Микроорганизмы для получения путресцина или орнитина и способ получения путресцина или орнитина с использованием указанных микроорганизмов - RU2708165C2
Код документа: RU2708165C2
Чертежи

Описание
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к рекомбинантному микроорганизму, продуцирующему путресцин или орнитин, и к способу получения путресцина или орнитина с использованием указанного микроорганизма.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Биогенные амины (БА) представляют собой соединения азота, получаемые, главным образом, посредством декарбоксилирования аминокислот или аминирования и трансаминирования альдегидов и кетонов. Эти биогенные амины имеют малую молекулярную массу и синтезируются во время метаболических процессов у микроорганизмов, растений и животных, поэтому они известны как составные элементы, часто обнаруживаемые в их клетках.
Среди них, путресцин обнаруживают у грамотрицательных бактерий или грибов, и он присутствует в высокой концентрации у различных видов, и поэтому полагают, что путресцин играет важную роль в метаболизме микроорганизмов. В целом, путресцин является важным сырьем для синтеза полиамина нейлона-4,6, и его получают преимущественно химическим синтезом. Химический синтез представляет собой 3-хстадийный процесс, включающий реакцию каталитического окисления, реакцию с использованием цианидного соединения и реакцию гидрогенизации с использованием высокого давления водорода. Соответственно, существует потребность в разработке более экологически безопасного и энергоэффективного способа, включающего использование биомассы.
С учетом этих обстоятельств были описаны различные способы получения путресцина в высокой концентрации трансформацией Е. coli и микроорганизма рода Corynebacterium (публикация международной заявки на патент № WO 06/005603; публикация международной заявки на патент № WO 09/125924; Qian ZD et al., Biotechnol. Bioeng. 104 (4): 651-662, 2009; Schneider et al., Appl. Microbiol. Biotechnol. 88 (4): 859-868, 2010; Schneider et al., Appl. Microbiol. Biotechnol. 95: 169-178, 2012).
С другой стороны орнитин представляет собой вещество, часто обнаруживаемое у растений, животных и микроорганизмов, и является предшественником для биосинтеза аргинина, пролина и полиамина. Кроме того, орнитин играет важную роль в метаболическом пути образования мочевины из аминокислот или аммиака и ее выведения посредством орнитинового цикла в процессе метаболизма у высших организмов in vivo. Орнитин эффективен при наращивании мышечной массы и уменьшении жировой массы тела, и поэтому его используют в качестве пищевой добавки, а также в качестве фармацевтического лекарственного средства для лечения цирроза печени и печеночной недостаточности. Способы получения орнитина включают способ с использованием казеина молока и пищеварительного фермента и способ с использованием Е. coli или микроорганизма рода Corynebacterium (патент Кореи №10-1372635; Т. Gotoh et al., Bioprocess Biosyst. Eng., 33: 773-777, 2010).
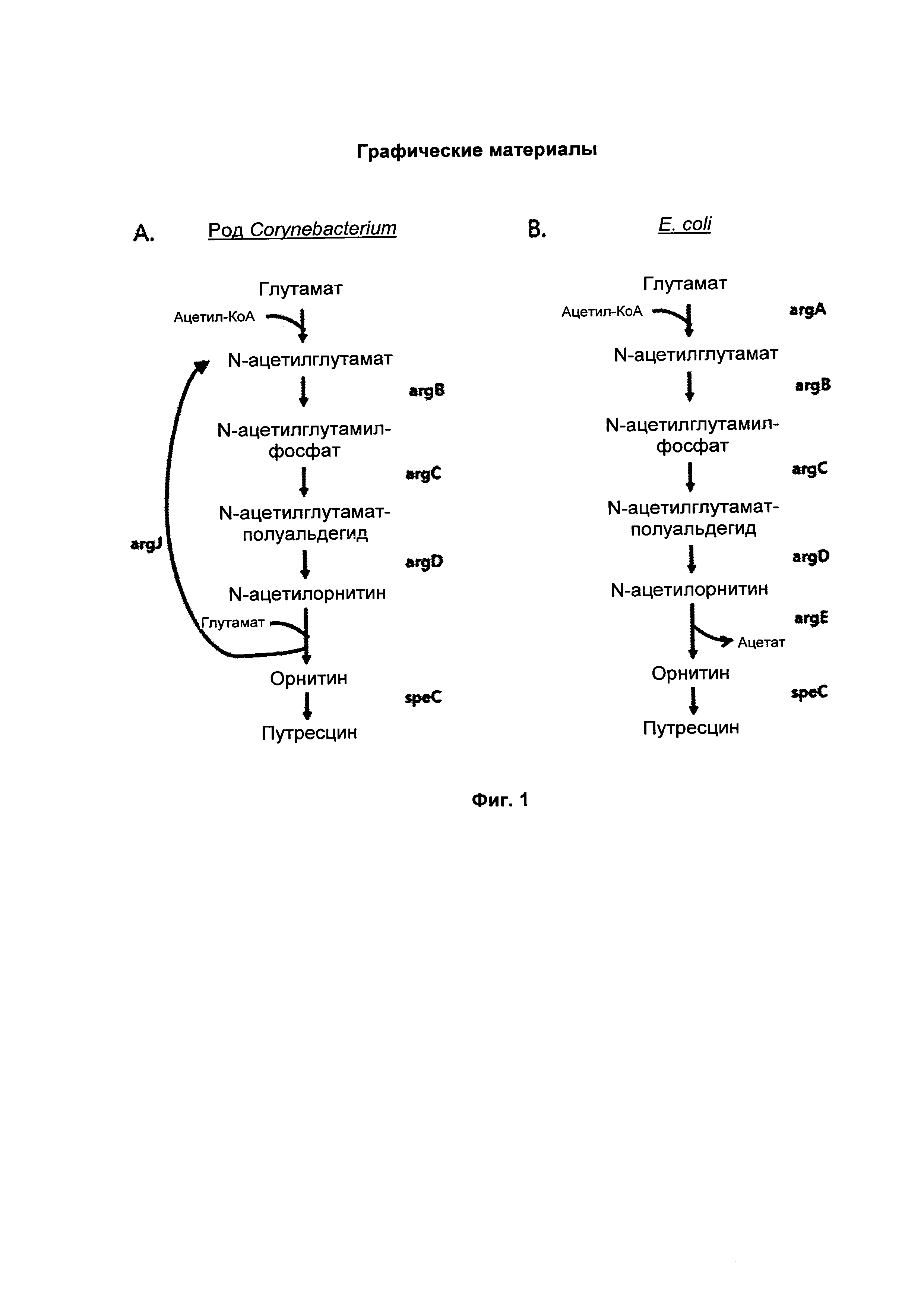
Биосинтетические пути образования путресцина и орнитина у Е. coli и микроорганизма рода Corynebacterium сходны, но также имеют следующие различия. Во-первых, микроорганизм рода Corynebacterium имеет «циклический путь» с превращением глутаминовой кислоты в N-ацетил-L-глутаминовую кислоту и превращением N-ацетил-L-орнитина в L-орнитин посредством argJ (бифункциональной орнитинацетилтрансферазы/N-ацетилглутаматсинтазы, ЕС 2.3.1.35). В отличие от этого, у Е. coli биосинтез путресцина или орнитина проходит по «линейному пути», где функцию argJ микроорганизма рода Corynebacterium выполняют argA (N-ацетилглутаматсинтаза, ЕС 2.3.1.1) и argE (ацетилорнитиндезацетилаза, ЕС 3.5.1.16).
Известно, что в ArgJ микроорганизма рода Corynebacterium происходит повторный обмен ацетильной группой между орнитином и глутаминовой кислотой. Однако у Е. coli ArgA присоединяет ацетильную группу ацетил-КоА к глутамату с образованием N-ацетилглутамата и ArgE обеспечивает распад N-ацетилорнитина с образованием орнитина и ацетата (Schneider et al., Appl. Microbiol. Biotechnol. 91, 17-30, 2011).
В частности, известно, что оперон pta-ackA (pta, фосфотрансацетилаза; ackA, ацетаткиназа) и ацетилкоэнзим-А-синтетаза (acs) являются генами, обеспечивающими синтез ацетил-КоА с использованием ацетата.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Техническая проблема
Авторы настоящего изобретения приложили значительные усилия для улучшения способности микроорганизма рода Corynebacterium продуцировать орнитин и путресцин и, в результате, обнаружили, что введение argA и argE, имеющих происхождение от Е. coli, в микроорганизм рода Corynebacterium может улучшить его способность продуцировать орнитин и путресцин, завершив посредством этого настоящее изобретение.
Техническое решение
Задачей настоящего изобретения является обеспечение рекомбинантного микроорганизма, продуцирующего путресцин или орнитин с высоким выходом.
Другой задачей настоящего изобретения является обеспечение способа получения путресцина или орнитина с использованием указанного выше микроорганизма.
Полезные эффекты изобретения
Была подтверждена возможность увеличения количества путресцина или орнитина, продуцируемого микроорганизмом рода Corynebacterium по настоящему изобретению, продуцирующим путресцин или орнитин, при введении в микроорганизм argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от Е. coli, а также при усилении пути утилизации ацетата. Соответственно, микроорганизм по настоящему изобретению может быть широко использован для получения путресцина или орнитина, а также может быть широко использован в качестве эффективного и желательного, с экономической и экологической точек зрения, средства получения сырья для получения различных полимерных продуктов, где путресцин или орнитин используют в качестве сырья.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На ФИГ. 1 представлена схематическая диаграмма, на которой показан биосинтетический путь (циклический путь) образования путресцина и орнитина у микроорганизма рода Corynebacterium (А) и биосинтетический путь (линейный путь) образования путресцина и орнитина у Е. coli (В).
На ФИГ. 2 представлена схематическая диаграмма, на которой показан биосинтетический путь с улучшением способности к продукции путрецина и орнитина посредством введения argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от Е. coli, в микроорганизм рода Corynebacterium, находящийся в состоянии экспрессии argJ.
НАИЛУЧШИЙ ВАРИАНТ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Согласно одному аспекту настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, в который введена активность N-ацетилглутаматсинтазы из Е. coli и ацетилорнитиндезацетилазы из Е. coli.
Согласно типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где N-ацетилглутаматсинтаза, имеющая происхождение от Е. coli, состоит из аминокислотной последовательности SEQ ID NO: 1.
Согласно другому типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где ацетилорнитиндезацетилаза, имеющая происхождение от Е. coli, состоит из аминокислотной последовательности SEQ ID NO: 3.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, выбранный из группы, состоящей из Corynebacterium glutamicum, Corynebacterium ammoniagenes, Corynebacterium thermoaminogenes, Brevibacterium flavum и Brevibacterium lactofermentum.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность фосфотрансацетилазного и ацетаткиназного оперона (оперона pta-ackA) дополнительно усилена по сравнению с его эндогенной активностью.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где фосфотрансацетилазный и ацетаткиназный оперон состоит из аминокислотной последовательности SEQ ID NO: 5 или 7.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, в который дополнительно введена активность ацетил-КоА-синтетазы (acs), имеющей происхождение от Е. coli.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где ацетил-КоА-синтетаза (acs), имеющая происхождение от Е. coli, состоит из аминокислотной последовательности SEQ ID NO: 9.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, в который дополнительно введена активность орнитиндекарбоксилазы (ODC).
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность (1) орнитинкарбамоилтрансферазы (ArgF), (2) экспортера глутамата или (3) орнитинкарбамоилтрансферазы и экспортера глутамата дополнительно ослаблена по сравнению с их эндогенной активностью.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность по меньшей мере одного, выбранного из группы, состоящей из ацетил-гамма-глутамилфосфатредуктазы (ArgC), ацетилглутаматсинтазы или орнитинацетилтрансферазы (ArgJ), ацетилглутаматкиназы (ArgB) и ацетилорнитинаминотрансферазы (ArgD), дополнительно усилена по сравнению с их эндогенной активностью.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность ацетилтрансферазы дополнительно ослаблена по сравнению с ее эндогенной активностью.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где ацетилтрансфераза состоит из аминокислотной последовательности SEQ ID NO: 30 или 31.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность экспортера путресцина дополнительно усилена по сравнению с его эндогенной активностью.
Согласно еще одному типичному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где экспортер путресцина состоит из аминокислотной последовательности SEQ ID NO: 26 или 28.
Согласно другому аспекту настоящего изобретения предложен способ получения путресцина или орнитина, включающий:
(1) культивирование модифицированного микроорганизма рода Corynebacterium, продуцирующего путресцин или орнитин, в среде; и
(2) выделение путресцина или орнитина из культивированного микроорганизма или среды.
В типичном воплощении настоящего изобретения модифицированный микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
Далее настоящее изобретение будет описано подробно.
Согласно одному воплощению настоящего изобретения предложен модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, в который введены активности N-ацетилглутаматсинтазы, имеющей происхождение от Е. coli, и ацетилорнитиндезацетилазы, имеющей происхождение от Е. coli.
При использовании здесь термин «N-ацетилглутаматсинтаза» относится к ферменту, опосредующему реакцию образования N-ацетилглутамата из глутамата и ацетил-КоА, и образованный таким образом N-ацетилглутамат может быть использован как предшественник орнитина и аргинина.
В настоящем изобретении N-ацетилглутаматсинтаза может включать, например, без ограничения, белок, имеющий аминокислотную последовательность SEQ ID NO: 1, и любой белок, на 70% или более, конкретно, на 80% или более, конкретнее, на 90% или более, еще конкретнее, на 95% или более, еще конкретнее, на 98% или более, и, наиболее конкретно, на 99% или более гомологичный указанной выше аминокислотной последовательности, при условии, что белок обладает существенной N-ацетилглутаматсинтазной активностью.
Кроме того, аминокислотные последовательности белков, демонстрирующих указанную выше активность, могут различаться у разных видов и штаммов микроорганизмов. Соответственно, N-ацетилглутаматсинтаза по настоящему изобретению может представлять собой, например, без ограничения, N-ацетилглутаматсинтазу из Е. coli.
Очевидно, что в качестве последовательностей, гомологичных указанной выше последовательности, в объем настоящего изобретения также следует включать аминокислотные последовательности с делецией, модификацией, заменой или добавлением части последовательности, если биологическая активность этих аминокислотных последовательностей идентична или соответствует биологической активности белка по SEQ ID NO: 1.
Полинуклеотид, кодирующий N-ацетилглутаматсинтазу по настоящему изобретению, может включать, без ограничения, полинуклеотид, кодирующий белок, имеющий аминокислотную последовательность SEQ ID NO: 1, и любой белок, на 70% или более, конкретно, на 80% или более, конкретнее, на 90% или более, еще конкретнее, на 95% или более, еще конкретнее, на 98% или более, и, наиболее конкретно, на 99% или более гомологичный указанной выше аминокислотной последовательности, при условии, что полинуклеотид обладает активностью, сходной с активностью N-ацетилглутаматсинтазы, и, например, может быть включена полинуклеотидная последовательность SEQ ID NO: 2.
При использовании здесь термин «ацетилорнитиндезацетилаза» относится к ферменту, опосредующему реакцию, вовлеченную в образование уксусной кислоты и орнитина, путем опосредования гидролиза ацетилорнитина.
В настоящем изобретении ацетилорнитиндезацетилаза может включать, без ограничения, белок, имеющий аминокислотную последовательность SEQ ID NO: 3, и любой белок, на 70% или более, конкретно, на 80% или более, конкретнее, на 90% или более, еще конкретнее, на 95% или более, еще конкретнее, на 98% или более, и, наиболее конкретно, на 99% или более гомологичный указанной выше аминокислотной последовательности, при условии, что белок обладает существенной активностью по разделению ацетилорнитина на ацетильную группу и орнитин.
Кроме того, аминокислотные последовательности белков, демонстрирующих указанную выше активность, могут различаться у разных видов и штаммов микроорганизмов. Соответственно, ацетилорнитиндезацетилаза по настоящему изобретению может представлять собой, без ограничения, ацетилорнитиндезацетилазу из Е. coli. Очевидно, что в качестве гомологичных последовательностей в объем настоящего изобретения также следует включать аминокислотные последовательности с делецией, модификацией, заменой или добавлением части последовательности, если биологическая активность этих аминокислотных последовательностей идентична или соответствует биологической активности белка по SEQ ID NO: 3.
Полинуклеотид, кодирующий ацетилорнитиндезацетилазу по настоящему изобретению, может включать, при условии, что полинуклеотид обладает активностью, сходной с активностью белка ацетилорнитиндезацетилазы, белок, имеющий аминокислотную последовательность SEQ ID NO: 3, или полинуклеотид, кодирующий белок, на 70% или более, конкретно, на 80% или более, конкретнее, на 90% или более, еще конкретнее, на 95% или более, еще конкретнее, на 98% или более, и, наиболее конкретно, на 99% или более гомологичный указанной выше аминокислотной последовательности, например, полинуклеотидную последовательность SEQ ID NO: 4.
Кроме того, полинуклеотид, кодирующий N-ацетилглутаматсинтазу или ацетилорнитиндезацетилазу по настоящему изобретению, можно гибридизовать с полинуклеотидной последовательностью SEQ ID NO: 2 или SEQ ID NO: 4 или зондом, имеющим происхождение от указанной полинуклеотидной последовательности, в жестких условиях, и он может представлять собой модифицированный тип N-ацетилглутаматсинтазы или ацетилорнитиндезацетилазы, обладающий нормальной функцией. Использованный выше термин «жесткие условия» относится к условиям, обеспечивающим специфичность гибридизации полинуклеотидов. Например, конкретные жесткие условия описаны в литературе (например, J. Sambrook et al., supra).
Использованный выше термин «гомология» относится к степени идентичности данной аминокислотной последовательности или полинуклеотидной последовательности и может быть указан в процентах. При использовании здесь гомологичная последовательность, обладающая такой же активностью, что данная полипептидная последовательность или полинуклеотидная последовательность, или сходной активностью, может быть обозначена посредством термина «% гомологии». Например, % гомологии может быть подтвержден с применением стандартного программного обеспечения, то есть BLAST 2.0, для расчета таких параметров, как показатель, идентичность и сходство, или сравнением последовательностей посредством гибридизационных экспериментов по Саузерну, и подходящие условия гибридизации могут быть определены способом, известным специалисту в данной области (например, J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York).
С другой стороны, микроорганизм по настоящему изобретению может включать как микроорганизмы естественного типа, так и микроорганизмы модифицированного типа, например, микроорганизмы, принадлежащие к роду Escherichia, роду Shigella, роду Citrobacter, роду Salmonella, роду Enterobacter, роду Yersinia, роду Klebsiella, роду Erwinia, роду Corynebacterium, роду Brevibacterium, роду Lactobacillus, роду Selenomanas, роду Vibrio, роду Pseudomonas, роду Streptomyces, роду Arcanobacterium, роду Alcaligenes и так далее. Конкретно, микроорганизм по настоящему изобретению может представлять собой, без ограничения, микроорганизм, принадлежащий к роду Corynebacterium, конкретнее, микроорганизм, выбранный из группы, состоящей из Corynebacterium glutamicum, Corynebacterium ammoniagenes, Corynebacterium thermoaminogenes, Brevibacterium flavum и Brevibacterium lactofermentum, и, конкретнее, Corynebacterium glutamicum.
Конкретно, при использовании здесь термин микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, относится к микроорганизму рода Corynebacterium, продуцирующему путресцин или орнитин в естественном состоянии, или микроорганизму рода Corynebacterium, продуцирующему путресцин или орнитин, полученному приданием способности продуцировать путресцин или орнитин его исходному штамму, неспособному продуцировать путресцин или орнитин.
Микроорганизм, которому придана способность продуцировать путресцин или орнитин, или способный продуцировать путресцин или орнитин, может обладать улучшенной способностью продуцировать орнитин, используемый в качестве исходного вещества для биосинтеза путресцина, благодаря, без ограничения, модификации активности ацетилглутаматсинтазы (превращающей глутамат в N-ацетилглутамат), орнитинацетилтрансферазы (ArgJ, превращающей ацетилорнитин в орнитин), ацетилглутаматкиназы (ArgB, превращающей ацетилглутамат в N-ацетилглутамилфосфат), гамма-глутамилфосфатредуктазы (ArgC, превращающей N-ацетилглутамилфосфат в N-ацетилглутаматполуальдегид) и ацетилорнитинаминотрансферазы (ArgD, превращающей ацетилглутаматполуальдегид в N-ацетилорнитин) в сторону повышения, по сравнению с их эндогенной активностью, для усиления биосинтетического пути от глутамата до орнитина.
Кроме того, микроорганизм может быть модифицирован для ослабления активности орнитинкарбамоилтрансферазы (ArgF, вовлеченной в синтез аргинина из орнитина), белка (белков), вовлеченного в экспорт глутамата, и/или белка (белков), ацетилирующего путресцин, по сравнению с их эндогенной активностью, и/или введения активности орнитиндекарбоксилазы (ODC).
При использовании здесь термин «введение активности» может относиться к активности белка, отсутствующей или слабой у микроорганизма, вводимой впервые или усиленной у соответствующего микроорганизма. Конкретно, он может включать, без ограничения, введение или доставку гена, кодирующего белок, отсутствующий у микроорганизма, в микроорганизм для экспрессии в нем или введение модификации белка для усиления экспрессии белка, не экспрессируемого или почти не экспрессируемого у микроорганизма.
С другой стороны, в настоящем изобретении модификации, такие как введение активности, усиление активности, ослабление активности и так далее, могут быть проведены способом, называемым трансформацией. При использовании здесь термин «трансформация» относится к процессу введения вектора, содержащего полинуклеотид, кодирующий определенный белок, или промоторную последовательность с сильной или слабой активностью и так далее в клетку-хозяина, что обеспечивает экспрессию белка, кодируемого полинуклеотидом, или приводит к модификации хромосомы клетки-хозяина. Кроме того, полинуклеотид включает ДНК и РНК, кодирующие целевой белок. Полинуклеотид может быть введен в любой форме, при условии, что он может быть введен в клетку-хозяина и обеспечить экспрессию или привести к модификации в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, представляющей собой генную конструкцию, содержащую все обязательные элементы, необходимые для самостоятельной экспрессии. Экспрессионная кассета может обычно содержать промотор, функционально связанный с полинуклеотидом, сигнал терминации транскрипции, домен связывания рибосом и сигнал терминации трансляции и может иметь форму вектора экспрессии, способного к самостоятельной репликации. Кроме того, полинуклеотид может, без ограничения, быть введен в клетку-хозяина сам по себе и функционально связан с последовательностью, необходимой для его экспрессии в клетке-хозяине.
Кроме того, при использовании здесь термин «функционально связанный» относится к функциональной связи промоторной последовательности, инициирующей и опосредующей транскрипцию полинуклеотида, кодирующего определенный белок по настоящему изобретению, с последовательностью гена.
При использовании здесь термин «вектор» относится к ДНК-конструкции, содержащей нуклеотидную последовательность полинуклеотида, кодирующего интересующий белок, где интересующий белок функционально связан с подходящей регуляторной последовательностью, таким образом, что интересующий белок может быть экспрессирован у подходящего хозяина. Регуляторная последовательность включает промотор, способный инициировать транскрипцию, любую операторную последовательность для регуляции транскрипции, последовательность, кодирующую подходящий домен связывания рибосом на мРНК, и последовательность для регуляции транскрипции и трансляции. После трансформации в подходящую клетку-хозяина вектор может быть реплицирован или функционировать независимо от генома хозяина, или может быть интегрирован в сам геном хозяина.
Относительно вектора, используемого в настоящем изобретении, может не быть существенных ограничений, при условии, что вектор реплицирован в клетке-хозяине, и может быть использован любой вектор, известный в данной области. Примеры вектора могут включать естественные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора могут быть использованы pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A и так далее; и в качестве плазмидного вектора могут быть использованы векторы на основе pBR, pUC, pBluescriptll, pGEM, pTZ, pCL, pET и так далее. Относительно вектора для использования в настоящем изобретении, может не быть существенных ограничений, и может быть использован любой вектор, известный в данной области. Конкретно, могут быть использованы векторы pDZTn, pACYC177, PACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCCIBAC и так далее.
Как таковой, полинуклеотид, кодирующий белок-мишень, может быть заменен модифицированным полинуклеотидом с использованием вектора для введения в хромосомы бактерий. Введение полинуклеотида в хромосому может быть проведено, без ограничения, любым способом, известным в данной области, например, гомологичной рекомбинацией. Поскольку вектор по настоящему изобретению может быть введен в хромосому гомологичной рекомбинацией, может быть дополнительно включен селективный маркер для подтверждения введения в хромосому. Селективный маркер используют для отбора трансформированных клеток, то есть для подтверждения введения целевого полинуклеотида, и могут быть использованы маркеры, обеспечивающие селектируемые фенотипы, такие как резистентность к лекарственным средствам, потребность в питательных веществах, резистентность к цитотоксическим агентам и экспрессия поверхностных белков. При обработке селективными агентами только клетки, экспрессирующие селективные маркеры, могут выжить или проявлять другие фенотипические признаки, что позволяет легко отбирать трансформированные клетки.
Микроорганизм рода Corynebacterium по настоящему изобретению может представлять собой модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность фосфотрансацетилазного и ацетаткиназного оперона (оперона pta-ackA) дополнительно усилена по сравнению с его эндогенными ферментами.
В настоящем изобретении фосфотрансацетилазный и ацетаткиназный оперон (оперон pta-ackA) представляет собой опероны, содержащие гены, обратно опосредующие метаболический путь, в котором происходит превращение ацетил-КоА, полученного из глюкозы или пирувата, в уксусную кислоту через ацетилфосфат, и метаболический путь противоположного направления.
В настоящем изобретении фосфотрансацетилазный и ацетаткиназный оперон может включать, без ограничения, белки, содержащие аминокислотную последовательность SEQ ID NO: 5 или SEQ ID NO: 7, или любой белок, на 70% или более, конкретно, на 80% или более, конкретнее, на 90% или более, еще конкретнее, на 95% или более, еще конкретнее, на 98% или более, или, наиболее конкретно, на 99% или более гомологичный указанным выше аминокислотным последовательностям, при условии, что белок существенным образом опосредует реакцию образования ацетил-КоА из уксусной кислоты.
Кроме того, поскольку аминокислотные последовательности белков, демонстрирующих указанные активности, могут различаться у разных видов или штаммов данного микроорганизма, фосфотрансацетилазный и ацетаткиназный оперон в настоящем изобретении может не быть ограничен теми источниками, от которых он имеет происхождение. Очевидно, что объем настоящего изобретения может также включать любую аминокислотную последовательность, гомологичную указанным выше последовательностям и обладающую биологической активностью, по существу идентичной или соответствующей белку по SEQ ID NO: 5 или SEQ ID NO: 7, несмотря на то, что аминокислотная последовательность может иметь делецию, модификацию, замену или добавление части последовательности.
Полинуклеотид, кодирующий фосфотрансацетилазный и ацетаткиназный оперон по настоящему изобретению, может включать полинуклеотид, кодирующий аминокислотную последовательность SEQ ID NO: 5 или SEQ ID NO: 7, или полинуклеотид, кодирующий белок, на 70% или более, конкретно, на 80% или более, конкретнее, на 90% или более, еще конкретнее, на 95% или более, еще конкретнее, на 98% или более, и, наиболее конкретно, на 99% или более гомологичный указанным выше аминокислотным последовательностям, и, наиболее конкретно, может включать полинуклеотидную последовательность SEQ ID NO: 6 или SEQ ID NO: 8.
При использовании здесь термин «усиление активности» включает не только оказание эффекта, превосходящего исходную функцию, благодаря введению новой активности или повышению активности самого белка, но также включает повышение его активности посредством повышения активности эндогенного гена, амплификации эндогенного гена под действием внутреннего или внешнего фактора (факторов), устранения регуляторного фактора (факторов), ингибирующего экспрессию гена, увеличения числа копий гена, введения гена извне, модификации последовательности регуляции экспрессии и, конкретно, повышение ферментативной активности, благодаря замене или модификации промотора и мутации гена, и так далее.
Конкретно, в настоящем изобретении повышение активности может быть осуществлено посредством:
1) увеличения числа копий полинуклеотида, кодирующего фермент;
2) модификации последовательности регуляции экспрессии для повышения экспрессии полинуклеотида;
3) модификации полинуклеотидной последовательности в хромосоме для усиления активности фермента; или
4) модификации комбинацией указанного выше;
но способ повышения активности не ограничен указанными вариантами.
Увеличение числа копий полинуклеотида (способ 1) может быть осуществлено в форме функционального связывания полинуклеотида с вектором или введением полинуклеотида в хромосому клетки-хозяина, однако способ не ограничен указанными вариантами. Конкретно, число копий полинуклеотида в хромосоме клетки-хозяина может быть увеличено введением вектора, способного проходить репликацию и функционировать независимо от клетки-хозяина, с которым функционально связан полинуклеотид, кодирующий белок по настоящему изобретению; или оно может быть увеличено введением в вектора, способного интегрировать полинуклеотид в хромосому клетки-хозяина, с которым функционально связан полинуклеотид, в клетку-хозяина.
Далее, модификация последовательности регуляции экспрессии для повышения экспрессии полинуклеотида (способ 2) может быть осуществлена модификацией последовательности посредством делеции, вставки, неконсервативной или консервативной замены в полинуклеотидной последовательности или их комбинации для дополнительного усиления активности последовательности регуляции экспрессии или заменой полинуклеотидной последовательности на полинуклеотидную последовательность, обладающую более сильной активностью, однако способ не ограничен указанными вариантами. Последовательность регуляции экспрессии включает, без ограничения, промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосом, и последовательность, регулирующую терминацию транскрипции и трансляции.
Вместо исходного промотора с начальной областью экспрессионной единицы полинуклеотида может быть связан сильный экзогенный промотор. Примерами сильных промоторов могут быть, без ограничения, промотор CJ7, промотор lysCP1, промотор EF-Tu, промотор groEL, промотор асеА или промотор асеВ и так далее, и, конкретнее, интенсивность экспрессии может быть улучшена функциональным связыванием с промотором lysCP1 (WO 2009/096689) или промотором CJ7 (патент Кореи №10-0620092 и WO 2006/065095), имеющими происхождение от Corynebacterium.
Кроме того, модификация полинуклеотидной последовательности в хромосоме (способ 3) может быть осуществлена модификацией последовательности регуляции экспрессии посредством делеции, вставки, неконсервативной или консервативной замены в полинуклеотидной последовательности или их комбинации для дополнительного усиления активности полинуклеотидной последовательности или заменой полинуклеотидной последовательности на улучшенную полинуклеотидную последовательность, обладающую более сильной активностью, однако способ не ограничен указанными вариантами.
Конкретно, в настоящем изобретении активность фосфотрансацетилазного и ацетаткиназного оперона (оперона pta-ackA) может быть усилена по сравнению с его эндогенной активностью любым способом, выбранным из группы, состоящей из способа увеличения числа копий оперона в клетке, способа модификации последовательности регуляции экспрессии оперона, способа замены последовательности регуляции экспрессии гена в опероне на последовательность, обладающую более сильной активностью, способа замены генов, кодирующих ферменты в хромосоме, мутантными генами для повышения активности ферментов, составляющих оперон, и способа модификации гена в хромосоме для повышения активности ферментов, составляющих оперон. Конкретно, способ замены последовательности регуляции экспрессии гена в опероне на последовательность, обладающую более сильной активностью, может быть осуществлен, без ограничения, заменой эндогенного промотора ацетилазного и ацетаткиназного оперона на промотор CJ7, промотор lysCP1, промотор EF-Tu, промотор groEL, промотор асеА или промотор асеВ и так далее.
При использовании здесь термин «эндогенная активность» относится к активному состоянию фермента в немодифицированном состоянии, исходно присутствующего у микроорганизма, и термин «усиление по сравнению с эндогенной активностью» относится к состоянию повышенной активности фермента, присутствующего у микроорганизма после манипуляции, такой как введение гена, демонстрирующего активность, или увеличение числа копий соответствующего гена, делеция регуляторного фактора, ингибирующего экспрессию гена, или модификация последовательности регуляции экспрессии, например, использование промотора, улучшенного по сравнению с активностью у микроорганизма до манипуляции.
В настоящем изобретении в микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, может быть дополнительно введена активность ацетил-КоА-синтетазы (acs), имеющей происхождение от Е. coli.
В настоящем изобретении ацетил-КоА-синтетаза (acs) представляет собой фермент, опосредующий реакцию образования ацетил-КоА из АТФ, уксусной кислоты и КоА.
В настоящем изобретении ацетил-КоА-синтетаза может включать, без ограничения, белки, имеющие аминокислотную последовательность SEQ ID NO: 9, или любой белок, на 70% или более, конкретно, на 80% или более, конкретнее, на 90% или более, еще конкретнее, на 95% или более, еще конкретнее, на 98% или более, и, наиболее конкретно, на 99% или более гомологичный указанной выше аминокислотной последовательности, при условии, что белок обладает существенной активностью по опосредованию синтеза ацетил-КоА.
Кроме того, поскольку аминокислотные последовательности белков, демонстрирующих указанную активность, могут различаться у разных видов или штаммов данного микроорганизма, ацетил-КоА-синтетаза (acs) в настоящем изобретении может не быть ограничена источником, от которого она имеет происхождение, и, например, она может иметь происхождение от Е. coli. Очевидно, что объем настоящего изобретения может также включать любую аминокислотную последовательность, гомологичную указанной выше последовательности и обладающую биологической активностью, по существу идентичной или соответствующей белку по SEQ ID NO: 9, несмотря на то, что аминокислотная последовательность может иметь делецию, модификацию, замену или добавление части последовательности.
Полинуклеотид, кодирующий ацетил-КоА-синтетазу (acs) по настоящему изобретению, может включать полинуклеотид, кодирующий белок, содержащий аминокислотную последовательность SEQ ID NO: 9, или любой белок, на 70% или более, конкретно, на 80% или более, конкретнее, на 90% или более, еще конкретнее, на 95% или более, еще конкретнее, на 98% или более, и, наиболее конкретно, на 99% или более гомологичный указанной выше аминокислотной последовательности, и, наиболее конкретно, он может включать полинуклеотидную последовательность SEQ ID NO: 10.
Микроорганизм рода Corynebacterium по настоящему изобретению может представлять собой модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, в который дополнительно введена активность орнитиндекарбоксилазы (ODC), имеющей происхождение от Е. coli.
При использовании здесь термин «орнитиндекарбоксилаза» относится к ферменту, обеспечивающему образование путресцина, опосредуя декарбоксилирование орнитина. Несмотря на то, что у микроорганизма рода Corynebacterium нет фермента биосинтеза путресцина, при введении орнитиндекарбоксилазы (ODC) извне происходит экспорт путресцина из клеток по мере его синтеза. Орнитиндекарбоксилаза, которая может быть введена извне, может быть использована в настоящем изобретении при условии наличия у нее указанной выше активности, независимо от источника, от которого микроорганизм имеет происхождение, и, конкретно, может быть введена орнитиндекарбоксилаза из Е. coli.
Микроорганизм рода Corynebacterium по настоящему изобретению может представлять собой модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность (1) орнитинкарбамоилтрансферазы (ArgF), (2) экспортера глутамата или (3) орнитинкарбамоилтрансферазы и экспортера глутамата дополнительно ослаблена по сравнению с их эндогенной активностью. Экспортер глутамата рода Corynebacterium может представлять собой NCgl1221.
Микроорганизм рода Corynebacterium по настоящему изобретению может представлять собой модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность по меньшей мере одного, выбранного из группы, состоящей из ацетил-гамма-глутамилфосфатредуктазы (ArgC), ацетилглутаматсинтазы или орнитинацетилтрансферазы (ArgJ), ацетилглутаматкиназы (ArgB) и ацетилорнитинаминотрансферазы (ArgD), дополнительно усилена по сравнению с их эндогенной активностью.
Кроме того, микроорганизм рода Corynebacterium может представлять собой модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность ацетилтрансферазы, конкретно, активность NCgl1469, дополнительно ослаблена по сравнению с ее эндогенной активностью.
В заключение, микроорганизм рода Corynebacterium может представлять собой модифицированный микроорганизм рода Corynebacterium, продуцирующий путресцин или орнитин, где активность экспортера путресцина, конкретно, активность NCgl2522, дополнительно усилена по сравнению с его эндогенной активностью.
При использовании здесь «ослабление активности» включает не только оказание эффекта, уступающего исходной функции, ввиду снижения или устранения активности самого белка, но также включает снижение его активности посредством снижения активности эндогенного гена, активации регуляторного фактора (факторов), ингибирующего экспрессию гена, уменьшения числа копий гена, модификации последовательности регуляции экспрессии и, конкретно, устранение или снижение ферментативной активности, ввиду замены или модификации промотора и мутации гена, и так далее.
Конкретно, в настоящем изобретении ослабление активности может быть осуществлено посредством:
1) частичного или полного удаления полинуклеотида, кодирующего белок;
2) модификации последовательности регуляции экспрессии для снижения экспрессии полинуклеотида;
3) модификации полинуклеотидной последовательности в хромосомах для ослабления активности белка; и
4) способа, выбранного из комбинации указанного выше;
но способ ослабления активности не ограничен указанными вариантами.
Конкретно, способ частичного или полного удаления полинуклеотида, кодирующего белок, может быть осуществлен заменой полинуклеотида, кодирующего эндогенный целевой белок, в хромосоме на полинуклеотид с частичной делецией полинуклеотидной последовательности или маркерный ген с использованием вектора для введения в хромосомы бактерий. При использовании здесь термин «часть» может варьировать в зависимости от типа полинуклеотида и, конкретно, может относиться к 1-300, конкретнее, 1-100, и, еще конкретнее, 1-50.
Кроме того, способ модификации последовательности регуляции экспрессии может быть осуществлен модификацией последовательности регуляции экспрессии посредством делеции, вставки, неконсервативной или консервативной замены в полинуклеотидной последовательности или их комбинации для дополнительного ослабления активности последовательности регуляции экспрессии или заменой полинуклеотидной последовательности на полинуклеотидную последовательность, обладающую более слабой активностью. Последовательность регуляции экспрессии включает промотор, последовательность оператора, последовательность, кодирующую сайт связывания рибосом, и последовательность, регулирующую терминацию транскрипции и трансляции.
Кроме того, способ модификации полинуклеотидной последовательности в хромосоме может быть осуществлен модификацией последовательности посредством делеции, вставки, неконсервативной или консервативной замены в полинуклеотидной последовательности или их комбинации для дополнительного ослабления активности фермента или заменой полинуклеотидной последовательности на улучшенную полинуклеотидную последовательность, обладающую более сильной активностью.
Кроме того, способ удаления регуляторного фактора, ингибирующего экспрессию полинуклеотида фермента, может быть осуществлен заменой полинуклеотида фактора, ингибирующего экспрессию, на полинуклеотид с частичной делецией полинуклеотидной последовательности или маркерный ген. При использовании здесь термин «часть» может варьировать в зависимости от типа полинуклеотида и, конкретно, может относиться к 1-300, конкретнее, 1-100, и, еще конкретнее, 1-50.
В частности, ацетил-гамма-глутамилфосфатредуктаза (ArgC), ацетилглутаматсинтаза или орнитинацетилтрансфераза (ArgJ), ацетилглутаматкиназа (ArgB), ацетилорнитинаминотрансфераза (ArgD), орнитинкарбамоилтрансфераза (ArgF), белки, вовлеченные в экспорт глутамата, и орнитиндекарбоксилаза (ODC) могут, соответственно, без ограничения, содержать аминокислотную последовательность SEQ ID NO: 32, 33, 34, 35, 36, 37 или 38 или, конкретно, любую аминокислотную последовательность, на 70% или более, конкретнее, на 80% или более, и, еще конкретнее, на 90% или более гомологичную указанным выше аминокислотным последовательностям. Кроме того, белок, ацетилирующий путресцин, может, без ограничения, содержать аминокислотную последовательность SEQ ID NO: 30 или 31 или, конкретно, любую аминокислотную последовательность, на 70% или более, конкретнее, на 80% или более, и, еще конкретнее, на 90% или более гомологичную указанным выше аминокислотным последовательностям.
Кроме того, в настоящем изобретении экспортер путресцина может содержать аминокислотную последовательность SEQ ID NO: 26 или 28 или, конкретно, любую аминокислотную последовательность, на 70% или более, конкретнее, на 80% или более, и, еще конкретнее, на 90% или более гомологичную указанным выше аминокислотным последовательностям.
Среди описанных выше белков, усиление активности ацетил-гамма-глутамилфосфатредуктазы (ArgC), ацетилглутаматсинтазы или орнитинацетилтрансферазы (ArgJ), ацетилглутаматкиназы (ArgB), ацетилорнитинаминотрансферазы (ArgD), орнитиндекарбоксилазы (ODC) и экспортера путресцина может быть осуществлено, например, способом, выбранным из увеличения числа копий полинуклеотидов, кодирующих белки, модификации последовательности регуляции экспрессии для повышения экспрессии полинуклеотидов, модификации полинуклеотидных последовательностей в хромосоме для усиления активности указанных выше ферментов, удаления регуляторного фактора (факторов), ингибирующего экспрессию полинуклеотидов указанных выше ферментов, или их комбинаций.
Кроме того, ослабление активности орнитинкарбамоилтрансферазы (ArgF), белков, вовлеченных в экспорт глутамата, и белков, ацетилирующих путресцин, может быть осуществлено способом, выбранным из частичного или полного удаления полинуклеотидов, кодирующих белки, модификации последовательности регуляции экспрессии для снижения экспрессии полинуклеотидов, модификации полинуклеотидных последовательностей в хромосоме для ослабления активности белков и их комбинаций.
Согласно другому аспекту настоящего изобретения предложен способ получения путресцина или орнитина, включающий:
(1) культивирование микроорганизма рода Corynebacterium, продуцирующего путресцин или орнитин, в среде; и
(2) выделение путресцина или орнитина из культивированного микроорганизма или среды.
В указанном выше способе микроорганизм можно культивировать, без ограничения, в периодической культуре, непрерывной культуре, подпитываемой культуре и так далее, как известно в данной области. В частности, применительно к условиям культивирования, надлежащий рН (то есть оптимальный рН от 5 до 9, конкретно, рН 6-8, и, наиболее конкретно, рН 6,8) можно поддерживать с использованием, без ограничения, основного соединения (например, гидроксид натрия, гидроксид калия или аммиак) или кислого соединения (например, фосфорная кислота или серная кислота). Кроме того, можно поддерживать аэробные условия, добавляя в клеточную культуру кислород или кислород-содержащую смесь газов. В культуре можно поддерживать температуру от 20°С до 45°С и, конкретно, от 25°С до 40°С, и микроорганизм можно культивировать в течение приблизительно 10-160 часов. Путресцин или орнитин, продуцируемый посредством описанного выше культивировании, может секретироваться в культуральную среду или оставаться внутри клеток.
Кроме того, в культуральной среде могут быть использованы источники углерода, такие как, без ограничения, сахара и углеводы (например, глюкоза, сахароза, лактоза, фруктоза, мальтоза, меласса, крахмал и целлюлоза), масла и жиры (например, соевое масло, подсолнечное масло, арахисовое масло и кокосовое масло), жирные кислоты (например, пальмитиновая кислота, стеариновая кислота и линолевая кислота), спирты (например, глицерин и этанол) и органические кислоты (например, уксусная кислота), по отдельности или в комбинации; источники азота, такие как, без ограничения, азотсодержащие органические соединения (например, пептон, дрожжевой экстракт, мясной сок, солодовый экстракт, жидкий кукурузный экстракт, соевая мука и мочевина) или неорганические соединения (например, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония), по отдельности или в комбинации; и источники калия, такие как, без ограничения, дигидрофосфат калия, гидрофосфат калия или соответствующие натрийсодержащие соли, по отдельности или в комбинации. Кроме того, среда может дополнительно содержать, без ограничения, другие необходимые ростостимулирующие вещества, включая соли металлов (например, сульфат магния или сульфат железа), аминокислоты и витамины.
Способ выделения путресцина или орнитина, полученного при культивировании по настоящему изобретению, может быть осуществлен подходящим методом культивирования, известным в данной области, например, таким как периодическая культура, непрерывная культура или подпитываемая культура, и, посредством этого, целевая аминокислота может быть выделена из культуры.
ВАРИАНТ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Далее настоящее изобретение будет описано более подробно со ссылкой на следующие Примеры. Тем не менее, эти Примеры приведены лишь в целях наглядности, и изобретение не следует ограничивать этими Примерами.
Пример 1: Введение argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от Е. coli, в штамм, продуцирующий путресцин, и подтверждение способности полученного штамма продуцировать путресцин
1-1. Получение штамма с одновременным введением argA, имеющего происхождение от E. coli, и argE, имеющего происхождение от Е. coli, в ген транспозона штамма, продуцирующего путресцин, на основе АТСС13032
Для подтверждения того, что введение гена argA, имеющего происхождение от Е. coli, и гена argE, имеющего происхождение от Е. coli, в штамм, продуцирующий путресцин, на основе АТСС13032 может улучшить способность продуцировать путресцин, гены argA и argE вводили в ген транспозона указанного штамма.
В качестве вектора для трансформации, позволяющего осуществлять введение генной области транспозона микроорганизма рода Corynebacterium в хромосому, использовали pDZTn (WO 2009/125992) и в качестве промотора использовали промотор lysCPI (публикация международной заявки на патент №WO 2009/096689, SEQ ID NO: 39).
Конкретно, на основе полинуклеотидной последовательности (SEQ ID NO: 2) гена argA, имеющего происхождение от Е. coli, кодирующего N-ацетилглутаматсинтазу, получали пару праймеров SEQ ID NO: 11 и 12 для получения гомологичных рекомбинантных фрагментов открытой рамки считывания (ORF) argA. Кроме того, на основе полинуклеотидной последовательности (SEQ ID NO: 4) гена argE, имеющего происхождение от Е. coli, кодирующего ацетилорнитиндезацетилазу, получали пару праймеров SEQ ID NO: 15 и 16 для получения гомологичных рекомбинантных фрагментов ORF argE, и на основе полинуклеотидной последовательности (SEQ ID NO: 39) lysCP1 получали пару праймеров SEQ ID NO: 13 и 14 для получения гомологичных рекомбинантных фрагментов области lysCP1 (Таблица 1).

Сначала для получения гена argA амплифицировали фрагмент гена размером приблизительно 1,6 тысячи пар оснований (т.п.о.) с использованием хромосомы штамма E. coli W3110 в качестве матрицы с парой праймеров SEQ ID NO: 11 и 12. В частности, полимеразную цепную реакцию (ПЦР) проводили, повторяя 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и удлинения при 72°С в течение 1 минуты и 30 секунд. Полученные таким образом фрагменты подвергали электрофорезу в 0,8%-м агарозном геле и проводили элюирование и очистку полос желаемого размера.
Кроме того, амплифицировали промоторную область lysCP1, проводя ПЦР с использованием хромосомы штамма КССМ10919Р (публикация международной заявки на патент №WO 2009/096689) в качестве матрицы с парой праймеров SEQ ID NO: 13 и 14, повторяя 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и удлинения при 72°С в течение 30 секунд.
Вектор pDZTn обрабатывали Xnol, после чего каждый из полученных ПЦР-продуктов подвергали клонированию посредством слияния. Клонирование посредством слияния проводили с использованием набора In-Fusion® HD Cloning Kit (Clontech), и полученная таким образом плазмида была названа pDZTn-lysCP1-argA.
Затем получали ПЦР-продукты для получения гена argE, амплифицируя фрагмент гена размером приблизительно 1,4 т.п.о. таким же образом, как описано выше, с использованием хромосомы штамма Е. coli W3110 в качестве матрицы с парой праймеров SEQ ID NO: 15 и 16, и подвергали их клонированию посредством слияния вместе с промоторной областью lysCP1. Полученная таким образом плазмида была названа pDZTn-lysCP1-argE.
Затем плазмиду pDZTn-lysCP1-argA вводили в штамм КССМ11240Р (публикация заявки на патент Кореи №10-2013-0082478) электропорацией с получением трансформантов и трансформантов высевали на твердую среду BHIS (сердечно-мозговой экстракт (37 г/л), сорбит (91 г/л) и агар (2%)), содержащую канамицин (25 мкг/мл) и X-gal (5-бром-4-хлор-3-индолин-D-галактозид), и культивировали с образованием колоний. Из полученных колоний отбирали синие колонии, проводя посредством этого отбор трансформированных штаммов с введенной плазмидой pDZTn-lysCP1-argA.
Отобранные штаммы культивировали с покачиванием (30°С, 8 часов) в среде СМ (глюкоза (10 г/л), полипептон (10 г/л), дрожжевой экстракт (5 г/л), мясной экстракт (5 г/л), NaCl (2,5 г/л), мочевина (2 г/л), рН 6,8) и последовательно разводили от 10-4 до 10-10, высевали на твердую среду, содержащую X-gal, и культивировали с образованием колоний. Из полученных таким образом колоний отбирали белые колонии, встречавшиеся со сравнительно низкой частотой, проводя окончательный отбор штамма с геном, кодирующим argA, введенным посредством вторичного кроссинговера. Отобранный в итоге штамм подвергали ПЦР с использованием пары праймеров SEQ ID NO: 12 и 13, подтвердив введение гена, кодирующего argA, и модифицированный штамм Corynebacterium glutamicum был назван КССМ11240Р Tn:lysCP1-argA.
Для введения в штамм с введенным argA, полученный выше, КССМ11240Р Tn:lysCP1-argA трансформировали полученной выше pDZTn-lysCP1-argE таким же образом, как описано выше, и введение argE в транспозон отобранного в итоге штамма было подтверждено проведением ПЦР с использованием пары праймеров SEQ ID NO: 13 и 16. Отобранный таким образом модифицированный штамм Corynebacterium glutamicum был назван КССМ11240Р Tn:lysCP1-argA Tn:lysCP1-argE.
1-2. Получение штамма с одновременным введением argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от Е. coli, в ген транспозона штамма, продуцирующего путресцин, на основе АТСС13869
DAB12-a ΔNCgl1469 (публикация заявки на патент Кореи №10-2013-0082478), представляющий собой штамм, продуцирующий путресцин, на основе Corynebacterium glutamicum АТСС13869, был назван DAB12-b, и в ген его транспозона вводили argA и argE для подтверждения того, что введение генов argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от Е. coli, может быть связано с улучшением способности полученного штамма продуцировать путресцин.
Сначала pDZTn-lysCP1-argA, полученной ранее, трансформировали Corynebacterium glutamicum DAB12-b таким же образом, как в Примере 1-1, и подтверждали введение argA в транспозон. Отобранный таким образом модифицированный штамм Corynebacterium glutamicum был назван DAB12-b Tn: lysCP1-argA.
Затем, для введения argE в штамм с уже введенным argA, DAB12-b Tn:lysCP1-argA трансформировали полученной ранее pDZTn-lysCP1-argE таким же образом, как в Примере 1-1, и подтверждали введение argE в транспозон. Отобранный таким образом модифицированный штамм Corynebacterium glutamicum был назван DAB12-b Tn: lysCPI-argE.
1-3. Оценка способности штамма Corynebacterium, продуцирующего путресцин, с введенным геном argA, имеющим происхождение от Е. coli, и геном argE, имеющим происхождение от Е. coli, продуцировать путресцин
Проводили сравнение способности модифицированных штаммов Corynebacterium glutamicum, полученных в Примерах 1-1 и 1-2, продуцировать путресцин для подтверждения эффекта введения argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от Е. coli, в штамм, продуцирующий путресцин, на продукцию путресцина.
Конкретно, модифицированные штаммы Corynebacterium glutamicum двух разных типов (то есть КССМ11240Р Tn:lysCP1-argA Tn:lysCP1-argE; DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE), полученные в Примерах 1-1 и 1-2, и исходные штаммы двух разных типов (то есть КССМ11240Р и DAB12-b), соответственно, высевали на твердую среду СМ (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-го NaOH, 2% агара, рН 6,8, на основе 1 л), содержащую 1 мМ аргинина, и культивировали при 30°С в.течение 24 часов.
Каждый из культивированных штаммов в количестве приблизительно одной платиновой петли засевали в 25 мл титрационной среды (8% глюкозы, 0,25% соевого белка, 0,50% сухого кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, биотин (100 мг), тиамин-HCl (3 мг), пантотенат кальция (3 мг), никотинамид (3 мг), 5% СаСО3, на основе 1 л) и культивировали с покачиванием при 30°С и частоте 200 об/мин в течение 98 часов. Во всех культурах штаммов в среду добавляли 1 мМ аргинина. По завершении культивирования в каждой культуральной среде измеряли концентрацию полученного путресцина, и результаты показаны в Таблице 2 ниже.

Как показано в Таблице 2 выше, оба модифицированных штамма Corynebacterium glutamicum с одновременным введением генов argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от E. coli, продемонстрировали повышение продукции путресцина на 9,8% или более.
Пример 2: Усиление pta-ackA у штамма, продуцирующего путресцин, с введенными argA, имеющим происхождение от Е. coli, и argE, имеющим происхождение от Е. coli, и подтверждение способности полученного штамма продуцировать путресцин
2-1. Получение штамма с заменой промотора pta-ackA из штамма Corynebacterium, продуцирующего путресцин, на основе АТСС13032
У штамма, продуцирующего путресцин, с введенными генами argA, имеющим происхождение от Е. coli, и argE, имеющим происхождение от Е. coli, полученного в Примере 1, проводили дополнительное усиление активности фосфотрансацетилазы и ацетаткиназы (pta-ackA) и оценивали эффект такого усиления на способность штамма продуцировать путресцин.
С этой целью промотор оперона pta-ackA в хромосоме заменяли на промотор, обладающий более сильной активностью, по сравнению с его эндогенным промотором, конкретно перед инициирующим кодоном оперона pta-ackA вводили промотор lysCP1 (публикация международной заявки на патент № WO 2009/096689).
Сначала получали гомологичный рекомбинантный фрагмент, содержащий промотор lysCP1, где оба конца промотора имеют оригинальную хромосомную последовательность pta-ackA. Конкретно, 5'-концевую область промотора lysCP1 получали, проводя ПЦР с использованием геномной ДНК Corynebacterium glutamicum АТСС13032 с парой праймеров SEQ ID NO: 17 и 18. В частности, ПЦР проводили, повторяя 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и удлинения при 72°С в течение 30 секунд.
Кроме того, промоторную область lysCP1 получали, проводя ПЦР в таких же условиях с использованием пары праймеров SEQ ID NO: 14 и 19, и 3'-концевую область промотора lysCP1 получали, проводя ПЦР с использованием геномной ДНК Corynebacterium glutamicum АТСС13032 в качестве матрицы с парой праймеров SEQ ID NO: 20 и 21. Праймеры, использованные при получении промотора lysCP1, показаны в Таблице 1 выше и в Таблице 3 ниже.

Каждый из полученных выше ПЦР-продуктов подвергали клонированию посредством слияния с использованием вектора pDZ, обработанного Xbal. Клонирование посредством слияния проводили с использованием набора In-Fusion® HD Cloning Kit (Clontech), и полученная таким образом плазмида была названа pDZ-lysCP1-1'pta-ackA.
Полученную выше плазмиду pDZ-lysCP1-1'pta-ackA вводили, соответственно, в штаммы КССМ11240Р и КССМ11240Р Tn:lysCP1-argA Tn:lysCP1-argE, являющийся модифицированным штаммом Corynebacterium glutamicum, полученным в Примере 1-1, электропорацией с получением трансформантов и трансформантов высевали на твердую среду BHIS (сердечно-мозговой экстракт (37 г/л), сорбит (91 г/л) и агар (2%)), содержащую канамицин (25 мкг/мл) и X-gal (5-бром-4-хлор-3-индолин-D-галактозид), и культивировали с образованием колоний. Из полученных колоний отбирали синие колонии, проводя посредством этого отбор трансформированных штаммов с введенной плазмидой pDZ-lysCP1-1'pta-ackA.
Отобранные штаммы культивировали с покачиванием (30°С, 8 часов) в среде СМ (глюкоза (10 г/л), полипептон (10 г/л), дрожжевой экстракт (5 г/л), мясной экстракт (5 г/л), NaCl (2,5 г/л), мочевина (2 г/л), рН 6,8) и последовательно разводили от 10-4 до 10-10, высевали на твердую среду, содержащую X-gal, и культивировали с образованием колоний. Из полученных таким образом колоний отбирали белые колонии, встречавшиеся со сравнительно низкой частотой, проводя окончательный отбор штамма, в котором промотор pta-ackA был заменен на промотор lysCP1 посредством вторичного кроссинговера.
Отобранный в итоге штамм подвергали ПЦР с использованием пары праймеров SEQ ID NO: 19 и 21, подтвердив введение промотора lysCP1 в хромосому перед инициирующим кодоном pta-ackA. В частности, ПЦР проводили, повторяя 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и удлинения при 72°С в течение 1 минуты.
Отобранные таким образом модифицированные штаммы Corynebacterium glutamicum были названы КССМ11240Р lysCP1-1'pta-ackA и КССМ11240Р Tn:lysCP1-argA Tn:lysCP1-argE lysCP1-1'pta-ackA, соответственно.
2-2. Получение штамма с заменой промотора pta-ackA из штамма Corynebacterium, продуцирующего путресцин, на основе АТСС13869
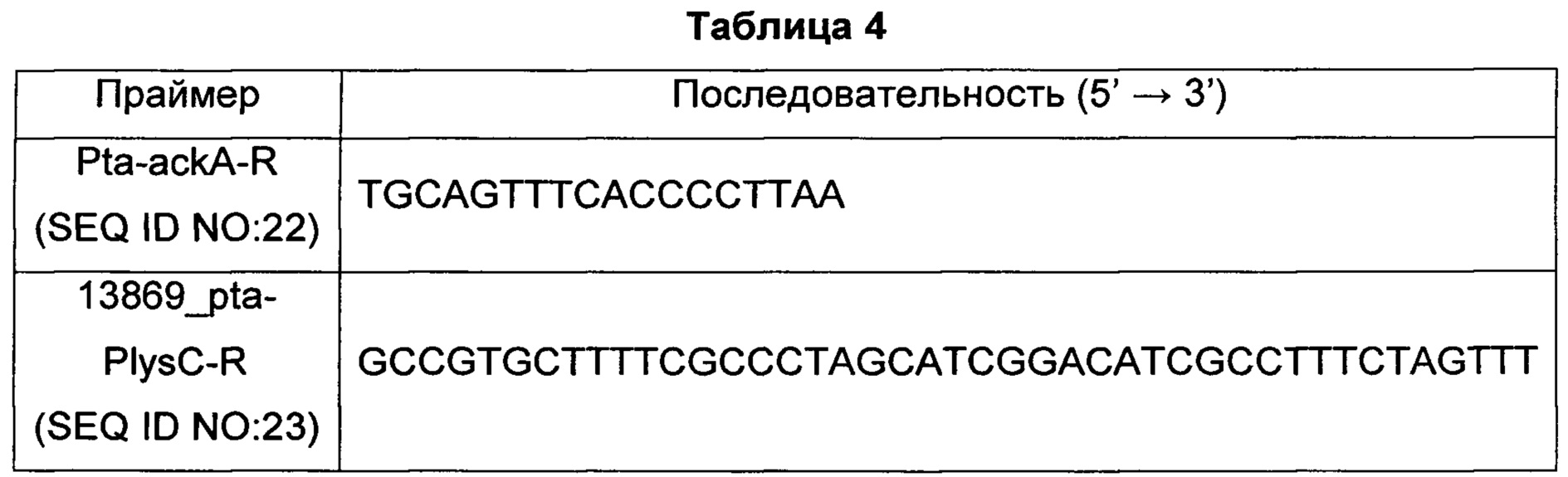
Для подтверждения последовательности гена, кодирующего pta-ackA, имеющего происхождение от Corynebacterium glutamicum АТСС13869, и экспрессированного с него белка, полученного методом, раскрытым в Примере 2-1, проводили ПЦР с использованием геномной ДНК Corynebacterium glutamicum АТСС13869 в качестве матрицы с парой праймеров SEQ ID NO: 17 и 22 (Таблицы 3 и 4). В частности, ПЦР проводили, повторяя 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и удлинения при 72°С в течение 3 минут.
Полученные таким образом ПЦР-продукты разделяли электрофорезом и анализировали их последовательности. В результате, было подтверждено, что ген, кодирующий pta-ackA, имеющий происхождение от Corynebacterium glutamicum АТСС13869, содержит полинуклеотидную последовательность, описываемую SEQ ID NO: 8, и что белок, кодируемый этим геном, содержит аминокислотную последовательность, описываемую SEQ ID NO: 7.
С другой стороны, в результате сравнения аминокислотной последовательности pta-ackA, имеющей происхождение от Corynebacterium glutamicum АТСС13032 (SEQ ID NO: 5), и аминокислотной последовательности pta-ackA, имеющей происхождение от Corynebacterium glutamicum АТСС13869, было подтверждено, что эти последовательности гомологичны на 99,4%.
Сначала получали гомологичный рекомбинантный фрагмент, содержащий промотор lysCP1, где оба конца промотора имеют оригинальную хромосомную последовательность pta-ackA. Конкретно, 5'-концевую область промотора lysCP1 получали, проводя ПЦР с использованием геномной ДНК Corynebacterium glutamicum АТСС13869 с парой праймеров SEQ ID NO: 17 и 23. В частности, ПЦР проводили, повторяя 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и удлинения при 72°С в течение 30 секунд. Кроме того, промоторную область lysCP1 получали, проводя ПЦР в таких же условиях с использованием пары праймеров SEQ ID NO: 14 и 19, и 3'-концевую область промотора lysCP1 получали, проводя ПЦР с использованием геномной ДНК Corynebacterium glutamicum АТСС13869 в качестве матрицы с парой праймеров SEQ ID NO: 20 и 21. Праймеры, использованные при замене промотора, показаны в Таблицах 1, 3 и 4.
Каждый из полученных ПЦР-продуктов подвергали клонированию посредством слияния с использованием вектора pDZTn, обработанного Xhol. Клонирование посредством слияния проводили с использованием набора In-Fusion® HD Cloning Kit (Clontech), и полученная таким образом плазмида была названа pDZ-lysCP1-2'pta-ackA.
Плазмидой pDZ-lysCP1-2'pta-ackA, полученной выше, трансформировали, соответственно, DAB12-b и DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE, являющийся модифицированным штаммом Corynebacterium glutamicum, полученным в Примере 1-2, таким же образом, как в Примере 2-1. В результате, было подтверждено, что промотор lysCP1 был введен в хромосому перед инициирующим кодоном pta-ackA. Модифицированные штаммы Corynebacterium glutamicum были названы DAB12-b lysCP1-2'pta-ackA и DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE lysCP1-2'pta-ackA, соответственно. 2-3. Оценка способности штамма с усилением pta-ackA продуцировать путресцин
Для подтверждения эффекта усиления pta-ackA у штамма, продуцирующего путресцин, с введенными argA, имеющим происхождение от Е. coli, и argE, имеющим происхождение от Е. coli, проводили сравнение способности модифицированных штаммов Corynebacterium glutamicum, полученных в Примерах 2-1 и 2-2, продуцировать путресцин.
Конкретно, модифицированные штаммы Corynebacterium glutamicum четырех типов (KCCM11240P lysCP1-1'pta-ackA; KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE lysCP1-1'pta-ackA; DAB12-b lysCP1-2'pta-ackA; и DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE lysCP1-2'pta-ackA) и исходные штаммы четырех типов (KCCM11240P; KCCM11240P Tn:lysCP1-argA Tn: lysCP1-argE; DAB12-b; и DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE), соответственно, высевали на твердую среду СМ (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-го NaOH, 2% агара, рН 6,8, на основе 1 л), содержащую 1 мМ аргинина, и культивировали при 30°С в течение 24 часов. Каждый из культивированных штаммов в количестве приблизительно одной платиновой петли засевали в 25 мл титрационной среды (8% глюкозы, 0,25% соевого белка, 0,50% сухого кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, биотин (100 мг), тиамин-HCl (3 мг), пантотенат кальция (3 мг), никотинамид (3 мг), 5% CaCO3, на основе 1 л) и культивировали с покачиванием при 30°С и частоте 200 об/мин в течение 98 часов. Во всех культурах штаммов в среду добавляли 1 мМ аргинина. По завершении культивирования в каждой культуральной среде измеряли концентрацию полученного путресцина, и результаты показаны в Таблице 5 ниже.


Как показано в Таблице 5, при усилении pta-ackA у KCCM11240P и DAB12-b, соответственно, количество продуцируемого путресцина оставалось на том же уровне. Тем не менее, при усилении pta-ackA у модифицированных штаммов Corynebacterium glutamicum двух разных типов с одновременным введением генов argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от Е. coli (KCCM11240P Tn: lysCPI-argA Tn: lysCPI-argE; DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE), соответственно, количество продуцируемого путресцина увеличивалось на 14,3% или более по сравнению с исходным штаммом. Кроме того, количество продуцируемого путресцина увеличивалось на 4% или более по сравнению с модифицированными штаммами.
Как таковой, полученный микроорганизм рода Corynebacterium (Corynebacterium glutamicum KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE:lysCP1-1'pta-ackA), обладающий улучшенной способностью продуцировать путресцин, полученный из штамма Corynebacterium glutamicum KCCM11240P, продуцирующего путресцин, введением активности argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от Е. coli и усилением активности pta-ackA у штамма Corynebacterium glutamicum KCCM11240P, был назван авторами настоящего изобретения СС01-1145 и депонирован в Корейском центре культур микроорганизмов (Korean Culture Center of Microorganisms, KCCM) 21 ноября 2014 г. под регистрационным номером KCCM11606P в соответствии с Будапештским соглашением.
Пример 3: Введение acs, имеющего происхождение от Е. coli, в штамм, продуцирующий путресцин. с введенными argA, имеющим происхождение от Е. coli, и argE, имеющим происхождение от Е. coli, и подтверждение способности полученного штамма продуцировать путресцин 3-1. Получение штамма с введением acs, имеющего происхождение от Е. coli, в ген транспозона штамма, продуцирующего путресцин, на основе АТСС13032
acs вводили в ген транспозона с использованием промотора lysCP1 для подтверждения того, что введение гена ацетил-КоА-синтетазы (acs), имеющего происхождение от Е. coli, в штамм, продуцирующий путресцин, на основе АТСС13032 с уже введенными argA, имеющим происхождение от Е. coli, и argE, имеющим происхождение от Е. coli, может улучшить способность продуцировать путресцин.
Конкретно, на основе полинуклеотидной последовательности гена, кодирующего acs, описываемой SEQ ID NO: 10, получали пару праймеров SEQ ID NO: 24 и 25 для получения гомологичного рекомбинантного фрагмента вокруг ORF-области acs и пару праймеров SEQ ID NO: 13 и 14 для получения гомологичного рекомбинантного фрагмента вокруг промоторной области lysCP1, как показано в Таблице 1 выше и в Таблице 6 ниже.
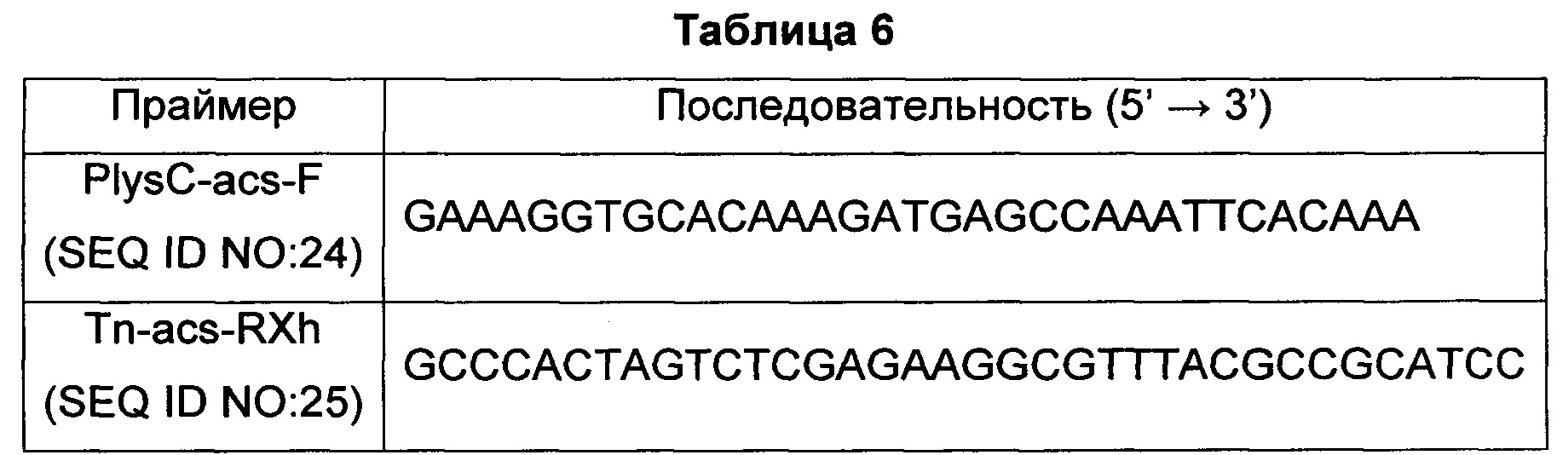
Конкретно, для получения гена acs амплифицировали фрагмент гена размером приблизительно 2 т.п.о. с использованием хромосомы штамма Е. coli W3110 в качестве матрицы с парой праймеров SEQ ID NO: 24 и 25. В частности, ПЦР проводили, повторяя 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и удлинения при 72°С в течение 1 минуты и 30 секунд. Затем полученные таким образом ПЦР-продукты подвергали электрофорезу в 0,8%-м агарозном геле и проводили элюирование и очистку полос желаемого размера.
Кроме того, получали промоторную область lysCP1, проводя ПЦР с использованием хромосомы штамма КССМ10919Р (публикация международной заявки на патент № WO 2009/096689) в качестве матрицы с парой праймеров SEQ ID NO: 13 и 14, повторяя 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и удлинения при 72°С в течение 30 секунд.
Вектор pDZ обрабатывали Xhol и каждый из полученных таким образом ПЦР-продуктов подвергали клонированию посредством слияния. Клонирование посредством слияния проводили с использованием набора In-Fusion® HD Cloning Kit (Clontech). Полученная таким образом плазмида была названа pDZTn-lysCP1-acs.
Затем плазмиду pDZTn-lysCP1-acs вводили, соответственно, в KCCM11240P и KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE, являющийся модифицированным штаммом Corynebacterium glutamicum, полученным в Примере 1-1, электропорацией с получением трансформантов и трансформантов высевали на твердую среду BHIS (сердечно-мозговой экстракт (37 г/л), сорбит (91 г/л) и агар (2%)), содержащую канамицин (25 мкг/мл) и X-gal (5-бром-4-хлор-3-индолин-D-галактозид), и культивировали с образованием колоний. Из полученных колоний отбирали синие колонии, проводя посредством этого отбор трансформированных штаммов с введенной плазмидой pDZTn-lysCP1-acs.
Отобранные штаммы культивировали с покачиванием (30°С, 8 часов) в среде СМ (глюкоза (10 г/л), полипептон (10 г/л), дрожжевой экстракт (5 г/л), мясной экстракт (5 г/л), NaCl (2,5 г/л), мочевина (2 г/л), рН 6,8) и последовательно разводили от 10-4 до 10-10, высевали на твердую среду, содержащую X-gal, и культивировали с образованием колоний. Из полученных таким образом колоний отбирали белые колонии, встречавшиеся со сравнительно низкой частотой, проводя окончательный отбор штаммов с геном, кодирующим acs, введенным посредством вторичного кроссинговера. Отобранные в итоге штаммы подвергали ПЦР с использованием пары праймеров SEQ ID NO: 13 и 25, подтвердив введение гена, кодирующего acs, и модифицированные штаммы Corynebacterium glutamicum были названы КССМ11240Р Tn:lysCP1-acs и KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE Tn:lysCP1-acs, соответственно.
3-2. Получение штамма с введением acs, имеющего происхождение от Е. coli, в ген транспозона штамма, продуцирующего путресцин, на основе АТСС13869
Как в Примере 3-1, pDZTn-lysCP1-acs, полученной выше, трансформировали DAB12-b и DAB12-b Tn: lysCP1-argA Tn: lysCP1-argE, являющийся модифицированным штаммом Corynebacterium glutamicum, полученным в Примере 1-2, соответственно, таким же образом, как в Примере 3-1, подтвердив введение acs в ген транспозона.
Отобранные таким образом модифицированные штаммы Corynebacterium glutamicum были названы DAB12-b Tn:lysCP1-acs и DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE Tn:lysCP1-acs, соответственно.
3-3. Оценка способности штамма с введенным acs, имеющим происхождение от Е. coli, продуцировать путресцин
Для подтверждения эффекта введения acs в штамм, продуцирующий путресцин, с уже введенными argA, имеющим происхождение от Е. coli, и argE, имеющим происхождение от Е. coli, проводили сравнение способности модифицированных штаммов Corynebacterium glutamicum, полученных в Примерах 3-1 и 3-2, продуцировать путресцин.
Конкретно, модифицированные штаммы Corynebacterium glutamicum четырех типов (KCCM11240P Tn: lysCP1-acs; KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE Tn:lysCP1-acs; DAB12-b Tn:lysCP1-acs; и DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE Tn:lysCP1-acs) и исходные штаммы четырех типов (KCCM11240P; КССМ11240Р Tn:lysCP1-argA Tn:lysCP1-argE; DAB12-b; и DAB12-b Tn: lysCPI-argA Tn:lysCP1-argE), соответственно, высевали на твердую среду СМ (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-го NaOH, 2% агара, рН 6,8, на основе 1 л), содержащую 1 мМ аргинина, и культивировали при 30°С в течение 24 часов. Каждый из культивированных штаммов в количестве приблизительно одной платиновой петли засевали в 25 мл титрационной среды (8% глюкозы, 0,25% соевого белка, 0,50% сухого кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, биотин (100 мг), тиамин-HCl (3 мг), пантотенат кальция (3 мг), никотинамид (3 мг), 5% СаСО3, на основе 1 л) и культивировали с покачиванием при 30°С и частоте 200 об/мин в течение 98 часов. Во всех культурах штаммов в среду добавляли 1 мМ аргинина. По завершении культивирования в каждой культуральной среде измеряли концентрацию полученного путресцина, и результаты показаны в Таблице 7 ниже.

Как показано в Таблице 7, при введении acs в KCCM11240P и DAB12-b, соответственно, количество продуцируемого путресцина оставалось на том же уровне. Тем не менее, при введении acs в модифицированные штаммы Corynebacterium glutamicum двух разных типов с одновременным введением генов argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от Е. coli (KCCM11240P Tn:lysCP1-argA Tn:lysCP1-argE; DAB12-b Tn:lysCP1-argA Tn:lysCP1-argE), соответственно, количество продуцируемого путресцина увеличивалось на 13,5% или более по сравнению с исходным штаммом. Кроме того, количество продуцируемого путресцина увеличивалось на 3,4% или более по сравнению с указанными выше модифицированными штаммами.
Пример 4: Штамм с введением argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от Е. coli, и заменой промотора pta-ackA из штамма, продуцирующего путресцин, с улучшенной способностью к экспорту путресцина и способность штамма продуцировать путресцин 4-1. Получение штамма с введением argA и argE, имеющих происхождение от Е. coli, и заменой промотора pta-ackA из штамма с улучшенной способностью к экспорту путресцина
На основе штамма KCCM11401P (публикация заявки на патент Кореи №10-2014-0115244) с улучшенной способностью к экспорту путресцина получали штамм для определения того, может ли введение argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от Е. coli, и усиление активности pta-ackA Corynebacterium улучшить способность продуцировать путресцин.
Конкретно, pDZTn-lysCP1-argA, полученной в Примере 1-1, трансформировали KCCM11401P таким же образом, как в Примере 1-1, и, в результате, введение argA в ген транспозона было подтверждено. Отобранный таким образом модифицированный штамм Corynebacterium glutamicum был назван KCCM11401P Tn:lysCP1-argA.
Кроме того, для введения argE в штамм с уже введенным argA, как в Примере 1-1, pDZTn-lysCP1-argE, полученной в Примере 1-1, трансформировали KCCM11401P Tn:lysCP1-argA таким же образом, как в Примере 1-1, и введение argE в ген транспозона было подтверждено. Отобранный таким образом модифицированный штамм был назван KCCM11401P Tn:lysCP1-argA Tn:lysCP1-argE.
Затем pDZ-lysCP1-1'pta-ackA, полученной в Примере 2-1, трансформировали KCCM11401P Tn:lysCP1-argA Tn: lysCP1-argE таким же образом, как в Примере 2-1, и было подтверждено, что промотор lysCPI был введен в хромосому перед инициирующим кодоном pta-ackA. Полученный выше модифицированный штамм Corynebacterium glutamicum был назван KCCM11401P Tn:lysCP1-argA Tn:lysCP1-argE lysCP1-1'pta-ackA.
4-2. Оценка штамма с введением argA, имеющего происхождение от Е. coli, argE, имеющего происхождение от Е. coli, и заменой промотора pta-ackA из штамма с улучшенной способностью к экспорту путресцина
Для подтверждения эффекта введения argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от Е. coli, и усиления активности pta-ackA у штамма Corynebacterium glutamicum, продуцирующего путресцин, с улучшенной способностью к экспорту путресцина проводили сравнение способности модифицированных штаммов Corynebacterium glutamicum, полученных в Примере 4-1, продуцировать путресцин.
Конкретно, модифицированные штаммы Corynebacterium glutamicum (KCCM11401P Tn:lysCP1-argA Tn:lysCP1-argE, KCCM11401P Tn:lysCP1-argA Tn:lysCP1-argE lysCP1-1'pta-ackA) и исходный штамм (KCCM11401P), соответственно, высевали на твердую среду СМ (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-го NaOH, 2% агара, рН 6,8, на основе 1 л), содержащую 1 мМ аргинина, и культивировали при 30°С в течение 24 часов. Каждый из культивированных штаммов в количестве приблизительно одной платиновой петли засевали в 25 мл титрационной среды (8% глюкозы, 0,25% соевого белка, 0,50% сухого кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, биотин (100 мг), тиамин-HCl (3 мг), пантотенат кальция (3 мг), никотинамид (3 мг), 5% СаСО3, на основе 1 л) и культивировали с покачиванием при 30°С и частоте 200 об/мин в течение 98 часов. Во всех культурах штаммов в среду добавляли 1 мМ аргинина. По завершении культивирования в каждой культуральной среде измеряли концентрацию полученного путресцина, и результаты показаны в Таблице 8 ниже.
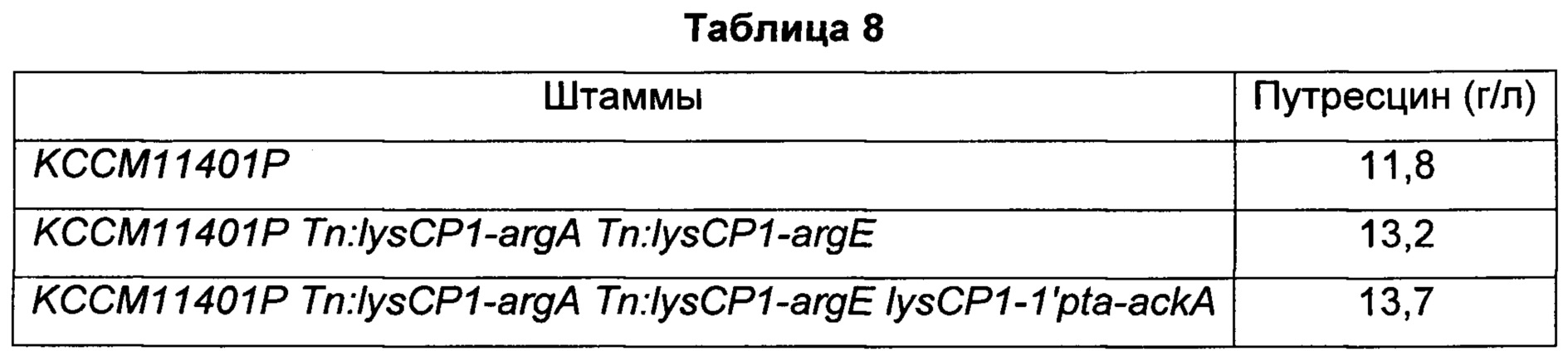
Как показано в Таблице 8, было подтверждено, что при введении гена argA, имеющего происхождение от Е. coli, и гена argE, имеющего происхождение от Е. coli, в KCCM11401P с улучшенной способностью к экспорту путресцина количество продуцируемого путресцина увеличивалось на 11,9% по сравнению с исходным штаммом, а при дополнительном усилении штамма посредством pta-ackA количество продуцируемого путресцина увеличивалось на 16,1% по сравнению с исходным штаммом.
Пример 5: Введение argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от Е. coli, в штамм, продуцирующий орнитин, и подтверждение способности штамма продуцировать орнитин 5-1. Получение штамма с одновременным введением argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от Е. coli, в ген транспозона штамма, продуцирующего орнитин, на основе KCCM11137P
Для подтверждения того, что введение гена argA, имеющего происхождение от Е. coli, и гена argE, имеющего происхождение от Е. coli, в штамм KCCM11137P (публикация заявки на патент Кореи №10-1372635), представляющий собой штамм, продуцирующий орнитин, на основе Corynebacterium glutamicum АТСС13032, может улучшать способность продуцировать орнитин, ген argA и ген argE вводили в ген транспозона указанного штамма с использованием вектора, полученного в Примере 1-1.
Сначала плазмиду pDZTn-lysCP1-argA вводили в штамм KCCM11137P электропорацией с получением трансформантов и трансформантов высевали на твердую среду BHIS (сердечно-мозговой экстракт (37 г/л), сорбит (91 г/л) и агар (2%)), содержащую канамицин (25 мкг/мл) и X-gal (5-бром-4-хлор-3-индолин-О-галактозид), и культивировали с образованием колоний. Из полученных колоний отбирали синие колонии, проводя посредством этого отбор штаммов с введенной плазмидой pDZTn-lysCP1-argA.
Отобранные штаммы культивировали с покачиванием (30°С, 8 часов) в среде СМ (глюкоза (10 г/л), полипептон (10 г/л), дрожжевой экстракт (5 г/л), мясной экстракт (5 г/л), NaCl (2,5 г/л), мочевина (2 г/л), рН 6,8) и последовательно разводили от 10-4 до 10-10, высевали на твердую среду, содержащую X-gal, и культивировали с образованием колоний. Из полученных таким образом колоний отбирали белые колонии, встречавшиеся со сравнительно низкой частотой, проводя окончательный отбор штамма с геном, кодирующим argA, введенным посредством вторичного кроссинговера. Отобранный в итоге штамм подвергали ПЦР с использованием пары праймеров SEQ ID NO: 12 и 13, подтвердив введение гена, кодирующего argA, и модифицированный штамм Corynebacterium glutamicum был назван KCCM11137P Tn:lysCP1-argA.
Для введения argE в штамм с уже введенным argA, полученный выше, pDZTn-lysCP1-argE, полученной в Примере 1-1, трансформировали KCCM11137P Tn:lysCP1-argA таким же образом, как в Примере 1-1, и, таким образом, введение argE в ген транспозона было подтверждено.
Отобранный таким образом модифицированный штамм Corynebacterium glutamicum был назван KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE.
5-2. Оценка способности штамма Corynebacterium, продуцирующего орнитин, с введенным argA, имеющим происхождение от Е. coli, и argE, имеющим происхождение от Е. coli, продуцировать орнитин
Для подтверждения эффекта введения argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от Е. coli, на продукцию орнитина штаммом, продуцирующим орнитин, проводили сравнение способности модифицированных штаммов Corynebacterium glutamicum, полученных в Примере 5-1, продуцировать орнитин.
Конкретно, модифицированный штамм Corynebacterium glutamicum одного типа (KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE) и исходный штамм одного типа (KCCM11137P), соответственно, высевали на твердую среду СМ (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-го NaOH, 2% агара, рН 6,8, на основе 1 л), содержащую 1 мМ аргинина, и культивировали при 30°С в течение 24 часов. Каждый из культивированных штаммов в количестве приблизительно одной платиновой петли засевали в 25 мл титрационной среды (8% глюкозы, 0,25% соевого белка, 0,50% сухого кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, биотин (100 мг), тиамин-HCl (3 мг), пантотенат кальция (3 мг), никотинамид (3 мг), 5% СаСО3, на основе 1 л) и культивировали с покачиванием при 30°С и частоте 200 об/мин в течение 98 часов. Во всех культурах штаммов в среду добавляли 1 мМ аргинина. По завершении культивирования в каждой культуральной среде измеряли концентрацию полученного путресцина, и результаты показаны в Таблице 9 ниже.
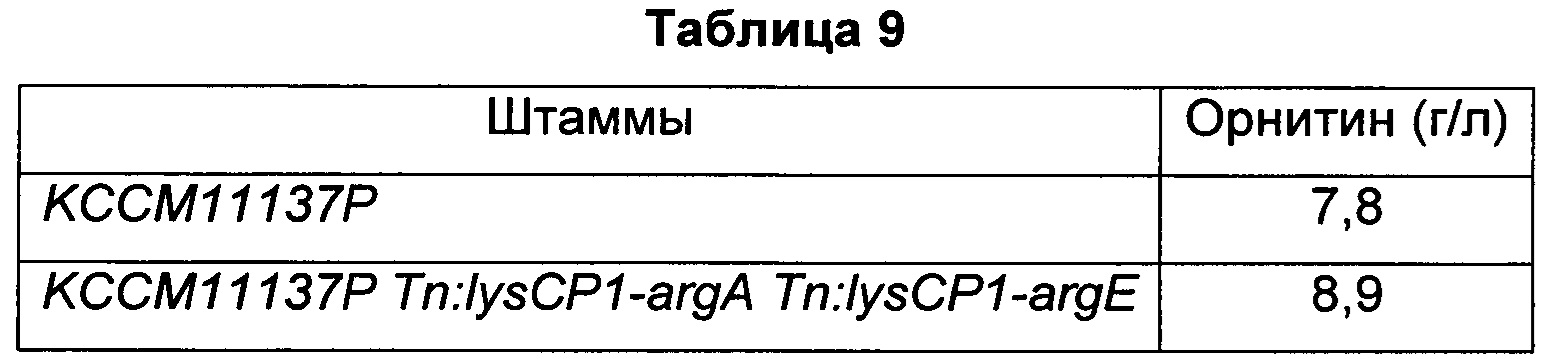
Как показано в Таблице 9, было подтверждено, что при введение гена argA, имеющего происхождение от Е. coli, и гена argE, имеющего происхождение от Е. coli, в модифицированный штамм Corynebacterium glutamicum продемонстрировало увеличение количества продуцируемого орнитина на 14,1% по сравнению с исходным штаммом.
Пример 6: Усиление pta-ackA у штамма с введенными argA, имеющим происхождение от Е. coli, и argE, имеющим происхождение от Е. coli, и подтверждение способности штамма продуцировать орнитин 6-1. Получение штамма с заменой промотора pta-ackA из штамма, продуцирующего орнитин, на основе АТСС13032
Для подтверждения того, что усиление pta-ackA у штамма, продуцирующего орнитин, на основе АТСС13032 с введенными argA, имеющим происхождение от Е. coli, и argE, имеющим происхождение от Е. coli, может улучшить способность продуцировать орнитин, перед инициирующим кодоном оперона pta-ackA в хромосому вводили промотор lysCPI (WO 2009/096689).
Сначала плазмиду pDZ-lysCP1-1'pta-ackA, полученную в Примере 2-1, вводили в штаммы KCCM11137P и KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE, соответственно, электропорацией с получением трансформантов и трансформантов высевали на твердую среду BHIS (сердечно-мозговой экстракт (37 г/л), сорбит (91 г/л) и агар (2%)), содержащую канамицин (25 мкг/мл) и X-gal (5-бром-4-хлор-3-индолин-D-галактозид), и культивировали с образованием колоний. Из полученных колоний отбирали синие колонии, проводя посредством этого отбор трансформированных штаммов с введенной плазмидой pDZ-lysCP1-1'pta-ackA.
Отобранные штаммы культивировали с покачиванием (30°С, 8 часов) в среде СМ (глюкоза (Юг/л), полипептон (10 г/л), дрожжевой экстракт (5 г/л), мясной экстракт (5 г/л), NaCl (2,5 г/л), мочевина (2 г/л), рН 6,8) и последовательно разводили от 10-4 до 10-10, высевали на твердую среду, содержащую X-gal, и культивировали с образованием колоний. Из полученных таким образом колоний отбирали белые колонии, встречавшиеся со сравнительно низкой частотой, проводя окончательный отбор штамма, в котором промотор pta-ackA был заменен на промотор lysCP1 посредством вторичного кроссинговера. Отобранный в итоге штамм подвергали ПЦР с использованием пары праймеров SEQ ID NO: 19 и 21, подтвердив введение промотора lysCP1 в хромосому перед инициирующим кодоном оперона pta-ackA. В частности, ПЦР проводили, повторяя 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и удлинения при 72°С в течение 1 минуты.
Отобранные таким образом модифицированные штаммы Corynebacterium glutamicum были названы KCCM11137P lysCP1-1'pta-ackA и KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE lysCP1-1'pta-ackA, соответственно.
6-2. Оценка способности штамма с усилением активности pta-ackA продуцировать орнитин
Для подтверждения эффекта усиления активности pta-ackA у штамма, продуцирующего орнитин, с введенными argA, имеющим происхождение от Е. coli, и argE, имеющим происхождение от Е. coli, проводили сравнение способности модифицированных штаммов Corynebacterium glutamicum, полученных в Примере 6-1, продуцировать орнитин.
Конкретно, модифицированные штаммы Corynebacterium glutamicum двух разных типов (KCCM11137P lysCP1-1'pta-ackA; KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE lysCP1-1'pta-ackA) и исходные штаммы двух разных типов (KCCM11137P; KCCM11137P Tn: lysCP1-argA Tn:lysCP1-argE), соответственно, высевали на твердую среду СМ (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-го NaOH, 2% агара, рН 6,8, на основе 1 л), содержащую 1 мМ аргинина, и культивировали при 30°С в течение 24 часов. Каждый из культивированных штаммов в количестве приблизительно одной платиновой петли засевали в 25 мл титрационной среды (8% глюкозы, 0,25% соевого белка, 0,50% сухого кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, биотин (100 мг), тиамин-HCl (3 мг), пантотенат кальция (3 мг), никотинамид (3 мг), 5% СаСО3, на основе 1 л) и культивировали с покачиванием при 30°С и частоте 200 об/мин в течение 98 часов. Во всех культурах штаммов в среду добавляли 1 мМ аргинина. По завершении культивирования в каждой культуральной среде измеряли концентрацию полученного орнитина, и результаты показаны в Таблице 10 ниже.
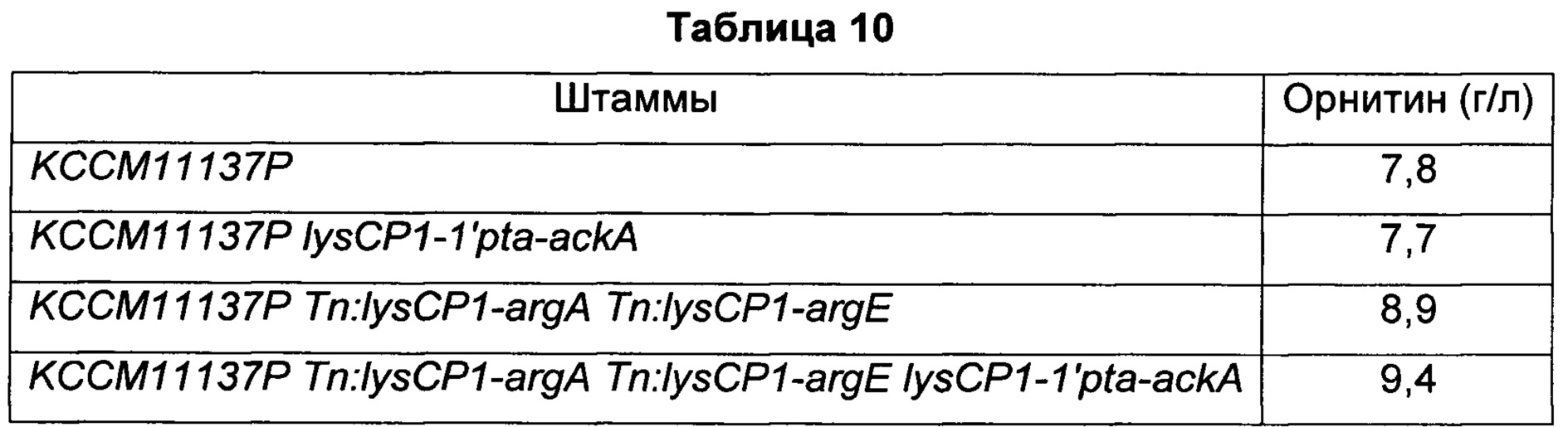
Как показано в Таблице 10, было подтверждено, что при усилении активности pta-ackA у штамма KCCM11137P количество продуцируемого орнитина не увеличивалось, в то время как в случае штамма KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE, представляющего собой модифицированный штамм Corynebacterium glutamicum с одновременным введением гена argA, имеющего происхождение от Е. coli, и гена argE, имеющего происхождение от Е. coli, количество продуцируемого орнитина увеличивалось на 20,5% по сравнению со штаммом KCCM11137P и на 5,6% по сравнению со штаммом KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE.
Пример 7: Введение acs, имеющего происхождение от Е. coli, в штамм с введенными argA, имеющим происхождение от Е. coli, и argE. имеющим происхождение от Е. coli, и подтверждение способности штамма продуцировать орнитин
7-1. Получение штамма с введением acs, имеющего происхождение от Е. coli, в ген транспозона штамма, продуцирующего орнитин, на основе КССМ11137
Для подтверждения того, что введение acs, имеющего происхождение от Е. coli, в штамм КССМ11137Р (патент Кореи №10-1372635), представляющий собой штамм, продуцирующий орнитин, на основе Corynebacterium glutamicum АТСС13032, может улучшить способность продуцировать орнитин, acs вводили в ген транспозона с использованием промотора lysCP1.
Сначала плазмиду pDZTn-lysCP1-acs, полученную в Примере 3-1, вводили в штаммы KCCM11137P и KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE, соответственно, электропорацией с получением трансформантов и трансформантов высевали на твердую среду BHIS (сердечно-мозговой экстракт (37 г/л), сорбит (91 г/л) и агар (2%)), содержащую канамицин (25 мкг/мл) и X-gal (5-бром-4-хлор-3-индолин-D-галактозид), и культивировали с образованием колоний. Из полученных колоний отбирали синие колонии, проводя посредством этого отбор трансформированных штаммов с введенной плазмидой pDZTn-lysCP1-acs.
Отобранные штаммы культивировали с покачиванием (30°С, 8 часов) в среде СМ (глюкоза (10 г/л), полипептон (10 г/л), дрожжевой экстракт (5 г/л), мясной экстракт (5 г/л), NaCl (2,5 г/л), мочевина (2 г/л), рН 6,8) и последовательно разводили от 10-4 до 10-10, высевали на твердую среду, содержащую X-gal, и культивировали с образованием колоний. Из полученных таким образом колоний отбирали белые колонии, встречавшиеся со сравнительно низкой частотой, проводя окончательный отбор штамма с геном, кодирующим acs, введенным посредством вторичного кроссинговера.
Отобранные в итоге штаммы подвергали ПЦР с использованием пары праймеров SEQ ID NO: 13 и 25, подтвердив введение гена, кодирующего acs. Отобранные таким образом модифицированные штаммы Corynebacterium glutamicum были названы KCCM11137P Tn:lysCP1-acs и KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE Tn:lysCP1-acs, соответственно.
7-2. Оценка способности штамма с введенным acs, имеющим происхождение от Е. coli, продуцировать орнитин
Для подтверждения эффекта введения acs в штамм, продуцирующий орнитин, с введенными argA, имеющим происхождение от Е. coli, и argE, имеющим происхождение от Е. coli, проводили сравнение способности модифицированных штаммов Corynebacterium glutamicum, полученных в Примере 7-1, продуцировать орнитин.
Конкретно, модифицированные штаммы Corynebacterium glutamicum двух разных типов (KCCM11137P Tn:lysCP1-acs; KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE Tn:lysCP1-acs) и исходные штаммы двух разных типов (KCCM11137P; KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE), соответственно, высевали на твердую среду СМ (1% глюкозы, 1% полипептона, 0,5% дрожжевого экстракта, 0,5% мясного экстракта, 0,25% NaCl, 0,2% мочевины, 100 мкл 50%-го NaOH, 2% агара, рН 6,8, на основе 1 л), содержащую 1 мМ аргинина, и культивировали при 30°С в течение 24 часов. Каждый из культивированных штаммов в количестве приблизительно одной платиновой петли засевали в 25 мл титрационной среды (8% глюкозы, 0,25% соевого белка, 0,50% сухого кукурузного экстракта, 4% (NH4)2SO4, 0,1% KH2PO4, 0,05% MgSO4⋅7H2O, 0,15% мочевины, биотин (100 мг), тиамин-HCl (3 мг), пантотенат кальция (3 мг), никотинамид (3 мг), 5% СаСО3, на основе 1 л) и культивировали с покачиванием при 30°С и частоте 200 об/мин в течение 98 часов. Во всех культурах штаммов в среду добавляли 1 мМ аргинина. По завершении культивирования в каждой культуральной среде измеряли концентрацию полученного орнитина, и результаты показаны в Таблице 11 ниже.

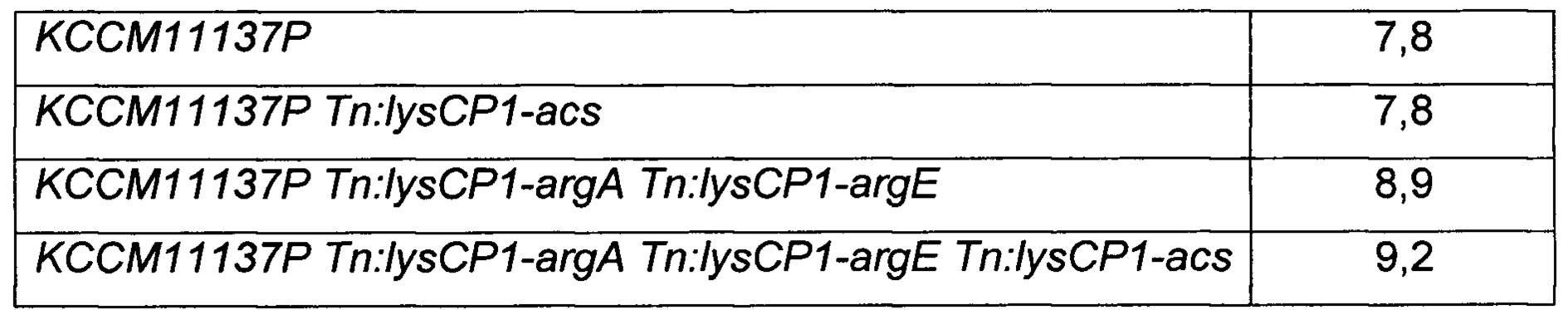
Как показано в Таблице 11, было подтверждено, что при введении acs в штамм KCCM11137P количество продуцируемого орнитина не увеличивалось, в то время как в случае штамма KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE, представляющего собой модифицированный штамм Corynebacterium glutamicum с одновременным введением гена argA, имеющего происхождение от Е. coli, и гена argE, имеющего происхождение от Е. coli, количество продуцируемого орнитина увеличивалось на 17,9% по сравнению со штаммом KCCM11137P и на 3,4% по сравнению со штаммом KCCM11137P Tn:lysCP1-argA Tn:lysCP1-argE.
Суммируя вышеизложенное, было подтверждено, что введение argA, имеющего происхождение от Е. coli, и argE, имеющего происхождение от Е. coli, в штамм Corynebacterium может увеличить количество продуцируемого путресцина и орнитина, и, кроме того, было подтверждено, что усиление активности гена pta-ackA у штамма Corynebacterium или введение acs, имеющего происхождение от Е. coli, может дополнительно увеличить количество продуцируемого путресцина и орнитина.
Исходя из изложенного выше, специалисту в области, к которой относится настоящее изобретение, будет ясно, что настоящее изобретение может быть воплощено в других конкретных формах без изменения технической концепции или существенных характеристик настоящего изобретения. В этом отношении, типичные воплощения раскрыты здесь исключительно в целях наглядности, и их не следует толковать как ограничение объема настоящего изобретения. Наоборот, подразумевают, что настоящее изобретение охватывает не только типичные воплощения, но также различные альтернативы, модификации, эквиваленты и другие воплощения, которые могут быть включены в сущность и объем настоящего изобретения, как определено приложенной формулой изобретения.
Реферат
Группа изобретений относится к модифицированному микроорганизму рода Corynebacterium, продуцирующему путресцин, и способу получения путресцина с использованием указанного микроорганизма. Предложен модифицированный микроорганизм рода Corynebacterium, в котором активности N-ацетилглутаматсинтазы из E. coli и ацетилорнитиндезацетилазы из E. coli введены в указанный микроорганизм рода Corynebacterium, обладающий активностью ацетилглутаматсинтазы или орнитинацетилтрансферазы (ArgJ). Также предложен способ получения путресцина, включающий культивирование указанного модифицированного микроорганизма рода Corynebacterium в среде и выделение путресцина из культивированного микроорганизма или среды. Группа изобретений позволяет получить путресцин с увеличенным выходом. 2 н. и 15 з.п. ф-лы, 2 ил., 11 табл., 7 пр.
Формула
Документы, цитированные в отчёте о поиске
Способ получения l-аргинина с использованием бактерии семейства enterobacteriaceae, содержащей n-ацетилорнитиндеацетилазу с нарушенной активностью
Комментарии