 171, 5314-5321
29. Koncz, C. and Schell, J. (1986) Mol Gen Genet 204, 383-396
30. Clough, S.J. and Bent, A.F. (1998) Plant J. 16, 735-743
31. Jefferson, R.A. (1987) Plant Mol. Biol. Rep. 5, 387-405
32. Wolfger, H., Mamnun, Y.M. and Kuchler, K. (2001) Res Microbiol. 152, 375-89
33. Li, Y., Baldauf, S., Lim, E.-K., and Bowles, D.J. (2001) J.Biol Chem 276, 4338-4343
34. Fraissinet-Tachet, L., Baltz, R., Chong, J., Kauffmann, S., Fritig, B. and Saindrenan, P. (1998) FEBS 437, 319-323
35. Horvath, D.M. and Chua, N.-H. (1996) Plant Mol Biol 31, 1061-1072
36. Truesdale, M.R., Doherty, H.M., Loake, G.J., McPherson, M.J., Roberts, M.R. and Bowles, D.J. (1996) Plant Physiol 112, 446
37. Horvath, D., Huang, D.J. and Chua, N.-H. (1998) Mol. Plant-Microbe Interact. 11, 895-905
38. Vogt, T., Grimm, R. and Strack, D. (1999) Plant J 19, 509-519
39. Nganje, W.E., Johnson, D.D., Wilson, W.W., Leistritz, F.L., Bangsund, D.A. and Tiapo, N.M. (2001) Agribusiness and Applied Economics Report 464
40. Jenczmionka, N.J., Maier, F.J., Losch, A.P. and Schafer, W. (2003) Curr. Genet. 43, 87-95
41. Hou, Z., Xue, C, Peng, Y., Katan, T., Kistler, H.C. and Xu, J.R. (2002) Mol Plant Microbe Interact. 15, 1119-1127
42. Okubara, P.A., Blechl, A.E., McCormick, S.P., Alexander, N.J., Dill-Macky, R. and Hohn, T.M. (2002) Theor Appl Genet. 106, 74-83
43. Jones, P. and Vogt, T. (2001) Planta 213, 164-174
44. Engelhardt, G., Ruhland, M. and Wallnofer, P.R. (1999) Adv. Food Sci. 21, 71-78
45. Lim, E.-K., Baldauf, S., Li, Y., Elias, L., Worrall, D., Spencer, S.P., Jackson, R.G., Taguchi, G., Ross, J. and Bowles, D. (2003) glucobiol. 13, 139-145
46. Wetzel, A. and Sandermann, H. (1994) Arch. Biochem. Biophys. 314, 323-328
47. Leah, J.M., Worral, T.L. and Cobb, A.H. (1992) Pest. Science 34, 81-87
48. Mebner, B., Thulke, O. and Schäffner, A.R. (2003) Planta 217, 138-146
49. Jackson, R.G., Lim, E.-K., Li, Y., Kowalczyk, M., Sandberg, G., Hoggett, J., Ashford, D.A. and Bowles, D. (2001) J. Biol. Chem. 276, 4350-4356
50. Reymond, P. and Farmer, E.E. (1998) Curr Opin Plant Biol. 1, 404-411
51. Laskin, J.D., Heck, D.E. and Laskin, D.L. (2002) Toxicol Sci. 69, 289-291
52. Meyers, B.C., Kozik, A.., Griego, A., Kuang, H. and Michel-more, R.W. (2003) Plant Cell 15, 809-834
53. Eudes F., Comeau, A., Rioux S. and Collin J. (2000) Can. J. Plant Path. 22, 286-292
54. Potrykus, I. and Spangenberg, G. (Eds.): Gene Transfer to Plants. Springer Verlag Berlin (1995), ISBN 3-54058406-4
Сокращения:
FHB - фузариоз колосьев; DON - дезоксиниваленол; 15-ADON - 15-ацетил-дезоксиниваленол; NIV - ниваленол; UGT - УДФ-глюкозилтрансфераза; ABC - АТФ-связывающая кассета; DOGT1 - DON-глюкозилтрансфераза; ORF - открытая рамка считывания; GUS - β-глюкуронидаза; СК (SA) - салициловая кислота; ЖК (JA) - жасмоновая кислота; АЦС (ACC) - 1-аминоциклопропилкарбоновая кислота; TDI - переносимое дневное поступление; ИУК (IAA) - индол-3-уксусная кислота; DAG - дни после прорастания.
Реферат
Изобретение относится к биотехнологии и представляет собой способ детоксикации микотоксинов, согласно которому микотоксин подвергают взаимодействию с глюкозилтрансферазой в присутствии активированной глюкозы. Изобретение позволяет эффективно снизить токсичность микотоксинов. 17 з.п. ф-лы, 6 ил.
Комментарии